
ヘリコバクターピロリに対する高親和性レセプターおよびその用途
本発明はヘリコバクター ピロリに結合するオリゴ糖物質またはレセプター、ならびにその用途を開示する。用途しては、例えば、該オリゴ糖物質またはレセプターを用いた、ヘリコバクター ピロリの存在によって生じる病態の治療用の医薬組成物または栄養補助組成物が挙げられる。本発明はまた、該レセプターを用いた、ヘリコバクター ピロリの診断方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヘリコバクター ピロリに対する新規な高親和性レセプターを開示する。このレセプターは、シアリル化ポリ−N−アセチルラクトサミン構造を含むものである。本発明は、主要な胃病原体であるヘリコバクター ピロリに対して上記物質を用いた治療方法および予防方法に関する。また本発明は、上記構造を用いた分析方法および診断方法に関する。本発明はさらに、上記構造を用いた、ヘリコバクター ピロリ感染を防止するための機能性食品の製造方法に関する。
【背景技術】
【0002】
糖タンパク質や糖脂質に提示されているシアリルラクトースおよびシアル酸は、ヘリコバクター ピロリに対するレセプターとして知られている。これらレセプターの構造は、NeuNAcα3Gal- や NeuNAcα3Galβ4Glc(シアリル−ラクトース)(US Patent Nos. 5,883,079、5,753,630および5,514,660、ならびにMysore et al., 1999)または NeuNAcα3Galβ4GlcNAc(Johansson and Miller-Podraza, 1998、Miller-Podraza et al., 1997)であると考えられている。さらに最近では、フコシル化された異型、例えば、シアリル−ルイスx(NeuNAcα3Galβ4(Fucα3)GlcNAc;sLex)、特にジフコシル化構造であるシアリル−2量体−ルイスx(sialyl-dimeric Lewis x)(NeuNAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4(Fucα3)GlcNAc;sdiLex)(Mahdavi et al., 2002)が、ヘリコバクターに対するリガンドであると考えられている。これらの構造が糖脂質に提示された場合、sdiLexはsLexよりもヘリコバクター ピロリに対して何十倍も優れたレセプターである(Mahdavi et al., 2002)。本発明は、他のより大きな非フコシル化ポリ−N−アセチルラクトサミン構造にも関する。フコシル化およびシアリル化されたポリラクトサミンは、ヘリコバクター ピロリに対して優れたレセプターである。しかしながら、比較的高い親和性を示すエピトープが非フコシル化ポリ−N−アセチルラクトサミンに提示されることもある。このような構造の利点は、5〜10工程もの追加工程を経る化学合成によってエピトープにフコースを付加する必要がない点である。フコシル化グリカンの酵素学的または発酵学的な合成は未だ商業レベルに達していないので、フコースの付加は工業的にも困難であると考えられる。
【0003】
本発明はさらに、NeuNAcα3Galβ4GlcNAc-構造がヘリコバクター ピロリを効果的に結合するためには、-β4Glc(NAc)構造を含む必要があることを示す。一般的に、他の結合基を介して結合したNeuNAcα3Galβ3GlcNAcやNeuNAcα3Galβ3GalNAcを含む類似の糖脂質エピトープは結合構造ではないことから、この知見は、末端NeuNAcα3Gal-エピトープについて示唆された結果と対照的である。
【0004】
七糖糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCerは、ヘリコバクター ピロリと結合することが報告されている(Roche. et al. 2001、Johansson and Miller-Podraza, 1998、Miller-Podraza et al., 1997)。本発明は、非フコシル化構造が、より短いNeuNAcα3Galβ4Glc(NAc)エピトープやsLexエピトープよりもヘリコバクター ピロリに対して実際により有効なレセプターであることを示し、ヘリコバクター ピロリに対するより低コストの高親和性阻害剤や診断剤が設計できる可能性を提供する。本発明は、公知のシアル酸含有レセプターの中からシアリル化ポリラクトサミンを高親和性レセプターとして見出すことを可能にする。さらに本発明は、糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCerに代表されるより長いポリラクトサミン鎖、および糖脂質であるNeuNAcα3Galβ4GlcNAcβ3(NeuNAcα3Galβ4GlcNAcβ6)Galβ4GlcNAcβ3Galβ4GlcβCerなどに代表される分岐ポリラクトサミン構造に関する。
【0005】
すでに本発明者らは、ヘリコバクター ピロリに対する2型ラクトサミン結合エピトープである「ネオラクト結合エピトープ」(-GlcNAcβ3Galβ4GlcNAcβ-)も開示している。この結合エピトープはより長い直鎖ポリ−N−アセチルラクトサミン(例えばNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer)において活性を示す。しかしながら、(シアル酸非結合種においては)脱シアリル化構造のみならず、本発明におけるシアル酸結合特異性によって認識されない、N−グリコリル−ノイラミン酸含有構造であるNeuNGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer(FI 20010118)も活性を示すことから、結合特異性にはシアル酸の存在は必要ない。一般的なネオラクトエピトープをシアル酸非結合ヘリコバクター ピロリ種によって特徴付けたときには、NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerに対する結合も見られなかった(FI20010118)。シアル酸結合特異性は、ネオラクト結合特異性よりも一般的ではないが、シアル酸結合特異性はおそらく炎症に関連し、新鮮な病理学的単離物中に存在する(Mahdavi et al., 2002)。ポリラクトサミン鎖の分岐によってネオラクトエピトープへの結合を阻害することができるが、本発明による高親和性シアル酸レセプターへの結合を阻害することはできない(FI20010118)。
【0006】
従来技術には、HPNAPと命名されたヘリコバクター ピロリ結合タンパク質も開示されており、このタンパク質は好中球の活性化に関する潜在的な機能を有すると考えられる。本願のデータは、シアル酸結合活性を示すNAP−タンパク質が、シアリル化構造に結合するヘリコバクター ピロリの阻害に関与しないことを示す。より長いポリラクトサミンを含有するNeuNAcα3Galβ4GlcNAcβ3Gal-型構造への結合も含む、NAP−タンパク質のシアル酸結合作用の潜在的な機能は、ヘリコバクター ピロリと好中球との相互作用に関与すると考えられるが、その特異性や生物学的意義は明らかではない(Teneberg et al., 1997、Teneberg et al., 2000)。
【0007】
さらに従来技術には、次に挙げる非シアリル化1型エピトープも開示されている:ラクトシルセラミドであるGalβ4GlcβCer、ガングリオテトラオシルセラミドであるGalβ3GalNAcβ4LacβCer(Lingwood et al., 1992)、末端Galβ3GlcNAcβ-を含有するラクト−構造(Teneberg et al., 2002)およびルイスbであるFucα2Galβ3(Fucα4)GlcNAcβ-(Boren et al., 1993)。これらのエピトープは種々の炭水化物主鎖、即ちラクトース、1型アセチルラクトサミンまたはガングリオコア、をベースとし、本発明によるシアリル化されたより長い2型N−アセチルラクトサミンとは一次構造と三次構造が明確に異なるものである。さらに、本願明細書で挙げた参考文献によると、種々のレセプターに対して明らかに特異的な種が存在し、シアル酸(sLex)結合特異性(Mahdavi et al., 2002)およびルイスb結合の場合には、非常に特異的なレセプタータンパク質が実際に特徴付けられている(Ilver D. et al., 1998)。
【0008】
発明の詳細の説明
本発明は、複数の直鎖状および分岐状のNeuNAcα3−ポリ−N−アセチルラクトサミン構造がヘリコバクター ピロリに対する高親和性リガンドとして機能し得ることを開示する。このような結合は、2型ラクトサミンであるGalβ4GlcNAcに結合したNeuNAcα3に特異的である。本発明は、末端三糖エピトープよりも高い結合能を有するより大きなポリラクトサミンに関する。直鎖ポリラクトサミン構造である NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer および NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCerを考慮すると、β3Galβ4GlcNAcおよびより大きなポリラクトサミンであるβ3Galβ4GlcNAcβ3Galβ4GlcNAcに提示された末端NeuNAcα3Galβ4GlcNAcが有効であることは明確である。このような結果は、Galまたはラクトースにβ3結合した末端構造が有用な最小エピトープであることも示した。
【0009】
ヘリコバクター ピロリに対する新規な高親和性阻害剤の一般式
具体的には、本発明は、下記式(1)で表される高親和性ヘリコバクター ピロリ結合オリゴ糖配列に関する:
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal[β4GlcNAcβ3Gal]s2−
{β4Glc[NAc]s3}s4 (1)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4GlcNAcβ3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;そして
側鎖として存在し得るものを( )内に示す。)
本発明はまた、ヘリコバクター ピロリに対する結合能を有する該オリゴ糖配列の構造類似体、特に立体配座類似体、または誘導体にも関する。
【0010】
非シアリル化分岐構造と考えられるものの構造
上記式中のR1またはR2がNeuNAcα3ではない非シアリル化置換基である場合には、R1またはR2は、Galの2位および3位の少なくとも1種に結合した、下記式(2)で表される末端置換基であることが好ましい。
Hex[NAc]t1α/β3[(DeoxyHexα2)]t2 (2)
(式中、HexはGalまたはGlcであることが好ましく、
t1およびt2は各々独立に0または1であり、そして
α/βはα結合またはβ結合である。)
【0011】
さらに非シアリル化置換基であるR1またはR2が、下記式からなる群より選ばれる構造であることが好ましい:
Galα3、GalNAcα3、Fucα2、Galα3(Fucα2)、GalNAcα3(Fucα2)、
NeuNAcα3Galβ4GlcNAcβ3、Galβ4GlcNAcβ3、GlcNAcβ3Galβ4GlcNAcβ3、GlcNAcα3、GlcNAcβ3、GalNAcβ3、Galβ3、Glcβ3、そしてGlcα3。さらに好ましくは、下記式からなる群より選ばれる血液型抗原型の構造であることが好ましい:Galα3、GalNAcα3、
Fucα2、Galα3(Fucα2)、そしてGalNAcα3(Fucα2)。
【0012】
好ましい分岐構造
本発明の好ましい態様においては、式中のs1が1であり、R1およびR2が共にNeuNAcα3およびNeuNAcα3Galβ4GlcNAcβ3からなる群より選ばれるものである。好ましいポリ−N−アセチルラクトサミン構造としては、下記の配列で表されるオリゴ糖配列が挙げられる:
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、および
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Gal 。
【0013】
本発明によるオリゴ糖配列は、下記式で表されるような分岐したポリ−N−アセチルラクトサミンでもよい。
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc
(式中、LacNAcはN−アセチルラクトサミン、即ちGalβ4GlcNAcであり、Lacはラクトース、即ちGalβ4Glcである。従って、下記の2種の配列は同じ構造を表す:
NeuNAcα3Galβ4GlcNAcβ3(NeuNAcα3Galβ4GlcNAcβ6)Galβ4Glc と
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac 。)
【0014】
好ましい長鎖ラクトサミンエピトープ
上記式(1)においてs1が0である場合、式(1)は、例えば、下記式のいずれかで表される直鎖シアリルポリラクトサミンを表す:
NeuNAcα3LacNAcβ3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3Lac、
NeuNAcα3LacNAcβ3LacNAcβ3Gal、または
NeuNAcα3LacNAcβ3LacNAc;あるいは
より短い、最小エピトープである、
NeuNAcα3LacNAcβ3Lac、または
NeuNAcα3LacNAcβ3Gal 。
【0015】
好ましい類似体構造
本発明は、NeuNAc残基のグリセロール尾部は、酸化や還元によって生じる修飾を許容しないことを示す。NeuNAcのアセチル基は多くの変更を許容しないので、グリコリル類似体、プロピル類似体または脱アセチル化アミン類似体は、低活性であるか全く活性を示さない。また、シアル酸のカルボキシル基が還元されてアルコールになった異型も活性を示さない。同様の特異性が小さな五糖エピトープと分岐した大きなポリグリコシルセラミドでも見られた。シアル酸残基の位置も重要であり、ジシアル酸構造やNeuNAcα6-構造は活性を示さなかった。さらに、NeuNAc-残基がα3結合した末端N−アセチルラクトサミンは、2型N−アセチルラクトサミン(即ちGalβ4GlcNAc)であるはずであり、1型アセチルラクトサミン末端(即ちGalβ3GlcNAc)やアシアロ−ガングリオシド末端(即ちGalβ3GalNAc)は受け入れられなかった。このようなデータによって、不活性構造の特徴を回避した許容可能な類似体および誘導体の効果的な設計が可能となる。
【0016】
本発明は特に、式(1)で表される構造の類似体に関する。具体的には、式(1)中のN−アセチルラクトサミン残基の少なくとも1つが2型N−アセチルラクトサミン類似体構造、好ましくはラクトース残基、で置換されている、下記式(3)で表される類似体である。
R1Galβ4Glc[NAc]u1β3{(R2Galβ4Glc[NAc]u2β6)}s1Gal{β4Glc[NAc]u3β3Gal}s2−
{β4Glc[NAc] s3}s4 (3)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;そして
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0である。)
本願明細書で開示する2型N−アセチルラクトサミン類似体構造にはGalβ4Glc2-X構造が含まれ、この構造においてはGlc-環の2位の炭素が置換基Xに結合している。Xとしては、-NH2基(GalβGlcN−類似体となる)、N−アルキル基(Galβ4GlcN-アルキル基類似体となる)またはN−アルカノイル基が好ましい。好ましいアルカノイル基はプロパノイル基である。2位の炭素は、O−アセチル基などのO−エステルまたはO−メチル基などのO−エーテルによって誘導されていてもよい。特に、以下のジシアリル化された直鎖モノシアリル化構造が好ましい:
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β6)Lac[NAc]u3β3Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3(NeuNAcα3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、そして
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3Gal{β4Glc[NAc]s3}s4 。
【0017】
本発明はまた、下記式で表される類似体に関する。
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2
(但し、u2が0である場合にはu1も0である。)
【0018】
フコシル化類似体
上述のように、sLexやsdiLexなどのフコシル化構造についても、別途、研究している。本発明は、フコシル化および非フコシル化したシアリルα3−N−アセチルラクトサミンの両方が同じ結合特異性に関与することを示す。上記したポリ−N−アセチルラクトサミンのラクトサミン類似体(特にラクトース含有類似体)もまた、Glc残基および(存在する場合には)GlcNAc残基がα3−フコシル化されている場合に活性を示すであろうことがわかる。このような類似体は、より安価な主鎖構造と、フコース残基によって誘導されたより高い親和性とを兼ね備える。本発明は、このようなシアリル化およびフコシル化構造に関する。
【0019】
最も好ましいフコシル化構造は下記式(4)で表される構造である。
R1Galβ4[(Fucα3)]t1Glc[NAc]u1β3{(R2Galβ4[(Fucα3)]t2Glc[NAc]u2β6)}s1−
Gal{β4[(Fucα3)]t3Glc[NAc]u3β3Gal}s2{β4[(Fucα3)]t4Glc[NAc] s3}s4 (4)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4[(Fucα3)]t5Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0であり;そして
t1、t2、t3、t4およびt5は各々独立に0または1であり、[ ]内のFucα3側鎖構造の存在または不在を示す。)
式(4)で表される構造としては、特に、ジシアリル化構造および直鎖モノシアリル化構造が好ましい。
【0020】
本発明はまた、非還元位置にN−アセチルラクトサミン類似体を有する、非フコシル化類似体構造およびフコシル化類似体構造にも関する。特に、ラクトースを有する類似体、即ち、式中のu1、u2およびu3からなる群より選ばれる少なくとも1種の変数が0であるものが好ましく、u1、u2およびu3からなる群より選ばれる2種が0であることがより好ましく、u1、u2およびu3のすべてが0であることが最も好ましい。好ましい態様においては、類似体は分岐しており、さらに好ましくはジシアリル化したものである。別の態様においては、五糖構造、六糖構造および七糖構造が本発明による類似体として好ましい。
【0021】
NeuNAcα3Galβ4Glc(NAc) がより効果的に提示された最小エピトープ
セラミドの近くに位置する構造は、TLC結合アッセイにおいて他の位置の構造よりも立体構造に制限がある。遊離の一価阻害剤や多価の結合体として用いるオリゴ糖配列としては、NeuNAcα3Galβ4GlcNAcβ3Galβ?およびNeuNAcα3Galβ4GlcNAcβ3Galβ4Glc、特にNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcが、シアリルラクトースやNeuNAcα3Galβ4GlcNAcよりも高い活性を示す。ネオ糖脂質構造の使用により、還元的アミノ化構造であるNeuNAcα3Galβ4GlcNAcβ3Galβ4Glc-ヘキサデシルアニリンは、還元的結合後の還元末端グルコースが環状ではないにもかかわらず、活性を有することが示された。この活性は、三糖糖脂質であるNeuNAcα3Galβ4GlcβCer(即ち、GM3)が不活性であるにもかかわらず、糖脂質構造であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerが示す活性に類似していた。
【0022】
本発明は具体的には、機能性食品、特に乳幼児用ミルクなどの乳幼児用食品および食品添加剤に関し、機能性食品は、外部からの添加または濃縮によって、ヒトおよび動物の乳に存在する本発明のシアリル化ポリラクトサミンの濃度が高められたものである。下記のオリゴ糖からなる群より選ばれる少なくとも1種が食品用途に特に好ましい構造である:
NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc、
NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc、および
NeuNAcα3Galβ4GlcNAcβ3(NeuNAcα3Galβ4GlcNAcβ6)Galβ4Glc 。
機能性食品用の構造としては、NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc および/または NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcがさらに好ましく、NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc が最も好ましい。本発明はさらに、実質的に純粋な状態で(炭水化物質量の少なくとも80%、さらに好ましくは少なくとも90%、最も好ましくは少なくとも95%が所望のオリゴ糖である実質的に純粋なオリゴ糖画分として)上記3種のオリゴ糖全て、または上記群より2種のオリゴ糖を選択してなるいずれかの組み合わせを包含するオリゴ糖組成物、およびオリゴ糖組成物を用いた機能性食品、医薬組成物または治療用組成物の製造方法に関する。本発明はさらに、チューインガムや種々の消費製品に用いるオリゴ糖配列に関する。
【0023】
本発明の他の一つの態様においては、上記群より選ばれるシアリル化オリゴ糖あるいは2種または3種のシアリル化オリゴ糖からなる実質的に純粋な混合物を、ヒトの乳および動物の乳に含まれるオリゴ糖(例えば、シアリルラクトース類、ラクトース、ラクト−N−ネオテトラオース、パラ−ラクト−N−ネオテトラオース、ラクト−N−ネオヘキサオース、およびそれらのα3−フコシル化誘導体)からなる群より選ばれる1種または複数種と共に含有するものも好ましい。このようなシアル化オリゴ糖を含む混合物は、NeuNAcα3Lac、NeuNacα3LacNAcβ3LacおよびNeuNAcα3(LacNAcβ3)2Lacを含むものが好ましく、これら3種の糖の1番目および2番目の糖のみを含むものがさらに好ましい。
【0024】
本発明は特に、下記式(5)で表される四糖からなるより高い親和性の最小レセプターオリゴ糖配列に関するが、但し、この構造は五糖の糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerとしては提示されない。
NeuNAcα3Galβ4GlcNAcβ3Gal (5)
四糖からなる上記オリゴ糖配列は、炭素数が22を超えるセラミド、疎水性アグリコンまたは疎水性スペーサーに結合していないことがより好ましく、疎水性構造から得られた炭素数8未満のアグリコンまたはスペーサーにカップリングしていることがさらに好ましい。
【0025】
本発明はまた、上記構造の立体配座類似体および誘導体にも関する。構造類似体または誘導体構造は、ヘリコバクター ピロリに対して同様のまたはより優れた親和性を示すことが好ましい。類似体の好ましい分類においては、類似体は、ラクトースを含んでいるかまたは2型N−アセチルラクトサミンに類似した立体配座を有する他の二糖エピトープを含んでいる。好ましい類似体構造としては、オリゴ糖配列であるNeuNAcα3Galβ4Glcβ3Galが挙げられる。本発明は、具体的には、末端オリゴ糖配列としてNeuNAcα3Galβ4Glcβ3Galを含む物質に関する。ラクトースをベースとした構造は、N−アセチルラクトサミン構造よりも化学的に製造する際にずっと安価である。
【0026】
本発明は特に、下記式(6)で表されるより高い親和性の最小レセプターオリゴ糖配列に関するが、但し、この構造は五糖の糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerとしては提示されない。
NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc(NAc)m (6)
(式中、mは0または1である。)
上記五糖からなるオリゴ糖配列は、炭素数が22を超えるセラミド、疎水性アグリコンまたは疎水性スペーサーに結合していないことがさらに好ましく、疎水性構造から得られた炭素数8未満のアグリコンまたはスペーサーにカップリングしていることがさらに好ましい。高い親和性の最小レセプターは多価の結合体および遊離の糖として特に有用である。
【0027】
本発明はまた、上記構造の立体配座類似体および誘導体にも関する。構造類似体または誘導体構造は、ヘリコバクター ピロリに対して同様のまたはより優れた親和性を示すことが好ましい。類似体の好ましい分類においては、類似体は、ラクトースを含んでいるかまたは2型N−アセチルラクトサミンと類似の立体配座を有する他の二糖エピトープを含んでいる。好ましい類似体構造としては、オリゴ糖配列であるNeuNAcα3Galβ4Glcβ3Galβ4Glc(NAc)m(式中、mは0または1である)が挙げられる。本発明は、具体的には、末端オリゴ糖配列としてNeuNAcα3Galβ4Glcβ3Galβ4GlcまたはNeuNAcα3Galβ4Glcβ3Galβ4GlcNAcを含む物質に関する。ラクトースをベースとした構造は、N−アセチルラクトサミン構造よりも化学的に製造する際にずっと安価である。
【0028】
親水性オリゴ糖物質
本発明は、具体的には、親水性オリゴ糖物質に関する。本発明者らは、本発明の新規な高親和性オリゴ糖配列への結合は、そこに存在するセラミド構造に依存しないことを見出した。すでに、糖脂質への細菌の結合、特にヘリコバクター ピロリの結合の一部はセラミド構造に依存しており、他の一部はセラミド構造に依存しないことが知られている。本発明は、脂質部分に依存しない、有用な高親和性オリゴ糖結合に関する。本願実施例のデータは、ヘリコバクター ピロリがセラミド構造に依存することなく、特異的な高親和性オリゴ糖配列に結合することを示す。
【0029】
すでに、三糖であるより短いシアリル−ラクトース構造/シアリルラクトサミン構造や、より大きなフコシル化シアリルラクトサミンについて、セラミドに依存しない効果が観察されている。より短いエピトープに対する特異性は特定の条件下で培養したヘリコバクター ピロリには見られないことから、出版物に記載されているより大きなシアリル−ポリラクトサミンとの結合特異性は、三糖に対する特異性とは異なると考えられる。このような高親和性の結合特異性またはヘリコバクター ピロリの結合様式(mode of H. pylori)に関する本発明は、小さな糖による研究とは明らかに異なるものである。本発明はさらに、本発明の物質を用いたヘリコバクター ピロリの結合の阻害、および本発明の物質を用いた、患者からヘリコバクター ピロリを除去する方法に関する。従来技術はまた、フコシル化シアリルラクトサミンやシアリル−ルイスx型構造に特異的に関連する種々の高親和性効果も開示している。
【0030】
セラミドは細胞膜と会合する傾向をもともと示すため、セラミドをベースとした構造は、一般的には単一分子をベースとした治療に有用ではない。付着阻害を目的としたセラミド化合物を付着阻害に用いると、脂質が組織に付着する危険性、即ち、付着を阻害する代わりに感染組織への結合が増加する危険性がある。さらに、セラミドをベースとした構造をいかなる医療用途で用いる場合にも、その合成、製造および処方が困難であり、且つ比較的費用がかかる。糖脂質物質はさらに、抗原性があることが知られている。
【0031】
したがって、セラミド非依存性の高親和性オリゴ糖配列に関する本発明は、ヘリコバクター ピロリ感染の治療または予防のための種々の治療用アプローチに特に有用である。糖脂質の使用に代わる、本発明は親水性オリゴ糖配列の用途に関する。本発明の親水性オリゴ糖配列は、本発明で開示する高親和性オリゴ糖配列、および所望により親水性構造を含む。具体的な態様においては、オリゴ糖配列はその配列からなるオリゴ糖である。具体的な態様においては、オリゴ糖配列は機能性食品の分野に用いることが好ましく、特に本願に記載する乳幼児用食品分野に用いることが好ましい。ヒトまたは動物の乳の天然オリゴ糖を用いることのさらに有用な点は、優れた安全性を有する天然オリゴ糖の使用が可能になったことと、感染症の治療において数々の有益な側面が考えられるということである。親水性構造はオリゴ糖配列の還元末端に結合していることが好ましく、このような構造は下記(i)または(ii)で定義される構造である。
(i)オリゴ糖配列の一価の結合を増加させる非疎水性アグリコン構造、あるいは
(ii)オリゴ糖配列を多価(polyvalent or multivalent)の担体または親水性の一価の担体に結合するスペーサーである非疎水性アグリコン構造。
【0032】
非疎水性アグリコン構造とは、天然のセラミド構造よりも疎水性が実質的に低い構造を意味する。このような構造は、有機炭素をベースとした炭素数の少ない構造や、親水性基(例えば、脂肪族構造中の−CH2−と関連した酸素原子または窒素原子)を天然の糖脂質よりも実質的に大きな比率で含有する構造により得られる。親水性基の−CH2−構造に対する割合は少なくとも1:4であることが好ましく、より好ましくは少なくとも1:3であり、さらに好ましくは少なくとも1:2であり、最も好ましくは約1:1であり、特定のより好ましい態様においては、1:1未満である。アグリコンは、3を越える数の互いに結合した−CH2−構造を含有する脂肪族構造を1つ以上有していないことが好ましい。アグリコン構造は炭素数が23未満であることが好ましく、さらに好ましくは炭素数は15未満であり、さらに好ましくは10未満である。好ましい態様においては、アグリコン構造は環構造(より好ましくは六員環構造または五員環構造、さらに好ましくは六員環構造)、および/または少なくとも1つの水酸基(好ましくは少なくとも2つの水酸基、より好ましい態様においては、少なくとも3つの水酸基)、および/または少なくとも1つのアミド構造[好ましくはアセトアミド(N−アセチルともいう)−構造またはGlcNAc-残基のアセトアミド−構造を模倣する構造]を含むことが好ましい。アグリコンは、環構造と水酸基または環構造とアミド構造を含むことがさらに好ましい。好ましい六員環としては、シクロヘキサンやベンゼン環およびそれらの置換誘導体、並びに5個の炭素原子と共に窒素へテロ原子、酸素へテロ原子または硫黄へテロ原子を含む複素環構造が挙げられる。
【0033】
非疎水性アグリコン物質がオリゴ糖配列の還元末端に結合している場合には、その目的がオリゴ糖配列の親和性を増加させることによってオリゴ糖の親和性を増加させることであることが好ましい。特に、一価の物質の親和性を増加させるのには非疎水性アグリコン物質が好ましく、このようなアグリコン物質は、好ましい態様においてはオリゴ糖配列の親和性をさらに増加させる構造を含むスペーサーである。アグリコン構造は、長鎖である本発明の高親和性オリゴ糖配列の還元末端オリゴ糖構造を模倣する五員環構造または六員環構造を少なくとも1つ含むことが好ましく、さらに好ましくは、アグリコンは、本発明のオリゴ糖配列よりも単糖1つ分長いオリゴ糖配列の還元末端単糖残基を模倣する六員環構造中の5つの原子を1組含むものである。さらに具体的な態様においては、アグリコンは六炭糖構造の模倣物(mimic)であり、より好ましくは、オリゴ糖配列の還元末端単糖残基がGlcNAcまたはGlcである場合には、アグリコンはD−ガラクトピラノシル構造の模倣物であり、オリゴ糖配列の還元末端単糖残基がGalである場合には、アグリコンはD−グルコピラノシルまたはD−N−アセチルグルコピラノシルの模倣物である。さらに好ましくは、アグリコンは、単糖1つ分長いオリゴ糖配列中の単糖間に存在するグリコシド結合構造もさらに模倣するように、付加されている還元末端単糖を模倣し、この時還元末端構造は、オリゴ糖が3位に(好ましくはβ3結合で)結合したGal、あるいはオリゴ糖が4位に(好ましくはβ4結合で)結合したGlcまたはGlcNAcのいずれかを模倣することが好ましい。
【0034】
オリゴ糖が遊離オリゴ糖であるか、またはオリゴ糖がその還元末端から本願で説明したアグリコンに結合している場合は、本発明は、特に親水性オリゴ糖に関する。アグリコンは炭素数が23未満の親水性物質であることが好ましい。特定の態様においては、アグリコンは担体(好ましくは多価またはオリゴ価の担体)へのリンカーであり、これはオリゴ糖配列がオリゴ価または多価の形態で担体構造に結合することができることを意味する。
【0035】
好ましいアグリコン構造としては、炭素数が1〜約8、より好ましくは1〜3、最も好ましくは1または2、の小さな構造がさらに挙げられる。このような小さな構造には、例えば、還元単糖単位などのポリオール物質、グリコシド類、メタノール、エタノール、プロパノール、イソプロパノールおよびグリコールなどの単一アルコール類(simple alcohols)、グリコール酸、グリセロール、グリセラート、カルボン酸アルコールおよびアミノ酸残基が挙げられる。アミノ酸残基は、天然のL−アミノ酸残基であることが好ましく、さらに好ましくはセリンまたはスレオニンのグリコシドである。好ましい小さなアグリコン構造としては、還元末端に結合したカルボン酸、より好ましくはグリコシルアミドに誘導化したカルボン酸、がさらに挙げられる。好ましいカルボン酸はC1〜C8のカルボン酸であり、より好ましくはC1〜C4カルボン酸であり、さらに好ましくは酢酸、最も好ましくはオリゴ糖配列の還元末端に結合した酢酸アミドである。このような好ましい小さな構造はスペーサーとしても用いることもでき、2つの官能基を有する小さなスペーサー構造をスペーサーとして用いることが好ましい。
【0036】
本発明における好ましいスペーサー構造は、例えば、本発明で用いる親水性スペーサー構造として記載されている。好ましくは、多価の担体は多糖であり、オリゴ糖は、オリゴ糖の還元アルデヒドと特異的に反応する官能基によって、オリゴ糖の化学的保護を必要とすることなく、多糖に結合している。担体は、親水性分子および水溶性分子であることが好ましく、最も好ましくは、キトサン、グリコーゲン、デンプンの重合体またはオリゴマー、シクロデキストリン、コンドロイチン、コンドロイチン硫酸、ヘパリンおよびヒアルロン酸などの多糖である。
【0037】
オリゴ糖構造はアグリコン構造にβ結合していることが好ましい。
【0038】
具体的な態様においては、アグリコン構造は還元末端の単糖の結合相互作用を模倣する。単糖残基の結合相互作用を模倣するためには、アグリコン物質は、単糖残基の水酸基または環構造に含まれる酸素(ring oxygens)の結合を模倣する水素結合構造、または単糖残基とタンパク質表面の親水性アミノ酸側鎖との親水性相互作用を模倣する親水性構造を含んでいてもよい。好ましい態様においては、芳香環構造、さらに好ましくは六員環である芳香環構造を、還元末端単糖残基の結合を模倣するのに用いる。
【0039】
本発明は、具体的には、本発明のオリゴ糖配列のヘリコバクター ピロリへの結合を用いた、本発明において好ましいアグリコン構造をスクリーニングするための方法およびアッセイに関する。好ましい方法においては、シアル酸結合付着因子(sialic acid binding adhesin)のタンパク質構造をアグリコン構造の設計に用いる。本発明はさらに、還元末端単糖残基を本発明で開示する還元末端単糖残基を模倣するアグリコンによって置換することによる、本発明において最も短いオリゴ糖配列の類似体の探索および設計に関する。好ましくは、本発明は、NeuNAcα3Galβ4Glcβ3GalおよびNeuNAcα3Galβ4GlcNAcβ3Galの類似体のスクリーニングに関し、ここでは3Galを、この単糖残基を模倣するアグリコンまたはヘリコバクター ピロリと3Galとの結合を模倣する構造で置換する。さらに好ましくは、本発明は、NeuNAcα3Galβ4Glcβ3Galβ4Glc(NAc)0または1およびNeuNAcα3Galβ4GlcNAcβ3Galβ4Glc(NAc)0または1の類似体のスクリーニングに関し、ここでは4Glc(NAc)0または1を、この単糖残基またはヘリコバクター ピロリと単糖残基との結合を模倣するアグリコンで置換する。したがって、本発明は、好ましくは下記式で表される本発明のオリゴ糖配列の類似体のスクリーニングに関する。
NeuNAcα3Galβ4Glc(NAc)s1{β3Gal}s2- R
(式中、s1およびs2は各々独立に0または1であり、そして
s2が0である場合は、Rはβ3Galまたはヘリコバクター ピロリへのβ3Galの結合を模倣するアグリコンであり、s2が1である場合は、Rはβ4GlcNAcまたはβ4Glcあるいはβ4GlcNAcまたはβ4Glcの結合を模倣するアグリコンである。)
具体的な態様においては、オリゴ糖部分を安価な天然材料、例えばウシ初乳およびチーズ製造工程で生じる廃棄物(waste stream of cheese production)から製造することができるので、s2が0である式で表される構造が好ましい。
【0040】
好ましい大きなポリ−N−アセチルラクトサミン類およびその好ましい用途
具体的な態様においては、本発明は、医薬として用いることを特徴とする、非還元末端配列となる下記式で表される構造、あるいはヘリコバクター ピロリに対する結合能を有する該構造の類似体または誘導体に関する。
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal{β4Glc[NAc]s3}s4
(式中、R1およびR2は末端単糖置換基または末端オリゴ糖置換基であり、該置換基の少なくとも1種はNeuNAcα3であり;そして
s1、s3およびs4は各々独立に0または1であり、{ }または[ ]の中の構造の存在または不在を示す。)
【0041】
上記のようなより大きなポリ−N−アセチルラクトサミン構造は、天然材料から製造する場合に特に好ましい。このような大きなポリ−N−アセチルラクトサミンは、オリゴ糖の遊離に用いられている方法によって天然の糖タンパク質または糖脂質から製造することができ、このような方法としては、(例えばタンパク質であるN−グリコシダーゼを用いた)N−グリカンを遊離するためのエンド−グリコシダーゼ法や、糖タンパク質をプロテアーゼで対応の糖ペプチドに消化するO−結合グリカンを遊離するための方法が挙げられる。食品分野で許容可能な方法、特にプロテアーゼ法、による糖ペプチドの製造が好ましい。また、タンパク質結合N−グリカンは、所望によりオリゴ糖の還元末端の還元を行うβ−脱離法(beta-elimination methods)や、例えば当業界で知られているヒドラジンに基づく遊離法(hydrazine based release methods)により遊離することができる。音波分解法(otsonolysis)やエンド−グリコシルセラミドによって糖脂質からオリゴ糖を遊離することもできる。具体的な態様においては、本発明は酵素学的に遊離したタンパク質結合オリゴ糖や糖ペプチド、化学的に遊離したタンパク質結合オリゴ糖または糖ペプチド、ならびにそれらの混合物に関する。本発明の他の態様は、化学的または酵素学的な手段で糖脂質から製造したオリゴ糖に関する。このような構造は、天然材料から製造した混合物であることが特に好ましい。本発明はさらに、本発明の大きなポリ−N−アセチルラクトサミンに富んだ、天然または遊離のオリゴ糖または糖ペプチドに関する。
【0042】
本発明はさらに、本発明の大きなポリ−N−アセチルラクトサミンに富んだ天然の糖タンパク質画分に関する。本発明はさらに、ポリ−N−アセチルラクトサミン前駆体を酵素学的にリモデリングして本発明の構造とするための、天然のポリ−N−アセチルラクトサミン前駆体画分に関する。ポリ−N−アセチルラクトサミン構造を含む、種々の動物由来のタンパク質材料および脂質材料について報告されており、当業界で知られている。本発明は特に、食品分野で許容可能な材料に関する。
【0043】
具体的な態様においては、本発明は、本発明の大きなポリ−N−アセチルラクトサミンとして好ましいものを糖脂質としてミセルの形状で用いた、本発明の治療方法またはヘリコバクター ピロリ結合性の試験方法に関する。このような物質は、高親和性リガンドを探索するために、ヘリコバクター ピロリ結合性をスクリーニングするのに特に好ましい。
【0044】
天然グリカンのリモデリングによる機能性オリゴ糖の製造
具体的な態様においては、本発明は天然グリカン、好ましくは食物材料から得たポリ−N−アセチルラクトサミン、の機能性食品炭水化物へのリモデリングに関する。グリカンのリモデリング方法は、Neose-companyによる最近の米国特許出願および国際特許出願に記載されている。本発明は特に、天然の食物の糖タンパク質やオリゴ糖、または遊離オリゴ糖の脱シアリル化および/または脱フコシル化に関する。最も好ましくは、オリゴ糖や糖ペプチドを穏やかな酸処理により脱シアリル化する。好ましい態様においては、脱シアリル化は、当業界で知られているように、食品分野で許容されるカルボン酸による処理後に約80〜100℃に加熱することで行い、好ましくは酢酸により処理する。他の一つの態様においては、熱処理により除去することが可能な揮発性の酸(例えば塩酸)で食物材料を脱シアリル化する。オリゴ糖構造の主要部分はそのまま残るように行う化学的な脱フコシル化法、例えば硫酸を用いる方法、も知られている。本発明はさらに、天然材料から得たオリゴ糖または糖ペプチドを含む糖を本発明による構造にするためのリモデリングに関する。糖はタンパク質材料に比べて安定性が高いので、精製または単離した糖混合物をリモデリングに用いることが好ましい。本発明は具体的には、α3−シアリル化による天然の糖のリモデリング、好ましくは脱シアリル化および/またはフコシル化後に行うリモデリングに関する。他の目的のためには、本発明はさらに、α3−ガラクトシル化、α4−ガラクトシル化、β4−GalNAc−転移、β3−GalNAc−転移、β3−GlcNAc−転移、α6−シアリル化、α3−フコシル化、α2−フコシル化またはα6−フコシル化、好ましくはα3−ガラクトシル化、α4−ガラクトシル化、β3−GlcNAc−転移またはα6−シアリル化による遊離オリゴ糖のリモデリングに関し、好ましい態様においては、β3−GlcNAc−転移またはα6−シアリル化でリモデリングを行う。このようなグリコシル化反応は、グリコシルトランスフェラーゼ酵素、トランスグリコシル化グリコシダーゼまたはS. Whithersらのグループが発表した変異トランスグリコシル化酵素を用いて、公知の方法で行うことができる。
【0045】
本発明は、下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料由来の遊離の糖または単離した糖のリモデリング方法に関する。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) 該原料オリゴ糖からなる画分、必要な場合には所望の糖、好ましくはポリ−N−アセチルラクトサミン、に富んだ画分を単離し、
(3) 該単離した画分を処理、好ましくは穏やかな酸処理に付して、末端単糖、好ましくはフコースとシアル酸の少なくとも1種、さらに好ましくはシアル酸、を遊離させて末端単糖除去オリゴ糖を含む糖混合物を得、そして
(4) 糖混合物の末端単糖除去オリゴ糖に、単糖、好ましくはα3結合するシアル酸をグリコシルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【0046】
本発明は、下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料由来の遊離の糖または単離した糖のリモデリング方法に関する。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) 該原料オリゴ糖からなる画分、必要な場合には所望の糖、好ましくはポリ−N−アセチルラクトサミン、に富んだ画分を単離し、
(3) 所望により末端単糖、好ましくはフコースとシアル酸の少なくとも1種、さらに好ましくはシアル酸、を遊離させて末端単糖除去オリゴ糖を含む糖混合物を得、そして
(4) 糖混合物の末端単糖除去オリゴ糖に、単糖、好ましくはα3結合するシアル酸をα3−シアリルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【0047】
好ましい態様においては、上記リモデリング方法は種々の数のシアル酸を含有するオリゴ糖を精製する工程をさらに包含し、1つ、2つ、3つまたは4つのシアル酸を含有するオリゴ糖あるいはそれらの混合物を含む画分を得ることが好ましく、精製はイオン交換クロマトグラフィーまたはアフィニティークロマトグラフィーで行うことが好ましい。
【0048】
好ましい態様においては、ポリ−N−アセチルラクトサミン画分をサイズ排除クロマトグラフィーまたはアフィニティークロマトグラフィーにより単離する。単離にはポリ−N−アセチルラクトサミン結合性のレクチンを用いることが好ましく、ポリ−N−アセチルラクトサミン結合性の食物由来レクチン(好ましい例としてはトマトレクチン、ジャガイモレクチンおよび小麦麦芽凝集素)を用いることがより好ましい。
【0049】
好ましいポリ−N−アセチルラクトサミングリカンは、少なくとも1つのGalβ4Glcβ3Galβ4Glc(NAc)0または1を含有し、Galβ4Glcβ3Galβ構造はそのものの形で含有されいてもよいし、より大きなグリカン構造の一部として含有されていてもよい。
【0050】
具体的な態様においては、本発明は、上記リモデリング方法の工程(4)が2つの工程からなる方法に関し、この方法では、Galβ4を転移させる工程と、次いで転移したGalβ4に末端単糖、好ましくはα3−結合するシアル酸をα3−シアリルトランスフェラーゼによって転移する工程を包含する。このリモデリング方法に用いる原料は、末端GlcNAc残基を有するものである。
【0051】
トランスシアリダーゼ酵素は、例えばトリパノソーマ寄生虫より調製し、クローニングした。
【0052】
ヘリコバクター ピロリに対する結合能を有する、炭素修飾した中性または酸性の新規なシアル酸誘導体
本発明はさらに、ヘリコバクター ピロリに対する結合親和性を有する新規なシアル酸誘導体を開示する。このような誘導体は下記式で表される。
SA(X−R)
(式中、SAはシアル酸であり;
Xはシアル酸のC1位に結合している結合原子または結合基、および/またはシアル酸の7位、8位および8位の炭素原子からなる群より選ばれる少なくとも1種であって、この場合にシアル酸は8位の炭素原子または8位と9位の炭素原子を欠失しているシアル酸誘導体でもよく;
特に、Rで表される置換基がシアル酸のC7位またはC8位に結合している時、シアル酸はC7位とC8位を含む構造またはC9位を含む構造をそれぞれ欠失した、短いシアル酸誘導体であることが好ましく;そして
Rは水素原子または炭素数が3を超える有機ラジカルである。)
上記式中のXは、シアル酸残基のカルボキシル基と共にアミド構造を形成する−NHであることが好ましい。また、上記式中のRは、水素原子あるいは環構造および脂肪鎖の少なくとも1種を含むC4〜C30有機ラジカルであることが好ましく、C6〜C24有機ラジカルであることがより好ましく、C6〜C24脂肪族アルキル鎖であることが最も好ましい。SAは、NeuNAcであることが好ましい。
【0053】
シアル酸は、そのカルボン酸および/またはC7位〜C9位のグリセロール尾部構造で修飾可能であることを見出した。グリセロール尾部構造を短縮(好ましくはC7位まで短縮)した後に、疎水性構造でグリセロール尾部を修飾することが、特に効果的な修飾であることが判明した。好ましい態様においては、修飾に用いる基は直鎖状の脂肪族アルキル鎖である。さらに、修飾したシアル酸構造は本発明の他の構造と共に用いることができることも判明した。特に、C7位〜C9位が修飾された構造を本発明のポリラクトサミン型構造と共に用いることが好ましい。このような構造をヘリコバクター ピロリに対する他の阻害剤のスクリーニングおよび設計に用いることがさらに好ましい。
【0054】
本発明はさらに、修飾したシアル酸構造の類似体または誘導体に関する。
【0055】
さらに好ましくは、このようなシアル酸構造は、ラクトースまたはN−アセチルラクトサミン構造に結合している。好ましい態様においては、シアル酸誘導体は下記式で表されるように、2型N−アセチルラクトサミン配列にα3結合を介して結合している。
SA(X-R)αxGal{β4GlcNAc[β3Gal(β4Glc)p1]p2}p3
(式中、xはシアル酸誘導体の結合位置を示し;そして
p1、p2およびp3は各々独立に0または1であり、{ }、[ ]または( )の中の構造全体の存在または不在を示す。)
【0056】
本願明細書においては、「オリゴ糖配列(OSs)」という用語には、修飾したシアル酸構造も含まれる。
【0057】
本発明はまた、本発明で開示するヘリコバクター ピロリ結合性のオリゴ糖配列またはシアル酸修飾構造、あるいはその類似体または誘導体の多価の結合体に関し、多価の結合体は、例えば下記式で表される構造からなる。
[OS−(X)n−L−Y]m−Z
(式中、mは1以上の整数であり、好ましくは1を超える整数であり、nは0または1であり;
OSはオリゴ糖配列であり;
Lは酸素原子、窒素原子、硫黄原子または炭素原子であり;
Xは単糖残基またはオリゴ糖残基、好ましくはラクトシル残基、ガラクトシル残基、ポリ−N−アセチル−ラクトサミニル残基、あるいはO−グリカンオリゴ糖配列またはN−グリカンオリゴ糖配列の一部、あるいは少なくとも1つのマンノシル残基またはN−アセチルガラクトサミニル残基を含むものであり;
Yはアグリコン、スペーサー、または該LとZの間の結合であり;
Zはオリゴ価または多価の担体であり、多糖などの炭水化物材料を含み;そして
該OSの還元末端の1位の炭素原子または本発明のヘリコバクター ピロリ結合性末端配列を含む修飾シアル酸のR基によって、該Yおよび所望により該Xを介して該Zに該Lから結合している。)
【0058】
さらに、新規なシアル酸誘導体を、NeuNAcに関連して記載した本発明のポリ−N−アセチルラクトサミンや、ヘリコバクター ピロリに対する結合能を有する他の糖結合体に提示できることが明かとなった。
【0059】
中性のシアル酸エピトープは、酸性のNeuNAcが結合するのとは異なる、ヘリコバクター ピロリの表面に存在するレセプターに結合し得ることが判明した。本発明は特に、このようなエピトープを新規なレセプターとして用いた、該レセプターに対応する付着因子をヘリコバクター ピロリの表面から同定する方法に関する。本発明はさらに、ヘリコバクター ピロリのヒトおよび動物の細胞や組織への結合の阻害、またはヘリコバクター ピロリの凝集において有用であると考えられる新規なリガンドの設計に関する。このような新規なヘリコバクター ピロリ結合性物質の設計または合成は、ヘリコバクター ピロリの同じレセプターに結合する他の炭水化物構造を用いたモデリング方法にも関する。
【0060】
本発明において、「類似体」及び「誘導体」は次のように定義される。本発明によると、ヘリコバクター ピロリ結合性オリゴ糖配列の構造類似体や誘導体を設計することができる。従って、本発明は、本発明の結合性物質の構造類似体にも関する。本発明における構造類似体は、ヘリコバクター ピロリのオリゴ糖配列に対する結合に重要な構造因子を包含する。効果的な構造類似体を設計するには、ヘリコバクター ピロリと糖類との結合に重要な構造因子を知ることが必要である。重要な構造因子は、修飾されていないか、重要な構造因子との類似性が非常に高い模倣物となるように修飾されていることが好ましい。
【0061】
