
プラス一本鎖RNAウイルスへの感染の予防又は治療のためのヒト大型2’,5’−オリゴアデニル酸合成酵素OAS3
プラス一本鎖RNAウイルスへの感染の診断、予防及び治療のため、並びにプラス一本鎖RNAウイルス関連疾患へのヒトの遺伝的感受性の予測のためのヒト大型2',5'-オリゴアデニル酸合成酵素(OAS3)の使用。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、プラス一本鎖RNAウイルスへの感染の予防又は治療のための医薬品としてのヒト大型(large form) 2',5'-オリゴアデニル酸合成酵素(OAS3)に関する。
本発明は、プラス一本鎖RNAウイルスへの感染に対する遺伝的感受性を決定するためのマーカーとしてのヒト大型2',5'-オリゴアデニル酸合成酵素(OAS3)にも関する。
【背景技術】
【0002】
フラビウイルス科(Flaviviridae)及びトガウイルス科(Togaviridae)は、世界的にヒト及び動物の健康に影響し得る病原体を含む2つのプラス一本鎖(ss)RNAウイルスの科である。これらのウイルスの例は、フラビウイルス(Flavivirus)属(フラビウイルス科)のデング、黄熱(YF)、日本脳炎(JE)及びダニ媒介脳炎(TBE)の抗原性群(antigenic complexes)のメンバー、アルファウイルス(Alphavirus)属(トガウイルス科)の東部ウマ脳炎(EEE)/ベネズエラウマ脳炎(VEE)、セムリキ森林(SF)及びシンドビス(SIN)群のメンバー、並びにヘパシウイルス(Hepacivirus)属(フラビウイルス科)のメンバー、例えばC型肝炎(HCV)及びG型肝炎(HGV)ウイルスを含む。
【0003】
デングウイルス(DV; フラビウイルス属のDEN抗原性群)は、熱帯のほとんどの都市中心において流行性である。なぜなら、都市化の劇的な増加が、蚊媒介デング疾患の伝達の増加に理想的な条件を作り出したからである。デングウイルスの4つの血清型(DV-1〜DV-4)は、媒介体である蚊のネッタイシマカ(Ae. aegypti)によりヒトに伝達される。DV感染は、流感様疾患(デング熱、DF)から、デングショック症候群(DSS)及び死に発展し得るデング出血熱(DHF)までにわたる連続する疾病をもたらす。現在までに、デング疾病は、推定100百万件のヒトにおける最も重要なアルボウイルス症であり、各年に500,000件を超えるDHF/DSSが発生し、約25,000件の死に至る件を含み、これは主に15歳未満の子供である。DHF/DSSの高い頻度の流行病は、アジア及び南アメリカにおいて地理的に拡大し続けている。健康及び経済的な影響の増加にもかかわらず、デング疾患の病因は、現在のところほとんど理解されていない。抗デングワクチン又はデングウイルス感染の治療のための特定の療法は利用可能でない。
【0004】
西ナイルウイルス(WNV; フラビウイルス属のJE抗原性群)は、蚊(クレクス(Culex)種)及び鳥類を含む天然の伝達サイクルで循環し、ウマ及びヒトが偶生宿主である。人畜共通感染症のWNVは、1998年にイスラエルで高度に神経浸潤性の株(WNウイルスI型系統のクレードIaからの変種Isr98/NY99)が出現したので(Ceccaldiら, FEMS Microbiol. Lett., 2004, 233, 1〜6)、北米、中東及び欧州で主な健康問題になった。WNVは、中枢神経系に感染し、広い範囲の動物種においてウイルス性脳炎を引き起こす。ヒトにおいては、臨床感染は、無併発性の西ナイル熱から致死的な髄膜脳炎までの範囲の重症度にわたり得る。WNVの出現は、ヒトにおける感染の重症度の劇的な増加に関連し、ウイルス性脳炎が公衆衛生の問題として注目を集めることとなった。過去10年間に、WNVは、西半球及びカリブ海諸国に広がった。WNVの米国での大発生は、数千人の患者を伴い、重篤な神経疾患(髄膜脳炎及び灰白髄炎様症候群)及び数百名の関連する死者の原因となった。WNVの蚊媒介伝達は主な形態であるが、輸血、臓器提供及び胎児への経胎盤伝達により伝達されるWNV感染も認められた。WNV関連疾患のためのワクチン又は抗ウイルス療法は利用可能でない。
【0005】
チクングンヤウイルス(CHIKV; アルファウイルスのSF群)は、アフリカ、東南アジア、インド及び西太平洋に広まっており、多数の流行病がこれらの地域で報告されている。臨床的には、CHIKVへの感染は、発熱、発疹及び強く、無気力にさせ、ときに持続的な関節痛をもたらす。2005〜06年に、CHIKウイルスは、インド洋の南、特にレユニオン諸島(フランス)に広がり、ここでの大発生は、数十万の患者を伴った。より最近では、このウイルスはイタリアの東部で出現し、ここでは200名を超える人が感染した。
【0006】
C型肝炎ウイルス(HCV)感染は、世界的に一般的である。世界の人口の約3%がHCVを有し、欧州だけで約4百万のキャリアがいると見積もられる。これらの個体の20〜30%が肝硬変、及び肝細胞癌のような長期の続発症を発症する。リバビリンと組み合わせたPEG化インターフェロンアルファ(IFN-α)を用いるHCV感染の治療は、HCVウイルス遺伝子型2又は3に感染した患者において持続した応答を達成できる(80%)が、HCVウイルス遺伝子型1に感染した患者においては、応答率がかなり低い(42%)。
【0007】
I型インターフェロン(IFN-α/β)により媒介される先天的な抗ウイルス機構は、ウイルス複製を制限する宿主細胞防御の最も重要な経路の可能性がある。実際に、IFN-α/βは、特異的シグナル伝達経路の活性化を引き起こして、IFN-活性化遺伝子(ISG)の誘導を導くことができ、これは抗ウイルス状態の確立を担う。単細胞においてRNAウイルスの複製に影響すると考えられるISGは、RNA特異的アデノシンデアミナーゼ(ADAR)、ミクソウイルス耐性(Mx)ファミリーのタンパク質、2本鎖RNA依存性タンパク質キナーゼ(PKR)、及びエンドリボヌクレアーゼRNase Lと関連する2',5'-オリゴアデニル酸合成酵素(2',5'-OAS又はOAS)ファミリーである。
【0008】
OAS/RNase L系は、確立された内因性抗ウイルス経路において重要な役割を有することが知られているRNA分解経路である。酵素的に活性なOASが、アクチベーターである2本鎖(ds)ウイルスRNAに結合すると、2'-5'結合オリゴアデニル酸(2-5A)の生成をもたらす。潜在性の単量体RNase Lは、2-5Aオリゴマーへの結合により誘導されるホモ二量体形成により酵素的に活性化される。一旦活性化されると、RNase Lは、mRNA及びウイルスRNAを含む1本鎖RNA分子を分解し、ウイルス複製を抑制する(Silverman, J. Virol. 81: 12720, 2007)。
【0009】
ヒトOASは、以下の順序での染色体12q24.2上の3つの密接に結合した遺伝子によりコードされる酵素のファミリーである:小型(OAS1, p40/46)、中型(OAS2, p69/71)及び大型(OAS3, p100)のOASアイソフォーム(Hovnanianら, Genomics, 1998, 52, 267〜277; Rebouillat, D.及びHovanessian, A.G., Journal of Interferon and Cytokine Research, 1999, 19, 295〜308; Rebouillatら, Genomics, 2000, 70, 232〜240; Justesenら, Cellular and Molecular Life Sciences, 2000, 57, 1593〜1612; Rebouillat; Hovanessian, A.G., Cytokine and Growth Factor Reviews, 2007, 18, 351〜361)。各OAS遺伝子は、5つの翻訳されるエキソン(エキソンA〜E)で構成される保存OASユニットからなる。OAS1は1つのユニットを有するが、OAS2及びOAS3はそれぞれ2つ及び3つのユニットを有し、3つ全ての遺伝子は、活性型2',5'-オリゴアデニル酸合成酵素をコードする。別の遺伝子であるOASL (OAS様)は、単独のユニットのOAS様タンパク質をコードするが、これは2'-5'合成酵素活性を欠く(Hartmannら, Nucleic Acids Res., 1998, 26, 4121〜4128; Rebouillatら, Eur. J. Biochem., 1998; 257, 319〜330)。それぞれのサイズクラス内で、最初の転写産物の選択的スプライシングの結果として、複数のメンバーが生じる。OASタンパク質は、約350アミノ酸の保存ユニット/ドメイン(OASユニット)を共有する;OAS1 (p40/p46)、OAS2 (p69/71)及びOAS3 (p100)は、OASユニットのそれぞれ1つ、2つ及び3つのタンデムコピーを有する。各OASタンパク質は、細胞で異なる位置に蓄積し、活性化されるために異なる量のdsRNAを必要とし、異なるサイズの2-5A生成物の形成を触媒する。OAS1は四量体として機能するが、OAS2は二量体としてのみ活性であり、OAS3は単量体としてのみ観察されている。さらに、大型のヒトOASは、おそらく、RNase L活性化に関与しない(概説として、Rebouillat, D.及びHovanessian, A.G., Journal of Interferon and Cytokine Research, 1999, 19, 295〜308)。
【0010】
OASファミリーが、IFNにより示される抗ウイルス効果に関与することについての最初の直接的な証拠は、2',5'-オリゴアデニル酸合成酵素(OAS) cDNAを細胞に形質移入することにより得られた。OAS1又はOAS2の過剰発現は、ピコルナウイルス複製に対する細胞の耐性を導く(Hovanessian, A.G., Cytokine and Growth Factor Reviews, 2007, 18, 351〜361)。インビボにおいてWNV感染を除去するためのOAS1の重要性は、ヒトOAS1のオルソロガス遺伝子であるマウスOas1bが、WNV誘導性脳炎に対するマウスの感受性/耐性表現型において重要な役割を有し得るとの知見によっても支持された(Mashimo, T.ら, Proc. Natl. Acad. Sci. USA, 2002, 99, 11311〜11316; Lucasら, Immunol. Cell. Biol., 2003, 81, 230〜236; Kajaste-Rudnitskiら, Journal of Biological Chemistry, 2006, 281, 46244637; 国際PCT出願WO 02/081741)。ヒトにおけるOAS遺伝子多型の分析は、OAS遺伝子中の遺伝マーカーが酵素活性と最も強く関連することを示した。OAS1が、ウイルス感染に対する宿主の感受性に影響するヒト遺伝子としての優れた候補であるとすると(Bonnevie-Nielsenら, Am. J. Hum. Genet., 2005, 76, 623〜633)、ヒトOAS1及びOASL遺伝子における遺伝子的変動は、ウイルス性脳炎、1型DM、HCV関連疾患及びその他のウイルス感染の危険性と関連した。HCV疾患について特に強調すると、HCV感染の結果と関係する一連のOAS1遺伝子型が報告されている(国際PCT出願WO 03/089003及びWO 2005/040428)。
【0011】
OASファミリーの抗ウイルス活性がプラス鎖ssRNAウイルスに対して選択的であるかは、まだ調べられていない重要な問題である。OAS1の異所発現が、脳心筋炎ウイルス(ピコルナウイルス)複製に対する耐性を導くが、VSVウイルス複製に対しては耐性を導かないことが報告されている。しかし、OASは、IFN誘導性RNA依存性プロテインキナーゼ(PKR)及びミクソウイルス耐性1 (MxA)遺伝子のようなその他のISGとは反対に、IFN-αで処置された患者でのHCVの除去において役割を示さないと考えられている。OAS、PKR及びMxの転写レベルは、非ヒト霊長類モデルにおいてDV感染の後に上方制御された(Sariolら, Clinical and Vaccine Immunology, 14, 2007, 756〜766)。デングショック症候群に関連する遺伝子転写パターンの分析は、OAS3遺伝子転写産物が、非DSSのデング患者におけるよりもDSSのデング患者においてより少ないことを示した (Simmonsら, J.Infect.Dis., 195:1097, 2007)。しかし、DV感染の病因におけるこれらのISGの役割は、ほとんど理解されていない。
【0012】
CHIKVは、I型インターフェロン(IFN-α/β)の抗ウイルス作用に対して感度が高いことが報告されている(Coudercら, PloS Pathogens, 2007, 4,e29)。アルファウイルスの場合、ウイルス成長のIFN媒介性阻害はRNase Lを必要としないことを示唆する一連の証拠が存在する(Rymanら, J. Virol., 2005, 79, 1487〜1499; 概説としてSilverman, J. Virol., 2007, 81, 12720〜9)。OASファミリーのようなヒトISGのメンバーのいずれかが、抗アルファウイルス活性を示し得るかが、まだ調べられていない重要な問題である。
【発明の概要】
【発明が解決しようとする課題】
【0013】
アルファウイルス(CHIKV、SINV及びSFV)のようなプラス鎖ssRNAウイルスに対する確立された内因性抗ウイルス経路におけるOAS3の役割を、本発明者らが初めて立証する。本発明者らは、OAS3が、感染ヒト上皮細胞の内部でウイルスタンパク質合成及びウイルスRNA複製を遮断してCHIKVの成長の段階に対して作用することを示す。アルファウイルス感染に対するヒトの遺伝的感受性について、ほとんど情報は得られていない。健康な白人個体における多型についてOAS3遺伝子をスクリーニングすることにより、CをTに置換することによりナンセンス変異が生じるコドンCGA-844の最初の位置にて1塩基多型(SNP)が同定された(OAS3.R844X)。OAS3.R844X位でのSNPは、カルボキシ末端からの約20%を欠くOAS3タンパク質の切断型をもたらすと予測される。変異体OAS3の異所発現は、全長OAS3タンパク質と比較してより低い効率のCHIKV阻害をもたらした。OAS3の遺伝子多型がその抗アルファウイルス活性を制御できるという見解は、チクングンヤ熱のようなアルファウイルス関連疾患の病因におけるヒトOAS遺伝子の役割を示唆する。
【0014】
本発明者らは、DV (肝細胞癌細胞)及びWNV (肝細胞癌細胞及び上皮細胞)に感染したヒト細胞において、OAS3が抗フラビウイルス活性を示すことを初めて立証する。DSS患者及び非DSSデング患者における完全OAS3遺伝子の検査により、SNP rs2285993が、GのCへの置換がArgからSerへのアミノ酸の変更をもたらすコドンAGG-381の3番目の位置にて同定された。我々の遺伝子データは、バリアントSer-381が、タイ人のデング患者におけるDSSの危険性に対する優勢な保護(donimanto protection)と関連することを示唆する。本発明者らは、今回、Ser-381を有するOAS3が、ヒト肝細胞におけるDV成長の潜在的な阻害物質であることを立証する。
【0015】
本発明者らは、YFVの生弱毒化ワクチン株17D-204 (STAMARIL, Sanofi-Pasteur)が、感染ヒト上皮細胞及び肝細胞癌細胞においてOAS3媒介性抗ウイルス経路に対する固有の耐性を有することも示す。しかし、IFN-αは、ヒト細胞においてYFVのワクチン株17D-204に対する抗ウイルス状態を確立することができた。
【0016】
これらの知見は、CHIKV、WNV及びDVを含む医療的に重要性が高いプラス鎖ssRNAウイルスに対するOAS3に基づく予防及び治療の発展にとって有効である。これらは、アルファウイルス、フラビウイルス及び医療的に重要性が高いその他のプラス鎖ssRNAウイルスへの感染に対するヒトの感受性の予測のため、特にデング、西ナイル及びチクングンヤの患者における疾患の重症形態の予測のためのOAS3に基づく新しい分子ツールの開発のためにも有効である。
【課題を解決するための手段】
【0017】
本発明の主題は、医薬品としての単離2',5'-オリゴアデニル酸合成酵素3タンパク質又は該2'-5'-オリゴアデニル酸合成酵素3タンパク質をコードする単離ポリヌクレオチドである。
【発明を実施するための形態】
【0018】
定義
- 「ポリヌクレオチド」とは、ゲノムDNAフラグメント、cDNAフラグメント又はRNA分子のことである。
- 「2'-5'-オリゴアデニル酸合成酵素3」、「OAS3」、「OAS 3」、「2'-5'-オリゴアデニル酸合成酵素3 (100 kD)」、「(2-5')オリゴ(A)合成酵素3」、「オリゴアデニル酸合成酵素p100」、「p100 OAS」又は「p100OAS」とは、哺乳動物のOAS3遺伝子によりコードされ、2'-5'-オリゴアデニル酸合成酵素活性を有するタンパク質、並びにOAS3遺伝子の多型に起因する天然のバリアントと、OAS3遺伝子/オープンリーディングフレーム(ORF)配列における1つ以上のヌクレオチドの変異(挿入、欠失、置換)に起因する人工的バリアント(但し、バリアントが2'-5'-オリゴアデニル酸合成酵素活性を有し、プラス一本鎖RNAウイルス複製を阻害できることを条件とする)とを含む派生バリアントのことをいう。好ましくは、バリアントは3つのOASドメインを含む。種々の哺乳動物のOAS3遺伝子、並びに推定OAS3 ORF及びアミノ酸配列は、配列データベースで入手可能であり、その他のOAS3遺伝子/ORF配列は、当業者に知られる標準的なクローニング及び配列決定の技術により決定してよい。
【0019】
- 「ヒトOAS3遺伝子」は、GenBank配列受入番号NC_000012の111860632位〜111895437位に相当する34806 bpの配列である。ヒトOAS3遺伝子は、第12染色体(12q24.2)上にあり、16個のエキソンを含む:エキソン1:1〜264;エキソン2:3127〜3409;エキソン3:6033〜6208;エキソン4:8300〜8538;エキソン5:9503〜9656;エキソン6:10418〜10762;エキソン7:12250〜12532;エキソン8:22628〜22803;エキソン9:24209〜24459;エキソン10:24870〜25014;エキソン11:25792〜25965;エキソン12:27301〜27586;エキソン13:28975〜29150;エキソン14:29493〜29731;エキソン15:31165〜31312;エキソン16:31519〜34806。
【0020】
- 「ヒトOAS3オープンリーディングフレーム(ORF)」は、6646 bpのヒトcDNA配列であるGenBank受入番号NM_006187 (配列番号27)上の88位〜3351位に相当する配列番号1の配列である。cDNA配列NM_006187上のエキソン1〜16の位置は、それぞれ1〜264、265〜547、548〜723、724〜962、963〜1116、1117〜1461、1462〜1744、1745〜1920、1921〜2171、2172〜2316、2317〜2490、2491〜2776、2777〜2952、2953〜3191、3192〜3339、3340〜6627である。
- 「ヒトOAS3タンパク質」は、GenBank受入番号NP_006178又は配列番号2に相当する1087アミノ酸の配列である。3つのOASドメインは、6位〜343位(配列番号3)、411位〜742位(配列番号4)及び750位〜1084位(配列番号5)にそれぞれ相当する。
【0021】
- 「OAS3活性」とは、.2'-5'-オリゴアデニル酸合成酵素活性及びプラス一本鎖RNAウイルス複製阻害活性の両方のことをいう。
本発明のOAS3タンパク質の2',5'-オリゴアデニル酸合成酵素活性は、クロマトグラフィー法又は電気泳動法によりアッセイして、形成されたオリゴアデニル酸の終点の量を決定できる(St Laurentら, Cell, 1983, 33, 95〜102; Interferon 3: Mechanisms of Production and Action, 1984, 189〜298, Friedman, R.M.編, Elsevier, Amsterdam中のJohnstonら; Justesenら, Proc. Natl. Acad. Sci., USA, 1980, 77, 4618〜4622; Justesenら, Nucleic Acids Res., 1980, 8, 3073〜3085; Justesen, J.及びKjelgaard, N.O., Anal. Biochem., 1992, 207, 90〜93)。
【0022】
- 「本発明のOAS3タンパク質によるプラス鎖 ssRNAウイルス複製の阻害」とは、ウイルス感染細胞内に外因性OAS3タンパク質(細胞のゲノムによりコードされていない;例えば組換えOAS3タンパク質)が存在する場合のウイルス成長(ウイルス複製)の部分的又は完全な低減のことをいう。この阻害は、OAS3タンパク質を発現する適切な組換え株化細胞に、プラス一本鎖RNAウイルスを感染させることにより決定できる。ウイルスに感染した同じタイプの非組み換え細胞を、対照として用いる。次いで、ウイルス感染細胞の上清中の子孫ウイルス生成を、任意の公知のウイルス力価決定アッセイにより測定できる。代わりに、ウイルスタンパク質の生成を、ウェスタンブロット若しくはウイルス抗原の免疫標識により分析できるか、又はウイルスのゲノム及びサブゲノムRNA生成を、ノザンブロット若しくはRT-PCRにより分析できる。
- 「プラス鎖ssRNAウイルス」とは、プラス一本鎖リボ核酸(ssRNA)をその遺伝的材料として有し、DNA中間体を用いて複製しないウイルスのことをいう。プラス鎖ssRNAウイルスは、ウイルス分類のボルティモア分類方式の第IV群に属する。
【0023】
- アミノ酸配列及び核酸配列の両方に関する「同一性」とは、整列させるアミノ酸残基若しくはヌクレオチド同士の同一性を最大にする配列のアラインメントに基づく2つの配列間の同一性の程度の指標のことであり、これは、同一残基若しくはヌクレオチド、残基(本発明の場合に1087残基)若しくはヌクレオチド(本発明の場合に3261ヌクレオチド)の合計数、並びに配列アラインメント中のギャップの存在及び長さの関数である。GCG配列分析パッケージ(University of Wisconsin, Madison, Wis.)の一部分として利用可能であり、例えばデフォルト設定で用いることができるFASTA又はBLASTを含む種々のアラインメントアルゴリズム及び/又はコンピュータプログラムが、標準的なパラメータを用いて配列同一性を決定するために利用可能である。
配列番号2の残基1〜1087と75%の同一性を有するタンパク質は、タンパク質配列を配列番号2の残基1〜1087と整列させて比較したときに、その配列が、(25×10.87 = 271)までの変更を含み得るタンパク質である。1つの変更とは、配列番号2の残基1〜1087と比較して1つのアミノ酸の欠失、置換又は挿入のことをいう。例えば、配列番号2の最初の1080残基から50の変更を有する1080アミノ酸の配列は、配列番号2と1080-50/10.87= 94.75%の同一性を有する。例えば、配列番号2の残基1〜1087から50の変更を有する1090アミノ酸の配列は、配列番号2と1087-50/10.87= 95.4%の同一性を有する。
【0024】
- 「類似性」とは、整列させるアミノ酸残基同士の類似性を最大にする配列のアラインメントに基づく2つのアミノ酸配列間の類似性の程度の指標のことをいい、これは、同一若しくは類似の残基の数、残基の合計数(本発明の場合に1087残基)、並びに配列アラインメント中のギャップの存在及び長さの関数である。GCG配列分析パッケージ(University of Wisconsin, Madison, Wis.)の一部分として利用可能であり、例えばデフォルト設定で用いることができるFASTA又はBLASTを含む種々のアラインメントアルゴリズム及び/又はコンピュータプログラムが、標準的なパラメータを用いて配列類似性を決定するために利用可能である。類似の残基とは、同等の化学的特性(サイズ、電荷(中性、塩基性、酸性)、親水性/疎水性)を有する残基のことを言う。
【0025】
-「個体」とは、哺乳動物、及びその他の脊椎動物(例えば鳥類、魚類及び爬虫類)を含む。本明細書において用いる場合、「哺乳動物」及び「哺乳類」の用語は、それらの子に授乳し、かつ生きている子を産む(真獣類または胎盤哺乳類)か、又は産卵する(後獣亜綱または無胎盤哺乳類)、単孔類、有袋類及び胎盤哺乳類(placental)を含む任意の脊椎動物のことをいう。哺乳動物種の例は、ヒト及びその他の霊長類(例えばサル、チンパンジー)、げっ歯類(例えばラット、マウス、モルモット)、及びウシ、ブタ及びウマのようなその他のものを含む。
- 「変異」により、ポリヌクレオチド(cDNA、遺伝子)又はポリペプチド配列中の1つ以上のヌクレオチド/アミノ酸の置換、欠失、挿入を意図する。この置換は、遺伝子のコード配列又はその調節配列に影響し得る。これは、ゲノム配列の構造、又はコードされるmRNAの構造/安定性にも影響し得る。
【0026】
本発明は、OAS3アミノ酸配列中の1つ以上のアミノ酸の変異(挿入、欠失、置換)、アミノ酸融合部分の付加、非天然アミノ酸(Dアミノ酸又は非アミノ酸アナログ)でのアミノ酸の置換、ペプチド結合の改変、環化、側鎖への化学基の付加(脂質、オリゴ糖又は多糖)、並びに適切な担体へのカップリングからなる群より選択される1つ以上の改変を含む改変OAS3タンパク質を包含する。当該技術において公知の手順により導入されるこれらの改変は、2'-5'-オリゴアデニル酸合成酵素活性及びプラス一本鎖RNAウイルス複製阻害活性についてまだ活性な改変OAS3タンパク質をもたらす。
【0027】
本発明の好ましい実施形態によると、上記の2'-5'-オリゴアデニル酸合成酵素3 (OAS3)は、ヒト2'-5'-オリゴアデニル酸合成酵素3である。
本発明の別の好ましい実施形態によると、上記のOAS3タンパク質は、配列番号2の残基1〜1087と少なくとも70%のアミノ酸配列同一性又は80%のアミノ酸配列類似性、好ましくは少なくとも80%のアミノ酸配列同一性又は90%のアミノ酸配列類似性を有する。
【0028】
本発明のより好ましい実施形態によると、上記のOAS3タンパク質は、配列番号2の381位にセリンを含む。好ましくは、上記のOAS3タンパク質は、配列番号2及び配列番号7からなる群より選択されるアミノ酸配列を含むか又はそれからなる。
本発明の別の好ましい実施形態によると、上記のOAS3ポリヌクレオチドは、上記で規定されるタンパク質をコードし、より好ましくは、これは、配列番号2のタンパク質をコードする配列番号1、及び配列番号7のタンパク質をコードする配列番号6からなる群より選択されるヌクレオチド配列を含むか又はそれからなる。
【0029】
本発明の別の好ましい実施形態によると、上記のポリヌクレオチドは、発現ベクターに挿入される。
ベクターとは、それが連結された別の核酸を輸送できる核酸分子のことをいう。本発明において用いることができるベクターは、限定されないが、ウイルスベクター、プラスミド、RNAベクター、又は染色体、非染色体、半合成若しくは合成の核酸分子からなり得る直鎖状若しくは環状のDNA又はRNA分子を含む。好ましいベクターは、自己複製できるか(エピソームベクター)及び/又はそれらが連結された核酸を発現できる(発現ベクター)ものである。多数の適切なベクターが当業者に知られ、市販されている。
【0030】
