
ペプチド翻訳合成におけるRAPIDディスプレイ法
【課題】遺伝子型と表現型の対応付け技術(ディスプレイ法)において、核酸とその翻訳産物であるペプチドとの連結体を再構成型の無細胞翻訳系内で作製する際に好適なリンカーを提供すること。
【解決手段】一方の端に、mRNAの3'末端側の塩基と対合する側鎖塩基を有する一本鎖構造領域を含み、もう一方の端に、塩基配列ACCAからなるオリゴRNAにアミノ酸がエステル結合した構造からなるペプチドアクセプター領域を含み、無細胞翻訳反応液中でリンカーとmRNAが結合可能であることを特徴とするリンカー。
【解決手段】一方の端に、mRNAの3'末端側の塩基と対合する側鎖塩基を有する一本鎖構造領域を含み、もう一方の端に、塩基配列ACCAからなるオリゴRNAにアミノ酸がエステル結合した構造からなるペプチドアクセプター領域を含み、無細胞翻訳反応液中でリンカーとmRNAが結合可能であることを特徴とするリンカー。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、遺伝子型と表現型の対応付け技術(ディスプレイ系)において、cDNAまたはmRNAとその翻訳産物であるペプチドまたはタンパク質との連結体を作製する際に用いられる新規な方法に関する。本発明者らは、この方法をRAPIDディスプレイ法と命名した。この方法は、先に報告されたフレキシザイムシステム(あるいは、RAPIDシステム:Random Peptide Integrated Discovery system)を用いて構築される特殊ペプチドライブラリーからの、薬剤候補となり得るペプチドアプタマーのスクリーニングに好適である。
【背景技術】
【0002】
進化分子工学のツールとして誕生した遺伝子型と表現型の対応付け技術は、ディスプレイ法とも称され、mRNAディスプレイ法 ("In vitro virus"、Nemoto N, et al. FEBS Lett. 414, 405-408 (1997)、国際公開WO98/16636;又は"RNA-peptide fusions"、Roberts, R. W. & Szostak, J. W. , Proc. Natl. Acad. Sci. USA., 94, 12297-12302 (1997)、国際公開WO98/31700)、STABLE法(非共有結合DNAディスプレイ)、マイクロビーズドロップレット法、共有結合DNAディスプレイ法、ファージディスプレイ法、リボソーム・ディスプレイ法などが知られている。ディスプレイ法では、ライブラリから機能のあるペプチドやタンパク質の分子を選択した際に、それに対応する遺伝子が連結しているのでその配列を容易に読み取ることができ、特定の機能を有するポリペプチドの遺伝情報を選択する際に有用である。
【0003】
mRNAディスプレイ法は無細胞翻訳系(in vitroタンパク質合成系)を用いて、遺伝子型としてのmRNAと表現型としてのペプチド分子を共有結合させることにより、遺伝子型と表現型を一体化する技術であり、現在、チロシルtRNA3’末端部分のアナログであるピューロマイシンを介して、合成されたペプチド分子とこれをコードするmRNAとを連結する方法がとられている。
【0004】
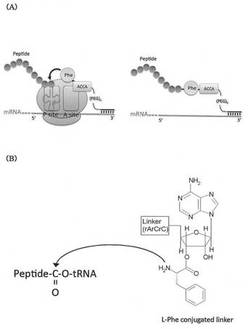
mRNAディスプレイ法においては、ピューロマイシンを適当なリンカーを介してmRNAの3’末端に結合させておき、これを無細胞翻訳系に導入してmRNAからペプチドを合成すると、ピューロマイシンがリボソームにおけるペプチド転移反応の基質として伸長中のペプチド鎖のC末端に連結し、翻訳産物であるペプチド分子がピューロマイシンを介してmRNAと連結する(図1A)。リンカーは、主として、ピューロマイシンをリボソームのAサイトに効率良く取り込ませるために、mRNAとピューロマイシンの間に挿入される。ピューロマイシンの特徴として、アミノアシルtRNAの3’末端とは異なり、アデノシン様部分とアミノ酸(チロシン)様部分がエステル結合ではなくアミド結合を形成していることが挙げられる(図1B)。そのため、リボソーム上で連結したピューロマイシンとペプチドとの結合体は、加水分解されにくく安定である。
【0005】
mRNAディスプレイ法においては、ピューロマイシンを介してmRNAと翻訳産物を連結するために、無細胞翻訳系の外で、予め、ピューロマイシンをmRNAの3’末端に連結しておくことが必要である。この連結方法として、まずピューロマイシンから5’末端側へ直鎖高分子からなるスペーサーを合成したピューロマイシン結合リンカー(Puromycin conjugated linker)を作成してから、当該リンカーとmRNAの3’末端を結合する方法と、mRNAの3’末端側にスペーサーを結合してから、当該結合体にピューロマイシンを結合する方法がある。いずれの方法でも、スペーサーである直鎖高分子の端は、典型的には、リン酸基またはヌクレオチドであり、mRNAの3’末端とリンカーの5’末端側との結合は、リン酸基を介した共有結合となっている。この共有結合は、RNAリガーゼまたはDNAリガーゼを用いた反応、または一般的な有機化学反応によって形成される。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特許第3683282号広報(国際公開WO98/16636)
【特許文献2】特許第3683902号広報
【特許文献3】特許第3692542号広報(国際公開WO98/31700)
【非特許文献】
【0007】
【非特許文献1】Nemoto N, et al. FEBS Lett. 414, 405-408 (1997)
【非特許文献2】Roberts, R. W. & Szostak, J. W. , Proc. Natl. Acad. Sci. USA., 94, 12297-12302 (1997)
【発明の概要】
【発明が解決しようとする課題】
【0008】
公知のmRNAディスプレイ法においては、3’末端にピューロマイシンを持つmRNAを鋳型として、これを小麦胚芽やウサギ網状赤血球の抽出液を使用する無細胞翻訳系に加えてペプチドへの翻訳を行う。従って、DNAからmRNAへの転写、及びmRNAとピューロマイシンとの連結反応を、予め翻訳系外で行っておく必要がある。
【0009】
近年、大腸菌のリボソームを用いる系で、翻訳に必要な因子をそれぞれ精製して混ぜ合わせた、再構成型の無細胞翻訳系が開発された(H. F. Kung, B. Redfield, B. V. Treadwell, B. Eskin, C. Spears and H. Weissbach (1977) “DNA-directed in vitrosynthesis of beta-galactosidase. Studies with purified factors” The Journal of Biological Chemistry Vol. 252, No. 19, 6889-6894 ; M. C. Gonza, C. Cunningham and R. M. Green (1985) “Isolation and point of action of a factor from Escherichia coli required to reconstruct translation” Proceeding of National Academy of Sciences of the United States of America Vol. 82, 1648-1652 ; M. Y. Pavlov and M. Ehrenberg (1996) “Rate of translation of natural mRNAs in an optimized in Vitrosystem” Archives of Biochemistry and Biophysics Vol. 328, No. 1, 9-16 ; Y. Shimizu, A. Inoue, Y. Tomari, T. Suzuki, T. Yokogawa, K. Nishikawa and T. Ueda (2001) “Cell-free translation reconstituted with purified components” Nature Biotechnology Vol. 19, No. 8, 751-755;H. Ohashi, Y. Shimizu, B. W. Ying, and T. Ueda (2007) “Efficient protein selection based on ribosome display system with purified components” Biochemical and Biophysical Research Communications Vol. 352, No. 1, 270-276)。再構成型の無細胞翻訳系とは、主に大腸菌抽出液を使用する無細胞翻訳系を細分化し、各因子を再構成することにより、翻訳と関係の無い成分が除かれたシステムであり、従来の細胞抽出液を使用する無細胞翻訳系よりもヌクレアーゼやプロテアーゼなどの阻害物質の混入を容易に防ぐことができる。転写反応に必要な因子を加えることにより、DNAからの転写を翻訳と同時に行うこともできる。
【0010】
このような転写カップル型の無細胞翻訳系を用いれば、翻訳反応と同一の系内で、鋳型DNAからの転写によるmRNAの合成を行うことができる。さらに、mRNAとリンカー分子との複合体形成もその同じ系内で行うことができれば、従来のmRNAディスプレイ法とは異なり、cDNAの転写からmRNA−ペプチド連結体の作製までをワンポット(同一反応容器内)で調製することが可能になる。そのような、反応系成分の制御が可能な再構成型の無細胞翻訳系の利点を生かしたディスプレイ法を提供することが、本発明の第一の課題である。
【0011】
さらに、このようなディスプレイ法を、先に報告された、アシル化tRNA合成触媒能を有するリボザイム(フレキシザイム)を用いた特殊ペプチド合成技術と組み合わせることで、特殊ペプチドが提示されたmRNAまたはcDNAのライブラリーの構築を可能とすることが、本発明の第二の課題である。
【課題を解決するための手段】
【0012】
本発明のRAPIDディスプレイ法では、mRNAディスプレイ法を改変して、ピューロマイシン結合リンカーの代わりに本発明者らが今回新たに開発したリンカー分子を利用し、さらに再構成型の無細胞翻訳系を最適化することにより、転写、翻訳、およびリンカーとmRNAとの複合体形成、それに続くペプチドとリンカーとの結合までを、一つの翻訳系内で完結させることが可能となった。
【0013】
従来のmRNAディスプレイ法と比較した、RAPIDディスプレイ法の主な特徴は以下の通りである。
(ア)リンカー分子の3’末端はピューロマイシンではなく、アミノ酸がエステルを介してアデノシンに結合(アミノアシル化)した構造となっている。
(イ)アミノアシル化反応は、人工RNA触媒(リボザイム)により行う。
(ウ)再構成型の無細胞翻訳系を利用する。
(エ)リンカーとmRNAとの結合は、ライゲーションではなく、翻訳系内でのハイブリダイゼーションによる複合体形成で行う。
(オ)転写、翻訳およびリンカーとの複合体形成を単一の翻訳反応容器中で行うことができる。
【0014】
さらに、同じ翻訳系において、特殊ペプチドの翻訳合成技術を適用することにより、表現型として特殊ペプチドを提示することもできる。特殊ペプチドの構成単位である、非タンパク質性のアミノ酸やヒドロキシ酸をtRNAに担持させるアシル化反応も、人工RNA触媒(リボザイム)を使用して行う。
【0015】
本発明の概略は以下の通りである。
(1)再構成型のin vitroタンパク質合成系において、mRNAと、その翻訳産物であるペプチドとが、リンカーを介して結合した連結体を作製するために使用されるリンカーであって、
リンカーの一方の端に、mRNAの3'末端側の塩基と対合する側鎖塩基を有する一本鎖構造領域を含み、
リンカーのもう一方の端に、ペプチド転移反応によって翻訳産物と結合可能な基を含むペプチドアクセプター領域を含み、
ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにアミノ酸がエステル結合した構造からなることを特徴とするリンカー。
(1−1)当該エステル結合が、人工RNA触媒を用いるアミノアシル化反応により形成されたことを特徴とする、前記リンカー。
(1−2)無細胞翻訳反応液中でリンカーとmRNAが結合可能であることを特徴とする、前記リンカー。
(1−3)ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにフェニルアラニンがエステル結合した構造からなる、前記リンカー。
(1−4)再構成型のin vitroタンパク質合成系において、DNAの転写からmRNA−ペプチド連結体の調製までを翻訳反応と同一の系内で行うために使用される、前記リンカー。
(2)一本鎖構造領域とペプチドアクセプター領域の間が、オリゴヌクレオチド、ポリアルキレン、ポリアルキレングリコール、ポリスチレン、多糖類、又はこれらの組合わせから選択される直鎖状物質で連結されている、前記リンカー。
(2−1)一本鎖構造領域とペプチドアクセプター領域の間がポリエチレングリコールで連結されている、前記リンカー。
(3)一本鎖構造領域が一本鎖DNAからなる、前記リンカー。
(4)アミノアシル化反応に使用される人工RNA触媒が、以下のいずれかのRNA配列
GGAUCGAAAGAUUUCCGCAGGCCCGAAAGGGUAUUGGCGUUAGGU (配列番号3)
GGAUCGAAAGAUUUCCGCGGCCCCGAAAGGGGAUUAGCGUUAGGU (配列番号4)
GGAUCGAAAGAUUUCCGCAUCCCCGAAAGGGUACAUGGCGUUAGGU (配列番号5)
GGAUCGAAAGAUUUCCGCACCCCCGAAAGGGGUAAGUGGCGUUAGGU (配列番号19)
からなる化学構造を有する、前記(1−1)のリンカー。
(5)mRNAと、その翻訳産物であるペプチドが、リンカーを介して結合した〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製する方法であって、
前記のいずれかのリンカーを調製する工程、
リンカーの一本鎖構造領域の塩基配列にハイブリダイズできる配列を、ペプチドをコードする配列の下流に有するmRNAを合成する工程、
再構成型のin vitroタンパク質合成反応液中で、リンカーとmRNAとを接触させるとともに、mRNAからペプチドの翻訳を行う工程
を含む、前記方法。
(6)mRNAと、その翻訳産物であるペプチドが、リンカーを介して結合した〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製する方法であって、
前記のいずれかのリンカーを調製する工程、
リンカーの一本鎖構造領域の塩基配列にハイブリダイズできる配列を、ペプチドをコードする配列の下流に有するmRNAの鋳型となるDNAを合成する工程、
再構成型のin vitroタンパク質合成反応液中にリンカーとDNAを供することにより、DNAからmRNAへの転写およびペプチドへの翻訳ならびにリンカーとmRNAとの複合体形成を行う工程
を含む、前記方法。
(7)再構成型のin vitroタンパク質合成反応液中に、非タンパク質性アミノ酸またはヒドロキシ酸でチャージされたtRNAを含むことにより、翻訳産物であるペプチドが特殊ペプチドとなる、(5)または(6)に記載の方法。
(8)前記(7)に記載の方法で作製された、〔mRNA〕−〔リンカー〕−〔特殊ペプチド〕連結体からなるライブラリー。
【発明の効果】
【0016】
再構成型の無細胞翻訳反応液中でmRNAと結合可能なリンカー分子を利用することで、単一の反応容器中にリンカーとcDNAまたはmRNAを加えて翻訳反応を行うだけで、〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製することができる。
【0017】
さらに、同じ再構成型の無細胞翻訳反応液中に、非タンパク質性のアミノ酸やヒドロキシ酸を担持させたtRNAを導入することにより、鋳型である核酸分子の配列情報から翻訳により合成された特殊ペプチドを提示した、〔mRNA〕−〔リンカー〕−〔特殊ペプチド〕連結体を得ることができる。
【0018】
このように、RAPIDディスプレイ法を、特殊ペプチドの翻訳合成技術と組み合わせることにより、生体内での安定性や標的タンパク質への結合力の向上が期待される人工的なペプチドアプタマーの遺伝子ライブラリーを簡便に構築することが可能となる。
【図面の簡単な説明】
【0019】
【図1】図1Aは、mRNAディスプレイ法において、翻訳産物であるペプチド分子がピューロマイシン結合リンカー(Puromycin conjugated linker)を介してmRNAと連結する様子を模式的に示す。図1Bは、リンカー(Puromycin conjugated linker)のペプチドアクセプター部分の構造を示す。
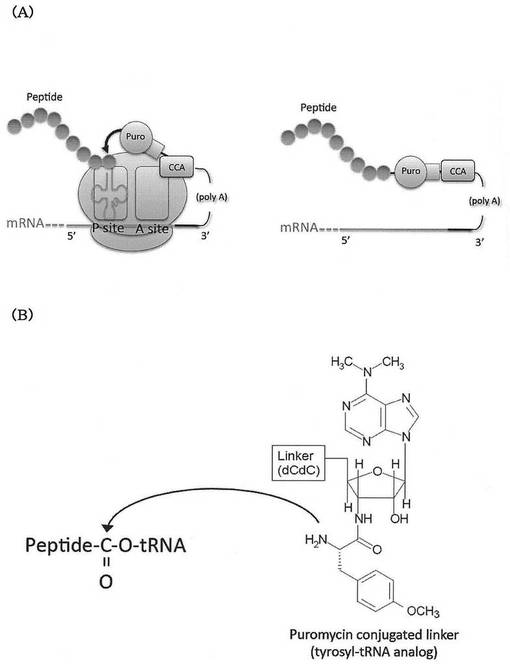
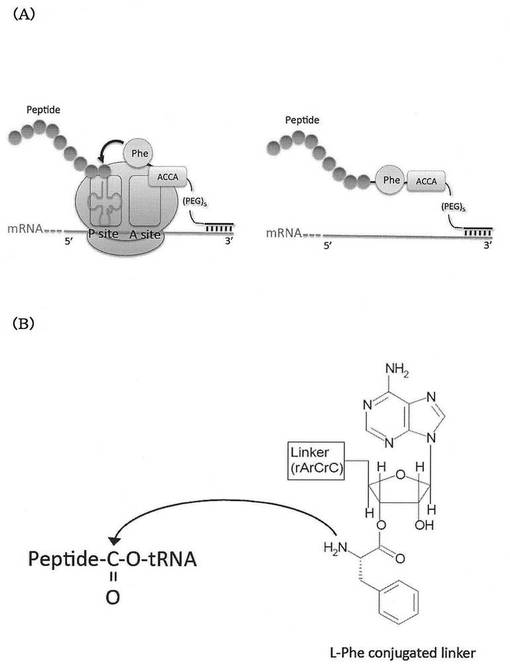
【図2】図2Aは、本発明のRAPIDディスプレイ法において、翻訳産物であるペプチド分子がRAPIDリンカー(L-Phe conjugated linkerが例示されている)を介してmRNAと連結する様子を模式的に示す。図2Bは、リンカー(L-Phe conjugated linker)のペプチドアクセプター部分の構造を示す。
【図3】リンカー分子(an21-ACCA)がフレキシザイムの触媒する反応によりアミノアシル化されていることを示す。(実施例1)
【図4】反応液にmRNAを添加した場合の、ペプチドアプタマー(TNF-α-DW)とmRNAの複合体の回収率を示す。(実施例1)
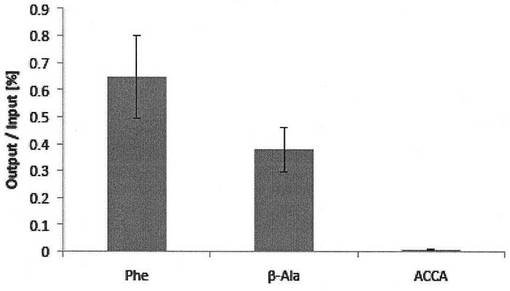
【図5】反応液にcDNAを添加した場合の、ペプチドアプタマー(TNF-α-DW)とmRNAの複合体の回収率を示す。(実施例1)
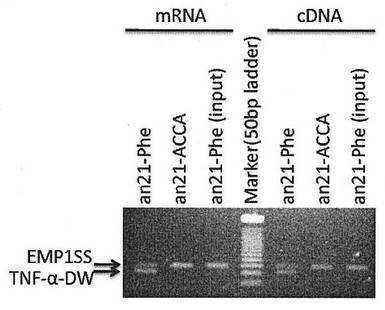
【図6】TNF-α-DWを提示したペプチド-mRNA複合体が、ターゲットに結合しない対照(EMP1SS)の複合体よりも効率よく回収されていることを確認した結果である。(実施例2)
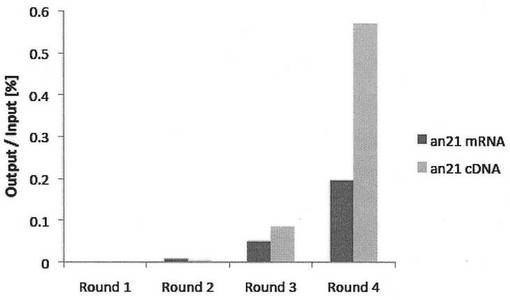
【図7】TNF-α-DWスパイクドライブラリーとan21-Pheリンカーを用いて、複数回のセレクションを繰り返すことによりペプチドアプタマーを選別した。ラウンド毎の結合率の変化を示した。(実施例3)
【発明を実施するための形態】
【0020】
1.リンカー
リンカーは、本発明のRAPIDディスプレイ法においても公知のmRNAディスプレイ法と同様に、一方の端でmRNAの3’末端側と、もう一方の端でペプチドのC末端側と結合することにより、mRNAとその翻訳産物であるペプチドを連結する。
