
ペルオキシソーム生合成および機能に関連する障害を治療および診断するための医薬組成物
【課題】本発明は、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む、細胞におけるペルオキシソームの生成および機能に関連する障害を処置するための医薬組成物を提供することを目的とする。さらに本発明は、細胞内におけるペルオキシソームの生成および/または機能に関連する疾患または症候群に診断された被検体を処置する方法であり、該被検体にプロテアーゼ活性を有しそしてペルオキシソーム酵素をプロセッシングするペプチドを投与する工程を含む方法を提供する。
【解決手段】本発明は、細胞におけるペルオキシソームの生成に関連する障害を治療または診断するための医薬組成物であって、システインプロテアーゼ活性を有し、PTS1またはPTS2シグナルによって標的化されるペルオキシソーム酵素を直接にプロセッシングするポリペプチドを含む医薬組成物に関する。ポリペプチドは好ましくは、Tysnd1によってコードされる。
【解決手段】本発明は、細胞におけるペルオキシソームの生成に関連する障害を治療または診断するための医薬組成物であって、システインプロテアーゼ活性を有し、PTS1またはPTS2シグナルによって標的化されるペルオキシソーム酵素を直接にプロセッシングするポリペプチドを含む医薬組成物に関する。ポリペプチドは好ましくは、Tysnd1によってコードされる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ペルオキシソームを標的とするポリペプチドをコードする核酸配列に関し、該ポリペプチドはプロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングする。特定の実施形態において、本発明は、ペルオキシソームプロテアーゼTysnd1が、ペルオキシソームβ酸化およびα酸化に関与するPTS1およびPTS2シグナルを含有する酵素Acaa1、Acox1、Scp2およびHsd17b4を、プロセッシング可能であるという証拠を提供する。本発明はさらに、ペルオキシソームプロテアーゼをコードするこれらの哺乳類核酸のいずれか1つが関与する障害の診断、処置、および予防のための治療、診断および調査方法に関する。
【背景技術】
【0002】
ペルオキシソームは、これまで研究されて来た全ての真核生物に存在するオルガネラである。ペルオキシソームはDNAおよびタンパク質合成能力を欠いているため、全てのペルオキシソームタンパク質は、細胞質ゾルコンパートメント内で合成され、翻訳後にペルオキシソームに仕分けられる(非特許文献1および2)。2つの異なるペルオキシソームシグナル標的化配列(PTS)およびその変異体、C末端PTS1およびN末端PTS2が明らかにされている。ほぼ全てのペルオキシソーム酵素が、その後[STAGQCN]−[KRH]−[LIVMAFY]に拡張されるPTS1シグナル[SA]−K−Lを有し(非特許文献3)、このPTS1シグナルはサイトゾル溶解性レセプターPex5pによって認識される。数個のペルオキシソームタンパク質のみがN末端に位置するPTS2モチーフ[RK]−[LVI]−[X5]−[HQ]−[LAF]を介して標的化される(非特許文献4−6)。PTS1およびPTS2シグナルの両方を欠いている少数のペルオキシソームマトリクスタンパク質は、十分に明らかにされていない内部PTSによってオルガネラに対して標的化されている(非特許文献7)。
【0003】
ペルオキシソームの機能は極めて多様であり、細胞タイプおよび外部刺激に依存している。ヒトにおいては、ペルオキシソームは、多様な同化および異化経路(例えばコレステロール生合成、脂肪酸酸化、プリン代謝、過酸化水素解毒、胆汁酸合成、プラスマロゲン合成、アミノ酸代謝)(非特許文献8および9)、ヒト免疫不全ウイルスおよびロタウイルスの感染性(非特許文献10)および代謝状態とは独立したある種の発生プロセス(非特許文献11)に関与している。酵母においては、ペルオキシソームは、普通でない炭素源、例えばオレイン酸、一級アミン、プリン、D−アミノ酸、およびメタノールの代謝には不可欠である(非特許文献12および13)。植物においては、ペルオキシソームは光呼吸に関与し(非特許文献14)、トリパノソーマにおいては、ペルオキシソームは解糖に関与し(非特許文献15および16)、真菌類においては、それらは二次代謝産物、例えばβ−ラクタムペニシリンの合成に関与している(非特許文献17)。
【0004】
