
プロテアソーム干渉に関連する避妊法および組成物
本発明は、動物、特にほ乳類の受精能を調節または評価する組成物および方法の使用に関する。本発明の種々の態様において、避妊方法は、配偶子、特に精子のプロテアソーム活性を阻害する段階を含む。プロテアソーム活性はインビトロまたはインビボにおいて阻害することができる。プロテアソーム経路の阻害剤には、プロテアソーム経路の種々の成分に結合する小分子、ペプチド、ポリペプチド(例えば、抗体等)および親和性物質(例えば、アプタマー等)が挙げられるが、これらに限定されない。いくつかの態様において、プロテアソーム経路の活性またはプロテアソーム経路の成分の活性は、プロテアソーム経路の1つ以上の成分に結合し、阻害する抗体によって阻害される。
【発明の詳細な説明】
【技術分野】
【0001】
A. 発明の分野
本発明は、一般に、繁殖生物学、薬物開発、免疫学および分子生物学の分野に関する。より具体的には、本発明は、受精能の調整および評価または不妊の診断のための、ユビキチン-プロテアソーム経路およびユビキチン-プロテアソーム経路の成分由来の抗原の阻害剤に関する。なお、本願は、全体の開示内容が具体的に参照として本明細書に組み入れられている、2003年2月14日提出の米国仮特許出願第60/447,675号の優先権を主張する。政府は、米国農務省からの付与番号1999-3520-11743に準じて本発明に権利を有しうる。
【背景技術】
【0002】
B. 関連技術の説明
ユビキチン化は広範で、進化的に保存されているが、真核細胞におけるタンパク質分解の基質特異性が高い経路である(Pickart, 1998;LaneyおよびHochstrasser, 1999)。ユビキチンによる翻訳後タンパク質修飾は、ユビキチンのC-末端グリシン(Gly)残基によって76アミノ酸ユビキチン残基が基質アミノ酸配列内のリジン(Lys)残基に共有結合することによって生ずる。ユビキチン分子は自身の7つのLys-残基を保有するので、この最初のモノ-ユビキチン化の次に追加のユビキチン分子が第1の基質-結合ユビキチンモノマーにGly-Lys結合することが多く、ジ-、トリ-テトラ-またはポリ-ユビキチン鎖の形成を誘導する(HershkoおよびCiechanover, 1998)。モノ-、ジ-およびトリ-ユビキチン化は、リソソームタンパク質分解、膜受容体のエンドサイトーシス、信号伝達および転写調節を含むが、これらに限定されない種々の目的を果たす(Conawayら, 2002;GlickmanおよびCiechanover, 2002により総説が報告されている)。テトラおよびポリ-ユビキチン化は、ユビキチン化タンパク質に特異的な活性を有するマルチ-サブユニットプロテアーゼである26-Sプロテアソームによるタンパク質ドッキングおよび分解のコンセンサスシグナルである(TanakaおよびTsurumi, 1997;Hochstrasser, 2002)。バレル型プロテアソームのルーメン内では、基質タンパク質が小ペプチドに切断されて細胞質に放出され、細胞質エンドペプチダーゼセットによりタンパク質分解が完了する(Hochstrasser, 2002;Kohlerら, 2001によって総説が示されている)。遊離されたポリ-ユビキチン鎖はプロテアソームによって分解されるのではなく、細胞質に戻り、他のタンパク質の修飾に再使用される(Chenら, 2002;Vogesら, 1999)。
【0003】
種々の基質のユビキチン化およびプロテアソーム分解は細胞質および核内で実証されている(GlickmanおよびCiechanover, 2002)。興味深いことに、ほ乳類の生殖の研究は、ユビキチン化は細胞表面および細胞外空間でも生じうるという証拠をもたらした。ユビキチンは、卵巣の卵胞液(Einspanierら, 1993)、精漿(Lippertら, 1993)、精巣上体液(HermoおよびJacks, 2002;Sutovskyら, 2001b)および精巣上体を通過する欠陥精子の表面(Sutovskyら, 2001a, b;2002, 2003)において検出された。卵子の卵黄(viteline)膜表面の精子受容体のユビキチン化およびプロテアソーム分解はホヤの受精中に生じる(Sawadaら, 2002a, b)。
【0004】
ほ乳類の精子が透明帯(「帯」またはZP)に結合して通過する能力は、ZP基質のZP3-精子受容体への結合の結果、エキソサイトーシス(Gerton, 2002)を受ける精子先体帽に存在する精子表面受容体およびタンパク質分解トリプシン様酵素セット(PrimakoffおよびMyles, 2002によって総説が報告されている)に帰されている(WassarmanおよびLitscher, 1995)。いくつかの候補先体タンパク質および精子表面受容体の標的突然変異およびノックアウトは、マウス精子がZPに結合し、透過/消化する能力を排除しなかった(Talbotら, 2003;Evans, 2001によって総説が報告されている)。セリンプロテアーゼ以外のタンパク質分解系が精子先体に存在し、受精中のZPの消化に寄与することをこれは示唆している。
【0005】
現在の避妊方法には、米国で最も一般的な避妊方法である女性の外科的不妊化および女性の経口避妊薬の使用が挙げられる。ホルモン調節避妊薬には、配合注射用避妊薬、配合経口避妊薬などの配合(エストロゲン/プロゲスチン)避妊薬;およびノルプラントインプラント、プロゲスチン-単独注射用避妊薬またはプロゲスチン-単独ピルなどのプロゲスチン-単独避妊薬が挙げられる。しかし、配合エストロゲン/プロゲスチン経口避妊薬は、血栓塞栓障害、脳血管事故、冠動脈疾患、肝異常、エストロゲン依存性の癌および妊娠を生じるまたはそれらに至ることがある。他の避妊方法には、子宮内避妊器具;子宮頸管キャップ;男性のコンドーム、女性のコンドーム、ペッサリー、殺精子剤または避妊用スポンジなどの遮断方法;および関与する個人への依存性が高い周期避妊法が挙げられる。これらの方法の多くは、生理的および物理的に広範囲の副作用を有することがある。従って、ヒト並びに種々の家畜および野生動物の受精を調節する改善された方法および受精を診断する方法の必要性がある。
【発明の開示】
【0006】
発明の概要
本発明は、ユビキチン-プロテアソーム経路の成分を阻害または検出することによって、ヒトまたは動物、特にほ乳類の受精を調節または評価する方法を含む。本発明の種々の態様において、避妊方法は精子のプロテアソーム活性を阻害する段階を含む。本明細書において使用する「避妊薬」は、投与したとき、意図的に、直接または間接的に受精または妊娠を妨害する薬剤を意味する。同様に、本明細書において使用する「避妊」は、直接または間接的に受精または妊娠を意図的に妨害することを意味する。本発明の目的のためには、プロテアソーム経路の成分に対する免疫応答を誘導する組成物を含む、プロテアソーム経路の成分の活性を阻害する薬剤は避妊薬と考えることができる。いくつかの態様において、精子のプロテアソーム活性は、インビトロ、エクスビボまたはインビボにおいて阻害することができる。プロテアソーム経路の阻害剤には、ユビキチン-プロテアソーム経路の種々の成分に結合する小分子、ペプチド、ポリペプチド(例えば、抗体等)および親和性物質(例えば、アプタマー等)が挙げられるが、これらに限定されない。いくつかの態様において、ユビキチン-プロテアソーム経路の活性またはユビキチン-プロテアソーム経路の成分の活性は、1つ以上のポリペプチドに結合し、その活性を阻害する抗体によって阻害される。ユビキチン-プロテアソーム経路に関連する種々のポリペプチドの説明は、国立バイオテクノロジー情報センター(National Center for Biotechnology Information(NCBI))が管理するヒト遺伝子変異と遺伝子疾患に関わる情報の統合データベース(Online Mendelian Inheritance in Man(OMIM)データベース)において見つけることができる。
【0007】
ある態様において、方法は、ユビキチン-プロテアソーム経路のタンパク質成分由来の少なくとも1つのペプチドまたは抗原で被験者または動物を免疫化することによって抗体を誘導する段階を含む。抗原は、プロテアソームサブユニット、ユビキチン、ユビキチン関連酵素またはプロテアソーム経路に関連するポリペプチド由来のペプチドであってもよい。特定の態様において、ペプチドは20Sプロテアソームコアのα-型および/またはβ-型サブユニット由来またはある種のプロテアソームに存在する19S調節キャップ(regulatory cap)もしくは11Sアクチベーター複合体のサブユニット由来である。本発明の1つ以上のペプチドまたは抗原はワクチン組成物に含まれてもよい。本発明のペプチドは、その例が配列番号:1、2、4および6に記載されているアミノ酸配列を有するプロテアソーム-由来のペプチドであってもよい。本発明のペプチドまたは抗原は融合タンパク質に関連してもよい。いくつかの態様において、本発明のペプチドまたは抗原は、種々のポリペプチド由来であってもよい多重抗原エピトープを単一ポリペプチドまたは組成物に含んでもよい。ある態様において、抗体は、プロテアソーム経路の成分由来の少なくとも1つのペプチドまたは抗原をコードする少なくとも1つのポリヌクレオチドで免疫化することによって誘導される。少なくとも1つのペプチドまたは抗原をコードするポリヌクレオチドは、プラスミド、鎖状発現要素またはウイルス発現ベクターなどの発現ベクターに含まれてもよい。特定の態様において、ポリヌクレオチドがコードするペプチドまたは抗原は、α-型またはβ-型プロテアソームサブユニットポリペプチドなどのプロテアソームタンパク質由来であってもよい。
【0008】
本発明の一態様において、プロテアソームの活性は、精子もしくは卵母細胞または両方にプロテアソーム阻害剤を接触させることによって阻害される。プロテアソーム阻害剤は薬学的に許容されうる製剤に含まれてもよい。ある態様において、プロテアソーム阻害剤またはプロテアソーム阻害剤組成物はMG132および/またはラクタシスチンを含む。薬学的に許容されうる製剤は、溶液、懸濁液、錠剤、ピル、カプセル、徐放性製剤、粉末、クリーム、軟膏(ointment)、軟膏(salve)、スプレー、ポンプ、リポソーム、坐剤、吸入剤、パッチまたは他の送達系に製剤化されてもよい。プロテアソーム経路の活性を阻害する組成物は、静脈内、腹腔内、皮内、筋肉内、子宮内、皮膚、鼻、口腔、膣、吸入または局所的投与経路を含むが、これらに限定されない経路によって投与することができる。ある態様において、投与経路は子宮内または膣である。
【0009】
本発明の種々の態様において、ほ乳類において抗プロテアソーム抗体を誘導する方法は、プロテアソーム経路の成分由来の1つ以上のペプチドまたは抗原をほ乳類に投与する段階を含む。本発明の1つ以上のペプチドまたは抗原を投与する段階は、ほ乳類のユビキチン-プロテアソーム経路に結合して、その活性を阻害する抗体の産生を誘導することができる。ある態様において、ほ乳類はヒト、家畜(farm animal)、家畜(domestic animal)、野生動物または害獣、例えば、ラット、スカンク等である。動物には、ブタ、ウシ、ウマ、ネコ、げっ歯類、ウサギ、シカ、アライグマ、オポッサム、スカンク等が挙げられるが、これらに限定されない。1つ以上のペプチドまたは抗原は、1つ以上のペプチドまたは抗原をコードするポリヌクレオチドを発現することによって投与することができる。ポリヌクレオチドは、筋肉内、静脈内、皮下、腹腔内、皮内、経口、吸入または他の既知の投与経路によって投与することができる。1つ以上のペプチドまたは抗原は、筋肉内、静脈内、皮下、腹腔内、皮内、経口、吸入または他の既知の投与経路によって投与することができる。
【0010】
本発明のいくつかの態様において、方法は、精子、卵母細胞または動物もしくはヒトの受精に寄与する他の生殖組織(例えば、精巣、精巣上体、卵巣、前立腺、精嚢および他の性付属腺)に関連するユビキチン-プロテアソーム経路の成分の有無を検出することによって不妊の診断および/または受精能の評価/推定を含む。方法は、ELISA、タンパク質ブロット法、免疫組織化学法等を含んでもよいが、これらに限定されない。
【0011】
本発明の方法は、インビトロにおける受精に使用する精液試料にプロテアソーム活性の阻害剤を添加する段階を含むインビトロにおける受精方法であって、阻害剤は、受精を妨害するには不十分であるが、多精受精を妨害するには十分な濃度が添加される方法をさらに提供する。本発明の方法では、受精の約25%未満が多精受精であってもよく、受精の10%未満を含む。阻害剤は、本明細書に記載する任意の方法によってインビトロまたはインビボにおいて添加することができる。任意の阻害剤を、本明細書に記載する方法に使用することができる。
【0012】
本明細書に記載する任意の方法または組成物は、本明細書に記載する任意の他の方法または組成物に関して実施することができることが考慮される。
【0013】
特許請求の範囲および/または明細書において「含む」という用語と共に使用する場合の「1つのもの」という用語の使用は「1つ」を意味することができるが、「1つ以上」、「少なくとも1つ」および「1つまたは1つ以上」の意味にも一致する。
【0014】
本発明の他の目的、特徴および利点は、以下の詳細な説明から明らかになる。しかし、本発明の精神および範囲内の種々の変更および改良は、この詳細な説明から当業者に明らかになるので、詳細な説明および具体的な実施例は、本発明の具体的な態様を示すが、例示のためだけに提供されていることが理解されるべきである。
【0015】
例示的な態様の説明
上記に記載するように、受精の調節、受精能の評価または不妊の診断の改善された方法が必要とされている。受精能を調節する際の大半の努力は、配偶子形成または局在化/移動性を阻害する化学的または機械的方法の使用に絞られている。受精能調節のための改善された組成物および方法が必要とされている。本明細書に記載する発明は、受精能を調節および診断する組成物および関連の方法を記載する。本発明は、受精能を調節するためのユビキチン-プロテアソーム経路の成分の阻害剤または結合剤を含む。本発明は、卵母細胞の透明帯の透過性はプロテアソームおよびプロテアソームに関連する経路に関与し、抗プロテアソーム抗体および阻害剤によって阻害することができるという驚くべき観察を利用している。本明細書に記載するように、本発明は、受精能および、いくつかの例においては多精受精をインビボおよびインビトロにおいて低下または阻害する改善された方法を提供する。
【0016】
本発明者らは、プロテアソーム阻害剤、特に化合物ラクタシスチンおよびMG132並びに、プロテアソームに結合してプロテアソーム活性を阻害するまたはユビキチン依存性タンパク質分解を遮断する抗体の受精能調節特性を同定した。ほ乳類のユビキチン-プロテアソーム経路の阻害は、精子による透明帯(ZP)の透過性を遮断する。本発明者らは、接合体の細胞質内の父系ミトコンドリアの分解を可逆的に遮断する試みの成功においてこれらの新規特性を観察した。父系ミトコンドリア分解を生じるためには、精子がZPを通過して、卵母細胞の細胞質に侵入直後に受精培地にプロテアソーム阻害剤を添加しなければならなかった。ZP透過性はセリンプロテアーゼ活性に関連すると考えられており、プロテアソーム阻害剤はセリンプロテアーゼの活性を遮断しないという事実からこの観察は予想されなかった(Fenteanyら,1995;Goldbergら, 1995)。さらに、本発明者らは、ユビキチンおよびプロテアソームサブユニット以外に、ほ乳類の精子アクロソームは、ユビキチン-基質結合に必要な結合酵素であるユビキチンキャリアE2を含有し、おそらく他の脱ユビキチン化、ユビキチン結合およびユビキチン活性化酵素も含有すると思われることを示した。
【0017】
本明細書に開示するように、本発明は、ほ乳類の受精およびZP透過性は精子アクロソームのプロテアソームのタンパク質分解作用に依存していることを実証している。精子が卵子表面を活発にユビキチン化すると思われるホヤとは異なり、ユビキチン化タンパク質は受精前または卵形成中にほ乳類の卵子ZPによって獲得されうる。このようなデータは、ほ乳類の受精の謎並びに新規避妊標的および不妊マーカーにおける重要な点を提供する。
【0018】
I. ユビキチン-プロテアソーム経路
本発明の組成物は、避妊薬としてまたは避妊方法において並びに受精能を診断する際に使用することができる。ユビキチン-プロテアソーム経路に関連するポリペプチドの阻害または検出は避妊またはプロテアソーム成分が無傷/欠損しているもしくは存在する/存在しない配偶子の検出にそれぞれ使用することができる。
【0019】
プロテアソームは、ATP依存的タンパク質分解経路の必須成分である非リソソームタンパク質分解の中心酵素複合体として広く認識されている。この経路は多くの律速酵素、転写調節因子および重要な調節タンパク質の迅速な分解を触媒する。この経路は、突然変異または翻訳後損傷によって生じる異常性の高いタンパク質の迅速な排除に必須であり、ほ乳類細胞のタンパク質の大部分の迅速でない分解の主要な役割を果たす。それはまた、高等な真核生物における抗原プロセシングに大きく関与している。
【0020】
26Sプロテアソームは、ユビキチン/ATP依存的タンパク質分解経路の主要酵素複合体である。触媒中心は、電子顕微鏡によって示されるように、バレル型構造の2000 kDa複合体である20Sプロテアソームによって形成される。触媒中心は、各々7つのサブユニット(サブユニットα1〜7、β1〜7)を含む4つの環から構成される。
【0021】
配列の類似性に基づいて考慮すると、全ての20Sプロテアソームサブユニット配列は、各グループが別個の構造的および機能的役割を有する2つのグループ、αおよびβに分類することができる。7つのα-サブユニットは20Sプロテアソームの外側リングを含み、7つのβ-サブユニットは20Sプロテアソームの内側リングを含む。各サブユニットはα-またはβ-リングの独自の位置に位置する。
【0022】
典型的な26S複合体は、20S複合体と19S調節複合体と呼ばれる別個の複合体に位置する25kDa〜10 kDaの分子量範囲の20を超える追加のタンパク質を含む。26S複合体は、基質特異性を決定し、タンパク質分解および存続度に必要な複数の酵素機能を提供する。サブユニット成分の系統的な分析は、ATP結合タンパク質の新規ファミリーに属するATPaseである少なくとも6つのメンバーと、ATPに結合する能力のないさらに別の15のサブユニット、20Sプロテアソームのタンパク質分解中心への挿入前にタンパク質基質の変性を担当すると考えられるイソペプチダーゼおよびいくつかの他のタンパク質を明らかにした。いくつかのプロテアソームにおいて、19S調節複合体は11Sアクチベーター複合体と置換されている。
【0023】
26S複合体はATPに結合し、ユビキチンと結合することによって分解の標的となったタンパク質の分解を担当する。ユビキチンは、上記のように、一連のユビキチン活性化(UBA)およびユビキチン結合(UBC)酵素、例えば、酵素E1、E2およびE3によって、標的のリジンのε-アミノ基とユビキチンのC-末端グリシン残基の間に形成されるイソペプチド結合によって標的タンパク質に結合する。ユビキチン活性化酵素および結合酵素は、ユビキチン分子を1つの酵素から次の酵素に移動させ、次に活性化されたユビキチン分子をE2酵素から標的タンパク質に移動させることによって連続的に作用する。次いで、モノ-ユビキチン化タンパク質が作用されて、同じ酵素が先の分子に追加のユビキチン分子を結合する。ユビキチン結合が継続し、高分子量タンパク質複合体が生じる。次いで、このポリ-ユビキチン化産物が26Sプロテアソームによる迅速な分解の標的となり、同時にイソペプチダーゼによって触媒されるユビキチンが再利用される。
【0024】
複数のペプチダーゼ活性を有する複合体として最初に記載されたにもかかわらず、その後の研究により、20Sプロテアソームの5つの活性:大きい疎水性残基の後方を切断するキモトリプシン様活性;塩基性残基の後方を切断するトリプシン様活性;酸性残基の後方を切断するポストグルタミルヒドロラーゼ;分岐鎖アミノ酸の後方を主に切断するもの;および小型の中性アミノ酸の後方を切断するものが規定された。
【0025】
ATPが存在しない場合に20Sプロテアソームと関連することができる第2のアクチベーターは11Sレギュレーターとして既知である。純粋なPA28アクチベーターは、約50%の相同性を有する2つの交互のサブユニット、PA28αおよびPA28βの複合体である。電子顕微鏡による研究により、PA28は、19S複合体のように、両方またはどちらかの末端においてα-リングに結合することによって20Sプロテアソームにキャップをするリング状粒子であることが示された。PA28は、MHCクラスI分子の免疫系に提示される抗原ペプチドのプロテアソーム-触媒産生を調節するという所見は、抗原プロセシングにおけるこのアクチベーターの細胞機能を示している。詳細な総説は、SchmidおよびBriand, 1997;Bogyoら, 1997を参照されたい。
【0026】
ある態様において、ユビキチン-プロテアソーム経路に関連する種々のポリペプチドおよびそれらの断片を使用することができる。例えば、ユビキチン-プロテアソーム経路関連ポリペプチドには、ヒト遺伝子変異と遺伝子疾患に関わる情報の統合データベース(Online Mendelian Inheritance in Man(OMIM)データベース)において同定されている化合物が挙げられるが、これらに限定されず、その例には、OMIM第600307プロテアソームサブユニット、β-型、6;Psmb6(Genbank アクセッション番号第 NM_002798);176844 プロテアソームサブユニット, α-型, 5; Psma5 (Genbank アクセッション番号第 NM_002790) ; 176844 プロテアソームサブユニット、α-型、5;Psma5(Genbankアクセッション番号第 NM_002790);177045 プロテアソームサブユニット, β-型,9 ; Psmb9 (Genbank アクセッション番号第 NM_002800 および NM_148954) ; 604030 プロテアソームサブユニット、 β-型、7 ; Psmb7 (Genbank アクセッション番号第 NM_002799) ; 600654 プロテアソーム アクチベーター サブユニット 1 ; Psmel (Genbank アクセッション番号第 NM_006263) ; 606223 プロテアソーム 26S サブユニット、 非ATPase、 2; Psmd2 (Genbank アクセッション番号第 NM_002808) ; 176847 プロテアソームサブユニット、β-型、 10; PsmblO (Genbank アクセッション番号第 NM_002801) ; 177046 プロテアソームサブユニット、 β-型、 8; Psmb8 (Genbank アクセッション番号第 NM_004159 および NM_148919) ; 602855 プロテアソームサブユニット、α-型、 6; Psma6 (Genbank アクセッション番号第 NM_002791) ; 602854 プロテアソームサブユニット, α-型、 1; Psmal (Genbank アクセッション番号第 NM_002786 およびNM_ 148976) ; 602161 プロテアソーム アクチベーター サブユニット 2; Psme2 (Genbank アクセッション番号第 NM_002818) ; 600306 プロテアソームサブユニット、 β-型、 5; Psmb5 (Genbank アクセッション番号第 NM_002797) ; 605129 プロテアソーム アクチベーター サブユニット 3; Psme3 (Genbank アクセッション番号第 NM_005789) ; 602175 プロテアソームサブユニット、β-型、 2; Psmb2 (Genbank アクセッション番号第 NM_002794) ; 603146 プロテアソーム 26s サブユニット、非ATPase、 9; Psmd9 (Genbank アクセッション番号第 NM_002813) ; 154365 プロテアソーム 26s サブユニット、 ATPase, 2; Psmc2 (Genbank アクセッション番号第 NM_002803) ; 604449 プロテアソーム 26s サブユニット、非ATPase、11; Psmdll (Genbank アクセッション番号第 NM_002815) ; 606607 プロテアソームサブユニット、 α-型、 7; Psma7 (Genbank アクセッション番号第 NM_002792 および NM_152255) ; 176843 プロテアソームサブユニット、α-型、 3; Psma3 (Genbank アクセッション番号第 NM_002788 およびNM_ 152132) ; 607173 Padl ; (Genbank アクセッション番号第 NM_005805) ; 604450 プロテアソーム 26s サブユニット、非ATPase、 12; Psmdl2 (Genbank アクセッション番号第 NM_002816) ; 602706 プロテアソーム 26s サブユニット、 ATPase, 1; Psmcl (Genbank アクセッション番号第 NM_002802) ; 602708 プロテアソーム 26s サブユニット、 ATPase、 6; Psmc6 (Genbank アクセッション番号第 NM_002806) ; 186852 プロテアソーム 26s サブユニット、 ATPase、 3; Psmc3 (Genbank アクセッション番号第 NM_002804) ; 176842 プロテアソームサブユニット、α-型、 2; Psma2 (Genbank アクセッション番号第 NM_002787) ; 603481 プロテアソーム 26s サブユニット、非ATPase、 13; Psmdl3 (Genbank アクセッション番号第 NM_002817) ; 603480 プロテアソーム 26s サブユニット、非ATPase、 10;PsmdlO (Genbank アクセッション番号第 NM_002814) ; 602017 プロテアソームサブユニット、β-型、 1 ; Psmbl (Genbank アクセッション番号第 NM_002793) ; 604452 プロテアソーム 26s サブユニット、非ATPase、 5; Psmd5 (Genbank アクセッション番号第 NM_005047) ; 601648 プロテアソーム 26s サブユニット、 非ATPase、 4; Psmd4 (Genbank アクセッション番号第 NM_002810 および NM_153822) ; 602177 プロテアソームサブユニット、 β-型、 4; Psmb4 (Genbank アクセッション番号第 NM_002796) ; 602176 プロテアソームサブユニット、β-型、 3; Psmb3 (Genbank アクセッション番号第 NM_002795) ; 602707 プロテアソーム 26s サブユニット、 ATPase、 4; Psmc4 (Genbank アクセッション番号第 NM_153001 およびNM_006503) ; 601681 プロテアソーム 26s サブユニット、 ATPase、 5; Psmc5 (Genbank アクセッション番号第 NM_002805) ; 157970 プロテアソーム 26s サブユニット、非ATPase、 7; Psmd7 (Genbank アクセッション番号第 NM_002811) ; 602163 ユビキチン結合酵素 E2e 2; Ube2e2 (Genbank アクセッション番号第 Z44894) ; 605046 ユビキリン(Ubiquilin) 1 ; Ubqln1 (Genbank アクセッション番号第 NM_013438 および NM_053067) ; 602544 Parkin ; Park2 (Genbank アクセッション番号第 NM_004562、 NM_013987 および NM_013988) ; 314370ユビキチン活性化酵素 1; Ubel (Genbank アクセッション番号第 NM_003334 および NM_153280) ; 300264 ユビキリン(Ubiquilin) 2; Ubqln2 (Genbank アクセッション番号第 NM_013444) ; 180470 リボフォリン(Ribophorin) I;Rpnl (Genbank アクセッション番号第 NM_002950) ; 191342 ユビキチンカルボキシル-ターミナルエステラーゼ LI (Genbank アクセッション番号第 NM_004181) ; 118888 キモトリプシン様プロテアーゼ; Ctrl (Genbank アクセッション番号第 NM_001907) ; 605532 Smad ユビキチン化調節因子 2 (Genbank アクセッション番号第 NM_022739) ; 607119 ダブルリングフィンガータンパク質(Double Ring Finger Protein) (Genbank アクセッション番号第 NM_015435) ; 605624 アリアドネ(Ariadne)、ショウジョウバエ(Drosophila)、ホモログ Of、 1; Arih1 (Genbank アクセッション番号第 NM_005744) ; 605568 Smad ユビキチン化調節因子 1 (Genbank アクセッション番号第 AF199364); 603124 ユビキチン結合酵素 E2g 2; Ube2g2 (Genbank アクセッション番号第 NM_003343) ; 603091 ユビキチン特異的プロテアーゼ 12; Uspl2 (Genbank アクセッション番号第 AF022789); 602995 ユビキチン結合酵素 E2 変種1;Ube2vl (Genbank アクセッション番号第 NM_003349、 NM021988 および NM-022442) ; 602961 ユビキチン結合酵素 E2d 1; Ube2dl (Genbank アクセッション番号第 NM_003338) ; 602916 ユビキチン結合酵素 E2e 1; Ube2el (Genbank アクセッション番号第 NM_003341) ; および 300420 Praja 1; Pjal (Genbank アクセッション番号第 NM_022368)が挙げられ、その各々は参照として本明細書に組み入れられている。Genbankなどの同様のデータベースを使用してユビキチン-プロテアソーム関連ポリペプチドを同定することもできる。ユビキチン-プロテアソーム関連ポリペプチドは、複合体もしくは経路の一部である、または経路の活性もしくは組成物に影響を与える任意のポリペプチドをいう。
【0027】
II. ポリペプチドおよびタンパク様組成物
本発明の種々の局面において、ペプチド、ポリペプチドまたはタンパク質を含むタンパク様組成物を、抗原、抗体、ワクチン、遮断剤、治療剤としてまたは本明細書に記載する組成物および方法の成分として使用することができる。ペプチド、ポリペプチドおよび/またはタンパク質は単離/精製された組換えまたは合成ペプチド、ポリペプチドおよび/またはタンパク質であってもよい。
【0028】
本発明のポリペプチドおよびペプチドは、免疫原性ペプチド、抗原もしくはエピトープ、またはほ乳類のユビキチン-プロテアソームおよびそれに関連する経路の1つ以上のタンパク質成分に対する抗体等を含んでもよい。このようなペプチドは、例えば、ウサギ、雌ウシ、ブタ、ウマ、ヒヒおよびヒトを含む種々のほ乳類種のプロテアソームタンパク質に対する免疫応答を誘発するのに有用となりうる。プロテアソームタンパク質に対する免疫応答を誘発することによって、本発明のペプチドは避妊剤として有用である。他の態様において、抗体は単離されて、避妊または診断組成物に使用してもよい。
【0029】
ある態様において、プロテアソームポリペプチドまたは抗原は合成ペプチドであってもよい。さらに他の態様において、ペプチドは、分子工学的手法によって作製された組換えペプチドであってもよい。本セクションは、本発明に使用するためのタンパク性組成物を作製する際に関係する方法および組成物を記載する。
【0030】
A. ポリペプチド
ユビキチン-プロテアソーム経路の1つ以上のタンパク質成分由来のポリペプチドは、すなわち当業者に既知のタンパク質抽出技法を使用して抽出した天然型ポリペプチドであってもよい。特定の態様において、プロテアソーム抗原を同定して、ほ乳類をワクチン化するための薬学的に許容されうる担体中で作製してもよい。
【0031】
ポリペプチドの配列変種を作製してもよい。ポリペプチド配列変種は、集団内の天然の変更によって生ずるまたは他の動物に見られる相同物であってもよいポリペプチドのわずかな配列変種であってもよい。それらは、天然には生じないが、天然型のポリペプチドと同様に機能するおよび/または天然型のポリペプチドと交差反応する免疫応答を誘発する程度に十分に同様である配列であってもよい。配列変種は、Sambrookら2001に記載されているものなどの標準的な部位特異的突然変異方法によって作製することができる。
【0032】
抗原性プロテアソームポリペプチドの別の合成または組換え変更は、プロテアソームタンパク質に天然に見られるエピトープ決定基の反復を含むポリペプチド部分である。このような合成ポリペプチドタンパク質は、任意の1つのプロテアソームタンパク質エピトープのいくつかのホモマー反復から作製されても;または同じもしくは異なるポリペプチド由来の2つ以上のエピトープを含んでもよい。
【0033】
ポリペプチドのアミノ酸配列変種は置換、挿入または欠失変種であってもよい。欠失変種は、機能または免疫原性活性に必須でない未変性のタンパク質の1つ以上の残基を欠損している。別の一般的な種類の欠失変異は、分泌シグナル配列または細胞の特定の部分にタンパク質を結合させるシグナル配列を欠損するものである。
【0034】
置換変種は、典型的には、タンパク質内の1つ以上の部位における1つのアミノ酸と別のアミノ酸の交換を含み、タンパク質分解に対する安定性などのポリペプチドの1つ以上の特性を調節するように設計することができる。置換は、好ましくは、同類置換であり、すなわち、1つのアミノ酸が同様の形状および電荷を有するアミノ酸と交換される。同類置換は当技術分野において既知であり、例えば、以下の変更を含む:アラニンとセリン;アルギニンとリジン;アスパラギンとグルタミンまたはヒスチジン;アスパラギン酸とグルタミン酸;システインとセリン;グルタミンとアスパラギン;グルタミンとアスパラギン;グリシンとプロリン;ヒスチジンとアスパラギンまたはグルタミン;イソロイシンとロイシンまたはバリン;ロイシンとバリンまたはイソロイシン;リジンとアルギニン;メチオニンとロイシンまたはイソロイシン;フェニルアラニンとチロシン、ロイシンまたはメチオニン;セリンとスレオニン;スレオニンとセリン;トリプトファンとチロシン;チロシンとトリプトファンまたはフェニルアラニン;およびバリンとイソロイシンまたはロイシン。