【0021】
しかしながら、本発明のRAPIDディスプレイ法では、リンカーの両端の構造が公知のmRNAディスプレイ法と相違する。本明細書では、本発明のRAPIDディスプレイ法で使用されるリンカーのことを、「RAPIDリンカー」と称することもある。
【0022】
まず、リンカーの一方の端である、ペプチドのC末端と結合する部分について説明する。本明細書においては、この部分を「ペプチドアクセプター(peptidyl acceptor)」あるいは、単に「アクセプター」とも称する。すなわち、ペプチドアクセプターとは、リボソームにおけるペプチド転移反応によって伸長中のペプチド(ペプチジルtRNA)と結合できる構造を持った分子を意味する。ペプチドアクセプターは、リンカーの末端に位置する部分のことを指す場合もあるし、リンカーを含んだ全体の構造を指す場合もある。例えば、公知のmRNAディスプレイ法におけるペプチドアクセプターは、リンカーの末端に位置するピューロマイシン、またはリンカーを含んだ全体の構造であるピューロマイシン結合リンカーである。
【0023】
本発明のRAPIDリンカーでは、ペプチドアクセプターの構造とその作製方法に特徴がある。
RAPIDディスプレイ法では、3’末端側に4残基のリボヌクレオチドからなるACCA配列を有するリンカーを合成してから、その3’末端のアデノシンに任意のアミノ酸を結合させることにより、リンカーにペプチドアクセプターとしての構造を付与する。リボソーム上のペプチド伸長反応において、リンカー末端に結合したアミノ酸が、ペプチジルtRNAのペプチドC末端を受け取ってペプチドと結合する。本明細書において、RNA配列ACCAにアミノ酸がエステル結合した構造を「ペプチドアクセプター領域」と称する。
【0024】
公知のmRNAディスプレイ法におけるペプチドアクセプターであるピューロマイシンは、アデノシン様部分のリボースとアミノ酸とがアミド結合したアミノヌクレオシド構造を有する。これに対して、本発明のRAPIDディスプレイ法では、リボースの3’-Oにアミノ酸がエステル結合した構造となっている。つまり、本発明のRAPIDディスプレイにおけるペプチドアクセプターは、天然のアミノアシルtRNAと同様のヌクレオシド構造を有する。このようなペプチドアクセプターの一例として、L−フェニルアラニンが結合したリンカーの構造について、図2B(L-Phe conjugated linker)を参照し、図1B(Puromycin conjugated linker)と比較されたい。本発明では、ペプチドアクセプターが、より天然に近い構造を用いることでピューロマイシンと同等あるいはそれ以上の取り込み効率を示す。
【0025】
ペプチドアクセプターとペプチドC末端との結合の形成はリボソーム内で起こる通常のペプチド転移反応と同様に、P部位のペプチジルtRNAに付いたペプチドのC末端のエステル結合に、A部位に取り込まれたペプチジルアクセプターのアミノ基が近接することにより起こると思われる。従って、当該ペプチド鎖C末端との間で形成される共有結合は、mRNAディスプレイ法の場合と同様に、典型的にはアミド結合である。なお、本発明のRAPIDディスプレイでは、リンカーの合成に後述する人工RNA触媒(フレキシザイム)を用いることで天然型(非タンパク質性)のアミノ酸とは異なるD-アミノ酸あるいはβ(beta)-アミノ酸を有するものを用いることも可能である。
【0026】
次に、リンカーのもう一方の端におけるmRNAとの結合について説明する。
本発明のRAPIDディスプレイ法においては、リンカーの5’末端側とmRNA分子の3’末端側とが、塩基対形成に基くハイブリダイゼーションにより複合体を形成する。従って、リンカーの5’末端側は核酸塩基を側鎖に持つ一本鎖構造を取る。RAPIDリンカーにおける、この部分を、本明細書では「一本鎖構造領域」と称する。核酸塩基を側鎖に持つ一本鎖構造の具体的な例は、一本鎖DNA、一本鎖RNA、一本鎖PNA(ペプチド核酸)などである。形成された複合体は、ペプチドのセレクションの間も安定的に保持される必要がある。リンカーの一本鎖構造領域の塩基配列と、mRNA分子の3’末端側配列との、相補性が高いほど二本鎖形成の効率が高くなり、安定性も高くなる。安定性は、GC含量、反応溶液の塩濃度、反応温度にも依存する。特に、高いGC含量を有することが望ましく、具体的には80%以上、好ましくは85%以上のGC含量を有することが望ましい。そのようなリンカーの5’末端構造の具体例を挙げて説明するならば、決して限定されるものではないが、実施例で使用された
5'-CTCCCGCCCCCCGTCC-3’(配列番号1)
5'-CCCGCCTCCCGCCCCCCGTCC-3’(配列番号2)
といった塩基配列を有する、13〜21ヌクレオチドからなる一本鎖DNAが挙げられる。
【0027】
リンカーの両端以外の部分は、公知のmRNAディスプレイ法で使用されるリンカーの構造と同様に、全体として、柔軟性があり、親水性で、側鎖の少ない単純な直鎖構造を有するように設計される。従って、例えば、一本鎖又は二本鎖DNAやRNA等のオリゴヌクレオチド、ポリエチレン等のポリアルキレン、ポリエチレングリコール等のポリアルキレングリコール、ポリスチレン、多糖類等の直鎖状物質又はこれらの組合わせを適宜選択して用いることができる。リンカーの長さは、100Å以上あることが好ましく、100〜1000Å程度であることがさらに好ましい。
【0028】
本発明で使用可能なリンカーの非限定的な具体例としては、一本鎖構造領域がGC含量の高い配列を有する一本鎖DNAであり、3’末端がACCA配列からなるRNAであり、DNAとRNAの間がポリエチレングリコール(PEGリンカー)で連結されたDNA/RNAキメラオリゴヌクレオチドが上げられる。例えば、実施例1で合成された、〔DNA〕−〔Spacer18〕n−rArCrCrA(但し、Spacer18はヘキサエチレングリコールであり、nは4〜8の整数である)が典型的な例として挙げられる。
【0029】
2.リンカーのアミノ酸修飾
上述の通り、リンカーにペプチドアクセプターとしての構造を付与するためには、リンカー分子末端のオリゴRNA(ACCA配列)部分に、アミノ酸を結合させる必要がある。このアミノ酸の結合を、人工RNA触媒を用いて行うことが、本発明の特徴である。
【0030】
固相合成法などの通常の化学合成を用いて合成したオリゴRNAの3'-O部位にエステルを介してアミノ酸を結合させることは理論上可能ではあるが、生産物は反応性が高く安定性を欠くため、これを合成反応後の後処理を行った後に無細胞翻訳系に供することは事実上不可能である。本発明においては、この問題点を、アミノアシルtRNA合成酵素として開発された人工RNA触媒である「フレキシザイム」を用いたオリゴRNAのアミノアシル化反応により解決している。この方法では、アミノアシル化反応は穏和な条件で行うことができ、容易な後処理を行うだけで翻訳系に導入して用いることができる。詳細は以下の文献を参照されたい。
H. Murakami, H. Saito, and H. Suga, (2003), Chemistry & Biology, Vol. 10, 655-662;
H. Murakami, D. Kourouklis, and H. Suga, (2003), Chemistry & Biology, Vol. 10, 1077-1084;
H. Murakami, A. Ohta, H. Ashigai, H. Suga (2006) Nature Methods 3, 357-359;
N. Niwa, Y. Yamagishi, H. Murakami, H. Suga (2009) Bioorganic & Medicinal Chemistry Letters 19, 3892-3894; および
特開2008-125396またはWO2007/066627。
【0031】
フレキシザイムは、ジニトロベンジルフレキシザイム(dFx)、エンハンスドフレキシザイム(eFx)、アミノフレキシザイム (aFx) 等の呼称でも知られる。フレキシザイムは、弱く活性化されたアミノ酸を基質として、アミノ酸の反応点であるカルボニル基、およびアミノ酸側鎖あるいは脱離基である芳香環、並びにリンカーの3’末端に存在するACC-3'配列部分を認識して、3’末端のアデノシンにアミノアシル化する触媒能を有する。RAPIDリンカーの端にACCA-3'の配列からなるオリゴRNA構造が必須なのはこのためである。フレキシザイムによるアミノアシル化反応は、アミノ酸基質と、結合相手であるオリゴRNA部分を有するリンカー分子を、フレキシザイム存在下、2時間程度氷上に置くだけで進行する。
【0032】
本発明においては、以下の式で示される配列を有するフレキシザイムが好適に使用される。
原型のフレキシザイム Fx
[GGAUCGAAAGAUUUCCGCAGGCCCGAAAGGGUAUUGGCGUUAGGU-3’, 45nt] (配列番号3)
エンハンスドフレキシザイム eFx
[5'-GGAUCGAAAGAUUUCCGCGGCCCCGAAAGGGGAUUAGCGUUAGGU-3’,45nt]) (配列番号4)
ジニトロベンジルフレキシザイム dFx
[5'-GGAUCGAAAGAUUUCCGCAUCCCCGAAAGGGUACAUGGCGUUAGGU-3’,46nt] (配列番号5)
アミノフレキシザイム aFx
[5'-GGAUCGAAAGAUUUCCGCACCCCCGAAAGGGGUAAGUGGCGUUAGGU-3’,47nt]) (配列番号19)
フレキシザイム Fxは、脱離基としてシアノメチル基を、側鎖として芳香環をもつアミノ酸(例えば、フェニルアラニン、チロシンなどのシアノメチルエステル)、フレキシザイム eFx, dFx, aFxはフレキシザイム Fxで利用可能な脱離基とアミノ酸に加え、脱離基として4−クロロベンジルチオールを、側鎖としては芳香環以外の側鎖をもつアミノ酸を基質としたアミノアシル化を触媒することができる。詳細は、前述の文献を参照されたい。したがって、このような構造を持つアミノ酸とリンカーをフレキシザイムの存在下で反応させることにより、リンカー3’末端のアデノシン内のリボース環上の3’位の水酸基に、アミノ酸がエステル結合した分子を得ることができる。アミノ酸基質としては、任意の構造を持つものを結合させることができ、天然の翻訳で使用されるアミノ酸のみならず、D-アミノ酸あるいはβ(beta)-アミノ酸などの非タンパク質性アミノ酸(すなわち、天然に存在するタンパク質中に通常見出されるL-アミノ酸以外のもの)も利用できる。さらに、アミノ酸の代わりにヒドロキシカルボン酸をリンカー3’末端に結合させることもでき、ペプチドアクセプターとしてリボソームへの取り込みも可能である。
【0033】
3.翻訳系内での反応
RAPIDディスプレイ法では、リンカーをcDNAあるいはmRNAと共に無細胞翻訳系(「in vitroタンパク質合成系」ともいう)に加えて一定時間反応させることで、mRNAからの翻訳およびリンカー分子との複合体形成、それに続くペプチドとリンカーとの連結を行うことができる。
【0034】
無細胞翻訳系としては、慣用の再構成型の無細胞翻訳系を適宜改変して利用でき、DNA依存性RNAポリメラーゼ(好ましくはT7RNAポリメラーゼ)を加えた系であればDNAからの転写も翻訳と同じ系で行うことができる。
【0035】
転写・翻訳型の合成系であれば、以下の(ア)から(エ)の反応を一つの反応容器(one-pot)内で行うことが可能である。
(ア)DNAをmRNAに転写する反応
(イ)mRNAの3’末端側が、リンカー末端の一本鎖構造領域とハイブリダイゼーションにより複合体を形成する反応
(ウ)mRNAからペプチドに翻訳する反応
(エ)翻訳が完了したペプチドのC末端がリンカーのもう一方の末端のペプチドアクセプター領域にアミド結合する反応
(オ)〔ペプチド〕−〔リンカー〕−〔mRNA〕複合体が、リボソームから解離する反応。
【0036】
あるいは、予め調製されたmRNAを、リンカーと一緒に翻訳系に加える場合は、(イ)から始まる一連の反応を再構成型の無細胞翻訳系内で行う。
再構成型の無細胞翻訳系には精製したリボソーム、翻訳開始因子、翻訳伸長因子、mRNA、アミノアシルtRNA、基質としてATPやGTP等が必要である(M. H. Schreier, B. Erni and T. Staehelin (1977) “Initiation of mammalian protein synthesis. I. Purification and characterization of seven initiation factors.” Journal of Molecular Biology, Vol. 116, No. 4, 727-53. H. Trachsel, B. Emi, M. H. Schreier and T. Staehelin (1977)“Initiation of mammalian protein synthesis. II. The assembly of the initiation complex with purified initiation factors.” Journal of Molecular Biology, Vol. 116, No. 4, 755-67.)。これらのうち、アミノアシルtRNAは同一反応液中にtRNAとアミノアシル-tRNA合成酵素およびその基質を加えることで代替が可能である。また、一般の無細胞翻訳系において行われているように、翻訳反応の効率や忠実性を上げるため、翻訳終止因子、リボソーム再生因子、クレアチンキナーゼ、ミオキナーゼ、ヌクレオチド二リン酸キナーゼ、ピロフォスファターゼなどのタンパク質や酵素およびその基質を添加することも可能である(P.C. Jelenc and C.G. Kurland (1979) “Nucleoside triphosphate regeneration decreases the frequency of translation errors” Proceedings of the Natural Academy Science of the United States of America Vol. 76, No. 7, 3174-3178)。また、翻訳と同時に転写反応を行う場合は、mRNAの代わりにcDNAおよびT7 RNAポリメラーゼとその基質を添加することができる。
【0037】
本発明においては、目的に合わせて最適化された構成因子からなる無細胞翻訳系に、必要な配列を有するDNAまたはRNAが供される。DNAまたはRNAの配列については、使用する合成系で、cDNAがmRNAに転写され、当該mRNAの翻訳が開始されるための配列が必要である。ペプチドのアミノ酸配列をコードする部分の全長が、最後まで翻訳される必要があり、さらに、翻訳されたアミノ酸のC末端側にも柔軟性を持たせるためのペプチドからなるスペーサー配列が連結し、その直後に停止コドンが来る。例えば、ペプチド配列(Cys-(Gly-Ser-)x3)と、その直後にアンバーコドン(停止コドン)がコードされる。また、mRNAの3’末端側は、リンカーの一本鎖構造領域とハイブリダイズして二本鎖を形成できるような構造を有するので、コード領域の下流(停止コドンの直後)は、一本鎖構造領域の配列に相補的な配列を有する必要がある。この部分(二本鎖形成部)の配列を、本明細書中ではリンカーハイブリダイゼーション配列と称する。
【0038】
cDNAまたはmRNAは、具体的には、以下のような配列を含むことが望ましい。
(1)cDNAにおいて、使用するRNAポリメラーゼに対応したプロモーター配列。
T7プロモーターの場合、TAATACGACTCACTATA (配列番号6)
(2)開始コドン上流の適切な配列をコードする配列。
無細胞翻訳系で大腸菌由来のリボソームを使用する場合、SD配列遺伝子を含む。これは通常のタンパク質合成と同様である。例えば、GGGTTAACTTTAA GAAGGA GATATACAT (配列番号7): T7ファージのgene 10 proteinの上流配列を改変したもの。下線部はSD配列。
【0039】
(3)開始コドン(ATG)から始まる、変異遺伝子ライブラリーのORF〔ペプチドアプタマーのC末端側にスペーサーが融合したアミノ酸配列をコードする配列〕を構成する配列。
【0040】
ここで使用される停止コドンにより、無細胞翻訳系から対応するrelease factor(解離因子)を除く必要がある。TAG(アンバーコドン)を用いた場合はRF1を、TGA(オパールコドン)を用いた場合はRF2を、TAA(オーカーコドン)を用いた場合はRF1とRF2の両方を除いて無細胞翻訳反応液を調製する。
【0041】
(4)二本鎖形成部をコードする配列(リンカーハイブリダイゼーション配列)。
本発明で使用される再構成型の無細胞翻訳反応液としては、構成因子を目的に合わせて改変して利用することが好ましい。例えば、実施例においては、アンバーコドンに対応するrelease factor、ペプチジル-tRNAを分解する酵素であるペプチジル-tRNAヒドロキシダーゼ(PTH)などを除いている。
【0042】
また、再構成無細胞翻訳系に、限られた天然アミノ酸だけを与えた系に、予め所望の非タンパク質性アミノ酸(又はヒドロキシ酸)でチャージされた(つまり、活性化されたアミノ酸が結合した)アシル化tRNAを添加することも可能である。添加しない天然アミノ酸のコドンを、非タンパク質性アミノ酸(又はヒドロキシ酸)でアシル化されたtRNAのアンチコドンに対応させることにより、非タンパク質性アミノ酸(又はヒドロキシ酸)を含むペプチドをmRNAの遺伝情報に基づき、リボソーム上で翻訳合成することが可能である。あるいは、天然アミノ酸を含まない再構成無細胞翻訳系に、非タンパク質性アミノ酸(又はヒドロキシ酸)でチャージされたアシル化tRNAを加えることにより、天然アミノ酸を全く含まない特殊ペプチドを翻訳合成することも可能である。
【0043】
非タンパク質性アミノ酸(又はヒドロキシ酸)を有するアシル化tRNAは、アミノアシルtRNA合成触媒能を有する上述の人工RNA触媒「フレキシザイム」を用いて調製することができる。上述したように、これらの人工RNA触媒は、任意の側鎖を持つアミノ酸をチャージさせることができ、また、tRNAの3'末端に存在する共通配列である5'-RCC-3'配列部分(R = A or G)のみを認識し、tRNAの3'末端にアシル化する機能を有するので、アンチコドンの異なるどのようなtRNAにも対応することが可能である。さらに、アミノ酸の認識部位にα位の置換基が含まれていないため、Lアミノ酸に限らず、ヒドロキシ酸(α位が水酸基)、N-メチルアミノ酸(α位がN-メチルアミノ酸)、N-アシルアミノ酸(α位がN-アシルアミノ基)、D-アミノ酸なども基質とすることができる。詳細については、上述のフレキシザイムに関する文献に加え、Y. Goto, H. Suga (2009) "Translation initiation with initiator tRNA charged with exotic peptides" Journal of the American Chemical Society, Vol. 131, No. 14, 5040-5041、WO2008/059823「N末端に非天然骨格をもつポリペプチドの翻訳合成とその応用」、Goto et al., ACS Chem. Biol., 2008, 3, 120-129、WO2008/117833「環状ペプチド化合物の合成方法」などに記載されている。