本発明における構造誘導体は、ヘリコバクター ピロリに対する結合能を保持するか向上するように本発明のオリゴ糖配列を化学的に修飾したものである。本発明によると、オリゴ糖配列の水酸基またはアセトアミド基の1つまたは複数を誘導化することが好ましい。本発明は、類似体や誘導体を調製する際に変えることのできる分子上の位置を複数開示する。少なくともある種の修飾を許容することが好ましい水酸基またはアセトアミド基は、本願に記載した化学式から当業者には明かである。
【0062】
嵩高い置換基や酸性の置換基および他の構造(例えば単糖残基)は許容されないが、オリゴ糖類似体、例えばレクチン結合性のオリゴ糖類似体、を製造する方法はよく知られている。例えば、シアリル−ルイスxオリゴ糖の類似体は複数製造されており、これらは異なる骨格の活性官能基である(Sears and Wong 1996、12090ページを参照)。同様に、へパリンオリゴ糖の類似体もサノフィ社によって製造されており、シアリダーゼ酵素に対するシアル酸模倣型阻害剤、例えばZanamivirやTamiflu(Relenza)、が複数のグループによって製造されている。オリゴ糖類似体は少なくとも1つの六員環または五員環構造を含む分子に基づいて構築されていることが好ましく、類似体は6個または5個の原子を含む少なくとも2つの環構造を含有することがより好ましい。模倣構造においては、単糖環はシクロヘキサンやシクロペンタンなどの環状分子、ベンゼン環を含む芳香環によって置換されていてもよく、複素環構造は酸素原子の他に、例えば窒素原子や硫黄原子を含んでいてもよい。活性環状立体構造を固定するためには、環構造を許容できる結合基で繋ぐこともできる。典型的な模倣構造は、オリゴ糖配列またはその一部に対するペプチド類似体構造をさらに含んでいてもよい。
【0063】
活性基が結合能に与える影響は累積するので、1つの活性基が失われても、分子の異なる場所に活性基を付加することで補うことができる。分子モデリング、好ましくはコンピューターを用いた分子モデリングを本発明のヘリコバクター ピロリ結合性オリゴ糖配列の類似構造を製造するのに用いることができる。種々のオリゴ糖配列の分子モデリングの結果が実施例に示されており、NMRとX線結晶回折に加えてこれらと同じまたは類似した方法を用いて本発明の他のオリゴ糖配列に対する類似体を得ることができる。オリゴ糖配列の類似体を見つけるために、オリゴ糖配列をヘリコバクター ピロリの炭水化物結合分子、最も可能性が高いのはヘリコバクター ピロリのレクチン、と合体させることで、さらなる潜在的な結合相互作用を探索することができる。
【0064】
コンビナトリアルケミストリーによってオリゴ糖の誘導体を調製することで、レクチンに対してより高い活性を示すように1価、オリゴ価または多価のオリゴ糖を活性化することができることにも注目されたい。オリゴ糖配列の1つまたは複数の残基を置換することでライブラリーを構築した場合には、それは誘導体ライブラリーと考えることができ、また、本発明で開示するオリゴ糖配列の類似体からライブラリーを構築することもできる。コンビナトリアルケミストリーによるライブラリーは本発明のオリゴ糖、その前駆体または糖結合体に基づいて構築することができる。例えば、種々の還元末端を有するオリゴ糖をカーボハイブリッド技術(carbohydrid technology)と呼ばれる技法によって製造することができる。
【0065】
好ましい態様においては、コンビナトリアルケミストリーによるライブラリーは、本発明のヘリコバクター ピロリ結合性物質に結合されている。さらに好ましい態様においては、ライブラリーは少なくとも6種の異なる分子を包含する。このようなライブラリーは、本発明のオリゴ糖配列に対する微生物による結合を検出するために用いるのが好ましい。コンビナトリアルケミストリーによるライブラリーから高親和性のバインダーを同定することは、例えば阻害アッセイによって可能であり、ここでは本発明で開示する糖脂質または糖結合体に対する細菌の結合を阻害するために、ライブラリー中の化合物を用いる。本発明において好ましい構造類似体や誘導体は、本発明のヘリコバクター ピロリ結合性オリゴ糖配列がヘリコバクター ピロリに結合するのを阻害することができる。
【0066】
以下の記載においては、ヘリコバクター ピロリ結合性配列を単に「オリゴ糖配列」と記載する。本願で定義するオリゴ糖配列は、天然のまたは合成した糖結合体の一部、遊離オリゴ糖または遊離オリゴ糖の一部であってもよい。このようなオリゴ糖配列は、多糖鎖に結合した種々の単糖、オリゴ糖または多糖に結合させることができ、オリゴ糖配列が細菌の多糖の一部として発現している場合などが例示できる。さらに、O−アセチル化誘導体または硫酸化誘導体に代表される、単糖の種々の天然の修飾が知られている。本願で定義するヘリコバクター ピロリ結合性物質には、天然のまたは合成した糖結合体の一部、それに相当する遊離オリゴ糖、または遊離オリゴ糖の一部として記載したオリゴ糖配列が含まれる。ヘリコバクター ピロリ結合性物質には、ヘリコバクター ピロリ結合性オリゴ糖配列の混合物も含まれる。
【0067】
本発明のヘリコバクター ピロリ結合性物質は、糖鎖または糖結合体の一部、あるいは他の公知のヘリコバクター ピロリ結合性エピトープを含有する複数の糖化合物との混合物であってもよく、上記した公知のヘリコバクター ピロリ結合性エピトープとは、本発明のヘリコバクター ピロリ結合性物質とは異なる糖配列や立体構造を有するもの、例えばルイスb、Fucα2Galβ3(Fucα4)GlcNAc、Neu5Acα3Galβ4Glc/GlcNAcなどである。尚、本発明のヘリコバクター ピロリ結合性物質を治療のために用いる場合、複数の結合性物質との併用は有効手段となり得る。
【0068】
本発明は特に、本発明のオリゴ糖構造の類似体の設計に関し、類似体はオリゴ糖残基中の単糖残基に類似した環構造を含む。さらに好ましくは、ヘリコバクター ピロリの結合または阻害について類似体を試験し、製品開発のために最適な結合性配列を選択する。他の一つの態様においては、本発明の分子あるいはその類似体または誘導体を、他の微生物またはウイルスの結合、好ましくはClostridium difficileのトキシンAに対する結合について試験する。オリゴ糖類似体は少なくとも1つの六員環または五員環構造に基づいて構築されていることが好ましく、類似体は6個または5個の原子を含む少なくとも2つの環構造を含有することがより好ましい。オリゴ糖の好ましい類似体の型は末端ウロン酸アミドを包含するか、あるいは類似体または誘導体がGal/GalNAcβ4GlcNAc−糖模倣構造に結合したものである。本発明によると、末端単糖残基の2位または4位の水酸基は結合に重要ではなく、6位の水酸基は分子の親和性を実際に上昇させるような構造になるように修飾することができ、親和性が高い類似体はこのような位置を修飾することで製造することができる。本願のデータは、末端単糖残基のすべての水酸基を含有しない類似体の設計が可能であることを示している。本発明は特に、末端Gal/GalNAcの3位を修飾するための種々の有機誘導化分子(例えば芳香族または脂肪族環状有機残基)を付加することで行う、機能的類似体の設計に関する。誘導体は、Gal/GalNAcβ4の3位に環状有機残基を結合させ得る特別な結合化学的手法(linker chemistry)によって製造することができる。結合構造の幾何構造、さらにその長さでさえもが、グリコシド結合構造とは異なっていてもよいが、環状有機残基は、対応する末端単糖残基、特に末端単糖残基の6位の水酸基/カルボキシル基/アミド基の近傍で見られる正の結合相互作用の少なくとも一部を示さなければならない。
【0069】
具体的な態様においては、類似体はGal/GalNAcβ4の3位の水酸基の置換によってアミン基で製造する。本発明は特に、ヘリコバクター ピロリに結合する三糖エピトープ中の末端Gal/GalNAcの3位に末端環状分子を有する類似体のスクリーニングに関する。末端環状分子は有機六員環残基であることが好ましく、オリゴ糖構造の末端および/または本願で開示する化学式中に見られる内部ヘキスロン酸と同様のカルボン酸構造、アミド構造またはアルキルアミド構造をさらに含有することがより好ましい。
【0070】
また、末端ウロン酸アミドあるいはその類似体または誘導体が、Galに1−3結合しており、該GalがGlcNAc模倣構造に結合しているものも挙げられる。模倣構造においては、単糖環はシクロヘキサンやシクロペンタンなどの環状分子、ベンゼン環を含む芳香族環によって置換されていてもよく、そのような複素環構造は酸素原子の他に、例えば窒素原子や硫黄原子を含んでいてもよい。活性環状立体構造を固定するためには、環構造に許容される結合基で繋ぐこともできる。典型的な模倣構造は、オリゴ糖配列またはその一部に対するペプチド類似体構造をさらに含んでいてもよい。本発明は、オリゴ糖配列のためのペプチド類似体の設計および/またはスクリーニングにも関する。さらに本発明は、本発明のオリゴ糖配列に対する、DNAまたはRNAに基づく類似体(例えば、いわゆるアプタマー)のスクリーニングに関する。活性基が結合能に与える影響は累積するので、1つの活性基が失われても、分子の異なる場所に活性基を付加することで補うことができる。
【0071】
分子モデリング、好ましくはコンピューターを用いた分子モデリングを本発明のヘリコバクター ピロリ結合性オリゴ糖配列の類似体構造を製造するのに用いることができる。種々のオリゴ糖配列の分子モデリングの結果が実施例に示されており、NMRとX線結晶回折に加えてこれらと同じまたは類似した方法を用いて本発明の他の結合性オリゴ糖配列あるいはその類似体または誘導体の構造を決定することができる。好ましい態様においては、本発明のオリゴ糖と同じまたは類似の立体構造を有するオリゴ糖構造の類似体または誘導体は、コンピューター支援分子モデリング(computer assisted molecular modeling)で得られた結果または分子の三次元構造を含むスクリーニング用データベースから選択するが、このような方法を電算化スクリーニング方法(computerized screening method)と称する。
【0072】
本発明はさらに、電算化スクリーニング方法によって選択したオリゴ糖構造あるいはその類似体または誘導体の試験に関し、該試験においては、他の病原性の微生物、ウイルスまたは毒素に対して、本発明の一つまたは複数のオリゴ糖配列とヘリコバクター ピロリとの結合特異性に類似した結合特異性を示すものを検出する。好ましい態様においては、電算化スクリーニング方法によって選択されたオリゴ糖構造あるいはその類似体または誘導体をClostridium difficileのトキシンAに対する結合について試験する。
【0073】
類似体分子は、合成によって製造するか天然材料から得ることができる。分子は、コンピューター上で仮想的に作成することも可能であり、活性分子のスクリーニングの一部をin silicoで行うこともできる。本発明は、炭水化物構造、類似体または誘導体をコンピューター上でヘリコバクター ピロリに存在する炭水化物結合部位に適合させることによって行う、本発明のオリゴ糖配列に対するヘリコバクター ピロリ結合性および/または阻害性の類似体および/または誘導体の探索に関する。
【0074】
ヘリコバクター ピロリ結合性オリゴ糖配列あるいはその類似体または誘導体を、ヘリコバクター ピロリの炭水化物結合分子、最も可能性が高いのはヘリコバクター ピロリのレクチン、と分子モデリングの方法によって合体させることで、さらなる結合相互作用を探索することができる。オリゴ糖配列の三次元構造を炭水化物結合部位の三次元モデルとコンピューター上で合体させることは、結合相互作用および考えられるさらなる結合相互作用部位の決定を可能にするので、結合活性類似体の設計にさらに貢献する。この方法は、電算化合体方法(computerized docking method)によるオリゴ糖構造あるいはその類似体または誘導体の結合性の比較にも関する。
【0075】
本発明はさらに、電算化合体方法によるオリゴ糖構造あるいはその類似体または誘導体の試験に関し、該試験においては、他の病原性の微生物、ウイルスまたは毒素に対して、本発明の一つまたは複数のオリゴ糖配列とヘリコバクター ピロリとの結合特異性に類似した結合特異性を示すものを検出する。好ましい態様においては、オリゴ糖構造あるいはその類似体または誘導体のClostridium difficileのトキシンAに対する結合性を、電算化合体方法で試験する。
【0076】
コンビナトリアルケミストリーによってオリゴ糖の誘導体を調製することで、レクチンに対してより高い活性を示すように1価、オリゴ価または多価のオリゴ糖を活性化することもできることにも注目されたい。オリゴ糖配列の1つまたは複数の残基を置換することでライブラリーを構築した場合には、それは誘導体ライブラリーと考えることができる。また、本発明のオリゴ糖配列の類似体からライブラリーを構築した場合には、それは類似体ライブラリーと考えることができる。コンビナトリアルケミストリーによるライブラリーは本発明のオリゴ糖、その前駆体または糖結合体に基づいて構築することができる。例えば、種々の還元末端を有するオリゴ糖をカーボハイブリッド技術と呼ばれる技法によって製造することができる。本発明は、コンビナトリアルケミストリーによるライブラリー、即ち、本発明のオリゴ糖構造の多種多様な化学的な類似体および/または誘導体の設計および構築、ならびにこれらのヘリコバクター ピロリに対する結合またはヘリコバクター ピロリの阻害についての試験に関する。本発明はさらに、コンビナトリアルケミストリーによるライブラリーの試験に関し、該試験においては、他の病原性の微生物、ウイルスまたは毒素に対して、本発明の一つまたは複数のオリゴ糖配列とヘリコバクター ピロリとの結合特異性に類似した結合特異性を示すものを検出する。好ましい態様においては、コンビナトリアルケミストリーによるライブラリーのClostridium difficileのトキシンAに対する結合性を試験する。
【0077】
本発明の好ましい態様においては、コンビナトリアルケミストリーによるライブラリーは、本発明のヘリコバクター ピロリ結合性物質に結合されている。より好ましい態様においては、ライブラリーは少なくとも6種の異なる分子を包含する。コンビナトリアルケミストリーによる修飾は、式(9)のR8基またはR9基であるカルボキシル基を種々のアミド基に変化させることで得られるものが好ましい。R8基を修飾する置換基は、アルデヒド、アミンまたはその他の反応性基でもよい。このようなライブラリーは、本発明のオリゴ糖配列に対する微生物性の結合を検出するために用いるのが好ましい。アミノ酸または有機アミド類のコレクションは市販されており、このような物質はウロン酸アミドのコンビナトリアルケミストリーによるライブラリーを合成するために用いることができる。コンビナトリアルケミストリーによるライブラリーから高親和性のバインダーを同定することは、例えば阻害アッセイによって可能であり、ここでは本発明で開示する糖脂質または糖結合体に対する細菌の結合を阻害するために、ライブラリー中の化合物を用いる。本発明おいて好ましい構造類似体や誘導体は、本発明のヘリコバクター ピロリ結合性オリゴ糖配列がヘリコバクター ピロリに結合するのを阻害することができる。
【0078】
以下の記載においては、ヘリコバクター ピロリ結合性配列を単に「オリゴ糖配列」と記載する。本願で定義するオリゴ糖配列は、天然のまたは合成した糖結合体の一部、遊離オリゴ糖または遊離オリゴ糖の一部であってもよい。このようなオリゴ糖配列は、多糖鎖に結合した種々の単糖、オリゴ糖または多糖に結合させることができ、オリゴ糖配列が細菌の多糖の一部として発現している場合などが例示できる。さらに、単糖に対する天然の修飾も多数知られており、オリゴ糖配列のO−アセチル化誘導体や硫酸化誘導体が例示できる。特定の態様においては、オリゴ糖配列とは、所望により還元末端が修飾されていることを除いては、他のいかなる単糖残基によっても修飾されていない非還元末端オリゴ糖配列を意味する。広義においては、オリゴ糖配列という用語には、本発明のオリゴ糖構造の構造類似体や誘導体が含まれることが好ましく、本願で開示するように、ヘリコバクター ピロリに対して同一または同様の結合能を有することが好ましい。本願で定義するヘリコバクター ピロリ結合性物質には、天然のまたは合成した糖結合体またはその一部、それに相当する遊離オリゴ糖、または遊離オリゴ糖の一部として記載したオリゴ糖配列が含まれてもよい。ヘリコバクター ピロリ結合性物質には、ヘリコバクター ピロリ結合性オリゴ糖配列の混合物も含まれてもよい。
【0079】
ヘリコバクター ピロリ結合性オリゴ糖配列は、グリコシルトランスフェラーゼによって酵素学的に合成するか、あるいはグリコシダーゼ酵素またはトランスグリコシダーゼ酵素によって触媒されるトランスグリコシル化反応で合成することができる(Ernst et al., 2000)。これらの酵素の特異性および補因子の必要性については改良することができる。修飾された特定の酵素を用いてより効率よく合成することもでき、例えば、グリコシンセターゼをトランスグリコシル化のみを触媒するよう修飾することができる。本発明で開示する糖および糖結合体またはそれらと類似した化合物の有機合成も知られている(Ernst et al., 2000)。糖物質は天然の原料から単離して、化学的または酵素学的に修飾してヘリコバクター ピロリ結合性化合物とすることができる。種々の反芻動物の乳から天然のオリゴ糖を単離することができる。グリコシル化酵素を発現するトランスジェニックな生物、例えばウシや微生物も糖の製造に用いることができる。
【0080】
細菌結合性物質は、クラスター、即ち、細胞膜上の糖脂質、ミセルまたはリポソームとして提示されるか、あるいはアッセイに用いるTCLプレートのような固相に提示されていることが好ましい。正しい空間配置によって提示されたクラスターは高い結合親和性を示す。
【0081】
本発明においては、ヘリコバクター ピロリ結合性エピトープあるいはヘリコバクター ピロリに対して同様または改善された結合能を有する、該エピトープの天然のまたは合成した類似体や誘導体を用いることもできる。さらに、細菌結合性物質を含有する物質、例えば、本願で開示するレセプター活性ガングリオシドやヘリコバクター ピロリに対して同様または改善された結合能を有する、該ガングリオシドの類似体や誘導体などを用いることもできる。細菌結合性物質は、オリゴ糖鎖のグリコシル結合した末端エピトープでもよい。また、細菌結合性エピトープはオリゴ糖鎖の側鎖、好ましくはポリラクトサミン鎖でもよい。
【0082】
ヘリコバクター ピロリ結合性物質は、抗生物質、好ましくはペニシリン系抗生物質、に結合していてもよい。ヘリコバクター ピロリ結合性物質は抗生物質をヘリコバクター ピロリにターゲティングする。このような結合体は、ヘリコバクター ピロリに対する処置や治療に必要な抗生物質が少量となるので治療に有利であり、その結果、抗生物質による副作用を低減させる。結合体の抗生物質部分は細菌を殺すか弱めるためのものであるが、後述するように、結合体には付着防止効果があってもよい。
【0083】
細菌結合性物質、好ましくはオリゴ価またはクラスター状のものは、ヘリコバクター ピロリの存在によって生じる疾患または病態の治療に用いることができる。これはヘリコバクター ピロリ結合性物質を付着防止に用いることで達成される、即ち、ヘリコバクター ピロリがターゲットとなる細胞や組織のレセプター エピトープに結合するのを阻害する。ヘリコバクター ピロリ結合性物質または医薬組成物を投与すると、それは細菌の結合に対してターゲット細胞上のレセプター糖結合体と競合する。その結果、細菌の一部またはすべてがターゲットとなる細胞や組織のレセプターの代わりにヘリコバクター ピロリ結合性物質に結合する。ヘリコバクター ピロリ結合性物質に結合した細菌は患者から(例えば、胃腸器系における液体の流入によって)除去され、その結果、患者の健康に細菌が与える影響を低減させる。使用する結合性物質は、好ましくはヘリコバクター ピロリ結合性物質を包含する可溶性の組成物である。結合性物質は、担体、好ましくはタンパク質以外の担体に結合させてもよい。担体分子を用いる場合には、ヘリコバクター ピロリ結合性物質の複数の分子を1つの担体に結合し、阻害効率を改善することもできる。
【0084】
ターゲット細胞は主としてターゲット組織(特に胃腸器系)またはターゲット組織と考えられるその他の組織(肝臓や膵臓)の上皮細胞である。ターゲット組織のグリコシル化は病原体による感染によって変化すると考えられる(Karlsson et al., 2000)。ターゲット細胞はターゲット組織中の悪性細胞、形質転換細胞または癌化/腫瘍細胞でもよい。形質転換した細胞または組織は異なる形のグリコシル化反応を発現し、細菌に対するレセプターを提供する場合がある。レクチンまたは糖類が糖タンパク質または糖脂質からなるレセプターの糖質と結合すること(炭水化物−炭水化物間相互作用)によって細胞が活性化され、癌化/悪性細胞の場合には、これは癌の増殖または転移につながる。オリゴ糖エピトープのいくつか、ならびに悪性細胞由来のシアリル化ポリラクトサミン(Stroud et al., 1996)は、癌関連抗原または癌抗原として報告されている。ヘリコバクター ピロリは胃リンパ腫とも関連する。本明細書に記載する結合性物質は前癌状態または悪性の細胞に対するヘリコバクター ピロリの結合ならびに癌の増殖および転移の活性化を防止するために用いることができる。結合阻害は胃がん、特にリンパ腫を治療する可能性がある。
【0085】
ターゲット細胞としては血液細胞、特に白血球が挙げられる。消化性潰瘍と関連するヘリコバクター ピロリ株、例えば、本発明で主として用いた株は、オプソニン作用を受けていないにもかかわらず、顆粒球による炎症反応を刺激した(Rautelin et al., 1994a,b)。細菌の食作用において最初に起こる事柄には特定のレクチン様相互作用が含まれると考えられ、その結果、顆粒球の凝集が生じる(Ofek and Sharon, 1988)。食作用に続いて酸化的バースト反応が生じるが、これはヘリコバクター ピロリ関連疾患の病因を導くものとも考えられる(Babior, 1978)。N−アセチルラクトサミン繰り返し単位を有するいくつかのシアリル化および非酸性グリコスフィンゴ脂質は顆粒球から単離されて、特徴付けられており(Fukuda et al., 1985; Stroud et al., 1996)、白血球表面のヘリコバクター ピロリに対する潜在的レセプターとして機能し得る。さらに、x2グリコスフィンゴ脂質も同じ材料から単離されている(Teneberg, S.、未発表)。本発明は、N−アセチルラクトサミン特異的レクチンおよびモノクローナル抗体(X2、GalNAcβ3Galβ4GlcNAc-)によって認識される、ヒト赤血球および顆粒球に存在するレセプター糖質の存在を立証する。ヘリコバクター ピロリ結合性物質は、ヘリコバクター ピロリに対する白血球の結合の阻害ならびに白血球の活性化に続く酸化的バーストおよび/または炎症の防止に有用である。
【0086】
ヘリコバクター ピロリは種々のオリゴ糖配列に結合し得ることが知られている。特定の株による結合の中にはより共生的な相互作用を示すものもあり、癌や重度の病態に進行することはない。癌型糖エピトープへの結合に関する本願のデータはヘリコバクター ピロリ結合性物質がより病原性の高い相互作用も防止することが可能であり、その際、他のレセプター構造に結合した病原性の低いヘリコバクター ピロリ株がいくらか残る。従って、細菌の完全な除去はヘリコバクター ピロリ関連疾患の予防には必ずしも必要ない。病原性の低い細菌は、病原性の高いヘリコバクター ピロリ株による感染の予防においてプロバイオティックな効果を発揮することも考えられる。
【0087】
さらに、ヘリコバクター ピロリは大きなポリラクトサミンオリゴ糖類をその表面に有し、少なくともいくつかの株は、本願で説明するように細菌が結合する未フコシル化エピトープを含有する。本願に記載した結合性物質はヘリコバクター ピロリ同士の結合を防止し、そうすることによって、例えば、細菌のコロニー形成を阻害することができる。
【0088】
本発明によると、ヘリコバクター ピロリ結合性物質を、所望により担体と共に医薬組成物に導入することができる。このような医薬組成物は、ヘリコバクター ピロリが患者に存在することによって生じる病態の治療またはヘリコバクター ピロリ結合性物質を用いたこのような病態の治療方法に適している。本発明によって治療することのできる病態の具体例としては、慢性表在性胃炎、胃潰瘍、十二指腸潰瘍、ヒトの胃における非ホジキン悪性リンパ腫、胃腺癌、ならびにある種の膵臓病、皮膚病、肝臓病、心臓病、乳幼児突然死症候群、自己免疫疾患(自己免疫性胃炎、悪性貧血および非ステロイド性抗炎症薬(NSAID)関連胃疾患を含む)であり、すべてまたは少なくとも一部がヘリコバクター ピロリ感染によって生じる病態である。
【0089】
ヘリコバクター ピロリ結合性物質を包含する医薬組成物は、さらに他の物質、例えば不活性なベヒクルまたは医薬的に許容される担体や保存剤などの公知の物質を包含してもよい。ヘリコバクター ピロリ結合性物質は他の薬剤、例えばヘリコバクター ピロリに対する抗生物質、と共に投与してもよい。
【0090】
ヘリコバクター ピロリ結合性物質またはこのような結合性物質を包含する医薬組成物は、適当な方法で投与すればよく、経口投与が好ましい。
【0091】
本願において「治療」とは、疾患または病態を治癒または改善するための治療と、疾患または病態の発症を予防するための治療の両方を意味する。治療は短期間または継続的に行うことができる。
【0092】
本願において「患者」とは、本発明による治療を必要とするヒトまたはヒト以外の哺乳類である。
【0093】
ヘリコバクター ピロリ結合性物質は、ヘリコバクター ピロリ結合性物質に結合するタンパク質または(炭水化物−炭水化物相互作用によって結合する)炭水化物をスクリーニングすることで1つ以上の付着因子の同定に用いることもできる。炭水化物結合性タンパク質はレクチンまたは炭水化物結合性酵素である。スクリーニングは、例えば、アフィニティークロマトグラフィーやアフィニティー交差結合法で行うことができる(Ilver et al., 1998)。
【0094】
さらに、ヒト組織に提示されているヘリコバクター ピロリ結合性物質に特異的に結合するか不活化する物質を用い、ヘリコバクター ピロリの結合を防止することもできる。このような物質の例としては、Erythrina cristagalliおよびErythrina corallodendronなどの植物性レクチン(Teneberg et al., 1994)、ジャガイモレクチンなどのポリラクトサミン結合性レクチン、およびSambucus nigra凝集素などのNeuNAcα3特異的レクチンなどが挙げられる。ヒトに用いる場合には、このような結合性物質は、その用途に適したものでなければならず、例えば、ヒト化抗体またはヒト由来の組み換えグリコシダーゼなどの非免疫原性であり、ヘリコバクター ピロリ結合性物質の末端単糖残基を開裂させることのできる物質である。しかし、胃腸器系においては、例えば食品に由来する、多種の天然のレクチンやグリコシダーゼが許容されている。
【0095】
また、ヘリコバクター ピロリ結合性物質を食品や飼料を含む栄養補助組成物の一部として用いることができる。ヘリコバクター ピロリ結合性物質を機能性食品または機能化食品の一部として用いることが好ましい。機能性食品は、ヘリコバクター ピロリがターゲットとなる細胞または組織に結合するのを阻害するか防止することで、ヒトまたは動物の健康に正の影響を与える。ヘリコバクター ピロリ結合性物質は既に知られている食品や機能性食品組成物の一部でもよい。機能性食品は、専門機関、例えば米国FDA、によって許可されている他の許容される食品材料を含有してもよい。ヘリコバクター ピロリ結合性物質は栄養補助剤、好ましくは食品添加剤や飲料物添加剤として機能性食品または機能性飲料を製造するために用いることができる。食品または食品添加剤は、例えば、ウシなどの家畜やその他の動物が多量のヘリコバクター ピロリ結合性物質を天然にその乳に産生するようにすることで得ることもできる。これは、動物がその乳に適切なグリコシルトランスフェラーゼを過剰発現させることで達成される。家畜の特定の品種または種を選択し、ヘリコバクター ピロリ結合性物質を多量に産生するように交配することができる。栄養補助組成物または栄養補助剤のためのヘリコバクター ピロリ結合性物質は、細菌や酵母などの微生物によって製造することもできる。
【0096】
ヘリコバクター ピロリ結合性物質を乳幼児用の食品の一部、好ましくは乳児用ミルクの一部として用いることは特に有用である。多くの乳児は、ヒトの天然乳の代わりとして特別な処方のミルクを与えられている。ヘリコバクター ピロリは特に乳児および幼児に高い感染性を示し、後に発症することを考慮すると、この段階で感染を予防することは妥当である。ヘリコバクター ピロリは乳幼児突然死症候群の原因となることも知られているが、細菌を一掃するための強い抗生物質による治療は、特に幼児や乳児には不適切である。
【0097】
さらに、ヘリコバクター ピロリ結合性物質をヘリコバクター ピロリの感染によって生じる病態の診断に用いることができる。診断用途には、ヘリコバクター ピロリ結合性物質を用いたヘリコバクター ピロリの型の同定方法も含まれる。結合性物質を診断または型同定に用いた場合、例えば、結合性物質はプローブやテストスティックに含まれていてもよく、所望により、テストキットの一部を構成してもよい。このようなプローブやテストスティックがヘリコバクター ピロリを含有するサンプルと接触すると、細菌はプローブまたはテストスティックに結合し、サンプルから取り出されてさらに分析される。
【0098】
末端残基は、好ましくはβ3結合したグルクロン酸であり、より好ましくはそのメチルアミドなどの6位にアミド基を有するグルクロン酸である。従って、配列の類似体や誘導体は三糖エピトープの末端6位を変化させるか誘導化することによって製造することができる。
【0099】
好ましいヘリコバクター ピロリ結合性物質
本発明のオリゴ糖配列は、予想に反して、薄層表面に提示された時に効果的なバインダーとなることが判明した。この方法は、糖脂質配列を多価の状態で提示することを可能にした。多価の状態で提示されたオリゴ糖配列の活性は驚くほど高いので、多価の状態こそが本発明のオリゴ糖配列を提示するのに好ましい方法である。
【0100】
糖脂質構造は、天然では多価の状態で細胞膜に提示されている。このような提示方法は後述する固相アッセイ法や糖脂質またはネオ糖脂質のリポソームを形成することで模倣することができる。
【0101】
疎水性ヘキサデシルアニリンの還元的アミノ化反応によって製造される本発明の新規なネオ糖脂質は、オリゴ糖を効果的に提示することができた。従来の細菌結合に用いられるネオ糖脂質結合体の大部分は、フォスファチジル エタノールアミン ネオ糖脂質のフォスフォロエステルなどの負の電荷を帯びた基を有している。このような化合物の問題点は、この物質の負の電荷とリン脂質構造に関連した天然の生物学的結合である。負の電荷を帯びた分子はタンパク質やその他の生物学的物質との多数の非特異的結合に関与することが知られている。さらに多くのこのような構造は不安定であり、酵素学的または化学的に分解されることがある。本発明は、オリゴ糖配列の非酸性結合体に関するものであり、これはオリゴ糖配列が非酸性化学構造に結合していることを意味する。非酸性結合体は中性の物質であることが好ましく、これはオリゴ糖配列が中性の、電荷を帯びていない化学構造体に結合していることを意味する。本発明において好ましい結合体は多価の物質である。
【0102】
従来技術においては、生物活性を有するオリゴ糖配列は、レセプター活性型オリゴ糖構造の一部を還元することで担体構造に結合していることが多い。アルキル鎖(-CH2-)nおよび/またはベンゼン環を含有する疎水性のスペーサーが用いられている。しかし、疎水性構造は、一般的にタンパク質やその他の生物活性分子との非特異的相互作用に関与することが知られている。
【0103】
後述する実施例で得られたネオ糖脂質に関するデータによると、アッセイに用いた実験条件においてネオ糖脂質化合物のヘキサデシニルアニリン部分は、研究に用いた細菌に対する非特異的結合を起こさなかった。ネオ糖脂質においては、糖結合体のヘキサデシニルアニリン部分は脂質層状の構造を形成すると考えられ、結合のためには存在していない。本発明は結合エピトープに属する単糖残基を還元すると結合が破壊されることを示す。さらに、還元された単糖は、レセプターエピトープと多価に提示された構造体とを繋ぐ親水性スペーサーとして使えることを見出した。本発明によると、生物活性を有するオリゴ糖を親水性スペーサーを介して多価またはオリゴ価の担体分子に結合し、多価またはオリゴ価の構造体を形成することが好ましい。本願においては、多価構造体(10個を超えるオリゴ糖残基を含むもの)およびオリゴ価構造体(2〜10個のオリゴ糖残基を含むもの)を全て多価構造体というが、用途によっては大きな多価構造体よりもオリゴ価の構造体のほうが好ましい場合がある。親水性スペーサーは少なくとも1つの水酸基を含有することが好ましい。さらに好ましいスペーサーは少なくとも2つ、最も好ましくは少なくとも3つの水酸基を含有する。
【0104】
本発明によると、親水性スペーサーは、1つまたは複数の−CHOH−基および/またはアセトアミド基である−NHCOCH3やアルキルアミド基などのアミド側鎖を含有する可撓性鎖であることが好ましい。水酸基および/またはアセトアミド基はin vivoにおける酵素的加水分解からスペーサーの保護も行う。可撓性という用語は、スペーサーが可撓性の結合を有し、可撓性ではない環構造を形成しないことを意味する。本発明において(例えば、還元的アミノ化反応によって得られた)還元単糖残基は、可撓性親水性スペーサーの一例である。可撓性親水性スペーサーはネオ糖脂質や多価結合体の非特異的結合を防止するために最適である。これは、例えば、医薬や機能性食品におけるバイオアッセイおよび生物活性において必須な最適活性である。
【0105】
可撓性親水性リンカーを有する結合体は下記式IVで表される。
[OS-O-(X)n-L1-CH(H/{CH1-2OH}p1)-{CH1OH}p2-{CH(NH-R)}p3-{CH1OH}p4-L2]m-Z (IV)
(式中、
OSはヘリコバクター ピロリ結合性オリゴ糖配列であり;
Xは炭素(−C−)、窒素(−N−)、または硫黄(−S−)で置換可能な糖残基であり;
L1およびL2は結合基あって、各々独立に酸素原子、窒素原子、硫黄原子または炭素原子である結合原子あるいは置換基を形成する結合に含まれる2個の原子を包含し、該結合原子および該置換基を形成する結合は−O−、−S−、−CH2−、−N−、−N(COCH3)−、−CO−NH−または−NH−CO−で表されるアミド基、−N−N−(ヒドラジン誘導体)、あるいは−O−N−または−N−O−で表されるアミノオキシ結合であり;
L1は該Xの還元末端単糖のC1位に結合した結合基であり、nが0の場合には、L1は−O−を置換して、該OSの還元末端のC1位に直接結合しており;
p1、p2、p3およびp4は各々独立に0〜7の整数であるが、但し、p1、p2、p3およびp4の少なくとも1種が1以上の整数であり;
{CH1-2OH}p1が側鎖構造を示す場合には、末端基となるCH1-2OHはCH2OHであり、p1が1を超える場合には、該末端基をスペーサーである該Yの残りの部分に結合させるための第二級アルコール基である−CHOH−が存在し;
Rは好ましくはアセチル基(−COCH3)、Zに連結する結合、類似体形成基、水素原子または式中でアミンを形成するC1-4アルキル基であり、Rが結合である場合にはL2は糖鎖末端基の1個または2個の原子であり、該類似体形成基は、アミド構造を有するC1-4アシル基(好ましくは親水性の基、例えばヒドロキシアルキル基)を含み、;
mは1を超える整数であり;
Zは多価の担体である。)
【0106】
式(IV)に示した可撓性親水性スペーサーを有する好ましい多価構造体としては、ヘリコバクター ピロリ結合性オリゴ糖配列(OS)が Galβ4Glc(red)-Zにβ1−3結合したもの、OSβ6GlcNAc(red)-Z および OSβ6GalNAc(red)-Z(式中、“(red)”は還元末端単糖および多価担体Zのアミノ基の還元的アミノ化によって得られたアミン結合構造である)が挙げられる。
【0107】
本発明においてオリゴ糖基は、抗原性やアレルギー反応の出現を防止するために、多価またはオリゴ価の形でタンパク質またはペプチドではない担体に結合していることが好ましく、より好ましくは、担体の骨格は天然の非抗原性多糖である。
【0108】
本発明のex vivoにおける用途
本発明により、生体外(ex vivo)における病原体、特に下痢発症性大腸菌の阻害に用いることができることが判明した。また、このような方法は、消毒および保存のために用いることもできる。好ましくは単一成分物質の部分または単一成分物質そのものである本発明で開示するレセプターオリゴ糖配列、さらに好ましくは、少なくとも2種の本発明で開示するレセプターオリゴ糖配列を包含する組成物を、病原体、好ましくは大腸菌を阻害するためにex vivoで用いる。本発明で開示する多価結合体、特に、病原体、好ましくは下痢原性大腸菌を凝集することができる可溶性多価結合体が、ex vivoでの使用には好ましい。本発明のex vivoでの使用の特殊な態様として、病院や準病院環境などで使用するテーブル、医療器具および包装容器などの表面の洗浄や消毒において、本発明で開示するレセプターオリゴ糖配列を含有する洗浄剤や消毒剤を用いる方法が挙げられる。本発明で開示するレセプター糖類はまた、病院や準病院環境などにおいて患者の洗浄用または入浴用の石鹸や洗剤の成分としても有用である。
【0109】
口腔感染症および口腔衛生製品
本発明が標的とする感染症は経口経路で、またおそらく鼻から口腔への経路でも広がるものと考えられる。本発明は、ヒトの口に存在している感染症の阻害に関する。本発明は、少なくとも2種の異なるオリゴ糖配列による口腔感染症の治療に関し、該オリゴ糖配列は、病原体(好ましくは経口的に感染する細菌、さらに好ましくは下痢発症性細菌)の結合特異性の少なくとも2種を阻害するものである。口腔感染症や鼻腔感染症の予防のために、単一成分物質の部分または単一成分物質そのものとして本発明で開示するレセプターオリゴ糖配列を用いるか、本発明の異なる種類のレセプターオリゴ糖配列を少なくとも2種包含する組成物として用いることが好ましい。本発明によると、本発明で開示したレセプターオリゴ糖配列を、組成物または単一成分物質として、ヒトの口腔中の病原体を阻害する製品(ここでは「口腔衛生製品」と称する)に用いる。
【0110】
ヒトの口腔には、ヒトの腸と同様のレセプターが存在すると考えられ、特にタンパク質上には、少なくともネオラクトレセプター、マンノースレセプター、および本発明のフコースレセプターに類似したオリゴ糖レセプターが存在するものと考えられる。別の態様においては、本発明の物質および組成物は、カリエスを発症させる病原体を阻害するのにも有用である。具体的な態様においては、本発明は、経口的に広がる他の感染症、例えば、中耳炎を併発する感染症やインフルエンザ、気管支炎または肺炎を含む肺の感染症を治療するための組成物にも関する。本発明の口腔衛生製品は、カリエス、中耳炎、気管支炎および肺炎の治療を目的とすることもできる。本発明の具体的態様の一例として、Neu5Acα3Galβ4GlcNAc および Neu5Acα3Galβ4Glcからなる群より選ばれる少なくとも1種のオリゴ糖配列、より好ましくは Neu5Acα6Galβ4GlcNAc および Neu5Acα6Galβ4Glcからなる群より選ばれる少なくとも1種のオリゴ糖配列を包含する、口腔衛生製品の製造または経口的に感染する病原体の阻害のために用いる組成物がある。そのような組成物は、少なくともヒト インフルエンザウイルスの処置を目的とし、好ましくはインフルエンザウイルスに対する予防を目的としたものである。
【0111】
特に本発明は、病原体阻害性オリゴ糖配列、特に本発明で開示するオリゴ糖配列、を含む物質または組成物を含有する口腔衛生製品に関する。口腔衛生製品は、練り歯磨、洗口薬、口腔用タブレット、咀嚼錠(chewing tablets)およびチューインガムからなる群より選ばれることが好ましい。一価のレセプターオリゴ糖配列または多価のレセプターオリゴ糖配列のいずれかを用いることが好ましい。他の一つの好ましい態様においては、口腔衛生製品は、本発明で開示する多価のオリゴ糖配列を含有する。ヒトの口腔の大きさおよび口腔表面に存在する唾液の容量を考慮すると、口腔において病原体阻害性レセプターの飽和濃度を達成するには比較的少量のオリゴ糖で十分である。レセプター活性を有する一価エピトープの典型的な量は、レセプター活性オリゴ糖として約100nmol〜100μmol(分子量が1,000Daの場合、100μg〜100mgに相当する)の範囲内である。より一般的な量は、約1〜10μmolと推定される。別の態様においては、本発明の治療用医薬組成物は、病原体を阻害するスプレー式点鼻薬にも関する。スプレー式点鼻薬は、中耳炎または肺の感染症の治療を目的としたものである。
【0112】
局所用製品、洗剤および化粧品
一般的な病原体は、ヒトの肌、性器上皮、毛髪、日用品の表面およびヒトの環境中に存在する他の表面に広がりうると考えられる。本発明のオリゴ糖配列は、このような環境中に存在する病原体の阻害にも有用である。したがって、局所用製品または化粧品(例えば、クリーム、ローションまたはジェル)に、単一成分物質の部分または単一成分物質そのものとして本発明のオリゴ糖配列を用いるか、本発明の異なる種類のレセプターオリゴ糖配列を少なくとも2種包含する組成物を用いることも好ましい。また、本発明の物質または組成物を、ヒトの肌、毛髪または性器上皮を洗浄するための製品(個人用衛生製品とも言う)や、日用品の表面、皿または衣服を洗浄するための製品に用いることも好ましい。従来の抗生物質は、日用品用の洗浄液に用いることを目的としているが、本発明の物質とは違って耐性の問題があるため有用ではない。好ましい態様においては、多価オリゴ糖配列を洗浄液に用い、他の一つの好ましい態様においては、一価オリゴ糖配列を洗浄液に用いる。
【0113】
食品または飼料、飲料、酒類および水に用いる食品安全性向上製品
ヒトまたは動物の治療方法の他に、本発明は、本発明で開示するレセプターおよび組成物を用いた感染症の予防方法、即ち、本発明を用いて食品製品の内部または表面上の病原体や細菌を中和する方法に関する。例えば、病原体の蔓延を防止するために、食肉生産の際に本発明の炭水化物を食肉製品や動物の体またはその部分の表面に塗布することができる。細菌を被覆し凝集させるために可溶性多価結合体および他の多価結合体を用いることが好ましい。固体または半固体の食品製品の表面に用いる具体的な方法には、細菌に本発明の炭水化物レセプターを接触させ、所望により病原体と炭水化物との複合体を洗い流すことが含まれる。この種の方法には、従来の抗生物質を適用することはできない。本発明の炭水化物は、液状食品製品、濃縮物またはそれらの元となる粉末、例えば、ミルク、液状ミルク様製品およびジュース、ソフトドリンク、スポーツドリンク、アルコール飲料などの種々の飲料に対して用いることもできる。
【0114】
具体的な態様においては、本発明の炭水化物を多価の型で液状食品製品または飲料製品に対して使用し、潜在的な病原体をこのような多価結合体によって凝集させ、凝集複合体をその大きさや溶解性に基づいた方法によって除去する。不溶性の凝集物は、沈殿物として標準的な方法、例えば、凝集物の上に存在する溶液をデカンテーションする方法や、より一般的且つ効果的なろ過法で除去することができる。ろ過法は、より大きな凝集複合体を除去するのに用いることができる。
【0115】
本発明の炭水化物で処理する好ましい食品としては、種々の動物性食品製品、特に食肉製品および加工途中の製品が挙げられる。下痢発症性大腸菌菌体を含む多くの病原体は、正しく洗浄されていない野菜、果物、サラダおよび他の植物性食品から伝播する。洗浄が必要であるにもかかわらず、正しく洗浄されていないか汚染された水で洗浄された食物は、発展途上国で特に問題となっている。本発明は、洗浄することが必要な植物性食品や他の食品の安全性を向上させるための方法であって、病原体、特に食品製品中の病原性大腸菌の菌体量を制御するための方法に関する。特に本発明は、食品からの感染を防止するための、家庭用製品および食品工業用製品にも関する。このような製品は、加工中の食品に利用することができる、粉末状または錠剤状の固体製品か、本発明で開示するレセプターの溶液を内包するカプセルであることが好ましい。このような製品は、発展途上国で発生する下痢、特に子供の下痢の予防に用いることができる。食品安全性向上製品は、旅行者の下痢の予防も目的とする。食品安全性向上製品および以下に述べる飼料安全性向上製品は、新規な安全性向上保存料であると考えられる。
【0116】
飲料および水を精製するためのフィルター製品
汚染された飲料および水は胃腸器系の疾患、特に下痢の主要な病因である。本発明で開示するレセプターは、液状食品、飲料および水(特に、飲料および食品を作るための水)に存在する病原体、特に細菌を除去するためのフィルターの作製に用いることもできる。少なくとも2種のレセプター構造を用いることが好ましい。フィルターを得るために炭水化物配列を結合させる固相材料を、例えばセルロース、プラスチック、アガロースおよびそれに類似した材料から製造する方法は知られている。本発明のフィルターとしては、公知技術であるアフィニティークロマトグラフィー用の材料も挙げられる。このようなフィルターから結合物を除去するための方法は公知であり、具体的な態様においては、フィルターは、汚染物質を除去し、所望により熱または他の滅菌方法により滅菌することにより、再生する。
【0117】
飼料安全性向上製品
上記の食品安全性向上製品は、動物用の固体および液状の飼料ならびに動物用の飲料水に用いることもできる。保護する動物として好ましいものとしては愛玩動物、特にネコとイヌ、および家畜または農場動物(例えばウシや他の反芻動物、ブタ、ヒツジ、ウマ、家禽(例えばメンドリ、アヒルや七面鳥)およびウサギ)が挙げられる。
【0118】
水、食品および飼料の安全性の分析
本発明のレセプター炭水化物を組み合わせた標準的な分析および診断方法を用いて水、飲料、食品および飼料を分析し、レセプター炭水化物に結合する病原体の存在量を測定することができる。汚染病原体の結合特異性に関する知見は、感染患者の治療計画をたてる場合や、上記の病原体を除去または制御して食品の安全性を向上させるための方法に適用することができる。
【0119】
本発明によって阻害することのできる、炭水化物に基づく他の相互作用
異なる種類の付着因子の阻害の他にも、本発明は、炭水化物−炭水化物相互作用および炭水化物−レクチン相互作用の阻害に用いることもできる。
【0120】
本発明の炭水化物含有組成物および炭水化物含有物質はオリゴ糖配列を包含している。オリゴ糖配列は、少なくとも1種の病原体および/または少なくとも1種の病原体に対するレセプターに結合することによって、少なくとも1種の病原体を阻害する。少なくとも2種の病原体阻害性オリゴ糖配列を用いることが好ましく、少なくとも3種の病原体阻害性オリゴ糖配列を用いることがより好ましい。他の好ましい態様においては、少なくとも4種、5種、6種または7種の病原体阻害性オリゴ糖配列を用いる。
【0121】
具体的な治療においては、1つまたは複数のオリゴ糖配列を異なる時期に別々に投与する。これは、全種類のオリゴ糖配列の同時投与が正常細菌叢に負の影響を与える場合に特に有効である。治療用組成物を別々に投与することは、胃腸器系における本発明のオリゴ糖配列の安定性が胃腸器系の栄養状態の影響を受けることからも有効である。
【0122】
本発明とプロバイオティック細菌を一緒に用いる方法
本発明を細菌の結合、特に複数の細菌の結合を阻害するために用いる場合、いくつかの有益な細菌の結合も阻害されることが考えられる。正常細菌叢は、例えばヒト胃腸器系において、多くの重要な機能を有する。しかし、従来の抗生物質を用いた場合に生じる正常細菌叢の破壊は、より大きな問題となる。
【0123】
他の態様においては、少なくとも2種の病原体阻害性オリゴ糖配列を、プロバイオティック微生物および/またはプレバイオティック化合物と共に投与する。本発明においてプロバイオティック微生物とは、例えば食物の消化、栄養およびビタミンの提供または組織表面の被覆による病原性細菌からの保護といった有益な機能を有する無害な細菌を意味する。プロバイオティック細菌には、正常細菌叢から選ばれる1種、数種または多種の細菌が含まれることが好ましい。好ましい態様においては、プロバイオティック細菌には、少なくとも1種類または1株または1種(species)の乳酸菌が含まれる。
【0124】
プレバイオティック化合物とは、正常細菌叢またはプロバイオティック微生物を支える物質である。好ましいプレバイオティック化合物としては、プレバイオティック炭水化物(例えば、プレバイオティック化合物として用いられるガラクトースオリゴ糖、キシロースオリゴ糖またはフルクトースオリゴ糖)が挙げられ、プレバイオティック活性を有する多糖および繊維(例えばイヌリンまたは修飾した澱粉)も含まれる。本発明は、食品または栄養補助的目的のために用いられる他の多糖、例えばキトサンまたはβ−グルカン(例えばオート麦のグルカン)を用いてコレステロールおよび脂肪を減少させる方法にも関する。好ましい態様においては、非還元末端にβ−結合したガラクトース残基を有する炭水化物のような、プレバイオティック化合物としても機能する少なくとも1種の病原体阻害性炭水化物を選択する。好ましい治療方法においては、
a)病原体および正常細菌叢と考えられるものの一部を、1種、さらに好ましくは少なくとも2種の本発明の炭水化物を用いて初めに除去し、そして
b)プロバイオティック微生物および/またはプレバイオティック化合物を投与する。
上記の第一および第二の工程は逆の順番で行ってもよく、好ましくは大量のプロバイオティック微生物および/またはプレバイオティック化合物を用いてから、第一の工程を行う。本発明によると、工程の順番を変えながら、第一および/または第二の工程を数回繰り返すこともできる。第一および第二の工程は同時に行ってもよい。本発明の物質は、プロバイオティック微生物および/またはプレバイオティック化合物と共に投与することができるが、プロバイオティック微生物および/またはプレバイオティック化合物を本発明の組成物に含ませることもでき、その場合、第一および第二の工程は同時に行うことができる。
【0125】
本発明のオリゴ糖配列の中には、プレバイオティック効果を有することが知られているものがある。そのようなものの例としては、N−アセチル−ラクトサミン型オリゴ糖配列や、特にヒト乳オリゴ糖などのフコシル化オリゴ糖が挙げられる。また、プロバイオティック微生物および/またはプレバイオティック化合物と共にヒト乳オリゴ糖、特に、例えばラクト−N−ネオテトラオース、ラクト−N−テトラオース、ラクト−N−ヘキサオース、ラクト−N−ネオヘキサオース、パララクト−N−ヘキサオース、およびパララクト−N−ネオヘキサオースからなる群より選ばれる少なくとも1種を含有するN−アセチル−ラクトサミン、および/またはそれらより誘導されるフコシル化オリゴ糖、および/または、例えば、モノフコシル化、ジフコシル化またはトリフコシル化したラクト−N−テトラオース(LNT)やラクト−N−ネオテトラオース(LNnT)、および/またはフコシルラクトースであるオリゴ糖(例えば2'フコシルラクトース、3−フコシルラクトースやジフコシルラクトース)を投与する方法がある。
【0126】
本発明の物質および/または組成物と共に用いられるその他の有用な物質
本発明によると、病原体阻害性(pathogenesis preventing)炭水化物を、グリコシダーゼ阻害剤、好ましくはシアリダーゼ阻害剤と併用することも有用である。
【0127】
本発明によると、病原体阻害性炭水化物を、レクチンまたはそれ以外の炭水化物結合性タンパク質と併用することも有用である。レクチンは、炭水化物レセプター、例えば、菌体外多糖類をブロックするために用いることができる。
【0128】
糖脂質および炭水化物の命名は、IUPAC−IUB生化学命名委員会の推奨する命名法(Carbohydrate Res. 1998, 312, 167; Carbohydrate Res. 1997, 297, 1; Eur. J. Biochem. 1998, 257, 29)に従った。
【0129】
