
プロヒビチン2(PHB2)のミトコンドリア機能
【課題】PHB2遺伝子調節剤、PHB2タンパク質機能調節剤及びミトコンドリア機能関連疾患治療薬を提供することを目的とする。
【解決手段】PHB2遺伝子に対する特定のsiRNAを含有するPHB2遺伝子発現調節剤、特定の核レセプターのアンタゴニスト又はアゴニストを含有するPHB2タンパク質機能調節剤、及びこれらを含有するミトコンドリア機能関連疾患治療薬。
【解決手段】PHB2遺伝子に対する特定のsiRNAを含有するPHB2遺伝子発現調節剤、特定の核レセプターのアンタゴニスト又はアゴニストを含有するPHB2タンパク質機能調節剤、及びこれらを含有するミトコンドリア機能関連疾患治療薬。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、例えばPHB2遺伝子調節剤のスクリーニング方法に関する。
【背景技術】
【0002】
ミトコンドリアは、真核生物において多くの反応を行う。特に電子伝達系によるATP合成は生物にとって重要であり、細胞におけるほとんどのATPがミトコンドリアから供給される。ミトコンドリアにおけるその他の反応系としては、TCA回路、ヘム合成系、脂肪酸のβ酸化回路、アミノ酸代謝回路などが挙げられる。さらに、Ca恒常性の維持機能、活性酸素の産生系、並びに代謝物質、イオン及びタンパク質などの輸送系がミトコンドリアに存在する。このように、ミトコンドリアは真核生物の異化作用及び同化作用に重要な働きを持つ細胞内小器官である。
【0003】
ヒトミトコンドリアに存在するタンパク質は、1000〜1500種類存在すると予測されている。そのうちの13種類は、ミトコンドリアDNAにコードされるタンパク質で、電子伝達系のサブユニットである。その他の約99%のタンパク質は、核DNAにコードされるタンパク質であり、細胞質でタンパク質が合成された後、ミトコンドリアに移行する。プロテオーム解析によれば、ヒトミトコンドリアに存在するタンパク質のうち約544種類のタンパク質が同定されている(非特許文献1)が、未だ未知のタンパク質が多く存在すると予測される。
【0004】
上述したように、ミトコンドリアDNAは電子伝達系の複合体I、III、IV、Vの一部のサブユニットをコードする。即ち、ミトコンドリアDNAの変異は、電子伝達系の機能異常を引き起こす。電子伝達系の機能異常に関連する疾患としては、例えばMELAS、MERRF、心筋症、LHON、Leigh脳症などが挙げられる。また、ミトコンドリアDNAの塩基変異が、肝臓、前立腺、膀胱及び頭頸部の早期癌、原発性肺癌、Barrett's esophagusにおいても認められている(非特許文献2)。
【0005】
一方、核DNAにコードされたミトコンドリアタンパク質の異常も、例えば、以下のように多くの疾患の原因に成っている。(i)Friedreich's ataxiaは、ミトコンドリアにおいてFe-Sタンパク質の生合成に関与するfrataxinタンパク質の異常によって生じる。(ii)ミトコンドリアへのタンパク質移行に関与する因子Deafness dystonia peptide 1(DDP1)の異常は、Mohr-Tranebjaerg syndromeに関与する。(iii)常染色体優性遺伝を示す網膜萎縮症は、ミトコンドリア膜融合を生じさせるOPA1タンパク質の異常によって生じる。(iv)ミトコンドリア膜融合に関与する他の因子Mfn2の異常により、Charcot-Marie-Tooth neuropathy type 2が生じる。(v)また、ミトコンドリアのチミジンホスホリラーゼの異常により、常染色体劣性遺伝で、消化器症状の強いMNGIE(mitochondrial neurogastrointestinal encephalomyopathy)が生じる。
【0006】
その他に、ミトコンドリア機能異常は、より一般的な疾患への関与も示されている。例えば、ミトコンドリア機能異常による糖代謝や脂質代謝の異常は、肥満や糖尿病などの原因となる。また、ミトコンドリア機能異常による細胞内ATPの低下は、パーキンソン病やアルツハイマー病の病因の主要因子である。近年、アルツハイマー病において蓄積物質のアミロイドβタンパク質が、細胞内でミトコンドリアタンパク質であるABADタンパク質と結合し、ミトコンドリア機能を傷害するという報告もなされている(非特許文献3)。
【0007】
また、ミトコンドリアの消費する酸素の0.4〜4%は、電子伝達系で活性酸素となることが知られている。この活性酸素がDNAやタンパク質などに障害を与え、細胞障害や細胞数の減少などを生じさせることにより、細胞の機能低下や個体の老化を促進すると考えられている。
【0008】
さらに、ミトコンドリアはアポトーシス誘導に関与しており、これらの経路は細胞増殖や癌化と関連があると考えられている。
【0009】
従って、ミトコンドリアの機能を正常に保ち、当該機能をうまく制御することが、老化の防止だけでなく、個体の恒常性を維持していく上で極めて重要である。
【0010】
ところで、プロヒビチン(以下、「PHB」という)と呼ばれるタンパク質が、細胞増殖抑制因子として最初に哺乳動物で単離された。PHBは酵母から哺乳動物に至るまで高度に保存されたタンパク質である。PHBタンパク質には、アミノ酸の一次構造が類似した2種類のタンパク質PHB1及びPHB2が存在し、互いに複合体を形成し、ミトコンドリアの内膜に局在することが知られている。酵母PHBタンパク質に関しては、細胞周期制御や新規に合成されたミトコンドリアタンパク質の安定化を担うシャペロン様の機能を果たすことが明らかにされている(非特許文献4〜6)。また、線虫では、PHB1の老化や初期発生への関与が報告されている(非特許文献7)。
【0011】
一方、哺乳動物では、PHB1及びPHB2による転写制御等の多様な機能が示唆されているものの、ミトコンドリアにおける生理的機能は明らかにされていない(非特許文献8及び9)。
【先行技術文献】
【非特許文献】
【0012】
【非特許文献1】Reichert AS及びNeupert W., 「Trends in Genetics」, 2004年, 第20巻, 第11号, p.556-562
【非特許文献2】Verma M.ら, 「Nature reviews cancer」, 2003年, 第3巻, 第10号, p.789-795
【非特許文献3】Lustbader, J.W.ら, 「Science」, 2004年, 第304巻, 第5669号, p.448-452
【非特許文献4】Berger, K.H.及びYaffe, M.P., 「Mol Cell Biol.」, 1998年, 第18巻, 第7号, p.4043-4052
【非特許文献5】Nijtmans, L.G.ら, 「EMBO J.」, 2000年, 第19巻, 第11号, p.2444-2451
【非特許文献6】Piper P.W.及びBringloe, D., 「Mech Ageing Dev.」, 2002年, 第123巻, 第4号, p.287-295
【非特許文献7】Artal-Sanz M.ら, 「J Biol Chem.」, 2003年, 第278巻, 第34号, p.32091-32099
【非特許文献8】Delage-Mourroux R.ら, 「J Biol Chem.」, 2000年, 第275巻, 第46号, p.35848-35856
【非特許文献9】Sun L.ら, 「J Cell Sci.」, 2004年, 第117巻, p.3021-3029
【発明の概要】
【発明が解決しようとする課題】
【0013】
上述したように、ミトコンドリアの機能を正常に保ち、当該機能をうまく制御することが、老化の防止だけでなく、個体の恒常性を維持していく上で極めて重要である。
【0014】
そこで、本発明は、上述した実情に鑑み、例えばミトコンドリア機能調節剤のスクリーニング方法を提供することを目的とする。
【課題を解決するための手段】
【0015】
上記課題を解決するため鋭意研究を行った結果、ミトコンドリアにおけるヒトPHB2タンパク質は、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持の機能を有することを見出し、本発明を完成するに至った。
【0016】
本発明は以下を包含する。
(1)PHB2遺伝子発現細胞をその培養下で候補物質と接触させる工程と、候補物質非存在下での該PHB2遺伝子発現細胞と比較して、該候補物質と接触した該PHB2遺伝子発現細胞においてPHB2遺伝子発現又はPHB2タンパク質機能が調節されていることにより、該候補物質がPHB2遺伝子調節剤であると判定する工程とを含む、PHB2遺伝子調節剤のスクリーニング方法。
(2)上記PHB2タンパク質機能が、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持から成る群から選択されるものであることを特徴とする、(1)記載の方法。
(3)上記PHB2タンパク質機能が、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質との相互作用能であることを特徴とする、(1)又は(2)記載の方法。
(4)上記相互作用能が複合体形成能であることを特徴とする、(3)記載の方法。
(5)上記PHB2タンパク質機能が、PHB2タンパク質の核-ミトコンドリア移行機能であることを特徴とする、(1)又は(2)記載の方法。
【0017】
(6)上記PHB2遺伝子発現細胞がERα、PPARα及びPPARγ2から成る群から選択される核レセプターをさらに発現していることを特徴とする、(5)記載の方法。
(7)上記PHB2遺伝子発現細胞の培養が、エストラジオール存在下で行われることを特徴とする、(6)記載の方法。
(8)(1)〜(7)のいずれか1記載の方法により得られたPHB2遺伝子調節剤。
(9)以下の(a)又は(b)記載のsiRNAを含有することを特徴とするPHB2遺伝子調節剤。
(a)配列番号1〜20記載のいずれか1つに示される塩基配列から成るsiRNA。
(b)上記(a)記載のsiRNAにおいて、1又は数個の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2遺伝子発現阻害活性を有するsiRNA。
(10)ERα、PPARα及びPPARγ2から成る群から選択される核レセプターのアンタゴニスト又はアゴニストを含有することを特徴とするPHB2遺伝子調節剤。
【0018】
(11)上記PHB2遺伝子調節剤が抗アポトーシス作用剤、ミトコンドリア膜電位調節剤、及びミトコンドリア膜形態調節剤から成る群から選択されるものである、(8)〜(10)のいずれか1記載のPHB2遺伝子調節剤。
(12)(8)〜(11)のいずれか1記載のPHB2遺伝子調節剤を含有するミトコンドリア機能関連疾患治療薬。
(13)被験者由来の生物学的サンプルにおいて、PHB2遺伝子発現又はPHB2タンパク質機能を測定する工程と、該PHB2遺伝子発現又はPHB2タンパク質機能が異常であることを指標として、該被験者がミトコンドリア機能関連疾患を有するか又はそれらを有する疑いがあると決定する工程とを含む、ミトコンドリア機能関連疾患の検出方法。
(14)上記PHB2タンパク質機能が、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持から成る群から選択されるものであることを特徴とする、(13)記載の方法。
(15)上記PHB2タンパク質機能が、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質との相互作用能であることを特徴とする、(13)又は(14)記載の方法。
【0019】
(16)上記相互作用能が複合体形成能であることを特徴とする、(15)記載の方法。
(17)上記PHB2タンパク質機能が、PHB2タンパク質の核-ミトコンドリア移行機能であることを特徴とする、(13)又は(14)記載の方法。
(18)PHB2遺伝子発現可能なミトコンドリア機能異常細胞をその培養下で候補物質と接触させる工程と、候補物質非存在下での該ミトコンドリア機能異常細胞と比較して、該候補物質と接触した該ミトコンドリア機能異常細胞において、PHB2遺伝子発現又はPHB2タンパク質機能が調節される結果としてミトコンドリア機能が回復又は正常化することを指標として、上記候補物質がミトコンドリア機能調節剤であると判定する工程とを含む、ミトコンドリア機能調節剤のスクリーニング方法。
(19)上記PHB2タンパク質機能が、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持から成る群から選択されるものであることを特徴とする、(18)記載の方法。
(20)上記PHB2タンパク質機能が、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質との相互作用能であることを特徴とする、(18)又は(19)記載の方法。
【0020】
(21)上記相互作用能が複合体形成能であることを特徴とする、(20)記載の方法。
(22)上記PHB2タンパク質機能が、PHB2タンパク質の核-ミトコンドリア移行機能であることを特徴とする、(18)又は(19)記載の方法。
(23)上記ミトコンドリア機能異常細胞がERα、PPARα及びPPARγ2から成る群から選択される核レセプターをさらに発現していることを特徴とする、(22)記載の方法。
(24)上記ミトコンドリア機能異常細胞の培養が、エストラジオール存在下で行われることを特徴とする、(23)記載の方法。
【発明の効果】
【0021】
本発明によれば、PHB2遺伝子調節剤、ミトコンドリア機能関連疾患治療薬及びミトコンドリア機能調節剤を提供することができる。さらに、本発明によれば、ミトコンドリア機能関連疾患を検出することができる。
【図面の簡単な説明】
【0022】
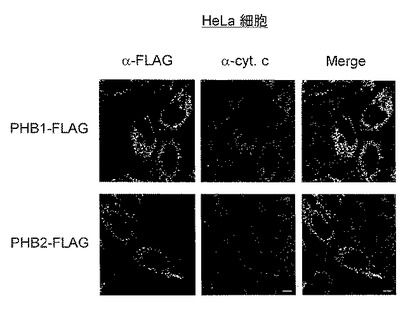
【図1】図1は、実施例1におけるPHB1-FLAG又はPHB2-FLAGを発現するHeLa細胞の共焦点レーザー顕微鏡写真を示す。
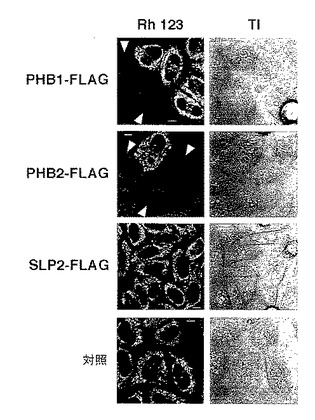
【図2】図2は、実施例2における各遺伝子を発現するHeLa細胞をRh123で染色した際の写真を示す。
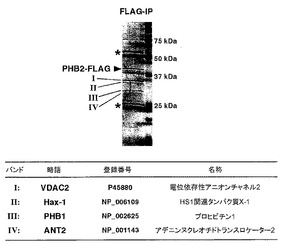
【図3】図3は、実施例3におけるIP解析及びMASS解析の結果を示す。
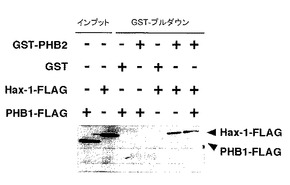
【図4】図4は、実施例4におけるPHB2タンパク質とHax-1タンパク質との相互作用に関するin vitro結合実験の結果を示す。
【図5】図5は、実施例5におけるRNAi法によるPHB2ノックダウンの結果を示す。
【図6】図6は、実施例6での、PHB1又はPHB2のノックダウンに供したHeLa細胞におけるミトコンドリア膜電位及びカスパーゼ依存性細胞死(アポトーシス)についての検討結果を示す。
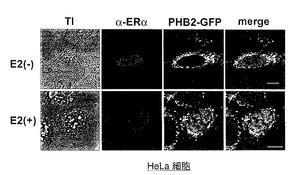
【図7】図7は、実施例7におけるRNAi法によるHax-1のノックダウンの結果を示す。
【図8】図8は、実施例8におけるMitoTracker Redでミトコンドリアを染色したPHB1、PHB2又はHax-1ノックダウン細胞の写真を示す。
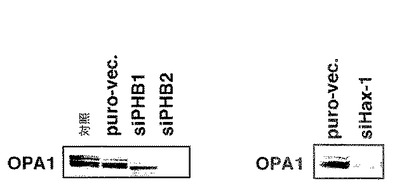
【図9】図9は、実施例8におけるOPA1タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。
【図10】図10は、実施例9におけるPHB1-GFP又はPHB2-GFPを発現するMCF-7細胞及びHeLa細胞の顕微鏡写真を示す。
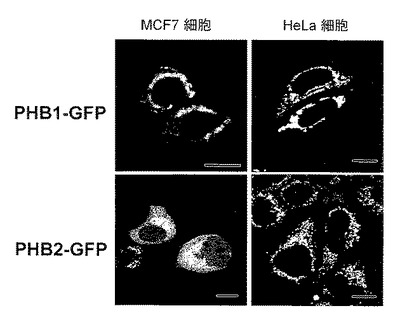
【図11】図11は、実施例10での、HeLa細胞においてPHB2-GFPとERαとを共発現させた際の共焦点レーザー顕微鏡写真を示す。
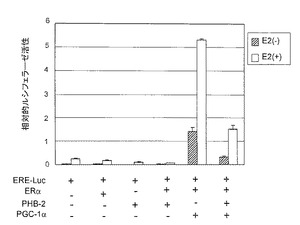
【図12−1】図12は、実施例11におけるPHB2タンパク質の構造分析の結果を示す。
【図12−2】図12−1の続きである。
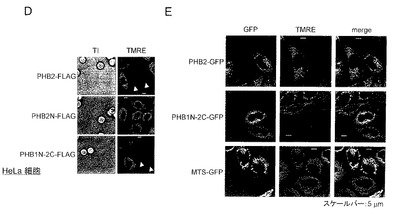
【図13】図13は、実施例12におけるPHB1-GFP又はPHB2-GFPとPPARα又はPPARγ2とを発現するHeLa細胞の顕微鏡写真を示す。
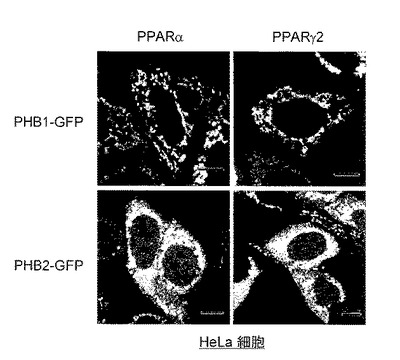
【図14】図14は、実施例13での、各遺伝子を導入したHeLa細胞における相対的ルシフェラーゼ活性を示す。