ウイルスベクターは、レトロウイルス、アデノウイルス、パルボウイルス(例えばアデノ随伴ウイルス又はAAV)、コロナウイルス、マイナス鎖RNAウイルス、例えばオルトミクソウイルス(例えばインフルエンザウイルス)、ラブドウイルス(例えば狂犬病及び水疱性口内炎ウイルス)、パラミクソウイルス(例えば麻疹及びセンダイ)、プラス鎖RNAウイルス、例えばピコルナウイルス及びアルファウイルス、並びにアデノウイルス、ヘルペスウイルス(例えば単純ヘルペスウイルス1及び2型、エプスタイン-バーウイルス、サイトメガロウイルス)及びポックスウイルス(例えばワクシニア、鶏痘及びカナリア痘)を含む二本鎖DNAウイルスを含む。その他のウイルスは、例えば、ノーウォークウイルス、トガウイルス、フラビウイルス、レオウイルス、パポバウイルス、ヘパドナウイルス及び肝炎ウイルスを含む。レトロウイルスの例は、トリ白血病肉腫、哺乳類C型、B型ウイルス、D型ウイルス、HTLV-BLV群、レンチウイルス、スプマウイルスを含む(Coffin, J. M., Retroviridae: The viruses and their replication, In Fundamental Virology, 第3版, B. N. Fieldsら編, Lippincott-Raven Publishers, Philadelphia, 1996)。
【0031】
好ましくは、上記のベクターは、本発明のOAS3タンパク質をコードする配列が、適切な転写及び翻訳制御要素の制御下に位置して、該タンパク質の生成又は合成を許容する発現ベクターである。よって、上記のポリヌクレオチドは、発現カセットに含まれる。より具体的には、該ベクターは、複製起点、該コードポリヌクレオチドに機能可能に連結されるプロモーター、リボソーム結合部位、RNAスプライシング部位(ゲノムDNAを用いる場合)、ポリアデニル化部位、及び転写終結部位を含む。これは、エンハンサーも含み得る。プロモーターの選択は、ポリペプチドが発現される細胞に依存する。適切なプロモーターは、組織特異的及び/又は誘導性プロモーターを含む。誘導性プロモーターの例は、重金属のレベルの増加により誘導される真核メタロチオネインプロモーター、温度の増加により誘導される熱ショックプロモーターである。組織特異的プロモーターの例は、骨格筋クレアチンキナーゼ、前立腺特異的抗原(PSA)、α-アンチトリプシンプロテアーゼ、ヒトサーファクタント(SP)タンパク質A及びB、並びにβ-カゼインである。ベクターは、選択マーカー、例えば真核細胞培養についてネオマイシンホスホトランスフェラーゼ、ヒスチジノールデヒドロゲナーゼ、ジヒドロ葉酸レダクターゼ、ハイグロマイシンホスホトランスフェラーゼ、単純ヘルペスウイルスチミジンキナーゼ、アデノシンデアミナーゼ、グルタミンシンセターゼ及びヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ;エス・セレビシエ(S. cerevisiae)についてTRP1、URA3及びLEU2;大腸菌(E. coli)においてテトラサイクリン、リファンピシン又はアンピシリン耐性を含み得る。
【0032】
ベクターの選択は、それらの使用(安定発現又は一過性発現)又は及び宿主細胞に依存する。ウイルスベクター及び「裸の」核酸ベクターは、哺乳動物細胞(ヒト及び動物)における発現のために好ましいベクターである。なかでも、興味対象の配列が予め挿入された、アデノウイルス、レトロウイルス、レンチウイルス及びAAVのようなウイルスベクターを用いることができる。
【0033】
本発明の主題は、上記で規定されるような少なくとも1つのOAS3タンパク質、又は好ましくは発現ベクター中に挿入された少なくとも1つのOAS3ポリヌクレオチドと、少なくとも1つの許容され得るビヒクル、担体、添加剤及び/又は免疫賦活剤とを含むことを特徴とする医薬組成物でもある。
当業者に知られる任意の適切な担体を、本発明の医薬組成物に用いることができ、担体のタイプは、投与の様式に依存して変動する。皮下注射のような非経口投与について、担体は、好ましくは水、塩水バッファー、ラクトース、マンニトール、グルタメート、脂肪又はワックスを含み、注射可能医薬組成物は、好ましくは等張溶液である(300〜320 m osmole程度)。経口投与について、マンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、タルク、セルロース、グルコース、スクロース及び炭酸マグネシウムのような上記の担体又は固体担体のいずれも用いることができる。生分解性マイクロスフェア(例えばポリ乳酸ガラクチド)を、本発明の医薬組成物の担体として用いることもできる。適切な生分解性マイクロスフェアは、例えば米国特許第4,897,268号及び第5,075,109号に開示されている。添加剤は、凝集防止剤、抗酸化剤、色素、風味増強剤、又は平滑剤、集合剤(assembling agent)若しくは単離剤(isolating agent)から、そして一般的に製薬産業で通常用いられる任意の賦形剤から選択できる。任意の種類の免疫賦活剤を本発明の組成物において用いて、免疫応答を増強し得る。
【0034】
医薬組成物は、経口投与に適する形態であり得る。例えば、組成物は、経口投与用の錠剤、通常のカプセル剤、ゼラチンカプセル剤又はシロップ剤の形態である。これらのゼラチンカプセル剤、通常のカプセル剤及び錠剤の形態は、デンプン、ガム及びゼラチンのようなアジュバント若しくは結合剤、リン酸カルシウムのようなアジュバント、トウモロコシデンプン若しくはアルギン酸のような崩壊剤、ステアリン酸マグネシウムのような滑沢剤、甘味料又は香料のような医薬製剤において通常用いられる賦形剤を含み得る。液剤又は懸濁剤は、薬理学的に適合し得る溶剤を加えることにより水性又は非水性の媒体中で調製できる。これらは、グリコール、ポリグリコール、プロピレングリコール、ポリグリコールエーテル、DMSO及びエタノールを含む。
【0035】
OAS3タンパク質又はOAS3ポリヌクレオチド(単離されているか又はベクターに挿入されている)は、細胞に、インビトロ、エクスビボ又はインビボで、特定の細胞タイプに適する当該技術において公知の任意の簡便な手段により、単独で、又は少なくとも適切なビヒクル及び/又は担体のいずれかとともに導入される。例えば、OAS3タンパク質/ポリヌクレオチドは、生体中で該配列への保護を提供できるか、又はそれが宿主細胞の膜を通過することを可能にする物質と組み合わせ得る。OAS3タンパク質は、有利には、リポソーム、ポリエチレンイミン(PEI)及び/又は膜輸送ペプチド(Bonetta, The Scientist, 2002, 16, 38; Fordら, Gene Ther., 2001, 8, 1〜4 ; Wadia及びDowdy, Curr. Opin. Biotechnol., 2002, 13, 52〜56; Handbook of cell penetrating peptides (第2版), 2006, Lavoisier, FRANCE中のLangel, U.)と組み合わせ得る。後者の場合、OAS3タンパク質の配列を、膜輸送ペプチドの配列と融合させる(融合ペプチド)。OAS3をコードするポリヌクレオチド(単離されているか又はベクターに挿入されている)は、種々の方法により細胞に導入できる(例えば注入、直接取り込み、発射衝撃、リポソーム、エレクトロポレーション)。OAS3タンパク質は、上記で規定される適切な発現ベクターを用いて細胞において安定的に又は一過的に発現され得る。
【0036】
本発明のある実施形態において、OAS3タンパク質/ポリヌクレオチドは、実質的に非免疫原性であり、すなわち、ほとんど又は全く有害な免疫応答を生じない。この種の有害な免疫反応を緩和又は消去するための種々の方法を、本発明に従って用いることができる。好ましい実施形態において、OAS3タンパク質は、N-ホルミルメチオニンを実質的に含まない。望ましくない免疫応答を回避する別の方法は、タンパク質/ポリヌクレオチドを、ポリエチレングリコール(「PEG」)又はポリプロピレングリコール(「PPG」) (好ましくは平均分子量(MW)が500〜20,000ダルトンのもの)と複合させることである。
【0037】
本発明の別の主題は、プラス一本鎖RNAウイルスへの感染を予防又は治療するための上記で規定されるOAS3タンパク質又は該OAS3タンパク質をコードするポリヌクレオチドである。
より好ましい実施形態によると、上記のウイルスは、アルファウイルス属のものである。より好ましくは、これは、チクングンヤ(CHIK)、シンドビス(SIN)、セムリキ森林(SF)、東部ウマ脳炎(EEE)、西部ウマ脳炎(WEE)、ベネズエラウマ脳炎(VEE)、ロスリバー(RR)、オニョンニョン(ONN)及びバーマフォレスト(BF)ウイルスからなる群より選択される。
【0038】
別のより好ましい実施形態によると、上記のウイルスは、フラビウイルス属のものである。より好ましくは、上記のウイルスは、デング、日本脳炎、キャサヌール森林病、マレー渓谷脳炎、セントルイス脳炎、ダニ媒介脳炎、西ナイル、黄熱及びオムスク出血熱(OHF)ウイルスからなる群より選択される。
【0039】
別のより好ましい実施形態によると、上記のウイルスは、ヘパシウイルス属のものである。より好ましくは、上記のウイルスは、C型肝炎ウイルスである。
【0040】
本発明の主題は、プラス一本鎖RNAウイルス感染の予防又は治療用の同時、別々又は逐次的な使用のための組み合わせ調製物としての、少なくとも、上記で規定されるOAS3タンパク質、又は好ましくは発現ベクターに挿入されたOAS3ポリヌクレオチドと、抗ウイルス薬、抗炎症薬及び免疫調節薬からなる群より選択される、第1の製品とは異なる第2の製品とを含む製品でもある。
【0041】
本発明の主題は、必要とする個体に、上記で規定される組成物を任意の手段により投与するステップを含む、必要とする個体におけるプラス一本鎖RNAウイルス感染を予防又は治癒するための方法でもある。
一般的に、組成物は、非経口注射(例えば皮内、筋肉内、静脈内及び皮下)、鼻腔内(例えば吸入又は噴霧)、経口、舌下又は局所的に、皮膚又は直腸を通すことにより投与できる。
【0042】
本発明の組成物中に存在するOAS3 (タンパク質/ポリペプチド)の量は、治療有効量である。OAS3 (タンパク質/ポリペプチド)の治療有効量は、組成物が投与される対象者において過度に負の影響を引き起こすことなく、OAS3タンパク質が、プラス一本鎖RNAウイルス複製を阻害するというその役割を働かせるのに必要な量である。用いられるOAS3 (タンパク質/ポリペプチド)及び投与される組成物の正確な量は、プラス一本鎖RNAウイルスの種、及び治療される個体の種(ヒト、動物)、投与の様式、投与頻度及び組成物中のその他の成分のような因子によって変動する。
好ましくは、組成物は、約10μg〜約10 mg、より好ましくは約100μg〜約1 mgのOAS3 (タンパク質/ポリペプチド)で構成される。「約」により、そこで述べたOAS3の量の値(μg又はmg)が、そのような量を評価するために用いた方法の誤差限界に依存するある範囲内で変動できることを意味する。
例えば、本発明の組成物の経口投与中に、治療される個体は、連続する3日間に1日当たり約10μg〜約10 mgのOAS3 (タンパク質/ポリペプチド)の1用量スケジュールに供することができる。治療は、1週間後に1回反復できる。
皮下注射のような非経口投与について、治療される個体は、約10μg〜約10 mg、より好ましくは約100μg〜約1 mgのOAS3 (タンパク質/ポリペプチド)の1用量に供することができる。治療は、1週間後に1回反復できる。
【0043】
本発明の主題は、個体から得られる核酸試料中のOAS3遺伝子における多型を検出するステップを含む、上記で規定されるプラス一本鎖RNAウイルスへの感染に対する個体の感受性をインビトロで評価する方法でもある。
核酸試料は、ゲノムDNA、トータルmRNA又はcDNAであり得る。
多型は、核酸配列中の変異の検出を可能にする、例えばIn Current Protocols in Human Genetics, 2008, John Wiley & Sons, Incに記載されるような当該技術において知られる任意の方法により検出される。遺伝子型同定アッセイの例は、限定されないが:RAPD、RFLP、AFLP、配列特異的オリゴヌクレオチドハイブリッド形成、SnapShot PCR、リガーゼ検出反応、PCR及びMaldi-TOF、ピロシーケンスを含む。このアッセイは、表II及びIIIのOAS3特異的プライマー、特に配列番号11、12、62〜86及び118〜142のプライマーを用いることができる。
【0044】
上記の方法の好ましい実施形態によると、上記のプラス一本鎖RNAウイルスは、チクングンヤウイルスのようなアルファウイルス、又はデングウイルスのようなフラビウイルスである。
【0045】
上記の方法の別の好ましい実施形態によると、上記の多型は、Arg844コドンの停止コドンへの変異(R844X)である。白人個体群において検出されるこの多型は、プラス一本鎖RNAウイルス感染、特にチクングンヤウイルス感染への感受性の増加と関連する。R844X変異は、プライマー対(配列番号11及び配列番号12)を用いるPCR-RFLPと、その後の183 bpのPCR生成物のBglIIでの消化とにより検出できる。115 bp及び68 bpの2つのフラグメントの存在は、OAS3.R844X変異の存在を示す。
【0046】
上記の方法の別の好ましい実施形態によると、上記の多型は、コドン381の3番目の位置での1塩基多型(SNP)である。好ましくは、このSNPは、アミノ酸をArgからSerに変更するコドンAGG-381のGからCへの置換である(R381S)。遺伝的データは、バリアントSer-381が、デング患者におけるDSSの危険性に対する優勢な保護と関連することを示唆する(表VI)。このSNPは、上記で規定される任意の適切な遺伝子型同定アッセイにより検出できる。例えば、このアッセイは、表IIIに示すOAS3 エキソン6に特異的なPCRプライマー対(配列番号70及び126)のようなOAS3特異的プライマー対を用いて増幅されたゲノムDNAの直接配列決定を含み得る。
【0047】
本発明の主題は、配列番号7又は配列番号30の配列を含むか又はそれからなる単離OAS3タンパク質でもある。
【0048】
本発明の主題は、配列番号9の配列を含むか又はそれからなる単離OAS3タンパク質フラグメントでもある。配列番号7の残基1〜843を含むこのOAS3フラグメントは、OAS3の第1及び第2のOASドメインを含むが、第3の(C末端) OASドメインの大部分を欠く。
【0049】
本発明の主題は:
- 配列番号7のOAS3タンパク質をコードする配列番号6の配列を含むか又はそれからなる単離ポリヌクレオチド、
-配列番号9のOAS3タンパク質フラグメントをコードする配列番号8の配列を含むか又はそれからなる単離ポリヌクレオチド、
- 配列番号30のOAS3タンパク質バリアントをコードする配列番号28又は29の配列を含むか又はそれからなる単離ポリヌクレオチド
でもある。
【0050】
本発明の主題は、配列番号6、配列番号8、配列番号28及び配列番号29からなる群より選択される配列を有するポリヌクレオチドを含む組換えベクター、好ましくは発現ベクターでもある。
【0051】
本発明の主題は、配列番号6、配列番号8、配列番号28及び配列番号29からなる群より選択される配列を含むか又はそれからなるポリヌクレオチドで形質移入又は形質転換された宿主細胞でもある。
本発明の好ましい実施形態によると、これは、HeLa-Tet-Off/OAS3#C417-1と命名され、2008年2月26日にCollection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3927の下で寄託された、組換えヒトOAS3を発現するHeLa-Tet-Off株化細胞である。
本発明の別の好ましい実施形態によると、これは、HeLa-Tet-Off/OAS3/delta/1Cと命名され、2008年4月17日にCollection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3968の下で寄託された、切断型組換えヒトOAS3を発現するa-Tet-Off株化細胞である。
本発明の好ましい実施形態によると、これは、HeLa-Tet-Off/OAS3#F8と命名され、2009年5月15日にCollection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-4158の下で寄託された、組換えヒトOAS3を発現するHepG2-Tet-Off株化細胞である。
種々のウイルスが感染できるHeLa又はHepG2細胞に由来するこれらの株化細胞は、OAS3媒介抗ウイルス活性に対するウイルスの感受性をアッセイするために有用である。
【0052】
本発明の主題は、上記で規定されるポリヌクレオチドを含む非ヒトトランスジェニック動物でもある。
本発明の主題は、上記で規定されるポリヌクレオチドを含むトランスジェニック植物でもある。
【0053】
本発明のOAS3タンパク質/ポリヌクレオチドは、公知の組換えDNA及び遺伝子工学技術を用いて調製される。例えば、OAS3のORFを含む配列は、DNA鋳型から、特異的プライマーを用いるポリメラーゼ連鎖反応により増幅される。PCRフラグメントは、次いで、適切な制限部位を用いることにより発現ベクターにクローニングされる。OAS3タンパク質は、発現ベクターで改変された宿主細胞又はトランスジェニック動物/植物において、OAS3タンパク質の発現に適する条件下で発現され、OAS3タンパク質は、宿主細胞培養物又はトランスジェニック動物/植物から回収される。
【0054】
本発明の実行は、そうでないと記載しない限り、当該技術の範囲内である細胞生物学、細胞培養、分子生物学、トランスジェニック生物学、微生物学、組換えDNA及び免疫学の通常の技術を用いる。このような技術は、文献に十分に説明されている。例えば、Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, 第3版, (Sambrookら, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); Oligonucleotide Synthesis (M. J. Gait編, 1984); Mullisら、米国特許第4,683,195号; Nucleic Acid Hybridization (B. D. Harries及びS. J. Higgins編1984); Transcription And Translation (B. D. Hames及びS. J. Higgins編1984); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); Methods In ENZYMOLOGYのシリーズ(J. Abelson及びM. Simon編, Academic Press, Inc., New York)、特に第154巻及び第155巻(Wuら編)並びに第185巻「Gene Expression Technology」(D. Goeddel編); Gene Transfer Vectors For Mammalian Cells (J. H. Miller及びM. P. Calos編, 1987, Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (Mayer及びWalker編, Academic Press, London, 1987); Handbook Of Experimental Immunology, 第I〜IV巻(D. M. Weir及びC. C. Blackwell編, 1986); 並びにManipulating the Mouse Embryo, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1986)、Current Protocols in Human Genetics (John Wiley & Sons, Inc, 2008)、特に第12章「Vectors For Gene Therapy」及び第13章「Delivery Systems for Gene Therapy」を参照されたい。
【図面の簡単な説明】
【0055】
【図1】c-mycエピトープをタグ付加されたヒト組換えOAS3 (クローンOAS3C 17.1)のヌクレオチド(A)及びアミノ酸(B)の配列を表す。これらのヌクレオチド及びアミノ酸配列は、それぞれ配列番号23及び24に相当する。OAS3 mRNA参照配列(配列番号27)と、クローニングされたOAS3 cDNAとの間の変更は、同義であれば下線を付し、非同義であれば太字で示す。翻訳開始及び停止コドンは、太字で示す。C末端c-mycエピトープに相当する付加ヌクレオチド及びアミノ酸配列は、下線を付す。5'のNotI及び3'のEcoRVの制限部位は、斜体で示す。
【図2】組換えヒトOAS3を発現する誘導性HeLa-Tet-Off/OAS3#C417-1株化細胞の樹立を説明する。
【図3】HeLa細胞におけるOAS3の検出を示す。(A)において、HeLa.Tet-Off細胞に、CHIKウイルス06-49株(CHIKV 06-49)を、細胞あたり異なるフォーカス形成単位(AP61FFU)で感染させた。感染後(p.i.) 20時間で、感染細胞の上清中に生成されるウイルス粒子を、蚊エーデス・シュードスクテラリス(Aedes pseudoscutellaris) AP61細胞でのフォーカス免疫アッセイにより力価決定した。(B)において、細胞あたり1AP61FFUの感染多重度(1 MOI)でのCHIKVの投入の5時間前に、HeLa.Tet-Off細胞を、1,000 IU/mlのヒトIFN-αで処理したか、又は模擬処理(対照)した。ウイルス子孫の生成を、上記のようにして18時間p.iにて決定した。(C)において、グラフは、全長及び切断型のOAS3の構造を描く。(D)において、誘導(- Tet)又は非誘導(+ Tet) Tet-Off/OAS3細胞におけるOAS3の異所発現を、OAS3特異的抗体を用いるイムノブロット分析により分析した。対照として、HeLa.Tet-Off細胞を、1,000 IUのIFN-α(+ IFN)で処理したか、又は模擬処理した(模擬)。β-アクチンは、ハウスキーピングタンパク質対照であった。
【図4】OAS3発現HeLa細胞におけるCHIKVの成長の阻害を示す。(A)において、細胞に18時間、1MOIでCHIKVを感染させ、抗CHIK.E2 MAb 3E4を用いてフローサイトメトリーにより分析した。模擬感染細胞(ウイルスなし)、ウイルス投入の5時間前に1,000 IU/mlのヒトIFN-αとインキュベートした(+ IFN)か、又は模擬処理した(対照)、CHIKVに感染したHeLa.Tet-off細胞、及びテトラサイクリンの存在下(+ tet)又は非存在下で(- tet)のCHIKVに感染したTet-Off/OAS3細胞におけるCHIKV E2タンパク質生成の分析。(B)において、細胞に、種々のMOIでCHIKVを感染させ、ウイルス子孫の生成を18及び24時間のp.i.で決定した。(C)において、CHIKV成長の阻害を、種々のp.i.時間でモニターした。(D)において、細胞に1MOIにてSINV又はSFVを感染させ、ウイルス子孫の生成を18時間p.i.で決定した。
【図5】OAS3の抗ウイルス活性に対するWNウイルスの感度を示す。HeLa/Tet-off及び誘導HeLa.Tet-Off/OAS3#C417-1細胞に、細胞あたり0.1、1.0又は10 AP61FFUのWNウイルスIS-98-ST1株を感染させた。感染した細胞の上清中に生成される感染性ウイルス粒子を、感染後48時間で、蚊エーデス・シュードスクテラリスAP61細胞でのフォーカス免疫アッセイにより力価決定した。
【図6】OAS3発現細胞でのWNウイルス成長の阻害を示す。HeLa/Tet-Off及び誘導HeLa.Tet-Off/OAS3#C417-1細胞に、細胞あたり0.1 AP61FFUのWNウイルスIS-98-ST1株を感染させ、感染した細胞の上清中に生成される感染性ウイルス粒子を、感染後種々の時間にて(24、48及び72時間)、蚊エーデス・シュードスクテラリスAP61細胞でのフォーカス免疫アッセイにより力価決定した。
【図7】CHIKV複製に対するOAS3発現の意義を示す。(A)において、イムノブロットアッセイを、CHIKV (ウイルス)に感染させたか又は模擬感染させた(m.i.) HeLa.Tet-off細胞(対照)及びHeLa.Tet-Off/OAS3 (OAS3)細胞からの細胞抽出物について、抗CHIK HMAF (左)又は抗CHIKV E2 MAb 3E4 (右)を用いて行った。βアクチンンは、タンパク質対照であった。(B)において、1MOIでのCHIKVに感染させたHeLa.Tet-Off細胞(対照)及びHeLa.Tet-Off/OAS3細胞(OAS3)におけるウイルスRNA生成のリアルタイムRT-PCR分析を、8時間p.i.にて行った。
【図8】切断型OAS3の抗アルファウイルス活性を示す。HeLa.Tet-off (対照)、HeLa.Tet-Off/OAS3 (OAS3)及びHeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)細胞に、1MOIにてCHIKVを感染させ、ウイルス子孫の生成を、18時間p.i.にて決定した。値は、スチューデントのt検定に従って統計学的に比較した(***:P<0.001)。
【図9】切断型OAS3の抗フラビウイルス活性を示す。HeLa.Tet-off (対照)、HeLa.Tet-Off/OAS3 (OAS3)及びHeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)細胞に、1MOIにてWNVを感染させ、ウイルス子孫の生成を、18時間p.i.にて決定した。値は、スチューデントのt検定に従って統計学的に比較した(***:P<0.001)。
【図10】I型IFN経路に対するDV-1ウイルスの感度を示す。A:DV感染に対するHepG2.Tet-Off株化細胞の感受性。細胞に、漸増投入量(細胞あたりのAP61FFU)のDV-1ウイルスFGA/NA d1d株を感染させた。感染後40時間にて、細胞を、DV E糖タンパク質に対して反応性のMAb 4E11を用いるフローサイトメトリーにより分析した。B:I型IFN経路に対するDV-1ウイルスの感度。HepG2.Tet-off細胞を、DV感染(10 MOI)の5時間前に、1,000 IU.mL-1のヒトIFN-α(+IFN)で前処理したか又は処理しなかった(模擬処理)。40時間p.i.にて、ウイルス子孫の生成を、以前に記載されたようにして決定した(Duarte dos Santosら, Virology, 2000, 274, 292〜)。
【図11】OAS3発現HepG2細胞におけるデングウイルス成長の阻害。2μg.mL-1のテトラサイクリンの存在下(非誘導)又は非存在下(誘導)での親のHepG2.Tet-Off細胞及びHepG2.Tet-Off/OAS3#F8細胞クローンに、デングウイルス1型FGA/NA d1d株を種々の感染多重度で感染させた。感染後40時間にて、細胞を、-20℃にて20分間、メタノール/アセトンで固定した。免疫蛍光アッセイを、FITC-結合抗デングE mAb 4E1を用いて行った。核をDAPIで染色した。ウイルス抗原について陽性のHepG2細胞のパーセンテージを、3重の実験にて決定した。ウイルス子孫の生成を、25の感染多重度にて決定した。ウイルス力価決定を、以前に記載されたようにして行った(Duarte dos Santosら, Virology, 2000, 274, 292〜)。
