
ペプチドアクセプターの連結法
【課題】ペプチドアクセプターをRNA分子に結合させる方法ならびにRNA-ペプチドアクセプター産物を提供する。
【解決手段】(a)ヘアピン構造を形成する3’配列を有するRNA分子を提供する段階;(b)核酸リンカー分子に共有結合したペプチドアクセプターを提供する段階;および(c)ペプチドアクセプターとRNA分子間における共有結合の形成が可能な条件下で、該RNA分子と該核酸リンカー分子のハイブリッドを形成させる段階を含むペプチドアクセプターをRNA分子に結合させる方法。
【解決手段】(a)ヘアピン構造を形成する3’配列を有するRNA分子を提供する段階;(b)核酸リンカー分子に共有結合したペプチドアクセプターを提供する段階;および(c)ペプチドアクセプターとRNA分子間における共有結合の形成が可能な条件下で、該RNA分子と該核酸リンカー分子のハイブリッドを形成させる段階を含むペプチドアクセプターをRNA分子に結合させる方法。
【発明の詳細な説明】
【背景技術】
【0001】
発明の背景
一般に本発明は連結法、特にペプチドアクセプターを核酸に連結させる方法に関する。
【0002】
RNA-タンパク質融合体を調製する方法は現在いくつか存在する。RNA-タンパク質融合体は、ペプチドアクセプターをRNA分子の3’端に結合させた後に、そのRNAをインビトロまたはインサイチューで翻訳することで作製される。生じる産物は、ペプチドであり、このペプチドはこれをコードするRNAの3’端で結合している。このようなRNA-タンパク質融合体を作ることで、部分的または全体的にランダムなアミノ酸配列の大規模プールから所望の特徴をもつタンパク質を容易に単離することが可能となり、またRNAのコード配列をそれに対応するタンパク質分子に共有結合を介して結合させることで、タンパク質配列の情報を回収して増幅する際の問題が解決される。
【発明の概要】
【0003】
本発明は、ペプチドアクセプターをRNA分子に結合させる方法ならびにRNA-ペプチドアクセプター産物を特徴とする。この方法により例えば、部分的または全体的にランダムなアミノ酸配列または核酸配列の大規模プールから所望の特徴を有するタンパク質や核酸を単離する際に使用可能なRNA-タンパク質融合体の産生を容易にする。本発明の方法は、ペプチドアクセプターを核酸分子に結合させる種々のストラテジーで実施することができる。このような多様なアプローチは、ペプチドと核酸の結合のタイプ、および結合に使用する試薬によって異なる。
【0004】
したがって本発明の第一の局面は、ペプチドアクセプターとRNA分子との結合法を特徴とする。この結合法は、ヘアピン構造を形成する3’配列を有するRNA分子を提供する段階、ペプチドアクセプターを核酸リンカー分子に共有結合させる段階、およびペプチドアクセプターとRNA分子間に共有結合が形成可能な条件下でRNA分子と核酸リンカー分子のハイブリッドを形成させる段階を含む。
【0005】
本発明の第二の局面は、ペプチドアクセプターをRNA分子に結合させる方法を特徴とする。この方法は、ヘアピンを形成する5’配列をもつリンカーを有するペプチドアクセプターを提供する段階、ペプチドアクセプターとRNA分子のハイブリッドを形成させる段階、およびペプチドアクセプターとRNAを共有結合させる段階を含む。本発明の上記の局面の一つの態様においては、T4 DNAリガーゼを用いてペプチドアクセプターをRNA分子に結合させる。
【0006】
本発明の第三の局面は、デオキシヌクレオチド三リン酸またはジデオキシヌクレオチド三リン酸から始まるリンカー分子に共有結合させたRNA分子およびペプチドアクセプターを提供する段階、ならびにRNA分子およびペプチドアクセプターを末端デオキシヌクレオチジルトランスフェラーゼにより接触させて、ペプチドアクセプターをRNA分子に共有結合させる段階により、ペプチドアクセプターをRNA分子に結合させる方法を特徴とする。
【0007】
本発明の第四の局面は、RNA分子をペプチドアクセプターに化学的に連結することでペプチドアクセプターをRNA分子に結合させる方法を特徴とする。
【0008】
この局面の一つの態様においては、ペプチドアクセプターをソラレン部分に結合し、ソラレン部分を介してRNA分子に架橋する。ソラレン部分は、ペプチドアクセプターに結合させたリンカー分子の5’端または3’端に結合させることができる。またはソラレン部分は、リンカー分子の内部に位置させることができる。この方法では、ペプチドアクセプターを、UV照射によってRNA分子に架橋する。この特定の局面の別の態様においては、ソラレンをC6アルキル鎖を介してペプチドアクセプターに結合させ、および/またはRNA分子は、その3’端の近傍に停止コドンを有する。好ましくは、このようなリンカーの長さは25〜40ヌクレオチド単位である。さらに、ペプチドアクセプターとRNA分子を架橋させる前に、RNAと、光切断可能(photocleavable)な部分をさらに含むリンカーとのハイブリッドを形成させることができる。ハイブリッドを形成したRNAは次に光切断可能な部分を介して固相支持体に固定化することができる。好ましくは、光切断可能な部分はビオチンである。
【0009】
本発明の第四の局面の別の態様においては、RNA分子に官能基を導入し、ペプチドアクセプターとRNA分子間の化学結合形成が可能となるようにあらかじめ適切に修飾しておいたペプチドに結合させる。好ましくは、このようなRNA分子はIO4-酸化により官能基を導入する。ペプチドアクセプターには、アミン、ヒドラジン、(チオ)ヒドラジド、および(チオ)セミカルバゾンからなる群より選択されるペプチドアクセプターに分子を結合させることで官能基を導入することができる。
【0010】
本発明の第四の局面のさらに別の態様においては、外部鋳型を添加することなく化学的連結を行う。または、この化学的連結反応は、外部鋳型を添加することで進行させることができる。後者の方法は、鋳型の5’端がペプチドアクセプターのリンカー部分とハイブリッドを形成し、鋳型の3’端がRNA分子とハイブリッドを形成するように、RNA分子とペプチドアクセプターのリンカー部分を鋳型を用いて整列させる段階を含む。RNA分子のペプチドアクセプターへの化学的連結はまた、ペプチドアクセプターに共有結合させたリンカー分子そのものとRNA分子のハイブリッドを形成させることで、外部鋳型の非存在下で行うこともできる。ハイブリッドを形成させることで、ペプチドアクセプターとRNA分子は、連結可能な程度に近接する。好ましくは、官能基は、ペプチドアクセプターのリンカー領域の5’端にあるか、または片側がハイブリッド形成ドメインに隣接し、もう片側がペプチドアクセプターに隣接する。
【0011】
本発明の第四の局面のさらなる態様においては、ペプチドアクセプターとRNA分子間の化学的連結は、RNAの還元的アミノ化を介した官能基とRNA分子の結合段階と、それに続く、RNA分子と反応するペプチドアクセプターの修飾が含まれる。両分子は次に共有結合を形成して結合する。好ましくは、RNA分子に結合させる官能基は、チオール、マレイミド、またはアミンである。
【0012】
本発明の第五の局面は、非共有結合を介してペプチドアクセプターをRNA分子に結合させる方法を特徴とする。一つの態様においては、このような結合は、ペプチド核酸(PNA)とペプチドアクセプター間の共有結合の生成、およびPNAを介したペプチドアクセプターとRNA分子との非共有結合の生成によって生じる。この態様においては、RNA分子は停止コドンを含むことができる。
【0013】
本発明のさらに別の局面は、ペプチドアクセプターならびにこれらのRNA分子の転写および翻訳(および必要に応じて逆転写および/または増幅)により生じた核酸-タンパク質融合体に、化学的または非共有結合的に連結させたRNA分子を特徴とする。一つの態様においては、ペプチドアクセプターは、RNA分子の3’端に連結される。
【0014】
本発明のさらに別の局面は、本発明のRNA-ペプチドアクセプター分子を用いて、所望のタンパク質または核酸を選択する方法を特徴とする。この選択法では、RNA-タンパク質融合体を形成する本発明の分子を用いて、対象タンパク質または核酸を選択する。このような選択方法は例えば、参照として本明細書に組み入れられる、ショステク(Szostak)らによる国際公開公報第98/31700号、およびショステクらによる米国特許出願第09/247,190号に記載された任意のアプローチで実施することができる。
【0015】
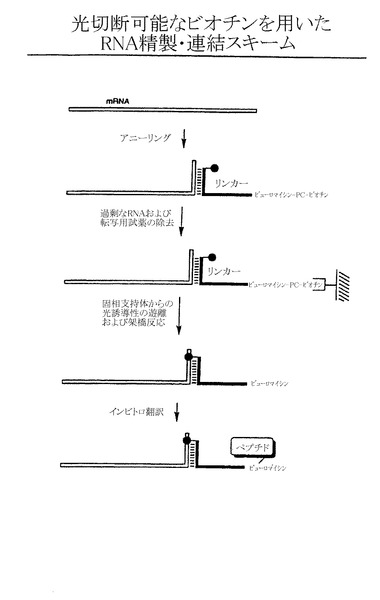
本発明の最後の局面は、RNA-タンパク質融合体を生成する方法を特徴とする。この方法は、光切断可能な部分、ソラレン部分、およびペプチドアクセプターを含むリンカーとハイブリッドを形成するRNA分子を提供する段階;固定されていない状態のRNAを支持体から実質的に除去する条件下でRNAを固相支持体に固定化させる段階;ソラレン部分を介してペプチドアクセプターを該RNAに架橋させ、この架橋形成が、架橋RNAを固相支持体から同時に遊離させる段階;および、架橋RNAを翻訳してRNA融合タンパク質を形成させる段階を含む。一つの態様においては、光切断可能な部分はビオチンである。
【0016】
上述した本発明のすべての局面におけるRNA分子は、候補タンパク質をコードする配列に使用可能に連結させた翻訳開始配列および開始コドンを含むことができる。さらに、好ましいペプチドアクセプターの一つは、伸長するペプチド鎖のC末端に付加して翻訳を停止させるヌクレオシド類似体であるピューロマイシンである。一つの態様においては、ペプチドアクセプターは、例えばヌクレオチドリンカーなどのリンカーに結合させたピューロマイシンを含む。このようなリンカーを使用することで、ペプチドアクセプターと結合対象RNA分子が整列しやすくなる。別の態様においては、ペプチドアクセプターのリンカー領域は例えばPEGなどの非ヌクレオチド成分を含む。アクセプターの他の選択肢には、RNAの3’端におけるtRNA様の構造、ならびにピューロマイシンと同様に作用する他の化合物が含まれる。このような化合物は、アデニンまたはアミノ酸ヌクレオチド、フェニルアラニル-アデノシン(A-Phe)、チロシルアデノシン(A-Tyr)、およびアラニルアデノシン(A-Ala)などのアデニン様化合物、ならびにフェニルアラニル3’デオキシ3’アミノアデノシン、アラニル3’デオキシ3’アミノアデノシン、およびチロシル3’デオキシ3’アミノアデノシンなどのアミド結合した構造に連結させたアミノ酸を有する任意の化合物を含むが、これらに限定されない。上記化合物についてはいずれも、天然のL-アミノ酸またはその類似体を任意に使用することができる。さらに、混合型のtRNA様3’構造-ピューロマイシン複合体を本発明に使用することができる。
【0017】
本発明の好ましい一つの設計において、DNA配列はメッセージの末端とペプチドアクセプターの間に含まれる。この配列は、読み枠の末端でリボソームを一時停止させて、ペプチドアクセプター(例えばピューロマイシン)が、新生ペプチド鎖を受け入れて、ペプチジル-tRNA結合が加水分解されるまでの時間を稼ぐように設計される。インビトロ翻訳の間には、リボソームを特にソラレン架橋部位またはPNAクランプなどの化学結合部位において一時停止させることも可能である。
【0018】
本発明の別の好ましい設計においては、大部分が非ヌクレオチドのリンカー部分が、ペプチドアクセプターに結合させたヌクレオチドリンカーに代わって使用される。このように設計することで、ペプチドアクセプターとRNA分子が連結しやすくなる。このリンカーは例えば、トリエチレングリコールのスペーサーを含むことができる。リンカーはまた、2’-OMe-RNAホスホラミダイトを含むことができる。化学的または酵素的な連結にハイブリッド形成が必要な場合は、連結部位に隣接するリンカーの十分な部分が、核酸を含む必要がある。
【0019】
さらに本発明のRNAまたはリンカーは、例えばRNA、またはRNAもしくはリンカーから形成されるRNA-タンパク質融合体分子のアフィニティー精製法などに用いる配列(例えばポリ(A)配列)を含んでもよい。
【0020】
さらに、上述した本発明のすべての局面においては、ペプチドアクセプターに結合させたRNA分子をインビトロまたはインサイチューで翻訳してRNA-タンパク質融合分子を作ることができる。このRNA-タンパク質融合分子を次に、高塩濃度下でインキュベートするか、および/またはショステク(Szostak)ら(09/247,190)の方法に基づいて低温でインキュベートする(例えば-20℃で一晩)。RNA-タンパク質融合分子はまた、例えば標準的なポリ(A)精製法で精製することもできる。
【0021】
本発明で用いる「タンパク質」とは、2種またはそれ以上の天然のアミノ酸、または一つまたは複数のペプチド結合で連結された修飾アミノ酸を意味する。「タンパク質」と「ペプチド」とは同じ意味で用いられる。
【0022】
「RNA」とは、共有結合で結合された天然または修飾型リボヌクレオチドの2種またはそれ以上の配列を意味する。この用語に含まれる修飾型RNAの一例が、ホスホロチオエート型RNAである。
【0023】
「翻訳開始配列」とは、機能性のリボソームエントリー部位を提供可能な配列を意味する。細菌系では、この領域はシャイン・ダルガルノ配列と呼ばれることがある。
【0024】
「開始コドン」とは、タンパク質コード配列の開始シグナルとなる3塩基を意味する。「停止コドン」とは、タンパク質コード配列の停止シグナルとなる3塩基を意味する。一般に開始コドンはAUG(すなわちATG)であり、停止コドンはUAA(すなわちTAA)、UAG(すなわちTAG)、またはUGA(すなわちTGA)であるが、開始コドンまたは停止コドンとして使用可能な他の3塩基配列で代用される場合がある。
【0025】
「共有結合で結合される」という用語は、共有結合を介して直接に、または別の共有結合で結合された配列(例えば一時停止部位に対応するDNA)を介して間接的に連結されることを意味する。
【0026】
「非共有結合で結合される」という用語は、共有結合以外の方法で連結されることを意味する
【0027】
「ヘアピン構造」とは、一本鎖の核酸によって形成される二本鎖領域を意味する。好ましくは、そのようなヘアピン構造の長さは少なくとも8塩基対であり、さらに好ましくは8〜15塩基対である。
【0028】
「化学的に連結する」という用語は、酵素を使用せずに2分子を連結することを意味する。化学的に連結することで、非共有結合ならびに共有結合が生じる。
【0029】
「ペプチドアクセプター」とは、リボソームのペプチジルトランスフェラーゼ機能の触媒活性により、伸長するタンパク質鎖のC末端へ付加されうる任意の分子を意味する。通常、このような分子は、(i)例えばアデノシンまたはアデノシン類似体(N6位のアミノ基におけるジメチル化が許容される)などのヌクレオチドまたはヌクレオチド様の成分、(ii)20種類のD型またはL型のアミノ酸や、エールマン(Ellman)らによる文献(Meth. Enzymol. 202:301、1991)に記載されているO-メチルチロシンを始めとする任意の類似体を含む、アミノ酸またはアミノ酸様成分、および(iii)(3’位、またはそれほど好ましくないものの2’位における2種の成分(例えばエステル、アミド、またはケトン結合など)の結合を含む。好ましくは、これらの結合は、天然のリボヌクレオチドコンホメーションに基づく環の歪みを大きく変えないことである。ペプチドアクセプターはまた求核基を有する場合がある。求核基には、アミノ基、ヒドロキシル基、またはスルフヒドリル基などが含まれるが、これらに限定されない。さらにペプチドアクセプターは、ヌクレオチド模倣体、アミノ酸模倣体、または結合型ヌクレオチド-アミノ酸構造模倣体から構成されることもある。
【0030】
「リンカー」または「リンカー分子」とは、デオキシリボヌクレオチド、リボヌクレオチド、またはそれらの類似体を含む配列を意味する。
【0031】
「官能基を導入する」という用語は、官能基(または官能性を有する部分)が結合するように修飾することを意味する。例えばRNA分子には、IO4-酸化またはアミノ化によって官能基を導入することができ、ペプチドアクセプターには、アミン、ヒドラジン、(チオ)ヒドラジド、または(チオ)セミカルバゾン基を結合することで官能基を導入することができる。
【0032】
「外部鋳型」とは、連結反応混合物に添加されるものの連結反応の最終産物の一部とはならない核酸配列を意味する。
【0033】
「高塩濃度(high salt)」とは、一価の陽イオン濃度が少なくとも200 mM、好ましくは少なくとも500 mM、またはさらには1 Mであること、および/または二価または多価の陽イオン濃度が少なくとも25 mM、好ましくは少なくとも50 mM、最も好ましくは少なくとも100 mMであることを意味する。
【0034】
「アフィニティー精製用の配列」とは、核酸または核酸-タンパク質融合体分子の精製に使用されるヌクレオチド配列を意味する。アフィニティー精製用の配列は例えば、オリゴdTセルロース上における核酸または融合体分子の精製に使用可能なA8-20などのポリ(A)配列である。アフィニティー精製用の配列はまた、核酸-タンパク質融合体分子の精製に使用されるポリペプチド配列の場合もある。他の例示的な精製法は、ショステク(Szostak)らによる米国特許出願第09/247,190号に記載されている。
【0035】
本発明には多数の利点がある。本明細書に記載された方法は例えば、ペプチドアクセプターとRNA分子間の効率のよい連結を、RNAとペプチドアクセプターを結合させるための外部鋳型を必要とすることなく、いくつかの局面において容易にする。本発明ではまた、RNA-タンパク質融合体の生成にかかるコストも低い。
【0036】
本発明の他の特徴および利点は、以下の詳細な説明および特許請求の範囲から明らかになると考えられる。
【図面の簡単な説明】
【0037】
図面について簡単に説明する。
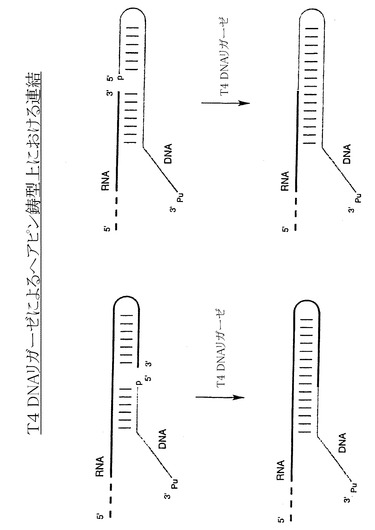
【図1】ヘアピン鋳型上における、T4 DNAリガーゼを用いたペプチドアクセプターリンカーとRNAとの連結の例示的な段階を示す略図である。RNAの3’側の配列またはペプチドアクセプターリンカーのいずれかを、ヘアピン構造を形成するように設計し、ペプチドアクセプターリンカーとRNAのハイブリッドを形成させることで、RNAとペプチドアクセプターが近接する。