【発明を実施するための形態】
【0023】
以下、本発明を詳細に説明する。
ミトコンドリアにおけるヒトPHB2タンパク質の相互作用因子の探索を、HeLa細胞由来の精製ミトコンドリア画分を用いた免疫沈降解析及び質量分析により行ったところ、抗アポトーシス作用を有するミトコンドリアタンパク質Hax-1(NCBI 登録番号:NP_006109)(Cilenti L.ら, J Biol Chem., 279:50295-50301, 2004)、PTP(permeability transition pore)の構成因子であるVDAC2タンパク質(Swiss-Prot 登録番号:P45880)、ANT2タンパク質(NCBI 登録番号:NP_001143)及びPHB1タンパク質(NCBI 登録番号:NP_002625)の各タンパク質を相互作用因子として同定した。また、in vitro結合実験によれば、PHB2タンパク質とHax-1タンパク質との相互作用が直接的であることが明らかになった。また、RNA干渉法を用いて、HeLa細胞をPHB2ノックダウンに供したところ、PHB1タンパク質及びHax-1タンパク質の発現量減少、ミトコンドリア膜電位の減少、カスパーゼ依存性細胞死の誘導、ミトコンドリア膜形態調節因子OPA1タンパク質(NCBI 登録番号:NP_056375)量の減少及びミトコンドリア形態の断片化が観察された。
【0024】
一方、Hax-1ノックダウン細胞では、PHB2タンパク質発現量に影響を与えないものの、PHB2ノックダウンと同様のアポトーシスが誘導されることから、PHB2ノックダウンによるアポトーシス誘導は、Hax-1タンパク質量の減少によるものと考えられた。また、ミトコンドリアの形態は、Hax-1ノックダウン細胞ではミトコンドリアの断片化は観察できなかった。
【0025】
以上の知見によれば、PHB2タンパク質は、(1)ミトコンドリアにおいて相互作用因子Hax-1タンパク質の安定化、(2)それを介した抗アポトーシス作用、(3)PTP複合体を形成するVDAC2タンパク質及びANT2タンパク質との結合を介すると考えられるミトコンドリア膜電位形成、(4)OPA1タンパク質を介したミトコンドリア形態調節に関与していることが示された。
【0026】
従って、ミトコンドリアにおいて、PHB2タンパク質は、Hax-1タンパク質、VDAC2タンパク質、ANT2タンパク質、PHB1タンパク質、OPA1タンパク質等と物理的又は生理的な相互作用を示し、抗アポトーシス作用、ミトコンドリア膜電位形成作用及びミトコンドリア形態維持作用を有する。
【0027】
一方、PHB2タンパク質は、これまで、哺乳動物では核に存在し、転写抑制に働くこと、ヒストン脱アセチル化酵素(HDAC)をリクルートすることが報告されている(J. Biol. Chem. 279 (23), 24834-24843 (2004))。ヒト乳癌由来細胞であるMCF-7細胞では、PHB2タンパク質の核への局在が報告されている(J. Biol. Chem. 279 (23), 24834-24843 (2004))。本願では、PHB2タンパク質をミトコンドリア及び核に局在化させる機構を明らかにした。HeLa細胞では、MCF-7細胞と異なり、PHB2タンパク質はミトコンドリアにのみ局在し、核には局在化しない。しかしながら、PHB2タンパク質がエストロゲンレセプター(以下、「ER」という)存在下でエストラジオール(以下、「E2」という)依存的にミトコンドリアから核に移行することを明らかにした。このPHB2の核移行に必要な領域は、PHB2タンパク質のカルボキシル末端に依存する。また、PHB2タンパク質のアミノ末端にはミトコンドリア標的シグナルと弱い膜貫通ドメインが存在し、この部分のみでミトコンドリアに移行する。PHB2の核移行は、ER以外の核レセプターであるPPARαやPPARγ2存在下でも生じる。
【0028】
このように、本願では、PHB2タンパク質はミトコンドリア及び核の双方に局在するタンパク質であり、核では転写抑制に、ミトコンドリアでは各種タンパク質を安定化することにより、ミトコンドリア膜電位形成、抗アポトーシス作用、ミトコンドリア形態維持等に働いていることを明らかにした。
以上に説明した新規のPHB2タンパク質の機能に基づき、本発明を説明する。
【0029】
本発明に係るPHB2遺伝子調節剤のスクリーニング方法は、PHB2遺伝子発現細胞をその培養下で候補物質と接触させ、候補物質非存在下での該PHB2遺伝子発現細胞と比較して、該候補物質と接触した該PHB2遺伝子発現細胞においてPHB2遺伝子発現又はPHB2タンパク質機能が調節されていることにより、該候補物質がPHB2遺伝子調節剤であると判定するものである。
【0030】
ヒトPHB2遺伝子は、NCBI登録番号NM_007273としてNCBIに登録されており、配列番号21記載の塩基配列から成るDNAである。なお、CDSは配列番号21記載の塩基配列において、186〜1085番目の塩基配列である。当該ヒトPHB2タンパク質は、NCBI登録番号NP_009204としてNCBIに登録されており、配列番号22記載のアミノ酸配列から成るタンパク質である。ヒト以外の生物由来のPHB2遺伝子又はPHB2タンパク質としては、例えば、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)由来(PHB2_YEAST(swissprot:accession P50085))、線虫(Caenorhabditis elegans)由来(PHB2_CAEEL(swissprot:accession P50093))、シロイヌナズナ(Arabidopsis thaliana)由来(ATPHB2 (PROHIBITIN 2) [Arabidopsis thaliana](NCBI登録番号 NP_973755))、アフリカツメガエル(Xenopus tropicalis)由来(prohibitin 2 [Xenopus tropicalis](NCBI登録番号NP_001016551))、マウス由来(PHB2_MOUSE(swissprot:accession O35129))及びラット由来(PHB2_RAT(swissprot:accession Q5XIH7))のものが挙げられる。本発明では、PHB2遺伝子には、上述した配列番号又は登録番号に記載の塩基配列から成るDNAに限らず、これらDNAにおいて、1又は複数(例えば、1〜10個、好ましくは1〜5個)の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2タンパク質機能を有するタンパク質をコードするDNAも含まれる。また、PHB2遺伝子には、上述した配列番号又は登録番号に記載の塩基配列から成るDNAに90%以上、好ましくは95%以上、97%以上、より好ましくは98%以上、特に好ましくは99%以上の同一性を有し、且つPHB2タンパク質機能を有するタンパク質をコードするDNAも含まれる。あるいは、上述した配列番号又は登録番号に記載のアミノ酸配列をコードするDNAに限らず、これらアミノ酸配列において、1又は複数(例えば、1〜10個、好ましくは1〜5個)のアミノ酸が欠失、置換又は付加されたアミノ酸配列から成り、且つPHB2タンパク質機能を有するタンパク質をコードするDNAもPHB2遺伝子に含まれる。
【0031】
ここで、PHB2タンパク質機能としては、例えば、抗アポトーシス作用、ミトコンドリア膜電位形成、ミトコンドリア形態維持、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質(以下では、各タンパク質をそれぞれ「VDAC2タンパク質、Hax-1タンパク質、PHB1タンパク質、ANT2タンパク質及びOPA1タンパク質」という)との相互作用能(例えば、複合体形成能、シグナル伝達能、タンパク質安定化能、タンパク質立体構造・機能調節能などを指し、生理的相互作用を含む)、並びに核-ミトコンドリア移行機能が挙げられる。本明細書中で使用される「複合体形成能」とは、ミトコンドリアや核、または細胞質などで直接的な結合や間接的な結合を介して形成される複合体の形成能を指し、その複合体の構成成分としては、タンパク質や核酸、またはある種の脂質などが含まれる。また、「核-ミトコンドリア移行機能」とは、ミトコンドリアと核との間の移行機能を指し、ミトコンドリアと細胞質との間、または細胞質と核との間の移行機能を含む。また、当該核-ミトコンドリア移行機構には、これらの移行を介在する輸送体やこれを刺激するリガンドを含めた輸送機構が含まれる。
【0032】
PHB2遺伝子発現細胞とは、PHB2遺伝子を発現する細胞であればいずれのものであってよいが、例えばHeLa細胞(ヒト子宮頸癌由来)、MCF-7細胞(ヒト乳癌由来)、U-2OS細胞(ヒト骨肉腫由来)及びヒト線維芽細胞等のヒト由来の細胞系、並びにB6.1細胞(マウスミエローマ由来)、マウス胚性幹細胞及びSWISS 3T3細胞(マウス線維芽細胞由来)が挙げられる。さらに、上述したPHB2遺伝子を導入した(又は過剰発現させた)細胞もPHB2遺伝子発現細胞に含まれる。例えば、PCR産物やベクターに含まれるPHB2遺伝子を、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等により細胞へ導入することができる。PHB2遺伝子発現細胞において、PHB2遺伝子が発現していることの確認方法としては、mRNAレベルでは、例えばPHB2遺伝子に特異的なプライマーやプローブを用いたRT-PCR、定量PCRやノーザンブロッティングによって確認する方法が挙げられる。また、タンパク質レベルでは、例えばPHB2タンパク質に特異的な抗体を用いたELISA、フローサイトメトリー、ウエスタンブロッテイング等の免疫学的方法を用いて、PHB2遺伝子の発現を確認することができる。
【0033】
一方、本発明において、候補物質としては、例えば、核酸、ペプチド、タンパク質、合成化合物、微生物の培養上清、植物や海洋生物由来の天然成分、植物抽出物、動物組織抽出物等が挙げられる。
【0034】
本発明に係るPHB2遺伝子調節剤のスクリーニング方法においては、先ずPHB2遺伝子発現細胞をその培養下で候補物質と接触させる。PHB2遺伝子発現細胞の培養は、各細胞に応じて、培地、培養条件(例えば、温度、pH)を適宜選択して行うことができる。ここで、接触とは、PHB2遺伝子発現細胞に候補物質が影響を及ぼす状態を意味する。例えば、PHB2遺伝子発現細胞の培地に候補物質を添加するだけでもよく、あるいは、候補物質をリポソームに抱埋または結合させて培地に添加してもよく、候補物質をある種のキャリアー物質(タンパク質や脂質など)とともに培地に添加してもよい。また、PHB2遺伝子発現細胞にマイクロインジェクションなどで候補物質を直接導入してもよい。培養時間は、候補物質がPHB2遺伝子発現細胞に影響を及ぼすのに十分な時間であればよいが、例えば1〜72時間、好ましくは12〜24時間とすることができる。
【0035】
次いで、本発明に係るPHB2遺伝子調節剤のスクリーニング方法においては、候補物質非存在下でのPHB2遺伝子発現細胞と比較して、該候補物質と接触した該PHB2遺伝子発現細胞においてPHB2遺伝子発現又はPHB2タンパク質機能が調節されている否かを決定する。ここで、PHB2遺伝子発現の調節とは、mRNAレベル又はタンパク質レベルでのPHB2遺伝子発現の低下又は増加を意味する。さらに、PHB2タンパク質機能の調節とは、上述したPHB2タンパク質機能の上昇を意味する。
【0036】
PHB2遺伝子発現の調節の評価方法では、培養後、当該PHB2遺伝子発現細胞からmRNA又はタンパク質を抽出する。次いで、得られたmRNA又はタンパク質中のPHB2遺伝子発現量を、候補物質不在下で培養したPHB2遺伝子発現細胞におけるPHB2遺伝子発現量と比較する。なお、mRNAレベル又はタンパク質レベルでのPHB2遺伝子の発現量の測定は、上記で説明した方法に準じて行うことができる。
【0037】
候補物質不在下で培養したPHB2遺伝子発現細胞に比べて、候補物質を接触させたPHB2遺伝子発現細胞において、mRNAレベル又はタンパク質レベルで有意(例えば、1.5〜100倍、好ましくは2〜5倍)にPHB2遺伝子発現量が増加した場合に、あるいは、有意(例えば、1/2〜1/1000、好ましくは1/4〜1/10)にPHB2遺伝子発現量が低下した場合に、候補物質がPHB2遺伝子調節剤であると判定することができる。
一方、PHB2タンパク質機能の調節の評価は、各機能に応じて個別に行うことができる。
【0038】
抗アポトーシス作用は、例えば、DNA断片化、Annexin Vの結合による細胞膜構造変化、ミトコンドリア膜電位消失、カスパーゼ等のアポトーシス関連酵素の活性、cytochrome c のミトコンドリアから細胞質への転移等を検出することによって評価することができる。
【0039】
ミトコンドリア膜電位形成については、例えば、ミトコンドリア膜電位に依存したミトコンドリア染色マーカーRh123を用いた染色にPHB2遺伝子発現細胞を供し、顕微鏡による観察により染色度合いによって評価することができる。
【0040】
ミトコンドリア形態維持については、例えば、MitoTracker Redを用いたミトコンドリア染色にPHB2遺伝子発現細胞を供し、顕微鏡観察によりミトコンドリア形態を観察することにより評価することができる。
【0041】
また、PHB2タンパク質とVDAC2タンパク質、Hax-1タンパク質、PHB1タンパク質、ANT2タンパク質又はOPA1タンパク質との相互作用能(例えば、複合体形成能)については、例えばPHB2遺伝子発現細胞由来のタンパク質を各タンパク質に特異的な抗体を用いた免疫学的解析に供することで、タンパク質間の会合を観察することで評価することができる。
【0042】
さらに、核-ミトコンドリア移行機能については、ERα、PPARα又はPPARγ2を発現するPHB2遺伝子発現細胞を(ERαについてはE2存在下で)培養することで、ミトコンドリアから核へのPHB2タンパク質の移行を、例えばPHB2タンパク質に特異的な抗体を用いた染色により評価することができる。培地に対するE2添加量は、例えば、1x10-12〜1x10-4M、好ましくは1x10-7〜1x10-6Mとする。
【0043】
候補物質不在下で培養したPHB2遺伝子発現細胞に比べて、候補物質を接触させたPHB2遺伝子発現細胞において、有意(例えば、1.5〜100倍、好ましくは2〜10倍)にPHB2タンパク質機能が上昇した場合に、候補物質がPHB2遺伝子調節剤であると判定することができる。
【0044】
一方、上述した本発明に係るPHB2遺伝子調節剤のスクリーニング方法に準じて、ミトコンドリア機能調節剤をスクリーニングすることができる。本発明に係るミトコンドリア機能調節剤のスクリーニング方法は、PHB2遺伝子発現可能なミトコンドリア機能異常細胞をその培養下で候補物質と接触させ、候補物質非存在下での該ミトコンドリア機能異常細胞と比較して、該候補物質と接触した該ミトコンドリア機能異常細胞において、PHB2遺伝子発現又はPHB2タンパク質機能が調節される結果としてミトコンドリア機能が回復又は正常化することを指標として、上記候補物質がミトコンドリア機能調節剤であると判定するものである。
【0045】
ここで、PHB2遺伝子発現可能なミトコンドリア機能異常細胞(以下、単に「ミトコンドリア機能異常細胞」という)とは、PHB2遺伝子を発現し、且つ上述したミトコンドリア機能に異常を有する細胞である。このような細胞としては、例えば、PHB2をノックダウンした細胞に別の変異PHB2を発現させた細胞などが挙げられる。
【0046】
本発明に係るミトコンドリア機能調節剤のスクリーニング方法では、上述した本発明に係るPHB2遺伝子調節剤のスクリーニング方法に準じて、候補物質非存在下でのミトコンドリア機能異常細胞と比較して、該候補物質と接触したミトコンドリア機能異常細胞において、PHB2遺伝子発現又はPHB2タンパク質機能が調節され、ミトコンドリア機能が回復又は正常化するか否かを決定する。そのような指標により、上記候補物質の中からミトコンドリア機能調節剤を選択することができる。
【0047】
本発明に係るPHB2遺伝子調節剤としては、例えば、上述したPHB2遺伝子調節剤のスクリーニング方法により得られたもの、以下の(a)又は(b)記載のsiRNAを含有するもの:(a)配列番号1〜20記載のいずれか1つに示される塩基配列から成るsiRNA:(b)上記(a)記載のsiRNAにおいて、1又は数個(例えば1〜5個、好ましくは1〜3個)の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2遺伝子発現阻害活性を有するsiRNA、並びにERα、PPARα又はPPARγ2の核レセプターのアンタゴニスト又はアゴニストを含有するものが挙げられる。また、上記(a)記載のsiRNAに90%以上、好ましくは95%以上、97%以上、より好ましくは98%以上、特に好ましくは99%以上の同一性を有し、且つPHB2遺伝子発現阻害活性を有するsiRNAも本発明に係るPHB2遺伝子調節剤とすることができる。
