
ペプチドおよびタンパク質を加水分解するためのプロリン特異的エンドプロテアーゼの使用
【課題】セリアック病関連エピトープを欠く食物を製造するためのプロリン特異的エンドプロテアーゼの使用法を提供する。
【解決手段】セリアック病関連エピトープを含むコムギまたはオオムギのグルテンをアスペルギルス(Aspergillus)の酵素であるプロリン特異的エンドプロテアーゼで加水分解し、得られたペプチドをセリアック病患者のためのビールおよびパンの製造に用いる。
【解決手段】セリアック病関連エピトープを含むコムギまたはオオムギのグルテンをアスペルギルス(Aspergillus)の酵素であるプロリン特異的エンドプロテアーゼで加水分解し、得られたペプチドをセリアック病患者のためのビールおよびパンの製造に用いる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ペプチドまたはポリペプチドのタンパク質加水分解に関する。
【背景技術】
【0002】
プロリンリッチ食餌タンパク質、例えば、牛乳中カゼインまたは穀物中グルテンは、ヒト消化管におけるタンパク質分解に耐性であることが公知である。その結果、プロリンリッチペプチドが構成され得、そして特定のグループの個体において所望されない影響をもたらし得る。これらの影響のいくつかは、プロリンリッチペプチドが末梢組織および中枢神経系の受容体に結合するオピオイドとして作用するという事実によるとされている。例えば、自閉症および統合失調症患者に見られる症状は、プロリンリッチ食餌タンパク質の体内摂取と関連付けられている。他の影響は、プロリンリッチペプチドに対する不耐症の結果である。例えば、特定のプロリンリッチ配列は、セリアック病において観察されるグルテンの毒性を担う。セリアック病は小腸の広範に認められる自己免疫疾患であり、生涯にわたるグルテンを含まない食餌によってでしか処置することができない。セリアック病は時にはまた、プロリンリッチペプチドの代謝障害が有し得る遠大な結果を例示する精神的および神経学的症状を伴う。
【0003】
牛乳中のタンパク質は成長および健康に関連し、ヒト食餌における重要な成分を形成する。カゼインは牛乳中の全タンパク質の約80%を構成し、アミノ酸、カルシウムおよびリン酸塩の重要な供給源である。カゼインは、概ね50%のα−カゼイン、35%のβ−カゼイン、13%のκ−カゼインおよび3%のγ−カゼインよりなる。ヒト乳汁には、一般にα−カゼイン画分は存在しない。
【0004】
カゼインの代謝においては、多くの新たな生体活性ペプチドが形成されることが公知である。αおよびβ−カゼイン画分からは、それぞれ、α−カゾモルフィンおよびβ−カゾモルフィンと呼ばれるオピオイドペプチドが同定され、単離されている。特に、β−カゾモルフィンの薬理学的効果については、広範に研究されている。配列Tyr−Pro−Phe−Pro−Gly−Pro−Ileを伴うβ−カゾモルフィンは牛乳中の主要なオピオイドペプチドであり、BCM−7と呼ばれる(β−カゾモルフィン(1−7);チャン(Chang)ら(1985)Journal of Biological Chemistry,260,9706−9712)。β−カゼイン分子のアミノ酸位置60〜66におけるこのBCM−7フラグメントの他に、それぞれ、アミノ酸位置60〜63および60〜64におけるTyr−Pro−Phe−Pro(β−カゾモルフィン(1−4))およびTyr−Pro−Phe−Pro−Gly(β−カゾモルフィン(1−5))のようなBCM−7のより小さなフラグメントならびに(アミノ酸位置60〜70における)11アミノ酸の鎖長までのすべてのより大きなBCM−7関連ペプチドは少なくともいくらかの程度のオピオイド活性を示す。BCM−7のN末端トリペプチド、即ち、位置60〜62における配列Tyr−Pro−Pheはオピオイド活性を有さない。A1と呼ばれる遺伝的β−カゼイン変異体(アミノ酸位置67においてA2β−カゼインのプロリン残基ではなくヒスチジンを有する)は、高いレベルのBCM−7分子の形成をもたらすことが要求される。
【0005】
多様なβ−カゾモルフィンの作製のための基本的な理由は、それらのアミノ酸配列が比較的プロリン残基に富むことである。プロリン残基に関与するペプチド結合はタンパク質分解に耐性であるため、β−カゾモルフィン配列は、胃および腸管腔における消化管プロテアーゼへの暴露後も残存する傾向がある。同じ理由により、これらのβ−カゾモルフィン配列は、他のプロテアーゼ、例えば、タンパク質加水分解物の産業的生産において一般に使用されるそれらのプロテアーゼとのインキュベーション後にも残存する傾向があることが想定され得る。この仮定は、一般に利用可能なタンパク質加水分解物またはこれらのタンパク質加水分解物を含有する産物がすべてBCM−7もしくは緊密に関連するペプチドを含有することを含意する。BCM−7ペプチドフラグメントおよびその関連分子は所定の疾患と関連付けられているため、幼児、高齢者および患者のような脆弱なグループの食餌においてかなり頻繁に使用されるタンパク質加水分解物にそのような分子が存在することは、所望されない状況である。ヒトおよびウシβ−カゾモルフィンのオピオイド受容体結合アッセイの結果は、オピオイド活性を伴うフラグメントがラット脳膜のオピオイド受容体に結合することを示す。β−カゼインは、δおよびκ受容体サブタイプにほとんど親和性を伴わないμ−リガンドに対してより選択的であることが示されている。これらおよび他の研究に従えば、β−カゾモルフィンは、多様な消化管、鎮痛性、呼吸器、循環器、内分泌および免疫調節効果を有することが要求される。プロリン残基を組み入れているオピオイドペプチドに共通の構造的特徴は、Tyr−Pro−Phe/Trpモチーフである(オカダ(Okada)ら、Vitamins and Hormones2002,65,257−279)。
【0006】
健康な個体では、腸上皮層および血液におけるペプチダーゼは、β−カゾモルフィンを処理することができるが、これは、統合失調症、自閉症、ADHDまたは他の気分障害を患う患者の場合では必ずしもそうではないようである。例えば、プロリンリッチペプチドの不完全な分解をもたらす血漿中ジペプチジルペプチダーゼIV(DPP IV)酵素活性の遺伝的変更は、これらの疾患の発症と関連付けられている。さらに、高ペプチド尿症、即ち、尿中における高い濃度のカゼインまたはグルテン由来のペプチドが、一定の頻度で認められる(ライヘルト、W.H.(Reichelt,W.H.)ら;(1997)Dev.Brain Dysfunct;10:44−55)。最近の科学的文献は、プロリンリッチペプチドの不完全な分解がそのような疾患の発達および重症度に寄与し得るという説得力のある証拠を提供している。カゼイン(caseine)由来BCM−7フラグメントの他にも、これに関して、グルテン由来プロテアーゼ耐性ペプチドについて述べられている。すでに1979年には、パンクセップ(Panksepp)(Trends in Neuroscience1979;2:174−177)はオピオイド過剰理論を提唱し、その中で彼はオピオイド代謝障害が自閉症の病因の部分であることを示唆した。現在では、本発明者らは、多くのプロリンリッチペプチドが胃および膵臓のペプチダーゼ、例えば、ペプシン、トリプシン、キモトリプシンなどによる切断に高度に耐性であり、なかでも消化管の刷子縁上皮層に存在するような特異的酵素だけが、プロリンに関与するペプチド結合を加水分解することが可能であることを理解した。
【0007】
グルテンはデンプンおよび水溶性成分を除去するための洗浄後に残留するコムギ、ライムギ、カラスムギ、オオムギ、トウモロコシおよびイネのような穀物の不溶性タンパク質画分である。グルテンは、4つの主要な可溶性画分、即ち、アルブミン、グロブリン、プロラミンおよびグルテリンに再分割することができる。これらのなかでも、特に、コムギ、トウモロコシ、オオムギならびにカラスムギのプロラミンおよびグルテリン画分は、比較的高い含有量のアミノ酸、プロリンおよびグルタミンによって特徴付けられる。最近の証拠は、セリアック病の発達における主要な因子として、プロリンリッチグルテン配列を含意している。セリアックスプルーとしても公知であるセリアック病は、グルテンタンパク質の摂取によって生じる小腸の自己免疫疾患である。それは、慢性の下痢および腹部膨満のような重度の症状を伴う幼児において一般に認められ;その後の生涯では、症状は、疲労、吸収不良による体重減少および神経学的症状を含む。多様な穀物のプロリンリッチ画分のなかでも、コムギ由来のα−グリアジン、オオムギ由来のホルデイン、ライムギ由来のセカリンおよびカラスムギ由来のアベニンは、最も毒性が高いようである(Schuppan,D.;Gastroenterology2000;119:234−242)。生涯にわたるグルテンを含まない食餌のみが、セリアック病患者に有効な処置である。セリアック病の患者のなかでも、高い罹病率の多様な自己免疫障害、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎が観察されている。これは、未知の機構によって、非処置のセリアック病が他の器官に対する自己免疫を起こしやすいことを示す(シュパン、D.(Schuppan,D.)2000Gasteroenterology119:234−242)。さらに、軽度の形態のセリアック病が、過敏性腸症候群(IBS)を患うヒトのグループに存在する徴候がある。IBSは大腸の正常な機能を妨害する障害であり、差し込むような腹痛、便秘および下痢によって特徴付けられる。IBSは、通常、約20歳で始まり、多大な不快感および苦痛を生じる。コムギ、オオムギ、ライムギまたは乳製品の摂食は、IBSの症状の悪化と関係付けられている。
【0008】
最近、シャン(Shan)ら(Science;第297巻、2002年9月27日:2275−2279)は、1組の主要なセリアック病の患者特異的T細胞のエピトープの供給源と考えられるグリアジン由来、プロリンリッチ、33アミノ酸長ペプチドを同定した。小腸刷子縁細胞から調製された酵素抽出物はこの33マーを加水分解することができなかったが、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)由来の細菌性プロリルオリゴペプチダーゼの補充は、関連T細胞クローンの刺激の付随的な顕著な減少を伴う迅速な消化をもたらす。パパインの経口投与に関する早期の研究(メッセル、M.(Messer,M.)およびボーメ、P.E.(Baume,P.E.);Lancet1976;2:1022)を模倣すると、該論文より、酵素療法によるグルテンの無毒化における食餌中酵素としてのプロリルオリゴペプチダーゼの能力が示される。
【0009】
プロリルオリゴペプチダーゼ(EC3.4.21.26)は、プロリン残基のカルボキシル側でペプチドを好適に切断する独特な可能性を有する。哺乳動物供給源から単離されるプロリルオリゴペプチダーゼならびにフラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から単離されるプロリルオリゴペプチダーゼにおいて、酵素の活性部位から構造決定された大きなタンパク質を含まない独特なペプチダーゼドメインが同定されている。事実、これらの酵素は、約30を超えるアミノ酸残基を含有するタンパク質を分解することができず、そのため、これらの酵素は、現在、「プロリルオリゴペプチダーゼ」と称される(フロプ(Fulop)ら:Cell、第94巻、161−170、1998年7月24日)。すべての既知のプロリルオリゴペプチダーゼは、中性付近で至適pH値を示すサイトゾルの酵素であり、それらが約30を超えるアミノ酸残基を含有する分子を効率的に分解することができないという事実によって特徴付けられる。これらの酵素が、消化管のより遠位部において示現するpH値に対応する至適pH値を示すという事実は、それらを、食餌中グルテンの腸での消化を支持する栄養補助食品として理想的に安定にする。
【0010】
毒性のプロリンリッチペプチドの不活化において利益を有することができるもう1つの酵素は、酵素ジペプチジルペプチダーゼIVである(米国公開特許第2002/0041871A号明細書)。Xaa−Pro−ジペプチジル−アミノペプチダーゼ(EC3.4.14.5)とも呼ばれるジペプチジルペプチダーゼIVは、好ましくは、Xbbがプロリンであるが、但し、Xccがプロリンでない場合、N末端配列Xaa−Xbb−Xcc−を伴うペプチドからのN末端ジペプチド、Xaa−Xbbの放出を触媒する。ジペプチジル−ペプチダーゼIVが、多くの哺乳動物供給源、例えば、酵素の豊富な供給源由来の腸刷子縁膜から単離されている。さらに、酵素は、微生物供給源、例えば、食物用微生物、サッカロミセス(Saccharomyces)、ラクトコッカス(Lactococcus)およびアスペルギルス(Aspergillus)から単離されている。プロリルオリゴペプチダーゼのように、すべての既知のジペプチジル−ペプチダーゼIVは、中性付近の至適pH値を伴う酵素であり、従って、腸での消化プロセスを支えるのに適している。
【0011】
セリアック病もしくは統合失調症、自閉症または他の気分障害の処置におけるプロリン特異的オリゴペプチダーゼおよびジペプチジル−ペプチダーゼIVの可能な関連のため、これらのデータから、この事柄の多様な局面を取り扱う多くの特許出願が生じている。例えば、米国特許第6,447,772号明細書および国際公開第01/24816号パンフレットはジペプチジルペプチダーゼIVを含有する組成物について記載し、国際公開第03/068170号パンフレットは場合により、ジペプチジル−ペプチダーゼIVと組み合わされたプロリン特異的オリゴペプチダーゼを含有する組成物について記載し、国際公開第02/45523号パンフレットはプロリン特異的エンドプロテアーゼにより調製される低アレルギー性タンパク質加水分解物について記載し、国際公開第03/028745号パンフレットは腸内の毒性プロリンリッチペプチドの濃度を低下することができる細菌株を含んでなる組成物について記載している。国際公開第96/36239号パンフレットは、β−カゼインA1対立遺伝子を実質的に含まないウシから誘導される産物の利点について記載している。
【発明の概要】
【課題を解決するための手段】
【0012】
本発明は、ペプチドまたはポリペプチドのタンパク質加水分解のための方法であって、前記ペプチドまたはポリペプチドは4〜40、好ましくは5〜35アミノ酸残基を含んでなり、前記ペプチドまたはポリペプチドはスブチリシンによって加水分解され得ず、それによって、前記ペプチドまたはポリペプチドは、前記ペプチドまたはポリペプチドを加水分解するための6.5もしくはそれ以下、好ましくは5.5もしくはそれ以下、およびより好ましくは5.0もしくはそれ以下のpHでプロリン特異的エンドプロテアーゼにより加水分解される、方法に関する。好ましくは、少なくとも70%、より好ましくは、少なくとも80%、および最も好ましくは、少なくとも90%のペプチドまたはポリペプチドがこの方法において加水分解される。
【0013】
別の実施態様に従えば、本発明は、ペプチドまたはポリペプチドのタンパク質加水分解のための方法であって、前記ペプチドまたはポリペプチドは4〜40、好ましくは5〜35アミノ酸残基を含んでなり、そしてトリペプチドモチーフGlu−Xxx−Pro、Gln−Xxx−Pro、Tyr−Pro−PheまたはTyr−Pro−Trpを含んでなり、それによって、前記ペプチドまたはポリペプチドは、前記ペプチドまたはポリペプチドを加水分解するための6.5もしくはそれ以下、好ましくは5.5もしくはそれ以下およびより好ましくは5.0もしくはそれ以下のpHでプロリン特異的エンドプロテアーゼにより加水分解される、方法に関する。好ましくは、少なくとも70%、より好ましくは、少なくとも80%、および最も好ましくは、少なくとも90%のペプチドまたはポリペプチドがこの方法において加水分解される。
【0014】
さらに、本発明は、ペプチドまたはポリペプチドのタンパク質加水分解のための方法であって、前記ペプチドまたはポリペプチドは4〜40、好ましくは5〜35アミノ酸残基を含んでなり、それによって、ペプチドまたはポリペプチドのアミノ酸残基は少なくとも30%、好ましくは少なくとも40%のプロリンおよび/またはグルタミン残基を含んでなり、それによって、前記ペプチドまたはポリペプチドは、前記ペプチドまたはポリペプチドを加水分解するための6.5もしくはそれ以下、好ましくは5.5もしくはそれ以下およびより好ましくは5.0もしくはそれ以下のpHでプロリン特異的エンドプロテアーゼにより加水分解されるが、但し、ペプチドまたはポリペプチドは少なくとも10%のプロリン残基を含んでなる、方法を提供する。好ましくは、少なくとも70%、より好ましくは、少なくとも80%、および最も好ましくは、少なくとも90%のペプチドまたはポリペプチドがこの方法において加水分解される。
【0015】
好ましくは、本発明の方法におけるペプチドまたはポリペプチドは、モチーフGln−Xxx−ProまたはGlu−Xxx−Proを含んでなり、9もしくはそれ以上のアミノ酸残基を含有する。このペプチドまたはポリペプチドは、有利なことに、8もしくはそれ以下のアミノ酸残基を含有するペプチドに加水分解される。ペプチドまたはポリペプチドがモチーフTyr−Pro−PheまたはTyr−Pro−Trpを有する場合、好ましくは、ProとPheとの間またはProとTrpとの間の結合が加水分解される。
【0016】
本発明の方法において使用される好適なエンドプロリン特異的エンドプロテアーゼは、好ましくは、アスペルギルス(Aspergillus)から誘導されるか、またはセリンプロテアーゼのS28ファミリーに属するプロリン特異的エンドプロテアーゼである。この酵素は、好ましくは、スブチリシンによって加水分解され得ない4〜40、好ましくは5〜35アミノ酸残基を含んでなるペプチドまたはポリペプチドを加水分解するための6.5未満、好ましくは5.5未満、より好ましくは5.0未満の至適pH値を有する。
【0017】
プロリン特異的エンドプロテアーゼは、自閉症、統合失調症、ADHD、双極性気分障害およびうつを含む精神障害ならびに自己免疫障害、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎のようなセリアック病に関連する障害ならびにIBSに関連するプロリンリッチペプチドを5.5未満のpHで加水分解するために使用することができる。
【0018】
有利なことに、この酵素は、セリアック病関連エピトープ、好ましくはグルテンのエピトープ、より好ましくはコムギまたはオオムギのエピトープを欠く食物、例えば、ビールもしくはパンを生成するために使用される。
【0019】
本発明はまた、医薬品としての使用のためかまたは医薬品の製造における使用のためのプロリン特異的エンドプロテアーゼに関し、該プロリン特異的エンドプロテアーゼは、好ましくは、アスペルギルス(Aspergillus)、より好ましくは、アスペルギルス・ニガー(Aspergillus niger)の酵素である。
【0020】
本発明の別の実施態様に従えば、プロリン特異的エンドプロテアーゼは、自閉症、統合失調症、ADHD、双極性気分障害およびうつを含む精神障害ならびに自己免疫障害(autoimmne disorder)、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎のようなセリアック病に関連する障害ならびにIBSの処置もしくは防止のための栄養補助食品または医薬品の製造のために使用される。
【0021】
有利なことに、プロリン特異的エンドプロテアーゼは、年齢25歳未満の個体のための栄養補助食品もしくは医薬品の製造のために使用される。
【0022】
本発明に従えば、プロリン特異的エンドプロテアーゼはまた、自閉症、統合失調症、ADHD、双極性気分障害およびうつを含む精神障害ならびに自己免疫障害(autoimmne disorder)、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎のようなセリアック病に関連する障害ならびにIBSの処置もしくは防止のための栄養補助食品または医薬品のために使用される。
【0023】
さらに、本発明は、30を超えるアミノ酸残基を有するタンパク質またはペプチドを加水分解するためのプロリン特異的エンドプロテアーゼの使用に関する。本発明に従う使用では、好ましくは、アスペルギルス(Aspergillus)、より好ましくは、A.niger(A.ニガー)のプロリン特異的エンドプロテアーゼが適用される。
【0024】
本発明はまた、ペプシンの存在下、5もしくは5以下のpHで活性であるプロリン特異的プロテアーゼの使用に関する。
【0025】
有利なことに、本発明は、β−カゼインA1またはβ−カゼインA2対立遺伝子を担持するウシから得られる乳タンパク質中に存在する前記ペプチドもしくはタンパク質のタンパク質加水分解の方法に関する。
【0026】
タンパク質はまた、本発明の方法においても使用することができる。第1に、タンパク質は、4〜40アミノ酸残基を含んでなるペプチドに加水分解されなければならない。タンパク質の加水分解は、本発明の方法の前、または同時に行うことができる。
【0027】
さらに、本発明は、非ヒト動物、好ましくは哺乳動物を意図した栄養補助食品の生成のため、医薬品の生成のためまたはペットフードを含む食物の生成のための栄養補助食品として、医薬品としての6.5未満、好ましくは5.5未満、より好ましくは5.0未満の至適pH値を有するプロリン特異的エンドプロテアーゼの使用に関する。
【0028】
一般に、先行技術は、体内摂取の前の食物から毒性のプロリンリッチペプチドを取り出すこと、または不適切な腸内消化プロセスを補整するための調整酵素の経口的供給を目的とする。正常な消化管消化プロセスでは、酵素ペプシンによる胃内のタンパク質分解は、食餌タンパク質の分解における第1の工程であると理解される。第2の工程は、小腸、即ち、胃のすぐ下流に位置する十二指腸および空腸において生じる。十二指腸では、胃内容物の酸性pHは、様々なエンド−およびカルボキシペプチダーゼをも含有する膵液の添加によって生じる。後者の酵素によって触媒されるペプシン分解食餌タンパク質のさらなる分解は、5を超えるpH値で十二指腸および空腸の内腔において達成される。腸壁を超える輸送の前に、第3の工程は、腸(intestestinal)上皮の刷子縁表面膜におけるさらなるペプチド加水分解に関与する。後者の工程は、これらの上皮細胞の膜に位置するDPP IVを含む多くのプロテアーゼによって達成される。先行技術は、いくらかの疾患を患う個体の腸において、本発明は、いくらかのこれらの上皮酵素を目的としており、とりわけ、DPP IVは部分的に不活性であるかまたは存在さえしないことを示唆している。セリアック病の患者では、これらの上皮酵素のレベルはおそらく正常であるが、このグループの個体内では、低レベルの所定のプロリンリッチペプチドであっても強力な炎症性T細胞応答を発揮することができる。先行技術において示される解決方法は、消化器官の管腔に存在する毒性のプロリンリッチペプチドのレベルを最小限にするために、経口用調整酵素をヒト消化器官に導入することである。このアプローチでは、先行技術は、できるだけ天然のヒト酵素を模倣する選択された酵素を有し、即ち、酵素は、pH5を超える条件下の消化器官において活性である。これらの先行技術の酵素は酸性条件下では活性ではないか、またはこれらの先行技術の酵素を含有するオーラル・プレゼンテーション形態は、酸性条件下でのそれらの活性を防止するために適切に被覆される。
【0029】
本発明者らは、胃における酸性条件下でその主要な活性を発達する酵素が、不十分に消化されたプロリンリッチペプチドの問題に対し、より優れた解決を提供することを見出した。このアプローチは新規であり、問題を解決するために文献に記載された酵素以外の酵素を使用する可能性を開く。
【0030】
上記で考察したように、いくらかの刊行物は、オピオイド代謝障害の防止におけるジペプチジルペプチダーゼIVおよびプロリルオリゴペプチダーゼのような酵素を使用する可能性について指摘している。さらに、中性付近のpH条件下で典型的に活性であるジペプチジルペプチダーゼIVならびにプロリルオリゴペプチダーゼ酵素が選択される。ここで意味することは、これらの酵素のみが、pH5を超える条件下で効率的にプロリンリッチタンパク質を加水分解することである。ヒト消化管のpHプロフィールを考慮すると、これらの先行技術の酵素は、十二指腸の遠位部、即ち、胃よりかなり先方に到達する前に、活性になることはない。しかし、十二指腸の遠位部は、タンパク質吸収の主要部位であり、また、セリアック病患者では影響を受けることが公知である。そのため、事実上、これらの先行技術の酵素は、障害の部分を受けている場所で作用し始める。
【0031】
さらに、先行技術の酵素は、プロリンリッチタンパク質の有意なプレ消化後にのみ活性になる。膵臓の酵素によるこのようなプレ消化を達成するのに必要な時間は、経口的に適用された酵素がプロリンリッチ配列の完全な加水分解を達成するのに利用可能な期間をさらに制限する。本発明に従えば、食物が小腸に侵入する前にほとんどのプロリンリッチおよび/またはグルタミンリッチタンパク質配列を分解する酵素が使用される。胃における正常な食物滞留時間は、プロリンリッチまたはグルタミンリッチタンパク質の適切な加水分解に必要な期間を提供する。さらに、本発明に従えば、好ましくは、プロリン残基のカルボキシル側でペプチドを切断することが可能であり、また、プロリン残基のカルボキシル側でかつペプシンの存在下であっても無傷なタンパク質を切断することが可能である酵素が使用される。先行技術の酵素は、胃の低いpH条件下で、かつ胃に分泌されるタンパク質分解性ペプシン酵素に暴露される場合、加水分解される。しかし、これまで、そのような酸性/ペプシン安定性酵素の使用は、現在の方法では知られていない。本アプローチは、腸においてプロリンリッチペプチドを加水分解する天然のプロセスを模倣することを提唱する先行技術のアプローチの代わりに、腸に到達する前にプロリンリッチペプチド、ポリペプチドおよびタンパク質の実質的な分解を生じる。本発明の方法において使用される好適な酵素は、5未満のpH値、ペプシンの存在下で活性である酵素であり、食餌タンパク質またはポリペプチドならびにペプチドを分解することが可能である。先行技術の解決方法は、常に、食餌タンパク質をペプチドにプレ加水分解する補酵素を必要とし、その後にのみ、さらなるプロリン特異的酵素がそれらの活性を展開し始めることができる。
【0032】
「ペプチド」または「オリゴペプチド」は、本明細書において、ペプチド結合を介して連結される2〜30アミノ酸残基の鎖として規定される。用語「ペプチド」および「オリゴペプチド」は、同義(共通認識される)とみなされ、各用語は、状況に応じて交換可能に使用することができる。「ポリペプチド」は、本明細書において、30を超えるアミノ酸残基を含有する鎖として規定される。
【0033】
スブチリシン(EC3.4.21.62)、好ましくは、スブチリシンCarlsbergによって加水分解され得ない4〜40アミノ酸残基を有するペプチドまたはポリペプチドは、20g/lタンパク質および1グラムのタンパク質あたり0.12AU−A(アンソン単位アルカラーゼ(Alcalase))プロテアーゼ単位の酵素対基質を含有する懸濁液または溶液において、pH8.0ならびに60℃で2時間のインキュベーション後にも無傷(intact)を保持するペプチドまたはポリペプチドであると理解される。AU−Aプロテアーゼ単位は、Novozymes(丁国)により提出された分析方法LUNA番号2003−32153−01において特定されるように規定される。無傷な(intact)とは、インキュベーション後、本来のペプチドまたはポリペプチドが80%を超え、好ましくは90%を超え、より好ましくは、95%を超えて得られる酵素反応産物を形成することを意味する。実際には、酵素消化は、示される条件下、および存在する1グラムの基質タンパク質あたりの40マイクロリットルのアルカラーゼ(Alcalase)を使用して、行われる。そのような非加水分解性ペプチドの例は、実施例4に記載のβ−カゼイン加水分解から得られるVYPFPGPIPNペプチドである。そのような非加水分解性ポリペプチドのもう1つの例は、実施例6に記載の33マーである。
【0034】
オリゴペプチダーゼは、EC3.4.21.26として分類される酵素であり、セリンプロテアーゼclanSCのファミリー、ファミリーS9に属する。
【0035】
本明細書に記載のすべての(オリゴ)ペプチドおよびポリペプチドの式または配列は、定法に従い、アミノ末端からカルボキシ末端の方向で、左から右に記載される。本明細書において使用するアミノ酸の1文字コードは、当該分野において一般に公知であり、サンブルック(Sambrook)ら(Molecular Cloning:A Laboratory Manual、第2版Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor、ニューヨーク州、1989年)において見出すことができる。アミノ酸Xxx、Xaa、XbbまたはXccは、任意のアミノ酸であることが意図される。モチーフとは、ペプチド、ポリペプチドまたはタンパク質の部分であるアミノ酸配列を意味する。ここで、グルタミン(QもしくはGln)は、グルタミンまたはグルタミン酸(EもしくはGlu)であることが理解される。例えば、化学的に脱アミド化されたグルテンでは、存在する天然のグルタミン残基の大きな部分が、グルタミン酸に変換される。そのため、モチーフGln−Xxx−Proはまた、モチーフGlu−Xxx−Proも含んでなっている。また、他のペプチドでは、ポリペプチドまたはタンパク質のグルタミン(glumine)を、例えば、pH条件、温度によりまたは例えば、トランスグルタミナーゼによる酵素変換によって、グルタミン酸に変換することができる。
【0036】
プロリンリッチペプチドまたはポリペプチドとは、4〜40アミノ酸残基を含んでなるペプチドまたはポリペプチドを意味し、ここで、ペプチドまたはポリペプチドのアミノ酸残基は、少なくとも30%、好ましくは、少なくとも40%のプロリンおよび/またはグルタミン残基を含んでなるが、但し、ペプチドまたはポリペプチドは少なくとも10%のプロリン残基を含んでなる。栄養補助食品は、DSHEA、以下の食餌成分の1つもしくはそれ以上を有するかまたは含有する食餌を補助することを目的とした製品(タバコ以外)の適応された規定に従う:酵素を含むタンパク質、ポリペプチド、ペプチド、ビタミン、ミネラル、ハーブまたは他の植物性薬品、アミノ酸、全摂取量を増加することによって食餌を補助するためのヒトによる使用のための食餌物質、または濃縮物、代謝物、構成物、抽出物、あるいはこれらの成分の組み合わせ。
【0037】
さらに、栄養補助食品は、
・丸剤、カプセル剤、錠剤、または液体剤形での摂取を目的とする。
・従来の食物としてまたは食事もしくは食餌の単独の品目としての使用には該当しない。
・一般に、「栄養補助食品」として標識される。
・承認された新規の薬物、保証された抗生物質、または承認、保証、もしくは認可前に食物の栄養補助食品として市販された認可された生物工学製品のような製品を含む(但し、Secretary of Health and Human Servicesがこの条項を放棄する場合を除く)。
【0038】
毒性のプロリンリッチペプチドまたはポリペプチドは、オピオイド受容体への結合または精神もしくはセリアック障害の発達もしくは重症度において関連するペプチドまたはポリペプチドである。
【0039】
精神障害としては、自閉症、統合失調症、ADHD、双極性気分障害ならびにうつが挙げられる。
【0040】
セリアック病に関連する障害は、自己免疫障害、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎である。
【0041】
IUMB由来のすべての酵素の分類および命名法についての国際的に認められたスキームは、プロテアーゼを含む。プロテアーゼEC番号の更新されたIUMBテキストは、インターネットサイト:http://www.chem.qmw/ac.uk/iubmb/enzyme/EC3/4/11/で見出すことができる。このシステムでは、酵素は、それらが単一の反応を触媒するという事実によって規定される。該システムは、プロテアーゼをエンドおよびエキソプロテアーゼを分類する。エンドペプチダーゼは、内部のペプチド結合を加水分解するそれらの酵素であり、エキソプロテアーゼは、末端のα−アミノ基に隣接するペプチド結合(「アミノペプチダーゼ」)、または末端のカルボキシル基と最後から2番目のアミノ酸との間のペプチド結合(「カルボキシペプチダーゼ」)を加水分解する。エンドペプチダーゼは、触媒機構に基づいてサブ‐サブクラスに分けられる。セリンエンドペプチダーゼ(EC3.4.21)、システインエンドペプチダーゼ(EC3.4.22)、アスパラギン酸エンドプロテアーゼ(EC3.4.23)、メタロエンドプロテアーゼ(EC3.4.24)およびスレオニンエンドプロテアーゼ(EC3.4.25)のサブ‐サブクラスが存在する。
