
プリオン病治療薬及びそのスクリーニング方法
【課題】 迅速、簡便に効率良くプリオン病に対する低分子治療候補化合物を探索するためのスクリーニング方法、及びプリオン病治療薬を提供する。
【解決手段】 プリオン病治療薬のスクリーニング方法は、硫酸化多糖を構成する二糖構造を含む糖と、異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列を少なくとも含む正常型プリオン蛋白質断片、或は、全長正常型プリオン蛋白質との結合の阻害を指標とする。プリオン病治療薬は、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上を有効成分として含んでなる。
【解決手段】 プリオン病治療薬のスクリーニング方法は、硫酸化多糖を構成する二糖構造を含む糖と、異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列を少なくとも含む正常型プリオン蛋白質断片、或は、全長正常型プリオン蛋白質との結合の阻害を指標とする。プリオン病治療薬は、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上を有効成分として含んでなる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、プリオン病治療薬及びそのスクリーニング方法に関する。
【背景技術】
【0002】
プリオン病は感染因子プリオンにより中枢神経が侵される疾患であり、ヒトでは全身の不随意運動と急速に進行する痴呆を主な特徴とするクロイツフェルト・ヤコブ病(CJD)、ゲルストマン・ストロイスラー・シャインカー症候群や致死性家族性不眠症などがある。ヒト以外の哺乳動物のプリオン病については、例えば、牛海綿状脳症(BSE)、スクレイピー、鹿慢性消耗性疾患(CWD)、伝達性ミンク脳症(TME)など、ウシ、ヤギ、ヒツジ、シカ、オオジカ、ミンク、ブタ、サル、マウス、ラット、ハムスター、ネコにおける各種プリオン病が公知である。これまでにプリオン病に対する実用性に優れた治療薬は発見されていないものの、硫酸化多糖が薬効のある化合物群の一つとして知られている。硫酸化多糖の一種であるペントサンポリサルフェートは実際にプリオン病患者に使用され、脳室内投与による臨床研究が進行している。このような硫酸化多糖は様々な部分構造と分子量分布を含み不均一性を有する高分子であり、プリオン蛋白質との結合は定性的な評価に限られている。高分子ゆえに脳血液関門を越えてプリオン病の病変の主座である脳実質内にまで移行しないこと、薬理活性の作用機序が明らかになっていないこと、強力な抗凝血作用を有するものがあること等が問題となっており、脳実質内への移行が期待できる低分子化合物で硫酸化多糖と同程度の抗プリオン活性を有する治療薬の開発が望まれている。
【0003】
プリオン病治療薬のスクリーニング技術としては、異常型プリオン蛋白と正常型プリオン蛋白の相互作用を阻害する化合物をスクリーニングする方法[例えば非特許文献1参照]、プリオン感染培養細胞を用いて異常型プリオン蛋白の産生を阻害する化合物をスクリーニングする方法[例えば非特許文献2参照]、酵母を用いて酵母プリオンの産生を阻害する化合物をスクリーニングする方法[例えば非特許文献3参照]、正常型プリオン蛋白との相互作用の強い化合物をスクリーニングする方法[例えば非特許文献4参照]、予想される正常型プリオン蛋白構造よりインシリコで正常型プリオン蛋白と相互作用する化合物をスクリーニングする方法[例えば非特許文献5参照]、また、イムノアッセイによりプリオンペプチド106−126に結合する候補物質を選択し、選択候補物質につき分子間相互作用解析にて結合選択性評価し、ウェスタンブロットによって異常型プリオンへの変換阻害効果を検定する方法[例えば特許文献1参照]などが提案されている。しかしながら、非特許文献1乃至5、及び特許文献1のようなプリオン病治療薬のスクリーニング技術については、アッセイにかなりの時間を要したり、十分な科学的エビデンスがなく闇雲に探索するものであり、治療効果を有する化合物が発見される効率が必ずしも高くないという問題があった。
【0004】
【非特許文献1】ギース A.ら(Giese A.,et al)、ジャーナル オブ ヴァイロロジー(Journal of Virology)2005年、79巻、7785−7791頁
【非特許文献2】レイス R.E.ら(Race R.E.,et al)、ジャーナル オブ ジェネラル ヴァイロロジー(Journal of General Virology)1987年、68巻、1391−1399頁
【非特許文献3】バック S.ら(Bach S.,et al)、ネイチャー バイオテクノロジー(Nature Biotechnology)2003年、21巻、1075−1081頁
【非特許文献4】カワタケ S.、ドウウラ K.ら(Kawatake S.,Doh−ura K.,et al)、バイオロジカル アンド ファーマスーティカル ブレチン(Biological & Pharmaceutical Bulletin)2006年、29巻、927−932頁
【非特許文献5】ローレンツェン S.ら(Lorenzen S.,et al)、バイオシステムス(Biosystems)2005年、80巻、117−122頁
【特許文献1】特開2003−149237号公報
【発明の開示】
【発明が解決しようとする課題】
【0005】
かかる問題点に鑑みてなされた本発明の目的は、迅速、簡便に効率良くプリオン病に対する低分子治療候補化合物を探索するための方法、及びプリオン病治療薬を提供することにある。
【課題を解決するための手段】
【0006】
上記目的を達成するため、本発明によるプリオン病治療薬のスクリーニング方法は、a)硫酸化多糖を構成する二糖構造を含む糖と、b)異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列を少なくとも含む正常型プリオン蛋白質断片、或は、全長正常型プリオン蛋白質と、の結合の阻害を指標とすることを特徴とする。このようなプリオン病治療薬のスクリーニング方法において、糖が、硫酸化多糖を酸化剤により10糖以下に低分子化したヘテロ多糖又はヘテロオリゴ糖、或は化学合成したヘテロオリゴ糖であることが好ましく、また、硫酸化多糖がヘパリンであることが良く、酸化剤が過ヨウ素酸又は過塩素酸又はこれらの塩であることが好適である。そして、糖が、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれる1種以上であることが好適であり、また、正常型プリオン蛋白質は配列番号1のアミノ酸配列を有するマウスプリオン蛋白質(23−231)とし、プリオン蛋白質断片は、N末端側に少なくとも配列番号2のアミノ酸配列(マウスプリオン蛋白質23−89)を含むマウスプリオン蛋白質断片とすることが好ましい。なお、指標は、表面プラズモン共鳴法によって評価されることが好適である。
【0007】
また、上記目的を達成するため、本発明のプリオン病治療薬のスクリーニング方法は、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上を有効成分として含んでなることを特徴とする。
【発明の効果】
【0008】
本発明のプリオン病治療薬のスクリーニング方法によれば、プリオン病治療効果発現の分子メカニズムに基づいた直接的な作用によって合理的にプリオン病治療薬を探索できることから、迅速に、高い効率(確度)でプリオン病治療薬候補化合物を探り当てることが可能である。また、プリオン病感染材料を扱うことなく簡便にスクリーニングを行うことができ、安全衛生面においても優れている。糖として、硫酸化多糖を酸化剤によって10糖以下に低分子化したヘテロ多糖(6〜10糖)又はヘテロオリゴ糖(2〜5糖)、或は化学合成した(N−硫酸化及び/又はO−硫酸化)ヘテロオリゴ糖を用いる場合、また、例えばマウスプリオン蛋白質(23−89)のような異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域部に相当する正常型プリオン蛋白質断片を用いる場合には、より高い効率でプリオン病治療薬候補化合物のスクリーニングができる。糖として、例えばグルコサミン6−硫酸とイズロン酸2−硫酸とがα1−4結合したへテロ二糖、特に「GlcNS6Sα4IdoA2Sα」を用いる場合、また、マウスプリオン蛋白質(23−89)と組み合わせて用いる場合には、さらに低分子のプリオン病治療薬候補化合物スクリーニングの確度が増す。また、本発明のプリオン病治療薬は、強力な抗プリオン活性を持つヘパリンと同様の作用機序による抗プリオン作用を有しており、且つ、ヘパリンに比して抗凝血作用が極めて弱い有効成分を含有することから有用である。
【発明を実施するための最良の形態】
【0009】
以下、本発明によるプリオン病治療薬及びそのスクリーニング方法を実施するための最良の形態について、詳細に説明する。
【0010】
本実施形態によるプリオン病治療薬のスクリーニング方法は、硫酸化多糖を構成する二糖構造を含む糖と、全長正常型プリオン蛋白質或は正常型プリオン蛋白質断片との結合の阻害を指標とするものである。ここで、硫酸化多糖は、硫酸化された二糖(単位)構造を有していれば特に限定されず、好ましくは、例えば、ヘキソサミンとヘキスロン酸(ウロン酸)とのヘテロ二糖構造を有するヘパリン、ヘパラン硫酸、コンドロイチン硫酸、デルマタン硫酸、また、ヘキソサミンとヘキソースとのヘテロ二糖構造を有するケラタン硫酸や、ヘキスロン酸とペントースとのヘテロ二糖構造及びペントースからなるホモ二糖構造を有するペントサンポリサルフェートなどを挙げることができる。これらの硫酸化多糖の内でも、特に、二糖構造として、D−グルクロン酸(GlcA)又はL−イズロン酸(IdoA)のヘキスロン酸と、N−スルホ−D−グルコサミン(GlcNS)又はN−アセチル−D−グルコサミン(GlcNAc)のヘキソサミンとがβ1−4或はα1−4結合した構造を有し、2位、3位、6位の一部又は全部が硫酸化(N−硫酸化、O−硫酸化)されたヘパリン又はヘパラン硫酸が、低分子化した場合にも抗プリオン活性を有する場合がある点でより好ましい。強力な抗プリオン活性を持ち、例えば10糖にまで低分子化した場合にも抗プリオン活性が殆ど低下しない場合がある点でヘパリンがさらに好適に使用できる。ヘパリンは、如何なる動物種の如何なる臓器由来のものであっても良く特に限定されないが、安全衛生面からプリオン病発症例を認めない点でブタ由来のヘパリンが好ましい。
【0011】
このような硫酸化多糖は、液相において各種分解酵素や分解剤によって10糖以下に低分子化することができる。例えば、硫酸化多糖としてヘパリンを低分子化する場合には、上述の二糖単位構造を保持して低分子化可能であることから、酸化剤によって又は酸化剤及びアルカリによって低分子化することが好ましい。酸化剤として、例えば、過ヨウ素酸、過塩素酸、亜硝酸又はこれらの塩を使用して、10糖以下のヘテロ多糖(10糖〜6糖)又はヘテロオリゴ糖(5糖〜2糖)とすることができる。例えば、スダ、ソーベル(Suda, Sobel)らの方法[スダ Y.、ソーベル M.ら、スロンボシス リサーチ(Thrombosis Research)1993年、69巻、501−508頁]に準じて低分子化することができる。また、上述の二糖単位構造を含むヘテロオリゴ糖を化学合成しても良く、スダ(Suda)らの方法[スダ Y.ら、テトラへドロン レターズ(Tetrahedron Letters)1996年、37巻、1053−1056頁]に準じて合成することができる。ヘテロオリゴ糖の内でも、プリオン蛋白質との結合における必須の中心構造となることから、二糖単位構造が好ましい。N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルファミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上がより好適である。ヘテロ二糖の金属塩としては、アルカリ金属塩が好ましく、アルカリ金属塩の中でもNa塩、K塩がより好ましい。
【0012】
また、上記ヘテロ二糖、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上の糖は、強力な抗プリオン活性を持つヘパリンと同様の作用機序による抗プリオン作用を有しており、且つ、ヘパリンに比して抗凝血作用が極めて弱いことから、プリオン病治療薬の有効成分として有用である。
【0013】
上述のN−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖(単位)構造は、以下の式(1)で表される。
【0014】
【化1】

【0015】
また、上記スダ(Suda)らの方法[スダ Y.ら、テトラへドロン レターズ(Tetrahedron Letters)1996年、37巻、1053−1056頁]に準じた化学合成により得られるヘテロ二糖は、以下の一般式(2)で表されるものとすることができる〔式(2)中、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す〕。一般式(2)におけるR1がNa又はK、R2が水素又はメチル基であるヘテロ二糖が好ましい。
【0016】
【化2】
【0017】
また、硫酸化多糖を構成する二糖構造を含む糖として、一般式(2)で表されるヘテロ二糖におけるR2にグルコース残基を導入した下記一般式(3)で表されるヘテロ三糖も好適に使用することができる〔式(3)中、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す〕。一般式(3)におけるR1がNa又はK、R2が水素又はメチル基であるヘテロ三糖が好ましい。このようなヘテロ三糖は、例えば、コシダ(S Koshida)、スダ(Y Suda)らの方法[コシダ S.、スダ Y.ら、テトラへドロン レターズ(Tetrahedron Letters)2001年、42巻、1289−1292頁]に準じて化学合成することができる。一般式(3)中のR2にグルコース残基を導入してヘテロ四糖としても良い。
【0018】
【化3】
【0019】
正常型プリオン蛋白質は、上述のような硫酸化多糖を構成する二糖構造を含む糖との結合阻害を指標としてプリオン病治療薬をスクリーニングする際に用いるものであり、如何なる動物種のものであっても良いが、動物種間における相同性が高いことから、哺乳動物のプリオン蛋白質が好ましい。特に、前駆体プリオン蛋白質におけるN末端のシグナル配列部分、及びC末端疎水性領域(GPIアンカー付加)が成熟、分泌過程において除かれた哺乳動物の正常型プリオン蛋白質では、動物種によるアミノ酸配列の差異がより小さく、例えば、ヒトとマウスやハムスターのようなげっ歯類とのホモロジーは95%以上である。本実施形態においては、マウスプリオン蛋白質全長に相当する配列番号1に示すアミノ酸配列を有するプリオン蛋白質(23−231)を哺乳動物全般を対象とするスクリーニングに適用可能な全長正常型プリオン蛋白質として用いることができる。また、上述の糖との結合における必須領域(配列)はN末端側部分にあり、プリオン蛋白質(23−231)のN末端からアミノ酸残基数67の配列に相当する配列番号2に示すアミノ酸配列に含まれることが後述の実施例により確認された。このプリオン蛋白質(23−231)のN末端からアミノ酸残基数67のマウスプリオン蛋白質(23−89)は、哺乳動物異常型プリオン蛋白質のプロテアーゼ(プロテイナーゼK)処理により消化除去されるN末端領域部に相当する。したがって、本実施形態のスクリーニング方法における正常型プリオン蛋白質断片として、異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列をN末端に少なくとも含むプリオン蛋白質断片が好適に使用できる。異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域部のみからなる正常型プリオン蛋白質断片が、硫酸化二糖構造との結合に必須の最小構造であり、さらに低分子量のプリオン病治療薬候補化合物を探索できる場合があることからより好ましい。このような正常型プリオン蛋白質断片のうち、マウスプリオン蛋白質(23−89)を哺乳動物全般を対象とするスクリーニングに用いることができ、少なくとも配列番号2のアミノ酸配列を含んでなるプリオン蛋白質断片であればいずれも好適に使用できる。マウスプリオン蛋白質(23−231)の前駆体蛋白質におけるアミノ酸配列は、配列番号3に示すとおりである。