【0048】
上述した(a)又は(b)記載のsiRNAは、例えば、公知の核酸合成方法により化学的に合成することができる。
【0049】
ERα、PPARα又はPPARγ2の核レセプターのアンタゴニスト又はアゴニストとしては、例えばERのアンタゴニストであるICI182,780、DDE及びTamoxifenなど、ERのアゴニストであるDES(Diethylstilbestrol)、ゲニステイン、ノニルフェノール及びビスフェノールA(Bisphenol A)など、PPARαのアゴニストであるWY-14643、ETYA、Benzafibrate及びLY171883など、PPARαのアンタゴニストであるGW6471など、PPARγのアゴニストであるGW9662及びTroglitazoneなど、並びにPPARγのアンタゴニストであるチアゾリジンジオンなどが挙げられる。
【0050】
更なるPHB2遺伝子調節剤としては、例えばTR、PR及びRXR等の他の核レセプターのアゴニストやアンタゴニストが挙げられる。
【0051】
また、PHB2遺伝子調節剤として、例えばPHB1タンパク質のミトコンドリア標的シグナル及び膜貫通ドメイン(ヒトPHB1タンパク質では、N末端より1番目〜50番目のアミノ酸残基)とPHB2タンパク質のC末端(ヒトPHB2タンパク質では、N末端より51番目〜299番目のアミノ酸残基)とのキメラタンパク質を用いることができる。
【0052】
なお、PHB2タンパク質の機能に基づき、PHB2遺伝子調節剤は、抗アポトーシス作用剤、ミトコンドリア膜電位調節剤又はミトコンドリア膜形態調節剤として使用することができる。
【0053】
本発明に係るミトコンドリア機能関連疾患治療薬は、本発明に係るPHB2遺伝子調節剤を含有することを特徴とする。ここで、ミトコンドリア機能関連疾患とは、ミトコンドリア機能の異常に関連する疾患を意味し、例えば、肥満、糖尿病、パーキンソン病、アルツハイマー病、乳癌等の癌が挙げられるが、これらに限定されない。
【0054】
本発明に係るミトコンドリア機能関連疾患治療薬は、本発明に係るPHB2遺伝子調節剤それ自体であっても、あるいは通常用いられる固体または液体の担体、乳化分散剤等により錠剤、粉剤、乳剤、カプセル剤等の剤形に製剤化されたものであってもよい。上記担体としては、水、ゼラチン、澱粉、ステアリン酸マグネシウム、ラクトース、植物油等が挙げられる。本発明に係るミトコンドリア機能関連疾患治療薬におけるPHB2遺伝子調節剤の含有量及び当該治療薬の投与量は、投与目的、投与経路、剤形等によって適宜変更しうる。
【0055】
本発明に係るミトコンドリア機能関連疾患治療薬の薬理評価は、例えば、上述したミトコンドリア機能異常細胞を使用したin vitroレベル、アルツハイマー病等のミトコンドリア機能関連疾患動物モデルを使用したin vivoレベルで評価することができる。例えば、本発明に係るミトコンドリア機能関連疾患治療薬非存在下に比べて、当該治療薬存在下で培養したミトコンドリア機能異常細胞において、ミトコンドリア機能異常が回復又は正常化するか否かにより薬理評価を行うことができる。あるいは、ミトコンドリア機能関連疾患動物モデルに本発明に係るミトコンドリア機能関連疾患治療薬を投与しない場合と比較して、当該治療薬を投与した場合にミトコンドリア機能関連疾患が治療又は改善するか否かにより薬理評価を行うことができる。
【0056】
本発明に係るミトコンドリア機能関連疾患の検出方法は、被験者由来の生物学的サンプルにおいて、PHB2遺伝子発現又はPHB2タンパク質機能を測定し、該PHB2遺伝子発現又はPHB2タンパク質機能が異常であることを指標として、該被験者がミトコンドリア機能関連疾患を有するか又はそれらを有する疑いがあると決定するものである。
【0057】
生物学的サンプルとしては、例えば、被験者由来の線維芽細胞、筋芽細胞、白血球細胞、精子細胞や卵子細胞等の細胞、これら細胞を含む組織や器官、血液や唾液等の体液、排出物が挙げられる。
【0058】
本発明に係るミトコンドリア機能関連疾患の検出方法では、上述した本発明に係るPHB2遺伝子調節剤のスクリーニング方法に準じて行うことができる。即ち、先ず生物学的サンプル中の細胞におけるPHB2遺伝子発現又はPHB2タンパク質機能を測定する。次いで、PHB2遺伝子発現又はPHB2タンパク質機能が異常であるか否かを正常者由来の細胞と比較して判定する。ここで、PHB2遺伝子発現又はPHB2タンパク質機能の異常とは、例えば、PHB2遺伝子の発現増加若しくは低下又はPHB2タンパク質機能の低下を意味する。PHB2遺伝子発現又はPHB2タンパク質機能に異常が見られた場合には、生物学的サンプルが由来する被験者がミトコンドリア機能関連疾患を有するか又はそれらを有する疑いがあると決定することができる。
【0059】
以上に説明したように、新規PHB2タンパク質の機能に基づけば、PHB2遺伝子調節剤を同定することができ、またミトコンドリア機能調節剤やミトコンドリア機能関連疾患治療薬を提供することができる。
【0060】
本明細書は本願の優先権の基礎である日本国特許出願2005-339354号の明細書及び/又は図面に記載される内容を包含する。
【実施例】
【0061】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲はこれら実施例に限定されるものではない。
【0062】
なお、本実施例で示すタンパク質は、標識やレポーターとして使用したタンパク質(例えば、FLAGタグ、GST、GFP及びルシフェラーゼ等)以外はいずれもヒト由来のタンパク質である。
【0063】
〔実施例1〕HeLa細胞におけるPHB1タンパク質又はPHB2タンパク質の細胞内局在
カルボキシル末端にFLAGタグを融合したPHB1タンパク質(以下、「PHB1-FLAG」という)又はPHB2タンパク質(以下、「PHB2-FLAG」という)をコードする遺伝子をそれぞれHeLa細胞(ヒト子宮頸癌由来)に一過性に発現させた。
【0064】
方法としては、哺乳細胞発現ベクターpCMV-SPORTにPHB1-FLAG又はPHB2-FLAGをコードするcDNAを挿入し、これをHeLa細胞に遺伝子導入し、約12時間後、これらの細胞を固定し、免疫染色に供した。抗体は抗FLAGウサギポリクローナル抗体およびミトコンドリアを染色するコントロール抗体として抗チトクロームC(cyt.c)マウスモノクローナル抗体を一次抗体として、二次抗体にはAlexa488標識抗ウサギ抗体およびCy3標識抗マウス抗体を用いた。これらの細胞は共焦点レーザー顕微鏡にて観察した。
【0065】
結果を図1に示す。図1は、PHB1-FLAG又はPHB2-FLAGを発現するHeLa細胞の共焦点レーザー顕微鏡写真である。図1において、α-FLAGは、抗FLAG抗体の染色像(緑色)を示し、α-cyt.cは、抗チトクロームC抗体の染色像(赤色)を示す。また、Mergeは、両者の重ね合わせ像を示す。
【0066】
図1に示すように、PHB1-FLAG及びPHB2-FLAGをそれぞれHeLa細胞に一過性に発現させると、PHB1-FLAG及びPHB2-FLAGはミトコンドリアに局在した。HeLa細胞におけるPHB1-FLAG及びPHB2-FLAGは、ミトコンドリアマーカーであるcyt.cと共局在を示した。
【0067】
〔実施例2〕PHB1タンパク質又はPHB2タンパク質発現によるミトコンドリア膜電位消失
実施例1に示すように、PHB1-FLAG及びPHB2-FLAGをそれぞれHeLa細胞に一過性に発現させると、PHB1-FLAG及びPHB2-FLAGはミトコンドリアに局在した。
【0068】
そこで、PHB1-FLAG又はPHB2-FLAGを発現するHeLa細胞を、ミトコンドリア膜電位に依存したミトコンドリア染色マーカーであるRh123で染色した。同様に、SLP2タンパク質(NCBI登録番号:NP_038470)(J Biol Chem. 2000 Mar 17;275(11):8062-71)のカルボキシル末端にFLAGタグを融合した融合タンパク質(以下、「SLP2-FLAG」という)をコードする遺伝子をHeLa細胞に一過性に発現させたものを、Rh123で染色した。また、対照として、いずれの遺伝子も導入されていないHeLa細胞をRh123で染色した。
【0069】
結果を図2に示す。図2は、各遺伝子を発現するHeLa細胞をRh123で染色した際の写真を示す。図2において、TIは、透過像の写真を示す。
【0070】
図2に示すように、PHB1-FLAG又はPHB2-FLAGを過剰発現するHeLa細胞は、Rh123で染色されないことが明らかとなった。このことは、PHB1-FLAG又はPHB2-FLAGの強制発現によってミトコンドリア膜電位が消失したことを示し、PHBタンパク質がミトコンドリアにおいて膜電位の調節に関与している可能性を示唆するものである。
【0071】
〔実施例3〕PHB2タンパク質の分子機能分析
PHB2タンパク質の分子機能を明らかにするために、ミトコンドリアにおけるPHB2タンパク質の相互作用因子の探索を行った。
【0072】
まず、実施例1で作製したPHB2-FLAGを発現するHeLa細胞由来の精製ミトコンドリア画分を用いた免疫沈降(IP)解析及び質量分析(MASS解析)を行った。
【0073】
IP解析では、抗FLAG抗体を用いて免疫沈降法を行い、この沈降物を12%SDS-PAGEに供した後、タンパク質染色を行い、PHB2-FLAGタンパク質と共沈するタンパク質を検出した。
【0074】
一方、MASS解析では、IP解析にて得たタンパク質のバンドを切り出し、トリプシンによるゲル内消化を行った後、消化されたペプチドを抽出後、ナノスケール高速液体クロマトグラフィーおよびタンデム型質量分析装置(nano LC-MS/MS)にて、ペプチド断片を同定した。次いで、データベース解析にてタンパク質を同定した。
【0075】
IP解析及びMASS解析の結果を図3に示す。
図3から判るように、抗アポトーシス作用を有するミトコンドリアタンパク質Hax-1(NCBI登録番号:NP_006109)、PTP(permeability transition pore)の構成因子であるVDAC2タンパク質(Swiss-Prot登録番号:P45880)及びANT2タンパク質(NCBI登録番号:NP_001143)、並びにPHB2タンパク質と複合体を形成することが知られている(Curr. Biol. 7 (8), 607-610 (1997))PHB1タンパク質(NCBI登録番号:NP_002625)が、PHB2タンパク質の相互作用因子として得られた。
【0076】
〔実施例4〕PHB2タンパク質とHax-1タンパク質との相互作用
実施例3に記載のIP解析、並びにPHB2タンパク質、Hax-1タンパク質、VDAC2タンパク質及びPHB1タンパク質に対する特異的抗体を用いたウエスタンブロット分析により、PHB2タンパク質とHax-1タンパク質、VDAC2タンパク質及びPHB1タンパク質との相互作用が確認された。
【0077】
そこで、in vitro結合実験により、PHB2タンパク質とHax-1タンパク質との相互作用を検討した。
【0078】
方法としては、大腸菌を用いて合成したグルタチオンS-トランスフェラーゼ(GST)-PHB2融合タンパク質(PHB2タンパク質のN末端にGSTタンパク質を融合したタンパク質)と、網状赤血球溶解液を用いて試験管内で翻訳したPHB1-FLAGおよびHax-1-FLAGタンパク質(Hax-1タンパク質のC末端にFLAGタグを融合したタンパク質)を、試験管内で混合し、Glutathione-Sepharoseビーズを用いてGST融合タンパク質をPull downした沈降産物を得た。次いで、この沈降産物をSDS-PAGEに供した後、抗FLAG抗体にてウエスタンブロット法を行い、PHB2タンパク質に各々のFLAG融合タンパク質が結合したか否かを検討した。
【0079】
結果を図4に示す。図4において、「インプット」は、混入したFLAG融合タンパク質を示す。一方、「GST-プルダウン」は、Glutathione-Sepharoseビーズで沈降させた沈降物を示す。さらに、「-」は反応系に混入していないことを示し、「+」は反応系に混入していることを示す。
【0080】
図4に示すように、PHB2タンパク質とHax-1タンパク質との相互作用は直接的であることが明らかとなった。
【0081】
〔実施例5〕RNAi法によるPHB2のノックダウン
RNAi法を用いて、HeLa細胞においてPHB2のノックダウンを行った。siRNAとしては、配列番号1記載の塩基配列から成るものを用いた。また、PHB1に対するsiRNA(配列番号23)を用いて、PHB1のノックダウンも行った。
【0082】
方法としては、PHB2およびPHB1のcDNAの一部に一致したshort hairpin RNAを発現させる塩基配列を、ピューロマイシン耐性遺伝子を有するpScilencer 3.1-H1 puro vectorに挿入したプラスミドを作製し、これを培養細胞に遺伝子導入した。その後、ピューロマイシンを培地に添加することで、目的遺伝子をノックダウンした細胞を得ることが可能であった。
【0083】
結果を図5A〜Cに示す。図5Aは、PHB1又はPHB2をノックダウンした際の各タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。図5Bは、PHB1又はPHB2をノックダウンした際のHax-1タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。図5Cは、PHB1又はPHB2をノックダウンした際のHax-1 mRNA量を測定したRT-PCRの結果を示す。図5A〜Cにおいて、siPHB1又はsiPHB2は、それぞれRNAi法によりPHB1又はPHB2をノックダウンした際の結果を示す。対照は遺伝子を導入していない細胞を示し、puro-vec.は、ピューロマイシン耐性遺伝子を導入し、その後、薬剤選択を行った細胞を示す。
【0084】
図5A〜Cに示すように、RNAi法を用いて、HeLa細胞においてPHB2のノックダウンを行ったところ、相互作用因子であるHax-1タンパク質やPHB1タンパク質の発現量が減少していることがわかった。しかしながら、PHB1及びPHB2のmRNAは減少していなかった。酵母PHBが新規に合成されたミトコンドリアタンパク質と相互作用し、その安定化を行うとの報告(Mol. Cell. Biol. 19, 3435-3442(1999).及びEMBO J. 19, 2444-2451 (2000).)があることから、ヒトPHB2タンパク質も相互作用因子Hax-1タンパク質の安定化を行っている可能性が考えられる。
【0085】
〔実施例6〕PHB2ノックダウンHeLa細胞におけるミトコンドリア膜電位低下及びカスパーゼ依存性細胞死(アポトーシス)の誘導
本実施例では、実施例5においてPHB1又はPHB2のノックダウンに供したHeLa細胞において、ミトコンドリア膜電位及びカスパーゼ依存性細胞死(アポトーシス)について検討した。
【0086】
方法としては、実施例5で行ったノックダウンと同様の方法を用いた。
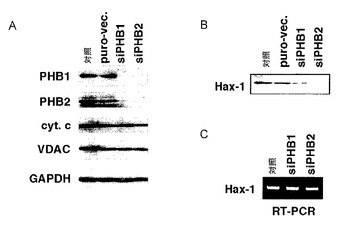
結果を図6A〜Cに示す。図6A及びBは、PHB1又はPHB2のノックダウンに供したHeLa細胞における相対的細胞数を示す。図6Bにおいて、Z-VAD-FMKは、アポトーシスで活性化されるカスパーゼの阻害剤である。Z-VAD-FMKに関する+又は-は、培地への添加又は非添加を示す。図6Cは、PHB1又はPHB2のノックダウンに供したHeLa細胞におけるRh123染色の結果を示す。図6A〜Cにおいて、siPHB1又はsiPHB2は、それぞれRNAi法によりPHB1又はPHB2をノックダウンした際の結果を示す。puro-vec.は、ピューロマイシン耐性遺伝子を有するプラスミドベクターを導入した際の結果を示す。
【0087】
図6A及びBに示すように、PHB1又はPHB2をノックダウンしたHeLa細胞では、カスパーゼ依存性細胞死(アポトーシス)が誘導されていることが明らかとなった。また、図6Cに示すように、PHB1又はPHB2をノックダウンしたHeLa細胞では、一部の細胞でミトコンドリア膜電位が低下していることが明らかとなった。
【0088】
