
プロオキシダント抗ガン化合物
本発明は、抗ガン化合物およびガンの治療または予防方法に関する。さらに詳しくは、本発明は、ガン性細胞のミトコンドリア呼吸鎖の複合体IIと選択的に相互作用し、反応性酸素種を生成し、それらの細胞のアポトーシスを誘導する、ビタミンEのプロオキシダント型などのプロオキシダント抗ガン化合物に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗ガン化合物およびガンの治療または予防方法に関する。さらに詳しくは、本発明は、ガン性細胞のミトコンドリア呼吸鎖の複合体IIと選択的に相互作用し、反応性酸素種を生成し、それらの細胞のアポトーシスを誘導する、ビタミンEのプロオキシダント型などのプロオキシダント抗ガン化合物に関する。
【背景技術】
【0002】
理想的抗ガン薬は、ガン細胞の内部においてのみ活性であり、ガン細胞の本質的な成分を標的とするか、またはその本質的な過程を破壊する薬物である。細胞エネルギーの大部分およびアポトーシスの重要な制御因子を供給するミトコンドリアは、抗ガン療法に望まれる細胞選択性を提供しうる有効な標的として浮かび上がっている(Don and Hogg、2004、Armstrong、2006)。
【0003】
動物実験において報告されているように(Koら、2004)、ガン細胞内のミトコンドリアに選択的に影響を及ぼすことによって作用する細胞毒性薬物である「マイトカン(マイトカン)(抗ガン成分)」は、副作用が少ないか、あるいは副作用のない強力な抗ガン剤として働くので、ガンの治療にとって非常に興味をそそるものであることがわかっている。マイトカンは、ガン細胞ミトコンドリアのエネルギー生成システムを破壊し、ガン細胞内部での活性酸素種(ROS)の増加およびミトコンドリア依存性細胞死シグナル伝達経路の活性化をもたらす。
【0004】
マイトカンとして、以下のミトコンドリア関連活性に影響を及ぼす薬物が挙げられる:ヘキソキナーゼインヒビター;電子輸送/呼吸鎖ブロッカー;ミトコンドリア膜透過性転移孔標的化構成タンパク質サブユニット、電位依存性アニオン選択的チャネル(VDAC)またはアデニンヌクレオチドトランスポーター(ANT)のいずれかのアクチベーター;Bcl-2抗アポトーシスファミリータンパク質およびBax/Bidアポトーシス促進性模倣体のインヒビター。副作用が少ないか、あるいは副作用のないマイトカンの2つの主な例は、3-ブロモピルビン酸エステル(3-BP)およびα-トコフェリルコハク酸エステル(α-TOS)であり、それらの両方が、非常に大きい有効性で、ガン細胞においてアポトーシスを誘導する(Koら、2004、Geschwindら、2002、Xuら、2005、Neuzilら、2001、2004)。
【0005】
マイトカンの1つのグループとして、ビタミンEのロオキシダント類縁体が挙げられる(Wangら 2006)。α-TOSによって代表されるプロオキシダントビタミンE類縁体の抗ガン剤としての有望性は、それらが悪性腫瘍を抑制することを明らかにした、ヌードマウスで増殖するヒト意趣移植片などの実験的に作成されたガンを用いた研究に起因する(Neuzilら、2004に概説)。このような研究として、結腸直腸(Neuzilら、2001、Weberら、2002)および肺ガン(Quinら、2005)、黒色腫(Malafaら、2002)ならびに中皮腫(Tomasettiら、2004、Stapelbergら、2005)が挙げられる。α-TOSが、乳ガンの休止状態を促進すること(Malafaら、2000)および結腸ガンのへの転移を抑制することも明らかにされている(Barnettら、2002)。
【0006】
ビタミンE(α-トコフェロール、α-TOH)は、細胞において強力な抗酸化剤として作用するけれども、ビタミンEのエステル化された酸化還元サイレントおよびプロオキシダント類縁体であるα-TOSは、特徴的性質を有する。α-TOHとは対照的に、α-TOSは、種々のガン細胞系の範囲においてROSの急速な生成を引き起こす強力な細胞ストレッサーとして作用する(Neuzilら、2004、Weberら、2003、Wangら、2005、Swettenhamら、2005、Stapelbergら、2005)。α-TOSは、Bcl-2/Bcl-xLに結合し、それらを阻害する能力を有する(Dongら、2006)。今までの証拠は、正常な細胞は、より大きい抗酸化防御に恵まれており(Allen and Balin、2003、Saffordら、1994、Churchら、1993)、および/またはコハク酸エステル部分を放出することにより酸化還元活性、非アポトーシス性α-TOHを産生することによってα-TOSを不活性化する高濃度のエステラーゼを含む(Farissら、2001、Neuzilら、2004、Neuzil and Massa、2005)ので、α-TOSのガン細胞特異的性質および正常な細胞における毒性作用の欠如が起こることを示唆している。
【0007】
天然のビタミンEは、クロマノール環のメチル化パターン(α-、β-、γ-、δ-トコフェロール)およびフィチル側鎖の二重結合の数(α-、β-、γ-、δ-トコトリエノール)によって相異する8種の化合物の混合物からなる。インビトロおよびインビボでの親油性抗酸化剤としてのこれらの分子の役割は、広く受け入れられている。さらに、VEファミリーのメンバーの非抗酸化特性も研究されている(Azziら、2002)。
【0008】
ビタミンE分子は、3つの異なるドメインに分割することができる。機能的ドメイン(I)は、クロマノール環のC6位における置換パターンから生じる。遊離のヒドロキシ基は、ビタミンEが抗酸化剤として機能するために必須であるので、この位置は、分子が酸化還元活性または酸化還元サイレントとして振る舞うかどうかを決定する。文書で十分に立証された、4種のトコフェロール異性体の抗酸化特性は、ガン臨床試験への適用をもたらした。これらの研究は、ガンの予防における遊離トコフェロールの使用に関して肯定的な結果を明らかにしなかった(Pham and Plakogiannis、2005)。しかし、C6における特定の化学的修飾は、強力な抗腫瘍剤であること立証されたエーテル(RO-)、エステル(RCOO-)およびアミド(RCONH-)をもたらした。下記第1表を参照。
【0009】
第1表
ビタミンE類縁体の抗増殖活性
化合物は、シグナル伝達ドメインによって分類される。
【表1】

【0010】
【表2】
a無効;b細胞増殖の阻害;cα-TOSよりも細胞毒性が高い;d54よりも高価が低い;eエーテル類縁体がα-TOS自体よりも効果が低い;fα-TOSと同程度;gEC50 [μg/ml];hα-TOSよりも効率的。
【0011】
第2表
修飾疎水性ドメインを有するビタミンE類縁体の抗増殖活性
【表3】
a無効;b50 μMにおいて弱い阻害;c10 μMにおいて82%阻害。
【0012】
第3表
ビタミンE類縁体の抗増殖活性
化合物は、シグナル伝達ドメインによって分類される。
【表4】
【0013】
【表5】
a0-40 μMの範囲において細胞毒性;b非常に強力;c完全阻害;dα-TOSと同程度;eγ-トコトリエノールよりも2倍強力;f細胞増殖の阻害。
【0014】
シグナル伝達ドメイン(II)と称される第2のドメインは、トコフェロールの抗酸化特性とは無関係である活性を示す。これらの特性は、芳香環のメチル化パターンに由来する。たとえば、α-トコフェロールは、ジアシルグリセロール(DAG)レベルを低下させることによってタンパク質キナーゼC(PKC)を阻害することが報告されているが、同様の抗酸化能力を有する他のトコフェロール(たとえば、β-トコフェロール)は、PKCを阻害しない。したがって、α-トコフェロールのPKC阻害活性は、その鉱産か能力とは無関係である(Tasinatoら、1995;Kunisakiら、1995)。しかし、いくつかの場合、種々のトコフェロールの生物活性は、実際に特定の種に対する抗ガン活性において重大な影響を有する、シグナル伝達ドメインにおける構造的差異によって影響を及ぼされる。たとえば、γ-トコフェロールは、α-トコフェロールよりも非常によい活性酸化窒素種(たとえば、ペルオキシ亜硝酸)のスカベンジャーである。したがって、C5にメチル基を欠いているγ-分子は、その部位において容易にニトロ化される(Mortonら、2002;Christenら、1997)。
【0015】
ビタミンE異性体の親油性側鎖は、飽和イソプレニルユニットを有するトコフェロールと飽和イソプレニルユニットを有するトコトリエノールを区別する。疎水性ドメイン(III)は、分子がリポタンパク質および膜にそれぞれ結合することができるかどうか、または第1相酵素によって分解されうるかどうかを決定する(Birringerら、2002;Neuzil and Massa、2005)。
【0016】
修飾ヒドロキシル基を有する多くのトコフェロール誘導体が、それらのアポトーシス促進活性について試験されている(第1表)。試験されたもののうち最も優れた誘導体は、クロマノール環のC6位にスクシニルエステルを有するα-TOS(エントリー1)である。その低いpKa(<6)により、α-TOSは、生理的条件下で完全に脱プロトン化され、ミトコンドリア膜を不安定にし、複合体IIにおける効果を有する界面活性剤様分子をもたらす。トコフェロールのジカルボン酸エステルは、構造と活性の関係(SAR)について最もよく研究された化合物を提供する。強いアポトーシス性化合物として、コハク酸α-トコフェロール(1)、シュウ酸トコフェロール(10)、およびマロン酸トコフェロール(11)が挙げられ、後の2つは、黒色腫細胞を接種されたマウスにおいて非選択的細胞毒性を誘導する(Kogureら、2005)。さらに大きいアポトーシス促進活性が、不飽和ジカルボン酸様α-トコフェリルマレエート(3)(Birringerら、2003)およびα-トコフェリルフマレートに観察されている。ジカルボン酸の鎖長の増加は、グルタミン酸(5)、メチル化グルタル酸(6、7、8)(Birringerら、2003)において明らかにされたように活性の低下をもたらし、ピメリン酸(24)(Kogureら、2004)はまったく活性を示さかった。
【0017】
全α-TOS分子が、その完全なアポトーシス誘導活性に必要であることが確立されている(Birringerら、2003)。遊離カルボキシル基のエステル化は、アポトーシス促進活性のない非荷電誘導体(9、25)をもたらす。酢酸トコフェリルおよびプロピオン酸トコフェリル(19)などの脂肪族カルボン酸は、それぞれ、メチルエーテル(18)と同様に不活性であった。α-TOSは腸内エステラーゼによって切断されるので、この化合物の経口投与は効果的ではない(Wuら、2004b;Cheesemanら、1995)。エステル結合切断の問題を解消するために、エステル結合を加水分解に耐性があるのでエーテル結合と置き換えて、化合物(20、21)および側鎖切断型(truncated)誘導体(42)が合成されている(Wuら、2004b;Nishikawaら、2003;Shunら、2004;Shiauら、2006)。メチレン基によるエーテル結合の置き換えは、アポトーシスを促進するのに十分であることに留意すべきである(22)(Sandersら、2001)。
【0018】
エステル結合をアミド結合と置き換える場合、さらに増強されたアポトーシス促進活性が観察された(12、13、37、38)(Tomic-Vaticら、2005)。また、不飽和アミド(13、38)は、飽和アミドよりも優れていた。エステル結合の代わりにアミド結合を導入する理論的根拠は、アニリンアミドは、対応するフェノールエステルよりも加水分解する傾向がほとんどないという十分に確立された事実に基づいていた。これらのトコフェリルエステル誘導体の安定性を増強することは、インビボにおいてこれらの分子を保護し、より長く無傷でいることを可能にし、そのことによってそれらのバイオアベイラビリティが増加する。アミドによるエステルのイソステリック置換は、その連結を酵素加水分解しにくくもする。いつかの非特異的エステラーゼは、腸内粘膜細胞および血液中に存在する。対照的に、ペプチドは、より狭い特異性を示す。たとえば、アミド結合にアミノ酸を有するプロドラッグは、腸および血液内で、それらの対応するエステル類縁体よりも安定である(Sugawaraら 2000)。
【0019】
化合物の最後のグループは、一連のプラスに荷電したN-末端を有するリシンα-トコフェリルエステルからなった(15-17)。親水性アンモニウム官能基は、そのカルボキシレート対応物と同様のアポトーシス促進効果を発揮したが、一般的モチーフが、親油性側鎖および親水性頭部基からなる活性にとって必要であることを示唆する。しかし、長鎖脂肪族アルコールのスクシニルエステル(たとえば、フィトールおよびオレオール)は、どのような活性も示さなかった(Birringerら、2003)。
【0020】
一般的な構造と活性の関係は、第1表に示すデータから引き出すことができる。
1.絶大なプロオキシダントおよびアポトーシス促進活性を得るために、機能的ドメインIの修飾は、解離した酸または荷電したアンモニウム基からなる親水性頭部基を要求した。
2.機能的ドメインの鎖長および不飽和の程度が、アポトーシス活性を決定した。コンホメーション的制限が、活性を増強するように見えた。
3.機能的ドメインの化学結合は、エステルに限定されず、他の官能基は、誘導体の酵素的分解を防止した。
【0021】
クロマノール環の置換パターンは、トコフェロールの抗酸化特性に関係するのみではないことが多い(Azziら、2002)。異なる生化学的観察が、シグナル伝達および代謝過程におけるα-トコフェロールの役割を強調している。したがって、α-トコフェロールは、肝臓内で、他のトコフェロールおよびトコトリエノールと比べてα-トコフェロールに高い親和性をもつ32 kDaのタンパク質であるα-トコフェロールトランスファータンパク質(α-TTP)によって選択的に認識される。α-TTPへの相対的親和性は、クロマノール環のメチルバイオアベイラビリティの損失とともに減少する (α-トコフェロール 100 %、β-トコフェロール 38%、γ-トコフェロール 9%およびδ-トコフェロール 2%)(Hosomiら、1997)。近年発見されたトコフェロール関連タンパク質(TAPs)は、トコフェロール結合に同様の優先傾向を示している(Yamauchiら、2001)。内皮細胞において、トロンビン誘発PKC活性化およびエンドセリン分泌は、α-トコフェロールによって阻害されるが、β-トコフェロールによっては阻害されない(Martin-Nizardら、1998)。転写レベルにおいて、α-トコフェロールは、α-トロポミオシン発現のアップレギュレーション(Aratriら、1999)ならびにLDLスカベンジャー受容体SR-AおよびCD36のダウンレギュレーションを引き起こすが、β-トコフェロールは、効果はない(Ricciarelliら、2000;Devarajら、2001)。さらに、細胞培養において、γ-およびδ-トコフェロールは、α-またはβ-トコフェロールよりも、より速く分解されるので(Birringerら、2001)、置換パターンは、側鎖分解の速度に関与すると思われる。4つのトコフェロール異性体のスクシニル化は、化合物1、32、33および35を生成する。これらのα-TOS(1)が、最も高いアポトーシス活性を有し、β-TOS(32)、γ-TOS(33)と続き、δ-TOS(35)が最も効果が低いことは驚くには当たらない(Birringerら、2001)。一般に、トコフェロールファミリーのメチル化の程度の高いメンバーが、最も強力であるが、この傾向は、トコトリエノールでは逆である(後記参照)。
【0022】
短縮された側鎖を有する水溶性ビタミンE誘導体であるトロロックスのスクシニル化は、アポトーシス促進活性の完全な損失をもたらした。前立腺ガン細胞において、最高レベルのアポトーシス活性を示した、切断されたフィトール側鎖を有する種々のコハク酸トコフェロールの構造と活性の関係の実験(第2表、43、44、45)は、側鎖長が2つのイソプレニルユニットである誘導体から得られた。コンピューターを使った分子モデリングおよび免疫共沈降実験は、Bcl-xLおよびBcl-2へのBak BH3ペプチドの結合が、トコフェロール類縁体によって阻害されたことを示した(Shiauら、2006)。抗腫瘍活性のための中心的要求は、クロマノール環のスクシニル化および1つのイソプレニルユニットの最短の鎖長であった(42、46)。ドメインIII側鎖に結合したエーテル/エステルを有する一連のトコフェリルリシンエステルは、鎖長とIC50(47-50)との間に負の相関関係を示した(Aryaら、1998)。
【0023】
トコトリエノールは、効率的な抗ガン剤であり、そのアポトーシス促進特性は、タンパク質のRasファミリーの不活性化に関連する。トコトリエノールは、機能的ドメインの修飾することなく、そのアポトーシス促進活性を示す。シグナル伝達ドメインにおける階層も逆転され、δ-トコトリエノール(59)をマウスB16-F10黒色腫細胞モデルにおける最も強力な作用剤にし、次に、γ-(56)およびα-トコトリエノール(53)が続く(第3表;Heら、1997)。興味深いことに、すべての芳香族メチル基を欠いているデスメチルトコトリエノール(60)は、IC50が0.9 μMという幾分高い活性を示す。この化合物は、米ぬかから単離されている(Qureshiら、2000)。Rasタンパク質の膜係留システインが、通例の構造エレメントであるファルネシル鎖によって修飾されるので、トコトリエノールの直接阻害作用が提案されている。したがって、活性化しているras突然変異を含むヒト肺腺ガン細胞系であるA549細胞において、トコトリエノールによって、Rasファルネシル化およびRhoAプレニル化が阻害された(Yanoら、2005)。トコトリエノールの短いインビボ半減期を延ばすために、機能的ドメインが導入されている。これらの修飾は、分子の抗増殖活性も増強した(54、57、58)。側鎖の切断もまた、化合物55について見られたのと同様に、活性を改良した。
【0024】
機能的ドメインに修飾が行われた化合物の多くは、抗増殖活性を示し、さらなる特殊化された特性を提供した。たとえば、α-トコフェリルポリエチレングリコールコハク酸エステル(23)は、ドラッグデリバリーシステムのためのビヒクルとして用いられている。この化合物は、ヌードマウスに移植されたヒト肺ガン細胞に対する抗ガン活性を有することが明らかにされた。この化合物のアポトーシス誘導効果は、その細胞への取り込みの増加によるものではなく、むしろ、活性酸素種を生成する能力の増大によるものであった(Youkら、2005)。α-トコフェリルホスフェート(30)は、トコフェロール関連シグナル伝達中に起こる代謝から得られると考えられている(Negisら、2005)。30およびジ-α-トコフェリルホスフェート(31)の混合物は、ラット大動脈平滑筋細胞およびヒトTHP-1単球白血病細胞の増殖を阻害した(Munteanuら、2004)。著者らは、コハク酸トコフェリルおよびマレイン酸トコフェリルが、リン酸トコフェリルを模倣し、置換することによって、ガンにおいて作用し、それによって細胞シグナル伝達の持続的活性化を引き起こすことを提案した。
【0025】
すべての-transレチノイン酸(28)および9-cisレチノイン酸(29)という2つの実験的α-トコフェリルエステルをそれぞれ用いて、急性前骨髄球性白血病細胞の増殖を減少させている(Makishimaら、1998)。レチノイド受容体反応性リポーター構築物を用いるトランス活性化実験は、これらの化合物の両方が、レチノイン酸受容体(RARs)に対するアゴニストとして作用したことを明らかにした。尿中に分泌されているのが見出されることが多いγ-トコフェロールの分解産物であるカルボキシエチルヒドロキシクロマン(52)は、サイクリンD1発現を阻害することによってPC-3前立腺ガン細胞の細胞増殖を低下させることができる(Galliら、2004)。
【発明の概要】
【発明が解決しようとする課題】
【0026】
本発明者らは、特に、呼吸鎖の複合体II(コハク酸ユビキノン酸化還元酵素)を標的化することを手段として、ガン性細胞のミトコンドリア呼吸鎖に沿った電子の伝達を選択的に破壊することが、ROSの生成およびこれらの細胞の死をもたらしうることを見出した。さらに詳しくは、本発明者らは、ビタミンEのプロオキシダント型が、ガン性細胞の複合体IIのユビキノン結合部位からユビキノンを選択的に置き換え、電子の正常な伝達を遮断する能力がある有効な抗ガン化合物であることを見出した。
【課題を解決するための手段】
【0027】
(発明の詳細な記載)
本発明の第1の態様にしたがって、ガン性細胞のミトコンドリア呼吸鎖の複合体II(コハク酸ユビキノン酸化還元酵素)と相互作用して、細胞内に活性酸素種を生成する能力がある、ガン性細胞の死を選択的に誘導するためのプロオキシダント化合物を提供する。
【0028】
本発明の第2の態様にしたがって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力がある、ガンを予防または治療するためのプロオキシダント化合物を提供する。
【0029】
本発明の第3の態様にしたがって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者においてガン性細胞の死を誘導する方法を提供する。
【0030】
本発明の第4の態様にしたがって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者におけるガンを予防または治療する方法を提供する。
【0031】
本発明の第5の態様にしたがって、ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0032】
本発明の第6の態様にしたがって、ガンの予防または治療のための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0033】
本発明の第7の態様にしたがって、ガン性細胞の存在を検出し、ガン性細胞の死を選択的に誘導するプロオキシダント化合物であって、該化合物が、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有する化合物を提供する。
【0034】
本発明の第8の態様にしたがって、患者においてガン性細胞の存在を検出し、ガン性細胞の死を選択的に誘導するための方法であって、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有するプロオキシダント化合物の治療有効量を患者に投与するステップを含む方法を提供する。
【0035】
いすれかの適当なタイプのプロオキシダント化合物を用いることができ、該化合物は、ミトコンドリア複合体IIといずれかの適当な手段で相互作用することができる。好ましくは、化合物は、複合体IIのユビキノン結合部位に結合し、複合体IIと優先的に相互作用する天然の基質ユビキノン、ユビセミキノンまたはユビキノール(コエンザイムQ)またはその他のユビキノンもしくは関連化合物を容易に置き換えることができる。このような基質は、たとえば、Briereら、2004、Tanら、1993、and Espostiら、1996において特定化されている。
【0036】
本発明の第9の態様にしたがって、ガン性細胞の死を選択的に誘導するための、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物を提供する。
【0037】
本発明の第10の態様にしたがって、ガンを予防または治療するための、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物を提供する。
【0038】
本発明の第11の態様にしたがって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者においてガン性細胞の死を誘導する方法を提供する。
【0039】
本発明の第12の態様にしたがって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者におけるガンを予防または治療する方法を提供する。
【0040】
本発明の第13の態様にしたがって、ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0041】
本発明の第14の態様にしたがって、ガンの予防または治療のための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0042】
本発明の第15の態様にしたがって、本発明の第1、第2、第9または第10の態様のプロオキシダント化合物またはその生理的に許容しうる塩および生理的に許容しうる担体を含む医薬組成物または獣医用組成物を提供する。
【0043】
アポトーシスは、ガン性細胞のミトコンドリアにおける活性酸素種のレベルの増加の結果として単独に起こってもよく、または該化合物は、細胞内のミトコンドリア依存性細胞死シグナル伝達経路の活性化を目的として、さらにアポトーシス促進性であってもよい。好ましくは、該化合物は、複合体IIへの結合を目的として活性酸素種を生成し、さらにミトコンドリア依存性細胞死シグナル伝達経路の活性化を目的としてアポトーシス促進性である。
【0044】
好ましくは、プロオキシダント化合物は、プロオキシダント活性を欠いている無害な形体へと、非ガン性細胞内で切断、処置または代謝される。
【0045】
好ましくは、化合物は、プロオキシダントビタミンE類縁体である。本発明者らは、プロオキシダントビタミンE類縁体が、複合体IIを結合し、ユビキノンへの電子伝達を中断させることができることを見出している。本発明者らは、プロオキシダントビタミンE類縁体が、アポトーシス促進性であることも見出している。本発明者らは、have further found thatプロオキシダントビタミンE類縁体が、非ガン性細胞において、無害な抗酸化剤に処置されうることをさらに見出している。
【0046】
対照的に、これも複合体IIに結合するテノイルトリフルオロアセトン(TTFA) (Sunら、2005)は、すべての細胞にとって毒性である(Zhangら、2001b)。3-BPもまた、複合体IIに結合するが、プロオキシダント活性を有すると特徴づけられっていない。
【0047】
「プロオキシダントビタミンE類縁体」は、ガン性細胞のミトコンドリアに位置する場合、酸化還元サイレントであり、複合体IIのユビキノン結合部位に結合して、細胞にダメージを引き起こす代謝の酸素副産物の生成の引き金をひく能力があるビタミンE類縁体として定義づけられる。プロオキシダントビタミンE類縁体の例は、α-トコフェリルコハク酸エステル(α-TOS)である。
【0048】
一方、「抗酸化剤ビタミンE類縁体」は、ガン性細胞のミトコンドリアに位置
する場合、たとえば、α-トコフェロール(α-TOH)などの抗酸化(酸化還元)活性があるビタミンE類縁体である。したがって、プロオキシダントビタミンE類縁体およびビタミンE類縁体の生物活性は、正反対である。
【0049】
特に好ましいプロオキシダントビタミンE類縁体の例を上記第1−3表、下記第4−5表に示し、本明細書の従来技術の欄で議論する。
あるいは、化合物は、以下から選ぶことができる:
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
Gu LQら、1990;He DYら、1994、Yang Fら、1991、Yankovskayaら、1996、およびYabunakaら、2002に記載のユビキノン誘導体(これらは、全体として相互参照することにより本発明に援用される)。
【0050】
化合物は、たとえば、肺、肝臓、腎臓、脳、前立腺、乳房、卵巣、リンパ、皮膚、眼、結腸、胃、口腔扁平および造血系などの患者におけるいずれかの型のガン性細胞の死を誘導するために用いることができる。
【0051】
驚いたことに、化合物α-TOSは、erbB2-低または高ガン細胞を効率的に殺すことが本発明者らによって見出されている。化合物α-TOSは、中皮腫を治療することも本発明者らによって見出されている。
【0052】
驚いたことに、化合物α-TOSは、正常酸素圧および低酸素性の両方のガン性細胞の死を誘導することが本発明者らによって見出されている。したがって、化合物は、患者における早期および後期段階の両方の腫瘍の死を誘導することができるという利点を有する。
【0053】
治療の対象は、ヒト、哺乳類または動物であってよい。好ましくは、患者は、ヒトまたは他の哺乳類である。
【0054】
化合物は、組成物中に、その医薬的にまたは獣医学的に許容しうる誘導体として含まれてもよい。本明細書で用いる本発明化合物の「誘導体」は、塩、Mn2+およびZn2+などの金属イオンとの配位錯体、インビボ加水分解可能エステルなどのエステル、遊離酸もしくは塩基、水和物またはプロドラッグが包含される。リン酸または硫酸などの酸性基を有する化合物は、Na、K、MgおよびCaなどのアルカリまたはアルカリ土類金属およびトリエチルアミンおよびトリス(2-ヒドロキシエチル)アミンなどの有機アミンとの塩を形成することができる。塩は、アミンなどの塩基性基と、塩酸、リン酸または硫酸などの無機酸と、または酢酸、クエン酸、安息香酸、フマル酸または酒石酸などの有機酸を有する化合物との間に形成されることができる。酸性および塩基性基を有する化合物は、内部塩を形成することができる。
【0055】
エステルは、当業者に公知の技術を用いて、化合物内に存在する水酸基またはカルボン酸基と適当なカルボン酸またはアルコール反応パートナーとの間に形成されることができる。
【0056】
組成物は、考慮中である特定の状況に必要な予防有効量または治療有効量のいずれかで患者に投与することができる。組成物中の化合物の実際の量および組成物の投与の速度および経時経過は、治療または予防を必要とするガンの性質および重篤度に応じて変わる。用量の決定などの治療の処方は、患者の介護に責任を負うべき医師または獣医によってなされる。しかし、典型的には、ヒトの患者に投与するための組成物は、約0.01〜100 mgの化合物/体重kg、より好ましくは、約0.1〜10 mg/体重kgである。α-トコフェリルスクシネートまたは他の類縁体が、ヒト患者に経皮適用される場合、化合物の血清レベルは、IC50値の近辺、約40-50 μMであるのが好ましい。
【0057】
組成物は、非経口、局所、経口、吸入スプレー、直腸内、経鼻、バッカル、膣内または埋め込み貯蔵器などの適当な経路で患者に投与することができる。本明細書で用いる用語「非経口」は、皮下、静脈内、筋肉内、動脈内、滑液内、胸骨内、髄腔内、肝内、病巣内および頭蓋内注射または輸液技術を包含する。
【0058】
担体は、いずれかの適当な希釈剤、アジュバント、賦形剤、緩衝剤、安定化剤、等張剤、保存剤または抗酸化剤を含むことができる。担体が、非毒性であり、本発明化合物の有効性を妨げないべきであるのは当然のことである。担体または組成物へのいずれかの他の添加剤の正確な性質は、投与経路および必要とされる治療の型に応じて変わる。たとえば、Alfonso R.Gennaro.Remington:The Science and Practice of Pharmacy、20th Edition.Baltimore、MD:Lippincott Williams & Wilkins、2000、and Goodman and Gilman’s The Pharmaceutical Basis of Therapeutics、Pergamon Press、New York、NYを参照のこと(これらは、全体として参照することにより本発明に援用される)。医薬組成物は、たとえば、従来の混合、溶解、造粒、糖衣形成、磨り潰し、乳化、カプセル化、封入または凍結乾燥処置によって製造することができる。
【0059】
組成物の滅菌注射剤形は、水性または油性懸濁液であってよい。このような剤形は、当業者には公知である。処置するのが望ましい部位での静脈内、皮膚または皮下注射用には、組成物は、適当なpH、等張性および安定性を有する非経口的に許容しうる水性溶液の剤形であってよい。
【0060】
組成物の経口的に許容しうる投与剤形として、カプセル剤、錠剤、丸剤、散剤、リポソーム、顆粒剤、スフィア、糖衣錠、液剤、ゲル剤、シロップ剤、スラリー剤、懸濁剤などが挙げられる。適当な経口剤形は、当業者には公知である。錠剤は、ゼラチンなどの固体担体、アジュバントまたは不活性希釈剤を含むことができる。液体医薬組成物は、一般に、水、石油、動物または植物油、鉱物油または合成油などの液体担体を含む。生理食塩水、またはエチレングリコール、プロピレングリコールまたはポリエチレングリコールなどのグリコールを含んでもよい。このような組成物および製剤は、与えられた担体におけるその溶解度に応じて、一般に、少なくとも、0.1 重量%の化合物、好ましくは、約25 重量%の化合物を含む。
【0061】
組成物は、特に治療の標的が、眼、皮膚または下部消化管のガンなどの局所適用によって容易に到達可能である領域または器官である場合に、局所投与されてよい。組成物は、液体、懸濁液、エマルジョン、軟膏、クリーム、ローション、ペースト、ゲル、泡またはエアロゾルの剤形で適用されてよい。適当な局所剤形は、当業者には公知である。
【0062】
組成物は、生物学的効力を損失することなく、化合物を特定の器官、組織またはガンの型にデリバリーするため、および/または、化合物が、たとえば皮膚を通して吸収されるか、または消化管を介して摂取されることができるのを確実にするために、デリバリービヒクルを含んでもよい。デリバリービヒクルは、たとえば、脂質、ポリマー、りぽそーむ、エマルジョン、抗体および/またはタンパク質であってよい。たとえば、中皮腫を治療するために皮膚を介して化合物をデリバリーするには、リポソームが特に好ましい。
【0063】
化合物を含む固体疎水性ポリマー半透性物質などの持続放出システムを用いて、組成物をデリバリーすることができる。様々な持続放出物質が、入手可能であり、当業者には周知である。持続放出カプセル剤は、約1〜20週間化合物を放出する。
【0064】
化合物は、プロドラッグの形態であってもよい。プロドラッグは、組成物がたとえば経口などで摂取される場合に化合物の活性が損なわれないように保護基を有することができる。プロドラッグは、特定の器官または細胞型に活性化合物をデリバリーすることができる。適当なプロドラッグ剤形および保護基は、当業者には公知であろう。受容体であるチロシンキナーゼerbB2を過剰発現しているガン細胞を標的化するために、α-TOSのアダクトが、ヘプタペプチドLTVSPWYに結合するのが好ましい。
【0065】
患者は、化合物とともに最適予防または治療効果を達成するために1種以上の他の活性成分を含む組成物を投与されてもよい。活性成分は、たとえば、アルキル化剤、血管新生抑制薬、抗アンドロゲン薬、抗エストロゲン薬、代謝拮抗剤、アポトーシス剤、アロマターゼインヒビター、細胞周期制御薬、細胞ストレッサー、細胞障害剤、細胞保護剤、ホルモン剤、免疫療法薬、キナーゼインヒビター、モノクローナル抗体、白金薬、呼吸抑制剤、レチノイド、シグナル伝達阻害剤、タキサンおよびトポイソメラーゼインヒビターであってよい。特に好ましい作用剤として、2-デオキシグルコースおよび3-BPなどの解糖インヒビターが挙げられる。他の特に好ましい作用剤として、TRAILおよびAkt1インヒビターならびにRalph SJら、2006、Neuzil J、Tomasettiら、2007およびNeuzil J、Dongら、2007(これらは、全体として相互参照することにより本発明に援用されるに概説される)他のマイトカンが挙げられる。
【0066】
本発明者らは、たとえば、ガン細胞が、α-TOSと3-BPとの併用(ならびに他の複合薬)によって、いずれかの薬物を単独で用いる場合と比べて、殺傷に対してより感受性が高くなることを見出している。
【0067】
組成物は、非経口または局所投与されるのが好ましい。特に好ましいプロオキシダントビタミンE類縁体は、α-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステル、α-トコフェリルマレイルアミドおよび2,5,7,8-テトラメチル-2R-(4R,8R,12-トリメチルトリデシル)-クロマン-6-イルオキシ酢酸(α-トコフェリルオキシ酢酸)である。エステルであるα-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルおよびα-トコフェリルマレイルアミドのための好ましい担体は、リポソームベースクリーム「Lipoderm」などの経皮適用可能なクリームである。非加水分解性エーテル類縁体であるα-トコフェリルオキシ酢酸が、経口デリバリーに好ましい。
【0068】
検出可能な部分のいずれかの適当なタイプを用いることができ、化合物をいずれかの適当な方法で検出することができる。検出可能な部分は、蛍光色素(たとえば、ローダミン123など)または放射性標識カチオンである。このような検出可能な部分は、たとえば、光線力学療法などの付加的抗ガン療法の同時実践を可能にする。
【0069】
本発明の第16の態様にしたがって、ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)第1のガン性細胞のミトコンドリアに試験化合物を導入し、第2のガン性細胞にガン細胞死を誘導しないことがわかっているコントロール化合物を導入し;次いで、
(ii)ミトコンドリア複合体IIへの化合物の結合および第1および第2のガン性細胞内での活性酸素種の生成をアッセイし、試験化合物およびコントロール化合物によるガン性細胞の死ををアッセイする;
[ここで、コントロール化合物と比較しての試験化合物による複合体II結合、活性酸素種の生成および第1のガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法を提供する。
【0070】
好ましくは、化学物質は、複合体IIのユビキノン結合部位に結合するプロオキシダントビタミンE類縁体である。
【0071】
好ましくは、試験化合物を同時にスクリーニングすることができるハイスループットスクリーニングを用いる。
【0072】
本発明の第17の態様にしたがって、本発明の第16の態様の方法によって同定される化学物質を提供する。
【0073】
本発明の第18の態様にしたがって、ミトコンドリア呼吸鎖複合体IIまたは1種以上のそのサブユニットに結合した本発明の第1、第2、第9または第10の態様の化合物を含む単離または精製された複合体を提供する。
【0074】
好ましくは、化合物が、α-TOSなどのプロオキシダントビタミンE類縁体である。好ましくは、ミトコンドリア呼吸鎖複合体IIが、哺乳類由来であり、より好ましくは、ヒト由来である。
【0075】
本発明の第19の態様にしたがって、ミトコンドリア呼吸鎖複合体IIまたは1種以上のそのサブユニットに結合した本発明の第1、第2、第9または第10の態様の化合物を含む結晶または結晶化可能な組成物を提供する。
【0076】
好ましくは、化合物が、α-TOSなどのプロオキシダントビタミンE類縁体である。好ましくは、ミトコンドリア呼吸鎖複合体IIが、哺乳類由来であり、より好ましくは、ヒト由来である。
【0077】
本発明の第20の態様にしたがって、ミトコンドリア呼吸鎖複合体IIのユビキノン結合部位に結合する能力がある化学物質を同定または設計する方法であって、
(i)呼吸鎖複合体IIと、複合体IIに結合してユビキノンへの電子流を妨げる少なくとも1つのビタミンE類縁体との間の相互作用をコンピューターモデリングし;次いで、
(ii)コンピューターモデリングによって作成されたデータを用いて、複合体IIに結合してユビキノンへの電子流を妨げる能力を有する化学物質を同定または設計する;
ステップを含む方法を提供する。
【0078】
好ましくは、α-TOSなどの少なくとも1つのビタミンE類縁体が、プロオキシダント活性を有し、活性酸素種の生成を促進する。より好ましくは、複合体IIに結合してユビキノンへの電子流を妨げる複数のビタミンE類縁体をステップ(i)で用いる。
【0079】
好ましくは、ミトコンドリア呼吸鎖複合体IIが、哺乳類由来であり、より好ましくは、ヒト由来である。
【0080】
本発明の第21の態様にしたがって、本発明の第20の態様によって同定または設計された化学物質を提供する。
【0081】
好ましくは、化学物質が、ユビキノンに等しいか、またはより大きい複合体II結合親和性を有するプロオキシダントビタミンE類縁体であるか、または以下から選ばれる化学物質である:
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