【0020】
以上のような、全長正常型プリオン蛋白質や正常型プリオン蛋白質断片は、公知の遺伝子組み換え蛋白質合成法に準じて調製することができる。ここで形質転換する宿主細胞株は動物や酵母由来の真核細胞を用いても良いが、糖鎖が結合した糖蛋白質として分泌される場合があり、得られたリコンビナントプリオン蛋白質と上述の硫酸化多糖を構成する二糖構造を含む糖との結合、結合阻害の評価において影響を与える可能性がある点では、大腸菌株のような原核細胞を用いることが好ましい。具体的には、例えば、ホーネマンらの方法[ホーネマン(Hornemann S)ら、Journal of Molcular Biology,1996年、261巻、614−619頁]、及び[ホーネマン(S.Hornemann)ら、FEBS Letters,1997年、413巻、277−281頁]、リーマンらの方法[リーマン(Liemann S)ら、Biochemistry,1999年、38巻、3258−3267頁]などに準じて、リコンビナントプリオン蛋白質及びその断片を調製することができる。
【0021】
上述のような糖と、プリオン蛋白質断片又は全長正常型プリオン蛋白質との結合の阻害を指標として評価する方法としては、糖と蛋白質との相互作用(反応、構造や重量などの状態変化)を検出可能な方法であれば公知の如何なる探索、評価方法であっても適用できる。このような探索、評価方法(装置)として、例えば、表面プラズモン共鳴法(SPR法)、水晶発振子の周波数変化を測定する方法、吸発熱を測定するカロリメトリー法、蛍光偏光測定法、核磁気共鳴法(NMR)、原子間力顕微鏡法(AFM)、透過型電子顕微鏡法(TEM)、走査型電子顕微鏡法(SEM)などの他、ウェスタンブロッティング法、ELISA法を挙げることができる。抗体の調製や標識体の調製が不要である点では、蛍光偏光測定法、ウェスタンブロッティング法、ELISA法以外の方法、装置が好ましく、夫々、適用する方法、装置のインストラクションマニュアル等に基づき実施することができる。例えば、表面プラズモン共鳴法を適用する場合には、[永田知宏、半田宏編集、シュプリンガー・フェアラーク東京株式会社1998年発行、生体物質相互作用のリアルタイム解析実験法 ]、[スダ Y.、ソーベル M.ら、バイオコンジュゲート ケミストリー (Bioconjugate Chemistry)2006年、17巻、1125−1135頁]、特開2003−83969号公報などの記載に準じ、市販のSPRセンサー(装置)を用いて行なうことができる。本実施形態では、上述の糖(硫酸化多糖、硫酸化オリゴ糖)又はプリオン蛋白質(全長正常型プリオン蛋白質、或はその断片)の内の一方をセンサーチップ上に固定し、他方を結合させた系において、両者の結合を競合阻害する化合物を共存させ表面プラズモン共鳴角度の変化をリアルタイムに測定する。この測定結果から両者の結合や解離の相互作用を反応速度論的、化学平衡論的に解析、評価することによってプリオン病治療薬の候補化合物を探索することができる。また、AFM、TEM、SEMなどを適用する場合には、例えば、上述の糖又はプリオン蛋白質(或はその断片)の内の一方をSPRのセンサーチップに固定する代わりに、金などの金属ナノ粒子に固定し、他方を結合させた系において、両者の結合を競合阻害する化合物を共存させ各装置を用いて相互作用を測定、解析、評価することによってプリオン病治療薬の候補化合物を探索することができる。
【0022】
プリオン蛋白質断片又は全長正常型プリオン蛋白質をリガンドとしてセンサーチップ上に固定する場合には、金表面にカルボキシメチルデキストラン層を設けたセンサーチップ(CM5:Biacore社製)に全長正常型プリオン蛋白質(マウスプリオン蛋白質(23−231))をアミンカップリングにより固定化するとN末端部分が分析中の条件化において変質・変性してしまうことがある(非特許文献4)ことから、全長正常型プリオン蛋白質よりマウスプリオン蛋白質断片(23−89)を用いることが好ましい。また、センサーチップ上へ固定化するリガンドとしては、以上のようなプリオン蛋白質やその断片に比較して上述のような糖を用いることがより好ましく、糖の内でも一般式(1)で表される二糖単位構造からなるヘテロ二糖、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれる1種以上がさらに好ましい。一般式(2)で表される化学合成のヘテロ二糖も一般式(1)の糖と同様好適に使用できる。一般式(1)、(2)で表されるような(硫酸化)糖をリガンドとしてセンサーチップ上に固定化する場合には、例えば、オクタンチオールのような末端にチオール基を有する長鎖アルキル化合物をセンサーチップ金表面にAu−S結合によって固定、アルキル鎖が露出する疎水性層を設け、糖の還元末端にも疎水性基を導入して疎水吸着によって糖を固定することもできるが、糖固定化後に添加するプリオン蛋白質が糖との結合のみならずチップ表面の疎水性層に吸着するなど好ましくない非特異的な相互作用を生じることがある点では、リガンドである糖鎖とリンカー化合物を結合させたリガンド・リンカー複合体(以下リガンド複合体)を調製してセンサーチップ金表面に直接結合、固定化することが好適である。一般式(1)で表される二糖構造を有する糖や、一般式(2)で表される合成へテロ二糖をリンカーと結合する際に、プリオン蛋白質との結合に必須の最小二糖構造が失われることがある点では、一般式(3)で表されるヘテロ三糖を用いてリガンド複合体を調製することが好ましい。硫酸化多糖を酸化剤によって10糖以下に低分子化したヘテロ多糖、ヘテロオリゴ糖を用いてリガンド複合体を調製する場合には、酸化によって生じるアルデヒド基を利用し、一般式(3)の合成法に準じてリンカーと結合することが好適である。リガンド複合体の調製は、[スダ Y.、ソーベル M.ら、バイオコンジュゲート ケミストリー (Bioconjugate Chemistry)2006年、17巻、1125−1135頁]、特開2003−83969号公報などに準じて行なうことができる。一般式(3)で表される合成へテロ三糖をリガンドとするリガンド複合体として、例えば、一般式(4)で表される(芳香族アミンと環状SS結合を持つチオクト酸をリンカー部とする)リガンド複合体を挙げることができる〔式(4)中、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す〕。一般式(4)におけるR1がNa又はK、R2が水素又はメチル基であるリガンド複合体が好ましい。このような、リガンド複合体のセンサーチップ金表面への結合は、1μM〜100μM濃度で水溶性アルコール溶液中に溶解した液にセンサーチップを室温、6〜24時間浸漬することによって行なうことができる。式(4)におけるR1がNa、R2がメチル基であるリガンド複合体:GlcNS6Sα4IdoA2Sα6Glc−monoをセンサーチップ金表面に結合する場合には、例えば、5〜50μM濃度でメタノール・水混液(1:1)に溶解した液に浸漬すれば良い。
【0023】
【化4】
【0024】
また、リガンド複合体として、一般式(5)、(6)、(7)で表される(ヘテロ三糖を集積した)GlcNS6Sα4IdoA2Sα6Glc−Tri、GlcNS6Sα4IdoA2Sα6Glc−Tetra、GlcNS6Sα4IdoA2Sα6Glc−Tetra−Longについても、GlcNS6Sα4IdoA2Sα6Glc−monoと同様に好適に使用できる。さらに、一般式(8)で表されるビオチン・アビジン結合によるリガンド複合体も使用可能であるが、金表面にアビジンを結合したセンサーチップを用いるためプリオン蛋白質などとの非特異的相互作用が残る。一般式(5)〜(8)の夫々において、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す。一般式(5)〜(8)の夫々において、R1がNa又はK、R2が水素又はメチル基であるリガンド複合体が好ましい。
【0025】
【化5】
【0026】
【化6】
【0027】
【化7】
【0028】
【化8】
【実施例】
【0029】
以下、本発明によるプリオン病治療薬及びそのスクリーニング方法について、実施例を示して具体的に説明するが、これによって本発明を限定するものではない。
【0030】
〔実施例1〕
ヘパリンを構成する糖は数種類あり、また、それらの糖修飾の程度、モノマー間の結合様式などには多様性がある。ヘパリンはプリオン持続感染細胞を用いた薬剤スクリーニングで抗プリオン活性があることが示されている。そこで、この活性を示すヘパリンの部分構造を推定する実験を行った。
〔方法〕
プリオン持続感染細胞としては、スクレイピープリオンであるRML株がマウス神経芽細胞腫細胞(Neuro2a)に持続感染したScN2a細胞を用い、以下の処理を行ったヘパリンと共存下で培養した。培養液(10%胎児牛血清加MEM(Opti−MEM、インビトロジェン社製))に添加するヘパリン処理物の一つは、過ヨウ素酸による、炭素−炭素結合で隣接した2個の水酸基の酸化と引き続くベータ脱離反応による分解、もう一つは、酵素(heparinase I)による酵素消化である。これらのヘパリンの低分子化処理は、スダ、ソーベル(Suda, Sobel)らの方法[スダ Y.、ソーベル M.ら、スロンボシス リサーチ(Thrombosis Research)1993年、69巻、501−508頁]に準じて行った。これら低分子量化(低分子化処理)されたヘパリンのうち、10糖に相当する鎖長の分画を用いた。コントロールとして、処理を施していないヘパリン(ナカライテスク社製、分子量17600)の計3種類のサンプルを用いた。これらを上記の細胞培養時、培養液に添加した。終濃度は、未処理ヘパリンに対して、0, 0.1, 0.5, 1, 5, 10μg/mL、過ヨウ素酸処理したヘパリンについて、0, 0.2, 0.6, 1, 10, 50μg/mL、ヘパリナーゼ処理(HeparinaseI(EC4.2.2.7)、シグマ社製)したヘパリンに対して、0, 1, 5, 10, 25, 50μg/mLとした。細胞培養は、細胞培養フラスコ(底面積9.7cm2 )にて培養し、細胞密度がコンフルエントとなった時点で培養上清を除き、リン酸バッファー生理食塩水[PBS、pH6.5]で細胞を洗浄した。洗浄液を十分に除き、リシス液[lysis液:0.5%デオキシコール酸ナトリウム、0.5% NP−40(商品名、和光純薬工業(株)オクチルフェニルエーテル)、残部PBS]0.5mLを用いて細胞を溶解した。軽く遠心分離して、高分子核酸を除いた後、細胞溶解液にトリチラキウム アルブム(Tritirachium album)由来のプロテイナーゼK(PK:Proteinase K)を終濃度10μg/mLとなるように加えて37℃で30分反応(蛋白質分解)させた後、フェニルメタンスルホニルフロライド(phenylmethan sulfonylfluoride: PMSF)を終濃度1mMとなるように加えて蛋白質分解反応を停止させた。
【0031】
蛋白質分解処理した細胞溶解液0.5mLに核酸分離用シリカとして市販されているFOG((商品名)、Qbiogene社製)を滅菌水(オートクレーブ滅菌蒸留水)で1000倍希釈したシリカ懸濁液5μL(固形分(シリカ 約1.5μg含有))を加え、2〜3分間室温で混和した後、軽く遠心(9,000×g、 1分間)してシリカを沈殿させ、上清を除いた。この沈殿全量に対し、SDSポリアクリルアミドゲル電気泳動(SDS−PAGE)用バッファー[1% SDS(ソディウムドデシルサルフェート)、0.05% ブロモフェノールブルー、4% グリセロール、1% 2−メルカプトエタノール、25mM トリス塩酸 pH6.8] 20μLを加えて懸濁し、95℃、5分間の熱変性後に、イシカワ、ドウウラ(Ishikawa, Doh−ura)らの方法[イシカワ K.、ドウウラ K.ら、ジャーナル オブ ジェネラル ヴァイロロジー(Journal of General Virology)2004年、85巻、1785−1790頁]に準じ、15% トリス・グリシンSDS−PAGEゲルで電気泳動し、ウエスタンブロット法でプロテアーゼ抵抗性の異常型プリオン蛋白質を検出した。ウエスタンブロット法においては、電気泳動した蛋白質をポリビニリデンジフルオライド(PVDF)膜(ミリポア社製、PVDFメンブレン)にブロッティングし、プリオン蛋白質に対するマウスモノクローナル抗体(SPI−BIO社製、SAF83(5000倍希釈))、及び、アルカリフォスファターゼ・コンジュゲートヤギ抗マウス免疫グロブリン抗体(プロメガ社製、(20000倍希釈))、化学発光検出試薬(アマーシャム社製、CDP−Star detection reagent(商品名))を使用してプリオン蛋白質の検出を行った。
【0032】
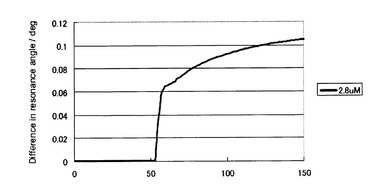
[結果]
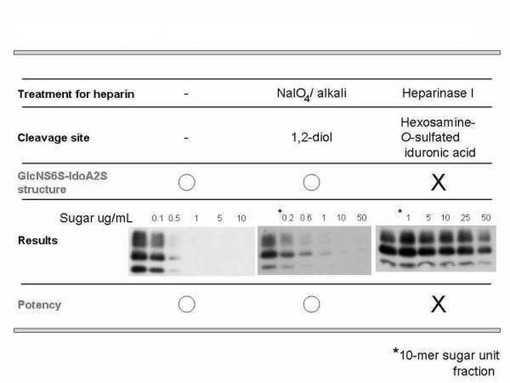
結果を図1に示す。図中の最上段はヘパリンに施した処理法を示している。“−”、“NaIO4 /alkali”、“HeparinaseI”はそれぞれ、未処理、過ヨウ素酸酸化、ヘパリナーゼ消化(酵素消化)を示している。二段目には、ヘパリンの過ヨウ素酸酸化とアルカリ処理、酵素消化処理によって切断される化学結合の種類を示している。三段目にそれぞれの処理を行ったヘパリンを添加した時の検出される変異型プリオン蛋白質の添加に伴う容量依存性を示している。四段目には、それぞれの容量依存性からプリオン持続感染細胞、ScN2a、を用いた薬剤の効能評価の結果を示している。過ヨウ素酸処理ヘパリンでは未処理ヘパリンと同程度の濃度において薬理活性を示した。すなわち、ヘパリンのもっている抗プリオン活性を担う微細構造(二糖単位構造)は過ヨウ素酸処理によって構造変化を受けないことを示している。一方、ヘパリナーゼ処理を行ったヘパリンに関しては、より高濃度の薬剤を添加しているにもかかわらず、ヘパリンによる治療効果が消失していることがわかる。以上の結果から、ヘパリナーゼ処理では、ヘパリンのもっている抗プリオン活性を消失するような反応が起きている。以上の結果から、ScN2aでの抗プリオン活性を担っているのは、GlcNS6Sα4IdoA2Sαという二糖構造(二糖単位構造)(図2)であると推定した。
【0033】
〔実施例2〕
実施例1の結果を踏まえ、二糖構造GlcNS6Sα4IdoA2Sαがプリオン蛋白質と相互作用するかどうかを、リコンビナントプリオン蛋白質を用いて表面プラズモン共鳴法で検討した。
【0034】
〔方法〕
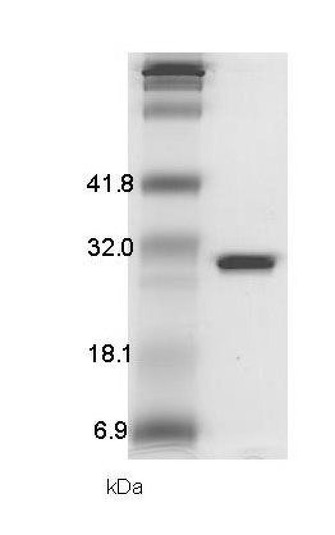