【図12】OAS3発現HepG2細胞は、デングウイルス感染に対する耐性を示す。親のHepG2.Tet-Off細胞及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、デングウイルス1型FGA/NA d1d株(Duarte dos Santosら, Virology, 2000, 274, 292〜)を、感染多重度30 AP61FFU/細胞で感染させたか、又は模擬感染させた。感染後40時間にて、免疫蛍光アッセイを、図11の説明に記載したようにして行った。
【図13】OAS3発現HepG2細胞は、西ナイルウイルス感染に対する耐性を示す。親のHepG2.Tet-Off細胞及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、西ナイルウイルスIS-98-ST1株(Lucasら, Virology J., 2004, 1, 9〜)を、感染多重度1AP61FFU/細胞で感染させたか、又は模擬感染させた。感染後40時間にて、細胞を-20℃にて20分間、メタノール/アセトンで固定した。免疫蛍光アッセイを、cy3-結合抗西ナイルE mAb E24を用いて行った。核をDAPIで染色した。
【図14】IFN-αが、YFV 17Dに対する抗ウイルス状態を、ヒト上皮HeLa細胞において確立できることを示す。HeLa細胞に、YFV 17Dを1PFU/細胞にて感染させ、次いで、感染後の種々の時点にて、1,000 IU/mlのヒトIFN-αで処理したか、又は模擬処理した(対照)。ウイルス子孫の生成を、感染後72時間にて決定した。
【図15】YFVの17D生弱毒化株が、ヒト細胞において、感染後の時間又は感染多重度のいずれにも関係なく、OAS3媒介抗ウイルス効果に対する予期せぬ耐性を示すことを示す。HeLa (対照)及び誘導HeLa.Tet-Off/OAS3 (OAS3)細胞を、低ウイルス投入量(パネルA; 1PFU/細胞)又は高ウイルス投入量(パネルB; 10 PFU/細胞)でYFV 17D株に曝露した。
【図16】OAS3発現HepG2細胞での黄熱ワクチン17D-204の阻害はない。親のHepG2.Tet-Off及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、Vero E6細胞で一旦成長させた黄熱の生弱毒化STAMARILワクチン(Sanofi-Pasteur)を感染させたか、又は模擬感染させた。感染後40時間にて、細胞を-20℃にて20分間、メタノール/アセトンで固定した。免疫蛍光アッセイを、抗フランス神経親和性ウイルスHMAF及びFITC-結合ヤギ抗マウスIgを用いて行った。核をDAPIで染色した。
【図17】切断型ヒト組換えOAS3 (OAS3 1-843)のヌクレオチド(A)及びアミノ酸(B)配列を示す。これらのヌクレオチド及びアミノ酸配列は、それぞれ配列番号25及び配列番号26に相当する。OAS3 mRNA参照配列と、クローニングされたOAS3 cDNAフラグメントとの間の変更は、同義であれば下線を付し、非同義であれば太字で示す。翻訳開始及び停止コドンは、太字で示す。C末端c-mycエピトープに相当する付加ヌクレオチド及びアミノ酸配列は、下線を付す。5'のNotI及び3'のEcoRVの制限部位は、斜体で示す。
【実施例】
【0056】
実施例1:HeLaTet-Off/OAS3株化細胞の確立及び評価
1) 材料及び方法
a) 細胞培養
HeLa.Tet-Off株化細胞を、BD BIOSCIENCES CLONTECHから購入した。HeLa.Tet-Off細胞は、5% CO2中で37℃にて、10%熱不活化胎児ウシ血清(FCS)、4 mM L-グルタミン、100 UI/mlペニシリン、10μg/mlストレプトマイシン及び200μg.mL-1 G418 (INVITROGEN)を補ったDMEM (INVITROGEN)中で維持する。
HeLa.Tet-Off/OAS3株化細胞(CNCM I-3927)は、DMEM、10% FCS、4 mM L-グルタミン、100 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB (BD BIOSCIENCES CLONTECH)及び2μg.mL-1テトラサイクリン(SIGMA-ALDRICH)中で維持する。細胞を、単層で成長させる。予測される細胞密度は、80%〜100%である。集団倍加時間は、約2日である。細胞を、トリプシン処理により採集する。これを、毎週1/10で継代培養する。細胞は、制限された寿命を有する(15〜20継代)。細胞を、20% FCS、10% DMSO、4 mMグルタミン、100 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB及び2μg.mL-1テトラサイクリンを補ったDMEM中で凍結させる。
【0057】
HeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)株化細胞(CNCM I-3968)は、DMEM、10% FCS、20 mM L-グルタミン、10,000 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB (BD BIOSCIENCES CLONTECH)及び10μg.mL-1テトラサイクリン(SIGMA-ALDRICH)中で維持する。細胞を、単層で成長させる。予測される細胞密度は90%である。集団倍加時間は、約2日である。細胞を、トリプシン処理により採集する。これを、毎週1/10で継代培養する。細胞は、制限された寿命を有する(25継代)。細胞を、20% FCS、10% DMSO、4 mMグルタミン、10,000 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB及び10μg.mL-1テトラサイクリンを補ったDMEM中で凍結させる。
【0058】
b) OAS3タンパク質を発現するHeLa.Tet-Off株化細胞の樹立
PCR4-TOPOベクター(INVITROGEN)中で別々にクローニングしたヒトOAS3の2つのオーバーラップcDNAフラグメントを、全長OAS3タンパク質(aa 1〜1,087;配列番号24及び図1)、又は切断型OAS3ΔC-term若しくはOAS3[1-843] (aa 1〜843; 配列番号26及び図3C、9及び17)の発現のための鋳型として用いた。参照OAS3配列(GenBank受入番号NM-006187; 配列番号27)と、本研究で用いたOAS3 cDNA (図1)との間のヌクレオチド及びアミノ酸の違いを、表Iに列挙する。
【0059】
【表1】

【0060】
OAS3配列は、3'オープンリーディングフレーム末端で、追加の10残基の配列EQKLISKEDL (配列番号10)及びその後の停止コドンと接するように、OAS3についてプライマー対pTet-OAS3-univ1及びpTet-OAS3-rev2を、そしてOAS3[1-843]についてプライマー対pTet-OAS3-univ1bis及びpTet-OAS3-rev2bis (表II)を用いてPCRにより改変した。
【0061】
【表2】
【0062】
酵素認識部位に下線を付す。停止コドンに相補的な配列を太字で示す。OAS3配列(配列番号23〜25)は、NotI及びEcoRV制限酵素部位で、それぞれ下流の端及び上流の端にて挟まれている。PCR生成物は、NotI及びEcoRVを用いて消化し、pTRE2hyg発現ベクター(BD BIOSCIENCES CLONTECH)のユニークNotI及びEcoRV部位に挿入して、pTRE2hyg-OAS3 (図1)及びpTRE2hyg/OAS3CΔC-term (OAS3 1-843)を作製した。この構成において、OAS3挿入断片は、Tet-Off発現系の制御下にある。Tet-Off系は、リプレッサーであるテトラサイクリン(Tet)の除去により外来遺伝子発現の誘導を可能にする。HeLa.Tet-Off細胞(BD BIOSCIENCES CLONTECH)に、pTRE2hyg-OAS3又はpTRE2hyg/OAS3CΔC-termを、形質移入試薬Fugene 6 (ROCHE)を製造業者の推奨する手順に従って用いて、形質移入した。Tet-Off発現系を、2μg/mlのTetを培養培地に加えることにより抑圧した。形質移入された細胞を、阻害剤G418及びハイグロマイシンを含む成長培地上で選択し、次いで、10μg.mL-1のリプレッサーであるテトラサイクリン(Tet)の存在下で限界希釈により単一細胞からクローニングした。Tet除去のために、細胞単層をトリプシン処理し、何も補充していないDMEMで少なくとも5回洗浄した後に、遺伝毒性薬剤のみを補充したDMEM/10 % FBSに置き換えた。非誘導細胞(+ Tet)でのものに対する誘導細胞(- Tet)での組換えOAS3 mRNA生成のレベルを、RT-PCR分析により、プライマー対OAS3-For及びOAS3-Rev (表II)を用いて決定した。これらの実験に基づいて、誘導性のHeLa.Tet-Off/OAS3#C417-1及びHeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)クローンを選択した。HeLa.Tet-Off/OAS3#C417-1株化細胞は、2008年2月26日に、Collection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3927の下で寄託した。HeLa-Tet-Off/OAS3/delta/1C株化細胞は、2008年4月17日に、Collection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3968の下で寄託した。両方のHeLa.Tet-Off/OAS3株化細胞は、2μg.mL-1 Tetの存在下の抑圧条件下で維持した。
【0063】
c) ウイルス
臨床単離株であるチクングンヤウイルス06-49株(CHIKV 06-49; Schuffeneckerら, PLoS Medicine 3, 2006, e263)の、蚊エーデス・シュードスクテラリス(AP61)細胞単層上での生成、及びフォーカス免疫検出アッセイによるウイルス力価決定(Despresら, Virology, 1993, 196, 209〜219)は、以前に記載されたようにして行った(Brehinら, Virology, 2008, 371, 185〜195)。感染性の力価は、AP61細胞でのフォーカス形成単位(FFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。
【0064】
d) イムノブロットアッセイ
細胞性タンパク質を、以前に記載されたようにしてイムノブロット分析に供した(Brehinら, Virology, 2008, 371, 185〜195)。ウイルスタンパク質発現を、抗CHIKV HMAF又は抗CHIK.E2 MAb 3E4を用いて検出した(Brehinら, Virology, 2008, 371, 185〜195)。OAS3タンパク質発現を、抗OAS3 N-term (Santa-Cruz)又はC-term (ABGENT)抗体を用いて検出した。
【0065】
e) フローサイトメトリー分析
細胞を剥離し、次いで、PBS中の3.2%パラホルムアルデヒドを用いて固定した。固定された細胞を、以前に記載されたようにして(Brehinら, Virology, 2008, 371, 185〜195)、透過にし、抗CHIKV HMAFで染色し、フローサイトメトリーにより分析した。
【0066】
2) 結果
まず、親のHeLa.Tet-Off株化細胞の、CHIKV感染に対する感受性を評価した(図3A)。1感染多重度(MOI)で感染させたこれらの細胞におけるCHIKV複製の分析は、子孫ウイルスの生成が、18時間p.i.にておよそ7.0 log FFU.mL-1に達したことを示した。CHIKV E2糖タンパク質に対して反応性のMAb 3E4を用いるフローサイトメトリー分析により(Brehinら, Virology, 2008, 371, 185〜195)、約50%のCHIKV感染HeLa.Tet-Off細胞が、ウイルス抗原について陽性であった。次いで、IFN-αがHeLa.Tet-Off細胞において抗ウイルス状態を確立する能力について調べた。HeLa.Tet-off細胞を、1,000 IU.mL-1ヒトIFN-αで、CHIK曝露(1MOI)の5時間前に前処理することにより、18時間p.i.にて、約1.5 logのウイルス力価の低減がもたらされた(図3B)。よって、IFN-依存的抗ウイルス経路はHeLa.Tet-Off細胞において機能的であり、細胞レベルにてCHIKVに対する保護を提供する。しかし、CHIKV感染細胞は、感染HeLa.Tet-Off細胞内部でウイルス複製が一旦確立されると、5時間p.i.にてIFN-αに対する完全な耐性を示した。
【0067】
OAS3タンパク質発現をTet-Off発現系の制御下で上方制御する安定HeLa.Tet-Off/OAS3#C417-1細胞クローンを選択して、大型のOASの抗ウイルス活性を評価した。組換えOAS3タンパク質は、3つの潜在的活性触媒部位を含む3つの近接するOASユニットで構成される(ドメインI、II及びIII) (図3C)。組換えOAS3タンパク質の発現を、ヒトOAS3タンパク質のC末端領域に対して指向されたポリクローナル免疫血清を用いる、細胞溶解物のイムノブロッティングにより分析した(図3D)。1,000 IU.mL-1のIFN-αと5時間インキュベートした細胞は、陽性対照であった。IFN-α処理の際に、内因性OAS3分子の生成がHeLa.Tet-Off細胞において明確に検出されたが、休止細胞においては、OAS3の基底発現は観察されなかった。よって、OAS3遺伝子発現は、IFN-α/βにより能動的に上方制御され得る。組換えOAS3タンパク質の基底レベルは、非誘導HeLa.Tet-Off/OAS3細胞(2μg.mL-1 Tet)において観察され、このことは、遮断が不完全であったことを示す(図3D)。リプレッサーであるTetを培地から24時間除去したときに、非誘導細胞で見出されたものと比較して、組換えOAS3タンパク質の生成レベルが著しく増加した。
【0068】
実施例2:CHIKVに対するOAS3の抗ウイルス活性
1) 材料及び方法
実験手順は、実施例1に記載されるとおりである。
【0069】
2) 結果
ヒト上皮細胞におけるCHIKV成長に対するOAS3タンパク質の効果を調べるために、HeLa.Tet-Off/OAS3細胞に、CHIKV.06-49を1MOIにて18時間感染させた。MAb 3E4を用いるフローサイトメトリーにより決定されるように、非誘導HeLa.Tet-Off/OAS3及びHeLa.Tet-Off細胞は、CHIKV感染に対して同様の感受性を示した(図4A)。ウイルス曝露の24時間前のHeLa.Tet-Off/OAS3細胞の誘導は、CHIKV感染細胞を少なくとも20%減少させた。陽性対照として、IFN-α処理は、感染HeLa.Tet-Off細胞のパーセンテージを75%減少させた。よって、CHIKVは、ヒト上皮細胞におけるOAS3の異所発現に対して感受性である。
【0070】
HeLa.Tet-Off/OAS3細胞がCHIKV成長を阻害する効率をさらに評価するために、誘導した細胞を、漸増投入量のCHIKV-06.49に曝露した(図4B)。18時間p.i.にて、組換えOAS3タンパク質の発現は、子孫ウイルス生成を、試験したMOIに関係なく1.5〜2.0 log減少させた。24時間p.i.で、試験したより低いMOIにて、ウイルス力価は約3.0 log減少した。よって、OAS3の発現は、CHIKVの細胞から細胞への広がりを、ヒト線維芽細胞において抑制できる。中程度のMOIにて、ウイルス子孫は、少なくとも2.0 log、まだ減少した。10 MOIにて、ウイルス成長は約0.5 logしか減少せず、このことは、感染が高いウイルス投入量で行われたときに、CHIKVが、OAS3の抗ウイルス活性を回避する能力を有することを示唆した。
【0071】
動態研究は、CHIKVのOAS3により媒介される阻害が、ウイルス生活環の間に有効であったことを示した(図4C)。12時間p.i.にて、HeLa.Tet-Off/OAS3細胞から回収されたウイルスの力価は、HeLa.Tet-Off細胞と比較して、2.0 log減少した。OAS3タンパク質発現の誘導は、子孫ウイルス生成を、24時間p.i.にて少なくとも1.5 log、まだ減少させることができた。
細胞溶解の際に培養培地中に放出されるので膜の完全性の指標である細胞質酵素ラクテートデヒドロゲナーゼ(LDH)の測定は、感染させた親の細胞と比較して、感染の最初の24時間以内の感染HeLa/Tet-Off/OAS3細胞の生存性の著しい喪失を示さなかった。よって、CHIKV感染に対する耐性は、アポトーシス誘導によるウイルス感染細胞の排除よりもむしろ、OAS3の抗ウイルス活性に直接関連した。
【0072】
実施例3:ヒトOAS3を発現するHeLa.Tet-Off組換え株化細胞における、その他のプラス一本鎖RNAウイルスに対するOAS3の抗ウイルス作用
1) 材料及び方法
実験手順は、実施例1に記載されるとおりである。さらに、ウイルス生成及び力価決定は、以下に記載するようにして行った。
【0073】
ウイルス
継代数が低い(low passaged)西ナイルウイルス(WNV) IS-98-ST1株(GenBank受入番号AF 481864)の、蚊エーデス・シュードスクテラリス(AP61)細胞単層上での生成と、フォーカス免疫検出アッセイによるウイルス力価決定(Despresら, Virology, 1993, 196, 209〜219)は、以前に記載されたようにして行った(Brehinら, Virology, 2008, 371, 185〜195; 国際出願WO 02/081741)。感染性の力価は、AP61細胞でのフォーカス形成単位(FFU)として表した。シンドビスウイルスAR339株(SINV AR339)及びセムリキ森林ウイルスSF 64株(SFV 64)を、ミドリザル腎臓(VERO)株化細胞で繁殖させ、感染性の力価を、VERO細胞でのプラーク形成単位(PFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。
【0074】
2) 結果
大型のOASが、ヒト上皮細胞においてSINV AR339株、SFV SF 64株及びWNV IS-98-ST1株の成長を阻害する能力を調べた(図4D、5及び6)。HeLa.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞に、アルファウイルスを1MOIにて感染させた。OAS3タンパク質発現に応答して、18時間p.iにて、SINVとSFVとの間で、子孫ウイルスの生成が、同程度に(およそ2 log)阻害された(図4D)。HeLa.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞に、フラビウイルス(WNV)を0.1、1又は10 MOIで48時間感染させた(図5及び6)。ウイルス力価の測定に基づいて、ヒト上皮細胞が、OAS3発現に対して、WNウイルス複製を効率的に阻害することにより応答することが示された。0.1 MOIにて、HeLa.Tet-Off/OAS3から回収されたウイルス力価が、HeLa.Tet-Off細胞と比較して、1.5 log減少した(図6)。OAS3タンパク質発現の誘導は、子孫ウイルス生成を、感染後72時間で少なくとも1.0 log減少させることができた。よって、OAS3発現HeLa細胞は、親の細胞と比較して、WNウイルス感染に対するより低い感受性を示す。結論として、このデータは、OAS3が、ヒト細胞の内部でアルファウイルス及びフラビウイルスのようなRNAウイルスに対して抗ウイルス作用を有することを示す。
【0075】
実施例4:OAS3媒介CHIKV阻害の機構
1) 材料及び方法
実験手順は、実施例1に記載されるとおりである。さらに、定量的RT-PCRを、以下に記載するようにして行った。
【0076】
定量RT-PCR
ウイルスRNA蓄積のリアルタイムRT-PCR分析を、SYBR (登録商標) Green PCRを用いるABI Prism 7700配列検出で、実質的に以前に記載されたようにして行った(Kajaste-Rudnistkiら, J. Biol. Chem., 2006, 281, 4624〜4637)。CHIKV E2遺伝子についてのプライマーはChik/E2/9018/+及びChik/E2/9235/-である(表II)。GAPDH mRNAを、各試料の標準化のための内因性配列対照として用いた。プライマーGAPDH-For及びGAPDH-Revは、表IIに列挙する。ウイルスRNAコピーの定量のために、E2のN末端領域をコードするインビトロCHIK RNA転写産物を行って、以前に記載されたようにして標準曲線を構築した(Vazeilleら, PLoS One, 2007, 2(11): e1168)。
【0077】
2) 結果
OAS3の抗ウイルス作用の分子的な基礎を解明するための努力において、CHIKV成長の阻害が、ウイルス構成タンパク質の蓄積の欠如によるものかどうかを決定した。全タンパク質をHela.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞から、感染後18時間の時点で抽出し、カプシドタンパク質(C)、エンベロープ糖タンパク質pE2 (E2の前駆体)、E2及びE1を、抗CHIKV HMAFを用いるイムノブロットにより検出したが、エンベロープ糖タンパク質pE2 (E2の前駆体)及びE2は、抗CHIKV E2 MAb 3E4を用いるイムノブロットにより特異的に検出した。図7Aに示すように、OAS3タンパク質発現に応答して、バックグラウンドレベルでのウイルス構造タンパク質が存在した。よって、CHIKVが、OAS3発現HeLa細胞に生産的に感染できないことは、ウイルスタンパク質合成における著しい減少と関連する。
【0078】
ウイルスタンパク質合成の無能性が、ウイルスRNAの蓄積の欠如によるものかどうかを決定した。トータルRNAをHela.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞から、感染後8時間の時点で抽出し、ゲノム及びサブゲノムのウイルスRNAの生成を、CHIKV E2遺伝子に基づいて設計したプライマーを用いるRT-PCRを用いて分析した。リアルタイムRT-PCR分析により決定されるように、親の細胞で見出されたものと比較すると、OAS3発現細胞から回収されたウイルスRNAレベルは2 log減少した(図7B)。つまり、OAS3発現は、感染細胞の内部でウイルスRNAの蓄積を妨げる。これらの結果に基づいて、OAS3発現が、感染ヒト細胞の内部でアルファウイルスの複製に影響すると結論付けることができる。
【0079】
実施例5:OAS3抗ウイルス効果に対する健常個体でのOAS3遺伝子多型の影響
1) 材料及び方法
実験手順は、実施例1に記載したとおりである。さらに、OAS3多型の遺伝子型同定は、以下に記載するようにしてアッセイした。
【0080】
遺伝子型同定
SNP OAS3.R844Xは、表IIに示すプライマー対OAS3.R844X-F(配列番号11)及びOAS3.R844X-R (配列番号12)を用いるPCR-RFLPアッセイにより遺伝子型同定した。183 bp長のPCR生成物を、Bgl IIを用いる消化に付し、115及び68 bpの2つのフラグメントの検出が、OAS3.R844Xの存在を示した。
【0081】
2) 結果
OAS3遺伝子を、多型についてスクリーニングした。48名の健常白人個体における合計で16個のエキソン、フランキングイントロン及びOAS3遺伝子の5'側2 kbの再配列決定は、コドンCGA-844の最初の位置での1塩基多型(SNP)を同定し、これは、CのTへの置換がナンセンス変異をもたらす(OAS3.R844X) (表I)。このSNPを、180名の健常白人個体においてさらにスクリーニングし、0.5%の対立遺伝子頻度を与える2つのヘテロ接合体が同定された。OAS3.R844XはOAS3タンパク質のカルボキシ末端から約20%がおそらく切断されているので(図3C)、OAS3-1〜OAS3-843配列を含む安定HeLa.Tet-off/OAS3ΔC-term細胞クローンを作製した。OAS3ΔC-termは、OAS3の第3ドメインの活性触媒部位を規定する三連構造を構成するD889残基を欠くことに注目することが興味深い(Sarkarら, 1999; Rambouillatら, 2000)。変異OAS3がCHIKV及びWNVの成長を阻害する能力を評価するために、誘導HeLa.Tet-Off/OAS3及びHeLa.Tet-off/OAS3ΔC-term細胞に、1MOIにてCHIKV-06-49又は西ナイルウイルス(WNV) IS-98-ST1株を感染させた。図8及び9に示すように、OAS3ΔC-termを発現する細胞は、CHIKV及びWNV感染に対する耐性を示し、このことは、OAS-1〜OAS3-843配列がまだ保護を提供できることを示す。ウイルス成長の分析は、OAS3ΔC-termの異所発現が、全長OAS3と比較してCHIKV及びWNVの阻害のより低い効率をもたらしたことを示した。このような結果は、コドンCGA-844でのSNPが抗ウイルス活性に対して影響を有し得ることを示唆する。
【0082】
実施例6:ヒトOAS3を発現するHep.G2.Tet-Off株化細胞におけるDV及びWNVに対するOAS3の抗ウイルス作用
1) 材料及び方法
a) 細胞培養
HepG2.Tet-Off株化細胞を、CLONTECH (#632106)から購入した。HepG2.Tet-Off細胞は、5% CO2中で37℃にて、10%熱不活化胎児ウシ血清(FCS)、4 mM L-グルタミン、100 UI/mlペニシリンGナトリウム、100μg/mlストレプトマイシンサルフェート(GIBCO, #15140-122)、0.1 mM非必須アミノ酸、100μg.mL-1 G418 (GIBCO, #10131-019)を補ったDMEM (GIBCO, #41965)中で維持する。
HepG2.Tet-Off/OAS3#F8株化細胞(CNCM I-4158)は、DMEM、10% FCS、4 mM L-グルタミン、100 UI/mlペニシリン、100μg/mlストレプトマイシン、100μg.mL-1 G418、100μg.mL-1ハイグロマイシンB (CLONTECH, #631309)、及び2μg.mL-1テトラサイクリン(SIGMA-ALDRICH, # T-7660)中で維持する。
【0083】
b) OAS3タンパク質を発現するHepG2.Tet-Off株化細胞の樹立
全長ヒトOAS3タンパク質(配列番号24)を発現する誘導性HepG2.Tet-Off/OAS3#F8クローンを、誘導性HeLa.Tet-Off/OAS3#C417-1について実施例1に記載するようにして選択した。HepG2.Tet-Off/OAS3#F8株化細胞は、2009年5月15日に、Collection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-4158の下で寄託した。
この株化細胞を、2μg.mL-1 Tetの存在下の抑圧条件下で維持する。
【0084】
c) ウイルス
