
ペプチド生成酵素、その遺伝子及びペプチドの製造方法
【課題】プロリン残基を含有するペプチドを高純度で製造することが可能なペプチド生成酵素、その遺伝子及びペプチドの製造方法の提供。
【解決手段】Streptomyces aureofaciens TH−3株(FERM P−21343)のプロリンアミノペプチダーゼ(PAP TH−3)のアミノ酸配列、およびそれと類似のアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させてプロリン残基を含有するペプチドを生成する活性を有するペプチド生成酵素。
【解決手段】Streptomyces aureofaciens TH−3株(FERM P−21343)のプロリンアミノペプチダーゼ(PAP TH−3)のアミノ酸配列、およびそれと類似のアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させてプロリン残基を含有するペプチドを生成する活性を有するペプチド生成酵素。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ペプチド生成酵素、その遺伝子及びペプチドの製造方法に関する。
【背景技術】
【0002】
コラーゲンは、3本のポリペプチド鎖からなる三重らせん構造を有しており、ポリペプチド鎖は、トリペプチド配列Gly−X−Yを繰り返し単位とする繰り返し構造を有する。このX及びYの位置のアミノ酸は、プロリン(Pro)及び翻訳後修飾によって生成する4−ヒドロキシプロリン(L−ヒドロキシプロリン、Hyp)であることが多い。コラーゲンは、4−ヒドロキシプロリンによってその三重らせんが非常に安定化されていることから、水に不溶であり、ほとんどのプロテアーゼに対して抵抗性を有する。
【0003】
コラーゲンの分解物であるコラーゲンペプチドは、工業及び医療の分野において、種々の有用な生物学的活性を有することが報告されており、急速にそして広範に適用されている。例えば、骨粗鬆症、胃潰瘍、変形性関節症、褥瘡及び高血圧用の薬剤、皮膚保湿剤、並びに、保存剤の用途が挙げられる。
【0004】
近年、コラーゲンペプチドのなかでも、プロリン及び4−ヒドロキシプロリンから構成されるPro−Hyp等のジペプチドが上記生物学的活性に寄与していることが明らかとなり、このようなジペプチドが注目されている(特許文献1)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2010−106003号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
特許文献1では、Pro−Hyp等のジペプチドを生成する方法として、(1)コラゲナーゼ等の酵素の存在下、コラーゲン又はゼラチンを分解することによって得る方法、並びに、(2)固相合成法及び液相合成法等の化学合成法によって、アミノ酸から合成する方法が挙げられている。
【0007】
しかしながら、上記(1)の方法では、原料であるコラーゲン又はゼラチン、並びに、反応中間体であるコラーゲンペプチド等のポリペプチドが不純物としてPro−Hypと共に含まれるため、Pro−Hypを高純度の状態で得るためには、更に精製する工程が必要となる。上記(2)の方法では、トリフルオロ酢酸等を使用することから、生成したジペプチドを医薬品等の目的で使用することは困難な場合が多い。
【0008】
本発明は、上記事情に鑑みてなされたものであり、N末端にプロリン残基を有するペプチドを高純度で製造することが可能なペプチド生成酵素、その遺伝子及びペプチドの製造方法を提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者等は、鋭意研究を行った結果、ある特定のアミノ酸配列を有するタンパク質が、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有していることを見出し、本願発明を完成するに至った。
【0010】
すなわち、本発明は、(A)配列表の配列番号10に記載のアミノ酸配列、(B)配列表の配列番号10に記載のアミノ酸配列において1又は数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列、及び(C)配列表の配列番号10に記載のアミノ酸配列に対して80%以上の配列同一性を有するアミノ酸配列、からなる群から選択されるアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させて、N末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素を提供する。
【0011】
本発明に係るペプチド生成酵素は、このようなアミノ酸配列を有することで、プロリン誘導体とアミノ酸とを縮合させることによって、N末端にプロリン残基を有するペプチドを生成することが可能となる。その結果、N末端にプロリン残基を有するペプチドを高純度の状態で提供することが可能となる。
【0012】
本発明は、(a)上記ジペプチド生成酵素をコードする塩基配列、及び(b)配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖とストリンジェントな条件下にハイブリダイズし、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素をコードする塩基配列、からなる群より選択される塩基配列を有する遺伝子を提供する。
【0013】
本発明は、上記ペプチド生成酵素の存在下で、プロリン誘導体とアミノ酸とを縮合させる工程を含む、N末端にプロリン残基を有するペプチドの製造方法を提供する。
【0014】
上記プロリン誘導体は、プロリンエステル又はプロリンアミドであることが好ましい。プロリン誘導体をこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0015】
上記プロリンエステルは、ベンジルエステル又はメチルエステルであることが好ましい。プロリンエステルをこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0016】
上記アミノ酸は、α−アミノ酸であることが好ましい。アミノ酸をこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0017】
上記アミノ酸は、L−ヒドロキシプロリン、L−プロリン、L−バリン、L−アラニン、グリシン、L−システイン、L−リジン、L−セリン、L−トレオニン、L−アルギニン、(R)−α−アリルグリシン、(S)−α−プロパギルグリシン、(R)−α−プロパギルグリシン、(S)−α−メチルロイシン、(R)−α−メチルロイシン、又は(S)−α−メチルフェニルアラニンであることが好ましい。アミノ酸をこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0018】
pH9.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させることが好ましい。このようにすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【発明の効果】
【0019】
本発明によれば、N末端にプロリン残基を有するペプチドを高純度で製造することが可能なペプチド生成酵素、その遺伝子及びペプチドの製造方法が提供される。
【図面の簡単な説明】
【0020】
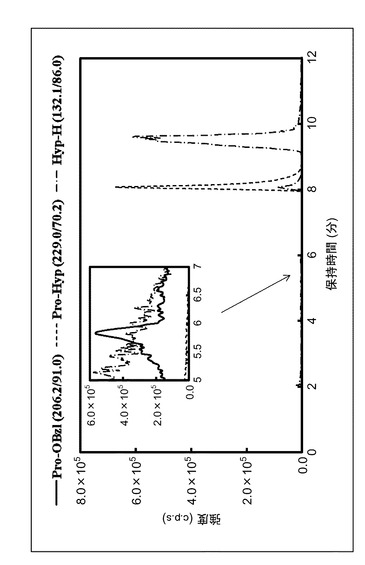
【図1】反応混合物を多重反応モニタリングクロマトグラフィーで分析した結果を示すグラフである。
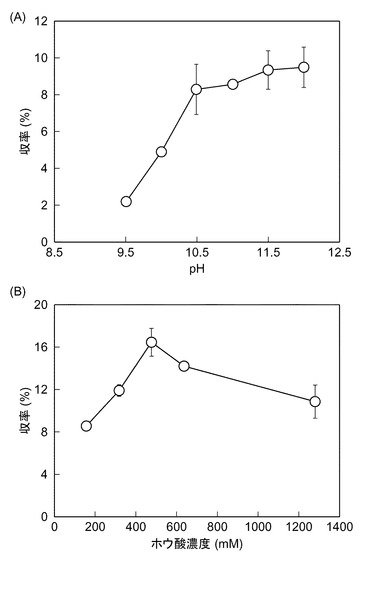
【図2】Pro−Hypジペプチドの生成反応における、(A)pH依存性及び(B)緩衝水溶液の濃度の依存性を示すグラフである。
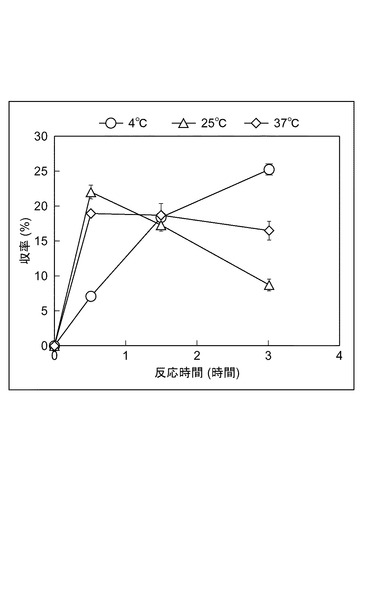
【図3】Pro−Hypジペプチドの生成反応における、温度依存性を示すグラフである。
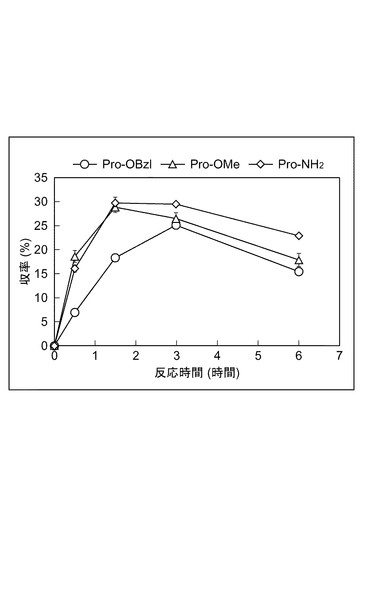
【図4】Pro−Hypジペプチドの生成反応における、プロリン誘導体の影響を示すグラフである。
【図5】Pro−Hypジペプチドの生成反応における、プロリン誘導体を連続して添加したときの影響を示すグラフである。
【発明を実施するための形態】
【0021】
本明細書において、アミノ酸の表記を、3文字又は1文字で表記する場合がある。すなわち、アラニンは、Ala又はA、アルギニンは、Arg又はR、アスパラギンは、Asn又はN、アスパラギン酸は、Asp又はD、システインは、Cys又はC、グルタミン酸は、Glu又はE、グルタミンは、Gln又はQ、グリシンは、Gly又はG、ヒスチジンは、His又はH、イソロイシンは、Ile又はI、ロイシンは、Leu又はL、リジンは、Lys又はK、メチオニンは、Met又はM、フェニルアラニンは、Phe又はF、プロリンは、Pro又はP、セリンは、Ser又はS、トレオニンは、Thr又はT、トリプトファンは、Trp又はW、チロシンは、Tyr又はY、並びに、バリンは、Val又はVを表記する。
【0022】
本実施形態に係るペプチド生成酵素は、
(A)配列表の配列番号10に記載のアミノ酸配列、
(B)配列表の配列番号10に記載のアミノ酸配列において1又は数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列、及び
(C)配列表の配列番号10に記載のアミノ酸配列に対して80%以上の配列同一性を有するアミノ酸配列、からなる群から選択されるアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有する。
【0023】
配列表の配列番号10に記載のアミノ酸配列は、Streptomyces aureofaciens TH−3株(FERM P−21343)のプロリンアミノペプチダーゼ(PAP TH−3)のアミノ酸配列である。
【0024】
上記(B)のアミノ酸配列について、配列表の配列番号10のアミノ酸残基の置換、欠失、挿入又は付加されたアミノ酸(変異されたアミノ酸)の数は、アミノ酸配列が後述するプロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性(以下、単にペプチド生成活性と呼ぶ場合がある。)の測定法の条件下に同様の活性を示す範囲であればよい。アミノ酸の変異の数は、少なくとも1個、好ましくは、1個〜117個、より好ましくは、1個〜12個である。変異されたアミノ酸配列は、天然に存在するアミノ酸配列、すなわち、自然発生による変異を有するアミノ酸配列であってもよく、人為的に所望の部位又はアミノ酸残基に変異を導入したアミノ酸配列であってもよい。人為的に所望の部位又はアミノ酸残基に変異を導入する場合、天然に存在する配列表の配列番号10のアミノ酸配列の変異体に準じて人為的に変異を導入してもよい。人為的な変異の導入は、例えば、PCR法を用いた部位特異的変異導入法等によって行われる。
【0025】
変異されたアミノ酸配列としては、例えば、配列表の配列番号10のアミノ酸配列において少なくとも1個のアミノ酸残基が類似した物理学的性質を有する他のアミノ酸残基に置換(保存的置換)されたアミノ酸配列が挙げられる。保存的置換は、例えば、疎水性、電荷、pK、又は立体構造上類似したアミノ酸残基同士の置換、並びに、本来のポリペプチドの生理活性を維持する程度に該ポリペプチドの立体構造や折り畳み構造が維持されるようなアミノ酸残基の置換である。より具体的には、上記(B)のアミノ酸配列は、配列表の配列番号10のアミノ酸配列において、少なくとも1個のアミノ酸残基が、以下のアミノ酸群(1)〜(6):
(1)グリシン及びアラニン、
(2)バリン、イソロイシン及びロイシン、
(3)アスパラギン酸、グルタミン酸、アスパラギン及びグルタミン、
(4)セリン及びスレオニン、
(5)リジン及びアルギニン、並びに
(6)フェニルアラニン及びチロシン
における、同一のアミノ酸群に属するアミノ酸残基同士の置換により他のアミノ酸と保存的置換されたものであることが好ましい。
【0026】