【0035】
挿入変種は、ポリペプチドの迅速な精製を可能にするために使用されるものなどの融合タンパク質を含み、ポリペプチドの相同物である他のタンパク質またはポリペプチド由来の配列を含むハイブリッドタンパク質を含んでもよい。例えば、挿入変種は、1つの種由来のポリペプチドのアミノ酸配列の一部と、別の種由来の相同ポリペプチドの一部を含んでもよい。「キメラタンパク質」という用語は、相同または非相同タンパク質である2つの異なるタンパク質由来のセグメントを含むタンパク質をいうために使用される。セグメントは天然に隣接するセグメントでなくても、または任意のセグメントが任意の特定の位置もしくは順序を維持しなくてもよい。他の挿入変種は、追加のアミノ酸がポリペプチドのコード配列に導入されているものを含んでもよい。これらは、典型的には、上記の融合タンパク質より短い挿入であり、例えば、プロテアーゼ切断部位に導入される。
【0036】
一つの態様において、ポリペプチドの主要抗原決定基は、ポリペプチドをコードする遺伝子の一部が組換え宿主において発現され、得られたタンパク質は、免疫応答を誘発する能力について試験される実験方法によって同定することができる。例えば、ポリメラーゼ連鎖反応(PCR)を使用して、タンパク質のC-末端の連続する長い断片を欠損するペプチドをコードする範囲のcDNAを作製することができる。次いで、これらのペプチドの各々の免疫原性活性は、この活性に必須のポリペプチドの断片またはドメインを同定する。次いで、ごく少数のアミノ酸が各反復において除去または付加されるさらに別の実験によって、ポリペプチドの他の抗原性決定基を位置づけることができる。従って、熱安定性DNAポリメラーゼ、デオキシリボヌクレオチドおよびプライマー配列を使用する多数サイクルの変性-再生によってDNAの特定のセグメントを増幅する技法であるポリメラーゼ連鎖反応が本発明において考慮される(Mullis, 1990;Mullisら, 1992)。
【0037】
本発明によるポリペプチドを作製するための別の態様は、ペプチド模倣物の使用である。模倣物は、タンパク質の二次構造の要素を模倣する分子である。多数のタンパク質は、折りたたみ面の比較的小さい領域によって生物作用を発揮するので、それらの作用は、生理活性面を保持し、薬物動態学的/動力学的特性が改善されていると思われるかなり小型の設計(模倣)分子によって再現することができる(Fairlieら, 1998)。二次構造(ヘリックス、ターン、ストランド、シート)の個々の要素を模倣し、それらの組み合わせを三次構造(ヘリックスバンドル、マルチプルループ、ヘリックス-ループ-ヘリックスモチーフ)に集成する方法は総説が報告されている(Fairlieら, 1998;Moore, 1994)。模倣ペプチドを予測し、作製し、改変し、スクリーニングする方法は米国特許第5,933,819号および米国特許第5,869,451号(各々が具体的に参照として本明細書に組み入れられている)に記載されている。
【0038】
以下は、同等物または場合によっては改善された第二世代の分子を作製するためにタンパク質またはポリペプチドのアミノ酸を変更することに基づいた考察である。アミノ酸の変更は、以下の実施例により、DNA配列のコドンを変更することによってまたは化学的なペプチド合成によって実施することができる。
【0039】
例えば、抗体の抗原結合領域または基質分子の結合部位などの構造の相互作用的な結合能力をあまり損失しないで、例えば、ポリペプチド構造のあるアミノ酸を他のアミノ酸と置換することができる。それは、生物活性を規定するポリペプチドの相互作用的な能力および性質であるので、あるアミノ酸置換はポリペプチド配列およびポリペプチドをコードする根本的なDNA内で行うことができ、それでもなお、同様の特性または改善された特性を有するポリペプチドを得ることができる。表1は、特定のアミノ酸をコードするコドンを示す。
【0040】
このような変更を行う際には、アミノ酸の疎水性親水性指数を考慮することができる。タンパク質に相互作用的な生物機能を与える際のアミノ酸の疎水性親水性指数の重要性は、一般に、当技術分野において理解されている(KyteおよびDoolittle, 1982)。
【0041】
あるアミノ酸を同様の疎水性親水性指数またはスコアを有する他のアミノ酸によって置換しても、同様の生物作用を有するタンパク質またはポリペプチドを生じることができることは当技術分野において既知である。同様のアミノ酸の置換は親水性に基づいて効果的に実施されうることも当技術分野において理解されている。参照として本明細書に組み入れられている米国特許第4,554,101号は、隣接アミノ酸の親水性によって支配されるタンパク質の最も大きい局所的な平均親水性はタンパク質の生物学的特性に関連することを記載している。
【0042】
アミノ酸置換は、一般に、アミノ酸側鎖の置換基の相対的な類似性、例えば、疎水性、親水性、電荷、サイズ等に基づいている。上記の種々の特徴を考慮している例示的な置換は当業者に既知であり、以下を含む:アルギニンとリジン;グルタミン酸とアスパラギン酸;セリンとスレオニン;グルタミンとアスパラギン;およびバリン、ロイシンとイソロイシン等。
【0043】
本発明の有用なペプチドは、配列番号:1、2、4および6またはその断片の1つ以上を含むアミノ酸配列を含む。好ましくは、本発明のペプチドは30以下のアミノ酸の配列を含む。
【0044】
本発明の免疫原性ペプチドは、配列番号:1、2、4および6またはその断片の1つ以上並びに任意に非プロテアソームアミノ酸配列を有する複合ペプチドであってもよい。
【0045】
本発明の有用な組成物は、本発明の1つ以上のペプチドおよび/または融合タンパク質を、薬学的に許容されうる担体および/または他の薬学的添加剤成分を合わせて含む。組成物は、本発明のペプチドまたは融合タンパク質のエピトープに対する免疫応答を増強することができるアジュバントを含んでもよい。
【0046】
本発明はまたポリペプチドの断片にも関係する場合もある。分子のN-末端を含む断片は、コード領域内の翻訳停止部位の遺伝子操作によって作製することができる。または、ポリペプチドをプロテアーゼとして既知のタンパク質分解酵素で処理すると、種々のN-末端、C-末端および内部断片を作製することができる。ある態様において、ペプチドは既知の方法によって合成することができる。断片の例は、長さ

またはそれ以上のアミノ酸の連続残基を含んでもよい。これらの断片は、沈殿(例えば、硫酸アンモニウム)、HPLC、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー(イムノアフィニティークロマトグラフィーを含む)または種々のサイズ分離(沈降、電気泳動、ゲルろ過)などの既知の方法によって精製することができる。
【0047】
B. 合成ペプチド
本発明の種々の態様は、受精能を調節および/または診断するための方法および組成物に使用するためのペプチドまたはポリペプチドを記載している。いくつかの態様において、動物の受精能を阻止または低下および検出する際に使用するための抗プロテアソームまたは抗ユビキチン抗体の作製が考慮されている。本発明のペプチドは、従来技法により溶液または固相支持体でも合成することができる。種々の自動合成装置が市販されており、既知のプロトコールにより使用することができる。例えば、おのおの参照として本明細書に組み入れられている、StewartおよびYoung(1984);Tamら(1983);Meerifield(1986)並びにBaranyおよびMerrifield(1979)を参照されたい。本明細書に記載するペプチドに相当する通常約6から35〜50アミノ酸のオーバーラップペプチドの短いペプチド配列またはライブラリーを容易に合成することができる。いくつかの態様において、本発明のペプチドまたはポリペプチドをコードするヌクレオチド配列を発現ベクターに挿入し、適当な宿主細胞に形質転換またはトランスフェクションし、発現に好適な条件下において培養する組換えDNA技術を使用することができる。
【0048】
C. 融合ペプチドまたはポリペプチド
特殊な種類の挿入変種は融合タンパク質である。この分子は、一般に、第1の分子の全てまたは実質的な部分が第2のポリペプチドの全てまたは一部のN-末端またはC-末端に結合している。例えば、融合は、典型的には、異種宿主におけるタンパク質の組換え発現を可能にするために他種のリーダー配列を使用する。別の有用な融合は、融合タンパク質の精製を容易にするために、抗体エピトープなどの免疫学的に活性なドメインの追加を含む。融合接合部または接合部付近に切断部位を導入すると、精製後の外来ポリペプチドの除去が容易になる。他の有用な融合には、酵素の活性部位、グリコシル化ドメイン、細胞標的シグナルまたは膜貫通領域などの機能的ドメインの結合が挙げられる。本発明の他の融合には、2つ以上のプロテアソーム抗原の融合が挙げられる。ある態様において、2つ以上のプロテアソーム抗原が可逆的または非可逆的に互いに結合される。
【0049】
D. ペプチドまたはポリペプチドの精製
ある態様において、ペプチドまたはポリペプチドを精製することが望ましい場合がある。タンパク質精製技法は当業者に既知である。これらの技法は、一つのレベルにおいて、ポリペプチドおよび非ポリペプチド分画への細胞環境の粗分画化に関係する。ポリペプチドを他の分子から分離したら、クロマトグラフィー技法および電気泳動技法を使用して関心対象のポリペプチドをさらに精製して、部分または完全精製(または、均質までの精製)することができる。純粋なペプチドの作製に特に好適な分析方法はイオン交換クロマトグラフィー、排除クロマトグラフィー;ポリアクリルアミドゲル電気泳動;等電点電気泳動法である。特に効率的なペプチド精製方法は高速タンパク質液体クロマトグラフィーまたは場合によってはHPLCである。
【0050】
タンパク質精製に使用するのに好適な種々の技法は当業者に既知である。これらには、例えば、硫酸アンモニウムによる沈殿、PEG、抗体等または熱変性後の遠心分離;イオン交換、ゲルろ過、逆相、ヒドロキシアパタイトおよびアフィニティークロマトグラフィーなどのクロマトグラフィー段階;等電点電気泳動法;ゲル電気泳動並びにこのような技法および他の技法の組み合わせが挙げられる。当技術分野において一般に既知であるように、種々の精製段階を実施する順序を変更することができること、またはある段階を省略しても実質的に精製されたタンパク質もしくはペプチドの作製に好適な方法になることが考えられる。
【0051】
本発明のある局面はペプチドの精製に関し、特定の態様においてペプチドの実質的な精製に関する。本明細書において使用する「精製されたペプチド、ポリペプチドまたはタンパク質」という用語は、他の成分から単離可能な組成をいうことが意図されており、ペプチドは天然に入手可能な状態に対して任意の程度まで精製される。従って、精製されたタンパク質またはペプチドは、それが天然に生じうる環境を含まないタンパク質またはペプチドのこともいう。
【0052】
一般に、「精製された」は、種々の他の成分を除去するために分画化が実施されており、実質的に活性を保持するタンパク質またはペプチド組成物をいう。「実質的に精製された」という用語を使用する場合には、この意味は、タンパク質またはペプチドが組成物中で約50%、約60%、約70%、約80%、約90%、約95%またはそれ以上のペプチド、ポリペプチドまたはタンパク質を構成するなどの、組成物の主要な成分を形成する組成物をいう。タンパク質またはペプチドの精製程度を定量化する種々の方法は、本発明の開示内容を考慮して当業者に既知である。
【0053】
E. 抗原
本発明の抗原は、典型的には、ほ乳類のプロテアソームの1つ以上のタンパク質成分から単離または誘導され、総説は、例えば、SchmidおよびBriand, 1997またはBogyoら, 1997を参照されたい。本発明の種々の態様において、動物の受精能を調節するためまたは受精能の調節に使用するための抗体を作製するために抗原を動物に投与する。特定の態様において、本発明による脊椎動物の免疫化は、1つ以上のプロテアソームペプチドをコードするポリヌクレオチドの投与を含んでもよい。
【0054】
いくつかの態様において、本発明は:(a)プロテアソーム抗原を同定する段階;(b)本明細書に記載する抗原で脊椎動物に免疫応答を形成する段階;(c)動物細胞または免疫した動物から回収した細胞と不死化癌細胞の融合によって作製したハイブリドーマ細胞系統において作製された抗体を入手する段階とを含むプロテアソーム抗原に対する抗体を作製する方法に関する。
【0055】
本発明はまた、免疫原性であるが、ワクチンとして必ずしも防御的ではないプロテアソームポリペプチドに対する抗体を作製する方法に関する。例えば、プロテアソーム特異的抗体は、受精能の評価、不妊の診断および検討または抗体治療に有用である可能性がある。抗原または抗原をコードするポリヌクレオチドで動物を免疫化することを使用して、抗プロテアソーム抗体を作製することができる。抗プロテアソーム抗体を作製する他の方法において、同定された抗原は、ファージライブラリーをパニングするために使用されてもよい。この手法はインビトロにおいて抗体を単離するために使用してもよい。
【0056】
F. 抗体
別の局面において、本発明は、プロテアソーム経路またはその任意の部分の成分に免疫反応性である抗体組成物を含む。さらに他の態様において、本発明の抗原は、抗体および/または抗体組成物を作製するために使用することができる。抗体は、特異的または優先的に、プロテアソームもしくはユビキチンポリペプチドまたはユビキチン-プロテアソーム経路の酵素のポリペプチドに反応性となりうる。例示的な抗体は、配列番号:1、2、4および6に記載する配列、それらの断片、変種もしくは模倣物または密接に関連する配列を有する抗原に反応性の抗体を含むプロテアソーム経路の成分もしくは関連ポリペプチドに反応性のものを含む。抗体は、ポリクローナルまたはモノクローナルであってもよく、当技術分野において既知の方法によって作製することができる。抗体はまた1価または2価であってもよい。抗体は、種々の生物学的または化学的手段によって分割されてもよい。抗体の半分は各々1つの抗原に結合することができ、従って、1価と規定される。抗体を作製し、特徴づける手段は当技術分野において既知である(例えば、参照として本明細書に組み入れられている、HarlowおよびLane, 1988を参照されたい)。
【0057】
抗体を作製するために、本発明のポリペプチドの1つ以上の抗原決定基に対応するペプチドを作製することができる。このようなペプチドは、一般に、長さ少なくとも5または6アミノ酸残基である必要があり、好ましくは、長さ約10、15、20、25または約30アミノ酸残基であり、最高約35残基〜50残基ほどを含んでもよい。合成ペプチドは、一般に、Applied Biosystems(Foster City, CA)社製のものなどの、自動ペプチド合成機械のほぼ上限の長さである、長さ約35残基である。例えば、組換え手段によってさらに長いペプチドも作製することができる。他の方法において、完全長または実質的に完全長のポリペプチドを本発明の抗体を作製するために使用することができる。
【0058】
少なくとも1つ以上の抗原決定基を含むペプチドが作製されたら、ポリペプチドに対する抗血清の作製にペプチドを使用する。例えば、PCRクローニング方法を使用する標準的な方法によって、これらの決定基をコードするミニ遺伝子または遺伝子融合を構築し、発現ベクターに挿入することもできる。抗体形成またはワクチン化のためのペプチド使用は、典型的には、B型肝炎表面抗原、キーホールリンペットヘモシアニンまたはウシ血清アルブミンなどの免疫原性キャリアタンパク質にペプチドを結合することが必要である。この結合を実施する方法は、種々のアジュバントと同様に、当技術分野において既知である。
【0059】
本発明の方法に使用する抗体は、すなわち、抗体に任意の種類の分子を共有結合することによって修飾されている誘導体を含む。例えば、抗体誘導体には、例えば、グリコシル化、アセチル化、PEG付化、リン酸化、アミド化、既知の保護/ブロック基による誘導体化、タンパク質分解的切断、細胞リガンド(他のタンパク質)への結合等によって修飾されている抗体を含むが、これらに限定されない。数多くの化学的修飾のいずれかを既知の技法によって実施することができる。また、誘導体は1つ以上の非古典的アミノ酸を含んでもよい。
【0060】
ヒトにおける抗体のインビボにおける使用およびインビトロにおける検出アッセイを含むいくつかの使用のためには、キメラ、ヒト化またはヒト抗体を使用することが好ましい場合がある。キメラ抗体は、マウスモノクローナル抗体由来の可変領域とヒト免疫グロブリン由来の定常領域を有する抗体などの、抗体の異なる部分が異なる動物種または分子由来である分子である。キメラ抗体を作製する方法は当技術分野において既知である。例えば、全体の内容が参照として本明細書に組み入れられている、Morrison, 1985;Gilliesら, 1989;米国特許第5,807,715号;同第4,816,567号および同第4,816,397号を参照されたい。ヒト化抗体は、非ヒト種の1つ以上の相補性決定領域(CDR)およびヒト免疫グロブリン分子のフレームワーク領域を有する望ましい抗原に結合する非ヒト種由来の抗体分子である。抗体は、例えば、全て全体の内容が参照として本明細書に組み入れられている、CDR-グラフティング
、ベニアリング(veneering)またはリサーフェイシング(resurfacing)(欧州特許第592,106号;欧州特許第519,596号;Padlan, 1991;Studnickaら, 1994;Roguskaら, 1994)およびチェーンシャフリング(米国特許第5,565,332号)を含む当技術分野において既知の種々の技法を使用してヒト化することができる。
【0061】
完全ヒト抗体は、患者の治療的療法に特に望ましい。ヒト抗体は、ヒト免疫グロブリン配列由来の抗体ライブラリーを使用する上記のファージディスプレイ方法を含む当技術分野において既知の種々の方法によって作製することができる。各々全体の内容が参照として本明細書に組み入れられている、
を参照されたい。
【0062】
ヒト抗体は、機能的な内因性免疫グロブリンを発現することができないが、ヒト免疫グロブリン遺伝子を発現することができる遺伝子組換えマウスを使用しても作製することができる。ヒト抗体を作製するためのこの技術の概略は、LonbergおよびHuszar, 1995を参照されたい。ヒト抗体およびヒトモノクローナル抗体を作製するためのこの技術並びにこのような抗体を作製するためのプロトコールの詳細な考察は、例えば、全体の内容が参照として本明細書に組み入れられている、
を参照されたい。また、Abgenix, Inc.(Freemont、CA)、Kirin, Inc.(日本)、Medarex(NJ)およびGenpharm(San Jose、CA)などの会社が、上記のものと同様の技術を使用して、選択した抗原に対するヒト抗体を提供することに関与することがある。
【0063】
本発明の抗体は、免疫原性応答を実質的に全く生じないで、ほ乳類の循環系に注射することができる組成物を提供するために、Davisら(米国特許第4,179,337号)によって記載されている方法およびカップリング剤によって修飾することもできる。
【0064】
III. ポリヌクレオチド
本発明は、ポリヌクレオチドを含む組成物と、被験者における受精能を調節および診断するためにこれらの組成物を使用する方法を提供する。いくつかの態様において、本発明の方法および組成物は、プロテアソーム、ユビキチンまたはユビキチン-プロテアソーム経路の関連酵素に対する動物または被験者の免疫応答を誘導することに関してもよい。
【0065】
本発明の種々の態様において、プロテアソームタンパク質、モノマーユビキチンまたはユビキチンポリペプチドおよびそれらの断片をコードするポリヌクレオチドが提供される。プロテアソームタンパク質は、プロテアソームのα1-7およびβ1-7サブユニット、特にMECL1サブユニットを含むが、これらに限定されない。プロテアソームポリペプチドまたはポリペプチド断片をコードするポリヌクレオチドは原核細胞または真核細胞において発現することができる。発現されたポリペプチドまたはポリペプチド断片は、脊椎動物に投与して、プロテアソームポリペプチドまたはポリペプチド断片と免疫反応する抗体を作製するためのプロテアソーム抗原として使用するために精製することができる。
【0066】
本発明は、任意の特定のほ乳類のプロテアソームポリヌクレオチドまたはポリペプチドに範囲が限定されない。当業者は、本明細書に記載する核酸を使用して、他のほ乳類において関連する相同物を容易に同定することができると思われる。また、本発明は、本明細書に開示されている特定の核酸に限定されないことが明らかにされるべきである。以下に考察するように、特定の「プロテアソームタンパク質」遺伝子またはポリヌクレオチド断片は、種々の異なる塩基を含有し、本明細書に開示するポリヌクレオチドと機能的に識別できない、および場合によっては、構造的に識別できない対応するポリペプチドを作製することができる。種々の動物由来の種々のプロテアソームまたはユビキチン配列の核酸配列はGenBankおよび他の公的なデータベースにおいて見つけることができる。
【0067】
A. ペプチドまたは抗原をコードする核酸
本発明のいくつかの態様において、脊椎動物において免疫応答を誘導することができるおよび抗プロテアソームまたは抗ユビキチン抗体を作製するための抗原として使用するための抗原性ポリペプチドをコードするポリヌクレオチドが考慮される。ある例において、ワクチンとして使用するまたは受動免疫のための抗プロテアソーム抗体を作製する際に使用する特定の抗原性プロテアソームポリペプチドドメインをコードするプロテアソームポリヌクレオチドを発現することが望ましい場合がある。本発明の核酸は、プロテアソームタンパク質全体または任意の他の断片をコードしてもよい。核酸は、特定の生物のPCR増幅DNAから誘導することができる。しかし、他の態様において、核酸は、ゲノムDNA、相補的DNA(cDNA)または合成DNAを含んでもよい。タンパク質は、ワクチンに使用するため、または抗体を単離する方法に使用するために指定された配列から誘導することができる。抗原性プロテアソームポリペプチドを作製するために、プロテアソームポリペプチドをコードする遺伝子または対応するcDNAを適当な発現ベクターに挿入することができる。
【0068】
「cDNA」という用語は、鋳型としてメッセンジャーRNA(mRNA)を使用して作製されるDNAをいうことが意図されている。ゲノムDNA鋳型または非プロセッシングもしくは部分プロセッシングRNA鋳型から増幅または合成されるDNAと違って、cDNAを使用する利点は、cDNAは、主に、対応するタンパク質のオープンリーディングフレーム(ORF)を含むコード配列を含むということである。非コード領域が最適な発現に必要とされる場合などの、完全長または部分的なゲノム配列が好ましい場合もある。
【0069】
さらに別の態様において、所定の動物のポリヌクレオチドは、わずかに異なる核酸配列を有するが、それでもなお、同じポリペプチドをコードする天然の変種によって提供されてもよい(以下の表1を参照されたい)。また、異なる核酸配列を生じるが、同じポリペプチドをコードする別のコドンを使用して、動物の所定のプロテアソームポリペプチドを作製することができることが考慮される。
【0070】
本願において使用する「プロテアソームポリヌクレオチドをコードする核酸」という用語は、細胞の総核酸を含まない単離されている核酸分子をいう。「機能的に同等なコドン」という用語は、アルギニンまたはセリンの6つのコドン(表1、以下)などの同じアミノ酸をコードするコドンをいうために本明細書において使用され、また以下のページに考察するように、生物学的に同等なアミノ酸をコードするコドンもいう。
【0071】
遺伝子コードの縮重を考慮すると、配列は、所定のプロテアソーム遺伝子またはポリヌクレオチドのヌクレオチドと同一のヌクレオチドの少なくとも約50%、通常少なくとも約60%、さらに通常約70%、最も通常には約80%、好ましくは少なくとも約90%、および最も好ましくは約95%を有するプロテアソーム遺伝子またはポリヌクレオチドに記載されているものと本質的に同じであると考えられる。プロテアソーム遺伝子またはポリヌクレオチドに記載されているものと本質的に同じである配列はまた、標準的な条件においてプロテアソームポリヌクレオチドの相補鎖を含む核酸セグメントにハイブリダイゼーションすることができる配列として機能的に規定することもできる。密接に関連する配列という用語は、実質的な配列の類似性を有する配列または本明細書に記載するものと同様の抗原応答を実施または誘発するタンパク質をコードする配列をいう。密接に関連する配列という用語は、比較しているポリヌクレオチドまたはポリペプチドと最小または50%の類似性を有する配列をいうために本明細書において使用される。
【0072】
本発明のDNAセグメントは、上記のように、生物学的に機能的な同等なプロテアソームタンパク質およびペプチドをコードするものを含む。このような配列は、核酸配列内に天然に生じることが既知のコドン冗長性およびアミノ酸の機能的同等性の結果並びにこのようにコードされるタンパク質として生じうる。または、機能的に同等なタンパク質またはペプチドは、タンパク質構造の変更が、交換されるアミノ酸の特性の考慮に基づいて工作することができる組換えDNA技術の適用によって作製することができる。変更は、以下に記載するように、部位特異的突然変異技法の適用によって工作しても、または無作為に導入して、望ましい機能について後にスクリーニングしてもよい。
【0073】
当然のことながら、本発明は、プロテアソームポリヌクレオチドの配列に相補的であるまたは本質的に相補的であるオリゴヌクレオチドも含む。「相補的」である核酸配列は、標準的なワトソン-クリック相補則により塩基対形成することができるものである。本明細書において使用する「相補的な配列」という用語は、上記と同じヌクレオチド比較によって評価するとき、または本明細書に記載するものなどの比較的ストリンジェントな条件下において、プロテアソームポリヌクレオチドの核酸セグメントとハイブリダイゼーションすることができると規定するとき、実質的に相補的である核酸配列を意味する。
【0074】
または、ハイブリダイゼーションセグメントは短いオリゴヌクレオチドであってもよい。長さ17塩基の配列がヒトゲノムにおいて1回だけ生じるはずであるので、独自の標的配列を指定するのに十分である。短いオリゴマーは作製が容易であり、インビボにおけるアクセスしやすさを高めるが、ハイブリダイゼーションの特異性の決定には数多くの他の因子が関与する。相補的な標的へのオリゴヌクレオチドの結合親和性および配列特異性は共に長さの増加と共に増加する。
またはそれ以上の塩基対の例示的なオリゴヌクレオチドが使用されることが考慮されるが、他も考慮される。250、500、1000、1212、1500、2000、2500、3000または3500塩基およびそれ以上の塩基をコードする長いポリヌクレオチドも考慮される。このようなオリゴヌクレオチドまたはポリヌクレオチドは、典型的には、例えば、サザンブロットおよびノーザンブロットのプローブとして、並びに増幅反応のプライマーとして、またはワクチンのための用途を見出している。
【0075】
(表1)
【0076】
C. 非細菌的に増幅した核酸
本発明の核酸またはポリヌクレオチドは、例えば、化学的合成または酵素的作製などの当業者に既知の任意の技法によって作製することができる。本発明の方法において、1つ以上のオリゴヌクレオチドまたはポリヌクレオチドを使用することができる。種々の異なる機序のオリゴヌクレオチド合成が、例えば、各々参照として本明細書に組み入れられている、
に開示されている。
【0077】
酵素的に作製される核酸またはポリヌクレオチドの限定するものではない例は、PCR(商標)などの増幅反応において酵素によって作製されるもの(例えば、各々参照として本明細書に組み入れられている、米国特許第4,683,202号および米国特許第4,682,195号を参照されたい)または参照として本明細書に組み入れられている米国特許第5,645,897号に記載されているオリゴヌクレオチドの合成によって作製されるものを含む。
【0078】
核酸またはポリヌクレオチド増幅の別の方法は、全体の内容が参照として本明細書に組み入れられている、欧州特許第320 308号に開示されているリガーゼ連鎖反応(「LCR」)である。米国特許第4,883,750号は、標的配列にプローブ塩基対を結合するための、LCRに類似した方法を記載している。結果として得られる「ジ-オリゴヌクレオチド」の配列を有し、それによってジ-オリゴヌクレオチドを増幅する核酸の存在下における2つ(または2つ以上)のオリゴヌクレオチドのライゲーションに基づいた方法も本発明の増幅段階に使用することができる。Wuら, (1989)は、全体の内容が参照として本明細書に組み入れられている。
【0079】
D. ポリヌクレオチド送達
本発明のある態様において、真核細胞において機能することができるプロモーターの制御下においてプロテアソームポリヌクレオチドまたはポリヌクレオチドセグメントを含む発現構築物を提供する。本発明のある局面における一般的な方法は、特定のタンパク質、ポリペプチドまたはペプチド断片をコードする発現構築物を細胞に提供し、それによって細胞において抗原性タンパク質、ポリペプチドまたはペプチドの発現を可能にすることである。発現構築物の送達後、発現構築物によってコードされるタンパク質、ポリペプチドまたはペプチド断片は、細胞の転写および翻訳機構によって合成される。種々のポリヌクレオチド送達組成物および方法が既知である(各々参照として本明細書に組み入れられている、Sambrookら, 2001;LiuおよびHuang, 2002;Ravidら, 1998;BalickiおよびBeutler, 2002を参照されたい)。
【0080】
ウイルスおよび非ウイルス送達システムは、抗原性タンパク質、ポリペプチド、ポリペプチド断片をコードする発現構築物の送達のための種々の送達システムのうちの2つである。両方の種類の送達系が当技術分野において既知であり、以下に簡単に記載する。遺伝子による免疫化のために発現構築物の送達に使用される主な方法も2つある;間接的なエクスビボ方法または直接的なインビボ方法。エクスビボ遺伝子導入は、培養中の(宿主)細胞のベクター改変およびベクターで改変した細胞の被験者への投与または移植を含む。インビボにおける遺伝子導入は、免疫化対象被験者へのワクチンベクターの直接導入を含む。
【0081】
種々の態様において、発現される核酸は、典型的には、遺伝子発現成分の完全なセット(プロモーター、コード配列およびターミネーター)を含む、直鎖状発現要素(「LEE」)および/または環状発現要素(「CEE」)に関連してもよい。これらのLEEおよびCEEは細胞または無傷の生物に直接導入されて、発現されて、標準的な高次コイル複製プラスミドによるものに匹敵する発現レベルを生じることができる(SykesおよびJohnston, 1999)。
【0082】
1. 非ウイルスポリヌクレオチド送達
本発明の一態様において、ポリヌクレオチド発現構築物は、組換えによって作製されるDNAプラスミドまたはインビトロにおいて作製されるDNAを含んでもよい。本発明の種々の態様において、例えば、プロテアソームまたはユビキチンポリヌクレオチドを含む発現構築物は、注射および/または微粒子銃(例えば、遺伝子銃)によって生物または被験者に投与される(Kleinら, 1987およびSanfordら, 1991)。いくつかの態様において、ポリヌクレオチドの投与は、筋肉内、静脈内、皮下、皮内または腹腔内注射によってもよい。
【0083】
本発明のプロテアソームまたは同様のポリヌクレオチドを含む発現構築物の導入は、細胞膜を物理的または化学的に透過性にする方法のいずれか(例えば、リン酸カルシウム沈殿、DEAE-デキストラン、エレクトロポレーション、直接マイクロインジェクション、DNA負荷リポソームおよびリポフェクタミンDNA複合体、細胞の超音波処理、高速微粒子を使用する遺伝子銃並びに受容体媒介性形質移入)によって実施することができる。ある態様において、脂質製剤および/またはナノカプセルの使用が、プロテアソームポリヌクレオチド、プロテアソームポリペプチドまたはプロテアソームポリヌクレオチドを含む発現ベクターを宿主細胞に導入するために考慮される(各々参照として本明細書に組み入れられている、Banghamら(1965)、Gregoriadis(1979)、DeamerおよびUster(1983)、SzokaおよびPapahadjopoulos(1978)、Nicolauら, 1987およびWattら, 1986の例示的な方法および組成物を参照されたい)。本発明の別の態様において、発現構築物は、単純に、裸の(naked)組換えDNA、発現カセットまたはプラスミドからなってもよい。
【0084】
2. ウイルスベクター
ある態様において、本発明による免疫応答を与えるポリヌクレオチドまたはコードされるポリペプチドはウイルスベクターによって送達することができることが考慮される。特定の態様において、免疫応答はプロテアソームまたはユビキチンペプチドまたはポリペプチドであってもよい。細胞に効率的に感染して侵入し、宿主細胞ゲノムに組込み、ウイルス遺伝子を安定して発現するある種のウイルスベクターの能力によって、数多くの異なるウイルスベクター系が開発され、適用されている(Robbinsら, 1998)。ウイルスシステムは、現在、エクスビボおよびインビボにおける遺伝子導入のためのベクターとして使用するために開発されている。例えば、アデノウイルス、単純ヘルペスウイルス、レトロウイルスおよびアデノ随伴ウイルスベクターが、現在、癌、嚢胞性線維症、ゴーシェ病、腎疾患および関節炎の治療のために評価されている(RobbinsおよびGhivizzani, 1998;Imaiら, 1998;米国特許第5,670,488号)。
【0085】
特定の態様において、アデノウイルス(各々具体的に参照として本明細書に組み入れられている、米国特許第6,383,795号、同第6,328,958号および同第6,287,571号)、レトロウイルス(各々具体的に参照として本明細書に組み入れられている、米国特許第5,955,331号;同第5,888,502号、同第5,830,725号)、単純ヘルペスウイルス(各々具体的に参照として本明細書に組み入れられている、米国特許第5,879,934号;同第5,851,826号)、アデノ随伴ウイルス(AAV)、ポックスウイルス;例えば、ワクシニアウイルス(Gnantら,1999a;Gnantら, 1999b)、αウイルス;例えば、シンドビスウイルス、セムリキ森林熱ウイルス(Lundstorm, 1999)、レトロウイルス(Coffeyら, 1998)およびインフルエンザAウイルス(Neumannら, 1999)、ポックスウイルス/レトロウイルスキメラベクター(Holzerら, 1999)、アデノウイルス/レトロウイルスベクター(Fengら, 1997;Bilbaoら, 1997;Caplenら, 1999)およびアデノウイルス/アデノ随伴ウイルスベクター(Fisherら, 1996;米国特許第5,871,982号)発現ベクターが発現構築物の送達のために考慮される。「ウイルス発現ベクター」は、(a)構築物のパッケージングを支持し、(b)クローニングされている組織または細胞特異的構築物を最終的に発現するのに十分なウイルス配列を含むような構築物を含むことを意味している。ウイルスの増殖および操作は当技術分野において既知である。