【0044】
本出願人らは、「フレキシザイム」を用いて非タンパク質性アミノ酸やヒドロキシ酸をアシル化したtRNAを用いた特殊ペプチドの合成系(キットまたは合成方法の両方を含む概念)をコア技術としてペプチドの翻訳合成・修飾・スクリーニングを行う総合技術をRAPIDシステム(Random Peptide Integrated Discovery system)と呼んでいる。RAPIDシステムでは、適当な配列の鋳型mRNAに基くin vitro翻訳産物として、様々な特殊ペプチドを翻訳合成することができる。これまでの説明から自明であるが、本明細書において、特殊ペプチドとは、RAPIDシステムで合成可能なあらゆる翻訳産物を含み、20種類の天然アミノ酸以外に、様々な側鎖を有するアミノ酸、β(beta)-アミノ酸、γ(gamma)-アミノ酸及びδ(delta)-アミノ酸、D-アミノ酸、アミノ酸骨格上のアミノ基やカルボキシル基が置換された構造を有する誘導体を含む、上述した様々な基質を構成要素とする重合体である。さらに、特殊ペプチドは、主鎖骨格として通常のアミド結合とは異なる構造も有し得るものである。例えば、アミノ酸とヒドロキシ酸から構成されるデプシペプチド、ヒドロキシ酸が連続して縮合したポリエステル、N-メチルアミノ酸の導入によりアミド結合の窒素がメチル化されたペプチド、N-末端に様々なアシル基(アセチル基、ピログルタミン酸、脂肪酸など)を有するペプチドも、特殊ペプチドに含まれる。また、両端に互いの間で結合形成反応が可能な一組の官能基を配置したアミノ酸配列からなる非環状ペプチドを適切な反応条件下で環化させて得られる環状ペプチド(N-メチルペプチドが導入された場合は、環状N-メチルペプチド)もRAPIDシステムで合成可能である。一組の官能基の種類によっては無細胞翻訳系の条件下において環化することがあり、後述の実施例のように、両端にそれぞれクロロアセチル基とシステインを配置したペプチド配列を翻訳合成して得られるチオエーテル結合で環化した環状ペプチドはその一例である。
【0045】
RAPIDシステムでは、リボソームによる翻訳で特殊ペプチドを合成するため、鋳型であるmRNAを代えるだけで様々な構造を有するペプチドを合成できる。ランダム配列を含むmRNA(または対応するDNA)を用いて翻訳合成を行えば、ランダムペプチドライブラリーを容易に構築することができる。本発明のRAPIDディスプレイ法は、RAPIDシステムにより合成される特殊ペプチドを、その遺伝子型であるmRNAと対応付けるために使用するのに、好適である。特殊ペプチドの合成用に最適化された無細胞翻訳系に、鋳型cDNAあるいはmRNAと共に、本発明のRAPIDリンカーを加えて、一定時間反応させることで、翻訳産物である特殊ペプチドがリンカーを介してmRNAと連結して提示される。
【0046】
次に、無細胞翻訳系内で起こる、リンカーとmRNAの結合について説明する。従来技術の項で述べたように、公知のmRNAディスプレイ法では、翻訳反応の前段階に、リンカーとmRNAの連結(ライゲーション)を翻訳系外で行っておくことが必要であった。これに対し、本発明のRAPIDディスプレイ法ではリンカーとmRNAのハイブリダイゼーションによる複合体形成を翻訳系内で行うことができることが特徴である。
【0047】
図2を参照しながら、mRNAをRAPIDリンカーと共に無細胞翻訳系に供することにより、mRNAとリンカーとのハイブリダイゼーションおよびペプチド分子の合成、そしてペプチドC末端とリンカーとの連結が起こる過程について説明する。
【0048】
上述したように、mRNAとリンカーとの結合は、リンカーの一本鎖構造領域の塩基配列とmRNAの3’末端の相補的な塩基配列が水素結合で二本鎖を形成することによる。この結合は、mRNAと翻訳されたペプチド分子とを対応付けるためのものであるから、リボソーム上で翻訳が行われている間に形成された〔mRNA〕−〔リンカー〕−〔ペプチド〕複合体が、翻訳産物であるペプチドのセレクションの間も安定に維持される必要がある。
【0049】
mRNAをRAPIDリンカーと共に無細胞翻訳系に供することにより、リボソームがmRNA上に配置されて翻訳反応が開始し、ペプチド鎖が伸長し、そして、終結したペプチド鎖のC末端が、リンカーの〔rACCA-アミノ酸〕からなるペプチドアクセプター領域のアミノ酸と結合してtRNAから切り離される。本発明の概念を限定する意図ではなく、あくまでも説明のためであるが、mRNAをRAPIDリンカーと同時に無細胞翻訳系に供する場合、mRNAとRAPIDリンカーとのハイブリダイゼーションは、ペプチド鎖の伸長反応開始よりも前の段階で起こり得ると本発明者らは考えている。あるいは、cDNAからの転写および翻訳反応を先に行ってから、終結因子およびPTHを除いた翻訳反応液中にリンカーを後から添加することによっても、リボソーム上のmRNAとRAPIDリンカーとのハイブリダイゼーションによる複合体形成は正しく起こる。そして、リンカー末端の〔rACCA-アミノ酸〕とペプチドC末端との共有結合は、翻訳終了時に偶然にこの部分がリボソームAサイトに入った場合に起こると考えられる。翻訳終了時には既にmRNAとリンカーとのハイブリダイゼーションが起こっているので、この〔rACCA-アミノ酸〕基質はリンカーでリボソーム上のmRNAとつながっており局所的な濃度が非常に高くなっているため、反応は高い効率で起こる。
【0050】
このようにしてリボソーム上で形成された〔mRNA〕−〔リンカー〕−〔ペプチド〕複合体が、ペプチド鎖がリボソームから解離した後も安定に維持されていることは、後述の実施例においても裏付けられている。
【0051】
4.ペプチドアプタマーの選択
進化分子工学では、所望の機能や性質を持つタンパク質やペプチドを創製することを目的として、可能性のある遺伝子を大規模に準備し、その中から狙った表現型を有するクローンを選択する。
【0052】
基本的には、最初にDNA集団を調製し、in vitro転写産物としてRNA集団を得て、in vitro翻訳産物としてペプチド集団を得る。このペプチド集団から、所望の機能や性質を持つものを何らかのスクリーニング系で選択することになる。例えば、特定のタンパク質に結合するペプチド分子を得たい場合は、標的タンパク質を固相化したカラムにペプチド集団を流し込み、カラムに結合したペプチド分子の混合物を回収することができる。このとき、各ペプチド分子には、その鋳型であるmRNAがタグのように付加されているので、回収したペプチド−mRNA複合体の集団から逆転写酵素でDNAに戻し、PCRで増幅して狙った表現型を有するクローンが多く含まれるバイアスのかかったライブラリーを得た後に、再度同じような選択実験を行う。あるいは、RNAアプタマーを回収してしまう可能性を回避するため、核酸部分を2本鎖にする目的で、選択前に逆転写反応を行うことも可能である。この操作を繰返すことで、世代の経過とともに所望の表現型を有するクローンが集団中で濃縮されていく。
【0053】
ペプチドアプタマーを同定する場合、対応付け分子のライブラリーと標的物質とを混合し、標的物質に結合したペプチドを提示する対応付け分子(活性種)を選択し、選択された対応付け分子の核酸部分からPCRにより核酸ライブラリーを調製する工程を繰り返すことで、標的物質に結合するペプチドアプタマーの遺伝子をクローニングできる。標的物質に結合した対応付け分子を選択する工程は、〔RNA(あるいはDNA/RNAハイブリッド)〕−〔リンカー〕−〔ペプチド〕からなる複合体を標的物質と結合させ、適当な方法で他の複合体から分離して、目的の結合特性を有するペプチドを同定することにより行うことができる。
【0054】
標的物質としては、タンパク質、核酸、糖質、脂質、その他どのような化合物でもよい。標的物質と結合する活性種である複合体を、他の複合体から分離するために、標的物質に、固相への結合により回収可能な修飾を施しておくと便利である。例えば、後述の実施例では、標的物質をビオチン修飾しておき、固相化されたビオチン結合タンパク質への特異的な結合を利用して回収している。このような特異的な結合としては、ビオチン結合タンパク質(アビジン、ストレプトアビジンなど)/ビオチンの組み合わせの他にも、マルトース結合タンパク質/マルトース、ポリヒスチジンペプチド/金属イオン(ニッケル、コバルトなど)、グルタチオン-S-トランスフェラーゼ/グルタチオン、抗体/抗原(エピトープ)などが、利用可能であるが、これらに限定されるものでは勿論ない。
【0055】
進化分子工学を用いることで、遺伝子ライブラリーとして、A、T、G、Cの4つの塩基がランダムに結合したDNA配列から、天然には全く存在しないアミノ酸配列を有するペプチドを得ることが原理的に可能である。また、翻訳系に非タンパク質性アミノ酸(またはヒドロキシ酸)でアシル化されたtRNAを導入することにより、in vitro翻訳産物として非タンパク質性アミノ酸(またはヒドロキシ酸)が導入された特殊ペプチドの翻訳による合成も可能である。特殊ペプチドとmRNA(あるいはcDNA)の複合体の集団から、所望の結合特性を有するペプチドが提示された活性種を選択し、対応付けされた遺伝子部分を増幅、再び翻訳するという作業を繰返すことで、特殊ペプチドのライブラリーを効率的に得ることができる。
【0056】
これまでの説明および後述の実施例の内容に関し、分子生物学的手法の詳細は、例えば、Sambrook, Molecular Cloning: A Laboratory Manual, 3rd edition, Cold Spring Harbor Laboratory Press, 2001; Golemis, Protein-Protein Interactions: A Molecular Cloning Manual, 2nd edition, Cold Spring Laboratory Press, 2005などを参照されたい。
【0057】
以下、本発明を実施例によって具体的に説明する。なお、これらの実施例は、本発明を説明するためのものであって、本発明の範囲を限定するものではない。
【実施例1】
【0058】
〔リンカーの合成〕
リンカーとしてDNAとRNA間をポリエチレングリコール(5ユニットのSpacer18)で連結したDNA/RNAキメラオリゴヌクレオチドを使用した。種々リンカーはBEX社(東京)より購入した。以下に示す配列の*Aおよび*CはRNA、SPC18はSpacer18(ヘキサエチレングリコール)に相当する。
【0059】
an21-ACCA: 5'-CCCGCCTCCCGCCCCCCGTCC-[SPC18]5-A*-C*-C*-A*-3'
〔リンカーのアミノアシル化〕
フレキシザイムが触媒する反応により、an21-ACCAリンカー分子の3’末端側にエステル結合を介してL-フェニルアラニンもしくはβ-L-アラニンを結合させた。反応産物は精製した反応産物溶液を酸性条件下でのアクリルアミド電気泳動分析により確認した。リンカー分子に由来するバンドがアミノアシル化されると移動度が少なくなる。そのため、未反応物由来のバンドと反応産物由来のバンドの強度を比較することでアミノアシル化効率を決定することができる。
【0060】
アシル化反応は、L-フェニルアラニンを結合させる場合は、0.1 M HEPES-カリウムバッファー(pH 7.5)、600 mM 塩化マグネシウム中で、20 μMのフレキシザイムeFx 5 μL、20 μMのan21-ACCAリンカー、および基質(L- フェニルアラニンシアノメチルエステル)を20 %ジメチルスルホキシドに加えて、氷上で2時間反応させた。詳細な手順としては、まず、純水に溶解した40 μMのリンカー分子を0.2 M HEPES-カリウムバッファー(pH 7.5)およびフレキシザイムeFx(200 μM, 0.5 μL)を加え、サーモブロック(ND-MD1, 日伸理化)上で95 ℃で2分間加熱し、室温で5分間静置した。その後、氷上で塩化マグネシウム(3 M, 1 μL) および基質(ジメチルスルホキシド中25 mM, 1 μL)を加えることによりリンカー分子のアシル化反応を開始し、氷上で2時間静置した。β-L-アラニンを結合させる場合は、同様の条件にて、pHを8.0とし、基質としてβ-L-アラニンp-クロロベンジルチオエーテルを用いて行った。反応は0.3 M 酢酸ナトリウム(pH 5.0)を40 μL加えることにより停止した。反応産物をエタノール沈殿し、70% エタノールで洗浄したペレットを10 μLの1 mM酢酸ナトリウムに溶解した。反応後の溶液を酸性条件下、20 %の変性ポリアクリルアミド電気泳動(50 mM 酢酸ナトリウム(pH 5.0)、6 M 尿素)で分離し、泳動後のゲルをSYBR Green II (Invitrogen、SYBRはMolecular Probes Inc.の登録商標)を用いた蛍光染色により解析した。
【0061】
結果は図3で示したようになり、対照となる未反応のリンカー分子よりも、反応産物のリンカー分子の移動度が小さいことが確認され、リンカー分子がフレキシザイムの触媒する反応によりアミノアシル化されていることが確認された。
【0062】
〔cDNAの合成〕
ペプチド-mRNA複合体を形成するために用いるcDNAは、合成オリゴヌクレオチドをPCRにより連結することにより調製した。以下に配列を示す合成DNAはオペロン社(東京)より購入した。これらのオリゴDNAを連結することにより得られる各々のcDNAは、T7プロモーター配列、リボソーム結合配列、開始コドン、ペプチドアプタマー配列、スペーサーペプチド配列(CGSGSGS)、アンバーコドンおよびリンカーハイブリダイゼーション配列より構成される。
【0063】
TNF-a_D-Trp.R66:
GCCGCTGCCGCTGCCGCAATGCTTCAGATACAGACAATGCAGACGTTGCATATGTATATCTCCTTC
EMP1SS.F63: GAAGGAGATATACATATGGCAGCAGGTGGTACCTATTCTTCTCATTTTGGTCCGCTGACCTGG
EMP1SS.R63: GCCGCTGCCGCTGCCGCATGCTGCACCACCTTGCGGCTTAGAAACCCAGGTCAGCGGACCAAA
T7g10M.F48: TAATACGACTCACTATAGGGTTAACTTTAAGAAGGAGATATACATATG
CGS3an13.R39: TTTCCGCCCCCCGTCCTAGCTGCCGCTGCCGCTGCCGCA
CGS3an21.R44: CCCGCCTCCCGCCCCCCGTCCTAGCTGCCGCTGCCGCTGCCGCA
PCRによる連結は以下の手順にて行った。
【0064】
T7g10M.F48およびTNF-a_D-Trp.R66(もしくはEMP1SS.F63とEMP1SS.R63の組み合わせ)を250 nM含むPCR反応液(10 mM Tris-HCl (pH 9.0), 50 mM 塩化カリウム, 2.5 mM 塩化マグネシウム, 250 μM dNTPs, 0.2 % Triton X-100(トリトンX-100、ナカライテスク)およびTaq ポリメラーゼ) 20 uLを調製し、サーマルサイクラー(TC-3000, Techne社)を用いて94 ℃ 1 min, {50 ℃ 30 sec, 72 ℃ 30 sec}5サイクルによる反応を行った。次に各々500 nM のプライマー、T7g10M.F48およびGCSan21.R44(もしくはGCSan13.R39)を含むPCR反応液100 μLを調製し、これに先のPCR反応後液1 μLを加え、温度サイクル{94 ℃ 40 sec, 50 ℃ 40 sec, 72 ℃ 40 sec}10サイクルで反応を行った。反応産物は常法に従いフェノール・クロロホルム抽出、クロロホルム抽出、さらにエタノール沈殿により精製を行った。
【0065】
〔mRNAの調製〕
mRNAは前述の方法で作成したcDNA(TNF-α-DW(an21))を元にT7 RNA ポリメラーゼによる転写反応によりRNAを合成し、得られた反応産物は常法に従いフェノール・クロロホルム抽出、クロロホルム抽出、さらに2-プロパノール沈殿により精製を行った。精製後のmRNAの濃度を260 nmにおけるUVの吸収から求め、10 μMになるように希釈した。
【0066】
〔転写・翻訳、リンカーのハイブリダイゼーション、ペプチドアクセプターとの連結〕
mRNAからの翻訳およびリンカー分子との複合体形成もしくはcDNAからの転写、翻訳およびリンカー分子との複合体形成は再構成型の無細胞翻訳系を用いて行った。
【0067】
本実施例で使用した再構成型の無細胞翻訳系には以下の生体高分子が含まれる:
70Sリボソーム(1.2uM)、翻訳開始因子(IF1(0.7uM), IF2(0.4uM), IF3(1.5uM))、
伸長因子(EF-G(0.26uM), EF-Tu/EF-Ts複合体(10uM))、翻訳終止因子(RF2(0.25uM), RF3(0.17uM))メチオニルtRNAトランスフォルミラーゼ(MTF(0.6uM))、リボソーム再生因子(RRF(0.5uM))、アミノアシルtRNA合成酵素(AlaRS(0.73uM), ArgRS(0.03uM), AsnRS(0.38uM), CysRS(0.02uM), GlnRS(0.06uM), GluRS(0.23uM), GlyRS(0.09uM), HisRS(0.02uM), IleRS(0.4uM), LeuRS(0.04uM), MetRS(0.03uM), PheRS(0.68uM), ProRS(0.16uM), SerRS(0.04uM), ThrRS(0.09uM), TrpRS(0.03uM), ValRS(0.02uM), AspRS(0.13uM), LysRS(0.11uM), TyrRS(0.02uM))、クレアチンキナーゼ(ロシュ社製CK(4ug/mL)、ミオキナーゼ(ロシュ社製、MK(3ug/mL))、ピロフォスファターゼ(PPa(0.1uM))、
ヌクレオチド二リン酸キナーゼ(NDK(0.1uM))、T7 RNAポリメラーゼ(T7ファージ遺伝子由来の組み換え体、0.1uM)、大腸菌tRNA(ロシュ社製、1.5mg/mL)。リボソームは対数増殖期の大腸菌から精製したもの、リボソーム以外の各種タンパク質は、特に指示がある場合を除き、大腸菌の遺伝子をクローニングし、それを元に発現、精製した組み換えタンパク質である。
【0068】
また、生体高分子以外の成分として、以下の物が含まれる:
50mM HEPES-KOH (pH7.6), 2mM NTPs, 20mM リン酸クレアチン, 100mM 酢酸カリウム, 2mM スペルミジン, 1mM ジチオスレイトール, 6mM 酢酸マグネシウム, 0.1mM 10-ホルミル-5, 6, 7, 8-テトラヒドロ葉酸。
【0069】
これらに加えて、mRNA 1.5 μMもしくはcDNA 0.15 μM、アシル基としてN-クロロアセチル-D-トリプトファン(ClAc-D-Trp)を担持したアミノアシルイニシエーションtRNA(特開2008-125396で開示の方法により調製)、メチオニンを除く天然タンパク質を構成する19種類のアミノ酸(各々5 mM)を含む転写・翻訳反応溶液2.5 μLを調製した。
【0070】
あらかじめ転写反応を行いmRNAを調製し、これを元に翻訳およびペプチドとリンカーとの結合を行う場合は、以下の手順によりペプチド-mRNA複合体を調製した。まず、4 mM酢酸ナトリウム pH 5.0とmRNAを1:3で混合し、これをサーモブロック上で95 ℃で1分間加熱したのちに室温下で5分静置した。1 mM酢酸ナトリウムに溶解した25 μM のリンカー溶液を混合し10分間静置することでmRNA-リンカーの複合体を形成させた。これに翻訳系の他のコンポーネントを加え、恒温水槽(NT-202D, 日伸理化)中で37 ℃、30分間、続いて室温で12分インキュベートし、pH 7.5に調製した0.1 M EDTA(エチレンジアミン4酢酸、分子生物学実験用、ナカライテスク)を終濃度が20 mMになるように添加し、さらに恒温水槽中で37 ℃で30分間インキュベートした。
【0071】
反応溶液中にcDNAを加え、この反応溶液中で転写、翻訳およびリンカーとの結合を行う場合は、以下の手順によりペプチド-mRNA複合体を調製した。まず始めにリンカー以外の物を含む溶液を調製し、これを恒温水槽中で37 ℃で30分間反応させることにより、転写および翻訳反応を行った。これに25 μMのリンカー溶液0.25 μLを加え、さらに37 ℃で30分、続いて室温で12分インキュベートした。その後、pH 7.5に調製した0.1 M EDTA(エチレンジアミン4酢酸、分子生物学実験用、ナカライテスク)を終濃度が20 mMになるように添加し、さらに恒温水槽中で37 ℃ で30分間インキュベートした。
【0072】
〔逆転写〕
ペプチド−mRNA複合体のmRNA部分の安定性を向上させるため、逆転写反応を行うことにより、RNA-DNAハイブリッド鎖を形成させた。具体的には以下の操作を行った。前述の翻訳反応産物に、12 mM Tris-HCl (pH 8.3), 5 μM 逆転写プライマー(CGS3an13.R21), 0.5 mM dNTPs, 18 mM Mg(OAc)2, 10 mM KOH(各々の濃度は終濃度)を添加した。これにM-MLV Reverse Transcriptase (RNaseH Minus, Point Mutant, Promega) 1 unitを添加し恒温水槽中で42 ℃で10分間反応させた。その後、終濃度が10 mMおよび18 mMになるようにEDTAと塩酸を添加した。
【0073】
〔セレクション〕
このようにして作成したペプチド−mRNA複合体分子が、提示ペプチドとターゲットとなるタンパク質との相互作用により回収されるかどうかを以下に示す方法により確認した。
【0074】
ターゲットとなるタンパク質として、ヒトの腫瘍壊死因子アルファ(以下、TNF-α)を選択した。
ペプチド-mRNA複合体分子を含む無細胞翻訳反応産物液6 μLに対し、ビオチン修飾されたTNF-αタンパク質を終濃度250 nMになるように添加し、この溶液を低吸着処理を施した0.6 mLエッペンドルフチューブ(プラチナ(超低吸着)チューブ、ビーエム機器)に移し、ローテーター(RT-30 mini、タイテック)の上で4 ℃、1 hr 穏やかに攪拌し結合させた。この溶液に対して、ストレプトアビジンを固相化した磁性ビーズ(Dynabeads 登録商標Streptavidin M-280, Invitrogen社)の懸濁液3 μLを加え、さらに10分攪拌した。その後、遠心分離およびマグネットホルダーを使用して磁性ビーズを分離し上清を除き、再度0.05 % のTween20(ポリオキシエチレンソルビタンモノラウレート、ナカライテスク)を含むTBS (Tris Buffered Saline, 50 mM Tris-HCl(トリスヒドロキシメチルアミノメタン、ナカライテスク) pH 7.5, 150 mM 塩化ナトリウム) 50 μL に懸濁した。この洗浄操作を4回繰り返した。
【0075】
続いて磁性ビーズにPCR(taq-)バッファー(10 mM Tris-HCl (pH 9.0), 50 mM 塩化カリウム, 2.5 mM 塩化マグネシウム, 250 μM dNTPs, 0.25 μM T7g10m.F48, 0.25 μM CGS3an13.R21, 0.2 % Triton X-100)を25 μL加え、サーモブロック上で95 ℃で5分間加熱後に上清を回収することにより、磁性ビーズより逆転写後のDNAを回収した。
【0076】
〔リアルタイムPCRによる定量〕
ターゲットタンパク質添加前、および磁性ビーズより回収後のDNAの量をリアルタイムPCR法により定量した。リアルタイムPCR測定装置はLightCycler(登録商標) 1.5 (Roche Applied Science社)を使用し、前述のPCR(taq-)バッファーにTaqポリメラーゼ、SYBR(登録商標) Green I (100,000希釈、Invitrogen社)、および測定サンプル溶液を添加した反応液を測定に供した。
【0077】
TNF-αを標的としたモデル実験
リンカーの機能を確認するために用いたターゲットとなるタンパク質およびペプチドアプタマーとして、TNF-αおよびTNF-αに結合するペプチドアプタマーであるTNF-α-DWを選択した。
【0078】
本実施例で用いたTNF-αタンパク質として大腸菌により発現した組換え体である可溶型TNF-αを使用した。組換えTNF-αは野生型TNF-αの77番目から233番目のアミノ酸配列の部分、そのN末端側にAviTag配列(GLNDIFEAQKIEWHE)およびHis×6 tag配列を融合したものである。このタンパク質とAviTagを基質とするビオチンリガーゼ(BirA)を大腸菌内で共発現させることによりAviTagのリジン残基の側鎖がビオチン化修飾されるため、本実施例で用いているTNF-αはストレプトアビジン固相化ビーズにより容易に分離することができる。
【0079】
ペプチドアプタマーとして用いたTNF-α-DWは、非タンパク質性アミノ酸であるN-クロロアセチル-D-トリプトファン(ClAc-D-Trp)のクロロアセチル基とペプチド配列中のシステインの側鎖のチオール基が反応し、チオエーテルを形成することにより環化したペプチドXQRLHCLYLKH(X: Ac-D-Trp)である。このペプチドのC末端側にスペーサーアミノ酸配列CGSGSGSを融合した配列をコードするcDNAを前述の方法にて作成し、無細胞転写・翻訳系にてペプチドとmRNA複合体分子を作成した。
【0080】
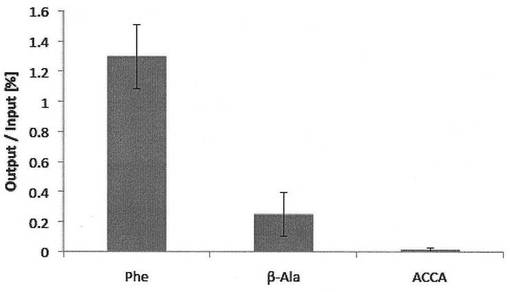
このペプチドアプタマーと標的タンパク質の組み合わせを用いた場合のペプチド-mRNA複合体の回収率は図4および図5に示すとおりであった。L-フェニルアラニンを3’末端にエステルを介して結合させたan21- Phe、an21-β-Alaの回収率は翻訳反応液にmRNAを添加した場合(図4)は各々1.30 %, 0.25 %、cDNAを添加した場合(図5)は0.65 %, 0.38 %であった。一方、3’末端が修飾されていないan21-ACCAリンカーでの回収率はそれらのものと比較して数十倍程度低く(mRNAの場合0.015%, cDNAの場合0.007 %)、L-フェニルアラニン、β-L-アラニンが翻訳合成されたペプチドのC末端側に共有結合を介して連結することにより、ペプチド-mRNA複合体分子として効率よく回収されることが示された。
【実施例2】
【0081】
提示ペプチドとmRNAとの対応付けの確認
前述までの実験に於いては、提示しているペプチドと、その配列をコードしているmRNAは単一の種類であるため、mRNAから翻訳合成されたペプチドに対して、別のリンカー分子がアクセプターとして結合している可能性や、2本鎖を形成しているリンカー分子とmRNA分子が操作の途中で別のリンカー分子やmRNAと交換している可能性を排除できない。そのため、回収されるペプチド-mRNA-リンカー複合分子の提示ペプチドとmRNAとの対応付けが正確に行われていることを確認するために以下に示す実験を行った。
【0082】
まず、TNF-α-DWの対照としてターゲットとするタンパク質に結合しないペプチド配列(EMP1SS:XAAGGTYSSHFGPLTWVSKPQGGAA、XはTNF-α-DWと同様にAc-D-Trpである)をコードするmRNAもしくはcDNAを実施例1に示す方法で調製し、これとTNF-α-DWをコードするmRNAもしくはcDNAをモル比100:1で混合したものを用意した。これを元に前述と同様の実験を行い、回収したDNAをPCRにて増幅した。その際の条件として、PCR(taq-)バッファーにTaqポリメラーゼおよび測定サンプル溶液を添加した反応液20 μLを用意し、サーマルサイクラーを用いて、温度サイクル{94 ℃ 40 sec, 61 ℃ 40 sec, 72 ℃ 40 sec}(61 ℃から72 ℃への昇温は0.5 ℃/sec)を30サイクル行い反応させた。反応産物液2.5 μLをTAE(40 mM Tris酢酸、1 mM EDTA)および3 % アガロース(低電気浸透度、ナカライテスク)からなるゲルを用いた電気泳動により分離し、エチジウムブロミドによる染色およびトランスイルミネーターを用いて泳動像を得た。その結果を図6に示す。
【0083】
TNF-α-DWをコードするmRNAに由来するバンドは約90 bpの位置に確認され、対照とするEMP1SSをコードするmRNAに由来するバンドは約150 bpの位置に観察される。ペプチドアクセプターを含まないan21-ACCAリンカーを用いた場合は、はじめに2種類のcDNAを混合した際のモル比を反映しEMP1SSのmRNAに由来するバンドのみが確認された。それに対して、an21-Pheにおいて、TNF-α-DWのmRNAに由来するバンドが確認され、その蛍光強度はEMP1SSと同程度もしくはそれ以上であった。したがって、ターゲットタンパク質に結合するTNF-α-DWを提示したペプチド-mRNA複合体が、ターゲットに結合しないEMP1SSを提示したペプチド-mRNA複合体よりも効率よく回収されていることが確認された。このことは、ペプチド-mRNA-リンカー複合分子の提示ペプチドとmRNAとの対応付けが実験の途中で変化しないことを示している。また、この結果は翻訳反応液中にmRNAを添加してペプチド-mRNA複合体を形成させた場合、およびcDNAを添加してペプチド-mRNA複合体を形成させた場合ともに同様の結果が得えられた。
【実施例3】
【0084】
TNF-α-DWスパイクドライブラリーのセレクション
〔ランダムペプチドライブラリー〕
本手法によりランダムペプチド配列をコードするmRNAライブラリー中に含まれる低コピー数のペプチドアプタマーをコードするmRNAもしくはcDNAを、複数回のセレクションを繰り返すことにより選別することが可能かどうかを検討した。
【0085】
始めに、8-12アミノ酸のランダム配列を提示するペプチドアプタマーライブラリを調製した。このライブラリーはランダム配列をコードするmRNAとして調製したものである。このライブラリーの構造は前述のTNF-α-DWやEMP1SSの提示配列の部分がランダム化された物であり、それ以外の部分は同様の構造を有しているが、リンカーがハイブリダイズし2本鎖を形成する領域の長さは13塩基となっている。
【0086】
〔TNF-α-DWスパイクドライブラリーの調製〕
このランダムペプチドライブラリーに対して、TNF-α-DWをコードするmRNA(TNF-α-DW(an13): リンカーとの2本鎖形成部の長さが13塩基)をモル比で1,000,000 : 1となるように混合したスパイクドライブラリーを用意した。
【0087】
〔ラウンド1〕
このスパイクドライブラリーmRNAおよびan21-Pheリンカーを用いて、前述の実施例1および2に示した手順に従い翻訳とペプチド−mRNA複合体の形成を行った(ラウンド1)。
【0088】
セレクションの際には、ネガティブセレクションとしてペプチド-mRNA複合体溶液を何も結合させていないストレプトアビジン固相化磁性ビーズと4 ℃,10分間混合することにより、磁性ビーズに結合するペプチド-mRNA複合体を除いた。この溶液にTNF-αを終濃度250 nMになるように添加し、4 ℃ で1時間混和した。これにストレプトアビジン固相化磁性ビーズを加え、4 ℃, 10 分で結合させた後にTBSTで4回洗浄した。
【0089】
次に、洗浄後の磁性ビーズに逆転写反応液(5 units / μL M-MLV Reverse Transcriptase (Promega), 2 μM CGS3an21.R44, 0.5 mM dNTP, 10 mM Tris-HCl (pH 8.3), 15 mM 塩化カリウム, 0.6 mM 塩化マグネシウム, 2 mM ジチオスレイトール)10 μLに懸濁し、42 ℃で1 hr加温し逆転写反応を行った。続いて磁性ビーズ懸濁液にPCR(taq-)バッファー(プライマーとして0.25 μM T7g10m.F48および 0.25 μM CGS3an21.R44を含む)を15 μL加え、これをサーモブロック上で95 ℃で5分間加熱後に上清を回収することにより、磁性ビーズより逆転写後のDNAを回収した。
【0090】
翻訳直後のmRNAおよび回収したDNAの一部を取り、そのコピー数を実施例1と同様にリアルタイムPCRを用いて定量した。また、実施例2と同様にPCRによる増幅を行い、PCRによる増幅の際には、反応産物同士の再アニーリングを避けるため、増幅反応が飽和する前で終了した。
【0091】
〔ラウンド2、3および4〕
ラウンド2以降に於いては、前ラウンドのPCR産物から合成したmRNAを翻訳反応に用いた場合と、前ラウンドのPCR産物をcDNAとして転写・翻訳反応を同時に行った場合とを比較した。実験の手順は実施例1および2と同様に行った。その際のネガティブセレクションの操作を複数回行った。具体的にはラウンド毎に1回ずつネガティブセレクションの回数を増やしている。ラウンド2以降もPCRは増幅反応が飽和する前に増幅を終了した。
【0092】
〔選択された配列の確認〕
ラウンド2の操作を複数回繰り返した結果、ラウンド4において結合率の大幅な増大を認めた。ラウンド毎の結合率の変化を図7に示した。PCR産物をpGEM-T easy Vector System I (Promega社)を用い常法に従いクローニングし、各々5クローンをDNAシーケンシングにより解析した。その結果、得られたクローンは全てTNF-α-DW(an21)と同一の配列であった。
【0093】
本手法によってランダム配列からなるペプチドライブラリーよりペプチドアプタマーを選別しうることが示された。
【配列表フリーテキスト】
【0094】
配列番号1 合成オリゴヌクレオチド(synthetic oligonucleotide)
配列番号2 合成オリゴヌクレオチド
配列番号3 フレキシザイムFx (Flexizyme Fx)
配列番号4 フレキシザイムeFX (Flexizyme eFx)
配列番号5 フレキシザイムdFx (Flexizyme dFx)
配列番号6 合成オリゴヌクレオチド
配列番号7 合成オリゴヌクレオチド
配列番号8 合成オリゴヌクレオチド TNF-a_D-Trp.R66
配列番号9 合成オリゴヌクレオチド EMP1SS.F63
配列番号10 合成オリゴヌクレオチド EMP1SS.R63
配列番号11 合成オリゴヌクレオチド T7g10M.F48
配列番号12 合成オリゴヌクレオチド CGS3an13.R39
配列番号13 合成オリゴヌクレオチド CGS3an21.R44
配列番号14 合成オリゴヌクレオチド CGSan13.R21
配列番号15 TNF-alpha-DW(an21)
配列番号16 TNF-alpha-DW(an13)
配列番号17 EMP1SS (an21)
配列番号18 TNF-alpha
配列番号19 フレキシザイムaFx (Flexizyme aFx)
(注)配列番号15〜17のDNAでは、メチオニンのコドンが非タンパク質性アミノ酸(ClAc-D-Trp)に割り当てられている(遺伝暗号のリプログラミング)。
【技術分野】
【0001】
本発明は、遺伝子型と表現型の対応付け技術(ディスプレイ系)において、cDNAまたはmRNAとその翻訳産物であるペプチドまたはタンパク質との連結体を作製する際に用いられる新規な方法に関する。本発明者らは、この方法をRAPIDディスプレイ法と命名した。この方法は、先に報告されたフレキシザイムシステム(あるいは、RAPIDシステム:Random Peptide Integrated Discovery system)を用いて構築される特殊ペプチドライブラリーからの、薬剤候補となり得るペプチドアプタマーのスクリーニングに好適である。
【背景技術】
【0002】
進化分子工学のツールとして誕生した遺伝子型と表現型の対応付け技術は、ディスプレイ法とも称され、mRNAディスプレイ法 ("In vitro virus"、Nemoto N, et al. FEBS Lett. 414, 405-408 (1997)、国際公開WO98/16636;又は"RNA-peptide fusions"、Roberts, R. W. & Szostak, J. W. , Proc. Natl. Acad. Sci. USA., 94, 12297-12302 (1997)、国際公開WO98/31700)、STABLE法(非共有結合DNAディスプレイ)、マイクロビーズドロップレット法、共有結合DNAディスプレイ法、ファージディスプレイ法、リボソーム・ディスプレイ法などが知られている。ディスプレイ法では、ライブラリから機能のあるペプチドやタンパク質の分子を選択した際に、それに対応する遺伝子が連結しているのでその配列を容易に読み取ることができ、特定の機能を有するポリペプチドの遺伝情報を選択する際に有用である。
【0003】
mRNAディスプレイ法は無細胞翻訳系(in vitroタンパク質合成系)を用いて、遺伝子型としてのmRNAと表現型としてのペプチド分子を共有結合させることにより、遺伝子型と表現型を一体化する技術であり、現在、チロシルtRNA3’末端部分のアナログであるピューロマイシンを介して、合成されたペプチド分子とこれをコードするmRNAとを連結する方法がとられている。
【0004】
