
ヘテロクリティックアナログおよび関連方法
【課題】本発明は,対応する類似した野生型エピトープと比べて、免疫応答をもたらす増強された能力を有するエピトープを含むペプチドを、調製する方法を提供する。
【解決手段】クラスIエピトープのヘテロクリティックアナログは、これらのエピトープの3および/または5および/または7位で保存的または半保存的アミノ酸置換を提供することで調製される。このアナログは、対応する野生型エピトープに関連する免疫応答の誘発において有用である。本発明はまた、エピトープを含むペプチドの免疫原性を高める方法を提供する。本発明はまた、上記の方法によって得られ得るクラスIエピトープを含むペプチドを少なくとも1つ含む組成物を提供する。
【解決手段】クラスIエピトープのヘテロクリティックアナログは、これらのエピトープの3および/または5および/または7位で保存的または半保存的アミノ酸置換を提供することで調製される。このアナログは、対応する野生型エピトープに関連する免疫応答の誘発において有用である。本発明はまた、エピトープを含むペプチドの免疫原性を高める方法を提供する。本発明はまた、上記の方法によって得られ得るクラスIエピトープを含むペプチドを少なくとも1つ含む組成物を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の引用)
本出願は、米国35 U.S.C.§119(e)の下、特許仮出願番号60/166,529(1999年11月18日に出願)および同60/239,008(2000年10月6日に出願)に対する優先権を主張する。これらはその全体が参照として援用される。
【0002】
(発明の分野)
本発明は所定のT細胞に対する増加した刺激能力を有する、元のペプチドのヘテロクリティック(heteroclitic)アナログを生成するための方法に関係する。
【背景技術】
【0003】
(発明の背景)
数々の研究は、細胞障害性リンパ球(CTL)が、免疫系による感染性の疾患およびガンの根絶に中心的な役割を果たすことを示唆する(Byrneら、J.Immumol.51:682(1984)、McMichealら、N.Englnd J.Med.,309:13(1983))。CTLはエピトープを含むペプチドにより刺激されると、CTL応答を刺激するエピトープに基づくワクチンの開発においてかなりの努力が進められている。ヘテロクリティックアナログと命名したエピトープの一つのクラスは、ワクチン成分としての利点を提供する。なぜならこれらのアナログは、ネイティブエピトープによって誘導されるT細胞応答よりも強力なT細胞応答を誘導するからである。ヘテロクリティックアナログは、所定の用量に対する応答の増加もしくは同じ応答を達成するためのより少ない必要量によって測定されるような、特定のT細胞に対する増加した刺激能力もしくは刺激効力を有するペプチドと定義される。
【0004】
臨床適応におけるヘテロクリティックアナログの使用と関連する利点は、以下のとおりである。第一に、ヘテロクリティックアナログは、T細胞アネルギーの状態を逆転することによってか、T細胞の非寛容化交差反応クローンを活性化することによってか、または、「免疫偏向」(すなわち、産生されるCTLの型(例えば、Th1またはTh2))を媒介することによって、寛容を崩壊/克服する能力を有する。最近の研究は、ヘテロクリティックアナログが内因的にプロセシングされたエピトープを認識するCTLを誘導し得るという点で、ヘテロクリティックアナログが免疫原であることを示す(Zarembaら、Cancer Research、57:4570(1997);Rivoltoniら、Cancer Research、59:301(1999);Selbyら、162(2):669(1999))。これは異なる免疫系における研究によって確認される(Zugelら、J.Immunol.、161:1705(1998)、Wangら、J.Exp.Med.、190:983(1999)、Menら、J.Immunol.、162:3566(1999)。例えば、Zugelらの研究(Zugelら、前出)は、成体マウスにおいて免疫優性のT細胞エピトープに対するT細胞寛容が、ペプチドのそのヘテロクリティック交差反応性ペプチドアナログで免疫することによって克服され得ることを示す。
【0005】
この事は、ガンのワクチンの分野で特に有意であり、ここで、CTLエピトープの大部分が、自己抗原から誘導される。ガン関連抗体が、しばしば、自己抗原であるという事実に起因して、これらの抗原に対する既存の寛容が存在し得るという対応した現象が存在し、それによって、そのようなエピトープに対するT細胞応答の発生が課題である。ヘテロクリティックアナログによる寛容の崩壊は、マウスクラスII系における最近の研究で示されている(Wangら、J.Exp.Med.190:983(1999)。この研究では、寛容の崩壊に関与するメカニズムがアネルギーの逆転よりむしろ、寛容化の低親和性クローンの刺激であった。本明細書中で実証されるヘテロクリシティーは、高いアビディティのCTLの誘導に関連しており、これは重要な差異を表す。
【0006】
第二に、ペプチドアナログは、T細胞からのサイトカイン産生を調節することが示されている(Pfeifferら、J.Exp.Med.、181:1569(1995)、Taoら、J.Immunol.、158:4237(1997)、Salazarら、Int.J.Cancer 85(6):829−38(2000)、Nicholsonら、Int.Immunol.12(2):205−13(2000))。そのようなアナログによって誘導される免疫偏向は、Th細胞応答の特定のサブセットの発生が腫瘍退行に相関する、(Zitvogelら、J.Exp.Med.、183:87(1996)、Celluzziら、J.Exp.Med.、183:283(1996))または、自己免疫疾患もしくは感染性の疾患の臨床結果に影響を与える(Romagnaniら、Annu.Rev.Immumol.、12:227−57(1994))数々の疾患状態と関連を有する。従って、ヘテロクリティックアナログを伴う免疫は、エフェクターT細胞の特定のサブセットの誘導によってサイトカイン産生を調節する能力を提供し、それによって、疾患の経過を変化させる。
【0007】
第三に、ヘテロクリティックアナログは、薬剤の開発において利点を有する。なぜなら、それらは強力な生物学的効力により、有意により少量のペプチドが、処置用量に必要とされるからである。この特徴は、特定の製造および毒性の懸念を克服する。この事については、黒色腫患者において抗原特異的T細胞を生じた、MART−1ペプチドのヘテロクリティックアナログが、ネイティブエピトープよりはるかに低濃度で活性であることが示されている(Rivoltiniら、Cancer Research 59:301(1999)。同様の結果が、CEA由来CAP1エピトープのヘテロクリティックアナログに関して、Schlomおよび共同研究者によって報告された。(Zarembaら、Cancer Research 57:4570(1997))。しかし、野生型ペプチドに対する並行したprecursor frequency分析もしくはTCRアビディティ分析は、行われなかった。
【0008】
従って、それらの生物学的関連性もために、所定のエピトープにヘテロクリティック活性を与えるアミノ酸置換を予測することが非常に有用である。しかしながら、本開示以前では、そのような置換を予想するための簡単な方法は存在しない。しかし、以前の研究では(非特許文献1、非特許文献2)、黒色腫細胞から天然に存在する変異ペプチドを溶出することによってか、または、エピトープ内のほとんど全ての位置での変異からなる多くのアナログを系統的にスクリーニングすることによって、ヘテロクリティックエピトープが、偶発的に同定された(非特許文献3、非特許文献4、非特許文献5)。あるいは、ヘテロクリティックアナログは、ランダムコンビナトリアムペプチドライブラリーをスクリーニングすることによって同定された。このライブラリーはまた、多くのペプチドの困難な合成およびスクリーニングを必要であった(非特許文献6)。DNA発現ライブラリーのスクリーニングのような、遺伝学的なアプローチが、CTLエピトープおよびアナログの生成のための別の方法を提供する(非特許文献7、非特許文献8)。しかし、このアプローチは、生成されるエピトープの潜在的な少ない量および複雑性が問題になり得る。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Selbyら、J.Immunol.、162(2):669(1999)
【非特許文献2】Skipperら、J.Exp.Med.183:527(1996)
【非特許文献3】Zarembaら、Cancer Research 57:4570(1997)
【非特許文献4】Loftusら、Cancer Research 58:2433(1998)
【非特許文献5】Blakeら、J.Exp.Med.18:121(1996)
【非特許文献6】Pinillaら、Current Opinion in Immunology 11:193−202(1999)
【非特許文献7】Boonら、Annu.Rev.Immunol.12:337−65(1994)
【非特許文献8】Gavinら、Eur.J.Immunol.24(9):2124−33(1994)
【発明の概要】
【課題を解決するための手段】
【0010】
(発明の開示)
本発明は,対応する類似した野生型エピトープと比べて、免疫応答をもたらす増強された能力を有するエピトープを含むペプチドを、調製する方法を提供する。この得られた「ヘテロクリティックアナログ」は、ウイルス性疾患、ガン、および標的細胞上の提示された抗原によって特徴付けられる他の疾患の処置のための、免疫学的組成物として有用である。また、本発明は以下を提供する:
(項目1) エピトープを含むペプチドの免疫原性を高める方法であって、該方法が、以下:
i)第一のクラスIエピトープを含むペプチドを提供する工程であって、ここで該エピトープが、本質的にN末端およびC末端ならびに少なくとも1つの一次アンカー残基を有するアミノ酸配列からなり、ここで該アミノ酸残基が、連続して数えられ、そして該エピトープのN末端に最も近い一次アンカー残基が、2位または3位である、工程;および
ii)該エピトープのN末端とC末端との間の3位および/もしくは5位および/もしくは7位での1つ以上の保存的、または半保存的置換を誘導する工程であって、ただし、該位が一次アンカー残基でない、工程、
それにより第二のクラスIエピトープを含むペプチドを構築する工程であって、該エピトープが、該第一のクラスIエピトープと比較して高められた免疫原性を示す、工程、
を包含する、方法。
【0011】
(項目2) 前記第二のクラスIエピトープが、前記第一のクラスIエピトープと比較して、特定のT細胞について少なくとも約50%増加した効力を示す、項目1に記載の方法。
【0012】
(項目3) 1つの置換のみが導入される、項目1に記載の方法。
【0013】
(項目4) 前記置換が保存的置換である、項目1に記載の方法。
【0014】
(項目5) 前記置換が半保存的置換である、項目1に記載の方法。
【0015】
(項目6) 項目1に記載の方法であって、前記ペプチドがHLAクラスI分子によって結合されそして細胞傷害性のT細胞と接触する場合、前記第二のクラスIエピトープを含むペプチドが、Th1およびTh2サイトカインの両方を誘導する、方法。
【0016】
(項目7) 前記第一のクラスIエピトープが、A1、A2、A3、A24、B7、B27、B44、B58およびB62からなる群より選択されるスーパーモチーフを含む、項目1に記載の方法。
【0017】
(項目8) 項目1に記載の方法であって、ここで前記第一のクラスIエピトープが、ウイルス抗原、腫瘍関連抗原、寄生生物抗原、細菌抗原または真菌抗原由来である、方法。
【0018】
(項目9) 項目1の方法によって調製された第二のクラスIエピトープを含むペプチド。
【0019】
(項目10) 免疫応答を誘発する方法であって、該方法が細胞傷害性Tリンパ球(CTL)と項目9に記載のペプチドとを接触させる工程を包含する、方法。
【0020】
(項目11) 前記接触させる工程がインビトロで、抗原提示細胞の存在下で実施される、項目10に記載の方法。
【0021】
(項目12) 前記接触工程が、被験体に前記ペプチドをコードするヌクレオチド配列を含む核酸分子を投与する工程によって実施される、項目10に記載の方法。
【0022】
(項目13) 少なくとも1つのペプチドを含む組成物であって、該ペプチドが、項目1に記載の方法によって得られ得るクラスIエピトープを含む、組成物。
【0023】
(項目14) 前記ペプチドが、9個〜15個のアミノ酸を含む、項目13に記載の組成物。
【0024】
(項目15) 項目13に記載の組成物であって、前記ペプチドが、配列番号2、配列番号3、配列番号5、配列番号6、配列番号8、配列番号9、配列番号11、配列番号12、配列番号14、配列番号15、配列番号17、配列番号19および配列番号20からなる群より選択されるアミノ酸配列を含む、組成物。
【0025】
(項目16) 前記ペプチドがCTLエピトープと混ぜられるか、または連結される、項目13に記載の組成物。
【0026】
(項目17) 前記ペプチドが、HTLエピトープと混ぜられるか、または連結される、項目13に記載の組成物。
【0027】
(項目18) 前記HTLエピトープが、pan−DR結合分子である、項目17に記載の組成物。
【0028】
(項目19) リポソームをさらに含む、項目13に記載の組成物。
【0029】
(項目20) 前記エピトープが、脂質に結合される、項目13に記載の組成物。
【0030】
(項目21) 前記エピトープが、ヘテロポリマーに含まれる、項目13に記載の組成物。
【0031】
(項目22) 前記エピトープが、ホモポリマーに含まれる、項目13に記載の組成物。
【0032】
(項目23) 前記エピトープが、HLA重鎖、β2ミクログロブリン、およびストレプトアビジン複合体に結合され、これによりトリマーが形成される、項目13に記載の組成物。
【0033】
(項目24) 項目13に記載の組成物であって、抗原提示細胞をさらに含み、ここで、前記エピトープが、該抗原提示細胞上または抗原提示細胞内にある、組成物。
【0034】
(項目25) 項目24に記載の組成物であって、前記エピトープが前記抗原提示細胞上のHLA分子に結合し、これにより、該HLA分子に制限される細胞傷害性リンパ球(CTL)が存在する場合、該CTLのレセプターが該HLA分子と該エピトープとの複合体に結合する、組成物。
【0035】
(項目26) 前記抗原提示細胞が、樹状細胞である、項目25に記載の組成物。
【0036】
(項目27) 項目13に記載の組成物であって、HLA分子をさらに含み、ここで、前記ペプチドが、該HLA分子により結合される、組成物。
【0037】
(項目28) 標識をさらに含む、項目13に記載の組成物。
【0038】
(項目29) 項目28に記載の組成物であって、ここで、前記標識が、ビオチン、蛍光部分、非哺乳動物糖、放射性標識、またはモノクローナル抗体が結合する低分子である、項目28に記載の組成物。
【0039】
(項目30) 項目13に記載の組成物であって、以下:
前記ペプチドの単位投薬量、および
薬学的賦形剤、
を含む、ワクチンである、組成物。
【0040】
(項目31) 項目1に記載の方法により得られ得る第二のクラスIエピトープを含む、9〜15個のアミノ酸のペプチドをコードするヌクレオチド配列を含む核酸分子。
【0041】
(項目32) 項目31に記載の核酸分子であって、前記ペプチドが、配列番号2、配列番号3、配列番号5、配列番号6、配列番号8、配列番号9、配列番号11、配列番号12、配列番号14、配列番号15、配列番号17、配列番号19、および配列番号20、からなる群より選択されるアミノ酸配列からなるエピトープを含む、核酸分子。
【0042】
(項目33) 前記ヌクレオチド配列の発現のための制御配列をさらに含む、項目32に記載の核酸分子。
【0043】
(項目34) 項目31に記載の核酸分子を、活性成分として含む、薬学的組成物。
【0044】
従って、一つの局面において、本発明は、エピトープを含むペプチドの免疫原性を増強するための方法に関する。この方法は、以下の工程を包含する:I)第一のクラスIエピトープを含むペプチドを提供する工程であって、ここで、このエピトープは、本質的に、N末端およびC末端ならびに少なくとも一つの一次アンカー残基を有するアミノ酸配列からなり、ここで、このエピトープのアミノ酸残基は連続的に番号付けられ、そして、このエピトープのN末端に最も近い一次アンカー残基は、位置2または位置3である工程;II)一次アンカー残基を含まない位置3および/または5および/または7で、このエピト−プのN末端とC末端との間に一つ以上の保存的または半保存的な置換を導入する工程で、これにより、増強された第一のクラスIエピトープと比較される抗原原性を示す第二のクラスIエピトープを含むペプチドを構成する工程。
【0045】
上記の第二のクラスIエピトープは、一般に「ヘテロクリティックアナログ」と称する。
【0046】
好ましい実施形態において、このヘテロクリティックアナログは、対応する野生型クラスIエピトープに比較して、特定のT細胞に対する少なくとも約50%増加した効力を示す。このアナログは、1つの置換のみを含み得るか、または2もしくは3つの置換を含み得、そしてこの置換は、保存的または半保存的であり得る。このヘテロクリティックアナログは、HLAクラスI分子に結合され、そして関連する細胞傷害性T細胞と接触された場合、Th1およびTh2サイトカインの両方を誘導し得る。好ましくは、クラスIエピトープは、A1、A2、A3、A24、B7、B27、B44、B58およびB62からなる群から選択されたスーパーモチーフを含み、より好ましくは、クラスIエピトープは、A2スーパーモチーフ、最も好ましくは、A2.1モチーフを含む。
【0047】
本発明はまた、事前に選択したクラスIペプチドエピトープに対してヒト細胞傷害性T細胞応答を誘導する方法を提供する。この方法は、以下の工程を包含する:上記のヘテロクリティックアナログを提供する工程;および、ヒトCTLにこのヘテロクリティックアナログを接触させる工程。
【0048】
いくつかの局面において、この接触の工程は、インビトロで実施される。いくつかの局面において、この接触の工程は、このヘテロクリティックアナログペプチドエピトープをコードする配列を含む核酸分子を、被験者に投与することによって実施される。
【0049】
本発明はまた、上記の方法によって入手可能であるヘテロクリティックアナログエピトープを含むペプチドに関する。特に、そして、好ましくは、そのようなペプチドとしては、そのエピトープが、配列番号2、配列番号3、配列番号5、配列番号6、配列番号8、配列番号9、配列番号11、配列番号12、配列番号14、配列番号15、配列番号17、配列番号19、および配列番号20からなる群から選択されるアミノ酸の配列からなる、ペプチドが挙げられる。このペプチドは、9〜20アミノ酸、好ましくは、9〜16アミノ酸、より好ましくは9〜15アミノ酸を含み得るが、また、全部で9、10、11、12、13または14アミノ酸のみを含み得る。定義されたヘテロクリティックアナログエピトープは、より長いポリペプチドまたはタンパク質に含まれ得、このタンパク質またポリペプチドは、同じエピトープのホモポリマーであるか、あるいは、種々のこのようなエピトープを含むかまたは野生型ペプチドと組み合わせてヘテロクリティックアナログエピトープを含む、ヘテロポリマーである。これらペプチドおよびタンパク質は、薬学的使用のために設計される組成物に含まれ得る。
【0050】
ヘテロクリティックアナログエピトープを含むペプチドまたはヘテロポリマーもしくはホモポリマーは、他の成分を結合させ、免疫応答の誘発におけるそれらの活性をさらに増強し得るか、または調節し得る。これらさらなる変更物は、共有結合的に結合され得るか、または混合物中に非共有結合的に含まれ得る。従って、ヘテロクリティックアナログエピトープは、CTLエピトープまたはこのHTLエピトープに混合され得るか、または投与され得る。特にHTLエピトープは、pan−DR結合分子である。ヘテロクリティックアナログエピトープを含む組成物は、さらにリポソームを含み得る。ここで、このエピトープは、リポソーム上もしくはリポソーム中に存在するか、またはこのエピトープは、脂質に結合され得る。このヘテロクリティックエピトープは、HLA重鎖、β2−ミクログロブリンおよびストレプトアビジン(strepavidin)複合体に結合され得、それによって、4量体が形成される。さらに、このヘテロクリティックエピトープは、抗原提示細胞を含む組成物において改変され得る。ここで、このエピトープは、抗原提示細胞上または抗原提示細胞中にあり、ここで、このエピトープは、抗原提示細胞上のHLA分子に結合される。従って、HLA分子に制限される細胞傷害性リンパ球(CTL)が存在する場合、CTLのレセプターは、HLA分子とそのエピトープの複合体に結合する。抗原提示細胞は、樹状細胞であり得る。この組成物はまた、単にHLA分子を含み得る。ここで、このエピトープを含むペプチドは、HLA分子に結合される。この組成物は、モノクローナル抗体が結合する標識(例えば、ビオチン)、蛍光部分、非哺乳動物性の糖、放射標識または小さい分子をまた含み得る。
【0051】
記載される組成物は、対応する野生型エピトープに対する免疫応答の誘発において有用である。代表的には、ヘテロクリティックアナログは、適切な賦形剤をさらに含むそのような組成物に含まれる。活性な成分であるヘテロクリティックエピトープは、投薬形態単位で存在し得る。被験体の処置に有用な組成物はまた、これらの核酸分子は、上記のペプチドをコードする核酸分子を含み得、それらの発現のための制御配列を必要に応じて含む。
【図面の簡単な説明】
【0052】
【図1−1】図1Aおよび1Bは、CEA.691およびMAGE3.112のそれぞれのアナログのパネルを、その対応するCTLにおいてIFNγ産生を誘導する能力について、試験した結果を示す。
【図1−2】図1Cおよび1Dは、それぞれCEA.691およびMAGE3.112ヘテロクリティックアナログについてその対応する用量応答曲線である。
【図2−1】図2Aは、MAGE2.157、HIVPol.476、およびHBVPol.455エピトープアナログのアナログのパネルを、対応するCTLにおいてIFNγ産生を誘導するこれらのアナログの能力に関して、試験した結果を示す。
【図2−2】図2Bは、MAGE2.157、HIVPol.476、およびHBVPol.455エピトープアナログのアナログのパネルを、対応するCTLにおいてIFNγ産生を誘導するこれらのアナログの能力に関して、試験した結果を示す。
【図2−3】図2Cは、MAGE2.157、HIVPol.476、およびHBVPol.455エピトープアナログのアナログのパネルを、対応するCTLにおいてIFNγ産生を誘導するこれらのアナログの能力に関して、試験した結果を示す。
【図2−4】図2Dは、首尾よいHIVPol.476アナログについての関連した用量応答曲線である。
【図3】図3Aおよび3Bは、適切なCTLからIFNγ産生またはIL10産生を誘導する能力に関して、野生型との比較におけるMAGE2.157のヘテロクリティックアナログの用量応答曲線の結果を示す。
【図4】図4Aおよび4Bは、適切なCTLにおいてIFNγおよびIL10の産生に対する、野生型およびHIVPol.476のヘテロクリティックアナログについての用量応答曲線である。
【図5】図5は、エピトープp53.149M2の潜在的なヘテロクリティックアナログのパネルを、適切なCTLからのIFNγの産生に関して、試験した結果である。
【図6】図6Aおよび6Bは、p53.149M2の首尾よいヘテロクリティックアナログによるIFNγおよびIL10の産生に対する、対応する用量応答曲線である。
【図7】図7は、CTLにおけるIFNγ産生に対するp53.Mu184エピト−プの潜在的なアナログのパネルを試験した結果である。
【図8】図8は、IFNγ産生に関する、野生型、およびp53.Mu184の2つ首尾よいヘテロクリティックアナログについての用量応答曲線である。
【図9】図9A〜Dは、対応する野生型エピトープに関する、ヘテロクリティックアナログの反応交差性を示す。図9Aおよび9Bにおいて、IFNγ産生は、免疫ペプチドによる刺激を使用して、濃度の関数としてプロットされる。図9Cおよび9Dは、野生型エピトープが、CTLの初期誘導のために使用されるヘテロクリティックアナログとは対抗的に、刺激剤として使用される場合の、対応する結果である。
【図10】図10は、p53.261およびそのヘテロクリティックアナログによる誘導に関する、IFNγの放出である。
【図11】図11は、種々のヘテロクリティックアナログに関するElispotの結果である。
【図12】図12は、種々のヘテロクリティックアナログを使用する内因性のペプチドに対するCTL活性の刺激の結果である。
【図101−1】図101は、実施例で用いられるペプチド、および腫瘍抗原およびウイルス抗原から同定されたヘテロクリティックアナログの特徴を示す、表1である。
【図101−2】図101は、実施例で用いられるペプチド、および腫瘍抗原およびウイルス抗原から同定されたヘテロクリティックアナログの特徴を示す、表1である。
【図102−1】図102は、所定のアミノ酸置換を、保存的置換、半保存的置換、または非保存的置換であるとして特徴付けし得るように、任意の所定のアミノ酸対の間の類似性の割り当てを示す、表2である。
【図102−2】図102は、所定のアミノ酸置換を、保存的置換、半保存的置換、または非保存的置換であるとして特徴付けし得るように、任意の所定のアミノ酸対の間の類似性の割り当てを示す、表2である。
【図103−1】図103は、表3である。
【図103−2】図103は、表3である。
【図104−1】図104は、表4である。
【図104−2】図104は、表4である。
【発明を実施するための形態】
【0053】
(発明を実施するための形態)
(1.概要)
本発明は、HLAクラスI分子に結合するヘテロクリティックアナログの設計の方法に関連する。本明細書中に記載される「ヘテロクリティックアナログ」は、特定のT細胞に対する増強された効力を持つエピトープを含むペプチドである。それは、所定の投与量に対する増強された応答によって、または相同性のあるクラスIペプチドとして同じ応答を達成するより少ないの必要量によって測定される。本発明の方法は、特に、ヒトの癌および前癌性の状態に関連する、ウイルス、細菌、真菌および原生動物の寄生虫のような感染性の病原体に由来する、ならびに任意のクラスIペプチドの改変に有用である。
【0054】
重要なことに、ヘテロクリシティーの現象は、個々のクラスIエピトープに結合するHLA分子にわたってあてはまる。例えば、A2スーパーモチーフを有するヘテロクリティックアナログペプチドは、HLA−スーパータイプ(例えば、A0201、A0202、A0203、A0204、A0205、A0206、A0207など)における全てのHLA分子にわたって、ヘテロクリティック(すなわち、より高い効力を有する)である。同様に、異なる配列モチーフ(例えば、A1、A3、A24、B7、B27、B44、B58、B62など)を有するヘテロクリティックアナログペプチドは、その特定のHLA−スーパーファミリーにおける全てのHLA分子にわたって、免疫応答をより強力にする。
【0055】
出願者は、対応する野生型エピトープに対する、免疫応答を高めるヘテロクリティックアナログを設計する特定の法則を見つけた。これらの法則は、任意のクラスI対立遺伝子によってコードされるHLA分子に結合するモチーフ、またはス−パーモチーフを有するエピトープに関して適応できる。従って、これらの法則の使用によって、任意の「野生型」または「ネイティブ」クラスIエピトープの免疫原性を高めることが可能である。
【0056】
簡略には、この法則は、野生型クラスIエピトープが、エピトープの位置3および/または位置5および/または位置7として保存的または半保存的なアミノ酸の置換によって改変されることを述べる。置換されるべき保存的または半保存的アミノ酸の性質は、本明細書以下の、調製Bの記載によって定義され、この結果は表2に要約される。従って、表2を参照すれば、これらの位置での置換のための適した候補を決め得る。表2に示されるように、表の最上段にわたって示されるそれぞれのアミノ酸は、残存する19の遺伝的にコードされるアミノ酸と数値的に規定された関係を有する。指標がより低いほど、保存性はより高く;同じアミノ酸は、1.0の類似性の割当を有し;最も大きく異なるアミノ酸は、20に近い類似割当を有する。調製B中で示す方法を使用すると、遺伝子コードされないアミノ酸もまた、類似性の指標を割り当てられ得、そして保存的または半保存的(または非保存的)として、任意に天然に存在するアミノ酸について、分類され得る。
【0057】
本発明のヘテロクリティックアナログペプチドは、被験体の免疫系が耐性となる抗原に対する免疫応答を誘導するのに特に有用である。ヒト被験体は、特に好ましいが、この方法はまた、これらの被験体に関して、対応するHLAモチーフを考慮にいれることにより、他の哺乳動物、例えば実験室マウスにも適用し得る。耐性は、先立つ抗体への曝露によって誘導される特定の免疫学的な非反応性をいう。耐性は、患者が耐性である特定のクラスIペプチドエピトープを同定することによって、本発明の方法に従ってペプチドエピトープ配列を改変することによって、および寛容化エピトープ(抗原)に対する交差反応をする免疫反応を誘導することによって、克服され得る。例えば、被験体の免疫系がウイルス抗原または腫瘍関連抗原に耐性である場合、耐性の克服は特に望ましく、後者の抗原は、細胞の形質転換の結果として、しばしば過剰発現する自己タンパク質である。
【0058】
ヘテロクリティックを設計するための法則を決定するため、いくつかの種々のCTL株がアナログパネルに対する反応性によりスクリーニングされた。T細胞刺激容量の改変は一次MHCアンカーの変更なしに達成された。
【0059】
その野生型エピトープとしては、上皮細胞の癌において特異的に上方制御され、そして免疫原性であることが示された自己抗原に由来する腫瘍エピトープが挙げられる。HIVおよびHBVのポリメラーゼ遺伝子由来のエピトープのような使用されるウイルスのエピトープは、同様に免疫原性であることが示された。
【0060】
本明細書中に記載した法則は、ヘテロクリティックなアナログを設計するための基盤を提供し、そうでなければ要求されるスクリーニングを劇的に減少させ、そして癌および感染性疾患のためのエピトープを基礎としたワクチンの設計に非常に有用である。
【0061】
以下に示す実施例において、スクリーニングされた(本明細書に開示されるヘテロクリティックの法則に適合する)総アナログの17%がヘテロクリティックであった(16/95)。これは、二つの理由において有意である:第1に、ヘテロクリティック置換の法則に従ったアナログを使用することでヘテロクリティック検出の効率が2.2%から17%まで上昇した;第2に、合成されることが必要なペプチドの数が一つのエピトープあたり約100アナログから約15アナログに劇的に減少され、この工程費用を効果的にし、高い処理能力に耐えられるようにする。本発明のヘテロクリティック置換の法則の適用を通して、ヘテロクリティックアナログが生じる効率は、100から1000倍近く、0.2%(233のCEA.691およびMAGE3.112アナログのスクリーニングから4つが同定された)から30%(9つの予測されたアナログのスクリーニングから3つが同定された)まで増加した。後者の頻度は、最初のアッセイにおいて潜在的なヘテロクリティック活性を示した6つのアナログのうちの4つのみをさらなる分析に供したので、ひどい過小評価でありうる。
【0062】
以前の研究は、ヘテロクリティックアナログによるT細胞応答の調節が、TCR接触残基を含むことを示した(Byrneら、 J. Immunol. 51:682(1984)、 McMichaelら、 N. England. J. Med. 309:13(1983)、 Zugelら、 J. Immunol. 161:1705(1998)、 Rivoltiniら、 Cancer Research 59:301(1999)、 Parhurstら、 J. Immunol. 157:2539(1996))が、本研究ではこれを発見しなかった。例えば、CEA.691エピトープに対しては、TCR接触残基が8位であり、一方で、ヘテロクリティックは、3および5位でアナログ置換が認められた。いかなる仮定によっても束縛されることを意図しないが、MHC結合の変更は機構となり得る。このアナログに対して実施された結合分析は、大多数の場合(80%)において好むと好まざるとにかかわらずMHC結合における変更があることを示した。HLA−A2結合について試験された13アナログのうち10アナログはMHC結合の変更を持ち、6アナログは、野生型のペプチドおよび野生型より結合の悪い4アナログよりも良好に結合するが、まだ実質的に増加した生物学的応答を生じた。いくつかの研究ではMHC結合を増強するために一次のMHCアンカー残基を修飾する(この手段は、アナログを生じるためにいくつかのグループによって用いられた(Pfeifferら、 J. Exp. Med. 181:1569(1995)、 Valmoriら、J.Immunol.160:1750−1758(1998))。一次のTCRの接触残基あるいは一次のMHCアンカー残基の変化無しに生物学的応答の増加は本研究においては認められなかった。応答の増加がMHC結合における変更にともない媒介されるので、その効果は二次アンカーの位置の変化により媒介しうることを前提とする。これを支持するさらなる証拠は、ヘテロクリティックの置換がそのペプチドの中間における奇数位置(3、5、7)で起こるという発見から生じる。3、5および7のこれらの位置全ては、HLA−A2分子に対する結合にとって二次アンカー位置であると示された(Boonら、 Annu. Rev. Immunol. 12:337−65 (1994)、 Ishiokaら、 J. Immunol. 162(7):3915−25 (1999) )。
【0063】
これらの位置のうちの2つ(3および7)は、HLA−A2.1分子に結合するための二次アンカー位置であることがいくつかのグループにより示された(Ruppertら、 Cell 74:929(1993)、 Madden、 Annu. Rev. Immunol. 13:587−622(1995))。そのような二次アンカー位置の変更は、T細胞認識差異に転換しうる(Valmoriら、 J. Immnol. 160:1750(1998); Davisら、 Annu. Rev. Immnol. 16:523 (1998))が、これらの研究では、T細胞の認識差異は、MHC結合における変化に付随し、ヘテロクリティックの獲得に関与するアミノ酸置換の種類に関する法則は規定されなかった。二次アンカーの変化からT細胞の認識の変化へのこのような転換が生じる機構は現在不明瞭である。しかし、いくつかのモデルは、二次アンカーの位置の残基がMHCと結合する様式の変化がTCR接触残基の配向における変更あるいは増加した可撓性を引き起こし得ることを示唆しており、それは、TCRに対するこれらのアナログの結合の増加に帰着する(Kershら、 J. Exp. Med. 184:1259(1996)、 Evavoldら、 J. Immunol. 148:347(1992)、 Alamら、 Immunity 10:227(1999)、 Hamplら、 Immunity 7:379−85(1997))。また、いくつかの以前の研究では、ヘテロクリティックアナログによるT細胞応答の調節が主なTCR接触残基に直接関与することを意味していた(Zarembaら、 Cancer Research 57:4570(1997)、 Loftusら、 Cancer Research 58:2433(1998)、 Dresselら、 J. Immunol. 159:4943(1997))。けれどもこの発見は、現在の体系的解析により実証されない。本研究において同定されたアナログに対して増加したT細胞認識は、おそらくMHC結合容量の増加に起因しないが、増加される結合は、おそらく一次アンカーの位置が最適化されたアナログの場合において、重要な役割を果たす。本研究は、ヘテロクリティックアナログが、おそらくTCRあるいはMHCの結合容量の大きな変更によるよりもむしろ構造における微妙な変更により生じることを示唆する。
【0064】
Th1あるいはTh2サイトカイン産生の調節差異は認められなかった。それどころか、本データは、ヘテロクリティックアナログがTh1およびTh2両応答の産生を増加することを示唆したが、その増加の大きさおよび動力学が異なり得る。事実、いくつかのグループ(Nicholsonら、 Int. Immunol. 12(2):205−13(2000)、 Parhurstら、 J. Immunol. 157:2539(1996))は、最近ペプチドアナログによるそのような全体刺激を報告した。これは、アナログによって誘起されるより強いTCRのシグナルに起因するが、そのような全体刺激の機構は、今後解明されるべきである。
【0065】
関連する腫瘍モデルあるいは自己抗原に対する寛容が存在するモデルを使用するインビボでのヘテロクリティックアナログの効力は評価される。従って、効果的なCTLを生じさせることが攻撃であるとこれまで判明している腫瘍に対するワクチン化として、ヘテロクリティックアナログでの免疫化がより効果的で有効な戦略であることが見出される。
【0066】
要約すると、本出願人らが癌およびウイルス起源のHLA−A2.1制限CTLエピトープのいくつもの異なるヘテロクリティックアナログを同定した。その適切な野生型エピトープは表1に示される。全てのこれらのエピトープは本発明者らのより早い報告において免疫原性であることが示された(Kawashimaら、 Human Immunology 59:1−14(1998)、 Ishiokaら、 J. Immunol. 162(7):3915−25(1999))。最初の実験の中で、CEA.691およびMAGE3.112CTLエピトープの233のアナログの抗原性が調査された。同定された4つのヘテロクリティックアナログの性質は、ヘテロクリティック置換が3、5および7の位置で保存的な置換を含むことを示した。この仮説は、3つのさらなるエピトープであるMAGE2.157、HIVPol.476およびHBVPol.455が関与する引き続きの研究において試験された。このように同定された全てのヘテロクリティックアナログは、提案した本法則、すなわちヘテロクリティックアナログが3、5および/または7の位置での保存的あるいは半保存的な置換に付随されることに一致した。
【0067】
癌免疫治療におけるヘテロクリティックアナログの臨床的適用をより綿密に模倣するため、マウスのエピトープ、p53.261もまた修飾された。T細胞寛容の部分的な状態はこのエピトープに関して報告された(Theobaldら、 Proc.Natl. Acad. Sci. 92:11993−11997(1995)、 Theobaldら、 J. Exp. Med., 185(5):833−841(1997))。予想された9つのp53.261アナログのうち4つは、未変性ペプチドにより誘導されるCTL応答に比べてインビボでのアナログ特異的CTLの応答をより強く誘導することが発見された。より顕著に、ヘテロクリティックアナログによる免疫化により生じたCTLの交差反応性が解析された場合に、3つのp53.261アナログが未変性のp53.261エピトープに対して活発に応答するCTLを誘導した。最終的に、ヒトCTLについてこれらの発見の妥当性はMAGE3.112エピトープのヘテロクリティックアナログがインビトロでのヒトT細胞に対して免疫原性であることを実証することにより考慮された。帰着するCTLは腫瘍細胞株の形態において自然に作用される野生型抗原を認識しうる。
【0068】
ヘテロクリティックアナログが、研究されたすべてのエピトープに対して同定されたので、本明細書に示した研究は、ヘテロクリシティーが全世界的な現象であることを証明する。加えて、本願は、クローンのT細胞集団(以前の研究で明示されたような)およびインビボでの免疫化の後の大部分のT細胞集団の両方におけるヘテロクリティックアナログを検出することが可能であることを示す。さらに、本明細書において、ヘテロクリシティー(HLA A2.1系におけるものと他のクラスIスーパーモチーフに関するものとの両方)が、合理的なヘテロクリシティーの予測を見込む別々の構造上の特徴に付随することが実証される。
【0069】
さらに、p53.261ヘテロクリティックアナログがより高いアビディティによりCTLを誘導し、また、野生型ペプチドにより誘導されるより多数の(前駆体出現率)これらの細胞を誘導することが実証される;インビボでのヘテロクリティックCTLの誘導およびT細胞の寛容を中断するためのその適用は実証される。
【0070】
ヘテロクリティックアナログは、インビボでの免疫化に続いて大部分の特異的なT細胞集団を生じるのに効果的であった。多数のTCR遺伝子由来のTCRを提示するポリクローナルの応答は臨床背景で疾患状態を治療するのにより有効である。最終的に、野生型エピトープに対する強い交差反応性のアビディティを有するCTLの高い前駆体出現率を生じるための能力が、通常は免疫系に対して寛容であるエピトープに対する効果的なCTL応答が要求される例において重要である。
【0071】
(2.定義)
特定のアミノ酸配列に関して、「エピトープ」は、特定の免疫グロブリンによる認識に関与するアミノ酸残基のセットであるか、またはT細胞の状況においては、主要組織適合遺伝子複合体(MHC)によりコードされるHLAの状況に提示される場合にT細胞レセプタータンパク質による認識にとって必要な残基である。インビトロまたはインビボでの免疫系の設定において、エピトープは、一次、二次および三次ペプチド構造、ならびに電荷のような分子の集団的特徴であり、これらは一緒になって、免疫グロブリン、T細胞レセプターまたはHLA分子によりコードされる部位を形成する。本開示を通してエピトープおよびペプチドは、しばしば交換可能に使用される。しかし、本発明のエピトープより大きくかつ本エピトープを含む、分離または精製されたタンパク質またはペプチド分子が、まだ本発明の範囲内にあることは、評価されることである。
【0072】
「クラスIエピトープ」は、クラスI HLA分子に対して結合するペプチドをいう。本明細書において記載されたように、クラスIエピトープは代表的に約8から約13アミノ酸長である。HLA分子に対する結合は、2つの一次アンカー残基により主に調節され、このうちの一方はエピトープのC末端にあり、そして他方は、2または3位にある。結合は、また、1つ以上の二次アンカー残基により補助されうる。読み手の利便性のために、様々な一次HLAクラスI結合アンカーは、表4にて示される。アンカーのパターンは、「モチーフ」と称される。「スーパーモチーフ」は、2つ以上のHLA対立遺伝子によりコードされるHLA分子により共有されるペプチド結合の特異性である。好ましくは、スーパーモチーフ保有ペプチドは、高いまたは中間の親和性(本明細書で定義されるような)で2つ以上のHLA抗原により認識される。クラスIスーパーモチーフの例としては、例えば、A1、A2、A3、A24、B7、B27、B44、B58およびB62が挙げられる。
【0073】
本開示をとおして、「結合データ」の結果を、しばしば「IC50」に換算して表す。IC50は、参照ペプチドの結合の50%阻害が認められる、結合アッセイにおけるペプチドの濃度である。アッセイを行う条件(すなわち、制限HLAタンパク質濃度および標識ペプチド濃度)が付与されると、これらの値は、Kd値を概算する。結合を測定するためのアッセイは、例えば、参考として本明細書に援用される、PCT公開WO94/20127およびWO94/03205に、詳細に記載される。IC50値が、アッセイ条件が変化する場合、そして使用する特定の試薬(例えば、HLA調製物など)に依存して、(しばしば、劇的に)変化し得ることに留意する。例えば、過剰濃度のHLA分子は、所定のリガンドの見かけの測定されたIC50を増加する。あるいは、結合は、参照ペプチドと関連して表される。特定のアッセイが、より高感度またはより低感度になるにつれて、試験するペプチドのIC50は、幾分変化し得るが、参照ペプチドに関連する結合は、有意には変化しない。例えば、参照ペプチドのIC50が10倍増加するような条件下で行うアッセイにおいて、試験ペプチドのIC50値もまた、約10倍シフトする。従って、あいまい性を回避するために、ペプチドが良好、中程度、弱いあるいはネガティブな結合因子であるかの評価は、一般に、標準ペプチドのIC50に対する、そのペプチドのIC50に基づく。結合はまた、当該分野において公知の他のアッセイ系を使用して測定され得る。
【0074】
「カルボキシルあるいはC末端」としてのエピトープ中の残基の位置の指定は、ペプチドのカルボキシル末端に最も近いエピトープの末端の残基の位置をいい、これは以下に定義したような従来の命名法を使用して指定される。エピトープの「C末端」は、ペプチドあるいはポリペプチドの末端に実際に対応してもよいし、対応しなくてもよい。
【0075】
「N末端」または「アミノの末端の位置」としてのエピトープ中の残基の位置の指定は、ペプチドのN末端に最も近いエピトープの末端の残基の位置をいい、これは以下に定義したような従来の命名法を使用して指定される。エピトープの「N末端」は、ペプチドあるいはポリペプチドの末端に実際に対応してもよいし、対応しなくてもよい。
【0076】
「コンピューター」または「コンピューターシステム」は、一般に、以下を備える:プロセッサー;少なくとも1つの情報記憶/検索装置、例えば、ハードドライブ、ディスクドライブまたはテープドライブなど;少なくとも1つの入力装置、例えば、キーボード、マウス、タッチスクリーン、またはマイクロホンなど;およびディスプレイ構造。さらに、コンピューターは、ネットワークと連絡する通信チャネルを備え得る。このようなコンピューターは、多かれ少なかれ、上記に列挙したものを備え得る。
【0077】
本明細書で使用される場合、「保存性」または「保存的」および「半保存性」または「半保存的」であるアミノ酸は、調製Bに従って定義され、表2に示される。
【0078】
本明細書中で使用される場合、HLAクラスI分子に関する「高い親和性」とは、50nM以下のIC50またはKD値での結合として定義され;「中程度の親和性」とは、約50nMと約500nMとの間のIC50またはKD値での結合である。HLAクラスII分子への結合に関する「高い親和性」とは、100nM以下のIC50またはKD値での結合として定義され;「中程度の親和性」とは、約100nMと約1000nMとの間のIC50またはKD値での結合である。
【0079】
「免疫原性ペプチド」または「ペプチドエピトープ」とは、そのペプチドがHLA分子を結合し、そしてCTL応答および/またはHTL応答を誘導するような、対立遺伝子特異的モチーフまたはスーパーモチーフを含むペプチドである。従って、本発明の免疫原性ペプチドは、適切なHLA分子に結合し、そしてその後、免疫原性ペプチドが由来する抗原に対する、細胞傷害性T細胞応答またはヘルパーT細胞応答を誘導し得る。
【0080】
句「単離された」または「生物学的に純粋」とは、そのネイティブ状態において見い出されるような通常その材料に付随する成分を、実質的または本質的に含まない材料をいう。従って、本発明に従う単離されたペプチドは、好ましくは、そのインサイチュ環境下でそのペプチドに通常会合する物質を含まない。