Gal、Glc、GlcNAc および Neu5AcはD型であり、FucはL型であり、全ての単糖単位はピラノース環構造であるものとする。グルコサミンはGlcNまたはGlcNH2と示し、ガラクトサミンはGalNまたはGalNH2と示す。グリコシド結合は略記で示す場合と正式名称で示す場合とあり、Neu5Ac-残基のα3結合およびα6結合はそれぞれα2−3結合およびα2−6結合と同じであり、他の単糖残基については、α1−3結合、β1−3結合、β1−4結合およびβ1−6結合はそれぞれα3、β3、β4およびβ6と略すことができる。ラクトサミンはN−アセチルラクトサミン(Galβ4GlcNAc)を示し、シアル酸はN−アセチルノイラミン酸(Neu5Ac, NeuNAc または NeuAc)、N−グリコリルノイラミン酸(Neu5Gc)またはその他の天然のシアル酸を示す。本願においてグリカンという用語は、ヒトまたは動物に存在する糖結合体、特に糖脂質または糖タンパクに含まれるオリゴ糖や多糖鎖を広く包含する。脂肪酸と塩基の略記命名法においては、コロンの前の数字が分子の炭素鎖の長さを表し、コロンの後の数字が炭化水素鎖の二重結合の合計を示す。GSLという略語はグリコスフィンゴ脂質を表す。グリコスフィンゴ脂質の略語、略称または記号は表1および2に記載した。ヘリコバクター ピロリとは、ヘリコバクター ピロリに類似した細菌も意味する。
【0130】
「末端オリゴ糖配列」または「末端配列」という表現は、オリゴ糖配列またはシアル酸誘導体配列が、所望によりオリゴ糖配列の還元末端から結合している場合を除いては、他の単糖構造またはオリゴ糖構造に結合していないことを表す。オリゴ糖配列が結合体として存在している場合は、オリゴ糖配列はその還元末端から結合していることが好ましいが、病原体結合性に影響しない他の結合位置を用いることもできる。さらに具体的な態様においては、本発明のオリゴ糖配列とは、天然のグリコシド結合によって他の単糖構造またはオリゴ糖構造に結合していない、上記配列に相当するオリゴ糖残基を意味する。このようなオリゴ糖残基は、遊離オリゴ糖、結合体を形成するもの、またはオリゴ糖残基の還元末端を誘導化したものであることが好ましい。
【0131】
「α3/β3」は、オリゴ糖配列において隣り合った糖残基は互いにα3−またはβ3−結合していることを表す。
【発明を実施するための最良の形態】
【0132】
本発明を以下の実施例によってさらに説明するが、これらは本発明の範囲を限定するものではない。
【0133】
ガングリオシドの調製 − ガングリオシドを単離するために、複合ガングリオシドを含有することが知られている種々の組織、例えばヒト赤血球、ウシ赤血球、ウサギ胸腺、ヒト胎便およびヒト癌(Stults et al., 1989)を収集した。総酸性グリコスフィンゴ脂質画分の単離は公知の方法(Karlsson, 1987)で行った。端的に記すと、各組織を凍結乾燥し、次いでソックスレー抽出器でクロロフォルム/メタノール混合溶媒(2:1と9:1、容量比)を用いた2段階の抽出を行った。抽出物をプールし、穏やかなアルカリ加水分解および透析に付した後、ケイ酸カラムによる分離に付した。酸性グリコスフィンゴ脂質と非酸性グリコスフィンゴ脂質はDEAEカラムで分離した。
【0134】
酸性グリコスフィンゴ脂質画分をDEAE−セファロ−スクロマトグラフィーにより分離し、さらにケイ酸クロマトグラフィーに繰り返し付し、最終的な分離をHPLCにより行った。HPLCは、長さ250mm、内径10mmで粒径が5μmのKromasil 5 シリカカラム(米国、カリフォルニア州、トランス、Phenomenex製)を用い、クロロフォルム/メタノール/水の直線濃度勾配(60:35:8〜40:40:12または65:25:4〜40:40:12、容量比)で、流速2ml/分で180分間行った。回収した各2mlの画分をそれぞれ薄層クロマトグラフィーおよびアニスアルデヒド染色(下記参照)により分析し、ヘリコバクター ピロリ結合能をクロマトグラム結合アッセイ(下記参照)により評価した。薄層クロマトグラム上の移動度およびヘリコバクター ピロリ結合能に基づいてHPLC画分をプールした。
【0135】
参照用グリコスフィンゴ脂質 − 参照用グリコスフィンゴ脂質は、スウェーデン国、イェーテボリ大学、医薬生化学研究所(Institute of Medical Biochemistry)にて単離し、特徴付けた。構造の特徴付けはプロトンNMR法(Koerner et al., 1983)、質量分析法(Samuelsson et al., 1990)および分解的手法(degradation studies)(Yang and Hakomori, 1971、Stellner, 1973)により行った。シアリル−Lex ヘキサグリコシルセラミドはARC社(カナダ国、エドモントン)より購入した。
【0136】
細菌株、生育条件および標識 − ヘリコバクター ピロリ CCUG 17874株はイェーテボリ大学カルチャーコレクション(Culture Collection University of Goteborg(CCUG))より入手した。J99株はTim Cover博士、John Atherton博士およびMartin Blaser博士より寄贈されたものである。
【0137】
細菌の培養は、10%の56℃で不活性化したウシ胎児血清(英国、ラフバラ、Harlan Sera-Lab製)とBBL IsoVitale X エンリッチメント(ニュージャージー州、フランクリン・レイクス、Becton Dickson Microbiology Systems製)を含有するブルセラ培地(カリフォルニア州、アーヴィン、Difco Laboratories製)で行った。細菌の放射線標識は、0.5mlのリン酸緩衝化生理食塩水(PBS)(pH7.3)で希釈した50μCiの35S−メチオニン(英国、リトル・チャルフォント、Amersham Pharmacia Biotech製)を、固体培養中の菌に添加することにより行った。微好気性条件下、37℃で12〜72時間インキュベートした後に細菌を回収し、回収した細菌をPBS中、3,500rpmで10分間の遠心分離に3回付した。
【0138】
放射線標識のもう一つの方法においては、10%の熱不活性化ウシ胎児血清と50μCiの35S−メチオニンを添加したハムF12培地(英国、カリフォルニア州、カールズバッド、Invitrogen Corp.製)に(1×105CFU/mlとなるように)コロニーを接種した。この培地の入った培養用ボトルを微好気性条件下、37℃で24時間、振とうさせながらインキュベートした。菌体は遠心分離によって回収し、PBSで3回洗浄した。
【0139】
上記いずれの方法においても、標識した細菌の濃度が約1×108CFU/mlになるように、最後に2%(w/v)ウシ血清アルブミン含有PBS(PBS/BSA)に再懸濁した。いずれの標識方法によっても、比活性がヘリコバクター ピロリ100個あたり約1cpmの懸濁液が得られた。
【0140】
薄層クロマトグラフィー − 総酸性グリコスフィンゴ脂質画分(40μg)または純粋なガングリオシド(0.0002〜4μg)を、アルミニウムで裏打ちしたシリカゲル60HPTLCプレート(ドイツ国、ダルムシュタット、Merck製)上で、クロロフォルム/メタノール/水(60:35:8、容量比)、クロロフォルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)または1−プロパノール/水/25%NH3(7:3:1、容量比)を溶媒系として分離した。薄層クロマトグラム上のグリコスフィンゴ脂質の化学的検出は、アニスアルデヒド試薬(Waldi, 1962)またはレゾルシノール試薬(Svennerholm, 1963)を用いて行った。
【0141】
クロマトグラム結合アッセイ − クロマトグラム結合アッセイは、公知の方法(Angstrom et al., 1994)と実質的に同様に行った。分離したグリコスフィンゴ脂質を含む乾燥薄層クロマトグラムを、0.5%(w/v)ポリイソブチルメタクリレート(ウィスコンシン州、ミルウォーキー、Aldrich Chemical Company Inc.製)のジエチルエーテル/n−ヘキサン(1:5、容量比)溶液で1分間処理し、風乾した。非特異的結合を減少させるために、プレートを0.1%NaN3(w/v)と0.1%Tween20(容量比)を含有するPBS/BSA中で、室温で2時間インキュベートした。次いでプレートをPBS/BSAで希釈した35S−標識ヘリコバクター ピロリと共に室温で2時間インキュベートした。また、薄層クロマトグラム上のGM1ガングリオシド希釈系列に対して125I−標識コレラトキシンB−サブユニットを公知の方法(Karlsson and Stromberg, 1987)で結合させた。最後の洗浄と乾燥を行った後、Biomaxフィルム(米国、ニューヨーク州、Eastman Kodak Company製)を用いてオートラジオグラフィーを一晩行った。他のガングリオシドに対する、1つの純粋なガングリオシドへの細菌の結合量を定量するために、薄層クロマトグラム上のガングリオシド希釈液系列に対する細菌の結合と、それに続くオートラジオグラフィーの後に、得られたオートラジオグラムをデンシトメトリーに付した。ガングリオシドの濃度範囲は、飽和濃度を求めた予備実験に基づいて選択した。オートラジオグラムはCCDカメラ(インディアナ州、ミシガンシティ、Dage-MTI, Inc.製)で撮影し、得られたイメージをパブリックドメインプログラムであるNIHイメージプログラム(米国国立衛生研究所(U.S. National Institute of Health)によって開発され、http://rsb.info.nih.gov/nih-image/より入手可能)を用いて解析した。ここに示したデータは、バックグラウンド値を引いた値である。
【0142】
ガングリオシドの誘導化 − ガングリオシドは、水酸化ナトリウムとヨウ化メチルのジメチルスルホキシド溶液を用いて公知の方法(Larsson et al., 1987)によりパーメチル化した。パーメチル化した試料はLiAlH4のジエチルエーテル溶液を用いて還元した(Karlsson, 1974)。
【0143】
高速原子衝突質量分析および電子イオン化質量分析 − 陰イオンFABおよびEI質量スペクトルをJEOL SX 102A質量分析計(日本国、東京、日本電子株式会社製)を用いて得た。誘導化前のガングリオシドの陰イオンFAB質量スペクトルは、キセノン原子衝突(6eV)、加速電圧(〜8kV)およびトリエタノールアミンをマトリックスとする条件下で得た。誘導化したグリコスフィンゴ脂質のEIスペクトルは、イオン化電圧70eV、イオン化電流300μAおよび加速電圧8kVの条件下で得た。温度は、150℃から410℃に昇温速度10℃/分で昇温した。FABスペクトルおよびEIスペクトルのいずれのスペクトルも取得する際の解像度は1000とした。
【0144】
エレクトロスプレーイオン化質量分析法および衝突誘起解離法 − ヒト赤血球由来の、パーメチル化ヘリコバクター ピロリ結合性ガングリオシドを10%(容量比)アセトニトリル水溶液に溶解した。エレクトロスプレーイオン化は、Q−TOF質量分析計(英国、Micromass製)でナノフローエレクトロスプレーオプションを用いて、約15〜40nL/分の一定流速で行った。噴霧はナノスプレーニードルに900Vの電圧をかけることにより行った。イオン源温度は80℃とした。流速150リットル/時間の窒素を乾燥用ガスとして用いた。質量スペクトルにおいてm/z1594.4で観察された二価荷電ピークを、衝突誘起解離を行うために選択した。突誘起解離は、アルゴンを30〜80Vの衝突エネルギーで用いることにより達成した。データは50〜2600質量単位に渡って回収した。ここでは、ドモン(Domon)とコステロ(Costello)の命名法(Domon and Costello, 1988)を使用した。
【0145】
プロトンNMRスペクトル法 − 1H NMRスペクトルは、Varian 500MHz NMR分光計およびVarian 600MHz NMR分光計を30℃で用いて得た。試料は、重水素置換後に、ジメチルスルホキシド/D2O(98:2、容量比)に溶解した。
【0146】
詳細なエピトープ分析実験に用いる特別な材料および方法
天然糖脂質の入手源
PGC(ヒト赤血球由来)は、パーアセチル化法(Miller-Podraza et al., 1993)に従って本発明者らが単離した。S−3−PG(ヒト赤血球由来およびヒト白血球由来)、ジシアリルパラグロボシド(ヒト赤血球由来)、S−6−PG(ヒト白血球由来)、七糖ネオラクトガングリオシド(ヒト赤血球由来およびヒト白血球由来)およびグロボシド(ヒト赤血球由来)も本研究室で調製した(Karlsson, 1987)。ヒト脳のGQ1bは、イェーテボリ大学神経化学科(Department of Neurochemistry of Goteborg University)より入手した(Miller-Podraza et al., 1992)。ウシ脳由来のガングリオシドであるGM1、GD1a、GD1bおよびGT1bは、Calbiochem(米国)から購入した。
【0147】
炭水化物の入手源
NeuAcα3Gαβ4GlcNAcβ3Galβ4Glcは、S−3−PG(ヒト赤血球由来)をセラミドグリカナーゼ(ヒル由来;ドイツ国、Boehringer Mannheim GmBH製)により消化し(Ito and Yamagata, 1989)、クロロフォルム/メタノール/水(2:1:0.6)を用いた相分離に付すことにより、本研究室で調製した。上記五糖を上相から回収した。NeuAcα3Galβ3GlcNAcβ3Galβ4Glc、NeuAcα6Galβ4GlcNAcβ3Galβ4GlcおよびGalβ3(NeuAcα6)GlcNAcβ3Galβ4Glcは、IsoSep(スウェーデン国、トウリンゲ)より入手した。
【0148】
その他の試薬
4−ヘキサデシルアニリン、メチルアミン、エチルアミン、プロピルアミン、ブチルアミンおよびベンジルアミンは、Aldrich Chemical Company(米国、ミルウォーキー)製を用いた。グリコール酸、エタノールアミンおよびオクタデシルアミンは、Sigma-Aldrich(ドイツ国)から購入した。Sephadex LH 20は、Pharmacia(スウェーデン国、ウプサラ)製であり、エチレングリコールはFluka(スウェーデン国)製であった。
【0149】
シアル酸のグリセロール尾部の化学的修飾
A.過ヨウ素酸による穏やかな酸化(Veh et al., 1977)とそれに続く還元反応(R-CHOH-CHOH-CH2OH → R-CHOH-CH2OH/R-CH2OH):
反応材料(0.5〜1μmol)を、1〜2mMのNaIO4を含有する0.05mM酢酸緩衝液(pH5.5)500μl中で、氷上で40分間インキュベートすることにより反応を行った。反応を過剰量のエチレングリコールにより停止し、反応混合物を試料とした。次いで、試料を凍結乾燥により濃縮し(約5倍)、室温で一晩、過剰量のNaBH4により還元反応に付した。最後にこのような処理を施した試料を蒸留水に対して2日間透析し、次いで凍結乾燥した。
【0150】
B.S−3−PGの過ヨウ素酸による穏やかな酸化とそれに続くメチルアミンまたはエタノールアミンとのカップリング(R-CHOH-CHOH-CH2OH → R-CH2-NH-CH3/R-CH2-NH-CH2-OH):
ヨウ素酸を用いたS−3−PGの穏やかな酸化(上記参照)とエチレングリコールの添加の後、得られた反応混合物を蒸留水に対して2日間透析し、凍結乾燥して酸化S−3−PGとした。以下の条件下で酸化S−3−PGとメチルアミンまたはエチルアミンとのカップリングを行った:糖脂質(0.5mg)を200μlのM/C(3:1)に溶解し、50μlのアミン、200μlの、NaBH3CNのメタノール溶液(62mg/ml)および200μlのグリコール酸水溶液(136mg/ml)と混合し、試料を得た。メチルアミンを用いた場合には、溶解性を高めるために50μlのテトラヒドロフランをさらに添加して混合した。得られた試料を30℃で4時間インキュベートした後、窒素雰囲気下で水分を蒸発した。得られた残渣をC/M/水(60:30:4.5)に懸濁し、メタノールで飽和させたSephadex LH-20カラムを用いて脱塩した。試料(約0.6ml/0.5×15cmのカラム)をカラムにアプライした後、カラムをメタノールによる溶出に付し、糖に対して陽性を示した画分を収集することにより糖脂質を回収した(TLCとアニスアルデヒドによりモニターした)。分子種(上記式参照)の最終的な精製を行うために、C/M/水(60:35:8)を展開溶媒とした分取TLCにより、目的材料を分離した。(プレートからストリップを切り出した後にアニスアルデヒドで検出した)主要なバンドをかき取り、上記と同じ溶媒で抽出した。
【0151】
カルボキシル基の修飾(Lanne et al., 1995)
S−3−PG(0.5〜5mg)をまず、ヨウ化メチル(100μl)のジメチルスルホキシド(DMSO、0.5ml)溶液と共に室温で1時間インキュベートすることでメチルエステルに転化した(R-COOH → R-COCH3)。反応産物はSephadex LH-20を用いて上記と同様に精製し、メチルエステルを得た。S−3−PGのアルコール誘導体(R-COCH3 → R-CH2OH)を調製するために、上記のメチルエステル(0.5mg)を0.5mlのメタノールに溶解し、そこに5mgのNaBH4を添加した。室温で1時間還元反応に付した後、還元したS−3−PGをSephadex LH-20を用いて脱塩した(上記参照)。
【0152】
S−3−PGのアミドならびにS−3−PGのメチルアミド、エチルアミド、プロピルアミド、ベンジルアミドおよびステアリルアミドを合成するために、上記メチルエステルの溶液(0.5mlのメタノール中に0.5mg)を次の溶液のそれぞれと混合した:0.2mlの30%NH3水溶液、0.5mlの40%メチルアミン水溶液、1mlの70%エチルアミン水溶液、100μlのプロピルアミン、100μlのベンジルアミンおよび200μlの、ステアリルアミンのTHF溶液(飽和溶液)。室温で一晩インキュベートした後、反応産物から水分を窒素雰囲気下で蒸発し、アミド類とした。ベンジルアミドおよびステアリルアミドはさらに、ヘキサン/アセトン(1:1、容量比)による抽出により精製した。各アミドを過剰量の上記混合溶媒で洗浄し、遠心分離に付し、上澄みを廃棄した。
【0153】
PGC誘導体を、以下の2つの条件を変えた以外は上記と同様に調製した。
1.還元したPGC調製物をDEAE-Sephadexカラムで分離し、中性画分のみをさらに分析した、そして
2.酸化/還元工程を2回行った。
【0154】
遊離の糖(30)へのヘキサデシルアニリンのカップリング
糖(0.5mg)を100μlのメタノールに溶解し、NaBH3CNのメタノール溶液(62mg/ml)100μl、ヘキサデシルアニリンのテトラヒドロフラン溶液(40mg/ml)100μlおよび1.8Mのグリコール酸水溶液(136mg/ml)100μlと混合した。さらに、沈殿しているヘキサデシルアニリンの溶解性を高めるために、100〜200μlのテトラヒドロフランを添加した。このようにして得られた試料を30℃で一晩インキュベートして反応し、得られた反応産物をSephadex LHクロマトグラフィーにより精製した(上記「カルボキシル基の修飾」の項を参照)。この反応の収率は90%を超えた。
【0155】
分岐した脂質部分の合成(Magnusson et al., 1994、Read et al., 1977)
分岐した脂質部分を有するネオ糖脂質を得るために、HDA−誘導化糖をN−アシル化によりさらに修飾した。500μlの、p−ニトロフェニルパルミチン酸の脱水DMSO溶液を、200〜300μgの乾燥させたHDA−糖に添加して試料とした。窒素雰囲気下で37℃で3日間インキュベートした試料に、トリエチルアミンを4滴添加した。糖脂質をSephadex LH-20カラムクロマトグラフィー(上記参照)および分取TLCにより精製した。収率は約20%であった。
【0156】
ラクトン類の調製
S−3−PGを、公知の方法(Laferriere and Roy(33))によりそのラクトン型に転換した。1mgのS−3−PGを濃縮酢酸に溶解し、室温で2日間反応に付した。その後酢酸を蒸発で除去し、残渣をC/M/水(60:35:8、容量比)(0.5ml)に溶解した。この反応の収率を、0.1mmのシリカゲル60(ドイツ国、Merck製)で被覆した、アルミニウムで裏打ちしたHPTLCプレート上の薄層クロマトグラフィーにより検査したところ、約50%であった。
【0157】
ガングリオシドGM1bの合成と調製
ガングリオシドGM1の脱シアリル化により調製したガングリオテトラオシルセラミドの末端ガラクトースを、α−2,3−シアリルトランスフェラーゼ(EC番号:2.4.99.4)を用いてシアリル化した。反応条件としては、公知の条件(Lee et al., 1994)に次の若干の変更を加えて用いた。ガングリオテトラオシルセラミド(100μg)を、Triton CF−54(2%)を含有する10μlの500mM MES(4−モルホリンエタン−スルホン酸)緩衝液(pH6.0)に溶解し、74.1μlの水で希釈して反応系とした。そこに10μlのCMP-NeuAc(15mMの水溶液)および6μlのα−2,3−シアリルトランスフェラーゼ(338mU/ml)を添加して、室温で一晩静置した。反応の進行は、C/M/0.25% KCl水溶液(50:40:10、容量比)を溶出液としたTLCで分析した。次いで反応混合液から水分を窒素気流下で蒸発し、残渣をC/M/H2O混合溶媒(60:30:4.5、容量比)(2ml)に溶解し、Sephadex G-25を充填した小さいカラム(約2.5×0.6cm)(5mlの上記の溶媒系で予め洗浄したもの)にアプライした。糖脂質材料を2.5mlの上記の混合溶媒および2.5mlのC/M(2:1、容量比)で溶出した。最後に、溶出液から水分を蒸発し、少量のC/M/H2O(60:30:4.5)に溶解した。
【0158】
ヘリコバクター ピロリ株
ヘリコバクター ピロリCCGU 17874株はスウェーデン国、イェーテボリ大学カルチャーコレクション(Culture Collection University of Goteborg)から入手したものであり、ヘリコバクター ピロリ032株はスウェーデン国、ルンド大学医療微生物学科(Department of Medical Microbiology, Lund University)のT. Wadstrom教授より寄贈されたものである。ハムF12液体培地およびブルセラ寒天培地での細菌の培養は、それぞれ公知の条件(Miller-Podraza et al., 1996)で行った。本明細書においては、S−3−PG(直鎖構造)に対する結合を示す菌体として、寒天プレートで培養したCCGU 17874株を用い、PGC(分岐構造)に対する結合を示す菌体として、液体培地で培養した032株を用いた。
【0159】
結果
調製したガングリオシド類 − ヘリコバクター ピロリのガングリオシド結合選択性の分析を可能にするため、ガングリオシドライブラリーを構築した(構築したガングリオシドライブラリーを表1にまとめた)。各ガングリオシドを質量分析法およびプロトンNMRにより特徴付けた。分析方法は、ヒト赤血球のヘリコバクター ピロリ結合性ガングリオシドの1種(表1のNo.19)の単離と特徴付けに関する以下の記載によって詳細に説明する。
【0160】
総酸性グリコスフィンゴ脂質を、血液型Bの赤血球のプール500リットルから標準的な手順(Karlsson, 1987)で単離し、5.5gの総酸性グリコスフィンゴ脂質を得た。そこから得た副画分390mgを、700mlのDEAE-Sepharoseカラムと、酢酸アンモニウムのメタノール溶液2100mlを用いた酢酸アンモニウムの直線濃度勾配(0.05M〜0.45M)によって分離した。各10mlの画分を回収し、レゾルシノール試薬を用いた薄層クロマトグラフィーにより分析した。主要化合物の移動度に基づいて画分をプールした。画分62〜67をプールすると15.4mgの酸性グリコスフィンゴ脂質が得られ、このプール画分はシアリル−ネオラクトテトラオシルセラミドの領域に移動する主要化合物を含んでいた。しかし、ヘリコバクター ピロリ結合能をクロマトグラム結合アッセイを用いて試験した時に、結合能を示す遅移動性の化合物が検出された。この15.4mgの酸性グリコスフィンゴ脂質を、クロロフォルム/メタノール/水(60:35:8〜40:40:12、容量比)の直線濃度勾配を用いたHPLCによりさらに分離した。画分39〜56にヘリコバクター ピロリ結合性化合物が溶出し、これらをプールして0.9mgの化合物を得た。
【0161】
糖脂質構造の特徴を図1〜5に示すが、さらなる詳細は今後、別途発表する。
【0162】
ガングリオシドライブラリーに対するヘリコバクター ピロリの結合
クロマトグラム結合実験で用いたヘリコバクター ピロリ CCUG 17874株およびJ99株は、両方ともシアル酸結合性である(Mahdavi et al., 2002)。
【0163】
I.結合性および非結合性のガングリオシド − 単離したガングリオシドに対するヘリコバクター ピロリ株の結合実験の結果を図6〜10に示し、そのまとめを表1に示した。4μgのガングリオシドを薄層プレートにアプライしても結合が見られなかった場合、このガングリオシドを非結合性に分類した。図6に示したように、シアル酸結合性の野生株であるCCUG 17874株およびJ99株は、以下のNeuAc−末端ガングリオシドを認識した:NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−ネオラクトヘキサオシルセラミド;レーン1)、NeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−G−10ガングリオシド;レーン3)、Galα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド;レーン6)。しかし、これらに対応するNeuGc−末端異性体構造(レーン2、4および5)は認識しなかった。さらに認識されたガングリオシドは、NeuAcα3(Galβ4GlcNAcβ3)3Galβ4Glcβ1Cer(NeuAc−ネオラクトオクタアオシルセラミド、図8、レーン1〜7)、NeuAcα3Galβ4GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(VIM−2ガングリオシド、図9、レーン6〜10)およびNeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(シアリル−2量体−Lexガングリオシド、図9、レーン11〜14)であった。
【0164】
NeuAcα3−ネオラクトテトラオシルセラミド(表1のNo.2;図8、レーン1〜7)への結合も時折検出されたが、NeuAcα6−ネオラクトテトラオシルセラミド(No.3)は非結合性であり、これは過去の報告(Miller-Podraza et al., 1997、Johansson and Miller-Podraza, 1998)と一致した。NeuAcα6−を有するガングリオシドであるGalβ4GlcNAcβ6(NeuAcα6Galβ4GlcNAcβ3)Galβ4Glcβ1Cer(No.14)およびGalβ4GlcNAcβ6(NeuAcα6Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(No.15)もまた非結合性であった。NeuGcα3−ネオラクトテトラオシルセラミド(No.4)またはジシアリル−ネオラクトテトラオシルセラミド(No.5)への結合性は得られなかった。シアリル−Lexヘキサグリコシルセラミド(No.7)への結合が時折観察された。シアリル−Leaヘキサグリコシルセラミド(No.6)はCCUG 17874株には認識されなかったが、J99株はこの化合物に時折結合した。
【0165】
II.相対的結合親和性の比較 − 初めに、マイクロタイターウェル中の希釈したグリコスフィンゴ脂質類への放射線標識したヘリコバクター ピロリの結合を、結合能を示す種々のガングリオシドに関する相対的結合親和性を評価するために試みた。しかし、ここで得られた結果には再現性がなかった。そこでガングリオシド類の希釈物を用いた結合アッセイを薄層プレート上で行った。初期実験では、NeuAcα3−ネオラクトヘキサオシルセラミドおよびNeuAc−2量体−Lexガングリオシドへの結合は、約100pmolで飽和に達したことを発見したので、結合曲線を得るためにより低い濃度範囲(1〜100pmol)を用いた。図8〜10に示した結果は、多数の結合アッセイのうちの代表的なものである。放射線標識バクテリアのバッチによって結合のレベルは多少変化したが、結合能を示すガングリオシドとの関係性については同じ結果が繰り返し得られた。十分に特徴付けられたガングリオシドの認識系と比較するために、薄層クロマトグラム上のGM1ガングリオシドの希釈系列に対する125I−標識コレラトキシンB−サブユニットの結合の結果を図10に示した。
【0166】
A.炭水化物鎖の長さの影響 − NeuAcα3−ネオラクトテトラオシルセラミド(表1のNo.2)、NeuAcα3−ネオラクトヘキサオシルセラミド(No.8)およびNeuAcα3−ネオラクトオクタオシルセラミド(No.10)の希釈系列に対するヘリコバクター ピロリ CCUG 17874株の結合結果には、NeuAcα3−ネオラクトオクタオシルセラミドに対する明確な選択性が見られた(図8)。
【0167】
B.分岐の影響 − 炭水化物鎖の分岐の影響を評価するために、NeuAcα3−ネオラクトヘキサオシルセラミド(No.8)、NeuAc−G−10ガングリオシド(No.16)およびG9−Bガングリオシド(No.19)(以上、表1に記載)に対するヘリコバクター ピロリCCUG 17874株の結合性を比較した。図9に示すように、G9−Bガングリオシドのβ6−結合側鎖上に存在する血液型Bの2型エピトープは、直鎖状のNeuAcα3−ネオラクトヘキサオシルセラミドと比べて結合を減少させた。一方、NeuAc−G−10ガングリオシドは好ましいリガンドであったため、この結果はβ6−結合した側鎖上のNeuAcα3Galβ4GlcNAc配列が結合親和性を増加させることを示す。
【0168】
C.フコース残基の影響 − フコース側鎖の影響を調べるために、NeuAcα3−ネオラクトヘキサオシルセラミド(No.8)、VIM−2ガングリオシド(No.12)およびシアリル−2量体−Lexガングリオシド(No.13)に対するヘリコバクター ピロリCCUG 17874株の相対的な結合性を評価した。図10に示すように、この細菌は、NeuAcα3-ネオラクトヘキサオシルセラミドと比較して、VIM−2ガングリオシドおよびシアリル−2量体−Lexガングリオシドより高い親和性で結合した。
【0169】
末端NeuNAcα3LacNAc−構造の詳細なエピトープ分析
1.直鎖状炭水化物に関連した結合エピトープ
S−3−PG
化学誘導体のモデル化合物として選択したS−3−PGは、オーバーレイアッセイにおいてヘリコバクター ピロリと結合した、ヒト好中球の最も単純なガングリオシドであった(図11)。図に示したように、ヒト好中球には細菌に対する結合親和性を有するガングリオシドの混合物が含まれており、より複雑な移動度の遅い種ほど高い結合強度を示すようである。TLCプレート上の典型的な実験で陽性反応を得るのに必要なS−3−PGの最小量は、1×7mmのスポットあたり20〜30pmol(2.8〜4.2pmol/mm2に相当)であった。ヘリコバクター ピロリのいくつかのバッチにおいては、S−3−PG検出の感度はさらに高く、0.3〜0.4pmol/mm2のレベルに達した。
【0170】
化学的に修飾されたS−3−PG
ヘリコバクター ピロリとの相互作用における、グリセロール尾部およびNeuAcのカルボキシル基の重要性を試験するために、S−3−PGを種々の方法で化学的に修飾した。具体的には、次の誘導化を行った:(a)シアル酸グリセロール尾部の過ヨウ素酸による穏やかな酸化とそれに続く還元、あるいは過ヨウ素酸による穏やかな酸化とそれに続くメチルアミンまたはエタノールアミンとのカップリング、(b)カルボキシル基の第一アルコールへの還元、(C)カルボキシル基の種々のアミドへの転化、および(d)ラクトンの合成。こうして得られた誘導体を、その構造の特性を確認するために陰イオンFAB MSで分析した(図12参照)。全ての分子イオンの質量が予想質量と一致し、変更はシアル酸残基に限られていた(表2)。後者は、糖鎖主鎖(core chain)中の糖配列に帰属するm/z1339、1176、973および811に観測されるフラグメントイオン(現行の命名法によるとYシリーズイオン;Domon and Costello, 1988, Harvey, 1999)に変化がないことによって示された。破壊されていないセラミド部分の存在はm/z649に観測されたフラグメントイオン(Y0イオン)(18:1−24:0)により示された。いずれの分子イオンもサテライトイオン(−28質量単位)と共に出現し、サテライトイオンの出現は、d18:1−22:0で表されるセラミドがS−3−PG調製物中に少量存在するためである(例えば、図2A中のm/z1629と1601や、図2C中のm/z1582と1554)。唯一の例外は図2Bであり、m/z1599と1569に観測されるイオンは、m/z1629に観測される主成分(図2A)に由来する2種の異なる誘導体を表している。図2Bにおいては、d18:1−22:0成分に帰属されるイオンはバックグラウンドに溶け込んでいた。
【0171】
放射線標識したヘリコバクター ピロリでTLCプレートをオーバーレイすることにより、S−3−PGから得た全ての誘導体について、ヘリコバクター ピロリによる結合性を試験した。得られた結果を表2にまとめた。結合性の実験の一例を図13に示す。この図に見られるように、シアル酸残基に加えた修飾の大部分は、S−3−PGに対するヘリコバクター ピロリの結合性を喪失させるか、あるいは劇的に減少させた。S−3−PGのアミド誘導体およびベンジルアミド誘導体に対する結合は若干見られた。これらの結合の一部はより多くの糖脂質材料を必要とした。いくつかのプレート上では、酸化/還元SPGとの相互作用も見られたが(結果は示さない)、これはおそらくこのような調製物中に誘導化されていないSPGが痕跡量存在するためであろう。さらに、グリセロール尾部の重要性を分析するために、S−3−PGを酸化し、メチルアミンまたはエタノールアミンとカップリングした。得られた2種の主要な誘導体はR-CH2-NH-CH3 および R-CH2-NH-CH2-CH2OHであった。この2種の誘導体はいずれもTLCプレート上の結合分子としては不活性であることが判明した。対照的に、S−3−PGのオクタデシルアミドには強い結合性が見られ、TLCプレート上でより低いpmolレベルでヘリコバクター ピロリと相互作用した(図14)。さらに、シアル酸を過ヨウ素酸でアルデヒドに酸化し、オクタデシルアミンで還元的にアミノ化したシアル酸の7位の炭素の誘導体は、ヘリコバクター ピロリに非常に効果的に結合することが分かった。さらに、修飾を過ヨウ素酸による穏やかな酸化およびアルデヒドの還元により行った場合には、1位の炭素のアミド誘導体は、シアル酸のグリセロール尾部の短縮を許容することが分かった。このデータは、より高い活性を示す誘導体や類似体を、シアル酸よりも単純な環状構造(好ましくは6員環であるピラノース様環状構造)から製造または設計可能であることを示している。
【0172】
他の糖脂質およびネオ糖脂質
種々の天然ガングリオシドおよびネオガングリオシドのパネルをヘリコバクター ピロリとの相互作用に関して試験した(表3)。ガングリオ系のガングリオシド、S−6−PG、NeuAcα8NeuAcα3−PG、またはラクト(Galβ3GlcNAcβ3Galβ4Glc)主鎖に基づくシアリル化オリゴ糖から調製したネオガングリオシドに対するヘリコバクター ピロリの結合は見られなかった。一方、糖がHDAまたは分岐脂質鎖とカップリングしたS−3−PGは、同じオーバーレイ条件下で活性を示した。本発明者らの研究室で合成したネオ糖脂質へのヘリコバクター ピロリの結合を図15に示す。主要なバンドに含まれる物質の構造は、プレートからバンドに相当する部分をかき取り、質量分析により確認した。レーン2の矢印はそれぞれHDA誘導体(下のバンド)および分岐状誘導体(上の2重のバンド)に相当するバンドを示す。これらの画分のFABスペクトルを図16に示す。分子イオンおよびフラグメントイオンは予想通りだった。上記バンドに対応する、図15のレーン3〜5の画分についても、同様のFABスペクトルを得た。結合系列(binding series)に含まれる他のシアリル化分子を少量含有するSPG調製物から、糖であるNeuAcα3Galβ4GlcNAcβ3Galβ4Glcを得た(図15のレーン1、図11も参照)。このような微量の結合画分が誘導化後でさえ観測された(図15のレーン2)。実験を繰り返すことにより、TLCプレート上のHDAネオ糖脂質に対するヘリコバクター ピロリの結合の再現性は、分岐脂質鎖を有する誘導体よりも低いことが分かった。
【0173】
2.PGCの分岐ポリ−N−アセチルラクトサミン鎖に関連した結合エピトープ
上記で説明したとおり、この選択的な結合性は、液体培地で培養したヘリコバクター ピロリの数株が発現した。例として、PGCへは結合するがS−3−PGへは結合しないことを示す図18を参照されたい。典型的な実験において、TLCプレート上で陽性反応を得るのに必要なPGCの最小量は0.16pmol NeuAc/mm2であった。PGCのNeuAcのグリセロール尾部を過ヨウ素酸による穏やかな酸化とそれに続く還元(R-CHOH-CHOH-CH2OH → R-CHOH-CH2OH/R-CH2OH)またはエタノールアミンとのカップリング(R-CHOH-CHOH-CH2OH → R-CH2-NH-CH2-OH)により修飾し、カルボキシル基を還元(R-COOH → CH2OH)により修飾した。修飾したPGCをパーメチルしてEI MSにより分析した結果を図17に例示した。図17には、還元前(図6A)および還元後(図6B)の末端シアリル酸残基に対応するイオンが観察された。予想通り、NeuAcに帰属するm/z376および344に観測されたフラグメントイオンは、還元したNeuAcに帰属する、m/z362および330に観測されるイオンで置換されていた。
【0174】
放射線標識細菌によるTLCプレートのオーバーレイを用いた結合試験は、修飾がPGCとヘリコバクター ピロリとの相互作用に負の影響を与えることを示した。修飾したPGCへのヘリコバクター ピロリの結合の一例を図18に示し、その結果を表4にまとめた。
【0175】
考察
ある種のヘリコバクター ピロリ株によるシアル酸含有糖結合体の認識はこれまで繰り返し報告されている(Roche et al., 2001、Mahdavi, et al., 2002、Miller-Podraza et al, 1997、Johansson and Miller-Podraza 1998)。本願の実験では、ガングリオシドのライブラリーを収集し、代表的なシアル酸認識ヘリコバクター ピロリを用いてヘリコバクター ピロリの結合選択性を分析した。
【0176】
シアリル−Leaヘキサグリコシルセラミドに対するJ99株の結合が時折観察されたが、CCUG17874株はこのガングリオシドを認識しなかった。また、シアリル−Leaネオ糖タンパク質およびシアリル−Lexネオ糖タンパク質の両方に対するJ99株の結合も報告されている(Mahdavi, 2002)。このことは、J99株とCCUG17874株ではSabA炭水化物結合部位が同じではないことを示している。しかし、その他の全ての点において、CCUG17874株とJ99株はガングリオシドに同一の様式で結合し、いずれの株も、末端NeuAcα3を有する、N−アセチルラクトサミンをベースにしたガングリオシドを認識したが、末端NeuAcα6を有するものは認識しなかった。この結果は従来の報告(Miller-Podraza et al, 1997、Johansson and Miller-Podraza, 1998)と一致していた。また、末端NeuGcα3または末端NeuAcα8NeuAcα3を有するガングリオシドは認識されなかった。
【0177】
結合親和性に影響を及ぼす要因として、i) N−アセチルラクトサミン炭水化物鎖の長さ、ii) 炭水化物鎖の分岐、およびiii) N−アセチルラクトサミン主鎖のフコース置換、の3つを明らかにした。
【0178】
1.N−アセチルラクトサミン主鎖の長さ:NeuAcα3−ネオラクトヘキサオシルセラミドおよびNeuAcα3−ネオラクトテトラオシルセラミドよりも、NeuAcα3−ネオラクトオクタオシルセラミドに対してヘリコバクター ピロリが選択的に結合することを観察した。このような効果は、炭水化物の頭部基(carbohydrate head group)がより長い主鎖に提示された場合、その基に対する接触性が向上するからである可能性が最も高い。
【0179】
2.二価性:2つのNeuAcα3Galβ4GlcNAcβ側鎖を有するNeuAc−G−10ガングリオシドに対して観察される、直鎖状のNeuAcα3−ネオラクトヘキサオシルセラミドに対するよりも高い親和性は、共同的結合によると考えられる。このことは、シアリルラクトース(NeuAcα3Galβ4Glc)の多価アルブミン結合体が、一価のシアリル−ラクトースよりも上皮単層へのヘリコバクター ピロリの付着を効果的に阻害することを実証したSimon et al. (1997)の報告と合致する。本発明は、シアリルラクトサミンがオリゴ価または多価に提示されることによって、このエピトープが効果的に提示されることを示す。
【0180】
NeuAcα3−ネオラクトヘキサオシルセラミドに比べ、G9−Bガングリオシドに対する結合親和性がより低いことは、β6結合した側鎖上の血液型B決定因子が結合作用に干渉したことを示す。G9−Bガングリオシドの検知レベルは約100pmolだったため、完全な障害ではない。このことは、ヘリコバクター ピロリ結合決定因子は、Galα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer構造体のβ3軸上に主として露出されていることを示唆している。
【0181】
3.N−アセチルラクトサミン主鎖に提示されたフコース側鎖:NeuAcα3−ネオラクトヘキサオシルセラミドに比べ、VIM−2ガングリオシドに対する結合親和性がより高いことは、最も内側に位置するGlcNAcにα3結合しているフコースが、シアリル−2量体−Lexの高親和性結合に関与することを示唆する。このフコース残基は、SabA付着因子の炭水化物結合部位と相互作用するか、あるいはガングリオシドの立体構造に影響を及ぼして炭水化物の頭部基の提示を最適化すると考えられる。しかし、この問題を解明するには、SabA付着因子の発現と結晶化を待たねばならない。
【0182】
SabA付着因子の発現は、Leb結合性のBabA付着因子とは異なり、相変異にさらされる(Mahdavi et al., 2002)。結合アッセイに用いた菌体群の中でSabA付着因子を発現する細菌の数が変化することが、あるガングリオシドに対する結合性の絶対的な親和性を測定することが困難な理由であると考えられる。しかし、繰り返し行った結合アッセイにおいては、結合能を有するガングリオシド間で同じ関係性が認められた。しかし、最適条件下では、NeuAcα3−ネオラクトヘキサオシルセラミドに対するヘリコバクター ピロリの結合が、GM1ガングリオシドに対するコレラトキシンB−サブユニットの結合に匹敵することは注目すべき知見である。コレラトキシンの結合は、非常に強力なタンパク質−炭水化物相互作用の一例である。
【0183】
上記知見の生物学的重要性は、さらなる研究が必要である。ヘリコバクター ピロリの主要ターゲット組織、即ちヒトの胃上皮組織、のシアル酸含有量は非常に低い(Madrid et al., 1990)。しかしながら、最近になって、胃に炎症が起きるとシアル酸含有糖結合体の発現が上昇するように調節されることが発表された(Mahdavi et al., 2002)。さらに、数種類のヘリコバクター ピロリ結合性ガングリオシドもヒトの好中球に存在し(Miller-Podraza et al., 1999、Stroud et al., 1996a, 1996b)、最近の研究によると、レクチノファゴサイトーシス(lectinophagocytosis)によってヒトの好中球の非オプソニン性ヘリコバクター ピロリ誘導活性化(即ち、細菌性付着因子による好中球細胞表面のシアリル化糖結合体の認識)が生じると、食作用、そして反応性酸素代謝産物の産生を伴う酸化的バーストが起こる(Teneberg, et al., 2001)。このように、ヘリコバクター ピロリのシアル酸結合能力には2つの役割がある。即ち、一方ではすでに病理に冒された胃の上皮への細菌の付着を仲介し、もう一方では好中球を活性化して反応性酸素代謝産物の産生を伴う酸化的バーストを起こし、生物学的活性を有する酵素を放出して組織破壊にさらなる拍車をかけることである。
【0184】
1.炭水化物鎖に提示された末端NeuNAcα3LacNAcに対するヘリコバクター ピロリの結合
調製が比較的容易な糖脂質であり、十分に定義された構造を有することから、本研究のモデル化合物としてS−3−PGを選んだ。In vitroにおける数種のヘリコバクター ピロリ株によるシアル酸含有糖結合体の認識は、種々の文献で議論されており(Karlsson, 1998、Karlsson, 2000、Miller-Podraza et al., 1997a、Johansson and Miller-Podraza, 1998、Miller-Podraza et al., 1996、Miller-Podraza et al., 1997b、Evans et al., 1988、Hirmo et al., 1996、Simon et al., 1997)、本願に示す結果は、これらの発表内容と合致し、α3結合したNeuAcとβ4結合したGalに対するヘリコバクター ピロリの選択性をはっきりと明示した。NeuAcα3Galβ3GlcNAc-R と NeuAcα3Galβ3GalNAc-Rが不活性であることから、ネオラクト構造が明らかに必要であった。しかしながら、シアリルラクトースであるNeuAcα3Galβ4Glcがシアリル化構造とヘリコバクター ピロリとの相互作用に対して弱い阻害性を示すことが他のグループによって報告されているので(Evans et al., 1988、Hirmo et al., 1996、Simon et al., 1997)、GlcNAc自体は結合に絶対的に必要なものではないと考えられる。また、(シアリル−ルイスxオリゴ糖および糖脂質を用いて試験した)GlcNAcに結合したα3Fucおよび脱−N−アセチル化などのGlcNAcの他の修飾(Johansson and Miller-Podraza、出版準備中)は結合を喪失させることはない。従って本発明は、より長いヘリコバクター ピロリ結合性オリゴ糖においては、その活性を損失させることなくGlcまたはGlcNAcの2位の炭素を修飾することも可能であることを示す。
【0185】
S−3−PGを誘導化とそれに続く結合試験により、ヘリコバクター ピロリの結合はグリセロール尾部とカルボキシル基に提示されたシアリル化エピトープに依存することが分かった。カルボキシル基の重要性はCOOHの還元およびアミドの合成によって実証された。遊離アミドおよびベンジルアミドは、TLCプレート上でヘリコバクター ピロリに対する結合能をいくらか示したが、これらの反応は常に再現性があるわけではなく、親和性の減少を示した。ベンジルアミド中の大きな疎水性部分と遊離アミド中の正の電荷を帯びたNH2基が非特異的結合を引き起こすことができる。一方、SPGのオクタデシルアミドへの結合性はより低いピコモルレベルであり(図4)、この結合が特異的な相互作用であることを示した。未修飾カルボキシル基はオクタデシルアミド中のアミドによって置換されてもよいという事実は、相互作用にはカルボキシル基の1つの酸素原子しか必要ではないことを示す。
【0186】
ヒトや動物の種々の糖結合体の一部であるNeuAcα3Galβ4GlcNAcが、天然のシアリル化ヘリコバクター ピロリ結合性糖の最適末端部を表していると考えられる。重要なのは、ヘリコバクター ピロリ関連感染症に積極的に関与するヒトの好中球が(Rautelin et al., 1993, Fiocca et al., 1994)、特にS−3−PGと他のネオラクトガングリオシドを豊富に含んでいる(Fukuda et al., 1985, Muthing 1996, Stroud et al., 1996)ということである。上で述べたように、S−3−PGは、TLCプレート上でヘリコバクター ピロリを結合した、ヒト好中球由来のガングリオシドの中で最も単純なものである。図11は、より複雑な構造のガングリオシドに対して比較的強い結合が生じることを示す。TLC表面でエピトープがより効果的に提示されていることがこの強い相互作用の最も大きな理由であると考えられる。研究段階で、糖鎖長および脂質部分の構造が結合に影響を及ぼすかもしれないことに気が付いた。例えば、ヘキサデシルアニリンを有するネオ糖脂質へのヘリコバクター ピロリの結合は、分岐脂質鎖を有するネオ糖脂質への結合よりも再現性が低く、また、TLCプレート上の結果(表11参照)から判断すると、シアロネオヘキサオシルセラミドに対する結合はシアロネオテトラオシルセラミドに対する結合よりも強い。しかし、他の要因、例えば繰返しエピトープ単位、フコース側鎖または他の置換基なども考慮に入れる必要がある。より大きな種に強く結合するのは、長く伸びた主鎖の末端領域と内部領域の両方を認識する様々な細菌付着因子の結合の組み合わせによるものであると考えられる。ヘリコバクター ピロリは、シアリル化糖鎖および中性糖鎖の両者に関与する様々な結合特異性を示すことが分かっている(Karlsson 1998)。近年、Roche N. et al. によって、ヒトの胃癌細胞から調製した繰返しラクトサミン単位を有するガングリオシドへのヘリコバクター ピロリの結合が報告されている(Roche et al., 2001)。
【0187】
2.分岐ポリ−N−アセチルラクトサミン鎖に関与する結合エピトープ
本発明者らは以前に、ヘリコバクター ピロリ結合性PGC中のNeuAcは1つのラクトサミン単位(NeuAc-Hex-HexNAc)に基づく短い側鎖に関係があることを明かにした。このことは、エンド−β−ガラクトシダーゼでPGCを消化し、遊離したオリゴ糖断片を様々な質量分析技術を用いて分析することにより証明された(Karlsson et al., 2000)。分岐したPGC中にNeuAcα3Galβ4GlcNAcが存在することは、複数の異種の糖の間で水素結合を形成するための好的な条件を提供するので、NeuAcのグリセロール尾部の9位の炭素と、隣接する2本の側鎖中の各GlcNAcとの間に相互作用に基づいて新しい結合エピトープが発生するという仮説が立てられた(Angstrom et al., 投稿済み)。このような仮説は、相互作用におけるグリセロール尾部の重要性にも合致する上、シアリル化構造に対するヘリコバクター ピロリの結合様式には異なる2種類の様式が明かに存在することの説明にもなる。本明細書は、グリセロール基の重要性に関する本発明者らの過去の知見(Miller-Podraza et al., 1996)を確立させると同時に、カルボキシル基さえも結合にとって非常に重要であることを示す。COOHの存在によって、ラクトン化または他の可逆的修飾によって、in vivoで素早く結合性のオンとオフを切り替えられる可能性が示されることから、この知見は非常に興味深い。
【0188】
【表1】

【0189】
a)結合強度は以下のように定義した:「+++」は薄層クロマトグラムにグリコスフィンゴ脂質を0.5μg未満アプライした場合に結合が検出されたことを示し;「+」は0.5μgアプライした場合に時折結合が検出されたことを示し;「−」は4μgアプライした場合でも結合が検出されないことを示す。
【0190】
【表2】
【0191】
【表3】
【0192】
【表4】
【0193】
参考文献
Boren, T., Falk, P., Roth, K. A. and Normark, S. (1993) Science 262, 1892-1895.