【0042】
国際公開第02/45524号パンフレットは、アスペルギルス・ニガー(Aspergillus niger)から入手可能なプロリン特異的エンドプロテアーゼについて説明している。驚くべきことに、本発明者らは、今回、このアスペルギルス(Aspergillus)の酵素は、胃の酸性条件下で本方法において有利に使用され、これらの条件下でも無傷な食餌タンパク質、ポリペプチドならびにより小さなペプチド分子を加水分解することができることを見出した。さらに、この酵素は、酸性条件で酵素ペプシンの存在下でも残存し、十二指腸通じてその活性を継続するようである。本発明者らは、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが、活性ならびに進化の観点から既知のプロリン特異的プロテアーゼおよび国際公開第03/068170号パンフレットにおいて特定されるグルテナーゼとはまったく異なることを実証している。後者の特徴は、国際公開第03/068170号パンフレットにおいて特定されるグルテナーゼとA.niger(A.ニガー)由来酵素との間のアミノ酸配列相同性が、グローバルアラインメントアルゴリズム解析を使用して典型的に20%未満であるという事実により十分に実証される。この結果は、プロリルオリゴペプチダーゼが、本発明に従うプロリン特異的エンドプロテアーゼが単離されるアスペルギルス・ニガー(Aspergillus niger)のような真菌では発生しないという現在の観点(ヴェナライネンJ.I.(Venaelaeinen,J.I.)ら、Eur J Biochem271,2705−2715(2004))と一致する。
【0043】
本発明に従うかまたは本発明に従って使用されるプロリン特異的エンドプロテアーゼとは、例えば、本明細書において参考として援用される国際公開第02/45524号パンフレットの請求項1〜5、11および13に記載のポリペプチドを意味する。従って、このプロリン特異的エンドプロテアーゼは、以下よりなる群から選択されるプロリン特異的エンド型プロテアーゼ活性を有するポリペプチドである:
(a)配列番号2のアミノ酸1〜526またはそのフラグメントと少なくとも40%アミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド;
(b)低いストリンジェンシー条件で、(i)少なくとも80%もしくは90%同一な60を超える、好ましくは100を超えるヌクレオチド、より好ましくは少なくとも90%同一な200を超えるヌクレオチドである配列番号1の核酸配列またはそのフラグメント、あるいは(ii)配列番号1の核酸配列に相補的な核酸配列、とハイブリダイズするポリヌクレオチドによってコードされるポリペプチド。配列番号1および配列番号2は国際公開第02/45524号パンフレットに示される通りである。好ましくは、ポリペプチドは単離された形態である。
【0044】
好適なポリペプチドは、配列番号2のアミノ酸1〜526と少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも65%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、およびさらに最も好ましくは少なくとも約97%同一性を有するかまたは配列番号2のアミノ酸配列を含んでなる。
【0045】
好ましくは、ポリペプチドは、低いストリンジェンシー条件、より好ましくは中等度のストリンジェンシー条件、および最も好ましくは高いストリンジェンシー条件下、(i)配列番号1の核酸配列もしくはそのフラグメント、または(ii)配列番号1の核酸配列に相補的な核酸配列とハイブリダイズするポリヌクレオチドによってコードされる。
【0046】
用語「ハイブリダイズすることが可能」は、本発明の標的ポリヌクレオチドが、プローブとして使用される核酸(例えば、配列番号1のヌクレオチド配列、もしくはそのフラグメント、または配列番号1の相補物)に、バックグランドよりも有意に高いレベルでハイブリダイズすることができることを意味する。本発明はまた、本発明のプロリン特異的エンドプロテアーゼをコードするポリヌクレオチド、ならびにそれに相補的であるヌクレオチド配列を含む。ヌクレオチド配列は、RNAまたはゲノムDNA、合成DNAもしくはcDNAを含むDNAであってもよい。好ましくは、ヌクレオチド配列は、DNAおよび最も好ましくは、ゲノムDNA配列である。典型的に、本発明のポリヌクレオチドは、選択的条件下で、コーディング配列または配列番号1のコーディング配列の相補物にハイブリダイズすることが可能であるヌクレオチドの連続配列を含んでなる。そのようなヌクレオチドは、当該分野において周知の方法に従って合成することができる。
【0047】
本発明のポリヌクレオチドは、コーディング配列または配列番号1のコーディング配列の相補物に、バックグランドよりも有意に高いレベルでハイブリダイズすることができる。例えば、cDNAライブラリーに存在する他のcDNAのため、バックグランドのハイブリダイゼーションが生じ得る。本発明のポリヌクレオチドとコーディング配列または配列番号1のコーディング配列の相補物との間の相互作用によって作製されるシグナルレベルは、典型的に、他のポリヌクレオチドと配列番号1のコーディング配列との間の相互作用の少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも50倍、およびさらにより好ましくは少なくとも100倍の強度である。相互作用の強度は、例えば、プローブを、例えば、32Pで放射性標識することによって、測定することができる。選択的ハイブリダイゼーションは、典型的に、低いストリンジェンシー(0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約40℃)、中等度のストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約50℃)または高いストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約60℃)の条件を使用して、達成することができる。
【0048】
UWGCGパッケージは、(例えば、そのデフォルト設定値に対して使用される)同一性を算出するために使用することができるBESTFITプログラムを提供する。
【0049】
PILEUPおよびBLAST Nアルゴリズムを使用して、配列同一性を算出するかまたは(例えば、それらのデフォルト設定値に対する等価もしくは対応する配列を同定するような)配列を列挙することができる。
【0050】
BLAST解析を実施するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に入手することができる。このアルゴリズムは、データベース配列において同じ長さのワードと共に整列される場合、いくつかのポジティブ評価された閾値スコアTに一致または満たすかのいずれかである問い合わせ配列における長さWの短いワードを同定することによって、ハイスコアリング配列対(high scoring sequence pair)(HSP)を第1に同定することに関与する。Tは隣接ワードスコア閾値と称される。これらの初期の隣接ワードのヒットは、それらを含有するHSPを見出すための検索を初期化するためのシードとして作用する。ワードヒットは、累積アラインメントスコアが増加し得る限り、各配列に沿って両方向において拡張される。累積アラインメントスコアがその最大到達値から量Xだけ低下する場合;1つもしくはそれ以上のネガティブスコアリング残基アラインメントの累積により累積スコアが0もしくはそれ以下となる場合;またはいずれかの配列の末端に到達する場合、各方向におけるワードヒットの拡張は停止する。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプログラムは、デフォルトとして、11のワード長(W)、50のBLOSUM62スコアリングマトリックスアラインメント(B)、10の期待値(E)、M=5、N=4、および両鎖の比較を使用する。
【0051】
BLASTアルゴリズムは、2つの配列間の類似性の統計解析を実施する。BLASTアルゴリズムによって提供される類似性の1つの測定は、2つのヌクレオチドまたはアミノ酸配列間の一致が偶然生じる確率の示度を提供する最小の全確率(P(N))である。例えば、配列は、第1の配列と第2の配列との比較における最小全確率が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、および最も好ましくは約0.001未満である場合、もう1つの配列に類似であるとみなされる。
【0052】
本発明に従う酵素は、好ましくは、真菌、好ましくは、アスペルギルス(Aspergillus)、より好ましくはアスペルギルス・ニガー(Aspergillus niger)から得られる。そのため、アスペルギルス(Aspergillus)由来酵素は、酸性安定であり、好ましくは、胃において示現するpH条件下でのペプシンの存在の影響を受けない真のエンドペプチダーゼとして見出される。フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から得ることができる酵素を一般に含むプロリン特異的オリゴペプチダーゼは、pH5以下でほとんど活性を有さず、低pHおよび酵素ペプシンの存在の組み合わせにより不活化される。さらに、プロリン特異的プロテアーゼは、一般に、無傷なタンパク質を分解することができず、より小さなペプチドのみ、即ち、約30アミノ酸残基の長さまでのペプチドを効率的に加水分解することが見出された。
【0053】
本発明は、毒性のプロリンリッチペプチドもしくはオピオイド代謝障害の現象を延期または最小限にする経済的な食品用組成物を提供する。組成物は、食物的、薬学的および獣医学的使用に適した経口用酵素処方ならびに有意に低減されたレベルのオピオイドまたは毒性プロリンリッチペプチドを伴うタンパク質加水分解物および食品の生成に適した酵素処方を含む。
【0054】
本発明は、精神障害もしくはセリアック病またはオピオイド代謝障害の発達に関連するプロリンリッチペプチドまたはポリペプチドを加水分解するための方法を開示する。本発明はまた、幼児におけるかもしくは一般に年齢25歳未満の個体のそのような障害の発達を防止または遅延することができる食物を生成する方法についても開示する。また、セリアック病およびIBSに関係する胃の症状を患う人により良好に忍容性である食物は、そのような方法によって調製することができる。本発明の実施形態は、体内摂取前のこれらのプロリンリッチペプチドまたはポリペプチドの分解に関連付けられ、それによって、身体の毒性プロリンリッチペプチドへの暴露が防止または最小限にされる。幼児では、これによりオピオイドへの早期の暴露が回避され、未成熟な免疫系は、そのような毒性のプロリンリッチペプチドに感作されない。また、十代および成人については、低いレベルの毒性プロリンリッチペプチドを含有する食餌は、例えば、看過され得るセリアック病またはIBSを患うヒトに対し予防的利益を有する。本発明はまた、胃、即ち、酸性pH条件下、好ましくはpH5未満の条件下で、これらの毒性のプロリンリッチペプチドまたはペプチドの身体(ヒトもしくは動物)における分解に安定な酵素の補充に関する。後者の実施態様に従えば、セリアック病、セリアック病の発症に関係する疾患またはこれらのペプチドまたはポリペプチドの分解に必要な低いレベルの身体中プロリン特異的プロテアーゼによって生じる疾患を患うヒトが、胃および十二指腸の近接部において関連のプロリンリッチペプチドまたはポリペプチドを分解することも可能である。さらに、本発明に従う酵素は、保護被覆を必要としない。
【0055】
好ましくは、中性pH条件下でのペプチドもしくはポリペプチドまたはタンパク質とスブチリシン(EC3.4.21.62)、好ましくは、バチルス・リケニフォルミス(Bacillus licheniformis)のスブチリシン(またはスブチリシンCarlsberg)とのインキュベーション時に形成される少なくとも80%の毒性プロリンリッチペプチドまたはポリペプチドが、本発明に従うプロリン特異的エンドプロテアーゼによって加水分解される。そのようなスブチリシン耐性ペプチドの形成を、本出願の実施例4に例示する。そのようなスブチリシン耐性プロリンリッチペプチドは、しばしば、上記の疾患と関連付けられる。これらのペプチドの例には、BCM−7、BCM−7関連ペプチド、即ち、β−カゼイン分子の位置60〜63に存在するアミノ酸配列YPFPを含んでなるペプチドがある。さらに、モチーフGln−Xxx−Pro(Q−X−P)を含んでなるグリアジン由来ペプチド、例えば、PYPQPQLPYエピトープ、ならびにグリアジン、ホルデイン、セカリンまたはアベニンから得ることができるこのQ−X−PまたはE−X−Pモチーフを含んでなる他のスブチリシン耐性分子がその例である。スブチリシンによる加水分解によって形成され得るプロリンリッチペプチドのうち、より好ましくは、少なくとも90%、なおより好ましくは、少なくとも95%および最も好ましくは、少なくとも99%が、本発明に従うプロリン特異的エンドプロテアーゼを使用することによって分解されるか、または形成されない。最も好ましくは、これらのプロリンリッチペプチドは、pH5.5未満の条件下で本発明の方法に従って分解することができる。
【0056】
必要とする酵素を供給する1つの方法は、例えば、食餌中プロリンリッチペプチドまたはポリペプチドの胃腸消化を援助するために食物と共に同時摂取される安定化された酵素処方としての消化剤の形態であり得る。もう1つの方法は、例えばアスペルギルス(Aspergillus)の酵素によって「予め消化された」タンパク質食物を使用することによって、問題のある毒性のプロリンリッチ配列の摂取を防止または制限することである。そのようなタンパク質食物は、加水分解物、例えば、グルテンまたは乳タンパク質加水分解物の形態で供給され得、多量の遊離アミノ酸を遊離するためのエンド−ならびにエキソプロテアーゼによって広範に消化されている加水分解物を含む。後者のアプリケーションの典型的な例は、とりわけセイボリーアプリケーションのためのグルタミン酸に富むグルテン加水分解物の作製であり得る。加水分解物は、そのままの形で体内摂取され得るか、または食物成分として役立ち得る。別の方法では、本発明は、組成物が本発明に従う酸性に安定なプロリン特異的エンドペプチダーゼを含んでなることを特徴とする食品の忍容性を改善するための組成物の使用を提供する。さらに別の方法では、本発明は、食品の改善された忍容性のための酸性に安定なプロリン特異的エンドペプチダーゼの添加を含んでなる食品を調製するための方法を提供する。すべてのそのようなアプリケーションにおいて、プロリン特異的酵素は、いわゆる加工助剤として使用され得る。
【0057】
アスペルギルス(Aspergillus)属の株は食物用として位置付けられており、これらの微生物由来の酵素は、思いも寄らない食物用供給源を形成することが公知である。別の好適な実施態様に従えば、酵素は、非分泌型のいわゆるサイトゾルまたはペリプラズム酵素を提示するのではなく、その産生細胞によって分泌される。この方法では、酵素は、高価な精製工程を伴わずに、本質的に純粋な状態で細胞ブロスから回収することができる。好ましくは、酵素は、示現するpHおよび温度条件下でその基質に対して高い親和性を有する。好ましくは、酵素は、消化管の当該関連部位のpHで、消化管タンパク質分解酵素、例えば、ペプシンまたはトリプシン、エラスターゼおよびキモトリプシンによって不活化されない。より好ましくは、酵素は、胃ならびに十二指腸において活性であり、保護被覆を必要としない。
【0058】
ヒトの消化管は、異なるコンパートメントの連続である。食物が摂取され、嚥下された後、それは、胃に到達し、ここで、それは酸およびエンドペプチダーゼのペプシンと混合される。胃における固形食物の典型的な残留時間は、1〜数時間の範囲である。幽門が時々開放することにより、酸性化されかつ部分的に加水分解された食物を小腸に流動させることが可能である。小腸の第1の部分、即ち、十二指腸では、胆汁ならびに膵液が添加される。膵液は、胃の内容物を部分的に中和するための重炭酸塩を含有する。膵液はまた、プロテアーゼのさらなる組、即ち、エンドペプチダーゼ、トリプシン、キモトリプシンおよびエラスターゼならびに胃においてペプシンにより形成されるペプチドおよびポリペプチドをさらに分解するためのカルボキシペプチダーゼAおよびBを含有する。十二指腸後、消化物は空腸に到達する。十二指腸および空腸は、消化管におけるタンパク質吸収の主要な部位である。このような吸収プロセスは、腸管上皮の刷子縁細胞に固定された異なるプロテアーゼによる食餌タンパク質のさらなるタンパク質分解に関与する。後者の加水分解は、腸壁の上での小さなペプチドならびに遊離のアミノ酸の促進された輸送によって達成される。小腸の最後の部分は回腸によって形成され、その後、消化物は大腸(直腸)に進入する。直腸では、集中的な発酵が存在するが、アミノ酸またはペプチドの吸収は認められない。
【0059】
セリアック病患者によるグルテンの摂取により、近位小腸の病巣がもたらされる。腸絨毛の萎縮は、そのような病巣の1つの最も特徴的な特徴である。この絨毛の萎縮は、空腸に限定されず、遠位の十二指腸においても実証され得る(メイジャー,J.W.R.(Meijer,J.W.R.)ら;Virchows Arch.2003年2月;442:124−128)。このような観察は、セリアック病の患者における毒性のプロリンリッチペプチドの損傷の影響は、胃のすぐ下流の領域において常に明らかであることを示す。本発明者らは、セリアック病の症状の防止を目的とする酵素療法が、消化管のより遠位部ではなく、胃における関連のプロリンリッチペプチドを加水分解することを意図すべきであることを見出した。有利なことに、酵素療法において適用される酵素の活性は、胃において作用し始め、十二指腸においても継続して活性であるほどである。本発明者らは、精神、呼吸器および循環器障害の発達において役割を果たすオピオイドペプチドもまた、胃において最も良好に破壊されることを見出した。十二指腸および空腸はタンパク質吸収の主要部位であるため、小腸のこの部位における毒性ペプチドのレベルは、これらの毒性ペプチドの存在に関係する症状を最小限にするためにできるだけ低くあるべきである。
【0060】
消化剤としての使用では、調整酵素は、37℃の温度で十分に活性あるべきであり、好ましくは、胃における酸性条件下で残存するために低い至適pH値を有するべきである。公開されたデータに従えば、摂取された食物の酸性度は、摂取の30分後、初期値のpH5からpH3.5に減少し、続いて、摂取60分後にさらにpH2に減少する(論文マンス・ミンクス(Mans Minkus);University of Utrecht;蘭国;ISBN:90−393−1666−X)。そのため、酵素療法を意図する理想的な酵素は、2のような低いpH値下で活性であるべきである。胃内容排出に関する研究では、摂取45分後、固形食物のほぼ90%がなお胃に存在することが示されている(R.ノチボル(R.Notivol)ら、1984年、Scand J.Gastroenterol.19:1107−1113)。胃より先では、食物のpHは緩徐に上昇して、十二指腸の遠位部、即ち、幽門部より約50cm先で5のpHに到達する(Handbook of Physiology,American Physiological Society、ワシントン市、1968年、ベルナー・ハイデル(Werner Heidel)編;セクション6:Alimentary Canal、第111巻、1457−1490頁)。国際公開第03/068170号パンフレットでは、患者による摂取の前または後のいずれかで、食材中の毒性グルテンオリゴペプチドのレベルを減少することができる異なる「グルテナーゼ」が特定されている。用語「グルテナーゼ」は、プロテアーゼまたはペプチダーゼ酵素、より具体的には、グルテンタンパク質の毒性オリゴペプチドを切断して非毒性のフラグメントにすることが可能であるプロピル特異的プロテアーゼを指す。これらのすべてのグルタナーゼのすべては、典型的に腸において活性であることが意図されるため、それらは、中性付近のpH条件下で、最適に活性である。国際公開第03/068170号パンフレットは、胃または十二指腸の近接部ではなく、十二指腸中もしくはそれより先における毒性のプロリンリッチペプチドの分解について教示している。さらに、国際公開第03/068170号パンフレットは、存在する酵素が胃を通過し、中性pH条件下の腸においてのみ活性となるように、低いpHで酵素活性を遮蔽するいわゆる腸溶コーティングによって保護される酵素調製物を目的とする。
【0061】
タンパク質加水分解物の生成における産業用加工助剤として使用される場合、酵素は、非滅菌産業的条件下での微生物学的に安全なインキュベーションを可能にする条件下で、十分に活性であるべきである。少なくとも50℃のプロセシング温度およびpH5.5よりも十分に低いpH値での適切な酵素活性は、これらの要件を満たす。
【0062】
担子菌アガリクス・ビスポラス(Agaricus bisporus)(サッタル(Sattar)ら、J.Biochem.107,256−261(1990))および非関連の子嚢菌アスペルギルス・ニガー(Aspergillus niger)(国際公開第02/45524号パンフレット)は、両方とも、細胞外プロリルエンドペプチダーゼを産生することが明らかにされている。しかし、担子菌から得られる酵素は、5未満のpH値では残存せず、従って、あまり興味を引かない。好ましくは、酸性の至適pH値を有するA.niger(A.ニガー)由来のプロリルエンドペプチダーゼが使用される。
【0063】
本発明は、低コスト、法的許容性とプロリンリッチペプチド配列に対する酸性pH条件下で証明された有効性とを組み合わせた酵素調製物を提供する。好ましくは、同酵素を使用して、A1ならびにA2型β−カゾモルフィンだけではなく、多様なグルテンのエピトープをも分解することができる。
【0064】
このアスペルギルス(Aspergillus)由来プロリン特異的エンドプロテアーゼは、プロリンリッチペプチドまたはポリペプチドまたはさらにタンパク質の分解において極めて活性であることが見出されている。有利なことに、この酵素は、産生微生物によって発酵ブロスに分泌され、酸性の至適pH値を有し、食物用かつ経済的方法で産生させることができる。関連のβ−カゾモルフィンペプチドは、分子中に4個までのプロリン残基を含有し、さらに、A1およびA2β−カゾモルフィンは異なるアミノ酸配列を有する。かなり驚くべきことに、本発明者らは、アスペルギルス(Aspergillus)の酵素が位置61のプロリンのC末端側でβ−カゾモルフィンを加水分解することが可能であり、従って、A1またはA2β−カゼインの両方について、すべてのBCM−7およびBCM−7関連ペプチドを効率的に不活化することを見出した。本発明者らは、例えば、アルカラーゼ(Alcalase)のように市販のスブチリシン(EC3.4.21.64)のような広い基質特異性を伴う広範に使用されかつ高度に強度のエンドペプチダーゼが、BCM−7を分解することができないことを見出したため、これはかなり注目に値する。同様に、産業上利用可能なプロテアーゼは、BCM−7を分解することができない。事実、アスペルギルス(Aspergillus)由来プロリン特異的エンドプロテアーゼは、β−カゾモルフィン(beta−casomophine)を、但し、かなり驚くべきことに、BCM−7に存在する4個のプロリン残基のうちの1箇所のみで加水分解することができる。それにもかかわらず、この特定のプロリン残基に対するこの特定のアスペルギルス(Aspergillus)の酵素の特異性のため、BCM−7ならびにすべてのBCM−7関連分子は、Tyr−Pro−Pheモチーフが酵素によって切断されるため、この酵素とのインキュベーションによって効率的に分解される。さらに、β−カゼイン由来のA1ならびにA2から誘導されるBCM−7分子は、アスペルギルス(Aspergillus)プロリン特異的エンドプロテアーゼとのインキュベーションによって不活化される。
【0065】
真のプロリン特異的エンドプロテアーゼの利点は、プロリン特異的エンドプロテアーゼは、酵素とタンパク質とを接触させるときに直ちにプロリンリッチ配列を加水分解し始めることができることである。プロリルオリゴペプチダーゼは、例えば、他の消化管エンドペプチダーゼによって、グルテンまたはカゼイン分子の有意なプレ消化後にのみ活性になることができる。必要とされる広範な分解および食物が小腸に進入する前に利用することができる限られた時間の観点から、真のプロリン特異的エンドプロテアーゼは、既知のオリゴペプチダーゼを超える重要なアプリケーションの利点を有する。真のプロリン特異的エンドペプチダーゼの別の利点は、グルテン構造の全体的な破壊を伴わずに、グルテン中の毒性プロリンリッチペプチドのレベルを減少するために、それを産業的に使用することができることである。例えば、アスペルギルス(Aspergillus)由来酵素と共にコムギグルテンのペーストを押し出し形成すると、いくつかの残留する基本構造特性を伴うが、強度に減少したレベルの毒性プロリンリッチペプチドを伴う生成物を生じる。プロリルオリゴペプチダーゼによる毒性プロリンリッチペプチドの同じような減少を達成するために、他のプロテアーゼによるほとんどすべてのプレ加水分解が、30アミノ酸残基未満の所望されるペプチド長を達成するのに必要であり得る。これにより、グルテンのすべての関連する物理化学的特性の完全な消失、即ち、生地の固さが消失し、許容可能な形状および容積でパンを焼くことができなくなることは言うまでもない。ディ・カグノ(Di Cagno)ら(Appl.Environ.Microbiol.、第70(2)巻、1088−1096、2004年)は、セリアックスプルー患者によって良好に忍容されるサワー種のパンの作製について報告している。彼らのアプローチでは、高い割合の非毒性小麦粉で調製した生地を使用し、これを選択されたラクトバチルス(lactobacilli)で24時間の期間、発酵させることによって、毒性のプロリンリッチエピトープのレベルを最小限にした。彼らの結果は科学的には確かに興味深いが、24時間もの長い産業環境での発酵期間は、経済的に容易ではないことは明白である。さらに、彼らの高い割合の非毒性小麦粉は、「規格通りの」グルテンが不足し、そのため最終的なパンの容積が制限されることを含意する。従って、現存の産業的手順を使用するが、毒性プロリンリッチペプチドのレベルが制限される100%コムギパンの生産を可能にする経済的な方法の役に立つことは有利である。そのようなパンは、毒性ペプチドの日々の摂取量を減少することを目的とする食餌の重要な成分となることができる。そのような食餌と特別な関連性がある消費者グループとして、幼児、IBSを患う若年者、高齢者ならびに記載の疾患および症状を患う個体が挙げられる。セリアック病の患者では、50ミリグラム未満のグルテンの日々の摂取は安全とみなされるが、平均的な欧米での消費量である1日あたり13グラムのグルテンに存在する量より有意に少ない毒性プロリンリッチペプチドを含有する任意の食餌は、上記の消費者グループにとって有益であるようである。
【0066】
総合的に見ると、これらの考慮から、先行技術において記載の酵素よりもA.niger(A.ニガー)由来酵素は新規であり、かなり有利であることが示される。本発明に従う酵素を含有する組成物は、グルテンの感作またはオピオイド代謝障害の現象もしくはIBSの現象を抑制あるいは遅延させるために有利に使用される。そのような組成物を消化剤として適用し、毒性プロリンリッチペプチドの消化管でのインサイチュ減少を達成することができる。あるいは、そのような組成物を加工助剤として適用し、そのような毒性のプロリンリッチペプチドを伴わないタンパク質加水分解物を生成させることができる。タンパク質加水分解物の分野内では、ビールの醸造は、A.niger(A.ニガー)由来、低いpHのプロリン特異的エンドプロテアーゼの適用が驚くべき数の利点を付与する特別な場合を提供する。国際公開第02/046381号パンフレットは、ビール糖化工程中またはビールのろ過前もしくはビールのラガーリングの前のいずれかに適用されるプロリル特異的エンドペプチダーゼは、ビールの濁りの形成を減少することを教示している。本発明者らの示すデータは、低いpH酵素との適切なインキュベーションもまた、穀物由来毒性プロリンリッチペプチドのレベルを減少することを例示する。驚くべきことに、本発明者らは、穀物タンパク質を含有するビールまたは他の飲料をプロリン特異的エンドペプチダーゼで処置すると、濁りのレベルが減少するだけではなく、セリアック病を患うヒトに安全な製品がもたらされることを見出した。
【0067】
本出願の実施例1は、アスペルギルス(Aspergillus)由来のプロリン特異的エンドプロテアーゼの酸性の至適pH値および理想的な至適温度を示している。実施例2および3では、本発明者らは、アスペルギルス・ニガー(Aspergillus niger)より産生可能なプロリン特異的エンドプロテアーゼが、より小さなペプチドまたはポリペプチドと同じ効率で大きな無傷なタンパク質を切断することができる真のプロリン特異的エンドプロテアーゼであることを示している。事実、本発明者らのデータは、アスペルギルス(Aspergillus)の酵素が、既知のオリゴペプチダーゼが属するS9ファミリー(N.D.ローリングス(N.D.Rawlings)およびA.J.バレット(A.J.Barrett)、Methods in Enzymology、第244巻、19−61頁、1994年;N.D.ローリングス(N.D.Rawlings)およびA.J.バレット(A.J.Barrett)、Biochimica&Biophysica Acta1298(1996年)1−3)ではなく、S28ファミリーの新規のメンバーであることを示している。本発明者らは、既知のプロリルオリゴペプチダーゼとは対照的に、アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼが、胃における酸性条件下で活性であり、大きなプロリンリッチタンパク質フラグメントおよびポリペプチドの加水分解に対して高い有効性を示すことを見出した。そのような高い効率は、実施例4、5および6において例示される。実施例4では、本発明者らは、アルカラーゼ(Alcalase)(タンパク質加水分解物の生成において頻繁に使用される強力な広域スペクトルのプロテアーゼ)によるミルクタンパク質加水分解物の生成中に、BCM−7配列を組み入れたいくらかのペプチドが、加水分解方法において残存することを実証している。しかし、これらのペプチドは、アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼとの酸性条件下でのインキュベーション時に迅速に消滅する。実施例5に提供されるデータは、アスペルギルス(Aspergillus)の酵素が、β−A2カゼイン由来BCM−7分子において利用可能な4個のプロリン残基のうちの1つでしか切断しないという驚くべき事実を表している。これは、プロリンリッチ基質と任意のプロリン特異的プロテアーゼとのインキュベーションが、劇的に増加した酵素/基質比の条件下であっても、プロリン残基を含むすべてのペプチド結合の切断を自動的に含意するわけではないことを示す。実施例6では、本発明者らは、セリアック病の患者において主要なエピトープであることが要求されるグリアジン由来33マーに対するアスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼの有効性について実証している。また、広域スペクトルのアルカラーゼ(Alcalase)は、アルカリ性または酸性条件下のいずれでも、この分子を切断することができないが、アスペルギルス(Aspergillus)由来酵素は、頻繁に、酸性条件下で分子を切断し、最大で6アミノ酸残基の長さを伴う99.5%のペプチドが作製される。そのため、酸性条件下でのプロリンリッチペプチドに対するその高い有効性にもかかわらず、アスペルギルス(Aspergillus)由来酵素でさえ、アミノ酸配列YPQPQLPを伴う少なくとも0.5%のヘプタマーを遊離する。配列PYPQPQLPYが既知のセリアック病の患者に特異的なT細胞エピトープであるため、この所見は、最適とは言えないプロリン特異的酵素、例えば、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から誘導される酵素を含む既知のプロリン特異的オリゴペプチダーゼでは、グルテン分子由来の毒性ペプチドの形成を防止するための現実的なインビボアプリケーションが、実現不可能であることを証明することを例示している。後者の結論は、実施例7および8において示される実験によって確認される。実施例7では、本発明者らは、pH2〜12の間で、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼとF.メニンゴゼプチクム(F.meningosepticum)由来プロリン特異的オリゴペプチダーゼとの活性プロフィールを比較している。A.niger(A.ニガー)由来酵素はpH2.5〜7.0の間で活性を示すが、F.メニンゴゼプチクム(F.meningosepticum)の酵素は、活性になるために、5.0を超えるpHを必要とする。実施例8では、本発明者らは、両方のプロリン特異的酵素を、胃の酵素ペプシンの非存在下および存在下で低いpH条件に暴露することによって、胃における状況を模倣している。個々の酵素に至適なpH条件下でのその後の活性測定は、F.メニンゴゼプチクム(F.meningosepticum)の酵素がpH2またはpH3条件下でも残存することができることを示している。ペプシンの存在下では、F.メニンゴゼプチクム(F.meningosepticum)の酵素は、pH4で既に不可逆的な損傷を受ける。対照的に、A.niger(A.ニガー)由来酵素は、pH2の低いpH条件下およびペプシンの存在下でも残存し、それによって、胃において毒性ペプチドを加水分解するその価値が強調される。実施例9では、本発明者らは、A.niger(A.ニガー)由来酵素が多数の既知のHLA−DQ2グルテンのエピトープを切断することができることを例示している。興味深いことに、観察される切断部位の位置から、文献から公知のすべてのT細胞のエピトープが破壊されることが推定される。実施例10では、本発明者らは、100%麦芽ビールおよび100%コムギパンからグルテンのエピトープを回収し、本発明者らが抗体アッセイにおいてこれらのエピトープを検出することができることを実証している。実施例11では、本発明者らは、A.niger(A.ニガー)由来酵素を生産プロセスに組み入れることによって生成されるビールが、認知されうる程度に低減されたレベルのグルテンのエピトープを生じることを実証している。実施例12では、本発明者らは、酵素をコムギ生地に組み入れ、低減されたレベルのグルテンのエピトープでダッチ(Dutch)型焼きパンを生成することによって、類似の効果を得ることができることを示している。