〔実施例7〕RNAi法によるHax-1のノックダウン
本実施例では、HeLa細胞においてPHB2の相互作用因子であるHax-1のノックダウンを行った。Hax-1に対するsiRNAとして、配列番号24記載の塩基配列から成るものを用いた。
【0089】
方法としては、Hax-1のcDNAの一部に一致したshort hairpin RNAを発現させる塩基配列を、ピューロマイシン耐性遺伝子を有するpScilencer 3.1-H1 puro vectorに挿入したプラスミドを作製し、実施例5の方法に準じた。
【0090】
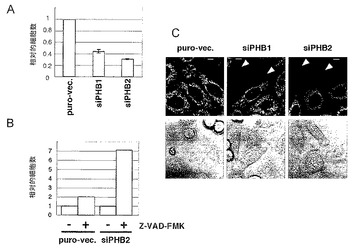
結果を図7A〜Cに示す。図7Aは、Hax-1をノックダウンした際の各タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。図7B及びCは、Hax-1のノックダウンに供したHeLa細胞における相対的細胞数を示す。図7におけるZ-VAD-FMK及びZ-VAD-FMKに関する+又は-の意味は、図6と同様である。図7A〜Cにおいて、siHax-1は、RNAi法によりHax-1をノックダウンした際の結果を示す。puro-vec.は、ピューロマイシン耐性遺伝子を有するプラスミドベクターを導入した際の結果を示す。
【0091】
図7A〜Cに示すように、Hax-1のノックダウンが、PHB1タンパク質及びPHB2タンパク質の発現量には影響を与えずに、PHB2のノックダウンと同様のアポトーシスを誘導することから、Hax-1タンパク質の発現量減少がPHB2ノックダウンによって誘導されるアポトーシスの一因であると考えられた。従って、PHB2タンパク質はHax-1タンパク質の発現量を維持することにより抗アポトーシス作用を持つことが示された。
【0092】
〔実施例8〕PHB2のノックダウンによるミトコンドリア形態変化
本実施例では、実施例5及び7に記載するPHB1、PHB2又はHax-1のノックダウンに供したHeLa細胞において、ミトコンドリア形態の変化を検討した。
【0093】
方法としては、実施例5及び7で作製したノックダウン細胞をMitoTracker Redで染色して、共焦点レーザー顕微鏡でミトコンドリアの形態を観察した。
【0094】
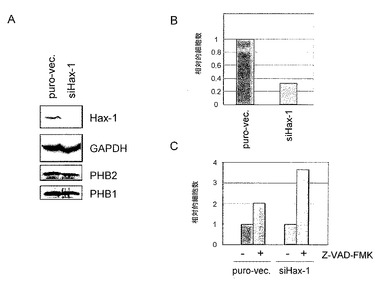
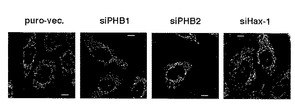
結果を図8に示す。図8において、puro-vec.は、ピューロマイシン耐性遺伝子を有するプラスミドベクターを導入した際の結果を示す。また、siPHB1、siPHB2、siHax-1は、それぞれPHB1、PHB2、Hax-1のノックダウンに供したHeLa細胞の結果を示す。
【0095】
図8に示すように、MitoTracker Redでミトコンドリアを染色したPHB1又はPHB2ノックダウン細胞では、ミトコンドリア形態の断片化が観察された。こうしたミトコンドリアの断片化はHax-1ノックダウン細胞では見られず、PHBタンパク質ノックダウン特異的で、且つHax-1を介在しない形態変化であると考えられた。
【0096】
また、上述したPHB1、PHB2又はHax-1ノックダウン細胞を、ミトコンドリア融合因子OPA1タンパク質に対する抗体を用いたウエスタンブロット分析に供した。
【0097】
方法としては、ノックダウンした細胞から細胞抽出液を作製し、これをウエスタンブロット法に供した。一次抗体は抗OPA1ポリクローナルウサギ抗体を用いた。
【0098】
結果を図9に示す。図9において、対照は、遺伝子非導入細胞である。また、puro-vec.、siPHB1、siPHB2、siHax-1は図8と同様の細胞の結果である。
【0099】
図9に示すように、PHB2ノックダウン細胞では、OPA1タンパク質の著明な発現減少が見られたため、OPA1タンパク質の減少によるミトコンドリアの断片化が生じたと考えられた。一方、Hax-1ノックダウン細胞においてもOPA1タンパク質は減少していた。また、PHB1のノックダウンによるOPA1タンパク質の発現減少は、PHB2のノックダウンと比較して顕著ではないが、PHB2ノックダウンと同等のミトコンドリア断片化を引き起こす(図8)。
【0100】
OPA1タンパク質は、他のミトコンドリア形態を調節する因子Drp1等とのバランスによりミトコンドリア形態を維持していることが知られている(Cell. 2004 Dec 17;119(6):873-87.)。PHB1ノックダウン細胞におけるミトコンドリアの断片化は、OPA1の発現量以外にも何らかの因子の関与があるものと考えられる。
【0101】
〔実施例9〕MCF-7細胞におけるPHB1タンパク質又はPHB2タンパク質の細胞内局在
実施例1で示すように、HeLa細胞では、PHB1タンパク質及びPHB2タンパク質が、ミトコンドリアに局在した。
【0102】
本実施例では、HeLa細胞(ヒト子宮頸癌由来)に加えて、MCF-7細胞(ヒト乳癌由来)でもPHB1タンパク質及びPHB2タンパク質の細胞内局在を検討した。
【0103】
方法としては、緑色蛍光タンパク質(GFP)をC末端に連結したPHB2又はPHB1タンパク質(各融合タンパク質を、「PHB2-GFP」、「PHB1-GFP」という)をコードするcDNAを連結した哺乳細胞発現ベクターを、培養細胞に導入し、これを共焦点レーザー顕微鏡にて、GFP融合タンパク質の細胞内局在を観察した。
【0104】
結果を図10に示す。図10は、PHB1-GFP又はPHB2-GFPを発現するMCF-7細胞及びHeLa細胞の顕微鏡写真を示す。図10に示すように、PHB1-GFPをコードする遺伝子をMCF-7細胞及びHeLa細胞に導入すると、PHB1-GFPはいずれにおいてもミトコンドリアに局在した。一方、PHB2-GFPはHeLa細胞ではミトコンドリアに局在するものの、MCF-7細胞では、PHB2-GFPの一部は核にも局在した。
【0105】
〔実施例10〕PHB2タンパク質と核レセプターERαとの相互作用
本実施例ではPHB2タンパク質と核レセプターERαとの相互作用について検討した。
方法としては、発現ベクターに組み込んだERα遺伝子とPHB2-GFP遺伝子をHeLa細胞に遺伝子導入後、この細胞を固定し、抗ERαウサギポリクローナル抗体を一次抗体として、Cy3標識抗ウサギ抗体を二次抗体として用いて細胞免疫染色を行った。
【0106】
結果を図11に示す。図11は、HeLa細胞においてPHB2-GFPとERαとを共発現させた際の共焦点レーザー顕微鏡写真を示す。図11において、TIは、透過像を示し、α-ERαは、抗ERα抗体染色像(赤色)を示す。また、PHB2-GFPは、GFP像(緑色)を示し、mergeは、重ね合わせ像を示す。E2(-)はE2非存在下、E2(+)はE2存在下で培養した細胞である。
【0107】
図11に示すように、PHB2-GFPは、HeLa細胞でERαと共発現させると、E2存在下で核に局在化した。これまでの報告から、PHB2タンパク質と核レセプターERαとが結合することが知られている(J. Biol. Chem. Vol. 275, No. 46, pp. 35848-35856, 2000)。従って、PHB2はE2依存的にERαと結合し、核に移行するのではないかと考えられた。
【0108】
〔実施例11〕PHB2タンパク質の構造分析
上記実施例で示すように、PHB2タンパク質はミトコンドリア及び核に局在化することが明らかになった。この機構を明らかにするために、PHB2タンパク質の欠失変異体及びPHB2タンパク質のC末端を有するキメラタンパク質等を作製した。同様に、PHB1タンパク質の欠失変異体を作製した。
【0109】
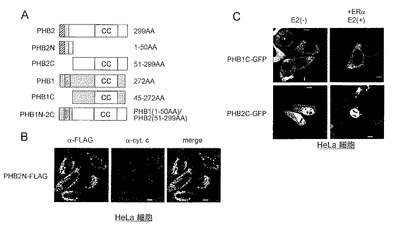
図12Aには、作製したPHB2タンパク質の欠失変異体及びキメラタンパク質、並びにPHB1タンパク質の欠失変異体を示す。PHB2Nは、PHB2タンパク質のN末端より1番目〜50番目のアミノ酸残基から成る欠失変異体である。PHB2Cは、PHB2タンパク質のN末端より51番目〜299番目のアミノ酸残基から成る欠失変異体である。また、PHB1Cは、PHB1タンパク質のN末端より45番目〜272番目のアミノ酸残基から成る欠失変異体である。さらにキメラタンパク質PHB1N-2Cは、PHB1タンパク質のN末端より1番目〜50番目のアミノ酸残基(内膜に局在化させるシグナルを有する)とPHB2タンパク質のN末端より51番目〜299番目のアミノ酸残基とを連結した変異体である。図12Aにおいて、ccは、コイルドコイルドメインを示す。
【0110】
欠失変異体等をコードする遺伝子の作製方法では、任意の塩基部位に相補的なプライマーを作製し、PCRにて任意の領域を増幅し、発現ベクターに組み込むことで各変異体をコードする遺伝子を含む発現ベクターを作製した。
【0111】
結果を図12B〜Dに示す。
【0112】
図12Bは、PHB2N-FLAG(PHB2Nタンパク質のC末端にFLAGタグを融合したタンパク質)を発現するHeLa細胞を用いた細胞免疫染色の結果を示す。図12Bにおいて、α-FLAGは、抗FLAG抗体による染色像を示し、α-cyt.cは抗チトクロームC抗体による染色像を示す。また、mergeは重ねあわせ像を示す。
【0113】
図12Cは、HeLa細胞におけるPHB1C-GFP(PHB1Cタンパク質のC末端にGFPタンパク質を融合したタンパク質)又はPHB2C-GFP(PHB2Cタンパク質のC末端にGFPタンパク質を融合したタンパク質)の細胞内局在を示す。E2(-)はE2非存在下で培養したHeLa細胞であり、+ERα E2(+)はE2存在下で培養したERα発現HeLa細胞である。
【0114】
さらに、図12Dは、HeLa細胞において、PHB2全長(PHB2-FLAG)、PHB2N末端(PHB2N-FLAG)、およびPHB1N-2Cキメラタンパク質(PHB1N-2C-FLAG)のミトコンドリア膜電位に与える影響を検討した結果を示す。発現させた各タンパク質は、C末端にFLAGタグを有する融合タンパク質である。なお、ミトコンドリア膜電位の検出には、膜電位に依存してミトコンドリアを染色するTMREを用いた染色を利用した。図12Dにおいて、「TI」は透過像を示す。
【0115】
図12Bに示すように、PHB2N-FLAGがミトコンドリアで発現することから、PHB2タンパク質のアミノ末端にミトコンドリア標的シグナルが存在することが明らかになった(図12AのPHB2タンパク質及びPHB2Nタンパク質において、斜線で示すボックスがミトコンドリア標的シグナルであり、それに隣接するボックスが膜貫通ドメインである)。
【0116】
また、図12Cに示すように、PHB2C-GFPがE2存在下で核に移行していることから、PHB2タンパク質のカルボキシル末端に、E2依存的な核移行シグナルが存在することが明らかになった。一方、PHB1C-GFPはE2存在下でも核に移行しないことから、PHB1タンパク質のカルボキシル末端にはこの核移行シグナルが存在せず、このシグナル(又は核移行機能)はPHB2タンパク質特異的であることが明らかになった。
【0117】
さらに、図12Dに示すように、PHB2のC末端のキメラタンパク質(PHB1N-2C-FLAG)がミトコンドリアの膜電位を調節することが明らかになった。PHB1のミトコンドリア標的シグナルと膜貫通ドメインを持ったN末端にPHB2のC末端を融合させたキメラタンパク質をコードする遺伝子を遺伝子導入すると、PHB2N末端のみ(PHB2N-FLAG)ではミトコンドリア機能になんら変化は認められないが、上述したキメラタンパク質を発現させると、PHB2と同様にミトコンドリア膜電位が低下することが明らかになった。なお、PHB1やPHB2のC末端のみでは、膜電位の低下は認められない。
【0118】
さらに、PHB2のC末端のキメラタンパク質がミトコンドリア膜電位を消失させることについて、以下の実験より確認した。
【0119】
陽性対照としてPHB2-GFP(PHB2タンパク質全長のC末端にGFPタンパク質を融合したタンパク質)、陰性対照としてMTS-GFP(他のタンパク質(ヒトチトクロムC酸化酵素サブユニット8A, 1-29a.a. (NP_004065))のミトコンドリア標的シグナル(MTS)のC末端にGFPタンパク質を融合したタンパク質)及びPHB1N-2C-GFP(PHB1N-2Cキメラタンパク質のC末端にGFPタンパク質を融合したタンパク質)を、それぞれHeLa細胞に発現させ、ミトコンドリアの膜電位の有無をTMRE(tetramethylrhodamine ethyl ester)で染色することによって共焦点レーザー顕微鏡で観察した。
【0120】
結果を図12Eに示す。図12Eにおいて、「GFP」はGFPタンパク質による蛍光像を示し、「TMRE」はTMRE染色像を示し、「merge」は重ねあわせ像を示す。
【0121】
図12Eに示すように、陽性対照及びキメラタンパク質(PHB1N-2C-GFP)が発現した細胞では、有意に膜電位が低下した。しかし、陰性対照が発現しているミトコンドリアでは膜電位の低下は認められなかった。即ち、PHB2のキメラタンパク質が、ミトコンドリア機能調節剤の例として示された。
【0122】
〔実施例12〕PHB2タンパク質と核レセプターPPARα又はPPARγ2との相互作用
実施例10では、PHB2はE2依存的にERαと結合し、核に移行するのではないかと考えられた。
本実施例では、PHB2タンパク質と、ERαと同様に核レセプターであるPPARα又はPPARγ2との相互作用について検討した。
【0123】
方法としては、発現ベクターに組み込んだPPARα又はPPARγ2をコードする遺伝子とPHB2-GFP又はPHB1-GFPをコードする遺伝子とを共発現させた細胞を、共焦点レーザー顕微鏡で観察した。
【0124】
結果を図13に示す。図13は、PHB1-GFP又はPHB2-GFPとPPARα又はPPARγ2とを発現するHeLa細胞の顕微鏡写真を示す。図13に示すように、PHB2タンパク質の核移行は、ERαだけでなく、同様の核レセプターPPARα又はPPARγ2存在下においても生じ、且つPHB2タンパク質特異的に核への局在化が生じることを、HeLa細胞において確認した。
【0125】
〔実施例13〕PHB2タンパク質の転写抑制活性の検討
本実施例では、PHB2タンパク質の転写抑制活性を検討するために、エストロゲン応答性エレメントの下流にルシフェラーゼ遺伝子を連結したレポーター遺伝子(ERE-Luc)を用いて、デュアルルシフェラーゼアッセイを行った。
【0126】
方法としては、HeLa細胞に、ERE-Luc遺伝子、各々発現ベクターに挿入したERα遺伝子、PHB-2遺伝子、PGC-1α遺伝子を導入した。さらにコントロールとしてCMVプロモーターを持つウミシイタケルシフェラーゼを同時に導入し、これを内部コントロールとしてルシフェラーゼ活性を測定した。
【0127】
結果を図14に示す。図14は、各遺伝子を導入したHeLa細胞における相対的ルシフェラーゼ活性を示す。E2(-)はE2非存在下で培養した場合、E2(+)はE2存在下で培養した場合を示す。図14における、+又は-は、各遺伝子の導入の有無を示す。なお、図14では、各バーは独立して3回行った実験の平均値と1SD(標準偏差)で示す。
【0128】
図14に示すように、ERα遺伝子及びERαのコアクチベーターであるPGC-1α遺伝子をHeLa細胞に導入すると、転写活性が増強し、特にE2存在下では転写活性がさらに増強した。これに、PHB2タンパク質を共発現させると、転写活性の抑制効果が検出できた。従って、PHB2タンパク質はERαのコアクチベーターであるPGC-1αタンパク質の転写調節に抑制的に働くことが明らかになった。
【0129】
本明細書で引用した全ての刊行物、特許及び特許出願をそのまま参考として本明細書にとり入れるものとする。
【技術分野】
【0001】
本発明は、例えばPHB2遺伝子調節剤のスクリーニング方法に関する。
【背景技術】
【0002】
ミトコンドリアは、真核生物において多くの反応を行う。特に電子伝達系によるATP合成は生物にとって重要であり、細胞におけるほとんどのATPがミトコンドリアから供給される。ミトコンドリアにおけるその他の反応系としては、TCA回路、ヘム合成系、脂肪酸のβ酸化回路、アミノ酸代謝回路などが挙げられる。さらに、Ca恒常性の維持機能、活性酸素の産生系、並びに代謝物質、イオン及びタンパク質などの輸送系がミトコンドリアに存在する。このように、ミトコンドリアは真核生物の異化作用及び同化作用に重要な働きを持つ細胞内小器官である。
【0003】