【0081】
「PanDR結合ペプチド」は、1つより多くのHLAクラスIIDR分子(例えば、PADRETMペプチド、Epimmune Inc.、San Diego、CA)を結合する分子ファミリーのメンバーである。PADRETMファミリーの分子を定義するパターンは、HLAクラスIIスーパーモチーフとして考えられ得る。PADRETM分子に見い出されたパターンを含むペプチドは、大部分HLA−DR分子に結合し、インビトロおよびインビボでヒトヘルパーTリンパ球(HTL)応答を刺激する。
【0082】
「薬学的に受容可能な」は、一般に、非毒性の、不活性な、および/または生理学的に適合性の組成物をいう。
【0083】
(3.本発明のペプチド)
本発明に従うペプチドは、組み換えDNA技術または化学合成によるか、あるいは天然腫瘍あるいは病原性生物のような天然源から合成的に調製されうる。ペプチドエピトープは、個々にまたはポリエピトープペプチドとして合成され得る。ペプチドは、好ましくは、他の天然に存在する宿主細胞のタンパク質およびそのフラグメントを実質的に有しないが、いくつかの実施形態において、ペプチドは、天然フラグメントまたは粒子に合成的に結合体化され得る。
【0084】
HLAクラスIペプチドは、当該分野において周知であり、MHCクラスI分子に結合するペプチドとして定義される。本発明に従うペプチドは、様々な長さであり、そしてそれらの中性(非荷電)形態または塩である形態のいずれかであり得る。本発明に従うペプチドは、グリコシル化、側鎖酸化、もしくはリン酸化反応のような修飾を有しないか、またはそれらは修飾が本明細書中に記載されるようなペプチドの生物学的活性を破壊しない条件を前提としてこれらの修飾を含むかのいずれかである。
【0085】
対応する「野生型」として役目を果たすクラスIエピトープは、任意のタンパク質源に由来し得る。例えば、クラスIペプチドは、ウイルス性抗原、腫瘍付随抗原、寄生物抗原、細菌抗原、または真菌抗原由来であり得る。本発明のいくつかの好ましい局面において、クラスIペプチドは、被験体の免疫系が寛容(すなわち先の抗原に対する曝露により引き起こされる特異的免疫学的非応答性)を発現する抗原に由来する。
【0086】
従って、いくつもの潜在的標的エピトープに基するヘテロクリティックアナログは、本発明にて利用され得る。適合した腫瘍付随抗原の例としては、前立腺特異的抗原(PSA)、黒色腫抗原MAGE1、MAGE2、MAGE3、MAGE−11、MAGE−A10、並びにBAGE、GAGE、RAGE、MAGE−C1、LAGE−1、CAG−3、DAM、MUC1、MUC2、MUC18、NY−ESO−1、CDK4、BRCA2、NY−LU−1、NY−LU−7、NY−LU−12、CASP8、RAS、KIAA−2−5、SCCs、p53、p73、CEA、Her2/neu、Melan−A、gp100、チロシナーゼ、TRP2、gp75/TRP1、カリクレイン、前立腺特異的膜抗原(PSM)、前立腺酸性ホスファターゼ(PAP)、前立腺特異的抗原(PSA)、PT1−1、B−カテニン、PRAME、テロメラーゼ、FAK、サイクリンD1タンパク質、NOEY2、EGF−R、SART−1、CAPB、HPVE7、p15、葉酸レセプターCDC27、PAGE−1、そしてPAGE−4が挙げられる。適合した感染症付随抗原の例としては、B型肝炎コア抗原および表面抗原(HBVc、HBVs)、C型肝炎抗原、エプスタイン−バーウイルス抗原、ヒト免疫不全症ウイルス(HIV)抗原、そしてヒトパピローマウイルス(HPV)抗原、Mycobacterium tuberculosisおよびChlamydiaが挙げられる。適合する真菌抗原の例としては、Candida albicans、Cryptococcus neoformans、Coccidoides spp.、Histoplasma spp.およびAspergillus fumigatisに由来する抗原が挙げられる。適合する原性動物の寄生物抗原の例としては、P. falciparum、Trypanosoma spp.、Schistosoma spp.、Leishmania sppなどを含む、Plasmodium spp.由来のものが挙げられる。
【0087】
対応するヘテロクリティックアナログを構築するために本発明の法則が適用される野生型の配列として用いられ得るエピトープは、任意のクラスIエピトープに対応することが見出され得る。上に記載されるもののような任意の抗原に対して、特定のクラスI対立遺伝子に付随するモチーフは、このようなエピトープが備わっている抗原のアミノ酸配列中の位置を決定する指針として利用され得る。この決定は、視覚的にかまたは好ましくはコンピューター技術および関連のソフトウェアを使用して行われ得る。従って、例えば2位のバリンおよびC末端のアルギニンを含むような、例えばA3スーパーモチーフの認識により、要求される任意の抗原のアミノ酸配列は、このモチーフを保有するエピトープについて調査され得る。次いで、そのエピトープは、要求されるアナログを獲得するために本発明に記載される法則に従って修飾され得る。
【0088】
可能ならば、本発明のHLAクラスI結合エピトープを最適化することが所望され得、例えばこれは、約8から約13、しばしば8から11、好ましくは9から10アミノ酸残基長に対するポリエピトープの構築に利用され得る。好ましくは、ペプチドエピトープは、内在的に処理される関連するHLA分子に対して結合される病原体由来ペプチドまたは腫瘍細胞ペプチドとサイズにおいて同一基準であるが、本発明のエピトープを含むペプチドの同定および調製は、また、本明細書に記載された技術を利用して行われ得る。
【0089】
代替の実施形態において、本発明のエピトープは、ポリエピトープペプチドとして、または、ポリエピトープペプチドをコードするミニ遺伝子として関連づけられ得る。
【0090】
別の実施形態において、高濃度のクラスIエピトープおよび/またはクラスIIエピトープを含む天然のペプチド領域を同定することが好ましい。そのような配列は、1つのアミノ酸長に対して最大のエピトープ数を含むことを基として通常選択される。エピトープが、ネスティングされるか、もしくは重なる様式で存在し得ることは、評価されることであり、例えば、10アミノ酸長ペプチドは、2つの9アミノ酸長エピトープならびに1つの10アミノ酸長エピトープを含み得た;細胞内のプロセシングにおいて、それぞれのエピトープは、そのようなペプチドの投与の際にHLA分子により曝露および結合されうる。このより大きく、好ましいマルチ−エピトープのペプチドは、合成的、組換え、あるいは天然素材からの切断を経て生じ得る。
【0091】
本発明のペプチドは、広く多様な方法から調製され得る。好ましい比較的短いサイズに関しては、ペプチドは、従来の技術に従って溶液中あるいは固体支持体上で合成され得る。種々の自動合成機は、市販されており、公知のプロトコールに従って使用され得る。(例えば、Stewart & Young、SOLID PHASE PEPTIDE SYNTHESIS、第2版、Pierce Chemical Co.、1984年を参照)。さらに、個々のペプチドエピトープは、いまだ本発明の範囲内であるより大きなペプチドを作製するために化学連結を使用して結合され得る。
【0092】
あるいは、目的の免疫原性ペプチドをコードするヌクレオチドの配列が、発現ベクターに挿入され、適切な宿主細胞に形質転換またはトランスフェクトされ、そして発現のために適切な条件下で培養される、組換えDNAの技術が利用され得る。これらの製法は、Sambrookら、Molecular Cloning、A LABORATORY MANUAL、Cold Spring Harbor Press、Cold Spring Harbor、 New York(1989)に一般に記載されるように、当該分野において一般的に公知である。従って、1つ以上の本発明のペプチド配列を含む組換えポリペプチドは、適切なT細胞エピトープを提示するために使用され得る。
【0093】
本明細書において企図される好ましい長さのペプチドエピトープに対応するヌクレオチドがコードする配列は、例えば、Matteucciら、J.Am.Chem.Soc.103:3185(1981)のホスホトリエステル法のような化学技術により合成され得る。ペプチドアナログは、天然ペプチド配列をコードする核酸塩基を、適切で、また要求される核酸塩基で置換することにより簡単に作製され得る;典型的な核酸の置換は、本明細書において、モチーフ/スーパーモチーフにより定義されるアミノ酸をコードするものである。次いで、コード配列は、適切なリンカーに供され、また当該分野において通例利用可能な発現ベクター中に連結され得、そしてそのベクターは、要求される融合タンパク質を作製するために適切な宿主を形質転換するために使用され得る。いくつものそのようなベクターおよび適切な宿主系は現在使用可能である。融合タンパク質の発現のために、コードする配列は、作動可能に連結される開始および終止コドン、プロモーターおよびターミネータ領域、ならびに通例要求される細胞の宿主での発現のための発現ベクターを提供する複製系を提供される。例えば、細菌の宿主との適合性を持つプロモーター配列は、要求されるコード配列の挿入のための便利な制限部位を含むプラスミドに提供される。得られる発現ベクターは、適切な細菌の宿主に形質転換される。もちろん、酵母、昆虫あるいは哺乳動物の細胞の宿主はまた、適切なベクター、ならびに制御配列を用いて使用され得る。
【0094】
本発明のアナログは、生じるペプチドの物理的特性(例えば、安定性または溶解性)を修飾するための置換を含むペプチドが含まれ得る。例えば、ペプチドはシステイン(C)のα−アミノ酪酸での置換によって修飾され得る。この化学的特質に起因して、システインはジスルフィド架橋を形成する傾向を有し、そして結合能力を減らすようにペプチド構造を十分に変化させる。Cの代わりにα−アミノ酪酸を用いることは、この問題を緩和させるだけでなく、実際に特定の例において結合および交差結合能力を改善する。システインをα−アミノ酪酸と置換することは、ペプチドエピトープの任意の残基で、すなわちアンカー位置または非アンカー位置のいずれかで起こり得る。
【0095】
種々のアミノ酸模倣物または非天然のアミノ酸を有する修飾されたペプチドは特に有用である。なぜならそれらはインビボにおいて増加した安定性を表す傾向があるからである。このようなアナログはまた、改善された貯蔵寿命または製造の特性を有し得る。より詳細には、重要ではないアミノ酸はタンパク質中の天然に存在するアミノ酸(例えばL−α−アミノ酸またはそれらのD−異性体)に制限される必要はないが、非天然のアミノ酸、例えばアミノ酸の模倣物(例えばD−ナフチルアラニン(naphylalanin)またはL−ナフチルアラニン;D−フェニルグリシンまたはL−フェニルグリシン;D−2−チエニルアラニン(thieneylalanine)またはL−2−チエニルアラニン;D−1、−2、3−もしくは4−ピレネイルアラニン(pyreneylalanin)またはL−1、−2、3−もしくは4−ピレネイルアラニン;D−3−チエニルアラニンまたはL−3チエニルアラニン;D−(2−ピリジニル)−アラニンまたはL−(2−ピリジニル)−アラニン;D−(3−ピリジニル)−アラニンまたはL−(3−ピリジニル)−アラニン;D−(2−ピラジニル)−アラニンまたはL−(2−ピラジニル)−アラニン;D−(4−イソプロピル)−フェニルグリシンまたはL−(4−イソプロピル)−フェニルグリシン;D−(トリフルオロメチル)−フェニルグリシン;D−(トリフルオロメチル)−フェニルアラニン;D−ρ−フルオロフェニルアラニン;D−ρ−ビフェニルフェニルアラニンまたはL−ρ−ビフェニルフェニルアラニン;D−ρ−メトキシビフェニルフェニルアラニンまたはL−ρ−メトキシビフェニルフェニルアラニン;D−インドール(アルキル)アラニンまたはL−2−インドール(アルキル)アラニンおよびD−2−アルキルアラニンまたはL−アルキルアラニン(ここでアルキル基はメチル、エチル、プロピル、ヘキシル、ブチル、ペンチル、イソプロピル、イソブチル、sec−イソチル、イソ−ペンチルまたは酸性ではないアミノ酸に置換され得るかまたは置換されない)が挙げられ得る。非天然のアミノ酸の芳香環として、例えばチアゾリル、チオフェニル、ピラゾリル、ベンジミダゾリル、ナフチル、フラニル、ピロリル、およびピリジル芳香環が挙げられる。
【0096】
ペプチドの安定性は多数の方法でアッセイされ得る。例えば、ペプチダーゼおよびヒト血漿および血清のような種々の生物学的培地は安定性を試験するために用いられている。例えば、Verhoefら、Eur.J.Drug Metab.Pharmacokinetics 11:291(1986)を参照のこと。本発明のペプチドの半減期は25%ヒト血清(v/v)アッセイを用いて簡便に決定される。そのプルトコールは一般的に以下のようである:プールしたヒト血清(AB型、熱で不活性化されていない)を使用する前に遠心分離により脱脂する。次いで血清をRPMI−1640または別の適切な組織培養培地で25%に希釈する。予め決定された時間間隔で、少量の反応溶液が取り出され、そして6%のトリクロロ酢酸水溶液(TCA)またはエタノールのいずれかを加えられる。濁った反応サンプルは4℃に15分間冷却され、その後スピンして沈殿した血清タンパク質をペレットにする。次いでペプチドの存在は安定性特異的なクロマトグラフィ条件を用いて逆相HPLCによって決定される。
【0097】
(4.クラスIモチーフ)
過去数年間で、HLAクラスI分子の巨大な割合が比較的少ないスーパータイプに分類され得、それぞれは広く重複するペプチド結合のレパートリーおよび主なペプチド結合ポケットのコンセンサス構造によって特徴付ける、ということを実証する証拠が集まってきている。従って本発明のペプチドは、いくつかのHLA特異的アミノ酸モチーフのうちのいずれか1つによってか(例えば表3〜4を参照のこと)、またはモチーフの存在がいくつかの対立遺伝子特異的HLA抗原に結合する能力に対応する場合はスーパーモチーフによって同定されている。特定のアミノ酸スーパーモチーフを有するペプチドに結合するHLA分子は、集合的にHLA「スーパータイプ」という。
【0098】
読者に便利にするため、以下に記載され表3〜4に要約されるペプチドのモチーフおよびスーパーモチーフは同定のための手引きおよび発明に従うペプチドエピトープの用途を提供する。これは、アナログを構築するための本明細書中に示される法則を適用するために、以下の実施例に示されるものとは異なる種々のクラスIモチーフに対応する候補野生型エピトープまたは以下に示されるものを保有するが異なる抗原の中にあるエピトープの同定を可能にする。
【0099】
ヘテロクリティックアナログは、ペプチドが属するモチーフまたはスーパーモチーフに関わり無く、そのペプチドから発明の方法に従って設計され得る。下記に描いたHLAクラスIペプチドエピトープのスーパーモチーフおよびモチーフの一次アンカー残基を表3に要約する。表3(a)で述べたHLAクラスIモチーフは、ここで主張される本発明に最も著しい関連のモチーフである。HLAクラスIスーパータイプファミリーを含む対立遺伝子特異的なHLA分子を表4に列挙する。いくつかの例において、ペプチドエピトープはモチーフおよびスーパーモチーフの両方に列挙され得る。特定のモチーフとそれぞれのスーパーモチーフとの関係は、個々のモチーフの説明中に示される。
【0100】
(i.HLA−A1スーパーモチーフ)
HLA−A1スーパーモチーフは、2位おける小さな(TまたはS)または疎水性(L、I、VまたはM)の一次アンカー残基のペプチドリガンド中の存在、そしてエピトープのC末端位置における芳香族(Y、FまたはW)の一次アンカー残基の存在により特徴付けられる。A1スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、HLA−A1スーパータイプ)は、少なくともA*0101、A*2601、A*2602、A*2501、およびA*3201を含む(例えば、DiBrino,M.ら、J.Immunol.151:5930,1993;DiBrino,M.ら、J.Immunol.152:620,1994;Kondo,A.ら、Immunogenetics 45:249,1997を参照のこと)。A1スーパーファミリーのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表4に示される。
【0101】
(ii.HLA−A2スーパーモチーフ)
対立遺伝子特異的HLA−A2.1分子に対する一次アンカーの特異性(例えば、Falkら、Nature 351:290−296,1991;Huntら、Science 255:1261−1263,1992;Parkerら、J.Immunol.149:3580−3587,1992;Ruppertら、Cell 74:929−937,1993を参照のこと)およびHLA−A2およびHLA−A28分子の間での交差反応性結合が記載される(例えば、関連データの総説については、Fruciら、Human Immunol.38:187−192,1993;Tanigakiら、Human Immunol.39:155−162,1994;Del Guercioら、J.Immunol.154:685−693,1995;Kastら、J.Immunol.152:3904−3912,1994を参照のこと)。これらの一次アンカー残基は、HLA−A2スーパーモチーフを規定し;ペプチドリガンド中の存在は、いくつかの異なるHLA−A2およびHLA−A28分子を結合する能力に対応する。HLA−A2スーパーモチーフは、2位に一次アンカー残基としてL、I、V、M、A、TまたはQを有するペプチドリガンド、およびエピトープのC末端位置に一次アンカー残基としてL、I、V、M、AまたはTを有するペプチドリガンドを含む。
【0102】
HLA分子の対応するファミリー(すなわち、これらのペプチドを結合するHLA−A2スーパータイプ)は、少なくとも以下を含む:A*0201、A*0202、A*0203、A*0204、A*0205、A*0206、A*0207、A*0209、A*0214、A*6802およびA*6901。A2スーパーファミリーのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表4に示される。
【0103】
(iii.HLA−A3スーパーモチーフ)
HLA−A3スーパーモチーフは、2位における一次アンカーとしてのA、L、I、V、M、SまたはTのペプチドリガンド中の存在、そしてエピトープのC末端位置における(例えば、9マーの9位に)正に荷電した残基RまたはKの存在により特徴付けられる(例えば、Sidneyら、Hum.Immunol.45:79,1996を参照のこと)。A3スーパーモチーフを結合するHLA分子の対応するファミリー(HLA−A3スーパータイプ)の例示的メンバーとしては、少なくとも:A*0301、A*1101、A*3101、A*3301およびA*6801が挙げられる。A3スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0104】
(iv.HLA−A24スーパーモチーフ)
HLA−A24スーパーモチーフは、2位における一次アンカーとして芳香族残基(F、W、またはY)または疎水性脂肪族残基(L、I、V、M、またはT)がペプチドリガンド中に存在し、そしてエピトープのC末端位置に一次アンカーとしてのY、F、W、L、IまたはMの存在により特徴付けられる(例えば、SetteおよびSidney,Immunogenetics(印刷中)1999を参照のこと)。A24スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、A24スーパータイプ)としては、少なくともA*2402、A*3001およびA*2301が挙げられる。A24スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0105】
(v.HLA−B7スーパーモチーフ)
HLA−B7スーパーモチーフは、一次アンカーとして2位にプロリンを有するペプチド、およびエピトープのC末端位置における一次アンカーとして疎水性アミノ酸または脂肪族アミノ酸(L、I、V、M、A、F、W、またはY)により特徴付けられる。B7スーパーモチーフを結合するHLA分子の対応するファミリー(すなわち、HLA−B7スーパータイプ)は、少なくとも26のHLA−Bタンパク質を含み、これらとしては以下が挙げられる:B*0702、B*0703、B*0704、B*0705、B*1508、B*3501、B*3502、B*3503、B*3504、B*3505、B*3506、B*3507、B*3508、B*5101、B*5102、B*5103、B*5104、B*5105、B*5301、B*5401、B*5501、B*5502、B*5601、B*5602、B*6701、およびB*7801(例えば、関連データの総説については、Sidneyら、J.Immunol.154:247,1995;Barberら、Curr.Biol.5:179,1995;Hillら、Nature 360:434,1992;Rammenseeら、Immunogenetics 41:178,1995を参照のこと)。B7スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0106】
(vi.HLA−B27スーパーモチーフ)
HLA−B27スーパーモチーフは、2位における一次アンカーとしての正に荷電した残基(R、H、またはK)のペプチドリガンド中の存在、そしてエピトープのC末端位置における一次アンカーとして疎水性残基(F、Y、L、W、M、I、A、またはV)の存在により特徴付けられる(例えば、SidneyおよびSette,Immunogenetics(印刷中)1999を参照のこと)。B27スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B27スーパータイプ)の例示的なメンバーとしては、少なくとも:B*1401、B*1402、B*1509、B*2702、B*2703、B*2704、B*2705、B*2706、B*3801、B*3901、B*3902、およびB*7301が挙げられる。B27スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0107】
(vii.HLA−B44スーパーモチーフ)
HLA−B44スーパーモチーフは、2位における一次アンカーとしての負に荷電した残基(DまたはE)のペプチドリガンド中の存在、そしてエピトープのC末端位置における一次アンカーとしての疎水性残基(F、W、Y、L、I、M、V、またはA)の存在により特徴付けられる(例えば、Sidneyら、Immunol.Today 17:261,1996参照のこと)。B44スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B44スーパータイプ)の例示的なメンバーとしては、少なくとも以下が挙げられる:B*1801、B*1802、B*3701、B*4001、B*4002、B*4006、B*4402、B*4403、およびB*4006。
【0108】
(viii.HLA−B58スーパーモチーフ)
HLA−B58スーパーモチーフは、2位での一次アンカー残基としての小さな脂肪族残基(A、S、またはT)のペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としての芳香族または疎水性の残基(F、W、Y、L、I、V、M、またはA)の存在によって、特徴付けられる(例えば、関連するデータの総説についてはSidneyおよびSette,Immunogenetics,印刷中、1999を参照のこと)。B58スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B58スーパータイプ)の例示的なメンバーとしては、少なくとも以下が挙げられる:B*1516、B*1517、B*5701、B*5702、およびB*5801。B58スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0109】
(ix.HLA−B62スーパーモチーフ)
HLA−B62スーパーモチーフは、2位での一次アンカーとしての極性脂肪族残基Qまたは疎水性脂肪族残基(L、V、M、I、またはP)のペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカーとして疎水性残基(F、W、Y、M、I、V、L、またはA)の存在によって、特徴付けられる(例えば、SidneyおよびSette,Immunogenetics,印刷中、1999を参照のこと)。B62スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B62スーパータイプ)の例示的なメンバーとしては、少なくとも以下が挙げられる:B*1501、B*1502、B*1513、およびB5201。B62スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0110】
(x.HLA−A1モチーフ)
HLA−A1モチーフは、2位での一次アンカー残基としてのT、S、またはMのペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としてのYの存在によって、特徴付けられる。代替の対立遺伝子特異的A1モチーフは、2位よりむしろ3位における、一次アンカー残基によって特徴付けられる。このモチーフは、D、E、A、またはSの一次アンカー残基としての3位における存在、および、エピトープのC末端位置における一次アンカー残基としてのYの存在によって、特徴付けられる(例えば、関連するデータの総説については、DiBrinoら、J.Immunol.,152:620、1994;Kondoら、Immunogenetics 45:249、1997;およびKuboら、J.Immunol.152:3913、1994を参照のこと)。
【0111】
(xi.HLA−A*0201モチーフ)
HLA−A2*0201モチーフは、2位での一次アンカー残基としての、LまたはMのペプチドリガンドにおける存在、およびLまたはVの9残基ペプチドのC末端位置での一次アンカー残基としての存在によって特徴付けられることが決定され(例えば、Falkら、Nature 351:290−296、1991を参照のこと)、そしてさらに、9アミノ酸ペプチドの2位にIを含み、そしてC末端位置にIまたはAを含むことが、見出された(例えば、Huntら、Science 255:1261−1263、3月6日、1992;Parkerら、J.Immunol.149:3580−3587、1992を参照のこと)。A*0201対立遺伝子特異的モチーフはまた、本発明者らによって、さらにV、A、T、またはQを、一次アンカー残基としてこのエピトープの2位に含み、そしてMまたはTを、一次アンカー残基としてこのエピトープのC末端位置に含むことが規定された(例えば、Kastら、J.Immunol.152:3904−3912、1994を参照のこと)。従って、HLA−A*0201モチーフは、2位に一次アンカー残基としてL、I、V、M、A、T、またはQを有し、そしてこのエピトープのC末端位置に一次アンカー残基としてL、I、V、M、A、またはTを有する、ペプチドリガンドを含む。HLA−A*0201モチーフの一次アンカー位置を特徴付ける、好ましい許容される残基は、A2スーパーモチーフを記載する残基と同一である。
【0112】
(xii.HLA−A3モチーフ)
HLA−A3モチーフは、2位での一次アンカー残基としてのL、M、V、I、S、A、T、F、C、G、またはDのペプチドリガンドにおける存在、およびエピトープのC末端位置における一次アンカー残基としてのK、Y、R、H、F、またはAの、存在によって、特徴付けられる(例えば、DiBrinoら、Proc.Natl.Acad.Sci.USA 90:1508、1993;およびKuboら、J.Immunol.152:3913−3924、1994を参照のこと)。
【0113】
(xiii.HLA−A11モチーフ)
HLA−A11モチーフは、2位での一次アンカー残基としてのV、T、M、L、I、S、A、G、N、C、D、またはFのペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としてのK、R、Y、またはHの存在によって、特徴付けられる(例えば、Zhangら、Proc.Natl.Acad.Sci.USA 90:2217−2221、1993;およびKuboら、J.Immunol.152:3913−3924、1994を参照のこと)。
【0114】
(xiv.HLA−A24モチーフ)
HLA−A24モチーフは、2位での一次アンカー残基としてのY、F、W、またはMのペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としてのF、L、I、またはWの存在によって、特徴付けられる(例えば、Kondoら、J.Immunol.155:4307−4312、1995;およびKuboら、J.Immunol.152:3913−3924、1994を参照のこと)。
【0115】
(5.T細胞応答を検出するためのアッセイ)
一旦、本発明のヘテロクリティックアナログが合成されると、それらは、T細胞応答を誘発する能力について試験され得る。モチーフ保有ペプチド(例えば、ヘテロクリティックアナログ)の調製および評価は、PCT公報WO94/20127およびWO94/03205に記載される。簡単に述べると、特定の抗原由来のエピトープを含むペプチドが合成され、適切なHLAタンパク質に結合するそれらの能力について試験される。これらのアッセイは、放射性ヨウ素標識した参照ペプチドの結合に関して、精製されるHLAクラスI分子への本発明のペプチドの結合を評価する工程を包含し得る。あるいは、空のクラスI分子を発現する細胞(すなわち、その中にペプチドを含まない)は、免疫蛍光染色およびフローマイクロ蛍光定量法によってペプチド結合について評価され得る。ペプチド結合を評価するために使用され得る他のアッセイは、ペプチド依存クラスIアセンブリアッセイおよび/またはペプチド競合によるCTL認識の阻害を含む。代表的に、500nM以下の親和性でクラスI分子に結合するそれらのペプチドは、感染された個体または免疫化された個体から誘導体化されたCTLに対する標的として作用する能力について、および、疾患に関連する選択された標的細胞と反応し得るCTL集団を引き起こし得る一次インビトロCTL応答または一次インビボCTL応答を誘発する能力について、さらに評価される。
【0116】
T細胞応答を検出するために使用される従来のアッセイとしては、増殖アッセイ、リンホカイン分泌アッセイ、直接細胞傷害性アッセイ、および限界希釈アッセイが挙げられる。このようなアッセイは、ヘテロクリティックアナログのペプチドによる免疫応答の誘導を、非ヘテロクリティックアナログのクラスIペプチド(例えば、ここからヘテロクリティックアナログ配列が基礎にされた)により誘導される応答と比較する際に有用である。例えば、ペプチドを用いてインキュベートされた抗原提示細胞は、応答細胞集団(responder cell population)においてCTL応答を誘導する能力についてアッセイされ得る。抗原提示細胞は、末梢血単核細胞または樹状細胞のような正常細胞であり得る。あるいは、内部的にプロセスされたペプチドを有するクラスI分子を装填するそれらの能力が欠乏し、そして適切なヒトクラスI遺伝子でトランスフェクトされた変異体非ヒト哺乳動物細胞株が、インビトロで一次CTL応答を誘発するペプチドの能力について試験するために使用され得る。
【0117】
末梢血単核細胞(PBMC)は、CTL前躯体の応答細胞源として使用され得る。適切な抗原提示細胞は、ペプチドと共にインキュベートされ、次いでこの後、ペプチド装填した抗原提示細胞が、最適な培養条件下で応答細胞集団と共にインキュベートされる。陽性CTL活性化は、放射性標識した標的細胞(特定のペプチドをパルスされた標的およびペプチド配列が誘導される内因的にプロセスされた形態の抗原を発現する標的細胞)を殺傷するCTLの存在について培養物をアッセイすることによって決定され得る。
【0118】
さらに、フルオレセイン標識したHLA四量体複合体で染色することによって抗原特異的T細胞の直接的定量を可能にする方法が、もたらされた(Altman,J.D.ら、Proc.Natl.Acad.Sci.USA 90:10330、1993;Altman,J.D.ら、Science 274:94,1996)。他の比較的最近の技術的開発としては、細胞内リンホカインの染色、およびインターフェロン−γ放出アッセイまたはElispotアッセイが挙げられる。四量体染色、細胞内リンホカイン染色およびElispotアッセイは全て、さらなる従来のアッセイより少なくとも10倍より感度が高いようである(Lalvani,Aら、J.Exp.Med.186:859、1997;Dunbar、P.R.ら、Curr.Biol.8:413、1998;Murali−Krishna,K.ら、Immunity 8:177、1998)。
【0119】
所望される場合、HTLの活性化はまた、当該分野で公知の技術(例えば、T細胞増殖およびリンホカイン(例えば、IL−2)の分泌)を使用して評価され得る(例えば、Alexanderら、Immunity 1:751〜761、1994を参照のこと)。
【0120】
あるいは、HLAトランスジェニックマウスの免疫化は、ペプチドエピトープの免疫原性を決定するために使用され得る。いくつかのトランスジェニックマウスモデル(ヒトA2.1、A11(これはHLA−A3エピトープを分析するためにさらに使用され得る)、およびB7対立遺伝子を有するマウスを含む)が特徴付けられ、そして他のマウス(例えば、HLA−A1およびA24に対するトランスジェニックマウス)が開発されている。HLA−DR1およびHLA−DR3マウスモデルもまた開発されている。他のHLA対立遺伝子を有するさらなるトランスジェニックマウスモデルが、必要な場合作製され得る。マウスは、不完全フロイトアジュバント中の乳化したペプチドで免疫化され得、そして得られるT細胞は、ペプチドをパルスされた標的細胞および適切な遺伝子でトランスフェクトされた標的細胞を認識するそれらの能力について試験される。CTL応答は、上記の細胞傷害性アッセイを使用して分析され得る。同様に、HTL応答は、T細胞増殖またはリンホカインの分泌のようなアッセイを使用して分析され得る。
【0121】
本発明のヘテロクリティックアナログは、しばしばTh1サイトカイン応答およびTh2サイトカイン応答の両方を誘導する。それゆえ、ヘテロクリティックな候補物を予め選択されたクラスIペプチドと比較するための1つの方法は、Th1サイトカインおよびTh2サイトカインの誘導を試験するための方法である。予め選抜したクラスIペプチドは、代表的にヘテロクリティックアナログに誘導されるペプチド、または、そのようなペプチドが存在しない場合は候補物に対して最も高い類似性を有するクラスIペプチドである。本発明のヘテロクリティックアナログは、代表的にTh1サイトカイン応答およびTh2サイトカイン応答の両方を誘導するが、これはアナログが誘導されたクラス1ペプチドに比較して非常に増強されたレベルにおいてである。例えば、所定のヘテロクリティックアナログは、等しいレベルのTh1サイトカインまたはTh2サイトカイン(50〜100pg/ml)を、このアナログが誘導された野生型ペプチドと比較して10倍以下の用量で刺激する。さらに、クラスIペプチドがTh1応答またはTh2応答のどちらかのみ、またはいずれかを主に誘導する場合、ヘテロクリティックアナログはTh1応答およびTh2応答の両方を誘導し得る。Th1サイトカインとしては、例えばIFNγ、IL−2およびIL3が挙げられる。Th2サイトカインとしては、例えばIL−4、IL−5、IL−6およびIL−10が挙げられる。サイトカインの産生は、例えばELISA法または他の免疫学的定量法を用いて測定される。(例えば、McKinneyら、Journal of Immunological Methods 237:105〜117、2000を参照のこと)
(6.診断薬としておよび免疫応答を評価するためのペプチドエピトープの使用)
本発明の一実施形態において、本明細書中に記載されるようなヘテロクリティックアナログペプチドが、免疫応答を評価するための試薬として使用される。評価される免疫応答は、免疫原として任意の因子を使用することによって、誘導され、この因子は、試薬として用いられるペプチドエピトープを認識し、そしてこれに結合する抗原特異性CTLまたはHTLの誘導を生成し得る。ペプチド試薬は、免疫原として使用される必要はない。このような分析のために使用され得るアッセイシステムとしては、比較的最近の技術開発(例えば、細胞内リンホカインに関する四量体染色およびインターフェロン放出アッセイ、またはElispotアッセイ)が挙げられる。
【0122】
例えば、本発明のペプチドは、腫瘍細胞抗原または免疫原の曝露後の抗原特異的CTLの存在について末梢血単核細胞を評価するために、四量体染色アッセイにおいて使用される。HLA四量体複合体を使用して、抗原特異性CTLを直接可視化し(例えば、Oggら、Science 279:2103〜2106、1998;およびAltmanら、Science 174:94〜96、1996を参照のこと)、そして末梢血単核細胞のサンプル中の抗原特異的CTL集団の頻度を決定する。本発明のペプチドを使用する四量体試薬は、以下のように生成され得る:HLA分子に結合するペプチドは、三分子複合体を生成するために、対応するHLA重鎖およびβ2−ミクログロブリンの存在下で再折り畳まれる。この複合体は、先にタンパク質中の操作された部位において重鎖のカルボキシル末端でビオチン化される。四量体形成は、次いで、ストレプトアビジンの付加によって誘導される。蛍光標識されたストレプトアビジンを用いて、抗原特異的細胞を染色するために四量体が使用され得る。これらの細胞は、次いで、例えば、フローサイトメトリーによって同定され得る。このような分析は、診断目的または予後目的のために使用され得る。この手順によって同定された細胞はまた、治療目的のために使用され得る。
【0123】
本発明のペプチドはまた、免疫リコール応答を評価するための試薬として使用される(例えば、Bertoniら、J.Clin.Invest.100:503〜513、1997、およびPennaら、J.Exp.Med.174:1565〜1570、1991を参照のこと)。例えば、癌を有する個体由来の患者のPBMCサンプルは、特定のペプチドを使用して、抗原特異的CTLまたはHTLの存在について分析され得る。単核細胞を含む血液サンプルは、PBMCを培養し、そして本発明のペプチドでその細胞を刺激することによって評価され得る。適切な培養期間の後、拡大された細胞集団は、例えば、CTLまたはHTL活性について分析され得る。
【0124】
ペプチドはまた、ワクチンの効力を評価するための試薬として使用される。免疫原をワクチン接種された患者から得たPBMCは、例えば、上記の方法のいずれかを使用して分析され得る。患者は、HLA型であり、そしてその患者に存在する対立遺伝子特異的分子を認識するペプチドエピトープ試薬が、分析のために選択される。ワクチンの免疫原性は、PBMCサンプル中のエピトープ特異的CTLおよび/またはHTLの存在によって示される。
【0125】
本発明のペプチドはまた、当該分野で周知の技術を使用して、抗体を作製するために使用され(例えば、CURRENT PROTOCOLS IN IMMUNOLOGY、Wiley/Greene、NY;およびAntibodies A Laboratory Manual 、Harlow and Lane、Cold Spring Harbor Laboratory Press、1989を参照のこと)、これは癌を診断またはモニターするための試薬として有用であり得る。このような抗体としては、HLA分子の状況においてペプチドを認識する抗体(すなわち、ペプチド−MHC複合体に結合する抗体)が挙げられる。
【0126】
(7.ワクチン組成物)
免疫学的に有効な量の本明細書中に述べるような1つ以上のペプチドを含むワクチンおよびそのワクチンを準備する方法は、本発明のさらなる実施形態である。一旦、適切な免疫原性エピトープが規定されると、それらは様々な手段により分類され送達され得るが、本明細書中ではこれらを「ワクチン」組成物と呼ぶ。このようなワクチン組成物は、例えば以下のようなものが挙げられる:リポペプチド(例えば、Vitiello、A.ら、J.Clin.Invest.95:341,1995)、ポリ(DL−ラクチド−co−グリコリド)(「PLG」)ミクロスフィアに封入されたペプチド組成物(例えば、Eldridgeら、Molec.Immunol.28:287−294,1991:Alonsoら、Vaccine 12:299−306,1994;Jonesら、Vaccine 13:675−681,1995)、免疫刺激複合体(ISCOMS)により包まれたペプチド組成物(例えば、Takahashiら、Nature, 344:873−875,1990;Huら、Clin Exp Immunol.113:235−243,1998)、多抗原ペプチドシステム(MAPs)(例えば、Tam,J.P.、Proc.Natl.Acad.Sci.USA 85:5409−5413,1988;Tam、J.P.,J. Immunol.Methods 196:17−32,1996)、多価のペプチドとして処方されるペプチド;銃式送達システム(ballistic delivery system)に処方されるペプチド、代表的には結晶化されたペプチド、ウイルス送達ベクター(Perkus,M.E.ら、In:Concepts in vaccine development,Kaufman,S.H.E.,編 p.379,1996;Chakrabarti,S.ら、Nature 320:535,1986、Hu,S.L.ら、Nature 320:537,1986;Kiney,M.−P.ら、AIDS Bio/Technology 4:790,1986;Top,F.H.ら、J.Infect.Dis. 124:148,1971;Chanda,P.K.ら、Virology 175;535,1990),ウイルス性もしくは合成起源の粒子(例えば、Kofler,N.ら、J.Immunol.Methods 192:25,1996;Eldridge、J.H.ら、Sem.Hematol.30:16,1993;Falo,L.D.,Jr.ら、Nature Med.7:649,1995)、アジュバント(Warren,H.S.,Vogel,F.R.および Chedid,L.A.Annu.Rev.Immunol.4:369,1986;Gupta,R.K.ら、Vaccine 11:293,1993)、リポソーム(Reddy,R.ら、J.Immunol.148:1585,1992;Rock,K.L.,Immunol.Today 17:131,1996)、あるいは裸または粒子吸着されたcDNA(Ulmer,J.B.ら、Science 259:1745,1993;Robinson,H.L.,Hunt,L.A.,およびWebster,R.G.,Vaccine 11:957,1993;Shiver,J.W.ら、Concepts in vaccine development, Kaufmann,S.H.E.編、p.423,1996;Cease, K.B.,およびBerzofsky,J.A.,Annu.Rev.Immunol.12:923,1994 および Eldridge, J. H. ら、Sem. Hematol.30:16,1993)。Avant Immunotherapeutics,Inc(Needham,Massachusetts)の技術のようなレセプター媒介標的化(recepter mediated targeting)として公知の毒素標的とする送達技術もまた使用され得る。
【0127】
本発明のワクチンは核酸が介在する様式を含む。一つ以上の本発明のペプチドをコードしているDNAまたはRNAはまた患者に投与され得る。このアプローチは、例えば、Wolff ら、Science 247:1465 (1990)ならびに米国特許第5,580,859号;同第5,589,466号;同第5,804,566号;同第5,739,118号;同第5,736,524号;同第5,679,647号;WO 98/04720に記載され、さらに詳細は以下に記載される。DNAに基づく送達技術の例としては、「裸のDNA」、促進(ブピバカイン、ポリマー、ペプチド介在)送達、陽イオン性液体複合体、粒子媒介(「遺伝子銃」)もしくは圧力媒介送達(例えば米国特許第5,922,687号を参照のこと)が挙げられる。
【0128】
治療的または予防的免疫処置の目的で、本発明のペプチドはまた、ウイルスベクターまたは細菌ベクターによって発現され得る。発現ベクターの例としては、痘疹や鶏痘などの弱毒化したウイルス宿主が挙げられる。このアプローチの例のように、ワクシニアウイルスは本発明のペプチドをコードするヌクレオチド配列を発現するベクターとして使用される。腫瘍を保有する宿主に導入する際、組換え型ワクシニアウイルスは免疫原性のペプチドを発現し、それによってCTLおよび/またはHTLを誘発する。ワクシニアベクターおよび免疫処置プロコトルにおいて有用な方法は、例えば米国特許第4,722,848号に記載されている。別のベクターは、BCG(Bacille Calmette Guerin)である。BCGベクターについてはStover ら、Nature 351:456−460(1991)に記載されている。アデノウイルスベクターおよびアデノ随伴ウイルスベクター、レトロウイルスベクター、Salmonella typhiベクター、無毒化した炭疽毒素ベクターなど発明のペプチドの治療的投与または免疫処置に有用な幅広い種々の他のベクターは、本明細書中の記載から当業者にとっては明白である。
【0129】
さらに、本発明に従ったワクチンは、一つ以上の特許請求されるペプチドの組成物を包含する。ペプチドは個々にワクチン中に存在し得る。あるいは、このペプチドは同一のペプチドの複数のコピーから構成されるホモポリマーまたは様々なペプチドのヘテロポリマーとして存在し得る。ポリマーは免疫反応を増大するという利点を有し、そしてこのポリマーをつくるために異種のペプチドエピトープが使われる場合、免疫応答の標的となる病原体生物または腫瘍関連ペプチドの異なる抗原性決定因子と反応する抗体および/またはCTLを誘導する更なる能力という利点を持つ。本組成物は抗原の天然に存在する領域であり得るか、あるいは例えば組換えまたは化学合成によって調製され得る。
【0130】
本発明のワクチンとともに使われる担体は当該分野において周知であり、例えば、サイログロブリン、ヒト血清アルブミンのようなアルブミン、破傷風トキソイド、ポリL−ロイシン、ポリL−グルタミン酸のようなポリアミノ酸、インフルエンザ、B型肝炎ウイルスコアタンパク質などが挙げられる。本ワクチンは水または食塩水、好ましくはリン酸緩衝化食塩水のような生理学的に耐性な(すなわち、受容されうる)希釈剤を含み得る。本ワクチンは、代表的なものにまたアジュバントを含む。フロイントの不完全アジュバント、リン酸アルミニウム、水酸化アルミニウム、またはミョウバンのようなアジュバントは当該分野で周知の材料例である。加えて、本明細書中で開示したように、CTL応答はトリパルミトイル−S−グリセリルシステイニルセリル−セリン(P3CSS)のような本発明のペプチドの脂質への結合によって開始され得る。
【0131】
