
ペプチジルアルギニンデイミナーゼ、およびシトルリン化タンパク質およびペプチド生成におけるその使用
本出願は、ジベレラ・ゼアエ(Gibberella zeae)(フザリウム・グラミネアルム(Fusarium graminearum)とも称される)からのペプチジルアルギニンデイミナーゼ酵素(EC3.5.3.15)の単離に関する。本出願は、前記アルギニンデイミナーゼを使用して酵素的に生成されたシトルリン化タンパク質およびペプチド、およびこのようなシトルリン化タンパク質およびペプチドの食材中での使用を開示する。本発明は、タンパク質、ペプチドまたはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換された、修飾タンパク質、ペプチドまたはタンパク質加水分解産物に関する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、シトルリン残基を含んでなるタンパク質およびペプチドに関する。
【0002】
[背景技術]
アルギニンは、哺乳類の生理機能において重要な役割を果たす準必須アミノ酸である。アルギニンの代謝経路については、詳しく述べられている。その食餌摂取に際して、アルギニンは肝臓によって肝臓の門脈から取り込まれ、酵素アルギナーゼによって迅速にオルニチンに転換する。後者の過程で尿素が形成される。次にアルギニンから発生したオルニチンはシトルリンに転換され、またはアミノ酸グルタメートおよびプロリンに代謝されることができる。代案としては形成されたオルニチンは、プトレシンなどのポリアミン化合物に組み込まれる。オルニチンに代謝されなかった食餌性アルギニンは、タンパク質合成の目的で、例えば酸化窒素またはアルギニル−tRNAに加工されることができる。アルギニンに向けた内在性合成経路もまた存在する。後者の過程は、アルギニンがオルニチンおよびシトルリン前駆物質から合成される腎臓で主に起きる。
【0003】
シトルリンは、スイカ、カボチャ、およびキュウリのようなウリ類中で遊離アミノ酸として存在すると述べられている天然アミノ酸である。遊離アミノ酸のその他の起源は、プルーン果汁、いくつかのブドウの変異株、および醤油およびワインなどの発酵食品である。自然界では、シトルリンがその他のアミノ酸に結合して存在することは稀である。食用キノコでは、ジペプチドピログルタメート−シトルリンが、アイリッシュコケではジペプチドシトルリン−アルギニンの存在が示されている。哺乳類では免疫化学的技術を使用して、シトルリンを取り込んだ低レベルのペプチドまたはタンパク質の存在が示されている。
【0004】
哺乳類では、シトルリンは腸内でグルタミンから合成され、血中に放出されて腎臓内で転換されてアルギニンに戻る。健康な成人では、腎臓によって転換されるシトルリンは、身体の全アルギニン所要量を提供するのに十分である。しかし新生児では、腎臓内でのこのシトルリンからアルギニンへの反応は不十分であり、追加的機序が関与する。様々な生理学的過程におけるアルギニンに代わるものとしてのシトルリンの重要な役割は、最近の研究によって明らかにされている。肝臓による食餌性アルギニンの捕捉は非常に効率的であるので、肝臓下流の血中アルギニン濃度は比較的低い。それゆえ例えば迅速な生育期間中に、栄養失調またはアミノ酸代謝変化の結果として、または外傷性または病的侵襲に応えて、全ての臓器におけるアルギニン需要が完全に満たされないかもしれない状態が生じる場合がある。このような状況では、シトルリンはアルギニンに代わるものとして機能するかもしれない。食餌性アルギニンとは対照的に、食餌性シトルリンは肝臓によって門脈血から取り除かれない。それゆえシトルリンは、筋肉をはじめとする末梢組織が利用できる自由に循環するアルギニン源の代替えに相当するが、より効率的である(Curisら著、Amino Acids(2005年)29:177〜205頁)。食餌性シトルリンはまた、アルギニンがオルニチンに転換するアルギナーゼ反応について述べられているような肝臓内における尿素生成をもたらさない。したがってシトルリンを摂取することにより、この尿素生成および免疫学的状態の低下の結果としての腎臓損傷などの補給されるアルギニンのネガティブな副作用を回避できる。シトルリンのさらに別の利点は、それが過剰なアンモニアを捕らえることができ、すなわちそれがいわゆる血中アンモニア低下剤として作用して、特定の酵素機能障害を患っている患者、てんかんを患っている個人に、そして健康な個人では、長期の高強度筋肉運動に起因する疲労を予防する利点を提供できることである。
【0005】
多くの国々で、食物への遊離アミノ酸の添加を規制する法規がある。結果として、遊離アミノ酸は、臨床栄養物でのみ使用でき、上述の補給されるアルギニンまたはシトルリンの生理学的利点を一般の消費者に提供することはできない。さらに遊離アルギニンの非常に苦い味は、臨床および非臨床用途の重要な欠点となる。重病患者は、味の悪い食物または栄養補給剤を単に避け、高齢者またはスポーツ選手などの医療ニーズがない消費者も同様である。したがって食物中での遊離アミノ酸の使用は、特に推奨アミノ酸投薬量を考慮した場合、重大な美味性の問題を引き起こすことが予期できる。これらの結論の意味合いは、遊離アミノ酸として存在しない形態であり、改善された美味性を有するシトルリンに対する明らかな必要性が存在することである。
【0006】
[発明の概要]
本発明は、タンパク質、ペプチドまたはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換する、修飾タンパク質、ペプチドまたはタンパク質加水分解産物に関する。したがって本発明のタンパク質、ペプチドまたはタンパク質加水分解産物は、好ましくは少なくとも0.15、好ましくは少なくとも0.30、より好ましくは少なくとも0.5、なおもより好ましくは少なくとも1.0、さらになおもより好ましくは2.0、最も好ましくは少なくとも4のシトルリン対アルギニン(タンパク質、ペプチドまたはタンパク質加水分解産物中に存在する)のモル比を有する。加水分解産物の場合、アルギニンとして存在する遊離アミノ酸の量は、シトルリン形成の測定に使用せず、シトルリン対アルギニン比の考慮にも入れない。それゆえ本発明は、高比率の結合シトルリン残基を有する、タンパク質、ペプチドまたは加水分解産物に関する。結合シトルリンまたはペプチド結合シトルリンとは、遊離アミノ酸である遊離シトルリンとは対照的に、ペプチドまたはタンパク質の一部であるシトルリン残基を意味する。
【0007】
さらに本発明は、タンパク質、ペプチドまたはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換する、タンパク質、ペプチドまたはタンパク質加水分解産物を酵素的に生成する方法に関する。本発明のタンパク質、ペプチドまたはタンパク質を得るために、開始タンパク質、ペプチドまたはタンパク質加水分解産物基質をタンパク質アルギニンデイミナーゼと共にインキュベートする。
【0008】
さらに食物と、食物成分と、飼料と、飼料成分と、栄養補助食品または薬物などの栄養補給食品と、または栄養補助食品または薬物などの栄養補給食品の成分として使用でき、または食物と、飼料と、または栄養補助食品または薬物などの栄養補給食品の生産において成分として使用できる、タンパク質、タンパク質加水分解産物、ペプチドまたはペプチド混合物を提供することが本発明の目的である。
【0009】
本発明の別の態様に従って、生産宿主によって培養液中に能動的に分泌される、タンパク質アルギニンデイミナーゼが開示されている。本発明はまた、このようなタンパク質アルギニンデイミナーゼの生成および使用にも関する。
【0010】
したがって本発明は、
(a)配列番号6、8、9、10、13または14の1〜640番目のアミノ酸と少なくとも30%のアミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド、
(b)低ストリンジェンシー条件下で、(i)60個を超える、好ましくは100個を超えるヌクレオチドと少なくとも80%または90%同一の、より好ましくは200個を超えるヌクレオチドと少なくとも90%同一の配列番号3の核酸配列またはその断片、または(ii)配列番号3の核酸配列に相補的な核酸配列とハイブリダイズするポリヌクレオチドによってコードされるポリペプチド
からなる群から選択される、タンパク質アルギニンデイミナーゼ活性を有する単離されたポリペプチドに関する。
【0011】
[発明の詳細な説明]
シトルリンはアミノ酸でありながらDNAによってコードされず、タンパク質合成中にタンパク質に組み立てられない。それにもかかわらず、いくつかの哺乳類の組織内では、免疫化学的技術を使用して微少量のペプチド結合シトルリンが検出されている。例は、滑液、滑膜組織、造血性細胞、および活性化マクロファージである。シトルリン残基を含有するタンパク質は、ペプチド結合アルギニン残基のいわゆる翻訳後修飾中に発生する。この特定の修飾は、シトルリン化または脱イミノ化と称される過程で、ペプチドまたはタンパク質結合アルギニンをペプチドまたはタンパク質結合シトルリンに転換する、タンパク質またはペプチジルアルギニンデイミナーゼ(EC3.5.3.15)と称される酵素ファミリーによって触媒される。タンパク質アルギニンデイミナーゼおよびペプチジルアルギニンデイミナーゼという用語は、ここで同義的に使用される。アルギニンからシトルリンへの反応において、アルギニン側鎖の末端窒素原子の1つは酸素によって置換される。反応は1個の水分子を使用して、副生物としてアンモニアを生じる(http://en.wikipedia.org/wiki/Citrullination)。中性pHにおいてアルギニンが正に帯電する一方、シトルリンは非荷電である。したがってシトルリン化はペプチドまたはタンパク質の疎水性を増大させ、これはタンパク質またはペプチドの特性を変化させることができ、究極的にタンパク質のアンフォールディングをもたらすかもしれない過程である。シトルリン残基を含有することが知られている哺乳類タンパク質としては、ミエリン塩基性タンパク質(MBP)、フラグリンおよびいくつかのヒストンタンパク質が挙げられる一方、線維素およびビメンチンのようなその他のタンパク質は、細胞死および組織炎症においてシトルリン化されることができる。タンパク質のシトルリン化は、尿素回路の一環としての、または酸化窒素シンターゼファミリーの酵素副産物としての遊離アミノ酸シトルリンの形成とは異なることに留意されたい。
【0012】
アルギニン含有タンパク質またはペプチドとは対照的に、シトルリン化タンパク質またはペプチドは、工業規模では市販されない。意外にも我々は、タンパク質またはペプチド結合シトルリンの生成を含んでなる、食品等級で工業的に応用可能な方法を同定した。この方法では、タンパク質またはペプチド結合アルギニン残基をタンパク質またはペプチド結合シトルリン残基に効率的に転換する、酵素タンパク質アルギニンデイミナーゼまたはペプチジルアルギニンデイミナーゼ(下文においてPADと称される)を利用する。
【0013】
主に哺乳類組織からのいくつかのタイプのPADが知られているが、これらの酵素のいずれも産業上の利用には適さない。多くの微生物をスクリーニングすることで、我々はかなり意外なことに、発酵ブロス中にPAD酵素を能動的に分泌できるいくつかの微生物に行き当たった。目下記述されている全ての哺乳類タイプのPADは培養液中に能動的に分泌されないので、このような能動的に分泌されるPADが自然界に見いだされることは予期されていなかった。能動的分泌は、ここで生物が生育または培養液中にポリペプチドを蓄積する能力と定義される。ポリペプチドの能動的分泌は宿主生物からのエネルギー、および宿主生物からの専用分泌経路を必要とする。一般に能動的に分泌されるポリペプチドは、シグナル配列またはシグナルペプチドとも称されるアミノ−末端プレ配列を含有する。能動的分泌は発酵ブロスへの酵素の輸送のために、必ずしも細胞壁の破壊を伴わない。一般にグラム陰性細菌は、能動的分泌を有さないことが知られている。
【0014】
哺乳類のPADは互いに強いつながりがあり、異なるイソ型が多様な異なる臓器中で発現される。実際、酵素的PAD活性は、細胞の溶解後にのみ検出でき、それはPADが哺乳類の組織中で細胞内酵素であることを強力に示唆し(Vossenaarらによるレビュー(2003年)Bioessays 25:1106〜1118頁)、酵素は能動的に分泌されない。さらに全ての哺乳類PADは、常態において効率的分泌のために必要とされる明白なシグナル配列を欠いている。したがって能動的に分泌されるPAD酵素は明白な工業利点を有するであろうが、微生物中のこのような酵素についてスクリーンすることは論理的でないようである。
【0015】
能動的分泌は、煩わしい精製工程を経ることなくほぼ純粋な形態での酵素の回収ができるようにするので、経済的製造工程に最も重要である。アスペルギルス(Aspergillus)などの食品等級真菌宿主によるこのような能動的に分泌されるPADの過剰発現は、食品等級酵素と、シトルリン化タンパク質またはペプチドに向けた費用効率が高い製造工程を生じる。
【0016】
我々が知る限りでは、我々はアスペルギルス(Aspergillus)のような食品等級宿主生物によって発酵ブロス中に効率的に分泌されるPADについて初めて述べ、シトルリン化タンパク質またはペプチドに向けた費用効率が高い製造工程を初めて報告する。
【0017】
酵素アルギニンデイミナーゼ(EC3.5.3.6)はよく知られており、遊離アルギニンの遊離シトルリンへの転換におけるその使用については、幅広く述べられている。しかしPAD(EC3.5.3.15)は、タンパク質またはペプチド結合アルギニン残基の脱イミノ化を触媒してタンパク質またはペプチド結合シトルリン残基を生じ、それよってアンモニアを放出する比較的新しい酵素ファミリーを形成する。今までに多岐にわたる哺乳類の組織内に酵素が同定されている。ヒトでは、例えば4つの異なるPAD酵素が同定されており、例えば皮膚、子宮、筋肉、脳、膵臓、脾臓、胃、胸線、脊髄、およびマクロファージなどの造血性細胞に見られる。これらのCa2+−依存性PAD酵素は、例えば表皮分化、神経軸索の髄鞘形成、および毛嚢の角質化に関与する。既知のいずれの哺乳類PADも細胞により能動的に分泌されない。哺乳類組織中に広範に存在しながら、微生物からの活性PADに関する報告は非常に限られている。唯一の例外は、ヒトグラム陰性病原体ポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)からの細菌酵素である(McGrawら(1999年)Infect Immun.67(7):3248〜3256頁)。この酵素はペリプラズム間隙に輸送されて、この細菌外のペリプラズム間隙から漏出するのみである。成人発症型歯周炎の発症および進行は、ポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)による歯肉溝の感染と関連付けられている。このポルフィロモナス(Porphyromonas)PADは、脊椎動物PADと進化的に関連していないが、いくつかのアルギニンデイミナーゼと配列相同性を共有する(Shiraiら(2001年)Trends Biochem Sci.;26(8):465〜468頁)。しかし酵素は、哺乳類のPADとは対照的に、ペプチド−結合ならびに遊離L−アルギニンをシトルリンに転換でき、カルシウムイオンに依存しない。したがって酵素は、能動的に分泌されず高度に不安定であり、毒性因子であると述べられているので、食品等級のこのP.ジンジバリス(gingivalis)由来酵素の産業上の利用は、全くありそうにない。したがってポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)からのPADおよびその使用は、本発明の一部ではない。
【0018】
ここで我々は、我々の知る限りでは、初めて記述された分泌PADである、真菌フザリウム・グラミネアルム(Fusarium graminearum)から分泌PADを単離して同定した。PADアッセイに基づいて、我々はフザリウム(Fusarium)株中で、分泌PADのいくつかの変異株を単離および特性決定できた。これらの真菌からのPADをコードする遺伝子は、単離および特性決定された(配列番号3および4)。真菌アスペルギルス・ニガー(Aspergillus niger)中でのこのPADの過剰発現は、培養液中へのPADの効率的な分泌をもたらす。これらの全ての新しい変異株は、分泌PADの特徴を含む。さらにこの知識に基づいて、我々は、そのDNA配列が公共データベース中に存在するその他の微生物中で、分泌PADの遺伝子を同定して正しくアノテートできた。潜在的分泌PADは、真菌ケトミウム・グロボスム(Chaetomium globosum)、ファエオスフェリア・ノドルム(Phaeosphaeria nodorum)、および細菌ストレプトミセス・スキャビエス(Streptomyces scabies)およびストレプトミセス・クラブリゲルス(Streptomyces clavuligerus)に見られる。これらのタンパク質の分泌された特徴に関する我々の知識に基づいて、我々は正確なタンパク質配列が配列番号8〜10、13、および14で示されることを提案する。これらの全配列は、これまで分泌されるペプチジルアルギニンデイミナーゼをコーディングすると述べられたことがないが、フザリウム・グラミネアルム(Fusarium graminearum)の分泌PADの遺伝子構造に関する我々の知識から、それとしてのみ同定できる。
【0019】
異なる培養コレクションからの真菌の大規模なスクリーニングの後、我々は、発酵ブロス中に能動的に分泌されるPAD様酵素源として、フザリウム(Fusarium)種を同定することができた。このフザリウム(Fusarium)由来PAD様酵素は哺乳類のPADに対してかなりの相同性を示すが、P.ジンジバリス(gingivalis)PADに対しては示さない。クロマトグラフィー精製によるフザリウム(Fusarium)発酵ブロスからのPADの単離は、この分泌PADの分子量が約55kDaであり、最適活性がpH8前後で、最適温度は40〜50℃の間であることを明らかにした。フザリウム(Fusarium)酵素の分子量は哺乳類酵素の分子量よりも顕著に低いが、我々は最適pHおよび温度が同様であることを見いだした。
【0020】
経済的観点から、PADを大量にかつ比較的純粋な形態で生成する、改善された手段に対する明白な必要がある。これを行う好ましい方法は、組み換えDNA技術を使用したこのようなPADの過剰産生を通じたものである。これを行う特に好ましい方法は、フザリウム(Fusarium)由来PADの過剰産生を通じたものであり、これを行う最も好ましい方法は、フザリウム・グラミネアルム(Fusarium graminearum)由来PADの過剰産生を通じたものである。後者の生成経路を可能にするために、フザリウム(Fusarium)由来ペプチジルアルギニンデイミナーゼのユニークな配列情報が必須である。より好ましくは、コードする遺伝子の全ヌクレオチド配列を利用できなくてはならない。
【0021】
新たに同定された分泌PADを大量にかつ比較的純粋な形態で生成する改善された手段は、組み換えDNA技術を使用して、フザリウム(Fusarium)がコードする酵素の過剰産生を通じたものである。これを行う好ましい方法は、食品等級宿主微生物中におけるこのような分泌PADの過剰産生を通じたものである。よく知られている食品等級微生物としては、アスペルギルス(Aspergilli)、トリコデルマ(Trichoderma)、ストレプトミセス(Streptomyces)、桿菌(Bacilli)、およびサッカロミセス(Saccharomyces)およびクリヴェロミセス(Kluyveromyces)などの酵母が挙げられる。これを行うさらにより好ましい方法は、アスペルギルス(Aspergillus)などの食品等級真菌中における、フザリウム(Fusarium)由来分泌PADの過剰産生を通じたものである。最も好ましいのは、その中でPADコード遺伝子のコドン使用が、使用される食品等級発現宿主のために最適化されている、食品等級真菌中における分泌PADの過剰産生である。一般に後者の最適化経路を可能にするために、分泌PADのユニークな配列情報が望ましい。より好ましいのは、PADコード遺伝子の全ヌクレオチド配列が利用できることである。ひとたび新しい酵素が大量にかつ比較的純粋な形態で利用できるようになれば、シトルリン含有食物タンパク質またはこのようなシトルリン含有食物タンパク質の加水分解産物が、食品等級で経済的なやり方によって生産できる。好ましくはこのようなシトルリン含有食物タンパク質は、高率のタンパク質結合アルギニンを含有する食物タンパク質、またはこのような食物タンパク質の加水分解産物から得られる。本発明に従ったPADのための好ましい基質タンパク質は、少なくとも10:1(mol/mol)のアルギニン対シトルリン比を有し、好ましくはこのような基質タンパク質は、少なくとも100:1(mol/mol)のアルギニン対シトルリン比を有し、最も好ましくは基質タンパク質はシトルリンを全く含有しない。本発明に従ったPADのための好ましい基質タンパク質は、少なくとも3mol%のタンパク質結合アルギニンを含有し、より好ましくはそれらは少なくとも6mol%のタンパク質結合アルギニンを含有する。このような基質タンパク質の例は、(脱脂)乳タンパク質、乳清タンパク質、カゼインまたは卵タンパク質などの動物起源からの市販食物タンパク質である。その他のこのような基質タンパク質の例は、穀物タンパク質、ジャガイモタンパク質、ダイズタンパク質、エンドウマメタンパク質、米タンパク質、エンドウマメタンパク質などの植物起源からの市販食物タンパク質、ならびにルピナス、ゴマ、ヤシの実などのアルギニンに富むことが知られているその他の植物起源からのタンパク質である。穀物タンパクの例は、小麦またはトウモロコシまたは例えば小麦グルテンなどのその画分である。微生物タンパク質の例は、肉代替品のための酵母抽出物または単細胞タンパク質である。これらのアルギニンに富むタンパク質またはペプチド中における特異的アミノ酸の相対的欠乏を代償するために、シトルリン含有タンパク質、ペプチドまたは加水分解産物の栄養組成物は、シトルリン化材料中で不足するアミノ酸に比較的富む、選択された遊離アミノ酸またはタンパク質、ペプチドまたは加水分解産物を添加することにより最適化できる。シトルリン含有タンパク質、ペプチドまたは加水分解産物の栄養価を増強するための好ましいアミノ酸は、システイン、ヒスチジン、イソロイシン、グルタミン、およびリジンである。代案としてはシトルリン含有タンパク質、ペプチドまたは加水分解産物をカゼイン、ジャガイモ、小麦またはダイズタンパク質などの比較的これらのアミノ酸に富むタンパク質源から得られたタンパク質、ペプチドまたは加水分解産物と混合できる。特に本発明の方法は酵素を修飾するのに有用であり、活性または安定性などの特徴が変更された酵素がもたらされる。
【0022】
哺乳類の組織を使用する場合、この組織は好ましくは非ヒト組織である。哺乳類のタンパク質を使用する場合、このタンパク質は好ましくは血液タンパク質、神経組織、脳、臓器、筋肉または毛髪ではない。好ましくは本発明に従って使用される基質タンパク質は、植物タンパク質、脱脂(乳)タンパク質、乳清タンパク質、カゼインタンパク質、ゼラチンタンパク質、卵タンパク質または微生物のタンパク質である。
【0023】
別の応用は、このようなシトルリン含有食物タンパク質の加水分解産物の創出である。これらの加水分解産物は、当該技術分野で知られている方法に従って生成できる。代案としては動物または植物起源のタンパク質を最初に加水分解して、タンパク質加水分解産物を得ることができ、引き続いてこの加水分解産物を本発明に従ったPADと共にインキュベートできる。
【0024】
より濃縮された形態でペプチド結合シトルリンを提供するために、PAD酵素のための基質として、アルギニン濃縮加水分解産物を使用できる。このアプローチでは、例えば米タンパク質またはエンドウマメタンパク質などのアルギニンに富むタンパク質源を、最初にサブチリシン(EC3.4.21.62)またはケカビペプシン(mucorpepsin)(EC3.4.23.23)またはプロリン−特異的プロテアーゼ(EC3.4.21.26)などの適切なエンドプロテアーゼによって加水分解し、次にクロマトグラフィーを使用して、得られた加水分解産物をアルギニン含有ペプチドに濃縮する。このようなクロマトグラフィー分離技術では、特定のpH条件下のアルギニン残基の正電荷を利用する。ペプチド精製におけるこれらの特徴の使用の実用的背景は、例えば「タンパク質精製ハンドブック(Protein Purification Handbook)」(発行元:アマシャム・ファーマシア・バイオテック(Amersham Pharmacia Biotech)、現在はベルギー国ディーゲムのGEヘルスケア・バイオサイエンシズ(GE Healthcare Bio−Sciences(Diegem,Belgium))にある。次に得られたアルギニン濃縮ペプチドをPAD酵素と共にインキュベートして、本発明に従ったシトルリンを含んでなる加水分解産物を得る。アルギニン濃縮加水分解産物を得ることに向けた別のアプローチでは、アルギニンに富むタンパク質源を最初にアルギニン−特異的エンドプロテアーゼトリプシン(EC3.4.21.4)と共にインキュベートする。加水分解後、より大きな非加水分解ペプチドが沈殿してより小さいアルギニンに富むペプチドが溶液中に残るように、インキュベーションのpHをタンパク質基質の水溶性が最小となる値に調節する。例えば中性に近いpHでは、エンドウマメタンパク質の溶解度はかなり低い。エンドウマメタンパク質の懸濁液を中性pHでトリプシンと共にインキュベートすることで(最大活性のためにはトリプシンは中性に近いpHを必要とする)、アルギニン残基に富むエンドウマメタンパク質の部分のみが溶液中に入り、引き続くデカンテーションまたは濾過ステップによって、それぞれ上清または濾液中にアルギニンに富むペプチドが生じる。この上清または濾液に本発明に従ったPADを添加することで、所望の高濃度シトルリンを含有するペプチドが生じる。
【0025】
本発明は、本タンパク質またはタンパク質加水分解産物を含んでなる新規な栄養補給食品組成物に関する。栄養補給食品組成物は、このような治療タンパク質を必要とする対象に、本発明のペプチドまたはタンパク質加水分解産物を投与するステップを含んでなる、例えば高血圧予防のため、および栄養失調からまたは腸管疾患から回復するための活性成分として、タンパク質加水分解産物を含んでなる。
【0026】
シトルリンを含んでなる本タンパク質、タンパク質加水分解産物、ペプチドまたはペプチド混合物は、固形製品、半固形製品(ペースト)または飲料などの液体製品などのあらゆる適切な形態で使用できる。例えば高レベルのシトルリンを含んでなる製品は、栄養補助食品としてまたは食物、飲料、飼料またはペットフード成分として使用される。高レベルのシトルリンを含んでなる製品はまた、ローション、ゲルまたはエマルジョンの形態の局所塗布をはじめとするパーソナルケア用途の形態で使用できる。これらの全ての用途で、タンパク質、ペプチドまたは加水分解産物は、ビタミンAまたはビタミンCなどのビタミンを含んでなるマルチビタミン製剤と共に、亜鉛などの微量元素と共に、および正常な代謝機能の維持に必須であるが体内で合成されないミネラルと共に、共調合されてもよい。さらにシトルリン化タンパク質、ペプチドまたは加水分解産物は、特定脂肪酸、グルタミンなどの特定アミノ酸、または特定アミノ酸に富むタンパク質またはタンパク質加水分解産物と組み合わせてもよい。またレスベラトロールまたはEGCGなどのポリフェノール、グリコシル化および脱グリコシル化ダイズイソフラボン、プレバイオティクスまたはプロバイオティクスとの組み合わせが予見される。
【0027】
栄養補給食品という用語は、本明細書での用法では、栄養および製薬分野用途の双方における有用性を意味する。したがって新規な栄養補給食品組成物は、食物および飲料への栄養補給剤としての、およびカプセルまたは錠剤などの固形製剤、または溶液または懸濁液などの液体製剤であってもよい、経腸的または非経口用途のための医薬製剤または薬物としての用途がある。上記から明らかなように、栄養補給食品組成物という用語は、本ペプチド含有組成物と、場合により炭水化物、ならびに例えば上記シトルリン化タンパク質、ペプチドまたは加水分解産物を含んでなる栄養補助食品などの栄養補助組成物を含んでなる、食物および飲料もまた含んでなる。
【0028】
栄養補助食品という用語は、本明細書での用法では、食餌への栄養補給を意図する「食餌性成分」を含有する、口から摂取される製品を意味する。これらの製品中の「食餌性成分」は、ビタミンと、ミネラルと、薬草またはその他の植物と、アミノ酸と、酵素、臓器組織、小腺、および代謝産物などの物質とを含んでもよい。栄養補助食品はまた、抽出または濃縮でき、錠剤、カプセル、ソフトジェル、ジェルキャップ、液体、または粉末などの多くの形態であってもよい。それらはまた、バーなどのその他の形態であることができるが、その場合、栄養補助食品のラベルの情報は、一般に製品を従来の食物として、または食事または食餌の唯一のアイテムとして表示しない。加水分解産物、タンパク質加水分解産物または加水分解タンパク質とは、基質タンパク質のタンパク質分解加水分解によって形成された生成物を意味する。好ましくは加水分解は、酵素的加水分解である。可溶性加水分解産物は、タンパク質加水分解産物の(水)可溶性画分であり、それはまた本明細書で、可溶性ペプチド含有組成物、または可溶性ペプチドを含んでなる組成物、またはタンパク質加水分解産物と可溶性加水分解産物の混合物としても述べられている。
【0029】
「ペプチド」または「オリゴペプチド」は、本明細書においてペプチド結合を通じて連結された少なくとも2つのアミノ酸の鎖と定義される。「ペプチド」および「オリゴペプチド」という用語は、(一般に認識されるように)同義と見なされ、各用語は文脈の必要に応じて同義的に使用できる。「ポリペプチド」または「タンパク質」は、本明細書において30個を超えるアミノ酸残基を含んでなる鎖と定義される。全ての(オリゴ)ペプチドおよびポリペプチドの式または配列は、本明細書では慣行に従ってアミノ−末端からカルボキシ−末端の方向で左から右に記述される。本明細書において使用されるアミノ酸の1文字コードについては、一般に当該技術分野で知られており、Sambrookら著、「分子クローニング実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、第2版、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press、Cold Spring Harbor,NY、1989年、にある。
【0030】
タンパク質生合成において細胞によって使用される20個の「標準」アミノ酸があり、それらは一般遺伝コードによって規定される。本明細書では、アミノ酸とはこれらの20個の「標準」アミノ酸およびシトルリンを意味する。
【0031】
好ましくは単離されたジペプチドシトルリン−アルギニンは、本発明の一部ではない。しかしこのジペプチドを含んでなる加水分解産物、およびこのような加水分解産物の使用は、本発明のさらなる実施態様である。栄養失調からまたは腸管疾患から回復するための活性成分としての単離されたジペプチドシトルリン−アルギニンの使用もまた、本発明の目的である。その中でアイリッシュモスまたは抽出物、またはアイリッシュモスから得られたその他の製品が使用される製品は、本発明の一部ではない。
【0032】
大量のシトルリンを組み入れたタンパク質またはペプチドは、顕著な利点を提供できる。最も重要には、それらは特殊食品、幼児栄養、または特別な消費者グループのための栄養補給剤などの非医療用途のために、ペプチド結合シトルリンを利用できるようにする可能性を初めて提供する。さらにシトルリン残基を含んでなるタンパク質およびタンパク質加水分解産物は苦味を呈さないので、このようなタンパク質またはペプチドは機能的利点を提供する。さらに本発明に従ったシトルリンは、国の法令によっては許容されないかもしれない遊離シトルリンとは対照的に、ペプチド結合シトルリンとして存在する。その結果、本発明に従った方法は、シトルリンを含有する許容可能な味覚プロフィールがある食物、栄養補給剤、および臨床製品の生産を初めて可能にする。
【0033】
本発明に従ったタンパク質またはペプチドの別の重要な利点は、それらがより低いアレルゲン性を示すかもしれないことである。ミルクおよびそのカゼインおよび乳清タンパク質画分、ならびに例えばダイズ単離物、米タンパク質および小麦グルテンから得られる植物タンパク質画分などの全ての主要な食物タンパク質は、重要な抗原化合物と見なされる。通常タンパク質抗原性は、タンパク質を8〜10個未満のアミノ酸残基を有するペプチドに加水分解することで克服される。しかしこのような高度なタンパク質分解消化によって作り出された加水分解産物は、苦さ、肉汁のような異臭、および浸透圧値増大をはじめとする不都合を示す。タンパク質抗原性の経済的重要性は、幼児および小児における食物アレルギーおよび喘息の罹患率が上昇しているという事実によって例示される。例えば牛乳アレルギーは、3才未満の小児の約2.5%が罹患している。牛乳アレルギーは、生後1ヶ月以内および牛乳の導入1週間以内に遭遇することが多い。異なる程度に加水分解された牛乳タンパク質または牛乳画分が組み込まれた様々な乳児用調製粉乳製品には牛乳アレルギーが予期される。我々は、食物タンパク質を本発明に従ったPADと共にインキュベートすることで、得られたシトルリン化タンパク質またはタンパク質加水分解産物の抗原性またはアレルゲン性が低下することを見いだした。最も重要には、この効果は、損なわれた味または乳化力低下などの高度なタンパク質分解に結びついた悪影響なしに得られた。運動愛好家、高齢者、および食物アレルギーを患っている人々が増えていることを鑑みて、本発明に従った製品には大きな経済的重要性がある。これらの全ての標的グループにとって、改善された味は顕著な心理学的重要性がある側面である。今までに工業規模で経済的な方法で、このような食物または栄養補給剤を生産することは不可能だった。目下工業的に入手できる食物タンパク質および/または食物タンパク質加水分解産物は、タンパク質−またはペプチド結合シトルリン残基を組み込んでいないので、シトルリン残基に富むタンパク質またはタンパク質加水分解産物またはペプチドは新しい。「濃縮」という用語は、タンパク質またはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換することを意味することが意図される。したがって本方法の生成物、すなわちシトルリン化タンパク質、シトルリン化タンパク質の加水分解産物またはシトルリン化加水分解産物中では、存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%はシトルリン残基に転換している。存在するアルギニンまたはシトルリン残基量を確立するために使用されるアミノ酸分析法は、本明細書の材料と方法セクションで特定されている。アミノ酸分析中に典型的に使用されて、遊離アミノ酸を解放する酸加水分解過程の人工産物として新たに発生したシトルリン残基の一部がオルニチン残基に転換することに留意することが重要である。存在するシトルリン残基のレベルを計算するために、存在するシトルリンおよびオルニチン残基のレベルを合計しなくてはならない。
【0034】
シトルリン含有タンパク質加水分解産物を得るために、シトルリン含有タンパク質を酵素的に加水分解できる。代案としては既存のタンパク質加水分解産物、すなわちペプチド結合シトルリンを含まない加水分解産物を本発明に従ったPADと共にインキュベートして、ペプチド結合シトルリンを含んでなるタンパク質加水分解産物を得ることができる。タンパク質加水分解産物は、当該技術分野で知られている加水分解法を使用して生成できる。好ましくはこのような加水分解産物は、5〜50の間、より好ましくは10〜35の間の加水分解度(DH)を有する。DH値を確立するための方法は、本明細書の材料と方法セクションで特定されている。本発明に従ったシトルリンを含んでなるタンパク質加水分解産物は、幼児および臨床栄養物、治療用食餌ならびに民生用食餌、およびスポーツ栄養において使用できる。このようなシトルリン化タンパク質、タンパク質加水分解産物またはペプチドを組み込んだ食物または食餌または臨床製品もまた新しい。さらにタンパク質−またはペプチド結合シトルリンを含んでなるタンパク質またはタンパク質加水分解産物は、パーソナルケア用途をはじめとする様々な局所用途において、および動物およびペットのための栄養製品において使用できる。
【0035】