Domon, B. and Costello, C. E. (1988) Glycoconj. J. 5, 397-409
Evans, D.G., Evans, D.J., Jr., Moulds, J.J. and Graham, D.Y. (1988) Infect. Immun. 56(11): 2896-2906.
Fiocca, R., Luinetti, O., Villani, L., Chiaravalli, A.M., Capella, C. and Solcia, E. (1994) Scand. J. Gastroenterol. 205(29S):11-21.
Fukuda, M.N., Dell, A., Oates, J.E., Wu, P., Klock, J.C. and Fukuda, M. (1985) J. Biol. Chem. 260:1067-1082.
Harvey, D.J. (1999) Mass Spectrometry Reviews 18: 349-450.
Hirmo, S., Kelm, S., Schauer, R., Nilsson, B. and Wadstrom, T. (1996) Glycoconjugate J. 13:1005-1011.
Ilver, D. et al. (1998) Science 279, 373-.
Ito, M. and Yamagata, T. (1989) Methods Enzymol. 179:488-496.
Johansson, L., Miller-Podraza, H. (1998) Anal. Biochem. 265, 260-268.
Karlsson, K.-A. (1987) Meth. Enzymol. 138, 212-220
Karlsson, K.-A. (1974) Biochemistry 13, 3643-3647
Karlsson, K.-A. (1998) Mol. Microbiol. 29:1-11.
Karlsson, K.-A. (2000) Glycobiology 10(8): 761-771.
Karlsson, H., Larsson, T., Karlsson, K.-A. and Miller-Podraza, H. (2000) Glycobiology 10(12):1291-1309.
Karlsson, K.-A. and Stromberg, N. (1987) Meth. Enzymol. 138, 220-231
Koerner, T., A.,W., Jr., Prestegard, J., H., Demou, P. C. and Yu, R. K. (1983) Biochemistry 22, 2676-2687
Larsson, G., Karlsson, H., Hansson, G. C. and Pimlott, W. (1987) Carbohydr. Res. 161, 281-290
Lanne, B., Uggla, L., Stenhagen, G. and Karlsson, K.-A. (1995) Biochemistry 34:1845-1850.
Lee, Y.-C., Kojima, N., Wada, E., Kurosawa, N., Nakaoka, T., Hamamoto, T. and Tsuji, S. (1994) J. Biol. Chem. 269(13): 10028-10033.
Lingwood, C. A., Huesca, M. and Kuksis, A. (1992) Infect. Immun. 60, 2470-2474
Madrid, J. F., Ballesta, J., Castells, M. T. and Hernandez, F. (1990) Histochemistry 95, 179-187
Magnusson, G., Chernyak, A.Y., Kihlberg, J. and Kononov, L.O. Synthesis of neoglycoconjugates. In: Neoglycoconjugates, Preparation and applications, (Y. C. Lee and Reiko, T. Lee, eds.), Academic Press, Inc., A Division of Harcourt Brace & Company, San Diego 1994, pp. 53-117.
Mahdavi, J., Sonden, B., Forsberg, L., Hurtig, M., Olfat, F. O., Forsberg, L., Roche, N., ・ngstrom, J., Larsson, T., Teneberg, S., Karlsson, K.-A., Altraja, S., Wadstrom, T., Kersulyte, D., Berg, D. E., Dubois, A., Petterson, C., Magnusson, K.-E,. Norberg, T., Lindh, F., Lundskog, B. B., Arnqvist, A., Hammarstrom, L., Boren, T. (2002) Science 297, 573-578
Miller-Podraza, H., Mansson, J.-E. and Svennerholm, L. (1992) Biochim. Biophys. Acta 1124:45-51.
Miller-Podraza, H., Andersson, C. and Karlsson, K.-A. (1993) Biochim. Biophys. Acta 1168: 330-339
Miller-Podraza, H., Abul Milh, M., Bergstrom, J. and Karlsson, K.-A. (1996) Glycoconjugate J. 13: 453-460.
Miller-Podraza, H., Abul Milh, M., Teneberg, S. and Karlsson, K.-A. (1997a) Infect. Immun. 65, 2480-2482
Miller-Podraza, H., Bergstrom, J., Abul Milh, M. and Karlsson, K.-A. (1997b) Glycoconjugate J. 14:467-471.
Miller-Podraza, H., Bergstrom, J., Teneberg, S., Abul Milh, M., Longard, M., Olsson, B.-M., Uggla, L. and Karlsson, K.-A. (1999) Infect. Immun. 67, 6309-6313
Muthing, J. (1996) Carbohydrate Research.290:217-224.
Mysore, J.V., Wiggington, T., Simon, P.M., Zopf, D., Heman-Ackah, L.M. and Dubois, A.
(1999) Gastroenterology, 117, 1316-1325
Rautelin, H., Blomberg, B., Fredlund, H., Jarnerot, G. and Danielsson, D. (1993) Gut 34:599-603.
Read, B., Demel, R.A., Wiegandt, H. and Van Deenen, L.L.M. (1977) Biochim. Biophys. Acta, 470:325-330.
Roche, N., Angstom, J., Larsson, T. and Teneberg, S. (2001) Glycobiology 11, 935-944
Samuelsson, B. E., Pimlott, W. and Karlsson, K.-A. (1990) Meth. Enzymol. 193, 623-646
Simon, P. M., Goode, P. L., Mobasseri, A. and Zopf, D. (1997) Infect. Immun. 65, 750-757
Stellner, K., Saito, H. and Hakomori, S.-i. (1973) Arch. Biochem. Biophys. 155, 464-472
Stroud, M. R., Handa, K., Salyan, M. E. K., Ito, K., Levery, S. B., Hakomori, S.-i., Reinhold, B. B. and Reinhold, V. N. (1996a) Biochemistry 35, 758-769
Stroud, M. R., Handa, K., Salyan, M. E. K., Ito, K., Levery, S. B., Hakomori, S.-i., Reinhold, B. B. and Reinhold, V. N. (1996b) Biochemistry 35, 770-778
Stults, C. L. M., Sweeley, C. C. and Macher, B. A. (1989) Meth. Enzymol. 179, 167-214
Svennerholm, L. (1963) J. Neurochem. 10, 613-623
Teneberg, S., Jurstrand, M., Karlsson, K.-A. and Danielsson, D. (2000) Glycobiology 10, 1171-1181
Teneberg, S., Miller-Podraza, H., Lampert, H. C., Evans Jr., D. J., Evans, D. G., Danielsson, D., and Karlsson, K.-A. (1997) J. Biol. Chem. 272, 19067-19071
Teneberg, S., Leonardsson, I., Karlsson, H., Jovall, P.-A., Angstrom, J., Danielsson, D., Naslund, I., Ljungh, A., Wadstrom, T., and Karlsson, K.-A. (2002) J. Biol. Chem. 277, 19709-19719
Veh, R.W., Cornfield, A.P., Sander, M. and Schauer R. (1977) Biochim. Biophys. Acta 486:145-160.
Waldi, D. (1962) in Dunnschicht-Chromatographie (Stahl, E., ed.) pp. 496-515. Springer-Verlag, Berlin
Yang, H. and Hakomori, S.-i. (1971) J. Biol. Chem. 246, 1192-1200
Angstrom, J., Teneberg, S. and Karlsson, K.-A. (1994) Proc. Natl. Acad. Sci. USA 91, 11859-11863.
【図面の簡単な説明】
【0194】
【図1】ヒト赤血球由来ガングリオシドの陰イオンFAB MSスペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。2種類のスペクトルAとB、具体的には、加速電圧を10kVとしたスペクトルAと、加圧電圧を8kVとしたスペクトルBをそれぞれ図1Aおよび図1Bに示す。スペクトルAにおいては、100〜2400質量単位に渡ってスキャニングし、一方のスペクトルBにおいては、2000〜3200質量単位に渡ってスキャニングし、2.4分間のデータを回収した。
【図2】パーメチル化したヒト赤血球由来ガングリオシドの、電子衝突イオン化マススペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。スペクトルの記録温度は380℃とした。m/z 354に観測されたピークは混入物に帰属されるものである。
【図3】パーメチル化したヒト赤血球由来ガングリオシドの、エレクトロスプレーイオン化質量分析および衝突誘起解離によって得たマススペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。
【図4】パーメチル化と還元処理を行ったヒト赤血球由来ガングリオシドの、電子衝突イオン化マススペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。スペクトルの記録温度は340℃とした。
【図5】ヒト赤血球由来ガングリオシド(30℃)の、600MHzにおけるプロトンNMRスペクトル。試料は、重水素置換後に、ジメチルスルホキシド/D2O(98:2、容量比)に溶解した。約4.8ppmを中心とする幅広のピーク(記号*で示した)は、未同定の混入物に帰属されるものである。
【図6】末端NeuAcを有するガングリオシド、および末端NeuGcを有するガングリオシドに対するヘリコバクター ピロリの結合。Aはアニスアルデヒドによる化学的検出結果であり、BとCはそれぞれ、35Sで標識したヘリコバクター ピロリのCCUG 17874株とJ 99株の結合によって得られたオートラジオグラムである。ガングリオシドは、アルミニウムで裏打ちしたシリカゲルプレート上で、クロロホルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)を溶媒系として分離し、「実施例」に記載の方法で結合アッセイを行った。レーンは以下の通りである: ヒト肝細胞癌由来のNeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-ネオラクトヘキサオシルセラミド)、2μg(レーン1); ウサギ胸腺由来のNeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuGc-ネオラクトヘキサオシルセラミド)、2μg(レーン2); ヒト赤血球由来のNeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-G-10型ガングリオシド)、2μg(レーン3); ウシ赤血球由来のNeuGcα3Galβ4GlcNAcβ6(NeuGcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuGc-G-10型ガングリオシド)、2μg(レーン4); ウシ赤血球由来のGalα3Galβ4GlcNAcβ6(NeuGcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer、2μg(レーン5); ヒト赤血球由来のGalα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド)、2μg(レーン6)。オートラジオグラフィーは12時間行った。
【図7】ヘリコバクター ピロリのCCUG 17874株とJ 99株の結合の比較。Aはアニスアルデヒドによる化学的検出結果であり、BおよびCはそれぞれ、35Sで標識したヘリコバクター ピロリのCCUG 17874株およびJ 99株の結合によって得られたオートラジオグラムである。ガングリオシドは、アルミニウムで裏打ちしたシリカゲルプレート上で、クロロホルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)を溶媒系として分離し、「実施例」に記載の方法で結合アッセイを行った。各レーンは以下の通りである: ヒト好中球由来のガングリオシド、20μg(レーン1); ウサギ胸腺由来のNeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuGc-ネオラクトヘキサオシルセラミド)、2μg(レーン2); ヒト肝細胞癌由来のNeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-ネオラクトヘキサオシルセラミド)、1μg(レーン3); ヒト赤血球由来のNeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−G−10ガングリオシド)、1μg(レーン4); ヒト赤血球由来の Galα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド)、1μg(レーン5); ヒト赤血球由来のGalα3(Fucα2)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(B6 2型ヘキサグリコシルセラミド)、4μg(レーン6); マウス糞由来のGalβ3GalNAcβ4Galβ4Glcβ1Cer(ガングリオテトラシルセラミド)、4μg(レーン7)。オートラジオグラフィーは12〜24時間行った。
【図8】段階希釈したガングリオシドに対するヘリコバクター ピロリの結合。Aは、クロマトグラム結合アッセイを用いた、ヘリコバクター ピロリ CCUG 17874株の結合によって得られたオートラジオグラムである。レーン1〜7はそれぞれ、以下の糖の段階希釈系列(1〜100pmol)である: NeuAcα3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトテトラオシルセラミド)、 NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトヘキサオシルセラミド)および NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトオクタオシルセラミド)。レーン8は、1mmolのNeuAcα3Galβ3(Fucα4)GlcNAcβ3Galβ4Glcβ1Cer(シアリルLeaヘキサグリコシルセラミド)である。結合アッセイは、「実施例」に記載の方法で行った。3回の実験のうち、最も代表的な実験結果をAに示す。Bは,デンシトメトリーによって結合を定量化した結果である。Aに示したオートラジオグラムはNIHイメージプログラムで分析した。
【図9】段階希釈したガングリオシドに対するヘリコバクター ピロリの結合。Aは、クロマトグラム結合アッセイを用いた、ヘリコバクター ピロリ 17874株の結合によって得られたオートラジオグラムであり、レーンは以下の段階希釈系列である: NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-ネオラクトヘキサオシルセラミド)、(10〜100pmole)(レーン1〜5); NeuAcα3Galβ4GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(VIM−2ガングリオシド)、(10〜100pmole)(レーン6〜10); NeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-2量体-Lexガングリオシド)、(10〜100pmole)(レーン11〜14); NeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(シアリルLexヘキサグリコシルセラミド)、1mmole(レーン15)。結合アッセイは、「実施例」に記載の方法で行った。3回の実験のうち、最も代表的な実験結果をAに示す。Bは,デンシトメトリーによって結合を定量化した結果である。Aに示したオートラジオグラムはNIHイメージプログラムで分析した。
【図10】段階希釈したガングリオシドに対するヘリコバクター ピロリの結合。Aは、クロマトグラム結合アッセイを用いた、ヘリコバクター ピロリ CCUG 17874株の結合によって得られたのオートラジオグラムであり、以下の糖の段階希釈系列(1〜100pmol)を使用した: NeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-Lex)、 NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトヘキサオシルセラミド)、 NeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−G−10ガングリオシド)およびGalα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド)。結合アッセイは、「実施例」に記載の方法で行った。3回の実験のうち、最も代表的な実験結果をAに示す。Bは,デンシトメトリーによって結合を定量化した結果である。Aに示したオートラジオグラムはNIHイメージプログラムで分析した。比較のために、薄層クロマトグラム上の段階希釈したGM1ガングリオシドに対する、125Iで標識したコレラトキシンB−サブユニット(CTB)の結合に関するデンシトメトリーの結果もAに示した。
【図11】放射線標識したヘリコバクター ピロリを用いた、TLCプレート上の顆粒球由来ガングリオシドの検出レベル。プレートは、クロロホルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)で展開し、アニスアルデヒドを噴霧するか、または35Sで標識したヘリコバクター ピロリ CCGU 17874株をオーバーレイして視覚化した。レーン1〜10は、顆粒球由来ガングリオシドの2倍希釈系列(レーン1には合計12μgをアプライした)であり;レーン11は、ウシ脳由来ガングリオシド、2μg(GM1、GD1a、GD1bおよびGT1bの混合物)である。図中のS−3−PGとS−6−PGはそれぞれ、NeuAcα3Galβ4GlcNAcβ3Galβ4GlcCer と NeuAcα6Galβ4GlcNAcβ3Galβ4GlcCerを表す。「7s」および「8s」という注釈は、それぞれ1分子中に7個か8個の単糖単位を含むガングリオシドに対応するクロマトグラフィー領域を示す。
【図12】シアル酸の -CH2CH2CH2OH 基および -COOH 基が誘導化されたS−3−PGの陰イオンFAB MSスペクトル。Aは未修飾のS−3−PG、Bは酸化還元されたS−3−PG、Cは酸化後メチルアミンにより誘導化したS−3−PG、Dは酸化後エタノールアミンにより誘導化したS−3−PG、Eは-COOH基を還元したS−3−PG、Fは遊離アミド、Gはメチルアミド、Hはエチルアミド、Iはプロピルアミド、Jはベンジルアミド、Kはオクタデシルアミドの結果である。
【図13】修飾したS−3−PGに対するヘリコバクター ピロリ(CCGU 17874株)の結合の一例。糖脂質をTLCプレート上で分離し、アニスアルデヒド(左側のプレート)または35Sで標識した細菌(ヘリコバクター ピロリ、右側のプレート)で視覚化した。クロマトグラフィーの条件に関しては図1を参照のこと。レーン1はS−3−PGだが、結合系列に属するより長い物質が痕跡量含まれている;レーン2はS−3−PGのエチルアミドであり;レーン3はS−3−PGのプロピルアミドであり;レーン4はS−3−PGのベンジルアミドであり;レーン5は酸化還元したS−3−PGであり(表3を参照);レーン6はヘリコバクター ピロリの正の対照であり;レーン7はウシ脳由来ガングリオシドであり(上から:GM1、GD1a、GD1b、GT1b);レーン8はスルファチド混合物であり;レーン9はウサギ胸腺由来の五糖含有ガングリオシドの混合物であり;レーン10はヒト顆粒球から調製した複合ガングリオシド混合物である。
【図14】TLCプレート上のネオ糖脂質に対するヘリコバクター ピロリ(CCGU 17874株)の結合。クロマトグラフィーの条件に関しては図1を参照のこと。左側のプレートは、アニスアルデヒドを用いて炭水化物を染色したものであり、右側のプレートは35Sで標識したヘリコバクター ピロリをオーバーレイしたものである。レーン1はS−3−PGだが、図3と同様に、SPG調製物には結合系列に属するより長い物質が痕跡量含まれている;レーン2はNeuAcα3Galβ4GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン3はNeuAcα3Galβ3GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン4はNeuAcα6Galβ4GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン5はGalβ3(NeuAcα6)GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン6はウシ脳由来ガングリオシドである(上から:GM1、GD1a、GD1b、GT1b)。レーン2に示した二つの矢印は、それぞれ、ヘキサデシルアニリン誘導体(下側のバンド)および分岐した脂質鎖を有するネオ糖脂質(上側の2重のバンド)を示す。レーン2の矢印で示した2つの画分と、それぞれに相当するレーン3〜5の画分をかき出し、マススペクトロメトリーで分析した(図6参照)。
【図15】NeuAcα3Galβ4GlcNAcβ4Galβ4Glcから誘導化したネオ糖脂質の陰イオンFAB MSスペクトル。Aはヘキサデシルアニリン誘導体の結果であり;Bは分岐した脂質部を有するネオ糖脂質の結果である。
【図16】パーメチル化ポリグリコシルセラミドのEI質量スペクトルの一例であり、NeuAcと還元NeuAcに相当するフラグメントイオンを示す。
【図17】TLCプレート上の誘導化PGCに対する、ヘリコバクター ピロリ(032株)の結合。左側のプレートは、アニスアルデヒドを用いて炭水化物を染色したものであり、右側のプレートは35Sで標識したヘリコバクター ピロリをオーバーレイしたものである。レーン1と2はヒト赤血球由来の未誘導化PGCであり;レーン3は還元PGC(COOH → CH2OH)であり;レーン4はウシ脳由来のガングリオシド(GM1、GD1a、GD1b、GT1bの混合物)であり;レーン5はS−3−PGである。(液体培養した032株は、直鎖構造を表すS−3−PGには結合しないことに注意されたい。)
【図1A】
【図1B】
【技術分野】
【0001】
本発明は、ヘリコバクター ピロリに対する新規な高親和性レセプターを開示する。このレセプターは、シアリル化ポリ−N−アセチルラクトサミン構造を含むものである。本発明は、主要な胃病原体であるヘリコバクター ピロリに対して上記物質を用いた治療方法および予防方法に関する。また本発明は、上記構造を用いた分析方法および診断方法に関する。本発明はさらに、上記構造を用いた、ヘリコバクター ピロリ感染を防止するための機能性食品の製造方法に関する。
【背景技術】
【0002】
糖タンパク質や糖脂質に提示されているシアリルラクトースおよびシアル酸は、ヘリコバクター ピロリに対するレセプターとして知られている。これらレセプターの構造は、NeuNAcα3Gal- や NeuNAcα3Galβ4Glc(シアリル−ラクトース)(US Patent Nos. 5,883,079、5,753,630および5,514,660、ならびにMysore et al., 1999)または NeuNAcα3Galβ4GlcNAc(Johansson and Miller-Podraza, 1998、Miller-Podraza et al., 1997)であると考えられている。さらに最近では、フコシル化された異型、例えば、シアリル−ルイスx(NeuNAcα3Galβ4(Fucα3)GlcNAc;sLex)、特にジフコシル化構造であるシアリル−2量体−ルイスx(sialyl-dimeric Lewis x)(NeuNAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4(Fucα3)GlcNAc;sdiLex)(Mahdavi et al., 2002)が、ヘリコバクターに対するリガンドであると考えられている。これらの構造が糖脂質に提示された場合、sdiLexはsLexよりもヘリコバクター ピロリに対して何十倍も優れたレセプターである(Mahdavi et al., 2002)。本発明は、他のより大きな非フコシル化ポリ−N−アセチルラクトサミン構造にも関する。フコシル化およびシアリル化されたポリラクトサミンは、ヘリコバクター ピロリに対して優れたレセプターである。しかしながら、比較的高い親和性を示すエピトープが非フコシル化ポリ−N−アセチルラクトサミンに提示されることもある。このような構造の利点は、5〜10工程もの追加工程を経る化学合成によってエピトープにフコースを付加する必要がない点である。フコシル化グリカンの酵素学的または発酵学的な合成は未だ商業レベルに達していないので、フコースの付加は工業的にも困難であると考えられる。
【0003】
本発明はさらに、NeuNAcα3Galβ4GlcNAc-構造がヘリコバクター ピロリを効果的に結合するためには、-β4Glc(NAc)構造を含む必要があることを示す。一般的に、他の結合基を介して結合したNeuNAcα3Galβ3GlcNAcやNeuNAcα3Galβ3GalNAcを含む類似の糖脂質エピトープは結合構造ではないことから、この知見は、末端NeuNAcα3Gal-エピトープについて示唆された結果と対照的である。
【0004】
七糖糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCerは、ヘリコバクター ピロリと結合することが報告されている(Roche. et al. 2001、Johansson and Miller-Podraza, 1998、Miller-Podraza et al., 1997)。本発明は、非フコシル化構造が、より短いNeuNAcα3Galβ4Glc(NAc)エピトープやsLexエピトープよりもヘリコバクター ピロリに対して実際により有効なレセプターであることを示し、ヘリコバクター ピロリに対するより低コストの高親和性阻害剤や診断剤が設計できる可能性を提供する。本発明は、公知のシアル酸含有レセプターの中からシアリル化ポリラクトサミンを高親和性レセプターとして見出すことを可能にする。さらに本発明は、糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCerに代表されるより長いポリラクトサミン鎖、および糖脂質であるNeuNAcα3Galβ4GlcNAcβ3(NeuNAcα3Galβ4GlcNAcβ6)Galβ4GlcNAcβ3Galβ4GlcβCerなどに代表される分岐ポリラクトサミン構造に関する。
【0005】
すでに本発明者らは、ヘリコバクター ピロリに対する2型ラクトサミン結合エピトープである「ネオラクト結合エピトープ」(-GlcNAcβ3Galβ4GlcNAcβ-)も開示している。この結合エピトープはより長い直鎖ポリ−N−アセチルラクトサミン(例えばNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer)において活性を示す。しかしながら、(シアル酸非結合種においては)脱シアリル化構造のみならず、本発明におけるシアル酸結合特異性によって認識されない、N−グリコリル−ノイラミン酸含有構造であるNeuNGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer(FI 20010118)も活性を示すことから、結合特異性にはシアル酸の存在は必要ない。一般的なネオラクトエピトープをシアル酸非結合ヘリコバクター ピロリ種によって特徴付けたときには、NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerに対する結合も見られなかった(FI20010118)。シアル酸結合特異性は、ネオラクト結合特異性よりも一般的ではないが、シアル酸結合特異性はおそらく炎症に関連し、新鮮な病理学的単離物中に存在する(Mahdavi et al., 2002)。ポリラクトサミン鎖の分岐によってネオラクトエピトープへの結合を阻害することができるが、本発明による高親和性シアル酸レセプターへの結合を阻害することはできない(FI20010118)。
【0006】
従来技術には、HPNAPと命名されたヘリコバクター ピロリ結合タンパク質も開示されており、このタンパク質は好中球の活性化に関する潜在的な機能を有すると考えられる。本願のデータは、シアル酸結合活性を示すNAP−タンパク質が、シアリル化構造に結合するヘリコバクター ピロリの阻害に関与しないことを示す。より長いポリラクトサミンを含有するNeuNAcα3Galβ4GlcNAcβ3Gal-型構造への結合も含む、NAP−タンパク質のシアル酸結合作用の潜在的な機能は、ヘリコバクター ピロリと好中球との相互作用に関与すると考えられるが、その特異性や生物学的意義は明らかではない(Teneberg et al., 1997、Teneberg et al., 2000)。
【0007】
さらに従来技術には、次に挙げる非シアリル化1型エピトープも開示されている:ラクトシルセラミドであるGalβ4GlcβCer、ガングリオテトラオシルセラミドであるGalβ3GalNAcβ4LacβCer(Lingwood et al., 1992)、末端Galβ3GlcNAcβ-を含有するラクト−構造(Teneberg et al., 2002)およびルイスbであるFucα2Galβ3(Fucα4)GlcNAcβ-(Boren et al., 1993)。これらのエピトープは種々の炭水化物主鎖、即ちラクトース、1型アセチルラクトサミンまたはガングリオコア、をベースとし、本発明によるシアリル化されたより長い2型N−アセチルラクトサミンとは一次構造と三次構造が明確に異なるものである。さらに、本願明細書で挙げた参考文献によると、種々のレセプターに対して明らかに特異的な種が存在し、シアル酸(sLex)結合特異性(Mahdavi et al., 2002)およびルイスb結合の場合には、非常に特異的なレセプタータンパク質が実際に特徴付けられている(Ilver D. et al., 1998)。
【0008】
発明の詳細の説明
本発明は、複数の直鎖状および分岐状のNeuNAcα3−ポリ−N−アセチルラクトサミン構造がヘリコバクター ピロリに対する高親和性リガンドとして機能し得ることを開示する。このような結合は、2型ラクトサミンであるGalβ4GlcNAcに結合したNeuNAcα3に特異的である。本発明は、末端三糖エピトープよりも高い結合能を有するより大きなポリラクトサミンに関する。直鎖ポリラクトサミン構造である NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCer および NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcβCerを考慮すると、β3Galβ4GlcNAcおよびより大きなポリラクトサミンであるβ3Galβ4GlcNAcβ3Galβ4GlcNAcに提示された末端NeuNAcα3Galβ4GlcNAcが有効であることは明確である。このような結果は、Galまたはラクトースにβ3結合した末端構造が有用な最小エピトープであることも示した。
【0009】
ヘリコバクター ピロリに対する新規な高親和性阻害剤の一般式
具体的には、本発明は、下記式(1)で表される高親和性ヘリコバクター ピロリ結合オリゴ糖配列に関する:
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal[β4GlcNAcβ3Gal]s2−
{β4Glc[NAc]s3}s4 (1)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4GlcNAcβ3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;そして
側鎖として存在し得るものを( )内に示す。)
本発明はまた、ヘリコバクター ピロリに対する結合能を有する該オリゴ糖配列の構造類似体、特に立体配座類似体、または誘導体にも関する。
【0010】
非シアリル化分岐構造と考えられるものの構造
上記式中のR1またはR2がNeuNAcα3ではない非シアリル化置換基である場合には、R1またはR2は、Galの2位および3位の少なくとも1種に結合した、下記式(2)で表される末端置換基であることが好ましい。
Hex[NAc]t1α/β3[(DeoxyHexα2)]t2 (2)
(式中、HexはGalまたはGlcであることが好ましく、
t1およびt2は各々独立に0または1であり、そして
α/βはα結合またはβ結合である。)
【0011】
さらに非シアリル化置換基であるR1またはR2が、下記式からなる群より選ばれる構造であることが好ましい:
Galα3、GalNAcα3、Fucα2、Galα3(Fucα2)、GalNAcα3(Fucα2)、
NeuNAcα3Galβ4GlcNAcβ3、Galβ4GlcNAcβ3、GlcNAcβ3Galβ4GlcNAcβ3、GlcNAcα3、GlcNAcβ3、GalNAcβ3、Galβ3、Glcβ3、そしてGlcα3。さらに好ましくは、下記式からなる群より選ばれる血液型抗原型の構造であることが好ましい:Galα3、GalNAcα3、
Fucα2、Galα3(Fucα2)、そしてGalNAcα3(Fucα2)。
【0012】
好ましい分岐構造
本発明の好ましい態様においては、式中のs1が1であり、R1およびR2が共にNeuNAcα3およびNeuNAcα3Galβ4GlcNAcβ3からなる群より選ばれるものである。好ましいポリ−N−アセチルラクトサミン構造としては、下記の配列で表されるオリゴ糖配列が挙げられる:
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、および
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Gal 。
【0013】
本発明によるオリゴ糖配列は、下記式で表されるような分岐したポリ−N−アセチルラクトサミンでもよい。
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc
(式中、LacNAcはN−アセチルラクトサミン、即ちGalβ4GlcNAcであり、Lacはラクトース、即ちGalβ4Glcである。従って、下記の2種の配列は同じ構造を表す:
NeuNAcα3Galβ4GlcNAcβ3(NeuNAcα3Galβ4GlcNAcβ6)Galβ4Glc と
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac 。)
【0014】
好ましい長鎖ラクトサミンエピトープ
上記式(1)においてs1が0である場合、式(1)は、例えば、下記式のいずれかで表される直鎖シアリルポリラクトサミンを表す:
NeuNAcα3LacNAcβ3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3Lac、
NeuNAcα3LacNAcβ3LacNAcβ3Gal、または
NeuNAcα3LacNAcβ3LacNAc;あるいは
より短い、最小エピトープである、
NeuNAcα3LacNAcβ3Lac、または
NeuNAcα3LacNAcβ3Gal 。
【0015】
好ましい類似体構造
本発明は、NeuNAc残基のグリセロール尾部は、酸化や還元によって生じる修飾を許容しないことを示す。NeuNAcのアセチル基は多くの変更を許容しないので、グリコリル類似体、プロピル類似体または脱アセチル化アミン類似体は、低活性であるか全く活性を示さない。また、シアル酸のカルボキシル基が還元されてアルコールになった異型も活性を示さない。同様の特異性が小さな五糖エピトープと分岐した大きなポリグリコシルセラミドでも見られた。シアル酸残基の位置も重要であり、ジシアル酸構造やNeuNAcα6-構造は活性を示さなかった。さらに、NeuNAc-残基がα3結合した末端N−アセチルラクトサミンは、2型N−アセチルラクトサミン(即ちGalβ4GlcNAc)であるはずであり、1型アセチルラクトサミン末端(即ちGalβ3GlcNAc)やアシアロ−ガングリオシド末端(即ちGalβ3GalNAc)は受け入れられなかった。このようなデータによって、不活性構造の特徴を回避した許容可能な類似体および誘導体の効果的な設計が可能となる。
【0016】
本発明は特に、式(1)で表される構造の類似体に関する。具体的には、式(1)中のN−アセチルラクトサミン残基の少なくとも1つが2型N−アセチルラクトサミン類似体構造、好ましくはラクトース残基、で置換されている、下記式(3)で表される類似体である。
R1Galβ4Glc[NAc]u1β3{(R2Galβ4Glc[NAc]u2β6)}s1Gal{β4Glc[NAc]u3β3Gal}s2−
{β4Glc[NAc] s3}s4 (3)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;そして
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0である。)
本願明細書で開示する2型N−アセチルラクトサミン類似体構造にはGalβ4Glc2-X構造が含まれ、この構造においてはGlc-環の2位の炭素が置換基Xに結合している。Xとしては、-NH2基(GalβGlcN−類似体となる)、N−アルキル基(Galβ4GlcN-アルキル基類似体となる)またはN−アルカノイル基が好ましい。好ましいアルカノイル基はプロパノイル基である。2位の炭素は、O−アセチル基などのO−エステルまたはO−メチル基などのO−エーテルによって誘導されていてもよい。特に、以下のジシアリル化された直鎖モノシアリル化構造が好ましい:
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β6)Lac[NAc]u3β3Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3(NeuNAcα3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、そして
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3Gal{β4Glc[NAc]s3}s4 。
【0017】
本発明はまた、下記式で表される類似体に関する。
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2
(但し、u2が0である場合にはu1も0である。)
【0018】
フコシル化類似体
上述のように、sLexやsdiLexなどのフコシル化構造についても、別途、研究している。本発明は、フコシル化および非フコシル化したシアリルα3−N−アセチルラクトサミンの両方が同じ結合特異性に関与することを示す。上記したポリ−N−アセチルラクトサミンのラクトサミン類似体(特にラクトース含有類似体)もまた、Glc残基および(存在する場合には)GlcNAc残基がα3−フコシル化されている場合に活性を示すであろうことがわかる。このような類似体は、より安価な主鎖構造と、フコース残基によって誘導されたより高い親和性とを兼ね備える。本発明は、このようなシアリル化およびフコシル化構造に関する。
【0019】
最も好ましいフコシル化構造は下記式(4)で表される構造である。
R1Galβ4[(Fucα3)]t1Glc[NAc]u1β3{(R2Galβ4[(Fucα3)]t2Glc[NAc]u2β6)}s1−
Gal{β4[(Fucα3)]t3Glc[NAc]u3β3Gal}s2{β4[(Fucα3)]t4Glc[NAc] s3}s4 (4)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4[(Fucα3)]t5Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0であり;そして
t1、t2、t3、t4およびt5は各々独立に0または1であり、[ ]内のFucα3側鎖構造の存在または不在を示す。)
式(4)で表される構造としては、特に、ジシアリル化構造および直鎖モノシアリル化構造が好ましい。
【0020】
本発明はまた、非還元位置にN−アセチルラクトサミン類似体を有する、非フコシル化類似体構造およびフコシル化類似体構造にも関する。特に、ラクトースを有する類似体、即ち、式中のu1、u2およびu3からなる群より選ばれる少なくとも1種の変数が0であるものが好ましく、u1、u2およびu3からなる群より選ばれる2種が0であることがより好ましく、u1、u2およびu3のすべてが0であることが最も好ましい。好ましい態様においては、類似体は分岐しており、さらに好ましくはジシアリル化したものである。別の態様においては、五糖構造、六糖構造および七糖構造が本発明による類似体として好ましい。
【0021】
NeuNAcα3Galβ4Glc(NAc) がより効果的に提示された最小エピトープ
セラミドの近くに位置する構造は、TLC結合アッセイにおいて他の位置の構造よりも立体構造に制限がある。遊離の一価阻害剤や多価の結合体として用いるオリゴ糖配列としては、NeuNAcα3Galβ4GlcNAcβ3Galβ?およびNeuNAcα3Galβ4GlcNAcβ3Galβ4Glc、特にNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcが、シアリルラクトースやNeuNAcα3Galβ4GlcNAcよりも高い活性を示す。ネオ糖脂質構造の使用により、還元的アミノ化構造であるNeuNAcα3Galβ4GlcNAcβ3Galβ4Glc-ヘキサデシルアニリンは、還元的結合後の還元末端グルコースが環状ではないにもかかわらず、活性を有することが示された。この活性は、三糖糖脂質であるNeuNAcα3Galβ4GlcβCer(即ち、GM3)が不活性であるにもかかわらず、糖脂質構造であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerが示す活性に類似していた。
【0022】
本発明は具体的には、機能性食品、特に乳幼児用ミルクなどの乳幼児用食品および食品添加剤に関し、機能性食品は、外部からの添加または濃縮によって、ヒトおよび動物の乳に存在する本発明のシアリル化ポリラクトサミンの濃度が高められたものである。下記のオリゴ糖からなる群より選ばれる少なくとも1種が食品用途に特に好ましい構造である:
NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc、
NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glc、および
NeuNAcα3Galβ4GlcNAcβ3(NeuNAcα3Galβ4GlcNAcβ6)Galβ4Glc 。
機能性食品用の構造としては、NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc および/または NeuNAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcがさらに好ましく、NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc が最も好ましい。本発明はさらに、実質的に純粋な状態で(炭水化物質量の少なくとも80%、さらに好ましくは少なくとも90%、最も好ましくは少なくとも95%が所望のオリゴ糖である実質的に純粋なオリゴ糖画分として)上記3種のオリゴ糖全て、または上記群より2種のオリゴ糖を選択してなるいずれかの組み合わせを包含するオリゴ糖組成物、およびオリゴ糖組成物を用いた機能性食品、医薬組成物または治療用組成物の製造方法に関する。本発明はさらに、チューインガムや種々の消費製品に用いるオリゴ糖配列に関する。
【0023】
本発明の他の一つの態様においては、上記群より選ばれるシアリル化オリゴ糖あるいは2種または3種のシアリル化オリゴ糖からなる実質的に純粋な混合物を、ヒトの乳および動物の乳に含まれるオリゴ糖(例えば、シアリルラクトース類、ラクトース、ラクト−N−ネオテトラオース、パラ−ラクト−N−ネオテトラオース、ラクト−N−ネオヘキサオース、およびそれらのα3−フコシル化誘導体)からなる群より選ばれる1種または複数種と共に含有するものも好ましい。このようなシアル化オリゴ糖を含む混合物は、NeuNAcα3Lac、NeuNacα3LacNAcβ3LacおよびNeuNAcα3(LacNAcβ3)2Lacを含むものが好ましく、これら3種の糖の1番目および2番目の糖のみを含むものがさらに好ましい。
【0024】
本発明は特に、下記式(5)で表される四糖からなるより高い親和性の最小レセプターオリゴ糖配列に関するが、但し、この構造は五糖の糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerとしては提示されない。
NeuNAcα3Galβ4GlcNAcβ3Gal (5)
四糖からなる上記オリゴ糖配列は、炭素数が22を超えるセラミド、疎水性アグリコンまたは疎水性スペーサーに結合していないことがより好ましく、疎水性構造から得られた炭素数8未満のアグリコンまたはスペーサーにカップリングしていることがさらに好ましい。
【0025】
本発明はまた、上記構造の立体配座類似体および誘導体にも関する。構造類似体または誘導体構造は、ヘリコバクター ピロリに対して同様のまたはより優れた親和性を示すことが好ましい。類似体の好ましい分類においては、類似体は、ラクトースを含んでいるかまたは2型N−アセチルラクトサミンに類似した立体配座を有する他の二糖エピトープを含んでいる。好ましい類似体構造としては、オリゴ糖配列であるNeuNAcα3Galβ4Glcβ3Galが挙げられる。本発明は、具体的には、末端オリゴ糖配列としてNeuNAcα3Galβ4Glcβ3Galを含む物質に関する。ラクトースをベースとした構造は、N−アセチルラクトサミン構造よりも化学的に製造する際にずっと安価である。
【0026】
本発明は特に、下記式(6)で表されるより高い親和性の最小レセプターオリゴ糖配列に関するが、但し、この構造は五糖の糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerとしては提示されない。
NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc(NAc)m (6)
(式中、mは0または1である。)
上記五糖からなるオリゴ糖配列は、炭素数が22を超えるセラミド、疎水性アグリコンまたは疎水性スペーサーに結合していないことがさらに好ましく、疎水性構造から得られた炭素数8未満のアグリコンまたはスペーサーにカップリングしていることがさらに好ましい。高い親和性の最小レセプターは多価の結合体および遊離の糖として特に有用である。
【0027】
本発明はまた、上記構造の立体配座類似体および誘導体にも関する。構造類似体または誘導体構造は、ヘリコバクター ピロリに対して同様のまたはより優れた親和性を示すことが好ましい。類似体の好ましい分類においては、類似体は、ラクトースを含んでいるかまたは2型N−アセチルラクトサミンと類似の立体配座を有する他の二糖エピトープを含んでいる。好ましい類似体構造としては、オリゴ糖配列であるNeuNAcα3Galβ4Glcβ3Galβ4Glc(NAc)m(式中、mは0または1である)が挙げられる。本発明は、具体的には、末端オリゴ糖配列としてNeuNAcα3Galβ4Glcβ3Galβ4GlcまたはNeuNAcα3Galβ4Glcβ3Galβ4GlcNAcを含む物質に関する。ラクトースをベースとした構造は、N−アセチルラクトサミン構造よりも化学的に製造する際にずっと安価である。
【0028】
親水性オリゴ糖物質
本発明は、具体的には、親水性オリゴ糖物質に関する。本発明者らは、本発明の新規な高親和性オリゴ糖配列への結合は、そこに存在するセラミド構造に依存しないことを見出した。すでに、糖脂質への細菌の結合、特にヘリコバクター ピロリの結合の一部はセラミド構造に依存しており、他の一部はセラミド構造に依存しないことが知られている。本発明は、脂質部分に依存しない、有用な高親和性オリゴ糖結合に関する。本願実施例のデータは、ヘリコバクター ピロリがセラミド構造に依存することなく、特異的な高親和性オリゴ糖配列に結合することを示す。
【0029】
すでに、三糖であるより短いシアリル−ラクトース構造/シアリルラクトサミン構造や、より大きなフコシル化シアリルラクトサミンについて、セラミドに依存しない効果が観察されている。より短いエピトープに対する特異性は特定の条件下で培養したヘリコバクター ピロリには見られないことから、出版物に記載されているより大きなシアリル−ポリラクトサミンとの結合特異性は、三糖に対する特異性とは異なると考えられる。このような高親和性の結合特異性またはヘリコバクター ピロリの結合様式(mode of H. pylori)に関する本発明は、小さな糖による研究とは明らかに異なるものである。本発明はさらに、本発明の物質を用いたヘリコバクター ピロリの結合の阻害、および本発明の物質を用いた、患者からヘリコバクター ピロリを除去する方法に関する。従来技術はまた、フコシル化シアリルラクトサミンやシアリル−ルイスx型構造に特異的に関連する種々の高親和性効果も開示している。
【0030】
セラミドは細胞膜と会合する傾向をもともと示すため、セラミドをベースとした構造は、一般的には単一分子をベースとした治療に有用ではない。付着阻害を目的としたセラミド化合物を付着阻害に用いると、脂質が組織に付着する危険性、即ち、付着を阻害する代わりに感染組織への結合が増加する危険性がある。さらに、セラミドをベースとした構造をいかなる医療用途で用いる場合にも、その合成、製造および処方が困難であり、且つ比較的費用がかかる。糖脂質物質はさらに、抗原性があることが知られている。
【0031】