本発明に従ってペプチド組成物を用いて免疫処置を行う際、注入的、エアロゾル的、経口的、経皮的、経粘膜的、胸膜内腔的、クモ膜下腔的、またはその他の適切な経路を介して、宿主の免疫系は、所望の抗原に特異的な多量のCTLおよび/またはHTLを産生することにより本ワクチンに応答する。結果として、宿主はその後の感染に対して少なくとも部分的に免疫されるか、または現在進行中の慢性的な感染が進展することに対して少なくとも部分的に耐性をもつか、あるいは抗原が腫瘍関連の場合は少なくとも治療的利益を導き出す。
【0132】
いくつかの実施形態において、本発明のヘテロクリティックアナログペプチドを、中和抗体およびまたはヘルパーT細胞の興味ある標的抗原に対する応答を誘導するか、あるいは促進する成分と結合することは望ましいことであり得る。このような組成物の好ましい実施形態は本発明に従って、クラスIエピトープとクラスIIエピトープを包含する。このような組成物の代替の実施形態は、PADRETM(Epimmune,San Diego,CA)分子(例えば、米国特許第5,736,142号に記載されている)のようなpan−DR結合ペプチドに加え、本発明に従ったクラスIエピトープおよび/またはクラスIIエピトープを包含する。
【0133】
本発明のワクチンはまた、樹状細胞(DC)のような抗原提示細胞(APC)を本発明のペプチドを提供するための媒体として包含する。ワクチン組成物は樹状細胞の動員および採取につづいてインビトロでつくられ得、これによって樹状細胞の負荷がインビトロで生じる。例えば、樹状細胞は例えば、本発明に従ったミニ遺伝子(minigene)を伴ってトランスフェクションされるかまたはペプチドとともにパルスされる。それから、樹状細胞は、インビボで免疫応答を誘発するために患者に投与され得る。
【0134】
DNAまたはペプチドのいずれかに基づくワクチン組成物は、また樹状細胞の動員と組み合わせてインビボに投与され得、これによって樹状細胞の負荷がインビボで生じる。
【0135】
抗原性のペプチドは同様にエキソビボでCTLおよび/またはHTL応答を誘発するために用いられる。結果として生じるCTLまたはHTL細胞は、他の従来の治療形式では応答しないか、あるいは本発明の治療的なワクチンペプチドまたは核酸に応答しない患者の腫瘍を処置するのに使われ得る。エキソビボでのある特定の腫瘍関連する抗原に対するCTLまたはHTL応答は、患者または遺伝的に適合性のあるCTLもしくはHTL前駆細胞を樹状細胞のような抗原提示細胞の供給源と適切な免疫原性のペプチドとともに組織培養中でインキュベートすることにより誘導される。適切なインキュベーション時間(代表的には約7〜28日)経過後、この期間中この前駆細胞は活性化され効果細胞へと発展するが、この細胞は患者に再注入され、ここでこれらは特定の標的細胞(感染した細胞または腫瘍細胞)を破壊するか(CTL)またはその破壊の促進を行う(HTL)。トランスフェクションされた樹状細胞はまた抗原提示細胞として使われ得る。
【0136】
本発明のワクチン組成物は、またIL−2、IL−12、GM−CSFなどのような免疫アジュバントと組み合わせる使用法を含む、癌に対する他の処置と組み合わせて使用され得る。
【0137】
好ましくは、ワクチンの用途でポリエピトープ組成物においてアレイのエピトープを封入のために選択する場合に、またはワクチンに取り込むため、および/またはミニ遺伝子のように核酸にコードされるために別個のエピトープを選択する場合、次の原則が利用される。選択を行うために、以下の原則のそれぞれのバランスを保つことが好ましい。所定のワクチン組成物に組み込まれた複数のエピトープは、エピトープが誘導される天然の抗原の配列と隣接し得るが、しかしながらこれは隣接する必要はない。
【0138】
1)投与されると、腫瘍クリアランスに相関することが観察された免疫応答を模倣するエピトープが選択される。HLA I型について、これは少なくとも1つの腫瘍関連抗原(TTA)に由来する3〜4個のエピトープを含む。HLA II型について、類似の原理が採用される;再び、3〜4個のエピトープは、少なくとも1つのTTAから選択される(例えば、Rosenbergら、Science 278:1447〜1450を参照のこと)。一つのTAAからのエピトープは一つ以上のさらなるTAAからのエピトープとの組み合わせで使用され得、頻出発現TAA(frequently−expressed TAA)の様々な発現パターンを持つ腫瘍を標的とするワクチンを産生する。
【0139】
2)免疫原性と相関することが確立された必須結合親和性を有するエピトープが選択される:HLA I型は、500nM以下しばしば200nM以下のIC50を有し、II型については、1000nM以下のIC50を有する。
【0140】
3)十分なスーパーモチーフ保有ペプチドまたは対立遺伝子特異的モチーフ保有ペプチドの十分なアレイは、広い集団適用範囲を与えるように選択される。例えば、少なくとも80%の集団適用範囲を有することが好ましい。モンテカルロ解析(当該分野で公知の統計的評価)を使用して、集団適用範囲の広さまたは冗長性を評価し得る。
【0141】
4)癌関連性抗原からエピトープを選択する場合、患者は天然のエピトープに対して寛容性を発達し得ているため、アナログを選択することがしばしば有用である。感染疾患関連性抗原についてエピトープを選択する場合、天然のエピトープまたはアナログのエピトープのいずれかを選択することが好ましい。
【0142】
5)「入れ子状(nested)エピトープ」と称されるエピトープは特定の関連性を有する。少なくとも2つのエピトープが所定のペプチド配列において重複している場合、入れ子状エピトープが生じる。入れ子状ペプチド配列は、HLAクラスIとHLAクラスIIエピトープの両方を包含する。入れ子状エピトープを提供する場合、一般的な目的は、一配列あたりの最多数のエピトープを提供することである。従って、1つの局面は、ペプチド中のアミノ末端エピトープのアミノ末端およびカルボキシル末端エピトープのカルボキシル末端より長いペプチドを提供しないようにすることである。多重エピトープ性配列(例えば、入れ子状エピトープを含む配列)を提供する場合、一般的にこれが病理学的または他の有害な生物学的特性を有さないことを保証するために、この配列をスクリーニングすることが重要である。
【0143】
6)ポリエピトープタンパク質が作製される場合、またはミニ遺伝子を作製する場合、目的のエピトープを含む最小のペプチドを生成することが目的である。入れ子状エピトープを含むペプチドを選択する場合に用いられるものと同じではない場合、この原理は類似している。しかし、人工ポリエピトープペプチドを用いて、サイズ最小化の目的は、ポリエピトープタンパク質中のエピトープ間の任意のスペーサー配列を組み込む必要性に対してバランスを保たれる。スペーサーアミノ酸残基は、例えば、接合部エピトープ(免疫系により認識されるエピトープであって、標的抗原には存在せず、そしてエピトープの人工並列によってのみ作製される)を回避するため、またはエピトープ間の切断を促進し、それによりエピトープ提示を増大するために導入され得る。接合部エピトープは一般的に回避されるべきである。なぜなら、レシピエントは、非ネイティブのエピトープに対する免疫応答を生成し得るからである。「優性エピトープ」である接合部エピトープは特に関心をもたれる。優性エピトープは、他のエピトープに対する免疫応答が減少または抑制されるこのような強力な応答を導き得る。
【0144】
(8.ミニ遺伝子ワクチン)
複数のエピトープを同時に送達し得る多数の異なるアプローチが利用可能である。本発明のペプチドをコードする核酸は、本発明の特に有用な実施形態である。ミニ遺伝子に含まれるエピトープは、好ましくは、前の章に記載されるガイドラインに従って選択される。本発明のペプチドをコードする核酸を投与する好ましい手段は、本発明の1つまたは複数のエピトープを含むペプチドをコードするミニ遺伝子構築物(minigene construct)を使用する。
【0145】
多重エピトープ性ミニ遺伝子の使用は、以下に記載される(例えば、同時係属出願U.S.S.N.09/311,784;Ishioka ら、J.Immunol.162:3915−3925,1999;An,L.およびWhitton,J.L.,J.Virol.71:2292,1997;Thomson,S.A.ら、J.Immunol.157:822,1996;Whitton,J.L.ら、J.Virol.67:348,1993;Hanke,R ら、Vaccine 16:426,1998)。例えば、TAA、PADRETM汎用ヘルパーT細胞エピトープのようなpan_DR結合ペプチドおよび小胞体トランスロケイティングシグナル(endoplasmic reticulum−translocating signal)配列の複数の領域に由来する、スーパーモチーフおよび/またはモチーフを保有するエピトープ(例えば、PSA、PSM、PAP、およびhK2)をコードする多重エピトープ性DNAプラスミドが操作され得る。ワクチンはまた、他のTAA由来のエピトープを包含し得る。
【0146】
多重エピトープ性ミニ遺伝子の免疫原性をトランスジェニックマウスで試験して、この試験したエピトープに対するCTL誘導応答の大きさを評価し得る。さらに、インビボにおいてDNAでコードされたエピトープの免疫原性を、DNAプラスミドでトランスフェクションした標的細胞に対する特異的CTL株のインビトロ応答と相関させ得る。従って、これらの実験は、このミニ遺伝子が以下の両方に有用であることを示し得る:1)CTL応答を発生させること、および2)誘導CTLがコードされたエピトープを発現する細胞を認識すること。
【0147】
例えば、ヒトの細胞中での発現のために選択したエピトープ(ミニ遺伝子)をコードするDNA配列を産生するために、このエピトープのアミノ酸配列は、逆翻訳され得る。ヒトのコドン使用表を利用して、各アミノ酸に対するコドン選択を誘導し得る。これらのエピトープをコードするDNA配列は、直接隣接されてもよく、その結果、翻訳される場合、連続するポリペプチド配列を産生する。発現および/または免疫原性を最適にするために、さらなるエレメントがミニ遺伝子の設計に取り込まれ得る。逆翻訳され得、そしてミニ遺伝子配列に含まれ得るアミノ酸配列の例としては、以下が挙げられる:HLAクラスIエピトープ、HLAクラスIIエピトープ、ユビキチン結合シグナル配列、および/または小胞体標的シグナル。さらに、CTLおよびHTLエピトープのHLA提示は、合成(例えば、ポリ−アラニン)または天然に存在する、CTLエピトープまたはHTLエピトープに隣接する隣接配列(flanking sequences)を含むことによって改良され得;エピトープ(単数または複数)を含むこれらのより大きなペプチドは、本発明の範囲内である。
【0148】
ミニ遺伝子配列は、ミニ遺伝子のプラス鎖およびマイナス鎖をコードするオリゴヌクレオチドを構築することによってDNAに変換され得る。重複性オリゴヌクレオチド(30〜100の塩基長)を、周知の技術を使用する適切な条件下で、合成、リン酸化、精製およびアニールし得る。オリゴヌクレオチドの末端を、例えば、T4 DNAリガーゼを使用して結合し得る。この合成ミニ遺伝子(エピトープポリペプチドをコードする)を、所望の発現ベクターにクローニングし得る。
【0149】
当業者に周知の標準調節配列は、好ましくは、標的細胞での発現を確実にするためにベクター中に含まれる。いくつかのベクターエレメントが所望される:ミニ遺伝子の挿入のための下流クローニング部位を有するプロモーター;効果的な転写終結のためのポリアデニル化シグナル;E.coliの複製起点;およびE.coli 選択マーカー(例えば、アンピシリンまたはカナマイシン耐性)。多数のプロモーター(例えば、ヒトサイトメガロウイルス(hCMV)プロモーター)が、この目的のために使用され得る。他の適切なプロモーター配列について、例えば、米国特許第5,580,859号および同第5,589,466号を参照のこと。
【0150】
さらなるベクターの改変は、ミニ遺伝子の発現および免疫原性を最適化するために所望され得る。いくつかの場合において、イントロンは、効果的な遺伝子発現のために必要とされ、そして1つ以上の合成イントロンまたは天然に存在するイントロンは、ミニ遺伝子の転写領域に取り込まれ得る。ミニ遺伝子の発現を増加させるために、mRNA安定化配列(mRNA stabilization
sequence)および哺乳動物細胞中での複製のための配列の封入がまた考慮され得る。
【0151】
一旦、発現ベクターが選択されると、ミニ遺伝子は、プロモーターの下流のポリリンカー領域にクローニングされる。このプラスミドを、適切なE.coli株に形質転換し、そしてDNAを標準的な技術を使用して調製する。ミニ遺伝子の方向およびDNA配列、ならびにベクター中に含まれる他の全エレメントは、制限マップおよびDNA配列解析を使用して確認される。正確なプラスミドを有する細菌細胞は、マスター細胞バンク(master cell bank)および作業用細胞バンク(working cell bank)として保存され得る。
【0152】
さらに、免疫刺激配列(ISSまたはCpG)は、DNAワクチンの免疫原性の役割を果たすようである。これらの配列は、所望の場合、免疫原性を増強するために、ミニ遺伝子コード配列の外側のベクター中に含まれ得る。
【0153】
いくつかの実施形態において、ミニ遺伝子コードエピトープと第二タンパク質(免疫原性を増強または減少させるために含まれる)との両方の産生を可能にするbi−シストロン(bi−cistronic)発現ベクターを使用し得る。同時発現する場合、免疫応答を有利に増強し得るタンパク質またはポリペプチドの例としては、サイトカイン(例えば、IL−2、IL−12、GM−CSF)、サイトカイン誘導分子(例えば、LeIF)、同時刺激分子(costimulatory molecule)、またはHTL応答、pan−DR結合タンパク質(例えば、PADRETM,Epimmune,San Diego,CA)が挙げられる。ヘルパー(HTL)エピトープを細胞内標的シグナルに結合し、そして発現したCTLエピトープから別個に発現し得る。これにより、HTLエピトープをCTLエピトープとは異なる細胞画分に指向させることを可能にする。必要である場合、これにより、HTLエピトープのHLAクラスII経路へのより効果的な参加を可能にし、これによりHTL誘導を改良する。HTLまたはCTL誘導に対して、免疫抑制性の分子(例えば、TGF−β)の同時発現による免疫応答を特異的に減少させることは、特定の疾患において有用であり得る。
【0154】
治療量のプラスミドDNAは、例えば、E.coliでの発酵、続く精製により産生され得る。作業用細胞バンクからのアリコートを、増殖培地に接種するために使用し、周知の技術に従って、振盪フラスコまたはバイオリアクター中で飽和まで増殖させる。プラスミドDNAを、標準的なバイオ分離技術(例えば、QIAGEN,Inc.(Valencia,California)によって供給される固相陰イオン交換樹脂)を使用して精製し得る。必要とされる場合、スーパーコイルDNAを、ゲル電気泳動または他の方法を使用して開環形態および線状形態から単離し得る。
【0155】
精製したプラスミドDNAを、種々の処方物を使用して注入のために調製し得る。これらの最も単純なものは、滅菌リン酸緩衝化生理食塩水(PBS)中の凍結乾燥したDNAの再構成物である。「裸のDNA」として公知のこのアプローチは、現在、臨床試験において筋内(IM)投与のために使用されている。ミニ遺伝子のDNAワクチンの免疫療法的効果を最大にするために、精製したプラスミドDNAを処方するための代替方法が所望され得る。種々の方法が記載され、そして新規の技術が利用可能になり得る。カチオン性脂質、糖脂質、およびフソジェニック(fusogenic)リポソームがまた処方物中で使用され得る(例えば、WO93/24640;Mannino & Gould−Fogerite,BioTechnisques6(7):682(1988);米国特許第5,279,833号;WO91/06309号;およびFelgerら、Proc.Nat’l Acad.Sci.USA 84:7413(1987)を参照のこと)。さらに、保護的、相互作用的、非縮合(non−condensing)化合物(PINC)として集合的に言及されるペプチドおよび化合物はまた、精製されたプラスミドDNAに複合体化されて、安定性、筋内分散、または特定の器官もしくは細胞型への輸送のような変数に影響を与える。
【0156】
標的細胞の感作は、ミニ遺伝子コード化CTLエピトープの発現およびHLAクラスI提示のための機能的アッセイとして使用され得る。例えば、プラスミドDNAは、標準的CTLのクロム放出アッセイ(standard CTL chromium release assay)に対する標的として適切である哺乳動物細胞株に導入される。使用されるトランスフェクションの方法は、最終の処方物に依存する。エレクトロポレーションが「裸の」DNAのために使用され、一方、カチオン性脂質は、直接的なインビトロでのトランスフェクションを可能にする。緑色蛍光タンパク質(GFP)を発現するプラスミドは同時にトランスフェクションされて、蛍光活性化細胞分類(FACS)を使用してトランスフェクションされた細胞の濃縮を可能にし得る。次いで、これらの細胞は、クロミウム−51(51Cr)で標識され、エピトープ特異的CTL株のための標的細胞として使用される;51Cr放出によって検出された細胞溶解は、ミニ遺伝子コード化CTLエピトープの産生とミニ遺伝子コード化CTLエピトープのHLA提示との両方を示す。HTLエピトープの発現は、HTL活性を評価するためのアッセイを使用して、類似様式で評価され得る。
【0157】
インビボでの免疫原性は、ミニ遺伝子のDNA処方物の機能的試験についての第2のアプローチである。適切なヒトHLAタンパク質を発現するトランスジェニックマウスは、DNA産物で免疫される。投与の用量および経路は、処方物に依存する(例えば、PBS中でのDNAに対するIM、脂質複合体化DNAに対する腹腔内(IP))。免疫の21日後、脾細胞を収集し、そして試験される各エピトープをコードするペプチドの存在下で1週間、再刺激した。その後、CTLエフェクター細胞について、標準的な技術を使用してペプチド負荷した51Crで標識した標的細胞の細胞溶解についてアッセイを行う。ミニ遺伝子コード化エピトープに対応する、ペプチドエピトープで負荷したHLAによって感作された標的細胞の溶解は、CTLのインビボでの誘導についてDNAワクチン機能を証明する。HTLエピトープの免疫原性を、類似の様式でトランスジェニックマウスにおいて評価する。
【0158】
あるいは、例えば、注入または例えば米国特許第5,204,253号に記載されるような銃式送達によって、核酸を皮内に投与し得る。この技術を使用して、単独でDNAを含む粒子を投与する。さらなる代替の実施形態において、DNAは、粒子(例えば、金粒子)に接着され得る。
【0159】
ミニ遺伝子はまた、当該分野で他の周知の細菌性またはウイルス性の送達システムを使用して送達され得る。例えば本発明のエピトープをコードしている発現構築物はワクシニアのようなウイルスベクターに組み込まれ得る。
【0160】
(9. ヘルパーペプチドとCTLペプチドの組み合わせ)
本発明のペプチドを含むワクチンの組成物は、血清中における改良された半減期のような所望の性質を与えるため、または免疫原性を増大させるために改変され得る。
【0161】
例えば、Tヘルパー細胞の応答を誘導し得るエピトープを少なくとも1つ含むペプチドを結合させることにより、CTL活性を誘導するペプチドの能力は増大され得る。免疫原性を増大させるための、CTLエピトープと組み合わせたTヘルパーエピトープの使用の例が、例えば以下の同時係属中出願、U.S.S.N.08/820,360、U.S.S.N.08/197,484、およびU.S.S.N.08/464,234に記載されている。
【0162】
CTLペプチドはTヘルパーペプチドと直接結合し得るが、しばしばCTLエピトープ/HTLエピトープ結合体はスペーサー分子により連結される。スペーサーは、代表的に生理的条件下において、実質的に荷電を帯びていないアミノ酸またはアミノ酸ミメティックのような、比較的小さな中性の分子を含む。スペーサーは、代表的に、例えばAla、Gly、またはその他の中性の非極性アミノ酸および中性の極性アミノ酸の中性スペーサーから選択される。必要に応じて存在するスペーサーは同じ残基から構成される必要はなく、ヘテロオリゴマーまたはホモオリゴマーであり得ることと理解される。スペーサーが存在する場合、通常、スペーサーは少なくとも1つまたは2つの残基であるが、さらに通常には3〜6つの残基、時には10残基以上になる。CTLペプチドエピトープはTヘルパーペプチドエピトープと直接またはスペーサーを介してのどちらかで、CTLペプチドのアミノ末端またはカルボキシ末端のいずれかで結合され得る。免疫原性ペプチドまたはTヘルパーペプチドのいずれかのアミノ末端はアシル化され得る。
【0163】
特定の実施形態において、Tヘルパーペプチドは、集団の大部分に存在するTヘルパー細胞により認識されるものである。これは、多数の、ほとんどの、あるいは全てのHLA クラスII分子と結合するアミノ酸配列の選択によって達成され得る。これらのことは「ゆるいHLA制限」、または「乱雑な」Tヘルパー配列として、公知である。乱雑なペプチドの例としては、破傷風毒の位置830〜843(QYIKANSKFIGITE)、Plasmodium falciparum circumsporozoite(CS)タンパク質の位置378〜398(DIEKKIAKMEKASSVFNVVNS)およびStreptococcus 18kD タンパク質の位置116(GAVDSILGGVATYGAA)のような抗原由来の配列が挙げられる。その他の例としては、DR1−4−7スーパーモチーフ、もしくはDR3モチーフのいずれかを保有するペプチドが挙げられる。
【0164】
あるいは、ゆるいHLA制限様式において、天然では見出されないアミノ酸配列を使用して、Tヘルパーリンパ球を刺激し得る合成ペプチドを調製することが可能である(例えば、PCT刊行物WO95/07707を参照)。Pan−DR結合エピトープ(例えば、PADRETM、Epimmune,Inc.,San Diego,CA)と呼ばれるこれらの合成化合物はほとんどのHLA−DR(ヒトHLAクラスII)分子に、最も優先的に結合するよう設計されている。例えば、式:aKXVAAWTLKAAa(ここで、「X」はシクロヘキシルアラニン、フェニルアラニン、またはチロシンのいずれかであり、「a」はD−アラニンまたはL−アラニンのいずれかである)を有するpan−DR結合エピトープペプチドは、それらのHLA型に関わらずほとんどのHLA−DR対立遺伝子(allele)と結合すること、およびほとんどの個体由来のTヘルパーリンパ球の応答を刺激することが見出されている。Pan−DR結合エピトープの代替物は、全て「L」の天然アミノ酸を含み、エピトープをコードする核酸の形態で提供され得る。
【0165】
HTLペプチドエピトープはまた、それらの生物学的性質を変えるように修飾され得る。例えば、それらはD−アミノ酸を含むように修飾されて、それらのプロテアーゼ耐性を増大させ、そしてそれらの血清中半減期を延長させ得、または、それらを例えば、脂質、タンパク質、炭水化物などのような他の分子と結合させて、それらの生物学的活性を増大し得る。例えば、Tヘルパーペプチドは、アミノ末端またはカルボキシ末端のいずれかにおいて1以上のパルミチン酸鎖と結合され得る。
【0166】
(10.T細胞刺激(priming)因子とCTLペプチドの組み合わせ)
いくつかの実施形態において、少なくとも1つの、細胞傷害性Tリンパ球を刺激する成分が、本発明の薬学的組成物に含まれることが所望され得る。脂質は、インビボでウイルス抗原に対し、CTL刺激をなし得る因子として同定されている。例えば、パルミチン酸残基は、リジン残基のε−アミノ基およびα−アミノ基に結合され得、次いで、例えば1つ以上の連結した残基(例えばGly、Gly−Gly−、Ser、Ser−Serなど)を介して免疫原性ペプチドに結合され得る。次いで、脂質化ペプチドは、直接にミセルまたは微粒子のいずれかで投与され得るか、リポソームへ組み込まれ得るか、またはアジュバント(例えば不完全フロイントアジュバント)へ乳化され得る。好ましい免疫原性組成物は、Lysのε−アミノ基およびα−アミノ基と結合したパルミチン酸を含み、このLysは、例えばSer−Serのような結合を介して、免疫原性ペプチドのアミノ末端と結合している。
【0167】
CTL応答を刺激する脂質のもう1つの例として、E.coliのリポタンパク質、例えばトリパルミトイル−S−グリセリルシステイニルセリル−セリン(tripalmitoyl−S−glycerylcysteinlyseryl−serine:P3CSS)は、ある適切なペプチドに共有結合した場合に、ウイルス特異的CTLを刺激することに使用され得る(例えば、Deres,ら、Nature 342:561,1989を参照)。本発明のペプチドは、例えば、P3CSSに結合され得、標的抗原に対するCTL応答を特異的に刺激するため、個体に投与され得る。さらに、中和抗体の誘導もまた、P3CSS結合エピトープにより刺激され得るため、2つのそのような組成物は、組み合わされて、体液性および細胞媒介性応答の両方をさらに効果的に誘発し得る。
【0168】
CTLおよび/あるいはHTLペプチドは、ペプチドの末端へのアミノ酸の付加によっても修飾され、あるペプチドをもう1つのペプチドへ結合することの容易さ、キャリア支持体またはより大きなペプチドへの結合、ペプチド、オリゴペプチドなどの物理学的または化学的特性の修飾を提供し得る。アミノ酸(例えばチロシン、システイン、リジン、グルタミン酸、アスパラギン酸など)は、ペプチドまたはオリゴペプチド、特にクラスIペプチドのC−末端またはN−末端に導入され得る。しかしながら、CTLエピトープのカルボキシ末端においての修飾が、いくつかの場合において、ペプチドへの結合特性を変化させ得ることは注意されるべきである。さらに、ペプチド配列またはオリゴペプチド配列は末端のNH2アシル化(例えば、アルカノイル(C1〜C20)またはチオグリコリルアセチル化)による、末端カルボキシルアミド化(例えば、アンモニア、メチルアミンなど)によって修飾されることによって天然の配列と異なり得る。いくつかの例において、これらの修飾は、支持体または他の分子との結合のための部位を提供し得る。
【0169】
(11.CTLおよび/またはHTLペプチドでパルス化(pulsed)されたDCを含むワクチン組成物)
本発明に従うワクチン組成物の実施形態は、患者の血液由来のPBMCへのエピトープ保有ペプチドのカクテル、またはPBMCから単離したDCへのエピトープ保有ペプチドのカクテルのエキソビボ投与を包含する。DCの採取を容易にする薬(例えばProgenipoietinTM(Monsanto,St.Louis,MO)またはGM−CSF/IL−4)が使用され得る。ペプチドでDCをパルス化した後でかつ、患者に再注入する前に、DCは結合していないペプチドを除くため洗浄される。この実施形態において、ワクチンは、HLA分子とその表面で複合体になった、パルス化されたペプチドエピトープを提示するペプチドパルス化DCを包含する。
【0170】
DCは、エキソビボでペプチドカクテル(それらのうちのいくつかは、目的の1つ以上の抗原に対するCTL応答を刺激する)でパルス化され得る。必要に応じて、ヘルパーT細胞ペプチド(例えば、PADRETMファミリー分子)は、CTL応答を容易にするするため、含有され得る。
【0171】
(12.治療目的または予防目的のためのワクチンの投与)
本発明のペプチド、本発明の薬学的組成物およびワクチン組成物は、代表的に癌の処置のために治療的に使用される。本発明のペプチドを含有したワクチン組成物は、代表的に、1つ以上の抗原の発現に関連した悪影響を有する癌患者へ投与される。あるいは、ワクチン組成物は、癌の進行に対して影響を受けやすい、さもなくば癌の進行の危険にある個人に対し投与され得る。
【0172】
治療適用において、ペプチド組成物および/もしくは核酸組成物は、腫瘍抗原に対する効果的なCTL応答および/もしくはHTL応答を誘発するため、そして症状および/もしくは合併症を治癒するか、少なくとも部分的に止める、もしくは遅らせるために十分な量で患者に投与される。これを満たすのに十分な量は、「治療的有効用量」と定義される。この使用に十分な量は、例えば、投与される特定の組成物、投与方法、治療される疾患の段階および重篤度、患者の体重および全般的な健康状態、ならびに処方医の判断に依存する。
【0173】
上に記載したように、本発明のCTLエピトープおよび/もしくはHTLエピトープを含むペプチドは、CTLまたはHTL特異的にHLA分子によって提示される場合、およびこのペプチドに含まれるエピトープに特異的なCTLまたはHTLと接触される場合に、免疫応答を誘導する。ペプチド(またはそれらをコードするDNA)は、個別に、または1つ以上のペプチド配列の融合物として投与され得る。ペプチドがCTLまたはHTLと接触される方法は、本発明には重要でない。例えば、ペプチドは、インビボもしくはインビトロのいずれかで、CTLまたはHTLと接触し得る。接触が、インビボで起きる場合、ペプチド自体が患者に投与され得るか、または他のビーイクル(例えば、1つ以上のペプチドをコードしたDNAベクター、ペプチドをコードしたウイルスベクター、リポソームなど)が、本明細書中に記載されるように使用され得る。
【0174】
ペプチドが、インビトロで接触する場合、ワクチン因子は、細胞の集団(例えば、ペプチドパルス化樹状細胞)、または抗原提示細胞をペプチドでパルス化すること、もしくは本発明品のミニ遺伝子を抗原提示細胞にトランスフェクトすることによって誘導されたTAA特異的CTLを含有し得る。そのような細胞集団は、続いて、治療的有効用量で患者に投与される。
【0175】
治療に用いるために、投与は、一般的に、最初の癌の診断の時点で始まるべきである。これに続いて、少なくとも実質的に症状がやわらげられるまで、そしてその後ある期間にわたって追加抗原投与量がブーストされる。患者に送達されるワクチン組成物の実施形態(すなわち、例えばペプチドカクテル、ポリエピトープ性ポリペプチド、ミニ遺伝子、またはTAA特異的CTL、またはパルス化された樹状突起細胞のような実施形態を含むが、これらに限定されない)は、疾患の段階、あるいは患者の健康状態によって変り得る。例えば、TAA特異的CTLを含むワクチンは、進行した疾患を有する患者中において、腫瘍細胞を殺傷する際に、代変の実施形態より効果的であり得る。
【0176】
本発明のワクチン組成物はまた、外科手術などの処置と組み合わせても、治療的に用いられ得る。1つの例は、患者が、原発性腫瘍を取り除くための手術を受けており、次いで、ワクチンが、再発および/もしくは転移を、遅らせるか、または予防するために使用される状況である。
【0177】
感受性の個体(例えば、前立腺腫瘍を発病するような遺伝的素因があると診断され得る個体)が、癌の診断に先立って特定される場合、組成物が、彼らに対して標的化され得、したがってより大きな集団への投与の必要性を最小にす得る。
【0178】
最初の、治療的免疫のための用量決定は、一般的に、下限の値が、約1、5、50、500、または1,000μgであり、そして上限の値が、約10,000;20,000;30,000;または50,000μgの単位投薬範囲にある。ヒトについての投薬量の値は、典型的に、70キログラムの患者あたり約500μg〜約50,000μgの範囲である。確立された間隔(例えば、4週間から6ヶ月)で、ブースト投薬前の、最初の投薬は、患者を効果的に処置するため、ことによると長期の時間の間、必要とされ得る。ブーストレジメンに従って、数週間から数ヶ月にわたる、ペプチドの約1.0μg〜約50,000μgの間のブースト投薬は、患者の血液から得られるCTLおよびHTLの特異的活性を測定することによって決定される患者の応答および状態に依存して投与され得る。
【0179】
少なくとも臨床的症状または実験的試験が、腫瘍が除去されたかまたは腫瘍細胞荷重が実質的に減少したことを示すまでそしてその後のある期間の間、投薬が続けられるべきである。投薬、投与の経路、および投薬スケジュールは、当該分野で公知の方法論に従って調節される。
【0180】
ある実施形態において、本発明のペプチドおよび組成物は、重症な疾患状態、すなわち、生命を危うくする状況または潜在的に生命を危うくする状況で使用される。このような場合において、最小量の外来の物質および本発明の好ましい組成物におけるペプチドの比較的非毒性の性質の結果として、これらの記述された投薬量に対して実質的に過剰のこれらのペプチド組成物を投与することが、可能であり、そして処置する医師によって望ましいと感じられ得る。
【0181】
本発明のワクチン組成物は、予防因子としても使用され得る。例えば、組成物は、前立腺癌進行の危険性を有する個体に投与され得る。一般的に、最初の予防的免疫のための一般的な投与量は、下限の値が約1、5、50、500、または1000μgであり、そして上限の値が約10,000;20,000;30,000;または50,000μgである単位投薬範囲にある。ヒトについての投薬量の値は、典型的に、70キログラムの患者あたり約500μg〜約50,000μgの範囲である。これに続いて、最初のワクチン投与後、約4週間〜約6ヶ月の規定された間隔で投与される、約1.0μg〜約50,000μgの間のペプチドがブースト投薬される。ワクチンの免疫原性は、患者の血液のサンプルから得られたCTLおよびHTLの特異的活性を測定することによって評価され得る。
【0182】
治療的処置のための薬学的組成物は、非経口的投与、局所的投与、経口投与、くも膜下投与、または局所投与に意図される。好ましくは、薬学的組成物は、非経口的に、例えば、静脈内に、皮下に、皮内に、または筋内に投与される。従って、本発明は、受容可能なキャリア(好ましくは、水性キャリア)に溶解されるかまたは懸濁される免疫原性ペプチドの溶液を含む非経口投与のための組成物を提供する。種々の水性キャリアは、例えば、水、緩衝化水、0.8%生理食塩水、0.3%グリシン、ヒアルロン酸などが使用され得る。これらの組成物は、従来の周知の滅菌化技術によって滅菌化され得るか、または滅菌濾過され得る。得られる水溶液は、そのままでの使用のためにパッケージングされ得るか、または凍結乾燥され得、この凍結乾燥された調製物は、投与の前に滅菌溶液と組み合わせられる。この組成物は、およそ生理学的条件に必要とされる薬学的に受容可能な補助物質、例えば、pH調節剤、緩衝化剤、張性調節剤、湿潤剤、保存剤など(例えば、酢酸ナトリウム、乳酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、ソルビタンモノラウレート、オレイン酸トリエタノールアミンなど)を含み得る。
【0183】
薬学的処方物中の本発明のペプチドの濃度は、幅広く、すなわち、約0.1重量%未満、通常、約2重量%または少なくとも約2重量%から、20重量%〜50重量%以上の多さまでで変化し得、主に、選択される投与の特定の様式に従って、流体容積、粘度などによって選択される。
【0184】
ペプチド組成物のヒト単位用量形態は、代表的に、ヒト単位用量の受容可能なキャリア(好ましくは、水性キャリア)を含む薬学的組成物中に含まれ、そしてこのような組成物のヒトへの投与に使用されるための当業者によって公知の流体の容積で投与される(例えば、Remington’s Pharmaceutical Sciences、17th Edition、A.Gennaro、Editor、Mack Publishing Co.、Easton、Pennsylvania、1985を参照)。
【0185】
本発明のペプチドはまた、リポソームを介して投与され得、リポソームは、ペプチドを特定の組織(例えば、リンパ組織)に標的化させるために、または感染した細胞に選択的に標的化させるために、そしてペプチド組成物の半減期を増加させるために役立つ。リポソームには、エマルジョン、泡状物、ミセル、不溶性単層、液晶、リン脂質分散物、層状層などが挙げられる。この調製物において、送達されるべきペプチドは、リポソームの一部として、単独であるいはリンパ系細胞に広く行き渡るレセプターに結合する分子(例えば、CD45抗原に結合するモノクローナル抗体)とともに、または他の治療組成物もしくは免疫原性組成物とともに組み込まれる。従って、本発明の所望のペプチドで負荷されるかまたは装飾(decorated)されるかのいずれかのリポソームが、リンパ系細胞の部位に指向され得、ここで、そのときリポソームがペプチド組成物を送達する。本発明に従う使用のためのリポソームは、標準的な小胞形成脂質から形成され、これには、一般的に、中性および陰性に荷電されたリン脂質およびステロール(例えば、コレステロール)が挙げられる。脂質の選択は、一般的に、例えば、リポソームサイズ、酸不安定性および血流中のリポソームの安定性を考慮してガイドされる。種々の方法が、リポソームを調製するために利用可能であり、例えば、Szokaら、Ann.Rev.Biophys.Bioeng.9:467(1980)および米国特許第4,235,871号、同第4,501,728号、同第4,837,028号、および同第5,019,369号に記載される。
【0186】
免疫系の細胞を標的化するために、リポソームに組み込まれるリガンドには、例えば、所望の免疫系細胞の細胞表面決定基に特異的な抗体またはそのフラグメントが挙げられ得る。ペプチドを含むリポソーム懸濁物は、静脈内、局所的(locally、topically)などで、特に投与の方法、送達されるペプチド、および処置される疾患の段階に従って変化する用量で投与され得る。
【0187】
固体組成物について、従来の非毒性固体キャリアが使用され得、これには、例えば、薬学的等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、タルク、セルロース、グルコース、スクロース、炭酸マグネシウムなどが挙げられる。経口投与のために、薬学的に受容可能な非毒性組成物は、任意の通常使用される賦形剤(例えば、先に列挙されたもの)および一般的に10〜95%の活性成分(すなわち、本発明の1つ以上のペプチド)、より好ましくは、25%〜75%の濃度で組み込むことによって形成される。
【0188】
エアロゾル投与のために、免疫原性ペプチドは、好ましくは、界面活性剤および噴霧剤とともに細かく分割された形態で供給される。ペプチドの代表的な割合は、0.01重量%〜20重量%、好ましくは、1重量%〜10重量%である。界面活性剤は、もちろん非毒性でなければならず、そして好ましくは噴霧剤に可溶である。このような薬剤の代表は、6〜22個の炭素原子を含む脂肪酸のエステルまたは部分エステル、例えば、カプロン酸、オクタン酸、ラウリン酸、パルミチン酸、ステアリン酸、リノール酸、リノレン酸、オレステリン酸(olesteric acid)およびオレイン酸と、脂肪族多水酸基アルコールまたはその環式無水物である。混合エステル(例えば、混合または中性グリセリド)が使用され得る。界面活性剤は、組成物の0.1重量%〜20重量%、好ましくは、0.25〜5重量%を構成し得る。組成物の残りは、通常、噴霧剤である。キャリアはまた、望ましいならば、例えば、鼻腔内送達のためのレシチンとともに含まれ得る。
【0189】
(13.キット)
本発明のペプチドおよび核酸組成物は、ワクチン投与のための指示書とともにキットの形態で提供され得る。代表的なキットは、容器内に、好ましくは、単位用量形態の所望のペプチド組成物および投与のための指示書を含む。代替的なキットは、投与のための指示書とともに、容器内に、好ましくは単位用量形態で、本発明の所望の核酸を伴うミニ遺伝子構築物を含む。IL−2またはIL−12のようなリンホカインもまた、キットに含まれ得る。所望であり得る他のキット構成要素には、例えば、滅菌シリンジ、ブースター投薬、および他の所望の賦形剤が挙げられる。
【0190】
本発明に従うエピトープは、免疫応答を誘導するために首尾良く使用された。これらのエピトープを用いる免疫応答は、種々の形態においてエピトープを投与することによって導入された。エピトープは、ペプチドとして、核酸として、そして本発明のエピトープをコードする核酸を含むウイルスベクターとして投与された。ペプチドに基づくエピトープ形態の投与において、免疫応答は、エピトープを細胞で発現される空のHLA分子上に直接負荷することによって、そしてエピトープのインターナリゼーションを介して、そしてHLA I型経路を介してプロセスすることによって導入された;いずれの場合においても、エピトープを発現するHLA分子は、次いで、相互作用し得、そしてCTL応答を誘導し得た。ペプチドは、直接的に、またはリポソームのような薬剤を使用して送達され得る。これらは、さらに、銃式送達(ballistic delivery)(ここで、ペプチドは、代表的には、結晶形態である)を使用して送達され得る。DNAが免疫応答を誘導するために使用される場合、裸のDNAとして(一般的に、約1〜5mgの投薬範囲)または銃式「遺伝子銃」送達を介して(代表的に、約10〜100μgの投薬範囲)のいずれかで投与される。DNAは、種々のコンホメーション(例えば、線状、環状など)で送達され得る。種々のウイルスベクターもまた、本発明に従ってエピトープをコードする核酸を含んで首尾良く使用されている。
【0191】
従って、本発明に従う組成物は、いくつかの形態で存在する。本発明に従うこれらの組成物形態のそれぞれの実施形態は、免疫応答を誘導するために首尾良く使用されている。
【0192】
本発明に従う1つの組成物は、複数のペプチドを含む。この複数またはカクテルのペプチドは、一般的に、1つ以上の薬学的に受容可能な賦形剤と混合される。ペプチドカクテルは、同じペプチドの複数のコピーを含み得るか、またはペプチドの混合物を含み得る。ペプチドは、天然のエピトープのアナログであり得る。ペプチドは、人工のアミノ酸および/または化学的改変(例えば、脂質化;アセチル化、グリコシル化、ビオチニル化、リン酸化のような表面活性分子の添加)を含み得る。ペプチドは、CTLまたはHTLエピトープであり得る。好ましい実施形態において、ペプチドカクテルは、複数の異なるCTLエピトープおよび少なくとも1つのHTLエピトープを含む。HTLエピトープは、天然または非天然(例えば、PADRE(登録商標)、Epimmune Inc.,San Diego、CA)であり得る。本発明の実施形態における異なるエピトープの数は、一般的に、1〜150の整数(whole unit integer)である(例えば、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、・・・、150)。
【0193】
本発明に従う組成物のさらなる実施形態は、ポリペプチド多重エピトープ性構築物(すなわち、ポリエピトープペプチド)を含む。本発明に従うポリエピトープペプチドは、当該分野において周知の技術の使用によって調製される。これらの公知の技術の使用によって、本発明に従うエピトープは、互いに接続される。ポリエピトープペプチドは、線状または非線状(例えば、多価)であり得る。これらのポリエピトープ構築物は、人工アミノ酸、スペーシングまたはスペーサーアミノ酸、隣接アミノ酸(flanking amino acids)、または隣接エピトープユニット間の化学改変を含み得る。ポリエピトープ構築物は、ヘテロポリマーまたはホモポリマーであり得る。ポリエピトープ構築物は、一般的に、2〜150の間の任意の整数の量(例えば、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、・・・、150)でエピトープを含む。ポリエピトープ構築物は、CTLおよび/またはHTLエピトープを含み得る。構築物中の1つ以上のエピトープは、例えば、表面活性物質(例えば、脂質)の付加によって改変され得るか、または化学的に改変され得る(例えば、アセチル化など)。さらに、多重エピトープ性構築物における結合は、ペプチド結合以外(例えば、共有結合、エステルまたはエーテル結合、ジスルフィド結合、水素結合、イオン結合など)であり得る。
【0194】
あるいは、本発明に従う組成物は、ネイティブな配列に対する相同性を有する(すなわち、対応するかまたは接触(contiguous)する)アミノ酸の系列、配列、ストレッチ(stretch)を含む構築物を含む。このアミノ酸のストレッチは、より長い系列のアミノ酸から切断されるかまたは単離される場合、本発明に従って、HLA I型エピトープまたはHLA II型エピトープとして機能するアミノ酸の少なくとも1つのサブ配列(subsequence)を含む。この実施形態において、ペプチド配列は、当該分野で公知であるかまたは提供される多数の技術を使用することによって、本明細書中に規定されるような構築物になるように改変される。ポリエピトープ構築物は、70〜100%(例えば、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、または100%)の任意の整数の増分でネイティブな配列に対して相同性を含み得る。
【0195】
本発明に従う組成物のさらなる実施形態は、本発明に従う1つ以上のエピトープを含む抗原提示細胞である。抗原提示細胞は、「専門的」抗原提示細胞(例えば、樹状細胞)であり得る。抗原提示細胞は、当該分野において公知の任意の手段または当該分野において決定された任意の手段によって本発明のエピトープを含み得る。このような手段には、樹状細胞を1つ以上の個々のエピトープまたは複数のエピトープを含む1つ以上のペプチドで、銃式核酸送達のような核酸投与によって、または核酸の投与について当該分野の他の技術(ベクターに基づく(例えば、ウイルスベクター)核酸の送達を含む)によって適用することを含む。
【0196】
本発明に従う組成物のさらなる実施形態は、本発明の1つ以上のペプチドをコードする核酸、または本発明に従うポリエピトープペプチドをコードする核酸を含む。当業者によって理解されるように、種々の核酸組成物が、遺伝子コードの冗長性に起因して同じペプチドをコードする。これらの核酸組成物のそれぞれは、本発明の範囲内に含まれる。本発明のこの実施形態は、DNAまたはRNAを含み、特定の実施形態において、DNAおよびRNAの組み合わせを含む。本発明に従うペプチドをコードする核酸を含む任意の組成物または本発明に従う任意の他のペプチドに基づく組成物が、本発明の範囲内であることが理解される。
【実施例】
【0197】
(実施例)
(調製A)
(ペプチド合成およびペプチドアナログの生成)
これらの実施例で用いられるペプチドを表1に示す。腫瘍関連抗原に由来している全ての野生型ヒトCTLエピトープならびにHIVおよびB型肝炎ウイルス(HBV2)のポリメラーゼ遺伝子に由来している野生型ウイルスエピトープは、ヒトおよびトランスジェニックマウスの系における免疫原性が示されている(Kawashima,I.ら、Human Immunol.(1998)59:1;Ishioka,G.ら、J.Immunol.(1999)162:3915)。
【0198】
ヘテロクリティックな活性について最初に試験されたペプチドは、Chiron Technologies(Victor、Australia)によって合成された。さらなる生物学的特徴付けを必要としているペプチドは、(Ruppert,J.ら、Cell(1993)74:929)の方法を用いてEpimmuneにて合成され、分析用逆相HPLCによって決定された場合に、これらのペプチドの純度は通常95%より高かった。後者のペプチドの正体を質量スペクトル分析によって確認した。
【0199】
(調製B)
(単一アミノ酸置換の選択についてのスキーム)