【0086】
E. 発現ベクター
プロテアソームペプチドをコードするポリヌクレオチドは、遺伝的免疫化ベクターまたは任意の他の好適な発現構築物にクローニングすることができる。ベクターは、真核細胞において機能可能なプロモーター、例えば、CMVプロモーターまたは任意の他の好適なプロモーターを含むことができる。このような方法では、ポリヌクレオチドは、筋肉内注射、皮内注射または表皮注射または微粒子銃によって投与することができる。ポリヌクレオチドは、静脈内、皮下、病巣内、腹腔内、経口、他の粘膜または吸入投与経路によっても投与することができる。いくつかの具体的な例示的な態様において、投与は、少なくとも0.005μg〜5.0μgのポリヌクレオチドの表皮注射/微粒子銃によってもよい。いくつかの例において、第1の投与後少なくとも約2週間以上経過してから、第2の投与、例えば、筋肉内注射および/または表皮注射を実施してもよい。これらの方法では、ポリヌクレオチドは、ウイルス発現ベクター、例えば、アデノウイルス、単純ヘルペスウイルス、レトロウイルスまたはアデノ随伴ウイルスベクターを含むウイルス発現ベクターにクローニングすることができるが、必ずしも必要はない。ポリヌクレオチドは、本明細書に開示するまたは当業者に既知の任意の他の方法においても投与することができる。1つ以上のポリヌクレオチドを1つ以上の発現ベクターに含ませることができる。
【0087】
発現に好適なベクターは既知であり、細菌発現ベクター、酵母発現ベクターおよびバキュロウイルスベクターを含むが、これらに限定されない。宿主細胞が好適な条件下で培養されると、プロテアソームペプチドは宿主細胞内で発現される。宿主細胞は、カイコ幼虫、CHO細胞、大腸菌(E.coli)および酵母を挙げることができるが、これらに限定されない。宿主細胞において発現されるペプチドの回収方法は既知である。ベクター、宿主、ベクターを発現する方法およびペプチドを回収する方法は既知である(例えば、Sambrookら, 2001およびO'Reilleyら, 1994)。
【0088】
IV. 免疫化
プロテアソームおよびユビキチンペプチドまたはポリペプチドを含むが、これらに限定されない、本明細書に記載する種々のペプチドまたはポリペプチドに対して免疫応答を誘発することができる。本発明のプロテアソームまたはユビキチンペプチド由来の免疫原性ペプチドは、好ましくは、抗プロテアソームまたは抗ユビキチン抗体をそれぞれ誘導するために使用される。例えば、1つ以上のプロテアソームエピトープを含む融合タンパク質を、非プロテアソーム免疫原と共にまたは伴わないで、例えば、注射によって動物に投与する。免疫原は、当技術分野において既知のアジュバントと共に送達してもよい。投与経過は、抗プロテアソーム抗体価を誘導するための必要性に応じて、1回または多数回の追加投与であってもよい。従って、十分な抗体価を保持する必要性に応じて、維持投与を実施してもよい。本発明のペプチド、ペプチド組成物またはワクチンの適当な用量および免疫法は、本明細書および以下の実施例に提供されている情報を利用して、当業者が決定することができる。
【0089】
ワクチン化/免疫化の概念は、免疫系の2つの基本的な特徴、すなわち免疫系成分の特異性および記憶に基づいている。ワクチン化/免疫化は、被験者に投与した抗原に特異的に向かう応答を開始する。さらに、メモリーBおよびTリンパ球集団が誘導されうる。抗原に再度接触すると、免疫系はさらに迅速に且つさらにより激しく応答を開始し、それによってワクチン化/免疫化した被験者に抗原に対する免疫学的防御を与える。同じ抗原もしくは異なる抗原を被験者に反復投与することによってまたは被験者にワクチン組成物を追加免疫投与することによって免疫応答を増強することができる。
【0090】
ワクチン化は、抗原の全てまたは一部を投与することによる、能動的に獲得される免疫の人工的な誘導である。能動的に獲得される免疫以外に、受動免疫方法を使用して被験者に治療的恩恵を提供することもできる、以下を参照されたい。
【0091】
特に、DNA免疫化としても既知の遺伝的ワクチン化は、抗原をコードする発現ベクターをインビボ、インビトロまたはエクスビボにおいて投与して、適当な生物、組織、細胞または標的細胞内に適切に折りたたまれた抗原の産生を誘導することに関係する。発現されたタンパク質またはペプチドは、典型的には、正常な細胞の主要組織適合性(MHC)抗原複合体と共に、形質移入した細胞の細胞表面で提示される。MHC抗原と関連したこれらの抗原決定基の提示は、決定基に特異的な細胞障害性T-リンパ球クローンの増殖を誘発することを意図している。さらに、発現中の形質移入細胞によって放出されるタンパク質は、抗原提示細胞によってピックアップ、内部移行または発現されて、全身の液性抗体応答を誘発することもできる。
【0092】
ワクチンは、非病原性で、ワクチン化に使用するのに好適であるように改変されている免疫原性物質の全てもしくは一部またはそれらの模倣物由来の抗原を含む組成物である。ワクチンの種類には、遺伝子ワクチン、ビロソーム、弱毒化または不活性化した生物体そのものを使用したワクチン、組換えタンパク質ワクチン、結合ワクチン、遺伝子組換え植物ワクチン、トキソイドワクチン、精製サブユニットワクチン、遺伝子組換え複合ワクチン、抗イディオタイプワクチンおよび当技術分野において既知の他のワクチンが挙げられるが、これらに限定されない。
【0093】
免疫応答は能動または受動免疫応答であってもよい。能動免疫は、生体が種々の抗原に接触すると形成する。それは、典型的には、BまたはTリンパ球に関係する。Bリンパ球(B細胞とも呼ばれる)は抗体を産生する。抗体は特異的な抗原に結合して、貪食細胞が抗原を破壊しやすくする。典型的には、Tリンパ球(T細胞)が抗原を直接攻撃して、免疫応答に対してある種のコントロールを提供することができる。特定の抗原または抗原種に特異的なB細胞およびT細胞が発生する。受動免疫化は、抗原に結合する事前に形成された抗体または他の結合物質の投与をいう。
【0094】
ある場合では、免疫応答は養子免疫治療の結果であってもよい。養子免疫治療では、リンパ球を被験者から入手し、抗原組成物と接触または抗原組成物で標識(pulsed)する。例示的な方法または組成物は、
を参照されたい。
【0095】
本発明は、プロテアソーム抗原またはプロテアソーム抗原、ユビキチン抗原もしくはユビキチン-プロテアソーム経路において活性な酵素由来の抗原をコードするポリヌクレオチドを含む抗原組成物を被験者に接触させることによって、被験者を免疫化、治療またはワクチン化する方法を含む。抗原組成物は、核酸;ポリペプチド;プロテアソーム抗原を発現してもよいウイルス、細菌、真菌または寄生虫などの弱毒化病原体;プロテアソーム抗原を発現する原核細胞;プロテアソーム抗原を発現する真核細胞;ビロソーム等またはそれらの組み合わせを含んでもよい。本明細書において使用する「抗原組成物」は、典型的には、薬学的に許容されうる製剤中に抗原を含む。
【0096】
抗原は、特に、抗原に反応性の特異的な抗体または細胞種の形態の、生物が免疫応答を誘発する任意の基質、分子または基質をコードする分子をいう。抗原組成物は、本明細書に記載するアジュバント、免疫調節物質、ワクチン基剤および/または他の賦形剤をさらに含んでもよく、当技術分野において既知である(例えば、Remington's Pharmaceutical Scienceを参照されたい)。
【0097】
抗原または抗原組成物を被験者に導入する種々の方法が当技術分野において既知である。ワクチン化方法には、DNAワクチンまたは遺伝子免疫化(例えば、各々参照として本明細書に組み入れられている、
を参照されたい)、遺伝子組換え食用植物ワクチン(例えば、各々参照として本明細書に組み入れられている
を参照されたい)、経皮的免疫化(各々参照として本明細書に組み入れられている、Glennら, 1999および米国特許第5,980,898号)、経鼻的または粘膜免疫化(例えば、各々参照として本明細書に組み入れられている、米国特許第4,512,972号、同第5,429,599号、同第5,707,644号、同第5,942,242号を参照されたい);ビロソーム(各々参照として本明細書に組み入れられている、
)、生ベクター等が挙げられるが、これらに限定されない。抗原送達方法は1つ以上のワクチン法と組合わせてもよい。
【0098】
抗原、ポリペプチドまたは抗原をコードするポリヌクレオチドを含むワクチンは、免疫応答を刺激するために種々の状況において抗原を提示することができる。種々のワクチン状況のいくつかは弱毒化病原体、不活性化病原体、トキソイド、結合物、組換えベクター等を含む。本発明のポリペプチドは、種々のワクチン組成物によって発現されてもまたは種々のワクチン組成物に結合してもよい。種々のワクチン組成物は、抗原または抗原をコードする分子を産生または発現する細胞に、抗原を直接提供してもまたは抗原を産生する組成物、例えば、発現構築物を送達してもよい。
【0099】
本発明のワクチンは、抗プロテアソームまたは抗ユビキチン抗体の産生を誘導するのに十分な有効な治療量の免疫原性プロテアソームまたはユビキチンペプチドを含む。得られる免疫によって避妊効果を生ずることができる。好ましくは、ワクチンは1種以上のほ乳類において避妊効果を生ずるのに有効である。最も好ましくは、ワクチンは、げっ歯類またはウサギ、ブタ、ウマ、サル、イヌ、ネコ、雌ウシおよびヒトの1種以上を含む非げっ歯類に投与すると、抗プロテアソーム抗体を産生する。
【0100】
他の態様において、タンパク性組成物またはポリペプチド、断片または模倣物を使用して、ワクチンに使用するための抗イディオタイプ抗体を作製することができる。抗イディオタイプワクチンでは、免疫原は、病原体の抗原分子に対して形成される第2の抗体のFab末端に対する抗体である。第1の抗体のFab末端は、病原体の抗原分子と同じ抗原形状を有し、抗原として使用することができる(例示的な米国特許第5,614,610号、同第5,766,588号を参照されたい)。本明細書において使用するための「ヒト化」抗体は、分泌される1つ以上のアミノ酸が、ヒト抗体においてより一般的に見られるアミノ酸と交換されている非ヒト種の抗体であってもよい。これは、通常の組換え技術、特に部位特異的突然変異を使用することによって容易に実施することができる。ヒト化抗体は、以下に記載するように、受動免疫剤としても使用することができる。
【0101】
V. 薬学的組成物
本発明の薬学的水性組成物は、薬学的に許容されうる担体または水性媒体に溶解または分散させた1つ以上のプロテアソーム阻害剤の有効量を含む。「薬学的または薬理学的に許容されうる」というフレーズは、そのような応答を誘発することが具体的に示されていない限り、ヒトに投与したとき有害反応、アレルギー反応または他の望ましくない反応を生じない分子的実態および組成物をいう。本明細書において使用する「薬学的に許容されうる担体」は、溶媒、分散媒体、抗菌剤および抗真菌剤、等張剤および吸収遅延剤等のいずれかおよび全てを含む。薬学的に活性な物質のためのこのような媒体および物質の使用は当技術分野において既知である。任意の従来の媒体または物質が作用成分と不適合である場合を除いて、治療用組成物におけるその使用が考慮される。追加の作用成分も本発明の組成物に組み入れることができる。
【0102】
被験者に投与される本発明の組成物の実際の投与量は、体重などの物理的および生理学的因子および投与経路によって決定される。これらを考慮すると、特定の被験者および/または治療経過のためのペプチド、ポリペプチド、ポリヌクレオチド、免疫グロブリンまたはプロテアソーム阻害剤組成物の用量を容易に決定することができる。
【0103】
本発明に使用することができるプロテアソーム阻害剤は当技術分野において既知であり、例えば、BIOMOL Research Laboratories, Inc.、Plymouth Meeting, PAから購入可能である。本発明において用途を見出すことができるこのような阻害剤の例には、N-アセチル-Leu-Leu-Nle-H、N-アセチル-Leu-Leu-Met-H、アクラシノマイシンA(アクラルビシン)、Ada-(Ahx)3-(Leu)3-ビニルスルホン、Ada-Lys(ビオチニル)-(Ahx)3-(Leu)3-ビニルスルホン、Ada-Tyr-(Ahx)3-(Leu)3-ビニルスルホン、バクテネシン-5、ブレフェルジンA、クルクミン、没食子酸(-)エピガロカテキン(EGCG)、エポキソマイシン、グリオトキシン、ラクタシスチン、クラスト(clasto)-ラクタシスチン a-ラクトン、N-トシル-Lys-クロロメチルケトン(TLCK)、N-トシル-Phe-クロロメチルケトン(TPCK)、NIP-(Leu)3-ビニルスルホン、フェプロペプチン(Phepropeptin) Aキモトリプシン、フェプロペプチン(Phepropeptin) Bキモトリプシン、フェプロペプチン(Phepropeptin) Cキモトリプシン、フェプロペプチン(Phepropeptin) Dキモトリプシン、PR11、PR39、ユビキチン5+1、Z-Il2-Glu(OBut)-Ala-Leu-H(PSI)、Z-Leu-Leu-Leu-ビニルスルホン、Z-Leu-Leu-Nva-H(MG115)、Z-Leu-Leu-Leu-H(MG132)、Z-Leu-Leu-Leu-B(OH)2(MG262)およびZ-Leu-Leu-Tyr-COCHOが挙げられる。
【0104】
本発明の薬学的組成物は、静脈内、皮内、動脈内、腹腔内、関節内、胸膜内、気管内、鼻腔内、膣内、局所的、筋肉内、腹腔内、皮下、小胞内、粘膜、経口、局所的、エアゾールを使用して局所的、注射、注入、持続注入、標的細胞を直接つける局所的灌流またはカテーテルもしくは洗浄投与することができる。典型的には、このような組成物は溶液または懸濁液として注射剤として調製され;注射前に液体を添加して溶液または懸濁液を調製するのに好適な固体剤を作製してもよく;製剤を乳化してもよい。組成物は滅菌されており、容易なシリンガビリティー(syringability)が存在する程度に流動体であり、製造および保存条件下において安定であり、細菌および真菌などの微生物の汚染作用から保護されている。内毒素汚染は安全レベル、例えば、0.5 ng/mgタンパク質未満に最小に保たれるべきであることが考慮される。
【0105】
本発明の組成物は、他の非作用成分を含有する滅菌水中で調製されて、注射に好適にされることが最も好ましいが、このような作用成分の溶液は、適宜、ヒドロキシプロピルセルロースなどの界面活性剤と好適に混合された水中でも調製することができる。分散剤も、液体ポリエチレングリコールおよびその混合物並びにオイル中で調製することができる。担体は、例えば、水、エタノール、ポリオール(例えば、プロピレングリコールおよび液体ポリエチレングリコール等)、それらの好適な混合物および植物油を含有する溶媒または分散媒体であってもよい。適切な流動性は、例えば、レシチンなどのコーティングを使用することによって、分散剤の場合には必要な粒子サイズを維持することによっておよび界面活性剤を使用することによって維持することができる。
【0106】
微生物の作用は、種々の抗菌および抗真菌剤、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チメロサール等によって防止することができる。多数の例において、等張剤、例えば、糖または塩化ナトリウムを加えることが好ましい。注射用組成物の吸収の遅延は、吸収遅延剤、例えば、モノステアリン酸アルミニウムおよびゼラチンを組成物中に使用することによって生じることができる。
【0107】
製剤化したら、投与製剤に適合性した方法で、避妊作用を誘導するのに有効である量の溶液を投与する。水溶液での非経口投与のためには、例えば、溶液を適宜好適に緩衝化するべきであり、最初に希釈液を十分量の生理食塩液またはグルコースで等張にするべきである。これらの特定の水溶液は、静脈内、筋肉内、皮下および腹腔内投与に特に好適である。これに関連して、使用することができる滅菌水性媒体は、本発明の開示内容を考慮して当業者に既知である。治療する被験者の状態に応じて、用量の変更を必ず行う。投与担当者は、個々の被験者に適当な用量を任意に決定する。
【0108】
VI. 実施例
以下の実施例は、本発明の好ましい態様を実証するために含まれる。以下の実施例に開示する技術は、本発明を実施する際に十分に機能するために本発明者らが発見した技術を提供しており、従って本発明の好ましい実施様式を構成すると考えることができることが当業者に理解されるべきである。しかし、本発明の精神および範囲から逸脱することなく、多数の変更を開示されている具体的な態様に加えることができ、それでも同様または類似の結果を得ることができることを当業者は、本発明の開示内容に照らして理解するべきである。
【0109】
実施例1
材料と方法
A. 抗体
プロテアソームサブユニットサンプラーパックおよびプロテアソームサブユニットに対する以下のウサギ血清は、Affinity Research products. Ltd.、Mammahead、UKから購入した:
のマウスMECL-1-アミノ酸配列に基づいて組換えペプチドに対して形成された抗-MECL1サブユニット血清(カタログ番号#PW8150)は、免疫蛍光およびイムノゴールド透過型電子顕微鏡(TEM)には1/200で、ウェスタンブロットには1/4000で、インビトロにおける受精の阻止には1/2000〜1/8000で使用した。この抗体は、29 kDa MECL-1サブユニットおよびこのサブユニットを含むいくつかの他のさらに大きいタンパク質複合体を認識することが製造業者によって示された。抗-α/β-サブユニット血清(カタログ番号#PW 8155)は、ヒト赤血球細胞から単離したプロテアソーム調製物に対して形成され(TanakaおよびTsurumi, 1997)、ヒト赤血球溶解液の25kDa〜30 kDaのバンドセット並びにいくつかの別個のα-型およびβ-型サブユニットに相当する、他の細胞溶解液の別のさらに高いMWバンドを認識する。抗β1iサブユニット血清(カタログ番号#PW 8205)はマウスβ1iサブユニットの組換え完全長配列に対して形成され(Groettrupら, 1996)、26 kDa、36 kDaのタンパク質およびおそらく他のさらに高いMWタンパク質複合体を認識する。α/βおよびβ1i抗血清は、免疫蛍光およびイムノゴールドTEMでは1/200の希釈で、ウェスタンブロットでは1/4000で使用した。
【0110】
ブタ透明帯の外側面のユビキチンは、組換えヒトユビキチンの全分子に対して形成された抗体KM691(免疫蛍光では1/200の希釈、ウェスタンでは1:1000の希釈;Kamiya Biomedical Company、Seattle、WAから購入)によって検出された。前核は、ブタ胎仔核ユビキチン-CEP 52テイル融合タンパク質を認識し(McCauleyら, 2002)、Chemicon International Inc.、Temecula、CAから購入したウサギポリクローナル抗体AB 1690(1/100の希釈)を使用して対比染色した。この抗体は、卵巣組織切片のZPの外側層のユビキチン化タンパク質も認識した(図3Fを参照されたい)。
【0111】
アクロソームエキソサイトーシスのマーカーであるアクロソームチロシンキナーゼc-Yes(LeclercおよびGoupil, 2002)は、Santa Cruz Biotechnology, Inc.、Santa Cruz、CAから購入した、ヒトc-Yes(希釈、1/100)の進化的に保存されているC-末端配列に相当する組換えペプチドに対するウサギ血清を使用して検出した。ウサギ抗プロヒビチン抗体Ab-2(希釈、1/100;Thompsonら, 1997)はNeomarkers Inc.、Union City、CAから購入した。
【0112】
免疫蛍光検討に使用した二次抗体は、Zymed Inc.、South San Francisco、CA社製で、1/80に希釈した、赤色蛍光のTRITC-結合および緑色蛍光のFITC結合ヤギ抗マウスIgG、ヤギ抗マウスIgMおよびヤギ抗ウサギIgGを含んだ。DNAは、青色蛍光染料DAPI(Molecular Probes Inc.、Eugene、OR)で対比染色した。コロイド金TEMは、10 nm金結合ヤギ抗ウサギIgGをElectron Microscopy Sciences、Washington、PAから購入し、1/10希釈で使用した。
【0113】
B. 配偶子およびインビトロ受精
思春期前の雌ブタの卵巣を地元の食肉処理場から回収して、実験室に運び、3mm〜6mmの大きい胞状卵胞から卵母細胞を単離した。多層卵丘が存在するために卵丘細胞-卵母細胞複合体(COC)を解剖顕微鏡下で選択し、0.1%(w/v)ポリビニルアルコール(PVA)を含有するHEPES-緩衝タイロードの乳酸(TL-HEPES)培地で繰り返し洗浄した。3.05 mMグルコース、0.91 mMピルビン酸ナトリウム、0.57 mMシステイン、10 ng/ml上皮細胞増殖因子、0.5μg/ml FSH、0.5μg/ml LH、0.1%(w/v)PVA、75μg/mlぺニシリン-Gおよび50μg/ml硫酸ストレプトマイシン(AbeydeeraおよびDay, 1997)を添加した数滴の無血清改良組織培養培地(TCM 199;Gibco、Grand Island、NY)中で39℃において5%CO2雰囲気下で50のCOC群を22時間成熟させた。この後、FSHまたはLHを含有しない改良TCM 199でCOCを洗浄し、さらに20時間成熟させた。
【0114】
受精前に、0.1%(w/v)ヒアルロニダーゼを含有するTL-HEPES培地中でCOCをボルテックスして卵丘細胞を取り出し、113.1 mM NaCl、3 mM KCl、7.5 mM CaCl2、20 mM Tris、11 mMグルコース、5 mMピルビン酸ナトリウム、2 mM カフェインおよび0.2%(w/v)BSAからなる50μlの改良Tris緩衝培地(mTBM)中で洗浄した。受精は50μlのmTBM培地液滴中で実施した。0.1%(w/v)BSAを添加した10 mlのダルベッコのPBS(Gibco、Grand Island、NY) 中で凍結保存精子を解凍した。受精卵内に精子尾部および前核が存在することによって受精を判定するために、以前に記載されているように(Sutovskyら, 2003)、精子を生体ミトコンドリアプローブMitoTracker CMTM Rosで事前標識し、得られた接合体を、以下に記載するように、DNA染料DAPIで対比染色した。精子を遠心分離によって2回洗浄し、最終濃度5×105精子/mlまで受精液に添加した。IVFの6時間後に、接合体と思われるものを無血清TALP Hepes中で3回洗浄し、免疫蛍光、イムノゴールドTEMまたはウェスタンブロット用に処理した。ウェスタンブロット用の透明帯を含まない卵母細胞を入手するために、接合体を、0.5%w/vプロナーゼ(Protease、Sigma)を含有し、タンパク質を含有しないTALP-HEPESで30秒間処理した。
【0115】
C. プロテアソーム阻害剤およびIVF中の抗体処理
精子-透明帯相互作用におけるユビキチン-プロテアソーム経路の役割を調査するために、抗プロテアソーム抗体MECL-1およびプロテアソームのタンパク質作用の特異的な阻害剤、MG132(Z-Leu-Leu-Leu-CHO;可逆的阻害剤;Biomol Research Labs Inc、Plymouth Meeting、PA)およびラクタシスチン(C15H24N2O7S;非可逆的阻害剤;Biomol社製)の影響を評価した。IVFは、受精培地にMG132(10μM)またはラクタシスチン(10μMまたは100μM)または抗体MECL-1(希釈1/2000、1/4000、1/8000;Affinity Research、Mamhead、英国)を添加して上記のように実施した。阻害剤を含有しない対照は、IVF中阻害剤と同等な希釈液量の適当な溶媒(MG132:100%エタノール;ラクタシスチン:H2O)を添加した。抗体ブロックの対照は、抗体を添加せず、等量の免疫前のウサギ血清を添加した。
【0116】
MG132、ラクタシスチンまたはMECL-1抗体の封入は、透明帯で囲まれた卵母細胞の受精を完全に妨害したので、ブロックが生じた正確な受精段階を同定するために追加の実験を実施した。MG132および抗体MECL-1の存在下または非存在下において透明帯で囲まれたおよび透明帯を含まない(ZFまたはZP-)卵母細胞を使用してIVFを実施した。透明帯の除去は、卵母細胞を0.5%プロナーゼ(Sigma、St. Louis、MO)に短時間(30秒)接触させて実施した。卵母細胞を十分に洗浄し、ZF卵母細胞を、精子注入の1時間前に酵素処理から回収した。IVFのためには、ZF卵母細胞を1×105精子/mlと共にインキュベーションしたが、透明帯で囲まれた/透明帯が無傷(ZIまたはZP+)の卵母細胞は上記のように処理した。卵母細胞は、精子注入後(p.i.)12時間経過時に25%酢酸/エタノールで固定し、受精の頻度を求めるために1%オルセインで染色した。いくつかの実験において、卵母細胞を免疫蛍光顕微鏡で評価し、接合体の細胞質中の前核および精子尾部の存在を検出した。透明帯で囲まれたおよび透明帯を含まない胚も精子注入後6時間または20時間経過時に固定し、MG132によるプロテアソーム活性の阻害が精子-透明帯相互作用を特異的に遮断したかどうか、精子-卵細胞膜融合を阻害したかどうかまたは精子の移動性、生存性もしくは精子がアクロソームエキソサイトーシスを受ける能力に影響を与えたかどうかを判定するために免疫蛍光分析用に処理した。
【0117】
精子-透明帯結合および精子のアクロソーム状態を評価するために、透明帯で囲まれた卵母細胞を固定し、無傷のアクロソームを有する精子の核周囲卵胞膜(perinuclear theca)のサブアクロソーム(subacrosomal)および外側アクロソーム層に存在することが既知のチロシンキナーゼc-Yes(LeclercおよびGoupil, 2002)を免疫蛍光によって評価した。透明帯に結合した精子の数は、DAPI染色後に求めた。群あたり5つの卵を、アクロソームチロシンキナーゼc-Yes(免疫蛍光を参照されたい)に対する抗体およびDAPIで対比染色し、60×の倍率で写真撮影した。ZP結合アクロソーム反応性およびアクロソーム無傷精子を計数し、値をアクロソーム無傷精子/卵のパーセント(%)として表した。処理の結果をX2検定およびSAS8.2の一般的な線形モデル手法によって比較した。
【0118】
D. 卵巣組織切片、単離精子および接合体の全載試料の免疫蛍光
従来の組織化学的プロトコールに準じて卵巣組織片を4%パラホルムアルデヒドで固定し、洗浄して、パラフィンに包埋した。厚さ4ミクロンのパラフィン組織切片を切断し、顕微鏡スライドにのせ、キシレンで脱ろうし、100%〜70%エタノールシリーズおよび水で再水和し、スチーマーを使用してクエン酸緩衝液(pH 6.0)中で20分間前処理し、水に浸した組織を入れた生物トレイ上で抗体で処理した。遠心分離によって精子を精液試料からペレット化し、ポリ-L-リジンコーティングしたカバースリップに結合し、2%ホルムアルデヒド中で40分間固定し、4ウェルペトリ皿中でウェルからウェルに通過させることによって処理した。透明帯を含まないおよび透明帯が無傷の卵母細胞を、スライドまたはカバースリップに結合しないで、2%ホルムアルデヒドのPBS溶液中で固定し、1 ccシリンジに取り付けたアナペット(Unapette)ピペット(Beckton Dickinson)を使用して取り扱い、9ウェルパイレックスガラスプレート(Fischer Scientific)の個々のウェルを通過させることによって処理した。全ての試料は、上記および結果のセクションに記載する抗体で処理した。
【0119】
一般的なプロトコールにおいて、カバースリップ上の精子および未結合の卵母細胞は0.1%Triton-X100中で40分間または終夜透過性にしたが、パラフィン切片のスライドは透過性にしなかった。全ての試料は、0.1% TX-100を添加した5%正常ヤギ血清(NGS;Sigma)のPBS溶液中でブロックした。1%NGSおよび0.1% TX-100で強化した中性PBSの標識溶液を使用して、その後のインキュベーションおよび洗浄を実施した。カバースリップに、上記および結果のセクションに掲載し、標識溶液で希釈した一時抗体を積層し、40分(カバースリップに全細胞をのせたもの)または2時間(スライドに組織切片をのせたもの)おいた。標識溶液で手早く洗浄してから、蛍光を結合した適当な抗マウスIgGまたは抗マウスIgMを、異なる蛍光の抗ウサギIgGおよび2.5μg/ml DAPI(Molecular probes Inc.、Eugene、OR)と混合し、試料と共に30分〜40分インキュベーションした。標識溶液中で洗浄後、カバースリップを顕微鏡スライドにのせ、透明マニキュアで密封し、適当なフィルターセットおよびDIC光学を使用したNikon Eclipse 800エピ蛍光顕微鏡で観察した。画像はCoolSnap CCD HQカメラ(Roper Scientific、Tucson、AZ)およびMetaMorphソフトウェア(Universal Imaging Corp.、Downington、PA)で獲得し、記録可能なCDに保管し、Adobe Photoshop 5.5(Adobe Systems、Mountain View、CA)を使用して編集し、Epson Stylus 1280フォトプリンターで印刷した。
【0120】
E. コロイド金イムノラベリングおよび透過型電子顕微鏡
免疫蛍光について以下に記載するように、透明帯が無傷の接合体を、精子注入後6時間経過時に4%ホルムアルデヒド中で固定し、洗浄し、透過性にし、ブロックし、プロテアソームサブユニットに対するウサギ血清およびc-Yesチロシンキナーゼに対する血清と共に2時間インキュベーションした。洗浄後、接合体をヤギ抗ウサギIgG(希釈1% NGSおよび0.1% TX-100を添加したPBSで1/10)と共に1時間インキュベーションし、再度洗浄し、電子顕微鏡用に再固定し、パラホルムアルデヒドおよびグルタルアルデヒドに包埋した(以下を参照されたい)。微細構造の検討のための接合体は、免疫蛍光固定および処理しないで、TEM用に固定した。
【0121】
電子顕微鏡用の包埋のためには、標識および未標識接合体を、2%パラホルムアルデヒドおよび0.6%グルタルアルデヒドのカコジル酸緩衝液混合物中で固定し、1%四酸化オスミウム中で後固定し、上昇エタノールシリーズ(30%〜100%)で脱水し、PolyBed 812樹脂に包埋した。極薄切片をLeica Ultracut UCTウルトラミクロトームで作製し、100 MESH銅グリッド上に置き、酢酸ウラニルおよびクエン酸鉛により2段階で染色した。連続切片を調査し、Jeol 1200 EX電子顕微鏡で撮影した。各抗体および未処理/未標識接合体の微細構造検討のために10の卵母細胞を処理した。ネガフィルムをUmax Magic Scanフラットベッドスキャナーによってスキャンし、CD-Rに記録し、Adobe Photoshop 5.5ソフトウェアを使用してEpson Stylus 1280フォトプリンターで印刷した。
【0122】
F. SDS-PAGEおよびウェスタンブロット法
SDS-PAGEおよびウェスタンブロット法は、一般にSambrookら(2001)に記載されているように実施した。卵母細胞および接合体を所定の経過時点において培養から取り出し、タンパク質を含有しないTALP HEPES中で繰り返し洗浄し、群あたり350の接合体を、Tris(50 mM)、β-メルカプトエタノール(5%)、グリセロール(10%)、SDS(2%)およびPMSF(100 mM)を含有する10μlのロード緩衝液中で5分間煮沸した。雄ブタ精子タンパク質は、抽出緩衝液(50 mM Tris、20 mMイミダゾール、1 mM EDTA、5 mMベンズアミジンHCl、5μg/mlロイペプチン、1μg/mlペプスタチンA、100 mM PMSFおよび0.1% SDS)中で粉砕することによって抽出し、17000×gで30分間遠心分離した。β-メルカプトエタノール、グリセロール、SDSを上清に添加し、ロード緩衝液と同じ最終濃度を達成した。ヒト精子をロード緩衝液と共に煮沸し、次に25000×gで30分間遠心分離して、DNAペレットを除去した。電気泳動分離を10%ゲル上で実施し、Mini-Tank Electroblotter(Owl Scientific、Woburn、MA)を使用してタンパク質をPVDF膜に移した。膜を1%カゼイン溶液でブロックした。一次抗ユビキチン抗体と共に終夜インキュベーションし、次いで適当なビオチン結合二次抗体と共にインキュベーションした後、Vectastain ABC-Ampキット(Vector Lab Inc.、Burlingame、CA)を使用してアルカリ性ホスファターゼ反応を用いてシグナルを検出した。
【0123】
実施例2
プロテアソーム阻害剤による、精子-透明帯結合およびアクロソーム反応に影響を与えない、雄ブタ精子による透明帯の透過性の遮断
プロテアソーム阻害剤ラクタシスチンおよびMG132は、ブタ接合体内の父系的な精子ミトコンドリアのプロテアソーム分解経路を与える目的の検討経過において受精を妨害することを最初に観察した(Sutovskyら, 2003)。この検討は、受精のどの段階において、阻害剤が遮断を発揮するかを決定するために設計された実験によって追跡した。MG132およびラクタシスチンは共に、精子注入時に添加すると、受精を妨害した。実験を4回繰り返し、一致した結果が得られた。これらの試験において、MG132は卵母細胞の100%の受精を妨害した。MG132ほど強力でなく、非可逆的であることが既知のラクタシスチン(Goldbergら, 1995)は、受精速度に用量依存的に作用し、100μm濃度において100%遮断に達した。既知のプロテアソーム特異性および可逆性(LeeおよびGoldberg, 1998)、並びにラクタシスチンは、アクロソームに存在すると思われる非プロテアソームプロテアーゼの一部の活性、すなわちカテプシンAの活性を部分的に阻害する可能性があるという懸念(Ostrowskaら, 2000)により、さらに別の検討をMG132だけを用いて実施した。MG132は精子の生存性、精子-透明帯結合またはアクロソームエキソサイトーシスに影響を与えるということを排除するために、10μM MG132の存在下において受精した卵母細胞を精子注入後20時間経過時に透明帯を無傷のままで固定した。ZPに結合した精子頭部はDAPIで検出し、無傷のアクロソームは、アクロソーム-無傷精子の核周囲卵胞膜(perinuclear theca)のサブアクロソームおよび外側アクロソーム層に存在することが既知のチロシンキナーゼc-Yesに対する抗体によって検出された(図1 A〜D)。この実験の精子はMitoTracker CMTM Rosで事前に標識されたので、精子ミトコンドリアの鞘は、受精した卵母細胞内で検出可能になった。MG132を用いた以前の実験と同様に、阻害剤は受精を完全に妨害した(MG132群の受精0%対対照の受精77%)。しかし、卵あたりの結合精子の数(MG132の99.3対対照の93.3)またはアクロソーム無傷、透明帯結合精子の割合(MG132の2.01%対対照の2.14%)には有意な差はなかった。