上記(C)において、配列同一性は、参照配列(例えば、配列表の配列番号10のアミノ酸配列)に対して、クエリー配列(評価対象のアミノ酸配列)を、適切にアラインメントし、算出された値である。具体的には、本実施形態においては、配列同一性は、CLUSTALアルゴリズムで算出された値である。
【0027】
本実施形態においては、配列同一性は、高いほど好ましく、具体的には、80%以上、85%以上、90%以上、95%以上、又は98%以上である。
【0028】
本実施形態において、ペプチド生成酵素のペプチド生成活性は、任意の方法で測定され得る。例えば、プロリン誘導体とアミノ酸とを基質として、上記ペプチド生成酵素によって生成されたN末端にプロリン残基を有するペプチドの生成量を多重反応モニタリングクロマトグラフィー等の方法を用いて測定する方法が挙げられる。
【0029】
本実施形態に係るペプチド生成酵素は、また、以下に詳述する本実施形態に係る遺伝子を用いて、遺伝子工学的に製造することができる。例えば、Streptomyces aureofaciens TH−3株とは異なる異種細胞において、本実施形態に係るペプチド生成酵素を発現させることができる。
【0030】
本実施形態に係るペプチド生成酵素をコードする遺伝子は、
(a)上記ペプチド生成酵素をコードする塩基配列、及び
(b)配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖と、ストリンジェントな条件下にハイブリダイズし、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素をコードする塩基配列、
からなる群より選択される塩基配列を有する。
【0031】
このうち(a)の塩基配列は、それぞれ、上記(A)〜(C)のアミノ酸配列をコードする塩基配列であり、そして(b)の塩基配列は、上記(A)〜(C)のアミノ酸配列のいずれかをコードする塩基配列であり得る。この遺伝子によってコードされるペプチド生成酵素は、ペプチド生成活性を有する。
【0032】
上記(b)の「ストリンジェントな条件」とは、6×SSC(1×SSCの組成:0.15MのNaCl、0.015Mのクエン酸ナトリウム、pH7.0)と0.5%SDSと5×デンハルトと100μg/mLの変性サケ精子DNAと50%(v/v)ホルムアミドとを含む溶液中、室温にて12時間インキュベートし、更に0.5×SSCで50℃以上の温度で洗浄する条件をいう。さらに、よりストリンジェントな条件、例えば、45℃又は60℃にて12時間インキュベートすること、0.2×SSC又は0.1×SSCで洗浄すること、洗浄に際し60℃又は65℃以上の温度条件で洗浄すること等の、より厳しい条件も含む。また、本実施形態においては、配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖とストリンジェントな条件下でハイブリダイズする塩基配列は、該塩基配列によりコードされるアミノ酸配列に関して、配列表の配列番号10のアミノ酸配列との配列同一性が、高いほど好ましい。具体的には、アミノ酸の配列同一性は80%以上、85%以上、90%以上、95%以上、又は98%以上である。また、該塩基配列に関して、配列表の配列番号10のアミノ酸配列をコードする塩基配列との配列同一性が、高いほど好ましい。具体的には、塩基配列の配列同一性は80%以上、85%以上、90%以上、又は95%以上である塩基配列との特異的なハイブリダイゼーションが達成され得る条件であることが好ましい。本実施形態においては、塩基配列の配列同一性は、CLUSTALアルゴリズムで算出された値である。
【0033】
本実施形態に係る遺伝子に基づき、本実施形態に係るペプチド生成酵素と同等のペプチド生成活性を示すアイソザイムをスクリーニングするための、プローブ及びプライマーセットも提供できる。
【0034】
本実施形態に係る遺伝子を含むペプチド生成酵素の発現用担体を用いて、本実施形態に係る遺伝子によってコードされるペプチド生成酵素の組換えタンパク質(組換えペプチド生成酵素)を製造することもできる。本実施形態において、「発現用担体」とは、ベクター等を基本骨格として含有し、かつ適切な位置に本実施形態に係る遺伝子が作動可能に連結された核酸の構築物、並びに、金粒子、リポソーム、デキストラン、及びリン酸カルシウム等の担体に、細胞内での発現に適したエレメント等を有する本実施形態に係る遺伝子を担持させた構築物等を意味する。
【0035】
本実施形態に係るペプチド生成酵素の発現用担体を用いて、本実施形態に係るペプチド生成酵素をコードする遺伝子をStreptomyces aureofaciens TH−3株以外の細胞に導入することができ、それによって、異種細胞において本実施形態に係るペプチド生成酵素を発現させることができる。したがって、本実施形態に係るペプチド生成酵素の工業的な生産のための手段が提供される。
【0036】
上記ベクターとしては、pUC118、pUC119、pBR322、pCR3、pCMVSPORT、pETkmS2(Mishima, N.ら、Biotechnol. Prog.、第13巻、864-868頁、1997年)、pOMal(New England Biolabs)、及びpET−28a(+)(ノバジェン社製)等の大腸菌用プラスミドベクター、λZAPII、及びλgt11等の大腸菌用ファージベクター、pYES2、pYEUra3、及びpPICZα(インビトロジェン株式会社)等の酵母用ベクター、pIJ486(Mol. Gen. Genet.、203巻、468-478頁、1986年)、pKC1064(Gene、103巻、97-99頁、1991年)、pUWL−KS(Gene、165巻、149-150頁、1995年)、pIJ702(Katz, E.ら、J. Gen. Microbiol.、129巻、2703-2714頁、1983年)、及びpIJ8600(Sun, J.ら、Microbiology、145巻、2221-2227頁、1999年)等の放線菌用ベクター、pAcSGHisNT−A等の昆虫細胞用ベクター、pKCR、pEFBOS、cDM8、及びpCEV4等の動物細胞用ベクター等が挙げられる。このようなベクターには、適切なプロモーター(例えば、lacプロモーター、tacプロモーター、trcプロモーター、trpプロモーター、CMVプロモーター、及びSV40初期プロモーター等)、選択マーカー遺伝子、ターミネーター等のエレメントを適宜有していてもよい。
【0037】
上記ベクターは、本実施形態に係るペプチド生成酵素を、Hisタグ等のタグ、又はグルタチオンS−トランスフェラーゼ等の可溶性のタンパク質と融合した状態で発現させ得るベクターであってもよい。
【0038】
このようなベクターの調製に用いられる菌としては、例えば、当該技術分野で通常用いられる宿主菌が挙げられ、具体的には、放線菌、及び大腸菌が挙げられる。より具体的には、放線菌としては、ストレプトマイセス リビダンス1326、及びストレプトマイセス リビダンスTK−23等、そして大腸菌としては、BL21(DE3)、BL21−Gold (DE3)、及びJM109等が挙げられる。これらの宿主菌のうち、精製の容易化、大量調製、及び安全性等の観点から、大腸菌BL21(DE3)、BL21−Gold (DE3)、及びJM109が好ましい。
【0039】
本実施形態に係るペプチド生成酵素の発現用担体は、適切な宿主細胞へ導入されて、宿主細胞を形質転換して、形質転換細胞を作製する。形質転換法として、例えば、リン酸カルシウム法、DEAE−デキストラン法、エレクトロポレーション法、及びパーティクル銃によるボンバードメント法等、当業者に周知の方法が採用される。
【0040】
本実施形態に係るペプチド生成酵素の発現用の宿主細胞は、好適には微生物(宿主菌)が用いられる。宿主菌としては、放線菌、大腸菌、バシラス属菌、シュードモナス属菌、サーマス属菌、アグロバクテリウム属菌、及び酵母(例えば、メタノール資化性の酵母であるPichia)等が挙げられる。中でも、本発明においては、大腸菌が好適に用いられる。
【0041】
宿主細胞へ上記発現用担体の導入によって得られた細胞が、目的の形質転換細胞であることは、細胞又は該細胞の培養上清について、上記ペプチド生成酵素の活性の測定法により、ペプチド生成活性が検出されることを指標として確認され得る。
【0042】
上記形質転換細胞を培養して増殖させることによって、本実施形態に係るペプチド生成酵素を遺伝子工学的に容易に精製できる状態で発現させることができ、それによって、本実施形態に係るペプチド生成酵素を工業的に製造することができる。
【0043】
本実施形態に係る組換えペプチド生成酵素の製造においては、培地組成、培地のpH、培養温度、培養時間の他、インデューサーの使用量、及び使用時間等について、組換えペプチド生成酵素の発現の最適な条件を決定することによって、効率よく組換えペプチド生成酵素を生産させることができる。
【0044】
本実施形態に係る形質転換細胞の培養に通常用いられる培地としては、天然培地及び合成培地のいずれを用いてもよい。液体培地又は固体培地を使用することができる。例えば、宿主菌が資化し得る炭素源、窒素源、及び無機塩類等を含有し、形質転換体の培養を効率的に行うことができる培地であればよい。炭素源としては、グルコース、ガラクトース、フラクトース、スクロース、ラフィノース、及びデンプン等の炭水化物、酢酸、及びプロピオン酸等の有機酸、並びに、エタノール、及びプロパノール等のアルコール類が挙げられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、及びリン酸アンモニウム等の無機酸、若しくは有機酸のアンモニウム塩又はその他の含窒素化合物が挙げられる。その他、ペプトン、肉エキス、コーンスティープリカー、及び各種アミノ酸等を含んでいてもよい。その他に、無機塩として、微量のカリウム、ナトリウム、及び鉄等のリン酸塩、塩酸塩、硝酸塩、並びに、酢酸塩等を含んでいてもよい。また、必要に応じて植物油、界面活性剤、及びシリコン等の消泡剤を添加してもよい。市販の培地としては、Overnight Express(登録商標) Instant TB培地(ノバジェン社製)等が挙げられる。
【0045】
培養条件は、培地の種類、培養方法等により適宜選択すればよく、宿主細胞が増殖し、目的のタンパク質を産生できる条件であれば特に制限はない。通常、液体培地中で振とう培養又は攪拌培養等の好気的条件下で、10℃〜40℃、好ましくは25℃で12時間〜120時間行われる。pHは、4〜10、好ましくは6〜8に調節される。pHの調整は、無機酸又は有機酸、並びに、アルカリ溶液等を用いて行う。
【0046】
形質転換細胞が産生する組換えペプチド生成酵素は、それが細胞内に産生されるときは細胞内の他のタンパク質、又は他のポリペプチド等が共存するが、これらは発現されるペプチド生成酵素の量に比べて微量にすぎないため、その精製は極めて容易であるという優れた利点がある。また、ベクターとして菌体外分泌型のベクターを用いた場合、ペプチド生成酵素が菌体外に分泌される。
【0047】
形質転換細胞の培養物からペプチド生成酵素を精製するには通常の方法が用いられる。形質転換細胞が大腸菌のように細胞内にペプチド生成酵素が蓄積する場合には、培養終了後、遠心分離によって形質転換細胞を集め、得られた細胞を超音波処理等によって破砕した後、遠心分離等によって無細胞抽出液を得る。これを出発材料とし、塩析法、並びに、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、疎水クロマトグラフィー、及びアフィニティークロマトグラフィー等の各種クロマトグラフィー等の一般的なタンパク質精製法によって精製することができる。用いる形質転換細胞によっては、発現した組換えタンパク質が細胞外に分泌される場合がある。このような場合、培養上清から同様に精製を行えばよい。
【0048】
以下、本実施形態に係るペプチド生成酵素を用いた、N末端にプロリン残基を有するペプチドの製造方法について説明する。本実施形態に係る製造方法は、本実施形態に係るペプチド生成酵素の存在下で、プロリン誘導体とアミノ酸とを縮合させる工程を含む。
【0049】
プロリン誘導体としては、プロリンエステル又はプロリンアミドであることが好ましい。プロリンエステルとしては、例えば、ベンジルエステル、メチルエステル等が挙げられる。特にベンジルエステル及びメチルエステルが好ましい。プロリン誘導体をこのような化合物とすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0050】