二糖構造GlcNS6Sα4IdoA2Sαにグルコースのリンカーを介して還元末端にチオクト酸が結合した分子(GlcNS6Sα4IdoA2Sα6Glc−mono)がそのチオクト酸の2個の硫黄原子で金表面に固定化された「糖チップ」をスダらの方法[スダ Y.、ソーベル M.ら、バイオコンジュゲート ケミストリー (Bioconjugate Chemistry)2006年、17巻、1125−1135頁]に準じて作製した。この「糖チップ」は、簡易型表面プラズモン共鳴(SPR)測定装置(モリテックス社製、NanoSensor)に適合するように作出されたものである。リコンビナントプリオン蛋白質(マウスの配列、23−231)は、アンピシリン耐性遺伝子とT7プロモータ支配下にクローニングサイトを有する大腸菌用蛋白質発現ベクター(pETシステム、ノバジェン社製)のクローニングサイトにマウスプリオン蛋白質遺伝子(配列は、NCBI(米国の生物情報研究所)における、NM−011170のうち上記配列に相当する部分)を導入した発現プラスミドで、大腸菌(BL21(DE3)株)を形質転換した。プラスミドへの遺伝子導入は、制限酵素として、プリオン蛋白質(23−231)のセンス鎖の5′末端側にNdeI、3′末端側にBamHIを利用した。形質転換した大腸菌をアンピシリン存在下で液体培養し、イソプロピルチオガラクトシド(isopropylthiogalactoside(IPTG), ナカライテスク社)によって、プロモーター支配の蛋白質発現誘導を行い、一晩培養後、遠心分離によって培養上清を除き、プリオン蛋白質(23−231)を含む大腸菌ペレットを得た。得られたプリオン蛋白質(23−231)を含む大腸菌ペレットをリシス液[lysis液:20mMトリスヒドロキシアミノメタン、pH7.5、300mM塩化ナトリウム 水溶液]に懸濁し、超音波によって、大腸菌細胞膜を破砕した。得られた大腸菌破砕物懸濁液を遠心分離(5000×g, 15分間)し、プリオン蛋白質(23−231)を含む沈殿物を回収した。得られた沈殿物を8M尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清をリシス液に対して透析(透析膜;分子量カット3,500、スペクトラ社製)し、一晩放置して得られた沈殿を再び8M尿素を含むリシス液に溶解させ、Ni−NTAカラム(商品名、GEヘルスケアバイオサイエンス社製)に蛋白質溶液を添加し、8M尿素を含むリシス液にて洗浄後、600mMイミダゾール、8M尿素を含むリシス液によって、カラムに結合した蛋白質を溶出させた。得られた蛋白質溶液は、水に対して透析(透析膜;分子量カット3,500、スペクトラ社製)することによって尿素や、塩化ナトリウムなどの低分子化合物の除去を行った。この一連の工程は、15%トリス・グリシンSDS−PAGEゲルで電気泳動をおこない、電気泳動後蛋白質を酢酸によって固定し、固定後、蛋白質をクーマーシーブリリアントブルー(ナカライテスク社製)で染色することによって追跡した。プリオン蛋白質(23−231)は以下のナノセンサーによるチップとの結合解析実験に十分な精製度であることがわかる(図3)。
【0035】
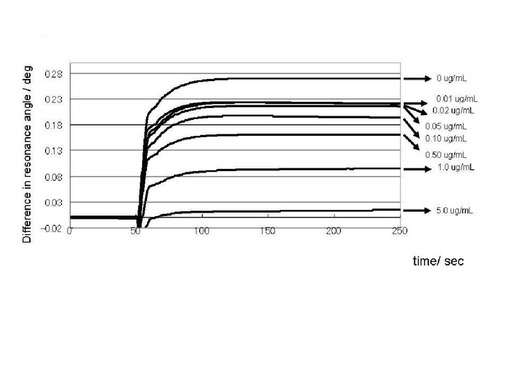
プリオン蛋白質(23−231)溶液を糖チップ測定用の平衡化バッファー(0.05% Tween 20(ポリオキシエチレンソルビタンモノラウレート、ナカライテスク社製)、残部PBS)で希釈することにより、18, 11, 7.0, 5.5, 4.6, 3.7, 2.8μM の濃度のプリオン蛋白質(23−231)溶液を200μLずつ調製した。GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置へ、調製した各濃度のプリオン蛋白質(23−231)溶液を添加し、共鳴プラズモン効果の共鳴角の変化を観測した。図4に代表的なセンサーグラムの変化のプロファイルを示した。横軸は、時間を表しており、数値の単位は秒である。縦軸は、表面プラズモン共鳴角の変化角度を表している。測定開始後、100秒間ベースラインが安定していることを確認後、プリオン蛋白質(23−231)溶液を添加した。添加後、シグナルの上昇が観測された(プリオン蛋白質(23−231)の糖チップへの結合カーブ)。200秒間、結合カーブのデータを収集後、平衡化バッファーで洗浄した。緩やかなシグナル現象のカーブ(プリオン蛋白質(23−231)の解離カーブ)得られた。観測後、チップに洗浄液1(300mM 水酸化ナトリウム水溶液)、引き続き、洗浄液2(2.0M 塩化ナトリウム水溶液)を添加して、チップ上の洗浄を行ない、各濃度のプリオン蛋白質(23−231)溶液を添加して、同様の結合カーブのデータを収集した。得られたデータは、デジタルデータとして、表計算ソフト、エクセル(商品名、マイクロソフト社製)へ移し、化学反応速度論的、化学平衡論的な解析を行った。すなわち、得られた各プリオン蛋白質(23−231)濃度における結合カーブのシグナル値を横軸とする結合カーブの時間微分に対して、近似直線を求めその切片と傾きから、一次反応結合速度定数と一次反応解離速度定数を算出した。プリオン蛋白質(23−231)濃度に対するこれらの一次反応速度定数の値から、平衡解離定数を算出した。ここではセンサーグラムを1:1の化学量論比での結合を仮定して解析したことになり、みかけの解離定数(K(app))が算出される。
【0036】
〔結果〕
調製したリコンビナントプリオン蛋白(23−231)は十分な精製度であり、SDS−PAGE上で一本のバンドしか観測されず、またその移動度はプリオン蛋白質のそれと合致した(図3)。プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合が観測され(図4)、平衡論的解離定数を見積もったところ7.1μMであった。速度論的定数は、解離〔kd((μMプリオン蛋白(23−231))−1・sec−1)〕、結合〔ka((μMプリオン蛋白(23−231))−1・sec−1)〕がそれぞれ、kd=0.015、ka=0.0021であった。実施例1において推察したヘパリンの抗プリオン活性を担う構造(図2)と、プリオン蛋白質(23−231)との相互作用が観測された。
【0037】
〔実施例3〕
実施例2において観測されたプリオン蛋白質(23−231)とGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合の特性を調べた。
〔方法〕
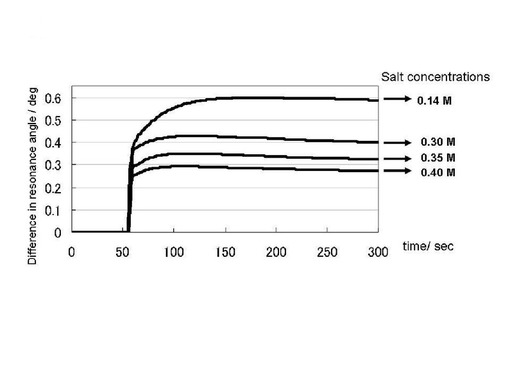
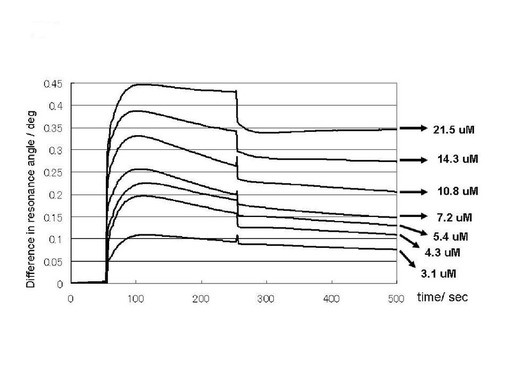
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈、および、塩化ナトリウムを添加することにより、プリオン蛋白質溶液(23−231)の濃度が11μM、塩化ナトリウム濃度が、0.14, 0.30, 0.35, 0,40Mの試料を200μLずつ調製した。上記のサンプルをGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置で測定した。測定条件は実施例2に準じた。ただし、ベースラインの確認は50秒、結合カーブのデータを収集は、250秒間とした。
【0038】
〔結果〕
図5に塩化ナトリウム濃度を変化させたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。結合は、高い塩濃度においても認められた。
【0039】
〔実施例4〕
実施例2および実施例3において観測されたプリオン蛋白質(23−231)添加によるセンサーグラムの上昇が、プリオン蛋白質(23−231)の結合であることを確認した。
〔方法〕
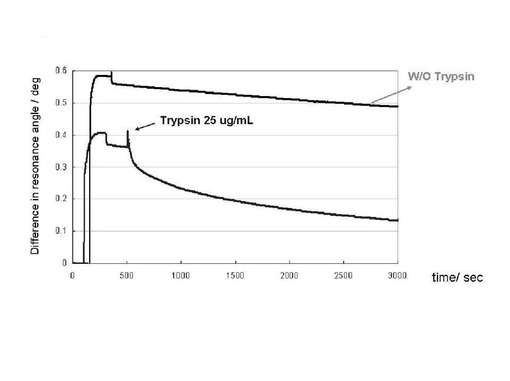
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈し、5.6μMの溶液を200μL調製した。2糖構造GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置へ添加した。200秒間、結合カーブのデータを収集後、PKを終濃度20μg/mLとなるように加えた平衡化バッファーでチップ上の溶液を置換した。実施例2に準じて、系の洗浄、データの解析を行った。次に、実施例2において調製を行ったプリオン蛋白質(23−231)をトリプシン反応バッファー(10mM 炭酸水素アンモニウム、pH7.0)で希釈し、11μMの濃度のプリオン蛋白質溶液(23−231)を200μL調製し、糖チップへ結合させた。200秒間、結合カーブのデータを収集後、トリプシン反応バッファーでチップを洗浄した。200秒間後、トリプシン(プロメガ社製)を終濃度25μg/mLとなるように加えたトリプシン反応バッファーでチップ上の溶液を置換した。2400秒間放置した後、チップ上のトリプシン消化反応液を75μLのトリプシン反応バッファー、25μLの空気、50μLのトリプシン反応バッファー、50μLの空気で溶出、押し出しを行い回収した。得られた反応溶液に5%のトリフルオロ酢酸水溶液、25μL添加し、攪拌したのち溶液を凍結させた。凍結乾燥させて得られた白色の粉を、5%アセトニトリル、0.1%トリフルオロ酢酸水溶液100μLにて溶解させた。その100μLの溶解液のうち、50μLを、ZipTip C18(商品名、ミリポア社製)で吸引、排出を3回繰り返した。その後、0.1%トリフルオロ酢酸水溶液の吸引、排出を行うことにより、ZipTipを洗浄した。その後、5mg/mLの濃度でα−シアノ−4−ヒドロキシケイヒ酸(ブルカー・ダルトニクス社製)と0.1%トリフルオロ酢酸を含むアセトニトリル2μLでZipTipへの結合物をマトリックス支援レーザー脱離イオン化−飛行時間型質量分析計用の試料台へと溶出した。風乾後、試料をマトリックス支援レーザー脱離イオン化−飛行時間型質量分析計(商品名、VoyagerSTR、アプライドバイオシステム社製)にて測定し、得られたデータをソフトウエア(dataexplore, アプライドバイオシステム社製)で表示した。得られた質量分析値のセットをウエブ上での解析サイト(http://www.matrixscience.com/)にて解析した。
【0040】
〔結果〕
図6にプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合、それに引き続くチップ上でのプロテイナーゼK処理によるセンサーグラムの変化のプロファイルを示した。実施例2において観測した場合と同様に、プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合に伴うシグナルの上昇が観測された。その後プロテイナーゼKの添加により、シグナル強度の減少が観測された。このことから、プリオン蛋白質(23−231)の糖チップ上での分解が可能であることがわかる。上記の条件下での速度論的解析の結果、糖チップ上でのプリオン蛋白質(23−231)の見かけの一次分解速度(プロテイナーゼK 1μg/mLの濃度条件下において)は、2.0×10−4 sec−1 であった。図7にチップ上でのトリプシン処理によるセンサーグラムの変化のプロファイルを示した。プロテイナーゼKによる処理のときと同様なセンサーグラムの変化が観測された。回収したトリプシン消化産物の脱塩を行い、質量分析を行った。図8にその測定結果を示す。ペプチド・マス・フィンガープリント法によって、プリオン蛋白質(23−231)が同定された(図9)。同定の確かさでは、種の異なる動物のプリオン蛋白質がヒットしたものの、マウスプリオン蛋白質(23−231)が確かに同定された。同定に利用されたペプチド断片は、プリオン蛋白質(23−231)のうち、25−37, 38−48, 148−155, 194−203, 220−228, 220−229に相当する断片計6本であった。これらのことから、糖チップ上での蛋白質分解によって得られるペプチド断片を用いてチップ結合蛋白質が、ペプチド・マス・フィンガープリント法によって同定できることがわかった。
【0041】
〔実施例5〕
実施例1においてGlcNS6Sα4IdoA2Sαという二糖構造が抗プリオン活性に重要であることを示し、実施例2〜実施例4においてリコンビナントプリオン蛋白質(23−231)はGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」と結合することを示した。そこで、この結合はプリオン蛋白質(23−231)ではどのような部位にその結合サイトが存在するかを明らかにした。
〔方法〕
プリオン蛋白質のN末端側ドメインを欠損した以下のプリオン蛋白質断片、プリオン蛋白質(121−231)、プリオン蛋白質(89−231)の調製を行った。プリオン蛋白質(121−231)においては、実施例2で得た発現ベクターをテンプレートとして、プリオン蛋白質(121−231)に相当するDNAフラグメントを増幅、発現ベクターに導入しクローニングした。実施例2に準じた培養、処理を行い、プリオン蛋白質(121−231)を含む沈殿物を回収した。得られた沈殿物を、8M尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清を透析バッファー1(5mMトリスヒドロキシアミノメタン、pH8.5)に対して透析(透析膜;分子量カット3,500、スペクトラ社製)し、プリオン蛋白質(121−231)を含む溶液をHi−trap DEAEカラム(GEヘルスケアバイオサイエンス社製)に添加し、透析バッファー1から200mM 塩化ナトリウムを含む透析バッファー1への塩濃度勾配により精製した。分離されたプリオン蛋白質(121−231)を含む溶液を、水に対して透析(透析膜;分子量カット3,500、スペクトラ社製)を行った。プリオン蛋白質(89−231)においては、実施例2で得た発現ベクターをテンプレートとして、プリオン蛋白質の23に相当する部位から、3′末端側方向へのプライマーとプリオン蛋白質の89に相当する部位から、5′末端側方向へのプライマーのセットで、プリオン蛋白質(23−88)に相当するDNAフラグメントを欠損した直鎖状の二本鎖DNAを増幅した。この直鎖状の二本鎖DNAの環化反応を行いプリオン蛋白質(89−231)の発現プラスミドとした。実施例2に準じた培養、処理を行い、プリオン蛋白質(89−231)を含む沈殿物を回収した。得られた沈殿物を、8M 尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清をリシス液に対して透析(透析膜;分子量カット3,500、スペクトラ社製)し、一晩放置して得られた沈殿を遠心分離によって除き、その上清を0.1%ギ酸水溶液に透析し、一晩放置した。遠心分離の上清を回収し、C4カラム(Cosmosil、10mm×250mm、ナカライテスク社製)に蛋白質溶液を添加し、0.1%トリフルオロ酢酸水溶液から、0.1%トリフルオロ酢酸を含むアセトニトリルへの直線勾配で精製した。プリオン蛋白質(89−231)を含むピークを分取し、凍結乾燥させた。一連の工程は、SDS−PAGEにより確認を行った。プリオン蛋白質(121−231)溶液、プリオン蛋白質(89−231)溶液のそれぞれを糖チップ測定用の平衡化バッファーで希釈することにより10μM の濃度に調整したプリオン蛋白質溶液を200μLずつ調製し、実施例2に準じた条件によりSPR測定装置で測定した。
【0042】
〔結果〕
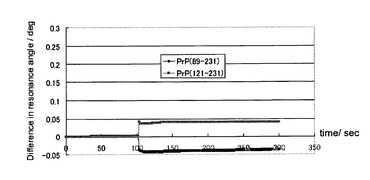
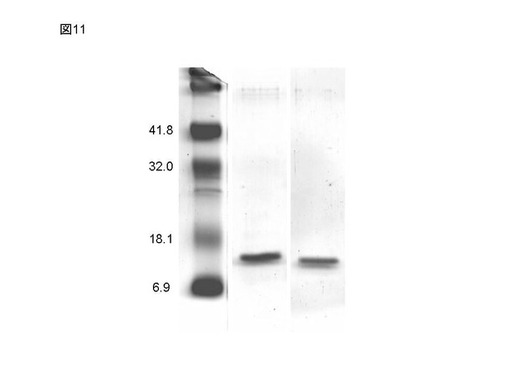
ここで調製したプリオン蛋白質(121−231)およびプリオン蛋白質(89−231)は、十分な精製度であり、SDS−PAGE上で一本のバンドしか観測されず、またその移動度はプリオン蛋白質のそれと合致した(図11)。図10にこれらのプリオン蛋白質の測定結果を示す。有意な結合は観測されず、プリオン蛋白(23−231)のうち、23−89の領域が結合に関与していることが示唆された。
【0043】
〔実施例6〕
実施例2にて観測されたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合は、実施例5においてプリオン蛋白(23−231)のうち、23−89の領域が結合に関与していることが示唆された。この点をさらに詳しく調べて、N末端部位を欠損させたことによるアーティファクトではないことを検証した。
〔方法〕