表2は、所定のアミノ酸置換を、保存的置換、半保存的置換、または非保存的置換であるとして特徴付けし得るように、任意の所定のアミノ酸対の間の類似性の割り当てを示す。
【0200】
アミノ酸対の間の類似性の程度を、各アミノ酸対について、PAM250に関する順位係数スコア(rank coefficient score)、疎水性度および以下に述べるような側鎖の体積を平均化することによって定量した。これらの複合した順位の平均値に基づいて、この表は各対が保存されているか、半保存されているか、または保存されていないかを示す。
【0201】
Dayhoff PAM250スコア(Dayhoff,M.O.ら、Atlas of Protein Sequence and Structure,第5巻,補遺3.(1978)M.O.Dayhoff編.National
Biomedical Research Foundation,Washington DC,345頁;Creighton,T.E.,Proteins:structures and molecular properties(1993)(第2版)W.H.Freeman and Company,NY;http://prowl.rockefeller.edu/aainfo/pam250.html)は、規定された時間枠の範囲内で受容可能な点変異(PAM)の百分率を測定する、一般的に利用されるタンパク質アラインメントスコア付けマトリクスである。これらの変異の頻度はランダム変異の確率から予想されるものとは異なり、恐らく、置換に関与するアミノ酸対の物理的および化学的類似性の程度に起因する偏向を反映する。類似性の他の尺度を用いて標準化され得るアミノ酸類似性のスコアを得るために、PAM250スコアを順位値(rank value)に変換した。ここで、1は受容された変異であるという最も高い確率を示している。
【0202】
20個の天然に存在するアミノ酸の相対的疎水性度を表すために、最もよく一般的に利用されるスケール(Cornette,J.ら、J.Mol.Biol(1987)195:659)は、KyteおよびDoolittle(Kyte,J.およびR.F.Doolittle,J.Mol.Biol.(1982)157:105)、ならびにFauchereおよびPliska(Fauchere,J.およびV.Pliska,Eur.J.Med.Chem.(1983)18:369)による実験データを基礎として発展したものである。Kyte/Doolittleスケールは個々のアミノ酸のH2O/有機溶媒分配を測定する。このスケールは折り畳まれたタンパク質のアミノ酸の位置を考慮するので、このスケールは、タンパク質の状況下におけるネイティブな疎水性度を最も正確に反映し得る。Fauchere/PliskaスケールはN−アセチルアミノ酸アミドのオクタノール/H2O分配を測定し、変性タンパク質および/または、低分子合成ペプチドの状況下における疎水性度を最も正確に反映する。疎水性度についてのスコアを得るために、各アミノ酸残基をKyte/DoolittleおよびFauchere/Pliska疎水性度スケールの両方において順位付けした。2つのスケール間の平均順位を計算し、各対についての疎水性度の差の平均を計算した。
【0203】
最後に、アミノ酸側鎖の体積を計算するために、1分子または1グラムのいずれかのアミノ酸残基を添加した後の水の体積の増加を留意することによって得られる溶液中の分体積を考慮した。(Zamyatnin,A.A.,Ann.Rev.Biophys.Bioeng.(1984)13:145;Zamyatnin,A.A.,Prog.Biophys.Mol.Biol.(1972)24:107)。20個の天然に存在するアミノ酸に関して可能な各対の分体積の絶対的差異を計算し、そして順位付けた。ここで、1は最も類似する体積を有する残基、そして20は最も異なる体積を有する残基を示した。
【0204】
(調製C)
(アッセイのための材料)
(1.APC株)
HLA−A2.1の状況下でぺプチドを提示する細胞株は以下のように調製された:
.221A2.1細胞株は、HLA−A2.1遺伝子をHLA−A、−B、−C−ヌル変異体EBV−形質転換ヒトB−リンパ芽球細胞株3A4−721.221(Kawashima,I.ら、Human Immunol.(1998)59:1)にトランスフェクションすることによって生成した。
【0205】
腫瘍細胞株はメチルコラントレン(methylcholanthrene)誘導性肉腫であるMeth A細胞のトランスフェクションによって調製した。そして、Jurkat細胞株ではHLA−A2.1またはHLA−A2.1/Kb導入遺伝子のトランスフェクションを、他(Vitiello,A.らJ.Exp.Med.(1991)173:1007)で述べた方法を用いて行った。HLA型黒色腫細胞株624mel(A2.1+,MAGE+)および888mel(A2.1-,MAGE-)の組み合わせはY.KawakamiおよびS.Rosenberg(National Cancer Institute)の御好意により提供され、そして内因的にプロセシングされたMAGE3エピトープの提示を測定するために用いられた。(Boon,T.ら,Ann.Rev.Immunol.(1994)12:337)。その黒色腫細胞株をAPCとして使用する前に、100IU/mlヒトIFNγ(Genzyme,Cambridge,MA)で48時間、37℃で処理した。
【0206】
本研究で用いた全ての細胞は、抗生物質、ピルビン酸ナトリウム、非必須アミノ酸および10%(v/v)熱不活化FBSを補充したRPMI−1640培地で増殖させた。
【0207】
(2.インビトロでのヒトPBMCからのCTLの誘導およびヒトCTL株の誘導体化)
MAGE3.112および癌胎児抗原(CEA)エピトープであるCEA.691に対するペプチド特異的なCTL株を生成するために、インビトロにおいて正常な被験体からのPBMCを、(Kawashima,I.らHuman Immunol.(1998)59:1)に記載のようなペプチドを用いて繰り返し刺激した。簡潔には、ペプチドを適用した(pulsed)樹状細胞(GM−CSFおよびIL4中で培養することによって接着性PBMCから区別した)を、抗体でコーティングしたビーズ(Dynal A.S.,Oslo,Noway)を用いたポジティブ選択によって得られた自己CD8+T細胞とともに、48ウェルプレート中で共存培養した。IL2、IL7およびIL10の存在下での培養の7日目後、インビトロにおいて、ペプチドを適用した(pulsed)接着性PBMCを用いて各PBMC培養物(ウェル)を再刺激した。次いで、ペプチドの存在下または非存在下において、腫瘍APC.221A2.1を用いて刺激した後のIFNγ生成を測定することによって、CTL活性について培養物を試験した。CTL株はペプチド特異的なIFNγ応答を示したPBMC培養物から、ペプチドを適用した接着性PBMCを用いたさらなるインビトロ刺激よって増殖した。
【0208】
(3.マウスCTL株)
エピトープHBV Pol.455およびHIV Pol.476ペプチドに対するCTL株を、他(Ishioka、G.らJ.Immunol.(1999)162:3915)に記載されているようなDNA免疫によって、HLA−A2.1/KbXSトランスジェニックマウス中で産生した。HLA−A2.1/KbXSおよびHLA−A2.1/KbXdトランスジェニックマウスはEpimmuneで飼育された。これらの株は、C57BL/6のバックグラウンド(Vitiello,A.らJ.Exp.Med.(1991)173:1007)において作製されたHLA−A2.1/Kbトランスジェニック株と、それぞれSJLまたはBALB/cマウス(Jackson Laboratories、Bar Harbor、ME)との間の交雑のF1世代を表す。MAGE2.157エピトープに対するCTL株は、8〜12週齢HLA−A2.1/KbXSマウスの尾部の基部に、IFA中で乳化された50μgのペプチドおよび140μgのHBV Core.128ThエピトープであるTPPAYRPPNAPIL(配列番号30)を免疫し、そしてインビトロでペプチドを用いて繰り返し初回刺激した脾細胞を再刺激することよって生じた。
【0209】
(調製D)
(アッセイ方法)
(1.HLA−A2.1分子に対するペプチド結合親和性の測定)
放射性標識された標準ペプチドのHLA−A2.1への結合について、所定の試験ペプチドによって誘導される競合のレベルを決定することによって、試験ペプチドのHLA−A2.1への結合を測定した。ゲル濾過によってMHC結合性放射能の百分率を決定し、そして、標識された標準ペプチドの結合の50%を阻害した試験ペプチドの濃度(IC50)を計算した(Ruppert,J.ら、Cell(1993)74:929;Sette,A.ら、Mol.Immunol.(1994)31:813)。標準ペプチドはHBV Core.18エピトープであった(配列FLPSDFFPSV)。
【0210】
(2.CTLによるマウスおよびヒトのIFNγ、IL5およびIL10産生の測定)
インサイチュの捕捉ELISAを、CTLからのIFNγ放出を測定するために使用した(McKinney,D.ら、J.Immunol.Methods(2000)237:105)。簡潔には、CTLを抗マウスIFNγ(クローンR4−6A2,Pharmingen,San Diego,CA)または抗ヒトIFNγ mAb(クローンNIB42,Pharmigen)のいずれかをプレコーティングしたELISA等級の96ウェル平底ウェル中においてAPCおよびペプチドを用いて刺激した。細胞培養後、ウェルを洗浄し、ビオチン化した抗マウスIFNγ(クローンXMG1.2,Pharmingen)mAbまたは抗ヒトIFNγ(クローン4S.B3,Pharmingen)mAbを加えた後、酵素結合体化ストレプトアビジン(Zymed,South San Francisco,CA)および3,3',5,5'テトラメチルベンジジン基質(ImmunoPure TMB基質キット、Pierce,Rockford,IL)を加えることによって現像した。各ウェルの吸光度は450nmでLabsystems Multiskan RC ELISAプレートリーダーにおいて測定した。各ウェルで生成されたIFNγのレベルを、同じアッセイ方法で確立されたマウスまたはヒトのIFNγの検量線から外挿することによって決定づけた。
【0211】
マウスおよびヒトのIL5およびIL10をELISAキット(R&D Biosystems、Minneapolis、MN)を用いて培養上清中で測定した。これらのアッセイを定量的サンドイッチELISA技術を用い、製造業者のプロトコールに従って行った。
【0212】
(3.エキソビボCTL応答を測定するための酵素結合免疫スポット(Enzyme−liked immunospot)(Elispot)アッセイ)
Elispotアッセイを標準プロトコール(Murali−Krishna,K.ら、Immunity(1998)8:177;Lewis,J.J.ら、Int.J.Cancer(2000)87:391)に従って行った。簡潔には、平底96ウェルニトロセルロースプレート(Immobilon−Pメンブレン,Millipore,Bedford,MA)を抗IFNγmAb(10μg/ml,クローンR4−6A2)でコーティングし、4℃で一晩中インキュベートした。PBSで洗浄した後、プレートを10%FBSを含むRPMI培地で37℃で1時間でブロックした。磁気ビーズ(Miltenyi,Auburn,CA)によって単離された4×105個の脾性CD8+細胞および10μg/mlのペプチドを適用された5×104個のJurkat−A2.1/Kb細胞を各ウェルに加え、細胞を10%FBSを含むRPMI培地で20時間インキュベートした。インキュベーション後、PBS/0.05%Tweenを用いてプレートを徹底的に洗浄した後、ビオチン化した抗IFNγ mAb(2μg/ml,クローン XMG1.2)を各プレートに加え、プレートを37℃で4時間インキュベートした。次いで、プレートをPBS(0.1%Tween−20を含む)およびVectastain ABCペルオキシダーゼ(Vectastain Elite キット;Vector Laboratories,Burlingame,CA)で4回洗浄した。室温で1時間インキュベートした後、プレートを1×PBS/0.05%Tweenを用いて3回洗浄し、次いでさらに3回1×PBSで洗浄した。スポットを現像するために100μlのAEC溶液(Sigma Chemical,St.Louis,MO)を加えた。反応は流水している水道水のもとで4−6分後に停止した。スポットをコンピューター支援画像分析(Zeiss KS Elispot Reader,Jena,Germany)を用いてカウントした。106個のCD8+細胞あたりの正味のスポット数を、以下[(関連ペプチドに対するスポット数)−(非関連ペプチドに対するスポット数)]×2.5として計算した。
【0213】
(実施例1)
(ヘテロクリティックな活性についてのペプチドアナログのスクリーニング)
(A.IFNγ放出の増加に関与しているCEA.691およびMAGE3.112アナログの同定)
アナログのスクリーニングの前に、広範囲な野生型ペプチド用量にわたってCTL株からのIFNγ生成のペプチド用量滴定を、行った。.221A2.1腫瘍細胞に異なる用量のペプチドを適用し、次いで、105個のペプチド負荷細胞を同数のマウスまたはヒトCTLと共に培養した。37℃で24時間(マウス)または48時間(ヒト)のインキュベーション後、CTLによって放出されたIFNγのレベルをインサイチュ捕捉ELISAアッセイによって測定した。用量滴定曲線を決定した後、ペプチドアナログのパネルの抗原性をスクリーニングするために、野生型ペプチドに対する活性がほとんど検出可能でない最適以下ペプチド用量を選択した。全てのマウスおよびヒトCTL株について、この最適以下用量の範囲は0.1〜1μg/mlであった。マウスCTL株は第3ドメイン内のマウスH−2Kb配列を有しているHLA分子を発現するHLA−A2.1/Kbxsトランスジェニックマウスにおいて生成されたが、すべてがネイティブなHLA−A2.1分子を発現するAPC上に提示されているペプチドに反応したということに留意すべきである。
【0214】
ペプチドアナログのスクリーニングについて、.221A2.1細胞に選択された最適以下用量の各アナログを適用し、前述したようにペプチド負荷APCをCTLと共に培養した。次いで、さらなる特徴づけのために、野生型ペプチドと比較して増強されたCTL応答を誘導するアナログを選択した。これらのアナログを、前述したのと同じ条件のもとで野生型エピトープと並行してペプチド用量滴定を行うことによって特徴付けた。
【0215】
HLA−A2.1拘束CEA.691およびMAGE3.112エピトープに特異的なCTL株を、調製Cで記載したように、インビトロにおいてペプチド負荷樹状細胞または接着性単球を用いてヒトPBMCの再刺激を繰り返すことによって、誘導した。
【0216】
17個の異なる単一アミノ酸で各残基を体系的に置換することによって合計117CEA.691および116個のMAGE3.112アナログを作製した。CEA.691はIMIGVLVGV(配列番号1);MAGE3.122はKVAELVHFL(配列番号5)である。Cys、TrpおよびMet残基は、それらが保存的変化に対応しない限り、一般に回避した。主なMHCアンカー位置、位置2およびC末端以外のペプチドの全ての位置で置換を導入した。
【0217】
次いで、インビトロでこれらのアナログのを、その抗原性について試験した。野生型ペプチドに応答するIFNγ生成がほとんど検出可能でない抗原濃度を規定にするために、前述したように、予備的な用量滴定実験を各CTL株について行った。ついで、T細胞刺激の許容量の増加に関連したアナログを同定するために、引き続きこの最適以下濃度をアナログペプチドの各エピトープについてのすべての抗原性分析に用いた。このような抗原性分析の結果を図1に示す。図1Aに示すように、最適以下100ng/ml用量では、野生型CEA.691ペプチドは最低限のIFNγ生成(50pg/ウェル未満)しか生じなかった。対照的に、いくつかのCEA.691アナログ(M3、L4、P4、H5、L5、H6、T6およびI7)は、同じ用量で、150〜350pg/ウェルの範囲内で検出可能なレベルのIFNγの産生を、誘導した。図1Bに示したように、野生型ペプチドのMAGE3.112特異的CTL株100ng/mlが100pg/mlのIFNγの放出を誘導したが、2つのアナログ(I5およびW7)は300pg/ウェル以上のIFNγレベルの誘導に関連していた。
【0218】
さらなる特徴付けのために、100pg/ウェルを超えるIFNγを刺激したCEA.691およびMAGE3.112の全てのアナログを選択し、完全な用量滴定はヘテロクリティックなアナログを同定するために実施した。ヘテロクリティックなアナログは野生型ペプチドの10倍以下のペプチド濃度で、有意なIFNγ放出(100pg/ウェルより大きい)を刺激するアナログである。CEA.691エピトープについては2つの異なるアナログ、M3(配列番号2)およびH5(配列番号3)を同定した。図1Cで見られるように、エピトープCEA691について、アナログM3およびH5が0.01ng/ml程度の少ないペプチドを用いて有意な放出を刺激したが、野生型ペプチドは1〜100μg/mlの用量範囲で有意に検出可能なIFNγシグナルを産出した。これらの基準によると、これらの2つのCEA.691アナログは、モル濃度に基づき、IFNγ放出の点において、その未改変野生型対応物よりも100,000倍以上強力である。
【0219】
同様に、MAGE3.112エピトープについて、2つのヘテロクリティックなアナログ、I5およびW7を同定した。図1Dに示すように、等価な応答を刺激するためには、0.1ng/mlのI5(配列番号6)またはW7(配列番号7)アナログのいずれかが必要であったが、1μg/mlの野生型ペプチド濃度が有意なIFNγの放出に必要とされた。これは、野生型ペプチドと比較して100,000倍より高い生物学的な活性の増加に対応する。
【0220】
一般的に、ヘテロクリティックなアナログは用量応答変化を誘導するのみならず、野生型ペプチドと比較してより高いレベルのIFNγを産出するためのCTL応答を刺激した。その結果、アナログに応答して達成される最大用量応答(プラトー)は未改変抗原に応答して得られる応答よりはるかに高かった。
【0221】
(実施例2)
(さらなるヘテロクリティックなアナログの同定)
3つのさらなるA2.1拘束エピトープ、MAGE2.157 YLQLVFGIEV、配列番号7腫瘍エピトープ、およびウイルス抗原からの2つのエピトープ、HBV Pol.455、GLSRYVARL(配列番号16)およびHIV pol.476 ILKEPVHGF(配列番号18)を分析した。これらのエピトープは全て、CTLに対して免疫原性であることが以前に示されている。
【0222】
調製Bおよび表2で述べたアミノ酸保存性割り当て(assignment)を使って、240個の異なるアナログのパネルを、3つのエピトープのそれぞれにおいてエピトープ位置3、5、7、およびエピトープ位置1、4、6に、5つの保存的アミノ酸置換および5つの非保存的アミノ酸置換を含むように合成した。これらのアナログを、HLA−A2.1/Kbxsトランスジェニックマウスで生じたマウスTCL株を使用し、そしてCEA.691およびMAGE3.112エピトープについての実施例1で述べたものと類似した実験ストラテジーに従い、へテロクリシティーについて試験した。マウス株の生成および維持に関連した技術的容易さに起因して、HLAトランスジェニックマウス由来のマウスCTL株をヒトCTL株の代わりに用いた。
【0223】
この結果を図2DのHIV pol.467について対応する用量滴定プロフィールと共に図2A(MAGE2.157)、2B(HBV pol.455)、および2C(HIV pol.476)に示した(MAGE2.157およびHIV Pol.455について、実施例3を参照のこと)。
【0224】
試験されたMAGE2.157エピトープの総計85個の異なるアナログの分析は、野生型ペプチドより1/100〜1/100,000倍の用量でIFNγ応答を刺激した2つのヘテロクリティックなアナログ、I5(配列番号8)およびF5(配列番号9)の同定を生じた(図1);これらのアナログは両方ともペプチドの中央部の奇数位置(5位)において天然に存在している保存的または半保存的な置換を有していた。
【0225】
HIV pol.476エピトープについては、スクリーニングされた78個の異なるアナログの中から、2つ(H3(配列番号19)およびL3(配列番号20))が、ヘテロクリティックな活性を有するとして同定された(表1);両方のアナログはペプチド中央部の奇数位置に保存的または半保存的置換のいずれかを保有していた。77個の試験されたものの中から、HIV pol.455エピトープの1つのヘテロクリティックなアナログの正体が同定された。このアナログはペプチド(配列番号17)の位置7に保存的置換基(P)を有していた(表1)。
【0226】
従って、腫瘍およびウイルス起源の3つのさらなるエピトープ(MAGE2.157、HIV pol.476およびHBV Pol.455)についての240個アナログから得られたデータは、実施例1で示したようなMAGE3.112およびCEA.691エピトープの分析と一致する。
【0227】
ヘテロクリシティーな分析をまた、2つのp53エピトープにおいて行った。1つのエピトープP53.149M2、SMPPPGTRV(配列番号10)はMHC結合を増強するメチオニン残基置換を有するヒトp53エピトープの固定アンカーアナログ(fixed anchor analog)を表す。第2のエピトープ、P53Mu.184、GLAPPQHLIRV(配列番号13)はマウスとヒトとの間で完全に保存された配列を有している(Theobaldら、92(26):11993(1995))。
【0228】
p53.149M2において行った用量滴定分析によって、1μg/mlおよび0.1μg/ml用量範囲における最適および最適以下応答が明らかになった。p53.149M2(各位置で5つの保存的および5つの半保存的置換)についての76個のアナログのパネルがスクリーニングされ、わずか2つのアナログ、C1(配列番号11)およびP7(配列番号12)が同定された。この両方とも、最適以下用量で100pg/ウェルのIFNγ放出を与える、図5。さらに分析すると、両方のアナログは野生型ペプチドの1/10倍の濃度で有意なIFNγ産生を誘導した。さらに、C1アナログもまた1/100倍のペプチド濃度で有意なIL10レベルを誘導した、図6。
【0229】
p53mu.184エピトープに関して、用量滴定分析を行った後、ペプチドの最適および最適以下レベルは、それぞれ500ng/mlおよび10ng/mlであると決定した。63個の保存的および半保存的置換アナログのパネルを免疫原性について試験した。増強された免疫原性を有する2つのアナログが見い出された−T3(配列番号14)およびT3、E6(配列番号15)。図7および8を参照のこと。
【0230】
(実施例3)
(ヘテロクリティックなアナログによって誘導されるリンホカインプロフィ
ール)
ヘテロクリティックなアナログはT細胞からのサイトカインの生成を示差的に活性化することが以前に示されており、それにより、いくつかのアナログがTh1サイトカインを生成するために特異的にT細胞を活性化するが、他は優先的にTh2サイトカインの生成を活性化する。本発明のヘテロクリティックなアナログに関連するリンホカイン放出のパターンを研究するため、CTL株からのTh2サイトカインであるIL5および/またはIL10の生成をIFNγの生成と比較した。2つの異なるエピトープからの代表的なデータを図3および4に示した。
【0231】
図3はMAGE2.157アナログによって誘導されたリンホカインのプロフィールを示す。アナログI5もしくはF5、または野生型(WT)ペプチドを適用した.221A2.1標的に応答して、MAGE2.157特異的CTL株によって生成されたIFNγ(A)およびIL10(B)をいくつかの異なる用量に対して測定した。点線はIFNγ(100pg/ウェル)またはIL10(50pg/ml)の有意なレベルを示している。図3Aで見られるように、MAGE2.157のF5およびI5アナログは、それぞれ野生型ペプチドの1/100倍または1/10,000倍の濃度でIFNγ生成の有意なレベルを誘導した。さらに、同じアナログはまた、野生型ペプチドの1/10倍または1/100倍のペプチド濃度で、有意なIL10生成誘導した。
【0232】
同じ傾向を示す、別のエピトープHBV Pol.455からのデータを、図4Aおよび4Bに示した。いくつかの異なるペプチド用量に対して、アナログP7または野生型(WT)ペプチドに応答してHBV Pol.455CTL株によって放出されたIFNγ(A)またはIL10(B)を示した。再度、HBV
Pol.455のP7アナログは、有意なレベルのIFNγ(図4A)およびIL10(図4B)を野生型ペプチドの1/100倍のペプチド濃度において誘導した。Th2サイトカイン(表1)の誘導について試験した全てのヘテロクリティックなアナログをまとめたデータを総合すると、ほとんどのヘテロクリティックなアナログは増加したTh1およびTh2サイトカインの両方の生成を刺激することが示される。
【0233】
(実施例4)
(ヘテロクリティックアナログのHLA−A2.1との結合親和力)
観察されたCTL株によって向上した認識が、アナログエピトープとMHCとの結合能力における偶然の増加に起因しないことを実証するために、精製したHLA−A2.1分子を、インビトロで利用して全てのヘテロクリティックアナログとMHCとの結合親和力を測定し、そして調製Dに記載されているように未改変の野生型対照物と比較した。
【0234】
表1に概説されているように、3つのアナログ(MAGE3.112W7、HIV Pol.476H3、およびHIVPol.476L3)は、HLA−A2.1と野生型ペプチドよりも4倍以上高い親和力で結合し、そして2つのアナログ(MAGE2.157I5、MAGE2.157F5)は低い親和力で結合した。残る4つのヘテロクリティックアナログであるMAGE3.112I5、CEA.691M3、CEA.691H5、およびHBV Pol.455P7は、HLA−A2.1との結合能力において、ほとんどあるいは全く変化なしで結合した。これらのデータからひとまとめにして、増加した結合性とヘテロクリシティーとの間に相関はないことが示唆される。
【0235】
(実施例5)
(マウスp53.261エピトープに関するアナログの予測および免疫原性)
インビボで免疫原性に関して試験するために、HLA−A2.1を制限したマウスp53.261エピトープを使用した。この理由は、このエピトープに対するCTL応答は、HLA−A2.1/Kbトランスジェニックマウスにおいて局部的に寛容化されることが示されているためである。これはインビボでT細胞の耐性を破壊すると予測されたヘテロクリティックアナログの能力の分析を可能にする。従来まで、ヘテロクリティックアナログは野生型エピトープに対して引き起こされるCTL株とともにインビトロでのスクリーニングを通じて検出されてきたが、置換法によって同定したアナログは野生型エピトープに対してヘテロクリティックであるCTLをインビトロで潜在的に誘導し得り、寛容な腫瘍関連エピトープに対するワクチンを設計するための目的の適応である。
【0236】
p53.261の予測されるアナログに関する免疫原性を、HLA−A2.1/Kbxdトランスジェニックマウスにおいて、p53.261エピトープ(LLGRDSFEV)50μg、または予測されるアナログおよびIFAにおけるHBV Core.128ヘルパーエピトープ140μgとを用いてマウスを共同免疫することで試験した。11日後、プライム化脾臓細胞を収集し、そして10μg/mlのペプチドでパルスした照射同系LPS活性化脾臓細胞とともにインビトロで培養した。培養から10日後、CTLをIL2の供給源としてCon Aの馴化培地の存在下で、ペプチドでパルスしたLPS芽細胞で再刺激した(Ishioka,G.ら、J.Immunol.(1999)162:3915)。
予測アナログによって免疫化されたマウス由来の脾臓細胞を、野生型ペプチド(交差反応性、アビディティおよび野生型抗原に応答するCTLの前駆体度数を決定するため)とそれぞれの免疫化アナログ(アビディティとアナログに応答するCTLの前駆体度数を決定するため)の両方に対してインビトロで刺激した。APCとしてJurkat−A2.1腫瘍細胞を用いることで、インビトロでの2回目の再刺激の5日後でのIFNγのインサイチュのELISAアッセイによるペプチドの特異性に関して、CTLのバルク集団を全ての短い周期で試験した。あるいは、CTL応答を、Elispotアッセイを用いて免疫化動物から新たに単離した脾臓細胞上で実施した。
【0237】
9マーペプチドの3、5および7の位置における3つの保存的、または半保存的置換からなるp53.261エピトープの9つのアナログのパネルを、HLA−A2.1/Kbxdトランスジェニックマウスにおける免疫原性について試験した。その結果、それぞれ9つのアナログでのマウスの免疫、およびそれぞれの免疫化アナログでのプライム化脾細胞のインビトロでの膨張により6つのアナログ(L7、D3、H7、H3、N5、およびG5)を同定し、これは野生型ペプチドを用いてインビボで誘導され、そしてインビトロで膨張されたCLTと比較して、かなりより低いペプチド濃度で、100pg/ウェルのIFNγ産生によって特徴づけられたCTL応答が得られた。
【0238】
WTペプチドまたは適応したアナログのいずれかによって免疫化されたマウス由来の脾臓細胞を、インビトロで対応する免疫ペプチド(図9AおよびB)またはWTペプチド(図9CおよびD)を用いて刺激した。次いで、これらのCTLによるIFNγの放出を、免疫ペプチド(図9AおよびB)またはWTペプチド(図9CおよびD)を用いてパルス化した標的に対して用量範囲にわたって測定した。100pg/ウェルにおけるIFNγの放出を点線で示す。これらの結果より、アナログの有意なパーセンテージは、野生型ペプチドそれ自体で誘導されたアビディティよりも高いアビディティのCTLを誘導することが示される。
【0239】
野生型ペプチドに対するヘテロクリティックアナログでプライム化されたCTLの交差反応性を、図9Cおよび図9Dに示す。野生型ペプチドで免疫化し、そして再刺激した動物由来のCTLは、0.1から10μg/mlの間のペプチド用量において100pg/ウェルのIFNγを誘導した。もう一方で、L7、H3、およびD3アナログで免疫化し、そして野生型ペプチドを用いてインビトロで刺激し、そして試験した動物から得られたCTLは、100pg/ウェルのIFNγを誘導するために、それぞれ10倍、100倍または1000倍低い用量の野生型ペプチドが必要であった(図4C)。これにより、6つのケースの内3つのケースにおいて、予測されるヘテロクリティックアナログは、野生型抗原に対して寛容である一部のCTLが存在する条件において、野生型ペプチドと応答するCTLを誘導するのに10〜1000倍の活性および能力であったことが示唆される。
【0240】
(実施例6)
(野生型との交差反応性)
D3およびH3アナログにより誘導されたCTLの交差反応性もまた、自然処理された野生型エピトープに対してHLA−A2.1/Kbを用いてトランスフェクトされたp−53発現Meth A腫瘍細胞クローンによって試験した。野生型エピトープに対してヘテロクリティックであるp−53.261アナログによって産生したCTLは、Meth A/A2.1Kb腫瘍細胞によって出現した内因的に処理されたp−53.261エピトープと応答することが明らかとなった。
【0241】
CTL集団(105/ウェル)は、HLA−A2.1/Kbと共にトランスフェクトされたMeth Aクローン、または2.5×104のMeth A腫瘍細胞とともに培養し、IFNγの放出はインサイチュELISAアッセイによって測定した。図10に示しているように、p−53.261エピトープのD3とH3アナログの両方に対して増加したCTL株は、Meth A/A2.1Kb腫瘍細胞クローンによって発現された内因性エピトープと応答し、親HLA−A2.1ネガティブMeth A腫瘍細胞株とは応答しなかった。
【0242】
(実施例7)
(Elispotアッセイを用いた前駆体度数の分析)
野生型ペプチドに対する交差反応性CTLが、アナログで免疫化されたマウスにおいて産生されることを確証するために、インビトロでのさらなるCTL膨張なしでアナログまたは野生型ペプチドで免疫化されたマウスの脾臓細胞からCD8+細胞を単離し、そして野生型またはアナログのどちらか一方に対するCTL反応性の前駆体度数をElispotアッセイを用いて測定した。
【0243】
WTエピトープ、またはD3、H3、L7およびH7アナログの何れか一方で免疫化されたマウスから単離したCD8+細胞を、ElispotアッセイにおいてWTエピトープで刺激した場合のIFNγの放出する能力について分析した。図11に示すように、野生型ペプチド反応性CTLの前駆体度数は、野生型ペプチドで免疫化されたマウスにおいて1/66,000(15点/106)であったのに対して、予測アナログで免疫化されたマウスにおける野生型ペプチド反応性細胞の前駆体度数は、D3、H3およびL7アナログに対してほぼ1/15,000(60〜75点/106細胞)であり、そしてH7アナログに対して1/83,000(12点/106)であった。これより、野生型反応性細胞は、天然ペプチドで免疫化されたマウスに比べて4つのアナログのうち3つのアナログで免疫化されたマウスにおいて4倍高い度数で存在していたことが示される。この発見は、インビボ−プライム化CTLのインビトロでの膨張を回避する、より直接的な分析システムを用いたヘテロクリティックアナログでのインビボ免疫化は、野生型ペプチドに対して実際により複数のCTL反応性を誘導することを示すので重要である。
【0244】
(実施例8)
(ヘテロクリティックアナログはインビトロで腫瘍細胞を認識し得るヒトCTLを誘導する)
調製Cに記載されているように、MAGE3.112のヘテロクリティックアナログの免疫原性もまた、MAGE3.112ペプチド、またはこのエピトープのI5およびW7アナログのどちらか一方に対するPBMCから1次CTLを誘導することによって試験した。インビトロでの2回目の刺激の後、48ウェルのPBMC培養は、正味のIFNγ産物が100pg/ウェルより多い場合にはCTL誘導に対して陽性を記録し、そして産物はペプチドの存在下、または非存在下で.221−A2.1 APCで刺激した後ではバックグラウンドより少なくとも2倍以上となった。
【0245】
ヒト腫瘍抗原に対する本発明者らの所見の生理学的な関連性を強調するために、本発明者らはインビトロでの1次誘導システムにおいて、MAGE3.112エピトープのヘテロクリティックアナログがヒトCTLを誘導し得るかどうかを試験した。正常のドナー由来の新しい未処理のヒトPBMCを、前述しているように野生型またはアナログのどちらか一方を用いてインビトロで繰り返し刺激した(Kawashima,I.ら、Human Immunol.(1998)59:1)。ペプチド特異的CTL応答を、野生型ペプチド(図12A)またはI5アナログ(図12B)およびW7アナログ(図12C)のいずれかで刺激された培地中で検出した。手短に言えば、.221−A2.1細胞をWTペプチド(図12A)、I5アナログ(図12B)またはW7アナログ(図12C)10μg/mlで一晩パルスした。48ウェルプレートの個々のウェルに成長しているCTLによるIFNγ産物を、ペプチドの存在下、または非存在下で.221−A2.1細胞に対して、あるいは内因性ネガティブエピトープ888メル腫瘍細胞株および内因性ポジティブエピトープ624メル腫瘍細胞株に対して試験した。ポジティブペプチド特異的CTL応答を示すウェルのみが示される。
【0246】
さらに重要なことに、これらのアナログとともに誘導された培地により、内因的に処理し、そして野生型配列を示す624メル腫瘍細胞株が認識された。これは、ヘテロクリティックアナログは腫瘍細胞によって示される内因的に産生された野生型エピトープを認識する生理学的に関連性のあるヒトCTLを誘導し得ることの証拠であり、そしてその現象は、ヒトとトランスジェニックマウスシステムの両方に関連性があることの証拠である。
【0247】
(実施例9)
(HLA A2スーパーファミリーメンバーにおけるHLA−A2.1スーパーモチーフから誘導されたヘテロクリティックアナログの合成および分析)
A2スーパーファミリー内部の他のHLA分子におけるヘテロクリティック置換法則をさらに確認するために、3、5および7の位置における3つの保存的、または半保存的置換よりなる、以前に議論したp53.261エピトープの9つのアナログのパネルを、以下に示すヒトHLA分子(A*0202、A*0203、A*0204、A*0205、A*0206、A*0207、A*0209、A*0214、A*6802、A*6901)の1つを発現しているトランスジェニックマウスにおける免疫原性についてインビボで試験した。
【0248】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、p53.261エピトープ)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型エピトープ自体に対するよりも、天然野生型エピトープに対してより高いアビディティCTLを誘導するのにより強力であることを示す。
【0249】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0250】
(実施例10)
(HLA B7スーパーファミリーメンバーにおけるHLA−B7スーパーモチーフから誘導されたヘテロクリティックアナログの合成および分析)
ヘテロクリティック置換法則をさらに確認するために、HLA−B7スーパーモチーフの内部に配列を持っているペプチドから誘導されたヘテロクリティックアナログを用いて追加研究を行った。例えば、アナログをインビボで免疫原性について試験し得る。
【0251】
本研究では、ペプチドを持ったHLA−B7スーパーモチーフであるAPRTLVYLLエピトープを選択し、そして合成した。9マーペプチドの3、5および7の位置における3つの保存的、または半保存的置換よりなるアナログのパネルを、HLA−B*0702/Kbトランスジェニックマウスにおける免疫原性について試験した。パネルはAPETLVYLL、APRTWVYLLおよびAPRTLVPLLを含み、それぞれ3、5および7番目の位置における半保存的置換と対応している。
【0252】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγの誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、APRTLVYLL)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体よりも高いアビディティCTLを誘導するのにより強力であることを示す。
【0253】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0254】
B7スーパーファミリー内部の他のHLA分子についてヘテロクリティック置換法則をさらに確認するために、APETLVYLL、APRTWVYLLおよびAPRTLVPLLペプチドを、以下のヒトHLA分子(B*0702、B*0703、B*0704、B*0705、B*1508、B*3501、B*3502、B*3503、B*3503、B*3504、B*3505、B*3506、B*3507、B*3508、B*5101、B*5102、B*5103、B*5104、B*5105、B*5301、B*5401、B*5501、B*5502、B*5601、B*5602、B*6701、およびB*7801)の1つを発現しているトランスジェニックマウスにおける免疫原性についてインビボで試験した。
【0255】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、APRTLVYLL)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体より高いアビディティCTLを誘導するのにより強力であることを示す。
【0256】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0257】
(Elispotアッセイを用いた前駆体度数の分析)
野生型ペプチドに対する交差反応性CTLが、アナログで免疫化されたマウスにおいて産生されることを確証するために、インビトロでのさらなるCTL膨張なしでアナログまたは野生型ペプチドで免疫化された脾臓からCD8+細胞を単離した。この材料より、野生型、またはアナログのどちらか一方に対するCTL反応性の前駆体度数をElispotアッセイを用いて測定した。代表的に、野生型ペプチドの反応性CTLの前駆体度数は、アナログの前駆体度数よりもかなり低い。
【0258】
(ヘテロクリティックアナログによってインビトロでエピトープを認識し得るヒトCTLを誘導し得る)
ヘテロクリティックアナログを、インビトロでの1次誘導システムにおけるCTLの誘導について分析し得る。前記のように、48ウェルプレートにおいて正常のドナーからの新しい未処理のヒトPBMCを、野生型またはアナログのどちらか一方を用いてインビトロで繰り返し刺激した。次に、ペプチド特異的CTL応答を、野生型ペプチドまたはヘテロクリティックアナログのどちらか一方で刺激された培地中で検出する。これらのアナログで誘導された培地において、内因的に処理され、そして野生型配列を示す標的を認識し得る。これは、ヘテロクリティックアナログが内因的に産生された細胞上で発現した野生型ペプチドを認識する生理学的に関連性のあるヒトCTLを誘導し得ることの証拠であり、そしてその現象はヒトとトランスジェニックマウスシステムの両方に関連性があることの証拠である。
【0259】
(実施例11)
(HLA A3スーパーファミリーメンバーにおけるHLA−A3スーパーモチーフから誘導されたヘテロクリティックアナログの合成および分析)
ヘテロクリティック置換法則をさらに進んで確認するために、HLA−A3スーパーモチーフの内部に配列を有するペプチドから誘導されたヘテロクリティックアナログを用いて追加研究を行う。例えば、インビボで免疫原性についてアナログを試験し得る。
【0260】
本研究では、ペプチドを有するHLA−A3スーパーモチーフであるKVFPYALINK(配列番号22)エピトープを選択し、そして合成する。9マーペプチドの3、5および7の位置における3つの保存的/半保存的置換よりなる配列番号22のアナログのパネルを、HLA−A*3101/Kbトランスジェニックマウスにおける免疫原性について試験した。パネルはKVHPYALINK、KVFPQALINKおよびKVFPYAKINKを含み、それぞれ3、5および7番目の位置における半保存的変化と対応している。
【0261】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、KVFPYALINK)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体より野生型に対して高いアビディティCTLを誘導するのにより強力であることを示す。
【0262】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0263】
A3スーパーファミリー内部の他のHLA分子についてヘテロクリティック置換法則をさらに確認するために、KVHPYALINK、KVFPQALINKおよびKVFPYAKINKペプチドを、以下のヒトHLA分子(A*0301、A*1101、A*3101、A*3301、A*6801)の1つを発現しているトランスジェニックマウスにおけるインビボでの免疫原性について試験する。
【0264】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、KVFPYALINK)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体より高いアビディティCTLを誘導するのにより強力であることを示す。
【0265】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0266】
(Elispotアッセイを用いた前駆体度数の分析)
野生型ペプチドに対する交差反応性CTLが、アナログで免疫化されたマウスにおいて産生することを確証するために、インビトロでのさらなるCTL膨張なしでアナログまたは野生型ペプチドで免疫化された脾臓からCD8+細胞を単離した。この材料より、野生型、またはアナログのどちらか一方に対するCTL反応性の前駆体度数をElispotアッセイを用いて測定した。代表的に、野生型ペプチドの反応性CTLの前駆体度数は、アナログの前駆体度数よりもかなり低い。
【0267】
(ヘテロクリティックアナログはインビトロでエピトープを認識し得るヒトCTLを誘導し得る。)
ヘテロクリティックアナログを、インビトロでの1次誘導システムにおけるCTLの誘導について分析する。前記したように、48ウェルプレートにおいて正常のドナー由来の新しい未処理のヒトPBMCを、野生型またはアナログのどちらか一方を用いてインビトロで繰り返し刺激した。次に、ペプチド特異的CTL応答を、野生型ペプチドまたはヘテロクリティックアナログのどちらか一方で刺激された培地中で検出する。これらのアナログで誘導された培地において、内因的に処理し、そして野生型配列を示す標的を認知する。これは、ヘテロクリティックアナログが内因的に産生される細胞上に発現した野生型ペプチドを認識する生理学的に関連性のあるヒトCTLを誘導することの証拠であり、そしてその現象はヒトとトランスジェニックマウスシステムの両方に関連性があることの証拠である。
【0268】
本明細書中に記載される実施例および実施形態は、例示の目的のためだけであり、本発明に照らして、様々な改良または変更は当業者に示唆され、そして本出願の意図および範囲ならびに添付の特許請求の範囲内に含まれることが理解される。この結果、本明細書中で引用される全ての刊行物、特許、および特許出願は、全ての目的のためにその全体が参考として援用される。
【技術分野】
【0001】
(関連出願の引用)
本出願は、米国35 U.S.C.§119(e)の下、特許仮出願番号60/166,529(1999年11月18日に出願)および同60/239,008(2000年10月6日に出願)に対する優先権を主張する。これらはその全体が参照として援用される。
【0002】
(発明の分野)
本発明は所定のT細胞に対する増加した刺激能力を有する、元のペプチドのヘテロクリティック(heteroclitic)アナログを生成するための方法に関係する。
【背景技術】
【0003】
(発明の背景)
数々の研究は、細胞障害性リンパ球(CTL)が、免疫系による感染性の疾患およびガンの根絶に中心的な役割を果たすことを示唆する(Byrneら、J.Immumol.51:682(1984)、McMichealら、N.Englnd J.Med.,309:13(1983))。CTLはエピトープを含むペプチドにより刺激されると、CTL応答を刺激するエピトープに基づくワクチンの開発においてかなりの努力が進められている。ヘテロクリティックアナログと命名したエピトープの一つのクラスは、ワクチン成分としての利点を提供する。なぜならこれらのアナログは、ネイティブエピトープによって誘導されるT細胞応答よりも強力なT細胞応答を誘導するからである。ヘテロクリティックアナログは、所定の用量に対する応答の増加もしくは同じ応答を達成するためのより少ない必要量によって測定されるような、特定のT細胞に対する増加した刺激能力もしくは刺激効力を有するペプチドと定義される。