アミノ酸は、α−アミノ酸であることが好ましく、炭素数2〜10のα−アミノ酸であることがより好ましく、L−ヒドロキシプロリン、L−プロリン、L−バリン、L−アラニン、グリシン、L−システイン、L−リジン、L−セリン、L−トレオニン、L−アルギニン、(R)−α−アリルグリシン、(S)−α−プロパギルグリシン、(R)−α−プロパギルグリシン、(S)−α−メチルロイシン、(R)−α−メチルロイシン、又は(S)−α−メチルフェニルアラニンであることがさらに好ましい。アミノ酸をこのような化合物とすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0051】
pH9.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させることが好ましく、pH10.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させることがより好ましい。pHが9.5未満の場合、酵素のペプチド生成活性が低下する傾向がある。pHの上限は特に設けないが、取り扱い性の観点からpH12未満が好ましい。
【0052】
上記反応液は、酵素を失活させなければどの様な溶媒でも用いることが可能であるが、通常は緩衝水溶液が好ましく、ホウ酸緩衝水溶液がより好ましい。緩衝水溶液の濃度は特に制限はないが、終濃度で、80mM〜1280mMが好ましく、480mM〜640mMがより好ましい。
【0053】
上記プロリン誘導体とアミノ酸とを縮合させる工程は、反応時間が0.5時間〜3時間であることが好ましく、1.5時間〜3時間であることがより好ましい。
【0054】
反応時間が1時間未満の場合、反応温度は、4℃〜37℃が好ましく、25℃〜37℃がより好ましく、25℃が特に好ましい。一方反応時間が2時間を超える場合、反応温度は、4℃〜37℃が好ましく、4℃がより好ましい。
【0055】
本実施形態に係る製造方法によって製造されたペプチドは、高純度の状態で得ることが可能である。また、得られたペプチドの純度をさらに上げるため、塩析法、並びに、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、疎水クロマトグラフィー、及びアフィニティークロマトグラフィー等の各種クロマトグラフィー等の一般的なタンパク質精製法により精製してもよい。
【実施例】
【0056】
以下、実施例により本発明をより詳細に説明するが、本発明は、実施例により限定されるものではない。特に明記しない限り、各種操作は、モレキュラークローニング ア ラボラトリー マニュアル(Molecular Cloning A Laboratory Manual)第2版(ザンブルーク(Sambrook, J)ら、1989年)に従って行った。なお、実施例において、%は、特に明記しない限り、w/v%を表す。
【0057】
(PAP遺伝子のクローニング)
ストレプトマイセス サーモグリセウス(Streptomyces thermogriseus) NBRC100772株からゲノムDNAを、Hopwoodらの方法(D.A. Hopwoodら、A Laboratory Manual, The John Ines Foundation,Norwich,1985年,70-84頁)を用いて調製した。次に、Streptomyces coelicolor (gene ID: SCO5122)、Streptomyces avermitilis (gene ID: SAV3144)及びStreptomyces griseus (gene ID: SGR2407)由来のPAPの一次配列から縮重PCRプライマー(フォワードプライマー:5’−CAGWSSTTCGGSGGSTTCTGC−3’(配列番号13)及びリバースプライマー:5’−ACGTACATGTCGTCGTGGTA−3’(配列番号14))を設計した。上記ゲノムDNAを鋳型鎖とし、上記縮重PCRプライマーセット及び、LA Taq DNAポリメラーゼ(宝ホールディングス株式会社製)を用いて、PCR法によってNBRC100772株由来のPAP(PAP10)のゲノムDNAの内部配列を増幅した。PCRは、94℃にて1分のインキュベーションの後、94℃にて30秒、60℃にて30秒、72℃にて90秒を30サイクル行い、次いで72℃にて5分のインキュベーションで行った。PCR産物と、上記PCRプライマーを用いてPAP10をコードする遺伝子(pap10遺伝子)の部分内部配列を決定した。得られた部分内部配列を基に、pap10遺伝子の全長を得るためのPCRプライマー(センスプライマー:5’−ACCGACTGGCGGCCCCTGTACGAC−3’(配列番号15)及びアンチセンスプライマー:5’−GACCTTGCGCTCGATGCGCGGGTA−3’(配列番号16))を設計した。一方、上記ゲノムDNAを制限酵素CpoI(37℃、一晩)で消化し、消化DNAを自己ライゲートさせた。ライゲーション産物を上述したpap10遺伝子の部分内部配列から設計したプライマーセット(配列番号15及び配列番号16)を用いてPCR法によって増幅した。PCR産物をクローニングして塩基配列を決定した。PAP14(Streptomyces thermoluteus subsp. fuscus strain NBRC14270株由来のPAP)の遺伝子(pap14遺伝子)及びPAP TH−3(Streptomyces aureofaciens TH−3株由来のPAP)の遺伝子(pap th−3遺伝子)の全長塩基配列も同様の方法で取得した。配列表の配列番号との対応を表1に示す。
【0058】
【表1】

【0059】
(組換えPAP発現ベクターの構築)
大腸菌を宿主とする場合のPAPを発現するためのベクター(組換えPAP発現ベクター)を以下のようにして構築した。まず、各遺伝子に対応するNdeI部位を含むセンスプライマーと停止コドンの下流にHindIII部位を含むアンチセンスプライマーとのセット及びPrime STAR(登録商標) GXL DNA ポリメラーゼ (タカラバイオ社製)を用いてPCR法により各PAP遺伝子を、増幅した。各PAP遺伝子に対応するプライマーセットは、表2に示した。得られたDNAフラグメントを、Zero Blunt II TOPO PCRクローニングキット(インビトロジェン社製)を用いてクローニングして塩基配列を決定した。PAPをコードするDNAフラグメントをNdeI及びHindIIIの制限酵素で切断し、そしてpET−28a(+)(ノバジェン社製)のNdeI−HindIIIギャップに連結し、N末端側にHisタグを有する組換えPAP発現ベクターを得た。
【0060】
【表2】
【0061】
(組換えPAPの発現及び精製)
上述のようにして得られた組換えPAPの発現ベクターを、GenePulseXcell(登録商標)(バイオラッド社製)を用いて、それぞれ大腸菌BL21−Gold (DE3)(アジレント テクノロジー株式会社製)に導入して形質転換した。形質転換体の単一コロニーを、50mlのOvernight Express(登録商標) Instant TB培地(ノバジェン社製)中で、overnight express system 1(ノバジェン社製)を用いて培養した。25℃、180rpmの条件で48時間培養した後、遠心分離によって培養液から菌体を回収した。得られた菌体は、Elestin NP035SP(ネッパジーン社製、商品名)を用いて超音波破砕した。破砕条件は、出力最大、5分間超音波破砕した後に1分間のインターバルを開けて、トータルで45分間超音波破砕するように設定した。得られた無細胞抽出液から、コバルトアフィニティ樹脂(クロンテック社製、商品名TALON(登録商標))を用いて組換えPAPを精製した。精製して得られた各PAPは、SDS−PAGE(還元条件、クマシーブルー染色)で解析した。各PAPの濃度は、Bio−Rad protein assay reagent(バイオラッド社製)を用いて定量した。
【0062】
(プロリン残基を加水分解する活性(プロリン残基加水分解活性)の評価)
水320μl、200mM緩衝水溶液(PAP14の場合、リン酸緩衝水溶液 pH7;PAP10の場合、リン酸緩衝水溶液 pH7.5;PAP TH−3の場合、トリスヒドロキシメチルアミノメタン緩衝水溶液(トリス緩衝水溶液) pH8)320μl、L−プロリン−p−ニトロアニリド(SIGMA社製、Pro−pNA)のジメチルスルホキシド(ナカライテスク社製、DMSO)溶液(0.0625mM〜8.0mM)80μlの混合液を用意し、そこに20μlのPAP水溶液を加えて37℃で反応させた。加水分解反応によって遊離したpNAに起因する405nmの吸光度の変化を分光光度計(日立製作所製、商品名U2800)で1分間モニターした。405nmにおける吸光度のプロファイル(ε405nm=10600M−1cm−1)を基に、上記加水分解反応における速度論的なパラメーターを求めた。結果を表3に示す。なお、酵素活性(プロリン残基加水分解活性)の1ユニット(U)は、各反応条件において、1分間に1μmolのpNAを遊離させるのに必要な酵素量として定義した。
【0063】
【表3】
【0064】
(PAPの生化学的性質の評価)
PAPの組成は、Superdex 200 10/300カラム(GE ヘルスケア社製)を用いたゲル濾過クロマトグラフィーによって評価した。プロリン残基加水分解活性の至適pHは、終濃度80mMのリン酸緩衝水溶液(pH 6.0〜8.0)又はトリス緩衝水溶液(pH 7.5〜9.0)を用いて決定した。またプロリン残基加水分解活性の至適温度は、35.0℃〜65.0℃の間で決定した。熱安定性は、酵素を至適pHで40.0℃〜80.0℃の間で30分間インキュベートした後の残存するプロリン加水分解活性と、酵素を4℃で保存した時のプロリン残基加水分解活性とを比較することで決定した。pH安定性は、酵素をpH6〜10.5で、30℃で30分間インキュベートした後の残存するプロリン残基加水分解活性と、至適pHにおけるプロリン残基加水分解活性とを比較することで決定した。結果を表4に示す。
【0065】
【表4】
【0066】
表3及び表4の結果から、PAP14、PAP10及びPAP TH−3のプロリン残基加水分解活性、及び生化学的性質に大きな差は認められないことが明らかとなった。
【0067】
(Pro−Hypジペプチドを生成する活性(ペプチド生成活性)の評価)
以下に示す手順にしたがって、Pro−Hypジペプチドの生成反応を行った。400mMホウ酸緩衝水溶液(pH11.0)40μl中に、アシル供与体(プロリン誘導体)として、1MのL−プロリンベンジル塩酸塩(Bachem社製、Pro−OBzl)のDMSO溶液を5μl、アシル受容体(アミノ酸)として、2MのL−ヒドロキシプロリン(和光純薬工業株式会社製、Hyp−H)の水溶液を50μl、及び720mUのPAPを含む、酵素溶液を5μlそれぞれ加えて、900rpm、37℃で3時間反応させた。その後、200μlのメタノールを反応液に加えることで、反応を終了させた。反応液を10000rpmで遠心した後、得られた上精を33倍希釈した溶液2.5μlを用いて液体クロマトグラフィー/マススペクトル(LC/MS)分析を行った。
【0068】
LC/MS/MS分析は、TUV検出器、バイナリーソルベントマネージャー、及びAPI2000 LC/MS/MSシステム(AB SCIEX社製)を備えたACQUITY(登録商標)ウルトラパフォーマンス液体クロマトグラフィー(UPLC)システム(ウォーターズ社製)を用いて行った。カラムは、XBridge(登録商標)アミドカラム(3.5μm、2.1×50mm、ウォーターズ社製)を用いた。移動層は、(A)液として、水(0.1%ギ酸)、(B)液としてアセトニトリル(0.1%ギ酸)を用いた。グラジェントプロファイルは、以下のように設定した、流速 0.3ml/min、0〜2分 2%(A)液、2〜13.5分 2〜30%(A)液。Pro−Hypと基質を含む反応混合液を検出するため、イオンクロマトグラフィーから溶出した時に現れる各ピークエリアをAPI2000のデバイスシステム(マルチリアクションモニターモード(MRM))によって計算した。Pro−OBzl、Pro−NH2、Pro−Hyp、及びHyp−Hの各フラグメンテーションのパターンは、それぞれ、m/z=206.2/91.0、114.7/70、229.0/131.9及び132.1/86.0であった。