【0068】
グルテンは、複雑な3次元構造を伴う水に不溶な化合物である。そのプロリンリッチアミノ酸組成物との組み合わせにおけるこれらの特性により、胃および腸でのタンパク質分解に対し、グルテン分子を耐性にする。消化管腔に分泌される天然のタンパク質分解活性のうち、プロリンに関与するペプチド結合を切断することが可能である活性は存在しないため、共同作用性の内因性プロリン特異的酵素を使用することが理解される。しかし、セリアック病を患うヒトは、グルテンに存在する多くのエピトープに対して極めて感受性であることができる。本発明に従えば、天然の消化プロテアーゼの効果は、アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼで改善することができ、このタンパク質分解混合物の加水分解能のさらなる増強についてさえも本明細書胃おいて開示する。
【0069】
負に荷電した残基、例えば、Glu(E)およびAsp(D)を含むペプチド結合が、プロテアーゼに対して不十分な基質を形成することは周知である。また、天然の消化管タンパク質分解酵素は、これらの残基を処理することができず、これは、グリアジンから胃および膵プロテアーゼ耐性ペプチドWQIPEQSRを単離することによって証明される(シャン(Shan)らを参照のこと)後者の刊行物はまた、グルテンに普遍的に存在するグルタミン残基(Q)を、組織トランスグルタミナーゼによって、グルタミン酸残基(E)に脱アミド化することができることを明らかにしている。不幸なことに、グリアジンペプチドのこの位置特異的脱アミノ化は、それらの潜在的免疫原性をさらに増加する。このような背景に対し、本発明者らは、アスペルギルス(Aspergillus)由来プロリン特異的エンドプロテアーゼと補助的なエンドペプチダーゼとの有効な酵素の組み合わせを生じさせ、プロリンリッチ毒性プロリンリッチペプチドの形成を防止することを可能にしている。本発明に従えば、グルタミン酸特異的エンドペプチダーゼ(EC3.4.21.19)、例えば、バチルス(Bacillus)およびストレプトミセス(Streptomyces)のような多くの食物用微生物によって過剰分泌される該グルタミン酸特異的エンドペプチダーゼを使用することができる。これらの酵素は、経済的かつ食物用の方法で産生させることができる。胃を安全に通過する酵素が、それらの酵素活性に関して、好適である。一般に、該酵素は、酸性または中性の至適pH値を有する。アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼとの組み合わせで、グルタミン酸特異的エンドペプチダーゼのこのカテゴリーは、低いレベルの毒性プロリンリッチペプチドを伴うタンパク質加水分解物の産生において、有用であるとみなされる。
【0070】
かなり驚くべきことに、本発明者らの示す研究は、グルタミン酸特異的エンドペプチダーゼとは別に、アスペルギルス(Aspergillus)由来のプロリン特異的エンドプロテアーゼとのインキュベーションに対して相乗効果を有する他のエンドペプチダーゼが存在することを実証している。本発明者らは、アミノ酸残基Q(グルタミン)とL(ロイシン)との間を切断することが可能なエンドプロテアーゼ(EC3.4.21−99)は、アスペルギルス(Aspergillus)由来のプロリン特異的エンドペプチダーゼと有利に組み合わせられると結論している。具体的には、pH5.0未満の至適pH値を有し、基質のP1またはP1’位置においてグルタミンまたはロイシン残基のいずれかを選好するエンドプロテアーゼ、例えば、アスペルギロペプシン(EC3.4.23.18および19)ならびにムコールペプシン(EC3.4.23.23)が有利に使用される。
【0071】
本発明に従う酵素の1つのアプリケーションは消化剤としてのそれらの使用である。本出願では、本発明の組成物は、好ましくは経口的に投与されるが、他の直接的経路を介してとよしてもよい。組成物は、典型的に、ヒトに投与されるが、増加したグルテン感受性またはカゼインもしくはグルテンの代謝障害あるいはIBSに典型的な症状を緩和するために、動物、好ましくは哺乳動物に投与してもよい。消化剤としてのそれらのアプリケーションでは、本発明に従う酵素は、例えば、丸剤、錠剤、顆粒剤、サシェ(sachet)またはカプセル剤中の乾燥粉末として処方してもよい。あるいは、本発明に従う酵素は、例えば、シロップ剤もしくはカプセル剤中の液体として処方してもよく、または0.85未満の水分活性(Aw)を伴う食品に組み入れてもよい。多様な処方物において使用され、かつ本発明に従う酵素を含有する組成物はまた、生理学的に許容可能なキャリア、アジュバント、賦形剤、安定剤、緩衝液および希釈剤よりなる群の少なくとも1つの化合物を組み入れてもよく、該用語は、包装、送達、吸収、安定化、またはアジュバントの場合には、酵素の生理学的効果を増強することを援助する物質を示すためにそれらの常識的な意味で使用される。粉末化された形態において本発明に従う酵素と組み合わせて使用することができる多様な化合物に関する関連背景は、「Pharmaceutical Dosage Forms」、第2版、第1、2および3巻、ISBN0−8247−8044−2 Marcel Dekker,Incに見出すことができる。乾燥粉末として処方される本発明に従う酵素は、幾分長期間保存することができるが、水分または多湿な空気との接触は、例えば、アルミニウムブリスターのような適切な包装を選択することによって、回避すべきである。液体形態で処方する場合、酵素活性を安定化するためおよび防微生物のために使用される化合物は重要な役割を果たす。酵素活性の安定化は、ポリオール、例えば、グリセロールまたは多様な糖の使用によって得ることができる低減された水分活性を必要とし得る。さらに、Ca2+またはMg2+のような2価のカチオンは、それらの安定化効果について公知であり、ならびにイオウ含有アミノ酸のような還元剤およびBHTまたは没食子酸プロピルのようなフェノール化合物も公知である。食物用防微生物は、低pH条件または低い水分活性とソルビン酸塩もしくは安息香酸塩もしくはパラベンとの周知の組み合わせを使用して、達成することができる。さらに、ハイドロコロイドのような食物用増粘財も必要とし得る。比較的新規の経口アプリケーション形態は、液体を含有するゼラチンカプセルの使用である。本出願では、液体は、本発明の乾燥させた酵素を懸濁することができる典型的にオイルまたはポリエチレングリコールまたはレシチンである。改善された酵素安定性を伴う錠剤処方物の例は、米国公開特許第2002/0136800号明細書において提供される。
【図面の簡単な説明】
【0072】
【図1】基質として合成ペプチドZ−Gly−Pro−pNAを使用するA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの至適pH値のグラフ図を表す(実施例1を参照のこと)。
【図2】クロマトグラフィー精製されたA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼとのインキュベーション後の無傷なオボアルブミン(ovalbumine)および合成27マーペプチドのSDS−PAGEを表す(実施例3を参照のこと)。
【図3】A.niger(A.ニガー)およびF.メニンゴゼプチクム(F.meningosepticum)由来プロリン特異的エンドプロテアーゼの至適pH値のグラフ図を表す(実施例7を参照のこと)。
【発明を実施するための形態】
【0073】
材料および方法
材料
次の酵素は、Sigmaから入手した:アスペルギルス・ニガー(Aspergillus niger)由来のアミログルコシダーゼ、300U/ml、SigmaA−7095;ブタ胃粘膜由来のペプシン、2331U/mg、SigmaP−7012;モルモット由来のトランスグルタミナーゼ、SigmaT−5398。トリプシン溶液2.5%はGibco(BRL25090−028)から入手し、Sep−Pak Plus tC18カートリッジ、Waters No.036810はWatersから入手した。2.6AU(A)(アンソン単位アルカラーゼ(Alcalase))単位/グラム製品の活性を有するアルカラーゼ(Alcalase)(登録商標)AF2.4Lは、NovozymesA/S(Bagsvaerd、丁国)から入手した。Novozymesに従い、この活性測定の詳細については、Novozymes分析方法LUNA番号2003−32153−01/SOP番号:EB−SM−0218.02/02)に見出すことができる。
【0074】
合成ペプチドは、PepscanSystemsB.V.(Lelystad、蘭国)から入手した。発色ペプチド基質は、PepscanSystemsまたはBachem、瑞国のいずれか一方から入手した。
【0075】
A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼ
アスペルギルス・ニガー(Aspergillus niger)由来のプロリン特異的エンドプロテアーゼの過剰発現およびクロマトグラフィー精製は、国際公開第02/45524号パンフレットに記載の通りに達成させた。酵素(1単位/10mgのタンパク質)の活性は、37℃で、クエン酸/リン酸二ナトリウム緩衝液pH4.6中合成ペプチドZ−Gly−Pro−pNAに対して試験した。反応産物は、分光光度法により405nMでモニターした。市販のプロリルオリゴペプチダーゼ酵素(ICN Biomedicals/MP Biomedicals、Aurora、オハイオ州、米国から購入)の活性は、1mgの製品あたり35単位であり、30℃で、pH7.0緩衝液中Z−Gly−Pro−pNAに対し試験した。反応産物は、分光光度法により405nMでモニターした。両酵素について、単位は、指定された条件下、1分間あたり1μmolのp−ニトロアニリドの遊離を誘発する酵素の量として規定される。
【0076】
LC/MS分析
P4000ポンプ(Thermoquest(登録商標)、Breda、蘭国)に連結されたイオントラップ質量分析計(Thermo Electront(登録商標)、Breda、蘭国)を使用するHPLCを、本発明の酵素混合物によって産生される酵素によるタンパク質加水分解物を特徴付けるのに使用した。溶出のためのMilliQウォーター中0.1%ギ酸(Millipore、Bedford、マサチューセッツ州、米国;溶液A)およびアセトニトリル中0.1%ギ酸(溶液B)の勾配と組み合わせたPEPMAP C18 300A(MIC−15−03−C18−PM,LC Packings、Amsterdam、蘭国)カラムを使用して、ペプチドを分離した。勾配は95%の溶液Aで開始し、140分間で40%の溶液Bにまで増加させ、さらに5分間、後者の割合で保持した。使用した注入容積は50マイクロリットルであり、流速は1分間あたり50マイクロリットルであり、カラム温度は30℃に維持した。注入したサンプルのタンパク質濃度は、約50マイクログラム/ミリリットルであった。
【0077】
個々のペプチドに関する詳細な情報については、イオントラップ質量分析計のための特徴的なアルゴリズムである「スキャン依存的」MS/MSアルゴリズムを使用することによって、入手した。
【0078】
フルスキャン分析に続いて、フルスキャンの質量範囲において最も高い強度のイオンの荷電状態の決定のためのズームスキャン分析を行った。後者のイオンのその後のMS/MS分析により部分的なペプチド配列情報が得られ、これは、Xcalibur Bioworks(Thermoquest(登録商標)、Breda、蘭国)由来のSEQUESTアプリケーションを使用するデータベース検索のために使用し得る。使用したデータベースは、使用したアプリケーションのための目的のタンパク質を含有するNCBI(National Centre for Biotechnology informatics)において利用可能なOWL.ファスタ・データバンクから抽出した。乳清タンパク質またはカゼインのような良好に特徴付けられたタンパク質基質が測定される該実験では、分析技術の正確さは、配列適合性が50%未満であるそれらのMS/MSスペクトルを省略することによって、増加する。
【0079】
アンギオテンシン(M=1295.6)を使用して、MSモードにおける至適感受性およびMS/MSモードにおける至適フラグメント化のために同調し、60μg/mlの一定の輸注を実施し、MSモードでは主にダブルおよびトリプルチャージ種が、およびMS/MSモードでは約35%の至適衝突エネルギーが生じた。
【0080】
MALDI−TOF
MALDI−TOFは、Voyager De−Pro(Applied Biosystems)質量分析計を使用して実施した。適切なマトリックスと混合後、ペプチドサンプルをリニアモードで測定した。MassLynxソフトウェアを介して見出される質量から、Post Source Decay(PSD)によって配列決定し、提唱されるペプチドのアミノ酸配列を決定した。
【0081】
食物サンプルにおけるグルテンペプチドの定量化
T細胞刺激α−グリアジン、γ−グリアジンおよびLMW−グルテニンペプチドに特異的なモノクローナル抗体が利用可能であり、食物サンプルにおけるこれらのペプチドの検出のための競合アッセイに組み入れた(E.H.A.スペイニ−デッキング(E.H.A.Spaenij−Dekking)ら、GUT,53:1267−1273(2004)。
【0082】
抗体に基づくアッセイ
抗体に基づくアッセイの作製のために、既知のT細胞刺激α、γ−グリアジンおよびLMWグルテニンペプチドに対し、BalbCマウスにおいてモノクローナル抗体を惹起させた。マウスの脾臓とマウス骨髄腫細胞系統との融合後、抗体産生ハイブリドーマを得た。これらを制限希釈によってクローニングし、これらの細胞によって分泌されるモノクローナル抗体を、モノクローナル抗体競合アッセイにおけるそれらの使用のために試験した。それぞれの特異性について、1つもしくは2つの適切なモノクローナル抗体を選択し、異なるモノクローナル抗体によって認識されるエピトープを決定した(以下の表を参照のこと)。
【0083】
【表1】

【実施例】
【0084】
実施例1
A.niger(A.ニガー)から得られるプロリン特異的エンドプロテアーゼの至適pHおよび温度
A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼをA.niger(A.ニガー)宿主において過剰産生させ、単離し、国際公開第02/45524号パンフレットに記載の材料および方法を使用してクロマトグラフィー精製した。このようにして得られた酵素の至適pH値を確立するために、異なるpH値を伴う緩衝液を調製した。pH4.0−4.5−4.8−5.0−5.5および6.0の緩衝液は、0.05mol/l酢酸Naおよび0.02M CaCl2を使用して作製し;pH7.0および8.0の緩衝液は、0.02M CaCl2を含有する0.05M Tris/HCl緩衝液を使用して作製した。pH値は、それぞれ、酢酸およびHClを使用して調整した。発色合成ペプチドZ−Gly−Pro−pNAを基質として使用した。緩衝溶液、基質溶液およびプロリルエンドペプチダーゼプレ希釈液(0.1U/mLの活性)を、水浴中で正確に37.0℃にまで加熱した。混合後、反応物を、分光光度的に405nm、37.0℃で3.5分間追跡し、0.5分間毎に測定した。図1に示す結果から、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが約4の至適pH値を有することが明らかである。
【0085】
また、プロリルエンドプロテアーゼの至適温度を確立した。そのために、基質としてCaseine Resorufine(Rocheバージョン3)を使用し、精製された酵素調製物を、pH5.0で0.02mol/l CaCl2を含有する0.1mol/l酢酸Naにおいて、2時間、異なる温度でインキュベートし、574nmで測定することによって酵素活性を定量した。得られる結果に従えば、A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼは約50℃の至適温度を有する。
【0086】
該酵素は産業アプリケーションに好適な条件ならびに胃および小腸の初期部分において示現する条件下で至適に活性であるため、極めて強い酸性の至適pH値は、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが産業アプリケーションならびに経口的体内摂取に理想的な特性を有することを強く示唆する。また、酵素の至適温度は、酵素を両アプリケーションに理想的に適切にする。
【0087】
実施例2
A.niger(A.ニガー)から得られる酵素は新規のクラスのプロリン特異的酵素を提示する。
国際公開第02/45524号パンフレットにおいて提供されるA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼのコーディング配列全体から、526アミノ酸のタンパク質配列を決定することができる。酵素の新規性は、SwissProt,PIRおよびtrEMBLのようなデータベースのBLAST検索によって確認した。驚くべきことに、A.niger(A.ニガー)の酵素と既知のプロリルオリゴペプチダーゼとの間に明確な相同性を検出することができなかった。しかし、アミノ酸配列をより緊密に調べたところ、Pro−Xカルボキシペプチダーゼ(EC3.4.16.2)、ジペプチジルアミノペプチダーゼI(EC3.4.14.2)、および胸腺特異的セリンプロテアーゼに対して低いが有意な相同性を示した。これらのすべての酵素は、セリンペプチダーゼのファミリーS28に割り当てられている。また、活性部位セリンの周りのGxSYxGの立体配置は、これらの酵素とA.niger(A.ニガー)由来エンドペプチダーゼとの間で保存される。さらに、ファミリーS28のメンバーは、酸性の至適pH値を有し、プロリン残基のカルボキシ末端側で切断するための特異性を有し、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼと同様のシグナル配列およびプロペプチドで合成される。また、A.niger(A.ニガー)酵素のサイズは、ファミリーS28のメンバーのサイズに類似する。従って、A.niger(A.ニガー)プロリン特異的エンドプロテアーゼは、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から得られる酵素を含むほとんどのサイトゾルプロリルオリゴペプチダーゼが分類されているS9ファミリーではなく、セリンプロテアーゼのファミリーS28のメンバーであるようである。これらの構造的および生理学的特徴に基づいて、本発明者らは、A.niger(A.ニガー)の酵素がセリンプロテアーゼのS9ファミリーではなく、S28に属すると結論されている。S9ファミリーに属するプロリルオリゴペプチダーゼ由来のA.niger(A.ニガー)由来酵素を区別するさらなる特徴は、後者のファミリーに属するサイトゾルプロリルエンドペプチダーゼとは異なり、新たに同定されたA.niger(A.ニガー)の酵素は増殖培地に分泌されるという事実である。これまでのところ、担子菌アガリクス・ビスポラス(Agaricus bisporus)(サッタル(Sattar)ら、J.Biochem.107,256−261(1990))および非関連の子嚢菌アスペルギルス・ニガー(Aspergillus niger)(国際公開第02/45524号パンフレット)のみが、細胞外プロリルエンドペプチダーゼを産生することが明らかにされている。しかし、担子菌から得られる酵素は、5未満のpH値では残存せず、従って、産業アプリケーションならびに経口的体内摂取に適切とは言い難い。
【0088】
これは、下等真核生物由来のファミリーS28のメンバーの単離および特徴付けに関する最初の報告である。
【0089】
実施例3
A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼは、大きなタンパク質ならびに小さなペプチドを加水分解することができ、従って、真のエンドプロテアーゼである。
特定の構造的特徴のため、S9ファミリーに属するプロリルオリゴペプチダーゼは30アミノ酸を超えるペプチドを消化することができない。この制限は、できるだけ迅速かつ効率的にすべての潜在的なプロリンリッチ毒性プロリンリッチペプチドを加水分解することが意図される酵素に明らかに不利である。A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが基質分子のサイズに関して同じ制限を示すかどうかを調べるために、本発明者らは、A.niger(A.ニガー)由来のクロマトグラフィー精製したプロリルエンドペプチダーゼを、小さな合成ペプチドおよび大きなオボアルブミン(ovalbumine)分子と共にインキュベートし、SDS−PAGEによって、形成された加水分解産物を分析した。
【0090】
使用した合成ペプチドは、27マーの配列NH2−FRASDNDRVIDPGKVETLTIRRLHIPR−COOHであり、Pepscan company(Lelystad、蘭国)より贈与された。
【0091】
そのアミノ酸配列から示されるように、このペプチドは、2個のプロリン残基を含有し、1つはペプチドの中央にあり、1つは極めて末端にある。無傷なオボアルブミン(ovalbumine)分子(Pierce Imject、20mgの凍結乾燥材料を含有するバイアル)は、42750Daの分子量を伴う385アミノ酸よりなる。この分子は、14個のプロリン残基を含有し、このうち1つは分子の究極的C末端に位置し、プロリン特異的エンドプロテアーゼによって切断することができない。
【0092】
オボアルブミンおよびオリゴペプチドを、個別に、50℃で、精製されたA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼと共にインキュベートした。いくらかの時間間隔で、サンプルを採取し、SDS−PAGEを使用して分析した。
【0093】
4.5単位/mlの活性を伴うクロマトグラフィー精製されたA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼを、20mM CaCl2を含有する0.1M酢酸緩衝液pH4で100倍希釈した。オボアルブミン(ovalbumine)を、1mg/mlの濃度(22μM)で酢酸緩衝液pH4に溶解した。27マーを同じ緩衝液に溶解し、0.48mg/mlの濃度(152μM)に到達させた。オボアルブミン(ovalbumine)および27マー溶液のモル濃度は、両溶液が切断可能なプロリン残基において同じモル濃度を含有するような方法で、選択した。オボアルブミン(ovalbumine)は13個の潜在的プロリン切断部位を含有する一方、27マーペプチドは2個しか有さない。両基質溶液のうち、0.5mlを、エッペンドルフ(Eppendorf)サーモミキサー中、50℃で10μl(0.45ミリU)の酵素溶液と共にインキュベートした。いくらかの時間間隔で、10μlサンプルをインキュベーション混合物から抜き取り、SDS−PAGEまで20℃で保持した。SDS−PAGEおよび染色のために使用されるすべての材料は、Invitrogenから購入した。サンプルは、製造者の指示に従い、LDS緩衝液を使用して調製し、製造者の指示に従い、MES−SDS緩衝液システムを使用して12%Bis−Trisゲル上で分離した。染色は、Simply Blue Safe Stain(Collodial Coomassie G250)を使用して実施した。
【0094】
図2に認められ得るように、オボアルブミン(ovalbumine)は、アスペルギルス(Aspergillus)由来酵素によって、インキュベーションの最初の4.75時間において約35〜36kDの個別のバンドに切断される(レーン3)。インキュベーション期間を延長すると、多様な分子量のより小さな産物へのさらなる分解が生じる(レーン7)。
【0095】
27マーペプチドもまた分解され、レーン2と比較して、レーン4、6および8におけるより微弱なバンドによって判定される。産物の極めて小さな分子量のシフト(レーン9および8を比較すること)は、ペプチドのカルボキシ末端でのアルギニン残基の切断によるためである可能性が最も高い。差異は約200Dであり(AlphaImager2000システムの上のAlphaImager3.3dソフトウェアを使用して測定される)、アルギニンは174のMWを有する。この小さな分子量のシフトは、恐らく、ペプチドの分解の第1の工程である。
【0096】
産物のさらなる崩壊は、SDSゲル上のバンドの強度の減少によってのみ、見ることができる。約1000のMWを伴う成分のゲル染色はクマシー・ブリリアント・ブルー(Coomassie Brillant Blue)では可能ではないため、さらなる産物の崩壊は認められない。
【0097】
この実験から、S9ファミリーに属する既知のプロリルオリゴペプチダーゼとは異なり、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼは、極めてより大きなタンパク質よりも小型のペプチドの方を特に選好して切断するわけではないと結論付けることができる。A.niger(A.ニガー)由来酵素は、それ自体で、真のエンドプロテアーゼおよび潜在的な毒性のプロリンリッチプロリンリッチペプチドを切断するのに好適な酵素を提示する。
【0098】
実施例4
アルカラーゼ(Alcalase)およびアルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせとのインキュベーション後に形成される加水分解物におけるβ−カゾモルフィン
消化管タンパク質分解中のプロテアーゼ耐性β−カゾモルフィンの形成と同様に、本発明者らは、ミルクタンパク質加水分解物の産業的生産中にも、BCM−7関連ペプチドフラグメントの類似の蓄積が生じるのであろうかと考えた。そのために、本発明者らは、牛乳から単離されるA2β−カゼインを、産業上頻繁に使用されるサブチリシンアルカラーゼ(Alcalase)およびアルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼと共にインキュベートした。LC/MS/MS分析を使用して、そのようにして形成されるペプチドを分析した。
【0099】
牛乳は、1kgの乳汁あたりほぼ10グラムのβ−カゼイン(caseine)を含有し、存在するすべてのタンパク質のうち28%を提示する。BCM−7関連アミノ酸配列の分析を容易にするために、この実験では、本発明者らは、最低でも90%の(A2)β−カゼインを含有する濃縮調製物(Sigma由来)を使用した。後者の産物を、1リットルあたり20グラムの濃度で水に溶解し、その後、NaOHを使用してpHを8に調整し、1リットルのカゼイン溶液あたり800マイクロリットルの酵素濃縮物の量でアルカラーゼ(Alcalase)を添加した。インキュベーションを、2時間、60℃で行った。次いで、クエン酸を使用して、溶液のpHを4.5まで低下させた。次いで、溶液を2つの部に分けた:一方の部は、5分間、90℃で加熱してアルカラーゼ(Alcalase)を不活化し、他方の部には、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼを添加して、存在するカゼインの1グラムあたり1単位の酵素濃度(単位の定義に関する材料および方法のセクションを参照のこと)を得た。A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼとのインキュベーションを、16時間、55℃で継続し、続いて、プロリン特異的エンドプロテアーゼを不活化するために、さらに加熱処置を施した。最後に、20mg/mlの推定β−カゼイン濃度を伴う2つのサンプルを、LC/MS/MS分析のために供給した。2つのサンプルを、10分間、13000rpmで遠心分離し、LC/MS/MS分析の前にMilliQ中で20倍希釈した。材料および方法のセクションに記載の通りに、LC/MS/MS分析を行った。
【0100】
β−カゼイン分子のアミノ酸位置60〜66におけるBCM−7配列Tyr−Pro−Phe−Pro−Gly−Pro−Ileの他に、それぞれ、アミノ酸位置60〜63および60〜64におけるTyr−Pro−Phe−Pro(β−カゾモルフィン(1−4))およびTyr−Pro−Phe−Pro−Gly(β−カゾモルフィン(1−5))のようなより小さなフラグメントならびに(アミノ酸位置60〜70における)11アミノ酸の鎖長までのすべてのより大きなペプチドは、少なくともいくらかの程度のオピオイド活性を示す。位置60〜62のトリペプチドTyr−Pro−Pheは、オピオイド活性を有さないことが報告されている。
【0101】
ペプチド同定のために、プロトン化分子のダイレクトLC/MS/MSを使用した。関連の可能性のあるペプチドのプロトン化された質量を表1に提供する。すべてのスペクトルを、35%の衝突の大きさおよび3Daのピーク幅で得た。
【0102】
すべての実験データを、いわゆるBおよびYイオンに基づく理論的フラグメント化パターンと比較する。これは、普通、自動データ処理を使用して実施される方法である。
【0103】
【表2】
【0104】
βカゾモルフィン(Betacasomorphine)アミノ酸配列は、チロシン(Y;β−カゼインペプチド鎖の残基番号60)で開始し、太字で示す。m/z値は、他の関連する可能性のあるペプチドのプロトン化分子を提示する。位置67のプロリン残基の検出は、使用した基質がA2β−カゼインを提示することを示す。
【0105】
アルカラーゼ(Alcalase)かまたはアルカラーゼ(Alcalase)+プロリン特異的エンドプロテアーゼのいずれか一方とのインキュベーション後に得られるβ−カゼイン加水分解物のLC/MS/MS分析に対して得られる結果を表2に示すが、これらは、以下のようにまとめることができる:
・正確なβ−カゾモルフィン(casomorphine)配列、即ち、YPFPGPIおよび誘導体は、アルカラーゼ(Alcalase)かまたはアルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせのいずれか一方で処置した(A2)β−カゼインには存在しない。
・しかし、β−カゾモルフィン配列を含有する2つのペプチドは、アルカラーゼ(Alcalase)処置サンプル、即ち、LVYPFPGPIPNおよびVYPFPGPIPNに存在する。
・これらのβ−カゾモルフィン含有配列の強度は、A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼによる処置時に劇的に減少する。