ヒトミトコンドリアに存在するタンパク質は、1000〜1500種類存在すると予測されている。そのうちの13種類は、ミトコンドリアDNAにコードされるタンパク質で、電子伝達系のサブユニットである。その他の約99%のタンパク質は、核DNAにコードされるタンパク質であり、細胞質でタンパク質が合成された後、ミトコンドリアに移行する。プロテオーム解析によれば、ヒトミトコンドリアに存在するタンパク質のうち約544種類のタンパク質が同定されている(非特許文献1)が、未だ未知のタンパク質が多く存在すると予測される。
【0004】
上述したように、ミトコンドリアDNAは電子伝達系の複合体I、III、IV、Vの一部のサブユニットをコードする。即ち、ミトコンドリアDNAの変異は、電子伝達系の機能異常を引き起こす。電子伝達系の機能異常に関連する疾患としては、例えばMELAS、MERRF、心筋症、LHON、Leigh脳症などが挙げられる。また、ミトコンドリアDNAの塩基変異が、肝臓、前立腺、膀胱及び頭頸部の早期癌、原発性肺癌、Barrett's esophagusにおいても認められている(非特許文献2)。
【0005】
一方、核DNAにコードされたミトコンドリアタンパク質の異常も、例えば、以下のように多くの疾患の原因に成っている。(i)Friedreich's ataxiaは、ミトコンドリアにおいてFe-Sタンパク質の生合成に関与するfrataxinタンパク質の異常によって生じる。(ii)ミトコンドリアへのタンパク質移行に関与する因子Deafness dystonia peptide 1(DDP1)の異常は、Mohr-Tranebjaerg syndromeに関与する。(iii)常染色体優性遺伝を示す網膜萎縮症は、ミトコンドリア膜融合を生じさせるOPA1タンパク質の異常によって生じる。(iv)ミトコンドリア膜融合に関与する他の因子Mfn2の異常により、Charcot-Marie-Tooth neuropathy type 2が生じる。(v)また、ミトコンドリアのチミジンホスホリラーゼの異常により、常染色体劣性遺伝で、消化器症状の強いMNGIE(mitochondrial neurogastrointestinal encephalomyopathy)が生じる。
【0006】
その他に、ミトコンドリア機能異常は、より一般的な疾患への関与も示されている。例えば、ミトコンドリア機能異常による糖代謝や脂質代謝の異常は、肥満や糖尿病などの原因となる。また、ミトコンドリア機能異常による細胞内ATPの低下は、パーキンソン病やアルツハイマー病の病因の主要因子である。近年、アルツハイマー病において蓄積物質のアミロイドβタンパク質が、細胞内でミトコンドリアタンパク質であるABADタンパク質と結合し、ミトコンドリア機能を傷害するという報告もなされている(非特許文献3)。
【0007】
また、ミトコンドリアの消費する酸素の0.4〜4%は、電子伝達系で活性酸素となることが知られている。この活性酸素がDNAやタンパク質などに障害を与え、細胞障害や細胞数の減少などを生じさせることにより、細胞の機能低下や個体の老化を促進すると考えられている。
【0008】
さらに、ミトコンドリアはアポトーシス誘導に関与しており、これらの経路は細胞増殖や癌化と関連があると考えられている。
【0009】
従って、ミトコンドリアの機能を正常に保ち、当該機能をうまく制御することが、老化の防止だけでなく、個体の恒常性を維持していく上で極めて重要である。
【0010】
ところで、プロヒビチン(以下、「PHB」という)と呼ばれるタンパク質が、細胞増殖抑制因子として最初に哺乳動物で単離された。PHBは酵母から哺乳動物に至るまで高度に保存されたタンパク質である。PHBタンパク質には、アミノ酸の一次構造が類似した2種類のタンパク質PHB1及びPHB2が存在し、互いに複合体を形成し、ミトコンドリアの内膜に局在することが知られている。酵母PHBタンパク質に関しては、細胞周期制御や新規に合成されたミトコンドリアタンパク質の安定化を担うシャペロン様の機能を果たすことが明らかにされている(非特許文献4〜6)。また、線虫では、PHB1の老化や初期発生への関与が報告されている(非特許文献7)。
【0011】
一方、哺乳動物では、PHB1及びPHB2による転写制御等の多様な機能が示唆されているものの、ミトコンドリアにおける生理的機能は明らかにされていない(非特許文献8及び9)。
【先行技術文献】
【非特許文献】
【0012】
【非特許文献1】Reichert AS及びNeupert W., 「Trends in Genetics」, 2004年, 第20巻, 第11号, p.556-562
【非特許文献2】Verma M.ら, 「Nature reviews cancer」, 2003年, 第3巻, 第10号, p.789-795
【非特許文献3】Lustbader, J.W.ら, 「Science」, 2004年, 第304巻, 第5669号, p.448-452
【非特許文献4】Berger, K.H.及びYaffe, M.P., 「Mol Cell Biol.」, 1998年, 第18巻, 第7号, p.4043-4052
【非特許文献5】Nijtmans, L.G.ら, 「EMBO J.」, 2000年, 第19巻, 第11号, p.2444-2451
【非特許文献6】Piper P.W.及びBringloe, D., 「Mech Ageing Dev.」, 2002年, 第123巻, 第4号, p.287-295
【非特許文献7】Artal-Sanz M.ら, 「J Biol Chem.」, 2003年, 第278巻, 第34号, p.32091-32099
【非特許文献8】Delage-Mourroux R.ら, 「J Biol Chem.」, 2000年, 第275巻, 第46号, p.35848-35856
【非特許文献9】Sun L.ら, 「J Cell Sci.」, 2004年, 第117巻, p.3021-3029
【発明の概要】
【発明が解決しようとする課題】
【0013】
上述したように、ミトコンドリアの機能を正常に保ち、当該機能をうまく制御することが、老化の防止だけでなく、個体の恒常性を維持していく上で極めて重要である。
【0014】
そこで、本発明は、上述した実情に鑑み、例えばミトコンドリア機能調節剤のスクリーニング方法を提供することを目的とする。
【課題を解決するための手段】
【0015】
上記課題を解決するため鋭意研究を行った結果、ミトコンドリアにおけるヒトPHB2タンパク質は、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持の機能を有することを見出し、本発明を完成するに至った。
【0016】
本発明は以下を包含する。
(1)PHB2遺伝子発現細胞をその培養下で候補物質と接触させる工程と、候補物質非存在下での該PHB2遺伝子発現細胞と比較して、該候補物質と接触した該PHB2遺伝子発現細胞においてPHB2遺伝子発現又はPHB2タンパク質機能が調節されていることにより、該候補物質がPHB2遺伝子調節剤であると判定する工程とを含む、PHB2遺伝子調節剤のスクリーニング方法。
(2)上記PHB2タンパク質機能が、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持から成る群から選択されるものであることを特徴とする、(1)記載の方法。
(3)上記PHB2タンパク質機能が、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質との相互作用能であることを特徴とする、(1)又は(2)記載の方法。
(4)上記相互作用能が複合体形成能であることを特徴とする、(3)記載の方法。
(5)上記PHB2タンパク質機能が、PHB2タンパク質の核-ミトコンドリア移行機能であることを特徴とする、(1)又は(2)記載の方法。
【0017】
(6)上記PHB2遺伝子発現細胞がERα、PPARα及びPPARγ2から成る群から選択される核レセプターをさらに発現していることを特徴とする、(5)記載の方法。
(7)上記PHB2遺伝子発現細胞の培養が、エストラジオール存在下で行われることを特徴とする、(6)記載の方法。
(8)(1)〜(7)のいずれか1記載の方法により得られたPHB2遺伝子調節剤。
(9)以下の(a)又は(b)記載のsiRNAを含有することを特徴とするPHB2遺伝子調節剤。
(a)配列番号1〜20記載のいずれか1つに示される塩基配列から成るsiRNA。
(b)上記(a)記載のsiRNAにおいて、1又は数個の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2遺伝子発現阻害活性を有するsiRNA。
(10)ERα、PPARα及びPPARγ2から成る群から選択される核レセプターのアンタゴニスト又はアゴニストを含有することを特徴とするPHB2遺伝子調節剤。
【0018】
(11)上記PHB2遺伝子調節剤が抗アポトーシス作用剤、ミトコンドリア膜電位調節剤、及びミトコンドリア膜形態調節剤から成る群から選択されるものである、(8)〜(10)のいずれか1記載のPHB2遺伝子調節剤。
(12)(8)〜(11)のいずれか1記載のPHB2遺伝子調節剤を含有するミトコンドリア機能関連疾患治療薬。
(13)被験者由来の生物学的サンプルにおいて、PHB2遺伝子発現又はPHB2タンパク質機能を測定する工程と、該PHB2遺伝子発現又はPHB2タンパク質機能が異常であることを指標として、該被験者がミトコンドリア機能関連疾患を有するか又はそれらを有する疑いがあると決定する工程とを含む、ミトコンドリア機能関連疾患の検出方法。
(14)上記PHB2タンパク質機能が、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持から成る群から選択されるものであることを特徴とする、(13)記載の方法。
(15)上記PHB2タンパク質機能が、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質との相互作用能であることを特徴とする、(13)又は(14)記載の方法。
【0019】
(16)上記相互作用能が複合体形成能であることを特徴とする、(15)記載の方法。
(17)上記PHB2タンパク質機能が、PHB2タンパク質の核-ミトコンドリア移行機能であることを特徴とする、(13)又は(14)記載の方法。
(18)PHB2遺伝子発現可能なミトコンドリア機能異常細胞をその培養下で候補物質と接触させる工程と、候補物質非存在下での該ミトコンドリア機能異常細胞と比較して、該候補物質と接触した該ミトコンドリア機能異常細胞において、PHB2遺伝子発現又はPHB2タンパク質機能が調節される結果としてミトコンドリア機能が回復又は正常化することを指標として、上記候補物質がミトコンドリア機能調節剤であると判定する工程とを含む、ミトコンドリア機能調節剤のスクリーニング方法。
(19)上記PHB2タンパク質機能が、抗アポトーシス作用、ミトコンドリア膜電位形成及びミトコンドリア形態維持から成る群から選択されるものであることを特徴とする、(18)記載の方法。
(20)上記PHB2タンパク質機能が、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質との相互作用能であることを特徴とする、(18)又は(19)記載の方法。
【0020】
(21)上記相互作用能が複合体形成能であることを特徴とする、(20)記載の方法。
(22)上記PHB2タンパク質機能が、PHB2タンパク質の核-ミトコンドリア移行機能であることを特徴とする、(18)又は(19)記載の方法。
(23)上記ミトコンドリア機能異常細胞がERα、PPARα及びPPARγ2から成る群から選択される核レセプターをさらに発現していることを特徴とする、(22)記載の方法。
(24)上記ミトコンドリア機能異常細胞の培養が、エストラジオール存在下で行われることを特徴とする、(23)記載の方法。
【発明の効果】
【0021】
本発明によれば、PHB2遺伝子調節剤、ミトコンドリア機能関連疾患治療薬及びミトコンドリア機能調節剤を提供することができる。さらに、本発明によれば、ミトコンドリア機能関連疾患を検出することができる。
【図面の簡単な説明】
【0022】
【図1】図1は、実施例1におけるPHB1-FLAG又はPHB2-FLAGを発現するHeLa細胞の共焦点レーザー顕微鏡写真を示す。
【図2】図2は、実施例2における各遺伝子を発現するHeLa細胞をRh123で染色した際の写真を示す。
【図3】図3は、実施例3におけるIP解析及びMASS解析の結果を示す。
【図4】図4は、実施例4におけるPHB2タンパク質とHax-1タンパク質との相互作用に関するin vitro結合実験の結果を示す。
【図5】図5は、実施例5におけるRNAi法によるPHB2ノックダウンの結果を示す。
【図6】図6は、実施例6での、PHB1又はPHB2のノックダウンに供したHeLa細胞におけるミトコンドリア膜電位及びカスパーゼ依存性細胞死(アポトーシス)についての検討結果を示す。
【図7】図7は、実施例7におけるRNAi法によるHax-1のノックダウンの結果を示す。
【図8】図8は、実施例8におけるMitoTracker Redでミトコンドリアを染色したPHB1、PHB2又はHax-1ノックダウン細胞の写真を示す。
【図9】図9は、実施例8におけるOPA1タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。
【図10】図10は、実施例9におけるPHB1-GFP又はPHB2-GFPを発現するMCF-7細胞及びHeLa細胞の顕微鏡写真を示す。
【図11】図11は、実施例10での、HeLa細胞においてPHB2-GFPとERαとを共発現させた際の共焦点レーザー顕微鏡写真を示す。
【図12−1】図12は、実施例11におけるPHB2タンパク質の構造分析の結果を示す。
【図12−2】図12−1の続きである。
【図13】図13は、実施例12におけるPHB1-GFP又はPHB2-GFPとPPARα又はPPARγ2とを発現するHeLa細胞の顕微鏡写真を示す。
【図14】図14は、実施例13での、各遺伝子を導入したHeLa細胞における相対的ルシフェラーゼ活性を示す。
【発明を実施するための形態】
【0023】
以下、本発明を詳細に説明する。
ミトコンドリアにおけるヒトPHB2タンパク質の相互作用因子の探索を、HeLa細胞由来の精製ミトコンドリア画分を用いた免疫沈降解析及び質量分析により行ったところ、抗アポトーシス作用を有するミトコンドリアタンパク質Hax-1(NCBI 登録番号:NP_006109)(Cilenti L.ら, J Biol Chem., 279:50295-50301, 2004)、PTP(permeability transition pore)の構成因子であるVDAC2タンパク質(Swiss-Prot 登録番号:P45880)、ANT2タンパク質(NCBI 登録番号:NP_001143)及びPHB1タンパク質(NCBI 登録番号:NP_002625)の各タンパク質を相互作用因子として同定した。また、in vitro結合実験によれば、PHB2タンパク質とHax-1タンパク質との相互作用が直接的であることが明らかになった。また、RNA干渉法を用いて、HeLa細胞をPHB2ノックダウンに供したところ、PHB1タンパク質及びHax-1タンパク質の発現量減少、ミトコンドリア膜電位の減少、カスパーゼ依存性細胞死の誘導、ミトコンドリア膜形態調節因子OPA1タンパク質(NCBI 登録番号:NP_056375)量の減少及びミトコンドリア形態の断片化が観察された。
【0024】
一方、Hax-1ノックダウン細胞では、PHB2タンパク質発現量に影響を与えないものの、PHB2ノックダウンと同様のアポトーシスが誘導されることから、PHB2ノックダウンによるアポトーシス誘導は、Hax-1タンパク質量の減少によるものと考えられた。また、ミトコンドリアの形態は、Hax-1ノックダウン細胞ではミトコンドリアの断片化は観察できなかった。
【0025】
以上の知見によれば、PHB2タンパク質は、(1)ミトコンドリアにおいて相互作用因子Hax-1タンパク質の安定化、(2)それを介した抗アポトーシス作用、(3)PTP複合体を形成するVDAC2タンパク質及びANT2タンパク質との結合を介すると考えられるミトコンドリア膜電位形成、(4)OPA1タンパク質を介したミトコンドリア形態調節に関与していることが示された。
【0026】
従って、ミトコンドリアにおいて、PHB2タンパク質は、Hax-1タンパク質、VDAC2タンパク質、ANT2タンパク質、PHB1タンパク質、OPA1タンパク質等と物理的又は生理的な相互作用を示し、抗アポトーシス作用、ミトコンドリア膜電位形成作用及びミトコンドリア形態維持作用を有する。
【0027】