【0069】
ペプチド生成酵素としてPAP TH−3(実施例1)を、プロリン誘導体としてPro−OBzl、及びアミノ酸としてHyp−Hを用いて、上記反応手順によって得られた反応混合物を、多重反応モニタリングクロマトグラフィーで分析した結果を図1に示す。Pro−Hypを示すピークが保持時間8分のところに現れていることから、PAP TH−3のペプチド生成活性によってPro−OBzl及びHyp−Hから、Pro−Hypが合成されたことが明らかとなった。また、Pro−Hypを示すピーク以外には、過剰に用いたHyp−Hのピーク(保持時間9.7分)しかみられなかったことから、高純度のPro−Hypを得ることが可能な反応であることが分かった。同様の実験をPAP14(比較例1)及びPAP10(比較例2)でも行ったが、Pro−Hypは検出できなかった(不図示)。以下、PAP TH−3のペプチド生成活性を詳細に検討した。
【0070】
(ペプチド生成活性のpH依存性及び緩衝水溶液濃度依存性)
図2は、Pro−Hypジペプチドの生成反応における、(A)pH依存性及び(B)緩衝水溶液の濃度の依存性を示すグラフである。(A)pHを9.5〜12.0の範囲で変化させたこと又は、(B)ホウ酸緩衝水溶液の濃度を80mM〜1280mMの範囲で変化させたこと以外は、上記(Pro−Hypジペプチド生成活性の評価)の手順に従って収率を求めた。pH9.5から収率が上昇し始め、pH10.5〜12.0の範囲において、約9%になることが明らかとなった(図2(A))。またpH11.0において、ホウ酸濃度480mMで収率が約17%になることが明らかとなった(図2(B))。
【0071】
(ペプチド生成活性の温度依存性)
図3は、Pro−Hypジペプチドの生成反応における、温度依存性を示すグラフである。緩衝水溶液を1200mMのホウ酸緩衝水溶液(pH 11.0)にして、反応温度と反応時間を変化させたこと以外は、上記(Pro−Hypジペプチド生成活性の評価)の手順に従って収率を求めた。反応温度37℃においては、反応時間30分で収率が最大となり、その後徐々に収率は減少した。反応温度25℃においては、反応時間30分で収率が最大となり、その後収率は、減少した。反応温度4℃においては、反応時間の増加に伴い収率も徐々に上昇し、反応時間3時間で収率25%を示した。
【0072】
(ペプチド生成活性のプロリン誘導体の影響)
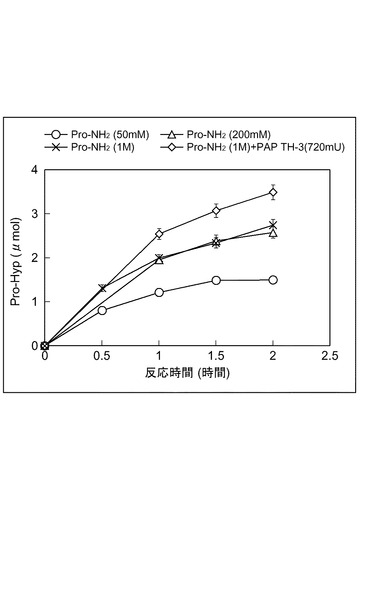
図4は、Pro−Hypジペプチドの生成反応における、プロリン誘導体の影響を示すグラフである。プロリン誘導体を変えたこと、反応温度を4℃にしたこと以外は、上記(ペプチド生成活性の温度依存性)の手順に従って収率を求めた。プロリン誘導体としては、Pro−OBzlの他に、L−プロリンメチル塩酸塩(ALDRICHI社製、Pro−OMe)及び、L−プロリンアミド塩酸塩(SIGMA社製、Pro−NH2)を用いた。いずれの反応時間においても、Pro−NH2、Pro−OMe、Pro−OBzlの順に収率が高いことが明らかになった。
【0073】
(Pro−Hypジペプチドの生成反応における、プロリン誘導体を連続して添加したときの影響)
Pro−Hypジペプチドの生成反応における、プロリン誘導体を連続して添加したときの影響を調べた。まず、1200mMホウ酸緩衝水溶液(pH11.0)40μl中に、プロリン誘導体として、1MのPro−NH2のDMSO溶液を5μl、アミノ酸として、2MのHyp−Hの水溶液を50μl、及び720mUのPAP TH−3を含む酵素溶液を5μlを、それぞれ加えて、900rpm、4℃で反応させた。上記反応混合物に、50mMのPro−NH2、200mMのPro−NH2、若しくは1MのPro−NH2のDMSO溶液を5μl、又は、1MのPro−NH2のDMSO溶液を5μlと720mUのPAP TH−3溶液を1.6μlとの混合液を終容量100μlとなるように調整して、30分おきに加えた。結果を図5に示す。いずれの場合もプロリン誘導体を添加するたびに、Pro−Hypの収量は、増加した。特に、Pro−NH2とPAP TH−3を同時に加えたとき収量がもっとも増加しており、50mM Pro−NH2を添加したときと比べて、反応時間2時間の時点で2.3倍であった。
【0074】
(アミノ酸の検討)
Hyp−Hの他にも、アシル受容体として有効なアミノ酸があるのかどうかを式(1)〜式(9)で表わされる天然アミノ酸及び、式(10)〜式(18)で表わされる非天然アミノ酸について検討した。
【0075】
【化1】
【0076】
【化2】
【0077】
まず、1200mMホウ酸緩衝水溶液(pH11.0)40μl中に、プロリン誘導体として、1MのPro−NH2のDMSO溶液を5μl、アミノ酸として、上記式(1)〜式(18)で表される化合物の水溶液を、50μl、及び720mUのPAP TH−3を含む酵素溶液を5μlそれぞれ加えて、4℃で3時間反応させた。なお、アミノ酸は、終濃度が下記表5に示す濃度となるように加えた。反応混合物を液体クロマトグラフィー/マススペクトル(LC/MS)分析することで、反応生成物を検出した。結果を表5に示す。
【0078】
【表5】
【0079】
表5に示される結果から、PAP TH−3のペプチド生成活性は、基質として用いるアミノ酸として、天然アミノ酸のみならず、非天然アミノ酸でも有効であることが明らかになった。さらに、式(11)と式(12)の化合物、及び式(13)と式(14)の化合物は、互いに光学異性体の関係にあることから、PAP TH−3のペプチド生成活性は、基質となるアミノ酸の光学活性に依存しないことが明らかになった。
【0080】
(参考例1)
(C末端の12アミノ酸残基が欠損したPAP TH−3の発現ベクターの構築)
大腸菌を宿主とする場合の、C末端の12アミノ酸残基が欠損したPAP TH−3を発現するためのベクター(組換えC末端12アミノ酸残基欠損PAP TH−3発現ベクター)を以下のようにして構築した。まず、PAP TH−3遺伝子に対応するNdeI部位を含むセンスプライマー:5’−CATATGTCCACCGTCAGCCGGCTG−3’(配列番号17)と停止コドンの下流にHindIII部位を含むアンチセンスプライマー:5’−AAGCTTAGGCCATCTTGATCAGCCGGTC−3’(配列番号18)及びPrime STAR(登録商標) GXL DNA ポリメラーゼ (タカラバイオ社製)を用いてPCR法によりC末端の12アミノ酸残基が欠損したPAP TH−3をコードする遺伝子を増幅した。得られたDNAフラグメントを、Zero Blunt II TOPO PCRクローニングキット(インビトロジェン社製)を用いてクローニングして塩基配列を決定した。PAPをコードするDNAフラグメントをNdeI及びHindIIIの制限酵素で切断し、そしてpET−28a(+)(ノバジェン社製)のNdeI−HindIIIギャップに連結し、N末端側にHisタグを有する組換えC末端12アミノ酸残基欠損PAP TH−3発現ベクターを得た。
【0081】
(組換えC末端12アミノ酸残基欠損PAP TH−3の発現及び精製)
上記(組換えPAPの発現及び精製)に記載の方法と同様の方法で発現及び精製を行った。
【0082】
(ペプチド生成活性の評価)
上記(Pro−Hypジペプチドを生成する活性(ペプチド生成活性)の評価)に記載の方法と同様の方法で組換えC末端12アミノ酸残基欠損PAP TH−3のペプチド生成活性評価を行い、Pro−Hypジペプチドの生成を確認することができた。
【0083】
(参考例2)
(PAP10とPAP TH−3とのキメラPAP遺伝子のクローニング及びそのキメラPAP発現ベクターの構築)
まず、PAP TH−3遺伝子にNsbI部位を導入するため、センスプライマー:5’−GGTGCGCGGCCTGCGCACGTGGATCACCAA−3’(配列番号19)とアンチセンスプライマー:5’−TTGGTGATCCACGTGCGCAGGCCGCGCACC−3’(配列番号20)及びクイックチェンジキット(Stratagene製)を用いて、PAP TH−3遺伝子にNsbI部位が導入された遺伝子を増幅した。得られたDNAフラグメントは、PAP TH−3遺伝子にNsbI部位が導入されていることを確認した。
【0084】
PAP TH−3遺伝子にNsbI部位が導入されたDNAフラグメントをNdeI及びHindIIIの制限酵素で切断し、そしてpET−28a(+)(ノバジェン社製)のNdeI−HindIIIギャップに連結した。得られたプラスミドを、NsbI及びClaIで切断した。一方、PAP−10遺伝子が挿入されているプラスミドを、NsbI及びClaIの制限酵素で切断してPAP10遺伝子のフラグメントを得た。得られたPAP10遺伝子のフラグメントを、上述のPAP TH−3遺伝子が挿入されたpET−28a(+)のNsbI−ClaIギャップに挿入して、PAP−10とPAP TH−3とのキメラPAP遺伝子(配列番号21)の発現ベクターを得た。このキメラPAPは、PAP TH−3のN末側402個のアミノ酸残基がPAP10のものに置き換わっている。また、このキメラPAPは、PAP TH−3と比較して、117アミノ酸残基が異なっていることが確認された。
【0085】
(PAP−10とPAP TH−3とのキメラPAPの発現及び精製)
上記(組換えPAPの発現及び精製)に記載の方法と同様の方法で発現及び精製を行った。
【0086】
(ペプチド生成活性の評価)
上記(Pro−Hypジペプチドを生成する活性(ペプチド生成活性)の評価)に記載の方法と同様の方法で組換えPAP−10とPAP TH−3とのキメラPAPのペプチド生成活性評価を行い、Pro−Hypジペプチドの生成を確認することができた。
【産業上の利用可能性】
【0087】
本発明のタンパク質(ペプチド生成酵素)は、Pro−Hyp等のN末端にプロリン残基を有するペプチドを、高純度で製造することができるため、工業及び医療の分野において応用が可能である。例えば、骨粗鬆症、胃潰瘍、変形性関節症、褥瘡及び高血圧用の薬剤、皮膚保湿剤、並びに、保存剤の用途が挙げられる。
【受託番号】
【0088】
Streptomyces aureofaciens TH−3として命名及び表示される微生物は、受託番号 FERM P−21343(寄託日:2007年8月17日)として、独立行政法人産業技術総合研究所 特許生物寄託センター(日本国茨城県つくば市東1丁目1−1)に寄託されている。
【技術分野】
【0001】
本発明は、ペプチド生成酵素、その遺伝子及びペプチドの製造方法に関する。
【背景技術】
【0002】
コラーゲンは、3本のポリペプチド鎖からなる三重らせん構造を有しており、ポリペプチド鎖は、トリペプチド配列Gly−X−Yを繰り返し単位とする繰り返し構造を有する。このX及びYの位置のアミノ酸は、プロリン(Pro)及び翻訳後修飾によって生成する4−ヒドロキシプロリン(L−ヒドロキシプロリン、Hyp)であることが多い。コラーゲンは、4−ヒドロキシプロリンによってその三重らせんが非常に安定化されていることから、水に不溶であり、ほとんどのプロテアーゼに対して抵抗性を有する。
【0003】
コラーゲンの分解物であるコラーゲンペプチドは、工業及び医療の分野において、種々の有用な生物学的活性を有することが報告されており、急速にそして広範に適用されている。例えば、骨粗鬆症、胃潰瘍、変形性関節症、褥瘡及び高血圧用の薬剤、皮膚保湿剤、並びに、保存剤の用途が挙げられる。
【0004】
近年、コラーゲンペプチドのなかでも、プロリン及び4−ヒドロキシプロリンから構成されるPro−Hyp等のジペプチドが上記生物学的活性に寄与していることが明らかとなり、このようなジペプチドが注目されている(特許文献1)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2010−106003号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
特許文献1では、Pro−Hyp等のジペプチドを生成する方法として、(1)コラゲナーゼ等の酵素の存在下、コラーゲン又はゼラチンを分解することによって得る方法、並びに、(2)固相合成法及び液相合成法等の化学合成法によって、アミノ酸から合成する方法が挙げられている。