継代数が低い西ナイルウイルス(WNV) IS-98-ST1株(GenBank受入番号AF 481864; Lucasら, Virology J., 2004, 1, 9〜)の、蚊エーデス・シュードスクテラリス(AP61)細胞単層上での生成、及びフォーカス免疫検出アッセイによるウイルス力価決定(Despresら, Virology, 1993, 196, 209〜219)は、以前に記載されたようにして行った(Brehinら, Virology, 2008, 371, 185〜195)。DV1 FGA/NA d1d株の生成及びウイルス力価決定は、以前に記載されたようにして行った(Duarte dos Santosら, Virology, 2000, 274, 292〜)。感染性の力価は、AP61細胞上でのフォーカス形成単位(FFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。
【0085】
d) 免疫蛍光アッセイ
ウイルス感染細胞を、メタノール/アセトンを用いて-20℃にて20分間固定した。免疫蛍光アッセイを、FITC-結合抗デングE mAb 4E1又はcy3-結合抗WNV.E MAb E24を用いて行った。核をDAPIで染色した。
【0086】
2) 結果
まず、親のHepG2.Tet-Off株化細胞のDV感染に対する感受性を評価した(図10A)。細胞を、細胞あたり異なるフォーカス形成単位(AP61FFU)のDV-1ウイルスFGA/NA d1d株に感染させた。感染後40時間にて、細胞を、DV E糖タンパク質に反応性のMAb 4E11を用いるフローサイトメトリーにより分析した。細胞あたり30 AP61FFUのDV-1ウイルス投入量で、約50%のHepG2.Tet-Off細胞が、デングE糖タンパク質についての陽性シグナルにより示されるように、DV-1ウイルスに感染した。
次いで、IFN-αがHepG2.Tet-Off細胞において抗ウイルス状態を確立する能力を調べた。HepG2.Tet-off細胞を1,000 IU.mL-1のヒトIFN-αでDV感染(10 MOI)の5時間前に前処理すると、40時間p.i.にて約0.7 logのウイルス力価が減少した(図10B)。つまり、IFN依存性抗ウイルス経路はHepG2.Tet-Off細胞において機能的であり、細胞レベルにてDVに対する保護を提供する。
【0087】
Hep.G2細胞におけるDV成長に対するOAS3タンパク質の効果を調べるために、誘導(-Tet)及び非誘導(+Tet)のHepG2.Tet-Off/OAS3細胞、並びに親のHepG2.Tet-Off細胞に、漸増投入量のDV-1ウイルスFGA/NA d1d株を感染させた。40時間p.i.にて、抗デングE mAbを用いる免疫蛍光アッセイを行って、ウイルス抗原について陽性のHepG2細胞のパーセンテージを決定した(図11及び12)。子孫ウイルスの生成を、25の感染多重度にて決定した(図11)。組換えOAS3タンパク質の発現は、DV感染細胞のパーセンテージを少なくとも75% (図11及び12)、及び子孫ウイルスの生成を2 log (図11)減少させた。
つまり、OAS3発現HepG2細胞は、DV感染に対する耐性を示す。
【0088】
Hep.G2細胞におけるWNV成長に対するOAS3タンパク質の効果も、免疫蛍光アッセイにより分析した。親のHepG2.Tet-Off及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、西ナイルウイルスIS-98-ST1株を1 AP61FFU/細胞の感染多重度で感染させたか、又は模擬感染させた。感染後40時間にて、ウイルス抗原(WNV E糖タンパク質)を、cy3-結合mAb E24を用いる免疫蛍光により決定した。結果(図13)は、HeLa細胞において得られた結果(実施例3)を確認し、OAS3発現ヒト細胞がWNV感染に対する耐性を示すことを示す。
【0089】
実施例7:黄熱ウイルスの生弱毒化17D-204ワクチン株は、ヒト細胞において、ヒト大型2',5'-オリゴアデニル酸合成酵素(OAS3)により媒介される抗ウイルス効果に対して驚くべき耐性を示す
1) 材料及び方法
弱毒化黄熱ウイルス(YFV)ワクチン株(STAMARIL (登録商標)、AVENTIS-PASTEUR)を、ミドリザル腎臓(VERO)株化細胞にて繁殖させ、感染性の力価を、VERO細胞でのプラーク形成単位(PFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。免疫蛍光アッセイを、抗フランス神経親和性ウイルスHMAF及びFITC-結合ヤギ抗マウスIgを用いて実施例6に記載されるようにして行った。
【0090】
2) 結果
実施例2、3、4及び6は、IFN-α/βにより媒介されるOAS3依存性抗ウイルス活性が、チクングンヤウイルスのようなアルファウイルス並びに西ナイルウイルス及びデングウイルスのようなフラビウイルスに対する主なヒト細胞防御ストラテジーをヒト細胞において示すことの初めての証拠を提供した。
OAS3が黄熱ウイルス(YFV)に対する抗ウイルス活性を示すかを調べた。このために、YFVの生弱毒化17D-204ワクチン株(STAMARIL, Aventis-Pasteur)を、ミドリザル腎臓(VERO)株化細胞上で2回繁殖させ、感染性の力価を、VERO細胞でのプラーク形成単位(PFU)として表した。HeLa細胞に、YFV 17Dを1PFU/細胞にて感染させ、次いで、感染後の種々の時点にて1,000 IU/mlのヒトIFN-αで処理したか又は模擬処理した(対照)。ウイルス子孫生成を、感染後72時間にて決定した。図14に示すように、IFN-αは、ヒト上皮HeLa細胞においてYFV 17Dに対する抗ウイルス状態を確立できる。OAS3がウイルス成長を阻害する能力を評価するために、HeLa (対照)及び誘導HeLa.Tet-Off/OAS3 (OAS3)細胞を、YFV 17D株に、低(1PFU/細胞; 図15A)又は高(10 PFU/細胞; 図15B)ウイルス投入量にて曝露した。両方の細胞集団の間で、YFV複製についての明確な違いはなかった(図15)。動態研究は、17Dが、ヒト細胞におけるOAS3により媒介される抗ウイルス効果に対して、感染後の時間又は感染多重度のいずれにも関係なく、耐性であることを示した。
【0091】
YFV 17DがOAS-3により媒介される抗ウイルス効果に対して耐性であることは、YFV 17Dに感染させた親のHepG2.Tet-Off及びHepG2.Tet-Off/OAS3#F8細胞クローンにおける免疫蛍光アッセイによって確認された(図16)。
YFVの生弱毒化株が、OAS3の抗フラビウイルス作用に対して予期せぬ耐性を示すとの知見は、フラビウイルス弱毒化の機構を解明する新たな道を開く。
【0092】
実施例8:OAS3の非同義バリアントは、デングショック症候群の病因と関連する。
1) 材料及び方法
a) 患者及び対照
Bangkokの2つの医療センターRamathibodi病院及びSiriraj病院、Mahidol University、並びにタイのKhon-Kaen州の1つの病院からの3年間(2000年6月〜2003年)の症候性デングウイルス(DV)感染の750名の患者(男性:女性の比率=0.99)を、研究に含めた。患者の年齢は、1〜25歳の範囲であり、平均は9.6歳であった。高熱、重度の頭痛、後眼窩痛、筋肉痛、関節痛、悪心及び嘔吐、並びに発疹を含む臨床上の特徴に基づいてデングウイルス感染が疑われる患者を、臨床観察及び治療のために病院に収容した。デングウイルス感染の診断は、のちに、後期急性期及び/又は回復期の血清に対する比較IgG及びIgM酵素結合免疫吸着アッセイ力価決定により確認した。DF (デング熱)及びDHF (デング出血熱)を区別する診断は、血液濃縮又は胸水貯留により表される血管透過性の増加についての証拠の不在(DF)又は存在(DHF)に基づいて確立した。具体的に、DHFの診断は、以下の4つの特徴の全てに基づいて行った:1) 2〜7日間連続する高熱、2) 駆血帯試験の陽性、点状出血(petechii)、紫斑、又は吐血のような出血性の傾向、3) 血小板減少(血小板計数<100,000/μl)及び4)血液濃縮(20%以上のヘマトクリットの増加)又は胸水貯留により表される血管透過性の増加による血漿漏出の証拠。臨床症状が明確でないのでDF又はDHFと分類できなかったいくらかのデング患者を、未定DF/DHF状態に割り当てた。DHFの重症度は、WHO基準に従う4つのグレードにより分類した。グレードIII及びIVは、特徴的な拡張期圧の上昇から重度のショックまでを有する脈圧の狭小化を伴うDHFであった。二次感染は、デング特異的IgM/IgG比<1.8と規定した。計画のプロトコル及び研究の目的は、患者及びその両親又は親族に注意深く説明した。インフォームドコンセントは、全ての対象者から個別に得た。プロトコルは、各病院の倫理委員会により承認されている。
【0093】
対照は、Ramathibodi病院からの296名の血液提供者及びSiriraj病院からの216名の血液提供者、並びにKhon-Kaen地域からの184名の人種適合健常対象者(男性:女性の比率=1)からなった。症例及び対照群はともに、タイのBangkok及び中央部に由来した。全血試料を患者及び対照からEDTA上に回収し、DNAを、標準的なフェノール/クロロホルム抽出法を用いて抽出した。
2004〜2006年の間に、さらに254名の患者(男性:女性の比率=1.07)を、Ramathibodi及びKhon-Kaen病院から同じ臨床的及びウイルス診断基準を用いて含めた。
【0094】
b) 多型の同定及び遺伝子型同定
多型は、個別のDNA試料(24名のデング患者及び32名のタイ人対照)に対するPCR増幅ゲノムDNAの直接配列決定により同定した。OASのエキソン、イントロンの一部分、並びに5'及び3'領域を配列決定するために用いたプライマーを、表IIIに示す。
【0095】
【表3−1】
【0096】
【表3−2】
【0097】
配列決定の結果は、Genalysソフトウェア(Takahashiら, J. Bioinform. Comput Biol., 2003, 1, 253〜265)により分析した。OAS3-R381Sを、推奨プロトコルを用いてABI Prism 7000配列決定システムを用いるTaqManアッセイにより遺伝子型同定した。OAS3配列のより一般的なバリアントのコドン381は、アルギニンに相当するAGGである。rs2285933と命名したOAS3-R381Sの多型は、GからCへの置換を、OAS3のコドン381の3番目のヌクレオチドにて有し(AGGからAGC)、これはアミノ酸をアルギニンからセリンに変える。OAS3の2つのバリアントであるOAS3-(R381及びS381)のmRNA、ORF及びアミノ酸配列は、それぞれ配列番号28、29、30 (R381)並びに配列番号27、1及び2(S381)に相当する。
【0098】
c) 統計分析
関連の研究を、Haploviewプログラム(Barrettら, Bioinformatics, 2005, 21, 263〜265)に組み込まれるPearsonのカイ二乗検定により、対立遺伝子カウントを用いて行った。このプログラムは、並べ替え検定を行って、経験的P値を得ることを可能にする。遺伝子型の結果をコードするロジスティック回帰分析を、STATAバージョン9を用いて試験した遺伝子モデルに従って行った。
【0099】
2) 結果
2000年〜2003年の間に、580名のデング患者をBangkokの2つの病院(Siriraj, SI及びRamathibodi, RA)から、及び170名をタイ北東部のKhonkaen州(KK)の別の病院から採用した。Bangkok の同じ病院からの512名の血液提供者、及びタイの北東地域からの184名の健常ボランティアからDNA試料を回収し、一般集団における遺伝子頻度を代表した。これらの患者及び対照の一部の遺伝子研究は、以前に報告された(Sakuntabhaiら, Nature Genetics, 2005, 37, 507〜513)。ウイルス診断は、血清試験により確認され、患者は、WHO基準に従って4つの群に分類された:DF (血漿漏出の証拠なし)及びDHF1 (血漿漏出の証拠)、DHF2 (自発的出血を伴う)及びDSS (ショックを伴う) (表IV)。
【0100】
【表4】
【0101】
WHO基準に従ってDF又はDHFを有すると分類できなかった患者は、51名であった(未分類;UC)。それぞれの重症度カテゴリーの患者の割合は、病院間で有意に異なっていた(P =1.5×10-8)。このことは、病院のタイプ(KKについて二次看護、SI及びRAについて医科学校)及び採用基準(KKにおいて3日未満の発熱、SI及びRAにおいてデング症例の疑い)における違いを反映する。
デング感染に対してIgM応答が優勢な患者と比較して、IgG応答が優勢であった患者において、血漿漏出(DHF1/DHF2/DSS対DF)及び自発的出血(DHF2/DSS対DF/UC/DHF1)の危険性の非常に有意な増加があった(それぞれP = 1.1×10-7及びP = 7.6×10-5)が、ショックについては限界有意(marginal significance)のみであった(DSS対残り、P = 0.038) (表V)。
【0102】
【表5】
【0103】
これらの知見は、DVに対するIgG応答を有する個体が、疾患のより重症な形態にかかりやすくなるという見解と一貫する。
タグ付加された多型のスクリーニングについて、関連についての試験を、血漿漏出の危険性(DHF1/DHF2/DSS対DF)、自発的出血の危険性(DHF2/DSS対DF/UC/DHF1)、ショックの危険性(DSS対非ショックDV患者)についての対立遺伝子カウントを、Bangkokの2つの病院からの患者について行った(表VI)。
【0104】
【表6】
【0105】
表VIに示すように、いずれの多型も疾患の危険性との関連の証拠を示さなかった。しかし、1つのOAS3の非同義多型が、疾患の重症度に依存する非常に著しく異なる対立遺伝子カウント分布を有した。このバリアント(OAS3-R381S: rs2285933)は、OAS3のコドン381の3番目のヌクレオチドでのGからCへの置換であり、これはアミノ酸をアルギニンからセリンに変更する。両方の病院において、この多型のC対立遺伝子は、DF、DHF1及びDHF2において、DSSよりも頻度が高かった。2つの病院間の患者の各群と対照との間に対立遺伝子頻度の著しい差はなかった。組み合わせ分析において、C対立遺伝子は、一般集団において14%、DFにおいて20%、DHF1及びDHF2において15〜16%、DSSにおいて7%の頻度を有した。DSSと非ショックDV症例との間の違いは、非常に有意であった(P = 7.10-4)。我々は10,000の並べ替えを行い、0.0032の経験的P値を得た。
【0106】
遺伝の様式
OAS3-R381Sの遺伝の様式を見出すために、C対立遺伝子の相加、優性及び劣性の3つの異なる遺伝様式に基づくモデルの尤度を、DSSの危険性に対するロジスティック回帰を用いて比較した。C対立遺伝子の劣性モデルは著しい関連を示さなかったが、C対立遺伝子の優性(<10-5 P<10-5, OR = 0.30 95CI = 0.17-0.53)及び相加(17.93, P<10-5)モデルを用いて高度に有意なレベルが得られた。log尤度比検定により、優性モデルは、相加モデルよりも有意により適切である(P = 0.04)。よって、我々は、残りの我々の分析のために優性モデルを用いた。
【0107】
第3コホートにおける再現
我々は、同じ流行(2001〜2003年)中の3番目の病院(KK)からの患者におけるOAS3-R381Sの再現研究を行った。C対立遺伝子は、以前の結果と同様に、DSS群において、ショックのないDV患者と比較してより低い頻度を示した(C対立遺伝子頻度について8%対17%、CG+GG頻度について17%対32%)。この差は、患者が少数であるので統計学的に有意でなかったが、3つの病院からの全ての患者を組み合わせたときに、より高い有意性の関連が得られた(C対立遺伝子の優性モデルについてOR = 0.48, 95 CI = 0.31-0.75, P= 7×10-4)。
【0108】
OAS3及び免疫応答のタイプ
我々は、OAS3-R381Sの影響が、ウイルスに対する免疫応答のタイプに依存するかをさらに調べた。患者における多型及びDSSの、優勢IgM免疫応答との著しい関連はなかった。しかし、各病院において及び組み合わせた場合に、優勢IgG応答を有する患者において、有意及びほぼ有意の関連が得られた(Chi2 = 19.41, OR = 0.27 95 CI = 0.14-0.53, P<10-5) (表VII)。
【0109】
【表7】
【0110】
2004〜2006年流行中の患者の関連研究
2004〜2006年の間に、我々は、以前の2つの病院(RA及びKK、表IV)からさらなる患者を得た。OAS3-R381Sの関連研究は、研究した全ての仮定との有意な関連を明らかにしなかったが、これらの患者を元の症例/対照研究に加えた場合に、有意な関連が残っていた(OR = 0.48, 95 CI = 0.31-0.75, P= 7×10-4)。この結果は、2つの期間の結果の間の不一致を調べる動機となった。
デング疾患の重症度に貢献し得る1つの因子は、ウイルス因子である。我々は、我々が471名の患者のデータを有するSiriraj病院及びKhonkaen病院からの患者のデータを用いて、2001〜2006年の間のDV-4血清型の罹患率を調べた(表VIII)。
【0111】
【表8】
【0112】
DV1及びDV2血清型は2001〜2003年の間に優勢であり、2004年においてDV1は減少し、DV4の流行が2004年に開始し、次いで、2006年まで優勢であった(表5A)。デング疾患の重症度は、ウイルスの異なる血清型の間で有意に異なっており(P = 0.012、表5B)、DV2血清型はその他のDV血清型よりもより重症の症例があった。驚くべきことに、ウイルスの血清型は、免疫応答のタイプによって有意に異なっており(フィッシャー正確P<10-4、表5C)、DV2及びDV4は、優勢IgGを示した。これらの結果は、Ministry of Public Healthからのデータと同様であった。我々は、OAS3-R381Sの影響が、いくつかのウイルス血清型について特異的であり得ると仮定した。我々は、DSSに対するOAS3-R381Sの有意に優勢な保護効果を、DV2でのみ見出した(OR = 0.24, 95 CI = 0.08-0.74, P = 0.0045, 表IX)。
【0113】
【表9】
【0114】
DF/DHF/DSS患者及び対照におけるOAS遺伝子の系統的スクリーニングに基づいて、我々は、OAS3遺伝子の多型とショックの危険性(DSS)、及びデングウイルスに感染した患者における疾患の重症度との間に関連を示した。バリアント(OAS3-R381S: rs2285933)は、OAS3のコドン381の3番目のヌクレオチドのGからCへの置換であり、これは、アミノ酸をアルギニンからセリンに変更する。Cバリアントは、非ショックデング疾患と比較してDSSに対する優勢な保護と関連し、頻度の比は0.13:0.32である(OR = 0.48, 95 CI = 0.31-0.75, P= 7×10-4)。さらに、優勢なIgG応答を有する患者(OR = 0.46, 95 CI = 0.28-0.74, P = 8×10-4)及びDV血清型2に感染した患者(OR = 0.24, 95 CI = 0.08-0.74, P = 0.0045)においてのみ、著しい関連が見出された。
【0115】
これらの結果は、非同義多型OAS3-R381Sの機能的役割を支持し、デング病因におけるOAS3の役割と、OAS3のバリアントとウイルスの血清型との間の相互関係とを証明する。
我々の知見は、DV感染患者における疾患重症度の予測、並びにデングウイルス及び医療的に重要性が高いその他のプラス一本鎖RNAウイルスによる感染に対するOAS3に基づく予防及び治療の開発について重要な影響を有するであろう。
【受託番号】
【0116】
CNCM I−3927
CNCM I−3968
CNCM I−4158
【図1−1】
【図1−2】
【図17−1】
【図17−2】
【技術分野】
【0001】
本発明は、プラス一本鎖RNAウイルスへの感染の予防又は治療のための医薬品としてのヒト大型(large form) 2',5'-オリゴアデニル酸合成酵素(OAS3)に関する。
本発明は、プラス一本鎖RNAウイルスへの感染に対する遺伝的感受性を決定するためのマーカーとしてのヒト大型2',5'-オリゴアデニル酸合成酵素(OAS3)にも関する。
【背景技術】
【0002】
フラビウイルス科(Flaviviridae)及びトガウイルス科(Togaviridae)は、世界的にヒト及び動物の健康に影響し得る病原体を含む2つのプラス一本鎖(ss)RNAウイルスの科である。これらのウイルスの例は、フラビウイルス(Flavivirus)属(フラビウイルス科)のデング、黄熱(YF)、日本脳炎(JE)及びダニ媒介脳炎(TBE)の抗原性群(antigenic complexes)のメンバー、アルファウイルス(Alphavirus)属(トガウイルス科)の東部ウマ脳炎(EEE)/ベネズエラウマ脳炎(VEE)、セムリキ森林(SF)及びシンドビス(SIN)群のメンバー、並びにヘパシウイルス(Hepacivirus)属(フラビウイルス科)のメンバー、例えばC型肝炎(HCV)及びG型肝炎(HGV)ウイルスを含む。
【0003】
デングウイルス(DV; フラビウイルス属のDEN抗原性群)は、熱帯のほとんどの都市中心において流行性である。なぜなら、都市化の劇的な増加が、蚊媒介デング疾患の伝達の増加に理想的な条件を作り出したからである。デングウイルスの4つの血清型(DV-1〜DV-4)は、媒介体である蚊のネッタイシマカ(Ae. aegypti)によりヒトに伝達される。DV感染は、流感様疾患(デング熱、DF)から、デングショック症候群(DSS)及び死に発展し得るデング出血熱(DHF)までにわたる連続する疾病をもたらす。現在までに、デング疾病は、推定100百万件のヒトにおける最も重要なアルボウイルス症であり、各年に500,000件を超えるDHF/DSSが発生し、約25,000件の死に至る件を含み、これは主に15歳未満の子供である。DHF/DSSの高い頻度の流行病は、アジア及び南アメリカにおいて地理的に拡大し続けている。健康及び経済的な影響の増加にもかかわらず、デング疾患の病因は、現在のところほとんど理解されていない。抗デングワクチン又はデングウイルス感染の治療のための特定の療法は利用可能でない。
【0004】
西ナイルウイルス(WNV; フラビウイルス属のJE抗原性群)は、蚊(クレクス(Culex)種)及び鳥類を含む天然の伝達サイクルで循環し、ウマ及びヒトが偶生宿主である。人畜共通感染症のWNVは、1998年にイスラエルで高度に神経浸潤性の株(WNウイルスI型系統のクレードIaからの変種Isr98/NY99)が出現したので(Ceccaldiら, FEMS Microbiol. Lett., 2004, 233, 1〜6)、北米、中東及び欧州で主な健康問題になった。WNVは、中枢神経系に感染し、広い範囲の動物種においてウイルス性脳炎を引き起こす。ヒトにおいては、臨床感染は、無併発性の西ナイル熱から致死的な髄膜脳炎までの範囲の重症度にわたり得る。WNVの出現は、ヒトにおける感染の重症度の劇的な増加に関連し、ウイルス性脳炎が公衆衛生の問題として注目を集めることとなった。過去10年間に、WNVは、西半球及びカリブ海諸国に広がった。WNVの米国での大発生は、数千人の患者を伴い、重篤な神経疾患(髄膜脳炎及び灰白髄炎様症候群)及び数百名の関連する死者の原因となった。WNVの蚊媒介伝達は主な形態であるが、輸血、臓器提供及び胎児への経胎盤伝達により伝達されるWNV感染も認められた。WNV関連疾患のためのワクチン又は抗ウイルス療法は利用可能でない。
【0005】
チクングンヤウイルス(CHIKV; アルファウイルスのSF群)は、アフリカ、東南アジア、インド及び西太平洋に広まっており、多数の流行病がこれらの地域で報告されている。臨床的には、CHIKVへの感染は、発熱、発疹及び強く、無気力にさせ、ときに持続的な関節痛をもたらす。2005〜06年に、CHIKウイルスは、インド洋の南、特にレユニオン諸島(フランス)に広がり、ここでの大発生は、数十万の患者を伴った。より最近では、このウイルスはイタリアの東部で出現し、ここでは200名を超える人が感染した。
【0006】
C型肝炎ウイルス(HCV)感染は、世界的に一般的である。世界の人口の約3%がHCVを有し、欧州だけで約4百万のキャリアがいると見積もられる。これらの個体の20〜30%が肝硬変、及び肝細胞癌のような長期の続発症を発症する。リバビリンと組み合わせたPEG化インターフェロンアルファ(IFN-α)を用いるHCV感染の治療は、HCVウイルス遺伝子型2又は3に感染した患者において持続した応答を達成できる(80%)が、HCVウイルス遺伝子型1に感染した患者においては、応答率がかなり低い(42%)。
【0007】
I型インターフェロン(IFN-α/β)により媒介される先天的な抗ウイルス機構は、ウイルス複製を制限する宿主細胞防御の最も重要な経路の可能性がある。実際に、IFN-α/βは、特異的シグナル伝達経路の活性化を引き起こして、IFN-活性化遺伝子(ISG)の誘導を導くことができ、これは抗ウイルス状態の確立を担う。単細胞においてRNAウイルスの複製に影響すると考えられるISGは、RNA特異的アデノシンデアミナーゼ(ADAR)、ミクソウイルス耐性(Mx)ファミリーのタンパク質、2本鎖RNA依存性タンパク質キナーゼ(PKR)、及びエンドリボヌクレアーゼRNase Lと関連する2',5'-オリゴアデニル酸合成酵素(2',5'-OAS又はOAS)ファミリーである。
【0008】
OAS/RNase L系は、確立された内因性抗ウイルス経路において重要な役割を有することが知られているRNA分解経路である。酵素的に活性なOASが、アクチベーターである2本鎖(ds)ウイルスRNAに結合すると、2'-5'結合オリゴアデニル酸(2-5A)の生成をもたらす。潜在性の単量体RNase Lは、2-5Aオリゴマーへの結合により誘導されるホモ二量体形成により酵素的に活性化される。一旦活性化されると、RNase Lは、mRNA及びウイルスRNAを含む1本鎖RNA分子を分解し、ウイルス複製を抑制する(Silverman, J. Virol. 81: 12720, 2007)。
【0009】
ヒトOASは、以下の順序での染色体12q24.2上の3つの密接に結合した遺伝子によりコードされる酵素のファミリーである:小型(OAS1, p40/46)、中型(OAS2, p69/71)及び大型(OAS3, p100)のOASアイソフォーム(Hovnanianら, Genomics, 1998, 52, 267〜277; Rebouillat, D.及びHovanessian, A.G., Journal of Interferon and Cytokine Research, 1999, 19, 295〜308; Rebouillatら, Genomics, 2000, 70, 232〜240; Justesenら, Cellular and Molecular Life Sciences, 2000, 57, 1593〜1612; Rebouillat; Hovanessian, A.G., Cytokine and Growth Factor Reviews, 2007, 18, 351〜361)。各OAS遺伝子は、5つの翻訳されるエキソン(エキソンA〜E)で構成される保存OASユニットからなる。OAS1は1つのユニットを有するが、OAS2及びOAS3はそれぞれ2つ及び3つのユニットを有し、3つ全ての遺伝子は、活性型2',5'-オリゴアデニル酸合成酵素をコードする。別の遺伝子であるOASL (OAS様)は、単独のユニットのOAS様タンパク質をコードするが、これは2'-5'合成酵素活性を欠く(Hartmannら, Nucleic Acids Res., 1998, 26, 4121〜4128; Rebouillatら, Eur. J. Biochem., 1998; 257, 319〜330)。それぞれのサイズクラス内で、最初の転写産物の選択的スプライシングの結果として、複数のメンバーが生じる。OASタンパク質は、約350アミノ酸の保存ユニット/ドメイン(OASユニット)を共有する;OAS1 (p40/p46)、OAS2 (p69/71)及びOAS3 (p100)は、OASユニットのそれぞれ1つ、2つ及び3つのタンデムコピーを有する。各OASタンパク質は、細胞で異なる位置に蓄積し、活性化されるために異なる量のdsRNAを必要とし、異なるサイズの2-5A生成物の形成を触媒する。OAS1は四量体として機能するが、OAS2は二量体としてのみ活性であり、OAS3は単量体としてのみ観察されている。さらに、大型のヒトOASは、おそらく、RNase L活性化に関与しない(概説として、Rebouillat, D.及びHovanessian, A.G., Journal of Interferon and Cytokine Research, 1999, 19, 295〜308)。