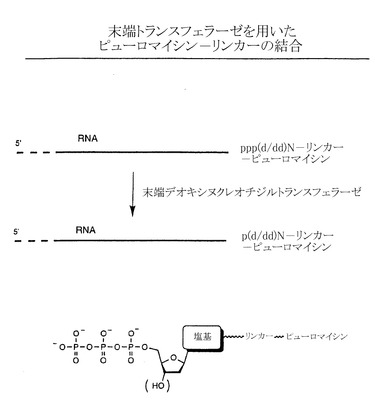
【図2】末端デオキシヌクレオチジルトランスフェラーゼを用いたペプチドアクセプターリンカーとRNAとの連結の例示的な段階を示す略図である。
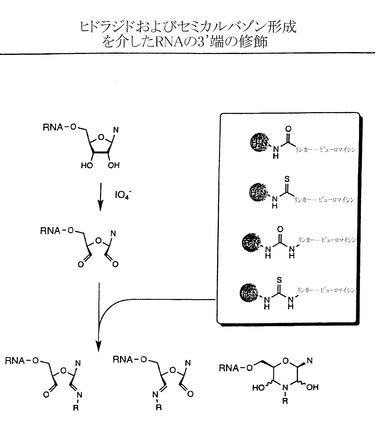
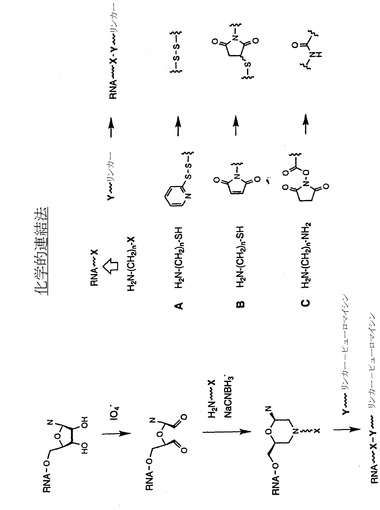
【図3A】ヒドラジド形成およびセミカルバゾン形成を介したRNAの3’端における修飾の例示的な段階を示す略図である。
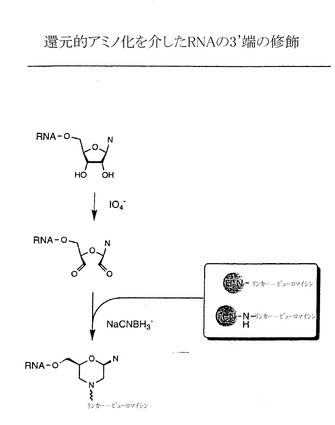
【図3B】還元的アミノ化によるRNAの3’端における修飾の例示的な段階を示す略図である。
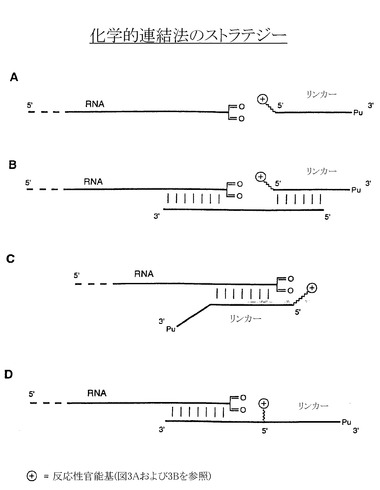
【図4】官能基を導入したピューロマイシンリンカーを、官能基を導入したRNAに化学的に連結させる例示的なストラテジーを示す一連の図である。一つのストラテジーは鋳型に依存しない(図4A)。他のストラテジーでは、外部オリゴ鋳型を使用して、RNAとピューロマイシンリンカーを整列させる。外部オリゴ鋳型が関与するストラテジーでは、オリゴ鋳型は、RNAとピューロマイシンリンカーの両方とハイブリッドを形成するか(図4B)、またはピューロマイシンリンカーと結合してRNAとハイブリッドを形成する(図4D)。さらに官能基は、ピューロマイシンリンカーの5’端(図4Bおよび4C)または内部(図4D)に導入することができる。
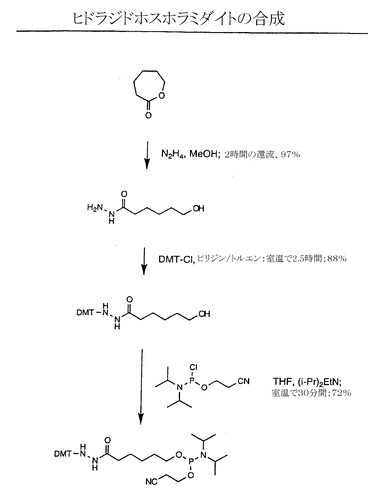
【図5】修飾型ピューロマイシンリンカーに使用する十分に保護されたカルボヒドラジドホスホラミダイトの合成の例示的な段階を示す略図である。
【図6】官能基のRNAの3’端への結合と、それに続く化学的結合による、ペプチドアクセプターリンカーとRNAとの連結の例示的な段階を示す略図である。官能基は次に、適切に修飾されたリンカー分子と反応し、チオール基(A)、マレイミド基(B)、またはアミン基(C)を介してRNAとピューロマイシンリンカーを共有結合させることができる。
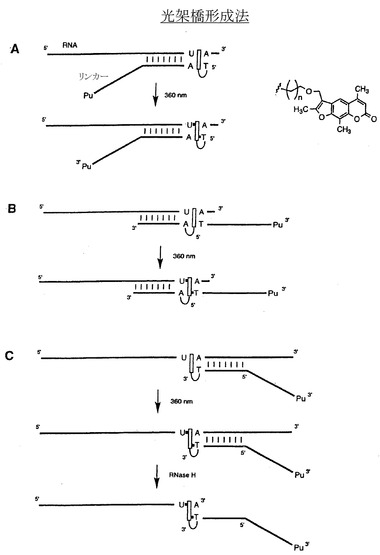
【図7A−C】光架橋法によるペプチドアクセプターとRNAとの連結の例示的な段階を示す一連の略図である。この一般的な方法では、ピューロマイシンリンカーに結合させたソラレン部分が、UV照射を受けてピューロマイシンリンカーとRNA間を光架橋する。ソラレン部分は、ピューロマイシンリンカーの5’端(図7A)、内部(図7B)、または3’端(図7C)に位置することができる。
【図7D】光架橋形成に使用されるmRNAを示す略図である。
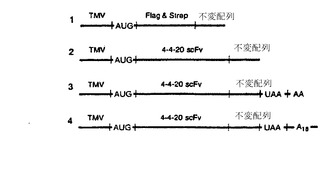
【図7E】mRNAの不変の3’配列(図7DにおけるmRNAの不変領域)と、推定ソラレン挿入部位を示す光リンカー間に形成される二本鎖を示す略図である。C6アルキル鎖を介してリンカーの5’リン酸に結合させたソラレンの構造も示す。変数xおよびyは、リンカーに含まれるdAヌクレオチド数とトリエチレングリコール(TEG)単位数をそれぞれ示す。左側に示したオートラジオグラフは、mRNA 1とリンカーB間の光架橋反応をゲル電気泳動の解析結果を示す。
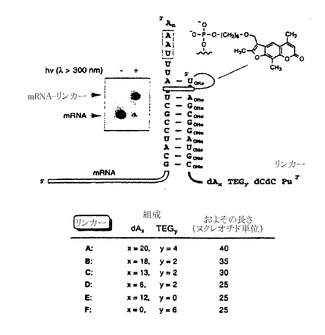
【図7F】インビトロ翻訳および融合体形成を示すゲル像である。mRNA鋳型2および鋳型3に由来するタンパク質合成をそれぞれレーン1とレーン3に示す。リンカーBへ光架橋したmRNA鋳型の翻訳により、mRNA-タンパク質融合体が生じた(レーン2と4)。鋳型3は、リンカー架橋部位に続いて停止コドンを有する。レーン5は、オリゴdTセルロースで精製した後の融合産物を示す。停止コドンを含むmRNA分子を用いた融合体形成を示す略図をゲル像の上に示す。
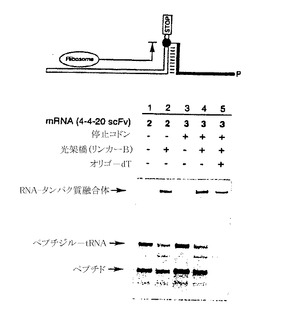
【図7G】mRNA-タンパク質融合体分子の収率が、リンカー組成と塩濃度に依存することを示すゲル像である。
【図7H】mRNAと光切断可能なビオチンを用いたハイブリッド形成用リンカーとの連結および、それに続く融合体形成にかかわる段階を示す略図である。
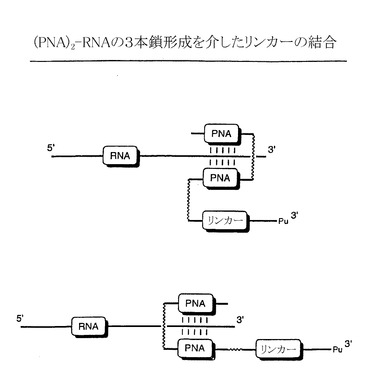
【図8A】(PNA)2-RNAの三本鎖形成を介したピューロマイシンリンカーとRNAとの結合を示す略図である。ピューロマイシンリンカーは、強固な非共有結合を介して三本鎖を形成するRNAに結合する2個のPNA分子と結合する。
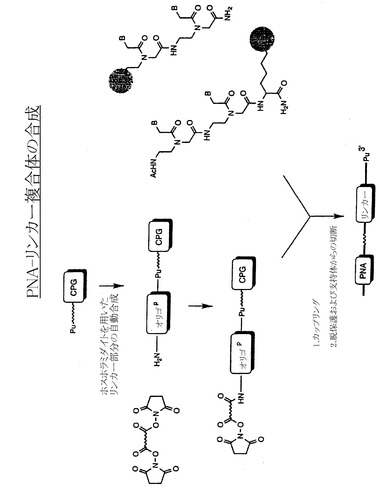
【図8B】PNA-リンカー複合体の合成段階を示す略図である。リンカーを固相支持体に結合し、PNAと結合するように修飾する。次にPNAを所望のリンカーと結合させ、PNAリンカー分子を脱保護して固相支持体から分離する。
【発明を実施するための形態】
【0038】
発明の詳細な説明
本明細書では、ペプチドアクセプターをRNA分子に結合させる種々の方法を記載する。RNAは、通常の細胞を用いた合成法、組換え技術、および化学合成を含む標準的なアプローチで生成することができ、RNAには、細胞のRNAライブラリー、mRNAライブラリー、およびランダム合成RNAライブラリーが含まれるがこれらに限定されない。ペプチドアクセプター(例えばピューロマイシン)をDNAリンカーまたはRNAリンカーに通常結合させる。このようなペプチドアクセプター分子は、任意の標準的な方法で生成することができる。これには例えば、ロバーツ(Roberts)およびショステク(Szostak)の論文(Proc. Natl. Acad. Sci. USA 94:12297、1997)、ショステクら(国際公開公報第98/31700号)、およびショステクらによる米国特許出願第09/247,190号に記載される手法がある。本発明の各方法を実施する方法について、特定の実施例を用いて詳細に説明する。特定の例は本発明を説明する目的で提供するものであって、本発明を制約するものと解釈すべきではない。
【実施例】
【0039】
実施例1:ヘアピン鋳型上でT4 DNAリガーゼが関与する酵素的連結法
本発明の特定のアプローチの一つにおいては、T4 DNAリガーゼを使用して、例えば図1に示すヘアピンを含む鋳型を用いてペプチドアクセプターをRNA分子に結合させる。連結反応は、自己鋳型的に進み、副子オリゴ(splint oligo)を使用しない。RNAの3’端の配列、またはペプチドアクセプターのリンカー領域の5’端のいずれかを、ヘアピン構造を形成するように設計する。ペプチドアクセプターのリンカー領域の5’端およびヘアピン構造を形成するRNAの3’端が近接することで、T4 DNAリガーゼによる酵素的連結が促される。これは例えば、サムブロック(Sambrook)、フリッシュ(Fritsch)およびマニアティス(Maniatis)の「分子クローニング(Molecular Cloning)」、Cold Spring Harbor、New York、Cold Spring Harbor Laboratory Press、1989に記載されている。反応物の比はおよそ1:1であるが、いずれかの反応物が若干過剰(1:1.2)であってもよい。最適な連結条件は、反応によって若干変動する場合があるので、当業者に周知の方法で実験的に決定してもよい。
【0040】
実施例2:末端トランスフェラーゼが関与する酵素的連結法
ペプチドアクセプターをRNA分子に結合させる別の酵素的方法では、RNAの3’端を修飾してから、例えば図2に示すように末端デオキシヌクレオチジルトランスフェラーゼ(TdT)を用いて連結を行う。TdTによる反応は、サムブロック(Sambrook)、フリッシュ(Fritsch)およびマニアティス(Maniatis)の書籍(前記)に一般的に記述されている方法で行う。この酵素は、核酸の3’端にデオキシ(またはジデオキシ)ヌクレオチド三リン酸(NTP)を用いて伸長させる。このような(d/dd)NTPは、塩基成分を化学的に修飾することで所望のリンカー構造をもたせることができる(例えばMeyer、「分子生物学における方法(Methods in Molecular Biology)」、 Agrawal、ed.、第26巻、Totowa:Humana Press、1994、73〜91ページ;Kumarら、Anal. Biochem. 169:376、1988;Rileyら、DNA 5:333、1986;およびSchmitzら、Anal. Biochem. 192:222、1991に記載されている)。図2に示すように、リンカー分子は(d/dd)NTPで開始しなければならない。しかし残りのリンカー組成は変化してもよい。この末端トランスフェラーゼ連結法の特徴は、ペプチドアクセプターのDNAリンカー領域が最短で1残基の(d/dd)NTPでもよいことである。
【0041】
実施例3:IO4-酸化とそれに続く化学的連結によるRNAの3’端への官能基導入が関与する化学的連結法
本発明の方法では、図3に示すように、ペプチドアクセプターはIO4-酸化を介したRNAの3’端への官能基導入と、それに続くペプチドアクセプターとRNAの化学的連結によってRNA分子に結合させることもできる。IO4-酸化は下記の方法で、またアグラワル(Agrawal)(「分子生物学における方法(Methods in Molecular Biology)」、 Agrawal編、第26巻、Totowa:Humana Press、1994、93〜120ページ)、プロウドニコフ(Proudnikov)およびミルザベコフ(Mirzabekov)、Nucleic Acids Res. 24:4535、1996;ゴーシュ(Gosh)ら、Anal. Biochem. 178:43、1989;バウマン(Bauman)ら、J. Histochem. Cytochem. 29:227、1981;およびウー(Wu)ら、Nucleic Acids Res. 24:3472、1996、の各論文に記載された方法で実施する。IO4-酸化段階は反応に1,2-ジオールを厳格に必要とするので、末端ヌクレオチドのみを修飾し、RNA分子の内部にある残基は影響を受けない。結果として生じるジアルデヒドはさらに、アミン、ヒドラジン、カルボ(チオ)ヒドラジド、または(チオ)セミカルバジドなど様々な求核基による反応を受けてシッフ塩基様の構造を生じる。これらの反応は例えばアグラワル(前記)、プロウドニコフおよびミルザベコフ(前記)、ゴーシュら(前記)、バウマンら(前記)、およびウーら(前記)の各論文に記載された通りに進む。それぞれカルボ(チオ)ヒドラジドおよび(チオ)セミカルバジドとの反応後にそれぞれ得られる(チオ)ヒドラジドおよび(チオ)セミカルバゾンは極めて安定であり、アミンまたはヒドラジンを含む初期付加物は通常、その後に図3Bおよびアグラワル(前記)、プロウドニコフおよびミルザベコフ(前記)、ゴーシュら(前記)、バウマンら(前記)、およびウーら(前記)の論文に記載されるように、新規合成された結合を加水分解に対して安定なものとする、その後の還元的アミノ化などの還元段階を必要とする。
【0042】
上述のカップリング反応は、図4A-4Dに示す多種多様なストラテジーで行うことができる。例えば連結は図4Aに示すように、外部鋳型に依存せずに行うことができる。このストラテジーには、RNA分子との連結を成功させるために大過剰の修飾型ペプチドアクセプター(例えば100〜1000倍過剰量)が必要である。RNAの連結効率を上げるために図4Bに示すように、外部鋳型オリゴを使用して基質を整列させることができる。好ましくは、配列がRNA配列およびペプチドアクセプターリンカー配列と相補的なこのようなオリゴの長さは、少なくとも約10ヌクレオチドである。このようなオリゴは通常、RNA分子に相補的な少なくとも約10ヌクレオチド、およびペプチドアクセプターリンカーに相補的な約10ヌクレオチドを含む。または反応部位は、図4C〜4Dに示すようにリンカーとRNAドメインが直接ハイブリッドを形成することで近接する場合がある。この場合、ハイブリッドを形成する配列の長さは、少なくとも10〜15ヌクレオチドである。
【0043】
架橋形成によってペプチドアクセプターをRNAに結合させるために多くの多様なコンストラクトを使用することができる。例えば、例示的なコンストラクトのタイプの一つには、図4A〜4Cに示すように5’端を修飾したリンカーに結合させるペプチドアクセプターがある。別のコンストラクトは、片側がハイブリダイゼーションドメインに隣接し、もう片側がピューロマイシンリンカー部分に隣接する内部官能基を含む(図4D)。
【0044】
このような修飾型リンカーの合成では、ヌクレオチドの本体またはスペーサー部分を集合させる市販のホスホラミダイト(Glen Research、 スターリング、バージニア州)を用いた標準的な自動DNA合成を含む。3’端のピューロマイシンは、合成用の固相支持体としてピューロマイシン-CPG(Glen Research、 スターリング、バージニア州)を用いて導入することができる。反応性官能基は、アミノ末端修飾物質(Glen Research、スターリング、バージニア州)やuni-linkアミノ修飾物質(Clontech、パロアルト、カリフォルニア州)など市販の試薬を用いて結合することができる。適切なホスホラミダイトを用いることで他の官能基を組み入れることが可能である。カルボヒドラジドホスホラミダイトを生成する1つの例示的な方法の一つを図5に示す。カルボヒドラジドホスホラミダイトは、ラクトンにヒドラジンおよびメタノールを結合させ、反応物を2時間還流してカルボヒドラジド成分をつくることで生成した。この合成段階の収率は97%であった。結果として生じた産物を次にジメトキシトリチル基およびピリジン/トルエンの塩と室温で2.5時間反応させて、保護基のついたカルボヒドラジド成分を得た。この段階の産物収率は88%であった。この産物をさらに、テトラヒドロフランおよびジイソプロピルアミンの存在下でホスホラミダイト成分と室温で30分間反応させ、十分に保護されたカルボヒドラジドホスホラミダイトの反応産物を得た(72%)。
【0045】
さらに、IO4-で酸化されたRNAに対するペプチドアクセプターの反応性は、複数コピーの反応基を導入することでさらに高まる。
【0046】
例示的な連結反応の一つは以下のように進んだ。flagエピトープおよびstrepタグをコードする転写産物を含む1 nmoleのRNA(配列:

)を、20 μlの500 mM NaOAc(pH 5.2)、10 μlの5 mM NaIO4と混合し、水を加えて最終容量を100 μlとした。この反応混合物を室温で15分間インキュベートした。次に、10 μlの10 mM Na2SO3を添加し、反応混合物を再び15分間室温でインキュベートした。40 μlの1 Mリン酸緩衝液(pH 8.0)、1.5 nmoleのペプチドアクセプターリンカーUni-A1/8(配列:
(XはUni-linkアミノ修飾物質[Clontech]、またPuはピューロマイシン-CPG[Glen Research]を意味する)、および20 μlのNaCNBH3を反応混合物に添加した。この混合物を次に18時間室温でインキュベートして沈澱を生じさせ、6%のTBE-尿素ゲルで精製し、一晩破砕・浸漬してRNA-タンパク質融合分子を得た。この反応で230 pmoleの産物が得られた。
【0047】
実施例4: RNA分子の3’端への官能基の結合と、それに続く化学的連結が関与する化学的連結法
ペプチドアクセプターは図6に示すように、RNAの3’端に官能基を結合させた後に化学的に連結させることで、RNA分子に結合させることもできる。上記のプロセスの変法では、還元的アミノ化および関連反応によってRNAの3’端に官能基を結合させる。新しく導入したこれらの官能基を次に、ペプチドアクセプターの適切に修飾したリンカーとさらに反応させて、RNAとペプチドアクセプター間に共有結合を形成させる。例示的な反応基には、チオール(ジスルフィド形成、またはピリジルジスルフィドなどのチオール親和性(thiolphilic)試薬との反応用;図6の反応A)またはマレイミド(図6の反応B)などがある。
【0048】
RNAの3’端への可能な官能基の導入とそれに続くペプチドアクセプターの結合に用いられる他の反応基がアミン類である。例えば、N-ヒドロキシスクシンイミド-エステル(NHS-エステル)は、ジスクシンイミジルグルタレート(DSG)または関連試薬と反応させることで、ペプチドアクセプターの5’側のアミノ修飾リンカー上で生成する。これはコックス(Cox)ら(J. Immunol. 145:1719、1990)およびhttp://www.piercenet.com/Products/linksearch.cfm(Pierce、ロックフォード、イリノイ州;図6の反応C)に記載されている。次にこの修飾型リンカーは、修飾型RNAのアミノ官能基と反応させることができる。
【0049】
このタイプの連結反応は、上述の外来鋳型に依存しない方法、または依存した方法のいずれかにより、同様の一般的なアプローチで行うことができる。
【0050】
実施例5:光化学的な方法を含む化学的連結法
ペプチドアクセプターは、図7A〜7Hに示すような光化学的な方法でRNA分子に結合させることもできる。ソラレン基を有するペプチドアクセプターリンカー分子は、長波長のUV光照射を受けて相補的RNA鎖に架橋を導入することができる。この手法は通常、ピーレス(Pieles)およびイングリッシュ(Englisch)の論文(Nucleic Acids Res. 17:285、1989);およびゴダール(Godard)ら(Nucleic Acids Res. 22:4789、1994)の記述にしたがって実施される。ペプチドアクセプターのリンカー領域の5’端へのソラレン部分の結合は、市販のソラレンアミダイト、2’-OMe-RNA ホスホラミダイト、ソラレンC6ホスホラミダイト、およびトリエチレングリコール(TEG)ホスホラミダイト(Glen Research、スターリング、バージニア州)を用いて、標準的なDNA合成装置を用いて達成できる。
【0051】
例示的なアプローチの一つにおいては、この方法は以下のように実施された。flagエピトープ、strepタグ、および光化学的な標的部位(配列:
)をコードするRNA転写物からなる1 nmoleのRNA、標準的な製造業者プロトコル通りに合成された1.2 nmoleのPhotolinker 30/10(配列:
)(PsoはソラレンC2アミダイト[Glen Research]、Puはピューロマイシン-CPG [Glen Research]を意味する)、または光リンカー30/15(配列:
)(PsoはソラレンC2アミダイト、Puはピューロマイシン-CPGを意味する)、10×緩衝液(250 mM Tris pH 7.0;1 M NaCl)、および水(最終容量を360 μlにする)を混合して80℃で2分間加熱した。次に反応混合物を緩やかに室温まで冷却した。次にこの反応混合物に、石英浸漬用ウェル(ACE Glass、カタログ番号7854-25)に入れたPyrex製吸収用スリーブ(ACE Glass、カタログ番号7835-44)を備えた450 Wの浸漬型ランプ(中圧;ACE Glass、カタログ番号7825-34番)を用いて、0℃で15分間、310 nm以上の波長で照射した。この際、試料は浸漬用ウェルに固定化した微小遠心用チューブに入れて氷中で冷却した。次に試料に40 μlの3 M NaOAcと1000 μlのエタノールを添加して沈澱を生じさせ、75 μlの水に再懸濁した。次いで75 μlの2×ローディング緩衝液(Novex)を試料に添加し、プレキャストタイプの6% TBE尿素ゲル(Novex)で試料を精製した。産物を破砕・浸漬法(0.3 M NaOAc、室温で一晩)で回収した後にエタノールで沈澱させた。この光架橋法により、Photolinker 10/30を用いて272 pmoleのRNA-タンパク質融合産物を得た。および、Photolinker 15/30を用いて227 pmoleのRNA-タンパク質融合産物を得た。
【0052】
上記の光架橋法による化学的連結法では、種々の反応パラメータの評価を行った。最初に光架橋形成の塩依存性を検討した。100〜1000 mMのNaClを含む緩衝液を用いた一連の架橋実験を行った。種々の反応間で連結効率に差は認められなかった。さらにRNA標的配列を、
(下線部の配列がソラレンリンカーの標的となる)に変えたところ、産物の収率は大きく減少した(15〜20%の減少)。このことから、RNA標的配列が重要であることがわかった。次に産物の収率が、反応過程にあらかじめ不活性化しておいたソラレンリンカーを繰り返し置換することで上昇しうることを確認した。この実験は以下の手順で実施した。RNA、リンカー、および10×緩衝液を混合して80℃で2分間加熱した。次に反応混合物を緩やかに室温に冷却し、上述の通りに紫外線を照射した。次にさらに1 nmoleのリンカーと1 μlの緩衝液を添加した。リンカーをRNAとアニーリングさせて紫外線を照射した。このプロセスを、2 nmoleのリンカーと2 μlの緩衝液を添加して紫外線照射を繰り返し行った。この手順を採用することで、産物の収率を、ある特定の配列について20%から40%を超えて増大させることができた。
【0053】
光化学的架橋法で生成させる連結産物の能力についても評価を行った。種々の長さのリンカー(ソラレン+15塩基対の標的ハイブリッド形成用ドメイン+dAnCCPu[n=7、12、17、または22])を対象とした実験では以下の結果が得られた。長いリンカーは、高塩濃度条件(500 mM KCl+50 mM MgCl2)でRNA-タンパク質融合体の収率が最も高かった。低塩濃度の緩衝液(250 mM KCl+10 mM MgCl2、または250 mM KCl)中では、短いリンカーの方が長いリンカーよりも収率が高かったが、全体的な収率は概して低値であった。一般に収率は、酵素的に連結させたRNA鋳型と同等であると考えられた。
【0054】
別の例示的な方法においては、種々のmRNAおよびピューロマイシンリンカーを、ペプチドアクセプターがリンカーの3’端に位置するように合成した。リンカーを標的mRNAとアニーリングさせ、mRNA-タンパク質融合体分子の形成効率をインビトロ翻訳法で評価した。リンカーの長さと組成が融合体分子の収率に及ぼす作用についても決定した。
【0055】
図7Dは、架橋反応に使用したmRNAの設計を示す。各RNA分子は、良好な開始コドン組成を有するタバコモザイクウイルス(TMV)の5’-UTR(Gallieら、Nucleic Acids Res.、16:883〜93、1988;およびKozak、Microbiol. Rev.、47:1〜45、1983)、および翻訳開始コドン(AUG)を含む。すべてのmRNAはまた、3’末端に10ヌクレオチドのリンカーハイブリッド形成配列
(アミノ酸配列ASAをコードする)、およびソラレン光架橋用のジヌクレオチド配列5’-UA(不変)を末端に有する。これらは、シンデン(Sinden)およびヘイガーマン(Hagerman)(Biochemistry、23:6299〜6303、198)、およびガンパー(Gamper)ら(Photochem. Photobiol.、40:29〜34、1984)の論文に記載されている。さらに一種のmRNA(mRNA 1)は、FlagエピトープDYKDDDDK(Hoppら、Biotechnology、6:1205〜1210、1988)と、それに続くStrep-Tag II配列WSHPQFEK(Schmidtら、J. Mol. Biol.、255:753〜66、1996)を含む。他のmRNA分子(mRNA 2、3、および4)は、4-4-20 scFv、抗フルオレセイン一本鎖抗体鋳型を含む。これらは、ベドジク(Bedzyk)ら(J. Biol. Chem.、265:18615〜20、1990)、およびマレンデール(Mallender)らの論文(J. Biol. Chem.、271:5338〜46、1996)に記載されている。さらにmRNA 3および4は、残存する非架橋鋳型の翻訳後にリボソームからのmRNAの遊離を誘導する下流の停止コドン(UAA)を含む(下記参照)。オリゴdTによる精製用のポリAテールもmRNA 4に結合させた。
【0056】
以上の検討に使用したmRNA(図7D)は、PCR DNA鋳型を対象に、メリガン(Milligan)らの論文(Nucleic Acids Res.、15:8783〜8798、1987)に記載されている、T7 RNAポリメラーゼのラン・オフ転写法(Megashortscript Transcription Kit、Ambion、テキサス州)で調製した。転写後に全RNAを6%のTBE-尿素ポリアクリルアミドゲル(Novex、カリフォルニア州)を用いた電気泳動で精製した。産物のバンドをUVシャドウイング法で可視化し、ゲルから切り出して破砕し、0.3 M NaOAcに一晩浸漬した。エタノールで沈澱させた後に、RNAを水に再懸濁して保存した。同じ手法で、放射標識したRNAを[α-32P]UTP(Amersham、イリノイ州)を転写用緩衝液に含めることで合成した。
【0057】
以上の検討に使用したピューロマイシンリンカー(図7E)は、Expedite Synthesizer モデル8909(PerSeptive Biosystems、マサチューセッツ州)を用いて、従来の固相支持体ホスホラミダイト化学法で調製した。ピューロマイシン-CPG、DNA ホスホラミダイト、2’-OMe-RNA ホスホラミダイト、ソラレンC6ホスホラミダイト、およびトリエチレングリコール(TEG)ホスホラミダイト(スペーサー9)を、推奨プロトコルに基づいて使用した(Glen Research)。ソラレン部分は、C6アルキル鎖を介してリンカーの5’-リン酸に結合させた。柔軟性のあるリン酸トリエチレングリコール(TEG)のスペーサー、および種々の長さのポリヌクレオチド配列を用いて5’-dCdC-ピューロマイシンを3’端に連結させた(図7Eの表を参照)。リンカーのハイブリッド形成配列は、幹状構造の対形成時の安定性を強化するために2’-OMe-RNA ホスホラミダイトから調製した(Inoueら、Nucleic Acids Res.、15:6131〜6148、1987;およびMajlessiら、Nucleic Acids Res.、26:2224〜2229、1998)。濃水酸化アンモニウム中で55℃で8時間かけて保護基を外した後に、逆相HPLC法でC18 Spheri-5カラム(Perkin Elmer、カリフォルニア州)と、緩衝液Aに50 mM酢酸トリエチレンアンモニウム(溶媒:5% v/v アセトニトリル)を、および緩衝液Bに50 mM酢酸トリエチルアンモニウム(溶媒:70% v/v アセトニトリル)を用いて1.5 ml/分の流速でリンカーを精製した。15〜60%の緩衝液Bの直線性勾配を用いて45分間にわたる溶出を行った。リンカーは乾燥後に水に再懸濁して保存した。
【0058】
リンカー(5 μM)を標的mRNA(2.5 μM)と、25 mM Tris HCl緩衝液(pH 7)および100 mM NaCl中で、85℃で30秒間加熱してアニーリングさせた後に、5分間かけて4℃に冷却した。この反応混合物をホウケイ酸ガラス製バイアル(Kimble/Kontes、ニュージャージー州)に入れ、手持ち式マルチ波長UVランプモデルUVGL-25(UVP、カリフォルニア州)を長波長(波長λ>300 nm)に設定したもので15分間室温で紫外線を照射した。放射標識したmRNA 1とリンカーB間の光架橋反応の産物混合物を、変性6% TBE-尿素ゲル(Novex)で解析し、ホスホイメージングシステム(Molecular Dynamics、カリフォルニア州)で可視化した(図7E)。光架橋産物混合物は一般に、20%未満の未反応mRNAと80%を超える光架橋mRNAを含んでおり、これをさらに精製することなくインビトロ翻訳とそれに続くmRNA-タンパク質合体形成に直接使用した。より長いmRNA基質2、3、および4(>800ヌクレオチド)については、mRNAと架橋されたmRNA間の相対サイズの差は極めて小さくなり、ゲル上で分離することはできなかった。このような場合は、粗光架橋反応混合物を、さらに精製することなく融合体形成用の溶解液に直接添加した。
【0059】
mRNA融合体分子の翻訳および融合体形成は、mRNA 2を用いて以下の実験で最初に検討した。インビトロ翻訳反応は、ウサギ網状赤血球溶解液(Ambion)を用いて30分間30℃で行った。この反応物は、100 pmoleの光架橋mRNA(上述)、10 mMのクレアチンリン酸、150 mMのKOAc、0.5 mMのMgCl2、0.1 mMの各アミノ酸(メチオニンを除く)、150 μCiの[35S]メチオニン(Amersham)、および67%(v/v)の溶解液を最終容量300 μlとして含んでいた。mRNA-タンパク質融合体の形成は、ロバーツ(Roberts)およびショステク(Szostak)の方法ならびにショステクらの方法(上記)通りにKClとMgCl2を最終濃度がそれぞれ590 mMと50 mMとなるように、かつ容量が500 μlになるように添加して促進した。インキュベーションは20℃でさらに60分間継続した。融合体形成の塩依存性を探るためにさまざまな濃度のKClとMgCl2も検討した。
【0060】
インビトロ翻訳産物は、溶解液を10 mlの結合用緩衝液(100 mM Tris HCl、pH 8.0、10 mM EDTA、1 M NaCl、0.25% v/v Triton X-100)に希釈し、混合物に10 mgのオリゴ-dTセルロース7型(Pharmacia、ニュージャージー州)を添加して単離した。このサンプルを60分間4℃で回転した。次に固相支持体を5 mlの氷冷した結合用緩衝液で洗浄した後に100 μlの脱イオン水のアリコートで溶出した。単離したmRNA-タンパク質融合体の量は、取り込まれた[35S]メチオニンのシンチレーションを測定して決定した。この産物を4〜12%のMESランニング緩衝液(Novex)を用いたNuPageゲル電気泳動で解析した。ゲルを十分に洗浄して過剰な[35S]メチオニンを除去した後に乾燥させ、バンドをホスホイメージャーシステム(Molecular Dynamics)で可視化した。
【0061】
ゲル解析から、ペプチジル-tRNAおよび遊離ペプチドに対応する二本のバンドの存在が認められた(図7Fのレーン1)。光架橋したmRNA 2の翻訳を実施したところ、緩やかに移動する第三のバンドが現われた。これはmRNA-タンパク質融合体が良好に形成されていることを意味する(図7Fのレーン2)。遊離タンパク質および融合体(約20%多い)の収率は、コード配列がmRNA 2と同一であるが光架橋部位の下流に停止コドンを有するmRNA 3では上昇した(図7Fのレーン3および4)。相対的なバンドの強度から、合成タンパク質の総量の30%がmRNA-タンパク質融合体に変換されたことが判明した(図7Fのレーン4)。
【0062】
図7EのmRNA 4から調製したmRNA-scFvの融合体分子は、ロバーツ(Roberts)およびショステク(Szostak)ならびにショステクら(上記)の方法で、A18リンカー領域をオリゴdTセルロースに結合させた後に結合用緩衝液で洗浄することで精製した(図7Eのレーン5)。融合体産物は、光架橋した入力mRNA量を元にした場合、1.3%の収率で単離することができた。光架橋したmRNAから調製した融合体の物理的性質(ゲル移動度、オリゴdTセルロースに対する結合性、親和性試薬に対する選択的なペプチド結合性)は、酵素を用いて連結させたmRNA鋳型から得られた融合体産物と同一であることが判明した。
【0063】
融合体分子のペプチド部分の組成を確認するために、図7EのリンカーBと架橋させた、FlagおよびStrep-Tag IIエピトープをコードするmRNA鋳型1から調製した融合体のタンパク質結合性を検討した。35Sで標識したmRNA-ペプチド融合体(mRNA 1からリンカーBで調製したもの)溶液10 μlを、50 mM Tris HCl、pH 7.4、1% NP 40、150 mM NaCl、1 mM EDTA、1 mM Na3VO4、および1 mM NaFを含む300 μlの緩衝液に溶解した20 μlのAnti-Flagアフィニティーゲル(Sigma、ミズーリ州)に添加した。同じ融合体産物を、100 mM Tris-HCl、pH 7.1、1 mM EDTA、および0.5 mg/mlの酵母tRNAを含む300 μlの緩衝液に溶解した20 μlのStrepTactin Sepharose(Genosys、テキサス州)に添加して二回目の沈澱実験を行った。両沈澱混合物とも、同一の条件で並行して処理した。混合物を1時間4℃で回転させた後に、Ultrafree-MCフィルターユニット(0.45 μm;Millipore、マサチューセッツ州)上に移した。遠心して緩衝液を除き、残渣を5×300 μlの氷冷緩衝液で洗浄した。残渣のシンチレーションを測定して解析し、融合体結合をAnti-Flag M2マトリックスとStrepTactinマトリックスについてそれぞれ54%と62%であると判定した。プロテインAアガロース(Sigma)を対象とした対照反応では、マトリックスに対する結合は検出されなかった。
【0064】
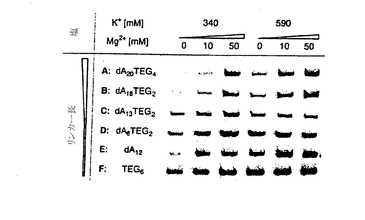
リンカーの長さと組成が、種々の塩濃度で融合体形成に及ぼす影響を調べるために、mRNA 3を図7EのリンカーA〜Fに光架橋した後に、各鋳型の融合体形成を検討した(図7G)。30℃で30分間インキュベートした後に試料をアリコートに分け、各アリコートにKClおよびMgCl2を量を変えて添加した。この試料をさらに60分間20℃でインキュベートした後に、融合体産物をゲル電気泳動で解析した。mRNA-タンパク質融合体の最高収率は、高塩濃度下で長いリンカーAおよびB(それぞれ40ヌクレオチドおよび35ヌクレオチド)で得られた。低い塩濃度では、リンカーAおよびBでは融合体形成の有意な低下が認められた。一方、より短いリンカーC〜Fの融合体の収率は、それほど塩濃度に依存していなかった。さらに、融合体分子の収率は一般に、柔軟性のあるTEGスペーサーの数とともに上昇した。粗mRNA-タンパク質融合体分子溶解液を解析したところ、総タンパク質の最大45%がmRNA-タンパク質融合体として存在することが判明した。リンカーFには、オリゴdTによる精製に必要なオリゴdAトラックがないので、mRNA 4は、その3’端にある、オリゴdTセルロース上における融合体精製を可能とするA18の連続を使用して調製した。
【0065】
リンカーの5’端にソラレンを結合させる上記手法の別法では、ソラレン部分を、図7B、およびピールス(Pieles)らの論文(Nucleic Acids Res. 17:8967、1989)に示されるように、ペプチドアクセプターのリンカー領域の内部に組み込まれる場合もあるほか、リンカー配列内部における分枝状ホスホラミダイト(Clontech、パロアルト、カリフォルニア州)を組み込んだ後にソラレンホスホラミダイト(Glen Research、スターリング、バージニア州)を添加する場合がある。例えばソラレン部分は、参照として本明細書に組み入れられる米国特許出願第09/453,190号に記載されているように分枝状リンカーを介してRNAに架橋される。
【0066】
例示的なアプローチの一つでは、配列が
(Puはピューロマイシン-CPG[Glen Research];CおよびAは標準的な3’-アミダイト[Glen Research];a、t、c、およびgは5’-ホスホラミダイト[Glen Research];Xは非対称性の分枝状アミダイト[Clontech]を意味する)であるリンカーを、標準的な製造業者プロトコル通りに合成した後に、分枝点Xの選択的脱保護(Clontechの指示書による)と、それに続くソラレンC6アミダイト(Glen Research)のカップリングを行った。次にこのリンカーは、標的配列
をもつRNAに、上述の光架橋法で光架橋した。このRNA-リンカーコンストラクトを次に、RNA-タンパク質融合体の合成に問題なく使用することができた。
【0067】
さらに別の方法においては、標的ハイブリッド形成ドメインの3’端に結合させたソラレンを含むリンカーを構築する場合もある(図7C)。この方法では、ソラレンを含むリンカーの5’端が、別のリンカーにより伸長し、鎖の方向が逆転した後に3’ピューロマイシンで終結する。続くRNase HによるRNA標的ドメインの切断は任意選択で行うが、こうすることでリンカーコンストラクトの柔軟性が上昇する。さらに、このアプローチを採用することで、かなり長いRNA分子の内部にリンカーを結合させることが可能となり、続いてリンカー部位の下流にある未翻訳領域を切断した後に、翻訳を行うことでRNA-タンパク質融合体が形成する。