Gu LQら、1990;He DYら、1994、Yang Fら、1991、Yankovskayaら、1996、およびYabunakaら、2002に記載のユビキノン誘導体。
【0082】
本発明の第22の態様にしたがって、ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)ガン性細胞のミトコンドリアに試験化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位の天然のユビキノン基質の置換をアッセイし、ガン性細胞の死をアッセイする;
[ここで、試験化合物による天然のユビキノン基質の置換およガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法を提供する。
【0083】
本発明の第23の態様にしたがって、本発明の第22の態様によって同定された化学物質を提供する。
【0084】
ヒト神経悪性腫瘍、褐色細胞腫および傍神経節腫、ならびに平滑筋腫および腎細胞ガンの特定の型の研究は、それによってガン細胞がそれら自身の生存を促進する1つの保護装置への洞察力を提供した。この過程は、ガンの低酸素性の発達のための通常の経路とは区別して、「偽性低酸素」として知られるようになっている。これは、これらのガンが、酸素のレベルが正常であっても細胞があたかも低酸素性であるかのように振る舞うようにする突然変異を示すからである。したがって、偽性低酸素性ガンにおいて、体細胞変異は、細胞質ゾルにおけるグルコース代謝とミトコンドリアにおける酸化的ホスホリル化との間のトリカルボン酸(TCA)回路連鎖の一部である酵素であるコハク酸デヒドロゲナーゼ(SDH)またはフマル酸ヒドラターゼ(FHまたはフマラーゼ)のいずれかにおいて起こる。SDHAは、ユビキノン酸化還元酵素複合体IIの3つの他のサブユニットであるSDHB、SDHCおよびSDHDに関連する膜結合タンパク質として内部ミトコンドリア膜のマトリッククス側に位置する酵素サブユニットをコードする。SDHB、SDHCまたはSDHD遺伝子における遺伝性または体細胞突然変異は、褐色細胞腫または傍神経節腫に関連するが、FH遺伝子におけるそのような変異は、平滑筋腫、平滑筋肉腫または人材棒ガンを引き起こす(Gottlieb EおよびTomlinson IP、2005;King Aら、2006;Astuti Dら、2003;Bayley JPら、2006)。
【0085】
SDHまたはFHをコードする遺伝子における突然変異の結果としてSDHまたはFHの酵素活性が低下するので、代謝中間体であるスクシネートおよびフマレートが、これらのガン細胞の内側で上昇することになる。SDHの酵素活性の低下の結果として、このようなガンは、α-TOSなどのプロオキシダント化合物による治療への感受性が高まるか、または低くなる。SDHまたはFHサブユニットをコードする遺伝子に突然変異を含むことが疑われる褐色細胞腫または傍神経節腫、平滑筋腫または腎細胞ガンのタイプのガンは、それらのSDHまたはFH酵素活性のレベルについての診断試験によって分析することができる。SDHの酵素活性は、600 nmにて測定された、細胞または肝臓ミトコンドリア調合液の存在下におけるCII基質であるDCPIPの測定に基づいて、King、TS、1967に記載されるような標準的手順に基づくことができる。反応混合物は、0.5 mM NADH、5 mMスクシネート、10 mM KCN、50 μM DCPIPおよび50 μM PMSを含む。別法として、たとえば、Berridge MVおよびTan AS、1993に概略された、そして、Berridge MVら、2005に概説された、複合体IIのSDH活性を分析するためのいずれかのテトラゾリウム色素還元アッセイがある。
【0086】
別法として、このような異常を検出するために、SDH突然変異の分子または免疫組織化学解析を用いることができる。方法は、PCRベースDNA配列決定、RFLP解析、遺伝子アレイ発現解析およびvan Nederveen FHら、 2007、Pollard PJら、2005およびBayley JPら、2006に記載の例のような同様の技術を包含する。
【0087】
本発明の第24の態様にしたがって、ガンを有する患者が、本発明の第1または第2の態様のプロオキシダント化合物の投与を含む療法に反応する可能性があるかどうかを試験する方法であって、
(i)該患者由来のガン性細胞のミトコンドリアにプロオキシダント化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位へのプロオキシダント化合物の結合をアッセイし、ガン性細胞の死をアッセイする;
[ここで、プロオキシダント化合物の結合およびプロオキシダント化合物によるガン性細胞の死は、患者が、療法に反応する可能性があることを示す]
ステップを含む方法を提供する。
【図面の簡単な説明】
【0088】
【図1】α-TOS、3-BPおよびTRAILは、乳ガン細胞においてATPの還元を引き起こす。示した濃度および示した期間において、α-TOS、3-BPまたはTRIALによって、MDA-MB-453、MCF-7およびNeuTL乳ガン細胞を処置した。次いで、「一般的材料および方法」に記載のATP生物発光アッセイキットを用いて、ATPレベルを評価した。示したデータは、平均値±S.D.(n=5)を表す。記号「*」は、非処置グループと比較しての細胞のATPのレベルにおける有意な差(p<0.05)を意味する。
【図2】α-TOSおよび3-BPによる乳ガン細胞系におけるアポトーシスの誘導。正常酸素圧または低酸素のいずれかにおいて、示した濃度で24時間、α-TOSまたは3-BPに、MCF-7(A、B)およびMDA-MB-453細胞(C、D)を曝露し、アネキシンV-FITC法を用いてアポトーシスを評価した。示したデータは、平均値±S.D.(n=3)を表す。記号「*」は、非処置グループと比較しての処置された細胞のアポトーシスのレベルにおける有意な差(p<0.05)を意味する。
【図3】MDA-MB-453細胞でのアポトーシス誘導におけるα-TOSおよび3-BPの併用効果。正常酸素圧または低酸素のいずれかにおいて、示した濃度にて、α-TOSまたは3-BP単独で、あるいはα-TOS+3-BPの併用で、MDA-MB-453細胞を24時間処置した。時間、α-TOSまたは3-BPに、MCF-7(A、B)およびMDA-MB-453細胞(C、D)を曝露し、アネキシンV-FITC法を用いてアポトーシスを評価した。示したデータは、平均値±S.D.(n=3)を表す。記号「*」は、単独薬物処置と比較してのα-TOS+3-BPの併用によって処置された細胞のアポトーシスのレベルにおける有意な差(p<0.05)を意味する。
【図4】Akt1ノックダウンは、MDA-MB-453細胞をα-TOS、3-BPおよびTRAILに対して感受性にする。60 pM Akt1 siRNAで、50%コンフルエントにおけるMDA-MB-453細胞を24時間前処置した後、示した濃度にて、α-TOS、TRAILまたは3-BPで、さらに24時間処置した。アネキシンV-FITC法を用いてアポトーシスを評価した。2つの独立した実験を行った。結果を平均値±S.D.で示す。
【図5】NeuTL細胞のMTTを還元する能力における3NPA、BP、TTFAおよびα-TOSの効果。(A)示した濃度にて用いた3NPAまたは3-BPの存在下、4時間の共インキュベーション後、PBS中でのMTT還元を評価した。(B)MitoQで細胞を60分間プレインキュベートした後、示した濃度にて3-BPを添加し、2時間インキュベートした後、PBS中でMTTを還元する能力を評価した。20 mMコハク酸(pH 7.4)を含むRPMI中でのMTTを還元する能力について、MitoQで60分間前処置した細胞を、示した濃度の(C)TTFAまたは(D)α-TOSの存在下で2時間インキュベートした後に評価した。結果を、コントロール(非処置)±S.D.と比較してのMTTの平均還元%で表す。記号「*」は、有意な差(p<0.05)を意味する。
【図6】単離されたラット肝臓ミトコンドリア(A、C)またはパラコッカス・デニトリフィカンス(B、D)における、α-TOSによるSDH/複合体II活性の阻害。「一般的材料および方法」に概略したように細胞を分画し、ラット肝臓由来のミトコンドリアまたはパラコッカス・デニトリフィカンス由来の膜の調合液を、ミトコンドリアSDH/複合体II活性を促進し、インジケーター色素DCIP+PMSを含む反応体積中でインキュベートした。AおよびB中のサンプルは、スクシネートを含み、示したように100または300 μMのα-TOSのいずれかで処置した。コントロール反応物は、希釈液DMSO単独の等価の添加を受けた。波長600 nmにおける吸光度を測定し、酵素活性(μmol/分/ml)における変化として表した。棒グラフAおよびBにおける結果は、平均値±S.D.(n=3)を表す。記号「*」は、コントロールからの有意な差(p<0.05)を意味する。パネルCおよびDは、100または300 μMのα-TOSの不在または存在下で、示されたスクシネートの異なる濃度下で測定された反応速度(μmol/分)における変化を示す。
【図7】α-TOSによるアポトーシス誘導は、CII機能不全細胞において抑制される。ヒトCybL(B9rec)でのトランスフェクションによる複合体II再構築の後に、親(B1)、CI-機能不全(B10)、CII-機能不全(CybL-突然変異;B9)およびCybL-突然変異細胞を、示した濃度および時間にてα-TOSに曝露し、収穫し、ROS蓄積(A)、基質としてのスクシネートによるMTT還元に基づいて全細胞において評価されたSDH活性(B)およびアポトーシス(C)について評価した。CybLまたは非特異的(NS)siRNAでMCF7細胞を前処置し、示したようにα-TOSに曝露し、ROS蓄積(D)、SDH活性(E)およびアポトーシス誘導(F)について評価した。パネルGは、ヒトSDHCSDHCプライマーを用いるB1、B10、B9またはB9rec細胞ならびにsiRNA処置MCF細胞のRT-PCR解析の結果を示す。パネルHは、ヒトモノクローナル抗-SDHC IgGB1を用いる、B10、B9およびB9rec細胞のウエスタンブロッティングの結果を示す。ウエスタンブロッティングは、異なるSDHC siRNA二本鎖およびNS siRNA(I)で処置したMCF7細胞におけるSDHCのレベルを記録するためにも示し、アクチンバンド(J)と比較して評価した。結果は、平均値±S.D.(n=3)を表し、画像は、3つの独立した実験の代表である。記号「*」は、コントロールからの有意な差(p<0.05)を意味する。
【図8A】分子モデリング。複合体II結晶構造におけるQP結合部位に結合したα-TOS構造と比較してのUbQ分子の最良のドッキングポーズ。画像は、棒線画として表した結合した薬物を有する空間充填モードにおける複合体IIの横断面図/浮き彫りとして示す。同定のために様々な成分をラベル付けする。
【図8B】分子モデリング。棒線画様式での複合体II結晶構造から引き出されたこの部位のための相対的UbQ構造と比較してのQP結合部位へのα-TOSの最良のドッキングポーズ。結合に関わる複合体IIの重要なアミノ酸残基を同定し、それぞれについてそれらのサブユニット鎖および残基数にしたがってコード付する。α-TOS構をハイライトし、より暗い棒線画としてラベル付する。
【図8C】分子モデリング。重要なアミノ酸残基および複合体II QP結合ポケットとα-TOSの相互作用に関与するそれらの側基の空間分布の三次元視覚化を提供する(B)から誘導された立体画像。驚いたことに、α-TOSが、UbQ分子よりもポケットにより深く貫通し、水素結合を提供するSDHC(CybL)サブユニットのセリン42への結合を調和させるのを見ることができる。
【図8D】分子モデリング。α-TOSの最良結合コンホメーションと結合にとって重要なQP結合部位アミノ酸残基との間の主な相互作用を示すLigplot図。
【図8E】ポケットの正面を横切って伸びる複合体IIの一部によって形成された橋(半透明の形式)を示す、分子モデリング。QD結合部位へのα-TOSの最良ドッキングポーズ;ポケットの内部に包まれているUbQと比較してのα-TOSの結合。
【図8F】分子モデリング。α-TOSの最良結合コンホメーションと結合にとって重要なQP結合部位アミノ酸残基との間の主な相互作用を示すLigplot図。複合体IIのQdポケットにおけるα-TOSの結合相互作用に関与する重大な意味を持つ空間分布。注:TOSのスクシニル基と複合体II SDHDサブユニットのリシン残基D128、D135との間の水素結合(点線)。
【図8G】分子モデリング。UbQ5およびα-TOSの化学構造ならびに複合体IIのQPおよびQD部位に結合する構造のそれぞれについて計算された相互作用エネルギー。注:パネルAおよびEは、Astex Viewer(Hartshorn、2002)を用いて作製したが、パネルDおよびFは、Ligplot(Wallaceら、1995)を用いて作製した。
【図9】第4表および第5表(後記)からの異なるTOS類縁体の生物活性と、複合体IIのQp部位へのそれらの計算された結合親和性との相関関係を示すグラフ。IC50:インビトロにおいてガン細胞の50%死亡を誘導するために特定の薬物について必要なマイクロモルで表す濃度。−log Kd:特定の薬物の結合親和性の逆対数。
【図10】分子モデリング−複合体II活性部位および他の化学種。ミトコンドリアの複合体II Qp活性部位に強力に結合する分子モデリングによって同定された10個の最良化学種の詳細。
【図11】共役ペプチドLTVSPWY-α-TOSは、乳ガンを抑制する。超音波画像化(USI)によって検出される乳ガンの7ヶ月齢以上のFVB/N202 c-neu雌性マウスを、示した時点でビヒクルまたはトウモロコシ油/エタノール(100:5、v/v)に可溶化したLTVSPWY-α-TOSの5 μmol/マウス/投与の腹腔内注射で処置した。USIを用いて腫瘍を視覚化し、定量し、処置(A)の開始時におけるそれらの体積と比較して表す。パネルBは、実験の第15日におけるコントロールマウス(左)および処置動物(5 μmol LTVSPWY-α-TOS /投与/マウス)(右)由来の腫瘍の画像を表し、それぞれ402.1および34.8 mm3の腫瘍体積を示す。体積は、両方の腫瘍について、実験の開始時において〜40 mm3であった、示したデータは、平均値±SD(n=4-6)であるUSI画像は、各グループ由来の腫瘍の代表である。
【図12】α-TOSによるマウスモデルにおける乳ガンの阻害。(A)ヌードマウスにMCF-7細胞を接種し、腫瘍が確立された後、3日に1回、腹腔内(i.p.)注射により動物を10 μモル/マウスのDMSOに溶解したα-TOSまたはDMSO単独で処置した。キャリパーを用いて腫瘍サイズを測定し、治療の開始時のガンのサイズと相関させた。各グループにおいて4匹の動物を用いた。示した結果は、平均値±S.E.である。記号「*」は、有意な差(p<0.05)を意味する。(B)触知可能な腫瘍を有する雌性FVB/N erbB2マウス(「一般的材料および方法」を参照)に、3日に1回、腹腔内注射により、トウモロコシ油/4%エタノールに可溶化した10 μモルのα-TOS (n=11)もしくはトウモロコシ油/4%エタノール単独(コントロール、n=9)か、または腫瘍部位に毎日、0.2 μモルの3-BP(n=8)/PBSのいずれかを与えた。キャリパーを用いて、腫瘍サイズを記録した。2つの独立した実験を行った。結果は、平均値±S.E.で表す。記号「*」は、3-BPおよびα-TOS曲線の間の分散解析における有意な差(p<0.05)を意味する。
【発明を実施するための形態】
【0089】
本発をより良く理解することができるように、以下の実施例によって説明する。これらの実施例は、説明のみを目的とするものであり、いかなる方法においても本発明の範囲を限定するものと解釈されるべきではない。
【0090】
一般的材料および方法
細胞培養および処置
erbB2を低レベルで発現するヒト乳ガン細胞系MCF-7および高レベルで発現するヒト乳ガン細胞系MDA-MB-453、ならびにFVB/N202ラットc-neuトランスジェニックマウス株由来のerbB2を高レベルで発現するマウス乳ガン細胞系NeuTL(Guyら、1992)をこの実験に用いた。湿気のある5% CO2環境において、37℃にて、10% FCS、100 U/mlのペニシリンおよび100 μg/mlのストレプトマイシンを加えたDMEM中で、細胞を培養した。10% FCS、10 g/mlのグルコースおよび非必須アミノ酸を加えたDMEM中で、ミトコンドリア複合体I欠損(BreenおよびScheffler 1979;DeFrancescoら 1976;Schefflerら 2004)および複合体II欠損細胞(Albayrakら 2003;Oostveenら 1995)を成長させた。
【0091】
約70%のコンフルエントにて、正常酸素圧または低酸素条件下で50 μM以下のα-TOS(Sigma)、100 μM以下の3-BP(Aldrich)または30 ng/mlのヒト組換え腫瘍壊死因子関連アポトーシス誘導リガンド(TRAIL)(Weberら、2002)で細胞を処置した。低酸素のために、窒素パージ培地で覆った細胞を、大量の窒素を予め流した低酸素室に置いた。次いで、細胞を入れた低酸素室を、要求に応じてインキュベーター内に置いた。α-TOSをエタノールに溶解し、細胞培養培地における最終エタノール濃度が≦0.1%(v/v)になるようにして用いた。使用前に3-BPの貯蔵溶液をPBS中で調製した。
【0092】
ATP測定
1.2x105個/mlのMDA-MB-453およびNeuTL細胞を、96ウエル組織培養マイクロタイタープレートに播き、使用前に一夜回復させた。上述のように、正常酸素圧または低酸素下で、α-TOSまたは3-BPとともに6または24時間、細胞をインキュベートした。次いで、使用説明書にしたがって生物発光アッセイキットHS11(Roche Applied Science)を用いて、ATPレベルを測定した。簡単に述べると、室温にて細胞溶解試薬とともに5分間インキュベートした後に、コントロールまたは処置細胞を黒いマイクロタイタープレートに移した。ルシフェラーゼ試薬を加え、Flurostar Optimaプレートリーダー(BMG Labtech)を用いてシグナルを迅速に検出した。各条件について三組みウエルを読み、既知の濃度のATPで作成した標準的較正曲線を用いてデータを評価した。
【0093】
ROS蓄積の評価
フローサイトメトリーにより、プローブとしてジヒドロエチジニウム(DHE)(Molecular Probes)(Weberら、2003)を用いて、あるいは、電子常磁性共鳴(EPR)分光分析(Weberら、2003)を用いる5,5-ジメチル-1-ピロリン N-オキシド(DMPO;Sigma)でのトラッピングによって、細胞ROSを検出した。いくつかの実験において、2 μMのMitoQ(Jamesら、2005 )で細胞を1時間前処置するか、またはSOD(PEG-SOD、750 units/ml;Sigma)で共インキュベートした。
【0094】
アポトーシス評価
アポトーシスの早期に外面化されたホスファチジルセリンを検出するV-FITC法を用いて、アポトーシスを定量した(Boersmaら、1996)。簡単に述べると、細胞を一夜療養し、α-TOSもしくは3-BP単独または併用で処置した後、24ウエルプレートに105個/ウエルにて細胞を播く。浮いている細胞および付着した細胞を採集し、PBSで洗浄し、0.2 mlの結合緩衝液(10 mM Hepes、140 mM NaCl、5 mM CaCl2、pH 7.4)に再懸濁し、2 μlのアネキシンV-FITC(PharMingen)とともに4℃にて30分間インキュベートし、50 μg/mlのヨウ化プロピジウム(PI;Sigma)200 μlを補足し、アネキシンV-FITC 結合についてチャネル1を用い、PI染色についてチャネル2を用いるフローサイトメトリー(FACScalibur、Becton Dickinson)によって分析した。アネキシンV結合およびPI染色の増加した細胞のパーセンテージとして、細胞死を定量した。
【0095】
RNA干渉(RNAi)
Akt1に特異的な3’-dTdTオーバーハングをもつ短い干渉RNA(siRNA)オリゴヌクレオチドをQiagenによって合成した。配列は、以下のとおりである:標的配列:CCT GCC CTT CTA CAA CCA GGA;siRNA二本鎖配列:センス - r(UGC CCU UCU ACA ACC AGG A)dTdT、アンチセンス - r(UCC UGG UUG UAG AAG GGC A)dTdT。使用説明書にしたがって、OligofectAmine(Invitrogen)およびOptiMEMを用いて、培養した細胞のsiRNAによるトランスフェクションを行った。簡単に述べると、細胞を50%コンフルエントに到らせ、OligofectAmineとともに前インキュベートし、OptiMEMで覆った60 pMのsiRNAを補足する。24時間後に細胞をPBSで洗浄し、完全DMEMで覆い、さらに24時間後、要求に応じて処置した。
【0096】
MTTアッセイ
2.5 mg/mlの3-(4,5-ジメチル-2,5-ジフェニル 2H-テトラゾリウムブロミド(MTT)(Sigma)をPBS単独または20 mMコハク酸、pH 7.4を含むフェノールレッド-フリーRPMI培地に溶解することによって、MTT溶液を使用前に調製した。エタノール中の3-ニトロプロピオン酸(3NPA)、DMSO中のテノイルトリフルオロアセトン(TTFA)、PBS中の3-BP、DMSO中のα-TOS、およびエタノール中のMitoQ(Allevaら、2003、Jamesら、2005)の貯蔵溶液を調製した。アッセイした希釈液ごとに4-8組みウエルを用い、96ウエルマイクロタイタープレートにて指数増殖期に培養したNeuTL細胞において薬物を試験した。MitoQのMTT還元を回復する能力を評価するために、2または5 μM MitoQとともに細胞を60分間前インキュベートした。培養物中のエタノールまたはDMSOの最終濃度は、≦0.1%であった。処置およびコントロール細胞は、37℃および5% CO2にて、2〜4時間、MTTを還元してブルーホルマザン結晶にすることができた。インキュベーション後、30 μl以外の培養上清を除去した後、150 μlのDMSOを加えることによってホルマザン結晶を溶解し、Fluorostar Optimaプレートリーダーを用い、570 nmにて吸光度の値を測定した。
【0097】
単離されたラット肝臓からのサブミトコンドリア粒子(SMPs)の調製
RiceおよびLindsay、1997の変更法にしたがって、分画遠心法によってラット肝臓ミトコンドリアを調製した。2 mlのミトコンドリアペレットを15 mlの20 mM KH2PO4 pH 7.4に再懸濁し、80 Wattで3x15秒間超音波処置した。次いで、超音波処置溶液を6,500xgにて7分間遠心分離した。上清画分を氷上に保持し、残留ペレットを再懸濁し、再度超音波処置した。超音波処置溶液を前述のように遠心分離し、第2の遠心分離ステップから除去した上清を第1の遠心分離からのものと合わせた。合わせた上清を108,000xgにて1時間再遠心分離した。SMPsのペレットを5 mlの20 mM KH2PO4 pH 7.4に再懸濁し、使用するまで−20℃にて凍結保存した。
【0098】
パラコッカス・デニトリフィカンスからの膜小胞の調製
パラコッカス・デニトリフィカンスCCM 982(NCIB 8944)を、炭素源として50 mMスクシネートおよび最終受容体として10 mMナイトレートを含む培地中で30℃にて嫌気的に増殖させた。Burnellら(1975)にしたがって、リゾチームで処置することによって採集した細胞から膜を調製した。約20 mg/mlの最終濃度になるまで0.1 M リン酸Na、pH 7.3中に希釈した後、使用するまで1 mlのアリコートにて膜を−20℃にて保存した。ビシンコニン酸法およびiEMSプレートリーダー(Labsystem、Finland)を用いて、タンパク質濃度を決定した。
【0099】
ミトコンドリア複合体IIおよび複合体I活性の測定
1 mlの反応体積を有する1 cmキュベットにおける600 nmでの吸光度を測定することによって、ミトコンドリア調製物による複合体II基質2,6-ジクロロフェノールインドフェノール(DCIP)の還元のための経時変化を追跡した(ε600=21x103 M-1cm-1)。反応成分は、NADH、0.5 mM;スクシネート、5 mM;KCN、10 mM;DCIP、50 μM;PMS、50 μMを含んだ。各アッセイポイントにおいて、0.5 mgのサンプルタンパク質を用い、示したように100または300 μMにてα-TOSを加えた。分光光度計(UVIKON XL、Secomam)を用いて、DCIPの吸光度における変化を測定し、繰り返しサンプルをアッセイした(n=3)。複合体I(NADHデヒドロゲナーゼ活性)を測定する場合、PMSを除外した。α-TOSを含まないコントロール反応について、採集濃度<0.1%(v/v)であるように希釈液DMSOを加えた。
【0100】
分子モデリング−複合体IIおよびビタミンE類縁体
Brookhaven Protein Databank(code 1ZOY)(Sunら、2005)から、ブタ心臓由来のミトコンドリア呼吸膜タンパク質複合体IIの結晶構造を入手した。複合体は、4個のタンパク質を含む。この複合体における3個のサブユニットである鉄イオウタンパク質(鎖B)、大(鎖C)および小(鎖D)膜貫通タンパク質は、UbQへの結合に関与する。NCBIウェブサイトからのBLAST調査は、ブタおよびヒト複合体IIの間の配列同一性が、鉄イオウタンパク質について97%、大膜貫通タンパク質について90%および小膜貫通タンパク質について94%と非常に高いことを示した。
【0101】
最初に除去されるヘテロ原子で、AutoDock Tools(Sanner、1999)を用いて、ドッキングのために、タンパク質構造を作成した。該構造に極性水素を付加し、タンパク質原子のためにKollman United Atomチャージを用いた。InsightII (Accelrys、2001)を用いて、結合したUbQ(1ZOY)の結晶構造配位からUbQ5を構築した。再びInsightIIを用い、α-TOSの環系についてのサブ構造調査によって、Cambridge Structural Database(Allen、2002)から検索された結晶構造MOPHLB01からα-TOSを構築した。次いで、非極性水素を融合し、Gasteigerチャージを割り当て、回転可能な結合を決定することを含む、AutoDock Toolsによって、ドッキングのために、両方のリガンドを作成した。
【0102】
Autodock 3.0.5.(Morrisら、1998)において実施されたように、Lamarckian Genetic Algorithmを用いて、ドッキングを行った;2つのドッキンググリッドを作成した。両方は、0.375オングストロームのグリッドペーシングである126x126x126ポイントであり、第1のものは、QP部位におけるTyr173(鎖B)を中心とし、第2のものは、QD部位におけるTrp134(鎖D)を中心とした。以下のもの以外は、対象のリガンドに存在する回転可能な結合の相対的に高い数(UbQ5=16、α-TOS =17)によって増加する、デフォルトパラメーターを用いた:- ga_run=250、ga_pop_size=250、ga_num_evals=10,000,000。また、計算の分析相中に、より管理可能なクラスターを生み出すために、パラメーターrmstolを、2.5に増加した。各ドッキング計算は、2 GHz G5 PowerPC Macintoshを用いて、49時間を僅かに越えた。AutoDockで提供されたスクリプトを用いて、結果の分析を行い、Astex Viewer(Hartshorn、2002)を用いて、ドッキングした構造を視覚化した。
第4表に示す他のビタミンE類縁体のAutodockingも、上述のとおり行った。
【0103】
分子モデリング−複合体II Qp部位および他の化学物質
ZINC Database project(http://blaster.docking.org/zinc/)から65,000個の商業上入手可能な化合物のバーチャルライブラリーをダウンロードした。これらの化合物を、計算された物理的特性に基づくさらに大きなデータベースから検索条件を追加したので、それらは、大変似ているドラッグリード様候補である(Teague、SJら、1999)。次いで、最終的データベースを作成するために、それらを多様性解析に付した。バーチャルスクリーニング実験中のDOCK 6.1(http://dock.compbio.ucsf.edu/DOCK_6/index.htm)を用いて、これらの化合物のそれぞれをブタ複合体II(1ZOY)の結晶構造にドッキングさせ、相互作用エネルギーに基づいてトップ1000個の化合物を同定した。次いで、再度DOCK 6.1を用いて、これらの最終化合物をより厳密なドッキング実験に付し、視覚化プログラムChimera (http://www.rbvi.ucsf.edu/chimera/)を用いて、最もブタ複合体IIと相互作用する可能性のある化合物として10個の化合物を同定した。
【0104】
マウス腫瘍実験
MCF-7細胞(2x106細胞/マウス)をヌードマウスに皮下接種した。腫瘍が発達した後(直径3-7 mm)、DMSO中の10 μモルのα-TOSを3日毎に、マウスに腹腔内注射した。コントロールマウスには、等量(100 μl)のDMSOのみを注射した。デジタルキャリパーで測定することによって、腫瘍サイズを評価した。H-2q FVB/Nバックグラウンド上のMMTVプロモーターによって誘発されたラットHER-2/neu プロトオンコジーンを有するトランスジェニックFVB/N202ラットc-neuマウスのコロニー(Guyら、1992)を、Griffith University Animal Facilityにて確立し、厳密な同系交配条件下に維持した。PCRによって、トランスジーンの存在をルーチン的に確認した。約70%の雌性マウスが、平均潜伏時間10ヶ月で同時の乳ガンを発達させた。平均直径10±5 mmの徐々に増殖する腫瘍を有する雌性トランスジェニックFVB/N202ラットc-neuマウスをコントロールまたは処置グループにランダムに割り当てた。デジタルキャリパーで腫瘍サイズを測定した。α-TOS療法のために、マウスは、3日毎に腹腔内投与で投与されるトウモロコシ油/4%エタノール単独での処置(コントロール)、またはトウモロコシ油/4%エタノール中の10 μモルのα-TOSでの処置を受けた。3-BP療法のために、1日当たり、0.2 μモルの薬物が腫瘍内に注射された。使用した3-BPの薬物用量は、以前の報告(Koら、2004)に基づいたものであり、この薬物をラット(平均体重〜300 g)に適用し、マウスに対して、低体重(〜30 g)を考慮した。
【0105】
統計的分析
グループ比較間の対応のないスチューデントt検定を用いて、アポトーシスの平均パーセンテージ、ATPおよびMTT還元±SDを比較した。共変動として日数を用いる共分散分析(ANCOVA)を用いて、平均相対腫瘍サイズ±SEMにおける差異を審査した。SPSS(登録商標) 10.0分析ソフトウェア(SPSS、Chicago、USA)を用いて統計的分析を行った。差異は、p<0.05である場合に統計的に有意であるとみなした。
【実施例1】
【0106】
3-BP、α-TOSまたはTRAILは、乳ガン細胞のATPレベルを低下させる。
3-BPは、従来、使用した薬物の特定の用量に応じて、処置の数時間後に起こる、細胞ATPレベルを枯渇させる活性の高い作用剤として報告されていた(Koら、2004、Xuら、2005)。さらに、細胞ATPレベルの減少が、3-BP薬物誘発性ガン細胞死の出現の前に起こることが示された(Xuら、2005)。低下したATPレベルは、次いで、アポトーシス促進性Bax分子がミトコンドリアに移行するのを可能にしてアポトーシスシグナル伝達経路を活性化させる、pBadホスホリル化の損失と相関した(Xuら、2005)。
したがって、本発明者らは、ヒト乳ガンMDA-MB-453(erbB2-高)およびMCF-7(erbB2-低)細胞系における細胞ATPの産生における3つの薬物α-TOS、3-BPおよびTRAILの効果を比較するために、それらの比較に着手した。さらに、FVB/Nラットc-neuトランスジェニックマウス由来のマウス乳ガン細胞系NeuTLにおいて、薬物を試験した。
【0107】
結果
(図1A)は、6時間以内に、MDA-MB-453細胞が、ATPレベルを有意に低下したことを示す。24時間までに、すべての3つの細胞系は、ATPレベルの低下を示したP(図1B-D)。3-BPおよびTRAILは、試験した異なる濃度の範囲にわたって、α-TOSよりも強力なインヒビターであることが分かった。α-TOSのみが、50 μMという高濃度で付加した場合に、細胞ATPレベルを有意に低下させた(図1)。細胞は、低酸素または正常酸素圧下のいずれかでインキュベートした場合に、細胞ATPを枯渇させることにおいて、3つの薬物の作用に対して、同じように反応性であった。
【実施例2】
【0108】
高度にerbB2-陽性である乳ガン細胞は、3-BPによって誘導されたアポトーシスに耐性があるが、α-TOSによって誘導されたアポトーシスには耐性がない。
高レベルのerbB2発現を示している乳ガン細胞は、いくつかのガン治療によって誘導されたアポトーシスに対する関連するより大きい耐性を有する(Burstein、2005)。これらの発見と一致して、本発明者らは、近年、高度にerbB2-陽性であるMDA-MB-453細胞系が、弱くerbB2-陽性であるMCF-7細胞系におけるその効果と比較した場合に、死受容体アクチベーターTRAILによって誘導されるアポトーシスに対してより耐性があることを観察している。しかし、MDA-MB-453細胞は、α-TOS誘導アポトーシスに反応性であることが分かった(Wangら、2005)。この実験において、本発明者らは、2つのミトコンドリア影響性薬物、α-TOSおよび3-BPを、高および低erbB2-発現乳ガン細胞系における、そして異なる酸素条件下でのアポトーシスインデューサーとしてのそれらの相対的効力について比較した。
【0109】
結果(図2)は、MDA-MB-453およびMCF-7細胞系の両方が、正常酸素圧および低酸素下の両方で、α-TOSによって誘導されるアポトーシスに対して感受性があり、MCF-7細胞がわずかにより影響を受けやすいことを示した。MDA-MB-453細胞は、100 μMという高濃度でさえも、3-BPによる殺傷に著しく耐性があることが見出された。α-TOSに関しては、反応性MCF-7細胞が、正常酸素圧よりも低酸素下において、3-BPによる殺傷に対して、より感受性があることが見出された(図2)。
【実施例3】
【0110】
3-BPおよびα-TOSの併用は、高erbB2-陽性細胞において、アポトーシスを増強する。
この実験において、本発明者らは、α-TOSおよび3-BPが、オーバーラップするかどうか、またはそれらが3-BP-耐性MDA-MB-453乳ガン細胞の殺傷を増強することができるかどうかを決定した。この実験は、これらの薬物が、同様のメカニズムで働くか、あるいは他方とは独立しているかどうかを理解する手がかりとなる。3-BPが、スクシネートであるデヒドロゲナーゼの活性部位に結合することによって、複合体IIのインヒビターとして作用することが知られている(Sanbornら 1971)。したがって、それらの両方が違いにオーバーラップする可能性があった。
【0111】
図3は、MDA-MB-453細胞においてアポトーシスを誘導するためのα-TOSおよび3-BPの併用効果の結果を示す。事実、MDA-MB-453細胞は、驚いたことに、それぞれの薬物単独での使用と比較して、α-TOSおよび3-BPの併用による殺傷に対して、感受性を高め、薬物併用を用いた場合のガン細胞アポトーシスの程度は、有意(p<0.05)に増強された。さらに、併用で増強された効果は、細胞培養条件が正常酸素圧か低酸素かどうかにかかわらず起こった。したがって、これらの結果から、2つの異なる薬物が、互いに独立して作用し、α-TOS処置が、3-BP耐性MDA-MB-453細胞を、3-BPの細胞毒性効果に対して感受性があるようにするという並はずれた発見を提供することが明らかとなった。
【実施例4】
【0112】
Akt siRNAは、3-BPへのMDA-MB-453細胞の反応性を回復し、α-TOSおよびTRAILへの感受性を増加する。
過去の研究は、高レベルのerbB2発現が、Bad(Zhaoら、2004)およびカスパーゼ-9(Cardoneら、1998)などの多くの異なる細胞標的をホスホリル化するAktの活性化をもたらすことを明らかにしている。このように、ガン細胞は生存の増加を示し、薬物治療によって誘導されるアポトーシスに対して耐性ができる(Westら、2002、Osakiら、2004、Fresnoら、2004、Grandageら、2005)。実際、アンチセンスRNAアプローチを用いるAktの細胞発現を阻害することが、薬物療法へのガン細胞の感受性を回復させるための一般的方法として提案されている(Kimら、2004)。さらに、近年、Aktに対してsiRNAが、ガン細胞においてAkt発現をダウンレギュレートすることのガン細胞の生存に及ぼす効果を調査するいくつかの研究において用いられている(Takeuchiら、2005、Yoeli-Lernerら、2005)。したがって、本発明者らは、Akt1発現を低下させて、このことが、erbB2-陽性 3-BP-およびTRAIL-耐性MDA-MB-453細胞を2つの作用剤に対して反応性にするかどうか、ならびに、このことが、α-TOS媒介性アポトーシスに対する該細胞の感受性を高めるかどうかを決定する手段として、RNAi技術を活用している。
【0113】
OligofectAmineおよびOptiMEMを用いて、siRNAsによるMDA-MB-453細胞のトランスフェクションを行った。簡単に述べると、細胞を〜50%コンフルエントに到達させ、OligofectAmineとともにプレインキュベートした60 pmol/l siRNAを補足し、OptiMEMで覆った。24時間後にPBSで細胞を洗浄し、完全DMEMで覆い、さらに24時間、48時間培養し、ウエスタンブロッティングを用いてトランスジーン発現を確認した後、次の実験に用いた。
【0114】
結果(図4)は、Akt1 siRNAによるMDA-MB-453細胞の前処置が、試験する3つの薬物それぞれに対する感受性を増加させることにおいて有効であり、3-BPに対する細胞毒性反応において特に目覚ましい増加を生み出すことを示した。この証拠は、高レベルAkt活性をもつ乳ガンが、3-BP治療への耐性をもつ傾向があり、このことが、分子操作によって効率的に調節されうることを示唆する。
【0115】
3-BPへの細胞反応性におけるAkt siRNAの効果およびα-TOS/3-BP併用治療によるアポトーシスの増加は、α-TOSが、Akt活性にも影響を及ぼし、そのことによって、Akt siRNAと同様の、3-BPへのガン細胞の感受性の増加を提供するという可能性を引き起こす。
【実施例5】
【0116】
α-TOSは、複合体II上のUbQ-結合ポケットを標的とする。
本発明者らによる過去の研究(Neuzilら、2004に概説)は、ミトコンドリアが、ガン細胞におけるα-TOSの作用のメカニズムの中心をなし、ミトコンドリアアポトーシスシグナル伝達経路の活性化をもたらすことを明らかにした。近年そのBcl-2/Bcl-xL阻害活性の発見まで、α-TOSは「オーファンドラッグ」状態を保持している。しかし、ガン細胞へのα-TOSの付加の早くも1時間後にROS生成をもたらすこの薬物のための特異的ミトコンドリア標的およびアポトーシス促進性経路の次の開始(Weberら、2002、Stapelbergら、2005)は、明確ではない(Zhangら、2001)。
【0117】
この目的を達成するために、本発明者らは、α-TOSが、複合体IIにおいてUbQ結合を妨げるかどうかを詳細に調べた。全細胞における複合体II活性を特に測定するために、テトラゾリウム塩MTTの還元に基づく短時間(1-2時間)アッセイを用いた(Maeharaら、1988)。したがって、ミトコンドリア呼吸が複合体IIを介して全細胞において最適に進行特するのを確実にする特定の条件を用いた。このために、マウスNeuTL乳ガン細胞の酸化呼吸を高レベルのスクシネート(20 mM)で持続させ、それによって、複合体IIのSDH活性を促進した(Maeharaら、1988)。次いで、高スクシネート条件下で複合体II/SDH依存性活性の特異的インジケーターになる、MTTを酵素的に還元する能力について、1-2時間以内に細胞をアッセイした(Maeharaら、1988、Berridge and Tan、1993)。