質量分析法(MS)は、配列データベースの急速な発展と相まって、モデル生物におけるペルオキシソームタンパク質組成の特徴づけを世界的に著しく向上させてきた。MSに基づく方法はラット肝臓から、34個の既知の、そして5個の推定のペルオキシソームタンパク質の同定を可能にした(非特許文献18(Kikuchi et al 2004))。複数の遺伝子が、ペルオキシソーム生合成とも呼ばれる、細胞内でのペルオキシソームの生成に関与している。ペルオキシンをコードするいわゆるPEX遺伝子が、機能性ペルオキシソームを欠いた酵母変異株の機能相補性によってクローニングされている(非特許文献7)。配列データベースのスクリーニングによって13個のヒトPEX相同体が同定され、そのうち11個は、ペルオキシソーム障害を有する患者の細胞系にてペルオキシソーム生合成を回復することが証明された(非特許文献19)。ペルオキシソーム生合成の欠陥は、複数の遺伝性ヒト障害、例えば代謝性疾患の中でもレフサム病(非特許文献20)、X染色体連鎖副腎白質ジストロフィー(X−ALD)、メバロン酸尿症(非特許文献21)の原因となっている。
【0005】
ペルオキシソームタンパク質の移入には、著しいタンパク質の修飾が関与していないらしい(非特許文献22)。折畳みポリペプチドおよびタンパク質ダイマーは、ペルオキシソームマトリクス中に取り込ませることができる(非特許文献23および24)。アルコールオキシダーゼモノマーは、酵素的に活性なオクタマーのアセンブリ前に、マトリクス中に取り込まれ(非特許文献25および26)、アラニン:グリオキシル酸アミノトランスフェラーゼ1は、ダイマーとしてでもモノマーとしてでも、同じ効率で取り込ませることができる(非特許文献24)。2つのペルオキシソーム移入モデルが提案されている。1つのモデルでは、ペルオキシソーム移入レセプターがサイトゾルとペルオキシソームとの間で往復することを提案している(非特許文献27−29)。もう一方のモデルでは、ペルオキシソームレセプターがタンパク質をペルオキシソームに引き込むことが示唆されている(非特許文献30−31)。
【0006】
しかしながら、そのプロセッシング、活性化、分解および関連する疾患に関するタンパク質の運命については、タンパク質がいったんペルオキシソーム内に入るとほとんど不明である。これまで2つのプロテアーゼ、すなわちインスリン分解酵素(IDE)(非特許文献32)およびLonプロテアーゼのペルオキシソーム特異的な型(非特許文献18)が、ペルオキシソーム中で実験によって検出されている。IDEは、酸化ペルオキシソームリゾチームの分解で役割を果たし得、一方新規なLONプロテアーゼはペルオキシソーム生合成に関与する可能性がある。
【0007】
Kurochkinら(非特許文献33−35)は、GenBank 139.0のげっ歯類および霊長類mRNA配列の130629個の推定翻訳物を探索した中から、29個の新規なペルオキシソームPTS1標的化タンパク質候補をコンピュータにより同定した。現在Tysnd1またはトリプシンドメイン含有1(GenBankおよびGenPeptアクセッションAK005069およびBAB23793)と呼ばれるそれらの候補の内の一つ1300019N10Rik、ならびにラット(XM_345106およびXP_345107)およびヒト(NM_173555およびNP_775826)でのそのオルソログは、シロイヌナズナに由来するプロテアーゼ関連タンパク質にわずかに似ている。それは、2つのプロテアーゼ関連ドメインであるグルタミルエンドペプチダーゼI(IPR008256)ならびにトリプシン様セリンおよびシステインプロテアーゼ(IPR009003)も含有する。プロテアーゼのグルタミルエンドペプチダーゼIファミリーのメンバーは、セリンタイプのペプチダーゼ活性を保有している。その触媒活動においてセリンを利用するタンパク質分解酵素は遍在的であり、ウイルス、細菌および真核生物に見られる。ペルオキシソームは、それぞれが独自の特異性を備えた、広範囲に渡るプロテアーゼを採用して(recruit)、オルガネラ中のタンパク質の効率的な分解を達成する可能性が高い。
【0008】
Tysnd1は、マウス染色体10に位置する。そのヒトオルソログは、染色体10に対するシンテニック位置に位置する。Tysnd1は脂肪組織、大動脈、肝臓、腎臓および肺で発現される(表1を参照)。同時発現遺伝子は、ペルオキシソーム標的化遺伝子産物をコードすることが知られているPeci、Pex6、Pex16を含む。他の同時発現遺伝子(例えばFsp27およびCas1)は、脂質代謝に関連している。公開されている遺伝子発現リソース(GNF U74A、GNF Atlas 2、マウス遺伝子予測データベース)に由来するデータは、Tysnd1がペルオキシソーム調節脂質代謝に関与することを示唆している。
【0009】
【表1】

【0010】
【非特許文献1】Subramani S, Koller A, Snyder WB. Import of peroxisomal matrix and membrane proteins. Annu Rev Biochem. 2000;69:399-418.
