
プロテオームチップを用いるタンパク質活性の包括的分析
【課題】単一の種において発現された全タンパク質の大半を有するアレイからなるプロテオームチップに関し、プロテオームチップの作製方法に関する。さらに、プロテオームチップを使ってハイスループット方式で1種における全てのタンパク質相互作用を系統的にアッセイする方法を提供する。
【解決手段】基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種における既知遺伝子のうち少なくとも50%によりコードされる少なくとも1つのタンパク質を含むアレイ。
【解決手段】基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種における既知遺伝子のうち少なくとも50%によりコードされる少なくとも1つのタンパク質を含むアレイ。
【発明の詳細な説明】
【技術分野】
【0001】
1. 発明の分野
本発明は、単一の種において発現される全タンパク質の大部分を含んでなるプロテオームチップに関する。本発明はまた、プロテオームチップの製造方法に関する。本発明はさらに、ハイスループット方式で単一の種における全タンパク質の相互作用を系統的にアッセイするためのプロテオームチップの使用方法に関する。
【0002】
加えて、本発明は、高密度アレイフォーマットで真核生物タンパク質を製造し精製するための方法に関する。本発明はまた、二重タグ付けされた融合タンパク質を基板に結合させることによるタンパク質アレイの製造方法に関する。本発明はさらに、シグナルが陽性であるかどうかを確認する方法に関する。
【背景技術】
【0003】
2. 発明の背景
ポストゲノム配列決定時代の厄介なタスクは、ゲノムによりコードされるあらゆるタンパク質の機能、修飾および調節を理解することである (Fieldsら, 1999, Proc Natl Acad Sci. 96: 8825; Goffeauら, 1996, Science 274: 563)。現在、mRNA発現プロファイル、遺伝子破壊表現型、ツーハイブリッド相互作用、およびタンパク質細胞内局在を分析することによって遺伝子(それゆえにタンパク質)の機能を研究することに多くの努力が払われている (Ross-Macdonaldら, 1999, Nature 402: 413; DeRisiら, 1997, Science 278: 680; Winzelerら, 1999, Science 285: 901; Uetzら, 2000, Nature 403: 623; Itoら, 2000, Proc. Natl. Acad. Sci. U.S.A. 97: 1143)。こうした研究は有用であるけれども、転写プロファイルは必ずしも細胞のタンパク質レベルと相関しない。したがって、生化学的活性の分析がタンパク質機能に関する情報をもたらし、かかる情報はゲノム解析を補完して細胞の働きのより完全な全体図を提供することができる (Zhuら, 2001, Curr. Opin. Chem. Biol. 5: 40; Martzenら, 1999, Science 286: 1153; Zhuら, 2000, Nat. Genet. 26 : 283; MacBeath, 2000, Science 289: 1760; Caveman, 2000, J. Cell Sci. 113: 3543)。
【0004】
近年、幾つかのグループは、タンパク質活性をスクリーニングするためのマイクロアレイフォーマットを記載している (Zhuら, 2000, Nat. Genet. 26: 283; MacBeathら, 2000, Science 289: 1763; Arenkovら, 2000, Anal. Biochem 278: 123)。さらに、酵母タンパク質の過剰発現クローンの集合体が作製されて、生化学的活性に関してスクリーニングされた (Martzenら, 1999, Science 286: 1153)。しかしながら、ほぼ全プロテオームにあたる何千もの個々のタンパク質はまだ調製もアレイ化もされておらず、また、多数の活性に関するスクリーニングもされていない (Caveman, 2000, J. Cell Sci. 113:3543)。
【0005】
全プロテオームのスクリーニングには、ハイスループット方式で生産されたタンパク質の生化学的活性を系統的に精査し、並行して、何千何百というタンパク質試料の機能を分析することが必要となろう (Zhuら, 2000, Nat. Genet. 26: 283; MacBeathら, 2000, Science 289 : 1763; Arenkovら, 2000, Anal. Biochem 278: 123)。全プロテオームアレイをスクリーニングする試みは、必要な発現クローンの作製や、発現タンパク質のハイスループット方式での発現および精製が可能でないことを含めて、大きな障害に遭遇してきた。これまで、in vitroアッセイはランダム発現ライブラリーまたはプーリング(pooling)ストラテジーを用いて行われているが、これらはいずれも欠点を有する (Martzenら, 1999, Science 286: 1153; Bussowら, 2000, Genomics 65: 1)。特に、ランダム発現ライブラリーはスクリーニングに時間がかかり、また、しばしば完全長ではないクローンを含む。最近のもう一つのアプローチは、規定されたアレイを作製して、そのアレイをプーリングストラテジーによりスクリーニングすることであった (Martzenら, 1999, Science 286: 1153)。しかし、プーリングストラテジーはスクリーニングされるタンパク質の実際の数を覆い隠してしまい、また、このストラテジーは多数の陽性試料を同定する場合には面倒である。
【0006】
タンパク質-タンパク質相互作用を検出するのに有用な別の方法は、ツーハイブリッド法である (Uetzら, 2000, Nature 403: 623; Itoら, 2000, Proc. Natl. Acad. Sci. U.S.A. 97: 1143)。しかし、この方法を使って検出することができる相互作用のタイプは限られている。なぜなら、相互作用は核内で検出されるのが一般的だからである。
【0007】
したがって、当技術分野では、生化学的機能の大規模分析の必要性が存在しており、それには、ハイスループット方式で、種のゲノムによりコードされる広範囲にわたるタンパク質のセットを調製し、スクリーニングすることが必要となるだろう。
【0008】
セクション2または本明細書の他のセクションでの文献の引用または確認は、そのような文献が本発明の先行技術として利用可能であることを容認するものとして考えられるべきでない。
【発明の開示】
【0009】
3. 発明の概要
本発明は、ある種のタンパク質相互作用をハイスループット方式で包括的に研究するのに有用なプロテオームチップを提供する。本発明の方法および組成物は、本発明者らが、新たに、また予期せざることに、ゲノムのタンパク質コード配列を含む広範囲の発現構築物のセットを作製し、ハイスループット様式で宿主細胞にタンパク質産物を産生させ、マイクロアレイを使って複数のタンパク質の機能をハイスループット方式で分析する手段を見出したことにより、可能となったものである。
【0010】
本発明はプロテオームチップに関し、かかるチップは、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイであり、ここで、該複数のタンパク質は単一の種において発現された全タンパク質の大部分に相当し、その際、1つのオープンリーディングフレームの翻訳産物が1つのタンパク質とみなされる。
【0011】
アッセイを1回ずつ行うことに比べて、アレイを用いることの利点は、多くのタンパク質−プローブ相互作用を同時に同定して特徴づけられる点である。さらに、複雑なプローブ混合物をプロテオームチップと接触させることにより、例えば、細胞の環境を代表する環境における相互作用を検出し、多くの潜在的結合化合物を迅速に評価することができる。
本発明は、より具体的には、以下の特徴を有する:
〔1〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種における既知遺伝子のうち少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記アレイ。
〔2〕前記複数のタンパク質が単一の種における既知遺伝子のうち少なくとも70%によりコードされる少なくとも1つのタンパク質を含む、上記〔1〕に記載のアレイ。
〔3〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される全タンパク質のうち少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記アレイ。
〔4〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記アレイ。
〔5〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記アレイ。
〔6〕前記タンパク質がタンパク質の分類に従って前記アレイ上に組織化されている、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔7〕前記分類が存在量、機能、酵素活性、相同性、タンパク質ファミリー、特定の代謝経路との関連性、または翻訳後修飾によるものである、上記〔6〕に記載のアレイ。
〔8〕前記タンパク質がHisタグを介して基板に結合されている、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔9〕前記基板がニッケルを含む、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔10〕前記基板がニッケルをコーティングしたスライドガラスからなる、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔11〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記方法。
〔12〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される全タンパク質の少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記方法。
〔13〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記方法。
〔14〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記方法。
〔15〕基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
〔16〕前記結合ステップに先立って、前記タンパク質を第1タグの結合パートナーと接触させることにより前記タンパク質を精製するステップを行い、前記結合ステップにおいて第2タグを用いて基板に前記タンパク質を結合させる、上記〔15〕に記載の方法。
〔17〕第1タグがGSTタグであり、第2タグがHisタグである、上記〔16〕に記載の方法。
〔18〕第1タグと第2タグが前記タンパク質のアミノ末端に存在する、上記〔15〕に記載の方法。
〔19〕第1タグと第2タグが前記タンパク質のカルボキシ末端に存在する、上記〔15〕に記載の方法。
〔20〕複数の精製されたタンパク質サンプルを調製して単離する方法であって、マルチサイトアレイの複数サイトの各サイトにおいて、
(a) 調節配列と機能的に連結された異種ヌクレオチド配列を有する真核細胞を生育させ、
(b) 該調節配列を、異種ヌクレオチド配列によりコードされるタンパク質の発現を高める誘導物質と接触させ、
(c) 該細胞を溶解して細胞溶解物を形成し、
(d) 該細胞溶解物またはそれから得られたタンパク質含有サンプルを結合物質と接触させて、結果的に該タンパク質と結合物質との複合体を形成させ、そして
(e) 該複合体から該タンパク質を単離する、
各ステップを含んでなり、その際、各ステップがマルチアレイフォーマットで実施される、上記方法。
〔21〕各サイトがウェルである、上記〔20〕に記載の方法。
〔22〕前記タンパク質がアフィニティータグを含む融合タンパク質であり、該アフィニティータグに前記結合物質が結合する、上記〔20〕に記載の方法。
〔23〕前記細胞が酵母細胞である、上記〔20〕に記載の方法。
〔24〕前記溶解ステップがペイントシェーカーを使って実施される、上記〔20〕に記載の方法。
〔25〕脂質結合タンパク質を検出する方法であって、
(a) 脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置に該相互作用が検出されれば、その位置に脂質結合タンパク質が存在することを示す、上記方法。
〔26〕前記脂質がリン脂質である、上記〔25〕に記載の方法。
〔27〕前記リン脂質がホスファチジルコリンまたはホスファチジルイノシトールである、上記〔26〕に記載の方法。
〔28〕前記プローブがリポソームからなる、上記〔25〕に記載の方法。
〔29〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔30〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも50%を含むものであり、その際、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなし、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔31〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される少なくとも1000のタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔32〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔33〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数の融合タンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該融合タンパク質は第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔34〕前記プローブが核酸、タンパク質、小分子、候補薬物または脂質を含むものである、上記〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔35〕前記核酸がRNAまたはDNAからなる、上記〔34〕に記載の方法。
〔36〕前記プローブが酵母タンパク質である、上記〔34〕に記載の方法。
〔37〕酵母タンパク質がMyo2、Rho1、Rho2、Rho3、Rho4、Cdc11、Cdc12またはHs17である、上記〔36〕に記載の方法。
〔38〕前記プローブが抗体である、上記〔34〕に記載の方法。
〔39〕前記抗体がサイクリン、キナーゼ、GST、Clb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2、またはCdc11に特異的である、上記〔38〕に記載の方法。
〔40〕前記プローブがカルモジュリンである、上記〔34〕に記載の方法。
〔41〕前記プローブがATP、GTP、cAMP、ホスホチロシン、ホスホセリン、およびホスホトレオニンからなる群より選択される小分子を含むものである、上記〔34〕に記載の方法。
〔42〕前記プローブがホスファチジルコリンまたはホスファチジルイノシトールを含むものである、上記〔34〕に記載の方法。
〔43〕前記プローブがリポソームからなる、上記〔34〕に記載の方法。
〔44〕前記プローブが哺乳動物由来のものである、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔45〕前記哺乳動物がヒトである、上記〔44〕に記載の方法。
〔46〕前記複数のタンパク質がヒト以外のものである、上記〔44〕に記載の方法。
〔47〕前記複数のタンパク質がHisタグを介して基板に結合される、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔48〕前記基板がニッケルを含む、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔49〕前記基板がニッケルをコーティングしたスライドガラスからなる、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔50〕前記検出ステップでタンパク質との相互作用が検出されたプローブの正体を同定するステップをさらに含んでなる、上記〔34〕に記載の方法。
〔51〕前記相互作用は、前記同定されたプローブが抗菌性、抗真菌性、または抗ウイルス性タンパク質であることを示す、上記〔50〕に記載の方法。
〔52〕結合アッセイで使用するためのタンパク質を標識する方法であって、
(a) 該タンパク質の個々のアリコートを、ビオチン転移化合物と、アリコートごとに異なる程度にビオチン化されたタンパク質を生成する条件下および時間にわたり接触させ、そして
(b) 個々のアリコートを一緒に合わせて、異なる程度にビオチン化されたタンパク質のサンプルを得る、
各ステップを含んでなる、上記方法。
〔53〕結合タンパク質を検出する方法であって、
(a) 上記〔52〕に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じているアレイ上の位置を検出する、
各ステップを含んでなる、上記方法。
〔54〕結合タンパク質を検出する方法であって、
(a) 上記〔52〕に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) 該アレイに、蛍光物質とコンジュゲートさせたストレプトアビジンを接触させ、そして
(c) 蛍光が発生しているアレイ上の位置を検出する、
各ステップを含んでなり、その際、蛍光は、ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じていることを示す、上記方法。
〔55〕タンパク質がホスファチジルコリンに比べてホスファチジルイノシトールと優先的に結合するかどうかを判定する方法であって、
(a) ホスファチジルイノシトールを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルイノシトール結合タンパク質の存在を示すものであり、
(c) ホスファチジルコリンを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該タンパク質はステップ(a)におけると同じタンパク質を少なくとも一部含んでなり、
(d) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルコリン結合タンパク質の存在を示すものであり、そして
(e) 複数のタンパク質のそれぞれについて、ステップ(b)とステップ(d)の結果を比較する、
各ステップを含んでなる、上記方法。
〔56〕リン脂質が細胞における代謝経路またはシグナル伝達経路を調節するのかどうか、あるいは該代謝経路またはシグナル伝達経路が膜表面で起こるのかどうかを判定する方法であって、
(a) リン脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該複数のタンパク質は該経路の少なくとも一部を構成する1以上のタンパク質を含んでなり、そして
(b) 該プローブと該経路中のタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、該相互作用が生じれば、該プローブが代謝経路またはシグナル伝達経路を調節すること、あるいは、該経路が膜表面で起こることを示す、上記方法。
〔57〕カルモジュリンと結合する非天然タンパク質を製造する方法であって、下記の配列:
I/L-Q-X-X-K-K/X-G-B (配列番号1)
(ここで、Xは任意のアミノ酸であり、Bは塩基性アミノ酸である)
を含んでなる非天然タンパク質を製造することからなる上記方法。
〔58〕タンパク質における翻訳後修飾の有無を判定する方法であって、
(a) 翻訳後修飾と結合するプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 該プローブとタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置で該相互作用が生じれば、その位置のタンパク質が翻訳後修飾を有することを示す、上記方法。
〔59〕前記翻訳後修飾がメチル化、リン酸化、ビオチン化、アセチル化、ペグ化、グリコシル化、脂質修飾、ユビキチン化、またはSUMO化である、上記〔58〕に記載の方法。
〔60〕酵母細胞の培養物を調製する方法であって、
(a) 複数の酵母細胞を増殖培地中で、OD600が0.3〜1.0となるまで増殖させ、ここで、該複数の酵母細胞は調節配列に機能的に連結された異種ヌクレオチド配列を含むものであり、
(b) 該細胞を、該異種ヌクレオチド配列によりコードされるタンパク質の発現を増強する誘導物質と接触させ、
(c) 該細胞を該培地から分離し、
(d) 該細胞を冷水に接触させ、
(e) 該細胞を該冷水から分離し、
(f) 該細胞を冷溶解バッファーに接触させ、
(g) 該細胞を該溶解バッファーから分離し、そして
(h) 保存のために該細胞を半乾燥状態で凍結する、
各ステップを含んでなる上記方法。
〔61〕細胞からタンパク質を精製する方法であって、
(a) 複数の細胞サンプルのそれぞれについて、各サンプルの細胞を溶解して細胞溶解物を調製し、ここで、該細胞はアフィニティータグをもつ融合タンパク質を含み、また、該溶解ステップはペイントシェーカーを使って実施し、
(b) 各溶解物を可溶性画分と非可溶性画分とに分離し、
(c) 各可溶性画分をマルチサイトアレイの異なるサイトに移動させ、その際、該移動ステップはワイドオープンチップを使って実施し、
(d) 各可溶性画分を結合物質と接触させて、該融合タンパク質と結合物質との複合体が形成されるようにし、
(e) 該複合体から各融合タンパク質を単離し、そして
(f) 各融合タンパク質を高粘性のバッファー中に保存する、
各ステップを含んでなる上記方法。
〔62〕シグナルが陽性であるかどうかを判定する方法であって、
(a) 各スポットについて局所的にフォアグラウンドシグナルおよびバックグラウンドシグナルを測定し、該フォアグラウンドシグナルとバックグラウンドシグナルの差から正味のシグナルを決定し、
(b) 第1および第2の正味シグナル分布に関する下四分位値、中位値、および上四分位値を決定し、
(c) 第1の正味シグナル分布から第1の中位値を減じ、さらに第2の正味シグナル分布から第2の中位値を減じて、それぞれ第1および第2の減算値を得、
(d) 第1の減算値を、第1の正味シグナル分布に関する上四分位値と下四分位値との差で割り算し、さらに第2の減算値を、第2の正味シグナル分布の上四分位値と下四分位値との差で割り算して、それぞれ第1および第2の基準化された値を得、
(e) 近傍領域の基準化されたシグナル分布に関する局所中位値を算定し、ただし、該近傍領域はその領域内に複数のサイトを含んでなり、そして
(f) 基準化されたシグナルから局所中位値を減じて、基準化された過剰値を得る、
各ステップを含んでなる、上記方法。
〔63〕陽性シグナルがタンパク質−プローブ相互作用を示す、上記〔62〕に記載の方法。
〔64〕前記近傍領域が前記シグナルの上2列、下2列、左2列、および右2列である、上記〔62〕に記載の方法。
〔65〕サンプル値の1つとサンプル値の平均との差が基準化された過剰値の誤差の3標準偏差より大きい場合には、基準化された過剰値の並列サンプルを除外するステップをさらに含む、上記〔62〕に記載の方法。
〔66〕複数の異なるアレイを用いて測定したシグナルの中から陽性シグナルを確認する方法であって、
(a) 異なるアレイを用いて測定したシグナルを変換して変換シグナルを得、
(b) 各変換シグナルを、該変換シグナルから局所中位値シグナルを減じて修正された変換シグナルを得る方法によって修正し、ここで、該局所中位値シグナルは近傍領域におけるシグナルの中位値であり、該近傍領域は該変換シグナルのサイトのまわりの1以上のサイトを含んでなり、そして
(c) 修正された各変換シグナルを閾値と比較して、該修正された変換シグナルが該閾値より大きい場合には、該修正された変換シグナルを陽性と判定する、
各ステップを含んでなり、
ここで、該アレイは基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイからなるものである、上記方法。
〔67〕前記変換ステップが、
(a) 異なるアレイのそれぞれを用いて測定したシグナルについて、シグナル分布の下四分位値、中位値、および上四分位値を決定し、
(b) 異なるアレイのそれぞれを用いて測定したシグナルから該中位値を減じて、該アレイについての翻訳シグナルを得、
(c) 該翻訳シグナルを、該アレイの上四分位値と下四分位値との差で割り算することによって変換シグナルを得る、
各ステップを含んでなる、上記〔66〕に記載の方法。
〔68〕前記近傍領域が変換シグナルの上2列、下2列、左2列、および右2列の領域内の複数のサイトからなる、上記〔66〕に記載の方法。
〔69〕陽性シグナルがタンパク質−プローブ相互作用を示す、上記〔66〕、〔67〕または〔68〕に記載の方法。
〔70〕データポイントを捨てるステップをさらに含み、ただし、該データポイントはアレイ上の二重反復サイトで測定され、該二重反復サイト間の変化が3標準偏差より大きいものである、上記〔66〕、〔67〕または〔68〕に記載の方法。
〔71〕修正された変換シグナルを次式:
r+εr = G+εG/R+εR
(式中、Gは修正された変換シグナルであり、RはGSTシグナルであり、εGはGの誤差であり、εRはRの誤差であり、そしてεrはrの誤差である)
を用いて正規化するステップをさらに含む、上記〔66〕、〔67〕または〔68〕に記載の方法。
〔72〕マルチサイトアレイが96サイトアレイである、上記〔20〕に記載の方法。
〔73〕基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
〔74〕(a) 2つの二重反復スポットのシグナル値を平均して平均値を出し、(b) 該平均値が基準化された過剰値の誤差の3標準偏差より大きいかどうかを調べる、各ステップをさらに含んでなり、その際、該平均値が基準化された過剰値の誤差の3標準偏差より大きければ、該スポットのシグナルを陽性とする、上記〔62〕または〔65〕に記載の方法。
〔75〕タンパク質における酵素活性の有無を判定する方法であって、
(a) 該酵素活性の基質であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素活性をもつことを示す、上記方法。
〔76〕タンパク質における酵素基質の有無を判定する方法であって、
(a) 該酵素基質のための酵素であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素基質を含むことを示す、上記方法。
〔77〕前記タンパク質が基板にビオチンタグを介して結合される、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔78〕第1タグが前記タンパク質のカルボキシ末端に存在し、第2タグが前記タンパク質のアミノ末端に存在する、上記〔15〕に記載の方法。
〔79〕アフィニティータグがビオチンである、上記〔22〕に記載の方法。
〔80〕前記基板がニトロセルロースをコーティングしたスライドガラスからなる、上記〔1、3、4または5〕に記載のアレイ。
【0012】
したがって、一つの実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%を包含する。
【0013】
別の実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、または50%を包含し、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。特定の実施形態では、複数のタンパク質は単一の種において発現される全タンパク質の少なくとも50%を包含し、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0014】
別の実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000のタンパク質を包含する。
【0015】
別の実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0016】
さらなる実施形態において、前記タンパク質はタンパク質分類に従ってアレイ上に組織化される。こうした分類は、存在量、機能、機能上のクラス、酵素活性、相同性、タンパク質ファミリー、特定の代謝もしくはシグナル伝達経路との関連、関係した代謝もしくはシグナル伝達経路との関連、または翻訳後修飾によるものであってよい。
【0017】
別の実施形態において、本発明は、基板がガラス、セラミック、ニトロセルロース、無定形シリコンカーバイド、注型可能な酸化物、ポリイミド、ポリメチルメタクリレート、ポリスチレン、またはシリコーンエラストマーで構成されている、上記の位置指定可能なアレイを提供する。
【0018】
さらなる実施形態において、基板は複数のタンパク質を基板に結合させるのに役立つ物質を含んでなる。例えば、基板に、各タンパク質のアフィニティータグと結合する物質をコーティングすることができる。特定の実施形態では、基板がグルタチオンを含む。別の特定の実施形態では、基板のコーティングがニッケルまたはニトロセルロースを含んでなる。別の特定の実施形態では、基板のコーティングがグルタチオンとニッケルを含む。ある実施形態では、基板がニッケルをコーティングしたスライドガラスである。好ましい実施形態では、基板がニトロセルロースをコーティングしたスライドガラスである。タンパク質(およびDNA)マイクロアレイを作製するためのニトロセルロースコーティングスライドガラスは市販されている(例えば、Schleicher & Schuell社(Keene, NH)は、ニトロセルロース系のポリマーをコーティングしたスライドガラスを販売している(カタログ番号10 484 182))。特定の実施形態では、各タンパク質がOMNIGRIDTM (GeneMachines, San Carlos, CA)を用いてニトロセルロースコーティングスライドガラス上にスポットされる。
【0019】
プロテオームチップ上のタンパク質は、好ましくは、該タンパク質を精製したり、かつ/またはプロテオームチップに結合させたりするのに有用なアフィニティータグを少なくとも1つ含む融合タンパク質である。
【0020】
本発明はまた、プロテオームチップの作製方法を提供する。したがって、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの構築方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%を包含する。
【0021】
一つの実施形態において、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの作製方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、または50%を包含し、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0022】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの構築方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000のタンパク質を包含する。
【0023】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの作製方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0024】
本発明はさらに、ハイスループット分析に適合しやすく、容易に評価できるフォーマットで、ウイルス、原核生物または真核生物のタンパク質を調製して単離する方法を提供する。好ましい方法は、自動化技術に適合しうるアレイフォーマットでタンパク質を合成し精製することを含む。したがって、一実施形態において、本発明は、ウイルス、原核生物または真核生物のタンパク質を調製して単離する方法を提供し、該方法は、調節配列と機能的に連結された異種配列を含むベクターにより形質転換された真核細胞を増殖させ、該調節配列を、該異種配列によりコードされたタンパク質の発現を高める誘導物質と接触させ、該細胞を溶解し、該タンパク質を結合物質と接触させて該タンパク質と結合物質との複合体が形成されるようにし、該複合体を細胞破砕物から単離し、そして該タンパク質を該複合体から分離するステップを含んでなり、ここで、各ステップは96ウェルフォーマットで実施される。
【0025】
上記タンパク質は、異種配列が目的タンパク質のコード領域とアフィニティータグなどのタグをコードする配列とを含むような融合タンパク質であることが好ましい。かかるタグは、そのタンパク質をモニタリングしたり、融合タンパク質を細胞破砕物や混入試薬から分離したり、かつ/または該タンパク質を本発明のプロテオームチップに結合させたりするのに有用でありうる。
【0026】
本発明はさらに、複数の融合タンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの作製方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該タンパク質は第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである。特定の実施形態では、該タンパク質のアミノ末端には1つのタグが付いており、また、そのカルボキシ末端には異なる第2タグが付いている。他の実施形態では、該タンパク質のアミノ末端に2つのタグが付いているか、あるいはカルボキシ末端に2つのタグが付いている。さらに別の実施形態では、該タンパク質のアミノまたはカルボキシ末端以外の部位に1つ以上のタグが付いている。二重タグ付けされたタンパク質を使用することの利点としては、高度に精製されたタンパク質を得ることができるだけでなく、細胞破砕物からタンパク質を精製してそのタンパク質を基板に結合させる簡便な方法を提供できることにある。
【0027】
したがって、特定の実施形態においては、第1タグがグルタチオン-S-トランスフェラーゼタグ(GSTタグ)であり、第2タグがポリヒスチジンタグ(Hisタグ)である。別の実施形態では、GSTタグとHisタグをタンパク質のアミノ末端に結合させる。これとは別に、GSTタグとHisタグをタンパク質のカルボキシ末端に結合させることもできる。GSTタグおよびHisタグはタンパク質のアミノ末端またはカルボキシ末端のどちらに存在してもよい。特定の実施形態においては、GSTタグをタンパク質のアミノ末端に結合させ、Hisタグをカルボキシ末端に結合させる。その他の実施形態では、Hisタグをタンパク質のアミノ末端に結合させ、GSTタグをカルボキシ末端に結合させる。
【0028】
融合タンパク質のアフィニティータグの配置を変えると、機能的な二次構造、細胞外ドメインの適切な折りたたみ、ならびに、適切なタンパク質の輸送、局在化および/または分泌がもたらされる可能性がある。例えば、GSTタグおよびHisタグのカルボキシ末端への融合は、不適切な折りたたみまたは翻訳開始コドンの上流の領域が遮断される場合の発現を未然に防ぐことができる。
【0029】
本発明はさらに、タンパク質アレイを用いて脂質結合タンパク質をスクリーニングする方法を提供する。したがって、一実施形態において、本発明は、各タンパク質が基板の異なる位置にある、複数のタンパク質を含んでなる位置指定可能なアレイの使用方法であり、該方法は、プローブをアレイに接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなり、ここで、該プローブは脂質を含むものである。特定の実施形態では、かかる脂質はリン脂質を含み、例えば、ホスファチジルコリンおよびホスファチジルイノシトールであるが、これらに限らない。別の特定の実施形態では、プローブは対象のリン脂質を含むリポソームの形態をしている。
【0030】
さらに、プロテオームチップを使用する方法も提供される。本発明のプロテオームチップを使用すると、1つの種または細胞における本質的に全てのタンパク質−タンパク質相互作用をアッセイすることができる。また、試験化合物と相互作用する1つの種の全タンパク質を体系的にアッセイするために、プロテオームチップを使用することも可能である。したがって、本発明のプロテオームチップを使用すると、多数の活性をアッセイして豊富な情報を手に入れることができる。かかる情報として、制限するものではないが、刺激に応答した細胞もしくは生物の「フィンガープリント」(指紋)もしくは「サイン」を規定すること、対象のプローブと相互作用する1つの種における全タンパク質を特徴づけること、対象のプローブと相互作用する2以上の種における全タンパク質を特徴づけること、生物学的経路(例えば、代謝もしくはシグナル伝達経路)または関連した生物学的経路に関与する全タンパク質を特徴づけること、対象の酵素活性(例えば、キナーゼ活性、プロテアーゼ活性、ホスファターゼ活性、グリコシダーゼ、アセチラーゼ活性、およびその他の化学基転移酵素活性)を有する1つの種における全タンパク質を特徴づけること、対象の翻訳後修飾を伴う1つの種における全タンパク質を特徴づけること、および薬物標的を同定することがある。特定の実施形態においては、本発明のプロテオームチップを用いて、対象の薬物または候補薬物と相互作用する1つの種における全タンパク質(例えば、薬物標的)を特徴づける。
【0031】
かくして、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は単一の種における既知遺伝子の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%によりコードされる少なくとも1つのタンパク質を含むものである。
【0032】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%を含んでなり、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0033】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000の既知タンパク質を含むものである。
【0034】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は全体として、単一の種における少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000または50000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0035】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は全体として、少なくとも2つの種における少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000または50000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0036】
別の実施形態において、本発明は、プローブを、複数の融合タンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該融合タンパク質は第1タグ、第2タグ、および生物のゲノム核酸によりコードされるタンパク質配列を含むものである。先に記載したように、特定の実施形態では、2つのタグはHisおよびGSTでありうる。
【0037】
本発明はさらに、結合アッセイにおいて使用するためのタンパク質の標識方法を提供し、該方法は、該タンパク質の個々のアリコートを、ビオチン転移化合物と、アリコートごとに異なる程度にビオチン化されたタンパク質を生成する条件下および時間にわたり接触させ、個々のアリコートを一緒に合わせて、異なる程度にビオチン化されたタンパク質のサンプルを得るステップを含んでなる。
【0038】
本発明はまた、結合タンパク質の検出方法を提供し、該方法は、上記方法により得られたビオチン化タンパク質のサンプルを、複数のタンパク質(各タンパク質が基板上の異なる位置にある)を含んでなる位置指定可能なアレイと接触させ、アレイタンパク質−プローブ相互作用(ビオチン化タンパク質とアレイ上のタンパク質との相互作用が起こる)の位置を検出するステップを含んでなる。
【0039】
本発明はまた、結合タンパク質の検出方法を提供し、該方法は、上記方法により得られたビオチン化タンパク質のサンプルを、複数のタンパク質(各タンパク質が基板上の異なる位置にある)を含んでなる位置指定可能なアレイと接触させ、該アレイを蛍光物質にコンジュゲートさせたストレプトアビジンと接触させ、蛍光を発するアレイ上の位置を検出するステップを含んでなり、その際、蛍光が発せられれば、ビオチン化タンパク質とアレイ上のタンパク質との間で相互作用が起こったことを示す。
【0040】
本発明はさらに、シグナルが陽性であるかどうかを確認する方法を提供する。したがって、本発明の一実施形態は、タンパク質マイクロアレイを用いるアッセイで得られたシグナルが陽性である(相互作用物質へのプローブの結合を示す)かどうかを確認する方法を提供する。
【0041】
3.1 定義
本明細書中で用いる「タンパク質」という語は、全長タンパク質、タンパク質の一部分、またはペプチドをさす。タンパク質はより大きなタンパク質の断片化によって生成することも、化学合成することもできる。好ましくは、細菌、酵母、昆虫細胞、哺乳動物細胞など(しかし、これらに限らない)の種における組換え体の過剰発現によりタンパク質を調製する。本発明のタンパク質マイクロアレイに配置されるタンパク質は融合タンパク質であることが好ましく、精製および/または固定化を助けるアフィニティータグを少なくとも1つもつものがより好ましい。
【0042】
本明細書中で用いる「相互作用物質」(interactor)という語は、プローブと相互作用するタンパク質マイクロアレイ上のタンパク質をさす。
【0043】
本明細書中で用いる「プローブ」という語は、マイクロアレイのタンパク質との相互作用に関してアッセイするために本発明のタンパク質マイクロアレイに適用することができる、タンパク質、核酸(例えば、DNA、RNA、オリゴヌクレオチド、ポリヌクレオチド)、小分子、基質、インヒビター、薬物もしくは候補薬物、受容体、抗原、ホルモン、ステロイド、脂質、リン脂質、リポソーム、抗体、補因子、サイトカイン、グルタチオン、免疫グロブリンドメイン、炭水化物、マルトース、ニッケル、ジヒドロトリプシン、カルモジュリン、ビオチン、レクチン、および重金属など(しかし、これらに限らない)のいずれかの化学試薬をさす。
【0044】
4. 図面の説明
図1A〜1B. プロテインチップ技術を用いた酵母プロテオーム解析の手順。
【0045】
A. 酵母ORFを、二重タグ付けされた酵母GAL1発現ベクターに組換え法によりクローニングし、配列決定により正体を確認した。次に、純粋なプラスミド構築物のそれぞれを、大規模タンパク質精製用の酵母株に再導入した。酵母培養物を96ウェルフォーマットで増殖させ、ガラクトースを添加して誘導した。ハイスループット精製ステップの後、精製されたタンパク質をアリコートに分けて、プリンティングに先立ってグリセロールバッファー中に-80℃で保存した。高精度のマイクロアレイヤー(microarrayer)を用いると、1つの実験で80枚のスライドガラス上に6566のタンパク質サンプルを二重反復してスポットすることができる。
【0046】
B. 3mlの酵母培養物から精製されたタンパク質のイムノブロット分析。
【0047】
図2A〜2C. 抗GST抗体を用いてプローブしたプロテオームチップ。
【0048】
60のサンプルを、抗GST抗体を用いるイムノブロット分析により検査した;19の代表的な例を示してある。80%を超える調製物から高収量の融合タンパク質が得られる。
【0049】
A. GST::酵母融合タンパク質を96ウェルフォーマットで精製した。5800のユニークなタンパク質を代表する6566のタンパク質サンプルを、1枚のニッケルコーティングを施した顕微鏡用スライドガラスの上に二重反復してスポットした。このスライドガラスを抗GST抗体でプローブした。
【0050】
B. 48ブロックのうちの1つの拡大図をプロテオームチップの右側に示す。文字は二重反復タンパク質サンプルを示し、数字は起源プレート番号を表す。すぐれた分解能と高いシグナル対ノイズ比に留意されたい。
【0051】
C. 各ORF産物の二重反復スポット間の誤差分布。εの単位はAxonスキャナーにより測定したシグナル強度である。GST対照により測定されるように、10単位はおよそ32fgのタンパク質に等しい。ε分布の中心は1.2単位であり、95%のサンプルは中心から10単位以内のところにある。
【0052】
図3A〜3B. プロテオームチップでの様々なアッセイの例。
【0053】
A. 6566の酵母タンパク質を含むプロテオームチップは二重反復してスポットし、図示したビオチン化プローブと共にインキュベートした。陽性の相互作用物質(ボックスで示す)を6つのタンパク質−リポソーム相互作用アッセイ(「PI(3)P」、「PI(4,5)P2」、「PI(4)P」、「PI(3,4)P2」、「PI(3,4,5)P3」、「PC」)、カルモジュリン結合アッセイ(「カルモジュリン」)、およびDNA−タンパク質相互作用アッセイ(「ゲノムDNA」)で同定した。各ブロックは16X18のタンパク質スポットを含む。二重反復陽性シグナルは下のパネルに水平な対として現れる。二重反復スポッティングは内部対照として役立ち、これはシグナルがバックグラウンドに対して弱いときに重要である。上のパネルは、抗GST抗体(「α-GST」)でプローブした対照プロテオームチップの同じ酵母タンパク質調製物を示す。図面から実証されるように、比較的低レベルのGST融合タンパク質(「プローブ」)を含有するサンプルにおいて強いシグナルがしばしば観察されており、このことは、結合が高感度かつ特異的であることを示している。PI、ホスファチジルイノシトール。PC、ホスファチジルコリン。
【0054】
B. 様々なカルモジュリン標的が共有するアミノ酸配列をサーチすることにより同定された推定上のカルモジュリン結合モチーフ(Zhuら 2000, Nat. Genet. 26:283)。39の陽性タンパク質のうち14は、コンセンサスがI/L-Q-X-K-K/X-G-B (配列番号1)(ここで、Xは任意の残基であり、Bは塩基性残基である)であるモチーフを共有している。アラインメントの上に示した文字の大きさは、表示したアミノ酸の相対頻度を示す。YFL003C/MSH4 (配列番号2); YJR073C/OPI3 (配列番号3); YBR050C/REG2 (配列番号4); YNL202W/SPS19 (配列番号5); YOL016C/CMK2 (配列番号6); BR011C/IPP1 (配列番号7); YGR034W/RPL26B (配列番号8);YFR004W/RPN11 (配列番号9); YIL021W/RPB3 (配列番号10); YGL063W/PUS2 (配列番号11); YDR292C/SRP101 (配列番号12); YFR014C/CMK1 (配列番号13); YBR213W/MET8 (配列番号14); YAL029C/MY04 (配列番号15)。
【0055】
図4A〜4D. ホスファチジルイノシトール結合タンパク質の分析。150の陽性タンパク質のホスファチジルイノシトール(「PI」)結合特異性を調べるために、それらの結合シグナルをホスファチジルコリン(「PC」)の対応する結合シグナルに対して正規化した。比率(「PI/PC」)に基づいて、タンパク質を4つのカテゴリーに分類した。すなわち、(A) 30の強くかつ特異的、(B) 43の強くかつ非特異的、(C) 19の弱くかつ特異的、および、(D) 58の弱くかつ非特異的、なホスファチジルイノシトール結合タンパク質である。強度はPI/PCシグナル比(図面のスケールで示される)を表す。PI/PC結合比の右に「10n」と表記される欄は、最大結合シグナル強度(白抜きのボックス)およびその信頼区間(水平の実線)を示す。数字はその値のlogを示す。信頼区間欄の右側の3つの欄のボックスは、それぞれ膜結合タンパク質(「膜」)、キナーゼ(「キナーゼ」)、および特性決定されなかったORF(「未知」)を示す。
【0056】
図5A〜5C. プロテオームマイクロアレイにより検出されたタンパク質−脂質相互作用を慣用方法で確認する(Casamayorら, 1999, Curr. Biol. 9: 186 ; Guerraら, 2000, Biosci. Rep. 20: 41)。
【0057】
A. 最初に、PI(4,5)P2リポソームを、BSAでブロックしたニトロセルロース膜に付着させた。Rim15p、Eno2pおよびHxk1p、ならびにGST対照の連続希釈物を用いて膜をプローブした。結合したタンパク質は抗GST抗体および増強ケミルミネッセンス(「ECL」)キットを用いて検出した。
【0058】
B. タンパク質−脂質相互作用を試験するためにリバースアッセイを実施した。タンパク質を調製して連続希釈でニトロセルロースフィルターにスポットし、6種のリポソームでプローブした。対照として、6種のリポソームもBSAブロック膜に添加した。十分な洗浄後、結合したリポソームを、HRPコンジュゲート化ストレプトアビジンおよびECLキットを用いて検出した。
【0059】
C. 膜アッセイにおける結合シグナルとRim15pの量との線形相関。膜アッセイ(図4B)からのRim15pのリポソーム結合シグナルを、スポットしたRim15pの濃度勾配に対してプロットしたとき、PI(4)P、PI(3,4)P2、PI(3)P、およびPI(4,5)P2の結合シグナルはRim15pの量と線形に相関した。PI(4)PとRim15pの相互作用は最高の親和性を示し、対照PCとRim15pとの親和性より少なくとも3倍高かった。
【0060】
5. 発明の詳細な説明
本発明は、本発明者らが酵母で発現される全タンパク質の約80%を含有する酵母プロテオームマイクロアレイを構築したことに、部分的に基づいており、種にかかわらずプロテオームアレイに関する最初の記述となる。本発明のプロテオームチップは、ある1つの種においてタンパク質の相互作用および活性を包括的に分析するために使用することができる。プロテオームチップの利用は、既存のアプローチよりも顕著な利点を有する。本明細書で提示されるプロテオームマイクロアレイ技術の利点の1つは、個々のタンパク質からなる大きなセットを、何百もの生化学的活性について、あるいは何千でさえ、同時に、直接、ハイスループット様式でスクリーニングすることができることである。例えば、プロテオームチップ法の利点は、タンパク質-薬物相互作用(例えば、薬物標的-薬物相互作用)、タンパク質-脂質相互作用および酵素的アッセイを包含するがそれに限定されない多種多様な活性についてタンパク質を直接in vitroでスクリーニングできることである。それに加えて、広範囲のin vitro条件を容易に試験することができる。さらに、ひとたびタンパク質が調製されれば、プロテオームスクリーニングは極めて速く、安価であり、さらに、多くのマイクロアレイフォーマットによる迅速なデータ分析は、既存の装置および分析用ソフトウェアに適合する。
【0061】
5.1 プロテオームアレイ
本発明は、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は、ある単一の種における既知遺伝子のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%によってコードされる、少なくとも1つのタンパク質を含んでなる。すなわち、1つの遺伝子に由来するすべてのタンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0062】
本発明は、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は、単一の種において発現される全タンパク質のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%を含んでなり、この場合、タンパク質アイソフォームおよびスプライス変異体は単一タンパク質とみなされる。ある実施形態において、前記の複数のタンパク質は、ある種において発現される全タンパク質の約90%、95%または99%を含んでなる。特定の実施形態において、この複数のタンパク質はある種において発現される全タンパク質の約93.5%を含んでなる。
【0063】
本発明はまた、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は、単一の種において発現される、少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000のタンパク質を含んでなる。
【0064】
本発明はまた、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は全体として、単一の種における、少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000、または50000の異なる既知の遺伝子を含んでなる。
【0065】
本発明はまた、それぞれの融合タンパク質が基板上の異なる位置に存在する、基板表面に融合した複数の融合タンパク質を含んでなる位置指定可能なアレイを包含するが、ここでこの融合タンパク質は、第1タグ、第2タグ、およびある生物のゲノム核酸によってコードされるタンパク質配列を含んでなる。別の実施形態において、前記融合タンパク質のタンパク質配列は、生物のゲノム核酸にコードされる必要はないが、結合タンパク質の機能および/または活性を特定することが必要とされる配列である。
【0066】
位置指定可能なアレイは、対象とする各プローブもしくはタンパク質が基板上の既知の場所に存在するような配置を提供し、したがって、各プローブもしくはタンパク質の正体をアレイ上のその位置から決定することが可能である。したがって、アレイ上のそれぞれのタンパク質は、各タンパク質の正体を基板上のその位置から決定することができるように、基板上の既知の所定の場所にあることが望ましい。
【0067】
ある実施形態において、前記の種はウイルスである。別の実施形態において、前記の種とは原核生物である。また別の実施形態において、前記の種は真核生物である。別の実施形態において、前記の種は脊椎動物である。さらにまた別の実施形態において前記の種は、哺乳動物である。特定の実施形態において、前記の種は、動物であって、昆虫、霊長類、および齧歯類を含めるがこれらに限定されない。特定の実施形態において、前記の種はサル、ショウジョウバエ、ウシ、ウマ、ヒツジ、ブタ、ニワトリ、シチメンチョウ、ウズラ、ネコ、イヌ、マウス、ラット、ウサギ、線虫もしくは魚である。望ましい実施形態において、前記の種はヒトである。別の望ましい実施形態において、前記の種は酵母である。
【0068】
本発明のプロテオームチップのタンパク質は、全長タンパク質、全長タンパク質の一部分、およびペプチドを包含するが、これらを組換え体の過剰発現、大きいタンパク質の断片化、もしくは化学合成によって調製することができる。例えば酵母、細菌、昆虫、ヒト、もしくはヒト以外の哺乳動物、例えばマウス、ラット、ネコ、イヌ、ブタ、ウシ、およびウマ、に由来する細胞においてタンパク質を過剰発現させることができる。さらに、天然の、または合成のタンパク質に連結された規定のドメインを含んでなる融合タンパク質を使用することができる。プロテオームチップのタンパク質を、チップの基板に付着させる前に、精製することができる。また、プロテオームチップのタンパク質をプロテオームチップに付着させている間にこれを精製したり、さらに追加して精製したりしてもよい。
【0069】