プリオン蛋白質(23−89)については、実施例5のプリオン蛋白質(121−231)の方法に準じて発現ベクターを調製した。実施例2に準じた培養、処理を行い、プリオン蛋白質(23−89)を含む沈殿物を回収した。得られた沈殿物を8M尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清をNi−NTAカラム(GEヘルスケアバイオサイエンス社製)に蛋白質溶液を添加し、8M尿素を含むリシス液にて洗浄後、600mMイミダゾール、8M尿素を含むリシス液によって、カラムに結合した蛋白質を溶出させた。得られた蛋白質溶液を、水に対して透析(透析膜;分子量カット3,500、スペクトラ社製)した。一晩放置した透析内容物を遠心分離し、その上清を回収し、C18カラム(Cosmosil、10mm×250mm、ナカライテスク社製)に蛋白質溶液を添加し、0.1%トリフルオロ酢酸水溶液から、0.1%トリフルオロ酢酸を含むアセトニトリルへの直線勾配で精製した。プリオン蛋白質(23−89)を含むピークを分取し、凍結乾燥させた。この一連の工程は、SDS−PAGEで追跡した。試料の一部をマトリックス支援レーザー脱離イオン化−飛行時間型質量分析計(商品名、VoyagerSTR、アプライドバイオシステム社製)にて測定した。プリオン蛋白質(23−89)溶液を糖チップ測定用の平衡化バッファーで希釈することにより、22, 14, 11, 7.2, 5.4, 4.3, 3.1μM の濃度のプリオン蛋白質(23−89)溶液を200μLずつ調製した。各濃度のプリオン蛋白質(23−89)溶液を実施例2に準じて測定した。結合カーブは200秒間データを収集した。洗浄後、緩やかなプリオン蛋白質(23−231)の解離カーブが得られた。観測後、実施例2に準じた処理と解析を行った。
【0044】
〔結果〕
調製したリコンビナントプリオン蛋白(23−89)は、図13に精製時におけるHPLC(C18カラム)のクロマトグラムを示すとおり十分な純度であり(SDS−PAGEによっても一本のバンドしか観測されず)、またその質量分析値はプリオン蛋白質(23−89)のそれと合致した(図13)。プリオン蛋白質(23−89)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合が観測され(図12)、平衡論的解離定数を見積もったところ16μMであった。速度論的定数は、解離〔kd((μMプリオン蛋白(23−89))−1・sec−1)〕、結合〔ka((μMプリオン蛋白(23−89))−1・sec−1)〕がそれぞれ、kd=0.061、ka=0.0037であった。実施例2において解析したプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合の平衡論的解離定数および速度論学的結合・解離定数とよい一致が見られた。プリオン蛋白質(23−89)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への速度論学的結合・解離定数はプリオン蛋白質(23−231)のそれらと比べて、若干大きな値を示したが、これはプリオン蛋白質(23−89)の分子量がプリオン蛋白質(23−231)にくらべて小さく、分子の並進拡散速度が大きいためであると考察される。実施例2、実施例5および実施例6により、プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合は、23−89の領域が結合を担っていることが明らかになった。
【0045】
〔実施例7〕
プリオン蛋白質の51−91の領域は銅イオンとの結合が報告されている[ホルムショウ MP.ら、バイオケミカル アンド バイオフィジカル リサーチ コミュニケーションズ(Biochemical & Biophysical Research Communications)1995年、214巻、993−999頁]。実施例6において明らかとなったプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合は、この銅結合領域を含んでいる。そのため、プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合が銅イオンの存在下で影響されるかどうかを検証する必要があり、以下の実験を実施した。
〔方法〕
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈、さらに硫酸銅水溶液を添加することにより、プリオン蛋白質溶液(23−231)の濃度が10μM、硫酸銅濃度が0, 50, 100, 200, 400, 1000μMの試料を200μLずつ調製した。実施例2に準じてSPR測定をおこなった。450秒間、結合カーブのデータを収集し、実施例2に準じて解析した。
【0046】
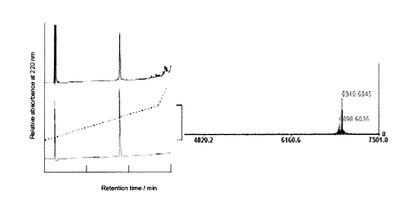
〔結果〕
図14に硫酸銅濃度を変化させたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。プリオン蛋白質(23−231)に対して化学量論的に大過剰に硫酸銅が存在する条件下(400, 1000μM)では、蛋白質性の沈殿物が混合後、すぐに目視で確認できた。硫酸銅50μMの添加では、硫酸銅を添加しない条件での結合プロファイルと差は見られなかった。50μMを超える高濃度の硫酸銅存在下では、硫酸銅の濃度の上昇に伴いプリオン蛋白質(23−231)の結合量は低下した。これらの結果から、10μMのプリオン蛋白質(23−231)に対して、50μMまでの硫酸銅は結合を阻害しないが、それ以上の濃度の硫酸銅ではプリオン蛋白質(23−231)の沈殿を生じさせ、有効な結合を阻害するため、プリオン蛋白質(23−231)とGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との相互作用を検出する上で不都合であることがわかった。
【0047】
〔実施例8〕
プリオン蛋白質(23−231)とGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合の阻害を利用した治療薬スクリーニングが可能であるかどうかを検証した。
〔方法〕
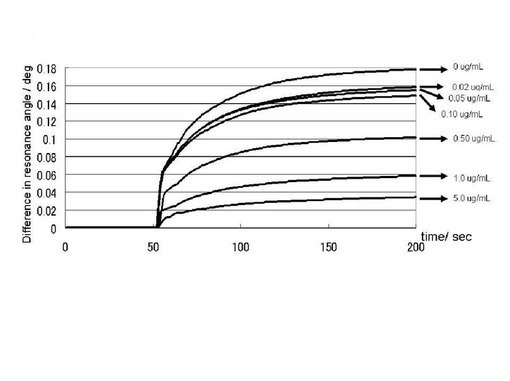
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈、さらにヘパリン(ナカライテスク社製、分子量17600)水溶液を添加することにより、プリオン蛋白質溶液(23−231)の濃度が5.5μM、ヘパリン濃度が、0, 0.02, 0.05, 0.10, 0.50, 1.0, 5.0μg/mLの試料を200μLずつ調製し、また、ペントサンポリサルフェートについても同様の試料調製を行った。両薬剤のシリーズについて実施例2に準じてSPR測定を行った。プリオン蛋白質(23−231)の糖チップへの結合カーブについて150秒間データの収集を行った。
【0048】
〔結果〕
図15にヘパリン濃度を変化させたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。ヘパリンの濃度依存的にプリオン蛋白質(23−231)の結合の低下が観測された。糖チップへと添加したヘパリンを含むプリオン蛋白質(23−231)の溶液には、沈殿は観測されなかった。したがって、このプリオン蛋白質(23−231)の GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合量の低下は、ヘパリンによる競合的阻害であることが明らかである。ヘパリンは実施例1において示したように抗プリオン活性を有する化合物であり、本実施例は、抗プリオン活性を有する化合物がプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合を競合的に阻害することを示すものである。さらに、現在臨床応用もなされているペントサンポリサルフェートについても、ヘパリンの場合と同様、濃度依存的なプリオン蛋白質(23−231)の結合の低下が観測された(図16)。
【0049】
〔実施例9〕
シグナルを誘起したプリオン蛋白質(23−231)が、GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」から回収可能であるかどうかを調べた。
〔方法〕
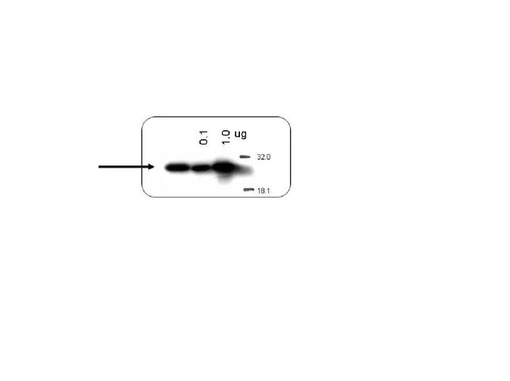
実施例2において調製を行ったプリオン蛋白質(23−231)を実施例1におけるリシス液で希釈することにより、2.8μMプリオン蛋白質(23−231)溶液の試料を200μL調製した。このサンプルをGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置にて測定した。100秒間結合カーブのデータを収集した。得られたデータは、実施例2に準じて図示した。その後、1mLの脱イオン水を用いて経路を洗浄し、0.1%トリフルオロ酢酸、50%アセトニトリルを含む液300μLを用いてGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」から結合蛋白質を溶出した。溶出液を凍結乾燥させ、この乾燥粉末全量に対しSDSポリアクリルアミドゲル電気泳動(SDS−PAGE)用バッファーを加え、実施例1に準じてウエスタンブロット法でプリオン蛋白質(23−231)を検出した。コントロールとして0.1μgプリオン蛋白質(23−231)を含む試料を並行して泳動した。検出したプリオン蛋白質(23−231)のシグナル強度を、コントールと比較することによって、表面プラズモン共鳴角の変化を誘起したGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」上のプリオン蛋白質(23−231) を定量した。
【0050】
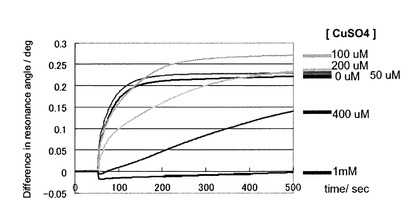
〔結果〕
図17に2.8μMの濃度のプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。図18に糖チップ上から回収したプリオン蛋白質(23−231) (左)と、0.10μgのプリオン蛋白質(23−231)(右)のウエスタンブロットの結果を示している。回収したプリオン蛋白質(23−231)の量は、コントロールとの比較により、120ngであることが明らかになった。また、これらの結果は、GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」に結合したプリオン蛋白質を回収することが可能であることを示している。
【図面の簡単な説明】
【0051】
【図1】本発明における硫酸化多糖の一例であるヘパリンについて、抗プリオン活性を担う部分構造を推定する試験を行なった結果の例として、ウエスタンブロットの泳動像を示す図面代用写真を含む説明図である(実施例1)。
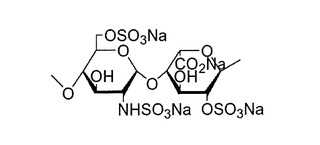
【図2】本発明における糖(二糖単位構造)の一例を示す構造式である。
【図3】本発明における正常型プリオン蛋白質についてSDS−PAGEにより試験を行なった結果の一例として、泳動像を示す図面代用写真である(実施例2)。
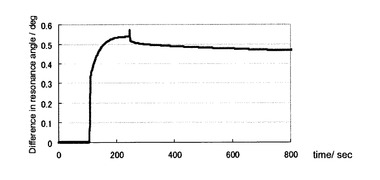
【図4】本発明における糖と正常型プリオン蛋白質との相互作用による表面プラズモン共鳴角の変化角度(Difference in resonance angle /deg)の経時的な変化プロファイルの一例を示すセンサーグラムである(実施例2)。
【図5】同、糖と正常型プリオン蛋白質との相互作用において正常型プリオン蛋白質溶液の塩濃度を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例3)。
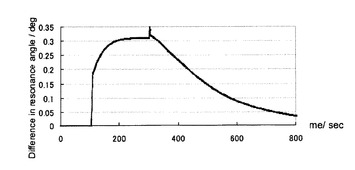
【図6】同、糖と正常型プリオン蛋白質との相互作用において、両者結合後、プロテイナーゼKの添加によってチップ上で正常型プリオン蛋白質が分解することによる変化プロファイルの一例を示すセンサーグラムである(実施例4)。
【図7】同、糖と正常型プリオン蛋白質との相互作用において、両者結合後、トリプシンの添加によってチップ上で正常型プリオン蛋白質が消化することによる変化プロファイルの一例を示すセンサーグラムである(実施例4)。
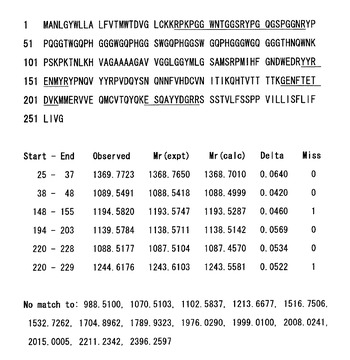
【図8】(図7で確認のトリプシン添加によって消化された)正常型プリオン蛋白質のトリプシン消化産物(回収、脱塩)について質量分析を行なった結果の一例を示す解析図であり、各行「Start − End」の数値は、上方に示されたアミノ酸配列(1−254:配列番号3)におけるN末端側からのアミノ酸残基番号であり、同定された各行ペプチド断片のアミノ酸配列(範囲)を表している(実施例4)。
【図9】図8に示した質量分析測定値に基づくペプチド・マス・フィンガープリント法による同定結果の一例を示す解析図である(実施例4)。
【図10】本発明における糖と比較試験例に相当するプリオン蛋白質断片(マウスプリオン蛋白質断片(121−231)、(89−231))との相互作用による表面プラズモン共鳴角の変化角度の経時的な変化プロファイルの一例を示すセンサーグラムである(実施例5)。
【図11】図10に示した試験に用いた比較試験例に相当するプリオン蛋白質断片について、SDS−PAGEにより試験を行なった結果の一例として、泳動像を示す図面代用写真であり、左側がマウスプリオン蛋白質断片(89−231)、右側がマウスプリオン蛋白質断片(121−231)である(実施例5)。
【図12】本発明における糖とプリオン蛋白質断片(マウスプリオン蛋白質断片(23−89))との相互作用による表面プラズモン共鳴角の変化角度の経時的な変化プロファイルの一例を示すセンサーグラムである(実施例6)。
【図13】図12に示した試験に用いたプリオン蛋白質断片について、その調製精製時のHPLCクロマトグラム及び質量分析の試験結果の一例を併せて示す図である(実施例6)。
【図14】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液の銅イオン濃度(硫酸銅添加濃度)を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例7)。
【図15】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液へのヘパリン添加濃度を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例8)。
【図16】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液へのペントサンポリサルフェート添加濃度を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例8)。
【図17】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液にリシス液を添加した場合における変化プロファイルの一例を示すセンサーグラムである(実施例9)。
【図18】本発明における糖と正常型プリオン蛋白質との相互作用(結合)測定の後、チップ上から回収した正常型プリオン蛋白質(左端)について、コントロールとの比較から定量する試験を行なった結果の一例として、ウエスタンブロットの泳動像を示す図面代用写真である(実施例9)。
【技術分野】
【0001】
本発明は、プリオン病治療薬及びそのスクリーニング方法に関する。
【背景技術】
【0002】
プリオン病は感染因子プリオンにより中枢神経が侵される疾患であり、ヒトでは全身の不随意運動と急速に進行する痴呆を主な特徴とするクロイツフェルト・ヤコブ病(CJD)、ゲルストマン・ストロイスラー・シャインカー症候群や致死性家族性不眠症などがある。ヒト以外の哺乳動物のプリオン病については、例えば、牛海綿状脳症(BSE)、スクレイピー、鹿慢性消耗性疾患(CWD)、伝達性ミンク脳症(TME)など、ウシ、ヤギ、ヒツジ、シカ、オオジカ、ミンク、ブタ、サル、マウス、ラット、ハムスター、ネコにおける各種プリオン病が公知である。これまでにプリオン病に対する実用性に優れた治療薬は発見されていないものの、硫酸化多糖が薬効のある化合物群の一つとして知られている。硫酸化多糖の一種であるペントサンポリサルフェートは実際にプリオン病患者に使用され、脳室内投与による臨床研究が進行している。このような硫酸化多糖は様々な部分構造と分子量分布を含み不均一性を有する高分子であり、プリオン蛋白質との結合は定性的な評価に限られている。高分子ゆえに脳血液関門を越えてプリオン病の病変の主座である脳実質内にまで移行しないこと、薬理活性の作用機序が明らかになっていないこと、強力な抗凝血作用を有するものがあること等が問題となっており、脳実質内への移行が期待できる低分子化合物で硫酸化多糖と同程度の抗プリオン活性を有する治療薬の開発が望まれている。