【0007】
しかしながら、上記(1)の方法では、原料であるコラーゲン又はゼラチン、並びに、反応中間体であるコラーゲンペプチド等のポリペプチドが不純物としてPro−Hypと共に含まれるため、Pro−Hypを高純度の状態で得るためには、更に精製する工程が必要となる。上記(2)の方法では、トリフルオロ酢酸等を使用することから、生成したジペプチドを医薬品等の目的で使用することは困難な場合が多い。
【0008】
本発明は、上記事情に鑑みてなされたものであり、N末端にプロリン残基を有するペプチドを高純度で製造することが可能なペプチド生成酵素、その遺伝子及びペプチドの製造方法を提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者等は、鋭意研究を行った結果、ある特定のアミノ酸配列を有するタンパク質が、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有していることを見出し、本願発明を完成するに至った。
【0010】
すなわち、本発明は、(A)配列表の配列番号10に記載のアミノ酸配列、(B)配列表の配列番号10に記載のアミノ酸配列において1又は数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列、及び(C)配列表の配列番号10に記載のアミノ酸配列に対して80%以上の配列同一性を有するアミノ酸配列、からなる群から選択されるアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させて、N末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素を提供する。
【0011】
本発明に係るペプチド生成酵素は、このようなアミノ酸配列を有することで、プロリン誘導体とアミノ酸とを縮合させることによって、N末端にプロリン残基を有するペプチドを生成することが可能となる。その結果、N末端にプロリン残基を有するペプチドを高純度の状態で提供することが可能となる。
【0012】
本発明は、(a)上記ジペプチド生成酵素をコードする塩基配列、及び(b)配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖とストリンジェントな条件下にハイブリダイズし、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素をコードする塩基配列、からなる群より選択される塩基配列を有する遺伝子を提供する。
【0013】
本発明は、上記ペプチド生成酵素の存在下で、プロリン誘導体とアミノ酸とを縮合させる工程を含む、N末端にプロリン残基を有するペプチドの製造方法を提供する。
【0014】
上記プロリン誘導体は、プロリンエステル又はプロリンアミドであることが好ましい。プロリン誘導体をこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0015】
上記プロリンエステルは、ベンジルエステル又はメチルエステルであることが好ましい。プロリンエステルをこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0016】
上記アミノ酸は、α−アミノ酸であることが好ましい。アミノ酸をこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0017】
上記アミノ酸は、L−ヒドロキシプロリン、L−プロリン、L−バリン、L−アラニン、グリシン、L−システイン、L−リジン、L−セリン、L−トレオニン、L−アルギニン、(R)−α−アリルグリシン、(S)−α−プロパギルグリシン、(R)−α−プロパギルグリシン、(S)−α−メチルロイシン、(R)−α−メチルロイシン、又は(S)−α−メチルフェニルアラニンであることが好ましい。アミノ酸をこのような化合物にすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0018】
pH9.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させることが好ましい。このようにすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【発明の効果】
【0019】
本発明によれば、N末端にプロリン残基を有するペプチドを高純度で製造することが可能なペプチド生成酵素、その遺伝子及びペプチドの製造方法が提供される。
【図面の簡単な説明】
【0020】
【図1】反応混合物を多重反応モニタリングクロマトグラフィーで分析した結果を示すグラフである。
【図2】Pro−Hypジペプチドの生成反応における、(A)pH依存性及び(B)緩衝水溶液の濃度の依存性を示すグラフである。
【図3】Pro−Hypジペプチドの生成反応における、温度依存性を示すグラフである。
【図4】Pro−Hypジペプチドの生成反応における、プロリン誘導体の影響を示すグラフである。
【図5】Pro−Hypジペプチドの生成反応における、プロリン誘導体を連続して添加したときの影響を示すグラフである。
【発明を実施するための形態】
【0021】
本明細書において、アミノ酸の表記を、3文字又は1文字で表記する場合がある。すなわち、アラニンは、Ala又はA、アルギニンは、Arg又はR、アスパラギンは、Asn又はN、アスパラギン酸は、Asp又はD、システインは、Cys又はC、グルタミン酸は、Glu又はE、グルタミンは、Gln又はQ、グリシンは、Gly又はG、ヒスチジンは、His又はH、イソロイシンは、Ile又はI、ロイシンは、Leu又はL、リジンは、Lys又はK、メチオニンは、Met又はM、フェニルアラニンは、Phe又はF、プロリンは、Pro又はP、セリンは、Ser又はS、トレオニンは、Thr又はT、トリプトファンは、Trp又はW、チロシンは、Tyr又はY、並びに、バリンは、Val又はVを表記する。
【0022】
本実施形態に係るペプチド生成酵素は、
(A)配列表の配列番号10に記載のアミノ酸配列、
(B)配列表の配列番号10に記載のアミノ酸配列において1又は数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列、及び
(C)配列表の配列番号10に記載のアミノ酸配列に対して80%以上の配列同一性を有するアミノ酸配列、からなる群から選択されるアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有する。
【0023】
配列表の配列番号10に記載のアミノ酸配列は、Streptomyces aureofaciens TH−3株(FERM P−21343)のプロリンアミノペプチダーゼ(PAP TH−3)のアミノ酸配列である。
【0024】
上記(B)のアミノ酸配列について、配列表の配列番号10のアミノ酸残基の置換、欠失、挿入又は付加されたアミノ酸(変異されたアミノ酸)の数は、アミノ酸配列が後述するプロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性(以下、単にペプチド生成活性と呼ぶ場合がある。)の測定法の条件下に同様の活性を示す範囲であればよい。アミノ酸の変異の数は、少なくとも1個、好ましくは、1個〜117個、より好ましくは、1個〜12個である。変異されたアミノ酸配列は、天然に存在するアミノ酸配列、すなわち、自然発生による変異を有するアミノ酸配列であってもよく、人為的に所望の部位又はアミノ酸残基に変異を導入したアミノ酸配列であってもよい。人為的に所望の部位又はアミノ酸残基に変異を導入する場合、天然に存在する配列表の配列番号10のアミノ酸配列の変異体に準じて人為的に変異を導入してもよい。人為的な変異の導入は、例えば、PCR法を用いた部位特異的変異導入法等によって行われる。
【0025】
変異されたアミノ酸配列としては、例えば、配列表の配列番号10のアミノ酸配列において少なくとも1個のアミノ酸残基が類似した物理学的性質を有する他のアミノ酸残基に置換(保存的置換)されたアミノ酸配列が挙げられる。保存的置換は、例えば、疎水性、電荷、pK、又は立体構造上類似したアミノ酸残基同士の置換、並びに、本来のポリペプチドの生理活性を維持する程度に該ポリペプチドの立体構造や折り畳み構造が維持されるようなアミノ酸残基の置換である。より具体的には、上記(B)のアミノ酸配列は、配列表の配列番号10のアミノ酸配列において、少なくとも1個のアミノ酸残基が、以下のアミノ酸群(1)〜(6):
(1)グリシン及びアラニン、
(2)バリン、イソロイシン及びロイシン、
(3)アスパラギン酸、グルタミン酸、アスパラギン及びグルタミン、
(4)セリン及びスレオニン、
(5)リジン及びアルギニン、並びに
(6)フェニルアラニン及びチロシン
における、同一のアミノ酸群に属するアミノ酸残基同士の置換により他のアミノ酸と保存的置換されたものであることが好ましい。
【0026】
上記(C)において、配列同一性は、参照配列(例えば、配列表の配列番号10のアミノ酸配列)に対して、クエリー配列(評価対象のアミノ酸配列)を、適切にアラインメントし、算出された値である。具体的には、本実施形態においては、配列同一性は、CLUSTALアルゴリズムで算出された値である。
【0027】
本実施形態においては、配列同一性は、高いほど好ましく、具体的には、80%以上、85%以上、90%以上、95%以上、又は98%以上である。
【0028】
本実施形態において、ペプチド生成酵素のペプチド生成活性は、任意の方法で測定され得る。例えば、プロリン誘導体とアミノ酸とを基質として、上記ペプチド生成酵素によって生成されたN末端にプロリン残基を有するペプチドの生成量を多重反応モニタリングクロマトグラフィー等の方法を用いて測定する方法が挙げられる。
【0029】
本実施形態に係るペプチド生成酵素は、また、以下に詳述する本実施形態に係る遺伝子を用いて、遺伝子工学的に製造することができる。例えば、Streptomyces aureofaciens TH−3株とは異なる異種細胞において、本実施形態に係るペプチド生成酵素を発現させることができる。
【0030】
本実施形態に係るペプチド生成酵素をコードする遺伝子は、
(a)上記ペプチド生成酵素をコードする塩基配列、及び
(b)配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖と、ストリンジェントな条件下にハイブリダイズし、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素をコードする塩基配列、
からなる群より選択される塩基配列を有する。
【0031】
このうち(a)の塩基配列は、それぞれ、上記(A)〜(C)のアミノ酸配列をコードする塩基配列であり、そして(b)の塩基配列は、上記(A)〜(C)のアミノ酸配列のいずれかをコードする塩基配列であり得る。この遺伝子によってコードされるペプチド生成酵素は、ペプチド生成活性を有する。
【0032】
上記(b)の「ストリンジェントな条件」とは、6×SSC(1×SSCの組成:0.15MのNaCl、0.015Mのクエン酸ナトリウム、pH7.0)と0.5%SDSと5×デンハルトと100μg/mLの変性サケ精子DNAと50%(v/v)ホルムアミドとを含む溶液中、室温にて12時間インキュベートし、更に0.5×SSCで50℃以上の温度で洗浄する条件をいう。さらに、よりストリンジェントな条件、例えば、45℃又は60℃にて12時間インキュベートすること、0.2×SSC又は0.1×SSCで洗浄すること、洗浄に際し60℃又は65℃以上の温度条件で洗浄すること等の、より厳しい条件も含む。また、本実施形態においては、配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖とストリンジェントな条件下でハイブリダイズする塩基配列は、該塩基配列によりコードされるアミノ酸配列に関して、配列表の配列番号10のアミノ酸配列との配列同一性が、高いほど好ましい。具体的には、アミノ酸の配列同一性は80%以上、85%以上、90%以上、95%以上、又は98%以上である。また、該塩基配列に関して、配列表の配列番号10のアミノ酸配列をコードする塩基配列との配列同一性が、高いほど好ましい。具体的には、塩基配列の配列同一性は80%以上、85%以上、90%以上、又は95%以上である塩基配列との特異的なハイブリダイゼーションが達成され得る条件であることが好ましい。本実施形態においては、塩基配列の配列同一性は、CLUSTALアルゴリズムで算出された値である。