プロテインチップに付着させる前、または付着させるときにタンパク質を人工膜もしくは天然の膜(例えば、リポソーム、膜小胞)に埋め込むことができる。実際、特定のタンパク質合成を人工膜もしくは天然の膜の存在下で行うことが望ましく、例えばタンパク質の折りたたみ、タンパク質のプロセシングを促進し、活性を維持し、および/またはタンパク質の沈澱を防止することができる。
【0070】
さらに、タンパク質をプロテオームチップの基板に付着させることができる。あるいはまた、タンパク質をプロテオームチップのウェル内に送達することができるが、この場合、このタンパク質はプロテオームチップの基板には結合しない状態で存在する。
【0071】
本発明は、また、本発明のプロテオームチップのための基板として有用な、化合物に関する。例えば、シリコン、ガラス、石英、ポリイミド、アクリル、ポリメタクリル酸メチル(LUCITE(登録商標))、セラミック、ニトロセルロース、無定形シリコンカーバイド、ポリスチレン、および/またはマイクロファブリケーション、マイクロリソグラフィーもしくはキャスティングに適したあらゆる他の材料、といった材料から基板を作製することができるが、これらに限定されない。例えば、基板は、親水性マイクロタイタープレート(例、MILLIPORETM)もしくはニトロセルロースコーティングスライドガラスとすることができる。好ましい実施形態において、基板はニトロセルロースコーティングスライドガラスである。プロテイン(およびDNA)マイクロアレイを作製するためのニトロセルロースコーティングスライドガラスは、市販されており、入手可能である(例えば、Schleicher & Schuell(Keene, NH)から、この会社はニトロセルロース系のポリマーでコーティングされたスライドガラス(カタログ番号10 484 182)を販売する)。特定の実施形態において、それぞれのタンパク質を、OMNIGRIDTM(GeneMachines, San Carlos, CA)を用いてニトロセルロースコーティングスライドガラス上にスポットする。本発明は、プロテインチップを作製するために有用な他の基板を検討するが、これらの一部は例えば、2001年5月4日出願の同時係属中の米国特許出願第09/849,781号において開示されており、この出願はその全体を参考として本明細書に含めるものとする。
【0072】
特定の実施形態において、基板はシリコーンエラストマー素材(例えばポリジメチルシロキサン(”PDMS”)であるが、それに限らない)を含んでなる。シリコーンエラストマー素材の利点はその柔軟性である。
【0073】
もう1つの特定の実施形態において、基板はシリコンウェハーである。シリコンウェハーは、パターンを形成してエッチングすることができる(例えば、G. Kovacs, 1998, Micromachined Transducers Sourcebook, Academic Press; M. Madou, 1997, Fundamentals of Microfabrication, CRC Pressを参照されたい)。エッチングされたウェハーを用いて、本発明のプロテオームチップを成形することができる。
【0074】
ある実施形態において、本発明は、スライドガラスのような、ただしそれに限定されないが、平らな表面を有する基板を含んでなるプロテオームチップを提供する。高密度プロテインアレイを、例えばスライドガラス上に作成することができるが、その結果、タンパク質の存在、量、および/または機能性に関するアッセイをハイスループット方式で行うことができる。
【0075】
したがって、ある実施形態において、プロテオームチップは基板表面に付着させた複数のタンパク質を含んでなるが、ここでタンパク質が付けられるサイトの密度は、少なくとも100サイト/cm2、10,000サイト/cm2、1,000,00サイト/cm2、10,000,000サイト/cm2、25,000,000サイト/cm2、10,000,000,000サイト/cm2、または10,000,000,000,000サイト/cm2である。個々のタンパク質試料をそれぞれチップ上の別々のサイトに付着させることが望ましい。チップ上のそれぞれの位置にあるタンパク質の正体は既知である。
【0076】
別の実施形態において、基板は整列したウェルを有する。マイクロリソグラフィー技術およびマイクロマシン加工技術(例えば、同時係属中の米国出願第09/849,781号、2001年5月4日出願、これは参考としてその全体を本明細書に含めるものとする)によって、数100ミクロンから100nmもしくはさらに小さいものにまで及ぶ種々多様な寸法で、深さは同じ程度の、整列したウェルを製造することができる。ある実施形態において、シリコンウェハーをマイクロマシンで微細加工して、マスター鋳型として使用し、直径400μmのウェルを200μm離した間隔をおいて成形するが、ウェルの密度は約277ウェル/cm2で、個々のウェルの容積は深さ100μmのウェルについて約30nlである。
【0077】
別の実施形態において、マイクロリソグラフィーによる微細加工によって、直径500nmおよび275nmのウェルを1μmの間隔をおいて作製し、その結果ウェル密度はそれぞれ4400万および6100万ウェル/cm2以上となる。直径が小さいことはもちろん、間隔が狭いほど、密度を高くすることができる。
【0078】
また別の実施形態において、精密マイクロレーザー加工技術によって、アクリルを材料として、1.5mmを超えるものから下は500μmまでの範囲の大きさで、約500μm間隔のウェルを有する鋳型構造を直接製作することができる。このようなウェルの容積は50〜500nlの範囲である。
【0079】
したがって、ある実施形態において、プロテオームチップは基板の表面上に複数のウェルを含んでなり、ここで、ウェルの密度は少なくとも100ウェル/cm2、1000ウェル/cm2、10,000ウェル/cm2、100,000ウェル/cm2、1,000,000ウェル/cm2、10,000,000ウェル/cm2、25,000,000ウェル/cm2、10,000,000,000ウェル/cm2、または10,000,000,000,000ウェル/cm2である。本発明は複数のウェルを含んでなるプロテインチップの変更を意図するが、これらは例えば同時係属中の米国出願第09/849,781号、出願日2001年5月4日、において開示されており、これは参考としてその全体を本明細書に含めるのもとする。
【0080】
また、本発明は、プロテオームチップにおいてウェルの形、幅対深さの比、および容積が変化したものを包含するが、これらは例えば同時係属中の米国出願第09/849,781号、出願日2001年5月4日、において開示されており、これは参考としてその全体を本明細書に含めるのもとする。このような形としては、円、楕円、長方形、正方形などが挙げられるがそれに限定されない。こうしたウェルの底面は、例えば、方形、丸みのあるV型、またはU型とすることもできる。
【0081】
ある実施形態において、基板は金を含んでなる。好ましい実施形態において、基板は金をコーティングしたスライドガラスを含んでなる。別の実施形態において、基板はニッケルを含んでなる。別の好ましい実施形態において、基板はニッケルコーティングしたスライドガラスを含んでなる。ニッケルを含んでなる基板はポリヒスチジンタグ(「Hisタグ」)を有する融合タンパク質の精製および付着に有利である。もう1つの実施形態において、基板はニトロセルロースを含んでなる。さらに別の好ましい実施形態において、基板はニトロセルロースをコーティングしたスライドガラスを含んでなる。
【0082】
本発明はさらに、プロテオームチップ基板の誘導体化に有用な化合物に関する。タンパク質を直接基板に結合することができるが、リンカー分子または化合物を介して基板に付着させることもできる。このリンカーは、基板の表面を誘導体化して基板表面にタンパク質が付着しやすくする、あらゆる分子もしくは化合物とすることができる。リンカーは、共有結合もしくは非共有結合によって、タンパク質またはプローブを基板表面に結合させることができる。さらに、リンカーは無機または有機分子とすることができる。ある実施形態において、リンカーはシラン、例えばシアノシラン、チオシラン、アミノシランなどとすることができる。本発明はプロテインチップの誘導体化に有用な化合物を意図するが、その一部は、例えば同時係属中の米国出願第09/849,781号、出願日2001年5月4日、において開示されており、これは参考としてその全体を本明細書に含めるのもとする。
【0083】
したがって、ある実施形態において、プロテオームチップのタンパク質は非共有結合によって(例えば吸着によって)基板に結合する。非共有結合によって基板に結合するタンパク質を、例えば、水素結合、ファンデルワールス結合、静電結合、もしくは金属-キレート配位結合のような様々な分子相互作用によって、基板の表面に付着させることができる。特定の実施形態において、タンパク質はポリリシンでコーティングされた基板の表面に結合する。さらに、上記のように、ある実施形態において、タンパク質はシラン(シアノシラン、チオシラン、アミノシランなど)でコーティングされた基板表面に結合する。
【0084】
その他に、当技術分野でよく知られた架橋化合物、例えば、同一の、または異なる官能基の架橋化合物(例、ビス[スルホスクシンイミジル]スベリン酸、N-[ガンマ-マレイミドブチルオキシル]スクシンイミド エステル、もしくは1-エチル-3-[3-ジメチルアミノプロピル]カルボジイミド)を用いて、共有結合性または非共有結合性相互作用によってタンパク質を基板に付けることができる。
【0085】
別の実施形態において、プロテオームチップのタンパク質は共有結合によって基板に結合する。例えば、タンパク質は、受容体-リガンド相互作用によって基板に結合することができるが、この相互作用には、抗体と抗原、DNA結合タンパク質とDNA、酵素と基質、アビジン(もしくはストレプトアビジン)とビオチン(もしくはビオチン化分子)の間の相互作用、および脂質結合タンパク質とリン脂質(または、リン脂質を含んでなる、膜、小胞、もしくはリポソーム)の間の相互作用を含める。
【0086】
当業界において公知の様々な方法によって、精製されたタンパク質をアレイ上に置くことができる。ある実施形態において、タンパク質を基板上にプリントすることができる。もう1つの実施形態においては、タンパク質をアフィニティータグによって基板に付着させる。タンパク質の精製に用いたものと異なるアフィニティータグを使用することが好ましいが、これはプロテインアレイを作るときにさらなる精製が達成されるためである。
【0087】
したがって、好ましい実施形態において、プロテオームチップのタンパク質は、基板表面に付着する化合物に親和性を示す、少なくとも1つの異種ドメインを含有する融合タンパク質として発現される。融合タンパク質を基板上に結合するために役立つ適当な化合物(すなわち結合パートナーとして機能する)には、トリプシン/アンヒドロトリプシン、グルタチオン、免疫グロブリンドメイン、マルトース、ニッケル、またはビオチンおよびその誘導体があるが、それらに限定されない。前記化合物が結合するのは、それぞれ、ウシ膵臓トリプシンインヒビター、グルタチオン-S-トランスフェラーゼ、プロテインAもしくは抗原、マルトース結合タンパク質、ポリヒスチジン(例えば、HisX6タグ)およびアビジン/ストレプトアビジンである。例えば、プロテインA、プロテインGおよびプロテインA/Gは、哺乳類の免疫グロブリン分子、特にIgG、のFc部分に結合できるタンパク質である。前記タンパク質を例えばSepharose(登録商標)支持体に共有結合で連結して、Fcドメインを含んでなるタグを有する融合タンパク質を精製する効率的な方法を提供することができる。
【0088】
別の実施形態では、タンパク質を基板に直接結合させる。さらに別の実施形態では、タンパク質をリンカーによって基板に結合させる。特定の実施形態では、タンパク質をHisタグを介して基板に付着させる。別の特定の実施形態では、タンパク質を3-グリシドオキシプロピルトリメトキシシラン(”GPTS”)リンカーを介して基板に付着させる。特定の実施形態では、タンパク質をHisタグを介して基板に結合させるが、ここで基板は平坦な表面を含んでなる。好ましい実施形態では、タンパク質をHisタグを介して基板に結合させるが、ここで、基板はニッケル-コーティングされたスライドガラスからなる。
【0089】
本発明のプロテオームチップは、その物理的大きさに限定されず、使用できるあらゆる大きさをとることができる。プロテオームチップは、自動化技術に適合したアレイフォーマットを有することが望ましく、それによって迅速なデータ分析が可能となる。したがって、ある実施形態において、プロテオームマイクロアレイフォーマットは、実験装置および/または分析用ソフトウェアに適合する。好ましい実施形態において、プロテオームチップは標準の顕微鏡スライドガラスの大きさである。別の好ましい実施形態において、プロテインチップは質量分析計の試料チャンバーにはまるようにデザインされる。
【0090】
5.2 高密度アレイフォーマットにおいてタンパク質を調製し精製する方法
本発明は、ウイルス、原核生物もしくは真核生物のタンパク質を、ハイスループット分析に供しやすく、容易に測定できるフォーマットで作製し、単離する方法に関する。好ましい方法は、自動化技術に適合したアレイフォーマットで、タンパク質を合成し、精製することを包含する。したがって、ある実施形態において、本発明は、適切に機能するように調節配列に連結された異種配列を有するベクターで形質転換された真核細胞を増殖させるステップ、異種配列によってコードされたタンパク質の発現を増強する誘導物質と前記調節配列を接触させるステップ、細胞を溶解するステップ、結合物質を前記タンパク質と接触させてタンパク質と結合物質との複合体を形成させるステップ、細胞破砕物から前記複合体を分離するステップ、およびこの複合体からタンパク質を単離するステップを含んでなり、これらの各ステップが96-ウェルフォーマットにおいて実施される、真核生物タンパク質を調製し単離する方法を提供する。
【0091】
ある実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は、単一の種における既知遺伝子の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%によってコードされる、少なくとも1つのタンパク質を含んでなる。
【0092】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は、単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%を含んでなる。ここでタンパク質のアイソフォームおよびスプライス変異体は単一タンパク質とみなされる。
【0093】
もう1つの実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は、単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000もしくは1,000,000のタンパク質を含んでなる。
【0094】
さらに別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は全体として、単一の種における少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000、もしくは50000の異なる既知の遺伝子を含んでなる。
【0095】
また別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記タンパク質は、第1タグ、第2タグ、および、生物のゲノム核酸によってコードされるタンパク質を含んでなる融合タンパク質である。
【0096】
ある実施形態において、合成および精製手順の各ステップは、迅速な自動化を行いやすいアレイにおいて実施される。このようなアレイは、基板表面上の複数のウェルを含んでなるが、ここでウェルの密度は、例えば、少なくとも10、20、30、40、50、100、1000、10,000、100,000、もしくは1,000,000ウェル/cm2である。あるいはまた、こうしたアレイは、基板表面上の複数のサイトを含んでなり、こうしたサイトの密度は、例えば、少なくとも10、20、30、40、50、100、1000、10,000、100,000、もしくは1,000,000サイト/cm2である。
【0097】
特別の実施形態において、真核生物のタンパク質は、96アレイフォーマット(すなわち、プロセシングが起こる基板上の各サイトは96サイトのうちの1つである)、例えば96ウェルマイクロタイタープレートで調製され、精製される。好ましい実施形態において、基板はタンパク質と結合しない(例えば、タンパク質非結合性マイクロタイタープレート)。
【0098】
特定の実施形態において、タンパク質は当技術分野で広く知られている方法にしたがって、in vitro翻訳によって合成される。
【0099】
タンパク質合成を進める誘導プロモーターを有するあらゆる発現構築物を、本発明の方法にしたがって使用することができる。発現構築物は、形質転換に使用すべき細胞の種類に合わせて作製することが望ましい。発現構築物と宿主細胞との適合性は当業界で知られており、その変異体を使用することも本発明に包含される。
【0100】
培養下で増殖可能な宿主細胞を用いて、対象のタンパク質を合成することができる。対象のタンパク質を過剰生産することが可能で、結果として、タンパク質の適正な合成、折りたたみ、および翻訳後修飾をもたらす宿主細胞を使用することが望ましい。こうしたタンパク質のプロセシングによって、in vivoに対応するin vitro分子相互作用を特徴づけるアッセイに有用なエピトープ、活性部位、結合部位などが形成されることが望ましい。
【0101】
したがって、真核細胞(例えば、酵母、ヒト細胞)を用いて真核生物タンパク質を合成することが望ましい。さらに、安定な形質転換がしやすく、対象の形質転換体を含有する細胞を同定および単離するための選択マーカーを有する真核細胞が好ましい。あるいはまた、ある遺伝子産物を欠損した真核宿主細胞を、その欠損を補完する発現構築物で形質転換する。改変されたウイルス、原核生物、もしくは真核生物タンパク質の発現に好都合な細胞は当業界で公知であり、そうした細胞の変異体を当業者であれば正しく認識することができる。
【0102】
例えば、Invitrogen社(Carlsbad, CA, カタログ番号K800-01)製のInsectSelect系は、非細胞溶解性、単一ベクター昆虫発現系であって、高品質のタンパク質の発現を単純化し、ウイルスストックを調製および増幅する必要のない系であるが、これを使用することができる。このシステムにおいて好ましいベクターは、pIB/V5-His TOPO TAベクター(カタログ番号K980-20)である。製造業者の記載した手順によって、ポリメラーゼ連鎖反応法(“PCR”)産物を直接このベクターにクローニングすることが可能であり、さらに発現されたタンパク質の精製に役立つN-末端ヒスチジンタグを有するタンパク質を発現させることができる。
【0103】
昆虫細胞における別の真核生物発現系として、BAC-TO-BACTM系(LIFETECHTM, Rockville, MD)を用いることもできる。相同的組換えを利用するのではなくて、BAC-TO-BACTM系は、大腸菌における部位特異的遺伝子転位に基づいて、組換えバキュロウイルスを生成する。遺伝子発現は非常に強力なポリヘドリンプロモーターによって駆動されるので、感染した昆虫細胞の細胞タンパク質の25%にまで相当することがある。
【0104】
特定の実施形態においては、酵母培養物を用いて真核生物融合タンパク質を合成する。タンパク質合成を効率よく誘導するために、特に小量の培地で実施する場合には、新しい培養物を使用することが望ましい。また、酵母培養物が増殖しすぎないよう注意することが望ましい。さらに、約3ml以下の酵母培養物が精製に十分なタンパク質を生産することが望ましい。培養物の通気を改善するために、全体量をいくつかの小量に分けることができる(例えば、4本の0.75ml培養物を調製して、全体量を3mlとすることができる)。
【0105】
次に、細胞を誘導物質(例えば、ガラクトース)と接触させ、その後回収する。誘導された細胞をそれぞれ、冷水(すなわち、4℃から約15℃まで)で洗浄して、これ以上の細胞増殖を止めた後、冷(すなわち、4℃から約15℃まで)細胞溶解バッファーで洗浄して培地を除去し、誘導した細胞をタンパク質精製のためにプレコンディショニングする。タンパク質精製の前に、誘導細胞を冷凍保存して、タンパク質の分解を防ぐことができる。特定の実施形態においては、誘導細胞を−80℃にて半乾燥状態で保存して、タンパク質の分解を防止または抑制する。
【0106】
細胞をアレイから別のアレイへ、なんらかの適当な機械装置によって移すことができる。例えば、自動操作システム(例えば自動ピペット)を用いて、培地を含有するアレイに対象の細胞を接種することができる。特別の実施形態において、寒天を含む増殖培地を含有する96ウェルアレイに96-prongerを用いて酵母細胞を植菌することができる。同様に、自動液体取扱装置(例えば、Q-FILLTM、Genetix, UK)を用いて、アレイからアレイへ液体(例えば、試薬)の移動を行うことができる。
【0107】
細胞周期のいかなる時点の細胞からもタンパク質を回収することができるが、タンパク質合成が高まっている対数増殖期に細胞を分離することが望ましい。例えば、酵母細胞は、OD600=0.3からOD600=1.5の間で、望ましくはOD600=0.5からOD600=1.5の間で、集めることができる。特定の実施形態において、対数増殖期中期後のある時点の細胞からタンパク質を回収する。集めた細胞を将来の操作に備えて凍結保存することができる。
【0108】
集めた細胞を当業界で公知の様々な方法によって溶解することができるが、これには、例えば機械的な力、酵素消化、化学的処理などがある。細胞溶解の方法は、宿主細胞のタイプに適合していなくてはならない。例えば、新鮮なプロテアーゼインヒビターを含有する溶解バッファーを、酵母細胞に、細胞壁を破壊する物質(例えば、砂、ガラスビーズ、ジルコニアビーズ)とともに加え、その後、この混合物を振盪機(例えば、ボルテックスミキサー、ペイントシェーカー)によって激しく振盪する。
【0109】
特定の実施形態では、ジルコニアビーズを酵母細胞に接触させ、ボルテックス混合による機械的破壊によって細胞を溶解する。もう1つの実施形態では、高密度アレイフォーマットでの酵母細胞の溶解は、ペイントシェーカーによって達成される。ペイントシェーカーには、少なくとも18個の96-ウェルボックスを3層にしっかりと保持できる台があり、それによって、培養物の高速大量処理が可能になる。さらに、ペイントシェーカーは、培養物が完全に融解する前でも、これを激しく撹拌するため、結果としてタンパク質の分解を最小限に抑えつつ細胞の効率的な破壊をもたらす。実際、顕微鏡観察によって判定されるように、90%を超える酵母細胞を2分足らずの振盪で溶解することができる。
【0110】
結果として得られた細胞破砕片を対象のタンパク質および/または他の分子から遠心によって分離することができる。また、ハイスループット様式でタンパク質試料の純度を高めるために、タンパク質濃縮上清を、望ましくはタンパク質非結合性基板上のフィルターを用いて、濾過することができる。対象のタンパク質を含有する可溶性画分を不溶性画分から分離するためにフィルタープレートを使用することは、タンパク質の分解を減らし、または回避するために非常に望ましい。さらに、細胞破砕片を含有する画分について、これらのステップを繰り返すことが望ましく、結果としてタンパク質の収率が向上する。
【0111】
次に、このタンパク質濃縮上清から、当業界で公知の様々なアフィニティー精製法によってタンパク質を精製することができる。融合タンパク質調製物をアフィニティータグに対する結合パートナーと接触させることによる、融合タンパク質のアフィニティー精製に有用なアフィニティータグは、カルモジュリン、トリプシン/アンヒドロトリプシン、グルタチオン、免疫グロブリンドメイン、マルトース、ニッケル、またはビオチンおよびその誘導体を包含するがそれに限定されない。これらはそれぞれ、カルモジュリン結合タンパク質、ウシ膵臓トリプシンインヒビター、グルタチオン-S-トランスフェラーゼ(「GSTタグ」)、抗原もしくはプロテインA、マルトース結合タンパク質、ポリヒスチジン(「Hisタグ」)、およびアビジン/ストレプトアビジンと結合する。他のアフィニティータグとしては、例えば、mycまたはFLAGが挙げられる。適当な結合化合物(すなわち、グルタチオンビーズのような結合パートナー)を用いて、融合タンパク質をアフィニティー精製して、単離することができるが、これは、例えば、タンパク質非結合性フィルター上に結合タンパク質を含有する複合体を捕捉することによる。アフィニティータグをタンパク質の一方の末端(例えば、カルボキシ末端)に、第2のアフィニティータグをタンパク質の他方の末端(例えば、アミノ末端)に配置することは、全長タンパク質の精製に役立つと考えられる。
【0112】
特別の実施形態において、融合タンパク質はGSTタグを有しており、このタンパク質をグルタチオンビーズと接触させることによってアフィニティー精製される。もう1つの実施形態においては、融合タンパク質の付着したグルタチオンビーズを、96ウェルボックス内でフィルタープレートを使用することなく洗浄して、試料の取り扱いを容易にし、試料の相互汚染を防止することができる。
【0113】
さらに、融合タンパク質を、結合化合物(例えば、グルタチオンビーズ)から、所望のタンパク質濃度を与えるように、溶出バッファーで溶出することができる。特定の実施形態において、融合タンパク質はグルタチオンビーズから30μlの溶出バッファーで溶出されて、好ましいタンパク質濃度を与える。
【0114】
最終的に顕微鏡スライドガラス上にスポットされる精製タンパク質の場合は、グルタチオンビーズをこの精製タンパク質から分離する。すべてのグルタチオンビーズを除去して、基板上に精製タンパク質をスポットするために使用されるマイクロアレイピンが塞がるのを避けることが望ましい。好ましい実施形態において、グルタチオンビーズは、フィルタープレートを用いて、精製タンパク質から分離されるが、このプレートはタンパク質非結合性基板を含んでなることが望ましい。精製タンパク質を含有する溶出液の濾過によって、結果としてタンパク質の回収率は90%を超えるはずである。
【0115】
溶出バッファーは、例えば、15%から50%までのグリセロール、望ましくは約40%グリセロールのような粘性の高い液体を含んでなることが望ましい。グリセロール溶液は溶液中のタンパク質を安定化し、マイクロアレイヤーを用いたプリンティングステップの間、タンパク質溶液の乾燥を防ぐ。
【0116】
精製タンパク質は、タンパク質を安定化し、試料の乾燥を防ぐ媒質中で保存することが望ましい。例えば、15%から50%までのグリセロール、望ましくは約40%グリセロールのような粘性の高い液体中で、精製タンパク質を保存することができる。凍結/融解サイクルによって引き起こされるタンパク質活性の低下を避けるために、精製タンパク質を含有する試料をアリコートに分割することが望ましい。
【0117】
精製プロトコルを調整してタンパク質の純度を望ましいレベルに制御することができることを、当業者は正しく認識することができる。ある場合には、対象のタンパク質と会合する分子の分離が求められる。例えば、過剰生産された対象のタンパク質を含んでなる、二量体、三量体、もしくはそれ以上のホモタイプもしくはヘテロタイプ複合体を、本明細書に記載される精製方法、もしくはその変法によって単離することができる。さらに、結合した分子を当業界で公知の方法によって個別に単離して同定することができる(例えば、質量分析)。
【0118】
5.3 プロテオームアレイの作製方法
本発明は、また、プロテオームチップを作製する方法に関する。したがって、本発明は、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法を提供し、ここで、前記の複数のタンパク質は、ある単一の種における既知遺伝子のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%に相当する、少なくとも1つの代表的なタンパク質を含んでなり、この場合、前記タンパク質は、1つの遺伝子に由来するすべてのタンパク質アイソフォームおよびスプライス変異体とする。
【0119】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法を提供し、ここで、前記の複数のタンパク質は、単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%を含んでなる。
【0120】
また別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法を提供し、ここで、前記の複数のタンパク質は、単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000もしくは1,000,000のタンパク質を含んでなる。
【0121】
本発明はまた、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、次のような、位置指定可能なアレイを作製する方法に関する。すなわち、前記基板が微細加工された型から成形され、さらに、前記の複数のタンパク質が、単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%を含んでなるか、または、単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000もしくは1,000,000のタンパク質を含んでなるか、または、ある単一の種における既知遺伝子のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%に相当する、少なくとも1つの代表的なタンパク質を含んでなり、この場合、前記タンパク質は、1つの遺伝子に由来するすべてのタンパク質アイソフォームおよびスプライス変異体である。本発明は、微細加工された型から成形された様々な基板を包含するが、その一部は、例えば、2001年5月4日出願の同時係属中の米国特許出願第09/849,781号において開示されており、この出願はその全体を参考として本明細書に含めるものとする。
【0122】
本発明はまた、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法に関するが、ここで、前記タンパク質は第1のタグおよび第2のタグを含んでなる。二重タグ標識タンパク質を用いる利点としては、高度に精製されたタンパク質が得られること、ならびに、細胞破砕片からタンパク質を精製してそのタンパク質を基板に付着させる能率的な方法を提供することがある。特定の実施形態において、第1タグはグルタチオン-S-トランスフェラーゼタグ(「GSTタグ」)であり、第2タグはポリヒスチジンタグ(「Hisタグ」)である。もう1つの実施形態において、GSTタグおよびHisタグはタンパク質のアミノ末端に結合される。あるいはまた、GSTタグおよびHisタグはタンパク質のカルボキシ末端に結合される。
【0123】
さらに別の実施形態において、GSTタグはタンパク質のアミノ末端に結合される。また別の実施形態において、Hisタグはタンパク質のカルボキシ末端に結合される。もう1つの実施形態において、Hisタグはタンパク質のアミノ末端に結合される。また別の実施形態において、GSTタグはタンパク質のカルボキシ末端に結合される。
【0124】
別の実施形態において、タンパク質はGSTタグおよびHisタグを含んでなるが、GSTタグもHisタグも、タンパク質のアミノ末端もしくはカルボキシ末端には存在しない。特定の実施形態において、GSTタグおよびHisタグは、当該タンパク質のコード領域の中に存在する;対象とする結合ドメインに影響を及ぼさないタンパク質の領域にあることが望ましい。
【0125】
ある実施形態において、第1タグは融合タンパク質を精製するために用いられる。別の実施形態において、第2タグは融合タンパク質を基板に付着させるために使用される。別の特定の実施形態において、第1タグはGSTタグであり、第2タグはHisタグである。
【0126】
タンパク質は融合タンパク質であって、その異種配列が当該タンパク質のコード配列およびアフィニティータグのようなタグをコードする配列を含んでなる融合タンパク質であることが望ましい。このようなタグは、タンパク質をモニタリングし、細胞破砕片および混入試薬から融合タンパク質を分離し、および/または、タンパク質を本発明のプロテオームチップに付着させるために有用であると考えられる。
【0127】
誘導物質の例には、ガラクトース、エンハンサー結合タンパク質および他の転写因子があるが、それらに限定されない。ある実施形態においては、ガラクトースを、ガラクトース誘導性GAL1プロモーターを含む調節配列と接触させる。
【0128】
本発明にしたがって使用することができる結合物質としては、グルタチオンビーズ、ニッケルコーティングされた基板、および抗体があるが、それらに限定されない。ある実施形態において、複合体はグルタチオンビーズに結合したGSTタグを有する融合タンパク質を含んでなる。別の実施形態において。複合体はニッケルコーティングされた基板に結合したHisタグを有する融合タンパク質を含んでなる。さらに別の実施形態において、複合体は、抗体および、場合によっては、二次抗体に結合した対象のタンパク質を含んでなる。
【0129】
5.4 プロテオームアレイを使用する方法
本発明はまた、プロテオームチップを用いて、少なくとも1つの試料中に存在するタンパク質の存在、量、および/または機能性をアッセイする方法に関する。本発明のプロテオームチップを用いて、化学反応およびアッセイを大規模並行的な分析において実施して、生物学的状態または生物学的応答の特徴を明らかにし、タンパク質の存在、量、および/または生物活性を決定することができる。したがって、本発明のプロテオームチップを用いて、細胞、組織、器官、系、もしくは生物体における本質的にすべてのタンパク質-タンパク質相互作用をアッセイすることができる。
【0130】
さらに、本発明のプロテオームチップを用いて、宿主細胞(すなわち、融合タンパク質を調製するために使用した細胞)に加えられた特定の刺激に対する生物学的応答をアッセイすることができる。例えば、融合タンパク質をコードする発現ベクターで形質転換された酵母細胞に刺激を与えることができるが、その後、その融合細胞を精製してアレイ化する。アレイ化したタンパク質を、次に、例えば、何らかの結合物質を用いてプロービングすることによって特徴づけることができる。その結果得られた結合パターンを、次に、前記刺激を受けない、または異なる刺激を受けた酵母細胞から作製された同一のアレイと比較する。結合パターンの相違は、生物学的応答の特性を示していると考えられ、生物学的応答に関して特異的な相互作用物質を同定することができる。
【0131】
ある実施形態において、宿主細胞から作製されたプロテオームチップ(各チップは異なる刺激に曝された宿主細胞に対応する)を、標識レクチン(例えば、コンカナバリンA)でスクリーニングする。タンパク質-プローブ相互作用のパターンはグリコシル化タンパク質の存在を示唆するが、これを比較して、プロテオームチップのタンパク質のグリコシル化状態に関するそれぞれの刺激の影響を判定する。
【0132】
本発明のプロテオームチップを用いて測定できる生物活性は、酵素活性(例えば、キナーゼ活性、プロテアーゼ活性、ホスファターゼ活性、グリコシダーゼ、アセチラーゼ活性、および他の化学基転移酵素活性)、核酸の結合、ホルモンの結合、などを包含するが、それに限定されない。高密度および小容量の化学反応は、本発明のプロテオームチップの使用に関わる方法に好都合であると考えられる。
【0133】
生物学的状態もしくは応答についてこれ以上の情報を、本発明のプロテオームチップを用いて得ることができるが、この場合、チップ上のタンパク質はタンパク質の分類にしたがって組織化される。この分類は、存在量、機能、機能的クラス、酵素活性、相同性、タンパク質ファミリー、特定の代謝経路もしくはシグナル伝達経路との関連性、関連した代謝経路もしくはシグナル伝達経路との関連性、または翻訳後修飾によることができる。
【0134】
本発明のプロテオームチップを1以上のプローブと接触させる際に、当業界で公知の様々な技法を用いて、タンパク質-プローブ相互作用をアッセイすることができる。例えば、化学発光もしくは蛍光を生じる標準的な酵素測定法によって、プロテオームチップをアッセイすることができる。非タンパク質基質、酵素的発色、質量分析のサインマーカー、もしくはオリゴヌクレオチドタグの増幅を用いて、例えば、フォトルミネセンス、化学発光、もしくは蛍光によって、様々なタンパク質の修飾を検出することができる。
【0135】
当技術分野で広く知られている方法によって直接的もしくは間接的にその結合を検出できるように、プローブをマーカーで標識し、もしくはタグを付ける。当技術分野で公知のあらゆるマーカーを使用することが可能であって、こうしたマーカーは、それがプロテオームチップのタンパク質をチップの基板に付けるために使用されるアフィニティータグもしくは試薬と同一ではないという条件で、エピトープタグ、ハプテンおよびアフィニティータグといったタグ、抗体、標識などを包含するが、それらに限定されない。例えば、タンパク質をプロテオームチップアレイに付着させるリンカーとしてビオチンを使用するならば、このプロテオームチップのタンパク質中に存在しない別のタグ、例えばHisもしくはGSTを用いてプローブを標識し、タンパク質-プローブ相互作用を検出する。特定の実施形態においては、フォトルミネセンス、化学発光、蛍光、もしくは酵素タグを使用する。他の実施形態においては、質量分析のサインマーカーを使用する。さらに他の実施形態では、増幅可能なオリゴヌクレオチド、ペプチド、もしくは分子量標識を使用する。
【0136】
特定の実施形態において、本発明は、標識プローブ(例えば、標識タンパク質)試料を、複数のタンパク質を含んでなる位置指定可能なアレイ(それぞれのタンパク質は基板上の異なる位置に存在する)と接触させるステップ、およびアレイ上で標識プローブとタンパク質との相互作用が生じるアレイ上の位置を検出するステップを含んでなる、タンパク質-プローブ相互作用を検出する方法を提供する。
【0137】
したがって、タンパク質-プローブ相互作用を例えば下記によって検出することができる:すなわち、1)放射性標識リガンドを使用し、それに続いてオートラジオグラフィーおよび/またはホスホルイメージャー(PhosphorImager)分析を行うこと;2)ハプテンの結合、これを次に蛍光標識したもしくは酵素標識した抗体または高親和性ハプテンリガンド、例えばビオチンもしくはストレプトアビジン、によって検出する;3)質量分析;4)原子間力顕微鏡法;5)蛍光偏光法;6)赤外線赤色標識化合物もしくはタンパク質;7)増幅可能なオリゴヌクレオチド、ペプチドもしくは分子量標識;8)タンパク質の酵素活性の刺激もしくは阻害;9)ローリングサークル増幅-検出法(Hatchら、1999、「固体表面に固定化されたDNAのローリングサークル増幅および多重突然変異検出へのその応用」(”Rolling circle amplification of DNA immobilized on solid surfaces and its application to multiplex mutation detection”)、Genet. Anal. 15:35-40);10)競合的PCR(Finiら、1999、「マイクロプレート照度計を用いてパルボウイルスB19 DNAを検出および定量するための化学発光競合PCRの開発」(”Development of a chemiluminescence competitive PCR for the detection and quantification of parvovirus B19 DNA using a microplate luminometer”)、Clin. Chem. 45:1391-6; Kruseら、1999、「セミネステッド競合PCRアッセイによるトランスフォーミング増殖因子-β1(TGF-β1)遺伝子発現の検出および定量」(”Detection and quantitative measurement of transforming growth factor-beta 1 (TGF-beta 1) gene expression using a semi-nested competitive PCR assay”)、Cytokine 11: 179-85; GuenthnerおよびHart, 1998、「マイクロプレートを基本とする検出システムによるHIV-1のための定量的競合PCRアッセイ」(”Quantitative, competitive PCR assay for HIV-1 using a microplate-based detection system”)、Biotechniques 24: 810-6);11)比色定量法;および12)生物学的アッセイ(例えば、ウイルス力価)。
【0138】
特別の実施形態において、タンパク質-プローブ相互作用は、直接質量分析によって検出される。さらなる実施形態において、タンパク質および/またはプローブの正体は質量分析を用いて決定される。例えば、プロテオームチップ上のタンパク質と結合している1以上のプローブを、アレイから解離させて質量分析によって同定することができる(例えば、WO 98/59361を参照されたい)。別の実施形態において、質量分析によって、プロテオームチップ上のタンパク質の酵素的切断を検出し、切断されたタンパク質断片もしくは他の放出された化合物を同定することができる。
【0139】
ある実施形態において、プロテオームチップ上の各タンパク質をプローブと接触させてタンパク質-プローブ相互作用を検出し、定量する。別の実施形態において、プロテオームチップ上の各タンパク質を複数のプローブと接触させ、タンパク質-プローブ相互作用を検出して定量する。たとえば、プロテオームチップを同時に複数のプローブを用いてスクリーニングすることができる。このプローブは、複雑な混合物(例えば、細胞抽出物)、そのままの細胞成分(例えば、細胞小器官)、細胞全体、およびいくつかの起源から集められたプローブを包含するが、それらに限定されない。次に、タンパク質-プローブ相互作用を検出し、定量する。1つには、本発明のアレイの位置指定可能な性質によって、すなわち、プロテインチップ上の既知の位置にタンパク質を配置することによって、プローブの混合物を用いたアッセイから有用な情報が得られ、プローブが結合するタンパク質(「相互作用物質」)を特徴づけることができる。
【0140】
当業者であれば、プローブを用いて本発明のプロテオームチップをスクリーニングすることによって、様々な細胞相互作用をアッセイすることを目的とした数多くの様々な実施形態を正しく認識することができる。例えば、様々なプローブを用いて、プロテオームチップを多数回連続してスクリーニングすることによって、特定のシグナル伝達経路もしくは特定の代謝経路に関わるすべてのタンパク質を明確にすることができる。さらに、こうしたアッセイは、診断、予後判定および/または治療目的に役立てることができる。
【0141】
本発明の方法によれば、プローブは、細胞、細胞膜、細胞内小器官、タンパク質を含有する細胞材料、タンパク質、オリゴヌクレオチド、ポリヌクレオチド、DNA、RNA、小分子(すなわち、分子量が500より小さい化合物)、基質、薬物もしくは候補薬物、受容体、抗原、ステロイド、抗体、免疫グロブリンドメイン、グルタチオン、マルトース、ニッケル、ジヒドロトリプシン、レクチン、またはビオチンとすることができる。
【0142】
タンパク質-プローブ相互作用を検出するために、プロテインアレイと接触させる際に使用するプローブはビオチン化することができる。弱くビオチン化したタンパク質の方が、対象の生物活性を維持する可能性が大きい。このように、タンパク質の結合活性もしくは他の生物活性を保存するためには、より穏やかなビオチン化法が好ましい。したがって、特別の実施形態において、プローブタンパク質は、ビオチン転移化合物(例えば、Sulfo-NHS-LC-LC-Biotin; PIERCETM カタログ番号21338、USA)によって様々な程度にビオチン化される。
【0143】
さらに、酵素基質もしくは阻害剤をプローブとすることができる。例えば、プローブは次のような酵素の基質もしくは阻害剤とすることができる。すなわち、キナーゼ、ホスファターゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、および他の化学基転移酵素といった酵素であるが、これらに限定されない。核酸もしくはタンパク質プローブの組合せとチップ上のタンパク質をインキュベートした後、例えば、質量分析によって結合した核酸またはタンパク質プローブを同定することができる(Lakeyら、1998、「タンパク質-タンパク質相互作用の測定」(“Measuring protein-protein interactions”)、Curr Opin Struct Biol. 8: 119-23)。
【0144】
したがって、プロテオームチップ上のタンパク質との相互作用に対する様々な細胞性応答を、ホールセル(whole cell)をプローブとして探索することによってアッセイすることができる。例えば、プロテオームチップをリンパ球と接触させて、リンパ球の活性化を様々な方法によってアッセイすることができる。すなわち、前記方法には、抗体合成を検出すること、3H-チミジンの取り込みを検出もしくは測定すること、細胞表面の分子を抗体で標識して抗原認識および活性化によって誘導もしくは抑制される分子を同定すること(例えば、CD23、CD38、IgD、C3b受容体、IL-2受容体、トランスフェリン受容体、膜クラスII MHC分子、PCA-1分子、HLA-DR)、および、発現されたおよび/または分泌されたサイトカインを同定することがあるが、それらに限定されない。
【0145】
別の例において、細胞をプロテオームチップとともにインキュベートすることによって、特定の細胞型に対するマイトジェンを決定することができる。有糸分裂活性を、例えば、細胞による3H-チミジンの取り込みを検出もしくは測定することによって、測定することができる。細胞は同一の細胞型(すなわち、均一集団)であってもよく、異なる細胞型とすることもできる。
【0146】
また別の例において、細胞をプロテオームチップとともにインキュベートすることによって、特定の細胞型に対する分化因子を決定することができる。細胞の分化はたとえば、目視検査、マーカー特異的抗体による細胞表面分化マーカーの検出、もしくは分泌された分化マーカーの同定によって判定することができる。
【0147】
別の例において、細胞をプロテオームチップとともにインキュベートすることによって、特定の細胞型に対するアポトーシス因子を決定することができる。アポトーシスは、例えば、目視検査、マーカー特異的抗体による細胞表面アポトーシスマーカーの検出、または、培地中に放出された分泌マーカーもしくは他の細胞成分の同定によって判定することができる。
【0148】
プロテオームチップ上のタンパク質に対する細胞の分泌応答を、細胞を本発明のプロテオームチップとともにインキュベートすることによって、アッセイすることができる。分泌されたタンパク質および他の細胞性化合物を、例えば、培地中に放出された化合物を検出することで、アッセイすることができる。
【0149】
別の例において、例えば、1以上の細胞を本発明のプロテオームチップとともにインキュベートし、細胞凝集をアッセイすることによって、プロテオームチップ上のタンパク質が細胞凝集を媒介する能力についてアッセイすることができる。また、例えば、細胞および細胞外マトリックス成分をプロテオームチップとともにインキュベートし、チップ上のタンパク質による細胞もしくは細胞外マトリックス成分の親和性の増加をアッセイすることによって、細胞外マトリックスに対する親和性を媒介するタンパク質の能力をアッセイすることができる。こうしたアッセイで同定される相互作用物質は、例えば、癌、細胞移動、シナプス形成、樹状細胞の成長、突起の伸長、もしくは軸索の伸長において何らかの役割を担うと考えられる。
【0150】
さらに別の例において、本発明のプロテオームチップのタンパク質がイオン輸送もしくは他の小分子輸送(例えばATP)に及ぼす影響を測定することができる。例えば、プローブ細胞に放射性標識したイオンもしくは他の小分子を前もってロードして、本発明のプロテオームチップとインキュベートする。細胞をプロテオームアレイのタンパク質と接触させた後、様々な時点で、放射性標識の保持および放出を測定することができる。あるいはまた、当技術分野で公知の電気生理学的技法を用いてイオン輸送を検出し、特徴づけることができる。
【0151】
さらに別の例において、プロテオームチップ上のタンパク質の細胞による取り込みおよび/またはプロセシングを、例えば、チップ上に放射性もしくは蛍光標識したタンパク質を有するプロテオームチップとともに細胞をインキュベートし、プロテオームチップ上のシグナルの増加もしくは減少を測定するか、または細胞による標識タンパク質の取り込みを測定することによって、アッセイすることができる。
【0152】
あるいはまた、本発明のプロテオームチップを細胞および対象の標識化合物とともにインキュベートし、その結果として、細胞による化合物の取り込みおよび/またはプロセシングを検出および/または測定する。
【0153】
無細胞系において小分子をプローブとして探査することによって、小分子(すなわちMW=500より小さい化合物)とプロテオームチップ上のタンパク質との相互作用をアッセイすることができる。この小分子としては、例えば、ATP、GTP、cAMP、ホスホチロシン、ホスホセリンおよびホスホトレオニンがあるが、それらに限定されない。このようなアッセイは、対象の小分子と相互作用する、ある生物種におけるすべてのタンパク質を同定することができる。対象の小分子は、医薬品、候補薬物、殺真菌薬、殺虫薬、発癌物質、および汚染物質を包含するが、これらに限定されない。本発明の方法にしたがってプローブとして使用される小分子は、タンパク質以外の有機化合物であることが望ましい。
【0154】
別の実施形態において、ある生物種における特定のリガンドもしくは特定のクラスのリガンド群に対する本質的にすべての受容体は、対象の受容体を本発明のプロテオームチップと接触させることによって同定することができる。あるいはまた、ある生物種における本質的にすべてのリガンドが対象の特定の受容体もしくは受容体ファミリーによって同定されるが、こうしたリガンドは対象の受容体を本発明のプロテオームチップと接触させることによって同定可能である。別の実施形態において、特定の受容体-リガンド複合体の形成を阻害もしくは阻止できる、1つの生物種の本質的にすべてのタンパク質は、受容体およびそのリガンドを本発明のプロテオームチップと接触させて、受容体-リガンド相互作用がチップ上のタンパク質の不在下での受容体-リガンド相互作用の程度と比較して阻害されるかどうかを判定することによって、同定可能である。受容体-リガンド相互作用の検出およびリガンド相互作用物質の同定は、当技術分野でよく知られた方法によって実施することができる。
【0155】
別の実施形態において、1つの種の本質的にすべてのキナーゼ標的は、例えば、キナーゼを本発明のプロテオームチップと接触させ、標識リン酸の存在下で、当業界で公知の方法によってリン酸化された相互作用物質を検出することによって同定することができる。あるいはまた、リン酸化される可能性のある基質を本発明のプロテオームチップと接触させ、リン酸化された基質の存在および/またはレベルを、リン酸化アミノ酸に特異的な抗体を用いてアッセイすることによって、1つの種の本質的にすべてのキナーゼを同定することができる。別の実施形態において、キナーゼおよびその基質を本発明のプロテオームチップに接触させ、チップ上のタンパク質の不在下でのリン酸化レベルと比較して、基質のリン酸化が減少したかどうかを判定することによって、1つの種の本質的にすべてのキナーゼ阻害剤を同定することができる。
【0156】
キナーゼ活性を検出する方法は、当技術分野で公知であり、放射性標識(例えば、32P-ATPおよび35S-γ-ATP)もしくはリン酸化アミノ酸と結合する蛍光抗体プローブの使用を包含するがそれに限定されない。
【0157】
同様に、アッセイを行って、1つの種における全ホスファターゼおよびホスファターゼ阻害剤を同定することができる。例えば、あるアッセイにおいて放射性標識リンのタンパク質への取り込みはキナーゼ活性を示すが、別のアッセイを用いて放射性標識リンの培地中への放出を測定することが可能であり、それがホスファターゼ活性を示す。
【0158】
また、本発明のプロテオームチップは、例えば、プロテオームチップを、異なる細胞集団に対応する細胞もしくは細胞抽出物と接触させ、プロテオームチップ上のタンパク質-プローブ相互作用のパターンを比較することによって、異なる細胞型(形態的もしくは機能的のいずれか)を識別するために使用することができる。この方法を用いて、例えば、細胞周期の異なる時期、病気の状態、生理学的状態の変化(例えば、低酸素)、治療前後の生理学的状態(例えば、薬物療法)、代謝状態、分化の段階、発生の段階、環境刺激(例えば、光、熱)に対する応答、環境毒物(例えば、殺虫剤、除草剤、汚染)への応答、細胞-細胞相互作用、細胞特異的タンパク質の発現、および疾病特異的タンパク質の発現を特徴づけることもできる。
【0159】
タンパク質-タンパク質相互作用の発生プロファイルを用いて、発生の各段階で関与するシグナル伝達経路、代謝経路などを特徴づけて、発生段階間の推移を解明することができる。こうした研究によって得られる豊富な情報を用いて、各段階の薬物標的を同定したり、かつ/または疾病の経過中に治療計画を適合するように変更することができる。
【0160】
本発明のプロテオームチップを細胞抽出物とともにインキュベートして、特定の細胞型、刺激への応答、もしくは生理学的状態を特徴づけることができる。したがって、典型的な実施形態では、本発明のプロテオームチップを、化合物(例えば、薬物)で処理した細胞から、細胞分化の特定の段階(例えば多能性段階)にある細胞から、または特別な代謝状態(例えば、分裂状態)にある細胞から得られた細胞抽出物と接触させて、例えば、キナーゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、ホスファターゼ、および/または他の転移酵素活性をアッセイすることができる。
【0161】
プロテオームチップ上のタンパク質-プローブ相互作用のパターンは、したがって、生物学的状態に特徴的な「サイン」や「フィンガープリント」を提供することができる。例えば、薬物の存在下もしくは非存在下の細胞、いくつかの分化段階にある細胞、もしくは異なる代謝状態にある細胞を比較する、そのようなアッセイから得られた結果は、それぞれの状態の典型的特徴を提供することが可能であり、さらに異なる条件下での細胞の生理学的変化に関する情報を提供することができる。
【0162】
複数のプローブ(例えば、既知のプローブ混合物、細胞抽出物、細胞内小器官、細胞膜調製物、ホールセルなど)を用いて1つの種のプロテオームをスクリーニングすることによって、得られたタンパク質-プローブ相互作用の分析結果は、明らかに、細胞型、または細胞、組織、器官もしくは系の生理学的状態の「フィンガープリント」もしくは「サイン」を同定する土台を形成すると考えられる。こうした情報は、例えば、診断、予後判定、薬物試験、および薬物探索に役立てることができる。
【0163】
したがって、本発明のプロテオームチップを用いて、チップ上のタンパク質との薬物の相互作用を測定することができる。あるいはまた、本発明のプロテオームチップを使用して、例えば、ホールセル、細胞抽出物、もしくは組織ホモジネートのような複雑なタンパク質混合物への薬物の影響を特徴づけることができる。例えば、プロテオームチップを、複雑なタンパク質混合物と混合し、薬物が存在する状態もしくは存在しない状態で比較すれば、チップ上のタンパク質とのタンパク質混合物の相互作用の変化をアッセイすることができる。
【0164】
これにより、薬物の実際の効果は、1以上のプロテオームチップを、薬物処理した細胞、組織、もしくは抽出物によってスクリーニングすることにより分析できるが、このことは、ひいては、薬物処理状態に関する「サイン」を提供することを可能にし、さらに、未処理状態の「サイン」と比較するならば、例えば、効能、毒性および副作用に関する予測値を示すことができる。さらに、薬物の時間依存的効果は、例えば、細胞、細胞抽出物、組織ホモジネート、もしくは生物体そのものに薬物を添加し、処理の様々な時点で調製された薬物処理細胞もしくは抽出物をプロテオームチップに加えることによって、アッセイすることができる。こうしたアッセイは、疾病の診断もしくは予後判定に有用であると考えられる。