【0068】
この方法の例示的なアプローチの一つでは、5’ Pso atg cga gaa agt gat aaa aaa aaa aaa CC Pu(Puはピューロマイシン-CPG[Glen Research];Cは標準的な3’-アミダイト[Glen Research];a、t、c、およびgは5’-ホスホラミダイト[Glen Research];およびPsoはソラレンC6ホスホラミダイト[Glen Research]を意味する)の配列をもつリンカーを、標準的な製造業者プロトコル通りに構築した。次にリンカーを、
の標的配列をもつRNAに、上述の光架橋法で光架橋した。このRNA-リンカーコンストラクト(事前にRNase Hによる処理を任意で行う)は後に、RNA-タンパク質融合体の合成に問題なく使用することができた。
【0069】
ペプチドアクセプターをRNAに結合させる光化学的手法の利点の一つは、この方法が、連結前にRNAを化学的に修飾する必要がない点である。この利点があるため、方法が極めて確実かつ選択的となり、粗T7転写反応に由来するRNAを、化学的連結の基質として使用することができる。
【0070】
実施例6:光切断可能なビオチンを用いたRNAの精製および連結
必要に応じて、親和性を利用したRNA精製段階を、上述の光化学的連結法と組み合わせる(図7H)。適切なリンカー分子を、ピューロマイシン末端で光切断可能なビオチン成分(例えば、EZ-Link(商標) NHS-PC-LC-Biotin、Pierce、ロックフォード、イリノイ州)で修飾する。次に標的RNA(例えば粗転写反応で得られる)と一定量のリンカーとの間でハイブリッドを形成させ、得られた二本鎖を、ストレプトアビジン(または関連する)樹脂などの固相支持体上に補足する。次に、十分な洗浄を行って過剰なRNAならびに転写反応成分を除去する。長波長の紫外光を同時に照射することで光架橋が形成され、産物が樹脂から遊離する。連結したRNAは次に、さらに精製を行うことなく翻訳および融合形成反応に直接使用することができる。
【0071】
この方法には、従来のRNA精製スキームにはない利点がある。例えば転写後のRNA量は、使用するリンカーの量を上回ることを推定するだけである。記載の手順をふむことで、その量は使用リンカー量を超えない程度に自動的に減少する。このため、さらにRNA(または連結したRNA)を、例えばA260のUV値で定量することなく次の工程に進めることが可能となり、RNA量を別の方法で調整する必要はない。したがって核酸-タンパク質の融合体分子の調製法は自動化に適している。
【0072】
例示的な方法の一つでは、このようなビオチンを用いたRNA精製・連結プロトコルは以下の手順で実施される。光切断可能なビオチンを用いたRNA精製および連結法では、最初にリンカーをビオチン化する。このリンカー(C6-ソラレン-2-OMe[U AGC GGA UGC]dA18 TEG2 dCdC-ピューロマイシン)は、100 μlの100 μMリンカー(総量10 nmol)、50 μlの1 μmole EZ-Link(商標) PC-LC-Biotin(溶媒DMSO)、20 μlの10×PBS、pH 7.4、および30 μlのH2Oを混合することでビオチン化する。この混合物を室温で2時間インキュベートすると、定量可能な収量が得られる。次にこの混合物に2回エタノールを添加して沈澱を生じさせると、PC-ビオチン化リンカーの予想回収率は90%を超える。これを200 μlのH2Oに再懸濁する。
【0073】
次にRNAを、例えばT7 Megashortscriptキット(Ambion)を用いて転写する。転写には、250 μlの反応につき、10 pmoleのDNA鋳型(3’末端に配列
を含む)を使用する。この転写反応で約2〜5 nmolの粗RNA産物が得られる。次の工程に進む前に精製(フェノール抽出、NAP-5カラム、またはRNeasyカラム)は必要ない。
【0074】
この精製・連結反応に続く段階では、ビオチン化したリンカー(10 μlの50 pmol/μl ビオチン化リンカー;総量500 pmol)を、62.5〜250 μlの転写混合物(推定最小量として500 pmoleのRNAを含む)と、例えばPCR装置(80℃で30秒間加熱した後に0.3℃/秒で4℃に冷却する)で、15 μlの5 M NaCl(300 μlの反応について最終濃度を0.25 Mとする)およびH2Oを、最終反応容量が300 μlになるように添加してアニーリングを行う。次に反応混合物のRNAを、100 μlのNeutravidinビーズ(Pierce、ロックフォード、イリノイ州)に、反応混合物を4℃で30分間かけて緩やかに振盪しながら固定化する。次にこのビーズを洗浄して300 μlのH2O中に再懸濁する。
【0075】
このビーズを次に遠心して沈降させ、100 μlの緩衝液(25 mM Tris pH 7.0および0.25 M NaCl)で3回洗浄する。次にビーズに紫外線を15分間室温で(手持ち式UVランプUVGL-25を用いてビーズを含む微量遠心用チューブをランプ上に直接置いて)照射し、リンカーをRNAに化学的に連結し、かつ連結した分子をビーズから光化学的に遊離させる。250 pmolの連結RNAが、光化学的に遊離すると予測される。75 μlのH2Oを同チューブに添加し、チューブをボルテックスミキサーで30秒間撹拌して、ビーズを遠心して沈降させる。連結したRNA/リンカーを含む75 μlの上清を翻訳および核酸-タンパク質融合体分子の形成に使用する。
【0076】
核酸-タンパク質融合体分子は、前工程で得られた75 μlの上清を225 μlの緩衝液成分およびウサギ網状赤血球溶解物キット(Ambion)と混合して、その混合物を30分間30℃でインキュベートすることで形成される。次にKClおよびMgCl2を最終濃度がそれぞれ500 mMおよび50 mMになるように添加し、室温で60分間インキュベートして反応を続けて、核酸-タンパク質融合体分子を作製する。
【0077】
実施例7:強固な非共有結合を介したペプチドアクセプターとRNAとの結合
ペプチドアクセプターをRNA分子に結合させる共有結合形成法の別法として、強固な複合体形成にのみ依存する方法も可能であり、これもまた本発明の一部である。一つの方法では、図8Aに示すように、ペプチド核酸(PNA)を使用してRNAの認識および結合を行う。PNAはDNAの模倣体であり、アキラルおよび無電荷のN-(2-アミノエチル)グリシンのユニットから構成される主鎖をもつ(KnudsenおよびNielsen、Nucleic Acids Res. 24:494、1996)。PNAは、配列特異的かつ高い親和力で相補的な一本鎖DNAおよびRNAとハイブリッドを形成することが知られている。特に、RNAに結合してクランプを形成する2個のPNA分子を含む三本鎖形成コンストラクトは、RNAに強固に結合する効率のよい手法を提供することができる。というのは、このようなコンストラクトは、熱変性および、インビトロ翻訳の条件に対して極めて耐性が強いためである(図8)(Hanveyら、Science、258:1481、1992)。
【0078】
偽イソシトシン(Pseudoisocytosine)塩基を使用することで、中性pHおよび塩基性pHにおける安定性がさらに強化される(Egholmら、Nucleic Acids Res. 23:217、1995)。このようなPNA-クランプは、インビトロ翻訳条件下でmRNAと結合したまま維持され、リボソームで置換されないことが報告されている(KnudsenおよびNielsen、Nucleic Acids Res. 24:494、1996)。この特性のため、対応するRNA-タンパク質融合体コンストラクトの安定性は極めて大きくなる。
【0079】
核酸リンカー-PNAコンストラクトは、図8Bおよびウルマン(Uhlmann)らの論文(Angew. Chem. Int. Ed. Engl. 35:2633、1996)に記載されているように、ピューロマイシン-CPGで始まる固相支持体を用いた合成法で調製することができる。標準的な自動合成法による所望の核酸部分(または必要に応じてPEG-スペーサー)のアセンブリー後に、PNAドメインを適切な試薬(例えばウルマンらの論文(上記)に記載)に結合させて固相合成を続ける。またはPNAを個別の成分として事前に合成しておき(PE Biosystems、フォスターシティー、カリフォルニア州)、その後に所望のリンカー部分と化学的に結合させることができる。
【0080】
一つの特定の例においては、ピューロマイシン-DNAリンカーの5’末端をアミノ基で修飾して、化学的に活性のあるエステル(例えば、ジスクシンイミジルグルタレートまたは関連試薬との反応を介したNHS-エステル;この手法は例えばコックス(Cox)ら(J. Immunol. 145:1719、1990);およびhttp://www.piercenet.com/Products/linksearch.cfm;Pierce、ロックフォード、イリノイ州、に記載されている)にさらに変換することができる。その後の(非保護状態のアミノ末端またはカルボキシ末端のリジンのいずれかを有する)PNA成分との反応により、ドメインが共有結合で連結される。この工程は、最終的なDNA-リンカー産物を含む均質溶液中で、または保護基を有し、かつ固相樹脂に結合させた状態のままのDNAを用いることで行なわれる。
【0081】
実施例8:リンカーの長さと組成の最適化
上述のすべての方法では、リンカーコンストラクトを最適化することが好ましい。考慮対象となる諸因子には例えば、鋳型/標的認識エレメントを含めること、および結合させた官能基の空間的な接近のしやすさがある。特に、核酸のハイブリッド形成を介した鋳型または標的の認識が関与する場合は、好ましくはリンカーの標的配列および化学的性質を含む諸因子を最適化する。例えばRNAのハイブリッド形成強度およびその結果としての連結効率は、DNAではなく2-OMe RNAまたはプロピンで修飾した核酸塩基を使用することで上昇することが知られている(例えばInoue ら、Nucleic Acids Res. 15:6131、1987;Kibler-Herzogら、Nucleic Acids Res. 19:2979、1991;およびWagnerら、Science 260:1510、1993に記載)。
【0082】
リンカーは、RNA-タンパク質融合体反応における効率に対しても最適化することができる。これには一般に、種々の長さのリンカーが関与するが、RNA-タンパク質の結合に用いる種々の構築用ブロックを使用することもある。一つの特定の例においては、リンカーのデオキシヌクレオチドは、PEGスペーサーまたは2-OMe-RNAユニットと置換することができる(いずれもGlen Research、スターリング、バージニア州)。
【0083】
その他の態様は特許請求の範囲に含まれる。
【0084】
本明細書で説明したすべての特許および特許出願は、参照として組み入れられる。
【背景技術】
【0001】
発明の背景
一般に本発明は連結法、特にペプチドアクセプターを核酸に連結させる方法に関する。
【0002】
RNA-タンパク質融合体を調製する方法は現在いくつか存在する。RNA-タンパク質融合体は、ペプチドアクセプターをRNA分子の3’端に結合させた後に、そのRNAをインビトロまたはインサイチューで翻訳することで作製される。生じる産物は、ペプチドであり、このペプチドはこれをコードするRNAの3’端で結合している。このようなRNA-タンパク質融合体を作ることで、部分的または全体的にランダムなアミノ酸配列の大規模プールから所望の特徴をもつタンパク質を容易に単離することが可能となり、またRNAのコード配列をそれに対応するタンパク質分子に共有結合を介して結合させることで、タンパク質配列の情報を回収して増幅する際の問題が解決される。
【発明の概要】
【0003】
本発明は、ペプチドアクセプターをRNA分子に結合させる方法ならびにRNA-ペプチドアクセプター産物を特徴とする。この方法により例えば、部分的または全体的にランダムなアミノ酸配列または核酸配列の大規模プールから所望の特徴を有するタンパク質や核酸を単離する際に使用可能なRNA-タンパク質融合体の産生を容易にする。本発明の方法は、ペプチドアクセプターを核酸分子に結合させる種々のストラテジーで実施することができる。このような多様なアプローチは、ペプチドと核酸の結合のタイプ、および結合に使用する試薬によって異なる。
【0004】
したがって本発明の第一の局面は、ペプチドアクセプターとRNA分子との結合法を特徴とする。この結合法は、ヘアピン構造を形成する3’配列を有するRNA分子を提供する段階、ペプチドアクセプターを核酸リンカー分子に共有結合させる段階、およびペプチドアクセプターとRNA分子間に共有結合が形成可能な条件下でRNA分子と核酸リンカー分子のハイブリッドを形成させる段階を含む。
【0005】
本発明の第二の局面は、ペプチドアクセプターをRNA分子に結合させる方法を特徴とする。この方法は、ヘアピンを形成する5’配列をもつリンカーを有するペプチドアクセプターを提供する段階、ペプチドアクセプターとRNA分子のハイブリッドを形成させる段階、およびペプチドアクセプターとRNAを共有結合させる段階を含む。本発明の上記の局面の一つの態様においては、T4 DNAリガーゼを用いてペプチドアクセプターをRNA分子に結合させる。
【0006】
本発明の第三の局面は、デオキシヌクレオチド三リン酸またはジデオキシヌクレオチド三リン酸から始まるリンカー分子に共有結合させたRNA分子およびペプチドアクセプターを提供する段階、ならびにRNA分子およびペプチドアクセプターを末端デオキシヌクレオチジルトランスフェラーゼにより接触させて、ペプチドアクセプターをRNA分子に共有結合させる段階により、ペプチドアクセプターをRNA分子に結合させる方法を特徴とする。
【0007】
本発明の第四の局面は、RNA分子をペプチドアクセプターに化学的に連結することでペプチドアクセプターをRNA分子に結合させる方法を特徴とする。
【0008】
この局面の一つの態様においては、ペプチドアクセプターをソラレン部分に結合し、ソラレン部分を介してRNA分子に架橋する。ソラレン部分は、ペプチドアクセプターに結合させたリンカー分子の5’端または3’端に結合させることができる。またはソラレン部分は、リンカー分子の内部に位置させることができる。この方法では、ペプチドアクセプターを、UV照射によってRNA分子に架橋する。この特定の局面の別の態様においては、ソラレンをC6アルキル鎖を介してペプチドアクセプターに結合させ、および/またはRNA分子は、その3’端の近傍に停止コドンを有する。好ましくは、このようなリンカーの長さは25〜40ヌクレオチド単位である。さらに、ペプチドアクセプターとRNA分子を架橋させる前に、RNAと、光切断可能(photocleavable)な部分をさらに含むリンカーとのハイブリッドを形成させることができる。ハイブリッドを形成したRNAは次に光切断可能な部分を介して固相支持体に固定化することができる。好ましくは、光切断可能な部分はビオチンである。
【0009】
本発明の第四の局面の別の態様においては、RNA分子に官能基を導入し、ペプチドアクセプターとRNA分子間の化学結合形成が可能となるようにあらかじめ適切に修飾しておいたペプチドに結合させる。好ましくは、このようなRNA分子はIO4-酸化により官能基を導入する。ペプチドアクセプターには、アミン、ヒドラジン、(チオ)ヒドラジド、および(チオ)セミカルバゾンからなる群より選択されるペプチドアクセプターに分子を結合させることで官能基を導入することができる。
【0010】
本発明の第四の局面のさらに別の態様においては、外部鋳型を添加することなく化学的連結を行う。または、この化学的連結反応は、外部鋳型を添加することで進行させることができる。後者の方法は、鋳型の5’端がペプチドアクセプターのリンカー部分とハイブリッドを形成し、鋳型の3’端がRNA分子とハイブリッドを形成するように、RNA分子とペプチドアクセプターのリンカー部分を鋳型を用いて整列させる段階を含む。RNA分子のペプチドアクセプターへの化学的連結はまた、ペプチドアクセプターに共有結合させたリンカー分子そのものとRNA分子のハイブリッドを形成させることで、外部鋳型の非存在下で行うこともできる。ハイブリッドを形成させることで、ペプチドアクセプターとRNA分子は、連結可能な程度に近接する。好ましくは、官能基は、ペプチドアクセプターのリンカー領域の5’端にあるか、または片側がハイブリッド形成ドメインに隣接し、もう片側がペプチドアクセプターに隣接する。
【0011】
本発明の第四の局面のさらなる態様においては、ペプチドアクセプターとRNA分子間の化学的連結は、RNAの還元的アミノ化を介した官能基とRNA分子の結合段階と、それに続く、RNA分子と反応するペプチドアクセプターの修飾が含まれる。両分子は次に共有結合を形成して結合する。好ましくは、RNA分子に結合させる官能基は、チオール、マレイミド、またはアミンである。
【0012】
本発明の第五の局面は、非共有結合を介してペプチドアクセプターをRNA分子に結合させる方法を特徴とする。一つの態様においては、このような結合は、ペプチド核酸(PNA)とペプチドアクセプター間の共有結合の生成、およびPNAを介したペプチドアクセプターとRNA分子との非共有結合の生成によって生じる。この態様においては、RNA分子は停止コドンを含むことができる。
【0013】
本発明のさらに別の局面は、ペプチドアクセプターならびにこれらのRNA分子の転写および翻訳(および必要に応じて逆転写および/または増幅)により生じた核酸-タンパク質融合体に、化学的または非共有結合的に連結させたRNA分子を特徴とする。一つの態様においては、ペプチドアクセプターは、RNA分子の3’端に連結される。
【0014】
本発明のさらに別の局面は、本発明のRNA-ペプチドアクセプター分子を用いて、所望のタンパク質または核酸を選択する方法を特徴とする。この選択法では、RNA-タンパク質融合体を形成する本発明の分子を用いて、対象タンパク質または核酸を選択する。このような選択方法は例えば、参照として本明細書に組み入れられる、ショステク(Szostak)らによる国際公開公報第98/31700号、およびショステクらによる米国特許出願第09/247,190号に記載された任意のアプローチで実施することができる。
【0015】