【0106】
【表3】
【0107】
結果は、明らかに、アルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせがすべての潜在的なβ−カゾモルフィン配列を高い効率で破壊し、それによって、潜在的に毒性のプロリンリッチプロリンリッチ配列を伴わない加水分解物を付与することを示す。
【0108】
形成されるペプチドの間でのより正確な検索後、本発明者らは、2つの酵素の組み合わせによって形成される加水分解物においてペプチドVYPの存在を実証することができた。このペプチドは公正なアルカラーゼ(Alcalase)を使用することによって形成される加水分解物において追跡できなかったため、この所見は、アスペルギルス(Aspergillus)由来酵素によるβ−カゼイン分子の位置61におけるプロリン残基のC末端切断を示唆する。
【0109】
実施例5
アルカラーゼ(Alcalase)形成ペプチドVYPFPGPIPNとA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼとのインキュベーションにおいて形成されるペプチド
実施例4に見られるように、アルカラーゼ(Alcalase)およびA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせによるA2β−カゼインの加水分解は、すべての潜在的β−カゾモルフィン配列を効率的に取り出す。しかし、作製されたペプチドの複雑さのため、本発明者らは、アスペルギルス(Aspergillus)由来酵素がアルカラーゼ(Alcalase)形成ペプチドVYPFPGPIPNを切断する位置を確立することができなかった。そのために、この特定の配列を伴うペプチドを合成し、2種の濃度のプロリン特異的エンドプロテアーゼと共にインキュベートした。形成されたペプチドのその後のLC/MS/MS分析は、酵素の正確な切断部位を示した。
【0110】
凍結乾燥された10マー(Pepscan Systems;Lelystad、蘭国)を、2mg/mlの濃度で、クエン酸−リン酸緩衝液pH4.5に溶解した。溶液に、1グラムのペプチドあたり1および10単位の濃度でプロリン特異的エンドプロテアーゼを添加した。4時間、55℃でインキュベーションし、その後、5分間、90℃での加熱処置を使用して、酵素を不活化した。次いで、2つのサンプルを、10分間、13000rpmで遠心分離し、LC/MSおよびLC/MS/MS分析の前にMilliQウォーター中で20倍希釈した。サンプルを、まず、LC/MSモードで分析して、異なる量の酵素を使用して、10マーの強度の減少を観察し、酵素切断後のLC/MSイオンクロマトグラムにおいてどのペプチド質量が出現したかを観察した。
【0111】
次いで、LC/MS稼動において見出されるペプチドのプロトン化分子のダイレクトLC/MS/MSを実施した。すべてのスペクトルを、35%の衝突の大きさおよび3Daのピーク幅で得た。すべての実験データを、いわゆるBおよびYイオンに基づく理論的フラグメント化パターンと比較する。これは、普通、ペプチド、ポリペプチドおよびタンパク質の同定のための自動データ処理を使用して実施される方法である。
【0112】
1単位/gのタンパク質による10マーVYPFPGPIPN(M=1099.5)の処置は、既に、10マーのいくらかのペプチドへの全体的分解を生じた。m/z1100.5でのプロトン化分子の強度は、3桁の大きさだけ低下する。10単位/gによる10マーの処置では、プロトン化分子の強度のさらなる減少は生じず、また、他のペプチド質量も認められなかった。1単位/gによる酵素処置においては、4つのペプチドが形成され、VYP(M=377.2)を伴い、最も豊富に認められるm/z378.2によって特徴付けられる(ほぼ98%;表3を参照のこと)。4つのすべてのペプチドをLC/MS/MSモードで分析し、上記の基準に基づいて正確であることを見出した。表3:10マーVYPFPGPIPN M=1099.5のLC/MSおよびLC/MS/MSモードで分析したプロトン化ペプチド質量。第2のカラムはプロトン化分子のm/z値、第3のカラムはLC/MSモードにおいて観察されるプロトン化分子の強度、第4のカラムはプロトン化分子のピーク面積に基づく百分率、そして第5のカラムは10マーの全アミノ酸配列において見出されるペプチドの位置を示す。ペプチドのプロトン化分子のピーク面積を使用することには、イオン化効率の差異の影響を含まないことを強調すべきである。
【0113】
【表4】
【0114】
しかし、A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼは、位置61(この特定のデカペプチドでは位置3)のプロリンのC末端側でほとんど排他的に切断すると結論することができる。切断の性能は、酵素/基質比の増加による影響を受けない。
【0115】
オピオイド活性を伴うすべての既知のβ−カゾモルフィン(casomorhin)分子はN末端配列YPFを共有するため、プロリン特異的エンドプロテアーゼによるPとFとの間(即ち、位置61でのプロリン残基のカルボキシ末端)のこの配列の効率的な切断は、すべてのBCM−7およびβ−カゼインのA1またはA2遺伝子変異体由来であるBCM−7関連ペプチドを有効に不活化することが明らかである。
【0116】
μ受容体との相互作用における位置61のプロリン残基の極めて重要な役割もまた、最近のインターネット上での公開物において確認された(the Dipartimento di Scienza degli Alimenti Universita di Napoli「Frederico II」Facolta di AgrariaによるSintesi e affinita ai recettori oppiodi di analoghi delle beta−casomorphine contenenti beta−omo amminoacidi」)。
【0117】
実施例6
グリアジン由来33マーLQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPFとA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼとのインキュベーションにおいて形成されるペプチド
pH8またはpH5のいずれか一方でのアルカラーゼ(Alcalase)によるシャン(Shan)ら(Science、第297号、2002年9月27日)に記載の胃および膵液耐性グリアジン由来33マーLQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF(M=3911)の処置は、何ら分子の切断を生じなかった。しかし、β−カゼイン由来10マーを伴う状況に類似して、pH5における1単位のA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼとのインキュベーションにより、いくらかのペプチドへの分子の全体的な分解が生じた。m/z1304.4でのトリプルプロトン化(triple protonated)分子の強度は、3桁の大きさだけ低下する。プロトン化分子の強度のさらなる減少もおよびまた他のペプチド質量も、1グラムのタンパク質あたり10酵素単位による33マーの処置では観察されなかった。
【0118】
酵素処置後、約6つの主要なペプチドおよびいくらかの少数のペプチドが形成され、最も豊富に認められるm/z565.2によって特徴付けられるペプチドを伴う。6つのすべてのペプチドを、LC/MS/MSモードにおいて分析し、それらはすべてC末端でプロリンを含有することが見出され、酵素の特異性が確認された。形成された主要なペプチドは、m/z565.2によって特徴付けられる(表4の配列..QLP)。C末端配列「LP」は、このペプチドについて明確に実証することができたが、理論的には、同定される質量は、33マーのエンド型タンパク質分解では形成させることができないため、ペプチドの正確なN末端組成物に関しては、いくつか不確かな点が残る。ほぼ間違いなく、m/z565.2はQPQLP(M=581.3)のN−ピログルタミル変異体であるが、これについてさらに調べなかった。
【0119】
QPQLPの外観:LQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF
QPQLPYPの外観:LQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF
【0120】
m/z679を伴うペプチドのLC/MS/MSスペクトルは、PQPQLPであることが解明できた。m/z565.2を伴うペプチドの性質が十分に理解さなかったという事実にもかかわらず、得られたデータは、この特定の33マーの位置12、19および26でのプロリン残基のC末端側(即ち、プロリンおよびチロシン残基の間で排他的に)アスペルギルス(Aspergillus)由来のプロリン特異的エンドプロテアーゼの好適な切断を明確に実証する。この切断パターンは、より高い酵素/基質比による影響を受けない。表4に、すべての関連する情報をまとめる。
【0121】
第1のカラムは、誘導されるペプチド配列を指定し、使用したドット(またカラム5においても)は、説明されない質量の矛盾点のため、ペプチドの正確な開始位置を示すことができなかったことを示す。第2のカラムはプロトン化分子のm/z値、第3のカラムはLC/MSモードにおいて観察されるペプチドの強度、第4のカラムはプロトン化分子のピーク面積に基づく百分率、そして第5のカラムは全アミノ酸配列において同定されるペプチドの位置を示す。
【0122】
【表5】
【0123】
セリアック病の患者において主要なエピトープであることが要求される33マーの切断は、胃もしくは膵液によっても、あるいはアルカリまたは酸性条件下のいずれの強力な広域スペクトルのプロテアーゼアルカラーゼ(Alcalase)とのインキュベーションによっても、達成することができない。それにもかかわらず、本発明者らの結果は、酸性条件下でのプロリン特異的A.niger(A.ニガー)由来エンドプロテアーゼによる効率的な切断を示す。後者の切断は、分子のプロリンおよびチロシン残基との間で排他的に生じ、6以下のアミノ酸残基長を伴う99.5%のペプチドを作製する。そのため、酸性条件下でのプロリンリッチペプチドに対するその高い有効性にもかかわらず、アスペルギルス(Aspergillus)由来酵素でさえ、アミノ酸配列YPQPQLPを伴う少なくとも0.5%のヘプタマーを遊離する。配列PYPQPQLPYが既知のセリアック病の患者に特異的なT細胞エピトープであるため、この所見は、中性付近の至適pH値を伴う最適とは言えないプロリン特異的酵素、例えば、既知のプロリン特異的オリゴペプチダーゼ、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から誘導される酵素では、グルテン分子由来の毒性ペプチドの形成を防止するための現実的なインビボアプリケーションが、実現不可能であることを証明することを再度強調している。
【0124】
実施例7
A.niger(A.ニガー)プロリン特異的エンドプロテアーゼおよびF.メニンゴゼプチクム(F.meningosepticum)由来のプロリン特異的オリゴペプチダーゼのpH活性スペクトル
実施例1において実証されるように、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの至適pH値は約4.2である。この第1の試験では、極めて酸性のpH値を考慮しなかったため、胃に存在することができるpHの極値における酵素の挙動については、完全には明らかにされていない。国際公開第03/068170号パンフレットは、プロリン特異的プロテアーゼ、例えば、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)由来のプロリルオリゴペプチダーゼまたはアスペルギルス・フミガーツス(Aspergillus fumigatus)由来のDPPIVまたはストレプトミセス(Streptomyces)種由来のペプチジルジペプチダーゼは、毒性のプロリンリッチペプチドのインサイチュ分解のための好適な候補を示すことを教示している。典型的に、後者のプロテアーゼのすべては、胃において何ら加水分解活性を含まないようである中性付近の至適pH値を特徴とする。本発明のプロリン特異的エンドプロテアーゼと先行技術の酵素との間に存在する至適pH値の差異を実証するために、本発明者らは、アスペルギルス(Aspergillus)由来エンドペプチダーゼおよびフラボバクテリウム(Flavobacterium)由来オリゴペプチダーゼのpH活性スペクトルを比較した。実施例1に記載のように、アスペルギルス(Aspergillus)由来エンドペプチダーゼを得た。フラボバクテリウム(Flavobacterium)由来オリゴペプチダーゼは、ICN Biomedicalsから購入した(35単位/mg;カタログ番号32082;オハイオ州、米国)。
【0125】
2つの酵素のpH活性スペクトルを確立するために、異なるpH値を伴う緩衝液を調製した。0.1mol/lクエン酸を使用してpH2.0〜7.0の範囲の緩衝液を調製し、0.1mol/lトリスを使用してpH6.0〜9.0の範囲の緩衝液を調製し、0.2mol/lグリシンを使用してpH8.0〜12.0の範囲の緩衝液を作製した。必要とされるpH値は、HClまたはNaOHのいずれか一方を使用して調整した。発色合成ペプチドZ−Gly−Pro−AMC(Bachem、瑞国)を、両酵素の基質として使用した。各ウェル(Costar番号3631プレート)において、85μLの緩衝液、10μLの酵素溶液および5μLの基質(60%メタノール中4mM Z−Gly−Pro−AMC)を導入した。A.niger(A.ニガー)酵素の最終濃度は32μg/ml(3.2ミリ単位/ml)であり、F.メニンゴゼプチクム(F.meningosepticum)酵素の最終濃度は0.21μg/ml(7.4ミリ単位/ml)であった。混合後、反応を30分間、37.0℃で進行させ、その後、PerSeptive BiosciencesのCytoFluorマルチウェルプレートリーダーにおいて、蛍光を測定した。得られた関連データを図3に示す。これらの僅かに異なる試験条件下で得られたデータにより、実施例1において確立したA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの酸性至適pH値が確認される。データは、A.niger(A.ニガー)酵素がpH2.2および7.5で約20%の残存活性を有することを示す。図3に示されるデータによってもまた、F.メニンゴゼプチクム(F.meningosepticum)の約pH7.0の公開された至適pH値が確認される。本特許出願では、pH5.0未満において、F.メニンゴゼプチクム(F.meningosepticum)酵素が活性を有さないことが最も重要である。
【0126】
これらのデータは、F.メニンゴゼプチクム(F.menigosepticum)酵素とは対照的に、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが、胃および十二指腸の部分において示現する酸性条件下でその十分な活性を展開するために理想的に適切であることを示す。
【0127】
実施例8
胃に存在する条件下におけるA.niger(A.ニガー)プロリン特異的エンドプロテアーゼおよびF.メニンゴゼプチクム(F.meningosepticum)プロリン特異的オリゴペプチダーゼの安定性
首尾よい酵素療法のための必須条件は、そのようなペプチドが十二指腸の遠位部に到達する前の毒性プロリンリッチペプチドの効率的な消化管分解である。これは、酵素療法に使用すべき外因性酵素が、胃におけるおよび場合により胃より先における食餌タンパク質の滞留時間中に活性であるプロリン特異的プロテアーゼであることを必要とする。そのような「胃様」条件下で、A.niger(A.ニガー)およびF.メニンゴゼプチクム(F.meningosepticum)由来プロリン特異的プロテアーゼの活性を比較するために、本発明者らは、異なるpH条件下ならびにプロテアーゼペプシンの存在および非存在下で、37℃、異なる期間におけるインキュベーション後のそれらの残留活性をアッセイした。残留酵素活性は、関連酵素に至適なpH値および発色ペプチド、即ち、F.メニンゴゼプチクム(F.meningosepticum)酵素については7.0のpHおよびZ−Gly−Pro−pNA基質ならびにA.niger(A.ニガー)酵素については4.0のpHおよびAla−Ala−Pro−pNA基質を使用して、アッセイした。使用されるA.niger(A.ニガー)酵素は、Ala−Ala−Pro−pNAに対して高い活性およびAla−Ala−Ala−pNA基質に対して若干の活性(Ala−Ala−Pro−pNAに対するその活性の10%未満)を示す。使用されるA.niger(A.ニガー)酵素は、他のAla−Ala−X−pNA基質に対して有意な活性を示さない。
【0128】
0.2mol/lのクエン酸/HCl緩衝液は、必要とされる酸性pH条件を得るために使用した。A.niger(A.ニガー)由来酵素の用量は、1.5単位/ml、F.メニンゴゼプチクム(F.meningosepticum)酵素(MP Biochemicals、オハイオ州、米国)3.3単位/ml(単位の定義に関する材料および方法のセクションを参照のこと)の用量であった。ペプシン(Sigma)は、180マイクログラム/mlの濃度で添加した。ペプシンを不活化するために、サンプリング後、ペプスタチン(Sigma)を1.67マイクログラム/mlの濃度で添加した。これらの条件下で、ペプスタチンは、2種のプロリン特異的プロテアーゼに対し、阻害効果を有さなかった。
【0129】
それぞれ合成基質Ala−Ala−Pro−pNAおよびZ−Gly−Pro−pNAを利用して、2種のプロリン特異的プロテアーゼの残留活性を、動力学的に405nmで測定した。そのために、200μLの基質溶液(0.05mol/lリン酸緩衝液pH7 0中2mmol/l Z−Gly−Pro−pNAまたは0.05mol/l酢酸緩衝液pH4.0中1.5mmol/l Ala−Ala−Pro−pNA)を、MTPウェルにおける50マイクロリットル(10〜100×プレ希釈した)の酸性/ペプシン処置サンプルと混合した。TECAN Genios MTP Reader(Salzburg、Vienna)を利用して、10分間、405nm、30℃で、吸収を動力学的に測定した。
【0130】
表5に示される結果は、pH4が下限を提示することを示し、ここで、F.メニンゴゼプチクム(F.meningosepticum)酵素が残存することができることを示す。実施例7に記載のデータに従えば、酵素はこれらのpH条件下で活性を有さないことに留意していただきたい。pH4.0で2時間後、酵素は、至適条件下で試験する場合、約25%の残留活性を示す。しかし、ペプシンが存在するや否や、このpHで15分後に、酵素は完全に不活化された。対照的に、これらの結果はA.niger(A.ニガー)酵素により得られるデータである。後者の酵素は、pH2の低いpH値およびペプシンとの組み合わせ下でもその十分な活性を維持する。これらの結果は、5.5未満の至適pH値を伴う酵素のみが胃において活性であるようであり、そのため、酵素クラスEC3.4.21.26(プロリルオリゴペプチダーゼ)、EC3.4.14.5(ジペプチジル−ペプチダーゼIV)およびEC3.4.15.1(ペプチジルジペプチダーゼA)に属する既知のプロリン特異的プロテアーゼは、胃においてプロリンリッチペプチドを分解するのに適格でないことを強く示唆する。
【0131】
【表6】
【0132】
実施例9
多様なグルテンのエピトープに対するA.niger(A.ニガー)プロリン特異的エンドプロテアーゼの活性
多くの刊行物が、セリアック病患者由来の小腸T細胞によって認識されるコムギグルテン由来ペプチドについて詳記している。これに関して、HLA−DQ2ハプロタイプのみを担持するヒトは、セリアック病を発達する可能性が断然高い(マキ(Maki)ら;N Engl J Med.2003年6月19日;348(25):2517−24)ため、HLA−DQ2分子に対するペプチド結合は特に興味深い。検査したすべての成人HLA−DQ2患者は、α−GLIA配列Glia−α2およびGlia−α9に応答する(アレンツ−ハンセン(Arentz−Hansen)ら、J.Exp.Med.2000;6:337−342)。セリアック病を発症したばかりの小児は、ペプチドGlia−α20、Glia−γ30、Glt−17、Glt−156およびGlu−21に対して応答する(ヴァーデル(Vader)ら、Gastroenterology2002;122:1729−1737)。実施例6において、本発明者らは、既に、A.niger(A.ニガー)由来酵素によるGlia−α2ペプチド(PQPQLPYPQPQLPY)の首尾よい切断について実証した。ここでは、本発明者らは、A.niger(A.ニガー)酵素もまた、他の関連のコムギグルテン由来ペプチドならびに他の穀物から誘導されるこれらのペプチドの相同物を切断するかどうかについて試験する。
【0133】
そのために、関連ペプチドを合成し、その後、それらの純度を、材料および方法のセクションに記載の手順に従うrpHPLCおよび質量分析(MALDI−TOF)によって確認した。ペプチド消化は、50マイクロリットルのペプチド溶液(1mg/ml)と、50ミリモル/l酢酸アンモニウム緩衝液pH4.5中0.43マイクログラム/ml(0.043ミリ単位)を含有する50マイクロリットルのA.niger(A.ニガー)酵素溶液とを混合することによって行った。室温での多様なインキュベーション期間後、0.5マイクロリットルのサンプルを採取し、9.5マイクロリットルのマトリックス溶液(0.2%TFAを含有する1:1H2OおよびCH3CN)に添加した。脱塩後、Dowexイオン樹脂ビーズを使用して、液体を、いくつかの関連対照ペプチドと共にMALDI−プレート上のスポットし、その後、すべてのペプチドを分析した。得られたデータから、表6に示す切断部位を推測し得る。ペプチドGlt156およびGliaγ−2は10マーであり;Glt17は8マーであるが、至適結合のためにPおよびL残基によるC末端伸長を必要とする。明白な切断部位を二重縦線で示し、あまり明白ではない切断部位を一重縦線で示す。各ペプチド上の既知の最小のT細胞のエピトープに下線を付して示す。Glu−5において使用されるUは、ロイシンまたはイソロイシン残基のいずれか一方を提示する。
【0134】
得られるデータから、Glia−α2だけではなく、若年者ならびに成人HLA−DQ2患者に関連の事実上すべてのグルテンのエピトープが、A.niger(A.ニガー)由来プロリン特異的プロテアーゼによって効率的に加水分解されることが明らかである。ほとんどの切断部位は、既知の最小T細胞エピトープ結合部位に対し精密に位置を特定することができるため、グルテンのエピトープの酵素への暴露は、認識を廃止し、従って、究極的にセリアック病患者に特徴的な症状をもたらす以後のT細胞増殖を効率的に防止する可能性がある。
【0135】
【表7】
【0136】
類似のアプローチにおいて、コムギグルテンのエピトープの多くのカラスムギ、オオムギおよびライムギ由来グルテン相同物におけるA.niger(A.ニガー)由来酵素の切断部位を確立した(ヴェーデル,W(Vader,W)ら、Gastroenterology2003;125:1105−1113)。得られた結果(表7を参照のこと)は、A.niger(A.ニガー)由来酵素がセリアック病を担うコムギグルテンのエピトープだけではなく、カラスムギ、オオムギおよびライムギに存在するこれらのペプチドの相同物をも切断することができることを例示する。表6におけるように、明白な切断部位を二重線で示す。
【0137】
【表8】
【0138】
実施例10
100%麦芽ビールおよび100%コムギパンからのグルテンのエピトープの回収の試験
提案されている抽出手順(材料および方法のセクションを参照のこと)を、抗体に基づくアッセイと組み合わせて、PVPP処置した100%麦芽ビール(実施例11を参照のこと)および100%コムギパンサンプル(実施例12を参照のこと)に対して試験した。得られた結果(表8を参照のこと)に従って、抗体アッセイに組み合わせた抽出手順は、ビールならびにパンにおける抗−αグリアジン、抗−γグリアジンならびに抗−グルテニンエピトープを検出することができる。
【0139】
総合すると、得られたデータは、適用した抽出手順がビールおよびパンサンプルの両方におけるグルテンの検出に適切であることを強く示唆する。これらの結果に基づき、それらのプロセシング中に異なる濃度のA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼに供したビールおよびパンサンプルを、実施例11および12においてさらに緊密に調べた。
【0140】
【表9】
【0141】
実施例11
A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼに関与するビール生産は、より低いレベルのグルテンのエピトープをもたらす。
ビールの濁りは、グルテン由来プロリンリッチタンパク質、ポリペプチドおよびペプチドと、ビール生産に使用される穀物(大部分は、オオムギ)から抽出されるポリフェノールとの会合によって形成される。国際公開第02/046381号パンフレットに記載されているように、ビールの濁りの形成は、ビール生産の糖化、発酵またはラガーリング相のいずれかの間に、酸性安定なプロリン特異的エンドプロテアーゼを組み入れることによって、減少または防止することができる。従来のビールの醸造方法では、濁りの形成は、PVPP(存在する多様なポリフェノールに結合するが、濁り活性なプロリンリッチペプチドのレベルに対しては極少数の効果しか伴わない化合物)での処置によって防止される。従来の醸造方法は毒性のプロリンリッチペプチドを除去しないため、セリアック病の患者がビールを飲むことは認められていない。
【0142】
本研究の目的は、ビールの作製方法中のA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの添加により、完成品のビールにおいて低減されたレベルの毒性プロリンリッチペプチドがもたらされるかどうかを確立することである。そうであるならば、そのようなビールは、セリアック病の患者でも飲むことができる。
【0143】
ビール生産は、IFBM(Nancy、仏国)での20hlパイロットプラントにおいて行った。使用したA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼサンプルは50%グリセロール(w/w)中で安定化され、5単位/グラム液体の最終活性を有した(単位の定義に関する材料および方法セクションを参照のこと)。
【0144】
独立した生産稼動において、5つの100%麦芽ビールを、PVPP(対照)または異なる量のプロリン特異的エンドプロテアーゼのいずれかを使用して醸造し、濁りの形成を防止した。すべての実験において、使用した糖化プロトコルはまったく同じである。実験に依存して、プロリン特異的プロテアーゼを、糖化方法の開始かまたはビール発酵直前の糖化方法後のいずれか一方で添加した。糖化方法では、異なる3つの酵素用量、即ち、使用した麦芽1kgあたり2.5、5.0および7.5酵素単位を使用した。発酵では、単回の酵素用量のみ、即ち、添加された麦芽1kgあたり0.75酵素単位を試験した。対照ビールを、ビールろ過前に添加した30グラム/hlのPVPPで安定化させた。
【0145】
各醸造は、300kgのオオムギ麦芽およびホップペレットから生産した。3:1(vol/wt)の液体/グリストの糖化条件およびpH5.6を使用した。糖化ダイアグラムは、45℃で20分間の第1の工程、64℃で15分間の第2の工程および76℃で25分間の第3の工程ならびに最終的に78℃までの加熱を含む。工程間の加熱速度は1分間あたり1℃である。麦汁を90分間、煮沸した。良好なトラブ分離をWhirlpool上で実施した。発酵は、麦汁の17 106細胞/mlのピッチング速度および約97%のピッチングでの生存能で底部酵母株により行った。12℃+/−1℃での10日間の発酵期間に続いて、−1℃+/−1℃で5日間の低温成熟を行った。ビールに5.2g/lの圧力で炭酸ガスを注入し、ボトリング後、60℃で、20分間、低温殺菌した。
【0146】
【表10】
【0147】
結果に従えば、抗−αグリアジンおよび抗−γグリアジン抗体は同等のデータを生じる。このアッセイにおける抗−グルテニン抗体では結果に至っていない。糖化段階後の酵素の特定の適用により、ビールにおいて極めて低い毒性のプロリンリッチペプチドがもたらされることが明白である。抗体認識部位は、最低でも5アミノ酸残基の長さを有するが、T細胞認識部位では少なくとも9アミノ酸残基を必要とすることを考慮すると、T細胞が、発酵アプローチにおいて作製される短いペプチドを認識し得る可能性はさらにより低くなる。事実、これらのデータは、上記の酵素アプローチに従って、セリアック病の患者に安全なビールを醸造することができることを強く示唆する。
【0148】
実施例11
A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼを生地に組み入れることによって生成されるパンは、より低いレベルのグルテンのエピトープを生じる
パン作製のために、3500gの小麦粉(80%KolibriTMおよび20%IbisTM)、1990mlの水(56%)、77gの圧搾酵母(2.2%)、70gの塩(2%)、140mgのアスコルビン酸(40ppm)ならびに表10に記載の多様な量のA.niger(A.ニガー)由来酵素から生地を調製した。添加する酵素の量は、より少量の水を生地に添加することによって補正した。
【0149】
Diosnar(登録商標)スパイラルミキサーを2分間、スピード1で使用し、続いて、6分間、スピード2で混合することによって、成分を生地に混合した。875gの生地片を丸め、35分間、34℃および85%RHで発酵させ、型抜きし、成形し、パンニングし、75分間、38℃および87%RHで発酵させ、20分間、220℃で焼成した。生地および完成品のパンの評価は、パン製造の職人が行った。表10の結果から、パン容積および堅さの値は、1kg小麦粉あたり225単位もしくはそれ以下の酵素濃度によって影響を受けなかったため、プロリン特異的エンドプロテアーゼの添加は、グルテンネットワークおよび生地の気体保持能には明らかに影響を及ぼさないことが明白である。事実、より高い酵素用量を添加だけでは、生地にネガティブな影響を与え、ほとんどの消費者に許容され得ないと思われるパンが作製される。後者の観察のため、最も高い酵素濃度で生産されるパンに関する抗体アッセイデータは作製されなかった。
【0150】
表10において表されたデータは、1kgあたり約200単位の酵素濃度で生成させるパンに存在する毒性プロリンリッチペプチドの減少を示す。この程度の減少では、セリアック病の患者によるそのようなパンの消費を可能にするのに不十分であると証明される可能性が非常に高い。しかし、該減少は、看過され得るセリアック病もしくはIBSを患うヒトまたは未成熟な免疫系を伴う幼児に対してさえも予防的利益を伴う食物を調製するのにかなり十分であり得る。
【0151】
【表11】
【技術分野】
【0001】
本発明は、ペプチドまたはポリペプチドのタンパク質加水分解に関する。
【背景技術】
【0002】
プロリンリッチ食餌タンパク質、例えば、牛乳中カゼインまたは穀物中グルテンは、ヒト消化管におけるタンパク質分解に耐性であることが公知である。その結果、プロリンリッチペプチドが構成され得、そして特定のグループの個体において所望されない影響をもたらし得る。これらの影響のいくつかは、プロリンリッチペプチドが末梢組織および中枢神経系の受容体に結合するオピオイドとして作用するという事実によるとされている。例えば、自閉症および統合失調症患者に見られる症状は、プロリンリッチ食餌タンパク質の体内摂取と関連付けられている。他の影響は、プロリンリッチペプチドに対する不耐症の結果である。例えば、特定のプロリンリッチ配列は、セリアック病において観察されるグルテンの毒性を担う。セリアック病は小腸の広範に認められる自己免疫疾患であり、生涯にわたるグルテンを含まない食餌によってでしか処置することができない。セリアック病は時にはまた、プロリンリッチペプチドの代謝障害が有し得る遠大な結果を例示する精神的および神経学的症状を伴う。
【0003】
牛乳中のタンパク質は成長および健康に関連し、ヒト食餌における重要な成分を形成する。カゼインは牛乳中の全タンパク質の約80%を構成し、アミノ酸、カルシウムおよびリン酸塩の重要な供給源である。カゼインは、概ね50%のα−カゼイン、35%のβ−カゼイン、13%のκ−カゼインおよび3%のγ−カゼインよりなる。ヒト乳汁には、一般にα−カゼイン画分は存在しない。