【0165】
特に、本発明のプロテオームチップは、薬物の作用方式を特徴づけ、薬物の特異性を決定し、薬物の毒性を予測するのに役立ち、さらに薬物の探索に有用であると考えられる。例えば、種々のアッセイ条件下で薬物または薬物候補とともにプロテオームチップをインキュベートし、アレイ上のどの場所に薬物が結合するかを判定することによって薬物の特異性を決定し、それぞれ異なるタンパク質に結合した薬物の量を測定することによって、薬物と結合するタンパク質の正体およびそれらの相対的親和性をアッセイすることができる。
【0166】
本発明のプロテオームチップは、例えば、プロテオームチップを罹病した細胞、細胞抽出物、もしくは罹病組織から得られた組織ホモジネート、または病気にかかった患者から得られた体液と接触させ、プロテオームチップ上でのタンパク質-プローブ相互作用のパターンを、健康な対応部分のそれと比較することによって、疾病状態を判定するために使用することができる。こうしたアッセイは疾病の状態に関する「サイン」を提供することができる。さらに、健康な状態の「サイン」と比較するならば、疾病の診断もしくは予後判定に関する予測値を示すことができる。さらに、例えば、様々な病期でプロテオームチップ上にて生物学的調製物をアッセイすることによって病期を特徴づけることができる。
【0167】
結合アッセイではなくて、生物活性をアッセイするバイオアッセイも、同じプロテオームチップ上で、もしくは同一の第2のチップ上で実施することができる。したがって、本発明のプロテインチップによるこのようなタイプのアッセイは、薬物の特異性を研究したり、薬物の潜在的副作用を予測したり、薬物を分類したりするために有用である。
【0168】
さらに、本発明のプロテオームチップは、薬物候補の複雑なライブラリーのスクリーニングに適している。具体的には、チップ上のタンパク質を、薬物候補のライブラリーとともにインキュベートした後、結合した化合物を、例えば、質量分析によって同定することができる。このことにより、特定の一部のタンパク質に優先的に結合するか、もしくはチップ上のタンパク質のうちいくつかと結合する、すべてのライブラリー成分を同時に同定することが可能となる。さらに、アレイに含まれる様々なタンパク質に対する薬物候補の相対的親和性を測定することもできる。
【0169】
さらに、観察された相互作用、酵素活性もしくは生物学的応答の潜在的な阻害剤、触媒、モジュレーターもしくはエンハンサーの存在下で、本発明のプロテインチップをプローブすることができる。本発明のプロテオームチップを用いると、このような戦略的なスクリーニングは、1つの種で発現されたタンパク質であって、例えば、薬物の結合を阻止する、ウイルス感染を阻害する、静菌活性を示す、抗真菌活性を示す、寄生虫感染を改善するタンパク質を同定し、また、特定のカテゴリーのタンパク質に対する生理的エフェクターを同定することができる。
【0170】
酵素反応を行って、酵素活性を本発明のプロテオームチップにより測定することができる。特定の実施形態では、チップ上の1つもしくは複数のタンパク質の酵素活性を調節する化合物を同定することができる。例えば、1つの化合物もしくは化合物類の混合物を酵素反応混合物とともにインキュベートし、それによってシグナル(例えば、酵素活性により蛍光性となる基質からのシグナル)を生じることにより、酵素活性のレベルの変化を検出して定量することができる。試験化合物の存在と非存在との差異を特徴づけることができる。さらに、化合物が酵素活性に及ぼす影響の差異は、サンプルに対するそれらの相対的影響をプロテオームチップ内で、およびチップ間で、比較することによって検出することができる。
【0171】
本発明のプロテオームチップを使用する様々なストラテジーは、タンパク質の様々な物理学的および機能的特性を決定するために利用することができる。例えば、プロテインチップを用いて、抗体でプローブすることによりタンパク質の存在および存在量を評価することができる。ルミネセンス、化学ルミネセンス、蛍光、化学蛍光、もしくは質量分析のような標準的な検出アッセイによってタンパク質を検出することができる。例えば、対象のタンパク質に対する一次抗体を、蛍光標識した二次抗体によって認識させ、これを次に、蛍光産物を光源によって励起して、その結果生じた蛍光を検出する機器(例えば、Molecular Dynamicsスキャナー)によって測定する。感度をいっそう高めるために、対象のタンパク質に対する一次抗体を、アルカリホスファターゼもしくは西洋ワサビペルオキシダーゼのような酵素と結合した二次抗体によって認識させる。発光基質(化学発光のための)もしくは蛍光原基質(化学蛍光のための)の存在下で、酵素的切断は強い発光産物もしくは蛍光産物をもたらすが、これは例えば、Molecular Dynamicsスキャナーを使用することによって検出および定量することができる。あるいはまた、蛍光標識した二次抗体のシグナルを、アルカリホスファターゼと結合した、もしくは西洋ワサビペルオキシダーゼと結合した三次抗体を用いて増幅することができる。
【0172】
ある実施形態において、本発明のプロテオームチップは、1つの種における既知タンパク質に対する抗体でプローブすることが可能であるが、その結果、認識されたエピトープを有する相同タンパク質を別の種のプロテオームにおいて同定することができる。たとえば、相互作用物質の同種のホモログは、相同タンパク質のDNA配列情報を用いて他の種におけるホモログを同定することによって得ることができる。特定の実施形態において、抗体は、サイクリン、キナーゼ、GST、CIb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2もしくはCdc11に対するものである。
【0173】
別の実施形態において、第1の種のタンパク質を含有する本発明のプロテオームチップを、第2の種から得られたタンパク質を用いてプローブして、相互作用物質を同定することができる。次に、第1の種から得られた相互作用物質の第2の種におけるホモログは、例えば、ヌクレオチド配列相同性によって同定し、特徴づけることができる。したがって、特定の種のプロテオームが利用できない場合は、このストラテジーを用いて、対象のタンパク質と相互作用する同じ種のタンパク質を見つけ出すことができる。
【0174】
本発明のプロテオームチップは、1つの種に存在する各酵素に対する、1つの種における本質的にすべての基質を同定するために使用することができる。したがって、本発明のプロテオームチップ上で、プロテインキナーゼ、ホスファターゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、もしくは他の化学基転移酵素の基質の同定を行うことができる。
【0175】
ある実施形態において、プロテアーゼ活性は、標準的なアッセイ(例えば、質量分析、ペプチド断片に対する蛍光標識抗体、もしくは蛍光タグ付けした基質からの蛍光シグナルの減少)を用いて、プロテアーゼ活性によって生成し、媒質中に放出されたペプチド断片を同定することによって検出することができる。このようにして、化学基-転移酵素の活性は、任意の検出手段を用いたいくつかの方法(当業者は理解できるだろう)によって容易にアッセイすることができる。
【0176】
本発明のプロテオームチップは、任意の化合物についての、1つの種における本質的にすべての結合タンパク質を同定するために使用することができる。したがって、プロテオームチップを用いて、例えば、キナーゼ、プロテアーゼ、ホルモン、DNA、RNA、ホスファターゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、もしくは他の化学基転移酵素と結合する本質的にすべてのタンパク質を同定して特徴づけることができる。このように、チップをプローブによって探索して、タンパク質-プローブ相互作用をアッセイすることができ、かつ/または、望ましい活性をアッセイすることができる。例えば、もしRNA結合が関心のある活性であるならば、RNAをプローブとしてプロテオームチップを探索して、タンパク質-RNA複合体を同定する。
【0177】
例えば、本発明のプロテオームチップは、膜結合タンパク質もしくは他の膜結合化合物と結合する、1つの種における本質的にすべてのタンパク質を同定するために、このチップをプローブ(例えば、ホールセル、細胞膜調製物、膜小胞、もしくは対象の膜成分を含むリポソーム)と接触させ、タンパク質-プローブ相互作用を検出することによって、使用することができる。特定の実施形態において、プローブは1以上の対象のリン脂質を含むリポソームの形をとる。タンパク質-プローブ相互作用は当技術分野で公知の技術によって検出可能である。相互作用物質および/またはプローブの正体(素性)は当技術分野で公知の技術を用いて同定することができる。さらに、生物活性(例えば、酵素活性、細胞活性化)も当技術分野で公知の技術によって検出することもできる。
【0178】
本発明のプロテオームチップを使用することによって、回復期の患者もしくは回復していない患者の免疫応答において抗原として作用する病原体(例えば、ウイルス、細菌、真菌、または寄生虫のような感染症の病因)由来の標的タンパク質、もしくは異常細胞(例えば、新生物細胞、病変細胞、もしくは損傷細胞)由来の標的タンパク質の正体(素性)を同定することができる。例えば、患者から分離したリンパ球を用いて、病原体のタンパク質のすべてを含んでなるチップをスクリーニングすることができる。一般に、こうしたスクリーニングは、プロテオームチップを複数のリンパ球と接触させ(ただし、このプロテオームチップ上のタンパク質は可能性のある複数の抗原を含んでなる)、リンパ球の活性化が生じたチップ上の位置を検出することを含んでなる。特定の実施形態では、リンパ球を、プロテオームチップ上の病原体のタンパク質と接触させた後、抗原もしくは抗原混合物によるB細胞またはT細胞の活性化をアッセイし、それによって病原体に由来する標的抗原を同定する。
【0179】
あるいはまた、本発明のプロテオームチップを用いて、例えば、患者のリンパ球で感染性生物のプロテオームをスクリーニングして患者のB細胞および/またはT細胞の標的を同定することによって、免疫応答を特徴づけることができる。例えば、B細胞を本発明のプロテオームチップとともにインキュベートして、体液性免疫に対する抗原標的を同定することができる。
【0180】
別の実施形態において、本発明のプロテオームチップを使用して、自己免疫の基質もしくはアレルギー原因タンパク質を検出し、特徴付けることができる。例えば、患者のリンパ球または患者の循環抗体を用いて、ヒトタンパク質のプロテオームをスクリーニングし、患者のB細胞および/またはT細胞の標的を同定することができる。こうしたスクリーニングは、自己免疫またはアレルギー反応を特徴付けて、可能性のある標的薬物候補を同定することができる。
【0181】
ある実施形態において、本発明のプロテオームチップはB細胞もしくはT細胞を活性化できる物質を同定するために使用することができる。例えば、リンパ球をプロテオームチップに接触させてリンパ球活性化をアッセイし、それによってB細胞もしくはT細胞またはリンパ球の亜集団(例えば、細胞傷害性T細胞)を活性化する全般的な能力を有する物質を同定する。
【0182】
抗原認識によるB細胞活性化の誘導は、抗体合成の検出もしくは測定、3H-チミジンの取り込み、新たに発現もしくは抑制された細胞表面分子への標識抗体の結合、およびB細胞活性化を示す因子(例えば、サイトカイン)の分泌を含むがこれらに限らない様々な方法によってアッセイ可能である。同様に、本発明のプロテインチップを用いたスクリーニングでのT細胞活性化は様々なアッセイにより測定できる。例えば、クロム(51Cr)放出アッセイは、抗原の認識と、それに続く細胞傷害性T細胞の活性化を検出することができる(例えば、Palladinoら、1987, Cancer Res. 47: 5074-9; Blachereら、1993, J. Immunotherapy 14: 352-6を参照されたい)。
【0183】
本発明のプロテオームチップの使用によって、抗体調製物の特異性を決定することができるが、これは、チップを抗体調製物と接触させ、抗体調製物中の抗体による結合が生じた基板上の位置を検出することを含んでなる。抗体調製物は、Fabフラグメント、抗血清、およびポリクローナル、モノクローナル、キメラ、一本鎖、ヒト化、もしくは合成抗体であってよいが、それらに限定されない。ある実施形態では、モノクローナル抗体をプローブとして使用してプロテオームチップを探索し、その結合の強さおよび/またはその特異性を特徴づける。特定の実施形態において、抗体は、サイクリン、キナーゼ、GST、Clb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2もしくはCdc11に対するものである。
【0184】
本発明のプロテオームチップは、生物学的に関心のある特定の分子(例えば、潜在的リガンド分子の受容体、ウイルス受容体、およびオーファン受容体のリガンドを包含するがこれらに限定されない)と結合するタンパク質を同定するために有用である。
【0185】
本発明のプロテオームチップは、チップ上のタンパク質へのDNAの結合もしくはRNAの結合の検出、ならびにその結合特異性および強度の評価にも有用である。DNAは、一本鎖もしくは二本鎖とすることができる。RNAはmRNA、hnRNA、ポリA+RNA、もしくは全RNAであってよい。
【0186】
本発明のプロテオームチップは、翻訳後修飾されたタンパク質の同定に役立つ。本発明の方法によって検出可能な翻訳後修飾には、メチル化、アセチル化、ファルネシル化、ビオチン化、ステアロイル化、ホルミル化、リポ酸、ミリストイル化、パルミトイル化、ゲラニルゲラニル化、ペグ(PEG)化、リン酸化、硫酸化、グリコシル化、糖による修飾、脂質化、脂質による修飾、ユビキチン化、SUMO化、ジスルフィド結合、システイニル化、酸化、グルタチオニル化、ピログルタミン酸、カルボキシル化、および脱アミノ化があるが、これらに限定されない。さらに、ペントース、ヘキソサミン、N-アセチルヘキソサミン、デオキシヘキソース、ヘキソース、およびシアル酸で、タンパク質を翻訳後修飾することができる。
【0187】
ある実施形態において、ホスホチロシン、ホスホセリンもしくはホスホトレオニンをプローブとして用いて本発明のプロテオームチップを探索し、リン酸化相互作用物質を同定することができる。別の実施形態において、レクチン(例えば、小麦胚芽凝集素)をプローブとして用いて本発明のプロテオームチップを探索し、グリコシル化相互作用物質(例えば、N-アセチルグルコサミン)を同定することができる。リン酸化もしくはグリコシル化相互作用物質は、当技術分野で公知の方法によって検出可能である。
【0188】
また、本発明のプロテオームチップは、機能、リガンド結合、もしくは酵素活性の点で相違するタンパク質アイソフォームを同定し特徴づけるために有用である。特別の実施形態では、本発明のプロテオームチップを用いて、異なる対立遺伝子から誘導されたタンパク質アイソフォームによる様々な結合親和性を、それらの活性を他と比較してアッセイすることによって、特徴づける。
【0189】
ある実施形態において、本発明のプロテオームチップを用いてコンセンサス配列を決定することができる。例えば、プロテオームチップを結合物質(チップ上のタンパク質とは異なる種に由来するものであってよい)によってスクリーニングした後、相互作用物質のアミノ酸配列をアラインメントし、相互作用ドメインのコンセンサス配列を構築することができる。こうした情報は、例えば、結合物質-相互作用物質の相互作用の阻害剤をデザインするために、あるいは特定の相互作用物質もしくは特定クラスの相互作用物質に対する新規および/または改良型結合物質をデザインするために有用であると考えられる。
【0190】
特定の実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、カルモジュリンを用いたプロテオームチップのスクリーニングによって決定される。もうひとつの実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、アミノ酸配列I/L-Q-X-K-K/X-G-B(配列番号1)からなる。別の実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、アミノ酸配列I/L-Q-X-K-K/X-G-B(配列番号1)を含んでなる。さらに別の実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、アミノ酸配列I/L-Q-X-K-K/X-G-B(配列番号1)を含んでなり、その長さは10、15、30、50または100アミノ酸残基より小さい。
【0191】
本発明のプロテオームチップは、抗菌性、抗真菌性、抗寄生虫性もしくは抗ウイルス性が見込まれる一連の化合物を同定するためにも有用である。例えば、細胞溶解物、または他の細菌、真菌、寄生虫もしくはウイルスの調製物をプロテオームアレイに接触させ、タンパク質-プローブ相互作用を検出して同定することができる。さらに、感染性生物から得られた感染段階と非感染段階での相互作用パターンを比較することによって、宿主の感染に関わる相互作用を同定することができる。
【0192】
ファージディスプレイライブラリーのスクリーニングは、ライブラリーを本発明のプロテオームチップとともにインキュベートすることによって行うことができる。チップ上のタンパク質に結合するクローンの検出は当技術分野で公知の様々な方法(例えば、質量分析)によって実施することができるが、それによって対象のクローンを同定し、その後、対象クローンをコードするDNAを標準的な方法によって同定することができる(例えば、Amesら、1995, J. Immunol. Methods 184: 177-86; Kettleboroughら、1994, Eur. J. Immunol. 24: 952-8; Persicら、1997, Gene 187: 9-18を参照されたい)。このように、チップは、チップ上の特定のタンパク質と結合する表面成分を有する細胞を選択するうえで有用である。
【0193】
5.5 キット
本発明は、また、本発明のアッセイ計画を実施するためのキットを提供する。ある実施形態において、本発明のキットは1以上の本発明のプロテオームチップを含んでなる。こうしたキットは、さらに、1以上の容器内に、タンパク質もしくは分子の生物活性をアッセイするのに有用な試薬、タンパク質-プローブ相互作用をアッセイするのに役立つ試薬、および/または1以上のプローブ、タンパク質もしくは他の分子を含んでいてもよい。タンパク質もしくは他の分子の生物活性をアッセイするために、またはプローブとタンパク質もしくは他の分子との間の相互作用をアッセイするために有用な試薬を、プローブとともに用いて、プロテオームチップに付着させたり、またはプロテオームチップ上の1以上のウェルに入れることができる。かかる試薬は溶液または固体の状態であってよい。試薬は、当該アッセイを実施するために必要なタンパク質もしくは他の分子とプローブのいずれか一方または両方を含むことができる。
【0194】
プロテオームチップのタンパク質は、平坦な基板の表面に付着させたり、基板上のウェルに入れたり、または基板上のウェルの表面に付着させたりすることができる。ある実施形態において、キットのプロテオームチップは、基板上にすでに付着したタンパク質および/またはプローブを有する。別の実施形態において、キットのプロテオームチップは、タンパク質や他の分子の生物活性のアッセイに有用な、またはプローブとタンパク質もしくは他の分子との相互作用のアッセイに有用な、基板上のウェルにすでに付着した試薬または反応混合物を有する。さらに別の実施形態において、試薬は基板のウェルに付着していないが、ウェル内に含まれる。また別の実施形態において、試薬は基板のウェルに付着していないが、1以上の容器に入っており、これを基板上のウェルに加えることができる。別の実施形態において、キットはさらに、タンパク質または分子の生物活性をアッセイするための反応混合液を入れた1以上の容器を含んでなる。また別の実施形態において、キットは担体(例えば、ビーズ)を提供する。この担体に対象のプローブ、タンパク質もしくは分子、および/または1以上のアッセイを実施するために役立つ試薬を付着させ、その後、付着したプローブ、タンパク質もしくは試薬を有する担体をチップのウェル内に入れることができる。
【0195】
5.6 陽性判定アルゴリズムの設計
本発明はまた、シグナルが陽性であるかどうかを判定する方法を提供する。シグナルは、可視光、紫外線、赤外線、X線、蛍光、および比色可視化を包含するあらゆる測定可能な形をとることができるが、それらに限定されない。
【0196】
好ましい実施形態においては、シグナルを質量分析によって検出する。
【0197】
さらに、シグナルはどのような配置であってもよく、アレイ、ブロット、ゲル、およびスクリーンのような任意の形をとることができる。シグナルは定位に配置されたスポットであることが望ましい。
【0198】
別の好ましい実施形態において、シグナルは蛍光によって生じ、アレイ上に格子パターンで配置される。このように、シグナルは横列および縦列に関する位置座標を割り当てられる。横列および縦列はどのような幅の列であってもよい。
【0199】
シグナルを選別する第1のステップは、各スポットに関する局所フォアグラウンドおよびバックグラウンドシグナルを算定することである。局所フォアグラウンドシグナルはスポットそのものから発せられるが、局所バックグラウンドシグナルはそのスポットに直に接している領域から発せられる。正味のシグナルは、局所フォアグラウンドシグナルから局所バックグラウンドシグナルを差し引いたものであるが、これを、以後のすべての計算に使用する。局所フォアグラウンドおよびバックグラウンドシグナルは、GENEPIXTMのようなソフトウェアによって確認される。
【0200】
しかしながら、チップ間のばらつき(例えば、様々な脂質結合実験に相当する)、およびチップ上の局所ばらつき(例えば、担体の不均一な拡散による)によって、結果的に正味のシグナル強度に一層のばらつきが生じ、チップごとに正味のシグナル分布が異なることがある。チップ間のばらつきを補正するために、異なるチップから得られた正味のシグナルを共通の値域に基準化する必要がある。数個のチップのうち1個を基準として選び、各チップの正味のシグナル分布を、基準チップの正味のシグナル分布の値域および形状に基準化することを目標とする。
【0201】
例えば、各チップの正味シグナル分布の下四分位値、中位値、および上四分位値を計算する。次に、それぞれのチップについて、正味シグナルの中位値を、各スポットの正味シグナルから差し引く。さらに、各チップにつき基準化倍率を算定するが、これは特定のチップの上四分位値と下四分位値の差と、基準チップの上四分位値と下四分位値の差との比率に等しい。これは、基準チップに対する倍率が1に等しいことを意味する。次いで、各チップに関する正味のシグナルに、そのチップに特異的な倍率を乗じて、基準化された正味のシグナルを算定する。
【0202】
アレイ上の局所ばらつきを補正するために、それぞれのスポットについて「近傍減法」(neighborhood subtraction)を行うことができる。例えば、近傍領域はある単一のスポットの上下2列および左右2列の範囲と定義することができる。次に、この領域の中位値シグナルを、そのスポットのシグナルから減じて、そのスポットの近傍と比較した過剰シグナルを算出する。好ましくは、中位値があまり影響を受けず、その近傍領域のバックグラウンドシグナルを良好に表すように、いずれの近傍領域においても高いシグナル強度を示すスポットの数が十分少なくなるようにする。
【0203】
基準化された正味シグナルに近傍減法を適用して、基準化された過剰シグナルを得る。次のステップで、並列サンプルの基準化過剰シグナルを比較する。2つの並列サンプルの基準化過剰シグナルの平均値と、並列サンプルの一方の基準化過剰シグナルとの差が、基準化過剰シグナルの誤差の標準偏差の3倍より大きいならば、その2つの並列サンプルに属するスポットは、更なる分析から除外される。その後、残りのスポットおよびそれらの基準化過剰シグナルが、選別されたシグナルの集合に相当する。
【0204】
基準化された過剰シグナルの誤差の分布およびその標準偏差は、次のようにして算出可能である。並列サンプルの完全な集合に対して線形回帰を行い、並列サンプル間の一般線形関係を決定する。次に、それぞれの並列サンプルについて誤差の値を算出する:
εG = |G2−Gm|
ここで、G1およびG2は2つの並列サンプルの、2つの基準化された過剰シグナルを表し、Gm = (G1 + G2)/2であり、
そして関数f(G1) = a*G2 + b は、線形回帰によって決定されるパラメーターaおよびbを伴う、2つの並列サンプルG1およびG2の間の一般線形関係である。
【0205】
誤差の値の完全な集合は、すべての並列サンプルに関する誤差の値の集合であって、基準化された過剰シグナルに関する誤差の値の分布を表す。そこで、このような分布の標準偏差を算定することができる。
【0206】
最終的に、並列サンプルの選別されたシグナルの平均値が、基準化された過剰値の誤差よりも3標準偏差大きいならば、この1対の並列サンプルは陽性であるといえる。(ここで、留意すべきは、この閾値は、並列サンプルが選別されたシグナルの集合から除外されるべきかどうかを決定する閾値とは無関係であるということである。)
このような選別手順の後、選別されたシグナル(G)は、GSTシグナル(R)によって正規化され、比r = G/Rをもたらし、これはタンパク質量当たりの結合の尺度となり、異なるタンパク質間での結合シグナルの比較を可能にする。特異的結合の比rは、GおよびRシグナルの両者の誤差εGおよびεRに影響される。モンテカルロ(Monte Carlo)法によって、この比について90%および95%の信頼区間を算出することができる。
【0207】
r値の誤差eGは誤差εGおよびεRに関連している:
r + εR = (G + εG)/(R +εR)
ここで、εRは比rの誤差を表す。
【0208】
モンテカルロ法の場合は、εGおよびεRの両者の分布が明らかでなくてはならない。eGの分布は、上記の説明のように計算することができる。eRの分布は、入力として並列サンプルのGSTシグナル対を使用することによってeGと同様の方法で算出することができる。
【0209】
モンテカルロ法は次の通りである:
r + εRに対する信頼区間を決定するために、r + eRの無作為サンプルの母集団を算定する必要がある。これらは、εGおよびεRの無作為サンプルから導くことができる。εGの無作為サンプルは、次のように算定される。0から1の間に一様に分布する乱数のサンプルを、標準乱数発生器によって計算する。εGの分布から、eGの逆累積分布関数を標準法によって決定する。この関数の独立変数として乱数のサンプルを使用することで、εGに対する乱数サンプルの集合を得る。同様に、εGに対する無作為サンプルの集合を計算することができる。このようなεGおよびεRの無作為サンプルを式r +εR= (G +εG)/(R +εR)に結合させて、r +εRに対する無作為サンプルの母集団を作ることができる。
【0210】
ある実施形態において、本発明は、シグナルが陽性であるかどうかを判定する方法を提供するが、これは、次のステップを含んでなる。すなわち、それぞれのスポットについて局所的にフォアグラウンドおよびバックグラウンドシグナルを測定して、それらの差から正味のシグナルを決定し;第1および第2の正味シグナル分布に関する下四分位値、中位値、および上四分位値を決定し;第1の正味シグナル分布から第1の中位値を減じ、さらに第2の正味シグナル分布から第2の中位値を減じて、それぞれ第1および第2の減算値を得;第1の減算値を、第1のシグナル分布に関する上四分位値と下四分位値との差で割り算し、さらに第2の減算値を、上四分位値と下四分位値との差で割り算して、それぞれ第1および第2の規準化された値を得;近傍領域の規準化されたシグナル分布に関する局所中位値を算定し、ただし、前記近傍領域はその領域内に複数のサイトを含んでなり;規準化されたシグナルから局所中位値を減じて、規準化された過剰値を得;そして、サンプル値の1つとそれらの平均値との差が、規準化された過剰値の誤差の3標準偏差より大きいならば、規準化された過剰値の並列サンプルを除外する。
【0211】
選別された値を用いて、シグナルが陽性であるかどうかを判定することができる。並列サンプルの選別された値の平均が、規準化された過剰値の誤差の標準偏差の3倍より大きいならば、並列サンプルは「陽性」と称される。選別された陽性シグナルを、次の式によって正規化することができる:
r = G/R
ここで、Gは選別された値であり、RはGSTシグナルであり、そしてrは、タンパク質の量当たりのシグナルを表し、様々なスポットの間で比較することができる。比rはGおよびRの誤差に影響されやすい。次の式の信頼区間を計算することによって、このような影響されやすさを評価することができる:
r + εr = G + εG/R + εR
εGはGの誤差、εRはRの誤差、そしてεrはrの誤差である。
【0212】
特定に実施形態において、陽性シグナルはタンパク質-プローブ相互作用を示す。別の特定の実施形態において、近傍領域は、シグナルの上2列、下2列、左2列、右2列である。もう1つの特定の実施形態において、二つの並列サンプルから得られるデータポイントが更なる分析から除外されるが、この場合、前記サンプルの規準化された過剰シグナルとそれらの平均値との差が、規準化された過剰シグナルの誤差の3標準偏差より大きい。データポイントは、それが明らかに人為的結果である場合にも、除外される。
【0213】
6. 実施例
ハイスループット技術によって、出芽酵母由来の5800以上のタンパク質からなる確定した集団を調製し、タンパク質-タンパク質、タンパク質-DNA、タンパク質-RNA、およびタンパク質-リポソーム相互作用を含めた多くの活性についてスクリーニングした。多数の新規活性が同定され、以前には性質の明らかになっていなかった既知遺伝子に関する新たな情報が得られた。
【0214】
酵母プロテオームの研究を進めるために、5800のオープンリーディングフレームをクローニングして、過剰発現させ、それらに対応するタンパク質を精製した。このタンパク質をスライドガラス上に高い空間密度でプリントして、酵母プロテオームマイクロアレイを作製し、タンパク質およびリン脂質と相互作用するそれらの能力についてスクリーニングした。多数の新規なカルモジュリンおよびリン脂質-相互作用タンパク質が同定された;カルモジュリン結合タンパク質の多くに共通する推定結合モチーフを同定した。これらの研究から、完全な真核生物プロテオームのマイクロアレイを作製して、多数の生化学的活性についてスクリーニングすることが可能であり、結果として多くの新規タンパク質機能/相互作用の同定に至ることが実証される。こうしたマイクロアレイをスクリーニングして、タンパク質の翻訳後修飾を検出することもできる。
【0215】
6.1 酵母培養物の調製
1. −80℃にて96ウェルプレートで保存された酵母グリセロールストックを、96-プロンガーを用いてURA寒天平板(Omni, 米国)に接種した。
【0216】
2. 培養物を寒天上で30℃にて48時間増殖させると、目に見えるコロニー(直径2 mm)が認められる。
【0217】
3. 96-プロンガーを用いて、酵母細胞を寒天平板から96ウェル2 mlボックスに接種したが、このボックスには、それぞれのウェルにいずれも300μl URA-/ラフィノース液体培地および直径2 mmのガラス玉が入っており、これは均一な増殖を促進する。
【0218】
4. 激しく撹拌(300 rpm)しながら、30℃にて、およそ16時間で、培養物のO.D.600が4.0に達した後、3 mlの培養物を得るために、同じ菌株15μlを、4つの別々のボックスに入っている750μlのURA-/ラフィノース液体培地に接種した。再び、通気および均一な増殖を達成するために、各ウェルに同じガラス玉を入れた。激しく撹拌しながら30℃にて細胞を増殖させた。
【0219】
5. 12〜16時間増殖させた後、培養物のO.D.は0.6から0.8に達するはずである。O.D.が1.0を超えていれば、培養物は廃棄した。細胞を誘導するために、自動化液体操作装置(Q-Fill, Genetix, UK)を用いて、40%ガラクトースストックを各ウェルに最終濃度2%となるように添加した。この培養物を撹拌しながら4時間30℃にて誘導した。
【0220】
6. 3000 rpmで2〜10分間回転させることによって細胞を集め、細胞ペレットをボルテックス混合によって100〜1000μlの冷水に再懸濁した。次に、同じ菌株の細胞を4ウェルから1つに併せた。細胞を遠心によって集め、氷上でプロテアーゼインヒビターを含まない100〜1500μlの冷溶解バッファーに再懸濁した。洗浄した細胞を短時間の遠心によって集め、溶解バッファーを捨てた。洗浄した半乾燥の培養物を直ちに−80℃の冷凍庫で保存した。この培養物は何週間も保存することができる。
【0221】
6.2. 96ウェル方式によるタンパク質の精製
1. 96ウェルボックスに入れた凍結培養物を−80℃から氷に移し、100〜300μlのジルコニアビーズ(直径0.5 mm、BSP、ドイツ)を各ウェルに加えた。培養物がまだ凍っている間に、新調製のプロテアーゼインヒビターを含有する100〜500μlの溶解バッファーを添加した。各ウェルを密閉するためにキャップマットを使用した。氷上で5〜25分間培養物を融解させた後、96ウェルボックス内の細胞を氷上で1〜5分間の間隔をおいて3〜6回、20〜60秒ボルテックス混合した。酵母の細胞壁を効果的に破壊するために、さらに多くのプレートを同時に処理するために、ペイントシェーカー(HARBILTM 5G HG、最大積載量36 kg、圧力および振盪時間は調節可能、速度は200回/分に固定)を使用して試料を激しく撹拌した。
【0222】
2. 3000rpmで2〜10分間回転させた後、ワイドオープンチップ(wide-open tip)(Fisher, USA)を用いて上清を集め、96ウェルボックスの上に置いた96ウェルフィルタープレート(Whatman, 米国、Whatman UNIFILTERTM、カタログ番号7700-1801、容量はウェル当たり800μlで、親水性PVDFフィルターを有する)に移し入れた。
【0223】
3. より多くのタンパク質を得るために、新調製のプロテアーゼインヒビターを含有する100〜500μlの溶解バッファーを細胞破砕片に加え、ステップ1および2を繰り返した。
【0224】
4. 併せた細胞溶解物を3000 rpmで10〜30分間回転させて、フィルタープレートを通して清潔な冷却96ウェルボックスに入れた。それぞれのウェル内の濾過された溶解液の容量は、ざっと200〜1000μlであった。
【0225】
5. 一方で、必要量のグルタチオンビーズ(試料当たり約10〜50μlのビーズ)(Amersham、米国)を4回、プロテアーゼインヒビターを含まない冷溶解バッファーで洗浄し、最終的に元の容量の5倍となるように新調製のプロテアーゼインヒビターを含有する溶解バッファーで再懸濁した。
【0226】
6. 100μlの洗浄したグルタチオンビーズを各ウェルに加え、キャップマットでしっかりと密閉した。このビーズを溶解物とともにローラードラム上で1時間4℃にてインキュベートした。最適な混合を得るために、ボックスをローラードラム上で360度回転させた。
【0227】
7. 3000 rpmで10〜60秒間回転させてビーズを集め、上清を捨てた。ビーズを200〜800μlのプロテアーゼインヒビター含有洗浄バッファーIで1回、インヒビターなしで2回洗浄した。
【0228】
8. 次に、ビーズを200〜800μlの洗浄バッファーIIで3回洗浄した。バッファーを完全に除去した後、20〜50μlの溶出バッファーを各ウェルに添加した。溶出ステップで使用するフィルタープレートは、タンパク質に対する親和性が低い材料から構成されていた(MILLIPORE MULTISCREENTM、カタログ番号MADVN6550、容量は200μl/ウェルで、親水性PVDFフィルターを有する)。ボックスを短時間ボルテックス混合してビーズを再懸濁し、4℃にて1時間ローラードラム上でインキュベートした。
【0229】
9. 溶出液/ビーズスラリーを冷フィルタープレート(Millipore、米国)に移し、4℃にて3000 rpmで0.5〜2分間フィルタープレートを通して回転させることによって、溶出液を96ウェルPCRプレートに集めた。
【0230】
10. それぞれの精製タンパク質を3つの96ウェルPCRプレートに分注し、直ちに−80℃冷凍庫に保存した。
【0231】
溶解バッファー
30-300 mM Tris pH 7.5
50-300 mM NaCl
0.1-10mM EGTA
0.01-1.0% TritonX-100
0.01-1% β-メルカプトエタノール(BME)
0.1-3 mM フッ化フェニルメチルスルホニル(PMSF)
Rocheプロテアーゼインヒビター錠剤(EDTA含有)
BME, PMSFおよびインヒビター錠剤は新たに添加した。
【0232】
洗浄バッファーI:
30-300 mM Tris pH 7.5
300-600mM NaCl
0.1-10mM EGTA
0.01-1.0% TritonX-100
0.01-1% β-メルカプトエタノール(BME)
0.1-3 mM PMSF
Rocheプロテアーゼインヒビター錠剤(EDTA含有)
BME, PMSFおよびインヒビター錠剤は新たに添加した。
【0233】
洗浄バッファーII:
50-200 mM HEPES pH 7.5
50-300 mM NaCl
1-15% グリセロール
【0234】
溶出バッファー:
50-200 mM HEPES pH 7.5
50-200 mM NaCl
20-40% グリセロール
5-40 mM グルタチオン(還元型)
溶出バッファーは約pH=7.5とすべきである。
【0235】
6.3 ハイスループット96アレイフォーマットでのタンパク質の製造方法
ほぼすべての酵母タンパク質を含有する酵母プロテオームマイクロアレイを作製し、多数の生化学的活性についてスクリーニングした。5800の酵母ORF(全体の93.5%)からなる高品質のコレクションを、組換えクローニング(Mitchellら、1993, Yeast 9:715)によって酵母の高コピー数発現ベクターにクローニングした。酵母タンパク質をそのアミノ末端でGST-HisX6と融合させ、ガラクトース誘導性GAL1プロモーター(Zhuら、2000, Nat. Genet. 26: 283; Mitchellら、1993, Yeast 9:715)の制御下で酵母において発現させた。酵母発現菌株は、正しい酵母ORFがインフレームでGSTに適正に融合していることがDNA配列決定によって明らかにされた個別のプラスミドを含有する。手短に言えば、酵母ORFをPCRによって増殖し、酵母細胞にベクターとともに同時形質転換して発現クローンを生成した。このプラスミドを大腸菌(E. coli)でレスキューし、ベクター-インサート結合部分の配列を決定した(図1A)。クローニングされたORFが目的のORFでない場合、もしくはフレームシフトが検出されたならば、クローニングサイクルを繰り返した。ひとたび構築物が確認されたならば、プラスミドDNAを酵母および大腸菌に再導入して、今後の分析のための永久ストックを作製した(Zhuら、2000, Nat. Genet. 26: 283)。クローニングサイクルを繰り返すことによって、5800のユニークな酵母ORFをクローニングすることに成功したが、これは全体の93.5%に当たる。
【0236】
生化学的分析を目的として精製タンパク質を生成するために、96ウェルフォーマットでタンパク質を調製するための強力なハイスループット精製法を開発して、最適化した。グルタチオン-アガロースビーズを用いて、酵母抽出物を調製し、融合タンパク質を精製した(96ウェルフォーマットタンパク質精製プロトコール、すべての実験から得られた結果の完全な一覧、および陽性判定アルゴリズムの設計の詳細については、公開ウェブサイトspine.mbb.yale.edu/protein_chips/にアクセスされたい)。溶解バッファーおよび最初の洗浄液は、精製されたタンパク質が確実に脂質を含まないようにするため、0.1%のTritonを含有していた。本発明の方法を用いると、少なくとも1152タンパク質試料を10時間足らずのうちに細胞から調製することができる。
【0237】
精製されたタンパク質の質および量は、60の無作為抽出試料のイムノブロット分析によってモニターされた(図1B)。80%を上回る菌株が、予想分子量をもつ融合タンパク質を検出可能な量で生産した。手動プリンティング装置を用いて、3 nlの1152の精製タンパク質を二重反復でスライドガラス(19)上にスポットし、ポリクローナル抗GST抗体を用いてタンパク質を検出した。85%を超える試料が、バックグラウンドに対して目視可能なシグナルを示し、イムノブロット分析と符合した。本発明の方法を用いると、6144(64×96ウェルボックス)の酵母菌株から得られた融合タンパク質を2週間足らずで精製することができる。
【0238】
6.4 プロテオームマイクロアレイの作製方法
プロテオームチップを作製するために、5800の異なる酵母タンパク質に対応する6566のタンパク質調製物を、市販のマイクロアレイヤーを用いてスライドガラス上に二重反復でプリントした。対照として、既知量のGSTもプリントした。2つのタイプのスライドガラスを使用した。最初の実験ではアルデヒド処理済みの顕微鏡用スライドガラスを使用したが(MacBeathら、2000, Science 289: 1760)、この場合、融合タンパク質のN末端もしくは他の残基の1級アミンを介して融合タンパク質をスライドガラス表面に付着させたため、結果として、表面上のタンパク質は比較的ランダムな配向となった。その後の実験では、ニッケルコーティングスライドガラス上にタンパク質をスポットした。この場合、融合タンパク質は、そのHisX6タグを介して付着するので、クローニングされた融合タンパク質の酵母部分は、必然的に表面から離れて一様に方向付けられる。両タイプのスライドガラスはいずれも成功裏に使用できたが、本発明者らの特定のタンパク質調製物に関しては、ニッケルコーティングスライドガラスの方が有意に優れたシグナルを与えた(図2A)。
【0239】
どのくらいの量の融合タンパク質がそれぞれのガラス表面に共有結合しているかを判定するために、また、タンパク質結合の再現性を評価するために、抗GST抗体でチップをプローブした。93.5%を超えるタンパク質試料が、バックグラウンドを有意に上回るシグナルを与えた(すなわち、タンパク質は約10 fgより多い)。スライドガラス上にプリントされた既知量のGSTと比較して、約90%のスポットが約10 fgから950 fgまでのタンパク質を含有することが明らかになった。図2Aに示すように、プロテオームチップ上での蛍光標識抗体によるタンパク質の検出は極めて高感度であり、すなわち、スライドガラス上にスポットされるのは3 ml培養物から精製された精製タンパク質のわずか1/10,000であるにもかかわらず、シグナル対ノイズ比が高かった。この結果は、標準的な顕微鏡用スライドガラス(2.5cm×7.5cm)の半分の面積に、優れた分解能で、13,000のタンパク質試料をスポットすることが実現可能であることを実証する(図2Aおよび2B)。タンパク質のスポッティングの再現性を試験するために、それぞれ対をなす二重反復スポットから得られるシグナルを相互に比較した。図2Cの鋭いスパイクが実証するように、シグナルの95%が平均の5%以内に存在した(10)。
【0240】
6.5 プロテオームマイクロアレイの使用方法
いくつかの典型的な生物活性、すなわち、タンパク質-タンパク質相互作用、タンパク質-核酸相互作用、およびタンパク質-脂質相互作用をプローブすることによってプロテオームチップを試験した。
【0241】
一般的には、アッセイのためにプロテオームチップを次のように調製した。プロテオームチップをブロックするため、プリントされたスライドガラスを、BSA(PBSバッファー中の1〜3%(W/W) BSA;SIGMATM, 米国)もしくはグリシンブロッキングバッファー(30〜300 mMグリシン;50〜300 mM Tris, pH 6.5〜8.5; 50〜300 mM NaCl; SIGMATM, 米国)のいずれかにタンパク質側を上にしてゆっくりと浸漬した。粒子を除去するために2μmフィルターユニットを通してバッファーを濾過した。炭水化物結合タンパク質をプローブする場合にはグリシンの方が好ましい。このスライドガラスをブロッキングバッファー中で4℃にて一晩、振盪せずにインキュベートした(ブロッキングバッファーの妨害により、結果的にガラス表面にタンパク質線条(streaks)が生じる可能性がある)。
【0242】
プローブタンパク質は、一般的に、次のように調製した。グルタチオンビーズを用いたアフィニティーカラムによって、50 ml培養物から標準法を用いてビーズからの溶出なしに酵母タンパク質を精製した。タンパク質ビーズを冷PBSバッファー(pH8.0)(SIGMATM, 米国)で3〜5回洗浄した。約1 mlのスルホ-NHS-LC-LC-ビオチン(PIERCETM カタログ番号21338、米国)を0.1〜50mg/mlの濃度となるようにPBS(pH 8.0)に溶解して、それをグルタチオンビーズに添加し、4℃にて2時間インキュベートした。ビーズを冷PBSバッファー(pH 8.0)で5回洗浄し、100〜500 mlの溶出バッファー(50〜200 mM, HEPES pH7.5; 50〜200 mM NaCl; 20〜40%グリセロール;5〜40 mMグルタチオン)で溶出した。比較的弱くビオチン化されたタンパク質をもたらすようなプロトコールが望ましい。様々な程度にビオチン化されたタンパク質のバッチを、以後使用するためにプールした。
【0243】
6.5.1 カルモジュリン相互作用タンパク質の同定
タンパク質-タンパク質相互作用を試験するために、カルモジュリンで酵母プロテオームをプローブした(11)。カルモジュリンは、カルシウムによって調節される多くの細胞プロセスに関与する高度に保存されたカルシウム結合タンパク質であって、多くの既知の物理的パートナーを有する(Hookら、2001, Annu. Rev. Pharmacol. Toxicol. 41: 471)。カルモジュリンプローブをビオチン化し、結合したプローブをCy3標識ストレプトアビジンによって検出した。対照として、Cy3標識ストレプトアビジンのみを用いて、酵母プロテオームをプローブした。
【0244】
一般に、タンパク質-タンパク質相互作用は、次のようにアッセイすることができる。ブロックされたプロテオームチップをPBSバッファーで3〜5回洗浄し、ガラス表面上の余分な液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去する。200μlのビオチン化プローブをプロテオームチップに添加し、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。閉じこめられた気泡を取り除いた後、チップを加湿チャンバー内で室温(RT)にて1時間インキュベートする。カバーガラスを取り外すために、チップを多量のPBSバッファー(>50ml)中に浸すと、カバーガラスは浮き上がって離れるはずである。次にチップを第2のPBS槽(>50ml)に移動し、室温で振盪しながら、3×5分間、洗浄した。チップ表面上の余分な液体を除去した後、少なくとも150μlのCy3-もしくはCy5-結合ストレプトアビジン(PIERCETM, 米国;希釈率1:2000から1:4000)をその表面に加え、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。チップを暗所でRTにて、30分より長時間インキュベートする。次に、このチップを上記のように洗浄する。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥する。
【0245】
プロテオームチップを抗体でスクリーニングしようとする場合は、タンパク質-抗体相互作用を次のように検出することができる。ブロックされたプロテオームチップをPBSバッファーで3〜5回洗浄し、ガラス表面上の余分の液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去する。200μlの一次抗体(1〜3% BSAおよび0.1% TritonX-100を含有するPBSで適当に希釈する)をプロテオームチップに添加して、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。閉じこめられた気泡を取り除いた後、チップを加湿チャンバー内で室温(RT)にて1時間インキュベートする。カバーガラスを取り外すために、チップを多量のPBSバッファー(>50ml)中に浸すと、カバーガラスは浮き上がって離れるだろう。次にチップを第2のPBS槽(>50ml)に移動し、室温で振盪しながら、3×5分間、洗浄した。チップ表面上の余分な液体を除去した後、>150μlのCy3-もしくはCy5-結合二次抗体(1〜3% BSAおよび0.1% TritonX-100を含有するPBSで適当に希釈する)をその表面に加え、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。チップを暗所でRTにて30分より長時間インキュベートする。次に、このチップを上記のように洗浄する。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥する。
【0246】
ビオチン化カルモジュリンを用いて、カルシウム存在下でプロテオームチップをプローブしたところ、6つの既知カルモジュリン標的物質、すなわちCmk1p、Cmk2p、Cmp2p、Dst1p、Myo4pおよびArc35pが同定された(図3A)。Cmk1pおよびCmk2pはI型およびII型カルシウム/カルモジュリン依存性セリン/トレオニンプロテインキナーゼであり、これらはいずれも接合応答(mating response)でのシグナル伝達経路に関与する(Hookら、2001, Annu. Rev. Pharmacol. Toxicol. 41)。Cmp2p(Cna2p)は、2つの酵母カルシニュリンのうちの1つであって、in vivoでカルモジュリンと相互作用することが実証されている(Cyertら、1991, Proc. Natl. Acad. Sci. USA 88:7376)、Dst1pは転写伸長に関与する(Stirling ら、1994, EMBO J. 13:4329)、Myo4pは、ASH1転写物の適正な局在化のために必要とされるクラスVミオシン重鎖である(Bohlら、2000, EMBO J. 19:5514; Bertrandら、1998, Mol. Cell 2:437)。Arc35pは、Arp2/3アクチン構築複合体の成分であり、この複合体はアクチンの構築および機能、ならびにエンドサイトーシスに関与する(Winterら、1999, Proc. Natl. Acad. Sci. USA 96: 7288)。Arc35pは、ツーハイブリッド研究において、カルモジュリンと相互作用することが最近明らかになったが(Schaerer-Brodbeckら、2000, Mol. Biol. Cell 11:1113)、これは、Arc35pおよびカルモジュリンがin vitroで相互作用することを実証する本明細書のデータを確認するものである。検出されなかった6つの既知カルモジュリン標的物質のうち、2つはこのコレクションに集められておらず、残りの4つはGSTプロービング実験では検出不可能であった。既知の相互作用物質に加えて、このカルモジュリンプローブはさらに33の相互作用物質を同定した。これらの相互作用物質は、種々様々な異なるタイプのタンパク質を包含し(表1;公開ウェブサイトbioinfo.mbb.yale.edu/proteinchipを参照されたい)、多岐にわたる細胞プロセスにおけるカルモジュリンの役割と合致する。
【0247】
カルモジュリン結合標的物質の他に、Cy3標識ストレプトアビジンと結合する1つのタンパク質、Pyc1p、も同定された。Pyc1pは、高度に保存されたビオチン結合領域を含有するピルビン酸カルボキシラーゼ1ホモログをコードする(Menendezら、1998, Yeast 14:647)。したがって、その配列から予想されるように、Pyc1pはin vivoでビオチン化されており、これはストレプトアビジン検出法を用いたすべての実験で同定された。このように、プロテオームマイクロアレイを使用して、タンパク質の翻訳後修飾を同定することができる。
【0248】
6.5.2 カルモジュリン結合モチーフの同定
推定カルモジュリン結合ドメインを同定するために、異なるカルモジュリン結合標的物質(すなわち相互作用物質)の間で共通するアミノ酸配列を決定した(Zhuら、2000, Nat. Genet. 26:283)。39のカルモジュリン結合タンパク質のうち14が、あるモチーフを含有し、そのコンセンサス配列はI/L-Q-X-X-K-K/X-G-B(配列番号1)であるが、ここでXは任意の残基であり、Bは塩基性残基である(図3B)。ミオシンにおける関連配列、I-Q-X-X-X-X-K-X-X-X-R(配列番号16)は、カルモジュリンと結合することがすでに明らかになっている(Hommaら、2000, J. Biol. Chem. 275)。この結果は、モチーフ、I-Q-X-X-X-X-K-X-X-X-R(配列番号16)が多くのカルモジュリン結合タンパク質に存在することを実証する。このモチーフを持たない他のカルモジュリン結合相互作用物質は、他のカルモジュリン結合配列を保有すると考えられる。
【0249】
6.5.3 ATP/GTP結合タンパク質の同定
1. ブロックされたプロテオームチップを冷PBSバッファーで3〜5回洗浄し、ガラス表面の余分な液体を、KIMWIPETMの上でスライドガラスを縦にして軽くたたいて取り除く。
【0250】
2. ATPおよびGTP溶液(0.5〜5 mM ATP-Cy3, 0.5〜5 mM GTP-Cy5, 100〜400mM NaCl, 1〜30 mM MgCl2, 50-300 mM Tris, pH 7〜8.5)100μlをプロテオームチップに加え、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, OR, 米国)で覆う。閉じ込められた気泡を追い出した後、チップを加湿チャンバー内で4℃にて1時間インキュベートする。
【0251】
3. カバーガラスを取り外すために、チップを多量(>30 ml)の氷冷洗浄バッファー(100〜400mM NaCl, 1〜30 mM MgCl2, 50〜300 mM Tris, pH 7〜8.5)中に浸漬すると、カバーガラスは浮き上がって離れるだろう。次にチップを第2の洗浄槽(>50 ml)に移動して、4℃にて振盪しながら3×5分間洗浄する。
【0252】
4. チップ表面の余分な液体を除去した後、チップを4℃にて1500〜2000 rpmで5〜10分間回転させて乾燥する。
【0253】
6.5.4 DNA結合タンパク質の同定
タンパク質-DNAおよびタンパク質-RNA相互作用は、転写調節、染色体の分離および維持、ならびに、RNA輸送およびプロセシングといった、多くの基本的な生物学的機能にとって重要である(Williamson, 2000, Nat. Struct. Biol. 7(10):834-7)。プロテオームチップを用いてDNAもしくはRNA分子と結合するタンパク質を同定する可能性を探るために、酵母の全ゲノムDNAをCy3-CTPおよびKlenow断片を用いて標識し、また、ポリA+をビオチン化プソラレン誘導体(31)によって標識した。それぞれのプローブを、異なるプロテオームチップとともにインキュベートした(図3A)。
【0254】
タンパク質-核酸相互作用を次のようにアッセイした。核酸プローブをWinzelerら(1999, Science 285:901)に記載のように標識した。ブロックされたプロテオームチップをPBSバッファー中で3〜5回洗浄し、ガラス表面上の余分の液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去する。200μlの標識核酸プローブ(50〜200 mM ポリ(dC/dG);100〜300 mM NaCl; 50〜300 mM HEPES, pH 7.0〜8.5; 10 mM MgCl2)をプロテオームチップに添加して、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。閉じ込められた気泡を追い出した後、チップを加湿チャンバー内暗所で室温(RT)にて1時間インキュベートする。カバーガラスを取り外すために、チップを多量のPBSバッファー(>50ml)中に浸すと、カバーガラスは浮き上がって離れるだろう。次にチップを第2のPBS槽(>50ml)に移動し、室温にて暗所で振盪しながら、3×5分間、洗浄した。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥させた。
【0255】
目視検査によって、ゲノムDNAプローブが41種のDNA結合タンパク質を同定したことが明らかになった(表1を参照されたい)。これらは、8種の既知DNA関連タンパク質および2種の予想されるDNA結合タンパク質を含んでいた。既知のDNA結合タンパク質のうち、本発明者らが見いだしたのは、3種のDNA修復タンパク質(すなわち、xrs2p、Cdc1pおよびRad51p(Costanzoら、2001, Nucleic Acids Res. 29:75))、SWI-SNF包括的転写活性化因子複合体成分(すなわち、Snfl1p(Costanzoら、2001, Nucleic Acids Res. 29:75))、クラスII転写Bur6pのNC2(Dr1/Drap1)リプレッサーのαサブユニット(Sternbergら、1987, Cell 48:567)、転写因子Spt2p(HO遺伝子転写の負の制御因子(Sternbergら、1987, Cell 48:567))、および5つのWDリピートを含有し、特定のプロモーター領域と結合するMet30p(Thomasら、1995, Mol. Cell Biol. 15:6526)である。本発明者らはTrm2pも同定したが、これはDNAおよびRNAのいずれとも結合し、DNA修復およびRNAプロセシング/修飾に関与する(Sadekovaら、1996, Curr. Genet. 30(1):50-5)。
【0256】
8種のタンパク質が核酸結合タンパク質として同定されたが、これらは機能が不明である。これらのうち3種は、アミノ酸配列および本アッセイの結果に基づいて、DNA結合活性を有する可能性が高い。Ycr087cは共通のDNA結合モチーフであるジンクフィンガードメインを含む。Yhr54cは転写因子Rsc3pとの配列相同性を共有する(Costanzoら、2001, Nucleic Acids Res. 29:75)。Ybr025cは推定ヌクレオチド結合活性を有すると考えられる(Costanzoら、2001, Nucleic Acids Res. 29:75)。本アッセイは、これら8種のタンパク質がin vitroでDNAに結合し、おそらくin vivoで核酸に結合することを示す。
【表1】

【0257】
【0258】
【0259】
【0260】
【0261】
【0262】
【0263】
6.5.5 RNA結合タンパク質の同定
Trm2pに加えて、RNA結合活性を有する他の相互作用物質が同定された。例えば、イソロイシル-tRNA合成酵素(Ils1p)およびリボソームタンパク質(Rpl41B)はいずれもRNA分子と結合することが明らかになっている(Lankerら、1992, Cell 70:647; Suzukiら、1990, Curr. Genet 17:185)。翻訳に関与する他のタンパク質が、Rpl41b、Sqt1p、およびGcd11pのように、RNA結合タンパク質のスクリーニングで陽性と判定された。Rpl41bはリボソーム大サブユニットの成分である。Sqt1pはRpl10pと相互作用し、Rpl10pはRNA結合タンパク質である(Costanzo, 2001, Nucleic Acids Res. 29:75)。Gcd11pは翻訳開始因子eIF2のγサブユニットであるが、GTP結合活性を有し、ツーハイブリッド分析によって実証されるように、Lsm1-7複合体(U6特異的snRNP)に結合する(FromontRacineら、2000, Yeast 17:95)。DNAプローブと結合する他の2つのタンパク質標的物質もヌクレオチドと結合する。すなわち、Thi13pはヌクレオチド代謝に関与し、Ypt31pはGTP結合活性を有する(Costanzo, 2001, Nucleic Acids Res. 29:75)。DNAプローブによって同定されたRNA結合およびヌクレオチド結合タンパク質は、DNAとの低親和性相互作用を反映している可能性があり、あるいはまた、これまでに認識されていないDNAに対して標準の親和性を示すかもしれない。
【0264】
対数増殖期の細胞から得られたRNA-ポリA+をビオチンで標識し、プロテオームマイクロアレイをプローブするために使用した。Cy3-ストレプトアビジンを用いて陽性シグナルを検出した。19の標的相互作用物質が同定されたが(表1を参照)、これは、U1 snRNP成分(Smd1p)、mRNA輸送タンパク質と結合することが判明しているタンパク質(Sxm8p)、C2-H2ジンクフィンガータンパク質(Azf1p)、およびTos8pを包含し、Tos8pは、その配列から、RNAスプライシングおよびプロセシングに関与することが示唆された(Schwikowskiら、2000, Nat. Biot. 18:1257)。おそらく直接もしくは間接的にRNAと相互作用するタンパク質に加えて、本発明者らは2つの転写因子(Bur6pおよびTos8p)、1つの転写サイレンサ(Hst3p)、既知ORFによってコードされる6つの他のタンパク質、および特徴づけられていないORFに由来する7つのタンパク質を発見した。未知のORFの1つは最高のシグナルを示した(Yer152c)。ただ1つの相互作用物質であるBur6pのみがDNAおよびRNAプローブの両者と結合したことに注目することが重要である。他の相互作用物質は、二本鎖DNAプローブもしくはRNAプローブのいずれか一方でのみ検出されたが、これは、大半のタンパク質がDNAもしくはRNAのいずれかと特異的に結合する可能性が高いことを示唆する。
【0265】
要約すれば、41種の標的相互作用物質のうち、18種は核酸相互作用タンパク質である;11種は既知もしくは推定DNA結合タンパク質であり、3種はRNA結合タンパク質であり、そして4種はヌクレオチドと結合する。
【0266】
6.5.6 脂質結合タンパク質の同定
本発明のプロテオームチップは、例えばタンパク質-薬物相互作用およびタンパク質-脂質相互作用のような、他の実験手法が利用しにくいと思われる活性を同定するのに有用である。実際、リン脂質と結合するタンパク質に関するゲノム全体にわたる分析は、いかなる生物種についてもこれまで検討されたことがない。ホスファチジルイノシトール(PI)と相互作用するタンパク質を研究するために本発明のプロテオームチップを使用した。ホスファチジルイノシトールは、細胞膜の構成成分としての役割に加えて、成長、分化、細胞骨格再編成、および膜輸送を含めて、様々な細胞プロセスを制御する重要な第2メッセンジャーであって、核、液胞、および原形質膜に存在している(Odorizziら、2000, TIBS 25: 229; Frumanら、1998, Annu. Rev. Biochem. 67: 481; Martin, 2000, Annu. Rev. Cell Dev. Bio. 14:231; Weraら、2001, FEMS Yeast Res. 1406:17)。ホスファチジルイノシトールは細胞内に一時的に、しかも小量で存在することが多いため、酵母で広範に特徴づけられたことはなく、したがって、特定のリン脂質と結合するタンパク質についてはほとんど知られていない(Odorizziら、2000, TIBS 25: 229; Frumanら、1998, Annu. Rev. Biochem. 67: 229; Martin, 2000, Annu. Rev. Cell Dev. Bio. 14:231; Weraら、2001, FEMS Yeast Res. 1406:1)。
【0267】
酵母でPI結合タンパク質を同定するために、リポソームをプローブとして使用したが、これは、リポソームが相互作用物質をアッセイするために最適の生理的状態を提供するためである。6つのタイプのリポソームを使用した。それぞれが、1%(w/w)N-(ビオチノイル)-1,2-ジヘキサデカノイル-sn-グリセロ-3-ホスホエタノールアミン、トリエチルアンモニウム塩(ビオチンDHPE)とともに、ホスファチジルコリン(PC)を含有する。ビオチン化脂質はCy3-ストレプトアビジンによって検出可能な標識として機能する(21)。PCに加えて、5つのタイプの他のリポソームは、5%(w/w) PI(3)P、PI(4)、PI(3,4)P2、PI(4,5)P2もしくはPI(3,4,5)P3のいずれかを含有する(図3A)。それぞれの上記リン脂質は、PI(3,4,5)P3を除いて、酵母で発見されたものである(Odorizziら、2000, TIBS 25: 229; Frumanら、1998, Annu. Rev. Biochem. 67: 481; Martin, 2000, Annu. Rev. Cell Dev. Bio. 14:231; Weraら、2001, FEMS Yeast Res. 1406:1)。
【0268】
タンパク質-リポソーム相互作用を下記のようにアッセイした。クロロホルム中の適当量の各脂質を、窒素ガス下で混合して乾燥させた。この脂質混合物をボルテックス混合によってTBSバッファーに再懸濁した。超音波処理によってリポソームを形成した。ブロックされたプロテオームチップをPBSバッファーで3〜5回洗浄し、ガラス表面上の余分の液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去した。200μlのリポソーム溶液をプロテオームチップに添加して、直ちに、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)で覆った。閉じこめられた気泡を追い出した後、チップを加湿チャンバー内でRTにて1時間インキュベートした。カバーガラスを取り外すために、チップを多量のPBSバッファー(50ml)中に浸すと、カバーガラスは浮いて外れることになる。次にチップを第2のPBS槽(>50ml)に移動し、室温で振盪しながら、3×5分間、洗浄した。チップ表面上の余分な液体を除去した後、約100μlのCy3-もしくはCy5-結合ストレプトアビジン(PIERCETM, USA; 1:2000から1:4000希釈)をその表面に加え、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)で覆った。チップを、30分より長時間、暗所で、RTにてインキュベートした。次に、このチップを上記のように洗浄した。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥した。
【0269】
6つのタイプのリポソームが、バックグラウンドより有意に高いシグナルを発生した合計150の異なる相互作用物質を同定した。陽性シグナルの識別に役立つように、アルゴリズムが考案された(42)。52(35%)の相互作用物質(すなわち、脂質結合タンパク質)が特性決定されていないタンパク質に相当し、それゆえ、これらのタンパク質にこの最初の生化学的活性を帰属させた。上記の結果はまた、これまで性質が明らかにされていない多くのタンパク質が、潜在的に重要な生化学的活性を有することを示すものである。
【0270】
52の特性決定されていないタンパク質のうち、13(25%)は膜に結合していると予想される(Gerstein, 1998, Proteins 33:518)。他の多くは、負電荷を帯びたホスファチジルイノシトールとの静電相互作用を媒介することができる塩基性ストレッチを含有する。
【0271】
98の以前に記載された相互作用物質の中で、これらの81の細胞内局在が研究されている(Costanzoら、2001, Nucleic Acids Res. 29:75)。大半は膜結合タンパク質であるか、もしくは脂質代謝に関与するかのいずれかである。より詳細には、45のタンパク質が膜結合型であり、既知のもしくは予想される膜貫通領域を有する(Gerstein, 1998, Proteins 33:518)。これらの同定された相互作用物質には、内在性膜タンパク質、脂質修飾を有するタンパク質(例えば、グリコシルホスファチジルイノシトール(GPI)アンカータンパク質Tos6pおよびSps2p(Costanzoら、2001, Nucleic Acids Res. 29:75))、プレニル化タンパク質、Gpa2pおよび接合フェロモンaファクター(Ansariら、1999, J. Bio. Chem. 274:30052)、ならびに、周辺結合タンパク質(例えば、芽頸部(bud neck)および/または細胞周辺に見いだされるKcc4pおよびMyo4p(Bohl, 2000, EMBO J. 19: 5514; Bertrandら、1998, Mol. Cell 2:437))がある。別の8個は、脂質代謝(例えば、Bpl1p)、イノシトール環のリン酸化(例えば、Kcs1p)に関わり、もしくは膜/脂質機能(例えば、トリアシルグリセロールリパーゼに対して相同性を示すYlr020cp)に関係していると予想される。
【0272】
相互作用物質(すなわち、リン脂質結合タンパク質)を、幾通りかの方法でグループ分けした(42)。最初に、相互作用物質を、それらが脂質に強く結合するか、弱く結合するかにしたがって分類したが、これはGSTの量に対するリン脂質結合シグナルに基づく(図4)。やや多くの(72%)強い脂質結合タンパク質(図4Aおよび4B)が、弱い結合タンパク質(54%)(図4Cおよび4D)と比較して特徴付けられた。強い脂質結合タンパク質の多くが膜結合型であるか、または膜に結合していると予想されるのに対して、膜に結合していると思われる弱い結合タンパク質はより少数である(図4、「膜」の列)。驚くべきことに、19の脂質結合タンパク質がキナーゼであり、このうち17はプロテインキナーゼである。さらに、この17のプロテインキナーゼのうち13は非常に強くリン脂質に結合する。
【0273】
また、ホスファチジルコリンと比較して1以上のホスファチジルイノシトールと優先的に結合するかどうかによって、タンパク質をグループ分けした。101のタンパク質が、試験したホスファチジルイノシトール脂質と同様に、もしくはほぼ同様にホスファチジルコリンと結合した(PI/PC<1.3)(図4Bおよび4D)。しかしながら、49のタンパク質が1以上のホスファチジルイノシトールに優先的に結合した(PI/PC>1.3)(図4Aおよび4C)。強い相互作用物質(すなわち、ホスファチジルイノシトール結合タンパク質)の分析によって、多くが特定のホスファチジルイノシトール脂質に特異的に結合することが明らかになった。例えば、Stp22pは、Ste2pおよびCan1pのように、温度感受性原形質膜タンパク質の液胞ターゲティングに必要であるが、優先的にPI(4)Pと結合する(Liら、1999, Mol. Cell Biol. 19:3588)。9のプロテインキナーゼは特異的にPI(4)PおよびPI(3,4)P2と強く結合するが、1つは前記脂質とは弱く結合する。Atp1pは、ミトコンドリア内膜のF1-ATPシンターゼのαサブユニットであるが、やはり優先的にPI(3,4)P2に結合する(Arnold ら、1999, J. Biol. Chem.)。Sps2pは、胞子形成の中/後期に関与し、原生胞子膜に局在するが(Chuら、1998, Science 282:699)、PI(3)Pと優先的に結合する。興味深い一例は、優先的にPI(4,5)P2と結合するMyo4pであるが、おそらくこの脂質の結合は、細胞表皮でのその相互作用および/またはその調節における重要な部分である。PI(3,4,5)P3と特異的に結合する強い脂質結合標的物質はまったく見いだされなかったが、一部のタンパク質はこの脂質と他の脂質の両者と結合した(図4)。上記の結果は、内在性膜タンパク質および周辺結合タンパク質を含めて、多くの膜結合タンパク質がin vivoで特定のリン脂質と優先的に結合することを実証する。
【0274】
予想に反して、グルコース代謝の調節因子を含めて、グルコース代謝に関与する多くのタンパク質がリン脂質に結合する。具体的には、解糖の一連のステップに関わる3つのグルコース代謝酵素、すなわちホスホグリセリン酸ムターゼ(Gpm3p)、エノラーゼ(Eno2p)、およびピルビン酸キナーゼ(Cdc19p/Pyk1p)が同定された。グルコースをグルコース6リン酸に変換するヘキソキナーゼ(Hxk1p)、およびグルコース代謝の調節因子であることが明らかになっている2つのプロテインキナーゼ(すなわち、Snf1pおよびRim15p)も、脂質結合相互作用物質として同定された。HXk1pは、その活性を促進する双性イオンのミセルと結合することが最近明らかにされた(30);Eno2pは分泌されることが明らかになり、膜とも相互作用する可能性が示唆された(31)。したがって、ある実施形態において、リン脂質を用いて、グルコース代謝に関与するステップを調節することができる。別の実施形態において、例えば、細胞膜のようなリン脂質足場上でグルコース代謝のステップを行うことによって、解糖ステップを高めることができる。
【0275】
驚くべきことに、リン脂質を含むプローブは、膜機能もしくは脂質シグナル伝達に関与すると予想されない多くのタンパク質を認識した。したがって、6つのタンパク質、すなわち、Rim15p、Eno2p、Hxk1p、Sps1p、Ygl059wpおよびGcn2pはホスホイノシトール結合について2種類の標準アッセイによってさらに検討した(Williamson, 2000, Nat. Struct Biol. 7(10):834-7)。Rim15p、Eno2pおよびHxk1pに関しては、PI(4,5)P2リポソームを最初にニトロセルロース膜に付着させ、次にこれをBSAでブロックした;異なる量のGST融合タンパク質およびGST対照を用いてこの膜をプローブし、結合したタンパク質を抗GST抗体によって検出した。図5Aに示すように、それぞれの酵母融合タンパク質はしっかりとPI(4,5)P2に結合し、量的効果を示した;GST単独では結合しなかった。本発明者らは、4つのGST融合タンパク質、Rim15p、Sps1p、Ygl059wpおよびGcn2pについて、リバースアッセイも行った(Guerraら、2000, Biosci. Rep. 20:41)。異なる量の前記精製タンパク質をニトロセルロースフィルター上にスポットし、6つの異なるリポソームによってプローブした(図5B)。結合したリポソームを西洋ワサビペルオキシダーゼ(HRP)結合ストレプトアビジンによって検出した。プロテオームチップを用いた実験と同様に、リポソームはそれぞれのタンパク質に結合し、BSA対照とは結合しなかった。Sps1pは、5種すべてのホスホイノシトール含有リポソームとほぼ同等に結合した。Rim15p、Gcn2pおよびYgl059wpは、異なるリポソームに対して、異なる親和性を示した(Rim15pについては図5Bを参照されたい)。3種のタンパク質のすべてが、PI(3)PおよびPI(4)PおよびPI(3,4)P2にもっとも強く結合した。Rim15Pに関しては、結合シグナルとRim15Pレベルの間に直線的相関関係が明らかになった(図5C)。
【0276】
さらに、未知の機能を有するいくつかのタンパク質も、強い脂質結合活性を示したが、このことは、これらが細胞内でリン脂質と結合しやすいことを示す。当業者に公知の日常的なアッセイを利用して、プロテオームチップによって同定される脂質結合相互作用物質を迅速にスクリーニングし、in vivoで脂質結合活性を測定することができる。さらに、本発明にしたがって、プロテオームチップを使用して、これらの相互作用物質がリン脂質代謝経路、もしくは情報伝達経路に関与するかどうかを判定することができる。
【0277】
要約すると、上記の結果は、本発明のプロテオームチップを使用して、脂質と結合することが以前に知られていなかったタンパク質を含めて、多くの脂質結合タンパク質を検出し、特徴づけることができることを明白に実証する。さらに、本発明のプロテオームチップによって同定されたリン脂質結合相互作用物質は、従来のアッセイで真正な脂質結合タンパク質であることが示される。
【0278】
さらに加えて、酵母ツーハイブリッド系またはDNA/オリゴマイクロアレイ技術のいずれかを使用することによって、このような相互作用物質を研究することは不可能であるため、本発明のプロテオームチップおよびその使用方法は、分子-タンパク質相互作用を探求する先駆的方法となる。
【0279】
本研究は、真核生物に関するプロテオームマイクロアレイを作製しスクリーニングすることが実現可能であることを示す。いくつかの技術的特徴を新たに非自明的に組み合わせることが、本発明を実行するうえで不可欠である。第1に、本研究で分析されるタンパク質は、DNA塩基配列決定法によって確認され、かつ純粋なプラスミドを含有する、酵母発現クローンから調製されたものである。第2に、規定されたクローンおよびタンパク質をハイスループット様式で生産することが不可欠である。本明細書に記載の方法は、5,000プロテインチップに十分なタンパク質を提供する。第3に、タンパク質は真核生物宿主において生産される。そのようにして、正しく折り畳まれて翻訳後修飾を受け、かつ/または天然のパートナーとの複合体を形成した、多数の全長タンパク質を作製することができる。イムノブロット分析によって実証されるように、精製された融合タンパク質の少なくとも80%が全長タンパク質として発現される。
【0280】
多くの相互作用物質は、直接的にではなく、結合タンパク質を介してプローブと結合することが可能である。ある場合には、相互作用物質は多量体の一部であってよい。しかしながら、このような結合タンパク質は本発明の方法によって検出し同定することもできる。そうではあるが、本発明の方法で検出された相互作用物質は、おそらく直接的にプローブと相互作用し、もしくはプローブと相互作用するタンパク質と最終的にしっかりと結合する。本発明のプロテオームチップのタンパク質は、ストリンジェントな条件(例えば、0.5M NaClによる洗浄)によって調製され、クーマシー染色は抗GST抗体によって検出可能なタンパク質のバンドのみを明らかにしたが、このことは他のタンパク質による混入が最小であることを示す。
【0281】
構築されたタンパク質のコレクションは、正しく折り畳まれた細胞外ドメインを有する分泌タンパク質を十分に代表していない可能性が高い。GSTタグおよびHisX6タグを翻訳開始コドンの上流に融合すると、シグナルペプチドを有する膜タンパク質は、分泌経路に運ばれず、適切な折り畳みもしくは修飾を受けることができない。シグナルペプチドを有する3つのタンパク質が脂質結合タンパク質のスクリーニングで同定されたが、このことは、少なくとも一部の分泌タンパク質が生産されて機能ドメインを含有することを示唆する。
【0282】
さらに、実施したハイスループット方法を用いても、すべてのタンパク質が容易に過剰生産および精製されるわけではないので、必ずしもすべての相互作用が検出されたわけではない。しかしながら、採用したプロテインアレイは、スクリーニングのために合理的なレベルで、約80%の全長酵母タンパク質を含有すると見積もられ、したがって、タンパク質相互作用物質の大半が検出されることが期待できる。
【0283】
これらの結果は、プロテオームチップによって生化学的活性をスクリーニングすることができること、それによって包括的なプロテオーム分析が可能となることを実証する。同様の方法を用いて、ヒトおよび他の真核生物における包括的ハイスループットプロテオーム分析のために、10〜100,000タンパク質からなるプロテインアレイを作製することができる。
【0284】
参考文献および注釈:
1. S. Fields, Y. Kohara, D. J. Lockhart, Proc. Natl. Acad. Sci. 96, 8825(1999); A. Goffeauら、Science 274, 563(1996).
2. P. Ross-Macdonaldら、Nature 402, 413(1999); J. L. DeRisi, V. R. Iyer, P. O. Brown, Science 278, 680(1997); E. A. Winzelerら、Science 285, 901(1999).P. Uetzら、Nature 403, 623 (2000); T. Itoら、Proc. Natl. Acad. Sci. USA 97, 1143(2000)
3. H. Zhu, M. Snyder, Curr. Opin. Chem. Biol. 5, 40(2000).
4. M. R. Martzenら、Science 286, 1153(1999).
5. H. Zhuら、Nat. Genet. 26, 283(2000).
6. G. MacBeath, S. L. Schreiber, Science 289, 1760(2000).
7. A. Caveman, J. Cell Sci. 113, 3543(2000).
8. P. Arenkovら、Anal. Biochem. 278, 123(2000).
9. D. A. Mitchell, T. K. Marshall, R. J. Deschenes, Yeast 9, 715(1993). 発現ベクターpEGHは、RGS-HisX6エピトープタグを、pEG(KG)のGST遺伝子とポリクローニング部位の間に挿入することによって作製された。前記(5)に記載のストラテジーによって酵母ORFをクローニングしたが、ただし、96ウェルフォーマットで各ステップを行った。DNA配列決定によって確認されたプラスミドDNAを酵母(Y258)および大腸菌(DH5a)の両方に再導入した。ライブラリーは5800の固有のORFを含有する。
【0285】
10. 96ウェルフォーマットタンパク質精製プロトコールの詳細、すべての実験から得られた結果の完全な一覧、およびポジティブ識別アルゴリズムのデザインは、公開ウェブサイトbioinfo.mbb.yale.edu/proteinchipで見ることができる。
【0286】
11. ビオチン化カルモジュリン(CalBiochem, 米国)をPBSバッファー中0.02 mg/mlとして0.1 mMカルシウムとともにプロテオームチップに添加し、加湿チャンバー内で室温にて1時間インキュベートした。0.1 mMカルシウムはその後のすべてのステップでバッファー中に存在した。チップをRTにてPBSによって3回洗浄した。Cy3-結合ストレプトアビジン(Jackson IR, 米国)(1:5000希釈)をチップに添加し、RTにて30分間インキュベートした。広範に洗浄した後、チップを脱水し、マイクロアレイスキャナーを用いてスキャンした;次に、GENEPIXTMアレイ デンシトメトリー ソフトウェアによってデータを取得した(Axon, 米国)。
【0287】
12. S. S. Hook, A. R. Means, Annu. Rev. Pharmacol. Toxicol. 41, 471(2001).
13. M. S. Cyert, R. Kunisawa, D. Kaim, J. Thorner, Proc. Natl. Acad. Sci. USA 88, 7376(1991).
14. D. A. Stirling, K. A. Welch, M. J. Stark, EMBO J. 13, 4329(1994).
15. F. Bohl, Kruse, EMBO J. 19, 5514(2000); E. Bertrandら、Mol, Cell 2, 437(1998).
16. D. C. Winter, E. Y. Choe, R. Li, Proc. Natl. Acad. Sci. USA 96, 7288(1999).
17. C. Schaerer-Brodbeck, H. Riezman, Mol. Biol. Cell 11, 1113(2000).
18. K. Homma, J. Saito, R. Ikebe, J. Biol. Chem. 275, 34766(2000).
19. J. Menendez, J. Delgado, C. Gancedo, Yeast 14, 647(1998).
20. G. Odorizzi, M. Babst, S. D. Emr, TIBS 25, 229(2000); D. A. Frumanら、Annu. Rev. Biochem. 67, 481(1998); T. F. Martin, Annu. Rev. Cell Dev. Biol. 14, 231(2000); S. Wera, J. C. T. Bergsma, FEMS Yeast Res. 1406, 1(2001).
21. リポソームは標準的方法によって調製された(52)。簡単に述べると、適当な量の、クロロホルム中の各脂質を混合し、窒素ガス下で乾燥させた。この脂質混合物を、ボルテックス混合によってTBSバッファー中に再懸濁した。超音波処理によってリポソームが作られた。プロテオームチップを探索するために、異なるリポソーム60mlを異なるチップ上に添加した。このチップをRTにて1時間、加湿チャンバー内でインキュベートした。TBSバッファーで3回洗浄した後、Cy-3結合ストレプトアビジン(1:5000希釈)をチップに添加し、RTにて30分間インキュベートした。
【0288】
22. それぞれのスポットについて局所強度バックグラウンドを算定するGenePixソフトウェア、および本発明者らが開発した一連のアルゴリズムを組み合わせて使用して陽性を判定した。詳細は、公開ウェブサイトbioinfo.mbb.yale.edu/proteinchipで見ることができる。
【0289】
23. M. C. Costanzoら、Nucleic Acids Res. 29, 75(2001).
24. M. Gerstein, Proteins 33, 518(1998).
25. K. Ansariら、J. Biol. Chem. 274, 30052(1999).
26. Y. Barral, M. Parra, S. Bidlinmaier, M. Snyder, Genes Dev. 13, 176(1999).
27. Y. Li, T. Kane, C. Tipper, P. Spatrick. D. D. Jennes, Mol. Cell Biol. 19, 3588 (1999).
28. I. Arnoldら、J. Biol. Chem. 274, 36(1999).
29. S. Chuら、Science 282, 699(1998).
30. A. Casamayorら、Curr. Biol. 9, 186(1999); R. Guerra, M. L. Bianconi. Biosci. Rep. 20, 41(2000).
31. M. Pardoら、Yeast 15, 459(1999).
【0290】
7. 引用文献
本明細書のすべての引用文献は、それぞれ個別の出版物または特許もしくは特許出願がそのまま全体をあらゆる目的のために参考として取り入れられることが具体的かつ個別的に示されたのと同程度に、そのまま全体をあらゆる目的のために参考として本明細書に含めるものとする。
【0291】
8. 均等物
本発明の精神および範囲から逸脱することなく、本発明の多くの修正および変更を行うことができる。当業者は、本明細書に記載された本発明の特定の実施形態に対する様々な代替、改作、および修正を認識し、もしくはルーチンの実験操作を通して確認することができるであろう。ここで、前記はいずれも本発明の範囲に含めるものとする。したがって、特許請求の範囲に記載の本発明は、こうした均等物のすべてを包含することを意味する。このため、本明細書に記載の具体的な実施形態は、ほんの一例として提供されるのであって、本発明は、添付の特許請求の範囲が権利を有する均等物の全範囲とともに、添付の特許請求の範囲によってのみ限定されるべきものである。
【図面の簡単な説明】
【0292】
【図1A】プロテインチップ技術を用いた酵母プロテオーム解析の手順を示す。
【図1B】3mlの酵母培養物から精製されたタンパク質のイムノブロット分析を示す。
【図2A】抗GST抗体を用いてプローブしたプロテオームチップを示す。
【図2B】図2Aに示したプロテオームチップの48ブロックのうちの1つの拡大図を示す。
【図2C】各ORF産物の二重反復スポット間の誤差分布を示す。
【図3A】プロテオームチップでの様々なアッセイの例を示す。
【図3B】様々なカルモジュリン標的が共有するアミノ酸配列をサーチすることにより同定された推定上のカルモジュリン結合モチーフを示す。
【図4A】30の強くかつ特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図4B】43の強くかつ非特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図4C】19の弱くかつ特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図4D】58の弱くかつ非特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図5A−B】プロテオームマイクロアレイにより検出されたタンパク質−脂質相互作用の慣用方法を用いた確認を示す。
【図5C】膜アッセイにおける結合シグナルとRim15pの量との線形相関を示す。
【技術分野】
【0001】
1. 発明の分野
本発明は、単一の種において発現される全タンパク質の大部分を含んでなるプロテオームチップに関する。本発明はまた、プロテオームチップの製造方法に関する。本発明はさらに、ハイスループット方式で単一の種における全タンパク質の相互作用を系統的にアッセイするためのプロテオームチップの使用方法に関する。
【0002】
加えて、本発明は、高密度アレイフォーマットで真核生物タンパク質を製造し精製するための方法に関する。本発明はまた、二重タグ付けされた融合タンパク質を基板に結合させることによるタンパク質アレイの製造方法に関する。本発明はさらに、シグナルが陽性であるかどうかを確認する方法に関する。
【背景技術】
【0003】
2. 発明の背景
ポストゲノム配列決定時代の厄介なタスクは、ゲノムによりコードされるあらゆるタンパク質の機能、修飾および調節を理解することである (Fieldsら, 1999, Proc Natl Acad Sci. 96: 8825; Goffeauら, 1996, Science 274: 563)。現在、mRNA発現プロファイル、遺伝子破壊表現型、ツーハイブリッド相互作用、およびタンパク質細胞内局在を分析することによって遺伝子(それゆえにタンパク質)の機能を研究することに多くの努力が払われている (Ross-Macdonaldら, 1999, Nature 402: 413; DeRisiら, 1997, Science 278: 680; Winzelerら, 1999, Science 285: 901; Uetzら, 2000, Nature 403: 623; Itoら, 2000, Proc. Natl. Acad. Sci. U.S.A. 97: 1143)。こうした研究は有用であるけれども、転写プロファイルは必ずしも細胞のタンパク質レベルと相関しない。したがって、生化学的活性の分析がタンパク質機能に関する情報をもたらし、かかる情報はゲノム解析を補完して細胞の働きのより完全な全体図を提供することができる (Zhuら, 2001, Curr. Opin. Chem. Biol. 5: 40; Martzenら, 1999, Science 286: 1153; Zhuら, 2000, Nat. Genet. 26 : 283; MacBeath, 2000, Science 289: 1760; Caveman, 2000, J. Cell Sci. 113: 3543)。
【0004】
近年、幾つかのグループは、タンパク質活性をスクリーニングするためのマイクロアレイフォーマットを記載している (Zhuら, 2000, Nat. Genet. 26: 283; MacBeathら, 2000, Science 289: 1763; Arenkovら, 2000, Anal. Biochem 278: 123)。さらに、酵母タンパク質の過剰発現クローンの集合体が作製されて、生化学的活性に関してスクリーニングされた (Martzenら, 1999, Science 286: 1153)。しかしながら、ほぼ全プロテオームにあたる何千もの個々のタンパク質はまだ調製もアレイ化もされておらず、また、多数の活性に関するスクリーニングもされていない (Caveman, 2000, J. Cell Sci. 113:3543)。
【0005】
全プロテオームのスクリーニングには、ハイスループット方式で生産されたタンパク質の生化学的活性を系統的に精査し、並行して、何千何百というタンパク質試料の機能を分析することが必要となろう (Zhuら, 2000, Nat. Genet. 26: 283; MacBeathら, 2000, Science 289 : 1763; Arenkovら, 2000, Anal. Biochem 278: 123)。全プロテオームアレイをスクリーニングする試みは、必要な発現クローンの作製や、発現タンパク質のハイスループット方式での発現および精製が可能でないことを含めて、大きな障害に遭遇してきた。これまで、in vitroアッセイはランダム発現ライブラリーまたはプーリング(pooling)ストラテジーを用いて行われているが、これらはいずれも欠点を有する (Martzenら, 1999, Science 286: 1153; Bussowら, 2000, Genomics 65: 1)。特に、ランダム発現ライブラリーはスクリーニングに時間がかかり、また、しばしば完全長ではないクローンを含む。最近のもう一つのアプローチは、規定されたアレイを作製して、そのアレイをプーリングストラテジーによりスクリーニングすることであった (Martzenら, 1999, Science 286: 1153)。しかし、プーリングストラテジーはスクリーニングされるタンパク質の実際の数を覆い隠してしまい、また、このストラテジーは多数の陽性試料を同定する場合には面倒である。
【0006】
タンパク質-タンパク質相互作用を検出するのに有用な別の方法は、ツーハイブリッド法である (Uetzら, 2000, Nature 403: 623; Itoら, 2000, Proc. Natl. Acad. Sci. U.S.A. 97: 1143)。しかし、この方法を使って検出することができる相互作用のタイプは限られている。なぜなら、相互作用は核内で検出されるのが一般的だからである。
【0007】
したがって、当技術分野では、生化学的機能の大規模分析の必要性が存在しており、それには、ハイスループット方式で、種のゲノムによりコードされる広範囲にわたるタンパク質のセットを調製し、スクリーニングすることが必要となるだろう。
【0008】
セクション2または本明細書の他のセクションでの文献の引用または確認は、そのような文献が本発明の先行技術として利用可能であることを容認するものとして考えられるべきでない。
【発明の開示】
【0009】
3. 発明の概要
本発明は、ある種のタンパク質相互作用をハイスループット方式で包括的に研究するのに有用なプロテオームチップを提供する。本発明の方法および組成物は、本発明者らが、新たに、また予期せざることに、ゲノムのタンパク質コード配列を含む広範囲の発現構築物のセットを作製し、ハイスループット様式で宿主細胞にタンパク質産物を産生させ、マイクロアレイを使って複数のタンパク質の機能をハイスループット方式で分析する手段を見出したことにより、可能となったものである。
【0010】
本発明はプロテオームチップに関し、かかるチップは、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイであり、ここで、該複数のタンパク質は単一の種において発現された全タンパク質の大部分に相当し、その際、1つのオープンリーディングフレームの翻訳産物が1つのタンパク質とみなされる。
【0011】
アッセイを1回ずつ行うことに比べて、アレイを用いることの利点は、多くのタンパク質−プローブ相互作用を同時に同定して特徴づけられる点である。さらに、複雑なプローブ混合物をプロテオームチップと接触させることにより、例えば、細胞の環境を代表する環境における相互作用を検出し、多くの潜在的結合化合物を迅速に評価することができる。
本発明は、より具体的には、以下の特徴を有する:
〔1〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種における既知遺伝子のうち少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記アレイ。
〔2〕前記複数のタンパク質が単一の種における既知遺伝子のうち少なくとも70%によりコードされる少なくとも1つのタンパク質を含む、上記〔1〕に記載のアレイ。
〔3〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される全タンパク質のうち少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記アレイ。
〔4〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記アレイ。
〔5〕基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記アレイ。
〔6〕前記タンパク質がタンパク質の分類に従って前記アレイ上に組織化されている、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔7〕前記分類が存在量、機能、酵素活性、相同性、タンパク質ファミリー、特定の代謝経路との関連性、または翻訳後修飾によるものである、上記〔6〕に記載のアレイ。
〔8〕前記タンパク質がHisタグを介して基板に結合されている、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔9〕前記基板がニッケルを含む、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔10〕前記基板がニッケルをコーティングしたスライドガラスからなる、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔11〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記方法。
〔12〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される全タンパク質の少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記方法。
〔13〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記方法。
〔14〕基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記方法。
〔15〕基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
〔16〕前記結合ステップに先立って、前記タンパク質を第1タグの結合パートナーと接触させることにより前記タンパク質を精製するステップを行い、前記結合ステップにおいて第2タグを用いて基板に前記タンパク質を結合させる、上記〔15〕に記載の方法。
〔17〕第1タグがGSTタグであり、第2タグがHisタグである、上記〔16〕に記載の方法。
〔18〕第1タグと第2タグが前記タンパク質のアミノ末端に存在する、上記〔15〕に記載の方法。
〔19〕第1タグと第2タグが前記タンパク質のカルボキシ末端に存在する、上記〔15〕に記載の方法。
〔20〕複数の精製されたタンパク質サンプルを調製して単離する方法であって、マルチサイトアレイの複数サイトの各サイトにおいて、
(a) 調節配列と機能的に連結された異種ヌクレオチド配列を有する真核細胞を生育させ、
(b) 該調節配列を、異種ヌクレオチド配列によりコードされるタンパク質の発現を高める誘導物質と接触させ、
(c) 該細胞を溶解して細胞溶解物を形成し、
(d) 該細胞溶解物またはそれから得られたタンパク質含有サンプルを結合物質と接触させて、結果的に該タンパク質と結合物質との複合体を形成させ、そして
(e) 該複合体から該タンパク質を単離する、
各ステップを含んでなり、その際、各ステップがマルチアレイフォーマットで実施される、上記方法。
〔21〕各サイトがウェルである、上記〔20〕に記載の方法。
〔22〕前記タンパク質がアフィニティータグを含む融合タンパク質であり、該アフィニティータグに前記結合物質が結合する、上記〔20〕に記載の方法。
〔23〕前記細胞が酵母細胞である、上記〔20〕に記載の方法。
〔24〕前記溶解ステップがペイントシェーカーを使って実施される、上記〔20〕に記載の方法。
〔25〕脂質結合タンパク質を検出する方法であって、
(a) 脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置に該相互作用が検出されれば、その位置に脂質結合タンパク質が存在することを示す、上記方法。
〔26〕前記脂質がリン脂質である、上記〔25〕に記載の方法。
〔27〕前記リン脂質がホスファチジルコリンまたはホスファチジルイノシトールである、上記〔26〕に記載の方法。
〔28〕前記プローブがリポソームからなる、上記〔25〕に記載の方法。
〔29〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔30〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも50%を含むものであり、その際、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなし、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔31〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される少なくとも1000のタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔32〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔33〕結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数の融合タンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該融合タンパク質は第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
〔34〕前記プローブが核酸、タンパク質、小分子、候補薬物または脂質を含むものである、上記〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔35〕前記核酸がRNAまたはDNAからなる、上記〔34〕に記載の方法。
〔36〕前記プローブが酵母タンパク質である、上記〔34〕に記載の方法。
〔37〕酵母タンパク質がMyo2、Rho1、Rho2、Rho3、Rho4、Cdc11、Cdc12またはHs17である、上記〔36〕に記載の方法。
〔38〕前記プローブが抗体である、上記〔34〕に記載の方法。
〔39〕前記抗体がサイクリン、キナーゼ、GST、Clb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2、またはCdc11に特異的である、上記〔38〕に記載の方法。
〔40〕前記プローブがカルモジュリンである、上記〔34〕に記載の方法。
〔41〕前記プローブがATP、GTP、cAMP、ホスホチロシン、ホスホセリン、およびホスホトレオニンからなる群より選択される小分子を含むものである、上記〔34〕に記載の方法。
〔42〕前記プローブがホスファチジルコリンまたはホスファチジルイノシトールを含むものである、上記〔34〕に記載の方法。
〔43〕前記プローブがリポソームからなる、上記〔34〕に記載の方法。
〔44〕前記プローブが哺乳動物由来のものである、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔45〕前記哺乳動物がヒトである、上記〔44〕に記載の方法。
〔46〕前記複数のタンパク質がヒト以外のものである、上記〔44〕に記載の方法。
〔47〕前記複数のタンパク質がHisタグを介して基板に結合される、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔48〕前記基板がニッケルを含む、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔49〕前記基板がニッケルをコーティングしたスライドガラスからなる、上記〔25〕、〔29〕、〔30〕、〔31〕、〔32〕または〔33〕に記載の方法。
〔50〕前記検出ステップでタンパク質との相互作用が検出されたプローブの正体を同定するステップをさらに含んでなる、上記〔34〕に記載の方法。
〔51〕前記相互作用は、前記同定されたプローブが抗菌性、抗真菌性、または抗ウイルス性タンパク質であることを示す、上記〔50〕に記載の方法。
〔52〕結合アッセイで使用するためのタンパク質を標識する方法であって、
(a) 該タンパク質の個々のアリコートを、ビオチン転移化合物と、アリコートごとに異なる程度にビオチン化されたタンパク質を生成する条件下および時間にわたり接触させ、そして
(b) 個々のアリコートを一緒に合わせて、異なる程度にビオチン化されたタンパク質のサンプルを得る、
各ステップを含んでなる、上記方法。
〔53〕結合タンパク質を検出する方法であって、
(a) 上記〔52〕に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じているアレイ上の位置を検出する、
各ステップを含んでなる、上記方法。
〔54〕結合タンパク質を検出する方法であって、
(a) 上記〔52〕に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) 該アレイに、蛍光物質とコンジュゲートさせたストレプトアビジンを接触させ、そして
(c) 蛍光が発生しているアレイ上の位置を検出する、
各ステップを含んでなり、その際、蛍光は、ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じていることを示す、上記方法。
〔55〕タンパク質がホスファチジルコリンに比べてホスファチジルイノシトールと優先的に結合するかどうかを判定する方法であって、
(a) ホスファチジルイノシトールを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルイノシトール結合タンパク質の存在を示すものであり、
(c) ホスファチジルコリンを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該タンパク質はステップ(a)におけると同じタンパク質を少なくとも一部含んでなり、
(d) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルコリン結合タンパク質の存在を示すものであり、そして
(e) 複数のタンパク質のそれぞれについて、ステップ(b)とステップ(d)の結果を比較する、
各ステップを含んでなる、上記方法。
〔56〕リン脂質が細胞における代謝経路またはシグナル伝達経路を調節するのかどうか、あるいは該代謝経路またはシグナル伝達経路が膜表面で起こるのかどうかを判定する方法であって、
(a) リン脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該複数のタンパク質は該経路の少なくとも一部を構成する1以上のタンパク質を含んでなり、そして
(b) 該プローブと該経路中のタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、該相互作用が生じれば、該プローブが代謝経路またはシグナル伝達経路を調節すること、あるいは、該経路が膜表面で起こることを示す、上記方法。
〔57〕カルモジュリンと結合する非天然タンパク質を製造する方法であって、下記の配列:
I/L-Q-X-X-K-K/X-G-B (配列番号1)
(ここで、Xは任意のアミノ酸であり、Bは塩基性アミノ酸である)
を含んでなる非天然タンパク質を製造することからなる上記方法。
〔58〕タンパク質における翻訳後修飾の有無を判定する方法であって、
(a) 翻訳後修飾と結合するプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 該プローブとタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置で該相互作用が生じれば、その位置のタンパク質が翻訳後修飾を有することを示す、上記方法。
〔59〕前記翻訳後修飾がメチル化、リン酸化、ビオチン化、アセチル化、ペグ化、グリコシル化、脂質修飾、ユビキチン化、またはSUMO化である、上記〔58〕に記載の方法。
〔60〕酵母細胞の培養物を調製する方法であって、
(a) 複数の酵母細胞を増殖培地中で、OD600が0.3〜1.0となるまで増殖させ、ここで、該複数の酵母細胞は調節配列に機能的に連結された異種ヌクレオチド配列を含むものであり、
(b) 該細胞を、該異種ヌクレオチド配列によりコードされるタンパク質の発現を増強する誘導物質と接触させ、
(c) 該細胞を該培地から分離し、
(d) 該細胞を冷水に接触させ、
(e) 該細胞を該冷水から分離し、
(f) 該細胞を冷溶解バッファーに接触させ、
(g) 該細胞を該溶解バッファーから分離し、そして
(h) 保存のために該細胞を半乾燥状態で凍結する、
各ステップを含んでなる上記方法。
〔61〕細胞からタンパク質を精製する方法であって、
(a) 複数の細胞サンプルのそれぞれについて、各サンプルの細胞を溶解して細胞溶解物を調製し、ここで、該細胞はアフィニティータグをもつ融合タンパク質を含み、また、該溶解ステップはペイントシェーカーを使って実施し、
(b) 各溶解物を可溶性画分と非可溶性画分とに分離し、
(c) 各可溶性画分をマルチサイトアレイの異なるサイトに移動させ、その際、該移動ステップはワイドオープンチップを使って実施し、
(d) 各可溶性画分を結合物質と接触させて、該融合タンパク質と結合物質との複合体が形成されるようにし、
(e) 該複合体から各融合タンパク質を単離し、そして
(f) 各融合タンパク質を高粘性のバッファー中に保存する、
各ステップを含んでなる上記方法。
〔62〕シグナルが陽性であるかどうかを判定する方法であって、
(a) 各スポットについて局所的にフォアグラウンドシグナルおよびバックグラウンドシグナルを測定し、該フォアグラウンドシグナルとバックグラウンドシグナルの差から正味のシグナルを決定し、
(b) 第1および第2の正味シグナル分布に関する下四分位値、中位値、および上四分位値を決定し、
(c) 第1の正味シグナル分布から第1の中位値を減じ、さらに第2の正味シグナル分布から第2の中位値を減じて、それぞれ第1および第2の減算値を得、
(d) 第1の減算値を、第1の正味シグナル分布に関する上四分位値と下四分位値との差で割り算し、さらに第2の減算値を、第2の正味シグナル分布の上四分位値と下四分位値との差で割り算して、それぞれ第1および第2の基準化された値を得、
(e) 近傍領域の基準化されたシグナル分布に関する局所中位値を算定し、ただし、該近傍領域はその領域内に複数のサイトを含んでなり、そして
(f) 基準化されたシグナルから局所中位値を減じて、基準化された過剰値を得る、
各ステップを含んでなる、上記方法。
〔63〕陽性シグナルがタンパク質−プローブ相互作用を示す、上記〔62〕に記載の方法。
〔64〕前記近傍領域が前記シグナルの上2列、下2列、左2列、および右2列である、上記〔62〕に記載の方法。
〔65〕サンプル値の1つとサンプル値の平均との差が基準化された過剰値の誤差の3標準偏差より大きい場合には、基準化された過剰値の並列サンプルを除外するステップをさらに含む、上記〔62〕に記載の方法。
〔66〕複数の異なるアレイを用いて測定したシグナルの中から陽性シグナルを確認する方法であって、
(a) 異なるアレイを用いて測定したシグナルを変換して変換シグナルを得、