一方、PHB2タンパク質は、これまで、哺乳動物では核に存在し、転写抑制に働くこと、ヒストン脱アセチル化酵素(HDAC)をリクルートすることが報告されている(J. Biol. Chem. 279 (23), 24834-24843 (2004))。ヒト乳癌由来細胞であるMCF-7細胞では、PHB2タンパク質の核への局在が報告されている(J. Biol. Chem. 279 (23), 24834-24843 (2004))。本願では、PHB2タンパク質をミトコンドリア及び核に局在化させる機構を明らかにした。HeLa細胞では、MCF-7細胞と異なり、PHB2タンパク質はミトコンドリアにのみ局在し、核には局在化しない。しかしながら、PHB2タンパク質がエストロゲンレセプター(以下、「ER」という)存在下でエストラジオール(以下、「E2」という)依存的にミトコンドリアから核に移行することを明らかにした。このPHB2の核移行に必要な領域は、PHB2タンパク質のカルボキシル末端に依存する。また、PHB2タンパク質のアミノ末端にはミトコンドリア標的シグナルと弱い膜貫通ドメインが存在し、この部分のみでミトコンドリアに移行する。PHB2の核移行は、ER以外の核レセプターであるPPARαやPPARγ2存在下でも生じる。
【0028】
このように、本願では、PHB2タンパク質はミトコンドリア及び核の双方に局在するタンパク質であり、核では転写抑制に、ミトコンドリアでは各種タンパク質を安定化することにより、ミトコンドリア膜電位形成、抗アポトーシス作用、ミトコンドリア形態維持等に働いていることを明らかにした。
以上に説明した新規のPHB2タンパク質の機能に基づき、本発明を説明する。
【0029】
本発明に係るPHB2遺伝子調節剤のスクリーニング方法は、PHB2遺伝子発現細胞をその培養下で候補物質と接触させ、候補物質非存在下での該PHB2遺伝子発現細胞と比較して、該候補物質と接触した該PHB2遺伝子発現細胞においてPHB2遺伝子発現又はPHB2タンパク質機能が調節されていることにより、該候補物質がPHB2遺伝子調節剤であると判定するものである。
【0030】
ヒトPHB2遺伝子は、NCBI登録番号NM_007273としてNCBIに登録されており、配列番号21記載の塩基配列から成るDNAである。なお、CDSは配列番号21記載の塩基配列において、186〜1085番目の塩基配列である。当該ヒトPHB2タンパク質は、NCBI登録番号NP_009204としてNCBIに登録されており、配列番号22記載のアミノ酸配列から成るタンパク質である。ヒト以外の生物由来のPHB2遺伝子又はPHB2タンパク質としては、例えば、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)由来(PHB2_YEAST(swissprot:accession P50085))、線虫(Caenorhabditis elegans)由来(PHB2_CAEEL(swissprot:accession P50093))、シロイヌナズナ(Arabidopsis thaliana)由来(ATPHB2 (PROHIBITIN 2) [Arabidopsis thaliana](NCBI登録番号 NP_973755))、アフリカツメガエル(Xenopus tropicalis)由来(prohibitin 2 [Xenopus tropicalis](NCBI登録番号NP_001016551))、マウス由来(PHB2_MOUSE(swissprot:accession O35129))及びラット由来(PHB2_RAT(swissprot:accession Q5XIH7))のものが挙げられる。本発明では、PHB2遺伝子には、上述した配列番号又は登録番号に記載の塩基配列から成るDNAに限らず、これらDNAにおいて、1又は複数(例えば、1〜10個、好ましくは1〜5個)の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2タンパク質機能を有するタンパク質をコードするDNAも含まれる。また、PHB2遺伝子には、上述した配列番号又は登録番号に記載の塩基配列から成るDNAに90%以上、好ましくは95%以上、97%以上、より好ましくは98%以上、特に好ましくは99%以上の同一性を有し、且つPHB2タンパク質機能を有するタンパク質をコードするDNAも含まれる。あるいは、上述した配列番号又は登録番号に記載のアミノ酸配列をコードするDNAに限らず、これらアミノ酸配列において、1又は複数(例えば、1〜10個、好ましくは1〜5個)のアミノ酸が欠失、置換又は付加されたアミノ酸配列から成り、且つPHB2タンパク質機能を有するタンパク質をコードするDNAもPHB2遺伝子に含まれる。
【0031】
ここで、PHB2タンパク質機能としては、例えば、抗アポトーシス作用、ミトコンドリア膜電位形成、ミトコンドリア形態維持、PHB2タンパク質とVDAC2遺伝子、Hax-1遺伝子、PHB1遺伝子、ANT2遺伝子及びOPA1遺伝子から成る群から選択される遺伝子によりコードされるタンパク質(以下では、各タンパク質をそれぞれ「VDAC2タンパク質、Hax-1タンパク質、PHB1タンパク質、ANT2タンパク質及びOPA1タンパク質」という)との相互作用能(例えば、複合体形成能、シグナル伝達能、タンパク質安定化能、タンパク質立体構造・機能調節能などを指し、生理的相互作用を含む)、並びに核-ミトコンドリア移行機能が挙げられる。本明細書中で使用される「複合体形成能」とは、ミトコンドリアや核、または細胞質などで直接的な結合や間接的な結合を介して形成される複合体の形成能を指し、その複合体の構成成分としては、タンパク質や核酸、またはある種の脂質などが含まれる。また、「核-ミトコンドリア移行機能」とは、ミトコンドリアと核との間の移行機能を指し、ミトコンドリアと細胞質との間、または細胞質と核との間の移行機能を含む。また、当該核-ミトコンドリア移行機構には、これらの移行を介在する輸送体やこれを刺激するリガンドを含めた輸送機構が含まれる。
【0032】
PHB2遺伝子発現細胞とは、PHB2遺伝子を発現する細胞であればいずれのものであってよいが、例えばHeLa細胞(ヒト子宮頸癌由来)、MCF-7細胞(ヒト乳癌由来)、U-2OS細胞(ヒト骨肉腫由来)及びヒト線維芽細胞等のヒト由来の細胞系、並びにB6.1細胞(マウスミエローマ由来)、マウス胚性幹細胞及びSWISS 3T3細胞(マウス線維芽細胞由来)が挙げられる。さらに、上述したPHB2遺伝子を導入した(又は過剰発現させた)細胞もPHB2遺伝子発現細胞に含まれる。例えば、PCR産物やベクターに含まれるPHB2遺伝子を、エレクトロポレーション法、リン酸カルシウム法、リポフェクション法等により細胞へ導入することができる。PHB2遺伝子発現細胞において、PHB2遺伝子が発現していることの確認方法としては、mRNAレベルでは、例えばPHB2遺伝子に特異的なプライマーやプローブを用いたRT-PCR、定量PCRやノーザンブロッティングによって確認する方法が挙げられる。また、タンパク質レベルでは、例えばPHB2タンパク質に特異的な抗体を用いたELISA、フローサイトメトリー、ウエスタンブロッテイング等の免疫学的方法を用いて、PHB2遺伝子の発現を確認することができる。
【0033】
一方、本発明において、候補物質としては、例えば、核酸、ペプチド、タンパク質、合成化合物、微生物の培養上清、植物や海洋生物由来の天然成分、植物抽出物、動物組織抽出物等が挙げられる。
【0034】
本発明に係るPHB2遺伝子調節剤のスクリーニング方法においては、先ずPHB2遺伝子発現細胞をその培養下で候補物質と接触させる。PHB2遺伝子発現細胞の培養は、各細胞に応じて、培地、培養条件(例えば、温度、pH)を適宜選択して行うことができる。ここで、接触とは、PHB2遺伝子発現細胞に候補物質が影響を及ぼす状態を意味する。例えば、PHB2遺伝子発現細胞の培地に候補物質を添加するだけでもよく、あるいは、候補物質をリポソームに抱埋または結合させて培地に添加してもよく、候補物質をある種のキャリアー物質(タンパク質や脂質など)とともに培地に添加してもよい。また、PHB2遺伝子発現細胞にマイクロインジェクションなどで候補物質を直接導入してもよい。培養時間は、候補物質がPHB2遺伝子発現細胞に影響を及ぼすのに十分な時間であればよいが、例えば1〜72時間、好ましくは12〜24時間とすることができる。
【0035】
次いで、本発明に係るPHB2遺伝子調節剤のスクリーニング方法においては、候補物質非存在下でのPHB2遺伝子発現細胞と比較して、該候補物質と接触した該PHB2遺伝子発現細胞においてPHB2遺伝子発現又はPHB2タンパク質機能が調節されている否かを決定する。ここで、PHB2遺伝子発現の調節とは、mRNAレベル又はタンパク質レベルでのPHB2遺伝子発現の低下又は増加を意味する。さらに、PHB2タンパク質機能の調節とは、上述したPHB2タンパク質機能の上昇を意味する。
【0036】
PHB2遺伝子発現の調節の評価方法では、培養後、当該PHB2遺伝子発現細胞からmRNA又はタンパク質を抽出する。次いで、得られたmRNA又はタンパク質中のPHB2遺伝子発現量を、候補物質不在下で培養したPHB2遺伝子発現細胞におけるPHB2遺伝子発現量と比較する。なお、mRNAレベル又はタンパク質レベルでのPHB2遺伝子の発現量の測定は、上記で説明した方法に準じて行うことができる。
【0037】
候補物質不在下で培養したPHB2遺伝子発現細胞に比べて、候補物質を接触させたPHB2遺伝子発現細胞において、mRNAレベル又はタンパク質レベルで有意(例えば、1.5〜100倍、好ましくは2〜5倍)にPHB2遺伝子発現量が増加した場合に、あるいは、有意(例えば、1/2〜1/1000、好ましくは1/4〜1/10)にPHB2遺伝子発現量が低下した場合に、候補物質がPHB2遺伝子調節剤であると判定することができる。
一方、PHB2タンパク質機能の調節の評価は、各機能に応じて個別に行うことができる。
【0038】
抗アポトーシス作用は、例えば、DNA断片化、Annexin Vの結合による細胞膜構造変化、ミトコンドリア膜電位消失、カスパーゼ等のアポトーシス関連酵素の活性、cytochrome c のミトコンドリアから細胞質への転移等を検出することによって評価することができる。
【0039】
ミトコンドリア膜電位形成については、例えば、ミトコンドリア膜電位に依存したミトコンドリア染色マーカーRh123を用いた染色にPHB2遺伝子発現細胞を供し、顕微鏡による観察により染色度合いによって評価することができる。
【0040】
ミトコンドリア形態維持については、例えば、MitoTracker Redを用いたミトコンドリア染色にPHB2遺伝子発現細胞を供し、顕微鏡観察によりミトコンドリア形態を観察することにより評価することができる。
【0041】
また、PHB2タンパク質とVDAC2タンパク質、Hax-1タンパク質、PHB1タンパク質、ANT2タンパク質又はOPA1タンパク質との相互作用能(例えば、複合体形成能)については、例えばPHB2遺伝子発現細胞由来のタンパク質を各タンパク質に特異的な抗体を用いた免疫学的解析に供することで、タンパク質間の会合を観察することで評価することができる。
【0042】
さらに、核-ミトコンドリア移行機能については、ERα、PPARα又はPPARγ2を発現するPHB2遺伝子発現細胞を(ERαについてはE2存在下で)培養することで、ミトコンドリアから核へのPHB2タンパク質の移行を、例えばPHB2タンパク質に特異的な抗体を用いた染色により評価することができる。培地に対するE2添加量は、例えば、1x10-12〜1x10-4M、好ましくは1x10-7〜1x10-6Mとする。
【0043】
候補物質不在下で培養したPHB2遺伝子発現細胞に比べて、候補物質を接触させたPHB2遺伝子発現細胞において、有意(例えば、1.5〜100倍、好ましくは2〜10倍)にPHB2タンパク質機能が上昇した場合に、候補物質がPHB2遺伝子調節剤であると判定することができる。
【0044】
一方、上述した本発明に係るPHB2遺伝子調節剤のスクリーニング方法に準じて、ミトコンドリア機能調節剤をスクリーニングすることができる。本発明に係るミトコンドリア機能調節剤のスクリーニング方法は、PHB2遺伝子発現可能なミトコンドリア機能異常細胞をその培養下で候補物質と接触させ、候補物質非存在下での該ミトコンドリア機能異常細胞と比較して、該候補物質と接触した該ミトコンドリア機能異常細胞において、PHB2遺伝子発現又はPHB2タンパク質機能が調節される結果としてミトコンドリア機能が回復又は正常化することを指標として、上記候補物質がミトコンドリア機能調節剤であると判定するものである。
【0045】
ここで、PHB2遺伝子発現可能なミトコンドリア機能異常細胞(以下、単に「ミトコンドリア機能異常細胞」という)とは、PHB2遺伝子を発現し、且つ上述したミトコンドリア機能に異常を有する細胞である。このような細胞としては、例えば、PHB2をノックダウンした細胞に別の変異PHB2を発現させた細胞などが挙げられる。
【0046】
本発明に係るミトコンドリア機能調節剤のスクリーニング方法では、上述した本発明に係るPHB2遺伝子調節剤のスクリーニング方法に準じて、候補物質非存在下でのミトコンドリア機能異常細胞と比較して、該候補物質と接触したミトコンドリア機能異常細胞において、PHB2遺伝子発現又はPHB2タンパク質機能が調節され、ミトコンドリア機能が回復又は正常化するか否かを決定する。そのような指標により、上記候補物質の中からミトコンドリア機能調節剤を選択することができる。
【0047】
本発明に係るPHB2遺伝子調節剤としては、例えば、上述したPHB2遺伝子調節剤のスクリーニング方法により得られたもの、以下の(a)又は(b)記載のsiRNAを含有するもの:(a)配列番号1〜20記載のいずれか1つに示される塩基配列から成るsiRNA:(b)上記(a)記載のsiRNAにおいて、1又は数個(例えば1〜5個、好ましくは1〜3個)の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2遺伝子発現阻害活性を有するsiRNA、並びにERα、PPARα又はPPARγ2の核レセプターのアンタゴニスト又はアゴニストを含有するものが挙げられる。また、上記(a)記載のsiRNAに90%以上、好ましくは95%以上、97%以上、より好ましくは98%以上、特に好ましくは99%以上の同一性を有し、且つPHB2遺伝子発現阻害活性を有するsiRNAも本発明に係るPHB2遺伝子調節剤とすることができる。
【0048】
上述した(a)又は(b)記載のsiRNAは、例えば、公知の核酸合成方法により化学的に合成することができる。
【0049】
ERα、PPARα又はPPARγ2の核レセプターのアンタゴニスト又はアゴニストとしては、例えばERのアンタゴニストであるICI182,780、DDE及びTamoxifenなど、ERのアゴニストであるDES(Diethylstilbestrol)、ゲニステイン、ノニルフェノール及びビスフェノールA(Bisphenol A)など、PPARαのアゴニストであるWY-14643、ETYA、Benzafibrate及びLY171883など、PPARαのアンタゴニストであるGW6471など、PPARγのアゴニストであるGW9662及びTroglitazoneなど、並びにPPARγのアンタゴニストであるチアゾリジンジオンなどが挙げられる。
【0050】
更なるPHB2遺伝子調節剤としては、例えばTR、PR及びRXR等の他の核レセプターのアゴニストやアンタゴニストが挙げられる。
【0051】
また、PHB2遺伝子調節剤として、例えばPHB1タンパク質のミトコンドリア標的シグナル及び膜貫通ドメイン(ヒトPHB1タンパク質では、N末端より1番目〜50番目のアミノ酸残基)とPHB2タンパク質のC末端(ヒトPHB2タンパク質では、N末端より51番目〜299番目のアミノ酸残基)とのキメラタンパク質を用いることができる。
【0052】
なお、PHB2タンパク質の機能に基づき、PHB2遺伝子調節剤は、抗アポトーシス作用剤、ミトコンドリア膜電位調節剤又はミトコンドリア膜形態調節剤として使用することができる。
【0053】
本発明に係るミトコンドリア機能関連疾患治療薬は、本発明に係るPHB2遺伝子調節剤を含有することを特徴とする。ここで、ミトコンドリア機能関連疾患とは、ミトコンドリア機能の異常に関連する疾患を意味し、例えば、肥満、糖尿病、パーキンソン病、アルツハイマー病、乳癌等の癌が挙げられるが、これらに限定されない。
【0054】
