
プラスミドDNA醗酵プロセス

【課題】
【解決手段】
プラスミドDNA生成技術の改良は、将来のDNAワクチン及びDNA治療の経済的実現可能性を保証するために必要である。一般的な方法が、記載されており、これによって醗酵層中でのプラスミドDNA生産性を劇的に増加することができる。これらのプロセスは、新規の増殖と誘導期の温度シフトと組み合わせて、流加培養醗酵戦略を特徴付ける。
【解決手段】
プラスミドDNA生成技術の改良は、将来のDNAワクチン及びDNA治療の経済的実現可能性を保証するために必要である。一般的な方法が、記載されており、これによって醗酵層中でのプラスミドDNA生産性を劇的に増加することができる。これらのプロセスは、新規の増殖と誘導期の温度シフトと組み合わせて、流加培養醗酵戦略を特徴付ける。
【発明の詳細な説明】
【技術分野】
【0001】
連邦政府後援研究又は開発に関する陳述
適用されない。
【0002】
本出願は、2004年8月19日に出願された、米国暫定特許出願第60/603000号の利益を主張する。
【0003】
本発明の分野
本発明は、プラスミド、コスミド、バクテリア人工染色体(BACs)、バクテリオファージ、ウイルス性ベクタ及びこれらのハイブリッド等の閉環状(covalently closed circular:ccc)組み換え型DNA分子の生産に関し、より具体的には、醗酵培養における高レベルでのDNA分子の生産方法である。
【0004】
本発明の背景
本発明は、閉環状(ccc)組み換え型DNA分子の生産に関する。このような分子は、バイオテクノロジ、遺伝子組み換え生物、遺伝子治療、治療上のワクチン接種、農業及びDNAワクチンに有益である。
【0005】
本発明を念頭に置いて、先行技術の調査を実施した。大腸菌(E.coli)プラスミドは、長い間、研究者及び企業によって用いられた組み換え型DNA分子のただ1つの最も重要な源であった。今日、プラスミドDNAは、次世代のバイオテクノロジ生産物(遺伝子医薬品及びDNAワクチン)が、臨床試験及び、医薬市場への道を作るに当たって、ますます重要性が高まっている。プラスミドDNAワクチンは、ウイルス性、細菌性、又は寄生性疾患に対する予防的ワクチン;高力価免疫グロブリン生産物の生産用免疫剤;伝染性疾患用のワクチン;又は癌ワクチン;としての適用を見出すことができる。プラスミドは、また、遺伝子治療又は遺伝子置換用途に利用され、患者に投与した後に所望の遺伝子生産物が、プラスミドから発現する。
【0006】
今日、FDA(アメリカ食品医薬品局)基準は、暫定段階にあるもの以外には規定されていない(FDA Points to Consider on Plasmid DNA Vaccines for Preventive Infectious Disease Indications,1996参照)。しかしながら、将来は、プラスミドDNA純度についての国際基準が、大腸菌(E.coli)醗酵から同様に作られる組み換えタンパク生産物に使用される基準と同じか、又は非常に類似するものになりそうであり、このような基準は、確立された方法から達成できる現行の純度を超える。最も明らかなことには、投与量当たり100pg未満の主宿遺伝子DNAの許容された基準は(FDA Points to consider in the characterization of cell lines used to produce biologies,1993参照)、精製プラスミド製剤について現在達成できるレベルより遙かに低い(投与量1mg当たり100pgは、1000万分の1に等価である)。
【0007】
プラスミド(細菌醗酵によって)、及びこれらの純度(例えば、アルカリ溶菌法(alkaline lysis method)(Birnboim,HC,DoIy J.1979,Nucleic Acids Res.7:1513−1523)によって)を得る基本的な方法は公知である。最初に、醗酵した細菌性細胞ペーストを、再懸濁して、(水酸化ナトリウム及びドデシル硫酸ナトリウムの組み合わせを使用して)溶解し、その後溶液を、酸性塩(例えば、酢酸カリウム)の添加によって中和し、細菌性DNA及び細胞崩壊堆積物の大部分を沈殿させる。大量のスーパーコイル状のプラスミドDNAは、汚染細菌性RNA、DNA及びタンパク質、又は大腸菌内毒素(リポポリサッカライド、又はLPS)と共に溶液中に残る。次いで、可溶性画分をろ過によって分離し、種々の精製工程を受けさせる。この工程には、RNase(リボヌクレアーゼ)消化;クロマトグラフィ(イオン交換ゲルろ過、ハイドロキシアパタイト、ゲルろ過、疎水性相互作用、逆相、HPLC、等);膜分離;有機抽出(organic extraction);選択的沈殿、等が含まれる。
【0008】
明らかに、開始材料の純度が増し、より良い下流純度を達成することは、工業規模の臨床段階のDNAを生産するために不可欠な目標である。
【0009】
醗酵媒体の考察
安定した媒体の型は、細胞のエネルギィ所要量及び基本的な組成に基づいている。一般的に、栄養学的な所要量は、最少培地又は半合成培地によって満たされる。
【0010】
半合成培地は、酵母エキス、カザミノ酸、及びペプトン等の複合体成分を含む。複合体成分の追加は、増殖因子、アミノ酸、プリン類及びピリミジンを供給し、しばしばより高い細胞密度をサポートする。
【0011】
炭素は、細胞組成の半数を数える。従って、炭素は、最も多量に含まれている。炭素源は、エネルギィ及びバイオマスを提供し、制限栄養素として通常使用される。グルコースは、通常の炭素源である。グルコースは非常に効果的に代謝され、従って、細胞収率がより高い。しかしながら、高グルコース濃度は、(クラブトリー効果(Crabtree effect)として知られる)代謝過剰に起因して、望ましくない産生を引き起こす。グリセロールも用いられ、しばしば回文培養の好適な炭素源となる。グリセロールからの細胞収率は、グルコース由来のものよりも僅かに小さいが、グリセロールは、高レベルの酢酸産物を作らず、阻害されることなく、より高濃度で使用することができる。グリセロールも、最大比増殖濃度を減少させる。
【0012】
窒素の所要量には、無機又は有機窒素源によって応ずることができる。アンモニア及びアンモニア塩(例えば、NH4Cl、(NH4)2SO4)は、最少培地で使用される。半合成培地は、イースト抽出物、ペプトン、及びカザミノ酸を含有する複合体成分から部分的に又は全体的に窒素を供給する。
【0013】
鉱物は、増殖、代謝、及び酵素反応に必要である。マグネシウム、リン、カリウム、及び硫黄は、個別の媒質成分として一般的に加えられる。リン酸ジカリウム又はリン酸モノカリウムは、カリウム及びリンを提供し、ある比率で緩衝剤としても機能する。硫酸マグネシウム・7水和物は、マグネシウム及び硫黄の源として、しばしば利用される。その他の必須鉱物は、カルシウム、銅、コバルト、鉄、マンガン、モリブデン及び亜鉛を含む。これらは主要成分中に不純物として通常存在するが、これらは、極少量必要とされ、微量鉱物溶液の添加によって、しばしば補給される。浸透圧は、塩化ナトリウムで調節する。
【0014】
動物由来の生成物、特にプラスミド生産におけるウシ生成物の使用は、プリオン又はウイルス汚染物質の危険性に起因して許容できない。全培地成分は、認定された動物由来生成物を含まないものであるべきである。植物由来の代用物は、動物に起源を有する多くの成分に使用可能である(例えば、植物グリセロール、大豆ペプトン)。
【0015】
プラスミド醗酵プロセスの考察
増殖速度
減少した増殖速度の使用が、高品質、高収率プラスミド醗酵の統一原理である。高増殖速度は、酢酸生成物、プラスミド不安定性、及びスーパーコイル状のプラスミドのより低い割合と関連している。減少した増殖速度は、細胞分裂と同期したプラスミド複製のための時間を提供することによって、増殖速度依存プラスミド不安定性を緩和する。
【0016】
増殖状態
醗酵は、プラスミドの質及び収率に影響する多くのパラメータを制御及び監視する可能性を与える。スーパーコイル形成は、酸素及び温度によって影響されることが知られている(Dorman CJ et al.1988 J.Bacteriol.179:2816−2826)、(Goldstein E,Drlica K.1984 Proc Natl Acad Sci USA.81:4046−4050)。酸素は、プラスミド安定性において重要な役割を演じることが示されている。ある研究(Hopkins DJ,Betenbaugh MJ,Dhurjati P.1987 Biotechnol Bioeng.29:85−91)では、5%の空気飽和率に対する溶存酸素濃度中の1滴が、プラスミド安定化の急速な減少を導くことを発見した。別の研究(Namdev PK,Irwin N,Thompson BG,Gray MR.1993 Biotechnol Bioeng.41:666−670)では、酸素導入の変動が、プラスミドの不安定化をまねくことを示した。更に、ニックプラスミド及び多重体の形成は、温度、pH、溶存酸素、栄養素濃度、及び増殖速度を含む多くのパラメータによって影響を受け得る(Durland RH,Eastman EM.1998 Adv Drug Deliver Rev.30:33−48)。大腸菌増殖の最適温度は、37℃である。しかしながら、回分醗酵では、より低い温度(30から48℃)を用いて、最大比増殖速度を低くすることができる。また、より高い温度を用いて、pUC、及びpMM1(Wong EM,Muesing MA,Polisky,B.1982 Proc Natl Acad Sci USA.79:3570−3574)、(Lin−Chao S,Chen WT,Wong TT.1992 MoI Microbio.6:3385−3393)及びランアウェイレプリコンRプラスミド等の複製起源を伴う選択的なプラスミド増幅を誘発することもできる。Hamann et.al.2000(Hamann CW,Nielsen J,Ingerslev E.2000 World Patent Application WO0028048)は、プラスミド生産を(低温度によって)低レベルに維持して、プラスミドDNA合成による増殖の遅延を避けるRプラスミドの生産方法を報告している。ここでは、宿主細胞集団が高くなると、温度変化によってプラスミド生産が誘発される。
【0017】
回分醗酵
回分醗酵は、簡単であるという主な利点を有する。培養期間を通して細胞増殖及びプラスミド生産に用いられる全栄養素が、植菌時に存在する。回分醗酵は、誘導期、指数増殖期、及び静止期を有する。好適な植菌材料(1から5%の培養液量)は、誘導期の長さを低減させる。指数増殖期間には、全栄養素が過剰であり;従って、この比増殖速度は、本質的にモノ−キネティックスによって予測される最大比増殖速度、μmaxになる。上述したように、増殖速度が減少することは、プラスミド生産にとって望ましい。回分醗酵において、増殖速度は、μmaxを減少させることによってのみ低減する。これは、より低い温度での増殖によって、及びグルコースの代わりにグリセロール上で増殖することによって達成される。グリセロールを用いた30℃での回分醗酵は、一般的にμmax<0.3h−1となり、このことは、有害な酢酸蓄積及びプラスミド不安定に関する増殖速度を妨ぐのに十分である(Thatcher DR,Hitchcock A,Hanak JAJ,Varley DL.2003 U.S Patent No6503738)。また、阻害されることのなくグルコースよりずっと高い濃度でグリセロールを、使用することができ、より高いバイオマス収率にすることができる。一般的に、60g/L DCW以上のバイオマス収率を、回分醗酵で得ることができる。
【0018】
流加培養醗酵
流加培養醗酵は、プラスミド生産に特に有益である。制限栄養素を制御して加えることは、率<μmaxでの増殖速度の制御を可能にする。また、流加培養醗酵は、収率をより高くする結果となる。流加培養醗酵の鍵は、完全に消費されるような比率で基質を供給することである。結果として、残留基質濃度は、ほぼ0であり、基質の最大転換が得られる。過剰の基質からの代謝過剰を回避し、阻害酢酸の形成を低減する。
【0019】
流加培養醗酵は、回分期で開始する。全ての非制限栄養素を含有する開始量の培地及び開始濃度の制限基質内に細胞を植菌する。細胞がこの開始量の基質を消費したら、制限栄養素を制御して与え始める。
【0020】
最も簡単で、最も効果的な供給戦略の一つは、指数関数的供給である。この方法によれば、フィードバック制御を必要とすることなくμmax未満の所定の比率で培地が増殖することを可能である。醗酵は、非阻害濃度の基質を含有する回分モードで開始する。細胞は、この基質がなくなるまでμmaxで増殖し、この時点で、栄養素供給が始まる。
【0021】
DO−stat法及びpH−stat法を実施するのは実に簡単である。というのも、最も標準的な醗酵装置システムは、溶存酸素とpHの監視を含むからである。溶存酸素(DO)及びpHの傾向は、基質が細胞に利用できるかどうかを示す。基質の消耗は、酸素摂取量を低減させ、培地中のDO濃度を上げる。また、pHは、代謝性の酸の消費によって上昇する。DO又はpHが設定閾値以上に上昇すると、供給が誘発される。増殖速度は、DO又はpH閾値を変更することによって調節することができる。
【0022】
例示的プラスミド醗酵プロセス
現行の収率の試験により、一般的なラボの振盪フラスコ培養は1から5mgのプラスミドDNA/Lの培地を作るが、コンピュータ制御の醗酵装置は、一般的に、10から250mg/LのプラスミドDNA/Lの培地を作ることが明らかである。
【0023】
Lahijani et al.(Lahijani R,Hulley G,Soriano G,Horn NA,Marquet M.1996 Human Gene Therapy 7:1971−1980)は、指数関数的供給と、37℃から42−45℃の温度シフトで醗酵における温度感受性単一ポイント突然変異(pUC起源)を伴うpBR322−誘導プラスミドを用いることを報告している。このことは、10Lの醗酵装置中220mg/Lのプラスミド収率を達成した。30℃での回分醗酵(pBR322誘導起源)中の突然変異のない同じプラスミドは、3mg/Lプラスミドを得たのみである。Friehs et al.(Friehs K,Flaschel E,Schleef M,Schmidt T.2003 US Patent No 6664078)は、DO−statフィードバック制御供給で、グリセロールイースト抽出培地を用いる流加培養プロセスを記載している。醗酵は、7.5Lの開始回分体積で開始した。攪拌を増して30%以上のDOを保った。DOが閾値設定点の45%に達した時に供給培地を投入した。培養は、41時間後に静止期に達し、60g/L DCW及び230mg/Lのプラスミドを得た。Chen(Chen,W.1999 U.S.Patent No 5955323)は、DO−statとpH−statフィードバック制御を組み合わせた半合成培地の流加培養プロセスを用いた。DOとpH閾値設定点は、それぞれ50%と7.2であった。DOが30%以下に落ちると、高い代謝活性から攪拌速度が以前の速度の割合まで増加した。7Lの培養装置中で、この戦略は、0.13h−1の比増殖速度、及び82から98mg/Lのプラスミド収率を導いた。Durland and Eastman,Supra,1998は、専売培地中で37℃での回分醗酵を報告している。彼らのプロセスによれば、一般的には130mg/Lの収率が得られ、250mg/Lの高い収率が得られた。
【0024】
先行技術を考慮しても、高純度プラスミドDNA生産についての費用対効果の大きい方法の必要性がある。上記の醗酵培地及びプロセスは、現在技術的に知られているものを組み込んでおり、増殖速度の減少及び高温でのプラスミド複製数誘発等のプラスミド生産性を改善している。これらのプロセスは、約200から250mgのプラスミドDNA/Lで横ばいになる。この低い収率は、プラスミドDNA生産プロセスを商品化する上でのコスト及び純度に負担を与える。スケールメリットは将来著しくDNAのコストを下げるが、所望のコストを達成するためには、この問題に対する更なる経済的解決策が必要である。その上、プラスミドDNA純度の国際基準は、大腸菌(E.coli)醗酵から同様に生産される組み換えタンパク質生産物について使用される基準と同じ又は非常に似ている。この基準は、確立された方法から得られる現在の純度を超えている。醗酵における収率(細胞ペーストのDNA/グラムのmg)の増加は、コストを減少させ、DNAの純度を上げるであろう(処理される材料の量を低減するからである)。