(b) 各変換シグナルを、該変換シグナルから局所中位値シグナルを減じて修正された変換シグナルを得る方法によって修正し、ここで、該局所中位値シグナルは近傍領域におけるシグナルの中位値であり、該近傍領域は該変換シグナルのサイトのまわりの1以上のサイトを含んでなり、そして
(c) 修正された各変換シグナルを閾値と比較して、該修正された変換シグナルが該閾値より大きい場合には、該修正された変換シグナルを陽性と判定する、
各ステップを含んでなり、
ここで、該アレイは基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイからなるものである、上記方法。
〔67〕前記変換ステップが、
(a) 異なるアレイのそれぞれを用いて測定したシグナルについて、シグナル分布の下四分位値、中位値、および上四分位値を決定し、
(b) 異なるアレイのそれぞれを用いて測定したシグナルから該中位値を減じて、該アレイについての翻訳シグナルを得、
(c) 該翻訳シグナルを、該アレイの上四分位値と下四分位値との差で割り算することによって変換シグナルを得る、
各ステップを含んでなる、上記〔66〕に記載の方法。
〔68〕前記近傍領域が変換シグナルの上2列、下2列、左2列、および右2列の領域内の複数のサイトからなる、上記〔66〕に記載の方法。
〔69〕陽性シグナルがタンパク質−プローブ相互作用を示す、上記〔66〕、〔67〕または〔68〕に記載の方法。
〔70〕データポイントを捨てるステップをさらに含み、ただし、該データポイントはアレイ上の二重反復サイトで測定され、該二重反復サイト間の変化が3標準偏差より大きいものである、上記〔66〕、〔67〕または〔68〕に記載の方法。
〔71〕修正された変換シグナルを次式:
r+εr = G+εG/R+εR
(式中、Gは修正された変換シグナルであり、RはGSTシグナルであり、εGはGの誤差であり、εRはRの誤差であり、そしてεrはrの誤差である)
を用いて正規化するステップをさらに含む、上記〔66〕、〔67〕または〔68〕に記載の方法。
〔72〕マルチサイトアレイが96サイトアレイである、上記〔20〕に記載の方法。
〔73〕基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
〔74〕(a) 2つの二重反復スポットのシグナル値を平均して平均値を出し、(b) 該平均値が基準化された過剰値の誤差の3標準偏差より大きいかどうかを調べる、各ステップをさらに含んでなり、その際、該平均値が基準化された過剰値の誤差の3標準偏差より大きければ、該スポットのシグナルを陽性とする、上記〔62〕または〔65〕に記載の方法。
〔75〕タンパク質における酵素活性の有無を判定する方法であって、
(a) 該酵素活性の基質であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素活性をもつことを示す、上記方法。
〔76〕タンパク質における酵素基質の有無を判定する方法であって、
(a) 該酵素基質のための酵素であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素基質を含むことを示す、上記方法。
〔77〕前記タンパク質が基板にビオチンタグを介して結合される、上記〔1〕、〔3〕、〔4〕または〔5〕に記載のアレイ。
〔78〕第1タグが前記タンパク質のカルボキシ末端に存在し、第2タグが前記タンパク質のアミノ末端に存在する、上記〔15〕に記載の方法。
〔79〕アフィニティータグがビオチンである、上記〔22〕に記載の方法。
〔80〕前記基板がニトロセルロースをコーティングしたスライドガラスからなる、上記〔1、3、4または5〕に記載のアレイ。
【0012】
したがって、一つの実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%を包含する。
【0013】
別の実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、または50%を包含し、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。特定の実施形態では、複数のタンパク質は単一の種において発現される全タンパク質の少なくとも50%を包含し、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0014】
別の実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000のタンパク質を包含する。
【0015】
別の実施形態において、本発明は、各タンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを提供し、ここで、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0016】
さらなる実施形態において、前記タンパク質はタンパク質分類に従ってアレイ上に組織化される。こうした分類は、存在量、機能、機能上のクラス、酵素活性、相同性、タンパク質ファミリー、特定の代謝もしくはシグナル伝達経路との関連、関係した代謝もしくはシグナル伝達経路との関連、または翻訳後修飾によるものであってよい。
【0017】
別の実施形態において、本発明は、基板がガラス、セラミック、ニトロセルロース、無定形シリコンカーバイド、注型可能な酸化物、ポリイミド、ポリメチルメタクリレート、ポリスチレン、またはシリコーンエラストマーで構成されている、上記の位置指定可能なアレイを提供する。
【0018】
さらなる実施形態において、基板は複数のタンパク質を基板に結合させるのに役立つ物質を含んでなる。例えば、基板に、各タンパク質のアフィニティータグと結合する物質をコーティングすることができる。特定の実施形態では、基板がグルタチオンを含む。別の特定の実施形態では、基板のコーティングがニッケルまたはニトロセルロースを含んでなる。別の特定の実施形態では、基板のコーティングがグルタチオンとニッケルを含む。ある実施形態では、基板がニッケルをコーティングしたスライドガラスである。好ましい実施形態では、基板がニトロセルロースをコーティングしたスライドガラスである。タンパク質(およびDNA)マイクロアレイを作製するためのニトロセルロースコーティングスライドガラスは市販されている(例えば、Schleicher & Schuell社(Keene, NH)は、ニトロセルロース系のポリマーをコーティングしたスライドガラスを販売している(カタログ番号10 484 182))。特定の実施形態では、各タンパク質がOMNIGRIDTM (GeneMachines, San Carlos, CA)を用いてニトロセルロースコーティングスライドガラス上にスポットされる。
【0019】
プロテオームチップ上のタンパク質は、好ましくは、該タンパク質を精製したり、かつ/またはプロテオームチップに結合させたりするのに有用なアフィニティータグを少なくとも1つ含む融合タンパク質である。
【0020】
本発明はまた、プロテオームチップの作製方法を提供する。したがって、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの構築方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%を包含する。
【0021】
一つの実施形態において、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの作製方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、または50%を包含し、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0022】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの構築方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000のタンパク質を包含する。
【0023】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの作製方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0024】
本発明はさらに、ハイスループット分析に適合しやすく、容易に評価できるフォーマットで、ウイルス、原核生物または真核生物のタンパク質を調製して単離する方法を提供する。好ましい方法は、自動化技術に適合しうるアレイフォーマットでタンパク質を合成し精製することを含む。したがって、一実施形態において、本発明は、ウイルス、原核生物または真核生物のタンパク質を調製して単離する方法を提供し、該方法は、調節配列と機能的に連結された異種配列を含むベクターにより形質転換された真核細胞を増殖させ、該調節配列を、該異種配列によりコードされたタンパク質の発現を高める誘導物質と接触させ、該細胞を溶解し、該タンパク質を結合物質と接触させて該タンパク質と結合物質との複合体が形成されるようにし、該複合体を細胞破砕物から単離し、そして該タンパク質を該複合体から分離するステップを含んでなり、ここで、各ステップは96ウェルフォーマットで実施される。
【0025】
上記タンパク質は、異種配列が目的タンパク質のコード領域とアフィニティータグなどのタグをコードする配列とを含むような融合タンパク質であることが好ましい。かかるタグは、そのタンパク質をモニタリングしたり、融合タンパク質を細胞破砕物や混入試薬から分離したり、かつ/または該タンパク質を本発明のプロテオームチップに結合させたりするのに有用でありうる。
【0026】
本発明はさらに、複数の融合タンパク質を基板の表面に結合させるステップを含んでなる、位置指定可能なアレイの作製方法を提供し、ここで、各タンパク質は基板上の異なる位置に存在し、該タンパク質は第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである。特定の実施形態では、該タンパク質のアミノ末端には1つのタグが付いており、また、そのカルボキシ末端には異なる第2タグが付いている。他の実施形態では、該タンパク質のアミノ末端に2つのタグが付いているか、あるいはカルボキシ末端に2つのタグが付いている。さらに別の実施形態では、該タンパク質のアミノまたはカルボキシ末端以外の部位に1つ以上のタグが付いている。二重タグ付けされたタンパク質を使用することの利点としては、高度に精製されたタンパク質を得ることができるだけでなく、細胞破砕物からタンパク質を精製してそのタンパク質を基板に結合させる簡便な方法を提供できることにある。
【0027】
したがって、特定の実施形態においては、第1タグがグルタチオン-S-トランスフェラーゼタグ(GSTタグ)であり、第2タグがポリヒスチジンタグ(Hisタグ)である。別の実施形態では、GSTタグとHisタグをタンパク質のアミノ末端に結合させる。これとは別に、GSTタグとHisタグをタンパク質のカルボキシ末端に結合させることもできる。GSTタグおよびHisタグはタンパク質のアミノ末端またはカルボキシ末端のどちらに存在してもよい。特定の実施形態においては、GSTタグをタンパク質のアミノ末端に結合させ、Hisタグをカルボキシ末端に結合させる。その他の実施形態では、Hisタグをタンパク質のアミノ末端に結合させ、GSTタグをカルボキシ末端に結合させる。
【0028】
融合タンパク質のアフィニティータグの配置を変えると、機能的な二次構造、細胞外ドメインの適切な折りたたみ、ならびに、適切なタンパク質の輸送、局在化および/または分泌がもたらされる可能性がある。例えば、GSTタグおよびHisタグのカルボキシ末端への融合は、不適切な折りたたみまたは翻訳開始コドンの上流の領域が遮断される場合の発現を未然に防ぐことができる。
【0029】
本発明はさらに、タンパク質アレイを用いて脂質結合タンパク質をスクリーニングする方法を提供する。したがって、一実施形態において、本発明は、各タンパク質が基板の異なる位置にある、複数のタンパク質を含んでなる位置指定可能なアレイの使用方法であり、該方法は、プローブをアレイに接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなり、ここで、該プローブは脂質を含むものである。特定の実施形態では、かかる脂質はリン脂質を含み、例えば、ホスファチジルコリンおよびホスファチジルイノシトールであるが、これらに限らない。別の特定の実施形態では、プローブは対象のリン脂質を含むリポソームの形態をしている。
【0030】
さらに、プロテオームチップを使用する方法も提供される。本発明のプロテオームチップを使用すると、1つの種または細胞における本質的に全てのタンパク質−タンパク質相互作用をアッセイすることができる。また、試験化合物と相互作用する1つの種の全タンパク質を体系的にアッセイするために、プロテオームチップを使用することも可能である。したがって、本発明のプロテオームチップを使用すると、多数の活性をアッセイして豊富な情報を手に入れることができる。かかる情報として、制限するものではないが、刺激に応答した細胞もしくは生物の「フィンガープリント」(指紋)もしくは「サイン」を規定すること、対象のプローブと相互作用する1つの種における全タンパク質を特徴づけること、対象のプローブと相互作用する2以上の種における全タンパク質を特徴づけること、生物学的経路(例えば、代謝もしくはシグナル伝達経路)または関連した生物学的経路に関与する全タンパク質を特徴づけること、対象の酵素活性(例えば、キナーゼ活性、プロテアーゼ活性、ホスファターゼ活性、グリコシダーゼ、アセチラーゼ活性、およびその他の化学基転移酵素活性)を有する1つの種における全タンパク質を特徴づけること、対象の翻訳後修飾を伴う1つの種における全タンパク質を特徴づけること、および薬物標的を同定することがある。特定の実施形態においては、本発明のプロテオームチップを用いて、対象の薬物または候補薬物と相互作用する1つの種における全タンパク質(例えば、薬物標的)を特徴づける。
【0031】
かくして、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は単一の種における既知遺伝子の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%によりコードされる少なくとも1つのタンパク質を含むものである。
【0032】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、または99%を含んでなり、その際、タンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0033】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000の既知タンパク質を含むものである。
【0034】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は全体として、単一の種における少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000または50000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0035】
別の実施形態において、本発明は、プローブを、複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該複数のタンパク質は全体として、少なくとも2つの種における少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000または50000の異なる既知遺伝子によりコードされるタンパク質を包含する。
【0036】
別の実施形態において、本発明は、プローブを、複数の融合タンパク質を含んでなる位置指定可能なアレイと接触させ、タンパク質−プローブ相互作用を検出するステップを含んでなる結合タンパク質の検出方法を包含し、上記アレイにおいて、各タンパク質は基板上の異なる位置にあり、該融合タンパク質は第1タグ、第2タグ、および生物のゲノム核酸によりコードされるタンパク質配列を含むものである。先に記載したように、特定の実施形態では、2つのタグはHisおよびGSTでありうる。
【0037】
本発明はさらに、結合アッセイにおいて使用するためのタンパク質の標識方法を提供し、該方法は、該タンパク質の個々のアリコートを、ビオチン転移化合物と、アリコートごとに異なる程度にビオチン化されたタンパク質を生成する条件下および時間にわたり接触させ、個々のアリコートを一緒に合わせて、異なる程度にビオチン化されたタンパク質のサンプルを得るステップを含んでなる。
【0038】
本発明はまた、結合タンパク質の検出方法を提供し、該方法は、上記方法により得られたビオチン化タンパク質のサンプルを、複数のタンパク質(各タンパク質が基板上の異なる位置にある)を含んでなる位置指定可能なアレイと接触させ、アレイタンパク質−プローブ相互作用(ビオチン化タンパク質とアレイ上のタンパク質との相互作用が起こる)の位置を検出するステップを含んでなる。
【0039】
本発明はまた、結合タンパク質の検出方法を提供し、該方法は、上記方法により得られたビオチン化タンパク質のサンプルを、複数のタンパク質(各タンパク質が基板上の異なる位置にある)を含んでなる位置指定可能なアレイと接触させ、該アレイを蛍光物質にコンジュゲートさせたストレプトアビジンと接触させ、蛍光を発するアレイ上の位置を検出するステップを含んでなり、その際、蛍光が発せられれば、ビオチン化タンパク質とアレイ上のタンパク質との間で相互作用が起こったことを示す。
【0040】
本発明はさらに、シグナルが陽性であるかどうかを確認する方法を提供する。したがって、本発明の一実施形態は、タンパク質マイクロアレイを用いるアッセイで得られたシグナルが陽性である(相互作用物質へのプローブの結合を示す)かどうかを確認する方法を提供する。
【0041】
3.1 定義
本明細書中で用いる「タンパク質」という語は、全長タンパク質、タンパク質の一部分、またはペプチドをさす。タンパク質はより大きなタンパク質の断片化によって生成することも、化学合成することもできる。好ましくは、細菌、酵母、昆虫細胞、哺乳動物細胞など(しかし、これらに限らない)の種における組換え体の過剰発現によりタンパク質を調製する。本発明のタンパク質マイクロアレイに配置されるタンパク質は融合タンパク質であることが好ましく、精製および/または固定化を助けるアフィニティータグを少なくとも1つもつものがより好ましい。
【0042】
本明細書中で用いる「相互作用物質」(interactor)という語は、プローブと相互作用するタンパク質マイクロアレイ上のタンパク質をさす。
【0043】
本明細書中で用いる「プローブ」という語は、マイクロアレイのタンパク質との相互作用に関してアッセイするために本発明のタンパク質マイクロアレイに適用することができる、タンパク質、核酸(例えば、DNA、RNA、オリゴヌクレオチド、ポリヌクレオチド)、小分子、基質、インヒビター、薬物もしくは候補薬物、受容体、抗原、ホルモン、ステロイド、脂質、リン脂質、リポソーム、抗体、補因子、サイトカイン、グルタチオン、免疫グロブリンドメイン、炭水化物、マルトース、ニッケル、ジヒドロトリプシン、カルモジュリン、ビオチン、レクチン、および重金属など(しかし、これらに限らない)のいずれかの化学試薬をさす。
【0044】
4. 図面の説明
図1A〜1B. プロテインチップ技術を用いた酵母プロテオーム解析の手順。
【0045】
A. 酵母ORFを、二重タグ付けされた酵母GAL1発現ベクターに組換え法によりクローニングし、配列決定により正体を確認した。次に、純粋なプラスミド構築物のそれぞれを、大規模タンパク質精製用の酵母株に再導入した。酵母培養物を96ウェルフォーマットで増殖させ、ガラクトースを添加して誘導した。ハイスループット精製ステップの後、精製されたタンパク質をアリコートに分けて、プリンティングに先立ってグリセロールバッファー中に-80℃で保存した。高精度のマイクロアレイヤー(microarrayer)を用いると、1つの実験で80枚のスライドガラス上に6566のタンパク質サンプルを二重反復してスポットすることができる。
【0046】
B. 3mlの酵母培養物から精製されたタンパク質のイムノブロット分析。
【0047】
図2A〜2C. 抗GST抗体を用いてプローブしたプロテオームチップ。
【0048】
60のサンプルを、抗GST抗体を用いるイムノブロット分析により検査した;19の代表的な例を示してある。80%を超える調製物から高収量の融合タンパク質が得られる。
【0049】
A. GST::酵母融合タンパク質を96ウェルフォーマットで精製した。5800のユニークなタンパク質を代表する6566のタンパク質サンプルを、1枚のニッケルコーティングを施した顕微鏡用スライドガラスの上に二重反復してスポットした。このスライドガラスを抗GST抗体でプローブした。
【0050】
B. 48ブロックのうちの1つの拡大図をプロテオームチップの右側に示す。文字は二重反復タンパク質サンプルを示し、数字は起源プレート番号を表す。すぐれた分解能と高いシグナル対ノイズ比に留意されたい。
【0051】
C. 各ORF産物の二重反復スポット間の誤差分布。εの単位はAxonスキャナーにより測定したシグナル強度である。GST対照により測定されるように、10単位はおよそ32fgのタンパク質に等しい。ε分布の中心は1.2単位であり、95%のサンプルは中心から10単位以内のところにある。
【0052】
図3A〜3B. プロテオームチップでの様々なアッセイの例。
【0053】
A. 6566の酵母タンパク質を含むプロテオームチップは二重反復してスポットし、図示したビオチン化プローブと共にインキュベートした。陽性の相互作用物質(ボックスで示す)を6つのタンパク質−リポソーム相互作用アッセイ(「PI(3)P」、「PI(4,5)P2」、「PI(4)P」、「PI(3,4)P2」、「PI(3,4,5)P3」、「PC」)、カルモジュリン結合アッセイ(「カルモジュリン」)、およびDNA−タンパク質相互作用アッセイ(「ゲノムDNA」)で同定した。各ブロックは16X18のタンパク質スポットを含む。二重反復陽性シグナルは下のパネルに水平な対として現れる。二重反復スポッティングは内部対照として役立ち、これはシグナルがバックグラウンドに対して弱いときに重要である。上のパネルは、抗GST抗体(「α-GST」)でプローブした対照プロテオームチップの同じ酵母タンパク質調製物を示す。図面から実証されるように、比較的低レベルのGST融合タンパク質(「プローブ」)を含有するサンプルにおいて強いシグナルがしばしば観察されており、このことは、結合が高感度かつ特異的であることを示している。PI、ホスファチジルイノシトール。PC、ホスファチジルコリン。
【0054】
B. 様々なカルモジュリン標的が共有するアミノ酸配列をサーチすることにより同定された推定上のカルモジュリン結合モチーフ(Zhuら 2000, Nat. Genet. 26:283)。39の陽性タンパク質のうち14は、コンセンサスがI/L-Q-X-K-K/X-G-B (配列番号1)(ここで、Xは任意の残基であり、Bは塩基性残基である)であるモチーフを共有している。アラインメントの上に示した文字の大きさは、表示したアミノ酸の相対頻度を示す。YFL003C/MSH4 (配列番号2); YJR073C/OPI3 (配列番号3); YBR050C/REG2 (配列番号4); YNL202W/SPS19 (配列番号5); YOL016C/CMK2 (配列番号6); BR011C/IPP1 (配列番号7); YGR034W/RPL26B (配列番号8);YFR004W/RPN11 (配列番号9); YIL021W/RPB3 (配列番号10); YGL063W/PUS2 (配列番号11); YDR292C/SRP101 (配列番号12); YFR014C/CMK1 (配列番号13); YBR213W/MET8 (配列番号14); YAL029C/MY04 (配列番号15)。
【0055】
図4A〜4D. ホスファチジルイノシトール結合タンパク質の分析。150の陽性タンパク質のホスファチジルイノシトール(「PI」)結合特異性を調べるために、それらの結合シグナルをホスファチジルコリン(「PC」)の対応する結合シグナルに対して正規化した。比率(「PI/PC」)に基づいて、タンパク質を4つのカテゴリーに分類した。すなわち、(A) 30の強くかつ特異的、(B) 43の強くかつ非特異的、(C) 19の弱くかつ特異的、および、(D) 58の弱くかつ非特異的、なホスファチジルイノシトール結合タンパク質である。強度はPI/PCシグナル比(図面のスケールで示される)を表す。PI/PC結合比の右に「10n」と表記される欄は、最大結合シグナル強度(白抜きのボックス)およびその信頼区間(水平の実線)を示す。数字はその値のlogを示す。信頼区間欄の右側の3つの欄のボックスは、それぞれ膜結合タンパク質(「膜」)、キナーゼ(「キナーゼ」)、および特性決定されなかったORF(「未知」)を示す。
【0056】
図5A〜5C. プロテオームマイクロアレイにより検出されたタンパク質−脂質相互作用を慣用方法で確認する(Casamayorら, 1999, Curr. Biol. 9: 186 ; Guerraら, 2000, Biosci. Rep. 20: 41)。
【0057】
A. 最初に、PI(4,5)P2リポソームを、BSAでブロックしたニトロセルロース膜に付着させた。Rim15p、Eno2pおよびHxk1p、ならびにGST対照の連続希釈物を用いて膜をプローブした。結合したタンパク質は抗GST抗体および増強ケミルミネッセンス(「ECL」)キットを用いて検出した。
【0058】
B. タンパク質−脂質相互作用を試験するためにリバースアッセイを実施した。タンパク質を調製して連続希釈でニトロセルロースフィルターにスポットし、6種のリポソームでプローブした。対照として、6種のリポソームもBSAブロック膜に添加した。十分な洗浄後、結合したリポソームを、HRPコンジュゲート化ストレプトアビジンおよびECLキットを用いて検出した。
【0059】
C. 膜アッセイにおける結合シグナルとRim15pの量との線形相関。膜アッセイ(図4B)からのRim15pのリポソーム結合シグナルを、スポットしたRim15pの濃度勾配に対してプロットしたとき、PI(4)P、PI(3,4)P2、PI(3)P、およびPI(4,5)P2の結合シグナルはRim15pの量と線形に相関した。PI(4)PとRim15pの相互作用は最高の親和性を示し、対照PCとRim15pとの親和性より少なくとも3倍高かった。
【0060】
5. 発明の詳細な説明
本発明は、本発明者らが酵母で発現される全タンパク質の約80%を含有する酵母プロテオームマイクロアレイを構築したことに、部分的に基づいており、種にかかわらずプロテオームアレイに関する最初の記述となる。本発明のプロテオームチップは、ある1つの種においてタンパク質の相互作用および活性を包括的に分析するために使用することができる。プロテオームチップの利用は、既存のアプローチよりも顕著な利点を有する。本明細書で提示されるプロテオームマイクロアレイ技術の利点の1つは、個々のタンパク質からなる大きなセットを、何百もの生化学的活性について、あるいは何千でさえ、同時に、直接、ハイスループット様式でスクリーニングすることができることである。例えば、プロテオームチップ法の利点は、タンパク質-薬物相互作用(例えば、薬物標的-薬物相互作用)、タンパク質-脂質相互作用および酵素的アッセイを包含するがそれに限定されない多種多様な活性についてタンパク質を直接in vitroでスクリーニングできることである。それに加えて、広範囲のin vitro条件を容易に試験することができる。さらに、ひとたびタンパク質が調製されれば、プロテオームスクリーニングは極めて速く、安価であり、さらに、多くのマイクロアレイフォーマットによる迅速なデータ分析は、既存の装置および分析用ソフトウェアに適合する。
【0061】
5.1 プロテオームアレイ
本発明は、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は、ある単一の種における既知遺伝子のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%によってコードされる、少なくとも1つのタンパク質を含んでなる。すなわち、1つの遺伝子に由来するすべてのタンパク質アイソフォームおよびスプライス変異体は1つのタンパク質とみなされる。
【0062】
本発明は、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は、単一の種において発現される全タンパク質のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%を含んでなり、この場合、タンパク質アイソフォームおよびスプライス変異体は単一タンパク質とみなされる。ある実施形態において、前記の複数のタンパク質は、ある種において発現される全タンパク質の約90%、95%または99%を含んでなる。特定の実施形態において、この複数のタンパク質はある種において発現される全タンパク質の約93.5%を含んでなる。
【0063】
本発明はまた、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は、単一の種において発現される、少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000または1,000,000のタンパク質を含んでなる。
【0064】
本発明はまた、それぞれのタンパク質が基板上の異なる位置に存在する、複数のタンパク質を含んでなる位置指定可能なアレイを包含するが、ここで、前記の複数のタンパク質は全体として、単一の種における、少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000、または50000の異なる既知の遺伝子を含んでなる。
【0065】
本発明はまた、それぞれの融合タンパク質が基板上の異なる位置に存在する、基板表面に融合した複数の融合タンパク質を含んでなる位置指定可能なアレイを包含するが、ここでこの融合タンパク質は、第1タグ、第2タグ、およびある生物のゲノム核酸によってコードされるタンパク質配列を含んでなる。別の実施形態において、前記融合タンパク質のタンパク質配列は、生物のゲノム核酸にコードされる必要はないが、結合タンパク質の機能および/または活性を特定することが必要とされる配列である。
【0066】
位置指定可能なアレイは、対象とする各プローブもしくはタンパク質が基板上の既知の場所に存在するような配置を提供し、したがって、各プローブもしくはタンパク質の正体をアレイ上のその位置から決定することが可能である。したがって、アレイ上のそれぞれのタンパク質は、各タンパク質の正体を基板上のその位置から決定することができるように、基板上の既知の所定の場所にあることが望ましい。
【0067】
ある実施形態において、前記の種はウイルスである。別の実施形態において、前記の種とは原核生物である。また別の実施形態において、前記の種は真核生物である。別の実施形態において、前記の種は脊椎動物である。さらにまた別の実施形態において前記の種は、哺乳動物である。特定の実施形態において、前記の種は、動物であって、昆虫、霊長類、および齧歯類を含めるがこれらに限定されない。特定の実施形態において、前記の種はサル、ショウジョウバエ、ウシ、ウマ、ヒツジ、ブタ、ニワトリ、シチメンチョウ、ウズラ、ネコ、イヌ、マウス、ラット、ウサギ、線虫もしくは魚である。望ましい実施形態において、前記の種はヒトである。別の望ましい実施形態において、前記の種は酵母である。
【0068】
本発明のプロテオームチップのタンパク質は、全長タンパク質、全長タンパク質の一部分、およびペプチドを包含するが、これらを組換え体の過剰発現、大きいタンパク質の断片化、もしくは化学合成によって調製することができる。例えば酵母、細菌、昆虫、ヒト、もしくはヒト以外の哺乳動物、例えばマウス、ラット、ネコ、イヌ、ブタ、ウシ、およびウマ、に由来する細胞においてタンパク質を過剰発現させることができる。さらに、天然の、または合成のタンパク質に連結された規定のドメインを含んでなる融合タンパク質を使用することができる。プロテオームチップのタンパク質を、チップの基板に付着させる前に、精製することができる。また、プロテオームチップのタンパク質をプロテオームチップに付着させている間にこれを精製したり、さらに追加して精製したりしてもよい。
【0069】
プロテインチップに付着させる前、または付着させるときにタンパク質を人工膜もしくは天然の膜(例えば、リポソーム、膜小胞)に埋め込むことができる。実際、特定のタンパク質合成を人工膜もしくは天然の膜の存在下で行うことが望ましく、例えばタンパク質の折りたたみ、タンパク質のプロセシングを促進し、活性を維持し、および/またはタンパク質の沈澱を防止することができる。
【0070】
さらに、タンパク質をプロテオームチップの基板に付着させることができる。あるいはまた、タンパク質をプロテオームチップのウェル内に送達することができるが、この場合、このタンパク質はプロテオームチップの基板には結合しない状態で存在する。
【0071】
本発明は、また、本発明のプロテオームチップのための基板として有用な、化合物に関する。例えば、シリコン、ガラス、石英、ポリイミド、アクリル、ポリメタクリル酸メチル(LUCITE(登録商標))、セラミック、ニトロセルロース、無定形シリコンカーバイド、ポリスチレン、および/またはマイクロファブリケーション、マイクロリソグラフィーもしくはキャスティングに適したあらゆる他の材料、といった材料から基板を作製することができるが、これらに限定されない。例えば、基板は、親水性マイクロタイタープレート(例、MILLIPORETM)もしくはニトロセルロースコーティングスライドガラスとすることができる。好ましい実施形態において、基板はニトロセルロースコーティングスライドガラスである。プロテイン(およびDNA)マイクロアレイを作製するためのニトロセルロースコーティングスライドガラスは、市販されており、入手可能である(例えば、Schleicher & Schuell(Keene, NH)から、この会社はニトロセルロース系のポリマーでコーティングされたスライドガラス(カタログ番号10 484 182)を販売する)。特定の実施形態において、それぞれのタンパク質を、OMNIGRIDTM(GeneMachines, San Carlos, CA)を用いてニトロセルロースコーティングスライドガラス上にスポットする。本発明は、プロテインチップを作製するために有用な他の基板を検討するが、これらの一部は例えば、2001年5月4日出願の同時係属中の米国特許出願第09/849,781号において開示されており、この出願はその全体を参考として本明細書に含めるものとする。
【0072】
特定の実施形態において、基板はシリコーンエラストマー素材(例えばポリジメチルシロキサン(”PDMS”)であるが、それに限らない)を含んでなる。シリコーンエラストマー素材の利点はその柔軟性である。
【0073】
もう1つの特定の実施形態において、基板はシリコンウェハーである。シリコンウェハーは、パターンを形成してエッチングすることができる(例えば、G. Kovacs, 1998, Micromachined Transducers Sourcebook, Academic Press; M. Madou, 1997, Fundamentals of Microfabrication, CRC Pressを参照されたい)。エッチングされたウェハーを用いて、本発明のプロテオームチップを成形することができる。
【0074】
ある実施形態において、本発明は、スライドガラスのような、ただしそれに限定されないが、平らな表面を有する基板を含んでなるプロテオームチップを提供する。高密度プロテインアレイを、例えばスライドガラス上に作成することができるが、その結果、タンパク質の存在、量、および/または機能性に関するアッセイをハイスループット方式で行うことができる。
【0075】
したがって、ある実施形態において、プロテオームチップは基板表面に付着させた複数のタンパク質を含んでなるが、ここでタンパク質が付けられるサイトの密度は、少なくとも100サイト/cm2、10,000サイト/cm2、1,000,00サイト/cm2、10,000,000サイト/cm2、25,000,000サイト/cm2、10,000,000,000サイト/cm2、または10,000,000,000,000サイト/cm2である。個々のタンパク質試料をそれぞれチップ上の別々のサイトに付着させることが望ましい。チップ上のそれぞれの位置にあるタンパク質の正体は既知である。
【0076】
別の実施形態において、基板は整列したウェルを有する。マイクロリソグラフィー技術およびマイクロマシン加工技術(例えば、同時係属中の米国出願第09/849,781号、2001年5月4日出願、これは参考としてその全体を本明細書に含めるものとする)によって、数100ミクロンから100nmもしくはさらに小さいものにまで及ぶ種々多様な寸法で、深さは同じ程度の、整列したウェルを製造することができる。ある実施形態において、シリコンウェハーをマイクロマシンで微細加工して、マスター鋳型として使用し、直径400μmのウェルを200μm離した間隔をおいて成形するが、ウェルの密度は約277ウェル/cm2で、個々のウェルの容積は深さ100μmのウェルについて約30nlである。
【0077】
別の実施形態において、マイクロリソグラフィーによる微細加工によって、直径500nmおよび275nmのウェルを1μmの間隔をおいて作製し、その結果ウェル密度はそれぞれ4400万および6100万ウェル/cm2以上となる。直径が小さいことはもちろん、間隔が狭いほど、密度を高くすることができる。
【0078】
また別の実施形態において、精密マイクロレーザー加工技術によって、アクリルを材料として、1.5mmを超えるものから下は500μmまでの範囲の大きさで、約500μm間隔のウェルを有する鋳型構造を直接製作することができる。このようなウェルの容積は50〜500nlの範囲である。
【0079】
したがって、ある実施形態において、プロテオームチップは基板の表面上に複数のウェルを含んでなり、ここで、ウェルの密度は少なくとも100ウェル/cm2、1000ウェル/cm2、10,000ウェル/cm2、100,000ウェル/cm2、1,000,000ウェル/cm2、10,000,000ウェル/cm2、25,000,000ウェル/cm2、10,000,000,000ウェル/cm2、または10,000,000,000,000ウェル/cm2である。本発明は複数のウェルを含んでなるプロテインチップの変更を意図するが、これらは例えば同時係属中の米国出願第09/849,781号、出願日2001年5月4日、において開示されており、これは参考としてその全体を本明細書に含めるのもとする。
【0080】
また、本発明は、プロテオームチップにおいてウェルの形、幅対深さの比、および容積が変化したものを包含するが、これらは例えば同時係属中の米国出願第09/849,781号、出願日2001年5月4日、において開示されており、これは参考としてその全体を本明細書に含めるのもとする。このような形としては、円、楕円、長方形、正方形などが挙げられるがそれに限定されない。こうしたウェルの底面は、例えば、方形、丸みのあるV型、またはU型とすることもできる。
【0081】
ある実施形態において、基板は金を含んでなる。好ましい実施形態において、基板は金をコーティングしたスライドガラスを含んでなる。別の実施形態において、基板はニッケルを含んでなる。別の好ましい実施形態において、基板はニッケルコーティングしたスライドガラスを含んでなる。ニッケルを含んでなる基板はポリヒスチジンタグ(「Hisタグ」)を有する融合タンパク質の精製および付着に有利である。もう1つの実施形態において、基板はニトロセルロースを含んでなる。さらに別の好ましい実施形態において、基板はニトロセルロースをコーティングしたスライドガラスを含んでなる。
【0082】
本発明はさらに、プロテオームチップ基板の誘導体化に有用な化合物に関する。タンパク質を直接基板に結合することができるが、リンカー分子または化合物を介して基板に付着させることもできる。このリンカーは、基板の表面を誘導体化して基板表面にタンパク質が付着しやすくする、あらゆる分子もしくは化合物とすることができる。リンカーは、共有結合もしくは非共有結合によって、タンパク質またはプローブを基板表面に結合させることができる。さらに、リンカーは無機または有機分子とすることができる。ある実施形態において、リンカーはシラン、例えばシアノシラン、チオシラン、アミノシランなどとすることができる。本発明はプロテインチップの誘導体化に有用な化合物を意図するが、その一部は、例えば同時係属中の米国出願第09/849,781号、出願日2001年5月4日、において開示されており、これは参考としてその全体を本明細書に含めるのもとする。
【0083】
したがって、ある実施形態において、プロテオームチップのタンパク質は非共有結合によって(例えば吸着によって)基板に結合する。非共有結合によって基板に結合するタンパク質を、例えば、水素結合、ファンデルワールス結合、静電結合、もしくは金属-キレート配位結合のような様々な分子相互作用によって、基板の表面に付着させることができる。特定の実施形態において、タンパク質はポリリシンでコーティングされた基板の表面に結合する。さらに、上記のように、ある実施形態において、タンパク質はシラン(シアノシラン、チオシラン、アミノシランなど)でコーティングされた基板表面に結合する。
【0084】
その他に、当技術分野でよく知られた架橋化合物、例えば、同一の、または異なる官能基の架橋化合物(例、ビス[スルホスクシンイミジル]スベリン酸、N-[ガンマ-マレイミドブチルオキシル]スクシンイミド エステル、もしくは1-エチル-3-[3-ジメチルアミノプロピル]カルボジイミド)を用いて、共有結合性または非共有結合性相互作用によってタンパク質を基板に付けることができる。
【0085】
別の実施形態において、プロテオームチップのタンパク質は共有結合によって基板に結合する。例えば、タンパク質は、受容体-リガンド相互作用によって基板に結合することができるが、この相互作用には、抗体と抗原、DNA結合タンパク質とDNA、酵素と基質、アビジン(もしくはストレプトアビジン)とビオチン(もしくはビオチン化分子)の間の相互作用、および脂質結合タンパク質とリン脂質(または、リン脂質を含んでなる、膜、小胞、もしくはリポソーム)の間の相互作用を含める。
【0086】
当業界において公知の様々な方法によって、精製されたタンパク質をアレイ上に置くことができる。ある実施形態において、タンパク質を基板上にプリントすることができる。もう1つの実施形態においては、タンパク質をアフィニティータグによって基板に付着させる。タンパク質の精製に用いたものと異なるアフィニティータグを使用することが好ましいが、これはプロテインアレイを作るときにさらなる精製が達成されるためである。
【0087】
したがって、好ましい実施形態において、プロテオームチップのタンパク質は、基板表面に付着する化合物に親和性を示す、少なくとも1つの異種ドメインを含有する融合タンパク質として発現される。融合タンパク質を基板上に結合するために役立つ適当な化合物(すなわち結合パートナーとして機能する)には、トリプシン/アンヒドロトリプシン、グルタチオン、免疫グロブリンドメイン、マルトース、ニッケル、またはビオチンおよびその誘導体があるが、それらに限定されない。前記化合物が結合するのは、それぞれ、ウシ膵臓トリプシンインヒビター、グルタチオン-S-トランスフェラーゼ、プロテインAもしくは抗原、マルトース結合タンパク質、ポリヒスチジン(例えば、HisX6タグ)およびアビジン/ストレプトアビジンである。例えば、プロテインA、プロテインGおよびプロテインA/Gは、哺乳類の免疫グロブリン分子、特にIgG、のFc部分に結合できるタンパク質である。前記タンパク質を例えばSepharose(登録商標)支持体に共有結合で連結して、Fcドメインを含んでなるタグを有する融合タンパク質を精製する効率的な方法を提供することができる。
【0088】
別の実施形態では、タンパク質を基板に直接結合させる。さらに別の実施形態では、タンパク質をリンカーによって基板に結合させる。特定の実施形態では、タンパク質をHisタグを介して基板に付着させる。別の特定の実施形態では、タンパク質を3-グリシドオキシプロピルトリメトキシシラン(”GPTS”)リンカーを介して基板に付着させる。特定の実施形態では、タンパク質をHisタグを介して基板に結合させるが、ここで基板は平坦な表面を含んでなる。好ましい実施形態では、タンパク質をHisタグを介して基板に結合させるが、ここで、基板はニッケル-コーティングされたスライドガラスからなる。
【0089】
本発明のプロテオームチップは、その物理的大きさに限定されず、使用できるあらゆる大きさをとることができる。プロテオームチップは、自動化技術に適合したアレイフォーマットを有することが望ましく、それによって迅速なデータ分析が可能となる。したがって、ある実施形態において、プロテオームマイクロアレイフォーマットは、実験装置および/または分析用ソフトウェアに適合する。好ましい実施形態において、プロテオームチップは標準の顕微鏡スライドガラスの大きさである。別の好ましい実施形態において、プロテインチップは質量分析計の試料チャンバーにはまるようにデザインされる。
【0090】
5.2 高密度アレイフォーマットにおいてタンパク質を調製し精製する方法
本発明は、ウイルス、原核生物もしくは真核生物のタンパク質を、ハイスループット分析に供しやすく、容易に測定できるフォーマットで作製し、単離する方法に関する。好ましい方法は、自動化技術に適合したアレイフォーマットで、タンパク質を合成し、精製することを包含する。したがって、ある実施形態において、本発明は、適切に機能するように調節配列に連結された異種配列を有するベクターで形質転換された真核細胞を増殖させるステップ、異種配列によってコードされたタンパク質の発現を増強する誘導物質と前記調節配列を接触させるステップ、細胞を溶解するステップ、結合物質を前記タンパク質と接触させてタンパク質と結合物質との複合体を形成させるステップ、細胞破砕物から前記複合体を分離するステップ、およびこの複合体からタンパク質を単離するステップを含んでなり、これらの各ステップが96-ウェルフォーマットにおいて実施される、真核生物タンパク質を調製し単離する方法を提供する。
【0091】
ある実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は、単一の種における既知遺伝子の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%によってコードされる、少なくとも1つのタンパク質を含んでなる。
【0092】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は、単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%を含んでなる。ここでタンパク質のアイソフォームおよびスプライス変異体は単一タンパク質とみなされる。
【0093】
もう1つの実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は、単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000もしくは1,000,000のタンパク質を含んでなる。
【0094】
さらに別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記の複数のタンパク質は全体として、単一の種における少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、5000、10000、20000、30000、40000、もしくは50000の異なる既知の遺伝子を含んでなる。
【0095】
また別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップを含んでなり、それぞれのタンパク質が前記基板上の異なる位置に存在する、位置指定可能なアレイを作製する方法を提供するが、ここで前記タンパク質は、第1タグ、第2タグ、および、生物のゲノム核酸によってコードされるタンパク質を含んでなる融合タンパク質である。
【0096】
ある実施形態において、合成および精製手順の各ステップは、迅速な自動化を行いやすいアレイにおいて実施される。このようなアレイは、基板表面上の複数のウェルを含んでなるが、ここでウェルの密度は、例えば、少なくとも10、20、30、40、50、100、1000、10,000、100,000、もしくは1,000,000ウェル/cm2である。あるいはまた、こうしたアレイは、基板表面上の複数のサイトを含んでなり、こうしたサイトの密度は、例えば、少なくとも10、20、30、40、50、100、1000、10,000、100,000、もしくは1,000,000サイト/cm2である。
【0097】
特別の実施形態において、真核生物のタンパク質は、96アレイフォーマット(すなわち、プロセシングが起こる基板上の各サイトは96サイトのうちの1つである)、例えば96ウェルマイクロタイタープレートで調製され、精製される。好ましい実施形態において、基板はタンパク質と結合しない(例えば、タンパク質非結合性マイクロタイタープレート)。
【0098】
特定の実施形態において、タンパク質は当技術分野で広く知られている方法にしたがって、in vitro翻訳によって合成される。
【0099】
タンパク質合成を進める誘導プロモーターを有するあらゆる発現構築物を、本発明の方法にしたがって使用することができる。発現構築物は、形質転換に使用すべき細胞の種類に合わせて作製することが望ましい。発現構築物と宿主細胞との適合性は当業界で知られており、その変異体を使用することも本発明に包含される。
【0100】
培養下で増殖可能な宿主細胞を用いて、対象のタンパク質を合成することができる。対象のタンパク質を過剰生産することが可能で、結果として、タンパク質の適正な合成、折りたたみ、および翻訳後修飾をもたらす宿主細胞を使用することが望ましい。こうしたタンパク質のプロセシングによって、in vivoに対応するin vitro分子相互作用を特徴づけるアッセイに有用なエピトープ、活性部位、結合部位などが形成されることが望ましい。
【0101】
したがって、真核細胞(例えば、酵母、ヒト細胞)を用いて真核生物タンパク質を合成することが望ましい。さらに、安定な形質転換がしやすく、対象の形質転換体を含有する細胞を同定および単離するための選択マーカーを有する真核細胞が好ましい。あるいはまた、ある遺伝子産物を欠損した真核宿主細胞を、その欠損を補完する発現構築物で形質転換する。改変されたウイルス、原核生物、もしくは真核生物タンパク質の発現に好都合な細胞は当業界で公知であり、そうした細胞の変異体を当業者であれば正しく認識することができる。
【0102】
例えば、Invitrogen社(Carlsbad, CA, カタログ番号K800-01)製のInsectSelect系は、非細胞溶解性、単一ベクター昆虫発現系であって、高品質のタンパク質の発現を単純化し、ウイルスストックを調製および増幅する必要のない系であるが、これを使用することができる。このシステムにおいて好ましいベクターは、pIB/V5-His TOPO TAベクター(カタログ番号K980-20)である。製造業者の記載した手順によって、ポリメラーゼ連鎖反応法(“PCR”)産物を直接このベクターにクローニングすることが可能であり、さらに発現されたタンパク質の精製に役立つN-末端ヒスチジンタグを有するタンパク質を発現させることができる。
【0103】
昆虫細胞における別の真核生物発現系として、BAC-TO-BACTM系(LIFETECHTM, Rockville, MD)を用いることもできる。相同的組換えを利用するのではなくて、BAC-TO-BACTM系は、大腸菌における部位特異的遺伝子転位に基づいて、組換えバキュロウイルスを生成する。遺伝子発現は非常に強力なポリヘドリンプロモーターによって駆動されるので、感染した昆虫細胞の細胞タンパク質の25%にまで相当することがある。
【0104】
特定の実施形態においては、酵母培養物を用いて真核生物融合タンパク質を合成する。タンパク質合成を効率よく誘導するために、特に小量の培地で実施する場合には、新しい培養物を使用することが望ましい。また、酵母培養物が増殖しすぎないよう注意することが望ましい。さらに、約3ml以下の酵母培養物が精製に十分なタンパク質を生産することが望ましい。培養物の通気を改善するために、全体量をいくつかの小量に分けることができる(例えば、4本の0.75ml培養物を調製して、全体量を3mlとすることができる)。
【0105】
次に、細胞を誘導物質(例えば、ガラクトース)と接触させ、その後回収する。誘導された細胞をそれぞれ、冷水(すなわち、4℃から約15℃まで)で洗浄して、これ以上の細胞増殖を止めた後、冷(すなわち、4℃から約15℃まで)細胞溶解バッファーで洗浄して培地を除去し、誘導した細胞をタンパク質精製のためにプレコンディショニングする。タンパク質精製の前に、誘導細胞を冷凍保存して、タンパク質の分解を防ぐことができる。特定の実施形態においては、誘導細胞を−80℃にて半乾燥状態で保存して、タンパク質の分解を防止または抑制する。
【0106】
細胞をアレイから別のアレイへ、なんらかの適当な機械装置によって移すことができる。例えば、自動操作システム(例えば自動ピペット)を用いて、培地を含有するアレイに対象の細胞を接種することができる。特別の実施形態において、寒天を含む増殖培地を含有する96ウェルアレイに96-prongerを用いて酵母細胞を植菌することができる。同様に、自動液体取扱装置(例えば、Q-FILLTM、Genetix, UK)を用いて、アレイからアレイへ液体(例えば、試薬)の移動を行うことができる。
【0107】
細胞周期のいかなる時点の細胞からもタンパク質を回収することができるが、タンパク質合成が高まっている対数増殖期に細胞を分離することが望ましい。例えば、酵母細胞は、OD600=0.3からOD600=1.5の間で、望ましくはOD600=0.5からOD600=1.5の間で、集めることができる。特定の実施形態において、対数増殖期中期後のある時点の細胞からタンパク質を回収する。集めた細胞を将来の操作に備えて凍結保存することができる。
【0108】
集めた細胞を当業界で公知の様々な方法によって溶解することができるが、これには、例えば機械的な力、酵素消化、化学的処理などがある。細胞溶解の方法は、宿主細胞のタイプに適合していなくてはならない。例えば、新鮮なプロテアーゼインヒビターを含有する溶解バッファーを、酵母細胞に、細胞壁を破壊する物質(例えば、砂、ガラスビーズ、ジルコニアビーズ)とともに加え、その後、この混合物を振盪機(例えば、ボルテックスミキサー、ペイントシェーカー)によって激しく振盪する。
【0109】
特定の実施形態では、ジルコニアビーズを酵母細胞に接触させ、ボルテックス混合による機械的破壊によって細胞を溶解する。もう1つの実施形態では、高密度アレイフォーマットでの酵母細胞の溶解は、ペイントシェーカーによって達成される。ペイントシェーカーには、少なくとも18個の96-ウェルボックスを3層にしっかりと保持できる台があり、それによって、培養物の高速大量処理が可能になる。さらに、ペイントシェーカーは、培養物が完全に融解する前でも、これを激しく撹拌するため、結果としてタンパク質の分解を最小限に抑えつつ細胞の効率的な破壊をもたらす。実際、顕微鏡観察によって判定されるように、90%を超える酵母細胞を2分足らずの振盪で溶解することができる。
【0110】
結果として得られた細胞破砕片を対象のタンパク質および/または他の分子から遠心によって分離することができる。また、ハイスループット様式でタンパク質試料の純度を高めるために、タンパク質濃縮上清を、望ましくはタンパク質非結合性基板上のフィルターを用いて、濾過することができる。対象のタンパク質を含有する可溶性画分を不溶性画分から分離するためにフィルタープレートを使用することは、タンパク質の分解を減らし、または回避するために非常に望ましい。さらに、細胞破砕片を含有する画分について、これらのステップを繰り返すことが望ましく、結果としてタンパク質の収率が向上する。
【0111】