【0118】
過去の研究は、3-BP(Sanbornら、1971)および3NPA(Scalletら、2003)の両方が、SDH活性の強力なインヒビターとして作用することを明らかにしており、3NPAは、細胞への付加の4時間以内と比較的迅速にMTT還元を阻害する能力がある(Scalletら、2003)、SDHの特異的自殺型インヒビターである(Huangら、2006)ことを近年明らかにされている。2つのSDHインヒビター、3-BPおよび3NPAがNeuTL細胞において直接MTT還元を害するかどうかを決定するために、3-BPおよび3NPAを用い、別な方法でこれらのインヒビターと競合する追加のスクシネートの不在下にて初期実験を行った。結果(図5A)は、3-BPおよび3NPAが両方とも、複合体II-媒介性MTT還元の強力なインヒビターであり、したがって、アッセイが、SDH活性の効果を定量するという我々の目的に適していることを確認した。
【0119】
複合体IIの電子伝達流に関与するUbQ結合部位を標的とする特異的インヒビターとして、テノイルトリフルオロアセトン(TTFA)が確立されている(Berridge and Tan、1993、Sunら、2005)。これらの部位は、電子を複合体IIIに送るために、UbQに電子を伝達する状態にある、複合体IIの膜貫通領域におけるSDH反応中心の下に存在する。興味深いことに、TTFAは、すべての細胞に対して毒性である(Zhangら、2001b)。SDH酵素インヒビターである3-BPおよび3NPAと同様に、TTFAもまた、2,3時間以内に、乳ガン細胞によるMTTの酵素的還元を有意に阻害した(図5C)。事実、3-BP、α-TOSまたはTTFAのいずれかによる処置は、処置の早くも2,3時間後に、これらの細胞によるMTT還元のレベルにおいて生じる有意な減少をもって用量依存的変化を示した(図5B-D)。しかし、UbQのミトコンドリア的に標的化された形態であるMitoQとともに細胞を1時間プレインキュベートすること(Kelsoら、2001、Jamesら、2005)は、MitoQが、TTFA(図5C)またはα-TOS(図5D)のいずれかによって引き起こされたMTT還元における阻害に打ち勝つことができるが、3-BP(図5B)によって引き起こされた阻害には打ち勝つことができないことを示した。ここでの理論的解釈は、もし、これらの薬物のいずれかが、複合体IIのUbQ部位への結合によって活性化しているならば、増加したレベルのUbQ活性類縁体であるMitoQで細胞を充填することは、複合体IIによるMTT還元の薬物媒介性阻害の程度を低下させるであろうということであった。MTT還元における3-BPの阻害効果は、SDH頭部基を阻害することによって作用し、複合体IIの下流膜貫通UbQ部位を介さない3-BPと一致して、異なる濃度のMitoQ(図5B)の存在または不在下で細胞を処置した場合、有意には異ならなかった。しかし、TTFA(図5C)およびα-TOS(図5D)の付加によって引き起こされる阻害は、MitoQと細胞とのプレインキュベーションによって有意に打ち勝たれた。特に、α-TOSは、MitoQの存在下でのスクシネート駆動MTT還元を阻害することにおいて、有効性は低かったが、これも、近年、ミトコンドリア酸化還元鎖の複合体IIへの結合に対して特異的であることが明らかにされている(Jamesら、2005)。
【実施例6】
【0120】
α-TOSが、ミトコンドリア呼吸複合体II活性を阻害することの検証。
ミトコンドリア 呼吸複合体II活性を阻害することにおけるα-TOSの役割のためのさらなる裏付けは、ラット肝臓から細胞内フラクションを含んでいるミトコンドリ調合液、ならびにパラコッカス・デニトリフィカンスから膜フラクションを単離することによって探求された。これらの研究にとって、SDH/複合体II活性のアッセイ法は、フェナジンメト硫酸塩(PMS)およびインジケーター色素として末端電子受容体である2,6-ジクロロインドフェノール(DCIP)を用い、それが高スクシネート条件下で駆動されるSDHによって還元されるための600 nmにおける吸光度の減少を用いるKing(1967)の研究に基づいたものであった。結果(図6A,B)は、ラット肝臓ミトコンドリアおよびパラコッカス・デニトリフィカンス調合液の両方において、コントロールサンプルと比較して、DCIPの還元の速度が、α-TOS処置後に、急速および有意に低下したことを示した。したがって、結果は、複合体IIのUbQ部位で直接作用し、PMSおよびDCIPへの電子流を妨げるα-TOSを強く裏付ける。SDH活性についてのα-TOSの選択性は、NADHデヒドロゲナーゼ(複合体I)活性においてビタミンE類縁体の効果がないことを示す結果によって裏付けられる(データ示さず)。
【実施例7】
【0121】
複合体Iおよび複合体II欠損細胞を用いる、α-TOSがミトコンドリア呼吸複合体II活性を阻害することの検証。
複合体Iおよび複合体II欠損細胞を上記実施例6のインビトロの結果を確認するためにも用いた。CI機能不全細胞(B10細胞)(Seo、B.B.ら、1998)、CII機能不全細胞(突然変異チトクロームb大を有するB9細胞、CybL)および親チャイニーズハムスター肺線維芽細胞(B1細胞)(Oostveen、F.G.ら、1995)を、10% FCS、抗生物質、10 mg/mlのグルコースおおよび非必須アミノ酸を補足したDMEM中で増殖させた。α-TOSは、親チャイニーズハムスター肺線維芽細胞(B1細胞)、CI機能不全細胞(B10細胞)およびCII機能不全(SDHCサブユニットをコードする遺伝子において突然変異有するB9細胞、CybL)においてアポトーシスを開始した。B9細胞は、親(B1)またはB10細胞と比較して、α-TOSへの反応性が低く、ROS蓄積のレベルが低く(図7A)、SDH活性が減少した(図7B)。これらの発見に一致して、B9細胞は、VE類縁体によって誘導されるアポトーシスに対して相対的に耐性があった(図7C)。CybL突然変異(B9)細胞におけるCIIの差異構築は、SDH活性を正常化し(図7B)、α-TOS誘導殺傷への細胞感受性を回復させた(図7C)。
【0122】
CII欠損(B9)および再構築B9細胞により得られたデータを、CybLに対する4つの異なる短い干渉RNAs(siRNA)でのMCF7細胞の処置により、独立して検証した。二本鎖1、3および4は、実質的にCII活性を抑制した(図7D)。CybLにおける突然変異は、ROS蓄積およびSDH活性を阻害し、α-TOSによって媒介されるアポトーシスのレベルを低下させた(図7D-F)。
【0123】
ヒトモノクローナルSDHC抗体、B1、B9およびB10、ならびにCII再構築チャイニーズハムスター肺線維芽細胞(B9rec)の使用により、SDHCの存在を探った。ウエスタンブロッティング分析(図7H、I)は、B1、B9およびB10細胞におけるヒトSDHCの不在、ならびにSDHC siRNAsで処置したMCF-7細胞においてそのレベルが低いことを示した。再構築CIIを有するB9細胞は、SDHCの再出現を示した。デンシトメトリーの評価によって示されたように、MCF7細胞のsiRNA処置は、SDHCタンパク質のレベルを50-80%まで低下させた(図7J)。これらのデータは、CII再構築細胞におけるヒトSDHC mRNAの存在、ならびにSDHC siRNAで処置されたMCF7細胞における転写物の低レベルを示すRT-PCRの結果と一致した(図7G)。SDHC処置MCF7細胞におけるSDHBサブユニットタンパク質のレベルにおけるいずれかの変化が、観察されなかったので、siRNAアプローチはSDHCに対して特異的であった(データ示さず)。
【実施例8】
【0124】
分子モデリングは、UbQ自体に対してよりも、複合体II活性部位へのα-TOSのより強力な結合を示した。
α-TOSがUbQ結合部位を介してミトコンドリア複合体IIと相互作用することを示している上記の結果を理論的に説明するのを助けるために、本発明者らは、AutoDock(Morrisら、1998)を用いて、このシステムの分子モデリング研究を企てた。近年、ブタ心臓ミトコンドリア複合体IIの結晶構造が報告されており(Sunら、2005)、ヒトミトコンドリア複合体IIとの高い配列同一性ゆえに、本発明者らは、それらのAutoDock研究のための基準としてこの構造(1ZOY)を用いることを決定した。構造は、近位のUbQ結合部位(QP)を明確に示し、結合したインヒビターTTFAをもつ関連構造(1ZP0)は、提案された遠位のUbQ結合部位(QD)の位置を示唆する。
【0125】
このシステムを研究するためにAutoDockを用いることの実現可能性を試験するために、最初に彼らは、QPおよび提案されたQD結合部位の両方にUbQ5をドッキングさせるためにそれを用いた。UbQ5は、α-TOSと同様のサイズであり(図8)、同様の数の回転可能な結合を含む(それぞれ16および17)ので、選択された。UbQの部分について観察される位置よりわずかに深い位置へQPにドッキングしたUbQ5は、Sunら(2005)によって公開された結晶構造で解決した。興味深いことに、このより深いドッキング位置は、大腸菌由来の複合体IIの結晶構造にUbQ2をドッキングさせるためにドッキングプログラムGOLDが用いられる近年の研究において観察された(Horsefieldら、2006)。結合部位の正面に位置しているUbQ環および該部位の内側に位置する疎水性尾部をもつUbQ5もまた、提案されたQD部位にドッキングすることが見出された。AutoDock(図8)によって計算された相互作用エネルギーは、UbQ5が、提案されたQD部位よりも強くQP部位と相互作用することを示唆する。このことは、提案されたQD部位においてUbQ結合が観察されないという事実とよく相関し、QP部位が、UbQ5との最も強い相互作用を提供することを示唆した。
【0126】
図8A-Dに見られるように、AutoDockは、QPおよびQD部位の両方においてα-TOS結合を示した。QP部位において、α-TOSの環系は、UbQ環と同じ結合ポケットに位置するが、反対側に傾斜する。驚いたことに、コハク酸エステル部位が、結合ポケット内により深く伸び、補欠分子ヘム基の位置に向かって下方に伸びることが見出された。カルボキシル基は、このポケットに適切にフィットし、Ser42(C)との二座水素結合に関与する(図8B,C,D)。Ser42(C)もまた、α-TOSのエステル酸素と相互作用する。α-TOS分子の残部の疎水性性質に適しているので、該タンパク質とのすべての他の相互作用は、疎水性である。炭化水素側鎖は、周囲で輪になり、bQ5のイソプレノイド側鎖がドッキングするのが示される同じチャネルに沿ってQP部位からU伸びる。QD部位において、α-TOSの環系は、結晶構造において認識できるリン脂質の頭部基に観察される様式と同じ様式で該部位の底部まで伸びているコハク酸エステル部分をもち、結合部位の底部に向かって位置する。コハク酸部分は、Lys135(D)and Lys128(D)に水素結合するのが見られる(図8E)。
炭化水素側鎖は、結合部位の内側の周りで輪になる。
【0127】
α-TOSのドッキングしたコンホメーション(図8F)についての相互作用の計算されたエネルギーは、それが、QPまたはQD部位のいずれかにおいて結合することができるが、QD部位において結合が優先されることを示唆した。QP部位におけるα-TOSの結合エネルギーは、UbQ5よりもわずかに小さいが、α-TOSは、UbQ5と確実に競合することができる。QD部位において、α-TOSは、UbQ5よりも良好な結合エネルギーを示し、この結合部位から置き換えることができるはずである。
【0128】
まとめると、これらの発見は、α-TOSが、ユビキノン(UbQ)結合と相互作用し、呼吸鎖における複合体IIの膜ドメインによってUbQの還元を遮断することを示す。結果として、電子は、もはやFAD上のコハク酸デヒドロゲナーゼ親水性頭部の下に伝達されず、[4Fe-4S]を介してUbQへ中継されるが、代わりに、ミトコンドリア中に漏出し、最終的にガン細胞のアポトーシスを誘導する増加したレベルのROSを生成する。
【0129】
上記と同じプロトコルを用いて、下記の第表および第5表に示すのビタミンE類縁体のオートドッキングを行い、予測した相互作用エネルギーを各表の最後の欄に提示した。
【0130】
第4表
近位コエンザイムQ結合部位(Qp)におけるビタミンE類縁体の相互作用エネルギー。化合物は、シグナル伝達ドメインによって分類される。
【表6】
【表7】
a最初の値は、cis-、第2の値は、trans-についてである。
b2つの値は、キラル炭素における2つのエナンチオマーを意味する。
【0131】
第5表
修飾された疎水性ドメインとの、遠位コエンザイムQ結合部位(Qp)におけるビタミンE類縁体の相互作用エネルギー。
【表8】
【0132】
上記ビタミンE類縁体は、AutoDock 3.0.5および前に述べたものと同じプロトコルを用い、ブタ複合体IIの結晶構造(近位ユビキノン結合部位)にドッキングする。相互作用の予測されたエネルギーおよび生成したコンホメーションの数を比較することによって、結合において重要であるα-TOSテンプレートへの変化に関する以下の観察が得られる:
1.末端電荷。α-TOSテンプレートは、その末端カルボン酸による負の電荷を有する。この電荷の消滅は、相互作用エネルギーを減少したが、正の電荷とそれを置き換えることが、同様の相互作用エネルギーを維持した。この荷電した基がドッキングする領域は、極性アミノ酸残基(ヒスチジンおよびセリン)によって囲まれているが、荷電した残基はない。最も近い電荷は、近くのヘム基のカルボキシ基から来る。このことは、正または負の荷電した基のいずれかの蓄積を可能にする。
2.エステル結合。スクシネート部分を元のビタミンE骨格に連結するエステル結合は、相互作用エネルギーにおいてほとんど変化がないアミド結合と置き換えることができる。
3.スクシネート部分は、(二重結合における立体配置がtransに維持される限り)フマレート部分と置き換えることができ、良好な相互作用エネルギーをなお維持することができる。
4.α-TOSの芳香環上のメチル基は、近位ユビキノン結合部位内で物質を位置決定することにとって重要である。それらを除去すると、相互作用のエネルギーが低下する。
【0133】
図9において、本発明者らは、ユビキノン部位へのビタミンE類縁体の結合親和性と、ガン細胞のアポトーシスを引き起こすことにおけるそれらの生物活性との間の相関関係を解析した。結果は、該部位への結合親和性と、ガン細胞を殺傷する能力との間の直接的関係を示す。ドッキングと、結果のリストとの間に観察される際は、複合体II標的部位に到達する化合物の生物学的近接可能性を反映する。したがって、IC50は、部位に結合する類縁体の能力のみならず、細胞膜を横切って進入し、ミトコンドリアマトリックスへの接近を獲得する能力も反映する。IC50測定のために、本発明者らは、細胞ベースアッセイを用いたので、いくつかの化合物が、複合体IIと相互作用するためにミトコンドリア内に入るかどうかを評価することができない。最良の比較は、Autodockの結果と、市販の精製複合体IIキット(MS201 複合体II IMMUNOCAPTURE KIT MitoScience LLC、1850 Millrace Drive、Suite 3A、Eugene、OR 97403、USA、http://www.mitosciences.com) を用いる酵素アッセイとを比較することによってなされる。
【0134】
このキットを用いて、ヒト、マウス、ラットおよびウシの組織および細胞系から精製複合体IIを単離し、次いで、King TE、1967に概略されたDCPIPおよびPMSアッセイ反応を用いるか、または競合放射性ユビキノン結合アッセイを用いて、類縁体による活性および阻害についてスクリーニングすることができる。
【実施例9】
【0135】
分子モデリングは、複合体II Qp部位に結合する能力がある化学物質を示す。
この実施例において、コンピューターモデリング研究を用いて、ユビキノンQp結合部位に大きい親和性で結合し、電子伝達を中断させる能力を潜在的に有する化学物質を同定した。このような物質は、ガン性細胞を殺傷する能力を潜在的に有する。
【0136】
ZINC Database projectから65,000個の商業上入手可能な化合物のバーチャルライブラリーをダウンロードし、ブタ複合体II(1ZOY)の結晶構造に対して、望ましい特性を有するサブセットをドッキングさせた。相互作用エネルギーに基づいたトップ1000個の化合物をより厳密なドッキング実験に付し、最もブタ複合体IIと相互作用する可能性のある化合物として、最終的な10個の化合物を同定した。このような化合物を図10に示す。
【0137】
分子モデリングおよび生物アッセイからの化合物のリストおよび結合データに基づいて、複合体IIのQ部位への最良の結合に望ましい/有利な化学物質は、酸素がイオン化されるようになる能力があるかもしれないか、またはないかもしれない酸素含有側基をもつ疎水性化合物を包含する。コアの疎水性基として、トコフェロールおよびその同族体、ピリドキサール誘導体、ユビキノン誘導体、キニノリン誘導体、ベンゾスルホンアミドなどのベンジル基、ピリジル/ピリジン、プリンおよびピリミジン、パルテノライド、レチナミド、セスキテルペンラクトンが挙げられる。疎水性コア化合物の側基への置換修飾として、スルフェート、ホスフェート、ホスフィン酸、スルホンアミド、スルホニル基、プロパン酸、カルボキシレートおよびメトキシ、ジオキシまたはプロポキシ置換が挙げられる。
【実施例10】
【0138】
共役ペプチドLTVSPWY-α-TOSは、乳ガンを抑制する。
この実施例は、受容体チロシンキナーゼerbB2を過剰発現しているガン細胞へのα-TOSプロドラッグの標的化を記載する。α-TOSのアダクトを、erbB2を過剰発現しているガン細胞を標的化することが知られているヘプタペプチドLTVSPWYに結合した。受容体への結合に続いて、プロドラッグが取り込まれ、酸性ペプチダーゼによって加水分解されて、アポトーシス性α-TOSが得られることが見出されている。図11に見られるように、アダクトは、erbB2の高発現の結果として自然発生した乳腫瘍を特徴とするFVB/N MMTVラットc-neuマウスにおいて乳ガンを効率的に抑制した。事実、処置された腫瘍のサイズは、非処置コントロールと比較して、予想外の90%の相対的減少があった。したがって、このプロドラッグ/アダクトは、ガンの治療に特に有用である。
【実施例11】
【0139】
α-TOSは、erbB2の状態にかかわりなく腫瘍の増殖を阻害する。
インビボ研究のために、ヒト乳ガンの三分の一までが、高レベルのこの受容体チロシンキナーゼを発現することを考えれば(Slamonら、1989)、本発明者らは、α-TOSが、erbB2を低発現している乳ガンの動物モデルにおいて腫瘍増殖を抑制することができるかどうかを最初に決定した。この研究のために、ヌードマウスに低発現MCF-7細胞異種移植し、次いで、直径約5mmの腫瘍として確立されるようにした後、α-TOSで処置した(図12A)。結果は、コントロールマウスにおける腫瘍は、安楽死が必要になる時点までに、その初期のサイズの5倍まで増殖したが、ことを示したが、α-TOSは、マウスの処置グループにおいて腫瘍の増殖を抑制し、腫瘍のサイズは減少した。
【0140】
次に、本発明者らは、7ヶ月齢以上で、直径〜10 mmの認識できる乳ガンを明確に提示している自然発生トランスジェニックマウスモデルであるFVB/N MMTV-rat c-neuを用いるerbB2高ポジティブ乳ガンの治療に対する反応に興味をもった(Guyら、1992)。2つの薬物、3-BPおよびα-TOSを、腫瘍増殖のリプレッサーとしての、その相対的効力について比較した。結果(図12B)は、コントロール動物と比較して、処置マウスにおける腫瘍は、増殖停止または部分退行のいずれかを示すことが明らかになった。α-TOSは、これらの動物における乳腫瘍の増殖の退行を誘導することにおいて、3-BPと比べて、統計的により効果的であり、処置2週間で約30-40%まで腫瘍の平均サイズの低下をもたらすること(共分散分析によりp<0.05)が見出された。
【0141】
異種移植およびトランスジェニックマウスの乳腫瘍の増殖におけるα-TOSおよび3-BPの効果(図12参照)は、ヒト患者での臨床試験におけるそれらの提案された適用を裏付ける。しかし、本研究は、erbB2高発現細胞の3-BPへの耐性を示した。受容体は、ヒト乳ガンにおいて、通例、上昇するレベルで発現されるので(Slamonら、1989)、このことは、erbB2高ポジティブ患者の処置について問題を提供する。この問題は、Akt siRNAの同時投与、または3-BPとα-TOSの併用によって克服することができるかもしれないということが提案される。
【0142】
乳ガンを治療するためにα-TOSおよび類似する作用剤を用いることの利点は、以下のとおりである:α-TOSは、HER2-低(乳ガン患者の70%)およびHER2-高乳腫瘍(乳ガン患者の30%)の両方に対して、すなわち、HER-2発現に無関係に効率的である。このステージにおいて、HER2-低乳ガン患者を治療するための現在のアプローチは非常に効果的とはいえず、上記および本願発明を通して記載したように、α-TOSはガンの前臨床モデルにおいて選択的で、非常に効率がよい。HER2-高ガン患者に関して、現在の化学療法レジメンを補完するただ1つの治療のみが存在し、これは、HER2に対するヒト化抗体、ヘルセプチンの適用である。再度、α-TOSは、いくつかの理由から大いに融合である:ヘルセプチンのみが、非常に高レベルのHER2発現を有する乳ガンに対して有効である(蛍光インシトゥハイブリダイゼーションによる3+)が、α-TOSは、HER2発現が高レベルであろうと低レベルであろうと、乳腫瘍細胞を殺傷し、実験的乳ガンの有意な低下を引き起こす;ヘルセプチンは、心臓毒性であるが、心筋細胞(心臓筋肉細胞)は、α-TOSをアポトーシスサイレントビタミンEに加水分解するという傾向を有するので、α-TOSは毒性でない;ヘルセプチンは、はなはだしく高価:患者1名の治療の年間費用はAU$50,000であるが、α-TOSは比較的安価である;最後に、ヘルセプチンは、広範囲の乳ガン細胞のアポトーシス引き起こさず、むしろ、ヘルセプチンによる治療の中止後腫瘍が再発することができるように腫瘍細胞増殖を停止するにすぎない。対照的に、乳ガンの前臨床モデルを用いるα-TOSによる実験は、有意な縮小をを示し、続いて、TOS類縁体の適切な製剤での長期処置および最適投与レジメンの使用によって、乳ガンの完全な療法を達成する可能性があることを期待することができる。
【実施例12】
【0143】
中皮腫の治療におけるα-TOSの使用
α-TOSを用いて、ヒトガン患者において中皮腫を治療した。様々な化学療法およびその他の薬物による過去の失敗した試みにもかかわらず、α-TOSは、非常に効果的に働き、現在、患者はその生活の質を取り戻している。血漿レベルをモニターしたところ、血漿中のα-TOSのレベルは、長期間にわたって50 μMを達成した。これは、インビトロ分析に基づいてガン細胞を殺傷するのに十分すぎるほどである。
【0144】
臨床研究の概要:
これまでに5種の異なる細胞毒性作用剤を用いて治療を受けたが、改善を示すことができなかった末期中皮種を有する60歳の女性患者。患者は、最初は、高用量のアヘン剤投薬療法、神経障害性痛投薬療法、在宅酸素療法を受けており、余命6〜8週間と予測されていた。α-TOSで治療した後、患者は、もはや鎮痛または酸素を必要としなかった。α-TOを開始する前の患者は、触診可能な前胸壁腫瘤を有していたが、α-TOSでの療法により、劇的に縮小した。α-TOS療法は、目に付くような副作用がなく、非毒性であり、耐容性良好であった。
【0145】
病歴:
2001年7月に、患者は、最初に胸の痛みを発症し、2002年5月初旬に、生検によつ中皮腫であると診断された。患者は、重篤な痛みの管理のために苦痛緩和医療に付託され、2002年6月に、傍脊椎腫瘤に深部X線放射線療法を受けた。2002年7月に、シスプラチン、ゲムシタビンおよびタリドミドの併用による化学療法が開始された。
【0146】
重篤な末梢神経障害および毒性の問題および継続する吐き気により、2003年2月に、治療は中止された。2003年2月に、アリムタおよびカルボプラチンによる治療が開始された。骨髄抑制をともなう重篤な毒性ならびに継続する吐き気および嘔吐により、2003年9月に、治療は中止された。2003年9月に、経口定用量のフェノキソジオール(Novogen SAS薬物)による治療が開始され、2005年3月まで安定していた。しかし、フェノキソジオールにともなう継続する重篤な神経障害により、治療は再中止された。
【0147】
2005年3月に、中皮腫は、さらなる進行を明らかにし、再度前進した。患者は、ゲムシタビンの2用量による療法を開始したが、許容できない程度の毒性が再度生み出された。2005年8月に、患者は、ビノレルビンの2用量による療法を開始したが、末梢神経障害の重篤な症状の悪化が見られた。
【0148】
2005年12月に、患者は、基剤クリーム調製され、ジメチルスルホキシド(DMSO)で可溶化されたα-TOSの経皮デリバリーを受けることを開始した。患者は、約8週間の治療期間中、製剤の適用を続けたが、DMSOに続発する広範囲の皮膚反応問題(治癒に4週間を要するやけど)が発生した。中皮腫は、臨床反応を示したが、皮膚反応により、治療を中止しなければならなかった。
【0149】
2006年3月に、ガンは、神経根圧迫を引き起こす傍脊椎腫瘤とともに急速な進行を示した。患者は、コラムシン(SAS薬物、Soblec WA)による治療前に、さらなるDXRTを受けた。ガンは、6ヶ月間安定した反応を示したが、次いで、再度進行した。
【0150】
2006年11月に、患者は、アリムタ−アバスチン併用療法を再開したが、4ヶ月間療法を続けた後、吐き気、抑制されない下痢および神経障害の増加などの重篤な副作用をもつ合併症が現れ、治療を中止した。
【0151】
2007年4月に、リポダーマル(lipodermal)クリームと混合した変更製剤を用いるα-TOSによる療法を再開した。この時点で、中皮腫は、右肺を包み込む疾患の拡散を示し、両肺に多発性転移がCTスキャンによって視認された。さらに、触診可能な病変が、肋骨および右前胸壁の間で増殖するのが視認された。患者は、皮質脳波が低い非常に悪い状態であり、瀕死であったため、α-TOS治療を開始する時点でCTスキャンを行うことはできなかった。
【0152】
リポダーマル製剤を4週間適用した後、患者は、CTスキャンでは50%まで前胸壁腫瘤が減少し、非常に改善された回復を示し、継続中のα-TOS療法により、患者は、臨床的改善を続けた。
【0153】
α-TOS製剤:病院の薬局で混合された500gのリポダーマルクリーム+60gmsのα-TOS 化合物。2,3日ごとに約30gの混合された混合物を投与し、吸収を改善するために、上半身をプラスチックフィルム(サランラップ)で覆った。
【0154】
副作用:抗ヒスタミンで改善される軽いかゆみは別として、α-TOS療法には明らかな副作用はなかった。神経障害学的または他の薬害反応についての血液マーカーおよび他のサインのためのすべての実験室試験は、正常なままである。
【0155】
前記の実施態様は、本発明の原理の一例にすぎず、様々な変更および変化は、当業者にとって容易になしうることである。本発明は、様々な様式および他の実施態様において実践し、実行することができる。本明細書で用いた専門用語が、記載を目的とするためであり、限定とみなされるべきではないことも理解されよう。
【0156】
用語「含む」および「含んでいる」などのその変形は、本明細書において、述べられた完全体または述べられた完全体群の包含を意味するために用いられるものであり、文脈または用法において、用語の排他的解釈が必要でない限り、いずれかの他の完全体またはいずれかの他の完全体群を排除するものではない。
【0157】
本明細書に引用したいずれかの文献への言及は、開示がオーストラリアまたは他所における通例の一般的知識を構成するという承認ではない。
【0158】
参考文献のリスト
Albayrak T, Scherhammer V, Schoenfeld N, Braziulis E, Mund T, Bauer MK, Scheffler IE, Grimm S. (2003) The tumor suppressor cybL, a component of the respiratory chain, mediates apoptosis induction. Mol Biol Cell. Aug;14(8):3082-96.
Allen FH (2002) The Cambridge Structural Database: a quarter of a million crystal structures and rising. Acta Crystallogr B 58(Pt 3 Pt 1):380-388.
Allen RG, Balin AK (2003) Effects of oxygen on the antioxidant responses of normal and transformed cells. Exp Cell Res 289:307-316.
Alleva R, Tomasetti M, Andera L, Gellert N, Borghi B, Weber C, Murphy MP, Neuzil J (2003) Coenzyme Q blocks chemical but not receptor-mediated apoptosis by increasing mitochondrial antioxidant protection. FEBS Lett 503:46-50.
Aratri E, Spycher SE, Breyer I and Azzi A (1999) Modulation of α-tropomyosin expression by α-tocopherol in rat vascular smooth muscle cells. FEBS Lett 447:91-94.
Armstrong JS (2006) Mitochondria: a target for cancer therapy. Br J Pharmacol 147:239-48.
Arya P, Alibhai N, Qin H, Burton GW, Batist G, You SX and Alaoui-Jamali MA (1998) Design and synthesis of analogues of vitamin E: antiproliferative activity against human breast adenocarcinoma cells. Bioorg Med Chem Lett 8:2433-2438.
Astuti D, Hart-Holden N, Latif F, Lalloo F, Black GC, Lim C, Moran A, Grossman AB, Hodgson SV, Freemont A, Ramsden R, Eng C, Evans DG, Maher ER. (2003) Genetic analysis of mitochondrial complex II subunits SDHD, SDHB and SDHC in paraganglioma and phaeochromocytoma susceptibility.Clin Endocrinol (Oxf). Dec;59(6):728-33
Azzi A, Ricciarelli R and Zingg JM (2002) Non-antioxidant molecular functions of α-tocopherol (vitamin E). FEBS Lett 519:8-10.
Barnett KT, Fokum FD, Malafa MP (2002) Vitamin E succinate inhibits colon cancer liver metastases. J Surg Res 106:292-298.
Bayley JP, van Minderhout I, Weiss MM, Jansen JC, Oomen PH, Menko FH, Pasini B, Ferrando B, Wong N, Alpert LC, Williams R, Blair E, Devilee P, Taschner PE. (2006) Mutation analysis of SDHB and SDHC: novel germline mutations in sporadic head and neck paraganglioma and familial paraganglioma and/or pheochromocytoma. BMC Med Genet. Jan 11;7:1
Berridge MV, Herst PM, Tan AS. (2005) Tetrazolium dyes as tools in cell biology: new insights into their cellular reduction. Biotechnol Annu Rev.11:127-52
Berridge MV and Tan AS (1993) Characterization of the cellular reduction of 3-(4,5-dimethylthiazol- 2-yl)-2,5-diphenyltetrazolium bromide (MTT): subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch Biochem Biophys 303:474-478.
Birringer M, Drogan D and Brigelius-Flohe R (2001) Tocopherols are metabolized in HepG2 cells by side chain ω-oxidation and consecutive β-oxidation. Free Radic Biol Med 31:226-232.
Birringer M, EyTina JH, Salvatore BA and Neuzil J (2003) Vitamin E analogues as inducers of apoptosis: Structure-function relationship. Br J Cancer 88:1948-1955.
Birringer M, Pfluger P, Kluth D, Landes N and Brigelius-Flohe R (2002) Identities and differences in the metabolism of tocotrienols and tocopherols in HepG2 cells. J Nutr 132:3113-3118.
Boersma AW, Nooter K, Oostrum RG., Stoter G (1996) Quantification of apoptotic cells with fluorescein isothiocyanate-labeled annexin V in Chinese hamster ovary cell cultures with cisplatin. Cytometry 24:123-130
Breen GA, Scheffler IE. (1979) Respiration-deficient Chinese hamster cell mutants: biochemical characterization. Somatic Cell Genet. Jul;5(4):441-51.
Briere JJ, Schlemmer D, Chretien D, Rustin P. (2004) Quinone analogues regulate mitochondrial substrate competitive oxidation. Biochem Biophys Res Commun. Apr 16;316(4):1138-42.
Burstein HJ (2005) The distinctive nature of HER2-positive breast cancers. N Engl J Med 353:1652-1654.
Burnell JN, John P, Whatley FR (1975) The reversibility of active sulphate transport in membrane vesicles of Paracoccus denitrificans. Biochem J. 150: 527-536.
Cardone MH, Roy N, Stennicke HR, Salvesen GS, Franke TF, Stanbridge E, Frisch S, Reed JC (1998) Regulation of cell death protease caspase-9 by phosphorylation. Science 282:1318-1321.
Cheeseman KH, Holley AE, Kelly FJ, Wasil M, Hughes L and Burton G (1995) Biokinetics in humans of RRR-α-tocopherol: the free phenol, acetate ester, and succinate ester forms of vitamin E. Free Radic Biol Med 19:591-598
Choudhry ZM, Gavrikova EV, Kotlyar AB, Tushurashvili PR and Vinogradov AD. (1985) Pyridoxal phosphate-induced dissociation of the succinate: ubiquinone reductase.FEBS Lett. Mar 11;182(1):171-5.
Choudhry ZM, Kotlyar AB and Vinogradov AD. (1986) Studies on the succinate dehydrogenating system. Interaction of the mitochondrial succinate-ubiquinone reductase with pyridoxal phosphate.Biochim Biophys Acta. Jun 10;850(1):131-8.
Christen S, Woodall AA, Shigenaga MK, Southwell-Keely PT, Duncan MW, Ames BN. (1997) gamma-tocopherol traps mutagenic electrophiles such as NO(X) and complements alpha-tocopherol: physiological implications. Proc Natl Acad Sci U S A. Apr 1;94(7):3217-22.
Church SL, Grant JW, Ridnour LA, Oberley LW, Swanson PE, Meltzer PS, Trent JM (1993) Increased manganese superoxide dismutase expression supresses the malignant phenotype of human melanoma cells. Proc Natl Acad Sci USA 90:3113-3117.
DeFrancesco L, Scheffler IE, Bissell MJ. (1976) A respiration-deficient Chinese hamster cell line with a defect in NADH-coenzyme Q reductase. J Biol Chem. Aug 10;251(15):4588-95.