【0025】
本発明の開示
本発明は、プラスミド生産用の改善された回分及び流加培養プロセスを用いるDNAレプリコンの生産方法である。特に、流加培養醗酵方法が開示されており、ここでは、プラスミド含有大腸菌(E.coli)細胞が、流加培養期中に低い温度で増殖し、この増殖中は増殖速度が制限され、次いで、プラスミドを蓄積するために温度をアップシフト(up−shift)し、段階的に温度を高くして継続的に増殖させることによって、温度シフトと制限された増殖速度が、プラスミドの収率及び純度を改善する。好ましい実施例では、低減した増殖速度をほぼ2mg/L/OD以下のプラスミド収率を維持するように決定する。別の好ましい実施例では、この低減した温度はおよそ30℃である。別の好ましい実施例では、温度のアップシフトは、ほぼ36から45℃までである。別の好ましい実施例では、プラスミドはColE1誘導複製起源を含む。更にもう1つの好ましい実施例では、プラスミドはpUCGからpUCAまでの突然変異を含むpMB1複製起源を含む。更に別の実施例では、プラスミドは、VR1012バックボーンから誘導される。一の極めて好ましい実施例では、醗酵媒体が、実質的に半合成グリセロール媒体である。更に最後の好ましい実施例では、pBR322−誘導プラスミドは、増殖速度が流加培養期間で制限され、増殖がプラスミド生産物を継続して蓄積する間に、流加培養醗酵の大腸菌(E.coli)中で増殖する。要約すれば、これらのプロセスは、新規の増殖と誘発期の温度シフトとを組み合わせた、高い細胞密度培養戦略を特徴とする。DNAレプリコンを含有するpBR322−誘導起源の生産において、流加培養醗酵は、制限された細胞増殖速度で実施される。温度誘導DNAレプリコン(例えば、プラスミド含有pUC又はpMM1起源)の生産において、流加培養醗酵は、増殖期中の制限された細胞増殖速度と低減された温度で実施され;次いで、プラスミド生産は、温度をアップシフトすることによって誘発される。これらのプロセスは、プラスミドDNA醗酵収率を劇的に改善し、その一方で、従来技術のプロセスと比較して、プラスミドの完全性を維持又は改善する。
【0026】
本発明の簡単な概要
本発明の目的及び/又は対象は、プラスミドDNAの生産のための醗酵プロセスを提供することである。本発明のもう一つの対象及び/又は目的は、醗酵培養のプラスミドDNA生産収率を改善することである。本発明の更に別の対象及び/又は目的は、醗酵培養のプラスミドDNAの品質を改善することである。本発明の更に別の対象及び/又は目的は、精製プラスミドDNA中の不純物を低減することである。別の開示は、従来技術に記載されたプロセスと比較して改善された改善醗酵プロセスであり、バイオマス生産後のプラスミドレベルの誘導によってプラスミドの増加した収量;バイオマス生産中の低プラスミドレベルの維持による毒性又は不安定プラスミドと共に増加した収量及び完全性;プラスミドのニック(開環)又は直鎖バージョンの低減したレベルによってプラスミドの質を向上すること;プラスミドのパーセントモノマ(percent monomer)の増加によってプラスミドの質を向上すること;ロバスト、自動制御パラメータ及び供給を用いる簡略化した生産;増殖中に必要な増殖制御及び低減した酸素補給によって簡略化したスケーリング;ダウンストリームプロセシング内へのフィードストリームのプラスミドの濃縮レベルによるプラスミド精製後の不純物の低減したレベル;及び全ての動物性生成物誘導成分の除去によって、改善した標準コンプライアンス;によって改善される。
【0027】
本発明の更なる目的及び利点は、図面及び次の記載の考察から明らかになる。
【0028】
本発明の詳細な説明
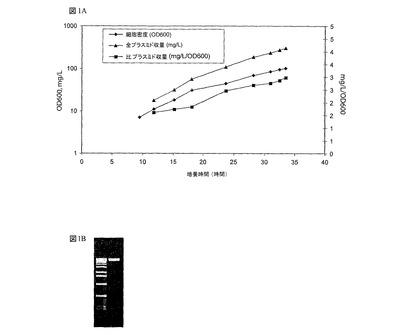
図面を参照すると、図1は、NTC3019培地を用いた大腸菌(E.coli)中のpBR322−誘導プラスミド流加培養醗酵を示し、これは(a)NTC3019培地を用いた流加培養中の、大腸菌(E.coli)中のpBR322−誘導プラスミドの一般的な増殖及びプラスミド生産性プロファイルと;(b)高スーパーコイル状であり、ギザギザで開環したアイソフォームを含まないNTC3019培地流加培養醗酵プロセスによって生産されたプラスミドDNAと;を明らかにしている。
【0029】
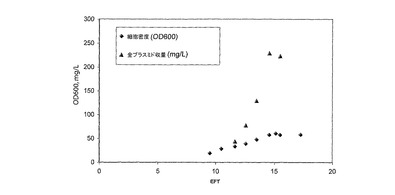
図2では、NTC3018培地を用いた大腸菌(E.coli)中のpUCプラスミド(pW2.0)回分醗酵のプラスミド増殖及びプラスミド生産性プロファイルが示されている。回分醗酵中のpW2.0プラスミドは、57OD600の細胞密度に達し、230mgプラスミド/Lの収率であった。
【0030】
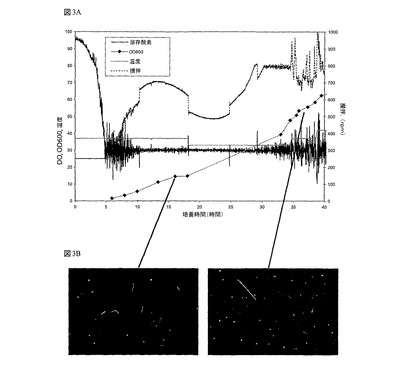
図3では、NTC3019培地を用いた大腸菌(E.coli)中のgWiz GFPプラスミド流加培養醗酵を示している:(a)gWiz GFPの流加培養醗酵プロファイルの増殖及び制御パラメータプロファイル(溶存酸素、温度、攪拌)と;(b)SYBR GREEN Iで染色された細胞の蛍光顕微鏡観察が、プラトウ(左)でフィラメンテーションを示すが、温度を33℃で低下させた後は、再開された増殖及びフィラメンテーションが低下した(右)。
【0031】
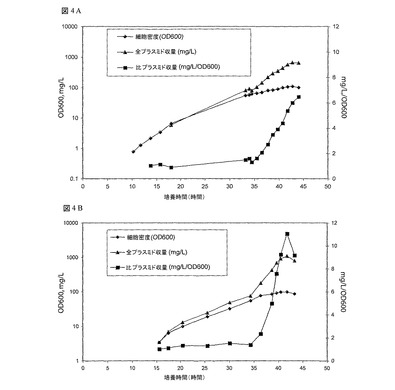
図4では、NTC3019培地(37℃又は42℃誘導)を用いた大腸菌(E.coli)中のgWiz GFP誘導プラスミド流加培養醗酵を示している:(a)35時間で、30→37℃の温度シフトを用いて、gWiz GFP/大腸菌(E.coli)DH5α醗酵の増殖及びプラスミド生産性プロファイルが明らかにされ、プラスミド収量が670mg/Lに達した;(b)35時間で、30→42℃の温度シフトを用いて、gWiz GFP/大腸菌(E.coli)DH5α醗酵の増殖及びプラスミド生産性プロファイルが明示されている。プラスミド収量は1100mg/Lに達した。
【0032】
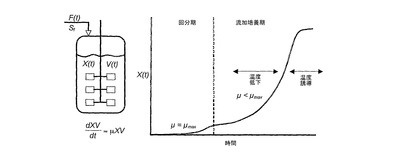
図5では、誘導流加培養醗酵プロセスを図示する。
【0033】
定義
【0034】
ccc:閉環状
【0035】
ColE1誘導起源:欠失(例えば、pBR322誘導起源)及び/又は塩基交換(例えば、pMB1、pMM1、ColE1等由来のpMM5由来のpUC)によるColE1型プラスミド(例えば、pMB1、ColE1)から誘導された複製された起源
【0036】
DNAレプリコン:プラスミド、コスミド、細菌性人工染色体(BACs)バクテリオファージ、ウイルス性ベクタ、及びこれらのハイブリッド
【0037】
NTC3018醗酵培地:グリセロール半合成回分醗酵培地
【0038】
NTC3019醗酵培地:グリセロール半合成流加培養培地
【0039】
pDNA:プラスミドDNA
【0040】
pBR322−誘導起源:rop(repressor of primer:プライマの抑制体)遺伝子が欠失した、pBR322由来のpMB1起源
【0041】
プラスミド:プラスミド、コスミド、細菌性人工染色体(BACs)、バクテリオファージ、ウイルス性ベクタ、及びこれらのハイブリッド
【0042】
pUC起源:段階的な温度でコピー数を増加するGからAへの転位を有する、pBR322−誘導起源
【0043】
半合成グリセロール培地:複合窒素源(例えば、イースト抽出物、大豆抽出物)及びグリセロール炭素源を含む醗酵培地
【0044】
本発明は、機械的醗酵容器を用いたプラスミド、コスミド、細菌性人工染色体(BACs)、バクテリオファージ、ウイルス性ベクタ及びこれらのハイブリッド(本明細書では、集合的にプラスミドと称される)等の閉環状(ccc)組み換えDNA分子の製造方法に関する。
【0045】
従来技術の醗酵プロセスは、次善のプラスミド収率、品質(例えば、プラスミドの切断又は線形化)、拡張性の無さ(例えば、過剰な酸素供給要求)、及び限られた応用範囲(例えば、不安定又は毒性シークエンスを含有するプラスミドで使用する能力がない)のために最適ではない。本発明は、醗酵培養中にプラスミドDNAの収率及び純度を改善する方法である。誘発性流加培養醗酵プロセスを用いる高収率醗酵に対する費用効率のアプローチを発展させて、プラスミド収率及び純度を改善する。
【0046】
プラスミド製造プロセス及び培地の好ましい実施例
pBR322−誘導プラスミドを生産する1つの好ましい実施例では、流加培養醗酵を制限された細胞増殖速度で実施する。このプロセスは、プラスミドDNA醗酵収率を劇的に改善するにも関わらず、従来技術に記載されているプロセスと比較して、プラスミドの完全性を維持又は改善する。
【0047】
適度の複製プラスミド(例えば、ここではpBR誘導プラスミドと呼ばれるrop遺伝子が欠損したpBR322複製起源)を使用したこのプロセスを用いて、0.12hr−1の比増殖速度を維持するように制御して供給をしながら、37℃で自動流加培養醗酵プロセスを行って、250から450mg/Lのプラスミド収率及び120OD600の細胞密度を達成した。これは、報告されている収率(Lahijani et al.,Supra,1996)の100倍の改良である。この新規で劇的な改良の分子的な根拠は、知られていない。この機構に関わらず、その他のプラスミドの生産への本発明を適用することで、プラスミドの質を下げることなく醗酵生産性は上がるであろう。
【0048】
温度誘発性DNAレプリコン(例えば、プラスミド含有pUC又はpMM1起源)の生産についての1つの好ましい実施例では、増殖期中に、制限された細胞増殖速度及び低減した温度で、流加培養醗酵を実施し;次いで、アップシフトした温度でプラスミド生産を誘発する。このプロセスは、従来技術に記載されているプロセスと比較すると、プラスミドの完全性を維持又は改善しつつ、プラスミドDNA醗酵収率を劇的に改善する。
【0049】
本明細書で開示されているプラスミド(例えば、pUC起源)を含む高コピー起源の高収率プラスミド生産についてのこの新規な戦略は、予想外に高いプラスミド生産性とプラスミドの品質をもたらす。加えて、流加培養期中に増殖速度が減少する。このプロセスは、図5に示されている。この要素の新規な組み合わせは、ColE1誘導起源プラスミドの生産には適用されず、試験してみると、新しく予想外の改善された生産性となった。従って、我々は、低減した増殖速度と、1)増殖及びバイオマス生産のための低減した温度、及び2)プラスミド生産の誘発のための上昇させた温度と、の組み合わせに対する新しい使用(改善された生産性)を教示する。
【0050】
初期の低減した温度増殖期と後の高温度生産期との組み合わせが、全体的な増殖、バイオマス、及びプラスミド収率の改善をもたらすという発見は、新規で予想外であり、従来技術には教示されていない。Hamann et al.,Supra,2000は、増殖中のRプラスミドに関連する代謝負担を特に低減するために温度誘発醗酵を用いた。この戦略は、Rプラスミドの生産に利用されたが、プラスミド含有ColE1起源については教示していない。同様に、Hamann et al.,Supra,2000は、この戦略を用いて、全体の生産性を改善することを意図していない。従って、この要素の組み合わせは、従来技術では、収率の改善のために提言されたものではない。改善された生産性によって明らかであるように、この組み合わせの相乗効果は、部分同士の総和より大きい。同様に、この高収量醗酵は、これまで要求されてきたが解決されなかった必要性を満たすものである。何故ならここ10年以上もの間、この分野で記載された様々なプロセスは、高収率醗酵を教示していないからである。要約すれば、我々は、プラスミド生産を含むColE1起源についての、新しい組み合わせを教示する。このことは、改善されたプラスミド生産性の、予想外で驚くべき新しい使用を示す温度誘発と組み合わせたゆっくりとした増殖を具える。
【0051】
高コピープラスミド生産用誘発性流加培養醗酵プロセスの適用は、1100mg/LプラスミドDNAの収率、及び100のOD600をもたらす。これは、従来技術に規定されている醗酵プロセスで得られる収率の5倍の改良である。これらの全プロセスから精製されたDNAは、高品質であり、事実上100%スーパーコイル状である。我々は、プラスミド生産性を改善するために本明細書に記載された回分及び流加培養醗酵プロセスの使用を意図する。
【0052】
本明細書に記載された誘発性流加培養プロセスは、このプロセスのバイオマス生産期の間、低プラスミドレベル(VR1012誘導ベクタに対して<2mg/L/OD600)を維持し、バイオマス生産後は予想外で前例のない超高プラスミド生産(VR1012誘導ベクタに対して>6mg/L/OD600)を容易にした。細菌1グラム当たりのプラスミド収率が増加することは、最終生産物のより高い純度に直接つながるので、高比産出量は非常に望ましい。我々は、本発明の温度シフトを用いて、増殖の間中、プラスミドレベルを低く保ち(VR1012誘導ベクタに対して<2mg/L/OD600)、バイオマス生産後のプラスミド生産を高いレベル(VR1012誘導ベクタ>3mg/L/OD600)に誘発する。これらのレベルは、VR1012誘導ベクタの生産に対するガイドラインである。その他のプラスミドは、増殖期中に2mg/L/OD600よりも高い又は低いレベルで許容される。許容できる代謝負担、プラスミド安定性、及び温度シフト後の高い生産性を維持する増殖期中の最大プラスミドレベルは、平均的な当業者によって、各新しいプラスミドについて実験的に決定することができる。
【0053】
醗酵培地の代替の実施例
本発明を実施する上で、例示的な動物由来成分を含まない醗酵培地配合、NTC3018(回分)、NTC3019(流加培養)を用いることができる。これらの培地は、グリセロール炭素源、イースト抽出窒素源、及び微量金属、塩及び緩衝液を含む半合成増殖培地である。従来技術に記載された培地を、この培地で代用することも本明細書に記載されているような醗酵プロセスを用いた改善されたプラスミド生産性をもたらす。
【0054】
NTC3018及びNTC3019培地用の代替の非動物源の窒素源が考えられる。複合培地成分として、イースト抽出物は、窒素、アミノ酸、ビタミン、及び炭素を提供する。プラスミド生産中の細胞代謝の鍵となる複合培地(例えば、イースト又は大豆抽出物)のあり得る成分は、制限アミノ酸、ビタミン、微量無機物及び代替の炭素源を含む。一例として、我々は、代替イースト抽出調製物、大豆調製物(例えば、BD Bioscience社製のソイトン、又はファイトンペプトンを選択する)又はNTC3018及びNTC3019培地中のその他の植物調製物(例えば、Oxoid社製のエンドウの花のペプトン)の使用を意図している。
【0055】
培地の規定された要素の変型も意図している。例えば、リン酸塩又はマグネシウムの増加は、回分培地成分の増加として、或いは流加培養醗酵における供給に対する追加として意図している。醗酵の分野における当業者は、系統だった成分測定による培地の更なる最適化を行うことができる。
【0056】
ある栄養要求性細胞株を利用する場合、培地への添加物を意図している。例えば、proAB欠失を含むStbl2(InVitrogen Corp.)等の細胞株は、最大増殖を達成するためには、アミノ酸補給、又はプロリンが豊富な代替窒素源を必要とする。このような修飾は、醗酵の分野における平均的な当業者によって決定することができる。
【0057】
また、我々は、従来技術に記載されているプラスミド醗酵培地を用いた本明細書に記載されている回分及び流加培養醗酵プロセスの使用を意図している。これは、Soubrier(Soubrier F.2004 US Patent application 2004/0142452)によって開示されているような規定された培地を含む。本発明の流加培養醗酵プロセスで使用する好ましい培地は、NTC3019や、Lahijani et al.,Supra,1996、Friehs et al.,Supra,2003、Chen Supra,1999.及びUrthaler et al(Urthaler J,Roman N,Ascher C,Woehrer H.2005 US Patent Application 2005/0026177)に開示されている培地等の半合成供給によるグリセロール培地配合である。
【0058】
醗酵プロセスの代替的実施例
改善されたプラスミド生産のための流加培養及び回分プロセスが、本明細書に開示されている。これらは、新規な増殖及び誘発期温度シフトと組み合わせて、指数関数的供給戦略を特徴付けるものである。