次に、このタンパク質濃縮上清から、当業界で公知の様々なアフィニティー精製法によってタンパク質を精製することができる。融合タンパク質調製物をアフィニティータグに対する結合パートナーと接触させることによる、融合タンパク質のアフィニティー精製に有用なアフィニティータグは、カルモジュリン、トリプシン/アンヒドロトリプシン、グルタチオン、免疫グロブリンドメイン、マルトース、ニッケル、またはビオチンおよびその誘導体を包含するがそれに限定されない。これらはそれぞれ、カルモジュリン結合タンパク質、ウシ膵臓トリプシンインヒビター、グルタチオン-S-トランスフェラーゼ(「GSTタグ」)、抗原もしくはプロテインA、マルトース結合タンパク質、ポリヒスチジン(「Hisタグ」)、およびアビジン/ストレプトアビジンと結合する。他のアフィニティータグとしては、例えば、mycまたはFLAGが挙げられる。適当な結合化合物(すなわち、グルタチオンビーズのような結合パートナー)を用いて、融合タンパク質をアフィニティー精製して、単離することができるが、これは、例えば、タンパク質非結合性フィルター上に結合タンパク質を含有する複合体を捕捉することによる。アフィニティータグをタンパク質の一方の末端(例えば、カルボキシ末端)に、第2のアフィニティータグをタンパク質の他方の末端(例えば、アミノ末端)に配置することは、全長タンパク質の精製に役立つと考えられる。
【0112】
特別の実施形態において、融合タンパク質はGSTタグを有しており、このタンパク質をグルタチオンビーズと接触させることによってアフィニティー精製される。もう1つの実施形態においては、融合タンパク質の付着したグルタチオンビーズを、96ウェルボックス内でフィルタープレートを使用することなく洗浄して、試料の取り扱いを容易にし、試料の相互汚染を防止することができる。
【0113】
さらに、融合タンパク質を、結合化合物(例えば、グルタチオンビーズ)から、所望のタンパク質濃度を与えるように、溶出バッファーで溶出することができる。特定の実施形態において、融合タンパク質はグルタチオンビーズから30μlの溶出バッファーで溶出されて、好ましいタンパク質濃度を与える。
【0114】
最終的に顕微鏡スライドガラス上にスポットされる精製タンパク質の場合は、グルタチオンビーズをこの精製タンパク質から分離する。すべてのグルタチオンビーズを除去して、基板上に精製タンパク質をスポットするために使用されるマイクロアレイピンが塞がるのを避けることが望ましい。好ましい実施形態において、グルタチオンビーズは、フィルタープレートを用いて、精製タンパク質から分離されるが、このプレートはタンパク質非結合性基板を含んでなることが望ましい。精製タンパク質を含有する溶出液の濾過によって、結果としてタンパク質の回収率は90%を超えるはずである。
【0115】
溶出バッファーは、例えば、15%から50%までのグリセロール、望ましくは約40%グリセロールのような粘性の高い液体を含んでなることが望ましい。グリセロール溶液は溶液中のタンパク質を安定化し、マイクロアレイヤーを用いたプリンティングステップの間、タンパク質溶液の乾燥を防ぐ。
【0116】
精製タンパク質は、タンパク質を安定化し、試料の乾燥を防ぐ媒質中で保存することが望ましい。例えば、15%から50%までのグリセロール、望ましくは約40%グリセロールのような粘性の高い液体中で、精製タンパク質を保存することができる。凍結/融解サイクルによって引き起こされるタンパク質活性の低下を避けるために、精製タンパク質を含有する試料をアリコートに分割することが望ましい。
【0117】
精製プロトコルを調整してタンパク質の純度を望ましいレベルに制御することができることを、当業者は正しく認識することができる。ある場合には、対象のタンパク質と会合する分子の分離が求められる。例えば、過剰生産された対象のタンパク質を含んでなる、二量体、三量体、もしくはそれ以上のホモタイプもしくはヘテロタイプ複合体を、本明細書に記載される精製方法、もしくはその変法によって単離することができる。さらに、結合した分子を当業界で公知の方法によって個別に単離して同定することができる(例えば、質量分析)。
【0118】
5.3 プロテオームアレイの作製方法
本発明は、また、プロテオームチップを作製する方法に関する。したがって、本発明は、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法を提供し、ここで、前記の複数のタンパク質は、ある単一の種における既知遺伝子のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%に相当する、少なくとも1つの代表的なタンパク質を含んでなり、この場合、前記タンパク質は、1つの遺伝子に由来するすべてのタンパク質アイソフォームおよびスプライス変異体とする。
【0119】
別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法を提供し、ここで、前記の複数のタンパク質は、単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%を含んでなる。
【0120】
また別の実施形態において、本発明は、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法を提供し、ここで、前記の複数のタンパク質は、単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000もしくは1,000,000のタンパク質を含んでなる。
【0121】
本発明はまた、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、次のような、位置指定可能なアレイを作製する方法に関する。すなわち、前記基板が微細加工された型から成形され、さらに、前記の複数のタンパク質が、単一の種において発現される全タンパク質の少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%もしくは99%を含んでなるか、または、単一の種において発現される少なくとも1、2、3、4、5、10、20、30、40、50、100、200、500、1000、1500、2000、2500、3000、4000、5000、6000、7000、8000、9000、10,000、100,000、500,000もしくは1,000,000のタンパク質を含んでなるか、または、ある単一の種における既知遺伝子のうち少なくとも1%、2%、3%、4%、5%、10%、20%、30%、40%、50%、60%、70%、80%、90%、95%または99%に相当する、少なくとも1つの代表的なタンパク質を含んでなり、この場合、前記タンパク質は、1つの遺伝子に由来するすべてのタンパク質アイソフォームおよびスプライス変異体である。本発明は、微細加工された型から成形された様々な基板を包含するが、その一部は、例えば、2001年5月4日出願の同時係属中の米国特許出願第09/849,781号において開示されており、この出願はその全体を参考として本明細書に含めるものとする。
【0122】
本発明はまた、複数のタンパク質を基板の表面に付着させるステップ(それぞれのタンパク質は基板上の異なる位置にある)を含んでなる、位置指定可能なアレイを作製する方法に関するが、ここで、前記タンパク質は第1のタグおよび第2のタグを含んでなる。二重タグ標識タンパク質を用いる利点としては、高度に精製されたタンパク質が得られること、ならびに、細胞破砕片からタンパク質を精製してそのタンパク質を基板に付着させる能率的な方法を提供することがある。特定の実施形態において、第1タグはグルタチオン-S-トランスフェラーゼタグ(「GSTタグ」)であり、第2タグはポリヒスチジンタグ(「Hisタグ」)である。もう1つの実施形態において、GSTタグおよびHisタグはタンパク質のアミノ末端に結合される。あるいはまた、GSTタグおよびHisタグはタンパク質のカルボキシ末端に結合される。
【0123】
さらに別の実施形態において、GSTタグはタンパク質のアミノ末端に結合される。また別の実施形態において、Hisタグはタンパク質のカルボキシ末端に結合される。もう1つの実施形態において、Hisタグはタンパク質のアミノ末端に結合される。また別の実施形態において、GSTタグはタンパク質のカルボキシ末端に結合される。
【0124】
別の実施形態において、タンパク質はGSTタグおよびHisタグを含んでなるが、GSTタグもHisタグも、タンパク質のアミノ末端もしくはカルボキシ末端には存在しない。特定の実施形態において、GSTタグおよびHisタグは、当該タンパク質のコード領域の中に存在する;対象とする結合ドメインに影響を及ぼさないタンパク質の領域にあることが望ましい。
【0125】
ある実施形態において、第1タグは融合タンパク質を精製するために用いられる。別の実施形態において、第2タグは融合タンパク質を基板に付着させるために使用される。別の特定の実施形態において、第1タグはGSTタグであり、第2タグはHisタグである。
【0126】
タンパク質は融合タンパク質であって、その異種配列が当該タンパク質のコード配列およびアフィニティータグのようなタグをコードする配列を含んでなる融合タンパク質であることが望ましい。このようなタグは、タンパク質をモニタリングし、細胞破砕片および混入試薬から融合タンパク質を分離し、および/または、タンパク質を本発明のプロテオームチップに付着させるために有用であると考えられる。
【0127】
誘導物質の例には、ガラクトース、エンハンサー結合タンパク質および他の転写因子があるが、それらに限定されない。ある実施形態においては、ガラクトースを、ガラクトース誘導性GAL1プロモーターを含む調節配列と接触させる。
【0128】
本発明にしたがって使用することができる結合物質としては、グルタチオンビーズ、ニッケルコーティングされた基板、および抗体があるが、それらに限定されない。ある実施形態において、複合体はグルタチオンビーズに結合したGSTタグを有する融合タンパク質を含んでなる。別の実施形態において。複合体はニッケルコーティングされた基板に結合したHisタグを有する融合タンパク質を含んでなる。さらに別の実施形態において、複合体は、抗体および、場合によっては、二次抗体に結合した対象のタンパク質を含んでなる。
【0129】
5.4 プロテオームアレイを使用する方法
本発明はまた、プロテオームチップを用いて、少なくとも1つの試料中に存在するタンパク質の存在、量、および/または機能性をアッセイする方法に関する。本発明のプロテオームチップを用いて、化学反応およびアッセイを大規模並行的な分析において実施して、生物学的状態または生物学的応答の特徴を明らかにし、タンパク質の存在、量、および/または生物活性を決定することができる。したがって、本発明のプロテオームチップを用いて、細胞、組織、器官、系、もしくは生物体における本質的にすべてのタンパク質-タンパク質相互作用をアッセイすることができる。
【0130】
さらに、本発明のプロテオームチップを用いて、宿主細胞(すなわち、融合タンパク質を調製するために使用した細胞)に加えられた特定の刺激に対する生物学的応答をアッセイすることができる。例えば、融合タンパク質をコードする発現ベクターで形質転換された酵母細胞に刺激を与えることができるが、その後、その融合細胞を精製してアレイ化する。アレイ化したタンパク質を、次に、例えば、何らかの結合物質を用いてプロービングすることによって特徴づけることができる。その結果得られた結合パターンを、次に、前記刺激を受けない、または異なる刺激を受けた酵母細胞から作製された同一のアレイと比較する。結合パターンの相違は、生物学的応答の特性を示していると考えられ、生物学的応答に関して特異的な相互作用物質を同定することができる。
【0131】
ある実施形態において、宿主細胞から作製されたプロテオームチップ(各チップは異なる刺激に曝された宿主細胞に対応する)を、標識レクチン(例えば、コンカナバリンA)でスクリーニングする。タンパク質-プローブ相互作用のパターンはグリコシル化タンパク質の存在を示唆するが、これを比較して、プロテオームチップのタンパク質のグリコシル化状態に関するそれぞれの刺激の影響を判定する。
【0132】
本発明のプロテオームチップを用いて測定できる生物活性は、酵素活性(例えば、キナーゼ活性、プロテアーゼ活性、ホスファターゼ活性、グリコシダーゼ、アセチラーゼ活性、および他の化学基転移酵素活性)、核酸の結合、ホルモンの結合、などを包含するが、それに限定されない。高密度および小容量の化学反応は、本発明のプロテオームチップの使用に関わる方法に好都合であると考えられる。
【0133】
生物学的状態もしくは応答についてこれ以上の情報を、本発明のプロテオームチップを用いて得ることができるが、この場合、チップ上のタンパク質はタンパク質の分類にしたがって組織化される。この分類は、存在量、機能、機能的クラス、酵素活性、相同性、タンパク質ファミリー、特定の代謝経路もしくはシグナル伝達経路との関連性、関連した代謝経路もしくはシグナル伝達経路との関連性、または翻訳後修飾によることができる。
【0134】
本発明のプロテオームチップを1以上のプローブと接触させる際に、当業界で公知の様々な技法を用いて、タンパク質-プローブ相互作用をアッセイすることができる。例えば、化学発光もしくは蛍光を生じる標準的な酵素測定法によって、プロテオームチップをアッセイすることができる。非タンパク質基質、酵素的発色、質量分析のサインマーカー、もしくはオリゴヌクレオチドタグの増幅を用いて、例えば、フォトルミネセンス、化学発光、もしくは蛍光によって、様々なタンパク質の修飾を検出することができる。
【0135】
当技術分野で広く知られている方法によって直接的もしくは間接的にその結合を検出できるように、プローブをマーカーで標識し、もしくはタグを付ける。当技術分野で公知のあらゆるマーカーを使用することが可能であって、こうしたマーカーは、それがプロテオームチップのタンパク質をチップの基板に付けるために使用されるアフィニティータグもしくは試薬と同一ではないという条件で、エピトープタグ、ハプテンおよびアフィニティータグといったタグ、抗体、標識などを包含するが、それらに限定されない。例えば、タンパク質をプロテオームチップアレイに付着させるリンカーとしてビオチンを使用するならば、このプロテオームチップのタンパク質中に存在しない別のタグ、例えばHisもしくはGSTを用いてプローブを標識し、タンパク質-プローブ相互作用を検出する。特定の実施形態においては、フォトルミネセンス、化学発光、蛍光、もしくは酵素タグを使用する。他の実施形態においては、質量分析のサインマーカーを使用する。さらに他の実施形態では、増幅可能なオリゴヌクレオチド、ペプチド、もしくは分子量標識を使用する。
【0136】
特定の実施形態において、本発明は、標識プローブ(例えば、標識タンパク質)試料を、複数のタンパク質を含んでなる位置指定可能なアレイ(それぞれのタンパク質は基板上の異なる位置に存在する)と接触させるステップ、およびアレイ上で標識プローブとタンパク質との相互作用が生じるアレイ上の位置を検出するステップを含んでなる、タンパク質-プローブ相互作用を検出する方法を提供する。
【0137】
したがって、タンパク質-プローブ相互作用を例えば下記によって検出することができる:すなわち、1)放射性標識リガンドを使用し、それに続いてオートラジオグラフィーおよび/またはホスホルイメージャー(PhosphorImager)分析を行うこと;2)ハプテンの結合、これを次に蛍光標識したもしくは酵素標識した抗体または高親和性ハプテンリガンド、例えばビオチンもしくはストレプトアビジン、によって検出する;3)質量分析;4)原子間力顕微鏡法;5)蛍光偏光法;6)赤外線赤色標識化合物もしくはタンパク質;7)増幅可能なオリゴヌクレオチド、ペプチドもしくは分子量標識;8)タンパク質の酵素活性の刺激もしくは阻害;9)ローリングサークル増幅-検出法(Hatchら、1999、「固体表面に固定化されたDNAのローリングサークル増幅および多重突然変異検出へのその応用」(”Rolling circle amplification of DNA immobilized on solid surfaces and its application to multiplex mutation detection”)、Genet. Anal. 15:35-40);10)競合的PCR(Finiら、1999、「マイクロプレート照度計を用いてパルボウイルスB19 DNAを検出および定量するための化学発光競合PCRの開発」(”Development of a chemiluminescence competitive PCR for the detection and quantification of parvovirus B19 DNA using a microplate luminometer”)、Clin. Chem. 45:1391-6; Kruseら、1999、「セミネステッド競合PCRアッセイによるトランスフォーミング増殖因子-β1(TGF-β1)遺伝子発現の検出および定量」(”Detection and quantitative measurement of transforming growth factor-beta 1 (TGF-beta 1) gene expression using a semi-nested competitive PCR assay”)、Cytokine 11: 179-85; GuenthnerおよびHart, 1998、「マイクロプレートを基本とする検出システムによるHIV-1のための定量的競合PCRアッセイ」(”Quantitative, competitive PCR assay for HIV-1 using a microplate-based detection system”)、Biotechniques 24: 810-6);11)比色定量法;および12)生物学的アッセイ(例えば、ウイルス力価)。
【0138】
特別の実施形態において、タンパク質-プローブ相互作用は、直接質量分析によって検出される。さらなる実施形態において、タンパク質および/またはプローブの正体は質量分析を用いて決定される。例えば、プロテオームチップ上のタンパク質と結合している1以上のプローブを、アレイから解離させて質量分析によって同定することができる(例えば、WO 98/59361を参照されたい)。別の実施形態において、質量分析によって、プロテオームチップ上のタンパク質の酵素的切断を検出し、切断されたタンパク質断片もしくは他の放出された化合物を同定することができる。
【0139】
ある実施形態において、プロテオームチップ上の各タンパク質をプローブと接触させてタンパク質-プローブ相互作用を検出し、定量する。別の実施形態において、プロテオームチップ上の各タンパク質を複数のプローブと接触させ、タンパク質-プローブ相互作用を検出して定量する。たとえば、プロテオームチップを同時に複数のプローブを用いてスクリーニングすることができる。このプローブは、複雑な混合物(例えば、細胞抽出物)、そのままの細胞成分(例えば、細胞小器官)、細胞全体、およびいくつかの起源から集められたプローブを包含するが、それらに限定されない。次に、タンパク質-プローブ相互作用を検出し、定量する。1つには、本発明のアレイの位置指定可能な性質によって、すなわち、プロテインチップ上の既知の位置にタンパク質を配置することによって、プローブの混合物を用いたアッセイから有用な情報が得られ、プローブが結合するタンパク質(「相互作用物質」)を特徴づけることができる。
【0140】
当業者であれば、プローブを用いて本発明のプロテオームチップをスクリーニングすることによって、様々な細胞相互作用をアッセイすることを目的とした数多くの様々な実施形態を正しく認識することができる。例えば、様々なプローブを用いて、プロテオームチップを多数回連続してスクリーニングすることによって、特定のシグナル伝達経路もしくは特定の代謝経路に関わるすべてのタンパク質を明確にすることができる。さらに、こうしたアッセイは、診断、予後判定および/または治療目的に役立てることができる。
【0141】
本発明の方法によれば、プローブは、細胞、細胞膜、細胞内小器官、タンパク質を含有する細胞材料、タンパク質、オリゴヌクレオチド、ポリヌクレオチド、DNA、RNA、小分子(すなわち、分子量が500より小さい化合物)、基質、薬物もしくは候補薬物、受容体、抗原、ステロイド、抗体、免疫グロブリンドメイン、グルタチオン、マルトース、ニッケル、ジヒドロトリプシン、レクチン、またはビオチンとすることができる。
【0142】
タンパク質-プローブ相互作用を検出するために、プロテインアレイと接触させる際に使用するプローブはビオチン化することができる。弱くビオチン化したタンパク質の方が、対象の生物活性を維持する可能性が大きい。このように、タンパク質の結合活性もしくは他の生物活性を保存するためには、より穏やかなビオチン化法が好ましい。したがって、特別の実施形態において、プローブタンパク質は、ビオチン転移化合物(例えば、Sulfo-NHS-LC-LC-Biotin; PIERCETM カタログ番号21338、USA)によって様々な程度にビオチン化される。
【0143】
さらに、酵素基質もしくは阻害剤をプローブとすることができる。例えば、プローブは次のような酵素の基質もしくは阻害剤とすることができる。すなわち、キナーゼ、ホスファターゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、および他の化学基転移酵素といった酵素であるが、これらに限定されない。核酸もしくはタンパク質プローブの組合せとチップ上のタンパク質をインキュベートした後、例えば、質量分析によって結合した核酸またはタンパク質プローブを同定することができる(Lakeyら、1998、「タンパク質-タンパク質相互作用の測定」(“Measuring protein-protein interactions”)、Curr Opin Struct Biol. 8: 119-23)。
【0144】
したがって、プロテオームチップ上のタンパク質との相互作用に対する様々な細胞性応答を、ホールセル(whole cell)をプローブとして探索することによってアッセイすることができる。例えば、プロテオームチップをリンパ球と接触させて、リンパ球の活性化を様々な方法によってアッセイすることができる。すなわち、前記方法には、抗体合成を検出すること、3H-チミジンの取り込みを検出もしくは測定すること、細胞表面の分子を抗体で標識して抗原認識および活性化によって誘導もしくは抑制される分子を同定すること(例えば、CD23、CD38、IgD、C3b受容体、IL-2受容体、トランスフェリン受容体、膜クラスII MHC分子、PCA-1分子、HLA-DR)、および、発現されたおよび/または分泌されたサイトカインを同定することがあるが、それらに限定されない。
【0145】
別の例において、細胞をプロテオームチップとともにインキュベートすることによって、特定の細胞型に対するマイトジェンを決定することができる。有糸分裂活性を、例えば、細胞による3H-チミジンの取り込みを検出もしくは測定することによって、測定することができる。細胞は同一の細胞型(すなわち、均一集団)であってもよく、異なる細胞型とすることもできる。
【0146】
また別の例において、細胞をプロテオームチップとともにインキュベートすることによって、特定の細胞型に対する分化因子を決定することができる。細胞の分化はたとえば、目視検査、マーカー特異的抗体による細胞表面分化マーカーの検出、もしくは分泌された分化マーカーの同定によって判定することができる。
【0147】
別の例において、細胞をプロテオームチップとともにインキュベートすることによって、特定の細胞型に対するアポトーシス因子を決定することができる。アポトーシスは、例えば、目視検査、マーカー特異的抗体による細胞表面アポトーシスマーカーの検出、または、培地中に放出された分泌マーカーもしくは他の細胞成分の同定によって判定することができる。
【0148】
プロテオームチップ上のタンパク質に対する細胞の分泌応答を、細胞を本発明のプロテオームチップとともにインキュベートすることによって、アッセイすることができる。分泌されたタンパク質および他の細胞性化合物を、例えば、培地中に放出された化合物を検出することで、アッセイすることができる。
【0149】
別の例において、例えば、1以上の細胞を本発明のプロテオームチップとともにインキュベートし、細胞凝集をアッセイすることによって、プロテオームチップ上のタンパク質が細胞凝集を媒介する能力についてアッセイすることができる。また、例えば、細胞および細胞外マトリックス成分をプロテオームチップとともにインキュベートし、チップ上のタンパク質による細胞もしくは細胞外マトリックス成分の親和性の増加をアッセイすることによって、細胞外マトリックスに対する親和性を媒介するタンパク質の能力をアッセイすることができる。こうしたアッセイで同定される相互作用物質は、例えば、癌、細胞移動、シナプス形成、樹状細胞の成長、突起の伸長、もしくは軸索の伸長において何らかの役割を担うと考えられる。
【0150】
さらに別の例において、本発明のプロテオームチップのタンパク質がイオン輸送もしくは他の小分子輸送(例えばATP)に及ぼす影響を測定することができる。例えば、プローブ細胞に放射性標識したイオンもしくは他の小分子を前もってロードして、本発明のプロテオームチップとインキュベートする。細胞をプロテオームアレイのタンパク質と接触させた後、様々な時点で、放射性標識の保持および放出を測定することができる。あるいはまた、当技術分野で公知の電気生理学的技法を用いてイオン輸送を検出し、特徴づけることができる。
【0151】
さらに別の例において、プロテオームチップ上のタンパク質の細胞による取り込みおよび/またはプロセシングを、例えば、チップ上に放射性もしくは蛍光標識したタンパク質を有するプロテオームチップとともに細胞をインキュベートし、プロテオームチップ上のシグナルの増加もしくは減少を測定するか、または細胞による標識タンパク質の取り込みを測定することによって、アッセイすることができる。
【0152】
あるいはまた、本発明のプロテオームチップを細胞および対象の標識化合物とともにインキュベートし、その結果として、細胞による化合物の取り込みおよび/またはプロセシングを検出および/または測定する。
【0153】
無細胞系において小分子をプローブとして探査することによって、小分子(すなわちMW=500より小さい化合物)とプロテオームチップ上のタンパク質との相互作用をアッセイすることができる。この小分子としては、例えば、ATP、GTP、cAMP、ホスホチロシン、ホスホセリンおよびホスホトレオニンがあるが、それらに限定されない。このようなアッセイは、対象の小分子と相互作用する、ある生物種におけるすべてのタンパク質を同定することができる。対象の小分子は、医薬品、候補薬物、殺真菌薬、殺虫薬、発癌物質、および汚染物質を包含するが、これらに限定されない。本発明の方法にしたがってプローブとして使用される小分子は、タンパク質以外の有機化合物であることが望ましい。
【0154】
別の実施形態において、ある生物種における特定のリガンドもしくは特定のクラスのリガンド群に対する本質的にすべての受容体は、対象の受容体を本発明のプロテオームチップと接触させることによって同定することができる。あるいはまた、ある生物種における本質的にすべてのリガンドが対象の特定の受容体もしくは受容体ファミリーによって同定されるが、こうしたリガンドは対象の受容体を本発明のプロテオームチップと接触させることによって同定可能である。別の実施形態において、特定の受容体-リガンド複合体の形成を阻害もしくは阻止できる、1つの生物種の本質的にすべてのタンパク質は、受容体およびそのリガンドを本発明のプロテオームチップと接触させて、受容体-リガンド相互作用がチップ上のタンパク質の不在下での受容体-リガンド相互作用の程度と比較して阻害されるかどうかを判定することによって、同定可能である。受容体-リガンド相互作用の検出およびリガンド相互作用物質の同定は、当技術分野でよく知られた方法によって実施することができる。
【0155】
別の実施形態において、1つの種の本質的にすべてのキナーゼ標的は、例えば、キナーゼを本発明のプロテオームチップと接触させ、標識リン酸の存在下で、当業界で公知の方法によってリン酸化された相互作用物質を検出することによって同定することができる。あるいはまた、リン酸化される可能性のある基質を本発明のプロテオームチップと接触させ、リン酸化された基質の存在および/またはレベルを、リン酸化アミノ酸に特異的な抗体を用いてアッセイすることによって、1つの種の本質的にすべてのキナーゼを同定することができる。別の実施形態において、キナーゼおよびその基質を本発明のプロテオームチップに接触させ、チップ上のタンパク質の不在下でのリン酸化レベルと比較して、基質のリン酸化が減少したかどうかを判定することによって、1つの種の本質的にすべてのキナーゼ阻害剤を同定することができる。
【0156】
キナーゼ活性を検出する方法は、当技術分野で公知であり、放射性標識(例えば、32P-ATPおよび35S-γ-ATP)もしくはリン酸化アミノ酸と結合する蛍光抗体プローブの使用を包含するがそれに限定されない。
【0157】
同様に、アッセイを行って、1つの種における全ホスファターゼおよびホスファターゼ阻害剤を同定することができる。例えば、あるアッセイにおいて放射性標識リンのタンパク質への取り込みはキナーゼ活性を示すが、別のアッセイを用いて放射性標識リンの培地中への放出を測定することが可能であり、それがホスファターゼ活性を示す。
【0158】
また、本発明のプロテオームチップは、例えば、プロテオームチップを、異なる細胞集団に対応する細胞もしくは細胞抽出物と接触させ、プロテオームチップ上のタンパク質-プローブ相互作用のパターンを比較することによって、異なる細胞型(形態的もしくは機能的のいずれか)を識別するために使用することができる。この方法を用いて、例えば、細胞周期の異なる時期、病気の状態、生理学的状態の変化(例えば、低酸素)、治療前後の生理学的状態(例えば、薬物療法)、代謝状態、分化の段階、発生の段階、環境刺激(例えば、光、熱)に対する応答、環境毒物(例えば、殺虫剤、除草剤、汚染)への応答、細胞-細胞相互作用、細胞特異的タンパク質の発現、および疾病特異的タンパク質の発現を特徴づけることもできる。
【0159】
タンパク質-タンパク質相互作用の発生プロファイルを用いて、発生の各段階で関与するシグナル伝達経路、代謝経路などを特徴づけて、発生段階間の推移を解明することができる。こうした研究によって得られる豊富な情報を用いて、各段階の薬物標的を同定したり、かつ/または疾病の経過中に治療計画を適合するように変更することができる。
【0160】
本発明のプロテオームチップを細胞抽出物とともにインキュベートして、特定の細胞型、刺激への応答、もしくは生理学的状態を特徴づけることができる。したがって、典型的な実施形態では、本発明のプロテオームチップを、化合物(例えば、薬物)で処理した細胞から、細胞分化の特定の段階(例えば多能性段階)にある細胞から、または特別な代謝状態(例えば、分裂状態)にある細胞から得られた細胞抽出物と接触させて、例えば、キナーゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、ホスファターゼ、および/または他の転移酵素活性をアッセイすることができる。
【0161】
プロテオームチップ上のタンパク質-プローブ相互作用のパターンは、したがって、生物学的状態に特徴的な「サイン」や「フィンガープリント」を提供することができる。例えば、薬物の存在下もしくは非存在下の細胞、いくつかの分化段階にある細胞、もしくは異なる代謝状態にある細胞を比較する、そのようなアッセイから得られた結果は、それぞれの状態の典型的特徴を提供することが可能であり、さらに異なる条件下での細胞の生理学的変化に関する情報を提供することができる。
【0162】
複数のプローブ(例えば、既知のプローブ混合物、細胞抽出物、細胞内小器官、細胞膜調製物、ホールセルなど)を用いて1つの種のプロテオームをスクリーニングすることによって、得られたタンパク質-プローブ相互作用の分析結果は、明らかに、細胞型、または細胞、組織、器官もしくは系の生理学的状態の「フィンガープリント」もしくは「サイン」を同定する土台を形成すると考えられる。こうした情報は、例えば、診断、予後判定、薬物試験、および薬物探索に役立てることができる。
【0163】
したがって、本発明のプロテオームチップを用いて、チップ上のタンパク質との薬物の相互作用を測定することができる。あるいはまた、本発明のプロテオームチップを使用して、例えば、ホールセル、細胞抽出物、もしくは組織ホモジネートのような複雑なタンパク質混合物への薬物の影響を特徴づけることができる。例えば、プロテオームチップを、複雑なタンパク質混合物と混合し、薬物が存在する状態もしくは存在しない状態で比較すれば、チップ上のタンパク質とのタンパク質混合物の相互作用の変化をアッセイすることができる。
【0164】
これにより、薬物の実際の効果は、1以上のプロテオームチップを、薬物処理した細胞、組織、もしくは抽出物によってスクリーニングすることにより分析できるが、このことは、ひいては、薬物処理状態に関する「サイン」を提供することを可能にし、さらに、未処理状態の「サイン」と比較するならば、例えば、効能、毒性および副作用に関する予測値を示すことができる。さらに、薬物の時間依存的効果は、例えば、細胞、細胞抽出物、組織ホモジネート、もしくは生物体そのものに薬物を添加し、処理の様々な時点で調製された薬物処理細胞もしくは抽出物をプロテオームチップに加えることによって、アッセイすることができる。こうしたアッセイは、疾病の診断もしくは予後判定に有用であると考えられる。
【0165】
特に、本発明のプロテオームチップは、薬物の作用方式を特徴づけ、薬物の特異性を決定し、薬物の毒性を予測するのに役立ち、さらに薬物の探索に有用であると考えられる。例えば、種々のアッセイ条件下で薬物または薬物候補とともにプロテオームチップをインキュベートし、アレイ上のどの場所に薬物が結合するかを判定することによって薬物の特異性を決定し、それぞれ異なるタンパク質に結合した薬物の量を測定することによって、薬物と結合するタンパク質の正体およびそれらの相対的親和性をアッセイすることができる。
【0166】
本発明のプロテオームチップは、例えば、プロテオームチップを罹病した細胞、細胞抽出物、もしくは罹病組織から得られた組織ホモジネート、または病気にかかった患者から得られた体液と接触させ、プロテオームチップ上でのタンパク質-プローブ相互作用のパターンを、健康な対応部分のそれと比較することによって、疾病状態を判定するために使用することができる。こうしたアッセイは疾病の状態に関する「サイン」を提供することができる。さらに、健康な状態の「サイン」と比較するならば、疾病の診断もしくは予後判定に関する予測値を示すことができる。さらに、例えば、様々な病期でプロテオームチップ上にて生物学的調製物をアッセイすることによって病期を特徴づけることができる。
【0167】
結合アッセイではなくて、生物活性をアッセイするバイオアッセイも、同じプロテオームチップ上で、もしくは同一の第2のチップ上で実施することができる。したがって、本発明のプロテインチップによるこのようなタイプのアッセイは、薬物の特異性を研究したり、薬物の潜在的副作用を予測したり、薬物を分類したりするために有用である。
【0168】
さらに、本発明のプロテオームチップは、薬物候補の複雑なライブラリーのスクリーニングに適している。具体的には、チップ上のタンパク質を、薬物候補のライブラリーとともにインキュベートした後、結合した化合物を、例えば、質量分析によって同定することができる。このことにより、特定の一部のタンパク質に優先的に結合するか、もしくはチップ上のタンパク質のうちいくつかと結合する、すべてのライブラリー成分を同時に同定することが可能となる。さらに、アレイに含まれる様々なタンパク質に対する薬物候補の相対的親和性を測定することもできる。
【0169】
さらに、観察された相互作用、酵素活性もしくは生物学的応答の潜在的な阻害剤、触媒、モジュレーターもしくはエンハンサーの存在下で、本発明のプロテインチップをプローブすることができる。本発明のプロテオームチップを用いると、このような戦略的なスクリーニングは、1つの種で発現されたタンパク質であって、例えば、薬物の結合を阻止する、ウイルス感染を阻害する、静菌活性を示す、抗真菌活性を示す、寄生虫感染を改善するタンパク質を同定し、また、特定のカテゴリーのタンパク質に対する生理的エフェクターを同定することができる。
【0170】
酵素反応を行って、酵素活性を本発明のプロテオームチップにより測定することができる。特定の実施形態では、チップ上の1つもしくは複数のタンパク質の酵素活性を調節する化合物を同定することができる。例えば、1つの化合物もしくは化合物類の混合物を酵素反応混合物とともにインキュベートし、それによってシグナル(例えば、酵素活性により蛍光性となる基質からのシグナル)を生じることにより、酵素活性のレベルの変化を検出して定量することができる。試験化合物の存在と非存在との差異を特徴づけることができる。さらに、化合物が酵素活性に及ぼす影響の差異は、サンプルに対するそれらの相対的影響をプロテオームチップ内で、およびチップ間で、比較することによって検出することができる。
【0171】
本発明のプロテオームチップを使用する様々なストラテジーは、タンパク質の様々な物理学的および機能的特性を決定するために利用することができる。例えば、プロテインチップを用いて、抗体でプローブすることによりタンパク質の存在および存在量を評価することができる。ルミネセンス、化学ルミネセンス、蛍光、化学蛍光、もしくは質量分析のような標準的な検出アッセイによってタンパク質を検出することができる。例えば、対象のタンパク質に対する一次抗体を、蛍光標識した二次抗体によって認識させ、これを次に、蛍光産物を光源によって励起して、その結果生じた蛍光を検出する機器(例えば、Molecular Dynamicsスキャナー)によって測定する。感度をいっそう高めるために、対象のタンパク質に対する一次抗体を、アルカリホスファターゼもしくは西洋ワサビペルオキシダーゼのような酵素と結合した二次抗体によって認識させる。発光基質(化学発光のための)もしくは蛍光原基質(化学蛍光のための)の存在下で、酵素的切断は強い発光産物もしくは蛍光産物をもたらすが、これは例えば、Molecular Dynamicsスキャナーを使用することによって検出および定量することができる。あるいはまた、蛍光標識した二次抗体のシグナルを、アルカリホスファターゼと結合した、もしくは西洋ワサビペルオキシダーゼと結合した三次抗体を用いて増幅することができる。
【0172】
ある実施形態において、本発明のプロテオームチップは、1つの種における既知タンパク質に対する抗体でプローブすることが可能であるが、その結果、認識されたエピトープを有する相同タンパク質を別の種のプロテオームにおいて同定することができる。たとえば、相互作用物質の同種のホモログは、相同タンパク質のDNA配列情報を用いて他の種におけるホモログを同定することによって得ることができる。特定の実施形態において、抗体は、サイクリン、キナーゼ、GST、CIb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2もしくはCdc11に対するものである。
【0173】
別の実施形態において、第1の種のタンパク質を含有する本発明のプロテオームチップを、第2の種から得られたタンパク質を用いてプローブして、相互作用物質を同定することができる。次に、第1の種から得られた相互作用物質の第2の種におけるホモログは、例えば、ヌクレオチド配列相同性によって同定し、特徴づけることができる。したがって、特定の種のプロテオームが利用できない場合は、このストラテジーを用いて、対象のタンパク質と相互作用する同じ種のタンパク質を見つけ出すことができる。
【0174】
本発明のプロテオームチップは、1つの種に存在する各酵素に対する、1つの種における本質的にすべての基質を同定するために使用することができる。したがって、本発明のプロテオームチップ上で、プロテインキナーゼ、ホスファターゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、もしくは他の化学基転移酵素の基質の同定を行うことができる。
【0175】
ある実施形態において、プロテアーゼ活性は、標準的なアッセイ(例えば、質量分析、ペプチド断片に対する蛍光標識抗体、もしくは蛍光タグ付けした基質からの蛍光シグナルの減少)を用いて、プロテアーゼ活性によって生成し、媒質中に放出されたペプチド断片を同定することによって検出することができる。このようにして、化学基-転移酵素の活性は、任意の検出手段を用いたいくつかの方法(当業者は理解できるだろう)によって容易にアッセイすることができる。
【0176】
本発明のプロテオームチップは、任意の化合物についての、1つの種における本質的にすべての結合タンパク質を同定するために使用することができる。したがって、プロテオームチップを用いて、例えば、キナーゼ、プロテアーゼ、ホルモン、DNA、RNA、ホスファターゼ、プロテアーゼ、グリコシダーゼ、アセチラーゼ、もしくは他の化学基転移酵素と結合する本質的にすべてのタンパク質を同定して特徴づけることができる。このように、チップをプローブによって探索して、タンパク質-プローブ相互作用をアッセイすることができ、かつ/または、望ましい活性をアッセイすることができる。例えば、もしRNA結合が関心のある活性であるならば、RNAをプローブとしてプロテオームチップを探索して、タンパク質-RNA複合体を同定する。
【0177】
例えば、本発明のプロテオームチップは、膜結合タンパク質もしくは他の膜結合化合物と結合する、1つの種における本質的にすべてのタンパク質を同定するために、このチップをプローブ(例えば、ホールセル、細胞膜調製物、膜小胞、もしくは対象の膜成分を含むリポソーム)と接触させ、タンパク質-プローブ相互作用を検出することによって、使用することができる。特定の実施形態において、プローブは1以上の対象のリン脂質を含むリポソームの形をとる。タンパク質-プローブ相互作用は当技術分野で公知の技術によって検出可能である。相互作用物質および/またはプローブの正体(素性)は当技術分野で公知の技術を用いて同定することができる。さらに、生物活性(例えば、酵素活性、細胞活性化)も当技術分野で公知の技術によって検出することもできる。
【0178】
本発明のプロテオームチップを使用することによって、回復期の患者もしくは回復していない患者の免疫応答において抗原として作用する病原体(例えば、ウイルス、細菌、真菌、または寄生虫のような感染症の病因)由来の標的タンパク質、もしくは異常細胞(例えば、新生物細胞、病変細胞、もしくは損傷細胞)由来の標的タンパク質の正体(素性)を同定することができる。例えば、患者から分離したリンパ球を用いて、病原体のタンパク質のすべてを含んでなるチップをスクリーニングすることができる。一般に、こうしたスクリーニングは、プロテオームチップを複数のリンパ球と接触させ(ただし、このプロテオームチップ上のタンパク質は可能性のある複数の抗原を含んでなる)、リンパ球の活性化が生じたチップ上の位置を検出することを含んでなる。特定の実施形態では、リンパ球を、プロテオームチップ上の病原体のタンパク質と接触させた後、抗原もしくは抗原混合物によるB細胞またはT細胞の活性化をアッセイし、それによって病原体に由来する標的抗原を同定する。
【0179】
あるいはまた、本発明のプロテオームチップを用いて、例えば、患者のリンパ球で感染性生物のプロテオームをスクリーニングして患者のB細胞および/またはT細胞の標的を同定することによって、免疫応答を特徴づけることができる。例えば、B細胞を本発明のプロテオームチップとともにインキュベートして、体液性免疫に対する抗原標的を同定することができる。
【0180】
別の実施形態において、本発明のプロテオームチップを使用して、自己免疫の基質もしくはアレルギー原因タンパク質を検出し、特徴付けることができる。例えば、患者のリンパ球または患者の循環抗体を用いて、ヒトタンパク質のプロテオームをスクリーニングし、患者のB細胞および/またはT細胞の標的を同定することができる。こうしたスクリーニングは、自己免疫またはアレルギー反応を特徴付けて、可能性のある標的薬物候補を同定することができる。
【0181】
ある実施形態において、本発明のプロテオームチップはB細胞もしくはT細胞を活性化できる物質を同定するために使用することができる。例えば、リンパ球をプロテオームチップに接触させてリンパ球活性化をアッセイし、それによってB細胞もしくはT細胞またはリンパ球の亜集団(例えば、細胞傷害性T細胞)を活性化する全般的な能力を有する物質を同定する。
【0182】
抗原認識によるB細胞活性化の誘導は、抗体合成の検出もしくは測定、3H-チミジンの取り込み、新たに発現もしくは抑制された細胞表面分子への標識抗体の結合、およびB細胞活性化を示す因子(例えば、サイトカイン)の分泌を含むがこれらに限らない様々な方法によってアッセイ可能である。同様に、本発明のプロテインチップを用いたスクリーニングでのT細胞活性化は様々なアッセイにより測定できる。例えば、クロム(51Cr)放出アッセイは、抗原の認識と、それに続く細胞傷害性T細胞の活性化を検出することができる(例えば、Palladinoら、1987, Cancer Res. 47: 5074-9; Blachereら、1993, J. Immunotherapy 14: 352-6を参照されたい)。
【0183】
本発明のプロテオームチップの使用によって、抗体調製物の特異性を決定することができるが、これは、チップを抗体調製物と接触させ、抗体調製物中の抗体による結合が生じた基板上の位置を検出することを含んでなる。抗体調製物は、Fabフラグメント、抗血清、およびポリクローナル、モノクローナル、キメラ、一本鎖、ヒト化、もしくは合成抗体であってよいが、それらに限定されない。ある実施形態では、モノクローナル抗体をプローブとして使用してプロテオームチップを探索し、その結合の強さおよび/またはその特異性を特徴づける。特定の実施形態において、抗体は、サイクリン、キナーゼ、GST、Clb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2もしくはCdc11に対するものである。
【0184】
本発明のプロテオームチップは、生物学的に関心のある特定の分子(例えば、潜在的リガンド分子の受容体、ウイルス受容体、およびオーファン受容体のリガンドを包含するがこれらに限定されない)と結合するタンパク質を同定するために有用である。
【0185】
本発明のプロテオームチップは、チップ上のタンパク質へのDNAの結合もしくはRNAの結合の検出、ならびにその結合特異性および強度の評価にも有用である。DNAは、一本鎖もしくは二本鎖とすることができる。RNAはmRNA、hnRNA、ポリA+RNA、もしくは全RNAであってよい。
【0186】
本発明のプロテオームチップは、翻訳後修飾されたタンパク質の同定に役立つ。本発明の方法によって検出可能な翻訳後修飾には、メチル化、アセチル化、ファルネシル化、ビオチン化、ステアロイル化、ホルミル化、リポ酸、ミリストイル化、パルミトイル化、ゲラニルゲラニル化、ペグ(PEG)化、リン酸化、硫酸化、グリコシル化、糖による修飾、脂質化、脂質による修飾、ユビキチン化、SUMO化、ジスルフィド結合、システイニル化、酸化、グルタチオニル化、ピログルタミン酸、カルボキシル化、および脱アミノ化があるが、これらに限定されない。さらに、ペントース、ヘキソサミン、N-アセチルヘキソサミン、デオキシヘキソース、ヘキソース、およびシアル酸で、タンパク質を翻訳後修飾することができる。
【0187】
ある実施形態において、ホスホチロシン、ホスホセリンもしくはホスホトレオニンをプローブとして用いて本発明のプロテオームチップを探索し、リン酸化相互作用物質を同定することができる。別の実施形態において、レクチン(例えば、小麦胚芽凝集素)をプローブとして用いて本発明のプロテオームチップを探索し、グリコシル化相互作用物質(例えば、N-アセチルグルコサミン)を同定することができる。リン酸化もしくはグリコシル化相互作用物質は、当技術分野で公知の方法によって検出可能である。
【0188】
また、本発明のプロテオームチップは、機能、リガンド結合、もしくは酵素活性の点で相違するタンパク質アイソフォームを同定し特徴づけるために有用である。特別の実施形態では、本発明のプロテオームチップを用いて、異なる対立遺伝子から誘導されたタンパク質アイソフォームによる様々な結合親和性を、それらの活性を他と比較してアッセイすることによって、特徴づける。
【0189】
ある実施形態において、本発明のプロテオームチップを用いてコンセンサス配列を決定することができる。例えば、プロテオームチップを結合物質(チップ上のタンパク質とは異なる種に由来するものであってよい)によってスクリーニングした後、相互作用物質のアミノ酸配列をアラインメントし、相互作用ドメインのコンセンサス配列を構築することができる。こうした情報は、例えば、結合物質-相互作用物質の相互作用の阻害剤をデザインするために、あるいは特定の相互作用物質もしくは特定クラスの相互作用物質に対する新規および/または改良型結合物質をデザインするために有用であると考えられる。
【0190】
特定の実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、カルモジュリンを用いたプロテオームチップのスクリーニングによって決定される。もうひとつの実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、アミノ酸配列I/L-Q-X-K-K/X-G-B(配列番号1)からなる。別の実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、アミノ酸配列I/L-Q-X-K-K/X-G-B(配列番号1)を含んでなる。さらに別の実施形態において、カルモジュリン結合タンパク質のコンセンサス配列は、アミノ酸配列I/L-Q-X-K-K/X-G-B(配列番号1)を含んでなり、その長さは10、15、30、50または100アミノ酸残基より小さい。
【0191】
本発明のプロテオームチップは、抗菌性、抗真菌性、抗寄生虫性もしくは抗ウイルス性が見込まれる一連の化合物を同定するためにも有用である。例えば、細胞溶解物、または他の細菌、真菌、寄生虫もしくはウイルスの調製物をプロテオームアレイに接触させ、タンパク質-プローブ相互作用を検出して同定することができる。さらに、感染性生物から得られた感染段階と非感染段階での相互作用パターンを比較することによって、宿主の感染に関わる相互作用を同定することができる。
【0192】
ファージディスプレイライブラリーのスクリーニングは、ライブラリーを本発明のプロテオームチップとともにインキュベートすることによって行うことができる。チップ上のタンパク質に結合するクローンの検出は当技術分野で公知の様々な方法(例えば、質量分析)によって実施することができるが、それによって対象のクローンを同定し、その後、対象クローンをコードするDNAを標準的な方法によって同定することができる(例えば、Amesら、1995, J. Immunol. Methods 184: 177-86; Kettleboroughら、1994, Eur. J. Immunol. 24: 952-8; Persicら、1997, Gene 187: 9-18を参照されたい)。このように、チップは、チップ上の特定のタンパク質と結合する表面成分を有する細胞を選択するうえで有用である。
【0193】
5.5 キット
本発明は、また、本発明のアッセイ計画を実施するためのキットを提供する。ある実施形態において、本発明のキットは1以上の本発明のプロテオームチップを含んでなる。こうしたキットは、さらに、1以上の容器内に、タンパク質もしくは分子の生物活性をアッセイするのに有用な試薬、タンパク質-プローブ相互作用をアッセイするのに役立つ試薬、および/または1以上のプローブ、タンパク質もしくは他の分子を含んでいてもよい。タンパク質もしくは他の分子の生物活性をアッセイするために、またはプローブとタンパク質もしくは他の分子との間の相互作用をアッセイするために有用な試薬を、プローブとともに用いて、プロテオームチップに付着させたり、またはプロテオームチップ上の1以上のウェルに入れることができる。かかる試薬は溶液または固体の状態であってよい。試薬は、当該アッセイを実施するために必要なタンパク質もしくは他の分子とプローブのいずれか一方または両方を含むことができる。
【0194】
プロテオームチップのタンパク質は、平坦な基板の表面に付着させたり、基板上のウェルに入れたり、または基板上のウェルの表面に付着させたりすることができる。ある実施形態において、キットのプロテオームチップは、基板上にすでに付着したタンパク質および/またはプローブを有する。別の実施形態において、キットのプロテオームチップは、タンパク質や他の分子の生物活性のアッセイに有用な、またはプローブとタンパク質もしくは他の分子との相互作用のアッセイに有用な、基板上のウェルにすでに付着した試薬または反応混合物を有する。さらに別の実施形態において、試薬は基板のウェルに付着していないが、ウェル内に含まれる。また別の実施形態において、試薬は基板のウェルに付着していないが、1以上の容器に入っており、これを基板上のウェルに加えることができる。別の実施形態において、キットはさらに、タンパク質または分子の生物活性をアッセイするための反応混合液を入れた1以上の容器を含んでなる。また別の実施形態において、キットは担体(例えば、ビーズ)を提供する。この担体に対象のプローブ、タンパク質もしくは分子、および/または1以上のアッセイを実施するために役立つ試薬を付着させ、その後、付着したプローブ、タンパク質もしくは試薬を有する担体をチップのウェル内に入れることができる。
【0195】
5.6 陽性判定アルゴリズムの設計
本発明はまた、シグナルが陽性であるかどうかを判定する方法を提供する。シグナルは、可視光、紫外線、赤外線、X線、蛍光、および比色可視化を包含するあらゆる測定可能な形をとることができるが、それらに限定されない。
【0196】
好ましい実施形態においては、シグナルを質量分析によって検出する。
【0197】
さらに、シグナルはどのような配置であってもよく、アレイ、ブロット、ゲル、およびスクリーンのような任意の形をとることができる。シグナルは定位に配置されたスポットであることが望ましい。
【0198】
別の好ましい実施形態において、シグナルは蛍光によって生じ、アレイ上に格子パターンで配置される。このように、シグナルは横列および縦列に関する位置座標を割り当てられる。横列および縦列はどのような幅の列であってもよい。
【0199】
シグナルを選別する第1のステップは、各スポットに関する局所フォアグラウンドおよびバックグラウンドシグナルを算定することである。局所フォアグラウンドシグナルはスポットそのものから発せられるが、局所バックグラウンドシグナルはそのスポットに直に接している領域から発せられる。正味のシグナルは、局所フォアグラウンドシグナルから局所バックグラウンドシグナルを差し引いたものであるが、これを、以後のすべての計算に使用する。局所フォアグラウンドおよびバックグラウンドシグナルは、GENEPIXTMのようなソフトウェアによって確認される。
【0200】
しかしながら、チップ間のばらつき(例えば、様々な脂質結合実験に相当する)、およびチップ上の局所ばらつき(例えば、担体の不均一な拡散による)によって、結果的に正味のシグナル強度に一層のばらつきが生じ、チップごとに正味のシグナル分布が異なることがある。チップ間のばらつきを補正するために、異なるチップから得られた正味のシグナルを共通の値域に基準化する必要がある。数個のチップのうち1個を基準として選び、各チップの正味のシグナル分布を、基準チップの正味のシグナル分布の値域および形状に基準化することを目標とする。
【0201】
例えば、各チップの正味シグナル分布の下四分位値、中位値、および上四分位値を計算する。次に、それぞれのチップについて、正味シグナルの中位値を、各スポットの正味シグナルから差し引く。さらに、各チップにつき基準化倍率を算定するが、これは特定のチップの上四分位値と下四分位値の差と、基準チップの上四分位値と下四分位値の差との比率に等しい。これは、基準チップに対する倍率が1に等しいことを意味する。次いで、各チップに関する正味のシグナルに、そのチップに特異的な倍率を乗じて、基準化された正味のシグナルを算定する。
【0202】
アレイ上の局所ばらつきを補正するために、それぞれのスポットについて「近傍減法」(neighborhood subtraction)を行うことができる。例えば、近傍領域はある単一のスポットの上下2列および左右2列の範囲と定義することができる。次に、この領域の中位値シグナルを、そのスポットのシグナルから減じて、そのスポットの近傍と比較した過剰シグナルを算出する。好ましくは、中位値があまり影響を受けず、その近傍領域のバックグラウンドシグナルを良好に表すように、いずれの近傍領域においても高いシグナル強度を示すスポットの数が十分少なくなるようにする。