【0124】
実施例3
雄ブタ精子アクロソームに存在するプロテアソームサブユニット
阻害剤のデータおよびプロテアソームサブユニットはほ乳類/ヒト精子アクロソーム内に存在することを確認している以前の報告(例えば、Bialyら, 2001)並びに無脊椎動物のアクロソーム活性におけるユビキチン-プロテアソーム系を示す証拠(MatsumuraおよびAketa, 1991;Sawadaら, 2001a、2001b)を考慮して、プロテアソームは雄ブタ精子アクロソームに存在するという可能性を検討することを決定した。コアサブユニットおよびサブユニットLMP-2(β1i)、MECL1およびMECL2に対するポリクローナル抗体はアクロソームにおける主要なシグナルを提供し、他の精子構造物におけるバックグラウンドはほとんどまたは全くなかった(図1 E〜I)。ユビキチンもアクロソームにおいて検出された(図1J)。同じプロテアソームサブユニットが、雄ブタ精子頭部および雄ブタ精子全体の抽出液中でウェスタンブロットによってさらに検出された(図2 A〜C)。透明帯透過性におけるプロテアソームのこの新規役割に一致して、マウスプロテアソームサブユニットMECL-1のAA配列(AA 218〜236)の組換えペプチドに対する抗体は低用量で、透明帯無傷卵母細胞における受精を確実に遮断したが、透明帯を含まない卵母細胞では遮断しなかった。
【0125】
実施例4
ユビキチン化タンパク質のブタ透明帯の外側面における存在
プロテアソーム阻害剤が透明帯侵入を遮断し、プロテアソームサブユニットおよびユビキチン経路の成分が共にアクロソームに存在する場合には(図1 E〜J)、ホヤ卵黄膜に見られるように(Sawadaら, 2002a)ユビキチン化感受性タンパク質/精子受容体はZPの外側層に存在するはずであり、または精子アクロソーム滲出液と接触する前にすでにユビキチン化されている、タンパク質/精子受容体がZPに存在するはずである。組換えヒトユビキチンに対する抗体KM 691を使用して、ブタ卵巣組織のパラフィン切片をスクリーニングし(図3 A〜D、F、G)、卵巣の胞状卵胞から単離したブタCOCの全載試料をスクリーニングした(図3E;図6A、Bも参照されたい)。
【0126】
ユビキチン交差反応性タンパク質の別個の濃密な層が、卵巣組織切片(図3 A〜D、E、F)および全載試料(図3E、図6A、B O)の前胞状および胞状卵胞のブタ卵母細胞の透明帯の外側層で検出された。これらの(ユビキチン免疫反応性)タンパク質の最も顕著な特徴は、それらが主にZPの外側面で検出されたということであり、これは、主要なZP成分、ZP1、2および3がほ乳類の透明帯に均一に分布することと異なる(ThalerおよびCardullo, 2002;WassarmanおよびLitscher 1995)。同様に、透明帯糖タンパク質に存在する糖残基はZPの内側面に存在するか、またはZPに均一に分散する(Avilesら, 1997;2000)。ユビキチン免疫反応性のこの局在化は、抗ユビキチン抗体のZPを通過する透過性が制限されているからではない。その理由は、このようなデータは組織切片の表面染色によって得られたからである。さらに、関連のないポリクローナル抗ユビキチン抗体AB1690もZPの外側面のユビキチン化タンパク質のこの層を認識し(図3F)、このような免疫反応性は、KM691を含有せず、免疫前ウサギ血清と共にインキュベーションし、次に適当な蛍光を結合した免疫グロブリンと共にインキュベーションした陰性対照では見られなかった(図3G)。
【0127】
次に、ユビキチン免疫反応性ZP基質の一部は、精子がZPに結合後分解を受けるかどうかおよびこのような分解はプロテアソーム阻害剤によって妨害されうるかどうかを判定した。卵母細胞はMG132の存在下/非存在下において受精し、精子注入後6時間経過時に回収し、レーンあたりの総数350の透明帯無傷(ZIまたはプロテアソーム+)または透明帯を含まない(ZFまたはZP-)をPAGEゲルにロードした。精子注入後6時間経過時のZF(図4A、レーン1)およびZI(図4A、レーン2)、受精後の卵母細胞の比較は、ZPが寄与する数多くのユビキチン免疫反応性バンドを示す。興味深いことに、123 kDaマーカーのわずかに下方に泳動するレーンセットはZP+/MG132-対照卵母細胞にはないが、ZP+MG132+卵母細胞には存在した(図4A、レーン3)。このようなバンドはヒト(図4A、レーン4)および雄ブタ(図4A、レーン5)精子の対照溶解液には存在しなかった。驚くべきことに、タンパク質移動後の卵母細胞ゲルのクーマシーブルー染色(図4A;右のパネル)は、このようなバンドの一部はゲルから膜に完全に移動しなかったことを示した。ユビキチンブロットと同様に、このようなバンドはZF卵母細胞レーンには全く見られず、ZP+/MG132-レーンでは劇的に低下した。ZP+/MG132+に存在するこのような大量のこのようなタンパク質は、このようなタンパク質はMG132が存在しない場合には対照受精中に消化されること、およびこの分解はMG132を受精培地に添加することによって効果的に妨害されることを示唆している。ポリ-ユビキチンなどのタンパク質の親水性AA残基含量である、高リジンがこの影響に寄与している可能性があるが、このようなタンパク質の一部があまり移動しない理由は不明である。
【0128】
接合体の総タンパク質含量に注目した上記の観察を、タンパク質移動前のゲルのクーマシーブルー染色によって評価した(図4B)。図4Aに示すゲルおよび膜と同様に、123 kDaの下方に泳動するバンドセットはZP+/MG132-接合体(レーン2)では劇的に低下しており、ZP+/MG132+接合体(レーン3)では優勢であり、ZP-/MG132+(レーン4)接合体、ZP-/MG132-接合体(示していない)および対照精子抽出液(レーン4および5)では見られなかった。
【0129】
その後の繰り返しでは(図4C;左のパネル)、ZP+未受精の卵母細胞のタンパク質(レーン2)、ZP+/MG132-接合体(レーン3)、ZP+/MG132+卵母細胞(レーン4)およびヒト(レーン5)および雄ブタ(レーン6)精子は分離し、移動した。膜を抗ユビキチンKM-691で探索し(図4C;左)、タンパク質移動後ゲルをクーマシーブルーで染色し(図4C;右)、ゲルと抗ユビキチン探索膜を重ねた(図4C;中央)。123 kDaマーカーの下方のバンドセットの移動は再度妨害され、ZP+/MG132+レーンにシングルバンドが存在し、M-IIおよびZP+/MG132-レーンには密度の低い、低MWバンドが見られた。ブロット膜では、約30 kDaの新たなユビキチン免疫反応性バンド(図3D、矢印)が、梯子上の密度の低いバンド(矢印)以外にZP+/MG132-接合体に出現したが、M-II卵母細胞またはZP+/MG132+接合体では出現しなかった。従って、このバンドは、受精後に生じ、MG132によって妨害されるタンパク質分解産物であった可能性がある。以前の試験と同様に、123 kDaマーカーの下方のバンドセットは受精後のZP+/MG132-対照接合体では低下していたが、M-II卵母細胞およびZP+/MG132+接合体では低下していなかった(図4E)。最後に、MG132が存在しない場合に、IVF後の約179 kDaレベルの2つのバンドの密度がわずかではあるが、識別できるほど低下した(図4F;矢印)。全体で、4つの試験を実施し、同様の結果が得られた。ZP+/MG132+およびZP+MG132-接合体は全ての試験に含め、対照卵母細胞レーンは異なった(M-II/ZP+;M-II/ZP-;IVF/ZP-/MG132+;IVF/ZP-/MG132-)。精子-卵母細胞結合は、ウェスタンブロットに使用した接合体の各バッチにおいて光学顕微鏡によって調査し、ZP+/MG132+接合体では、見かけ上正常で、MG132-処理に影響されなかった。従って、いくつかのZP-寄与卵母細胞タンパク質の分解はIVF後に生じ、選択的プロテアソーム阻害剤MG132によって妨害された。
【0130】
雄ブタ精子の受精能力は、精子の移動性または卵母細胞の細胞質膜と融合する精子の能力に対するプロテアソーム阻害剤の非特異的な影響によって低下されないことを確認するために、ブタ卵母細胞からZPを除去し、MG132の存在下または非存在下においてアクロソーム無傷精子を受精させた(図5 A〜H)。予測されたように、MG132は透明帯無傷卵母細胞の受精を阻害したが、透明帯を含まない卵母細胞には影響せず、その92%〜96%は受精し、そのほとんどは多精受精的であった(図5 A〜H)。卵母細胞または精子がMG132と共に事前にインキュベーションされ、十分に洗浄してから精子注入に使用すると、MG132は透明帯無傷卵母細胞の受精に影響を与えなかった。まとめると、これらのデータは、プロテアソーム阻害剤は、精子の移動性、精子-透明帯結合およびアクロソームエキソサイトーシスに影響を与えないで、雄ブタ精子による透明帯透過性を遮断することを納得させる証拠を提供する。
【0131】
次に、精子注入後6時間経過時の接合体の免疫蛍光および微分干渉コントラスト顕微鏡(DIC)画像によって、精子-アクロソームプロテアソームとユビキチン免疫反応性ZPタンパク質の相互作用を可視化することを試みた。ユビキチン標識のメッシュ様パターンが受精後の接合体のZP表面に見られ、ZP結合精子の下方および周囲の蛍光が低下していた(図6A)。後者の図では(図6 B〜B'')、ZP結合精子の下方のユビキチン免疫反応性層は消化されたと思われ(図6B';矢印)、精子頭部下方に空間を生じ、DIC光学的方法では明らかに識別可能である(図6B'';矢印)。無傷の精子アクロソームにおいて以前に検出されたプロテアソームサブユニットMECL-1、α/β-型サブユニットおよびβ1i(図1)は、無傷のアクロソームを有するZP結合精子(図6C、E、F;矢印)、エキソサイトーシスを受けたアクロソーム(図6C、F、G;矢印)および除去されたアクロソーム被覆(rejected acrosomal shrouds)(図6C、E;矢印)においても見られた。体細胞核のプロテアソームサブユニットおよびユビキチンシステムの活性の存在に一致して(Chenら, 2002;Rivett, 1998)、α-型およびβ-型のプロテアソームサブユニットが受精後の接合体内部の前核において検出された(図6D)。
【0132】
同じプロテアソームサブユニットは、アクロソームエキソサイトーシスおよび小胞形成を受けたZP結合精子のアクロソーム基質(図7 A〜C)および内側のアクロソーム膜(図7 D〜G)においてコロイド金TEMによって検出された(微細構造を図7 I〜Kに示す)。陽性対照として、抗c-Yes抗体(免疫蛍光の図1A、Bを参照されたい)は、主に外側アクロソーム膜および外側アクロソーム核周囲卵胞膜(perinuclear theca)の複合体を認識した(図7H)。
【0133】
実施例5
プロテアソーム阻害剤MG132および抗プロテアソーム血清MECL1によるインビトロにおけるウシ受精の阻害
雄ウシ精子がウシ卵子に受精する能力に対するプロテアソーム干渉の影響をインビトロにおいて分析した。特異的で、完全に可逆的なプロテアソーム阻害剤MG132(100μM)を、精子注入時に受精培地に添加した。これにより、精子注入後20時間経過時に調査した103の卵巣のうち3つにだけ受精が生じた(受精率2.8%;表2)。10μM MG132を使用した場合にはプロテアソーム干渉はウシ受精を一部遮断したが、100μM Mg132を適用した場合には完全に遮断することを分析は示した。MG132の溶媒である適量のエタノールを添加しても、受精または前核形成に影響を与えず、MG132の非存在下でのIVFの前の2時間の100μM MG132との未受精卵のプレインキュベーションも影響を与えなかった。同様に、プロテアソームサブユニットMECL1に対する免疫血清はウシIVFを遮断したが、適当な免疫前血清はしなかった。前核中のプロテアソームを、α-型およびβ-型プロテアソームサブユニットに対する抗体を用いて検出した。DNAはDAPIで可視化した(青色)。
【0134】
MG132の溶媒溶液である適当な量の100%エタノールの存在下において精子注入した卵は受精率85.9%を示した。ラクタシスチンなどの非可逆的なプロテアソーム阻害剤とは異なり、MG132はプロテアソームコアに永久的に結合しない。しかし、MG132は、精子アクロソーム媒介性プロテアソームに対する阻害作用とは異なり(Sutovskyら, 2003a、b)、透明帯卵細胞膜または卵質に対する直接作用によって受精を遮断または妨害することを排除するために、数群の卵を受精前に100μM MG132と共に2時間プレインキュベーションした。MG132の非存在下で受精させた、このようなプレインキュベーション卵において受精率97.5%が観察された(表2)。ブタIVFの上記の検討と同様に、20SプロテアソームコアサブユニットMECL1に対するウサギ血清は、1/200、1/500および場合によっては、1/1000の希釈時において、インビトロにおけるウシ卵の受精を効果的に遮断した(それぞれ、受精率15.4%、3.7%および32.1%;表1)。このような影響は、対応する希釈において免疫前のウサギ血清によっては誘発されなかった。
【0135】
MG132も抗-MECL1処理も、卵母細胞の形態の同定可能な異常を伴わなかった。これは、雄ウシ精子のプロテアソームサブユニットMECL1、LMP-2並びにα-型およびβ-型に実施された免疫局在化によって同定された。陰性対照は、適当な合成ペプチドで免疫飽和した抗MECL1抗体を使用して実施した。同じ細胞の微分干渉コントラスト画像を作製した。上記の観察は、上記の雄ブタの精子-アクロソームプロテアソームについて同定された局在化および機能並びに雄ウシ精子のMECL1、LMP2およびα/βのプロテアソームサブユニットの免疫局在化に一致した。
【0136】
(表2)プロテアソーム阻害剤MG132および抗プロテアソーム血清MECL1のインビトロにおけるウシ受精および前核形成の阻害
【0137】
実施例6
プロテアソームの接合体形成開始時からの前核への浸潤
ユビキチン-プロテアソーム経路は、最近まで、細胞質コンパートメントだけに存在するとされていた。体細胞の核コンパートメントにおけるプロテアソームの存在およびタンパク質分解作用が分析された。種々の前核発生段階のウシ前核接合体のプロテアソームサブユニットLMP2、α/βおよびMECL1の免疫局在化に特異的な免疫血清を使用した。卵質への導入直後、十分に凝縮された精子核は検出可能なプロテアソームを含まないことが示された。プロテアソームサブユニットは、精子核脱凝縮の開始段階において雄および雌の前核に浸潤を開始した。雄および雌の前核は、フルサイズおよび並置に達する前にプロテアソームに占められる。高密度プロテアソームが、単精受精および多精受精卵のフルサイズの並置された前核内並びに非同期的な前核発生の場合に観察された。受精前に卵を100μM MG132とプレインキュベーションしても、プロテアソームサブユニットの前核への流入に影響を与えず、精子および100μM MG132の受精培地への同時添加の20時間後に未受精のままであった卵ではプロテアソームと雌染色体の結合は観察されなかった。合成MECL1ペプチドを飽和させた抗MECL1抗体を使用する陰性対照は任意のシグナルを示さなかった。精子尾部は、受精前の精子を生体ミトコンドリア染料MitoTracker Green FM(緑色)と共にプレインキュベーションすることによって可視化した。
【0138】
ウシ卵質における上記プロテアソームサブユニットの発現はウェスタンブロットで確認した。プロテアソームサブユニットMECL1は、ウシ卵(図8、レーン1)、雄ウシ精子(図8、レーン2)およびウシ卵丘細胞(図8、レーン3)の溶解液中で検出された。卵質溶解液の予測された29 kDaレーン以外に、種々の細胞種の細胞溶解液に通常見られる高MWレーンは、おそらく他のプロテアソームサブユニットからのMECL1サブユニットの不完全な解離の結果として、精子および卵丘細胞溶解液中に存在した。
【0139】
実施例7
プロテアソーム干渉の正常な前核形成の妨害および、受精卵における未成熟染色体凝縮の発生
ウシ受精を阻害するための最適なMG132濃度を決定することを目的とした検討の経過において、低濃度(10μM)では、MG132処理は受精を部分的にしか阻害しないことが観察された。2つの別個の試験で得られた平均受精率は、10μM 132で38.2%(n=55)で、対照(n=37)では75.7%であった。低濃度MG132の存在下におけるこのような受精により、前核形成に対するプロテアソーム干渉の影響を観察することができた。受精卵中の前核は、Nup 153を含む核膜孔複合体(NPC)タンパク質群および他の関連するヌクレオポリンを認識するモノクローナル抗体mAb 414の組み合わせによって可視化した。DNAは青色蛍光DAPIで染色し、受精卵内の精子尾部は、精子のMitoTracker GreenFMとのプレインキュベーション中に精子ミトコンドリア鞘に蓄積する緑色蛍光の存在によって同定した。ヌクレオポリン標識は、ウシ(図12A;Sutovskyら, 1998)およびマウス(FitzHarrisら, 2003)の前核/接合体の早期形成段階における核エンベロープ強度および機能の適当なマーカーであるので、選択した。
【0140】
異常な前核形成および未成熟染色体凝縮(PCC)が、精子の透過性を一部可能にする条件である、10μM MG132の存在下で受精したウシ卵に観察された。前核は、核膜孔複合体(NPC;赤)タンパク質、ヌクレオポリンNup153に対するモノクローナル抗体およびDNA染料DAPI(青)の組み合わせで可視化した。同様に、このような卵におけるPN形成パターンは異常であった。未成熟染色体凝縮(PCC)および前核並置失敗の組み合わせは、単精受精および多精受精卵の最も一般的な異常であった。いくつかの場合において、精子核の不完全な脱凝縮が、雌クロマチンのPCCを伴って観察された。PCC中の染色体の散乱および凝縮されたクロマチン周囲への可溶性であると推定されるヌクレオポリンの集成が観察された。このようなヌクレオポリンの蓄積は父系的な寄与によるとは思われない:雄ウシ精子は、精子尾部接続片の冗長な核エンベロープ(HoおよびSuarez, 2003)に位置する残存量のヌクレオポリン(Sutovskyら, 1999)を保有する。
【0141】
実施例8
正常な前核形成の妨害機序の分析
以前の検討において、NPCおよびヌクレオポリンの環状の層状様配置が、自発的な活性化へと進行する成熟した中期-II卵において観察されることがあった。しかし、MG132処理した細胞質に接触した精子核も未成熟染色体凝縮を受けたことを考慮すべきである。プロテアソームの分解は精子クロマチンの最初の脱凝縮に必要でないが、雄前核の十分な形成に必要であることをこれは示唆している。このような影響は、接合体内の母系的および父系的クロマチンのプロテアソーム依存的なリモデリングによって直接的であっても、またはプロテアソーム依存的な細胞周期進行の遮断などの間接的であってもよい。減数分裂および前核形成の進行はラットではMG132によって阻害されると思われる(Josefsbergら, 2000)。この観察は、クローニングされた最初のラット子孫を作製するために使用され、研究者らはラット卵の未成熟な活性化を防ぐためにMG132を適用した(Zhouら, 2003)。
【0142】
PN並置の失敗は、精子-星状マイクロチューブが前核並置中に固定される、無傷の核エンベロープがないこと、または精子中心小体が、接合体中心小体再構成および精子星状形成の前に遊離されなければならない、精子尾部接続片におけるプロテアソーム機能の遮断によると思われる(Sutovskyら, 1996;SutovskyおよびSchatten, 1997;Wojcikら, 2000)。
【0143】
実施例9
胚移植のための胚盤胞を作製するためのブタインビトロにおける受精中の多精受精の低下
プロテアソーム干渉を使用して、全体的な受精率を低下させないで、IVF中に多精受精率を軽減または低下させることができる。その後、多精受精接合体由来の生存能力のない倍数体胚盤胞の数を低下させることによって、胚移植プログラムのための正常な2倍体胚盤胞の作製を増加させることができる受精媒体への新規添加剤を製剤化することができる。初期の検討では(表3)、ZPの透過性は、100 uM濃度の特異的なプロテアソーム阻害剤MG-132およびラクタシスチンによって完全に阻害された。閾値濃度10 uMでは、プロテアソーム阻害剤ラクタシスチンは受精を妨害しなかったが、多精受精率を10%まで低下すると同時に単精受精率を65%まで増加した。精子注入後20時間経過時の接合体内の蛍光標識された精子尾部の数によって評価したとき、この検討における対照受精は、多精受精および単精受精が、それぞれ、44%および38%であった。
【0144】
(表3)精子注入時の培養培地への10 uMラクタシスチンの添加によるブタIVF中の多精受精の低下
【0145】
本明細書に開示し、特許請求の範囲に記載する組成物および/または方法は全て、本発明の開示内容に照らして不要な実験を行うことなく作製することができ、実行することができる。本発明の組成物および方法は、好ましい態様に関して記載されているが、本発明の概念、精神および範囲から逸脱することなく、本発明の組成物および/または方法並びに本明細書に記載されている方法の段階および段階の順序に変更を適用することができることが当業者に明らかである。さらに具体的には、化学的および生理学的に関連するある一定の物質を本明細書に記載する物質と交換することができるが、同じまたは同様の結果が得られると思われることが明らかである。当業者に明らかな全てのこのような同様の置換および改良は、添付の特許請求の範囲によって規定される本発明の精神、範囲および概念の範囲内であると考えられる。
【0146】
参照文献
以下の参照文献は、本明細書に記載するものに補助的な例示的な手法または他の詳細を提供する程度に、参照として具体的に本明細書に組み入れられている。
【図面の簡単な説明】
【0147】
以下の図面は本明細書の一部を形成し、本発明のある局面をさらに証明するために含まれる。本発明は、本明細書に提供する具体的な態様の詳細な説明と合わせて、これらの図面の1つ以上を参照することによってさらに理解することができる。
【図1】図1A〜1Jは、プロテアソーム阻害剤処理した卵母細胞における精子-ZP結合の一例およびブタ配偶子におけるユビキチン-プロテアソーム系の成分を例示する。(図1A)常在性チロシンキナーゼc-Yesに対する抗体で標識した無傷のアクロソームを有する雄ブタ精子。(図1B)。表面帯(zona surface)へのアクロソームの分散後のc-Yes標識の残遺物。(図1C)10 μM MG132(プロテアソーム阻害剤)の存在下において受精した卵母細胞は、精子注入後(p.i.)20時間経過時に無傷のアクロソームを有するわずかに3つの精子を示す。これは、MG132は特異性高く受精を遮断するが、配偶子の生存性、精子-卵子結合および精子-アクロソーム反応自体を損なわないことを例示している。(図1D)精子注入後(p.i.)20時間経過時に3つのアクロソーム無傷精子を有する対照卵母細胞。図1 A〜Dの精子は生体ミトコンドリア染色色素MitoTracker CMTM Rosで事前に標識した。(図1E〜I)サブユニットLMP-2、MECL 1およびMECL 2に対する抗体並びにプロテアソームコアサブユニットに対する2つの異なる抗体による雄ブタ精子プロテアソームの検出。(図1J)Ab 1690によって検出されるユビキチン。
【図2】図2A〜2Cは、雄ブタ熱抽出(すなわち、アクロソームなし;レーン2)、TX-100抽出(レーン3)および無傷(レーン4)精子頭部の溶解物におけるプロテアソームサブユニットMECL-1(図2A)、β1i(図2B)並びに種々のα-型およびβ-型サブユニット(図2C)の免疫検出の一例を例示する。分子量マーカーはレーン1に示す。
【図3】図3A〜3Gは、ブタ透明帯の外側面のユビキチン免疫反応タンパク質の一例を例示する。ユビキチンは、ブタ前胞状卵胞(図3A、図3B、図3F、図3G)および胞状卵胞(図3C、図3D)内側並びに全載成熟卵母細胞表面(図3E、卵核胞段階の卵母細胞を示す)においてマウスモノクローナル抗体KM691によって(図3A〜D、図3F、図3E)またはウサギ血清AB1690(図3E;図3F)によって検出された。これらのパラフィン切片のいくつか(図3A、図3B)の卵胞顆粒膜細胞を、ミトコンドリアタンパク質プロヒビチンに対する抗体で対比染色した。図3Eでは、Ab1690による標識はこの卵母細胞の卵核胞内の核小体の輪郭を描き、表面帯の標識は抗体に対するZPの不透過性によるものではなかったことを示す。ユビキチン免疫反応性は、KM691を使用せず、免疫前ウサギ血清と共にインキュベーションし、次に蛍光的に結合した適当な免疫グロブリンと共にインキュベーションすることによって処理した陰性対照組織切片では見られない(図3G)。全ての画像のDNAはDAPIで対比染色した。
【図4】図4A〜4Fは、プロテアソーム阻害剤MG132が存在する場合または存在しない場合における受精前および受精後の透明帯無傷および透明帯除去ブタ卵母細胞の溶解物におけるユビキチン基質の免疫検出の一例を例示する。(図4A)左のパネル:抗-ユビキチンKM691で調査した、受精後6時間経過時の透明帯除去(レーン1)および透明帯無傷(レーン2)の受精卵母細胞の溶解物は、ZPによる数多くのユビキチン免疫反応バンドを示す。123 kDaマーカーのわずかに下に泳動するレーンセット(矢印)はZP+/MG132-対照卵母細胞には存在しないが、ZP+MG132+卵母細胞(レーン3)には存在する。このようなバンドは、ヒト(レーン4)および雄ブタ(レーン5)精子の対照溶解物には存在しない。右のパネル:タンパク質移動後の同じ卵母細胞ゲルのクーマシーブルー染色は、このような<123 kDaバンド(矢印)のいくつかはゲルから膜に完全に移動せず、ZP-卵母細胞レーン(レーン1)では存在せず、ZP+/MG132-レーン(レーン2)では低下していることを示している。存在するこのようなタンパク質の大部分はZP+/MG132+レーン(レーン3)に存在する。(図4B)タンパク質移動前のゲルのクーマシーブルー染色。(図4C)左のパネル:抗-ユビキチンKM-691で調査した、ZP+未受精卵母細胞(レーン2)、ZP+/MG132-接合体(レーン3)、ZP+/MG132+卵母細胞(レーン4)並びにヒト(レーン5)および雄ブタ(レーン6)精子のタンパク質。中央:右のパネルに示すように、移動後にクーマシーブルーで染色したゲルを左のパネルに重ねたもの。
【図5】図5A〜5Hは、プロテアソーム阻害剤による透明帯の透過性の遮断および透明帯除去卵母細胞ではこのような遮断が存在しないことの一例を例示する。MG132は透明帯無傷卵母細胞ではインビトロにおいて受精を阻害したが(図5C、図5D)、透明帯切除卵母細胞には影響を与えなかった(図5A、図5B)。適当な透明帯除去(図5E、図5F)および透明帯無傷(図5G、5H)対照を実施した。DNAをDAPIで対比染色し、受精卵母細胞内の前核および核小体を抗体Ab 1690で標識した。
【図6】精子注入後(p. i.)6時間経過時のZP表面の雄ブタ精子の微分干渉顕微鏡(DIC)画像と合わせて抗-ユビキチンおよび抗-プロテアソーム免疫蛍光によって可視化した、精子プロテアソームとユビキチン免疫反応性ZPタンパク質の相互作用の一例を例示する。
【図7】図7A〜7Kは、アクロソームエキソサイトーシス中のプロテアソームサブユニットの免疫電子顕微鏡検出の一例を例示する。(図7A〜G)サブユニットβ1i(図7A〜D)、α/β(図7E、F)およびMECL-1(図7G)のコロイド金標識はアクロソーム基質内および内側のアクロソーム膜に存在する。(図7H)アクロソームマーカー、チロシンキナーゼc-イエス(yes)は、主に外側のアクロソーム膜および外側-アクロソーム核周囲卵胞膜の複合体に限定される。(図7I〜K)従来の電子顕微鏡は、ブタ受精中のアクロソームエキソサイトーシスの超微細構造を示す。
【図8】100のウシ卵子(レーン1)、雄ウシ精子抽出物(レーン2)およびウシ卵丘細胞(レーン3)におけるプロテアソームサブユニットMECL1のウェスタンブロット法である。星印は予測される29 kDaレーンを表す。種々の細胞種の細胞溶解物に共通して見られる高MWレーン(参照文献 Groettrupら,1996)は、試料調製過程における他のプロテアソームサブユニット凝集の結果である。〜32 kDaのバンドは、未処理MECL-1前駆体タンパク質に相当する。
【技術分野】
【0001】
A. 発明の分野
本発明は、一般に、繁殖生物学、薬物開発、免疫学および分子生物学の分野に関する。より具体的には、本発明は、受精能の調整および評価または不妊の診断のための、ユビキチン-プロテアソーム経路およびユビキチン-プロテアソーム経路の成分由来の抗原の阻害剤に関する。なお、本願は、全体の開示内容が具体的に参照として本明細書に組み入れられている、2003年2月14日提出の米国仮特許出願第60/447,675号の優先権を主張する。政府は、米国農務省からの付与番号1999-3520-11743に準じて本発明に権利を有しうる。
【背景技術】
【0002】
B. 関連技術の説明
ユビキチン化は広範で、進化的に保存されているが、真核細胞におけるタンパク質分解の基質特異性が高い経路である(Pickart, 1998;LaneyおよびHochstrasser, 1999)。ユビキチンによる翻訳後タンパク質修飾は、ユビキチンのC-末端グリシン(Gly)残基によって76アミノ酸ユビキチン残基が基質アミノ酸配列内のリジン(Lys)残基に共有結合することによって生ずる。ユビキチン分子は自身の7つのLys-残基を保有するので、この最初のモノ-ユビキチン化の次に追加のユビキチン分子が第1の基質-結合ユビキチンモノマーにGly-Lys結合することが多く、ジ-、トリ-テトラ-またはポリ-ユビキチン鎖の形成を誘導する(HershkoおよびCiechanover, 1998)。モノ-、ジ-およびトリ-ユビキチン化は、リソソームタンパク質分解、膜受容体のエンドサイトーシス、信号伝達および転写調節を含むが、これらに限定されない種々の目的を果たす(Conawayら, 2002;GlickmanおよびCiechanover, 2002により総説が報告されている)。テトラおよびポリ-ユビキチン化は、ユビキチン化タンパク質に特異的な活性を有するマルチ-サブユニットプロテアーゼである26-Sプロテアソームによるタンパク質ドッキングおよび分解のコンセンサスシグナルである(TanakaおよびTsurumi, 1997;Hochstrasser, 2002)。バレル型プロテアソームのルーメン内では、基質タンパク質が小ペプチドに切断されて細胞質に放出され、細胞質エンドペプチダーゼセットによりタンパク質分解が完了する(Hochstrasser, 2002;Kohlerら, 2001によって総説が示されている)。遊離されたポリ-ユビキチン鎖はプロテアソームによって分解されるのではなく、細胞質に戻り、他のタンパク質の修飾に再使用される(Chenら, 2002;Vogesら, 1999)。
【0003】
種々の基質のユビキチン化およびプロテアソーム分解は細胞質および核内で実証されている(GlickmanおよびCiechanover, 2002)。興味深いことに、ほ乳類の生殖の研究は、ユビキチン化は細胞表面および細胞外空間でも生じうるという証拠をもたらした。ユビキチンは、卵巣の卵胞液(Einspanierら, 1993)、精漿(Lippertら, 1993)、精巣上体液(HermoおよびJacks, 2002;Sutovskyら, 2001b)および精巣上体を通過する欠陥精子の表面(Sutovskyら, 2001a, b;2002, 2003)において検出された。卵子の卵黄(viteline)膜表面の精子受容体のユビキチン化およびプロテアソーム分解はホヤの受精中に生じる(Sawadaら, 2002a, b)。
【0004】
ほ乳類の精子が透明帯(「帯」またはZP)に結合して通過する能力は、ZP基質のZP3-精子受容体への結合の結果、エキソサイトーシス(Gerton, 2002)を受ける精子先体帽に存在する精子表面受容体およびタンパク質分解トリプシン様酵素セット(PrimakoffおよびMyles, 2002によって総説が報告されている)に帰されている(WassarmanおよびLitscher, 1995)。いくつかの候補先体タンパク質および精子表面受容体の標的突然変異およびノックアウトは、マウス精子がZPに結合し、透過/消化する能力を排除しなかった(Talbotら, 2003;Evans, 2001によって総説が報告されている)。セリンプロテアーゼ以外のタンパク質分解系が精子先体に存在し、受精中のZPの消化に寄与することをこれは示唆している。
【0005】
現在の避妊方法には、米国で最も一般的な避妊方法である女性の外科的不妊化および女性の経口避妊薬の使用が挙げられる。ホルモン調節避妊薬には、配合注射用避妊薬、配合経口避妊薬などの配合(エストロゲン/プロゲスチン)避妊薬;およびノルプラントインプラント、プロゲスチン-単独注射用避妊薬またはプロゲスチン-単独ピルなどのプロゲスチン-単独避妊薬が挙げられる。しかし、配合エストロゲン/プロゲスチン経口避妊薬は、血栓塞栓障害、脳血管事故、冠動脈疾患、肝異常、エストロゲン依存性の癌および妊娠を生じるまたはそれらに至ることがある。他の避妊方法には、子宮内避妊器具;子宮頸管キャップ;男性のコンドーム、女性のコンドーム、ペッサリー、殺精子剤または避妊用スポンジなどの遮断方法;および関与する個人への依存性が高い周期避妊法が挙げられる。これらの方法の多くは、生理的および物理的に広範囲の副作用を有することがある。従って、ヒト並びに種々の家畜および野生動物の受精を調節する改善された方法および受精を診断する方法の必要性がある。
【発明の開示】
【0006】
発明の概要