【0004】
臨床適応におけるヘテロクリティックアナログの使用と関連する利点は、以下のとおりである。第一に、ヘテロクリティックアナログは、T細胞アネルギーの状態を逆転することによってか、T細胞の非寛容化交差反応クローンを活性化することによってか、または、「免疫偏向」(すなわち、産生されるCTLの型(例えば、Th1またはTh2))を媒介することによって、寛容を崩壊/克服する能力を有する。最近の研究は、ヘテロクリティックアナログが内因的にプロセシングされたエピトープを認識するCTLを誘導し得るという点で、ヘテロクリティックアナログが免疫原であることを示す(Zarembaら、Cancer Research、57:4570(1997);Rivoltoniら、Cancer Research、59:301(1999);Selbyら、162(2):669(1999))。これは異なる免疫系における研究によって確認される(Zugelら、J.Immunol.、161:1705(1998)、Wangら、J.Exp.Med.、190:983(1999)、Menら、J.Immunol.、162:3566(1999)。例えば、Zugelらの研究(Zugelら、前出)は、成体マウスにおいて免疫優性のT細胞エピトープに対するT細胞寛容が、ペプチドのそのヘテロクリティック交差反応性ペプチドアナログで免疫することによって克服され得ることを示す。
【0005】
この事は、ガンのワクチンの分野で特に有意であり、ここで、CTLエピトープの大部分が、自己抗原から誘導される。ガン関連抗体が、しばしば、自己抗原であるという事実に起因して、これらの抗原に対する既存の寛容が存在し得るという対応した現象が存在し、それによって、そのようなエピトープに対するT細胞応答の発生が課題である。ヘテロクリティックアナログによる寛容の崩壊は、マウスクラスII系における最近の研究で示されている(Wangら、J.Exp.Med.190:983(1999)。この研究では、寛容の崩壊に関与するメカニズムがアネルギーの逆転よりむしろ、寛容化の低親和性クローンの刺激であった。本明細書中で実証されるヘテロクリシティーは、高いアビディティのCTLの誘導に関連しており、これは重要な差異を表す。
【0006】
第二に、ペプチドアナログは、T細胞からのサイトカイン産生を調節することが示されている(Pfeifferら、J.Exp.Med.、181:1569(1995)、Taoら、J.Immunol.、158:4237(1997)、Salazarら、Int.J.Cancer 85(6):829−38(2000)、Nicholsonら、Int.Immunol.12(2):205−13(2000))。そのようなアナログによって誘導される免疫偏向は、Th細胞応答の特定のサブセットの発生が腫瘍退行に相関する、(Zitvogelら、J.Exp.Med.、183:87(1996)、Celluzziら、J.Exp.Med.、183:283(1996))または、自己免疫疾患もしくは感染性の疾患の臨床結果に影響を与える(Romagnaniら、Annu.Rev.Immumol.、12:227−57(1994))数々の疾患状態と関連を有する。従って、ヘテロクリティックアナログを伴う免疫は、エフェクターT細胞の特定のサブセットの誘導によってサイトカイン産生を調節する能力を提供し、それによって、疾患の経過を変化させる。
【0007】
第三に、ヘテロクリティックアナログは、薬剤の開発において利点を有する。なぜなら、それらは強力な生物学的効力により、有意により少量のペプチドが、処置用量に必要とされるからである。この特徴は、特定の製造および毒性の懸念を克服する。この事については、黒色腫患者において抗原特異的T細胞を生じた、MART−1ペプチドのヘテロクリティックアナログが、ネイティブエピトープよりはるかに低濃度で活性であることが示されている(Rivoltiniら、Cancer Research 59:301(1999)。同様の結果が、CEA由来CAP1エピトープのヘテロクリティックアナログに関して、Schlomおよび共同研究者によって報告された。(Zarembaら、Cancer Research 57:4570(1997))。しかし、野生型ペプチドに対する並行したprecursor frequency分析もしくはTCRアビディティ分析は、行われなかった。
【0008】
従って、それらの生物学的関連性もために、所定のエピトープにヘテロクリティック活性を与えるアミノ酸置換を予測することが非常に有用である。しかしながら、本開示以前では、そのような置換を予想するための簡単な方法は存在しない。しかし、以前の研究では(非特許文献1、非特許文献2)、黒色腫細胞から天然に存在する変異ペプチドを溶出することによってか、または、エピトープ内のほとんど全ての位置での変異からなる多くのアナログを系統的にスクリーニングすることによって、ヘテロクリティックエピトープが、偶発的に同定された(非特許文献3、非特許文献4、非特許文献5)。あるいは、ヘテロクリティックアナログは、ランダムコンビナトリアムペプチドライブラリーをスクリーニングすることによって同定された。このライブラリーはまた、多くのペプチドの困難な合成およびスクリーニングを必要であった(非特許文献6)。DNA発現ライブラリーのスクリーニングのような、遺伝学的なアプローチが、CTLエピトープおよびアナログの生成のための別の方法を提供する(非特許文献7、非特許文献8)。しかし、このアプローチは、生成されるエピトープの潜在的な少ない量および複雑性が問題になり得る。
【先行技術文献】
【非特許文献】
【0009】
【非特許文献1】Selbyら、J.Immunol.、162(2):669(1999)
【非特許文献2】Skipperら、J.Exp.Med.183:527(1996)
【非特許文献3】Zarembaら、Cancer Research 57:4570(1997)
【非特許文献4】Loftusら、Cancer Research 58:2433(1998)
【非特許文献5】Blakeら、J.Exp.Med.18:121(1996)
【非特許文献6】Pinillaら、Current Opinion in Immunology 11:193−202(1999)
【非特許文献7】Boonら、Annu.Rev.Immunol.12:337−65(1994)
【非特許文献8】Gavinら、Eur.J.Immunol.24(9):2124−33(1994)
【発明の概要】
【課題を解決するための手段】
【0010】
(発明の開示)
本発明は,対応する類似した野生型エピトープと比べて、免疫応答をもたらす増強された能力を有するエピトープを含むペプチドを、調製する方法を提供する。この得られた「ヘテロクリティックアナログ」は、ウイルス性疾患、ガン、および標的細胞上の提示された抗原によって特徴付けられる他の疾患の処置のための、免疫学的組成物として有用である。また、本発明は以下を提供する:
(項目1) エピトープを含むペプチドの免疫原性を高める方法であって、該方法が、以下:
i)第一のクラスIエピトープを含むペプチドを提供する工程であって、ここで該エピトープが、本質的にN末端およびC末端ならびに少なくとも1つの一次アンカー残基を有するアミノ酸配列からなり、ここで該アミノ酸残基が、連続して数えられ、そして該エピトープのN末端に最も近い一次アンカー残基が、2位または3位である、工程;および
ii)該エピトープのN末端とC末端との間の3位および/もしくは5位および/もしくは7位での1つ以上の保存的、または半保存的置換を誘導する工程であって、ただし、該位が一次アンカー残基でない、工程、
それにより第二のクラスIエピトープを含むペプチドを構築する工程であって、該エピトープが、該第一のクラスIエピトープと比較して高められた免疫原性を示す、工程、
を包含する、方法。
【0011】
(項目2) 前記第二のクラスIエピトープが、前記第一のクラスIエピトープと比較して、特定のT細胞について少なくとも約50%増加した効力を示す、項目1に記載の方法。
【0012】
(項目3) 1つの置換のみが導入される、項目1に記載の方法。
【0013】
(項目4) 前記置換が保存的置換である、項目1に記載の方法。
【0014】
(項目5) 前記置換が半保存的置換である、項目1に記載の方法。
【0015】
(項目6) 項目1に記載の方法であって、前記ペプチドがHLAクラスI分子によって結合されそして細胞傷害性のT細胞と接触する場合、前記第二のクラスIエピトープを含むペプチドが、Th1およびTh2サイトカインの両方を誘導する、方法。
【0016】
(項目7) 前記第一のクラスIエピトープが、A1、A2、A3、A24、B7、B27、B44、B58およびB62からなる群より選択されるスーパーモチーフを含む、項目1に記載の方法。
【0017】
(項目8) 項目1に記載の方法であって、ここで前記第一のクラスIエピトープが、ウイルス抗原、腫瘍関連抗原、寄生生物抗原、細菌抗原または真菌抗原由来である、方法。
【0018】
(項目9) 項目1の方法によって調製された第二のクラスIエピトープを含むペプチド。
【0019】
(項目10) 免疫応答を誘発する方法であって、該方法が細胞傷害性Tリンパ球(CTL)と項目9に記載のペプチドとを接触させる工程を包含する、方法。
【0020】
(項目11) 前記接触させる工程がインビトロで、抗原提示細胞の存在下で実施される、項目10に記載の方法。
【0021】
(項目12) 前記接触工程が、被験体に前記ペプチドをコードするヌクレオチド配列を含む核酸分子を投与する工程によって実施される、項目10に記載の方法。
【0022】
(項目13) 少なくとも1つのペプチドを含む組成物であって、該ペプチドが、項目1に記載の方法によって得られ得るクラスIエピトープを含む、組成物。
【0023】
(項目14) 前記ペプチドが、9個〜15個のアミノ酸を含む、項目13に記載の組成物。
【0024】
(項目15) 項目13に記載の組成物であって、前記ペプチドが、配列番号2、配列番号3、配列番号5、配列番号6、配列番号8、配列番号9、配列番号11、配列番号12、配列番号14、配列番号15、配列番号17、配列番号19および配列番号20からなる群より選択されるアミノ酸配列を含む、組成物。
【0025】
(項目16) 前記ペプチドがCTLエピトープと混ぜられるか、または連結される、項目13に記載の組成物。
【0026】
(項目17) 前記ペプチドが、HTLエピトープと混ぜられるか、または連結される、項目13に記載の組成物。
【0027】
(項目18) 前記HTLエピトープが、pan−DR結合分子である、項目17に記載の組成物。
【0028】
(項目19) リポソームをさらに含む、項目13に記載の組成物。
【0029】
(項目20) 前記エピトープが、脂質に結合される、項目13に記載の組成物。
【0030】
(項目21) 前記エピトープが、ヘテロポリマーに含まれる、項目13に記載の組成物。
【0031】
(項目22) 前記エピトープが、ホモポリマーに含まれる、項目13に記載の組成物。
【0032】
(項目23) 前記エピトープが、HLA重鎖、β2ミクログロブリン、およびストレプトアビジン複合体に結合され、これによりトリマーが形成される、項目13に記載の組成物。
【0033】
(項目24) 項目13に記載の組成物であって、抗原提示細胞をさらに含み、ここで、前記エピトープが、該抗原提示細胞上または抗原提示細胞内にある、組成物。
【0034】
(項目25) 項目24に記載の組成物であって、前記エピトープが前記抗原提示細胞上のHLA分子に結合し、これにより、該HLA分子に制限される細胞傷害性リンパ球(CTL)が存在する場合、該CTLのレセプターが該HLA分子と該エピトープとの複合体に結合する、組成物。
【0035】
(項目26) 前記抗原提示細胞が、樹状細胞である、項目25に記載の組成物。
【0036】
(項目27) 項目13に記載の組成物であって、HLA分子をさらに含み、ここで、前記ペプチドが、該HLA分子により結合される、組成物。
【0037】
(項目28) 標識をさらに含む、項目13に記載の組成物。
【0038】
(項目29) 項目28に記載の組成物であって、ここで、前記標識が、ビオチン、蛍光部分、非哺乳動物糖、放射性標識、またはモノクローナル抗体が結合する低分子である、項目28に記載の組成物。
【0039】
(項目30) 項目13に記載の組成物であって、以下:
前記ペプチドの単位投薬量、および
薬学的賦形剤、
を含む、ワクチンである、組成物。
【0040】
(項目31) 項目1に記載の方法により得られ得る第二のクラスIエピトープを含む、9〜15個のアミノ酸のペプチドをコードするヌクレオチド配列を含む核酸分子。
【0041】
(項目32) 項目31に記載の核酸分子であって、前記ペプチドが、配列番号2、配列番号3、配列番号5、配列番号6、配列番号8、配列番号9、配列番号11、配列番号12、配列番号14、配列番号15、配列番号17、配列番号19、および配列番号20、からなる群より選択されるアミノ酸配列からなるエピトープを含む、核酸分子。
【0042】
(項目33) 前記ヌクレオチド配列の発現のための制御配列をさらに含む、項目32に記載の核酸分子。
【0043】
(項目34) 項目31に記載の核酸分子を、活性成分として含む、薬学的組成物。
【0044】
従って、一つの局面において、本発明は、エピトープを含むペプチドの免疫原性を増強するための方法に関する。この方法は、以下の工程を包含する:I)第一のクラスIエピトープを含むペプチドを提供する工程であって、ここで、このエピトープは、本質的に、N末端およびC末端ならびに少なくとも一つの一次アンカー残基を有するアミノ酸配列からなり、ここで、このエピトープのアミノ酸残基は連続的に番号付けられ、そして、このエピトープのN末端に最も近い一次アンカー残基は、位置2または位置3である工程;II)一次アンカー残基を含まない位置3および/または5および/または7で、このエピト−プのN末端とC末端との間に一つ以上の保存的または半保存的な置換を導入する工程で、これにより、増強された第一のクラスIエピトープと比較される抗原原性を示す第二のクラスIエピトープを含むペプチドを構成する工程。
【0045】
上記の第二のクラスIエピトープは、一般に「ヘテロクリティックアナログ」と称する。
【0046】
好ましい実施形態において、このヘテロクリティックアナログは、対応する野生型クラスIエピトープに比較して、特定のT細胞に対する少なくとも約50%増加した効力を示す。このアナログは、1つの置換のみを含み得るか、または2もしくは3つの置換を含み得、そしてこの置換は、保存的または半保存的であり得る。このヘテロクリティックアナログは、HLAクラスI分子に結合され、そして関連する細胞傷害性T細胞と接触された場合、Th1およびTh2サイトカインの両方を誘導し得る。好ましくは、クラスIエピトープは、A1、A2、A3、A24、B7、B27、B44、B58およびB62からなる群から選択されたスーパーモチーフを含み、より好ましくは、クラスIエピトープは、A2スーパーモチーフ、最も好ましくは、A2.1モチーフを含む。
【0047】
本発明はまた、事前に選択したクラスIペプチドエピトープに対してヒト細胞傷害性T細胞応答を誘導する方法を提供する。この方法は、以下の工程を包含する:上記のヘテロクリティックアナログを提供する工程;および、ヒトCTLにこのヘテロクリティックアナログを接触させる工程。
【0048】
いくつかの局面において、この接触の工程は、インビトロで実施される。いくつかの局面において、この接触の工程は、このヘテロクリティックアナログペプチドエピトープをコードする配列を含む核酸分子を、被験者に投与することによって実施される。
【0049】
本発明はまた、上記の方法によって入手可能であるヘテロクリティックアナログエピトープを含むペプチドに関する。特に、そして、好ましくは、そのようなペプチドとしては、そのエピトープが、配列番号2、配列番号3、配列番号5、配列番号6、配列番号8、配列番号9、配列番号11、配列番号12、配列番号14、配列番号15、配列番号17、配列番号19、および配列番号20からなる群から選択されるアミノ酸の配列からなる、ペプチドが挙げられる。このペプチドは、9〜20アミノ酸、好ましくは、9〜16アミノ酸、より好ましくは9〜15アミノ酸を含み得るが、また、全部で9、10、11、12、13または14アミノ酸のみを含み得る。定義されたヘテロクリティックアナログエピトープは、より長いポリペプチドまたはタンパク質に含まれ得、このタンパク質またポリペプチドは、同じエピトープのホモポリマーであるか、あるいは、種々のこのようなエピトープを含むかまたは野生型ペプチドと組み合わせてヘテロクリティックアナログエピトープを含む、ヘテロポリマーである。これらペプチドおよびタンパク質は、薬学的使用のために設計される組成物に含まれ得る。
【0050】
ヘテロクリティックアナログエピトープを含むペプチドまたはヘテロポリマーもしくはホモポリマーは、他の成分を結合させ、免疫応答の誘発におけるそれらの活性をさらに増強し得るか、または調節し得る。これらさらなる変更物は、共有結合的に結合され得るか、または混合物中に非共有結合的に含まれ得る。従って、ヘテロクリティックアナログエピトープは、CTLエピトープまたはこのHTLエピトープに混合され得るか、または投与され得る。特にHTLエピトープは、pan−DR結合分子である。ヘテロクリティックアナログエピトープを含む組成物は、さらにリポソームを含み得る。ここで、このエピトープは、リポソーム上もしくはリポソーム中に存在するか、またはこのエピトープは、脂質に結合され得る。このヘテロクリティックエピトープは、HLA重鎖、β2−ミクログロブリンおよびストレプトアビジン(strepavidin)複合体に結合され得、それによって、4量体が形成される。さらに、このヘテロクリティックエピトープは、抗原提示細胞を含む組成物において改変され得る。ここで、このエピトープは、抗原提示細胞上または抗原提示細胞中にあり、ここで、このエピトープは、抗原提示細胞上のHLA分子に結合される。従って、HLA分子に制限される細胞傷害性リンパ球(CTL)が存在する場合、CTLのレセプターは、HLA分子とそのエピトープの複合体に結合する。抗原提示細胞は、樹状細胞であり得る。この組成物はまた、単にHLA分子を含み得る。ここで、このエピトープを含むペプチドは、HLA分子に結合される。この組成物は、モノクローナル抗体が結合する標識(例えば、ビオチン)、蛍光部分、非哺乳動物性の糖、放射標識または小さい分子をまた含み得る。
【0051】
記載される組成物は、対応する野生型エピトープに対する免疫応答の誘発において有用である。代表的には、ヘテロクリティックアナログは、適切な賦形剤をさらに含むそのような組成物に含まれる。活性な成分であるヘテロクリティックエピトープは、投薬形態単位で存在し得る。被験体の処置に有用な組成物はまた、これらの核酸分子は、上記のペプチドをコードする核酸分子を含み得、それらの発現のための制御配列を必要に応じて含む。
【図面の簡単な説明】
【0052】
【図1−1】図1Aおよび1Bは、CEA.691およびMAGE3.112のそれぞれのアナログのパネルを、その対応するCTLにおいてIFNγ産生を誘導する能力について、試験した結果を示す。
【図1−2】図1Cおよび1Dは、それぞれCEA.691およびMAGE3.112ヘテロクリティックアナログについてその対応する用量応答曲線である。
【図2−1】図2Aは、MAGE2.157、HIVPol.476、およびHBVPol.455エピトープアナログのアナログのパネルを、対応するCTLにおいてIFNγ産生を誘導するこれらのアナログの能力に関して、試験した結果を示す。
【図2−2】図2Bは、MAGE2.157、HIVPol.476、およびHBVPol.455エピトープアナログのアナログのパネルを、対応するCTLにおいてIFNγ産生を誘導するこれらのアナログの能力に関して、試験した結果を示す。
【図2−3】図2Cは、MAGE2.157、HIVPol.476、およびHBVPol.455エピトープアナログのアナログのパネルを、対応するCTLにおいてIFNγ産生を誘導するこれらのアナログの能力に関して、試験した結果を示す。
【図2−4】図2Dは、首尾よいHIVPol.476アナログについての関連した用量応答曲線である。
【図3】図3Aおよび3Bは、適切なCTLからIFNγ産生またはIL10産生を誘導する能力に関して、野生型との比較におけるMAGE2.157のヘテロクリティックアナログの用量応答曲線の結果を示す。
【図4】図4Aおよび4Bは、適切なCTLにおいてIFNγおよびIL10の産生に対する、野生型およびHIVPol.476のヘテロクリティックアナログについての用量応答曲線である。
【図5】図5は、エピトープp53.149M2の潜在的なヘテロクリティックアナログのパネルを、適切なCTLからのIFNγの産生に関して、試験した結果である。
【図6】図6Aおよび6Bは、p53.149M2の首尾よいヘテロクリティックアナログによるIFNγおよびIL10の産生に対する、対応する用量応答曲線である。
【図7】図7は、CTLにおけるIFNγ産生に対するp53.Mu184エピト−プの潜在的なアナログのパネルを試験した結果である。
【図8】図8は、IFNγ産生に関する、野生型、およびp53.Mu184の2つ首尾よいヘテロクリティックアナログについての用量応答曲線である。
【図9】図9A〜Dは、対応する野生型エピトープに関する、ヘテロクリティックアナログの反応交差性を示す。図9Aおよび9Bにおいて、IFNγ産生は、免疫ペプチドによる刺激を使用して、濃度の関数としてプロットされる。図9Cおよび9Dは、野生型エピトープが、CTLの初期誘導のために使用されるヘテロクリティックアナログとは対抗的に、刺激剤として使用される場合の、対応する結果である。
【図10】図10は、p53.261およびそのヘテロクリティックアナログによる誘導に関する、IFNγの放出である。
【図11】図11は、種々のヘテロクリティックアナログに関するElispotの結果である。
【図12】図12は、種々のヘテロクリティックアナログを使用する内因性のペプチドに対するCTL活性の刺激の結果である。
【図101−1】図101は、実施例で用いられるペプチド、および腫瘍抗原およびウイルス抗原から同定されたヘテロクリティックアナログの特徴を示す、表1である。
【図101−2】図101は、実施例で用いられるペプチド、および腫瘍抗原およびウイルス抗原から同定されたヘテロクリティックアナログの特徴を示す、表1である。
【図102−1】図102は、所定のアミノ酸置換を、保存的置換、半保存的置換、または非保存的置換であるとして特徴付けし得るように、任意の所定のアミノ酸対の間の類似性の割り当てを示す、表2である。
【図102−2】図102は、所定のアミノ酸置換を、保存的置換、半保存的置換、または非保存的置換であるとして特徴付けし得るように、任意の所定のアミノ酸対の間の類似性の割り当てを示す、表2である。
【図103−1】図103は、表3である。
【図103−2】図103は、表3である。
【図104−1】図104は、表4である。
【図104−2】図104は、表4である。
【発明を実施するための形態】
【0053】
(発明を実施するための形態)
(1.概要)
本発明は、HLAクラスI分子に結合するヘテロクリティックアナログの設計の方法に関連する。本明細書中に記載される「ヘテロクリティックアナログ」は、特定のT細胞に対する増強された効力を持つエピトープを含むペプチドである。それは、所定の投与量に対する増強された応答によって、または相同性のあるクラスIペプチドとして同じ応答を達成するより少ないの必要量によって測定される。本発明の方法は、特に、ヒトの癌および前癌性の状態に関連する、ウイルス、細菌、真菌および原生動物の寄生虫のような感染性の病原体に由来する、ならびに任意のクラスIペプチドの改変に有用である。
【0054】
重要なことに、ヘテロクリシティーの現象は、個々のクラスIエピトープに結合するHLA分子にわたってあてはまる。例えば、A2スーパーモチーフを有するヘテロクリティックアナログペプチドは、HLA−スーパータイプ(例えば、A0201、A0202、A0203、A0204、A0205、A0206、A0207など)における全てのHLA分子にわたって、ヘテロクリティック(すなわち、より高い効力を有する)である。同様に、異なる配列モチーフ(例えば、A1、A3、A24、B7、B27、B44、B58、B62など)を有するヘテロクリティックアナログペプチドは、その特定のHLA−スーパーファミリーにおける全てのHLA分子にわたって、免疫応答をより強力にする。
【0055】
出願者は、対応する野生型エピトープに対する、免疫応答を高めるヘテロクリティックアナログを設計する特定の法則を見つけた。これらの法則は、任意のクラスI対立遺伝子によってコードされるHLA分子に結合するモチーフ、またはス−パーモチーフを有するエピトープに関して適応できる。従って、これらの法則の使用によって、任意の「野生型」または「ネイティブ」クラスIエピトープの免疫原性を高めることが可能である。
【0056】
簡略には、この法則は、野生型クラスIエピトープが、エピトープの位置3および/または位置5および/または位置7として保存的または半保存的なアミノ酸の置換によって改変されることを述べる。置換されるべき保存的または半保存的アミノ酸の性質は、本明細書以下の、調製Bの記載によって定義され、この結果は表2に要約される。従って、表2を参照すれば、これらの位置での置換のための適した候補を決め得る。表2に示されるように、表の最上段にわたって示されるそれぞれのアミノ酸は、残存する19の遺伝的にコードされるアミノ酸と数値的に規定された関係を有する。指標がより低いほど、保存性はより高く;同じアミノ酸は、1.0の類似性の割当を有し;最も大きく異なるアミノ酸は、20に近い類似割当を有する。調製B中で示す方法を使用すると、遺伝子コードされないアミノ酸もまた、類似性の指標を割り当てられ得、そして保存的または半保存的(または非保存的)として、任意に天然に存在するアミノ酸について、分類され得る。
【0057】
本発明のヘテロクリティックアナログペプチドは、被験体の免疫系が耐性となる抗原に対する免疫応答を誘導するのに特に有用である。ヒト被験体は、特に好ましいが、この方法はまた、これらの被験体に関して、対応するHLAモチーフを考慮にいれることにより、他の哺乳動物、例えば実験室マウスにも適用し得る。耐性は、先立つ抗体への曝露によって誘導される特定の免疫学的な非反応性をいう。耐性は、患者が耐性である特定のクラスIペプチドエピトープを同定することによって、本発明の方法に従ってペプチドエピトープ配列を改変することによって、および寛容化エピトープ(抗原)に対する交差反応をする免疫反応を誘導することによって、克服され得る。例えば、被験体の免疫系がウイルス抗原または腫瘍関連抗原に耐性である場合、耐性の克服は特に望ましく、後者の抗原は、細胞の形質転換の結果として、しばしば過剰発現する自己タンパク質である。
【0058】
ヘテロクリティックを設計するための法則を決定するため、いくつかの種々のCTL株がアナログパネルに対する反応性によりスクリーニングされた。T細胞刺激容量の改変は一次MHCアンカーの変更なしに達成された。
【0059】
その野生型エピトープとしては、上皮細胞の癌において特異的に上方制御され、そして免疫原性であることが示された自己抗原に由来する腫瘍エピトープが挙げられる。HIVおよびHBVのポリメラーゼ遺伝子由来のエピトープのような使用されるウイルスのエピトープは、同様に免疫原性であることが示された。
【0060】
本明細書中に記載した法則は、ヘテロクリティックなアナログを設計するための基盤を提供し、そうでなければ要求されるスクリーニングを劇的に減少させ、そして癌および感染性疾患のためのエピトープを基礎としたワクチンの設計に非常に有用である。
【0061】
以下に示す実施例において、スクリーニングされた(本明細書に開示されるヘテロクリティックの法則に適合する)総アナログの17%がヘテロクリティックであった(16/95)。これは、二つの理由において有意である:第1に、ヘテロクリティック置換の法則に従ったアナログを使用することでヘテロクリティック検出の効率が2.2%から17%まで上昇した;第2に、合成されることが必要なペプチドの数が一つのエピトープあたり約100アナログから約15アナログに劇的に減少され、この工程費用を効果的にし、高い処理能力に耐えられるようにする。本発明のヘテロクリティック置換の法則の適用を通して、ヘテロクリティックアナログが生じる効率は、100から1000倍近く、0.2%(233のCEA.691およびMAGE3.112アナログのスクリーニングから4つが同定された)から30%(9つの予測されたアナログのスクリーニングから3つが同定された)まで増加した。後者の頻度は、最初のアッセイにおいて潜在的なヘテロクリティック活性を示した6つのアナログのうちの4つのみをさらなる分析に供したので、ひどい過小評価でありうる。
【0062】
以前の研究は、ヘテロクリティックアナログによるT細胞応答の調節が、TCR接触残基を含むことを示した(Byrneら、 J. Immunol. 51:682(1984)、 McMichaelら、 N. England. J. Med. 309:13(1983)、 Zugelら、 J. Immunol. 161:1705(1998)、 Rivoltiniら、 Cancer Research 59:301(1999)、 Parhurstら、 J. Immunol. 157:2539(1996))が、本研究ではこれを発見しなかった。例えば、CEA.691エピトープに対しては、TCR接触残基が8位であり、一方で、ヘテロクリティックは、3および5位でアナログ置換が認められた。いかなる仮定によっても束縛されることを意図しないが、MHC結合の変更は機構となり得る。このアナログに対して実施された結合分析は、大多数の場合(80%)において好むと好まざるとにかかわらずMHC結合における変更があることを示した。HLA−A2結合について試験された13アナログのうち10アナログはMHC結合の変更を持ち、6アナログは、野生型のペプチドおよび野生型より結合の悪い4アナログよりも良好に結合するが、まだ実質的に増加した生物学的応答を生じた。いくつかの研究ではMHC結合を増強するために一次のMHCアンカー残基を修飾する(この手段は、アナログを生じるためにいくつかのグループによって用いられた(Pfeifferら、 J. Exp. Med. 181:1569(1995)、 Valmoriら、J.Immunol.160:1750−1758(1998))。一次のTCRの接触残基あるいは一次のMHCアンカー残基の変化無しに生物学的応答の増加は本研究においては認められなかった。応答の増加がMHC結合における変更にともない媒介されるので、その効果は二次アンカーの位置の変化により媒介しうることを前提とする。これを支持するさらなる証拠は、ヘテロクリティックの置換がそのペプチドの中間における奇数位置(3、5、7)で起こるという発見から生じる。3、5および7のこれらの位置全ては、HLA−A2分子に対する結合にとって二次アンカー位置であると示された(Boonら、 Annu. Rev. Immunol. 12:337−65 (1994)、 Ishiokaら、 J. Immunol. 162(7):3915−25 (1999) )。
【0063】
これらの位置のうちの2つ(3および7)は、HLA−A2.1分子に結合するための二次アンカー位置であることがいくつかのグループにより示された(Ruppertら、 Cell 74:929(1993)、 Madden、 Annu. Rev. Immunol. 13:587−622(1995))。そのような二次アンカー位置の変更は、T細胞認識差異に転換しうる(Valmoriら、 J. Immnol. 160:1750(1998); Davisら、 Annu. Rev. Immnol. 16:523 (1998))が、これらの研究では、T細胞の認識差異は、MHC結合における変化に付随し、ヘテロクリティックの獲得に関与するアミノ酸置換の種類に関する法則は規定されなかった。二次アンカーの変化からT細胞の認識の変化へのこのような転換が生じる機構は現在不明瞭である。しかし、いくつかのモデルは、二次アンカーの位置の残基がMHCと結合する様式の変化がTCR接触残基の配向における変更あるいは増加した可撓性を引き起こし得ることを示唆しており、それは、TCRに対するこれらのアナログの結合の増加に帰着する(Kershら、 J. Exp. Med. 184:1259(1996)、 Evavoldら、 J. Immunol. 148:347(1992)、 Alamら、 Immunity 10:227(1999)、 Hamplら、 Immunity 7:379−85(1997))。また、いくつかの以前の研究では、ヘテロクリティックアナログによるT細胞応答の調節が主なTCR接触残基に直接関与することを意味していた(Zarembaら、 Cancer Research 57:4570(1997)、 Loftusら、 Cancer Research 58:2433(1998)、 Dresselら、 J. Immunol. 159:4943(1997))。けれどもこの発見は、現在の体系的解析により実証されない。本研究において同定されたアナログに対して増加したT細胞認識は、おそらくMHC結合容量の増加に起因しないが、増加される結合は、おそらく一次アンカーの位置が最適化されたアナログの場合において、重要な役割を果たす。本研究は、ヘテロクリティックアナログが、おそらくTCRあるいはMHCの結合容量の大きな変更によるよりもむしろ構造における微妙な変更により生じることを示唆する。
【0064】
Th1あるいはTh2サイトカイン産生の調節差異は認められなかった。それどころか、本データは、ヘテロクリティックアナログがTh1およびTh2両応答の産生を増加することを示唆したが、その増加の大きさおよび動力学が異なり得る。事実、いくつかのグループ(Nicholsonら、 Int. Immunol. 12(2):205−13(2000)、 Parhurstら、 J. Immunol. 157:2539(1996))は、最近ペプチドアナログによるそのような全体刺激を報告した。これは、アナログによって誘起されるより強いTCRのシグナルに起因するが、そのような全体刺激の機構は、今後解明されるべきである。
【0065】
関連する腫瘍モデルあるいは自己抗原に対する寛容が存在するモデルを使用するインビボでのヘテロクリティックアナログの効力は評価される。従って、効果的なCTLを生じさせることが攻撃であるとこれまで判明している腫瘍に対するワクチン化として、ヘテロクリティックアナログでの免疫化がより効果的で有効な戦略であることが見出される。
【0066】
要約すると、本出願人らが癌およびウイルス起源のHLA−A2.1制限CTLエピトープのいくつもの異なるヘテロクリティックアナログを同定した。その適切な野生型エピトープは表1に示される。全てのこれらのエピトープは本発明者らのより早い報告において免疫原性であることが示された(Kawashimaら、 Human Immunology 59:1−14(1998)、 Ishiokaら、 J. Immunol. 162(7):3915−25(1999))。最初の実験の中で、CEA.691およびMAGE3.112CTLエピトープの233のアナログの抗原性が調査された。同定された4つのヘテロクリティックアナログの性質は、ヘテロクリティック置換が3、5および7の位置で保存的な置換を含むことを示した。この仮説は、3つのさらなるエピトープであるMAGE2.157、HIVPol.476およびHBVPol.455が関与する引き続きの研究において試験された。このように同定された全てのヘテロクリティックアナログは、提案した本法則、すなわちヘテロクリティックアナログが3、5および/または7の位置での保存的あるいは半保存的な置換に付随されることに一致した。
【0067】
癌免疫治療におけるヘテロクリティックアナログの臨床的適用をより綿密に模倣するため、マウスのエピトープ、p53.261もまた修飾された。T細胞寛容の部分的な状態はこのエピトープに関して報告された(Theobaldら、 Proc.Natl. Acad. Sci. 92:11993−11997(1995)、 Theobaldら、 J. Exp. Med., 185(5):833−841(1997))。予想された9つのp53.261アナログのうち4つは、未変性ペプチドにより誘導されるCTL応答に比べてインビボでのアナログ特異的CTLの応答をより強く誘導することが発見された。より顕著に、ヘテロクリティックアナログによる免疫化により生じたCTLの交差反応性が解析された場合に、3つのp53.261アナログが未変性のp53.261エピトープに対して活発に応答するCTLを誘導した。最終的に、ヒトCTLについてこれらの発見の妥当性はMAGE3.112エピトープのヘテロクリティックアナログがインビトロでのヒトT細胞に対して免疫原性であることを実証することにより考慮された。帰着するCTLは腫瘍細胞株の形態において自然に作用される野生型抗原を認識しうる。
【0068】
ヘテロクリティックアナログが、研究されたすべてのエピトープに対して同定されたので、本明細書に示した研究は、ヘテロクリシティーが全世界的な現象であることを証明する。加えて、本願は、クローンのT細胞集団(以前の研究で明示されたような)およびインビボでの免疫化の後の大部分のT細胞集団の両方におけるヘテロクリティックアナログを検出することが可能であることを示す。さらに、本明細書において、ヘテロクリシティー(HLA A2.1系におけるものと他のクラスIスーパーモチーフに関するものとの両方)が、合理的なヘテロクリシティーの予測を見込む別々の構造上の特徴に付随することが実証される。
【0069】
さらに、p53.261ヘテロクリティックアナログがより高いアビディティによりCTLを誘導し、また、野生型ペプチドにより誘導されるより多数の(前駆体出現率)これらの細胞を誘導することが実証される;インビボでのヘテロクリティックCTLの誘導およびT細胞の寛容を中断するためのその適用は実証される。
【0070】
ヘテロクリティックアナログは、インビボでの免疫化に続いて大部分の特異的なT細胞集団を生じるのに効果的であった。多数のTCR遺伝子由来のTCRを提示するポリクローナルの応答は臨床背景で疾患状態を治療するのにより有効である。最終的に、野生型エピトープに対する強い交差反応性のアビディティを有するCTLの高い前駆体出現率を生じるための能力が、通常は免疫系に対して寛容であるエピトープに対する効果的なCTL応答が要求される例において重要である。
【0071】
(2.定義)
特定のアミノ酸配列に関して、「エピトープ」は、特定の免疫グロブリンによる認識に関与するアミノ酸残基のセットであるか、またはT細胞の状況においては、主要組織適合遺伝子複合体(MHC)によりコードされるHLAの状況に提示される場合にT細胞レセプタータンパク質による認識にとって必要な残基である。インビトロまたはインビボでの免疫系の設定において、エピトープは、一次、二次および三次ペプチド構造、ならびに電荷のような分子の集団的特徴であり、これらは一緒になって、免疫グロブリン、T細胞レセプターまたはHLA分子によりコードされる部位を形成する。本開示を通してエピトープおよびペプチドは、しばしば交換可能に使用される。しかし、本発明のエピトープより大きくかつ本エピトープを含む、分離または精製されたタンパク質またはペプチド分子が、まだ本発明の範囲内にあることは、評価されることである。
【0072】
「クラスIエピトープ」は、クラスI HLA分子に対して結合するペプチドをいう。本明細書において記載されたように、クラスIエピトープは代表的に約8から約13アミノ酸長である。HLA分子に対する結合は、2つの一次アンカー残基により主に調節され、このうちの一方はエピトープのC末端にあり、そして他方は、2または3位にある。結合は、また、1つ以上の二次アンカー残基により補助されうる。読み手の利便性のために、様々な一次HLAクラスI結合アンカーは、表4にて示される。アンカーのパターンは、「モチーフ」と称される。「スーパーモチーフ」は、2つ以上のHLA対立遺伝子によりコードされるHLA分子により共有されるペプチド結合の特異性である。好ましくは、スーパーモチーフ保有ペプチドは、高いまたは中間の親和性(本明細書で定義されるような)で2つ以上のHLA抗原により認識される。クラスIスーパーモチーフの例としては、例えば、A1、A2、A3、A24、B7、B27、B44、B58およびB62が挙げられる。
【0073】
本開示をとおして、「結合データ」の結果を、しばしば「IC50」に換算して表す。IC50は、参照ペプチドの結合の50%阻害が認められる、結合アッセイにおけるペプチドの濃度である。アッセイを行う条件(すなわち、制限HLAタンパク質濃度および標識ペプチド濃度)が付与されると、これらの値は、Kd値を概算する。結合を測定するためのアッセイは、例えば、参考として本明細書に援用される、PCT公開WO94/20127およびWO94/03205に、詳細に記載される。IC50値が、アッセイ条件が変化する場合、そして使用する特定の試薬(例えば、HLA調製物など)に依存して、(しばしば、劇的に)変化し得ることに留意する。例えば、過剰濃度のHLA分子は、所定のリガンドの見かけの測定されたIC50を増加する。あるいは、結合は、参照ペプチドと関連して表される。特定のアッセイが、より高感度またはより低感度になるにつれて、試験するペプチドのIC50は、幾分変化し得るが、参照ペプチドに関連する結合は、有意には変化しない。例えば、参照ペプチドのIC50が10倍増加するような条件下で行うアッセイにおいて、試験ペプチドのIC50値もまた、約10倍シフトする。従って、あいまい性を回避するために、ペプチドが良好、中程度、弱いあるいはネガティブな結合因子であるかの評価は、一般に、標準ペプチドのIC50に対する、そのペプチドのIC50に基づく。結合はまた、当該分野において公知の他のアッセイ系を使用して測定され得る。
【0074】
「カルボキシルあるいはC末端」としてのエピトープ中の残基の位置の指定は、ペプチドのカルボキシル末端に最も近いエピトープの末端の残基の位置をいい、これは以下に定義したような従来の命名法を使用して指定される。エピトープの「C末端」は、ペプチドあるいはポリペプチドの末端に実際に対応してもよいし、対応しなくてもよい。
【0075】
「N末端」または「アミノの末端の位置」としてのエピトープ中の残基の位置の指定は、ペプチドのN末端に最も近いエピトープの末端の残基の位置をいい、これは以下に定義したような従来の命名法を使用して指定される。エピトープの「N末端」は、ペプチドあるいはポリペプチドの末端に実際に対応してもよいし、対応しなくてもよい。
【0076】
「コンピューター」または「コンピューターシステム」は、一般に、以下を備える:プロセッサー;少なくとも1つの情報記憶/検索装置、例えば、ハードドライブ、ディスクドライブまたはテープドライブなど;少なくとも1つの入力装置、例えば、キーボード、マウス、タッチスクリーン、またはマイクロホンなど;およびディスプレイ構造。さらに、コンピューターは、ネットワークと連絡する通信チャネルを備え得る。このようなコンピューターは、多かれ少なかれ、上記に列挙したものを備え得る。
【0077】
本明細書で使用される場合、「保存性」または「保存的」および「半保存性」または「半保存的」であるアミノ酸は、調製Bに従って定義され、表2に示される。
【0078】
本明細書中で使用される場合、HLAクラスI分子に関する「高い親和性」とは、50nM以下のIC50またはKD値での結合として定義され;「中程度の親和性」とは、約50nMと約500nMとの間のIC50またはKD値での結合である。HLAクラスII分子への結合に関する「高い親和性」とは、100nM以下のIC50またはKD値での結合として定義され;「中程度の親和性」とは、約100nMと約1000nMとの間のIC50またはKD値での結合である。
【0079】
「免疫原性ペプチド」または「ペプチドエピトープ」とは、そのペプチドがHLA分子を結合し、そしてCTL応答および/またはHTL応答を誘導するような、対立遺伝子特異的モチーフまたはスーパーモチーフを含むペプチドである。従って、本発明の免疫原性ペプチドは、適切なHLA分子に結合し、そしてその後、免疫原性ペプチドが由来する抗原に対する、細胞傷害性T細胞応答またはヘルパーT細胞応答を誘導し得る。
【0080】
句「単離された」または「生物学的に純粋」とは、そのネイティブ状態において見い出されるような通常その材料に付随する成分を、実質的または本質的に含まない材料をいう。従って、本発明に従う単離されたペプチドは、好ましくは、そのインサイチュ環境下でそのペプチドに通常会合する物質を含まない。
【0081】
「PanDR結合ペプチド」は、1つより多くのHLAクラスIIDR分子(例えば、PADRETMペプチド、Epimmune Inc.、San Diego、CA)を結合する分子ファミリーのメンバーである。PADRETMファミリーの分子を定義するパターンは、HLAクラスIIスーパーモチーフとして考えられ得る。PADRETM分子に見い出されたパターンを含むペプチドは、大部分HLA−DR分子に結合し、インビトロおよびインビボでヒトヘルパーTリンパ球(HTL)応答を刺激する。
【0082】
「薬学的に受容可能な」は、一般に、非毒性の、不活性な、および/または生理学的に適合性の組成物をいう。
【0083】
(3.本発明のペプチド)
本発明に従うペプチドは、組み換えDNA技術または化学合成によるか、あるいは天然腫瘍あるいは病原性生物のような天然源から合成的に調製されうる。ペプチドエピトープは、個々にまたはポリエピトープペプチドとして合成され得る。ペプチドは、好ましくは、他の天然に存在する宿主細胞のタンパク質およびそのフラグメントを実質的に有しないが、いくつかの実施形態において、ペプチドは、天然フラグメントまたは粒子に合成的に結合体化され得る。
【0084】
HLAクラスIペプチドは、当該分野において周知であり、MHCクラスI分子に結合するペプチドとして定義される。本発明に従うペプチドは、様々な長さであり、そしてそれらの中性(非荷電)形態または塩である形態のいずれかであり得る。本発明に従うペプチドは、グリコシル化、側鎖酸化、もしくはリン酸化反応のような修飾を有しないか、またはそれらは修飾が本明細書中に記載されるようなペプチドの生物学的活性を破壊しない条件を前提としてこれらの修飾を含むかのいずれかである。
【0085】
対応する「野生型」として役目を果たすクラスIエピトープは、任意のタンパク質源に由来し得る。例えば、クラスIペプチドは、ウイルス性抗原、腫瘍付随抗原、寄生物抗原、細菌抗原、または真菌抗原由来であり得る。本発明のいくつかの好ましい局面において、クラスIペプチドは、被験体の免疫系が寛容(すなわち先の抗原に対する曝露により引き起こされる特異的免疫学的非応答性)を発現する抗原に由来する。
【0086】