【0033】
本実施形態に係る遺伝子に基づき、本実施形態に係るペプチド生成酵素と同等のペプチド生成活性を示すアイソザイムをスクリーニングするための、プローブ及びプライマーセットも提供できる。
【0034】
本実施形態に係る遺伝子を含むペプチド生成酵素の発現用担体を用いて、本実施形態に係る遺伝子によってコードされるペプチド生成酵素の組換えタンパク質(組換えペプチド生成酵素)を製造することもできる。本実施形態において、「発現用担体」とは、ベクター等を基本骨格として含有し、かつ適切な位置に本実施形態に係る遺伝子が作動可能に連結された核酸の構築物、並びに、金粒子、リポソーム、デキストラン、及びリン酸カルシウム等の担体に、細胞内での発現に適したエレメント等を有する本実施形態に係る遺伝子を担持させた構築物等を意味する。
【0035】
本実施形態に係るペプチド生成酵素の発現用担体を用いて、本実施形態に係るペプチド生成酵素をコードする遺伝子をStreptomyces aureofaciens TH−3株以外の細胞に導入することができ、それによって、異種細胞において本実施形態に係るペプチド生成酵素を発現させることができる。したがって、本実施形態に係るペプチド生成酵素の工業的な生産のための手段が提供される。
【0036】
上記ベクターとしては、pUC118、pUC119、pBR322、pCR3、pCMVSPORT、pETkmS2(Mishima, N.ら、Biotechnol. Prog.、第13巻、864-868頁、1997年)、pOMal(New England Biolabs)、及びpET−28a(+)(ノバジェン社製)等の大腸菌用プラスミドベクター、λZAPII、及びλgt11等の大腸菌用ファージベクター、pYES2、pYEUra3、及びpPICZα(インビトロジェン株式会社)等の酵母用ベクター、pIJ486(Mol. Gen. Genet.、203巻、468-478頁、1986年)、pKC1064(Gene、103巻、97-99頁、1991年)、pUWL−KS(Gene、165巻、149-150頁、1995年)、pIJ702(Katz, E.ら、J. Gen. Microbiol.、129巻、2703-2714頁、1983年)、及びpIJ8600(Sun, J.ら、Microbiology、145巻、2221-2227頁、1999年)等の放線菌用ベクター、pAcSGHisNT−A等の昆虫細胞用ベクター、pKCR、pEFBOS、cDM8、及びpCEV4等の動物細胞用ベクター等が挙げられる。このようなベクターには、適切なプロモーター(例えば、lacプロモーター、tacプロモーター、trcプロモーター、trpプロモーター、CMVプロモーター、及びSV40初期プロモーター等)、選択マーカー遺伝子、ターミネーター等のエレメントを適宜有していてもよい。
【0037】
上記ベクターは、本実施形態に係るペプチド生成酵素を、Hisタグ等のタグ、又はグルタチオンS−トランスフェラーゼ等の可溶性のタンパク質と融合した状態で発現させ得るベクターであってもよい。
【0038】
このようなベクターの調製に用いられる菌としては、例えば、当該技術分野で通常用いられる宿主菌が挙げられ、具体的には、放線菌、及び大腸菌が挙げられる。より具体的には、放線菌としては、ストレプトマイセス リビダンス1326、及びストレプトマイセス リビダンスTK−23等、そして大腸菌としては、BL21(DE3)、BL21−Gold (DE3)、及びJM109等が挙げられる。これらの宿主菌のうち、精製の容易化、大量調製、及び安全性等の観点から、大腸菌BL21(DE3)、BL21−Gold (DE3)、及びJM109が好ましい。
【0039】
本実施形態に係るペプチド生成酵素の発現用担体は、適切な宿主細胞へ導入されて、宿主細胞を形質転換して、形質転換細胞を作製する。形質転換法として、例えば、リン酸カルシウム法、DEAE−デキストラン法、エレクトロポレーション法、及びパーティクル銃によるボンバードメント法等、当業者に周知の方法が採用される。
【0040】
本実施形態に係るペプチド生成酵素の発現用の宿主細胞は、好適には微生物(宿主菌)が用いられる。宿主菌としては、放線菌、大腸菌、バシラス属菌、シュードモナス属菌、サーマス属菌、アグロバクテリウム属菌、及び酵母(例えば、メタノール資化性の酵母であるPichia)等が挙げられる。中でも、本発明においては、大腸菌が好適に用いられる。
【0041】
宿主細胞へ上記発現用担体の導入によって得られた細胞が、目的の形質転換細胞であることは、細胞又は該細胞の培養上清について、上記ペプチド生成酵素の活性の測定法により、ペプチド生成活性が検出されることを指標として確認され得る。
【0042】
上記形質転換細胞を培養して増殖させることによって、本実施形態に係るペプチド生成酵素を遺伝子工学的に容易に精製できる状態で発現させることができ、それによって、本実施形態に係るペプチド生成酵素を工業的に製造することができる。
【0043】
本実施形態に係る組換えペプチド生成酵素の製造においては、培地組成、培地のpH、培養温度、培養時間の他、インデューサーの使用量、及び使用時間等について、組換えペプチド生成酵素の発現の最適な条件を決定することによって、効率よく組換えペプチド生成酵素を生産させることができる。
【0044】
本実施形態に係る形質転換細胞の培養に通常用いられる培地としては、天然培地及び合成培地のいずれを用いてもよい。液体培地又は固体培地を使用することができる。例えば、宿主菌が資化し得る炭素源、窒素源、及び無機塩類等を含有し、形質転換体の培養を効率的に行うことができる培地であればよい。炭素源としては、グルコース、ガラクトース、フラクトース、スクロース、ラフィノース、及びデンプン等の炭水化物、酢酸、及びプロピオン酸等の有機酸、並びに、エタノール、及びプロパノール等のアルコール類が挙げられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、及びリン酸アンモニウム等の無機酸、若しくは有機酸のアンモニウム塩又はその他の含窒素化合物が挙げられる。その他、ペプトン、肉エキス、コーンスティープリカー、及び各種アミノ酸等を含んでいてもよい。その他に、無機塩として、微量のカリウム、ナトリウム、及び鉄等のリン酸塩、塩酸塩、硝酸塩、並びに、酢酸塩等を含んでいてもよい。また、必要に応じて植物油、界面活性剤、及びシリコン等の消泡剤を添加してもよい。市販の培地としては、Overnight Express(登録商標) Instant TB培地(ノバジェン社製)等が挙げられる。
【0045】
培養条件は、培地の種類、培養方法等により適宜選択すればよく、宿主細胞が増殖し、目的のタンパク質を産生できる条件であれば特に制限はない。通常、液体培地中で振とう培養又は攪拌培養等の好気的条件下で、10℃〜40℃、好ましくは25℃で12時間〜120時間行われる。pHは、4〜10、好ましくは6〜8に調節される。pHの調整は、無機酸又は有機酸、並びに、アルカリ溶液等を用いて行う。
【0046】
形質転換細胞が産生する組換えペプチド生成酵素は、それが細胞内に産生されるときは細胞内の他のタンパク質、又は他のポリペプチド等が共存するが、これらは発現されるペプチド生成酵素の量に比べて微量にすぎないため、その精製は極めて容易であるという優れた利点がある。また、ベクターとして菌体外分泌型のベクターを用いた場合、ペプチド生成酵素が菌体外に分泌される。
【0047】
形質転換細胞の培養物からペプチド生成酵素を精製するには通常の方法が用いられる。形質転換細胞が大腸菌のように細胞内にペプチド生成酵素が蓄積する場合には、培養終了後、遠心分離によって形質転換細胞を集め、得られた細胞を超音波処理等によって破砕した後、遠心分離等によって無細胞抽出液を得る。これを出発材料とし、塩析法、並びに、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、疎水クロマトグラフィー、及びアフィニティークロマトグラフィー等の各種クロマトグラフィー等の一般的なタンパク質精製法によって精製することができる。用いる形質転換細胞によっては、発現した組換えタンパク質が細胞外に分泌される場合がある。このような場合、培養上清から同様に精製を行えばよい。
【0048】
以下、本実施形態に係るペプチド生成酵素を用いた、N末端にプロリン残基を有するペプチドの製造方法について説明する。本実施形態に係る製造方法は、本実施形態に係るペプチド生成酵素の存在下で、プロリン誘導体とアミノ酸とを縮合させる工程を含む。
【0049】
プロリン誘導体としては、プロリンエステル又はプロリンアミドであることが好ましい。プロリンエステルとしては、例えば、ベンジルエステル、メチルエステル等が挙げられる。特にベンジルエステル及びメチルエステルが好ましい。プロリン誘導体をこのような化合物とすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0050】
アミノ酸は、α−アミノ酸であることが好ましく、炭素数2〜10のα−アミノ酸であることがより好ましく、L−ヒドロキシプロリン、L−プロリン、L−バリン、L−アラニン、グリシン、L−システイン、L−リジン、L−セリン、L−トレオニン、L−アルギニン、(R)−α−アリルグリシン、(S)−α−プロパギルグリシン、(R)−α−プロパギルグリシン、(S)−α−メチルロイシン、(R)−α−メチルロイシン、又は(S)−α−メチルフェニルアラニンであることがさらに好ましい。アミノ酸をこのような化合物とすることで、N末端にプロリン残基を有するペプチドの収率が向上する。
【0051】
pH9.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させることが好ましく、pH10.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させることがより好ましい。pHが9.5未満の場合、酵素のペプチド生成活性が低下する傾向がある。pHの上限は特に設けないが、取り扱い性の観点からpH12未満が好ましい。
【0052】
上記反応液は、酵素を失活させなければどの様な溶媒でも用いることが可能であるが、通常は緩衝水溶液が好ましく、ホウ酸緩衝水溶液がより好ましい。緩衝水溶液の濃度は特に制限はないが、終濃度で、80mM〜1280mMが好ましく、480mM〜640mMがより好ましい。
【0053】
上記プロリン誘導体とアミノ酸とを縮合させる工程は、反応時間が0.5時間〜3時間であることが好ましく、1.5時間〜3時間であることがより好ましい。
【0054】
反応時間が1時間未満の場合、反応温度は、4℃〜37℃が好ましく、25℃〜37℃がより好ましく、25℃が特に好ましい。一方反応時間が2時間を超える場合、反応温度は、4℃〜37℃が好ましく、4℃がより好ましい。
【0055】
本実施形態に係る製造方法によって製造されたペプチドは、高純度の状態で得ることが可能である。また、得られたペプチドの純度をさらに上げるため、塩析法、並びに、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、疎水クロマトグラフィー、及びアフィニティークロマトグラフィー等の各種クロマトグラフィー等の一般的なタンパク質精製法により精製してもよい。
【実施例】
【0056】
以下、実施例により本発明をより詳細に説明するが、本発明は、実施例により限定されるものではない。特に明記しない限り、各種操作は、モレキュラークローニング ア ラボラトリー マニュアル(Molecular Cloning A Laboratory Manual)第2版(ザンブルーク(Sambrook, J)ら、1989年)に従って行った。なお、実施例において、%は、特に明記しない限り、w/v%を表す。
【0057】
(PAP遺伝子のクローニング)