したがって、セラミド非依存性の高親和性オリゴ糖配列に関する本発明は、ヘリコバクター ピロリ感染の治療または予防のための種々の治療用アプローチに特に有用である。糖脂質の使用に代わる、本発明は親水性オリゴ糖配列の用途に関する。本発明の親水性オリゴ糖配列は、本発明で開示する高親和性オリゴ糖配列、および所望により親水性構造を含む。具体的な態様においては、オリゴ糖配列はその配列からなるオリゴ糖である。具体的な態様においては、オリゴ糖配列は機能性食品の分野に用いることが好ましく、特に本願に記載する乳幼児用食品分野に用いることが好ましい。ヒトまたは動物の乳の天然オリゴ糖を用いることのさらに有用な点は、優れた安全性を有する天然オリゴ糖の使用が可能になったことと、感染症の治療において数々の有益な側面が考えられるということである。親水性構造はオリゴ糖配列の還元末端に結合していることが好ましく、このような構造は下記(i)または(ii)で定義される構造である。
(i)オリゴ糖配列の一価の結合を増加させる非疎水性アグリコン構造、あるいは
(ii)オリゴ糖配列を多価(polyvalent or multivalent)の担体または親水性の一価の担体に結合するスペーサーである非疎水性アグリコン構造。
【0032】
非疎水性アグリコン構造とは、天然のセラミド構造よりも疎水性が実質的に低い構造を意味する。このような構造は、有機炭素をベースとした炭素数の少ない構造や、親水性基(例えば、脂肪族構造中の−CH2−と関連した酸素原子または窒素原子)を天然の糖脂質よりも実質的に大きな比率で含有する構造により得られる。親水性基の−CH2−構造に対する割合は少なくとも1:4であることが好ましく、より好ましくは少なくとも1:3であり、さらに好ましくは少なくとも1:2であり、最も好ましくは約1:1であり、特定のより好ましい態様においては、1:1未満である。アグリコンは、3を越える数の互いに結合した−CH2−構造を含有する脂肪族構造を1つ以上有していないことが好ましい。アグリコン構造は炭素数が23未満であることが好ましく、さらに好ましくは炭素数は15未満であり、さらに好ましくは10未満である。好ましい態様においては、アグリコン構造は環構造(より好ましくは六員環構造または五員環構造、さらに好ましくは六員環構造)、および/または少なくとも1つの水酸基(好ましくは少なくとも2つの水酸基、より好ましい態様においては、少なくとも3つの水酸基)、および/または少なくとも1つのアミド構造[好ましくはアセトアミド(N−アセチルともいう)−構造またはGlcNAc-残基のアセトアミド−構造を模倣する構造]を含むことが好ましい。アグリコンは、環構造と水酸基または環構造とアミド構造を含むことがさらに好ましい。好ましい六員環としては、シクロヘキサンやベンゼン環およびそれらの置換誘導体、並びに5個の炭素原子と共に窒素へテロ原子、酸素へテロ原子または硫黄へテロ原子を含む複素環構造が挙げられる。
【0033】
非疎水性アグリコン物質がオリゴ糖配列の還元末端に結合している場合には、その目的がオリゴ糖配列の親和性を増加させることによってオリゴ糖の親和性を増加させることであることが好ましい。特に、一価の物質の親和性を増加させるのには非疎水性アグリコン物質が好ましく、このようなアグリコン物質は、好ましい態様においてはオリゴ糖配列の親和性をさらに増加させる構造を含むスペーサーである。アグリコン構造は、長鎖である本発明の高親和性オリゴ糖配列の還元末端オリゴ糖構造を模倣する五員環構造または六員環構造を少なくとも1つ含むことが好ましく、さらに好ましくは、アグリコンは、本発明のオリゴ糖配列よりも単糖1つ分長いオリゴ糖配列の還元末端単糖残基を模倣する六員環構造中の5つの原子を1組含むものである。さらに具体的な態様においては、アグリコンは六炭糖構造の模倣物(mimic)であり、より好ましくは、オリゴ糖配列の還元末端単糖残基がGlcNAcまたはGlcである場合には、アグリコンはD−ガラクトピラノシル構造の模倣物であり、オリゴ糖配列の還元末端単糖残基がGalである場合には、アグリコンはD−グルコピラノシルまたはD−N−アセチルグルコピラノシルの模倣物である。さらに好ましくは、アグリコンは、単糖1つ分長いオリゴ糖配列中の単糖間に存在するグリコシド結合構造もさらに模倣するように、付加されている還元末端単糖を模倣し、この時還元末端構造は、オリゴ糖が3位に(好ましくはβ3結合で)結合したGal、あるいはオリゴ糖が4位に(好ましくはβ4結合で)結合したGlcまたはGlcNAcのいずれかを模倣することが好ましい。
【0034】
オリゴ糖が遊離オリゴ糖であるか、またはオリゴ糖がその還元末端から本願で説明したアグリコンに結合している場合は、本発明は、特に親水性オリゴ糖に関する。アグリコンは炭素数が23未満の親水性物質であることが好ましい。特定の態様においては、アグリコンは担体(好ましくは多価またはオリゴ価の担体)へのリンカーであり、これはオリゴ糖配列がオリゴ価または多価の形態で担体構造に結合することができることを意味する。
【0035】
好ましいアグリコン構造としては、炭素数が1〜約8、より好ましくは1〜3、最も好ましくは1または2、の小さな構造がさらに挙げられる。このような小さな構造には、例えば、還元単糖単位などのポリオール物質、グリコシド類、メタノール、エタノール、プロパノール、イソプロパノールおよびグリコールなどの単一アルコール類(simple alcohols)、グリコール酸、グリセロール、グリセラート、カルボン酸アルコールおよびアミノ酸残基が挙げられる。アミノ酸残基は、天然のL−アミノ酸残基であることが好ましく、さらに好ましくはセリンまたはスレオニンのグリコシドである。好ましい小さなアグリコン構造としては、還元末端に結合したカルボン酸、より好ましくはグリコシルアミドに誘導化したカルボン酸、がさらに挙げられる。好ましいカルボン酸はC1〜C8のカルボン酸であり、より好ましくはC1〜C4カルボン酸であり、さらに好ましくは酢酸、最も好ましくはオリゴ糖配列の還元末端に結合した酢酸アミドである。このような好ましい小さな構造はスペーサーとしても用いることもでき、2つの官能基を有する小さなスペーサー構造をスペーサーとして用いることが好ましい。
【0036】
本発明における好ましいスペーサー構造は、例えば、本発明で用いる親水性スペーサー構造として記載されている。好ましくは、多価の担体は多糖であり、オリゴ糖は、オリゴ糖の還元アルデヒドと特異的に反応する官能基によって、オリゴ糖の化学的保護を必要とすることなく、多糖に結合している。担体は、親水性分子および水溶性分子であることが好ましく、最も好ましくは、キトサン、グリコーゲン、デンプンの重合体またはオリゴマー、シクロデキストリン、コンドロイチン、コンドロイチン硫酸、ヘパリンおよびヒアルロン酸などの多糖である。
【0037】
オリゴ糖構造はアグリコン構造にβ結合していることが好ましい。
【0038】
具体的な態様においては、アグリコン構造は還元末端の単糖の結合相互作用を模倣する。単糖残基の結合相互作用を模倣するためには、アグリコン物質は、単糖残基の水酸基または環構造に含まれる酸素(ring oxygens)の結合を模倣する水素結合構造、または単糖残基とタンパク質表面の親水性アミノ酸側鎖との親水性相互作用を模倣する親水性構造を含んでいてもよい。好ましい態様においては、芳香環構造、さらに好ましくは六員環である芳香環構造を、還元末端単糖残基の結合を模倣するのに用いる。
【0039】
本発明は、具体的には、本発明のオリゴ糖配列のヘリコバクター ピロリへの結合を用いた、本発明において好ましいアグリコン構造をスクリーニングするための方法およびアッセイに関する。好ましい方法においては、シアル酸結合付着因子(sialic acid binding adhesin)のタンパク質構造をアグリコン構造の設計に用いる。本発明はさらに、還元末端単糖残基を本発明で開示する還元末端単糖残基を模倣するアグリコンによって置換することによる、本発明において最も短いオリゴ糖配列の類似体の探索および設計に関する。好ましくは、本発明は、NeuNAcα3Galβ4Glcβ3GalおよびNeuNAcα3Galβ4GlcNAcβ3Galの類似体のスクリーニングに関し、ここでは3Galを、この単糖残基を模倣するアグリコンまたはヘリコバクター ピロリと3Galとの結合を模倣する構造で置換する。さらに好ましくは、本発明は、NeuNAcα3Galβ4Glcβ3Galβ4Glc(NAc)0または1およびNeuNAcα3Galβ4GlcNAcβ3Galβ4Glc(NAc)0または1の類似体のスクリーニングに関し、ここでは4Glc(NAc)0または1を、この単糖残基またはヘリコバクター ピロリと単糖残基との結合を模倣するアグリコンで置換する。したがって、本発明は、好ましくは下記式で表される本発明のオリゴ糖配列の類似体のスクリーニングに関する。
NeuNAcα3Galβ4Glc(NAc)s1{β3Gal}s2- R
(式中、s1およびs2は各々独立に0または1であり、そして
s2が0である場合は、Rはβ3Galまたはヘリコバクター ピロリへのβ3Galの結合を模倣するアグリコンであり、s2が1である場合は、Rはβ4GlcNAcまたはβ4Glcあるいはβ4GlcNAcまたはβ4Glcの結合を模倣するアグリコンである。)
具体的な態様においては、オリゴ糖部分を安価な天然材料、例えばウシ初乳およびチーズ製造工程で生じる廃棄物(waste stream of cheese production)から製造することができるので、s2が0である式で表される構造が好ましい。
【0040】
好ましい大きなポリ−N−アセチルラクトサミン類およびその好ましい用途
具体的な態様においては、本発明は、医薬として用いることを特徴とする、非還元末端配列となる下記式で表される構造、あるいはヘリコバクター ピロリに対する結合能を有する該構造の類似体または誘導体に関する。
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal{β4Glc[NAc]s3}s4
(式中、R1およびR2は末端単糖置換基または末端オリゴ糖置換基であり、該置換基の少なくとも1種はNeuNAcα3であり;そして
s1、s3およびs4は各々独立に0または1であり、{ }または[ ]の中の構造の存在または不在を示す。)
【0041】
上記のようなより大きなポリ−N−アセチルラクトサミン構造は、天然材料から製造する場合に特に好ましい。このような大きなポリ−N−アセチルラクトサミンは、オリゴ糖の遊離に用いられている方法によって天然の糖タンパク質または糖脂質から製造することができ、このような方法としては、(例えばタンパク質であるN−グリコシダーゼを用いた)N−グリカンを遊離するためのエンド−グリコシダーゼ法や、糖タンパク質をプロテアーゼで対応の糖ペプチドに消化するO−結合グリカンを遊離するための方法が挙げられる。食品分野で許容可能な方法、特にプロテアーゼ法、による糖ペプチドの製造が好ましい。また、タンパク質結合N−グリカンは、所望によりオリゴ糖の還元末端の還元を行うβ−脱離法(beta-elimination methods)や、例えば当業界で知られているヒドラジンに基づく遊離法(hydrazine based release methods)により遊離することができる。音波分解法(otsonolysis)やエンド−グリコシルセラミドによって糖脂質からオリゴ糖を遊離することもできる。具体的な態様においては、本発明は酵素学的に遊離したタンパク質結合オリゴ糖や糖ペプチド、化学的に遊離したタンパク質結合オリゴ糖または糖ペプチド、ならびにそれらの混合物に関する。本発明の他の態様は、化学的または酵素学的な手段で糖脂質から製造したオリゴ糖に関する。このような構造は、天然材料から製造した混合物であることが特に好ましい。本発明はさらに、本発明の大きなポリ−N−アセチルラクトサミンに富んだ、天然または遊離のオリゴ糖または糖ペプチドに関する。
【0042】
本発明はさらに、本発明の大きなポリ−N−アセチルラクトサミンに富んだ天然の糖タンパク質画分に関する。本発明はさらに、ポリ−N−アセチルラクトサミン前駆体を酵素学的にリモデリングして本発明の構造とするための、天然のポリ−N−アセチルラクトサミン前駆体画分に関する。ポリ−N−アセチルラクトサミン構造を含む、種々の動物由来のタンパク質材料および脂質材料について報告されており、当業界で知られている。本発明は特に、食品分野で許容可能な材料に関する。
【0043】
具体的な態様においては、本発明は、本発明の大きなポリ−N−アセチルラクトサミンとして好ましいものを糖脂質としてミセルの形状で用いた、本発明の治療方法またはヘリコバクター ピロリ結合性の試験方法に関する。このような物質は、高親和性リガンドを探索するために、ヘリコバクター ピロリ結合性をスクリーニングするのに特に好ましい。
【0044】
天然グリカンのリモデリングによる機能性オリゴ糖の製造
具体的な態様においては、本発明は天然グリカン、好ましくは食物材料から得たポリ−N−アセチルラクトサミン、の機能性食品炭水化物へのリモデリングに関する。グリカンのリモデリング方法は、Neose-companyによる最近の米国特許出願および国際特許出願に記載されている。本発明は特に、天然の食物の糖タンパク質やオリゴ糖、または遊離オリゴ糖の脱シアリル化および/または脱フコシル化に関する。最も好ましくは、オリゴ糖や糖ペプチドを穏やかな酸処理により脱シアリル化する。好ましい態様においては、脱シアリル化は、当業界で知られているように、食品分野で許容されるカルボン酸による処理後に約80〜100℃に加熱することで行い、好ましくは酢酸により処理する。他の一つの態様においては、熱処理により除去することが可能な揮発性の酸(例えば塩酸)で食物材料を脱シアリル化する。オリゴ糖構造の主要部分はそのまま残るように行う化学的な脱フコシル化法、例えば硫酸を用いる方法、も知られている。本発明はさらに、天然材料から得たオリゴ糖または糖ペプチドを含む糖を本発明による構造にするためのリモデリングに関する。糖はタンパク質材料に比べて安定性が高いので、精製または単離した糖混合物をリモデリングに用いることが好ましい。本発明は具体的には、α3−シアリル化による天然の糖のリモデリング、好ましくは脱シアリル化および/またはフコシル化後に行うリモデリングに関する。他の目的のためには、本発明はさらに、α3−ガラクトシル化、α4−ガラクトシル化、β4−GalNAc−転移、β3−GalNAc−転移、β3−GlcNAc−転移、α6−シアリル化、α3−フコシル化、α2−フコシル化またはα6−フコシル化、好ましくはα3−ガラクトシル化、α4−ガラクトシル化、β3−GlcNAc−転移またはα6−シアリル化による遊離オリゴ糖のリモデリングに関し、好ましい態様においては、β3−GlcNAc−転移またはα6−シアリル化でリモデリングを行う。このようなグリコシル化反応は、グリコシルトランスフェラーゼ酵素、トランスグリコシル化グリコシダーゼまたはS. Whithersらのグループが発表した変異トランスグリコシル化酵素を用いて、公知の方法で行うことができる。
【0045】
本発明は、下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料由来の遊離の糖または単離した糖のリモデリング方法に関する。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) 該原料オリゴ糖からなる画分、必要な場合には所望の糖、好ましくはポリ−N−アセチルラクトサミン、に富んだ画分を単離し、
(3) 該単離した画分を処理、好ましくは穏やかな酸処理に付して、末端単糖、好ましくはフコースとシアル酸の少なくとも1種、さらに好ましくはシアル酸、を遊離させて末端単糖除去オリゴ糖を含む糖混合物を得、そして
(4) 糖混合物の末端単糖除去オリゴ糖に、単糖、好ましくはα3結合するシアル酸をグリコシルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【0046】
本発明は、下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料由来の遊離の糖または単離した糖のリモデリング方法に関する。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) 該原料オリゴ糖からなる画分、必要な場合には所望の糖、好ましくはポリ−N−アセチルラクトサミン、に富んだ画分を単離し、
(3) 所望により末端単糖、好ましくはフコースとシアル酸の少なくとも1種、さらに好ましくはシアル酸、を遊離させて末端単糖除去オリゴ糖を含む糖混合物を得、そして
(4) 糖混合物の末端単糖除去オリゴ糖に、単糖、好ましくはα3結合するシアル酸をα3−シアリルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【0047】
好ましい態様においては、上記リモデリング方法は種々の数のシアル酸を含有するオリゴ糖を精製する工程をさらに包含し、1つ、2つ、3つまたは4つのシアル酸を含有するオリゴ糖あるいはそれらの混合物を含む画分を得ることが好ましく、精製はイオン交換クロマトグラフィーまたはアフィニティークロマトグラフィーで行うことが好ましい。
【0048】
好ましい態様においては、ポリ−N−アセチルラクトサミン画分をサイズ排除クロマトグラフィーまたはアフィニティークロマトグラフィーにより単離する。単離にはポリ−N−アセチルラクトサミン結合性のレクチンを用いることが好ましく、ポリ−N−アセチルラクトサミン結合性の食物由来レクチン(好ましい例としてはトマトレクチン、ジャガイモレクチンおよび小麦麦芽凝集素)を用いることがより好ましい。
【0049】
好ましいポリ−N−アセチルラクトサミングリカンは、少なくとも1つのGalβ4Glcβ3Galβ4Glc(NAc)0または1を含有し、Galβ4Glcβ3Galβ構造はそのものの形で含有されいてもよいし、より大きなグリカン構造の一部として含有されていてもよい。
【0050】
具体的な態様においては、本発明は、上記リモデリング方法の工程(4)が2つの工程からなる方法に関し、この方法では、Galβ4を転移させる工程と、次いで転移したGalβ4に末端単糖、好ましくはα3−結合するシアル酸をα3−シアリルトランスフェラーゼによって転移する工程を包含する。このリモデリング方法に用いる原料は、末端GlcNAc残基を有するものである。
【0051】
トランスシアリダーゼ酵素は、例えばトリパノソーマ寄生虫より調製し、クローニングした。
【0052】
ヘリコバクター ピロリに対する結合能を有する、炭素修飾した中性または酸性の新規なシアル酸誘導体
本発明はさらに、ヘリコバクター ピロリに対する結合親和性を有する新規なシアル酸誘導体を開示する。このような誘導体は下記式で表される。
SA(X−R)
(式中、SAはシアル酸であり;
Xはシアル酸のC1位に結合している結合原子または結合基、および/またはシアル酸の7位、8位および8位の炭素原子からなる群より選ばれる少なくとも1種であって、この場合にシアル酸は8位の炭素原子または8位と9位の炭素原子を欠失しているシアル酸誘導体でもよく;
特に、Rで表される置換基がシアル酸のC7位またはC8位に結合している時、シアル酸はC7位とC8位を含む構造またはC9位を含む構造をそれぞれ欠失した、短いシアル酸誘導体であることが好ましく;そして
Rは水素原子または炭素数が3を超える有機ラジカルである。)
上記式中のXは、シアル酸残基のカルボキシル基と共にアミド構造を形成する−NHであることが好ましい。また、上記式中のRは、水素原子あるいは環構造および脂肪鎖の少なくとも1種を含むC4〜C30有機ラジカルであることが好ましく、C6〜C24有機ラジカルであることがより好ましく、C6〜C24脂肪族アルキル鎖であることが最も好ましい。SAは、NeuNAcであることが好ましい。
【0053】
シアル酸は、そのカルボン酸および/またはC7位〜C9位のグリセロール尾部構造で修飾可能であることを見出した。グリセロール尾部構造を短縮(好ましくはC7位まで短縮)した後に、疎水性構造でグリセロール尾部を修飾することが、特に効果的な修飾であることが判明した。好ましい態様においては、修飾に用いる基は直鎖状の脂肪族アルキル鎖である。さらに、修飾したシアル酸構造は本発明の他の構造と共に用いることができることも判明した。特に、C7位〜C9位が修飾された構造を本発明のポリラクトサミン型構造と共に用いることが好ましい。このような構造をヘリコバクター ピロリに対する他の阻害剤のスクリーニングおよび設計に用いることがさらに好ましい。
【0054】
本発明はさらに、修飾したシアル酸構造の類似体または誘導体に関する。
【0055】
さらに好ましくは、このようなシアル酸構造は、ラクトースまたはN−アセチルラクトサミン構造に結合している。好ましい態様においては、シアル酸誘導体は下記式で表されるように、2型N−アセチルラクトサミン配列にα3結合を介して結合している。
SA(X-R)αxGal{β4GlcNAc[β3Gal(β4Glc)p1]p2}p3
(式中、xはシアル酸誘導体の結合位置を示し;そして
p1、p2およびp3は各々独立に0または1であり、{ }、[ ]または( )の中の構造全体の存在または不在を示す。)
【0056】
本願明細書においては、「オリゴ糖配列(OSs)」という用語には、修飾したシアル酸構造も含まれる。
【0057】
本発明はまた、本発明で開示するヘリコバクター ピロリ結合性のオリゴ糖配列またはシアル酸修飾構造、あるいはその類似体または誘導体の多価の結合体に関し、多価の結合体は、例えば下記式で表される構造からなる。
[OS−(X)n−L−Y]m−Z
(式中、mは1以上の整数であり、好ましくは1を超える整数であり、nは0または1であり;
OSはオリゴ糖配列であり;
Lは酸素原子、窒素原子、硫黄原子または炭素原子であり;
Xは単糖残基またはオリゴ糖残基、好ましくはラクトシル残基、ガラクトシル残基、ポリ−N−アセチル−ラクトサミニル残基、あるいはO−グリカンオリゴ糖配列またはN−グリカンオリゴ糖配列の一部、あるいは少なくとも1つのマンノシル残基またはN−アセチルガラクトサミニル残基を含むものであり;
Yはアグリコン、スペーサー、または該LとZの間の結合であり;
Zはオリゴ価または多価の担体であり、多糖などの炭水化物材料を含み;そして
該OSの還元末端の1位の炭素原子または本発明のヘリコバクター ピロリ結合性末端配列を含む修飾シアル酸のR基によって、該Yおよび所望により該Xを介して該Zに該Lから結合している。)
【0058】
さらに、新規なシアル酸誘導体を、NeuNAcに関連して記載した本発明のポリ−N−アセチルラクトサミンや、ヘリコバクター ピロリに対する結合能を有する他の糖結合体に提示できることが明かとなった。
【0059】
中性のシアル酸エピトープは、酸性のNeuNAcが結合するのとは異なる、ヘリコバクター ピロリの表面に存在するレセプターに結合し得ることが判明した。本発明は特に、このようなエピトープを新規なレセプターとして用いた、該レセプターに対応する付着因子をヘリコバクター ピロリの表面から同定する方法に関する。本発明はさらに、ヘリコバクター ピロリのヒトおよび動物の細胞や組織への結合の阻害、またはヘリコバクター ピロリの凝集において有用であると考えられる新規なリガンドの設計に関する。このような新規なヘリコバクター ピロリ結合性物質の設計または合成は、ヘリコバクター ピロリの同じレセプターに結合する他の炭水化物構造を用いたモデリング方法にも関する。
【0060】
本発明において、「類似体」及び「誘導体」は次のように定義される。本発明によると、ヘリコバクター ピロリ結合性オリゴ糖配列の構造類似体や誘導体を設計することができる。従って、本発明は、本発明の結合性物質の構造類似体にも関する。本発明における構造類似体は、ヘリコバクター ピロリのオリゴ糖配列に対する結合に重要な構造因子を包含する。効果的な構造類似体を設計するには、ヘリコバクター ピロリと糖類との結合に重要な構造因子を知ることが必要である。重要な構造因子は、修飾されていないか、重要な構造因子との類似性が非常に高い模倣物となるように修飾されていることが好ましい。
【0061】
本発明における構造誘導体は、ヘリコバクター ピロリに対する結合能を保持するか向上するように本発明のオリゴ糖配列を化学的に修飾したものである。本発明によると、オリゴ糖配列の水酸基またはアセトアミド基の1つまたは複数を誘導化することが好ましい。本発明は、類似体や誘導体を調製する際に変えることのできる分子上の位置を複数開示する。少なくともある種の修飾を許容することが好ましい水酸基またはアセトアミド基は、本願に記載した化学式から当業者には明かである。
【0062】
嵩高い置換基や酸性の置換基および他の構造(例えば単糖残基)は許容されないが、オリゴ糖類似体、例えばレクチン結合性のオリゴ糖類似体、を製造する方法はよく知られている。例えば、シアリル−ルイスxオリゴ糖の類似体は複数製造されており、これらは異なる骨格の活性官能基である(Sears and Wong 1996、12090ページを参照)。同様に、へパリンオリゴ糖の類似体もサノフィ社によって製造されており、シアリダーゼ酵素に対するシアル酸模倣型阻害剤、例えばZanamivirやTamiflu(Relenza)、が複数のグループによって製造されている。オリゴ糖類似体は少なくとも1つの六員環または五員環構造を含む分子に基づいて構築されていることが好ましく、類似体は6個または5個の原子を含む少なくとも2つの環構造を含有することがより好ましい。模倣構造においては、単糖環はシクロヘキサンやシクロペンタンなどの環状分子、ベンゼン環を含む芳香環によって置換されていてもよく、複素環構造は酸素原子の他に、例えば窒素原子や硫黄原子を含んでいてもよい。活性環状立体構造を固定するためには、環構造を許容できる結合基で繋ぐこともできる。典型的な模倣構造は、オリゴ糖配列またはその一部に対するペプチド類似体構造をさらに含んでいてもよい。
【0063】
活性基が結合能に与える影響は累積するので、1つの活性基が失われても、分子の異なる場所に活性基を付加することで補うことができる。分子モデリング、好ましくはコンピューターを用いた分子モデリングを本発明のヘリコバクター ピロリ結合性オリゴ糖配列の類似構造を製造するのに用いることができる。種々のオリゴ糖配列の分子モデリングの結果が実施例に示されており、NMRとX線結晶回折に加えてこれらと同じまたは類似した方法を用いて本発明の他のオリゴ糖配列に対する類似体を得ることができる。オリゴ糖配列の類似体を見つけるために、オリゴ糖配列をヘリコバクター ピロリの炭水化物結合分子、最も可能性が高いのはヘリコバクター ピロリのレクチン、と合体させることで、さらなる潜在的な結合相互作用を探索することができる。
【0064】
コンビナトリアルケミストリーによってオリゴ糖の誘導体を調製することで、レクチンに対してより高い活性を示すように1価、オリゴ価または多価のオリゴ糖を活性化することができることにも注目されたい。オリゴ糖配列の1つまたは複数の残基を置換することでライブラリーを構築した場合には、それは誘導体ライブラリーと考えることができ、また、本発明で開示するオリゴ糖配列の類似体からライブラリーを構築することもできる。コンビナトリアルケミストリーによるライブラリーは本発明のオリゴ糖、その前駆体または糖結合体に基づいて構築することができる。例えば、種々の還元末端を有するオリゴ糖をカーボハイブリッド技術(carbohydrid technology)と呼ばれる技法によって製造することができる。
【0065】
好ましい態様においては、コンビナトリアルケミストリーによるライブラリーは、本発明のヘリコバクター ピロリ結合性物質に結合されている。さらに好ましい態様においては、ライブラリーは少なくとも6種の異なる分子を包含する。このようなライブラリーは、本発明のオリゴ糖配列に対する微生物による結合を検出するために用いるのが好ましい。コンビナトリアルケミストリーによるライブラリーから高親和性のバインダーを同定することは、例えば阻害アッセイによって可能であり、ここでは本発明で開示する糖脂質または糖結合体に対する細菌の結合を阻害するために、ライブラリー中の化合物を用いる。本発明において好ましい構造類似体や誘導体は、本発明のヘリコバクター ピロリ結合性オリゴ糖配列がヘリコバクター ピロリに結合するのを阻害することができる。
【0066】
以下の記載においては、ヘリコバクター ピロリ結合性配列を単に「オリゴ糖配列」と記載する。本願で定義するオリゴ糖配列は、天然のまたは合成した糖結合体の一部、遊離オリゴ糖または遊離オリゴ糖の一部であってもよい。このようなオリゴ糖配列は、多糖鎖に結合した種々の単糖、オリゴ糖または多糖に結合させることができ、オリゴ糖配列が細菌の多糖の一部として発現している場合などが例示できる。さらに、O−アセチル化誘導体または硫酸化誘導体に代表される、単糖の種々の天然の修飾が知られている。本願で定義するヘリコバクター ピロリ結合性物質には、天然のまたは合成した糖結合体の一部、それに相当する遊離オリゴ糖、または遊離オリゴ糖の一部として記載したオリゴ糖配列が含まれる。ヘリコバクター ピロリ結合性物質には、ヘリコバクター ピロリ結合性オリゴ糖配列の混合物も含まれる。
【0067】
本発明のヘリコバクター ピロリ結合性物質は、糖鎖または糖結合体の一部、あるいは他の公知のヘリコバクター ピロリ結合性エピトープを含有する複数の糖化合物との混合物であってもよく、上記した公知のヘリコバクター ピロリ結合性エピトープとは、本発明のヘリコバクター ピロリ結合性物質とは異なる糖配列や立体構造を有するもの、例えばルイスb、Fucα2Galβ3(Fucα4)GlcNAc、Neu5Acα3Galβ4Glc/GlcNAcなどである。尚、本発明のヘリコバクター ピロリ結合性物質を治療のために用いる場合、複数の結合性物質との併用は有効手段となり得る。
【0068】
本発明は特に、本発明のオリゴ糖構造の類似体の設計に関し、類似体はオリゴ糖残基中の単糖残基に類似した環構造を含む。さらに好ましくは、ヘリコバクター ピロリの結合または阻害について類似体を試験し、製品開発のために最適な結合性配列を選択する。他の一つの態様においては、本発明の分子あるいはその類似体または誘導体を、他の微生物またはウイルスの結合、好ましくはClostridium difficileのトキシンAに対する結合について試験する。オリゴ糖類似体は少なくとも1つの六員環または五員環構造に基づいて構築されていることが好ましく、類似体は6個または5個の原子を含む少なくとも2つの環構造を含有することがより好ましい。オリゴ糖の好ましい類似体の型は末端ウロン酸アミドを包含するか、あるいは類似体または誘導体がGal/GalNAcβ4GlcNAc−糖模倣構造に結合したものである。本発明によると、末端単糖残基の2位または4位の水酸基は結合に重要ではなく、6位の水酸基は分子の親和性を実際に上昇させるような構造になるように修飾することができ、親和性が高い類似体はこのような位置を修飾することで製造することができる。本願のデータは、末端単糖残基のすべての水酸基を含有しない類似体の設計が可能であることを示している。本発明は特に、末端Gal/GalNAcの3位を修飾するための種々の有機誘導化分子(例えば芳香族または脂肪族環状有機残基)を付加することで行う、機能的類似体の設計に関する。誘導体は、Gal/GalNAcβ4の3位に環状有機残基を結合させ得る特別な結合化学的手法(linker chemistry)によって製造することができる。結合構造の幾何構造、さらにその長さでさえもが、グリコシド結合構造とは異なっていてもよいが、環状有機残基は、対応する末端単糖残基、特に末端単糖残基の6位の水酸基/カルボキシル基/アミド基の近傍で見られる正の結合相互作用の少なくとも一部を示さなければならない。
【0069】
具体的な態様においては、類似体はGal/GalNAcβ4の3位の水酸基の置換によってアミン基で製造する。本発明は特に、ヘリコバクター ピロリに結合する三糖エピトープ中の末端Gal/GalNAcの3位に末端環状分子を有する類似体のスクリーニングに関する。末端環状分子は有機六員環残基であることが好ましく、オリゴ糖構造の末端および/または本願で開示する化学式中に見られる内部ヘキスロン酸と同様のカルボン酸構造、アミド構造またはアルキルアミド構造をさらに含有することがより好ましい。
【0070】
また、末端ウロン酸アミドあるいはその類似体または誘導体が、Galに1−3結合しており、該GalがGlcNAc模倣構造に結合しているものも挙げられる。模倣構造においては、単糖環はシクロヘキサンやシクロペンタンなどの環状分子、ベンゼン環を含む芳香族環によって置換されていてもよく、そのような複素環構造は酸素原子の他に、例えば窒素原子や硫黄原子を含んでいてもよい。活性環状立体構造を固定するためには、環構造に許容される結合基で繋ぐこともできる。典型的な模倣構造は、オリゴ糖配列またはその一部に対するペプチド類似体構造をさらに含んでいてもよい。本発明は、オリゴ糖配列のためのペプチド類似体の設計および/またはスクリーニングにも関する。さらに本発明は、本発明のオリゴ糖配列に対する、DNAまたはRNAに基づく類似体(例えば、いわゆるアプタマー)のスクリーニングに関する。活性基が結合能に与える影響は累積するので、1つの活性基が失われても、分子の異なる場所に活性基を付加することで補うことができる。
【0071】
分子モデリング、好ましくはコンピューターを用いた分子モデリングを本発明のヘリコバクター ピロリ結合性オリゴ糖配列の類似体構造を製造するのに用いることができる。種々のオリゴ糖配列の分子モデリングの結果が実施例に示されており、NMRとX線結晶回折に加えてこれらと同じまたは類似した方法を用いて本発明の他の結合性オリゴ糖配列あるいはその類似体または誘導体の構造を決定することができる。好ましい態様においては、本発明のオリゴ糖と同じまたは類似の立体構造を有するオリゴ糖構造の類似体または誘導体は、コンピューター支援分子モデリング(computer assisted molecular modeling)で得られた結果または分子の三次元構造を含むスクリーニング用データベースから選択するが、このような方法を電算化スクリーニング方法(computerized screening method)と称する。
【0072】
本発明はさらに、電算化スクリーニング方法によって選択したオリゴ糖構造あるいはその類似体または誘導体の試験に関し、該試験においては、他の病原性の微生物、ウイルスまたは毒素に対して、本発明の一つまたは複数のオリゴ糖配列とヘリコバクター ピロリとの結合特異性に類似した結合特異性を示すものを検出する。好ましい態様においては、電算化スクリーニング方法によって選択されたオリゴ糖構造あるいはその類似体または誘導体をClostridium difficileのトキシンAに対する結合について試験する。
【0073】
類似体分子は、合成によって製造するか天然材料から得ることができる。分子は、コンピューター上で仮想的に作成することも可能であり、活性分子のスクリーニングの一部をin silicoで行うこともできる。本発明は、炭水化物構造、類似体または誘導体をコンピューター上でヘリコバクター ピロリに存在する炭水化物結合部位に適合させることによって行う、本発明のオリゴ糖配列に対するヘリコバクター ピロリ結合性および/または阻害性の類似体および/または誘導体の探索に関する。
【0074】
ヘリコバクター ピロリ結合性オリゴ糖配列あるいはその類似体または誘導体を、ヘリコバクター ピロリの炭水化物結合分子、最も可能性が高いのはヘリコバクター ピロリのレクチン、と分子モデリングの方法によって合体させることで、さらなる結合相互作用を探索することができる。オリゴ糖配列の三次元構造を炭水化物結合部位の三次元モデルとコンピューター上で合体させることは、結合相互作用および考えられるさらなる結合相互作用部位の決定を可能にするので、結合活性類似体の設計にさらに貢献する。この方法は、電算化合体方法(computerized docking method)によるオリゴ糖構造あるいはその類似体または誘導体の結合性の比較にも関する。
【0075】
本発明はさらに、電算化合体方法によるオリゴ糖構造あるいはその類似体または誘導体の試験に関し、該試験においては、他の病原性の微生物、ウイルスまたは毒素に対して、本発明の一つまたは複数のオリゴ糖配列とヘリコバクター ピロリとの結合特異性に類似した結合特異性を示すものを検出する。好ましい態様においては、オリゴ糖構造あるいはその類似体または誘導体のClostridium difficileのトキシンAに対する結合性を、電算化合体方法で試験する。
【0076】
コンビナトリアルケミストリーによってオリゴ糖の誘導体を調製することで、レクチンに対してより高い活性を示すように1価、オリゴ価または多価のオリゴ糖を活性化することもできることにも注目されたい。オリゴ糖配列の1つまたは複数の残基を置換することでライブラリーを構築した場合には、それは誘導体ライブラリーと考えることができる。また、本発明のオリゴ糖配列の類似体からライブラリーを構築した場合には、それは類似体ライブラリーと考えることができる。コンビナトリアルケミストリーによるライブラリーは本発明のオリゴ糖、その前駆体または糖結合体に基づいて構築することができる。例えば、種々の還元末端を有するオリゴ糖をカーボハイブリッド技術と呼ばれる技法によって製造することができる。本発明は、コンビナトリアルケミストリーによるライブラリー、即ち、本発明のオリゴ糖構造の多種多様な化学的な類似体および/または誘導体の設計および構築、ならびにこれらのヘリコバクター ピロリに対する結合またはヘリコバクター ピロリの阻害についての試験に関する。本発明はさらに、コンビナトリアルケミストリーによるライブラリーの試験に関し、該試験においては、他の病原性の微生物、ウイルスまたは毒素に対して、本発明の一つまたは複数のオリゴ糖配列とヘリコバクター ピロリとの結合特異性に類似した結合特異性を示すものを検出する。好ましい態様においては、コンビナトリアルケミストリーによるライブラリーのClostridium difficileのトキシンAに対する結合性を試験する。
【0077】
本発明の好ましい態様においては、コンビナトリアルケミストリーによるライブラリーは、本発明のヘリコバクター ピロリ結合性物質に結合されている。より好ましい態様においては、ライブラリーは少なくとも6種の異なる分子を包含する。コンビナトリアルケミストリーによる修飾は、式(9)のR8基またはR9基であるカルボキシル基を種々のアミド基に変化させることで得られるものが好ましい。R8基を修飾する置換基は、アルデヒド、アミンまたはその他の反応性基でもよい。このようなライブラリーは、本発明のオリゴ糖配列に対する微生物性の結合を検出するために用いるのが好ましい。アミノ酸または有機アミド類のコレクションは市販されており、このような物質はウロン酸アミドのコンビナトリアルケミストリーによるライブラリーを合成するために用いることができる。コンビナトリアルケミストリーによるライブラリーから高親和性のバインダーを同定することは、例えば阻害アッセイによって可能であり、ここでは本発明で開示する糖脂質または糖結合体に対する細菌の結合を阻害するために、ライブラリー中の化合物を用いる。本発明おいて好ましい構造類似体や誘導体は、本発明のヘリコバクター ピロリ結合性オリゴ糖配列がヘリコバクター ピロリに結合するのを阻害することができる。
【0078】
以下の記載においては、ヘリコバクター ピロリ結合性配列を単に「オリゴ糖配列」と記載する。本願で定義するオリゴ糖配列は、天然のまたは合成した糖結合体の一部、遊離オリゴ糖または遊離オリゴ糖の一部であってもよい。このようなオリゴ糖配列は、多糖鎖に結合した種々の単糖、オリゴ糖または多糖に結合させることができ、オリゴ糖配列が細菌の多糖の一部として発現している場合などが例示できる。さらに、単糖に対する天然の修飾も多数知られており、オリゴ糖配列のO−アセチル化誘導体や硫酸化誘導体が例示できる。特定の態様においては、オリゴ糖配列とは、所望により還元末端が修飾されていることを除いては、他のいかなる単糖残基によっても修飾されていない非還元末端オリゴ糖配列を意味する。広義においては、オリゴ糖配列という用語には、本発明のオリゴ糖構造の構造類似体や誘導体が含まれることが好ましく、本願で開示するように、ヘリコバクター ピロリに対して同一または同様の結合能を有することが好ましい。本願で定義するヘリコバクター ピロリ結合性物質には、天然のまたは合成した糖結合体またはその一部、それに相当する遊離オリゴ糖、または遊離オリゴ糖の一部として記載したオリゴ糖配列が含まれてもよい。ヘリコバクター ピロリ結合性物質には、ヘリコバクター ピロリ結合性オリゴ糖配列の混合物も含まれてもよい。
【0079】
ヘリコバクター ピロリ結合性オリゴ糖配列は、グリコシルトランスフェラーゼによって酵素学的に合成するか、あるいはグリコシダーゼ酵素またはトランスグリコシダーゼ酵素によって触媒されるトランスグリコシル化反応で合成することができる(Ernst et al., 2000)。これらの酵素の特異性および補因子の必要性については改良することができる。修飾された特定の酵素を用いてより効率よく合成することもでき、例えば、グリコシンセターゼをトランスグリコシル化のみを触媒するよう修飾することができる。本発明で開示する糖および糖結合体またはそれらと類似した化合物の有機合成も知られている(Ernst et al., 2000)。糖物質は天然の原料から単離して、化学的または酵素学的に修飾してヘリコバクター ピロリ結合性化合物とすることができる。種々の反芻動物の乳から天然のオリゴ糖を単離することができる。グリコシル化酵素を発現するトランスジェニックな生物、例えばウシや微生物も糖の製造に用いることができる。
【0080】
細菌結合性物質は、クラスター、即ち、細胞膜上の糖脂質、ミセルまたはリポソームとして提示されるか、あるいはアッセイに用いるTCLプレートのような固相に提示されていることが好ましい。正しい空間配置によって提示されたクラスターは高い結合親和性を示す。
【0081】
本発明においては、ヘリコバクター ピロリ結合性エピトープあるいはヘリコバクター ピロリに対して同様または改善された結合能を有する、該エピトープの天然のまたは合成した類似体や誘導体を用いることもできる。さらに、細菌結合性物質を含有する物質、例えば、本願で開示するレセプター活性ガングリオシドやヘリコバクター ピロリに対して同様または改善された結合能を有する、該ガングリオシドの類似体や誘導体などを用いることもできる。細菌結合性物質は、オリゴ糖鎖のグリコシル結合した末端エピトープでもよい。また、細菌結合性エピトープはオリゴ糖鎖の側鎖、好ましくはポリラクトサミン鎖でもよい。
【0082】
ヘリコバクター ピロリ結合性物質は、抗生物質、好ましくはペニシリン系抗生物質、に結合していてもよい。ヘリコバクター ピロリ結合性物質は抗生物質をヘリコバクター ピロリにターゲティングする。このような結合体は、ヘリコバクター ピロリに対する処置や治療に必要な抗生物質が少量となるので治療に有利であり、その結果、抗生物質による副作用を低減させる。結合体の抗生物質部分は細菌を殺すか弱めるためのものであるが、後述するように、結合体には付着防止効果があってもよい。
【0083】
細菌結合性物質、好ましくはオリゴ価またはクラスター状のものは、ヘリコバクター ピロリの存在によって生じる疾患または病態の治療に用いることができる。これはヘリコバクター ピロリ結合性物質を付着防止に用いることで達成される、即ち、ヘリコバクター ピロリがターゲットとなる細胞や組織のレセプター エピトープに結合するのを阻害する。ヘリコバクター ピロリ結合性物質または医薬組成物を投与すると、それは細菌の結合に対してターゲット細胞上のレセプター糖結合体と競合する。その結果、細菌の一部またはすべてがターゲットとなる細胞や組織のレセプターの代わりにヘリコバクター ピロリ結合性物質に結合する。ヘリコバクター ピロリ結合性物質に結合した細菌は患者から(例えば、胃腸器系における液体の流入によって)除去され、その結果、患者の健康に細菌が与える影響を低減させる。使用する結合性物質は、好ましくはヘリコバクター ピロリ結合性物質を包含する可溶性の組成物である。結合性物質は、担体、好ましくはタンパク質以外の担体に結合させてもよい。担体分子を用いる場合には、ヘリコバクター ピロリ結合性物質の複数の分子を1つの担体に結合し、阻害効率を改善することもできる。
【0084】
ターゲット細胞は主としてターゲット組織(特に胃腸器系)またはターゲット組織と考えられるその他の組織(肝臓や膵臓)の上皮細胞である。ターゲット組織のグリコシル化は病原体による感染によって変化すると考えられる(Karlsson et al., 2000)。ターゲット細胞はターゲット組織中の悪性細胞、形質転換細胞または癌化/腫瘍細胞でもよい。形質転換した細胞または組織は異なる形のグリコシル化反応を発現し、細菌に対するレセプターを提供する場合がある。レクチンまたは糖類が糖タンパク質または糖脂質からなるレセプターの糖質と結合すること(炭水化物−炭水化物間相互作用)によって細胞が活性化され、癌化/悪性細胞の場合には、これは癌の増殖または転移につながる。オリゴ糖エピトープのいくつか、ならびに悪性細胞由来のシアリル化ポリラクトサミン(Stroud et al., 1996)は、癌関連抗原または癌抗原として報告されている。ヘリコバクター ピロリは胃リンパ腫とも関連する。本明細書に記載する結合性物質は前癌状態または悪性の細胞に対するヘリコバクター ピロリの結合ならびに癌の増殖および転移の活性化を防止するために用いることができる。結合阻害は胃がん、特にリンパ腫を治療する可能性がある。
【0085】
ターゲット細胞としては血液細胞、特に白血球が挙げられる。消化性潰瘍と関連するヘリコバクター ピロリ株、例えば、本発明で主として用いた株は、オプソニン作用を受けていないにもかかわらず、顆粒球による炎症反応を刺激した(Rautelin et al., 1994a,b)。細菌の食作用において最初に起こる事柄には特定のレクチン様相互作用が含まれると考えられ、その結果、顆粒球の凝集が生じる(Ofek and Sharon, 1988)。食作用に続いて酸化的バースト反応が生じるが、これはヘリコバクター ピロリ関連疾患の病因を導くものとも考えられる(Babior, 1978)。N−アセチルラクトサミン繰り返し単位を有するいくつかのシアリル化および非酸性グリコスフィンゴ脂質は顆粒球から単離されて、特徴付けられており(Fukuda et al., 1985; Stroud et al., 1996)、白血球表面のヘリコバクター ピロリに対する潜在的レセプターとして機能し得る。さらに、x2グリコスフィンゴ脂質も同じ材料から単離されている(Teneberg, S.、未発表)。本発明は、N−アセチルラクトサミン特異的レクチンおよびモノクローナル抗体(X2、GalNAcβ3Galβ4GlcNAc-)によって認識される、ヒト赤血球および顆粒球に存在するレセプター糖質の存在を立証する。ヘリコバクター ピロリ結合性物質は、ヘリコバクター ピロリに対する白血球の結合の阻害ならびに白血球の活性化に続く酸化的バーストおよび/または炎症の防止に有用である。
【0086】
ヘリコバクター ピロリは種々のオリゴ糖配列に結合し得ることが知られている。特定の株による結合の中にはより共生的な相互作用を示すものもあり、癌や重度の病態に進行することはない。癌型糖エピトープへの結合に関する本願のデータはヘリコバクター ピロリ結合性物質がより病原性の高い相互作用も防止することが可能であり、その際、他のレセプター構造に結合した病原性の低いヘリコバクター ピロリ株がいくらか残る。従って、細菌の完全な除去はヘリコバクター ピロリ関連疾患の予防には必ずしも必要ない。病原性の低い細菌は、病原性の高いヘリコバクター ピロリ株による感染の予防においてプロバイオティックな効果を発揮することも考えられる。
【0087】
さらに、ヘリコバクター ピロリは大きなポリラクトサミンオリゴ糖類をその表面に有し、少なくともいくつかの株は、本願で説明するように細菌が結合する未フコシル化エピトープを含有する。本願に記載した結合性物質はヘリコバクター ピロリ同士の結合を防止し、そうすることによって、例えば、細菌のコロニー形成を阻害することができる。
【0088】
本発明によると、ヘリコバクター ピロリ結合性物質を、所望により担体と共に医薬組成物に導入することができる。このような医薬組成物は、ヘリコバクター ピロリが患者に存在することによって生じる病態の治療またはヘリコバクター ピロリ結合性物質を用いたこのような病態の治療方法に適している。本発明によって治療することのできる病態の具体例としては、慢性表在性胃炎、胃潰瘍、十二指腸潰瘍、ヒトの胃における非ホジキン悪性リンパ腫、胃腺癌、ならびにある種の膵臓病、皮膚病、肝臓病、心臓病、乳幼児突然死症候群、自己免疫疾患(自己免疫性胃炎、悪性貧血および非ステロイド性抗炎症薬(NSAID)関連胃疾患を含む)であり、すべてまたは少なくとも一部がヘリコバクター ピロリ感染によって生じる病態である。
【0089】
ヘリコバクター ピロリ結合性物質を包含する医薬組成物は、さらに他の物質、例えば不活性なベヒクルまたは医薬的に許容される担体や保存剤などの公知の物質を包含してもよい。ヘリコバクター ピロリ結合性物質は他の薬剤、例えばヘリコバクター ピロリに対する抗生物質、と共に投与してもよい。
【0090】
ヘリコバクター ピロリ結合性物質またはこのような結合性物質を包含する医薬組成物は、適当な方法で投与すればよく、経口投与が好ましい。
【0091】
本願において「治療」とは、疾患または病態を治癒または改善するための治療と、疾患または病態の発症を予防するための治療の両方を意味する。治療は短期間または継続的に行うことができる。
【0092】
本願において「患者」とは、本発明による治療を必要とするヒトまたはヒト以外の哺乳類である。
【0093】
ヘリコバクター ピロリ結合性物質は、ヘリコバクター ピロリ結合性物質に結合するタンパク質または(炭水化物−炭水化物相互作用によって結合する)炭水化物をスクリーニングすることで1つ以上の付着因子の同定に用いることもできる。炭水化物結合性タンパク質はレクチンまたは炭水化物結合性酵素である。スクリーニングは、例えば、アフィニティークロマトグラフィーやアフィニティー交差結合法で行うことができる(Ilver et al., 1998)。
【0094】
さらに、ヒト組織に提示されているヘリコバクター ピロリ結合性物質に特異的に結合するか不活化する物質を用い、ヘリコバクター ピロリの結合を防止することもできる。このような物質の例としては、Erythrina cristagalliおよびErythrina corallodendronなどの植物性レクチン(Teneberg et al., 1994)、ジャガイモレクチンなどのポリラクトサミン結合性レクチン、およびSambucus nigra凝集素などのNeuNAcα3特異的レクチンなどが挙げられる。ヒトに用いる場合には、このような結合性物質は、その用途に適したものでなければならず、例えば、ヒト化抗体またはヒト由来の組み換えグリコシダーゼなどの非免疫原性であり、ヘリコバクター ピロリ結合性物質の末端単糖残基を開裂させることのできる物質である。しかし、胃腸器系においては、例えば食品に由来する、多種の天然のレクチンやグリコシダーゼが許容されている。
【0095】
また、ヘリコバクター ピロリ結合性物質を食品や飼料を含む栄養補助組成物の一部として用いることができる。ヘリコバクター ピロリ結合性物質を機能性食品または機能化食品の一部として用いることが好ましい。機能性食品は、ヘリコバクター ピロリがターゲットとなる細胞または組織に結合するのを阻害するか防止することで、ヒトまたは動物の健康に正の影響を与える。ヘリコバクター ピロリ結合性物質は既に知られている食品や機能性食品組成物の一部でもよい。機能性食品は、専門機関、例えば米国FDA、によって許可されている他の許容される食品材料を含有してもよい。ヘリコバクター ピロリ結合性物質は栄養補助剤、好ましくは食品添加剤や飲料物添加剤として機能性食品または機能性飲料を製造するために用いることができる。食品または食品添加剤は、例えば、ウシなどの家畜やその他の動物が多量のヘリコバクター ピロリ結合性物質を天然にその乳に産生するようにすることで得ることもできる。これは、動物がその乳に適切なグリコシルトランスフェラーゼを過剰発現させることで達成される。家畜の特定の品種または種を選択し、ヘリコバクター ピロリ結合性物質を多量に産生するように交配することができる。栄養補助組成物または栄養補助剤のためのヘリコバクター ピロリ結合性物質は、細菌や酵母などの微生物によって製造することもできる。
【0096】
ヘリコバクター ピロリ結合性物質を乳幼児用の食品の一部、好ましくは乳児用ミルクの一部として用いることは特に有用である。多くの乳児は、ヒトの天然乳の代わりとして特別な処方のミルクを与えられている。ヘリコバクター ピロリは特に乳児および幼児に高い感染性を示し、後に発症することを考慮すると、この段階で感染を予防することは妥当である。ヘリコバクター ピロリは乳幼児突然死症候群の原因となることも知られているが、細菌を一掃するための強い抗生物質による治療は、特に幼児や乳児には不適切である。
【0097】
さらに、ヘリコバクター ピロリ結合性物質をヘリコバクター ピロリの感染によって生じる病態の診断に用いることができる。診断用途には、ヘリコバクター ピロリ結合性物質を用いたヘリコバクター ピロリの型の同定方法も含まれる。結合性物質を診断または型同定に用いた場合、例えば、結合性物質はプローブやテストスティックに含まれていてもよく、所望により、テストキットの一部を構成してもよい。このようなプローブやテストスティックがヘリコバクター ピロリを含有するサンプルと接触すると、細菌はプローブまたはテストスティックに結合し、サンプルから取り出されてさらに分析される。
【0098】
末端残基は、好ましくはβ3結合したグルクロン酸であり、より好ましくはそのメチルアミドなどの6位にアミド基を有するグルクロン酸である。従って、配列の類似体や誘導体は三糖エピトープの末端6位を変化させるか誘導化することによって製造することができる。