【0010】
OASファミリーが、IFNにより示される抗ウイルス効果に関与することについての最初の直接的な証拠は、2',5'-オリゴアデニル酸合成酵素(OAS) cDNAを細胞に形質移入することにより得られた。OAS1又はOAS2の過剰発現は、ピコルナウイルス複製に対する細胞の耐性を導く(Hovanessian, A.G., Cytokine and Growth Factor Reviews, 2007, 18, 351〜361)。インビボにおいてWNV感染を除去するためのOAS1の重要性は、ヒトOAS1のオルソロガス遺伝子であるマウスOas1bが、WNV誘導性脳炎に対するマウスの感受性/耐性表現型において重要な役割を有し得るとの知見によっても支持された(Mashimo, T.ら, Proc. Natl. Acad. Sci. USA, 2002, 99, 11311〜11316; Lucasら, Immunol. Cell. Biol., 2003, 81, 230〜236; Kajaste-Rudnitskiら, Journal of Biological Chemistry, 2006, 281, 46244637; 国際PCT出願WO 02/081741)。ヒトにおけるOAS遺伝子多型の分析は、OAS遺伝子中の遺伝マーカーが酵素活性と最も強く関連することを示した。OAS1が、ウイルス感染に対する宿主の感受性に影響するヒト遺伝子としての優れた候補であるとすると(Bonnevie-Nielsenら, Am. J. Hum. Genet., 2005, 76, 623〜633)、ヒトOAS1及びOASL遺伝子における遺伝子的変動は、ウイルス性脳炎、1型DM、HCV関連疾患及びその他のウイルス感染の危険性と関連した。HCV疾患について特に強調すると、HCV感染の結果と関係する一連のOAS1遺伝子型が報告されている(国際PCT出願WO 03/089003及びWO 2005/040428)。
【0011】
OASファミリーの抗ウイルス活性がプラス鎖ssRNAウイルスに対して選択的であるかは、まだ調べられていない重要な問題である。OAS1の異所発現が、脳心筋炎ウイルス(ピコルナウイルス)複製に対する耐性を導くが、VSVウイルス複製に対しては耐性を導かないことが報告されている。しかし、OASは、IFN誘導性RNA依存性プロテインキナーゼ(PKR)及びミクソウイルス耐性1 (MxA)遺伝子のようなその他のISGとは反対に、IFN-αで処置された患者でのHCVの除去において役割を示さないと考えられている。OAS、PKR及びMxの転写レベルは、非ヒト霊長類モデルにおいてDV感染の後に上方制御された(Sariolら, Clinical and Vaccine Immunology, 14, 2007, 756〜766)。デングショック症候群に関連する遺伝子転写パターンの分析は、OAS3遺伝子転写産物が、非DSSのデング患者におけるよりもDSSのデング患者においてより少ないことを示した (Simmonsら, J.Infect.Dis., 195:1097, 2007)。しかし、DV感染の病因におけるこれらのISGの役割は、ほとんど理解されていない。
【0012】
CHIKVは、I型インターフェロン(IFN-α/β)の抗ウイルス作用に対して感度が高いことが報告されている(Coudercら, PloS Pathogens, 2007, 4,e29)。アルファウイルスの場合、ウイルス成長のIFN媒介性阻害はRNase Lを必要としないことを示唆する一連の証拠が存在する(Rymanら, J. Virol., 2005, 79, 1487〜1499; 概説としてSilverman, J. Virol., 2007, 81, 12720〜9)。OASファミリーのようなヒトISGのメンバーのいずれかが、抗アルファウイルス活性を示し得るかが、まだ調べられていない重要な問題である。
【発明の概要】
【発明が解決しようとする課題】
【0013】
アルファウイルス(CHIKV、SINV及びSFV)のようなプラス鎖ssRNAウイルスに対する確立された内因性抗ウイルス経路におけるOAS3の役割を、本発明者らが初めて立証する。本発明者らは、OAS3が、感染ヒト上皮細胞の内部でウイルスタンパク質合成及びウイルスRNA複製を遮断してCHIKVの成長の段階に対して作用することを示す。アルファウイルス感染に対するヒトの遺伝的感受性について、ほとんど情報は得られていない。健康な白人個体における多型についてOAS3遺伝子をスクリーニングすることにより、CをTに置換することによりナンセンス変異が生じるコドンCGA-844の最初の位置にて1塩基多型(SNP)が同定された(OAS3.R844X)。OAS3.R844X位でのSNPは、カルボキシ末端からの約20%を欠くOAS3タンパク質の切断型をもたらすと予測される。変異体OAS3の異所発現は、全長OAS3タンパク質と比較してより低い効率のCHIKV阻害をもたらした。OAS3の遺伝子多型がその抗アルファウイルス活性を制御できるという見解は、チクングンヤ熱のようなアルファウイルス関連疾患の病因におけるヒトOAS遺伝子の役割を示唆する。
【0014】
本発明者らは、DV (肝細胞癌細胞)及びWNV (肝細胞癌細胞及び上皮細胞)に感染したヒト細胞において、OAS3が抗フラビウイルス活性を示すことを初めて立証する。DSS患者及び非DSSデング患者における完全OAS3遺伝子の検査により、SNP rs2285993が、GのCへの置換がArgからSerへのアミノ酸の変更をもたらすコドンAGG-381の3番目の位置にて同定された。我々の遺伝子データは、バリアントSer-381が、タイ人のデング患者におけるDSSの危険性に対する優勢な保護(donimanto protection)と関連することを示唆する。本発明者らは、今回、Ser-381を有するOAS3が、ヒト肝細胞におけるDV成長の潜在的な阻害物質であることを立証する。
【0015】
本発明者らは、YFVの生弱毒化ワクチン株17D-204 (STAMARIL, Sanofi-Pasteur)が、感染ヒト上皮細胞及び肝細胞癌細胞においてOAS3媒介性抗ウイルス経路に対する固有の耐性を有することも示す。しかし、IFN-αは、ヒト細胞においてYFVのワクチン株17D-204に対する抗ウイルス状態を確立することができた。
【0016】
これらの知見は、CHIKV、WNV及びDVを含む医療的に重要性が高いプラス鎖ssRNAウイルスに対するOAS3に基づく予防及び治療の発展にとって有効である。これらは、アルファウイルス、フラビウイルス及び医療的に重要性が高いその他のプラス鎖ssRNAウイルスへの感染に対するヒトの感受性の予測のため、特にデング、西ナイル及びチクングンヤの患者における疾患の重症形態の予測のためのOAS3に基づく新しい分子ツールの開発のためにも有効である。
【課題を解決するための手段】
【0017】
本発明の主題は、医薬品としての単離2',5'-オリゴアデニル酸合成酵素3タンパク質又は該2'-5'-オリゴアデニル酸合成酵素3タンパク質をコードする単離ポリヌクレオチドである。
【発明を実施するための形態】
【0018】
定義
- 「ポリヌクレオチド」とは、ゲノムDNAフラグメント、cDNAフラグメント又はRNA分子のことである。
- 「2'-5'-オリゴアデニル酸合成酵素3」、「OAS3」、「OAS 3」、「2'-5'-オリゴアデニル酸合成酵素3 (100 kD)」、「(2-5')オリゴ(A)合成酵素3」、「オリゴアデニル酸合成酵素p100」、「p100 OAS」又は「p100OAS」とは、哺乳動物のOAS3遺伝子によりコードされ、2'-5'-オリゴアデニル酸合成酵素活性を有するタンパク質、並びにOAS3遺伝子の多型に起因する天然のバリアントと、OAS3遺伝子/オープンリーディングフレーム(ORF)配列における1つ以上のヌクレオチドの変異(挿入、欠失、置換)に起因する人工的バリアント(但し、バリアントが2'-5'-オリゴアデニル酸合成酵素活性を有し、プラス一本鎖RNAウイルス複製を阻害できることを条件とする)とを含む派生バリアントのことをいう。好ましくは、バリアントは3つのOASドメインを含む。種々の哺乳動物のOAS3遺伝子、並びに推定OAS3 ORF及びアミノ酸配列は、配列データベースで入手可能であり、その他のOAS3遺伝子/ORF配列は、当業者に知られる標準的なクローニング及び配列決定の技術により決定してよい。
【0019】
- 「ヒトOAS3遺伝子」は、GenBank配列受入番号NC_000012の111860632位〜111895437位に相当する34806 bpの配列である。ヒトOAS3遺伝子は、第12染色体(12q24.2)上にあり、16個のエキソンを含む:エキソン1:1〜264;エキソン2:3127〜3409;エキソン3:6033〜6208;エキソン4:8300〜8538;エキソン5:9503〜9656;エキソン6:10418〜10762;エキソン7:12250〜12532;エキソン8:22628〜22803;エキソン9:24209〜24459;エキソン10:24870〜25014;エキソン11:25792〜25965;エキソン12:27301〜27586;エキソン13:28975〜29150;エキソン14:29493〜29731;エキソン15:31165〜31312;エキソン16:31519〜34806。
【0020】
- 「ヒトOAS3オープンリーディングフレーム(ORF)」は、6646 bpのヒトcDNA配列であるGenBank受入番号NM_006187 (配列番号27)上の88位〜3351位に相当する配列番号1の配列である。cDNA配列NM_006187上のエキソン1〜16の位置は、それぞれ1〜264、265〜547、548〜723、724〜962、963〜1116、1117〜1461、1462〜1744、1745〜1920、1921〜2171、2172〜2316、2317〜2490、2491〜2776、2777〜2952、2953〜3191、3192〜3339、3340〜6627である。
- 「ヒトOAS3タンパク質」は、GenBank受入番号NP_006178又は配列番号2に相当する1087アミノ酸の配列である。3つのOASドメインは、6位〜343位(配列番号3)、411位〜742位(配列番号4)及び750位〜1084位(配列番号5)にそれぞれ相当する。
【0021】
- 「OAS3活性」とは、.2'-5'-オリゴアデニル酸合成酵素活性及びプラス一本鎖RNAウイルス複製阻害活性の両方のことをいう。
本発明のOAS3タンパク質の2',5'-オリゴアデニル酸合成酵素活性は、クロマトグラフィー法又は電気泳動法によりアッセイして、形成されたオリゴアデニル酸の終点の量を決定できる(St Laurentら, Cell, 1983, 33, 95〜102; Interferon 3: Mechanisms of Production and Action, 1984, 189〜298, Friedman, R.M.編, Elsevier, Amsterdam中のJohnstonら; Justesenら, Proc. Natl. Acad. Sci., USA, 1980, 77, 4618〜4622; Justesenら, Nucleic Acids Res., 1980, 8, 3073〜3085; Justesen, J.及びKjelgaard, N.O., Anal. Biochem., 1992, 207, 90〜93)。
【0022】
- 「本発明のOAS3タンパク質によるプラス鎖 ssRNAウイルス複製の阻害」とは、ウイルス感染細胞内に外因性OAS3タンパク質(細胞のゲノムによりコードされていない;例えば組換えOAS3タンパク質)が存在する場合のウイルス成長(ウイルス複製)の部分的又は完全な低減のことをいう。この阻害は、OAS3タンパク質を発現する適切な組換え株化細胞に、プラス一本鎖RNAウイルスを感染させることにより決定できる。ウイルスに感染した同じタイプの非組み換え細胞を、対照として用いる。次いで、ウイルス感染細胞の上清中の子孫ウイルス生成を、任意の公知のウイルス力価決定アッセイにより測定できる。代わりに、ウイルスタンパク質の生成を、ウェスタンブロット若しくはウイルス抗原の免疫標識により分析できるか、又はウイルスのゲノム及びサブゲノムRNA生成を、ノザンブロット若しくはRT-PCRにより分析できる。
- 「プラス鎖ssRNAウイルス」とは、プラス一本鎖リボ核酸(ssRNA)をその遺伝的材料として有し、DNA中間体を用いて複製しないウイルスのことをいう。プラス鎖ssRNAウイルスは、ウイルス分類のボルティモア分類方式の第IV群に属する。
【0023】
- アミノ酸配列及び核酸配列の両方に関する「同一性」とは、整列させるアミノ酸残基若しくはヌクレオチド同士の同一性を最大にする配列のアラインメントに基づく2つの配列間の同一性の程度の指標のことであり、これは、同一残基若しくはヌクレオチド、残基(本発明の場合に1087残基)若しくはヌクレオチド(本発明の場合に3261ヌクレオチド)の合計数、並びに配列アラインメント中のギャップの存在及び長さの関数である。GCG配列分析パッケージ(University of Wisconsin, Madison, Wis.)の一部分として利用可能であり、例えばデフォルト設定で用いることができるFASTA又はBLASTを含む種々のアラインメントアルゴリズム及び/又はコンピュータプログラムが、標準的なパラメータを用いて配列同一性を決定するために利用可能である。
配列番号2の残基1〜1087と75%の同一性を有するタンパク質は、タンパク質配列を配列番号2の残基1〜1087と整列させて比較したときに、その配列が、(25×10.87 = 271)までの変更を含み得るタンパク質である。1つの変更とは、配列番号2の残基1〜1087と比較して1つのアミノ酸の欠失、置換又は挿入のことをいう。例えば、配列番号2の最初の1080残基から50の変更を有する1080アミノ酸の配列は、配列番号2と1080-50/10.87= 94.75%の同一性を有する。例えば、配列番号2の残基1〜1087から50の変更を有する1090アミノ酸の配列は、配列番号2と1087-50/10.87= 95.4%の同一性を有する。
【0024】
- 「類似性」とは、整列させるアミノ酸残基同士の類似性を最大にする配列のアラインメントに基づく2つのアミノ酸配列間の類似性の程度の指標のことをいい、これは、同一若しくは類似の残基の数、残基の合計数(本発明の場合に1087残基)、並びに配列アラインメント中のギャップの存在及び長さの関数である。GCG配列分析パッケージ(University of Wisconsin, Madison, Wis.)の一部分として利用可能であり、例えばデフォルト設定で用いることができるFASTA又はBLASTを含む種々のアラインメントアルゴリズム及び/又はコンピュータプログラムが、標準的なパラメータを用いて配列類似性を決定するために利用可能である。類似の残基とは、同等の化学的特性(サイズ、電荷(中性、塩基性、酸性)、親水性/疎水性)を有する残基のことを言う。
【0025】
-「個体」とは、哺乳動物、及びその他の脊椎動物(例えば鳥類、魚類及び爬虫類)を含む。本明細書において用いる場合、「哺乳動物」及び「哺乳類」の用語は、それらの子に授乳し、かつ生きている子を産む(真獣類または胎盤哺乳類)か、又は産卵する(後獣亜綱または無胎盤哺乳類)、単孔類、有袋類及び胎盤哺乳類(placental)を含む任意の脊椎動物のことをいう。哺乳動物種の例は、ヒト及びその他の霊長類(例えばサル、チンパンジー)、げっ歯類(例えばラット、マウス、モルモット)、及びウシ、ブタ及びウマのようなその他のものを含む。
- 「変異」により、ポリヌクレオチド(cDNA、遺伝子)又はポリペプチド配列中の1つ以上のヌクレオチド/アミノ酸の置換、欠失、挿入を意図する。この置換は、遺伝子のコード配列又はその調節配列に影響し得る。これは、ゲノム配列の構造、又はコードされるmRNAの構造/安定性にも影響し得る。
【0026】
本発明は、OAS3アミノ酸配列中の1つ以上のアミノ酸の変異(挿入、欠失、置換)、アミノ酸融合部分の付加、非天然アミノ酸(Dアミノ酸又は非アミノ酸アナログ)でのアミノ酸の置換、ペプチド結合の改変、環化、側鎖への化学基の付加(脂質、オリゴ糖又は多糖)、並びに適切な担体へのカップリングからなる群より選択される1つ以上の改変を含む改変OAS3タンパク質を包含する。当該技術において公知の手順により導入されるこれらの改変は、2'-5'-オリゴアデニル酸合成酵素活性及びプラス一本鎖RNAウイルス複製阻害活性についてまだ活性な改変OAS3タンパク質をもたらす。
【0027】
本発明の好ましい実施形態によると、上記の2'-5'-オリゴアデニル酸合成酵素3 (OAS3)は、ヒト2'-5'-オリゴアデニル酸合成酵素3である。
本発明の別の好ましい実施形態によると、上記のOAS3タンパク質は、配列番号2の残基1〜1087と少なくとも70%のアミノ酸配列同一性又は80%のアミノ酸配列類似性、好ましくは少なくとも80%のアミノ酸配列同一性又は90%のアミノ酸配列類似性を有する。
【0028】
本発明のより好ましい実施形態によると、上記のOAS3タンパク質は、配列番号2の381位にセリンを含む。好ましくは、上記のOAS3タンパク質は、配列番号2及び配列番号7からなる群より選択されるアミノ酸配列を含むか又はそれからなる。
本発明の別の好ましい実施形態によると、上記のOAS3ポリヌクレオチドは、上記で規定されるタンパク質をコードし、より好ましくは、これは、配列番号2のタンパク質をコードする配列番号1、及び配列番号7のタンパク質をコードする配列番号6からなる群より選択されるヌクレオチド配列を含むか又はそれからなる。
【0029】
本発明の別の好ましい実施形態によると、上記のポリヌクレオチドは、発現ベクターに挿入される。
ベクターとは、それが連結された別の核酸を輸送できる核酸分子のことをいう。本発明において用いることができるベクターは、限定されないが、ウイルスベクター、プラスミド、RNAベクター、又は染色体、非染色体、半合成若しくは合成の核酸分子からなり得る直鎖状若しくは環状のDNA又はRNA分子を含む。好ましいベクターは、自己複製できるか(エピソームベクター)及び/又はそれらが連結された核酸を発現できる(発現ベクター)ものである。多数の適切なベクターが当業者に知られ、市販されている。
【0030】
ウイルスベクターは、レトロウイルス、アデノウイルス、パルボウイルス(例えばアデノ随伴ウイルス又はAAV)、コロナウイルス、マイナス鎖RNAウイルス、例えばオルトミクソウイルス(例えばインフルエンザウイルス)、ラブドウイルス(例えば狂犬病及び水疱性口内炎ウイルス)、パラミクソウイルス(例えば麻疹及びセンダイ)、プラス鎖RNAウイルス、例えばピコルナウイルス及びアルファウイルス、並びにアデノウイルス、ヘルペスウイルス(例えば単純ヘルペスウイルス1及び2型、エプスタイン-バーウイルス、サイトメガロウイルス)及びポックスウイルス(例えばワクシニア、鶏痘及びカナリア痘)を含む二本鎖DNAウイルスを含む。その他のウイルスは、例えば、ノーウォークウイルス、トガウイルス、フラビウイルス、レオウイルス、パポバウイルス、ヘパドナウイルス及び肝炎ウイルスを含む。レトロウイルスの例は、トリ白血病肉腫、哺乳類C型、B型ウイルス、D型ウイルス、HTLV-BLV群、レンチウイルス、スプマウイルスを含む(Coffin, J. M., Retroviridae: The viruses and their replication, In Fundamental Virology, 第3版, B. N. Fieldsら編, Lippincott-Raven Publishers, Philadelphia, 1996)。
【0031】
好ましくは、上記のベクターは、本発明のOAS3タンパク質をコードする配列が、適切な転写及び翻訳制御要素の制御下に位置して、該タンパク質の生成又は合成を許容する発現ベクターである。よって、上記のポリヌクレオチドは、発現カセットに含まれる。より具体的には、該ベクターは、複製起点、該コードポリヌクレオチドに機能可能に連結されるプロモーター、リボソーム結合部位、RNAスプライシング部位(ゲノムDNAを用いる場合)、ポリアデニル化部位、及び転写終結部位を含む。これは、エンハンサーも含み得る。プロモーターの選択は、ポリペプチドが発現される細胞に依存する。適切なプロモーターは、組織特異的及び/又は誘導性プロモーターを含む。誘導性プロモーターの例は、重金属のレベルの増加により誘導される真核メタロチオネインプロモーター、温度の増加により誘導される熱ショックプロモーターである。組織特異的プロモーターの例は、骨格筋クレアチンキナーゼ、前立腺特異的抗原(PSA)、α-アンチトリプシンプロテアーゼ、ヒトサーファクタント(SP)タンパク質A及びB、並びにβ-カゼインである。ベクターは、選択マーカー、例えば真核細胞培養についてネオマイシンホスホトランスフェラーゼ、ヒスチジノールデヒドロゲナーゼ、ジヒドロ葉酸レダクターゼ、ハイグロマイシンホスホトランスフェラーゼ、単純ヘルペスウイルスチミジンキナーゼ、アデノシンデアミナーゼ、グルタミンシンセターゼ及びヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ;エス・セレビシエ(S. cerevisiae)についてTRP1、URA3及びLEU2;大腸菌(E. coli)においてテトラサイクリン、リファンピシン又はアンピシリン耐性を含み得る。
【0032】
ベクターの選択は、それらの使用(安定発現又は一過性発現)又は及び宿主細胞に依存する。ウイルスベクター及び「裸の」核酸ベクターは、哺乳動物細胞(ヒト及び動物)における発現のために好ましいベクターである。なかでも、興味対象の配列が予め挿入された、アデノウイルス、レトロウイルス、レンチウイルス及びAAVのようなウイルスベクターを用いることができる。
【0033】
本発明の主題は、上記で規定されるような少なくとも1つのOAS3タンパク質、又は好ましくは発現ベクター中に挿入された少なくとも1つのOAS3ポリヌクレオチドと、少なくとも1つの許容され得るビヒクル、担体、添加剤及び/又は免疫賦活剤とを含むことを特徴とする医薬組成物でもある。
当業者に知られる任意の適切な担体を、本発明の医薬組成物に用いることができ、担体のタイプは、投与の様式に依存して変動する。皮下注射のような非経口投与について、担体は、好ましくは水、塩水バッファー、ラクトース、マンニトール、グルタメート、脂肪又はワックスを含み、注射可能医薬組成物は、好ましくは等張溶液である(300〜320 m osmole程度)。経口投与について、マンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、タルク、セルロース、グルコース、スクロース及び炭酸マグネシウムのような上記の担体又は固体担体のいずれも用いることができる。生分解性マイクロスフェア(例えばポリ乳酸ガラクチド)を、本発明の医薬組成物の担体として用いることもできる。適切な生分解性マイクロスフェアは、例えば米国特許第4,897,268号及び第5,075,109号に開示されている。添加剤は、凝集防止剤、抗酸化剤、色素、風味増強剤、又は平滑剤、集合剤(assembling agent)若しくは単離剤(isolating agent)から、そして一般的に製薬産業で通常用いられる任意の賦形剤から選択できる。任意の種類の免疫賦活剤を本発明の組成物において用いて、免疫応答を増強し得る。
【0034】
医薬組成物は、経口投与に適する形態であり得る。例えば、組成物は、経口投与用の錠剤、通常のカプセル剤、ゼラチンカプセル剤又はシロップ剤の形態である。これらのゼラチンカプセル剤、通常のカプセル剤及び錠剤の形態は、デンプン、ガム及びゼラチンのようなアジュバント若しくは結合剤、リン酸カルシウムのようなアジュバント、トウモロコシデンプン若しくはアルギン酸のような崩壊剤、ステアリン酸マグネシウムのような滑沢剤、甘味料又は香料のような医薬製剤において通常用いられる賦形剤を含み得る。液剤又は懸濁剤は、薬理学的に適合し得る溶剤を加えることにより水性又は非水性の媒体中で調製できる。これらは、グリコール、ポリグリコール、プロピレングリコール、ポリグリコールエーテル、DMSO及びエタノールを含む。
【0035】
OAS3タンパク質又はOAS3ポリヌクレオチド(単離されているか又はベクターに挿入されている)は、細胞に、インビトロ、エクスビボ又はインビボで、特定の細胞タイプに適する当該技術において公知の任意の簡便な手段により、単独で、又は少なくとも適切なビヒクル及び/又は担体のいずれかとともに導入される。例えば、OAS3タンパク質/ポリヌクレオチドは、生体中で該配列への保護を提供できるか、又はそれが宿主細胞の膜を通過することを可能にする物質と組み合わせ得る。OAS3タンパク質は、有利には、リポソーム、ポリエチレンイミン(PEI)及び/又は膜輸送ペプチド(Bonetta, The Scientist, 2002, 16, 38; Fordら, Gene Ther., 2001, 8, 1〜4 ; Wadia及びDowdy, Curr. Opin. Biotechnol., 2002, 13, 52〜56; Handbook of cell penetrating peptides (第2版), 2006, Lavoisier, FRANCE中のLangel, U.)と組み合わせ得る。後者の場合、OAS3タンパク質の配列を、膜輸送ペプチドの配列と融合させる(融合ペプチド)。OAS3をコードするポリヌクレオチド(単離されているか又はベクターに挿入されている)は、種々の方法により細胞に導入できる(例えば注入、直接取り込み、発射衝撃、リポソーム、エレクトロポレーション)。OAS3タンパク質は、上記で規定される適切な発現ベクターを用いて細胞において安定的に又は一過的に発現され得る。
【0036】
本発明のある実施形態において、OAS3タンパク質/ポリヌクレオチドは、実質的に非免疫原性であり、すなわち、ほとんど又は全く有害な免疫応答を生じない。この種の有害な免疫反応を緩和又は消去するための種々の方法を、本発明に従って用いることができる。好ましい実施形態において、OAS3タンパク質は、N-ホルミルメチオニンを実質的に含まない。望ましくない免疫応答を回避する別の方法は、タンパク質/ポリヌクレオチドを、ポリエチレングリコール(「PEG」)又はポリプロピレングリコール(「PPG」) (好ましくは平均分子量(MW)が500〜20,000ダルトンのもの)と複合させることである。
【0037】
本発明の別の主題は、プラス一本鎖RNAウイルスへの感染を予防又は治療するための上記で規定されるOAS3タンパク質又は該OAS3タンパク質をコードするポリヌクレオチドである。