本発明に従ったシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、多数の新しい驚くべき用途で使用できる。基本的にシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質水解物は、遊離アルギニンの補充が有益であることが示されている全ての用途で使用できる。したがってアルギニン欠乏状態によって特徴づけられる全ての病状において、本発明に従ったシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、このアルギニン欠乏状態を低下させることが予期される。一般にシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、栄養失調から、または短腸症候群などの腸管疾患から、または加齢の結果としてのタンパク質−エネルギー栄養失調から回復中の個人においてタンパク質代謝を維持する利点を提供する。さらにシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、胃腸管の健康を維持する上で、重要な役割を果たすことができる。アルギニンならびにシトルリンは酸化窒素生成、ひいては血管拡張の前駆物質であるので、これらのアミノ酸の適切な供給は、多様な血管疾患のリスクを予防する上で重要な役割を果たす。これは科学文献で、特に末梢動脈疾患、移植冠動脈疾患および喘息について実証されている。したがってシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、これらの疾患を予防する食餌への付加として、ならびにストレス性潰瘍の予防、またはストレス潰瘍治癒の改善において特に興味深い。さらに本発明に従ったタンパク質、ペプチド、およびタンパク質加水分解産物は、例えばアテローム動脈硬化、狭心症、高血圧、冠動脈心疾患の治療的措置において、II型糖尿病でインシュリンおよびグルコース濃度を低下させるのに、およびHIV感染症、火傷、外傷、および癌治療においても特に有用である。筋肉減少症の治療のためのシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物の使用は、特に注目に値する。さらに術前食餌中でのシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物の使用は、特にストレス条件下における免疫学的状態を改善して、患者の微小循環低灌流を回復させる。敗血症は高死亡率を伴う主要な健康問題であるので、敗血症の予防におけるシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物を組み込んだ食餌の有益な効果もまた言及に値する。シトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物はまた、それらの血中アンモニア低下効果のために、例えば遺伝性酵素欠陥を患っている患者において乱された尿素回路の症例で使用でき、または腎不全を患っている人々の腎臓の糸球体機能に対するストレスを低下させるのに使用できる。本発明に従ったタンパク質、ペプチド、およびタンパク質加水分解産物はまた、例えば運動選手または運動愛好家などの非医学的ニーズがある消費者向け製品でも使用できる。特にシトルリンは、eNOS NO産生を刺激することによって血流を改善することが予期されるので、筋肉血流改善を通じて運動性能が改善できることが予期される。より最近では、NO合成の阻害が、高脂血症および脂肪付着を引き起こすことを示唆する証拠が集まっている。したがって本発明に従ったペプチド結合シトルリンでの食餌補給は、肥満の人およびペットなどの動物における代謝症候群の予防および治療の助けになるかもしれない。このカテゴリーの製品としては、疲労感に対処して、身体持久力および長時間の高強度運動後の回復を増強する強化果汁およびスポーツドリンクが挙げられる。別の重要な用途は、例えばアレルギーの幼児のため、アレルギー反応を発症している乳児のため、または非アレルギーの幼児のための乳児用調製粉乳中での本発明に従ったタンパク質、ペプチド、およびタンパク質加水分解産物の使用であり、シトルリン化タンパク質またはペプチドは、牛乳感作を遅延させまたは予防する働きをする。局所用途では、シトルリン化タンパク質またはペプチドは意外にもヒドロキシルラジカルを除去し、活性酸素種を制御してそれを相殺するのに効果的である。さらに本発明に従った活性PADは直接的局所塗布のために使用でき、表皮の角質化を改善し、または乾癬の徴候に対処する。活性酵素を皮膚に塗布する有用なやり方は、米国特許第6,117,433号明細書で述べられている。本発明に従った活性PADの別の新しい用途はあらゆる種類の生地中への組み込みであり、観察されているように、これは得られたベークド製品の風味と匂いを増大させる。本発明に従った活性PADはまた、発酵食品、およびワイン、ビールおよび蒸留酒などの飲料中の尿素量を低下させることで、このような発酵食品および飲料中の発癌性物質エチルカルバメートのレベルを低下させるのに使用できる。
【0036】
ペプチジルアルギニンデイミナーゼ活性を有する本発明のポリペプチドは、単離形態であってもよい。本明細書において定義されるように、単離されたポリペプチドは、その他の非ペプチジルアルギニンデイミナーゼポリペプチドを本質的に含まない内因的に生成されたまたは組み換えされたポリペプチドであり、SDS−PAGEによる判定で、典型的に少なくとも約20%純粋、好ましくは少なくとも約40%純粋、より好ましくは少なくとも約60%純粋、さらにより好ましくは少なくとも約80%純粋、なおもより好ましくは約90%純粋、および最も好ましくは約95%純粋である。ポリペプチドは、遠心分離およびクロマトグラフィー法、または粗製溶液から純粋タンパク質を得るための当該技術分野で知られているあらゆるその他の技術によって単離されてもよい。ポリペプチドは、ポリペプチドの意図される目的を妨げないキャリアまたは希釈剤と混合されてもよく、したがってこの形態のポリペプチドは、なおも単離されていると見なされると理解される。それは一般に調製品中のポリペプチドを構成し、その中では調製品中のタンパク質重量の20%超、例えば30%、40%、50%、80%、90%、95%または99%超が本発明のポリペプチドである。
【0037】
好ましくは本発明のポリペプチドは、ペプチジルアルギニンデイミナーゼ活性がある酵素をコードする遺伝子を有する微生物から得られる。より好ましくは本発明のポリペプチドは、この微生物から分泌される。さらにより好ましくは微生物は真菌であり、および最適には糸状菌である。それゆえに好ましい生物は、フザリウム・グラミネアルム(Fusarium graminearum)種のものなどのフザリウム(Fusarium)属のものである。
【0038】
第1の実施態様では、本発明は、配列番号6の1〜640番目のアミノ酸(すなわちポリペプチド)に対して、少なくとも30%、好ましくは少なくとも40%、好ましくは少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、なおもより好ましくは少なくとも95%、および最も好ましくは少なくとも97%の程度のアミノ酸配列同一性を有するアミノ酸配列を有して、ペプチジルアルギニンデイミナーゼ活性を有する単離されたポリペプチドを提供する。
【0039】
本発明の目的で、2つ以上のアミノ酸配列間の同一性の程度は、BLAST Pタンパク質データベース検索プログラム(Altschulら著、1997年、Nucleic Acids Research 25:3389〜3402頁)によって、マトリックスBlosum 62および期待閾値10を使用して判定される。
【0040】
本発明のポリペプチドは、配列番号6に記載のアミノ酸配列、または実質的に相同的な配列、またはペプチジルアルギニンデイミナーゼ活性を有するどちらかの配列の断片を含んでなってもよい。一般に配列番号6に記載の天然アミノ酸配列が好ましい。
【0041】
本発明のポリペプチドはまた、配列番号6のポリペプチドの天然変異型、または種相同体を含んでなってもよい。
【0042】
変異型は、例えば真菌、細菌、酵母または植物細胞内で天然に生じるポリペプチドであり、変異型はペプチジルアルギニンデイミナーゼ活性および、配列番号6のタンパク質と実質的に類似の配列を有する。「変異型」という用語は、配列番号6のペプチジルアルギニンデイミナーゼと同じ本質的特性または基本的生物学的機能性を有するポリペプチドを指し、対立遺伝子多型を含む。好ましくは変異型ポリペプチドは、配列番号6のポリペプチドと少なくとも同一レベルのペプチジルアルギニンデイミナーゼ活性を有する。変異型は、配列番号6のポリペプチドと同じ株から、または同じ属または種の異なる株からのどちらかの対立遺伝子多型を含む。配列番号6のポリペプチドの変異型の例は、配列番号7に記載される。
【0043】
同様に発明のタンパク質の種相同体は、同様の配列の同等のタンパク質であり、それはペプチジルアルギニンデイミナーゼであり、別の種において天然に生じる。配列番号6のポリペプチド種相同体の例は、配列番号8〜10、13、および14に記載される。
【0044】
変異型および種相同体は、配列番号6のポリペプチドを単離するのに使用される本明細書で述べられている手順を使用して、このような手順を例えば細菌、酵母、真菌または植物細胞などの適切な細胞源に実施して単離できる。また配列番号6のポリペプチドの変異型または種相同体を発現するクローンを得るために、本発明のプローブを使用して、酵母、細菌、真菌または植物細胞からできたライブラリーを探索することも可能である。既知の遺伝子の変異型および種相同体を単離するのに使用できる方法は文献で詳細に述べられており、当業者に知られている。これらの遺伝子を従来の技術によって操作して、その後それ自体が既知である組み換えまたは合成技術によって生成されてもよい、本発明のポリペプチドを作り出すことができる。
【0045】
配列番号6のポリペプチドおよび変異型および種相同体の配列を修飾して、本発明のポリペプチドを提供することもできる。例えば1、2または3から10、20または30個の置換などのアミノ酸置換を行ってもよい。また同一数の欠失および挿入を行ってもよい。修飾されたポリペプチドがそのペプチジルアルギニンデイミナーゼ活性を維持するように、これらの変更をポリペプチドの機能に重要な領域の外で行ってもよい。
【0046】
本発明のポリペプチドは、配列番号6に記載の配列の断片をはじめとする、前述の全長ポリペプチドの断片、およびその変異型の断片を含む。このような断片は、典型的にペプチジルアルギニンデイミナーゼとしての活性を維持する。断片は、少なくとも50、100または200個のアミノ酸の長さであってもよく、またはこのアミノ酸数は、配列番号6に記載の全長配列に達していないくてもよい。
【0047】
通常、本発明のポリペプチドは後述するように組み換え的によって作られるが、それらは必要ならば合成手段によって生成できる。合成ポリペプチドは、例えばそれらの同定または精製を助けるヒスチジン残基またはT7 tagの添加によって、またはそれらの細胞からの分泌を促進するシグナル配列の添加によって修飾してもよい。
【0048】
したがって変異型配列は、配列番号6のポリペプチドが単離された株以外のフザリウム(Fusarium)株に由来するものを含んでなってもよい。変異型は、本明細書で述べられているように、ペプチジルアルギニンデイミナーゼ活性を調べることで、およびクローニングおよび配列決定によって、その他のフザリウム(Fusarium)株から識別できる。ペプチドが配列番号6のペプチジルアルギニンデイミナーゼの基本的生物学的機能性を維持しさえすれば、変異型は、欠失、修飾または単一アミノ酸またはアミノ酸群の付加をタンパク質配列内に含んでもよい。
【0049】
例えば1、2または3から10、20または30個の置換などのアミノ酸置換を行ってもよい。修飾ポリペプチドは、一般にペプチジルアルギニンデイミナーゼとしての活性を維持する。保存的置換を行ってもよく、このような置換については当該技術分野でよく知られている。
【0050】
より短いポリペプチド配列は、本発明の範囲内である。例えば長さが少なくとも50個のアミノ酸、または60、70、80、100、150または200個までのアミノ酸のペプチドは、それが配列番号6のペプチジルアルギニンデイミナーゼの基本的生物学的機能性を実証さえすれば、本発明の範囲内に入ると見なされる。特に、しかし排他的でなく、本発明のこの態様は、タンパク質が完全なタンパク質配列の断片である状況を包含する。
【0051】
本発明はまた、タンパク質アルギニンデイミナーゼ活性を有するポリペプチドをコードするポリヌクレオチドに関し、前記ポリヌクレオチドは、
(a)アミノ酸配列番号11をコードするポリヌクレオチド配列、および
(b)プレタンパク質シグナル配列をコードするポリヌクレオチド配列
を含んでなり、コードされたプレタンパク質シグナル配列は、コードされたプレポリペプチドのアミノ末端に位置し、好ましくは長さが15〜30個のアミノ酸である。
【0052】
本発明では、配列番号11のPAD共通配列を含有するポリペプチドは、本発明の範囲内である。好ましくは配列番号11のPAD共通配列の双方を含有して、シグナル配列を含有するプレタンパク質としてコードされるポリペプチドは、本発明の一部である。本発明では、関心のあるタンパク質が増殖培地中に能動的に分泌されることは、特に妥当である。分泌タンパク質は、常態では当初はプレタンパク質として合成され、引き続いて分泌過程においてプレ配列(シグナル配列)が除去される。分泌過程は、原核生物および真核生物において基本的に同様である。能動的に分泌されたプレタンパク質は膜を通り抜け、特異的シグナルペプチダーゼによってシグナル配列が除去され、成熟タンパク質が(再度)折りたたまれる。シグナル配列についてもまた、一般構造が認識できる。分泌のためのシグナル配列はプレタンパク質のアミノ−末端に位置し、一般に長さが15〜30個のアミノ酸である。アミノ−末端は、好ましくは正に帯電したアミノ酸を含有し、好ましくは非酸性アミノ酸を含有しない。この正に帯電した領域が、膜のリン脂質の負に帯電した頭部基と相互作用すると考えられる。この領域に疎水性の膜通過コア領域が続く。この領域は、一般に長さが10〜20個のアミノ酸であり、主に疎水性アミノ酸から成る。荷電したアミノ酸は、常態ではこの領域内に存在しない。膜貫通領域にシグナルペプチダーゼ認識部位が続く。認識部位は、small−X−smallを好むアミノ酸からなる。小型アミノ酸は、アラニン、グリシン、セリンまたはシステインであることができる。Xはあらゆるアミノ酸であることができる。このような規則を使用して、真核生物および原核生物からこのようなシグナル配列を認識できるアルゴリズムが記述されている(Bendtsen、Nielsen、von Heijne、およびBrunak著、(2004年)J.Mol.Biol.、340:783〜795頁)。タンパク質中のシグナル配列を計算および認識するSignalPプログラムは、一般に利用できる(http://www.cbs.dtu.dk/services/SignalP/)。
【0053】
本発明に関連があるのは、配列決定された遺伝子の推定タンパク質配列から、シグナル配列が認識できることである。SignalPプログラムを使用してシグナル配列が予測されるタンパク質を遺伝子がコードする場合、このタンパク質が分泌される確率は高い。したがって本発明の目的は、SignalPプログラムによって検出されるシグナル配列の存在と合わせて、配列番号11のコンセンサスを使用して、PAD活性を有する新しいタンパク質を発見する新しい方法を提供することである。
【0054】
第2の実施態様では、本発明は、ペプチジルアルギニンデイミナーゼ活性を有して、低ストリンジェンシー条件下、より好ましくは中度ストリンジェンシー条件、および最も好ましくは高ストリンジェンシー条件で、(I)配列番号3の核酸配列または少なくとも配列番号3のc−末端部分を含んでなるが、配列番号3の塩基の全てに満たない、または異なる塩基を有する核酸断片と、または(ii)鎖配列番号3に相補的な核酸ストランドとハイブリダイズするまたはハイブリダイズできるポリヌクレオチドによってコードされる、単離されたポリペプチドを提供する。
【0055】
「ハイブリダイズできる」という用語は、本発明の標的ポリヌクレオチドが、プローブとして使用される核酸(例えば配列番号3に記載のヌクレオチド配列、またはその断片、または配列番号3の相補体、またはその断片)と、バックグラウンドを顕著に超えるレベルでハイブリダイズできることを意味する。本発明はまた、本発明のペプチジルアルギニンデイミナーゼをコードするポリヌクレオチド、ならびにそれに相補的なヌクレオチド配列も含む。ヌクレオチド配列は、ゲノムDNA、合成DNAまたはcDNAをはじめとするRNAまたはDNAであってもよい。好ましくはヌクレオチド配列はDNA、最も好ましくはゲノムDNA配列である。典型的に本発明のポリヌクレオチドは、選択的条件下で、配列番号3のコード配列またはコード配列相補体とハイブリダイズできる近接するヌクレオチド配列を含んでなる。このようなヌクレオチドが、当該技術分野でよく知られている方法に従って合成できる。
【0056】
本発明のポリヌクレオチドは、配列番号3のコード配列またはコード配列相補体と、バックグラウンドを顕著に超えるレベルでハイブリダイズできる。バックグラウンドハイブリダイゼーションは、例えばcDNAライブラリー中に存在するその他のcDNAのために起こるかもしれない。本発明のポリヌクレオチドと配列番号3のコード配列またはコード配列相補体間の相互作用によって生じるシグナルレベルは、典型的に、その他のポリヌクレオチドと配列番号3のコード配列間の相互作用強度の少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも50倍、およびさらにより好ましくは少なくとも100倍である。相互作用強度は、例えば32Pを使用して、例えばプローブを放射標識して測定されてもよい。選択的ハイブリダイゼーションは、典型的に、低ストリンジェンシー(約40℃で0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム)、中程度ストリンジェンシー(例えば約50℃で0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム)または高ストリンジェンシー(例えば約60℃で0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム)条件を使用して達成されてもよい。
【0057】
本発明のポリヌクレオチドはまた、配列番号6のポリペプチドまたはその変異型をコードできる合成遺伝子を含む。遺伝子のコドン使用頻度を生産宿主中の好ましいバイアスに適応させることが好ましい場合もある。合成遺伝子をデザインおよび設計する技術は、一般に利用できる(例えばhttp://www.dnatwopointo.com/)。
【0058】
[修飾]
本発明のポリヌクレオチドは、DNAまたはRNAを含んでなってもよい。それらは一本鎖または二本鎖であってもよい。それらはまた、ペプチド核酸をはじめとする合成または修飾ヌクレオチドをその中に含むポリヌクレオチドであってもよい。いくつかの異なるタイプのポリヌクレオチド修飾が当該技術分野で知られている。これらとしては、メチルホスホン酸およびホスホロチオエート主鎖、および分子の3’および/または5’末端へのアクリジンまたはポリリジン鎖の付加が挙げられる。本発明の目的で、本明細書で述べられているポリヌクレオチドは、当該技術分野で利用できるあらゆる方法で修飾されてもよいものと理解される。
【0059】
当業者は、日常的技術を使用して、本発明のポリヌクレオチドによってコードされるポリペプチド配列に影響しないヌクレオチド置換を行い、その中で本発明のポリペプチドが発現される、あらゆる特定の宿主生物のコドン使用頻度を反映させてもよいものと理解される。
【0060】
配列番号3のコード配列は、例えば1、2または3から10、25、50、100個またはそれ以上の置換などのヌクレオチド置換によって修飾されてもよい。配列番号3のポリヌクレオチドは、1つ以上の挿入および/または欠失によって、および/または一端または両端の延長によって、代案としてまたは追加的に修飾されてもよい。修飾ポリヌクレオチドは、一般に、ペプチジルアルギニンデイミナーゼ活性を有するポリペプチドをコードする。例えば後でポリペプチドについて考察するように、縮重置換を行ってもよく、および/または修飾配列の翻訳時に保存的アミノ酸置換をもたらす置換を行ってもよい。
【0061】
[相同体]
配列番号3のDNAコード配列の相補体と選択的にハイブリダイズできるヌクレオチド配列は本発明に含まれ、少なくとも60、好ましくは少なくとも100、より好ましくは少なくとも200個の近接するヌクレオチドにわたり、または最も好ましくは配列番号3の全長の領域にわたって、一般に配列番号3のコード配列と少なくとも50%または60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%または少なくとも99%の配列同一性を有する。同様に能動的ペプチジルアルギニンデイミナーゼをコードして、配列番号3のDNAコード配列の相補体断片と選択的にハイブリダイズできるヌクレオチドもまた、本発明で受け入れられる。60個を超える、好ましくは100個を超えるヌクレオチドが少なくとも80%または90%同一であり、より好ましくは200個を超えるヌクレオチドが少なくとも90%同一である、配列番号3の核酸配列のC−末端断片は、本発明に包含される。
【0062】
前述の同一性程度および最小サイズのあらゆる組み合わせを使用して本発明のポリヌクレオチドを定義してもよく、よりストリンジェントな組み合わせ(すなわちより長い範囲にわたるより高い同一性)が好ましい。したがって例えば60個を超える、好ましくは100個を超えるヌクレオチドが少なくとも80%または90%同一であるポリヌクレオチドは本発明の一態様を形成し、200個を超えるヌクレオチドが少なくとも90%同一であるポリヌクレオチドについても同様である。
【0063】
UWGCGパッケージは、同一性を計算するのに使用してもよい(例えばそのデフォルト設定で使用する)BESTFITプログラムを提供する。
【0064】
PILEUPおよびBLAST Nアルゴリズムもまた、(例えばそれらのデフォルト設定で同等のまたは対応する配列を同定するなど)配列同一性を計算し、または配列を位置合わせするのに使用できる。
【0065】
BLAST分析を実施するためのソフトウェアは、国立バイオテクノロジー情報センター(http://www.ncbi.nlm.nih.gov/)を通じて公的に利用可能である。このアルゴリズムは最初に、データベース配列中の同じ長さのワードと配列比較すると、ある正値の閾値スコアTとマッチするかそれを満たすかのいずれかである、クエリー配列中の長さWの短いワードを同定することで、高採点配列対(HSP)を同定することを伴う。Tは近傍単語スコア閾値と称される。これらの初期近傍単語ヒットは、それらを含有するHSPを見つけるために検索を開始するシード(seed)として機能する。ワードヒットは、累積アラインメントスコアが増大できる限り、各配列に沿って両方向に延長される。各方向におけるワードヒットのための延長は、累積アラインメントスコアがその最大達成値からXの量だけ低下した場合、1つ以上の負の採点の残基アラインメントの蓄積のために累積スコアがゼロ以下になった場合、またはどちらかの配列の終わりに達した場合、停止する。BLASTアルゴリズムパラメーターのW、T、およびXは、配列比較の感受性およびスピード(speed)を決定する。BLASTプログラムは、デフォルトとしてワード長さ(W)11、BLOSUM 62採点マトリックス配列比較(B)50、期待値(E)10、M=5、N=4、および両ストランドの比較を使用する。
【0066】
BLASTアルゴリズムは、2つの配列間の類似性の統計学的分析を実施する。BLASTアルゴリズムによって提供される類似性の1つの尺度は、それによって2つのヌクレオチドまたはアミノ酸配列間のマッチが偶発する確率の指標を提供する、最少確率和(P(N))である。例えば第1の配列と第2の配列との比較における最少確率和が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、最も好ましくは約0.001未満であれば、配列は別の配列に類似していると見なされる。
【0067】
[プライマーおよびプローブ]
本発明のポリヌクレオチドはプライマーを含み、例えばポリメラーゼ連鎖反応(PCR)プライマーとして、代案の増幅反応のためのプライマーとして、または例えば放射性または非放射性標識を使用して従来の手段によって明示用標識で標識してプローブとして使用してもよく、またはポリヌクレオチドをベクター中にクローンしてもよい。このようなプライマー、プローブ、およびその他の断片は、長さが少なくとも15個、例えば少なくとも20、25、30または40個のヌクレオチドである。それらは長さが典型的に40、50、60、70、100、150、200または300個までのヌクレオチドであり、または配列番号5のコード配列よりも短い数個までのヌクレオチド(5または10個のヌクレオチドなど)でさえある。
【0068】
一般にプライマーは、ヌクレオチド1つずつの所望の核酸配列の段階的製造を伴う、合成手段によって生成される。これを達成するための技術およびプロトコールは、当該技術分野で容易に利用できる。より長いポリヌクレオチドは、例えばPCRクローニング技術を使用して、一般に組み換え手段を使用して生成される。これは(典型的に約15〜30個のヌクレオチドの)プライマー対を作成して、クローンするペプチジルアルギニンデイミナーゼの所望の領域を増幅し、プライマーを酵母、細菌、植物、原核生物または好ましくはフザリウム(Fusarium)株である真菌細胞から得られたmRNA、cDNAまたはゲノムDNAに接触させて、所望領域の増幅に適した条件下でポリメラーゼ連鎖反応を実施し、(例えば反応混合物をアガロースゲル上で精製することで)増幅された断片を単離して、増幅されたDNAを回収することを伴う。増幅されたDNAが適切なクローニングベクター中にクローンできるように、プライマーは適切な制限酵素認識部位を含有するようにデザインされてもよい。このような配列番号3のPAD遺伝子の変異型のPCRクローニングは本発明内で実施され、これらの遺伝子のDNA配列は配列番号4に記載され、推定タンパク質配列は配列番号7に記載される。
【0069】
代案としては、分泌されたペプチジルアルギニンデイミナーゼまたはその変異型のコード領域を包含する合成遺伝子を構築できる。これらの技術を使用して、多くの位置で修飾されながら、なおも同一タンパク質をコードするポリヌクレオチドを好都合にデザインして構築できる。これはコドン使用頻度を好ましい発現宿主に適応できるという利点を有し、したがってこの宿主中でのタンパク質生産性を改善できる。また遺伝子のポリヌクレオチド配列を変更して、mRNAの安定性を改善し、または回転率を低下させることができる。これは所望のタンパク質またはその変異型の改善された発現をもたらすことができる。さらに分泌効率、安定性、タンパク質分解敏感性、最適温度、比活性、またはタンパク質の工業生産または応用に妥当なその他の特性に対して好ましい効果を有する突然変異が、タンパク質配列中で生じるように、ポリヌクレオチド配列を合成遺伝子中で変更できる。合成遺伝子を構築してコドン使用頻度を最適化するサービスを提供する会社が、一般に利用できる。
【0070】
このような技術を使用して、本明細書で述べられているペプチジルアルギニンデイミナーゼ配列をコードするポリヌクレオチドの全部または一部を得てもよい。イントロン、プロモーター、およびトレーラー領域は本発明の範囲内であり、これらもまた真菌、酵母、細菌、植物または原核細胞からのゲノムDNAから開始して、類似様式で(例えば組み換え手段、PCRまたはクローニング技術によって)得てもよい。
【0071】
ポリヌクレオチドまたはプライマーは、明示用標識を保有しても良い。適切な標識としては、32Pまたは35Sなどの放射性同位体、蛍光性標識、酵素標識、またはビオチンなどのその他のタンパク質標識が挙げられる。このような標識を本発明のポリヌクレオチドまたはプライマーに添加してもよく、当業者に知られている技術を使用して検出してもよい。
【0072】
真菌サンプル中で、ペプチジルアルギニンデイミナーゼまたはその変異型を検出または配列決定するための核酸ベースの試験において、標識または非標識ポリヌクレオチドまたはプライマー(またはその断片)を使用してもよい。このような検出試験は、一般に、ハイブリダイズ条件下で、関心のあるDNAを含有することが疑われる真菌サンプルと、本発明のポリヌクレオチドまたはプライマーを含んでなるプローブとを接触させるステップと、サンプルのプローブと核酸との間に形成されたあらゆる二本鎖を検出するステップを含んでなる。検出は、PCRなどの技術を使用することで、または固体担体上にプローブを固定し、プローブとハイブリダイズしないサンプル中のあらゆる核酸を除去し、次にプローブとハイブリダイズするあらゆる核酸を検出することで達成してもよい。代案としてはサンプル核酸を固体担体上に固定化し、プローブをハイブリダイズして、あらゆる非結合プローブの除去後に、このような担体に結合したプローブ量を検出してもよい。
【0073】
本発明のプローブは、好都合には、適切な容器内に試験キットの形態で包装されてもよい。そのためにキットがそのためにデザインされたアッセイ形態がこのような結合を必要とする場合、このようなキット中でプローブが固体担体に結合していてもよい。キットはまた、プローブするサンプルを処理し、サンプル中の核酸にプローブをハイブリダイズするための適切な試薬、対照試薬、説明書なども含有する。プローブおよび本発明のポリヌクレオチドはまた、微量検定法で使用してもよい。
【0074】
好ましくは本発明のポリヌクレオチドは、ポリペプチドと同じ、真菌など、特にフザリウム(Fusarium)属の真菌である生物から得ることが可能である。
【0075】
[ポリヌクレオチドの生成]
配列番号3と100%の同一性を有さないが、本発明の範囲内に入るポリヌクレオチドをいくつかの方法で得ることができる。したがって本明細書で述べられているペプチジルアルギニンデイミナーゼ配列の変異型を、例えば本発明のポリペプチドの起源として考察されたものなどの一連の生物からできたゲノムDNAライブラリーを探査して得てもよい。さらにその他のペプチジルアルギニンデイミナーゼの真菌、植物または原核生物相同体を得てもよく、このような相同体およびその断片は、一般に配列番号3とハイブリダイズすることができる。このような配列は、その他の種からのcDNAライブラリーまたはゲノムDNAライブラリーを探査して得てもよく、このようなライブラリーを(先に述べたように)低、中程度から高ストリンジェンシー条件下で、配列番号3の全部または一部を含んでなるプローブで探査する。配列番号3の全部または一部を含んでなる核酸プローブを使用して、本発明のポリペプチドの起源として述べたものなどのその他の種からのcDNAまたはゲノムのライブラリーを探査してもよい。
【0076】
種相同体はまた、保存アミノ酸配列をコードする変異型および相同体中の配列を標的とするようにデザインされたプライマーを使用する縮重PCRを使用して得てもよい。プライマーは1つ以上の縮重位置を含有でき、既知の配列に対する単一配列プライマーがある配列のクローニングのために使用されるものよりも、低いストリンジェンシー条件で使用される。PADの種相同体を得るための好ましい方法は、配列番号11に記載の共通配列をコードする配列を標的とするプライマーをデザインすることである。
【0077】
代案としては、このようなポリヌクレオチドは、ペプチジルアルギニンデイミナーゼ配列またはその変異型の部位特異的変異誘発によって得てもよい。これは例えば、その中でポリヌクレオチド配列が発現される特定の宿主細胞のためにコドン選択を最適化するのに、配列に対するサイレントなコドン変化が必要とされる場合に有用かもしれない。制限酵素認識部位を導入するため、またはポリヌクレオチドによってコードされるポリペプチドの特質または機能を変更させるために、その他の配列変化を加えてもよい。
【0078】
本発明は、本発明のポリヌクレオチドおよびその相補体を含んでなる二本鎖ポリヌクレオチド含む。
【0079】
本発明はまた、上述の本発明のポリペプチドをコードするポリヌクレオチドを提供する。このようなポリヌクレオチドは、本発明のポリペプチドの組み換え生産のための配列として有用であるので、それらが配列番号3の配列とハイブリダイズできる必要はないが、これは一般に望ましい。さもなければこのようなポリヌクレオチドは、所望ならば標識されて、使用され、上述のように作成されてもよい。
【0080】
[組み換えポリヌクレオチド]
本発明はまた、クローニングおよび発現ベクターをはじめとする、本発明のポリヌクレオチドを含んでなるベクターも提供し、別の態様ではこのようなベクターを生育させ、例えば本発明のポリペプチド、または本発明の配列によってコードされるポリペプチドの発現が起きる条件下で、適切な宿主細胞に形質転換または形質移入する方法も提供する。本発明のポリヌクレオチドまたはベクターを含んでなる宿主細胞もまた提供され、ここでポリヌクレオチドは宿主細胞ゲノムにとって異種である。「異種」という用語は、通常宿主細胞に対し、ポリヌクレオチドが宿主細胞ゲノム中で自然発生しないこと、またはポリペプチドが細胞によって自然に生成されないことを意味する。好ましくは宿主細胞は、酵母細胞、例えばクリヴェロミセス(Kluyveromyces)、ピチア(Pichia)、ハンゼヌラ(Hansenula)またはサッカロミセス(Saccharomyces)属の酵母細胞であり、または、例えばアスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)またはフザリウム(Fusarium)属の糸状菌細胞である。
【0081】
[ベクター]
その中に本発明の発現カセットが挿入されるベクターは、好都合には組み換えDNA手順を施してもよい、あらゆるベクターであってもよく、ベクターの選択は、その中にそれが導入される宿主細胞に左右されることが多い。したがってベクターは自律的に複製するベクターであってもよく、すなわちプラスミドなど、その複製が染色体の複製から独立している、染色体外の実体として存在するベクターであってもよい。代案としてはベクターは、宿主細胞に導入されると宿主細胞ゲノムに組み込まれ、それが組み込まれた染色体と共に複製されるものであってもよい。
【0082】
好ましくは本発明のポリヌクレオチドがベクター中にある場合、それは宿主細胞によるコード配列の発現を提供できる制御配列と作動的に連結し、すなわちベクターは発現ベクターである。「作動的に連結する」という用語は、述べられている構成要素が、意図される様式でそれらが機能できるようになる関係にある並置位置を指す。コード配列と「作動的に連結する」プロモーター、エンハンサーまたはその他の発現調節シグナルなどの制御配列は、生成条件下でコード配列の発現が達成されるように配置される。
【0083】
例えばプラスミド、コスミド、ウイルスまたはファージベクターの場合、ベクターには複製起点、場合によりポリヌクレオチド発現のためのプロモーター、および場合によりエンハンサーおよび/またはプロモーターのレギュレーターが備わっていてもよい。ターミネーター配列が、ポリアデニル化配列として存在してもよい。ベクターは、例えば細菌プラスミドの場合はアンピシリン抵抗性遺伝子、または哺乳類ベクターではネオマイシン抵抗性遺伝子などの1つ以上の選択可能なマーカー遺伝子をを含有してもよい。ベクターは、例えばRNA生成のために生体外で使用されてもよく、または宿主細胞を形質移入または形質転換するのに使用できる。
【0084】
ポリペプチドをコードするDNA配列は、好ましくはその中でDNA配列と、宿主細胞内におけるDNA配列の発現を指示できる発現シグナルとが作動的に連結する発現コンストラクトの一部として、適切な宿主中に導入される。発現コンストラクトによる適切な宿主の形質転換のためには、当業者によく知られている形質転換手順が利用できる。発現コンストラクトは、選択可能なマーカーを保有するベクターの一部として、宿主の形質転換のために使用でき、または発現コンストラクトは、選択可能なマーカーを保有するベクターと共に、別個の分子として同時形質転換される。ベクターは、1つ以上の選択可能なマーカー遺伝子を含有してもよい。
【0085】
好ましい選択可能なマーカーとしては、宿主細胞内の欠損を補完するもの、または薬剤抵抗性を与えるものが挙げられるが、これらに限定されるものではない。それらとしては、例えばアセトアミダーゼ遺伝子またはcDNA(A.ニデュランス(A.nidulans)、コウジカビ(A.oryzae)、またはA.ニガー(A.niger)からのamdS、niaD、facA遺伝子またはcDNA)などのほとんどの糸状菌および酵母の形質転換のために使用できる用途の広いマーカー遺伝子、またはG418、ハイグロマイシン、ブレオマイシン、カナマイシン、フレオマイシンまたはベノミル抵抗性(benA)のような抗生物質に対する抵抗性を提供する遺伝子が挙げられる。代案としては、例えばURA3(S.セレヴィシエ(S.cerevisiae)からのまたはその他の酵母からの類似遺伝子)、pyrGまたはpyrA(A.ニデュランス(A.nidulans)またはA.ニガー(A.niger)からの)、argB(A.ニデュランス(A.nidulans)またはA.ニガー(A.niger)からの)またはtrpCなどの対応する突然変異宿主株を必要とする栄養要求性マーカーなどの特異的選択マーカーが使用できる。好ましい実施態様では、選択マーカー遺伝子を含まないポリペプチドを生成できる転換宿主細胞を得るように、発現コンストラクト導入後に選択マーカーが転換宿主細胞から除去される。
【0086】
その他のマーカーとしては、ATPシンセターゼサブユニット9(oliC)、オロチジン−5’−ホスフェート−デカルボキシラーゼ(pvrA)、細菌G418抵抗性遺伝子(酵母中で有用であるが糸状菌中では有用でない)、アンピシリン抵抗性遺伝子(大腸菌(E.coli))、ネオマイシン抵抗性遺伝子(バシラス(Bacillus))、およびグルクロニターゼ(GUS)をコードする大腸菌(E.coli)uidA遺伝子が挙げられる。ベクターは、例えばRNAの生成のために、または宿主細胞を形質移入または形質転換するために生体外で使用してもよい。
【0087】
ほとんどの糸状菌および酵母では、安定した形質転換体を得るために、発現コンストラクトは好ましくは宿主細胞のゲノム中に組み込まれる。しかし特定の酵母では、安定した高レベル発現のために、その中に発現コンストラクトを組み込むことができる、適切なエピソームベクターシステムもまた利用できる。その例としては、それぞれサッカロミセス(Saccharomyces)およびクリヴェロミセス(Kluyveromyces)の2μm CENおよびpKD1プラスミドに由来するベクター、またはAMA配列(例えばアスペルギルス(Aspergillus)からのAMA1)を含有するベクターが挙げられる。発現コンストラクトが宿主細胞ゲノムに組み込まれる場合、コンストラクトは、ゲノム中の無作為の遺伝子座に組み込まれるか、または相同的組換えを使用して所定の標的遺伝子座に組み込まれるかのどちらかであり、後者の場合、標的遺伝子座は好ましくは高度に発現する遺伝子を含んでなる。高度に発現される遺伝子とは、例えば誘導された条件下で、そのmRNAが総細胞mRNAの少なくとも0.01%(w/w)を構成できる遺伝子であり、または代案としては、その遺伝子産物が総細胞タンパク質の少なくとも0.2%(w/w)を構成できる遺伝子であり、または分泌遺伝子産物の場合は、少なくとも0.05g/lのレベルまで分泌されることができる。
【0088】
特定の宿主細胞のための発現コンストラクトは、通常、第1の態様のポリペプチドをコードする配列のコード鎖に相対して5’−末端から3’−末端までの連続的順序で、互いに作動的に連結する以下の要素を含有する。(1)特定の宿主細胞内でポリペプチドをコードするDNA配列の転写を指示できるプロモーター配列、(2)好ましくは5’−非翻訳領域(リーダー)、(3)場合により特定の宿主細胞から培養液中へのポリペプチドの分泌を指示できるシグナル配列、(4)成熟した好ましくは活性形態のポリペプチドをコードするDNA配列、および好ましくはまた(5)ポリペプチドをコードするDNA配列の転写下流を終結できる転写終結領域(ターミネーター)。
【0089】
ポリペプチドをコードするDNA配列の下流で、発現コンストラクトは、ターミネーターとも称される、1つ以上の転写終結部位を含有する3’非翻訳領域を好ましくは含有する。ターミネーターの起源はそれほど重要でない。ターミネーターは例えばポリペプチドをコードするDNA配列に天然であることができる。しかし好ましくは細菌ターミネーターが細菌宿主細胞内で使用され、酵母ターミネーターが酵母宿主細胞内で使用され、糸状菌ターミネーターが糸状菌宿主細胞内で使用される。より好ましくはターミネーターは、その中でポリペプチドをコードするDNA配列が発現される宿主細胞に対して内在性である。