【0099】
好ましいヘリコバクター ピロリ結合性物質
本発明のオリゴ糖配列は、予想に反して、薄層表面に提示された時に効果的なバインダーとなることが判明した。この方法は、糖脂質配列を多価の状態で提示することを可能にした。多価の状態で提示されたオリゴ糖配列の活性は驚くほど高いので、多価の状態こそが本発明のオリゴ糖配列を提示するのに好ましい方法である。
【0100】
糖脂質構造は、天然では多価の状態で細胞膜に提示されている。このような提示方法は後述する固相アッセイ法や糖脂質またはネオ糖脂質のリポソームを形成することで模倣することができる。
【0101】
疎水性ヘキサデシルアニリンの還元的アミノ化反応によって製造される本発明の新規なネオ糖脂質は、オリゴ糖を効果的に提示することができた。従来の細菌結合に用いられるネオ糖脂質結合体の大部分は、フォスファチジル エタノールアミン ネオ糖脂質のフォスフォロエステルなどの負の電荷を帯びた基を有している。このような化合物の問題点は、この物質の負の電荷とリン脂質構造に関連した天然の生物学的結合である。負の電荷を帯びた分子はタンパク質やその他の生物学的物質との多数の非特異的結合に関与することが知られている。さらに多くのこのような構造は不安定であり、酵素学的または化学的に分解されることがある。本発明は、オリゴ糖配列の非酸性結合体に関するものであり、これはオリゴ糖配列が非酸性化学構造に結合していることを意味する。非酸性結合体は中性の物質であることが好ましく、これはオリゴ糖配列が中性の、電荷を帯びていない化学構造体に結合していることを意味する。本発明において好ましい結合体は多価の物質である。
【0102】
従来技術においては、生物活性を有するオリゴ糖配列は、レセプター活性型オリゴ糖構造の一部を還元することで担体構造に結合していることが多い。アルキル鎖(-CH2-)nおよび/またはベンゼン環を含有する疎水性のスペーサーが用いられている。しかし、疎水性構造は、一般的にタンパク質やその他の生物活性分子との非特異的相互作用に関与することが知られている。
【0103】
後述する実施例で得られたネオ糖脂質に関するデータによると、アッセイに用いた実験条件においてネオ糖脂質化合物のヘキサデシニルアニリン部分は、研究に用いた細菌に対する非特異的結合を起こさなかった。ネオ糖脂質においては、糖結合体のヘキサデシニルアニリン部分は脂質層状の構造を形成すると考えられ、結合のためには存在していない。本発明は結合エピトープに属する単糖残基を還元すると結合が破壊されることを示す。さらに、還元された単糖は、レセプターエピトープと多価に提示された構造体とを繋ぐ親水性スペーサーとして使えることを見出した。本発明によると、生物活性を有するオリゴ糖を親水性スペーサーを介して多価またはオリゴ価の担体分子に結合し、多価またはオリゴ価の構造体を形成することが好ましい。本願においては、多価構造体(10個を超えるオリゴ糖残基を含むもの)およびオリゴ価構造体(2〜10個のオリゴ糖残基を含むもの)を全て多価構造体というが、用途によっては大きな多価構造体よりもオリゴ価の構造体のほうが好ましい場合がある。親水性スペーサーは少なくとも1つの水酸基を含有することが好ましい。さらに好ましいスペーサーは少なくとも2つ、最も好ましくは少なくとも3つの水酸基を含有する。
【0104】
本発明によると、親水性スペーサーは、1つまたは複数の−CHOH−基および/またはアセトアミド基である−NHCOCH3やアルキルアミド基などのアミド側鎖を含有する可撓性鎖であることが好ましい。水酸基および/またはアセトアミド基はin vivoにおける酵素的加水分解からスペーサーの保護も行う。可撓性という用語は、スペーサーが可撓性の結合を有し、可撓性ではない環構造を形成しないことを意味する。本発明において(例えば、還元的アミノ化反応によって得られた)還元単糖残基は、可撓性親水性スペーサーの一例である。可撓性親水性スペーサーはネオ糖脂質や多価結合体の非特異的結合を防止するために最適である。これは、例えば、医薬や機能性食品におけるバイオアッセイおよび生物活性において必須な最適活性である。
【0105】
可撓性親水性リンカーを有する結合体は下記式IVで表される。
[OS-O-(X)n-L1-CH(H/{CH1-2OH}p1)-{CH1OH}p2-{CH(NH-R)}p3-{CH1OH}p4-L2]m-Z (IV)
(式中、
OSはヘリコバクター ピロリ結合性オリゴ糖配列であり;
Xは炭素(−C−)、窒素(−N−)、または硫黄(−S−)で置換可能な糖残基であり;
L1およびL2は結合基あって、各々独立に酸素原子、窒素原子、硫黄原子または炭素原子である結合原子あるいは置換基を形成する結合に含まれる2個の原子を包含し、該結合原子および該置換基を形成する結合は−O−、−S−、−CH2−、−N−、−N(COCH3)−、−CO−NH−または−NH−CO−で表されるアミド基、−N−N−(ヒドラジン誘導体)、あるいは−O−N−または−N−O−で表されるアミノオキシ結合であり;
L1は該Xの還元末端単糖のC1位に結合した結合基であり、nが0の場合には、L1は−O−を置換して、該OSの還元末端のC1位に直接結合しており;
p1、p2、p3およびp4は各々独立に0〜7の整数であるが、但し、p1、p2、p3およびp4の少なくとも1種が1以上の整数であり;
{CH1-2OH}p1が側鎖構造を示す場合には、末端基となるCH1-2OHはCH2OHであり、p1が1を超える場合には、該末端基をスペーサーである該Yの残りの部分に結合させるための第二級アルコール基である−CHOH−が存在し;
Rは好ましくはアセチル基(−COCH3)、Zに連結する結合、類似体形成基、水素原子または式中でアミンを形成するC1-4アルキル基であり、Rが結合である場合にはL2は糖鎖末端基の1個または2個の原子であり、該類似体形成基は、アミド構造を有するC1-4アシル基(好ましくは親水性の基、例えばヒドロキシアルキル基)を含み、;
mは1を超える整数であり;
Zは多価の担体である。)
【0106】
式(IV)に示した可撓性親水性スペーサーを有する好ましい多価構造体としては、ヘリコバクター ピロリ結合性オリゴ糖配列(OS)が Galβ4Glc(red)-Zにβ1−3結合したもの、OSβ6GlcNAc(red)-Z および OSβ6GalNAc(red)-Z(式中、“(red)”は還元末端単糖および多価担体Zのアミノ基の還元的アミノ化によって得られたアミン結合構造である)が挙げられる。
【0107】
本発明においてオリゴ糖基は、抗原性やアレルギー反応の出現を防止するために、多価またはオリゴ価の形でタンパク質またはペプチドではない担体に結合していることが好ましく、より好ましくは、担体の骨格は天然の非抗原性多糖である。
【0108】
本発明のex vivoにおける用途
本発明により、生体外(ex vivo)における病原体、特に下痢発症性大腸菌の阻害に用いることができることが判明した。また、このような方法は、消毒および保存のために用いることもできる。好ましくは単一成分物質の部分または単一成分物質そのものである本発明で開示するレセプターオリゴ糖配列、さらに好ましくは、少なくとも2種の本発明で開示するレセプターオリゴ糖配列を包含する組成物を、病原体、好ましくは大腸菌を阻害するためにex vivoで用いる。本発明で開示する多価結合体、特に、病原体、好ましくは下痢原性大腸菌を凝集することができる可溶性多価結合体が、ex vivoでの使用には好ましい。本発明のex vivoでの使用の特殊な態様として、病院や準病院環境などで使用するテーブル、医療器具および包装容器などの表面の洗浄や消毒において、本発明で開示するレセプターオリゴ糖配列を含有する洗浄剤や消毒剤を用いる方法が挙げられる。本発明で開示するレセプター糖類はまた、病院や準病院環境などにおいて患者の洗浄用または入浴用の石鹸や洗剤の成分としても有用である。
【0109】
口腔感染症および口腔衛生製品
本発明が標的とする感染症は経口経路で、またおそらく鼻から口腔への経路でも広がるものと考えられる。本発明は、ヒトの口に存在している感染症の阻害に関する。本発明は、少なくとも2種の異なるオリゴ糖配列による口腔感染症の治療に関し、該オリゴ糖配列は、病原体(好ましくは経口的に感染する細菌、さらに好ましくは下痢発症性細菌)の結合特異性の少なくとも2種を阻害するものである。口腔感染症や鼻腔感染症の予防のために、単一成分物質の部分または単一成分物質そのものとして本発明で開示するレセプターオリゴ糖配列を用いるか、本発明の異なる種類のレセプターオリゴ糖配列を少なくとも2種包含する組成物として用いることが好ましい。本発明によると、本発明で開示したレセプターオリゴ糖配列を、組成物または単一成分物質として、ヒトの口腔中の病原体を阻害する製品(ここでは「口腔衛生製品」と称する)に用いる。
【0110】
ヒトの口腔には、ヒトの腸と同様のレセプターが存在すると考えられ、特にタンパク質上には、少なくともネオラクトレセプター、マンノースレセプター、および本発明のフコースレセプターに類似したオリゴ糖レセプターが存在するものと考えられる。別の態様においては、本発明の物質および組成物は、カリエスを発症させる病原体を阻害するのにも有用である。具体的な態様においては、本発明は、経口的に広がる他の感染症、例えば、中耳炎を併発する感染症やインフルエンザ、気管支炎または肺炎を含む肺の感染症を治療するための組成物にも関する。本発明の口腔衛生製品は、カリエス、中耳炎、気管支炎および肺炎の治療を目的とすることもできる。本発明の具体的態様の一例として、Neu5Acα3Galβ4GlcNAc および Neu5Acα3Galβ4Glcからなる群より選ばれる少なくとも1種のオリゴ糖配列、より好ましくは Neu5Acα6Galβ4GlcNAc および Neu5Acα6Galβ4Glcからなる群より選ばれる少なくとも1種のオリゴ糖配列を包含する、口腔衛生製品の製造または経口的に感染する病原体の阻害のために用いる組成物がある。そのような組成物は、少なくともヒト インフルエンザウイルスの処置を目的とし、好ましくはインフルエンザウイルスに対する予防を目的としたものである。
【0111】
特に本発明は、病原体阻害性オリゴ糖配列、特に本発明で開示するオリゴ糖配列、を含む物質または組成物を含有する口腔衛生製品に関する。口腔衛生製品は、練り歯磨、洗口薬、口腔用タブレット、咀嚼錠(chewing tablets)およびチューインガムからなる群より選ばれることが好ましい。一価のレセプターオリゴ糖配列または多価のレセプターオリゴ糖配列のいずれかを用いることが好ましい。他の一つの好ましい態様においては、口腔衛生製品は、本発明で開示する多価のオリゴ糖配列を含有する。ヒトの口腔の大きさおよび口腔表面に存在する唾液の容量を考慮すると、口腔において病原体阻害性レセプターの飽和濃度を達成するには比較的少量のオリゴ糖で十分である。レセプター活性を有する一価エピトープの典型的な量は、レセプター活性オリゴ糖として約100nmol〜100μmol(分子量が1,000Daの場合、100μg〜100mgに相当する)の範囲内である。より一般的な量は、約1〜10μmolと推定される。別の態様においては、本発明の治療用医薬組成物は、病原体を阻害するスプレー式点鼻薬にも関する。スプレー式点鼻薬は、中耳炎または肺の感染症の治療を目的としたものである。
【0112】
局所用製品、洗剤および化粧品
一般的な病原体は、ヒトの肌、性器上皮、毛髪、日用品の表面およびヒトの環境中に存在する他の表面に広がりうると考えられる。本発明のオリゴ糖配列は、このような環境中に存在する病原体の阻害にも有用である。したがって、局所用製品または化粧品(例えば、クリーム、ローションまたはジェル)に、単一成分物質の部分または単一成分物質そのものとして本発明のオリゴ糖配列を用いるか、本発明の異なる種類のレセプターオリゴ糖配列を少なくとも2種包含する組成物を用いることも好ましい。また、本発明の物質または組成物を、ヒトの肌、毛髪または性器上皮を洗浄するための製品(個人用衛生製品とも言う)や、日用品の表面、皿または衣服を洗浄するための製品に用いることも好ましい。従来の抗生物質は、日用品用の洗浄液に用いることを目的としているが、本発明の物質とは違って耐性の問題があるため有用ではない。好ましい態様においては、多価オリゴ糖配列を洗浄液に用い、他の一つの好ましい態様においては、一価オリゴ糖配列を洗浄液に用いる。
【0113】
食品または飼料、飲料、酒類および水に用いる食品安全性向上製品
ヒトまたは動物の治療方法の他に、本発明は、本発明で開示するレセプターおよび組成物を用いた感染症の予防方法、即ち、本発明を用いて食品製品の内部または表面上の病原体や細菌を中和する方法に関する。例えば、病原体の蔓延を防止するために、食肉生産の際に本発明の炭水化物を食肉製品や動物の体またはその部分の表面に塗布することができる。細菌を被覆し凝集させるために可溶性多価結合体および他の多価結合体を用いることが好ましい。固体または半固体の食品製品の表面に用いる具体的な方法には、細菌に本発明の炭水化物レセプターを接触させ、所望により病原体と炭水化物との複合体を洗い流すことが含まれる。この種の方法には、従来の抗生物質を適用することはできない。本発明の炭水化物は、液状食品製品、濃縮物またはそれらの元となる粉末、例えば、ミルク、液状ミルク様製品およびジュース、ソフトドリンク、スポーツドリンク、アルコール飲料などの種々の飲料に対して用いることもできる。
【0114】
具体的な態様においては、本発明の炭水化物を多価の型で液状食品製品または飲料製品に対して使用し、潜在的な病原体をこのような多価結合体によって凝集させ、凝集複合体をその大きさや溶解性に基づいた方法によって除去する。不溶性の凝集物は、沈殿物として標準的な方法、例えば、凝集物の上に存在する溶液をデカンテーションする方法や、より一般的且つ効果的なろ過法で除去することができる。ろ過法は、より大きな凝集複合体を除去するのに用いることができる。
【0115】
本発明の炭水化物で処理する好ましい食品としては、種々の動物性食品製品、特に食肉製品および加工途中の製品が挙げられる。下痢発症性大腸菌菌体を含む多くの病原体は、正しく洗浄されていない野菜、果物、サラダおよび他の植物性食品から伝播する。洗浄が必要であるにもかかわらず、正しく洗浄されていないか汚染された水で洗浄された食物は、発展途上国で特に問題となっている。本発明は、洗浄することが必要な植物性食品や他の食品の安全性を向上させるための方法であって、病原体、特に食品製品中の病原性大腸菌の菌体量を制御するための方法に関する。特に本発明は、食品からの感染を防止するための、家庭用製品および食品工業用製品にも関する。このような製品は、加工中の食品に利用することができる、粉末状または錠剤状の固体製品か、本発明で開示するレセプターの溶液を内包するカプセルであることが好ましい。このような製品は、発展途上国で発生する下痢、特に子供の下痢の予防に用いることができる。食品安全性向上製品は、旅行者の下痢の予防も目的とする。食品安全性向上製品および以下に述べる飼料安全性向上製品は、新規な安全性向上保存料であると考えられる。
【0116】
飲料および水を精製するためのフィルター製品
汚染された飲料および水は胃腸器系の疾患、特に下痢の主要な病因である。本発明で開示するレセプターは、液状食品、飲料および水(特に、飲料および食品を作るための水)に存在する病原体、特に細菌を除去するためのフィルターの作製に用いることもできる。少なくとも2種のレセプター構造を用いることが好ましい。フィルターを得るために炭水化物配列を結合させる固相材料を、例えばセルロース、プラスチック、アガロースおよびそれに類似した材料から製造する方法は知られている。本発明のフィルターとしては、公知技術であるアフィニティークロマトグラフィー用の材料も挙げられる。このようなフィルターから結合物を除去するための方法は公知であり、具体的な態様においては、フィルターは、汚染物質を除去し、所望により熱または他の滅菌方法により滅菌することにより、再生する。
【0117】
飼料安全性向上製品
上記の食品安全性向上製品は、動物用の固体および液状の飼料ならびに動物用の飲料水に用いることもできる。保護する動物として好ましいものとしては愛玩動物、特にネコとイヌ、および家畜または農場動物(例えばウシや他の反芻動物、ブタ、ヒツジ、ウマ、家禽(例えばメンドリ、アヒルや七面鳥)およびウサギ)が挙げられる。
【0118】
水、食品および飼料の安全性の分析
本発明のレセプター炭水化物を組み合わせた標準的な分析および診断方法を用いて水、飲料、食品および飼料を分析し、レセプター炭水化物に結合する病原体の存在量を測定することができる。汚染病原体の結合特異性に関する知見は、感染患者の治療計画をたてる場合や、上記の病原体を除去または制御して食品の安全性を向上させるための方法に適用することができる。
【0119】
本発明によって阻害することのできる、炭水化物に基づく他の相互作用
異なる種類の付着因子の阻害の他にも、本発明は、炭水化物−炭水化物相互作用および炭水化物−レクチン相互作用の阻害に用いることもできる。
【0120】
本発明の炭水化物含有組成物および炭水化物含有物質はオリゴ糖配列を包含している。オリゴ糖配列は、少なくとも1種の病原体および/または少なくとも1種の病原体に対するレセプターに結合することによって、少なくとも1種の病原体を阻害する。少なくとも2種の病原体阻害性オリゴ糖配列を用いることが好ましく、少なくとも3種の病原体阻害性オリゴ糖配列を用いることがより好ましい。他の好ましい態様においては、少なくとも4種、5種、6種または7種の病原体阻害性オリゴ糖配列を用いる。
【0121】
具体的な治療においては、1つまたは複数のオリゴ糖配列を異なる時期に別々に投与する。これは、全種類のオリゴ糖配列の同時投与が正常細菌叢に負の影響を与える場合に特に有効である。治療用組成物を別々に投与することは、胃腸器系における本発明のオリゴ糖配列の安定性が胃腸器系の栄養状態の影響を受けることからも有効である。
【0122】
本発明とプロバイオティック細菌を一緒に用いる方法
本発明を細菌の結合、特に複数の細菌の結合を阻害するために用いる場合、いくつかの有益な細菌の結合も阻害されることが考えられる。正常細菌叢は、例えばヒト胃腸器系において、多くの重要な機能を有する。しかし、従来の抗生物質を用いた場合に生じる正常細菌叢の破壊は、より大きな問題となる。
【0123】
他の態様においては、少なくとも2種の病原体阻害性オリゴ糖配列を、プロバイオティック微生物および/またはプレバイオティック化合物と共に投与する。本発明においてプロバイオティック微生物とは、例えば食物の消化、栄養およびビタミンの提供または組織表面の被覆による病原性細菌からの保護といった有益な機能を有する無害な細菌を意味する。プロバイオティック細菌には、正常細菌叢から選ばれる1種、数種または多種の細菌が含まれることが好ましい。好ましい態様においては、プロバイオティック細菌には、少なくとも1種類または1株または1種(species)の乳酸菌が含まれる。
【0124】
プレバイオティック化合物とは、正常細菌叢またはプロバイオティック微生物を支える物質である。好ましいプレバイオティック化合物としては、プレバイオティック炭水化物(例えば、プレバイオティック化合物として用いられるガラクトースオリゴ糖、キシロースオリゴ糖またはフルクトースオリゴ糖)が挙げられ、プレバイオティック活性を有する多糖および繊維(例えばイヌリンまたは修飾した澱粉)も含まれる。本発明は、食品または栄養補助的目的のために用いられる他の多糖、例えばキトサンまたはβ−グルカン(例えばオート麦のグルカン)を用いてコレステロールおよび脂肪を減少させる方法にも関する。好ましい態様においては、非還元末端にβ−結合したガラクトース残基を有する炭水化物のような、プレバイオティック化合物としても機能する少なくとも1種の病原体阻害性炭水化物を選択する。好ましい治療方法においては、
a)病原体および正常細菌叢と考えられるものの一部を、1種、さらに好ましくは少なくとも2種の本発明の炭水化物を用いて初めに除去し、そして
b)プロバイオティック微生物および/またはプレバイオティック化合物を投与する。
上記の第一および第二の工程は逆の順番で行ってもよく、好ましくは大量のプロバイオティック微生物および/またはプレバイオティック化合物を用いてから、第一の工程を行う。本発明によると、工程の順番を変えながら、第一および/または第二の工程を数回繰り返すこともできる。第一および第二の工程は同時に行ってもよい。本発明の物質は、プロバイオティック微生物および/またはプレバイオティック化合物と共に投与することができるが、プロバイオティック微生物および/またはプレバイオティック化合物を本発明の組成物に含ませることもでき、その場合、第一および第二の工程は同時に行うことができる。
【0125】
本発明のオリゴ糖配列の中には、プレバイオティック効果を有することが知られているものがある。そのようなものの例としては、N−アセチル−ラクトサミン型オリゴ糖配列や、特にヒト乳オリゴ糖などのフコシル化オリゴ糖が挙げられる。また、プロバイオティック微生物および/またはプレバイオティック化合物と共にヒト乳オリゴ糖、特に、例えばラクト−N−ネオテトラオース、ラクト−N−テトラオース、ラクト−N−ヘキサオース、ラクト−N−ネオヘキサオース、パララクト−N−ヘキサオース、およびパララクト−N−ネオヘキサオースからなる群より選ばれる少なくとも1種を含有するN−アセチル−ラクトサミン、および/またはそれらより誘導されるフコシル化オリゴ糖、および/または、例えば、モノフコシル化、ジフコシル化またはトリフコシル化したラクト−N−テトラオース(LNT)やラクト−N−ネオテトラオース(LNnT)、および/またはフコシルラクトースであるオリゴ糖(例えば2'フコシルラクトース、3−フコシルラクトースやジフコシルラクトース)を投与する方法がある。
【0126】
本発明の物質および/または組成物と共に用いられるその他の有用な物質
本発明によると、病原体阻害性(pathogenesis preventing)炭水化物を、グリコシダーゼ阻害剤、好ましくはシアリダーゼ阻害剤と併用することも有用である。
【0127】
本発明によると、病原体阻害性炭水化物を、レクチンまたはそれ以外の炭水化物結合性タンパク質と併用することも有用である。レクチンは、炭水化物レセプター、例えば、菌体外多糖類をブロックするために用いることができる。
【0128】
糖脂質および炭水化物の命名は、IUPAC−IUB生化学命名委員会の推奨する命名法(Carbohydrate Res. 1998, 312, 167; Carbohydrate Res. 1997, 297, 1; Eur. J. Biochem. 1998, 257, 29)に従った。
【0129】
Gal、Glc、GlcNAc および Neu5AcはD型であり、FucはL型であり、全ての単糖単位はピラノース環構造であるものとする。グルコサミンはGlcNまたはGlcNH2と示し、ガラクトサミンはGalNまたはGalNH2と示す。グリコシド結合は略記で示す場合と正式名称で示す場合とあり、Neu5Ac-残基のα3結合およびα6結合はそれぞれα2−3結合およびα2−6結合と同じであり、他の単糖残基については、α1−3結合、β1−3結合、β1−4結合およびβ1−6結合はそれぞれα3、β3、β4およびβ6と略すことができる。ラクトサミンはN−アセチルラクトサミン(Galβ4GlcNAc)を示し、シアル酸はN−アセチルノイラミン酸(Neu5Ac, NeuNAc または NeuAc)、N−グリコリルノイラミン酸(Neu5Gc)またはその他の天然のシアル酸を示す。本願においてグリカンという用語は、ヒトまたは動物に存在する糖結合体、特に糖脂質または糖タンパクに含まれるオリゴ糖や多糖鎖を広く包含する。脂肪酸と塩基の略記命名法においては、コロンの前の数字が分子の炭素鎖の長さを表し、コロンの後の数字が炭化水素鎖の二重結合の合計を示す。GSLという略語はグリコスフィンゴ脂質を表す。グリコスフィンゴ脂質の略語、略称または記号は表1および2に記載した。ヘリコバクター ピロリとは、ヘリコバクター ピロリに類似した細菌も意味する。
【0130】
「末端オリゴ糖配列」または「末端配列」という表現は、オリゴ糖配列またはシアル酸誘導体配列が、所望によりオリゴ糖配列の還元末端から結合している場合を除いては、他の単糖構造またはオリゴ糖構造に結合していないことを表す。オリゴ糖配列が結合体として存在している場合は、オリゴ糖配列はその還元末端から結合していることが好ましいが、病原体結合性に影響しない他の結合位置を用いることもできる。さらに具体的な態様においては、本発明のオリゴ糖配列とは、天然のグリコシド結合によって他の単糖構造またはオリゴ糖構造に結合していない、上記配列に相当するオリゴ糖残基を意味する。このようなオリゴ糖残基は、遊離オリゴ糖、結合体を形成するもの、またはオリゴ糖残基の還元末端を誘導化したものであることが好ましい。
【0131】
「α3/β3」は、オリゴ糖配列において隣り合った糖残基は互いにα3−またはβ3−結合していることを表す。
【発明を実施するための最良の形態】
【0132】
本発明を以下の実施例によってさらに説明するが、これらは本発明の範囲を限定するものではない。
【0133】
ガングリオシドの調製 − ガングリオシドを単離するために、複合ガングリオシドを含有することが知られている種々の組織、例えばヒト赤血球、ウシ赤血球、ウサギ胸腺、ヒト胎便およびヒト癌(Stults et al., 1989)を収集した。総酸性グリコスフィンゴ脂質画分の単離は公知の方法(Karlsson, 1987)で行った。端的に記すと、各組織を凍結乾燥し、次いでソックスレー抽出器でクロロフォルム/メタノール混合溶媒(2:1と9:1、容量比)を用いた2段階の抽出を行った。抽出物をプールし、穏やかなアルカリ加水分解および透析に付した後、ケイ酸カラムによる分離に付した。酸性グリコスフィンゴ脂質と非酸性グリコスフィンゴ脂質はDEAEカラムで分離した。
【0134】
酸性グリコスフィンゴ脂質画分をDEAE−セファロ−スクロマトグラフィーにより分離し、さらにケイ酸クロマトグラフィーに繰り返し付し、最終的な分離をHPLCにより行った。HPLCは、長さ250mm、内径10mmで粒径が5μmのKromasil 5 シリカカラム(米国、カリフォルニア州、トランス、Phenomenex製)を用い、クロロフォルム/メタノール/水の直線濃度勾配(60:35:8〜40:40:12または65:25:4〜40:40:12、容量比)で、流速2ml/分で180分間行った。回収した各2mlの画分をそれぞれ薄層クロマトグラフィーおよびアニスアルデヒド染色(下記参照)により分析し、ヘリコバクター ピロリ結合能をクロマトグラム結合アッセイ(下記参照)により評価した。薄層クロマトグラム上の移動度およびヘリコバクター ピロリ結合能に基づいてHPLC画分をプールした。
【0135】
参照用グリコスフィンゴ脂質 − 参照用グリコスフィンゴ脂質は、スウェーデン国、イェーテボリ大学、医薬生化学研究所(Institute of Medical Biochemistry)にて単離し、特徴付けた。構造の特徴付けはプロトンNMR法(Koerner et al., 1983)、質量分析法(Samuelsson et al., 1990)および分解的手法(degradation studies)(Yang and Hakomori, 1971、Stellner, 1973)により行った。シアリル−Lex ヘキサグリコシルセラミドはARC社(カナダ国、エドモントン)より購入した。
【0136】
細菌株、生育条件および標識 − ヘリコバクター ピロリ CCUG 17874株はイェーテボリ大学カルチャーコレクション(Culture Collection University of Goteborg(CCUG))より入手した。J99株はTim Cover博士、John Atherton博士およびMartin Blaser博士より寄贈されたものである。
【0137】
細菌の培養は、10%の56℃で不活性化したウシ胎児血清(英国、ラフバラ、Harlan Sera-Lab製)とBBL IsoVitale X エンリッチメント(ニュージャージー州、フランクリン・レイクス、Becton Dickson Microbiology Systems製)を含有するブルセラ培地(カリフォルニア州、アーヴィン、Difco Laboratories製)で行った。細菌の放射線標識は、0.5mlのリン酸緩衝化生理食塩水(PBS)(pH7.3)で希釈した50μCiの35S−メチオニン(英国、リトル・チャルフォント、Amersham Pharmacia Biotech製)を、固体培養中の菌に添加することにより行った。微好気性条件下、37℃で12〜72時間インキュベートした後に細菌を回収し、回収した細菌をPBS中、3,500rpmで10分間の遠心分離に3回付した。
【0138】
放射線標識のもう一つの方法においては、10%の熱不活性化ウシ胎児血清と50μCiの35S−メチオニンを添加したハムF12培地(英国、カリフォルニア州、カールズバッド、Invitrogen Corp.製)に(1×105CFU/mlとなるように)コロニーを接種した。この培地の入った培養用ボトルを微好気性条件下、37℃で24時間、振とうさせながらインキュベートした。菌体は遠心分離によって回収し、PBSで3回洗浄した。
【0139】
上記いずれの方法においても、標識した細菌の濃度が約1×108CFU/mlになるように、最後に2%(w/v)ウシ血清アルブミン含有PBS(PBS/BSA)に再懸濁した。いずれの標識方法によっても、比活性がヘリコバクター ピロリ100個あたり約1cpmの懸濁液が得られた。
【0140】
薄層クロマトグラフィー − 総酸性グリコスフィンゴ脂質画分(40μg)または純粋なガングリオシド(0.0002〜4μg)を、アルミニウムで裏打ちしたシリカゲル60HPTLCプレート(ドイツ国、ダルムシュタット、Merck製)上で、クロロフォルム/メタノール/水(60:35:8、容量比)、クロロフォルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)または1−プロパノール/水/25%NH3(7:3:1、容量比)を溶媒系として分離した。薄層クロマトグラム上のグリコスフィンゴ脂質の化学的検出は、アニスアルデヒド試薬(Waldi, 1962)またはレゾルシノール試薬(Svennerholm, 1963)を用いて行った。
【0141】
クロマトグラム結合アッセイ − クロマトグラム結合アッセイは、公知の方法(Angstrom et al., 1994)と実質的に同様に行った。分離したグリコスフィンゴ脂質を含む乾燥薄層クロマトグラムを、0.5%(w/v)ポリイソブチルメタクリレート(ウィスコンシン州、ミルウォーキー、Aldrich Chemical Company Inc.製)のジエチルエーテル/n−ヘキサン(1:5、容量比)溶液で1分間処理し、風乾した。非特異的結合を減少させるために、プレートを0.1%NaN3(w/v)と0.1%Tween20(容量比)を含有するPBS/BSA中で、室温で2時間インキュベートした。次いでプレートをPBS/BSAで希釈した35S−標識ヘリコバクター ピロリと共に室温で2時間インキュベートした。また、薄層クロマトグラム上のGM1ガングリオシド希釈系列に対して125I−標識コレラトキシンB−サブユニットを公知の方法(Karlsson and Stromberg, 1987)で結合させた。最後の洗浄と乾燥を行った後、Biomaxフィルム(米国、ニューヨーク州、Eastman Kodak Company製)を用いてオートラジオグラフィーを一晩行った。他のガングリオシドに対する、1つの純粋なガングリオシドへの細菌の結合量を定量するために、薄層クロマトグラム上のガングリオシド希釈液系列に対する細菌の結合と、それに続くオートラジオグラフィーの後に、得られたオートラジオグラムをデンシトメトリーに付した。ガングリオシドの濃度範囲は、飽和濃度を求めた予備実験に基づいて選択した。オートラジオグラムはCCDカメラ(インディアナ州、ミシガンシティ、Dage-MTI, Inc.製)で撮影し、得られたイメージをパブリックドメインプログラムであるNIHイメージプログラム(米国国立衛生研究所(U.S. National Institute of Health)によって開発され、http://rsb.info.nih.gov/nih-image/より入手可能)を用いて解析した。ここに示したデータは、バックグラウンド値を引いた値である。
【0142】
ガングリオシドの誘導化 − ガングリオシドは、水酸化ナトリウムとヨウ化メチルのジメチルスルホキシド溶液を用いて公知の方法(Larsson et al., 1987)によりパーメチル化した。パーメチル化した試料はLiAlH4のジエチルエーテル溶液を用いて還元した(Karlsson, 1974)。
【0143】
高速原子衝突質量分析および電子イオン化質量分析 − 陰イオンFABおよびEI質量スペクトルをJEOL SX 102A質量分析計(日本国、東京、日本電子株式会社製)を用いて得た。誘導化前のガングリオシドの陰イオンFAB質量スペクトルは、キセノン原子衝突(6eV)、加速電圧(〜8kV)およびトリエタノールアミンをマトリックスとする条件下で得た。誘導化したグリコスフィンゴ脂質のEIスペクトルは、イオン化電圧70eV、イオン化電流300μAおよび加速電圧8kVの条件下で得た。温度は、150℃から410℃に昇温速度10℃/分で昇温した。FABスペクトルおよびEIスペクトルのいずれのスペクトルも取得する際の解像度は1000とした。
【0144】
エレクトロスプレーイオン化質量分析法および衝突誘起解離法 − ヒト赤血球由来の、パーメチル化ヘリコバクター ピロリ結合性ガングリオシドを10%(容量比)アセトニトリル水溶液に溶解した。エレクトロスプレーイオン化は、Q−TOF質量分析計(英国、Micromass製)でナノフローエレクトロスプレーオプションを用いて、約15〜40nL/分の一定流速で行った。噴霧はナノスプレーニードルに900Vの電圧をかけることにより行った。イオン源温度は80℃とした。流速150リットル/時間の窒素を乾燥用ガスとして用いた。質量スペクトルにおいてm/z1594.4で観察された二価荷電ピークを、衝突誘起解離を行うために選択した。突誘起解離は、アルゴンを30〜80Vの衝突エネルギーで用いることにより達成した。データは50〜2600質量単位に渡って回収した。ここでは、ドモン(Domon)とコステロ(Costello)の命名法(Domon and Costello, 1988)を使用した。
【0145】
プロトンNMRスペクトル法 − 1H NMRスペクトルは、Varian 500MHz NMR分光計およびVarian 600MHz NMR分光計を30℃で用いて得た。試料は、重水素置換後に、ジメチルスルホキシド/D2O(98:2、容量比)に溶解した。
【0146】
詳細なエピトープ分析実験に用いる特別な材料および方法
天然糖脂質の入手源
PGC(ヒト赤血球由来)は、パーアセチル化法(Miller-Podraza et al., 1993)に従って本発明者らが単離した。S−3−PG(ヒト赤血球由来およびヒト白血球由来)、ジシアリルパラグロボシド(ヒト赤血球由来)、S−6−PG(ヒト白血球由来)、七糖ネオラクトガングリオシド(ヒト赤血球由来およびヒト白血球由来)およびグロボシド(ヒト赤血球由来)も本研究室で調製した(Karlsson, 1987)。ヒト脳のGQ1bは、イェーテボリ大学神経化学科(Department of Neurochemistry of Goteborg University)より入手した(Miller-Podraza et al., 1992)。ウシ脳由来のガングリオシドであるGM1、GD1a、GD1bおよびGT1bは、Calbiochem(米国)から購入した。
【0147】
炭水化物の入手源
NeuAcα3Gαβ4GlcNAcβ3Galβ4Glcは、S−3−PG(ヒト赤血球由来)をセラミドグリカナーゼ(ヒル由来;ドイツ国、Boehringer Mannheim GmBH製)により消化し(Ito and Yamagata, 1989)、クロロフォルム/メタノール/水(2:1:0.6)を用いた相分離に付すことにより、本研究室で調製した。上記五糖を上相から回収した。NeuAcα3Galβ3GlcNAcβ3Galβ4Glc、NeuAcα6Galβ4GlcNAcβ3Galβ4GlcおよびGalβ3(NeuAcα6)GlcNAcβ3Galβ4Glcは、IsoSep(スウェーデン国、トウリンゲ)より入手した。
【0148】
その他の試薬
4−ヘキサデシルアニリン、メチルアミン、エチルアミン、プロピルアミン、ブチルアミンおよびベンジルアミンは、Aldrich Chemical Company(米国、ミルウォーキー)製を用いた。グリコール酸、エタノールアミンおよびオクタデシルアミンは、Sigma-Aldrich(ドイツ国)から購入した。Sephadex LH 20は、Pharmacia(スウェーデン国、ウプサラ)製であり、エチレングリコールはFluka(スウェーデン国)製であった。
【0149】
シアル酸のグリセロール尾部の化学的修飾
A.過ヨウ素酸による穏やかな酸化(Veh et al., 1977)とそれに続く還元反応(R-CHOH-CHOH-CH2OH → R-CHOH-CH2OH/R-CH2OH):
反応材料(0.5〜1μmol)を、1〜2mMのNaIO4を含有する0.05mM酢酸緩衝液(pH5.5)500μl中で、氷上で40分間インキュベートすることにより反応を行った。反応を過剰量のエチレングリコールにより停止し、反応混合物を試料とした。次いで、試料を凍結乾燥により濃縮し(約5倍)、室温で一晩、過剰量のNaBH4により還元反応に付した。最後にこのような処理を施した試料を蒸留水に対して2日間透析し、次いで凍結乾燥した。
【0150】
B.S−3−PGの過ヨウ素酸による穏やかな酸化とそれに続くメチルアミンまたはエタノールアミンとのカップリング(R-CHOH-CHOH-CH2OH → R-CH2-NH-CH3/R-CH2-NH-CH2-OH):
ヨウ素酸を用いたS−3−PGの穏やかな酸化(上記参照)とエチレングリコールの添加の後、得られた反応混合物を蒸留水に対して2日間透析し、凍結乾燥して酸化S−3−PGとした。以下の条件下で酸化S−3−PGとメチルアミンまたはエチルアミンとのカップリングを行った:糖脂質(0.5mg)を200μlのM/C(3:1)に溶解し、50μlのアミン、200μlの、NaBH3CNのメタノール溶液(62mg/ml)および200μlのグリコール酸水溶液(136mg/ml)と混合し、試料を得た。メチルアミンを用いた場合には、溶解性を高めるために50μlのテトラヒドロフランをさらに添加して混合した。得られた試料を30℃で4時間インキュベートした後、窒素雰囲気下で水分を蒸発した。得られた残渣をC/M/水(60:30:4.5)に懸濁し、メタノールで飽和させたSephadex LH-20カラムを用いて脱塩した。試料(約0.6ml/0.5×15cmのカラム)をカラムにアプライした後、カラムをメタノールによる溶出に付し、糖に対して陽性を示した画分を収集することにより糖脂質を回収した(TLCとアニスアルデヒドによりモニターした)。分子種(上記式参照)の最終的な精製を行うために、C/M/水(60:35:8)を展開溶媒とした分取TLCにより、目的材料を分離した。(プレートからストリップを切り出した後にアニスアルデヒドで検出した)主要なバンドをかき取り、上記と同じ溶媒で抽出した。
【0151】
カルボキシル基の修飾(Lanne et al., 1995)
S−3−PG(0.5〜5mg)をまず、ヨウ化メチル(100μl)のジメチルスルホキシド(DMSO、0.5ml)溶液と共に室温で1時間インキュベートすることでメチルエステルに転化した(R-COOH → R-COCH3)。反応産物はSephadex LH-20を用いて上記と同様に精製し、メチルエステルを得た。S−3−PGのアルコール誘導体(R-COCH3 → R-CH2OH)を調製するために、上記のメチルエステル(0.5mg)を0.5mlのメタノールに溶解し、そこに5mgのNaBH4を添加した。室温で1時間還元反応に付した後、還元したS−3−PGをSephadex LH-20を用いて脱塩した(上記参照)。
【0152】
S−3−PGのアミドならびにS−3−PGのメチルアミド、エチルアミド、プロピルアミド、ベンジルアミドおよびステアリルアミドを合成するために、上記メチルエステルの溶液(0.5mlのメタノール中に0.5mg)を次の溶液のそれぞれと混合した:0.2mlの30%NH3水溶液、0.5mlの40%メチルアミン水溶液、1mlの70%エチルアミン水溶液、100μlのプロピルアミン、100μlのベンジルアミンおよび200μlの、ステアリルアミンのTHF溶液(飽和溶液)。室温で一晩インキュベートした後、反応産物から水分を窒素雰囲気下で蒸発し、アミド類とした。ベンジルアミドおよびステアリルアミドはさらに、ヘキサン/アセトン(1:1、容量比)による抽出により精製した。各アミドを過剰量の上記混合溶媒で洗浄し、遠心分離に付し、上澄みを廃棄した。
【0153】
PGC誘導体を、以下の2つの条件を変えた以外は上記と同様に調製した。
1.還元したPGC調製物をDEAE-Sephadexカラムで分離し、中性画分のみをさらに分析した、そして
2.酸化/還元工程を2回行った。
【0154】
遊離の糖(30)へのヘキサデシルアニリンのカップリング
糖(0.5mg)を100μlのメタノールに溶解し、NaBH3CNのメタノール溶液(62mg/ml)100μl、ヘキサデシルアニリンのテトラヒドロフラン溶液(40mg/ml)100μlおよび1.8Mのグリコール酸水溶液(136mg/ml)100μlと混合した。さらに、沈殿しているヘキサデシルアニリンの溶解性を高めるために、100〜200μlのテトラヒドロフランを添加した。このようにして得られた試料を30℃で一晩インキュベートして反応し、得られた反応産物をSephadex LHクロマトグラフィーにより精製した(上記「カルボキシル基の修飾」の項を参照)。この反応の収率は90%を超えた。
【0155】
分岐した脂質部分の合成(Magnusson et al., 1994、Read et al., 1977)
分岐した脂質部分を有するネオ糖脂質を得るために、HDA−誘導化糖をN−アシル化によりさらに修飾した。500μlの、p−ニトロフェニルパルミチン酸の脱水DMSO溶液を、200〜300μgの乾燥させたHDA−糖に添加して試料とした。窒素雰囲気下で37℃で3日間インキュベートした試料に、トリエチルアミンを4滴添加した。糖脂質をSephadex LH-20カラムクロマトグラフィー(上記参照)および分取TLCにより精製した。収率は約20%であった。
【0156】
ラクトン類の調製
S−3−PGを、公知の方法(Laferriere and Roy(33))によりそのラクトン型に転換した。1mgのS−3−PGを濃縮酢酸に溶解し、室温で2日間反応に付した。その後酢酸を蒸発で除去し、残渣をC/M/水(60:35:8、容量比)(0.5ml)に溶解した。この反応の収率を、0.1mmのシリカゲル60(ドイツ国、Merck製)で被覆した、アルミニウムで裏打ちしたHPTLCプレート上の薄層クロマトグラフィーにより検査したところ、約50%であった。
【0157】
ガングリオシドGM1bの合成と調製
ガングリオシドGM1の脱シアリル化により調製したガングリオテトラオシルセラミドの末端ガラクトースを、α−2,3−シアリルトランスフェラーゼ(EC番号:2.4.99.4)を用いてシアリル化した。反応条件としては、公知の条件(Lee et al., 1994)に次の若干の変更を加えて用いた。ガングリオテトラオシルセラミド(100μg)を、Triton CF−54(2%)を含有する10μlの500mM MES(4−モルホリンエタン−スルホン酸)緩衝液(pH6.0)に溶解し、74.1μlの水で希釈して反応系とした。そこに10μlのCMP-NeuAc(15mMの水溶液)および6μlのα−2,3−シアリルトランスフェラーゼ(338mU/ml)を添加して、室温で一晩静置した。反応の進行は、C/M/0.25% KCl水溶液(50:40:10、容量比)を溶出液としたTLCで分析した。次いで反応混合液から水分を窒素気流下で蒸発し、残渣をC/M/H2O混合溶媒(60:30:4.5、容量比)(2ml)に溶解し、Sephadex G-25を充填した小さいカラム(約2.5×0.6cm)(5mlの上記の溶媒系で予め洗浄したもの)にアプライした。糖脂質材料を2.5mlの上記の混合溶媒および2.5mlのC/M(2:1、容量比)で溶出した。最後に、溶出液から水分を蒸発し、少量のC/M/H2O(60:30:4.5)に溶解した。
【0158】
ヘリコバクター ピロリ株
ヘリコバクター ピロリCCGU 17874株はスウェーデン国、イェーテボリ大学カルチャーコレクション(Culture Collection University of Goteborg)から入手したものであり、ヘリコバクター ピロリ032株はスウェーデン国、ルンド大学医療微生物学科(Department of Medical Microbiology, Lund University)のT. Wadstrom教授より寄贈されたものである。ハムF12液体培地およびブルセラ寒天培地での細菌の培養は、それぞれ公知の条件(Miller-Podraza et al., 1996)で行った。本明細書においては、S−3−PG(直鎖構造)に対する結合を示す菌体として、寒天プレートで培養したCCGU 17874株を用い、PGC(分岐構造)に対する結合を示す菌体として、液体培地で培養した032株を用いた。
【0159】
結果
調製したガングリオシド類 − ヘリコバクター ピロリのガングリオシド結合選択性の分析を可能にするため、ガングリオシドライブラリーを構築した(構築したガングリオシドライブラリーを表1にまとめた)。各ガングリオシドを質量分析法およびプロトンNMRにより特徴付けた。分析方法は、ヒト赤血球のヘリコバクター ピロリ結合性ガングリオシドの1種(表1のNo.19)の単離と特徴付けに関する以下の記載によって詳細に説明する。
【0160】
総酸性グリコスフィンゴ脂質を、血液型Bの赤血球のプール500リットルから標準的な手順(Karlsson, 1987)で単離し、5.5gの総酸性グリコスフィンゴ脂質を得た。そこから得た副画分390mgを、700mlのDEAE-Sepharoseカラムと、酢酸アンモニウムのメタノール溶液2100mlを用いた酢酸アンモニウムの直線濃度勾配(0.05M〜0.45M)によって分離した。各10mlの画分を回収し、レゾルシノール試薬を用いた薄層クロマトグラフィーにより分析した。主要化合物の移動度に基づいて画分をプールした。画分62〜67をプールすると15.4mgの酸性グリコスフィンゴ脂質が得られ、このプール画分はシアリル−ネオラクトテトラオシルセラミドの領域に移動する主要化合物を含んでいた。しかし、ヘリコバクター ピロリ結合能をクロマトグラム結合アッセイを用いて試験した時に、結合能を示す遅移動性の化合物が検出された。この15.4mgの酸性グリコスフィンゴ脂質を、クロロフォルム/メタノール/水(60:35:8〜40:40:12、容量比)の直線濃度勾配を用いたHPLCによりさらに分離した。画分39〜56にヘリコバクター ピロリ結合性化合物が溶出し、これらをプールして0.9mgの化合物を得た。
【0161】
糖脂質構造の特徴を図1〜5に示すが、さらなる詳細は今後、別途発表する。
【0162】
ガングリオシドライブラリーに対するヘリコバクター ピロリの結合
クロマトグラム結合実験で用いたヘリコバクター ピロリ CCUG 17874株およびJ99株は、両方ともシアル酸結合性である(Mahdavi et al., 2002)。
【0163】
I.結合性および非結合性のガングリオシド − 単離したガングリオシドに対するヘリコバクター ピロリ株の結合実験の結果を図6〜10に示し、そのまとめを表1に示した。4μgのガングリオシドを薄層プレートにアプライしても結合が見られなかった場合、このガングリオシドを非結合性に分類した。図6に示したように、シアル酸結合性の野生株であるCCUG 17874株およびJ99株は、以下のNeuAc−末端ガングリオシドを認識した:NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−ネオラクトヘキサオシルセラミド;レーン1)、NeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−G−10ガングリオシド;レーン3)、Galα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド;レーン6)。しかし、これらに対応するNeuGc−末端異性体構造(レーン2、4および5)は認識しなかった。さらに認識されたガングリオシドは、NeuAcα3(Galβ4GlcNAcβ3)3Galβ4Glcβ1Cer(NeuAc−ネオラクトオクタアオシルセラミド、図8、レーン1〜7)、NeuAcα3Galβ4GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(VIM−2ガングリオシド、図9、レーン6〜10)およびNeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(シアリル−2量体−Lexガングリオシド、図9、レーン11〜14)であった。
【0164】
NeuAcα3−ネオラクトテトラオシルセラミド(表1のNo.2;図8、レーン1〜7)への結合も時折検出されたが、NeuAcα6−ネオラクトテトラオシルセラミド(No.3)は非結合性であり、これは過去の報告(Miller-Podraza et al., 1997、Johansson and Miller-Podraza, 1998)と一致した。NeuAcα6−を有するガングリオシドであるGalβ4GlcNAcβ6(NeuAcα6Galβ4GlcNAcβ3)Galβ4Glcβ1Cer(No.14)およびGalβ4GlcNAcβ6(NeuAcα6Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(No.15)もまた非結合性であった。NeuGcα3−ネオラクトテトラオシルセラミド(No.4)またはジシアリル−ネオラクトテトラオシルセラミド(No.5)への結合性は得られなかった。シアリル−Lexヘキサグリコシルセラミド(No.7)への結合が時折観察された。シアリル−Leaヘキサグリコシルセラミド(No.6)はCCUG 17874株には認識されなかったが、J99株はこの化合物に時折結合した。
【0165】
II.相対的結合親和性の比較 − 初めに、マイクロタイターウェル中の希釈したグリコスフィンゴ脂質類への放射線標識したヘリコバクター ピロリの結合を、結合能を示す種々のガングリオシドに関する相対的結合親和性を評価するために試みた。しかし、ここで得られた結果には再現性がなかった。そこでガングリオシド類の希釈物を用いた結合アッセイを薄層プレート上で行った。初期実験では、NeuAcα3−ネオラクトヘキサオシルセラミドおよびNeuAc−2量体−Lexガングリオシドへの結合は、約100pmolで飽和に達したことを発見したので、結合曲線を得るためにより低い濃度範囲(1〜100pmol)を用いた。図8〜10に示した結果は、多数の結合アッセイのうちの代表的なものである。放射線標識バクテリアのバッチによって結合のレベルは多少変化したが、結合能を示すガングリオシドとの関係性については同じ結果が繰り返し得られた。十分に特徴付けられたガングリオシドの認識系と比較するために、薄層クロマトグラム上のGM1ガングリオシドの希釈系列に対する125I−標識コレラトキシンB−サブユニットの結合の結果を図10に示した。
【0166】
A.炭水化物鎖の長さの影響 − NeuAcα3−ネオラクトテトラオシルセラミド(表1のNo.2)、NeuAcα3−ネオラクトヘキサオシルセラミド(No.8)およびNeuAcα3−ネオラクトオクタオシルセラミド(No.10)の希釈系列に対するヘリコバクター ピロリ CCUG 17874株の結合結果には、NeuAcα3−ネオラクトオクタオシルセラミドに対する明確な選択性が見られた(図8)。
【0167】
B.分岐の影響 − 炭水化物鎖の分岐の影響を評価するために、NeuAcα3−ネオラクトヘキサオシルセラミド(No.8)、NeuAc−G−10ガングリオシド(No.16)およびG9−Bガングリオシド(No.19)(以上、表1に記載)に対するヘリコバクター ピロリCCUG 17874株の結合性を比較した。図9に示すように、G9−Bガングリオシドのβ6−結合側鎖上に存在する血液型Bの2型エピトープは、直鎖状のNeuAcα3−ネオラクトヘキサオシルセラミドと比べて結合を減少させた。一方、NeuAc−G−10ガングリオシドは好ましいリガンドであったため、この結果はβ6−結合した側鎖上のNeuAcα3Galβ4GlcNAc配列が結合親和性を増加させることを示す。
【0168】
C.フコース残基の影響 − フコース側鎖の影響を調べるために、NeuAcα3−ネオラクトヘキサオシルセラミド(No.8)、VIM−2ガングリオシド(No.12)およびシアリル−2量体−Lexガングリオシド(No.13)に対するヘリコバクター ピロリCCUG 17874株の相対的な結合性を評価した。図10に示すように、この細菌は、NeuAcα3-ネオラクトヘキサオシルセラミドと比較して、VIM−2ガングリオシドおよびシアリル−2量体−Lexガングリオシドより高い親和性で結合した。
【0169】
末端NeuNAcα3LacNAc−構造の詳細なエピトープ分析
1.直鎖状炭水化物に関連した結合エピトープ
S−3−PG