【非特許文献2】Purdue PE, Lazarow PB. Peroxisome biogenesis. Annu Rev Cell Dev Biol. 2001;17:701-52.
【非特許文献3】Olivier LM, Kovacs W, Masuda K, Keller GA, Krisans SK. Identification of peroxisomal targeting signals in cholesterol biosynthetic enzymes. AA-CoA thiolase, hmg-coa synthase, MPPD, and FPP synthase. J Lipid Res. 2000 Dec;41(12):1921-35.
【非特許文献4】Osumi T, Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T. Amino-terminal presequence of the precursor of peroxisomal 3-ketoacyl-CoA thiolase is a cleavable signal peptide for peroxisomal targeting. Biochem Biophys Res Commun. 1991 Dec 31;181(3):947-54.
【非特許文献5】Swinkels BW, Gould SJ, Bodnar AG, Rachubinski RA, Subramani S. A novel, cleavable peroxisomal targeting signal at the amino-terminus of the rat 3-ketoacyl-CoA thiolase. EMBO J. 1991 Nov;10(11):3255-62.
【非特許文献6】Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T, Osumi T. Characterization of the signal peptide at the amino terminus of the rat peroxisomal 3-ketoacyl-CoA thiolase precursor. J Biol Chem. 1994 Feb 25;269(8):6001-10.
【非特許文献7】Hettema EH, Distel B, Tabak HF. Import of proteins into peroxisomes. Biochim Biophys Acta. 1999 Aug 12;1451(1):17-34.
【非特許文献8】Titorenko VI, Rachubinski RA. The life cycle of the peroxisome. Nat Rev Mol Cell Biol. 2001 May;2(5):357-68.
【非特許文献9】Titorenko VI, Rachubinski RA. The peroxisome: orchestrating important developmental decisions from inside the cell. J Cell Biol. 2004 Mar 1;164(5):641-5.
【非特許文献10】Cohen GB, Rangan VS, Chen BK, Smith S, Baltimore D. The human thioesterase II protein binds to a site on HIV-1 Nef critical for CD4 down-regulation. J Biol Chem. 2000 Jul 28;275(30):23097-105.
【非特許文献11】Gavva NR, Wen SC, Daftari P, Moniwa M, Yang WM,Yang-Feng LP, Seto E, Davie JR, Shen CK. NAPP2, a peroxisomal membrane protein, is also a transcriptional corepressor. Genomics. 2002 Mar;79(3):423-31.
【非特許文献12】Van der Klei IJ, Veenhuis M. Yeast peroxisomes: function and biogenesis of a versatile cell organelle. Trends Microbiol. 1997 Dec;5(12):502-9.
【非特許文献13】Veenhuis M, Kiel JA, Van Der Klei IJ. Peroxisome assembly in yeast. Microsc Res Tech. 2003 Jun 1;61(2):139-50.