【0090】
本発明のポリペプチドをコードするポリヌクレオチドの改善された発現はまた、関心のあるタンパク質の発現レベルそして所望ならば選択された発現宿主からの分泌レベルを増大させ、および/または本発明のポリペプチド発現の誘導性制御を提供する役割をする、例えばプロモーター、シグナル配列およびターミネーター領域などの異種の調節領域の選択によって達成されてもよい。
【0091】
本発明のポリペプチドコードする遺伝子に天然であるプロモーターの他に、その他のプロモーターを使用して、本発明のポリペプチドの発現を指示してもよい。プロモーターは、所望の発現宿主中で本発明のポリペプチドを、発現を指示するその効率によって選択してもよい。
【0092】
プロモーター/エンハンサーおよびその他の発現調節シグナルは、発現ベクターがそのためにデザインされた宿主細胞と適合性であるように選択してもよい。例えば原核生物プロモーター、特にE.coli株中で使用するのに適したものを使用してもよい。本発明のポリペプチドの発現を哺乳類細胞内で実施する場合、哺乳類プロモーターを使用してもよい。例えば肝細胞細胞−特異的プロモーターなどの組織特異的プロモーターもまた使用してもよい。例えばモロニーマウス白血病ウイルスの長い末端反復末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒト サイトメガロウイルス(CMV)IEプロモーター、単純ヘルペスウイルスプロモーターまたはアデノウィルスプロモーターなどのウイルス性プロモーターもまた使用してもよい。
【0093】
適切な酵母プロモーターとしては、S.セレヴィシエ(cerevisiae)GAL4およびADHプロモーター、およびS.ポンベ(S.pombe)nmt1およびadhプロモーターが挙げられる。哺乳類プロモーターとしては、カドミウムなどの重金属に応えて誘導されることができるメタロチオネインプロモーターが挙げられる。SV40大型T抗原プロモーターまたはアデノウィルスプロモーターなどのウイルス性プロモーターもまた使用してもよい。これらの全てのプロモーターは、当該技術分野で容易に入手できる。
【0094】
β−アクチンプロモーターなどの哺乳類プロモーターを使用してもよい。組織−特異的プロモーター、特に内皮またはニューロン細胞特異的プロモーター(例えばDDAHIおよびDDAHIIプロモーター)が特に好ましい。例えばモロニーマウス白血病ウイルスの長い末端反復末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒト サイトメガロウイルス(CMV)IEプロモーター、アデノウィルス、HSVプロモーター(HSV IEプロモーターなど)、またはHPVプロモーターなどのウイルス性プロモーターもまた使用してもよく、特にHPV上流調節領域(URR)ウイルス性プロモーターは、当該技術分野で容易に入手できる。
【0095】
本発明の宿主細胞内で転写を指示できる多様なプロモーターが使用できる。好ましくはプロモーター配列は、先に定義したように高度に発現される遺伝子に由来する。プロモーターが好ましくはそれに由来し、および/または発現コンストラクト組み込みのために好ましい所定の標的遺伝子座に含まれる、好ましい高度に発現する遺伝子の例としては、トリオース−ホスフェートイソメラーゼ(TPI)、グリセルアルデヒド−ホスフェートデヒドロゲナーゼ(GAPDH)、ホスホグリセリン酸キナーゼ(PGK)、ピルビン酸キナーゼ(PYK)、アルコールデヒドロゲナーゼ(ADH)などの解糖作用酵素をコードする遺伝子、ならびにアミラーゼ、グルコアミラーゼ、プロテアーゼ、キシラナーゼ、セロビオ加水分解酵素、β−ガラクトシダーゼ、アルコール(メタノール)オキシダーゼ、延長因子およびリボソームタンパク質をコードする遺伝子が挙げられるが、これに限定されるものではない。適切な高度に発現する遺伝子の特定例としては、例えばクリヴェロミセス(Kluyveromyces)種からのLAC4遺伝子、それぞれハンゼヌラ(Hansenula)およびピチア(Pichia)からのメタノールオキシダーゼ遺伝子(AOXおよびMOX)、A.ニガー(A.niger)およびA.アワモリ(A.awamori)からのグルコアミラーゼ(glaA)遺伝子、コウジカビ(A.oryzae)TAKA−アミラーゼ遺伝子、A.ニデュランス(A.nidulans)gpdA遺伝子、およびT.リーセイ(T.reesei)セロビオ加水分解酵素遺伝子が挙げられる。
【0096】
真菌発現宿主中で使用するのに好ましい強力な構成的および/または誘導プロモーターの例は、キシラナーゼ(xlnA)、フィターゼ、ATP−シンセターゼサブユニット9(oliC)、トリオースホスフェートイソメラーゼ(tpi)、アルコールデヒドロゲナーゼ(AdhA)、アミラーゼ(amy)、アミログルコシダーゼ(AG−glaA遺伝子から)、アセトアミダーゼ(amdS)、およびグリセルアルデヒド−3−リン酸デヒドロゲナーゼ(gpd)プロモーターの真菌遺伝子から得られるものである。
【0097】
使用してもよい強力な酵母プロモーターの例としては、アルコールデヒドロゲナーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ラクターゼ、3−ホスホグリセリン酸キナーゼ、原形質膜ATPアーゼ(PMA1)およびトリオースホスフェートイソメラーゼの遺伝子から得られるものが挙げられる。
【0098】
使用してもよい強力な細菌プロモーターの例としては、アミラーゼおよびSPo2プロモーターならびに細胞外プロテアーゼ遺伝子からのプロモーターが挙げられる。
【0099】
使用してもよい植物細胞に適したプロモーターとしては、ノパリンシンターゼ(nos)、オクトピンシンターゼ(ocs)、マンノピンシンターゼ(mas)、リブロース小サブユニット(rubisco ssu)、ヒストン、米アクチン、ファゼオリン、カリフラワーモザイクウイルス(CMV)35Sおよび19S、およびサーコウイルスプロモーターが挙げられる。
【0100】
ベクターはポリヌクレオチドを挟む配列をさらに含んで、真核生物ゲノムの配列、好ましくは真菌ゲノム配列、または酵母ゲノム配列からのものと相同的な配列を含んでなるRNAを生じてもよい。これは相同的組換えによる、真菌または酵母ゲノムへの本発明のポリヌクレオチドの導入を可能にする。特に真菌配列で挟まれた発現カセットを含んでなるプラスミドベクターを使用して、本発明のポリヌクレオチドを真菌細胞に送達するのに適したベクターを調製できる。これらの真菌ベクターを使用する形質転換技術は、当業者に知られている。
【0101】
ベクターは、アンチセンス方向を向いた本発明のポリヌクレオチドを含有して、アンチセンスRNAの生成を提供してもよい。これは所望ならば、ポリペプチドの発現レベルを低下させるのに使用してもよい。
【0102】
[宿主細胞および発現]
さらなる態様では、本発明は、ポリペプチドをコードするコード配列のベクターによる発現に適した条件下で、上述のように発現ベクターによって転換されたまたは形質移入された宿主細胞を培養するステップと、発現したポリペプチドを回収するステップを含んでなる、本発明のポリペプチドを調製する方法を提供する。本発明のポリヌクレオチドは、発現ベクターなどの組み換え複製可能なベクター中に組み込むことができる。ベクターを使用して適合性宿主細胞中で核酸を複製してもよい。したがってさらなる実施態様では、本発明は、本発明のポリヌクレオチドを複製可能なベクター中に導入し、ベクターを適合性宿主細胞内に導入し、ベクターの複製をもたらす条件下で宿主細胞を生育させることで、本発明のポリヌクレオチドを作成する方法を提供する。適切な宿主細胞としては、大腸菌(E.coli)などの細菌と、酵母と、哺乳類細胞系と、例えばSf9細胞などの昆虫細胞および(例えば糸状)真菌細胞などのその他の真核生物の細胞系とが挙げられる。
【0103】
好ましくはポリペプチドは分泌タンパク質として産生され、その場合、発現コンストラクト中でポリペプチドの成熟形態をコードするDNA配列は、シグナル配列をコードするDNA配列と作動的に連結してもよい。分泌タンパク質をコードする遺伝子が野性型株中でシグナル配列を有する場合、好ましくは使用されるシグナル配列は、ポリペプチドをコードするDNA配列に天然(相同的)である。代案としてはシグナル配列は、ポリペプチドをコードするDNA配列に外来性(異種)であり、その場合、シグナル配列は好ましくは、その中でDNA配列が発現される宿主細胞に内在性である。酵母宿主細胞に適したシグナル配列の例は、酵母MFα遺伝子に由来するシグナル配列である。同様に糸状菌宿主細胞に適したシグナル配列は、例えばA.ニガー(A.niger)glaA遺伝子などの例えば糸状菌アミログルコシダーゼ(AG)遺伝子に由来するシグナル配列である。シグナル配列はアミログルコシダーゼ((グルコ)アミラーゼとも称される)プロモーターそれ自体と組み合わせて、ならびにその他のプロモーターと組み合わせて使用してもよい。ハイブリッドシグナル配列もまた、本発明の文脈で使用してもよい。
【0104】
好ましい異種の分泌リーダー配列は、真菌アミログルコシダーゼ(AG)遺伝子(例えばアスペルギルス(Aspergillus)からなどのglaA−18および24個のアミノ酸バージョンの双方)、MFα伝子(例えばサッカロミセス(Saccharomyces)などの酵母およびクリヴェロミセス(Kluyveromyces))、またはαアミラーゼ遺伝子(バシラス(Bacillus))起源のものである。
【0105】
ベクターを上述のように適切な宿主細胞に形質転換または形質移入して、本発明のポリペプチドの発現を提供してもよい。この方法は、ポリペプチドの発現に適した条件下で、上述のように発現ベクターで転換された宿主細胞を培養するステップと、発現したポリペプチドを場合により回収するステップを含んでなってもよい。
【0106】
したがって本発明のさらなる態様は、本発明のポリヌクレオチドまたはベクターで形質転換または形質移入された、またはそれを含んでなる宿主細胞を提供する。好ましくはポリヌクレオチドは、ポリヌクレオチドの複製および発現を可能にするベクター中で輸送される。細胞は前記ベクターと適合性であるように選択され、例えば原核生物(例えば細菌)、または真核生物の真菌、酵母または植物細胞であってもよい。
【0107】
本発明は、ポリペプチドをコードするDNA配列の組み換え発現の手段による、本発明のポリペプチドの生成のための方法を包含する。この目的で、本発明のDNA配列は、適切な相同的または異種宿主細胞内におけるポリペプチドの経済的生産を可能にするために、遺伝子増幅および/またはプロモーター、分泌シグナル配列などの発現シグナルの交換のために使用できる。相同的宿主細胞とは、本明細書においてDNA配列が由来する種と同一種である、または同一種内の変異型である、宿主細胞と定義される。
【0108】
適切な宿主細胞は、好ましくは細菌などの原核微生物、またはより好ましくは例えば真菌、酵母または糸状菌、または植物細胞などの真核生物である。一般に、より容易に操作できることから、酵母細胞が糸状菌細胞よりも好ましい。しかしいくつかのタンパク質は、酵母からの分泌が不良であったり、または場合によっては適切に処理されない(例えば酵母中での過度のグリコシル化)。このような場合は糸状菌宿主生物を選択すべきである。
【0109】
バシラス(Bacillus)属からの細菌は、培養液中にタンパク質を分泌するそれらの能力のために、異種の宿主として非常に適している。宿主として適切なその他の細菌は、ストレプトミセス(Streptomyces)およびシュードモナス(Pseudomonas)属からのものである。ポリペプチドをコードするDNA配列の発現のために好ましい酵母宿主細胞は、サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、ハンゼヌラ(Hansenula)、ピチア(Pichia)、ヤロウイア(Yarrowia)、または分裂酵母(Schizosaccharomyces)属のものである。より好ましくは酵母宿主細胞は、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)、クリヴェロミセス・ラクチス(Kluyveromyces lactis)(クリヴェロミセス・マルキシアナス(Kluyveromyces marxianus)変種ラクチス(lactis)としてもまた知られている)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、ヤロウィア・リポリティカ(Yarrowia lipolytica)、およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)種からなる群から選択される。
【0110】
しかしポリペプチドをコードするDNA配列の発現のために最も好ましいのは、糸状菌宿主細胞である。好ましい糸状菌宿主細胞は、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、フザリウム(Fusarium)、ディスポロトリカム(Disporotrichum)、ペニシリウム(Penicillium)、アクレモニウム(Acremonium)、ニューロスポラ(Neurospora)、サーモアスクス(Thermoascus)、ミセリオフトラ(Myceliophtora)、スポロトリカム(Sporotrichum)、チエラビア(Thielavia)、およびタラロミセス(Talaromyces)属からなる群から選択される。より好ましくは糸状菌宿主細胞は、コウジカビ(Aspergillus oyzae)、ショウユコウジカビ(Aspergillus sojae)またはアスペルギルス・ニデュランス(Aspergillus nidulans)種のものまたはアスペルギルス・ニガー(Aspergillus niger)群(RaperおよびFennell著、「アスペルギルス(Aspergillus)属(The Genus Aspergillus)」、The Williams&Wilkins Company、Baltimore、pp293〜344頁、1965年、によって定義される)からの種のものである。これらとしては、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・ツビゲンシス(Aspergillus tubigensis)、アスペルギルス・アクレアツス(Aspergillus aculeatus)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ニデュランス(Aspergillus nidulans)、アスペルギルス・ジャポニカス(Aspergillus japonicus)、コウジカビ(Aspergillus oryzae)およびアスペルギルス・フィクウム(Aspergillus ficuum)、およびトリコデルマ・リーセイ(Trichoderma reesei)、フザリウム・グラミネアルム(Fusarium graminearum)、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)、アクレモニウム・アラバメンセ(Acremonium alabamense)、ニューロスポラ・クラッサ(Neurospora crassa)、ミセリオフトラ・サーモフィラ(Myceliophtora thermophilum)、スポロトリカム・セルロフィルム(Sporotrichum cellulophilum)、ディスポロトリカム・ディモルフォスポラム(Disporotrichum dimorphosporum)、およびチエラビア・テルレストリス(Thielavia terrestris)種のものが挙げられるがこれらに限定されるものではない。
【0111】
本発明の範囲内の好ましい発現宿主の例は、アスペルギルス(Aspergillus)種(特に欧州特許出願公開第A−184,438号明細書および欧州特許出願公開第A−284,603号明細書で述べられているもの)およびトリコデルマ(Trichoderma)種などの真菌と、特に枯草菌(Bacillus subtilis)、バシラス・リケニフォルミス(Bacillus licheniforms)、バシラス・アミロリケファシエンス(Bacillus amyloliquefaciens)であるバシラス(Bacillus)種(特に欧州特許出願公開第A−134,048号明細書および欧州特許出願公開第A−253,455号明細書で述べられているものなど)、シュードモナス(Pseudomonas)種などの細菌と、クリヴェロミセス(Kluyveromyces)種(特にクリヴェロミセス・ラクチス(Kluyveromyces lactis)などの欧州特許出願公開第A−096,430号明細書で述べられているもの、および欧州特許出願公開第A−301,670号明細書で述べられているもの)およびサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)などのサッカロミセス(Saccharomyces)種などの酵母である。
【0112】
本発明に従った宿主細胞としては植物細胞が挙げられ、したがって本発明は、1つ以上の本発明の細胞を含有する植物、およびその部分などの遺伝子導入生物にまで及ぶ。細胞は本発明のポリペプチドを異種性に発現してもよく、または1つ以上の本発明のポリヌクレオチドを異種性に含有してもよい。したがって遺伝子導入(または遺伝子改変)植物は、そのゲノム中に本発明のポリペプチドをコードする配列を(典型的に安定して)挿入されてもよい。植物細胞形質転換は、例えばアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)からのTiまたはRiプラスミドを使用して、公知の技術を使用して実施できる。したがってプラスミド(またはベクター)は、植物に感染するのに必要な配列を含有してもよく、Tiおよび/またはRiプラスミドの誘導体を用いてもよい。
【0113】
宿主細胞はポリペプチドを過剰に発現してもよく、過剰発現のための技術はよく知られており、本発明で使用できる。したがって宿主は、2つ以上のポリヌクレオチドのコピーをを有してもよい。
【0114】
代案としては、葉、根または茎などの植物部分の直接感染を実行できる。この技術では、例えば植物を剃刀で切断する、植物を針で突き刺す、または植物を研磨材で擦ることで、感染させる植物を傷つけることができる。次に創傷にアグロバクテリウム(Agrobacterium)を接種する。次に植物または植物部分を適切な培地上で培養液生育させて、成熟植物に発育させることができる。転換細胞の遺伝子改変植物への再生は、例えば抗生物質を使用して形質転換された苗条を選択することで、および苗条を適切な栄養素、植物ホルモンなどを含有する培地上で二次培養することで、公知の技術を使用して達成できる。
【0115】
[宿主細胞の培養および組み換え生産]
本発明はまた、修飾されてペプチジルアルギニンデイミナーゼまたはその変異型を発現する細胞も含む。このような細胞としては、哺乳類細胞または昆虫細胞などの一時的なまたは好ましくは安定した改変高等真核細胞系、酵母および糸状菌細胞などの下等真核細胞、または細菌細胞などの原核細胞が挙げられる。
【0116】
本発明のポリペプチドを例えばバキュロウイルス(baculovirus)発現系などの細胞系中または膜上で一過性に発現することもまた可能である。本発明に従ったタンパク質を発現するように適応された、このような系もまた本発明の範囲内に含まれる。
【0117】
本発明に従って、従来の栄養素発酵培地中で、1つ以上の本発明のポリヌクレオチドで転換された微生物発現宿主を培養することで、本発明のポリペプチドの生成を実行できる。
【0118】
本発明に従った組み換え宿主細胞は、当該技術分野で知られている手順を使用して培養してもよい。プロモーターと宿主細胞との各組み合わせについて、ポリペプチドをコードするDNA配列の発現に貢献する培養条件が利用できる。所望の細胞密度またはポリペプチド力価に達した後、培養を停止して公知の手順を使用してポリペプチドを回収する。
【0119】
発酵培地は、炭素源(例えばグルコース、マルトース、糖蜜など)、窒素源(例えば硫酸アンモニウム、硝酸アンモニウム、塩化アンモニウムなど)、有機窒素源(例えば酵母抽出物、麦芽抽出物、ペプトンなど)、および無機栄養素源(例えばホスフェート、マグネシウム、カリウム、亜鉛、鉄など)を含有する公知の培養液を含んでなることができる。場合により(使用する発現コンストラクト次第で)誘導物質を含めても、または引き続いて添加してもよい。
【0120】
適切な培地の選択は、発現宿主の選択に基づいてもおよび/または発現コンストラクトの調節要件に基づいてもよい。適切な培地は当業者によく知られている。培地は、所望ならば、その他の潜在的汚染微生物よりも転換された発現宿主に有利に働く追加的構成要素を含有してもよい。
【0121】
発酵は0.5〜30日間にわたり実施されてもよい。発酵は、0℃〜45℃の範囲内の適切な温度で、例えば2〜10pHにおける、バッチ、連続または流加工程であってもよい。好ましい発酵条件としては、20℃〜37℃の範囲内の温度、および/または3〜9のpHが挙げられる。適切な条件は、通常、発現宿主および発現されるタンパク質の選択に基づいて選択される。
【0122】
発酵後、必要ならば遠心分離または濾過の手段によって、発酵ブロスから細胞を除去できる。発酵停止後または細胞除去後、次に本発明のポリペプチドを回収して、所望ならば従来の手段によって精製および単離してもよい。本発明のペプチジルアルギニンデイミナーゼは、菌糸体から、または培養された真菌細胞によってその中にペプチジルアルギニンデイミナーゼが放出された培養ブロスから精製できる。
【0123】
好ましい実施態様では、ポリペプチドは真菌から、より好ましくはアスペルギルス(Aspergillus)から、最も好ましくはアスペルギルス・ニガー(Aspergillus niger)から生成される。
【0124】
[修飾]
本発明のポリペプチドは、化学的に修飾されてもよく、例えば翻訳後に修飾されてもよい。例えばそれらは、グリコシル化されて(1回以上)、または修飾アミノ酸残基を含んでなってもよい。それらはまた、ヒスチジン残基の添加によって修飾されて、それらの精製を助けてもよく、またはシグナル配列の添加によって細胞からの分泌を促進してもよい。ポリペプチドは、アミノ−末端メチオニン残基などのアミノ−またはカルボキシル−末端延長と、約20〜25個の残基までの小型リンカーペプチドと、または精製を容易にするポリ−ヒスチジントラクト、抗原エピトープまたは結合領域などの小型延長部とを有してもよい。
【0125】
本発明のポリペプチドは、明示用標識で標識してもよい。明示用標識は、ポリペプチドが検出できるようにするあらゆる適切なラベルであってもよい。適切な標識としては、例えば125I、35Sなどの放射性同位体と、酵素と、抗体と、ポリヌクレオチドと、ビオチンなどのリンカーとが挙げられる。
【0126】
ポリペプチドは修飾されて、非天然アミノ酸を含んでもよく、またはポリペプチドの安定性が増大してもよい。タンパク質またはペプチドが合成手段によって生成される間に、このようなアミノ酸が生成中に導入されてもよい。タンパク質またはペプチドはまた、合成または組み換え生成のどちらかに引き続いて、修飾されてもよい。
【0127】
本発明のポリペプチドはまた、D−アミノ酸を使用して生成されてもよい。このような場合、アミノ酸はCからN方向に逆順で連結される。これはこのようなタンパク質またはペプチドを生成するために、当該技術分野で慣習的である。
【0128】
いくつかの側鎖修飾が当該技術分野で知られており、本発明のタンパク質またはペプチドの側鎖に施してもよい。このような修飾としては、例えばアルデヒドとの反応とそれに続くNaBH4での還元、アセトイミド酸メチルでのアミジン化または無水酢酸でのアシル化による、還元的アルキル化によるアミノ酸修飾が挙げられる。
【0129】
本発明によって提供される配列はまた、「第二世代」酵素構築のための出発原料として使用してもよい。「第二世代」ペプチジルアルギニンデイミナーゼは、変異誘発技術(例えば部位特異的変異誘発または遺伝子シャフリング技術)によって改変されたペプチジルアルギニンデイミナーゼであり、野生型ペプチジルアルギニンデイミナーゼ、または本発明によって生成されるものなどの組み換えペプチジルアルギニンデイミナーゼとは異なる特性を有する。例えばそれらの温度または最適pH、比活性、基質親和性または熱安定性は、特定の工程での使用によりふさわしいようにように改変されてもよい。
【0130】
本発明のペプチジルアルギニンデイミナーゼ活性に必須であり、したがって好ましくは置換されるアミノ酸は、部位特異的変異誘発またはアラニンスキャニング変異誘発などの当該技術分野で知られている手順に従って同定されてもよい。後者の技術では、突然変異が分子内のあらゆる残基に導入され、得られた突然変異分子を生物学的活性(例えばペプチジルアルギニンデイミナーゼ活性)について試験して、分子活性に決定的なアミノ酸残基を同定する。酵素−基質相互作用部位もまた、核磁気共鳴、結晶構造解析または光親和性標識などの技術によって判定される結晶構造の分析によって判定できる。
【0131】
遺伝子シャフリング技術は、ポリヌクレオチド配列中に突然変異を導入するランダムな方法を提供する。発現後、最良の特性がある単離物を再単離して合わせ、再度シャフリングして遺伝的多様性を増大させる。この手順を数回繰り返すことにより、急速に改善されたタンパク質をコードする遺伝子を単離できる。好ましくは遺伝子シャフリング手順は、同様の機能があるタンパク質をコードする遺伝子ファミリーを用いて開始する。本発明で提供されるポリヌクレオチド配列は、分泌されたペプチジルアルギニンデイミナーゼの特性を改善するための遺伝子シャフリングによく適している。
【0132】
代案としては、NTG処置による変異誘発またはUV変異誘発などの古典的ランダム変異誘発技術および選択を使用して、タンパク質の特性を改善できる。変異誘発は、単離されたDNAに直接に、または関心のあるDNAで転換された細胞に実施できる。代案としては、突然変異は、当業者に知られているいくつかの技術によって単離されたDNA中に導入できる。これらの方法の例は、誤りがちなPCR、反復−欠損宿主細胞内のプラスミドDNAの増幅などである。
【0133】
酵母および糸状菌宿主細胞の使用は、本発明の組み換え発現産物に最適生物学的活性を与えるのに必要に応じて、翻訳後修飾(例えばタンパク質分解過程、ミリスチン化(myristilation)、グリコシル化、トランケーション、およびチロシン、セリンまたはスレオニンホスホリル化)を提供することが予期される。
【0134】
[調製]
本発明のポリペプチドは単離形態であってもよい。ポリペプチドは、ポリペプチドの意図される目的を妨げないキャリアまたは希釈剤と混合してもよく、なおも単離されていると見なされるものと理解される。本発明のポリペプチドはまた、実質的に精製された形態であってもよく、その場合、それは一般に調製品中にポリペプチドを含んでなり、その中で70%を超える、例えば80%、90%、95%、98%または99%を超える、調製品中のタンパク質が本発明のポリペプチドである。
【0135】
本発明のポリペプチドは、それらがそれらの天然の細胞環境外であるような形態で提供されてもよい。したがってそれらは上で考察したように実質的に単離または精製されていてもよく、または例えばその他の真菌種、動物、植物または細菌細胞などのその中でそれらが自然に生じない細胞内にあってもよい。
【0136】
[ペプチジルアルギニンデイミナーゼ活性の除去または低下]
本発明はまた、親細胞よりも少ないポリペプチドを生成する突然変異細胞をもたらす、ポリペプチドまたはその制御配列をコードする内在性核酸配列を中断または消去するステップを含んでなる、親細胞の突然変異細胞を生成する方法に関する。
【0137】
低下したペプチジルアルギニンデイミナーゼ活性を有する株の構築は、細胞内でのペプチジルアルギニンデイミナーゼの発現に必要な核酸配列の修飾または不活性化によって、好都合に達成されてもよい。修飾または不活性化される核酸配列は、例えばペプチジルアルギニンデイミナーゼ活性を示すのに必須なポリペプチドまたはその一部をコードする核酸配列であってもよく、または核酸配列は、核酸配列のコード配列からのポリペプチド発現に必要な調節機能を有してもよい。このような調節または制御配列の例としては、プロモーター配列またはその機能性部分、すなわちポリペプチドの発現に影響を与えるのに十分な部分が挙げられる。修飾可能なその他の制御配列としては、リーダー配列、ポリアデニル化配列、プロペプチド配列、シグナル配列、および終止配列が挙げられるが、これらに限定されるものではない。
【0138】
核酸配列の修飾または不活性化は、細胞を変異誘発して、その中でペプチジルアルギニンデイミナーゼ生成能力が低下し、または排除されている細胞を選択して、実施してもよい。特異的またはランダムであってもよい変異誘発は、例えば適切な物理的または化学的変異誘発剤の使用によって、適切なオリゴヌクレオチドの使用によって、またはDNA配列をPCR変異誘発して、実施してもよい。さらに変異誘発は、これらの変異誘発剤のあらゆる組み合わせの使用によって実施してもよい。
【0139】
本目的に適した物理的または化学的変異誘発剤の例としては、紫外線(UV)照射、ヒドロキシルアミン、N−メチル−N’−ニトロ−N−ニトロソグアニジン(NTG)、O−メチルヒドロキシルアミン、亜硝酸、エチルメタンスルホネート(EMS)、亜硫酸水素ナトリウム、ギ酸、およびヌクレオチド類似体が挙げられる。
【0140】
このような薬剤を使用する場合、変異誘発は典型的に、適切な条件下で選択された変異誘発剤存在下で、変異誘発させる細胞をインキュベートして、低下したまたは皆無のペプチジルアルギニンデイミナーゼ活性発現を示す細胞を選択して実施される。
【0141】
本発明のポリペプチドの生成の修飾または不活性化は、ポリペプチドまたはその転写または翻訳に必要とされる調節要素をコードする核酸配列中の1つ以上のヌクレオチドの導入、置換、または除去によって達成されてもよい。例えばヌクレオチドは、停止コドンの導入、開始コドンの除去、または読み取り枠の変更をもたらすように挿入または除去されてもよい。このような修飾または不活性化は、当該技術分野で知られている方法に従って、部位特異的変異誘発またはPCR変異誘発によって達成されてもよい。
【0142】
原則として修飾は、生体内で、すなわち修飾する核酸配列を発現する細胞に直接に実施してもよいが、下で例証するように修飾を生体外で実施することが好ましい。
【0143】
選択された宿主細胞によるペプチジルアルギニンデイミナーゼの生成を不活性化するまたは低下させる都合のよい方法の例は、遺伝子置換または遺伝子中断の技術に基づく。例えば遺伝子中断法では、関心のある内在性遺伝子または遺伝子断片に対応する核酸配列を生体外で変異誘発して欠損核酸配列を生じ、次にそれを宿主細胞に転換して欠損遺伝子を生成する。相同的組換えによって、欠損核酸配列を内在性遺伝子または遺伝子断片に置き換える。好ましくは欠損遺伝子または遺伝子断片は、その中でポリペプチドをコードする遺伝子が修飾または破壊された転換体を選択するのに使用してもよいマーカーもまたコードする。
【0144】
代案としては、本発明のポリペプチドをコードする核酸配列の修飾または不活性化は、ポリペプチドをコードする配列に相補的なヌクレオチド配列を使用して、確立されたアンチセンス技術によって達成してもよい。より具体的には、ポリペプチドをコードする核酸配列に相補的なヌクレオチド配列を導入することで、細胞によるポリペプチドの生成を低下させまたは排除してもよい。次にアンチセンスポリヌクレオチドは、典型的に細胞内で転写され、ペプチジルアルギニンデイミナーゼをコードするmRNAにハイブリダイズできる。細胞内で生成するペプチジルアルギニンデイミナーゼの量は、相補的アンチセンスヌクレオチド配列とmRNAがハイブリダイズできる条件下で低下し、または排除される。
【0145】
本発明の方法に従って修飾される細胞は、細胞に同種または異種のどちらかの、例えば所望のタンパク質製品の生成に適した真菌株などの微生物起源であることが好ましい。
【0146】
本発明はさらに、親細胞よりも少ないポリペプチドを産生する突然変異細胞をもたらす、ポリペプチドまたはその制御配列をコードする内在性核酸配列の中断または欠失を含んでなる、親細胞の突然変異細胞に関する。
【0147】
このように作り出されたポリペプチド−欠損突然変異細胞は、同種および/または異種のポリペプチド発現のための宿主細胞として特に有用である。したがって本発明はさらに、(a)突然変異細胞をポリペプチドの生成に貢献する条件下で培養するステップと、(b)ポリペプチドを回収するステップを含んでなる、同種または異種のポリペプチドを生成する方法に関する。本文脈で、「異種のポリペプチド」という用語は、本明細書において宿主細胞に天然でないポリペプチド、その中で修飾がなされて天然配列が改変された天然タンパク質、または組み換えDNA技術による宿主細胞の操作の結果としてその発現が定量的に改変された天然タンパク質と定義される。
【0148】
なおもさらなる態様では、本発明は、本発明のペプチジルアルギニンデイミナーゼポリペプチドならびに関心のあるタンパク質生成物の双方を産生する細胞の発酵によって、本質的にペプチジルアルギニンデイミナーゼ活性を含まない、タンパク質生成物を生成する方法を提供する。本方法は、発酵中または発酵完了後のどちらかに、ペプチジルアルギニンデイミナーゼ活性を阻害できる有効量の薬剤を発酵ブロスに添加するステップと、発酵ブロスから関心のある生成物を回収するステップと、場合により回収された生成物にさらなる精製を施すステップを含んでなる。代案としては、培養後に、ペプチジルアルギニンデイミナーゼ活性を実質的に低下させて、培養ブロスからの生成物の回収ができるように、得られた培養ブロスにpHまたは温度処理を施すことができる。pHまたは温度処理の併用は、培養ブロスから回収されたタンパク質調製品に実施してもよい。
【0149】
本質的にペプチジルアルギニンデイミナーゼを含まない生成物を生成する本発明の方法は、真核生物ポリペプチドの生成において、特に酵素などの真菌タンパク質の生成において特に興味深い。ペプチジルアルギニンデイミナーゼ−欠損細胞はまた、食品産業で関心がある、または薬学的関心のある異種タンパク質を発現するのに使用してもよい。
【0150】
ペプチジルアルギニンデイミナーゼの好ましい起源は、ペプチジルアルギニンデイミナーゼをコードする微生物遺伝子を微生物宿主生物にクローニングして得られる。ペプチジルアルギニンデイミナーゼのより好ましい起源は、ペプチジルアルギニンデイミナーゼ遺伝子を過剰発現できるアスペルギルス(Aspergillus)属に属する宿主に、ペプチジルアルギニンデイミナーゼをコードするフザリウム(Fusarium)由来遺伝子をクローニングして得られる。一般に、過剰発現のためには相同宿主生物が好ましい。同種とはここで同一種であることを意味する。
【0151】
[配列一覧]
配列番号1 合成DNA
配列番号2 合成DNA
配列番号3 フザリウム・グラミネアルム(Fusarium graminearum)のDNA
配列番号4 フザリウム・グラミネアルム(Fusarium graminearum)のDNA
配列番号5 フザリウム・グラミネアルム(Fusarium graminearum)のDNA
配列番号6 フザリウム・グラミネアルム(Fusarium graminearum)のアミノ酸配列
配列番号7 フザリウム・グラミネアルム(Fusarium graminearum)のアミノ酸配列
配列番号8 ケトミウム・グロボスム(Chaetomium globosum)のアミノ酸配列
配列番号9 ストレプトミセス・スキャビエス(Streptomyces scabies)のアミノ酸配列
配列番号10 ストレプトミセス・スキャビエス(Streptomyces scabies)のアミノ酸配列
配列番号11 PADコンセンサスアミノ酸配列
配列番号12 人工アミノ酸配列
配列番号13 ストレプトミセス・クラブリゲルス(Streptomyces clavuligerus)のアミノ酸配列
配列番号14 ファエオスフェリア・ノドルム(Phaeosphaeria nodorum)のアミノ酸配列
【図面の簡単な説明】
【0152】
【図1】フザリウム(Fusarium)PADのクローニング。
【図2A】分泌PADタンパク質の配列比較。
【図2B】分泌PADタンパク質の配列比較。
【図3】発現ベクターpGBFINPADの構築。
【図4】SDS−PAGE上でのBSAおよびPADタンパク質濃度の比較。染色は、シンプリー・ブルー・セーフ・ステイン(コロイド状クーマシーG250)を使用して実施した。レーン1〜6はBSA、レーン7〜10はPAD。
【0153】
[材料と方法]
[加水分解度]
使用された様々なタンパク質分解混合物の加水分解度(DH)は、迅速なOPA試験(Nielsen,P.M.;Petersen,D.;Dambmann,C.著「食物タンパク質の加水分解度を判定するための改善された方法(Improved method for determining food protein degree of hydrolysis)」、Journal of Food Science 2001年、66、642〜646頁)を使用して測定された。
【0154】
[PADアッセイ]
PAD様活性は、2つの異なる方法でモニターした。スクリーニングアッセイでは、シグマ・アルドリッチ(Sigma−Aldrich)によって提供される、この酵素のためのシグマ品質管理試験手順(1584頁;ウサギ骨格筋から)を使用した。この色素産生方法は、N−α−ベンゾイル−L−アルギニンエチルエステル塩酸塩の使用に基づいた(BAEE;タケハラら著、(1986年)Journal of Biochemistry、99、1477〜1424頁)。
【0155】
別のアッセイでは、遊離またはペプチド結合アルギニンのシトルリンへの転換を測定して、古典的アミノ酸分析によって酵素活性を測定した。この方法は「アミノ酸分析」の下で規定される。
【0156】
[アミノ酸分析]
アミノ酸分析は、米国マサチューセッツ州ミルフォードのウォーターズ(Waters(Milford MA,USA))からのアミノ酸分析システムの操作手引で規定される、PicoTag法に従って実施した。最終サンプルを乾燥させ、フェニルイソチオシアネートを使用して直接誘導体化した。その中でシグマからの純粋化合物を誘導体化してクロマトグラフィーにかける試行を行って、クロマトグラム中でのシトルリンおよびオルニチンの位置を確立した。HPLC法を使用して、存在する誘導体化アミノ酸を定量化した。その中でタンパク質またはペプチドがPADのための基質として使用されるインキュベーション混合物を、米国マサチューセッツ州ミルフォードのウォーターズからのアミノ酸分析システムの操作手引に従って最初に酸加水分解し、次に誘導体化して、分離および定量化した。遊離アルギニンのみを含有するインキュベーション混合物中では、酸加水分解は任意であった。実施例6で例証するように、酸加水分解はシトルリンを部分的にオルニチンに転換する。アルギニンから形成されたシトルリンの総量を計算するために、シトルリンおよびオルニチンのレベルを合計した。加水分解中に、TrpおよびCysは破壊され、したがってこれらのアミノ酸はさらなる計算中で排除された。さらにGlnおよびAsn残基は、酸加水分解中にGluおよびAspに転換するので、GluおよびGlnの値、およびAspおよびAsnの値もまた合計して、酸加水分解前に得られたデータと比較できるようにした。