従って、いくつもの潜在的標的エピトープに基するヘテロクリティックアナログは、本発明にて利用され得る。適合した腫瘍付随抗原の例としては、前立腺特異的抗原(PSA)、黒色腫抗原MAGE1、MAGE2、MAGE3、MAGE−11、MAGE−A10、並びにBAGE、GAGE、RAGE、MAGE−C1、LAGE−1、CAG−3、DAM、MUC1、MUC2、MUC18、NY−ESO−1、CDK4、BRCA2、NY−LU−1、NY−LU−7、NY−LU−12、CASP8、RAS、KIAA−2−5、SCCs、p53、p73、CEA、Her2/neu、Melan−A、gp100、チロシナーゼ、TRP2、gp75/TRP1、カリクレイン、前立腺特異的膜抗原(PSM)、前立腺酸性ホスファターゼ(PAP)、前立腺特異的抗原(PSA)、PT1−1、B−カテニン、PRAME、テロメラーゼ、FAK、サイクリンD1タンパク質、NOEY2、EGF−R、SART−1、CAPB、HPVE7、p15、葉酸レセプターCDC27、PAGE−1、そしてPAGE−4が挙げられる。適合した感染症付随抗原の例としては、B型肝炎コア抗原および表面抗原(HBVc、HBVs)、C型肝炎抗原、エプスタイン−バーウイルス抗原、ヒト免疫不全症ウイルス(HIV)抗原、そしてヒトパピローマウイルス(HPV)抗原、Mycobacterium tuberculosisおよびChlamydiaが挙げられる。適合する真菌抗原の例としては、Candida albicans、Cryptococcus neoformans、Coccidoides spp.、Histoplasma spp.およびAspergillus fumigatisに由来する抗原が挙げられる。適合する原性動物の寄生物抗原の例としては、P. falciparum、Trypanosoma spp.、Schistosoma spp.、Leishmania sppなどを含む、Plasmodium spp.由来のものが挙げられる。
【0087】
対応するヘテロクリティックアナログを構築するために本発明の法則が適用される野生型の配列として用いられ得るエピトープは、任意のクラスIエピトープに対応することが見出され得る。上に記載されるもののような任意の抗原に対して、特定のクラスI対立遺伝子に付随するモチーフは、このようなエピトープが備わっている抗原のアミノ酸配列中の位置を決定する指針として利用され得る。この決定は、視覚的にかまたは好ましくはコンピューター技術および関連のソフトウェアを使用して行われ得る。従って、例えば2位のバリンおよびC末端のアルギニンを含むような、例えばA3スーパーモチーフの認識により、要求される任意の抗原のアミノ酸配列は、このモチーフを保有するエピトープについて調査され得る。次いで、そのエピトープは、要求されるアナログを獲得するために本発明に記載される法則に従って修飾され得る。
【0088】
可能ならば、本発明のHLAクラスI結合エピトープを最適化することが所望され得、例えばこれは、約8から約13、しばしば8から11、好ましくは9から10アミノ酸残基長に対するポリエピトープの構築に利用され得る。好ましくは、ペプチドエピトープは、内在的に処理される関連するHLA分子に対して結合される病原体由来ペプチドまたは腫瘍細胞ペプチドとサイズにおいて同一基準であるが、本発明のエピトープを含むペプチドの同定および調製は、また、本明細書に記載された技術を利用して行われ得る。
【0089】
代替の実施形態において、本発明のエピトープは、ポリエピトープペプチドとして、または、ポリエピトープペプチドをコードするミニ遺伝子として関連づけられ得る。
【0090】
別の実施形態において、高濃度のクラスIエピトープおよび/またはクラスIIエピトープを含む天然のペプチド領域を同定することが好ましい。そのような配列は、1つのアミノ酸長に対して最大のエピトープ数を含むことを基として通常選択される。エピトープが、ネスティングされるか、もしくは重なる様式で存在し得ることは、評価されることであり、例えば、10アミノ酸長ペプチドは、2つの9アミノ酸長エピトープならびに1つの10アミノ酸長エピトープを含み得た;細胞内のプロセシングにおいて、それぞれのエピトープは、そのようなペプチドの投与の際にHLA分子により曝露および結合されうる。このより大きく、好ましいマルチ−エピトープのペプチドは、合成的、組換え、あるいは天然素材からの切断を経て生じ得る。
【0091】
本発明のペプチドは、広く多様な方法から調製され得る。好ましい比較的短いサイズに関しては、ペプチドは、従来の技術に従って溶液中あるいは固体支持体上で合成され得る。種々の自動合成機は、市販されており、公知のプロトコールに従って使用され得る。(例えば、Stewart & Young、SOLID PHASE PEPTIDE SYNTHESIS、第2版、Pierce Chemical Co.、1984年を参照)。さらに、個々のペプチドエピトープは、いまだ本発明の範囲内であるより大きなペプチドを作製するために化学連結を使用して結合され得る。
【0092】
あるいは、目的の免疫原性ペプチドをコードするヌクレオチドの配列が、発現ベクターに挿入され、適切な宿主細胞に形質転換またはトランスフェクトされ、そして発現のために適切な条件下で培養される、組換えDNAの技術が利用され得る。これらの製法は、Sambrookら、Molecular Cloning、A LABORATORY MANUAL、Cold Spring Harbor Press、Cold Spring Harbor、 New York(1989)に一般に記載されるように、当該分野において一般的に公知である。従って、1つ以上の本発明のペプチド配列を含む組換えポリペプチドは、適切なT細胞エピトープを提示するために使用され得る。
【0093】
本明細書において企図される好ましい長さのペプチドエピトープに対応するヌクレオチドがコードする配列は、例えば、Matteucciら、J.Am.Chem.Soc.103:3185(1981)のホスホトリエステル法のような化学技術により合成され得る。ペプチドアナログは、天然ペプチド配列をコードする核酸塩基を、適切で、また要求される核酸塩基で置換することにより簡単に作製され得る;典型的な核酸の置換は、本明細書において、モチーフ/スーパーモチーフにより定義されるアミノ酸をコードするものである。次いで、コード配列は、適切なリンカーに供され、また当該分野において通例利用可能な発現ベクター中に連結され得、そしてそのベクターは、要求される融合タンパク質を作製するために適切な宿主を形質転換するために使用され得る。いくつものそのようなベクターおよび適切な宿主系は現在使用可能である。融合タンパク質の発現のために、コードする配列は、作動可能に連結される開始および終止コドン、プロモーターおよびターミネータ領域、ならびに通例要求される細胞の宿主での発現のための発現ベクターを提供する複製系を提供される。例えば、細菌の宿主との適合性を持つプロモーター配列は、要求されるコード配列の挿入のための便利な制限部位を含むプラスミドに提供される。得られる発現ベクターは、適切な細菌の宿主に形質転換される。もちろん、酵母、昆虫あるいは哺乳動物の細胞の宿主はまた、適切なベクター、ならびに制御配列を用いて使用され得る。
【0094】
本発明のアナログは、生じるペプチドの物理的特性(例えば、安定性または溶解性)を修飾するための置換を含むペプチドが含まれ得る。例えば、ペプチドはシステイン(C)のα−アミノ酪酸での置換によって修飾され得る。この化学的特質に起因して、システインはジスルフィド架橋を形成する傾向を有し、そして結合能力を減らすようにペプチド構造を十分に変化させる。Cの代わりにα−アミノ酪酸を用いることは、この問題を緩和させるだけでなく、実際に特定の例において結合および交差結合能力を改善する。システインをα−アミノ酪酸と置換することは、ペプチドエピトープの任意の残基で、すなわちアンカー位置または非アンカー位置のいずれかで起こり得る。
【0095】
種々のアミノ酸模倣物または非天然のアミノ酸を有する修飾されたペプチドは特に有用である。なぜならそれらはインビボにおいて増加した安定性を表す傾向があるからである。このようなアナログはまた、改善された貯蔵寿命または製造の特性を有し得る。より詳細には、重要ではないアミノ酸はタンパク質中の天然に存在するアミノ酸(例えばL−α−アミノ酸またはそれらのD−異性体)に制限される必要はないが、非天然のアミノ酸、例えばアミノ酸の模倣物(例えばD−ナフチルアラニン(naphylalanin)またはL−ナフチルアラニン;D−フェニルグリシンまたはL−フェニルグリシン;D−2−チエニルアラニン(thieneylalanine)またはL−2−チエニルアラニン;D−1、−2、3−もしくは4−ピレネイルアラニン(pyreneylalanin)またはL−1、−2、3−もしくは4−ピレネイルアラニン;D−3−チエニルアラニンまたはL−3チエニルアラニン;D−(2−ピリジニル)−アラニンまたはL−(2−ピリジニル)−アラニン;D−(3−ピリジニル)−アラニンまたはL−(3−ピリジニル)−アラニン;D−(2−ピラジニル)−アラニンまたはL−(2−ピラジニル)−アラニン;D−(4−イソプロピル)−フェニルグリシンまたはL−(4−イソプロピル)−フェニルグリシン;D−(トリフルオロメチル)−フェニルグリシン;D−(トリフルオロメチル)−フェニルアラニン;D−ρ−フルオロフェニルアラニン;D−ρ−ビフェニルフェニルアラニンまたはL−ρ−ビフェニルフェニルアラニン;D−ρ−メトキシビフェニルフェニルアラニンまたはL−ρ−メトキシビフェニルフェニルアラニン;D−インドール(アルキル)アラニンまたはL−2−インドール(アルキル)アラニンおよびD−2−アルキルアラニンまたはL−アルキルアラニン(ここでアルキル基はメチル、エチル、プロピル、ヘキシル、ブチル、ペンチル、イソプロピル、イソブチル、sec−イソチル、イソ−ペンチルまたは酸性ではないアミノ酸に置換され得るかまたは置換されない)が挙げられ得る。非天然のアミノ酸の芳香環として、例えばチアゾリル、チオフェニル、ピラゾリル、ベンジミダゾリル、ナフチル、フラニル、ピロリル、およびピリジル芳香環が挙げられる。
【0096】
ペプチドの安定性は多数の方法でアッセイされ得る。例えば、ペプチダーゼおよびヒト血漿および血清のような種々の生物学的培地は安定性を試験するために用いられている。例えば、Verhoefら、Eur.J.Drug Metab.Pharmacokinetics 11:291(1986)を参照のこと。本発明のペプチドの半減期は25%ヒト血清(v/v)アッセイを用いて簡便に決定される。そのプルトコールは一般的に以下のようである:プールしたヒト血清(AB型、熱で不活性化されていない)を使用する前に遠心分離により脱脂する。次いで血清をRPMI−1640または別の適切な組織培養培地で25%に希釈する。予め決定された時間間隔で、少量の反応溶液が取り出され、そして6%のトリクロロ酢酸水溶液(TCA)またはエタノールのいずれかを加えられる。濁った反応サンプルは4℃に15分間冷却され、その後スピンして沈殿した血清タンパク質をペレットにする。次いでペプチドの存在は安定性特異的なクロマトグラフィ条件を用いて逆相HPLCによって決定される。
【0097】
(4.クラスIモチーフ)
過去数年間で、HLAクラスI分子の巨大な割合が比較的少ないスーパータイプに分類され得、それぞれは広く重複するペプチド結合のレパートリーおよび主なペプチド結合ポケットのコンセンサス構造によって特徴付ける、ということを実証する証拠が集まってきている。従って本発明のペプチドは、いくつかのHLA特異的アミノ酸モチーフのうちのいずれか1つによってか(例えば表3〜4を参照のこと)、またはモチーフの存在がいくつかの対立遺伝子特異的HLA抗原に結合する能力に対応する場合はスーパーモチーフによって同定されている。特定のアミノ酸スーパーモチーフを有するペプチドに結合するHLA分子は、集合的にHLA「スーパータイプ」という。
【0098】
読者に便利にするため、以下に記載され表3〜4に要約されるペプチドのモチーフおよびスーパーモチーフは同定のための手引きおよび発明に従うペプチドエピトープの用途を提供する。これは、アナログを構築するための本明細書中に示される法則を適用するために、以下の実施例に示されるものとは異なる種々のクラスIモチーフに対応する候補野生型エピトープまたは以下に示されるものを保有するが異なる抗原の中にあるエピトープの同定を可能にする。
【0099】
ヘテロクリティックアナログは、ペプチドが属するモチーフまたはスーパーモチーフに関わり無く、そのペプチドから発明の方法に従って設計され得る。下記に描いたHLAクラスIペプチドエピトープのスーパーモチーフおよびモチーフの一次アンカー残基を表3に要約する。表3(a)で述べたHLAクラスIモチーフは、ここで主張される本発明に最も著しい関連のモチーフである。HLAクラスIスーパータイプファミリーを含む対立遺伝子特異的なHLA分子を表4に列挙する。いくつかの例において、ペプチドエピトープはモチーフおよびスーパーモチーフの両方に列挙され得る。特定のモチーフとそれぞれのスーパーモチーフとの関係は、個々のモチーフの説明中に示される。
【0100】
(i.HLA−A1スーパーモチーフ)
HLA−A1スーパーモチーフは、2位おける小さな(TまたはS)または疎水性(L、I、VまたはM)の一次アンカー残基のペプチドリガンド中の存在、そしてエピトープのC末端位置における芳香族(Y、FまたはW)の一次アンカー残基の存在により特徴付けられる。A1スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、HLA−A1スーパータイプ)は、少なくともA*0101、A*2601、A*2602、A*2501、およびA*3201を含む(例えば、DiBrino,M.ら、J.Immunol.151:5930,1993;DiBrino,M.ら、J.Immunol.152:620,1994;Kondo,A.ら、Immunogenetics 45:249,1997を参照のこと)。A1スーパーファミリーのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表4に示される。
【0101】
(ii.HLA−A2スーパーモチーフ)
対立遺伝子特異的HLA−A2.1分子に対する一次アンカーの特異性(例えば、Falkら、Nature 351:290−296,1991;Huntら、Science 255:1261−1263,1992;Parkerら、J.Immunol.149:3580−3587,1992;Ruppertら、Cell 74:929−937,1993を参照のこと)およびHLA−A2およびHLA−A28分子の間での交差反応性結合が記載される(例えば、関連データの総説については、Fruciら、Human Immunol.38:187−192,1993;Tanigakiら、Human Immunol.39:155−162,1994;Del Guercioら、J.Immunol.154:685−693,1995;Kastら、J.Immunol.152:3904−3912,1994を参照のこと)。これらの一次アンカー残基は、HLA−A2スーパーモチーフを規定し;ペプチドリガンド中の存在は、いくつかの異なるHLA−A2およびHLA−A28分子を結合する能力に対応する。HLA−A2スーパーモチーフは、2位に一次アンカー残基としてL、I、V、M、A、TまたはQを有するペプチドリガンド、およびエピトープのC末端位置に一次アンカー残基としてL、I、V、M、AまたはTを有するペプチドリガンドを含む。
【0102】
HLA分子の対応するファミリー(すなわち、これらのペプチドを結合するHLA−A2スーパータイプ)は、少なくとも以下を含む:A*0201、A*0202、A*0203、A*0204、A*0205、A*0206、A*0207、A*0209、A*0214、A*6802およびA*6901。A2スーパーファミリーのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表4に示される。
【0103】
(iii.HLA−A3スーパーモチーフ)
HLA−A3スーパーモチーフは、2位における一次アンカーとしてのA、L、I、V、M、SまたはTのペプチドリガンド中の存在、そしてエピトープのC末端位置における(例えば、9マーの9位に)正に荷電した残基RまたはKの存在により特徴付けられる(例えば、Sidneyら、Hum.Immunol.45:79,1996を参照のこと)。A3スーパーモチーフを結合するHLA分子の対応するファミリー(HLA−A3スーパータイプ)の例示的メンバーとしては、少なくとも:A*0301、A*1101、A*3101、A*3301およびA*6801が挙げられる。A3スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0104】
(iv.HLA−A24スーパーモチーフ)
HLA−A24スーパーモチーフは、2位における一次アンカーとして芳香族残基(F、W、またはY)または疎水性脂肪族残基(L、I、V、M、またはT)がペプチドリガンド中に存在し、そしてエピトープのC末端位置に一次アンカーとしてのY、F、W、L、IまたはMの存在により特徴付けられる(例えば、SetteおよびSidney,Immunogenetics(印刷中)1999を参照のこと)。A24スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、A24スーパータイプ)としては、少なくともA*2402、A*3001およびA*2301が挙げられる。A24スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0105】
(v.HLA−B7スーパーモチーフ)
HLA−B7スーパーモチーフは、一次アンカーとして2位にプロリンを有するペプチド、およびエピトープのC末端位置における一次アンカーとして疎水性アミノ酸または脂肪族アミノ酸(L、I、V、M、A、F、W、またはY)により特徴付けられる。B7スーパーモチーフを結合するHLA分子の対応するファミリー(すなわち、HLA−B7スーパータイプ)は、少なくとも26のHLA−Bタンパク質を含み、これらとしては以下が挙げられる:B*0702、B*0703、B*0704、B*0705、B*1508、B*3501、B*3502、B*3503、B*3504、B*3505、B*3506、B*3507、B*3508、B*5101、B*5102、B*5103、B*5104、B*5105、B*5301、B*5401、B*5501、B*5502、B*5601、B*5602、B*6701、およびB*7801(例えば、関連データの総説については、Sidneyら、J.Immunol.154:247,1995;Barberら、Curr.Biol.5:179,1995;Hillら、Nature 360:434,1992;Rammenseeら、Immunogenetics 41:178,1995を参照のこと)。B7スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0106】
(vi.HLA−B27スーパーモチーフ)
HLA−B27スーパーモチーフは、2位における一次アンカーとしての正に荷電した残基(R、H、またはK)のペプチドリガンド中の存在、そしてエピトープのC末端位置における一次アンカーとして疎水性残基(F、Y、L、W、M、I、A、またはV)の存在により特徴付けられる(例えば、SidneyおよびSette,Immunogenetics(印刷中)1999を参照のこと)。B27スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B27スーパータイプ)の例示的なメンバーとしては、少なくとも:B*1401、B*1402、B*1509、B*2702、B*2703、B*2704、B*2705、B*2706、B*3801、B*3901、B*3902、およびB*7301が挙げられる。B27スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0107】
(vii.HLA−B44スーパーモチーフ)
HLA−B44スーパーモチーフは、2位における一次アンカーとしての負に荷電した残基(DまたはE)のペプチドリガンド中の存在、そしてエピトープのC末端位置における一次アンカーとしての疎水性残基(F、W、Y、L、I、M、V、またはA)の存在により特徴付けられる(例えば、Sidneyら、Immunol.Today 17:261,1996参照のこと)。B44スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B44スーパータイプ)の例示的なメンバーとしては、少なくとも以下が挙げられる:B*1801、B*1802、B*3701、B*4001、B*4002、B*4006、B*4402、B*4403、およびB*4006。
【0108】
(viii.HLA−B58スーパーモチーフ)
HLA−B58スーパーモチーフは、2位での一次アンカー残基としての小さな脂肪族残基(A、S、またはT)のペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としての芳香族または疎水性の残基(F、W、Y、L、I、V、M、またはA)の存在によって、特徴付けられる(例えば、関連するデータの総説についてはSidneyおよびSette,Immunogenetics,印刷中、1999を参照のこと)。B58スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B58スーパータイプ)の例示的なメンバーとしては、少なくとも以下が挙げられる:B*1516、B*1517、B*5701、B*5702、およびB*5801。B58スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0109】
(ix.HLA−B62スーパーモチーフ)
HLA−B62スーパーモチーフは、2位での一次アンカーとしての極性脂肪族残基Qまたは疎水性脂肪族残基(L、V、M、I、またはP)のペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカーとして疎水性残基(F、W、Y、M、I、V、L、またはA)の存在によって、特徴付けられる(例えば、SidneyおよびSette,Immunogenetics,印刷中、1999を参照のこと)。B62スーパーモチーフに結合するHLA分子の対応するファミリー(すなわち、B62スーパータイプ)の例示的なメンバーとしては、少なくとも以下が挙げられる:B*1501、B*1502、B*1513、およびB5201。B62スーパータイプのメンバーであると推定される他の対立遺伝子特異的HLA分子は、表3に示される。
【0110】
(x.HLA−A1モチーフ)
HLA−A1モチーフは、2位での一次アンカー残基としてのT、S、またはMのペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としてのYの存在によって、特徴付けられる。代替の対立遺伝子特異的A1モチーフは、2位よりむしろ3位における、一次アンカー残基によって特徴付けられる。このモチーフは、D、E、A、またはSの一次アンカー残基としての3位における存在、および、エピトープのC末端位置における一次アンカー残基としてのYの存在によって、特徴付けられる(例えば、関連するデータの総説については、DiBrinoら、J.Immunol.,152:620、1994;Kondoら、Immunogenetics 45:249、1997;およびKuboら、J.Immunol.152:3913、1994を参照のこと)。
【0111】
(xi.HLA−A*0201モチーフ)
HLA−A2*0201モチーフは、2位での一次アンカー残基としての、LまたはMのペプチドリガンドにおける存在、およびLまたはVの9残基ペプチドのC末端位置での一次アンカー残基としての存在によって特徴付けられることが決定され(例えば、Falkら、Nature 351:290−296、1991を参照のこと)、そしてさらに、9アミノ酸ペプチドの2位にIを含み、そしてC末端位置にIまたはAを含むことが、見出された(例えば、Huntら、Science 255:1261−1263、3月6日、1992;Parkerら、J.Immunol.149:3580−3587、1992を参照のこと)。A*0201対立遺伝子特異的モチーフはまた、本発明者らによって、さらにV、A、T、またはQを、一次アンカー残基としてこのエピトープの2位に含み、そしてMまたはTを、一次アンカー残基としてこのエピトープのC末端位置に含むことが規定された(例えば、Kastら、J.Immunol.152:3904−3912、1994を参照のこと)。従って、HLA−A*0201モチーフは、2位に一次アンカー残基としてL、I、V、M、A、T、またはQを有し、そしてこのエピトープのC末端位置に一次アンカー残基としてL、I、V、M、A、またはTを有する、ペプチドリガンドを含む。HLA−A*0201モチーフの一次アンカー位置を特徴付ける、好ましい許容される残基は、A2スーパーモチーフを記載する残基と同一である。
【0112】
(xii.HLA−A3モチーフ)
HLA−A3モチーフは、2位での一次アンカー残基としてのL、M、V、I、S、A、T、F、C、G、またはDのペプチドリガンドにおける存在、およびエピトープのC末端位置における一次アンカー残基としてのK、Y、R、H、F、またはAの、存在によって、特徴付けられる(例えば、DiBrinoら、Proc.Natl.Acad.Sci.USA 90:1508、1993;およびKuboら、J.Immunol.152:3913−3924、1994を参照のこと)。
【0113】
(xiii.HLA−A11モチーフ)
HLA−A11モチーフは、2位での一次アンカー残基としてのV、T、M、L、I、S、A、G、N、C、D、またはFのペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としてのK、R、Y、またはHの存在によって、特徴付けられる(例えば、Zhangら、Proc.Natl.Acad.Sci.USA 90:2217−2221、1993;およびKuboら、J.Immunol.152:3913−3924、1994を参照のこと)。
【0114】
(xiv.HLA−A24モチーフ)
HLA−A24モチーフは、2位での一次アンカー残基としてのY、F、W、またはMのペプチドリガンド中の存在、およびエピトープのC末端位置における一次アンカー残基としてのF、L、I、またはWの存在によって、特徴付けられる(例えば、Kondoら、J.Immunol.155:4307−4312、1995;およびKuboら、J.Immunol.152:3913−3924、1994を参照のこと)。
【0115】
(5.T細胞応答を検出するためのアッセイ)
一旦、本発明のヘテロクリティックアナログが合成されると、それらは、T細胞応答を誘発する能力について試験され得る。モチーフ保有ペプチド(例えば、ヘテロクリティックアナログ)の調製および評価は、PCT公報WO94/20127およびWO94/03205に記載される。簡単に述べると、特定の抗原由来のエピトープを含むペプチドが合成され、適切なHLAタンパク質に結合するそれらの能力について試験される。これらのアッセイは、放射性ヨウ素標識した参照ペプチドの結合に関して、精製されるHLAクラスI分子への本発明のペプチドの結合を評価する工程を包含し得る。あるいは、空のクラスI分子を発現する細胞(すなわち、その中にペプチドを含まない)は、免疫蛍光染色およびフローマイクロ蛍光定量法によってペプチド結合について評価され得る。ペプチド結合を評価するために使用され得る他のアッセイは、ペプチド依存クラスIアセンブリアッセイおよび/またはペプチド競合によるCTL認識の阻害を含む。代表的に、500nM以下の親和性でクラスI分子に結合するそれらのペプチドは、感染された個体または免疫化された個体から誘導体化されたCTLに対する標的として作用する能力について、および、疾患に関連する選択された標的細胞と反応し得るCTL集団を引き起こし得る一次インビトロCTL応答または一次インビボCTL応答を誘発する能力について、さらに評価される。
【0116】
T細胞応答を検出するために使用される従来のアッセイとしては、増殖アッセイ、リンホカイン分泌アッセイ、直接細胞傷害性アッセイ、および限界希釈アッセイが挙げられる。このようなアッセイは、ヘテロクリティックアナログのペプチドによる免疫応答の誘導を、非ヘテロクリティックアナログのクラスIペプチド(例えば、ここからヘテロクリティックアナログ配列が基礎にされた)により誘導される応答と比較する際に有用である。例えば、ペプチドを用いてインキュベートされた抗原提示細胞は、応答細胞集団(responder cell population)においてCTL応答を誘導する能力についてアッセイされ得る。抗原提示細胞は、末梢血単核細胞または樹状細胞のような正常細胞であり得る。あるいは、内部的にプロセスされたペプチドを有するクラスI分子を装填するそれらの能力が欠乏し、そして適切なヒトクラスI遺伝子でトランスフェクトされた変異体非ヒト哺乳動物細胞株が、インビトロで一次CTL応答を誘発するペプチドの能力について試験するために使用され得る。
【0117】
末梢血単核細胞(PBMC)は、CTL前躯体の応答細胞源として使用され得る。適切な抗原提示細胞は、ペプチドと共にインキュベートされ、次いでこの後、ペプチド装填した抗原提示細胞が、最適な培養条件下で応答細胞集団と共にインキュベートされる。陽性CTL活性化は、放射性標識した標的細胞(特定のペプチドをパルスされた標的およびペプチド配列が誘導される内因的にプロセスされた形態の抗原を発現する標的細胞)を殺傷するCTLの存在について培養物をアッセイすることによって決定され得る。
【0118】
さらに、フルオレセイン標識したHLA四量体複合体で染色することによって抗原特異的T細胞の直接的定量を可能にする方法が、もたらされた(Altman,J.D.ら、Proc.Natl.Acad.Sci.USA 90:10330、1993;Altman,J.D.ら、Science 274:94,1996)。他の比較的最近の技術的開発としては、細胞内リンホカインの染色、およびインターフェロン−γ放出アッセイまたはElispotアッセイが挙げられる。四量体染色、細胞内リンホカイン染色およびElispotアッセイは全て、さらなる従来のアッセイより少なくとも10倍より感度が高いようである(Lalvani,Aら、J.Exp.Med.186:859、1997;Dunbar、P.R.ら、Curr.Biol.8:413、1998;Murali−Krishna,K.ら、Immunity 8:177、1998)。
【0119】
所望される場合、HTLの活性化はまた、当該分野で公知の技術(例えば、T細胞増殖およびリンホカイン(例えば、IL−2)の分泌)を使用して評価され得る(例えば、Alexanderら、Immunity 1:751〜761、1994を参照のこと)。
【0120】
あるいは、HLAトランスジェニックマウスの免疫化は、ペプチドエピトープの免疫原性を決定するために使用され得る。いくつかのトランスジェニックマウスモデル(ヒトA2.1、A11(これはHLA−A3エピトープを分析するためにさらに使用され得る)、およびB7対立遺伝子を有するマウスを含む)が特徴付けられ、そして他のマウス(例えば、HLA−A1およびA24に対するトランスジェニックマウス)が開発されている。HLA−DR1およびHLA−DR3マウスモデルもまた開発されている。他のHLA対立遺伝子を有するさらなるトランスジェニックマウスモデルが、必要な場合作製され得る。マウスは、不完全フロイトアジュバント中の乳化したペプチドで免疫化され得、そして得られるT細胞は、ペプチドをパルスされた標的細胞および適切な遺伝子でトランスフェクトされた標的細胞を認識するそれらの能力について試験される。CTL応答は、上記の細胞傷害性アッセイを使用して分析され得る。同様に、HTL応答は、T細胞増殖またはリンホカインの分泌のようなアッセイを使用して分析され得る。
【0121】
本発明のヘテロクリティックアナログは、しばしばTh1サイトカイン応答およびTh2サイトカイン応答の両方を誘導する。それゆえ、ヘテロクリティックな候補物を予め選択されたクラスIペプチドと比較するための1つの方法は、Th1サイトカインおよびTh2サイトカインの誘導を試験するための方法である。予め選抜したクラスIペプチドは、代表的にヘテロクリティックアナログに誘導されるペプチド、または、そのようなペプチドが存在しない場合は候補物に対して最も高い類似性を有するクラスIペプチドである。本発明のヘテロクリティックアナログは、代表的にTh1サイトカイン応答およびTh2サイトカイン応答の両方を誘導するが、これはアナログが誘導されたクラス1ペプチドに比較して非常に増強されたレベルにおいてである。例えば、所定のヘテロクリティックアナログは、等しいレベルのTh1サイトカインまたはTh2サイトカイン(50〜100pg/ml)を、このアナログが誘導された野生型ペプチドと比較して10倍以下の用量で刺激する。さらに、クラスIペプチドがTh1応答またはTh2応答のどちらかのみ、またはいずれかを主に誘導する場合、ヘテロクリティックアナログはTh1応答およびTh2応答の両方を誘導し得る。Th1サイトカインとしては、例えばIFNγ、IL−2およびIL3が挙げられる。Th2サイトカインとしては、例えばIL−4、IL−5、IL−6およびIL−10が挙げられる。サイトカインの産生は、例えばELISA法または他の免疫学的定量法を用いて測定される。(例えば、McKinneyら、Journal of Immunological Methods 237:105〜117、2000を参照のこと)
(6.診断薬としておよび免疫応答を評価するためのペプチドエピトープの使用)
本発明の一実施形態において、本明細書中に記載されるようなヘテロクリティックアナログペプチドが、免疫応答を評価するための試薬として使用される。評価される免疫応答は、免疫原として任意の因子を使用することによって、誘導され、この因子は、試薬として用いられるペプチドエピトープを認識し、そしてこれに結合する抗原特異性CTLまたはHTLの誘導を生成し得る。ペプチド試薬は、免疫原として使用される必要はない。このような分析のために使用され得るアッセイシステムとしては、比較的最近の技術開発(例えば、細胞内リンホカインに関する四量体染色およびインターフェロン放出アッセイ、またはElispotアッセイ)が挙げられる。
【0122】
例えば、本発明のペプチドは、腫瘍細胞抗原または免疫原の曝露後の抗原特異的CTLの存在について末梢血単核細胞を評価するために、四量体染色アッセイにおいて使用される。HLA四量体複合体を使用して、抗原特異性CTLを直接可視化し(例えば、Oggら、Science 279:2103〜2106、1998;およびAltmanら、Science 174:94〜96、1996を参照のこと)、そして末梢血単核細胞のサンプル中の抗原特異的CTL集団の頻度を決定する。本発明のペプチドを使用する四量体試薬は、以下のように生成され得る:HLA分子に結合するペプチドは、三分子複合体を生成するために、対応するHLA重鎖およびβ2−ミクログロブリンの存在下で再折り畳まれる。この複合体は、先にタンパク質中の操作された部位において重鎖のカルボキシル末端でビオチン化される。四量体形成は、次いで、ストレプトアビジンの付加によって誘導される。蛍光標識されたストレプトアビジンを用いて、抗原特異的細胞を染色するために四量体が使用され得る。これらの細胞は、次いで、例えば、フローサイトメトリーによって同定され得る。このような分析は、診断目的または予後目的のために使用され得る。この手順によって同定された細胞はまた、治療目的のために使用され得る。
【0123】
本発明のペプチドはまた、免疫リコール応答を評価するための試薬として使用される(例えば、Bertoniら、J.Clin.Invest.100:503〜513、1997、およびPennaら、J.Exp.Med.174:1565〜1570、1991を参照のこと)。例えば、癌を有する個体由来の患者のPBMCサンプルは、特定のペプチドを使用して、抗原特異的CTLまたはHTLの存在について分析され得る。単核細胞を含む血液サンプルは、PBMCを培養し、そして本発明のペプチドでその細胞を刺激することによって評価され得る。適切な培養期間の後、拡大された細胞集団は、例えば、CTLまたはHTL活性について分析され得る。
【0124】
ペプチドはまた、ワクチンの効力を評価するための試薬として使用される。免疫原をワクチン接種された患者から得たPBMCは、例えば、上記の方法のいずれかを使用して分析され得る。患者は、HLA型であり、そしてその患者に存在する対立遺伝子特異的分子を認識するペプチドエピトープ試薬が、分析のために選択される。ワクチンの免疫原性は、PBMCサンプル中のエピトープ特異的CTLおよび/またはHTLの存在によって示される。
【0125】
本発明のペプチドはまた、当該分野で周知の技術を使用して、抗体を作製するために使用され(例えば、CURRENT PROTOCOLS IN IMMUNOLOGY、Wiley/Greene、NY;およびAntibodies A Laboratory Manual 、Harlow and Lane、Cold Spring Harbor Laboratory Press、1989を参照のこと)、これは癌を診断またはモニターするための試薬として有用であり得る。このような抗体としては、HLA分子の状況においてペプチドを認識する抗体(すなわち、ペプチド−MHC複合体に結合する抗体)が挙げられる。
【0126】
(7.ワクチン組成物)
免疫学的に有効な量の本明細書中に述べるような1つ以上のペプチドを含むワクチンおよびそのワクチンを準備する方法は、本発明のさらなる実施形態である。一旦、適切な免疫原性エピトープが規定されると、それらは様々な手段により分類され送達され得るが、本明細書中ではこれらを「ワクチン」組成物と呼ぶ。このようなワクチン組成物は、例えば以下のようなものが挙げられる:リポペプチド(例えば、Vitiello、A.ら、J.Clin.Invest.95:341,1995)、ポリ(DL−ラクチド−co−グリコリド)(「PLG」)ミクロスフィアに封入されたペプチド組成物(例えば、Eldridgeら、Molec.Immunol.28:287−294,1991:Alonsoら、Vaccine 12:299−306,1994;Jonesら、Vaccine 13:675−681,1995)、免疫刺激複合体(ISCOMS)により包まれたペプチド組成物(例えば、Takahashiら、Nature, 344:873−875,1990;Huら、Clin Exp Immunol.113:235−243,1998)、多抗原ペプチドシステム(MAPs)(例えば、Tam,J.P.、Proc.Natl.Acad.Sci.USA 85:5409−5413,1988;Tam、J.P.,J. Immunol.Methods 196:17−32,1996)、多価のペプチドとして処方されるペプチド;銃式送達システム(ballistic delivery system)に処方されるペプチド、代表的には結晶化されたペプチド、ウイルス送達ベクター(Perkus,M.E.ら、In:Concepts in vaccine development,Kaufman,S.H.E.,編 p.379,1996;Chakrabarti,S.ら、Nature 320:535,1986、Hu,S.L.ら、Nature 320:537,1986;Kiney,M.−P.ら、AIDS Bio/Technology 4:790,1986;Top,F.H.ら、J.Infect.Dis. 124:148,1971;Chanda,P.K.ら、Virology 175;535,1990),ウイルス性もしくは合成起源の粒子(例えば、Kofler,N.ら、J.Immunol.Methods 192:25,1996;Eldridge、J.H.ら、Sem.Hematol.30:16,1993;Falo,L.D.,Jr.ら、Nature Med.7:649,1995)、アジュバント(Warren,H.S.,Vogel,F.R.および Chedid,L.A.Annu.Rev.Immunol.4:369,1986;Gupta,R.K.ら、Vaccine 11:293,1993)、リポソーム(Reddy,R.ら、J.Immunol.148:1585,1992;Rock,K.L.,Immunol.Today 17:131,1996)、あるいは裸または粒子吸着されたcDNA(Ulmer,J.B.ら、Science 259:1745,1993;Robinson,H.L.,Hunt,L.A.,およびWebster,R.G.,Vaccine 11:957,1993;Shiver,J.W.ら、Concepts in vaccine development, Kaufmann,S.H.E.編、p.423,1996;Cease, K.B.,およびBerzofsky,J.A.,Annu.Rev.Immunol.12:923,1994 および Eldridge, J. H. ら、Sem. Hematol.30:16,1993)。Avant Immunotherapeutics,Inc(Needham,Massachusetts)の技術のようなレセプター媒介標的化(recepter mediated targeting)として公知の毒素標的とする送達技術もまた使用され得る。
【0127】
本発明のワクチンは核酸が介在する様式を含む。一つ以上の本発明のペプチドをコードしているDNAまたはRNAはまた患者に投与され得る。このアプローチは、例えば、Wolff ら、Science 247:1465 (1990)ならびに米国特許第5,580,859号;同第5,589,466号;同第5,804,566号;同第5,739,118号;同第5,736,524号;同第5,679,647号;WO 98/04720に記載され、さらに詳細は以下に記載される。DNAに基づく送達技術の例としては、「裸のDNA」、促進(ブピバカイン、ポリマー、ペプチド介在)送達、陽イオン性液体複合体、粒子媒介(「遺伝子銃」)もしくは圧力媒介送達(例えば米国特許第5,922,687号を参照のこと)が挙げられる。
【0128】
治療的または予防的免疫処置の目的で、本発明のペプチドはまた、ウイルスベクターまたは細菌ベクターによって発現され得る。発現ベクターの例としては、痘疹や鶏痘などの弱毒化したウイルス宿主が挙げられる。このアプローチの例のように、ワクシニアウイルスは本発明のペプチドをコードするヌクレオチド配列を発現するベクターとして使用される。腫瘍を保有する宿主に導入する際、組換え型ワクシニアウイルスは免疫原性のペプチドを発現し、それによってCTLおよび/またはHTLを誘発する。ワクシニアベクターおよび免疫処置プロコトルにおいて有用な方法は、例えば米国特許第4,722,848号に記載されている。別のベクターは、BCG(Bacille Calmette Guerin)である。BCGベクターについてはStover ら、Nature 351:456−460(1991)に記載されている。アデノウイルスベクターおよびアデノ随伴ウイルスベクター、レトロウイルスベクター、Salmonella typhiベクター、無毒化した炭疽毒素ベクターなど発明のペプチドの治療的投与または免疫処置に有用な幅広い種々の他のベクターは、本明細書中の記載から当業者にとっては明白である。
【0129】
さらに、本発明に従ったワクチンは、一つ以上の特許請求されるペプチドの組成物を包含する。ペプチドは個々にワクチン中に存在し得る。あるいは、このペプチドは同一のペプチドの複数のコピーから構成されるホモポリマーまたは様々なペプチドのヘテロポリマーとして存在し得る。ポリマーは免疫反応を増大するという利点を有し、そしてこのポリマーをつくるために異種のペプチドエピトープが使われる場合、免疫応答の標的となる病原体生物または腫瘍関連ペプチドの異なる抗原性決定因子と反応する抗体および/またはCTLを誘導する更なる能力という利点を持つ。本組成物は抗原の天然に存在する領域であり得るか、あるいは例えば組換えまたは化学合成によって調製され得る。
【0130】
本発明のワクチンとともに使われる担体は当該分野において周知であり、例えば、サイログロブリン、ヒト血清アルブミンのようなアルブミン、破傷風トキソイド、ポリL−ロイシン、ポリL−グルタミン酸のようなポリアミノ酸、インフルエンザ、B型肝炎ウイルスコアタンパク質などが挙げられる。本ワクチンは水または食塩水、好ましくはリン酸緩衝化食塩水のような生理学的に耐性な(すなわち、受容されうる)希釈剤を含み得る。本ワクチンは、代表的なものにまたアジュバントを含む。フロイントの不完全アジュバント、リン酸アルミニウム、水酸化アルミニウム、またはミョウバンのようなアジュバントは当該分野で周知の材料例である。加えて、本明細書中で開示したように、CTL応答はトリパルミトイル−S−グリセリルシステイニルセリル−セリン(P3CSS)のような本発明のペプチドの脂質への結合によって開始され得る。
【0131】
本発明に従ってペプチド組成物を用いて免疫処置を行う際、注入的、エアロゾル的、経口的、経皮的、経粘膜的、胸膜内腔的、クモ膜下腔的、またはその他の適切な経路を介して、宿主の免疫系は、所望の抗原に特異的な多量のCTLおよび/またはHTLを産生することにより本ワクチンに応答する。結果として、宿主はその後の感染に対して少なくとも部分的に免疫されるか、または現在進行中の慢性的な感染が進展することに対して少なくとも部分的に耐性をもつか、あるいは抗原が腫瘍関連の場合は少なくとも治療的利益を導き出す。
【0132】
いくつかの実施形態において、本発明のヘテロクリティックアナログペプチドを、中和抗体およびまたはヘルパーT細胞の興味ある標的抗原に対する応答を誘導するか、あるいは促進する成分と結合することは望ましいことであり得る。このような組成物の好ましい実施形態は本発明に従って、クラスIエピトープとクラスIIエピトープを包含する。このような組成物の代替の実施形態は、PADRETM(Epimmune,San Diego,CA)分子(例えば、米国特許第5,736,142号に記載されている)のようなpan−DR結合ペプチドに加え、本発明に従ったクラスIエピトープおよび/またはクラスIIエピトープを包含する。
【0133】
本発明のワクチンはまた、樹状細胞(DC)のような抗原提示細胞(APC)を本発明のペプチドを提供するための媒体として包含する。ワクチン組成物は樹状細胞の動員および採取につづいてインビトロでつくられ得、これによって樹状細胞の負荷がインビトロで生じる。例えば、樹状細胞は例えば、本発明に従ったミニ遺伝子(minigene)を伴ってトランスフェクションされるかまたはペプチドとともにパルスされる。それから、樹状細胞は、インビボで免疫応答を誘発するために患者に投与され得る。
【0134】
DNAまたはペプチドのいずれかに基づくワクチン組成物は、また樹状細胞の動員と組み合わせてインビボに投与され得、これによって樹状細胞の負荷がインビボで生じる。
【0135】
抗原性のペプチドは同様にエキソビボでCTLおよび/またはHTL応答を誘発するために用いられる。結果として生じるCTLまたはHTL細胞は、他の従来の治療形式では応答しないか、あるいは本発明の治療的なワクチンペプチドまたは核酸に応答しない患者の腫瘍を処置するのに使われ得る。エキソビボでのある特定の腫瘍関連する抗原に対するCTLまたはHTL応答は、患者または遺伝的に適合性のあるCTLもしくはHTL前駆細胞を樹状細胞のような抗原提示細胞の供給源と適切な免疫原性のペプチドとともに組織培養中でインキュベートすることにより誘導される。適切なインキュベーション時間(代表的には約7〜28日)経過後、この期間中この前駆細胞は活性化され効果細胞へと発展するが、この細胞は患者に再注入され、ここでこれらは特定の標的細胞(感染した細胞または腫瘍細胞)を破壊するか(CTL)またはその破壊の促進を行う(HTL)。トランスフェクションされた樹状細胞はまた抗原提示細胞として使われ得る。