Devaraj S, Hugou I and Jialal I (2001) α-Tocopherol decreases CD36 expression in human monocyte-derived macrophages. J Lipid Res 42:521-527.
Don AS, Hogg PJ (2004) Mitochondria as cancer drug targets. Trends Mol Med 10:372-378.
Dong LF, Wang XF, Zhao Y, Tomasetti M, Wu K, Neuzil J (2006) Vitamin E analogues as anti-cancer agents: the role of modulation of apoptosis signalling pathways. Cancer Therapy 4:35-46.
Esposti MD, Ngo A, Ghelli A, Benelli B, Carelli V, McLennan H, Linnane AW (1996) The interaction of Q analogs, particularly hydroxydecyl benzoquinone (idebenone), with the respiratory complexes of heart mitochondria. Arch Biochem Biophys. Jun 15;330(2):395-400.. Please specify the different types of ubiquinone
Fariss MW, Nicholls-Grzemski FA, Tirmenstein MA, Zhang JG (2001) Enhanced antioxidant and cytoprotective abilities of vitamin E succinate is associated with a rapid uptake advantage in rat hepatocytes and mitochondria. Free Radic Biol Med 31:530-541.
Fresno Vara JA, Casado E, de Castro J, Cejas P, Belda-Iniesta C, Gonzalez-Baron M (2004) PI3K/Akt signalling pathway and cancer. Cancer Treat Rev 30:193-204.
Galli F, Stabile AM, Betti M, Conte C, Pistilli A, Rende M, Floridi A and Azzi A (2004) The effect of α- and γ-tocopherol and their carboxyethyl hydroxychroman metabolites on prostate cancer cell proliferation. Arch Biochem Biophys 423:97-102.
Geschwind JF, Ko YH, Torbenson MS, Magee C, Pedersen PL (2002) Novel therapy for liver cancer: direct intraarterial injection of a potent inhibitor of ATP production. Cancer Res 62:3909-3913.
Gottlieb E, Tomlinson IP. (2005) Mitochondrial tumour suppressors: a genetic and biochemical update.Nat Rev Cancer. Nov;5(11):857-66
Grandage VL, Gale RE, Linch DC, Khwaja A (2005) PI3-kinase/Akt is constitutively active in primary acute myeloid leukaemia cells and regulates survival and chemoresistance via NF-κB, MAP kinase and p53 pathways. Leukemia 19:586-594.
Gu LQ, Yu L, Yu CA. (1990) Effect of substituents of the benzoquinone ring on electron-transfer activities of ubiquinone derivatives. Biochim Biophys Acta. Feb 22;1015(3):482-92.
Guthrie N, Gapor A, Chambers AF and Carroll KK (1997) Inhibition of proliferation of estrogen receptor-negative MDA-MB-435 and -positive MCF-7 human breast cancer cells by palm oil tocotrienols and tamoxifen, alone and in combination. J Nutr 127: 544S-548S.
Guy CT, Webster MA, Schaller M, Parsons TJ, Cardiff RD, Muller WJ (1992) Expression of the neu protooncogene in the mammary epithelium of transgenic mice induces metastatic disease. Proc Natl Acad Sci USA 89:10578-10582.
Hartshorn MJ (2002) AstexViewer(TM): An aid for structure-based drug design. J Computer Aided Mol Des 16: 871-881.
He L, Mo H, Hadisusilo S, Qureshi AA and Elson CE (1997) Isoprenoids suppress the growth of murine B16 melanomas in vitro and in vivo. J Nutr 127:668-674.
He DY, Yu L, Yu CA. (1994) Protein ubiquinone interaction. Synthesis and biological properties of 5-alkyl ubiquinone derivatives. J Biol Chem. Nov 11;269(45):27885-8.
Hosomi A, Arita M, Sato Y, Kiyose C, Ueda T, Igarashi O, Arai H and Inoue K (1997) Affinity for α-tocopherol transfer protein as a determinant of the biological activities of vitamin E analogs. FEBS Lett 409:105-108.
Huang LS, Sun G, Cobessi D, Wang AC, Shen JT, Tung EY, Anderson VE, Berry EA (2006) 3-nitropropionic acid is a suicide inhibitor of mitochondrial respiration that, upon oxidation by complex II, forms a covalent adduct with a catalytic base arginine in the active site of the enzyme. J Biol Chem 281:5965-5972
James AM, Cocheme HM, Smith RA, Murphy MP (2005) Interactions of mitochondria-targeted and untargeted ubiquinones with the mitochondrial respiratory chain and reactive oxygen species. Implications for the use of exogenous ubiquinones as therapies and experimental tools. J Biol Chem 280:21295-212312.
Kelso GF, Porteous CM, Coulter CV, Hughes G, Porteous WK, Ledgerwood EC, Smith RA, Murphy MP (2001) Selective targeting of a redox-active ubiquinone to mitochondria within cells: antioxidant and antiapoptotic properties. J Biol Chem 276:4588-4596.
Kim R, Tanabe K, Emi M, Uchida Y, Toge T (2004) Potential roles of antisense therapy in the molecular targeting of genes involved in cancer. Int J Oncol 24:5-17
King A, Selak MA, Gottlieb E. (2006) Succinate dehydrogenase and fumarate hydratase: linking mitochondrial dysfunction and cancer. Oncogene. Aug 7;25(34):4675-82
King TE (1967). Preparation of succinate dehydrdogenase and reconstitution of succinate oxidase. Methods Enzymol 10: 322-331.
King, T.S. (1967) Preparations of succinate-cytochrome c reductase and the cytochrome b-c1 particle, and reconstitution of succinate-cytochrome c reductase. Methods in Enzymology, 10, 216-225
Ko YH, Smith BL, Wang Y, Pomper MG, Rini DA, Torbenson MS, Hullihen J, Pedersen PL (2004) Advanced cancers: eradication in all cases using 3-bromopyruvate therapy to deplete ATP. Biochem Biophys Res Commun 324:269-275.
Kogure K, Hama S, Kisaki M, Takemasa H, Tokumura A, Suzuki I and Fukuzawa K (2004) Structural characteristic of terminal dicarboxylic moiety required for apoptogenic activity of α-tocopheryl esters. Biochim Biophys Acta 1672: 93-99.
Kogure K, Manabe S, Suzuki I, Tokumura A and Fukuzawa K (2005) Cytotoxicity of α-tocopheryl succinate, malonate and oxalate in normal and cancer cells in vitro and their anti-cancer effects on mouse melanoma in vivo. J Nutr Sci Vitaminol 51:392-397.
Kunisaki M, Bursell SE, Clermont AC, Ishii H, Ballas LM, Jirousek MR, Umeda F, Nawata H and King GL (1995) Vitamin E prevents diabetes-induced abnormal retinal blood flow via the diacylglycerol-protein kinase C pathway. Am J Physiol 269:E239-246.
Maehara Y, Kusumoto T, Kusumoto H, Anai H, Sugimachi K (1988) Sodium succinate enhances the colorimetric reaction of the in vitro chemosensitivity test: MTT assay. Oncology 5:434-436.
Makishima M, Umesono K, Shudo K, Naoe T, Kishi K and Honma Y (1998) Induction of differentiation in acute promyelocytic leukemia cells by 9-cis retinoic acid α-tocopherol ester (9-cis tretinoin tocoferil). Blood 91:4715-4726.
Malafa MP, Fokum FD, Mowlavi A, Abusief M, King M (2002) Vitamin E inhibits melanoma growth in mice. Surgery 131:85-91.
Malafa MP, Neitzel LT. Vitamin E succinate promotes breast cancer tumor dormancy. J Surg Res 93:163-170.
Martin-Nizard F, Boullier A, Fruchart JC and Duriez P (1998) α-Tocopherol but not β-tocopherol inhibits thrombin-induced PKC activation and endothelin secretion in endothelial cells. J Cardiovasc Risk 5:339-345.
Morris G M, Goodsell DS, Halliday RS, Huey R, Hart W E, Belew RK, Olson AJ (1998) Automated docking using a Lamarckian genetic algorithm and empirical binding free energy function. J Comp Chem 19: 1639-1662.
Morton LW, Ward NC, Croft KD, Puddey IB. (2002) Evidence for the nitration of gamma-tocopherol in vivo: 5-nitro-gamma-tocopherol is elevated in the plasma of subjects with coronary heart disease. Biochem J. Jun 15;364(Pt 3):625-8.
Munteanu A, Zingg JM, Ogru E, Libinaki R, Gianello R, West S, Negis Y and Azzi (2004) AModulation of cell proliferation and gene expression by α-tocopheryl phosphates: relevance to atherosclerosis and inflammation. Biochem Biophys Res Commun 318:311-316.
Negis Y, Zingg JM, Ogru E, Gianello R, Libinaki R and Azzi A (2005) On the existence of cellular tocopheryl phosphate, its synthesis, degradation and cellular roles: a hypothesis. IUBMB Life 57:23-25.
Nesaretnam K, Stephen R, Dils R and Darbre P (1998) Tocotrienols inhibit the growth of human breast cancer cells irrespective of estrogen receptor status. Lipids 33:461-469.
Neuzil J, Dong LF, Ramanathapuram L, Hahn T, Chladova M, Wang XF, Zobalova R, Prochazka L, Gold M, Freeman R, Turanek J, Akporiaye ET, Dyason JC, Ralph SJ. (2007) Vitamin E analogues as a novel group of mitocans: Anti-cancer agents that act by targeting mitochondria. Mol Aspects Med. Feb 23.
Neuzil J, Massa H (2005) Hepatic processing determines dual activity of vitamin E succinate. Biochem Biophys Res Commun 327:1024-1027.
Neuzil J, Tomasetti M, Mellick AS, Alleva R, Salvatore BA, Birringer M, Fariss MW (2004) Vitamin E analogues: A new class of inducers of apoptosis with selective anti-cancer effects. Curr Cancer Drug Targets 4:355-372.
Neuzil J, Tomasetti M, Zhao Y, Dong LF, Birringer M, Wang XF, Low P, Wu K, Salvatore BA, Ralph SJ. (2007) Vitamin E analogs, a novel group of "mitocans," as anticancer agents: the importance of being redox-silent. Mol Pharmacol. May;71(5):1185-99.
Neuzil J, Weber T, Gellert N, Weber C (2001) Selective cancer cell killing by α-tocopheryl succinate. Br J Cancer 84:87-89.
Neuzil J, Weber T, Schroder A, Lu M, Ostermann G, Gellert N, Mayne GC, Olejnicka B, Negre-Salvayre A, Sticha M, Coffey RJ, Weber C (2001) Induction of apoptosis in cancer cells by α-tocopheryl succinate: Molecular pathways and structural requirements. FASEB J 15:403-415.
Nishikawa K, Satoh H, Hirai A, Suzuzki K, Asano R, Kumadaki I, Hagiwara K and Yano T (2003) α-Tocopheryloxybutyric acid enhances necrotic cell death in breast cancer cells treated with chemotherapy agent. Cancer Lett 201:51-56
Oostveen FG, Au HC, Meijer PJ, Scheffler IE. (1995) A Chinese hamster mutant cell line with a defect in the integral membrane protein CII-3 of complex II of the mitochondrial electron transport chain. J Biol Chem. Nov 3;270(44):26104-8.
Osaki M, Oshimura M, Ito H (2004) PI3K-Akt pathway: its functions and alterations in human cancer. Apoptosis 9:667-676.
Pham DQ and Plakogiannis R (2005) Vitamin E supplementation in cardiovascular disease and cancer prevention: Part 1. Ann Pharmacother 39:1870-8
Pollard PJ, Briere JJ, Alam NA, Barwell J, Barclay E, Wortham NC, Hunt T, Mitchell M, Olpin S, Moat SJ, Hargreaves IP, Heales SJ, Chung YL, Griffiths JR, Dalgleish A, McGrath JA, Gleeson MJ, Hodgson SV, Poulsom R, Rustin P, Tomlinson IP. (2005) Accumulation of Krebs cycle intermediates and over-expression of HIF1alpha in tumours which result from germline FH and SDH mutations. Hum Mol Genet. Aug 1;14(15):2231-9.
Quin J, Engle D, Litwiller A, Peralta E, Grasch A, Boley T, Hazelrigg S (2005) Vitamin E succinate decreases lung cancer tumor growth in mice. J Surg Res 127:139-143.
Qureshi AA, Mo H, Packer L and Peterson DM (2000) Isolation and identification of novel tocotrienols from rice bran with hypocholesterolemic, antioxidant, and antitumor properties. J Agric Food Chem 48:3130-3140.
Ralph SJ, Dong LF, Low P, Lawen A, and Neuzil J (2006) Mitocans: mitochondria targeted anti-cancer drugs as improved therapies and related patents. Recent Pat Anticancer Drug Discov 1:305-326.
Ricciarelli R, Zingg JM and Azzi A (2000) Vitamin E reduces the uptake of oxidized LDL by inhibiting CD36 scavenger receptor expression in cultured aortic smooth muscle cells. Circulation 102:82-87.
Safford SE, Oberley TD, Urano M, St Clair DK (1994) Suppression of fibrosarcoma metastasis by elevated expression of manganese superoxide dismutase. Cancer Res 54:4261-4265.
Sanborn BM, Felberg NT, Hollocher TC (1971) The inactivation of succinate dehydrogenase by bromopyruvate. Biochim Biophys Acta 227:219-231.
Sanner MF (1999) Python: a programming language for software integration and development, J Mol Graphic Mod 17: 57-61.
Sanders G., et al (2001) Preparation of tocopherols, tocotrienols, other chroman and side chain derivatives that induce cell apoptosis for therapeutic use as antiproliferative agents. 2001: PCT Int. Appl. WO 2001058889. p. 120.
Scallet AC, Haley RL, Scallet DM, Duhart HM, Binienda ZK (2003) 3-nitropropionic acid inhibition of succinate dehydrogenase (complex II) activity in cultured Chinese hamster ovary cells: antagonism by L-carnitine. Ann N Y Acad Sci 993:305-312.
Scheffler IE, Yadava N, Potluri P. (2004) Molecular genetics of complex I-deficient Chinese hamster cell lines. Biochim Biophys Acta. Dec 6;1659(2-3):160-71. Review.
Seo, B.B., Kitajima-Ihara, T., Chan, E.K., Scheffler, I.E., Matsuno-Yagi, A., and Yagi, T. (1998) Proc. Natl. Acad. Sci. USA 95, 9167-9171
Shah SJ and Sylvester PW (2005) γ-Tocotrienol inhibits neoplastic mammary epithelial cell proliferation by decreasing Akt and nuclear factor κB activity. Exp Biol Med 230: 235-241.
Shiau CW, Huang JW, Wang DS, Weng JR, Yang CC, Lin CH, Li C, Chen CS (2006) alpha-Tocopheryl succinate induces apoptosis in prostate cancer cells in part through inhibition of Bcl-xL/Bcl-2 function. J Biol Chem 281:11819-11825.
Shun MC, Yu W, Gapor A, Parsons R, Atkinson J, Sanders BG and Kline K (2004) Pro-apoptotic mechanisms of action of a novel vitamin E analog (α-TEA) and a naturally occurring form of vitamin E (δ-tocotrienol) in MDA-MB-435 human breast cancer cells. Nutr Cancer 48:95-105.
Slamon DJ, Godolphin W, Jones LA, Holt JA, Wong SG, Keith DE, Levin WJ, Stuart SG, Udove J, Ullrich A, et al (1989) Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science 244:707-712.
Stapelberg M, Gellert N, Swettenham E, Tomasetti M, Witting PK, Procopio A, Neuzil J (2005) α-Tocopheryl succinate inhibits malignant mesothelioma by disruption of the FGF autocrine signaling loop: Mechanism and the role of oxidative stress. J Biol Chem 280:25369-25376.
Sugawara M, Huang W, Fei Y-J, Leibach FH, Ganapathy V and Ganapathy ME (2000) Transport of valganciclovir, a ganciclovir prodrug, via peptide transporters PEPT1 and PEPT2. J Pharm Sci 89:781-789.
Sun F, Huo X, Zhai Y, Wang A, Xu J, Su D, Bartlam M, Rao Z. Crystal structure of mitochondrial respiratory membrane protein complex II. Cell 121:1043-1057.
Swettenham E, Witting PK, Salvatore BA, Neuzil J (2005) α-Tocopheryl succinate selectively induces apoptosis in neuroblastoma cells: Potential therapy of malignancies of the nervous system? J Neurochem 94:1448-1456.
Takeuchi H, Kim J, Fujimoto A, Umetani N, Mori T, Bilchik A, Turner R, Tran A, Kuo C, Hoon DS (2005) X-Linked inhibitor of apoptosis protein expression level in colorectal cancer is regulated by hepatocyte growth factor/C-met pathway via Akt signaling. Clin Cancer Res 11:7621-7628.
Tan AK, Ramsay RR, Singer TP, Miyoshi H. (1993) Comparison of the structures of the quinone-binding sites in beef heart mitochondria. J Biol Chem. Sep 15;268(26):19328-33.
Tasinato A, Boscoboinik D, Bartoli GM, Maroni P and Azzi A (1995) d-α-Tocopherol inhibition of vascular smooth muscle cell proliferation occurs at physiological concentrations, correlates with protein kinase C inhibition, and is independent of its antioxidant properties. Proc Natl Acad Sci USA 92:12190-12194.
Teague, S. J.; Davis, A. M.; Leeson, P. D.; Oprea, T. (1999) The Design of Leadlike Combinatorial Libraries. Angew Chem Int Ed Engl, 38, 3743-3748
Tomasetti M, Gellert N, Procopio A, Neuzil J (2004) A vitamin E analogue suppresses malignant mesothelioma in a pre-clinical model: A prototype of a future drug against a fatal neoplastic disease? Int J Cancer 109:641-642.
Tomic-Vatic A, EyTina JH, Chapmann JM, Mahdavian E, Neuzil J and Salvatore BA (2005) Vitamin E amides, a new class of vitamin E analogues with enhanced pro-apoptotic activity. Int J Cancer 117:118-193.
van Nederveen FH, Korpershoek E, Lenders JW, de Krijger RR, Dinjens WN. (2007) Somatic SDHB mutation in an extraadrenal pheochromocytoma. N Engl J Med. Jul 19;357(3):306-8.
Vraka PS, Drouza C, Rikkou MP, Odysseos AD and Keramidas AD (2006) Synthesis and study of the cancer cell growth inhibitory properties of α-, γ-tocopheryl and γ-tocotrienyl 2-phenylselenyl succinates. Bioorg Med Chem 14: 2684-2696.
Wallace, A.C., Laskowski, R.A. and Thornton, J.M. (1995). LIGPLOT: a program to generate schematic diagrams of protein-ligand interactions. Protein Eng 8, 127-134.
Wang XF, Dong LF, Zhao Y, Tomasetti M, Wu K, Neuzil J (2006) Vitamin E analogues as anti-cancer agents: Lessons from studies with α-tocopheryl succinate. Mol Nutr Food Res 50:675-685.
Wang XF, Witting PK, Salvatore BA, Neuzil J (2005) α-Tocopheryl succinate induces apoptosis in HER2/erbB2-overexpressing breast cancer cells by signalling via the mitochondrial pathway. Biochem Biophys Res Commun 326:282-289.
Weber T, Dalen H, Andera L, Negre-Salvayre A, Auge N, Sticha M, Lloret A, Terman A, Witting PK, Higuchi M, Plasilova M, Zivny J, Gellert N, Weber C, Neuzil J (2003) Mitochondria play a central role in apoptosis induced by α-tocopheryl succinate, an agent with anticancer activity. Comparison with receptor-mediated pro-apoptotic signaling. Biochemistry 42:4277-4291.
Weber T, Lu M, Andera L, Lahm H, Gellert N, Fariss MW, Korinek V, Sattler W, Ucker DS, Terman A, Schroder A, Erl W, Brunk UT, Coffey RJ, Weber C, Neuzil J (2002) Vitamin E succinate is a potent novel anti-neoplastic agent with high tumor selectivity and cooperativity with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL, Apo2L) in vivo. Clin Cancer Res 8:863-869.
West KA, Castillo SS, Dennis PA (2002) Activation of the PI3K/Akt pathway and chemotherapeutic resistance. Drug Resist Updat 5:234-248.
Wu Y, Zu K, Ni J, Yeh S, Kasi D, James NS, Chemler S and Ip C (2004) Cellular and molecular effects of α-tocopheryloxybutyrate: lessons for the design of vitamin E analog for cancer prevention. Anticancer Res 24:3795-3802.
Xu RH, Pelicano H, Zhou Y, Carew JS, Feng L, Bhalla KN, Keating MJ, Huang P (2005) Inhibition of glycolysis in cancer cells: a novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia. Cancer Res 65:613-621.
Yabunaka H, Kenmochi A, Nakatogawa Y, Sakamoto K, Miyoshi H. (2002) Hybrid ubiquinone: novel inhibitor of mitochondrial complex I. Biochim Biophys Acta. Dec 2;1556(2-3):106-12.
Yamauchi J, Iwamoto T, Kida S, Masushige S, Yamada K and Esashi T (2001) Tocopherol-associated protein is a ligand-dependent transcriptional activator. Biochem Biophys Res Commun 285:295-299.
Yang F, Yu L, He DY, Yu CA. (1991) Protein-ubiquinone interaction in bovine heart mitochondrial succinate-cytochrome c reductase. Synthesis and biological properties of fluorine substituted ubiquinone derivatives. J Biol Chem. Nov 5;266(31):20863-9.
Yankovskaya V, Sablin SO, Ramsay RR, Singer TP, Ackrell BA, Cecchini G, Miyoshi H. (1996) Inhibitor probes of the quinone binding sites of mammalian complex II and Escherichia coli fumarate reductase. J Biol Chem. Aug 30;271(35):21020-4.
Yano Y, Satoh H, Fukumoto K, Kumadaki I, Ichikawa T, Yamada K, Hagiwara K and Yano T (2005) Induction of cytotoxicity in human lung adenocarcinoma cells by 6-O-carboxy-propyl-α-tocotrienol, a redox-silent derivative of α-tocotrienol. Int J Cancer 115:839-846.
Yoeli-Lerner M, Yiu GK, Rabinovitz I, Erhardt P, Jauliac S, Toker A (2005) Akt blocks breast cancer cell motility and invasion through the transcription factor NFAT. Mol Cell 20:539-550.
Youk HJ, Lee E, Choi MK, Lee YJ, Chung JH, Kim SH, Lee CH and Lim SJ (2005) Enhanced anticancer efficacy of α-tocopheryl succinate by conjugation with polyethylene glycol. J Control Release 107:43-52.
Zhang JG, Nicholls-Grzemski FA, Tirmenstein MA, Fariss MW (2001a) Vitamin E succinate protects hepatocytes against the toxic effect of reactive oxygen species generated at mitochondrial complexes I and III by alkylating agents. Chem Biol Interact 138:267-284.
Zhang JG, Tirmenstein MA, Nicholls-Grzemski FA, Fariss MW (2001b) Mitochondrial electron transport inhibitors cause lipid peroxidation-dependent and -independent cell death: protective role of antioxidants. Arch Biochem Biophys 393:87-96.
Zhao S, Konopleva M, Cabreira-Hansen M, Xie Z, Hu W, Milella M, Estrov Z, Mills GB, Andreeff M (2004) Inhibition of phosphatidylinositol 3-kinase dephosphorylates BAD and promotes apoptosis in myeloid leukemias. Leukemia 18:267-27.
【技術分野】
【0001】
本発明は、抗ガン化合物およびガンの治療または予防方法に関する。さらに詳しくは、本発明は、ガン性細胞のミトコンドリア呼吸鎖の複合体IIと選択的に相互作用し、反応性酸素種を生成し、それらの細胞のアポトーシスを誘導する、ビタミンEのプロオキシダント型などのプロオキシダント抗ガン化合物に関する。
【背景技術】
【0002】
理想的抗ガン薬は、ガン細胞の内部においてのみ活性であり、ガン細胞の本質的な成分を標的とするか、またはその本質的な過程を破壊する薬物である。細胞エネルギーの大部分およびアポトーシスの重要な制御因子を供給するミトコンドリアは、抗ガン療法に望まれる細胞選択性を提供しうる有効な標的として浮かび上がっている(Don and Hogg、2004、Armstrong、2006)。
【0003】
動物実験において報告されているように(Koら、2004)、ガン細胞内のミトコンドリアに選択的に影響を及ぼすことによって作用する細胞毒性薬物である「マイトカン(マイトカン)(抗ガン成分)」は、副作用が少ないか、あるいは副作用のない強力な抗ガン剤として働くので、ガンの治療にとって非常に興味をそそるものであることがわかっている。マイトカンは、ガン細胞ミトコンドリアのエネルギー生成システムを破壊し、ガン細胞内部での活性酸素種(ROS)の増加およびミトコンドリア依存性細胞死シグナル伝達経路の活性化をもたらす。
【0004】
マイトカンとして、以下のミトコンドリア関連活性に影響を及ぼす薬物が挙げられる:ヘキソキナーゼインヒビター;電子輸送/呼吸鎖ブロッカー;ミトコンドリア膜透過性転移孔標的化構成タンパク質サブユニット、電位依存性アニオン選択的チャネル(VDAC)またはアデニンヌクレオチドトランスポーター(ANT)のいずれかのアクチベーター;Bcl-2抗アポトーシスファミリータンパク質およびBax/Bidアポトーシス促進性模倣体のインヒビター。副作用が少ないか、あるいは副作用のないマイトカンの2つの主な例は、3-ブロモピルビン酸エステル(3-BP)およびα-トコフェリルコハク酸エステル(α-TOS)であり、それらの両方が、非常に大きい有効性で、ガン細胞においてアポトーシスを誘導する(Koら、2004、Geschwindら、2002、Xuら、2005、Neuzilら、2001、2004)。
【0005】
マイトカンの1つのグループとして、ビタミンEのロオキシダント類縁体が挙げられる(Wangら 2006)。α-TOSによって代表されるプロオキシダントビタミンE類縁体の抗ガン剤としての有望性は、それらが悪性腫瘍を抑制することを明らかにした、ヌードマウスで増殖するヒト意趣移植片などの実験的に作成されたガンを用いた研究に起因する(Neuzilら、2004に概説)。このような研究として、結腸直腸(Neuzilら、2001、Weberら、2002)および肺ガン(Quinら、2005)、黒色腫(Malafaら、2002)ならびに中皮腫(Tomasettiら、2004、Stapelbergら、2005)が挙げられる。α-TOSが、乳ガンの休止状態を促進すること(Malafaら、2000)および結腸ガンのへの転移を抑制することも明らかにされている(Barnettら、2002)。
【0006】