mRNAディスプレイ法においては、ピューロマイシンを適当なリンカーを介してmRNAの3’末端に結合させておき、これを無細胞翻訳系に導入してmRNAからペプチドを合成すると、ピューロマイシンがリボソームにおけるペプチド転移反応の基質として伸長中のペプチド鎖のC末端に連結し、翻訳産物であるペプチド分子がピューロマイシンを介してmRNAと連結する(図1A)。リンカーは、主として、ピューロマイシンをリボソームのAサイトに効率良く取り込ませるために、mRNAとピューロマイシンの間に挿入される。ピューロマイシンの特徴として、アミノアシルtRNAの3’末端とは異なり、アデノシン様部分とアミノ酸(チロシン)様部分がエステル結合ではなくアミド結合を形成していることが挙げられる(図1B)。そのため、リボソーム上で連結したピューロマイシンとペプチドとの結合体は、加水分解されにくく安定である。
【0005】
mRNAディスプレイ法においては、ピューロマイシンを介してmRNAと翻訳産物を連結するために、無細胞翻訳系の外で、予め、ピューロマイシンをmRNAの3’末端に連結しておくことが必要である。この連結方法として、まずピューロマイシンから5’末端側へ直鎖高分子からなるスペーサーを合成したピューロマイシン結合リンカー(Puromycin conjugated linker)を作成してから、当該リンカーとmRNAの3’末端を結合する方法と、mRNAの3’末端側にスペーサーを結合してから、当該結合体にピューロマイシンを結合する方法がある。いずれの方法でも、スペーサーである直鎖高分子の端は、典型的には、リン酸基またはヌクレオチドであり、mRNAの3’末端とリンカーの5’末端側との結合は、リン酸基を介した共有結合となっている。この共有結合は、RNAリガーゼまたはDNAリガーゼを用いた反応、または一般的な有機化学反応によって形成される。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特許第3683282号広報(国際公開WO98/16636)
【特許文献2】特許第3683902号広報
【特許文献3】特許第3692542号広報(国際公開WO98/31700)
【非特許文献】
【0007】
【非特許文献1】Nemoto N, et al. FEBS Lett. 414, 405-408 (1997)
【非特許文献2】Roberts, R. W. & Szostak, J. W. , Proc. Natl. Acad. Sci. USA., 94, 12297-12302 (1997)
【発明の概要】
【発明が解決しようとする課題】
【0008】
公知のmRNAディスプレイ法においては、3’末端にピューロマイシンを持つmRNAを鋳型として、これを小麦胚芽やウサギ網状赤血球の抽出液を使用する無細胞翻訳系に加えてペプチドへの翻訳を行う。従って、DNAからmRNAへの転写、及びmRNAとピューロマイシンとの連結反応を、予め翻訳系外で行っておく必要がある。
【0009】
近年、大腸菌のリボソームを用いる系で、翻訳に必要な因子をそれぞれ精製して混ぜ合わせた、再構成型の無細胞翻訳系が開発された(H. F. Kung, B. Redfield, B. V. Treadwell, B. Eskin, C. Spears and H. Weissbach (1977) “DNA-directed in vitrosynthesis of beta-galactosidase. Studies with purified factors” The Journal of Biological Chemistry Vol. 252, No. 19, 6889-6894 ; M. C. Gonza, C. Cunningham and R. M. Green (1985) “Isolation and point of action of a factor from Escherichia coli required to reconstruct translation” Proceeding of National Academy of Sciences of the United States of America Vol. 82, 1648-1652 ; M. Y. Pavlov and M. Ehrenberg (1996) “Rate of translation of natural mRNAs in an optimized in Vitrosystem” Archives of Biochemistry and Biophysics Vol. 328, No. 1, 9-16 ; Y. Shimizu, A. Inoue, Y. Tomari, T. Suzuki, T. Yokogawa, K. Nishikawa and T. Ueda (2001) “Cell-free translation reconstituted with purified components” Nature Biotechnology Vol. 19, No. 8, 751-755;H. Ohashi, Y. Shimizu, B. W. Ying, and T. Ueda (2007) “Efficient protein selection based on ribosome display system with purified components” Biochemical and Biophysical Research Communications Vol. 352, No. 1, 270-276)。再構成型の無細胞翻訳系とは、主に大腸菌抽出液を使用する無細胞翻訳系を細分化し、各因子を再構成することにより、翻訳と関係の無い成分が除かれたシステムであり、従来の細胞抽出液を使用する無細胞翻訳系よりもヌクレアーゼやプロテアーゼなどの阻害物質の混入を容易に防ぐことができる。転写反応に必要な因子を加えることにより、DNAからの転写を翻訳と同時に行うこともできる。
【0010】
このような転写カップル型の無細胞翻訳系を用いれば、翻訳反応と同一の系内で、鋳型DNAからの転写によるmRNAの合成を行うことができる。さらに、mRNAとリンカー分子との複合体形成もその同じ系内で行うことができれば、従来のmRNAディスプレイ法とは異なり、cDNAの転写からmRNA−ペプチド連結体の作製までをワンポット(同一反応容器内)で調製することが可能になる。そのような、反応系成分の制御が可能な再構成型の無細胞翻訳系の利点を生かしたディスプレイ法を提供することが、本発明の第一の課題である。
【0011】
さらに、このようなディスプレイ法を、先に報告された、アシル化tRNA合成触媒能を有するリボザイム(フレキシザイム)を用いた特殊ペプチド合成技術と組み合わせることで、特殊ペプチドが提示されたmRNAまたはcDNAのライブラリーの構築を可能とすることが、本発明の第二の課題である。
【課題を解決するための手段】
【0012】
本発明のRAPIDディスプレイ法では、mRNAディスプレイ法を改変して、ピューロマイシン結合リンカーの代わりに本発明者らが今回新たに開発したリンカー分子を利用し、さらに再構成型の無細胞翻訳系を最適化することにより、転写、翻訳、およびリンカーとmRNAとの複合体形成、それに続くペプチドとリンカーとの結合までを、一つの翻訳系内で完結させることが可能となった。
【0013】
従来のmRNAディスプレイ法と比較した、RAPIDディスプレイ法の主な特徴は以下の通りである。
(ア)リンカー分子の3’末端はピューロマイシンではなく、アミノ酸がエステルを介してアデノシンに結合(アミノアシル化)した構造となっている。
(イ)アミノアシル化反応は、人工RNA触媒(リボザイム)により行う。
(ウ)再構成型の無細胞翻訳系を利用する。
(エ)リンカーとmRNAとの結合は、ライゲーションではなく、翻訳系内でのハイブリダイゼーションによる複合体形成で行う。
(オ)転写、翻訳およびリンカーとの複合体形成を単一の翻訳反応容器中で行うことができる。
【0014】
さらに、同じ翻訳系において、特殊ペプチドの翻訳合成技術を適用することにより、表現型として特殊ペプチドを提示することもできる。特殊ペプチドの構成単位である、非タンパク質性のアミノ酸やヒドロキシ酸をtRNAに担持させるアシル化反応も、人工RNA触媒(リボザイム)を使用して行う。
【0015】
本発明の概略は以下の通りである。
(1)再構成型のin vitroタンパク質合成系において、mRNAと、その翻訳産物であるペプチドとが、リンカーを介して結合した連結体を作製するために使用されるリンカーであって、
リンカーの一方の端に、mRNAの3'末端側の塩基と対合する側鎖塩基を有する一本鎖構造領域を含み、
リンカーのもう一方の端に、ペプチド転移反応によって翻訳産物と結合可能な基を含むペプチドアクセプター領域を含み、
ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにアミノ酸がエステル結合した構造からなることを特徴とするリンカー。
(1−1)当該エステル結合が、人工RNA触媒を用いるアミノアシル化反応により形成されたことを特徴とする、前記リンカー。
(1−2)無細胞翻訳反応液中でリンカーとmRNAが結合可能であることを特徴とする、前記リンカー。
(1−3)ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにフェニルアラニンがエステル結合した構造からなる、前記リンカー。
(1−4)再構成型のin vitroタンパク質合成系において、DNAの転写からmRNA−ペプチド連結体の調製までを翻訳反応と同一の系内で行うために使用される、前記リンカー。
(2)一本鎖構造領域とペプチドアクセプター領域の間が、オリゴヌクレオチド、ポリアルキレン、ポリアルキレングリコール、ポリスチレン、多糖類、又はこれらの組合わせから選択される直鎖状物質で連結されている、前記リンカー。
(2−1)一本鎖構造領域とペプチドアクセプター領域の間がポリエチレングリコールで連結されている、前記リンカー。
(3)一本鎖構造領域が一本鎖DNAからなる、前記リンカー。
(4)アミノアシル化反応に使用される人工RNA触媒が、以下のいずれかのRNA配列
GGAUCGAAAGAUUUCCGCAGGCCCGAAAGGGUAUUGGCGUUAGGU (配列番号3)
GGAUCGAAAGAUUUCCGCGGCCCCGAAAGGGGAUUAGCGUUAGGU (配列番号4)
GGAUCGAAAGAUUUCCGCAUCCCCGAAAGGGUACAUGGCGUUAGGU (配列番号5)
GGAUCGAAAGAUUUCCGCACCCCCGAAAGGGGUAAGUGGCGUUAGGU (配列番号19)
からなる化学構造を有する、前記(1−1)のリンカー。
(5)mRNAと、その翻訳産物であるペプチドが、リンカーを介して結合した〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製する方法であって、
前記のいずれかのリンカーを調製する工程、
リンカーの一本鎖構造領域の塩基配列にハイブリダイズできる配列を、ペプチドをコードする配列の下流に有するmRNAを合成する工程、
再構成型のin vitroタンパク質合成反応液中で、リンカーとmRNAとを接触させるとともに、mRNAからペプチドの翻訳を行う工程
を含む、前記方法。
(6)mRNAと、その翻訳産物であるペプチドが、リンカーを介して結合した〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製する方法であって、
前記のいずれかのリンカーを調製する工程、
リンカーの一本鎖構造領域の塩基配列にハイブリダイズできる配列を、ペプチドをコードする配列の下流に有するmRNAの鋳型となるDNAを合成する工程、
再構成型のin vitroタンパク質合成反応液中にリンカーとDNAを供することにより、DNAからmRNAへの転写およびペプチドへの翻訳ならびにリンカーとmRNAとの複合体形成を行う工程
を含む、前記方法。
(7)再構成型のin vitroタンパク質合成反応液中に、非タンパク質性アミノ酸またはヒドロキシ酸でチャージされたtRNAを含むことにより、翻訳産物であるペプチドが特殊ペプチドとなる、(5)または(6)に記載の方法。
(8)前記(7)に記載の方法で作製された、〔mRNA〕−〔リンカー〕−〔特殊ペプチド〕連結体からなるライブラリー。
【発明の効果】
【0016】
再構成型の無細胞翻訳反応液中でmRNAと結合可能なリンカー分子を利用することで、単一の反応容器中にリンカーとcDNAまたはmRNAを加えて翻訳反応を行うだけで、〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製することができる。
【0017】
さらに、同じ再構成型の無細胞翻訳反応液中に、非タンパク質性のアミノ酸やヒドロキシ酸を担持させたtRNAを導入することにより、鋳型である核酸分子の配列情報から翻訳により合成された特殊ペプチドを提示した、〔mRNA〕−〔リンカー〕−〔特殊ペプチド〕連結体を得ることができる。
【0018】
このように、RAPIDディスプレイ法を、特殊ペプチドの翻訳合成技術と組み合わせることにより、生体内での安定性や標的タンパク質への結合力の向上が期待される人工的なペプチドアプタマーの遺伝子ライブラリーを簡便に構築することが可能となる。
【図面の簡単な説明】
【0019】
【図1】図1Aは、mRNAディスプレイ法において、翻訳産物であるペプチド分子がピューロマイシン結合リンカー(Puromycin conjugated linker)を介してmRNAと連結する様子を模式的に示す。図1Bは、リンカー(Puromycin conjugated linker)のペプチドアクセプター部分の構造を示す。
【図2】図2Aは、本発明のRAPIDディスプレイ法において、翻訳産物であるペプチド分子がRAPIDリンカー(L-Phe conjugated linkerが例示されている)を介してmRNAと連結する様子を模式的に示す。図2Bは、リンカー(L-Phe conjugated linker)のペプチドアクセプター部分の構造を示す。
【図3】リンカー分子(an21-ACCA)がフレキシザイムの触媒する反応によりアミノアシル化されていることを示す。(実施例1)
【図4】反応液にmRNAを添加した場合の、ペプチドアプタマー(TNF-α-DW)とmRNAの複合体の回収率を示す。(実施例1)
【図5】反応液にcDNAを添加した場合の、ペプチドアプタマー(TNF-α-DW)とmRNAの複合体の回収率を示す。(実施例1)
【図6】TNF-α-DWを提示したペプチド-mRNA複合体が、ターゲットに結合しない対照(EMP1SS)の複合体よりも効率よく回収されていることを確認した結果である。(実施例2)
【図7】TNF-α-DWスパイクドライブラリーとan21-Pheリンカーを用いて、複数回のセレクションを繰り返すことによりペプチドアプタマーを選別した。ラウンド毎の結合率の変化を示した。(実施例3)
【発明を実施するための形態】
【0020】
1.リンカー
リンカーは、本発明のRAPIDディスプレイ法においても公知のmRNAディスプレイ法と同様に、一方の端でmRNAの3’末端側と、もう一方の端でペプチドのC末端側と結合することにより、mRNAとその翻訳産物であるペプチドを連結する。
【0021】
しかしながら、本発明のRAPIDディスプレイ法では、リンカーの両端の構造が公知のmRNAディスプレイ法と相違する。本明細書では、本発明のRAPIDディスプレイ法で使用されるリンカーのことを、「RAPIDリンカー」と称することもある。
【0022】
まず、リンカーの一方の端である、ペプチドのC末端と結合する部分について説明する。本明細書においては、この部分を「ペプチドアクセプター(peptidyl acceptor)」あるいは、単に「アクセプター」とも称する。すなわち、ペプチドアクセプターとは、リボソームにおけるペプチド転移反応によって伸長中のペプチド(ペプチジルtRNA)と結合できる構造を持った分子を意味する。ペプチドアクセプターは、リンカーの末端に位置する部分のことを指す場合もあるし、リンカーを含んだ全体の構造を指す場合もある。例えば、公知のmRNAディスプレイ法におけるペプチドアクセプターは、リンカーの末端に位置するピューロマイシン、またはリンカーを含んだ全体の構造であるピューロマイシン結合リンカーである。
【0023】
本発明のRAPIDリンカーでは、ペプチドアクセプターの構造とその作製方法に特徴がある。
RAPIDディスプレイ法では、3’末端側に4残基のリボヌクレオチドからなるACCA配列を有するリンカーを合成してから、その3’末端のアデノシンに任意のアミノ酸を結合させることにより、リンカーにペプチドアクセプターとしての構造を付与する。リボソーム上のペプチド伸長反応において、リンカー末端に結合したアミノ酸が、ペプチジルtRNAのペプチドC末端を受け取ってペプチドと結合する。本明細書において、RNA配列ACCAにアミノ酸がエステル結合した構造を「ペプチドアクセプター領域」と称する。
【0024】
公知のmRNAディスプレイ法におけるペプチドアクセプターであるピューロマイシンは、アデノシン様部分のリボースとアミノ酸とがアミド結合したアミノヌクレオシド構造を有する。これに対して、本発明のRAPIDディスプレイ法では、リボースの3’-Oにアミノ酸がエステル結合した構造となっている。つまり、本発明のRAPIDディスプレイにおけるペプチドアクセプターは、天然のアミノアシルtRNAと同様のヌクレオシド構造を有する。このようなペプチドアクセプターの一例として、L−フェニルアラニンが結合したリンカーの構造について、図2B(L-Phe conjugated linker)を参照し、図1B(Puromycin conjugated linker)と比較されたい。本発明では、ペプチドアクセプターが、より天然に近い構造を用いることでピューロマイシンと同等あるいはそれ以上の取り込み効率を示す。
【0025】
ペプチドアクセプターとペプチドC末端との結合の形成はリボソーム内で起こる通常のペプチド転移反応と同様に、P部位のペプチジルtRNAに付いたペプチドのC末端のエステル結合に、A部位に取り込まれたペプチジルアクセプターのアミノ基が近接することにより起こると思われる。従って、当該ペプチド鎖C末端との間で形成される共有結合は、mRNAディスプレイ法の場合と同様に、典型的にはアミド結合である。なお、本発明のRAPIDディスプレイでは、リンカーの合成に後述する人工RNA触媒(フレキシザイム)を用いることで天然型(非タンパク質性)のアミノ酸とは異なるD-アミノ酸あるいはβ(beta)-アミノ酸を有するものを用いることも可能である。
【0026】
次に、リンカーのもう一方の端におけるmRNAとの結合について説明する。