【0003】
プリオン病治療薬のスクリーニング技術としては、異常型プリオン蛋白と正常型プリオン蛋白の相互作用を阻害する化合物をスクリーニングする方法[例えば非特許文献1参照]、プリオン感染培養細胞を用いて異常型プリオン蛋白の産生を阻害する化合物をスクリーニングする方法[例えば非特許文献2参照]、酵母を用いて酵母プリオンの産生を阻害する化合物をスクリーニングする方法[例えば非特許文献3参照]、正常型プリオン蛋白との相互作用の強い化合物をスクリーニングする方法[例えば非特許文献4参照]、予想される正常型プリオン蛋白構造よりインシリコで正常型プリオン蛋白と相互作用する化合物をスクリーニングする方法[例えば非特許文献5参照]、また、イムノアッセイによりプリオンペプチド106−126に結合する候補物質を選択し、選択候補物質につき分子間相互作用解析にて結合選択性評価し、ウェスタンブロットによって異常型プリオンへの変換阻害効果を検定する方法[例えば特許文献1参照]などが提案されている。しかしながら、非特許文献1乃至5、及び特許文献1のようなプリオン病治療薬のスクリーニング技術については、アッセイにかなりの時間を要したり、十分な科学的エビデンスがなく闇雲に探索するものであり、治療効果を有する化合物が発見される効率が必ずしも高くないという問題があった。
【0004】
【非特許文献1】ギース A.ら(Giese A.,et al)、ジャーナル オブ ヴァイロロジー(Journal of Virology)2005年、79巻、7785−7791頁
【非特許文献2】レイス R.E.ら(Race R.E.,et al)、ジャーナル オブ ジェネラル ヴァイロロジー(Journal of General Virology)1987年、68巻、1391−1399頁
【非特許文献3】バック S.ら(Bach S.,et al)、ネイチャー バイオテクノロジー(Nature Biotechnology)2003年、21巻、1075−1081頁
【非特許文献4】カワタケ S.、ドウウラ K.ら(Kawatake S.,Doh−ura K.,et al)、バイオロジカル アンド ファーマスーティカル ブレチン(Biological & Pharmaceutical Bulletin)2006年、29巻、927−932頁
【非特許文献5】ローレンツェン S.ら(Lorenzen S.,et al)、バイオシステムス(Biosystems)2005年、80巻、117−122頁
【特許文献1】特開2003−149237号公報
【発明の開示】
【発明が解決しようとする課題】
【0005】
かかる問題点に鑑みてなされた本発明の目的は、迅速、簡便に効率良くプリオン病に対する低分子治療候補化合物を探索するための方法、及びプリオン病治療薬を提供することにある。
【課題を解決するための手段】
【0006】
上記目的を達成するため、本発明によるプリオン病治療薬のスクリーニング方法は、a)硫酸化多糖を構成する二糖構造を含む糖と、b)異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列を少なくとも含む正常型プリオン蛋白質断片、或は、全長正常型プリオン蛋白質と、の結合の阻害を指標とすることを特徴とする。このようなプリオン病治療薬のスクリーニング方法において、糖が、硫酸化多糖を酸化剤により10糖以下に低分子化したヘテロ多糖又はヘテロオリゴ糖、或は化学合成したヘテロオリゴ糖であることが好ましく、また、硫酸化多糖がヘパリンであることが良く、酸化剤が過ヨウ素酸又は過塩素酸又はこれらの塩であることが好適である。そして、糖が、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれる1種以上であることが好適であり、また、正常型プリオン蛋白質は配列番号1のアミノ酸配列を有するマウスプリオン蛋白質(23−231)とし、プリオン蛋白質断片は、N末端側に少なくとも配列番号2のアミノ酸配列(マウスプリオン蛋白質23−89)を含むマウスプリオン蛋白質断片とすることが好ましい。なお、指標は、表面プラズモン共鳴法によって評価されることが好適である。
【0007】
また、上記目的を達成するため、本発明のプリオン病治療薬のスクリーニング方法は、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上を有効成分として含んでなることを特徴とする。
【発明の効果】
【0008】
本発明のプリオン病治療薬のスクリーニング方法によれば、プリオン病治療効果発現の分子メカニズムに基づいた直接的な作用によって合理的にプリオン病治療薬を探索できることから、迅速に、高い効率(確度)でプリオン病治療薬候補化合物を探り当てることが可能である。また、プリオン病感染材料を扱うことなく簡便にスクリーニングを行うことができ、安全衛生面においても優れている。糖として、硫酸化多糖を酸化剤によって10糖以下に低分子化したヘテロ多糖(6〜10糖)又はヘテロオリゴ糖(2〜5糖)、或は化学合成した(N−硫酸化及び/又はO−硫酸化)ヘテロオリゴ糖を用いる場合、また、例えばマウスプリオン蛋白質(23−89)のような異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域部に相当する正常型プリオン蛋白質断片を用いる場合には、より高い効率でプリオン病治療薬候補化合物のスクリーニングができる。糖として、例えばグルコサミン6−硫酸とイズロン酸2−硫酸とがα1−4結合したへテロ二糖、特に「GlcNS6Sα4IdoA2Sα」を用いる場合、また、マウスプリオン蛋白質(23−89)と組み合わせて用いる場合には、さらに低分子のプリオン病治療薬候補化合物スクリーニングの確度が増す。また、本発明のプリオン病治療薬は、強力な抗プリオン活性を持つヘパリンと同様の作用機序による抗プリオン作用を有しており、且つ、ヘパリンに比して抗凝血作用が極めて弱い有効成分を含有することから有用である。
【発明を実施するための最良の形態】
【0009】
以下、本発明によるプリオン病治療薬及びそのスクリーニング方法を実施するための最良の形態について、詳細に説明する。
【0010】
本実施形態によるプリオン病治療薬のスクリーニング方法は、硫酸化多糖を構成する二糖構造を含む糖と、全長正常型プリオン蛋白質或は正常型プリオン蛋白質断片との結合の阻害を指標とするものである。ここで、硫酸化多糖は、硫酸化された二糖(単位)構造を有していれば特に限定されず、好ましくは、例えば、ヘキソサミンとヘキスロン酸(ウロン酸)とのヘテロ二糖構造を有するヘパリン、ヘパラン硫酸、コンドロイチン硫酸、デルマタン硫酸、また、ヘキソサミンとヘキソースとのヘテロ二糖構造を有するケラタン硫酸や、ヘキスロン酸とペントースとのヘテロ二糖構造及びペントースからなるホモ二糖構造を有するペントサンポリサルフェートなどを挙げることができる。これらの硫酸化多糖の内でも、特に、二糖構造として、D−グルクロン酸(GlcA)又はL−イズロン酸(IdoA)のヘキスロン酸と、N−スルホ−D−グルコサミン(GlcNS)又はN−アセチル−D−グルコサミン(GlcNAc)のヘキソサミンとがβ1−4或はα1−4結合した構造を有し、2位、3位、6位の一部又は全部が硫酸化(N−硫酸化、O−硫酸化)されたヘパリン又はヘパラン硫酸が、低分子化した場合にも抗プリオン活性を有する場合がある点でより好ましい。強力な抗プリオン活性を持ち、例えば10糖にまで低分子化した場合にも抗プリオン活性が殆ど低下しない場合がある点でヘパリンがさらに好適に使用できる。ヘパリンは、如何なる動物種の如何なる臓器由来のものであっても良く特に限定されないが、安全衛生面からプリオン病発症例を認めない点でブタ由来のヘパリンが好ましい。
【0011】
このような硫酸化多糖は、液相において各種分解酵素や分解剤によって10糖以下に低分子化することができる。例えば、硫酸化多糖としてヘパリンを低分子化する場合には、上述の二糖単位構造を保持して低分子化可能であることから、酸化剤によって又は酸化剤及びアルカリによって低分子化することが好ましい。酸化剤として、例えば、過ヨウ素酸、過塩素酸、亜硝酸又はこれらの塩を使用して、10糖以下のヘテロ多糖(10糖〜6糖)又はヘテロオリゴ糖(5糖〜2糖)とすることができる。例えば、スダ、ソーベル(Suda, Sobel)らの方法[スダ Y.、ソーベル M.ら、スロンボシス リサーチ(Thrombosis Research)1993年、69巻、501−508頁]に準じて低分子化することができる。また、上述の二糖単位構造を含むヘテロオリゴ糖を化学合成しても良く、スダ(Suda)らの方法[スダ Y.ら、テトラへドロン レターズ(Tetrahedron Letters)1996年、37巻、1053−1056頁]に準じて合成することができる。ヘテロオリゴ糖の内でも、プリオン蛋白質との結合における必須の中心構造となることから、二糖単位構造が好ましい。N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルファミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上がより好適である。ヘテロ二糖の金属塩としては、アルカリ金属塩が好ましく、アルカリ金属塩の中でもNa塩、K塩がより好ましい。
【0012】
また、上記ヘテロ二糖、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上の糖は、強力な抗プリオン活性を持つヘパリンと同様の作用機序による抗プリオン作用を有しており、且つ、ヘパリンに比して抗凝血作用が極めて弱いことから、プリオン病治療薬の有効成分として有用である。
【0013】
上述のN−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖(単位)構造は、以下の式(1)で表される。
【0014】
【化1】
【0015】
また、上記スダ(Suda)らの方法[スダ Y.ら、テトラへドロン レターズ(Tetrahedron Letters)1996年、37巻、1053−1056頁]に準じた化学合成により得られるヘテロ二糖は、以下の一般式(2)で表されるものとすることができる〔式(2)中、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す〕。一般式(2)におけるR1がNa又はK、R2が水素又はメチル基であるヘテロ二糖が好ましい。
【0016】
【化2】
【0017】
また、硫酸化多糖を構成する二糖構造を含む糖として、一般式(2)で表されるヘテロ二糖におけるR2にグルコース残基を導入した下記一般式(3)で表されるヘテロ三糖も好適に使用することができる〔式(3)中、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す〕。一般式(3)におけるR1がNa又はK、R2が水素又はメチル基であるヘテロ三糖が好ましい。このようなヘテロ三糖は、例えば、コシダ(S Koshida)、スダ(Y Suda)らの方法[コシダ S.、スダ Y.ら、テトラへドロン レターズ(Tetrahedron Letters)2001年、42巻、1289−1292頁]に準じて化学合成することができる。一般式(3)中のR2にグルコース残基を導入してヘテロ四糖としても良い。
【0018】
【化3】
【0019】
正常型プリオン蛋白質は、上述のような硫酸化多糖を構成する二糖構造を含む糖との結合阻害を指標としてプリオン病治療薬をスクリーニングする際に用いるものであり、如何なる動物種のものであっても良いが、動物種間における相同性が高いことから、哺乳動物のプリオン蛋白質が好ましい。特に、前駆体プリオン蛋白質におけるN末端のシグナル配列部分、及びC末端疎水性領域(GPIアンカー付加)が成熟、分泌過程において除かれた哺乳動物の正常型プリオン蛋白質では、動物種によるアミノ酸配列の差異がより小さく、例えば、ヒトとマウスやハムスターのようなげっ歯類とのホモロジーは95%以上である。本実施形態においては、マウスプリオン蛋白質全長に相当する配列番号1に示すアミノ酸配列を有するプリオン蛋白質(23−231)を哺乳動物全般を対象とするスクリーニングに適用可能な全長正常型プリオン蛋白質として用いることができる。また、上述の糖との結合における必須領域(配列)はN末端側部分にあり、プリオン蛋白質(23−231)のN末端からアミノ酸残基数67の配列に相当する配列番号2に示すアミノ酸配列に含まれることが後述の実施例により確認された。このプリオン蛋白質(23−231)のN末端からアミノ酸残基数67のマウスプリオン蛋白質(23−89)は、哺乳動物異常型プリオン蛋白質のプロテアーゼ(プロテイナーゼK)処理により消化除去されるN末端領域部に相当する。したがって、本実施形態のスクリーニング方法における正常型プリオン蛋白質断片として、異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列をN末端に少なくとも含むプリオン蛋白質断片が好適に使用できる。異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域部のみからなる正常型プリオン蛋白質断片が、硫酸化二糖構造との結合に必須の最小構造であり、さらに低分子量のプリオン病治療薬候補化合物を探索できる場合があることからより好ましい。このような正常型プリオン蛋白質断片のうち、マウスプリオン蛋白質(23−89)を哺乳動物全般を対象とするスクリーニングに用いることができ、少なくとも配列番号2のアミノ酸配列を含んでなるプリオン蛋白質断片であればいずれも好適に使用できる。マウスプリオン蛋白質(23−231)の前駆体蛋白質におけるアミノ酸配列は、配列番号3に示すとおりである。
【0020】
以上のような、全長正常型プリオン蛋白質や正常型プリオン蛋白質断片は、公知の遺伝子組み換え蛋白質合成法に準じて調製することができる。ここで形質転換する宿主細胞株は動物や酵母由来の真核細胞を用いても良いが、糖鎖が結合した糖蛋白質として分泌される場合があり、得られたリコンビナントプリオン蛋白質と上述の硫酸化多糖を構成する二糖構造を含む糖との結合、結合阻害の評価において影響を与える可能性がある点では、大腸菌株のような原核細胞を用いることが好ましい。具体的には、例えば、ホーネマンらの方法[ホーネマン(Hornemann S)ら、Journal of Molcular Biology,1996年、261巻、614−619頁]、及び[ホーネマン(S.Hornemann)ら、FEBS Letters,1997年、413巻、277−281頁]、リーマンらの方法[リーマン(Liemann S)ら、Biochemistry,1999年、38巻、3258−3267頁]などに準じて、リコンビナントプリオン蛋白質及びその断片を調製することができる。
【0021】