【0136】
本発明のワクチン組成物は、またIL−2、IL−12、GM−CSFなどのような免疫アジュバントと組み合わせる使用法を含む、癌に対する他の処置と組み合わせて使用され得る。
【0137】
好ましくは、ワクチンの用途でポリエピトープ組成物においてアレイのエピトープを封入のために選択する場合に、またはワクチンに取り込むため、および/またはミニ遺伝子のように核酸にコードされるために別個のエピトープを選択する場合、次の原則が利用される。選択を行うために、以下の原則のそれぞれのバランスを保つことが好ましい。所定のワクチン組成物に組み込まれた複数のエピトープは、エピトープが誘導される天然の抗原の配列と隣接し得るが、しかしながらこれは隣接する必要はない。
【0138】
1)投与されると、腫瘍クリアランスに相関することが観察された免疫応答を模倣するエピトープが選択される。HLA I型について、これは少なくとも1つの腫瘍関連抗原(TTA)に由来する3〜4個のエピトープを含む。HLA II型について、類似の原理が採用される;再び、3〜4個のエピトープは、少なくとも1つのTTAから選択される(例えば、Rosenbergら、Science 278:1447〜1450を参照のこと)。一つのTAAからのエピトープは一つ以上のさらなるTAAからのエピトープとの組み合わせで使用され得、頻出発現TAA(frequently−expressed TAA)の様々な発現パターンを持つ腫瘍を標的とするワクチンを産生する。
【0139】
2)免疫原性と相関することが確立された必須結合親和性を有するエピトープが選択される:HLA I型は、500nM以下しばしば200nM以下のIC50を有し、II型については、1000nM以下のIC50を有する。
【0140】
3)十分なスーパーモチーフ保有ペプチドまたは対立遺伝子特異的モチーフ保有ペプチドの十分なアレイは、広い集団適用範囲を与えるように選択される。例えば、少なくとも80%の集団適用範囲を有することが好ましい。モンテカルロ解析(当該分野で公知の統計的評価)を使用して、集団適用範囲の広さまたは冗長性を評価し得る。
【0141】
4)癌関連性抗原からエピトープを選択する場合、患者は天然のエピトープに対して寛容性を発達し得ているため、アナログを選択することがしばしば有用である。感染疾患関連性抗原についてエピトープを選択する場合、天然のエピトープまたはアナログのエピトープのいずれかを選択することが好ましい。
【0142】
5)「入れ子状(nested)エピトープ」と称されるエピトープは特定の関連性を有する。少なくとも2つのエピトープが所定のペプチド配列において重複している場合、入れ子状エピトープが生じる。入れ子状ペプチド配列は、HLAクラスIとHLAクラスIIエピトープの両方を包含する。入れ子状エピトープを提供する場合、一般的な目的は、一配列あたりの最多数のエピトープを提供することである。従って、1つの局面は、ペプチド中のアミノ末端エピトープのアミノ末端およびカルボキシル末端エピトープのカルボキシル末端より長いペプチドを提供しないようにすることである。多重エピトープ性配列(例えば、入れ子状エピトープを含む配列)を提供する場合、一般的にこれが病理学的または他の有害な生物学的特性を有さないことを保証するために、この配列をスクリーニングすることが重要である。
【0143】
6)ポリエピトープタンパク質が作製される場合、またはミニ遺伝子を作製する場合、目的のエピトープを含む最小のペプチドを生成することが目的である。入れ子状エピトープを含むペプチドを選択する場合に用いられるものと同じではない場合、この原理は類似している。しかし、人工ポリエピトープペプチドを用いて、サイズ最小化の目的は、ポリエピトープタンパク質中のエピトープ間の任意のスペーサー配列を組み込む必要性に対してバランスを保たれる。スペーサーアミノ酸残基は、例えば、接合部エピトープ(免疫系により認識されるエピトープであって、標的抗原には存在せず、そしてエピトープの人工並列によってのみ作製される)を回避するため、またはエピトープ間の切断を促進し、それによりエピトープ提示を増大するために導入され得る。接合部エピトープは一般的に回避されるべきである。なぜなら、レシピエントは、非ネイティブのエピトープに対する免疫応答を生成し得るからである。「優性エピトープ」である接合部エピトープは特に関心をもたれる。優性エピトープは、他のエピトープに対する免疫応答が減少または抑制されるこのような強力な応答を導き得る。
【0144】
(8.ミニ遺伝子ワクチン)
複数のエピトープを同時に送達し得る多数の異なるアプローチが利用可能である。本発明のペプチドをコードする核酸は、本発明の特に有用な実施形態である。ミニ遺伝子に含まれるエピトープは、好ましくは、前の章に記載されるガイドラインに従って選択される。本発明のペプチドをコードする核酸を投与する好ましい手段は、本発明の1つまたは複数のエピトープを含むペプチドをコードするミニ遺伝子構築物(minigene construct)を使用する。
【0145】
多重エピトープ性ミニ遺伝子の使用は、以下に記載される(例えば、同時係属出願U.S.S.N.09/311,784;Ishioka ら、J.Immunol.162:3915−3925,1999;An,L.およびWhitton,J.L.,J.Virol.71:2292,1997;Thomson,S.A.ら、J.Immunol.157:822,1996;Whitton,J.L.ら、J.Virol.67:348,1993;Hanke,R ら、Vaccine 16:426,1998)。例えば、TAA、PADRETM汎用ヘルパーT細胞エピトープのようなpan_DR結合ペプチドおよび小胞体トランスロケイティングシグナル(endoplasmic reticulum−translocating signal)配列の複数の領域に由来する、スーパーモチーフおよび/またはモチーフを保有するエピトープ(例えば、PSA、PSM、PAP、およびhK2)をコードする多重エピトープ性DNAプラスミドが操作され得る。ワクチンはまた、他のTAA由来のエピトープを包含し得る。
【0146】
多重エピトープ性ミニ遺伝子の免疫原性をトランスジェニックマウスで試験して、この試験したエピトープに対するCTL誘導応答の大きさを評価し得る。さらに、インビボにおいてDNAでコードされたエピトープの免疫原性を、DNAプラスミドでトランスフェクションした標的細胞に対する特異的CTL株のインビトロ応答と相関させ得る。従って、これらの実験は、このミニ遺伝子が以下の両方に有用であることを示し得る:1)CTL応答を発生させること、および2)誘導CTLがコードされたエピトープを発現する細胞を認識すること。
【0147】
例えば、ヒトの細胞中での発現のために選択したエピトープ(ミニ遺伝子)をコードするDNA配列を産生するために、このエピトープのアミノ酸配列は、逆翻訳され得る。ヒトのコドン使用表を利用して、各アミノ酸に対するコドン選択を誘導し得る。これらのエピトープをコードするDNA配列は、直接隣接されてもよく、その結果、翻訳される場合、連続するポリペプチド配列を産生する。発現および/または免疫原性を最適にするために、さらなるエレメントがミニ遺伝子の設計に取り込まれ得る。逆翻訳され得、そしてミニ遺伝子配列に含まれ得るアミノ酸配列の例としては、以下が挙げられる:HLAクラスIエピトープ、HLAクラスIIエピトープ、ユビキチン結合シグナル配列、および/または小胞体標的シグナル。さらに、CTLおよびHTLエピトープのHLA提示は、合成(例えば、ポリ−アラニン)または天然に存在する、CTLエピトープまたはHTLエピトープに隣接する隣接配列(flanking sequences)を含むことによって改良され得;エピトープ(単数または複数)を含むこれらのより大きなペプチドは、本発明の範囲内である。
【0148】
ミニ遺伝子配列は、ミニ遺伝子のプラス鎖およびマイナス鎖をコードするオリゴヌクレオチドを構築することによってDNAに変換され得る。重複性オリゴヌクレオチド(30〜100の塩基長)を、周知の技術を使用する適切な条件下で、合成、リン酸化、精製およびアニールし得る。オリゴヌクレオチドの末端を、例えば、T4 DNAリガーゼを使用して結合し得る。この合成ミニ遺伝子(エピトープポリペプチドをコードする)を、所望の発現ベクターにクローニングし得る。
【0149】
当業者に周知の標準調節配列は、好ましくは、標的細胞での発現を確実にするためにベクター中に含まれる。いくつかのベクターエレメントが所望される:ミニ遺伝子の挿入のための下流クローニング部位を有するプロモーター;効果的な転写終結のためのポリアデニル化シグナル;E.coliの複製起点;およびE.coli 選択マーカー(例えば、アンピシリンまたはカナマイシン耐性)。多数のプロモーター(例えば、ヒトサイトメガロウイルス(hCMV)プロモーター)が、この目的のために使用され得る。他の適切なプロモーター配列について、例えば、米国特許第5,580,859号および同第5,589,466号を参照のこと。
【0150】
さらなるベクターの改変は、ミニ遺伝子の発現および免疫原性を最適化するために所望され得る。いくつかの場合において、イントロンは、効果的な遺伝子発現のために必要とされ、そして1つ以上の合成イントロンまたは天然に存在するイントロンは、ミニ遺伝子の転写領域に取り込まれ得る。ミニ遺伝子の発現を増加させるために、mRNA安定化配列(mRNA stabilization
sequence)および哺乳動物細胞中での複製のための配列の封入がまた考慮され得る。
【0151】
一旦、発現ベクターが選択されると、ミニ遺伝子は、プロモーターの下流のポリリンカー領域にクローニングされる。このプラスミドを、適切なE.coli株に形質転換し、そしてDNAを標準的な技術を使用して調製する。ミニ遺伝子の方向およびDNA配列、ならびにベクター中に含まれる他の全エレメントは、制限マップおよびDNA配列解析を使用して確認される。正確なプラスミドを有する細菌細胞は、マスター細胞バンク(master cell bank)および作業用細胞バンク(working cell bank)として保存され得る。
【0152】
さらに、免疫刺激配列(ISSまたはCpG)は、DNAワクチンの免疫原性の役割を果たすようである。これらの配列は、所望の場合、免疫原性を増強するために、ミニ遺伝子コード配列の外側のベクター中に含まれ得る。
【0153】
いくつかの実施形態において、ミニ遺伝子コードエピトープと第二タンパク質(免疫原性を増強または減少させるために含まれる)との両方の産生を可能にするbi−シストロン(bi−cistronic)発現ベクターを使用し得る。同時発現する場合、免疫応答を有利に増強し得るタンパク質またはポリペプチドの例としては、サイトカイン(例えば、IL−2、IL−12、GM−CSF)、サイトカイン誘導分子(例えば、LeIF)、同時刺激分子(costimulatory molecule)、またはHTL応答、pan−DR結合タンパク質(例えば、PADRETM,Epimmune,San Diego,CA)が挙げられる。ヘルパー(HTL)エピトープを細胞内標的シグナルに結合し、そして発現したCTLエピトープから別個に発現し得る。これにより、HTLエピトープをCTLエピトープとは異なる細胞画分に指向させることを可能にする。必要である場合、これにより、HTLエピトープのHLAクラスII経路へのより効果的な参加を可能にし、これによりHTL誘導を改良する。HTLまたはCTL誘導に対して、免疫抑制性の分子(例えば、TGF−β)の同時発現による免疫応答を特異的に減少させることは、特定の疾患において有用であり得る。
【0154】
治療量のプラスミドDNAは、例えば、E.coliでの発酵、続く精製により産生され得る。作業用細胞バンクからのアリコートを、増殖培地に接種するために使用し、周知の技術に従って、振盪フラスコまたはバイオリアクター中で飽和まで増殖させる。プラスミドDNAを、標準的なバイオ分離技術(例えば、QIAGEN,Inc.(Valencia,California)によって供給される固相陰イオン交換樹脂)を使用して精製し得る。必要とされる場合、スーパーコイルDNAを、ゲル電気泳動または他の方法を使用して開環形態および線状形態から単離し得る。
【0155】
精製したプラスミドDNAを、種々の処方物を使用して注入のために調製し得る。これらの最も単純なものは、滅菌リン酸緩衝化生理食塩水(PBS)中の凍結乾燥したDNAの再構成物である。「裸のDNA」として公知のこのアプローチは、現在、臨床試験において筋内(IM)投与のために使用されている。ミニ遺伝子のDNAワクチンの免疫療法的効果を最大にするために、精製したプラスミドDNAを処方するための代替方法が所望され得る。種々の方法が記載され、そして新規の技術が利用可能になり得る。カチオン性脂質、糖脂質、およびフソジェニック(fusogenic)リポソームがまた処方物中で使用され得る(例えば、WO93/24640;Mannino & Gould−Fogerite,BioTechnisques6(7):682(1988);米国特許第5,279,833号;WO91/06309号;およびFelgerら、Proc.Nat’l Acad.Sci.USA 84:7413(1987)を参照のこと)。さらに、保護的、相互作用的、非縮合(non−condensing)化合物(PINC)として集合的に言及されるペプチドおよび化合物はまた、精製されたプラスミドDNAに複合体化されて、安定性、筋内分散、または特定の器官もしくは細胞型への輸送のような変数に影響を与える。
【0156】
標的細胞の感作は、ミニ遺伝子コード化CTLエピトープの発現およびHLAクラスI提示のための機能的アッセイとして使用され得る。例えば、プラスミドDNAは、標準的CTLのクロム放出アッセイ(standard CTL chromium release assay)に対する標的として適切である哺乳動物細胞株に導入される。使用されるトランスフェクションの方法は、最終の処方物に依存する。エレクトロポレーションが「裸の」DNAのために使用され、一方、カチオン性脂質は、直接的なインビトロでのトランスフェクションを可能にする。緑色蛍光タンパク質(GFP)を発現するプラスミドは同時にトランスフェクションされて、蛍光活性化細胞分類(FACS)を使用してトランスフェクションされた細胞の濃縮を可能にし得る。次いで、これらの細胞は、クロミウム−51(51Cr)で標識され、エピトープ特異的CTL株のための標的細胞として使用される;51Cr放出によって検出された細胞溶解は、ミニ遺伝子コード化CTLエピトープの産生とミニ遺伝子コード化CTLエピトープのHLA提示との両方を示す。HTLエピトープの発現は、HTL活性を評価するためのアッセイを使用して、類似様式で評価され得る。
【0157】
インビボでの免疫原性は、ミニ遺伝子のDNA処方物の機能的試験についての第2のアプローチである。適切なヒトHLAタンパク質を発現するトランスジェニックマウスは、DNA産物で免疫される。投与の用量および経路は、処方物に依存する(例えば、PBS中でのDNAに対するIM、脂質複合体化DNAに対する腹腔内(IP))。免疫の21日後、脾細胞を収集し、そして試験される各エピトープをコードするペプチドの存在下で1週間、再刺激した。その後、CTLエフェクター細胞について、標準的な技術を使用してペプチド負荷した51Crで標識した標的細胞の細胞溶解についてアッセイを行う。ミニ遺伝子コード化エピトープに対応する、ペプチドエピトープで負荷したHLAによって感作された標的細胞の溶解は、CTLのインビボでの誘導についてDNAワクチン機能を証明する。HTLエピトープの免疫原性を、類似の様式でトランスジェニックマウスにおいて評価する。
【0158】
あるいは、例えば、注入または例えば米国特許第5,204,253号に記載されるような銃式送達によって、核酸を皮内に投与し得る。この技術を使用して、単独でDNAを含む粒子を投与する。さらなる代替の実施形態において、DNAは、粒子(例えば、金粒子)に接着され得る。
【0159】
ミニ遺伝子はまた、当該分野で他の周知の細菌性またはウイルス性の送達システムを使用して送達され得る。例えば本発明のエピトープをコードしている発現構築物はワクシニアのようなウイルスベクターに組み込まれ得る。
【0160】
(9. ヘルパーペプチドとCTLペプチドの組み合わせ)
本発明のペプチドを含むワクチンの組成物は、血清中における改良された半減期のような所望の性質を与えるため、または免疫原性を増大させるために改変され得る。
【0161】
例えば、Tヘルパー細胞の応答を誘導し得るエピトープを少なくとも1つ含むペプチドを結合させることにより、CTL活性を誘導するペプチドの能力は増大され得る。免疫原性を増大させるための、CTLエピトープと組み合わせたTヘルパーエピトープの使用の例が、例えば以下の同時係属中出願、U.S.S.N.08/820,360、U.S.S.N.08/197,484、およびU.S.S.N.08/464,234に記載されている。
【0162】
CTLペプチドはTヘルパーペプチドと直接結合し得るが、しばしばCTLエピトープ/HTLエピトープ結合体はスペーサー分子により連結される。スペーサーは、代表的に生理的条件下において、実質的に荷電を帯びていないアミノ酸またはアミノ酸ミメティックのような、比較的小さな中性の分子を含む。スペーサーは、代表的に、例えばAla、Gly、またはその他の中性の非極性アミノ酸および中性の極性アミノ酸の中性スペーサーから選択される。必要に応じて存在するスペーサーは同じ残基から構成される必要はなく、ヘテロオリゴマーまたはホモオリゴマーであり得ることと理解される。スペーサーが存在する場合、通常、スペーサーは少なくとも1つまたは2つの残基であるが、さらに通常には3〜6つの残基、時には10残基以上になる。CTLペプチドエピトープはTヘルパーペプチドエピトープと直接またはスペーサーを介してのどちらかで、CTLペプチドのアミノ末端またはカルボキシ末端のいずれかで結合され得る。免疫原性ペプチドまたはTヘルパーペプチドのいずれかのアミノ末端はアシル化され得る。
【0163】
特定の実施形態において、Tヘルパーペプチドは、集団の大部分に存在するTヘルパー細胞により認識されるものである。これは、多数の、ほとんどの、あるいは全てのHLA クラスII分子と結合するアミノ酸配列の選択によって達成され得る。これらのことは「ゆるいHLA制限」、または「乱雑な」Tヘルパー配列として、公知である。乱雑なペプチドの例としては、破傷風毒の位置830〜843(QYIKANSKFIGITE)、Plasmodium falciparum circumsporozoite(CS)タンパク質の位置378〜398(DIEKKIAKMEKASSVFNVVNS)およびStreptococcus 18kD タンパク質の位置116(GAVDSILGGVATYGAA)のような抗原由来の配列が挙げられる。その他の例としては、DR1−4−7スーパーモチーフ、もしくはDR3モチーフのいずれかを保有するペプチドが挙げられる。
【0164】
あるいは、ゆるいHLA制限様式において、天然では見出されないアミノ酸配列を使用して、Tヘルパーリンパ球を刺激し得る合成ペプチドを調製することが可能である(例えば、PCT刊行物WO95/07707を参照)。Pan−DR結合エピトープ(例えば、PADRETM、Epimmune,Inc.,San Diego,CA)と呼ばれるこれらの合成化合物はほとんどのHLA−DR(ヒトHLAクラスII)分子に、最も優先的に結合するよう設計されている。例えば、式:aKXVAAWTLKAAa(ここで、「X」はシクロヘキシルアラニン、フェニルアラニン、またはチロシンのいずれかであり、「a」はD−アラニンまたはL−アラニンのいずれかである)を有するpan−DR結合エピトープペプチドは、それらのHLA型に関わらずほとんどのHLA−DR対立遺伝子(allele)と結合すること、およびほとんどの個体由来のTヘルパーリンパ球の応答を刺激することが見出されている。Pan−DR結合エピトープの代替物は、全て「L」の天然アミノ酸を含み、エピトープをコードする核酸の形態で提供され得る。
【0165】
HTLペプチドエピトープはまた、それらの生物学的性質を変えるように修飾され得る。例えば、それらはD−アミノ酸を含むように修飾されて、それらのプロテアーゼ耐性を増大させ、そしてそれらの血清中半減期を延長させ得、または、それらを例えば、脂質、タンパク質、炭水化物などのような他の分子と結合させて、それらの生物学的活性を増大し得る。例えば、Tヘルパーペプチドは、アミノ末端またはカルボキシ末端のいずれかにおいて1以上のパルミチン酸鎖と結合され得る。
【0166】
(10.T細胞刺激(priming)因子とCTLペプチドの組み合わせ)
いくつかの実施形態において、少なくとも1つの、細胞傷害性Tリンパ球を刺激する成分が、本発明の薬学的組成物に含まれることが所望され得る。脂質は、インビボでウイルス抗原に対し、CTL刺激をなし得る因子として同定されている。例えば、パルミチン酸残基は、リジン残基のε−アミノ基およびα−アミノ基に結合され得、次いで、例えば1つ以上の連結した残基(例えばGly、Gly−Gly−、Ser、Ser−Serなど)を介して免疫原性ペプチドに結合され得る。次いで、脂質化ペプチドは、直接にミセルまたは微粒子のいずれかで投与され得るか、リポソームへ組み込まれ得るか、またはアジュバント(例えば不完全フロイントアジュバント)へ乳化され得る。好ましい免疫原性組成物は、Lysのε−アミノ基およびα−アミノ基と結合したパルミチン酸を含み、このLysは、例えばSer−Serのような結合を介して、免疫原性ペプチドのアミノ末端と結合している。
【0167】
CTL応答を刺激する脂質のもう1つの例として、E.coliのリポタンパク質、例えばトリパルミトイル−S−グリセリルシステイニルセリル−セリン(tripalmitoyl−S−glycerylcysteinlyseryl−serine:P3CSS)は、ある適切なペプチドに共有結合した場合に、ウイルス特異的CTLを刺激することに使用され得る(例えば、Deres,ら、Nature 342:561,1989を参照)。本発明のペプチドは、例えば、P3CSSに結合され得、標的抗原に対するCTL応答を特異的に刺激するため、個体に投与され得る。さらに、中和抗体の誘導もまた、P3CSS結合エピトープにより刺激され得るため、2つのそのような組成物は、組み合わされて、体液性および細胞媒介性応答の両方をさらに効果的に誘発し得る。
【0168】
CTLおよび/あるいはHTLペプチドは、ペプチドの末端へのアミノ酸の付加によっても修飾され、あるペプチドをもう1つのペプチドへ結合することの容易さ、キャリア支持体またはより大きなペプチドへの結合、ペプチド、オリゴペプチドなどの物理学的または化学的特性の修飾を提供し得る。アミノ酸(例えばチロシン、システイン、リジン、グルタミン酸、アスパラギン酸など)は、ペプチドまたはオリゴペプチド、特にクラスIペプチドのC−末端またはN−末端に導入され得る。しかしながら、CTLエピトープのカルボキシ末端においての修飾が、いくつかの場合において、ペプチドへの結合特性を変化させ得ることは注意されるべきである。さらに、ペプチド配列またはオリゴペプチド配列は末端のNH2アシル化(例えば、アルカノイル(C1〜C20)またはチオグリコリルアセチル化)による、末端カルボキシルアミド化(例えば、アンモニア、メチルアミンなど)によって修飾されることによって天然の配列と異なり得る。いくつかの例において、これらの修飾は、支持体または他の分子との結合のための部位を提供し得る。
【0169】
(11.CTLおよび/またはHTLペプチドでパルス化(pulsed)されたDCを含むワクチン組成物)
本発明に従うワクチン組成物の実施形態は、患者の血液由来のPBMCへのエピトープ保有ペプチドのカクテル、またはPBMCから単離したDCへのエピトープ保有ペプチドのカクテルのエキソビボ投与を包含する。DCの採取を容易にする薬(例えばProgenipoietinTM(Monsanto,St.Louis,MO)またはGM−CSF/IL−4)が使用され得る。ペプチドでDCをパルス化した後でかつ、患者に再注入する前に、DCは結合していないペプチドを除くため洗浄される。この実施形態において、ワクチンは、HLA分子とその表面で複合体になった、パルス化されたペプチドエピトープを提示するペプチドパルス化DCを包含する。
【0170】
DCは、エキソビボでペプチドカクテル(それらのうちのいくつかは、目的の1つ以上の抗原に対するCTL応答を刺激する)でパルス化され得る。必要に応じて、ヘルパーT細胞ペプチド(例えば、PADRETMファミリー分子)は、CTL応答を容易にするするため、含有され得る。
【0171】
(12.治療目的または予防目的のためのワクチンの投与)
本発明のペプチド、本発明の薬学的組成物およびワクチン組成物は、代表的に癌の処置のために治療的に使用される。本発明のペプチドを含有したワクチン組成物は、代表的に、1つ以上の抗原の発現に関連した悪影響を有する癌患者へ投与される。あるいは、ワクチン組成物は、癌の進行に対して影響を受けやすい、さもなくば癌の進行の危険にある個人に対し投与され得る。
【0172】
治療適用において、ペプチド組成物および/もしくは核酸組成物は、腫瘍抗原に対する効果的なCTL応答および/もしくはHTL応答を誘発するため、そして症状および/もしくは合併症を治癒するか、少なくとも部分的に止める、もしくは遅らせるために十分な量で患者に投与される。これを満たすのに十分な量は、「治療的有効用量」と定義される。この使用に十分な量は、例えば、投与される特定の組成物、投与方法、治療される疾患の段階および重篤度、患者の体重および全般的な健康状態、ならびに処方医の判断に依存する。
【0173】
上に記載したように、本発明のCTLエピトープおよび/もしくはHTLエピトープを含むペプチドは、CTLまたはHTL特異的にHLA分子によって提示される場合、およびこのペプチドに含まれるエピトープに特異的なCTLまたはHTLと接触される場合に、免疫応答を誘導する。ペプチド(またはそれらをコードするDNA)は、個別に、または1つ以上のペプチド配列の融合物として投与され得る。ペプチドがCTLまたはHTLと接触される方法は、本発明には重要でない。例えば、ペプチドは、インビボもしくはインビトロのいずれかで、CTLまたはHTLと接触し得る。接触が、インビボで起きる場合、ペプチド自体が患者に投与され得るか、または他のビーイクル(例えば、1つ以上のペプチドをコードしたDNAベクター、ペプチドをコードしたウイルスベクター、リポソームなど)が、本明細書中に記載されるように使用され得る。
【0174】
ペプチドが、インビトロで接触する場合、ワクチン因子は、細胞の集団(例えば、ペプチドパルス化樹状細胞)、または抗原提示細胞をペプチドでパルス化すること、もしくは本発明品のミニ遺伝子を抗原提示細胞にトランスフェクトすることによって誘導されたTAA特異的CTLを含有し得る。そのような細胞集団は、続いて、治療的有効用量で患者に投与される。
【0175】
治療に用いるために、投与は、一般的に、最初の癌の診断の時点で始まるべきである。これに続いて、少なくとも実質的に症状がやわらげられるまで、そしてその後ある期間にわたって追加抗原投与量がブーストされる。患者に送達されるワクチン組成物の実施形態(すなわち、例えばペプチドカクテル、ポリエピトープ性ポリペプチド、ミニ遺伝子、またはTAA特異的CTL、またはパルス化された樹状突起細胞のような実施形態を含むが、これらに限定されない)は、疾患の段階、あるいは患者の健康状態によって変り得る。例えば、TAA特異的CTLを含むワクチンは、進行した疾患を有する患者中において、腫瘍細胞を殺傷する際に、代変の実施形態より効果的であり得る。
【0176】
本発明のワクチン組成物はまた、外科手術などの処置と組み合わせても、治療的に用いられ得る。1つの例は、患者が、原発性腫瘍を取り除くための手術を受けており、次いで、ワクチンが、再発および/もしくは転移を、遅らせるか、または予防するために使用される状況である。
【0177】
感受性の個体(例えば、前立腺腫瘍を発病するような遺伝的素因があると診断され得る個体)が、癌の診断に先立って特定される場合、組成物が、彼らに対して標的化され得、したがってより大きな集団への投与の必要性を最小にす得る。
【0178】
最初の、治療的免疫のための用量決定は、一般的に、下限の値が、約1、5、50、500、または1,000μgであり、そして上限の値が、約10,000;20,000;30,000;または50,000μgの単位投薬範囲にある。ヒトについての投薬量の値は、典型的に、70キログラムの患者あたり約500μg〜約50,000μgの範囲である。確立された間隔(例えば、4週間から6ヶ月)で、ブースト投薬前の、最初の投薬は、患者を効果的に処置するため、ことによると長期の時間の間、必要とされ得る。ブーストレジメンに従って、数週間から数ヶ月にわたる、ペプチドの約1.0μg〜約50,000μgの間のブースト投薬は、患者の血液から得られるCTLおよびHTLの特異的活性を測定することによって決定される患者の応答および状態に依存して投与され得る。
【0179】
少なくとも臨床的症状または実験的試験が、腫瘍が除去されたかまたは腫瘍細胞荷重が実質的に減少したことを示すまでそしてその後のある期間の間、投薬が続けられるべきである。投薬、投与の経路、および投薬スケジュールは、当該分野で公知の方法論に従って調節される。
【0180】
ある実施形態において、本発明のペプチドおよび組成物は、重症な疾患状態、すなわち、生命を危うくする状況または潜在的に生命を危うくする状況で使用される。このような場合において、最小量の外来の物質および本発明の好ましい組成物におけるペプチドの比較的非毒性の性質の結果として、これらの記述された投薬量に対して実質的に過剰のこれらのペプチド組成物を投与することが、可能であり、そして処置する医師によって望ましいと感じられ得る。
【0181】
本発明のワクチン組成物は、予防因子としても使用され得る。例えば、組成物は、前立腺癌進行の危険性を有する個体に投与され得る。一般的に、最初の予防的免疫のための一般的な投与量は、下限の値が約1、5、50、500、または1000μgであり、そして上限の値が約10,000;20,000;30,000;または50,000μgである単位投薬範囲にある。ヒトについての投薬量の値は、典型的に、70キログラムの患者あたり約500μg〜約50,000μgの範囲である。これに続いて、最初のワクチン投与後、約4週間〜約6ヶ月の規定された間隔で投与される、約1.0μg〜約50,000μgの間のペプチドがブースト投薬される。ワクチンの免疫原性は、患者の血液のサンプルから得られたCTLおよびHTLの特異的活性を測定することによって評価され得る。
【0182】
治療的処置のための薬学的組成物は、非経口的投与、局所的投与、経口投与、くも膜下投与、または局所投与に意図される。好ましくは、薬学的組成物は、非経口的に、例えば、静脈内に、皮下に、皮内に、または筋内に投与される。従って、本発明は、受容可能なキャリア(好ましくは、水性キャリア)に溶解されるかまたは懸濁される免疫原性ペプチドの溶液を含む非経口投与のための組成物を提供する。種々の水性キャリアは、例えば、水、緩衝化水、0.8%生理食塩水、0.3%グリシン、ヒアルロン酸などが使用され得る。これらの組成物は、従来の周知の滅菌化技術によって滅菌化され得るか、または滅菌濾過され得る。得られる水溶液は、そのままでの使用のためにパッケージングされ得るか、または凍結乾燥され得、この凍結乾燥された調製物は、投与の前に滅菌溶液と組み合わせられる。この組成物は、およそ生理学的条件に必要とされる薬学的に受容可能な補助物質、例えば、pH調節剤、緩衝化剤、張性調節剤、湿潤剤、保存剤など(例えば、酢酸ナトリウム、乳酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、ソルビタンモノラウレート、オレイン酸トリエタノールアミンなど)を含み得る。
【0183】
薬学的処方物中の本発明のペプチドの濃度は、幅広く、すなわち、約0.1重量%未満、通常、約2重量%または少なくとも約2重量%から、20重量%〜50重量%以上の多さまでで変化し得、主に、選択される投与の特定の様式に従って、流体容積、粘度などによって選択される。
【0184】
ペプチド組成物のヒト単位用量形態は、代表的に、ヒト単位用量の受容可能なキャリア(好ましくは、水性キャリア)を含む薬学的組成物中に含まれ、そしてこのような組成物のヒトへの投与に使用されるための当業者によって公知の流体の容積で投与される(例えば、Remington’s Pharmaceutical Sciences、17th Edition、A.Gennaro、Editor、Mack Publishing Co.、Easton、Pennsylvania、1985を参照)。
【0185】
本発明のペプチドはまた、リポソームを介して投与され得、リポソームは、ペプチドを特定の組織(例えば、リンパ組織)に標的化させるために、または感染した細胞に選択的に標的化させるために、そしてペプチド組成物の半減期を増加させるために役立つ。リポソームには、エマルジョン、泡状物、ミセル、不溶性単層、液晶、リン脂質分散物、層状層などが挙げられる。この調製物において、送達されるべきペプチドは、リポソームの一部として、単独であるいはリンパ系細胞に広く行き渡るレセプターに結合する分子(例えば、CD45抗原に結合するモノクローナル抗体)とともに、または他の治療組成物もしくは免疫原性組成物とともに組み込まれる。従って、本発明の所望のペプチドで負荷されるかまたは装飾(decorated)されるかのいずれかのリポソームが、リンパ系細胞の部位に指向され得、ここで、そのときリポソームがペプチド組成物を送達する。本発明に従う使用のためのリポソームは、標準的な小胞形成脂質から形成され、これには、一般的に、中性および陰性に荷電されたリン脂質およびステロール(例えば、コレステロール)が挙げられる。脂質の選択は、一般的に、例えば、リポソームサイズ、酸不安定性および血流中のリポソームの安定性を考慮してガイドされる。種々の方法が、リポソームを調製するために利用可能であり、例えば、Szokaら、Ann.Rev.Biophys.Bioeng.9:467(1980)および米国特許第4,235,871号、同第4,501,728号、同第4,837,028号、および同第5,019,369号に記載される。
【0186】
免疫系の細胞を標的化するために、リポソームに組み込まれるリガンドには、例えば、所望の免疫系細胞の細胞表面決定基に特異的な抗体またはそのフラグメントが挙げられ得る。ペプチドを含むリポソーム懸濁物は、静脈内、局所的(locally、topically)などで、特に投与の方法、送達されるペプチド、および処置される疾患の段階に従って変化する用量で投与され得る。
【0187】
固体組成物について、従来の非毒性固体キャリアが使用され得、これには、例えば、薬学的等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、タルク、セルロース、グルコース、スクロース、炭酸マグネシウムなどが挙げられる。経口投与のために、薬学的に受容可能な非毒性組成物は、任意の通常使用される賦形剤(例えば、先に列挙されたもの)および一般的に10〜95%の活性成分(すなわち、本発明の1つ以上のペプチド)、より好ましくは、25%〜75%の濃度で組み込むことによって形成される。
【0188】
エアロゾル投与のために、免疫原性ペプチドは、好ましくは、界面活性剤および噴霧剤とともに細かく分割された形態で供給される。ペプチドの代表的な割合は、0.01重量%〜20重量%、好ましくは、1重量%〜10重量%である。界面活性剤は、もちろん非毒性でなければならず、そして好ましくは噴霧剤に可溶である。このような薬剤の代表は、6〜22個の炭素原子を含む脂肪酸のエステルまたは部分エステル、例えば、カプロン酸、オクタン酸、ラウリン酸、パルミチン酸、ステアリン酸、リノール酸、リノレン酸、オレステリン酸(olesteric acid)およびオレイン酸と、脂肪族多水酸基アルコールまたはその環式無水物である。混合エステル(例えば、混合または中性グリセリド)が使用され得る。界面活性剤は、組成物の0.1重量%〜20重量%、好ましくは、0.25〜5重量%を構成し得る。組成物の残りは、通常、噴霧剤である。キャリアはまた、望ましいならば、例えば、鼻腔内送達のためのレシチンとともに含まれ得る。
【0189】
(13.キット)
本発明のペプチドおよび核酸組成物は、ワクチン投与のための指示書とともにキットの形態で提供され得る。代表的なキットは、容器内に、好ましくは、単位用量形態の所望のペプチド組成物および投与のための指示書を含む。代替的なキットは、投与のための指示書とともに、容器内に、好ましくは単位用量形態で、本発明の所望の核酸を伴うミニ遺伝子構築物を含む。IL−2またはIL−12のようなリンホカインもまた、キットに含まれ得る。所望であり得る他のキット構成要素には、例えば、滅菌シリンジ、ブースター投薬、および他の所望の賦形剤が挙げられる。
【0190】
本発明に従うエピトープは、免疫応答を誘導するために首尾良く使用された。これらのエピトープを用いる免疫応答は、種々の形態においてエピトープを投与することによって導入された。エピトープは、ペプチドとして、核酸として、そして本発明のエピトープをコードする核酸を含むウイルスベクターとして投与された。ペプチドに基づくエピトープ形態の投与において、免疫応答は、エピトープを細胞で発現される空のHLA分子上に直接負荷することによって、そしてエピトープのインターナリゼーションを介して、そしてHLA I型経路を介してプロセスすることによって導入された;いずれの場合においても、エピトープを発現するHLA分子は、次いで、相互作用し得、そしてCTL応答を誘導し得た。ペプチドは、直接的に、またはリポソームのような薬剤を使用して送達され得る。これらは、さらに、銃式送達(ballistic delivery)(ここで、ペプチドは、代表的には、結晶形態である)を使用して送達され得る。DNAが免疫応答を誘導するために使用される場合、裸のDNAとして(一般的に、約1〜5mgの投薬範囲)または銃式「遺伝子銃」送達を介して(代表的に、約10〜100μgの投薬範囲)のいずれかで投与される。DNAは、種々のコンホメーション(例えば、線状、環状など)で送達され得る。種々のウイルスベクターもまた、本発明に従ってエピトープをコードする核酸を含んで首尾良く使用されている。
【0191】
従って、本発明に従う組成物は、いくつかの形態で存在する。本発明に従うこれらの組成物形態のそれぞれの実施形態は、免疫応答を誘導するために首尾良く使用されている。
【0192】
本発明に従う1つの組成物は、複数のペプチドを含む。この複数またはカクテルのペプチドは、一般的に、1つ以上の薬学的に受容可能な賦形剤と混合される。ペプチドカクテルは、同じペプチドの複数のコピーを含み得るか、またはペプチドの混合物を含み得る。ペプチドは、天然のエピトープのアナログであり得る。ペプチドは、人工のアミノ酸および/または化学的改変(例えば、脂質化;アセチル化、グリコシル化、ビオチニル化、リン酸化のような表面活性分子の添加)を含み得る。ペプチドは、CTLまたはHTLエピトープであり得る。好ましい実施形態において、ペプチドカクテルは、複数の異なるCTLエピトープおよび少なくとも1つのHTLエピトープを含む。HTLエピトープは、天然または非天然(例えば、PADRE(登録商標)、Epimmune Inc.,San Diego、CA)であり得る。本発明の実施形態における異なるエピトープの数は、一般的に、1〜150の整数(whole unit integer)である(例えば、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、・・・、150)。
【0193】
本発明に従う組成物のさらなる実施形態は、ポリペプチド多重エピトープ性構築物(すなわち、ポリエピトープペプチド)を含む。本発明に従うポリエピトープペプチドは、当該分野において周知の技術の使用によって調製される。これらの公知の技術の使用によって、本発明に従うエピトープは、互いに接続される。ポリエピトープペプチドは、線状または非線状(例えば、多価)であり得る。これらのポリエピトープ構築物は、人工アミノ酸、スペーシングまたはスペーサーアミノ酸、隣接アミノ酸(flanking amino acids)、または隣接エピトープユニット間の化学改変を含み得る。ポリエピトープ構築物は、ヘテロポリマーまたはホモポリマーであり得る。ポリエピトープ構築物は、一般的に、2〜150の間の任意の整数の量(例えば、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、・・・、150)でエピトープを含む。ポリエピトープ構築物は、CTLおよび/またはHTLエピトープを含み得る。構築物中の1つ以上のエピトープは、例えば、表面活性物質(例えば、脂質)の付加によって改変され得るか、または化学的に改変され得る(例えば、アセチル化など)。さらに、多重エピトープ性構築物における結合は、ペプチド結合以外(例えば、共有結合、エステルまたはエーテル結合、ジスルフィド結合、水素結合、イオン結合など)であり得る。
【0194】
あるいは、本発明に従う組成物は、ネイティブな配列に対する相同性を有する(すなわち、対応するかまたは接触(contiguous)する)アミノ酸の系列、配列、ストレッチ(stretch)を含む構築物を含む。このアミノ酸のストレッチは、より長い系列のアミノ酸から切断されるかまたは単離される場合、本発明に従って、HLA I型エピトープまたはHLA II型エピトープとして機能するアミノ酸の少なくとも1つのサブ配列(subsequence)を含む。この実施形態において、ペプチド配列は、当該分野で公知であるかまたは提供される多数の技術を使用することによって、本明細書中に規定されるような構築物になるように改変される。ポリエピトープ構築物は、70〜100%(例えば、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、または100%)の任意の整数の増分でネイティブな配列に対して相同性を含み得る。
【0195】
本発明に従う組成物のさらなる実施形態は、本発明に従う1つ以上のエピトープを含む抗原提示細胞である。抗原提示細胞は、「専門的」抗原提示細胞(例えば、樹状細胞)であり得る。抗原提示細胞は、当該分野において公知の任意の手段または当該分野において決定された任意の手段によって本発明のエピトープを含み得る。このような手段には、樹状細胞を1つ以上の個々のエピトープまたは複数のエピトープを含む1つ以上のペプチドで、銃式核酸送達のような核酸投与によって、または核酸の投与について当該分野の他の技術(ベクターに基づく(例えば、ウイルスベクター)核酸の送達を含む)によって適用することを含む。
【0196】
本発明に従う組成物のさらなる実施形態は、本発明の1つ以上のペプチドをコードする核酸、または本発明に従うポリエピトープペプチドをコードする核酸を含む。当業者によって理解されるように、種々の核酸組成物が、遺伝子コードの冗長性に起因して同じペプチドをコードする。これらの核酸組成物のそれぞれは、本発明の範囲内に含まれる。本発明のこの実施形態は、DNAまたはRNAを含み、特定の実施形態において、DNAおよびRNAの組み合わせを含む。本発明に従うペプチドをコードする核酸を含む任意の組成物または本発明に従う任意の他のペプチドに基づく組成物が、本発明の範囲内であることが理解される。
【実施例】
【0197】
(実施例)
(調製A)
(ペプチド合成およびペプチドアナログの生成)
これらの実施例で用いられるペプチドを表1に示す。腫瘍関連抗原に由来している全ての野生型ヒトCTLエピトープならびにHIVおよびB型肝炎ウイルス(HBV2)のポリメラーゼ遺伝子に由来している野生型ウイルスエピトープは、ヒトおよびトランスジェニックマウスの系における免疫原性が示されている(Kawashima,I.ら、Human Immunol.(1998)59:1;Ishioka,G.ら、J.Immunol.(1999)162:3915)。
【0198】
ヘテロクリティックな活性について最初に試験されたペプチドは、Chiron Technologies(Victor、Australia)によって合成された。さらなる生物学的特徴付けを必要としているペプチドは、(Ruppert,J.ら、Cell(1993)74:929)の方法を用いてEpimmuneにて合成され、分析用逆相HPLCによって決定された場合に、これらのペプチドの純度は通常95%より高かった。後者のペプチドの正体を質量スペクトル分析によって確認した。
【0199】
(調製B)
(単一アミノ酸置換の選択についてのスキーム)
表2は、所定のアミノ酸置換を、保存的置換、半保存的置換、または非保存的置換であるとして特徴付けし得るように、任意の所定のアミノ酸対の間の類似性の割り当てを示す。
【0200】
アミノ酸対の間の類似性の程度を、各アミノ酸対について、PAM250に関する順位係数スコア(rank coefficient score)、疎水性度および以下に述べるような側鎖の体積を平均化することによって定量した。これらの複合した順位の平均値に基づいて、この表は各対が保存されているか、半保存されているか、または保存されていないかを示す。
【0201】
Dayhoff PAM250スコア(Dayhoff,M.O.ら、Atlas of Protein Sequence and Structure,第5巻,補遺3.(1978)M.O.Dayhoff編.National
Biomedical Research Foundation,Washington DC,345頁;Creighton,T.E.,Proteins:structures and molecular properties(1993)(第2版)W.H.Freeman and Company,NY;http://prowl.rockefeller.edu/aainfo/pam250.html)は、規定された時間枠の範囲内で受容可能な点変異(PAM)の百分率を測定する、一般的に利用されるタンパク質アラインメントスコア付けマトリクスである。これらの変異の頻度はランダム変異の確率から予想されるものとは異なり、恐らく、置換に関与するアミノ酸対の物理的および化学的類似性の程度に起因する偏向を反映する。類似性の他の尺度を用いて標準化され得るアミノ酸類似性のスコアを得るために、PAM250スコアを順位値(rank value)に変換した。ここで、1は受容された変異であるという最も高い確率を示している。
【0202】
20個の天然に存在するアミノ酸の相対的疎水性度を表すために、最もよく一般的に利用されるスケール(Cornette,J.ら、J.Mol.Biol(1987)195:659)は、KyteおよびDoolittle(Kyte,J.およびR.F.Doolittle,J.Mol.Biol.(1982)157:105)、ならびにFauchereおよびPliska(Fauchere,J.およびV.Pliska,Eur.J.Med.Chem.(1983)18:369)による実験データを基礎として発展したものである。Kyte/Doolittleスケールは個々のアミノ酸のH2O/有機溶媒分配を測定する。このスケールは折り畳まれたタンパク質のアミノ酸の位置を考慮するので、このスケールは、タンパク質の状況下におけるネイティブな疎水性度を最も正確に反映し得る。Fauchere/PliskaスケールはN−アセチルアミノ酸アミドのオクタノール/H2O分配を測定し、変性タンパク質および/または、低分子合成ペプチドの状況下における疎水性度を最も正確に反映する。疎水性度についてのスコアを得るために、各アミノ酸残基をKyte/DoolittleおよびFauchere/Pliska疎水性度スケールの両方において順位付けした。2つのスケール間の平均順位を計算し、各対についての疎水性度の差の平均を計算した。