本発明のRAPIDディスプレイ法においては、リンカーの5’末端側とmRNA分子の3’末端側とが、塩基対形成に基くハイブリダイゼーションにより複合体を形成する。従って、リンカーの5’末端側は核酸塩基を側鎖に持つ一本鎖構造を取る。RAPIDリンカーにおける、この部分を、本明細書では「一本鎖構造領域」と称する。核酸塩基を側鎖に持つ一本鎖構造の具体的な例は、一本鎖DNA、一本鎖RNA、一本鎖PNA(ペプチド核酸)などである。形成された複合体は、ペプチドのセレクションの間も安定的に保持される必要がある。リンカーの一本鎖構造領域の塩基配列と、mRNA分子の3’末端側配列との、相補性が高いほど二本鎖形成の効率が高くなり、安定性も高くなる。安定性は、GC含量、反応溶液の塩濃度、反応温度にも依存する。特に、高いGC含量を有することが望ましく、具体的には80%以上、好ましくは85%以上のGC含量を有することが望ましい。そのようなリンカーの5’末端構造の具体例を挙げて説明するならば、決して限定されるものではないが、実施例で使用された
5'-CTCCCGCCCCCCGTCC-3’(配列番号1)
5'-CCCGCCTCCCGCCCCCCGTCC-3’(配列番号2)
といった塩基配列を有する、13〜21ヌクレオチドからなる一本鎖DNAが挙げられる。
【0027】
リンカーの両端以外の部分は、公知のmRNAディスプレイ法で使用されるリンカーの構造と同様に、全体として、柔軟性があり、親水性で、側鎖の少ない単純な直鎖構造を有するように設計される。従って、例えば、一本鎖又は二本鎖DNAやRNA等のオリゴヌクレオチド、ポリエチレン等のポリアルキレン、ポリエチレングリコール等のポリアルキレングリコール、ポリスチレン、多糖類等の直鎖状物質又はこれらの組合わせを適宜選択して用いることができる。リンカーの長さは、100Å以上あることが好ましく、100〜1000Å程度であることがさらに好ましい。
【0028】
本発明で使用可能なリンカーの非限定的な具体例としては、一本鎖構造領域がGC含量の高い配列を有する一本鎖DNAであり、3’末端がACCA配列からなるRNAであり、DNAとRNAの間がポリエチレングリコール(PEGリンカー)で連結されたDNA/RNAキメラオリゴヌクレオチドが上げられる。例えば、実施例1で合成された、〔DNA〕−〔Spacer18〕n−rArCrCrA(但し、Spacer18はヘキサエチレングリコールであり、nは4〜8の整数である)が典型的な例として挙げられる。
【0029】
2.リンカーのアミノ酸修飾
上述の通り、リンカーにペプチドアクセプターとしての構造を付与するためには、リンカー分子末端のオリゴRNA(ACCA配列)部分に、アミノ酸を結合させる必要がある。このアミノ酸の結合を、人工RNA触媒を用いて行うことが、本発明の特徴である。
【0030】
固相合成法などの通常の化学合成を用いて合成したオリゴRNAの3'-O部位にエステルを介してアミノ酸を結合させることは理論上可能ではあるが、生産物は反応性が高く安定性を欠くため、これを合成反応後の後処理を行った後に無細胞翻訳系に供することは事実上不可能である。本発明においては、この問題点を、アミノアシルtRNA合成酵素として開発された人工RNA触媒である「フレキシザイム」を用いたオリゴRNAのアミノアシル化反応により解決している。この方法では、アミノアシル化反応は穏和な条件で行うことができ、容易な後処理を行うだけで翻訳系に導入して用いることができる。詳細は以下の文献を参照されたい。
H. Murakami, H. Saito, and H. Suga, (2003), Chemistry & Biology, Vol. 10, 655-662;
H. Murakami, D. Kourouklis, and H. Suga, (2003), Chemistry & Biology, Vol. 10, 1077-1084;
H. Murakami, A. Ohta, H. Ashigai, H. Suga (2006) Nature Methods 3, 357-359;
N. Niwa, Y. Yamagishi, H. Murakami, H. Suga (2009) Bioorganic & Medicinal Chemistry Letters 19, 3892-3894; および
特開2008-125396またはWO2007/066627。
【0031】
フレキシザイムは、ジニトロベンジルフレキシザイム(dFx)、エンハンスドフレキシザイム(eFx)、アミノフレキシザイム (aFx) 等の呼称でも知られる。フレキシザイムは、弱く活性化されたアミノ酸を基質として、アミノ酸の反応点であるカルボニル基、およびアミノ酸側鎖あるいは脱離基である芳香環、並びにリンカーの3’末端に存在するACC-3'配列部分を認識して、3’末端のアデノシンにアミノアシル化する触媒能を有する。RAPIDリンカーの端にACCA-3'の配列からなるオリゴRNA構造が必須なのはこのためである。フレキシザイムによるアミノアシル化反応は、アミノ酸基質と、結合相手であるオリゴRNA部分を有するリンカー分子を、フレキシザイム存在下、2時間程度氷上に置くだけで進行する。
【0032】
本発明においては、以下の式で示される配列を有するフレキシザイムが好適に使用される。
原型のフレキシザイム Fx
[GGAUCGAAAGAUUUCCGCAGGCCCGAAAGGGUAUUGGCGUUAGGU-3’, 45nt] (配列番号3)
エンハンスドフレキシザイム eFx
[5'-GGAUCGAAAGAUUUCCGCGGCCCCGAAAGGGGAUUAGCGUUAGGU-3’,45nt]) (配列番号4)
ジニトロベンジルフレキシザイム dFx
[5'-GGAUCGAAAGAUUUCCGCAUCCCCGAAAGGGUACAUGGCGUUAGGU-3’,46nt] (配列番号5)
アミノフレキシザイム aFx
[5'-GGAUCGAAAGAUUUCCGCACCCCCGAAAGGGGUAAGUGGCGUUAGGU-3’,47nt]) (配列番号19)
フレキシザイム Fxは、脱離基としてシアノメチル基を、側鎖として芳香環をもつアミノ酸(例えば、フェニルアラニン、チロシンなどのシアノメチルエステル)、フレキシザイム eFx, dFx, aFxはフレキシザイム Fxで利用可能な脱離基とアミノ酸に加え、脱離基として4−クロロベンジルチオールを、側鎖としては芳香環以外の側鎖をもつアミノ酸を基質としたアミノアシル化を触媒することができる。詳細は、前述の文献を参照されたい。したがって、このような構造を持つアミノ酸とリンカーをフレキシザイムの存在下で反応させることにより、リンカー3’末端のアデノシン内のリボース環上の3’位の水酸基に、アミノ酸がエステル結合した分子を得ることができる。アミノ酸基質としては、任意の構造を持つものを結合させることができ、天然の翻訳で使用されるアミノ酸のみならず、D-アミノ酸あるいはβ(beta)-アミノ酸などの非タンパク質性アミノ酸(すなわち、天然に存在するタンパク質中に通常見出されるL-アミノ酸以外のもの)も利用できる。さらに、アミノ酸の代わりにヒドロキシカルボン酸をリンカー3’末端に結合させることもでき、ペプチドアクセプターとしてリボソームへの取り込みも可能である。
【0033】
3.翻訳系内での反応
RAPIDディスプレイ法では、リンカーをcDNAあるいはmRNAと共に無細胞翻訳系(「in vitroタンパク質合成系」ともいう)に加えて一定時間反応させることで、mRNAからの翻訳およびリンカー分子との複合体形成、それに続くペプチドとリンカーとの連結を行うことができる。
【0034】
無細胞翻訳系としては、慣用の再構成型の無細胞翻訳系を適宜改変して利用でき、DNA依存性RNAポリメラーゼ(好ましくはT7RNAポリメラーゼ)を加えた系であればDNAからの転写も翻訳と同じ系で行うことができる。
【0035】
転写・翻訳型の合成系であれば、以下の(ア)から(エ)の反応を一つの反応容器(one-pot)内で行うことが可能である。
(ア)DNAをmRNAに転写する反応
(イ)mRNAの3’末端側が、リンカー末端の一本鎖構造領域とハイブリダイゼーションにより複合体を形成する反応
(ウ)mRNAからペプチドに翻訳する反応
(エ)翻訳が完了したペプチドのC末端がリンカーのもう一方の末端のペプチドアクセプター領域にアミド結合する反応
(オ)〔ペプチド〕−〔リンカー〕−〔mRNA〕複合体が、リボソームから解離する反応。
【0036】
あるいは、予め調製されたmRNAを、リンカーと一緒に翻訳系に加える場合は、(イ)から始まる一連の反応を再構成型の無細胞翻訳系内で行う。
再構成型の無細胞翻訳系には精製したリボソーム、翻訳開始因子、翻訳伸長因子、mRNA、アミノアシルtRNA、基質としてATPやGTP等が必要である(M. H. Schreier, B. Erni and T. Staehelin (1977) “Initiation of mammalian protein synthesis. I. Purification and characterization of seven initiation factors.” Journal of Molecular Biology, Vol. 116, No. 4, 727-53. H. Trachsel, B. Emi, M. H. Schreier and T. Staehelin (1977)“Initiation of mammalian protein synthesis. II. The assembly of the initiation complex with purified initiation factors.” Journal of Molecular Biology, Vol. 116, No. 4, 755-67.)。これらのうち、アミノアシルtRNAは同一反応液中にtRNAとアミノアシル-tRNA合成酵素およびその基質を加えることで代替が可能である。また、一般の無細胞翻訳系において行われているように、翻訳反応の効率や忠実性を上げるため、翻訳終止因子、リボソーム再生因子、クレアチンキナーゼ、ミオキナーゼ、ヌクレオチド二リン酸キナーゼ、ピロフォスファターゼなどのタンパク質や酵素およびその基質を添加することも可能である(P.C. Jelenc and C.G. Kurland (1979) “Nucleoside triphosphate regeneration decreases the frequency of translation errors” Proceedings of the Natural Academy Science of the United States of America Vol. 76, No. 7, 3174-3178)。また、翻訳と同時に転写反応を行う場合は、mRNAの代わりにcDNAおよびT7 RNAポリメラーゼとその基質を添加することができる。
【0037】
本発明においては、目的に合わせて最適化された構成因子からなる無細胞翻訳系に、必要な配列を有するDNAまたはRNAが供される。DNAまたはRNAの配列については、使用する合成系で、cDNAがmRNAに転写され、当該mRNAの翻訳が開始されるための配列が必要である。ペプチドのアミノ酸配列をコードする部分の全長が、最後まで翻訳される必要があり、さらに、翻訳されたアミノ酸のC末端側にも柔軟性を持たせるためのペプチドからなるスペーサー配列が連結し、その直後に停止コドンが来る。例えば、ペプチド配列(Cys-(Gly-Ser-)x3)と、その直後にアンバーコドン(停止コドン)がコードされる。また、mRNAの3’末端側は、リンカーの一本鎖構造領域とハイブリダイズして二本鎖を形成できるような構造を有するので、コード領域の下流(停止コドンの直後)は、一本鎖構造領域の配列に相補的な配列を有する必要がある。この部分(二本鎖形成部)の配列を、本明細書中ではリンカーハイブリダイゼーション配列と称する。
【0038】
cDNAまたはmRNAは、具体的には、以下のような配列を含むことが望ましい。
(1)cDNAにおいて、使用するRNAポリメラーゼに対応したプロモーター配列。
T7プロモーターの場合、TAATACGACTCACTATA (配列番号6)
(2)開始コドン上流の適切な配列をコードする配列。
無細胞翻訳系で大腸菌由来のリボソームを使用する場合、SD配列遺伝子を含む。これは通常のタンパク質合成と同様である。例えば、GGGTTAACTTTAA GAAGGA GATATACAT (配列番号7): T7ファージのgene 10 proteinの上流配列を改変したもの。下線部はSD配列。
【0039】
(3)開始コドン(ATG)から始まる、変異遺伝子ライブラリーのORF〔ペプチドアプタマーのC末端側にスペーサーが融合したアミノ酸配列をコードする配列〕を構成する配列。
【0040】
ここで使用される停止コドンにより、無細胞翻訳系から対応するrelease factor(解離因子)を除く必要がある。TAG(アンバーコドン)を用いた場合はRF1を、TGA(オパールコドン)を用いた場合はRF2を、TAA(オーカーコドン)を用いた場合はRF1とRF2の両方を除いて無細胞翻訳反応液を調製する。
【0041】
(4)二本鎖形成部をコードする配列(リンカーハイブリダイゼーション配列)。
本発明で使用される再構成型の無細胞翻訳反応液としては、構成因子を目的に合わせて改変して利用することが好ましい。例えば、実施例においては、アンバーコドンに対応するrelease factor、ペプチジル-tRNAを分解する酵素であるペプチジル-tRNAヒドロキシダーゼ(PTH)などを除いている。
【0042】
また、再構成無細胞翻訳系に、限られた天然アミノ酸だけを与えた系に、予め所望の非タンパク質性アミノ酸(又はヒドロキシ酸)でチャージされた(つまり、活性化されたアミノ酸が結合した)アシル化tRNAを添加することも可能である。添加しない天然アミノ酸のコドンを、非タンパク質性アミノ酸(又はヒドロキシ酸)でアシル化されたtRNAのアンチコドンに対応させることにより、非タンパク質性アミノ酸(又はヒドロキシ酸)を含むペプチドをmRNAの遺伝情報に基づき、リボソーム上で翻訳合成することが可能である。あるいは、天然アミノ酸を含まない再構成無細胞翻訳系に、非タンパク質性アミノ酸(又はヒドロキシ酸)でチャージされたアシル化tRNAを加えることにより、天然アミノ酸を全く含まない特殊ペプチドを翻訳合成することも可能である。
【0043】
非タンパク質性アミノ酸(又はヒドロキシ酸)を有するアシル化tRNAは、アミノアシルtRNA合成触媒能を有する上述の人工RNA触媒「フレキシザイム」を用いて調製することができる。上述したように、これらの人工RNA触媒は、任意の側鎖を持つアミノ酸をチャージさせることができ、また、tRNAの3'末端に存在する共通配列である5'-RCC-3'配列部分(R = A or G)のみを認識し、tRNAの3'末端にアシル化する機能を有するので、アンチコドンの異なるどのようなtRNAにも対応することが可能である。さらに、アミノ酸の認識部位にα位の置換基が含まれていないため、Lアミノ酸に限らず、ヒドロキシ酸(α位が水酸基)、N-メチルアミノ酸(α位がN-メチルアミノ酸)、N-アシルアミノ酸(α位がN-アシルアミノ基)、D-アミノ酸なども基質とすることができる。詳細については、上述のフレキシザイムに関する文献に加え、Y. Goto, H. Suga (2009) "Translation initiation with initiator tRNA charged with exotic peptides" Journal of the American Chemical Society, Vol. 131, No. 14, 5040-5041、WO2008/059823「N末端に非天然骨格をもつポリペプチドの翻訳合成とその応用」、Goto et al., ACS Chem. Biol., 2008, 3, 120-129、WO2008/117833「環状ペプチド化合物の合成方法」などに記載されている。
【0044】
本出願人らは、「フレキシザイム」を用いて非タンパク質性アミノ酸やヒドロキシ酸をアシル化したtRNAを用いた特殊ペプチドの合成系(キットまたは合成方法の両方を含む概念)をコア技術としてペプチドの翻訳合成・修飾・スクリーニングを行う総合技術をRAPIDシステム(Random Peptide Integrated Discovery system)と呼んでいる。RAPIDシステムでは、適当な配列の鋳型mRNAに基くin vitro翻訳産物として、様々な特殊ペプチドを翻訳合成することができる。これまでの説明から自明であるが、本明細書において、特殊ペプチドとは、RAPIDシステムで合成可能なあらゆる翻訳産物を含み、20種類の天然アミノ酸以外に、様々な側鎖を有するアミノ酸、β(beta)-アミノ酸、γ(gamma)-アミノ酸及びδ(delta)-アミノ酸、D-アミノ酸、アミノ酸骨格上のアミノ基やカルボキシル基が置換された構造を有する誘導体を含む、上述した様々な基質を構成要素とする重合体である。さらに、特殊ペプチドは、主鎖骨格として通常のアミド結合とは異なる構造も有し得るものである。例えば、アミノ酸とヒドロキシ酸から構成されるデプシペプチド、ヒドロキシ酸が連続して縮合したポリエステル、N-メチルアミノ酸の導入によりアミド結合の窒素がメチル化されたペプチド、N-末端に様々なアシル基(アセチル基、ピログルタミン酸、脂肪酸など)を有するペプチドも、特殊ペプチドに含まれる。また、両端に互いの間で結合形成反応が可能な一組の官能基を配置したアミノ酸配列からなる非環状ペプチドを適切な反応条件下で環化させて得られる環状ペプチド(N-メチルペプチドが導入された場合は、環状N-メチルペプチド)もRAPIDシステムで合成可能である。一組の官能基の種類によっては無細胞翻訳系の条件下において環化することがあり、後述の実施例のように、両端にそれぞれクロロアセチル基とシステインを配置したペプチド配列を翻訳合成して得られるチオエーテル結合で環化した環状ペプチドはその一例である。
【0045】
RAPIDシステムでは、リボソームによる翻訳で特殊ペプチドを合成するため、鋳型であるmRNAを代えるだけで様々な構造を有するペプチドを合成できる。ランダム配列を含むmRNA(または対応するDNA)を用いて翻訳合成を行えば、ランダムペプチドライブラリーを容易に構築することができる。本発明のRAPIDディスプレイ法は、RAPIDシステムにより合成される特殊ペプチドを、その遺伝子型であるmRNAと対応付けるために使用するのに、好適である。特殊ペプチドの合成用に最適化された無細胞翻訳系に、鋳型cDNAあるいはmRNAと共に、本発明のRAPIDリンカーを加えて、一定時間反応させることで、翻訳産物である特殊ペプチドがリンカーを介してmRNAと連結して提示される。
【0046】
次に、無細胞翻訳系内で起こる、リンカーとmRNAの結合について説明する。従来技術の項で述べたように、公知のmRNAディスプレイ法では、翻訳反応の前段階に、リンカーとmRNAの連結(ライゲーション)を翻訳系外で行っておくことが必要であった。これに対し、本発明のRAPIDディスプレイ法ではリンカーとmRNAのハイブリダイゼーションによる複合体形成を翻訳系内で行うことができることが特徴である。