本発明に係るミトコンドリア機能関連疾患治療薬は、本発明に係るPHB2遺伝子調節剤それ自体であっても、あるいは通常用いられる固体または液体の担体、乳化分散剤等により錠剤、粉剤、乳剤、カプセル剤等の剤形に製剤化されたものであってもよい。上記担体としては、水、ゼラチン、澱粉、ステアリン酸マグネシウム、ラクトース、植物油等が挙げられる。本発明に係るミトコンドリア機能関連疾患治療薬におけるPHB2遺伝子調節剤の含有量及び当該治療薬の投与量は、投与目的、投与経路、剤形等によって適宜変更しうる。
【0055】
本発明に係るミトコンドリア機能関連疾患治療薬の薬理評価は、例えば、上述したミトコンドリア機能異常細胞を使用したin vitroレベル、アルツハイマー病等のミトコンドリア機能関連疾患動物モデルを使用したin vivoレベルで評価することができる。例えば、本発明に係るミトコンドリア機能関連疾患治療薬非存在下に比べて、当該治療薬存在下で培養したミトコンドリア機能異常細胞において、ミトコンドリア機能異常が回復又は正常化するか否かにより薬理評価を行うことができる。あるいは、ミトコンドリア機能関連疾患動物モデルに本発明に係るミトコンドリア機能関連疾患治療薬を投与しない場合と比較して、当該治療薬を投与した場合にミトコンドリア機能関連疾患が治療又は改善するか否かにより薬理評価を行うことができる。
【0056】
本発明に係るミトコンドリア機能関連疾患の検出方法は、被験者由来の生物学的サンプルにおいて、PHB2遺伝子発現又はPHB2タンパク質機能を測定し、該PHB2遺伝子発現又はPHB2タンパク質機能が異常であることを指標として、該被験者がミトコンドリア機能関連疾患を有するか又はそれらを有する疑いがあると決定するものである。
【0057】
生物学的サンプルとしては、例えば、被験者由来の線維芽細胞、筋芽細胞、白血球細胞、精子細胞や卵子細胞等の細胞、これら細胞を含む組織や器官、血液や唾液等の体液、排出物が挙げられる。
【0058】
本発明に係るミトコンドリア機能関連疾患の検出方法では、上述した本発明に係るPHB2遺伝子調節剤のスクリーニング方法に準じて行うことができる。即ち、先ず生物学的サンプル中の細胞におけるPHB2遺伝子発現又はPHB2タンパク質機能を測定する。次いで、PHB2遺伝子発現又はPHB2タンパク質機能が異常であるか否かを正常者由来の細胞と比較して判定する。ここで、PHB2遺伝子発現又はPHB2タンパク質機能の異常とは、例えば、PHB2遺伝子の発現増加若しくは低下又はPHB2タンパク質機能の低下を意味する。PHB2遺伝子発現又はPHB2タンパク質機能に異常が見られた場合には、生物学的サンプルが由来する被験者がミトコンドリア機能関連疾患を有するか又はそれらを有する疑いがあると決定することができる。
【0059】
以上に説明したように、新規PHB2タンパク質の機能に基づけば、PHB2遺伝子調節剤を同定することができ、またミトコンドリア機能調節剤やミトコンドリア機能関連疾患治療薬を提供することができる。
【0060】
本明細書は本願の優先権の基礎である日本国特許出願2005-339354号の明細書及び/又は図面に記載される内容を包含する。
【実施例】
【0061】
以下、実施例を用いて本発明をより詳細に説明するが、本発明の技術的範囲はこれら実施例に限定されるものではない。
【0062】
なお、本実施例で示すタンパク質は、標識やレポーターとして使用したタンパク質(例えば、FLAGタグ、GST、GFP及びルシフェラーゼ等)以外はいずれもヒト由来のタンパク質である。
【0063】
〔実施例1〕HeLa細胞におけるPHB1タンパク質又はPHB2タンパク質の細胞内局在
カルボキシル末端にFLAGタグを融合したPHB1タンパク質(以下、「PHB1-FLAG」という)又はPHB2タンパク質(以下、「PHB2-FLAG」という)をコードする遺伝子をそれぞれHeLa細胞(ヒト子宮頸癌由来)に一過性に発現させた。
【0064】
方法としては、哺乳細胞発現ベクターpCMV-SPORTにPHB1-FLAG又はPHB2-FLAGをコードするcDNAを挿入し、これをHeLa細胞に遺伝子導入し、約12時間後、これらの細胞を固定し、免疫染色に供した。抗体は抗FLAGウサギポリクローナル抗体およびミトコンドリアを染色するコントロール抗体として抗チトクロームC(cyt.c)マウスモノクローナル抗体を一次抗体として、二次抗体にはAlexa488標識抗ウサギ抗体およびCy3標識抗マウス抗体を用いた。これらの細胞は共焦点レーザー顕微鏡にて観察した。
【0065】
結果を図1に示す。図1は、PHB1-FLAG又はPHB2-FLAGを発現するHeLa細胞の共焦点レーザー顕微鏡写真である。図1において、α-FLAGは、抗FLAG抗体の染色像(緑色)を示し、α-cyt.cは、抗チトクロームC抗体の染色像(赤色)を示す。また、Mergeは、両者の重ね合わせ像を示す。
【0066】
図1に示すように、PHB1-FLAG及びPHB2-FLAGをそれぞれHeLa細胞に一過性に発現させると、PHB1-FLAG及びPHB2-FLAGはミトコンドリアに局在した。HeLa細胞におけるPHB1-FLAG及びPHB2-FLAGは、ミトコンドリアマーカーであるcyt.cと共局在を示した。
【0067】
〔実施例2〕PHB1タンパク質又はPHB2タンパク質発現によるミトコンドリア膜電位消失
実施例1に示すように、PHB1-FLAG及びPHB2-FLAGをそれぞれHeLa細胞に一過性に発現させると、PHB1-FLAG及びPHB2-FLAGはミトコンドリアに局在した。
【0068】
そこで、PHB1-FLAG又はPHB2-FLAGを発現するHeLa細胞を、ミトコンドリア膜電位に依存したミトコンドリア染色マーカーであるRh123で染色した。同様に、SLP2タンパク質(NCBI登録番号:NP_038470)(J Biol Chem. 2000 Mar 17;275(11):8062-71)のカルボキシル末端にFLAGタグを融合した融合タンパク質(以下、「SLP2-FLAG」という)をコードする遺伝子をHeLa細胞に一過性に発現させたものを、Rh123で染色した。また、対照として、いずれの遺伝子も導入されていないHeLa細胞をRh123で染色した。
【0069】
結果を図2に示す。図2は、各遺伝子を発現するHeLa細胞をRh123で染色した際の写真を示す。図2において、TIは、透過像の写真を示す。
【0070】
図2に示すように、PHB1-FLAG又はPHB2-FLAGを過剰発現するHeLa細胞は、Rh123で染色されないことが明らかとなった。このことは、PHB1-FLAG又はPHB2-FLAGの強制発現によってミトコンドリア膜電位が消失したことを示し、PHBタンパク質がミトコンドリアにおいて膜電位の調節に関与している可能性を示唆するものである。
【0071】
〔実施例3〕PHB2タンパク質の分子機能分析
PHB2タンパク質の分子機能を明らかにするために、ミトコンドリアにおけるPHB2タンパク質の相互作用因子の探索を行った。
【0072】
まず、実施例1で作製したPHB2-FLAGを発現するHeLa細胞由来の精製ミトコンドリア画分を用いた免疫沈降(IP)解析及び質量分析(MASS解析)を行った。
【0073】
IP解析では、抗FLAG抗体を用いて免疫沈降法を行い、この沈降物を12%SDS-PAGEに供した後、タンパク質染色を行い、PHB2-FLAGタンパク質と共沈するタンパク質を検出した。
【0074】
一方、MASS解析では、IP解析にて得たタンパク質のバンドを切り出し、トリプシンによるゲル内消化を行った後、消化されたペプチドを抽出後、ナノスケール高速液体クロマトグラフィーおよびタンデム型質量分析装置(nano LC-MS/MS)にて、ペプチド断片を同定した。次いで、データベース解析にてタンパク質を同定した。
【0075】
IP解析及びMASS解析の結果を図3に示す。
図3から判るように、抗アポトーシス作用を有するミトコンドリアタンパク質Hax-1(NCBI登録番号:NP_006109)、PTP(permeability transition pore)の構成因子であるVDAC2タンパク質(Swiss-Prot登録番号:P45880)及びANT2タンパク質(NCBI登録番号:NP_001143)、並びにPHB2タンパク質と複合体を形成することが知られている(Curr. Biol. 7 (8), 607-610 (1997))PHB1タンパク質(NCBI登録番号:NP_002625)が、PHB2タンパク質の相互作用因子として得られた。
【0076】
〔実施例4〕PHB2タンパク質とHax-1タンパク質との相互作用
実施例3に記載のIP解析、並びにPHB2タンパク質、Hax-1タンパク質、VDAC2タンパク質及びPHB1タンパク質に対する特異的抗体を用いたウエスタンブロット分析により、PHB2タンパク質とHax-1タンパク質、VDAC2タンパク質及びPHB1タンパク質との相互作用が確認された。
【0077】
そこで、in vitro結合実験により、PHB2タンパク質とHax-1タンパク質との相互作用を検討した。
【0078】
方法としては、大腸菌を用いて合成したグルタチオンS-トランスフェラーゼ(GST)-PHB2融合タンパク質(PHB2タンパク質のN末端にGSTタンパク質を融合したタンパク質)と、網状赤血球溶解液を用いて試験管内で翻訳したPHB1-FLAGおよびHax-1-FLAGタンパク質(Hax-1タンパク質のC末端にFLAGタグを融合したタンパク質)を、試験管内で混合し、Glutathione-Sepharoseビーズを用いてGST融合タンパク質をPull downした沈降産物を得た。次いで、この沈降産物をSDS-PAGEに供した後、抗FLAG抗体にてウエスタンブロット法を行い、PHB2タンパク質に各々のFLAG融合タンパク質が結合したか否かを検討した。
【0079】
結果を図4に示す。図4において、「インプット」は、混入したFLAG融合タンパク質を示す。一方、「GST-プルダウン」は、Glutathione-Sepharoseビーズで沈降させた沈降物を示す。さらに、「-」は反応系に混入していないことを示し、「+」は反応系に混入していることを示す。
【0080】
図4に示すように、PHB2タンパク質とHax-1タンパク質との相互作用は直接的であることが明らかとなった。
【0081】
〔実施例5〕RNAi法によるPHB2のノックダウン
RNAi法を用いて、HeLa細胞においてPHB2のノックダウンを行った。siRNAとしては、配列番号1記載の塩基配列から成るものを用いた。また、PHB1に対するsiRNA(配列番号23)を用いて、PHB1のノックダウンも行った。
【0082】
方法としては、PHB2およびPHB1のcDNAの一部に一致したshort hairpin RNAを発現させる塩基配列を、ピューロマイシン耐性遺伝子を有するpScilencer 3.1-H1 puro vectorに挿入したプラスミドを作製し、これを培養細胞に遺伝子導入した。その後、ピューロマイシンを培地に添加することで、目的遺伝子をノックダウンした細胞を得ることが可能であった。
【0083】
結果を図5A〜Cに示す。図5Aは、PHB1又はPHB2をノックダウンした際の各タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。図5Bは、PHB1又はPHB2をノックダウンした際のHax-1タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。図5Cは、PHB1又はPHB2をノックダウンした際のHax-1 mRNA量を測定したRT-PCRの結果を示す。図5A〜Cにおいて、siPHB1又はsiPHB2は、それぞれRNAi法によりPHB1又はPHB2をノックダウンした際の結果を示す。対照は遺伝子を導入していない細胞を示し、puro-vec.は、ピューロマイシン耐性遺伝子を導入し、その後、薬剤選択を行った細胞を示す。
【0084】
図5A〜Cに示すように、RNAi法を用いて、HeLa細胞においてPHB2のノックダウンを行ったところ、相互作用因子であるHax-1タンパク質やPHB1タンパク質の発現量が減少していることがわかった。しかしながら、PHB1及びPHB2のmRNAは減少していなかった。酵母PHBが新規に合成されたミトコンドリアタンパク質と相互作用し、その安定化を行うとの報告(Mol. Cell. Biol. 19, 3435-3442(1999).及びEMBO J. 19, 2444-2451 (2000).)があることから、ヒトPHB2タンパク質も相互作用因子Hax-1タンパク質の安定化を行っている可能性が考えられる。
【0085】
〔実施例6〕PHB2ノックダウンHeLa細胞におけるミトコンドリア膜電位低下及びカスパーゼ依存性細胞死(アポトーシス)の誘導
本実施例では、実施例5においてPHB1又はPHB2のノックダウンに供したHeLa細胞において、ミトコンドリア膜電位及びカスパーゼ依存性細胞死(アポトーシス)について検討した。
【0086】
方法としては、実施例5で行ったノックダウンと同様の方法を用いた。
結果を図6A〜Cに示す。図6A及びBは、PHB1又はPHB2のノックダウンに供したHeLa細胞における相対的細胞数を示す。図6Bにおいて、Z-VAD-FMKは、アポトーシスで活性化されるカスパーゼの阻害剤である。Z-VAD-FMKに関する+又は-は、培地への添加又は非添加を示す。図6Cは、PHB1又はPHB2のノックダウンに供したHeLa細胞におけるRh123染色の結果を示す。図6A〜Cにおいて、siPHB1又はsiPHB2は、それぞれRNAi法によりPHB1又はPHB2をノックダウンした際の結果を示す。puro-vec.は、ピューロマイシン耐性遺伝子を有するプラスミドベクターを導入した際の結果を示す。
【0087】
図6A及びBに示すように、PHB1又はPHB2をノックダウンしたHeLa細胞では、カスパーゼ依存性細胞死(アポトーシス)が誘導されていることが明らかとなった。また、図6Cに示すように、PHB1又はPHB2をノックダウンしたHeLa細胞では、一部の細胞でミトコンドリア膜電位が低下していることが明らかとなった。
【0088】
〔実施例7〕RNAi法によるHax-1のノックダウン
本実施例では、HeLa細胞においてPHB2の相互作用因子であるHax-1のノックダウンを行った。Hax-1に対するsiRNAとして、配列番号24記載の塩基配列から成るものを用いた。
【0089】
方法としては、Hax-1のcDNAの一部に一致したshort hairpin RNAを発現させる塩基配列を、ピューロマイシン耐性遺伝子を有するpScilencer 3.1-H1 puro vectorに挿入したプラスミドを作製し、実施例5の方法に準じた。
【0090】
結果を図7A〜Cに示す。図7Aは、Hax-1をノックダウンした際の各タンパク質に対する抗体を用いたウエスタンブロット分析の結果を示す。図7B及びCは、Hax-1のノックダウンに供したHeLa細胞における相対的細胞数を示す。図7におけるZ-VAD-FMK及びZ-VAD-FMKに関する+又は-の意味は、図6と同様である。図7A〜Cにおいて、siHax-1は、RNAi法によりHax-1をノックダウンした際の結果を示す。puro-vec.は、ピューロマイシン耐性遺伝子を有するプラスミドベクターを導入した際の結果を示す。
【0091】
図7A〜Cに示すように、Hax-1のノックダウンが、PHB1タンパク質及びPHB2タンパク質の発現量には影響を与えずに、PHB2のノックダウンと同様のアポトーシスを誘導することから、Hax-1タンパク質の発現量減少がPHB2ノックダウンによって誘導されるアポトーシスの一因であると考えられた。従って、PHB2タンパク質はHax-1タンパク質の発現量を維持することにより抗アポトーシス作用を持つことが示された。
【0092】
〔実施例8〕PHB2のノックダウンによるミトコンドリア形態変化
本実施例では、実施例5及び7に記載するPHB1、PHB2又はHax-1のノックダウンに供したHeLa細胞において、ミトコンドリア形態の変化を検討した。
【0093】
方法としては、実施例5及び7で作製したノックダウン細胞をMitoTracker Redで染色して、共焦点レーザー顕微鏡でミトコンドリアの形態を観察した。
【0094】
結果を図8に示す。図8において、puro-vec.は、ピューロマイシン耐性遺伝子を有するプラスミドベクターを導入した際の結果を示す。また、siPHB1、siPHB2、siHax-1は、それぞれPHB1、PHB2、Hax-1のノックダウンに供したHeLa細胞の結果を示す。
【0095】
図8に示すように、MitoTracker Redでミトコンドリアを染色したPHB1又はPHB2ノックダウン細胞では、ミトコンドリア形態の断片化が観察された。こうしたミトコンドリアの断片化はHax-1ノックダウン細胞では見られず、PHBタンパク質ノックダウン特異的で、且つHax-1を介在しない形態変化であると考えられた。