ビタミンE(α-トコフェロール、α-TOH)は、細胞において強力な抗酸化剤として作用するけれども、ビタミンEのエステル化された酸化還元サイレントおよびプロオキシダント類縁体であるα-TOSは、特徴的性質を有する。α-TOHとは対照的に、α-TOSは、種々のガン細胞系の範囲においてROSの急速な生成を引き起こす強力な細胞ストレッサーとして作用する(Neuzilら、2004、Weberら、2003、Wangら、2005、Swettenhamら、2005、Stapelbergら、2005)。α-TOSは、Bcl-2/Bcl-xLに結合し、それらを阻害する能力を有する(Dongら、2006)。今までの証拠は、正常な細胞は、より大きい抗酸化防御に恵まれており(Allen and Balin、2003、Saffordら、1994、Churchら、1993)、および/またはコハク酸エステル部分を放出することにより酸化還元活性、非アポトーシス性α-TOHを産生することによってα-TOSを不活性化する高濃度のエステラーゼを含む(Farissら、2001、Neuzilら、2004、Neuzil and Massa、2005)ので、α-TOSのガン細胞特異的性質および正常な細胞における毒性作用の欠如が起こることを示唆している。
【0007】
天然のビタミンEは、クロマノール環のメチル化パターン(α-、β-、γ-、δ-トコフェロール)およびフィチル側鎖の二重結合の数(α-、β-、γ-、δ-トコトリエノール)によって相異する8種の化合物の混合物からなる。インビトロおよびインビボでの親油性抗酸化剤としてのこれらの分子の役割は、広く受け入れられている。さらに、VEファミリーのメンバーの非抗酸化特性も研究されている(Azziら、2002)。
【0008】
ビタミンE分子は、3つの異なるドメインに分割することができる。機能的ドメイン(I)は、クロマノール環のC6位における置換パターンから生じる。遊離のヒドロキシ基は、ビタミンEが抗酸化剤として機能するために必須であるので、この位置は、分子が酸化還元活性または酸化還元サイレントとして振る舞うかどうかを決定する。文書で十分に立証された、4種のトコフェロール異性体の抗酸化特性は、ガン臨床試験への適用をもたらした。これらの研究は、ガンの予防における遊離トコフェロールの使用に関して肯定的な結果を明らかにしなかった(Pham and Plakogiannis、2005)。しかし、C6における特定の化学的修飾は、強力な抗腫瘍剤であること立証されたエーテル(RO-)、エステル(RCOO-)およびアミド(RCONH-)をもたらした。下記第1表を参照。
【0009】
第1表
ビタミンE類縁体の抗増殖活性
化合物は、シグナル伝達ドメインによって分類される。
【表1】
【0010】
【表2】
a無効;b細胞増殖の阻害;cα-TOSよりも細胞毒性が高い;d54よりも高価が低い;eエーテル類縁体がα-TOS自体よりも効果が低い;fα-TOSと同程度;gEC50 [μg/ml];hα-TOSよりも効率的。
【0011】
第2表
修飾疎水性ドメインを有するビタミンE類縁体の抗増殖活性
【表3】
a無効;b50 μMにおいて弱い阻害;c10 μMにおいて82%阻害。
【0012】
第3表
ビタミンE類縁体の抗増殖活性
化合物は、シグナル伝達ドメインによって分類される。
【表4】
【0013】
【表5】
a0-40 μMの範囲において細胞毒性;b非常に強力;c完全阻害;dα-TOSと同程度;eγ-トコトリエノールよりも2倍強力;f細胞増殖の阻害。
【0014】
シグナル伝達ドメイン(II)と称される第2のドメインは、トコフェロールの抗酸化特性とは無関係である活性を示す。これらの特性は、芳香環のメチル化パターンに由来する。たとえば、α-トコフェロールは、ジアシルグリセロール(DAG)レベルを低下させることによってタンパク質キナーゼC(PKC)を阻害することが報告されているが、同様の抗酸化能力を有する他のトコフェロール(たとえば、β-トコフェロール)は、PKCを阻害しない。したがって、α-トコフェロールのPKC阻害活性は、その鉱産か能力とは無関係である(Tasinatoら、1995;Kunisakiら、1995)。しかし、いくつかの場合、種々のトコフェロールの生物活性は、実際に特定の種に対する抗ガン活性において重大な影響を有する、シグナル伝達ドメインにおける構造的差異によって影響を及ぼされる。たとえば、γ-トコフェロールは、α-トコフェロールよりも非常によい活性酸化窒素種(たとえば、ペルオキシ亜硝酸)のスカベンジャーである。したがって、C5にメチル基を欠いているγ-分子は、その部位において容易にニトロ化される(Mortonら、2002;Christenら、1997)。
【0015】
ビタミンE異性体の親油性側鎖は、飽和イソプレニルユニットを有するトコフェロールと飽和イソプレニルユニットを有するトコトリエノールを区別する。疎水性ドメイン(III)は、分子がリポタンパク質および膜にそれぞれ結合することができるかどうか、または第1相酵素によって分解されうるかどうかを決定する(Birringerら、2002;Neuzil and Massa、2005)。
【0016】
修飾ヒドロキシル基を有する多くのトコフェロール誘導体が、それらのアポトーシス促進活性について試験されている(第1表)。試験されたもののうち最も優れた誘導体は、クロマノール環のC6位にスクシニルエステルを有するα-TOS(エントリー1)である。その低いpKa(<6)により、α-TOSは、生理的条件下で完全に脱プロトン化され、ミトコンドリア膜を不安定にし、複合体IIにおける効果を有する界面活性剤様分子をもたらす。トコフェロールのジカルボン酸エステルは、構造と活性の関係(SAR)について最もよく研究された化合物を提供する。強いアポトーシス性化合物として、コハク酸α-トコフェロール(1)、シュウ酸トコフェロール(10)、およびマロン酸トコフェロール(11)が挙げられ、後の2つは、黒色腫細胞を接種されたマウスにおいて非選択的細胞毒性を誘導する(Kogureら、2005)。さらに大きいアポトーシス促進活性が、不飽和ジカルボン酸様α-トコフェリルマレエート(3)(Birringerら、2003)およびα-トコフェリルフマレートに観察されている。ジカルボン酸の鎖長の増加は、グルタミン酸(5)、メチル化グルタル酸(6、7、8)(Birringerら、2003)において明らかにされたように活性の低下をもたらし、ピメリン酸(24)(Kogureら、2004)はまったく活性を示さかった。
【0017】
全α-TOS分子が、その完全なアポトーシス誘導活性に必要であることが確立されている(Birringerら、2003)。遊離カルボキシル基のエステル化は、アポトーシス促進活性のない非荷電誘導体(9、25)をもたらす。酢酸トコフェリルおよびプロピオン酸トコフェリル(19)などの脂肪族カルボン酸は、それぞれ、メチルエーテル(18)と同様に不活性であった。α-TOSは腸内エステラーゼによって切断されるので、この化合物の経口投与は効果的ではない(Wuら、2004b;Cheesemanら、1995)。エステル結合切断の問題を解消するために、エステル結合を加水分解に耐性があるのでエーテル結合と置き換えて、化合物(20、21)および側鎖切断型(truncated)誘導体(42)が合成されている(Wuら、2004b;Nishikawaら、2003;Shunら、2004;Shiauら、2006)。メチレン基によるエーテル結合の置き換えは、アポトーシスを促進するのに十分であることに留意すべきである(22)(Sandersら、2001)。
【0018】
エステル結合をアミド結合と置き換える場合、さらに増強されたアポトーシス促進活性が観察された(12、13、37、38)(Tomic-Vaticら、2005)。また、不飽和アミド(13、38)は、飽和アミドよりも優れていた。エステル結合の代わりにアミド結合を導入する理論的根拠は、アニリンアミドは、対応するフェノールエステルよりも加水分解する傾向がほとんどないという十分に確立された事実に基づいていた。これらのトコフェリルエステル誘導体の安定性を増強することは、インビボにおいてこれらの分子を保護し、より長く無傷でいることを可能にし、そのことによってそれらのバイオアベイラビリティが増加する。アミドによるエステルのイソステリック置換は、その連結を酵素加水分解しにくくもする。いつかの非特異的エステラーゼは、腸内粘膜細胞および血液中に存在する。対照的に、ペプチドは、より狭い特異性を示す。たとえば、アミド結合にアミノ酸を有するプロドラッグは、腸および血液内で、それらの対応するエステル類縁体よりも安定である(Sugawaraら 2000)。
【0019】
化合物の最後のグループは、一連のプラスに荷電したN-末端を有するリシンα-トコフェリルエステルからなった(15-17)。親水性アンモニウム官能基は、そのカルボキシレート対応物と同様のアポトーシス促進効果を発揮したが、一般的モチーフが、親油性側鎖および親水性頭部基からなる活性にとって必要であることを示唆する。しかし、長鎖脂肪族アルコールのスクシニルエステル(たとえば、フィトールおよびオレオール)は、どのような活性も示さなかった(Birringerら、2003)。
【0020】
一般的な構造と活性の関係は、第1表に示すデータから引き出すことができる。
1.絶大なプロオキシダントおよびアポトーシス促進活性を得るために、機能的ドメインIの修飾は、解離した酸または荷電したアンモニウム基からなる親水性頭部基を要求した。
2.機能的ドメインの鎖長および不飽和の程度が、アポトーシス活性を決定した。コンホメーション的制限が、活性を増強するように見えた。
3.機能的ドメインの化学結合は、エステルに限定されず、他の官能基は、誘導体の酵素的分解を防止した。
【0021】
クロマノール環の置換パターンは、トコフェロールの抗酸化特性に関係するのみではないことが多い(Azziら、2002)。異なる生化学的観察が、シグナル伝達および代謝過程におけるα-トコフェロールの役割を強調している。したがって、α-トコフェロールは、肝臓内で、他のトコフェロールおよびトコトリエノールと比べてα-トコフェロールに高い親和性をもつ32 kDaのタンパク質であるα-トコフェロールトランスファータンパク質(α-TTP)によって選択的に認識される。α-TTPへの相対的親和性は、クロマノール環のメチルバイオアベイラビリティの損失とともに減少する (α-トコフェロール 100 %、β-トコフェロール 38%、γ-トコフェロール 9%およびδ-トコフェロール 2%)(Hosomiら、1997)。近年発見されたトコフェロール関連タンパク質(TAPs)は、トコフェロール結合に同様の優先傾向を示している(Yamauchiら、2001)。内皮細胞において、トロンビン誘発PKC活性化およびエンドセリン分泌は、α-トコフェロールによって阻害されるが、β-トコフェロールによっては阻害されない(Martin-Nizardら、1998)。転写レベルにおいて、α-トコフェロールは、α-トロポミオシン発現のアップレギュレーション(Aratriら、1999)ならびにLDLスカベンジャー受容体SR-AおよびCD36のダウンレギュレーションを引き起こすが、β-トコフェロールは、効果はない(Ricciarelliら、2000;Devarajら、2001)。さらに、細胞培養において、γ-およびδ-トコフェロールは、α-またはβ-トコフェロールよりも、より速く分解されるので(Birringerら、2001)、置換パターンは、側鎖分解の速度に関与すると思われる。4つのトコフェロール異性体のスクシニル化は、化合物1、32、33および35を生成する。これらのα-TOS(1)が、最も高いアポトーシス活性を有し、β-TOS(32)、γ-TOS(33)と続き、δ-TOS(35)が最も効果が低いことは驚くには当たらない(Birringerら、2001)。一般に、トコフェロールファミリーのメチル化の程度の高いメンバーが、最も強力であるが、この傾向は、トコトリエノールでは逆である(後記参照)。
【0022】
短縮された側鎖を有する水溶性ビタミンE誘導体であるトロロックスのスクシニル化は、アポトーシス促進活性の完全な損失をもたらした。前立腺ガン細胞において、最高レベルのアポトーシス活性を示した、切断されたフィトール側鎖を有する種々のコハク酸トコフェロールの構造と活性の関係の実験(第2表、43、44、45)は、側鎖長が2つのイソプレニルユニットである誘導体から得られた。コンピューターを使った分子モデリングおよび免疫共沈降実験は、Bcl-xLおよびBcl-2へのBak BH3ペプチドの結合が、トコフェロール類縁体によって阻害されたことを示した(Shiauら、2006)。抗腫瘍活性のための中心的要求は、クロマノール環のスクシニル化および1つのイソプレニルユニットの最短の鎖長であった(42、46)。ドメインIII側鎖に結合したエーテル/エステルを有する一連のトコフェリルリシンエステルは、鎖長とIC50(47-50)との間に負の相関関係を示した(Aryaら、1998)。
【0023】
トコトリエノールは、効率的な抗ガン剤であり、そのアポトーシス促進特性は、タンパク質のRasファミリーの不活性化に関連する。トコトリエノールは、機能的ドメインの修飾することなく、そのアポトーシス促進活性を示す。シグナル伝達ドメインにおける階層も逆転され、δ-トコトリエノール(59)をマウスB16-F10黒色腫細胞モデルにおける最も強力な作用剤にし、次に、γ-(56)およびα-トコトリエノール(53)が続く(第3表;Heら、1997)。興味深いことに、すべての芳香族メチル基を欠いているデスメチルトコトリエノール(60)は、IC50が0.9 μMという幾分高い活性を示す。この化合物は、米ぬかから単離されている(Qureshiら、2000)。Rasタンパク質の膜係留システインが、通例の構造エレメントであるファルネシル鎖によって修飾されるので、トコトリエノールの直接阻害作用が提案されている。したがって、活性化しているras突然変異を含むヒト肺腺ガン細胞系であるA549細胞において、トコトリエノールによって、Rasファルネシル化およびRhoAプレニル化が阻害された(Yanoら、2005)。トコトリエノールの短いインビボ半減期を延ばすために、機能的ドメインが導入されている。これらの修飾は、分子の抗増殖活性も増強した(54、57、58)。側鎖の切断もまた、化合物55について見られたのと同様に、活性を改良した。
【0024】
機能的ドメインに修飾が行われた化合物の多くは、抗増殖活性を示し、さらなる特殊化された特性を提供した。たとえば、α-トコフェリルポリエチレングリコールコハク酸エステル(23)は、ドラッグデリバリーシステムのためのビヒクルとして用いられている。この化合物は、ヌードマウスに移植されたヒト肺ガン細胞に対する抗ガン活性を有することが明らかにされた。この化合物のアポトーシス誘導効果は、その細胞への取り込みの増加によるものではなく、むしろ、活性酸素種を生成する能力の増大によるものであった(Youkら、2005)。α-トコフェリルホスフェート(30)は、トコフェロール関連シグナル伝達中に起こる代謝から得られると考えられている(Negisら、2005)。30およびジ-α-トコフェリルホスフェート(31)の混合物は、ラット大動脈平滑筋細胞およびヒトTHP-1単球白血病細胞の増殖を阻害した(Munteanuら、2004)。著者らは、コハク酸トコフェリルおよびマレイン酸トコフェリルが、リン酸トコフェリルを模倣し、置換することによって、ガンにおいて作用し、それによって細胞シグナル伝達の持続的活性化を引き起こすことを提案した。
【0025】
すべての-transレチノイン酸(28)および9-cisレチノイン酸(29)という2つの実験的α-トコフェリルエステルをそれぞれ用いて、急性前骨髄球性白血病細胞の増殖を減少させている(Makishimaら、1998)。レチノイド受容体反応性リポーター構築物を用いるトランス活性化実験は、これらの化合物の両方が、レチノイン酸受容体(RARs)に対するアゴニストとして作用したことを明らかにした。尿中に分泌されているのが見出されることが多いγ-トコフェロールの分解産物であるカルボキシエチルヒドロキシクロマン(52)は、サイクリンD1発現を阻害することによってPC-3前立腺ガン細胞の細胞増殖を低下させることができる(Galliら、2004)。
【発明の概要】
【発明が解決しようとする課題】
【0026】
本発明者らは、特に、呼吸鎖の複合体II(コハク酸ユビキノン酸化還元酵素)を標的化することを手段として、ガン性細胞のミトコンドリア呼吸鎖に沿った電子の伝達を選択的に破壊することが、ROSの生成およびこれらの細胞の死をもたらしうることを見出した。さらに詳しくは、本発明者らは、ビタミンEのプロオキシダント型が、ガン性細胞の複合体IIのユビキノン結合部位からユビキノンを選択的に置き換え、電子の正常な伝達を遮断する能力がある有効な抗ガン化合物であることを見出した。
【課題を解決するための手段】
【0027】
(発明の詳細な記載)
本発明の第1の態様にしたがって、ガン性細胞のミトコンドリア呼吸鎖の複合体II(コハク酸ユビキノン酸化還元酵素)と相互作用して、細胞内に活性酸素種を生成する能力がある、ガン性細胞の死を選択的に誘導するためのプロオキシダント化合物を提供する。
【0028】
本発明の第2の態様にしたがって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力がある、ガンを予防または治療するためのプロオキシダント化合物を提供する。
【0029】
本発明の第3の態様にしたがって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者においてガン性細胞の死を誘導する方法を提供する。
【0030】
本発明の第4の態様にしたがって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者におけるガンを予防または治療する方法を提供する。
【0031】
本発明の第5の態様にしたがって、ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0032】
本発明の第6の態様にしたがって、ガンの予防または治療のための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0033】
本発明の第7の態様にしたがって、ガン性細胞の存在を検出し、ガン性細胞の死を選択的に誘導するプロオキシダント化合物であって、該化合物が、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有する化合物を提供する。
【0034】
本発明の第8の態様にしたがって、患者においてガン性細胞の存在を検出し、ガン性細胞の死を選択的に誘導するための方法であって、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有するプロオキシダント化合物の治療有効量を患者に投与するステップを含む方法を提供する。
【0035】
いすれかの適当なタイプのプロオキシダント化合物を用いることができ、該化合物は、ミトコンドリア複合体IIといずれかの適当な手段で相互作用することができる。好ましくは、化合物は、複合体IIのユビキノン結合部位に結合し、複合体IIと優先的に相互作用する天然の基質ユビキノン、ユビセミキノンまたはユビキノール(コエンザイムQ)またはその他のユビキノンもしくは関連化合物を容易に置き換えることができる。このような基質は、たとえば、Briereら、2004、Tanら、1993、and Espostiら、1996において特定化されている。
【0036】
本発明の第9の態様にしたがって、ガン性細胞の死を選択的に誘導するための、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物を提供する。
【0037】
本発明の第10の態様にしたがって、ガンを予防または治療するための、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物を提供する。
【0038】
本発明の第11の態様にしたがって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者においてガン性細胞の死を誘導する方法を提供する。
【0039】
本発明の第12の態様にしたがって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量を患者に投与するステップを含む、患者におけるガンを予防または治療する方法を提供する。
【0040】
本発明の第13の態様にしたがって、ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0041】
本発明の第14の態様にしたがって、ガンの予防または治療のための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用を提供する。
【0042】
本発明の第15の態様にしたがって、本発明の第1、第2、第9または第10の態様のプロオキシダント化合物またはその生理的に許容しうる塩および生理的に許容しうる担体を含む医薬組成物または獣医用組成物を提供する。
【0043】
アポトーシスは、ガン性細胞のミトコンドリアにおける活性酸素種のレベルの増加の結果として単独に起こってもよく、または該化合物は、細胞内のミトコンドリア依存性細胞死シグナル伝達経路の活性化を目的として、さらにアポトーシス促進性であってもよい。好ましくは、該化合物は、複合体IIへの結合を目的として活性酸素種を生成し、さらにミトコンドリア依存性細胞死シグナル伝達経路の活性化を目的としてアポトーシス促進性である。
【0044】
好ましくは、プロオキシダント化合物は、プロオキシダント活性を欠いている無害な形体へと、非ガン性細胞内で切断、処置または代謝される。
【0045】
好ましくは、化合物は、プロオキシダントビタミンE類縁体である。本発明者らは、プロオキシダントビタミンE類縁体が、複合体IIを結合し、ユビキノンへの電子伝達を中断させることができることを見出している。本発明者らは、プロオキシダントビタミンE類縁体が、アポトーシス促進性であることも見出している。本発明者らは、have further found thatプロオキシダントビタミンE類縁体が、非ガン性細胞において、無害な抗酸化剤に処置されうることをさらに見出している。
【0046】
対照的に、これも複合体IIに結合するテノイルトリフルオロアセトン(TTFA) (Sunら、2005)は、すべての細胞にとって毒性である(Zhangら、2001b)。3-BPもまた、複合体IIに結合するが、プロオキシダント活性を有すると特徴づけられっていない。
【0047】
「プロオキシダントビタミンE類縁体」は、ガン性細胞のミトコンドリアに位置する場合、酸化還元サイレントであり、複合体IIのユビキノン結合部位に結合して、細胞にダメージを引き起こす代謝の酸素副産物の生成の引き金をひく能力があるビタミンE類縁体として定義づけられる。プロオキシダントビタミンE類縁体の例は、α-トコフェリルコハク酸エステル(α-TOS)である。
【0048】
一方、「抗酸化剤ビタミンE類縁体」は、ガン性細胞のミトコンドリアに位置
する場合、たとえば、α-トコフェロール(α-TOH)などの抗酸化(酸化還元)活性があるビタミンE類縁体である。したがって、プロオキシダントビタミンE類縁体およびビタミンE類縁体の生物活性は、正反対である。
【0049】
特に好ましいプロオキシダントビタミンE類縁体の例を上記第1−3表、下記第4−5表に示し、本明細書の従来技術の欄で議論する。
あるいは、化合物は、以下から選ぶことができる:
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
Gu LQら、1990;He DYら、1994、Yang Fら、1991、Yankovskayaら、1996、およびYabunakaら、2002に記載のユビキノン誘導体(これらは、全体として相互参照することにより本発明に援用される)。
【0050】
化合物は、たとえば、肺、肝臓、腎臓、脳、前立腺、乳房、卵巣、リンパ、皮膚、眼、結腸、胃、口腔扁平および造血系などの患者におけるいずれかの型のガン性細胞の死を誘導するために用いることができる。
【0051】
驚いたことに、化合物α-TOSは、erbB2-低または高ガン細胞を効率的に殺すことが本発明者らによって見出されている。化合物α-TOSは、中皮腫を治療することも本発明者らによって見出されている。
【0052】
驚いたことに、化合物α-TOSは、正常酸素圧および低酸素性の両方のガン性細胞の死を誘導することが本発明者らによって見出されている。したがって、化合物は、患者における早期および後期段階の両方の腫瘍の死を誘導することができるという利点を有する。
【0053】
治療の対象は、ヒト、哺乳類または動物であってよい。好ましくは、患者は、ヒトまたは他の哺乳類である。
【0054】
化合物は、組成物中に、その医薬的にまたは獣医学的に許容しうる誘導体として含まれてもよい。本明細書で用いる本発明化合物の「誘導体」は、塩、Mn2+およびZn2+などの金属イオンとの配位錯体、インビボ加水分解可能エステルなどのエステル、遊離酸もしくは塩基、水和物またはプロドラッグが包含される。リン酸または硫酸などの酸性基を有する化合物は、Na、K、MgおよびCaなどのアルカリまたはアルカリ土類金属およびトリエチルアミンおよびトリス(2-ヒドロキシエチル)アミンなどの有機アミンとの塩を形成することができる。塩は、アミンなどの塩基性基と、塩酸、リン酸または硫酸などの無機酸と、または酢酸、クエン酸、安息香酸、フマル酸または酒石酸などの有機酸を有する化合物との間に形成されることができる。酸性および塩基性基を有する化合物は、内部塩を形成することができる。
【0055】
エステルは、当業者に公知の技術を用いて、化合物内に存在する水酸基またはカルボン酸基と適当なカルボン酸またはアルコール反応パートナーとの間に形成されることができる。
【0056】
組成物は、考慮中である特定の状況に必要な予防有効量または治療有効量のいずれかで患者に投与することができる。組成物中の化合物の実際の量および組成物の投与の速度および経時経過は、治療または予防を必要とするガンの性質および重篤度に応じて変わる。用量の決定などの治療の処方は、患者の介護に責任を負うべき医師または獣医によってなされる。しかし、典型的には、ヒトの患者に投与するための組成物は、約0.01〜100 mgの化合物/体重kg、より好ましくは、約0.1〜10 mg/体重kgである。α-トコフェリルスクシネートまたは他の類縁体が、ヒト患者に経皮適用される場合、化合物の血清レベルは、IC50値の近辺、約40-50 μMであるのが好ましい。
【0057】
組成物は、非経口、局所、経口、吸入スプレー、直腸内、経鼻、バッカル、膣内または埋め込み貯蔵器などの適当な経路で患者に投与することができる。本明細書で用いる用語「非経口」は、皮下、静脈内、筋肉内、動脈内、滑液内、胸骨内、髄腔内、肝内、病巣内および頭蓋内注射または輸液技術を包含する。
【0058】
担体は、いずれかの適当な希釈剤、アジュバント、賦形剤、緩衝剤、安定化剤、等張剤、保存剤または抗酸化剤を含むことができる。担体が、非毒性であり、本発明化合物の有効性を妨げないべきであるのは当然のことである。担体または組成物へのいずれかの他の添加剤の正確な性質は、投与経路および必要とされる治療の型に応じて変わる。たとえば、Alfonso R.Gennaro.Remington:The Science and Practice of Pharmacy、20th Edition.Baltimore、MD:Lippincott Williams & Wilkins、2000、and Goodman and Gilman’s The Pharmaceutical Basis of Therapeutics、Pergamon Press、New York、NYを参照のこと(これらは、全体として参照することにより本発明に援用される)。医薬組成物は、たとえば、従来の混合、溶解、造粒、糖衣形成、磨り潰し、乳化、カプセル化、封入または凍結乾燥処置によって製造することができる。
【0059】
組成物の滅菌注射剤形は、水性または油性懸濁液であってよい。このような剤形は、当業者には公知である。処置するのが望ましい部位での静脈内、皮膚または皮下注射用には、組成物は、適当なpH、等張性および安定性を有する非経口的に許容しうる水性溶液の剤形であってよい。
【0060】
組成物の経口的に許容しうる投与剤形として、カプセル剤、錠剤、丸剤、散剤、リポソーム、顆粒剤、スフィア、糖衣錠、液剤、ゲル剤、シロップ剤、スラリー剤、懸濁剤などが挙げられる。適当な経口剤形は、当業者には公知である。錠剤は、ゼラチンなどの固体担体、アジュバントまたは不活性希釈剤を含むことができる。液体医薬組成物は、一般に、水、石油、動物または植物油、鉱物油または合成油などの液体担体を含む。生理食塩水、またはエチレングリコール、プロピレングリコールまたはポリエチレングリコールなどのグリコールを含んでもよい。このような組成物および製剤は、与えられた担体におけるその溶解度に応じて、一般に、少なくとも、0.1 重量%の化合物、好ましくは、約25 重量%の化合物を含む。
【0061】
組成物は、特に治療の標的が、眼、皮膚または下部消化管のガンなどの局所適用によって容易に到達可能である領域または器官である場合に、局所投与されてよい。組成物は、液体、懸濁液、エマルジョン、軟膏、クリーム、ローション、ペースト、ゲル、泡またはエアロゾルの剤形で適用されてよい。適当な局所剤形は、当業者には公知である。
【0062】
組成物は、生物学的効力を損失することなく、化合物を特定の器官、組織またはガンの型にデリバリーするため、および/または、化合物が、たとえば皮膚を通して吸収されるか、または消化管を介して摂取されることができるのを確実にするために、デリバリービヒクルを含んでもよい。デリバリービヒクルは、たとえば、脂質、ポリマー、りぽそーむ、エマルジョン、抗体および/またはタンパク質であってよい。たとえば、中皮腫を治療するために皮膚を介して化合物をデリバリーするには、リポソームが特に好ましい。
【0063】
化合物を含む固体疎水性ポリマー半透性物質などの持続放出システムを用いて、組成物をデリバリーすることができる。様々な持続放出物質が、入手可能であり、当業者には周知である。持続放出カプセル剤は、約1〜20週間化合物を放出する。
【0064】
化合物は、プロドラッグの形態であってもよい。プロドラッグは、組成物がたとえば経口などで摂取される場合に化合物の活性が損なわれないように保護基を有することができる。プロドラッグは、特定の器官または細胞型に活性化合物をデリバリーすることができる。適当なプロドラッグ剤形および保護基は、当業者には公知であろう。受容体であるチロシンキナーゼerbB2を過剰発現しているガン細胞を標的化するために、α-TOSのアダクトが、ヘプタペプチドLTVSPWYに結合するのが好ましい。
【0065】
患者は、化合物とともに最適予防または治療効果を達成するために1種以上の他の活性成分を含む組成物を投与されてもよい。活性成分は、たとえば、アルキル化剤、血管新生抑制薬、抗アンドロゲン薬、抗エストロゲン薬、代謝拮抗剤、アポトーシス剤、アロマターゼインヒビター、細胞周期制御薬、細胞ストレッサー、細胞障害剤、細胞保護剤、ホルモン剤、免疫療法薬、キナーゼインヒビター、モノクローナル抗体、白金薬、呼吸抑制剤、レチノイド、シグナル伝達阻害剤、タキサンおよびトポイソメラーゼインヒビターであってよい。特に好ましい作用剤として、2-デオキシグルコースおよび3-BPなどの解糖インヒビターが挙げられる。他の特に好ましい作用剤として、TRAILおよびAkt1インヒビターならびにRalph SJら、2006、Neuzil J、Tomasettiら、2007およびNeuzil J、Dongら、2007(これらは、全体として相互参照することにより本発明に援用されるに概説される)他のマイトカンが挙げられる。
【0066】
本発明者らは、たとえば、ガン細胞が、α-TOSと3-BPとの併用(ならびに他の複合薬)によって、いずれかの薬物を単独で用いる場合と比べて、殺傷に対してより感受性が高くなることを見出している。
【0067】
組成物は、非経口または局所投与されるのが好ましい。特に好ましいプロオキシダントビタミンE類縁体は、α-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステル、α-トコフェリルマレイルアミドおよび2,5,7,8-テトラメチル-2R-(4R,8R,12-トリメチルトリデシル)-クロマン-6-イルオキシ酢酸(α-トコフェリルオキシ酢酸)である。エステルであるα-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルおよびα-トコフェリルマレイルアミドのための好ましい担体は、リポソームベースクリーム「Lipoderm」などの経皮適用可能なクリームである。非加水分解性エーテル類縁体であるα-トコフェリルオキシ酢酸が、経口デリバリーに好ましい。
【0068】
検出可能な部分のいずれかの適当なタイプを用いることができ、化合物をいずれかの適当な方法で検出することができる。検出可能な部分は、蛍光色素(たとえば、ローダミン123など)または放射性標識カチオンである。このような検出可能な部分は、たとえば、光線力学療法などの付加的抗ガン療法の同時実践を可能にする。
【0069】
本発明の第16の態様にしたがって、ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)第1のガン性細胞のミトコンドリアに試験化合物を導入し、第2のガン性細胞にガン細胞死を誘導しないことがわかっているコントロール化合物を導入し;次いで、
(ii)ミトコンドリア複合体IIへの化合物の結合および第1および第2のガン性細胞内での活性酸素種の生成をアッセイし、試験化合物およびコントロール化合物によるガン性細胞の死ををアッセイする;
[ここで、コントロール化合物と比較しての試験化合物による複合体II結合、活性酸素種の生成および第1のガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法を提供する。
【0070】
好ましくは、化学物質は、複合体IIのユビキノン結合部位に結合するプロオキシダントビタミンE類縁体である。
【0071】
好ましくは、試験化合物を同時にスクリーニングすることができるハイスループットスクリーニングを用いる。
【0072】
本発明の第17の態様にしたがって、本発明の第16の態様の方法によって同定される化学物質を提供する。
【0073】
本発明の第18の態様にしたがって、ミトコンドリア呼吸鎖複合体IIまたは1種以上のそのサブユニットに結合した本発明の第1、第2、第9または第10の態様の化合物を含む単離または精製された複合体を提供する。
【0074】
好ましくは、化合物が、α-TOSなどのプロオキシダントビタミンE類縁体である。好ましくは、ミトコンドリア呼吸鎖複合体IIが、哺乳類由来であり、より好ましくは、ヒト由来である。
【0075】
本発明の第19の態様にしたがって、ミトコンドリア呼吸鎖複合体IIまたは1種以上のそのサブユニットに結合した本発明の第1、第2、第9または第10の態様の化合物を含む結晶または結晶化可能な組成物を提供する。
【0076】
好ましくは、化合物が、α-TOSなどのプロオキシダントビタミンE類縁体である。好ましくは、ミトコンドリア呼吸鎖複合体IIが、哺乳類由来であり、より好ましくは、ヒト由来である。
【0077】
本発明の第20の態様にしたがって、ミトコンドリア呼吸鎖複合体IIのユビキノン結合部位に結合する能力がある化学物質を同定または設計する方法であって、
(i)呼吸鎖複合体IIと、複合体IIに結合してユビキノンへの電子流を妨げる少なくとも1つのビタミンE類縁体との間の相互作用をコンピューターモデリングし;次いで、
(ii)コンピューターモデリングによって作成されたデータを用いて、複合体IIに結合してユビキノンへの電子流を妨げる能力を有する化学物質を同定または設計する;
ステップを含む方法を提供する。
【0078】
好ましくは、α-TOSなどの少なくとも1つのビタミンE類縁体が、プロオキシダント活性を有し、活性酸素種の生成を促進する。より好ましくは、複合体IIに結合してユビキノンへの電子流を妨げる複数のビタミンE類縁体をステップ(i)で用いる。
【0079】
好ましくは、ミトコンドリア呼吸鎖複合体IIが、哺乳類由来であり、より好ましくは、ヒト由来である。
【0080】
本発明の第21の態様にしたがって、本発明の第20の態様によって同定または設計された化学物質を提供する。
【0081】
好ましくは、化学物質が、ユビキノンに等しいか、またはより大きい複合体II結合親和性を有するプロオキシダントビタミンE類縁体であるか、または以下から選ばれる化学物質である:
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
Gu LQら、1990;He DYら、1994、Yang Fら、1991、Yankovskayaら、1996、およびYabunakaら、2002に記載のユビキノン誘導体。
【0082】
本発明の第22の態様にしたがって、ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)ガン性細胞のミトコンドリアに試験化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位の天然のユビキノン基質の置換をアッセイし、ガン性細胞の死をアッセイする;
[ここで、試験化合物による天然のユビキノン基質の置換およガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法を提供する。
【0083】
本発明の第23の態様にしたがって、本発明の第22の態様によって同定された化学物質を提供する。
【0084】
ヒト神経悪性腫瘍、褐色細胞腫および傍神経節腫、ならびに平滑筋腫および腎細胞ガンの特定の型の研究は、それによってガン細胞がそれら自身の生存を促進する1つの保護装置への洞察力を提供した。この過程は、ガンの低酸素性の発達のための通常の経路とは区別して、「偽性低酸素」として知られるようになっている。これは、これらのガンが、酸素のレベルが正常であっても細胞があたかも低酸素性であるかのように振る舞うようにする突然変異を示すからである。したがって、偽性低酸素性ガンにおいて、体細胞変異は、細胞質ゾルにおけるグルコース代謝とミトコンドリアにおける酸化的ホスホリル化との間のトリカルボン酸(TCA)回路連鎖の一部である酵素であるコハク酸デヒドロゲナーゼ(SDH)またはフマル酸ヒドラターゼ(FHまたはフマラーゼ)のいずれかにおいて起こる。SDHAは、ユビキノン酸化還元酵素複合体IIの3つの他のサブユニットであるSDHB、SDHCおよびSDHDに関連する膜結合タンパク質として内部ミトコンドリア膜のマトリッククス側に位置する酵素サブユニットをコードする。SDHB、SDHCまたはSDHD遺伝子における遺伝性または体細胞突然変異は、褐色細胞腫または傍神経節腫に関連するが、FH遺伝子におけるそのような変異は、平滑筋腫、平滑筋肉腫または人材棒ガンを引き起こす(Gottlieb EおよびTomlinson IP、2005;King Aら、2006;Astuti Dら、2003;Bayley JPら、2006)。
【0085】
SDHまたはFHをコードする遺伝子における突然変異の結果としてSDHまたはFHの酵素活性が低下するので、代謝中間体であるスクシネートおよびフマレートが、これらのガン細胞の内側で上昇することになる。SDHの酵素活性の低下の結果として、このようなガンは、α-TOSなどのプロオキシダント化合物による治療への感受性が高まるか、または低くなる。SDHまたはFHサブユニットをコードする遺伝子に突然変異を含むことが疑われる褐色細胞腫または傍神経節腫、平滑筋腫または腎細胞ガンのタイプのガンは、それらのSDHまたはFH酵素活性のレベルについての診断試験によって分析することができる。SDHの酵素活性は、600 nmにて測定された、細胞または肝臓ミトコンドリア調合液の存在下におけるCII基質であるDCPIPの測定に基づいて、King、TS、1967に記載されるような標準的手順に基づくことができる。反応混合物は、0.5 mM NADH、5 mMスクシネート、10 mM KCN、50 μM DCPIPおよび50 μM PMSを含む。別法として、たとえば、Berridge MVおよびTan AS、1993に概略された、そして、Berridge MVら、2005に概説された、複合体IIのSDH活性を分析するためのいずれかのテトラゾリウム色素還元アッセイがある。
【0086】
別法として、このような異常を検出するために、SDH突然変異の分子または免疫組織化学解析を用いることができる。方法は、PCRベースDNA配列決定、RFLP解析、遺伝子アレイ発現解析およびvan Nederveen FHら、 2007、Pollard PJら、2005およびBayley JPら、2006に記載の例のような同様の技術を包含する。
【0087】
本発明の第24の態様にしたがって、ガンを有する患者が、本発明の第1または第2の態様のプロオキシダント化合物の投与を含む療法に反応する可能性があるかどうかを試験する方法であって、
(i)該患者由来のガン性細胞のミトコンドリアにプロオキシダント化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位へのプロオキシダント化合物の結合をアッセイし、ガン性細胞の死をアッセイする;
[ここで、プロオキシダント化合物の結合およびプロオキシダント化合物によるガン性細胞の死は、患者が、療法に反応する可能性があることを示す]
ステップを含む方法を提供する。
【図面の簡単な説明】
【0088】
【図1】α-TOS、3-BPおよびTRAILは、乳ガン細胞においてATPの還元を引き起こす。示した濃度および示した期間において、α-TOS、3-BPまたはTRIALによって、MDA-MB-453、MCF-7およびNeuTL乳ガン細胞を処置した。次いで、「一般的材料および方法」に記載のATP生物発光アッセイキットを用いて、ATPレベルを評価した。示したデータは、平均値±S.D.(n=5)を表す。記号「*」は、非処置グループと比較しての細胞のATPのレベルにおける有意な差(p<0.05)を意味する。
【図2】α-TOSおよび3-BPによる乳ガン細胞系におけるアポトーシスの誘導。正常酸素圧または低酸素のいずれかにおいて、示した濃度で24時間、α-TOSまたは3-BPに、MCF-7(A、B)およびMDA-MB-453細胞(C、D)を曝露し、アネキシンV-FITC法を用いてアポトーシスを評価した。示したデータは、平均値±S.D.(n=3)を表す。記号「*」は、非処置グループと比較しての処置された細胞のアポトーシスのレベルにおける有意な差(p<0.05)を意味する。
【図3】MDA-MB-453細胞でのアポトーシス誘導におけるα-TOSおよび3-BPの併用効果。正常酸素圧または低酸素のいずれかにおいて、示した濃度にて、α-TOSまたは3-BP単独で、あるいはα-TOS+3-BPの併用で、MDA-MB-453細胞を24時間処置した。時間、α-TOSまたは3-BPに、MCF-7(A、B)およびMDA-MB-453細胞(C、D)を曝露し、アネキシンV-FITC法を用いてアポトーシスを評価した。示したデータは、平均値±S.D.(n=3)を表す。記号「*」は、単独薬物処置と比較してのα-TOS+3-BPの併用によって処置された細胞のアポトーシスのレベルにおける有意な差(p<0.05)を意味する。
【図4】Akt1ノックダウンは、MDA-MB-453細胞をα-TOS、3-BPおよびTRAILに対して感受性にする。60 pM Akt1 siRNAで、50%コンフルエントにおけるMDA-MB-453細胞を24時間前処置した後、示した濃度にて、α-TOS、TRAILまたは3-BPで、さらに24時間処置した。アネキシンV-FITC法を用いてアポトーシスを評価した。2つの独立した実験を行った。結果を平均値±S.D.で示す。
【図5】NeuTL細胞のMTTを還元する能力における3NPA、BP、TTFAおよびα-TOSの効果。(A)示した濃度にて用いた3NPAまたは3-BPの存在下、4時間の共インキュベーション後、PBS中でのMTT還元を評価した。(B)MitoQで細胞を60分間プレインキュベートした後、示した濃度にて3-BPを添加し、2時間インキュベートした後、PBS中でMTTを還元する能力を評価した。20 mMコハク酸(pH 7.4)を含むRPMI中でのMTTを還元する能力について、MitoQで60分間前処置した細胞を、示した濃度の(C)TTFAまたは(D)α-TOSの存在下で2時間インキュベートした後に評価した。結果を、コントロール(非処置)±S.D.と比較してのMTTの平均還元%で表す。記号「*」は、有意な差(p<0.05)を意味する。
【図6】単離されたラット肝臓ミトコンドリア(A、C)またはパラコッカス・デニトリフィカンス(B、D)における、α-TOSによるSDH/複合体II活性の阻害。「一般的材料および方法」に概略したように細胞を分画し、ラット肝臓由来のミトコンドリアまたはパラコッカス・デニトリフィカンス由来の膜の調合液を、ミトコンドリアSDH/複合体II活性を促進し、インジケーター色素DCIP+PMSを含む反応体積中でインキュベートした。AおよびB中のサンプルは、スクシネートを含み、示したように100または300 μMのα-TOSのいずれかで処置した。コントロール反応物は、希釈液DMSO単独の等価の添加を受けた。波長600 nmにおける吸光度を測定し、酵素活性(μmol/分/ml)における変化として表した。棒グラフAおよびBにおける結果は、平均値±S.D.(n=3)を表す。記号「*」は、コントロールからの有意な差(p<0.05)を意味する。パネルCおよびDは、100または300 μMのα-TOSの不在または存在下で、示されたスクシネートの異なる濃度下で測定された反応速度(μmol/分)における変化を示す。
【図7】α-TOSによるアポトーシス誘導は、CII機能不全細胞において抑制される。ヒトCybL(B9rec)でのトランスフェクションによる複合体II再構築の後に、親(B1)、CI-機能不全(B10)、CII-機能不全(CybL-突然変異;B9)およびCybL-突然変異細胞を、示した濃度および時間にてα-TOSに曝露し、収穫し、ROS蓄積(A)、基質としてのスクシネートによるMTT還元に基づいて全細胞において評価されたSDH活性(B)およびアポトーシス(C)について評価した。CybLまたは非特異的(NS)siRNAでMCF7細胞を前処置し、示したようにα-TOSに曝露し、ROS蓄積(D)、SDH活性(E)およびアポトーシス誘導(F)について評価した。パネルGは、ヒトSDHCSDHCプライマーを用いるB1、B10、B9またはB9rec細胞ならびにsiRNA処置MCF細胞のRT-PCR解析の結果を示す。パネルHは、ヒトモノクローナル抗-SDHC IgGB1を用いる、B10、B9およびB9rec細胞のウエスタンブロッティングの結果を示す。ウエスタンブロッティングは、異なるSDHC siRNA二本鎖およびNS siRNA(I)で処置したMCF7細胞におけるSDHCのレベルを記録するためにも示し、アクチンバンド(J)と比較して評価した。結果は、平均値±S.D.(n=3)を表し、画像は、3つの独立した実験の代表である。記号「*」は、コントロールからの有意な差(p<0.05)を意味する。
【図8A】分子モデリング。複合体II結晶構造におけるQP結合部位に結合したα-TOS構造と比較してのUbQ分子の最良のドッキングポーズ。画像は、棒線画として表した結合した薬物を有する空間充填モードにおける複合体IIの横断面図/浮き彫りとして示す。同定のために様々な成分をラベル付けする。
【図8B】分子モデリング。棒線画様式での複合体II結晶構造から引き出されたこの部位のための相対的UbQ構造と比較してのQP結合部位へのα-TOSの最良のドッキングポーズ。結合に関わる複合体IIの重要なアミノ酸残基を同定し、それぞれについてそれらのサブユニット鎖および残基数にしたがってコード付する。α-TOS構をハイライトし、より暗い棒線画としてラベル付する。
【図8C】分子モデリング。重要なアミノ酸残基および複合体II QP結合ポケットとα-TOSの相互作用に関与するそれらの側基の空間分布の三次元視覚化を提供する(B)から誘導された立体画像。驚いたことに、α-TOSが、UbQ分子よりもポケットにより深く貫通し、水素結合を提供するSDHC(CybL)サブユニットのセリン42への結合を調和させるのを見ることができる。