【0047】
図2を参照しながら、mRNAをRAPIDリンカーと共に無細胞翻訳系に供することにより、mRNAとリンカーとのハイブリダイゼーションおよびペプチド分子の合成、そしてペプチドC末端とリンカーとの連結が起こる過程について説明する。
【0048】
上述したように、mRNAとリンカーとの結合は、リンカーの一本鎖構造領域の塩基配列とmRNAの3’末端の相補的な塩基配列が水素結合で二本鎖を形成することによる。この結合は、mRNAと翻訳されたペプチド分子とを対応付けるためのものであるから、リボソーム上で翻訳が行われている間に形成された〔mRNA〕−〔リンカー〕−〔ペプチド〕複合体が、翻訳産物であるペプチドのセレクションの間も安定に維持される必要がある。
【0049】
mRNAをRAPIDリンカーと共に無細胞翻訳系に供することにより、リボソームがmRNA上に配置されて翻訳反応が開始し、ペプチド鎖が伸長し、そして、終結したペプチド鎖のC末端が、リンカーの〔rACCA-アミノ酸〕からなるペプチドアクセプター領域のアミノ酸と結合してtRNAから切り離される。本発明の概念を限定する意図ではなく、あくまでも説明のためであるが、mRNAをRAPIDリンカーと同時に無細胞翻訳系に供する場合、mRNAとRAPIDリンカーとのハイブリダイゼーションは、ペプチド鎖の伸長反応開始よりも前の段階で起こり得ると本発明者らは考えている。あるいは、cDNAからの転写および翻訳反応を先に行ってから、終結因子およびPTHを除いた翻訳反応液中にリンカーを後から添加することによっても、リボソーム上のmRNAとRAPIDリンカーとのハイブリダイゼーションによる複合体形成は正しく起こる。そして、リンカー末端の〔rACCA-アミノ酸〕とペプチドC末端との共有結合は、翻訳終了時に偶然にこの部分がリボソームAサイトに入った場合に起こると考えられる。翻訳終了時には既にmRNAとリンカーとのハイブリダイゼーションが起こっているので、この〔rACCA-アミノ酸〕基質はリンカーでリボソーム上のmRNAとつながっており局所的な濃度が非常に高くなっているため、反応は高い効率で起こる。
【0050】
このようにしてリボソーム上で形成された〔mRNA〕−〔リンカー〕−〔ペプチド〕複合体が、ペプチド鎖がリボソームから解離した後も安定に維持されていることは、後述の実施例においても裏付けられている。
【0051】
4.ペプチドアプタマーの選択
進化分子工学では、所望の機能や性質を持つタンパク質やペプチドを創製することを目的として、可能性のある遺伝子を大規模に準備し、その中から狙った表現型を有するクローンを選択する。
【0052】
基本的には、最初にDNA集団を調製し、in vitro転写産物としてRNA集団を得て、in vitro翻訳産物としてペプチド集団を得る。このペプチド集団から、所望の機能や性質を持つものを何らかのスクリーニング系で選択することになる。例えば、特定のタンパク質に結合するペプチド分子を得たい場合は、標的タンパク質を固相化したカラムにペプチド集団を流し込み、カラムに結合したペプチド分子の混合物を回収することができる。このとき、各ペプチド分子には、その鋳型であるmRNAがタグのように付加されているので、回収したペプチド−mRNA複合体の集団から逆転写酵素でDNAに戻し、PCRで増幅して狙った表現型を有するクローンが多く含まれるバイアスのかかったライブラリーを得た後に、再度同じような選択実験を行う。あるいは、RNAアプタマーを回収してしまう可能性を回避するため、核酸部分を2本鎖にする目的で、選択前に逆転写反応を行うことも可能である。この操作を繰返すことで、世代の経過とともに所望の表現型を有するクローンが集団中で濃縮されていく。
【0053】
ペプチドアプタマーを同定する場合、対応付け分子のライブラリーと標的物質とを混合し、標的物質に結合したペプチドを提示する対応付け分子(活性種)を選択し、選択された対応付け分子の核酸部分からPCRにより核酸ライブラリーを調製する工程を繰り返すことで、標的物質に結合するペプチドアプタマーの遺伝子をクローニングできる。標的物質に結合した対応付け分子を選択する工程は、〔RNA(あるいはDNA/RNAハイブリッド)〕−〔リンカー〕−〔ペプチド〕からなる複合体を標的物質と結合させ、適当な方法で他の複合体から分離して、目的の結合特性を有するペプチドを同定することにより行うことができる。
【0054】
標的物質としては、タンパク質、核酸、糖質、脂質、その他どのような化合物でもよい。標的物質と結合する活性種である複合体を、他の複合体から分離するために、標的物質に、固相への結合により回収可能な修飾を施しておくと便利である。例えば、後述の実施例では、標的物質をビオチン修飾しておき、固相化されたビオチン結合タンパク質への特異的な結合を利用して回収している。このような特異的な結合としては、ビオチン結合タンパク質(アビジン、ストレプトアビジンなど)/ビオチンの組み合わせの他にも、マルトース結合タンパク質/マルトース、ポリヒスチジンペプチド/金属イオン(ニッケル、コバルトなど)、グルタチオン-S-トランスフェラーゼ/グルタチオン、抗体/抗原(エピトープ)などが、利用可能であるが、これらに限定されるものでは勿論ない。
【0055】
進化分子工学を用いることで、遺伝子ライブラリーとして、A、T、G、Cの4つの塩基がランダムに結合したDNA配列から、天然には全く存在しないアミノ酸配列を有するペプチドを得ることが原理的に可能である。また、翻訳系に非タンパク質性アミノ酸(またはヒドロキシ酸)でアシル化されたtRNAを導入することにより、in vitro翻訳産物として非タンパク質性アミノ酸(またはヒドロキシ酸)が導入された特殊ペプチドの翻訳による合成も可能である。特殊ペプチドとmRNA(あるいはcDNA)の複合体の集団から、所望の結合特性を有するペプチドが提示された活性種を選択し、対応付けされた遺伝子部分を増幅、再び翻訳するという作業を繰返すことで、特殊ペプチドのライブラリーを効率的に得ることができる。
【0056】
これまでの説明および後述の実施例の内容に関し、分子生物学的手法の詳細は、例えば、Sambrook, Molecular Cloning: A Laboratory Manual, 3rd edition, Cold Spring Harbor Laboratory Press, 2001; Golemis, Protein-Protein Interactions: A Molecular Cloning Manual, 2nd edition, Cold Spring Laboratory Press, 2005などを参照されたい。
【0057】
以下、本発明を実施例によって具体的に説明する。なお、これらの実施例は、本発明を説明するためのものであって、本発明の範囲を限定するものではない。
【実施例1】
【0058】
〔リンカーの合成〕
リンカーとしてDNAとRNA間をポリエチレングリコール(5ユニットのSpacer18)で連結したDNA/RNAキメラオリゴヌクレオチドを使用した。種々リンカーはBEX社(東京)より購入した。以下に示す配列の*Aおよび*CはRNA、SPC18はSpacer18(ヘキサエチレングリコール)に相当する。
【0059】
an21-ACCA: 5'-CCCGCCTCCCGCCCCCCGTCC-[SPC18]5-A*-C*-C*-A*-3'
〔リンカーのアミノアシル化〕
フレキシザイムが触媒する反応により、an21-ACCAリンカー分子の3’末端側にエステル結合を介してL-フェニルアラニンもしくはβ-L-アラニンを結合させた。反応産物は精製した反応産物溶液を酸性条件下でのアクリルアミド電気泳動分析により確認した。リンカー分子に由来するバンドがアミノアシル化されると移動度が少なくなる。そのため、未反応物由来のバンドと反応産物由来のバンドの強度を比較することでアミノアシル化効率を決定することができる。
【0060】
アシル化反応は、L-フェニルアラニンを結合させる場合は、0.1 M HEPES-カリウムバッファー(pH 7.5)、600 mM 塩化マグネシウム中で、20 μMのフレキシザイムeFx 5 μL、20 μMのan21-ACCAリンカー、および基質(L- フェニルアラニンシアノメチルエステル)を20 %ジメチルスルホキシドに加えて、氷上で2時間反応させた。詳細な手順としては、まず、純水に溶解した40 μMのリンカー分子を0.2 M HEPES-カリウムバッファー(pH 7.5)およびフレキシザイムeFx(200 μM, 0.5 μL)を加え、サーモブロック(ND-MD1, 日伸理化)上で95 ℃で2分間加熱し、室温で5分間静置した。その後、氷上で塩化マグネシウム(3 M, 1 μL) および基質(ジメチルスルホキシド中25 mM, 1 μL)を加えることによりリンカー分子のアシル化反応を開始し、氷上で2時間静置した。β-L-アラニンを結合させる場合は、同様の条件にて、pHを8.0とし、基質としてβ-L-アラニンp-クロロベンジルチオエーテルを用いて行った。反応は0.3 M 酢酸ナトリウム(pH 5.0)を40 μL加えることにより停止した。反応産物をエタノール沈殿し、70% エタノールで洗浄したペレットを10 μLの1 mM酢酸ナトリウムに溶解した。反応後の溶液を酸性条件下、20 %の変性ポリアクリルアミド電気泳動(50 mM 酢酸ナトリウム(pH 5.0)、6 M 尿素)で分離し、泳動後のゲルをSYBR Green II (Invitrogen、SYBRはMolecular Probes Inc.の登録商標)を用いた蛍光染色により解析した。
【0061】
結果は図3で示したようになり、対照となる未反応のリンカー分子よりも、反応産物のリンカー分子の移動度が小さいことが確認され、リンカー分子がフレキシザイムの触媒する反応によりアミノアシル化されていることが確認された。
【0062】
〔cDNAの合成〕
ペプチド-mRNA複合体を形成するために用いるcDNAは、合成オリゴヌクレオチドをPCRにより連結することにより調製した。以下に配列を示す合成DNAはオペロン社(東京)より購入した。これらのオリゴDNAを連結することにより得られる各々のcDNAは、T7プロモーター配列、リボソーム結合配列、開始コドン、ペプチドアプタマー配列、スペーサーペプチド配列(CGSGSGS)、アンバーコドンおよびリンカーハイブリダイゼーション配列より構成される。
【0063】
TNF-a_D-Trp.R66:
GCCGCTGCCGCTGCCGCAATGCTTCAGATACAGACAATGCAGACGTTGCATATGTATATCTCCTTC
EMP1SS.F63: GAAGGAGATATACATATGGCAGCAGGTGGTACCTATTCTTCTCATTTTGGTCCGCTGACCTGG
EMP1SS.R63: GCCGCTGCCGCTGCCGCATGCTGCACCACCTTGCGGCTTAGAAACCCAGGTCAGCGGACCAAA
T7g10M.F48: TAATACGACTCACTATAGGGTTAACTTTAAGAAGGAGATATACATATG
CGS3an13.R39: TTTCCGCCCCCCGTCCTAGCTGCCGCTGCCGCTGCCGCA
CGS3an21.R44: CCCGCCTCCCGCCCCCCGTCCTAGCTGCCGCTGCCGCTGCCGCA
PCRによる連結は以下の手順にて行った。
【0064】
T7g10M.F48およびTNF-a_D-Trp.R66(もしくはEMP1SS.F63とEMP1SS.R63の組み合わせ)を250 nM含むPCR反応液(10 mM Tris-HCl (pH 9.0), 50 mM 塩化カリウム, 2.5 mM 塩化マグネシウム, 250 μM dNTPs, 0.2 % Triton X-100(トリトンX-100、ナカライテスク)およびTaq ポリメラーゼ) 20 uLを調製し、サーマルサイクラー(TC-3000, Techne社)を用いて94 ℃ 1 min, {50 ℃ 30 sec, 72 ℃ 30 sec}5サイクルによる反応を行った。次に各々500 nM のプライマー、T7g10M.F48およびGCSan21.R44(もしくはGCSan13.R39)を含むPCR反応液100 μLを調製し、これに先のPCR反応後液1 μLを加え、温度サイクル{94 ℃ 40 sec, 50 ℃ 40 sec, 72 ℃ 40 sec}10サイクルで反応を行った。反応産物は常法に従いフェノール・クロロホルム抽出、クロロホルム抽出、さらにエタノール沈殿により精製を行った。
【0065】
〔mRNAの調製〕
mRNAは前述の方法で作成したcDNA(TNF-α-DW(an21))を元にT7 RNA ポリメラーゼによる転写反応によりRNAを合成し、得られた反応産物は常法に従いフェノール・クロロホルム抽出、クロロホルム抽出、さらに2-プロパノール沈殿により精製を行った。精製後のmRNAの濃度を260 nmにおけるUVの吸収から求め、10 μMになるように希釈した。
【0066】
〔転写・翻訳、リンカーのハイブリダイゼーション、ペプチドアクセプターとの連結〕
mRNAからの翻訳およびリンカー分子との複合体形成もしくはcDNAからの転写、翻訳およびリンカー分子との複合体形成は再構成型の無細胞翻訳系を用いて行った。
【0067】
本実施例で使用した再構成型の無細胞翻訳系には以下の生体高分子が含まれる:
70Sリボソーム(1.2uM)、翻訳開始因子(IF1(0.7uM), IF2(0.4uM), IF3(1.5uM))、
伸長因子(EF-G(0.26uM), EF-Tu/EF-Ts複合体(10uM))、翻訳終止因子(RF2(0.25uM), RF3(0.17uM))メチオニルtRNAトランスフォルミラーゼ(MTF(0.6uM))、リボソーム再生因子(RRF(0.5uM))、アミノアシルtRNA合成酵素(AlaRS(0.73uM), ArgRS(0.03uM), AsnRS(0.38uM), CysRS(0.02uM), GlnRS(0.06uM), GluRS(0.23uM), GlyRS(0.09uM), HisRS(0.02uM), IleRS(0.4uM), LeuRS(0.04uM), MetRS(0.03uM), PheRS(0.68uM), ProRS(0.16uM), SerRS(0.04uM), ThrRS(0.09uM), TrpRS(0.03uM), ValRS(0.02uM), AspRS(0.13uM), LysRS(0.11uM), TyrRS(0.02uM))、クレアチンキナーゼ(ロシュ社製CK(4ug/mL)、ミオキナーゼ(ロシュ社製、MK(3ug/mL))、ピロフォスファターゼ(PPa(0.1uM))、
ヌクレオチド二リン酸キナーゼ(NDK(0.1uM))、T7 RNAポリメラーゼ(T7ファージ遺伝子由来の組み換え体、0.1uM)、大腸菌tRNA(ロシュ社製、1.5mg/mL)。リボソームは対数増殖期の大腸菌から精製したもの、リボソーム以外の各種タンパク質は、特に指示がある場合を除き、大腸菌の遺伝子をクローニングし、それを元に発現、精製した組み換えタンパク質である。
【0068】
また、生体高分子以外の成分として、以下の物が含まれる:
50mM HEPES-KOH (pH7.6), 2mM NTPs, 20mM リン酸クレアチン, 100mM 酢酸カリウム, 2mM スペルミジン, 1mM ジチオスレイトール, 6mM 酢酸マグネシウム, 0.1mM 10-ホルミル-5, 6, 7, 8-テトラヒドロ葉酸。
【0069】
これらに加えて、mRNA 1.5 μMもしくはcDNA 0.15 μM、アシル基としてN-クロロアセチル-D-トリプトファン(ClAc-D-Trp)を担持したアミノアシルイニシエーションtRNA(特開2008-125396で開示の方法により調製)、メチオニンを除く天然タンパク質を構成する19種類のアミノ酸(各々5 mM)を含む転写・翻訳反応溶液2.5 μLを調製した。
【0070】
あらかじめ転写反応を行いmRNAを調製し、これを元に翻訳およびペプチドとリンカーとの結合を行う場合は、以下の手順によりペプチド-mRNA複合体を調製した。まず、4 mM酢酸ナトリウム pH 5.0とmRNAを1:3で混合し、これをサーモブロック上で95 ℃で1分間加熱したのちに室温下で5分静置した。1 mM酢酸ナトリウムに溶解した25 μM のリンカー溶液を混合し10分間静置することでmRNA-リンカーの複合体を形成させた。これに翻訳系の他のコンポーネントを加え、恒温水槽(NT-202D, 日伸理化)中で37 ℃、30分間、続いて室温で12分インキュベートし、pH 7.5に調製した0.1 M EDTA(エチレンジアミン4酢酸、分子生物学実験用、ナカライテスク)を終濃度が20 mMになるように添加し、さらに恒温水槽中で37 ℃で30分間インキュベートした。
【0071】
反応溶液中にcDNAを加え、この反応溶液中で転写、翻訳およびリンカーとの結合を行う場合は、以下の手順によりペプチド-mRNA複合体を調製した。まず始めにリンカー以外の物を含む溶液を調製し、これを恒温水槽中で37 ℃で30分間反応させることにより、転写および翻訳反応を行った。これに25 μMのリンカー溶液0.25 μLを加え、さらに37 ℃で30分、続いて室温で12分インキュベートした。その後、pH 7.5に調製した0.1 M EDTA(エチレンジアミン4酢酸、分子生物学実験用、ナカライテスク)を終濃度が20 mMになるように添加し、さらに恒温水槽中で37 ℃ で30分間インキュベートした。
【0072】
〔逆転写〕
ペプチド−mRNA複合体のmRNA部分の安定性を向上させるため、逆転写反応を行うことにより、RNA-DNAハイブリッド鎖を形成させた。具体的には以下の操作を行った。前述の翻訳反応産物に、12 mM Tris-HCl (pH 8.3), 5 μM 逆転写プライマー(CGS3an13.R21), 0.5 mM dNTPs, 18 mM Mg(OAc)2, 10 mM KOH(各々の濃度は終濃度)を添加した。これにM-MLV Reverse Transcriptase (RNaseH Minus, Point Mutant, Promega) 1 unitを添加し恒温水槽中で42 ℃で10分間反応させた。その後、終濃度が10 mMおよび18 mMになるようにEDTAと塩酸を添加した。
【0073】
〔セレクション〕
このようにして作成したペプチド−mRNA複合体分子が、提示ペプチドとターゲットとなるタンパク質との相互作用により回収されるかどうかを以下に示す方法により確認した。
【0074】
ターゲットとなるタンパク質として、ヒトの腫瘍壊死因子アルファ(以下、TNF-α)を選択した。