【0203】
最後に、アミノ酸側鎖の体積を計算するために、1分子または1グラムのいずれかのアミノ酸残基を添加した後の水の体積の増加を留意することによって得られる溶液中の分体積を考慮した。(Zamyatnin,A.A.,Ann.Rev.Biophys.Bioeng.(1984)13:145;Zamyatnin,A.A.,Prog.Biophys.Mol.Biol.(1972)24:107)。20個の天然に存在するアミノ酸に関して可能な各対の分体積の絶対的差異を計算し、そして順位付けた。ここで、1は最も類似する体積を有する残基、そして20は最も異なる体積を有する残基を示した。
【0204】
(調製C)
(アッセイのための材料)
(1.APC株)
HLA−A2.1の状況下でぺプチドを提示する細胞株は以下のように調製された:
.221A2.1細胞株は、HLA−A2.1遺伝子をHLA−A、−B、−C−ヌル変異体EBV−形質転換ヒトB−リンパ芽球細胞株3A4−721.221(Kawashima,I.ら、Human Immunol.(1998)59:1)にトランスフェクションすることによって生成した。
【0205】
腫瘍細胞株はメチルコラントレン(methylcholanthrene)誘導性肉腫であるMeth A細胞のトランスフェクションによって調製した。そして、Jurkat細胞株ではHLA−A2.1またはHLA−A2.1/Kb導入遺伝子のトランスフェクションを、他(Vitiello,A.らJ.Exp.Med.(1991)173:1007)で述べた方法を用いて行った。HLA型黒色腫細胞株624mel(A2.1+,MAGE+)および888mel(A2.1-,MAGE-)の組み合わせはY.KawakamiおよびS.Rosenberg(National Cancer Institute)の御好意により提供され、そして内因的にプロセシングされたMAGE3エピトープの提示を測定するために用いられた。(Boon,T.ら,Ann.Rev.Immunol.(1994)12:337)。その黒色腫細胞株をAPCとして使用する前に、100IU/mlヒトIFNγ(Genzyme,Cambridge,MA)で48時間、37℃で処理した。
【0206】
本研究で用いた全ての細胞は、抗生物質、ピルビン酸ナトリウム、非必須アミノ酸および10%(v/v)熱不活化FBSを補充したRPMI−1640培地で増殖させた。
【0207】
(2.インビトロでのヒトPBMCからのCTLの誘導およびヒトCTL株の誘導体化)
MAGE3.112および癌胎児抗原(CEA)エピトープであるCEA.691に対するペプチド特異的なCTL株を生成するために、インビトロにおいて正常な被験体からのPBMCを、(Kawashima,I.らHuman Immunol.(1998)59:1)に記載のようなペプチドを用いて繰り返し刺激した。簡潔には、ペプチドを適用した(pulsed)樹状細胞(GM−CSFおよびIL4中で培養することによって接着性PBMCから区別した)を、抗体でコーティングしたビーズ(Dynal A.S.,Oslo,Noway)を用いたポジティブ選択によって得られた自己CD8+T細胞とともに、48ウェルプレート中で共存培養した。IL2、IL7およびIL10の存在下での培養の7日目後、インビトロにおいて、ペプチドを適用した(pulsed)接着性PBMCを用いて各PBMC培養物(ウェル)を再刺激した。次いで、ペプチドの存在下または非存在下において、腫瘍APC.221A2.1を用いて刺激した後のIFNγ生成を測定することによって、CTL活性について培養物を試験した。CTL株はペプチド特異的なIFNγ応答を示したPBMC培養物から、ペプチドを適用した接着性PBMCを用いたさらなるインビトロ刺激よって増殖した。
【0208】
(3.マウスCTL株)
エピトープHBV Pol.455およびHIV Pol.476ペプチドに対するCTL株を、他(Ishioka、G.らJ.Immunol.(1999)162:3915)に記載されているようなDNA免疫によって、HLA−A2.1/KbXSトランスジェニックマウス中で産生した。HLA−A2.1/KbXSおよびHLA−A2.1/KbXdトランスジェニックマウスはEpimmuneで飼育された。これらの株は、C57BL/6のバックグラウンド(Vitiello,A.らJ.Exp.Med.(1991)173:1007)において作製されたHLA−A2.1/Kbトランスジェニック株と、それぞれSJLまたはBALB/cマウス(Jackson Laboratories、Bar Harbor、ME)との間の交雑のF1世代を表す。MAGE2.157エピトープに対するCTL株は、8〜12週齢HLA−A2.1/KbXSマウスの尾部の基部に、IFA中で乳化された50μgのペプチドおよび140μgのHBV Core.128ThエピトープであるTPPAYRPPNAPIL(配列番号30)を免疫し、そしてインビトロでペプチドを用いて繰り返し初回刺激した脾細胞を再刺激することよって生じた。
【0209】
(調製D)
(アッセイ方法)
(1.HLA−A2.1分子に対するペプチド結合親和性の測定)
放射性標識された標準ペプチドのHLA−A2.1への結合について、所定の試験ペプチドによって誘導される競合のレベルを決定することによって、試験ペプチドのHLA−A2.1への結合を測定した。ゲル濾過によってMHC結合性放射能の百分率を決定し、そして、標識された標準ペプチドの結合の50%を阻害した試験ペプチドの濃度(IC50)を計算した(Ruppert,J.ら、Cell(1993)74:929;Sette,A.ら、Mol.Immunol.(1994)31:813)。標準ペプチドはHBV Core.18エピトープであった(配列FLPSDFFPSV)。
【0210】
(2.CTLによるマウスおよびヒトのIFNγ、IL5およびIL10産生の測定)
インサイチュの捕捉ELISAを、CTLからのIFNγ放出を測定するために使用した(McKinney,D.ら、J.Immunol.Methods(2000)237:105)。簡潔には、CTLを抗マウスIFNγ(クローンR4−6A2,Pharmingen,San Diego,CA)または抗ヒトIFNγ mAb(クローンNIB42,Pharmigen)のいずれかをプレコーティングしたELISA等級の96ウェル平底ウェル中においてAPCおよびペプチドを用いて刺激した。細胞培養後、ウェルを洗浄し、ビオチン化した抗マウスIFNγ(クローンXMG1.2,Pharmingen)mAbまたは抗ヒトIFNγ(クローン4S.B3,Pharmingen)mAbを加えた後、酵素結合体化ストレプトアビジン(Zymed,South San Francisco,CA)および3,3',5,5'テトラメチルベンジジン基質(ImmunoPure TMB基質キット、Pierce,Rockford,IL)を加えることによって現像した。各ウェルの吸光度は450nmでLabsystems Multiskan RC ELISAプレートリーダーにおいて測定した。各ウェルで生成されたIFNγのレベルを、同じアッセイ方法で確立されたマウスまたはヒトのIFNγの検量線から外挿することによって決定づけた。
【0211】
マウスおよびヒトのIL5およびIL10をELISAキット(R&D Biosystems、Minneapolis、MN)を用いて培養上清中で測定した。これらのアッセイを定量的サンドイッチELISA技術を用い、製造業者のプロトコールに従って行った。
【0212】
(3.エキソビボCTL応答を測定するための酵素結合免疫スポット(Enzyme−liked immunospot)(Elispot)アッセイ)
Elispotアッセイを標準プロトコール(Murali−Krishna,K.ら、Immunity(1998)8:177;Lewis,J.J.ら、Int.J.Cancer(2000)87:391)に従って行った。簡潔には、平底96ウェルニトロセルロースプレート(Immobilon−Pメンブレン,Millipore,Bedford,MA)を抗IFNγmAb(10μg/ml,クローンR4−6A2)でコーティングし、4℃で一晩中インキュベートした。PBSで洗浄した後、プレートを10%FBSを含むRPMI培地で37℃で1時間でブロックした。磁気ビーズ(Miltenyi,Auburn,CA)によって単離された4×105個の脾性CD8+細胞および10μg/mlのペプチドを適用された5×104個のJurkat−A2.1/Kb細胞を各ウェルに加え、細胞を10%FBSを含むRPMI培地で20時間インキュベートした。インキュベーション後、PBS/0.05%Tweenを用いてプレートを徹底的に洗浄した後、ビオチン化した抗IFNγ mAb(2μg/ml,クローン XMG1.2)を各プレートに加え、プレートを37℃で4時間インキュベートした。次いで、プレートをPBS(0.1%Tween−20を含む)およびVectastain ABCペルオキシダーゼ(Vectastain Elite キット;Vector Laboratories,Burlingame,CA)で4回洗浄した。室温で1時間インキュベートした後、プレートを1×PBS/0.05%Tweenを用いて3回洗浄し、次いでさらに3回1×PBSで洗浄した。スポットを現像するために100μlのAEC溶液(Sigma Chemical,St.Louis,MO)を加えた。反応は流水している水道水のもとで4−6分後に停止した。スポットをコンピューター支援画像分析(Zeiss KS Elispot Reader,Jena,Germany)を用いてカウントした。106個のCD8+細胞あたりの正味のスポット数を、以下[(関連ペプチドに対するスポット数)−(非関連ペプチドに対するスポット数)]×2.5として計算した。
【0213】
(実施例1)
(ヘテロクリティックな活性についてのペプチドアナログのスクリーニング)
(A.IFNγ放出の増加に関与しているCEA.691およびMAGE3.112アナログの同定)
アナログのスクリーニングの前に、広範囲な野生型ペプチド用量にわたってCTL株からのIFNγ生成のペプチド用量滴定を、行った。.221A2.1腫瘍細胞に異なる用量のペプチドを適用し、次いで、105個のペプチド負荷細胞を同数のマウスまたはヒトCTLと共に培養した。37℃で24時間(マウス)または48時間(ヒト)のインキュベーション後、CTLによって放出されたIFNγのレベルをインサイチュ捕捉ELISAアッセイによって測定した。用量滴定曲線を決定した後、ペプチドアナログのパネルの抗原性をスクリーニングするために、野生型ペプチドに対する活性がほとんど検出可能でない最適以下ペプチド用量を選択した。全てのマウスおよびヒトCTL株について、この最適以下用量の範囲は0.1〜1μg/mlであった。マウスCTL株は第3ドメイン内のマウスH−2Kb配列を有しているHLA分子を発現するHLA−A2.1/Kbxsトランスジェニックマウスにおいて生成されたが、すべてがネイティブなHLA−A2.1分子を発現するAPC上に提示されているペプチドに反応したということに留意すべきである。
【0214】
ペプチドアナログのスクリーニングについて、.221A2.1細胞に選択された最適以下用量の各アナログを適用し、前述したようにペプチド負荷APCをCTLと共に培養した。次いで、さらなる特徴づけのために、野生型ペプチドと比較して増強されたCTL応答を誘導するアナログを選択した。これらのアナログを、前述したのと同じ条件のもとで野生型エピトープと並行してペプチド用量滴定を行うことによって特徴付けた。
【0215】
HLA−A2.1拘束CEA.691およびMAGE3.112エピトープに特異的なCTL株を、調製Cで記載したように、インビトロにおいてペプチド負荷樹状細胞または接着性単球を用いてヒトPBMCの再刺激を繰り返すことによって、誘導した。
【0216】
17個の異なる単一アミノ酸で各残基を体系的に置換することによって合計117CEA.691および116個のMAGE3.112アナログを作製した。CEA.691はIMIGVLVGV(配列番号1);MAGE3.122はKVAELVHFL(配列番号5)である。Cys、TrpおよびMet残基は、それらが保存的変化に対応しない限り、一般に回避した。主なMHCアンカー位置、位置2およびC末端以外のペプチドの全ての位置で置換を導入した。
【0217】
次いで、インビトロでこれらのアナログのを、その抗原性について試験した。野生型ペプチドに応答するIFNγ生成がほとんど検出可能でない抗原濃度を規定にするために、前述したように、予備的な用量滴定実験を各CTL株について行った。ついで、T細胞刺激の許容量の増加に関連したアナログを同定するために、引き続きこの最適以下濃度をアナログペプチドの各エピトープについてのすべての抗原性分析に用いた。このような抗原性分析の結果を図1に示す。図1Aに示すように、最適以下100ng/ml用量では、野生型CEA.691ペプチドは最低限のIFNγ生成(50pg/ウェル未満)しか生じなかった。対照的に、いくつかのCEA.691アナログ(M3、L4、P4、H5、L5、H6、T6およびI7)は、同じ用量で、150〜350pg/ウェルの範囲内で検出可能なレベルのIFNγの産生を、誘導した。図1Bに示したように、野生型ペプチドのMAGE3.112特異的CTL株100ng/mlが100pg/mlのIFNγの放出を誘導したが、2つのアナログ(I5およびW7)は300pg/ウェル以上のIFNγレベルの誘導に関連していた。
【0218】
さらなる特徴付けのために、100pg/ウェルを超えるIFNγを刺激したCEA.691およびMAGE3.112の全てのアナログを選択し、完全な用量滴定はヘテロクリティックなアナログを同定するために実施した。ヘテロクリティックなアナログは野生型ペプチドの10倍以下のペプチド濃度で、有意なIFNγ放出(100pg/ウェルより大きい)を刺激するアナログである。CEA.691エピトープについては2つの異なるアナログ、M3(配列番号2)およびH5(配列番号3)を同定した。図1Cで見られるように、エピトープCEA691について、アナログM3およびH5が0.01ng/ml程度の少ないペプチドを用いて有意な放出を刺激したが、野生型ペプチドは1〜100μg/mlの用量範囲で有意に検出可能なIFNγシグナルを産出した。これらの基準によると、これらの2つのCEA.691アナログは、モル濃度に基づき、IFNγ放出の点において、その未改変野生型対応物よりも100,000倍以上強力である。
【0219】
同様に、MAGE3.112エピトープについて、2つのヘテロクリティックなアナログ、I5およびW7を同定した。図1Dに示すように、等価な応答を刺激するためには、0.1ng/mlのI5(配列番号6)またはW7(配列番号7)アナログのいずれかが必要であったが、1μg/mlの野生型ペプチド濃度が有意なIFNγの放出に必要とされた。これは、野生型ペプチドと比較して100,000倍より高い生物学的な活性の増加に対応する。
【0220】
一般的に、ヘテロクリティックなアナログは用量応答変化を誘導するのみならず、野生型ペプチドと比較してより高いレベルのIFNγを産出するためのCTL応答を刺激した。その結果、アナログに応答して達成される最大用量応答(プラトー)は未改変抗原に応答して得られる応答よりはるかに高かった。
【0221】
(実施例2)
(さらなるヘテロクリティックなアナログの同定)
3つのさらなるA2.1拘束エピトープ、MAGE2.157 YLQLVFGIEV、配列番号7腫瘍エピトープ、およびウイルス抗原からの2つのエピトープ、HBV Pol.455、GLSRYVARL(配列番号16)およびHIV pol.476 ILKEPVHGF(配列番号18)を分析した。これらのエピトープは全て、CTLに対して免疫原性であることが以前に示されている。
【0222】
調製Bおよび表2で述べたアミノ酸保存性割り当て(assignment)を使って、240個の異なるアナログのパネルを、3つのエピトープのそれぞれにおいてエピトープ位置3、5、7、およびエピトープ位置1、4、6に、5つの保存的アミノ酸置換および5つの非保存的アミノ酸置換を含むように合成した。これらのアナログを、HLA−A2.1/Kbxsトランスジェニックマウスで生じたマウスTCL株を使用し、そしてCEA.691およびMAGE3.112エピトープについての実施例1で述べたものと類似した実験ストラテジーに従い、へテロクリシティーについて試験した。マウス株の生成および維持に関連した技術的容易さに起因して、HLAトランスジェニックマウス由来のマウスCTL株をヒトCTL株の代わりに用いた。
【0223】
この結果を図2DのHIV pol.467について対応する用量滴定プロフィールと共に図2A(MAGE2.157)、2B(HBV pol.455)、および2C(HIV pol.476)に示した(MAGE2.157およびHIV Pol.455について、実施例3を参照のこと)。
【0224】
試験されたMAGE2.157エピトープの総計85個の異なるアナログの分析は、野生型ペプチドより1/100〜1/100,000倍の用量でIFNγ応答を刺激した2つのヘテロクリティックなアナログ、I5(配列番号8)およびF5(配列番号9)の同定を生じた(図1);これらのアナログは両方ともペプチドの中央部の奇数位置(5位)において天然に存在している保存的または半保存的な置換を有していた。
【0225】
HIV pol.476エピトープについては、スクリーニングされた78個の異なるアナログの中から、2つ(H3(配列番号19)およびL3(配列番号20))が、ヘテロクリティックな活性を有するとして同定された(表1);両方のアナログはペプチド中央部の奇数位置に保存的または半保存的置換のいずれかを保有していた。77個の試験されたものの中から、HIV pol.455エピトープの1つのヘテロクリティックなアナログの正体が同定された。このアナログはペプチド(配列番号17)の位置7に保存的置換基(P)を有していた(表1)。
【0226】
従って、腫瘍およびウイルス起源の3つのさらなるエピトープ(MAGE2.157、HIV pol.476およびHBV Pol.455)についての240個アナログから得られたデータは、実施例1で示したようなMAGE3.112およびCEA.691エピトープの分析と一致する。
【0227】
ヘテロクリシティーな分析をまた、2つのp53エピトープにおいて行った。1つのエピトープP53.149M2、SMPPPGTRV(配列番号10)はMHC結合を増強するメチオニン残基置換を有するヒトp53エピトープの固定アンカーアナログ(fixed anchor analog)を表す。第2のエピトープ、P53Mu.184、GLAPPQHLIRV(配列番号13)はマウスとヒトとの間で完全に保存された配列を有している(Theobaldら、92(26):11993(1995))。
【0228】
p53.149M2において行った用量滴定分析によって、1μg/mlおよび0.1μg/ml用量範囲における最適および最適以下応答が明らかになった。p53.149M2(各位置で5つの保存的および5つの半保存的置換)についての76個のアナログのパネルがスクリーニングされ、わずか2つのアナログ、C1(配列番号11)およびP7(配列番号12)が同定された。この両方とも、最適以下用量で100pg/ウェルのIFNγ放出を与える、図5。さらに分析すると、両方のアナログは野生型ペプチドの1/10倍の濃度で有意なIFNγ産生を誘導した。さらに、C1アナログもまた1/100倍のペプチド濃度で有意なIL10レベルを誘導した、図6。
【0229】
p53mu.184エピトープに関して、用量滴定分析を行った後、ペプチドの最適および最適以下レベルは、それぞれ500ng/mlおよび10ng/mlであると決定した。63個の保存的および半保存的置換アナログのパネルを免疫原性について試験した。増強された免疫原性を有する2つのアナログが見い出された−T3(配列番号14)およびT3、E6(配列番号15)。図7および8を参照のこと。
【0230】
(実施例3)
(ヘテロクリティックなアナログによって誘導されるリンホカインプロフィ
ール)
ヘテロクリティックなアナログはT細胞からのサイトカインの生成を示差的に活性化することが以前に示されており、それにより、いくつかのアナログがTh1サイトカインを生成するために特異的にT細胞を活性化するが、他は優先的にTh2サイトカインの生成を活性化する。本発明のヘテロクリティックなアナログに関連するリンホカイン放出のパターンを研究するため、CTL株からのTh2サイトカインであるIL5および/またはIL10の生成をIFNγの生成と比較した。2つの異なるエピトープからの代表的なデータを図3および4に示した。
【0231】
図3はMAGE2.157アナログによって誘導されたリンホカインのプロフィールを示す。アナログI5もしくはF5、または野生型(WT)ペプチドを適用した.221A2.1標的に応答して、MAGE2.157特異的CTL株によって生成されたIFNγ(A)およびIL10(B)をいくつかの異なる用量に対して測定した。点線はIFNγ(100pg/ウェル)またはIL10(50pg/ml)の有意なレベルを示している。図3Aで見られるように、MAGE2.157のF5およびI5アナログは、それぞれ野生型ペプチドの1/100倍または1/10,000倍の濃度でIFNγ生成の有意なレベルを誘導した。さらに、同じアナログはまた、野生型ペプチドの1/10倍または1/100倍のペプチド濃度で、有意なIL10生成誘導した。
【0232】
同じ傾向を示す、別のエピトープHBV Pol.455からのデータを、図4Aおよび4Bに示した。いくつかの異なるペプチド用量に対して、アナログP7または野生型(WT)ペプチドに応答してHBV Pol.455CTL株によって放出されたIFNγ(A)またはIL10(B)を示した。再度、HBV
Pol.455のP7アナログは、有意なレベルのIFNγ(図4A)およびIL10(図4B)を野生型ペプチドの1/100倍のペプチド濃度において誘導した。Th2サイトカイン(表1)の誘導について試験した全てのヘテロクリティックなアナログをまとめたデータを総合すると、ほとんどのヘテロクリティックなアナログは増加したTh1およびTh2サイトカインの両方の生成を刺激することが示される。
【0233】
(実施例4)
(ヘテロクリティックアナログのHLA−A2.1との結合親和力)
観察されたCTL株によって向上した認識が、アナログエピトープとMHCとの結合能力における偶然の増加に起因しないことを実証するために、精製したHLA−A2.1分子を、インビトロで利用して全てのヘテロクリティックアナログとMHCとの結合親和力を測定し、そして調製Dに記載されているように未改変の野生型対照物と比較した。
【0234】
表1に概説されているように、3つのアナログ(MAGE3.112W7、HIV Pol.476H3、およびHIVPol.476L3)は、HLA−A2.1と野生型ペプチドよりも4倍以上高い親和力で結合し、そして2つのアナログ(MAGE2.157I5、MAGE2.157F5)は低い親和力で結合した。残る4つのヘテロクリティックアナログであるMAGE3.112I5、CEA.691M3、CEA.691H5、およびHBV Pol.455P7は、HLA−A2.1との結合能力において、ほとんどあるいは全く変化なしで結合した。これらのデータからひとまとめにして、増加した結合性とヘテロクリシティーとの間に相関はないことが示唆される。
【0235】
(実施例5)
(マウスp53.261エピトープに関するアナログの予測および免疫原性)
インビボで免疫原性に関して試験するために、HLA−A2.1を制限したマウスp53.261エピトープを使用した。この理由は、このエピトープに対するCTL応答は、HLA−A2.1/Kbトランスジェニックマウスにおいて局部的に寛容化されることが示されているためである。これはインビボでT細胞の耐性を破壊すると予測されたヘテロクリティックアナログの能力の分析を可能にする。従来まで、ヘテロクリティックアナログは野生型エピトープに対して引き起こされるCTL株とともにインビトロでのスクリーニングを通じて検出されてきたが、置換法によって同定したアナログは野生型エピトープに対してヘテロクリティックであるCTLをインビトロで潜在的に誘導し得り、寛容な腫瘍関連エピトープに対するワクチンを設計するための目的の適応である。
【0236】
p53.261の予測されるアナログに関する免疫原性を、HLA−A2.1/Kbxdトランスジェニックマウスにおいて、p53.261エピトープ(LLGRDSFEV)50μg、または予測されるアナログおよびIFAにおけるHBV Core.128ヘルパーエピトープ140μgとを用いてマウスを共同免疫することで試験した。11日後、プライム化脾臓細胞を収集し、そして10μg/mlのペプチドでパルスした照射同系LPS活性化脾臓細胞とともにインビトロで培養した。培養から10日後、CTLをIL2の供給源としてCon Aの馴化培地の存在下で、ペプチドでパルスしたLPS芽細胞で再刺激した(Ishioka,G.ら、J.Immunol.(1999)162:3915)。
予測アナログによって免疫化されたマウス由来の脾臓細胞を、野生型ペプチド(交差反応性、アビディティおよび野生型抗原に応答するCTLの前駆体度数を決定するため)とそれぞれの免疫化アナログ(アビディティとアナログに応答するCTLの前駆体度数を決定するため)の両方に対してインビトロで刺激した。APCとしてJurkat−A2.1腫瘍細胞を用いることで、インビトロでの2回目の再刺激の5日後でのIFNγのインサイチュのELISAアッセイによるペプチドの特異性に関して、CTLのバルク集団を全ての短い周期で試験した。あるいは、CTL応答を、Elispotアッセイを用いて免疫化動物から新たに単離した脾臓細胞上で実施した。
【0237】
9マーペプチドの3、5および7の位置における3つの保存的、または半保存的置換からなるp53.261エピトープの9つのアナログのパネルを、HLA−A2.1/Kbxdトランスジェニックマウスにおける免疫原性について試験した。その結果、それぞれ9つのアナログでのマウスの免疫、およびそれぞれの免疫化アナログでのプライム化脾細胞のインビトロでの膨張により6つのアナログ(L7、D3、H7、H3、N5、およびG5)を同定し、これは野生型ペプチドを用いてインビボで誘導され、そしてインビトロで膨張されたCLTと比較して、かなりより低いペプチド濃度で、100pg/ウェルのIFNγ産生によって特徴づけられたCTL応答が得られた。
【0238】
WTペプチドまたは適応したアナログのいずれかによって免疫化されたマウス由来の脾臓細胞を、インビトロで対応する免疫ペプチド(図9AおよびB)またはWTペプチド(図9CおよびD)を用いて刺激した。次いで、これらのCTLによるIFNγの放出を、免疫ペプチド(図9AおよびB)またはWTペプチド(図9CおよびD)を用いてパルス化した標的に対して用量範囲にわたって測定した。100pg/ウェルにおけるIFNγの放出を点線で示す。これらの結果より、アナログの有意なパーセンテージは、野生型ペプチドそれ自体で誘導されたアビディティよりも高いアビディティのCTLを誘導することが示される。
【0239】
野生型ペプチドに対するヘテロクリティックアナログでプライム化されたCTLの交差反応性を、図9Cおよび図9Dに示す。野生型ペプチドで免疫化し、そして再刺激した動物由来のCTLは、0.1から10μg/mlの間のペプチド用量において100pg/ウェルのIFNγを誘導した。もう一方で、L7、H3、およびD3アナログで免疫化し、そして野生型ペプチドを用いてインビトロで刺激し、そして試験した動物から得られたCTLは、100pg/ウェルのIFNγを誘導するために、それぞれ10倍、100倍または1000倍低い用量の野生型ペプチドが必要であった(図4C)。これにより、6つのケースの内3つのケースにおいて、予測されるヘテロクリティックアナログは、野生型抗原に対して寛容である一部のCTLが存在する条件において、野生型ペプチドと応答するCTLを誘導するのに10〜1000倍の活性および能力であったことが示唆される。
【0240】
(実施例6)
(野生型との交差反応性)
D3およびH3アナログにより誘導されたCTLの交差反応性もまた、自然処理された野生型エピトープに対してHLA−A2.1/Kbを用いてトランスフェクトされたp−53発現Meth A腫瘍細胞クローンによって試験した。野生型エピトープに対してヘテロクリティックであるp−53.261アナログによって産生したCTLは、Meth A/A2.1Kb腫瘍細胞によって出現した内因的に処理されたp−53.261エピトープと応答することが明らかとなった。
【0241】
CTL集団(105/ウェル)は、HLA−A2.1/Kbと共にトランスフェクトされたMeth Aクローン、または2.5×104のMeth A腫瘍細胞とともに培養し、IFNγの放出はインサイチュELISAアッセイによって測定した。図10に示しているように、p−53.261エピトープのD3とH3アナログの両方に対して増加したCTL株は、Meth A/A2.1Kb腫瘍細胞クローンによって発現された内因性エピトープと応答し、親HLA−A2.1ネガティブMeth A腫瘍細胞株とは応答しなかった。
【0242】
(実施例7)
(Elispotアッセイを用いた前駆体度数の分析)
野生型ペプチドに対する交差反応性CTLが、アナログで免疫化されたマウスにおいて産生されることを確証するために、インビトロでのさらなるCTL膨張なしでアナログまたは野生型ペプチドで免疫化されたマウスの脾臓細胞からCD8+細胞を単離し、そして野生型またはアナログのどちらか一方に対するCTL反応性の前駆体度数をElispotアッセイを用いて測定した。
【0243】
WTエピトープ、またはD3、H3、L7およびH7アナログの何れか一方で免疫化されたマウスから単離したCD8+細胞を、ElispotアッセイにおいてWTエピトープで刺激した場合のIFNγの放出する能力について分析した。図11に示すように、野生型ペプチド反応性CTLの前駆体度数は、野生型ペプチドで免疫化されたマウスにおいて1/66,000(15点/106)であったのに対して、予測アナログで免疫化されたマウスにおける野生型ペプチド反応性細胞の前駆体度数は、D3、H3およびL7アナログに対してほぼ1/15,000(60〜75点/106細胞)であり、そしてH7アナログに対して1/83,000(12点/106)であった。これより、野生型反応性細胞は、天然ペプチドで免疫化されたマウスに比べて4つのアナログのうち3つのアナログで免疫化されたマウスにおいて4倍高い度数で存在していたことが示される。この発見は、インビボ−プライム化CTLのインビトロでの膨張を回避する、より直接的な分析システムを用いたヘテロクリティックアナログでのインビボ免疫化は、野生型ペプチドに対して実際により複数のCTL反応性を誘導することを示すので重要である。
【0244】
(実施例8)
(ヘテロクリティックアナログはインビトロで腫瘍細胞を認識し得るヒトCTLを誘導する)
調製Cに記載されているように、MAGE3.112のヘテロクリティックアナログの免疫原性もまた、MAGE3.112ペプチド、またはこのエピトープのI5およびW7アナログのどちらか一方に対するPBMCから1次CTLを誘導することによって試験した。インビトロでの2回目の刺激の後、48ウェルのPBMC培養は、正味のIFNγ産物が100pg/ウェルより多い場合にはCTL誘導に対して陽性を記録し、そして産物はペプチドの存在下、または非存在下で.221−A2.1 APCで刺激した後ではバックグラウンドより少なくとも2倍以上となった。
【0245】
ヒト腫瘍抗原に対する本発明者らの所見の生理学的な関連性を強調するために、本発明者らはインビトロでの1次誘導システムにおいて、MAGE3.112エピトープのヘテロクリティックアナログがヒトCTLを誘導し得るかどうかを試験した。正常のドナー由来の新しい未処理のヒトPBMCを、前述しているように野生型またはアナログのどちらか一方を用いてインビトロで繰り返し刺激した(Kawashima,I.ら、Human Immunol.(1998)59:1)。ペプチド特異的CTL応答を、野生型ペプチド(図12A)またはI5アナログ(図12B)およびW7アナログ(図12C)のいずれかで刺激された培地中で検出した。手短に言えば、.221−A2.1細胞をWTペプチド(図12A)、I5アナログ(図12B)またはW7アナログ(図12C)10μg/mlで一晩パルスした。48ウェルプレートの個々のウェルに成長しているCTLによるIFNγ産物を、ペプチドの存在下、または非存在下で.221−A2.1細胞に対して、あるいは内因性ネガティブエピトープ888メル腫瘍細胞株および内因性ポジティブエピトープ624メル腫瘍細胞株に対して試験した。ポジティブペプチド特異的CTL応答を示すウェルのみが示される。
【0246】
さらに重要なことに、これらのアナログとともに誘導された培地により、内因的に処理し、そして野生型配列を示す624メル腫瘍細胞株が認識された。これは、ヘテロクリティックアナログは腫瘍細胞によって示される内因的に産生された野生型エピトープを認識する生理学的に関連性のあるヒトCTLを誘導し得ることの証拠であり、そしてその現象は、ヒトとトランスジェニックマウスシステムの両方に関連性があることの証拠である。
【0247】
(実施例9)
(HLA A2スーパーファミリーメンバーにおけるHLA−A2.1スーパーモチーフから誘導されたヘテロクリティックアナログの合成および分析)
A2スーパーファミリー内部の他のHLA分子におけるヘテロクリティック置換法則をさらに確認するために、3、5および7の位置における3つの保存的、または半保存的置換よりなる、以前に議論したp53.261エピトープの9つのアナログのパネルを、以下に示すヒトHLA分子(A*0202、A*0203、A*0204、A*0205、A*0206、A*0207、A*0209、A*0214、A*6802、A*6901)の1つを発現しているトランスジェニックマウスにおける免疫原性についてインビボで試験した。
【0248】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、p53.261エピトープ)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型エピトープ自体に対するよりも、天然野生型エピトープに対してより高いアビディティCTLを誘導するのにより強力であることを示す。
【0249】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0250】
(実施例10)
(HLA B7スーパーファミリーメンバーにおけるHLA−B7スーパーモチーフから誘導されたヘテロクリティックアナログの合成および分析)
ヘテロクリティック置換法則をさらに確認するために、HLA−B7スーパーモチーフの内部に配列を持っているペプチドから誘導されたヘテロクリティックアナログを用いて追加研究を行った。例えば、アナログをインビボで免疫原性について試験し得る。
【0251】
本研究では、ペプチドを持ったHLA−B7スーパーモチーフであるAPRTLVYLLエピトープを選択し、そして合成した。9マーペプチドの3、5および7の位置における3つの保存的、または半保存的置換よりなるアナログのパネルを、HLA−B*0702/Kbトランスジェニックマウスにおける免疫原性について試験した。パネルはAPETLVYLL、APRTWVYLLおよびAPRTLVPLLを含み、それぞれ3、5および7番目の位置における半保存的置換と対応している。
【0252】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγの誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、APRTLVYLL)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体よりも高いアビディティCTLを誘導するのにより強力であることを示す。
【0253】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0254】
B7スーパーファミリー内部の他のHLA分子についてヘテロクリティック置換法則をさらに確認するために、APETLVYLL、APRTWVYLLおよびAPRTLVPLLペプチドを、以下のヒトHLA分子(B*0702、B*0703、B*0704、B*0705、B*1508、B*3501、B*3502、B*3503、B*3503、B*3504、B*3505、B*3506、B*3507、B*3508、B*5101、B*5102、B*5103、B*5104、B*5105、B*5301、B*5401、B*5501、B*5502、B*5601、B*5602、B*6701、およびB*7801)の1つを発現しているトランスジェニックマウスにおける免疫原性についてインビボで試験した。
【0255】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、APRTLVYLL)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体より高いアビディティCTLを誘導するのにより強力であることを示す。
【0256】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0257】
(Elispotアッセイを用いた前駆体度数の分析)
野生型ペプチドに対する交差反応性CTLが、アナログで免疫化されたマウスにおいて産生されることを確証するために、インビトロでのさらなるCTL膨張なしでアナログまたは野生型ペプチドで免疫化された脾臓からCD8+細胞を単離した。この材料より、野生型、またはアナログのどちらか一方に対するCTL反応性の前駆体度数をElispotアッセイを用いて測定した。代表的に、野生型ペプチドの反応性CTLの前駆体度数は、アナログの前駆体度数よりもかなり低い。
【0258】
(ヘテロクリティックアナログによってインビトロでエピトープを認識し得るヒトCTLを誘導し得る)
ヘテロクリティックアナログを、インビトロでの1次誘導システムにおけるCTLの誘導について分析し得る。前記のように、48ウェルプレートにおいて正常のドナーからの新しい未処理のヒトPBMCを、野生型またはアナログのどちらか一方を用いてインビトロで繰り返し刺激した。次に、ペプチド特異的CTL応答を、野生型ペプチドまたはヘテロクリティックアナログのどちらか一方で刺激された培地中で検出する。これらのアナログで誘導された培地において、内因的に処理され、そして野生型配列を示す標的を認識し得る。これは、ヘテロクリティックアナログが内因的に産生された細胞上で発現した野生型ペプチドを認識する生理学的に関連性のあるヒトCTLを誘導し得ることの証拠であり、そしてその現象はヒトとトランスジェニックマウスシステムの両方に関連性があることの証拠である。
【0259】
(実施例11)
(HLA A3スーパーファミリーメンバーにおけるHLA−A3スーパーモチーフから誘導されたヘテロクリティックアナログの合成および分析)
ヘテロクリティック置換法則をさらに進んで確認するために、HLA−A3スーパーモチーフの内部に配列を有するペプチドから誘導されたヘテロクリティックアナログを用いて追加研究を行う。例えば、インビボで免疫原性についてアナログを試験し得る。
【0260】
本研究では、ペプチドを有するHLA−A3スーパーモチーフであるKVFPYALINK(配列番号22)エピトープを選択し、そして合成する。9マーペプチドの3、5および7の位置における3つの保存的/半保存的置換よりなる配列番号22のアナログのパネルを、HLA−A*3101/Kbトランスジェニックマウスにおける免疫原性について試験した。パネルはKVHPYALINK、KVFPQALINKおよびKVFPYAKINKを含み、それぞれ3、5および7番目の位置における半保存的変化と対応している。
【0261】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、KVFPYALINK)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体より野生型に対して高いアビディティCTLを誘導するのにより強力であることを示す。
【0262】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0263】
A3スーパーファミリー内部の他のHLA分子についてヘテロクリティック置換法則をさらに確認するために、KVHPYALINK、KVFPQALINKおよびKVFPYAKINKペプチドを、以下のヒトHLA分子(A*0301、A*1101、A*3101、A*3301、A*6801)の1つを発現しているトランスジェニックマウスにおけるインビボでの免疫原性について試験する。
【0264】
上述したアナログで免疫化されたマウス由来のCTLを、少なくとも100pg/ウェルのIFNγ産生の誘導について試験した。代表的に、このIFNγ産生は、野生型ペプチド(例えば、KVFPYALINK)で誘導され、そして再刺激されたペプチド濃度よりもさらに低いペプチド濃度で存在する。これらの結果は、本発明者らの予測したヘテロクリティックアナログは、野生型ペプチド自体より高いアビディティCTLを誘導するのにより強力であることを示す。
【0265】
代表的に、野生型ペプチドで免疫化され、そして再刺激された動物から得られたCTLは、5〜10μg/ml用量のペプチドにおいて100pg/ウェルのIFNγを誘導するが、上述したアナログで免疫化された動物から得られ、野生型ペプチドでインビトロで刺激し、そして試験したCTLは、100pg/ウェルのIFNγを誘導するために野生型ペプチドの用量よりもそれぞれ10倍、100倍または1000倍までも低い用量を必要とする。
【0266】
(Elispotアッセイを用いた前駆体度数の分析)
野生型ペプチドに対する交差反応性CTLが、アナログで免疫化されたマウスにおいて産生することを確証するために、インビトロでのさらなるCTL膨張なしでアナログまたは野生型ペプチドで免疫化された脾臓からCD8+細胞を単離した。この材料より、野生型、またはアナログのどちらか一方に対するCTL反応性の前駆体度数をElispotアッセイを用いて測定した。代表的に、野生型ペプチドの反応性CTLの前駆体度数は、アナログの前駆体度数よりもかなり低い。
【0267】
(ヘテロクリティックアナログはインビトロでエピトープを認識し得るヒトCTLを誘導し得る。)
ヘテロクリティックアナログを、インビトロでの1次誘導システムにおけるCTLの誘導について分析する。前記したように、48ウェルプレートにおいて正常のドナー由来の新しい未処理のヒトPBMCを、野生型またはアナログのどちらか一方を用いてインビトロで繰り返し刺激した。次に、ペプチド特異的CTL応答を、野生型ペプチドまたはヘテロクリティックアナログのどちらか一方で刺激された培地中で検出する。これらのアナログで誘導された培地において、内因的に処理し、そして野生型配列を示す標的を認知する。これは、ヘテロクリティックアナログが内因的に産生される細胞上に発現した野生型ペプチドを認識する生理学的に関連性のあるヒトCTLを誘導することの証拠であり、そしてその現象はヒトとトランスジェニックマウスシステムの両方に関連性があることの証拠である。
【0268】
本明細書中に記載される実施例および実施形態は、例示の目的のためだけであり、本発明に照らして、様々な改良または変更は当業者に示唆され、そして本出願の意図および範囲ならびに添付の特許請求の範囲内に含まれることが理解される。この結果、本明細書中で引用される全ての刊行物、特許、および特許出願は、全ての目的のためにその全体が参考として援用される。
【特許請求の範囲】
【請求項1】
本明細書中に記載される発明。
【請求項1】
本明細書中に記載される発明。
【図1−1】


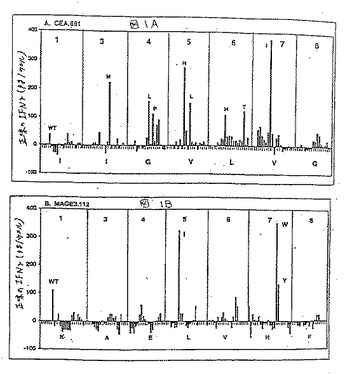
【図1−2】
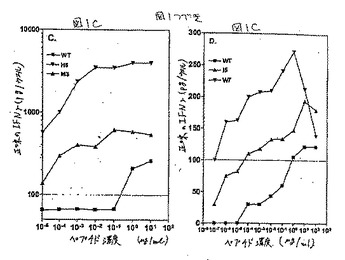
【図2−1】

【図2−2】

【図2−3】
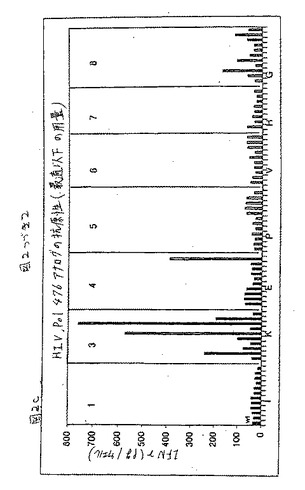
【図2−4】
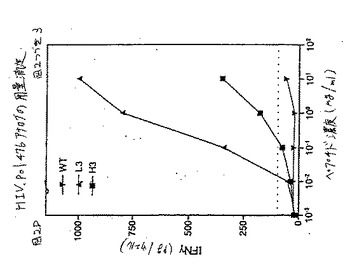
【図3】
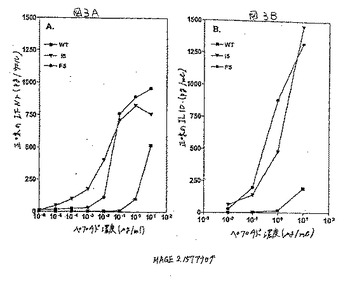
【図4】
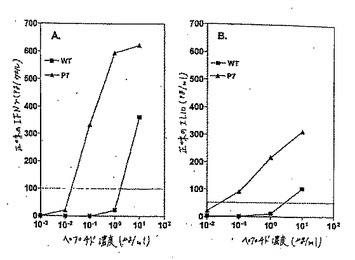
【図5】
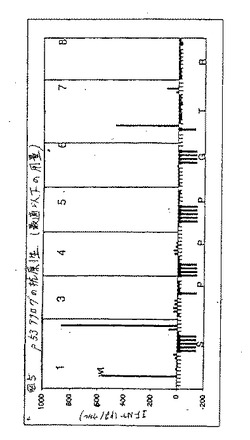
【図6】
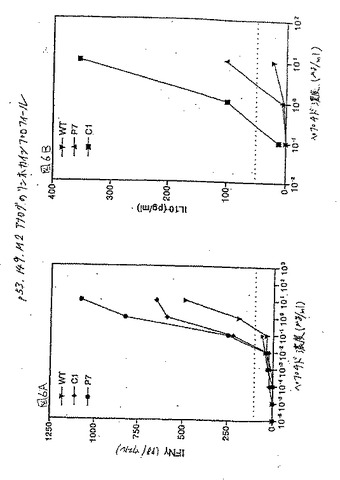
【図7】

【図8】
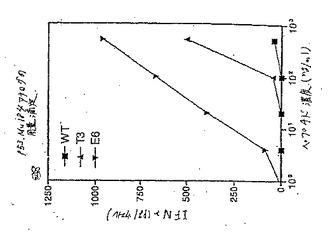
【図9】
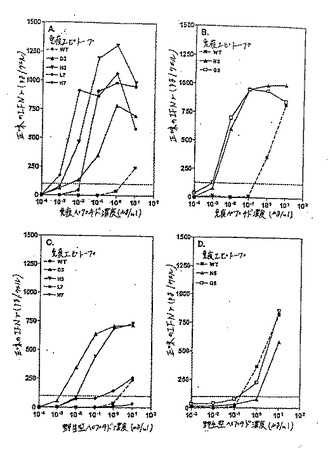
【図10】
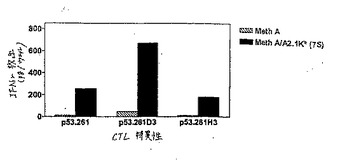
【図11】
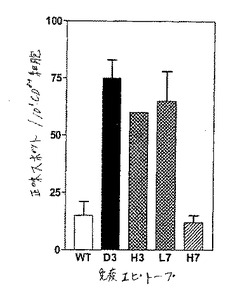
【図12】
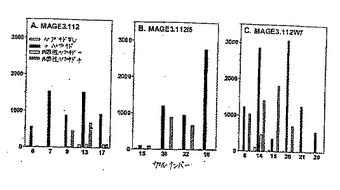
【図101−1】
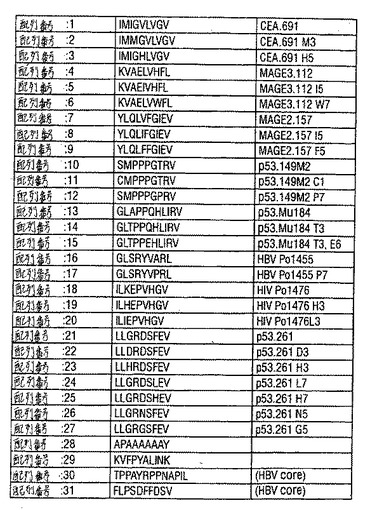
【図101−2】

【図102−1】

【図102−2】
【図103−1】
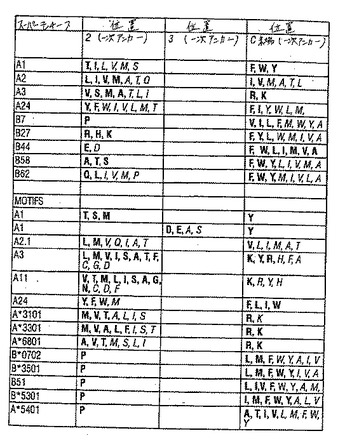
【図103−2】
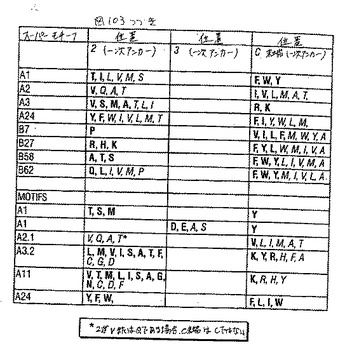
【図104−1】

【図104−2】
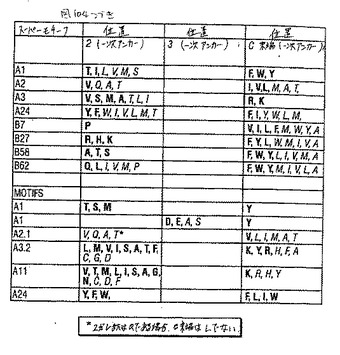
【図1−2】
【図2−1】
【図2−2】
【図2−3】
【図2−4】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図101−1】
【図101−2】
【図102−1】
【図102−2】
【図103−1】
【図103−2】
【図104−1】
【図104−2】
【公開番号】特開2011−101654(P2011−101654A)
【公開日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願番号】特願2011−1564(P2011−1564)
【出願日】平成23年1月6日(2011.1.6)
【分割の表示】特願2001−538941(P2001−538941)の分割
【原出願日】平成12年11月20日(2000.11.20)
【出願人】(500521038)エピミューン インコーポレイテッド (5)
【Fターム(参考)】
【公開日】平成23年5月26日(2011.5.26)
【国際特許分類】
【出願日】平成23年1月6日(2011.1.6)
【分割の表示】特願2001−538941(P2001−538941)の分割
【原出願日】平成12年11月20日(2000.11.20)
【出願人】(500521038)エピミューン インコーポレイテッド (5)
【Fターム(参考)】
[ Back to top ]