【図8D】分子モデリング。α-TOSの最良結合コンホメーションと結合にとって重要なQP結合部位アミノ酸残基との間の主な相互作用を示すLigplot図。
【図8E】ポケットの正面を横切って伸びる複合体IIの一部によって形成された橋(半透明の形式)を示す、分子モデリング。QD結合部位へのα-TOSの最良ドッキングポーズ;ポケットの内部に包まれているUbQと比較してのα-TOSの結合。
【図8F】分子モデリング。α-TOSの最良結合コンホメーションと結合にとって重要なQP結合部位アミノ酸残基との間の主な相互作用を示すLigplot図。複合体IIのQdポケットにおけるα-TOSの結合相互作用に関与する重大な意味を持つ空間分布。注:TOSのスクシニル基と複合体II SDHDサブユニットのリシン残基D128、D135との間の水素結合(点線)。
【図8G】分子モデリング。UbQ5およびα-TOSの化学構造ならびに複合体IIのQPおよびQD部位に結合する構造のそれぞれについて計算された相互作用エネルギー。注:パネルAおよびEは、Astex Viewer(Hartshorn、2002)を用いて作製したが、パネルDおよびFは、Ligplot(Wallaceら、1995)を用いて作製した。
【図9】第4表および第5表(後記)からの異なるTOS類縁体の生物活性と、複合体IIのQp部位へのそれらの計算された結合親和性との相関関係を示すグラフ。IC50:インビトロにおいてガン細胞の50%死亡を誘導するために特定の薬物について必要なマイクロモルで表す濃度。−log Kd:特定の薬物の結合親和性の逆対数。
【図10】分子モデリング−複合体II活性部位および他の化学種。ミトコンドリアの複合体II Qp活性部位に強力に結合する分子モデリングによって同定された10個の最良化学種の詳細。
【図11】共役ペプチドLTVSPWY-α-TOSは、乳ガンを抑制する。超音波画像化(USI)によって検出される乳ガンの7ヶ月齢以上のFVB/N202 c-neu雌性マウスを、示した時点でビヒクルまたはトウモロコシ油/エタノール(100:5、v/v)に可溶化したLTVSPWY-α-TOSの5 μmol/マウス/投与の腹腔内注射で処置した。USIを用いて腫瘍を視覚化し、定量し、処置(A)の開始時におけるそれらの体積と比較して表す。パネルBは、実験の第15日におけるコントロールマウス(左)および処置動物(5 μmol LTVSPWY-α-TOS /投与/マウス)(右)由来の腫瘍の画像を表し、それぞれ402.1および34.8 mm3の腫瘍体積を示す。体積は、両方の腫瘍について、実験の開始時において〜40 mm3であった、示したデータは、平均値±SD(n=4-6)であるUSI画像は、各グループ由来の腫瘍の代表である。
【図12】α-TOSによるマウスモデルにおける乳ガンの阻害。(A)ヌードマウスにMCF-7細胞を接種し、腫瘍が確立された後、3日に1回、腹腔内(i.p.)注射により動物を10 μモル/マウスのDMSOに溶解したα-TOSまたはDMSO単独で処置した。キャリパーを用いて腫瘍サイズを測定し、治療の開始時のガンのサイズと相関させた。各グループにおいて4匹の動物を用いた。示した結果は、平均値±S.E.である。記号「*」は、有意な差(p<0.05)を意味する。(B)触知可能な腫瘍を有する雌性FVB/N erbB2マウス(「一般的材料および方法」を参照)に、3日に1回、腹腔内注射により、トウモロコシ油/4%エタノールに可溶化した10 μモルのα-TOS (n=11)もしくはトウモロコシ油/4%エタノール単独(コントロール、n=9)か、または腫瘍部位に毎日、0.2 μモルの3-BP(n=8)/PBSのいずれかを与えた。キャリパーを用いて、腫瘍サイズを記録した。2つの独立した実験を行った。結果は、平均値±S.E.で表す。記号「*」は、3-BPおよびα-TOS曲線の間の分散解析における有意な差(p<0.05)を意味する。
【発明を実施するための形態】
【0089】
本発をより良く理解することができるように、以下の実施例によって説明する。これらの実施例は、説明のみを目的とするものであり、いかなる方法においても本発明の範囲を限定するものと解釈されるべきではない。
【0090】
一般的材料および方法
細胞培養および処置
erbB2を低レベルで発現するヒト乳ガン細胞系MCF-7および高レベルで発現するヒト乳ガン細胞系MDA-MB-453、ならびにFVB/N202ラットc-neuトランスジェニックマウス株由来のerbB2を高レベルで発現するマウス乳ガン細胞系NeuTL(Guyら、1992)をこの実験に用いた。湿気のある5% CO2環境において、37℃にて、10% FCS、100 U/mlのペニシリンおよび100 μg/mlのストレプトマイシンを加えたDMEM中で、細胞を培養した。10% FCS、10 g/mlのグルコースおよび非必須アミノ酸を加えたDMEM中で、ミトコンドリア複合体I欠損(BreenおよびScheffler 1979;DeFrancescoら 1976;Schefflerら 2004)および複合体II欠損細胞(Albayrakら 2003;Oostveenら 1995)を成長させた。
【0091】
約70%のコンフルエントにて、正常酸素圧または低酸素条件下で50 μM以下のα-TOS(Sigma)、100 μM以下の3-BP(Aldrich)または30 ng/mlのヒト組換え腫瘍壊死因子関連アポトーシス誘導リガンド(TRAIL)(Weberら、2002)で細胞を処置した。低酸素のために、窒素パージ培地で覆った細胞を、大量の窒素を予め流した低酸素室に置いた。次いで、細胞を入れた低酸素室を、要求に応じてインキュベーター内に置いた。α-TOSをエタノールに溶解し、細胞培養培地における最終エタノール濃度が≦0.1%(v/v)になるようにして用いた。使用前に3-BPの貯蔵溶液をPBS中で調製した。
【0092】
ATP測定
1.2x105個/mlのMDA-MB-453およびNeuTL細胞を、96ウエル組織培養マイクロタイタープレートに播き、使用前に一夜回復させた。上述のように、正常酸素圧または低酸素下で、α-TOSまたは3-BPとともに6または24時間、細胞をインキュベートした。次いで、使用説明書にしたがって生物発光アッセイキットHS11(Roche Applied Science)を用いて、ATPレベルを測定した。簡単に述べると、室温にて細胞溶解試薬とともに5分間インキュベートした後に、コントロールまたは処置細胞を黒いマイクロタイタープレートに移した。ルシフェラーゼ試薬を加え、Flurostar Optimaプレートリーダー(BMG Labtech)を用いてシグナルを迅速に検出した。各条件について三組みウエルを読み、既知の濃度のATPで作成した標準的較正曲線を用いてデータを評価した。
【0093】
ROS蓄積の評価
フローサイトメトリーにより、プローブとしてジヒドロエチジニウム(DHE)(Molecular Probes)(Weberら、2003)を用いて、あるいは、電子常磁性共鳴(EPR)分光分析(Weberら、2003)を用いる5,5-ジメチル-1-ピロリン N-オキシド(DMPO;Sigma)でのトラッピングによって、細胞ROSを検出した。いくつかの実験において、2 μMのMitoQ(Jamesら、2005 )で細胞を1時間前処置するか、またはSOD(PEG-SOD、750 units/ml;Sigma)で共インキュベートした。
【0094】
アポトーシス評価
アポトーシスの早期に外面化されたホスファチジルセリンを検出するV-FITC法を用いて、アポトーシスを定量した(Boersmaら、1996)。簡単に述べると、細胞を一夜療養し、α-TOSもしくは3-BP単独または併用で処置した後、24ウエルプレートに105個/ウエルにて細胞を播く。浮いている細胞および付着した細胞を採集し、PBSで洗浄し、0.2 mlの結合緩衝液(10 mM Hepes、140 mM NaCl、5 mM CaCl2、pH 7.4)に再懸濁し、2 μlのアネキシンV-FITC(PharMingen)とともに4℃にて30分間インキュベートし、50 μg/mlのヨウ化プロピジウム(PI;Sigma)200 μlを補足し、アネキシンV-FITC 結合についてチャネル1を用い、PI染色についてチャネル2を用いるフローサイトメトリー(FACScalibur、Becton Dickinson)によって分析した。アネキシンV結合およびPI染色の増加した細胞のパーセンテージとして、細胞死を定量した。
【0095】
RNA干渉(RNAi)
Akt1に特異的な3’-dTdTオーバーハングをもつ短い干渉RNA(siRNA)オリゴヌクレオチドをQiagenによって合成した。配列は、以下のとおりである:標的配列:CCT GCC CTT CTA CAA CCA GGA;siRNA二本鎖配列:センス - r(UGC CCU UCU ACA ACC AGG A)dTdT、アンチセンス - r(UCC UGG UUG UAG AAG GGC A)dTdT。使用説明書にしたがって、OligofectAmine(Invitrogen)およびOptiMEMを用いて、培養した細胞のsiRNAによるトランスフェクションを行った。簡単に述べると、細胞を50%コンフルエントに到らせ、OligofectAmineとともに前インキュベートし、OptiMEMで覆った60 pMのsiRNAを補足する。24時間後に細胞をPBSで洗浄し、完全DMEMで覆い、さらに24時間後、要求に応じて処置した。
【0096】
MTTアッセイ
2.5 mg/mlの3-(4,5-ジメチル-2,5-ジフェニル 2H-テトラゾリウムブロミド(MTT)(Sigma)をPBS単独または20 mMコハク酸、pH 7.4を含むフェノールレッド-フリーRPMI培地に溶解することによって、MTT溶液を使用前に調製した。エタノール中の3-ニトロプロピオン酸(3NPA)、DMSO中のテノイルトリフルオロアセトン(TTFA)、PBS中の3-BP、DMSO中のα-TOS、およびエタノール中のMitoQ(Allevaら、2003、Jamesら、2005)の貯蔵溶液を調製した。アッセイした希釈液ごとに4-8組みウエルを用い、96ウエルマイクロタイタープレートにて指数増殖期に培養したNeuTL細胞において薬物を試験した。MitoQのMTT還元を回復する能力を評価するために、2または5 μM MitoQとともに細胞を60分間前インキュベートした。培養物中のエタノールまたはDMSOの最終濃度は、≦0.1%であった。処置およびコントロール細胞は、37℃および5% CO2にて、2〜4時間、MTTを還元してブルーホルマザン結晶にすることができた。インキュベーション後、30 μl以外の培養上清を除去した後、150 μlのDMSOを加えることによってホルマザン結晶を溶解し、Fluorostar Optimaプレートリーダーを用い、570 nmにて吸光度の値を測定した。
【0097】
単離されたラット肝臓からのサブミトコンドリア粒子(SMPs)の調製
RiceおよびLindsay、1997の変更法にしたがって、分画遠心法によってラット肝臓ミトコンドリアを調製した。2 mlのミトコンドリアペレットを15 mlの20 mM KH2PO4 pH 7.4に再懸濁し、80 Wattで3x15秒間超音波処置した。次いで、超音波処置溶液を6,500xgにて7分間遠心分離した。上清画分を氷上に保持し、残留ペレットを再懸濁し、再度超音波処置した。超音波処置溶液を前述のように遠心分離し、第2の遠心分離ステップから除去した上清を第1の遠心分離からのものと合わせた。合わせた上清を108,000xgにて1時間再遠心分離した。SMPsのペレットを5 mlの20 mM KH2PO4 pH 7.4に再懸濁し、使用するまで−20℃にて凍結保存した。
【0098】
パラコッカス・デニトリフィカンスからの膜小胞の調製
パラコッカス・デニトリフィカンスCCM 982(NCIB 8944)を、炭素源として50 mMスクシネートおよび最終受容体として10 mMナイトレートを含む培地中で30℃にて嫌気的に増殖させた。Burnellら(1975)にしたがって、リゾチームで処置することによって採集した細胞から膜を調製した。約20 mg/mlの最終濃度になるまで0.1 M リン酸Na、pH 7.3中に希釈した後、使用するまで1 mlのアリコートにて膜を−20℃にて保存した。ビシンコニン酸法およびiEMSプレートリーダー(Labsystem、Finland)を用いて、タンパク質濃度を決定した。
【0099】
ミトコンドリア複合体IIおよび複合体I活性の測定
1 mlの反応体積を有する1 cmキュベットにおける600 nmでの吸光度を測定することによって、ミトコンドリア調製物による複合体II基質2,6-ジクロロフェノールインドフェノール(DCIP)の還元のための経時変化を追跡した(ε600=21x103 M-1cm-1)。反応成分は、NADH、0.5 mM;スクシネート、5 mM;KCN、10 mM;DCIP、50 μM;PMS、50 μMを含んだ。各アッセイポイントにおいて、0.5 mgのサンプルタンパク質を用い、示したように100または300 μMにてα-TOSを加えた。分光光度計(UVIKON XL、Secomam)を用いて、DCIPの吸光度における変化を測定し、繰り返しサンプルをアッセイした(n=3)。複合体I(NADHデヒドロゲナーゼ活性)を測定する場合、PMSを除外した。α-TOSを含まないコントロール反応について、採集濃度<0.1%(v/v)であるように希釈液DMSOを加えた。
【0100】
分子モデリング−複合体IIおよびビタミンE類縁体
Brookhaven Protein Databank(code 1ZOY)(Sunら、2005)から、ブタ心臓由来のミトコンドリア呼吸膜タンパク質複合体IIの結晶構造を入手した。複合体は、4個のタンパク質を含む。この複合体における3個のサブユニットである鉄イオウタンパク質(鎖B)、大(鎖C)および小(鎖D)膜貫通タンパク質は、UbQへの結合に関与する。NCBIウェブサイトからのBLAST調査は、ブタおよびヒト複合体IIの間の配列同一性が、鉄イオウタンパク質について97%、大膜貫通タンパク質について90%および小膜貫通タンパク質について94%と非常に高いことを示した。
【0101】
最初に除去されるヘテロ原子で、AutoDock Tools(Sanner、1999)を用いて、ドッキングのために、タンパク質構造を作成した。該構造に極性水素を付加し、タンパク質原子のためにKollman United Atomチャージを用いた。InsightII (Accelrys、2001)を用いて、結合したUbQ(1ZOY)の結晶構造配位からUbQ5を構築した。再びInsightIIを用い、α-TOSの環系についてのサブ構造調査によって、Cambridge Structural Database(Allen、2002)から検索された結晶構造MOPHLB01からα-TOSを構築した。次いで、非極性水素を融合し、Gasteigerチャージを割り当て、回転可能な結合を決定することを含む、AutoDock Toolsによって、ドッキングのために、両方のリガンドを作成した。
【0102】
Autodock 3.0.5.(Morrisら、1998)において実施されたように、Lamarckian Genetic Algorithmを用いて、ドッキングを行った;2つのドッキンググリッドを作成した。両方は、0.375オングストロームのグリッドペーシングである126x126x126ポイントであり、第1のものは、QP部位におけるTyr173(鎖B)を中心とし、第2のものは、QD部位におけるTrp134(鎖D)を中心とした。以下のもの以外は、対象のリガンドに存在する回転可能な結合の相対的に高い数(UbQ5=16、α-TOS =17)によって増加する、デフォルトパラメーターを用いた:- ga_run=250、ga_pop_size=250、ga_num_evals=10,000,000。また、計算の分析相中に、より管理可能なクラスターを生み出すために、パラメーターrmstolを、2.5に増加した。各ドッキング計算は、2 GHz G5 PowerPC Macintoshを用いて、49時間を僅かに越えた。AutoDockで提供されたスクリプトを用いて、結果の分析を行い、Astex Viewer(Hartshorn、2002)を用いて、ドッキングした構造を視覚化した。
第4表に示す他のビタミンE類縁体のAutodockingも、上述のとおり行った。
【0103】
分子モデリング−複合体II Qp部位および他の化学物質
ZINC Database project(http://blaster.docking.org/zinc/)から65,000個の商業上入手可能な化合物のバーチャルライブラリーをダウンロードした。これらの化合物を、計算された物理的特性に基づくさらに大きなデータベースから検索条件を追加したので、それらは、大変似ているドラッグリード様候補である(Teague、SJら、1999)。次いで、最終的データベースを作成するために、それらを多様性解析に付した。バーチャルスクリーニング実験中のDOCK 6.1(http://dock.compbio.ucsf.edu/DOCK_6/index.htm)を用いて、これらの化合物のそれぞれをブタ複合体II(1ZOY)の結晶構造にドッキングさせ、相互作用エネルギーに基づいてトップ1000個の化合物を同定した。次いで、再度DOCK 6.1を用いて、これらの最終化合物をより厳密なドッキング実験に付し、視覚化プログラムChimera (http://www.rbvi.ucsf.edu/chimera/)を用いて、最もブタ複合体IIと相互作用する可能性のある化合物として10個の化合物を同定した。
【0104】
マウス腫瘍実験
MCF-7細胞(2x106細胞/マウス)をヌードマウスに皮下接種した。腫瘍が発達した後(直径3-7 mm)、DMSO中の10 μモルのα-TOSを3日毎に、マウスに腹腔内注射した。コントロールマウスには、等量(100 μl)のDMSOのみを注射した。デジタルキャリパーで測定することによって、腫瘍サイズを評価した。H-2q FVB/Nバックグラウンド上のMMTVプロモーターによって誘発されたラットHER-2/neu プロトオンコジーンを有するトランスジェニックFVB/N202ラットc-neuマウスのコロニー(Guyら、1992)を、Griffith University Animal Facilityにて確立し、厳密な同系交配条件下に維持した。PCRによって、トランスジーンの存在をルーチン的に確認した。約70%の雌性マウスが、平均潜伏時間10ヶ月で同時の乳ガンを発達させた。平均直径10±5 mmの徐々に増殖する腫瘍を有する雌性トランスジェニックFVB/N202ラットc-neuマウスをコントロールまたは処置グループにランダムに割り当てた。デジタルキャリパーで腫瘍サイズを測定した。α-TOS療法のために、マウスは、3日毎に腹腔内投与で投与されるトウモロコシ油/4%エタノール単独での処置(コントロール)、またはトウモロコシ油/4%エタノール中の10 μモルのα-TOSでの処置を受けた。3-BP療法のために、1日当たり、0.2 μモルの薬物が腫瘍内に注射された。使用した3-BPの薬物用量は、以前の報告(Koら、2004)に基づいたものであり、この薬物をラット(平均体重〜300 g)に適用し、マウスに対して、低体重(〜30 g)を考慮した。
【0105】
統計的分析
グループ比較間の対応のないスチューデントt検定を用いて、アポトーシスの平均パーセンテージ、ATPおよびMTT還元±SDを比較した。共変動として日数を用いる共分散分析(ANCOVA)を用いて、平均相対腫瘍サイズ±SEMにおける差異を審査した。SPSS(登録商標) 10.0分析ソフトウェア(SPSS、Chicago、USA)を用いて統計的分析を行った。差異は、p<0.05である場合に統計的に有意であるとみなした。
【実施例1】
【0106】
3-BP、α-TOSまたはTRAILは、乳ガン細胞のATPレベルを低下させる。
3-BPは、従来、使用した薬物の特定の用量に応じて、処置の数時間後に起こる、細胞ATPレベルを枯渇させる活性の高い作用剤として報告されていた(Koら、2004、Xuら、2005)。さらに、細胞ATPレベルの減少が、3-BP薬物誘発性ガン細胞死の出現の前に起こることが示された(Xuら、2005)。低下したATPレベルは、次いで、アポトーシス促進性Bax分子がミトコンドリアに移行するのを可能にしてアポトーシスシグナル伝達経路を活性化させる、pBadホスホリル化の損失と相関した(Xuら、2005)。
したがって、本発明者らは、ヒト乳ガンMDA-MB-453(erbB2-高)およびMCF-7(erbB2-低)細胞系における細胞ATPの産生における3つの薬物α-TOS、3-BPおよびTRAILの効果を比較するために、それらの比較に着手した。さらに、FVB/Nラットc-neuトランスジェニックマウス由来のマウス乳ガン細胞系NeuTLにおいて、薬物を試験した。
【0107】
結果
(図1A)は、6時間以内に、MDA-MB-453細胞が、ATPレベルを有意に低下したことを示す。24時間までに、すべての3つの細胞系は、ATPレベルの低下を示したP(図1B-D)。3-BPおよびTRAILは、試験した異なる濃度の範囲にわたって、α-TOSよりも強力なインヒビターであることが分かった。α-TOSのみが、50 μMという高濃度で付加した場合に、細胞ATPレベルを有意に低下させた(図1)。細胞は、低酸素または正常酸素圧下のいずれかでインキュベートした場合に、細胞ATPを枯渇させることにおいて、3つの薬物の作用に対して、同じように反応性であった。
【実施例2】
【0108】
高度にerbB2-陽性である乳ガン細胞は、3-BPによって誘導されたアポトーシスに耐性があるが、α-TOSによって誘導されたアポトーシスには耐性がない。
高レベルのerbB2発現を示している乳ガン細胞は、いくつかのガン治療によって誘導されたアポトーシスに対する関連するより大きい耐性を有する(Burstein、2005)。これらの発見と一致して、本発明者らは、近年、高度にerbB2-陽性であるMDA-MB-453細胞系が、弱くerbB2-陽性であるMCF-7細胞系におけるその効果と比較した場合に、死受容体アクチベーターTRAILによって誘導されるアポトーシスに対してより耐性があることを観察している。しかし、MDA-MB-453細胞は、α-TOS誘導アポトーシスに反応性であることが分かった(Wangら、2005)。この実験において、本発明者らは、2つのミトコンドリア影響性薬物、α-TOSおよび3-BPを、高および低erbB2-発現乳ガン細胞系における、そして異なる酸素条件下でのアポトーシスインデューサーとしてのそれらの相対的効力について比較した。
【0109】
結果(図2)は、MDA-MB-453およびMCF-7細胞系の両方が、正常酸素圧および低酸素下の両方で、α-TOSによって誘導されるアポトーシスに対して感受性があり、MCF-7細胞がわずかにより影響を受けやすいことを示した。MDA-MB-453細胞は、100 μMという高濃度でさえも、3-BPによる殺傷に著しく耐性があることが見出された。α-TOSに関しては、反応性MCF-7細胞が、正常酸素圧よりも低酸素下において、3-BPによる殺傷に対して、より感受性があることが見出された(図2)。
【実施例3】
【0110】
3-BPおよびα-TOSの併用は、高erbB2-陽性細胞において、アポトーシスを増強する。
この実験において、本発明者らは、α-TOSおよび3-BPが、オーバーラップするかどうか、またはそれらが3-BP-耐性MDA-MB-453乳ガン細胞の殺傷を増強することができるかどうかを決定した。この実験は、これらの薬物が、同様のメカニズムで働くか、あるいは他方とは独立しているかどうかを理解する手がかりとなる。3-BPが、スクシネートであるデヒドロゲナーゼの活性部位に結合することによって、複合体IIのインヒビターとして作用することが知られている(Sanbornら 1971)。したがって、それらの両方が違いにオーバーラップする可能性があった。
【0111】
図3は、MDA-MB-453細胞においてアポトーシスを誘導するためのα-TOSおよび3-BPの併用効果の結果を示す。事実、MDA-MB-453細胞は、驚いたことに、それぞれの薬物単独での使用と比較して、α-TOSおよび3-BPの併用による殺傷に対して、感受性を高め、薬物併用を用いた場合のガン細胞アポトーシスの程度は、有意(p<0.05)に増強された。さらに、併用で増強された効果は、細胞培養条件が正常酸素圧か低酸素かどうかにかかわらず起こった。したがって、これらの結果から、2つの異なる薬物が、互いに独立して作用し、α-TOS処置が、3-BP耐性MDA-MB-453細胞を、3-BPの細胞毒性効果に対して感受性があるようにするという並はずれた発見を提供することが明らかとなった。
【実施例4】
【0112】
Akt siRNAは、3-BPへのMDA-MB-453細胞の反応性を回復し、α-TOSおよびTRAILへの感受性を増加する。
過去の研究は、高レベルのerbB2発現が、Bad(Zhaoら、2004)およびカスパーゼ-9(Cardoneら、1998)などの多くの異なる細胞標的をホスホリル化するAktの活性化をもたらすことを明らかにしている。このように、ガン細胞は生存の増加を示し、薬物治療によって誘導されるアポトーシスに対して耐性ができる(Westら、2002、Osakiら、2004、Fresnoら、2004、Grandageら、2005)。実際、アンチセンスRNAアプローチを用いるAktの細胞発現を阻害することが、薬物療法へのガン細胞の感受性を回復させるための一般的方法として提案されている(Kimら、2004)。さらに、近年、Aktに対してsiRNAが、ガン細胞においてAkt発現をダウンレギュレートすることのガン細胞の生存に及ぼす効果を調査するいくつかの研究において用いられている(Takeuchiら、2005、Yoeli-Lernerら、2005)。したがって、本発明者らは、Akt1発現を低下させて、このことが、erbB2-陽性 3-BP-およびTRAIL-耐性MDA-MB-453細胞を2つの作用剤に対して反応性にするかどうか、ならびに、このことが、α-TOS媒介性アポトーシスに対する該細胞の感受性を高めるかどうかを決定する手段として、RNAi技術を活用している。
【0113】
OligofectAmineおよびOptiMEMを用いて、siRNAsによるMDA-MB-453細胞のトランスフェクションを行った。簡単に述べると、細胞を〜50%コンフルエントに到達させ、OligofectAmineとともにプレインキュベートした60 pmol/l siRNAを補足し、OptiMEMで覆った。24時間後にPBSで細胞を洗浄し、完全DMEMで覆い、さらに24時間、48時間培養し、ウエスタンブロッティングを用いてトランスジーン発現を確認した後、次の実験に用いた。
【0114】
結果(図4)は、Akt1 siRNAによるMDA-MB-453細胞の前処置が、試験する3つの薬物それぞれに対する感受性を増加させることにおいて有効であり、3-BPに対する細胞毒性反応において特に目覚ましい増加を生み出すことを示した。この証拠は、高レベルAkt活性をもつ乳ガンが、3-BP治療への耐性をもつ傾向があり、このことが、分子操作によって効率的に調節されうることを示唆する。
【0115】
3-BPへの細胞反応性におけるAkt siRNAの効果およびα-TOS/3-BP併用治療によるアポトーシスの増加は、α-TOSが、Akt活性にも影響を及ぼし、そのことによって、Akt siRNAと同様の、3-BPへのガン細胞の感受性の増加を提供するという可能性を引き起こす。
【実施例5】
【0116】
α-TOSは、複合体II上のUbQ-結合ポケットを標的とする。
本発明者らによる過去の研究(Neuzilら、2004に概説)は、ミトコンドリアが、ガン細胞におけるα-TOSの作用のメカニズムの中心をなし、ミトコンドリアアポトーシスシグナル伝達経路の活性化をもたらすことを明らかにした。近年そのBcl-2/Bcl-xL阻害活性の発見まで、α-TOSは「オーファンドラッグ」状態を保持している。しかし、ガン細胞へのα-TOSの付加の早くも1時間後にROS生成をもたらすこの薬物のための特異的ミトコンドリア標的およびアポトーシス促進性経路の次の開始(Weberら、2002、Stapelbergら、2005)は、明確ではない(Zhangら、2001)。
【0117】
この目的を達成するために、本発明者らは、α-TOSが、複合体IIにおいてUbQ結合を妨げるかどうかを詳細に調べた。全細胞における複合体II活性を特に測定するために、テトラゾリウム塩MTTの還元に基づく短時間(1-2時間)アッセイを用いた(Maeharaら、1988)。したがって、ミトコンドリア呼吸が複合体IIを介して全細胞において最適に進行特するのを確実にする特定の条件を用いた。このために、マウスNeuTL乳ガン細胞の酸化呼吸を高レベルのスクシネート(20 mM)で持続させ、それによって、複合体IIのSDH活性を促進した(Maeharaら、1988)。次いで、高スクシネート条件下で複合体II/SDH依存性活性の特異的インジケーターになる、MTTを酵素的に還元する能力について、1-2時間以内に細胞をアッセイした(Maeharaら、1988、Berridge and Tan、1993)。
【0118】
過去の研究は、3-BP(Sanbornら、1971)および3NPA(Scalletら、2003)の両方が、SDH活性の強力なインヒビターとして作用することを明らかにしており、3NPAは、細胞への付加の4時間以内と比較的迅速にMTT還元を阻害する能力がある(Scalletら、2003)、SDHの特異的自殺型インヒビターである(Huangら、2006)ことを近年明らかにされている。2つのSDHインヒビター、3-BPおよび3NPAがNeuTL細胞において直接MTT還元を害するかどうかを決定するために、3-BPおよび3NPAを用い、別な方法でこれらのインヒビターと競合する追加のスクシネートの不在下にて初期実験を行った。結果(図5A)は、3-BPおよび3NPAが両方とも、複合体II-媒介性MTT還元の強力なインヒビターであり、したがって、アッセイが、SDH活性の効果を定量するという我々の目的に適していることを確認した。
【0119】
複合体IIの電子伝達流に関与するUbQ結合部位を標的とする特異的インヒビターとして、テノイルトリフルオロアセトン(TTFA)が確立されている(Berridge and Tan、1993、Sunら、2005)。これらの部位は、電子を複合体IIIに送るために、UbQに電子を伝達する状態にある、複合体IIの膜貫通領域におけるSDH反応中心の下に存在する。興味深いことに、TTFAは、すべての細胞に対して毒性である(Zhangら、2001b)。SDH酵素インヒビターである3-BPおよび3NPAと同様に、TTFAもまた、2,3時間以内に、乳ガン細胞によるMTTの酵素的還元を有意に阻害した(図5C)。事実、3-BP、α-TOSまたはTTFAのいずれかによる処置は、処置の早くも2,3時間後に、これらの細胞によるMTT還元のレベルにおいて生じる有意な減少をもって用量依存的変化を示した(図5B-D)。しかし、UbQのミトコンドリア的に標的化された形態であるMitoQとともに細胞を1時間プレインキュベートすること(Kelsoら、2001、Jamesら、2005)は、MitoQが、TTFA(図5C)またはα-TOS(図5D)のいずれかによって引き起こされたMTT還元における阻害に打ち勝つことができるが、3-BP(図5B)によって引き起こされた阻害には打ち勝つことができないことを示した。ここでの理論的解釈は、もし、これらの薬物のいずれかが、複合体IIのUbQ部位への結合によって活性化しているならば、増加したレベルのUbQ活性類縁体であるMitoQで細胞を充填することは、複合体IIによるMTT還元の薬物媒介性阻害の程度を低下させるであろうということであった。MTT還元における3-BPの阻害効果は、SDH頭部基を阻害することによって作用し、複合体IIの下流膜貫通UbQ部位を介さない3-BPと一致して、異なる濃度のMitoQ(図5B)の存在または不在下で細胞を処置した場合、有意には異ならなかった。しかし、TTFA(図5C)およびα-TOS(図5D)の付加によって引き起こされる阻害は、MitoQと細胞とのプレインキュベーションによって有意に打ち勝たれた。特に、α-TOSは、MitoQの存在下でのスクシネート駆動MTT還元を阻害することにおいて、有効性は低かったが、これも、近年、ミトコンドリア酸化還元鎖の複合体IIへの結合に対して特異的であることが明らかにされている(Jamesら、2005)。
【実施例6】
【0120】
α-TOSが、ミトコンドリア呼吸複合体II活性を阻害することの検証。
ミトコンドリア 呼吸複合体II活性を阻害することにおけるα-TOSの役割のためのさらなる裏付けは、ラット肝臓から細胞内フラクションを含んでいるミトコンドリ調合液、ならびにパラコッカス・デニトリフィカンスから膜フラクションを単離することによって探求された。これらの研究にとって、SDH/複合体II活性のアッセイ法は、フェナジンメト硫酸塩(PMS)およびインジケーター色素として末端電子受容体である2,6-ジクロロインドフェノール(DCIP)を用い、それが高スクシネート条件下で駆動されるSDHによって還元されるための600 nmにおける吸光度の減少を用いるKing(1967)の研究に基づいたものであった。結果(図6A,B)は、ラット肝臓ミトコンドリアおよびパラコッカス・デニトリフィカンス調合液の両方において、コントロールサンプルと比較して、DCIPの還元の速度が、α-TOS処置後に、急速および有意に低下したことを示した。したがって、結果は、複合体IIのUbQ部位で直接作用し、PMSおよびDCIPへの電子流を妨げるα-TOSを強く裏付ける。SDH活性についてのα-TOSの選択性は、NADHデヒドロゲナーゼ(複合体I)活性においてビタミンE類縁体の効果がないことを示す結果によって裏付けられる(データ示さず)。
【実施例7】
【0121】
複合体Iおよび複合体II欠損細胞を用いる、α-TOSがミトコンドリア呼吸複合体II活性を阻害することの検証。
複合体Iおよび複合体II欠損細胞を上記実施例6のインビトロの結果を確認するためにも用いた。CI機能不全細胞(B10細胞)(Seo、B.B.ら、1998)、CII機能不全細胞(突然変異チトクロームb大を有するB9細胞、CybL)および親チャイニーズハムスター肺線維芽細胞(B1細胞)(Oostveen、F.G.ら、1995)を、10% FCS、抗生物質、10 mg/mlのグルコースおおよび非必須アミノ酸を補足したDMEM中で増殖させた。α-TOSは、親チャイニーズハムスター肺線維芽細胞(B1細胞)、CI機能不全細胞(B10細胞)およびCII機能不全(SDHCサブユニットをコードする遺伝子において突然変異有するB9細胞、CybL)においてアポトーシスを開始した。B9細胞は、親(B1)またはB10細胞と比較して、α-TOSへの反応性が低く、ROS蓄積のレベルが低く(図7A)、SDH活性が減少した(図7B)。これらの発見に一致して、B9細胞は、VE類縁体によって誘導されるアポトーシスに対して相対的に耐性があった(図7C)。CybL突然変異(B9)細胞におけるCIIの差異構築は、SDH活性を正常化し(図7B)、α-TOS誘導殺傷への細胞感受性を回復させた(図7C)。
【0122】
CII欠損(B9)および再構築B9細胞により得られたデータを、CybLに対する4つの異なる短い干渉RNAs(siRNA)でのMCF7細胞の処置により、独立して検証した。二本鎖1、3および4は、実質的にCII活性を抑制した(図7D)。CybLにおける突然変異は、ROS蓄積およびSDH活性を阻害し、α-TOSによって媒介されるアポトーシスのレベルを低下させた(図7D-F)。
【0123】
ヒトモノクローナルSDHC抗体、B1、B9およびB10、ならびにCII再構築チャイニーズハムスター肺線維芽細胞(B9rec)の使用により、SDHCの存在を探った。ウエスタンブロッティング分析(図7H、I)は、B1、B9およびB10細胞におけるヒトSDHCの不在、ならびにSDHC siRNAsで処置したMCF-7細胞においてそのレベルが低いことを示した。再構築CIIを有するB9細胞は、SDHCの再出現を示した。デンシトメトリーの評価によって示されたように、MCF7細胞のsiRNA処置は、SDHCタンパク質のレベルを50-80%まで低下させた(図7J)。これらのデータは、CII再構築細胞におけるヒトSDHC mRNAの存在、ならびにSDHC siRNAで処置されたMCF7細胞における転写物の低レベルを示すRT-PCRの結果と一致した(図7G)。SDHC処置MCF7細胞におけるSDHBサブユニットタンパク質のレベルにおけるいずれかの変化が、観察されなかったので、siRNAアプローチはSDHCに対して特異的であった(データ示さず)。
【実施例8】
【0124】
分子モデリングは、UbQ自体に対してよりも、複合体II活性部位へのα-TOSのより強力な結合を示した。
α-TOSがUbQ結合部位を介してミトコンドリア複合体IIと相互作用することを示している上記の結果を理論的に説明するのを助けるために、本発明者らは、AutoDock(Morrisら、1998)を用いて、このシステムの分子モデリング研究を企てた。近年、ブタ心臓ミトコンドリア複合体IIの結晶構造が報告されており(Sunら、2005)、ヒトミトコンドリア複合体IIとの高い配列同一性ゆえに、本発明者らは、それらのAutoDock研究のための基準としてこの構造(1ZOY)を用いることを決定した。構造は、近位のUbQ結合部位(QP)を明確に示し、結合したインヒビターTTFAをもつ関連構造(1ZP0)は、提案された遠位のUbQ結合部位(QD)の位置を示唆する。
【0125】
このシステムを研究するためにAutoDockを用いることの実現可能性を試験するために、最初に彼らは、QPおよび提案されたQD結合部位の両方にUbQ5をドッキングさせるためにそれを用いた。UbQ5は、α-TOSと同様のサイズであり(図8)、同様の数の回転可能な結合を含む(それぞれ16および17)ので、選択された。UbQの部分について観察される位置よりわずかに深い位置へQPにドッキングしたUbQ5は、Sunら(2005)によって公開された結晶構造で解決した。興味深いことに、このより深いドッキング位置は、大腸菌由来の複合体IIの結晶構造にUbQ2をドッキングさせるためにドッキングプログラムGOLDが用いられる近年の研究において観察された(Horsefieldら、2006)。結合部位の正面に位置しているUbQ環および該部位の内側に位置する疎水性尾部をもつUbQ5もまた、提案されたQD部位にドッキングすることが見出された。AutoDock(図8)によって計算された相互作用エネルギーは、UbQ5が、提案されたQD部位よりも強くQP部位と相互作用することを示唆する。このことは、提案されたQD部位においてUbQ結合が観察されないという事実とよく相関し、QP部位が、UbQ5との最も強い相互作用を提供することを示唆した。
【0126】
図8A-Dに見られるように、AutoDockは、QPおよびQD部位の両方においてα-TOS結合を示した。QP部位において、α-TOSの環系は、UbQ環と同じ結合ポケットに位置するが、反対側に傾斜する。驚いたことに、コハク酸エステル部位が、結合ポケット内により深く伸び、補欠分子ヘム基の位置に向かって下方に伸びることが見出された。カルボキシル基は、このポケットに適切にフィットし、Ser42(C)との二座水素結合に関与する(図8B,C,D)。Ser42(C)もまた、α-TOSのエステル酸素と相互作用する。α-TOS分子の残部の疎水性性質に適しているので、該タンパク質とのすべての他の相互作用は、疎水性である。炭化水素側鎖は、周囲で輪になり、bQ5のイソプレノイド側鎖がドッキングするのが示される同じチャネルに沿ってQP部位からU伸びる。QD部位において、α-TOSの環系は、結晶構造において認識できるリン脂質の頭部基に観察される様式と同じ様式で該部位の底部まで伸びているコハク酸エステル部分をもち、結合部位の底部に向かって位置する。コハク酸部分は、Lys135(D)and Lys128(D)に水素結合するのが見られる(図8E)。
炭化水素側鎖は、結合部位の内側の周りで輪になる。
【0127】
α-TOSのドッキングしたコンホメーション(図8F)についての相互作用の計算されたエネルギーは、それが、QPまたはQD部位のいずれかにおいて結合することができるが、QD部位において結合が優先されることを示唆した。QP部位におけるα-TOSの結合エネルギーは、UbQ5よりもわずかに小さいが、α-TOSは、UbQ5と確実に競合することができる。QD部位において、α-TOSは、UbQ5よりも良好な結合エネルギーを示し、この結合部位から置き換えることができるはずである。
【0128】
まとめると、これらの発見は、α-TOSが、ユビキノン(UbQ)結合と相互作用し、呼吸鎖における複合体IIの膜ドメインによってUbQの還元を遮断することを示す。結果として、電子は、もはやFAD上のコハク酸デヒドロゲナーゼ親水性頭部の下に伝達されず、[4Fe-4S]を介してUbQへ中継されるが、代わりに、ミトコンドリア中に漏出し、最終的にガン細胞のアポトーシスを誘導する増加したレベルのROSを生成する。
【0129】
上記と同じプロトコルを用いて、下記の第表および第5表に示すのビタミンE類縁体のオートドッキングを行い、予測した相互作用エネルギーを各表の最後の欄に提示した。
【0130】
第4表
近位コエンザイムQ結合部位(Qp)におけるビタミンE類縁体の相互作用エネルギー。化合物は、シグナル伝達ドメインによって分類される。
【表6】
【表7】
a最初の値は、cis-、第2の値は、trans-についてである。
b2つの値は、キラル炭素における2つのエナンチオマーを意味する。
【0131】
第5表
修飾された疎水性ドメインとの、遠位コエンザイムQ結合部位(Qp)におけるビタミンE類縁体の相互作用エネルギー。
【表8】
【0132】
上記ビタミンE類縁体は、AutoDock 3.0.5および前に述べたものと同じプロトコルを用い、ブタ複合体IIの結晶構造(近位ユビキノン結合部位)にドッキングする。相互作用の予測されたエネルギーおよび生成したコンホメーションの数を比較することによって、結合において重要であるα-TOSテンプレートへの変化に関する以下の観察が得られる:
1.末端電荷。α-TOSテンプレートは、その末端カルボン酸による負の電荷を有する。この電荷の消滅は、相互作用エネルギーを減少したが、正の電荷とそれを置き換えることが、同様の相互作用エネルギーを維持した。この荷電した基がドッキングする領域は、極性アミノ酸残基(ヒスチジンおよびセリン)によって囲まれているが、荷電した残基はない。最も近い電荷は、近くのヘム基のカルボキシ基から来る。このことは、正または負の荷電した基のいずれかの蓄積を可能にする。
2.エステル結合。スクシネート部分を元のビタミンE骨格に連結するエステル結合は、相互作用エネルギーにおいてほとんど変化がないアミド結合と置き換えることができる。
3.スクシネート部分は、(二重結合における立体配置がtransに維持される限り)フマレート部分と置き換えることができ、良好な相互作用エネルギーをなお維持することができる。
4.α-TOSの芳香環上のメチル基は、近位ユビキノン結合部位内で物質を位置決定することにとって重要である。それらを除去すると、相互作用のエネルギーが低下する。
【0133】
図9において、本発明者らは、ユビキノン部位へのビタミンE類縁体の結合親和性と、ガン細胞のアポトーシスを引き起こすことにおけるそれらの生物活性との間の相関関係を解析した。結果は、該部位への結合親和性と、ガン細胞を殺傷する能力との間の直接的関係を示す。ドッキングと、結果のリストとの間に観察される際は、複合体II標的部位に到達する化合物の生物学的近接可能性を反映する。したがって、IC50は、部位に結合する類縁体の能力のみならず、細胞膜を横切って進入し、ミトコンドリアマトリックスへの接近を獲得する能力も反映する。IC50測定のために、本発明者らは、細胞ベースアッセイを用いたので、いくつかの化合物が、複合体IIと相互作用するためにミトコンドリア内に入るかどうかを評価することができない。最良の比較は、Autodockの結果と、市販の精製複合体IIキット(MS201 複合体II IMMUNOCAPTURE KIT MitoScience LLC、1850 Millrace Drive、Suite 3A、Eugene、OR 97403、USA、http://www.mitosciences.com) を用いる酵素アッセイとを比較することによってなされる。
【0134】
このキットを用いて、ヒト、マウス、ラットおよびウシの組織および細胞系から精製複合体IIを単離し、次いで、King TE、1967に概略されたDCPIPおよびPMSアッセイ反応を用いるか、または競合放射性ユビキノン結合アッセイを用いて、類縁体による活性および阻害についてスクリーニングすることができる。
【実施例9】
【0135】
分子モデリングは、複合体II Qp部位に結合する能力がある化学物質を示す。
この実施例において、コンピューターモデリング研究を用いて、ユビキノンQp結合部位に大きい親和性で結合し、電子伝達を中断させる能力を潜在的に有する化学物質を同定した。このような物質は、ガン性細胞を殺傷する能力を潜在的に有する。
【0136】
ZINC Database projectから65,000個の商業上入手可能な化合物のバーチャルライブラリーをダウンロードし、ブタ複合体II(1ZOY)の結晶構造に対して、望ましい特性を有するサブセットをドッキングさせた。相互作用エネルギーに基づいたトップ1000個の化合物をより厳密なドッキング実験に付し、最もブタ複合体IIと相互作用する可能性のある化合物として、最終的な10個の化合物を同定した。このような化合物を図10に示す。
【0137】
分子モデリングおよび生物アッセイからの化合物のリストおよび結合データに基づいて、複合体IIのQ部位への最良の結合に望ましい/有利な化学物質は、酸素がイオン化されるようになる能力があるかもしれないか、またはないかもしれない酸素含有側基をもつ疎水性化合物を包含する。コアの疎水性基として、トコフェロールおよびその同族体、ピリドキサール誘導体、ユビキノン誘導体、キニノリン誘導体、ベンゾスルホンアミドなどのベンジル基、ピリジル/ピリジン、プリンおよびピリミジン、パルテノライド、レチナミド、セスキテルペンラクトンが挙げられる。疎水性コア化合物の側基への置換修飾として、スルフェート、ホスフェート、ホスフィン酸、スルホンアミド、スルホニル基、プロパン酸、カルボキシレートおよびメトキシ、ジオキシまたはプロポキシ置換が挙げられる。
【実施例10】
【0138】
共役ペプチドLTVSPWY-α-TOSは、乳ガンを抑制する。
この実施例は、受容体チロシンキナーゼerbB2を過剰発現しているガン細胞へのα-TOSプロドラッグの標的化を記載する。α-TOSのアダクトを、erbB2を過剰発現しているガン細胞を標的化することが知られているヘプタペプチドLTVSPWYに結合した。受容体への結合に続いて、プロドラッグが取り込まれ、酸性ペプチダーゼによって加水分解されて、アポトーシス性α-TOSが得られることが見出されている。図11に見られるように、アダクトは、erbB2の高発現の結果として自然発生した乳腫瘍を特徴とするFVB/N MMTVラットc-neuマウスにおいて乳ガンを効率的に抑制した。事実、処置された腫瘍のサイズは、非処置コントロールと比較して、予想外の90%の相対的減少があった。したがって、このプロドラッグ/アダクトは、ガンの治療に特に有用である。
【実施例11】
【0139】
α-TOSは、erbB2の状態にかかわりなく腫瘍の増殖を阻害する。
インビボ研究のために、ヒト乳ガンの三分の一までが、高レベルのこの受容体チロシンキナーゼを発現することを考えれば(Slamonら、1989)、本発明者らは、α-TOSが、erbB2を低発現している乳ガンの動物モデルにおいて腫瘍増殖を抑制することができるかどうかを最初に決定した。この研究のために、ヌードマウスに低発現MCF-7細胞異種移植し、次いで、直径約5mmの腫瘍として確立されるようにした後、α-TOSで処置した(図12A)。結果は、コントロールマウスにおける腫瘍は、安楽死が必要になる時点までに、その初期のサイズの5倍まで増殖したが、ことを示したが、α-TOSは、マウスの処置グループにおいて腫瘍の増殖を抑制し、腫瘍のサイズは減少した。