【非特許文献14】Olsen LJ. The surprising complexity of peroxisome biogenesis. Plant Mol Biol. 1998 Sep;38(1-2):163-89.
【非特許文献15】Hannaert V, Michels PA. Structure, function, and biogenesis of glycosomes in kinetoplastida. J Bioenerg Biomembr. 1994 Apr;26(2):205-12.
【非特許文献16】Moyersoen J, Choe J, Fan E, Hol WG, Michels PA. Biogenesis of peroxisomes and glycosomes: trypanosomatid glycosome assembly is a promising new drug target. FEMS Microbiol Rev. 2004 Nov;28(5):603-43.
【非特許文献17】Van de Kamp M, Driessen AJ, Konings WN. Compartmentalization and transport in beta-lactam antibiotic biosynthesis by filamentous fungi. Antonie Van Leeuwenhoek. 1999 Jan-Feb;75(1-2):41-78.
【非特許文献18】Kikuchi M, Hatano N, Yokota S, Shimozawa N, Imanaka T, Taniguchi H. Proteomic analysis of rat liver peroxisome: presence of peroxisome-specific isozyme of Lon protease. J Biol Chem. 2004 Jan 2;279(1):421-8.
【非特許文献19】Gould SJ, Valle D. Peroxisome biogenesis disorders: genetics and cell biology. Trends Genet. 2000 Aug;16(8):340-5.
【非特許文献20】Jansen GA, Waterham HR, Wanders RJ. Molecular basis of Refsum disease: sequence variations in phytanoyl-CoA hydroxylase (PHYH) and the PTS2 receptor (PEX7). Hum Mutat. 2004 Mar;23(3):209-18.
【非特許文献21】Sacksteder KA, Gould SJ. The genetics of peroxisome biogenesis. Annu Rev Genet. 2000;34:623-652.
【非特許文献22】Titorenko VI, Smith JJ, Szilard RK, Rachubinski RA. Pex20p of the yeast Yarrowia lipolytica is required for the oligomerization of thiolase in the cytosol and for its targeting to the peroxisome. J Cell Biol. 1998 Jul 27;142(2):403-20.
【非特許文献23】Glover JR, Andrews DW, Rachubinski RA. Saccharomyces cerevisiae peroxisomal thiolase is imported as a dimer. Proc Natl Acad Sci U. S. A. 1994 Oct 25;91(22):10541-5.
【非特許文献24】Leiper JM, Oatey PB, Danpure CJ. Inhibition of alanine:glyoxylate aminotransferase 1 dimerization is a prerequisite for its peroxisome-to-mitochondrion mistargeting in primary hyperoxaluria type 1. J Cell Biol. 1996 Nov;135(4):939-51.
【非特許文献25】Bellion E, Goodman JM. Proton ionophores prevent assembly of a peroxisomal protein. Cell. 1987 Jan 16;48(1):165-73.
【非特許文献26】Waterham HR, Russell KA, Vries Y, Cregg JM. Peroxisomal targeting, import, and assembly of alcohol oxidase in Pichia pastoris. J Cell Biol. 1997 Dec 15;139(6):1419-31.
【非特許文献27】Marzioch M, Erdmann R, Veenhuis M, Kunau WH. PAS7 encodes a novel yeast member of the WD-40 protein family essential for import of 3-oxoacyl-CoA thiolase, a PTS2-containing protein, into peroxisomes. EMBO J. 1994 Oct 17;13(20):4908-18.
【非特許文献28】Albertini M, Rehling P, Erdmann R, Girzalsky W, Kiel JA, Veenhuis M, Kunau WH. Pex14p, a peroxisomal membrane protein binding both receptors of the two PTS-dependent import pathways. Cell. 1997 Apr 4;89(1):83-92.
【非特許文献29】Elgersma Y, Elgersma-Hooisma M, Wenzel T, McCafferyJM, Farquhar MG, Subramani S. A mobile PTS2 receptor for peroxisomal protein import in Pichia pastoris. J Cell Biol. 1998 Feb 23;140(4):807-20.