【0157】
[SDS−PAGE]
SDS−PAGEおよび染色のために使用した全ての材料は、米国カリフォルニア州カールスバッドのインヴィトロジェン(Invitrogen(Carlsbad,CA,US))から購入した。サンプルは、製造業者の説明書に従ってSDS緩衝液を使用して調製し、MES−SDS緩衝液系を使用して12%Bis−トリスゲル上で分離した。染色は、シンプリー・ブルー・セーフ・ステイン(コロイド状クーマシーG250)を使用して実施した。
【0158】
[LC/MS/MS分析]
PADインキュベーション後のペプチドQPRPFPFPRPRの分析では、オランダ国ブレダのサーモクエスト(Thermoquest)(登録商標)(Breda,the Netherlands))からのP4000ポンプと連結している、オランダ国ブレダのサーモクエスト(登録商標)からのイオントラップ質量分光計を使用したHPLCを使用した。溶出のための米国マサチューセッツ州ベッドフォードのミリポア(Millipore(Bedford,MA,USA)からのMilli Q水(溶液A)中の0.1%ギ酸、およびアセトニトリル中の0.1%ギ酸(溶液B)の勾配と組み合わせて、ベルギー国のVarian BelgiumからのInertsil 3 ODS3,3μm、150*2.1mmカラムを使用して、形成されたペプチドを分離した。勾配は100%の溶液Aで開始して5分間保ち、5%のBに直線的に10分間で増大させ、それに30分間での45%の溶液Bへの直線的増大が続き、即座に最初の条件にして、安定化のためにさらに15分間保った。使用した注入量は50μlであり、流速は分速200μlで、カラム温度は55℃に保たれた。注入サンプルのタンパク質濃度は、約50μgs/mlであった。約30%の最適衝突エネルギーを使用して、関心のあるペプチドのための専用MS/MSによって、インキュベーション後のペプチドQPRPFPFPRPR中の異なるアルギニン残基の反応および反応速度を経時的に追跡した。LC/MS/MSに先だって、インキュベーション混合物を周囲温度および13000rpmで10分間遠心分離し、0.22μmフィルターを通して濾過して上清をMilli Q水で1:100に希釈した。
【0159】
[クローニング技術]
核酸の単離および精製、核酸の電気泳動法、核酸の酵素修飾、切断および/または増幅、大腸菌(E.coli)などの形質転換などの標準分子クローニング技術は、Sambrookら(Sambrook,J.、Russell,D.W.著(2001年):分子クローニング;実験室マニュアル(Molecular cloning; a laboratory manual)(第3版)、Cold Spring Harbour laboratory press、Cold Spring Harbour,New York)または供給元の付属書に従って実施した。オランダ国ブレダのインヴィトロジェンは、合成オリゴヌクレオチドを供給する。DNA配列分析はオランダ国ライデン(Leiden,the Netherlands)のBaseClearで実施された。
【0160】
[実施例]
[実施例1]
[フザリウム(Fusarium)株はPAD様活性を分泌できる]
分泌されたPAD様活性を同定する目的で、カビの大きなコレクションをスクリーンした。そのために、株を30℃でジフコ(Difco)からのジャガイモデキストロースブロス(PDB)上で4〜5日間予備生育させた。次に培養を遠心分離によって収集して蒸留水で洗浄し、米タンパク質加水分解産物で強化された最少培地に移した。米タンパク質は比較的アルギニン残基に富み、その水溶性を増大させるために、ベルギー国ルーバン(Leuven,Belgium)のRemy Industriesからの米タンパク質をデンマーク国バウスヴェア(Bagsvaerd,Denmark)のNOVOからのアルカラーゼ(Alcalase)と共にpH7.5でプレインキュベートして、DHが約15の加水分解産物を得た。使用した最小増殖培地は、1リットルあたり0.52g KCl、1.52g KH2PO4、1.3ml 4M KOH、0.52g MgSO4・7H2O、22mg ZnSO4・7H2O、11mg H3BO3、5mg FeSO4・7H2O、1.7mg CoCl2・6H2O、1.6mg CuSO4・5H2O、5mg MnCl2・4H2O、1.5mg Na2MoO4・2H2O、50mg EDTA、40gグルコース、および5gの加水分解米タンパク質を含有した。真菌を後者の培地中でさらに2日間生育させた後、培養を再度遠心分離して、透明な上清のサンプルを凍結した。分泌されたPAD様活性が存在するかどうかを判定するために、材料と方法セクションで述べられているように、上清サンプルをシグマ品質管理試験手順比色分析にかけた。この試験では、0.2mlの実希釈上清を0.6mlの混合試薬に添加した。酵素インキュベーションを45℃で5時間行った。得られた結果次第で、フザリウム(Fusarium)株の全てではないがいくつかの上清は、いくらかの変色を示した。変色をもたらした上清は、オランダ国ユトレヒト(Utrecht,The Netherlands)のCBSから得られたフザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株から得られた。いくつかのその他のフザリウム(Fusarium)株、および例えば英国ウォリンフォード(Wallingford,UK)のCABIからのフザリウム・グラミネアルム(Fusarium graminearum)IMI145425などのその他の真菌の上清は、発色しなかった。得られた結果から、我々はいくつかのフザリウム(Fusarium)株は、PAD様活性を分泌すると結論した。我々の知る限り、これは微生物からのPAD分泌の最初の報告である。
【0161】
[実施例2]
[フザリウム(Fusarium)ゲノム配列中のPADコード遺伝子を同定する]
いくつかのフザリウム(Fusarium)株がPAD様活性を分泌できることが分かり、これらのPADをコードする遺伝子のゲノム配列を単離および分析した。これを行うために、フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株をジフコからのPDB(ジャガイモデキストロースブロス)中において30℃で3日間生育させ、供給元の説明書を使用して、オランダ国ブレダのOmnilabo International BVからのQ−Biogeneキット(カタログ番号6540−600)を使用して、菌糸から染色体のDNAを単離した。PCRを使用したPAD遺伝子コード配列の増幅のためにこの染色体DNAを使用した。
【0162】
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株の染色体DNAからのPAD遺伝子を特異的に増幅するために、2つのPCRプライマーをデザインした。ジベレラ・ゼアエ(Gibberella zeae)PH−1のゲノムDNA中に発見され、機能未知タンパク質(UNIPROT Q4IIR5_GIBZE)としてアノテートされた配列からプライマー配列を部分的に得た。我々はこの配列が、高等真核生物のPAD配列と相同性を有することを見いだした。第1のプライマーは、上流に12bpsのリーダー配列およびPacI制限部位(配列番号1)がある、ATG開始コドンに始まる24個のヌクレオチドのPADコード配列を含有した。第2のプライマーは、CTA停止コドン(配列番号2)のすぐ下流にAscI制限部位がある、PADコード配列に相補的な29個のヌクレオチドを含有した。これらのプライマーを使用して、テンプレートとしてフザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株からの染色体DNAを用いて、我々は2kbサイズの断片を増幅できた。これらの全例で、増幅された断片は同一サイズであった。このようにして得られた2kbサイズの断片を精製し、pCR−BluntII−TOPOベクター(Invitrogen)中にライゲートし、pGBPADシリーズのプラスミドを得た(図1参照)。PCR増幅された配列を配列分析によって分析した。興味深いことに、我々はこのサイズのゲノムDNA断片を例えばフザリウム・グラミネアルム(Fusarium graminearum)IMI145425から増幅できず、フザリウム・グラミネアルム(Fusarium graminearum)の分泌PAD活性の生成と関連がある2kb断片の存在が示唆された。
【0163】
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57およびCBS316.73株のPADコード領域のゲノム配列は、それぞれ配列番号3および配列番号4に記載される。フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57のPADを過剰発現する株から単離されたmRNAから、コード領域のcDNA配列を生じさせた(実施例3参照)。このcDNAは配列決定され、配列番号5に記載される。フザリウム・グラミネアルム(Fusarium graminearum)株CBS166.57およびCBS316.73によってコードされるPADの推定タンパク質配列は、それぞれ配列番号6および配列番号7に記載される。
【0164】
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57のPADゲノムDNA配列は、フザリウム・グラミネアルム(Fusarium graminearum)CBS316.73のPADゲノムDNA配列とは19箇所で異なる。推定タンパク質配列にとってこれは7個のアミノ酸が異なり、フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57およびCBS316.73株からの2つのPADが98.0%同一であることを意味する。興味深いことに、どちらの推定タンパク質配列も、タンパク質のアミノ−末端に配列シグナルを含有する。マトリックスBlosum 62および予期された閾値10でBlastPプログラムを使用して、これらの配列をタンパク質およびDNAデータベースと比較した(Altschulら著、1997年、Nucleic Acids Research 25:3389〜3402頁)。我々は、フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57のPADのDNAおよび推定タンパク質配列の双方が、我々がジベレラ・ゼアエ(Gibberella zeae)PH−1のゲノムDNA中に発見して、機能未知タンパク質(UNIPROT Q4IIR5_GIBZE)としてアノテートした配列と同一であることを見いだした。フザリウム(Fusarium)からのPADはまた、真菌タンパク質配列がシグナル配列を含有し、したがって細胞から能動的に分泌される見込みが高く、高等真核生物からのPADはこのようなシグナル配列を有さず、したがって能動的に分泌されないという本質的な違いを持って、高等真核生物のPADとの顕著な相同性も有した。フザリウム(Fusarium)PADは、ポルフィロモナス(Porphyromonas)からのPADとのいかなる相同性も示さなかった。
【0165】
さらにフザリウム(Fusarium)PADと、(フザリウム(Fusarium)のような)子嚢菌綱(ascomycetes)真菌であるが、異なる目(肉座菌目でなくフンタマカビ目)に属するケトミウム・グロボスム(Chaetomium globosum)CBS148.51のゲノムからの配列の間に相同性が見られた(http://www.broad.mit.edu/annotation/genome/chaetomium_globosum/Home.html;CHGG_01998.1)。フザリウム(Fusarium)とケトミウム(Chaetomium)のPAD配列間の相同性は、660個のアミノ酸中253個のアミノ酸である(38%同一)。ここでもまた、予測されたタンパク質は、機能未知タンパク質(UNIPROT Q2HCQ6_CHAGB)としてアノテートされた。さらにこの真菌中のPAD相同体のタンパク質配列のアノテーションは、おそらく不正確である。ケトミウム(Chaetomium)タンパク質と既知のPADとの相同性がタンパク質の最初の600個のアミノ酸にかけてのみに存在するのに対し、ケトミウム(Chaetomium)タンパク質は、1000個のアミノ酸のタンパク質としてアノテートされている。したがって我々は、UNIPROT Q2HCQ6_CHAGB中のケトミウム(Chaetomium)タンパク質のアノテーションは不正確と考える。ケトミウム・グロボスム(Chaetomium globosum)PADの正確なタンパク質配列を配列番号8に記載し、配列比較を図3に示す。SignalPプログラム(http://www.cbs.dtu.dk/services/SignalP/)を使用してこの配列をより綿密に調べると、タンパク質のアミノ−末端にシグナル配列を検出でき(図3参照)、このタンパク質もまた、我々がフザリウム(Fusarium)株からのPADについて発見したように、真菌から分泌される可能性が高いことを意味する。
【0166】
フザリウム(Fusarium)PADの別の相同体は、これもまた(フザリウム(Fusarium)およびケトミウム(Chaetomium)のような)子嚢菌綱(Ascomycetes)真菌であるが、さらに異なる綱(フンタマカビ綱(Sordariomycetes)でなくクロイボタケ綱(Dithideomycetes))に属するファエオスフェリア・ノドルム(Phaeosphaeria nodorum)SN15のゲノム中に見いだせる。この遺伝子によってコードされるタンパク質は、配列番号14に記載される。フザリウム(Fusarium)とファエオスフェリア(Phaeosphaeria)のPAD配列間の相同性は、629個のアミノ酸中231個のアミノ酸である(36%同一)。ここでもまた、予測されたタンパク質は機能未知タンパク質(機能未知タンパク質SNOG_13103;ジェンバンクEAT79430)としてアノテートされ、シグナル配列が認識できる。ここで示されるデータは、真菌界全体を通じて分泌PADが存在できるが、まれにしか生じないことを示唆する。ここで述べられる方法を使用すれば、真菌からの分泌PADを特異的に認識して単離できるであろう。
【0167】
フザリウム(Fusarium)PADの可能な相同体についてデータベースをさらに調べた際に、我々はジャガイモ瘡痂病の病原体であるストレプトミセス・スキャビエス(Streptomyces scabies)のゲノム中に2つの追加的配列見つけることができた(http://www.sanger.ac.uk/Projects/S_scabies/)。これらのPADのタンパク質配列は、配列番号9および配列番号10に記載される。またこれらの遺伝子でもタンパク質のアミノ−末端にシグナル配列が疑われるので、これらのタンパク質も分泌されるかもしれない。配列番号10に記載されたタンパク質のカルボキシル−末端は欠損しているが、これはこの遺伝子がcontigの終わりに位置し、コード領域の3’−末端が未だ配列決定されていないためである。
【0168】
ストレプトミセス・クラブリゲルス(Streptomyces clavuligerus)ATCC27064のゲノムを配列決定し分析した際に、SignalPプログラムを使用した検出で、我々は、これもまたシグナル配列を含有する別のPAD相同体を検出できた。アミノ酸配列は配列番号13に記載される。遺伝子が配列決定contigの終わりに位置し、したがって3’−末端が欠損していたことから、タンパク質はカルボキシ末端で短い。
【0169】
微生物からのこれらのPAD配列を高等真核生物のPADと比較すると、これらの配列間で検出可能な非常に相同的な領域がある。この共通配列はWLxVGHVDEであり、配列番号11に記載される。この共通配列を使用し、当業者が既知のクローニング技術を使用して、微生物からのPADをコードする遺伝子を同定および単離することは十分可能である。一つの可能性は、その中でPAD遺伝子を同定したい生物からの好ましいコドン使用頻度があるヌクレオチド配列への配列番号11の配列の逆翻訳に基づいて、オリゴヌクレオチドプライマーをデザインし、このオリゴヌクレオチドを遺伝子ライブラリーとのハイブリダイゼーションのために、または逆転写されたmRNAプール上のPCRプライマー中で使用することである。別の可能性は、Patscan(http://www−unix.mcs.anl.gov/compbio/PatScan/HTML/patscan.html)のようなプログラムを使用した、DNAデータバンクからの翻訳されたDNA配列における検索のために、配列番号11の配列を使用することである。これらの方法の1つを使用して同定された遺伝子は、次に当業者に知られているプログラムを使用してタンパク質配列に翻訳し、それらのアミノ−末端のシグナル配列の存在について調べることができる。シグナル配列を検出するためにSignalPのようなプログラムを使用できる(http://www.cbs.dtu.dk/services/SignalP/)。本発明では、我々は、配列番号11のコンセンサスと予測されたシグナル配列の双方を含有するタンパク質配列がPADを分泌する可能性があることを見いだした。これらの組み合わされた特性を探すことは、このような酵素の工業生産に大きな利点を与える。
【0170】
[実施例3]
[アスペルギルス・ニガー(Aspergillus niger)による推定上のフザリウム・グラミネアルム(Fusarium graminearum)PADの過剰発現]
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57からのゲノムPAD遺伝子を含有するpGBPADプラスミドから、PADコード配列を含んでなるPacI/AscI断片を単離して、pGBFIN−5(WO99/32617)中のPacI/AscIphyA断片と交換した。得られたプラスミドはpGBFINPADと命名されたPAD発現ベクターであった(図3参照)。全ての大腸菌(E.coli)由来配列を発現ベクターから除去するNotIでの消化によって、発現ベクターpGBFINPADを直線化した。フェノール:クロロホルム:イソアミルアルコール(24:23:1)抽出およびエタノール沈殿を使用して、消化されたDNAを精製した。これらのベクターを使用して、アスペルギルス・ニガー(Aspergillus niger)CBS513.88を形質転換した。アスペルギルス・ニガー(Aspergillus niger)形質転換手順は、国際公開第98/46772号パンフレットで詳細に述べられている。どのようにしてアセトアミドを含有する寒天プレート上で転換体を選択し、標的とされるマルチコピー組み込み体を選択するかについてもまた、述べられている。サンプル材料のさらなる産生のために、好ましくは発現カセットの複数コピーを含有するA.ニガー(A.niger)転換体が選択される。pGBFINPAD発現ベクター30のために、最初に個々の転換体を選択的培地プレートに播種し、それに続いて単一コロニーをPDAプレートに播種して、A.ニガー(A.niger)転換体を純化した。30℃で1週間の生育後に個々の転換体の胞子を採取し、胞子を冷蔵保存して液体培地の接種のために使用した。
【0171】
浸透フラスコ培養内での株の培養によって、発現カセットの複数コピーを含有するA.ニガー(A.niger)株をサンプル材料の産生のために使用した。A.ニガー(A.niger)株の培養および培養ブロスからの菌糸の分離のための有用な方法は、国際公開第98/46772号パンフレットで述べられている。培養媒体はCSM−MES(1リットルの培地あたり150gマルトース、60gのジフコからのソイトン(Soytone)、15g(NH4)2SO4、1g NaH2PO4・H2O、1g MgSO4・7H2O、1g L−アルギニン、80mg ツイーン−80、20g MES、pH6.2)中にあった。発酵の4〜8日目に5mlのサンプルを取って、Hereaus labofuge RF内で10分間5000rpmで遠心分離し、さらなる分析まで上清を−20℃で保存した。
【0172】
pGBFINPADベクターを含有する転換体は、SDS−PAGEで分析すると見かけ上の分子量がおよそ60kDaのタンパク質を産生することが明らかになった。これはタンパク質配列から予測された分子量よりもわずかに小さいので、我々は、フザリウム(Fusarium)PADをアスペルギルス・ニガー(Aspergillus niger)中で生産すると、シグナル配列の除去後に大規模なグリコシル化が起きないと推測する。
【0173】
選択された株は、発酵および後処理プロセスをスケールアップさせた際に、大量のPADの単離および精製のために使用できる。次にこの酵素をさらなる分析のために、そして多様な産業上の利用のために使用できる。
【0174】
[実施例4]
[A.ニガー(A.niger)上清からの過剰発現された推定上のPADの精製]
1リットルあたり、マルトース、40g H2O、30gソイトン(Difco)、15g (NH4)2SO4、1g NaH2PO4・H2O、1 MgSO4・7H2O、1g L−アルギニン、0.08gツイーン−80、70g Na−クエン酸を組み込んだ増殖培地を使用して、プラスミドpGBFINPADを組み込んだA.ニガー(A.niger)を10リットルの規模で生育させた。pHを6.2に調節した。30℃で6日間の生育後、3.5g/lの安息香酸ナトリウムを添加して細胞を死滅させ、さらに6時間インキュベーションを延長した。次に10g/lのCaCl2およびベルギー国ゲント(Gent,Belgium)のDicalite BFからの45g/lの濾過助剤をブロスに添加した。最初に布帛での濾過によって菌糸を除去し、Z−2000およびZ−200フィルター(Pall)を通した濾過がそれに続いた。最後に、ミリポアからの0.22μm GP Express PLUS膜を使用して滅菌濾過を実施した。限外濾過は、ミリポアからのPelliconシステムを使用して実施した。
【0175】
緩衝液条件を25mM Na−クエン酸、pH5.0(緩衝液A)に変更後、同一緩衝液で平衡化したベルギー国ディーゲムのGEヘルスケアからのSP−セファロースXKカラムに溶液を入れた。カラムを緩衝液Aから緩衝液A+1M NaClへの直線濃度勾配で溶出した。得られた画分をSDS−PAGEにかけて、染色した。見かけ上のMWが60kDaのバンドを明らかに示した画分をプールした。グリセロール(50%w/w最終濃度)、CaCl2(0.02%w/w)、および安息香酸ナトリウム(0.1%w/w)を添加して、得られた酵素溶液を安定化した。この最終調製品中のPADタンパク質濃度を推定するために、異なる量の最終調製品を異なる量のシグマからのBSA溶液(Fraction V)と共にSDS−PAGEにかけて、シンプリー・ブルー・セーフ・ステイン(Simply Blue Safe Stain)で染色した。得られた結果により(図4参照)、最終調製品中のPAD濃度は3.8mgタンパク質/ml液体である。
【0176】
[実施例5]
[過剰発現されて精製された推定上のPADはペプチド結合アルギニンをシトルリンに転換できる]
PADとして単離された酵素の性質を確認するために、オランダ国アルメレ(Almere,The Netherlands)のPepscanからの合成ペプチドQPRPFPFPRPRと共に、クロマトグラフ的に精製された酵素をインキュベートし、LC/MSを使用して得られた生成物を分析した。LC/MS分析の目的は、アルギニン残基のシトルリン残基への転換を確認することであった。このために、pH6.5および50℃で、合成ペプチド(10mM)と共に様々な量の精製された酵素をインキュベートした。0、1、および4時間のインキュベーション後に、サンプルをインキュベーション混合物から取り除いた。全てのサンプルを95℃で10分間加熱して、あらゆる残留酵素活性を不活性化し、引き続いて遠心分離した。ペプチドをインキュベーション混合物に添加した直後に、標準サンプル(0時間のインキュベーション)を加熱処理して遠心分離した。材料と方法セクションに規定する条件下で、透明な上清をLC/MS分析にかけた。
【0177】
MSを最適化するために、5μg/mlのQPRPFPFPRPR溶液を使用した。(major)m/z697.8=[M+2H]2+および(minor)m/z465.6=[M+3H]3+のESI/posモードで、ウンデカ−ペプチドを特性決定した。QPRPFPFPRPRの滞留時間を判定するために、LC/MSを実施した。アルギニン(R)のプロトン化質量は175であり、シトルリン(R=O)への転換後、プロトン化質量は1Daから176に増大する。質量軸の定義は質量電荷比(m/z)であるので、ペプチドは[M+H]+でなく二重荷電の[M+2H]2+によって特徴づけられるという事実のために、質量の差は転換したアルギニンあたり0.5Daの違いによって特徴づけられる。インキュベート0時間サンプルのマスクロマトグラムは、9.72分でペプチドQPRPFPFPRPR(m/z697.8)に相当する大きなピークを示す。しかし11.36分に溶出した小さなピークは、このサンプル中でさえ、いくらかのアルギニン残基がシトルリンに転換されていることを示す。1時間インキュベートしたサンプルのマスクロマトグラム中では、3つのピークが明らかである。インキュベーション0時間後に出現したピークの他に、13.36分(2つのR’がR=Oに転換されたQPRPFPFPRPR)および15.94分(3つのR’がR=Oに転換されたQPRPFPFPRPR)にピークが検出された。4時間インキュベートしたサンプル中では、後者の2つのピークのみが見られた。これらのデータは、確かに単離された酵素画分が、PAD活性がある酵素に相当することの強力な示唆を形成する。かなり意外なことに、得られたデータはまた、ペプチドのN−末端寄りのアルギニン残基が最初に判定されるので、PADがペプチドのN−末端に始まると、我々が結論できるようにした。ペプチドのC−末端寄りのアルギニン残基が後に従う。
【0178】
[実施例6]
[タンパク質結合アルギニン、ペプチド結合アルギニンおよび遊離アルギニンは、過剰発現されるフザリウム(Fusarium)PADのための適切な基質を形成する]
大量のPADの可用性は、我々が古典的なアミノ酸分析によって、アルギニンからシトルリンへの転換を調べられるようにした。この目的で我々は、使用したクロマトグラム中におけるシトルリンの位置を確立して、存在する遊離アミノ酸のレベルを定量化しなくてはならなかった。アミノ酸分析で使用した酸加水分解はシトルリンをオルニチンに分解することが予期されたので、どちらもシグマからの純粋なシトルリンおよびオルニチンを誘導体化して、PicoTag法に従ってクロマトグラフィーにかけた。試行は双方の誘導体化合物が、クロマトグラム中の個々のピークとして、その他の誘導体化アミノ酸中で追跡できることを実証した。
【0179】
新しい酵素の試験では、0、5、50、および500μlの過剰発現され精製されたPADと共に、3つの異なる基質、すなわちカゼイン酸ナトリウム、カゼイン加水分解産物、および遊離アルギニンの10%(w/w)溶液をインキュベートした(実施例4参照)。インキュベーションをpH6.5および45℃で3時間実施した。カゼイン酸ナトリウムはオランダ国フェーヘルのDMVインターナショナル(DMV International(Veghel,The Netherlands)から得られ、カゼイン加水分解産物は、サブチリシンおよびプロリン特異的エンドプロテアーゼでのカゼイン酸ナトリウムの消化によって調製した(Edensら著、JAFC 53(20)、7950〜7957、2005年)。L−アルギニンはシグマから得られた。
【0180】
インキュベーション後、様々な混合物をアミノ酸分析によって分析した。表1に示すデータから結論できるように、新たに同定されたPADの相当量を含有する全てのインキュベーション中で、顕著な量のアルギニンがシトルリンに転換された。予期されたように、クロマトグラムはまた、オルニチンの存在も示唆した。これらの所見はフザリウム(Fusarium)PADが、タンパク質結合およびペプチド結合アルギニンを転換できることを暗示する。かなり意外にも、遊離アルギニンもまた基質として受容される。結果によれば、アルギニンからシトルリンへの転換において、基質が小さいほど酵素はより効果的である。後者の観察は、遊離アルギニンよりもペプチジルアルギニン基質への好みを示すと報告されている、ポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)酵素について公開されるデータ(McGrawら著、Infection and immunity,1999年7月、3248〜3256頁)と対照的なようである。
【0181】
オルニチンの存在がシトルリンの酸加水分解の結果であることを確認するために、遊離アルギニンを基質とする実験を実施したが、今度は酸加水分解を省いた。表2に示すように、この場合、形成されたオルニチンの量はごくわずかであり、存在するオルニチンが酵素によって形成された生成物ではないことが実証された。
【0182】
【表1】

【0183】
【表2】
【0184】
[実施例7]
[過剰発現されたPADの最適pHおよび温度]
実施例6で述べられているのと非常によく似た一組の実験で、フザリウム(Fusarium)PADの最適pHおよび温度を判定した。これらの結果によれば、過剰発現された酵素は、その最適pHを8.0前後に、その最適温度を40〜50℃の間に有する。
【0185】
[実施例8]
[過剰発現されたPADのカルシウム依存性]
文献に従って、真核生物のペプチジルアルギニンデイミナーゼは、Ca2+−依存性酵素のファミリーを代表する。フザリウム(Fusarium)PADもまたCa2+を必要とするかどうかを試験するために、EDTA存在下でインキュベーションを実施した。この目的で、イリノイ州ロックフォード(Rockford,IL)のPierceからのDeSalt Spinカラムを使用してカゼイン酸ナトリウムを最初に脱塩し、次に10mM(最終濃度)のEDTAの存在および不在下で、5%カゼイネート溶液をPADと共に45℃およびpH7.2で4時間インキュベートした。実施したアミノ酸分析によれば、どちらのインキュベーションも酸加水分解カゼイネート中に遊離アルギニンの同一の減少(約20%)を示し、フザリウム(Fusarium)PADはCa2+非依存性であることが示唆された。
【0186】
[実施例9]
30%果汁入り清涼飲料
典型的な1食分:240ml
活性成分:
カゼイン加水分解産物(アルギニンの50%はシトルリンに転換されている)および炭水化物源としてマルトデキストリンをこの食品に組み込む:
カゼイン加水分解産物:1.5〜15g/1食当たり
マルトデキストリン:3〜30g/1食当たり
【0187】
[I.以下の成分から清涼飲料化合物を調製する:]
[果汁濃縮物および水溶性香料]
【0188】
【表3】
【0189】
[1.6 活性成分]
上述の濃度の活性成分(これは上述の活性成分を意味する)タンパク質加水分解産物およびマルトデキストリン。
【0190】
果汁濃縮物および水溶性香料を空気を入れずに混合する。着色剤を脱イオン水に溶解する。アスコルビン酸およびクエン酸を水に溶解する。安息香酸ナトリウムを水に溶解する。ペクチンを撹拌しながら添加して沸騰させながら溶解する。溶液を冷却する。オレンジ油および油溶性香料を予混合する。1.6で言及される活性成分を乾燥混合し、次に好ましくは果汁濃縮物混合物(1.1)中で撹拌する。
【0191】
清涼飲料化合物を調製するために、3.1.1〜3.1.6の全ての部分をTurrax、次に高圧ホモジナイザー(p1=200bar、p2=50bar)を使用して、均質化するまで共に混合する。
【0192】
[II.以下の成分から瓶詰シロップを調製する:]
【0193】
【表4】
【0194】
瓶詰めシロップの成分を混ぜ合わせる。瓶詰めシロップを水で希釈して、1リットルのすぐに飲める飲料にする。
【0195】
[バリエーション:]
安息香酸ナトリウムを使用する代わりに、飲料を低温殺菌してもよい。飲料はまた、炭酸ガスを添加してもよい。
【発明の詳細な説明】
【0001】
[技術分野]
本発明は、シトルリン残基を含んでなるタンパク質およびペプチドに関する。
【0002】
[背景技術]
アルギニンは、哺乳類の生理機能において重要な役割を果たす準必須アミノ酸である。アルギニンの代謝経路については、詳しく述べられている。その食餌摂取に際して、アルギニンは肝臓によって肝臓の門脈から取り込まれ、酵素アルギナーゼによって迅速にオルニチンに転換する。後者の過程で尿素が形成される。次にアルギニンから発生したオルニチンはシトルリンに転換され、またはアミノ酸グルタメートおよびプロリンに代謝されることができる。代案としては形成されたオルニチンは、プトレシンなどのポリアミン化合物に組み込まれる。オルニチンに代謝されなかった食餌性アルギニンは、タンパク質合成の目的で、例えば酸化窒素またはアルギニル−tRNAに加工されることができる。アルギニンに向けた内在性合成経路もまた存在する。後者の過程は、アルギニンがオルニチンおよびシトルリン前駆物質から合成される腎臓で主に起きる。
【0003】
シトルリンは、スイカ、カボチャ、およびキュウリのようなウリ類中で遊離アミノ酸として存在すると述べられている天然アミノ酸である。遊離アミノ酸のその他の起源は、プルーン果汁、いくつかのブドウの変異株、および醤油およびワインなどの発酵食品である。自然界では、シトルリンがその他のアミノ酸に結合して存在することは稀である。食用キノコでは、ジペプチドピログルタメート−シトルリンが、アイリッシュコケではジペプチドシトルリン−アルギニンの存在が示されている。哺乳類では免疫化学的技術を使用して、シトルリンを取り込んだ低レベルのペプチドまたはタンパク質の存在が示されている。
【0004】
哺乳類では、シトルリンは腸内でグルタミンから合成され、血中に放出されて腎臓内で転換されてアルギニンに戻る。健康な成人では、腎臓によって転換されるシトルリンは、身体の全アルギニン所要量を提供するのに十分である。しかし新生児では、腎臓内でのこのシトルリンからアルギニンへの反応は不十分であり、追加的機序が関与する。様々な生理学的過程におけるアルギニンに代わるものとしてのシトルリンの重要な役割は、最近の研究によって明らかにされている。肝臓による食餌性アルギニンの捕捉は非常に効率的であるので、肝臓下流の血中アルギニン濃度は比較的低い。それゆえ例えば迅速な生育期間中に、栄養失調またはアミノ酸代謝変化の結果として、または外傷性または病的侵襲に応えて、全ての臓器におけるアルギニン需要が完全に満たされないかもしれない状態が生じる場合がある。このような状況では、シトルリンはアルギニンに代わるものとして機能するかもしれない。食餌性アルギニンとは対照的に、食餌性シトルリンは肝臓によって門脈血から取り除かれない。それゆえシトルリンは、筋肉をはじめとする末梢組織が利用できる自由に循環するアルギニン源の代替えに相当するが、より効率的である(Curisら著、Amino Acids(2005年)29:177〜205頁)。食餌性シトルリンはまた、アルギニンがオルニチンに転換するアルギナーゼ反応について述べられているような肝臓内における尿素生成をもたらさない。したがってシトルリンを摂取することにより、この尿素生成および免疫学的状態の低下の結果としての腎臓損傷などの補給されるアルギニンのネガティブな副作用を回避できる。シトルリンのさらに別の利点は、それが過剰なアンモニアを捕らえることができ、すなわちそれがいわゆる血中アンモニア低下剤として作用して、特定の酵素機能障害を患っている患者、てんかんを患っている個人に、そして健康な個人では、長期の高強度筋肉運動に起因する疲労を予防する利点を提供できることである。
【0005】
多くの国々で、食物への遊離アミノ酸の添加を規制する法規がある。結果として、遊離アミノ酸は、臨床栄養物でのみ使用でき、上述の補給されるアルギニンまたはシトルリンの生理学的利点を一般の消費者に提供することはできない。さらに遊離アルギニンの非常に苦い味は、臨床および非臨床用途の重要な欠点となる。重病患者は、味の悪い食物または栄養補給剤を単に避け、高齢者またはスポーツ選手などの医療ニーズがない消費者も同様である。したがって食物中での遊離アミノ酸の使用は、特に推奨アミノ酸投薬量を考慮した場合、重大な美味性の問題を引き起こすことが予期できる。これらの結論の意味合いは、遊離アミノ酸として存在しない形態であり、改善された美味性を有するシトルリンに対する明らかな必要性が存在することである。
【0006】
[発明の概要]
本発明は、タンパク質、ペプチドまたはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換する、修飾タンパク質、ペプチドまたはタンパク質加水分解産物に関する。したがって本発明のタンパク質、ペプチドまたはタンパク質加水分解産物は、好ましくは少なくとも0.15、好ましくは少なくとも0.30、より好ましくは少なくとも0.5、なおもより好ましくは少なくとも1.0、さらになおもより好ましくは2.0、最も好ましくは少なくとも4のシトルリン対アルギニン(タンパク質、ペプチドまたはタンパク質加水分解産物中に存在する)のモル比を有する。加水分解産物の場合、アルギニンとして存在する遊離アミノ酸の量は、シトルリン形成の測定に使用せず、シトルリン対アルギニン比の考慮にも入れない。それゆえ本発明は、高比率の結合シトルリン残基を有する、タンパク質、ペプチドまたは加水分解産物に関する。結合シトルリンまたはペプチド結合シトルリンとは、遊離アミノ酸である遊離シトルリンとは対照的に、ペプチドまたはタンパク質の一部であるシトルリン残基を意味する。
【0007】
さらに本発明は、タンパク質、ペプチドまたはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換する、タンパク質、ペプチドまたはタンパク質加水分解産物を酵素的に生成する方法に関する。本発明のタンパク質、ペプチドまたはタンパク質を得るために、開始タンパク質、ペプチドまたはタンパク質加水分解産物基質をタンパク質アルギニンデイミナーゼと共にインキュベートする。
【0008】