本発明は、ユビキチン-プロテアソーム経路の成分を阻害または検出することによって、ヒトまたは動物、特にほ乳類の受精を調節または評価する方法を含む。本発明の種々の態様において、避妊方法は精子のプロテアソーム活性を阻害する段階を含む。本明細書において使用する「避妊薬」は、投与したとき、意図的に、直接または間接的に受精または妊娠を妨害する薬剤を意味する。同様に、本明細書において使用する「避妊」は、直接または間接的に受精または妊娠を意図的に妨害することを意味する。本発明の目的のためには、プロテアソーム経路の成分に対する免疫応答を誘導する組成物を含む、プロテアソーム経路の成分の活性を阻害する薬剤は避妊薬と考えることができる。いくつかの態様において、精子のプロテアソーム活性は、インビトロ、エクスビボまたはインビボにおいて阻害することができる。プロテアソーム経路の阻害剤には、ユビキチン-プロテアソーム経路の種々の成分に結合する小分子、ペプチド、ポリペプチド(例えば、抗体等)および親和性物質(例えば、アプタマー等)が挙げられるが、これらに限定されない。いくつかの態様において、ユビキチン-プロテアソーム経路の活性またはユビキチン-プロテアソーム経路の成分の活性は、1つ以上のポリペプチドに結合し、その活性を阻害する抗体によって阻害される。ユビキチン-プロテアソーム経路に関連する種々のポリペプチドの説明は、国立バイオテクノロジー情報センター(National Center for Biotechnology Information(NCBI))が管理するヒト遺伝子変異と遺伝子疾患に関わる情報の統合データベース(Online Mendelian Inheritance in Man(OMIM)データベース)において見つけることができる。
【0007】
ある態様において、方法は、ユビキチン-プロテアソーム経路のタンパク質成分由来の少なくとも1つのペプチドまたは抗原で被験者または動物を免疫化することによって抗体を誘導する段階を含む。抗原は、プロテアソームサブユニット、ユビキチン、ユビキチン関連酵素またはプロテアソーム経路に関連するポリペプチド由来のペプチドであってもよい。特定の態様において、ペプチドは20Sプロテアソームコアのα-型および/またはβ-型サブユニット由来またはある種のプロテアソームに存在する19S調節キャップ(regulatory cap)もしくは11Sアクチベーター複合体のサブユニット由来である。本発明の1つ以上のペプチドまたは抗原はワクチン組成物に含まれてもよい。本発明のペプチドは、その例が配列番号:1、2、4および6に記載されているアミノ酸配列を有するプロテアソーム-由来のペプチドであってもよい。本発明のペプチドまたは抗原は融合タンパク質に関連してもよい。いくつかの態様において、本発明のペプチドまたは抗原は、種々のポリペプチド由来であってもよい多重抗原エピトープを単一ポリペプチドまたは組成物に含んでもよい。ある態様において、抗体は、プロテアソーム経路の成分由来の少なくとも1つのペプチドまたは抗原をコードする少なくとも1つのポリヌクレオチドで免疫化することによって誘導される。少なくとも1つのペプチドまたは抗原をコードするポリヌクレオチドは、プラスミド、鎖状発現要素またはウイルス発現ベクターなどの発現ベクターに含まれてもよい。特定の態様において、ポリヌクレオチドがコードするペプチドまたは抗原は、α-型またはβ-型プロテアソームサブユニットポリペプチドなどのプロテアソームタンパク質由来であってもよい。
【0008】
本発明の一態様において、プロテアソームの活性は、精子もしくは卵母細胞または両方にプロテアソーム阻害剤を接触させることによって阻害される。プロテアソーム阻害剤は薬学的に許容されうる製剤に含まれてもよい。ある態様において、プロテアソーム阻害剤またはプロテアソーム阻害剤組成物はMG132および/またはラクタシスチンを含む。薬学的に許容されうる製剤は、溶液、懸濁液、錠剤、ピル、カプセル、徐放性製剤、粉末、クリーム、軟膏(ointment)、軟膏(salve)、スプレー、ポンプ、リポソーム、坐剤、吸入剤、パッチまたは他の送達系に製剤化されてもよい。プロテアソーム経路の活性を阻害する組成物は、静脈内、腹腔内、皮内、筋肉内、子宮内、皮膚、鼻、口腔、膣、吸入または局所的投与経路を含むが、これらに限定されない経路によって投与することができる。ある態様において、投与経路は子宮内または膣である。
【0009】
本発明の種々の態様において、ほ乳類において抗プロテアソーム抗体を誘導する方法は、プロテアソーム経路の成分由来の1つ以上のペプチドまたは抗原をほ乳類に投与する段階を含む。本発明の1つ以上のペプチドまたは抗原を投与する段階は、ほ乳類のユビキチン-プロテアソーム経路に結合して、その活性を阻害する抗体の産生を誘導することができる。ある態様において、ほ乳類はヒト、家畜(farm animal)、家畜(domestic animal)、野生動物または害獣、例えば、ラット、スカンク等である。動物には、ブタ、ウシ、ウマ、ネコ、げっ歯類、ウサギ、シカ、アライグマ、オポッサム、スカンク等が挙げられるが、これらに限定されない。1つ以上のペプチドまたは抗原は、1つ以上のペプチドまたは抗原をコードするポリヌクレオチドを発現することによって投与することができる。ポリヌクレオチドは、筋肉内、静脈内、皮下、腹腔内、皮内、経口、吸入または他の既知の投与経路によって投与することができる。1つ以上のペプチドまたは抗原は、筋肉内、静脈内、皮下、腹腔内、皮内、経口、吸入または他の既知の投与経路によって投与することができる。
【0010】
本発明のいくつかの態様において、方法は、精子、卵母細胞または動物もしくはヒトの受精に寄与する他の生殖組織(例えば、精巣、精巣上体、卵巣、前立腺、精嚢および他の性付属腺)に関連するユビキチン-プロテアソーム経路の成分の有無を検出することによって不妊の診断および/または受精能の評価/推定を含む。方法は、ELISA、タンパク質ブロット法、免疫組織化学法等を含んでもよいが、これらに限定されない。
【0011】
本発明の方法は、インビトロにおける受精に使用する精液試料にプロテアソーム活性の阻害剤を添加する段階を含むインビトロにおける受精方法であって、阻害剤は、受精を妨害するには不十分であるが、多精受精を妨害するには十分な濃度が添加される方法をさらに提供する。本発明の方法では、受精の約25%未満が多精受精であってもよく、受精の10%未満を含む。阻害剤は、本明細書に記載する任意の方法によってインビトロまたはインビボにおいて添加することができる。任意の阻害剤を、本明細書に記載する方法に使用することができる。
【0012】
本明細書に記載する任意の方法または組成物は、本明細書に記載する任意の他の方法または組成物に関して実施することができることが考慮される。
【0013】
特許請求の範囲および/または明細書において「含む」という用語と共に使用する場合の「1つのもの」という用語の使用は「1つ」を意味することができるが、「1つ以上」、「少なくとも1つ」および「1つまたは1つ以上」の意味にも一致する。
【0014】
本発明の他の目的、特徴および利点は、以下の詳細な説明から明らかになる。しかし、本発明の精神および範囲内の種々の変更および改良は、この詳細な説明から当業者に明らかになるので、詳細な説明および具体的な実施例は、本発明の具体的な態様を示すが、例示のためだけに提供されていることが理解されるべきである。
【0015】
例示的な態様の説明
上記に記載するように、受精の調節、受精能の評価または不妊の診断の改善された方法が必要とされている。受精能を調節する際の大半の努力は、配偶子形成または局在化/移動性を阻害する化学的または機械的方法の使用に絞られている。受精能調節のための改善された組成物および方法が必要とされている。本明細書に記載する発明は、受精能を調節および診断する組成物および関連の方法を記載する。本発明は、受精能を調節するためのユビキチン-プロテアソーム経路の成分の阻害剤または結合剤を含む。本発明は、卵母細胞の透明帯の透過性はプロテアソームおよびプロテアソームに関連する経路に関与し、抗プロテアソーム抗体および阻害剤によって阻害することができるという驚くべき観察を利用している。本明細書に記載するように、本発明は、受精能および、いくつかの例においては多精受精をインビボおよびインビトロにおいて低下または阻害する改善された方法を提供する。
【0016】
本発明者らは、プロテアソーム阻害剤、特に化合物ラクタシスチンおよびMG132並びに、プロテアソームに結合してプロテアソーム活性を阻害するまたはユビキチン依存性タンパク質分解を遮断する抗体の受精能調節特性を同定した。ほ乳類のユビキチン-プロテアソーム経路の阻害は、精子による透明帯(ZP)の透過性を遮断する。本発明者らは、接合体の細胞質内の父系ミトコンドリアの分解を可逆的に遮断する試みの成功においてこれらの新規特性を観察した。父系ミトコンドリア分解を生じるためには、精子がZPを通過して、卵母細胞の細胞質に侵入直後に受精培地にプロテアソーム阻害剤を添加しなければならなかった。ZP透過性はセリンプロテアーゼ活性に関連すると考えられており、プロテアソーム阻害剤はセリンプロテアーゼの活性を遮断しないという事実からこの観察は予想されなかった(Fenteanyら,1995;Goldbergら, 1995)。さらに、本発明者らは、ユビキチンおよびプロテアソームサブユニット以外に、ほ乳類の精子アクロソームは、ユビキチン-基質結合に必要な結合酵素であるユビキチンキャリアE2を含有し、おそらく他の脱ユビキチン化、ユビキチン結合およびユビキチン活性化酵素も含有すると思われることを示した。
【0017】
本明細書に開示するように、本発明は、ほ乳類の受精およびZP透過性は精子アクロソームのプロテアソームのタンパク質分解作用に依存していることを実証している。精子が卵子表面を活発にユビキチン化すると思われるホヤとは異なり、ユビキチン化タンパク質は受精前または卵形成中にほ乳類の卵子ZPによって獲得されうる。このようなデータは、ほ乳類の受精の謎並びに新規避妊標的および不妊マーカーにおける重要な点を提供する。
【0018】
I. ユビキチン-プロテアソーム経路
本発明の組成物は、避妊薬としてまたは避妊方法において並びに受精能を診断する際に使用することができる。ユビキチン-プロテアソーム経路に関連するポリペプチドの阻害または検出は避妊またはプロテアソーム成分が無傷/欠損しているもしくは存在する/存在しない配偶子の検出にそれぞれ使用することができる。
【0019】
プロテアソームは、ATP依存的タンパク質分解経路の必須成分である非リソソームタンパク質分解の中心酵素複合体として広く認識されている。この経路は多くの律速酵素、転写調節因子および重要な調節タンパク質の迅速な分解を触媒する。この経路は、突然変異または翻訳後損傷によって生じる異常性の高いタンパク質の迅速な排除に必須であり、ほ乳類細胞のタンパク質の大部分の迅速でない分解の主要な役割を果たす。それはまた、高等な真核生物における抗原プロセシングに大きく関与している。
【0020】
26Sプロテアソームは、ユビキチン/ATP依存的タンパク質分解経路の主要酵素複合体である。触媒中心は、電子顕微鏡によって示されるように、バレル型構造の2000 kDa複合体である20Sプロテアソームによって形成される。触媒中心は、各々7つのサブユニット(サブユニットα1〜7、β1〜7)を含む4つの環から構成される。
【0021】
配列の類似性に基づいて考慮すると、全ての20Sプロテアソームサブユニット配列は、各グループが別個の構造的および機能的役割を有する2つのグループ、αおよびβに分類することができる。7つのα-サブユニットは20Sプロテアソームの外側リングを含み、7つのβ-サブユニットは20Sプロテアソームの内側リングを含む。各サブユニットはα-またはβ-リングの独自の位置に位置する。
【0022】
典型的な26S複合体は、20S複合体と19S調節複合体と呼ばれる別個の複合体に位置する25kDa〜10 kDaの分子量範囲の20を超える追加のタンパク質を含む。26S複合体は、基質特異性を決定し、タンパク質分解および存続度に必要な複数の酵素機能を提供する。サブユニット成分の系統的な分析は、ATP結合タンパク質の新規ファミリーに属するATPaseである少なくとも6つのメンバーと、ATPに結合する能力のないさらに別の15のサブユニット、20Sプロテアソームのタンパク質分解中心への挿入前にタンパク質基質の変性を担当すると考えられるイソペプチダーゼおよびいくつかの他のタンパク質を明らかにした。いくつかのプロテアソームにおいて、19S調節複合体は11Sアクチベーター複合体と置換されている。
【0023】
26S複合体はATPに結合し、ユビキチンと結合することによって分解の標的となったタンパク質の分解を担当する。ユビキチンは、上記のように、一連のユビキチン活性化(UBA)およびユビキチン結合(UBC)酵素、例えば、酵素E1、E2およびE3によって、標的のリジンのε-アミノ基とユビキチンのC-末端グリシン残基の間に形成されるイソペプチド結合によって標的タンパク質に結合する。ユビキチン活性化酵素および結合酵素は、ユビキチン分子を1つの酵素から次の酵素に移動させ、次に活性化されたユビキチン分子をE2酵素から標的タンパク質に移動させることによって連続的に作用する。次いで、モノ-ユビキチン化タンパク質が作用されて、同じ酵素が先の分子に追加のユビキチン分子を結合する。ユビキチン結合が継続し、高分子量タンパク質複合体が生じる。次いで、このポリ-ユビキチン化産物が26Sプロテアソームによる迅速な分解の標的となり、同時にイソペプチダーゼによって触媒されるユビキチンが再利用される。
【0024】
複数のペプチダーゼ活性を有する複合体として最初に記載されたにもかかわらず、その後の研究により、20Sプロテアソームの5つの活性:大きい疎水性残基の後方を切断するキモトリプシン様活性;塩基性残基の後方を切断するトリプシン様活性;酸性残基の後方を切断するポストグルタミルヒドロラーゼ;分岐鎖アミノ酸の後方を主に切断するもの;および小型の中性アミノ酸の後方を切断するものが規定された。
【0025】
ATPが存在しない場合に20Sプロテアソームと関連することができる第2のアクチベーターは11Sレギュレーターとして既知である。純粋なPA28アクチベーターは、約50%の相同性を有する2つの交互のサブユニット、PA28αおよびPA28βの複合体である。電子顕微鏡による研究により、PA28は、19S複合体のように、両方またはどちらかの末端においてα-リングに結合することによって20Sプロテアソームにキャップをするリング状粒子であることが示された。PA28は、MHCクラスI分子の免疫系に提示される抗原ペプチドのプロテアソーム-触媒産生を調節するという所見は、抗原プロセシングにおけるこのアクチベーターの細胞機能を示している。詳細な総説は、SchmidおよびBriand, 1997;Bogyoら, 1997を参照されたい。
【0026】
ある態様において、ユビキチン-プロテアソーム経路に関連する種々のポリペプチドおよびそれらの断片を使用することができる。例えば、ユビキチン-プロテアソーム経路関連ポリペプチドには、ヒト遺伝子変異と遺伝子疾患に関わる情報の統合データベース(Online Mendelian Inheritance in Man(OMIM)データベース)において同定されている化合物が挙げられるが、これらに限定されず、その例には、OMIM第600307プロテアソームサブユニット、β-型、6;Psmb6(Genbank アクセッション番号第 NM_002798);176844 プロテアソームサブユニット, α-型, 5; Psma5 (Genbank アクセッション番号第 NM_002790) ; 176844 プロテアソームサブユニット、α-型、5;Psma5(Genbankアクセッション番号第 NM_002790);177045 プロテアソームサブユニット, β-型,9 ; Psmb9 (Genbank アクセッション番号第 NM_002800 および NM_148954) ; 604030 プロテアソームサブユニット、 β-型、7 ; Psmb7 (Genbank アクセッション番号第 NM_002799) ; 600654 プロテアソーム アクチベーター サブユニット 1 ; Psmel (Genbank アクセッション番号第 NM_006263) ; 606223 プロテアソーム 26S サブユニット、 非ATPase、 2; Psmd2 (Genbank アクセッション番号第 NM_002808) ; 176847 プロテアソームサブユニット、β-型、 10; PsmblO (Genbank アクセッション番号第 NM_002801) ; 177046 プロテアソームサブユニット、 β-型、 8; Psmb8 (Genbank アクセッション番号第 NM_004159 および NM_148919) ; 602855 プロテアソームサブユニット、α-型、 6; Psma6 (Genbank アクセッション番号第 NM_002791) ; 602854 プロテアソームサブユニット, α-型、 1; Psmal (Genbank アクセッション番号第 NM_002786 およびNM_ 148976) ; 602161 プロテアソーム アクチベーター サブユニット 2; Psme2 (Genbank アクセッション番号第 NM_002818) ; 600306 プロテアソームサブユニット、 β-型、 5; Psmb5 (Genbank アクセッション番号第 NM_002797) ; 605129 プロテアソーム アクチベーター サブユニット 3; Psme3 (Genbank アクセッション番号第 NM_005789) ; 602175 プロテアソームサブユニット、β-型、 2; Psmb2 (Genbank アクセッション番号第 NM_002794) ; 603146 プロテアソーム 26s サブユニット、非ATPase、 9; Psmd9 (Genbank アクセッション番号第 NM_002813) ; 154365 プロテアソーム 26s サブユニット、 ATPase, 2; Psmc2 (Genbank アクセッション番号第 NM_002803) ; 604449 プロテアソーム 26s サブユニット、非ATPase、11; Psmdll (Genbank アクセッション番号第 NM_002815) ; 606607 プロテアソームサブユニット、 α-型、 7; Psma7 (Genbank アクセッション番号第 NM_002792 および NM_152255) ; 176843 プロテアソームサブユニット、α-型、 3; Psma3 (Genbank アクセッション番号第 NM_002788 およびNM_ 152132) ; 607173 Padl ; (Genbank アクセッション番号第 NM_005805) ; 604450 プロテアソーム 26s サブユニット、非ATPase、 12; Psmdl2 (Genbank アクセッション番号第 NM_002816) ; 602706 プロテアソーム 26s サブユニット、 ATPase, 1; Psmcl (Genbank アクセッション番号第 NM_002802) ; 602708 プロテアソーム 26s サブユニット、 ATPase、 6; Psmc6 (Genbank アクセッション番号第 NM_002806) ; 186852 プロテアソーム 26s サブユニット、 ATPase、 3; Psmc3 (Genbank アクセッション番号第 NM_002804) ; 176842 プロテアソームサブユニット、α-型、 2; Psma2 (Genbank アクセッション番号第 NM_002787) ; 603481 プロテアソーム 26s サブユニット、非ATPase、 13; Psmdl3 (Genbank アクセッション番号第 NM_002817) ; 603480 プロテアソーム 26s サブユニット、非ATPase、 10;PsmdlO (Genbank アクセッション番号第 NM_002814) ; 602017 プロテアソームサブユニット、β-型、 1 ; Psmbl (Genbank アクセッション番号第 NM_002793) ; 604452 プロテアソーム 26s サブユニット、非ATPase、 5; Psmd5 (Genbank アクセッション番号第 NM_005047) ; 601648 プロテアソーム 26s サブユニット、 非ATPase、 4; Psmd4 (Genbank アクセッション番号第 NM_002810 および NM_153822) ; 602177 プロテアソームサブユニット、 β-型、 4; Psmb4 (Genbank アクセッション番号第 NM_002796) ; 602176 プロテアソームサブユニット、β-型、 3; Psmb3 (Genbank アクセッション番号第 NM_002795) ; 602707 プロテアソーム 26s サブユニット、 ATPase、 4; Psmc4 (Genbank アクセッション番号第 NM_153001 およびNM_006503) ; 601681 プロテアソーム 26s サブユニット、 ATPase、 5; Psmc5 (Genbank アクセッション番号第 NM_002805) ; 157970 プロテアソーム 26s サブユニット、非ATPase、 7; Psmd7 (Genbank アクセッション番号第 NM_002811) ; 602163 ユビキチン結合酵素 E2e 2; Ube2e2 (Genbank アクセッション番号第 Z44894) ; 605046 ユビキリン(Ubiquilin) 1 ; Ubqln1 (Genbank アクセッション番号第 NM_013438 および NM_053067) ; 602544 Parkin ; Park2 (Genbank アクセッション番号第 NM_004562、 NM_013987 および NM_013988) ; 314370ユビキチン活性化酵素 1; Ubel (Genbank アクセッション番号第 NM_003334 および NM_153280) ; 300264 ユビキリン(Ubiquilin) 2; Ubqln2 (Genbank アクセッション番号第 NM_013444) ; 180470 リボフォリン(Ribophorin) I;Rpnl (Genbank アクセッション番号第 NM_002950) ; 191342 ユビキチンカルボキシル-ターミナルエステラーゼ LI (Genbank アクセッション番号第 NM_004181) ; 118888 キモトリプシン様プロテアーゼ; Ctrl (Genbank アクセッション番号第 NM_001907) ; 605532 Smad ユビキチン化調節因子 2 (Genbank アクセッション番号第 NM_022739) ; 607119 ダブルリングフィンガータンパク質(Double Ring Finger Protein) (Genbank アクセッション番号第 NM_015435) ; 605624 アリアドネ(Ariadne)、ショウジョウバエ(Drosophila)、ホモログ Of、 1; Arih1 (Genbank アクセッション番号第 NM_005744) ; 605568 Smad ユビキチン化調節因子 1 (Genbank アクセッション番号第 AF199364); 603124 ユビキチン結合酵素 E2g 2; Ube2g2 (Genbank アクセッション番号第 NM_003343) ; 603091 ユビキチン特異的プロテアーゼ 12; Uspl2 (Genbank アクセッション番号第 AF022789); 602995 ユビキチン結合酵素 E2 変種1;Ube2vl (Genbank アクセッション番号第 NM_003349、 NM021988 および NM-022442) ; 602961 ユビキチン結合酵素 E2d 1; Ube2dl (Genbank アクセッション番号第 NM_003338) ; 602916 ユビキチン結合酵素 E2e 1; Ube2el (Genbank アクセッション番号第 NM_003341) ; および 300420 Praja 1; Pjal (Genbank アクセッション番号第 NM_022368)が挙げられ、その各々は参照として本明細書に組み入れられている。Genbankなどの同様のデータベースを使用してユビキチン-プロテアソーム関連ポリペプチドを同定することもできる。ユビキチン-プロテアソーム関連ポリペプチドは、複合体もしくは経路の一部である、または経路の活性もしくは組成物に影響を与える任意のポリペプチドをいう。
【0027】
II. ポリペプチドおよびタンパク様組成物
本発明の種々の局面において、ペプチド、ポリペプチドまたはタンパク質を含むタンパク様組成物を、抗原、抗体、ワクチン、遮断剤、治療剤としてまたは本明細書に記載する組成物および方法の成分として使用することができる。ペプチド、ポリペプチドおよび/またはタンパク質は単離/精製された組換えまたは合成ペプチド、ポリペプチドおよび/またはタンパク質であってもよい。
【0028】
本発明のポリペプチドおよびペプチドは、免疫原性ペプチド、抗原もしくはエピトープ、またはほ乳類のユビキチン-プロテアソームおよびそれに関連する経路の1つ以上のタンパク質成分に対する抗体等を含んでもよい。このようなペプチドは、例えば、ウサギ、雌ウシ、ブタ、ウマ、ヒヒおよびヒトを含む種々のほ乳類種のプロテアソームタンパク質に対する免疫応答を誘発するのに有用となりうる。プロテアソームタンパク質に対する免疫応答を誘発することによって、本発明のペプチドは避妊剤として有用である。他の態様において、抗体は単離されて、避妊または診断組成物に使用してもよい。
【0029】
ある態様において、プロテアソームポリペプチドまたは抗原は合成ペプチドであってもよい。さらに他の態様において、ペプチドは、分子工学的手法によって作製された組換えペプチドであってもよい。本セクションは、本発明に使用するためのタンパク性組成物を作製する際に関係する方法および組成物を記載する。
【0030】
A. ポリペプチド
ユビキチン-プロテアソーム経路の1つ以上のタンパク質成分由来のポリペプチドは、すなわち当業者に既知のタンパク質抽出技法を使用して抽出した天然型ポリペプチドであってもよい。特定の態様において、プロテアソーム抗原を同定して、ほ乳類をワクチン化するための薬学的に許容されうる担体中で作製してもよい。
【0031】
ポリペプチドの配列変種を作製してもよい。ポリペプチド配列変種は、集団内の天然の変更によって生ずるまたは他の動物に見られる相同物であってもよいポリペプチドのわずかな配列変種であってもよい。それらは、天然には生じないが、天然型のポリペプチドと同様に機能するおよび/または天然型のポリペプチドと交差反応する免疫応答を誘発する程度に十分に同様である配列であってもよい。配列変種は、Sambrookら2001に記載されているものなどの標準的な部位特異的突然変異方法によって作製することができる。
【0032】
抗原性プロテアソームポリペプチドの別の合成または組換え変更は、プロテアソームタンパク質に天然に見られるエピトープ決定基の反復を含むポリペプチド部分である。このような合成ポリペプチドタンパク質は、任意の1つのプロテアソームタンパク質エピトープのいくつかのホモマー反復から作製されても;または同じもしくは異なるポリペプチド由来の2つ以上のエピトープを含んでもよい。
【0033】
ポリペプチドのアミノ酸配列変種は置換、挿入または欠失変種であってもよい。欠失変種は、機能または免疫原性活性に必須でない未変性のタンパク質の1つ以上の残基を欠損している。別の一般的な種類の欠失変異は、分泌シグナル配列または細胞の特定の部分にタンパク質を結合させるシグナル配列を欠損するものである。
【0034】
置換変種は、典型的には、タンパク質内の1つ以上の部位における1つのアミノ酸と別のアミノ酸の交換を含み、タンパク質分解に対する安定性などのポリペプチドの1つ以上の特性を調節するように設計することができる。置換は、好ましくは、同類置換であり、すなわち、1つのアミノ酸が同様の形状および電荷を有するアミノ酸と交換される。同類置換は当技術分野において既知であり、例えば、以下の変更を含む:アラニンとセリン;アルギニンとリジン;アスパラギンとグルタミンまたはヒスチジン;アスパラギン酸とグルタミン酸;システインとセリン;グルタミンとアスパラギン;グルタミンとアスパラギン;グリシンとプロリン;ヒスチジンとアスパラギンまたはグルタミン;イソロイシンとロイシンまたはバリン;ロイシンとバリンまたはイソロイシン;リジンとアルギニン;メチオニンとロイシンまたはイソロイシン;フェニルアラニンとチロシン、ロイシンまたはメチオニン;セリンとスレオニン;スレオニンとセリン;トリプトファンとチロシン;チロシンとトリプトファンまたはフェニルアラニン;およびバリンとイソロイシンまたはロイシン。
【0035】
挿入変種は、ポリペプチドの迅速な精製を可能にするために使用されるものなどの融合タンパク質を含み、ポリペプチドの相同物である他のタンパク質またはポリペプチド由来の配列を含むハイブリッドタンパク質を含んでもよい。例えば、挿入変種は、1つの種由来のポリペプチドのアミノ酸配列の一部と、別の種由来の相同ポリペプチドの一部を含んでもよい。「キメラタンパク質」という用語は、相同または非相同タンパク質である2つの異なるタンパク質由来のセグメントを含むタンパク質をいうために使用される。セグメントは天然に隣接するセグメントでなくても、または任意のセグメントが任意の特定の位置もしくは順序を維持しなくてもよい。他の挿入変種は、追加のアミノ酸がポリペプチドのコード配列に導入されているものを含んでもよい。これらは、典型的には、上記の融合タンパク質より短い挿入であり、例えば、プロテアーゼ切断部位に導入される。
【0036】
一つの態様において、ポリペプチドの主要抗原決定基は、ポリペプチドをコードする遺伝子の一部が組換え宿主において発現され、得られたタンパク質は、免疫応答を誘発する能力について試験される実験方法によって同定することができる。例えば、ポリメラーゼ連鎖反応(PCR)を使用して、タンパク質のC-末端の連続する長い断片を欠損するペプチドをコードする範囲のcDNAを作製することができる。次いで、これらのペプチドの各々の免疫原性活性は、この活性に必須のポリペプチドの断片またはドメインを同定する。次いで、ごく少数のアミノ酸が各反復において除去または付加されるさらに別の実験によって、ポリペプチドの他の抗原性決定基を位置づけることができる。従って、熱安定性DNAポリメラーゼ、デオキシリボヌクレオチドおよびプライマー配列を使用する多数サイクルの変性-再生によってDNAの特定のセグメントを増幅する技法であるポリメラーゼ連鎖反応が本発明において考慮される(Mullis, 1990;Mullisら, 1992)。
【0037】
本発明によるポリペプチドを作製するための別の態様は、ペプチド模倣物の使用である。模倣物は、タンパク質の二次構造の要素を模倣する分子である。多数のタンパク質は、折りたたみ面の比較的小さい領域によって生物作用を発揮するので、それらの作用は、生理活性面を保持し、薬物動態学的/動力学的特性が改善されていると思われるかなり小型の設計(模倣)分子によって再現することができる(Fairlieら, 1998)。二次構造(ヘリックス、ターン、ストランド、シート)の個々の要素を模倣し、それらの組み合わせを三次構造(ヘリックスバンドル、マルチプルループ、ヘリックス-ループ-ヘリックスモチーフ)に集成する方法は総説が報告されている(Fairlieら, 1998;Moore, 1994)。模倣ペプチドを予測し、作製し、改変し、スクリーニングする方法は米国特許第5,933,819号および米国特許第5,869,451号(各々が具体的に参照として本明細書に組み入れられている)に記載されている。
【0038】
以下は、同等物または場合によっては改善された第二世代の分子を作製するためにタンパク質またはポリペプチドのアミノ酸を変更することに基づいた考察である。アミノ酸の変更は、以下の実施例により、DNA配列のコドンを変更することによってまたは化学的なペプチド合成によって実施することができる。
【0039】
例えば、抗体の抗原結合領域または基質分子の結合部位などの構造の相互作用的な結合能力をあまり損失しないで、例えば、ポリペプチド構造のあるアミノ酸を他のアミノ酸と置換することができる。それは、生物活性を規定するポリペプチドの相互作用的な能力および性質であるので、あるアミノ酸置換はポリペプチド配列およびポリペプチドをコードする根本的なDNA内で行うことができ、それでもなお、同様の特性または改善された特性を有するポリペプチドを得ることができる。表1は、特定のアミノ酸をコードするコドンを示す。
【0040】
このような変更を行う際には、アミノ酸の疎水性親水性指数を考慮することができる。タンパク質に相互作用的な生物機能を与える際のアミノ酸の疎水性親水性指数の重要性は、一般に、当技術分野において理解されている(KyteおよびDoolittle, 1982)。
【0041】
あるアミノ酸を同様の疎水性親水性指数またはスコアを有する他のアミノ酸によって置換しても、同様の生物作用を有するタンパク質またはポリペプチドを生じることができることは当技術分野において既知である。同様のアミノ酸の置換は親水性に基づいて効果的に実施されうることも当技術分野において理解されている。参照として本明細書に組み入れられている米国特許第4,554,101号は、隣接アミノ酸の親水性によって支配されるタンパク質の最も大きい局所的な平均親水性はタンパク質の生物学的特性に関連することを記載している。
【0042】
アミノ酸置換は、一般に、アミノ酸側鎖の置換基の相対的な類似性、例えば、疎水性、親水性、電荷、サイズ等に基づいている。上記の種々の特徴を考慮している例示的な置換は当業者に既知であり、以下を含む:アルギニンとリジン;グルタミン酸とアスパラギン酸;セリンとスレオニン;グルタミンとアスパラギン;およびバリン、ロイシンとイソロイシン等。
【0043】
本発明の有用なペプチドは、配列番号:1、2、4および6またはその断片の1つ以上を含むアミノ酸配列を含む。好ましくは、本発明のペプチドは30以下のアミノ酸の配列を含む。
【0044】
本発明の免疫原性ペプチドは、配列番号:1、2、4および6またはその断片の1つ以上並びに任意に非プロテアソームアミノ酸配列を有する複合ペプチドであってもよい。
【0045】
本発明の有用な組成物は、本発明の1つ以上のペプチドおよび/または融合タンパク質を、薬学的に許容されうる担体および/または他の薬学的添加剤成分を合わせて含む。組成物は、本発明のペプチドまたは融合タンパク質のエピトープに対する免疫応答を増強することができるアジュバントを含んでもよい。
【0046】
本発明はまたポリペプチドの断片にも関係する場合もある。分子のN-末端を含む断片は、コード領域内の翻訳停止部位の遺伝子操作によって作製することができる。または、ポリペプチドをプロテアーゼとして既知のタンパク質分解酵素で処理すると、種々のN-末端、C-末端および内部断片を作製することができる。ある態様において、ペプチドは既知の方法によって合成することができる。断片の例は、長さ