【0059】
本発明の流加培養プロセスの実施において、我々は、フィードバック、フィードフォワード、及び栄養素供給の所定の制御を含む、増殖速度を低減するための種々の供給戦略を意図している。例えば、炭素制御した指数関数的供給戦略によって、栄養素供給を加えてもよい。我々は、許容できる範囲内で増殖速度を制御するための、この供給戦略の変型を予想している。好ましい増殖速度の範囲は、μ=0.05から0.3h−1である。好ましいターゲット増殖速度は、μ=0.12hr−1である。許容される増殖速度の範囲は、プラスミド特異的であり、当業者によって実験的に決定することができる。
【0060】
本発明の流加培養プロセスの実施において、我々は、生産時間を低減するための種々の回分戦略を意図している。回分期に含まれる半合成栄養素を調節して、回分期をより高い濃度又はバイオマス濃度にすることができる。より高いバイオマス濃度の場合に、流加培養期と比較して回分期で増殖がより早く進むので、全体的な発酵時間が低減される。我々は、プロセスの流加培養の開始を制御するための、この回分戦略の変型を予想している。開始のための流加培養期の好ましいOD600の範囲は、1から60である。許容できる流加培養開始OD600の範囲はプラスミド特異的であり、当業者によって実験的に決定することができる。
【0061】
本発明の誘導性流加培養及び回分プロセスの実施において、我々は、増殖期から誘導期への種々の温度シフト戦略を意図している。増殖期は25から37℃、好ましくは30から37℃の温度で実施することができる。高コピープラスミドについては、増殖期は、最も好ましくは30から32℃である。誘導期を33℃から45℃、好ましくは37から42℃の温度で実施することができる。
【0062】
温度シフト戦略を伴う誘導性プロセスの実施において、我々は、増殖期から誘導期へ移る時の指標として栄養素濃度、バイオマス濃度、又は光学密度を使用することを意図している。醗酵培養の規則的なサンプリングは、バイオマス又は光学密度測定を得るための材料を提供することができ、温度シフトは、あるバイオマス濃度で実施することができる。また、オンラインセンサを使用して、バイオマス濃度の継続的なモニタリングを行い、醗酵装置を設定して特定のバイオマス濃度で温度シフトを自動的に実施することができる。温度シフト戦略を伴う誘導流加培養プロセスの実施において、我々は、また、ある量の供給栄養素を加えると温度シフトが実行されることを意図している。
【0063】
温度シフト戦略を伴う誘導性プロセスの実施において、我々は、生産回数を最小限にするために増殖期中により高い温度を用いることを意図している。本明細書に記載されている誘導性流加培養プロセスは、このプロセスの増殖期の間中、低い(<2mg/L/OD600)プラスミドレベルを維持した。温度シフト後に、許容できる代謝負担、プラスミド安定性、及び生産性の向上を維持する増殖期中に利用できる最大温度は、プラスミド特異的であり、様々なプラスミドバックボーン間で異なる。例えば、いくつかのプラスミドは、2mg/L/OD600よりずっと高いレベルに許容できる。同様に、プラスミド生産性は、低い温度で変化し、従って、いくつかのプラスミドは、高いレベルのプラスミドとしては生産されない。これらの場合、我々は、増殖期中に、より高い温度を使用することを意図している。加えて、上昇した最大温度は、無制限の増殖とこれに相当するプラスミドコピー数の低下に起因して、開始回分期中で許容される(図5)。増殖期(回分及び流加培養成分)中に用いることができ、温度シフト後に許容できる代謝負担、プラスミド安定性、及び生産性の向上を維持する最大温度は、この分野の平均的な当業者によって実験的に決定することができる。
【0064】
プラスミド及び宿主菌株
我々は、高コピー、低コピー及び中程度のコピーのいずれかであり、温度誘導性であるか温度誘導性ではない種々の起源の複製を伴うプラスミドの生産において本発明を使用することを意図している。いくつかの好ましい複製の起源、及びこれらを組み込んだプラスミドが、表1に記されている。これらの起源の変更はこの分野において公知であり、また、使用を意図している。
【0065】
表1:複製起源

【0066】
代替宿主菌株が意図されている。大腸菌(E.coli)株DH5αは、プラスミド生産用に広く使用されている宿主である。その鍵となる品質は、クローンDNAの非特異的組み換えを最小限にするrecA突然変異、及びエンドヌクレアーゼIによってプラスミドの非特異的消化を除去するendA1突然変異を含む。DH5αに加えて、その他の様々な菌株が、プラスミド生産に好適である;例示的な大腸菌(E.coli)宿主菌株の非限定的リストを表2に示す。
【0067】
表2:宿主菌株
【0068】
DH5α、XL1−Blue、DH10B、JM109及びTop10は、プラスミド生産菌株として、非常に良く確立されている。Mach1、及びECOS101は、最近開発されたものであり、望ましいプラスミド生産宿主である。Stb12、GT116及びSure細胞はプラスミド含有不安定DNAの生産に利用されている。不安定DNAは、直接(例えば、レトロウイルスの長い末端反復)又は逆方向反復(例えば、shRNAパリンドローム)ZDNA、その他のような構造を含む。GT116のdcm遺伝子の削除は、免疫刺激であるダムメチル化(dam methylation)を解消する。このため、GT116の生産は、プラスミドDNAの免疫原性を低減する。類似の免疫原性の低減は、CpGメチラーゼを発現する菌株を使用して観察される。
【0069】
不安定プラスミドの生産
【0070】
我々は、また、不安定なシークエンスを含むプラスミドの生産に本発明を使用することを意図している。パリンドロームシークエンス、定方向又は逆方向反復塩基配列、及びシークエンスを形成するZDNAが不安定であり、大腸菌(E,coli)宿主によって除去、又は再編成される。いくつかの場合には、治療に使用するプラスミドは、不安定なシークエンス(AAV及びHIV等のウイルス性ベクタについての逆方向又は定方向反復塩基配列、ある治療用遺伝子についてセグメント又は三塩基反復を形成するZDNA)を含んでいなければならない。不安定シークエンスを含むプラスミドを維持するための現在行われている戦略は、安定化突然変異を用いた宿主細胞株を使用することである。いくつかの宿主は、これらのプラスミド、例えば、Sure細胞(Stratagene)、GT115(Invivogen)又はStb12及びStb14(Invitrogen)の増殖用に商業的に入手できる。Stb12及びStb14細胞株は、レトロウイルスベクタ等のベクタを含む定方向反復塩基配列の安定性を増進する未公表の突然変異を用い;この効果は、恐らくは減少したコピー数に起因して低下した温度で強化される。修復突然変異の特異的組み合わせは、プラスミド増殖を、特に低温で安定化することができる。Sure及びSure2細胞株は、UV修復(uvrC)及びSOS修復(umuC)欠損(LTRsを安定化するため)と関連した相同性組み換え欠損(recB、recJ)、及びZDNAを安定化するためのSbcC(及びrecJ)を有するこのような組み合わせを用いる。GT116細胞株は、パリンドロームシークエンスを安定化するためにSbcC及びSbcDを用いる。これらの菌株は、恐らくはプラスミドコピー数の低下に起因して、低温(即ち、30℃)でのみプラスミドを安定化するように機能する。この戦略は明らかに生産コストを上げる。本明細書に記載されている誘導性醗酵プロセスの使用は、集菌前の短期間にのみコピー数が増加する前に、安定化細胞株中の30℃の不安定プラスミドを増殖することができる。これは、不安定プラスミドの収量及び安定性(即ち、質)を最大にするであろう。
【0071】
DNA及びプラスミド生産
誘導性流加培養プロセスにおけるプラスミドDNA(>6mg/L/OD600)の実際の増加した収量についての根底にある機構は知られていない。遅い増殖及び低下した温度でのバイオマス生産中に、DNA凝縮剤(例えば、dps遺伝子生産物等のヒストン状タンパク質、又はその他のクロマチン結合タンパク質)に潜在的に起因する。
【0072】
誘導期中にDNA凝縮を変化させることで、許容プラスミドレベル又はコピー数を増加することによって、プラスミド収量を増やすことができる。DNAの凝縮度合は、2つの対立因子;クロマチンタンパク質の凝縮と、転写複合体の脱凝縮と、によって設定される。プラスミド凝縮は、プラスミドプロモータからの転写レベルによって影響を受けることがある。転写が少ないほど、より高い凝縮となり、潜在的により高い運搬能力となる。
【0073】
E.coli中で、DNA凝縮に含まれる多数のクロマチンタンパク質を同定した。これらの遺伝子生産物は、プラスミドとゲノムDNAを結合させる。ゲノムDNAの場合には、核様体内にDNAを凝縮する((Robinow C,Kellenberger E.1994 Microbiol Review 58:211−232)参照)。核様体の主要成分は、ヒストン状タンパク質HU、IHF、及びHN−S、StpA(HN−Sに関連して、約1/10レベルで発現される)及びDpsであり、これらは核様体内に均一に分配され、一方で、SeqA、CbpA、CbpB、Fis及びIciA等のその他のタンパク質はより少ない量であり、核様体内に不均一に分配され、調整機能を有する。遊離Rプラスミドタンパク質複合体は、3つの主要なタンパク質、23%のHN−S、23%のRNAポリメラーゼ、及び5%のHUを含む。これは、クロマチン関連遺伝子生産物が異なる培地、異なる細胞密度で、増殖期及び静止期中に様々に調整されるので、増殖期に依存して、恐らく変化するであろう。Fis、HU、HF−1は、一般的に、対数期により高度に発現し、一方で、IHF及びDpsは、静止期により高いレベルにある。Dpsは、静止期に液状生物結晶性複合体にDNAを凝縮して、ストレス耐性を改善する。増殖期中のHN−Sの過剰発現は、DNA凝縮と生存度の減少を引き起こす。細胞は、DNAが高度に凝縮される場合、より高度なプラスミド能力を有する。NTC誘導性醗酵プロセスにおいて、増殖期細胞は、誘導期細胞よりもプラスミドDNAに対する全体的な能力が低い。これは、許容プラスミドを増殖期よりも高いレベルにする、誘導期に存在するクロマチンタンパク質の組み合わせの相違に起因する。誘導期中のクロマチンタンパク質の比率の変化が、プラスミド凝縮、及び運搬能力を上げる。
【0074】
また、誘導期中のクロマチンタンパク質の比率の変化は、プラスミド複製率を上げる。例えば、p15A起源RNAIIプロモータからの発現であって、pMB1(pBR322)RNAIIプロモータからではない発現は、IHFによって表される;p15A RNAII転写は、IHF突然変異中で増加する。Dps及びHUは、非特異的DNA結合剤、HN−S、CbpA及びCbpBであり、湾曲したDNAと結合する。StpAは、HN−Sに関連し、より高い親和性でDNAと結合し、また、湾曲DNAと結合する。Fis、IHF、IciA及びseqAは、シークエンス特異的である。HN−Sは、湾曲DNAを含む多数のプロモータからの転写を抑制する。pMB1(pUC及びpBR322)のRNAIIプロモータは、ポリA及びポリTトラックを含み、これらのシークエンスは、湾曲DNAを形成する。静止期の転写(例えば、HN−S)を抑制する減少したレベルの湾曲DNA結合性クロマチンタンパク質は、RNAIIのRNAIへの転写の増加した比率に関連し、文書化静止期は、プラスミドコピー数を増やす。NTC醗酵プロセスの誘導期中に存在するクロマチンタンパク質の組成の変化(例えば、HN−Sの更なる減少)は、pUC等のpMB1を有するプラスミドコピー数の増加をもたらす。
【0075】
大腸菌(E.coli)中で発現する場合、異種DNA凝縮体、例えば、バシラス(Bacillus)属の酸可溶性胞子タンパク質は、プラスミド収量を上げるための有益なDNA凝縮体でもある。例えば、大腸菌(E.coli)中の枯草菌(B.subtilis)酸可溶性小タンパク質の発現は、DNAに起因する(Setlow B,Hand AR,and Setlow P.JBacteriol.173:1642−1653)。
【0076】
また、プロセス変化は、DNA凝縮の効果を通じて収量を改善する。Dpsは、マグネシウム(Mg++)濃度;Dpsの存在は、DNA凝縮をもたらさない;密集した結晶性DNA:Mg++濃度が閾値以下に下がると、Dps複合体を形成する[(Frenkiel−Krispin D,Levin−Zaidman S,Shimoni E,Wolf SG,Wachtel EJ,Arad T,Finkel SE,Kolter R,Minsky A.2001 EMBO J.20:1184−1191)参照];によって調整される。形態学的に、この複合体は、静止期中にクロラムフェニコールを添加することによって誘導されるのに似ている。増殖する培地への0.2mMのスペルミジンの添加は、Dpsの不存在下でDNA凝縮を加速する。リン酸塩欠乏は、恐らくスペルミジンに対するスレオニン及びアルギニンの分解の促進を介して同様の効果を有する。2価カチオンのレベルを変更(例えば、Mg++、外因性添加又は減少による)するための、又は正電荷ポリアミン(例えば、スペルミジン、細菌合成の外因性添加又は制御による)の変化を変更するための誘導期中の醗酵組成又は状態への変化によって、プラスミド収量を増やすことができ、この変更は、培地の交換を意図する。
【0077】
我々は、更なる収量の増加が、プラスミドDNAの更なる凝縮によって得られることを意図している。これは、醗酵プロセス中に、スペルミン生産又はdpsタンパク質生産等の宿主菌株DNA凝縮剤の生産を増やす、供給(例えば、ポリエチレンイミン、スペルミジン、スペルミン)又は菌株組み換えへのDNA凝縮剤の添加によって達成される。このような菌株組み換えは、関連する遺伝子生産物が、醗酵プロセス中に減少することを可能にする変更になり得る。
【0078】
誘導性プロセスの実施において、我々は、代替戦略を用いて、増殖期中に低レベルでプラスミドコピー数を維持することを意図している。例えば、低温での増殖に加えて、増殖期中に組み込むことができるコピー数を減らすためのその他の機構が存在する。例えば、醗酵中の溶存酸素の減少は、プラスミドコピー数を減少させるために示される (Carnes AE,2005 BioProcess International 3:9,in press)。
【0079】
最終産物純度の改善
我々は、例示的なプラスミド精製プロセス中に記載された醗酵培地からのプラスミド濃縮供給ストリームを用いることを意図している。このようなプロセスは、技術的によく知られている。高収量醗酵と例示的な精製プロセスの組み合わせは、費用対効果のある方法論を提供し、ゲノミックDNAを更に遺伝子治療及びDNAワクチン接種適用に許容されるレベルにまで減らす。
【0080】
例
本発明の方法を、以下の例において更に説明する。これらは、例示によって提供され、本発明の範囲を限定するものではない。
【0081】
例1:NTC3018及びNTC3019醗酵培地
次の基準はプラスミドDNAを製造するための最適化醗酵プロセスの評価として確立された:
1) プラスミドの高い比収量(g細胞質量当たりのmgプラスミドDNA);
2) 高バイオマス収量;
3) 高度のスーパーコイル状のプラスミドを保つであろう;
4) ダウンストリームプロセシング中に生じる問題が最小になる;
5) 調整要求条件を満足する;
6) プラスミド構造を保持する(例えば、遺伝子欠失がない、又はその他の再配列);
【0082】
高い比プラスミド収量、高バイオマス収量、及び高プラスミド品質をサポートするように、培養培地を調整した。回分醗酵培地は、比増殖速度を減らすように構成した。低減した増殖速度の使用は、より高いプラスミドコピー数とより良いプラスミド安定性と関連していた。流加培養醗酵中に、制限栄養素の供給によって、増殖速度を0.12hr−1に制御した。加えて、培地に使用した全組成は、良く特徴付けられており、認定された動物由来成分が含まれていなかった。
【0083】
NTC3018(回分)及びNTC3019(流加培養)培地は、回分及び流加培養プロセス培地の双方の多くの成分用に最適化されている。例えば、グリセロールを、炭素源として使用して、増殖速度を低減している。イースト抽出物は、窒素源として使用される。大腸菌(E.coli)生産菌株の所定の条件に基づいて、微量金属及びMgSO4濃度を最適化した。
【0084】
例2:pBR322−誘導プラスミドを用いたNTC3019培地流加培養