さらに食物と、食物成分と、飼料と、飼料成分と、栄養補助食品または薬物などの栄養補給食品と、または栄養補助食品または薬物などの栄養補給食品の成分として使用でき、または食物と、飼料と、または栄養補助食品または薬物などの栄養補給食品の生産において成分として使用できる、タンパク質、タンパク質加水分解産物、ペプチドまたはペプチド混合物を提供することが本発明の目的である。
【0009】
本発明の別の態様に従って、生産宿主によって培養液中に能動的に分泌される、タンパク質アルギニンデイミナーゼが開示されている。本発明はまた、このようなタンパク質アルギニンデイミナーゼの生成および使用にも関する。
【0010】
したがって本発明は、
(a)配列番号6、8、9、10、13または14の1〜640番目のアミノ酸と少なくとも30%のアミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド、
(b)低ストリンジェンシー条件下で、(i)60個を超える、好ましくは100個を超えるヌクレオチドと少なくとも80%または90%同一の、より好ましくは200個を超えるヌクレオチドと少なくとも90%同一の配列番号3の核酸配列またはその断片、または(ii)配列番号3の核酸配列に相補的な核酸配列とハイブリダイズするポリヌクレオチドによってコードされるポリペプチド
からなる群から選択される、タンパク質アルギニンデイミナーゼ活性を有する単離されたポリペプチドに関する。
【0011】
[発明の詳細な説明]
シトルリンはアミノ酸でありながらDNAによってコードされず、タンパク質合成中にタンパク質に組み立てられない。それにもかかわらず、いくつかの哺乳類の組織内では、免疫化学的技術を使用して微少量のペプチド結合シトルリンが検出されている。例は、滑液、滑膜組織、造血性細胞、および活性化マクロファージである。シトルリン残基を含有するタンパク質は、ペプチド結合アルギニン残基のいわゆる翻訳後修飾中に発生する。この特定の修飾は、シトルリン化または脱イミノ化と称される過程で、ペプチドまたはタンパク質結合アルギニンをペプチドまたはタンパク質結合シトルリンに転換する、タンパク質またはペプチジルアルギニンデイミナーゼ(EC3.5.3.15)と称される酵素ファミリーによって触媒される。タンパク質アルギニンデイミナーゼおよびペプチジルアルギニンデイミナーゼという用語は、ここで同義的に使用される。アルギニンからシトルリンへの反応において、アルギニン側鎖の末端窒素原子の1つは酸素によって置換される。反応は1個の水分子を使用して、副生物としてアンモニアを生じる(http://en.wikipedia.org/wiki/Citrullination)。中性pHにおいてアルギニンが正に帯電する一方、シトルリンは非荷電である。したがってシトルリン化はペプチドまたはタンパク質の疎水性を増大させ、これはタンパク質またはペプチドの特性を変化させることができ、究極的にタンパク質のアンフォールディングをもたらすかもしれない過程である。シトルリン残基を含有することが知られている哺乳類タンパク質としては、ミエリン塩基性タンパク質(MBP)、フラグリンおよびいくつかのヒストンタンパク質が挙げられる一方、線維素およびビメンチンのようなその他のタンパク質は、細胞死および組織炎症においてシトルリン化されることができる。タンパク質のシトルリン化は、尿素回路の一環としての、または酸化窒素シンターゼファミリーの酵素副産物としての遊離アミノ酸シトルリンの形成とは異なることに留意されたい。
【0012】
アルギニン含有タンパク質またはペプチドとは対照的に、シトルリン化タンパク質またはペプチドは、工業規模では市販されない。意外にも我々は、タンパク質またはペプチド結合シトルリンの生成を含んでなる、食品等級で工業的に応用可能な方法を同定した。この方法では、タンパク質またはペプチド結合アルギニン残基をタンパク質またはペプチド結合シトルリン残基に効率的に転換する、酵素タンパク質アルギニンデイミナーゼまたはペプチジルアルギニンデイミナーゼ(下文においてPADと称される)を利用する。
【0013】
主に哺乳類組織からのいくつかのタイプのPADが知られているが、これらの酵素のいずれも産業上の利用には適さない。多くの微生物をスクリーニングすることで、我々はかなり意外なことに、発酵ブロス中にPAD酵素を能動的に分泌できるいくつかの微生物に行き当たった。目下記述されている全ての哺乳類タイプのPADは培養液中に能動的に分泌されないので、このような能動的に分泌されるPADが自然界に見いだされることは予期されていなかった。能動的分泌は、ここで生物が生育または培養液中にポリペプチドを蓄積する能力と定義される。ポリペプチドの能動的分泌は宿主生物からのエネルギー、および宿主生物からの専用分泌経路を必要とする。一般に能動的に分泌されるポリペプチドは、シグナル配列またはシグナルペプチドとも称されるアミノ−末端プレ配列を含有する。能動的分泌は発酵ブロスへの酵素の輸送のために、必ずしも細胞壁の破壊を伴わない。一般にグラム陰性細菌は、能動的分泌を有さないことが知られている。
【0014】
哺乳類のPADは互いに強いつながりがあり、異なるイソ型が多様な異なる臓器中で発現される。実際、酵素的PAD活性は、細胞の溶解後にのみ検出でき、それはPADが哺乳類の組織中で細胞内酵素であることを強力に示唆し(Vossenaarらによるレビュー(2003年)Bioessays 25:1106〜1118頁)、酵素は能動的に分泌されない。さらに全ての哺乳類PADは、常態において効率的分泌のために必要とされる明白なシグナル配列を欠いている。したがって能動的に分泌されるPAD酵素は明白な工業利点を有するであろうが、微生物中のこのような酵素についてスクリーンすることは論理的でないようである。
【0015】
能動的分泌は、煩わしい精製工程を経ることなくほぼ純粋な形態での酵素の回収ができるようにするので、経済的製造工程に最も重要である。アスペルギルス(Aspergillus)などの食品等級真菌宿主によるこのような能動的に分泌されるPADの過剰発現は、食品等級酵素と、シトルリン化タンパク質またはペプチドに向けた費用効率が高い製造工程を生じる。
【0016】
我々が知る限りでは、我々はアスペルギルス(Aspergillus)のような食品等級宿主生物によって発酵ブロス中に効率的に分泌されるPADについて初めて述べ、シトルリン化タンパク質またはペプチドに向けた費用効率が高い製造工程を初めて報告する。
【0017】
酵素アルギニンデイミナーゼ(EC3.5.3.6)はよく知られており、遊離アルギニンの遊離シトルリンへの転換におけるその使用については、幅広く述べられている。しかしPAD(EC3.5.3.15)は、タンパク質またはペプチド結合アルギニン残基の脱イミノ化を触媒してタンパク質またはペプチド結合シトルリン残基を生じ、それよってアンモニアを放出する比較的新しい酵素ファミリーを形成する。今までに多岐にわたる哺乳類の組織内に酵素が同定されている。ヒトでは、例えば4つの異なるPAD酵素が同定されており、例えば皮膚、子宮、筋肉、脳、膵臓、脾臓、胃、胸線、脊髄、およびマクロファージなどの造血性細胞に見られる。これらのCa2+−依存性PAD酵素は、例えば表皮分化、神経軸索の髄鞘形成、および毛嚢の角質化に関与する。既知のいずれの哺乳類PADも細胞により能動的に分泌されない。哺乳類組織中に広範に存在しながら、微生物からの活性PADに関する報告は非常に限られている。唯一の例外は、ヒトグラム陰性病原体ポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)からの細菌酵素である(McGrawら(1999年)Infect Immun.67(7):3248〜3256頁)。この酵素はペリプラズム間隙に輸送されて、この細菌外のペリプラズム間隙から漏出するのみである。成人発症型歯周炎の発症および進行は、ポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)による歯肉溝の感染と関連付けられている。このポルフィロモナス(Porphyromonas)PADは、脊椎動物PADと進化的に関連していないが、いくつかのアルギニンデイミナーゼと配列相同性を共有する(Shiraiら(2001年)Trends Biochem Sci.;26(8):465〜468頁)。しかし酵素は、哺乳類のPADとは対照的に、ペプチド−結合ならびに遊離L−アルギニンをシトルリンに転換でき、カルシウムイオンに依存しない。したがって酵素は、能動的に分泌されず高度に不安定であり、毒性因子であると述べられているので、食品等級のこのP.ジンジバリス(gingivalis)由来酵素の産業上の利用は、全くありそうにない。したがってポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)からのPADおよびその使用は、本発明の一部ではない。
【0018】
ここで我々は、我々の知る限りでは、初めて記述された分泌PADである、真菌フザリウム・グラミネアルム(Fusarium graminearum)から分泌PADを単離して同定した。PADアッセイに基づいて、我々はフザリウム(Fusarium)株中で、分泌PADのいくつかの変異株を単離および特性決定できた。これらの真菌からのPADをコードする遺伝子は、単離および特性決定された(配列番号3および4)。真菌アスペルギルス・ニガー(Aspergillus niger)中でのこのPADの過剰発現は、培養液中へのPADの効率的な分泌をもたらす。これらの全ての新しい変異株は、分泌PADの特徴を含む。さらにこの知識に基づいて、我々は、そのDNA配列が公共データベース中に存在するその他の微生物中で、分泌PADの遺伝子を同定して正しくアノテートできた。潜在的分泌PADは、真菌ケトミウム・グロボスム(Chaetomium globosum)、ファエオスフェリア・ノドルム(Phaeosphaeria nodorum)、および細菌ストレプトミセス・スキャビエス(Streptomyces scabies)およびストレプトミセス・クラブリゲルス(Streptomyces clavuligerus)に見られる。これらのタンパク質の分泌された特徴に関する我々の知識に基づいて、我々は正確なタンパク質配列が配列番号8〜10、13、および14で示されることを提案する。これらの全配列は、これまで分泌されるペプチジルアルギニンデイミナーゼをコーディングすると述べられたことがないが、フザリウム・グラミネアルム(Fusarium graminearum)の分泌PADの遺伝子構造に関する我々の知識から、それとしてのみ同定できる。
【0019】
異なる培養コレクションからの真菌の大規模なスクリーニングの後、我々は、発酵ブロス中に能動的に分泌されるPAD様酵素源として、フザリウム(Fusarium)種を同定することができた。このフザリウム(Fusarium)由来PAD様酵素は哺乳類のPADに対してかなりの相同性を示すが、P.ジンジバリス(gingivalis)PADに対しては示さない。クロマトグラフィー精製によるフザリウム(Fusarium)発酵ブロスからのPADの単離は、この分泌PADの分子量が約55kDaであり、最適活性がpH8前後で、最適温度は40〜50℃の間であることを明らかにした。フザリウム(Fusarium)酵素の分子量は哺乳類酵素の分子量よりも顕著に低いが、我々は最適pHおよび温度が同様であることを見いだした。
【0020】
経済的観点から、PADを大量にかつ比較的純粋な形態で生成する、改善された手段に対する明白な必要がある。これを行う好ましい方法は、組み換えDNA技術を使用したこのようなPADの過剰産生を通じたものである。これを行う特に好ましい方法は、フザリウム(Fusarium)由来PADの過剰産生を通じたものであり、これを行う最も好ましい方法は、フザリウム・グラミネアルム(Fusarium graminearum)由来PADの過剰産生を通じたものである。後者の生成経路を可能にするために、フザリウム(Fusarium)由来ペプチジルアルギニンデイミナーゼのユニークな配列情報が必須である。より好ましくは、コードする遺伝子の全ヌクレオチド配列を利用できなくてはならない。
【0021】
新たに同定された分泌PADを大量にかつ比較的純粋な形態で生成する改善された手段は、組み換えDNA技術を使用して、フザリウム(Fusarium)がコードする酵素の過剰産生を通じたものである。これを行う好ましい方法は、食品等級宿主微生物中におけるこのような分泌PADの過剰産生を通じたものである。よく知られている食品等級微生物としては、アスペルギルス(Aspergilli)、トリコデルマ(Trichoderma)、ストレプトミセス(Streptomyces)、桿菌(Bacilli)、およびサッカロミセス(Saccharomyces)およびクリヴェロミセス(Kluyveromyces)などの酵母が挙げられる。これを行うさらにより好ましい方法は、アスペルギルス(Aspergillus)などの食品等級真菌中における、フザリウム(Fusarium)由来分泌PADの過剰産生を通じたものである。最も好ましいのは、その中でPADコード遺伝子のコドン使用が、使用される食品等級発現宿主のために最適化されている、食品等級真菌中における分泌PADの過剰産生である。一般に後者の最適化経路を可能にするために、分泌PADのユニークな配列情報が望ましい。より好ましいのは、PADコード遺伝子の全ヌクレオチド配列が利用できることである。ひとたび新しい酵素が大量にかつ比較的純粋な形態で利用できるようになれば、シトルリン含有食物タンパク質またはこのようなシトルリン含有食物タンパク質の加水分解産物が、食品等級で経済的なやり方によって生産できる。好ましくはこのようなシトルリン含有食物タンパク質は、高率のタンパク質結合アルギニンを含有する食物タンパク質、またはこのような食物タンパク質の加水分解産物から得られる。本発明に従ったPADのための好ましい基質タンパク質は、少なくとも10:1(mol/mol)のアルギニン対シトルリン比を有し、好ましくはこのような基質タンパク質は、少なくとも100:1(mol/mol)のアルギニン対シトルリン比を有し、最も好ましくは基質タンパク質はシトルリンを全く含有しない。本発明に従ったPADのための好ましい基質タンパク質は、少なくとも3mol%のタンパク質結合アルギニンを含有し、より好ましくはそれらは少なくとも6mol%のタンパク質結合アルギニンを含有する。このような基質タンパク質の例は、(脱脂)乳タンパク質、乳清タンパク質、カゼインまたは卵タンパク質などの動物起源からの市販食物タンパク質である。その他のこのような基質タンパク質の例は、穀物タンパク質、ジャガイモタンパク質、ダイズタンパク質、エンドウマメタンパク質、米タンパク質、エンドウマメタンパク質などの植物起源からの市販食物タンパク質、ならびにルピナス、ゴマ、ヤシの実などのアルギニンに富むことが知られているその他の植物起源からのタンパク質である。穀物タンパクの例は、小麦またはトウモロコシまたは例えば小麦グルテンなどのその画分である。微生物タンパク質の例は、肉代替品のための酵母抽出物または単細胞タンパク質である。これらのアルギニンに富むタンパク質またはペプチド中における特異的アミノ酸の相対的欠乏を代償するために、シトルリン含有タンパク質、ペプチドまたは加水分解産物の栄養組成物は、シトルリン化材料中で不足するアミノ酸に比較的富む、選択された遊離アミノ酸またはタンパク質、ペプチドまたは加水分解産物を添加することにより最適化できる。シトルリン含有タンパク質、ペプチドまたは加水分解産物の栄養価を増強するための好ましいアミノ酸は、システイン、ヒスチジン、イソロイシン、グルタミン、およびリジンである。代案としてはシトルリン含有タンパク質、ペプチドまたは加水分解産物をカゼイン、ジャガイモ、小麦またはダイズタンパク質などの比較的これらのアミノ酸に富むタンパク質源から得られたタンパク質、ペプチドまたは加水分解産物と混合できる。特に本発明の方法は酵素を修飾するのに有用であり、活性または安定性などの特徴が変更された酵素がもたらされる。
【0022】
哺乳類の組織を使用する場合、この組織は好ましくは非ヒト組織である。哺乳類のタンパク質を使用する場合、このタンパク質は好ましくは血液タンパク質、神経組織、脳、臓器、筋肉または毛髪ではない。好ましくは本発明に従って使用される基質タンパク質は、植物タンパク質、脱脂(乳)タンパク質、乳清タンパク質、カゼインタンパク質、ゼラチンタンパク質、卵タンパク質または微生物のタンパク質である。
【0023】
別の応用は、このようなシトルリン含有食物タンパク質の加水分解産物の創出である。これらの加水分解産物は、当該技術分野で知られている方法に従って生成できる。代案としては動物または植物起源のタンパク質を最初に加水分解して、タンパク質加水分解産物を得ることができ、引き続いてこの加水分解産物を本発明に従ったPADと共にインキュベートできる。
【0024】
より濃縮された形態でペプチド結合シトルリンを提供するために、PAD酵素のための基質として、アルギニン濃縮加水分解産物を使用できる。このアプローチでは、例えば米タンパク質またはエンドウマメタンパク質などのアルギニンに富むタンパク質源を、最初にサブチリシン(EC3.4.21.62)またはケカビペプシン(mucorpepsin)(EC3.4.23.23)またはプロリン−特異的プロテアーゼ(EC3.4.21.26)などの適切なエンドプロテアーゼによって加水分解し、次にクロマトグラフィーを使用して、得られた加水分解産物をアルギニン含有ペプチドに濃縮する。このようなクロマトグラフィー分離技術では、特定のpH条件下のアルギニン残基の正電荷を利用する。ペプチド精製におけるこれらの特徴の使用の実用的背景は、例えば「タンパク質精製ハンドブック(Protein Purification Handbook)」(発行元:アマシャム・ファーマシア・バイオテック(Amersham Pharmacia Biotech)、現在はベルギー国ディーゲムのGEヘルスケア・バイオサイエンシズ(GE Healthcare Bio−Sciences(Diegem,Belgium))にある。次に得られたアルギニン濃縮ペプチドをPAD酵素と共にインキュベートして、本発明に従ったシトルリンを含んでなる加水分解産物を得る。アルギニン濃縮加水分解産物を得ることに向けた別のアプローチでは、アルギニンに富むタンパク質源を最初にアルギニン−特異的エンドプロテアーゼトリプシン(EC3.4.21.4)と共にインキュベートする。加水分解後、より大きな非加水分解ペプチドが沈殿してより小さいアルギニンに富むペプチドが溶液中に残るように、インキュベーションのpHをタンパク質基質の水溶性が最小となる値に調節する。例えば中性に近いpHでは、エンドウマメタンパク質の溶解度はかなり低い。エンドウマメタンパク質の懸濁液を中性pHでトリプシンと共にインキュベートすることで(最大活性のためにはトリプシンは中性に近いpHを必要とする)、アルギニン残基に富むエンドウマメタンパク質の部分のみが溶液中に入り、引き続くデカンテーションまたは濾過ステップによって、それぞれ上清または濾液中にアルギニンに富むペプチドが生じる。この上清または濾液に本発明に従ったPADを添加することで、所望の高濃度シトルリンを含有するペプチドが生じる。
【0025】
本発明は、本タンパク質またはタンパク質加水分解産物を含んでなる新規な栄養補給食品組成物に関する。栄養補給食品組成物は、このような治療タンパク質を必要とする対象に、本発明のペプチドまたはタンパク質加水分解産物を投与するステップを含んでなる、例えば高血圧予防のため、および栄養失調からまたは腸管疾患から回復するための活性成分として、タンパク質加水分解産物を含んでなる。
【0026】
シトルリンを含んでなる本タンパク質、タンパク質加水分解産物、ペプチドまたはペプチド混合物は、固形製品、半固形製品(ペースト)または飲料などの液体製品などのあらゆる適切な形態で使用できる。例えば高レベルのシトルリンを含んでなる製品は、栄養補助食品としてまたは食物、飲料、飼料またはペットフード成分として使用される。高レベルのシトルリンを含んでなる製品はまた、ローション、ゲルまたはエマルジョンの形態の局所塗布をはじめとするパーソナルケア用途の形態で使用できる。これらの全ての用途で、タンパク質、ペプチドまたは加水分解産物は、ビタミンAまたはビタミンCなどのビタミンを含んでなるマルチビタミン製剤と共に、亜鉛などの微量元素と共に、および正常な代謝機能の維持に必須であるが体内で合成されないミネラルと共に、共調合されてもよい。さらにシトルリン化タンパク質、ペプチドまたは加水分解産物は、特定脂肪酸、グルタミンなどの特定アミノ酸、または特定アミノ酸に富むタンパク質またはタンパク質加水分解産物と組み合わせてもよい。またレスベラトロールまたはEGCGなどのポリフェノール、グリコシル化および脱グリコシル化ダイズイソフラボン、プレバイオティクスまたはプロバイオティクスとの組み合わせが予見される。
【0027】
栄養補給食品という用語は、本明細書での用法では、栄養および製薬分野用途の双方における有用性を意味する。したがって新規な栄養補給食品組成物は、食物および飲料への栄養補給剤としての、およびカプセルまたは錠剤などの固形製剤、または溶液または懸濁液などの液体製剤であってもよい、経腸的または非経口用途のための医薬製剤または薬物としての用途がある。上記から明らかなように、栄養補給食品組成物という用語は、本ペプチド含有組成物と、場合により炭水化物、ならびに例えば上記シトルリン化タンパク質、ペプチドまたは加水分解産物を含んでなる栄養補助食品などの栄養補助組成物を含んでなる、食物および飲料もまた含んでなる。
【0028】
栄養補助食品という用語は、本明細書での用法では、食餌への栄養補給を意図する「食餌性成分」を含有する、口から摂取される製品を意味する。これらの製品中の「食餌性成分」は、ビタミンと、ミネラルと、薬草またはその他の植物と、アミノ酸と、酵素、臓器組織、小腺、および代謝産物などの物質とを含んでもよい。栄養補助食品はまた、抽出または濃縮でき、錠剤、カプセル、ソフトジェル、ジェルキャップ、液体、または粉末などの多くの形態であってもよい。それらはまた、バーなどのその他の形態であることができるが、その場合、栄養補助食品のラベルの情報は、一般に製品を従来の食物として、または食事または食餌の唯一のアイテムとして表示しない。加水分解産物、タンパク質加水分解産物または加水分解タンパク質とは、基質タンパク質のタンパク質分解加水分解によって形成された生成物を意味する。好ましくは加水分解は、酵素的加水分解である。可溶性加水分解産物は、タンパク質加水分解産物の(水)可溶性画分であり、それはまた本明細書で、可溶性ペプチド含有組成物、または可溶性ペプチドを含んでなる組成物、またはタンパク質加水分解産物と可溶性加水分解産物の混合物としても述べられている。
【0029】
「ペプチド」または「オリゴペプチド」は、本明細書においてペプチド結合を通じて連結された少なくとも2つのアミノ酸の鎖と定義される。「ペプチド」および「オリゴペプチド」という用語は、(一般に認識されるように)同義と見なされ、各用語は文脈の必要に応じて同義的に使用できる。「ポリペプチド」または「タンパク質」は、本明細書において30個を超えるアミノ酸残基を含んでなる鎖と定義される。全ての(オリゴ)ペプチドおよびポリペプチドの式または配列は、本明細書では慣行に従ってアミノ−末端からカルボキシ−末端の方向で左から右に記述される。本明細書において使用されるアミノ酸の1文字コードについては、一般に当該技術分野で知られており、Sambrookら著、「分子クローニング実験室マニュアル(Molecular Cloning:A Laboratory Manual)」、第2版、Cold Spring Harbor Laboratory、Cold Spring Harbor Laboratory Press、Cold Spring Harbor,NY、1989年、にある。
【0030】
タンパク質生合成において細胞によって使用される20個の「標準」アミノ酸があり、それらは一般遺伝コードによって規定される。本明細書では、アミノ酸とはこれらの20個の「標準」アミノ酸およびシトルリンを意味する。
【0031】
好ましくは単離されたジペプチドシトルリン−アルギニンは、本発明の一部ではない。しかしこのジペプチドを含んでなる加水分解産物、およびこのような加水分解産物の使用は、本発明のさらなる実施態様である。栄養失調からまたは腸管疾患から回復するための活性成分としての単離されたジペプチドシトルリン−アルギニンの使用もまた、本発明の目的である。その中でアイリッシュモスまたは抽出物、またはアイリッシュモスから得られたその他の製品が使用される製品は、本発明の一部ではない。
【0032】
大量のシトルリンを組み入れたタンパク質またはペプチドは、顕著な利点を提供できる。最も重要には、それらは特殊食品、幼児栄養、または特別な消費者グループのための栄養補給剤などの非医療用途のために、ペプチド結合シトルリンを利用できるようにする可能性を初めて提供する。さらにシトルリン残基を含んでなるタンパク質およびタンパク質加水分解産物は苦味を呈さないので、このようなタンパク質またはペプチドは機能的利点を提供する。さらに本発明に従ったシトルリンは、国の法令によっては許容されないかもしれない遊離シトルリンとは対照的に、ペプチド結合シトルリンとして存在する。その結果、本発明に従った方法は、シトルリンを含有する許容可能な味覚プロフィールがある食物、栄養補給剤、および臨床製品の生産を初めて可能にする。
【0033】
本発明に従ったタンパク質またはペプチドの別の重要な利点は、それらがより低いアレルゲン性を示すかもしれないことである。ミルクおよびそのカゼインおよび乳清タンパク質画分、ならびに例えばダイズ単離物、米タンパク質および小麦グルテンから得られる植物タンパク質画分などの全ての主要な食物タンパク質は、重要な抗原化合物と見なされる。通常タンパク質抗原性は、タンパク質を8〜10個未満のアミノ酸残基を有するペプチドに加水分解することで克服される。しかしこのような高度なタンパク質分解消化によって作り出された加水分解産物は、苦さ、肉汁のような異臭、および浸透圧値増大をはじめとする不都合を示す。タンパク質抗原性の経済的重要性は、幼児および小児における食物アレルギーおよび喘息の罹患率が上昇しているという事実によって例示される。例えば牛乳アレルギーは、3才未満の小児の約2.5%が罹患している。牛乳アレルギーは、生後1ヶ月以内および牛乳の導入1週間以内に遭遇することが多い。異なる程度に加水分解された牛乳タンパク質または牛乳画分が組み込まれた様々な乳児用調製粉乳製品には牛乳アレルギーが予期される。我々は、食物タンパク質を本発明に従ったPADと共にインキュベートすることで、得られたシトルリン化タンパク質またはタンパク質加水分解産物の抗原性またはアレルゲン性が低下することを見いだした。最も重要には、この効果は、損なわれた味または乳化力低下などの高度なタンパク質分解に結びついた悪影響なしに得られた。運動愛好家、高齢者、および食物アレルギーを患っている人々が増えていることを鑑みて、本発明に従った製品には大きな経済的重要性がある。これらの全ての標的グループにとって、改善された味は顕著な心理学的重要性がある側面である。今までに工業規模で経済的な方法で、このような食物または栄養補給剤を生産することは不可能だった。目下工業的に入手できる食物タンパク質および/または食物タンパク質加水分解産物は、タンパク質−またはペプチド結合シトルリン残基を組み込んでいないので、シトルリン残基に富むタンパク質またはタンパク質加水分解産物またはペプチドは新しい。「濃縮」という用語は、タンパク質またはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換することを意味することが意図される。したがって本方法の生成物、すなわちシトルリン化タンパク質、シトルリン化タンパク質の加水分解産物またはシトルリン化加水分解産物中では、存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%はシトルリン残基に転換している。存在するアルギニンまたはシトルリン残基量を確立するために使用されるアミノ酸分析法は、本明細書の材料と方法セクションで特定されている。アミノ酸分析中に典型的に使用されて、遊離アミノ酸を解放する酸加水分解過程の人工産物として新たに発生したシトルリン残基の一部がオルニチン残基に転換することに留意することが重要である。存在するシトルリン残基のレベルを計算するために、存在するシトルリンおよびオルニチン残基のレベルを合計しなくてはならない。
【0034】
シトルリン含有タンパク質加水分解産物を得るために、シトルリン含有タンパク質を酵素的に加水分解できる。代案としては既存のタンパク質加水分解産物、すなわちペプチド結合シトルリンを含まない加水分解産物を本発明に従ったPADと共にインキュベートして、ペプチド結合シトルリンを含んでなるタンパク質加水分解産物を得ることができる。タンパク質加水分解産物は、当該技術分野で知られている加水分解法を使用して生成できる。好ましくはこのような加水分解産物は、5〜50の間、より好ましくは10〜35の間の加水分解度(DH)を有する。DH値を確立するための方法は、本明細書の材料と方法セクションで特定されている。本発明に従ったシトルリンを含んでなるタンパク質加水分解産物は、幼児および臨床栄養物、治療用食餌ならびに民生用食餌、およびスポーツ栄養において使用できる。このようなシトルリン化タンパク質、タンパク質加水分解産物またはペプチドを組み込んだ食物または食餌または臨床製品もまた新しい。さらにタンパク質−またはペプチド結合シトルリンを含んでなるタンパク質またはタンパク質加水分解産物は、パーソナルケア用途をはじめとする様々な局所用途において、および動物およびペットのための栄養製品において使用できる。
【0035】
本発明に従ったシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、多数の新しい驚くべき用途で使用できる。基本的にシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質水解物は、遊離アルギニンの補充が有益であることが示されている全ての用途で使用できる。したがってアルギニン欠乏状態によって特徴づけられる全ての病状において、本発明に従ったシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、このアルギニン欠乏状態を低下させることが予期される。一般にシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、栄養失調から、または短腸症候群などの腸管疾患から、または加齢の結果としてのタンパク質−エネルギー栄養失調から回復中の個人においてタンパク質代謝を維持する利点を提供する。さらにシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、胃腸管の健康を維持する上で、重要な役割を果たすことができる。アルギニンならびにシトルリンは酸化窒素生成、ひいては血管拡張の前駆物質であるので、これらのアミノ酸の適切な供給は、多様な血管疾患のリスクを予防する上で重要な役割を果たす。これは科学文献で、特に末梢動脈疾患、移植冠動脈疾患および喘息について実証されている。したがってシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物は、これらの疾患を予防する食餌への付加として、ならびにストレス性潰瘍の予防、またはストレス潰瘍治癒の改善において特に興味深い。さらに本発明に従ったタンパク質、ペプチド、およびタンパク質加水分解産物は、例えばアテローム動脈硬化、狭心症、高血圧、冠動脈心疾患の治療的措置において、II型糖尿病でインシュリンおよびグルコース濃度を低下させるのに、およびHIV感染症、火傷、外傷、および癌治療においても特に有用である。筋肉減少症の治療のためのシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物の使用は、特に注目に値する。さらに術前食餌中でのシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物の使用は、特にストレス条件下における免疫学的状態を改善して、患者の微小循環低灌流を回復させる。敗血症は高死亡率を伴う主要な健康問題であるので、敗血症の予防におけるシトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物を組み込んだ食餌の有益な効果もまた言及に値する。シトルリンを含んでなるタンパク質、ペプチドまたはタンパク質加水分解産物はまた、それらの血中アンモニア低下効果のために、例えば遺伝性酵素欠陥を患っている患者において乱された尿素回路の症例で使用でき、または腎不全を患っている人々の腎臓の糸球体機能に対するストレスを低下させるのに使用できる。本発明に従ったタンパク質、ペプチド、およびタンパク質加水分解産物はまた、例えば運動選手または運動愛好家などの非医学的ニーズがある消費者向け製品でも使用できる。特にシトルリンは、eNOS NO産生を刺激することによって血流を改善することが予期されるので、筋肉血流改善を通じて運動性能が改善できることが予期される。より最近では、NO合成の阻害が、高脂血症および脂肪付着を引き起こすことを示唆する証拠が集まっている。したがって本発明に従ったペプチド結合シトルリンでの食餌補給は、肥満の人およびペットなどの動物における代謝症候群の予防および治療の助けになるかもしれない。このカテゴリーの製品としては、疲労感に対処して、身体持久力および長時間の高強度運動後の回復を増強する強化果汁およびスポーツドリンクが挙げられる。別の重要な用途は、例えばアレルギーの幼児のため、アレルギー反応を発症している乳児のため、または非アレルギーの幼児のための乳児用調製粉乳中での本発明に従ったタンパク質、ペプチド、およびタンパク質加水分解産物の使用であり、シトルリン化タンパク質またはペプチドは、牛乳感作を遅延させまたは予防する働きをする。局所用途では、シトルリン化タンパク質またはペプチドは意外にもヒドロキシルラジカルを除去し、活性酸素種を制御してそれを相殺するのに効果的である。さらに本発明に従った活性PADは直接的局所塗布のために使用でき、表皮の角質化を改善し、または乾癬の徴候に対処する。活性酵素を皮膚に塗布する有用なやり方は、米国特許第6,117,433号明細書で述べられている。本発明に従った活性PADの別の新しい用途はあらゆる種類の生地中への組み込みであり、観察されているように、これは得られたベークド製品の風味と匂いを増大させる。本発明に従った活性PADはまた、発酵食品、およびワイン、ビールおよび蒸留酒などの飲料中の尿素量を低下させることで、このような発酵食品および飲料中の発癌性物質エチルカルバメートのレベルを低下させるのに使用できる。
【0036】
ペプチジルアルギニンデイミナーゼ活性を有する本発明のポリペプチドは、単離形態であってもよい。本明細書において定義されるように、単離されたポリペプチドは、その他の非ペプチジルアルギニンデイミナーゼポリペプチドを本質的に含まない内因的に生成されたまたは組み換えされたポリペプチドであり、SDS−PAGEによる判定で、典型的に少なくとも約20%純粋、好ましくは少なくとも約40%純粋、より好ましくは少なくとも約60%純粋、さらにより好ましくは少なくとも約80%純粋、なおもより好ましくは約90%純粋、および最も好ましくは約95%純粋である。ポリペプチドは、遠心分離およびクロマトグラフィー法、または粗製溶液から純粋タンパク質を得るための当該技術分野で知られているあらゆるその他の技術によって単離されてもよい。ポリペプチドは、ポリペプチドの意図される目的を妨げないキャリアまたは希釈剤と混合されてもよく、したがってこの形態のポリペプチドは、なおも単離されていると見なされると理解される。それは一般に調製品中のポリペプチドを構成し、その中では調製品中のタンパク質重量の20%超、例えば30%、40%、50%、80%、90%、95%または99%超が本発明のポリペプチドである。
【0037】
好ましくは本発明のポリペプチドは、ペプチジルアルギニンデイミナーゼ活性がある酵素をコードする遺伝子を有する微生物から得られる。より好ましくは本発明のポリペプチドは、この微生物から分泌される。さらにより好ましくは微生物は真菌であり、および最適には糸状菌である。それゆえに好ましい生物は、フザリウム・グラミネアルム(Fusarium graminearum)種のものなどのフザリウム(Fusarium)属のものである。
【0038】
第1の実施態様では、本発明は、配列番号6の1〜640番目のアミノ酸(すなわちポリペプチド)に対して、少なくとも30%、好ましくは少なくとも40%、好ましくは少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、なおもより好ましくは少なくとも95%、および最も好ましくは少なくとも97%の程度のアミノ酸配列同一性を有するアミノ酸配列を有して、ペプチジルアルギニンデイミナーゼ活性を有する単離されたポリペプチドを提供する。
【0039】
本発明の目的で、2つ以上のアミノ酸配列間の同一性の程度は、BLAST Pタンパク質データベース検索プログラム(Altschulら著、1997年、Nucleic Acids Research 25:3389〜3402頁)によって、マトリックスBlosum 62および期待閾値10を使用して判定される。
【0040】
本発明のポリペプチドは、配列番号6に記載のアミノ酸配列、または実質的に相同的な配列、またはペプチジルアルギニンデイミナーゼ活性を有するどちらかの配列の断片を含んでなってもよい。一般に配列番号6に記載の天然アミノ酸配列が好ましい。
【0041】
本発明のポリペプチドはまた、配列番号6のポリペプチドの天然変異型、または種相同体を含んでなってもよい。
【0042】
変異型は、例えば真菌、細菌、酵母または植物細胞内で天然に生じるポリペプチドであり、変異型はペプチジルアルギニンデイミナーゼ活性および、配列番号6のタンパク質と実質的に類似の配列を有する。「変異型」という用語は、配列番号6のペプチジルアルギニンデイミナーゼと同じ本質的特性または基本的生物学的機能性を有するポリペプチドを指し、対立遺伝子多型を含む。好ましくは変異型ポリペプチドは、配列番号6のポリペプチドと少なくとも同一レベルのペプチジルアルギニンデイミナーゼ活性を有する。変異型は、配列番号6のポリペプチドと同じ株から、または同じ属または種の異なる株からのどちらかの対立遺伝子多型を含む。配列番号6のポリペプチドの変異型の例は、配列番号7に記載される。
【0043】
同様に発明のタンパク質の種相同体は、同様の配列の同等のタンパク質であり、それはペプチジルアルギニンデイミナーゼであり、別の種において天然に生じる。配列番号6のポリペプチド種相同体の例は、配列番号8〜10、13、および14に記載される。
【0044】