【0004】
カゼインの代謝においては、多くの新たな生体活性ペプチドが形成されることが公知である。αおよびβ−カゼイン画分からは、それぞれ、α−カゾモルフィンおよびβ−カゾモルフィンと呼ばれるオピオイドペプチドが同定され、単離されている。特に、β−カゾモルフィンの薬理学的効果については、広範に研究されている。配列Tyr−Pro−Phe−Pro−Gly−Pro−Ileを伴うβ−カゾモルフィンは牛乳中の主要なオピオイドペプチドであり、BCM−7と呼ばれる(β−カゾモルフィン(1−7);チャン(Chang)ら(1985)Journal of Biological Chemistry,260,9706−9712)。β−カゼイン分子のアミノ酸位置60〜66におけるこのBCM−7フラグメントの他に、それぞれ、アミノ酸位置60〜63および60〜64におけるTyr−Pro−Phe−Pro(β−カゾモルフィン(1−4))およびTyr−Pro−Phe−Pro−Gly(β−カゾモルフィン(1−5))のようなBCM−7のより小さなフラグメントならびに(アミノ酸位置60〜70における)11アミノ酸の鎖長までのすべてのより大きなBCM−7関連ペプチドは少なくともいくらかの程度のオピオイド活性を示す。BCM−7のN末端トリペプチド、即ち、位置60〜62における配列Tyr−Pro−Pheはオピオイド活性を有さない。A1と呼ばれる遺伝的β−カゼイン変異体(アミノ酸位置67においてA2β−カゼインのプロリン残基ではなくヒスチジンを有する)は、高いレベルのBCM−7分子の形成をもたらすことが要求される。
【0005】
多様なβ−カゾモルフィンの作製のための基本的な理由は、それらのアミノ酸配列が比較的プロリン残基に富むことである。プロリン残基に関与するペプチド結合はタンパク質分解に耐性であるため、β−カゾモルフィン配列は、胃および腸管腔における消化管プロテアーゼへの暴露後も残存する傾向がある。同じ理由により、これらのβ−カゾモルフィン配列は、他のプロテアーゼ、例えば、タンパク質加水分解物の産業的生産において一般に使用されるそれらのプロテアーゼとのインキュベーション後にも残存する傾向があることが想定され得る。この仮定は、一般に利用可能なタンパク質加水分解物またはこれらのタンパク質加水分解物を含有する産物がすべてBCM−7もしくは緊密に関連するペプチドを含有することを含意する。BCM−7ペプチドフラグメントおよびその関連分子は所定の疾患と関連付けられているため、幼児、高齢者および患者のような脆弱なグループの食餌においてかなり頻繁に使用されるタンパク質加水分解物にそのような分子が存在することは、所望されない状況である。ヒトおよびウシβ−カゾモルフィンのオピオイド受容体結合アッセイの結果は、オピオイド活性を伴うフラグメントがラット脳膜のオピオイド受容体に結合することを示す。β−カゼインは、δおよびκ受容体サブタイプにほとんど親和性を伴わないμ−リガンドに対してより選択的であることが示されている。これらおよび他の研究に従えば、β−カゾモルフィンは、多様な消化管、鎮痛性、呼吸器、循環器、内分泌および免疫調節効果を有することが要求される。プロリン残基を組み入れているオピオイドペプチドに共通の構造的特徴は、Tyr−Pro−Phe/Trpモチーフである(オカダ(Okada)ら、Vitamins and Hormones2002,65,257−279)。
【0006】
健康な個体では、腸上皮層および血液におけるペプチダーゼは、β−カゾモルフィンを処理することができるが、これは、統合失調症、自閉症、ADHDまたは他の気分障害を患う患者の場合では必ずしもそうではないようである。例えば、プロリンリッチペプチドの不完全な分解をもたらす血漿中ジペプチジルペプチダーゼIV(DPP IV)酵素活性の遺伝的変更は、これらの疾患の発症と関連付けられている。さらに、高ペプチド尿症、即ち、尿中における高い濃度のカゼインまたはグルテン由来のペプチドが、一定の頻度で認められる(ライヘルト、W.H.(Reichelt,W.H.)ら;(1997)Dev.Brain Dysfunct;10:44−55)。最近の科学的文献は、プロリンリッチペプチドの不完全な分解がそのような疾患の発達および重症度に寄与し得るという説得力のある証拠を提供している。カゼイン(caseine)由来BCM−7フラグメントの他にも、これに関して、グルテン由来プロテアーゼ耐性ペプチドについて述べられている。すでに1979年には、パンクセップ(Panksepp)(Trends in Neuroscience1979;2:174−177)はオピオイド過剰理論を提唱し、その中で彼はオピオイド代謝障害が自閉症の病因の部分であることを示唆した。現在では、本発明者らは、多くのプロリンリッチペプチドが胃および膵臓のペプチダーゼ、例えば、ペプシン、トリプシン、キモトリプシンなどによる切断に高度に耐性であり、なかでも消化管の刷子縁上皮層に存在するような特異的酵素だけが、プロリンに関与するペプチド結合を加水分解することが可能であることを理解した。
【0007】
グルテンはデンプンおよび水溶性成分を除去するための洗浄後に残留するコムギ、ライムギ、カラスムギ、オオムギ、トウモロコシおよびイネのような穀物の不溶性タンパク質画分である。グルテンは、4つの主要な可溶性画分、即ち、アルブミン、グロブリン、プロラミンおよびグルテリンに再分割することができる。これらのなかでも、特に、コムギ、トウモロコシ、オオムギならびにカラスムギのプロラミンおよびグルテリン画分は、比較的高い含有量のアミノ酸、プロリンおよびグルタミンによって特徴付けられる。最近の証拠は、セリアック病の発達における主要な因子として、プロリンリッチグルテン配列を含意している。セリアックスプルーとしても公知であるセリアック病は、グルテンタンパク質の摂取によって生じる小腸の自己免疫疾患である。それは、慢性の下痢および腹部膨満のような重度の症状を伴う幼児において一般に認められ;その後の生涯では、症状は、疲労、吸収不良による体重減少および神経学的症状を含む。多様な穀物のプロリンリッチ画分のなかでも、コムギ由来のα−グリアジン、オオムギ由来のホルデイン、ライムギ由来のセカリンおよびカラスムギ由来のアベニンは、最も毒性が高いようである(Schuppan,D.;Gastroenterology2000;119:234−242)。生涯にわたるグルテンを含まない食餌のみが、セリアック病患者に有効な処置である。セリアック病の患者のなかでも、高い罹病率の多様な自己免疫障害、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎が観察されている。これは、未知の機構によって、非処置のセリアック病が他の器官に対する自己免疫を起こしやすいことを示す(シュパン、D.(Schuppan,D.)2000Gasteroenterology119:234−242)。さらに、軽度の形態のセリアック病が、過敏性腸症候群(IBS)を患うヒトのグループに存在する徴候がある。IBSは大腸の正常な機能を妨害する障害であり、差し込むような腹痛、便秘および下痢によって特徴付けられる。IBSは、通常、約20歳で始まり、多大な不快感および苦痛を生じる。コムギ、オオムギ、ライムギまたは乳製品の摂食は、IBSの症状の悪化と関係付けられている。
【0008】
最近、シャン(Shan)ら(Science;第297巻、2002年9月27日:2275−2279)は、1組の主要なセリアック病の患者特異的T細胞のエピトープの供給源と考えられるグリアジン由来、プロリンリッチ、33アミノ酸長ペプチドを同定した。小腸刷子縁細胞から調製された酵素抽出物はこの33マーを加水分解することができなかったが、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)由来の細菌性プロリルオリゴペプチダーゼの補充は、関連T細胞クローンの刺激の付随的な顕著な減少を伴う迅速な消化をもたらす。パパインの経口投与に関する早期の研究(メッセル、M.(Messer,M.)およびボーメ、P.E.(Baume,P.E.);Lancet1976;2:1022)を模倣すると、該論文より、酵素療法によるグルテンの無毒化における食餌中酵素としてのプロリルオリゴペプチダーゼの能力が示される。
【0009】
プロリルオリゴペプチダーゼ(EC3.4.21.26)は、プロリン残基のカルボキシル側でペプチドを好適に切断する独特な可能性を有する。哺乳動物供給源から単離されるプロリルオリゴペプチダーゼならびにフラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から単離されるプロリルオリゴペプチダーゼにおいて、酵素の活性部位から構造決定された大きなタンパク質を含まない独特なペプチダーゼドメインが同定されている。事実、これらの酵素は、約30を超えるアミノ酸残基を含有するタンパク質を分解することができず、そのため、これらの酵素は、現在、「プロリルオリゴペプチダーゼ」と称される(フロプ(Fulop)ら:Cell、第94巻、161−170、1998年7月24日)。すべての既知のプロリルオリゴペプチダーゼは、中性付近で至適pH値を示すサイトゾルの酵素であり、それらが約30を超えるアミノ酸残基を含有する分子を効率的に分解することができないという事実によって特徴付けられる。これらの酵素が、消化管のより遠位部において示現するpH値に対応する至適pH値を示すという事実は、それらを、食餌中グルテンの腸での消化を支持する栄養補助食品として理想的に安定にする。
【0010】
毒性のプロリンリッチペプチドの不活化において利益を有することができるもう1つの酵素は、酵素ジペプチジルペプチダーゼIVである(米国公開特許第2002/0041871A号明細書)。Xaa−Pro−ジペプチジル−アミノペプチダーゼ(EC3.4.14.5)とも呼ばれるジペプチジルペプチダーゼIVは、好ましくは、Xbbがプロリンであるが、但し、Xccがプロリンでない場合、N末端配列Xaa−Xbb−Xcc−を伴うペプチドからのN末端ジペプチド、Xaa−Xbbの放出を触媒する。ジペプチジル−ペプチダーゼIVが、多くの哺乳動物供給源、例えば、酵素の豊富な供給源由来の腸刷子縁膜から単離されている。さらに、酵素は、微生物供給源、例えば、食物用微生物、サッカロミセス(Saccharomyces)、ラクトコッカス(Lactococcus)およびアスペルギルス(Aspergillus)から単離されている。プロリルオリゴペプチダーゼのように、すべての既知のジペプチジル−ペプチダーゼIVは、中性付近の至適pH値を伴う酵素であり、従って、腸での消化プロセスを支えるのに適している。
【0011】
セリアック病もしくは統合失調症、自閉症または他の気分障害の処置におけるプロリン特異的オリゴペプチダーゼおよびジペプチジル−ペプチダーゼIVの可能な関連のため、これらのデータから、この事柄の多様な局面を取り扱う多くの特許出願が生じている。例えば、米国特許第6,447,772号明細書および国際公開第01/24816号パンフレットはジペプチジルペプチダーゼIVを含有する組成物について記載し、国際公開第03/068170号パンフレットは場合により、ジペプチジル−ペプチダーゼIVと組み合わされたプロリン特異的オリゴペプチダーゼを含有する組成物について記載し、国際公開第02/45523号パンフレットはプロリン特異的エンドプロテアーゼにより調製される低アレルギー性タンパク質加水分解物について記載し、国際公開第03/028745号パンフレットは腸内の毒性プロリンリッチペプチドの濃度を低下することができる細菌株を含んでなる組成物について記載している。国際公開第96/36239号パンフレットは、β−カゼインA1対立遺伝子を実質的に含まないウシから誘導される産物の利点について記載している。
【発明の概要】
【課題を解決するための手段】
【0012】
本発明は、ペプチドまたはポリペプチドのタンパク質加水分解のための方法であって、前記ペプチドまたはポリペプチドは4〜40、好ましくは5〜35アミノ酸残基を含んでなり、前記ペプチドまたはポリペプチドはスブチリシンによって加水分解され得ず、それによって、前記ペプチドまたはポリペプチドは、前記ペプチドまたはポリペプチドを加水分解するための6.5もしくはそれ以下、好ましくは5.5もしくはそれ以下、およびより好ましくは5.0もしくはそれ以下のpHでプロリン特異的エンドプロテアーゼにより加水分解される、方法に関する。好ましくは、少なくとも70%、より好ましくは、少なくとも80%、および最も好ましくは、少なくとも90%のペプチドまたはポリペプチドがこの方法において加水分解される。
【0013】
別の実施態様に従えば、本発明は、ペプチドまたはポリペプチドのタンパク質加水分解のための方法であって、前記ペプチドまたはポリペプチドは4〜40、好ましくは5〜35アミノ酸残基を含んでなり、そしてトリペプチドモチーフGlu−Xxx−Pro、Gln−Xxx−Pro、Tyr−Pro−PheまたはTyr−Pro−Trpを含んでなり、それによって、前記ペプチドまたはポリペプチドは、前記ペプチドまたはポリペプチドを加水分解するための6.5もしくはそれ以下、好ましくは5.5もしくはそれ以下およびより好ましくは5.0もしくはそれ以下のpHでプロリン特異的エンドプロテアーゼにより加水分解される、方法に関する。好ましくは、少なくとも70%、より好ましくは、少なくとも80%、および最も好ましくは、少なくとも90%のペプチドまたはポリペプチドがこの方法において加水分解される。
【0014】
さらに、本発明は、ペプチドまたはポリペプチドのタンパク質加水分解のための方法であって、前記ペプチドまたはポリペプチドは4〜40、好ましくは5〜35アミノ酸残基を含んでなり、それによって、ペプチドまたはポリペプチドのアミノ酸残基は少なくとも30%、好ましくは少なくとも40%のプロリンおよび/またはグルタミン残基を含んでなり、それによって、前記ペプチドまたはポリペプチドは、前記ペプチドまたはポリペプチドを加水分解するための6.5もしくはそれ以下、好ましくは5.5もしくはそれ以下およびより好ましくは5.0もしくはそれ以下のpHでプロリン特異的エンドプロテアーゼにより加水分解されるが、但し、ペプチドまたはポリペプチドは少なくとも10%のプロリン残基を含んでなる、方法を提供する。好ましくは、少なくとも70%、より好ましくは、少なくとも80%、および最も好ましくは、少なくとも90%のペプチドまたはポリペプチドがこの方法において加水分解される。
【0015】
好ましくは、本発明の方法におけるペプチドまたはポリペプチドは、モチーフGln−Xxx−ProまたはGlu−Xxx−Proを含んでなり、9もしくはそれ以上のアミノ酸残基を含有する。このペプチドまたはポリペプチドは、有利なことに、8もしくはそれ以下のアミノ酸残基を含有するペプチドに加水分解される。ペプチドまたはポリペプチドがモチーフTyr−Pro−PheまたはTyr−Pro−Trpを有する場合、好ましくは、ProとPheとの間またはProとTrpとの間の結合が加水分解される。
【0016】
本発明の方法において使用される好適なエンドプロリン特異的エンドプロテアーゼは、好ましくは、アスペルギルス(Aspergillus)から誘導されるか、またはセリンプロテアーゼのS28ファミリーに属するプロリン特異的エンドプロテアーゼである。この酵素は、好ましくは、スブチリシンによって加水分解され得ない4〜40、好ましくは5〜35アミノ酸残基を含んでなるペプチドまたはポリペプチドを加水分解するための6.5未満、好ましくは5.5未満、より好ましくは5.0未満の至適pH値を有する。
【0017】
プロリン特異的エンドプロテアーゼは、自閉症、統合失調症、ADHD、双極性気分障害およびうつを含む精神障害ならびに自己免疫障害、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎のようなセリアック病に関連する障害ならびにIBSに関連するプロリンリッチペプチドを5.5未満のpHで加水分解するために使用することができる。
【0018】
有利なことに、この酵素は、セリアック病関連エピトープ、好ましくはグルテンのエピトープ、より好ましくはコムギまたはオオムギのエピトープを欠く食物、例えば、ビールもしくはパンを生成するために使用される。
【0019】
本発明はまた、医薬品としての使用のためかまたは医薬品の製造における使用のためのプロリン特異的エンドプロテアーゼに関し、該プロリン特異的エンドプロテアーゼは、好ましくは、アスペルギルス(Aspergillus)、より好ましくは、アスペルギルス・ニガー(Aspergillus niger)の酵素である。
【0020】
本発明の別の実施態様に従えば、プロリン特異的エンドプロテアーゼは、自閉症、統合失調症、ADHD、双極性気分障害およびうつを含む精神障害ならびに自己免疫障害(autoimmne disorder)、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎のようなセリアック病に関連する障害ならびにIBSの処置もしくは防止のための栄養補助食品または医薬品の製造のために使用される。
【0021】
有利なことに、プロリン特異的エンドプロテアーゼは、年齢25歳未満の個体のための栄養補助食品もしくは医薬品の製造のために使用される。
【0022】
本発明に従えば、プロリン特異的エンドプロテアーゼはまた、自閉症、統合失調症、ADHD、双極性気分障害およびうつを含む精神障害ならびに自己免疫障害(autoimmne disorder)、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎のようなセリアック病に関連する障害ならびにIBSの処置もしくは防止のための栄養補助食品または医薬品のために使用される。
【0023】
さらに、本発明は、30を超えるアミノ酸残基を有するタンパク質またはペプチドを加水分解するためのプロリン特異的エンドプロテアーゼの使用に関する。本発明に従う使用では、好ましくは、アスペルギルス(Aspergillus)、より好ましくは、A.niger(A.ニガー)のプロリン特異的エンドプロテアーゼが適用される。
【0024】
本発明はまた、ペプシンの存在下、5もしくは5以下のpHで活性であるプロリン特異的プロテアーゼの使用に関する。
【0025】
有利なことに、本発明は、β−カゼインA1またはβ−カゼインA2対立遺伝子を担持するウシから得られる乳タンパク質中に存在する前記ペプチドもしくはタンパク質のタンパク質加水分解の方法に関する。
【0026】
タンパク質はまた、本発明の方法においても使用することができる。第1に、タンパク質は、4〜40アミノ酸残基を含んでなるペプチドに加水分解されなければならない。タンパク質の加水分解は、本発明の方法の前、または同時に行うことができる。
【0027】
さらに、本発明は、非ヒト動物、好ましくは哺乳動物を意図した栄養補助食品の生成のため、医薬品の生成のためまたはペットフードを含む食物の生成のための栄養補助食品として、医薬品としての6.5未満、好ましくは5.5未満、より好ましくは5.0未満の至適pH値を有するプロリン特異的エンドプロテアーゼの使用に関する。
【0028】
一般に、先行技術は、体内摂取の前の食物から毒性のプロリンリッチペプチドを取り出すこと、または不適切な腸内消化プロセスを補整するための調整酵素の経口的供給を目的とする。正常な消化管消化プロセスでは、酵素ペプシンによる胃内のタンパク質分解は、食餌タンパク質の分解における第1の工程であると理解される。第2の工程は、小腸、即ち、胃のすぐ下流に位置する十二指腸および空腸において生じる。十二指腸では、胃内容物の酸性pHは、様々なエンド−およびカルボキシペプチダーゼをも含有する膵液の添加によって生じる。後者の酵素によって触媒されるペプシン分解食餌タンパク質のさらなる分解は、5を超えるpH値で十二指腸および空腸の内腔において達成される。腸壁を超える輸送の前に、第3の工程は、腸(intestestinal)上皮の刷子縁表面膜におけるさらなるペプチド加水分解に関与する。後者の工程は、これらの上皮細胞の膜に位置するDPP IVを含む多くのプロテアーゼによって達成される。先行技術は、いくらかの疾患を患う個体の腸において、本発明は、いくらかのこれらの上皮酵素を目的としており、とりわけ、DPP IVは部分的に不活性であるかまたは存在さえしないことを示唆している。セリアック病の患者では、これらの上皮酵素のレベルはおそらく正常であるが、このグループの個体内では、低レベルの所定のプロリンリッチペプチドであっても強力な炎症性T細胞応答を発揮することができる。先行技術において示される解決方法は、消化器官の管腔に存在する毒性のプロリンリッチペプチドのレベルを最小限にするために、経口用調整酵素をヒト消化器官に導入することである。このアプローチでは、先行技術は、できるだけ天然のヒト酵素を模倣する選択された酵素を有し、即ち、酵素は、pH5を超える条件下の消化器官において活性である。これらの先行技術の酵素は酸性条件下では活性ではないか、またはこれらの先行技術の酵素を含有するオーラル・プレゼンテーション形態は、酸性条件下でのそれらの活性を防止するために適切に被覆される。
【0029】
本発明者らは、胃における酸性条件下でその主要な活性を発達する酵素が、不十分に消化されたプロリンリッチペプチドの問題に対し、より優れた解決を提供することを見出した。このアプローチは新規であり、問題を解決するために文献に記載された酵素以外の酵素を使用する可能性を開く。
【0030】
上記で考察したように、いくらかの刊行物は、オピオイド代謝障害の防止におけるジペプチジルペプチダーゼIVおよびプロリルオリゴペプチダーゼのような酵素を使用する可能性について指摘している。さらに、中性付近のpH条件下で典型的に活性であるジペプチジルペプチダーゼIVならびにプロリルオリゴペプチダーゼ酵素が選択される。ここで意味することは、これらの酵素のみが、pH5を超える条件下で効率的にプロリンリッチタンパク質を加水分解することである。ヒト消化管のpHプロフィールを考慮すると、これらの先行技術の酵素は、十二指腸の遠位部、即ち、胃よりかなり先方に到達する前に、活性になることはない。しかし、十二指腸の遠位部は、タンパク質吸収の主要部位であり、また、セリアック病患者では影響を受けることが公知である。そのため、事実上、これらの先行技術の酵素は、障害の部分を受けている場所で作用し始める。
【0031】
さらに、先行技術の酵素は、プロリンリッチタンパク質の有意なプレ消化後にのみ活性になる。膵臓の酵素によるこのようなプレ消化を達成するのに必要な時間は、経口的に適用された酵素がプロリンリッチ配列の完全な加水分解を達成するのに利用可能な期間をさらに制限する。本発明に従えば、食物が小腸に侵入する前にほとんどのプロリンリッチおよび/またはグルタミンリッチタンパク質配列を分解する酵素が使用される。胃における正常な食物滞留時間は、プロリンリッチまたはグルタミンリッチタンパク質の適切な加水分解に必要な期間を提供する。さらに、本発明に従えば、好ましくは、プロリン残基のカルボキシル側でペプチドを切断することが可能であり、また、プロリン残基のカルボキシル側でかつペプシンの存在下であっても無傷なタンパク質を切断することが可能である酵素が使用される。先行技術の酵素は、胃の低いpH条件下で、かつ胃に分泌されるタンパク質分解性ペプシン酵素に暴露される場合、加水分解される。しかし、これまで、そのような酸性/ペプシン安定性酵素の使用は、現在の方法では知られていない。本アプローチは、腸においてプロリンリッチペプチドを加水分解する天然のプロセスを模倣することを提唱する先行技術のアプローチの代わりに、腸に到達する前にプロリンリッチペプチド、ポリペプチドおよびタンパク質の実質的な分解を生じる。本発明の方法において使用される好適な酵素は、5未満のpH値、ペプシンの存在下で活性である酵素であり、食餌タンパク質またはポリペプチドならびにペプチドを分解することが可能である。先行技術の解決方法は、常に、食餌タンパク質をペプチドにプレ加水分解する補酵素を必要とし、その後にのみ、さらなるプロリン特異的酵素がそれらの活性を展開し始めることができる。
【0032】
「ペプチド」または「オリゴペプチド」は、本明細書において、ペプチド結合を介して連結される2〜30アミノ酸残基の鎖として規定される。用語「ペプチド」および「オリゴペプチド」は、同義(共通認識される)とみなされ、各用語は、状況に応じて交換可能に使用することができる。「ポリペプチド」は、本明細書において、30を超えるアミノ酸残基を含有する鎖として規定される。
【0033】
スブチリシン(EC3.4.21.62)、好ましくは、スブチリシンCarlsbergによって加水分解され得ない4〜40アミノ酸残基を有するペプチドまたはポリペプチドは、20g/lタンパク質および1グラムのタンパク質あたり0.12AU−A(アンソン単位アルカラーゼ(Alcalase))プロテアーゼ単位の酵素対基質を含有する懸濁液または溶液において、pH8.0ならびに60℃で2時間のインキュベーション後にも無傷(intact)を保持するペプチドまたはポリペプチドであると理解される。AU−Aプロテアーゼ単位は、Novozymes(丁国)により提出された分析方法LUNA番号2003−32153−01において特定されるように規定される。無傷な(intact)とは、インキュベーション後、本来のペプチドまたはポリペプチドが80%を超え、好ましくは90%を超え、より好ましくは、95%を超えて得られる酵素反応産物を形成することを意味する。実際には、酵素消化は、示される条件下、および存在する1グラムの基質タンパク質あたりの40マイクロリットルのアルカラーゼ(Alcalase)を使用して、行われる。そのような非加水分解性ペプチドの例は、実施例4に記載のβ−カゼイン加水分解から得られるVYPFPGPIPNペプチドである。そのような非加水分解性ポリペプチドのもう1つの例は、実施例6に記載の33マーである。
【0034】
オリゴペプチダーゼは、EC3.4.21.26として分類される酵素であり、セリンプロテアーゼclanSCのファミリー、ファミリーS9に属する。
【0035】
本明細書に記載のすべての(オリゴ)ペプチドおよびポリペプチドの式または配列は、定法に従い、アミノ末端からカルボキシ末端の方向で、左から右に記載される。本明細書において使用するアミノ酸の1文字コードは、当該分野において一般に公知であり、サンブルック(Sambrook)ら(Molecular Cloning:A Laboratory Manual、第2版Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor、ニューヨーク州、1989年)において見出すことができる。アミノ酸Xxx、Xaa、XbbまたはXccは、任意のアミノ酸であることが意図される。モチーフとは、ペプチド、ポリペプチドまたはタンパク質の部分であるアミノ酸配列を意味する。ここで、グルタミン(QもしくはGln)は、グルタミンまたはグルタミン酸(EもしくはGlu)であることが理解される。例えば、化学的に脱アミド化されたグルテンでは、存在する天然のグルタミン残基の大きな部分が、グルタミン酸に変換される。そのため、モチーフGln−Xxx−Proはまた、モチーフGlu−Xxx−Proも含んでなっている。また、他のペプチドでは、ポリペプチドまたはタンパク質のグルタミン(glumine)を、例えば、pH条件、温度によりまたは例えば、トランスグルタミナーゼによる酵素変換によって、グルタミン酸に変換することができる。
【0036】
プロリンリッチペプチドまたはポリペプチドとは、4〜40アミノ酸残基を含んでなるペプチドまたはポリペプチドを意味し、ここで、ペプチドまたはポリペプチドのアミノ酸残基は、少なくとも30%、好ましくは、少なくとも40%のプロリンおよび/またはグルタミン残基を含んでなるが、但し、ペプチドまたはポリペプチドは少なくとも10%のプロリン残基を含んでなる。栄養補助食品は、DSHEA、以下の食餌成分の1つもしくはそれ以上を有するかまたは含有する食餌を補助することを目的とした製品(タバコ以外)の適応された規定に従う:酵素を含むタンパク質、ポリペプチド、ペプチド、ビタミン、ミネラル、ハーブまたは他の植物性薬品、アミノ酸、全摂取量を増加することによって食餌を補助するためのヒトによる使用のための食餌物質、または濃縮物、代謝物、構成物、抽出物、あるいはこれらの成分の組み合わせ。
【0037】
さらに、栄養補助食品は、
・丸剤、カプセル剤、錠剤、または液体剤形での摂取を目的とする。
・従来の食物としてまたは食事もしくは食餌の単独の品目としての使用には該当しない。
・一般に、「栄養補助食品」として標識される。
・承認された新規の薬物、保証された抗生物質、または承認、保証、もしくは認可前に食物の栄養補助食品として市販された認可された生物工学製品のような製品を含む(但し、Secretary of Health and Human Servicesがこの条項を放棄する場合を除く)。
【0038】
毒性のプロリンリッチペプチドまたはポリペプチドは、オピオイド受容体への結合または精神もしくはセリアック障害の発達もしくは重症度において関連するペプチドまたはポリペプチドである。
【0039】
精神障害としては、自閉症、統合失調症、ADHD、双極性気分障害ならびにうつが挙げられる。
【0040】
セリアック病に関連する障害は、自己免疫障害、具体的には、1型糖尿病、疱疹状皮膚炎、自己免疫性甲状腺炎、膠原病、自己免疫性脱毛および自己免疫性肝炎である。
【0041】
IUMB由来のすべての酵素の分類および命名法についての国際的に認められたスキームは、プロテアーゼを含む。プロテアーゼEC番号の更新されたIUMBテキストは、インターネットサイト:http://www.chem.qmw/ac.uk/iubmb/enzyme/EC3/4/11/で見出すことができる。このシステムでは、酵素は、それらが単一の反応を触媒するという事実によって規定される。該システムは、プロテアーゼをエンドおよびエキソプロテアーゼを分類する。エンドペプチダーゼは、内部のペプチド結合を加水分解するそれらの酵素であり、エキソプロテアーゼは、末端のα−アミノ基に隣接するペプチド結合(「アミノペプチダーゼ」)、または末端のカルボキシル基と最後から2番目のアミノ酸との間のペプチド結合(「カルボキシペプチダーゼ」)を加水分解する。エンドペプチダーゼは、触媒機構に基づいてサブ‐サブクラスに分けられる。セリンエンドペプチダーゼ(EC3.4.21)、システインエンドペプチダーゼ(EC3.4.22)、アスパラギン酸エンドプロテアーゼ(EC3.4.23)、メタロエンドプロテアーゼ(EC3.4.24)およびスレオニンエンドプロテアーゼ(EC3.4.25)のサブ‐サブクラスが存在する。
【0042】
国際公開第02/45524号パンフレットは、アスペルギルス・ニガー(Aspergillus niger)から入手可能なプロリン特異的エンドプロテアーゼについて説明している。驚くべきことに、本発明者らは、今回、このアスペルギルス(Aspergillus)の酵素は、胃の酸性条件下で本方法において有利に使用され、これらの条件下でも無傷な食餌タンパク質、ポリペプチドならびにより小さなペプチド分子を加水分解することができることを見出した。さらに、この酵素は、酸性条件で酵素ペプシンの存在下でも残存し、十二指腸通じてその活性を継続するようである。本発明者らは、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが、活性ならびに進化の観点から既知のプロリン特異的プロテアーゼおよび国際公開第03/068170号パンフレットにおいて特定されるグルテナーゼとはまったく異なることを実証している。後者の特徴は、国際公開第03/068170号パンフレットにおいて特定されるグルテナーゼとA.niger(A.ニガー)由来酵素との間のアミノ酸配列相同性が、グローバルアラインメントアルゴリズム解析を使用して典型的に20%未満であるという事実により十分に実証される。この結果は、プロリルオリゴペプチダーゼが、本発明に従うプロリン特異的エンドプロテアーゼが単離されるアスペルギルス・ニガー(Aspergillus niger)のような真菌では発生しないという現在の観点(ヴェナライネンJ.I.(Venaelaeinen,J.I.)ら、Eur J Biochem271,2705−2715(2004))と一致する。
【0043】
本発明に従うかまたは本発明に従って使用されるプロリン特異的エンドプロテアーゼとは、例えば、本明細書において参考として援用される国際公開第02/45524号パンフレットの請求項1〜5、11および13に記載のポリペプチドを意味する。従って、このプロリン特異的エンドプロテアーゼは、以下よりなる群から選択されるプロリン特異的エンド型プロテアーゼ活性を有するポリペプチドである:
(a)配列番号2のアミノ酸1〜526またはそのフラグメントと少なくとも40%アミノ酸配列同一性を有するアミノ酸配列を有するポリペプチド;