本発明の最後の局面は、RNA-タンパク質融合体を生成する方法を特徴とする。この方法は、光切断可能な部分、ソラレン部分、およびペプチドアクセプターを含むリンカーとハイブリッドを形成するRNA分子を提供する段階;固定されていない状態のRNAを支持体から実質的に除去する条件下でRNAを固相支持体に固定化させる段階;ソラレン部分を介してペプチドアクセプターを該RNAに架橋させ、この架橋形成が、架橋RNAを固相支持体から同時に遊離させる段階;および、架橋RNAを翻訳してRNA融合タンパク質を形成させる段階を含む。一つの態様においては、光切断可能な部分はビオチンである。
【0016】
上述した本発明のすべての局面におけるRNA分子は、候補タンパク質をコードする配列に使用可能に連結させた翻訳開始配列および開始コドンを含むことができる。さらに、好ましいペプチドアクセプターの一つは、伸長するペプチド鎖のC末端に付加して翻訳を停止させるヌクレオシド類似体であるピューロマイシンである。一つの態様においては、ペプチドアクセプターは、例えばヌクレオチドリンカーなどのリンカーに結合させたピューロマイシンを含む。このようなリンカーを使用することで、ペプチドアクセプターと結合対象RNA分子が整列しやすくなる。別の態様においては、ペプチドアクセプターのリンカー領域は例えばPEGなどの非ヌクレオチド成分を含む。アクセプターの他の選択肢には、RNAの3’端におけるtRNA様の構造、ならびにピューロマイシンと同様に作用する他の化合物が含まれる。このような化合物は、アデニンまたはアミノ酸ヌクレオチド、フェニルアラニル-アデノシン(A-Phe)、チロシルアデノシン(A-Tyr)、およびアラニルアデノシン(A-Ala)などのアデニン様化合物、ならびにフェニルアラニル3’デオキシ3’アミノアデノシン、アラニル3’デオキシ3’アミノアデノシン、およびチロシル3’デオキシ3’アミノアデノシンなどのアミド結合した構造に連結させたアミノ酸を有する任意の化合物を含むが、これらに限定されない。上記化合物についてはいずれも、天然のL-アミノ酸またはその類似体を任意に使用することができる。さらに、混合型のtRNA様3’構造-ピューロマイシン複合体を本発明に使用することができる。
【0017】
本発明の好ましい一つの設計において、DNA配列はメッセージの末端とペプチドアクセプターの間に含まれる。この配列は、読み枠の末端でリボソームを一時停止させて、ペプチドアクセプター(例えばピューロマイシン)が、新生ペプチド鎖を受け入れて、ペプチジル-tRNA結合が加水分解されるまでの時間を稼ぐように設計される。インビトロ翻訳の間には、リボソームを特にソラレン架橋部位またはPNAクランプなどの化学結合部位において一時停止させることも可能である。
【0018】
本発明の別の好ましい設計においては、大部分が非ヌクレオチドのリンカー部分が、ペプチドアクセプターに結合させたヌクレオチドリンカーに代わって使用される。このように設計することで、ペプチドアクセプターとRNA分子が連結しやすくなる。このリンカーは例えば、トリエチレングリコールのスペーサーを含むことができる。リンカーはまた、2’-OMe-RNAホスホラミダイトを含むことができる。化学的または酵素的な連結にハイブリッド形成が必要な場合は、連結部位に隣接するリンカーの十分な部分が、核酸を含む必要がある。
【0019】
さらに本発明のRNAまたはリンカーは、例えばRNA、またはRNAもしくはリンカーから形成されるRNA-タンパク質融合体分子のアフィニティー精製法などに用いる配列(例えばポリ(A)配列)を含んでもよい。
【0020】
さらに、上述した本発明のすべての局面においては、ペプチドアクセプターに結合させたRNA分子をインビトロまたはインサイチューで翻訳してRNA-タンパク質融合分子を作ることができる。このRNA-タンパク質融合分子を次に、高塩濃度下でインキュベートするか、および/またはショステク(Szostak)ら(09/247,190)の方法に基づいて低温でインキュベートする(例えば-20℃で一晩)。RNA-タンパク質融合分子はまた、例えば標準的なポリ(A)精製法で精製することもできる。
【0021】
本発明で用いる「タンパク質」とは、2種またはそれ以上の天然のアミノ酸、または一つまたは複数のペプチド結合で連結された修飾アミノ酸を意味する。「タンパク質」と「ペプチド」とは同じ意味で用いられる。
【0022】
「RNA」とは、共有結合で結合された天然または修飾型リボヌクレオチドの2種またはそれ以上の配列を意味する。この用語に含まれる修飾型RNAの一例が、ホスホロチオエート型RNAである。
【0023】
「翻訳開始配列」とは、機能性のリボソームエントリー部位を提供可能な配列を意味する。細菌系では、この領域はシャイン・ダルガルノ配列と呼ばれることがある。
【0024】
「開始コドン」とは、タンパク質コード配列の開始シグナルとなる3塩基を意味する。「停止コドン」とは、タンパク質コード配列の停止シグナルとなる3塩基を意味する。一般に開始コドンはAUG(すなわちATG)であり、停止コドンはUAA(すなわちTAA)、UAG(すなわちTAG)、またはUGA(すなわちTGA)であるが、開始コドンまたは停止コドンとして使用可能な他の3塩基配列で代用される場合がある。
【0025】
「共有結合で結合される」という用語は、共有結合を介して直接に、または別の共有結合で結合された配列(例えば一時停止部位に対応するDNA)を介して間接的に連結されることを意味する。
【0026】
「非共有結合で結合される」という用語は、共有結合以外の方法で連結されることを意味する
【0027】
「ヘアピン構造」とは、一本鎖の核酸によって形成される二本鎖領域を意味する。好ましくは、そのようなヘアピン構造の長さは少なくとも8塩基対であり、さらに好ましくは8〜15塩基対である。
【0028】
「化学的に連結する」という用語は、酵素を使用せずに2分子を連結することを意味する。化学的に連結することで、非共有結合ならびに共有結合が生じる。
【0029】
「ペプチドアクセプター」とは、リボソームのペプチジルトランスフェラーゼ機能の触媒活性により、伸長するタンパク質鎖のC末端へ付加されうる任意の分子を意味する。通常、このような分子は、(i)例えばアデノシンまたはアデノシン類似体(N6位のアミノ基におけるジメチル化が許容される)などのヌクレオチドまたはヌクレオチド様の成分、(ii)20種類のD型またはL型のアミノ酸や、エールマン(Ellman)らによる文献(Meth. Enzymol. 202:301、1991)に記載されているO-メチルチロシンを始めとする任意の類似体を含む、アミノ酸またはアミノ酸様成分、および(iii)(3’位、またはそれほど好ましくないものの2’位における2種の成分(例えばエステル、アミド、またはケトン結合など)の結合を含む。好ましくは、これらの結合は、天然のリボヌクレオチドコンホメーションに基づく環の歪みを大きく変えないことである。ペプチドアクセプターはまた求核基を有する場合がある。求核基には、アミノ基、ヒドロキシル基、またはスルフヒドリル基などが含まれるが、これらに限定されない。さらにペプチドアクセプターは、ヌクレオチド模倣体、アミノ酸模倣体、または結合型ヌクレオチド-アミノ酸構造模倣体から構成されることもある。
【0030】
「リンカー」または「リンカー分子」とは、デオキシリボヌクレオチド、リボヌクレオチド、またはそれらの類似体を含む配列を意味する。
【0031】
「官能基を導入する」という用語は、官能基(または官能性を有する部分)が結合するように修飾することを意味する。例えばRNA分子には、IO4-酸化またはアミノ化によって官能基を導入することができ、ペプチドアクセプターには、アミン、ヒドラジン、(チオ)ヒドラジド、または(チオ)セミカルバゾン基を結合することで官能基を導入することができる。
【0032】
「外部鋳型」とは、連結反応混合物に添加されるものの連結反応の最終産物の一部とはならない核酸配列を意味する。
【0033】
「高塩濃度(high salt)」とは、一価の陽イオン濃度が少なくとも200 mM、好ましくは少なくとも500 mM、またはさらには1 Mであること、および/または二価または多価の陽イオン濃度が少なくとも25 mM、好ましくは少なくとも50 mM、最も好ましくは少なくとも100 mMであることを意味する。
【0034】
「アフィニティー精製用の配列」とは、核酸または核酸-タンパク質融合体分子の精製に使用されるヌクレオチド配列を意味する。アフィニティー精製用の配列は例えば、オリゴdTセルロース上における核酸または融合体分子の精製に使用可能なA8-20などのポリ(A)配列である。アフィニティー精製用の配列はまた、核酸-タンパク質融合体分子の精製に使用されるポリペプチド配列の場合もある。他の例示的な精製法は、ショステク(Szostak)らによる米国特許出願第09/247,190号に記載されている。
【0035】
本発明には多数の利点がある。本明細書に記載された方法は例えば、ペプチドアクセプターとRNA分子間の効率のよい連結を、RNAとペプチドアクセプターを結合させるための外部鋳型を必要とすることなく、いくつかの局面において容易にする。本発明ではまた、RNA-タンパク質融合体の生成にかかるコストも低い。
【0036】
本発明の他の特徴および利点は、以下の詳細な説明および特許請求の範囲から明らかになると考えられる。
【図面の簡単な説明】
【0037】
図面について簡単に説明する。
【図1】ヘアピン鋳型上における、T4 DNAリガーゼを用いたペプチドアクセプターリンカーとRNAとの連結の例示的な段階を示す略図である。RNAの3’側の配列またはペプチドアクセプターリンカーのいずれかを、ヘアピン構造を形成するように設計し、ペプチドアクセプターリンカーとRNAのハイブリッドを形成させることで、RNAとペプチドアクセプターが近接する。
【図2】末端デオキシヌクレオチジルトランスフェラーゼを用いたペプチドアクセプターリンカーとRNAとの連結の例示的な段階を示す略図である。
【図3A】ヒドラジド形成およびセミカルバゾン形成を介したRNAの3’端における修飾の例示的な段階を示す略図である。
【図3B】還元的アミノ化によるRNAの3’端における修飾の例示的な段階を示す略図である。
【図4】官能基を導入したピューロマイシンリンカーを、官能基を導入したRNAに化学的に連結させる例示的なストラテジーを示す一連の図である。一つのストラテジーは鋳型に依存しない(図4A)。他のストラテジーでは、外部オリゴ鋳型を使用して、RNAとピューロマイシンリンカーを整列させる。外部オリゴ鋳型が関与するストラテジーでは、オリゴ鋳型は、RNAとピューロマイシンリンカーの両方とハイブリッドを形成するか(図4B)、またはピューロマイシンリンカーと結合してRNAとハイブリッドを形成する(図4D)。さらに官能基は、ピューロマイシンリンカーの5’端(図4Bおよび4C)または内部(図4D)に導入することができる。
【図5】修飾型ピューロマイシンリンカーに使用する十分に保護されたカルボヒドラジドホスホラミダイトの合成の例示的な段階を示す略図である。
【図6】官能基のRNAの3’端への結合と、それに続く化学的結合による、ペプチドアクセプターリンカーとRNAとの連結の例示的な段階を示す略図である。官能基は次に、適切に修飾されたリンカー分子と反応し、チオール基(A)、マレイミド基(B)、またはアミン基(C)を介してRNAとピューロマイシンリンカーを共有結合させることができる。
【図7A−C】光架橋法によるペプチドアクセプターとRNAとの連結の例示的な段階を示す一連の略図である。この一般的な方法では、ピューロマイシンリンカーに結合させたソラレン部分が、UV照射を受けてピューロマイシンリンカーとRNA間を光架橋する。ソラレン部分は、ピューロマイシンリンカーの5’端(図7A)、内部(図7B)、または3’端(図7C)に位置することができる。
【図7D】光架橋形成に使用されるmRNAを示す略図である。
【図7E】mRNAの不変の3’配列(図7DにおけるmRNAの不変領域)と、推定ソラレン挿入部位を示す光リンカー間に形成される二本鎖を示す略図である。C6アルキル鎖を介してリンカーの5’リン酸に結合させたソラレンの構造も示す。変数xおよびyは、リンカーに含まれるdAヌクレオチド数とトリエチレングリコール(TEG)単位数をそれぞれ示す。左側に示したオートラジオグラフは、mRNA 1とリンカーB間の光架橋反応をゲル電気泳動の解析結果を示す。
【図7F】インビトロ翻訳および融合体形成を示すゲル像である。mRNA鋳型2および鋳型3に由来するタンパク質合成をそれぞれレーン1とレーン3に示す。リンカーBへ光架橋したmRNA鋳型の翻訳により、mRNA-タンパク質融合体が生じた(レーン2と4)。鋳型3は、リンカー架橋部位に続いて停止コドンを有する。レーン5は、オリゴdTセルロースで精製した後の融合産物を示す。停止コドンを含むmRNA分子を用いた融合体形成を示す略図をゲル像の上に示す。
【図7G】mRNA-タンパク質融合体分子の収率が、リンカー組成と塩濃度に依存することを示すゲル像である。
【図7H】mRNAと光切断可能なビオチンを用いたハイブリッド形成用リンカーとの連結および、それに続く融合体形成にかかわる段階を示す略図である。
【図8A】(PNA)2-RNAの三本鎖形成を介したピューロマイシンリンカーとRNAとの結合を示す略図である。ピューロマイシンリンカーは、強固な非共有結合を介して三本鎖を形成するRNAに結合する2個のPNA分子と結合する。
【図8B】PNA-リンカー複合体の合成段階を示す略図である。リンカーを固相支持体に結合し、PNAと結合するように修飾する。次にPNAを所望のリンカーと結合させ、PNAリンカー分子を脱保護して固相支持体から分離する。
【発明を実施するための形態】
【0038】
発明の詳細な説明
本明細書では、ペプチドアクセプターをRNA分子に結合させる種々の方法を記載する。RNAは、通常の細胞を用いた合成法、組換え技術、および化学合成を含む標準的なアプローチで生成することができ、RNAには、細胞のRNAライブラリー、mRNAライブラリー、およびランダム合成RNAライブラリーが含まれるがこれらに限定されない。ペプチドアクセプター(例えばピューロマイシン)をDNAリンカーまたはRNAリンカーに通常結合させる。このようなペプチドアクセプター分子は、任意の標準的な方法で生成することができる。これには例えば、ロバーツ(Roberts)およびショステク(Szostak)の論文(Proc. Natl. Acad. Sci. USA 94:12297、1997)、ショステクら(国際公開公報第98/31700号)、およびショステクらによる米国特許出願第09/247,190号に記載される手法がある。本発明の各方法を実施する方法について、特定の実施例を用いて詳細に説明する。特定の例は本発明を説明する目的で提供するものであって、本発明を制約するものと解釈すべきではない。
【実施例】
【0039】
実施例1:ヘアピン鋳型上でT4 DNAリガーゼが関与する酵素的連結法
本発明の特定のアプローチの一つにおいては、T4 DNAリガーゼを使用して、例えば図1に示すヘアピンを含む鋳型を用いてペプチドアクセプターをRNA分子に結合させる。連結反応は、自己鋳型的に進み、副子オリゴ(splint oligo)を使用しない。RNAの3’端の配列、またはペプチドアクセプターのリンカー領域の5’端のいずれかを、ヘアピン構造を形成するように設計する。ペプチドアクセプターのリンカー領域の5’端およびヘアピン構造を形成するRNAの3’端が近接することで、T4 DNAリガーゼによる酵素的連結が促される。これは例えば、サムブロック(Sambrook)、フリッシュ(Fritsch)およびマニアティス(Maniatis)の「分子クローニング(Molecular Cloning)」、Cold Spring Harbor、New York、Cold Spring Harbor Laboratory Press、1989に記載されている。反応物の比はおよそ1:1であるが、いずれかの反応物が若干過剰(1:1.2)であってもよい。最適な連結条件は、反応によって若干変動する場合があるので、当業者に周知の方法で実験的に決定してもよい。
【0040】
実施例2:末端トランスフェラーゼが関与する酵素的連結法
ペプチドアクセプターをRNA分子に結合させる別の酵素的方法では、RNAの3’端を修飾してから、例えば図2に示すように末端デオキシヌクレオチジルトランスフェラーゼ(TdT)を用いて連結を行う。TdTによる反応は、サムブロック(Sambrook)、フリッシュ(Fritsch)およびマニアティス(Maniatis)の書籍(前記)に一般的に記述されている方法で行う。この酵素は、核酸の3’端にデオキシ(またはジデオキシ)ヌクレオチド三リン酸(NTP)を用いて伸長させる。このような(d/dd)NTPは、塩基成分を化学的に修飾することで所望のリンカー構造をもたせることができる(例えばMeyer、「分子生物学における方法(Methods in Molecular Biology)」、 Agrawal、ed.、第26巻、Totowa:Humana Press、1994、73〜91ページ;Kumarら、Anal. Biochem. 169:376、1988;Rileyら、DNA 5:333、1986;およびSchmitzら、Anal. Biochem. 192:222、1991に記載されている)。図2に示すように、リンカー分子は(d/dd)NTPで開始しなければならない。しかし残りのリンカー組成は変化してもよい。この末端トランスフェラーゼ連結法の特徴は、ペプチドアクセプターのDNAリンカー領域が最短で1残基の(d/dd)NTPでもよいことである。
【0041】
実施例3:IO4-酸化とそれに続く化学的連結によるRNAの3’端への官能基導入が関与する化学的連結法