【非特許文献30】Szilard RK, Titorenko VI, Veenhuis M, Rachubinski RA.Pay32p of the yeast Yarrowia lipolytica is an intraperoxisomal component of the matrix protein translocation machinery. J Cell Biol. 1995 Dec;131(6 Pt 1):1453-69.
【非特許文献31】Zhang JW, Lazarow PB. PEB1 (PAS7) in Saccharomyces cerevisiae encodes a hydrophilic, intra-peroxisomal protein that is a member of the WD repeat family and is essential for the import of thiolase into peroxisomes. J Cell Biol. 1995 Apr;129(1):65-80.
【非特許文献32】Morita M, Kurochkin IV, Motojima K, Goto S, TakanoT, Okamura S, Sato R, Yokota S, Imanaka T. Insulin-degrading enzyme exists inside of rat liver peroxisomes and degrades oxidized proteins. Cell Struct Funct. 2000 Oct;25(5):309-15.
【非特許文献33】Kurochkin IV, RIKEN GER Group, GSL Members, Konagaya A, Nagashima T, Schönbach C. Identification of Potential Peroxisomal Proteins in Mouse. Genome Res. 2003 Jun; 13(6b):1560. [http://www.genome.org/cgi/content/full/13/6b/1560]
【非特許文献34】Kurochkin IV, Schönbach C. Konagaya A. In Silico Prediction of Peroxisomal Proteins in Mouse. Genome Informatics 2003 Dec; 14:539-40 [http://www.jsbi.org/journal/GIW03/GIW03P068.pdf]
【非特許文献35】Kurochkin IV, Nagashima T, Konagaya A, Schönbach C.Sequence-based Discovery of the Human and Rodent Peroxisomal Proteome. Appl Bioinformatics; in press June 2005.
【非特許文献36】UCSC Genome browser; mouse Tysnd1 http://genome.ucsc.edu/cgi-bin/hgTracks?db=mm5&position=chr10:61457382-61464634&hgsid=39784308
【非特許文献37】UCSC Genome browser; human TYSND1 http://genome.ucsc.edu/cgi-bin/hgTracks?db=hg17&position=chr10:71568974-71575956&hgsid=39784308
【非特許文献38】Gene Entrez mouse Tysnd1 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene&cmd=Retrieve&dopt=Graphics&list_uids=71767
【非特許文献39】Gene Entrez human TYSND1 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene&cmd=Retrieve&dopt=Graphics&list_uids=219743
【非特許文献40】Swissprot/TrEMBL mouse Tysnd1 and human TYSND1 http://kr.expasy.org/cgi-bin/sprot-search-de?TYSND1
【非特許文献41】InterPro domain: Peptidase, trypsin-like serine and cysteine proteases http://www.ebi.ac.uk/interpro/ISpy?mode=single&ac=[SwissProt/TrEMBL accession number]
【非特許文献42】UCSC Genome browser; gene expression for mouse Tysnd1 and neighbors http://genome.ucsc.edu/cgi-bin/hgNear?hgsid=39784308&org=Mouse&db=mm5&near_search=AK005069&submit=Go%21&near.order=expGnfAtlas2&near.count=50