(b)低いストリンジェンシー条件で、(i)少なくとも80%もしくは90%同一な60を超える、好ましくは100を超えるヌクレオチド、より好ましくは少なくとも90%同一な200を超えるヌクレオチドである配列番号1の核酸配列またはそのフラグメント、あるいは(ii)配列番号1の核酸配列に相補的な核酸配列、とハイブリダイズするポリヌクレオチドによってコードされるポリペプチド。配列番号1および配列番号2は国際公開第02/45524号パンフレットに示される通りである。好ましくは、ポリペプチドは単離された形態である。
【0044】
好適なポリペプチドは、配列番号2のアミノ酸1〜526と少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも65%、好ましくは少なくとも70%、より好ましくは少なくとも80%、さらにより好ましくは少なくとも90%、最も好ましくは少なくとも95%、およびさらに最も好ましくは少なくとも約97%同一性を有するかまたは配列番号2のアミノ酸配列を含んでなる。
【0045】
好ましくは、ポリペプチドは、低いストリンジェンシー条件、より好ましくは中等度のストリンジェンシー条件、および最も好ましくは高いストリンジェンシー条件下、(i)配列番号1の核酸配列もしくはそのフラグメント、または(ii)配列番号1の核酸配列に相補的な核酸配列とハイブリダイズするポリヌクレオチドによってコードされる。
【0046】
用語「ハイブリダイズすることが可能」は、本発明の標的ポリヌクレオチドが、プローブとして使用される核酸(例えば、配列番号1のヌクレオチド配列、もしくはそのフラグメント、または配列番号1の相補物)に、バックグランドよりも有意に高いレベルでハイブリダイズすることができることを意味する。本発明はまた、本発明のプロリン特異的エンドプロテアーゼをコードするポリヌクレオチド、ならびにそれに相補的であるヌクレオチド配列を含む。ヌクレオチド配列は、RNAまたはゲノムDNA、合成DNAもしくはcDNAを含むDNAであってもよい。好ましくは、ヌクレオチド配列は、DNAおよび最も好ましくは、ゲノムDNA配列である。典型的に、本発明のポリヌクレオチドは、選択的条件下で、コーディング配列または配列番号1のコーディング配列の相補物にハイブリダイズすることが可能であるヌクレオチドの連続配列を含んでなる。そのようなヌクレオチドは、当該分野において周知の方法に従って合成することができる。
【0047】
本発明のポリヌクレオチドは、コーディング配列または配列番号1のコーディング配列の相補物に、バックグランドよりも有意に高いレベルでハイブリダイズすることができる。例えば、cDNAライブラリーに存在する他のcDNAのため、バックグランドのハイブリダイゼーションが生じ得る。本発明のポリヌクレオチドとコーディング配列または配列番号1のコーディング配列の相補物との間の相互作用によって作製されるシグナルレベルは、典型的に、他のポリヌクレオチドと配列番号1のコーディング配列との間の相互作用の少なくとも10倍、好ましくは少なくとも20倍、より好ましくは少なくとも50倍、およびさらにより好ましくは少なくとも100倍の強度である。相互作用の強度は、例えば、プローブを、例えば、32Pで放射性標識することによって、測定することができる。選択的ハイブリダイゼーションは、典型的に、低いストリンジェンシー(0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約40℃)、中等度のストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約50℃)または高いストリンジェンシー(例えば、0.3M塩化ナトリウムおよび0.03Mクエン酸ナトリウム、約60℃)の条件を使用して、達成することができる。
【0048】
UWGCGパッケージは、(例えば、そのデフォルト設定値に対して使用される)同一性を算出するために使用することができるBESTFITプログラムを提供する。
【0049】
PILEUPおよびBLAST Nアルゴリズムを使用して、配列同一性を算出するかまたは(例えば、それらのデフォルト設定値に対する等価もしくは対応する配列を同定するような)配列を列挙することができる。
【0050】
BLAST解析を実施するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を介して公的に入手することができる。このアルゴリズムは、データベース配列において同じ長さのワードと共に整列される場合、いくつかのポジティブ評価された閾値スコアTに一致または満たすかのいずれかである問い合わせ配列における長さWの短いワードを同定することによって、ハイスコアリング配列対(high scoring sequence pair)(HSP)を第1に同定することに関与する。Tは隣接ワードスコア閾値と称される。これらの初期の隣接ワードのヒットは、それらを含有するHSPを見出すための検索を初期化するためのシードとして作用する。ワードヒットは、累積アラインメントスコアが増加し得る限り、各配列に沿って両方向において拡張される。累積アラインメントスコアがその最大到達値から量Xだけ低下する場合;1つもしくはそれ以上のネガティブスコアリング残基アラインメントの累積により累積スコアが0もしくはそれ以下となる場合;またはいずれかの配列の末端に到達する場合、各方向におけるワードヒットの拡張は停止する。BLASTアルゴリズムパラメータW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプログラムは、デフォルトとして、11のワード長(W)、50のBLOSUM62スコアリングマトリックスアラインメント(B)、10の期待値(E)、M=5、N=4、および両鎖の比較を使用する。
【0051】
BLASTアルゴリズムは、2つの配列間の類似性の統計解析を実施する。BLASTアルゴリズムによって提供される類似性の1つの測定は、2つのヌクレオチドまたはアミノ酸配列間の一致が偶然生じる確率の示度を提供する最小の全確率(P(N))である。例えば、配列は、第1の配列と第2の配列との比較における最小全確率が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、および最も好ましくは約0.001未満である場合、もう1つの配列に類似であるとみなされる。
【0052】
本発明に従う酵素は、好ましくは、真菌、好ましくは、アスペルギルス(Aspergillus)、より好ましくはアスペルギルス・ニガー(Aspergillus niger)から得られる。そのため、アスペルギルス(Aspergillus)由来酵素は、酸性安定であり、好ましくは、胃において示現するpH条件下でのペプシンの存在の影響を受けない真のエンドペプチダーゼとして見出される。フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から得ることができる酵素を一般に含むプロリン特異的オリゴペプチダーゼは、pH5以下でほとんど活性を有さず、低pHおよび酵素ペプシンの存在の組み合わせにより不活化される。さらに、プロリン特異的プロテアーゼは、一般に、無傷なタンパク質を分解することができず、より小さなペプチドのみ、即ち、約30アミノ酸残基の長さまでのペプチドを効率的に加水分解することが見出された。
【0053】
本発明は、毒性のプロリンリッチペプチドもしくはオピオイド代謝障害の現象を延期または最小限にする経済的な食品用組成物を提供する。組成物は、食物的、薬学的および獣医学的使用に適した経口用酵素処方ならびに有意に低減されたレベルのオピオイドまたは毒性プロリンリッチペプチドを伴うタンパク質加水分解物および食品の生成に適した酵素処方を含む。
【0054】
本発明は、精神障害もしくはセリアック病またはオピオイド代謝障害の発達に関連するプロリンリッチペプチドまたはポリペプチドを加水分解するための方法を開示する。本発明はまた、幼児におけるかもしくは一般に年齢25歳未満の個体のそのような障害の発達を防止または遅延することができる食物を生成する方法についても開示する。また、セリアック病およびIBSに関係する胃の症状を患う人により良好に忍容性である食物は、そのような方法によって調製することができる。本発明の実施形態は、体内摂取前のこれらのプロリンリッチペプチドまたはポリペプチドの分解に関連付けられ、それによって、身体の毒性プロリンリッチペプチドへの暴露が防止または最小限にされる。幼児では、これによりオピオイドへの早期の暴露が回避され、未成熟な免疫系は、そのような毒性のプロリンリッチペプチドに感作されない。また、十代および成人については、低いレベルの毒性プロリンリッチペプチドを含有する食餌は、例えば、看過され得るセリアック病またはIBSを患うヒトに対し予防的利益を有する。本発明はまた、胃、即ち、酸性pH条件下、好ましくはpH5未満の条件下で、これらの毒性のプロリンリッチペプチドまたはペプチドの身体(ヒトもしくは動物)における分解に安定な酵素の補充に関する。後者の実施態様に従えば、セリアック病、セリアック病の発症に関係する疾患またはこれらのペプチドまたはポリペプチドの分解に必要な低いレベルの身体中プロリン特異的プロテアーゼによって生じる疾患を患うヒトが、胃および十二指腸の近接部において関連のプロリンリッチペプチドまたはポリペプチドを分解することも可能である。さらに、本発明に従う酵素は、保護被覆を必要としない。
【0055】
好ましくは、中性pH条件下でのペプチドもしくはポリペプチドまたはタンパク質とスブチリシン(EC3.4.21.62)、好ましくは、バチルス・リケニフォルミス(Bacillus licheniformis)のスブチリシン(またはスブチリシンCarlsberg)とのインキュベーション時に形成される少なくとも80%の毒性プロリンリッチペプチドまたはポリペプチドが、本発明に従うプロリン特異的エンドプロテアーゼによって加水分解される。そのようなスブチリシン耐性ペプチドの形成を、本出願の実施例4に例示する。そのようなスブチリシン耐性プロリンリッチペプチドは、しばしば、上記の疾患と関連付けられる。これらのペプチドの例には、BCM−7、BCM−7関連ペプチド、即ち、β−カゼイン分子の位置60〜63に存在するアミノ酸配列YPFPを含んでなるペプチドがある。さらに、モチーフGln−Xxx−Pro(Q−X−P)を含んでなるグリアジン由来ペプチド、例えば、PYPQPQLPYエピトープ、ならびにグリアジン、ホルデイン、セカリンまたはアベニンから得ることができるこのQ−X−PまたはE−X−Pモチーフを含んでなる他のスブチリシン耐性分子がその例である。スブチリシンによる加水分解によって形成され得るプロリンリッチペプチドのうち、より好ましくは、少なくとも90%、なおより好ましくは、少なくとも95%および最も好ましくは、少なくとも99%が、本発明に従うプロリン特異的エンドプロテアーゼを使用することによって分解されるか、または形成されない。最も好ましくは、これらのプロリンリッチペプチドは、pH5.5未満の条件下で本発明の方法に従って分解することができる。
【0056】
必要とする酵素を供給する1つの方法は、例えば、食餌中プロリンリッチペプチドまたはポリペプチドの胃腸消化を援助するために食物と共に同時摂取される安定化された酵素処方としての消化剤の形態であり得る。もう1つの方法は、例えばアスペルギルス(Aspergillus)の酵素によって「予め消化された」タンパク質食物を使用することによって、問題のある毒性のプロリンリッチ配列の摂取を防止または制限することである。そのようなタンパク質食物は、加水分解物、例えば、グルテンまたは乳タンパク質加水分解物の形態で供給され得、多量の遊離アミノ酸を遊離するためのエンド−ならびにエキソプロテアーゼによって広範に消化されている加水分解物を含む。後者のアプリケーションの典型的な例は、とりわけセイボリーアプリケーションのためのグルタミン酸に富むグルテン加水分解物の作製であり得る。加水分解物は、そのままの形で体内摂取され得るか、または食物成分として役立ち得る。別の方法では、本発明は、組成物が本発明に従う酸性に安定なプロリン特異的エンドペプチダーゼを含んでなることを特徴とする食品の忍容性を改善するための組成物の使用を提供する。さらに別の方法では、本発明は、食品の改善された忍容性のための酸性に安定なプロリン特異的エンドペプチダーゼの添加を含んでなる食品を調製するための方法を提供する。すべてのそのようなアプリケーションにおいて、プロリン特異的酵素は、いわゆる加工助剤として使用され得る。
【0057】
アスペルギルス(Aspergillus)属の株は食物用として位置付けられており、これらの微生物由来の酵素は、思いも寄らない食物用供給源を形成することが公知である。別の好適な実施態様に従えば、酵素は、非分泌型のいわゆるサイトゾルまたはペリプラズム酵素を提示するのではなく、その産生細胞によって分泌される。この方法では、酵素は、高価な精製工程を伴わずに、本質的に純粋な状態で細胞ブロスから回収することができる。好ましくは、酵素は、示現するpHおよび温度条件下でその基質に対して高い親和性を有する。好ましくは、酵素は、消化管の当該関連部位のpHで、消化管タンパク質分解酵素、例えば、ペプシンまたはトリプシン、エラスターゼおよびキモトリプシンによって不活化されない。より好ましくは、酵素は、胃ならびに十二指腸において活性であり、保護被覆を必要としない。
【0058】
ヒトの消化管は、異なるコンパートメントの連続である。食物が摂取され、嚥下された後、それは、胃に到達し、ここで、それは酸およびエンドペプチダーゼのペプシンと混合される。胃における固形食物の典型的な残留時間は、1〜数時間の範囲である。幽門が時々開放することにより、酸性化されかつ部分的に加水分解された食物を小腸に流動させることが可能である。小腸の第1の部分、即ち、十二指腸では、胆汁ならびに膵液が添加される。膵液は、胃の内容物を部分的に中和するための重炭酸塩を含有する。膵液はまた、プロテアーゼのさらなる組、即ち、エンドペプチダーゼ、トリプシン、キモトリプシンおよびエラスターゼならびに胃においてペプシンにより形成されるペプチドおよびポリペプチドをさらに分解するためのカルボキシペプチダーゼAおよびBを含有する。十二指腸後、消化物は空腸に到達する。十二指腸および空腸は、消化管におけるタンパク質吸収の主要な部位である。このような吸収プロセスは、腸管上皮の刷子縁細胞に固定された異なるプロテアーゼによる食餌タンパク質のさらなるタンパク質分解に関与する。後者の加水分解は、腸壁の上での小さなペプチドならびに遊離のアミノ酸の促進された輸送によって達成される。小腸の最後の部分は回腸によって形成され、その後、消化物は大腸(直腸)に進入する。直腸では、集中的な発酵が存在するが、アミノ酸またはペプチドの吸収は認められない。
【0059】
セリアック病患者によるグルテンの摂取により、近位小腸の病巣がもたらされる。腸絨毛の萎縮は、そのような病巣の1つの最も特徴的な特徴である。この絨毛の萎縮は、空腸に限定されず、遠位の十二指腸においても実証され得る(メイジャー,J.W.R.(Meijer,J.W.R.)ら;Virchows Arch.2003年2月;442:124−128)。このような観察は、セリアック病の患者における毒性のプロリンリッチペプチドの損傷の影響は、胃のすぐ下流の領域において常に明らかであることを示す。本発明者らは、セリアック病の症状の防止を目的とする酵素療法が、消化管のより遠位部ではなく、胃における関連のプロリンリッチペプチドを加水分解することを意図すべきであることを見出した。有利なことに、酵素療法において適用される酵素の活性は、胃において作用し始め、十二指腸においても継続して活性であるほどである。本発明者らは、精神、呼吸器および循環器障害の発達において役割を果たすオピオイドペプチドもまた、胃において最も良好に破壊されることを見出した。十二指腸および空腸はタンパク質吸収の主要部位であるため、小腸のこの部位における毒性ペプチドのレベルは、これらの毒性ペプチドの存在に関係する症状を最小限にするためにできるだけ低くあるべきである。
【0060】
消化剤としての使用では、調整酵素は、37℃の温度で十分に活性あるべきであり、好ましくは、胃における酸性条件下で残存するために低い至適pH値を有するべきである。公開されたデータに従えば、摂取された食物の酸性度は、摂取の30分後、初期値のpH5からpH3.5に減少し、続いて、摂取60分後にさらにpH2に減少する(論文マンス・ミンクス(Mans Minkus);University of Utrecht;蘭国;ISBN:90−393−1666−X)。そのため、酵素療法を意図する理想的な酵素は、2のような低いpH値下で活性であるべきである。胃内容排出に関する研究では、摂取45分後、固形食物のほぼ90%がなお胃に存在することが示されている(R.ノチボル(R.Notivol)ら、1984年、Scand J.Gastroenterol.19:1107−1113)。胃より先では、食物のpHは緩徐に上昇して、十二指腸の遠位部、即ち、幽門部より約50cm先で5のpHに到達する(Handbook of Physiology,American Physiological Society、ワシントン市、1968年、ベルナー・ハイデル(Werner Heidel)編;セクション6:Alimentary Canal、第111巻、1457−1490頁)。国際公開第03/068170号パンフレットでは、患者による摂取の前または後のいずれかで、食材中の毒性グルテンオリゴペプチドのレベルを減少することができる異なる「グルテナーゼ」が特定されている。用語「グルテナーゼ」は、プロテアーゼまたはペプチダーゼ酵素、より具体的には、グルテンタンパク質の毒性オリゴペプチドを切断して非毒性のフラグメントにすることが可能であるプロピル特異的プロテアーゼを指す。これらのすべてのグルタナーゼのすべては、典型的に腸において活性であることが意図されるため、それらは、中性付近のpH条件下で、最適に活性である。国際公開第03/068170号パンフレットは、胃または十二指腸の近接部ではなく、十二指腸中もしくはそれより先における毒性のプロリンリッチペプチドの分解について教示している。さらに、国際公開第03/068170号パンフレットは、存在する酵素が胃を通過し、中性pH条件下の腸においてのみ活性となるように、低いpHで酵素活性を遮蔽するいわゆる腸溶コーティングによって保護される酵素調製物を目的とする。
【0061】
タンパク質加水分解物の生成における産業用加工助剤として使用される場合、酵素は、非滅菌産業的条件下での微生物学的に安全なインキュベーションを可能にする条件下で、十分に活性であるべきである。少なくとも50℃のプロセシング温度およびpH5.5よりも十分に低いpH値での適切な酵素活性は、これらの要件を満たす。
【0062】
担子菌アガリクス・ビスポラス(Agaricus bisporus)(サッタル(Sattar)ら、J.Biochem.107,256−261(1990))および非関連の子嚢菌アスペルギルス・ニガー(Aspergillus niger)(国際公開第02/45524号パンフレット)は、両方とも、細胞外プロリルエンドペプチダーゼを産生することが明らかにされている。しかし、担子菌から得られる酵素は、5未満のpH値では残存せず、従って、あまり興味を引かない。好ましくは、酸性の至適pH値を有するA.niger(A.ニガー)由来のプロリルエンドペプチダーゼが使用される。
【0063】
本発明は、低コスト、法的許容性とプロリンリッチペプチド配列に対する酸性pH条件下で証明された有効性とを組み合わせた酵素調製物を提供する。好ましくは、同酵素を使用して、A1ならびにA2型β−カゾモルフィンだけではなく、多様なグルテンのエピトープをも分解することができる。
【0064】
このアスペルギルス(Aspergillus)由来プロリン特異的エンドプロテアーゼは、プロリンリッチペプチドまたはポリペプチドまたはさらにタンパク質の分解において極めて活性であることが見出されている。有利なことに、この酵素は、産生微生物によって発酵ブロスに分泌され、酸性の至適pH値を有し、食物用かつ経済的方法で産生させることができる。関連のβ−カゾモルフィンペプチドは、分子中に4個までのプロリン残基を含有し、さらに、A1およびA2β−カゾモルフィンは異なるアミノ酸配列を有する。かなり驚くべきことに、本発明者らは、アスペルギルス(Aspergillus)の酵素が位置61のプロリンのC末端側でβ−カゾモルフィンを加水分解することが可能であり、従って、A1またはA2β−カゼインの両方について、すべてのBCM−7およびBCM−7関連ペプチドを効率的に不活化することを見出した。本発明者らは、例えば、アルカラーゼ(Alcalase)のように市販のスブチリシン(EC3.4.21.64)のような広い基質特異性を伴う広範に使用されかつ高度に強度のエンドペプチダーゼが、BCM−7を分解することができないことを見出したため、これはかなり注目に値する。同様に、産業上利用可能なプロテアーゼは、BCM−7を分解することができない。事実、アスペルギルス(Aspergillus)由来プロリン特異的エンドプロテアーゼは、β−カゾモルフィン(beta−casomophine)を、但し、かなり驚くべきことに、BCM−7に存在する4個のプロリン残基のうちの1箇所のみで加水分解することができる。それにもかかわらず、この特定のプロリン残基に対するこの特定のアスペルギルス(Aspergillus)の酵素の特異性のため、BCM−7ならびにすべてのBCM−7関連分子は、Tyr−Pro−Pheモチーフが酵素によって切断されるため、この酵素とのインキュベーションによって効率的に分解される。さらに、β−カゼイン由来のA1ならびにA2から誘導されるBCM−7分子は、アスペルギルス(Aspergillus)プロリン特異的エンドプロテアーゼとのインキュベーションによって不活化される。
【0065】
真のプロリン特異的エンドプロテアーゼの利点は、プロリン特異的エンドプロテアーゼは、酵素とタンパク質とを接触させるときに直ちにプロリンリッチ配列を加水分解し始めることができることである。プロリルオリゴペプチダーゼは、例えば、他の消化管エンドペプチダーゼによって、グルテンまたはカゼイン分子の有意なプレ消化後にのみ活性になることができる。必要とされる広範な分解および食物が小腸に進入する前に利用することができる限られた時間の観点から、真のプロリン特異的エンドプロテアーゼは、既知のオリゴペプチダーゼを超える重要なアプリケーションの利点を有する。真のプロリン特異的エンドペプチダーゼの別の利点は、グルテン構造の全体的な破壊を伴わずに、グルテン中の毒性プロリンリッチペプチドのレベルを減少するために、それを産業的に使用することができることである。例えば、アスペルギルス(Aspergillus)由来酵素と共にコムギグルテンのペーストを押し出し形成すると、いくつかの残留する基本構造特性を伴うが、強度に減少したレベルの毒性プロリンリッチペプチドを伴う生成物を生じる。プロリルオリゴペプチダーゼによる毒性プロリンリッチペプチドの同じような減少を達成するために、他のプロテアーゼによるほとんどすべてのプレ加水分解が、30アミノ酸残基未満の所望されるペプチド長を達成するのに必要であり得る。これにより、グルテンのすべての関連する物理化学的特性の完全な消失、即ち、生地の固さが消失し、許容可能な形状および容積でパンを焼くことができなくなることは言うまでもない。ディ・カグノ(Di Cagno)ら(Appl.Environ.Microbiol.、第70(2)巻、1088−1096、2004年)は、セリアックスプルー患者によって良好に忍容されるサワー種のパンの作製について報告している。彼らのアプローチでは、高い割合の非毒性小麦粉で調製した生地を使用し、これを選択されたラクトバチルス(lactobacilli)で24時間の期間、発酵させることによって、毒性のプロリンリッチエピトープのレベルを最小限にした。彼らの結果は科学的には確かに興味深いが、24時間もの長い産業環境での発酵期間は、経済的に容易ではないことは明白である。さらに、彼らの高い割合の非毒性小麦粉は、「規格通りの」グルテンが不足し、そのため最終的なパンの容積が制限されることを含意する。従って、現存の産業的手順を使用するが、毒性プロリンリッチペプチドのレベルが制限される100%コムギパンの生産を可能にする経済的な方法の役に立つことは有利である。そのようなパンは、毒性ペプチドの日々の摂取量を減少することを目的とする食餌の重要な成分となることができる。そのような食餌と特別な関連性がある消費者グループとして、幼児、IBSを患う若年者、高齢者ならびに記載の疾患および症状を患う個体が挙げられる。セリアック病の患者では、50ミリグラム未満のグルテンの日々の摂取は安全とみなされるが、平均的な欧米での消費量である1日あたり13グラムのグルテンに存在する量より有意に少ない毒性プロリンリッチペプチドを含有する任意の食餌は、上記の消費者グループにとって有益であるようである。
【0066】
総合的に見ると、これらの考慮から、先行技術において記載の酵素よりもA.niger(A.ニガー)由来酵素は新規であり、かなり有利であることが示される。本発明に従う酵素を含有する組成物は、グルテンの感作またはオピオイド代謝障害の現象もしくはIBSの現象を抑制あるいは遅延させるために有利に使用される。そのような組成物を消化剤として適用し、毒性プロリンリッチペプチドの消化管でのインサイチュ減少を達成することができる。あるいは、そのような組成物を加工助剤として適用し、そのような毒性のプロリンリッチペプチドを伴わないタンパク質加水分解物を生成させることができる。タンパク質加水分解物の分野内では、ビールの醸造は、A.niger(A.ニガー)由来、低いpHのプロリン特異的エンドプロテアーゼの適用が驚くべき数の利点を付与する特別な場合を提供する。国際公開第02/046381号パンフレットは、ビール糖化工程中またはビールのろ過前もしくはビールのラガーリングの前のいずれかに適用されるプロリル特異的エンドペプチダーゼは、ビールの濁りの形成を減少することを教示している。本発明者らの示すデータは、低いpH酵素との適切なインキュベーションもまた、穀物由来毒性プロリンリッチペプチドのレベルを減少することを例示する。驚くべきことに、本発明者らは、穀物タンパク質を含有するビールまたは他の飲料をプロリン特異的エンドペプチダーゼで処置すると、濁りのレベルが減少するだけではなく、セリアック病を患うヒトに安全な製品がもたらされることを見出した。
【0067】
本出願の実施例1は、アスペルギルス(Aspergillus)由来のプロリン特異的エンドプロテアーゼの酸性の至適pH値および理想的な至適温度を示している。実施例2および3では、本発明者らは、アスペルギルス・ニガー(Aspergillus niger)より産生可能なプロリン特異的エンドプロテアーゼが、より小さなペプチドまたはポリペプチドと同じ効率で大きな無傷なタンパク質を切断することができる真のプロリン特異的エンドプロテアーゼであることを示している。事実、本発明者らのデータは、アスペルギルス(Aspergillus)の酵素が、既知のオリゴペプチダーゼが属するS9ファミリー(N.D.ローリングス(N.D.Rawlings)およびA.J.バレット(A.J.Barrett)、Methods in Enzymology、第244巻、19−61頁、1994年;N.D.ローリングス(N.D.Rawlings)およびA.J.バレット(A.J.Barrett)、Biochimica&Biophysica Acta1298(1996年)1−3)ではなく、S28ファミリーの新規のメンバーであることを示している。本発明者らは、既知のプロリルオリゴペプチダーゼとは対照的に、アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼが、胃における酸性条件下で活性であり、大きなプロリンリッチタンパク質フラグメントおよびポリペプチドの加水分解に対して高い有効性を示すことを見出した。そのような高い効率は、実施例4、5および6において例示される。実施例4では、本発明者らは、アルカラーゼ(Alcalase)(タンパク質加水分解物の生成において頻繁に使用される強力な広域スペクトルのプロテアーゼ)によるミルクタンパク質加水分解物の生成中に、BCM−7配列を組み入れたいくらかのペプチドが、加水分解方法において残存することを実証している。しかし、これらのペプチドは、アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼとの酸性条件下でのインキュベーション時に迅速に消滅する。実施例5に提供されるデータは、アスペルギルス(Aspergillus)の酵素が、β−A2カゼイン由来BCM−7分子において利用可能な4個のプロリン残基のうちの1つでしか切断しないという驚くべき事実を表している。これは、プロリンリッチ基質と任意のプロリン特異的プロテアーゼとのインキュベーションが、劇的に増加した酵素/基質比の条件下であっても、プロリン残基を含むすべてのペプチド結合の切断を自動的に含意するわけではないことを示す。実施例6では、本発明者らは、セリアック病の患者において主要なエピトープであることが要求されるグリアジン由来33マーに対するアスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼの有効性について実証している。また、広域スペクトルのアルカラーゼ(Alcalase)は、アルカリ性または酸性条件下のいずれでも、この分子を切断することができないが、アスペルギルス(Aspergillus)由来酵素は、頻繁に、酸性条件下で分子を切断し、最大で6アミノ酸残基の長さを伴う99.5%のペプチドが作製される。そのため、酸性条件下でのプロリンリッチペプチドに対するその高い有効性にもかかわらず、アスペルギルス(Aspergillus)由来酵素でさえ、アミノ酸配列YPQPQLPを伴う少なくとも0.5%のヘプタマーを遊離する。配列PYPQPQLPYが既知のセリアック病の患者に特異的なT細胞エピトープであるため、この所見は、最適とは言えないプロリン特異的酵素、例えば、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から誘導される酵素を含む既知のプロリン特異的オリゴペプチダーゼでは、グルテン分子由来の毒性ペプチドの形成を防止するための現実的なインビボアプリケーションが、実現不可能であることを証明することを例示している。後者の結論は、実施例7および8において示される実験によって確認される。実施例7では、本発明者らは、pH2〜12の間で、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼとF.メニンゴゼプチクム(F.meningosepticum)由来プロリン特異的オリゴペプチダーゼとの活性プロフィールを比較している。A.niger(A.ニガー)由来酵素はpH2.5〜7.0の間で活性を示すが、F.メニンゴゼプチクム(F.meningosepticum)の酵素は、活性になるために、5.0を超えるpHを必要とする。実施例8では、本発明者らは、両方のプロリン特異的酵素を、胃の酵素ペプシンの非存在下および存在下で低いpH条件に暴露することによって、胃における状況を模倣している。個々の酵素に至適なpH条件下でのその後の活性測定は、F.メニンゴゼプチクム(F.meningosepticum)の酵素がpH2またはpH3条件下でも残存することができることを示している。ペプシンの存在下では、F.メニンゴゼプチクム(F.meningosepticum)の酵素は、pH4で既に不可逆的な損傷を受ける。対照的に、A.niger(A.ニガー)由来酵素は、pH2の低いpH条件下およびペプシンの存在下でも残存し、それによって、胃において毒性ペプチドを加水分解するその価値が強調される。実施例9では、本発明者らは、A.niger(A.ニガー)由来酵素が多数の既知のHLA−DQ2グルテンのエピトープを切断することができることを例示している。興味深いことに、観察される切断部位の位置から、文献から公知のすべてのT細胞のエピトープが破壊されることが推定される。実施例10では、本発明者らは、100%麦芽ビールおよび100%コムギパンからグルテンのエピトープを回収し、本発明者らが抗体アッセイにおいてこれらのエピトープを検出することができることを実証している。実施例11では、本発明者らは、A.niger(A.ニガー)由来酵素を生産プロセスに組み入れることによって生成されるビールが、認知されうる程度に低減されたレベルのグルテンのエピトープを生じることを実証している。実施例12では、本発明者らは、酵素をコムギ生地に組み入れ、低減されたレベルのグルテンのエピトープでダッチ(Dutch)型焼きパンを生成することによって、類似の効果を得ることができることを示している。
【0068】