本発明の方法では、図3に示すように、ペプチドアクセプターはIO4-酸化を介したRNAの3’端への官能基導入と、それに続くペプチドアクセプターとRNAの化学的連結によってRNA分子に結合させることもできる。IO4-酸化は下記の方法で、またアグラワル(Agrawal)(「分子生物学における方法(Methods in Molecular Biology)」、 Agrawal編、第26巻、Totowa:Humana Press、1994、93〜120ページ)、プロウドニコフ(Proudnikov)およびミルザベコフ(Mirzabekov)、Nucleic Acids Res. 24:4535、1996;ゴーシュ(Gosh)ら、Anal. Biochem. 178:43、1989;バウマン(Bauman)ら、J. Histochem. Cytochem. 29:227、1981;およびウー(Wu)ら、Nucleic Acids Res. 24:3472、1996、の各論文に記載された方法で実施する。IO4-酸化段階は反応に1,2-ジオールを厳格に必要とするので、末端ヌクレオチドのみを修飾し、RNA分子の内部にある残基は影響を受けない。結果として生じるジアルデヒドはさらに、アミン、ヒドラジン、カルボ(チオ)ヒドラジド、または(チオ)セミカルバジドなど様々な求核基による反応を受けてシッフ塩基様の構造を生じる。これらの反応は例えばアグラワル(前記)、プロウドニコフおよびミルザベコフ(前記)、ゴーシュら(前記)、バウマンら(前記)、およびウーら(前記)の各論文に記載された通りに進む。それぞれカルボ(チオ)ヒドラジドおよび(チオ)セミカルバジドとの反応後にそれぞれ得られる(チオ)ヒドラジドおよび(チオ)セミカルバゾンは極めて安定であり、アミンまたはヒドラジンを含む初期付加物は通常、その後に図3Bおよびアグラワル(前記)、プロウドニコフおよびミルザベコフ(前記)、ゴーシュら(前記)、バウマンら(前記)、およびウーら(前記)の論文に記載されるように、新規合成された結合を加水分解に対して安定なものとする、その後の還元的アミノ化などの還元段階を必要とする。
【0042】
上述のカップリング反応は、図4A-4Dに示す多種多様なストラテジーで行うことができる。例えば連結は図4Aに示すように、外部鋳型に依存せずに行うことができる。このストラテジーには、RNA分子との連結を成功させるために大過剰の修飾型ペプチドアクセプター(例えば100〜1000倍過剰量)が必要である。RNAの連結効率を上げるために図4Bに示すように、外部鋳型オリゴを使用して基質を整列させることができる。好ましくは、配列がRNA配列およびペプチドアクセプターリンカー配列と相補的なこのようなオリゴの長さは、少なくとも約10ヌクレオチドである。このようなオリゴは通常、RNA分子に相補的な少なくとも約10ヌクレオチド、およびペプチドアクセプターリンカーに相補的な約10ヌクレオチドを含む。または反応部位は、図4C〜4Dに示すようにリンカーとRNAドメインが直接ハイブリッドを形成することで近接する場合がある。この場合、ハイブリッドを形成する配列の長さは、少なくとも10〜15ヌクレオチドである。
【0043】
架橋形成によってペプチドアクセプターをRNAに結合させるために多くの多様なコンストラクトを使用することができる。例えば、例示的なコンストラクトのタイプの一つには、図4A〜4Cに示すように5’端を修飾したリンカーに結合させるペプチドアクセプターがある。別のコンストラクトは、片側がハイブリダイゼーションドメインに隣接し、もう片側がピューロマイシンリンカー部分に隣接する内部官能基を含む(図4D)。
【0044】
このような修飾型リンカーの合成では、ヌクレオチドの本体またはスペーサー部分を集合させる市販のホスホラミダイト(Glen Research、 スターリング、バージニア州)を用いた標準的な自動DNA合成を含む。3’端のピューロマイシンは、合成用の固相支持体としてピューロマイシン-CPG(Glen Research、 スターリング、バージニア州)を用いて導入することができる。反応性官能基は、アミノ末端修飾物質(Glen Research、スターリング、バージニア州)やuni-linkアミノ修飾物質(Clontech、パロアルト、カリフォルニア州)など市販の試薬を用いて結合することができる。適切なホスホラミダイトを用いることで他の官能基を組み入れることが可能である。カルボヒドラジドホスホラミダイトを生成する1つの例示的な方法の一つを図5に示す。カルボヒドラジドホスホラミダイトは、ラクトンにヒドラジンおよびメタノールを結合させ、反応物を2時間還流してカルボヒドラジド成分をつくることで生成した。この合成段階の収率は97%であった。結果として生じた産物を次にジメトキシトリチル基およびピリジン/トルエンの塩と室温で2.5時間反応させて、保護基のついたカルボヒドラジド成分を得た。この段階の産物収率は88%であった。この産物をさらに、テトラヒドロフランおよびジイソプロピルアミンの存在下でホスホラミダイト成分と室温で30分間反応させ、十分に保護されたカルボヒドラジドホスホラミダイトの反応産物を得た(72%)。
【0045】
さらに、IO4-で酸化されたRNAに対するペプチドアクセプターの反応性は、複数コピーの反応基を導入することでさらに高まる。
【0046】
例示的な連結反応の一つは以下のように進んだ。flagエピトープおよびstrepタグをコードする転写産物を含む1 nmoleのRNA(配列:
)を、20 μlの500 mM NaOAc(pH 5.2)、10 μlの5 mM NaIO4と混合し、水を加えて最終容量を100 μlとした。この反応混合物を室温で15分間インキュベートした。次に、10 μlの10 mM Na2SO3を添加し、反応混合物を再び15分間室温でインキュベートした。40 μlの1 Mリン酸緩衝液(pH 8.0)、1.5 nmoleのペプチドアクセプターリンカーUni-A1/8(配列:
(XはUni-linkアミノ修飾物質[Clontech]、またPuはピューロマイシン-CPG[Glen Research]を意味する)、および20 μlのNaCNBH3を反応混合物に添加した。この混合物を次に18時間室温でインキュベートして沈澱を生じさせ、6%のTBE-尿素ゲルで精製し、一晩破砕・浸漬してRNA-タンパク質融合分子を得た。この反応で230 pmoleの産物が得られた。
【0047】
実施例4: RNA分子の3’端への官能基の結合と、それに続く化学的連結が関与する化学的連結法
ペプチドアクセプターは図6に示すように、RNAの3’端に官能基を結合させた後に化学的に連結させることで、RNA分子に結合させることもできる。上記のプロセスの変法では、還元的アミノ化および関連反応によってRNAの3’端に官能基を結合させる。新しく導入したこれらの官能基を次に、ペプチドアクセプターの適切に修飾したリンカーとさらに反応させて、RNAとペプチドアクセプター間に共有結合を形成させる。例示的な反応基には、チオール(ジスルフィド形成、またはピリジルジスルフィドなどのチオール親和性(thiolphilic)試薬との反応用;図6の反応A)またはマレイミド(図6の反応B)などがある。
【0048】
RNAの3’端への可能な官能基の導入とそれに続くペプチドアクセプターの結合に用いられる他の反応基がアミン類である。例えば、N-ヒドロキシスクシンイミド-エステル(NHS-エステル)は、ジスクシンイミジルグルタレート(DSG)または関連試薬と反応させることで、ペプチドアクセプターの5’側のアミノ修飾リンカー上で生成する。これはコックス(Cox)ら(J. Immunol. 145:1719、1990)およびhttp://www.piercenet.com/Products/linksearch.cfm(Pierce、ロックフォード、イリノイ州;図6の反応C)に記載されている。次にこの修飾型リンカーは、修飾型RNAのアミノ官能基と反応させることができる。
【0049】
このタイプの連結反応は、上述の外来鋳型に依存しない方法、または依存した方法のいずれかにより、同様の一般的なアプローチで行うことができる。
【0050】
実施例5:光化学的な方法を含む化学的連結法
ペプチドアクセプターは、図7A〜7Hに示すような光化学的な方法でRNA分子に結合させることもできる。ソラレン基を有するペプチドアクセプターリンカー分子は、長波長のUV光照射を受けて相補的RNA鎖に架橋を導入することができる。この手法は通常、ピーレス(Pieles)およびイングリッシュ(Englisch)の論文(Nucleic Acids Res. 17:285、1989);およびゴダール(Godard)ら(Nucleic Acids Res. 22:4789、1994)の記述にしたがって実施される。ペプチドアクセプターのリンカー領域の5’端へのソラレン部分の結合は、市販のソラレンアミダイト、2’-OMe-RNA ホスホラミダイト、ソラレンC6ホスホラミダイト、およびトリエチレングリコール(TEG)ホスホラミダイト(Glen Research、スターリング、バージニア州)を用いて、標準的なDNA合成装置を用いて達成できる。
【0051】
例示的なアプローチの一つにおいては、この方法は以下のように実施された。flagエピトープ、strepタグ、および光化学的な標的部位(配列:
)をコードするRNA転写物からなる1 nmoleのRNA、標準的な製造業者プロトコル通りに合成された1.2 nmoleのPhotolinker 30/10(配列:
)(PsoはソラレンC2アミダイト[Glen Research]、Puはピューロマイシン-CPG [Glen Research]を意味する)、または光リンカー30/15(配列:
)(PsoはソラレンC2アミダイト、Puはピューロマイシン-CPGを意味する)、10×緩衝液(250 mM Tris pH 7.0;1 M NaCl)、および水(最終容量を360 μlにする)を混合して80℃で2分間加熱した。次に反応混合物を緩やかに室温まで冷却した。次にこの反応混合物に、石英浸漬用ウェル(ACE Glass、カタログ番号7854-25)に入れたPyrex製吸収用スリーブ(ACE Glass、カタログ番号7835-44)を備えた450 Wの浸漬型ランプ(中圧;ACE Glass、カタログ番号7825-34番)を用いて、0℃で15分間、310 nm以上の波長で照射した。この際、試料は浸漬用ウェルに固定化した微小遠心用チューブに入れて氷中で冷却した。次に試料に40 μlの3 M NaOAcと1000 μlのエタノールを添加して沈澱を生じさせ、75 μlの水に再懸濁した。次いで75 μlの2×ローディング緩衝液(Novex)を試料に添加し、プレキャストタイプの6% TBE尿素ゲル(Novex)で試料を精製した。産物を破砕・浸漬法(0.3 M NaOAc、室温で一晩)で回収した後にエタノールで沈澱させた。この光架橋法により、Photolinker 10/30を用いて272 pmoleのRNA-タンパク質融合産物を得た。および、Photolinker 15/30を用いて227 pmoleのRNA-タンパク質融合産物を得た。
【0052】
上記の光架橋法による化学的連結法では、種々の反応パラメータの評価を行った。最初に光架橋形成の塩依存性を検討した。100〜1000 mMのNaClを含む緩衝液を用いた一連の架橋実験を行った。種々の反応間で連結効率に差は認められなかった。さらにRNA標的配列を、
(下線部の配列がソラレンリンカーの標的となる)に変えたところ、産物の収率は大きく減少した(15〜20%の減少)。このことから、RNA標的配列が重要であることがわかった。次に産物の収率が、反応過程にあらかじめ不活性化しておいたソラレンリンカーを繰り返し置換することで上昇しうることを確認した。この実験は以下の手順で実施した。RNA、リンカー、および10×緩衝液を混合して80℃で2分間加熱した。次に反応混合物を緩やかに室温に冷却し、上述の通りに紫外線を照射した。次にさらに1 nmoleのリンカーと1 μlの緩衝液を添加した。リンカーをRNAとアニーリングさせて紫外線を照射した。このプロセスを、2 nmoleのリンカーと2 μlの緩衝液を添加して紫外線照射を繰り返し行った。この手順を採用することで、産物の収率を、ある特定の配列について20%から40%を超えて増大させることができた。
【0053】
光化学的架橋法で生成させる連結産物の能力についても評価を行った。種々の長さのリンカー(ソラレン+15塩基対の標的ハイブリッド形成用ドメイン+dAnCCPu[n=7、12、17、または22])を対象とした実験では以下の結果が得られた。長いリンカーは、高塩濃度条件(500 mM KCl+50 mM MgCl2)でRNA-タンパク質融合体の収率が最も高かった。低塩濃度の緩衝液(250 mM KCl+10 mM MgCl2、または250 mM KCl)中では、短いリンカーの方が長いリンカーよりも収率が高かったが、全体的な収率は概して低値であった。一般に収率は、酵素的に連結させたRNA鋳型と同等であると考えられた。
【0054】
別の例示的な方法においては、種々のmRNAおよびピューロマイシンリンカーを、ペプチドアクセプターがリンカーの3’端に位置するように合成した。リンカーを標的mRNAとアニーリングさせ、mRNA-タンパク質融合体分子の形成効率をインビトロ翻訳法で評価した。リンカーの長さと組成が融合体分子の収率に及ぼす作用についても決定した。
【0055】
図7Dは、架橋反応に使用したmRNAの設計を示す。各RNA分子は、良好な開始コドン組成を有するタバコモザイクウイルス(TMV)の5’-UTR(Gallieら、Nucleic Acids Res.、16:883〜93、1988;およびKozak、Microbiol. Rev.、47:1〜45、1983)、および翻訳開始コドン(AUG)を含む。すべてのmRNAはまた、3’末端に10ヌクレオチドのリンカーハイブリッド形成配列
(アミノ酸配列ASAをコードする)、およびソラレン光架橋用のジヌクレオチド配列5’-UA(不変)を末端に有する。これらは、シンデン(Sinden)およびヘイガーマン(Hagerman)(Biochemistry、23:6299〜6303、198)、およびガンパー(Gamper)ら(Photochem. Photobiol.、40:29〜34、1984)の論文に記載されている。さらに一種のmRNA(mRNA 1)は、FlagエピトープDYKDDDDK(Hoppら、Biotechnology、6:1205〜1210、1988)と、それに続くStrep-Tag II配列WSHPQFEK(Schmidtら、J. Mol. Biol.、255:753〜66、1996)を含む。他のmRNA分子(mRNA 2、3、および4)は、4-4-20 scFv、抗フルオレセイン一本鎖抗体鋳型を含む。これらは、ベドジク(Bedzyk)ら(J. Biol. Chem.、265:18615〜20、1990)、およびマレンデール(Mallender)らの論文(J. Biol. Chem.、271:5338〜46、1996)に記載されている。さらにmRNA 3および4は、残存する非架橋鋳型の翻訳後にリボソームからのmRNAの遊離を誘導する下流の停止コドン(UAA)を含む(下記参照)。オリゴdTによる精製用のポリAテールもmRNA 4に結合させた。
【0056】
以上の検討に使用したmRNA(図7D)は、PCR DNA鋳型を対象に、メリガン(Milligan)らの論文(Nucleic Acids Res.、15:8783〜8798、1987)に記載されている、T7 RNAポリメラーゼのラン・オフ転写法(Megashortscript Transcription Kit、Ambion、テキサス州)で調製した。転写後に全RNAを6%のTBE-尿素ポリアクリルアミドゲル(Novex、カリフォルニア州)を用いた電気泳動で精製した。産物のバンドをUVシャドウイング法で可視化し、ゲルから切り出して破砕し、0.3 M NaOAcに一晩浸漬した。エタノールで沈澱させた後に、RNAを水に再懸濁して保存した。同じ手法で、放射標識したRNAを[α-32P]UTP(Amersham、イリノイ州)を転写用緩衝液に含めることで合成した。
【0057】
以上の検討に使用したピューロマイシンリンカー(図7E)は、Expedite Synthesizer モデル8909(PerSeptive Biosystems、マサチューセッツ州)を用いて、従来の固相支持体ホスホラミダイト化学法で調製した。ピューロマイシン-CPG、DNA ホスホラミダイト、2’-OMe-RNA ホスホラミダイト、ソラレンC6ホスホラミダイト、およびトリエチレングリコール(TEG)ホスホラミダイト(スペーサー9)を、推奨プロトコルに基づいて使用した(Glen Research)。ソラレン部分は、C6アルキル鎖を介してリンカーの5’-リン酸に結合させた。柔軟性のあるリン酸トリエチレングリコール(TEG)のスペーサー、および種々の長さのポリヌクレオチド配列を用いて5’-dCdC-ピューロマイシンを3’端に連結させた(図7Eの表を参照)。リンカーのハイブリッド形成配列は、幹状構造の対形成時の安定性を強化するために2’-OMe-RNA ホスホラミダイトから調製した(Inoueら、Nucleic Acids Res.、15:6131〜6148、1987;およびMajlessiら、Nucleic Acids Res.、26:2224〜2229、1998)。濃水酸化アンモニウム中で55℃で8時間かけて保護基を外した後に、逆相HPLC法でC18 Spheri-5カラム(Perkin Elmer、カリフォルニア州)と、緩衝液Aに50 mM酢酸トリエチレンアンモニウム(溶媒:5% v/v アセトニトリル)を、および緩衝液Bに50 mM酢酸トリエチルアンモニウム(溶媒:70% v/v アセトニトリル)を用いて1.5 ml/分の流速でリンカーを精製した。15〜60%の緩衝液Bの直線性勾配を用いて45分間にわたる溶出を行った。リンカーは乾燥後に水に再懸濁して保存した。
【0058】
リンカー(5 μM)を標的mRNA(2.5 μM)と、25 mM Tris HCl緩衝液(pH 7)および100 mM NaCl中で、85℃で30秒間加熱してアニーリングさせた後に、5分間かけて4℃に冷却した。この反応混合物をホウケイ酸ガラス製バイアル(Kimble/Kontes、ニュージャージー州)に入れ、手持ち式マルチ波長UVランプモデルUVGL-25(UVP、カリフォルニア州)を長波長(波長λ>300 nm)に設定したもので15分間室温で紫外線を照射した。放射標識したmRNA 1とリンカーB間の光架橋反応の産物混合物を、変性6% TBE-尿素ゲル(Novex)で解析し、ホスホイメージングシステム(Molecular Dynamics、カリフォルニア州)で可視化した(図7E)。光架橋産物混合物は一般に、20%未満の未反応mRNAと80%を超える光架橋mRNAを含んでおり、これをさらに精製することなくインビトロ翻訳とそれに続くmRNA-タンパク質合体形成に直接使用した。より長いmRNA基質2、3、および4(>800ヌクレオチド)については、mRNAと架橋されたmRNA間の相対サイズの差は極めて小さくなり、ゲル上で分離することはできなかった。このような場合は、粗光架橋反応混合物を、さらに精製することなく融合体形成用の溶解液に直接添加した。