【非特許文献43】Mouse Gene Prediction Database; Search Results for 1300019N10Rik (XM_125636.1) and top 100 correlated genes; http://mgpd.med.utoronto.ca/profile.php?nameofgene=XM_125636.1
【非特許文献44】UCSC Genome browser; gene expression for human TYSND1 and neighbors http://genome.ucsc.edu/cgi-bin/hgNear?hgsid=73232388&org=Human&db=hg18&near_search=TYSND1&submit=Go%21&near_order=expGnfAtlas2&near.count=50
【非特許文献45】Array Prospector gene expression for human TYSND1 (SwissProt/TrEBML accession Q96AR5) http://string.embl-heidelberg.de:8080/prophecies_cgi/new_prophecies/new_prophecies.pl?genename=Q96AR5&species=9606&colorcode=1&maxgenes=10&submit=SUBMIT
【発明の開示】
【発明が解決しようとする課題】
【0011】
発明の概要
【0012】
Tysnd1またはトリプシンドメイン含有1は最初に、仮説上のタンパク質として報告された(Kawai J, et al., RIKEN Genome Exploration Research Group Phase II Team and the FANTOM Consortium. Functional annotation of a full-length mouse cDNA collection. Nature. 2001 Feb 8;409(6821):685-90; Okazaki Y et al., FANTOM Consortium; RIKEN Genome Exploration Research Group Phase, I & II Team. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature. 2002 Dec 5;420(6915):563-73.)。公開データベースで利用可能なタンパク質レベルに対する機能情報(Gene Entrez(Gene Entrez human TYSND1 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene&cmd=Retrieve&dopt=Graphics&list_uids=219743)、SwissProt/TrEMBL(Swissprot/TrEMBL mouse Tysnd1 and human TYSND1 http://kr.expasy.org/cgi-bin/sprot-search-de?TYSND1))はコンピュータにより推測され、タンパク質モチーフおよび配列相同性検索に基づいている。本明細書において初めて、Tysnd1タンパク質機能のインビトロにおける実証について記載される。その機能としては、1)ペルオキシソームへの局在化、2)ペルオキシソームプロテアーゼ機能、3)PTS2含有ペルオキシソーム酵素のタンパク質分解開裂、4)PTS1含有ペルオキシソーム酵素のタンパク質分解開裂、5)前駆型Tysnd1の成熟Tysnd1への翻訳後のプロセッシングが挙げられる。機能2)〜4)は、Tysnd1の基質として、ラット3−ケトアシル−CoAチオラーゼB、ペルオキシソームの前駆体(RGD1562373_predicted)、ラットアシル補酵素Aオキシダーゼ1(Acox1)、マウスステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−b)デヒドロゲナーゼ4(Hsd17b4)を用いて証明された。
【0013】
1つの態様において、本発明は、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む、細胞におけるペルオキシソームの生成および機能に関連する障害を処置するための医薬組成物を提供する。
さらなる態様において、本発明は、細胞内におけるペルオキシソームの生成および/または機能に関連する疾患または症候群に診断された被検体(例えば、ヒト患者)を処置する方法であり、該被検体にプロテアーゼ活性を有しそしてペルオキシソーム酵素をプロセッシングするペプチドを投与する工程を含む方法を提供する。
【課題を解決するための手段】
【0014】
ポリペプチドは好ましくは、Tysnd1によってコードされる。ペルオキシソーム酵素は、3−オキソアシル補酵素Bチオラーゼ(以降、Acaa1と呼ぶ)、アシル補酵素Aオキシダーゼ1(Acox1)、パルミトイルステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ 4(Hsd17b4)からなる群より選択される少なくとも1つであってよい。疾患は、代謝性疾患の中でも、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、メバロン酸尿症、および男性不妊症からなる群より選択されてもよい。
【0015】
別の態様において、本発明は、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む、細胞におけるペルオキシソームの生成および機能に関連する障害を診断する医薬組成物を提供する。