またはそれ以上のアミノ酸の連続残基を含んでもよい。これらの断片は、沈殿(例えば、硫酸アンモニウム)、HPLC、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー(イムノアフィニティークロマトグラフィーを含む)または種々のサイズ分離(沈降、電気泳動、ゲルろ過)などの既知の方法によって精製することができる。
【0047】
B. 合成ペプチド
本発明の種々の態様は、受精能を調節および/または診断するための方法および組成物に使用するためのペプチドまたはポリペプチドを記載している。いくつかの態様において、動物の受精能を阻止または低下および検出する際に使用するための抗プロテアソームまたは抗ユビキチン抗体の作製が考慮されている。本発明のペプチドは、従来技法により溶液または固相支持体でも合成することができる。種々の自動合成装置が市販されており、既知のプロトコールにより使用することができる。例えば、おのおの参照として本明細書に組み入れられている、StewartおよびYoung(1984);Tamら(1983);Meerifield(1986)並びにBaranyおよびMerrifield(1979)を参照されたい。本明細書に記載するペプチドに相当する通常約6から35〜50アミノ酸のオーバーラップペプチドの短いペプチド配列またはライブラリーを容易に合成することができる。いくつかの態様において、本発明のペプチドまたはポリペプチドをコードするヌクレオチド配列を発現ベクターに挿入し、適当な宿主細胞に形質転換またはトランスフェクションし、発現に好適な条件下において培養する組換えDNA技術を使用することができる。
【0048】
C. 融合ペプチドまたはポリペプチド
特殊な種類の挿入変種は融合タンパク質である。この分子は、一般に、第1の分子の全てまたは実質的な部分が第2のポリペプチドの全てまたは一部のN-末端またはC-末端に結合している。例えば、融合は、典型的には、異種宿主におけるタンパク質の組換え発現を可能にするために他種のリーダー配列を使用する。別の有用な融合は、融合タンパク質の精製を容易にするために、抗体エピトープなどの免疫学的に活性なドメインの追加を含む。融合接合部または接合部付近に切断部位を導入すると、精製後の外来ポリペプチドの除去が容易になる。他の有用な融合には、酵素の活性部位、グリコシル化ドメイン、細胞標的シグナルまたは膜貫通領域などの機能的ドメインの結合が挙げられる。本発明の他の融合には、2つ以上のプロテアソーム抗原の融合が挙げられる。ある態様において、2つ以上のプロテアソーム抗原が可逆的または非可逆的に互いに結合される。
【0049】
D. ペプチドまたはポリペプチドの精製
ある態様において、ペプチドまたはポリペプチドを精製することが望ましい場合がある。タンパク質精製技法は当業者に既知である。これらの技法は、一つのレベルにおいて、ポリペプチドおよび非ポリペプチド分画への細胞環境の粗分画化に関係する。ポリペプチドを他の分子から分離したら、クロマトグラフィー技法および電気泳動技法を使用して関心対象のポリペプチドをさらに精製して、部分または完全精製(または、均質までの精製)することができる。純粋なペプチドの作製に特に好適な分析方法はイオン交換クロマトグラフィー、排除クロマトグラフィー;ポリアクリルアミドゲル電気泳動;等電点電気泳動法である。特に効率的なペプチド精製方法は高速タンパク質液体クロマトグラフィーまたは場合によってはHPLCである。
【0050】
タンパク質精製に使用するのに好適な種々の技法は当業者に既知である。これらには、例えば、硫酸アンモニウムによる沈殿、PEG、抗体等または熱変性後の遠心分離;イオン交換、ゲルろ過、逆相、ヒドロキシアパタイトおよびアフィニティークロマトグラフィーなどのクロマトグラフィー段階;等電点電気泳動法;ゲル電気泳動並びにこのような技法および他の技法の組み合わせが挙げられる。当技術分野において一般に既知であるように、種々の精製段階を実施する順序を変更することができること、またはある段階を省略しても実質的に精製されたタンパク質もしくはペプチドの作製に好適な方法になることが考えられる。
【0051】
本発明のある局面はペプチドの精製に関し、特定の態様においてペプチドの実質的な精製に関する。本明細書において使用する「精製されたペプチド、ポリペプチドまたはタンパク質」という用語は、他の成分から単離可能な組成をいうことが意図されており、ペプチドは天然に入手可能な状態に対して任意の程度まで精製される。従って、精製されたタンパク質またはペプチドは、それが天然に生じうる環境を含まないタンパク質またはペプチドのこともいう。
【0052】
一般に、「精製された」は、種々の他の成分を除去するために分画化が実施されており、実質的に活性を保持するタンパク質またはペプチド組成物をいう。「実質的に精製された」という用語を使用する場合には、この意味は、タンパク質またはペプチドが組成物中で約50%、約60%、約70%、約80%、約90%、約95%またはそれ以上のペプチド、ポリペプチドまたはタンパク質を構成するなどの、組成物の主要な成分を形成する組成物をいう。タンパク質またはペプチドの精製程度を定量化する種々の方法は、本発明の開示内容を考慮して当業者に既知である。
【0053】
E. 抗原
本発明の抗原は、典型的には、ほ乳類のプロテアソームの1つ以上のタンパク質成分から単離または誘導され、総説は、例えば、SchmidおよびBriand, 1997またはBogyoら, 1997を参照されたい。本発明の種々の態様において、動物の受精能を調節するためまたは受精能の調節に使用するための抗体を作製するために抗原を動物に投与する。特定の態様において、本発明による脊椎動物の免疫化は、1つ以上のプロテアソームペプチドをコードするポリヌクレオチドの投与を含んでもよい。
【0054】
いくつかの態様において、本発明は:(a)プロテアソーム抗原を同定する段階;(b)本明細書に記載する抗原で脊椎動物に免疫応答を形成する段階;(c)動物細胞または免疫した動物から回収した細胞と不死化癌細胞の融合によって作製したハイブリドーマ細胞系統において作製された抗体を入手する段階とを含むプロテアソーム抗原に対する抗体を作製する方法に関する。
【0055】
本発明はまた、免疫原性であるが、ワクチンとして必ずしも防御的ではないプロテアソームポリペプチドに対する抗体を作製する方法に関する。例えば、プロテアソーム特異的抗体は、受精能の評価、不妊の診断および検討または抗体治療に有用である可能性がある。抗原または抗原をコードするポリヌクレオチドで動物を免疫化することを使用して、抗プロテアソーム抗体を作製することができる。抗プロテアソーム抗体を作製する他の方法において、同定された抗原は、ファージライブラリーをパニングするために使用されてもよい。この手法はインビトロにおいて抗体を単離するために使用してもよい。
【0056】
F. 抗体
別の局面において、本発明は、プロテアソーム経路またはその任意の部分の成分に免疫反応性である抗体組成物を含む。さらに他の態様において、本発明の抗原は、抗体および/または抗体組成物を作製するために使用することができる。抗体は、特異的または優先的に、プロテアソームもしくはユビキチンポリペプチドまたはユビキチン-プロテアソーム経路の酵素のポリペプチドに反応性となりうる。例示的な抗体は、配列番号:1、2、4および6に記載する配列、それらの断片、変種もしくは模倣物または密接に関連する配列を有する抗原に反応性の抗体を含むプロテアソーム経路の成分もしくは関連ポリペプチドに反応性のものを含む。抗体は、ポリクローナルまたはモノクローナルであってもよく、当技術分野において既知の方法によって作製することができる。抗体はまた1価または2価であってもよい。抗体は、種々の生物学的または化学的手段によって分割されてもよい。抗体の半分は各々1つの抗原に結合することができ、従って、1価と規定される。抗体を作製し、特徴づける手段は当技術分野において既知である(例えば、参照として本明細書に組み入れられている、HarlowおよびLane, 1988を参照されたい)。
【0057】
抗体を作製するために、本発明のポリペプチドの1つ以上の抗原決定基に対応するペプチドを作製することができる。このようなペプチドは、一般に、長さ少なくとも5または6アミノ酸残基である必要があり、好ましくは、長さ約10、15、20、25または約30アミノ酸残基であり、最高約35残基〜50残基ほどを含んでもよい。合成ペプチドは、一般に、Applied Biosystems(Foster City, CA)社製のものなどの、自動ペプチド合成機械のほぼ上限の長さである、長さ約35残基である。例えば、組換え手段によってさらに長いペプチドも作製することができる。他の方法において、完全長または実質的に完全長のポリペプチドを本発明の抗体を作製するために使用することができる。
【0058】
少なくとも1つ以上の抗原決定基を含むペプチドが作製されたら、ポリペプチドに対する抗血清の作製にペプチドを使用する。例えば、PCRクローニング方法を使用する標準的な方法によって、これらの決定基をコードするミニ遺伝子または遺伝子融合を構築し、発現ベクターに挿入することもできる。抗体形成またはワクチン化のためのペプチド使用は、典型的には、B型肝炎表面抗原、キーホールリンペットヘモシアニンまたはウシ血清アルブミンなどの免疫原性キャリアタンパク質にペプチドを結合することが必要である。この結合を実施する方法は、種々のアジュバントと同様に、当技術分野において既知である。
【0059】
本発明の方法に使用する抗体は、すなわち、抗体に任意の種類の分子を共有結合することによって修飾されている誘導体を含む。例えば、抗体誘導体には、例えば、グリコシル化、アセチル化、PEG付化、リン酸化、アミド化、既知の保護/ブロック基による誘導体化、タンパク質分解的切断、細胞リガンド(他のタンパク質)への結合等によって修飾されている抗体を含むが、これらに限定されない。数多くの化学的修飾のいずれかを既知の技法によって実施することができる。また、誘導体は1つ以上の非古典的アミノ酸を含んでもよい。
【0060】
ヒトにおける抗体のインビボにおける使用およびインビトロにおける検出アッセイを含むいくつかの使用のためには、キメラ、ヒト化またはヒト抗体を使用することが好ましい場合がある。キメラ抗体は、マウスモノクローナル抗体由来の可変領域とヒト免疫グロブリン由来の定常領域を有する抗体などの、抗体の異なる部分が異なる動物種または分子由来である分子である。キメラ抗体を作製する方法は当技術分野において既知である。例えば、全体の内容が参照として本明細書に組み入れられている、Morrison, 1985;Gilliesら, 1989;米国特許第5,807,715号;同第4,816,567号および同第4,816,397号を参照されたい。ヒト化抗体は、非ヒト種の1つ以上の相補性決定領域(CDR)およびヒト免疫グロブリン分子のフレームワーク領域を有する望ましい抗原に結合する非ヒト種由来の抗体分子である。抗体は、例えば、全て全体の内容が参照として本明細書に組み入れられている、CDR-グラフティング
、ベニアリング(veneering)またはリサーフェイシング(resurfacing)(欧州特許第592,106号;欧州特許第519,596号;Padlan, 1991;Studnickaら, 1994;Roguskaら, 1994)およびチェーンシャフリング(米国特許第5,565,332号)を含む当技術分野において既知の種々の技法を使用してヒト化することができる。
【0061】
完全ヒト抗体は、患者の治療的療法に特に望ましい。ヒト抗体は、ヒト免疫グロブリン配列由来の抗体ライブラリーを使用する上記のファージディスプレイ方法を含む当技術分野において既知の種々の方法によって作製することができる。各々全体の内容が参照として本明細書に組み入れられている、
を参照されたい。
【0062】
ヒト抗体は、機能的な内因性免疫グロブリンを発現することができないが、ヒト免疫グロブリン遺伝子を発現することができる遺伝子組換えマウスを使用しても作製することができる。ヒト抗体を作製するためのこの技術の概略は、LonbergおよびHuszar, 1995を参照されたい。ヒト抗体およびヒトモノクローナル抗体を作製するためのこの技術並びにこのような抗体を作製するためのプロトコールの詳細な考察は、例えば、全体の内容が参照として本明細書に組み入れられている、
を参照されたい。また、Abgenix, Inc.(Freemont、CA)、Kirin, Inc.(日本)、Medarex(NJ)およびGenpharm(San Jose、CA)などの会社が、上記のものと同様の技術を使用して、選択した抗原に対するヒト抗体を提供することに関与することがある。
【0063】
本発明の抗体は、免疫原性応答を実質的に全く生じないで、ほ乳類の循環系に注射することができる組成物を提供するために、Davisら(米国特許第4,179,337号)によって記載されている方法およびカップリング剤によって修飾することもできる。
【0064】
III. ポリヌクレオチド
本発明は、ポリヌクレオチドを含む組成物と、被験者における受精能を調節および診断するためにこれらの組成物を使用する方法を提供する。いくつかの態様において、本発明の方法および組成物は、プロテアソーム、ユビキチンまたはユビキチン-プロテアソーム経路の関連酵素に対する動物または被験者の免疫応答を誘導することに関してもよい。
【0065】
本発明の種々の態様において、プロテアソームタンパク質、モノマーユビキチンまたはユビキチンポリペプチドおよびそれらの断片をコードするポリヌクレオチドが提供される。プロテアソームタンパク質は、プロテアソームのα1-7およびβ1-7サブユニット、特にMECL1サブユニットを含むが、これらに限定されない。プロテアソームポリペプチドまたはポリペプチド断片をコードするポリヌクレオチドは原核細胞または真核細胞において発現することができる。発現されたポリペプチドまたはポリペプチド断片は、脊椎動物に投与して、プロテアソームポリペプチドまたはポリペプチド断片と免疫反応する抗体を作製するためのプロテアソーム抗原として使用するために精製することができる。
【0066】
本発明は、任意の特定のほ乳類のプロテアソームポリヌクレオチドまたはポリペプチドに範囲が限定されない。当業者は、本明細書に記載する核酸を使用して、他のほ乳類において関連する相同物を容易に同定することができると思われる。また、本発明は、本明細書に開示されている特定の核酸に限定されないことが明らかにされるべきである。以下に考察するように、特定の「プロテアソームタンパク質」遺伝子またはポリヌクレオチド断片は、種々の異なる塩基を含有し、本明細書に開示するポリヌクレオチドと機能的に識別できない、および場合によっては、構造的に識別できない対応するポリペプチドを作製することができる。種々の動物由来の種々のプロテアソームまたはユビキチン配列の核酸配列はGenBankおよび他の公的なデータベースにおいて見つけることができる。
【0067】
A. ペプチドまたは抗原をコードする核酸
本発明のいくつかの態様において、脊椎動物において免疫応答を誘導することができるおよび抗プロテアソームまたは抗ユビキチン抗体を作製するための抗原として使用するための抗原性ポリペプチドをコードするポリヌクレオチドが考慮される。ある例において、ワクチンとして使用するまたは受動免疫のための抗プロテアソーム抗体を作製する際に使用する特定の抗原性プロテアソームポリペプチドドメインをコードするプロテアソームポリヌクレオチドを発現することが望ましい場合がある。本発明の核酸は、プロテアソームタンパク質全体または任意の他の断片をコードしてもよい。核酸は、特定の生物のPCR増幅DNAから誘導することができる。しかし、他の態様において、核酸は、ゲノムDNA、相補的DNA(cDNA)または合成DNAを含んでもよい。タンパク質は、ワクチンに使用するため、または抗体を単離する方法に使用するために指定された配列から誘導することができる。抗原性プロテアソームポリペプチドを作製するために、プロテアソームポリペプチドをコードする遺伝子または対応するcDNAを適当な発現ベクターに挿入することができる。
【0068】
「cDNA」という用語は、鋳型としてメッセンジャーRNA(mRNA)を使用して作製されるDNAをいうことが意図されている。ゲノムDNA鋳型または非プロセッシングもしくは部分プロセッシングRNA鋳型から増幅または合成されるDNAと違って、cDNAを使用する利点は、cDNAは、主に、対応するタンパク質のオープンリーディングフレーム(ORF)を含むコード配列を含むということである。非コード領域が最適な発現に必要とされる場合などの、完全長または部分的なゲノム配列が好ましい場合もある。
【0069】
さらに別の態様において、所定の動物のポリヌクレオチドは、わずかに異なる核酸配列を有するが、それでもなお、同じポリペプチドをコードする天然の変種によって提供されてもよい(以下の表1を参照されたい)。また、異なる核酸配列を生じるが、同じポリペプチドをコードする別のコドンを使用して、動物の所定のプロテアソームポリペプチドを作製することができることが考慮される。
【0070】
本願において使用する「プロテアソームポリヌクレオチドをコードする核酸」という用語は、細胞の総核酸を含まない単離されている核酸分子をいう。「機能的に同等なコドン」という用語は、アルギニンまたはセリンの6つのコドン(表1、以下)などの同じアミノ酸をコードするコドンをいうために本明細書において使用され、また以下のページに考察するように、生物学的に同等なアミノ酸をコードするコドンもいう。
【0071】
遺伝子コードの縮重を考慮すると、配列は、所定のプロテアソーム遺伝子またはポリヌクレオチドのヌクレオチドと同一のヌクレオチドの少なくとも約50%、通常少なくとも約60%、さらに通常約70%、最も通常には約80%、好ましくは少なくとも約90%、および最も好ましくは約95%を有するプロテアソーム遺伝子またはポリヌクレオチドに記載されているものと本質的に同じであると考えられる。プロテアソーム遺伝子またはポリヌクレオチドに記載されているものと本質的に同じである配列はまた、標準的な条件においてプロテアソームポリヌクレオチドの相補鎖を含む核酸セグメントにハイブリダイゼーションすることができる配列として機能的に規定することもできる。密接に関連する配列という用語は、実質的な配列の類似性を有する配列または本明細書に記載するものと同様の抗原応答を実施または誘発するタンパク質をコードする配列をいう。密接に関連する配列という用語は、比較しているポリヌクレオチドまたはポリペプチドと最小または50%の類似性を有する配列をいうために本明細書において使用される。
【0072】
本発明のDNAセグメントは、上記のように、生物学的に機能的な同等なプロテアソームタンパク質およびペプチドをコードするものを含む。このような配列は、核酸配列内に天然に生じることが既知のコドン冗長性およびアミノ酸の機能的同等性の結果並びにこのようにコードされるタンパク質として生じうる。または、機能的に同等なタンパク質またはペプチドは、タンパク質構造の変更が、交換されるアミノ酸の特性の考慮に基づいて工作することができる組換えDNA技術の適用によって作製することができる。変更は、以下に記載するように、部位特異的突然変異技法の適用によって工作しても、または無作為に導入して、望ましい機能について後にスクリーニングしてもよい。
【0073】
当然のことながら、本発明は、プロテアソームポリヌクレオチドの配列に相補的であるまたは本質的に相補的であるオリゴヌクレオチドも含む。「相補的」である核酸配列は、標準的なワトソン-クリック相補則により塩基対形成することができるものである。本明細書において使用する「相補的な配列」という用語は、上記と同じヌクレオチド比較によって評価するとき、または本明細書に記載するものなどの比較的ストリンジェントな条件下において、プロテアソームポリヌクレオチドの核酸セグメントとハイブリダイゼーションすることができると規定するとき、実質的に相補的である核酸配列を意味する。
【0074】
または、ハイブリダイゼーションセグメントは短いオリゴヌクレオチドであってもよい。長さ17塩基の配列がヒトゲノムにおいて1回だけ生じるはずであるので、独自の標的配列を指定するのに十分である。短いオリゴマーは作製が容易であり、インビボにおけるアクセスしやすさを高めるが、ハイブリダイゼーションの特異性の決定には数多くの他の因子が関与する。相補的な標的へのオリゴヌクレオチドの結合親和性および配列特異性は共に長さの増加と共に増加する。
またはそれ以上の塩基対の例示的なオリゴヌクレオチドが使用されることが考慮されるが、他も考慮される。250、500、1000、1212、1500、2000、2500、3000または3500塩基およびそれ以上の塩基をコードする長いポリヌクレオチドも考慮される。このようなオリゴヌクレオチドまたはポリヌクレオチドは、典型的には、例えば、サザンブロットおよびノーザンブロットのプローブとして、並びに増幅反応のプライマーとして、またはワクチンのための用途を見出している。
【0075】
(表1)
【0076】
C. 非細菌的に増幅した核酸
本発明の核酸またはポリヌクレオチドは、例えば、化学的合成または酵素的作製などの当業者に既知の任意の技法によって作製することができる。本発明の方法において、1つ以上のオリゴヌクレオチドまたはポリヌクレオチドを使用することができる。種々の異なる機序のオリゴヌクレオチド合成が、例えば、各々参照として本明細書に組み入れられている、
に開示されている。
【0077】
酵素的に作製される核酸またはポリヌクレオチドの限定するものではない例は、PCR(商標)などの増幅反応において酵素によって作製されるもの(例えば、各々参照として本明細書に組み入れられている、米国特許第4,683,202号および米国特許第4,682,195号を参照されたい)または参照として本明細書に組み入れられている米国特許第5,645,897号に記載されているオリゴヌクレオチドの合成によって作製されるものを含む。
【0078】
核酸またはポリヌクレオチド増幅の別の方法は、全体の内容が参照として本明細書に組み入れられている、欧州特許第320 308号に開示されているリガーゼ連鎖反応(「LCR」)である。米国特許第4,883,750号は、標的配列にプローブ塩基対を結合するための、LCRに類似した方法を記載している。結果として得られる「ジ-オリゴヌクレオチド」の配列を有し、それによってジ-オリゴヌクレオチドを増幅する核酸の存在下における2つ(または2つ以上)のオリゴヌクレオチドのライゲーションに基づいた方法も本発明の増幅段階に使用することができる。Wuら, (1989)は、全体の内容が参照として本明細書に組み入れられている。
【0079】
D. ポリヌクレオチド送達
本発明のある態様において、真核細胞において機能することができるプロモーターの制御下においてプロテアソームポリヌクレオチドまたはポリヌクレオチドセグメントを含む発現構築物を提供する。本発明のある局面における一般的な方法は、特定のタンパク質、ポリペプチドまたはペプチド断片をコードする発現構築物を細胞に提供し、それによって細胞において抗原性タンパク質、ポリペプチドまたはペプチドの発現を可能にすることである。発現構築物の送達後、発現構築物によってコードされるタンパク質、ポリペプチドまたはペプチド断片は、細胞の転写および翻訳機構によって合成される。種々のポリヌクレオチド送達組成物および方法が既知である(各々参照として本明細書に組み入れられている、Sambrookら, 2001;LiuおよびHuang, 2002;Ravidら, 1998;BalickiおよびBeutler, 2002を参照されたい)。
【0080】
ウイルスおよび非ウイルス送達システムは、抗原性タンパク質、ポリペプチド、ポリペプチド断片をコードする発現構築物の送達のための種々の送達システムのうちの2つである。両方の種類の送達系が当技術分野において既知であり、以下に簡単に記載する。遺伝子による免疫化のために発現構築物の送達に使用される主な方法も2つある;間接的なエクスビボ方法または直接的なインビボ方法。エクスビボ遺伝子導入は、培養中の(宿主)細胞のベクター改変およびベクターで改変した細胞の被験者への投与または移植を含む。インビボにおける遺伝子導入は、免疫化対象被験者へのワクチンベクターの直接導入を含む。
【0081】
種々の態様において、発現される核酸は、典型的には、遺伝子発現成分の完全なセット(プロモーター、コード配列およびターミネーター)を含む、直鎖状発現要素(「LEE」)および/または環状発現要素(「CEE」)に関連してもよい。これらのLEEおよびCEEは細胞または無傷の生物に直接導入されて、発現されて、標準的な高次コイル複製プラスミドによるものに匹敵する発現レベルを生じることができる(SykesおよびJohnston, 1999)。
【0082】
1. 非ウイルスポリヌクレオチド送達
本発明の一態様において、ポリヌクレオチド発現構築物は、組換えによって作製されるDNAプラスミドまたはインビトロにおいて作製されるDNAを含んでもよい。本発明の種々の態様において、例えば、プロテアソームまたはユビキチンポリヌクレオチドを含む発現構築物は、注射および/または微粒子銃(例えば、遺伝子銃)によって生物または被験者に投与される(Kleinら, 1987およびSanfordら, 1991)。いくつかの態様において、ポリヌクレオチドの投与は、筋肉内、静脈内、皮下、皮内または腹腔内注射によってもよい。
【0083】
本発明のプロテアソームまたは同様のポリヌクレオチドを含む発現構築物の導入は、細胞膜を物理的または化学的に透過性にする方法のいずれか(例えば、リン酸カルシウム沈殿、DEAE-デキストラン、エレクトロポレーション、直接マイクロインジェクション、DNA負荷リポソームおよびリポフェクタミンDNA複合体、細胞の超音波処理、高速微粒子を使用する遺伝子銃並びに受容体媒介性形質移入)によって実施することができる。ある態様において、脂質製剤および/またはナノカプセルの使用が、プロテアソームポリヌクレオチド、プロテアソームポリペプチドまたはプロテアソームポリヌクレオチドを含む発現ベクターを宿主細胞に導入するために考慮される(各々参照として本明細書に組み入れられている、Banghamら(1965)、Gregoriadis(1979)、DeamerおよびUster(1983)、SzokaおよびPapahadjopoulos(1978)、Nicolauら, 1987およびWattら, 1986の例示的な方法および組成物を参照されたい)。本発明の別の態様において、発現構築物は、単純に、裸の(naked)組換えDNA、発現カセットまたはプラスミドからなってもよい。
【0084】
2. ウイルスベクター
ある態様において、本発明による免疫応答を与えるポリヌクレオチドまたはコードされるポリペプチドはウイルスベクターによって送達することができることが考慮される。特定の態様において、免疫応答はプロテアソームまたはユビキチンペプチドまたはポリペプチドであってもよい。細胞に効率的に感染して侵入し、宿主細胞ゲノムに組込み、ウイルス遺伝子を安定して発現するある種のウイルスベクターの能力によって、数多くの異なるウイルスベクター系が開発され、適用されている(Robbinsら, 1998)。ウイルスシステムは、現在、エクスビボおよびインビボにおける遺伝子導入のためのベクターとして使用するために開発されている。例えば、アデノウイルス、単純ヘルペスウイルス、レトロウイルスおよびアデノ随伴ウイルスベクターが、現在、癌、嚢胞性線維症、ゴーシェ病、腎疾患および関節炎の治療のために評価されている(RobbinsおよびGhivizzani, 1998;Imaiら, 1998;米国特許第5,670,488号)。
【0085】
特定の態様において、アデノウイルス(各々具体的に参照として本明細書に組み入れられている、米国特許第6,383,795号、同第6,328,958号および同第6,287,571号)、レトロウイルス(各々具体的に参照として本明細書に組み入れられている、米国特許第5,955,331号;同第5,888,502号、同第5,830,725号)、単純ヘルペスウイルス(各々具体的に参照として本明細書に組み入れられている、米国特許第5,879,934号;同第5,851,826号)、アデノ随伴ウイルス(AAV)、ポックスウイルス;例えば、ワクシニアウイルス(Gnantら,1999a;Gnantら, 1999b)、αウイルス;例えば、シンドビスウイルス、セムリキ森林熱ウイルス(Lundstorm, 1999)、レトロウイルス(Coffeyら, 1998)およびインフルエンザAウイルス(Neumannら, 1999)、ポックスウイルス/レトロウイルスキメラベクター(Holzerら, 1999)、アデノウイルス/レトロウイルスベクター(Fengら, 1997;Bilbaoら, 1997;Caplenら, 1999)およびアデノウイルス/アデノ随伴ウイルスベクター(Fisherら, 1996;米国特許第5,871,982号)発現ベクターが発現構築物の送達のために考慮される。「ウイルス発現ベクター」は、(a)構築物のパッケージングを支持し、(b)クローニングされている組織または細胞特異的構築物を最終的に発現するのに十分なウイルス配列を含むような構築物を含むことを意味している。ウイルスの増殖および操作は当技術分野において既知である。
【0086】
E. 発現ベクター
プロテアソームペプチドをコードするポリヌクレオチドは、遺伝的免疫化ベクターまたは任意の他の好適な発現構築物にクローニングすることができる。ベクターは、真核細胞において機能可能なプロモーター、例えば、CMVプロモーターまたは任意の他の好適なプロモーターを含むことができる。このような方法では、ポリヌクレオチドは、筋肉内注射、皮内注射または表皮注射または微粒子銃によって投与することができる。ポリヌクレオチドは、静脈内、皮下、病巣内、腹腔内、経口、他の粘膜または吸入投与経路によっても投与することができる。いくつかの具体的な例示的な態様において、投与は、少なくとも0.005μg〜5.0μgのポリヌクレオチドの表皮注射/微粒子銃によってもよい。いくつかの例において、第1の投与後少なくとも約2週間以上経過してから、第2の投与、例えば、筋肉内注射および/または表皮注射を実施してもよい。これらの方法では、ポリヌクレオチドは、ウイルス発現ベクター、例えば、アデノウイルス、単純ヘルペスウイルス、レトロウイルスまたはアデノ随伴ウイルスベクターを含むウイルス発現ベクターにクローニングすることができるが、必ずしも必要はない。ポリヌクレオチドは、本明細書に開示するまたは当業者に既知の任意の他の方法においても投与することができる。1つ以上のポリヌクレオチドを1つ以上の発現ベクターに含ませることができる。
【0087】
発現に好適なベクターは既知であり、細菌発現ベクター、酵母発現ベクターおよびバキュロウイルスベクターを含むが、これらに限定されない。宿主細胞が好適な条件下で培養されると、プロテアソームペプチドは宿主細胞内で発現される。宿主細胞は、カイコ幼虫、CHO細胞、大腸菌(E.coli)および酵母を挙げることができるが、これらに限定されない。宿主細胞において発現されるペプチドの回収方法は既知である。ベクター、宿主、ベクターを発現する方法およびペプチドを回収する方法は既知である(例えば、Sambrookら, 2001およびO'Reilleyら, 1994)。
【0088】
IV. 免疫化
プロテアソームおよびユビキチンペプチドまたはポリペプチドを含むが、これらに限定されない、本明細書に記載する種々のペプチドまたはポリペプチドに対して免疫応答を誘発することができる。本発明のプロテアソームまたはユビキチンペプチド由来の免疫原性ペプチドは、好ましくは、抗プロテアソームまたは抗ユビキチン抗体をそれぞれ誘導するために使用される。例えば、1つ以上のプロテアソームエピトープを含む融合タンパク質を、非プロテアソーム免疫原と共にまたは伴わないで、例えば、注射によって動物に投与する。免疫原は、当技術分野において既知のアジュバントと共に送達してもよい。投与経過は、抗プロテアソーム抗体価を誘導するための必要性に応じて、1回または多数回の追加投与であってもよい。従って、十分な抗体価を保持する必要性に応じて、維持投与を実施してもよい。本発明のペプチド、ペプチド組成物またはワクチンの適当な用量および免疫法は、本明細書および以下の実施例に提供されている情報を利用して、当業者が決定することができる。
【0089】
ワクチン化/免疫化の概念は、免疫系の2つの基本的な特徴、すなわち免疫系成分の特異性および記憶に基づいている。ワクチン化/免疫化は、被験者に投与した抗原に特異的に向かう応答を開始する。さらに、メモリーBおよびTリンパ球集団が誘導されうる。抗原に再度接触すると、免疫系はさらに迅速に且つさらにより激しく応答を開始し、それによってワクチン化/免疫化した被験者に抗原に対する免疫学的防御を与える。同じ抗原もしくは異なる抗原を被験者に反復投与することによってまたは被験者にワクチン組成物を追加免疫投与することによって免疫応答を増強することができる。
【0090】
ワクチン化は、抗原の全てまたは一部を投与することによる、能動的に獲得される免疫の人工的な誘導である。能動的に獲得される免疫以外に、受動免疫方法を使用して被験者に治療的恩恵を提供することもできる、以下を参照されたい。
【0091】
特に、DNA免疫化としても既知の遺伝的ワクチン化は、抗原をコードする発現ベクターをインビボ、インビトロまたはエクスビボにおいて投与して、適当な生物、組織、細胞または標的細胞内に適切に折りたたまれた抗原の産生を誘導することに関係する。発現されたタンパク質またはペプチドは、典型的には、正常な細胞の主要組織適合性(MHC)抗原複合体と共に、形質移入した細胞の細胞表面で提示される。MHC抗原と関連したこれらの抗原決定基の提示は、決定基に特異的な細胞障害性T-リンパ球クローンの増殖を誘発することを意図している。さらに、発現中の形質移入細胞によって放出されるタンパク質は、抗原提示細胞によってピックアップ、内部移行または発現されて、全身の液性抗体応答を誘発することもできる。
【0092】
ワクチンは、非病原性で、ワクチン化に使用するのに好適であるように改変されている免疫原性物質の全てもしくは一部またはそれらの模倣物由来の抗原を含む組成物である。ワクチンの種類には、遺伝子ワクチン、ビロソーム、弱毒化または不活性化した生物体そのものを使用したワクチン、組換えタンパク質ワクチン、結合ワクチン、遺伝子組換え植物ワクチン、トキソイドワクチン、精製サブユニットワクチン、遺伝子組換え複合ワクチン、抗イディオタイプワクチンおよび当技術分野において既知の他のワクチンが挙げられるが、これらに限定されない。
【0093】
免疫応答は能動または受動免疫応答であってもよい。能動免疫は、生体が種々の抗原に接触すると形成する。それは、典型的には、BまたはTリンパ球に関係する。Bリンパ球(B細胞とも呼ばれる)は抗体を産生する。抗体は特異的な抗原に結合して、貪食細胞が抗原を破壊しやすくする。典型的には、Tリンパ球(T細胞)が抗原を直接攻撃して、免疫応答に対してある種のコントロールを提供することができる。特定の抗原または抗原種に特異的なB細胞およびT細胞が発生する。受動免疫化は、抗原に結合する事前に形成された抗体または他の結合物質の投与をいう。