グルテンは、複雑な3次元構造を伴う水に不溶な化合物である。そのプロリンリッチアミノ酸組成物との組み合わせにおけるこれらの特性により、胃および腸でのタンパク質分解に対し、グルテン分子を耐性にする。消化管腔に分泌される天然のタンパク質分解活性のうち、プロリンに関与するペプチド結合を切断することが可能である活性は存在しないため、共同作用性の内因性プロリン特異的酵素を使用することが理解される。しかし、セリアック病を患うヒトは、グルテンに存在する多くのエピトープに対して極めて感受性であることができる。本発明に従えば、天然の消化プロテアーゼの効果は、アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼで改善することができ、このタンパク質分解混合物の加水分解能のさらなる増強についてさえも本明細書胃おいて開示する。
【0069】
負に荷電した残基、例えば、Glu(E)およびAsp(D)を含むペプチド結合が、プロテアーゼに対して不十分な基質を形成することは周知である。また、天然の消化管タンパク質分解酵素は、これらの残基を処理することができず、これは、グリアジンから胃および膵プロテアーゼ耐性ペプチドWQIPEQSRを単離することによって証明される(シャン(Shan)らを参照のこと)後者の刊行物はまた、グルテンに普遍的に存在するグルタミン残基(Q)を、組織トランスグルタミナーゼによって、グルタミン酸残基(E)に脱アミド化することができることを明らかにしている。不幸なことに、グリアジンペプチドのこの位置特異的脱アミノ化は、それらの潜在的免疫原性をさらに増加する。このような背景に対し、本発明者らは、アスペルギルス(Aspergillus)由来プロリン特異的エンドプロテアーゼと補助的なエンドペプチダーゼとの有効な酵素の組み合わせを生じさせ、プロリンリッチ毒性プロリンリッチペプチドの形成を防止することを可能にしている。本発明に従えば、グルタミン酸特異的エンドペプチダーゼ(EC3.4.21.19)、例えば、バチルス(Bacillus)およびストレプトミセス(Streptomyces)のような多くの食物用微生物によって過剰分泌される該グルタミン酸特異的エンドペプチダーゼを使用することができる。これらの酵素は、経済的かつ食物用の方法で産生させることができる。胃を安全に通過する酵素が、それらの酵素活性に関して、好適である。一般に、該酵素は、酸性または中性の至適pH値を有する。アスペルギルス(Aspergillus)由来プロリルエンドペプチダーゼとの組み合わせで、グルタミン酸特異的エンドペプチダーゼのこのカテゴリーは、低いレベルの毒性プロリンリッチペプチドを伴うタンパク質加水分解物の産生において、有用であるとみなされる。
【0070】
かなり驚くべきことに、本発明者らの示す研究は、グルタミン酸特異的エンドペプチダーゼとは別に、アスペルギルス(Aspergillus)由来のプロリン特異的エンドプロテアーゼとのインキュベーションに対して相乗効果を有する他のエンドペプチダーゼが存在することを実証している。本発明者らは、アミノ酸残基Q(グルタミン)とL(ロイシン)との間を切断することが可能なエンドプロテアーゼ(EC3.4.21−99)は、アスペルギルス(Aspergillus)由来のプロリン特異的エンドペプチダーゼと有利に組み合わせられると結論している。具体的には、pH5.0未満の至適pH値を有し、基質のP1またはP1’位置においてグルタミンまたはロイシン残基のいずれかを選好するエンドプロテアーゼ、例えば、アスペルギロペプシン(EC3.4.23.18および19)ならびにムコールペプシン(EC3.4.23.23)が有利に使用される。
【0071】
本発明に従う酵素の1つのアプリケーションは消化剤としてのそれらの使用である。本出願では、本発明の組成物は、好ましくは経口的に投与されるが、他の直接的経路を介してとよしてもよい。組成物は、典型的に、ヒトに投与されるが、増加したグルテン感受性またはカゼインもしくはグルテンの代謝障害あるいはIBSに典型的な症状を緩和するために、動物、好ましくは哺乳動物に投与してもよい。消化剤としてのそれらのアプリケーションでは、本発明に従う酵素は、例えば、丸剤、錠剤、顆粒剤、サシェ(sachet)またはカプセル剤中の乾燥粉末として処方してもよい。あるいは、本発明に従う酵素は、例えば、シロップ剤もしくはカプセル剤中の液体として処方してもよく、または0.85未満の水分活性(Aw)を伴う食品に組み入れてもよい。多様な処方物において使用され、かつ本発明に従う酵素を含有する組成物はまた、生理学的に許容可能なキャリア、アジュバント、賦形剤、安定剤、緩衝液および希釈剤よりなる群の少なくとも1つの化合物を組み入れてもよく、該用語は、包装、送達、吸収、安定化、またはアジュバントの場合には、酵素の生理学的効果を増強することを援助する物質を示すためにそれらの常識的な意味で使用される。粉末化された形態において本発明に従う酵素と組み合わせて使用することができる多様な化合物に関する関連背景は、「Pharmaceutical Dosage Forms」、第2版、第1、2および3巻、ISBN0−8247−8044−2 Marcel Dekker,Incに見出すことができる。乾燥粉末として処方される本発明に従う酵素は、幾分長期間保存することができるが、水分または多湿な空気との接触は、例えば、アルミニウムブリスターのような適切な包装を選択することによって、回避すべきである。液体形態で処方する場合、酵素活性を安定化するためおよび防微生物のために使用される化合物は重要な役割を果たす。酵素活性の安定化は、ポリオール、例えば、グリセロールまたは多様な糖の使用によって得ることができる低減された水分活性を必要とし得る。さらに、Ca2+またはMg2+のような2価のカチオンは、それらの安定化効果について公知であり、ならびにイオウ含有アミノ酸のような還元剤およびBHTまたは没食子酸プロピルのようなフェノール化合物も公知である。食物用防微生物は、低pH条件または低い水分活性とソルビン酸塩もしくは安息香酸塩もしくはパラベンとの周知の組み合わせを使用して、達成することができる。さらに、ハイドロコロイドのような食物用増粘財も必要とし得る。比較的新規の経口アプリケーション形態は、液体を含有するゼラチンカプセルの使用である。本出願では、液体は、本発明の乾燥させた酵素を懸濁することができる典型的にオイルまたはポリエチレングリコールまたはレシチンである。改善された酵素安定性を伴う錠剤処方物の例は、米国公開特許第2002/0136800号明細書において提供される。
【図面の簡単な説明】
【0072】
【図1】基質として合成ペプチドZ−Gly−Pro−pNAを使用するA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの至適pH値のグラフ図を表す(実施例1を参照のこと)。
【図2】クロマトグラフィー精製されたA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼとのインキュベーション後の無傷なオボアルブミン(ovalbumine)および合成27マーペプチドのSDS−PAGEを表す(実施例3を参照のこと)。
【図3】A.niger(A.ニガー)およびF.メニンゴゼプチクム(F.meningosepticum)由来プロリン特異的エンドプロテアーゼの至適pH値のグラフ図を表す(実施例7を参照のこと)。
【発明を実施するための形態】
【0073】
材料および方法
材料
次の酵素は、Sigmaから入手した:アスペルギルス・ニガー(Aspergillus niger)由来のアミログルコシダーゼ、300U/ml、SigmaA−7095;ブタ胃粘膜由来のペプシン、2331U/mg、SigmaP−7012;モルモット由来のトランスグルタミナーゼ、SigmaT−5398。トリプシン溶液2.5%はGibco(BRL25090−028)から入手し、Sep−Pak Plus tC18カートリッジ、Waters No.036810はWatersから入手した。2.6AU(A)(アンソン単位アルカラーゼ(Alcalase))単位/グラム製品の活性を有するアルカラーゼ(Alcalase)(登録商標)AF2.4Lは、NovozymesA/S(Bagsvaerd、丁国)から入手した。Novozymesに従い、この活性測定の詳細については、Novozymes分析方法LUNA番号2003−32153−01/SOP番号:EB−SM−0218.02/02)に見出すことができる。
【0074】
合成ペプチドは、PepscanSystemsB.V.(Lelystad、蘭国)から入手した。発色ペプチド基質は、PepscanSystemsまたはBachem、瑞国のいずれか一方から入手した。
【0075】
A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼ
アスペルギルス・ニガー(Aspergillus niger)由来のプロリン特異的エンドプロテアーゼの過剰発現およびクロマトグラフィー精製は、国際公開第02/45524号パンフレットに記載の通りに達成させた。酵素(1単位/10mgのタンパク質)の活性は、37℃で、クエン酸/リン酸二ナトリウム緩衝液pH4.6中合成ペプチドZ−Gly−Pro−pNAに対して試験した。反応産物は、分光光度法により405nMでモニターした。市販のプロリルオリゴペプチダーゼ酵素(ICN Biomedicals/MP Biomedicals、Aurora、オハイオ州、米国から購入)の活性は、1mgの製品あたり35単位であり、30℃で、pH7.0緩衝液中Z−Gly−Pro−pNAに対し試験した。反応産物は、分光光度法により405nMでモニターした。両酵素について、単位は、指定された条件下、1分間あたり1μmolのp−ニトロアニリドの遊離を誘発する酵素の量として規定される。
【0076】
LC/MS分析
P4000ポンプ(Thermoquest(登録商標)、Breda、蘭国)に連結されたイオントラップ質量分析計(Thermo Electront(登録商標)、Breda、蘭国)を使用するHPLCを、本発明の酵素混合物によって産生される酵素によるタンパク質加水分解物を特徴付けるのに使用した。溶出のためのMilliQウォーター中0.1%ギ酸(Millipore、Bedford、マサチューセッツ州、米国;溶液A)およびアセトニトリル中0.1%ギ酸(溶液B)の勾配と組み合わせたPEPMAP C18 300A(MIC−15−03−C18−PM,LC Packings、Amsterdam、蘭国)カラムを使用して、ペプチドを分離した。勾配は95%の溶液Aで開始し、140分間で40%の溶液Bにまで増加させ、さらに5分間、後者の割合で保持した。使用した注入容積は50マイクロリットルであり、流速は1分間あたり50マイクロリットルであり、カラム温度は30℃に維持した。注入したサンプルのタンパク質濃度は、約50マイクログラム/ミリリットルであった。
【0077】
個々のペプチドに関する詳細な情報については、イオントラップ質量分析計のための特徴的なアルゴリズムである「スキャン依存的」MS/MSアルゴリズムを使用することによって、入手した。
【0078】
フルスキャン分析に続いて、フルスキャンの質量範囲において最も高い強度のイオンの荷電状態の決定のためのズームスキャン分析を行った。後者のイオンのその後のMS/MS分析により部分的なペプチド配列情報が得られ、これは、Xcalibur Bioworks(Thermoquest(登録商標)、Breda、蘭国)由来のSEQUESTアプリケーションを使用するデータベース検索のために使用し得る。使用したデータベースは、使用したアプリケーションのための目的のタンパク質を含有するNCBI(National Centre for Biotechnology informatics)において利用可能なOWL.ファスタ・データバンクから抽出した。乳清タンパク質またはカゼインのような良好に特徴付けられたタンパク質基質が測定される該実験では、分析技術の正確さは、配列適合性が50%未満であるそれらのMS/MSスペクトルを省略することによって、増加する。
【0079】
アンギオテンシン(M=1295.6)を使用して、MSモードにおける至適感受性およびMS/MSモードにおける至適フラグメント化のために同調し、60μg/mlの一定の輸注を実施し、MSモードでは主にダブルおよびトリプルチャージ種が、およびMS/MSモードでは約35%の至適衝突エネルギーが生じた。
【0080】
MALDI−TOF
MALDI−TOFは、Voyager De−Pro(Applied Biosystems)質量分析計を使用して実施した。適切なマトリックスと混合後、ペプチドサンプルをリニアモードで測定した。MassLynxソフトウェアを介して見出される質量から、Post Source Decay(PSD)によって配列決定し、提唱されるペプチドのアミノ酸配列を決定した。
【0081】
食物サンプルにおけるグルテンペプチドの定量化
T細胞刺激α−グリアジン、γ−グリアジンおよびLMW−グルテニンペプチドに特異的なモノクローナル抗体が利用可能であり、食物サンプルにおけるこれらのペプチドの検出のための競合アッセイに組み入れた(E.H.A.スペイニ−デッキング(E.H.A.Spaenij−Dekking)ら、GUT,53:1267−1273(2004)。
【0082】
抗体に基づくアッセイ
抗体に基づくアッセイの作製のために、既知のT細胞刺激α、γ−グリアジンおよびLMWグルテニンペプチドに対し、BalbCマウスにおいてモノクローナル抗体を惹起させた。マウスの脾臓とマウス骨髄腫細胞系統との融合後、抗体産生ハイブリドーマを得た。これらを制限希釈によってクローニングし、これらの細胞によって分泌されるモノクローナル抗体を、モノクローナル抗体競合アッセイにおけるそれらの使用のために試験した。それぞれの特異性について、1つもしくは2つの適切なモノクローナル抗体を選択し、異なるモノクローナル抗体によって認識されるエピトープを決定した(以下の表を参照のこと)。
【0083】
【表1】
【実施例】
【0084】
実施例1
A.niger(A.ニガー)から得られるプロリン特異的エンドプロテアーゼの至適pHおよび温度
A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼをA.niger(A.ニガー)宿主において過剰産生させ、単離し、国際公開第02/45524号パンフレットに記載の材料および方法を使用してクロマトグラフィー精製した。このようにして得られた酵素の至適pH値を確立するために、異なるpH値を伴う緩衝液を調製した。pH4.0−4.5−4.8−5.0−5.5および6.0の緩衝液は、0.05mol/l酢酸Naおよび0.02M CaCl2を使用して作製し;pH7.0および8.0の緩衝液は、0.02M CaCl2を含有する0.05M Tris/HCl緩衝液を使用して作製した。pH値は、それぞれ、酢酸およびHClを使用して調整した。発色合成ペプチドZ−Gly−Pro−pNAを基質として使用した。緩衝溶液、基質溶液およびプロリルエンドペプチダーゼプレ希釈液(0.1U/mLの活性)を、水浴中で正確に37.0℃にまで加熱した。混合後、反応物を、分光光度的に405nm、37.0℃で3.5分間追跡し、0.5分間毎に測定した。図1に示す結果から、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが約4の至適pH値を有することが明らかである。
【0085】
また、プロリルエンドプロテアーゼの至適温度を確立した。そのために、基質としてCaseine Resorufine(Rocheバージョン3)を使用し、精製された酵素調製物を、pH5.0で0.02mol/l CaCl2を含有する0.1mol/l酢酸Naにおいて、2時間、異なる温度でインキュベートし、574nmで測定することによって酵素活性を定量した。得られる結果に従えば、A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼは約50℃の至適温度を有する。
【0086】
該酵素は産業アプリケーションに好適な条件ならびに胃および小腸の初期部分において示現する条件下で至適に活性であるため、極めて強い酸性の至適pH値は、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが産業アプリケーションならびに経口的体内摂取に理想的な特性を有することを強く示唆する。また、酵素の至適温度は、酵素を両アプリケーションに理想的に適切にする。
【0087】
実施例2
A.niger(A.ニガー)から得られる酵素は新規のクラスのプロリン特異的酵素を提示する。
国際公開第02/45524号パンフレットにおいて提供されるA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼのコーディング配列全体から、526アミノ酸のタンパク質配列を決定することができる。酵素の新規性は、SwissProt,PIRおよびtrEMBLのようなデータベースのBLAST検索によって確認した。驚くべきことに、A.niger(A.ニガー)の酵素と既知のプロリルオリゴペプチダーゼとの間に明確な相同性を検出することができなかった。しかし、アミノ酸配列をより緊密に調べたところ、Pro−Xカルボキシペプチダーゼ(EC3.4.16.2)、ジペプチジルアミノペプチダーゼI(EC3.4.14.2)、および胸腺特異的セリンプロテアーゼに対して低いが有意な相同性を示した。これらのすべての酵素は、セリンペプチダーゼのファミリーS28に割り当てられている。また、活性部位セリンの周りのGxSYxGの立体配置は、これらの酵素とA.niger(A.ニガー)由来エンドペプチダーゼとの間で保存される。さらに、ファミリーS28のメンバーは、酸性の至適pH値を有し、プロリン残基のカルボキシ末端側で切断するための特異性を有し、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼと同様のシグナル配列およびプロペプチドで合成される。また、A.niger(A.ニガー)酵素のサイズは、ファミリーS28のメンバーのサイズに類似する。従って、A.niger(A.ニガー)プロリン特異的エンドプロテアーゼは、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から得られる酵素を含むほとんどのサイトゾルプロリルオリゴペプチダーゼが分類されているS9ファミリーではなく、セリンプロテアーゼのファミリーS28のメンバーであるようである。これらの構造的および生理学的特徴に基づいて、本発明者らは、A.niger(A.ニガー)の酵素がセリンプロテアーゼのS9ファミリーではなく、S28に属すると結論されている。S9ファミリーに属するプロリルオリゴペプチダーゼ由来のA.niger(A.ニガー)由来酵素を区別するさらなる特徴は、後者のファミリーに属するサイトゾルプロリルエンドペプチダーゼとは異なり、新たに同定されたA.niger(A.ニガー)の酵素は増殖培地に分泌されるという事実である。これまでのところ、担子菌アガリクス・ビスポラス(Agaricus bisporus)(サッタル(Sattar)ら、J.Biochem.107,256−261(1990))および非関連の子嚢菌アスペルギルス・ニガー(Aspergillus niger)(国際公開第02/45524号パンフレット)のみが、細胞外プロリルエンドペプチダーゼを産生することが明らかにされている。しかし、担子菌から得られる酵素は、5未満のpH値では残存せず、従って、産業アプリケーションならびに経口的体内摂取に適切とは言い難い。
【0088】
これは、下等真核生物由来のファミリーS28のメンバーの単離および特徴付けに関する最初の報告である。
【0089】
実施例3
A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼは、大きなタンパク質ならびに小さなペプチドを加水分解することができ、従って、真のエンドプロテアーゼである。
特定の構造的特徴のため、S9ファミリーに属するプロリルオリゴペプチダーゼは30アミノ酸を超えるペプチドを消化することができない。この制限は、できるだけ迅速かつ効率的にすべての潜在的なプロリンリッチ毒性プロリンリッチペプチドを加水分解することが意図される酵素に明らかに不利である。A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが基質分子のサイズに関して同じ制限を示すかどうかを調べるために、本発明者らは、A.niger(A.ニガー)由来のクロマトグラフィー精製したプロリルエンドペプチダーゼを、小さな合成ペプチドおよび大きなオボアルブミン(ovalbumine)分子と共にインキュベートし、SDS−PAGEによって、形成された加水分解産物を分析した。
【0090】
使用した合成ペプチドは、27マーの配列NH2−FRASDNDRVIDPGKVETLTIRRLHIPR−COOHであり、Pepscan company(Lelystad、蘭国)より贈与された。
【0091】
そのアミノ酸配列から示されるように、このペプチドは、2個のプロリン残基を含有し、1つはペプチドの中央にあり、1つは極めて末端にある。無傷なオボアルブミン(ovalbumine)分子(Pierce Imject、20mgの凍結乾燥材料を含有するバイアル)は、42750Daの分子量を伴う385アミノ酸よりなる。この分子は、14個のプロリン残基を含有し、このうち1つは分子の究極的C末端に位置し、プロリン特異的エンドプロテアーゼによって切断することができない。
【0092】
オボアルブミンおよびオリゴペプチドを、個別に、50℃で、精製されたA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼと共にインキュベートした。いくらかの時間間隔で、サンプルを採取し、SDS−PAGEを使用して分析した。
【0093】
4.5単位/mlの活性を伴うクロマトグラフィー精製されたA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼを、20mM CaCl2を含有する0.1M酢酸緩衝液pH4で100倍希釈した。オボアルブミン(ovalbumine)を、1mg/mlの濃度(22μM)で酢酸緩衝液pH4に溶解した。27マーを同じ緩衝液に溶解し、0.48mg/mlの濃度(152μM)に到達させた。オボアルブミン(ovalbumine)および27マー溶液のモル濃度は、両溶液が切断可能なプロリン残基において同じモル濃度を含有するような方法で、選択した。オボアルブミン(ovalbumine)は13個の潜在的プロリン切断部位を含有する一方、27マーペプチドは2個しか有さない。両基質溶液のうち、0.5mlを、エッペンドルフ(Eppendorf)サーモミキサー中、50℃で10μl(0.45ミリU)の酵素溶液と共にインキュベートした。いくらかの時間間隔で、10μlサンプルをインキュベーション混合物から抜き取り、SDS−PAGEまで20℃で保持した。SDS−PAGEおよび染色のために使用されるすべての材料は、Invitrogenから購入した。サンプルは、製造者の指示に従い、LDS緩衝液を使用して調製し、製造者の指示に従い、MES−SDS緩衝液システムを使用して12%Bis−Trisゲル上で分離した。染色は、Simply Blue Safe Stain(Collodial Coomassie G250)を使用して実施した。
【0094】
図2に認められ得るように、オボアルブミン(ovalbumine)は、アスペルギルス(Aspergillus)由来酵素によって、インキュベーションの最初の4.75時間において約35〜36kDの個別のバンドに切断される(レーン3)。インキュベーション期間を延長すると、多様な分子量のより小さな産物へのさらなる分解が生じる(レーン7)。
【0095】
27マーペプチドもまた分解され、レーン2と比較して、レーン4、6および8におけるより微弱なバンドによって判定される。産物の極めて小さな分子量のシフト(レーン9および8を比較すること)は、ペプチドのカルボキシ末端でのアルギニン残基の切断によるためである可能性が最も高い。差異は約200Dであり(AlphaImager2000システムの上のAlphaImager3.3dソフトウェアを使用して測定される)、アルギニンは174のMWを有する。この小さな分子量のシフトは、恐らく、ペプチドの分解の第1の工程である。
【0096】
産物のさらなる崩壊は、SDSゲル上のバンドの強度の減少によってのみ、見ることができる。約1000のMWを伴う成分のゲル染色はクマシー・ブリリアント・ブルー(Coomassie Brillant Blue)では可能ではないため、さらなる産物の崩壊は認められない。
【0097】
この実験から、S9ファミリーに属する既知のプロリルオリゴペプチダーゼとは異なり、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼは、極めてより大きなタンパク質よりも小型のペプチドの方を特に選好して切断するわけではないと結論付けることができる。A.niger(A.ニガー)由来酵素は、それ自体で、真のエンドプロテアーゼおよび潜在的な毒性のプロリンリッチプロリンリッチペプチドを切断するのに好適な酵素を提示する。
【0098】
実施例4
アルカラーゼ(Alcalase)およびアルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせとのインキュベーション後に形成される加水分解物におけるβ−カゾモルフィン
消化管タンパク質分解中のプロテアーゼ耐性β−カゾモルフィンの形成と同様に、本発明者らは、ミルクタンパク質加水分解物の産業的生産中にも、BCM−7関連ペプチドフラグメントの類似の蓄積が生じるのであろうかと考えた。そのために、本発明者らは、牛乳から単離されるA2β−カゼインを、産業上頻繁に使用されるサブチリシンアルカラーゼ(Alcalase)およびアルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼと共にインキュベートした。LC/MS/MS分析を使用して、そのようにして形成されるペプチドを分析した。
【0099】
牛乳は、1kgの乳汁あたりほぼ10グラムのβ−カゼイン(caseine)を含有し、存在するすべてのタンパク質のうち28%を提示する。BCM−7関連アミノ酸配列の分析を容易にするために、この実験では、本発明者らは、最低でも90%の(A2)β−カゼインを含有する濃縮調製物(Sigma由来)を使用した。後者の産物を、1リットルあたり20グラムの濃度で水に溶解し、その後、NaOHを使用してpHを8に調整し、1リットルのカゼイン溶液あたり800マイクロリットルの酵素濃縮物の量でアルカラーゼ(Alcalase)を添加した。インキュベーションを、2時間、60℃で行った。次いで、クエン酸を使用して、溶液のpHを4.5まで低下させた。次いで、溶液を2つの部に分けた:一方の部は、5分間、90℃で加熱してアルカラーゼ(Alcalase)を不活化し、他方の部には、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼを添加して、存在するカゼインの1グラムあたり1単位の酵素濃度(単位の定義に関する材料および方法のセクションを参照のこと)を得た。A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼとのインキュベーションを、16時間、55℃で継続し、続いて、プロリン特異的エンドプロテアーゼを不活化するために、さらに加熱処置を施した。最後に、20mg/mlの推定β−カゼイン濃度を伴う2つのサンプルを、LC/MS/MS分析のために供給した。2つのサンプルを、10分間、13000rpmで遠心分離し、LC/MS/MS分析の前にMilliQ中で20倍希釈した。材料および方法のセクションに記載の通りに、LC/MS/MS分析を行った。
【0100】
β−カゼイン分子のアミノ酸位置60〜66におけるBCM−7配列Tyr−Pro−Phe−Pro−Gly−Pro−Ileの他に、それぞれ、アミノ酸位置60〜63および60〜64におけるTyr−Pro−Phe−Pro(β−カゾモルフィン(1−4))およびTyr−Pro−Phe−Pro−Gly(β−カゾモルフィン(1−5))のようなより小さなフラグメントならびに(アミノ酸位置60〜70における)11アミノ酸の鎖長までのすべてのより大きなペプチドは、少なくともいくらかの程度のオピオイド活性を示す。位置60〜62のトリペプチドTyr−Pro−Pheは、オピオイド活性を有さないことが報告されている。
【0101】
ペプチド同定のために、プロトン化分子のダイレクトLC/MS/MSを使用した。関連の可能性のあるペプチドのプロトン化された質量を表1に提供する。すべてのスペクトルを、35%の衝突の大きさおよび3Daのピーク幅で得た。
【0102】
すべての実験データを、いわゆるBおよびYイオンに基づく理論的フラグメント化パターンと比較する。これは、普通、自動データ処理を使用して実施される方法である。
【0103】
【表2】
【0104】
βカゾモルフィン(Betacasomorphine)アミノ酸配列は、チロシン(Y;β−カゼインペプチド鎖の残基番号60)で開始し、太字で示す。m/z値は、他の関連する可能性のあるペプチドのプロトン化分子を提示する。位置67のプロリン残基の検出は、使用した基質がA2β−カゼインを提示することを示す。
【0105】
アルカラーゼ(Alcalase)かまたはアルカラーゼ(Alcalase)+プロリン特異的エンドプロテアーゼのいずれか一方とのインキュベーション後に得られるβ−カゼイン加水分解物のLC/MS/MS分析に対して得られる結果を表2に示すが、これらは、以下のようにまとめることができる:
・正確なβ−カゾモルフィン(casomorphine)配列、即ち、YPFPGPIおよび誘導体は、アルカラーゼ(Alcalase)かまたはアルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせのいずれか一方で処置した(A2)β−カゼインには存在しない。
・しかし、β−カゾモルフィン配列を含有する2つのペプチドは、アルカラーゼ(Alcalase)処置サンプル、即ち、LVYPFPGPIPNおよびVYPFPGPIPNに存在する。
・これらのβ−カゾモルフィン含有配列の強度は、A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼによる処置時に劇的に減少する。
【0106】
【表3】
【0107】
結果は、明らかに、アルカラーゼ(Alcalase)+A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせがすべての潜在的なβ−カゾモルフィン配列を高い効率で破壊し、それによって、潜在的に毒性のプロリンリッチプロリンリッチ配列を伴わない加水分解物を付与することを示す。
【0108】
形成されるペプチドの間でのより正確な検索後、本発明者らは、2つの酵素の組み合わせによって形成される加水分解物においてペプチドVYPの存在を実証することができた。このペプチドは公正なアルカラーゼ(Alcalase)を使用することによって形成される加水分解物において追跡できなかったため、この所見は、アスペルギルス(Aspergillus)由来酵素によるβ−カゼイン分子の位置61におけるプロリン残基のC末端切断を示唆する。
【0109】
実施例5
アルカラーゼ(Alcalase)形成ペプチドVYPFPGPIPNとA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼとのインキュベーションにおいて形成されるペプチド
実施例4に見られるように、アルカラーゼ(Alcalase)およびA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼの組み合わせによるA2β−カゼインの加水分解は、すべての潜在的β−カゾモルフィン配列を効率的に取り出す。しかし、作製されたペプチドの複雑さのため、本発明者らは、アスペルギルス(Aspergillus)由来酵素がアルカラーゼ(Alcalase)形成ペプチドVYPFPGPIPNを切断する位置を確立することができなかった。そのために、この特定の配列を伴うペプチドを合成し、2種の濃度のプロリン特異的エンドプロテアーゼと共にインキュベートした。形成されたペプチドのその後のLC/MS/MS分析は、酵素の正確な切断部位を示した。
【0110】