【0059】
mRNA融合体分子の翻訳および融合体形成は、mRNA 2を用いて以下の実験で最初に検討した。インビトロ翻訳反応は、ウサギ網状赤血球溶解液(Ambion)を用いて30分間30℃で行った。この反応物は、100 pmoleの光架橋mRNA(上述)、10 mMのクレアチンリン酸、150 mMのKOAc、0.5 mMのMgCl2、0.1 mMの各アミノ酸(メチオニンを除く)、150 μCiの[35S]メチオニン(Amersham)、および67%(v/v)の溶解液を最終容量300 μlとして含んでいた。mRNA-タンパク質融合体の形成は、ロバーツ(Roberts)およびショステク(Szostak)の方法ならびにショステクらの方法(上記)通りにKClとMgCl2を最終濃度がそれぞれ590 mMと50 mMとなるように、かつ容量が500 μlになるように添加して促進した。インキュベーションは20℃でさらに60分間継続した。融合体形成の塩依存性を探るためにさまざまな濃度のKClとMgCl2も検討した。
【0060】
インビトロ翻訳産物は、溶解液を10 mlの結合用緩衝液(100 mM Tris HCl、pH 8.0、10 mM EDTA、1 M NaCl、0.25% v/v Triton X-100)に希釈し、混合物に10 mgのオリゴ-dTセルロース7型(Pharmacia、ニュージャージー州)を添加して単離した。このサンプルを60分間4℃で回転した。次に固相支持体を5 mlの氷冷した結合用緩衝液で洗浄した後に100 μlの脱イオン水のアリコートで溶出した。単離したmRNA-タンパク質融合体の量は、取り込まれた[35S]メチオニンのシンチレーションを測定して決定した。この産物を4〜12%のMESランニング緩衝液(Novex)を用いたNuPageゲル電気泳動で解析した。ゲルを十分に洗浄して過剰な[35S]メチオニンを除去した後に乾燥させ、バンドをホスホイメージャーシステム(Molecular Dynamics)で可視化した。
【0061】
ゲル解析から、ペプチジル-tRNAおよび遊離ペプチドに対応する二本のバンドの存在が認められた(図7Fのレーン1)。光架橋したmRNA 2の翻訳を実施したところ、緩やかに移動する第三のバンドが現われた。これはmRNA-タンパク質融合体が良好に形成されていることを意味する(図7Fのレーン2)。遊離タンパク質および融合体(約20%多い)の収率は、コード配列がmRNA 2と同一であるが光架橋部位の下流に停止コドンを有するmRNA 3では上昇した(図7Fのレーン3および4)。相対的なバンドの強度から、合成タンパク質の総量の30%がmRNA-タンパク質融合体に変換されたことが判明した(図7Fのレーン4)。
【0062】
図7EのmRNA 4から調製したmRNA-scFvの融合体分子は、ロバーツ(Roberts)およびショステク(Szostak)ならびにショステクら(上記)の方法で、A18リンカー領域をオリゴdTセルロースに結合させた後に結合用緩衝液で洗浄することで精製した(図7Eのレーン5)。融合体産物は、光架橋した入力mRNA量を元にした場合、1.3%の収率で単離することができた。光架橋したmRNAから調製した融合体の物理的性質(ゲル移動度、オリゴdTセルロースに対する結合性、親和性試薬に対する選択的なペプチド結合性)は、酵素を用いて連結させたmRNA鋳型から得られた融合体産物と同一であることが判明した。
【0063】
融合体分子のペプチド部分の組成を確認するために、図7EのリンカーBと架橋させた、FlagおよびStrep-Tag IIエピトープをコードするmRNA鋳型1から調製した融合体のタンパク質結合性を検討した。35Sで標識したmRNA-ペプチド融合体(mRNA 1からリンカーBで調製したもの)溶液10 μlを、50 mM Tris HCl、pH 7.4、1% NP 40、150 mM NaCl、1 mM EDTA、1 mM Na3VO4、および1 mM NaFを含む300 μlの緩衝液に溶解した20 μlのAnti-Flagアフィニティーゲル(Sigma、ミズーリ州)に添加した。同じ融合体産物を、100 mM Tris-HCl、pH 7.1、1 mM EDTA、および0.5 mg/mlの酵母tRNAを含む300 μlの緩衝液に溶解した20 μlのStrepTactin Sepharose(Genosys、テキサス州)に添加して二回目の沈澱実験を行った。両沈澱混合物とも、同一の条件で並行して処理した。混合物を1時間4℃で回転させた後に、Ultrafree-MCフィルターユニット(0.45 μm;Millipore、マサチューセッツ州)上に移した。遠心して緩衝液を除き、残渣を5×300 μlの氷冷緩衝液で洗浄した。残渣のシンチレーションを測定して解析し、融合体結合をAnti-Flag M2マトリックスとStrepTactinマトリックスについてそれぞれ54%と62%であると判定した。プロテインAアガロース(Sigma)を対象とした対照反応では、マトリックスに対する結合は検出されなかった。
【0064】
リンカーの長さと組成が、種々の塩濃度で融合体形成に及ぼす影響を調べるために、mRNA 3を図7EのリンカーA〜Fに光架橋した後に、各鋳型の融合体形成を検討した(図7G)。30℃で30分間インキュベートした後に試料をアリコートに分け、各アリコートにKClおよびMgCl2を量を変えて添加した。この試料をさらに60分間20℃でインキュベートした後に、融合体産物をゲル電気泳動で解析した。mRNA-タンパク質融合体の最高収率は、高塩濃度下で長いリンカーAおよびB(それぞれ40ヌクレオチドおよび35ヌクレオチド)で得られた。低い塩濃度では、リンカーAおよびBでは融合体形成の有意な低下が認められた。一方、より短いリンカーC〜Fの融合体の収率は、それほど塩濃度に依存していなかった。さらに、融合体分子の収率は一般に、柔軟性のあるTEGスペーサーの数とともに上昇した。粗mRNA-タンパク質融合体分子溶解液を解析したところ、総タンパク質の最大45%がmRNA-タンパク質融合体として存在することが判明した。リンカーFには、オリゴdTによる精製に必要なオリゴdAトラックがないので、mRNA 4は、その3’端にある、オリゴdTセルロース上における融合体精製を可能とするA18の連続を使用して調製した。
【0065】
リンカーの5’端にソラレンを結合させる上記手法の別法では、ソラレン部分を、図7B、およびピールス(Pieles)らの論文(Nucleic Acids Res. 17:8967、1989)に示されるように、ペプチドアクセプターのリンカー領域の内部に組み込まれる場合もあるほか、リンカー配列内部における分枝状ホスホラミダイト(Clontech、パロアルト、カリフォルニア州)を組み込んだ後にソラレンホスホラミダイト(Glen Research、スターリング、バージニア州)を添加する場合がある。例えばソラレン部分は、参照として本明細書に組み入れられる米国特許出願第09/453,190号に記載されているように分枝状リンカーを介してRNAに架橋される。
【0066】
例示的なアプローチの一つでは、配列が
(Puはピューロマイシン-CPG[Glen Research];CおよびAは標準的な3’-アミダイト[Glen Research];a、t、c、およびgは5’-ホスホラミダイト[Glen Research];Xは非対称性の分枝状アミダイト[Clontech]を意味する)であるリンカーを、標準的な製造業者プロトコル通りに合成した後に、分枝点Xの選択的脱保護(Clontechの指示書による)と、それに続くソラレンC6アミダイト(Glen Research)のカップリングを行った。次にこのリンカーは、標的配列
をもつRNAに、上述の光架橋法で光架橋した。このRNA-リンカーコンストラクトを次に、RNA-タンパク質融合体の合成に問題なく使用することができた。
【0067】
さらに別の方法においては、標的ハイブリッド形成ドメインの3’端に結合させたソラレンを含むリンカーを構築する場合もある(図7C)。この方法では、ソラレンを含むリンカーの5’端が、別のリンカーにより伸長し、鎖の方向が逆転した後に3’ピューロマイシンで終結する。続くRNase HによるRNA標的ドメインの切断は任意選択で行うが、こうすることでリンカーコンストラクトの柔軟性が上昇する。さらに、このアプローチを採用することで、かなり長いRNA分子の内部にリンカーを結合させることが可能となり、続いてリンカー部位の下流にある未翻訳領域を切断した後に、翻訳を行うことでRNA-タンパク質融合体が形成する。
【0068】
この方法の例示的なアプローチの一つでは、5’ Pso atg cga gaa agt gat aaa aaa aaa aaa CC Pu(Puはピューロマイシン-CPG[Glen Research];Cは標準的な3’-アミダイト[Glen Research];a、t、c、およびgは5’-ホスホラミダイト[Glen Research];およびPsoはソラレンC6ホスホラミダイト[Glen Research]を意味する)の配列をもつリンカーを、標準的な製造業者プロトコル通りに構築した。次にリンカーを、
の標的配列をもつRNAに、上述の光架橋法で光架橋した。このRNA-リンカーコンストラクト(事前にRNase Hによる処理を任意で行う)は後に、RNA-タンパク質融合体の合成に問題なく使用することができた。
【0069】
ペプチドアクセプターをRNAに結合させる光化学的手法の利点の一つは、この方法が、連結前にRNAを化学的に修飾する必要がない点である。この利点があるため、方法が極めて確実かつ選択的となり、粗T7転写反応に由来するRNAを、化学的連結の基質として使用することができる。
【0070】
実施例6:光切断可能なビオチンを用いたRNAの精製および連結
必要に応じて、親和性を利用したRNA精製段階を、上述の光化学的連結法と組み合わせる(図7H)。適切なリンカー分子を、ピューロマイシン末端で光切断可能なビオチン成分(例えば、EZ-Link(商標) NHS-PC-LC-Biotin、Pierce、ロックフォード、イリノイ州)で修飾する。次に標的RNA(例えば粗転写反応で得られる)と一定量のリンカーとの間でハイブリッドを形成させ、得られた二本鎖を、ストレプトアビジン(または関連する)樹脂などの固相支持体上に補足する。次に、十分な洗浄を行って過剰なRNAならびに転写反応成分を除去する。長波長の紫外光を同時に照射することで光架橋が形成され、産物が樹脂から遊離する。連結したRNAは次に、さらに精製を行うことなく翻訳および融合形成反応に直接使用することができる。
【0071】
この方法には、従来のRNA精製スキームにはない利点がある。例えば転写後のRNA量は、使用するリンカーの量を上回ることを推定するだけである。記載の手順をふむことで、その量は使用リンカー量を超えない程度に自動的に減少する。このため、さらにRNA(または連結したRNA)を、例えばA260のUV値で定量することなく次の工程に進めることが可能となり、RNA量を別の方法で調整する必要はない。したがって核酸-タンパク質の融合体分子の調製法は自動化に適している。
【0072】
例示的な方法の一つでは、このようなビオチンを用いたRNA精製・連結プロトコルは以下の手順で実施される。光切断可能なビオチンを用いたRNA精製および連結法では、最初にリンカーをビオチン化する。このリンカー(C6-ソラレン-2-OMe[U AGC GGA UGC]dA18 TEG2 dCdC-ピューロマイシン)は、100 μlの100 μMリンカー(総量10 nmol)、50 μlの1 μmole EZ-Link(商標) PC-LC-Biotin(溶媒DMSO)、20 μlの10×PBS、pH 7.4、および30 μlのH2Oを混合することでビオチン化する。この混合物を室温で2時間インキュベートすると、定量可能な収量が得られる。次にこの混合物に2回エタノールを添加して沈澱を生じさせると、PC-ビオチン化リンカーの予想回収率は90%を超える。これを200 μlのH2Oに再懸濁する。
【0073】
次にRNAを、例えばT7 Megashortscriptキット(Ambion)を用いて転写する。転写には、250 μlの反応につき、10 pmoleのDNA鋳型(3’末端に配列
を含む)を使用する。この転写反応で約2〜5 nmolの粗RNA産物が得られる。次の工程に進む前に精製(フェノール抽出、NAP-5カラム、またはRNeasyカラム)は必要ない。
【0074】
この精製・連結反応に続く段階では、ビオチン化したリンカー(10 μlの50 pmol/μl ビオチン化リンカー;総量500 pmol)を、62.5〜250 μlの転写混合物(推定最小量として500 pmoleのRNAを含む)と、例えばPCR装置(80℃で30秒間加熱した後に0.3℃/秒で4℃に冷却する)で、15 μlの5 M NaCl(300 μlの反応について最終濃度を0.25 Mとする)およびH2Oを、最終反応容量が300 μlになるように添加してアニーリングを行う。次に反応混合物のRNAを、100 μlのNeutravidinビーズ(Pierce、ロックフォード、イリノイ州)に、反応混合物を4℃で30分間かけて緩やかに振盪しながら固定化する。次にこのビーズを洗浄して300 μlのH2O中に再懸濁する。
【0075】
このビーズを次に遠心して沈降させ、100 μlの緩衝液(25 mM Tris pH 7.0および0.25 M NaCl)で3回洗浄する。次にビーズに紫外線を15分間室温で(手持ち式UVランプUVGL-25を用いてビーズを含む微量遠心用チューブをランプ上に直接置いて)照射し、リンカーをRNAに化学的に連結し、かつ連結した分子をビーズから光化学的に遊離させる。250 pmolの連結RNAが、光化学的に遊離すると予測される。75 μlのH2Oを同チューブに添加し、チューブをボルテックスミキサーで30秒間撹拌して、ビーズを遠心して沈降させる。連結したRNA/リンカーを含む75 μlの上清を翻訳および核酸-タンパク質融合体分子の形成に使用する。
【0076】
核酸-タンパク質融合体分子は、前工程で得られた75 μlの上清を225 μlの緩衝液成分およびウサギ網状赤血球溶解物キット(Ambion)と混合して、その混合物を30分間30℃でインキュベートすることで形成される。次にKClおよびMgCl2を最終濃度がそれぞれ500 mMおよび50 mMになるように添加し、室温で60分間インキュベートして反応を続けて、核酸-タンパク質融合体分子を作製する。
【0077】
実施例7:強固な非共有結合を介したペプチドアクセプターとRNAとの結合
ペプチドアクセプターをRNA分子に結合させる共有結合形成法の別法として、強固な複合体形成にのみ依存する方法も可能であり、これもまた本発明の一部である。一つの方法では、図8Aに示すように、ペプチド核酸(PNA)を使用してRNAの認識および結合を行う。PNAはDNAの模倣体であり、アキラルおよび無電荷のN-(2-アミノエチル)グリシンのユニットから構成される主鎖をもつ(KnudsenおよびNielsen、Nucleic Acids Res. 24:494、1996)。PNAは、配列特異的かつ高い親和力で相補的な一本鎖DNAおよびRNAとハイブリッドを形成することが知られている。特に、RNAに結合してクランプを形成する2個のPNA分子を含む三本鎖形成コンストラクトは、RNAに強固に結合する効率のよい手法を提供することができる。というのは、このようなコンストラクトは、熱変性および、インビトロ翻訳の条件に対して極めて耐性が強いためである(図8)(Hanveyら、Science、258:1481、1992)。
【0078】
偽イソシトシン(Pseudoisocytosine)塩基を使用することで、中性pHおよび塩基性pHにおける安定性がさらに強化される(Egholmら、Nucleic Acids Res. 23:217、1995)。このようなPNA-クランプは、インビトロ翻訳条件下でmRNAと結合したまま維持され、リボソームで置換されないことが報告されている(KnudsenおよびNielsen、Nucleic Acids Res. 24:494、1996)。この特性のため、対応するRNA-タンパク質融合体コンストラクトの安定性は極めて大きくなる。
【0079】
核酸リンカー-PNAコンストラクトは、図8Bおよびウルマン(Uhlmann)らの論文(Angew. Chem. Int. Ed. Engl. 35:2633、1996)に記載されているように、ピューロマイシン-CPGで始まる固相支持体を用いた合成法で調製することができる。標準的な自動合成法による所望の核酸部分(または必要に応じてPEG-スペーサー)のアセンブリー後に、PNAドメインを適切な試薬(例えばウルマンらの論文(上記)に記載)に結合させて固相合成を続ける。またはPNAを個別の成分として事前に合成しておき(PE Biosystems、フォスターシティー、カリフォルニア州)、その後に所望のリンカー部分と化学的に結合させることができる。
【0080】
一つの特定の例においては、ピューロマイシン-DNAリンカーの5’末端をアミノ基で修飾して、化学的に活性のあるエステル(例えば、ジスクシンイミジルグルタレートまたは関連試薬との反応を介したNHS-エステル;この手法は例えばコックス(Cox)ら(J. Immunol. 145:1719、1990);およびhttp://www.piercenet.com/Products/linksearch.cfm;Pierce、ロックフォード、イリノイ州、に記載されている)にさらに変換することができる。その後の(非保護状態のアミノ末端またはカルボキシ末端のリジンのいずれかを有する)PNA成分との反応により、ドメインが共有結合で連結される。この工程は、最終的なDNA-リンカー産物を含む均質溶液中で、または保護基を有し、かつ固相樹脂に結合させた状態のままのDNAを用いることで行なわれる。
【0081】
実施例8:リンカーの長さと組成の最適化
上述のすべての方法では、リンカーコンストラクトを最適化することが好ましい。考慮対象となる諸因子には例えば、鋳型/標的認識エレメントを含めること、および結合させた官能基の空間的な接近のしやすさがある。特に、核酸のハイブリッド形成を介した鋳型または標的の認識が関与する場合は、好ましくはリンカーの標的配列および化学的性質を含む諸因子を最適化する。例えばRNAのハイブリッド形成強度およびその結果としての連結効率は、DNAではなく2-OMe RNAまたはプロピンで修飾した核酸塩基を使用することで上昇することが知られている(例えばInoue ら、Nucleic Acids Res. 15:6131、1987;Kibler-Herzogら、Nucleic Acids Res. 19:2979、1991;およびWagnerら、Science 260:1510、1993に記載)。