37℃での、New Brunswick BioFlo 110 fermentor中で流加培養醗酵を実施した。30%の水酸化アンモニウム又は10%のリン酸を自動的に添加することによって、pHを制御した。窒素ガス散布によって0%に、及び空気飽和で100%に溶存酸素プローブを調節した。容器を1VVMで通気し、溶存酸素を攪拌の比例積分制御によって30%に維持した。また、約20OD600以上の細胞密度で、30%の飽和を維持するためにO2補給を必要とした。
【0085】
LBプラス50μg/mlカナマイシン内に植菌した単一の遊離コロニーから種培養を開始し、37℃で培養した。中間指数関数期(0.5から1.5OD600)で、種培養を用いて、醗酵装置に対して1%の接種材料を提供した。
【0086】
流加培養中に、炭素の制限指数関数的供給戦略に応じて、半合成供給栄養素を添加した。要約すれば、回分期中にμmaxの比増殖速度で炭素基質の初期量が消費される。炭素基質が消耗すると、流加培養期が始まり、供給栄養素が、次の式によって決定される割合で、自動的に加えられる(Carnes,Supra,2005):
【0087】
【0088】
ここで、
μ=流加培養期中の所望の比増殖速度、
XB=回分期の終わりのバイオマス濃度、gDCW/L、
VB=培地の初期液体体積、L、
Sf=栄養素供給培地の制限基質濃度、g/L、
YX/S=基質からのバイオマスの収量係数、g/g
t=流加培養期の開始からの時間、
である。
【0089】
一般的に、いくつかの独立したカナマイシン耐性pBR322−誘導プラスミドを用いたNTC3019培地の流加培養醗酵は、100から120OD600の細胞密度、又は1リットル当たり55から65g乾燥細胞重量に達する(図1)。プラスミドは、平均260mg/Lであり、430mg/L程度に高かった。比較すると、pBR322−誘導プラスミドを用いた公開されている醗酵収量は、約3から4mg/Lである(Lahijani et ai,Supra,1996)。
【0090】
重要なことには、比プラスミド収量が非常に高く、一般的に2.5から3.8mg/L/OD600の間であり、かなり高いコピーpUC起源プラスミドを用いるその他の醗酵培地/プロセスで観察されたレベルを十分越えている(表3)。比収量mg/L/OD600に関する発現プラスミド収量は、全細胞質量と比較したプラスミドの量を示す。細菌1グラム当たりの増加したプラスミドの収量が、より高い最終産物純度を直接導くので、高い比収量は非常に望ましい。
【0091】
表3:公開高収量醗酵プロセスと比プラスミド収量の比較
【0092】
これは、NTC3019醗酵培地が、従来技術に記載された培地及びプロセスと比較して、中程度のコピー数のプラスミド(例えば、rop欠失を有するpBR322)の醗酵収量を、劇的に促進することを明示している。この効果は、プラスミド特異的ではない。
【0093】
これらのプロセスで精製したDNAは高品質であり、欠失又はその他の再配列を検出することなく、実質的に100%スーパーコイル状である。同様に、このプロセスで、細胞を使用して1グラムスケールでDNA精製を実施した。これは、NTC3019流加培養培地で実施された醗酵が、大きいスケールのダウンストリームプロセシングに敏感に反応することを明示している。
【0094】
例3:高コピー遺伝子治療プラスミドを用いたNTC3018培養回分醗酵
pUC起源プラスミドを用いたNTC3018培地回分培養を実施した。以下のプラスミド含有pUC起源を使用した:
1)pW2.0、変質ポリリンカーシークエンスを有するpUC19の誘導体
2)pMaxGFP
3)pEGFP−C1
【0095】
37℃の、New Brunswick BioFlo 110 fermentor中で回分醗酵を実施した。30%水酸化アンモニウム又は10%リン酸の自動添加によって、pHを制御した。窒素ガス散布によって0%に、及び空気飽和で100%に溶存酸素プローブを調節した。容器を1VVMで通気し、溶存酸素を攪拌の比例積分制御によって30%に維持した。また、約20OD600以上の細胞密度で、O2補給を30%の飽和を維持するためには、O2の補給が必要であった。
【0096】
LBプラス50μg/mlカナマイシン中に植菌した単一遊離コロニーから種培養を開始し、37℃で培養した。中間指数関数期(0.5から1.5OD600)で、種培養を用いて、醗酵装置に対して1%の接種材料を提供した。
【0097】
全ての醗酵を37℃で実施した。pW2.0について、プラスミドコピー数を醗酵の遅い42℃での増殖によって誘発した。pUCプラスミドを用いた回分醗酵は、16から57OD600の細胞密度に達したのみであった。しかしながら、比プラスミド収量の結果は有望である。pEGFP−C1についての最終収量は、細胞密度が56OD600であり、収量が163mgプラスミド/L(2.9mg/L/OD600)であり、pMaxGFPについては、細胞密度が16OD600であり、収量が84mgプラスミド/L(5.3mg/L/OD600)であり、pW2.0については、細胞密度が57OD600であり、収量が230mgプラスミド/L(4.0mg/L/OD600;図2)であった。
【0098】
これらのプロセスから精製したDNAは高品質であり、欠失又はその他の再配列を検出することなく、実質的に100%スーパーコイル状である。同様に、このプロセスで細胞を使用して実質0.5グラムスケールでDNA精製を実施した。これは、NTC3018回分培地で実施された醗酵が、大きいスケールのダウンストリームプロセシングに敏感に反応することを明示している。
【0099】
例4:高コピー遺伝子治療プラスミドを用いたNTC3019培地の流加培養醗酵
Vical VR1012ベクタの誘導体を含有するGFP遺伝子であるプラスミドgWiz GFP(Gene Therapy Systems)を、流加培養醗酵評価用に選択した。これは、5757bpの大きさの広く用いられているDNAワクチンプラスミド含有カナマイシン耐性(kanR)pUC起源である。このプラスミドgWiz GFP(Gene Therapy Systems)を、大腸菌(E.coli)DH5αに変質させた。また、pMaxGFPを、流加培養培地内で試験し;両方のプラスミドについて、同様の結果を得た。
【0100】
これらのプラスミドの生産にNTC3019培地を使用したときに、2つの問題に直面した。増殖がpBR322−誘導プラスミドを用いてうまく行われているときに、37℃でのgWiz GFPプラスミド培地の増殖時に第1の問題が発見された。pUCプラスミド培地では、細胞増殖は15OD600のあたりで止まった。蛍光顕微鏡は、細胞分裂の阻害を意味する広範なフィラメンテーションを示した。フィラメントが実質的に溶解するので、このことは致命的である(Arends SJR, Weiss,DS.2004 J Bacteriol 186:880−884)。例えば、図3(A)は、プラスミドgWiz GFPを用いた流加培養醗酵を示す。細胞増殖は遅くなり、15OD600の早い段階で静止期に入るように見えた。プラスミド収量分析は、細胞増殖が停止し始めたときに、2.7mg/L/OD600の上昇比プラスミド収量を示した。pUCプラスミドは、30℃の場合と比較すると、42℃でのコピー数の30から40倍の増加を示す温度感受性点突然変異を含む(Lin−Chao et al,1992)。次いで、プラスミドコピー数を減らし、細胞の代謝負荷を軽減しようとして、温度を33℃に下げた。温度の低下後に、プラスミドは1.6mg/L/OD600に下がり、細胞増殖が再開した。興味深いことに、比プラスミド収量は再び徐々に増加し、供給栄養素が加えられていたにも関わらず、増殖は、予想通りに>100OD600に増殖することなく、約60OD600で静止期に入った。
【0101】
図3(B)は、37℃で発生する細胞フィラメンテーションを示す。温度を33℃に下げた後に、増殖細胞のサンプルは、より少ないフィラメンテーションを示した。
【0102】
考えられる説明は、37℃で増殖が開始した時に、細胞母集団は、フィラメンテーション及び生存率の低下から完全に回復しないことであった。このことを試験するために、同じプラスミドで、引き続いて2つの流加培養醗酵を、33℃で完全に実施した。両醗酵において、細胞密度<60OD600で培養がピークに達した。
【0103】
これらの醗酵からのバイオマス及びプラスミドDNA収量データは、比増殖速度の低下と、細胞増殖阻害前の比プラスミド収量の急激な上昇を示す。このような高いレベルへのプラスミド含有量の急激な増加は予想されておらず、増殖速度の低下を招く細胞母集団上の代謝負担を認識する。しかしながら、増殖速度の低下は、しばしばプラスミドコピー数の増加を導くことも示されている(Satyagal VN,Agrawal P.1989 Biotechnol.Bioeng.33:1135−1144)、(Seo JH,Bailey JE.1985 Biotechnol.Bioeng.27:1668−1674)。予期しない増加比プラスミド収量が、増殖速度の低下を引き起こしているのか、又は予期しない低下した比プラスミド収量が、増殖速度の増加を引き起こすのか、或いは、それぞれが、複合してどちらかを引き起こすかは、明らかではない。
【0104】
例5:NTC3019培地を用いた高コピープラスミドの高収量生産のための誘導流加培養プロセス
【0105】
33℃及び37℃での醗酵の結果(例5)に基づいて、pUC起源プラスミドを用いた流加培養モードで観察される予期しないプラスミド増加を克服するように戦略を構成した。DH5α中のプラスミドgWiz GFPを、誘導流加培養プロセスで用いた。NTC3019流加培養醗酵は、30℃で60OD600まで培養することを除いて、例5で説明されるように実施した。この時に、温度を37℃にシフトした。驚くべき結果が、図4Aに示されている。60OD600まで30℃で増殖することで、増殖阻止問題がなくなり、この培養は、全プラスミド収量が670mg/Lの100OD600を最終的に上回った。このプロセスのサンプルから精製されたDNAは高品質であり、欠失又はその他の再配列を検出することなく、実質的に100%スーパーコイル状である。
【0106】
2mg/L/OD600以下に維持して、増殖期を通じて低く維持される温度シフトの前に、プラスミドが産生する。このことは、33℃又は37℃の醗酵の結果と対照的である。注目すべきことに、温度シフト後の比プラスミド収量は、6.5mg/L/OD600と非常に高くなり、その他の醗酵培地/プロセスを用いて観察されたレベルを十分に越えている(表3)。増殖期を通して30℃での醗酵させ、42℃へシフトすることは、gWiz GFPを伴う1.1gm/L(11mg/L/OD600)の生産量をもたらす結果となった(図4B)。生産性安定期は、細胞の大多数が生き残るので、大量の細胞死に関連していない。
【0107】
また、流加培養期間を低減する(回分期をより高いOD600にすることによる)ためのNTC3019培地(グリセロール、イースト抽出物、及び回分培地のマグネシウムの4倍の増加)の修飾も、42℃での誘導後に同様の高プラスミド収量を産生し、プラスミド誘導のロスなしで、より高いOD600で流加培養期を開始できることを明示している。
【0108】
様々な抗生物質抵抗性遺伝子及び原核生物要素の配向性を含む、種々のpUC起源バックボーンを有する多数の異なるプラスミドが、DH5α中での30℃から42℃の誘導性プロセスを用いて、NTC3019培地中の0.5gm/Lよりも大きい収量で生産された。これらの結果は、この誘導性プロセスが、特定のプラスミドに特異的でないことを明示している。
【0109】
また、gWiz−GFPプラスミドが、DH1細胞株中で、30℃から42℃での誘導性プロセスを用いて、NTC3019培地の0.5gm/Lよりも大きい収量で生産された。この結果は誘導性プロセスが、特定の大腸菌株(E.coli strain)に特異的でないことを明示している。
【0110】
同様に、このプロセスからの細胞を用いて、1グラムスケールでのDNA精製を実施した。このことは、NTC3019流加培養培地で実施された誘導性醗酵が、大きいスケールのダウンストリームプロセシングに敏感に反応することを明示している。
【0111】
比収量(mg/L/OD600)の用語でのプラスミド収量の表現は、全細胞質量に対するプラスミドの量を示す。本明細書に記載された誘導性流加培養プロセスは、このプロセスの増殖期を通してプラスミドレベルを低く(<2mg/L/OD600)維持し、バイオマス生産後の、前代未聞の非常に高いプラスミド生産(6から11mg/L/OD600)を容易なものにした。細菌の1グラム当たりのプラスミド収量の増加は、直接的に最終産物純度を高くするので、高い比収量は非常に望ましい。
【0112】
これらの結果は、プラスミドDNA生産性と品質を改善する、本発明の流加培養及び回分醗酵プロセスの一般的な使用を明示する。
【0113】
このように、本発明の生産プロセスが、改善されたプラスミド生産方法を提供するものであることを、読者は理解するであろう。
【0114】
上記記述が多くの特異性を含んでいる一方で、これらは、本発明の限定として解釈するべきではなく、むしろ本発明の好ましい一の実施例の例示として解釈するべきである。その他の多くの変形が可能である。例えば、誘導性流加培養プロセスを、回分プロセスと一体化して、栄養素がなくなるまでNTC3018培養培地中で醗酵を行うことができる。ここでは、NTC3019由来の流加培養培地及び誘導は同時に開始される。いくつかのプラスミドを用いて、この実施例においては、プラスミドコピー数をより高い増殖速度で低下するため、増殖期を37℃以上で実施することができる。増殖期中に細胞分裂を可能にし、硫化培養期中にプラスミド誘導性を保持する最適な温度は、当業者が決定できる。従って、本発明の範囲は、説明した実施例によってではなく、添付の特許請求の範囲及びこれらの法律的均等物によって決定されるべきである。
【図面の簡単な説明】
【0115】
【図1】図1は、NTC3019培地を用いた大腸菌(E.coli)内のpBR322−誘導プラスミド流加培養醗酵を示す。
【図2】図2は、NTC3018培地を用いた大腸菌(E.coli)内のpUCプラスミド回分醗酵を示す。
【図3】図3は、NTC3019培地を用いた大腸菌(E.coli)内のgWiz GFPプラスミド流加培養醗酵を示す。
【図4】図4は、NTC3019培地を用いた大腸菌(E.coli)内のgWiz GFP誘導流加培養醗酵を示す。
【図5】図5は、誘導性流加培養醗酵プロセスを示す。
【技術分野】
【0001】
連邦政府後援研究又は開発に関する陳述
適用されない。
【0002】
本出願は、2004年8月19日に出願された、米国暫定特許出願第60/603000号の利益を主張する。
【0003】
本発明の分野
本発明は、プラスミド、コスミド、バクテリア人工染色体(BACs)、バクテリオファージ、ウイルス性ベクタ及びこれらのハイブリッド等の閉環状(covalently closed circular:ccc)組み換え型DNA分子の生産に関し、より具体的には、醗酵培養における高レベルでのDNA分子の生産方法である。
【0004】
本発明の背景
本発明は、閉環状(ccc)組み換え型DNA分子の生産に関する。このような分子は、バイオテクノロジ、遺伝子組み換え生物、遺伝子治療、治療上のワクチン接種、農業及びDNAワクチンに有益である。
【0005】
本発明を念頭に置いて、先行技術の調査を実施した。大腸菌(E.coli)プラスミドは、長い間、研究者及び企業によって用いられた組み換え型DNA分子のただ1つの最も重要な源であった。今日、プラスミドDNAは、次世代のバイオテクノロジ生産物(遺伝子医薬品及びDNAワクチン)が、臨床試験及び、医薬市場への道を作るに当たって、ますます重要性が高まっている。プラスミドDNAワクチンは、ウイルス性、細菌性、又は寄生性疾患に対する予防的ワクチン;高力価免疫グロブリン生産物の生産用免疫剤;伝染性疾患用のワクチン;又は癌ワクチン;としての適用を見出すことができる。プラスミドは、また、遺伝子治療又は遺伝子置換用途に利用され、患者に投与した後に所望の遺伝子生産物が、プラスミドから発現する。
【0006】