より好ましい実施形態によると、上記のウイルスは、アルファウイルス属のものである。より好ましくは、これは、チクングンヤ(CHIK)、シンドビス(SIN)、セムリキ森林(SF)、東部ウマ脳炎(EEE)、西部ウマ脳炎(WEE)、ベネズエラウマ脳炎(VEE)、ロスリバー(RR)、オニョンニョン(ONN)及びバーマフォレスト(BF)ウイルスからなる群より選択される。
【0038】
別のより好ましい実施形態によると、上記のウイルスは、フラビウイルス属のものである。より好ましくは、上記のウイルスは、デング、日本脳炎、キャサヌール森林病、マレー渓谷脳炎、セントルイス脳炎、ダニ媒介脳炎、西ナイル、黄熱及びオムスク出血熱(OHF)ウイルスからなる群より選択される。
【0039】
別のより好ましい実施形態によると、上記のウイルスは、ヘパシウイルス属のものである。より好ましくは、上記のウイルスは、C型肝炎ウイルスである。
【0040】
本発明の主題は、プラス一本鎖RNAウイルス感染の予防又は治療用の同時、別々又は逐次的な使用のための組み合わせ調製物としての、少なくとも、上記で規定されるOAS3タンパク質、又は好ましくは発現ベクターに挿入されたOAS3ポリヌクレオチドと、抗ウイルス薬、抗炎症薬及び免疫調節薬からなる群より選択される、第1の製品とは異なる第2の製品とを含む製品でもある。
【0041】
本発明の主題は、必要とする個体に、上記で規定される組成物を任意の手段により投与するステップを含む、必要とする個体におけるプラス一本鎖RNAウイルス感染を予防又は治癒するための方法でもある。
一般的に、組成物は、非経口注射(例えば皮内、筋肉内、静脈内及び皮下)、鼻腔内(例えば吸入又は噴霧)、経口、舌下又は局所的に、皮膚又は直腸を通すことにより投与できる。
【0042】
本発明の組成物中に存在するOAS3 (タンパク質/ポリペプチド)の量は、治療有効量である。OAS3 (タンパク質/ポリペプチド)の治療有効量は、組成物が投与される対象者において過度に負の影響を引き起こすことなく、OAS3タンパク質が、プラス一本鎖RNAウイルス複製を阻害するというその役割を働かせるのに必要な量である。用いられるOAS3 (タンパク質/ポリペプチド)及び投与される組成物の正確な量は、プラス一本鎖RNAウイルスの種、及び治療される個体の種(ヒト、動物)、投与の様式、投与頻度及び組成物中のその他の成分のような因子によって変動する。
好ましくは、組成物は、約10μg〜約10 mg、より好ましくは約100μg〜約1 mgのOAS3 (タンパク質/ポリペプチド)で構成される。「約」により、そこで述べたOAS3の量の値(μg又はmg)が、そのような量を評価するために用いた方法の誤差限界に依存するある範囲内で変動できることを意味する。
例えば、本発明の組成物の経口投与中に、治療される個体は、連続する3日間に1日当たり約10μg〜約10 mgのOAS3 (タンパク質/ポリペプチド)の1用量スケジュールに供することができる。治療は、1週間後に1回反復できる。
皮下注射のような非経口投与について、治療される個体は、約10μg〜約10 mg、より好ましくは約100μg〜約1 mgのOAS3 (タンパク質/ポリペプチド)の1用量に供することができる。治療は、1週間後に1回反復できる。
【0043】
本発明の主題は、個体から得られる核酸試料中のOAS3遺伝子における多型を検出するステップを含む、上記で規定されるプラス一本鎖RNAウイルスへの感染に対する個体の感受性をインビトロで評価する方法でもある。
核酸試料は、ゲノムDNA、トータルmRNA又はcDNAであり得る。
多型は、核酸配列中の変異の検出を可能にする、例えばIn Current Protocols in Human Genetics, 2008, John Wiley & Sons, Incに記載されるような当該技術において知られる任意の方法により検出される。遺伝子型同定アッセイの例は、限定されないが:RAPD、RFLP、AFLP、配列特異的オリゴヌクレオチドハイブリッド形成、SnapShot PCR、リガーゼ検出反応、PCR及びMaldi-TOF、ピロシーケンスを含む。このアッセイは、表II及びIIIのOAS3特異的プライマー、特に配列番号11、12、62〜86及び118〜142のプライマーを用いることができる。
【0044】
上記の方法の好ましい実施形態によると、上記のプラス一本鎖RNAウイルスは、チクングンヤウイルスのようなアルファウイルス、又はデングウイルスのようなフラビウイルスである。
【0045】
上記の方法の別の好ましい実施形態によると、上記の多型は、Arg844コドンの停止コドンへの変異(R844X)である。白人個体群において検出されるこの多型は、プラス一本鎖RNAウイルス感染、特にチクングンヤウイルス感染への感受性の増加と関連する。R844X変異は、プライマー対(配列番号11及び配列番号12)を用いるPCR-RFLPと、その後の183 bpのPCR生成物のBglIIでの消化とにより検出できる。115 bp及び68 bpの2つのフラグメントの存在は、OAS3.R844X変異の存在を示す。
【0046】
上記の方法の別の好ましい実施形態によると、上記の多型は、コドン381の3番目の位置での1塩基多型(SNP)である。好ましくは、このSNPは、アミノ酸をArgからSerに変更するコドンAGG-381のGからCへの置換である(R381S)。遺伝的データは、バリアントSer-381が、デング患者におけるDSSの危険性に対する優勢な保護と関連することを示唆する(表VI)。このSNPは、上記で規定される任意の適切な遺伝子型同定アッセイにより検出できる。例えば、このアッセイは、表IIIに示すOAS3 エキソン6に特異的なPCRプライマー対(配列番号70及び126)のようなOAS3特異的プライマー対を用いて増幅されたゲノムDNAの直接配列決定を含み得る。
【0047】
本発明の主題は、配列番号7又は配列番号30の配列を含むか又はそれからなる単離OAS3タンパク質でもある。
【0048】
本発明の主題は、配列番号9の配列を含むか又はそれからなる単離OAS3タンパク質フラグメントでもある。配列番号7の残基1〜843を含むこのOAS3フラグメントは、OAS3の第1及び第2のOASドメインを含むが、第3の(C末端) OASドメインの大部分を欠く。
【0049】
本発明の主題は:
- 配列番号7のOAS3タンパク質をコードする配列番号6の配列を含むか又はそれからなる単離ポリヌクレオチド、
-配列番号9のOAS3タンパク質フラグメントをコードする配列番号8の配列を含むか又はそれからなる単離ポリヌクレオチド、
- 配列番号30のOAS3タンパク質バリアントをコードする配列番号28又は29の配列を含むか又はそれからなる単離ポリヌクレオチド
でもある。
【0050】
本発明の主題は、配列番号6、配列番号8、配列番号28及び配列番号29からなる群より選択される配列を有するポリヌクレオチドを含む組換えベクター、好ましくは発現ベクターでもある。
【0051】
本発明の主題は、配列番号6、配列番号8、配列番号28及び配列番号29からなる群より選択される配列を含むか又はそれからなるポリヌクレオチドで形質移入又は形質転換された宿主細胞でもある。
本発明の好ましい実施形態によると、これは、HeLa-Tet-Off/OAS3#C417-1と命名され、2008年2月26日にCollection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3927の下で寄託された、組換えヒトOAS3を発現するHeLa-Tet-Off株化細胞である。
本発明の別の好ましい実施形態によると、これは、HeLa-Tet-Off/OAS3/delta/1Cと命名され、2008年4月17日にCollection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3968の下で寄託された、切断型組換えヒトOAS3を発現するa-Tet-Off株化細胞である。
本発明の好ましい実施形態によると、これは、HeLa-Tet-Off/OAS3#F8と命名され、2009年5月15日にCollection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-4158の下で寄託された、組換えヒトOAS3を発現するHepG2-Tet-Off株化細胞である。
種々のウイルスが感染できるHeLa又はHepG2細胞に由来するこれらの株化細胞は、OAS3媒介抗ウイルス活性に対するウイルスの感受性をアッセイするために有用である。
【0052】
本発明の主題は、上記で規定されるポリヌクレオチドを含む非ヒトトランスジェニック動物でもある。
本発明の主題は、上記で規定されるポリヌクレオチドを含むトランスジェニック植物でもある。
【0053】
本発明のOAS3タンパク質/ポリヌクレオチドは、公知の組換えDNA及び遺伝子工学技術を用いて調製される。例えば、OAS3のORFを含む配列は、DNA鋳型から、特異的プライマーを用いるポリメラーゼ連鎖反応により増幅される。PCRフラグメントは、次いで、適切な制限部位を用いることにより発現ベクターにクローニングされる。OAS3タンパク質は、発現ベクターで改変された宿主細胞又はトランスジェニック動物/植物において、OAS3タンパク質の発現に適する条件下で発現され、OAS3タンパク質は、宿主細胞培養物又はトランスジェニック動物/植物から回収される。
【0054】
本発明の実行は、そうでないと記載しない限り、当該技術の範囲内である細胞生物学、細胞培養、分子生物学、トランスジェニック生物学、微生物学、組換えDNA及び免疫学の通常の技術を用いる。このような技術は、文献に十分に説明されている。例えば、Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, 第3版, (Sambrookら, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); Oligonucleotide Synthesis (M. J. Gait編, 1984); Mullisら、米国特許第4,683,195号; Nucleic Acid Hybridization (B. D. Harries及びS. J. Higgins編1984); Transcription And Translation (B. D. Hames及びS. J. Higgins編1984); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); Methods In ENZYMOLOGYのシリーズ(J. Abelson及びM. Simon編, Academic Press, Inc., New York)、特に第154巻及び第155巻(Wuら編)並びに第185巻「Gene Expression Technology」(D. Goeddel編); Gene Transfer Vectors For Mammalian Cells (J. H. Miller及びM. P. Calos編, 1987, Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (Mayer及びWalker編, Academic Press, London, 1987); Handbook Of Experimental Immunology, 第I〜IV巻(D. M. Weir及びC. C. Blackwell編, 1986); 並びにManipulating the Mouse Embryo, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1986)、Current Protocols in Human Genetics (John Wiley & Sons, Inc, 2008)、特に第12章「Vectors For Gene Therapy」及び第13章「Delivery Systems for Gene Therapy」を参照されたい。
【図面の簡単な説明】
【0055】
【図1】c-mycエピトープをタグ付加されたヒト組換えOAS3 (クローンOAS3C 17.1)のヌクレオチド(A)及びアミノ酸(B)の配列を表す。これらのヌクレオチド及びアミノ酸配列は、それぞれ配列番号23及び24に相当する。OAS3 mRNA参照配列(配列番号27)と、クローニングされたOAS3 cDNAとの間の変更は、同義であれば下線を付し、非同義であれば太字で示す。翻訳開始及び停止コドンは、太字で示す。C末端c-mycエピトープに相当する付加ヌクレオチド及びアミノ酸配列は、下線を付す。5'のNotI及び3'のEcoRVの制限部位は、斜体で示す。
【図2】組換えヒトOAS3を発現する誘導性HeLa-Tet-Off/OAS3#C417-1株化細胞の樹立を説明する。
【図3】HeLa細胞におけるOAS3の検出を示す。(A)において、HeLa.Tet-Off細胞に、CHIKウイルス06-49株(CHIKV 06-49)を、細胞あたり異なるフォーカス形成単位(AP61FFU)で感染させた。感染後(p.i.) 20時間で、感染細胞の上清中に生成されるウイルス粒子を、蚊エーデス・シュードスクテラリス(Aedes pseudoscutellaris) AP61細胞でのフォーカス免疫アッセイにより力価決定した。(B)において、細胞あたり1AP61FFUの感染多重度(1 MOI)でのCHIKVの投入の5時間前に、HeLa.Tet-Off細胞を、1,000 IU/mlのヒトIFN-αで処理したか、又は模擬処理(対照)した。ウイルス子孫の生成を、上記のようにして18時間p.iにて決定した。(C)において、グラフは、全長及び切断型のOAS3の構造を描く。(D)において、誘導(- Tet)又は非誘導(+ Tet) Tet-Off/OAS3細胞におけるOAS3の異所発現を、OAS3特異的抗体を用いるイムノブロット分析により分析した。対照として、HeLa.Tet-Off細胞を、1,000 IUのIFN-α(+ IFN)で処理したか、又は模擬処理した(模擬)。β-アクチンは、ハウスキーピングタンパク質対照であった。
【図4】OAS3発現HeLa細胞におけるCHIKVの成長の阻害を示す。(A)において、細胞に18時間、1MOIでCHIKVを感染させ、抗CHIK.E2 MAb 3E4を用いてフローサイトメトリーにより分析した。模擬感染細胞(ウイルスなし)、ウイルス投入の5時間前に1,000 IU/mlのヒトIFN-αとインキュベートした(+ IFN)か、又は模擬処理した(対照)、CHIKVに感染したHeLa.Tet-off細胞、及びテトラサイクリンの存在下(+ tet)又は非存在下で(- tet)のCHIKVに感染したTet-Off/OAS3細胞におけるCHIKV E2タンパク質生成の分析。(B)において、細胞に、種々のMOIでCHIKVを感染させ、ウイルス子孫の生成を18及び24時間のp.i.で決定した。(C)において、CHIKV成長の阻害を、種々のp.i.時間でモニターした。(D)において、細胞に1MOIにてSINV又はSFVを感染させ、ウイルス子孫の生成を18時間p.i.で決定した。
【図5】OAS3の抗ウイルス活性に対するWNウイルスの感度を示す。HeLa/Tet-off及び誘導HeLa.Tet-Off/OAS3#C417-1細胞に、細胞あたり0.1、1.0又は10 AP61FFUのWNウイルスIS-98-ST1株を感染させた。感染した細胞の上清中に生成される感染性ウイルス粒子を、感染後48時間で、蚊エーデス・シュードスクテラリスAP61細胞でのフォーカス免疫アッセイにより力価決定した。
【図6】OAS3発現細胞でのWNウイルス成長の阻害を示す。HeLa/Tet-Off及び誘導HeLa.Tet-Off/OAS3#C417-1細胞に、細胞あたり0.1 AP61FFUのWNウイルスIS-98-ST1株を感染させ、感染した細胞の上清中に生成される感染性ウイルス粒子を、感染後種々の時間にて(24、48及び72時間)、蚊エーデス・シュードスクテラリスAP61細胞でのフォーカス免疫アッセイにより力価決定した。
【図7】CHIKV複製に対するOAS3発現の意義を示す。(A)において、イムノブロットアッセイを、CHIKV (ウイルス)に感染させたか又は模擬感染させた(m.i.) HeLa.Tet-off細胞(対照)及びHeLa.Tet-Off/OAS3 (OAS3)細胞からの細胞抽出物について、抗CHIK HMAF (左)又は抗CHIKV E2 MAb 3E4 (右)を用いて行った。βアクチンンは、タンパク質対照であった。(B)において、1MOIでのCHIKVに感染させたHeLa.Tet-Off細胞(対照)及びHeLa.Tet-Off/OAS3細胞(OAS3)におけるウイルスRNA生成のリアルタイムRT-PCR分析を、8時間p.i.にて行った。
【図8】切断型OAS3の抗アルファウイルス活性を示す。HeLa.Tet-off (対照)、HeLa.Tet-Off/OAS3 (OAS3)及びHeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)細胞に、1MOIにてCHIKVを感染させ、ウイルス子孫の生成を、18時間p.i.にて決定した。値は、スチューデントのt検定に従って統計学的に比較した(***:P<0.001)。
【図9】切断型OAS3の抗フラビウイルス活性を示す。HeLa.Tet-off (対照)、HeLa.Tet-Off/OAS3 (OAS3)及びHeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)細胞に、1MOIにてWNVを感染させ、ウイルス子孫の生成を、18時間p.i.にて決定した。値は、スチューデントのt検定に従って統計学的に比較した(***:P<0.001)。
【図10】I型IFN経路に対するDV-1ウイルスの感度を示す。A:DV感染に対するHepG2.Tet-Off株化細胞の感受性。細胞に、漸増投入量(細胞あたりのAP61FFU)のDV-1ウイルスFGA/NA d1d株を感染させた。感染後40時間にて、細胞を、DV E糖タンパク質に対して反応性のMAb 4E11を用いるフローサイトメトリーにより分析した。B:I型IFN経路に対するDV-1ウイルスの感度。HepG2.Tet-off細胞を、DV感染(10 MOI)の5時間前に、1,000 IU.mL-1のヒトIFN-α(+IFN)で前処理したか又は処理しなかった(模擬処理)。40時間p.i.にて、ウイルス子孫の生成を、以前に記載されたようにして決定した(Duarte dos Santosら, Virology, 2000, 274, 292〜)。
【図11】OAS3発現HepG2細胞におけるデングウイルス成長の阻害。2μg.mL-1のテトラサイクリンの存在下(非誘導)又は非存在下(誘導)での親のHepG2.Tet-Off細胞及びHepG2.Tet-Off/OAS3#F8細胞クローンに、デングウイルス1型FGA/NA d1d株を種々の感染多重度で感染させた。感染後40時間にて、細胞を、-20℃にて20分間、メタノール/アセトンで固定した。免疫蛍光アッセイを、FITC-結合抗デングE mAb 4E1を用いて行った。核をDAPIで染色した。ウイルス抗原について陽性のHepG2細胞のパーセンテージを、3重の実験にて決定した。ウイルス子孫の生成を、25の感染多重度にて決定した。ウイルス力価決定を、以前に記載されたようにして行った(Duarte dos Santosら, Virology, 2000, 274, 292〜)。
【図12】OAS3発現HepG2細胞は、デングウイルス感染に対する耐性を示す。親のHepG2.Tet-Off細胞及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、デングウイルス1型FGA/NA d1d株(Duarte dos Santosら, Virology, 2000, 274, 292〜)を、感染多重度30 AP61FFU/細胞で感染させたか、又は模擬感染させた。感染後40時間にて、免疫蛍光アッセイを、図11の説明に記載したようにして行った。
【図13】OAS3発現HepG2細胞は、西ナイルウイルス感染に対する耐性を示す。親のHepG2.Tet-Off細胞及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、西ナイルウイルスIS-98-ST1株(Lucasら, Virology J., 2004, 1, 9〜)を、感染多重度1AP61FFU/細胞で感染させたか、又は模擬感染させた。感染後40時間にて、細胞を-20℃にて20分間、メタノール/アセトンで固定した。免疫蛍光アッセイを、cy3-結合抗西ナイルE mAb E24を用いて行った。核をDAPIで染色した。
【図14】IFN-αが、YFV 17Dに対する抗ウイルス状態を、ヒト上皮HeLa細胞において確立できることを示す。HeLa細胞に、YFV 17Dを1PFU/細胞にて感染させ、次いで、感染後の種々の時点にて、1,000 IU/mlのヒトIFN-αで処理したか、又は模擬処理した(対照)。ウイルス子孫の生成を、感染後72時間にて決定した。
【図15】YFVの17D生弱毒化株が、ヒト細胞において、感染後の時間又は感染多重度のいずれにも関係なく、OAS3媒介抗ウイルス効果に対する予期せぬ耐性を示すことを示す。HeLa (対照)及び誘導HeLa.Tet-Off/OAS3 (OAS3)細胞を、低ウイルス投入量(パネルA; 1PFU/細胞)又は高ウイルス投入量(パネルB; 10 PFU/細胞)でYFV 17D株に曝露した。
【図16】OAS3発現HepG2細胞での黄熱ワクチン17D-204の阻害はない。親のHepG2.Tet-Off及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、Vero E6細胞で一旦成長させた黄熱の生弱毒化STAMARILワクチン(Sanofi-Pasteur)を感染させたか、又は模擬感染させた。感染後40時間にて、細胞を-20℃にて20分間、メタノール/アセトンで固定した。免疫蛍光アッセイを、抗フランス神経親和性ウイルスHMAF及びFITC-結合ヤギ抗マウスIgを用いて行った。核をDAPIで染色した。
【図17】切断型ヒト組換えOAS3 (OAS3 1-843)のヌクレオチド(A)及びアミノ酸(B)配列を示す。これらのヌクレオチド及びアミノ酸配列は、それぞれ配列番号25及び配列番号26に相当する。OAS3 mRNA参照配列と、クローニングされたOAS3 cDNAフラグメントとの間の変更は、同義であれば下線を付し、非同義であれば太字で示す。翻訳開始及び停止コドンは、太字で示す。C末端c-mycエピトープに相当する付加ヌクレオチド及びアミノ酸配列は、下線を付す。5'のNotI及び3'のEcoRVの制限部位は、斜体で示す。
【実施例】
【0056】
実施例1:HeLaTet-Off/OAS3株化細胞の確立及び評価
1) 材料及び方法
a) 細胞培養
HeLa.Tet-Off株化細胞を、BD BIOSCIENCES CLONTECHから購入した。HeLa.Tet-Off細胞は、5% CO2中で37℃にて、10%熱不活化胎児ウシ血清(FCS)、4 mM L-グルタミン、100 UI/mlペニシリン、10μg/mlストレプトマイシン及び200μg.mL-1 G418 (INVITROGEN)を補ったDMEM (INVITROGEN)中で維持する。
HeLa.Tet-Off/OAS3株化細胞(CNCM I-3927)は、DMEM、10% FCS、4 mM L-グルタミン、100 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB (BD BIOSCIENCES CLONTECH)及び2μg.mL-1テトラサイクリン(SIGMA-ALDRICH)中で維持する。細胞を、単層で成長させる。予測される細胞密度は、80%〜100%である。集団倍加時間は、約2日である。細胞を、トリプシン処理により採集する。これを、毎週1/10で継代培養する。細胞は、制限された寿命を有する(15〜20継代)。細胞を、20% FCS、10% DMSO、4 mMグルタミン、100 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB及び2μg.mL-1テトラサイクリンを補ったDMEM中で凍結させる。
【0057】
HeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)株化細胞(CNCM I-3968)は、DMEM、10% FCS、20 mM L-グルタミン、10,000 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB (BD BIOSCIENCES CLONTECH)及び10μg.mL-1テトラサイクリン(SIGMA-ALDRICH)中で維持する。細胞を、単層で成長させる。予測される細胞密度は90%である。集団倍加時間は、約2日である。細胞を、トリプシン処理により採集する。これを、毎週1/10で継代培養する。細胞は、制限された寿命を有する(25継代)。細胞を、20% FCS、10% DMSO、4 mMグルタミン、10,000 UI/mlペニシリン、10μg/mlストレプトマイシン、200μg.mL-1 G418、100μg.mL-1ハイグロマイシンB及び10μg.mL-1テトラサイクリンを補ったDMEM中で凍結させる。
【0058】
b) OAS3タンパク質を発現するHeLa.Tet-Off株化細胞の樹立
PCR4-TOPOベクター(INVITROGEN)中で別々にクローニングしたヒトOAS3の2つのオーバーラップcDNAフラグメントを、全長OAS3タンパク質(aa 1〜1,087;配列番号24及び図1)、又は切断型OAS3ΔC-term若しくはOAS3[1-843] (aa 1〜843; 配列番号26及び図3C、9及び17)の発現のための鋳型として用いた。参照OAS3配列(GenBank受入番号NM-006187; 配列番号27)と、本研究で用いたOAS3 cDNA (図1)との間のヌクレオチド及びアミノ酸の違いを、表Iに列挙する。
【0059】
【表1】
【0060】
OAS3配列は、3'オープンリーディングフレーム末端で、追加の10残基の配列EQKLISKEDL (配列番号10)及びその後の停止コドンと接するように、OAS3についてプライマー対pTet-OAS3-univ1及びpTet-OAS3-rev2を、そしてOAS3[1-843]についてプライマー対pTet-OAS3-univ1bis及びpTet-OAS3-rev2bis (表II)を用いてPCRにより改変した。
【0061】
【表2】
【0062】