化学誘導体のモデル化合物として選択したS−3−PGは、オーバーレイアッセイにおいてヘリコバクター ピロリと結合した、ヒト好中球の最も単純なガングリオシドであった(図11)。図に示したように、ヒト好中球には細菌に対する結合親和性を有するガングリオシドの混合物が含まれており、より複雑な移動度の遅い種ほど高い結合強度を示すようである。TLCプレート上の典型的な実験で陽性反応を得るのに必要なS−3−PGの最小量は、1×7mmのスポットあたり20〜30pmol(2.8〜4.2pmol/mm2に相当)であった。ヘリコバクター ピロリのいくつかのバッチにおいては、S−3−PG検出の感度はさらに高く、0.3〜0.4pmol/mm2のレベルに達した。
【0170】
化学的に修飾されたS−3−PG
ヘリコバクター ピロリとの相互作用における、グリセロール尾部およびNeuAcのカルボキシル基の重要性を試験するために、S−3−PGを種々の方法で化学的に修飾した。具体的には、次の誘導化を行った:(a)シアル酸グリセロール尾部の過ヨウ素酸による穏やかな酸化とそれに続く還元、あるいは過ヨウ素酸による穏やかな酸化とそれに続くメチルアミンまたはエタノールアミンとのカップリング、(b)カルボキシル基の第一アルコールへの還元、(C)カルボキシル基の種々のアミドへの転化、および(d)ラクトンの合成。こうして得られた誘導体を、その構造の特性を確認するために陰イオンFAB MSで分析した(図12参照)。全ての分子イオンの質量が予想質量と一致し、変更はシアル酸残基に限られていた(表2)。後者は、糖鎖主鎖(core chain)中の糖配列に帰属するm/z1339、1176、973および811に観測されるフラグメントイオン(現行の命名法によるとYシリーズイオン;Domon and Costello, 1988, Harvey, 1999)に変化がないことによって示された。破壊されていないセラミド部分の存在はm/z649に観測されたフラグメントイオン(Y0イオン)(18:1−24:0)により示された。いずれの分子イオンもサテライトイオン(−28質量単位)と共に出現し、サテライトイオンの出現は、d18:1−22:0で表されるセラミドがS−3−PG調製物中に少量存在するためである(例えば、図2A中のm/z1629と1601や、図2C中のm/z1582と1554)。唯一の例外は図2Bであり、m/z1599と1569に観測されるイオンは、m/z1629に観測される主成分(図2A)に由来する2種の異なる誘導体を表している。図2Bにおいては、d18:1−22:0成分に帰属されるイオンはバックグラウンドに溶け込んでいた。
【0171】
放射線標識したヘリコバクター ピロリでTLCプレートをオーバーレイすることにより、S−3−PGから得た全ての誘導体について、ヘリコバクター ピロリによる結合性を試験した。得られた結果を表2にまとめた。結合性の実験の一例を図13に示す。この図に見られるように、シアル酸残基に加えた修飾の大部分は、S−3−PGに対するヘリコバクター ピロリの結合性を喪失させるか、あるいは劇的に減少させた。S−3−PGのアミド誘導体およびベンジルアミド誘導体に対する結合は若干見られた。これらの結合の一部はより多くの糖脂質材料を必要とした。いくつかのプレート上では、酸化/還元SPGとの相互作用も見られたが(結果は示さない)、これはおそらくこのような調製物中に誘導化されていないSPGが痕跡量存在するためであろう。さらに、グリセロール尾部の重要性を分析するために、S−3−PGを酸化し、メチルアミンまたはエタノールアミンとカップリングした。得られた2種の主要な誘導体はR-CH2-NH-CH3 および R-CH2-NH-CH2-CH2OHであった。この2種の誘導体はいずれもTLCプレート上の結合分子としては不活性であることが判明した。対照的に、S−3−PGのオクタデシルアミドには強い結合性が見られ、TLCプレート上でより低いpmolレベルでヘリコバクター ピロリと相互作用した(図14)。さらに、シアル酸を過ヨウ素酸でアルデヒドに酸化し、オクタデシルアミンで還元的にアミノ化したシアル酸の7位の炭素の誘導体は、ヘリコバクター ピロリに非常に効果的に結合することが分かった。さらに、修飾を過ヨウ素酸による穏やかな酸化およびアルデヒドの還元により行った場合には、1位の炭素のアミド誘導体は、シアル酸のグリセロール尾部の短縮を許容することが分かった。このデータは、より高い活性を示す誘導体や類似体を、シアル酸よりも単純な環状構造(好ましくは6員環であるピラノース様環状構造)から製造または設計可能であることを示している。
【0172】
他の糖脂質およびネオ糖脂質
種々の天然ガングリオシドおよびネオガングリオシドのパネルをヘリコバクター ピロリとの相互作用に関して試験した(表3)。ガングリオ系のガングリオシド、S−6−PG、NeuAcα8NeuAcα3−PG、またはラクト(Galβ3GlcNAcβ3Galβ4Glc)主鎖に基づくシアリル化オリゴ糖から調製したネオガングリオシドに対するヘリコバクター ピロリの結合は見られなかった。一方、糖がHDAまたは分岐脂質鎖とカップリングしたS−3−PGは、同じオーバーレイ条件下で活性を示した。本発明者らの研究室で合成したネオ糖脂質へのヘリコバクター ピロリの結合を図15に示す。主要なバンドに含まれる物質の構造は、プレートからバンドに相当する部分をかき取り、質量分析により確認した。レーン2の矢印はそれぞれHDA誘導体(下のバンド)および分岐状誘導体(上の2重のバンド)に相当するバンドを示す。これらの画分のFABスペクトルを図16に示す。分子イオンおよびフラグメントイオンは予想通りだった。上記バンドに対応する、図15のレーン3〜5の画分についても、同様のFABスペクトルを得た。結合系列(binding series)に含まれる他のシアリル化分子を少量含有するSPG調製物から、糖であるNeuAcα3Galβ4GlcNAcβ3Galβ4Glcを得た(図15のレーン1、図11も参照)。このような微量の結合画分が誘導化後でさえ観測された(図15のレーン2)。実験を繰り返すことにより、TLCプレート上のHDAネオ糖脂質に対するヘリコバクター ピロリの結合の再現性は、分岐脂質鎖を有する誘導体よりも低いことが分かった。
【0173】
2.PGCの分岐ポリ−N−アセチルラクトサミン鎖に関連した結合エピトープ
上記で説明したとおり、この選択的な結合性は、液体培地で培養したヘリコバクター ピロリの数株が発現した。例として、PGCへは結合するがS−3−PGへは結合しないことを示す図18を参照されたい。典型的な実験において、TLCプレート上で陽性反応を得るのに必要なPGCの最小量は0.16pmol NeuAc/mm2であった。PGCのNeuAcのグリセロール尾部を過ヨウ素酸による穏やかな酸化とそれに続く還元(R-CHOH-CHOH-CH2OH → R-CHOH-CH2OH/R-CH2OH)またはエタノールアミンとのカップリング(R-CHOH-CHOH-CH2OH → R-CH2-NH-CH2-OH)により修飾し、カルボキシル基を還元(R-COOH → CH2OH)により修飾した。修飾したPGCをパーメチルしてEI MSにより分析した結果を図17に例示した。図17には、還元前(図6A)および還元後(図6B)の末端シアリル酸残基に対応するイオンが観察された。予想通り、NeuAcに帰属するm/z376および344に観測されたフラグメントイオンは、還元したNeuAcに帰属する、m/z362および330に観測されるイオンで置換されていた。
【0174】
放射線標識細菌によるTLCプレートのオーバーレイを用いた結合試験は、修飾がPGCとヘリコバクター ピロリとの相互作用に負の影響を与えることを示した。修飾したPGCへのヘリコバクター ピロリの結合の一例を図18に示し、その結果を表4にまとめた。
【0175】
考察
ある種のヘリコバクター ピロリ株によるシアル酸含有糖結合体の認識はこれまで繰り返し報告されている(Roche et al., 2001、Mahdavi, et al., 2002、Miller-Podraza et al, 1997、Johansson and Miller-Podraza 1998)。本願の実験では、ガングリオシドのライブラリーを収集し、代表的なシアル酸認識ヘリコバクター ピロリを用いてヘリコバクター ピロリの結合選択性を分析した。
【0176】
シアリル−Leaヘキサグリコシルセラミドに対するJ99株の結合が時折観察されたが、CCUG17874株はこのガングリオシドを認識しなかった。また、シアリル−Leaネオ糖タンパク質およびシアリル−Lexネオ糖タンパク質の両方に対するJ99株の結合も報告されている(Mahdavi, 2002)。このことは、J99株とCCUG17874株ではSabA炭水化物結合部位が同じではないことを示している。しかし、その他の全ての点において、CCUG17874株とJ99株はガングリオシドに同一の様式で結合し、いずれの株も、末端NeuAcα3を有する、N−アセチルラクトサミンをベースにしたガングリオシドを認識したが、末端NeuAcα6を有するものは認識しなかった。この結果は従来の報告(Miller-Podraza et al, 1997、Johansson and Miller-Podraza, 1998)と一致していた。また、末端NeuGcα3または末端NeuAcα8NeuAcα3を有するガングリオシドは認識されなかった。
【0177】
結合親和性に影響を及ぼす要因として、i) N−アセチルラクトサミン炭水化物鎖の長さ、ii) 炭水化物鎖の分岐、およびiii) N−アセチルラクトサミン主鎖のフコース置換、の3つを明らかにした。
【0178】
1.N−アセチルラクトサミン主鎖の長さ:NeuAcα3−ネオラクトヘキサオシルセラミドおよびNeuAcα3−ネオラクトテトラオシルセラミドよりも、NeuAcα3−ネオラクトオクタオシルセラミドに対してヘリコバクター ピロリが選択的に結合することを観察した。このような効果は、炭水化物の頭部基(carbohydrate head group)がより長い主鎖に提示された場合、その基に対する接触性が向上するからである可能性が最も高い。
【0179】
2.二価性:2つのNeuAcα3Galβ4GlcNAcβ側鎖を有するNeuAc−G−10ガングリオシドに対して観察される、直鎖状のNeuAcα3−ネオラクトヘキサオシルセラミドに対するよりも高い親和性は、共同的結合によると考えられる。このことは、シアリルラクトース(NeuAcα3Galβ4Glc)の多価アルブミン結合体が、一価のシアリル−ラクトースよりも上皮単層へのヘリコバクター ピロリの付着を効果的に阻害することを実証したSimon et al. (1997)の報告と合致する。本発明は、シアリルラクトサミンがオリゴ価または多価に提示されることによって、このエピトープが効果的に提示されることを示す。
【0180】
NeuAcα3−ネオラクトヘキサオシルセラミドに比べ、G9−Bガングリオシドに対する結合親和性がより低いことは、β6結合した側鎖上の血液型B決定因子が結合作用に干渉したことを示す。G9−Bガングリオシドの検知レベルは約100pmolだったため、完全な障害ではない。このことは、ヘリコバクター ピロリ結合決定因子は、Galα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer構造体のβ3軸上に主として露出されていることを示唆している。
【0181】
3.N−アセチルラクトサミン主鎖に提示されたフコース側鎖:NeuAcα3−ネオラクトヘキサオシルセラミドに比べ、VIM−2ガングリオシドに対する結合親和性がより高いことは、最も内側に位置するGlcNAcにα3結合しているフコースが、シアリル−2量体−Lexの高親和性結合に関与することを示唆する。このフコース残基は、SabA付着因子の炭水化物結合部位と相互作用するか、あるいはガングリオシドの立体構造に影響を及ぼして炭水化物の頭部基の提示を最適化すると考えられる。しかし、この問題を解明するには、SabA付着因子の発現と結晶化を待たねばならない。
【0182】
SabA付着因子の発現は、Leb結合性のBabA付着因子とは異なり、相変異にさらされる(Mahdavi et al., 2002)。結合アッセイに用いた菌体群の中でSabA付着因子を発現する細菌の数が変化することが、あるガングリオシドに対する結合性の絶対的な親和性を測定することが困難な理由であると考えられる。しかし、繰り返し行った結合アッセイにおいては、結合能を有するガングリオシド間で同じ関係性が認められた。しかし、最適条件下では、NeuAcα3−ネオラクトヘキサオシルセラミドに対するヘリコバクター ピロリの結合が、GM1ガングリオシドに対するコレラトキシンB−サブユニットの結合に匹敵することは注目すべき知見である。コレラトキシンの結合は、非常に強力なタンパク質−炭水化物相互作用の一例である。
【0183】
上記知見の生物学的重要性は、さらなる研究が必要である。ヘリコバクター ピロリの主要ターゲット組織、即ちヒトの胃上皮組織、のシアル酸含有量は非常に低い(Madrid et al., 1990)。しかしながら、最近になって、胃に炎症が起きるとシアル酸含有糖結合体の発現が上昇するように調節されることが発表された(Mahdavi et al., 2002)。さらに、数種類のヘリコバクター ピロリ結合性ガングリオシドもヒトの好中球に存在し(Miller-Podraza et al., 1999、Stroud et al., 1996a, 1996b)、最近の研究によると、レクチノファゴサイトーシス(lectinophagocytosis)によってヒトの好中球の非オプソニン性ヘリコバクター ピロリ誘導活性化(即ち、細菌性付着因子による好中球細胞表面のシアリル化糖結合体の認識)が生じると、食作用、そして反応性酸素代謝産物の産生を伴う酸化的バーストが起こる(Teneberg, et al., 2001)。このように、ヘリコバクター ピロリのシアル酸結合能力には2つの役割がある。即ち、一方ではすでに病理に冒された胃の上皮への細菌の付着を仲介し、もう一方では好中球を活性化して反応性酸素代謝産物の産生を伴う酸化的バーストを起こし、生物学的活性を有する酵素を放出して組織破壊にさらなる拍車をかけることである。
【0184】
1.炭水化物鎖に提示された末端NeuNAcα3LacNAcに対するヘリコバクター ピロリの結合
調製が比較的容易な糖脂質であり、十分に定義された構造を有することから、本研究のモデル化合物としてS−3−PGを選んだ。In vitroにおける数種のヘリコバクター ピロリ株によるシアル酸含有糖結合体の認識は、種々の文献で議論されており(Karlsson, 1998、Karlsson, 2000、Miller-Podraza et al., 1997a、Johansson and Miller-Podraza, 1998、Miller-Podraza et al., 1996、Miller-Podraza et al., 1997b、Evans et al., 1988、Hirmo et al., 1996、Simon et al., 1997)、本願に示す結果は、これらの発表内容と合致し、α3結合したNeuAcとβ4結合したGalに対するヘリコバクター ピロリの選択性をはっきりと明示した。NeuAcα3Galβ3GlcNAc-R と NeuAcα3Galβ3GalNAc-Rが不活性であることから、ネオラクト構造が明らかに必要であった。しかしながら、シアリルラクトースであるNeuAcα3Galβ4Glcがシアリル化構造とヘリコバクター ピロリとの相互作用に対して弱い阻害性を示すことが他のグループによって報告されているので(Evans et al., 1988、Hirmo et al., 1996、Simon et al., 1997)、GlcNAc自体は結合に絶対的に必要なものではないと考えられる。また、(シアリル−ルイスxオリゴ糖および糖脂質を用いて試験した)GlcNAcに結合したα3Fucおよび脱−N−アセチル化などのGlcNAcの他の修飾(Johansson and Miller-Podraza、出版準備中)は結合を喪失させることはない。従って本発明は、より長いヘリコバクター ピロリ結合性オリゴ糖においては、その活性を損失させることなくGlcまたはGlcNAcの2位の炭素を修飾することも可能であることを示す。
【0185】
S−3−PGを誘導化とそれに続く結合試験により、ヘリコバクター ピロリの結合はグリセロール尾部とカルボキシル基に提示されたシアリル化エピトープに依存することが分かった。カルボキシル基の重要性はCOOHの還元およびアミドの合成によって実証された。遊離アミドおよびベンジルアミドは、TLCプレート上でヘリコバクター ピロリに対する結合能をいくらか示したが、これらの反応は常に再現性があるわけではなく、親和性の減少を示した。ベンジルアミド中の大きな疎水性部分と遊離アミド中の正の電荷を帯びたNH2基が非特異的結合を引き起こすことができる。一方、SPGのオクタデシルアミドへの結合性はより低いピコモルレベルであり(図4)、この結合が特異的な相互作用であることを示した。未修飾カルボキシル基はオクタデシルアミド中のアミドによって置換されてもよいという事実は、相互作用にはカルボキシル基の1つの酸素原子しか必要ではないことを示す。
【0186】
ヒトや動物の種々の糖結合体の一部であるNeuAcα3Galβ4GlcNAcが、天然のシアリル化ヘリコバクター ピロリ結合性糖の最適末端部を表していると考えられる。重要なのは、ヘリコバクター ピロリ関連感染症に積極的に関与するヒトの好中球が(Rautelin et al., 1993, Fiocca et al., 1994)、特にS−3−PGと他のネオラクトガングリオシドを豊富に含んでいる(Fukuda et al., 1985, Muthing 1996, Stroud et al., 1996)ということである。上で述べたように、S−3−PGは、TLCプレート上でヘリコバクター ピロリを結合した、ヒト好中球由来のガングリオシドの中で最も単純なものである。図11は、より複雑な構造のガングリオシドに対して比較的強い結合が生じることを示す。TLC表面でエピトープがより効果的に提示されていることがこの強い相互作用の最も大きな理由であると考えられる。研究段階で、糖鎖長および脂質部分の構造が結合に影響を及ぼすかもしれないことに気が付いた。例えば、ヘキサデシルアニリンを有するネオ糖脂質へのヘリコバクター ピロリの結合は、分岐脂質鎖を有するネオ糖脂質への結合よりも再現性が低く、また、TLCプレート上の結果(表11参照)から判断すると、シアロネオヘキサオシルセラミドに対する結合はシアロネオテトラオシルセラミドに対する結合よりも強い。しかし、他の要因、例えば繰返しエピトープ単位、フコース側鎖または他の置換基なども考慮に入れる必要がある。より大きな種に強く結合するのは、長く伸びた主鎖の末端領域と内部領域の両方を認識する様々な細菌付着因子の結合の組み合わせによるものであると考えられる。ヘリコバクター ピロリは、シアリル化糖鎖および中性糖鎖の両者に関与する様々な結合特異性を示すことが分かっている(Karlsson 1998)。近年、Roche N. et al. によって、ヒトの胃癌細胞から調製した繰返しラクトサミン単位を有するガングリオシドへのヘリコバクター ピロリの結合が報告されている(Roche et al., 2001)。
【0187】
2.分岐ポリ−N−アセチルラクトサミン鎖に関与する結合エピトープ
本発明者らは以前に、ヘリコバクター ピロリ結合性PGC中のNeuAcは1つのラクトサミン単位(NeuAc-Hex-HexNAc)に基づく短い側鎖に関係があることを明かにした。このことは、エンド−β−ガラクトシダーゼでPGCを消化し、遊離したオリゴ糖断片を様々な質量分析技術を用いて分析することにより証明された(Karlsson et al., 2000)。分岐したPGC中にNeuAcα3Galβ4GlcNAcが存在することは、複数の異種の糖の間で水素結合を形成するための好的な条件を提供するので、NeuAcのグリセロール尾部の9位の炭素と、隣接する2本の側鎖中の各GlcNAcとの間に相互作用に基づいて新しい結合エピトープが発生するという仮説が立てられた(Angstrom et al., 投稿済み)。このような仮説は、相互作用におけるグリセロール尾部の重要性にも合致する上、シアリル化構造に対するヘリコバクター ピロリの結合様式には異なる2種類の様式が明かに存在することの説明にもなる。本明細書は、グリセロール基の重要性に関する本発明者らの過去の知見(Miller-Podraza et al., 1996)を確立させると同時に、カルボキシル基さえも結合にとって非常に重要であることを示す。COOHの存在によって、ラクトン化または他の可逆的修飾によって、in vivoで素早く結合性のオンとオフを切り替えられる可能性が示されることから、この知見は非常に興味深い。
【0188】
【表1】
【0189】
a)結合強度は以下のように定義した:「+++」は薄層クロマトグラムにグリコスフィンゴ脂質を0.5μg未満アプライした場合に結合が検出されたことを示し;「+」は0.5μgアプライした場合に時折結合が検出されたことを示し;「−」は4μgアプライした場合でも結合が検出されないことを示す。
【0190】
【表2】
【0191】
【表3】
【0192】
【表4】
【0193】
参考文献
Boren, T., Falk, P., Roth, K. A. and Normark, S. (1993) Science 262, 1892-1895.
Domon, B. and Costello, C. E. (1988) Glycoconj. J. 5, 397-409
Evans, D.G., Evans, D.J., Jr., Moulds, J.J. and Graham, D.Y. (1988) Infect. Immun. 56(11): 2896-2906.
Fiocca, R., Luinetti, O., Villani, L., Chiaravalli, A.M., Capella, C. and Solcia, E. (1994) Scand. J. Gastroenterol. 205(29S):11-21.
Fukuda, M.N., Dell, A., Oates, J.E., Wu, P., Klock, J.C. and Fukuda, M. (1985) J. Biol. Chem. 260:1067-1082.
Harvey, D.J. (1999) Mass Spectrometry Reviews 18: 349-450.
Hirmo, S., Kelm, S., Schauer, R., Nilsson, B. and Wadstrom, T. (1996) Glycoconjugate J. 13:1005-1011.
Ilver, D. et al. (1998) Science 279, 373-.
Ito, M. and Yamagata, T. (1989) Methods Enzymol. 179:488-496.
Johansson, L., Miller-Podraza, H. (1998) Anal. Biochem. 265, 260-268.
Karlsson, K.-A. (1987) Meth. Enzymol. 138, 212-220
Karlsson, K.-A. (1974) Biochemistry 13, 3643-3647
Karlsson, K.-A. (1998) Mol. Microbiol. 29:1-11.
Karlsson, K.-A. (2000) Glycobiology 10(8): 761-771.
Karlsson, H., Larsson, T., Karlsson, K.-A. and Miller-Podraza, H. (2000) Glycobiology 10(12):1291-1309.
Karlsson, K.-A. and Stromberg, N. (1987) Meth. Enzymol. 138, 220-231
Koerner, T., A.,W., Jr., Prestegard, J., H., Demou, P. C. and Yu, R. K. (1983) Biochemistry 22, 2676-2687
Larsson, G., Karlsson, H., Hansson, G. C. and Pimlott, W. (1987) Carbohydr. Res. 161, 281-290
Lanne, B., Uggla, L., Stenhagen, G. and Karlsson, K.-A. (1995) Biochemistry 34:1845-1850.
Lee, Y.-C., Kojima, N., Wada, E., Kurosawa, N., Nakaoka, T., Hamamoto, T. and Tsuji, S. (1994) J. Biol. Chem. 269(13): 10028-10033.
Lingwood, C. A., Huesca, M. and Kuksis, A. (1992) Infect. Immun. 60, 2470-2474
Madrid, J. F., Ballesta, J., Castells, M. T. and Hernandez, F. (1990) Histochemistry 95, 179-187
Magnusson, G., Chernyak, A.Y., Kihlberg, J. and Kononov, L.O. Synthesis of neoglycoconjugates. In: Neoglycoconjugates, Preparation and applications, (Y. C. Lee and Reiko, T. Lee, eds.), Academic Press, Inc., A Division of Harcourt Brace & Company, San Diego 1994, pp. 53-117.
Mahdavi, J., Sonden, B., Forsberg, L., Hurtig, M., Olfat, F. O., Forsberg, L., Roche, N., ・ngstrom, J., Larsson, T., Teneberg, S., Karlsson, K.-A., Altraja, S., Wadstrom, T., Kersulyte, D., Berg, D. E., Dubois, A., Petterson, C., Magnusson, K.-E,. Norberg, T., Lindh, F., Lundskog, B. B., Arnqvist, A., Hammarstrom, L., Boren, T. (2002) Science 297, 573-578
Miller-Podraza, H., Mansson, J.-E. and Svennerholm, L. (1992) Biochim. Biophys. Acta 1124:45-51.
Miller-Podraza, H., Andersson, C. and Karlsson, K.-A. (1993) Biochim. Biophys. Acta 1168: 330-339
Miller-Podraza, H., Abul Milh, M., Bergstrom, J. and Karlsson, K.-A. (1996) Glycoconjugate J. 13: 453-460.
Miller-Podraza, H., Abul Milh, M., Teneberg, S. and Karlsson, K.-A. (1997a) Infect. Immun. 65, 2480-2482
Miller-Podraza, H., Bergstrom, J., Abul Milh, M. and Karlsson, K.-A. (1997b) Glycoconjugate J. 14:467-471.
Miller-Podraza, H., Bergstrom, J., Teneberg, S., Abul Milh, M., Longard, M., Olsson, B.-M., Uggla, L. and Karlsson, K.-A. (1999) Infect. Immun. 67, 6309-6313
Muthing, J. (1996) Carbohydrate Research.290:217-224.
Mysore, J.V., Wiggington, T., Simon, P.M., Zopf, D., Heman-Ackah, L.M. and Dubois, A.
(1999) Gastroenterology, 117, 1316-1325
Rautelin, H., Blomberg, B., Fredlund, H., Jarnerot, G. and Danielsson, D. (1993) Gut 34:599-603.
Read, B., Demel, R.A., Wiegandt, H. and Van Deenen, L.L.M. (1977) Biochim. Biophys. Acta, 470:325-330.
Roche, N., Angstom, J., Larsson, T. and Teneberg, S. (2001) Glycobiology 11, 935-944
Samuelsson, B. E., Pimlott, W. and Karlsson, K.-A. (1990) Meth. Enzymol. 193, 623-646
Simon, P. M., Goode, P. L., Mobasseri, A. and Zopf, D. (1997) Infect. Immun. 65, 750-757
Stellner, K., Saito, H. and Hakomori, S.-i. (1973) Arch. Biochem. Biophys. 155, 464-472
Stroud, M. R., Handa, K., Salyan, M. E. K., Ito, K., Levery, S. B., Hakomori, S.-i., Reinhold, B. B. and Reinhold, V. N. (1996a) Biochemistry 35, 758-769
Stroud, M. R., Handa, K., Salyan, M. E. K., Ito, K., Levery, S. B., Hakomori, S.-i., Reinhold, B. B. and Reinhold, V. N. (1996b) Biochemistry 35, 770-778
Stults, C. L. M., Sweeley, C. C. and Macher, B. A. (1989) Meth. Enzymol. 179, 167-214
Svennerholm, L. (1963) J. Neurochem. 10, 613-623
Teneberg, S., Jurstrand, M., Karlsson, K.-A. and Danielsson, D. (2000) Glycobiology 10, 1171-1181
Teneberg, S., Miller-Podraza, H., Lampert, H. C., Evans Jr., D. J., Evans, D. G., Danielsson, D., and Karlsson, K.-A. (1997) J. Biol. Chem. 272, 19067-19071
Teneberg, S., Leonardsson, I., Karlsson, H., Jovall, P.-A., Angstrom, J., Danielsson, D., Naslund, I., Ljungh, A., Wadstrom, T., and Karlsson, K.-A. (2002) J. Biol. Chem. 277, 19709-19719
Veh, R.W., Cornfield, A.P., Sander, M. and Schauer R. (1977) Biochim. Biophys. Acta 486:145-160.
Waldi, D. (1962) in Dunnschicht-Chromatographie (Stahl, E., ed.) pp. 496-515. Springer-Verlag, Berlin
Yang, H. and Hakomori, S.-i. (1971) J. Biol. Chem. 246, 1192-1200
Angstrom, J., Teneberg, S. and Karlsson, K.-A. (1994) Proc. Natl. Acad. Sci. USA 91, 11859-11863.
【図面の簡単な説明】
【0194】
【図1】ヒト赤血球由来ガングリオシドの陰イオンFAB MSスペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。2種類のスペクトルAとB、具体的には、加速電圧を10kVとしたスペクトルAと、加圧電圧を8kVとしたスペクトルBをそれぞれ図1Aおよび図1Bに示す。スペクトルAにおいては、100〜2400質量単位に渡ってスキャニングし、一方のスペクトルBにおいては、2000〜3200質量単位に渡ってスキャニングし、2.4分間のデータを回収した。
【図2】パーメチル化したヒト赤血球由来ガングリオシドの、電子衝突イオン化マススペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。スペクトルの記録温度は380℃とした。m/z 354に観測されたピークは混入物に帰属されるものである。
【図3】パーメチル化したヒト赤血球由来ガングリオシドの、エレクトロスプレーイオン化質量分析および衝突誘起解離によって得たマススペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。
【図4】パーメチル化と還元処理を行ったヒト赤血球由来ガングリオシドの、電子衝突イオン化マススペクトル。スペクトルの解読を容易にするために、スフィンゴシンと非ヒドロキシ脂肪酸(24:0)を有する化合物種の簡略化した式をスペクトルの上方に示した。分析は、「実施例」に記載の方法で行った。スペクトルの記録温度は340℃とした。
【図5】ヒト赤血球由来ガングリオシド(30℃)の、600MHzにおけるプロトンNMRスペクトル。試料は、重水素置換後に、ジメチルスルホキシド/D2O(98:2、容量比)に溶解した。約4.8ppmを中心とする幅広のピーク(記号*で示した)は、未同定の混入物に帰属されるものである。
【図6】末端NeuAcを有するガングリオシド、および末端NeuGcを有するガングリオシドに対するヘリコバクター ピロリの結合。Aはアニスアルデヒドによる化学的検出結果であり、BとCはそれぞれ、35Sで標識したヘリコバクター ピロリのCCUG 17874株とJ 99株の結合によって得られたオートラジオグラムである。ガングリオシドは、アルミニウムで裏打ちしたシリカゲルプレート上で、クロロホルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)を溶媒系として分離し、「実施例」に記載の方法で結合アッセイを行った。レーンは以下の通りである: ヒト肝細胞癌由来のNeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-ネオラクトヘキサオシルセラミド)、2μg(レーン1); ウサギ胸腺由来のNeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuGc-ネオラクトヘキサオシルセラミド)、2μg(レーン2); ヒト赤血球由来のNeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-G-10型ガングリオシド)、2μg(レーン3); ウシ赤血球由来のNeuGcα3Galβ4GlcNAcβ6(NeuGcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuGc-G-10型ガングリオシド)、2μg(レーン4); ウシ赤血球由来のGalα3Galβ4GlcNAcβ6(NeuGcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer、2μg(レーン5); ヒト赤血球由来のGalα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド)、2μg(レーン6)。オートラジオグラフィーは12時間行った。
【図7】ヘリコバクター ピロリのCCUG 17874株とJ 99株の結合の比較。Aはアニスアルデヒドによる化学的検出結果であり、BおよびCはそれぞれ、35Sで標識したヘリコバクター ピロリのCCUG 17874株およびJ 99株の結合によって得られたオートラジオグラムである。ガングリオシドは、アルミニウムで裏打ちしたシリカゲルプレート上で、クロロホルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)を溶媒系として分離し、「実施例」に記載の方法で結合アッセイを行った。各レーンは以下の通りである: ヒト好中球由来のガングリオシド、20μg(レーン1); ウサギ胸腺由来のNeuGcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuGc-ネオラクトヘキサオシルセラミド)、2μg(レーン2); ヒト肝細胞癌由来のNeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-ネオラクトヘキサオシルセラミド)、1μg(レーン3); ヒト赤血球由来のNeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−G−10ガングリオシド)、1μg(レーン4); ヒト赤血球由来の Galα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド)、1μg(レーン5); ヒト赤血球由来のGalα3(Fucα2)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(B6 2型ヘキサグリコシルセラミド)、4μg(レーン6); マウス糞由来のGalβ3GalNAcβ4Galβ4Glcβ1Cer(ガングリオテトラシルセラミド)、4μg(レーン7)。オートラジオグラフィーは12〜24時間行った。
【図8】段階希釈したガングリオシドに対するヘリコバクター ピロリの結合。Aは、クロマトグラム結合アッセイを用いた、ヘリコバクター ピロリ CCUG 17874株の結合によって得られたオートラジオグラムである。レーン1〜7はそれぞれ、以下の糖の段階希釈系列(1〜100pmol)である: NeuAcα3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトテトラオシルセラミド)、 NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトヘキサオシルセラミド)および NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトオクタオシルセラミド)。レーン8は、1mmolのNeuAcα3Galβ3(Fucα4)GlcNAcβ3Galβ4Glcβ1Cer(シアリルLeaヘキサグリコシルセラミド)である。結合アッセイは、「実施例」に記載の方法で行った。3回の実験のうち、最も代表的な実験結果をAに示す。Bは,デンシトメトリーによって結合を定量化した結果である。Aに示したオートラジオグラムはNIHイメージプログラムで分析した。
【図9】段階希釈したガングリオシドに対するヘリコバクター ピロリの結合。Aは、クロマトグラム結合アッセイを用いた、ヘリコバクター ピロリ 17874株の結合によって得られたオートラジオグラムであり、レーンは以下の段階希釈系列である: NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-ネオラクトヘキサオシルセラミド)、(10〜100pmole)(レーン1〜5); NeuAcα3Galβ4GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(VIM−2ガングリオシド)、(10〜100pmole)(レーン6〜10); NeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-2量体-Lexガングリオシド)、(10〜100pmole)(レーン11〜14); NeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(シアリルLexヘキサグリコシルセラミド)、1mmole(レーン15)。結合アッセイは、「実施例」に記載の方法で行った。3回の実験のうち、最も代表的な実験結果をAに示す。Bは,デンシトメトリーによって結合を定量化した結果である。Aに示したオートラジオグラムはNIHイメージプログラムで分析した。
【図10】段階希釈したガングリオシドに対するヘリコバクター ピロリの結合。Aは、クロマトグラム結合アッセイを用いた、ヘリコバクター ピロリ CCUG 17874株の結合によって得られたのオートラジオグラムであり、以下の糖の段階希釈系列(1〜100pmol)を使用した: NeuAcα3Galβ4(Fucα3)GlcNAcβ3Galβ4Glcβ1Cer(NeuAc-Lex)、 NeuAcα3Galβ4GlcNAcβ3Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAcα3-ネオラクトヘキサオシルセラミド)、 NeuAcα3Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(NeuAc−G−10ガングリオシド)およびGalα3(Fucα2)Galβ4GlcNAcβ6(NeuAcα3Galβ4GlcNAcβ3)Galβ4GlcNAcβ3Galβ4Glcβ1Cer(G9−Bガングリオシド)。結合アッセイは、「実施例」に記載の方法で行った。3回の実験のうち、最も代表的な実験結果をAに示す。Bは,デンシトメトリーによって結合を定量化した結果である。Aに示したオートラジオグラムはNIHイメージプログラムで分析した。比較のために、薄層クロマトグラム上の段階希釈したGM1ガングリオシドに対する、125Iで標識したコレラトキシンB−サブユニット(CTB)の結合に関するデンシトメトリーの結果もAに示した。
【図11】放射線標識したヘリコバクター ピロリを用いた、TLCプレート上の顆粒球由来ガングリオシドの検出レベル。プレートは、クロロホルム/メタノール/0.25%KCl水溶液(50:40:10、容量比)で展開し、アニスアルデヒドを噴霧するか、または35Sで標識したヘリコバクター ピロリ CCGU 17874株をオーバーレイして視覚化した。レーン1〜10は、顆粒球由来ガングリオシドの2倍希釈系列(レーン1には合計12μgをアプライした)であり;レーン11は、ウシ脳由来ガングリオシド、2μg(GM1、GD1a、GD1bおよびGT1bの混合物)である。図中のS−3−PGとS−6−PGはそれぞれ、NeuAcα3Galβ4GlcNAcβ3Galβ4GlcCer と NeuAcα6Galβ4GlcNAcβ3Galβ4GlcCerを表す。「7s」および「8s」という注釈は、それぞれ1分子中に7個か8個の単糖単位を含むガングリオシドに対応するクロマトグラフィー領域を示す。
【図12】シアル酸の -CH2CH2CH2OH 基および -COOH 基が誘導化されたS−3−PGの陰イオンFAB MSスペクトル。Aは未修飾のS−3−PG、Bは酸化還元されたS−3−PG、Cは酸化後メチルアミンにより誘導化したS−3−PG、Dは酸化後エタノールアミンにより誘導化したS−3−PG、Eは-COOH基を還元したS−3−PG、Fは遊離アミド、Gはメチルアミド、Hはエチルアミド、Iはプロピルアミド、Jはベンジルアミド、Kはオクタデシルアミドの結果である。
【図13】修飾したS−3−PGに対するヘリコバクター ピロリ(CCGU 17874株)の結合の一例。糖脂質をTLCプレート上で分離し、アニスアルデヒド(左側のプレート)または35Sで標識した細菌(ヘリコバクター ピロリ、右側のプレート)で視覚化した。クロマトグラフィーの条件に関しては図1を参照のこと。レーン1はS−3−PGだが、結合系列に属するより長い物質が痕跡量含まれている;レーン2はS−3−PGのエチルアミドであり;レーン3はS−3−PGのプロピルアミドであり;レーン4はS−3−PGのベンジルアミドであり;レーン5は酸化還元したS−3−PGであり(表3を参照);レーン6はヘリコバクター ピロリの正の対照であり;レーン7はウシ脳由来ガングリオシドであり(上から:GM1、GD1a、GD1b、GT1b);レーン8はスルファチド混合物であり;レーン9はウサギ胸腺由来の五糖含有ガングリオシドの混合物であり;レーン10はヒト顆粒球から調製した複合ガングリオシド混合物である。
【図14】TLCプレート上のネオ糖脂質に対するヘリコバクター ピロリ(CCGU 17874株)の結合。クロマトグラフィーの条件に関しては図1を参照のこと。左側のプレートは、アニスアルデヒドを用いて炭水化物を染色したものであり、右側のプレートは35Sで標識したヘリコバクター ピロリをオーバーレイしたものである。レーン1はS−3−PGだが、図3と同様に、SPG調製物には結合系列に属するより長い物質が痕跡量含まれている;レーン2はNeuAcα3Galβ4GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン3はNeuAcα3Galβ3GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン4はNeuAcα6Galβ4GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン5はGalβ3(NeuAcα6)GlcNAcβ3Galβ4Glcより得られたネオ糖脂質であり;レーン6はウシ脳由来ガングリオシドである(上から:GM1、GD1a、GD1b、GT1b)。レーン2に示した二つの矢印は、それぞれ、ヘキサデシルアニリン誘導体(下側のバンド)および分岐した脂質鎖を有するネオ糖脂質(上側の2重のバンド)を示す。レーン2の矢印で示した2つの画分と、それぞれに相当するレーン3〜5の画分をかき出し、マススペクトロメトリーで分析した(図6参照)。
【図15】NeuAcα3Galβ4GlcNAcβ4Galβ4Glcから誘導化したネオ糖脂質の陰イオンFAB MSスペクトル。Aはヘキサデシルアニリン誘導体の結果であり;Bは分岐した脂質部を有するネオ糖脂質の結果である。
【図16】パーメチル化ポリグリコシルセラミドのEI質量スペクトルの一例であり、NeuAcと還元NeuAcに相当するフラグメントイオンを示す。
【図17】TLCプレート上の誘導化PGCに対する、ヘリコバクター ピロリ(032株)の結合。左側のプレートは、アニスアルデヒドを用いて炭水化物を染色したものであり、右側のプレートは35Sで標識したヘリコバクター ピロリをオーバーレイしたものである。レーン1と2はヒト赤血球由来の未誘導化PGCであり;レーン3は還元PGC(COOH → CH2OH)であり;レーン4はウシ脳由来のガングリオシド(GM1、GD1a、GD1b、GT1bの混合物)であり;レーン5はS−3−PGである。(液体培養した032株は、直鎖構造を表すS−3−PGには結合しないことに注意されたい。)
【図1A】
【図1B】
【特許請求の範囲】
【請求項1】
下記式(1)で表される親水性オリゴ糖配列を非還元末端配列として含む、医薬として用いるヘリコバクター ピロリ結合性物質、あるいはヘリコバクター ピロリに対する結合能を有する該ヘリコバクター ピロリ結合性物質の類似体または誘導体。
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal{β4Glc[NAc]s3}s4 (1)
(式中、R1およびR2は末端単糖置換基または末端オリゴ糖置換基であり、該置換基の少なくとも1種はNeuNAcα3であり;そして
s1、s3およびs4は各々独立に0または1であり、{ }または[ ]の中の構造の存在または不在を示す。)
【請求項2】
該オリゴ糖配列がアグリコンに結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項3】
該アグリコンの炭素数が23未満であることを特徴とする、請求項2に記載のヘリコバクター ピロリ結合性物質。
【請求項4】
該アグリコンが、オリゴ糖配列とオリゴ価または多価の担体との間に位置するスペーサーであることを特徴とする、請求項2に記載のヘリコバクター ピロリ結合性物質。
【請求項5】
オリゴ糖またはオリゴ糖混合物であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項6】
式(1)中のR1またはR2がNeuNAcα3ではない非シアリル化置換基である場合には、R1またはR2はGalの2位および3位の少なくとも1種に結合した、下記式(2)で表される末端置換基であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
Hex[NAc]t1α/β3[(DeoxyHexα2)]t2 (2)
(式中、HexはGalまたはGlcであることが好ましく、
t1およびt2は各々独立に0または1であり、そして
α/βはα結合またはβ結合である。)
【請求項7】
非シアリル化置換基であるR1またはR2が、下記式からなる群より選ばれる構造であることを特徴とする、請求項6に記載のヘリコバクター ピロリ結合性物質。
Galα3、GalNAcα3、Fucα2、Galα3(Fucα2)、GalNAcα3(Fucα2)、
Galβ4GlcNAcβ3、GlcNAcβ3Galβ4GlcNAcβ3、Galβ4GlcNAcβ3Galβ4GlcNAcβ3、
GlcNAcα3、GlcNAcβ3、GalNAcβ3、Galβ3、Glcβ3、そしてGlcα3 。
【請求項8】
非シアリル化置換基であるR1またはR2が、下記式からなる群より選ばれる血液型抗原型の構造であることを特徴とする、請求項6に記載のヘリコバクター ピロリ結合性物質。
Galα3、GalNAcα3、Fucα2、Galα3(Fucα2)、そしてGalNAcα3(Fucα2) 。
【請求項9】
下記式:
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、または
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Gal
で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項10】
該オリゴ糖配列に、さらにポリ−N−アセチルラクトサミン側鎖が結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項11】
下記式で表される構造からなることを特徴とする、請求項10に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc
(式中、LacNAcはN−アセチルラクトサミン、即ちGalβ4GlcNAcであり、
Lacはラクトース、即ちGalβ4Glcである。)
【請求項12】
式(1)中のs1が0であり、下記式のいずれかで表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3LacNAcβ3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3Lac、
NeuNAcα3LacNAcβ3LacNAcβ3Gal、
NeuNAcα3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3Lac、または
NeuNAcα3LacNAcβ3Gal 。
【請求項13】
式(1)中のN−アセチルラクトサミン残基の少なくとも1つが2型N−アセチルラクトサミン類似体構造、好ましくはラクトース残基、で置換されており、下記式(3)で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
R1Galβ4Glc[NAc]u1β3{(R2Galβ4Glc[NAc]u2β6)}s1Gal{β4Glc[NAc]u3β3Gal}s2−
{β4Glc[NAc]s3}s4 (3)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;そして
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0である。)
【請求項14】
下記式:
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β6)Lac[NAc]u3β3Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3(NeuNAcα3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3Gal{β4Glc[NAc]s3}s4、または
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2
で表されることを特徴とする、請求項13に記載のヘリコバクター ピロリ結合性物質。
【請求項15】
フコシル化されており、下記式(4)で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
R1Galβ4[(Fucα3)]t1Glc[NAc]u1β3{(R2Galβ4[(Fucα3)]t2Glc[NAc]u2β6)}s1−