【0096】
また、上述したPHB1、PHB2又はHax-1ノックダウン細胞を、ミトコンドリア融合因子OPA1タンパク質に対する抗体を用いたウエスタンブロット分析に供した。
【0097】
方法としては、ノックダウンした細胞から細胞抽出液を作製し、これをウエスタンブロット法に供した。一次抗体は抗OPA1ポリクローナルウサギ抗体を用いた。
【0098】
結果を図9に示す。図9において、対照は、遺伝子非導入細胞である。また、puro-vec.、siPHB1、siPHB2、siHax-1は図8と同様の細胞の結果である。
【0099】
図9に示すように、PHB2ノックダウン細胞では、OPA1タンパク質の著明な発現減少が見られたため、OPA1タンパク質の減少によるミトコンドリアの断片化が生じたと考えられた。一方、Hax-1ノックダウン細胞においてもOPA1タンパク質は減少していた。また、PHB1のノックダウンによるOPA1タンパク質の発現減少は、PHB2のノックダウンと比較して顕著ではないが、PHB2ノックダウンと同等のミトコンドリア断片化を引き起こす(図8)。
【0100】
OPA1タンパク質は、他のミトコンドリア形態を調節する因子Drp1等とのバランスによりミトコンドリア形態を維持していることが知られている(Cell. 2004 Dec 17;119(6):873-87.)。PHB1ノックダウン細胞におけるミトコンドリアの断片化は、OPA1の発現量以外にも何らかの因子の関与があるものと考えられる。
【0101】
〔実施例9〕MCF-7細胞におけるPHB1タンパク質又はPHB2タンパク質の細胞内局在
実施例1で示すように、HeLa細胞では、PHB1タンパク質及びPHB2タンパク質が、ミトコンドリアに局在した。
【0102】
本実施例では、HeLa細胞(ヒト子宮頸癌由来)に加えて、MCF-7細胞(ヒト乳癌由来)でもPHB1タンパク質及びPHB2タンパク質の細胞内局在を検討した。
【0103】
方法としては、緑色蛍光タンパク質(GFP)をC末端に連結したPHB2又はPHB1タンパク質(各融合タンパク質を、「PHB2-GFP」、「PHB1-GFP」という)をコードするcDNAを連結した哺乳細胞発現ベクターを、培養細胞に導入し、これを共焦点レーザー顕微鏡にて、GFP融合タンパク質の細胞内局在を観察した。
【0104】
結果を図10に示す。図10は、PHB1-GFP又はPHB2-GFPを発現するMCF-7細胞及びHeLa細胞の顕微鏡写真を示す。図10に示すように、PHB1-GFPをコードする遺伝子をMCF-7細胞及びHeLa細胞に導入すると、PHB1-GFPはいずれにおいてもミトコンドリアに局在した。一方、PHB2-GFPはHeLa細胞ではミトコンドリアに局在するものの、MCF-7細胞では、PHB2-GFPの一部は核にも局在した。
【0105】
〔実施例10〕PHB2タンパク質と核レセプターERαとの相互作用
本実施例ではPHB2タンパク質と核レセプターERαとの相互作用について検討した。
方法としては、発現ベクターに組み込んだERα遺伝子とPHB2-GFP遺伝子をHeLa細胞に遺伝子導入後、この細胞を固定し、抗ERαウサギポリクローナル抗体を一次抗体として、Cy3標識抗ウサギ抗体を二次抗体として用いて細胞免疫染色を行った。
【0106】
結果を図11に示す。図11は、HeLa細胞においてPHB2-GFPとERαとを共発現させた際の共焦点レーザー顕微鏡写真を示す。図11において、TIは、透過像を示し、α-ERαは、抗ERα抗体染色像(赤色)を示す。また、PHB2-GFPは、GFP像(緑色)を示し、mergeは、重ね合わせ像を示す。E2(-)はE2非存在下、E2(+)はE2存在下で培養した細胞である。
【0107】
図11に示すように、PHB2-GFPは、HeLa細胞でERαと共発現させると、E2存在下で核に局在化した。これまでの報告から、PHB2タンパク質と核レセプターERαとが結合することが知られている(J. Biol. Chem. Vol. 275, No. 46, pp. 35848-35856, 2000)。従って、PHB2はE2依存的にERαと結合し、核に移行するのではないかと考えられた。
【0108】
〔実施例11〕PHB2タンパク質の構造分析
上記実施例で示すように、PHB2タンパク質はミトコンドリア及び核に局在化することが明らかになった。この機構を明らかにするために、PHB2タンパク質の欠失変異体及びPHB2タンパク質のC末端を有するキメラタンパク質等を作製した。同様に、PHB1タンパク質の欠失変異体を作製した。
【0109】
図12Aには、作製したPHB2タンパク質の欠失変異体及びキメラタンパク質、並びにPHB1タンパク質の欠失変異体を示す。PHB2Nは、PHB2タンパク質のN末端より1番目〜50番目のアミノ酸残基から成る欠失変異体である。PHB2Cは、PHB2タンパク質のN末端より51番目〜299番目のアミノ酸残基から成る欠失変異体である。また、PHB1Cは、PHB1タンパク質のN末端より45番目〜272番目のアミノ酸残基から成る欠失変異体である。さらにキメラタンパク質PHB1N-2Cは、PHB1タンパク質のN末端より1番目〜50番目のアミノ酸残基(内膜に局在化させるシグナルを有する)とPHB2タンパク質のN末端より51番目〜299番目のアミノ酸残基とを連結した変異体である。図12Aにおいて、ccは、コイルドコイルドメインを示す。
【0110】
欠失変異体等をコードする遺伝子の作製方法では、任意の塩基部位に相補的なプライマーを作製し、PCRにて任意の領域を増幅し、発現ベクターに組み込むことで各変異体をコードする遺伝子を含む発現ベクターを作製した。
【0111】
結果を図12B〜Dに示す。
【0112】
図12Bは、PHB2N-FLAG(PHB2Nタンパク質のC末端にFLAGタグを融合したタンパク質)を発現するHeLa細胞を用いた細胞免疫染色の結果を示す。図12Bにおいて、α-FLAGは、抗FLAG抗体による染色像を示し、α-cyt.cは抗チトクロームC抗体による染色像を示す。また、mergeは重ねあわせ像を示す。
【0113】
図12Cは、HeLa細胞におけるPHB1C-GFP(PHB1Cタンパク質のC末端にGFPタンパク質を融合したタンパク質)又はPHB2C-GFP(PHB2Cタンパク質のC末端にGFPタンパク質を融合したタンパク質)の細胞内局在を示す。E2(-)はE2非存在下で培養したHeLa細胞であり、+ERα E2(+)はE2存在下で培養したERα発現HeLa細胞である。
【0114】
さらに、図12Dは、HeLa細胞において、PHB2全長(PHB2-FLAG)、PHB2N末端(PHB2N-FLAG)、およびPHB1N-2Cキメラタンパク質(PHB1N-2C-FLAG)のミトコンドリア膜電位に与える影響を検討した結果を示す。発現させた各タンパク質は、C末端にFLAGタグを有する融合タンパク質である。なお、ミトコンドリア膜電位の検出には、膜電位に依存してミトコンドリアを染色するTMREを用いた染色を利用した。図12Dにおいて、「TI」は透過像を示す。
【0115】
図12Bに示すように、PHB2N-FLAGがミトコンドリアで発現することから、PHB2タンパク質のアミノ末端にミトコンドリア標的シグナルが存在することが明らかになった(図12AのPHB2タンパク質及びPHB2Nタンパク質において、斜線で示すボックスがミトコンドリア標的シグナルであり、それに隣接するボックスが膜貫通ドメインである)。
【0116】
また、図12Cに示すように、PHB2C-GFPがE2存在下で核に移行していることから、PHB2タンパク質のカルボキシル末端に、E2依存的な核移行シグナルが存在することが明らかになった。一方、PHB1C-GFPはE2存在下でも核に移行しないことから、PHB1タンパク質のカルボキシル末端にはこの核移行シグナルが存在せず、このシグナル(又は核移行機能)はPHB2タンパク質特異的であることが明らかになった。
【0117】
さらに、図12Dに示すように、PHB2のC末端のキメラタンパク質(PHB1N-2C-FLAG)がミトコンドリアの膜電位を調節することが明らかになった。PHB1のミトコンドリア標的シグナルと膜貫通ドメインを持ったN末端にPHB2のC末端を融合させたキメラタンパク質をコードする遺伝子を遺伝子導入すると、PHB2N末端のみ(PHB2N-FLAG)ではミトコンドリア機能になんら変化は認められないが、上述したキメラタンパク質を発現させると、PHB2と同様にミトコンドリア膜電位が低下することが明らかになった。なお、PHB1やPHB2のC末端のみでは、膜電位の低下は認められない。
【0118】
さらに、PHB2のC末端のキメラタンパク質がミトコンドリア膜電位を消失させることについて、以下の実験より確認した。
【0119】
陽性対照としてPHB2-GFP(PHB2タンパク質全長のC末端にGFPタンパク質を融合したタンパク質)、陰性対照としてMTS-GFP(他のタンパク質(ヒトチトクロムC酸化酵素サブユニット8A, 1-29a.a. (NP_004065))のミトコンドリア標的シグナル(MTS)のC末端にGFPタンパク質を融合したタンパク質)及びPHB1N-2C-GFP(PHB1N-2Cキメラタンパク質のC末端にGFPタンパク質を融合したタンパク質)を、それぞれHeLa細胞に発現させ、ミトコンドリアの膜電位の有無をTMRE(tetramethylrhodamine ethyl ester)で染色することによって共焦点レーザー顕微鏡で観察した。
【0120】
結果を図12Eに示す。図12Eにおいて、「GFP」はGFPタンパク質による蛍光像を示し、「TMRE」はTMRE染色像を示し、「merge」は重ねあわせ像を示す。
【0121】
図12Eに示すように、陽性対照及びキメラタンパク質(PHB1N-2C-GFP)が発現した細胞では、有意に膜電位が低下した。しかし、陰性対照が発現しているミトコンドリアでは膜電位の低下は認められなかった。即ち、PHB2のキメラタンパク質が、ミトコンドリア機能調節剤の例として示された。
【0122】
〔実施例12〕PHB2タンパク質と核レセプターPPARα又はPPARγ2との相互作用
実施例10では、PHB2はE2依存的にERαと結合し、核に移行するのではないかと考えられた。
本実施例では、PHB2タンパク質と、ERαと同様に核レセプターであるPPARα又はPPARγ2との相互作用について検討した。
【0123】
方法としては、発現ベクターに組み込んだPPARα又はPPARγ2をコードする遺伝子とPHB2-GFP又はPHB1-GFPをコードする遺伝子とを共発現させた細胞を、共焦点レーザー顕微鏡で観察した。
【0124】
結果を図13に示す。図13は、PHB1-GFP又はPHB2-GFPとPPARα又はPPARγ2とを発現するHeLa細胞の顕微鏡写真を示す。図13に示すように、PHB2タンパク質の核移行は、ERαだけでなく、同様の核レセプターPPARα又はPPARγ2存在下においても生じ、且つPHB2タンパク質特異的に核への局在化が生じることを、HeLa細胞において確認した。
【0125】
〔実施例13〕PHB2タンパク質の転写抑制活性の検討
本実施例では、PHB2タンパク質の転写抑制活性を検討するために、エストロゲン応答性エレメントの下流にルシフェラーゼ遺伝子を連結したレポーター遺伝子(ERE-Luc)を用いて、デュアルルシフェラーゼアッセイを行った。
【0126】
方法としては、HeLa細胞に、ERE-Luc遺伝子、各々発現ベクターに挿入したERα遺伝子、PHB-2遺伝子、PGC-1α遺伝子を導入した。さらにコントロールとしてCMVプロモーターを持つウミシイタケルシフェラーゼを同時に導入し、これを内部コントロールとしてルシフェラーゼ活性を測定した。
【0127】
結果を図14に示す。図14は、各遺伝子を導入したHeLa細胞における相対的ルシフェラーゼ活性を示す。E2(-)はE2非存在下で培養した場合、E2(+)はE2存在下で培養した場合を示す。図14における、+又は-は、各遺伝子の導入の有無を示す。なお、図14では、各バーは独立して3回行った実験の平均値と1SD(標準偏差)で示す。
【0128】
図14に示すように、ERα遺伝子及びERαのコアクチベーターであるPGC-1α遺伝子をHeLa細胞に導入すると、転写活性が増強し、特にE2存在下では転写活性がさらに増強した。これに、PHB2タンパク質を共発現させると、転写活性の抑制効果が検出できた。従って、PHB2タンパク質はERαのコアクチベーターであるPGC-1αタンパク質の転写調節に抑制的に働くことが明らかになった。
【0129】
本明細書で引用した全ての刊行物、特許及び特許出願をそのまま参考として本明細書にとり入れるものとする。
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)記載のsiRNAを含有することを特徴とするPHB2遺伝子発現調節剤。
(a)配列番号1〜20記載のいずれか1つに示される塩基配列から成るsiRNA。
(b)上記(a)記載のsiRNAにおいて、1又は数個の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2遺伝子発現阻害活性を有するsiRNA。
【請求項2】
上記PHB2遺伝子発現調節剤が抗アポトーシス作用剤、ミトコンドリア膜電位調節剤、及びミトコンドリア形態調節剤から成る群から選択されるものである、請求項1記載のPHB2遺伝子発現調節剤。
【請求項3】
請求項1又は2記載のPHB2遺伝子発現調節剤を含有するミトコンドリア機能関連疾患治療薬。
【請求項4】
ERα、PPARα及びPPARγ2から成る群から選択される核レセプターのアンタゴニスト又はアゴニストを含有することを特徴とするPHB2タンパク質機能調節剤。
【請求項5】
上記PHB2タンパク質機能調節剤が抗アポトーシス作用剤、ミトコンドリア膜電位調節剤、及びミトコンドリア形態調節剤から成る群から選択されるものである、請求項4記載のPHB2タンパク質機能調節剤。
【請求項6】
請求項4又は5記載のPHB2タンパク質機能調節剤を含有するミトコンドリア機能関連疾患治療薬。
【請求項1】
以下の(a)又は(b)記載のsiRNAを含有することを特徴とするPHB2遺伝子発現調節剤。
(a)配列番号1〜20記載のいずれか1つに示される塩基配列から成るsiRNA。
(b)上記(a)記載のsiRNAにおいて、1又は数個の塩基が欠失、置換又は付加された塩基配列から成り、且つPHB2遺伝子発現阻害活性を有するsiRNA。
【請求項2】
上記PHB2遺伝子発現調節剤が抗アポトーシス作用剤、ミトコンドリア膜電位調節剤、及びミトコンドリア形態調節剤から成る群から選択されるものである、請求項1記載のPHB2遺伝子発現調節剤。
【請求項3】
請求項1又は2記載のPHB2遺伝子発現調節剤を含有するミトコンドリア機能関連疾患治療薬。
【請求項4】
ERα、PPARα及びPPARγ2から成る群から選択される核レセプターのアンタゴニスト又はアゴニストを含有することを特徴とするPHB2タンパク質機能調節剤。
【請求項5】
上記PHB2タンパク質機能調節剤が抗アポトーシス作用剤、ミトコンドリア膜電位調節剤、及びミトコンドリア形態調節剤から成る群から選択されるものである、請求項4記載のPHB2タンパク質機能調節剤。
【請求項6】
請求項4又は5記載のPHB2タンパク質機能調節剤を含有するミトコンドリア機能関連疾患治療薬。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12−1】
【図12−2】
【図13】
【図14】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12−1】
【図12−2】
【図13】
【図14】
【公開番号】特開2012−45001(P2012−45001A)
【公開日】平成24年3月8日(2012.3.8)
【国際特許分類】
【出願番号】特願2011−254346(P2011−254346)
【出願日】平成23年11月21日(2011.11.21)
【分割の表示】特願2007−546485(P2007−546485)の分割
【原出願日】平成18年11月16日(2006.11.16)
【出願人】(505246789)学校法人自治医科大学 (49)
【Fターム(参考)】
【公開日】平成24年3月8日(2012.3.8)
【国際特許分類】
【出願日】平成23年11月21日(2011.11.21)
【分割の表示】特願2007−546485(P2007−546485)の分割
【原出願日】平成18年11月16日(2006.11.16)
【出願人】(505246789)学校法人自治医科大学 (49)
【Fターム(参考)】
[ Back to top ]