上述のような糖と、プリオン蛋白質断片又は全長正常型プリオン蛋白質との結合の阻害を指標として評価する方法としては、糖と蛋白質との相互作用(反応、構造や重量などの状態変化)を検出可能な方法であれば公知の如何なる探索、評価方法であっても適用できる。このような探索、評価方法(装置)として、例えば、表面プラズモン共鳴法(SPR法)、水晶発振子の周波数変化を測定する方法、吸発熱を測定するカロリメトリー法、蛍光偏光測定法、核磁気共鳴法(NMR)、原子間力顕微鏡法(AFM)、透過型電子顕微鏡法(TEM)、走査型電子顕微鏡法(SEM)などの他、ウェスタンブロッティング法、ELISA法を挙げることができる。抗体の調製や標識体の調製が不要である点では、蛍光偏光測定法、ウェスタンブロッティング法、ELISA法以外の方法、装置が好ましく、夫々、適用する方法、装置のインストラクションマニュアル等に基づき実施することができる。例えば、表面プラズモン共鳴法を適用する場合には、[永田知宏、半田宏編集、シュプリンガー・フェアラーク東京株式会社1998年発行、生体物質相互作用のリアルタイム解析実験法 ]、[スダ Y.、ソーベル M.ら、バイオコンジュゲート ケミストリー (Bioconjugate Chemistry)2006年、17巻、1125−1135頁]、特開2003−83969号公報などの記載に準じ、市販のSPRセンサー(装置)を用いて行なうことができる。本実施形態では、上述の糖(硫酸化多糖、硫酸化オリゴ糖)又はプリオン蛋白質(全長正常型プリオン蛋白質、或はその断片)の内の一方をセンサーチップ上に固定し、他方を結合させた系において、両者の結合を競合阻害する化合物を共存させ表面プラズモン共鳴角度の変化をリアルタイムに測定する。この測定結果から両者の結合や解離の相互作用を反応速度論的、化学平衡論的に解析、評価することによってプリオン病治療薬の候補化合物を探索することができる。また、AFM、TEM、SEMなどを適用する場合には、例えば、上述の糖又はプリオン蛋白質(或はその断片)の内の一方をSPRのセンサーチップに固定する代わりに、金などの金属ナノ粒子に固定し、他方を結合させた系において、両者の結合を競合阻害する化合物を共存させ各装置を用いて相互作用を測定、解析、評価することによってプリオン病治療薬の候補化合物を探索することができる。
【0022】
プリオン蛋白質断片又は全長正常型プリオン蛋白質をリガンドとしてセンサーチップ上に固定する場合には、金表面にカルボキシメチルデキストラン層を設けたセンサーチップ(CM5:Biacore社製)に全長正常型プリオン蛋白質(マウスプリオン蛋白質(23−231))をアミンカップリングにより固定化するとN末端部分が分析中の条件化において変質・変性してしまうことがある(非特許文献4)ことから、全長正常型プリオン蛋白質よりマウスプリオン蛋白質断片(23−89)を用いることが好ましい。また、センサーチップ上へ固定化するリガンドとしては、以上のようなプリオン蛋白質やその断片に比較して上述のような糖を用いることがより好ましく、糖の内でも一般式(1)で表される二糖単位構造からなるヘテロ二糖、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれる1種以上がさらに好ましい。一般式(2)で表される化学合成のヘテロ二糖も一般式(1)の糖と同様好適に使用できる。一般式(1)、(2)で表されるような(硫酸化)糖をリガンドとしてセンサーチップ上に固定化する場合には、例えば、オクタンチオールのような末端にチオール基を有する長鎖アルキル化合物をセンサーチップ金表面にAu−S結合によって固定、アルキル鎖が露出する疎水性層を設け、糖の還元末端にも疎水性基を導入して疎水吸着によって糖を固定することもできるが、糖固定化後に添加するプリオン蛋白質が糖との結合のみならずチップ表面の疎水性層に吸着するなど好ましくない非特異的な相互作用を生じることがある点では、リガンドである糖鎖とリンカー化合物を結合させたリガンド・リンカー複合体(以下リガンド複合体)を調製してセンサーチップ金表面に直接結合、固定化することが好適である。一般式(1)で表される二糖構造を有する糖や、一般式(2)で表される合成へテロ二糖をリンカーと結合する際に、プリオン蛋白質との結合に必須の最小二糖構造が失われることがある点では、一般式(3)で表されるヘテロ三糖を用いてリガンド複合体を調製することが好ましい。硫酸化多糖を酸化剤によって10糖以下に低分子化したヘテロ多糖、ヘテロオリゴ糖を用いてリガンド複合体を調製する場合には、酸化によって生じるアルデヒド基を利用し、一般式(3)の合成法に準じてリンカーと結合することが好適である。リガンド複合体の調製は、[スダ Y.、ソーベル M.ら、バイオコンジュゲート ケミストリー (Bioconjugate Chemistry)2006年、17巻、1125−1135頁]、特開2003−83969号公報などに準じて行なうことができる。一般式(3)で表される合成へテロ三糖をリガンドとするリガンド複合体として、例えば、一般式(4)で表される(芳香族アミンと環状SS結合を持つチオクト酸をリンカー部とする)リガンド複合体を挙げることができる〔式(4)中、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す〕。一般式(4)におけるR1がNa又はK、R2が水素又はメチル基であるリガンド複合体が好ましい。このような、リガンド複合体のセンサーチップ金表面への結合は、1μM〜100μM濃度で水溶性アルコール溶液中に溶解した液にセンサーチップを室温、6〜24時間浸漬することによって行なうことができる。式(4)におけるR1がNa、R2がメチル基であるリガンド複合体:GlcNS6Sα4IdoA2Sα6Glc−monoをセンサーチップ金表面に結合する場合には、例えば、5〜50μM濃度でメタノール・水混液(1:1)に溶解した液に浸漬すれば良い。
【0023】
【化4】
【0024】
また、リガンド複合体として、一般式(5)、(6)、(7)で表される(ヘテロ三糖を集積した)GlcNS6Sα4IdoA2Sα6Glc−Tri、GlcNS6Sα4IdoA2Sα6Glc−Tetra、GlcNS6Sα4IdoA2Sα6Glc−Tetra−Longについても、GlcNS6Sα4IdoA2Sα6Glc−monoと同様に好適に使用できる。さらに、一般式(8)で表されるビオチン・アビジン結合によるリガンド複合体も使用可能であるが、金表面にアビジンを結合したセンサーチップを用いるためプリオン蛋白質などとの非特異的相互作用が残る。一般式(5)〜(8)の夫々において、R1は水素、アルカリ金属、アルカリ土類金属から選ばれる元素を示し、R2は水素、又は炭素数1〜4のアルキルまたはアルケニル基を示す。一般式(5)〜(8)の夫々において、R1がNa又はK、R2が水素又はメチル基であるリガンド複合体が好ましい。
【0025】
【化5】
【0026】
【化6】
【0027】
【化7】
【0028】
【化8】
【実施例】
【0029】
以下、本発明によるプリオン病治療薬及びそのスクリーニング方法について、実施例を示して具体的に説明するが、これによって本発明を限定するものではない。
【0030】
〔実施例1〕
ヘパリンを構成する糖は数種類あり、また、それらの糖修飾の程度、モノマー間の結合様式などには多様性がある。ヘパリンはプリオン持続感染細胞を用いた薬剤スクリーニングで抗プリオン活性があることが示されている。そこで、この活性を示すヘパリンの部分構造を推定する実験を行った。
〔方法〕
プリオン持続感染細胞としては、スクレイピープリオンであるRML株がマウス神経芽細胞腫細胞(Neuro2a)に持続感染したScN2a細胞を用い、以下の処理を行ったヘパリンと共存下で培養した。培養液(10%胎児牛血清加MEM(Opti−MEM、インビトロジェン社製))に添加するヘパリン処理物の一つは、過ヨウ素酸による、炭素−炭素結合で隣接した2個の水酸基の酸化と引き続くベータ脱離反応による分解、もう一つは、酵素(heparinase I)による酵素消化である。これらのヘパリンの低分子化処理は、スダ、ソーベル(Suda, Sobel)らの方法[スダ Y.、ソーベル M.ら、スロンボシス リサーチ(Thrombosis Research)1993年、69巻、501−508頁]に準じて行った。これら低分子量化(低分子化処理)されたヘパリンのうち、10糖に相当する鎖長の分画を用いた。コントロールとして、処理を施していないヘパリン(ナカライテスク社製、分子量17600)の計3種類のサンプルを用いた。これらを上記の細胞培養時、培養液に添加した。終濃度は、未処理ヘパリンに対して、0, 0.1, 0.5, 1, 5, 10μg/mL、過ヨウ素酸処理したヘパリンについて、0, 0.2, 0.6, 1, 10, 50μg/mL、ヘパリナーゼ処理(HeparinaseI(EC4.2.2.7)、シグマ社製)したヘパリンに対して、0, 1, 5, 10, 25, 50μg/mLとした。細胞培養は、細胞培養フラスコ(底面積9.7cm2 )にて培養し、細胞密度がコンフルエントとなった時点で培養上清を除き、リン酸バッファー生理食塩水[PBS、pH6.5]で細胞を洗浄した。洗浄液を十分に除き、リシス液[lysis液:0.5%デオキシコール酸ナトリウム、0.5% NP−40(商品名、和光純薬工業(株)オクチルフェニルエーテル)、残部PBS]0.5mLを用いて細胞を溶解した。軽く遠心分離して、高分子核酸を除いた後、細胞溶解液にトリチラキウム アルブム(Tritirachium album)由来のプロテイナーゼK(PK:Proteinase K)を終濃度10μg/mLとなるように加えて37℃で30分反応(蛋白質分解)させた後、フェニルメタンスルホニルフロライド(phenylmethan sulfonylfluoride: PMSF)を終濃度1mMとなるように加えて蛋白質分解反応を停止させた。
【0031】
蛋白質分解処理した細胞溶解液0.5mLに核酸分離用シリカとして市販されているFOG((商品名)、Qbiogene社製)を滅菌水(オートクレーブ滅菌蒸留水)で1000倍希釈したシリカ懸濁液5μL(固形分(シリカ 約1.5μg含有))を加え、2〜3分間室温で混和した後、軽く遠心(9,000×g、 1分間)してシリカを沈殿させ、上清を除いた。この沈殿全量に対し、SDSポリアクリルアミドゲル電気泳動(SDS−PAGE)用バッファー[1% SDS(ソディウムドデシルサルフェート)、0.05% ブロモフェノールブルー、4% グリセロール、1% 2−メルカプトエタノール、25mM トリス塩酸 pH6.8] 20μLを加えて懸濁し、95℃、5分間の熱変性後に、イシカワ、ドウウラ(Ishikawa, Doh−ura)らの方法[イシカワ K.、ドウウラ K.ら、ジャーナル オブ ジェネラル ヴァイロロジー(Journal of General Virology)2004年、85巻、1785−1790頁]に準じ、15% トリス・グリシンSDS−PAGEゲルで電気泳動し、ウエスタンブロット法でプロテアーゼ抵抗性の異常型プリオン蛋白質を検出した。ウエスタンブロット法においては、電気泳動した蛋白質をポリビニリデンジフルオライド(PVDF)膜(ミリポア社製、PVDFメンブレン)にブロッティングし、プリオン蛋白質に対するマウスモノクローナル抗体(SPI−BIO社製、SAF83(5000倍希釈))、及び、アルカリフォスファターゼ・コンジュゲートヤギ抗マウス免疫グロブリン抗体(プロメガ社製、(20000倍希釈))、化学発光検出試薬(アマーシャム社製、CDP−Star detection reagent(商品名))を使用してプリオン蛋白質の検出を行った。
【0032】
[結果]
結果を図1に示す。図中の最上段はヘパリンに施した処理法を示している。“−”、“NaIO4 /alkali”、“HeparinaseI”はそれぞれ、未処理、過ヨウ素酸酸化、ヘパリナーゼ消化(酵素消化)を示している。二段目には、ヘパリンの過ヨウ素酸酸化とアルカリ処理、酵素消化処理によって切断される化学結合の種類を示している。三段目にそれぞれの処理を行ったヘパリンを添加した時の検出される変異型プリオン蛋白質の添加に伴う容量依存性を示している。四段目には、それぞれの容量依存性からプリオン持続感染細胞、ScN2a、を用いた薬剤の効能評価の結果を示している。過ヨウ素酸処理ヘパリンでは未処理ヘパリンと同程度の濃度において薬理活性を示した。すなわち、ヘパリンのもっている抗プリオン活性を担う微細構造(二糖単位構造)は過ヨウ素酸処理によって構造変化を受けないことを示している。一方、ヘパリナーゼ処理を行ったヘパリンに関しては、より高濃度の薬剤を添加しているにもかかわらず、ヘパリンによる治療効果が消失していることがわかる。以上の結果から、ヘパリナーゼ処理では、ヘパリンのもっている抗プリオン活性を消失するような反応が起きている。以上の結果から、ScN2aでの抗プリオン活性を担っているのは、GlcNS6Sα4IdoA2Sαという二糖構造(二糖単位構造)(図2)であると推定した。
【0033】
〔実施例2〕
実施例1の結果を踏まえ、二糖構造GlcNS6Sα4IdoA2Sαがプリオン蛋白質と相互作用するかどうかを、リコンビナントプリオン蛋白質を用いて表面プラズモン共鳴法で検討した。
【0034】
〔方法〕
二糖構造GlcNS6Sα4IdoA2Sαにグルコースのリンカーを介して還元末端にチオクト酸が結合した分子(GlcNS6Sα4IdoA2Sα6Glc−mono)がそのチオクト酸の2個の硫黄原子で金表面に固定化された「糖チップ」をスダらの方法[スダ Y.、ソーベル M.ら、バイオコンジュゲート ケミストリー (Bioconjugate Chemistry)2006年、17巻、1125−1135頁]に準じて作製した。この「糖チップ」は、簡易型表面プラズモン共鳴(SPR)測定装置(モリテックス社製、NanoSensor)に適合するように作出されたものである。リコンビナントプリオン蛋白質(マウスの配列、23−231)は、アンピシリン耐性遺伝子とT7プロモータ支配下にクローニングサイトを有する大腸菌用蛋白質発現ベクター(pETシステム、ノバジェン社製)のクローニングサイトにマウスプリオン蛋白質遺伝子(配列は、NCBI(米国の生物情報研究所)における、NM−011170のうち上記配列に相当する部分)を導入した発現プラスミドで、大腸菌(BL21(DE3)株)を形質転換した。プラスミドへの遺伝子導入は、制限酵素として、プリオン蛋白質(23−231)のセンス鎖の5′末端側にNdeI、3′末端側にBamHIを利用した。形質転換した大腸菌をアンピシリン存在下で液体培養し、イソプロピルチオガラクトシド(isopropylthiogalactoside(IPTG), ナカライテスク社)によって、プロモーター支配の蛋白質発現誘導を行い、一晩培養後、遠心分離によって培養上清を除き、プリオン蛋白質(23−231)を含む大腸菌ペレットを得た。得られたプリオン蛋白質(23−231)を含む大腸菌ペレットをリシス液[lysis液:20mMトリスヒドロキシアミノメタン、pH7.5、300mM塩化ナトリウム 水溶液]に懸濁し、超音波によって、大腸菌細胞膜を破砕した。得られた大腸菌破砕物懸濁液を遠心分離(5000×g, 15分間)し、プリオン蛋白質(23−231)を含む沈殿物を回収した。得られた沈殿物を8M尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清をリシス液に対して透析(透析膜;分子量カット3,500、スペクトラ社製)し、一晩放置して得られた沈殿を再び8M尿素を含むリシス液に溶解させ、Ni−NTAカラム(商品名、GEヘルスケアバイオサイエンス社製)に蛋白質溶液を添加し、8M尿素を含むリシス液にて洗浄後、600mMイミダゾール、8M尿素を含むリシス液によって、カラムに結合した蛋白質を溶出させた。得られた蛋白質溶液は、水に対して透析(透析膜;分子量カット3,500、スペクトラ社製)することによって尿素や、塩化ナトリウムなどの低分子化合物の除去を行った。この一連の工程は、15%トリス・グリシンSDS−PAGEゲルで電気泳動をおこない、電気泳動後蛋白質を酢酸によって固定し、固定後、蛋白質をクーマーシーブリリアントブルー(ナカライテスク社製)で染色することによって追跡した。プリオン蛋白質(23−231)は以下のナノセンサーによるチップとの結合解析実験に十分な精製度であることがわかる(図3)。
【0035】