酵素認識部位に下線を付す。停止コドンに相補的な配列を太字で示す。OAS3配列(配列番号23〜25)は、NotI及びEcoRV制限酵素部位で、それぞれ下流の端及び上流の端にて挟まれている。PCR生成物は、NotI及びEcoRVを用いて消化し、pTRE2hyg発現ベクター(BD BIOSCIENCES CLONTECH)のユニークNotI及びEcoRV部位に挿入して、pTRE2hyg-OAS3 (図1)及びpTRE2hyg/OAS3CΔC-term (OAS3 1-843)を作製した。この構成において、OAS3挿入断片は、Tet-Off発現系の制御下にある。Tet-Off系は、リプレッサーであるテトラサイクリン(Tet)の除去により外来遺伝子発現の誘導を可能にする。HeLa.Tet-Off細胞(BD BIOSCIENCES CLONTECH)に、pTRE2hyg-OAS3又はpTRE2hyg/OAS3CΔC-termを、形質移入試薬Fugene 6 (ROCHE)を製造業者の推奨する手順に従って用いて、形質移入した。Tet-Off発現系を、2μg/mlのTetを培養培地に加えることにより抑圧した。形質移入された細胞を、阻害剤G418及びハイグロマイシンを含む成長培地上で選択し、次いで、10μg.mL-1のリプレッサーであるテトラサイクリン(Tet)の存在下で限界希釈により単一細胞からクローニングした。Tet除去のために、細胞単層をトリプシン処理し、何も補充していないDMEMで少なくとも5回洗浄した後に、遺伝毒性薬剤のみを補充したDMEM/10 % FBSに置き換えた。非誘導細胞(+ Tet)でのものに対する誘導細胞(- Tet)での組換えOAS3 mRNA生成のレベルを、RT-PCR分析により、プライマー対OAS3-For及びOAS3-Rev (表II)を用いて決定した。これらの実験に基づいて、誘導性のHeLa.Tet-Off/OAS3#C417-1及びHeLa.Tet-Off/OAS3/delta/1C (OAS3ΔC-term)クローンを選択した。HeLa.Tet-Off/OAS3#C417-1株化細胞は、2008年2月26日に、Collection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3927の下で寄託した。HeLa-Tet-Off/OAS3/delta/1C株化細胞は、2008年4月17日に、Collection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-3968の下で寄託した。両方のHeLa.Tet-Off/OAS3株化細胞は、2μg.mL-1 Tetの存在下の抑圧条件下で維持した。
【0063】
c) ウイルス
臨床単離株であるチクングンヤウイルス06-49株(CHIKV 06-49; Schuffeneckerら, PLoS Medicine 3, 2006, e263)の、蚊エーデス・シュードスクテラリス(AP61)細胞単層上での生成、及びフォーカス免疫検出アッセイによるウイルス力価決定(Despresら, Virology, 1993, 196, 209〜219)は、以前に記載されたようにして行った(Brehinら, Virology, 2008, 371, 185〜195)。感染性の力価は、AP61細胞でのフォーカス形成単位(FFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。
【0064】
d) イムノブロットアッセイ
細胞性タンパク質を、以前に記載されたようにしてイムノブロット分析に供した(Brehinら, Virology, 2008, 371, 185〜195)。ウイルスタンパク質発現を、抗CHIKV HMAF又は抗CHIK.E2 MAb 3E4を用いて検出した(Brehinら, Virology, 2008, 371, 185〜195)。OAS3タンパク質発現を、抗OAS3 N-term (Santa-Cruz)又はC-term (ABGENT)抗体を用いて検出した。
【0065】
e) フローサイトメトリー分析
細胞を剥離し、次いで、PBS中の3.2%パラホルムアルデヒドを用いて固定した。固定された細胞を、以前に記載されたようにして(Brehinら, Virology, 2008, 371, 185〜195)、透過にし、抗CHIKV HMAFで染色し、フローサイトメトリーにより分析した。
【0066】
2) 結果
まず、親のHeLa.Tet-Off株化細胞の、CHIKV感染に対する感受性を評価した(図3A)。1感染多重度(MOI)で感染させたこれらの細胞におけるCHIKV複製の分析は、子孫ウイルスの生成が、18時間p.i.にておよそ7.0 log FFU.mL-1に達したことを示した。CHIKV E2糖タンパク質に対して反応性のMAb 3E4を用いるフローサイトメトリー分析により(Brehinら, Virology, 2008, 371, 185〜195)、約50%のCHIKV感染HeLa.Tet-Off細胞が、ウイルス抗原について陽性であった。次いで、IFN-αがHeLa.Tet-Off細胞において抗ウイルス状態を確立する能力について調べた。HeLa.Tet-off細胞を、1,000 IU.mL-1ヒトIFN-αで、CHIK曝露(1MOI)の5時間前に前処理することにより、18時間p.i.にて、約1.5 logのウイルス力価の低減がもたらされた(図3B)。よって、IFN-依存的抗ウイルス経路はHeLa.Tet-Off細胞において機能的であり、細胞レベルにてCHIKVに対する保護を提供する。しかし、CHIKV感染細胞は、感染HeLa.Tet-Off細胞内部でウイルス複製が一旦確立されると、5時間p.i.にてIFN-αに対する完全な耐性を示した。
【0067】
OAS3タンパク質発現をTet-Off発現系の制御下で上方制御する安定HeLa.Tet-Off/OAS3#C417-1細胞クローンを選択して、大型のOASの抗ウイルス活性を評価した。組換えOAS3タンパク質は、3つの潜在的活性触媒部位を含む3つの近接するOASユニットで構成される(ドメインI、II及びIII) (図3C)。組換えOAS3タンパク質の発現を、ヒトOAS3タンパク質のC末端領域に対して指向されたポリクローナル免疫血清を用いる、細胞溶解物のイムノブロッティングにより分析した(図3D)。1,000 IU.mL-1のIFN-αと5時間インキュベートした細胞は、陽性対照であった。IFN-α処理の際に、内因性OAS3分子の生成がHeLa.Tet-Off細胞において明確に検出されたが、休止細胞においては、OAS3の基底発現は観察されなかった。よって、OAS3遺伝子発現は、IFN-α/βにより能動的に上方制御され得る。組換えOAS3タンパク質の基底レベルは、非誘導HeLa.Tet-Off/OAS3細胞(2μg.mL-1 Tet)において観察され、このことは、遮断が不完全であったことを示す(図3D)。リプレッサーであるTetを培地から24時間除去したときに、非誘導細胞で見出されたものと比較して、組換えOAS3タンパク質の生成レベルが著しく増加した。
【0068】
実施例2:CHIKVに対するOAS3の抗ウイルス活性
1) 材料及び方法
実験手順は、実施例1に記載されるとおりである。
【0069】
2) 結果
ヒト上皮細胞におけるCHIKV成長に対するOAS3タンパク質の効果を調べるために、HeLa.Tet-Off/OAS3細胞に、CHIKV.06-49を1MOIにて18時間感染させた。MAb 3E4を用いるフローサイトメトリーにより決定されるように、非誘導HeLa.Tet-Off/OAS3及びHeLa.Tet-Off細胞は、CHIKV感染に対して同様の感受性を示した(図4A)。ウイルス曝露の24時間前のHeLa.Tet-Off/OAS3細胞の誘導は、CHIKV感染細胞を少なくとも20%減少させた。陽性対照として、IFN-α処理は、感染HeLa.Tet-Off細胞のパーセンテージを75%減少させた。よって、CHIKVは、ヒト上皮細胞におけるOAS3の異所発現に対して感受性である。
【0070】
HeLa.Tet-Off/OAS3細胞がCHIKV成長を阻害する効率をさらに評価するために、誘導した細胞を、漸増投入量のCHIKV-06.49に曝露した(図4B)。18時間p.i.にて、組換えOAS3タンパク質の発現は、子孫ウイルス生成を、試験したMOIに関係なく1.5〜2.0 log減少させた。24時間p.i.で、試験したより低いMOIにて、ウイルス力価は約3.0 log減少した。よって、OAS3の発現は、CHIKVの細胞から細胞への広がりを、ヒト線維芽細胞において抑制できる。中程度のMOIにて、ウイルス子孫は、少なくとも2.0 log、まだ減少した。10 MOIにて、ウイルス成長は約0.5 logしか減少せず、このことは、感染が高いウイルス投入量で行われたときに、CHIKVが、OAS3の抗ウイルス活性を回避する能力を有することを示唆した。
【0071】
動態研究は、CHIKVのOAS3により媒介される阻害が、ウイルス生活環の間に有効であったことを示した(図4C)。12時間p.i.にて、HeLa.Tet-Off/OAS3細胞から回収されたウイルスの力価は、HeLa.Tet-Off細胞と比較して、2.0 log減少した。OAS3タンパク質発現の誘導は、子孫ウイルス生成を、24時間p.i.にて少なくとも1.5 log、まだ減少させることができた。
細胞溶解の際に培養培地中に放出されるので膜の完全性の指標である細胞質酵素ラクテートデヒドロゲナーゼ(LDH)の測定は、感染させた親の細胞と比較して、感染の最初の24時間以内の感染HeLa/Tet-Off/OAS3細胞の生存性の著しい喪失を示さなかった。よって、CHIKV感染に対する耐性は、アポトーシス誘導によるウイルス感染細胞の排除よりもむしろ、OAS3の抗ウイルス活性に直接関連した。
【0072】
実施例3:ヒトOAS3を発現するHeLa.Tet-Off組換え株化細胞における、その他のプラス一本鎖RNAウイルスに対するOAS3の抗ウイルス作用
1) 材料及び方法
実験手順は、実施例1に記載されるとおりである。さらに、ウイルス生成及び力価決定は、以下に記載するようにして行った。
【0073】
ウイルス
継代数が低い(low passaged)西ナイルウイルス(WNV) IS-98-ST1株(GenBank受入番号AF 481864)の、蚊エーデス・シュードスクテラリス(AP61)細胞単層上での生成と、フォーカス免疫検出アッセイによるウイルス力価決定(Despresら, Virology, 1993, 196, 209〜219)は、以前に記載されたようにして行った(Brehinら, Virology, 2008, 371, 185〜195; 国際出願WO 02/081741)。感染性の力価は、AP61細胞でのフォーカス形成単位(FFU)として表した。シンドビスウイルスAR339株(SINV AR339)及びセムリキ森林ウイルスSF 64株(SFV 64)を、ミドリザル腎臓(VERO)株化細胞で繁殖させ、感染性の力価を、VERO細胞でのプラーク形成単位(PFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。
【0074】
2) 結果
大型のOASが、ヒト上皮細胞においてSINV AR339株、SFV SF 64株及びWNV IS-98-ST1株の成長を阻害する能力を調べた(図4D、5及び6)。HeLa.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞に、アルファウイルスを1MOIにて感染させた。OAS3タンパク質発現に応答して、18時間p.iにて、SINVとSFVとの間で、子孫ウイルスの生成が、同程度に(およそ2 log)阻害された(図4D)。HeLa.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞に、フラビウイルス(WNV)を0.1、1又は10 MOIで48時間感染させた(図5及び6)。ウイルス力価の測定に基づいて、ヒト上皮細胞が、OAS3発現に対して、WNウイルス複製を効率的に阻害することにより応答することが示された。0.1 MOIにて、HeLa.Tet-Off/OAS3から回収されたウイルス力価が、HeLa.Tet-Off細胞と比較して、1.5 log減少した(図6)。OAS3タンパク質発現の誘導は、子孫ウイルス生成を、感染後72時間で少なくとも1.0 log減少させることができた。よって、OAS3発現HeLa細胞は、親の細胞と比較して、WNウイルス感染に対するより低い感受性を示す。結論として、このデータは、OAS3が、ヒト細胞の内部でアルファウイルス及びフラビウイルスのようなRNAウイルスに対して抗ウイルス作用を有することを示す。
【0075】
実施例4:OAS3媒介CHIKV阻害の機構
1) 材料及び方法
実験手順は、実施例1に記載されるとおりである。さらに、定量的RT-PCRを、以下に記載するようにして行った。
【0076】
定量RT-PCR
ウイルスRNA蓄積のリアルタイムRT-PCR分析を、SYBR (登録商標) Green PCRを用いるABI Prism 7700配列検出で、実質的に以前に記載されたようにして行った(Kajaste-Rudnistkiら, J. Biol. Chem., 2006, 281, 4624〜4637)。CHIKV E2遺伝子についてのプライマーはChik/E2/9018/+及びChik/E2/9235/-である(表II)。GAPDH mRNAを、各試料の標準化のための内因性配列対照として用いた。プライマーGAPDH-For及びGAPDH-Revは、表IIに列挙する。ウイルスRNAコピーの定量のために、E2のN末端領域をコードするインビトロCHIK RNA転写産物を行って、以前に記載されたようにして標準曲線を構築した(Vazeilleら, PLoS One, 2007, 2(11): e1168)。
【0077】
2) 結果
OAS3の抗ウイルス作用の分子的な基礎を解明するための努力において、CHIKV成長の阻害が、ウイルス構成タンパク質の蓄積の欠如によるものかどうかを決定した。全タンパク質をHela.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞から、感染後18時間の時点で抽出し、カプシドタンパク質(C)、エンベロープ糖タンパク質pE2 (E2の前駆体)、E2及びE1を、抗CHIKV HMAFを用いるイムノブロットにより検出したが、エンベロープ糖タンパク質pE2 (E2の前駆体)及びE2は、抗CHIKV E2 MAb 3E4を用いるイムノブロットにより特異的に検出した。図7Aに示すように、OAS3タンパク質発現に応答して、バックグラウンドレベルでのウイルス構造タンパク質が存在した。よって、CHIKVが、OAS3発現HeLa細胞に生産的に感染できないことは、ウイルスタンパク質合成における著しい減少と関連する。
【0078】
ウイルスタンパク質合成の無能性が、ウイルスRNAの蓄積の欠如によるものかどうかを決定した。トータルRNAをHela.Tet-off及び誘導HeLa.Tet-Off/OAS3細胞から、感染後8時間の時点で抽出し、ゲノム及びサブゲノムのウイルスRNAの生成を、CHIKV E2遺伝子に基づいて設計したプライマーを用いるRT-PCRを用いて分析した。リアルタイムRT-PCR分析により決定されるように、親の細胞で見出されたものと比較すると、OAS3発現細胞から回収されたウイルスRNAレベルは2 log減少した(図7B)。つまり、OAS3発現は、感染細胞の内部でウイルスRNAの蓄積を妨げる。これらの結果に基づいて、OAS3発現が、感染ヒト細胞の内部でアルファウイルスの複製に影響すると結論付けることができる。
【0079】
実施例5:OAS3抗ウイルス効果に対する健常個体でのOAS3遺伝子多型の影響
1) 材料及び方法
実験手順は、実施例1に記載したとおりである。さらに、OAS3多型の遺伝子型同定は、以下に記載するようにしてアッセイした。
【0080】
遺伝子型同定
SNP OAS3.R844Xは、表IIに示すプライマー対OAS3.R844X-F(配列番号11)及びOAS3.R844X-R (配列番号12)を用いるPCR-RFLPアッセイにより遺伝子型同定した。183 bp長のPCR生成物を、Bgl IIを用いる消化に付し、115及び68 bpの2つのフラグメントの検出が、OAS3.R844Xの存在を示した。
【0081】
2) 結果
OAS3遺伝子を、多型についてスクリーニングした。48名の健常白人個体における合計で16個のエキソン、フランキングイントロン及びOAS3遺伝子の5'側2 kbの再配列決定は、コドンCGA-844の最初の位置での1塩基多型(SNP)を同定し、これは、CのTへの置換がナンセンス変異をもたらす(OAS3.R844X) (表I)。このSNPを、180名の健常白人個体においてさらにスクリーニングし、0.5%の対立遺伝子頻度を与える2つのヘテロ接合体が同定された。OAS3.R844XはOAS3タンパク質のカルボキシ末端から約20%がおそらく切断されているので(図3C)、OAS3-1〜OAS3-843配列を含む安定HeLa.Tet-off/OAS3ΔC-term細胞クローンを作製した。OAS3ΔC-termは、OAS3の第3ドメインの活性触媒部位を規定する三連構造を構成するD889残基を欠くことに注目することが興味深い(Sarkarら, 1999; Rambouillatら, 2000)。変異OAS3がCHIKV及びWNVの成長を阻害する能力を評価するために、誘導HeLa.Tet-Off/OAS3及びHeLa.Tet-off/OAS3ΔC-term細胞に、1MOIにてCHIKV-06-49又は西ナイルウイルス(WNV) IS-98-ST1株を感染させた。図8及び9に示すように、OAS3ΔC-termを発現する細胞は、CHIKV及びWNV感染に対する耐性を示し、このことは、OAS-1〜OAS3-843配列がまだ保護を提供できることを示す。ウイルス成長の分析は、OAS3ΔC-termの異所発現が、全長OAS3と比較してCHIKV及びWNVの阻害のより低い効率をもたらしたことを示した。このような結果は、コドンCGA-844でのSNPが抗ウイルス活性に対して影響を有し得ることを示唆する。
【0082】
実施例6:ヒトOAS3を発現するHep.G2.Tet-Off株化細胞におけるDV及びWNVに対するOAS3の抗ウイルス作用
1) 材料及び方法
a) 細胞培養
HepG2.Tet-Off株化細胞を、CLONTECH (#632106)から購入した。HepG2.Tet-Off細胞は、5% CO2中で37℃にて、10%熱不活化胎児ウシ血清(FCS)、4 mM L-グルタミン、100 UI/mlペニシリンGナトリウム、100μg/mlストレプトマイシンサルフェート(GIBCO, #15140-122)、0.1 mM非必須アミノ酸、100μg.mL-1 G418 (GIBCO, #10131-019)を補ったDMEM (GIBCO, #41965)中で維持する。
HepG2.Tet-Off/OAS3#F8株化細胞(CNCM I-4158)は、DMEM、10% FCS、4 mM L-グルタミン、100 UI/mlペニシリン、100μg/mlストレプトマイシン、100μg.mL-1 G418、100μg.mL-1ハイグロマイシンB (CLONTECH, #631309)、及び2μg.mL-1テトラサイクリン(SIGMA-ALDRICH, # T-7660)中で維持する。
【0083】
b) OAS3タンパク質を発現するHepG2.Tet-Off株化細胞の樹立
全長ヒトOAS3タンパク質(配列番号24)を発現する誘導性HepG2.Tet-Off/OAS3#F8クローンを、誘導性HeLa.Tet-Off/OAS3#C417-1について実施例1に記載するようにして選択した。HepG2.Tet-Off/OAS3#F8株化細胞は、2009年5月15日に、Collection Nationale de Cultures de Microorganismes, 25 rue du Docteur Roux, 75724 Paris Cedex 15に、受入番号I-4158の下で寄託した。
この株化細胞を、2μg.mL-1 Tetの存在下の抑圧条件下で維持する。
【0084】
c) ウイルス
継代数が低い西ナイルウイルス(WNV) IS-98-ST1株(GenBank受入番号AF 481864; Lucasら, Virology J., 2004, 1, 9〜)の、蚊エーデス・シュードスクテラリス(AP61)細胞単層上での生成、及びフォーカス免疫検出アッセイによるウイルス力価決定(Despresら, Virology, 1993, 196, 209〜219)は、以前に記載されたようにして行った(Brehinら, Virology, 2008, 371, 185〜195)。DV1 FGA/NA d1d株の生成及びウイルス力価決定は、以前に記載されたようにして行った(Duarte dos Santosら, Virology, 2000, 274, 292〜)。感染性の力価は、AP61細胞上でのフォーカス形成単位(FFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。
【0085】
d) 免疫蛍光アッセイ
ウイルス感染細胞を、メタノール/アセトンを用いて-20℃にて20分間固定した。免疫蛍光アッセイを、FITC-結合抗デングE mAb 4E1又はcy3-結合抗WNV.E MAb E24を用いて行った。核をDAPIで染色した。
【0086】
2) 結果
まず、親のHepG2.Tet-Off株化細胞のDV感染に対する感受性を評価した(図10A)。細胞を、細胞あたり異なるフォーカス形成単位(AP61FFU)のDV-1ウイルスFGA/NA d1d株に感染させた。感染後40時間にて、細胞を、DV E糖タンパク質に反応性のMAb 4E11を用いるフローサイトメトリーにより分析した。細胞あたり30 AP61FFUのDV-1ウイルス投入量で、約50%のHepG2.Tet-Off細胞が、デングE糖タンパク質についての陽性シグナルにより示されるように、DV-1ウイルスに感染した。
次いで、IFN-αがHepG2.Tet-Off細胞において抗ウイルス状態を確立する能力を調べた。HepG2.Tet-off細胞を1,000 IU.mL-1のヒトIFN-αでDV感染(10 MOI)の5時間前に前処理すると、40時間p.i.にて約0.7 logのウイルス力価が減少した(図10B)。つまり、IFN依存性抗ウイルス経路はHepG2.Tet-Off細胞において機能的であり、細胞レベルにてDVに対する保護を提供する。
【0087】
Hep.G2細胞におけるDV成長に対するOAS3タンパク質の効果を調べるために、誘導(-Tet)及び非誘導(+Tet)のHepG2.Tet-Off/OAS3細胞、並びに親のHepG2.Tet-Off細胞に、漸増投入量のDV-1ウイルスFGA/NA d1d株を感染させた。40時間p.i.にて、抗デングE mAbを用いる免疫蛍光アッセイを行って、ウイルス抗原について陽性のHepG2細胞のパーセンテージを決定した(図11及び12)。子孫ウイルスの生成を、25の感染多重度にて決定した(図11)。組換えOAS3タンパク質の発現は、DV感染細胞のパーセンテージを少なくとも75% (図11及び12)、及び子孫ウイルスの生成を2 log (図11)減少させた。
つまり、OAS3発現HepG2細胞は、DV感染に対する耐性を示す。
【0088】
Hep.G2細胞におけるWNV成長に対するOAS3タンパク質の効果も、免疫蛍光アッセイにより分析した。親のHepG2.Tet-Off及び誘導HepG2.Tet-Off/OAS3#F8細胞クローンに、西ナイルウイルスIS-98-ST1株を1 AP61FFU/細胞の感染多重度で感染させたか、又は模擬感染させた。感染後40時間にて、ウイルス抗原(WNV E糖タンパク質)を、cy3-結合mAb E24を用いる免疫蛍光により決定した。結果(図13)は、HeLa細胞において得られた結果(実施例3)を確認し、OAS3発現ヒト細胞がWNV感染に対する耐性を示すことを示す。
【0089】
実施例7:黄熱ウイルスの生弱毒化17D-204ワクチン株は、ヒト細胞において、ヒト大型2',5'-オリゴアデニル酸合成酵素(OAS3)により媒介される抗ウイルス効果に対して驚くべき耐性を示す
1) 材料及び方法
弱毒化黄熱ウイルス(YFV)ワクチン株(STAMARIL (登録商標)、AVENTIS-PASTEUR)を、ミドリザル腎臓(VERO)株化細胞にて繁殖させ、感染性の力価を、VERO細胞でのプラーク形成単位(PFU)として表した。その抗ウイルス効果をアッセイするために、ヒトIFN-α(BIOSOURCE)を、培養培地に1,000 IU.mL-1にて直接加えた。免疫蛍光アッセイを、抗フランス神経親和性ウイルスHMAF及びFITC-結合ヤギ抗マウスIgを用いて実施例6に記載されるようにして行った。
【0090】
2) 結果
実施例2、3、4及び6は、IFN-α/βにより媒介されるOAS3依存性抗ウイルス活性が、チクングンヤウイルスのようなアルファウイルス並びに西ナイルウイルス及びデングウイルスのようなフラビウイルスに対する主なヒト細胞防御ストラテジーをヒト細胞において示すことの初めての証拠を提供した。
OAS3が黄熱ウイルス(YFV)に対する抗ウイルス活性を示すかを調べた。このために、YFVの生弱毒化17D-204ワクチン株(STAMARIL, Aventis-Pasteur)を、ミドリザル腎臓(VERO)株化細胞上で2回繁殖させ、感染性の力価を、VERO細胞でのプラーク形成単位(PFU)として表した。HeLa細胞に、YFV 17Dを1PFU/細胞にて感染させ、次いで、感染後の種々の時点にて1,000 IU/mlのヒトIFN-αで処理したか又は模擬処理した(対照)。ウイルス子孫生成を、感染後72時間にて決定した。図14に示すように、IFN-αは、ヒト上皮HeLa細胞においてYFV 17Dに対する抗ウイルス状態を確立できる。OAS3がウイルス成長を阻害する能力を評価するために、HeLa (対照)及び誘導HeLa.Tet-Off/OAS3 (OAS3)細胞を、YFV 17D株に、低(1PFU/細胞; 図15A)又は高(10 PFU/細胞; 図15B)ウイルス投入量にて曝露した。両方の細胞集団の間で、YFV複製についての明確な違いはなかった(図15)。動態研究は、17Dが、ヒト細胞におけるOAS3により媒介される抗ウイルス効果に対して、感染後の時間又は感染多重度のいずれにも関係なく、耐性であることを示した。
【0091】
YFV 17DがOAS-3により媒介される抗ウイルス効果に対して耐性であることは、YFV 17Dに感染させた親のHepG2.Tet-Off及びHepG2.Tet-Off/OAS3#F8細胞クローンにおける免疫蛍光アッセイによって確認された(図16)。
YFVの生弱毒化株が、OAS3の抗フラビウイルス作用に対して予期せぬ耐性を示すとの知見は、フラビウイルス弱毒化の機構を解明する新たな道を開く。
【0092】
実施例8:OAS3の非同義バリアントは、デングショック症候群の病因と関連する。
1) 材料及び方法
a) 患者及び対照
Bangkokの2つの医療センターRamathibodi病院及びSiriraj病院、Mahidol University、並びにタイのKhon-Kaen州の1つの病院からの3年間(2000年6月〜2003年)の症候性デングウイルス(DV)感染の750名の患者(男性:女性の比率=0.99)を、研究に含めた。患者の年齢は、1〜25歳の範囲であり、平均は9.6歳であった。高熱、重度の頭痛、後眼窩痛、筋肉痛、関節痛、悪心及び嘔吐、並びに発疹を含む臨床上の特徴に基づいてデングウイルス感染が疑われる患者を、臨床観察及び治療のために病院に収容した。デングウイルス感染の診断は、のちに、後期急性期及び/又は回復期の血清に対する比較IgG及びIgM酵素結合免疫吸着アッセイ力価決定により確認した。DF (デング熱)及びDHF (デング出血熱)を区別する診断は、血液濃縮又は胸水貯留により表される血管透過性の増加についての証拠の不在(DF)又は存在(DHF)に基づいて確立した。具体的に、DHFの診断は、以下の4つの特徴の全てに基づいて行った:1) 2〜7日間連続する高熱、2) 駆血帯試験の陽性、点状出血(petechii)、紫斑、又は吐血のような出血性の傾向、3) 血小板減少(血小板計数<100,000/μl)及び4)血液濃縮(20%以上のヘマトクリットの増加)又は胸水貯留により表される血管透過性の増加による血漿漏出の証拠。臨床症状が明確でないのでDF又はDHFと分類できなかったいくらかのデング患者を、未定DF/DHF状態に割り当てた。DHFの重症度は、WHO基準に従う4つのグレードにより分類した。グレードIII及びIVは、特徴的な拡張期圧の上昇から重度のショックまでを有する脈圧の狭小化を伴うDHFであった。二次感染は、デング特異的IgM/IgG比<1.8と規定した。計画のプロトコル及び研究の目的は、患者及びその両親又は親族に注意深く説明した。インフォームドコンセントは、全ての対象者から個別に得た。プロトコルは、各病院の倫理委員会により承認されている。