ストレプトマイセス サーモグリセウス(Streptomyces thermogriseus) NBRC100772株からゲノムDNAを、Hopwoodらの方法(D.A. Hopwoodら、A Laboratory Manual, The John Ines Foundation,Norwich,1985年,70-84頁)を用いて調製した。次に、Streptomyces coelicolor (gene ID: SCO5122)、Streptomyces avermitilis (gene ID: SAV3144)及びStreptomyces griseus (gene ID: SGR2407)由来のPAPの一次配列から縮重PCRプライマー(フォワードプライマー:5’−CAGWSSTTCGGSGGSTTCTGC−3’(配列番号13)及びリバースプライマー:5’−ACGTACATGTCGTCGTGGTA−3’(配列番号14))を設計した。上記ゲノムDNAを鋳型鎖とし、上記縮重PCRプライマーセット及び、LA Taq DNAポリメラーゼ(宝ホールディングス株式会社製)を用いて、PCR法によってNBRC100772株由来のPAP(PAP10)のゲノムDNAの内部配列を増幅した。PCRは、94℃にて1分のインキュベーションの後、94℃にて30秒、60℃にて30秒、72℃にて90秒を30サイクル行い、次いで72℃にて5分のインキュベーションで行った。PCR産物と、上記PCRプライマーを用いてPAP10をコードする遺伝子(pap10遺伝子)の部分内部配列を決定した。得られた部分内部配列を基に、pap10遺伝子の全長を得るためのPCRプライマー(センスプライマー:5’−ACCGACTGGCGGCCCCTGTACGAC−3’(配列番号15)及びアンチセンスプライマー:5’−GACCTTGCGCTCGATGCGCGGGTA−3’(配列番号16))を設計した。一方、上記ゲノムDNAを制限酵素CpoI(37℃、一晩)で消化し、消化DNAを自己ライゲートさせた。ライゲーション産物を上述したpap10遺伝子の部分内部配列から設計したプライマーセット(配列番号15及び配列番号16)を用いてPCR法によって増幅した。PCR産物をクローニングして塩基配列を決定した。PAP14(Streptomyces thermoluteus subsp. fuscus strain NBRC14270株由来のPAP)の遺伝子(pap14遺伝子)及びPAP TH−3(Streptomyces aureofaciens TH−3株由来のPAP)の遺伝子(pap th−3遺伝子)の全長塩基配列も同様の方法で取得した。配列表の配列番号との対応を表1に示す。
【0058】
【表1】
【0059】
(組換えPAP発現ベクターの構築)
大腸菌を宿主とする場合のPAPを発現するためのベクター(組換えPAP発現ベクター)を以下のようにして構築した。まず、各遺伝子に対応するNdeI部位を含むセンスプライマーと停止コドンの下流にHindIII部位を含むアンチセンスプライマーとのセット及びPrime STAR(登録商標) GXL DNA ポリメラーゼ (タカラバイオ社製)を用いてPCR法により各PAP遺伝子を、増幅した。各PAP遺伝子に対応するプライマーセットは、表2に示した。得られたDNAフラグメントを、Zero Blunt II TOPO PCRクローニングキット(インビトロジェン社製)を用いてクローニングして塩基配列を決定した。PAPをコードするDNAフラグメントをNdeI及びHindIIIの制限酵素で切断し、そしてpET−28a(+)(ノバジェン社製)のNdeI−HindIIIギャップに連結し、N末端側にHisタグを有する組換えPAP発現ベクターを得た。
【0060】
【表2】
【0061】
(組換えPAPの発現及び精製)
上述のようにして得られた組換えPAPの発現ベクターを、GenePulseXcell(登録商標)(バイオラッド社製)を用いて、それぞれ大腸菌BL21−Gold (DE3)(アジレント テクノロジー株式会社製)に導入して形質転換した。形質転換体の単一コロニーを、50mlのOvernight Express(登録商標) Instant TB培地(ノバジェン社製)中で、overnight express system 1(ノバジェン社製)を用いて培養した。25℃、180rpmの条件で48時間培養した後、遠心分離によって培養液から菌体を回収した。得られた菌体は、Elestin NP035SP(ネッパジーン社製、商品名)を用いて超音波破砕した。破砕条件は、出力最大、5分間超音波破砕した後に1分間のインターバルを開けて、トータルで45分間超音波破砕するように設定した。得られた無細胞抽出液から、コバルトアフィニティ樹脂(クロンテック社製、商品名TALON(登録商標))を用いて組換えPAPを精製した。精製して得られた各PAPは、SDS−PAGE(還元条件、クマシーブルー染色)で解析した。各PAPの濃度は、Bio−Rad protein assay reagent(バイオラッド社製)を用いて定量した。
【0062】
(プロリン残基を加水分解する活性(プロリン残基加水分解活性)の評価)
水320μl、200mM緩衝水溶液(PAP14の場合、リン酸緩衝水溶液 pH7;PAP10の場合、リン酸緩衝水溶液 pH7.5;PAP TH−3の場合、トリスヒドロキシメチルアミノメタン緩衝水溶液(トリス緩衝水溶液) pH8)320μl、L−プロリン−p−ニトロアニリド(SIGMA社製、Pro−pNA)のジメチルスルホキシド(ナカライテスク社製、DMSO)溶液(0.0625mM〜8.0mM)80μlの混合液を用意し、そこに20μlのPAP水溶液を加えて37℃で反応させた。加水分解反応によって遊離したpNAに起因する405nmの吸光度の変化を分光光度計(日立製作所製、商品名U2800)で1分間モニターした。405nmにおける吸光度のプロファイル(ε405nm=10600M−1cm−1)を基に、上記加水分解反応における速度論的なパラメーターを求めた。結果を表3に示す。なお、酵素活性(プロリン残基加水分解活性)の1ユニット(U)は、各反応条件において、1分間に1μmolのpNAを遊離させるのに必要な酵素量として定義した。
【0063】
【表3】
【0064】
(PAPの生化学的性質の評価)
PAPの組成は、Superdex 200 10/300カラム(GE ヘルスケア社製)を用いたゲル濾過クロマトグラフィーによって評価した。プロリン残基加水分解活性の至適pHは、終濃度80mMのリン酸緩衝水溶液(pH 6.0〜8.0)又はトリス緩衝水溶液(pH 7.5〜9.0)を用いて決定した。またプロリン残基加水分解活性の至適温度は、35.0℃〜65.0℃の間で決定した。熱安定性は、酵素を至適pHで40.0℃〜80.0℃の間で30分間インキュベートした後の残存するプロリン加水分解活性と、酵素を4℃で保存した時のプロリン残基加水分解活性とを比較することで決定した。pH安定性は、酵素をpH6〜10.5で、30℃で30分間インキュベートした後の残存するプロリン残基加水分解活性と、至適pHにおけるプロリン残基加水分解活性とを比較することで決定した。結果を表4に示す。
【0065】
【表4】
【0066】
表3及び表4の結果から、PAP14、PAP10及びPAP TH−3のプロリン残基加水分解活性、及び生化学的性質に大きな差は認められないことが明らかとなった。
【0067】
(Pro−Hypジペプチドを生成する活性(ペプチド生成活性)の評価)
以下に示す手順にしたがって、Pro−Hypジペプチドの生成反応を行った。400mMホウ酸緩衝水溶液(pH11.0)40μl中に、アシル供与体(プロリン誘導体)として、1MのL−プロリンベンジル塩酸塩(Bachem社製、Pro−OBzl)のDMSO溶液を5μl、アシル受容体(アミノ酸)として、2MのL−ヒドロキシプロリン(和光純薬工業株式会社製、Hyp−H)の水溶液を50μl、及び720mUのPAPを含む、酵素溶液を5μlそれぞれ加えて、900rpm、37℃で3時間反応させた。その後、200μlのメタノールを反応液に加えることで、反応を終了させた。反応液を10000rpmで遠心した後、得られた上精を33倍希釈した溶液2.5μlを用いて液体クロマトグラフィー/マススペクトル(LC/MS)分析を行った。
【0068】
LC/MS/MS分析は、TUV検出器、バイナリーソルベントマネージャー、及びAPI2000 LC/MS/MSシステム(AB SCIEX社製)を備えたACQUITY(登録商標)ウルトラパフォーマンス液体クロマトグラフィー(UPLC)システム(ウォーターズ社製)を用いて行った。カラムは、XBridge(登録商標)アミドカラム(3.5μm、2.1×50mm、ウォーターズ社製)を用いた。移動層は、(A)液として、水(0.1%ギ酸)、(B)液としてアセトニトリル(0.1%ギ酸)を用いた。グラジェントプロファイルは、以下のように設定した、流速 0.3ml/min、0〜2分 2%(A)液、2〜13.5分 2〜30%(A)液。Pro−Hypと基質を含む反応混合液を検出するため、イオンクロマトグラフィーから溶出した時に現れる各ピークエリアをAPI2000のデバイスシステム(マルチリアクションモニターモード(MRM))によって計算した。Pro−OBzl、Pro−NH2、Pro−Hyp、及びHyp−Hの各フラグメンテーションのパターンは、それぞれ、m/z=206.2/91.0、114.7/70、229.0/131.9及び132.1/86.0であった。
【0069】
ペプチド生成酵素としてPAP TH−3(実施例1)を、プロリン誘導体としてPro−OBzl、及びアミノ酸としてHyp−Hを用いて、上記反応手順によって得られた反応混合物を、多重反応モニタリングクロマトグラフィーで分析した結果を図1に示す。Pro−Hypを示すピークが保持時間8分のところに現れていることから、PAP TH−3のペプチド生成活性によってPro−OBzl及びHyp−Hから、Pro−Hypが合成されたことが明らかとなった。また、Pro−Hypを示すピーク以外には、過剰に用いたHyp−Hのピーク(保持時間9.7分)しかみられなかったことから、高純度のPro−Hypを得ることが可能な反応であることが分かった。同様の実験をPAP14(比較例1)及びPAP10(比較例2)でも行ったが、Pro−Hypは検出できなかった(不図示)。以下、PAP TH−3のペプチド生成活性を詳細に検討した。
【0070】
(ペプチド生成活性のpH依存性及び緩衝水溶液濃度依存性)
図2は、Pro−Hypジペプチドの生成反応における、(A)pH依存性及び(B)緩衝水溶液の濃度の依存性を示すグラフである。(A)pHを9.5〜12.0の範囲で変化させたこと又は、(B)ホウ酸緩衝水溶液の濃度を80mM〜1280mMの範囲で変化させたこと以外は、上記(Pro−Hypジペプチド生成活性の評価)の手順に従って収率を求めた。pH9.5から収率が上昇し始め、pH10.5〜12.0の範囲において、約9%になることが明らかとなった(図2(A))。またpH11.0において、ホウ酸濃度480mMで収率が約17%になることが明らかとなった(図2(B))。
【0071】
(ペプチド生成活性の温度依存性)
図3は、Pro−Hypジペプチドの生成反応における、温度依存性を示すグラフである。緩衝水溶液を1200mMのホウ酸緩衝水溶液(pH 11.0)にして、反応温度と反応時間を変化させたこと以外は、上記(Pro−Hypジペプチド生成活性の評価)の手順に従って収率を求めた。反応温度37℃においては、反応時間30分で収率が最大となり、その後徐々に収率は減少した。反応温度25℃においては、反応時間30分で収率が最大となり、その後収率は、減少した。反応温度4℃においては、反応時間の増加に伴い収率も徐々に上昇し、反応時間3時間で収率25%を示した。
【0072】
(ペプチド生成活性のプロリン誘導体の影響)
図4は、Pro−Hypジペプチドの生成反応における、プロリン誘導体の影響を示すグラフである。プロリン誘導体を変えたこと、反応温度を4℃にしたこと以外は、上記(ペプチド生成活性の温度依存性)の手順に従って収率を求めた。プロリン誘導体としては、Pro−OBzlの他に、L−プロリンメチル塩酸塩(ALDRICHI社製、Pro−OMe)及び、L−プロリンアミド塩酸塩(SIGMA社製、Pro−NH2)を用いた。いずれの反応時間においても、Pro−NH2、Pro−OMe、Pro−OBzlの順に収率が高いことが明らかになった。
【0073】
(Pro−Hypジペプチドの生成反応における、プロリン誘導体を連続して添加したときの影響)