今日、FDA(アメリカ食品医薬品局)基準は、暫定段階にあるもの以外には規定されていない(FDA Points to Consider on Plasmid DNA Vaccines for Preventive Infectious Disease Indications,1996参照)。しかしながら、将来は、プラスミドDNA純度についての国際基準が、大腸菌(E.coli)醗酵から同様に作られる組み換えタンパク生産物に使用される基準と同じか、又は非常に類似するものになりそうであり、このような基準は、確立された方法から達成できる現行の純度を超える。最も明らかなことには、投与量当たり100pg未満の主宿遺伝子DNAの許容された基準は(FDA Points to consider in the characterization of cell lines used to produce biologies,1993参照)、精製プラスミド製剤について現在達成できるレベルより遙かに低い(投与量1mg当たり100pgは、1000万分の1に等価である)。
【0007】
プラスミド(細菌醗酵によって)、及びこれらの純度(例えば、アルカリ溶菌法(alkaline lysis method)(Birnboim,HC,DoIy J.1979,Nucleic Acids Res.7:1513−1523)によって)を得る基本的な方法は公知である。最初に、醗酵した細菌性細胞ペーストを、再懸濁して、(水酸化ナトリウム及びドデシル硫酸ナトリウムの組み合わせを使用して)溶解し、その後溶液を、酸性塩(例えば、酢酸カリウム)の添加によって中和し、細菌性DNA及び細胞崩壊堆積物の大部分を沈殿させる。大量のスーパーコイル状のプラスミドDNAは、汚染細菌性RNA、DNA及びタンパク質、又は大腸菌内毒素(リポポリサッカライド、又はLPS)と共に溶液中に残る。次いで、可溶性画分をろ過によって分離し、種々の精製工程を受けさせる。この工程には、RNase(リボヌクレアーゼ)消化;クロマトグラフィ(イオン交換ゲルろ過、ハイドロキシアパタイト、ゲルろ過、疎水性相互作用、逆相、HPLC、等);膜分離;有機抽出(organic extraction);選択的沈殿、等が含まれる。
【0008】
明らかに、開始材料の純度が増し、より良い下流純度を達成することは、工業規模の臨床段階のDNAを生産するために不可欠な目標である。
【0009】
醗酵媒体の考察
安定した媒体の型は、細胞のエネルギィ所要量及び基本的な組成に基づいている。一般的に、栄養学的な所要量は、最少培地又は半合成培地によって満たされる。
【0010】
半合成培地は、酵母エキス、カザミノ酸、及びペプトン等の複合体成分を含む。複合体成分の追加は、増殖因子、アミノ酸、プリン類及びピリミジンを供給し、しばしばより高い細胞密度をサポートする。
【0011】
炭素は、細胞組成の半数を数える。従って、炭素は、最も多量に含まれている。炭素源は、エネルギィ及びバイオマスを提供し、制限栄養素として通常使用される。グルコースは、通常の炭素源である。グルコースは非常に効果的に代謝され、従って、細胞収率がより高い。しかしながら、高グルコース濃度は、(クラブトリー効果(Crabtree effect)として知られる)代謝過剰に起因して、望ましくない産生を引き起こす。グリセロールも用いられ、しばしば回文培養の好適な炭素源となる。グリセロールからの細胞収率は、グルコース由来のものよりも僅かに小さいが、グリセロールは、高レベルの酢酸産物を作らず、阻害されることなく、より高濃度で使用することができる。グリセロールも、最大比増殖濃度を減少させる。
【0012】
窒素の所要量には、無機又は有機窒素源によって応ずることができる。アンモニア及びアンモニア塩(例えば、NH4Cl、(NH4)2SO4)は、最少培地で使用される。半合成培地は、イースト抽出物、ペプトン、及びカザミノ酸を含有する複合体成分から部分的に又は全体的に窒素を供給する。
【0013】
鉱物は、増殖、代謝、及び酵素反応に必要である。マグネシウム、リン、カリウム、及び硫黄は、個別の媒質成分として一般的に加えられる。リン酸ジカリウム又はリン酸モノカリウムは、カリウム及びリンを提供し、ある比率で緩衝剤としても機能する。硫酸マグネシウム・7水和物は、マグネシウム及び硫黄の源として、しばしば利用される。その他の必須鉱物は、カルシウム、銅、コバルト、鉄、マンガン、モリブデン及び亜鉛を含む。これらは主要成分中に不純物として通常存在するが、これらは、極少量必要とされ、微量鉱物溶液の添加によって、しばしば補給される。浸透圧は、塩化ナトリウムで調節する。
【0014】
動物由来の生成物、特にプラスミド生産におけるウシ生成物の使用は、プリオン又はウイルス汚染物質の危険性に起因して許容できない。全培地成分は、認定された動物由来生成物を含まないものであるべきである。植物由来の代用物は、動物に起源を有する多くの成分に使用可能である(例えば、植物グリセロール、大豆ペプトン)。
【0015】
プラスミド醗酵プロセスの考察
増殖速度
減少した増殖速度の使用が、高品質、高収率プラスミド醗酵の統一原理である。高増殖速度は、酢酸生成物、プラスミド不安定性、及びスーパーコイル状のプラスミドのより低い割合と関連している。減少した増殖速度は、細胞分裂と同期したプラスミド複製のための時間を提供することによって、増殖速度依存プラスミド不安定性を緩和する。
【0016】
増殖状態
醗酵は、プラスミドの質及び収率に影響する多くのパラメータを制御及び監視する可能性を与える。スーパーコイル形成は、酸素及び温度によって影響されることが知られている(Dorman CJ et al.1988 J.Bacteriol.179:2816−2826)、(Goldstein E,Drlica K.1984 Proc Natl Acad Sci USA.81:4046−4050)。酸素は、プラスミド安定性において重要な役割を演じることが示されている。ある研究(Hopkins DJ,Betenbaugh MJ,Dhurjati P.1987 Biotechnol Bioeng.29:85−91)では、5%の空気飽和率に対する溶存酸素濃度中の1滴が、プラスミド安定化の急速な減少を導くことを発見した。別の研究(Namdev PK,Irwin N,Thompson BG,Gray MR.1993 Biotechnol Bioeng.41:666−670)では、酸素導入の変動が、プラスミドの不安定化をまねくことを示した。更に、ニックプラスミド及び多重体の形成は、温度、pH、溶存酸素、栄養素濃度、及び増殖速度を含む多くのパラメータによって影響を受け得る(Durland RH,Eastman EM.1998 Adv Drug Deliver Rev.30:33−48)。大腸菌増殖の最適温度は、37℃である。しかしながら、回分醗酵では、より低い温度(30から48℃)を用いて、最大比増殖速度を低くすることができる。また、より高い温度を用いて、pUC、及びpMM1(Wong EM,Muesing MA,Polisky,B.1982 Proc Natl Acad Sci USA.79:3570−3574)、(Lin−Chao S,Chen WT,Wong TT.1992 MoI Microbio.6:3385−3393)及びランアウェイレプリコンRプラスミド等の複製起源を伴う選択的なプラスミド増幅を誘発することもできる。Hamann et.al.2000(Hamann CW,Nielsen J,Ingerslev E.2000 World Patent Application WO0028048)は、プラスミド生産を(低温度によって)低レベルに維持して、プラスミドDNA合成による増殖の遅延を避けるRプラスミドの生産方法を報告している。ここでは、宿主細胞集団が高くなると、温度変化によってプラスミド生産が誘発される。
【0017】
回分醗酵
回分醗酵は、簡単であるという主な利点を有する。培養期間を通して細胞増殖及びプラスミド生産に用いられる全栄養素が、植菌時に存在する。回分醗酵は、誘導期、指数増殖期、及び静止期を有する。好適な植菌材料(1から5%の培養液量)は、誘導期の長さを低減させる。指数増殖期間には、全栄養素が過剰であり;従って、この比増殖速度は、本質的にモノ−キネティックスによって予測される最大比増殖速度、μmaxになる。上述したように、増殖速度が減少することは、プラスミド生産にとって望ましい。回分醗酵において、増殖速度は、μmaxを減少させることによってのみ低減する。これは、より低い温度での増殖によって、及びグルコースの代わりにグリセロール上で増殖することによって達成される。グリセロールを用いた30℃での回分醗酵は、一般的にμmax<0.3h−1となり、このことは、有害な酢酸蓄積及びプラスミド不安定に関する増殖速度を妨ぐのに十分である(Thatcher DR,Hitchcock A,Hanak JAJ,Varley DL.2003 U.S Patent No6503738)。また、阻害されることのなくグルコースよりずっと高い濃度でグリセロールを、使用することができ、より高いバイオマス収率にすることができる。一般的に、60g/L DCW以上のバイオマス収率を、回分醗酵で得ることができる。
【0018】
流加培養醗酵
流加培養醗酵は、プラスミド生産に特に有益である。制限栄養素を制御して加えることは、率<μmaxでの増殖速度の制御を可能にする。また、流加培養醗酵は、収率をより高くする結果となる。流加培養醗酵の鍵は、完全に消費されるような比率で基質を供給することである。結果として、残留基質濃度は、ほぼ0であり、基質の最大転換が得られる。過剰の基質からの代謝過剰を回避し、阻害酢酸の形成を低減する。
【0019】
流加培養醗酵は、回分期で開始する。全ての非制限栄養素を含有する開始量の培地及び開始濃度の制限基質内に細胞を植菌する。細胞がこの開始量の基質を消費したら、制限栄養素を制御して与え始める。
【0020】
最も簡単で、最も効果的な供給戦略の一つは、指数関数的供給である。この方法によれば、フィードバック制御を必要とすることなくμmax未満の所定の比率で培地が増殖することを可能である。醗酵は、非阻害濃度の基質を含有する回分モードで開始する。細胞は、この基質がなくなるまでμmaxで増殖し、この時点で、栄養素供給が始まる。
【0021】
DO−stat法及びpH−stat法を実施するのは実に簡単である。というのも、最も標準的な醗酵装置システムは、溶存酸素とpHの監視を含むからである。溶存酸素(DO)及びpHの傾向は、基質が細胞に利用できるかどうかを示す。基質の消耗は、酸素摂取量を低減させ、培地中のDO濃度を上げる。また、pHは、代謝性の酸の消費によって上昇する。DO又はpHが設定閾値以上に上昇すると、供給が誘発される。増殖速度は、DO又はpH閾値を変更することによって調節することができる。
【0022】
例示的プラスミド醗酵プロセス
現行の収率の試験により、一般的なラボの振盪フラスコ培養は1から5mgのプラスミドDNA/Lの培地を作るが、コンピュータ制御の醗酵装置は、一般的に、10から250mg/LのプラスミドDNA/Lの培地を作ることが明らかである。
【0023】
Lahijani et al.(Lahijani R,Hulley G,Soriano G,Horn NA,Marquet M.1996 Human Gene Therapy 7:1971−1980)は、指数関数的供給と、37℃から42−45℃の温度シフトで醗酵における温度感受性単一ポイント突然変異(pUC起源)を伴うpBR322−誘導プラスミドを用いることを報告している。このことは、10Lの醗酵装置中220mg/Lのプラスミド収率を達成した。30℃での回分醗酵(pBR322誘導起源)中の突然変異のない同じプラスミドは、3mg/Lプラスミドを得たのみである。Friehs et al.(Friehs K,Flaschel E,Schleef M,Schmidt T.2003 US Patent No 6664078)は、DO−statフィードバック制御供給で、グリセロールイースト抽出培地を用いる流加培養プロセスを記載している。醗酵は、7.5Lの開始回分体積で開始した。攪拌を増して30%以上のDOを保った。DOが閾値設定点の45%に達した時に供給培地を投入した。培養は、41時間後に静止期に達し、60g/L DCW及び230mg/Lのプラスミドを得た。Chen(Chen,W.1999 U.S.Patent No 5955323)は、DO−statとpH−statフィードバック制御を組み合わせた半合成培地の流加培養プロセスを用いた。DOとpH閾値設定点は、それぞれ50%と7.2であった。DOが30%以下に落ちると、高い代謝活性から攪拌速度が以前の速度の割合まで増加した。7Lの培養装置中で、この戦略は、0.13h−1の比増殖速度、及び82から98mg/Lのプラスミド収率を導いた。Durland and Eastman,Supra,1998は、専売培地中で37℃での回分醗酵を報告している。彼らのプロセスによれば、一般的には130mg/Lの収率が得られ、250mg/Lの高い収率が得られた。
【0024】
先行技術を考慮しても、高純度プラスミドDNA生産についての費用対効果の大きい方法の必要性がある。上記の醗酵培地及びプロセスは、現在技術的に知られているものを組み込んでおり、増殖速度の減少及び高温でのプラスミド複製数誘発等のプラスミド生産性を改善している。これらのプロセスは、約200から250mgのプラスミドDNA/Lで横ばいになる。この低い収率は、プラスミドDNA生産プロセスを商品化する上でのコスト及び純度に負担を与える。スケールメリットは将来著しくDNAのコストを下げるが、所望のコストを達成するためには、この問題に対する更なる経済的解決策が必要である。その上、プラスミドDNA純度の国際基準は、大腸菌(E.coli)醗酵から同様に生産される組み換えタンパク質生産物について使用される基準と同じ又は非常に似ている。この基準は、確立された方法から得られる現在の純度を超えている。醗酵における収率(細胞ペーストのDNA/グラムのmg)の増加は、コストを減少させ、DNAの純度を上げるであろう(処理される材料の量を低減するからである)。
【0025】
本発明の開示
本発明は、プラスミド生産用の改善された回分及び流加培養プロセスを用いるDNAレプリコンの生産方法である。特に、流加培養醗酵方法が開示されており、ここでは、プラスミド含有大腸菌(E.coli)細胞が、流加培養期中に低い温度で増殖し、この増殖中は増殖速度が制限され、次いで、プラスミドを蓄積するために温度をアップシフト(up−shift)し、段階的に温度を高くして継続的に増殖させることによって、温度シフトと制限された増殖速度が、プラスミドの収率及び純度を改善する。好ましい実施例では、低減した増殖速度をほぼ2mg/L/OD以下のプラスミド収率を維持するように決定する。別の好ましい実施例では、この低減した温度はおよそ30℃である。別の好ましい実施例では、温度のアップシフトは、ほぼ36から45℃までである。別の好ましい実施例では、プラスミドはColE1誘導複製起源を含む。更にもう1つの好ましい実施例では、プラスミドはpUCGからpUCAまでの突然変異を含むpMB1複製起源を含む。更に別の実施例では、プラスミドは、VR1012バックボーンから誘導される。一の極めて好ましい実施例では、醗酵媒体が、実質的に半合成グリセロール媒体である。更に最後の好ましい実施例では、pBR322−誘導プラスミドは、増殖速度が流加培養期間で制限され、増殖がプラスミド生産物を継続して蓄積する間に、流加培養醗酵の大腸菌(E.coli)中で増殖する。要約すれば、これらのプロセスは、新規の増殖と誘発期の温度シフトとを組み合わせた、高い細胞密度培養戦略を特徴とする。DNAレプリコンを含有するpBR322−誘導起源の生産において、流加培養醗酵は、制限された細胞増殖速度で実施される。温度誘導DNAレプリコン(例えば、プラスミド含有pUC又はpMM1起源)の生産において、流加培養醗酵は、増殖期中の制限された細胞増殖速度と低減された温度で実施され;次いで、プラスミド生産は、温度をアップシフトすることによって誘発される。これらのプロセスは、プラスミドDNA醗酵収率を劇的に改善し、その一方で、従来技術のプロセスと比較して、プラスミドの完全性を維持又は改善する。
【0026】
本発明の簡単な概要