プリオン蛋白質(23−231)溶液を糖チップ測定用の平衡化バッファー(0.05% Tween 20(ポリオキシエチレンソルビタンモノラウレート、ナカライテスク社製)、残部PBS)で希釈することにより、18, 11, 7.0, 5.5, 4.6, 3.7, 2.8μM の濃度のプリオン蛋白質(23−231)溶液を200μLずつ調製した。GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置へ、調製した各濃度のプリオン蛋白質(23−231)溶液を添加し、共鳴プラズモン効果の共鳴角の変化を観測した。図4に代表的なセンサーグラムの変化のプロファイルを示した。横軸は、時間を表しており、数値の単位は秒である。縦軸は、表面プラズモン共鳴角の変化角度を表している。測定開始後、100秒間ベースラインが安定していることを確認後、プリオン蛋白質(23−231)溶液を添加した。添加後、シグナルの上昇が観測された(プリオン蛋白質(23−231)の糖チップへの結合カーブ)。200秒間、結合カーブのデータを収集後、平衡化バッファーで洗浄した。緩やかなシグナル現象のカーブ(プリオン蛋白質(23−231)の解離カーブ)得られた。観測後、チップに洗浄液1(300mM 水酸化ナトリウム水溶液)、引き続き、洗浄液2(2.0M 塩化ナトリウム水溶液)を添加して、チップ上の洗浄を行ない、各濃度のプリオン蛋白質(23−231)溶液を添加して、同様の結合カーブのデータを収集した。得られたデータは、デジタルデータとして、表計算ソフト、エクセル(商品名、マイクロソフト社製)へ移し、化学反応速度論的、化学平衡論的な解析を行った。すなわち、得られた各プリオン蛋白質(23−231)濃度における結合カーブのシグナル値を横軸とする結合カーブの時間微分に対して、近似直線を求めその切片と傾きから、一次反応結合速度定数と一次反応解離速度定数を算出した。プリオン蛋白質(23−231)濃度に対するこれらの一次反応速度定数の値から、平衡解離定数を算出した。ここではセンサーグラムを1:1の化学量論比での結合を仮定して解析したことになり、みかけの解離定数(K(app))が算出される。
【0036】
〔結果〕
調製したリコンビナントプリオン蛋白(23−231)は十分な精製度であり、SDS−PAGE上で一本のバンドしか観測されず、またその移動度はプリオン蛋白質のそれと合致した(図3)。プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合が観測され(図4)、平衡論的解離定数を見積もったところ7.1μMであった。速度論的定数は、解離〔kd((μMプリオン蛋白(23−231))−1・sec−1)〕、結合〔ka((μMプリオン蛋白(23−231))−1・sec−1)〕がそれぞれ、kd=0.015、ka=0.0021であった。実施例1において推察したヘパリンの抗プリオン活性を担う構造(図2)と、プリオン蛋白質(23−231)との相互作用が観測された。
【0037】
〔実施例3〕
実施例2において観測されたプリオン蛋白質(23−231)とGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合の特性を調べた。
〔方法〕
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈、および、塩化ナトリウムを添加することにより、プリオン蛋白質溶液(23−231)の濃度が11μM、塩化ナトリウム濃度が、0.14, 0.30, 0.35, 0,40Mの試料を200μLずつ調製した。上記のサンプルをGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置で測定した。測定条件は実施例2に準じた。ただし、ベースラインの確認は50秒、結合カーブのデータを収集は、250秒間とした。
【0038】
〔結果〕
図5に塩化ナトリウム濃度を変化させたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。結合は、高い塩濃度においても認められた。
【0039】
〔実施例4〕
実施例2および実施例3において観測されたプリオン蛋白質(23−231)添加によるセンサーグラムの上昇が、プリオン蛋白質(23−231)の結合であることを確認した。
〔方法〕
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈し、5.6μMの溶液を200μL調製した。2糖構造GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置へ添加した。200秒間、結合カーブのデータを収集後、PKを終濃度20μg/mLとなるように加えた平衡化バッファーでチップ上の溶液を置換した。実施例2に準じて、系の洗浄、データの解析を行った。次に、実施例2において調製を行ったプリオン蛋白質(23−231)をトリプシン反応バッファー(10mM 炭酸水素アンモニウム、pH7.0)で希釈し、11μMの濃度のプリオン蛋白質溶液(23−231)を200μL調製し、糖チップへ結合させた。200秒間、結合カーブのデータを収集後、トリプシン反応バッファーでチップを洗浄した。200秒間後、トリプシン(プロメガ社製)を終濃度25μg/mLとなるように加えたトリプシン反応バッファーでチップ上の溶液を置換した。2400秒間放置した後、チップ上のトリプシン消化反応液を75μLのトリプシン反応バッファー、25μLの空気、50μLのトリプシン反応バッファー、50μLの空気で溶出、押し出しを行い回収した。得られた反応溶液に5%のトリフルオロ酢酸水溶液、25μL添加し、攪拌したのち溶液を凍結させた。凍結乾燥させて得られた白色の粉を、5%アセトニトリル、0.1%トリフルオロ酢酸水溶液100μLにて溶解させた。その100μLの溶解液のうち、50μLを、ZipTip C18(商品名、ミリポア社製)で吸引、排出を3回繰り返した。その後、0.1%トリフルオロ酢酸水溶液の吸引、排出を行うことにより、ZipTipを洗浄した。その後、5mg/mLの濃度でα−シアノ−4−ヒドロキシケイヒ酸(ブルカー・ダルトニクス社製)と0.1%トリフルオロ酢酸を含むアセトニトリル2μLでZipTipへの結合物をマトリックス支援レーザー脱離イオン化−飛行時間型質量分析計用の試料台へと溶出した。風乾後、試料をマトリックス支援レーザー脱離イオン化−飛行時間型質量分析計(商品名、VoyagerSTR、アプライドバイオシステム社製)にて測定し、得られたデータをソフトウエア(dataexplore, アプライドバイオシステム社製)で表示した。得られた質量分析値のセットをウエブ上での解析サイト(http://www.matrixscience.com/)にて解析した。
【0040】
〔結果〕
図6にプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合、それに引き続くチップ上でのプロテイナーゼK処理によるセンサーグラムの変化のプロファイルを示した。実施例2において観測した場合と同様に、プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合に伴うシグナルの上昇が観測された。その後プロテイナーゼKの添加により、シグナル強度の減少が観測された。このことから、プリオン蛋白質(23−231)の糖チップ上での分解が可能であることがわかる。上記の条件下での速度論的解析の結果、糖チップ上でのプリオン蛋白質(23−231)の見かけの一次分解速度(プロテイナーゼK 1μg/mLの濃度条件下において)は、2.0×10−4 sec−1 であった。図7にチップ上でのトリプシン処理によるセンサーグラムの変化のプロファイルを示した。プロテイナーゼKによる処理のときと同様なセンサーグラムの変化が観測された。回収したトリプシン消化産物の脱塩を行い、質量分析を行った。図8にその測定結果を示す。ペプチド・マス・フィンガープリント法によって、プリオン蛋白質(23−231)が同定された(図9)。同定の確かさでは、種の異なる動物のプリオン蛋白質がヒットしたものの、マウスプリオン蛋白質(23−231)が確かに同定された。同定に利用されたペプチド断片は、プリオン蛋白質(23−231)のうち、25−37, 38−48, 148−155, 194−203, 220−228, 220−229に相当する断片計6本であった。これらのことから、糖チップ上での蛋白質分解によって得られるペプチド断片を用いてチップ結合蛋白質が、ペプチド・マス・フィンガープリント法によって同定できることがわかった。
【0041】
〔実施例5〕
実施例1においてGlcNS6Sα4IdoA2Sαという二糖構造が抗プリオン活性に重要であることを示し、実施例2〜実施例4においてリコンビナントプリオン蛋白質(23−231)はGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」と結合することを示した。そこで、この結合はプリオン蛋白質(23−231)ではどのような部位にその結合サイトが存在するかを明らかにした。
〔方法〕
プリオン蛋白質のN末端側ドメインを欠損した以下のプリオン蛋白質断片、プリオン蛋白質(121−231)、プリオン蛋白質(89−231)の調製を行った。プリオン蛋白質(121−231)においては、実施例2で得た発現ベクターをテンプレートとして、プリオン蛋白質(121−231)に相当するDNAフラグメントを増幅、発現ベクターに導入しクローニングした。実施例2に準じた培養、処理を行い、プリオン蛋白質(121−231)を含む沈殿物を回収した。得られた沈殿物を、8M尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清を透析バッファー1(5mMトリスヒドロキシアミノメタン、pH8.5)に対して透析(透析膜;分子量カット3,500、スペクトラ社製)し、プリオン蛋白質(121−231)を含む溶液をHi−trap DEAEカラム(GEヘルスケアバイオサイエンス社製)に添加し、透析バッファー1から200mM 塩化ナトリウムを含む透析バッファー1への塩濃度勾配により精製した。分離されたプリオン蛋白質(121−231)を含む溶液を、水に対して透析(透析膜;分子量カット3,500、スペクトラ社製)を行った。プリオン蛋白質(89−231)においては、実施例2で得た発現ベクターをテンプレートとして、プリオン蛋白質の23に相当する部位から、3′末端側方向へのプライマーとプリオン蛋白質の89に相当する部位から、5′末端側方向へのプライマーのセットで、プリオン蛋白質(23−88)に相当するDNAフラグメントを欠損した直鎖状の二本鎖DNAを増幅した。この直鎖状の二本鎖DNAの環化反応を行いプリオン蛋白質(89−231)の発現プラスミドとした。実施例2に準じた培養、処理を行い、プリオン蛋白質(89−231)を含む沈殿物を回収した。得られた沈殿物を、8M 尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清をリシス液に対して透析(透析膜;分子量カット3,500、スペクトラ社製)し、一晩放置して得られた沈殿を遠心分離によって除き、その上清を0.1%ギ酸水溶液に透析し、一晩放置した。遠心分離の上清を回収し、C4カラム(Cosmosil、10mm×250mm、ナカライテスク社製)に蛋白質溶液を添加し、0.1%トリフルオロ酢酸水溶液から、0.1%トリフルオロ酢酸を含むアセトニトリルへの直線勾配で精製した。プリオン蛋白質(89−231)を含むピークを分取し、凍結乾燥させた。一連の工程は、SDS−PAGEにより確認を行った。プリオン蛋白質(121−231)溶液、プリオン蛋白質(89−231)溶液のそれぞれを糖チップ測定用の平衡化バッファーで希釈することにより10μM の濃度に調整したプリオン蛋白質溶液を200μLずつ調製し、実施例2に準じた条件によりSPR測定装置で測定した。
【0042】
〔結果〕
ここで調製したプリオン蛋白質(121−231)およびプリオン蛋白質(89−231)は、十分な精製度であり、SDS−PAGE上で一本のバンドしか観測されず、またその移動度はプリオン蛋白質のそれと合致した(図11)。図10にこれらのプリオン蛋白質の測定結果を示す。有意な結合は観測されず、プリオン蛋白(23−231)のうち、23−89の領域が結合に関与していることが示唆された。
【0043】
〔実施例6〕
実施例2にて観測されたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合は、実施例5においてプリオン蛋白(23−231)のうち、23−89の領域が結合に関与していることが示唆された。この点をさらに詳しく調べて、N末端部位を欠損させたことによるアーティファクトではないことを検証した。
〔方法〕
プリオン蛋白質(23−89)については、実施例5のプリオン蛋白質(121−231)の方法に準じて発現ベクターを調製した。実施例2に準じた培養、処理を行い、プリオン蛋白質(23−89)を含む沈殿物を回収した。得られた沈殿物を8M尿素を含むリシス液に溶解させ、再び遠心分離した。得られた上清をNi−NTAカラム(GEヘルスケアバイオサイエンス社製)に蛋白質溶液を添加し、8M尿素を含むリシス液にて洗浄後、600mMイミダゾール、8M尿素を含むリシス液によって、カラムに結合した蛋白質を溶出させた。得られた蛋白質溶液を、水に対して透析(透析膜;分子量カット3,500、スペクトラ社製)した。一晩放置した透析内容物を遠心分離し、その上清を回収し、C18カラム(Cosmosil、10mm×250mm、ナカライテスク社製)に蛋白質溶液を添加し、0.1%トリフルオロ酢酸水溶液から、0.1%トリフルオロ酢酸を含むアセトニトリルへの直線勾配で精製した。プリオン蛋白質(23−89)を含むピークを分取し、凍結乾燥させた。この一連の工程は、SDS−PAGEで追跡した。試料の一部をマトリックス支援レーザー脱離イオン化−飛行時間型質量分析計(商品名、VoyagerSTR、アプライドバイオシステム社製)にて測定した。プリオン蛋白質(23−89)溶液を糖チップ測定用の平衡化バッファーで希釈することにより、22, 14, 11, 7.2, 5.4, 4.3, 3.1μM の濃度のプリオン蛋白質(23−89)溶液を200μLずつ調製した。各濃度のプリオン蛋白質(23−89)溶液を実施例2に準じて測定した。結合カーブは200秒間データを収集した。洗浄後、緩やかなプリオン蛋白質(23−231)の解離カーブが得られた。観測後、実施例2に準じた処理と解析を行った。
【0044】
〔結果〕
調製したリコンビナントプリオン蛋白(23−89)は、図13に精製時におけるHPLC(C18カラム)のクロマトグラムを示すとおり十分な純度であり(SDS−PAGEによっても一本のバンドしか観測されず)、またその質量分析値はプリオン蛋白質(23−89)のそれと合致した(図13)。プリオン蛋白質(23−89)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合が観測され(図12)、平衡論的解離定数を見積もったところ16μMであった。速度論的定数は、解離〔kd((μMプリオン蛋白(23−89))−1・sec−1)〕、結合〔ka((μMプリオン蛋白(23−89))−1・sec−1)〕がそれぞれ、kd=0.061、ka=0.0037であった。実施例2において解析したプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合の平衡論的解離定数および速度論学的結合・解離定数とよい一致が見られた。プリオン蛋白質(23−89)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への速度論学的結合・解離定数はプリオン蛋白質(23−231)のそれらと比べて、若干大きな値を示したが、これはプリオン蛋白質(23−89)の分子量がプリオン蛋白質(23−231)にくらべて小さく、分子の並進拡散速度が大きいためであると考察される。実施例2、実施例5および実施例6により、プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合は、23−89の領域が結合を担っていることが明らかになった。
【0045】
〔実施例7〕
プリオン蛋白質の51−91の領域は銅イオンとの結合が報告されている[ホルムショウ MP.ら、バイオケミカル アンド バイオフィジカル リサーチ コミュニケーションズ(Biochemical & Biophysical Research Communications)1995年、214巻、993−999頁]。実施例6において明らかとなったプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合は、この銅結合領域を含んでいる。そのため、プリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」への結合が銅イオンの存在下で影響されるかどうかを検証する必要があり、以下の実験を実施した。