変異型および種相同体は、配列番号6のポリペプチドを単離するのに使用される本明細書で述べられている手順を使用して、このような手順を例えば細菌、酵母、真菌または植物細胞などの適切な細胞源に実施して単離できる。また配列番号6のポリペプチドの変異型または種相同体を発現するクローンを得るために、本発明のプローブを使用して、酵母、細菌、真菌または植物細胞からできたライブラリーを探索することも可能である。既知の遺伝子の変異型および種相同体を単離するのに使用できる方法は文献で詳細に述べられており、当業者に知られている。これらの遺伝子を従来の技術によって操作して、その後それ自体が既知である組み換えまたは合成技術によって生成されてもよい、本発明のポリペプチドを作り出すことができる。
【0045】
配列番号6のポリペプチドおよび変異型および種相同体の配列を修飾して、本発明のポリペプチドを提供することもできる。例えば1、2または3から10、20または30個の置換などのアミノ酸置換を行ってもよい。また同一数の欠失および挿入を行ってもよい。修飾されたポリペプチドがそのペプチジルアルギニンデイミナーゼ活性を維持するように、これらの変更をポリペプチドの機能に重要な領域の外で行ってもよい。
【0046】
本発明のポリペプチドは、配列番号6に記載の配列の断片をはじめとする、前述の全長ポリペプチドの断片、およびその変異型の断片を含む。このような断片は、典型的にペプチジルアルギニンデイミナーゼとしての活性を維持する。断片は、少なくとも50、100または200個のアミノ酸の長さであってもよく、またはこのアミノ酸数は、配列番号6に記載の全長配列に達していないくてもよい。
【0047】
通常、本発明のポリペプチドは後述するように組み換え的によって作られるが、それらは必要ならば合成手段によって生成できる。合成ポリペプチドは、例えばそれらの同定または精製を助けるヒスチジン残基またはT7 tagの添加によって、またはそれらの細胞からの分泌を促進するシグナル配列の添加によって修飾してもよい。
【0048】
したがって変異型配列は、配列番号6のポリペプチドが単離された株以外のフザリウム(Fusarium)株に由来するものを含んでなってもよい。変異型は、本明細書で述べられているように、ペプチジルアルギニンデイミナーゼ活性を調べることで、およびクローニングおよび配列決定によって、その他のフザリウム(Fusarium)株から識別できる。ペプチドが配列番号6のペプチジルアルギニンデイミナーゼの基本的生物学的機能性を維持しさえすれば、変異型は、欠失、修飾または単一アミノ酸またはアミノ酸群の付加をタンパク質配列内に含んでもよい。
【0049】
例えば1、2または3から10、20または30個の置換などのアミノ酸置換を行ってもよい。修飾ポリペプチドは、一般にペプチジルアルギニンデイミナーゼとしての活性を維持する。保存的置換を行ってもよく、このような置換については当該技術分野でよく知られている。
【0050】
より短いポリペプチド配列は、本発明の範囲内である。例えば長さが少なくとも50個のアミノ酸、または60、70、80、100、150または200個までのアミノ酸のペプチドは、それが配列番号6のペプチジルアルギニンデイミナーゼの基本的生物学的機能性を実証さえすれば、本発明の範囲内に入ると見なされる。特に、しかし排他的でなく、本発明のこの態様は、タンパク質が完全なタンパク質配列の断片である状況を包含する。
【0051】
本発明はまた、タンパク質アルギニンデイミナーゼ活性を有するポリペプチドをコードするポリヌクレオチドに関し、前記ポリヌクレオチドは、
(a)アミノ酸配列番号11をコードするポリヌクレオチド配列、および
(b)プレタンパク質シグナル配列をコードするポリヌクレオチド配列
を含んでなり、コードされたプレタンパク質シグナル配列は、コードされたプレポリペプチドのアミノ末端に位置し、好ましくは長さが15〜30個のアミノ酸である。
【0052】
本発明では、配列番号11のPAD共通配列を含有するポリペプチドは、本発明の範囲内である。好ましくは配列番号11のPAD共通配列の双方を含有して、シグナル配列を含有するプレタンパク質としてコードされるポリペプチドは、本発明の一部である。本発明では、関心のあるタンパク質が増殖培地中に能動的に分泌されることは、特に妥当である。分泌タンパク質は、常態では当初はプレタンパク質として合成され、引き続いて分泌過程においてプレ配列(シグナル配列)が除去される。分泌過程は、原核生物および真核生物において基本的に同様である。能動的に分泌されたプレタンパク質は膜を通り抜け、特異的シグナルペプチダーゼによってシグナル配列が除去され、成熟タンパク質が(再度)折りたたまれる。シグナル配列についてもまた、一般構造が認識できる。分泌のためのシグナル配列はプレタンパク質のアミノ−末端に位置し、一般に長さが15〜30個のアミノ酸である。アミノ−末端は、好ましくは正に帯電したアミノ酸を含有し、好ましくは非酸性アミノ酸を含有しない。この正に帯電した領域が、膜のリン脂質の負に帯電した頭部基と相互作用すると考えられる。この領域に疎水性の膜通過コア領域が続く。この領域は、一般に長さが10〜20個のアミノ酸であり、主に疎水性アミノ酸から成る。荷電したアミノ酸は、常態ではこの領域内に存在しない。膜貫通領域にシグナルペプチダーゼ認識部位が続く。認識部位は、small−X−smallを好むアミノ酸からなる。小型アミノ酸は、アラニン、グリシン、セリンまたはシステインであることができる。Xはあらゆるアミノ酸であることができる。このような規則を使用して、真核生物および原核生物からこのようなシグナル配列を認識できるアルゴリズムが記述されている(Bendtsen、Nielsen、von Heijne、およびBrunak著、(2004年)J.Mol.Biol.、340:783〜795頁)。タンパク質中のシグナル配列を計算および認識するSignalPプログラムは、一般に利用できる(http://www.cbs.dtu.dk/services/SignalP/)。
【0053】
本発明に関連があるのは、配列決定された遺伝子の推定タンパク質配列から、シグナル配列が認識できることである。SignalPプログラムを使用してシグナル配列が予測されるタンパク質を遺伝子がコードする場合、このタンパク質が分泌される確率は高い。したがって本発明の目的は、SignalPプログラムによって検出されるシグナル配列の存在と合わせて、配列番号11のコンセンサスを使用して、PAD活性を有する新しいタンパク質を発見する新しい方法を提供することである。
【0054】
第2の実施態様では、本発明は、ペプチジルアルギニンデイミナーゼ活性を有して、低ストリンジェンシー条件下、より好ましくは中度ストリンジェンシー条件、および最も好ましくは高ストリンジェンシー条件で、(I)配列番号3の核酸配列または少なくとも配列番号3のc−末端部分を含んでなるが、配列番号3の塩基の全てに満たない、または異なる塩基を有する核酸断片と、または(ii)鎖配列番号3に相補的な核酸ストランドとハイブリダイズするまたはハイブリダイズできるポリヌクレオチドによってコードされる、単離されたポリペプチドを提供する。
【0055】
「ハイブリダイズできる」という用語は、本発明の標的ポリヌクレオチドが、プローブとして使用される核酸(例えば配列番号3に記載のヌクレオチド配列、またはその断片、または配列番号3の相補体、またはその断片)と、バックグラウンドを顕著に超えるレベルでハイブリダイズできることを意味する。本発明はまた、本発明のペプチジルアルギニンデイミナーゼをコードするポリヌクレオチド、ならびにそれに相補的なヌクレオチド配列も含む。ヌクレオチド配列は、ゲノムDNA、合成DNAまたはcDNAをはじめとするRNAまたはDNAであってもよい。好ましくはヌクレオチド配列はDNA、最も好ましくはゲノムDNA配列である。典型的に本発明のポリヌクレオチドは、選択的条件下で、配列番号3のコード配列またはコード配列相補体とハイブリダイズできる近接するヌクレオチド配列を含んでなる。このようなヌクレオチドが、当該技術分野でよく知られている方法に従って合成できる。
【0056】
本発明のポリヌクレオチドは、配列番号3のコード配列またはコード配列相補体と、バックグラウンドを顕著に超えるレベルでハイブリダイズできる。バックグラウンドハイブリダイゼーションは、例えばcDNAライブラリー中に存在するその他のcDNAのために起こるかもしれない。本発明のポリヌクレオチドと配列番号3のコード配列またはコード配列相補体間の相互作用によって生じるシグナルレベルは、典型的に、その他のポリヌクレオチドと配列番号3のコード配列間の相互作用強度の少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも50倍、およびさらにより好ましくは少なくとも100倍である。相互作用強度は、例えば32Pを使用して、例えばプローブを放射標識して測定されてもよい。選択的ハイブリダイゼーションは、典型的に、低ストリンジェンシー(約40℃で0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム)、中程度ストリンジェンシー(例えば約50℃で0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム)または高ストリンジェンシー(例えば約60℃で0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム)条件を使用して達成されてもよい。
【0057】
本発明のポリヌクレオチドはまた、配列番号6のポリペプチドまたはその変異型をコードできる合成遺伝子を含む。遺伝子のコドン使用頻度を生産宿主中の好ましいバイアスに適応させることが好ましい場合もある。合成遺伝子をデザインおよび設計する技術は、一般に利用できる(例えばhttp://www.dnatwopointo.com/)。
【0058】
[修飾]
本発明のポリヌクレオチドは、DNAまたはRNAを含んでなってもよい。それらは一本鎖または二本鎖であってもよい。それらはまた、ペプチド核酸をはじめとする合成または修飾ヌクレオチドをその中に含むポリヌクレオチドであってもよい。いくつかの異なるタイプのポリヌクレオチド修飾が当該技術分野で知られている。これらとしては、メチルホスホン酸およびホスホロチオエート主鎖、および分子の3’および/または5’末端へのアクリジンまたはポリリジン鎖の付加が挙げられる。本発明の目的で、本明細書で述べられているポリヌクレオチドは、当該技術分野で利用できるあらゆる方法で修飾されてもよいものと理解される。
【0059】
当業者は、日常的技術を使用して、本発明のポリヌクレオチドによってコードされるポリペプチド配列に影響しないヌクレオチド置換を行い、その中で本発明のポリペプチドが発現される、あらゆる特定の宿主生物のコドン使用頻度を反映させてもよいものと理解される。
【0060】
配列番号3のコード配列は、例えば1、2または3から10、25、50、100個またはそれ以上の置換などのヌクレオチド置換によって修飾されてもよい。配列番号3のポリヌクレオチドは、1つ以上の挿入および/または欠失によって、および/または一端または両端の延長によって、代案としてまたは追加的に修飾されてもよい。修飾ポリヌクレオチドは、一般に、ペプチジルアルギニンデイミナーゼ活性を有するポリペプチドをコードする。例えば後でポリペプチドについて考察するように、縮重置換を行ってもよく、および/または修飾配列の翻訳時に保存的アミノ酸置換をもたらす置換を行ってもよい。
【0061】
[相同体]
配列番号3のDNAコード配列の相補体と選択的にハイブリダイズできるヌクレオチド配列は本発明に含まれ、少なくとも60、好ましくは少なくとも100、より好ましくは少なくとも200個の近接するヌクレオチドにわたり、または最も好ましくは配列番号3の全長の領域にわたって、一般に配列番号3のコード配列と少なくとも50%または60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも98%または少なくとも99%の配列同一性を有する。同様に能動的ペプチジルアルギニンデイミナーゼをコードして、配列番号3のDNAコード配列の相補体断片と選択的にハイブリダイズできるヌクレオチドもまた、本発明で受け入れられる。60個を超える、好ましくは100個を超えるヌクレオチドが少なくとも80%または90%同一であり、より好ましくは200個を超えるヌクレオチドが少なくとも90%同一である、配列番号3の核酸配列のC−末端断片は、本発明に包含される。
【0062】
前述の同一性程度および最小サイズのあらゆる組み合わせを使用して本発明のポリヌクレオチドを定義してもよく、よりストリンジェントな組み合わせ(すなわちより長い範囲にわたるより高い同一性)が好ましい。したがって例えば60個を超える、好ましくは100個を超えるヌクレオチドが少なくとも80%または90%同一であるポリヌクレオチドは本発明の一態様を形成し、200個を超えるヌクレオチドが少なくとも90%同一であるポリヌクレオチドについても同様である。
【0063】
UWGCGパッケージは、同一性を計算するのに使用してもよい(例えばそのデフォルト設定で使用する)BESTFITプログラムを提供する。
【0064】
PILEUPおよびBLAST Nアルゴリズムもまた、(例えばそれらのデフォルト設定で同等のまたは対応する配列を同定するなど)配列同一性を計算し、または配列を位置合わせするのに使用できる。
【0065】
BLAST分析を実施するためのソフトウェアは、国立バイオテクノロジー情報センター(http://www.ncbi.nlm.nih.gov/)を通じて公的に利用可能である。このアルゴリズムは最初に、データベース配列中の同じ長さのワードと配列比較すると、ある正値の閾値スコアTとマッチするかそれを満たすかのいずれかである、クエリー配列中の長さWの短いワードを同定することで、高採点配列対(HSP)を同定することを伴う。Tは近傍単語スコア閾値と称される。これらの初期近傍単語ヒットは、それらを含有するHSPを見つけるために検索を開始するシード(seed)として機能する。ワードヒットは、累積アラインメントスコアが増大できる限り、各配列に沿って両方向に延長される。各方向におけるワードヒットのための延長は、累積アラインメントスコアがその最大達成値からXの量だけ低下した場合、1つ以上の負の採点の残基アラインメントの蓄積のために累積スコアがゼロ以下になった場合、またはどちらかの配列の終わりに達した場合、停止する。BLASTアルゴリズムパラメーターのW、T、およびXは、配列比較の感受性およびスピード(speed)を決定する。BLASTプログラムは、デフォルトとしてワード長さ(W)11、BLOSUM 62採点マトリックス配列比較(B)50、期待値(E)10、M=5、N=4、および両ストランドの比較を使用する。
【0066】
BLASTアルゴリズムは、2つの配列間の類似性の統計学的分析を実施する。BLASTアルゴリズムによって提供される類似性の1つの尺度は、それによって2つのヌクレオチドまたはアミノ酸配列間のマッチが偶発する確率の指標を提供する、最少確率和(P(N))である。例えば第1の配列と第2の配列との比較における最少確率和が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、最も好ましくは約0.001未満であれば、配列は別の配列に類似していると見なされる。
【0067】
[プライマーおよびプローブ]
本発明のポリヌクレオチドはプライマーを含み、例えばポリメラーゼ連鎖反応(PCR)プライマーとして、代案の増幅反応のためのプライマーとして、または例えば放射性または非放射性標識を使用して従来の手段によって明示用標識で標識してプローブとして使用してもよく、またはポリヌクレオチドをベクター中にクローンしてもよい。このようなプライマー、プローブ、およびその他の断片は、長さが少なくとも15個、例えば少なくとも20、25、30または40個のヌクレオチドである。それらは長さが典型的に40、50、60、70、100、150、200または300個までのヌクレオチドであり、または配列番号5のコード配列よりも短い数個までのヌクレオチド(5または10個のヌクレオチドなど)でさえある。
【0068】
一般にプライマーは、ヌクレオチド1つずつの所望の核酸配列の段階的製造を伴う、合成手段によって生成される。これを達成するための技術およびプロトコールは、当該技術分野で容易に利用できる。より長いポリヌクレオチドは、例えばPCRクローニング技術を使用して、一般に組み換え手段を使用して生成される。これは(典型的に約15〜30個のヌクレオチドの)プライマー対を作成して、クローンするペプチジルアルギニンデイミナーゼの所望の領域を増幅し、プライマーを酵母、細菌、植物、原核生物または好ましくはフザリウム(Fusarium)株である真菌細胞から得られたmRNA、cDNAまたはゲノムDNAに接触させて、所望領域の増幅に適した条件下でポリメラーゼ連鎖反応を実施し、(例えば反応混合物をアガロースゲル上で精製することで)増幅された断片を単離して、増幅されたDNAを回収することを伴う。増幅されたDNAが適切なクローニングベクター中にクローンできるように、プライマーは適切な制限酵素認識部位を含有するようにデザインされてもよい。このような配列番号3のPAD遺伝子の変異型のPCRクローニングは本発明内で実施され、これらの遺伝子のDNA配列は配列番号4に記載され、推定タンパク質配列は配列番号7に記載される。
【0069】
代案としては、分泌されたペプチジルアルギニンデイミナーゼまたはその変異型のコード領域を包含する合成遺伝子を構築できる。これらの技術を使用して、多くの位置で修飾されながら、なおも同一タンパク質をコードするポリヌクレオチドを好都合にデザインして構築できる。これはコドン使用頻度を好ましい発現宿主に適応できるという利点を有し、したがってこの宿主中でのタンパク質生産性を改善できる。また遺伝子のポリヌクレオチド配列を変更して、mRNAの安定性を改善し、または回転率を低下させることができる。これは所望のタンパク質またはその変異型の改善された発現をもたらすことができる。さらに分泌効率、安定性、タンパク質分解敏感性、最適温度、比活性、またはタンパク質の工業生産または応用に妥当なその他の特性に対して好ましい効果を有する突然変異が、タンパク質配列中で生じるように、ポリヌクレオチド配列を合成遺伝子中で変更できる。合成遺伝子を構築してコドン使用頻度を最適化するサービスを提供する会社が、一般に利用できる。
【0070】
このような技術を使用して、本明細書で述べられているペプチジルアルギニンデイミナーゼ配列をコードするポリヌクレオチドの全部または一部を得てもよい。イントロン、プロモーター、およびトレーラー領域は本発明の範囲内であり、これらもまた真菌、酵母、細菌、植物または原核細胞からのゲノムDNAから開始して、類似様式で(例えば組み換え手段、PCRまたはクローニング技術によって)得てもよい。
【0071】
ポリヌクレオチドまたはプライマーは、明示用標識を保有しても良い。適切な標識としては、32Pまたは35Sなどの放射性同位体、蛍光性標識、酵素標識、またはビオチンなどのその他のタンパク質標識が挙げられる。このような標識を本発明のポリヌクレオチドまたはプライマーに添加してもよく、当業者に知られている技術を使用して検出してもよい。
【0072】
真菌サンプル中で、ペプチジルアルギニンデイミナーゼまたはその変異型を検出または配列決定するための核酸ベースの試験において、標識または非標識ポリヌクレオチドまたはプライマー(またはその断片)を使用してもよい。このような検出試験は、一般に、ハイブリダイズ条件下で、関心のあるDNAを含有することが疑われる真菌サンプルと、本発明のポリヌクレオチドまたはプライマーを含んでなるプローブとを接触させるステップと、サンプルのプローブと核酸との間に形成されたあらゆる二本鎖を検出するステップを含んでなる。検出は、PCRなどの技術を使用することで、または固体担体上にプローブを固定し、プローブとハイブリダイズしないサンプル中のあらゆる核酸を除去し、次にプローブとハイブリダイズするあらゆる核酸を検出することで達成してもよい。代案としてはサンプル核酸を固体担体上に固定化し、プローブをハイブリダイズして、あらゆる非結合プローブの除去後に、このような担体に結合したプローブ量を検出してもよい。
【0073】
本発明のプローブは、好都合には、適切な容器内に試験キットの形態で包装されてもよい。そのためにキットがそのためにデザインされたアッセイ形態がこのような結合を必要とする場合、このようなキット中でプローブが固体担体に結合していてもよい。キットはまた、プローブするサンプルを処理し、サンプル中の核酸にプローブをハイブリダイズするための適切な試薬、対照試薬、説明書なども含有する。プローブおよび本発明のポリヌクレオチドはまた、微量検定法で使用してもよい。
【0074】
好ましくは本発明のポリヌクレオチドは、ポリペプチドと同じ、真菌など、特にフザリウム(Fusarium)属の真菌である生物から得ることが可能である。
【0075】
[ポリヌクレオチドの生成]
配列番号3と100%の同一性を有さないが、本発明の範囲内に入るポリヌクレオチドをいくつかの方法で得ることができる。したがって本明細書で述べられているペプチジルアルギニンデイミナーゼ配列の変異型を、例えば本発明のポリペプチドの起源として考察されたものなどの一連の生物からできたゲノムDNAライブラリーを探査して得てもよい。さらにその他のペプチジルアルギニンデイミナーゼの真菌、植物または原核生物相同体を得てもよく、このような相同体およびその断片は、一般に配列番号3とハイブリダイズすることができる。このような配列は、その他の種からのcDNAライブラリーまたはゲノムDNAライブラリーを探査して得てもよく、このようなライブラリーを(先に述べたように)低、中程度から高ストリンジェンシー条件下で、配列番号3の全部または一部を含んでなるプローブで探査する。配列番号3の全部または一部を含んでなる核酸プローブを使用して、本発明のポリペプチドの起源として述べたものなどのその他の種からのcDNAまたはゲノムのライブラリーを探査してもよい。
【0076】
種相同体はまた、保存アミノ酸配列をコードする変異型および相同体中の配列を標的とするようにデザインされたプライマーを使用する縮重PCRを使用して得てもよい。プライマーは1つ以上の縮重位置を含有でき、既知の配列に対する単一配列プライマーがある配列のクローニングのために使用されるものよりも、低いストリンジェンシー条件で使用される。PADの種相同体を得るための好ましい方法は、配列番号11に記載の共通配列をコードする配列を標的とするプライマーをデザインすることである。
【0077】
代案としては、このようなポリヌクレオチドは、ペプチジルアルギニンデイミナーゼ配列またはその変異型の部位特異的変異誘発によって得てもよい。これは例えば、その中でポリヌクレオチド配列が発現される特定の宿主細胞のためにコドン選択を最適化するのに、配列に対するサイレントなコドン変化が必要とされる場合に有用かもしれない。制限酵素認識部位を導入するため、またはポリヌクレオチドによってコードされるポリペプチドの特質または機能を変更させるために、その他の配列変化を加えてもよい。
【0078】
本発明は、本発明のポリヌクレオチドおよびその相補体を含んでなる二本鎖ポリヌクレオチド含む。
【0079】
本発明はまた、上述の本発明のポリペプチドをコードするポリヌクレオチドを提供する。このようなポリヌクレオチドは、本発明のポリペプチドの組み換え生産のための配列として有用であるので、それらが配列番号3の配列とハイブリダイズできる必要はないが、これは一般に望ましい。さもなければこのようなポリヌクレオチドは、所望ならば標識されて、使用され、上述のように作成されてもよい。
【0080】
[組み換えポリヌクレオチド]
本発明はまた、クローニングおよび発現ベクターをはじめとする、本発明のポリヌクレオチドを含んでなるベクターも提供し、別の態様ではこのようなベクターを生育させ、例えば本発明のポリペプチド、または本発明の配列によってコードされるポリペプチドの発現が起きる条件下で、適切な宿主細胞に形質転換または形質移入する方法も提供する。本発明のポリヌクレオチドまたはベクターを含んでなる宿主細胞もまた提供され、ここでポリヌクレオチドは宿主細胞ゲノムにとって異種である。「異種」という用語は、通常宿主細胞に対し、ポリヌクレオチドが宿主細胞ゲノム中で自然発生しないこと、またはポリペプチドが細胞によって自然に生成されないことを意味する。好ましくは宿主細胞は、酵母細胞、例えばクリヴェロミセス(Kluyveromyces)、ピチア(Pichia)、ハンゼヌラ(Hansenula)またはサッカロミセス(Saccharomyces)属の酵母細胞であり、または、例えばアスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)またはフザリウム(Fusarium)属の糸状菌細胞である。
【0081】
[ベクター]
その中に本発明の発現カセットが挿入されるベクターは、好都合には組み換えDNA手順を施してもよい、あらゆるベクターであってもよく、ベクターの選択は、その中にそれが導入される宿主細胞に左右されることが多い。したがってベクターは自律的に複製するベクターであってもよく、すなわちプラスミドなど、その複製が染色体の複製から独立している、染色体外の実体として存在するベクターであってもよい。代案としてはベクターは、宿主細胞に導入されると宿主細胞ゲノムに組み込まれ、それが組み込まれた染色体と共に複製されるものであってもよい。
【0082】
好ましくは本発明のポリヌクレオチドがベクター中にある場合、それは宿主細胞によるコード配列の発現を提供できる制御配列と作動的に連結し、すなわちベクターは発現ベクターである。「作動的に連結する」という用語は、述べられている構成要素が、意図される様式でそれらが機能できるようになる関係にある並置位置を指す。コード配列と「作動的に連結する」プロモーター、エンハンサーまたはその他の発現調節シグナルなどの制御配列は、生成条件下でコード配列の発現が達成されるように配置される。
【0083】
例えばプラスミド、コスミド、ウイルスまたはファージベクターの場合、ベクターには複製起点、場合によりポリヌクレオチド発現のためのプロモーター、および場合によりエンハンサーおよび/またはプロモーターのレギュレーターが備わっていてもよい。ターミネーター配列が、ポリアデニル化配列として存在してもよい。ベクターは、例えば細菌プラスミドの場合はアンピシリン抵抗性遺伝子、または哺乳類ベクターではネオマイシン抵抗性遺伝子などの1つ以上の選択可能なマーカー遺伝子をを含有してもよい。ベクターは、例えばRNA生成のために生体外で使用されてもよく、または宿主細胞を形質移入または形質転換するのに使用できる。
【0084】
ポリペプチドをコードするDNA配列は、好ましくはその中でDNA配列と、宿主細胞内におけるDNA配列の発現を指示できる発現シグナルとが作動的に連結する発現コンストラクトの一部として、適切な宿主中に導入される。発現コンストラクトによる適切な宿主の形質転換のためには、当業者によく知られている形質転換手順が利用できる。発現コンストラクトは、選択可能なマーカーを保有するベクターの一部として、宿主の形質転換のために使用でき、または発現コンストラクトは、選択可能なマーカーを保有するベクターと共に、別個の分子として同時形質転換される。ベクターは、1つ以上の選択可能なマーカー遺伝子を含有してもよい。
【0085】
好ましい選択可能なマーカーとしては、宿主細胞内の欠損を補完するもの、または薬剤抵抗性を与えるものが挙げられるが、これらに限定されるものではない。それらとしては、例えばアセトアミダーゼ遺伝子またはcDNA(A.ニデュランス(A.nidulans)、コウジカビ(A.oryzae)、またはA.ニガー(A.niger)からのamdS、niaD、facA遺伝子またはcDNA)などのほとんどの糸状菌および酵母の形質転換のために使用できる用途の広いマーカー遺伝子、またはG418、ハイグロマイシン、ブレオマイシン、カナマイシン、フレオマイシンまたはベノミル抵抗性(benA)のような抗生物質に対する抵抗性を提供する遺伝子が挙げられる。代案としては、例えばURA3(S.セレヴィシエ(S.cerevisiae)からのまたはその他の酵母からの類似遺伝子)、pyrGまたはpyrA(A.ニデュランス(A.nidulans)またはA.ニガー(A.niger)からの)、argB(A.ニデュランス(A.nidulans)またはA.ニガー(A.niger)からの)またはtrpCなどの対応する突然変異宿主株を必要とする栄養要求性マーカーなどの特異的選択マーカーが使用できる。好ましい実施態様では、選択マーカー遺伝子を含まないポリペプチドを生成できる転換宿主細胞を得るように、発現コンストラクト導入後に選択マーカーが転換宿主細胞から除去される。
【0086】
その他のマーカーとしては、ATPシンセターゼサブユニット9(oliC)、オロチジン−5’−ホスフェート−デカルボキシラーゼ(pvrA)、細菌G418抵抗性遺伝子(酵母中で有用であるが糸状菌中では有用でない)、アンピシリン抵抗性遺伝子(大腸菌(E.coli))、ネオマイシン抵抗性遺伝子(バシラス(Bacillus))、およびグルクロニターゼ(GUS)をコードする大腸菌(E.coli)uidA遺伝子が挙げられる。ベクターは、例えばRNAの生成のために、または宿主細胞を形質移入または形質転換するために生体外で使用してもよい。
【0087】
ほとんどの糸状菌および酵母では、安定した形質転換体を得るために、発現コンストラクトは好ましくは宿主細胞のゲノム中に組み込まれる。しかし特定の酵母では、安定した高レベル発現のために、その中に発現コンストラクトを組み込むことができる、適切なエピソームベクターシステムもまた利用できる。その例としては、それぞれサッカロミセス(Saccharomyces)およびクリヴェロミセス(Kluyveromyces)の2μm CENおよびpKD1プラスミドに由来するベクター、またはAMA配列(例えばアスペルギルス(Aspergillus)からのAMA1)を含有するベクターが挙げられる。発現コンストラクトが宿主細胞ゲノムに組み込まれる場合、コンストラクトは、ゲノム中の無作為の遺伝子座に組み込まれるか、または相同的組換えを使用して所定の標的遺伝子座に組み込まれるかのどちらかであり、後者の場合、標的遺伝子座は好ましくは高度に発現する遺伝子を含んでなる。高度に発現される遺伝子とは、例えば誘導された条件下で、そのmRNAが総細胞mRNAの少なくとも0.01%(w/w)を構成できる遺伝子であり、または代案としては、その遺伝子産物が総細胞タンパク質の少なくとも0.2%(w/w)を構成できる遺伝子であり、または分泌遺伝子産物の場合は、少なくとも0.05g/lのレベルまで分泌されることができる。
【0088】
特定の宿主細胞のための発現コンストラクトは、通常、第1の態様のポリペプチドをコードする配列のコード鎖に相対して5’−末端から3’−末端までの連続的順序で、互いに作動的に連結する以下の要素を含有する。(1)特定の宿主細胞内でポリペプチドをコードするDNA配列の転写を指示できるプロモーター配列、(2)好ましくは5’−非翻訳領域(リーダー)、(3)場合により特定の宿主細胞から培養液中へのポリペプチドの分泌を指示できるシグナル配列、(4)成熟した好ましくは活性形態のポリペプチドをコードするDNA配列、および好ましくはまた(5)ポリペプチドをコードするDNA配列の転写下流を終結できる転写終結領域(ターミネーター)。
【0089】
ポリペプチドをコードするDNA配列の下流で、発現コンストラクトは、ターミネーターとも称される、1つ以上の転写終結部位を含有する3’非翻訳領域を好ましくは含有する。ターミネーターの起源はそれほど重要でない。ターミネーターは例えばポリペプチドをコードするDNA配列に天然であることができる。しかし好ましくは細菌ターミネーターが細菌宿主細胞内で使用され、酵母ターミネーターが酵母宿主細胞内で使用され、糸状菌ターミネーターが糸状菌宿主細胞内で使用される。より好ましくはターミネーターは、その中でポリペプチドをコードするDNA配列が発現される宿主細胞に対して内在性である。
【0090】
本発明のポリペプチドをコードするポリヌクレオチドの改善された発現はまた、関心のあるタンパク質の発現レベルそして所望ならば選択された発現宿主からの分泌レベルを増大させ、および/または本発明のポリペプチド発現の誘導性制御を提供する役割をする、例えばプロモーター、シグナル配列およびターミネーター領域などの異種の調節領域の選択によって達成されてもよい。
【0091】
本発明のポリペプチドコードする遺伝子に天然であるプロモーターの他に、その他のプロモーターを使用して、本発明のポリペプチドの発現を指示してもよい。プロモーターは、所望の発現宿主中で本発明のポリペプチドを、発現を指示するその効率によって選択してもよい。
【0092】
プロモーター/エンハンサーおよびその他の発現調節シグナルは、発現ベクターがそのためにデザインされた宿主細胞と適合性であるように選択してもよい。例えば原核生物プロモーター、特にE.coli株中で使用するのに適したものを使用してもよい。本発明のポリペプチドの発現を哺乳類細胞内で実施する場合、哺乳類プロモーターを使用してもよい。例えば肝細胞細胞−特異的プロモーターなどの組織特異的プロモーターもまた使用してもよい。例えばモロニーマウス白血病ウイルスの長い末端反復末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒト サイトメガロウイルス(CMV)IEプロモーター、単純ヘルペスウイルスプロモーターまたはアデノウィルスプロモーターなどのウイルス性プロモーターもまた使用してもよい。
【0093】
適切な酵母プロモーターとしては、S.セレヴィシエ(cerevisiae)GAL4およびADHプロモーター、およびS.ポンベ(S.pombe)nmt1およびadhプロモーターが挙げられる。哺乳類プロモーターとしては、カドミウムなどの重金属に応えて誘導されることができるメタロチオネインプロモーターが挙げられる。SV40大型T抗原プロモーターまたはアデノウィルスプロモーターなどのウイルス性プロモーターもまた使用してもよい。これらの全てのプロモーターは、当該技術分野で容易に入手できる。
【0094】
β−アクチンプロモーターなどの哺乳類プロモーターを使用してもよい。組織−特異的プロモーター、特に内皮またはニューロン細胞特異的プロモーター(例えばDDAHIおよびDDAHIIプロモーター)が特に好ましい。例えばモロニーマウス白血病ウイルスの長い末端反復末端反復(MMLV LTR)、ラウス肉腫ウイルス(RSV)LTRプロモーター、SV40プロモーター、ヒト サイトメガロウイルス(CMV)IEプロモーター、アデノウィルス、HSVプロモーター(HSV IEプロモーターなど)、またはHPVプロモーターなどのウイルス性プロモーターもまた使用してもよく、特にHPV上流調節領域(URR)ウイルス性プロモーターは、当該技術分野で容易に入手できる。
【0095】
本発明の宿主細胞内で転写を指示できる多様なプロモーターが使用できる。好ましくはプロモーター配列は、先に定義したように高度に発現される遺伝子に由来する。プロモーターが好ましくはそれに由来し、および/または発現コンストラクト組み込みのために好ましい所定の標的遺伝子座に含まれる、好ましい高度に発現する遺伝子の例としては、トリオース−ホスフェートイソメラーゼ(TPI)、グリセルアルデヒド−ホスフェートデヒドロゲナーゼ(GAPDH)、ホスホグリセリン酸キナーゼ(PGK)、ピルビン酸キナーゼ(PYK)、アルコールデヒドロゲナーゼ(ADH)などの解糖作用酵素をコードする遺伝子、ならびにアミラーゼ、グルコアミラーゼ、プロテアーゼ、キシラナーゼ、セロビオ加水分解酵素、β−ガラクトシダーゼ、アルコール(メタノール)オキシダーゼ、延長因子およびリボソームタンパク質をコードする遺伝子が挙げられるが、これに限定されるものではない。適切な高度に発現する遺伝子の特定例としては、例えばクリヴェロミセス(Kluyveromyces)種からのLAC4遺伝子、それぞれハンゼヌラ(Hansenula)およびピチア(Pichia)からのメタノールオキシダーゼ遺伝子(AOXおよびMOX)、A.ニガー(A.niger)およびA.アワモリ(A.awamori)からのグルコアミラーゼ(glaA)遺伝子、コウジカビ(A.oryzae)TAKA−アミラーゼ遺伝子、A.ニデュランス(A.nidulans)gpdA遺伝子、およびT.リーセイ(T.reesei)セロビオ加水分解酵素遺伝子が挙げられる。
【0096】
真菌発現宿主中で使用するのに好ましい強力な構成的および/または誘導プロモーターの例は、キシラナーゼ(xlnA)、フィターゼ、ATP−シンセターゼサブユニット9(oliC)、トリオースホスフェートイソメラーゼ(tpi)、アルコールデヒドロゲナーゼ(AdhA)、アミラーゼ(amy)、アミログルコシダーゼ(AG−glaA遺伝子から)、アセトアミダーゼ(amdS)、およびグリセルアルデヒド−3−リン酸デヒドロゲナーゼ(gpd)プロモーターの真菌遺伝子から得られるものである。
【0097】
使用してもよい強力な酵母プロモーターの例としては、アルコールデヒドロゲナーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ラクターゼ、3−ホスホグリセリン酸キナーゼ、原形質膜ATPアーゼ(PMA1)およびトリオースホスフェートイソメラーゼの遺伝子から得られるものが挙げられる。
【0098】
使用してもよい強力な細菌プロモーターの例としては、アミラーゼおよびSPo2プロモーターならびに細胞外プロテアーゼ遺伝子からのプロモーターが挙げられる。
【0099】
使用してもよい植物細胞に適したプロモーターとしては、ノパリンシンターゼ(nos)、オクトピンシンターゼ(ocs)、マンノピンシンターゼ(mas)、リブロース小サブユニット(rubisco ssu)、ヒストン、米アクチン、ファゼオリン、カリフラワーモザイクウイルス(CMV)35Sおよび19S、およびサーコウイルスプロモーターが挙げられる。
【0100】
ベクターはポリヌクレオチドを挟む配列をさらに含んで、真核生物ゲノムの配列、好ましくは真菌ゲノム配列、または酵母ゲノム配列からのものと相同的な配列を含んでなるRNAを生じてもよい。これは相同的組換えによる、真菌または酵母ゲノムへの本発明のポリヌクレオチドの導入を可能にする。特に真菌配列で挟まれた発現カセットを含んでなるプラスミドベクターを使用して、本発明のポリヌクレオチドを真菌細胞に送達するのに適したベクターを調製できる。これらの真菌ベクターを使用する形質転換技術は、当業者に知られている。
【0101】
ベクターは、アンチセンス方向を向いた本発明のポリヌクレオチドを含有して、アンチセンスRNAの生成を提供してもよい。これは所望ならば、ポリペプチドの発現レベルを低下させるのに使用してもよい。
【0102】
[宿主細胞および発現]