本発明の目的及び/又は対象は、プラスミドDNAの生産のための醗酵プロセスを提供することである。本発明のもう一つの対象及び/又は目的は、醗酵培養のプラスミドDNA生産収率を改善することである。本発明の更に別の対象及び/又は目的は、醗酵培養のプラスミドDNAの品質を改善することである。本発明の更に別の対象及び/又は目的は、精製プラスミドDNA中の不純物を低減することである。別の開示は、従来技術に記載されたプロセスと比較して改善された改善醗酵プロセスであり、バイオマス生産後のプラスミドレベルの誘導によってプラスミドの増加した収量;バイオマス生産中の低プラスミドレベルの維持による毒性又は不安定プラスミドと共に増加した収量及び完全性;プラスミドのニック(開環)又は直鎖バージョンの低減したレベルによってプラスミドの質を向上すること;プラスミドのパーセントモノマ(percent monomer)の増加によってプラスミドの質を向上すること;ロバスト、自動制御パラメータ及び供給を用いる簡略化した生産;増殖中に必要な増殖制御及び低減した酸素補給によって簡略化したスケーリング;ダウンストリームプロセシング内へのフィードストリームのプラスミドの濃縮レベルによるプラスミド精製後の不純物の低減したレベル;及び全ての動物性生成物誘導成分の除去によって、改善した標準コンプライアンス;によって改善される。
【0027】
本発明の更なる目的及び利点は、図面及び次の記載の考察から明らかになる。
【0028】
本発明の詳細な説明
図面を参照すると、図1は、NTC3019培地を用いた大腸菌(E.coli)中のpBR322−誘導プラスミド流加培養醗酵を示し、これは(a)NTC3019培地を用いた流加培養中の、大腸菌(E.coli)中のpBR322−誘導プラスミドの一般的な増殖及びプラスミド生産性プロファイルと;(b)高スーパーコイル状であり、ギザギザで開環したアイソフォームを含まないNTC3019培地流加培養醗酵プロセスによって生産されたプラスミドDNAと;を明らかにしている。
【0029】
図2では、NTC3018培地を用いた大腸菌(E.coli)中のpUCプラスミド(pW2.0)回分醗酵のプラスミド増殖及びプラスミド生産性プロファイルが示されている。回分醗酵中のpW2.0プラスミドは、57OD600の細胞密度に達し、230mgプラスミド/Lの収率であった。
【0030】
図3では、NTC3019培地を用いた大腸菌(E.coli)中のgWiz GFPプラスミド流加培養醗酵を示している:(a)gWiz GFPの流加培養醗酵プロファイルの増殖及び制御パラメータプロファイル(溶存酸素、温度、攪拌)と;(b)SYBR GREEN Iで染色された細胞の蛍光顕微鏡観察が、プラトウ(左)でフィラメンテーションを示すが、温度を33℃で低下させた後は、再開された増殖及びフィラメンテーションが低下した(右)。
【0031】
図4では、NTC3019培地(37℃又は42℃誘導)を用いた大腸菌(E.coli)中のgWiz GFP誘導プラスミド流加培養醗酵を示している:(a)35時間で、30→37℃の温度シフトを用いて、gWiz GFP/大腸菌(E.coli)DH5α醗酵の増殖及びプラスミド生産性プロファイルが明らかにされ、プラスミド収量が670mg/Lに達した;(b)35時間で、30→42℃の温度シフトを用いて、gWiz GFP/大腸菌(E.coli)DH5α醗酵の増殖及びプラスミド生産性プロファイルが明示されている。プラスミド収量は1100mg/Lに達した。
【0032】
図5では、誘導流加培養醗酵プロセスを図示する。
【0033】
定義
【0034】
ccc:閉環状
【0035】
ColE1誘導起源:欠失(例えば、pBR322誘導起源)及び/又は塩基交換(例えば、pMB1、pMM1、ColE1等由来のpMM5由来のpUC)によるColE1型プラスミド(例えば、pMB1、ColE1)から誘導された複製された起源
【0036】
DNAレプリコン:プラスミド、コスミド、細菌性人工染色体(BACs)バクテリオファージ、ウイルス性ベクタ、及びこれらのハイブリッド
【0037】
NTC3018醗酵培地:グリセロール半合成回分醗酵培地
【0038】
NTC3019醗酵培地:グリセロール半合成流加培養培地
【0039】
pDNA:プラスミドDNA
【0040】
pBR322−誘導起源:rop(repressor of primer:プライマの抑制体)遺伝子が欠失した、pBR322由来のpMB1起源
【0041】
プラスミド:プラスミド、コスミド、細菌性人工染色体(BACs)、バクテリオファージ、ウイルス性ベクタ、及びこれらのハイブリッド
【0042】
pUC起源:段階的な温度でコピー数を増加するGからAへの転位を有する、pBR322−誘導起源
【0043】
半合成グリセロール培地:複合窒素源(例えば、イースト抽出物、大豆抽出物)及びグリセロール炭素源を含む醗酵培地
【0044】
本発明は、機械的醗酵容器を用いたプラスミド、コスミド、細菌性人工染色体(BACs)、バクテリオファージ、ウイルス性ベクタ及びこれらのハイブリッド(本明細書では、集合的にプラスミドと称される)等の閉環状(ccc)組み換えDNA分子の製造方法に関する。
【0045】
従来技術の醗酵プロセスは、次善のプラスミド収率、品質(例えば、プラスミドの切断又は線形化)、拡張性の無さ(例えば、過剰な酸素供給要求)、及び限られた応用範囲(例えば、不安定又は毒性シークエンスを含有するプラスミドで使用する能力がない)のために最適ではない。本発明は、醗酵培養中にプラスミドDNAの収率及び純度を改善する方法である。誘発性流加培養醗酵プロセスを用いる高収率醗酵に対する費用効率のアプローチを発展させて、プラスミド収率及び純度を改善する。
【0046】
プラスミド製造プロセス及び培地の好ましい実施例
pBR322−誘導プラスミドを生産する1つの好ましい実施例では、流加培養醗酵を制限された細胞増殖速度で実施する。このプロセスは、プラスミドDNA醗酵収率を劇的に改善するにも関わらず、従来技術に記載されているプロセスと比較して、プラスミドの完全性を維持又は改善する。
【0047】
適度の複製プラスミド(例えば、ここではpBR誘導プラスミドと呼ばれるrop遺伝子が欠損したpBR322複製起源)を使用したこのプロセスを用いて、0.12hr−1の比増殖速度を維持するように制御して供給をしながら、37℃で自動流加培養醗酵プロセスを行って、250から450mg/Lのプラスミド収率及び120OD600の細胞密度を達成した。これは、報告されている収率(Lahijani et al.,Supra,1996)の100倍の改良である。この新規で劇的な改良の分子的な根拠は、知られていない。この機構に関わらず、その他のプラスミドの生産への本発明を適用することで、プラスミドの質を下げることなく醗酵生産性は上がるであろう。
【0048】
温度誘発性DNAレプリコン(例えば、プラスミド含有pUC又はpMM1起源)の生産についての1つの好ましい実施例では、増殖期中に、制限された細胞増殖速度及び低減した温度で、流加培養醗酵を実施し;次いで、アップシフトした温度でプラスミド生産を誘発する。このプロセスは、従来技術に記載されているプロセスと比較すると、プラスミドの完全性を維持又は改善しつつ、プラスミドDNA醗酵収率を劇的に改善する。
【0049】
本明細書で開示されているプラスミド(例えば、pUC起源)を含む高コピー起源の高収率プラスミド生産についてのこの新規な戦略は、予想外に高いプラスミド生産性とプラスミドの品質をもたらす。加えて、流加培養期中に増殖速度が減少する。このプロセスは、図5に示されている。この要素の新規な組み合わせは、ColE1誘導起源プラスミドの生産には適用されず、試験してみると、新しく予想外の改善された生産性となった。従って、我々は、低減した増殖速度と、1)増殖及びバイオマス生産のための低減した温度、及び2)プラスミド生産の誘発のための上昇させた温度と、の組み合わせに対する新しい使用(改善された生産性)を教示する。
【0050】
初期の低減した温度増殖期と後の高温度生産期との組み合わせが、全体的な増殖、バイオマス、及びプラスミド収率の改善をもたらすという発見は、新規で予想外であり、従来技術には教示されていない。Hamann et al.,Supra,2000は、増殖中のRプラスミドに関連する代謝負担を特に低減するために温度誘発醗酵を用いた。この戦略は、Rプラスミドの生産に利用されたが、プラスミド含有ColE1起源については教示していない。同様に、Hamann et al.,Supra,2000は、この戦略を用いて、全体の生産性を改善することを意図していない。従って、この要素の組み合わせは、従来技術では、収率の改善のために提言されたものではない。改善された生産性によって明らかであるように、この組み合わせの相乗効果は、部分同士の総和より大きい。同様に、この高収量醗酵は、これまで要求されてきたが解決されなかった必要性を満たすものである。何故ならここ10年以上もの間、この分野で記載された様々なプロセスは、高収率醗酵を教示していないからである。要約すれば、我々は、プラスミド生産を含むColE1起源についての、新しい組み合わせを教示する。このことは、改善されたプラスミド生産性の、予想外で驚くべき新しい使用を示す温度誘発と組み合わせたゆっくりとした増殖を具える。
【0051】
高コピープラスミド生産用誘発性流加培養醗酵プロセスの適用は、1100mg/LプラスミドDNAの収率、及び100のOD600をもたらす。これは、従来技術に規定されている醗酵プロセスで得られる収率の5倍の改良である。これらの全プロセスから精製されたDNAは、高品質であり、事実上100%スーパーコイル状である。我々は、プラスミド生産性を改善するために本明細書に記載された回分及び流加培養醗酵プロセスの使用を意図する。
【0052】
本明細書に記載された誘発性流加培養プロセスは、このプロセスのバイオマス生産期の間、低プラスミドレベル(VR1012誘導ベクタに対して<2mg/L/OD600)を維持し、バイオマス生産後は予想外で前例のない超高プラスミド生産(VR1012誘導ベクタに対して>6mg/L/OD600)を容易にした。細菌1グラム当たりのプラスミド収率が増加することは、最終生産物のより高い純度に直接つながるので、高比産出量は非常に望ましい。我々は、本発明の温度シフトを用いて、増殖の間中、プラスミドレベルを低く保ち(VR1012誘導ベクタに対して<2mg/L/OD600)、バイオマス生産後のプラスミド生産を高いレベル(VR1012誘導ベクタ>3mg/L/OD600)に誘発する。これらのレベルは、VR1012誘導ベクタの生産に対するガイドラインである。その他のプラスミドは、増殖期中に2mg/L/OD600よりも高い又は低いレベルで許容される。許容できる代謝負担、プラスミド安定性、及び温度シフト後の高い生産性を維持する増殖期中の最大プラスミドレベルは、平均的な当業者によって、各新しいプラスミドについて実験的に決定することができる。
【0053】
醗酵培地の代替の実施例
本発明を実施する上で、例示的な動物由来成分を含まない醗酵培地配合、NTC3018(回分)、NTC3019(流加培養)を用いることができる。これらの培地は、グリセロール炭素源、イースト抽出窒素源、及び微量金属、塩及び緩衝液を含む半合成増殖培地である。従来技術に記載された培地を、この培地で代用することも本明細書に記載されているような醗酵プロセスを用いた改善されたプラスミド生産性をもたらす。
【0054】
NTC3018及びNTC3019培地用の代替の非動物源の窒素源が考えられる。複合培地成分として、イースト抽出物は、窒素、アミノ酸、ビタミン、及び炭素を提供する。プラスミド生産中の細胞代謝の鍵となる複合培地(例えば、イースト又は大豆抽出物)のあり得る成分は、制限アミノ酸、ビタミン、微量無機物及び代替の炭素源を含む。一例として、我々は、代替イースト抽出調製物、大豆調製物(例えば、BD Bioscience社製のソイトン、又はファイトンペプトンを選択する)又はNTC3018及びNTC3019培地中のその他の植物調製物(例えば、Oxoid社製のエンドウの花のペプトン)の使用を意図している。
【0055】
培地の規定された要素の変型も意図している。例えば、リン酸塩又はマグネシウムの増加は、回分培地成分の増加として、或いは流加培養醗酵における供給に対する追加として意図している。醗酵の分野における当業者は、系統だった成分測定による培地の更なる最適化を行うことができる。
【0056】
ある栄養要求性細胞株を利用する場合、培地への添加物を意図している。例えば、proAB欠失を含むStbl2(InVitrogen Corp.)等の細胞株は、最大増殖を達成するためには、アミノ酸補給、又はプロリンが豊富な代替窒素源を必要とする。このような修飾は、醗酵の分野における平均的な当業者によって決定することができる。
【0057】
また、我々は、従来技術に記載されているプラスミド醗酵培地を用いた本明細書に記載されている回分及び流加培養醗酵プロセスの使用を意図している。これは、Soubrier(Soubrier F.2004 US Patent application 2004/0142452)によって開示されているような規定された培地を含む。本発明の流加培養醗酵プロセスで使用する好ましい培地は、NTC3019や、Lahijani et al.,Supra,1996、Friehs et al.,Supra,2003、Chen Supra,1999.及びUrthaler et al(Urthaler J,Roman N,Ascher C,Woehrer H.2005 US Patent Application 2005/0026177)に開示されている培地等の半合成供給によるグリセロール培地配合である。
【0058】
醗酵プロセスの代替的実施例
改善されたプラスミド生産のための流加培養及び回分プロセスが、本明細書に開示されている。これらは、新規な増殖及び誘発期温度シフトと組み合わせて、指数関数的供給戦略を特徴付けるものである。
【0059】
本発明の流加培養プロセスの実施において、我々は、フィードバック、フィードフォワード、及び栄養素供給の所定の制御を含む、増殖速度を低減するための種々の供給戦略を意図している。例えば、炭素制御した指数関数的供給戦略によって、栄養素供給を加えてもよい。我々は、許容できる範囲内で増殖速度を制御するための、この供給戦略の変型を予想している。好ましい増殖速度の範囲は、μ=0.05から0.3h−1である。好ましいターゲット増殖速度は、μ=0.12hr−1である。許容される増殖速度の範囲は、プラスミド特異的であり、当業者によって実験的に決定することができる。
【0060】
本発明の流加培養プロセスの実施において、我々は、生産時間を低減するための種々の回分戦略を意図している。回分期に含まれる半合成栄養素を調節して、回分期をより高い濃度又はバイオマス濃度にすることができる。より高いバイオマス濃度の場合に、流加培養期と比較して回分期で増殖がより早く進むので、全体的な発酵時間が低減される。我々は、プロセスの流加培養の開始を制御するための、この回分戦略の変型を予想している。開始のための流加培養期の好ましいOD600の範囲は、1から60である。許容できる流加培養開始OD600の範囲はプラスミド特異的であり、当業者によって実験的に決定することができる。
【0061】
本発明の誘導性流加培養及び回分プロセスの実施において、我々は、増殖期から誘導期への種々の温度シフト戦略を意図している。増殖期は25から37℃、好ましくは30から37℃の温度で実施することができる。高コピープラスミドについては、増殖期は、最も好ましくは30から32℃である。誘導期を33℃から45℃、好ましくは37から42℃の温度で実施することができる。
【0062】
温度シフト戦略を伴う誘導性プロセスの実施において、我々は、増殖期から誘導期へ移る時の指標として栄養素濃度、バイオマス濃度、又は光学密度を使用することを意図している。醗酵培養の規則的なサンプリングは、バイオマス又は光学密度測定を得るための材料を提供することができ、温度シフトは、あるバイオマス濃度で実施することができる。また、オンラインセンサを使用して、バイオマス濃度の継続的なモニタリングを行い、醗酵装置を設定して特定のバイオマス濃度で温度シフトを自動的に実施することができる。温度シフト戦略を伴う誘導流加培養プロセスの実施において、我々は、また、ある量の供給栄養素を加えると温度シフトが実行されることを意図している。
【0063】
温度シフト戦略を伴う誘導性プロセスの実施において、我々は、生産回数を最小限にするために増殖期中により高い温度を用いることを意図している。本明細書に記載されている誘導性流加培養プロセスは、このプロセスの増殖期の間中、低い(<2mg/L/OD600)プラスミドレベルを維持した。温度シフト後に、許容できる代謝負担、プラスミド安定性、及び生産性の向上を維持する増殖期中に利用できる最大温度は、プラスミド特異的であり、様々なプラスミドバックボーン間で異なる。例えば、いくつかのプラスミドは、2mg/L/OD600よりずっと高いレベルに許容できる。同様に、プラスミド生産性は、低い温度で変化し、従って、いくつかのプラスミドは、高いレベルのプラスミドとしては生産されない。これらの場合、我々は、増殖期中に、より高い温度を使用することを意図している。加えて、上昇した最大温度は、無制限の増殖とこれに相当するプラスミドコピー数の低下に起因して、開始回分期中で許容される(図5)。増殖期(回分及び流加培養成分)中に用いることができ、温度シフト後に許容できる代謝負担、プラスミド安定性、及び生産性の向上を維持する最大温度は、この分野の平均的な当業者によって実験的に決定することができる。