【0094】
ある場合では、免疫応答は養子免疫治療の結果であってもよい。養子免疫治療では、リンパ球を被験者から入手し、抗原組成物と接触または抗原組成物で標識(pulsed)する。例示的な方法または組成物は、
を参照されたい。
【0095】
本発明は、プロテアソーム抗原またはプロテアソーム抗原、ユビキチン抗原もしくはユビキチン-プロテアソーム経路において活性な酵素由来の抗原をコードするポリヌクレオチドを含む抗原組成物を被験者に接触させることによって、被験者を免疫化、治療またはワクチン化する方法を含む。抗原組成物は、核酸;ポリペプチド;プロテアソーム抗原を発現してもよいウイルス、細菌、真菌または寄生虫などの弱毒化病原体;プロテアソーム抗原を発現する原核細胞;プロテアソーム抗原を発現する真核細胞;ビロソーム等またはそれらの組み合わせを含んでもよい。本明細書において使用する「抗原組成物」は、典型的には、薬学的に許容されうる製剤中に抗原を含む。
【0096】
抗原は、特に、抗原に反応性の特異的な抗体または細胞種の形態の、生物が免疫応答を誘発する任意の基質、分子または基質をコードする分子をいう。抗原組成物は、本明細書に記載するアジュバント、免疫調節物質、ワクチン基剤および/または他の賦形剤をさらに含んでもよく、当技術分野において既知である(例えば、Remington's Pharmaceutical Scienceを参照されたい)。
【0097】
抗原または抗原組成物を被験者に導入する種々の方法が当技術分野において既知である。ワクチン化方法には、DNAワクチンまたは遺伝子免疫化(例えば、各々参照として本明細書に組み入れられている、
を参照されたい)、遺伝子組換え食用植物ワクチン(例えば、各々参照として本明細書に組み入れられている
を参照されたい)、経皮的免疫化(各々参照として本明細書に組み入れられている、Glennら, 1999および米国特許第5,980,898号)、経鼻的または粘膜免疫化(例えば、各々参照として本明細書に組み入れられている、米国特許第4,512,972号、同第5,429,599号、同第5,707,644号、同第5,942,242号を参照されたい);ビロソーム(各々参照として本明細書に組み入れられている、
)、生ベクター等が挙げられるが、これらに限定されない。抗原送達方法は1つ以上のワクチン法と組合わせてもよい。
【0098】
抗原、ポリペプチドまたは抗原をコードするポリヌクレオチドを含むワクチンは、免疫応答を刺激するために種々の状況において抗原を提示することができる。種々のワクチン状況のいくつかは弱毒化病原体、不活性化病原体、トキソイド、結合物、組換えベクター等を含む。本発明のポリペプチドは、種々のワクチン組成物によって発現されてもまたは種々のワクチン組成物に結合してもよい。種々のワクチン組成物は、抗原または抗原をコードする分子を産生または発現する細胞に、抗原を直接提供してもまたは抗原を産生する組成物、例えば、発現構築物を送達してもよい。
【0099】
本発明のワクチンは、抗プロテアソームまたは抗ユビキチン抗体の産生を誘導するのに十分な有効な治療量の免疫原性プロテアソームまたはユビキチンペプチドを含む。得られる免疫によって避妊効果を生ずることができる。好ましくは、ワクチンは1種以上のほ乳類において避妊効果を生ずるのに有効である。最も好ましくは、ワクチンは、げっ歯類またはウサギ、ブタ、ウマ、サル、イヌ、ネコ、雌ウシおよびヒトの1種以上を含む非げっ歯類に投与すると、抗プロテアソーム抗体を産生する。
【0100】
他の態様において、タンパク性組成物またはポリペプチド、断片または模倣物を使用して、ワクチンに使用するための抗イディオタイプ抗体を作製することができる。抗イディオタイプワクチンでは、免疫原は、病原体の抗原分子に対して形成される第2の抗体のFab末端に対する抗体である。第1の抗体のFab末端は、病原体の抗原分子と同じ抗原形状を有し、抗原として使用することができる(例示的な米国特許第5,614,610号、同第5,766,588号を参照されたい)。本明細書において使用するための「ヒト化」抗体は、分泌される1つ以上のアミノ酸が、ヒト抗体においてより一般的に見られるアミノ酸と交換されている非ヒト種の抗体であってもよい。これは、通常の組換え技術、特に部位特異的突然変異を使用することによって容易に実施することができる。ヒト化抗体は、以下に記載するように、受動免疫剤としても使用することができる。
【0101】
V. 薬学的組成物
本発明の薬学的水性組成物は、薬学的に許容されうる担体または水性媒体に溶解または分散させた1つ以上のプロテアソーム阻害剤の有効量を含む。「薬学的または薬理学的に許容されうる」というフレーズは、そのような応答を誘発することが具体的に示されていない限り、ヒトに投与したとき有害反応、アレルギー反応または他の望ましくない反応を生じない分子的実態および組成物をいう。本明細書において使用する「薬学的に許容されうる担体」は、溶媒、分散媒体、抗菌剤および抗真菌剤、等張剤および吸収遅延剤等のいずれかおよび全てを含む。薬学的に活性な物質のためのこのような媒体および物質の使用は当技術分野において既知である。任意の従来の媒体または物質が作用成分と不適合である場合を除いて、治療用組成物におけるその使用が考慮される。追加の作用成分も本発明の組成物に組み入れることができる。
【0102】
被験者に投与される本発明の組成物の実際の投与量は、体重などの物理的および生理学的因子および投与経路によって決定される。これらを考慮すると、特定の被験者および/または治療経過のためのペプチド、ポリペプチド、ポリヌクレオチド、免疫グロブリンまたはプロテアソーム阻害剤組成物の用量を容易に決定することができる。
【0103】
本発明に使用することができるプロテアソーム阻害剤は当技術分野において既知であり、例えば、BIOMOL Research Laboratories, Inc.、Plymouth Meeting, PAから購入可能である。本発明において用途を見出すことができるこのような阻害剤の例には、N-アセチル-Leu-Leu-Nle-H、N-アセチル-Leu-Leu-Met-H、アクラシノマイシンA(アクラルビシン)、Ada-(Ahx)3-(Leu)3-ビニルスルホン、Ada-Lys(ビオチニル)-(Ahx)3-(Leu)3-ビニルスルホン、Ada-Tyr-(Ahx)3-(Leu)3-ビニルスルホン、バクテネシン-5、ブレフェルジンA、クルクミン、没食子酸(-)エピガロカテキン(EGCG)、エポキソマイシン、グリオトキシン、ラクタシスチン、クラスト(clasto)-ラクタシスチン a-ラクトン、N-トシル-Lys-クロロメチルケトン(TLCK)、N-トシル-Phe-クロロメチルケトン(TPCK)、NIP-(Leu)3-ビニルスルホン、フェプロペプチン(Phepropeptin) Aキモトリプシン、フェプロペプチン(Phepropeptin) Bキモトリプシン、フェプロペプチン(Phepropeptin) Cキモトリプシン、フェプロペプチン(Phepropeptin) Dキモトリプシン、PR11、PR39、ユビキチン5+1、Z-Il2-Glu(OBut)-Ala-Leu-H(PSI)、Z-Leu-Leu-Leu-ビニルスルホン、Z-Leu-Leu-Nva-H(MG115)、Z-Leu-Leu-Leu-H(MG132)、Z-Leu-Leu-Leu-B(OH)2(MG262)およびZ-Leu-Leu-Tyr-COCHOが挙げられる。
【0104】
本発明の薬学的組成物は、静脈内、皮内、動脈内、腹腔内、関節内、胸膜内、気管内、鼻腔内、膣内、局所的、筋肉内、腹腔内、皮下、小胞内、粘膜、経口、局所的、エアゾールを使用して局所的、注射、注入、持続注入、標的細胞を直接つける局所的灌流またはカテーテルもしくは洗浄投与することができる。典型的には、このような組成物は溶液または懸濁液として注射剤として調製され;注射前に液体を添加して溶液または懸濁液を調製するのに好適な固体剤を作製してもよく;製剤を乳化してもよい。組成物は滅菌されており、容易なシリンガビリティー(syringability)が存在する程度に流動体であり、製造および保存条件下において安定であり、細菌および真菌などの微生物の汚染作用から保護されている。内毒素汚染は安全レベル、例えば、0.5 ng/mgタンパク質未満に最小に保たれるべきであることが考慮される。
【0105】
本発明の組成物は、他の非作用成分を含有する滅菌水中で調製されて、注射に好適にされることが最も好ましいが、このような作用成分の溶液は、適宜、ヒドロキシプロピルセルロースなどの界面活性剤と好適に混合された水中でも調製することができる。分散剤も、液体ポリエチレングリコールおよびその混合物並びにオイル中で調製することができる。担体は、例えば、水、エタノール、ポリオール(例えば、プロピレングリコールおよび液体ポリエチレングリコール等)、それらの好適な混合物および植物油を含有する溶媒または分散媒体であってもよい。適切な流動性は、例えば、レシチンなどのコーティングを使用することによって、分散剤の場合には必要な粒子サイズを維持することによっておよび界面活性剤を使用することによって維持することができる。
【0106】
微生物の作用は、種々の抗菌および抗真菌剤、例えば、パラベン、クロロブタノール、フェノール、ソルビン酸、チメロサール等によって防止することができる。多数の例において、等張剤、例えば、糖または塩化ナトリウムを加えることが好ましい。注射用組成物の吸収の遅延は、吸収遅延剤、例えば、モノステアリン酸アルミニウムおよびゼラチンを組成物中に使用することによって生じることができる。
【0107】
製剤化したら、投与製剤に適合性した方法で、避妊作用を誘導するのに有効である量の溶液を投与する。水溶液での非経口投与のためには、例えば、溶液を適宜好適に緩衝化するべきであり、最初に希釈液を十分量の生理食塩液またはグルコースで等張にするべきである。これらの特定の水溶液は、静脈内、筋肉内、皮下および腹腔内投与に特に好適である。これに関連して、使用することができる滅菌水性媒体は、本発明の開示内容を考慮して当業者に既知である。治療する被験者の状態に応じて、用量の変更を必ず行う。投与担当者は、個々の被験者に適当な用量を任意に決定する。
【0108】
VI. 実施例
以下の実施例は、本発明の好ましい態様を実証するために含まれる。以下の実施例に開示する技術は、本発明を実施する際に十分に機能するために本発明者らが発見した技術を提供しており、従って本発明の好ましい実施様式を構成すると考えることができることが当業者に理解されるべきである。しかし、本発明の精神および範囲から逸脱することなく、多数の変更を開示されている具体的な態様に加えることができ、それでも同様または類似の結果を得ることができることを当業者は、本発明の開示内容に照らして理解するべきである。
【0109】
実施例1
材料と方法
A. 抗体
プロテアソームサブユニットサンプラーパックおよびプロテアソームサブユニットに対する以下のウサギ血清は、Affinity Research products. Ltd.、Mammahead、UKから購入した:
のマウスMECL-1-アミノ酸配列に基づいて組換えペプチドに対して形成された抗-MECL1サブユニット血清(カタログ番号#PW8150)は、免疫蛍光およびイムノゴールド透過型電子顕微鏡(TEM)には1/200で、ウェスタンブロットには1/4000で、インビトロにおける受精の阻止には1/2000〜1/8000で使用した。この抗体は、29 kDa MECL-1サブユニットおよびこのサブユニットを含むいくつかの他のさらに大きいタンパク質複合体を認識することが製造業者によって示された。抗-α/β-サブユニット血清(カタログ番号#PW 8155)は、ヒト赤血球細胞から単離したプロテアソーム調製物に対して形成され(TanakaおよびTsurumi, 1997)、ヒト赤血球溶解液の25kDa〜30 kDaのバンドセット並びにいくつかの別個のα-型およびβ-型サブユニットに相当する、他の細胞溶解液の別のさらに高いMWバンドを認識する。抗β1iサブユニット血清(カタログ番号#PW 8205)はマウスβ1iサブユニットの組換え完全長配列に対して形成され(Groettrupら, 1996)、26 kDa、36 kDaのタンパク質およびおそらく他のさらに高いMWタンパク質複合体を認識する。α/βおよびβ1i抗血清は、免疫蛍光およびイムノゴールドTEMでは1/200の希釈で、ウェスタンブロットでは1/4000で使用した。
【0110】
ブタ透明帯の外側面のユビキチンは、組換えヒトユビキチンの全分子に対して形成された抗体KM691(免疫蛍光では1/200の希釈、ウェスタンでは1:1000の希釈;Kamiya Biomedical Company、Seattle、WAから購入)によって検出された。前核は、ブタ胎仔核ユビキチン-CEP 52テイル融合タンパク質を認識し(McCauleyら, 2002)、Chemicon International Inc.、Temecula、CAから購入したウサギポリクローナル抗体AB 1690(1/100の希釈)を使用して対比染色した。この抗体は、卵巣組織切片のZPの外側層のユビキチン化タンパク質も認識した(図3Fを参照されたい)。
【0111】
アクロソームエキソサイトーシスのマーカーであるアクロソームチロシンキナーゼc-Yes(LeclercおよびGoupil, 2002)は、Santa Cruz Biotechnology, Inc.、Santa Cruz、CAから購入した、ヒトc-Yes(希釈、1/100)の進化的に保存されているC-末端配列に相当する組換えペプチドに対するウサギ血清を使用して検出した。ウサギ抗プロヒビチン抗体Ab-2(希釈、1/100;Thompsonら, 1997)はNeomarkers Inc.、Union City、CAから購入した。
【0112】
免疫蛍光検討に使用した二次抗体は、Zymed Inc.、South San Francisco、CA社製で、1/80に希釈した、赤色蛍光のTRITC-結合および緑色蛍光のFITC結合ヤギ抗マウスIgG、ヤギ抗マウスIgMおよびヤギ抗ウサギIgGを含んだ。DNAは、青色蛍光染料DAPI(Molecular Probes Inc.、Eugene、OR)で対比染色した。コロイド金TEMは、10 nm金結合ヤギ抗ウサギIgGをElectron Microscopy Sciences、Washington、PAから購入し、1/10希釈で使用した。
【0113】
B. 配偶子およびインビトロ受精
思春期前の雌ブタの卵巣を地元の食肉処理場から回収して、実験室に運び、3mm〜6mmの大きい胞状卵胞から卵母細胞を単離した。多層卵丘が存在するために卵丘細胞-卵母細胞複合体(COC)を解剖顕微鏡下で選択し、0.1%(w/v)ポリビニルアルコール(PVA)を含有するHEPES-緩衝タイロードの乳酸(TL-HEPES)培地で繰り返し洗浄した。3.05 mMグルコース、0.91 mMピルビン酸ナトリウム、0.57 mMシステイン、10 ng/ml上皮細胞増殖因子、0.5μg/ml FSH、0.5μg/ml LH、0.1%(w/v)PVA、75μg/mlぺニシリン-Gおよび50μg/ml硫酸ストレプトマイシン(AbeydeeraおよびDay, 1997)を添加した数滴の無血清改良組織培養培地(TCM 199;Gibco、Grand Island、NY)中で39℃において5%CO2雰囲気下で50のCOC群を22時間成熟させた。この後、FSHまたはLHを含有しない改良TCM 199でCOCを洗浄し、さらに20時間成熟させた。
【0114】
受精前に、0.1%(w/v)ヒアルロニダーゼを含有するTL-HEPES培地中でCOCをボルテックスして卵丘細胞を取り出し、113.1 mM NaCl、3 mM KCl、7.5 mM CaCl2、20 mM Tris、11 mMグルコース、5 mMピルビン酸ナトリウム、2 mM カフェインおよび0.2%(w/v)BSAからなる50μlの改良Tris緩衝培地(mTBM)中で洗浄した。受精は50μlのmTBM培地液滴中で実施した。0.1%(w/v)BSAを添加した10 mlのダルベッコのPBS(Gibco、Grand Island、NY) 中で凍結保存精子を解凍した。受精卵内に精子尾部および前核が存在することによって受精を判定するために、以前に記載されているように(Sutovskyら, 2003)、精子を生体ミトコンドリアプローブMitoTracker CMTM Rosで事前標識し、得られた接合体を、以下に記載するように、DNA染料DAPIで対比染色した。精子を遠心分離によって2回洗浄し、最終濃度5×105精子/mlまで受精液に添加した。IVFの6時間後に、接合体と思われるものを無血清TALP Hepes中で3回洗浄し、免疫蛍光、イムノゴールドTEMまたはウェスタンブロット用に処理した。ウェスタンブロット用の透明帯を含まない卵母細胞を入手するために、接合体を、0.5%w/vプロナーゼ(Protease、Sigma)を含有し、タンパク質を含有しないTALP-HEPESで30秒間処理した。
【0115】
C. プロテアソーム阻害剤およびIVF中の抗体処理
精子-透明帯相互作用におけるユビキチン-プロテアソーム経路の役割を調査するために、抗プロテアソーム抗体MECL-1およびプロテアソームのタンパク質作用の特異的な阻害剤、MG132(Z-Leu-Leu-Leu-CHO;可逆的阻害剤;Biomol Research Labs Inc、Plymouth Meeting、PA)およびラクタシスチン(C15H24N2O7S;非可逆的阻害剤;Biomol社製)の影響を評価した。IVFは、受精培地にMG132(10μM)またはラクタシスチン(10μMまたは100μM)または抗体MECL-1(希釈1/2000、1/4000、1/8000;Affinity Research、Mamhead、英国)を添加して上記のように実施した。阻害剤を含有しない対照は、IVF中阻害剤と同等な希釈液量の適当な溶媒(MG132:100%エタノール;ラクタシスチン:H2O)を添加した。抗体ブロックの対照は、抗体を添加せず、等量の免疫前のウサギ血清を添加した。
【0116】
MG132、ラクタシスチンまたはMECL-1抗体の封入は、透明帯で囲まれた卵母細胞の受精を完全に妨害したので、ブロックが生じた正確な受精段階を同定するために追加の実験を実施した。MG132および抗体MECL-1の存在下または非存在下において透明帯で囲まれたおよび透明帯を含まない(ZFまたはZP-)卵母細胞を使用してIVFを実施した。透明帯の除去は、卵母細胞を0.5%プロナーゼ(Sigma、St. Louis、MO)に短時間(30秒)接触させて実施した。卵母細胞を十分に洗浄し、ZF卵母細胞を、精子注入の1時間前に酵素処理から回収した。IVFのためには、ZF卵母細胞を1×105精子/mlと共にインキュベーションしたが、透明帯で囲まれた/透明帯が無傷(ZIまたはZP+)の卵母細胞は上記のように処理した。卵母細胞は、精子注入後(p.i.)12時間経過時に25%酢酸/エタノールで固定し、受精の頻度を求めるために1%オルセインで染色した。いくつかの実験において、卵母細胞を免疫蛍光顕微鏡で評価し、接合体の細胞質中の前核および精子尾部の存在を検出した。透明帯で囲まれたおよび透明帯を含まない胚も精子注入後6時間または20時間経過時に固定し、MG132によるプロテアソーム活性の阻害が精子-透明帯相互作用を特異的に遮断したかどうか、精子-卵細胞膜融合を阻害したかどうかまたは精子の移動性、生存性もしくは精子がアクロソームエキソサイトーシスを受ける能力に影響を与えたかどうかを判定するために免疫蛍光分析用に処理した。
【0117】
精子-透明帯結合および精子のアクロソーム状態を評価するために、透明帯で囲まれた卵母細胞を固定し、無傷のアクロソームを有する精子の核周囲卵胞膜(perinuclear theca)のサブアクロソーム(subacrosomal)および外側アクロソーム層に存在することが既知のチロシンキナーゼc-Yes(LeclercおよびGoupil, 2002)を免疫蛍光によって評価した。透明帯に結合した精子の数は、DAPI染色後に求めた。群あたり5つの卵を、アクロソームチロシンキナーゼc-Yes(免疫蛍光を参照されたい)に対する抗体およびDAPIで対比染色し、60×の倍率で写真撮影した。ZP結合アクロソーム反応性およびアクロソーム無傷精子を計数し、値をアクロソーム無傷精子/卵のパーセント(%)として表した。処理の結果をX2検定およびSAS8.2の一般的な線形モデル手法によって比較した。
【0118】
D. 卵巣組織切片、単離精子および接合体の全載試料の免疫蛍光
従来の組織化学的プロトコールに準じて卵巣組織片を4%パラホルムアルデヒドで固定し、洗浄して、パラフィンに包埋した。厚さ4ミクロンのパラフィン組織切片を切断し、顕微鏡スライドにのせ、キシレンで脱ろうし、100%〜70%エタノールシリーズおよび水で再水和し、スチーマーを使用してクエン酸緩衝液(pH 6.0)中で20分間前処理し、水に浸した組織を入れた生物トレイ上で抗体で処理した。遠心分離によって精子を精液試料からペレット化し、ポリ-L-リジンコーティングしたカバースリップに結合し、2%ホルムアルデヒド中で40分間固定し、4ウェルペトリ皿中でウェルからウェルに通過させることによって処理した。透明帯を含まないおよび透明帯が無傷の卵母細胞を、スライドまたはカバースリップに結合しないで、2%ホルムアルデヒドのPBS溶液中で固定し、1 ccシリンジに取り付けたアナペット(Unapette)ピペット(Beckton Dickinson)を使用して取り扱い、9ウェルパイレックスガラスプレート(Fischer Scientific)の個々のウェルを通過させることによって処理した。全ての試料は、上記および結果のセクションに記載する抗体で処理した。
【0119】
一般的なプロトコールにおいて、カバースリップ上の精子および未結合の卵母細胞は0.1%Triton-X100中で40分間または終夜透過性にしたが、パラフィン切片のスライドは透過性にしなかった。全ての試料は、0.1% TX-100を添加した5%正常ヤギ血清(NGS;Sigma)のPBS溶液中でブロックした。1%NGSおよび0.1% TX-100で強化した中性PBSの標識溶液を使用して、その後のインキュベーションおよび洗浄を実施した。カバースリップに、上記および結果のセクションに掲載し、標識溶液で希釈した一時抗体を積層し、40分(カバースリップに全細胞をのせたもの)または2時間(スライドに組織切片をのせたもの)おいた。標識溶液で手早く洗浄してから、蛍光を結合した適当な抗マウスIgGまたは抗マウスIgMを、異なる蛍光の抗ウサギIgGおよび2.5μg/ml DAPI(Molecular probes Inc.、Eugene、OR)と混合し、試料と共に30分〜40分インキュベーションした。標識溶液中で洗浄後、カバースリップを顕微鏡スライドにのせ、透明マニキュアで密封し、適当なフィルターセットおよびDIC光学を使用したNikon Eclipse 800エピ蛍光顕微鏡で観察した。画像はCoolSnap CCD HQカメラ(Roper Scientific、Tucson、AZ)およびMetaMorphソフトウェア(Universal Imaging Corp.、Downington、PA)で獲得し、記録可能なCDに保管し、Adobe Photoshop 5.5(Adobe Systems、Mountain View、CA)を使用して編集し、Epson Stylus 1280フォトプリンターで印刷した。
【0120】
E. コロイド金イムノラベリングおよび透過型電子顕微鏡
免疫蛍光について以下に記載するように、透明帯が無傷の接合体を、精子注入後6時間経過時に4%ホルムアルデヒド中で固定し、洗浄し、透過性にし、ブロックし、プロテアソームサブユニットに対するウサギ血清およびc-Yesチロシンキナーゼに対する血清と共に2時間インキュベーションした。洗浄後、接合体をヤギ抗ウサギIgG(希釈1% NGSおよび0.1% TX-100を添加したPBSで1/10)と共に1時間インキュベーションし、再度洗浄し、電子顕微鏡用に再固定し、パラホルムアルデヒドおよびグルタルアルデヒドに包埋した(以下を参照されたい)。微細構造の検討のための接合体は、免疫蛍光固定および処理しないで、TEM用に固定した。
【0121】
電子顕微鏡用の包埋のためには、標識および未標識接合体を、2%パラホルムアルデヒドおよび0.6%グルタルアルデヒドのカコジル酸緩衝液混合物中で固定し、1%四酸化オスミウム中で後固定し、上昇エタノールシリーズ(30%〜100%)で脱水し、PolyBed 812樹脂に包埋した。極薄切片をLeica Ultracut UCTウルトラミクロトームで作製し、100 MESH銅グリッド上に置き、酢酸ウラニルおよびクエン酸鉛により2段階で染色した。連続切片を調査し、Jeol 1200 EX電子顕微鏡で撮影した。各抗体および未処理/未標識接合体の微細構造検討のために10の卵母細胞を処理した。ネガフィルムをUmax Magic Scanフラットベッドスキャナーによってスキャンし、CD-Rに記録し、Adobe Photoshop 5.5ソフトウェアを使用してEpson Stylus 1280フォトプリンターで印刷した。
【0122】
F. SDS-PAGEおよびウェスタンブロット法
SDS-PAGEおよびウェスタンブロット法は、一般にSambrookら(2001)に記載されているように実施した。卵母細胞および接合体を所定の経過時点において培養から取り出し、タンパク質を含有しないTALP HEPES中で繰り返し洗浄し、群あたり350の接合体を、Tris(50 mM)、β-メルカプトエタノール(5%)、グリセロール(10%)、SDS(2%)およびPMSF(100 mM)を含有する10μlのロード緩衝液中で5分間煮沸した。雄ブタ精子タンパク質は、抽出緩衝液(50 mM Tris、20 mMイミダゾール、1 mM EDTA、5 mMベンズアミジンHCl、5μg/mlロイペプチン、1μg/mlペプスタチンA、100 mM PMSFおよび0.1% SDS)中で粉砕することによって抽出し、17000×gで30分間遠心分離した。β-メルカプトエタノール、グリセロール、SDSを上清に添加し、ロード緩衝液と同じ最終濃度を達成した。ヒト精子をロード緩衝液と共に煮沸し、次に25000×gで30分間遠心分離して、DNAペレットを除去した。電気泳動分離を10%ゲル上で実施し、Mini-Tank Electroblotter(Owl Scientific、Woburn、MA)を使用してタンパク質をPVDF膜に移した。膜を1%カゼイン溶液でブロックした。一次抗ユビキチン抗体と共に終夜インキュベーションし、次いで適当なビオチン結合二次抗体と共にインキュベーションした後、Vectastain ABC-Ampキット(Vector Lab Inc.、Burlingame、CA)を使用してアルカリ性ホスファターゼ反応を用いてシグナルを検出した。
【0123】
実施例2
プロテアソーム阻害剤による、精子-透明帯結合およびアクロソーム反応に影響を与えない、雄ブタ精子による透明帯の透過性の遮断
プロテアソーム阻害剤ラクタシスチンおよびMG132は、ブタ接合体内の父系的な精子ミトコンドリアのプロテアソーム分解経路を与える目的の検討経過において受精を妨害することを最初に観察した(Sutovskyら, 2003)。この検討は、受精のどの段階において、阻害剤が遮断を発揮するかを決定するために設計された実験によって追跡した。MG132およびラクタシスチンは共に、精子注入時に添加すると、受精を妨害した。実験を4回繰り返し、一致した結果が得られた。これらの試験において、MG132は卵母細胞の100%の受精を妨害した。MG132ほど強力でなく、非可逆的であることが既知のラクタシスチン(Goldbergら, 1995)は、受精速度に用量依存的に作用し、100μm濃度において100%遮断に達した。既知のプロテアソーム特異性および可逆性(LeeおよびGoldberg, 1998)、並びにラクタシスチンは、アクロソームに存在すると思われる非プロテアソームプロテアーゼの一部の活性、すなわちカテプシンAの活性を部分的に阻害する可能性があるという懸念(Ostrowskaら, 2000)により、さらに別の検討をMG132だけを用いて実施した。MG132は精子の生存性、精子-透明帯結合またはアクロソームエキソサイトーシスに影響を与えるということを排除するために、10μM MG132の存在下において受精した卵母細胞を精子注入後20時間経過時に透明帯を無傷のままで固定した。ZPに結合した精子頭部はDAPIで検出し、無傷のアクロソームは、アクロソーム-無傷精子の核周囲卵胞膜(perinuclear theca)のサブアクロソームおよび外側アクロソーム層に存在することが既知のチロシンキナーゼc-Yesに対する抗体によって検出された(図1 A〜D)。この実験の精子はMitoTracker CMTM Rosで事前に標識されたので、精子ミトコンドリアの鞘は、受精した卵母細胞内で検出可能になった。MG132を用いた以前の実験と同様に、阻害剤は受精を完全に妨害した(MG132群の受精0%対対照の受精77%)。しかし、卵あたりの結合精子の数(MG132の99.3対対照の93.3)またはアクロソーム無傷、透明帯結合精子の割合(MG132の2.01%対対照の2.14%)には有意な差はなかった。
【0124】
実施例3
雄ブタ精子アクロソームに存在するプロテアソームサブユニット
阻害剤のデータおよびプロテアソームサブユニットはほ乳類/ヒト精子アクロソーム内に存在することを確認している以前の報告(例えば、Bialyら, 2001)並びに無脊椎動物のアクロソーム活性におけるユビキチン-プロテアソーム系を示す証拠(MatsumuraおよびAketa, 1991;Sawadaら, 2001a、2001b)を考慮して、プロテアソームは雄ブタ精子アクロソームに存在するという可能性を検討することを決定した。コアサブユニットおよびサブユニットLMP-2(β1i)、MECL1およびMECL2に対するポリクローナル抗体はアクロソームにおける主要なシグナルを提供し、他の精子構造物におけるバックグラウンドはほとんどまたは全くなかった(図1 E〜I)。ユビキチンもアクロソームにおいて検出された(図1J)。同じプロテアソームサブユニットが、雄ブタ精子頭部および雄ブタ精子全体の抽出液中でウェスタンブロットによってさらに検出された(図2 A〜C)。透明帯透過性におけるプロテアソームのこの新規役割に一致して、マウスプロテアソームサブユニットMECL-1のAA配列(AA 218〜236)の組換えペプチドに対する抗体は低用量で、透明帯無傷卵母細胞における受精を確実に遮断したが、透明帯を含まない卵母細胞では遮断しなかった。
【0125】
実施例4
ユビキチン化タンパク質のブタ透明帯の外側面における存在
プロテアソーム阻害剤が透明帯侵入を遮断し、プロテアソームサブユニットおよびユビキチン経路の成分が共にアクロソームに存在する場合には(図1 E〜J)、ホヤ卵黄膜に見られるように(Sawadaら, 2002a)ユビキチン化感受性タンパク質/精子受容体はZPの外側層に存在するはずであり、または精子アクロソーム滲出液と接触する前にすでにユビキチン化されている、タンパク質/精子受容体がZPに存在するはずである。組換えヒトユビキチンに対する抗体KM 691を使用して、ブタ卵巣組織のパラフィン切片をスクリーニングし(図3 A〜D、F、G)、卵巣の胞状卵胞から単離したブタCOCの全載試料をスクリーニングした(図3E;図6A、Bも参照されたい)。
【0126】
ユビキチン交差反応性タンパク質の別個の濃密な層が、卵巣組織切片(図3 A〜D、E、F)および全載試料(図3E、図6A、B O)の前胞状および胞状卵胞のブタ卵母細胞の透明帯の外側層で検出された。これらの(ユビキチン免疫反応性)タンパク質の最も顕著な特徴は、それらが主にZPの外側面で検出されたということであり、これは、主要なZP成分、ZP1、2および3がほ乳類の透明帯に均一に分布することと異なる(ThalerおよびCardullo, 2002;WassarmanおよびLitscher 1995)。同様に、透明帯糖タンパク質に存在する糖残基はZPの内側面に存在するか、またはZPに均一に分散する(Avilesら, 1997;2000)。ユビキチン免疫反応性のこの局在化は、抗ユビキチン抗体のZPを通過する透過性が制限されているからではない。その理由は、このようなデータは組織切片の表面染色によって得られたからである。さらに、関連のないポリクローナル抗ユビキチン抗体AB1690もZPの外側面のユビキチン化タンパク質のこの層を認識し(図3F)、このような免疫反応性は、KM691を含有せず、免疫前ウサギ血清と共にインキュベーションし、次に適当な蛍光を結合した免疫グロブリンと共にインキュベーションした陰性対照では見られなかった(図3G)。
【0127】
次に、ユビキチン免疫反応性ZP基質の一部は、精子がZPに結合後分解を受けるかどうかおよびこのような分解はプロテアソーム阻害剤によって妨害されうるかどうかを判定した。卵母細胞はMG132の存在下/非存在下において受精し、精子注入後6時間経過時に回収し、レーンあたりの総数350の透明帯無傷(ZIまたはプロテアソーム+)または透明帯を含まない(ZFまたはZP-)をPAGEゲルにロードした。精子注入後6時間経過時のZF(図4A、レーン1)およびZI(図4A、レーン2)、受精後の卵母細胞の比較は、ZPが寄与する数多くのユビキチン免疫反応性バンドを示す。興味深いことに、123 kDaマーカーのわずかに下方に泳動するレーンセットはZP+/MG132-対照卵母細胞にはないが、ZP+MG132+卵母細胞には存在した(図4A、レーン3)。このようなバンドはヒト(図4A、レーン4)および雄ブタ(図4A、レーン5)精子の対照溶解液には存在しなかった。驚くべきことに、タンパク質移動後の卵母細胞ゲルのクーマシーブルー染色(図4A;右のパネル)は、このようなバンドの一部はゲルから膜に完全に移動しなかったことを示した。ユビキチンブロットと同様に、このようなバンドはZF卵母細胞レーンには全く見られず、ZP+/MG132-レーンでは劇的に低下した。ZP+/MG132+に存在するこのような大量のこのようなタンパク質は、このようなタンパク質はMG132が存在しない場合には対照受精中に消化されること、およびこの分解はMG132を受精培地に添加することによって効果的に妨害されることを示唆している。ポリ-ユビキチンなどのタンパク質の親水性AA残基含量である、高リジンがこの影響に寄与している可能性があるが、このようなタンパク質の一部があまり移動しない理由は不明である。
【0128】