【0093】
対照は、Ramathibodi病院からの296名の血液提供者及びSiriraj病院からの216名の血液提供者、並びにKhon-Kaen地域からの184名の人種適合健常対象者(男性:女性の比率=1)からなった。症例及び対照群はともに、タイのBangkok及び中央部に由来した。全血試料を患者及び対照からEDTA上に回収し、DNAを、標準的なフェノール/クロロホルム抽出法を用いて抽出した。
2004〜2006年の間に、さらに254名の患者(男性:女性の比率=1.07)を、Ramathibodi及びKhon-Kaen病院から同じ臨床的及びウイルス診断基準を用いて含めた。
【0094】
b) 多型の同定及び遺伝子型同定
多型は、個別のDNA試料(24名のデング患者及び32名のタイ人対照)に対するPCR増幅ゲノムDNAの直接配列決定により同定した。OASのエキソン、イントロンの一部分、並びに5'及び3'領域を配列決定するために用いたプライマーを、表IIIに示す。
【0095】
【表3−1】
【0096】
【表3−2】
【0097】
配列決定の結果は、Genalysソフトウェア(Takahashiら, J. Bioinform. Comput Biol., 2003, 1, 253〜265)により分析した。OAS3-R381Sを、推奨プロトコルを用いてABI Prism 7000配列決定システムを用いるTaqManアッセイにより遺伝子型同定した。OAS3配列のより一般的なバリアントのコドン381は、アルギニンに相当するAGGである。rs2285933と命名したOAS3-R381Sの多型は、GからCへの置換を、OAS3のコドン381の3番目のヌクレオチドにて有し(AGGからAGC)、これはアミノ酸をアルギニンからセリンに変える。OAS3の2つのバリアントであるOAS3-(R381及びS381)のmRNA、ORF及びアミノ酸配列は、それぞれ配列番号28、29、30 (R381)並びに配列番号27、1及び2(S381)に相当する。
【0098】
c) 統計分析
関連の研究を、Haploviewプログラム(Barrettら, Bioinformatics, 2005, 21, 263〜265)に組み込まれるPearsonのカイ二乗検定により、対立遺伝子カウントを用いて行った。このプログラムは、並べ替え検定を行って、経験的P値を得ることを可能にする。遺伝子型の結果をコードするロジスティック回帰分析を、STATAバージョン9を用いて試験した遺伝子モデルに従って行った。
【0099】
2) 結果
2000年〜2003年の間に、580名のデング患者をBangkokの2つの病院(Siriraj, SI及びRamathibodi, RA)から、及び170名をタイ北東部のKhonkaen州(KK)の別の病院から採用した。Bangkok の同じ病院からの512名の血液提供者、及びタイの北東地域からの184名の健常ボランティアからDNA試料を回収し、一般集団における遺伝子頻度を代表した。これらの患者及び対照の一部の遺伝子研究は、以前に報告された(Sakuntabhaiら, Nature Genetics, 2005, 37, 507〜513)。ウイルス診断は、血清試験により確認され、患者は、WHO基準に従って4つの群に分類された:DF (血漿漏出の証拠なし)及びDHF1 (血漿漏出の証拠)、DHF2 (自発的出血を伴う)及びDSS (ショックを伴う) (表IV)。
【0100】
【表4】
【0101】
WHO基準に従ってDF又はDHFを有すると分類できなかった患者は、51名であった(未分類;UC)。それぞれの重症度カテゴリーの患者の割合は、病院間で有意に異なっていた(P =1.5×10-8)。このことは、病院のタイプ(KKについて二次看護、SI及びRAについて医科学校)及び採用基準(KKにおいて3日未満の発熱、SI及びRAにおいてデング症例の疑い)における違いを反映する。
デング感染に対してIgM応答が優勢な患者と比較して、IgG応答が優勢であった患者において、血漿漏出(DHF1/DHF2/DSS対DF)及び自発的出血(DHF2/DSS対DF/UC/DHF1)の危険性の非常に有意な増加があった(それぞれP = 1.1×10-7及びP = 7.6×10-5)が、ショックについては限界有意(marginal significance)のみであった(DSS対残り、P = 0.038) (表V)。
【0102】
【表5】
【0103】
これらの知見は、DVに対するIgG応答を有する個体が、疾患のより重症な形態にかかりやすくなるという見解と一貫する。
タグ付加された多型のスクリーニングについて、関連についての試験を、血漿漏出の危険性(DHF1/DHF2/DSS対DF)、自発的出血の危険性(DHF2/DSS対DF/UC/DHF1)、ショックの危険性(DSS対非ショックDV患者)についての対立遺伝子カウントを、Bangkokの2つの病院からの患者について行った(表VI)。
【0104】
【表6】
【0105】
表VIに示すように、いずれの多型も疾患の危険性との関連の証拠を示さなかった。しかし、1つのOAS3の非同義多型が、疾患の重症度に依存する非常に著しく異なる対立遺伝子カウント分布を有した。このバリアント(OAS3-R381S: rs2285933)は、OAS3のコドン381の3番目のヌクレオチドでのGからCへの置換であり、これはアミノ酸をアルギニンからセリンに変更する。両方の病院において、この多型のC対立遺伝子は、DF、DHF1及びDHF2において、DSSよりも頻度が高かった。2つの病院間の患者の各群と対照との間に対立遺伝子頻度の著しい差はなかった。組み合わせ分析において、C対立遺伝子は、一般集団において14%、DFにおいて20%、DHF1及びDHF2において15〜16%、DSSにおいて7%の頻度を有した。DSSと非ショックDV症例との間の違いは、非常に有意であった(P = 7.10-4)。我々は10,000の並べ替えを行い、0.0032の経験的P値を得た。
【0106】
遺伝の様式
OAS3-R381Sの遺伝の様式を見出すために、C対立遺伝子の相加、優性及び劣性の3つの異なる遺伝様式に基づくモデルの尤度を、DSSの危険性に対するロジスティック回帰を用いて比較した。C対立遺伝子の劣性モデルは著しい関連を示さなかったが、C対立遺伝子の優性(<10-5 P<10-5, OR = 0.30 95CI = 0.17-0.53)及び相加(17.93, P<10-5)モデルを用いて高度に有意なレベルが得られた。log尤度比検定により、優性モデルは、相加モデルよりも有意により適切である(P = 0.04)。よって、我々は、残りの我々の分析のために優性モデルを用いた。
【0107】
第3コホートにおける再現
我々は、同じ流行(2001〜2003年)中の3番目の病院(KK)からの患者におけるOAS3-R381Sの再現研究を行った。C対立遺伝子は、以前の結果と同様に、DSS群において、ショックのないDV患者と比較してより低い頻度を示した(C対立遺伝子頻度について8%対17%、CG+GG頻度について17%対32%)。この差は、患者が少数であるので統計学的に有意でなかったが、3つの病院からの全ての患者を組み合わせたときに、より高い有意性の関連が得られた(C対立遺伝子の優性モデルについてOR = 0.48, 95 CI = 0.31-0.75, P= 7×10-4)。
【0108】
OAS3及び免疫応答のタイプ
我々は、OAS3-R381Sの影響が、ウイルスに対する免疫応答のタイプに依存するかをさらに調べた。患者における多型及びDSSの、優勢IgM免疫応答との著しい関連はなかった。しかし、各病院において及び組み合わせた場合に、優勢IgG応答を有する患者において、有意及びほぼ有意の関連が得られた(Chi2 = 19.41, OR = 0.27 95 CI = 0.14-0.53, P<10-5) (表VII)。
【0109】
【表7】
【0110】
2004〜2006年流行中の患者の関連研究
2004〜2006年の間に、我々は、以前の2つの病院(RA及びKK、表IV)からさらなる患者を得た。OAS3-R381Sの関連研究は、研究した全ての仮定との有意な関連を明らかにしなかったが、これらの患者を元の症例/対照研究に加えた場合に、有意な関連が残っていた(OR = 0.48, 95 CI = 0.31-0.75, P= 7×10-4)。この結果は、2つの期間の結果の間の不一致を調べる動機となった。
デング疾患の重症度に貢献し得る1つの因子は、ウイルス因子である。我々は、我々が471名の患者のデータを有するSiriraj病院及びKhonkaen病院からの患者のデータを用いて、2001〜2006年の間のDV-4血清型の罹患率を調べた(表VIII)。
【0111】
【表8】
【0112】
DV1及びDV2血清型は2001〜2003年の間に優勢であり、2004年においてDV1は減少し、DV4の流行が2004年に開始し、次いで、2006年まで優勢であった(表5A)。デング疾患の重症度は、ウイルスの異なる血清型の間で有意に異なっており(P = 0.012、表5B)、DV2血清型はその他のDV血清型よりもより重症の症例があった。驚くべきことに、ウイルスの血清型は、免疫応答のタイプによって有意に異なっており(フィッシャー正確P<10-4、表5C)、DV2及びDV4は、優勢IgGを示した。これらの結果は、Ministry of Public Healthからのデータと同様であった。我々は、OAS3-R381Sの影響が、いくつかのウイルス血清型について特異的であり得ると仮定した。我々は、DSSに対するOAS3-R381Sの有意に優勢な保護効果を、DV2でのみ見出した(OR = 0.24, 95 CI = 0.08-0.74, P = 0.0045, 表IX)。
【0113】
【表9】
【0114】
DF/DHF/DSS患者及び対照におけるOAS遺伝子の系統的スクリーニングに基づいて、我々は、OAS3遺伝子の多型とショックの危険性(DSS)、及びデングウイルスに感染した患者における疾患の重症度との間に関連を示した。バリアント(OAS3-R381S: rs2285933)は、OAS3のコドン381の3番目のヌクレオチドのGからCへの置換であり、これは、アミノ酸をアルギニンからセリンに変更する。Cバリアントは、非ショックデング疾患と比較してDSSに対する優勢な保護と関連し、頻度の比は0.13:0.32である(OR = 0.48, 95 CI = 0.31-0.75, P= 7×10-4)。さらに、優勢なIgG応答を有する患者(OR = 0.46, 95 CI = 0.28-0.74, P = 8×10-4)及びDV血清型2に感染した患者(OR = 0.24, 95 CI = 0.08-0.74, P = 0.0045)においてのみ、著しい関連が見出された。
【0115】
これらの結果は、非同義多型OAS3-R381Sの機能的役割を支持し、デング病因におけるOAS3の役割と、OAS3のバリアントとウイルスの血清型との間の相互関係とを証明する。
我々の知見は、DV感染患者における疾患重症度の予測、並びにデングウイルス及び医療的に重要性が高いその他のプラス一本鎖RNAウイルスによる感染に対するOAS3に基づく予防及び治療の開発について重要な影響を有するであろう。
【受託番号】
【0116】
CNCM I−3927
CNCM I−3968
CNCM I−4158
【図1−1】
【図1−2】
【図17−1】
【図17−2】
【特許請求の範囲】
【請求項1】
医薬品としての単離2'-5'-オリゴアデニル酸合成酵素3タンパク質又は2'-5'-オリゴアデニル酸合成酵素3タンパク質をコードする単離ポリヌクレオチド。
【請求項2】
ヒト2'-5'-オリゴアデニル酸合成酵素3の配列を有する請求項1に記載のタンパク質又はポリヌクレオチド。
【請求項3】
配列番号2の残基1〜1087と少なくとも70%のアミノ酸配列同一性又は80%のアミノ酸配列類似性を有する請求項1又は2に記載のタンパク質。
【請求項4】
配列番号2の381位にセリンを含む請求項1〜3のいずれか1項に記載のタンパク質。
【請求項5】
配列番号2及び配列番号7からなる群より選択されるアミノ酸配列を含む請求項4に記載のタンパク質。
【請求項6】
請求項3〜5のいずれか1項で規定されるタンパク質をコードする請求項1又は2に記載のポリヌクレオチド。
【請求項7】
配列番号1及び配列番号6からなる群より選択される配列を含む請求項6に記載のポリヌクレオチド。
【請求項8】
発現ベクターに挿入された請求項1、2、6及び7のいずれか1項に記載のポリヌクレオチド。
【請求項9】
プラス一本鎖RNAウイルス感染を予防又は治療するための請求項1〜8のいずれか1項で規定される単離2'-5'-オリゴアデニル酸合成酵素3タンパク質又はポリヌクレオチド。
【請求項10】
アルファウイルス属のウイルスへの感染を治療するための請求項9に記載のタンパク質又はポリヌクレオチド。
【請求項11】
前記ウイルスが、チクングンヤウイルス、シンドビスウイルス、東部ウマ脳炎ウイルス、西部ウマ脳炎ウイルス、ベネズエラウマ脳炎ウイルス、ロスリバーウイルス、セムリキ森林ウイルス、バーマフォレストウイルス及びオニョンニョンウイルスからなる群より選択される請求項10に記載のタンパク質又はポリヌクレオチド。
【請求項12】
フラビウイルス属のウイルスへの感染を治療するための請求項9に記載のタンパク質又はポリヌクレオチド。
【請求項13】
前記ウイルスが、デングウイルス、日本脳炎ウイルス、、キャサヌール森林病ウイルス、マレー渓谷脳炎ウイルス、セントルイス脳炎ウイルス、ダニ媒介脳炎ウイルス、西ナイル脳炎ウイルス、黄熱ウイルス及びオムスク出血熱ウイルスからなる群より選択される請求黄12に記載のタンパク質又はポリヌクレオチド。
【請求項14】
C型肝炎ウイルス感染を治療するための請求項9に記載のタンパク質又はポリヌクレオチド。
【請求項15】
請求項9〜14のいずれか1項で規定されるプラス一本鎖RNAウイルス感染の予防又は治療における同時、別々又は逐次的な使用のための組み合わせ調製物としての、少なくとも、請求項1〜8のいずれか1項で規定される2'-5'-オリゴアデニル酸合成酵素3と、第1の製品とは異なる第2の製品とを含有し、第2の製品が抗ウイルス薬、抗炎症薬及び免疫調節薬からなる群より選択される製品。
【請求項16】
個体から得られた核酸試料中の2'-5'-オリゴアデニル酸合成酵素3遺伝子中の多型を検出するステップを含む、請求項9〜14のいずれか1項で規定されるプラス一本鎖RNAウイルスに対する個体の感受性を評価するためのインビトロ方法。
【請求項17】
検出が、配列番号11、12、62〜86及び118〜142の配列からなる群より選択されるOAS3特異的オリゴヌクレオチドを用いる請求項16に記載の方法。
【請求項18】
前記多型が、Arg844コドンの停止コドンへの変異(R844X)、及びアルギニン381コドンのセリンコドンへの変更(R381S)からなる群より選択される請求項16又は17に記載の方法。
【請求項19】
R844X変異が:
a) 配列番号11及び12のプライマー対を用いるPCRによる183 bpの生成物の増幅と、
b) BglIIエンドヌクレアーゼを用いる前記PCR生成物の消化であって、115 bp及び68 bpの2つのフラグメントの存在がR844X変異の存在を示す消化と
を含むPCR-RFLPアッセイにより検出される請求項18に記載の方法。
【請求項20】
S381バリアントが、デングウイルス感染患者におけるデングショック症候群の危険性に対する優勢な保護と関連している請求項18に記載の方法。
【請求項21】
配列番号7及び配列番号30からなる群より選択される配列を含む単離OAS3タンパク質。
【請求項22】
配列番号9の配列からなる単離OAS3タンパク質フラグメント。
【請求項23】
配列番号7のOAS3タンパク質をコードする配列番号6の配列、配列番号30のOAS3タンパク質をコードする配列番号28又は29の配列、及び配列番号9のOAS3タンパク質フラグメントをコードする配列番号8の配列からなる群より選択される配列を含む単離ポリヌクレオチド。
【請求項24】
請求項23で規定されるポリヌクレオチド配列を含む組換えベクター。
【請求項25】
請求項23で規定されるポリヌクレオチド配列又は請求項24で規定される組換えベクターで形質移入又は形質転換された宿主細胞。
【請求項26】
HeLa-Tet-Off/OAS3#C417-1と命名され、2008年2月26日にCollection Nationale de Cultures de Microorganismes、25 rue du Docteur Roux, 75724 Paris Cedex 15に受入番号I-3927の下で寄託された、組換えヒト2'-5'-オリゴアデニル酸合成酵素3を発現するHeLa-Tet-Off株化細胞である請求項25に記載の宿主細胞。
【請求項27】
HeLa-Tet-Off/OAS3/delta/1Cと命名され、2008年4月17日にCollection Nationale de Cultures de Microorganismes、25 rue du Docteur Roux, 75724 Paris Cedex 15に受入番号I-3968の下で寄託された、切断型組換えヒト2'-5'-オリゴアデニル酸合成酵素3を発現するHeLa-Tet-Off株化細胞である請求項25に記載の宿主細胞。
【請求項28】
HepG2-Tet-Off/OAS3#F8と命名され、2009年5月15日にCollection Nationale de Cultures de Microorganismes、25 rue du Docteur Roux, 75724 Paris Cedex 15に受入番号I-4158の下で寄託された、組換えヒト2'-5'-オリゴアデニル酸合成酵素3を発現するHepG2-Tet-Off株化細胞である請求項25に記載の宿主細胞。
【請求項1】
医薬品としての単離2'-5'-オリゴアデニル酸合成酵素3タンパク質又は2'-5'-オリゴアデニル酸合成酵素3タンパク質をコードする単離ポリヌクレオチド。
【請求項2】
ヒト2'-5'-オリゴアデニル酸合成酵素3の配列を有する請求項1に記載のタンパク質又はポリヌクレオチド。
【請求項3】
配列番号2の残基1〜1087と少なくとも70%のアミノ酸配列同一性又は80%のアミノ酸配列類似性を有する請求項1又は2に記載のタンパク質。
【請求項4】
配列番号2の381位にセリンを含む請求項1〜3のいずれか1項に記載のタンパク質。
【請求項5】
配列番号2及び配列番号7からなる群より選択されるアミノ酸配列を含む請求項4に記載のタンパク質。
【請求項6】
請求項3〜5のいずれか1項で規定されるタンパク質をコードする請求項1又は2に記載のポリヌクレオチド。
【請求項7】
配列番号1及び配列番号6からなる群より選択される配列を含む請求項6に記載のポリヌクレオチド。
【請求項8】
発現ベクターに挿入された請求項1、2、6及び7のいずれか1項に記載のポリヌクレオチド。
【請求項9】
プラス一本鎖RNAウイルス感染を予防又は治療するための請求項1〜8のいずれか1項で規定される単離2'-5'-オリゴアデニル酸合成酵素3タンパク質又はポリヌクレオチド。
【請求項10】
アルファウイルス属のウイルスへの感染を治療するための請求項9に記載のタンパク質又はポリヌクレオチド。
【請求項11】
前記ウイルスが、チクングンヤウイルス、シンドビスウイルス、東部ウマ脳炎ウイルス、西部ウマ脳炎ウイルス、ベネズエラウマ脳炎ウイルス、ロスリバーウイルス、セムリキ森林ウイルス、バーマフォレストウイルス及びオニョンニョンウイルスからなる群より選択される請求項10に記載のタンパク質又はポリヌクレオチド。
【請求項12】
フラビウイルス属のウイルスへの感染を治療するための請求項9に記載のタンパク質又はポリヌクレオチド。
【請求項13】
前記ウイルスが、デングウイルス、日本脳炎ウイルス、、キャサヌール森林病ウイルス、マレー渓谷脳炎ウイルス、セントルイス脳炎ウイルス、ダニ媒介脳炎ウイルス、西ナイル脳炎ウイルス、黄熱ウイルス及びオムスク出血熱ウイルスからなる群より選択される請求黄12に記載のタンパク質又はポリヌクレオチド。
【請求項14】
C型肝炎ウイルス感染を治療するための請求項9に記載のタンパク質又はポリヌクレオチド。
【請求項15】
請求項9〜14のいずれか1項で規定されるプラス一本鎖RNAウイルス感染の予防又は治療における同時、別々又は逐次的な使用のための組み合わせ調製物としての、少なくとも、請求項1〜8のいずれか1項で規定される2'-5'-オリゴアデニル酸合成酵素3と、第1の製品とは異なる第2の製品とを含有し、第2の製品が抗ウイルス薬、抗炎症薬及び免疫調節薬からなる群より選択される製品。
【請求項16】
個体から得られた核酸試料中の2'-5'-オリゴアデニル酸合成酵素3遺伝子中の多型を検出するステップを含む、請求項9〜14のいずれか1項で規定されるプラス一本鎖RNAウイルスに対する個体の感受性を評価するためのインビトロ方法。
【請求項17】
検出が、配列番号11、12、62〜86及び118〜142の配列からなる群より選択されるOAS3特異的オリゴヌクレオチドを用いる請求項16に記載の方法。
【請求項18】
前記多型が、Arg844コドンの停止コドンへの変異(R844X)、及びアルギニン381コドンのセリンコドンへの変更(R381S)からなる群より選択される請求項16又は17に記載の方法。
【請求項19】
R844X変異が:
a) 配列番号11及び12のプライマー対を用いるPCRによる183 bpの生成物の増幅と、
b) BglIIエンドヌクレアーゼを用いる前記PCR生成物の消化であって、115 bp及び68 bpの2つのフラグメントの存在がR844X変異の存在を示す消化と
を含むPCR-RFLPアッセイにより検出される請求項18に記載の方法。
【請求項20】
S381バリアントが、デングウイルス感染患者におけるデングショック症候群の危険性に対する優勢な保護と関連している請求項18に記載の方法。
【請求項21】
配列番号7及び配列番号30からなる群より選択される配列を含む単離OAS3タンパク質。
【請求項22】
配列番号9の配列からなる単離OAS3タンパク質フラグメント。
【請求項23】
配列番号7のOAS3タンパク質をコードする配列番号6の配列、配列番号30のOAS3タンパク質をコードする配列番号28又は29の配列、及び配列番号9のOAS3タンパク質フラグメントをコードする配列番号8の配列からなる群より選択される配列を含む単離ポリヌクレオチド。
【請求項24】
請求項23で規定されるポリヌクレオチド配列を含む組換えベクター。
【請求項25】
請求項23で規定されるポリヌクレオチド配列又は請求項24で規定される組換えベクターで形質移入又は形質転換された宿主細胞。
【請求項26】
HeLa-Tet-Off/OAS3#C417-1と命名され、2008年2月26日にCollection Nationale de Cultures de Microorganismes、25 rue du Docteur Roux, 75724 Paris Cedex 15に受入番号I-3927の下で寄託された、組換えヒト2'-5'-オリゴアデニル酸合成酵素3を発現するHeLa-Tet-Off株化細胞である請求項25に記載の宿主細胞。
【請求項27】
HeLa-Tet-Off/OAS3/delta/1Cと命名され、2008年4月17日にCollection Nationale de Cultures de Microorganismes、25 rue du Docteur Roux, 75724 Paris Cedex 15に受入番号I-3968の下で寄託された、切断型組換えヒト2'-5'-オリゴアデニル酸合成酵素3を発現するHeLa-Tet-Off株化細胞である請求項25に記載の宿主細胞。
【請求項28】
HepG2-Tet-Off/OAS3#F8と命名され、2009年5月15日にCollection Nationale de Cultures de Microorganismes、25 rue du Docteur Roux, 75724 Paris Cedex 15に受入番号I-4158の下で寄託された、組換えヒト2'-5'-オリゴアデニル酸合成酵素3を発現するHepG2-Tet-Off株化細胞である請求項25に記載の宿主細胞。
【図2】

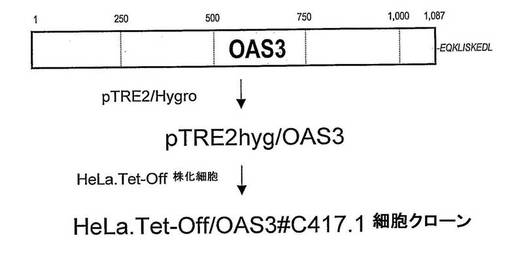
【図3−1】
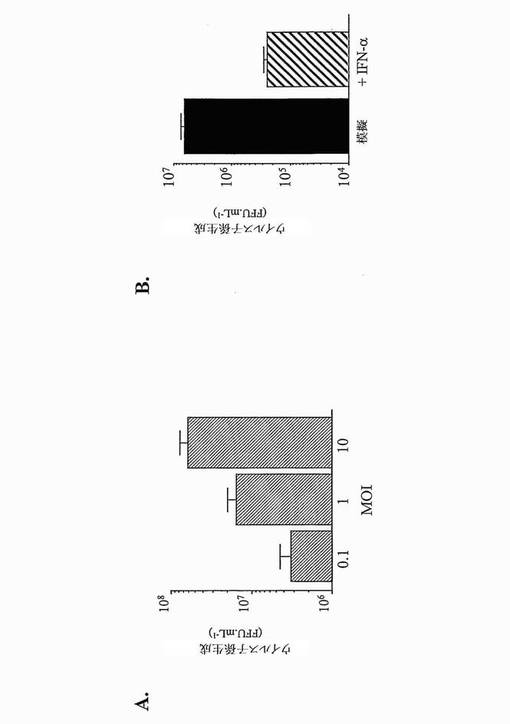
【図3−2】
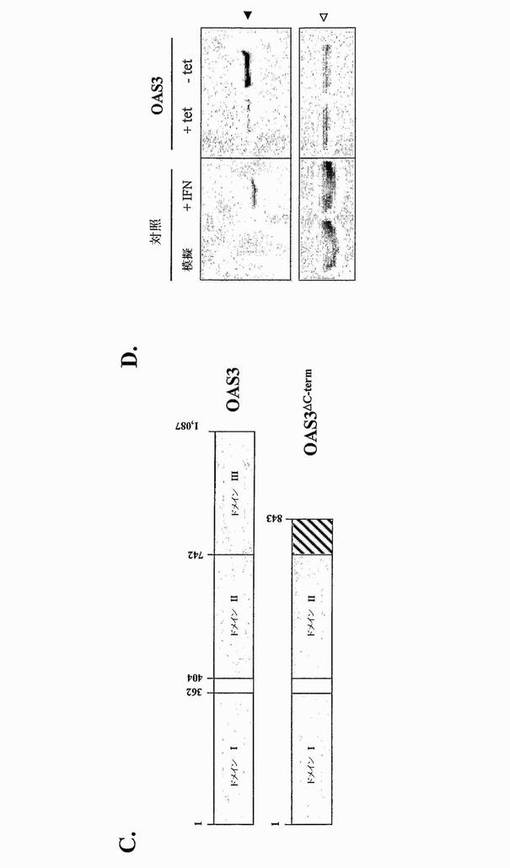
【図4−1】
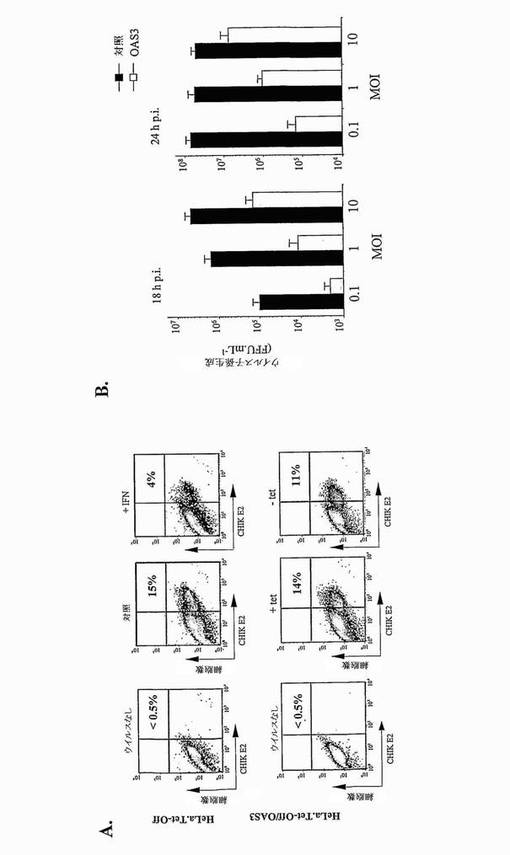
【図4−2】
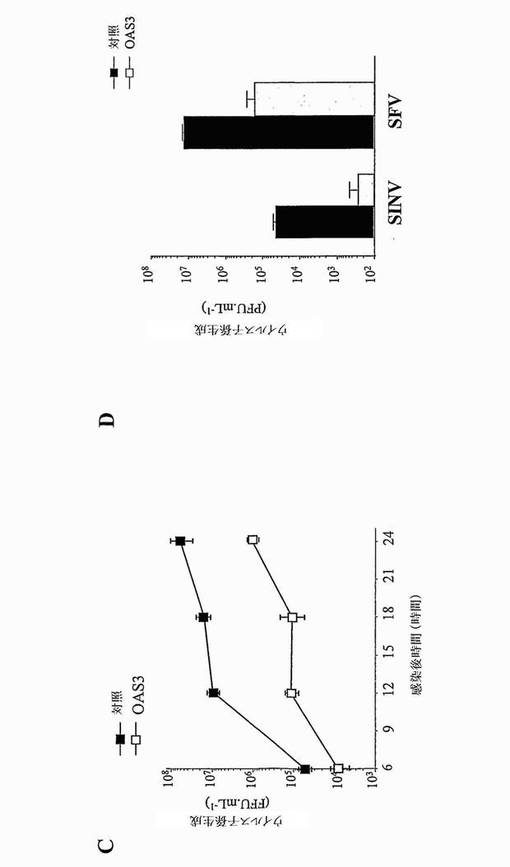
【図5】
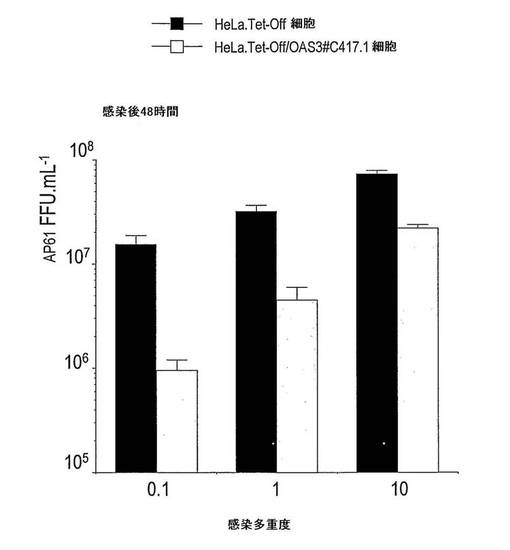
【図6】
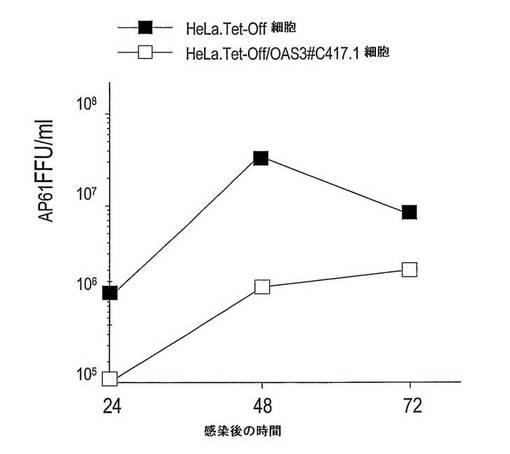
【図7】
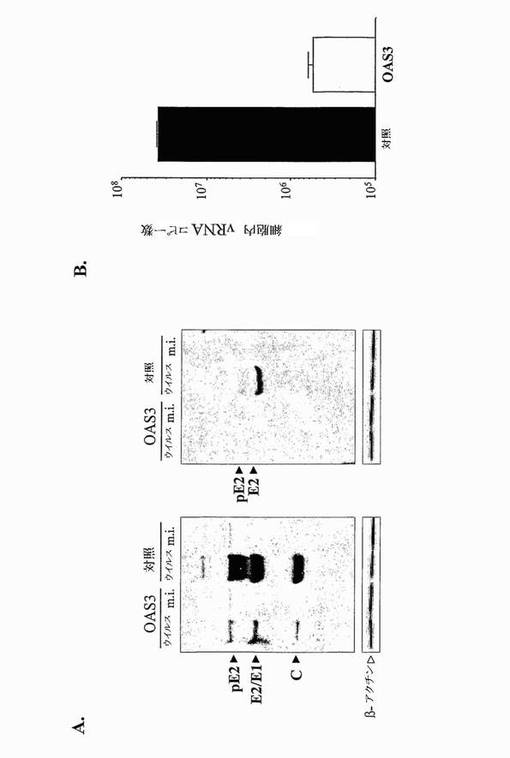
【図8】
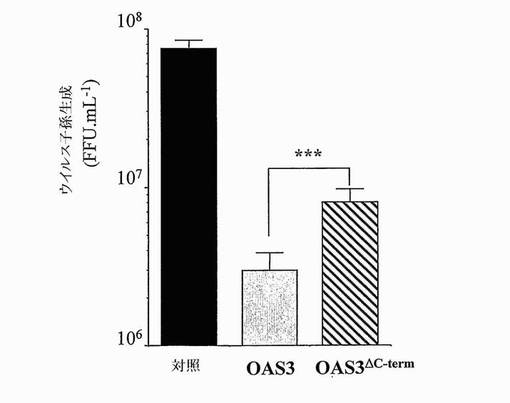
【図9】
【図10】
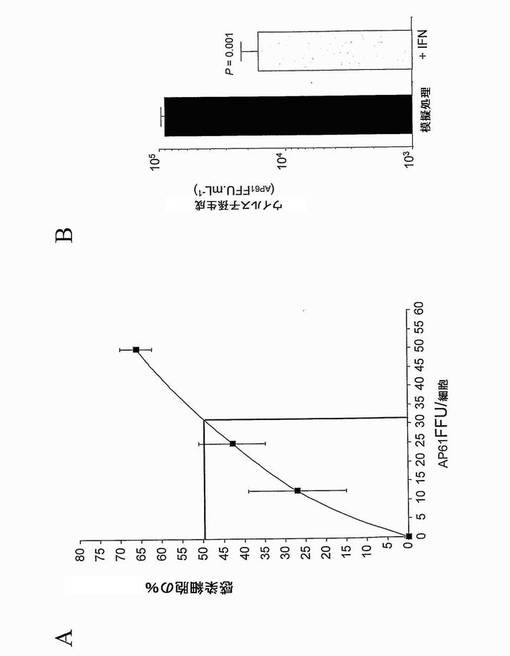
【図11】
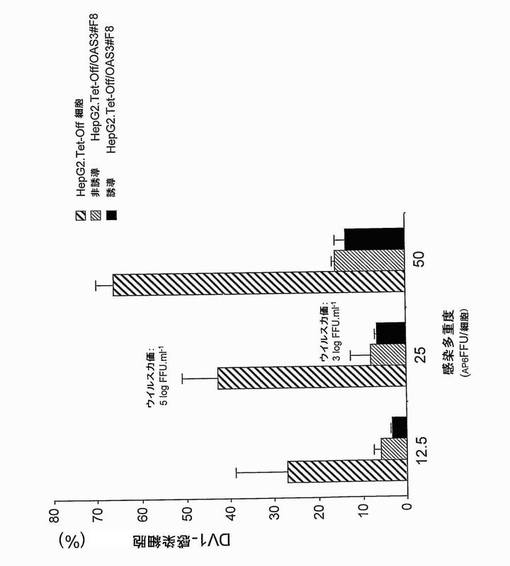
【図12】
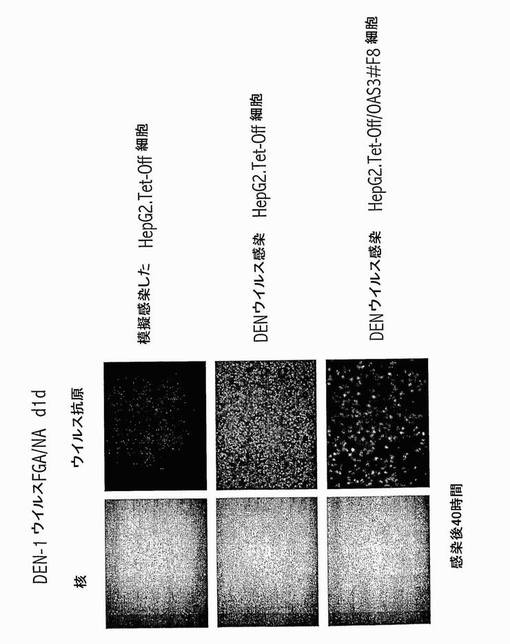
【図13】
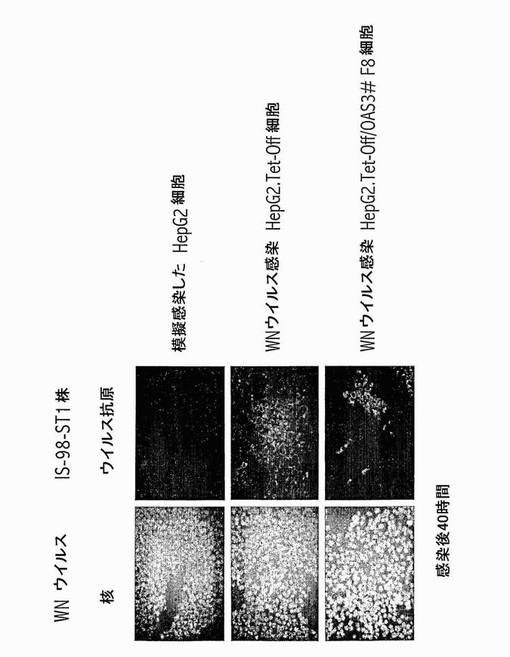
【図14】
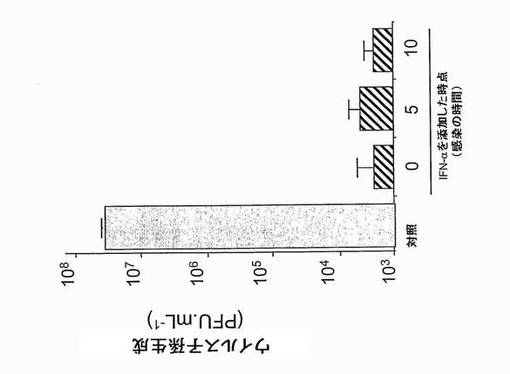
【図15】
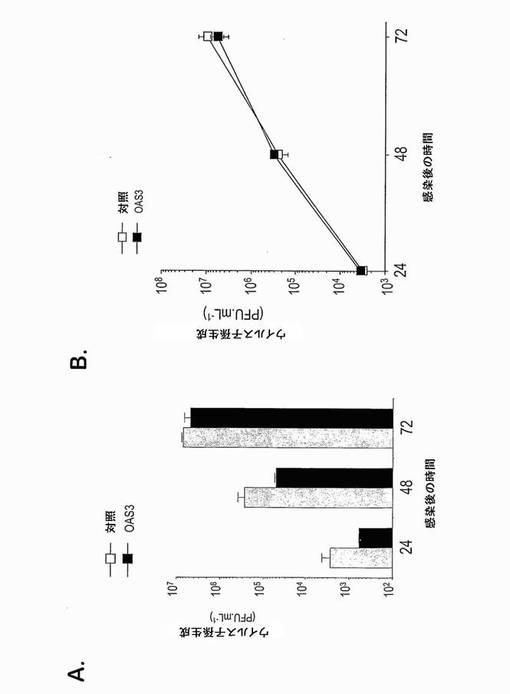
【図16】
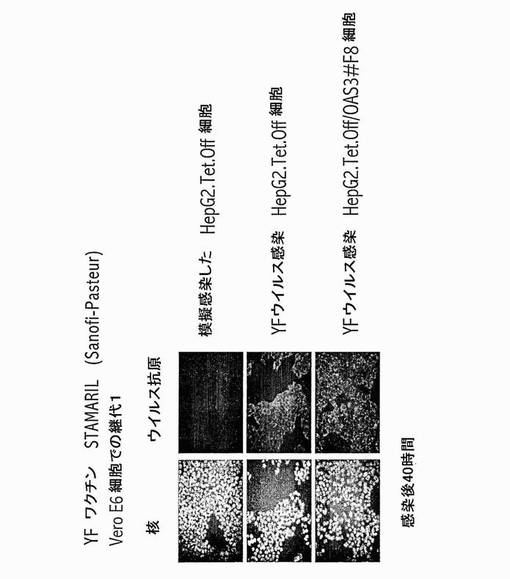
【図3−1】
【図3−2】
【図4−1】
【図4−2】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公表番号】特表2011−522527(P2011−522527A)
【公表日】平成23年8月4日(2011.8.4)
【国際特許分類】
【出願番号】特願2011−510066(P2011−510066)
【出願日】平成21年5月20日(2009.5.20)
【国際出願番号】PCT/IB2009/005956
【国際公開番号】WO2009/141731
【国際公開日】平成21年11月26日(2009.11.26)
【出願人】(501474748)インスティティ・パスツール (27)
【氏名又は名称原語表記】INSTITUT PASTEUR
【住所又は居所原語表記】28,rue du Docteur Roux,F−75724 Paris Cedex 15 FRANCE
【Fターム(参考)】
【公表日】平成23年8月4日(2011.8.4)
【国際特許分類】
【出願日】平成21年5月20日(2009.5.20)
【国際出願番号】PCT/IB2009/005956
【国際公開番号】WO2009/141731
【国際公開日】平成21年11月26日(2009.11.26)
【出願人】(501474748)インスティティ・パスツール (27)
【氏名又は名称原語表記】INSTITUT PASTEUR
【住所又は居所原語表記】28,rue du Docteur Roux,F−75724 Paris Cedex 15 FRANCE
【Fターム(参考)】
[ Back to top ]