【0064】
プラスミド及び宿主菌株
我々は、高コピー、低コピー及び中程度のコピーのいずれかであり、温度誘導性であるか温度誘導性ではない種々の起源の複製を伴うプラスミドの生産において本発明を使用することを意図している。いくつかの好ましい複製の起源、及びこれらを組み込んだプラスミドが、表1に記されている。これらの起源の変更はこの分野において公知であり、また、使用を意図している。
【0065】
表1:複製起源
【0066】
代替宿主菌株が意図されている。大腸菌(E.coli)株DH5αは、プラスミド生産用に広く使用されている宿主である。その鍵となる品質は、クローンDNAの非特異的組み換えを最小限にするrecA突然変異、及びエンドヌクレアーゼIによってプラスミドの非特異的消化を除去するendA1突然変異を含む。DH5αに加えて、その他の様々な菌株が、プラスミド生産に好適である;例示的な大腸菌(E.coli)宿主菌株の非限定的リストを表2に示す。
【0067】
表2:宿主菌株
【0068】
DH5α、XL1−Blue、DH10B、JM109及びTop10は、プラスミド生産菌株として、非常に良く確立されている。Mach1、及びECOS101は、最近開発されたものであり、望ましいプラスミド生産宿主である。Stb12、GT116及びSure細胞はプラスミド含有不安定DNAの生産に利用されている。不安定DNAは、直接(例えば、レトロウイルスの長い末端反復)又は逆方向反復(例えば、shRNAパリンドローム)ZDNA、その他のような構造を含む。GT116のdcm遺伝子の削除は、免疫刺激であるダムメチル化(dam methylation)を解消する。このため、GT116の生産は、プラスミドDNAの免疫原性を低減する。類似の免疫原性の低減は、CpGメチラーゼを発現する菌株を使用して観察される。
【0069】
不安定プラスミドの生産
【0070】
我々は、また、不安定なシークエンスを含むプラスミドの生産に本発明を使用することを意図している。パリンドロームシークエンス、定方向又は逆方向反復塩基配列、及びシークエンスを形成するZDNAが不安定であり、大腸菌(E,coli)宿主によって除去、又は再編成される。いくつかの場合には、治療に使用するプラスミドは、不安定なシークエンス(AAV及びHIV等のウイルス性ベクタについての逆方向又は定方向反復塩基配列、ある治療用遺伝子についてセグメント又は三塩基反復を形成するZDNA)を含んでいなければならない。不安定シークエンスを含むプラスミドを維持するための現在行われている戦略は、安定化突然変異を用いた宿主細胞株を使用することである。いくつかの宿主は、これらのプラスミド、例えば、Sure細胞(Stratagene)、GT115(Invivogen)又はStb12及びStb14(Invitrogen)の増殖用に商業的に入手できる。Stb12及びStb14細胞株は、レトロウイルスベクタ等のベクタを含む定方向反復塩基配列の安定性を増進する未公表の突然変異を用い;この効果は、恐らくは減少したコピー数に起因して低下した温度で強化される。修復突然変異の特異的組み合わせは、プラスミド増殖を、特に低温で安定化することができる。Sure及びSure2細胞株は、UV修復(uvrC)及びSOS修復(umuC)欠損(LTRsを安定化するため)と関連した相同性組み換え欠損(recB、recJ)、及びZDNAを安定化するためのSbcC(及びrecJ)を有するこのような組み合わせを用いる。GT116細胞株は、パリンドロームシークエンスを安定化するためにSbcC及びSbcDを用いる。これらの菌株は、恐らくはプラスミドコピー数の低下に起因して、低温(即ち、30℃)でのみプラスミドを安定化するように機能する。この戦略は明らかに生産コストを上げる。本明細書に記載されている誘導性醗酵プロセスの使用は、集菌前の短期間にのみコピー数が増加する前に、安定化細胞株中の30℃の不安定プラスミドを増殖することができる。これは、不安定プラスミドの収量及び安定性(即ち、質)を最大にするであろう。
【0071】
DNA及びプラスミド生産
誘導性流加培養プロセスにおけるプラスミドDNA(>6mg/L/OD600)の実際の増加した収量についての根底にある機構は知られていない。遅い増殖及び低下した温度でのバイオマス生産中に、DNA凝縮剤(例えば、dps遺伝子生産物等のヒストン状タンパク質、又はその他のクロマチン結合タンパク質)に潜在的に起因する。
【0072】
誘導期中にDNA凝縮を変化させることで、許容プラスミドレベル又はコピー数を増加することによって、プラスミド収量を増やすことができる。DNAの凝縮度合は、2つの対立因子;クロマチンタンパク質の凝縮と、転写複合体の脱凝縮と、によって設定される。プラスミド凝縮は、プラスミドプロモータからの転写レベルによって影響を受けることがある。転写が少ないほど、より高い凝縮となり、潜在的により高い運搬能力となる。
【0073】
E.coli中で、DNA凝縮に含まれる多数のクロマチンタンパク質を同定した。これらの遺伝子生産物は、プラスミドとゲノムDNAを結合させる。ゲノムDNAの場合には、核様体内にDNAを凝縮する((Robinow C,Kellenberger E.1994 Microbiol Review 58:211−232)参照)。核様体の主要成分は、ヒストン状タンパク質HU、IHF、及びHN−S、StpA(HN−Sに関連して、約1/10レベルで発現される)及びDpsであり、これらは核様体内に均一に分配され、一方で、SeqA、CbpA、CbpB、Fis及びIciA等のその他のタンパク質はより少ない量であり、核様体内に不均一に分配され、調整機能を有する。遊離Rプラスミドタンパク質複合体は、3つの主要なタンパク質、23%のHN−S、23%のRNAポリメラーゼ、及び5%のHUを含む。これは、クロマチン関連遺伝子生産物が異なる培地、異なる細胞密度で、増殖期及び静止期中に様々に調整されるので、増殖期に依存して、恐らく変化するであろう。Fis、HU、HF−1は、一般的に、対数期により高度に発現し、一方で、IHF及びDpsは、静止期により高いレベルにある。Dpsは、静止期に液状生物結晶性複合体にDNAを凝縮して、ストレス耐性を改善する。増殖期中のHN−Sの過剰発現は、DNA凝縮と生存度の減少を引き起こす。細胞は、DNAが高度に凝縮される場合、より高度なプラスミド能力を有する。NTC誘導性醗酵プロセスにおいて、増殖期細胞は、誘導期細胞よりもプラスミドDNAに対する全体的な能力が低い。これは、許容プラスミドを増殖期よりも高いレベルにする、誘導期に存在するクロマチンタンパク質の組み合わせの相違に起因する。誘導期中のクロマチンタンパク質の比率の変化が、プラスミド凝縮、及び運搬能力を上げる。
【0074】
また、誘導期中のクロマチンタンパク質の比率の変化は、プラスミド複製率を上げる。例えば、p15A起源RNAIIプロモータからの発現であって、pMB1(pBR322)RNAIIプロモータからではない発現は、IHFによって表される;p15A RNAII転写は、IHF突然変異中で増加する。Dps及びHUは、非特異的DNA結合剤、HN−S、CbpA及びCbpBであり、湾曲したDNAと結合する。StpAは、HN−Sに関連し、より高い親和性でDNAと結合し、また、湾曲DNAと結合する。Fis、IHF、IciA及びseqAは、シークエンス特異的である。HN−Sは、湾曲DNAを含む多数のプロモータからの転写を抑制する。pMB1(pUC及びpBR322)のRNAIIプロモータは、ポリA及びポリTトラックを含み、これらのシークエンスは、湾曲DNAを形成する。静止期の転写(例えば、HN−S)を抑制する減少したレベルの湾曲DNA結合性クロマチンタンパク質は、RNAIIのRNAIへの転写の増加した比率に関連し、文書化静止期は、プラスミドコピー数を増やす。NTC醗酵プロセスの誘導期中に存在するクロマチンタンパク質の組成の変化(例えば、HN−Sの更なる減少)は、pUC等のpMB1を有するプラスミドコピー数の増加をもたらす。
【0075】
大腸菌(E.coli)中で発現する場合、異種DNA凝縮体、例えば、バシラス(Bacillus)属の酸可溶性胞子タンパク質は、プラスミド収量を上げるための有益なDNA凝縮体でもある。例えば、大腸菌(E.coli)中の枯草菌(B.subtilis)酸可溶性小タンパク質の発現は、DNAに起因する(Setlow B,Hand AR,and Setlow P.JBacteriol.173:1642−1653)。
【0076】
また、プロセス変化は、DNA凝縮の効果を通じて収量を改善する。Dpsは、マグネシウム(Mg++)濃度;Dpsの存在は、DNA凝縮をもたらさない;密集した結晶性DNA:Mg++濃度が閾値以下に下がると、Dps複合体を形成する[(Frenkiel−Krispin D,Levin−Zaidman S,Shimoni E,Wolf SG,Wachtel EJ,Arad T,Finkel SE,Kolter R,Minsky A.2001 EMBO J.20:1184−1191)参照];によって調整される。形態学的に、この複合体は、静止期中にクロラムフェニコールを添加することによって誘導されるのに似ている。増殖する培地への0.2mMのスペルミジンの添加は、Dpsの不存在下でDNA凝縮を加速する。リン酸塩欠乏は、恐らくスペルミジンに対するスレオニン及びアルギニンの分解の促進を介して同様の効果を有する。2価カチオンのレベルを変更(例えば、Mg++、外因性添加又は減少による)するための、又は正電荷ポリアミン(例えば、スペルミジン、細菌合成の外因性添加又は制御による)の変化を変更するための誘導期中の醗酵組成又は状態への変化によって、プラスミド収量を増やすことができ、この変更は、培地の交換を意図する。
【0077】
我々は、更なる収量の増加が、プラスミドDNAの更なる凝縮によって得られることを意図している。これは、醗酵プロセス中に、スペルミン生産又はdpsタンパク質生産等の宿主菌株DNA凝縮剤の生産を増やす、供給(例えば、ポリエチレンイミン、スペルミジン、スペルミン)又は菌株組み換えへのDNA凝縮剤の添加によって達成される。このような菌株組み換えは、関連する遺伝子生産物が、醗酵プロセス中に減少することを可能にする変更になり得る。
【0078】
誘導性プロセスの実施において、我々は、代替戦略を用いて、増殖期中に低レベルでプラスミドコピー数を維持することを意図している。例えば、低温での増殖に加えて、増殖期中に組み込むことができるコピー数を減らすためのその他の機構が存在する。例えば、醗酵中の溶存酸素の減少は、プラスミドコピー数を減少させるために示される (Carnes AE,2005 BioProcess International 3:9,in press)。
【0079】
最終産物純度の改善
我々は、例示的なプラスミド精製プロセス中に記載された醗酵培地からのプラスミド濃縮供給ストリームを用いることを意図している。このようなプロセスは、技術的によく知られている。高収量醗酵と例示的な精製プロセスの組み合わせは、費用対効果のある方法論を提供し、ゲノミックDNAを更に遺伝子治療及びDNAワクチン接種適用に許容されるレベルにまで減らす。
【0080】
例
本発明の方法を、以下の例において更に説明する。これらは、例示によって提供され、本発明の範囲を限定するものではない。
【0081】
例1:NTC3018及びNTC3019醗酵培地
次の基準はプラスミドDNAを製造するための最適化醗酵プロセスの評価として確立された:
1) プラスミドの高い比収量(g細胞質量当たりのmgプラスミドDNA);
2) 高バイオマス収量;
3) 高度のスーパーコイル状のプラスミドを保つであろう;
4) ダウンストリームプロセシング中に生じる問題が最小になる;
5) 調整要求条件を満足する;
6) プラスミド構造を保持する(例えば、遺伝子欠失がない、又はその他の再配列);
【0082】
高い比プラスミド収量、高バイオマス収量、及び高プラスミド品質をサポートするように、培養培地を調整した。回分醗酵培地は、比増殖速度を減らすように構成した。低減した増殖速度の使用は、より高いプラスミドコピー数とより良いプラスミド安定性と関連していた。流加培養醗酵中に、制限栄養素の供給によって、増殖速度を0.12hr−1に制御した。加えて、培地に使用した全組成は、良く特徴付けられており、認定された動物由来成分が含まれていなかった。
【0083】
NTC3018(回分)及びNTC3019(流加培養)培地は、回分及び流加培養プロセス培地の双方の多くの成分用に最適化されている。例えば、グリセロールを、炭素源として使用して、増殖速度を低減している。イースト抽出物は、窒素源として使用される。大腸菌(E.coli)生産菌株の所定の条件に基づいて、微量金属及びMgSO4濃度を最適化した。
【0084】
例2:pBR322−誘導プラスミドを用いたNTC3019培地流加培養
37℃での、New Brunswick BioFlo 110 fermentor中で流加培養醗酵を実施した。30%の水酸化アンモニウム又は10%のリン酸を自動的に添加することによって、pHを制御した。窒素ガス散布によって0%に、及び空気飽和で100%に溶存酸素プローブを調節した。容器を1VVMで通気し、溶存酸素を攪拌の比例積分制御によって30%に維持した。また、約20OD600以上の細胞密度で、30%の飽和を維持するためにO2補給を必要とした。
【0085】
LBプラス50μg/mlカナマイシン内に植菌した単一の遊離コロニーから種培養を開始し、37℃で培養した。中間指数関数期(0.5から1.5OD600)で、種培養を用いて、醗酵装置に対して1%の接種材料を提供した。
【0086】
流加培養中に、炭素の制限指数関数的供給戦略に応じて、半合成供給栄養素を添加した。要約すれば、回分期中にμmaxの比増殖速度で炭素基質の初期量が消費される。炭素基質が消耗すると、流加培養期が始まり、供給栄養素が、次の式によって決定される割合で、自動的に加えられる(Carnes,Supra,2005):
【0087】
【0088】
ここで、
μ=流加培養期中の所望の比増殖速度、
XB=回分期の終わりのバイオマス濃度、gDCW/L、
VB=培地の初期液体体積、L、
Sf=栄養素供給培地の制限基質濃度、g/L、
YX/S=基質からのバイオマスの収量係数、g/g
t=流加培養期の開始からの時間、
である。
【0089】
一般的に、いくつかの独立したカナマイシン耐性pBR322−誘導プラスミドを用いたNTC3019培地の流加培養醗酵は、100から120OD600の細胞密度、又は1リットル当たり55から65g乾燥細胞重量に達する(図1)。プラスミドは、平均260mg/Lであり、430mg/L程度に高かった。比較すると、pBR322−誘導プラスミドを用いた公開されている醗酵収量は、約3から4mg/Lである(Lahijani et ai,Supra,1996)。
【0090】
重要なことには、比プラスミド収量が非常に高く、一般的に2.5から3.8mg/L/OD600の間であり、かなり高いコピーpUC起源プラスミドを用いるその他の醗酵培地/プロセスで観察されたレベルを十分越えている(表3)。比収量mg/L/OD600に関する発現プラスミド収量は、全細胞質量と比較したプラスミドの量を示す。細菌1グラム当たりの増加したプラスミドの収量が、より高い最終産物純度を直接導くので、高い比収量は非常に望ましい。
【0091】
表3:公開高収量醗酵プロセスと比プラスミド収量の比較
【0092】
これは、NTC3019醗酵培地が、従来技術に記載された培地及びプロセスと比較して、中程度のコピー数のプラスミド(例えば、rop欠失を有するpBR322)の醗酵収量を、劇的に促進することを明示している。この効果は、プラスミド特異的ではない。
【0093】
これらのプロセスで精製したDNAは高品質であり、欠失又はその他の再配列を検出することなく、実質的に100%スーパーコイル状である。同様に、このプロセスで、細胞を使用して1グラムスケールでDNA精製を実施した。これは、NTC3019流加培養培地で実施された醗酵が、大きいスケールのダウンストリームプロセシングに敏感に反応することを明示している。
【0094】
例3:高コピー遺伝子治療プラスミドを用いたNTC3018培養回分醗酵