ペプチド-mRNA複合体分子を含む無細胞翻訳反応産物液6 μLに対し、ビオチン修飾されたTNF-αタンパク質を終濃度250 nMになるように添加し、この溶液を低吸着処理を施した0.6 mLエッペンドルフチューブ(プラチナ(超低吸着)チューブ、ビーエム機器)に移し、ローテーター(RT-30 mini、タイテック)の上で4 ℃、1 hr 穏やかに攪拌し結合させた。この溶液に対して、ストレプトアビジンを固相化した磁性ビーズ(Dynabeads 登録商標Streptavidin M-280, Invitrogen社)の懸濁液3 μLを加え、さらに10分攪拌した。その後、遠心分離およびマグネットホルダーを使用して磁性ビーズを分離し上清を除き、再度0.05 % のTween20(ポリオキシエチレンソルビタンモノラウレート、ナカライテスク)を含むTBS (Tris Buffered Saline, 50 mM Tris-HCl(トリスヒドロキシメチルアミノメタン、ナカライテスク) pH 7.5, 150 mM 塩化ナトリウム) 50 μL に懸濁した。この洗浄操作を4回繰り返した。
【0075】
続いて磁性ビーズにPCR(taq-)バッファー(10 mM Tris-HCl (pH 9.0), 50 mM 塩化カリウム, 2.5 mM 塩化マグネシウム, 250 μM dNTPs, 0.25 μM T7g10m.F48, 0.25 μM CGS3an13.R21, 0.2 % Triton X-100)を25 μL加え、サーモブロック上で95 ℃で5分間加熱後に上清を回収することにより、磁性ビーズより逆転写後のDNAを回収した。
【0076】
〔リアルタイムPCRによる定量〕
ターゲットタンパク質添加前、および磁性ビーズより回収後のDNAの量をリアルタイムPCR法により定量した。リアルタイムPCR測定装置はLightCycler(登録商標) 1.5 (Roche Applied Science社)を使用し、前述のPCR(taq-)バッファーにTaqポリメラーゼ、SYBR(登録商標) Green I (100,000希釈、Invitrogen社)、および測定サンプル溶液を添加した反応液を測定に供した。
【0077】
TNF-αを標的としたモデル実験
リンカーの機能を確認するために用いたターゲットとなるタンパク質およびペプチドアプタマーとして、TNF-αおよびTNF-αに結合するペプチドアプタマーであるTNF-α-DWを選択した。
【0078】
本実施例で用いたTNF-αタンパク質として大腸菌により発現した組換え体である可溶型TNF-αを使用した。組換えTNF-αは野生型TNF-αの77番目から233番目のアミノ酸配列の部分、そのN末端側にAviTag配列(GLNDIFEAQKIEWHE)およびHis×6 tag配列を融合したものである。このタンパク質とAviTagを基質とするビオチンリガーゼ(BirA)を大腸菌内で共発現させることによりAviTagのリジン残基の側鎖がビオチン化修飾されるため、本実施例で用いているTNF-αはストレプトアビジン固相化ビーズにより容易に分離することができる。
【0079】
ペプチドアプタマーとして用いたTNF-α-DWは、非タンパク質性アミノ酸であるN-クロロアセチル-D-トリプトファン(ClAc-D-Trp)のクロロアセチル基とペプチド配列中のシステインの側鎖のチオール基が反応し、チオエーテルを形成することにより環化したペプチドXQRLHCLYLKH(X: Ac-D-Trp)である。このペプチドのC末端側にスペーサーアミノ酸配列CGSGSGSを融合した配列をコードするcDNAを前述の方法にて作成し、無細胞転写・翻訳系にてペプチドとmRNA複合体分子を作成した。
【0080】
このペプチドアプタマーと標的タンパク質の組み合わせを用いた場合のペプチド-mRNA複合体の回収率は図4および図5に示すとおりであった。L-フェニルアラニンを3’末端にエステルを介して結合させたan21- Phe、an21-β-Alaの回収率は翻訳反応液にmRNAを添加した場合(図4)は各々1.30 %, 0.25 %、cDNAを添加した場合(図5)は0.65 %, 0.38 %であった。一方、3’末端が修飾されていないan21-ACCAリンカーでの回収率はそれらのものと比較して数十倍程度低く(mRNAの場合0.015%, cDNAの場合0.007 %)、L-フェニルアラニン、β-L-アラニンが翻訳合成されたペプチドのC末端側に共有結合を介して連結することにより、ペプチド-mRNA複合体分子として効率よく回収されることが示された。
【実施例2】
【0081】
提示ペプチドとmRNAとの対応付けの確認
前述までの実験に於いては、提示しているペプチドと、その配列をコードしているmRNAは単一の種類であるため、mRNAから翻訳合成されたペプチドに対して、別のリンカー分子がアクセプターとして結合している可能性や、2本鎖を形成しているリンカー分子とmRNA分子が操作の途中で別のリンカー分子やmRNAと交換している可能性を排除できない。そのため、回収されるペプチド-mRNA-リンカー複合分子の提示ペプチドとmRNAとの対応付けが正確に行われていることを確認するために以下に示す実験を行った。
【0082】
まず、TNF-α-DWの対照としてターゲットとするタンパク質に結合しないペプチド配列(EMP1SS:XAAGGTYSSHFGPLTWVSKPQGGAA、XはTNF-α-DWと同様にAc-D-Trpである)をコードするmRNAもしくはcDNAを実施例1に示す方法で調製し、これとTNF-α-DWをコードするmRNAもしくはcDNAをモル比100:1で混合したものを用意した。これを元に前述と同様の実験を行い、回収したDNAをPCRにて増幅した。その際の条件として、PCR(taq-)バッファーにTaqポリメラーゼおよび測定サンプル溶液を添加した反応液20 μLを用意し、サーマルサイクラーを用いて、温度サイクル{94 ℃ 40 sec, 61 ℃ 40 sec, 72 ℃ 40 sec}(61 ℃から72 ℃への昇温は0.5 ℃/sec)を30サイクル行い反応させた。反応産物液2.5 μLをTAE(40 mM Tris酢酸、1 mM EDTA)および3 % アガロース(低電気浸透度、ナカライテスク)からなるゲルを用いた電気泳動により分離し、エチジウムブロミドによる染色およびトランスイルミネーターを用いて泳動像を得た。その結果を図6に示す。
【0083】
TNF-α-DWをコードするmRNAに由来するバンドは約90 bpの位置に確認され、対照とするEMP1SSをコードするmRNAに由来するバンドは約150 bpの位置に観察される。ペプチドアクセプターを含まないan21-ACCAリンカーを用いた場合は、はじめに2種類のcDNAを混合した際のモル比を反映しEMP1SSのmRNAに由来するバンドのみが確認された。それに対して、an21-Pheにおいて、TNF-α-DWのmRNAに由来するバンドが確認され、その蛍光強度はEMP1SSと同程度もしくはそれ以上であった。したがって、ターゲットタンパク質に結合するTNF-α-DWを提示したペプチド-mRNA複合体が、ターゲットに結合しないEMP1SSを提示したペプチド-mRNA複合体よりも効率よく回収されていることが確認された。このことは、ペプチド-mRNA-リンカー複合分子の提示ペプチドとmRNAとの対応付けが実験の途中で変化しないことを示している。また、この結果は翻訳反応液中にmRNAを添加してペプチド-mRNA複合体を形成させた場合、およびcDNAを添加してペプチド-mRNA複合体を形成させた場合ともに同様の結果が得えられた。
【実施例3】
【0084】
TNF-α-DWスパイクドライブラリーのセレクション
〔ランダムペプチドライブラリー〕
本手法によりランダムペプチド配列をコードするmRNAライブラリー中に含まれる低コピー数のペプチドアプタマーをコードするmRNAもしくはcDNAを、複数回のセレクションを繰り返すことにより選別することが可能かどうかを検討した。
【0085】
始めに、8-12アミノ酸のランダム配列を提示するペプチドアプタマーライブラリを調製した。このライブラリーはランダム配列をコードするmRNAとして調製したものである。このライブラリーの構造は前述のTNF-α-DWやEMP1SSの提示配列の部分がランダム化された物であり、それ以外の部分は同様の構造を有しているが、リンカーがハイブリダイズし2本鎖を形成する領域の長さは13塩基となっている。
【0086】
〔TNF-α-DWスパイクドライブラリーの調製〕
このランダムペプチドライブラリーに対して、TNF-α-DWをコードするmRNA(TNF-α-DW(an13): リンカーとの2本鎖形成部の長さが13塩基)をモル比で1,000,000 : 1となるように混合したスパイクドライブラリーを用意した。
【0087】
〔ラウンド1〕
このスパイクドライブラリーmRNAおよびan21-Pheリンカーを用いて、前述の実施例1および2に示した手順に従い翻訳とペプチド−mRNA複合体の形成を行った(ラウンド1)。
【0088】
セレクションの際には、ネガティブセレクションとしてペプチド-mRNA複合体溶液を何も結合させていないストレプトアビジン固相化磁性ビーズと4 ℃,10分間混合することにより、磁性ビーズに結合するペプチド-mRNA複合体を除いた。この溶液にTNF-αを終濃度250 nMになるように添加し、4 ℃ で1時間混和した。これにストレプトアビジン固相化磁性ビーズを加え、4 ℃, 10 分で結合させた後にTBSTで4回洗浄した。
【0089】
次に、洗浄後の磁性ビーズに逆転写反応液(5 units / μL M-MLV Reverse Transcriptase (Promega), 2 μM CGS3an21.R44, 0.5 mM dNTP, 10 mM Tris-HCl (pH 8.3), 15 mM 塩化カリウム, 0.6 mM 塩化マグネシウム, 2 mM ジチオスレイトール)10 μLに懸濁し、42 ℃で1 hr加温し逆転写反応を行った。続いて磁性ビーズ懸濁液にPCR(taq-)バッファー(プライマーとして0.25 μM T7g10m.F48および 0.25 μM CGS3an21.R44を含む)を15 μL加え、これをサーモブロック上で95 ℃で5分間加熱後に上清を回収することにより、磁性ビーズより逆転写後のDNAを回収した。
【0090】
翻訳直後のmRNAおよび回収したDNAの一部を取り、そのコピー数を実施例1と同様にリアルタイムPCRを用いて定量した。また、実施例2と同様にPCRによる増幅を行い、PCRによる増幅の際には、反応産物同士の再アニーリングを避けるため、増幅反応が飽和する前で終了した。
【0091】
〔ラウンド2、3および4〕
ラウンド2以降に於いては、前ラウンドのPCR産物から合成したmRNAを翻訳反応に用いた場合と、前ラウンドのPCR産物をcDNAとして転写・翻訳反応を同時に行った場合とを比較した。実験の手順は実施例1および2と同様に行った。その際のネガティブセレクションの操作を複数回行った。具体的にはラウンド毎に1回ずつネガティブセレクションの回数を増やしている。ラウンド2以降もPCRは増幅反応が飽和する前に増幅を終了した。
【0092】
〔選択された配列の確認〕
ラウンド2の操作を複数回繰り返した結果、ラウンド4において結合率の大幅な増大を認めた。ラウンド毎の結合率の変化を図7に示した。PCR産物をpGEM-T easy Vector System I (Promega社)を用い常法に従いクローニングし、各々5クローンをDNAシーケンシングにより解析した。その結果、得られたクローンは全てTNF-α-DW(an21)と同一の配列であった。
【0093】
本手法によってランダム配列からなるペプチドライブラリーよりペプチドアプタマーを選別しうることが示された。
【配列表フリーテキスト】
【0094】
配列番号1 合成オリゴヌクレオチド(synthetic oligonucleotide)
配列番号2 合成オリゴヌクレオチド
配列番号3 フレキシザイムFx (Flexizyme Fx)
配列番号4 フレキシザイムeFX (Flexizyme eFx)
配列番号5 フレキシザイムdFx (Flexizyme dFx)
配列番号6 合成オリゴヌクレオチド
配列番号7 合成オリゴヌクレオチド
配列番号8 合成オリゴヌクレオチド TNF-a_D-Trp.R66
配列番号9 合成オリゴヌクレオチド EMP1SS.F63
配列番号10 合成オリゴヌクレオチド EMP1SS.R63
配列番号11 合成オリゴヌクレオチド T7g10M.F48
配列番号12 合成オリゴヌクレオチド CGS3an13.R39
配列番号13 合成オリゴヌクレオチド CGS3an21.R44
配列番号14 合成オリゴヌクレオチド CGSan13.R21
配列番号15 TNF-alpha-DW(an21)
配列番号16 TNF-alpha-DW(an13)
配列番号17 EMP1SS (an21)
配列番号18 TNF-alpha
配列番号19 フレキシザイムaFx (Flexizyme aFx)
(注)配列番号15〜17のDNAでは、メチオニンのコドンが非タンパク質性アミノ酸(ClAc-D-Trp)に割り当てられている(遺伝暗号のリプログラミング)。
【特許請求の範囲】
【請求項1】
mRNAと、その翻訳産物であるペプチドとが、リンカーを介して結合した連結体を作製するために使用されるリンカーであって、
リンカーの一方の端に、mRNAの3'末端側の塩基と対合する側鎖塩基を有する一本鎖構造領域を含み、
リンカーのもう一方の端に、ペプチド転移反応によって翻訳産物と結合可能な基を含むペプチドアクセプター領域を含み、
ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにアミノ酸がエステル結合した構造からなり、
無細胞翻訳反応液中でリンカーとmRNAが結合可能である
ことを特徴とするリンカー。
【請求項2】
一本鎖構造領域とペプチドアクセプター領域の間が、オリゴヌクレオチド、ポリアルキレン、ポリアルキレングリコール、ポリスチレン、多糖類、又はこれらの組合わせから選択される直鎖状物質で連結されている、請求項1に記載のリンカー。
【請求項3】
一本鎖構造領域が一本鎖DNAからなる、請求項1または2に記載のリンカー。
【請求項4】
ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにフェニルアラニンがエステル結合した構造からなる、請求項1に記載のリンカー。
【請求項5】
再構成型のin vitroタンパク質合成系において、DNAの転写からmRNA−ペプチド連結体の調製までを翻訳反応と同一の系内で行うために使用される、請求項1のリンカー。
【請求項6】
mRNAと、その翻訳産物であるペプチドが、請求項1〜4のいずれかに記載のリンカーを介して結合した〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製する方法であって、
請求項1〜4のいずれかに記載のリンカーを調製する工程、
リンカーの一本鎖構造領域の塩基配列にハイブリダイズできる配列を、ペプチドをコードする配列の下流に有するmRNAの鋳型となるDNAを合成する工程、
再構成型のin vitroタンパク質合成反応液中にリンカーとDNAを供することにより、DNAからmRNAへの転写およびペプチドへの翻訳ならびにリンカーとmRNAとの複合体形成を行う工程
を含む、前記方法。
【請求項7】
再構成型のin vitroタンパク質合成反応液中に、非タンパク質性アミノ酸またはヒドロキシ酸でチャージされたtRNAを含むことにより、翻訳産物であるペプチドが特殊ペプチドとなる、請求項6に記載の方法。
【請求項8】
請求項7に記載の方法で作製された、〔mRNA〕−〔リンカー〕−〔特殊ペプチド〕連結体からなるライブラリー。
【請求項1】
mRNAと、その翻訳産物であるペプチドとが、リンカーを介して結合した連結体を作製するために使用されるリンカーであって、
リンカーの一方の端に、mRNAの3'末端側の塩基と対合する側鎖塩基を有する一本鎖構造領域を含み、
リンカーのもう一方の端に、ペプチド転移反応によって翻訳産物と結合可能な基を含むペプチドアクセプター領域を含み、
ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにアミノ酸がエステル結合した構造からなり、
無細胞翻訳反応液中でリンカーとmRNAが結合可能である
ことを特徴とするリンカー。
【請求項2】
一本鎖構造領域とペプチドアクセプター領域の間が、オリゴヌクレオチド、ポリアルキレン、ポリアルキレングリコール、ポリスチレン、多糖類、又はこれらの組合わせから選択される直鎖状物質で連結されている、請求項1に記載のリンカー。
【請求項3】
一本鎖構造領域が一本鎖DNAからなる、請求項1または2に記載のリンカー。
【請求項4】
ペプチドアクセプター領域は、塩基配列ACCAからなるオリゴRNAにフェニルアラニンがエステル結合した構造からなる、請求項1に記載のリンカー。
【請求項5】
再構成型のin vitroタンパク質合成系において、DNAの転写からmRNA−ペプチド連結体の調製までを翻訳反応と同一の系内で行うために使用される、請求項1のリンカー。
【請求項6】
mRNAと、その翻訳産物であるペプチドが、請求項1〜4のいずれかに記載のリンカーを介して結合した〔mRNA〕−〔リンカー〕−〔ペプチド〕連結体を作製する方法であって、
請求項1〜4のいずれかに記載のリンカーを調製する工程、
リンカーの一本鎖構造領域の塩基配列にハイブリダイズできる配列を、ペプチドをコードする配列の下流に有するmRNAの鋳型となるDNAを合成する工程、
再構成型のin vitroタンパク質合成反応液中にリンカーとDNAを供することにより、DNAからmRNAへの転写およびペプチドへの翻訳ならびにリンカーとmRNAとの複合体形成を行う工程
を含む、前記方法。
【請求項7】
再構成型のin vitroタンパク質合成反応液中に、非タンパク質性アミノ酸またはヒドロキシ酸でチャージされたtRNAを含むことにより、翻訳産物であるペプチドが特殊ペプチドとなる、請求項6に記載の方法。
【請求項8】
請求項7に記載の方法で作製された、〔mRNA〕−〔リンカー〕−〔特殊ペプチド〕連結体からなるライブラリー。
【図1】


【図2】
【図3】

【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2013−46637(P2013−46637A)
【公開日】平成25年3月7日(2013.3.7)
【国際特許分類】
【出願番号】特願2012−252252(P2012−252252)
【出願日】平成24年11月16日(2012.11.16)
【分割の表示】特願2011−537297(P2011−537297)の分割
【原出願日】平成22年10月21日(2010.10.21)
【出願人】(506269633)ペプチドリーム株式会社 (2)
【Fターム(参考)】
【公開日】平成25年3月7日(2013.3.7)
【国際特許分類】
【出願日】平成24年11月16日(2012.11.16)
【分割の表示】特願2011−537297(P2011−537297)の分割
【原出願日】平成22年10月21日(2010.10.21)
【出願人】(506269633)ペプチドリーム株式会社 (2)
【Fターム(参考)】
[ Back to top ]