接合体の総タンパク質含量に注目した上記の観察を、タンパク質移動前のゲルのクーマシーブルー染色によって評価した(図4B)。図4Aに示すゲルおよび膜と同様に、123 kDaの下方に泳動するバンドセットはZP+/MG132-接合体(レーン2)では劇的に低下しており、ZP+/MG132+接合体(レーン3)では優勢であり、ZP-/MG132+(レーン4)接合体、ZP-/MG132-接合体(示していない)および対照精子抽出液(レーン4および5)では見られなかった。
【0129】
その後の繰り返しでは(図4C;左のパネル)、ZP+未受精の卵母細胞のタンパク質(レーン2)、ZP+/MG132-接合体(レーン3)、ZP+/MG132+卵母細胞(レーン4)およびヒト(レーン5)および雄ブタ(レーン6)精子は分離し、移動した。膜を抗ユビキチンKM-691で探索し(図4C;左)、タンパク質移動後ゲルをクーマシーブルーで染色し(図4C;右)、ゲルと抗ユビキチン探索膜を重ねた(図4C;中央)。123 kDaマーカーの下方のバンドセットの移動は再度妨害され、ZP+/MG132+レーンにシングルバンドが存在し、M-IIおよびZP+/MG132-レーンには密度の低い、低MWバンドが見られた。ブロット膜では、約30 kDaの新たなユビキチン免疫反応性バンド(図3D、矢印)が、梯子上の密度の低いバンド(矢印)以外にZP+/MG132-接合体に出現したが、M-II卵母細胞またはZP+/MG132+接合体では出現しなかった。従って、このバンドは、受精後に生じ、MG132によって妨害されるタンパク質分解産物であった可能性がある。以前の試験と同様に、123 kDaマーカーの下方のバンドセットは受精後のZP+/MG132-対照接合体では低下していたが、M-II卵母細胞およびZP+/MG132+接合体では低下していなかった(図4E)。最後に、MG132が存在しない場合に、IVF後の約179 kDaレベルの2つのバンドの密度がわずかではあるが、識別できるほど低下した(図4F;矢印)。全体で、4つの試験を実施し、同様の結果が得られた。ZP+/MG132+およびZP+MG132-接合体は全ての試験に含め、対照卵母細胞レーンは異なった(M-II/ZP+;M-II/ZP-;IVF/ZP-/MG132+;IVF/ZP-/MG132-)。精子-卵母細胞結合は、ウェスタンブロットに使用した接合体の各バッチにおいて光学顕微鏡によって調査し、ZP+/MG132+接合体では、見かけ上正常で、MG132-処理に影響されなかった。従って、いくつかのZP-寄与卵母細胞タンパク質の分解はIVF後に生じ、選択的プロテアソーム阻害剤MG132によって妨害された。
【0130】
雄ブタ精子の受精能力は、精子の移動性または卵母細胞の細胞質膜と融合する精子の能力に対するプロテアソーム阻害剤の非特異的な影響によって低下されないことを確認するために、ブタ卵母細胞からZPを除去し、MG132の存在下または非存在下においてアクロソーム無傷精子を受精させた(図5 A〜H)。予測されたように、MG132は透明帯無傷卵母細胞の受精を阻害したが、透明帯を含まない卵母細胞には影響せず、その92%〜96%は受精し、そのほとんどは多精受精的であった(図5 A〜H)。卵母細胞または精子がMG132と共に事前にインキュベーションされ、十分に洗浄してから精子注入に使用すると、MG132は透明帯無傷卵母細胞の受精に影響を与えなかった。まとめると、これらのデータは、プロテアソーム阻害剤は、精子の移動性、精子-透明帯結合およびアクロソームエキソサイトーシスに影響を与えないで、雄ブタ精子による透明帯透過性を遮断することを納得させる証拠を提供する。
【0131】
次に、精子注入後6時間経過時の接合体の免疫蛍光および微分干渉コントラスト顕微鏡(DIC)画像によって、精子-アクロソームプロテアソームとユビキチン免疫反応性ZPタンパク質の相互作用を可視化することを試みた。ユビキチン標識のメッシュ様パターンが受精後の接合体のZP表面に見られ、ZP結合精子の下方および周囲の蛍光が低下していた(図6A)。後者の図では(図6 B〜B'')、ZP結合精子の下方のユビキチン免疫反応性層は消化されたと思われ(図6B';矢印)、精子頭部下方に空間を生じ、DIC光学的方法では明らかに識別可能である(図6B'';矢印)。無傷の精子アクロソームにおいて以前に検出されたプロテアソームサブユニットMECL-1、α/β-型サブユニットおよびβ1i(図1)は、無傷のアクロソームを有するZP結合精子(図6C、E、F;矢印)、エキソサイトーシスを受けたアクロソーム(図6C、F、G;矢印)および除去されたアクロソーム被覆(rejected acrosomal shrouds)(図6C、E;矢印)においても見られた。体細胞核のプロテアソームサブユニットおよびユビキチンシステムの活性の存在に一致して(Chenら, 2002;Rivett, 1998)、α-型およびβ-型のプロテアソームサブユニットが受精後の接合体内部の前核において検出された(図6D)。
【0132】
同じプロテアソームサブユニットは、アクロソームエキソサイトーシスおよび小胞形成を受けたZP結合精子のアクロソーム基質(図7 A〜C)および内側のアクロソーム膜(図7 D〜G)においてコロイド金TEMによって検出された(微細構造を図7 I〜Kに示す)。陽性対照として、抗c-Yes抗体(免疫蛍光の図1A、Bを参照されたい)は、主に外側アクロソーム膜および外側アクロソーム核周囲卵胞膜(perinuclear theca)の複合体を認識した(図7H)。
【0133】
実施例5
プロテアソーム阻害剤MG132および抗プロテアソーム血清MECL1によるインビトロにおけるウシ受精の阻害
雄ウシ精子がウシ卵子に受精する能力に対するプロテアソーム干渉の影響をインビトロにおいて分析した。特異的で、完全に可逆的なプロテアソーム阻害剤MG132(100μM)を、精子注入時に受精培地に添加した。これにより、精子注入後20時間経過時に調査した103の卵巣のうち3つにだけ受精が生じた(受精率2.8%;表2)。10μM MG132を使用した場合にはプロテアソーム干渉はウシ受精を一部遮断したが、100μM Mg132を適用した場合には完全に遮断することを分析は示した。MG132の溶媒である適量のエタノールを添加しても、受精または前核形成に影響を与えず、MG132の非存在下でのIVFの前の2時間の100μM MG132との未受精卵のプレインキュベーションも影響を与えなかった。同様に、プロテアソームサブユニットMECL1に対する免疫血清はウシIVFを遮断したが、適当な免疫前血清はしなかった。前核中のプロテアソームを、α-型およびβ-型プロテアソームサブユニットに対する抗体を用いて検出した。DNAはDAPIで可視化した(青色)。
【0134】
MG132の溶媒溶液である適当な量の100%エタノールの存在下において精子注入した卵は受精率85.9%を示した。ラクタシスチンなどの非可逆的なプロテアソーム阻害剤とは異なり、MG132はプロテアソームコアに永久的に結合しない。しかし、MG132は、精子アクロソーム媒介性プロテアソームに対する阻害作用とは異なり(Sutovskyら, 2003a、b)、透明帯卵細胞膜または卵質に対する直接作用によって受精を遮断または妨害することを排除するために、数群の卵を受精前に100μM MG132と共に2時間プレインキュベーションした。MG132の非存在下で受精させた、このようなプレインキュベーション卵において受精率97.5%が観察された(表2)。ブタIVFの上記の検討と同様に、20SプロテアソームコアサブユニットMECL1に対するウサギ血清は、1/200、1/500および場合によっては、1/1000の希釈時において、インビトロにおけるウシ卵の受精を効果的に遮断した(それぞれ、受精率15.4%、3.7%および32.1%;表1)。このような影響は、対応する希釈において免疫前のウサギ血清によっては誘発されなかった。
【0135】
MG132も抗-MECL1処理も、卵母細胞の形態の同定可能な異常を伴わなかった。これは、雄ウシ精子のプロテアソームサブユニットMECL1、LMP-2並びにα-型およびβ-型に実施された免疫局在化によって同定された。陰性対照は、適当な合成ペプチドで免疫飽和した抗MECL1抗体を使用して実施した。同じ細胞の微分干渉コントラスト画像を作製した。上記の観察は、上記の雄ブタの精子-アクロソームプロテアソームについて同定された局在化および機能並びに雄ウシ精子のMECL1、LMP2およびα/βのプロテアソームサブユニットの免疫局在化に一致した。
【0136】
(表2)プロテアソーム阻害剤MG132および抗プロテアソーム血清MECL1のインビトロにおけるウシ受精および前核形成の阻害
【0137】
実施例6
プロテアソームの接合体形成開始時からの前核への浸潤
ユビキチン-プロテアソーム経路は、最近まで、細胞質コンパートメントだけに存在するとされていた。体細胞の核コンパートメントにおけるプロテアソームの存在およびタンパク質分解作用が分析された。種々の前核発生段階のウシ前核接合体のプロテアソームサブユニットLMP2、α/βおよびMECL1の免疫局在化に特異的な免疫血清を使用した。卵質への導入直後、十分に凝縮された精子核は検出可能なプロテアソームを含まないことが示された。プロテアソームサブユニットは、精子核脱凝縮の開始段階において雄および雌の前核に浸潤を開始した。雄および雌の前核は、フルサイズおよび並置に達する前にプロテアソームに占められる。高密度プロテアソームが、単精受精および多精受精卵のフルサイズの並置された前核内並びに非同期的な前核発生の場合に観察された。受精前に卵を100μM MG132とプレインキュベーションしても、プロテアソームサブユニットの前核への流入に影響を与えず、精子および100μM MG132の受精培地への同時添加の20時間後に未受精のままであった卵ではプロテアソームと雌染色体の結合は観察されなかった。合成MECL1ペプチドを飽和させた抗MECL1抗体を使用する陰性対照は任意のシグナルを示さなかった。精子尾部は、受精前の精子を生体ミトコンドリア染料MitoTracker Green FM(緑色)と共にプレインキュベーションすることによって可視化した。
【0138】
ウシ卵質における上記プロテアソームサブユニットの発現はウェスタンブロットで確認した。プロテアソームサブユニットMECL1は、ウシ卵(図8、レーン1)、雄ウシ精子(図8、レーン2)およびウシ卵丘細胞(図8、レーン3)の溶解液中で検出された。卵質溶解液の予測された29 kDaレーン以外に、種々の細胞種の細胞溶解液に通常見られる高MWレーンは、おそらく他のプロテアソームサブユニットからのMECL1サブユニットの不完全な解離の結果として、精子および卵丘細胞溶解液中に存在した。
【0139】
実施例7
プロテアソーム干渉の正常な前核形成の妨害および、受精卵における未成熟染色体凝縮の発生
ウシ受精を阻害するための最適なMG132濃度を決定することを目的とした検討の経過において、低濃度(10μM)では、MG132処理は受精を部分的にしか阻害しないことが観察された。2つの別個の試験で得られた平均受精率は、10μM 132で38.2%(n=55)で、対照(n=37)では75.7%であった。低濃度MG132の存在下におけるこのような受精により、前核形成に対するプロテアソーム干渉の影響を観察することができた。受精卵中の前核は、Nup 153を含む核膜孔複合体(NPC)タンパク質群および他の関連するヌクレオポリンを認識するモノクローナル抗体mAb 414の組み合わせによって可視化した。DNAは青色蛍光DAPIで染色し、受精卵内の精子尾部は、精子のMitoTracker GreenFMとのプレインキュベーション中に精子ミトコンドリア鞘に蓄積する緑色蛍光の存在によって同定した。ヌクレオポリン標識は、ウシ(図12A;Sutovskyら, 1998)およびマウス(FitzHarrisら, 2003)の前核/接合体の早期形成段階における核エンベロープ強度および機能の適当なマーカーであるので、選択した。
【0140】
異常な前核形成および未成熟染色体凝縮(PCC)が、精子の透過性を一部可能にする条件である、10μM MG132の存在下で受精したウシ卵に観察された。前核は、核膜孔複合体(NPC;赤)タンパク質、ヌクレオポリンNup153に対するモノクローナル抗体およびDNA染料DAPI(青)の組み合わせで可視化した。同様に、このような卵におけるPN形成パターンは異常であった。未成熟染色体凝縮(PCC)および前核並置失敗の組み合わせは、単精受精および多精受精卵の最も一般的な異常であった。いくつかの場合において、精子核の不完全な脱凝縮が、雌クロマチンのPCCを伴って観察された。PCC中の染色体の散乱および凝縮されたクロマチン周囲への可溶性であると推定されるヌクレオポリンの集成が観察された。このようなヌクレオポリンの蓄積は父系的な寄与によるとは思われない:雄ウシ精子は、精子尾部接続片の冗長な核エンベロープ(HoおよびSuarez, 2003)に位置する残存量のヌクレオポリン(Sutovskyら, 1999)を保有する。
【0141】
実施例8
正常な前核形成の妨害機序の分析
以前の検討において、NPCおよびヌクレオポリンの環状の層状様配置が、自発的な活性化へと進行する成熟した中期-II卵において観察されることがあった。しかし、MG132処理した細胞質に接触した精子核も未成熟染色体凝縮を受けたことを考慮すべきである。プロテアソームの分解は精子クロマチンの最初の脱凝縮に必要でないが、雄前核の十分な形成に必要であることをこれは示唆している。このような影響は、接合体内の母系的および父系的クロマチンのプロテアソーム依存的なリモデリングによって直接的であっても、またはプロテアソーム依存的な細胞周期進行の遮断などの間接的であってもよい。減数分裂および前核形成の進行はラットではMG132によって阻害されると思われる(Josefsbergら, 2000)。この観察は、クローニングされた最初のラット子孫を作製するために使用され、研究者らはラット卵の未成熟な活性化を防ぐためにMG132を適用した(Zhouら, 2003)。
【0142】
PN並置の失敗は、精子-星状マイクロチューブが前核並置中に固定される、無傷の核エンベロープがないこと、または精子中心小体が、接合体中心小体再構成および精子星状形成の前に遊離されなければならない、精子尾部接続片におけるプロテアソーム機能の遮断によると思われる(Sutovskyら, 1996;SutovskyおよびSchatten, 1997;Wojcikら, 2000)。
【0143】
実施例9
胚移植のための胚盤胞を作製するためのブタインビトロにおける受精中の多精受精の低下
プロテアソーム干渉を使用して、全体的な受精率を低下させないで、IVF中に多精受精率を軽減または低下させることができる。その後、多精受精接合体由来の生存能力のない倍数体胚盤胞の数を低下させることによって、胚移植プログラムのための正常な2倍体胚盤胞の作製を増加させることができる受精媒体への新規添加剤を製剤化することができる。初期の検討では(表3)、ZPの透過性は、100 uM濃度の特異的なプロテアソーム阻害剤MG-132およびラクタシスチンによって完全に阻害された。閾値濃度10 uMでは、プロテアソーム阻害剤ラクタシスチンは受精を妨害しなかったが、多精受精率を10%まで低下すると同時に単精受精率を65%まで増加した。精子注入後20時間経過時の接合体内の蛍光標識された精子尾部の数によって評価したとき、この検討における対照受精は、多精受精および単精受精が、それぞれ、44%および38%であった。
【0144】
(表3)精子注入時の培養培地への10 uMラクタシスチンの添加によるブタIVF中の多精受精の低下
【0145】
本明細書に開示し、特許請求の範囲に記載する組成物および/または方法は全て、本発明の開示内容に照らして不要な実験を行うことなく作製することができ、実行することができる。本発明の組成物および方法は、好ましい態様に関して記載されているが、本発明の概念、精神および範囲から逸脱することなく、本発明の組成物および/または方法並びに本明細書に記載されている方法の段階および段階の順序に変更を適用することができることが当業者に明らかである。さらに具体的には、化学的および生理学的に関連するある一定の物質を本明細書に記載する物質と交換することができるが、同じまたは同様の結果が得られると思われることが明らかである。当業者に明らかな全てのこのような同様の置換および改良は、添付の特許請求の範囲によって規定される本発明の精神、範囲および概念の範囲内であると考えられる。
【0146】
参照文献
以下の参照文献は、本明細書に記載するものに補助的な例示的な手法または他の詳細を提供する程度に、参照として具体的に本明細書に組み入れられている。
【図面の簡単な説明】
【0147】
以下の図面は本明細書の一部を形成し、本発明のある局面をさらに証明するために含まれる。本発明は、本明細書に提供する具体的な態様の詳細な説明と合わせて、これらの図面の1つ以上を参照することによってさらに理解することができる。
【図1】図1A〜1Jは、プロテアソーム阻害剤処理した卵母細胞における精子-ZP結合の一例およびブタ配偶子におけるユビキチン-プロテアソーム系の成分を例示する。(図1A)常在性チロシンキナーゼc-Yesに対する抗体で標識した無傷のアクロソームを有する雄ブタ精子。(図1B)。表面帯(zona surface)へのアクロソームの分散後のc-Yes標識の残遺物。(図1C)10 μM MG132(プロテアソーム阻害剤)の存在下において受精した卵母細胞は、精子注入後(p.i.)20時間経過時に無傷のアクロソームを有するわずかに3つの精子を示す。これは、MG132は特異性高く受精を遮断するが、配偶子の生存性、精子-卵子結合および精子-アクロソーム反応自体を損なわないことを例示している。(図1D)精子注入後(p.i.)20時間経過時に3つのアクロソーム無傷精子を有する対照卵母細胞。図1 A〜Dの精子は生体ミトコンドリア染色色素MitoTracker CMTM Rosで事前に標識した。(図1E〜I)サブユニットLMP-2、MECL 1およびMECL 2に対する抗体並びにプロテアソームコアサブユニットに対する2つの異なる抗体による雄ブタ精子プロテアソームの検出。(図1J)Ab 1690によって検出されるユビキチン。
【図2】図2A〜2Cは、雄ブタ熱抽出(すなわち、アクロソームなし;レーン2)、TX-100抽出(レーン3)および無傷(レーン4)精子頭部の溶解物におけるプロテアソームサブユニットMECL-1(図2A)、β1i(図2B)並びに種々のα-型およびβ-型サブユニット(図2C)の免疫検出の一例を例示する。分子量マーカーはレーン1に示す。
【図3】図3A〜3Gは、ブタ透明帯の外側面のユビキチン免疫反応タンパク質の一例を例示する。ユビキチンは、ブタ前胞状卵胞(図3A、図3B、図3F、図3G)および胞状卵胞(図3C、図3D)内側並びに全載成熟卵母細胞表面(図3E、卵核胞段階の卵母細胞を示す)においてマウスモノクローナル抗体KM691によって(図3A〜D、図3F、図3E)またはウサギ血清AB1690(図3E;図3F)によって検出された。これらのパラフィン切片のいくつか(図3A、図3B)の卵胞顆粒膜細胞を、ミトコンドリアタンパク質プロヒビチンに対する抗体で対比染色した。図3Eでは、Ab1690による標識はこの卵母細胞の卵核胞内の核小体の輪郭を描き、表面帯の標識は抗体に対するZPの不透過性によるものではなかったことを示す。ユビキチン免疫反応性は、KM691を使用せず、免疫前ウサギ血清と共にインキュベーションし、次に蛍光的に結合した適当な免疫グロブリンと共にインキュベーションすることによって処理した陰性対照組織切片では見られない(図3G)。全ての画像のDNAはDAPIで対比染色した。
【図4】図4A〜4Fは、プロテアソーム阻害剤MG132が存在する場合または存在しない場合における受精前および受精後の透明帯無傷および透明帯除去ブタ卵母細胞の溶解物におけるユビキチン基質の免疫検出の一例を例示する。(図4A)左のパネル:抗-ユビキチンKM691で調査した、受精後6時間経過時の透明帯除去(レーン1)および透明帯無傷(レーン2)の受精卵母細胞の溶解物は、ZPによる数多くのユビキチン免疫反応バンドを示す。123 kDaマーカーのわずかに下に泳動するレーンセット(矢印)はZP+/MG132-対照卵母細胞には存在しないが、ZP+MG132+卵母細胞(レーン3)には存在する。このようなバンドは、ヒト(レーン4)および雄ブタ(レーン5)精子の対照溶解物には存在しない。右のパネル:タンパク質移動後の同じ卵母細胞ゲルのクーマシーブルー染色は、このような<123 kDaバンド(矢印)のいくつかはゲルから膜に完全に移動せず、ZP-卵母細胞レーン(レーン1)では存在せず、ZP+/MG132-レーン(レーン2)では低下していることを示している。存在するこのようなタンパク質の大部分はZP+/MG132+レーン(レーン3)に存在する。(図4B)タンパク質移動前のゲルのクーマシーブルー染色。(図4C)左のパネル:抗-ユビキチンKM-691で調査した、ZP+未受精卵母細胞(レーン2)、ZP+/MG132-接合体(レーン3)、ZP+/MG132+卵母細胞(レーン4)並びにヒト(レーン5)および雄ブタ(レーン6)精子のタンパク質。中央:右のパネルに示すように、移動後にクーマシーブルーで染色したゲルを左のパネルに重ねたもの。
【図5】図5A〜5Hは、プロテアソーム阻害剤による透明帯の透過性の遮断および透明帯除去卵母細胞ではこのような遮断が存在しないことの一例を例示する。MG132は透明帯無傷卵母細胞ではインビトロにおいて受精を阻害したが(図5C、図5D)、透明帯切除卵母細胞には影響を与えなかった(図5A、図5B)。適当な透明帯除去(図5E、図5F)および透明帯無傷(図5G、5H)対照を実施した。DNAをDAPIで対比染色し、受精卵母細胞内の前核および核小体を抗体Ab 1690で標識した。
【図6】精子注入後(p. i.)6時間経過時のZP表面の雄ブタ精子の微分干渉顕微鏡(DIC)画像と合わせて抗-ユビキチンおよび抗-プロテアソーム免疫蛍光によって可視化した、精子プロテアソームとユビキチン免疫反応性ZPタンパク質の相互作用の一例を例示する。
【図7】図7A〜7Kは、アクロソームエキソサイトーシス中のプロテアソームサブユニットの免疫電子顕微鏡検出の一例を例示する。(図7A〜G)サブユニットβ1i(図7A〜D)、α/β(図7E、F)およびMECL-1(図7G)のコロイド金標識はアクロソーム基質内および内側のアクロソーム膜に存在する。(図7H)アクロソームマーカー、チロシンキナーゼc-イエス(yes)は、主に外側のアクロソーム膜および外側-アクロソーム核周囲卵胞膜の複合体に限定される。(図7I〜K)従来の電子顕微鏡は、ブタ受精中のアクロソームエキソサイトーシスの超微細構造を示す。
【図8】100のウシ卵子(レーン1)、雄ウシ精子抽出物(レーン2)およびウシ卵丘細胞(レーン3)におけるプロテアソームサブユニットMECL1のウェスタンブロット法である。星印は予測される29 kDaレーンを表す。種々の細胞種の細胞溶解物に共通して見られる高MWレーン(参照文献 Groettrupら,1996)は、試料調製過程における他のプロテアソームサブユニット凝集の結果である。〜32 kDaのバンドは、未処理MECL-1前駆体タンパク質に相当する。
【特許請求の範囲】
【請求項1】
精子のプロテアソーム活性を阻害する段階を含む避妊方法。
【請求項2】
精子のプロテアソーム活性がインビトロにおいて阻害される、請求項1記載の方法。
【請求項3】
精子のプロテアソーム活性がインビボにおいて阻害される、請求項1記載の方法。
【請求項4】
プロテアソーム活性が、プロテアソームに結合してプロテアソーム活性を阻害する抗体によって阻害される、請求項1記載の方法。
【請求項5】
抗体がヒト化抗体を含む、請求項4記載の方法。
【請求項6】
プロテアソーム活性が、抗体による受動免疫によって阻害される、請求項1記載の方法。
【請求項7】
抗体が、プロテアソームペプチドの少なくとも1つの抗原で被験者を免疫することによって誘導される、請求項4記載の方法。
【請求項8】
抗原が抗原性プロテアソームペプチドである、請求項7記載の方法。
【請求項9】
少なくとも1つの抗原性ペプチドがワクチンに含まれる、請求項8記載の方法。
【請求項10】
1つ以上の抗原性プロテアソームペプチドが、配列番号:1または配列番号:2のアミノ酸配列を含む、請求項9記載の方法。
【請求項11】
抗原性プロテアソームペプチドが融合タンパク質である、請求項7記載の方法。
【請求項12】
多重抗原エピトープが単一タンパク質内に存在する、請求項7記載の方法。
【請求項13】
抗体が、少なくとも1つの抗原性プロテアソームペプチドをコードする少なくとも1つのポリヌクレオチドで免疫することによって誘導される、請求項4記載の方法。
【請求項14】
少なくとも1つの抗原性プロテアソームペプチドをコードするポリヌクレオチドが発現ベクターに含まれる、請求項13記載の方法。
【請求項15】
発現ベクターがプラスミド発現ベクターである、請求項14記載の方法。
【請求項16】
プロテアソーム活性が、精子にプロテアソーム阻害剤を接触させることによって阻害される、請求項1記載の方法。
【請求項17】
プロテアソーム阻害剤が、薬学的に許容されうる製剤に含まれる、請求項16記載の方法。
【請求項18】
プロテアソーム阻害剤がMG132またはラクタシスチンである、請求項16記載の方法。
【請求項19】
薬学的に許容されうる製剤が、溶液、懸濁液、錠剤、ピル、カプセル、徐放性製剤、粉末、クリーム、軟膏(ointment)、軟膏(salve)、スプレー、ポンプ、リポソーム、坐剤、吸入剤またはパッチの形態に製剤化される、請求項17記載の方法。
【請求項20】
薬学的に許容されうる製剤の投与経路が静脈内、腹腔内、皮内、筋肉内、子宮内、皮膚、鼻、口腔、膣、吸入または局所的である、請求項17記載の方法。
【請求項21】
投与経路が子宮内である、請求項20記載の方法。
【請求項22】
1つ以上のプロテアソームペプチドをほ乳類に投与する段階を含む、ほ乳類において抗プロテアソーム抗体を誘導する方法であって、投与する段階が、ほ乳類のプロテアソームに結合して、その活性を阻害する抗体の産生を誘導する方法。
【請求項23】
ほ乳類がヒトである、請求項22記載の方法。
【請求項24】
1つ以上のプロテアソームペプチドが、プロテアソームペプチドをコードするポリヌクレオチドの発現によって投与される、請求項22記載の方法。
【請求項25】
プロテアソームペプチドをコードするポリヌクレオチドが、筋肉内、静脈内、皮下、腹腔内、皮内、経口または吸入投与経路によって投与される、請求項24記載の方法。
【請求項26】
プロテアソームペプチドが、筋肉内、静脈内、皮下、腹腔内、皮内、経口または吸入投与経路によって投与される、請求項22記載の方法。
【請求項27】
卵母細胞に関連するユビキチンの分解を阻害する段階を含む避妊方法。
【請求項28】
ユビキチン分解の阻害が、ユビキチン、ユビキチン活性化酵素、ユビキチン-キャリア/結合酵素またはユビキチン-リガーゼに対する抗体の投与による、請求項27記載の方法。
【請求項29】
精子内または精子上のユビキチン-プロテアソーム経路に関連するタンパク質の有無を検出する段階を含む受精能を評価する方法。
【請求項30】
ユビキチン-プロテアソーム関連タンパク質の有無を検出する段階が、ユビキチン-プロテアソーム関連タンパク質に特異的に結合する抗体またはペプチドの結合を検出することによる、請求項29記載の方法。
【請求項31】
検出する段階が免疫蛍光法またはフローサイトメトリーによる、請求項30記載の方法。
【請求項32】
インビトロにおける受精に使用する精液試料にプロテアソーム活性の阻害剤を添加する段階を含むインビトロにおける受精方法であって、阻害剤は、受精を妨害するには不十分であるが、多精受精を妨害するには十分な濃度が添加される方法。
【請求項33】
受精の約25%未満が多精受精である、請求項32記載の方法。
【請求項34】
受精の約10%未満が多精受精である、請求項32記載の方法。
【請求項35】
阻害剤がインビトロにおいて添加される、請求項32記載の方法。
【請求項36】
阻害剤がインビボにおいて投与される、請求項32記載の方法。
【請求項1】
精子のプロテアソーム活性を阻害する段階を含む避妊方法。
【請求項2】
精子のプロテアソーム活性がインビトロにおいて阻害される、請求項1記載の方法。
【請求項3】
精子のプロテアソーム活性がインビボにおいて阻害される、請求項1記載の方法。
【請求項4】
プロテアソーム活性が、プロテアソームに結合してプロテアソーム活性を阻害する抗体によって阻害される、請求項1記載の方法。
【請求項5】
抗体がヒト化抗体を含む、請求項4記載の方法。
【請求項6】
プロテアソーム活性が、抗体による受動免疫によって阻害される、請求項1記載の方法。
【請求項7】
抗体が、プロテアソームペプチドの少なくとも1つの抗原で被験者を免疫することによって誘導される、請求項4記載の方法。
【請求項8】
抗原が抗原性プロテアソームペプチドである、請求項7記載の方法。
【請求項9】
少なくとも1つの抗原性ペプチドがワクチンに含まれる、請求項8記載の方法。
【請求項10】
1つ以上の抗原性プロテアソームペプチドが、配列番号:1または配列番号:2のアミノ酸配列を含む、請求項9記載の方法。
【請求項11】
抗原性プロテアソームペプチドが融合タンパク質である、請求項7記載の方法。
【請求項12】
多重抗原エピトープが単一タンパク質内に存在する、請求項7記載の方法。
【請求項13】
抗体が、少なくとも1つの抗原性プロテアソームペプチドをコードする少なくとも1つのポリヌクレオチドで免疫することによって誘導される、請求項4記載の方法。
【請求項14】
少なくとも1つの抗原性プロテアソームペプチドをコードするポリヌクレオチドが発現ベクターに含まれる、請求項13記載の方法。
【請求項15】
発現ベクターがプラスミド発現ベクターである、請求項14記載の方法。
【請求項16】
プロテアソーム活性が、精子にプロテアソーム阻害剤を接触させることによって阻害される、請求項1記載の方法。
【請求項17】
プロテアソーム阻害剤が、薬学的に許容されうる製剤に含まれる、請求項16記載の方法。
【請求項18】
プロテアソーム阻害剤がMG132またはラクタシスチンである、請求項16記載の方法。
【請求項19】
薬学的に許容されうる製剤が、溶液、懸濁液、錠剤、ピル、カプセル、徐放性製剤、粉末、クリーム、軟膏(ointment)、軟膏(salve)、スプレー、ポンプ、リポソーム、坐剤、吸入剤またはパッチの形態に製剤化される、請求項17記載の方法。
【請求項20】
薬学的に許容されうる製剤の投与経路が静脈内、腹腔内、皮内、筋肉内、子宮内、皮膚、鼻、口腔、膣、吸入または局所的である、請求項17記載の方法。
【請求項21】
投与経路が子宮内である、請求項20記載の方法。
【請求項22】
1つ以上のプロテアソームペプチドをほ乳類に投与する段階を含む、ほ乳類において抗プロテアソーム抗体を誘導する方法であって、投与する段階が、ほ乳類のプロテアソームに結合して、その活性を阻害する抗体の産生を誘導する方法。
【請求項23】
ほ乳類がヒトである、請求項22記載の方法。
【請求項24】
1つ以上のプロテアソームペプチドが、プロテアソームペプチドをコードするポリヌクレオチドの発現によって投与される、請求項22記載の方法。
【請求項25】
プロテアソームペプチドをコードするポリヌクレオチドが、筋肉内、静脈内、皮下、腹腔内、皮内、経口または吸入投与経路によって投与される、請求項24記載の方法。
【請求項26】
プロテアソームペプチドが、筋肉内、静脈内、皮下、腹腔内、皮内、経口または吸入投与経路によって投与される、請求項22記載の方法。
【請求項27】
卵母細胞に関連するユビキチンの分解を阻害する段階を含む避妊方法。
【請求項28】
ユビキチン分解の阻害が、ユビキチン、ユビキチン活性化酵素、ユビキチン-キャリア/結合酵素またはユビキチン-リガーゼに対する抗体の投与による、請求項27記載の方法。
【請求項29】
精子内または精子上のユビキチン-プロテアソーム経路に関連するタンパク質の有無を検出する段階を含む受精能を評価する方法。
【請求項30】
ユビキチン-プロテアソーム関連タンパク質の有無を検出する段階が、ユビキチン-プロテアソーム関連タンパク質に特異的に結合する抗体またはペプチドの結合を検出することによる、請求項29記載の方法。
【請求項31】
検出する段階が免疫蛍光法またはフローサイトメトリーによる、請求項30記載の方法。
【請求項32】
インビトロにおける受精に使用する精液試料にプロテアソーム活性の阻害剤を添加する段階を含むインビトロにおける受精方法であって、阻害剤は、受精を妨害するには不十分であるが、多精受精を妨害するには十分な濃度が添加される方法。
【請求項33】
受精の約25%未満が多精受精である、請求項32記載の方法。
【請求項34】
受精の約10%未満が多精受精である、請求項32記載の方法。
【請求項35】
阻害剤がインビトロにおいて添加される、請求項32記載の方法。
【請求項36】
阻害剤がインビボにおいて投与される、請求項32記載の方法。
【図1】

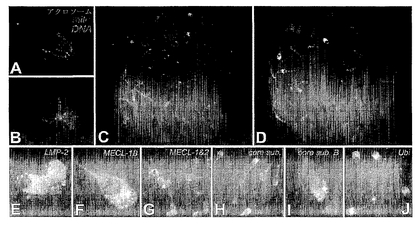
【図2】
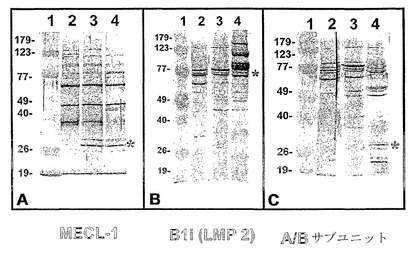
【図3】
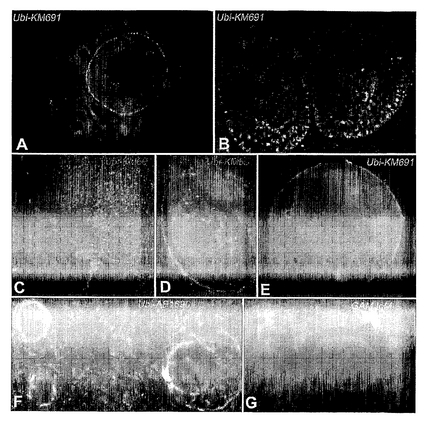
【図4】
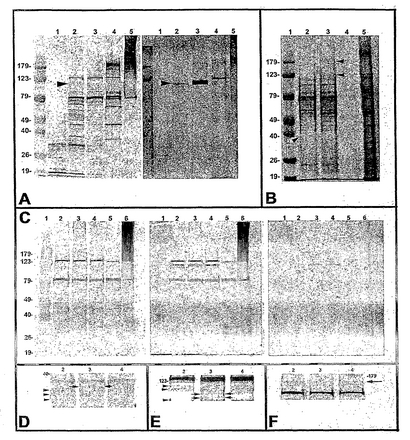
【図5】
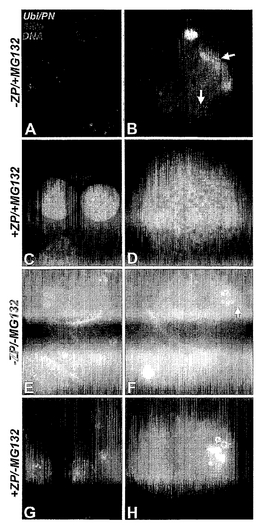
【図6】

【図7】

【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2006−522022(P2006−522022A)
【公表日】平成18年9月28日(2006.9.28)
【国際特許分類】
【出願番号】特願2006−503584(P2006−503584)
【出願日】平成16年2月13日(2004.2.13)
【国際出願番号】PCT/US2004/004378
【国際公開番号】WO2004/073624
【国際公開日】平成16年9月2日(2004.9.2)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
パイレックス
【出願人】(505101868)ザ キュレイターズ オブ ザ ユニバーシティー オブ ミズーリ (7)
【Fターム(参考)】
【公表日】平成18年9月28日(2006.9.28)
【国際特許分類】
【出願日】平成16年2月13日(2004.2.13)
【国際出願番号】PCT/US2004/004378
【国際公開番号】WO2004/073624
【国際公開日】平成16年9月2日(2004.9.2)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
パイレックス
【出願人】(505101868)ザ キュレイターズ オブ ザ ユニバーシティー オブ ミズーリ (7)
【Fターム(参考)】
[ Back to top ]