【0140】
次に、本発明者らは、7ヶ月齢以上で、直径〜10 mmの認識できる乳ガンを明確に提示している自然発生トランスジェニックマウスモデルであるFVB/N MMTV-rat c-neuを用いるerbB2高ポジティブ乳ガンの治療に対する反応に興味をもった(Guyら、1992)。2つの薬物、3-BPおよびα-TOSを、腫瘍増殖のリプレッサーとしての、その相対的効力について比較した。結果(図12B)は、コントロール動物と比較して、処置マウスにおける腫瘍は、増殖停止または部分退行のいずれかを示すことが明らかになった。α-TOSは、これらの動物における乳腫瘍の増殖の退行を誘導することにおいて、3-BPと比べて、統計的により効果的であり、処置2週間で約30-40%まで腫瘍の平均サイズの低下をもたらすること(共分散分析によりp<0.05)が見出された。
【0141】
異種移植およびトランスジェニックマウスの乳腫瘍の増殖におけるα-TOSおよび3-BPの効果(図12参照)は、ヒト患者での臨床試験におけるそれらの提案された適用を裏付ける。しかし、本研究は、erbB2高発現細胞の3-BPへの耐性を示した。受容体は、ヒト乳ガンにおいて、通例、上昇するレベルで発現されるので(Slamonら、1989)、このことは、erbB2高ポジティブ患者の処置について問題を提供する。この問題は、Akt siRNAの同時投与、または3-BPとα-TOSの併用によって克服することができるかもしれないということが提案される。
【0142】
乳ガンを治療するためにα-TOSおよび類似する作用剤を用いることの利点は、以下のとおりである:α-TOSは、HER2-低(乳ガン患者の70%)およびHER2-高乳腫瘍(乳ガン患者の30%)の両方に対して、すなわち、HER-2発現に無関係に効率的である。このステージにおいて、HER2-低乳ガン患者を治療するための現在のアプローチは非常に効果的とはいえず、上記および本願発明を通して記載したように、α-TOSはガンの前臨床モデルにおいて選択的で、非常に効率がよい。HER2-高ガン患者に関して、現在の化学療法レジメンを補完するただ1つの治療のみが存在し、これは、HER2に対するヒト化抗体、ヘルセプチンの適用である。再度、α-TOSは、いくつかの理由から大いに融合である:ヘルセプチンのみが、非常に高レベルのHER2発現を有する乳ガンに対して有効である(蛍光インシトゥハイブリダイゼーションによる3+)が、α-TOSは、HER2発現が高レベルであろうと低レベルであろうと、乳腫瘍細胞を殺傷し、実験的乳ガンの有意な低下を引き起こす;ヘルセプチンは、心臓毒性であるが、心筋細胞(心臓筋肉細胞)は、α-TOSをアポトーシスサイレントビタミンEに加水分解するという傾向を有するので、α-TOSは毒性でない;ヘルセプチンは、はなはだしく高価:患者1名の治療の年間費用はAU$50,000であるが、α-TOSは比較的安価である;最後に、ヘルセプチンは、広範囲の乳ガン細胞のアポトーシス引き起こさず、むしろ、ヘルセプチンによる治療の中止後腫瘍が再発することができるように腫瘍細胞増殖を停止するにすぎない。対照的に、乳ガンの前臨床モデルを用いるα-TOSによる実験は、有意な縮小をを示し、続いて、TOS類縁体の適切な製剤での長期処置および最適投与レジメンの使用によって、乳ガンの完全な療法を達成する可能性があることを期待することができる。
【実施例12】
【0143】
中皮腫の治療におけるα-TOSの使用
α-TOSを用いて、ヒトガン患者において中皮腫を治療した。様々な化学療法およびその他の薬物による過去の失敗した試みにもかかわらず、α-TOSは、非常に効果的に働き、現在、患者はその生活の質を取り戻している。血漿レベルをモニターしたところ、血漿中のα-TOSのレベルは、長期間にわたって50 μMを達成した。これは、インビトロ分析に基づいてガン細胞を殺傷するのに十分すぎるほどである。
【0144】
臨床研究の概要:
これまでに5種の異なる細胞毒性作用剤を用いて治療を受けたが、改善を示すことができなかった末期中皮種を有する60歳の女性患者。患者は、最初は、高用量のアヘン剤投薬療法、神経障害性痛投薬療法、在宅酸素療法を受けており、余命6〜8週間と予測されていた。α-TOSで治療した後、患者は、もはや鎮痛または酸素を必要としなかった。α-TOを開始する前の患者は、触診可能な前胸壁腫瘤を有していたが、α-TOSでの療法により、劇的に縮小した。α-TOS療法は、目に付くような副作用がなく、非毒性であり、耐容性良好であった。
【0145】
病歴:
2001年7月に、患者は、最初に胸の痛みを発症し、2002年5月初旬に、生検によつ中皮腫であると診断された。患者は、重篤な痛みの管理のために苦痛緩和医療に付託され、2002年6月に、傍脊椎腫瘤に深部X線放射線療法を受けた。2002年7月に、シスプラチン、ゲムシタビンおよびタリドミドの併用による化学療法が開始された。
【0146】
重篤な末梢神経障害および毒性の問題および継続する吐き気により、2003年2月に、治療は中止された。2003年2月に、アリムタおよびカルボプラチンによる治療が開始された。骨髄抑制をともなう重篤な毒性ならびに継続する吐き気および嘔吐により、2003年9月に、治療は中止された。2003年9月に、経口定用量のフェノキソジオール(Novogen SAS薬物)による治療が開始され、2005年3月まで安定していた。しかし、フェノキソジオールにともなう継続する重篤な神経障害により、治療は再中止された。
【0147】
2005年3月に、中皮腫は、さらなる進行を明らかにし、再度前進した。患者は、ゲムシタビンの2用量による療法を開始したが、許容できない程度の毒性が再度生み出された。2005年8月に、患者は、ビノレルビンの2用量による療法を開始したが、末梢神経障害の重篤な症状の悪化が見られた。
【0148】
2005年12月に、患者は、基剤クリーム調製され、ジメチルスルホキシド(DMSO)で可溶化されたα-TOSの経皮デリバリーを受けることを開始した。患者は、約8週間の治療期間中、製剤の適用を続けたが、DMSOに続発する広範囲の皮膚反応問題(治癒に4週間を要するやけど)が発生した。中皮腫は、臨床反応を示したが、皮膚反応により、治療を中止しなければならなかった。
【0149】
2006年3月に、ガンは、神経根圧迫を引き起こす傍脊椎腫瘤とともに急速な進行を示した。患者は、コラムシン(SAS薬物、Soblec WA)による治療前に、さらなるDXRTを受けた。ガンは、6ヶ月間安定した反応を示したが、次いで、再度進行した。
【0150】
2006年11月に、患者は、アリムタ−アバスチン併用療法を再開したが、4ヶ月間療法を続けた後、吐き気、抑制されない下痢および神経障害の増加などの重篤な副作用をもつ合併症が現れ、治療を中止した。
【0151】
2007年4月に、リポダーマル(lipodermal)クリームと混合した変更製剤を用いるα-TOSによる療法を再開した。この時点で、中皮腫は、右肺を包み込む疾患の拡散を示し、両肺に多発性転移がCTスキャンによって視認された。さらに、触診可能な病変が、肋骨および右前胸壁の間で増殖するのが視認された。患者は、皮質脳波が低い非常に悪い状態であり、瀕死であったため、α-TOS治療を開始する時点でCTスキャンを行うことはできなかった。
【0152】
リポダーマル製剤を4週間適用した後、患者は、CTスキャンでは50%まで前胸壁腫瘤が減少し、非常に改善された回復を示し、継続中のα-TOS療法により、患者は、臨床的改善を続けた。
【0153】
α-TOS製剤:病院の薬局で混合された500gのリポダーマルクリーム+60gmsのα-TOS 化合物。2,3日ごとに約30gの混合された混合物を投与し、吸収を改善するために、上半身をプラスチックフィルム(サランラップ)で覆った。
【0154】
副作用:抗ヒスタミンで改善される軽いかゆみは別として、α-TOS療法には明らかな副作用はなかった。神経障害学的または他の薬害反応についての血液マーカーおよび他のサインのためのすべての実験室試験は、正常なままである。
【0155】
前記の実施態様は、本発明の原理の一例にすぎず、様々な変更および変化は、当業者にとって容易になしうることである。本発明は、様々な様式および他の実施態様において実践し、実行することができる。本明細書で用いた専門用語が、記載を目的とするためであり、限定とみなされるべきではないことも理解されよう。
【0156】
用語「含む」および「含んでいる」などのその変形は、本明細書において、述べられた完全体または述べられた完全体群の包含を意味するために用いられるものであり、文脈または用法において、用語の排他的解釈が必要でない限り、いずれかの他の完全体またはいずれかの他の完全体群を排除するものではない。
【0157】
本明細書に引用したいずれかの文献への言及は、開示がオーストラリアまたは他所における通例の一般的知識を構成するという承認ではない。
【0158】
参考文献のリスト
Albayrak T, Scherhammer V, Schoenfeld N, Braziulis E, Mund T, Bauer MK, Scheffler IE, Grimm S. (2003) The tumor suppressor cybL, a component of the respiratory chain, mediates apoptosis induction. Mol Biol Cell. Aug;14(8):3082-96.
Allen FH (2002) The Cambridge Structural Database: a quarter of a million crystal structures and rising. Acta Crystallogr B 58(Pt 3 Pt 1):380-388.
Allen RG, Balin AK (2003) Effects of oxygen on the antioxidant responses of normal and transformed cells. Exp Cell Res 289:307-316.
Alleva R, Tomasetti M, Andera L, Gellert N, Borghi B, Weber C, Murphy MP, Neuzil J (2003) Coenzyme Q blocks chemical but not receptor-mediated apoptosis by increasing mitochondrial antioxidant protection. FEBS Lett 503:46-50.
Aratri E, Spycher SE, Breyer I and Azzi A (1999) Modulation of α-tropomyosin expression by α-tocopherol in rat vascular smooth muscle cells. FEBS Lett 447:91-94.
Armstrong JS (2006) Mitochondria: a target for cancer therapy. Br J Pharmacol 147:239-48.
Arya P, Alibhai N, Qin H, Burton GW, Batist G, You SX and Alaoui-Jamali MA (1998) Design and synthesis of analogues of vitamin E: antiproliferative activity against human breast adenocarcinoma cells. Bioorg Med Chem Lett 8:2433-2438.
Astuti D, Hart-Holden N, Latif F, Lalloo F, Black GC, Lim C, Moran A, Grossman AB, Hodgson SV, Freemont A, Ramsden R, Eng C, Evans DG, Maher ER. (2003) Genetic analysis of mitochondrial complex II subunits SDHD, SDHB and SDHC in paraganglioma and phaeochromocytoma susceptibility.Clin Endocrinol (Oxf). Dec;59(6):728-33
Azzi A, Ricciarelli R and Zingg JM (2002) Non-antioxidant molecular functions of α-tocopherol (vitamin E). FEBS Lett 519:8-10.
Barnett KT, Fokum FD, Malafa MP (2002) Vitamin E succinate inhibits colon cancer liver metastases. J Surg Res 106:292-298.
Bayley JP, van Minderhout I, Weiss MM, Jansen JC, Oomen PH, Menko FH, Pasini B, Ferrando B, Wong N, Alpert LC, Williams R, Blair E, Devilee P, Taschner PE. (2006) Mutation analysis of SDHB and SDHC: novel germline mutations in sporadic head and neck paraganglioma and familial paraganglioma and/or pheochromocytoma. BMC Med Genet. Jan 11;7:1
Berridge MV, Herst PM, Tan AS. (2005) Tetrazolium dyes as tools in cell biology: new insights into their cellular reduction. Biotechnol Annu Rev.11:127-52
Berridge MV and Tan AS (1993) Characterization of the cellular reduction of 3-(4,5-dimethylthiazol- 2-yl)-2,5-diphenyltetrazolium bromide (MTT): subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch Biochem Biophys 303:474-478.
Birringer M, Drogan D and Brigelius-Flohe R (2001) Tocopherols are metabolized in HepG2 cells by side chain ω-oxidation and consecutive β-oxidation. Free Radic Biol Med 31:226-232.
Birringer M, EyTina JH, Salvatore BA and Neuzil J (2003) Vitamin E analogues as inducers of apoptosis: Structure-function relationship. Br J Cancer 88:1948-1955.
Birringer M, Pfluger P, Kluth D, Landes N and Brigelius-Flohe R (2002) Identities and differences in the metabolism of tocotrienols and tocopherols in HepG2 cells. J Nutr 132:3113-3118.
Boersma AW, Nooter K, Oostrum RG., Stoter G (1996) Quantification of apoptotic cells with fluorescein isothiocyanate-labeled annexin V in Chinese hamster ovary cell cultures with cisplatin. Cytometry 24:123-130
Breen GA, Scheffler IE. (1979) Respiration-deficient Chinese hamster cell mutants: biochemical characterization. Somatic Cell Genet. Jul;5(4):441-51.
Briere JJ, Schlemmer D, Chretien D, Rustin P. (2004) Quinone analogues regulate mitochondrial substrate competitive oxidation. Biochem Biophys Res Commun. Apr 16;316(4):1138-42.
Burstein HJ (2005) The distinctive nature of HER2-positive breast cancers. N Engl J Med 353:1652-1654.
Burnell JN, John P, Whatley FR (1975) The reversibility of active sulphate transport in membrane vesicles of Paracoccus denitrificans. Biochem J. 150: 527-536.
Cardone MH, Roy N, Stennicke HR, Salvesen GS, Franke TF, Stanbridge E, Frisch S, Reed JC (1998) Regulation of cell death protease caspase-9 by phosphorylation. Science 282:1318-1321.
Cheeseman KH, Holley AE, Kelly FJ, Wasil M, Hughes L and Burton G (1995) Biokinetics in humans of RRR-α-tocopherol: the free phenol, acetate ester, and succinate ester forms of vitamin E. Free Radic Biol Med 19:591-598
Choudhry ZM, Gavrikova EV, Kotlyar AB, Tushurashvili PR and Vinogradov AD. (1985) Pyridoxal phosphate-induced dissociation of the succinate: ubiquinone reductase.FEBS Lett. Mar 11;182(1):171-5.
Choudhry ZM, Kotlyar AB and Vinogradov AD. (1986) Studies on the succinate dehydrogenating system. Interaction of the mitochondrial succinate-ubiquinone reductase with pyridoxal phosphate.Biochim Biophys Acta. Jun 10;850(1):131-8.
Christen S, Woodall AA, Shigenaga MK, Southwell-Keely PT, Duncan MW, Ames BN. (1997) gamma-tocopherol traps mutagenic electrophiles such as NO(X) and complements alpha-tocopherol: physiological implications. Proc Natl Acad Sci U S A. Apr 1;94(7):3217-22.
Church SL, Grant JW, Ridnour LA, Oberley LW, Swanson PE, Meltzer PS, Trent JM (1993) Increased manganese superoxide dismutase expression supresses the malignant phenotype of human melanoma cells. Proc Natl Acad Sci USA 90:3113-3117.
DeFrancesco L, Scheffler IE, Bissell MJ. (1976) A respiration-deficient Chinese hamster cell line with a defect in NADH-coenzyme Q reductase. J Biol Chem. Aug 10;251(15):4588-95.
Devaraj S, Hugou I and Jialal I (2001) α-Tocopherol decreases CD36 expression in human monocyte-derived macrophages. J Lipid Res 42:521-527.
Don AS, Hogg PJ (2004) Mitochondria as cancer drug targets. Trends Mol Med 10:372-378.
Dong LF, Wang XF, Zhao Y, Tomasetti M, Wu K, Neuzil J (2006) Vitamin E analogues as anti-cancer agents: the role of modulation of apoptosis signalling pathways. Cancer Therapy 4:35-46.
Esposti MD, Ngo A, Ghelli A, Benelli B, Carelli V, McLennan H, Linnane AW (1996) The interaction of Q analogs, particularly hydroxydecyl benzoquinone (idebenone), with the respiratory complexes of heart mitochondria. Arch Biochem Biophys. Jun 15;330(2):395-400.. Please specify the different types of ubiquinone
Fariss MW, Nicholls-Grzemski FA, Tirmenstein MA, Zhang JG (2001) Enhanced antioxidant and cytoprotective abilities of vitamin E succinate is associated with a rapid uptake advantage in rat hepatocytes and mitochondria. Free Radic Biol Med 31:530-541.
Fresno Vara JA, Casado E, de Castro J, Cejas P, Belda-Iniesta C, Gonzalez-Baron M (2004) PI3K/Akt signalling pathway and cancer. Cancer Treat Rev 30:193-204.
Galli F, Stabile AM, Betti M, Conte C, Pistilli A, Rende M, Floridi A and Azzi A (2004) The effect of α- and γ-tocopherol and their carboxyethyl hydroxychroman metabolites on prostate cancer cell proliferation. Arch Biochem Biophys 423:97-102.
Geschwind JF, Ko YH, Torbenson MS, Magee C, Pedersen PL (2002) Novel therapy for liver cancer: direct intraarterial injection of a potent inhibitor of ATP production. Cancer Res 62:3909-3913.
Gottlieb E, Tomlinson IP. (2005) Mitochondrial tumour suppressors: a genetic and biochemical update.Nat Rev Cancer. Nov;5(11):857-66
Grandage VL, Gale RE, Linch DC, Khwaja A (2005) PI3-kinase/Akt is constitutively active in primary acute myeloid leukaemia cells and regulates survival and chemoresistance via NF-κB, MAP kinase and p53 pathways. Leukemia 19:586-594.
Gu LQ, Yu L, Yu CA. (1990) Effect of substituents of the benzoquinone ring on electron-transfer activities of ubiquinone derivatives. Biochim Biophys Acta. Feb 22;1015(3):482-92.
Guthrie N, Gapor A, Chambers AF and Carroll KK (1997) Inhibition of proliferation of estrogen receptor-negative MDA-MB-435 and -positive MCF-7 human breast cancer cells by palm oil tocotrienols and tamoxifen, alone and in combination. J Nutr 127: 544S-548S.
Guy CT, Webster MA, Schaller M, Parsons TJ, Cardiff RD, Muller WJ (1992) Expression of the neu protooncogene in the mammary epithelium of transgenic mice induces metastatic disease. Proc Natl Acad Sci USA 89:10578-10582.
Hartshorn MJ (2002) AstexViewer(TM): An aid for structure-based drug design. J Computer Aided Mol Des 16: 871-881.
He L, Mo H, Hadisusilo S, Qureshi AA and Elson CE (1997) Isoprenoids suppress the growth of murine B16 melanomas in vitro and in vivo. J Nutr 127:668-674.
He DY, Yu L, Yu CA. (1994) Protein ubiquinone interaction. Synthesis and biological properties of 5-alkyl ubiquinone derivatives. J Biol Chem. Nov 11;269(45):27885-8.
Hosomi A, Arita M, Sato Y, Kiyose C, Ueda T, Igarashi O, Arai H and Inoue K (1997) Affinity for α-tocopherol transfer protein as a determinant of the biological activities of vitamin E analogs. FEBS Lett 409:105-108.
Huang LS, Sun G, Cobessi D, Wang AC, Shen JT, Tung EY, Anderson VE, Berry EA (2006) 3-nitropropionic acid is a suicide inhibitor of mitochondrial respiration that, upon oxidation by complex II, forms a covalent adduct with a catalytic base arginine in the active site of the enzyme. J Biol Chem 281:5965-5972
James AM, Cocheme HM, Smith RA, Murphy MP (2005) Interactions of mitochondria-targeted and untargeted ubiquinones with the mitochondrial respiratory chain and reactive oxygen species. Implications for the use of exogenous ubiquinones as therapies and experimental tools. J Biol Chem 280:21295-212312.
Kelso GF, Porteous CM, Coulter CV, Hughes G, Porteous WK, Ledgerwood EC, Smith RA, Murphy MP (2001) Selective targeting of a redox-active ubiquinone to mitochondria within cells: antioxidant and antiapoptotic properties. J Biol Chem 276:4588-4596.
Kim R, Tanabe K, Emi M, Uchida Y, Toge T (2004) Potential roles of antisense therapy in the molecular targeting of genes involved in cancer. Int J Oncol 24:5-17
King A, Selak MA, Gottlieb E. (2006) Succinate dehydrogenase and fumarate hydratase: linking mitochondrial dysfunction and cancer. Oncogene. Aug 7;25(34):4675-82
King TE (1967). Preparation of succinate dehydrdogenase and reconstitution of succinate oxidase. Methods Enzymol 10: 322-331.
King, T.S. (1967) Preparations of succinate-cytochrome c reductase and the cytochrome b-c1 particle, and reconstitution of succinate-cytochrome c reductase. Methods in Enzymology, 10, 216-225
Ko YH, Smith BL, Wang Y, Pomper MG, Rini DA, Torbenson MS, Hullihen J, Pedersen PL (2004) Advanced cancers: eradication in all cases using 3-bromopyruvate therapy to deplete ATP. Biochem Biophys Res Commun 324:269-275.
Kogure K, Hama S, Kisaki M, Takemasa H, Tokumura A, Suzuki I and Fukuzawa K (2004) Structural characteristic of terminal dicarboxylic moiety required for apoptogenic activity of α-tocopheryl esters. Biochim Biophys Acta 1672: 93-99.
Kogure K, Manabe S, Suzuki I, Tokumura A and Fukuzawa K (2005) Cytotoxicity of α-tocopheryl succinate, malonate and oxalate in normal and cancer cells in vitro and their anti-cancer effects on mouse melanoma in vivo. J Nutr Sci Vitaminol 51:392-397.
Kunisaki M, Bursell SE, Clermont AC, Ishii H, Ballas LM, Jirousek MR, Umeda F, Nawata H and King GL (1995) Vitamin E prevents diabetes-induced abnormal retinal blood flow via the diacylglycerol-protein kinase C pathway. Am J Physiol 269:E239-246.
Maehara Y, Kusumoto T, Kusumoto H, Anai H, Sugimachi K (1988) Sodium succinate enhances the colorimetric reaction of the in vitro chemosensitivity test: MTT assay. Oncology 5:434-436.
Makishima M, Umesono K, Shudo K, Naoe T, Kishi K and Honma Y (1998) Induction of differentiation in acute promyelocytic leukemia cells by 9-cis retinoic acid α-tocopherol ester (9-cis tretinoin tocoferil). Blood 91:4715-4726.
Malafa MP, Fokum FD, Mowlavi A, Abusief M, King M (2002) Vitamin E inhibits melanoma growth in mice. Surgery 131:85-91.
Malafa MP, Neitzel LT. Vitamin E succinate promotes breast cancer tumor dormancy. J Surg Res 93:163-170.
Martin-Nizard F, Boullier A, Fruchart JC and Duriez P (1998) α-Tocopherol but not β-tocopherol inhibits thrombin-induced PKC activation and endothelin secretion in endothelial cells. J Cardiovasc Risk 5:339-345.
Morris G M, Goodsell DS, Halliday RS, Huey R, Hart W E, Belew RK, Olson AJ (1998) Automated docking using a Lamarckian genetic algorithm and empirical binding free energy function. J Comp Chem 19: 1639-1662.
Morton LW, Ward NC, Croft KD, Puddey IB. (2002) Evidence for the nitration of gamma-tocopherol in vivo: 5-nitro-gamma-tocopherol is elevated in the plasma of subjects with coronary heart disease. Biochem J. Jun 15;364(Pt 3):625-8.
Munteanu A, Zingg JM, Ogru E, Libinaki R, Gianello R, West S, Negis Y and Azzi (2004) AModulation of cell proliferation and gene expression by α-tocopheryl phosphates: relevance to atherosclerosis and inflammation. Biochem Biophys Res Commun 318:311-316.
Negis Y, Zingg JM, Ogru E, Gianello R, Libinaki R and Azzi A (2005) On the existence of cellular tocopheryl phosphate, its synthesis, degradation and cellular roles: a hypothesis. IUBMB Life 57:23-25.
Nesaretnam K, Stephen R, Dils R and Darbre P (1998) Tocotrienols inhibit the growth of human breast cancer cells irrespective of estrogen receptor status. Lipids 33:461-469.
Neuzil J, Dong LF, Ramanathapuram L, Hahn T, Chladova M, Wang XF, Zobalova R, Prochazka L, Gold M, Freeman R, Turanek J, Akporiaye ET, Dyason JC, Ralph SJ. (2007) Vitamin E analogues as a novel group of mitocans: Anti-cancer agents that act by targeting mitochondria. Mol Aspects Med. Feb 23.
Neuzil J, Massa H (2005) Hepatic processing determines dual activity of vitamin E succinate. Biochem Biophys Res Commun 327:1024-1027.
Neuzil J, Tomasetti M, Mellick AS, Alleva R, Salvatore BA, Birringer M, Fariss MW (2004) Vitamin E analogues: A new class of inducers of apoptosis with selective anti-cancer effects. Curr Cancer Drug Targets 4:355-372.
Neuzil J, Tomasetti M, Zhao Y, Dong LF, Birringer M, Wang XF, Low P, Wu K, Salvatore BA, Ralph SJ. (2007) Vitamin E analogs, a novel group of "mitocans," as anticancer agents: the importance of being redox-silent. Mol Pharmacol. May;71(5):1185-99.
Neuzil J, Weber T, Gellert N, Weber C (2001) Selective cancer cell killing by α-tocopheryl succinate. Br J Cancer 84:87-89.
Neuzil J, Weber T, Schroder A, Lu M, Ostermann G, Gellert N, Mayne GC, Olejnicka B, Negre-Salvayre A, Sticha M, Coffey RJ, Weber C (2001) Induction of apoptosis in cancer cells by α-tocopheryl succinate: Molecular pathways and structural requirements. FASEB J 15:403-415.
Nishikawa K, Satoh H, Hirai A, Suzuzki K, Asano R, Kumadaki I, Hagiwara K and Yano T (2003) α-Tocopheryloxybutyric acid enhances necrotic cell death in breast cancer cells treated with chemotherapy agent. Cancer Lett 201:51-56
Oostveen FG, Au HC, Meijer PJ, Scheffler IE. (1995) A Chinese hamster mutant cell line with a defect in the integral membrane protein CII-3 of complex II of the mitochondrial electron transport chain. J Biol Chem. Nov 3;270(44):26104-8.
Osaki M, Oshimura M, Ito H (2004) PI3K-Akt pathway: its functions and alterations in human cancer. Apoptosis 9:667-676.
Pham DQ and Plakogiannis R (2005) Vitamin E supplementation in cardiovascular disease and cancer prevention: Part 1. Ann Pharmacother 39:1870-8
Pollard PJ, Briere JJ, Alam NA, Barwell J, Barclay E, Wortham NC, Hunt T, Mitchell M, Olpin S, Moat SJ, Hargreaves IP, Heales SJ, Chung YL, Griffiths JR, Dalgleish A, McGrath JA, Gleeson MJ, Hodgson SV, Poulsom R, Rustin P, Tomlinson IP. (2005) Accumulation of Krebs cycle intermediates and over-expression of HIF1alpha in tumours which result from germline FH and SDH mutations. Hum Mol Genet. Aug 1;14(15):2231-9.
Quin J, Engle D, Litwiller A, Peralta E, Grasch A, Boley T, Hazelrigg S (2005) Vitamin E succinate decreases lung cancer tumor growth in mice. J Surg Res 127:139-143.
Qureshi AA, Mo H, Packer L and Peterson DM (2000) Isolation and identification of novel tocotrienols from rice bran with hypocholesterolemic, antioxidant, and antitumor properties. J Agric Food Chem 48:3130-3140.
Ralph SJ, Dong LF, Low P, Lawen A, and Neuzil J (2006) Mitocans: mitochondria targeted anti-cancer drugs as improved therapies and related patents. Recent Pat Anticancer Drug Discov 1:305-326.
Ricciarelli R, Zingg JM and Azzi A (2000) Vitamin E reduces the uptake of oxidized LDL by inhibiting CD36 scavenger receptor expression in cultured aortic smooth muscle cells. Circulation 102:82-87.
Safford SE, Oberley TD, Urano M, St Clair DK (1994) Suppression of fibrosarcoma metastasis by elevated expression of manganese superoxide dismutase. Cancer Res 54:4261-4265.
Sanborn BM, Felberg NT, Hollocher TC (1971) The inactivation of succinate dehydrogenase by bromopyruvate. Biochim Biophys Acta 227:219-231.
Sanner MF (1999) Python: a programming language for software integration and development, J Mol Graphic Mod 17: 57-61.
Sanders G., et al (2001) Preparation of tocopherols, tocotrienols, other chroman and side chain derivatives that induce cell apoptosis for therapeutic use as antiproliferative agents. 2001: PCT Int. Appl. WO 2001058889. p. 120.
Scallet AC, Haley RL, Scallet DM, Duhart HM, Binienda ZK (2003) 3-nitropropionic acid inhibition of succinate dehydrogenase (complex II) activity in cultured Chinese hamster ovary cells: antagonism by L-carnitine. Ann N Y Acad Sci 993:305-312.
Scheffler IE, Yadava N, Potluri P. (2004) Molecular genetics of complex I-deficient Chinese hamster cell lines. Biochim Biophys Acta. Dec 6;1659(2-3):160-71. Review.