【0203】
基準化された正味シグナルに近傍減法を適用して、基準化された過剰シグナルを得る。次のステップで、並列サンプルの基準化過剰シグナルを比較する。2つの並列サンプルの基準化過剰シグナルの平均値と、並列サンプルの一方の基準化過剰シグナルとの差が、基準化過剰シグナルの誤差の標準偏差の3倍より大きいならば、その2つの並列サンプルに属するスポットは、更なる分析から除外される。その後、残りのスポットおよびそれらの基準化過剰シグナルが、選別されたシグナルの集合に相当する。
【0204】
基準化された過剰シグナルの誤差の分布およびその標準偏差は、次のようにして算出可能である。並列サンプルの完全な集合に対して線形回帰を行い、並列サンプル間の一般線形関係を決定する。次に、それぞれの並列サンプルについて誤差の値を算出する:
εG = |G2−Gm|
ここで、G1およびG2は2つの並列サンプルの、2つの基準化された過剰シグナルを表し、Gm = (G1 + G2)/2であり、
そして関数f(G1) = a*G2 + b は、線形回帰によって決定されるパラメーターaおよびbを伴う、2つの並列サンプルG1およびG2の間の一般線形関係である。
【0205】
誤差の値の完全な集合は、すべての並列サンプルに関する誤差の値の集合であって、基準化された過剰シグナルに関する誤差の値の分布を表す。そこで、このような分布の標準偏差を算定することができる。
【0206】
最終的に、並列サンプルの選別されたシグナルの平均値が、基準化された過剰値の誤差よりも3標準偏差大きいならば、この1対の並列サンプルは陽性であるといえる。(ここで、留意すべきは、この閾値は、並列サンプルが選別されたシグナルの集合から除外されるべきかどうかを決定する閾値とは無関係であるということである。)
このような選別手順の後、選別されたシグナル(G)は、GSTシグナル(R)によって正規化され、比r = G/Rをもたらし、これはタンパク質量当たりの結合の尺度となり、異なるタンパク質間での結合シグナルの比較を可能にする。特異的結合の比rは、GおよびRシグナルの両者の誤差εGおよびεRに影響される。モンテカルロ(Monte Carlo)法によって、この比について90%および95%の信頼区間を算出することができる。
【0207】
r値の誤差eGは誤差εGおよびεRに関連している:
r + εR = (G + εG)/(R +εR)
ここで、εRは比rの誤差を表す。
【0208】
モンテカルロ法の場合は、εGおよびεRの両者の分布が明らかでなくてはならない。eGの分布は、上記の説明のように計算することができる。eRの分布は、入力として並列サンプルのGSTシグナル対を使用することによってeGと同様の方法で算出することができる。
【0209】
モンテカルロ法は次の通りである:
r + εRに対する信頼区間を決定するために、r + eRの無作為サンプルの母集団を算定する必要がある。これらは、εGおよびεRの無作為サンプルから導くことができる。εGの無作為サンプルは、次のように算定される。0から1の間に一様に分布する乱数のサンプルを、標準乱数発生器によって計算する。εGの分布から、eGの逆累積分布関数を標準法によって決定する。この関数の独立変数として乱数のサンプルを使用することで、εGに対する乱数サンプルの集合を得る。同様に、εGに対する無作為サンプルの集合を計算することができる。このようなεGおよびεRの無作為サンプルを式r +εR= (G +εG)/(R +εR)に結合させて、r +εRに対する無作為サンプルの母集団を作ることができる。
【0210】
ある実施形態において、本発明は、シグナルが陽性であるかどうかを判定する方法を提供するが、これは、次のステップを含んでなる。すなわち、それぞれのスポットについて局所的にフォアグラウンドおよびバックグラウンドシグナルを測定して、それらの差から正味のシグナルを決定し;第1および第2の正味シグナル分布に関する下四分位値、中位値、および上四分位値を決定し;第1の正味シグナル分布から第1の中位値を減じ、さらに第2の正味シグナル分布から第2の中位値を減じて、それぞれ第1および第2の減算値を得;第1の減算値を、第1のシグナル分布に関する上四分位値と下四分位値との差で割り算し、さらに第2の減算値を、上四分位値と下四分位値との差で割り算して、それぞれ第1および第2の規準化された値を得;近傍領域の規準化されたシグナル分布に関する局所中位値を算定し、ただし、前記近傍領域はその領域内に複数のサイトを含んでなり;規準化されたシグナルから局所中位値を減じて、規準化された過剰値を得;そして、サンプル値の1つとそれらの平均値との差が、規準化された過剰値の誤差の3標準偏差より大きいならば、規準化された過剰値の並列サンプルを除外する。
【0211】
選別された値を用いて、シグナルが陽性であるかどうかを判定することができる。並列サンプルの選別された値の平均が、規準化された過剰値の誤差の標準偏差の3倍より大きいならば、並列サンプルは「陽性」と称される。選別された陽性シグナルを、次の式によって正規化することができる:
r = G/R
ここで、Gは選別された値であり、RはGSTシグナルであり、そしてrは、タンパク質の量当たりのシグナルを表し、様々なスポットの間で比較することができる。比rはGおよびRの誤差に影響されやすい。次の式の信頼区間を計算することによって、このような影響されやすさを評価することができる:
r + εr = G + εG/R + εR
εGはGの誤差、εRはRの誤差、そしてεrはrの誤差である。
【0212】
特定に実施形態において、陽性シグナルはタンパク質-プローブ相互作用を示す。別の特定の実施形態において、近傍領域は、シグナルの上2列、下2列、左2列、右2列である。もう1つの特定の実施形態において、二つの並列サンプルから得られるデータポイントが更なる分析から除外されるが、この場合、前記サンプルの規準化された過剰シグナルとそれらの平均値との差が、規準化された過剰シグナルの誤差の3標準偏差より大きい。データポイントは、それが明らかに人為的結果である場合にも、除外される。
【0213】
6. 実施例
ハイスループット技術によって、出芽酵母由来の5800以上のタンパク質からなる確定した集団を調製し、タンパク質-タンパク質、タンパク質-DNA、タンパク質-RNA、およびタンパク質-リポソーム相互作用を含めた多くの活性についてスクリーニングした。多数の新規活性が同定され、以前には性質の明らかになっていなかった既知遺伝子に関する新たな情報が得られた。
【0214】
酵母プロテオームの研究を進めるために、5800のオープンリーディングフレームをクローニングして、過剰発現させ、それらに対応するタンパク質を精製した。このタンパク質をスライドガラス上に高い空間密度でプリントして、酵母プロテオームマイクロアレイを作製し、タンパク質およびリン脂質と相互作用するそれらの能力についてスクリーニングした。多数の新規なカルモジュリンおよびリン脂質-相互作用タンパク質が同定された;カルモジュリン結合タンパク質の多くに共通する推定結合モチーフを同定した。これらの研究から、完全な真核生物プロテオームのマイクロアレイを作製して、多数の生化学的活性についてスクリーニングすることが可能であり、結果として多くの新規タンパク質機能/相互作用の同定に至ることが実証される。こうしたマイクロアレイをスクリーニングして、タンパク質の翻訳後修飾を検出することもできる。
【0215】
6.1 酵母培養物の調製
1. −80℃にて96ウェルプレートで保存された酵母グリセロールストックを、96-プロンガーを用いてURA寒天平板(Omni, 米国)に接種した。
【0216】
2. 培養物を寒天上で30℃にて48時間増殖させると、目に見えるコロニー(直径2 mm)が認められる。
【0217】
3. 96-プロンガーを用いて、酵母細胞を寒天平板から96ウェル2 mlボックスに接種したが、このボックスには、それぞれのウェルにいずれも300μl URA-/ラフィノース液体培地および直径2 mmのガラス玉が入っており、これは均一な増殖を促進する。
【0218】
4. 激しく撹拌(300 rpm)しながら、30℃にて、およそ16時間で、培養物のO.D.600が4.0に達した後、3 mlの培養物を得るために、同じ菌株15μlを、4つの別々のボックスに入っている750μlのURA-/ラフィノース液体培地に接種した。再び、通気および均一な増殖を達成するために、各ウェルに同じガラス玉を入れた。激しく撹拌しながら30℃にて細胞を増殖させた。
【0219】
5. 12〜16時間増殖させた後、培養物のO.D.は0.6から0.8に達するはずである。O.D.が1.0を超えていれば、培養物は廃棄した。細胞を誘導するために、自動化液体操作装置(Q-Fill, Genetix, UK)を用いて、40%ガラクトースストックを各ウェルに最終濃度2%となるように添加した。この培養物を撹拌しながら4時間30℃にて誘導した。
【0220】
6. 3000 rpmで2〜10分間回転させることによって細胞を集め、細胞ペレットをボルテックス混合によって100〜1000μlの冷水に再懸濁した。次に、同じ菌株の細胞を4ウェルから1つに併せた。細胞を遠心によって集め、氷上でプロテアーゼインヒビターを含まない100〜1500μlの冷溶解バッファーに再懸濁した。洗浄した細胞を短時間の遠心によって集め、溶解バッファーを捨てた。洗浄した半乾燥の培養物を直ちに−80℃の冷凍庫で保存した。この培養物は何週間も保存することができる。
【0221】
6.2. 96ウェル方式によるタンパク質の精製
1. 96ウェルボックスに入れた凍結培養物を−80℃から氷に移し、100〜300μlのジルコニアビーズ(直径0.5 mm、BSP、ドイツ)を各ウェルに加えた。培養物がまだ凍っている間に、新調製のプロテアーゼインヒビターを含有する100〜500μlの溶解バッファーを添加した。各ウェルを密閉するためにキャップマットを使用した。氷上で5〜25分間培養物を融解させた後、96ウェルボックス内の細胞を氷上で1〜5分間の間隔をおいて3〜6回、20〜60秒ボルテックス混合した。酵母の細胞壁を効果的に破壊するために、さらに多くのプレートを同時に処理するために、ペイントシェーカー(HARBILTM 5G HG、最大積載量36 kg、圧力および振盪時間は調節可能、速度は200回/分に固定)を使用して試料を激しく撹拌した。
【0222】
2. 3000rpmで2〜10分間回転させた後、ワイドオープンチップ(wide-open tip)(Fisher, USA)を用いて上清を集め、96ウェルボックスの上に置いた96ウェルフィルタープレート(Whatman, 米国、Whatman UNIFILTERTM、カタログ番号7700-1801、容量はウェル当たり800μlで、親水性PVDFフィルターを有する)に移し入れた。
【0223】
3. より多くのタンパク質を得るために、新調製のプロテアーゼインヒビターを含有する100〜500μlの溶解バッファーを細胞破砕片に加え、ステップ1および2を繰り返した。
【0224】
4. 併せた細胞溶解物を3000 rpmで10〜30分間回転させて、フィルタープレートを通して清潔な冷却96ウェルボックスに入れた。それぞれのウェル内の濾過された溶解液の容量は、ざっと200〜1000μlであった。
【0225】
5. 一方で、必要量のグルタチオンビーズ(試料当たり約10〜50μlのビーズ)(Amersham、米国)を4回、プロテアーゼインヒビターを含まない冷溶解バッファーで洗浄し、最終的に元の容量の5倍となるように新調製のプロテアーゼインヒビターを含有する溶解バッファーで再懸濁した。
【0226】
6. 100μlの洗浄したグルタチオンビーズを各ウェルに加え、キャップマットでしっかりと密閉した。このビーズを溶解物とともにローラードラム上で1時間4℃にてインキュベートした。最適な混合を得るために、ボックスをローラードラム上で360度回転させた。
【0227】
7. 3000 rpmで10〜60秒間回転させてビーズを集め、上清を捨てた。ビーズを200〜800μlのプロテアーゼインヒビター含有洗浄バッファーIで1回、インヒビターなしで2回洗浄した。
【0228】
8. 次に、ビーズを200〜800μlの洗浄バッファーIIで3回洗浄した。バッファーを完全に除去した後、20〜50μlの溶出バッファーを各ウェルに添加した。溶出ステップで使用するフィルタープレートは、タンパク質に対する親和性が低い材料から構成されていた(MILLIPORE MULTISCREENTM、カタログ番号MADVN6550、容量は200μl/ウェルで、親水性PVDFフィルターを有する)。ボックスを短時間ボルテックス混合してビーズを再懸濁し、4℃にて1時間ローラードラム上でインキュベートした。
【0229】
9. 溶出液/ビーズスラリーを冷フィルタープレート(Millipore、米国)に移し、4℃にて3000 rpmで0.5〜2分間フィルタープレートを通して回転させることによって、溶出液を96ウェルPCRプレートに集めた。
【0230】
10. それぞれの精製タンパク質を3つの96ウェルPCRプレートに分注し、直ちに−80℃冷凍庫に保存した。
【0231】
溶解バッファー
30-300 mM Tris pH 7.5
50-300 mM NaCl
0.1-10mM EGTA
0.01-1.0% TritonX-100
0.01-1% β-メルカプトエタノール(BME)
0.1-3 mM フッ化フェニルメチルスルホニル(PMSF)
Rocheプロテアーゼインヒビター錠剤(EDTA含有)
BME, PMSFおよびインヒビター錠剤は新たに添加した。
【0232】
洗浄バッファーI:
30-300 mM Tris pH 7.5
300-600mM NaCl
0.1-10mM EGTA
0.01-1.0% TritonX-100
0.01-1% β-メルカプトエタノール(BME)
0.1-3 mM PMSF
Rocheプロテアーゼインヒビター錠剤(EDTA含有)
BME, PMSFおよびインヒビター錠剤は新たに添加した。
【0233】
洗浄バッファーII:
50-200 mM HEPES pH 7.5
50-300 mM NaCl
1-15% グリセロール
【0234】
溶出バッファー:
50-200 mM HEPES pH 7.5
50-200 mM NaCl
20-40% グリセロール
5-40 mM グルタチオン(還元型)
溶出バッファーは約pH=7.5とすべきである。
【0235】
6.3 ハイスループット96アレイフォーマットでのタンパク質の製造方法
ほぼすべての酵母タンパク質を含有する酵母プロテオームマイクロアレイを作製し、多数の生化学的活性についてスクリーニングした。5800の酵母ORF(全体の93.5%)からなる高品質のコレクションを、組換えクローニング(Mitchellら、1993, Yeast 9:715)によって酵母の高コピー数発現ベクターにクローニングした。酵母タンパク質をそのアミノ末端でGST-HisX6と融合させ、ガラクトース誘導性GAL1プロモーター(Zhuら、2000, Nat. Genet. 26: 283; Mitchellら、1993, Yeast 9:715)の制御下で酵母において発現させた。酵母発現菌株は、正しい酵母ORFがインフレームでGSTに適正に融合していることがDNA配列決定によって明らかにされた個別のプラスミドを含有する。手短に言えば、酵母ORFをPCRによって増殖し、酵母細胞にベクターとともに同時形質転換して発現クローンを生成した。このプラスミドを大腸菌(E. coli)でレスキューし、ベクター-インサート結合部分の配列を決定した(図1A)。クローニングされたORFが目的のORFでない場合、もしくはフレームシフトが検出されたならば、クローニングサイクルを繰り返した。ひとたび構築物が確認されたならば、プラスミドDNAを酵母および大腸菌に再導入して、今後の分析のための永久ストックを作製した(Zhuら、2000, Nat. Genet. 26: 283)。クローニングサイクルを繰り返すことによって、5800のユニークな酵母ORFをクローニングすることに成功したが、これは全体の93.5%に当たる。
【0236】
生化学的分析を目的として精製タンパク質を生成するために、96ウェルフォーマットでタンパク質を調製するための強力なハイスループット精製法を開発して、最適化した。グルタチオン-アガロースビーズを用いて、酵母抽出物を調製し、融合タンパク質を精製した(96ウェルフォーマットタンパク質精製プロトコール、すべての実験から得られた結果の完全な一覧、および陽性判定アルゴリズムの設計の詳細については、公開ウェブサイトspine.mbb.yale.edu/protein_chips/にアクセスされたい)。溶解バッファーおよび最初の洗浄液は、精製されたタンパク質が確実に脂質を含まないようにするため、0.1%のTritonを含有していた。本発明の方法を用いると、少なくとも1152タンパク質試料を10時間足らずのうちに細胞から調製することができる。
【0237】
精製されたタンパク質の質および量は、60の無作為抽出試料のイムノブロット分析によってモニターされた(図1B)。80%を上回る菌株が、予想分子量をもつ融合タンパク質を検出可能な量で生産した。手動プリンティング装置を用いて、3 nlの1152の精製タンパク質を二重反復でスライドガラス(19)上にスポットし、ポリクローナル抗GST抗体を用いてタンパク質を検出した。85%を超える試料が、バックグラウンドに対して目視可能なシグナルを示し、イムノブロット分析と符合した。本発明の方法を用いると、6144(64×96ウェルボックス)の酵母菌株から得られた融合タンパク質を2週間足らずで精製することができる。
【0238】
6.4 プロテオームマイクロアレイの作製方法
プロテオームチップを作製するために、5800の異なる酵母タンパク質に対応する6566のタンパク質調製物を、市販のマイクロアレイヤーを用いてスライドガラス上に二重反復でプリントした。対照として、既知量のGSTもプリントした。2つのタイプのスライドガラスを使用した。最初の実験ではアルデヒド処理済みの顕微鏡用スライドガラスを使用したが(MacBeathら、2000, Science 289: 1760)、この場合、融合タンパク質のN末端もしくは他の残基の1級アミンを介して融合タンパク質をスライドガラス表面に付着させたため、結果として、表面上のタンパク質は比較的ランダムな配向となった。その後の実験では、ニッケルコーティングスライドガラス上にタンパク質をスポットした。この場合、融合タンパク質は、そのHisX6タグを介して付着するので、クローニングされた融合タンパク質の酵母部分は、必然的に表面から離れて一様に方向付けられる。両タイプのスライドガラスはいずれも成功裏に使用できたが、本発明者らの特定のタンパク質調製物に関しては、ニッケルコーティングスライドガラスの方が有意に優れたシグナルを与えた(図2A)。
【0239】
どのくらいの量の融合タンパク質がそれぞれのガラス表面に共有結合しているかを判定するために、また、タンパク質結合の再現性を評価するために、抗GST抗体でチップをプローブした。93.5%を超えるタンパク質試料が、バックグラウンドを有意に上回るシグナルを与えた(すなわち、タンパク質は約10 fgより多い)。スライドガラス上にプリントされた既知量のGSTと比較して、約90%のスポットが約10 fgから950 fgまでのタンパク質を含有することが明らかになった。図2Aに示すように、プロテオームチップ上での蛍光標識抗体によるタンパク質の検出は極めて高感度であり、すなわち、スライドガラス上にスポットされるのは3 ml培養物から精製された精製タンパク質のわずか1/10,000であるにもかかわらず、シグナル対ノイズ比が高かった。この結果は、標準的な顕微鏡用スライドガラス(2.5cm×7.5cm)の半分の面積に、優れた分解能で、13,000のタンパク質試料をスポットすることが実現可能であることを実証する(図2Aおよび2B)。タンパク質のスポッティングの再現性を試験するために、それぞれ対をなす二重反復スポットから得られるシグナルを相互に比較した。図2Cの鋭いスパイクが実証するように、シグナルの95%が平均の5%以内に存在した(10)。
【0240】
6.5 プロテオームマイクロアレイの使用方法
いくつかの典型的な生物活性、すなわち、タンパク質-タンパク質相互作用、タンパク質-核酸相互作用、およびタンパク質-脂質相互作用をプローブすることによってプロテオームチップを試験した。
【0241】
一般的には、アッセイのためにプロテオームチップを次のように調製した。プロテオームチップをブロックするため、プリントされたスライドガラスを、BSA(PBSバッファー中の1〜3%(W/W) BSA;SIGMATM, 米国)もしくはグリシンブロッキングバッファー(30〜300 mMグリシン;50〜300 mM Tris, pH 6.5〜8.5; 50〜300 mM NaCl; SIGMATM, 米国)のいずれかにタンパク質側を上にしてゆっくりと浸漬した。粒子を除去するために2μmフィルターユニットを通してバッファーを濾過した。炭水化物結合タンパク質をプローブする場合にはグリシンの方が好ましい。このスライドガラスをブロッキングバッファー中で4℃にて一晩、振盪せずにインキュベートした(ブロッキングバッファーの妨害により、結果的にガラス表面にタンパク質線条(streaks)が生じる可能性がある)。
【0242】
プローブタンパク質は、一般的に、次のように調製した。グルタチオンビーズを用いたアフィニティーカラムによって、50 ml培養物から標準法を用いてビーズからの溶出なしに酵母タンパク質を精製した。タンパク質ビーズを冷PBSバッファー(pH8.0)(SIGMATM, 米国)で3〜5回洗浄した。約1 mlのスルホ-NHS-LC-LC-ビオチン(PIERCETM カタログ番号21338、米国)を0.1〜50mg/mlの濃度となるようにPBS(pH 8.0)に溶解して、それをグルタチオンビーズに添加し、4℃にて2時間インキュベートした。ビーズを冷PBSバッファー(pH 8.0)で5回洗浄し、100〜500 mlの溶出バッファー(50〜200 mM, HEPES pH7.5; 50〜200 mM NaCl; 20〜40%グリセロール;5〜40 mMグルタチオン)で溶出した。比較的弱くビオチン化されたタンパク質をもたらすようなプロトコールが望ましい。様々な程度にビオチン化されたタンパク質のバッチを、以後使用するためにプールした。
【0243】
6.5.1 カルモジュリン相互作用タンパク質の同定
タンパク質-タンパク質相互作用を試験するために、カルモジュリンで酵母プロテオームをプローブした(11)。カルモジュリンは、カルシウムによって調節される多くの細胞プロセスに関与する高度に保存されたカルシウム結合タンパク質であって、多くの既知の物理的パートナーを有する(Hookら、2001, Annu. Rev. Pharmacol. Toxicol. 41: 471)。カルモジュリンプローブをビオチン化し、結合したプローブをCy3標識ストレプトアビジンによって検出した。対照として、Cy3標識ストレプトアビジンのみを用いて、酵母プロテオームをプローブした。
【0244】
一般に、タンパク質-タンパク質相互作用は、次のようにアッセイすることができる。ブロックされたプロテオームチップをPBSバッファーで3〜5回洗浄し、ガラス表面上の余分な液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去する。200μlのビオチン化プローブをプロテオームチップに添加し、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。閉じこめられた気泡を取り除いた後、チップを加湿チャンバー内で室温(RT)にて1時間インキュベートする。カバーガラスを取り外すために、チップを多量のPBSバッファー(>50ml)中に浸すと、カバーガラスは浮き上がって離れるはずである。次にチップを第2のPBS槽(>50ml)に移動し、室温で振盪しながら、3×5分間、洗浄した。チップ表面上の余分な液体を除去した後、少なくとも150μlのCy3-もしくはCy5-結合ストレプトアビジン(PIERCETM, 米国;希釈率1:2000から1:4000)をその表面に加え、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。チップを暗所でRTにて、30分より長時間インキュベートする。次に、このチップを上記のように洗浄する。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥する。
【0245】
プロテオームチップを抗体でスクリーニングしようとする場合は、タンパク質-抗体相互作用を次のように検出することができる。ブロックされたプロテオームチップをPBSバッファーで3〜5回洗浄し、ガラス表面上の余分の液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去する。200μlの一次抗体(1〜3% BSAおよび0.1% TritonX-100を含有するPBSで適当に希釈する)をプロテオームチップに添加して、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。閉じこめられた気泡を取り除いた後、チップを加湿チャンバー内で室温(RT)にて1時間インキュベートする。カバーガラスを取り外すために、チップを多量のPBSバッファー(>50ml)中に浸すと、カバーガラスは浮き上がって離れるだろう。次にチップを第2のPBS槽(>50ml)に移動し、室温で振盪しながら、3×5分間、洗浄した。チップ表面上の余分な液体を除去した後、>150μlのCy3-もしくはCy5-結合二次抗体(1〜3% BSAおよび0.1% TritonX-100を含有するPBSで適当に希釈する)をその表面に加え、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。チップを暗所でRTにて30分より長時間インキュベートする。次に、このチップを上記のように洗浄する。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥する。
【0246】
ビオチン化カルモジュリンを用いて、カルシウム存在下でプロテオームチップをプローブしたところ、6つの既知カルモジュリン標的物質、すなわちCmk1p、Cmk2p、Cmp2p、Dst1p、Myo4pおよびArc35pが同定された(図3A)。Cmk1pおよびCmk2pはI型およびII型カルシウム/カルモジュリン依存性セリン/トレオニンプロテインキナーゼであり、これらはいずれも接合応答(mating response)でのシグナル伝達経路に関与する(Hookら、2001, Annu. Rev. Pharmacol. Toxicol. 41)。Cmp2p(Cna2p)は、2つの酵母カルシニュリンのうちの1つであって、in vivoでカルモジュリンと相互作用することが実証されている(Cyertら、1991, Proc. Natl. Acad. Sci. USA 88:7376)、Dst1pは転写伸長に関与する(Stirling ら、1994, EMBO J. 13:4329)、Myo4pは、ASH1転写物の適正な局在化のために必要とされるクラスVミオシン重鎖である(Bohlら、2000, EMBO J. 19:5514; Bertrandら、1998, Mol. Cell 2:437)。Arc35pは、Arp2/3アクチン構築複合体の成分であり、この複合体はアクチンの構築および機能、ならびにエンドサイトーシスに関与する(Winterら、1999, Proc. Natl. Acad. Sci. USA 96: 7288)。Arc35pは、ツーハイブリッド研究において、カルモジュリンと相互作用することが最近明らかになったが(Schaerer-Brodbeckら、2000, Mol. Biol. Cell 11:1113)、これは、Arc35pおよびカルモジュリンがin vitroで相互作用することを実証する本明細書のデータを確認するものである。検出されなかった6つの既知カルモジュリン標的物質のうち、2つはこのコレクションに集められておらず、残りの4つはGSTプロービング実験では検出不可能であった。既知の相互作用物質に加えて、このカルモジュリンプローブはさらに33の相互作用物質を同定した。これらの相互作用物質は、種々様々な異なるタイプのタンパク質を包含し(表1;公開ウェブサイトbioinfo.mbb.yale.edu/proteinchipを参照されたい)、多岐にわたる細胞プロセスにおけるカルモジュリンの役割と合致する。
【0247】
カルモジュリン結合標的物質の他に、Cy3標識ストレプトアビジンと結合する1つのタンパク質、Pyc1p、も同定された。Pyc1pは、高度に保存されたビオチン結合領域を含有するピルビン酸カルボキシラーゼ1ホモログをコードする(Menendezら、1998, Yeast 14:647)。したがって、その配列から予想されるように、Pyc1pはin vivoでビオチン化されており、これはストレプトアビジン検出法を用いたすべての実験で同定された。このように、プロテオームマイクロアレイを使用して、タンパク質の翻訳後修飾を同定することができる。
【0248】
6.5.2 カルモジュリン結合モチーフの同定
推定カルモジュリン結合ドメインを同定するために、異なるカルモジュリン結合標的物質(すなわち相互作用物質)の間で共通するアミノ酸配列を決定した(Zhuら、2000, Nat. Genet. 26:283)。39のカルモジュリン結合タンパク質のうち14が、あるモチーフを含有し、そのコンセンサス配列はI/L-Q-X-X-K-K/X-G-B(配列番号1)であるが、ここでXは任意の残基であり、Bは塩基性残基である(図3B)。ミオシンにおける関連配列、I-Q-X-X-X-X-K-X-X-X-R(配列番号16)は、カルモジュリンと結合することがすでに明らかになっている(Hommaら、2000, J. Biol. Chem. 275)。この結果は、モチーフ、I-Q-X-X-X-X-K-X-X-X-R(配列番号16)が多くのカルモジュリン結合タンパク質に存在することを実証する。このモチーフを持たない他のカルモジュリン結合相互作用物質は、他のカルモジュリン結合配列を保有すると考えられる。
【0249】
6.5.3 ATP/GTP結合タンパク質の同定
1. ブロックされたプロテオームチップを冷PBSバッファーで3〜5回洗浄し、ガラス表面の余分な液体を、KIMWIPETMの上でスライドガラスを縦にして軽くたたいて取り除く。
【0250】
2. ATPおよびGTP溶液(0.5〜5 mM ATP-Cy3, 0.5〜5 mM GTP-Cy5, 100〜400mM NaCl, 1〜30 mM MgCl2, 50-300 mM Tris, pH 7〜8.5)100μlをプロテオームチップに加え、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, OR, 米国)で覆う。閉じ込められた気泡を追い出した後、チップを加湿チャンバー内で4℃にて1時間インキュベートする。
【0251】
3. カバーガラスを取り外すために、チップを多量(>30 ml)の氷冷洗浄バッファー(100〜400mM NaCl, 1〜30 mM MgCl2, 50〜300 mM Tris, pH 7〜8.5)中に浸漬すると、カバーガラスは浮き上がって離れるだろう。次にチップを第2の洗浄槽(>50 ml)に移動して、4℃にて振盪しながら3×5分間洗浄する。
【0252】
4. チップ表面の余分な液体を除去した後、チップを4℃にて1500〜2000 rpmで5〜10分間回転させて乾燥する。
【0253】
6.5.4 DNA結合タンパク質の同定
タンパク質-DNAおよびタンパク質-RNA相互作用は、転写調節、染色体の分離および維持、ならびに、RNA輸送およびプロセシングといった、多くの基本的な生物学的機能にとって重要である(Williamson, 2000, Nat. Struct. Biol. 7(10):834-7)。プロテオームチップを用いてDNAもしくはRNA分子と結合するタンパク質を同定する可能性を探るために、酵母の全ゲノムDNAをCy3-CTPおよびKlenow断片を用いて標識し、また、ポリA+をビオチン化プソラレン誘導体(31)によって標識した。それぞれのプローブを、異なるプロテオームチップとともにインキュベートした(図3A)。
【0254】
タンパク質-核酸相互作用を次のようにアッセイした。核酸プローブをWinzelerら(1999, Science 285:901)に記載のように標識した。ブロックされたプロテオームチップをPBSバッファー中で3〜5回洗浄し、ガラス表面上の余分の液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去する。200μlの標識核酸プローブ(50〜200 mM ポリ(dC/dG);100〜300 mM NaCl; 50〜300 mM HEPES, pH 7.0〜8.5; 10 mM MgCl2)をプロテオームチップに添加して、直ちに疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)によって覆う。閉じ込められた気泡を追い出した後、チップを加湿チャンバー内暗所で室温(RT)にて1時間インキュベートする。カバーガラスを取り外すために、チップを多量のPBSバッファー(>50ml)中に浸すと、カバーガラスは浮き上がって離れるだろう。次にチップを第2のPBS槽(>50ml)に移動し、室温にて暗所で振盪しながら、3×5分間、洗浄した。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥させた。
【0255】
目視検査によって、ゲノムDNAプローブが41種のDNA結合タンパク質を同定したことが明らかになった(表1を参照されたい)。これらは、8種の既知DNA関連タンパク質および2種の予想されるDNA結合タンパク質を含んでいた。既知のDNA結合タンパク質のうち、本発明者らが見いだしたのは、3種のDNA修復タンパク質(すなわち、xrs2p、Cdc1pおよびRad51p(Costanzoら、2001, Nucleic Acids Res. 29:75))、SWI-SNF包括的転写活性化因子複合体成分(すなわち、Snfl1p(Costanzoら、2001, Nucleic Acids Res. 29:75))、クラスII転写Bur6pのNC2(Dr1/Drap1)リプレッサーのαサブユニット(Sternbergら、1987, Cell 48:567)、転写因子Spt2p(HO遺伝子転写の負の制御因子(Sternbergら、1987, Cell 48:567))、および5つのWDリピートを含有し、特定のプロモーター領域と結合するMet30p(Thomasら、1995, Mol. Cell Biol. 15:6526)である。本発明者らはTrm2pも同定したが、これはDNAおよびRNAのいずれとも結合し、DNA修復およびRNAプロセシング/修飾に関与する(Sadekovaら、1996, Curr. Genet. 30(1):50-5)。
【0256】
8種のタンパク質が核酸結合タンパク質として同定されたが、これらは機能が不明である。これらのうち3種は、アミノ酸配列および本アッセイの結果に基づいて、DNA結合活性を有する可能性が高い。Ycr087cは共通のDNA結合モチーフであるジンクフィンガードメインを含む。Yhr54cは転写因子Rsc3pとの配列相同性を共有する(Costanzoら、2001, Nucleic Acids Res. 29:75)。Ybr025cは推定ヌクレオチド結合活性を有すると考えられる(Costanzoら、2001, Nucleic Acids Res. 29:75)。本アッセイは、これら8種のタンパク質がin vitroでDNAに結合し、おそらくin vivoで核酸に結合することを示す。
【表1】
【0257】
【0258】
【0259】
【0260】
【0261】
【0262】
【0263】
6.5.5 RNA結合タンパク質の同定
Trm2pに加えて、RNA結合活性を有する他の相互作用物質が同定された。例えば、イソロイシル-tRNA合成酵素(Ils1p)およびリボソームタンパク質(Rpl41B)はいずれもRNA分子と結合することが明らかになっている(Lankerら、1992, Cell 70:647; Suzukiら、1990, Curr. Genet 17:185)。翻訳に関与する他のタンパク質が、Rpl41b、Sqt1p、およびGcd11pのように、RNA結合タンパク質のスクリーニングで陽性と判定された。Rpl41bはリボソーム大サブユニットの成分である。Sqt1pはRpl10pと相互作用し、Rpl10pはRNA結合タンパク質である(Costanzo, 2001, Nucleic Acids Res. 29:75)。Gcd11pは翻訳開始因子eIF2のγサブユニットであるが、GTP結合活性を有し、ツーハイブリッド分析によって実証されるように、Lsm1-7複合体(U6特異的snRNP)に結合する(FromontRacineら、2000, Yeast 17:95)。DNAプローブと結合する他の2つのタンパク質標的物質もヌクレオチドと結合する。すなわち、Thi13pはヌクレオチド代謝に関与し、Ypt31pはGTP結合活性を有する(Costanzo, 2001, Nucleic Acids Res. 29:75)。DNAプローブによって同定されたRNA結合およびヌクレオチド結合タンパク質は、DNAとの低親和性相互作用を反映している可能性があり、あるいはまた、これまでに認識されていないDNAに対して標準の親和性を示すかもしれない。
【0264】
対数増殖期の細胞から得られたRNA-ポリA+をビオチンで標識し、プロテオームマイクロアレイをプローブするために使用した。Cy3-ストレプトアビジンを用いて陽性シグナルを検出した。19の標的相互作用物質が同定されたが(表1を参照)、これは、U1 snRNP成分(Smd1p)、mRNA輸送タンパク質と結合することが判明しているタンパク質(Sxm8p)、C2-H2ジンクフィンガータンパク質(Azf1p)、およびTos8pを包含し、Tos8pは、その配列から、RNAスプライシングおよびプロセシングに関与することが示唆された(Schwikowskiら、2000, Nat. Biot. 18:1257)。おそらく直接もしくは間接的にRNAと相互作用するタンパク質に加えて、本発明者らは2つの転写因子(Bur6pおよびTos8p)、1つの転写サイレンサ(Hst3p)、既知ORFによってコードされる6つの他のタンパク質、および特徴づけられていないORFに由来する7つのタンパク質を発見した。未知のORFの1つは最高のシグナルを示した(Yer152c)。ただ1つの相互作用物質であるBur6pのみがDNAおよびRNAプローブの両者と結合したことに注目することが重要である。他の相互作用物質は、二本鎖DNAプローブもしくはRNAプローブのいずれか一方でのみ検出されたが、これは、大半のタンパク質がDNAもしくはRNAのいずれかと特異的に結合する可能性が高いことを示唆する。
【0265】
要約すれば、41種の標的相互作用物質のうち、18種は核酸相互作用タンパク質である;11種は既知もしくは推定DNA結合タンパク質であり、3種はRNA結合タンパク質であり、そして4種はヌクレオチドと結合する。
【0266】
6.5.6 脂質結合タンパク質の同定
本発明のプロテオームチップは、例えばタンパク質-薬物相互作用およびタンパク質-脂質相互作用のような、他の実験手法が利用しにくいと思われる活性を同定するのに有用である。実際、リン脂質と結合するタンパク質に関するゲノム全体にわたる分析は、いかなる生物種についてもこれまで検討されたことがない。ホスファチジルイノシトール(PI)と相互作用するタンパク質を研究するために本発明のプロテオームチップを使用した。ホスファチジルイノシトールは、細胞膜の構成成分としての役割に加えて、成長、分化、細胞骨格再編成、および膜輸送を含めて、様々な細胞プロセスを制御する重要な第2メッセンジャーであって、核、液胞、および原形質膜に存在している(Odorizziら、2000, TIBS 25: 229; Frumanら、1998, Annu. Rev. Biochem. 67: 481; Martin, 2000, Annu. Rev. Cell Dev. Bio. 14:231; Weraら、2001, FEMS Yeast Res. 1406:17)。ホスファチジルイノシトールは細胞内に一時的に、しかも小量で存在することが多いため、酵母で広範に特徴づけられたことはなく、したがって、特定のリン脂質と結合するタンパク質についてはほとんど知られていない(Odorizziら、2000, TIBS 25: 229; Frumanら、1998, Annu. Rev. Biochem. 67: 229; Martin, 2000, Annu. Rev. Cell Dev. Bio. 14:231; Weraら、2001, FEMS Yeast Res. 1406:1)。
【0267】
酵母でPI結合タンパク質を同定するために、リポソームをプローブとして使用したが、これは、リポソームが相互作用物質をアッセイするために最適の生理的状態を提供するためである。6つのタイプのリポソームを使用した。それぞれが、1%(w/w)N-(ビオチノイル)-1,2-ジヘキサデカノイル-sn-グリセロ-3-ホスホエタノールアミン、トリエチルアンモニウム塩(ビオチンDHPE)とともに、ホスファチジルコリン(PC)を含有する。ビオチン化脂質はCy3-ストレプトアビジンによって検出可能な標識として機能する(21)。PCに加えて、5つのタイプの他のリポソームは、5%(w/w) PI(3)P、PI(4)、PI(3,4)P2、PI(4,5)P2もしくはPI(3,4,5)P3のいずれかを含有する(図3A)。それぞれの上記リン脂質は、PI(3,4,5)P3を除いて、酵母で発見されたものである(Odorizziら、2000, TIBS 25: 229; Frumanら、1998, Annu. Rev. Biochem. 67: 481; Martin, 2000, Annu. Rev. Cell Dev. Bio. 14:231; Weraら、2001, FEMS Yeast Res. 1406:1)。
【0268】
タンパク質-リポソーム相互作用を下記のようにアッセイした。クロロホルム中の適当量の各脂質を、窒素ガス下で混合して乾燥させた。この脂質混合物をボルテックス混合によってTBSバッファーに再懸濁した。超音波処理によってリポソームを形成した。ブロックされたプロテオームチップをPBSバッファーで3〜5回洗浄し、ガラス表面上の余分の液体を、KIMWIPETM上でスライドガラスを縦にして軽くたたくことによって除去した。200μlのリポソーム溶液をプロテオームチップに添加して、直ちに、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)で覆った。閉じこめられた気泡を追い出した後、チップを加湿チャンバー内でRTにて1時間インキュベートした。カバーガラスを取り外すために、チップを多量のPBSバッファー(50ml)中に浸すと、カバーガラスは浮いて外れることになる。次にチップを第2のPBS槽(>50ml)に移動し、室温で振盪しながら、3×5分間、洗浄した。チップ表面上の余分な液体を除去した後、約100μlのCy3-もしくはCy5-結合ストレプトアビジン(PIERCETM, USA; 1:2000から1:4000希釈)をその表面に加え、疎水性プラスチックカバーガラス(GRACE BIO-LABSTM, 米国)で覆った。チップを、30分より長時間、暗所で、RTにてインキュベートした。次に、このチップを上記のように洗浄した。チップ上の液体を完全に取り除くために、チップをRTにて5〜10分間1500〜2000 rpmで回転させて乾燥した。
【0269】
6つのタイプのリポソームが、バックグラウンドより有意に高いシグナルを発生した合計150の異なる相互作用物質を同定した。陽性シグナルの識別に役立つように、アルゴリズムが考案された(42)。52(35%)の相互作用物質(すなわち、脂質結合タンパク質)が特性決定されていないタンパク質に相当し、それゆえ、これらのタンパク質にこの最初の生化学的活性を帰属させた。上記の結果はまた、これまで性質が明らかにされていない多くのタンパク質が、潜在的に重要な生化学的活性を有することを示すものである。
【0270】
52の特性決定されていないタンパク質のうち、13(25%)は膜に結合していると予想される(Gerstein, 1998, Proteins 33:518)。他の多くは、負電荷を帯びたホスファチジルイノシトールとの静電相互作用を媒介することができる塩基性ストレッチを含有する。
【0271】
98の以前に記載された相互作用物質の中で、これらの81の細胞内局在が研究されている(Costanzoら、2001, Nucleic Acids Res. 29:75)。大半は膜結合タンパク質であるか、もしくは脂質代謝に関与するかのいずれかである。より詳細には、45のタンパク質が膜結合型であり、既知のもしくは予想される膜貫通領域を有する(Gerstein, 1998, Proteins 33:518)。これらの同定された相互作用物質には、内在性膜タンパク質、脂質修飾を有するタンパク質(例えば、グリコシルホスファチジルイノシトール(GPI)アンカータンパク質Tos6pおよびSps2p(Costanzoら、2001, Nucleic Acids Res. 29:75))、プレニル化タンパク質、Gpa2pおよび接合フェロモンaファクター(Ansariら、1999, J. Bio. Chem. 274:30052)、ならびに、周辺結合タンパク質(例えば、芽頸部(bud neck)および/または細胞周辺に見いだされるKcc4pおよびMyo4p(Bohl, 2000, EMBO J. 19: 5514; Bertrandら、1998, Mol. Cell 2:437))がある。別の8個は、脂質代謝(例えば、Bpl1p)、イノシトール環のリン酸化(例えば、Kcs1p)に関わり、もしくは膜/脂質機能(例えば、トリアシルグリセロールリパーゼに対して相同性を示すYlr020cp)に関係していると予想される。
【0272】
相互作用物質(すなわち、リン脂質結合タンパク質)を、幾通りかの方法でグループ分けした(42)。最初に、相互作用物質を、それらが脂質に強く結合するか、弱く結合するかにしたがって分類したが、これはGSTの量に対するリン脂質結合シグナルに基づく(図4)。やや多くの(72%)強い脂質結合タンパク質(図4Aおよび4B)が、弱い結合タンパク質(54%)(図4Cおよび4D)と比較して特徴付けられた。強い脂質結合タンパク質の多くが膜結合型であるか、または膜に結合していると予想されるのに対して、膜に結合していると思われる弱い結合タンパク質はより少数である(図4、「膜」の列)。驚くべきことに、19の脂質結合タンパク質がキナーゼであり、このうち17はプロテインキナーゼである。さらに、この17のプロテインキナーゼのうち13は非常に強くリン脂質に結合する。
【0273】
また、ホスファチジルコリンと比較して1以上のホスファチジルイノシトールと優先的に結合するかどうかによって、タンパク質をグループ分けした。101のタンパク質が、試験したホスファチジルイノシトール脂質と同様に、もしくはほぼ同様にホスファチジルコリンと結合した(PI/PC<1.3)(図4Bおよび4D)。しかしながら、49のタンパク質が1以上のホスファチジルイノシトールに優先的に結合した(PI/PC>1.3)(図4Aおよび4C)。強い相互作用物質(すなわち、ホスファチジルイノシトール結合タンパク質)の分析によって、多くが特定のホスファチジルイノシトール脂質に特異的に結合することが明らかになった。例えば、Stp22pは、Ste2pおよびCan1pのように、温度感受性原形質膜タンパク質の液胞ターゲティングに必要であるが、優先的にPI(4)Pと結合する(Liら、1999, Mol. Cell Biol. 19:3588)。9のプロテインキナーゼは特異的にPI(4)PおよびPI(3,4)P2と強く結合するが、1つは前記脂質とは弱く結合する。Atp1pは、ミトコンドリア内膜のF1-ATPシンターゼのαサブユニットであるが、やはり優先的にPI(3,4)P2に結合する(Arnold ら、1999, J. Biol. Chem.)。Sps2pは、胞子形成の中/後期に関与し、原生胞子膜に局在するが(Chuら、1998, Science 282:699)、PI(3)Pと優先的に結合する。興味深い一例は、優先的にPI(4,5)P2と結合するMyo4pであるが、おそらくこの脂質の結合は、細胞表皮でのその相互作用および/またはその調節における重要な部分である。PI(3,4,5)P3と特異的に結合する強い脂質結合標的物質はまったく見いだされなかったが、一部のタンパク質はこの脂質と他の脂質の両者と結合した(図4)。上記の結果は、内在性膜タンパク質および周辺結合タンパク質を含めて、多くの膜結合タンパク質がin vivoで特定のリン脂質と優先的に結合することを実証する。
【0274】
予想に反して、グルコース代謝の調節因子を含めて、グルコース代謝に関与する多くのタンパク質がリン脂質に結合する。具体的には、解糖の一連のステップに関わる3つのグルコース代謝酵素、すなわちホスホグリセリン酸ムターゼ(Gpm3p)、エノラーゼ(Eno2p)、およびピルビン酸キナーゼ(Cdc19p/Pyk1p)が同定された。グルコースをグルコース6リン酸に変換するヘキソキナーゼ(Hxk1p)、およびグルコース代謝の調節因子であることが明らかになっている2つのプロテインキナーゼ(すなわち、Snf1pおよびRim15p)も、脂質結合相互作用物質として同定された。HXk1pは、その活性を促進する双性イオンのミセルと結合することが最近明らかにされた(30);Eno2pは分泌されることが明らかになり、膜とも相互作用する可能性が示唆された(31)。したがって、ある実施形態において、リン脂質を用いて、グルコース代謝に関与するステップを調節することができる。別の実施形態において、例えば、細胞膜のようなリン脂質足場上でグルコース代謝のステップを行うことによって、解糖ステップを高めることができる。
【0275】
驚くべきことに、リン脂質を含むプローブは、膜機能もしくは脂質シグナル伝達に関与すると予想されない多くのタンパク質を認識した。したがって、6つのタンパク質、すなわち、Rim15p、Eno2p、Hxk1p、Sps1p、Ygl059wpおよびGcn2pはホスホイノシトール結合について2種類の標準アッセイによってさらに検討した(Williamson, 2000, Nat. Struct Biol. 7(10):834-7)。Rim15p、Eno2pおよびHxk1pに関しては、PI(4,5)P2リポソームを最初にニトロセルロース膜に付着させ、次にこれをBSAでブロックした;異なる量のGST融合タンパク質およびGST対照を用いてこの膜をプローブし、結合したタンパク質を抗GST抗体によって検出した。図5Aに示すように、それぞれの酵母融合タンパク質はしっかりとPI(4,5)P2に結合し、量的効果を示した;GST単独では結合しなかった。本発明者らは、4つのGST融合タンパク質、Rim15p、Sps1p、Ygl059wpおよびGcn2pについて、リバースアッセイも行った(Guerraら、2000, Biosci. Rep. 20:41)。異なる量の前記精製タンパク質をニトロセルロースフィルター上にスポットし、6つの異なるリポソームによってプローブした(図5B)。結合したリポソームを西洋ワサビペルオキシダーゼ(HRP)結合ストレプトアビジンによって検出した。プロテオームチップを用いた実験と同様に、リポソームはそれぞれのタンパク質に結合し、BSA対照とは結合しなかった。Sps1pは、5種すべてのホスホイノシトール含有リポソームとほぼ同等に結合した。Rim15p、Gcn2pおよびYgl059wpは、異なるリポソームに対して、異なる親和性を示した(Rim15pについては図5Bを参照されたい)。3種のタンパク質のすべてが、PI(3)PおよびPI(4)PおよびPI(3,4)P2にもっとも強く結合した。Rim15Pに関しては、結合シグナルとRim15Pレベルの間に直線的相関関係が明らかになった(図5C)。
【0276】
さらに、未知の機能を有するいくつかのタンパク質も、強い脂質結合活性を示したが、このことは、これらが細胞内でリン脂質と結合しやすいことを示す。当業者に公知の日常的なアッセイを利用して、プロテオームチップによって同定される脂質結合相互作用物質を迅速にスクリーニングし、in vivoで脂質結合活性を測定することができる。さらに、本発明にしたがって、プロテオームチップを使用して、これらの相互作用物質がリン脂質代謝経路、もしくは情報伝達経路に関与するかどうかを判定することができる。
【0277】
要約すると、上記の結果は、本発明のプロテオームチップを使用して、脂質と結合することが以前に知られていなかったタンパク質を含めて、多くの脂質結合タンパク質を検出し、特徴づけることができることを明白に実証する。さらに、本発明のプロテオームチップによって同定されたリン脂質結合相互作用物質は、従来のアッセイで真正な脂質結合タンパク質であることが示される。
【0278】
さらに加えて、酵母ツーハイブリッド系またはDNA/オリゴマイクロアレイ技術のいずれかを使用することによって、このような相互作用物質を研究することは不可能であるため、本発明のプロテオームチップおよびその使用方法は、分子-タンパク質相互作用を探求する先駆的方法となる。
【0279】
本研究は、真核生物に関するプロテオームマイクロアレイを作製しスクリーニングすることが実現可能であることを示す。いくつかの技術的特徴を新たに非自明的に組み合わせることが、本発明を実行するうえで不可欠である。第1に、本研究で分析されるタンパク質は、DNA塩基配列決定法によって確認され、かつ純粋なプラスミドを含有する、酵母発現クローンから調製されたものである。第2に、規定されたクローンおよびタンパク質をハイスループット様式で生産することが不可欠である。本明細書に記載の方法は、5,000プロテインチップに十分なタンパク質を提供する。第3に、タンパク質は真核生物宿主において生産される。そのようにして、正しく折り畳まれて翻訳後修飾を受け、かつ/または天然のパートナーとの複合体を形成した、多数の全長タンパク質を作製することができる。イムノブロット分析によって実証されるように、精製された融合タンパク質の少なくとも80%が全長タンパク質として発現される。
【0280】
多くの相互作用物質は、直接的にではなく、結合タンパク質を介してプローブと結合することが可能である。ある場合には、相互作用物質は多量体の一部であってよい。しかしながら、このような結合タンパク質は本発明の方法によって検出し同定することもできる。そうではあるが、本発明の方法で検出された相互作用物質は、おそらく直接的にプローブと相互作用し、もしくはプローブと相互作用するタンパク質と最終的にしっかりと結合する。本発明のプロテオームチップのタンパク質は、ストリンジェントな条件(例えば、0.5M NaClによる洗浄)によって調製され、クーマシー染色は抗GST抗体によって検出可能なタンパク質のバンドのみを明らかにしたが、このことは他のタンパク質による混入が最小であることを示す。
【0281】
構築されたタンパク質のコレクションは、正しく折り畳まれた細胞外ドメインを有する分泌タンパク質を十分に代表していない可能性が高い。GSTタグおよびHisX6タグを翻訳開始コドンの上流に融合すると、シグナルペプチドを有する膜タンパク質は、分泌経路に運ばれず、適切な折り畳みもしくは修飾を受けることができない。シグナルペプチドを有する3つのタンパク質が脂質結合タンパク質のスクリーニングで同定されたが、このことは、少なくとも一部の分泌タンパク質が生産されて機能ドメインを含有することを示唆する。
【0282】
さらに、実施したハイスループット方法を用いても、すべてのタンパク質が容易に過剰生産および精製されるわけではないので、必ずしもすべての相互作用が検出されたわけではない。しかしながら、採用したプロテインアレイは、スクリーニングのために合理的なレベルで、約80%の全長酵母タンパク質を含有すると見積もられ、したがって、タンパク質相互作用物質の大半が検出されることが期待できる。
【0283】
これらの結果は、プロテオームチップによって生化学的活性をスクリーニングすることができること、それによって包括的なプロテオーム分析が可能となることを実証する。同様の方法を用いて、ヒトおよび他の真核生物における包括的ハイスループットプロテオーム分析のために、10〜100,000タンパク質からなるプロテインアレイを作製することができる。
【0284】
参考文献および注釈:
1. S. Fields, Y. Kohara, D. J. Lockhart, Proc. Natl. Acad. Sci. 96, 8825(1999); A. Goffeauら、Science 274, 563(1996).
2. P. Ross-Macdonaldら、Nature 402, 413(1999); J. L. DeRisi, V. R. Iyer, P. O. Brown, Science 278, 680(1997); E. A. Winzelerら、Science 285, 901(1999).P. Uetzら、Nature 403, 623 (2000); T. Itoら、Proc. Natl. Acad. Sci. USA 97, 1143(2000)
3. H. Zhu, M. Snyder, Curr. Opin. Chem. Biol. 5, 40(2000).
4. M. R. Martzenら、Science 286, 1153(1999).
5. H. Zhuら、Nat. Genet. 26, 283(2000).
6. G. MacBeath, S. L. Schreiber, Science 289, 1760(2000).
7. A. Caveman, J. Cell Sci. 113, 3543(2000).
8. P. Arenkovら、Anal. Biochem. 278, 123(2000).
9. D. A. Mitchell, T. K. Marshall, R. J. Deschenes, Yeast 9, 715(1993). 発現ベクターpEGHは、RGS-HisX6エピトープタグを、pEG(KG)のGST遺伝子とポリクローニング部位の間に挿入することによって作製された。前記(5)に記載のストラテジーによって酵母ORFをクローニングしたが、ただし、96ウェルフォーマットで各ステップを行った。DNA配列決定によって確認されたプラスミドDNAを酵母(Y258)および大腸菌(DH5a)の両方に再導入した。ライブラリーは5800の固有のORFを含有する。
【0285】
10. 96ウェルフォーマットタンパク質精製プロトコールの詳細、すべての実験から得られた結果の完全な一覧、およびポジティブ識別アルゴリズムのデザインは、公開ウェブサイトbioinfo.mbb.yale.edu/proteinchipで見ることができる。
【0286】
11. ビオチン化カルモジュリン(CalBiochem, 米国)をPBSバッファー中0.02 mg/mlとして0.1 mMカルシウムとともにプロテオームチップに添加し、加湿チャンバー内で室温にて1時間インキュベートした。0.1 mMカルシウムはその後のすべてのステップでバッファー中に存在した。チップをRTにてPBSによって3回洗浄した。Cy3-結合ストレプトアビジン(Jackson IR, 米国)(1:5000希釈)をチップに添加し、RTにて30分間インキュベートした。広範に洗浄した後、チップを脱水し、マイクロアレイスキャナーを用いてスキャンした;次に、GENEPIXTMアレイ デンシトメトリー ソフトウェアによってデータを取得した(Axon, 米国)。
【0287】
12. S. S. Hook, A. R. Means, Annu. Rev. Pharmacol. Toxicol. 41, 471(2001).
13. M. S. Cyert, R. Kunisawa, D. Kaim, J. Thorner, Proc. Natl. Acad. Sci. USA 88, 7376(1991).
14. D. A. Stirling, K. A. Welch, M. J. Stark, EMBO J. 13, 4329(1994).
15. F. Bohl, Kruse, EMBO J. 19, 5514(2000); E. Bertrandら、Mol, Cell 2, 437(1998).
16. D. C. Winter, E. Y. Choe, R. Li, Proc. Natl. Acad. Sci. USA 96, 7288(1999).
17. C. Schaerer-Brodbeck, H. Riezman, Mol. Biol. Cell 11, 1113(2000).
18. K. Homma, J. Saito, R. Ikebe, J. Biol. Chem. 275, 34766(2000).
19. J. Menendez, J. Delgado, C. Gancedo, Yeast 14, 647(1998).
20. G. Odorizzi, M. Babst, S. D. Emr, TIBS 25, 229(2000); D. A. Frumanら、Annu. Rev. Biochem. 67, 481(1998); T. F. Martin, Annu. Rev. Cell Dev. Biol. 14, 231(2000); S. Wera, J. C. T. Bergsma, FEMS Yeast Res. 1406, 1(2001).
21. リポソームは標準的方法によって調製された(52)。簡単に述べると、適当な量の、クロロホルム中の各脂質を混合し、窒素ガス下で乾燥させた。この脂質混合物を、ボルテックス混合によってTBSバッファー中に再懸濁した。超音波処理によってリポソームが作られた。プロテオームチップを探索するために、異なるリポソーム60mlを異なるチップ上に添加した。このチップをRTにて1時間、加湿チャンバー内でインキュベートした。TBSバッファーで3回洗浄した後、Cy-3結合ストレプトアビジン(1:5000希釈)をチップに添加し、RTにて30分間インキュベートした。
【0288】
22. それぞれのスポットについて局所強度バックグラウンドを算定するGenePixソフトウェア、および本発明者らが開発した一連のアルゴリズムを組み合わせて使用して陽性を判定した。詳細は、公開ウェブサイトbioinfo.mbb.yale.edu/proteinchipで見ることができる。
【0289】
23. M. C. Costanzoら、Nucleic Acids Res. 29, 75(2001).
24. M. Gerstein, Proteins 33, 518(1998).
25. K. Ansariら、J. Biol. Chem. 274, 30052(1999).
26. Y. Barral, M. Parra, S. Bidlinmaier, M. Snyder, Genes Dev. 13, 176(1999).
27. Y. Li, T. Kane, C. Tipper, P. Spatrick. D. D. Jennes, Mol. Cell Biol. 19, 3588 (1999).
28. I. Arnoldら、J. Biol. Chem. 274, 36(1999).
29. S. Chuら、Science 282, 699(1998).
30. A. Casamayorら、Curr. Biol. 9, 186(1999); R. Guerra, M. L. Bianconi. Biosci. Rep. 20, 41(2000).
31. M. Pardoら、Yeast 15, 459(1999).
【0290】
7. 引用文献
本明細書のすべての引用文献は、それぞれ個別の出版物または特許もしくは特許出願がそのまま全体をあらゆる目的のために参考として取り入れられることが具体的かつ個別的に示されたのと同程度に、そのまま全体をあらゆる目的のために参考として本明細書に含めるものとする。
【0291】
8. 均等物
本発明の精神および範囲から逸脱することなく、本発明の多くの修正および変更を行うことができる。当業者は、本明細書に記載された本発明の特定の実施形態に対する様々な代替、改作、および修正を認識し、もしくはルーチンの実験操作を通して確認することができるであろう。ここで、前記はいずれも本発明の範囲に含めるものとする。したがって、特許請求の範囲に記載の本発明は、こうした均等物のすべてを包含することを意味する。このため、本明細書に記載の具体的な実施形態は、ほんの一例として提供されるのであって、本発明は、添付の特許請求の範囲が権利を有する均等物の全範囲とともに、添付の特許請求の範囲によってのみ限定されるべきものである。
【図面の簡単な説明】
【0292】
【図1A】プロテインチップ技術を用いた酵母プロテオーム解析の手順を示す。
【図1B】3mlの酵母培養物から精製されたタンパク質のイムノブロット分析を示す。
【図2A】抗GST抗体を用いてプローブしたプロテオームチップを示す。
【図2B】図2Aに示したプロテオームチップの48ブロックのうちの1つの拡大図を示す。
【図2C】各ORF産物の二重反復スポット間の誤差分布を示す。
【図3A】プロテオームチップでの様々なアッセイの例を示す。
【図3B】様々なカルモジュリン標的が共有するアミノ酸配列をサーチすることにより同定された推定上のカルモジュリン結合モチーフを示す。
【図4A】30の強くかつ特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図4B】43の強くかつ非特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図4C】19の弱くかつ特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図4D】58の弱くかつ非特異的なホスファチジルイノシトール結合タンパク質の分析を示す。
【図5A−B】プロテオームマイクロアレイにより検出されたタンパク質−脂質相互作用の慣用方法を用いた確認を示す。
【図5C】膜アッセイにおける結合シグナルとRim15pの量との線形相関を示す。
【特許請求の範囲】
【請求項1】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種における既知遺伝子のうち少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記アレイ。
【請求項2】
前記複数のタンパク質が単一の種における既知遺伝子のうち少なくとも70%によりコードされる少なくとも1つのタンパク質を含む、請求項1に記載のアレイ。
【請求項3】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される全タンパク質のうち少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記アレイ。
【請求項4】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記アレイ。
【請求項5】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記アレイ。
【請求項6】
前記タンパク質がタンパク質の分類に従って前記アレイ上に組織化されている、請求項1、3、4または5に記載のアレイ。
【請求項7】
前記分類が存在量、機能、酵素活性、相同性、タンパク質ファミリー、特定の代謝経路との関連性、または翻訳後修飾によるものである、請求項6に記載のアレイ。
【請求項8】
前記タンパク質がHisタグを介して基板に結合されている、請求項1、3、4または5に記載のアレイ。
【請求項9】
前記基板がニッケルを含む、請求項1、3、4または5に記載のアレイ。
【請求項10】
前記基板がニッケルをコーティングしたスライドガラスからなる、請求項1、3、4または5に記載のアレイ。
【請求項11】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記方法。
【請求項12】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される全タンパク質の少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記方法。
【請求項13】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記方法。
【請求項14】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記方法。
【請求項15】
基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
【請求項16】
前記結合ステップに先立って、前記タンパク質を第1タグの結合パートナーと接触させることにより前記タンパク質を精製するステップを行い、前記結合ステップにおいて第2タグを用いて基板に前記タンパク質を結合させる、請求項15に記載の方法。
【請求項17】
第1タグがGSTタグであり、第2タグがHisタグである、請求項16に記載の方法。
【請求項18】
第1タグと第2タグが前記タンパク質のアミノ末端に存在する、請求項15に記載の方法。
【請求項19】
第1タグと第2タグが前記タンパク質のカルボキシ末端に存在する、請求項15に記載の方法。
【請求項20】
複数の精製されたタンパク質サンプルを調製して単離する方法であって、マルチサイトアレイの複数サイトの各サイトにおいて、
(a) 調節配列と機能的に連結された異種ヌクレオチド配列を有する真核細胞を生育させ、
(b) 該調節配列を、異種ヌクレオチド配列によりコードされるタンパク質の発現を高める誘導物質と接触させ、
(c) 該細胞を溶解して細胞溶解物を形成し、
(d) 該細胞溶解物またはそれから得られたタンパク質含有サンプルを結合物質と接触させて、結果的に該タンパク質と結合物質との複合体を形成させ、そして
(e) 該複合体から該タンパク質を単離する、
各ステップを含んでなり、その際、各ステップがマルチアレイフォーマットで実施される、上記方法。
【請求項21】
各サイトがウェルである、請求項20に記載の方法。
【請求項22】
前記タンパク質がアフィニティータグを含む融合タンパク質であり、該アフィニティータグに前記結合物質が結合する、請求項20に記載の方法。
【請求項23】
前記細胞が酵母細胞である、請求項20に記載の方法。
【請求項24】
前記溶解ステップがペイントシェーカーを使って実施される、請求項20に記載の方法。
【請求項25】
脂質結合タンパク質を検出する方法であって、
(a) 脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置に該相互作用が検出されれば、その位置に脂質結合タンパク質が存在することを示す、上記方法。
【請求項26】
前記脂質がリン脂質である、請求項25に記載の方法。
【請求項27】
前記リン脂質がホスファチジルコリンまたはホスファチジルイノシトールである、請求項26に記載の方法。
【請求項28】
前記プローブがリポソームからなる、請求項25に記載の方法。
【請求項29】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項30】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも50%を含むものであり、その際、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなし、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項31】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される少なくとも1000のタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項32】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項33】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数の融合タンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該融合タンパク質は第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項34】
前記プローブが核酸、タンパク質、小分子、候補薬物または脂質を含むものである、請求項29、30、31、32または33に記載の方法。
【請求項35】
前記核酸がRNAまたはDNAからなる、請求項34に記載の方法。
【請求項36】
前記プローブが酵母タンパク質である、請求項34に記載の方法。
【請求項37】
酵母タンパク質がMyo2、Rho1、Rho2、Rho3、Rho4、Cdc11、Cdc12またはHs17である、請求項36に記載の方法。
【請求項38】
前記プローブが抗体である、請求項34に記載の方法。
【請求項39】
前記抗体がサイクリン、キナーゼ、GST、Clb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2、またはCdc11に特異的である、請求項38に記載の方法。
【請求項40】
前記プローブがカルモジュリンである、請求項34に記載の方法。
【請求項41】
前記プローブがATP、GTP、cAMP、ホスホチロシン、ホスホセリン、およびホスホトレオニンからなる群より選択される小分子を含むものである、請求項34に記載の方法。
【請求項42】
前記プローブがホスファチジルコリンまたはホスファチジルイノシトールを含むものである、請求項34に記載の方法。
【請求項43】
前記プローブがリポソームからなる、請求項34に記載の方法。
【請求項44】
前記プローブが哺乳動物由来のものである、請求項25、29、30、31、32または33に記載の方法。
【請求項45】
前記哺乳動物がヒトである、請求項44に記載の方法。
【請求項46】
前記複数のタンパク質がヒト以外のものである、請求項44に記載の方法。
【請求項47】
前記複数のタンパク質がHisタグを介して基板に結合される、請求項25、29、30、31、32または33に記載の方法。
【請求項48】
前記基板がニッケルを含む、請求項25、29、30、31、32または33に記載の方法。
【請求項49】
前記基板がニッケルをコーティングしたスライドガラスからなる、請求項25、29、30、31、32または33に記載の方法。
【請求項50】
前記検出ステップでタンパク質との相互作用が検出されたプローブの正体を同定するステップをさらに含んでなる、請求項34に記載の方法。
【請求項51】
前記相互作用は、前記同定されたプローブが抗菌性、抗真菌性、または抗ウイルス性タンパク質であることを示す、請求項50に記載の方法。
【請求項52】
結合アッセイで使用するためのタンパク質を標識する方法であって、
(a) 該タンパク質の個々のアリコートを、ビオチン転移化合物と、アリコートごとに異なる程度にビオチン化されたタンパク質を生成する条件下および時間にわたり接触させ、そして
(b) 個々のアリコートを一緒に合わせて、異なる程度にビオチン化されたタンパク質のサンプルを得る、
各ステップを含んでなる、上記方法。
【請求項53】
結合タンパク質を検出する方法であって、
(a) 請求項52に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じているアレイ上の位置を検出する、
各ステップを含んでなる、上記方法。
【請求項54】
結合タンパク質を検出する方法であって、
(a) 請求項52に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) 該アレイに、蛍光物質とコンジュゲートさせたストレプトアビジンを接触させ、そして
(c) 蛍光が発生しているアレイ上の位置を検出する、
各ステップを含んでなり、その際、蛍光は、ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じていることを示す、上記方法。
【請求項55】
タンパク質がホスファチジルコリンに比べてホスファチジルイノシトールと優先的に結合するかどうかを判定する方法であって、
(a) ホスファチジルイノシトールを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルイノシトール結合タンパク質の存在を示すものであり、
(c) ホスファチジルコリンを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該タンパク質はステップ(a)におけると同じタンパク質を少なくとも一部含んでなり、
(d) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルコリン結合タンパク質の存在を示すものであり、そして
(e) 複数のタンパク質のそれぞれについて、ステップ(b)とステップ(d)の結果を比較する、
各ステップを含んでなる、上記方法。
【請求項56】
リン脂質が細胞における代謝経路またはシグナル伝達経路を調節するのかどうか、あるいは該代謝経路またはシグナル伝達経路が膜表面で起こるのかどうかを判定する方法であって、
(a) リン脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該複数のタンパク質は該経路の少なくとも一部を構成する1以上のタンパク質を含んでなり、そして
(b) 該プローブと該経路中のタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、該相互作用が生じれば、該プローブが代謝経路またはシグナル伝達経路を調節すること、あるいは、該経路が膜表面で起こることを示す、上記方法。
【請求項57】
カルモジュリンと結合する非天然タンパク質を製造する方法であって、下記の配列:
I/L-Q-X-X-K-K/X-G-B (配列番号1)
(ここで、Xは任意のアミノ酸であり、Bは塩基性アミノ酸である)
を含んでなる非天然タンパク質を製造することからなる上記方法。
【請求項58】
タンパク質における翻訳後修飾の有無を判定する方法であって、
(a) 翻訳後修飾と結合するプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 該プローブとタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置で該相互作用が生じれば、その位置のタンパク質が翻訳後修飾を有することを示す、上記方法。
【請求項59】
前記翻訳後修飾がメチル化、リン酸化、ビオチン化、アセチル化、ペグ化、グリコシル化、脂質修飾、ユビキチン化、またはSUMO化である、請求項58に記載の方法。
【請求項60】
酵母細胞の培養物を調製する方法であって、
(a) 複数の酵母細胞を増殖培地中で、OD600が0.3〜1.0となるまで増殖させ、ここで、該複数の酵母細胞は調節配列に機能的に連結された異種ヌクレオチド配列を含むものであり、
(b) 該細胞を、該異種ヌクレオチド配列によりコードされるタンパク質の発現を増強する誘導物質と接触させ、
(c) 該細胞を該培地から分離し、
(d) 該細胞を冷水に接触させ、
(e) 該細胞を該冷水から分離し、
(f) 該細胞を冷溶解バッファーに接触させ、
(g) 該細胞を該溶解バッファーから分離し、そして
(h) 保存のために該細胞を半乾燥状態で凍結する、
各ステップを含んでなる上記方法。
【請求項61】
細胞からタンパク質を精製する方法であって、
(a) 複数の細胞サンプルのそれぞれについて、各サンプルの細胞を溶解して細胞溶解物を調製し、ここで、該細胞はアフィニティータグをもつ融合タンパク質を含み、また、該溶解ステップはペイントシェーカーを使って実施し、
(b) 各溶解物を可溶性画分と非可溶性画分とに分離し、
(c) 各可溶性画分をマルチサイトアレイの異なるサイトに移動させ、その際、該移動ステップはワイドオープンチップを使って実施し、
(d) 各可溶性画分を結合物質と接触させて、該融合タンパク質と結合物質との複合体が形成されるようにし、
(e) 該複合体から各融合タンパク質を単離し、そして
(f) 各融合タンパク質を高粘性のバッファー中に保存する、
各ステップを含んでなる上記方法。
【請求項62】
シグナルが陽性であるかどうかを判定する方法であって、
(a) 各スポットについて局所的にフォアグラウンドシグナルおよびバックグラウンドシグナルを測定し、該フォアグラウンドシグナルとバックグラウンドシグナルの差から正味のシグナルを決定し、
(b) 第1および第2の正味シグナル分布に関する下四分位値、中位値、および上四分位値を決定し、
(c) 第1の正味シグナル分布から第1の中位値を減じ、さらに第2の正味シグナル分布から第2の中位値を減じて、それぞれ第1および第2の減算値を得、
(d) 第1の減算値を、第1の正味シグナル分布に関する上四分位値と下四分位値との差で割り算し、さらに第2の減算値を、第2の正味シグナル分布の上四分位値と下四分位値との差で割り算して、それぞれ第1および第2の基準化された値を得、
(e) 近傍領域の基準化されたシグナル分布に関する局所中位値を算定し、ただし、該近傍領域はその領域内に複数のサイトを含んでなり、そして
(f) 基準化されたシグナルから局所中位値を減じて、基準化された過剰値を得る、
各ステップを含んでなる、上記方法。
【請求項63】
陽性シグナルがタンパク質−プローブ相互作用を示す、請求項62に記載の方法。
【請求項64】
前記近傍領域が前記シグナルの上2列、下2列、左2列、および右2列である、請求項62に記載の方法。
【請求項65】
サンプル値の1つとサンプル値の平均との差が基準化された過剰値の誤差の3標準偏差より大きい場合には、基準化された過剰値の並列サンプルを除外するステップをさらに含む、請求項62に記載の方法。
【請求項66】
複数の異なるアレイを用いて測定したシグナルの中から陽性シグナルを確認する方法であって、
(a) 異なるアレイを用いて測定したシグナルを変換して変換シグナルを得、
(b) 各変換シグナルを、該変換シグナルから局所中位値シグナルを減じて修正された変換シグナルを得る方法によって修正し、ここで、該局所中位値シグナルは近傍領域におけるシグナルの中位値であり、該近傍領域は該変換シグナルのサイトのまわりの1以上のサイトを含んでなり、そして
(c) 修正された各変換シグナルを閾値と比較して、該修正された変換シグナルが該閾値より大きい場合には、該修正された変換シグナルを陽性と判定する、
各ステップを含んでなり、
ここで、該アレイは基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイからなるものである、上記方法。
【請求項67】
前記変換ステップが、
(a) 異なるアレイのそれぞれを用いて測定したシグナルについて、シグナル分布の下四分位値、中位値、および上四分位値を決定し、
(b) 異なるアレイのそれぞれを用いて測定したシグナルから該中位値を減じて、該アレイについての翻訳シグナルを得、
(c) 該翻訳シグナルを、該アレイの上四分位値と下四分位値との差で割り算することによって変換シグナルを得る、
各ステップを含んでなる、請求項66に記載の方法。
【請求項68】
前記近傍領域が変換シグナルの上2列、下2列、左2列、および右2列の領域内の複数のサイトからなる、請求項66に記載の方法。
【請求項69】
陽性シグナルがタンパク質−プローブ相互作用を示す、請求項66、67または68に記載の方法。
【請求項70】
データポイントを捨てるステップをさらに含み、ただし、該データポイントはアレイ上の二重反復サイトで測定され、該二重反復サイト間の変化が3標準偏差より大きいものである、請求項66、67または68に記載の方法。
【請求項71】
修正された変換シグナルを次式:
r+εr = G+εG/R+εR
(式中、Gは修正された変換シグナルであり、RはGSTシグナルであり、εGはGの誤差であり、εRはRの誤差であり、そしてεrはrの誤差である)
を用いて正規化するステップをさらに含む、請求項66、67または68に記載の方法。
【請求項72】
マルチサイトアレイが96サイトアレイである、請求項20に記載の方法。
【請求項73】
基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
【請求項74】
(a) 2つの二重反復スポットのシグナル値を平均して平均値を出し、(b) 該平均値が基準化された過剰値の誤差の3標準偏差より大きいかどうかを調べる、各ステップをさらに含んでなり、その際、該平均値が基準化された過剰値の誤差の3標準偏差より大きければ、該スポットのシグナルを陽性とする、請求項62または65に記載の方法。
【請求項75】
タンパク質における酵素活性の有無を判定する方法であって、
(a) 該酵素活性の基質であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素活性をもつことを示す、上記方法。
【請求項76】
タンパク質における酵素基質の有無を判定する方法であって、
(a) 該酵素基質のための酵素であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素基質を含むことを示す、上記方法。
【請求項77】
前記タンパク質が基板にビオチンタグを介して結合される、請求項1、3、4または5に記載のアレイ。
【請求項78】
第1タグが前記タンパク質のカルボキシ末端に存在し、第2タグが前記タンパク質のアミノ末端に存在する、請求項15に記載の方法。
【請求項79】
アフィニティータグがビオチンである、請求項22に記載の方法。
【請求項80】
前記基板がニトロセルロースをコーティングしたスライドガラスからなる、請求項1、3、4または5に記載のアレイ。
【請求項1】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種における既知遺伝子のうち少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記アレイ。
【請求項2】
前記複数のタンパク質が単一の種における既知遺伝子のうち少なくとも70%によりコードされる少なくとも1つのタンパク質を含む、請求項1に記載のアレイ。
【請求項3】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される全タンパク質のうち少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記アレイ。
【請求項4】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記アレイ。
【請求項5】
基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる、位置指定可能なアレイであって、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記アレイ。
【請求項6】
前記タンパク質がタンパク質の分類に従って前記アレイ上に組織化されている、請求項1、3、4または5に記載のアレイ。
【請求項7】
前記分類が存在量、機能、酵素活性、相同性、タンパク質ファミリー、特定の代謝経路との関連性、または翻訳後修飾によるものである、請求項6に記載のアレイ。
【請求項8】
前記タンパク質がHisタグを介して基板に結合されている、請求項1、3、4または5に記載のアレイ。
【請求項9】
前記基板がニッケルを含む、請求項1、3、4または5に記載のアレイ。
【請求項10】
前記基板がニッケルをコーティングしたスライドガラスからなる、請求項1、3、4または5に記載のアレイ。
【請求項11】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものである、上記方法。
【請求項12】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される全タンパク質の少なくとも50%を含むものであり、ただし、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなす、上記方法。
【請求項13】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が単一の種において発現される少なくとも1000のタンパク質を含むものである、上記方法。
【請求項14】
基板の表面に複数のタンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各タンパク質は基板上の異なる位置に存在し、該複数のタンパク質が全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものである、上記方法。
【請求項15】
基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
【請求項16】
前記結合ステップに先立って、前記タンパク質を第1タグの結合パートナーと接触させることにより前記タンパク質を精製するステップを行い、前記結合ステップにおいて第2タグを用いて基板に前記タンパク質を結合させる、請求項15に記載の方法。
【請求項17】
第1タグがGSTタグであり、第2タグがHisタグである、請求項16に記載の方法。
【請求項18】
第1タグと第2タグが前記タンパク質のアミノ末端に存在する、請求項15に記載の方法。
【請求項19】
第1タグと第2タグが前記タンパク質のカルボキシ末端に存在する、請求項15に記載の方法。
【請求項20】
複数の精製されたタンパク質サンプルを調製して単離する方法であって、マルチサイトアレイの複数サイトの各サイトにおいて、
(a) 調節配列と機能的に連結された異種ヌクレオチド配列を有する真核細胞を生育させ、
(b) 該調節配列を、異種ヌクレオチド配列によりコードされるタンパク質の発現を高める誘導物質と接触させ、
(c) 該細胞を溶解して細胞溶解物を形成し、
(d) 該細胞溶解物またはそれから得られたタンパク質含有サンプルを結合物質と接触させて、結果的に該タンパク質と結合物質との複合体を形成させ、そして
(e) 該複合体から該タンパク質を単離する、
各ステップを含んでなり、その際、各ステップがマルチアレイフォーマットで実施される、上記方法。
【請求項21】
各サイトがウェルである、請求項20に記載の方法。
【請求項22】
前記タンパク質がアフィニティータグを含む融合タンパク質であり、該アフィニティータグに前記結合物質が結合する、請求項20に記載の方法。
【請求項23】
前記細胞が酵母細胞である、請求項20に記載の方法。
【請求項24】
前記溶解ステップがペイントシェーカーを使って実施される、請求項20に記載の方法。
【請求項25】
脂質結合タンパク質を検出する方法であって、
(a) 脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置に該相互作用が検出されれば、その位置に脂質結合タンパク質が存在することを示す、上記方法。
【請求項26】
前記脂質がリン脂質である、請求項25に記載の方法。
【請求項27】
前記リン脂質がホスファチジルコリンまたはホスファチジルイノシトールである、請求項26に記載の方法。
【請求項28】
前記プローブがリポソームからなる、請求項25に記載の方法。
【請求項29】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種における既知遺伝子の少なくとも50%によりコードされる少なくとも1つのタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項30】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される全タンパク質の少なくとも50%を含むものであり、その際、タンパク質アイソフォームおよびスプライス変異体は単一のタンパク質とみなし、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項31】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は単一の種において発現される少なくとも1000のタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項32】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該複数のタンパク質は全体として、単一の種における少なくとも1000の異なる既知遺伝子によりコードされるタンパク質を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項33】
結合タンパク質を検出する方法であって、
(a) プローブを、基板上の異なる位置にそれぞれ存在する複数の融合タンパク質を含んでなる位置指定可能なアレイと接触させ、ただし、該融合タンパク質は第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものであり、
(b) いずれかのタンパク質−プローブ相互作用を検出する、
各ステップを含んでなる、上記方法。
【請求項34】
前記プローブが核酸、タンパク質、小分子、候補薬物または脂質を含むものである、請求項29、30、31、32または33に記載の方法。
【請求項35】
前記核酸がRNAまたはDNAからなる、請求項34に記載の方法。
【請求項36】
前記プローブが酵母タンパク質である、請求項34に記載の方法。
【請求項37】
酵母タンパク質がMyo2、Rho1、Rho2、Rho3、Rho4、Cdc11、Cdc12またはHs17である、請求項36に記載の方法。
【請求項38】
前記プローブが抗体である、請求項34に記載の方法。
【請求項39】
前記抗体がサイクリン、キナーゼ、GST、Clb5、Cla4、Ste20、Cdc42、PI(3,4)P2、PI(4)P、SPA2、CLB1、CLB2、またはCdc11に特異的である、請求項38に記載の方法。
【請求項40】
前記プローブがカルモジュリンである、請求項34に記載の方法。
【請求項41】
前記プローブがATP、GTP、cAMP、ホスホチロシン、ホスホセリン、およびホスホトレオニンからなる群より選択される小分子を含むものである、請求項34に記載の方法。
【請求項42】
前記プローブがホスファチジルコリンまたはホスファチジルイノシトールを含むものである、請求項34に記載の方法。
【請求項43】
前記プローブがリポソームからなる、請求項34に記載の方法。
【請求項44】
前記プローブが哺乳動物由来のものである、請求項25、29、30、31、32または33に記載の方法。
【請求項45】
前記哺乳動物がヒトである、請求項44に記載の方法。
【請求項46】
前記複数のタンパク質がヒト以外のものである、請求項44に記載の方法。
【請求項47】
前記複数のタンパク質がHisタグを介して基板に結合される、請求項25、29、30、31、32または33に記載の方法。
【請求項48】
前記基板がニッケルを含む、請求項25、29、30、31、32または33に記載の方法。
【請求項49】
前記基板がニッケルをコーティングしたスライドガラスからなる、請求項25、29、30、31、32または33に記載の方法。
【請求項50】
前記検出ステップでタンパク質との相互作用が検出されたプローブの正体を同定するステップをさらに含んでなる、請求項34に記載の方法。
【請求項51】
前記相互作用は、前記同定されたプローブが抗菌性、抗真菌性、または抗ウイルス性タンパク質であることを示す、請求項50に記載の方法。
【請求項52】
結合アッセイで使用するためのタンパク質を標識する方法であって、
(a) 該タンパク質の個々のアリコートを、ビオチン転移化合物と、アリコートごとに異なる程度にビオチン化されたタンパク質を生成する条件下および時間にわたり接触させ、そして
(b) 個々のアリコートを一緒に合わせて、異なる程度にビオチン化されたタンパク質のサンプルを得る、
各ステップを含んでなる、上記方法。
【請求項53】
結合タンパク質を検出する方法であって、
(a) 請求項52に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じているアレイ上の位置を検出する、
各ステップを含んでなる、上記方法。
【請求項54】
結合タンパク質を検出する方法であって、
(a) 請求項52に記載の方法により得られたビオチン化タンパク質のサンプルを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) 該アレイに、蛍光物質とコンジュゲートさせたストレプトアビジンを接触させ、そして
(c) 蛍光が発生しているアレイ上の位置を検出する、
各ステップを含んでなり、その際、蛍光は、ビオチン化タンパク質とアレイ上のタンパク質との相互作用が生じていることを示す、上記方法。
【請求項55】
タンパク質がホスファチジルコリンに比べてホスファチジルイノシトールと優先的に結合するかどうかを判定する方法であって、
(a) ホスファチジルイノシトールを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、
(b) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルイノシトール結合タンパク質の存在を示すものであり、
(c) ホスファチジルコリンを含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該タンパク質はステップ(a)におけると同じタンパク質を少なくとも一部含んでなり、
(d) タンパク質−プローブ相互作用を検出し、ここで、基板上の位置での該相互作用はホスファチジルコリン結合タンパク質の存在を示すものであり、そして
(e) 複数のタンパク質のそれぞれについて、ステップ(b)とステップ(d)の結果を比較する、
各ステップを含んでなる、上記方法。
【請求項56】
リン脂質が細胞における代謝経路またはシグナル伝達経路を調節するのかどうか、あるいは該代謝経路またはシグナル伝達経路が膜表面で起こるのかどうかを判定する方法であって、
(a) リン脂質を含むプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、ここで、該複数のタンパク質は該経路の少なくとも一部を構成する1以上のタンパク質を含んでなり、そして
(b) 該プローブと該経路中のタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、該相互作用が生じれば、該プローブが代謝経路またはシグナル伝達経路を調節すること、あるいは、該経路が膜表面で起こることを示す、上記方法。
【請求項57】
カルモジュリンと結合する非天然タンパク質を製造する方法であって、下記の配列:
I/L-Q-X-X-K-K/X-G-B (配列番号1)
(ここで、Xは任意のアミノ酸であり、Bは塩基性アミノ酸である)
を含んでなる非天然タンパク質を製造することからなる上記方法。
【請求項58】
タンパク質における翻訳後修飾の有無を判定する方法であって、
(a) 翻訳後修飾と結合するプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 該プローブとタンパク質との相互作用を検出する、
各ステップを含んでなり、その際、基板上の位置で該相互作用が生じれば、その位置のタンパク質が翻訳後修飾を有することを示す、上記方法。
【請求項59】
前記翻訳後修飾がメチル化、リン酸化、ビオチン化、アセチル化、ペグ化、グリコシル化、脂質修飾、ユビキチン化、またはSUMO化である、請求項58に記載の方法。
【請求項60】
酵母細胞の培養物を調製する方法であって、
(a) 複数の酵母細胞を増殖培地中で、OD600が0.3〜1.0となるまで増殖させ、ここで、該複数の酵母細胞は調節配列に機能的に連結された異種ヌクレオチド配列を含むものであり、
(b) 該細胞を、該異種ヌクレオチド配列によりコードされるタンパク質の発現を増強する誘導物質と接触させ、
(c) 該細胞を該培地から分離し、
(d) 該細胞を冷水に接触させ、
(e) 該細胞を該冷水から分離し、
(f) 該細胞を冷溶解バッファーに接触させ、
(g) 該細胞を該溶解バッファーから分離し、そして
(h) 保存のために該細胞を半乾燥状態で凍結する、
各ステップを含んでなる上記方法。
【請求項61】
細胞からタンパク質を精製する方法であって、
(a) 複数の細胞サンプルのそれぞれについて、各サンプルの細胞を溶解して細胞溶解物を調製し、ここで、該細胞はアフィニティータグをもつ融合タンパク質を含み、また、該溶解ステップはペイントシェーカーを使って実施し、
(b) 各溶解物を可溶性画分と非可溶性画分とに分離し、
(c) 各可溶性画分をマルチサイトアレイの異なるサイトに移動させ、その際、該移動ステップはワイドオープンチップを使って実施し、
(d) 各可溶性画分を結合物質と接触させて、該融合タンパク質と結合物質との複合体が形成されるようにし、
(e) 該複合体から各融合タンパク質を単離し、そして
(f) 各融合タンパク質を高粘性のバッファー中に保存する、
各ステップを含んでなる上記方法。
【請求項62】
シグナルが陽性であるかどうかを判定する方法であって、
(a) 各スポットについて局所的にフォアグラウンドシグナルおよびバックグラウンドシグナルを測定し、該フォアグラウンドシグナルとバックグラウンドシグナルの差から正味のシグナルを決定し、
(b) 第1および第2の正味シグナル分布に関する下四分位値、中位値、および上四分位値を決定し、
(c) 第1の正味シグナル分布から第1の中位値を減じ、さらに第2の正味シグナル分布から第2の中位値を減じて、それぞれ第1および第2の減算値を得、
(d) 第1の減算値を、第1の正味シグナル分布に関する上四分位値と下四分位値との差で割り算し、さらに第2の減算値を、第2の正味シグナル分布の上四分位値と下四分位値との差で割り算して、それぞれ第1および第2の基準化された値を得、
(e) 近傍領域の基準化されたシグナル分布に関する局所中位値を算定し、ただし、該近傍領域はその領域内に複数のサイトを含んでなり、そして
(f) 基準化されたシグナルから局所中位値を減じて、基準化された過剰値を得る、
各ステップを含んでなる、上記方法。
【請求項63】
陽性シグナルがタンパク質−プローブ相互作用を示す、請求項62に記載の方法。
【請求項64】
前記近傍領域が前記シグナルの上2列、下2列、左2列、および右2列である、請求項62に記載の方法。
【請求項65】
サンプル値の1つとサンプル値の平均との差が基準化された過剰値の誤差の3標準偏差より大きい場合には、基準化された過剰値の並列サンプルを除外するステップをさらに含む、請求項62に記載の方法。
【請求項66】
複数の異なるアレイを用いて測定したシグナルの中から陽性シグナルを確認する方法であって、
(a) 異なるアレイを用いて測定したシグナルを変換して変換シグナルを得、
(b) 各変換シグナルを、該変換シグナルから局所中位値シグナルを減じて修正された変換シグナルを得る方法によって修正し、ここで、該局所中位値シグナルは近傍領域におけるシグナルの中位値であり、該近傍領域は該変換シグナルのサイトのまわりの1以上のサイトを含んでなり、そして
(c) 修正された各変換シグナルを閾値と比較して、該修正された変換シグナルが該閾値より大きい場合には、該修正された変換シグナルを陽性と判定する、
各ステップを含んでなり、
ここで、該アレイは基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイからなるものである、上記方法。
【請求項67】
前記変換ステップが、
(a) 異なるアレイのそれぞれを用いて測定したシグナルについて、シグナル分布の下四分位値、中位値、および上四分位値を決定し、
(b) 異なるアレイのそれぞれを用いて測定したシグナルから該中位値を減じて、該アレイについての翻訳シグナルを得、
(c) 該翻訳シグナルを、該アレイの上四分位値と下四分位値との差で割り算することによって変換シグナルを得る、
各ステップを含んでなる、請求項66に記載の方法。
【請求項68】
前記近傍領域が変換シグナルの上2列、下2列、左2列、および右2列の領域内の複数のサイトからなる、請求項66に記載の方法。
【請求項69】
陽性シグナルがタンパク質−プローブ相互作用を示す、請求項66、67または68に記載の方法。
【請求項70】
データポイントを捨てるステップをさらに含み、ただし、該データポイントはアレイ上の二重反復サイトで測定され、該二重反復サイト間の変化が3標準偏差より大きいものである、請求項66、67または68に記載の方法。
【請求項71】
修正された変換シグナルを次式:
r+εr = G+εG/R+εR
(式中、Gは修正された変換シグナルであり、RはGSTシグナルであり、εGはGの誤差であり、εRはRの誤差であり、そしてεrはrの誤差である)
を用いて正規化するステップをさらに含む、請求項66、67または68に記載の方法。
【請求項72】
マルチサイトアレイが96サイトアレイである、請求項20に記載の方法。
【請求項73】
基板の表面に複数の融合タンパク質を結合させるステップを含んでなる、位置指定可能なアレイの作製方法であって、各融合タンパク質は基板上の異なる位置に存在し、該融合タンパク質が第1タグ、第2タグ、および1つの生物のゲノム核酸によりコードされるタンパク質配列を含むものである、上記方法。
【請求項74】
(a) 2つの二重反復スポットのシグナル値を平均して平均値を出し、(b) 該平均値が基準化された過剰値の誤差の3標準偏差より大きいかどうかを調べる、各ステップをさらに含んでなり、その際、該平均値が基準化された過剰値の誤差の3標準偏差より大きければ、該スポットのシグナルを陽性とする、請求項62または65に記載の方法。
【請求項75】
タンパク質における酵素活性の有無を判定する方法であって、
(a) 該酵素活性の基質であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素活性をもつことを示す、上記方法。
【請求項76】
タンパク質における酵素基質の有無を判定する方法であって、
(a) 該酵素基質のための酵素であるプローブを、基板上の異なる位置にそれぞれ存在する複数のタンパク質を含んでなる位置指定可能なアレイと接触させ、そして
(b) 基板上の位置で該基質の触媒作用を検出する、
各ステップを含んでなり、その際、基板上の位置に触媒作用が検出されれば、該位置にあるタンパク質が酵素基質を含むことを示す、上記方法。
【請求項77】
前記タンパク質が基板にビオチンタグを介して結合される、請求項1、3、4または5に記載のアレイ。
【請求項78】
第1タグが前記タンパク質のカルボキシ末端に存在し、第2タグが前記タンパク質のアミノ末端に存在する、請求項15に記載の方法。
【請求項79】
アフィニティータグがビオチンである、請求項22に記載の方法。
【請求項80】
前記基板がニトロセルロースをコーティングしたスライドガラスからなる、請求項1、3、4または5に記載のアレイ。
【図1A】

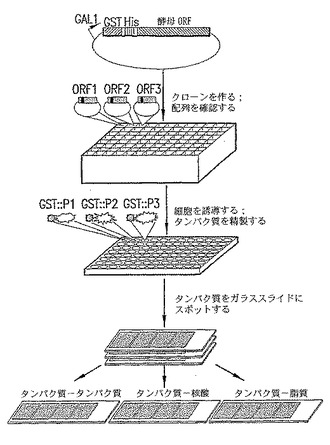
【図1B】
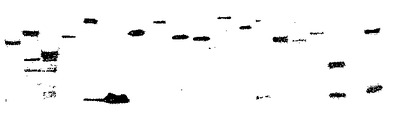
【図2A】
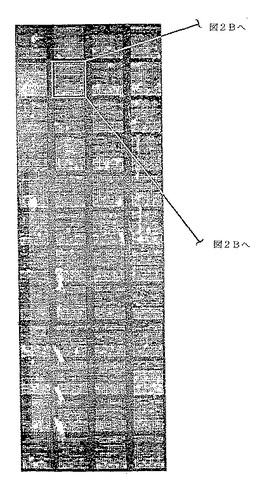
【図2B】
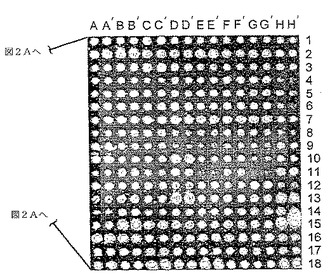
【図2C】

【図3A】

【図3B】

【図4A】

【図4B】

【図4C】
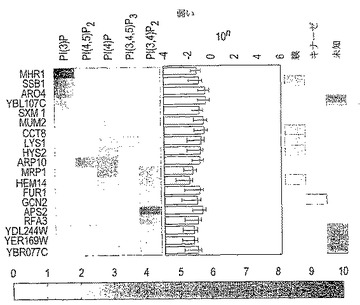
【図4D】

【図5A−B】
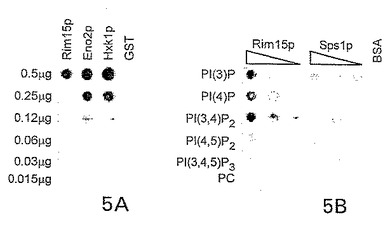
【図5C】
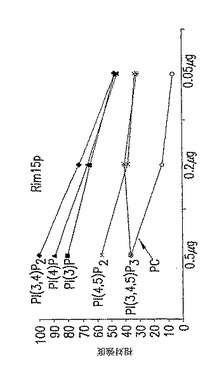
【図1B】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図4A】
【図4B】
【図4C】
【図4D】
【図5A−B】
【図5C】
【公開番号】特開2009−36772(P2009−36772A)
【公開日】平成21年2月19日(2009.2.19)
【国際特許分類】
【出願番号】特願2008−211919(P2008−211919)
【出願日】平成20年8月20日(2008.8.20)
【分割の表示】特願2002−589035(P2002−589035)の分割
【原出願日】平成14年5月13日(2002.5.13)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【Fターム(参考)】
【公開日】平成21年2月19日(2009.2.19)
【国際特許分類】
【出願日】平成20年8月20日(2008.8.20)
【分割の表示】特願2002−589035(P2002−589035)の分割
【原出願日】平成14年5月13日(2002.5.13)
【出願人】(392019352)イェール ユニバーシティー (38)
【氏名又は名称原語表記】YALE UNIVERSITY
【Fターム(参考)】
[ Back to top ]