Pro−Hypジペプチドの生成反応における、プロリン誘導体を連続して添加したときの影響を調べた。まず、1200mMホウ酸緩衝水溶液(pH11.0)40μl中に、プロリン誘導体として、1MのPro−NH2のDMSO溶液を5μl、アミノ酸として、2MのHyp−Hの水溶液を50μl、及び720mUのPAP TH−3を含む酵素溶液を5μlを、それぞれ加えて、900rpm、4℃で反応させた。上記反応混合物に、50mMのPro−NH2、200mMのPro−NH2、若しくは1MのPro−NH2のDMSO溶液を5μl、又は、1MのPro−NH2のDMSO溶液を5μlと720mUのPAP TH−3溶液を1.6μlとの混合液を終容量100μlとなるように調整して、30分おきに加えた。結果を図5に示す。いずれの場合もプロリン誘導体を添加するたびに、Pro−Hypの収量は、増加した。特に、Pro−NH2とPAP TH−3を同時に加えたとき収量がもっとも増加しており、50mM Pro−NH2を添加したときと比べて、反応時間2時間の時点で2.3倍であった。
【0074】
(アミノ酸の検討)
Hyp−Hの他にも、アシル受容体として有効なアミノ酸があるのかどうかを式(1)〜式(9)で表わされる天然アミノ酸及び、式(10)〜式(18)で表わされる非天然アミノ酸について検討した。
【0075】
【化1】
【0076】
【化2】
【0077】
まず、1200mMホウ酸緩衝水溶液(pH11.0)40μl中に、プロリン誘導体として、1MのPro−NH2のDMSO溶液を5μl、アミノ酸として、上記式(1)〜式(18)で表される化合物の水溶液を、50μl、及び720mUのPAP TH−3を含む酵素溶液を5μlそれぞれ加えて、4℃で3時間反応させた。なお、アミノ酸は、終濃度が下記表5に示す濃度となるように加えた。反応混合物を液体クロマトグラフィー/マススペクトル(LC/MS)分析することで、反応生成物を検出した。結果を表5に示す。
【0078】
【表5】
【0079】
表5に示される結果から、PAP TH−3のペプチド生成活性は、基質として用いるアミノ酸として、天然アミノ酸のみならず、非天然アミノ酸でも有効であることが明らかになった。さらに、式(11)と式(12)の化合物、及び式(13)と式(14)の化合物は、互いに光学異性体の関係にあることから、PAP TH−3のペプチド生成活性は、基質となるアミノ酸の光学活性に依存しないことが明らかになった。
【0080】
(参考例1)
(C末端の12アミノ酸残基が欠損したPAP TH−3の発現ベクターの構築)
大腸菌を宿主とする場合の、C末端の12アミノ酸残基が欠損したPAP TH−3を発現するためのベクター(組換えC末端12アミノ酸残基欠損PAP TH−3発現ベクター)を以下のようにして構築した。まず、PAP TH−3遺伝子に対応するNdeI部位を含むセンスプライマー:5’−CATATGTCCACCGTCAGCCGGCTG−3’(配列番号17)と停止コドンの下流にHindIII部位を含むアンチセンスプライマー:5’−AAGCTTAGGCCATCTTGATCAGCCGGTC−3’(配列番号18)及びPrime STAR(登録商標) GXL DNA ポリメラーゼ (タカラバイオ社製)を用いてPCR法によりC末端の12アミノ酸残基が欠損したPAP TH−3をコードする遺伝子を増幅した。得られたDNAフラグメントを、Zero Blunt II TOPO PCRクローニングキット(インビトロジェン社製)を用いてクローニングして塩基配列を決定した。PAPをコードするDNAフラグメントをNdeI及びHindIIIの制限酵素で切断し、そしてpET−28a(+)(ノバジェン社製)のNdeI−HindIIIギャップに連結し、N末端側にHisタグを有する組換えC末端12アミノ酸残基欠損PAP TH−3発現ベクターを得た。
【0081】
(組換えC末端12アミノ酸残基欠損PAP TH−3の発現及び精製)
上記(組換えPAPの発現及び精製)に記載の方法と同様の方法で発現及び精製を行った。
【0082】
(ペプチド生成活性の評価)
上記(Pro−Hypジペプチドを生成する活性(ペプチド生成活性)の評価)に記載の方法と同様の方法で組換えC末端12アミノ酸残基欠損PAP TH−3のペプチド生成活性評価を行い、Pro−Hypジペプチドの生成を確認することができた。
【0083】
(参考例2)
(PAP10とPAP TH−3とのキメラPAP遺伝子のクローニング及びそのキメラPAP発現ベクターの構築)
まず、PAP TH−3遺伝子にNsbI部位を導入するため、センスプライマー:5’−GGTGCGCGGCCTGCGCACGTGGATCACCAA−3’(配列番号19)とアンチセンスプライマー:5’−TTGGTGATCCACGTGCGCAGGCCGCGCACC−3’(配列番号20)及びクイックチェンジキット(Stratagene製)を用いて、PAP TH−3遺伝子にNsbI部位が導入された遺伝子を増幅した。得られたDNAフラグメントは、PAP TH−3遺伝子にNsbI部位が導入されていることを確認した。
【0084】
PAP TH−3遺伝子にNsbI部位が導入されたDNAフラグメントをNdeI及びHindIIIの制限酵素で切断し、そしてpET−28a(+)(ノバジェン社製)のNdeI−HindIIIギャップに連結した。得られたプラスミドを、NsbI及びClaIで切断した。一方、PAP−10遺伝子が挿入されているプラスミドを、NsbI及びClaIの制限酵素で切断してPAP10遺伝子のフラグメントを得た。得られたPAP10遺伝子のフラグメントを、上述のPAP TH−3遺伝子が挿入されたpET−28a(+)のNsbI−ClaIギャップに挿入して、PAP−10とPAP TH−3とのキメラPAP遺伝子(配列番号21)の発現ベクターを得た。このキメラPAPは、PAP TH−3のN末側402個のアミノ酸残基がPAP10のものに置き換わっている。また、このキメラPAPは、PAP TH−3と比較して、117アミノ酸残基が異なっていることが確認された。
【0085】
(PAP−10とPAP TH−3とのキメラPAPの発現及び精製)
上記(組換えPAPの発現及び精製)に記載の方法と同様の方法で発現及び精製を行った。
【0086】
(ペプチド生成活性の評価)
上記(Pro−Hypジペプチドを生成する活性(ペプチド生成活性)の評価)に記載の方法と同様の方法で組換えPAP−10とPAP TH−3とのキメラPAPのペプチド生成活性評価を行い、Pro−Hypジペプチドの生成を確認することができた。
【産業上の利用可能性】
【0087】
本発明のタンパク質(ペプチド生成酵素)は、Pro−Hyp等のN末端にプロリン残基を有するペプチドを、高純度で製造することができるため、工業及び医療の分野において応用が可能である。例えば、骨粗鬆症、胃潰瘍、変形性関節症、褥瘡及び高血圧用の薬剤、皮膚保湿剤、並びに、保存剤の用途が挙げられる。
【受託番号】
【0088】
Streptomyces aureofaciens TH−3として命名及び表示される微生物は、受託番号 FERM P−21343(寄託日:2007年8月17日)として、独立行政法人産業技術総合研究所 特許生物寄託センター(日本国茨城県つくば市東1丁目1−1)に寄託されている。
【特許請求の範囲】
【請求項1】
(A)配列表の配列番号10に記載のアミノ酸配列、
(B)配列表の配列番号10に記載のアミノ酸配列において1又は数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列、及び
(C)配列表の配列番号10に記載のアミノ酸配列に対して80%以上の配列同一性を有するアミノ酸配列、
からなる群から選択されるアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素。
【請求項2】
(a)請求項1に記載のペプチド生成酵素をコードする塩基配列、及び
(b)配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖とストリンジェントな条件下にハイブリダイズし、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素をコードする塩基配列、
からなる群より選択される塩基配列を有する遺伝子。
【請求項3】
請求項1に記載のペプチド生成酵素の存在下で、プロリン誘導体とアミノ酸とを縮合させる工程を含む、N末端にプロリン残基を有するペプチドの製造方法。
【請求項4】
前記プロリン誘導体がプロリンエステル又はプロリンアミドである請求項3に記載の製造方法。
【請求項5】
前記プロリンエステルがベンジルエステル又はメチルエステルである請求項4に記載の製造方法。
【請求項6】
前記アミノ酸がα−アミノ酸である請求項3〜5のいずれか一項に記載の製造方法。
【請求項7】
前記アミノ酸がL−ヒドロキシプロリン、L−プロリン、L−バリン、L−アラニン、グリシン、L−システイン、L−リジン、L−セリン、L−トレオニン、L−アルギニン、(R)−α−アリルグリシン、(S)−α−プロパギルグリシン、(R)−α−プロパギルグリシン、(S)−α−メチルロイシン、(R)−α−メチルロイシン、又は(S)−α−メチルフェニルアラニンである請求項3〜5のいずれか一項に記載の製造方法。
【請求項8】
pH9.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させる、請求項3〜7のいずれか一項に記載の製造方法。
【請求項1】
(A)配列表の配列番号10に記載のアミノ酸配列、
(B)配列表の配列番号10に記載のアミノ酸配列において1又は数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列、及び
(C)配列表の配列番号10に記載のアミノ酸配列に対して80%以上の配列同一性を有するアミノ酸配列、
からなる群から選択されるアミノ酸配列を有し、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素。
【請求項2】
(a)請求項1に記載のペプチド生成酵素をコードする塩基配列、及び
(b)配列表の配列番号10のアミノ酸配列をコードする塩基配列の相補鎖とストリンジェントな条件下にハイブリダイズし、プロリン誘導体とアミノ酸とを縮合させてN末端にプロリン残基を有するペプチドを生成する活性を有するペプチド生成酵素をコードする塩基配列、
からなる群より選択される塩基配列を有する遺伝子。
【請求項3】
請求項1に記載のペプチド生成酵素の存在下で、プロリン誘導体とアミノ酸とを縮合させる工程を含む、N末端にプロリン残基を有するペプチドの製造方法。
【請求項4】
前記プロリン誘導体がプロリンエステル又はプロリンアミドである請求項3に記載の製造方法。
【請求項5】
前記プロリンエステルがベンジルエステル又はメチルエステルである請求項4に記載の製造方法。
【請求項6】
前記アミノ酸がα−アミノ酸である請求項3〜5のいずれか一項に記載の製造方法。
【請求項7】
前記アミノ酸がL−ヒドロキシプロリン、L−プロリン、L−バリン、L−アラニン、グリシン、L−システイン、L−リジン、L−セリン、L−トレオニン、L−アルギニン、(R)−α−アリルグリシン、(S)−α−プロパギルグリシン、(R)−α−プロパギルグリシン、(S)−α−メチルロイシン、(R)−α−メチルロイシン、又は(S)−α−メチルフェニルアラニンである請求項3〜5のいずれか一項に記載の製造方法。
【請求項8】
pH9.5以上の反応液中でプロリン誘導体とアミノ酸とを縮合させる、請求項3〜7のいずれか一項に記載の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−147742(P2012−147742A)
【公開日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願番号】特願2011−10117(P2011−10117)
【出願日】平成23年1月20日(2011.1.20)
【出願人】(000214272)長瀬産業株式会社 (137)
【出願人】(591060980)岡山県 (96)
【Fターム(参考)】
【公開日】平成24年8月9日(2012.8.9)
【国際特許分類】
【出願日】平成23年1月20日(2011.1.20)
【出願人】(000214272)長瀬産業株式会社 (137)
【出願人】(591060980)岡山県 (96)
【Fターム(参考)】
[ Back to top ]