【0082】
リンカーは、RNA-タンパク質融合体反応における効率に対しても最適化することができる。これには一般に、種々の長さのリンカーが関与するが、RNA-タンパク質の結合に用いる種々の構築用ブロックを使用することもある。一つの特定の例においては、リンカーのデオキシヌクレオチドは、PEGスペーサーまたは2-OMe-RNAユニットと置換することができる(いずれもGlen Research、スターリング、バージニア州)。
【0083】
その他の態様は特許請求の範囲に含まれる。
【0084】
本明細書で説明したすべての特許および特許出願は、参照として組み入れられる。
【特許請求の範囲】
【請求項1】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)ヘアピン構造を形成する3’配列を有するRNA分子を提供する段階;
(b)核酸リンカー分子に共有結合したペプチドアクセプターを提供する段階;および
(c)ペプチドアクセプターとRNA分子間における共有結合の形成が可能な条件下で、該RNA分子と該核酸リンカー分子のハイブリッドを形成させる段階。
【請求項2】
共有結合形成がT4 DNAリガーゼで触媒される、請求項1記載の方法。
【請求項3】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA分子を提供する段階;
(b)5’配列がヘアピン構造を形成する核酸リンカー分子に、共有結合したペプチドアクセプターを提供する段階:および
(c)ペプチドアクセプターとRNA分子間における共有結合の形成が可能な条件下で、該RNA分子と該核酸リンカー分子のハイブリッドを形成させる段階。
【請求項4】
共有結合形成がT4 DNAリガーゼで触媒される、請求項3記載の方法。
【請求項5】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA分子を提供する段階;
(b)デオキシヌクレオチド三リン酸またはジデオキシヌクレオチド三リン酸から開始するリンカー分子に共有結合したペプチドアクセプターを提供する段階;および
(c)RNA分子およびペプチドアクセプターを末端デオキシヌクレオチジルトランスフェラーゼと接触させて、ペプチドアクセプターを該RNA分子に共有結合させる段階。
【請求項6】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA 分子を提供する段階;および
(b)ペプチドアクセプターをRNA分子に化学的に連結する段階。
【請求項7】
ペプチドアクセプターをソラレン部分に連結し、化学的連結段階が、ソラレン部分を介して該ペプチドアクセプターをRNA分子に架橋させる段階を含む、請求項6記載の方法。
【請求項8】
ソラレン部分をリンカーを介してペプチドアクセプターに結合させる、請求項7記載の方法。
【請求項9】
ソラレン部分がリンカーの5’末端に位置する、請求項8記載の方法。
【請求項10】
ソラレン部分がリンカーの3’末端に位置する、請求項8記載の方法。
【請求項11】
ソラレン部分がリンカーの内部に位置する、請求項8記載の方法。
【請求項12】
リンカーがC6アルキル鎖を含む、請求項8記載の方法。
【請求項13】
RNA分子が、その3’端の近傍に停止コドンを含む、請求項8記載の方法。
【請求項14】
リンカーの長さが25〜40ヌクレオチド単位である、請求項8記載の方法。
【請求項15】
架橋が紫外光の照射で達成される、請求項7記載の方法。
【請求項16】
架橋に先立ち、さらに光切断可能な部分を含むリンカーとRNAのハイブリッドを形成させ、該ハイブリッド形成RNAを光切断可能な部分を介して固相支持体に固定化する、請求項8記載の方法。
【請求項17】
光切断可能な部分がビオチンである、請求項16記載の方法。
【請求項18】
ペプチドアクセプターに結合する官能基をRNA分子に導入する、請求項6記載の方法。
【請求項19】
IO4-酸化によりRNA分子に官能基を導入する、請求項18記載の方法。
【請求項20】
化学的連結を外部鋳型の非存在下で行う、請求項18記載の方法。
【請求項21】
ペプチドアクセプターをリンカー分子に共有結合させ、化学的連結段階前に、外部鋳型、RNA分子の3’端およびリンカー分子の5’端とハイブリッドを形成する外部鋳型を用いて該RNA分子と該ペプチドアクセプターを整列させる、請求項18記載の方法。
【請求項22】
ペプチドアクセプターをリンカー分子と共有結合させ、化学的連結段階前に、リンカー分子とRNAの3’端のハイブリッドを形成させることで該RNA分子と該ペプチドアクセプターを整列させる、請求項18記載の方法。
【請求項23】
ペプチドアクセプターの官能基がリンカー分子の5’端に位置する、請求項22記載の方法。
【請求項24】
ペプチドアクセプターの官能基が、片側でRNA分子とハイブリッドを形成する領域に隣接し、もう片側で該ペプチドアクセプターに隣接する、請求項22記載の方法。
【請求項25】
ペプチドアクセプターの官能基が、アミン、ヒドラジン、(チオ)ヒドラジド、および(チオ)セミカルバゾンからなる群より選択される、請求項18記載の方法。
【請求項26】
化学的連結段階が以下の段階を含む、請求項6記載の方法:
(a)還元的アミノ化により、RNA分子に官能基を結合させる段階;
(b)RNA分子の官能基と反応するようにペプチドアクセプターを修飾する段階;および
(c)共有結合の形成を可能とする条件下で、ペプチドアクセプターを該RNA分子に接触させる段階。
【請求項27】
官能基がチオール、マレイミド、またはアミンである、請求項26記載の方法。
【請求項28】
以下の段階を含む、インビトロでペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA分子を提供する段階;および
(b)ペプチドアクセプターをRNA分子に非共有結合的に結合させる段階。
【請求項29】
ペプチドアクセプターをPNAに共有結合させ、該ペプチドアクセプターとRNA分子の非共有結合がPNAを介して生じる、請求項28記載の方法。
【請求項30】
RNA分子が停止コドンを含む、請求項29記載の方法。
【請求項31】
RNA分子が、タンパク質コード配列に使用可能に連結させた翻訳開始配列および開始コドンを含む、請求項1、3、5、6、または27記載の方法。
【請求項32】
ペプチドアクセプターがピューロマイシンである、請求項1、3、5、6、または27記載の方法。
【請求項33】
ペプチドアクセプターを非ヌクレオチドリンカー分子に共有結合させる、請求項1、3、5、6、または27記載の方法。
【請求項34】
リンカーがトリエチレングリコールスペーサーを含む、請求項1、3、5、または8記載の方法。
【請求項35】
リンカーが2’-OMe-RNAホスホラミダイトを含む、請求項1、3、5、または8記載の方法。
【請求項36】
RNAまたはリンカーが、アフィニティー精製用の配列を含み、該RNAを精製する段階をさらに含む、請求項1、3、5、6、8、または27記載の方法。
【請求項37】
アフィニティー精製用の配列がポリ(A)配列を含む、請求項36記載の方法。
【請求項38】
ペプチドアクセプターに化学的に連結させたRNA分子。
【請求項39】
RNAが、その光架橋部位に位置する停止コドンを含む、請求項38記載のRNA分子。
【請求項40】
非共有結合的にペプチドアクセプターに結合させるRNA分子であって、ペプチドアクセプターをリンカー分子およびPNA分子に結合させ、非共有結合がPNA分子とRNA分子間に形成されるRNA分子。
【請求項41】
ペプチドアクセプターをRNA分子の3’端に連結させる、請求項38または40記載の分子。
【請求項42】
RNA分子が、タンパク質コード配列に使用可能に連結させた翻訳開始配列および開始コドンを含む、請求項38または40記載の分子。
【請求項43】
ペプチドアクセプターがピューロマイシンである、請求項38または40記載の分子。
【請求項44】
以下の段階を含む、RNA-タンパク質融合体を作製する方法:
(a)光切断可能な部分、ソラレン部分、およびペプチドアクセプターを含むリンカーとハイブリッドを形成するRNA分子を提供する段階;
(b)固定化されていないRNAが固相支持体から実質的に除去される条件下で、RNAを固相支持体に固定化する段階;
(c)ペプチドアクセプターを該RNAに、ソラレン部分を介して架橋させ、それによって架橋が固相支持体から架橋RNAを同時に遊離させる段階;および
(d)段階(c)で形成された架橋RNAを翻訳して、RNA融合タンパク質を生成させる段階。
【請求項1】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)ヘアピン構造を形成する3’配列を有するRNA分子を提供する段階;
(b)核酸リンカー分子に共有結合したペプチドアクセプターを提供する段階;および
(c)ペプチドアクセプターとRNA分子間における共有結合の形成が可能な条件下で、該RNA分子と該核酸リンカー分子のハイブリッドを形成させる段階。
【請求項2】
共有結合形成がT4 DNAリガーゼで触媒される、請求項1記載の方法。
【請求項3】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA分子を提供する段階;
(b)5’配列がヘアピン構造を形成する核酸リンカー分子に、共有結合したペプチドアクセプターを提供する段階:および
(c)ペプチドアクセプターとRNA分子間における共有結合の形成が可能な条件下で、該RNA分子と該核酸リンカー分子のハイブリッドを形成させる段階。
【請求項4】
共有結合形成がT4 DNAリガーゼで触媒される、請求項3記載の方法。
【請求項5】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA分子を提供する段階;
(b)デオキシヌクレオチド三リン酸またはジデオキシヌクレオチド三リン酸から開始するリンカー分子に共有結合したペプチドアクセプターを提供する段階;および
(c)RNA分子およびペプチドアクセプターを末端デオキシヌクレオチジルトランスフェラーゼと接触させて、ペプチドアクセプターを該RNA分子に共有結合させる段階。
【請求項6】
以下の段階を含む、ペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA 分子を提供する段階;および
(b)ペプチドアクセプターをRNA分子に化学的に連結する段階。
【請求項7】
ペプチドアクセプターをソラレン部分に連結し、化学的連結段階が、ソラレン部分を介して該ペプチドアクセプターをRNA分子に架橋させる段階を含む、請求項6記載の方法。
【請求項8】
ソラレン部分をリンカーを介してペプチドアクセプターに結合させる、請求項7記載の方法。
【請求項9】
ソラレン部分がリンカーの5’末端に位置する、請求項8記載の方法。
【請求項10】
ソラレン部分がリンカーの3’末端に位置する、請求項8記載の方法。
【請求項11】
ソラレン部分がリンカーの内部に位置する、請求項8記載の方法。
【請求項12】
リンカーがC6アルキル鎖を含む、請求項8記載の方法。
【請求項13】
RNA分子が、その3’端の近傍に停止コドンを含む、請求項8記載の方法。
【請求項14】
リンカーの長さが25〜40ヌクレオチド単位である、請求項8記載の方法。
【請求項15】
架橋が紫外光の照射で達成される、請求項7記載の方法。
【請求項16】
架橋に先立ち、さらに光切断可能な部分を含むリンカーとRNAのハイブリッドを形成させ、該ハイブリッド形成RNAを光切断可能な部分を介して固相支持体に固定化する、請求項8記載の方法。
【請求項17】
光切断可能な部分がビオチンである、請求項16記載の方法。
【請求項18】
ペプチドアクセプターに結合する官能基をRNA分子に導入する、請求項6記載の方法。
【請求項19】
IO4-酸化によりRNA分子に官能基を導入する、請求項18記載の方法。
【請求項20】
化学的連結を外部鋳型の非存在下で行う、請求項18記載の方法。
【請求項21】
ペプチドアクセプターをリンカー分子に共有結合させ、化学的連結段階前に、外部鋳型、RNA分子の3’端およびリンカー分子の5’端とハイブリッドを形成する外部鋳型を用いて該RNA分子と該ペプチドアクセプターを整列させる、請求項18記載の方法。
【請求項22】
ペプチドアクセプターをリンカー分子と共有結合させ、化学的連結段階前に、リンカー分子とRNAの3’端のハイブリッドを形成させることで該RNA分子と該ペプチドアクセプターを整列させる、請求項18記載の方法。
【請求項23】
ペプチドアクセプターの官能基がリンカー分子の5’端に位置する、請求項22記載の方法。
【請求項24】
ペプチドアクセプターの官能基が、片側でRNA分子とハイブリッドを形成する領域に隣接し、もう片側で該ペプチドアクセプターに隣接する、請求項22記載の方法。
【請求項25】
ペプチドアクセプターの官能基が、アミン、ヒドラジン、(チオ)ヒドラジド、および(チオ)セミカルバゾンからなる群より選択される、請求項18記載の方法。
【請求項26】
化学的連結段階が以下の段階を含む、請求項6記載の方法:
(a)還元的アミノ化により、RNA分子に官能基を結合させる段階;
(b)RNA分子の官能基と反応するようにペプチドアクセプターを修飾する段階;および
(c)共有結合の形成を可能とする条件下で、ペプチドアクセプターを該RNA分子に接触させる段階。
【請求項27】
官能基がチオール、マレイミド、またはアミンである、請求項26記載の方法。
【請求項28】
以下の段階を含む、インビトロでペプチドアクセプターをRNA分子に結合させる方法:
(a)RNA分子を提供する段階;および
(b)ペプチドアクセプターをRNA分子に非共有結合的に結合させる段階。
【請求項29】
ペプチドアクセプターをPNAに共有結合させ、該ペプチドアクセプターとRNA分子の非共有結合がPNAを介して生じる、請求項28記載の方法。
【請求項30】
RNA分子が停止コドンを含む、請求項29記載の方法。
【請求項31】
RNA分子が、タンパク質コード配列に使用可能に連結させた翻訳開始配列および開始コドンを含む、請求項1、3、5、6、または27記載の方法。
【請求項32】
ペプチドアクセプターがピューロマイシンである、請求項1、3、5、6、または27記載の方法。
【請求項33】
ペプチドアクセプターを非ヌクレオチドリンカー分子に共有結合させる、請求項1、3、5、6、または27記載の方法。
【請求項34】
リンカーがトリエチレングリコールスペーサーを含む、請求項1、3、5、または8記載の方法。
【請求項35】
リンカーが2’-OMe-RNAホスホラミダイトを含む、請求項1、3、5、または8記載の方法。
【請求項36】
RNAまたはリンカーが、アフィニティー精製用の配列を含み、該RNAを精製する段階をさらに含む、請求項1、3、5、6、8、または27記載の方法。
【請求項37】
アフィニティー精製用の配列がポリ(A)配列を含む、請求項36記載の方法。
【請求項38】
ペプチドアクセプターに化学的に連結させたRNA分子。
【請求項39】
RNAが、その光架橋部位に位置する停止コドンを含む、請求項38記載のRNA分子。
【請求項40】
非共有結合的にペプチドアクセプターに結合させるRNA分子であって、ペプチドアクセプターをリンカー分子およびPNA分子に結合させ、非共有結合がPNA分子とRNA分子間に形成されるRNA分子。
【請求項41】
ペプチドアクセプターをRNA分子の3’端に連結させる、請求項38または40記載の分子。
【請求項42】
RNA分子が、タンパク質コード配列に使用可能に連結させた翻訳開始配列および開始コドンを含む、請求項38または40記載の分子。
【請求項43】
ペプチドアクセプターがピューロマイシンである、請求項38または40記載の分子。
【請求項44】
以下の段階を含む、RNA-タンパク質融合体を作製する方法:
(a)光切断可能な部分、ソラレン部分、およびペプチドアクセプターを含むリンカーとハイブリッドを形成するRNA分子を提供する段階;
(b)固定化されていないRNAが固相支持体から実質的に除去される条件下で、RNAを固相支持体に固定化する段階;
(c)ペプチドアクセプターを該RNAに、ソラレン部分を介して架橋させ、それによって架橋が固相支持体から架橋RNAを同時に遊離させる段階;および
(d)段階(c)で形成された架橋RNAを翻訳して、RNA融合タンパク質を生成させる段階。
【図1】

【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7A−C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図8A】
【図8B】
【図2】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7A−C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図8A】
【図8B】
【公開番号】特開2010−284172(P2010−284172A)
【公開日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願番号】特願2010−186725(P2010−186725)
【出願日】平成22年8月24日(2010.8.24)
【分割の表示】特願2001−512922(P2001−512922)の分割
【原出願日】平成12年7月19日(2000.7.19)
【出願人】(509054371)ブリストル−マイヤーズ スクウィブ カンパニー (10)
【Fターム(参考)】
【公開日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願日】平成22年8月24日(2010.8.24)
【分割の表示】特願2001−512922(P2001−512922)の分割
【原出願日】平成12年7月19日(2000.7.19)
【出願人】(509054371)ブリストル−マイヤーズ スクウィブ カンパニー (10)
【Fターム(参考)】
[ Back to top ]