さらなる態様では、本発明は、ポリペプチドをコードするコード配列のベクターによる発現に適した条件下で、上述のように発現ベクターによって転換されたまたは形質移入された宿主細胞を培養するステップと、発現したポリペプチドを回収するステップを含んでなる、本発明のポリペプチドを調製する方法を提供する。本発明のポリヌクレオチドは、発現ベクターなどの組み換え複製可能なベクター中に組み込むことができる。ベクターを使用して適合性宿主細胞中で核酸を複製してもよい。したがってさらなる実施態様では、本発明は、本発明のポリヌクレオチドを複製可能なベクター中に導入し、ベクターを適合性宿主細胞内に導入し、ベクターの複製をもたらす条件下で宿主細胞を生育させることで、本発明のポリヌクレオチドを作成する方法を提供する。適切な宿主細胞としては、大腸菌(E.coli)などの細菌と、酵母と、哺乳類細胞系と、例えばSf9細胞などの昆虫細胞および(例えば糸状)真菌細胞などのその他の真核生物の細胞系とが挙げられる。
【0103】
好ましくはポリペプチドは分泌タンパク質として産生され、その場合、発現コンストラクト中でポリペプチドの成熟形態をコードするDNA配列は、シグナル配列をコードするDNA配列と作動的に連結してもよい。分泌タンパク質をコードする遺伝子が野性型株中でシグナル配列を有する場合、好ましくは使用されるシグナル配列は、ポリペプチドをコードするDNA配列に天然(相同的)である。代案としてはシグナル配列は、ポリペプチドをコードするDNA配列に外来性(異種)であり、その場合、シグナル配列は好ましくは、その中でDNA配列が発現される宿主細胞に内在性である。酵母宿主細胞に適したシグナル配列の例は、酵母MFα遺伝子に由来するシグナル配列である。同様に糸状菌宿主細胞に適したシグナル配列は、例えばA.ニガー(A.niger)glaA遺伝子などの例えば糸状菌アミログルコシダーゼ(AG)遺伝子に由来するシグナル配列である。シグナル配列はアミログルコシダーゼ((グルコ)アミラーゼとも称される)プロモーターそれ自体と組み合わせて、ならびにその他のプロモーターと組み合わせて使用してもよい。ハイブリッドシグナル配列もまた、本発明の文脈で使用してもよい。
【0104】
好ましい異種の分泌リーダー配列は、真菌アミログルコシダーゼ(AG)遺伝子(例えばアスペルギルス(Aspergillus)からなどのglaA−18および24個のアミノ酸バージョンの双方)、MFα伝子(例えばサッカロミセス(Saccharomyces)などの酵母およびクリヴェロミセス(Kluyveromyces))、またはαアミラーゼ遺伝子(バシラス(Bacillus))起源のものである。
【0105】
ベクターを上述のように適切な宿主細胞に形質転換または形質移入して、本発明のポリペプチドの発現を提供してもよい。この方法は、ポリペプチドの発現に適した条件下で、上述のように発現ベクターで転換された宿主細胞を培養するステップと、発現したポリペプチドを場合により回収するステップを含んでなってもよい。
【0106】
したがって本発明のさらなる態様は、本発明のポリヌクレオチドまたはベクターで形質転換または形質移入された、またはそれを含んでなる宿主細胞を提供する。好ましくはポリヌクレオチドは、ポリヌクレオチドの複製および発現を可能にするベクター中で輸送される。細胞は前記ベクターと適合性であるように選択され、例えば原核生物(例えば細菌)、または真核生物の真菌、酵母または植物細胞であってもよい。
【0107】
本発明は、ポリペプチドをコードするDNA配列の組み換え発現の手段による、本発明のポリペプチドの生成のための方法を包含する。この目的で、本発明のDNA配列は、適切な相同的または異種宿主細胞内におけるポリペプチドの経済的生産を可能にするために、遺伝子増幅および/またはプロモーター、分泌シグナル配列などの発現シグナルの交換のために使用できる。相同的宿主細胞とは、本明細書においてDNA配列が由来する種と同一種である、または同一種内の変異型である、宿主細胞と定義される。
【0108】
適切な宿主細胞は、好ましくは細菌などの原核微生物、またはより好ましくは例えば真菌、酵母または糸状菌、または植物細胞などの真核生物である。一般に、より容易に操作できることから、酵母細胞が糸状菌細胞よりも好ましい。しかしいくつかのタンパク質は、酵母からの分泌が不良であったり、または場合によっては適切に処理されない(例えば酵母中での過度のグリコシル化)。このような場合は糸状菌宿主生物を選択すべきである。
【0109】
バシラス(Bacillus)属からの細菌は、培養液中にタンパク質を分泌するそれらの能力のために、異種の宿主として非常に適している。宿主として適切なその他の細菌は、ストレプトミセス(Streptomyces)およびシュードモナス(Pseudomonas)属からのものである。ポリペプチドをコードするDNA配列の発現のために好ましい酵母宿主細胞は、サッカロミセス(Saccharomyces)、クリヴェロミセス(Kluyveromyces)、ハンゼヌラ(Hansenula)、ピチア(Pichia)、ヤロウイア(Yarrowia)、または分裂酵母(Schizosaccharomyces)属のものである。より好ましくは酵母宿主細胞は、サッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)、クリヴェロミセス・ラクチス(Kluyveromyces lactis)(クリヴェロミセス・マルキシアナス(Kluyveromyces marxianus)変種ラクチス(lactis)としてもまた知られている)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ピチア・パストリス(Pichia pastoris)、ヤロウィア・リポリティカ(Yarrowia lipolytica)、およびシゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)種からなる群から選択される。
【0110】
しかしポリペプチドをコードするDNA配列の発現のために最も好ましいのは、糸状菌宿主細胞である。好ましい糸状菌宿主細胞は、アスペルギルス(Aspergillus)、トリコデルマ(Trichoderma)、フザリウム(Fusarium)、ディスポロトリカム(Disporotrichum)、ペニシリウム(Penicillium)、アクレモニウム(Acremonium)、ニューロスポラ(Neurospora)、サーモアスクス(Thermoascus)、ミセリオフトラ(Myceliophtora)、スポロトリカム(Sporotrichum)、チエラビア(Thielavia)、およびタラロミセス(Talaromyces)属からなる群から選択される。より好ましくは糸状菌宿主細胞は、コウジカビ(Aspergillus oyzae)、ショウユコウジカビ(Aspergillus sojae)またはアスペルギルス・ニデュランス(Aspergillus nidulans)種のものまたはアスペルギルス・ニガー(Aspergillus niger)群(RaperおよびFennell著、「アスペルギルス(Aspergillus)属(The Genus Aspergillus)」、The Williams&Wilkins Company、Baltimore、pp293〜344頁、1965年、によって定義される)からの種のものである。これらとしては、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・ツビゲンシス(Aspergillus tubigensis)、アスペルギルス・アクレアツス(Aspergillus aculeatus)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ニデュランス(Aspergillus nidulans)、アスペルギルス・ジャポニカス(Aspergillus japonicus)、コウジカビ(Aspergillus oryzae)およびアスペルギルス・フィクウム(Aspergillus ficuum)、およびトリコデルマ・リーセイ(Trichoderma reesei)、フザリウム・グラミネアルム(Fusarium graminearum)、ペニシリウム・クリソゲヌム(Penicillium chrysogenum)、アクレモニウム・アラバメンセ(Acremonium alabamense)、ニューロスポラ・クラッサ(Neurospora crassa)、ミセリオフトラ・サーモフィラ(Myceliophtora thermophilum)、スポロトリカム・セルロフィルム(Sporotrichum cellulophilum)、ディスポロトリカム・ディモルフォスポラム(Disporotrichum dimorphosporum)、およびチエラビア・テルレストリス(Thielavia terrestris)種のものが挙げられるがこれらに限定されるものではない。
【0111】
本発明の範囲内の好ましい発現宿主の例は、アスペルギルス(Aspergillus)種(特に欧州特許出願公開第A−184,438号明細書および欧州特許出願公開第A−284,603号明細書で述べられているもの)およびトリコデルマ(Trichoderma)種などの真菌と、特に枯草菌(Bacillus subtilis)、バシラス・リケニフォルミス(Bacillus licheniforms)、バシラス・アミロリケファシエンス(Bacillus amyloliquefaciens)であるバシラス(Bacillus)種(特に欧州特許出願公開第A−134,048号明細書および欧州特許出願公開第A−253,455号明細書で述べられているものなど)、シュードモナス(Pseudomonas)種などの細菌と、クリヴェロミセス(Kluyveromyces)種(特にクリヴェロミセス・ラクチス(Kluyveromyces lactis)などの欧州特許出願公開第A−096,430号明細書で述べられているもの、および欧州特許出願公開第A−301,670号明細書で述べられているもの)およびサッカロミセス・セレヴィシエ(Saccharomyces cerevisiae)などのサッカロミセス(Saccharomyces)種などの酵母である。
【0112】
本発明に従った宿主細胞としては植物細胞が挙げられ、したがって本発明は、1つ以上の本発明の細胞を含有する植物、およびその部分などの遺伝子導入生物にまで及ぶ。細胞は本発明のポリペプチドを異種性に発現してもよく、または1つ以上の本発明のポリヌクレオチドを異種性に含有してもよい。したがって遺伝子導入(または遺伝子改変)植物は、そのゲノム中に本発明のポリペプチドをコードする配列を(典型的に安定して)挿入されてもよい。植物細胞形質転換は、例えばアグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)からのTiまたはRiプラスミドを使用して、公知の技術を使用して実施できる。したがってプラスミド(またはベクター)は、植物に感染するのに必要な配列を含有してもよく、Tiおよび/またはRiプラスミドの誘導体を用いてもよい。
【0113】
宿主細胞はポリペプチドを過剰に発現してもよく、過剰発現のための技術はよく知られており、本発明で使用できる。したがって宿主は、2つ以上のポリヌクレオチドのコピーをを有してもよい。
【0114】
代案としては、葉、根または茎などの植物部分の直接感染を実行できる。この技術では、例えば植物を剃刀で切断する、植物を針で突き刺す、または植物を研磨材で擦ることで、感染させる植物を傷つけることができる。次に創傷にアグロバクテリウム(Agrobacterium)を接種する。次に植物または植物部分を適切な培地上で培養液生育させて、成熟植物に発育させることができる。転換細胞の遺伝子改変植物への再生は、例えば抗生物質を使用して形質転換された苗条を選択することで、および苗条を適切な栄養素、植物ホルモンなどを含有する培地上で二次培養することで、公知の技術を使用して達成できる。
【0115】
[宿主細胞の培養および組み換え生産]
本発明はまた、修飾されてペプチジルアルギニンデイミナーゼまたはその変異型を発現する細胞も含む。このような細胞としては、哺乳類細胞または昆虫細胞などの一時的なまたは好ましくは安定した改変高等真核細胞系、酵母および糸状菌細胞などの下等真核細胞、または細菌細胞などの原核細胞が挙げられる。
【0116】
本発明のポリペプチドを例えばバキュロウイルス(baculovirus)発現系などの細胞系中または膜上で一過性に発現することもまた可能である。本発明に従ったタンパク質を発現するように適応された、このような系もまた本発明の範囲内に含まれる。
【0117】
本発明に従って、従来の栄養素発酵培地中で、1つ以上の本発明のポリヌクレオチドで転換された微生物発現宿主を培養することで、本発明のポリペプチドの生成を実行できる。
【0118】
本発明に従った組み換え宿主細胞は、当該技術分野で知られている手順を使用して培養してもよい。プロモーターと宿主細胞との各組み合わせについて、ポリペプチドをコードするDNA配列の発現に貢献する培養条件が利用できる。所望の細胞密度またはポリペプチド力価に達した後、培養を停止して公知の手順を使用してポリペプチドを回収する。
【0119】
発酵培地は、炭素源(例えばグルコース、マルトース、糖蜜など)、窒素源(例えば硫酸アンモニウム、硝酸アンモニウム、塩化アンモニウムなど)、有機窒素源(例えば酵母抽出物、麦芽抽出物、ペプトンなど)、および無機栄養素源(例えばホスフェート、マグネシウム、カリウム、亜鉛、鉄など)を含有する公知の培養液を含んでなることができる。場合により(使用する発現コンストラクト次第で)誘導物質を含めても、または引き続いて添加してもよい。
【0120】
適切な培地の選択は、発現宿主の選択に基づいてもおよび/または発現コンストラクトの調節要件に基づいてもよい。適切な培地は当業者によく知られている。培地は、所望ならば、その他の潜在的汚染微生物よりも転換された発現宿主に有利に働く追加的構成要素を含有してもよい。
【0121】
発酵は0.5〜30日間にわたり実施されてもよい。発酵は、0℃〜45℃の範囲内の適切な温度で、例えば2〜10pHにおける、バッチ、連続または流加工程であってもよい。好ましい発酵条件としては、20℃〜37℃の範囲内の温度、および/または3〜9のpHが挙げられる。適切な条件は、通常、発現宿主および発現されるタンパク質の選択に基づいて選択される。
【0122】
発酵後、必要ならば遠心分離または濾過の手段によって、発酵ブロスから細胞を除去できる。発酵停止後または細胞除去後、次に本発明のポリペプチドを回収して、所望ならば従来の手段によって精製および単離してもよい。本発明のペプチジルアルギニンデイミナーゼは、菌糸体から、または培養された真菌細胞によってその中にペプチジルアルギニンデイミナーゼが放出された培養ブロスから精製できる。
【0123】
好ましい実施態様では、ポリペプチドは真菌から、より好ましくはアスペルギルス(Aspergillus)から、最も好ましくはアスペルギルス・ニガー(Aspergillus niger)から生成される。
【0124】
[修飾]
本発明のポリペプチドは、化学的に修飾されてもよく、例えば翻訳後に修飾されてもよい。例えばそれらは、グリコシル化されて(1回以上)、または修飾アミノ酸残基を含んでなってもよい。それらはまた、ヒスチジン残基の添加によって修飾されて、それらの精製を助けてもよく、またはシグナル配列の添加によって細胞からの分泌を促進してもよい。ポリペプチドは、アミノ−末端メチオニン残基などのアミノ−またはカルボキシル−末端延長と、約20〜25個の残基までの小型リンカーペプチドと、または精製を容易にするポリ−ヒスチジントラクト、抗原エピトープまたは結合領域などの小型延長部とを有してもよい。
【0125】
本発明のポリペプチドは、明示用標識で標識してもよい。明示用標識は、ポリペプチドが検出できるようにするあらゆる適切なラベルであってもよい。適切な標識としては、例えば125I、35Sなどの放射性同位体と、酵素と、抗体と、ポリヌクレオチドと、ビオチンなどのリンカーとが挙げられる。
【0126】
ポリペプチドは修飾されて、非天然アミノ酸を含んでもよく、またはポリペプチドの安定性が増大してもよい。タンパク質またはペプチドが合成手段によって生成される間に、このようなアミノ酸が生成中に導入されてもよい。タンパク質またはペプチドはまた、合成または組み換え生成のどちらかに引き続いて、修飾されてもよい。
【0127】
本発明のポリペプチドはまた、D−アミノ酸を使用して生成されてもよい。このような場合、アミノ酸はCからN方向に逆順で連結される。これはこのようなタンパク質またはペプチドを生成するために、当該技術分野で慣習的である。
【0128】
いくつかの側鎖修飾が当該技術分野で知られており、本発明のタンパク質またはペプチドの側鎖に施してもよい。このような修飾としては、例えばアルデヒドとの反応とそれに続くNaBH4での還元、アセトイミド酸メチルでのアミジン化または無水酢酸でのアシル化による、還元的アルキル化によるアミノ酸修飾が挙げられる。
【0129】
本発明によって提供される配列はまた、「第二世代」酵素構築のための出発原料として使用してもよい。「第二世代」ペプチジルアルギニンデイミナーゼは、変異誘発技術(例えば部位特異的変異誘発または遺伝子シャフリング技術)によって改変されたペプチジルアルギニンデイミナーゼであり、野生型ペプチジルアルギニンデイミナーゼ、または本発明によって生成されるものなどの組み換えペプチジルアルギニンデイミナーゼとは異なる特性を有する。例えばそれらの温度または最適pH、比活性、基質親和性または熱安定性は、特定の工程での使用によりふさわしいようにように改変されてもよい。
【0130】
本発明のペプチジルアルギニンデイミナーゼ活性に必須であり、したがって好ましくは置換されるアミノ酸は、部位特異的変異誘発またはアラニンスキャニング変異誘発などの当該技術分野で知られている手順に従って同定されてもよい。後者の技術では、突然変異が分子内のあらゆる残基に導入され、得られた突然変異分子を生物学的活性(例えばペプチジルアルギニンデイミナーゼ活性)について試験して、分子活性に決定的なアミノ酸残基を同定する。酵素−基質相互作用部位もまた、核磁気共鳴、結晶構造解析または光親和性標識などの技術によって判定される結晶構造の分析によって判定できる。
【0131】
遺伝子シャフリング技術は、ポリヌクレオチド配列中に突然変異を導入するランダムな方法を提供する。発現後、最良の特性がある単離物を再単離して合わせ、再度シャフリングして遺伝的多様性を増大させる。この手順を数回繰り返すことにより、急速に改善されたタンパク質をコードする遺伝子を単離できる。好ましくは遺伝子シャフリング手順は、同様の機能があるタンパク質をコードする遺伝子ファミリーを用いて開始する。本発明で提供されるポリヌクレオチド配列は、分泌されたペプチジルアルギニンデイミナーゼの特性を改善するための遺伝子シャフリングによく適している。
【0132】
代案としては、NTG処置による変異誘発またはUV変異誘発などの古典的ランダム変異誘発技術および選択を使用して、タンパク質の特性を改善できる。変異誘発は、単離されたDNAに直接に、または関心のあるDNAで転換された細胞に実施できる。代案としては、突然変異は、当業者に知られているいくつかの技術によって単離されたDNA中に導入できる。これらの方法の例は、誤りがちなPCR、反復−欠損宿主細胞内のプラスミドDNAの増幅などである。
【0133】
酵母および糸状菌宿主細胞の使用は、本発明の組み換え発現産物に最適生物学的活性を与えるのに必要に応じて、翻訳後修飾(例えばタンパク質分解過程、ミリスチン化(myristilation)、グリコシル化、トランケーション、およびチロシン、セリンまたはスレオニンホスホリル化)を提供することが予期される。
【0134】
[調製]
本発明のポリペプチドは単離形態であってもよい。ポリペプチドは、ポリペプチドの意図される目的を妨げないキャリアまたは希釈剤と混合してもよく、なおも単離されていると見なされるものと理解される。本発明のポリペプチドはまた、実質的に精製された形態であってもよく、その場合、それは一般に調製品中にポリペプチドを含んでなり、その中で70%を超える、例えば80%、90%、95%、98%または99%を超える、調製品中のタンパク質が本発明のポリペプチドである。
【0135】
本発明のポリペプチドは、それらがそれらの天然の細胞環境外であるような形態で提供されてもよい。したがってそれらは上で考察したように実質的に単離または精製されていてもよく、または例えばその他の真菌種、動物、植物または細菌細胞などのその中でそれらが自然に生じない細胞内にあってもよい。
【0136】
[ペプチジルアルギニンデイミナーゼ活性の除去または低下]
本発明はまた、親細胞よりも少ないポリペプチドを生成する突然変異細胞をもたらす、ポリペプチドまたはその制御配列をコードする内在性核酸配列を中断または消去するステップを含んでなる、親細胞の突然変異細胞を生成する方法に関する。
【0137】
低下したペプチジルアルギニンデイミナーゼ活性を有する株の構築は、細胞内でのペプチジルアルギニンデイミナーゼの発現に必要な核酸配列の修飾または不活性化によって、好都合に達成されてもよい。修飾または不活性化される核酸配列は、例えばペプチジルアルギニンデイミナーゼ活性を示すのに必須なポリペプチドまたはその一部をコードする核酸配列であってもよく、または核酸配列は、核酸配列のコード配列からのポリペプチド発現に必要な調節機能を有してもよい。このような調節または制御配列の例としては、プロモーター配列またはその機能性部分、すなわちポリペプチドの発現に影響を与えるのに十分な部分が挙げられる。修飾可能なその他の制御配列としては、リーダー配列、ポリアデニル化配列、プロペプチド配列、シグナル配列、および終止配列が挙げられるが、これらに限定されるものではない。
【0138】
核酸配列の修飾または不活性化は、細胞を変異誘発して、その中でペプチジルアルギニンデイミナーゼ生成能力が低下し、または排除されている細胞を選択して、実施してもよい。特異的またはランダムであってもよい変異誘発は、例えば適切な物理的または化学的変異誘発剤の使用によって、適切なオリゴヌクレオチドの使用によって、またはDNA配列をPCR変異誘発して、実施してもよい。さらに変異誘発は、これらの変異誘発剤のあらゆる組み合わせの使用によって実施してもよい。
【0139】
本目的に適した物理的または化学的変異誘発剤の例としては、紫外線(UV)照射、ヒドロキシルアミン、N−メチル−N’−ニトロ−N−ニトロソグアニジン(NTG)、O−メチルヒドロキシルアミン、亜硝酸、エチルメタンスルホネート(EMS)、亜硫酸水素ナトリウム、ギ酸、およびヌクレオチド類似体が挙げられる。
【0140】
このような薬剤を使用する場合、変異誘発は典型的に、適切な条件下で選択された変異誘発剤存在下で、変異誘発させる細胞をインキュベートして、低下したまたは皆無のペプチジルアルギニンデイミナーゼ活性発現を示す細胞を選択して実施される。
【0141】
本発明のポリペプチドの生成の修飾または不活性化は、ポリペプチドまたはその転写または翻訳に必要とされる調節要素をコードする核酸配列中の1つ以上のヌクレオチドの導入、置換、または除去によって達成されてもよい。例えばヌクレオチドは、停止コドンの導入、開始コドンの除去、または読み取り枠の変更をもたらすように挿入または除去されてもよい。このような修飾または不活性化は、当該技術分野で知られている方法に従って、部位特異的変異誘発またはPCR変異誘発によって達成されてもよい。
【0142】
原則として修飾は、生体内で、すなわち修飾する核酸配列を発現する細胞に直接に実施してもよいが、下で例証するように修飾を生体外で実施することが好ましい。
【0143】
選択された宿主細胞によるペプチジルアルギニンデイミナーゼの生成を不活性化するまたは低下させる都合のよい方法の例は、遺伝子置換または遺伝子中断の技術に基づく。例えば遺伝子中断法では、関心のある内在性遺伝子または遺伝子断片に対応する核酸配列を生体外で変異誘発して欠損核酸配列を生じ、次にそれを宿主細胞に転換して欠損遺伝子を生成する。相同的組換えによって、欠損核酸配列を内在性遺伝子または遺伝子断片に置き換える。好ましくは欠損遺伝子または遺伝子断片は、その中でポリペプチドをコードする遺伝子が修飾または破壊された転換体を選択するのに使用してもよいマーカーもまたコードする。
【0144】
代案としては、本発明のポリペプチドをコードする核酸配列の修飾または不活性化は、ポリペプチドをコードする配列に相補的なヌクレオチド配列を使用して、確立されたアンチセンス技術によって達成してもよい。より具体的には、ポリペプチドをコードする核酸配列に相補的なヌクレオチド配列を導入することで、細胞によるポリペプチドの生成を低下させまたは排除してもよい。次にアンチセンスポリヌクレオチドは、典型的に細胞内で転写され、ペプチジルアルギニンデイミナーゼをコードするmRNAにハイブリダイズできる。細胞内で生成するペプチジルアルギニンデイミナーゼの量は、相補的アンチセンスヌクレオチド配列とmRNAがハイブリダイズできる条件下で低下し、または排除される。
【0145】
本発明の方法に従って修飾される細胞は、細胞に同種または異種のどちらかの、例えば所望のタンパク質製品の生成に適した真菌株などの微生物起源であることが好ましい。
【0146】
本発明はさらに、親細胞よりも少ないポリペプチドを産生する突然変異細胞をもたらす、ポリペプチドまたはその制御配列をコードする内在性核酸配列の中断または欠失を含んでなる、親細胞の突然変異細胞に関する。
【0147】
このように作り出されたポリペプチド−欠損突然変異細胞は、同種および/または異種のポリペプチド発現のための宿主細胞として特に有用である。したがって本発明はさらに、(a)突然変異細胞をポリペプチドの生成に貢献する条件下で培養するステップと、(b)ポリペプチドを回収するステップを含んでなる、同種または異種のポリペプチドを生成する方法に関する。本文脈で、「異種のポリペプチド」という用語は、本明細書において宿主細胞に天然でないポリペプチド、その中で修飾がなされて天然配列が改変された天然タンパク質、または組み換えDNA技術による宿主細胞の操作の結果としてその発現が定量的に改変された天然タンパク質と定義される。
【0148】
なおもさらなる態様では、本発明は、本発明のペプチジルアルギニンデイミナーゼポリペプチドならびに関心のあるタンパク質生成物の双方を産生する細胞の発酵によって、本質的にペプチジルアルギニンデイミナーゼ活性を含まない、タンパク質生成物を生成する方法を提供する。本方法は、発酵中または発酵完了後のどちらかに、ペプチジルアルギニンデイミナーゼ活性を阻害できる有効量の薬剤を発酵ブロスに添加するステップと、発酵ブロスから関心のある生成物を回収するステップと、場合により回収された生成物にさらなる精製を施すステップを含んでなる。代案としては、培養後に、ペプチジルアルギニンデイミナーゼ活性を実質的に低下させて、培養ブロスからの生成物の回収ができるように、得られた培養ブロスにpHまたは温度処理を施すことができる。pHまたは温度処理の併用は、培養ブロスから回収されたタンパク質調製品に実施してもよい。
【0149】
本質的にペプチジルアルギニンデイミナーゼを含まない生成物を生成する本発明の方法は、真核生物ポリペプチドの生成において、特に酵素などの真菌タンパク質の生成において特に興味深い。ペプチジルアルギニンデイミナーゼ−欠損細胞はまた、食品産業で関心がある、または薬学的関心のある異種タンパク質を発現するのに使用してもよい。
【0150】
ペプチジルアルギニンデイミナーゼの好ましい起源は、ペプチジルアルギニンデイミナーゼをコードする微生物遺伝子を微生物宿主生物にクローニングして得られる。ペプチジルアルギニンデイミナーゼのより好ましい起源は、ペプチジルアルギニンデイミナーゼ遺伝子を過剰発現できるアスペルギルス(Aspergillus)属に属する宿主に、ペプチジルアルギニンデイミナーゼをコードするフザリウム(Fusarium)由来遺伝子をクローニングして得られる。一般に、過剰発現のためには相同宿主生物が好ましい。同種とはここで同一種であることを意味する。
【0151】
[配列一覧]
配列番号1 合成DNA
配列番号2 合成DNA
配列番号3 フザリウム・グラミネアルム(Fusarium graminearum)のDNA
配列番号4 フザリウム・グラミネアルム(Fusarium graminearum)のDNA
配列番号5 フザリウム・グラミネアルム(Fusarium graminearum)のDNA
配列番号6 フザリウム・グラミネアルム(Fusarium graminearum)のアミノ酸配列
配列番号7 フザリウム・グラミネアルム(Fusarium graminearum)のアミノ酸配列
配列番号8 ケトミウム・グロボスム(Chaetomium globosum)のアミノ酸配列
配列番号9 ストレプトミセス・スキャビエス(Streptomyces scabies)のアミノ酸配列
配列番号10 ストレプトミセス・スキャビエス(Streptomyces scabies)のアミノ酸配列
配列番号11 PADコンセンサスアミノ酸配列
配列番号12 人工アミノ酸配列
配列番号13 ストレプトミセス・クラブリゲルス(Streptomyces clavuligerus)のアミノ酸配列
配列番号14 ファエオスフェリア・ノドルム(Phaeosphaeria nodorum)のアミノ酸配列
【図面の簡単な説明】
【0152】
【図1】フザリウム(Fusarium)PADのクローニング。
【図2A】分泌PADタンパク質の配列比較。
【図2B】分泌PADタンパク質の配列比較。
【図3】発現ベクターpGBFINPADの構築。
【図4】SDS−PAGE上でのBSAおよびPADタンパク質濃度の比較。染色は、シンプリー・ブルー・セーフ・ステイン(コロイド状クーマシーG250)を使用して実施した。レーン1〜6はBSA、レーン7〜10はPAD。
【0153】
[材料と方法]
[加水分解度]
使用された様々なタンパク質分解混合物の加水分解度(DH)は、迅速なOPA試験(Nielsen,P.M.;Petersen,D.;Dambmann,C.著「食物タンパク質の加水分解度を判定するための改善された方法(Improved method for determining food protein degree of hydrolysis)」、Journal of Food Science 2001年、66、642〜646頁)を使用して測定された。
【0154】
[PADアッセイ]
PAD様活性は、2つの異なる方法でモニターした。スクリーニングアッセイでは、シグマ・アルドリッチ(Sigma−Aldrich)によって提供される、この酵素のためのシグマ品質管理試験手順(1584頁;ウサギ骨格筋から)を使用した。この色素産生方法は、N−α−ベンゾイル−L−アルギニンエチルエステル塩酸塩の使用に基づいた(BAEE;タケハラら著、(1986年)Journal of Biochemistry、99、1477〜1424頁)。
【0155】
別のアッセイでは、遊離またはペプチド結合アルギニンのシトルリンへの転換を測定して、古典的アミノ酸分析によって酵素活性を測定した。この方法は「アミノ酸分析」の下で規定される。
【0156】
[アミノ酸分析]
アミノ酸分析は、米国マサチューセッツ州ミルフォードのウォーターズ(Waters(Milford MA,USA))からのアミノ酸分析システムの操作手引で規定される、PicoTag法に従って実施した。最終サンプルを乾燥させ、フェニルイソチオシアネートを使用して直接誘導体化した。その中でシグマからの純粋化合物を誘導体化してクロマトグラフィーにかける試行を行って、クロマトグラム中でのシトルリンおよびオルニチンの位置を確立した。HPLC法を使用して、存在する誘導体化アミノ酸を定量化した。その中でタンパク質またはペプチドがPADのための基質として使用されるインキュベーション混合物を、米国マサチューセッツ州ミルフォードのウォーターズからのアミノ酸分析システムの操作手引に従って最初に酸加水分解し、次に誘導体化して、分離および定量化した。遊離アルギニンのみを含有するインキュベーション混合物中では、酸加水分解は任意であった。実施例6で例証するように、酸加水分解はシトルリンを部分的にオルニチンに転換する。アルギニンから形成されたシトルリンの総量を計算するために、シトルリンおよびオルニチンのレベルを合計した。加水分解中に、TrpおよびCysは破壊され、したがってこれらのアミノ酸はさらなる計算中で排除された。さらにGlnおよびAsn残基は、酸加水分解中にGluおよびAspに転換するので、GluおよびGlnの値、およびAspおよびAsnの値もまた合計して、酸加水分解前に得られたデータと比較できるようにした。
【0157】
[SDS−PAGE]
SDS−PAGEおよび染色のために使用した全ての材料は、米国カリフォルニア州カールスバッドのインヴィトロジェン(Invitrogen(Carlsbad,CA,US))から購入した。サンプルは、製造業者の説明書に従ってSDS緩衝液を使用して調製し、MES−SDS緩衝液系を使用して12%Bis−トリスゲル上で分離した。染色は、シンプリー・ブルー・セーフ・ステイン(コロイド状クーマシーG250)を使用して実施した。
【0158】
[LC/MS/MS分析]
PADインキュベーション後のペプチドQPRPFPFPRPRの分析では、オランダ国ブレダのサーモクエスト(Thermoquest)(登録商標)(Breda,the Netherlands))からのP4000ポンプと連結している、オランダ国ブレダのサーモクエスト(登録商標)からのイオントラップ質量分光計を使用したHPLCを使用した。溶出のための米国マサチューセッツ州ベッドフォードのミリポア(Millipore(Bedford,MA,USA)からのMilli Q水(溶液A)中の0.1%ギ酸、およびアセトニトリル中の0.1%ギ酸(溶液B)の勾配と組み合わせて、ベルギー国のVarian BelgiumからのInertsil 3 ODS3,3μm、150*2.1mmカラムを使用して、形成されたペプチドを分離した。勾配は100%の溶液Aで開始して5分間保ち、5%のBに直線的に10分間で増大させ、それに30分間での45%の溶液Bへの直線的増大が続き、即座に最初の条件にして、安定化のためにさらに15分間保った。使用した注入量は50μlであり、流速は分速200μlで、カラム温度は55℃に保たれた。注入サンプルのタンパク質濃度は、約50μgs/mlであった。約30%の最適衝突エネルギーを使用して、関心のあるペプチドのための専用MS/MSによって、インキュベーション後のペプチドQPRPFPFPRPR中の異なるアルギニン残基の反応および反応速度を経時的に追跡した。LC/MS/MSに先だって、インキュベーション混合物を周囲温度および13000rpmで10分間遠心分離し、0.22μmフィルターを通して濾過して上清をMilli Q水で1:100に希釈した。
【0159】
[クローニング技術]
核酸の単離および精製、核酸の電気泳動法、核酸の酵素修飾、切断および/または増幅、大腸菌(E.coli)などの形質転換などの標準分子クローニング技術は、Sambrookら(Sambrook,J.、Russell,D.W.著(2001年):分子クローニング;実験室マニュアル(Molecular cloning; a laboratory manual)(第3版)、Cold Spring Harbour laboratory press、Cold Spring Harbour,New York)または供給元の付属書に従って実施した。オランダ国ブレダのインヴィトロジェンは、合成オリゴヌクレオチドを供給する。DNA配列分析はオランダ国ライデン(Leiden,the Netherlands)のBaseClearで実施された。
【0160】
[実施例]
[実施例1]
[フザリウム(Fusarium)株はPAD様活性を分泌できる]
分泌されたPAD様活性を同定する目的で、カビの大きなコレクションをスクリーンした。そのために、株を30℃でジフコ(Difco)からのジャガイモデキストロースブロス(PDB)上で4〜5日間予備生育させた。次に培養を遠心分離によって収集して蒸留水で洗浄し、米タンパク質加水分解産物で強化された最少培地に移した。米タンパク質は比較的アルギニン残基に富み、その水溶性を増大させるために、ベルギー国ルーバン(Leuven,Belgium)のRemy Industriesからの米タンパク質をデンマーク国バウスヴェア(Bagsvaerd,Denmark)のNOVOからのアルカラーゼ(Alcalase)と共にpH7.5でプレインキュベートして、DHが約15の加水分解産物を得た。使用した最小増殖培地は、1リットルあたり0.52g KCl、1.52g KH2PO4、1.3ml 4M KOH、0.52g MgSO4・7H2O、22mg ZnSO4・7H2O、11mg H3BO3、5mg FeSO4・7H2O、1.7mg CoCl2・6H2O、1.6mg CuSO4・5H2O、5mg MnCl2・4H2O、1.5mg Na2MoO4・2H2O、50mg EDTA、40gグルコース、および5gの加水分解米タンパク質を含有した。真菌を後者の培地中でさらに2日間生育させた後、培養を再度遠心分離して、透明な上清のサンプルを凍結した。分泌されたPAD様活性が存在するかどうかを判定するために、材料と方法セクションで述べられているように、上清サンプルをシグマ品質管理試験手順比色分析にかけた。この試験では、0.2mlの実希釈上清を0.6mlの混合試薬に添加した。酵素インキュベーションを45℃で5時間行った。得られた結果次第で、フザリウム(Fusarium)株の全てではないがいくつかの上清は、いくらかの変色を示した。変色をもたらした上清は、オランダ国ユトレヒト(Utrecht,The Netherlands)のCBSから得られたフザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株から得られた。いくつかのその他のフザリウム(Fusarium)株、および例えば英国ウォリンフォード(Wallingford,UK)のCABIからのフザリウム・グラミネアルム(Fusarium graminearum)IMI145425などのその他の真菌の上清は、発色しなかった。得られた結果から、我々はいくつかのフザリウム(Fusarium)株は、PAD様活性を分泌すると結論した。我々の知る限り、これは微生物からのPAD分泌の最初の報告である。
【0161】
[実施例2]
[フザリウム(Fusarium)ゲノム配列中のPADコード遺伝子を同定する]
いくつかのフザリウム(Fusarium)株がPAD様活性を分泌できることが分かり、これらのPADをコードする遺伝子のゲノム配列を単離および分析した。これを行うために、フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株をジフコからのPDB(ジャガイモデキストロースブロス)中において30℃で3日間生育させ、供給元の説明書を使用して、オランダ国ブレダのOmnilabo International BVからのQ−Biogeneキット(カタログ番号6540−600)を使用して、菌糸から染色体のDNAを単離した。PCRを使用したPAD遺伝子コード配列の増幅のためにこの染色体DNAを使用した。