凍結乾燥された10マー(Pepscan Systems;Lelystad、蘭国)を、2mg/mlの濃度で、クエン酸−リン酸緩衝液pH4.5に溶解した。溶液に、1グラムのペプチドあたり1および10単位の濃度でプロリン特異的エンドプロテアーゼを添加した。4時間、55℃でインキュベーションし、その後、5分間、90℃での加熱処置を使用して、酵素を不活化した。次いで、2つのサンプルを、10分間、13000rpmで遠心分離し、LC/MSおよびLC/MS/MS分析の前にMilliQウォーター中で20倍希釈した。サンプルを、まず、LC/MSモードで分析して、異なる量の酵素を使用して、10マーの強度の減少を観察し、酵素切断後のLC/MSイオンクロマトグラムにおいてどのペプチド質量が出現したかを観察した。
【0111】
次いで、LC/MS稼動において見出されるペプチドのプロトン化分子のダイレクトLC/MS/MSを実施した。すべてのスペクトルを、35%の衝突の大きさおよび3Daのピーク幅で得た。すべての実験データを、いわゆるBおよびYイオンに基づく理論的フラグメント化パターンと比較する。これは、普通、ペプチド、ポリペプチドおよびタンパク質の同定のための自動データ処理を使用して実施される方法である。
【0112】
1単位/gのタンパク質による10マーVYPFPGPIPN(M=1099.5)の処置は、既に、10マーのいくらかのペプチドへの全体的分解を生じた。m/z1100.5でのプロトン化分子の強度は、3桁の大きさだけ低下する。10単位/gによる10マーの処置では、プロトン化分子の強度のさらなる減少は生じず、また、他のペプチド質量も認められなかった。1単位/gによる酵素処置においては、4つのペプチドが形成され、VYP(M=377.2)を伴い、最も豊富に認められるm/z378.2によって特徴付けられる(ほぼ98%;表3を参照のこと)。4つのすべてのペプチドをLC/MS/MSモードで分析し、上記の基準に基づいて正確であることを見出した。表3:10マーVYPFPGPIPN M=1099.5のLC/MSおよびLC/MS/MSモードで分析したプロトン化ペプチド質量。第2のカラムはプロトン化分子のm/z値、第3のカラムはLC/MSモードにおいて観察されるプロトン化分子の強度、第4のカラムはプロトン化分子のピーク面積に基づく百分率、そして第5のカラムは10マーの全アミノ酸配列において見出されるペプチドの位置を示す。ペプチドのプロトン化分子のピーク面積を使用することには、イオン化効率の差異の影響を含まないことを強調すべきである。
【0113】
【表4】
【0114】
しかし、A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼは、位置61(この特定のデカペプチドでは位置3)のプロリンのC末端側でほとんど排他的に切断すると結論することができる。切断の性能は、酵素/基質比の増加による影響を受けない。
【0115】
オピオイド活性を伴うすべての既知のβ−カゾモルフィン(casomorhin)分子はN末端配列YPFを共有するため、プロリン特異的エンドプロテアーゼによるPとFとの間(即ち、位置61でのプロリン残基のカルボキシ末端)のこの配列の効率的な切断は、すべてのBCM−7およびβ−カゼインのA1またはA2遺伝子変異体由来であるBCM−7関連ペプチドを有効に不活化することが明らかである。
【0116】
μ受容体との相互作用における位置61のプロリン残基の極めて重要な役割もまた、最近のインターネット上での公開物において確認された(the Dipartimento di Scienza degli Alimenti Universita di Napoli「Frederico II」Facolta di AgrariaによるSintesi e affinita ai recettori oppiodi di analoghi delle beta−casomorphine contenenti beta−omo amminoacidi」)。
【0117】
実施例6
グリアジン由来33マーLQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPFとA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼとのインキュベーションにおいて形成されるペプチド
pH8またはpH5のいずれか一方でのアルカラーゼ(Alcalase)によるシャン(Shan)ら(Science、第297号、2002年9月27日)に記載の胃および膵液耐性グリアジン由来33マーLQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF(M=3911)の処置は、何ら分子の切断を生じなかった。しかし、β−カゼイン由来10マーを伴う状況に類似して、pH5における1単位のA.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼとのインキュベーションにより、いくらかのペプチドへの分子の全体的な分解が生じた。m/z1304.4でのトリプルプロトン化(triple protonated)分子の強度は、3桁の大きさだけ低下する。プロトン化分子の強度のさらなる減少もおよびまた他のペプチド質量も、1グラムのタンパク質あたり10酵素単位による33マーの処置では観察されなかった。
【0118】
酵素処置後、約6つの主要なペプチドおよびいくらかの少数のペプチドが形成され、最も豊富に認められるm/z565.2によって特徴付けられるペプチドを伴う。6つのすべてのペプチドを、LC/MS/MSモードにおいて分析し、それらはすべてC末端でプロリンを含有することが見出され、酵素の特異性が確認された。形成された主要なペプチドは、m/z565.2によって特徴付けられる(表4の配列..QLP)。C末端配列「LP」は、このペプチドについて明確に実証することができたが、理論的には、同定される質量は、33マーのエンド型タンパク質分解では形成させることができないため、ペプチドの正確なN末端組成物に関しては、いくつか不確かな点が残る。ほぼ間違いなく、m/z565.2はQPQLP(M=581.3)のN−ピログルタミル変異体であるが、これについてさらに調べなかった。
【0119】
QPQLPの外観:LQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF
QPQLPYPの外観:LQLQPFPQPQLPYPQPQLPYPQPQLPYPQPQPF
【0120】
m/z679を伴うペプチドのLC/MS/MSスペクトルは、PQPQLPであることが解明できた。m/z565.2を伴うペプチドの性質が十分に理解さなかったという事実にもかかわらず、得られたデータは、この特定の33マーの位置12、19および26でのプロリン残基のC末端側(即ち、プロリンおよびチロシン残基の間で排他的に)アスペルギルス(Aspergillus)由来のプロリン特異的エンドプロテアーゼの好適な切断を明確に実証する。この切断パターンは、より高い酵素/基質比による影響を受けない。表4に、すべての関連する情報をまとめる。
【0121】
第1のカラムは、誘導されるペプチド配列を指定し、使用したドット(またカラム5においても)は、説明されない質量の矛盾点のため、ペプチドの正確な開始位置を示すことができなかったことを示す。第2のカラムはプロトン化分子のm/z値、第3のカラムはLC/MSモードにおいて観察されるペプチドの強度、第4のカラムはプロトン化分子のピーク面積に基づく百分率、そして第5のカラムは全アミノ酸配列において同定されるペプチドの位置を示す。
【0122】
【表5】
【0123】
セリアック病の患者において主要なエピトープであることが要求される33マーの切断は、胃もしくは膵液によっても、あるいはアルカリまたは酸性条件下のいずれの強力な広域スペクトルのプロテアーゼアルカラーゼ(Alcalase)とのインキュベーションによっても、達成することができない。それにもかかわらず、本発明者らの結果は、酸性条件下でのプロリン特異的A.niger(A.ニガー)由来エンドプロテアーゼによる効率的な切断を示す。後者の切断は、分子のプロリンおよびチロシン残基との間で排他的に生じ、6以下のアミノ酸残基長を伴う99.5%のペプチドを作製する。そのため、酸性条件下でのプロリンリッチペプチドに対するその高い有効性にもかかわらず、アスペルギルス(Aspergillus)由来酵素でさえ、アミノ酸配列YPQPQLPを伴う少なくとも0.5%のヘプタマーを遊離する。配列PYPQPQLPYが既知のセリアック病の患者に特異的なT細胞エピトープであるため、この所見は、中性付近の至適pH値を伴う最適とは言えないプロリン特異的酵素、例えば、既知のプロリン特異的オリゴペプチダーゼ、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)から誘導される酵素では、グルテン分子由来の毒性ペプチドの形成を防止するための現実的なインビボアプリケーションが、実現不可能であることを証明することを再度強調している。
【0124】
実施例7
A.niger(A.ニガー)プロリン特異的エンドプロテアーゼおよびF.メニンゴゼプチクム(F.meningosepticum)由来のプロリン特異的オリゴペプチダーゼのpH活性スペクトル
実施例1において実証されるように、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの至適pH値は約4.2である。この第1の試験では、極めて酸性のpH値を考慮しなかったため、胃に存在することができるpHの極値における酵素の挙動については、完全には明らかにされていない。国際公開第03/068170号パンフレットは、プロリン特異的プロテアーゼ、例えば、フラボバクテリウム・メニンゴセプチカム(Flavobacterium meningosepticum)由来のプロリルオリゴペプチダーゼまたはアスペルギルス・フミガーツス(Aspergillus fumigatus)由来のDPPIVまたはストレプトミセス(Streptomyces)種由来のペプチジルジペプチダーゼは、毒性のプロリンリッチペプチドのインサイチュ分解のための好適な候補を示すことを教示している。典型的に、後者のプロテアーゼのすべては、胃において何ら加水分解活性を含まないようである中性付近の至適pH値を特徴とする。本発明のプロリン特異的エンドプロテアーゼと先行技術の酵素との間に存在する至適pH値の差異を実証するために、本発明者らは、アスペルギルス(Aspergillus)由来エンドペプチダーゼおよびフラボバクテリウム(Flavobacterium)由来オリゴペプチダーゼのpH活性スペクトルを比較した。実施例1に記載のように、アスペルギルス(Aspergillus)由来エンドペプチダーゼを得た。フラボバクテリウム(Flavobacterium)由来オリゴペプチダーゼは、ICN Biomedicalsから購入した(35単位/mg;カタログ番号32082;オハイオ州、米国)。
【0125】
2つの酵素のpH活性スペクトルを確立するために、異なるpH値を伴う緩衝液を調製した。0.1mol/lクエン酸を使用してpH2.0〜7.0の範囲の緩衝液を調製し、0.1mol/lトリスを使用してpH6.0〜9.0の範囲の緩衝液を調製し、0.2mol/lグリシンを使用してpH8.0〜12.0の範囲の緩衝液を作製した。必要とされるpH値は、HClまたはNaOHのいずれか一方を使用して調整した。発色合成ペプチドZ−Gly−Pro−AMC(Bachem、瑞国)を、両酵素の基質として使用した。各ウェル(Costar番号3631プレート)において、85μLの緩衝液、10μLの酵素溶液および5μLの基質(60%メタノール中4mM Z−Gly−Pro−AMC)を導入した。A.niger(A.ニガー)酵素の最終濃度は32μg/ml(3.2ミリ単位/ml)であり、F.メニンゴゼプチクム(F.meningosepticum)酵素の最終濃度は0.21μg/ml(7.4ミリ単位/ml)であった。混合後、反応を30分間、37.0℃で進行させ、その後、PerSeptive BiosciencesのCytoFluorマルチウェルプレートリーダーにおいて、蛍光を測定した。得られた関連データを図3に示す。これらの僅かに異なる試験条件下で得られたデータにより、実施例1において確立したA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの酸性至適pH値が確認される。データは、A.niger(A.ニガー)酵素がpH2.2および7.5で約20%の残存活性を有することを示す。図3に示されるデータによってもまた、F.メニンゴゼプチクム(F.meningosepticum)の約pH7.0の公開された至適pH値が確認される。本特許出願では、pH5.0未満において、F.メニンゴゼプチクム(F.meningosepticum)酵素が活性を有さないことが最も重要である。
【0126】
これらのデータは、F.メニンゴゼプチクム(F.menigosepticum)酵素とは対照的に、A.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼが、胃および十二指腸の部分において示現する酸性条件下でその十分な活性を展開するために理想的に適切であることを示す。
【0127】
実施例8
胃に存在する条件下におけるA.niger(A.ニガー)プロリン特異的エンドプロテアーゼおよびF.メニンゴゼプチクム(F.meningosepticum)プロリン特異的オリゴペプチダーゼの安定性
首尾よい酵素療法のための必須条件は、そのようなペプチドが十二指腸の遠位部に到達する前の毒性プロリンリッチペプチドの効率的な消化管分解である。これは、酵素療法に使用すべき外因性酵素が、胃におけるおよび場合により胃より先における食餌タンパク質の滞留時間中に活性であるプロリン特異的プロテアーゼであることを必要とする。そのような「胃様」条件下で、A.niger(A.ニガー)およびF.メニンゴゼプチクム(F.meningosepticum)由来プロリン特異的プロテアーゼの活性を比較するために、本発明者らは、異なるpH条件下ならびにプロテアーゼペプシンの存在および非存在下で、37℃、異なる期間におけるインキュベーション後のそれらの残留活性をアッセイした。残留酵素活性は、関連酵素に至適なpH値および発色ペプチド、即ち、F.メニンゴゼプチクム(F.meningosepticum)酵素については7.0のpHおよびZ−Gly−Pro−pNA基質ならびにA.niger(A.ニガー)酵素については4.0のpHおよびAla−Ala−Pro−pNA基質を使用して、アッセイした。使用されるA.niger(A.ニガー)酵素は、Ala−Ala−Pro−pNAに対して高い活性およびAla−Ala−Ala−pNA基質に対して若干の活性(Ala−Ala−Pro−pNAに対するその活性の10%未満)を示す。使用されるA.niger(A.ニガー)酵素は、他のAla−Ala−X−pNA基質に対して有意な活性を示さない。
【0128】
0.2mol/lのクエン酸/HCl緩衝液は、必要とされる酸性pH条件を得るために使用した。A.niger(A.ニガー)由来酵素の用量は、1.5単位/ml、F.メニンゴゼプチクム(F.meningosepticum)酵素(MP Biochemicals、オハイオ州、米国)3.3単位/ml(単位の定義に関する材料および方法のセクションを参照のこと)の用量であった。ペプシン(Sigma)は、180マイクログラム/mlの濃度で添加した。ペプシンを不活化するために、サンプリング後、ペプスタチン(Sigma)を1.67マイクログラム/mlの濃度で添加した。これらの条件下で、ペプスタチンは、2種のプロリン特異的プロテアーゼに対し、阻害効果を有さなかった。
【0129】
それぞれ合成基質Ala−Ala−Pro−pNAおよびZ−Gly−Pro−pNAを利用して、2種のプロリン特異的プロテアーゼの残留活性を、動力学的に405nmで測定した。そのために、200μLの基質溶液(0.05mol/lリン酸緩衝液pH7 0中2mmol/l Z−Gly−Pro−pNAまたは0.05mol/l酢酸緩衝液pH4.0中1.5mmol/l Ala−Ala−Pro−pNA)を、MTPウェルにおける50マイクロリットル(10〜100×プレ希釈した)の酸性/ペプシン処置サンプルと混合した。TECAN Genios MTP Reader(Salzburg、Vienna)を利用して、10分間、405nm、30℃で、吸収を動力学的に測定した。
【0130】
表5に示される結果は、pH4が下限を提示することを示し、ここで、F.メニンゴゼプチクム(F.meningosepticum)酵素が残存することができることを示す。実施例7に記載のデータに従えば、酵素はこれらのpH条件下で活性を有さないことに留意していただきたい。pH4.0で2時間後、酵素は、至適条件下で試験する場合、約25%の残留活性を示す。しかし、ペプシンが存在するや否や、このpHで15分後に、酵素は完全に不活化された。対照的に、これらの結果はA.niger(A.ニガー)酵素により得られるデータである。後者の酵素は、pH2の低いpH値およびペプシンとの組み合わせ下でもその十分な活性を維持する。これらの結果は、5.5未満の至適pH値を伴う酵素のみが胃において活性であるようであり、そのため、酵素クラスEC3.4.21.26(プロリルオリゴペプチダーゼ)、EC3.4.14.5(ジペプチジル−ペプチダーゼIV)およびEC3.4.15.1(ペプチジルジペプチダーゼA)に属する既知のプロリン特異的プロテアーゼは、胃においてプロリンリッチペプチドを分解するのに適格でないことを強く示唆する。
【0131】
【表6】
【0132】
実施例9
多様なグルテンのエピトープに対するA.niger(A.ニガー)プロリン特異的エンドプロテアーゼの活性
多くの刊行物が、セリアック病患者由来の小腸T細胞によって認識されるコムギグルテン由来ペプチドについて詳記している。これに関して、HLA−DQ2ハプロタイプのみを担持するヒトは、セリアック病を発達する可能性が断然高い(マキ(Maki)ら;N Engl J Med.2003年6月19日;348(25):2517−24)ため、HLA−DQ2分子に対するペプチド結合は特に興味深い。検査したすべての成人HLA−DQ2患者は、α−GLIA配列Glia−α2およびGlia−α9に応答する(アレンツ−ハンセン(Arentz−Hansen)ら、J.Exp.Med.2000;6:337−342)。セリアック病を発症したばかりの小児は、ペプチドGlia−α20、Glia−γ30、Glt−17、Glt−156およびGlu−21に対して応答する(ヴァーデル(Vader)ら、Gastroenterology2002;122:1729−1737)。実施例6において、本発明者らは、既に、A.niger(A.ニガー)由来酵素によるGlia−α2ペプチド(PQPQLPYPQPQLPY)の首尾よい切断について実証した。ここでは、本発明者らは、A.niger(A.ニガー)酵素もまた、他の関連のコムギグルテン由来ペプチドならびに他の穀物から誘導されるこれらのペプチドの相同物を切断するかどうかについて試験する。
【0133】
そのために、関連ペプチドを合成し、その後、それらの純度を、材料および方法のセクションに記載の手順に従うrpHPLCおよび質量分析(MALDI−TOF)によって確認した。ペプチド消化は、50マイクロリットルのペプチド溶液(1mg/ml)と、50ミリモル/l酢酸アンモニウム緩衝液pH4.5中0.43マイクログラム/ml(0.043ミリ単位)を含有する50マイクロリットルのA.niger(A.ニガー)酵素溶液とを混合することによって行った。室温での多様なインキュベーション期間後、0.5マイクロリットルのサンプルを採取し、9.5マイクロリットルのマトリックス溶液(0.2%TFAを含有する1:1H2OおよびCH3CN)に添加した。脱塩後、Dowexイオン樹脂ビーズを使用して、液体を、いくつかの関連対照ペプチドと共にMALDI−プレート上のスポットし、その後、すべてのペプチドを分析した。得られたデータから、表6に示す切断部位を推測し得る。ペプチドGlt156およびGliaγ−2は10マーであり;Glt17は8マーであるが、至適結合のためにPおよびL残基によるC末端伸長を必要とする。明白な切断部位を二重縦線で示し、あまり明白ではない切断部位を一重縦線で示す。各ペプチド上の既知の最小のT細胞のエピトープに下線を付して示す。Glu−5において使用されるUは、ロイシンまたはイソロイシン残基のいずれか一方を提示する。
【0134】
得られるデータから、Glia−α2だけではなく、若年者ならびに成人HLA−DQ2患者に関連の事実上すべてのグルテンのエピトープが、A.niger(A.ニガー)由来プロリン特異的プロテアーゼによって効率的に加水分解されることが明らかである。ほとんどの切断部位は、既知の最小T細胞エピトープ結合部位に対し精密に位置を特定することができるため、グルテンのエピトープの酵素への暴露は、認識を廃止し、従って、究極的にセリアック病患者に特徴的な症状をもたらす以後のT細胞増殖を効率的に防止する可能性がある。
【0135】
【表7】
【0136】
類似のアプローチにおいて、コムギグルテンのエピトープの多くのカラスムギ、オオムギおよびライムギ由来グルテン相同物におけるA.niger(A.ニガー)由来酵素の切断部位を確立した(ヴェーデル,W(Vader,W)ら、Gastroenterology2003;125:1105−1113)。得られた結果(表7を参照のこと)は、A.niger(A.ニガー)由来酵素がセリアック病を担うコムギグルテンのエピトープだけではなく、カラスムギ、オオムギおよびライムギに存在するこれらのペプチドの相同物をも切断することができることを例示する。表6におけるように、明白な切断部位を二重線で示す。
【0137】
【表8】
【0138】
実施例10
100%麦芽ビールおよび100%コムギパンからのグルテンのエピトープの回収の試験
提案されている抽出手順(材料および方法のセクションを参照のこと)を、抗体に基づくアッセイと組み合わせて、PVPP処置した100%麦芽ビール(実施例11を参照のこと)および100%コムギパンサンプル(実施例12を参照のこと)に対して試験した。得られた結果(表8を参照のこと)に従って、抗体アッセイに組み合わせた抽出手順は、ビールならびにパンにおける抗−αグリアジン、抗−γグリアジンならびに抗−グルテニンエピトープを検出することができる。
【0139】
総合すると、得られたデータは、適用した抽出手順がビールおよびパンサンプルの両方におけるグルテンの検出に適切であることを強く示唆する。これらの結果に基づき、それらのプロセシング中に異なる濃度のA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼに供したビールおよびパンサンプルを、実施例11および12においてさらに緊密に調べた。
【0140】
【表9】
【0141】
実施例11
A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼに関与するビール生産は、より低いレベルのグルテンのエピトープをもたらす。
ビールの濁りは、グルテン由来プロリンリッチタンパク質、ポリペプチドおよびペプチドと、ビール生産に使用される穀物(大部分は、オオムギ)から抽出されるポリフェノールとの会合によって形成される。国際公開第02/046381号パンフレットに記載されているように、ビールの濁りの形成は、ビール生産の糖化、発酵またはラガーリング相のいずれかの間に、酸性安定なプロリン特異的エンドプロテアーゼを組み入れることによって、減少または防止することができる。従来のビールの醸造方法では、濁りの形成は、PVPP(存在する多様なポリフェノールに結合するが、濁り活性なプロリンリッチペプチドのレベルに対しては極少数の効果しか伴わない化合物)での処置によって防止される。従来の醸造方法は毒性のプロリンリッチペプチドを除去しないため、セリアック病の患者がビールを飲むことは認められていない。
【0142】
本研究の目的は、ビールの作製方法中のA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼの添加により、完成品のビールにおいて低減されたレベルの毒性プロリンリッチペプチドがもたらされるかどうかを確立することである。そうであるならば、そのようなビールは、セリアック病の患者でも飲むことができる。
【0143】
ビール生産は、IFBM(Nancy、仏国)での20hlパイロットプラントにおいて行った。使用したA.niger(A.ニガー)由来プロリン特異的エンドプロテアーゼサンプルは50%グリセロール(w/w)中で安定化され、5単位/グラム液体の最終活性を有した(単位の定義に関する材料および方法セクションを参照のこと)。
【0144】
独立した生産稼動において、5つの100%麦芽ビールを、PVPP(対照)または異なる量のプロリン特異的エンドプロテアーゼのいずれかを使用して醸造し、濁りの形成を防止した。すべての実験において、使用した糖化プロトコルはまったく同じである。実験に依存して、プロリン特異的プロテアーゼを、糖化方法の開始かまたはビール発酵直前の糖化方法後のいずれか一方で添加した。糖化方法では、異なる3つの酵素用量、即ち、使用した麦芽1kgあたり2.5、5.0および7.5酵素単位を使用した。発酵では、単回の酵素用量のみ、即ち、添加された麦芽1kgあたり0.75酵素単位を試験した。対照ビールを、ビールろ過前に添加した30グラム/hlのPVPPで安定化させた。
【0145】
各醸造は、300kgのオオムギ麦芽およびホップペレットから生産した。3:1(vol/wt)の液体/グリストの糖化条件およびpH5.6を使用した。糖化ダイアグラムは、45℃で20分間の第1の工程、64℃で15分間の第2の工程および76℃で25分間の第3の工程ならびに最終的に78℃までの加熱を含む。工程間の加熱速度は1分間あたり1℃である。麦汁を90分間、煮沸した。良好なトラブ分離をWhirlpool上で実施した。発酵は、麦汁の17 106細胞/mlのピッチング速度および約97%のピッチングでの生存能で底部酵母株により行った。12℃+/−1℃での10日間の発酵期間に続いて、−1℃+/−1℃で5日間の低温成熟を行った。ビールに5.2g/lの圧力で炭酸ガスを注入し、ボトリング後、60℃で、20分間、低温殺菌した。
【0146】
【表10】
【0147】
結果に従えば、抗−αグリアジンおよび抗−γグリアジン抗体は同等のデータを生じる。このアッセイにおける抗−グルテニン抗体では結果に至っていない。糖化段階後の酵素の特定の適用により、ビールにおいて極めて低い毒性のプロリンリッチペプチドがもたらされることが明白である。抗体認識部位は、最低でも5アミノ酸残基の長さを有するが、T細胞認識部位では少なくとも9アミノ酸残基を必要とすることを考慮すると、T細胞が、発酵アプローチにおいて作製される短いペプチドを認識し得る可能性はさらにより低くなる。事実、これらのデータは、上記の酵素アプローチに従って、セリアック病の患者に安全なビールを醸造することができることを強く示唆する。
【0148】
実施例11
A.niger(A.ニガー)由来のプロリン特異的エンドプロテアーゼを生地に組み入れることによって生成されるパンは、より低いレベルのグルテンのエピトープを生じる
パン作製のために、3500gの小麦粉(80%KolibriTMおよび20%IbisTM)、1990mlの水(56%)、77gの圧搾酵母(2.2%)、70gの塩(2%)、140mgのアスコルビン酸(40ppm)ならびに表10に記載の多様な量のA.niger(A.ニガー)由来酵素から生地を調製した。添加する酵素の量は、より少量の水を生地に添加することによって補正した。
【0149】
Diosnar(登録商標)スパイラルミキサーを2分間、スピード1で使用し、続いて、6分間、スピード2で混合することによって、成分を生地に混合した。875gの生地片を丸め、35分間、34℃および85%RHで発酵させ、型抜きし、成形し、パンニングし、75分間、38℃および87%RHで発酵させ、20分間、220℃で焼成した。生地および完成品のパンの評価は、パン製造の職人が行った。表10の結果から、パン容積および堅さの値は、1kg小麦粉あたり225単位もしくはそれ以下の酵素濃度によって影響を受けなかったため、プロリン特異的エンドプロテアーゼの添加は、グルテンネットワークおよび生地の気体保持能には明らかに影響を及ぼさないことが明白である。事実、より高い酵素用量を添加だけでは、生地にネガティブな影響を与え、ほとんどの消費者に許容され得ないと思われるパンが作製される。後者の観察のため、最も高い酵素濃度で生産されるパンに関する抗体アッセイデータは作製されなかった。
【0150】
表10において表されたデータは、1kgあたり約200単位の酵素濃度で生成させるパンに存在する毒性プロリンリッチペプチドの減少を示す。この程度の減少では、セリアック病の患者によるそのようなパンの消費を可能にするのに不十分であると証明される可能性が非常に高い。しかし、該減少は、看過され得るセリアック病もしくはIBSを患うヒトまたは未成熟な免疫系を伴う幼児に対してさえも予防的利益を伴う食物を調製するのにかなり十分であり得る。
【0151】
【表11】
【特許請求の範囲】
【請求項1】
セリアック病関連エピトープを欠く食物を製造するためのプロリン特異的エンドプロテアーゼの使用。
【請求項2】
セリアック病関連エピトープがグルテンのエピトープである、請求項1に記載の使用。
【請求項3】
グルテンのエピトープがコムギまたはオオムギのエピトープである、請求項2に記載の使用。
【請求項4】
食物がビールである、請求項1〜3のいずれか一項に記載の使用。
【請求項5】
ビール醸造工程において、糖化ステップ後発酵ステップ前にプロリン特異的エンドプロテアーゼを添加する、請求項4に記載の使用。
【請求項6】
食物がパンである、請求項1〜3のいずれか一項に記載の使用。
【請求項7】
プロリン特異的エンドプロテアーゼがアスペルギルス(Aspergillus)の酵素である、請求項1〜6のいずれか一項に記載の使用。
【請求項8】
プロリン特異的エンドプロテアーゼを使用するステップを含む、セリアック病関連エピトープを欠く食品の製造方法。
【請求項9】
穀物由来毒性プロリンリッチペプチドのレベルを減少するために、プロリン特異的エンドプロテアーゼを低いpHで使用する、請求項8に記載の方法。
【請求項10】
セリアック病を患うヒトに安全な、請求項4又は5に記載の使用によって得られるビール。
【請求項11】
セリアック病を患うヒトに安全な、請求項6に記載の使用によって得られるパン。
【請求項1】
セリアック病関連エピトープを欠く食物を製造するためのプロリン特異的エンドプロテアーゼの使用。
【請求項2】
セリアック病関連エピトープがグルテンのエピトープである、請求項1に記載の使用。
【請求項3】
グルテンのエピトープがコムギまたはオオムギのエピトープである、請求項2に記載の使用。
【請求項4】
食物がビールである、請求項1〜3のいずれか一項に記載の使用。
【請求項5】
ビール醸造工程において、糖化ステップ後発酵ステップ前にプロリン特異的エンドプロテアーゼを添加する、請求項4に記載の使用。
【請求項6】
食物がパンである、請求項1〜3のいずれか一項に記載の使用。
【請求項7】
プロリン特異的エンドプロテアーゼがアスペルギルス(Aspergillus)の酵素である、請求項1〜6のいずれか一項に記載の使用。
【請求項8】
プロリン特異的エンドプロテアーゼを使用するステップを含む、セリアック病関連エピトープを欠く食品の製造方法。
【請求項9】
穀物由来毒性プロリンリッチペプチドのレベルを減少するために、プロリン特異的エンドプロテアーゼを低いpHで使用する、請求項8に記載の方法。
【請求項10】
セリアック病を患うヒトに安全な、請求項4又は5に記載の使用によって得られるビール。
【請求項11】
セリアック病を患うヒトに安全な、請求項6に記載の使用によって得られるパン。
【図1】

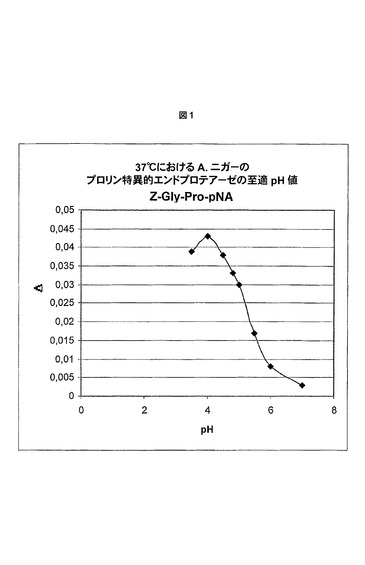
【図2】

【図3】
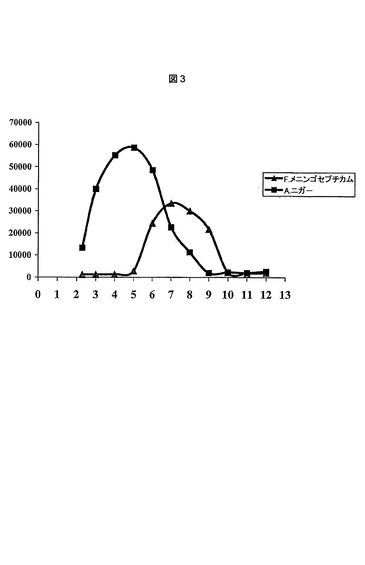
【図2】
【図3】
【公開番号】特開2011−120601(P2011−120601A)
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【出願番号】特願2011−23256(P2011−23256)
【出願日】平成23年2月4日(2011.2.4)
【分割の表示】特願2006−527369(P2006−527369)の分割
【原出願日】平成16年9月23日(2004.9.23)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公開日】平成23年6月23日(2011.6.23)
【国際特許分類】
【出願日】平成23年2月4日(2011.2.4)
【分割の表示】特願2006−527369(P2006−527369)の分割
【原出願日】平成16年9月23日(2004.9.23)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]