〔方法〕
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈、さらに硫酸銅水溶液を添加することにより、プリオン蛋白質溶液(23−231)の濃度が10μM、硫酸銅濃度が0, 50, 100, 200, 400, 1000μMの試料を200μLずつ調製した。実施例2に準じてSPR測定をおこなった。450秒間、結合カーブのデータを収集し、実施例2に準じて解析した。
【0046】
〔結果〕
図14に硫酸銅濃度を変化させたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。プリオン蛋白質(23−231)に対して化学量論的に大過剰に硫酸銅が存在する条件下(400, 1000μM)では、蛋白質性の沈殿物が混合後、すぐに目視で確認できた。硫酸銅50μMの添加では、硫酸銅を添加しない条件での結合プロファイルと差は見られなかった。50μMを超える高濃度の硫酸銅存在下では、硫酸銅の濃度の上昇に伴いプリオン蛋白質(23−231)の結合量は低下した。これらの結果から、10μMのプリオン蛋白質(23−231)に対して、50μMまでの硫酸銅は結合を阻害しないが、それ以上の濃度の硫酸銅ではプリオン蛋白質(23−231)の沈殿を生じさせ、有効な結合を阻害するため、プリオン蛋白質(23−231)とGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との相互作用を検出する上で不都合であることがわかった。
【0047】
〔実施例8〕
プリオン蛋白質(23−231)とGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合の阻害を利用した治療薬スクリーニングが可能であるかどうかを検証した。
〔方法〕
実施例2において調製を行ったプリオン蛋白質(23−231)を平衡化バッファーで希釈、さらにヘパリン(ナカライテスク社製、分子量17600)水溶液を添加することにより、プリオン蛋白質溶液(23−231)の濃度が5.5μM、ヘパリン濃度が、0, 0.02, 0.05, 0.10, 0.50, 1.0, 5.0μg/mLの試料を200μLずつ調製し、また、ペントサンポリサルフェートについても同様の試料調製を行った。両薬剤のシリーズについて実施例2に準じてSPR測定を行った。プリオン蛋白質(23−231)の糖チップへの結合カーブについて150秒間データの収集を行った。
【0048】
〔結果〕
図15にヘパリン濃度を変化させたプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。ヘパリンの濃度依存的にプリオン蛋白質(23−231)の結合の低下が観測された。糖チップへと添加したヘパリンを含むプリオン蛋白質(23−231)の溶液には、沈殿は観測されなかった。したがって、このプリオン蛋白質(23−231)の GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合量の低下は、ヘパリンによる競合的阻害であることが明らかである。ヘパリンは実施例1において示したように抗プリオン活性を有する化合物であり、本実施例は、抗プリオン活性を有する化合物がプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合を競合的に阻害することを示すものである。さらに、現在臨床応用もなされているペントサンポリサルフェートについても、ヘパリンの場合と同様、濃度依存的なプリオン蛋白質(23−231)の結合の低下が観測された(図16)。
【0049】
〔実施例9〕
シグナルを誘起したプリオン蛋白質(23−231)が、GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」から回収可能であるかどうかを調べた。
〔方法〕
実施例2において調製を行ったプリオン蛋白質(23−231)を実施例1におけるリシス液で希釈することにより、2.8μMプリオン蛋白質(23−231)溶液の試料を200μL調製した。このサンプルをGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」を搭載したSPR測定装置にて測定した。100秒間結合カーブのデータを収集した。得られたデータは、実施例2に準じて図示した。その後、1mLの脱イオン水を用いて経路を洗浄し、0.1%トリフルオロ酢酸、50%アセトニトリルを含む液300μLを用いてGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」から結合蛋白質を溶出した。溶出液を凍結乾燥させ、この乾燥粉末全量に対しSDSポリアクリルアミドゲル電気泳動(SDS−PAGE)用バッファーを加え、実施例1に準じてウエスタンブロット法でプリオン蛋白質(23−231)を検出した。コントロールとして0.1μgプリオン蛋白質(23−231)を含む試料を並行して泳動した。検出したプリオン蛋白質(23−231)のシグナル強度を、コントールと比較することによって、表面プラズモン共鳴角の変化を誘起したGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」上のプリオン蛋白質(23−231) を定量した。
【0050】
〔結果〕
図17に2.8μMの濃度のプリオン蛋白質(23−231)のGlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」との結合センサーグラムの変化のプロファイルを示した。図18に糖チップ上から回収したプリオン蛋白質(23−231) (左)と、0.10μgのプリオン蛋白質(23−231)(右)のウエスタンブロットの結果を示している。回収したプリオン蛋白質(23−231)の量は、コントロールとの比較により、120ngであることが明らかになった。また、これらの結果は、GlcNS6Sα4IdoA2Sα6Glc−mono固定化「糖チップ」に結合したプリオン蛋白質を回収することが可能であることを示している。
【図面の簡単な説明】
【0051】
【図1】本発明における硫酸化多糖の一例であるヘパリンについて、抗プリオン活性を担う部分構造を推定する試験を行なった結果の例として、ウエスタンブロットの泳動像を示す図面代用写真を含む説明図である(実施例1)。
【図2】本発明における糖(二糖単位構造)の一例を示す構造式である。
【図3】本発明における正常型プリオン蛋白質についてSDS−PAGEにより試験を行なった結果の一例として、泳動像を示す図面代用写真である(実施例2)。
【図4】本発明における糖と正常型プリオン蛋白質との相互作用による表面プラズモン共鳴角の変化角度(Difference in resonance angle /deg)の経時的な変化プロファイルの一例を示すセンサーグラムである(実施例2)。
【図5】同、糖と正常型プリオン蛋白質との相互作用において正常型プリオン蛋白質溶液の塩濃度を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例3)。
【図6】同、糖と正常型プリオン蛋白質との相互作用において、両者結合後、プロテイナーゼKの添加によってチップ上で正常型プリオン蛋白質が分解することによる変化プロファイルの一例を示すセンサーグラムである(実施例4)。
【図7】同、糖と正常型プリオン蛋白質との相互作用において、両者結合後、トリプシンの添加によってチップ上で正常型プリオン蛋白質が消化することによる変化プロファイルの一例を示すセンサーグラムである(実施例4)。
【図8】(図7で確認のトリプシン添加によって消化された)正常型プリオン蛋白質のトリプシン消化産物(回収、脱塩)について質量分析を行なった結果の一例を示す解析図であり、各行「Start − End」の数値は、上方に示されたアミノ酸配列(1−254:配列番号3)におけるN末端側からのアミノ酸残基番号であり、同定された各行ペプチド断片のアミノ酸配列(範囲)を表している(実施例4)。
【図9】図8に示した質量分析測定値に基づくペプチド・マス・フィンガープリント法による同定結果の一例を示す解析図である(実施例4)。
【図10】本発明における糖と比較試験例に相当するプリオン蛋白質断片(マウスプリオン蛋白質断片(121−231)、(89−231))との相互作用による表面プラズモン共鳴角の変化角度の経時的な変化プロファイルの一例を示すセンサーグラムである(実施例5)。
【図11】図10に示した試験に用いた比較試験例に相当するプリオン蛋白質断片について、SDS−PAGEにより試験を行なった結果の一例として、泳動像を示す図面代用写真であり、左側がマウスプリオン蛋白質断片(89−231)、右側がマウスプリオン蛋白質断片(121−231)である(実施例5)。
【図12】本発明における糖とプリオン蛋白質断片(マウスプリオン蛋白質断片(23−89))との相互作用による表面プラズモン共鳴角の変化角度の経時的な変化プロファイルの一例を示すセンサーグラムである(実施例6)。
【図13】図12に示した試験に用いたプリオン蛋白質断片について、その調製精製時のHPLCクロマトグラム及び質量分析の試験結果の一例を併せて示す図である(実施例6)。
【図14】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液の銅イオン濃度(硫酸銅添加濃度)を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例7)。
【図15】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液へのヘパリン添加濃度を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例8)。
【図16】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液へのペントサンポリサルフェート添加濃度を変えた場合における変化プロファイルの一例を示すセンサーグラムである(実施例8)。
【図17】本発明における糖と正常型プリオン蛋白質との相互作用において、正常型プリオン蛋白質溶液にリシス液を添加した場合における変化プロファイルの一例を示すセンサーグラムである(実施例9)。
【図18】本発明における糖と正常型プリオン蛋白質との相互作用(結合)測定の後、チップ上から回収した正常型プリオン蛋白質(左端)について、コントロールとの比較から定量する試験を行なった結果の一例として、ウエスタンブロットの泳動像を示す図面代用写真である(実施例9)。
【特許請求の範囲】
【請求項1】
a)硫酸化多糖を構成する二糖構造を含む糖と、
b)異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列を少なくとも含む正常型プリオン蛋白質断片、或は、全長正常型プリオン蛋白質と、の結合の阻害を指標とすることを特徴とするプリオン病治療薬のスクリーニング方法。
【請求項2】
前記糖が、硫酸化多糖を酸化剤により10糖以下に低分子化したヘテロ多糖又はヘテロオリゴ糖、或は化学合成したヘテロオリゴ糖であることを特徴とする請求項1に記載のプリオン病治療薬のスクリーニング方法。
【請求項3】
前記硫酸化多糖がヘパリンであることを特徴とする請求項1又は2に記載のプリオン病治療薬のスクリーニング方法。
【請求項4】
前記酸化剤が過ヨウ素酸又は過塩素酸又はこれらの塩であることを特徴とする請求項2又は3に記載のプリオン病治療薬のスクリーニング方法。
【請求項5】
前記糖が、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれる1種以上であることを特徴とする請求項1乃至4のいずれかに記載のプリオン病治療薬のスクリーニング方法。
【請求項6】
前記正常型プリオン蛋白質が、配列番号1のアミノ酸配列を有するマウスプリオン蛋白質(23−231)であり、
前記正常型プリオン蛋白質断片が、N末端側に少なくとも配列番号2のアミノ酸配列を含むマウスプリオン蛋白質断片であることを特徴とする請求項1乃至5のいずれかに記載のプリオン病治療薬のスクリーニング方法。
【請求項7】
前記指標は、表面プラズモン共鳴法によって評価されることを特徴とする請求項1乃至6のいずれかに記載のプリオン病治療薬のスクリーニング方法。
【請求項8】
N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上を有効成分として含んでなることを特徴とするプリオン病治療薬。
【請求項1】
a)硫酸化多糖を構成する二糖構造を含む糖と、
b)異常型プリオン蛋白質のプロテアーゼ処理により消化除去されるN末端領域のアミノ酸配列を少なくとも含む正常型プリオン蛋白質断片、或は、全長正常型プリオン蛋白質と、の結合の阻害を指標とすることを特徴とするプリオン病治療薬のスクリーニング方法。
【請求項2】
前記糖が、硫酸化多糖を酸化剤により10糖以下に低分子化したヘテロ多糖又はヘテロオリゴ糖、或は化学合成したヘテロオリゴ糖であることを特徴とする請求項1に記載のプリオン病治療薬のスクリーニング方法。
【請求項3】
前記硫酸化多糖がヘパリンであることを特徴とする請求項1又は2に記載のプリオン病治療薬のスクリーニング方法。
【請求項4】
前記酸化剤が過ヨウ素酸又は過塩素酸又はこれらの塩であることを特徴とする請求項2又は3に記載のプリオン病治療薬のスクリーニング方法。
【請求項5】
前記糖が、N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれる1種以上であることを特徴とする請求項1乃至4のいずれかに記載のプリオン病治療薬のスクリーニング方法。
【請求項6】
前記正常型プリオン蛋白質が、配列番号1のアミノ酸配列を有するマウスプリオン蛋白質(23−231)であり、
前記正常型プリオン蛋白質断片が、N末端側に少なくとも配列番号2のアミノ酸配列を含むマウスプリオン蛋白質断片であることを特徴とする請求項1乃至5のいずれかに記載のプリオン病治療薬のスクリーニング方法。
【請求項7】
前記指標は、表面プラズモン共鳴法によって評価されることを特徴とする請求項1乃至6のいずれかに記載のプリオン病治療薬のスクリーニング方法。
【請求項8】
N−スルホ−D−グルコサミン6−硫酸とL−イズロン酸2−硫酸とがα1−4結合したへテロ二糖〔GlcNS6Sα4IdoA2Sα:(2−デオキシ−2−スルホアミド−6−O−スルホ−α−D−グルコピラノシル)−(1−4)−O−(2−O−スルホ−α−L−イドピラノシルウロン酸)〕、そのアルカリ金属塩、そのアルカリ土類金属塩から選ばれた1種以上を有効成分として含んでなることを特徴とするプリオン病治療薬。
【図2】

【図4】
【図6】
【図8】
【図10】
【図13】
【図14】
【図17】
【図1】
【図3】
【図5】
【図7】
【図9】
【図11】
【図12】
【図15】
【図16】
【図18】
【図4】
【図6】
【図8】
【図10】
【図13】
【図14】
【図17】
【図1】
【図3】
【図5】
【図7】
【図9】
【図11】
【図12】
【図15】
【図16】
【図18】
【公開番号】特開2008−281557(P2008−281557A)
【公開日】平成20年11月20日(2008.11.20)
【国際特許分類】
【出願番号】特願2008−101561(P2008−101561)
【出願日】平成20年4月9日(2008.4.9)
【出願人】(504157024)国立大学法人東北大学 (2,297)
【出願人】(504258527)国立大学法人 鹿児島大学 (284)
【Fターム(参考)】
【公開日】平成20年11月20日(2008.11.20)
【国際特許分類】
【出願日】平成20年4月9日(2008.4.9)
【出願人】(504157024)国立大学法人東北大学 (2,297)
【出願人】(504258527)国立大学法人 鹿児島大学 (284)
【Fターム(参考)】
[ Back to top ]