【0162】
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株の染色体DNAからのPAD遺伝子を特異的に増幅するために、2つのPCRプライマーをデザインした。ジベレラ・ゼアエ(Gibberella zeae)PH−1のゲノムDNA中に発見され、機能未知タンパク質(UNIPROT Q4IIR5_GIBZE)としてアノテートされた配列からプライマー配列を部分的に得た。我々はこの配列が、高等真核生物のPAD配列と相同性を有することを見いだした。第1のプライマーは、上流に12bpsのリーダー配列およびPacI制限部位(配列番号1)がある、ATG開始コドンに始まる24個のヌクレオチドのPADコード配列を含有した。第2のプライマーは、CTA停止コドン(配列番号2)のすぐ下流にAscI制限部位がある、PADコード配列に相補的な29個のヌクレオチドを含有した。これらのプライマーを使用して、テンプレートとしてフザリウム・グラミネアルム(Fusarium graminearum)CBS166.57、CBS316.73、CBS11063、CBS18432、およびCBS792.70株からの染色体DNAを用いて、我々は2kbサイズの断片を増幅できた。これらの全例で、増幅された断片は同一サイズであった。このようにして得られた2kbサイズの断片を精製し、pCR−BluntII−TOPOベクター(Invitrogen)中にライゲートし、pGBPADシリーズのプラスミドを得た(図1参照)。PCR増幅された配列を配列分析によって分析した。興味深いことに、我々はこのサイズのゲノムDNA断片を例えばフザリウム・グラミネアルム(Fusarium graminearum)IMI145425から増幅できず、フザリウム・グラミネアルム(Fusarium graminearum)の分泌PAD活性の生成と関連がある2kb断片の存在が示唆された。
【0163】
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57およびCBS316.73株のPADコード領域のゲノム配列は、それぞれ配列番号3および配列番号4に記載される。フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57のPADを過剰発現する株から単離されたmRNAから、コード領域のcDNA配列を生じさせた(実施例3参照)。このcDNAは配列決定され、配列番号5に記載される。フザリウム・グラミネアルム(Fusarium graminearum)株CBS166.57およびCBS316.73によってコードされるPADの推定タンパク質配列は、それぞれ配列番号6および配列番号7に記載される。
【0164】
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57のPADゲノムDNA配列は、フザリウム・グラミネアルム(Fusarium graminearum)CBS316.73のPADゲノムDNA配列とは19箇所で異なる。推定タンパク質配列にとってこれは7個のアミノ酸が異なり、フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57およびCBS316.73株からの2つのPADが98.0%同一であることを意味する。興味深いことに、どちらの推定タンパク質配列も、タンパク質のアミノ−末端に配列シグナルを含有する。マトリックスBlosum 62および予期された閾値10でBlastPプログラムを使用して、これらの配列をタンパク質およびDNAデータベースと比較した(Altschulら著、1997年、Nucleic Acids Research 25:3389〜3402頁)。我々は、フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57のPADのDNAおよび推定タンパク質配列の双方が、我々がジベレラ・ゼアエ(Gibberella zeae)PH−1のゲノムDNA中に発見して、機能未知タンパク質(UNIPROT Q4IIR5_GIBZE)としてアノテートした配列と同一であることを見いだした。フザリウム(Fusarium)からのPADはまた、真菌タンパク質配列がシグナル配列を含有し、したがって細胞から能動的に分泌される見込みが高く、高等真核生物からのPADはこのようなシグナル配列を有さず、したがって能動的に分泌されないという本質的な違いを持って、高等真核生物のPADとの顕著な相同性も有した。フザリウム(Fusarium)PADは、ポルフィロモナス(Porphyromonas)からのPADとのいかなる相同性も示さなかった。
【0165】
さらにフザリウム(Fusarium)PADと、(フザリウム(Fusarium)のような)子嚢菌綱(ascomycetes)真菌であるが、異なる目(肉座菌目でなくフンタマカビ目)に属するケトミウム・グロボスム(Chaetomium globosum)CBS148.51のゲノムからの配列の間に相同性が見られた(http://www.broad.mit.edu/annotation/genome/chaetomium_globosum/Home.html;CHGG_01998.1)。フザリウム(Fusarium)とケトミウム(Chaetomium)のPAD配列間の相同性は、660個のアミノ酸中253個のアミノ酸である(38%同一)。ここでもまた、予測されたタンパク質は、機能未知タンパク質(UNIPROT Q2HCQ6_CHAGB)としてアノテートされた。さらにこの真菌中のPAD相同体のタンパク質配列のアノテーションは、おそらく不正確である。ケトミウム(Chaetomium)タンパク質と既知のPADとの相同性がタンパク質の最初の600個のアミノ酸にかけてのみに存在するのに対し、ケトミウム(Chaetomium)タンパク質は、1000個のアミノ酸のタンパク質としてアノテートされている。したがって我々は、UNIPROT Q2HCQ6_CHAGB中のケトミウム(Chaetomium)タンパク質のアノテーションは不正確と考える。ケトミウム・グロボスム(Chaetomium globosum)PADの正確なタンパク質配列を配列番号8に記載し、配列比較を図3に示す。SignalPプログラム(http://www.cbs.dtu.dk/services/SignalP/)を使用してこの配列をより綿密に調べると、タンパク質のアミノ−末端にシグナル配列を検出でき(図3参照)、このタンパク質もまた、我々がフザリウム(Fusarium)株からのPADについて発見したように、真菌から分泌される可能性が高いことを意味する。
【0166】
フザリウム(Fusarium)PADの別の相同体は、これもまた(フザリウム(Fusarium)およびケトミウム(Chaetomium)のような)子嚢菌綱(Ascomycetes)真菌であるが、さらに異なる綱(フンタマカビ綱(Sordariomycetes)でなくクロイボタケ綱(Dithideomycetes))に属するファエオスフェリア・ノドルム(Phaeosphaeria nodorum)SN15のゲノム中に見いだせる。この遺伝子によってコードされるタンパク質は、配列番号14に記載される。フザリウム(Fusarium)とファエオスフェリア(Phaeosphaeria)のPAD配列間の相同性は、629個のアミノ酸中231個のアミノ酸である(36%同一)。ここでもまた、予測されたタンパク質は機能未知タンパク質(機能未知タンパク質SNOG_13103;ジェンバンクEAT79430)としてアノテートされ、シグナル配列が認識できる。ここで示されるデータは、真菌界全体を通じて分泌PADが存在できるが、まれにしか生じないことを示唆する。ここで述べられる方法を使用すれば、真菌からの分泌PADを特異的に認識して単離できるであろう。
【0167】
フザリウム(Fusarium)PADの可能な相同体についてデータベースをさらに調べた際に、我々はジャガイモ瘡痂病の病原体であるストレプトミセス・スキャビエス(Streptomyces scabies)のゲノム中に2つの追加的配列見つけることができた(http://www.sanger.ac.uk/Projects/S_scabies/)。これらのPADのタンパク質配列は、配列番号9および配列番号10に記載される。またこれらの遺伝子でもタンパク質のアミノ−末端にシグナル配列が疑われるので、これらのタンパク質も分泌されるかもしれない。配列番号10に記載されたタンパク質のカルボキシル−末端は欠損しているが、これはこの遺伝子がcontigの終わりに位置し、コード領域の3’−末端が未だ配列決定されていないためである。
【0168】
ストレプトミセス・クラブリゲルス(Streptomyces clavuligerus)ATCC27064のゲノムを配列決定し分析した際に、SignalPプログラムを使用した検出で、我々は、これもまたシグナル配列を含有する別のPAD相同体を検出できた。アミノ酸配列は配列番号13に記載される。遺伝子が配列決定contigの終わりに位置し、したがって3’−末端が欠損していたことから、タンパク質はカルボキシ末端で短い。
【0169】
微生物からのこれらのPAD配列を高等真核生物のPADと比較すると、これらの配列間で検出可能な非常に相同的な領域がある。この共通配列はWLxVGHVDEであり、配列番号11に記載される。この共通配列を使用し、当業者が既知のクローニング技術を使用して、微生物からのPADをコードする遺伝子を同定および単離することは十分可能である。一つの可能性は、その中でPAD遺伝子を同定したい生物からの好ましいコドン使用頻度があるヌクレオチド配列への配列番号11の配列の逆翻訳に基づいて、オリゴヌクレオチドプライマーをデザインし、このオリゴヌクレオチドを遺伝子ライブラリーとのハイブリダイゼーションのために、または逆転写されたmRNAプール上のPCRプライマー中で使用することである。別の可能性は、Patscan(http://www−unix.mcs.anl.gov/compbio/PatScan/HTML/patscan.html)のようなプログラムを使用した、DNAデータバンクからの翻訳されたDNA配列における検索のために、配列番号11の配列を使用することである。これらの方法の1つを使用して同定された遺伝子は、次に当業者に知られているプログラムを使用してタンパク質配列に翻訳し、それらのアミノ−末端のシグナル配列の存在について調べることができる。シグナル配列を検出するためにSignalPのようなプログラムを使用できる(http://www.cbs.dtu.dk/services/SignalP/)。本発明では、我々は、配列番号11のコンセンサスと予測されたシグナル配列の双方を含有するタンパク質配列がPADを分泌する可能性があることを見いだした。これらの組み合わされた特性を探すことは、このような酵素の工業生産に大きな利点を与える。
【0170】
[実施例3]
[アスペルギルス・ニガー(Aspergillus niger)による推定上のフザリウム・グラミネアルム(Fusarium graminearum)PADの過剰発現]
フザリウム・グラミネアルム(Fusarium graminearum)CBS166.57からのゲノムPAD遺伝子を含有するpGBPADプラスミドから、PADコード配列を含んでなるPacI/AscI断片を単離して、pGBFIN−5(WO99/32617)中のPacI/AscIphyA断片と交換した。得られたプラスミドはpGBFINPADと命名されたPAD発現ベクターであった(図3参照)。全ての大腸菌(E.coli)由来配列を発現ベクターから除去するNotIでの消化によって、発現ベクターpGBFINPADを直線化した。フェノール:クロロホルム:イソアミルアルコール(24:23:1)抽出およびエタノール沈殿を使用して、消化されたDNAを精製した。これらのベクターを使用して、アスペルギルス・ニガー(Aspergillus niger)CBS513.88を形質転換した。アスペルギルス・ニガー(Aspergillus niger)形質転換手順は、国際公開第98/46772号パンフレットで詳細に述べられている。どのようにしてアセトアミドを含有する寒天プレート上で転換体を選択し、標的とされるマルチコピー組み込み体を選択するかについてもまた、述べられている。サンプル材料のさらなる産生のために、好ましくは発現カセットの複数コピーを含有するA.ニガー(A.niger)転換体が選択される。pGBFINPAD発現ベクター30のために、最初に個々の転換体を選択的培地プレートに播種し、それに続いて単一コロニーをPDAプレートに播種して、A.ニガー(A.niger)転換体を純化した。30℃で1週間の生育後に個々の転換体の胞子を採取し、胞子を冷蔵保存して液体培地の接種のために使用した。
【0171】
浸透フラスコ培養内での株の培養によって、発現カセットの複数コピーを含有するA.ニガー(A.niger)株をサンプル材料の産生のために使用した。A.ニガー(A.niger)株の培養および培養ブロスからの菌糸の分離のための有用な方法は、国際公開第98/46772号パンフレットで述べられている。培養媒体はCSM−MES(1リットルの培地あたり150gマルトース、60gのジフコからのソイトン(Soytone)、15g(NH4)2SO4、1g NaH2PO4・H2O、1g MgSO4・7H2O、1g L−アルギニン、80mg ツイーン−80、20g MES、pH6.2)中にあった。発酵の4〜8日目に5mlのサンプルを取って、Hereaus labofuge RF内で10分間5000rpmで遠心分離し、さらなる分析まで上清を−20℃で保存した。
【0172】
pGBFINPADベクターを含有する転換体は、SDS−PAGEで分析すると見かけ上の分子量がおよそ60kDaのタンパク質を産生することが明らかになった。これはタンパク質配列から予測された分子量よりもわずかに小さいので、我々は、フザリウム(Fusarium)PADをアスペルギルス・ニガー(Aspergillus niger)中で生産すると、シグナル配列の除去後に大規模なグリコシル化が起きないと推測する。
【0173】
選択された株は、発酵および後処理プロセスをスケールアップさせた際に、大量のPADの単離および精製のために使用できる。次にこの酵素をさらなる分析のために、そして多様な産業上の利用のために使用できる。
【0174】
[実施例4]
[A.ニガー(A.niger)上清からの過剰発現された推定上のPADの精製]
1リットルあたり、マルトース、40g H2O、30gソイトン(Difco)、15g (NH4)2SO4、1g NaH2PO4・H2O、1 MgSO4・7H2O、1g L−アルギニン、0.08gツイーン−80、70g Na−クエン酸を組み込んだ増殖培地を使用して、プラスミドpGBFINPADを組み込んだA.ニガー(A.niger)を10リットルの規模で生育させた。pHを6.2に調節した。30℃で6日間の生育後、3.5g/lの安息香酸ナトリウムを添加して細胞を死滅させ、さらに6時間インキュベーションを延長した。次に10g/lのCaCl2およびベルギー国ゲント(Gent,Belgium)のDicalite BFからの45g/lの濾過助剤をブロスに添加した。最初に布帛での濾過によって菌糸を除去し、Z−2000およびZ−200フィルター(Pall)を通した濾過がそれに続いた。最後に、ミリポアからの0.22μm GP Express PLUS膜を使用して滅菌濾過を実施した。限外濾過は、ミリポアからのPelliconシステムを使用して実施した。
【0175】
緩衝液条件を25mM Na−クエン酸、pH5.0(緩衝液A)に変更後、同一緩衝液で平衡化したベルギー国ディーゲムのGEヘルスケアからのSP−セファロースXKカラムに溶液を入れた。カラムを緩衝液Aから緩衝液A+1M NaClへの直線濃度勾配で溶出した。得られた画分をSDS−PAGEにかけて、染色した。見かけ上のMWが60kDaのバンドを明らかに示した画分をプールした。グリセロール(50%w/w最終濃度)、CaCl2(0.02%w/w)、および安息香酸ナトリウム(0.1%w/w)を添加して、得られた酵素溶液を安定化した。この最終調製品中のPADタンパク質濃度を推定するために、異なる量の最終調製品を異なる量のシグマからのBSA溶液(Fraction V)と共にSDS−PAGEにかけて、シンプリー・ブルー・セーフ・ステイン(Simply Blue Safe Stain)で染色した。得られた結果により(図4参照)、最終調製品中のPAD濃度は3.8mgタンパク質/ml液体である。
【0176】
[実施例5]
[過剰発現されて精製された推定上のPADはペプチド結合アルギニンをシトルリンに転換できる]
PADとして単離された酵素の性質を確認するために、オランダ国アルメレ(Almere,The Netherlands)のPepscanからの合成ペプチドQPRPFPFPRPRと共に、クロマトグラフ的に精製された酵素をインキュベートし、LC/MSを使用して得られた生成物を分析した。LC/MS分析の目的は、アルギニン残基のシトルリン残基への転換を確認することであった。このために、pH6.5および50℃で、合成ペプチド(10mM)と共に様々な量の精製された酵素をインキュベートした。0、1、および4時間のインキュベーション後に、サンプルをインキュベーション混合物から取り除いた。全てのサンプルを95℃で10分間加熱して、あらゆる残留酵素活性を不活性化し、引き続いて遠心分離した。ペプチドをインキュベーション混合物に添加した直後に、標準サンプル(0時間のインキュベーション)を加熱処理して遠心分離した。材料と方法セクションに規定する条件下で、透明な上清をLC/MS分析にかけた。
【0177】
MSを最適化するために、5μg/mlのQPRPFPFPRPR溶液を使用した。(major)m/z697.8=[M+2H]2+および(minor)m/z465.6=[M+3H]3+のESI/posモードで、ウンデカ−ペプチドを特性決定した。QPRPFPFPRPRの滞留時間を判定するために、LC/MSを実施した。アルギニン(R)のプロトン化質量は175であり、シトルリン(R=O)への転換後、プロトン化質量は1Daから176に増大する。質量軸の定義は質量電荷比(m/z)であるので、ペプチドは[M+H]+でなく二重荷電の[M+2H]2+によって特徴づけられるという事実のために、質量の差は転換したアルギニンあたり0.5Daの違いによって特徴づけられる。インキュベート0時間サンプルのマスクロマトグラムは、9.72分でペプチドQPRPFPFPRPR(m/z697.8)に相当する大きなピークを示す。しかし11.36分に溶出した小さなピークは、このサンプル中でさえ、いくらかのアルギニン残基がシトルリンに転換されていることを示す。1時間インキュベートしたサンプルのマスクロマトグラム中では、3つのピークが明らかである。インキュベーション0時間後に出現したピークの他に、13.36分(2つのR’がR=Oに転換されたQPRPFPFPRPR)および15.94分(3つのR’がR=Oに転換されたQPRPFPFPRPR)にピークが検出された。4時間インキュベートしたサンプル中では、後者の2つのピークのみが見られた。これらのデータは、確かに単離された酵素画分が、PAD活性がある酵素に相当することの強力な示唆を形成する。かなり意外なことに、得られたデータはまた、ペプチドのN−末端寄りのアルギニン残基が最初に判定されるので、PADがペプチドのN−末端に始まると、我々が結論できるようにした。ペプチドのC−末端寄りのアルギニン残基が後に従う。
【0178】
[実施例6]
[タンパク質結合アルギニン、ペプチド結合アルギニンおよび遊離アルギニンは、過剰発現されるフザリウム(Fusarium)PADのための適切な基質を形成する]
大量のPADの可用性は、我々が古典的なアミノ酸分析によって、アルギニンからシトルリンへの転換を調べられるようにした。この目的で我々は、使用したクロマトグラム中におけるシトルリンの位置を確立して、存在する遊離アミノ酸のレベルを定量化しなくてはならなかった。アミノ酸分析で使用した酸加水分解はシトルリンをオルニチンに分解することが予期されたので、どちらもシグマからの純粋なシトルリンおよびオルニチンを誘導体化して、PicoTag法に従ってクロマトグラフィーにかけた。試行は双方の誘導体化合物が、クロマトグラム中の個々のピークとして、その他の誘導体化アミノ酸中で追跡できることを実証した。
【0179】
新しい酵素の試験では、0、5、50、および500μlの過剰発現され精製されたPADと共に、3つの異なる基質、すなわちカゼイン酸ナトリウム、カゼイン加水分解産物、および遊離アルギニンの10%(w/w)溶液をインキュベートした(実施例4参照)。インキュベーションをpH6.5および45℃で3時間実施した。カゼイン酸ナトリウムはオランダ国フェーヘルのDMVインターナショナル(DMV International(Veghel,The Netherlands)から得られ、カゼイン加水分解産物は、サブチリシンおよびプロリン特異的エンドプロテアーゼでのカゼイン酸ナトリウムの消化によって調製した(Edensら著、JAFC 53(20)、7950〜7957、2005年)。L−アルギニンはシグマから得られた。
【0180】
インキュベーション後、様々な混合物をアミノ酸分析によって分析した。表1に示すデータから結論できるように、新たに同定されたPADの相当量を含有する全てのインキュベーション中で、顕著な量のアルギニンがシトルリンに転換された。予期されたように、クロマトグラムはまた、オルニチンの存在も示唆した。これらの所見はフザリウム(Fusarium)PADが、タンパク質結合およびペプチド結合アルギニンを転換できることを暗示する。かなり意外にも、遊離アルギニンもまた基質として受容される。結果によれば、アルギニンからシトルリンへの転換において、基質が小さいほど酵素はより効果的である。後者の観察は、遊離アルギニンよりもペプチジルアルギニン基質への好みを示すと報告されている、ポルフィロモナス・ジンジバリス(Porphyromonas gingivalis)酵素について公開されるデータ(McGrawら著、Infection and immunity,1999年7月、3248〜3256頁)と対照的なようである。
【0181】
オルニチンの存在がシトルリンの酸加水分解の結果であることを確認するために、遊離アルギニンを基質とする実験を実施したが、今度は酸加水分解を省いた。表2に示すように、この場合、形成されたオルニチンの量はごくわずかであり、存在するオルニチンが酵素によって形成された生成物ではないことが実証された。
【0182】
【表1】
【0183】
【表2】
【0184】
[実施例7]
[過剰発現されたPADの最適pHおよび温度]
実施例6で述べられているのと非常によく似た一組の実験で、フザリウム(Fusarium)PADの最適pHおよび温度を判定した。これらの結果によれば、過剰発現された酵素は、その最適pHを8.0前後に、その最適温度を40〜50℃の間に有する。
【0185】
[実施例8]
[過剰発現されたPADのカルシウム依存性]
文献に従って、真核生物のペプチジルアルギニンデイミナーゼは、Ca2+−依存性酵素のファミリーを代表する。フザリウム(Fusarium)PADもまたCa2+を必要とするかどうかを試験するために、EDTA存在下でインキュベーションを実施した。この目的で、イリノイ州ロックフォード(Rockford,IL)のPierceからのDeSalt Spinカラムを使用してカゼイン酸ナトリウムを最初に脱塩し、次に10mM(最終濃度)のEDTAの存在および不在下で、5%カゼイネート溶液をPADと共に45℃およびpH7.2で4時間インキュベートした。実施したアミノ酸分析によれば、どちらのインキュベーションも酸加水分解カゼイネート中に遊離アルギニンの同一の減少(約20%)を示し、フザリウム(Fusarium)PADはCa2+非依存性であることが示唆された。
【0186】
[実施例9]
30%果汁入り清涼飲料
典型的な1食分:240ml
活性成分:
カゼイン加水分解産物(アルギニンの50%はシトルリンに転換されている)および炭水化物源としてマルトデキストリンをこの食品に組み込む:
カゼイン加水分解産物:1.5〜15g/1食当たり
マルトデキストリン:3〜30g/1食当たり
【0187】
[I.以下の成分から清涼飲料化合物を調製する:]
[果汁濃縮物および水溶性香料]
【0188】
【表3】
【0189】
[1.6 活性成分]
上述の濃度の活性成分(これは上述の活性成分を意味する)タンパク質加水分解産物およびマルトデキストリン。
【0190】
果汁濃縮物および水溶性香料を空気を入れずに混合する。着色剤を脱イオン水に溶解する。アスコルビン酸およびクエン酸を水に溶解する。安息香酸ナトリウムを水に溶解する。ペクチンを撹拌しながら添加して沸騰させながら溶解する。溶液を冷却する。オレンジ油および油溶性香料を予混合する。1.6で言及される活性成分を乾燥混合し、次に好ましくは果汁濃縮物混合物(1.1)中で撹拌する。
【0191】
清涼飲料化合物を調製するために、3.1.1〜3.1.6の全ての部分をTurrax、次に高圧ホモジナイザー(p1=200bar、p2=50bar)を使用して、均質化するまで共に混合する。
【0192】
[II.以下の成分から瓶詰シロップを調製する:]
【0193】
【表4】
【0194】
瓶詰めシロップの成分を混ぜ合わせる。瓶詰めシロップを水で希釈して、1リットルのすぐに飲める飲料にする。
【0195】
[バリエーション:]
安息香酸ナトリウムを使用する代わりに、飲料を低温殺菌してもよい。飲料はまた、炭酸ガスを添加してもよい。
【特許請求の範囲】
【請求項1】
シトルリンと、タンパク質またはペプチドの一部であるアルギニン残基とのモル比が、少なくとも0.15、好ましくは少なくとも0.30、より好ましくは少なくとも0.5、なおもより好ましくは少なくとも1.0、さらになおもより好ましくは2.0、最も好ましくは少なくとも4である、タンパク質、ペプチドまたはタンパク質加水分解産物。
【請求項2】
植物タンパク質、脱脂乳タンパク質、乳タンパク質、小麦タンパク質、カゼインタンパク質、ゼラチンタンパク質、卵タンパク質、微生物タンパク質またはそれらの加水分解産物である、請求項1に記載の修飾タンパク質、ペプチドまたはタンパク質加水分解産物。
【請求項3】
平均ペプチド長が3〜9個のアミノ酸である、請求項1または2に記載のタンパク質加水分解産物。
【請求項4】
5〜50の間のDHを有する、請求項1〜3のいずれか一項に記載のタンパク質加水分解産物。
【請求項5】
栄養補助食品または薬物などの食物、飼料もしくは栄養補給食品中における、および栄養補助食品または薬物などの食物、飼料もしくは栄養補給食品の調製における、請求項1〜4のいずれか一項に記載のタンパク質、ペプチドまたはタンパク質加水分解産物の使用。
【請求項6】
タンパク質、ペプチドまたはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換され、タンパク質、ペプチドまたはタンパク質加水分解産物基質がタンパク質アルギニンデイミナーゼと共にインキュベートされる、タンパク質、ペプチドまたはタンパク質加水分解産物を酵素的に生成する方法。
【請求項7】
請求項1〜4のいずれか一項に記載の、または請求項6に記載の方法によって得られる、タンパク質、ペプチドまたはタンパク質加水分解産物を含んでなる食材。
【請求項8】
乳児用調製粉乳または臨床食物である、請求項7に記載の食材。
【請求項9】
(a)配列番号6、8、9、10、13または14の1〜640番目のアミノ酸と少なくとも30%のアミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド、
(b)(i)配列番号3の核酸配列、または60個を超える、好ましくは100個を超えるヌクレオチドと少なくとも80%または90%同一の、より好ましくは200個を超えるヌクレオチドと少なくとも90%同一のその断片、または(ii)配列番号3の核酸配列に相補的な核酸配列と、低ストリンジェンシー条件下でハイブリダイズするポリヌクレオチドによってコードされるポリペプチド
からなる群から選択される、タンパク質アルギニンデイミナーゼ活性を有する単離されたポリペプチド。
【請求項10】
配列番号6の1〜640番目のアミノ酸と、少なくとも40%、好ましくは少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸配列を有する、請求項9に記載のポリペプチド。
【請求項11】
配列番号6のアミノ酸配列を含んでなる、請求項9に記載のポリペプチド。
【請求項12】
(i)配列番号3の核酸配列またはその断片、または(ii)配列番号3の核酸配列に相補的な核酸配列と、低ストリンジェンシー条件、より好ましくは中度のストリンジェンシー条件、最も好ましくは高ストリンジェンシー条件下で、ハイブリダイズするポリヌクレオチドによってコードされる請求項9に記載のポリペプチド。
【請求項13】
真菌、好ましくはアスペルギルス(Aspergillus)、より好ましくはアスペルギルス・ニガー(Aspergillus niger)から得られる、請求項9に記載のポリペプチド。
【請求項14】
請求項9に記載のポリペプチドをコードする核酸配列を含んでなり、または低ストリンジェンシー条件、より好ましくは中度ストリンジェンシー条件、最も好ましくは高ストリンジェンシー条件下で、配列番号3とハイブリダイズする単離されたポリヌクレオチド。
【請求項15】
タンパク質アルギニンデイミナーゼ活性を有するポリペプチドをコードするポリヌクレオチドであって、
(c)アミノ酸配列番号11をコードするポリヌクレオチド配列、および
(d)プレタンパク質シグナル配列をコードするポリヌクレオチド配列を含んでなり、
コードされたプレタンパク質シグナル配列が、コードされたプレポリペプチドのアミノ末端に位置しかつ好ましくは15〜30個のアミノ酸の長さであるポリヌクレオチド。
【請求項16】
適切な発現宿主中でポリペプチドの生成を指示する1つ以上の制御配列と作動的に連結する、請求項14または15に記載のポリヌクレオチドを含んでなる核酸コンストラクト。
【請求項17】
請求項16に記載の核酸コンストラクトを含んでなる組み換え発現ベクター。
【請求項18】
請求項16に記載の核酸コンストラクトまたは請求項17に記載のベクターを含んでなる組み換え宿主細胞。
【請求項19】
タンパク質アルギニンデイミナーゼを分泌できる株または組み換え宿主を培養するステップと、タンパク質アルギニンデイミナーゼを回収するステップとを含んでなる、タンパク質アルギニンデイミナーゼを生成する方法。
【請求項20】
請求項9に記載の株/組み換え宿主細胞を培養して、ポリペプチドを含んでなる上清および/または細胞を生成するステップと、ポリペプチドを回収するステップとを含んでなる、請求項9〜13のいずれか一項に記載のポリペプチドを生成する方法。
【請求項21】
請求項19に記載の方法によって生成されるポリペプチド。
【請求項22】
ポリペプチドの生成に適した条件下で、ポリペプチドをコードするポリヌクレオチドを含んでなる核酸コンストラクトを含んでなる宿主細胞を培養するステップと、ポリペプチドを回収するステップとを含んでなる、請求項9〜13のいずれか一項に記載のポリペプチドを生成する方法。
【請求項23】
請求項20に記載の方法によって生成されるポリペプチド。
【請求項24】
請求項9に記載のタンパク質アルギニンデイミナーゼをコードするDNA分子。
【請求項1】
シトルリンと、タンパク質またはペプチドの一部であるアルギニン残基とのモル比が、少なくとも0.15、好ましくは少なくとも0.30、より好ましくは少なくとも0.5、なおもより好ましくは少なくとも1.0、さらになおもより好ましくは2.0、最も好ましくは少なくとも4である、タンパク質、ペプチドまたはタンパク質加水分解産物。
【請求項2】
植物タンパク質、脱脂乳タンパク質、乳タンパク質、小麦タンパク質、カゼインタンパク質、ゼラチンタンパク質、卵タンパク質、微生物タンパク質またはそれらの加水分解産物である、請求項1に記載の修飾タンパク質、ペプチドまたはタンパク質加水分解産物。
【請求項3】
平均ペプチド長が3〜9個のアミノ酸である、請求項1または2に記載のタンパク質加水分解産物。
【請求項4】
5〜50の間のDHを有する、請求項1〜3のいずれか一項に記載のタンパク質加水分解産物。
【請求項5】
栄養補助食品または薬物などの食物、飼料もしくは栄養補給食品中における、および栄養補助食品または薬物などの食物、飼料もしくは栄養補給食品の調製における、請求項1〜4のいずれか一項に記載のタンパク質、ペプチドまたはタンパク質加水分解産物の使用。
【請求項6】
タンパク質、ペプチドまたはタンパク質加水分解産物中に元々存在するアルギニン残基の少なくとも15%、好ましくは少なくとも30%、より好ましくは少なくとも45%、なおもより好ましくは少なくとも60%、最も好ましくは少なくとも80%がシトルリン残基に転換され、タンパク質、ペプチドまたはタンパク質加水分解産物基質がタンパク質アルギニンデイミナーゼと共にインキュベートされる、タンパク質、ペプチドまたはタンパク質加水分解産物を酵素的に生成する方法。
【請求項7】
請求項1〜4のいずれか一項に記載の、または請求項6に記載の方法によって得られる、タンパク質、ペプチドまたはタンパク質加水分解産物を含んでなる食材。
【請求項8】
乳児用調製粉乳または臨床食物である、請求項7に記載の食材。
【請求項9】
(a)配列番号6、8、9、10、13または14の1〜640番目のアミノ酸と少なくとも30%のアミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド、
(b)(i)配列番号3の核酸配列、または60個を超える、好ましくは100個を超えるヌクレオチドと少なくとも80%または90%同一の、より好ましくは200個を超えるヌクレオチドと少なくとも90%同一のその断片、または(ii)配列番号3の核酸配列に相補的な核酸配列と、低ストリンジェンシー条件下でハイブリダイズするポリヌクレオチドによってコードされるポリペプチド
からなる群から選択される、タンパク質アルギニンデイミナーゼ活性を有する単離されたポリペプチド。
【請求項10】
配列番号6の1〜640番目のアミノ酸と、少なくとも40%、好ましくは少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、さらに最も好ましくは少なくとも約97%の同一性を有するアミノ酸配列を有する、請求項9に記載のポリペプチド。
【請求項11】
配列番号6のアミノ酸配列を含んでなる、請求項9に記載のポリペプチド。
【請求項12】
(i)配列番号3の核酸配列またはその断片、または(ii)配列番号3の核酸配列に相補的な核酸配列と、低ストリンジェンシー条件、より好ましくは中度のストリンジェンシー条件、最も好ましくは高ストリンジェンシー条件下で、ハイブリダイズするポリヌクレオチドによってコードされる請求項9に記載のポリペプチド。
【請求項13】
真菌、好ましくはアスペルギルス(Aspergillus)、より好ましくはアスペルギルス・ニガー(Aspergillus niger)から得られる、請求項9に記載のポリペプチド。
【請求項14】
請求項9に記載のポリペプチドをコードする核酸配列を含んでなり、または低ストリンジェンシー条件、より好ましくは中度ストリンジェンシー条件、最も好ましくは高ストリンジェンシー条件下で、配列番号3とハイブリダイズする単離されたポリヌクレオチド。
【請求項15】
タンパク質アルギニンデイミナーゼ活性を有するポリペプチドをコードするポリヌクレオチドであって、
(c)アミノ酸配列番号11をコードするポリヌクレオチド配列、および
(d)プレタンパク質シグナル配列をコードするポリヌクレオチド配列を含んでなり、
コードされたプレタンパク質シグナル配列が、コードされたプレポリペプチドのアミノ末端に位置しかつ好ましくは15〜30個のアミノ酸の長さであるポリヌクレオチド。
【請求項16】
適切な発現宿主中でポリペプチドの生成を指示する1つ以上の制御配列と作動的に連結する、請求項14または15に記載のポリヌクレオチドを含んでなる核酸コンストラクト。
【請求項17】
請求項16に記載の核酸コンストラクトを含んでなる組み換え発現ベクター。
【請求項18】
請求項16に記載の核酸コンストラクトまたは請求項17に記載のベクターを含んでなる組み換え宿主細胞。
【請求項19】
タンパク質アルギニンデイミナーゼを分泌できる株または組み換え宿主を培養するステップと、タンパク質アルギニンデイミナーゼを回収するステップとを含んでなる、タンパク質アルギニンデイミナーゼを生成する方法。
【請求項20】
請求項9に記載の株/組み換え宿主細胞を培養して、ポリペプチドを含んでなる上清および/または細胞を生成するステップと、ポリペプチドを回収するステップとを含んでなる、請求項9〜13のいずれか一項に記載のポリペプチドを生成する方法。
【請求項21】
請求項19に記載の方法によって生成されるポリペプチド。
【請求項22】
ポリペプチドの生成に適した条件下で、ポリペプチドをコードするポリヌクレオチドを含んでなる核酸コンストラクトを含んでなる宿主細胞を培養するステップと、ポリペプチドを回収するステップとを含んでなる、請求項9〜13のいずれか一項に記載のポリペプチドを生成する方法。
【請求項23】
請求項20に記載の方法によって生成されるポリペプチド。
【請求項24】
請求項9に記載のタンパク質アルギニンデイミナーゼをコードするDNA分子。
【図2A】


【図2B】

【図4】
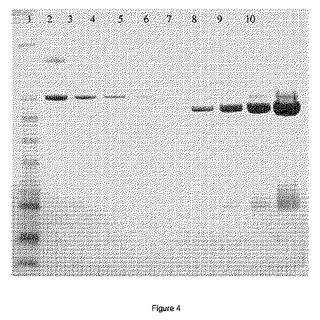
【図1】
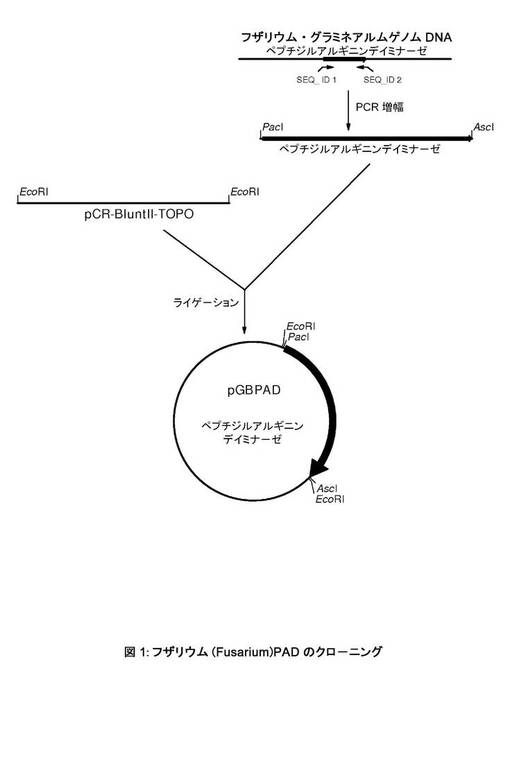
【図3】
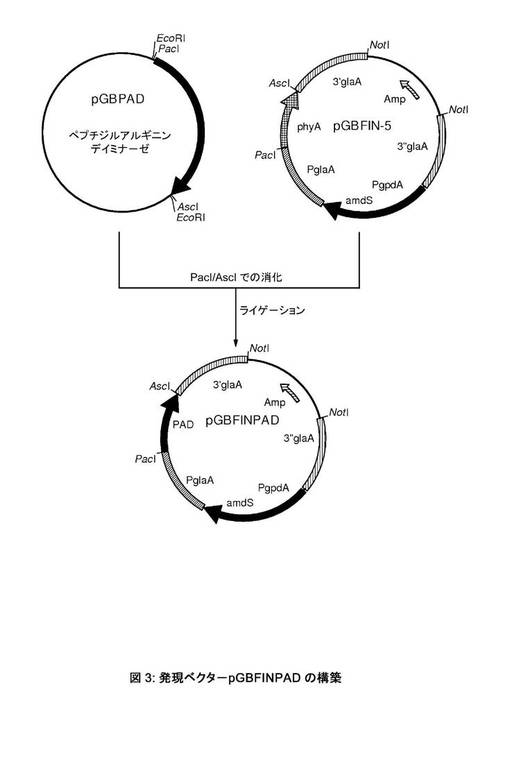
【図2B】
【図4】
【図1】
【図3】
【公表番号】特表2009−540847(P2009−540847A)
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願番号】特願2009−517153(P2009−517153)
【出願日】平成19年6月25日(2007.6.25)
【国際出願番号】PCT/EP2007/056310
【国際公開番号】WO2008/000714
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成21年11月26日(2009.11.26)
【国際特許分類】
【出願日】平成19年6月25日(2007.6.25)
【国際出願番号】PCT/EP2007/056310
【国際公開番号】WO2008/000714
【国際公開日】平成20年1月3日(2008.1.3)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]