Gal[β4[(Fucα3)]t3Glc[NAc]u3β3Gal]s2{β4[(Fucα3)]t4Glc[NAc]s3}s4 (4)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4[(Fucα3)]t5Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0であり;そして
t1、t2、t3、t4およびt5は各々独立に0または1であり、[ ]内のFucα3側鎖構造の存在または不在を示すので、t1、t2、t3、t4およびt5の少なくとも1種は1である。)
【請求項16】
下記式(5)で表される四糖からなるオリゴ糖配列を含むことを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3Galβ4GlcNAcβ3Gal (5)
【請求項17】
下記式(6)で表されるオリゴ糖配列を含むことを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc(NAc)m (6)
(式中、mは0または1である。)
【請求項18】
該オリゴ糖配列が、五糖糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerではないことを特徴とする、請求項16または17に記載のヘリコバクター ピロリ結合性物質。
【請求項19】
該オリゴ糖配列が、炭素数が22を超えるセラミド、疎水性アグリコンまたは疎水性スペーサーに結合していないことを特徴とする、請求項16または17に記載のヘリコバクター ピロリ結合性物質。
【請求項20】
該四糖からなるオリゴ糖配列が、疎水性構造から得られた炭素数8未満のアグリコンまたはスペーサーにカップリングしていることを特徴とする、請求項16または17に記載のヘリコバクター ピロリ結合性物質。
【請求項21】
多糖、好ましくはポリラクトサミン鎖、またはその結合体に結合していることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項22】
糖脂質であることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項23】
少なくとも2種のオリゴ糖配列を含有するオリゴマー分子であることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項24】
少なくとも1種の該結合性物質を包含するミセルである、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項25】
担体に結合していることを特徴とする、請求項1〜24のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項26】
ヘリコバクター ピロリに対して有効な抗生物質、好ましくはペニシリン系抗生物質、に共有結合していることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項27】
下記式で表される構造からなることを特徴とする請求項25に記載のヘリコバクター ピロリ結合性物質、またはヘリコバクター ピロリに対する結合能を有するその誘導体。
[OS−O−(X)n−Y]m−Z
(式中、mは1以上の整数であり、nは0または1であり;
OSはオリゴ糖配列であり;
Oは酸素原子であり;
Xは単糖残基、オリゴ糖残基またはそれらの誘導体、好ましくはラクトシル残基、ガラクトシル残基、ポリ−N−アセチル−ラクトサミニル残基、あるいはO−グリカンオリゴ糖配列またはN−グリカンオリゴ糖配列の一部であり;
Yはスペーサー、セラミド脂質成分などの末端結合基あるいは該Oまたは該XとZの間の結合であり;
Zはオリゴ価または多価の担体であり;そして
該OSのGal還元末端、Glc還元末端またはGlcNAc還元末端は、該末端のC1位の酸素原子によって、該Yおよび所望により該Xを介して該Zに結合している。)
【請求項28】
請求項1〜26のいずれかのヘリコバクター ピロリ結合性物質を包含する、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の医薬組成物。
【請求項29】
慢性表在性胃炎、胃潰瘍、十二指腸潰瘍、胃腺癌、ヒトの胃における非ホジキンリンパ腫、肝臓病、膵臓病、皮膚病、心臓病または自己免疫性胃炎、悪性貧血および非ステロイド性抗炎症薬(NSAID)関連胃疾患を含む自己免疫疾患の治療あるいは乳幼児突然死症候群の予防に用いることを特徴とする、請求項28に記載の医薬組成物。
【請求項30】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の医薬組成物または栄養補助組成物の製造方法。
【請求項31】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの感染によって生じる病態の診断方法。
【請求項32】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質または組成物を包含する、栄養補助剤、食物または飲料。
【請求項33】
乳幼児食品用であることを特徴とする、請求項32に記載の栄養補助剤。
【請求項34】
ヘリコバクター ピロリの存在によって生じる病態の治療方法であって、医薬的効果を生じる量の請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を対象物に投与することを包含してなる治療方法。
【請求項35】
該病態が、患者の消化器系におけるヘリコバクター ピロリの存在によって生じたものであることを特徴とする、請求項34に記載の治療方法。
【請求項36】
慢性表在性胃炎、胃潰瘍、十二指腸潰瘍、胃腺癌、ヒトの胃における非ホジキンリンパ腫、肝臓病、膵臓病、皮膚病、心臓病、または自己免疫性胃炎、悪性貧血および非ステロイド性抗炎症薬(NSAID)関連胃疾患を含む自己免疫疾患の治療用、あるいは乳幼児突然死症候群の予防用であることを特徴とする、請求項34に記載の治療方法。
【請求項37】
該結合性物質が、栄養補助剤であるか、または栄養補助組成物の一部であることを特徴とする、請求項34〜36のいずれかに記載の治療方法。
【請求項38】
ヘリコバクター ピロリに対する結合またはヘリコバクター ピロリの阻害に用いることを特徴とする、請求項1〜27のいずれかに記載のヘリコバクター ピロリ結合性物質または組成物。
【請求項39】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の栄養補助剤または栄養補助組成物の製造方法。
【請求項40】
該栄養補助剤または栄養補助組成物が乳幼児食品用であることを特徴とする、請求項39に記載の製造方法。
【請求項41】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、細菌性付着因子の同定方法。
【請求項42】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質または請求項38の方法によって同定された物質を用いた、ヘリコバクター ピロリに対するワクチンの製造方法。
【請求項43】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの型の同定方法。
【請求項44】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリ結合アッセイ。
【請求項45】
該構造が、L1-CH(H/{CH1-2OH}p1)-{CH1OH}p2-{CH(NH-R)}p3-{CH1OH}p4-L2をYとして用いる下記式で表され、非酸性多価物質であることを特徴とする、請求項27に記載のヘリコバクター ピロリ結合性物質。
[OS-O-(X)n-L1-CH(H/{CH1-2OH}p1)-{CH1OH}p2-{CH(NH-R)}p3-{CH1OH}p4-L2]m-Z
(式中、
OSおよびXは請求項16において定義した通りであり;
L1およびL2は結合基あって、各々独立に酸素原子、窒素原子、硫黄原子または炭素原子である結合原子あるいは置換基を形成する結合に含まれる2個の原子を包含し、該結合原子および該置換基を形成する結合は−O−、−S−、−CH2−、−N−、−N(COCH3)−、−CO−NH−または−NH−CO−で表されるアミド基、−N−N−(ヒドラジン誘導体)、あるいは−O−N−または−N−O−で表されるアミノオキシ結合であり;
L1は該Xの還元末端単糖のC1位に結合した結合基であり、nが0の場合には、L1は−O−を置換して、該OSの還元末端のC1位に直接結合しており;
p1、p2、p3およびp4は各々独立に0〜7の整数であるが、但し、p1、p2、p3およびp4の少なくとも1種が1以上の整数であり;
{CH1-2OH}p1が側鎖構造を示す場合には、末端基となるCH1-2OHはCH2OHであり、p1が1を超える場合には、該末端基をスペーサーである該Yの残りの部分に結合させるための第二級アルコール基である−CHOH−が存在し;
Rは好ましくはアセチル基(−COCH3)、Zに連結する結合、類似体形成基、水素原子または式中でアミンを形成するC1-4アルキル基であり、Rが結合である場合にはL2は糖鎖末端基の1個または2個の原子であり、該類似体形成基は、アミド構造を有するC1-4アシル基を含み、;
mは1を超える整数であり;
Zは多価の担体である。)
【請求項46】
下記式で表される、ヘリコバクター ピロリに対して結合親和性を示すシアル酸誘導体を、非還元末端配列として含むヘリコバクター ピロリ結合性物質。
SA(X−R)
(式中、SAはシアル酸であり;
Xはシアル酸のC1位に結合している結合原子または結合基、好ましくはシアル酸残基のカルボキシル基と共にアミド構造を形成する−NHであり;そして
Rは水素原子または炭素数が3を超える有機ラジカル、好ましくは水素原子あるいは環構造および脂肪鎖の少なくとも1種を含むC4〜C30有機ラジカル、さらに好ましくはC6〜C24有機ラジカル、最も好ましくはC6〜C24脂肪族アルキル鎖である。)
【請求項47】
該シアル酸がNeuNAcであることを特徴とする、請求項46に記載のヘリコバクター ピロリ結合性物質。
【請求項48】
該シアル酸が2型N−アセチルラクトサミン配列にα3結合を介して結合しており、下記式で表されることを特徴とする、請求項46に記載のヘリコバクター ピロリ結合性物質。
SA(X-R)αxGal{β4GlcNAc[β3Gal(β4Glc)p1]p2}p3
(式中、xはシアル酸誘導体の結合位置を示し;そして
p1、p2およびp3は各々独立に0または1であり、{ }、[ ]または( )の中の構造全体の存在または不在を示す。)
【請求項49】
医薬として用いることを特徴とする、請求項46〜48のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項50】
請求項13〜15のヘリコバクター ピロリ結合性物質。
【請求項51】
請求項1〜27および46〜48において定義されているオリゴ糖配列からなる群より選ばれる少なくとも2種の異なる種類のオリゴ糖配列を含有する可溶性多価物質。
【請求項52】
請求項1〜27および46〜48のいずれかにおいて定義されているオリゴ糖配列を少なくとも1種含有する食品保存剤。
【請求項53】
請求項1〜27および46〜48のいずれかにおいて定義されているオリゴ糖配列を少なくとも1種含有する口腔衛生製品。
【請求項54】
練り歯磨、洗口液、タブレット及びチューイングガムからなる群より選ばれることを特徴とする、請求項53に記載の口腔衛生製品。
【請求項55】
請求項1〜27および46〜48のいずれかにおいて定義されているオリゴ糖配列を少なくとも1種含有する局所用製品、洗剤または化粧品。
【請求項56】
練り歯磨き、洗口液、タブレット、清浄剤、消毒剤およびチューインガムからなる群より選ばれることを特徴とする、請求項55に記載の局所用製品、洗剤または化粧品。
【請求項57】
請求項1〜27および46〜48のいずれかの結合性物質を含む組成物を、ex vivoにおける病原体の非診断的な阻害または凝集に用いる方法。
【請求項58】
該病原体がヘリコバクター ピロリであることを特徴とする、請求項57に記載の方法。
【請求項59】
下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料のリモデリング方法。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) 該原料オリゴ糖からなる画分、必要な場合には所望の糖、好ましくはポリ−N−アセチルラクトサミン、に富んだ画分を単離し、
(3) 該単離した画分を処理、好ましくは穏やかな酸処理に付して、末端単糖、好ましくはフコースとシアル酸の少なくとも1種、さらに好ましくはシアル酸、を遊離させて末端単糖除去オリゴ糖を得、そして
(4) 該末端単糖除去オリゴ糖に、単糖、好ましくはα3結合するシアル酸をグリコシルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【特許請求の範囲】
【請求項1】
下記式(1)で表される親水性オリゴ糖配列を非還元末端配列として含む、医薬として用いるヘリコバクター ピロリ結合性物質、あるいはヘリコバクター ピロリに対する結合能を有する該ヘリコバクター ピロリ結合性物質の類似体または誘導体であって、該オリゴ糖配列は、遊離オリゴ糖または遊離オリゴ糖の一部であるか、炭素数が23未満のアグリコンに結合したオリゴ糖である。
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal{β4Glc[NAc]s3}s4 (1)
(式中、R1およびR2は末端単糖置換基または末端オリゴ糖置換基であり、該置換基の少なくとも1種はNeuNAcα3であり;そして
s1、s3およびs4は各々独立に0または1であり、{ }または[ ]の中の構造の存在または不在を示す。)
【請求項2】
式(1)中のR1またはR2がNeuNAcα3ではない非シアリル化置換基である場合には、R1またはR2は、Galの2位および3位の少なくとも1種に結合した、下記式(2)で表される末端置換基であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
Hex[NAc]t1α/β3[(DeoxyHexα2)]t2 (2)
(式中、HexはGalまたはGlcであり、
t1およびt2は各々独立に0または1であり、そして
α/βはα結合またはβ結合である。)
【請求項3】
下記式:
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3Lac、
NeuNAcα3LacNAcβ3LacNAcβ3Gal、
NeuNAcα3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3Lac、または
NeuNAcα3LacNAcβ3Gal
で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項4】
式(1)中のN−アセチルラクトサミン残基の少なくとも1つが2型N−アセチルラクトサミン類似体構造で置換されており、下記式(3)で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
R1Galβ4Glc[NAc]u1β3{(R2Galβ4Glc[NAc]u2β6)}s1Gal{β4Glc[NAc]u3β3Gal}s2−
{β4Glc[NAc]s3}s4 (3)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0であり;そして
Glc[NAc]単位はFucα3によって分岐していてもよい。)
【請求項5】
該オリゴ糖配列が、Glc、GlcNAcおよびGalからなる群より選ばれる1種の還元単糖残基を介してオリゴ価または多価の担体に結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項6】
多糖に結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項7】
少なくとも2種のオリゴ糖配列を含有するオリゴマー分子、少なくとも1種の結合性物質からなるミセル、または担体に結合した結合性物質のいずれかの形状であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項8】
下記式で表される構造からなることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質、またはヘリコバクター ピロリに対する結合能を有するその誘導体。
[OS−O−(X)n−Y]m−Z
(式中、mは1以上の整数であり、nは0または1であり;
OSはオリゴ糖配列であり;
Oは酸素原子であり;
Xはラクトシル残基、ガラクトシル残基、ポリ−N−アセチル−ラクトサミニル残基、ならびにO−グリカンオリゴ糖配列またはN−グリカンオリゴ糖配列の一部からなる群より選ばれる単糖残基、オリゴ糖残基またはそれらの誘導体であり;
Yはスペーサー、末端結合基あるいは該Oまたは該XとZの間の結合であり;
Zはオリゴ価または多価の担体であり;そして
該OSのGal還元末端、Glc還元末端またはGlcNAc還元末端は、該末端のC1位の酸素原子によって、該Yおよび該Xを介して該Zに結合している。)
【請求項9】
Yである該末端結合基がセラミド脂質成分であることを特徴とする、請求項8に記載のヘリコバクター ピロリ結合性物質。
【請求項10】
請求項1のヘリコバクター ピロリ結合性物質を包含する、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の医薬組成物または栄養補助組成物。
【請求項11】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの感染によって生じる病態の診断方法。
【請求項12】
請求項1のヘリコバクター ピロリ結合性物質を包含する、栄養補助剤、食物、食品保存剤または飲料。
【請求項13】
ヘリコバクター ピロリの存在によって生じる病態の治療方法であって、医薬的効果を生じる量の請求項1のヘリコバクター ピロリ結合性物質を対象物に投与することを包含してなる治療方法。
【請求項14】
該結合性物質が、栄養補助剤であるか、または栄養補助組成物の一部であることを特徴とする、請求項13に記載の治療方法。
【請求項15】
ヘリコバクター ピロリに対する結合またはヘリコバクター ピロリの阻害に用いることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項16】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の栄養補助剤または栄養補助組成物の製造方法。
【請求項17】
請求項1のヘリコバクター ピロリ結合性物質を用いた、細菌性付着因子の同定方法。
【請求項18】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの型の同定方法。
【請求項19】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリ結合アッセイ。
【請求項20】
ヘリコバクター ピロリに対して結合親和性を示すシアル酸誘導体を非還元末端配列として含む、下記式で表されるヘリコバクター ピロリ結合性物質。
NeuNAc(X-R)αxGalβ4GlcNAcβ3Gal(β4Glc)p1
Xはシアル酸のC1位に結合している結合原子または結合基であり;
Rは水素原子または炭素数が3を超える有機ラジカルであり;そして
p1は0または1であり、( )の中の構造全体の存在または不在を示す。)
【請求項21】
式中のXが、シアル酸残基のカルボキシル基と共にアミド構造を形成する−NHであることを特徴とする、請求項20に記載のヘリコバクター ピロリ結合性物質。
【請求項22】
式中のRが、水素原子、あるいは環構造および脂肪鎖の少なくとも1種を含むC4〜C30有機ラジカルであることを特徴とする、請求項20または21に記載のヘリコバクター ピロリ結合性物質。
【請求項23】
式中のRがC6〜C24有機ラジカルであることを特徴とする、請求項22に記載のヘリコバクター ピロリ結合性物質。
【請求項24】
式中のRがC6〜C24脂肪族アルキル鎖であることを特徴とする請求項23に記載のヘリコバクター ピロリ結合性物質。
【請求項25】
請求項1において定義されているオリゴ糖配列を少なくとも1種含有する、練り歯磨き、洗口液、タブレット、清浄剤、消毒剤およびチューインガムからなる群より選ばれる局所用製品、洗剤または化粧品。
【請求項26】
下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料のリモデリング方法。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) ポリ−N−アセチルラクトサミンを含む所望の糖に富んだ、該原料オリゴ糖からなる画分を単離し、
(3) 該単離した画分を処理することで、フコースおよびシアル酸からなる群より選ばれる少なくとも1種の末端単糖を該原料オリゴ糖から遊離させて、末端単糖除去オリゴ糖を得、そして
(4) 該末端単糖除去オリゴ糖にα3結合するシアル酸をグリコシルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【請求項27】
工程(3)で行う処理が穏やかな酸処理であることを特徴とする、請求項26に記載のリモデリング方法。
【請求項1】
下記式(1)で表される親水性オリゴ糖配列を非還元末端配列として含む、医薬として用いるヘリコバクター ピロリ結合性物質、あるいはヘリコバクター ピロリに対する結合能を有する該ヘリコバクター ピロリ結合性物質の類似体または誘導体。
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal{β4Glc[NAc]s3}s4 (1)
(式中、R1およびR2は末端単糖置換基または末端オリゴ糖置換基であり、該置換基の少なくとも1種はNeuNAcα3であり;そして
s1、s3およびs4は各々独立に0または1であり、{ }または[ ]の中の構造の存在または不在を示す。)
【請求項2】
該オリゴ糖配列がアグリコンに結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項3】
該アグリコンの炭素数が23未満であることを特徴とする、請求項2に記載のヘリコバクター ピロリ結合性物質。
【請求項4】
該アグリコンが、オリゴ糖配列とオリゴ価または多価の担体との間に位置するスペーサーであることを特徴とする、請求項2に記載のヘリコバクター ピロリ結合性物質。
【請求項5】
オリゴ糖またはオリゴ糖混合物であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項6】
式(1)中のR1またはR2がNeuNAcα3ではない非シアリル化置換基である場合には、R1またはR2はGalの2位および3位の少なくとも1種に結合した、下記式(2)で表される末端置換基であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
Hex[NAc]t1α/β3[(DeoxyHexα2)]t2 (2)
(式中、HexはGalまたはGlcであることが好ましく、
t1およびt2は各々独立に0または1であり、そして
α/βはα結合またはβ結合である。)
【請求項7】
非シアリル化置換基であるR1またはR2が、下記式からなる群より選ばれる構造であることを特徴とする、請求項6に記載のヘリコバクター ピロリ結合性物質。
Galα3、GalNAcα3、Fucα2、Galα3(Fucα2)、GalNAcα3(Fucα2)、
Galβ4GlcNAcβ3、GlcNAcβ3Galβ4GlcNAcβ3、Galβ4GlcNAcβ3Galβ4GlcNAcβ3、
GlcNAcα3、GlcNAcβ3、GalNAcβ3、Galβ3、Glcβ3、そしてGlcα3 。
【請求項8】
非シアリル化置換基であるR1またはR2が、下記式からなる群より選ばれる血液型抗原型の構造であることを特徴とする、請求項6に記載のヘリコバクター ピロリ結合性物質。
Galα3、GalNAcα3、Fucα2、Galα3(Fucα2)、そしてGalNAcα3(Fucα2) 。
【請求項9】
下記式:
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、または
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Gal
で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項10】
該オリゴ糖配列に、さらにポリ−N−アセチルラクトサミン側鎖が結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項11】
下記式で表される構造からなることを特徴とする、請求項10に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc
(式中、LacNAcはN−アセチルラクトサミン、即ちGalβ4GlcNAcであり、
Lacはラクトース、即ちGalβ4Glcである。)
【請求項12】
式(1)中のs1が0であり、下記式のいずれかで表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3LacNAcβ3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3Lac、
NeuNAcα3LacNAcβ3LacNAcβ3Gal、
NeuNAcα3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3Lac、または
NeuNAcα3LacNAcβ3Gal 。
【請求項13】
式(1)中のN−アセチルラクトサミン残基の少なくとも1つが2型N−アセチルラクトサミン類似体構造、好ましくはラクトース残基、で置換されており、下記式(3)で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
R1Galβ4Glc[NAc]u1β3{(R2Galβ4Glc[NAc]u2β6)}s1Gal{β4Glc[NAc]u3β3Gal}s2−
{β4Glc[NAc]s3}s4 (3)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;そして
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0である。)
【請求項14】
下記式:
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β6)Lac[NAc]u3β3Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3(NeuNAcα3Lac[NAc]u2β3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3(NeuNAcα3Lac[NAc]u3β6)Gal{β4Glc[NAc]s3}s4、
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2β3Gal{β4Glc[NAc]s3}s4、または
NeuNAcα3Lac[NAc]u1β3Lac[NAc]u2
で表されることを特徴とする、請求項13に記載のヘリコバクター ピロリ結合性物質。
【請求項15】
フコシル化されており、下記式(4)で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
R1Galβ4[(Fucα3)]t1Glc[NAc]u1β3{(R2Galβ4[(Fucα3)]t2Glc[NAc]u2β6)}s1−
Gal[β4[(Fucα3)]t3Glc[NAc]u3β3Gal]s2{β4[(Fucα3)]t4Glc[NAc]s3}s4 (4)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4[(Fucα3)]t5Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0であり;そして
t1、t2、t3、t4およびt5は各々独立に0または1であり、[ ]内のFucα3側鎖構造の存在または不在を示すので、t1、t2、t3、t4およびt5の少なくとも1種は1である。)
【請求項16】
下記式(5)で表される四糖からなるオリゴ糖配列を含むことを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3Galβ4GlcNAcβ3Gal (5)
【請求項17】
下記式(6)で表されるオリゴ糖配列を含むことを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
NeuNAcα3Galβ4GlcNAcβ3Galβ4Glc(NAc)m (6)
(式中、mは0または1である。)
【請求項18】
該オリゴ糖配列が、五糖糖脂質であるNeuNAcα3Galβ4GlcNAcβ3Galβ4GlcβCerではないことを特徴とする、請求項16または17に記載のヘリコバクター ピロリ結合性物質。
【請求項19】
該オリゴ糖配列が、炭素数が22を超えるセラミド、疎水性アグリコンまたは疎水性スペーサーに結合していないことを特徴とする、請求項16または17に記載のヘリコバクター ピロリ結合性物質。
【請求項20】
該四糖からなるオリゴ糖配列が、疎水性構造から得られた炭素数8未満のアグリコンまたはスペーサーにカップリングしていることを特徴とする、請求項16または17に記載のヘリコバクター ピロリ結合性物質。
【請求項21】
多糖、好ましくはポリラクトサミン鎖、またはその結合体に結合していることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項22】
糖脂質であることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項23】
少なくとも2種のオリゴ糖配列を含有するオリゴマー分子であることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項24】
少なくとも1種の該結合性物質を包含するミセルである、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項25】
担体に結合していることを特徴とする、請求項1〜24のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項26】
ヘリコバクター ピロリに対して有効な抗生物質、好ましくはペニシリン系抗生物質、に共有結合していることを特徴とする、請求項1〜20のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項27】
下記式で表される構造からなることを特徴とする請求項25に記載のヘリコバクター ピロリ結合性物質、またはヘリコバクター ピロリに対する結合能を有するその誘導体。
[OS−O−(X)n−Y]m−Z
(式中、mは1以上の整数であり、nは0または1であり;
OSはオリゴ糖配列であり;
Oは酸素原子であり;
Xは単糖残基、オリゴ糖残基またはそれらの誘導体、好ましくはラクトシル残基、ガラクトシル残基、ポリ−N−アセチル−ラクトサミニル残基、あるいはO−グリカンオリゴ糖配列またはN−グリカンオリゴ糖配列の一部であり;
Yはスペーサー、セラミド脂質成分などの末端結合基あるいは該Oまたは該XとZの間の結合であり;
Zはオリゴ価または多価の担体であり;そして
該OSのGal還元末端、Glc還元末端またはGlcNAc還元末端は、該末端のC1位の酸素原子によって、該Yおよび所望により該Xを介して該Zに結合している。)
【請求項28】
請求項1〜26のいずれかのヘリコバクター ピロリ結合性物質を包含する、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の医薬組成物。
【請求項29】
慢性表在性胃炎、胃潰瘍、十二指腸潰瘍、胃腺癌、ヒトの胃における非ホジキンリンパ腫、肝臓病、膵臓病、皮膚病、心臓病または自己免疫性胃炎、悪性貧血および非ステロイド性抗炎症薬(NSAID)関連胃疾患を含む自己免疫疾患の治療あるいは乳幼児突然死症候群の予防に用いることを特徴とする、請求項28に記載の医薬組成物。
【請求項30】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の医薬組成物または栄養補助組成物の製造方法。
【請求項31】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの感染によって生じる病態の診断方法。
【請求項32】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質または組成物を包含する、栄養補助剤、食物または飲料。
【請求項33】
乳幼児食品用であることを特徴とする、請求項32に記載の栄養補助剤。
【請求項34】
ヘリコバクター ピロリの存在によって生じる病態の治療方法であって、医薬的効果を生じる量の請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を対象物に投与することを包含してなる治療方法。
【請求項35】
該病態が、患者の消化器系におけるヘリコバクター ピロリの存在によって生じたものであることを特徴とする、請求項34に記載の治療方法。
【請求項36】
慢性表在性胃炎、胃潰瘍、十二指腸潰瘍、胃腺癌、ヒトの胃における非ホジキンリンパ腫、肝臓病、膵臓病、皮膚病、心臓病、または自己免疫性胃炎、悪性貧血および非ステロイド性抗炎症薬(NSAID)関連胃疾患を含む自己免疫疾患の治療用、あるいは乳幼児突然死症候群の予防用であることを特徴とする、請求項34に記載の治療方法。
【請求項37】
該結合性物質が、栄養補助剤であるか、または栄養補助組成物の一部であることを特徴とする、請求項34〜36のいずれかに記載の治療方法。
【請求項38】
ヘリコバクター ピロリに対する結合またはヘリコバクター ピロリの阻害に用いることを特徴とする、請求項1〜27のいずれかに記載のヘリコバクター ピロリ結合性物質または組成物。
【請求項39】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の栄養補助剤または栄養補助組成物の製造方法。
【請求項40】
該栄養補助剤または栄養補助組成物が乳幼児食品用であることを特徴とする、請求項39に記載の製造方法。
【請求項41】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、細菌性付着因子の同定方法。
【請求項42】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質または請求項38の方法によって同定された物質を用いた、ヘリコバクター ピロリに対するワクチンの製造方法。
【請求項43】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの型の同定方法。
【請求項44】
請求項1〜27のいずれかのヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリ結合アッセイ。
【請求項45】
該構造が、L1-CH(H/{CH1-2OH}p1)-{CH1OH}p2-{CH(NH-R)}p3-{CH1OH}p4-L2をYとして用いる下記式で表され、非酸性多価物質であることを特徴とする、請求項27に記載のヘリコバクター ピロリ結合性物質。
[OS-O-(X)n-L1-CH(H/{CH1-2OH}p1)-{CH1OH}p2-{CH(NH-R)}p3-{CH1OH}p4-L2]m-Z
(式中、
OSおよびXは請求項16において定義した通りであり;
L1およびL2は結合基あって、各々独立に酸素原子、窒素原子、硫黄原子または炭素原子である結合原子あるいは置換基を形成する結合に含まれる2個の原子を包含し、該結合原子および該置換基を形成する結合は−O−、−S−、−CH2−、−N−、−N(COCH3)−、−CO−NH−または−NH−CO−で表されるアミド基、−N−N−(ヒドラジン誘導体)、あるいは−O−N−または−N−O−で表されるアミノオキシ結合であり;
L1は該Xの還元末端単糖のC1位に結合した結合基であり、nが0の場合には、L1は−O−を置換して、該OSの還元末端のC1位に直接結合しており;
p1、p2、p3およびp4は各々独立に0〜7の整数であるが、但し、p1、p2、p3およびp4の少なくとも1種が1以上の整数であり;
{CH1-2OH}p1が側鎖構造を示す場合には、末端基となるCH1-2OHはCH2OHであり、p1が1を超える場合には、該末端基をスペーサーである該Yの残りの部分に結合させるための第二級アルコール基である−CHOH−が存在し;
Rは好ましくはアセチル基(−COCH3)、Zに連結する結合、類似体形成基、水素原子または式中でアミンを形成するC1-4アルキル基であり、Rが結合である場合にはL2は糖鎖末端基の1個または2個の原子であり、該類似体形成基は、アミド構造を有するC1-4アシル基を含み、;
mは1を超える整数であり;
Zは多価の担体である。)
【請求項46】
下記式で表される、ヘリコバクター ピロリに対して結合親和性を示すシアル酸誘導体を、非還元末端配列として含むヘリコバクター ピロリ結合性物質。
SA(X−R)
(式中、SAはシアル酸であり;
Xはシアル酸のC1位に結合している結合原子または結合基、好ましくはシアル酸残基のカルボキシル基と共にアミド構造を形成する−NHであり;そして
Rは水素原子または炭素数が3を超える有機ラジカル、好ましくは水素原子あるいは環構造および脂肪鎖の少なくとも1種を含むC4〜C30有機ラジカル、さらに好ましくはC6〜C24有機ラジカル、最も好ましくはC6〜C24脂肪族アルキル鎖である。)
【請求項47】
該シアル酸がNeuNAcであることを特徴とする、請求項46に記載のヘリコバクター ピロリ結合性物質。
【請求項48】
該シアル酸が2型N−アセチルラクトサミン配列にα3結合を介して結合しており、下記式で表されることを特徴とする、請求項46に記載のヘリコバクター ピロリ結合性物質。
SA(X-R)αxGal{β4GlcNAc[β3Gal(β4Glc)p1]p2}p3
(式中、xはシアル酸誘導体の結合位置を示し;そして
p1、p2およびp3は各々独立に0または1であり、{ }、[ ]または( )の中の構造全体の存在または不在を示す。)
【請求項49】
医薬として用いることを特徴とする、請求項46〜48のいずれかに記載のヘリコバクター ピロリ結合性物質。
【請求項50】
請求項13〜15のヘリコバクター ピロリ結合性物質。
【請求項51】
請求項1〜27および46〜48において定義されているオリゴ糖配列からなる群より選ばれる少なくとも2種の異なる種類のオリゴ糖配列を含有する可溶性多価物質。
【請求項52】
請求項1〜27および46〜48のいずれかにおいて定義されているオリゴ糖配列を少なくとも1種含有する食品保存剤。
【請求項53】
請求項1〜27および46〜48のいずれかにおいて定義されているオリゴ糖配列を少なくとも1種含有する口腔衛生製品。
【請求項54】
練り歯磨、洗口液、タブレット及びチューイングガムからなる群より選ばれることを特徴とする、請求項53に記載の口腔衛生製品。
【請求項55】
請求項1〜27および46〜48のいずれかにおいて定義されているオリゴ糖配列を少なくとも1種含有する局所用製品、洗剤または化粧品。
【請求項56】
練り歯磨き、洗口液、タブレット、清浄剤、消毒剤およびチューインガムからなる群より選ばれることを特徴とする、請求項55に記載の局所用製品、洗剤または化粧品。
【請求項57】
請求項1〜27および46〜48のいずれかの結合性物質を含む組成物を、ex vivoにおける病原体の非診断的な阻害または凝集に用いる方法。
【請求項58】
該病原体がヘリコバクター ピロリであることを特徴とする、請求項57に記載の方法。
【請求項59】
下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料のリモデリング方法。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) 該原料オリゴ糖からなる画分、必要な場合には所望の糖、好ましくはポリ−N−アセチルラクトサミン、に富んだ画分を単離し、
(3) 該単離した画分を処理、好ましくは穏やかな酸処理に付して、末端単糖、好ましくはフコースとシアル酸の少なくとも1種、さらに好ましくはシアル酸、を遊離させて末端単糖除去オリゴ糖を得、そして
(4) 該末端単糖除去オリゴ糖に、単糖、好ましくはα3結合するシアル酸をグリコシルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【特許請求の範囲】
【請求項1】
下記式(1)で表される親水性オリゴ糖配列を非還元末端配列として含む、医薬として用いるヘリコバクター ピロリ結合性物質、あるいはヘリコバクター ピロリに対する結合能を有する該ヘリコバクター ピロリ結合性物質の類似体または誘導体であって、該オリゴ糖配列は、遊離オリゴ糖または遊離オリゴ糖の一部であるか、炭素数が23未満のアグリコンに結合したオリゴ糖である。
R1Galβ4GlcNAcβ3{(R2Galβ4GlcNAcβ6)}s1Gal{β4Glc[NAc]s3}s4 (1)
(式中、R1およびR2は末端単糖置換基または末端オリゴ糖置換基であり、該置換基の少なくとも1種はNeuNAcα3であり;そして
s1、s3およびs4は各々独立に0または1であり、{ }または[ ]の中の構造の存在または不在を示す。)
【請求項2】
式(1)中のR1またはR2がNeuNAcα3ではない非シアリル化置換基である場合には、R1またはR2は、Galの2位および3位の少なくとも1種に結合した、下記式(2)で表される末端置換基であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
Hex[NAc]t1α/β3[(DeoxyHexα2)]t2 (2)
(式中、HexはGalまたはGlcであり、
t1およびt2は各々独立に0または1であり、そして
α/βはα結合またはβ結合である。)
【請求項3】
下記式:
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Lac、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)Gal、
NeuNAcα3LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAcβ3(NeuNAcα3LacNAcβ6)LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3LacNAcβ3Lac、
NeuNAcα3LacNAcβ3LacNAcβ3Gal、
NeuNAcα3LacNAcβ3LacNAc、
NeuNAcα3LacNAcβ3Lac、または
NeuNAcα3LacNAcβ3Gal
で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項4】
式(1)中のN−アセチルラクトサミン残基の少なくとも1つが2型N−アセチルラクトサミン類似体構造で置換されており、下記式(3)で表されることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
R1Galβ4Glc[NAc]u1β3{(R2Galβ4Glc[NAc]u2β6)}s1Gal{β4Glc[NAc]u3β3Gal}s2−
{β4Glc[NAc]s3}s4 (3)
(式中、R1およびR2は各々独立に末端単糖置換基または末端オリゴ糖置換基であるか、あるいは存在しないが、但し、R1およびR2の少なくとも1種はNeuNAcα3またはNeuNAcα3Galβ4Glc[NAc]u4β3であり;
s1、s2、s3およびs4は各々独立に0または1であり、[ ]または{ }の中の構造の存在または不在を示し;
u1、u2、u3およびu4は各々独立に0または1であり、非還元末端または鎖中のラクトサミン残基に結合したN−アセチル基の存在または不在を示すが、但し、u1、u2、u3およびu4の少なくとも1種は0であり;そして
Glc[NAc]単位はFucα3によって分岐していてもよい。)
【請求項5】
該オリゴ糖配列が、Glc、GlcNAcおよびGalからなる群より選ばれる1種の還元単糖残基を介してオリゴ価または多価の担体に結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項6】
多糖に結合していることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項7】
少なくとも2種のオリゴ糖配列を含有するオリゴマー分子、少なくとも1種の結合性物質からなるミセル、または担体に結合した結合性物質のいずれかの形状であることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項8】
下記式で表される構造からなることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質、またはヘリコバクター ピロリに対する結合能を有するその誘導体。
[OS−O−(X)n−Y]m−Z
(式中、mは1以上の整数であり、nは0または1であり;
OSはオリゴ糖配列であり;
Oは酸素原子であり;
Xはラクトシル残基、ガラクトシル残基、ポリ−N−アセチル−ラクトサミニル残基、ならびにO−グリカンオリゴ糖配列またはN−グリカンオリゴ糖配列の一部からなる群より選ばれる単糖残基、オリゴ糖残基またはそれらの誘導体であり;
Yはスペーサー、末端結合基あるいは該Oまたは該XとZの間の結合であり;
Zはオリゴ価または多価の担体であり;そして
該OSのGal還元末端、Glc還元末端またはGlcNAc還元末端は、該末端のC1位の酸素原子によって、該Yおよび該Xを介して該Zに結合している。)
【請求項9】
Yである該末端結合基がセラミド脂質成分であることを特徴とする、請求項8に記載のヘリコバクター ピロリ結合性物質。
【請求項10】
請求項1のヘリコバクター ピロリ結合性物質を包含する、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の医薬組成物または栄養補助組成物。
【請求項11】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの感染によって生じる病態の診断方法。
【請求項12】
請求項1のヘリコバクター ピロリ結合性物質を包含する、栄養補助剤、食物、食品保存剤または飲料。
【請求項13】
ヘリコバクター ピロリの存在によって生じる病態の治療方法であって、医薬的効果を生じる量の請求項1のヘリコバクター ピロリ結合性物質を対象物に投与することを包含してなる治療方法。
【請求項14】
該結合性物質が、栄養補助剤であるか、または栄養補助組成物の一部であることを特徴とする、請求項13に記載の治療方法。
【請求項15】
ヘリコバクター ピロリに対する結合またはヘリコバクター ピロリの阻害に用いることを特徴とする、請求項1に記載のヘリコバクター ピロリ結合性物質。
【請求項16】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの存在によって生じる病態の治療用または予防用の栄養補助剤または栄養補助組成物の製造方法。
【請求項17】
請求項1のヘリコバクター ピロリ結合性物質を用いた、細菌性付着因子の同定方法。
【請求項18】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリの型の同定方法。
【請求項19】
請求項1のヘリコバクター ピロリ結合性物質を用いた、ヘリコバクター ピロリ結合アッセイ。
【請求項20】
ヘリコバクター ピロリに対して結合親和性を示すシアル酸誘導体を非還元末端配列として含む、下記式で表されるヘリコバクター ピロリ結合性物質。
NeuNAc(X-R)αxGalβ4GlcNAcβ3Gal(β4Glc)p1
Xはシアル酸のC1位に結合している結合原子または結合基であり;
Rは水素原子または炭素数が3を超える有機ラジカルであり;そして
p1は0または1であり、( )の中の構造全体の存在または不在を示す。)
【請求項21】
式中のXが、シアル酸残基のカルボキシル基と共にアミド構造を形成する−NHであることを特徴とする、請求項20に記載のヘリコバクター ピロリ結合性物質。
【請求項22】
式中のRが、水素原子、あるいは環構造および脂肪鎖の少なくとも1種を含むC4〜C30有機ラジカルであることを特徴とする、請求項20または21に記載のヘリコバクター ピロリ結合性物質。
【請求項23】
式中のRがC6〜C24有機ラジカルであることを特徴とする、請求項22に記載のヘリコバクター ピロリ結合性物質。
【請求項24】
式中のRがC6〜C24脂肪族アルキル鎖であることを特徴とする請求項23に記載のヘリコバクター ピロリ結合性物質。
【請求項25】
請求項1において定義されているオリゴ糖配列を少なくとも1種含有する、練り歯磨き、洗口液、タブレット、清浄剤、消毒剤およびチューインガムからなる群より選ばれる局所用製品、洗剤または化粧品。
【請求項26】
下記の工程(1)〜(4)を包含することを特徴とする、天然食物材料のリモデリング方法。
(1) 化学的または酵素学的に食物材料から糖を遊離させて、原料オリゴ糖を得、
(2) ポリ−N−アセチルラクトサミンを含む所望の糖に富んだ、該原料オリゴ糖からなる画分を単離し、
(3) 該単離した画分を処理することで、フコースおよびシアル酸からなる群より選ばれる少なくとも1種の末端単糖を該原料オリゴ糖から遊離させて、末端単糖除去オリゴ糖を得、そして
(4) 該末端単糖除去オリゴ糖にα3結合するシアル酸をグリコシルトランスフェラーゼ酵素またはトランスシアリダーゼ酵素によって転移する。
【請求項27】
工程(3)で行う処理が穏やかな酸処理であることを特徴とする、請求項26に記載のリモデリング方法。
【図2】

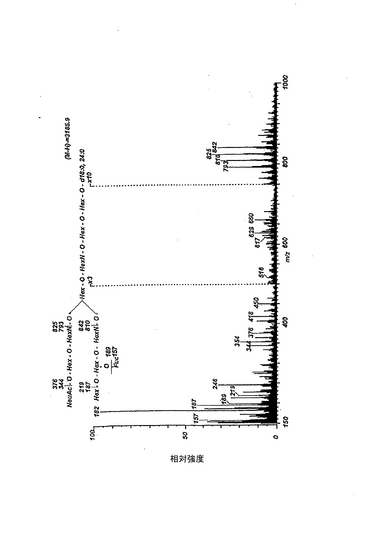
【図3】
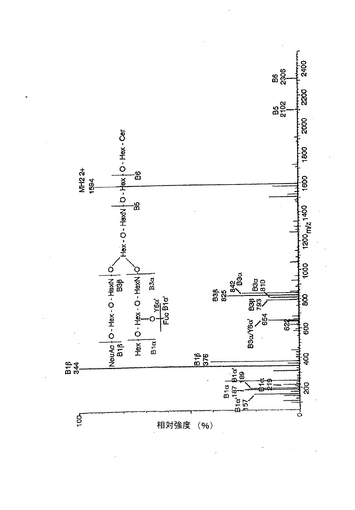
【図4】
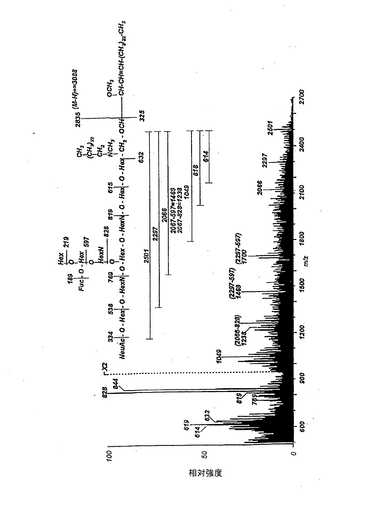
【図5A】
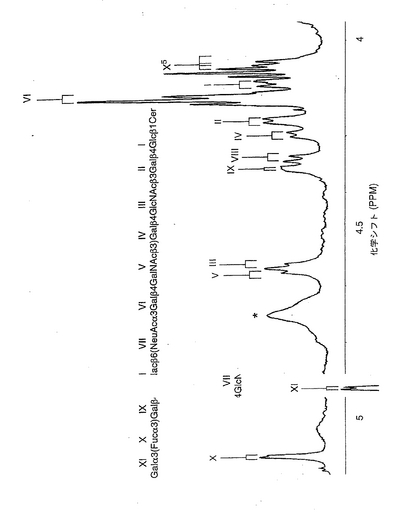
【図5B】
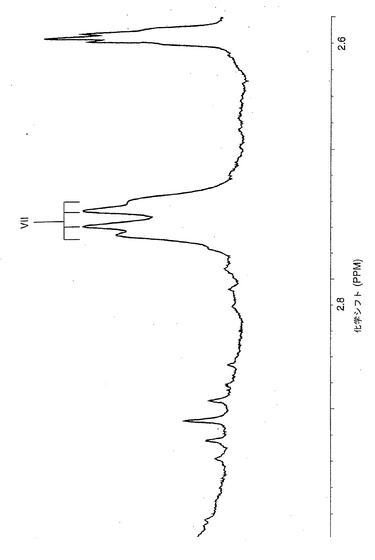
【図6】
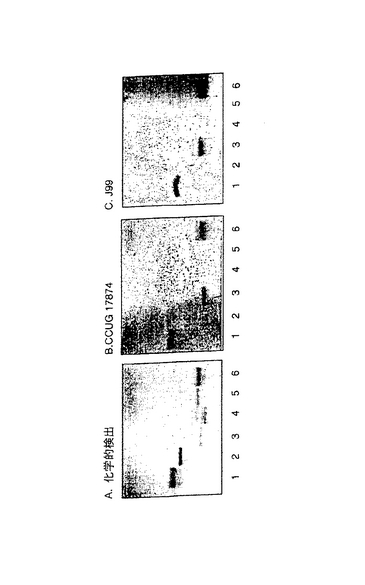
【図7】
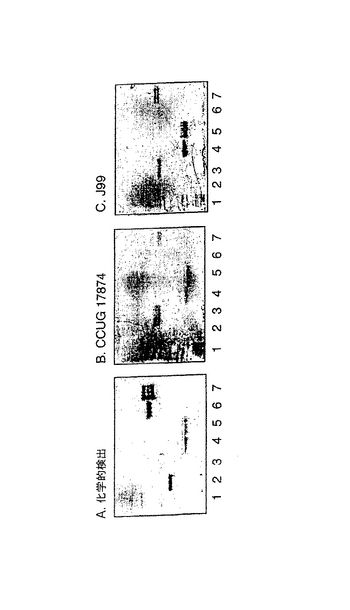
【図8】
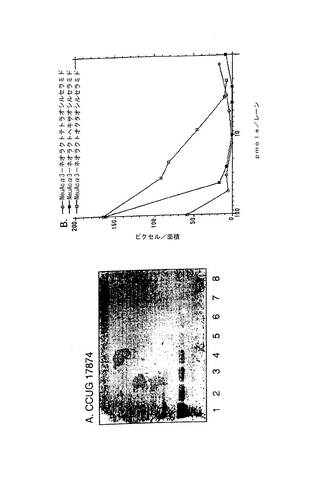
【図9】
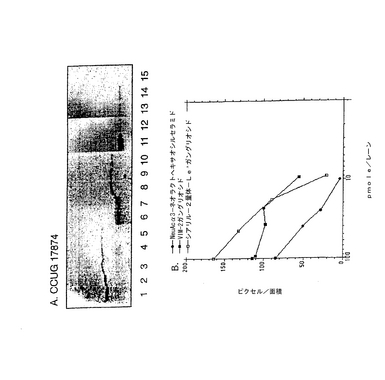
【図10】
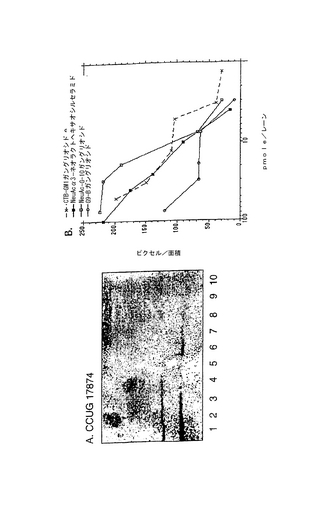
【図11】
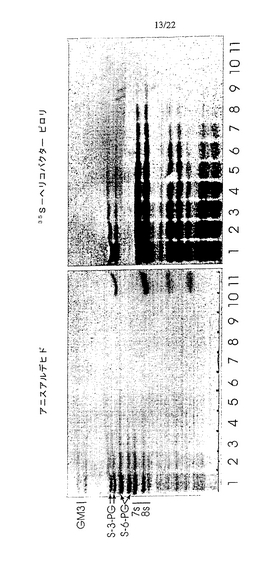
【図12−1】
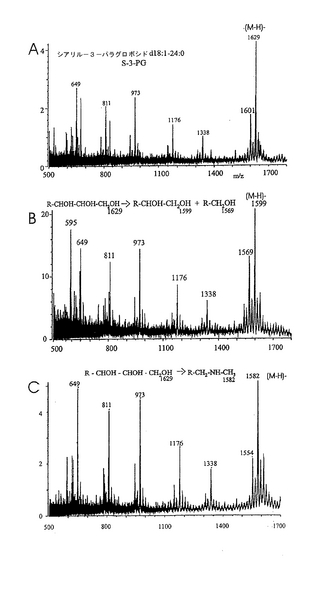
【図12−2】
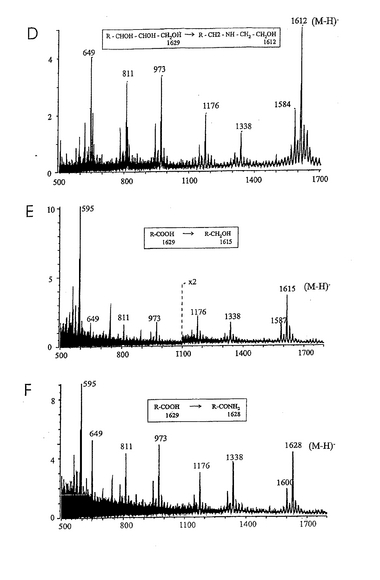
【図12−3】

【図12−4】
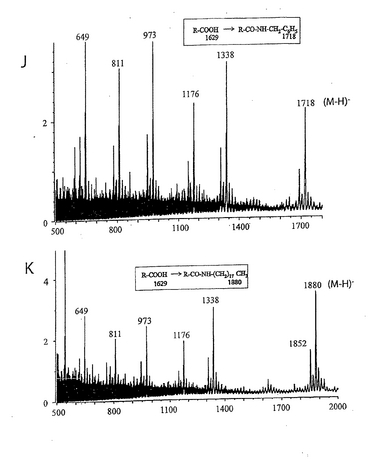
【図13】
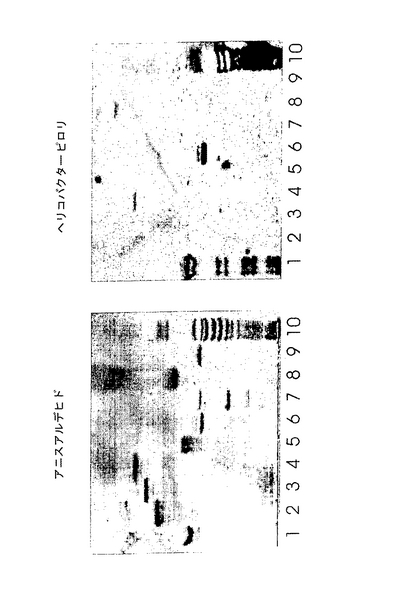
【図14】
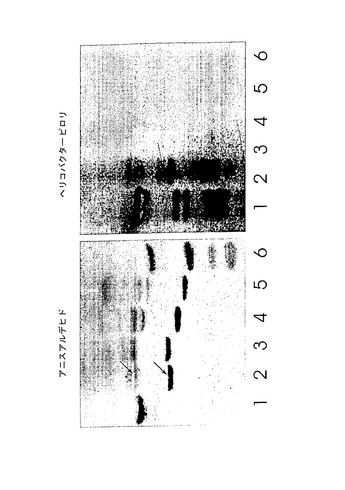
【図15】
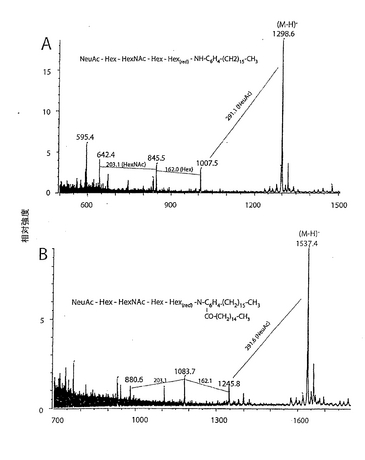
【図16】
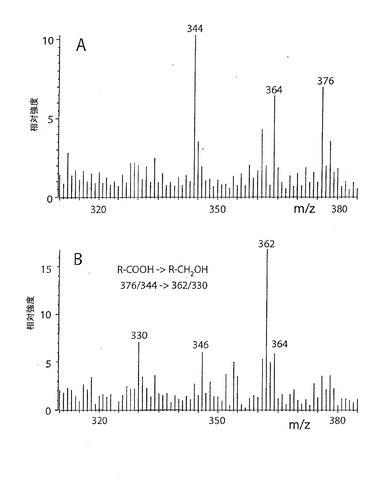
【図17】
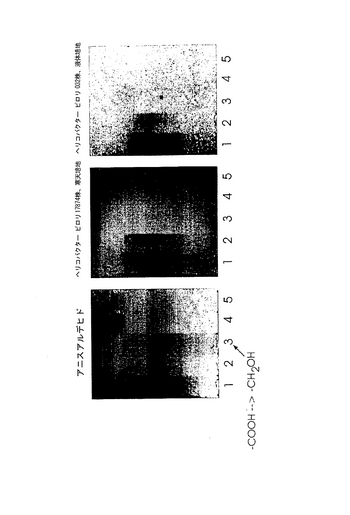
【図3】
【図4】
【図5A】
【図5B】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12−1】
【図12−2】
【図12−3】
【図12−4】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2006−511497(P2006−511497A)
【公表日】平成18年4月6日(2006.4.6)
【国際特許分類】
【出願番号】特願2004−549233(P2004−549233)
【出願日】平成15年11月6日(2003.11.6)
【国際出願番号】PCT/FI2003/000840
【国際公開番号】WO2004/041291
【国際公開日】平成16年5月21日(2004.5.21)
【出願人】(504472695)
【氏名又は名称原語表記】GLYKOS FINLAND OY
【Fターム(参考)】
【公表日】平成18年4月6日(2006.4.6)
【国際特許分類】
【出願日】平成15年11月6日(2003.11.6)
【国際出願番号】PCT/FI2003/000840
【国際公開番号】WO2004/041291
【国際公開日】平成16年5月21日(2004.5.21)
【出願人】(504472695)
【氏名又は名称原語表記】GLYKOS FINLAND OY
【Fターム(参考)】
[ Back to top ]