Seo, B.B., Kitajima-Ihara, T., Chan, E.K., Scheffler, I.E., Matsuno-Yagi, A., and Yagi, T. (1998) Proc. Natl. Acad. Sci. USA 95, 9167-9171
Shah SJ and Sylvester PW (2005) γ-Tocotrienol inhibits neoplastic mammary epithelial cell proliferation by decreasing Akt and nuclear factor κB activity. Exp Biol Med 230: 235-241.
Shiau CW, Huang JW, Wang DS, Weng JR, Yang CC, Lin CH, Li C, Chen CS (2006) alpha-Tocopheryl succinate induces apoptosis in prostate cancer cells in part through inhibition of Bcl-xL/Bcl-2 function. J Biol Chem 281:11819-11825.
Shun MC, Yu W, Gapor A, Parsons R, Atkinson J, Sanders BG and Kline K (2004) Pro-apoptotic mechanisms of action of a novel vitamin E analog (α-TEA) and a naturally occurring form of vitamin E (δ-tocotrienol) in MDA-MB-435 human breast cancer cells. Nutr Cancer 48:95-105.
Slamon DJ, Godolphin W, Jones LA, Holt JA, Wong SG, Keith DE, Levin WJ, Stuart SG, Udove J, Ullrich A, et al (1989) Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science 244:707-712.
Stapelberg M, Gellert N, Swettenham E, Tomasetti M, Witting PK, Procopio A, Neuzil J (2005) α-Tocopheryl succinate inhibits malignant mesothelioma by disruption of the FGF autocrine signaling loop: Mechanism and the role of oxidative stress. J Biol Chem 280:25369-25376.
Sugawara M, Huang W, Fei Y-J, Leibach FH, Ganapathy V and Ganapathy ME (2000) Transport of valganciclovir, a ganciclovir prodrug, via peptide transporters PEPT1 and PEPT2. J Pharm Sci 89:781-789.
Sun F, Huo X, Zhai Y, Wang A, Xu J, Su D, Bartlam M, Rao Z. Crystal structure of mitochondrial respiratory membrane protein complex II. Cell 121:1043-1057.
Swettenham E, Witting PK, Salvatore BA, Neuzil J (2005) α-Tocopheryl succinate selectively induces apoptosis in neuroblastoma cells: Potential therapy of malignancies of the nervous system? J Neurochem 94:1448-1456.
Takeuchi H, Kim J, Fujimoto A, Umetani N, Mori T, Bilchik A, Turner R, Tran A, Kuo C, Hoon DS (2005) X-Linked inhibitor of apoptosis protein expression level in colorectal cancer is regulated by hepatocyte growth factor/C-met pathway via Akt signaling. Clin Cancer Res 11:7621-7628.
Tan AK, Ramsay RR, Singer TP, Miyoshi H. (1993) Comparison of the structures of the quinone-binding sites in beef heart mitochondria. J Biol Chem. Sep 15;268(26):19328-33.
Tasinato A, Boscoboinik D, Bartoli GM, Maroni P and Azzi A (1995) d-α-Tocopherol inhibition of vascular smooth muscle cell proliferation occurs at physiological concentrations, correlates with protein kinase C inhibition, and is independent of its antioxidant properties. Proc Natl Acad Sci USA 92:12190-12194.
Teague, S. J.; Davis, A. M.; Leeson, P. D.; Oprea, T. (1999) The Design of Leadlike Combinatorial Libraries. Angew Chem Int Ed Engl, 38, 3743-3748
Tomasetti M, Gellert N, Procopio A, Neuzil J (2004) A vitamin E analogue suppresses malignant mesothelioma in a pre-clinical model: A prototype of a future drug against a fatal neoplastic disease? Int J Cancer 109:641-642.
Tomic-Vatic A, EyTina JH, Chapmann JM, Mahdavian E, Neuzil J and Salvatore BA (2005) Vitamin E amides, a new class of vitamin E analogues with enhanced pro-apoptotic activity. Int J Cancer 117:118-193.
van Nederveen FH, Korpershoek E, Lenders JW, de Krijger RR, Dinjens WN. (2007) Somatic SDHB mutation in an extraadrenal pheochromocytoma. N Engl J Med. Jul 19;357(3):306-8.
Vraka PS, Drouza C, Rikkou MP, Odysseos AD and Keramidas AD (2006) Synthesis and study of the cancer cell growth inhibitory properties of α-, γ-tocopheryl and γ-tocotrienyl 2-phenylselenyl succinates. Bioorg Med Chem 14: 2684-2696.
Wallace, A.C., Laskowski, R.A. and Thornton, J.M. (1995). LIGPLOT: a program to generate schematic diagrams of protein-ligand interactions. Protein Eng 8, 127-134.
Wang XF, Dong LF, Zhao Y, Tomasetti M, Wu K, Neuzil J (2006) Vitamin E analogues as anti-cancer agents: Lessons from studies with α-tocopheryl succinate. Mol Nutr Food Res 50:675-685.
Wang XF, Witting PK, Salvatore BA, Neuzil J (2005) α-Tocopheryl succinate induces apoptosis in HER2/erbB2-overexpressing breast cancer cells by signalling via the mitochondrial pathway. Biochem Biophys Res Commun 326:282-289.
Weber T, Dalen H, Andera L, Negre-Salvayre A, Auge N, Sticha M, Lloret A, Terman A, Witting PK, Higuchi M, Plasilova M, Zivny J, Gellert N, Weber C, Neuzil J (2003) Mitochondria play a central role in apoptosis induced by α-tocopheryl succinate, an agent with anticancer activity. Comparison with receptor-mediated pro-apoptotic signaling. Biochemistry 42:4277-4291.
Weber T, Lu M, Andera L, Lahm H, Gellert N, Fariss MW, Korinek V, Sattler W, Ucker DS, Terman A, Schroder A, Erl W, Brunk UT, Coffey RJ, Weber C, Neuzil J (2002) Vitamin E succinate is a potent novel anti-neoplastic agent with high tumor selectivity and cooperativity with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL, Apo2L) in vivo. Clin Cancer Res 8:863-869.
West KA, Castillo SS, Dennis PA (2002) Activation of the PI3K/Akt pathway and chemotherapeutic resistance. Drug Resist Updat 5:234-248.
Wu Y, Zu K, Ni J, Yeh S, Kasi D, James NS, Chemler S and Ip C (2004) Cellular and molecular effects of α-tocopheryloxybutyrate: lessons for the design of vitamin E analog for cancer prevention. Anticancer Res 24:3795-3802.
Xu RH, Pelicano H, Zhou Y, Carew JS, Feng L, Bhalla KN, Keating MJ, Huang P (2005) Inhibition of glycolysis in cancer cells: a novel strategy to overcome drug resistance associated with mitochondrial respiratory defect and hypoxia. Cancer Res 65:613-621.
Yabunaka H, Kenmochi A, Nakatogawa Y, Sakamoto K, Miyoshi H. (2002) Hybrid ubiquinone: novel inhibitor of mitochondrial complex I. Biochim Biophys Acta. Dec 2;1556(2-3):106-12.
Yamauchi J, Iwamoto T, Kida S, Masushige S, Yamada K and Esashi T (2001) Tocopherol-associated protein is a ligand-dependent transcriptional activator. Biochem Biophys Res Commun 285:295-299.
Yang F, Yu L, He DY, Yu CA. (1991) Protein-ubiquinone interaction in bovine heart mitochondrial succinate-cytochrome c reductase. Synthesis and biological properties of fluorine substituted ubiquinone derivatives. J Biol Chem. Nov 5;266(31):20863-9.
Yankovskaya V, Sablin SO, Ramsay RR, Singer TP, Ackrell BA, Cecchini G, Miyoshi H. (1996) Inhibitor probes of the quinone binding sites of mammalian complex II and Escherichia coli fumarate reductase. J Biol Chem. Aug 30;271(35):21020-4.
Yano Y, Satoh H, Fukumoto K, Kumadaki I, Ichikawa T, Yamada K, Hagiwara K and Yano T (2005) Induction of cytotoxicity in human lung adenocarcinoma cells by 6-O-carboxy-propyl-α-tocotrienol, a redox-silent derivative of α-tocotrienol. Int J Cancer 115:839-846.
Yoeli-Lerner M, Yiu GK, Rabinovitz I, Erhardt P, Jauliac S, Toker A (2005) Akt blocks breast cancer cell motility and invasion through the transcription factor NFAT. Mol Cell 20:539-550.
Youk HJ, Lee E, Choi MK, Lee YJ, Chung JH, Kim SH, Lee CH and Lim SJ (2005) Enhanced anticancer efficacy of α-tocopheryl succinate by conjugation with polyethylene glycol. J Control Release 107:43-52.
Zhang JG, Nicholls-Grzemski FA, Tirmenstein MA, Fariss MW (2001a) Vitamin E succinate protects hepatocytes against the toxic effect of reactive oxygen species generated at mitochondrial complexes I and III by alkylating agents. Chem Biol Interact 138:267-284.
Zhang JG, Tirmenstein MA, Nicholls-Grzemski FA, Fariss MW (2001b) Mitochondrial electron transport inhibitors cause lipid peroxidation-dependent and -independent cell death: protective role of antioxidants. Arch Biochem Biophys 393:87-96.
Zhao S, Konopleva M, Cabreira-Hansen M, Xie Z, Hu W, Milella M, Estrov Z, Mills GB, Andreeff M (2004) Inhibition of phosphatidylinositol 3-kinase dephosphorylates BAD and promotes apoptosis in myeloid leukemias. Leukemia 18:267-27.
【特許請求の範囲】
【請求項1】
ガン性細胞のミトコンドリア呼吸鎖の複合体II(コハク酸ユビキノン酸化還元酵素)と相互作用して、細胞内に活性酸素種を生成する能力がある、ガン性細胞の死を選択的に誘導するためのプロオキシダント化合物。
【請求項2】
ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力がある、ガンを予防または治療するためのプロオキシダント化合物。
【請求項3】
ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力がある、ガン性細胞の死を選択的に誘導するためのプロオキシダント化合物。
【請求項4】
ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力がある、ガンを予防または治療するためのプロオキシダント化合物。
【請求項5】
化合物が、プロオキシダントビタミンE類縁体である請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項6】
化合物が、α-トコフェリルコハク酸エステル(α-TOS)である請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項7】
化合物が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項8】
化合物が、ヘプタペプチドLTVSPWYに結合したα-TOSである請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項9】
請求項1〜8のいずれか1つに記載のプロオキシダント化合物またはその生理的に許容しうる塩および生理的に許容しうる担体を含む医薬組成物または獣医用組成物。
【請求項10】
さらに治療有効量の3-BPを含む請求項9に記載の医薬組成物または獣医用組成物。
【請求項11】
さらに治療有効量のTRAILを含む請求項9に記載の医薬組成物または獣医用組成物。
【請求項12】
さらに治療有効量のAkt1インヒビターを含む請求項9に記載の医薬組成物または獣医用組成物。
【請求項13】
組成物が、α-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルおよびα-トコフェリルマレイルアミドから選ばれるプロオキシダント化合物を含み、担体が、局所投与用のリポソームベースである請求項9に記載の医薬組成物または獣医用組成物。
【請求項14】
組成物が、プロオキシダント化合物である2,5,7,8-テトラメチル-2R-(4R,8R,12-トリメチルトリデシル)-クロマン-6-イルオキシ酢酸(α-トコフェリルオキシ酢酸)を含み、担体が、経口投与用に適するものである請求項9に記載の医薬組成物または獣医用組成物。
【請求項15】
患者においてガン性細胞の死を誘導する方法であって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項16】
患者においてガンを予防または治療する方法であって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項17】
患者においてガン性細胞の死を誘導する方法であって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項18】
患者においてガンを予防または治療する方法であって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項19】
化合物が、プロオキシダントビタミンE類縁体である請求項15〜18のいずれか1つに記載の方法。
【請求項20】
化合物が、α-TOSである請求項15〜18のいずれか1つに記載の方法。
【請求項21】
化合物が、ヘプタペプチドLTVSPWYに結合したα-TOSである請求項15〜18のいずれか1つに記載の方法。
【請求項22】
化合物が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項15〜18のいずれか1つに記載の方法。
【請求項23】
さらに治療有効量の3-BPを含む請求項15〜22のいずれか1つに記載の方法。
【請求項24】
さらに治療有効量のTRAILを含む請求項15〜22のいずれか1つに記載の方法。
【請求項25】
さらに治療有効量のAkt1インヒビターを含む請求項15〜22のいずれか1つに記載の方法。
【請求項26】
医薬組成物または獣医用組成物が、α-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルおよびα-トコフェリルマレイルアミドから選ばれるプロオキシダント化合物を含み、担体が、局所投与用のリポソームベースである請求項15〜18のいずれか1つに記載の方法。
【請求項27】
医薬組成物または獣医用組成物が、プロオキシダント化合物である2,5,7,8-テトラメチル-2R-(4R,8R,12-トリメチルトリデシル)-クロマン-6-イルオキシ酢酸(α-トコフェリルオキシ酢酸)を含み、担体が、経口投与用に適するものである請求項15〜18のいずれか1つに記載の方法。
【請求項28】
ガン性細胞が、肺、肝臓、腎臓、脳、前立腺、乳房、卵巣、リンパ、皮膚、眼、結腸、胃、口腔扁平または造血系由来である請求項15〜27のいずれか1つに記載の方法。
【請求項29】
ガン性細胞が、erbB2-低である請求項15〜21のいずれか1つに記載の方法。
【請求項30】
ガン性細胞が、erbB2-高である請求項15〜21のいずれか1つに記載の方法。
【請求項31】
ガン性細胞が、低酸素性である請求項15〜21のいずれか1つに記載の方法。
【請求項32】
患者における中皮腫を予防または治療する方法であって、方法が、治療有効量のα-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルまたはα-トコフェリルマレイルアミドおよびリポソームベース担体を含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項33】
ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項34】
ガンを予防または治療するための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項35】
ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項36】
ガンを予防または治療するための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項37】
ガン性細胞の存在を検出するため、およびガン性細胞の死を選択的に誘導するためのプロオキシダント化合物であって、該化合物が、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有する化合物。
【請求項38】
患者においてガン性細胞の存在を検出するため、およびガン性細胞の死を選択的に誘導するための方法であって、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有するプロオキシダント化合物の治療有効量を患者に投与するステップを含む方法。
【請求項39】
ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)第1のガン性細胞のミトコンドリアに試験化合物を導入し、第2のガン性細胞にガン細胞死を誘導しないことがわかっているコントロール化合物を導入し;次いで、
(ii)ミトコンドリア複合体IIへの化合物の結合および第1および第2のガン性細胞内での活性酸素種の生成をアッセイし、試験化合物およびコントロール化合物によるガン性細胞の死ををアッセイする;
[ここで、コントロール化合物と比較しての試験化合物による複合体II結合、活性酸素種の生成および第1のガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法。
【請求項40】
試験化合物が、プロオキシダントビタミンE類縁体である請求項39に記載の方法。
【請求項41】
試験化合物が、請求項7に記載の化合物である請求項40に記載の方法。
【請求項42】
請求項39〜41に記載の方法によって同定される化学物質。
【請求項43】
ミトコンドリア呼吸鎖複合体IIまたはその1つ以上のサブユニットに結合した請求項1〜8のいずれか1つに記載の化合物を含む単離または精製された複合体。
【請求項44】
ミトコンドリア呼吸鎖複合体IIに結合した請求項1〜8のいずれか1つに記載の化合物を含む結晶または結晶可能な組成物。
【請求項45】
ミトコンドリア呼吸鎖複合体IIのユビキノン結合部位に結合する能力がある化学物質を同定または設計する方法であって、
(i)呼吸鎖複合体IIと、複合体IIに結合してユビキノンへの電子流を妨げる少なくとも1つのビタミンE類縁体との間の相互作用をコンピューターモデリングし;次いで、
(ii)コンピューターモデリングによって作成されたデータを用いて、複合体IIに結合してユビキノンへの電子流を妨げる能力を有する化学物質を同定または設計する;
ステップを含む方法。
【請求項46】
ビタミンE類縁体が、プロオキシダントビタミンE類縁体である請求項45に記載の方法。
【請求項47】
ビタミンE類縁体が、α-TOSである請求項45に記載の方法。
【請求項48】
請求項45〜47のいずれか1つに記載の方法にしたがって同定または設計された化学物質。
【請求項49】
化学物質が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項48に記載の化学物質。
【請求項50】
ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)ガン性細胞のミトコンドリアに試験化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位の天然のユビキノン基質の置換をアッセイし、ガン性細胞の死をアッセイする;
[ここで、試験化合物による天然のユビキノン基質の置換およガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法。
【請求項51】
試験化合物が、プロオキシダントビタミンE類縁体である請求項50に記載の方法。
【請求項52】
試験化合物が、請求項7に記載の化合物である請求項51に記載の方法。
【請求項53】
請求項50〜52に記載の方法によって同定される化学物質。
【請求項54】
化学物質が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項53に記載の化学物質。
【請求項55】
ガンを有する患者が、本発明の第1または第2の態様のプロオキシダント化合物の投与を含む療法に反応する可能性があるかどうかを試験する方法であって、
(i)該患者由来のガン性細胞のミトコンドリアにプロオキシダント化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位へのプロオキシダント化合物の結合をアッセイし、ガン性細胞の死をアッセイする;
[ここで、プロオキシダント化合物の結合およびプロオキシダント化合物によるガン性細胞の死は、患者が、療法に反応する可能性があることを示す]
ステップを含む方法。
【請求項56】
プロオキシダント化合物が、プロオキシダントビタミンE類縁体である請求項55に記載の方法。
【請求項57】
プロオキシダントビタミンE類縁体が、α-TOSである請求項56に記載の方法。
【請求項1】
ガン性細胞のミトコンドリア呼吸鎖の複合体II(コハク酸ユビキノン酸化還元酵素)と相互作用して、細胞内に活性酸素種を生成する能力がある、ガン性細胞の死を選択的に誘導するためのプロオキシダント化合物。
【請求項2】
ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力がある、ガンを予防または治療するためのプロオキシダント化合物。
【請求項3】
ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力がある、ガン性細胞の死を選択的に誘導するためのプロオキシダント化合物。
【請求項4】
ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力がある、ガンを予防または治療するためのプロオキシダント化合物。
【請求項5】
化合物が、プロオキシダントビタミンE類縁体である請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項6】
化合物が、α-トコフェリルコハク酸エステル(α-TOS)である請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項7】
化合物が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項8】
化合物が、ヘプタペプチドLTVSPWYに結合したα-TOSである請求項1〜4のいずれか1つに記載のプロオキシダント化合物。
【請求項9】
請求項1〜8のいずれか1つに記載のプロオキシダント化合物またはその生理的に許容しうる塩および生理的に許容しうる担体を含む医薬組成物または獣医用組成物。
【請求項10】
さらに治療有効量の3-BPを含む請求項9に記載の医薬組成物または獣医用組成物。
【請求項11】
さらに治療有効量のTRAILを含む請求項9に記載の医薬組成物または獣医用組成物。
【請求項12】
さらに治療有効量のAkt1インヒビターを含む請求項9に記載の医薬組成物または獣医用組成物。
【請求項13】
組成物が、α-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルおよびα-トコフェリルマレイルアミドから選ばれるプロオキシダント化合物を含み、担体が、局所投与用のリポソームベースである請求項9に記載の医薬組成物または獣医用組成物。
【請求項14】
組成物が、プロオキシダント化合物である2,5,7,8-テトラメチル-2R-(4R,8R,12-トリメチルトリデシル)-クロマン-6-イルオキシ酢酸(α-トコフェリルオキシ酢酸)を含み、担体が、経口投与用に適するものである請求項9に記載の医薬組成物または獣医用組成物。
【請求項15】
患者においてガン性細胞の死を誘導する方法であって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項16】
患者においてガンを予防または治療する方法であって、ガン性細胞のミトコンドリア複合体IIと選択的に相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項17】
患者においてガン性細胞の死を誘導する方法であって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項18】
患者においてガンを予防または治療する方法であって、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位に結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の治療有効量含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項19】
化合物が、プロオキシダントビタミンE類縁体である請求項15〜18のいずれか1つに記載の方法。
【請求項20】
化合物が、α-TOSである請求項15〜18のいずれか1つに記載の方法。
【請求項21】
化合物が、ヘプタペプチドLTVSPWYに結合したα-TOSである請求項15〜18のいずれか1つに記載の方法。
【請求項22】
化合物が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項15〜18のいずれか1つに記載の方法。
【請求項23】
さらに治療有効量の3-BPを含む請求項15〜22のいずれか1つに記載の方法。
【請求項24】
さらに治療有効量のTRAILを含む請求項15〜22のいずれか1つに記載の方法。
【請求項25】
さらに治療有効量のAkt1インヒビターを含む請求項15〜22のいずれか1つに記載の方法。
【請求項26】
医薬組成物または獣医用組成物が、α-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルおよびα-トコフェリルマレイルアミドから選ばれるプロオキシダント化合物を含み、担体が、局所投与用のリポソームベースである請求項15〜18のいずれか1つに記載の方法。
【請求項27】
医薬組成物または獣医用組成物が、プロオキシダント化合物である2,5,7,8-テトラメチル-2R-(4R,8R,12-トリメチルトリデシル)-クロマン-6-イルオキシ酢酸(α-トコフェリルオキシ酢酸)を含み、担体が、経口投与用に適するものである請求項15〜18のいずれか1つに記載の方法。
【請求項28】
ガン性細胞が、肺、肝臓、腎臓、脳、前立腺、乳房、卵巣、リンパ、皮膚、眼、結腸、胃、口腔扁平または造血系由来である請求項15〜27のいずれか1つに記載の方法。
【請求項29】
ガン性細胞が、erbB2-低である請求項15〜21のいずれか1つに記載の方法。
【請求項30】
ガン性細胞が、erbB2-高である請求項15〜21のいずれか1つに記載の方法。
【請求項31】
ガン性細胞が、低酸素性である請求項15〜21のいずれか1つに記載の方法。
【請求項32】
患者における中皮腫を予防または治療する方法であって、方法が、治療有効量のα-トコフェリルコハク酸エステル、α-トコフェリルマレイン酸エステルまたはα-トコフェリルマレイルアミドおよびリポソームベース担体を含む医薬組成物または獣医用組成物を患者に投与するステップを含む方法。
【請求項33】
ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項34】
ガンを予防または治療するための医薬の製造における、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項35】
ガン性細胞の死を選択的に誘導するための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項36】
ガンを予防または治療するための医薬の製造における、ガン性細胞のミトコンドリア呼吸鎖の複合体IIのユビキノン結合部位と結合して、細胞内に活性酸素種を生成する能力があるプロオキシダント化合物の使用。
【請求項37】
ガン性細胞の存在を検出するため、およびガン性細胞の死を選択的に誘導するためのプロオキシダント化合物であって、該化合物が、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有する化合物。
【請求項38】
患者においてガン性細胞の存在を検出するため、およびガン性細胞の死を選択的に誘導するための方法であって、ガン性細胞のミトコンドリア複合体IIと相互作用して、細胞内に活性酸素種を生成する能力があり、ガン性細胞内で該化合物の検出を可能にする検出可能な部分を有するプロオキシダント化合物の治療有効量を患者に投与するステップを含む方法。
【請求項39】
ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)第1のガン性細胞のミトコンドリアに試験化合物を導入し、第2のガン性細胞にガン細胞死を誘導しないことがわかっているコントロール化合物を導入し;次いで、
(ii)ミトコンドリア複合体IIへの化合物の結合および第1および第2のガン性細胞内での活性酸素種の生成をアッセイし、試験化合物およびコントロール化合物によるガン性細胞の死ををアッセイする;
[ここで、コントロール化合物と比較しての試験化合物による複合体II結合、活性酸素種の生成および第1のガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法。
【請求項40】
試験化合物が、プロオキシダントビタミンE類縁体である請求項39に記載の方法。
【請求項41】
試験化合物が、請求項7に記載の化合物である請求項40に記載の方法。
【請求項42】
請求項39〜41に記載の方法によって同定される化学物質。
【請求項43】
ミトコンドリア呼吸鎖複合体IIまたはその1つ以上のサブユニットに結合した請求項1〜8のいずれか1つに記載の化合物を含む単離または精製された複合体。
【請求項44】
ミトコンドリア呼吸鎖複合体IIに結合した請求項1〜8のいずれか1つに記載の化合物を含む結晶または結晶可能な組成物。
【請求項45】
ミトコンドリア呼吸鎖複合体IIのユビキノン結合部位に結合する能力がある化学物質を同定または設計する方法であって、
(i)呼吸鎖複合体IIと、複合体IIに結合してユビキノンへの電子流を妨げる少なくとも1つのビタミンE類縁体との間の相互作用をコンピューターモデリングし;次いで、
(ii)コンピューターモデリングによって作成されたデータを用いて、複合体IIに結合してユビキノンへの電子流を妨げる能力を有する化学物質を同定または設計する;
ステップを含む方法。
【請求項46】
ビタミンE類縁体が、プロオキシダントビタミンE類縁体である請求項45に記載の方法。
【請求項47】
ビタミンE類縁体が、α-TOSである請求項45に記載の方法。
【請求項48】
請求項45〜47のいずれか1つに記載の方法にしたがって同定または設計された化学物質。
【請求項49】
化学物質が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項48に記載の化学物質。
【請求項50】
ガン性細胞の死を誘導する能力がある化学物質を同定する方法であって、
(i)ガン性細胞のミトコンドリアに試験化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位の天然のユビキノン基質の置換をアッセイし、ガン性細胞の死をアッセイする;
[ここで、試験化合物による天然のユビキノン基質の置換およガン性細胞の死は、細胞死を誘導する能力がある化学物質が同定されたことを示す]
ステップを含む方法。
【請求項51】
試験化合物が、プロオキシダントビタミンE類縁体である請求項50に記載の方法。
【請求項52】
試験化合物が、請求項7に記載の化合物である請求項51に記載の方法。
【請求項53】
請求項50〜52に記載の方法によって同定される化学物質。
【請求項54】
化学物質が、
[(N’-アミジノヒドラジノ)-(4-メトキシフェニル)-メチル]ホスフィン酸;
N-(4-メトキシ-6-メチル-ピリミジン-2-イル)ベンゼンスルホンアミド;
4,6-ジメチル-3-メチルスルホニル-2-プロピルスルホニル-ピリジン;
(4-ホルミル-5-ヒドロキシ-6-メチル-3-ピリジル)メトキシホスホン酸(ピリドキサルホスフェート);
3-(5-プロポキシ-2-スルフォフェニル)プロパン酸;
3-[ヒドロキシ-[(メチル-メチルスルホニル-アミノ)メチル]ホスホリル]プロパン酸;
(エトキシカルボニル-(2-エトキシカルボニルエチル)アミノ)メチルホスフィン酸;
1-[2-(3,7-ジメチル-2,6-ジオキソ-プリン-1-イル)エチリデンアミノ]グアニジン;
8-スルホキノリン-2,4-ジカルボン酸;
2-(2-カルボキシエチル-ヒドロキシ-ホスホリル)-3-(2-フリル)プロパン酸;
3-[2S,4S,5R)-5,6-ジクロロ-2,4-ジメチル-1-オキソヘキシル]-4-ヒドロキシ-5,6-ジメトキシ-2(1H)-ピリジノン(アツペニンA5);
2,3-ジメトキシ-5-ゲラニル-6-メチル-1,4-ベンゾキノン(ユビキノン-2);
2-(1-メチルヘキシル)-4,6-ジニトロフェノール(ジニトロフェノール-17);
5,6-ジヒドロ-2-メチル-1,4-オキサチイン-3-カルボキシアニリド(カルボキシン);
4,4,4-トリフルオロ-1-(2-チエニル)-1,3-ブタンジオン(2-テノイルトリフルオロアセトン);および
非天然ユビキノン基質;
から選ばれる請求項53に記載の化学物質。
【請求項55】
ガンを有する患者が、本発明の第1または第2の態様のプロオキシダント化合物の投与を含む療法に反応する可能性があるかどうかを試験する方法であって、
(i)該患者由来のガン性細胞のミトコンドリアにプロオキシダント化合物を導入し;次いで、
(ii)ミトコンドリアの複合体IIのユビキノン結合部位へのプロオキシダント化合物の結合をアッセイし、ガン性細胞の死をアッセイする;
[ここで、プロオキシダント化合物の結合およびプロオキシダント化合物によるガン性細胞の死は、患者が、療法に反応する可能性があることを示す]
ステップを含む方法。
【請求項56】
プロオキシダント化合物が、プロオキシダントビタミンE類縁体である請求項55に記載の方法。
【請求項57】
プロオキシダントビタミンE類縁体が、α-TOSである請求項56に記載の方法。
【図1】

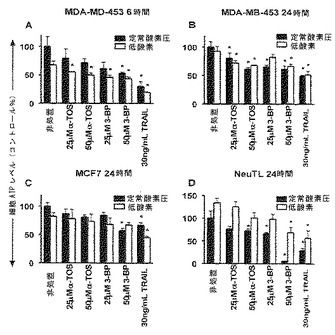
【図2】
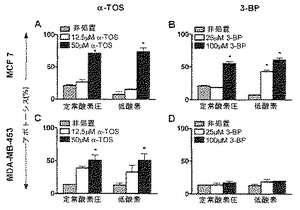
【図3】
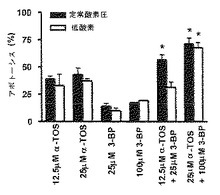
【図4】
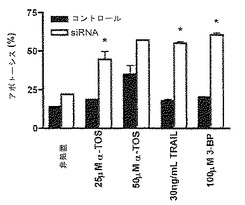
【図5】
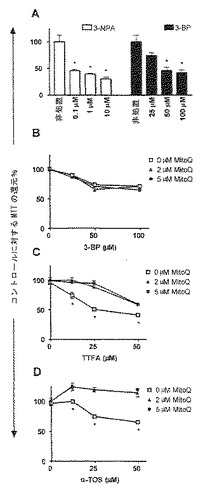
【図6】
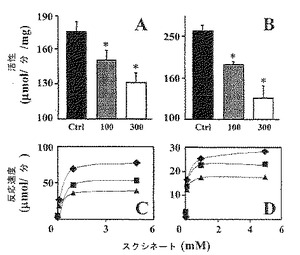
【図7】
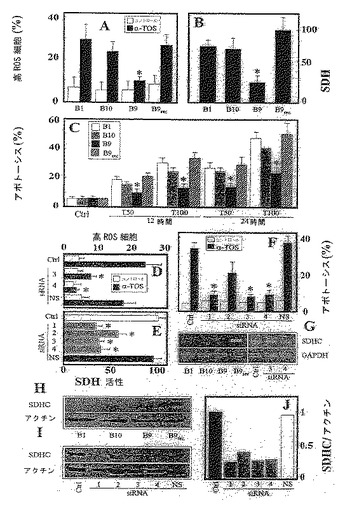
【図8A】
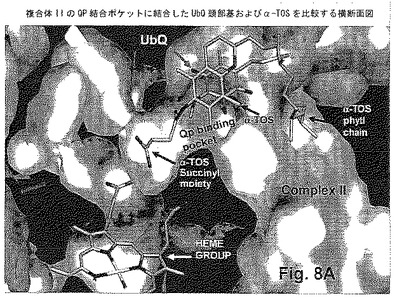
【図8B】

【図8C】
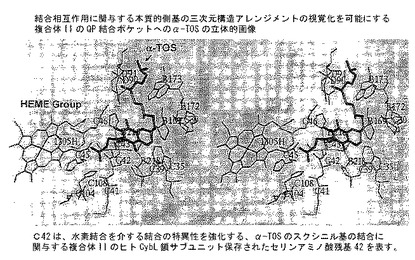
【図8D】
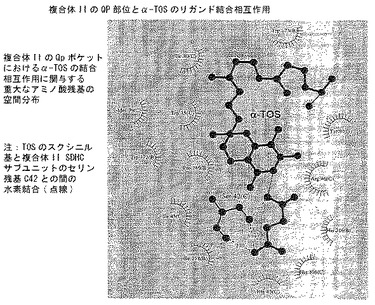
【図8E】
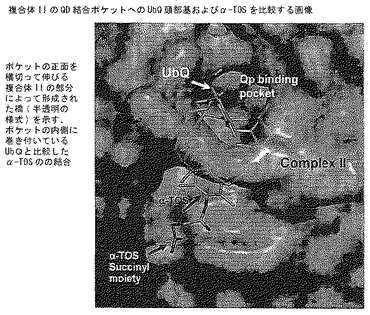
【図8F】
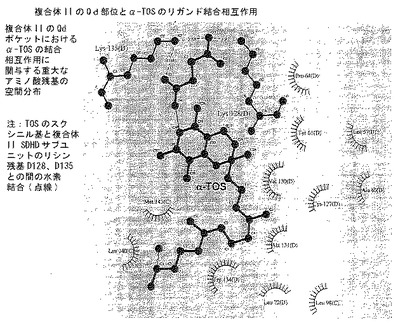
【図8G】
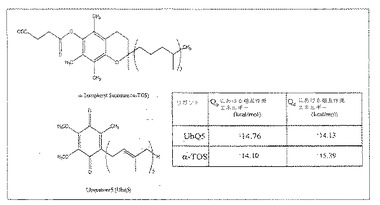
【図9】
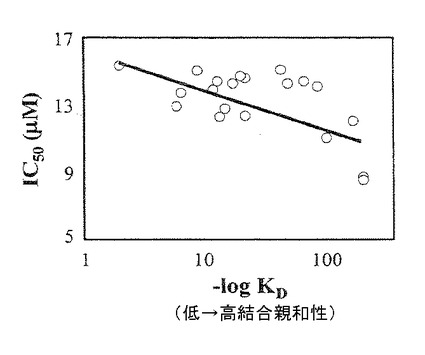
【図10−1】
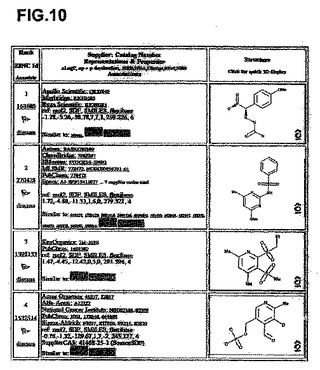
【図10−2】
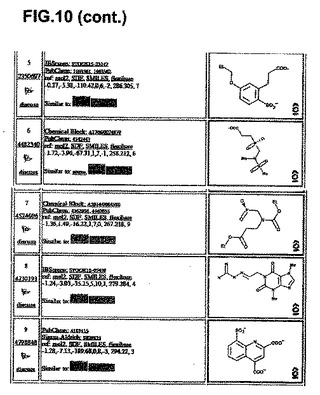
【図10−3】
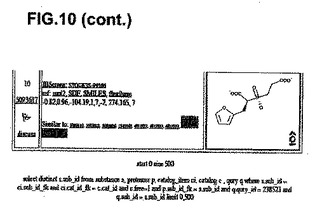
【図11】
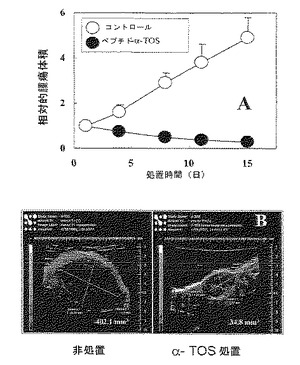
【図12】
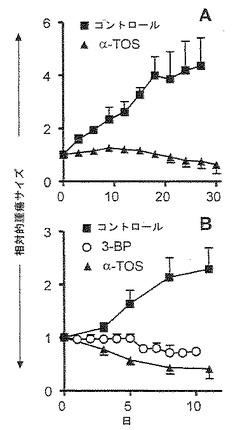
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図9】
【図10−1】
【図10−2】
【図10−3】
【図11】
【図12】
【公表番号】特表2010−508242(P2010−508242A)
【公表日】平成22年3月18日(2010.3.18)
【国際特許分類】
【出願番号】特願2009−527656(P2009−527656)
【出願日】平成19年9月17日(2007.9.17)
【国際出願番号】PCT/AU2007/001371
【国際公開番号】WO2008/031171
【国際公開日】平成20年3月20日(2008.3.20)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(509248095)
【氏名又は名称原語表記】Stephen John RALPH
【出願人】(509248109)
【氏名又は名称原語表記】Jiri NEUZIL
【Fターム(参考)】
【公表日】平成22年3月18日(2010.3.18)
【国際特許分類】
【出願日】平成19年9月17日(2007.9.17)
【国際出願番号】PCT/AU2007/001371
【国際公開番号】WO2008/031171
【国際公開日】平成20年3月20日(2008.3.20)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.サランラップ
【出願人】(509248095)
【氏名又は名称原語表記】Stephen John RALPH
【出願人】(509248109)
【氏名又は名称原語表記】Jiri NEUZIL
【Fターム(参考)】
[ Back to top ]