pUC起源プラスミドを用いたNTC3018培地回分培養を実施した。以下のプラスミド含有pUC起源を使用した:
1)pW2.0、変質ポリリンカーシークエンスを有するpUC19の誘導体
2)pMaxGFP
3)pEGFP−C1
【0095】
37℃の、New Brunswick BioFlo 110 fermentor中で回分醗酵を実施した。30%水酸化アンモニウム又は10%リン酸の自動添加によって、pHを制御した。窒素ガス散布によって0%に、及び空気飽和で100%に溶存酸素プローブを調節した。容器を1VVMで通気し、溶存酸素を攪拌の比例積分制御によって30%に維持した。また、約20OD600以上の細胞密度で、O2補給を30%の飽和を維持するためには、O2の補給が必要であった。
【0096】
LBプラス50μg/mlカナマイシン中に植菌した単一遊離コロニーから種培養を開始し、37℃で培養した。中間指数関数期(0.5から1.5OD600)で、種培養を用いて、醗酵装置に対して1%の接種材料を提供した。
【0097】
全ての醗酵を37℃で実施した。pW2.0について、プラスミドコピー数を醗酵の遅い42℃での増殖によって誘発した。pUCプラスミドを用いた回分醗酵は、16から57OD600の細胞密度に達したのみであった。しかしながら、比プラスミド収量の結果は有望である。pEGFP−C1についての最終収量は、細胞密度が56OD600であり、収量が163mgプラスミド/L(2.9mg/L/OD600)であり、pMaxGFPについては、細胞密度が16OD600であり、収量が84mgプラスミド/L(5.3mg/L/OD600)であり、pW2.0については、細胞密度が57OD600であり、収量が230mgプラスミド/L(4.0mg/L/OD600;図2)であった。
【0098】
これらのプロセスから精製したDNAは高品質であり、欠失又はその他の再配列を検出することなく、実質的に100%スーパーコイル状である。同様に、このプロセスで細胞を使用して実質0.5グラムスケールでDNA精製を実施した。これは、NTC3018回分培地で実施された醗酵が、大きいスケールのダウンストリームプロセシングに敏感に反応することを明示している。
【0099】
例4:高コピー遺伝子治療プラスミドを用いたNTC3019培地の流加培養醗酵
Vical VR1012ベクタの誘導体を含有するGFP遺伝子であるプラスミドgWiz GFP(Gene Therapy Systems)を、流加培養醗酵評価用に選択した。これは、5757bpの大きさの広く用いられているDNAワクチンプラスミド含有カナマイシン耐性(kanR)pUC起源である。このプラスミドgWiz GFP(Gene Therapy Systems)を、大腸菌(E.coli)DH5αに変質させた。また、pMaxGFPを、流加培養培地内で試験し;両方のプラスミドについて、同様の結果を得た。
【0100】
これらのプラスミドの生産にNTC3019培地を使用したときに、2つの問題に直面した。増殖がpBR322−誘導プラスミドを用いてうまく行われているときに、37℃でのgWiz GFPプラスミド培地の増殖時に第1の問題が発見された。pUCプラスミド培地では、細胞増殖は15OD600のあたりで止まった。蛍光顕微鏡は、細胞分裂の阻害を意味する広範なフィラメンテーションを示した。フィラメントが実質的に溶解するので、このことは致命的である(Arends SJR, Weiss,DS.2004 J Bacteriol 186:880−884)。例えば、図3(A)は、プラスミドgWiz GFPを用いた流加培養醗酵を示す。細胞増殖は遅くなり、15OD600の早い段階で静止期に入るように見えた。プラスミド収量分析は、細胞増殖が停止し始めたときに、2.7mg/L/OD600の上昇比プラスミド収量を示した。pUCプラスミドは、30℃の場合と比較すると、42℃でのコピー数の30から40倍の増加を示す温度感受性点突然変異を含む(Lin−Chao et al,1992)。次いで、プラスミドコピー数を減らし、細胞の代謝負荷を軽減しようとして、温度を33℃に下げた。温度の低下後に、プラスミドは1.6mg/L/OD600に下がり、細胞増殖が再開した。興味深いことに、比プラスミド収量は再び徐々に増加し、供給栄養素が加えられていたにも関わらず、増殖は、予想通りに>100OD600に増殖することなく、約60OD600で静止期に入った。
【0101】
図3(B)は、37℃で発生する細胞フィラメンテーションを示す。温度を33℃に下げた後に、増殖細胞のサンプルは、より少ないフィラメンテーションを示した。
【0102】
考えられる説明は、37℃で増殖が開始した時に、細胞母集団は、フィラメンテーション及び生存率の低下から完全に回復しないことであった。このことを試験するために、同じプラスミドで、引き続いて2つの流加培養醗酵を、33℃で完全に実施した。両醗酵において、細胞密度<60OD600で培養がピークに達した。
【0103】
これらの醗酵からのバイオマス及びプラスミドDNA収量データは、比増殖速度の低下と、細胞増殖阻害前の比プラスミド収量の急激な上昇を示す。このような高いレベルへのプラスミド含有量の急激な増加は予想されておらず、増殖速度の低下を招く細胞母集団上の代謝負担を認識する。しかしながら、増殖速度の低下は、しばしばプラスミドコピー数の増加を導くことも示されている(Satyagal VN,Agrawal P.1989 Biotechnol.Bioeng.33:1135−1144)、(Seo JH,Bailey JE.1985 Biotechnol.Bioeng.27:1668−1674)。予期しない増加比プラスミド収量が、増殖速度の低下を引き起こしているのか、又は予期しない低下した比プラスミド収量が、増殖速度の増加を引き起こすのか、或いは、それぞれが、複合してどちらかを引き起こすかは、明らかではない。
【0104】
例5:NTC3019培地を用いた高コピープラスミドの高収量生産のための誘導流加培養プロセス
【0105】
33℃及び37℃での醗酵の結果(例5)に基づいて、pUC起源プラスミドを用いた流加培養モードで観察される予期しないプラスミド増加を克服するように戦略を構成した。DH5α中のプラスミドgWiz GFPを、誘導流加培養プロセスで用いた。NTC3019流加培養醗酵は、30℃で60OD600まで培養することを除いて、例5で説明されるように実施した。この時に、温度を37℃にシフトした。驚くべき結果が、図4Aに示されている。60OD600まで30℃で増殖することで、増殖阻止問題がなくなり、この培養は、全プラスミド収量が670mg/Lの100OD600を最終的に上回った。このプロセスのサンプルから精製されたDNAは高品質であり、欠失又はその他の再配列を検出することなく、実質的に100%スーパーコイル状である。
【0106】
2mg/L/OD600以下に維持して、増殖期を通じて低く維持される温度シフトの前に、プラスミドが産生する。このことは、33℃又は37℃の醗酵の結果と対照的である。注目すべきことに、温度シフト後の比プラスミド収量は、6.5mg/L/OD600と非常に高くなり、その他の醗酵培地/プロセスを用いて観察されたレベルを十分に越えている(表3)。増殖期を通して30℃での醗酵させ、42℃へシフトすることは、gWiz GFPを伴う1.1gm/L(11mg/L/OD600)の生産量をもたらす結果となった(図4B)。生産性安定期は、細胞の大多数が生き残るので、大量の細胞死に関連していない。
【0107】
また、流加培養期間を低減する(回分期をより高いOD600にすることによる)ためのNTC3019培地(グリセロール、イースト抽出物、及び回分培地のマグネシウムの4倍の増加)の修飾も、42℃での誘導後に同様の高プラスミド収量を産生し、プラスミド誘導のロスなしで、より高いOD600で流加培養期を開始できることを明示している。
【0108】
様々な抗生物質抵抗性遺伝子及び原核生物要素の配向性を含む、種々のpUC起源バックボーンを有する多数の異なるプラスミドが、DH5α中での30℃から42℃の誘導性プロセスを用いて、NTC3019培地中の0.5gm/Lよりも大きい収量で生産された。これらの結果は、この誘導性プロセスが、特定のプラスミドに特異的でないことを明示している。
【0109】
また、gWiz−GFPプラスミドが、DH1細胞株中で、30℃から42℃での誘導性プロセスを用いて、NTC3019培地の0.5gm/Lよりも大きい収量で生産された。この結果は誘導性プロセスが、特定の大腸菌株(E.coli strain)に特異的でないことを明示している。
【0110】
同様に、このプロセスからの細胞を用いて、1グラムスケールでのDNA精製を実施した。このことは、NTC3019流加培養培地で実施された誘導性醗酵が、大きいスケールのダウンストリームプロセシングに敏感に反応することを明示している。
【0111】
比収量(mg/L/OD600)の用語でのプラスミド収量の表現は、全細胞質量に対するプラスミドの量を示す。本明細書に記載された誘導性流加培養プロセスは、このプロセスの増殖期を通してプラスミドレベルを低く(<2mg/L/OD600)維持し、バイオマス生産後の、前代未聞の非常に高いプラスミド生産(6から11mg/L/OD600)を容易なものにした。細菌の1グラム当たりのプラスミド収量の増加は、直接的に最終産物純度を高くするので、高い比収量は非常に望ましい。
【0112】
これらの結果は、プラスミドDNA生産性と品質を改善する、本発明の流加培養及び回分醗酵プロセスの一般的な使用を明示する。
【0113】
このように、本発明の生産プロセスが、改善されたプラスミド生産方法を提供するものであることを、読者は理解するであろう。
【0114】
上記記述が多くの特異性を含んでいる一方で、これらは、本発明の限定として解釈するべきではなく、むしろ本発明の好ましい一の実施例の例示として解釈するべきである。その他の多くの変形が可能である。例えば、誘導性流加培養プロセスを、回分プロセスと一体化して、栄養素がなくなるまでNTC3018培養培地中で醗酵を行うことができる。ここでは、NTC3019由来の流加培養培地及び誘導は同時に開始される。いくつかのプラスミドを用いて、この実施例においては、プラスミドコピー数をより高い増殖速度で低下するため、増殖期を37℃以上で実施することができる。増殖期中に細胞分裂を可能にし、硫化培養期中にプラスミド誘導性を保持する最適な温度は、当業者が決定できる。従って、本発明の範囲は、説明した実施例によってではなく、添付の特許請求の範囲及びこれらの法律的均等物によって決定されるべきである。
【図面の簡単な説明】
【0115】
【図1】図1は、NTC3019培地を用いた大腸菌(E.coli)内のpBR322−誘導プラスミド流加培養醗酵を示す。
【図2】図2は、NTC3018培地を用いた大腸菌(E.coli)内のpUCプラスミド回分醗酵を示す。
【図3】図3は、NTC3019培地を用いた大腸菌(E.coli)内のgWiz GFPプラスミド流加培養醗酵を示す。
【図4】図4は、NTC3019培地を用いた大腸菌(E.coli)内のgWiz GFP誘導流加培養醗酵を示す。
【図5】図5は、誘導性流加培養醗酵プロセスを示す。
【特許請求の範囲】
【請求項1】
閉環状スーパーコイル状プラスミドDNAの流加培養醗酵生産方法において:A)流加培養期中に、低下させた温度でプラスミド、コスミド、又は細菌性人工染色体複製を培養するステップと;B)前記流加培養期中に、増殖速度を制限するステップと;C)温度上昇シフトによってプラスミド生産を誘導するステップと;D)段階的温度で増殖を継続して、プラスミド産物を蓄積するステップと;を具え、
前記方法がプラスミド収量を増やすことを特徴とする方法。
【請求項2】
請求項1に記載の方法において、前記流加培養期中の低下した温度が、プラスミド収量をほぼ2mg/L/OD600以下に維持するように決定されることを特徴とする方法。
【請求項3】
請求項1に記載の方法において、前記流加培養期中の低下した温度が、ほぼ30℃であることを特徴とする方法。
【請求項4】
請求項1に記載の方法において、前記温度シフトが、36から45℃の範囲であることを特徴とする方法。
【請求項5】
請求項1に記載の方法において、前記プラスミドが、ColE1−誘導複製起源を含むことを特徴とする方法。
【請求項6】
請求項1に記載の方法において、前記プラスミドが、pUCGからpUCAの突然変異を含むpMB1複製起源を含むことを特徴とする方法。
【請求項7】
請求項1に記載の方法において、前記プラスミドが、VR1012バックボーンから誘導されることを特徴とする方法。
【請求項8】
請求項1に記載の方法において、前記醗酵培地が、実質的に半合成グリセロール培地であることを特徴とする方法。
【請求項9】
閉環状スーパーコイル状プラスミドDNAの醗酵生産方法において:A)流加培養醗酵培地中にpBR322−誘導プラスミド、コスミド、又は細菌性人工染色体複製を含む細菌性細胞を増殖するステップと;B)流加培養期中の増殖速度を制限するステップと;C)増殖を継続して、プラスミド産物を蓄積するステップと;を具え、
前記方法がプラスミド収量を増やすことを特徴とする方法。
【請求項10】
請求項9に記載の方法において、前記醗酵培地が、実質的に半合成グリセロール培地であることを特徴とする方法。
【請求項1】
閉環状スーパーコイル状プラスミドDNAの流加培養醗酵生産方法において:A)流加培養期中に、低下させた温度でプラスミド、コスミド、又は細菌性人工染色体複製を培養するステップと;B)前記流加培養期中に、増殖速度を制限するステップと;C)温度上昇シフトによってプラスミド生産を誘導するステップと;D)段階的温度で増殖を継続して、プラスミド産物を蓄積するステップと;を具え、
前記方法がプラスミド収量を増やすことを特徴とする方法。
【請求項2】
請求項1に記載の方法において、前記流加培養期中の低下した温度が、プラスミド収量をほぼ2mg/L/OD600以下に維持するように決定されることを特徴とする方法。
【請求項3】
請求項1に記載の方法において、前記流加培養期中の低下した温度が、ほぼ30℃であることを特徴とする方法。
【請求項4】
請求項1に記載の方法において、前記温度シフトが、36から45℃の範囲であることを特徴とする方法。
【請求項5】
請求項1に記載の方法において、前記プラスミドが、ColE1−誘導複製起源を含むことを特徴とする方法。
【請求項6】
請求項1に記載の方法において、前記プラスミドが、pUCGからpUCAの突然変異を含むpMB1複製起源を含むことを特徴とする方法。
【請求項7】
請求項1に記載の方法において、前記プラスミドが、VR1012バックボーンから誘導されることを特徴とする方法。
【請求項8】
請求項1に記載の方法において、前記醗酵培地が、実質的に半合成グリセロール培地であることを特徴とする方法。
【請求項9】
閉環状スーパーコイル状プラスミドDNAの醗酵生産方法において:A)流加培養醗酵培地中にpBR322−誘導プラスミド、コスミド、又は細菌性人工染色体複製を含む細菌性細胞を増殖するステップと;B)流加培養期中の増殖速度を制限するステップと;C)増殖を継続して、プラスミド産物を蓄積するステップと;を具え、
前記方法がプラスミド収量を増やすことを特徴とする方法。
【請求項10】
請求項9に記載の方法において、前記醗酵培地が、実質的に半合成グリセロール培地であることを特徴とする方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2008−510460(P2008−510460A)
【公表日】平成20年4月10日(2008.4.10)
【国際特許分類】
【出願番号】特願2007−527970(P2007−527970)
【出願日】平成17年8月16日(2005.8.16)
【国際出願番号】PCT/US2005/029238
【国際公開番号】WO2006/023546
【国際公開日】平成18年3月2日(2006.3.2)
【出願人】(507048765)ネイチャー テクノロジー コーポレーション (2)
【Fターム(参考)】
【公表日】平成20年4月10日(2008.4.10)
【国際特許分類】
【出願日】平成17年8月16日(2005.8.16)
【国際出願番号】PCT/US2005/029238
【国際公開番号】WO2006/023546
【国際公開日】平成18年3月2日(2006.3.2)
【出願人】(507048765)ネイチャー テクノロジー コーポレーション (2)
【Fターム(参考)】
[ Back to top ]