【0016】
ポリペプチドは好ましくは、Tysnd1によってコードされる。ペルオキシソーム酵素は、3−オキソアシル補酵素Bチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、パルミトイルステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ 4(Hsd17b4)からなる群より選択される少なくとも1つであってよい。疾患は、代謝性疾患の中でもレフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、メバロン酸尿症および男性不妊症からなる群より選択されてもよい。
発明の詳細な説明
【発明を実施するための最良の形態】
【0017】
定義
本発明の実施は、別途指示しない限り、当分野の技術範囲内である、分子生物学の従来技法を利用する。そのような技法は、文献に十分説明されている。
【0018】
以下の定義が、以下に文書化した説明で使用される具体的な用語について与えられている。
【0019】
「ペルオキシソーム」という用語は、本明細書で使用されるとき、真核細胞に見出される酵素を保持した、膜結合の小胞を指す。ペルオキシソームは時に、マイクロボディと呼ばれる。ペルオキシソームは、そのような酵素を残りの細胞部分から隔離する役割を果たし、細胞代謝を組織化する。
【0020】
ポリペプチドを「プロセッシングする」という用語は、本明細書で使用されるとき、成熟タンパク質またはペプチドを形成するための、細胞における合成されたポリペプチドの開裂(特に加水分解開裂)を指す。ポリペプチドは、他の基の付加などによってアミノ酸残基を修飾されるか、その活性な三次元コンホーメーションに折畳まれるか、あるいは分解のために標的化されてもよい。
【0021】
「Tysnd1」という用語は、第10染色体(マウス)に位置するゲノムDNA配列およびその遺伝子産物を指す。そのヒトオルソログは第10染色体に対するシンテニック位置に位置する。Tysnd1の特色付けは上の表1に示されている。
本発明はここで、本発明の実施形態が示されている添付図を参照して以下でより十分に説明される。
【0022】
[実施形態]
<材料および方法>
<発現ベクターの構築に使用するプライマー>
プライマーB1およびB2は、GenBankアクセッションAK005069のTysnd1ヌクレオチド配列に基づいている。B1およびB2は、構築プラスミドベクター、pEGFP−Tysnd1に使用した。
<B1>(フォワード)5’−GGATCCATGGGGCGGCAATGGGGAC−3’(配列番号1)は、BamHI部位(GGATCC)およびTysnd1配列位置59〜77を含有する。
<B2>(リバース)5’−GGATCCTCAGAGCTTGCTCCGTGGGAC−3’(配列番号2)は、BamHI部位(GGATCC)およびTysnd1配列位置1745〜1765を含有する。
プライマーAX1およびAX2は、GenBankアクセッションBC085743のラットAcox1ヌクレオチド配列に基づいている。
【0023】
<AX1>(フォワード)
(化1)
5’-ACCATGGGCTACCCTTACGACGTGCCTGACTACGCCAACCCCGACCTGCGCAAGGAGC-3’ (配列番号3)
は、ATG開始コドンを包囲する合成Kozak配列(太字)を含有する。HA−tagペプチドTyrAspValProAspTyrAlaは、下線の配列によってコードされる。プライマーの位置37〜58は、BC085743のAcox1配列位置94〜115に相当する。
<AX2>(リバース)5’−TCAAAGCTTGGACTGCAGGGGCTTC−3’(配列番号4)は、Acox1配列位置2052〜2076を含有する。停止コドンが含まれていた。
プライマーS5およびS2は、GenBankアクセッションBC034613のマウスScp2ヌクレオチド配列に基づいている。
【0024】
<S5>(フォワード)
(化2)
5’-ACCATGGGCTACCCTTACGACGTGCCTGACTACGCCCCTTCTGTCGCTTTGAA-3’
ATCTCC (配列番号5)
は、ATG開始コドンを包囲する合成Kozak配列(太字)を含有する。HA−tagペプチドTyrAspValProAspTyrAlaは、下線の配列によってコードされる。プライマーの位置37〜59は、Scp2配列位置25〜57に相当する。
<S2>(リバース)5’−CTCCTCACAGCTTAGCTTTGC−3’(配列番号6)は、Scp2配列位置1649〜1669に相当する。停止コドンが含まれていた。
プライマーHsd17b4−FwdおよびHsd17b4−Revは、GenBankアクセッションAK004866のマウスHsd17b4ヌクレオチド配列に基づいている。
【0025】
<Hsd17b4−Fwd>(フォワード)
5’−ACCATGGGCTACCCTTACGACGTGCCTGACTACGCCGCTTCGCCGCTGAGGTTCGAC−3’(配列番号17)
は、ATG開始コドンを包囲する合成Kozak配列(ACCATGG)を含有する。HA−tagペプチドTyrAspValProAspTyrAlaは、下線の配列によってコードされる。プライマーの位置37〜57は、Hsd17b4配列位置155〜175に相当する。
<Hsd17b4−Rev>(リバース)
5’−TCAGAGCTTGGCATAGTCTTTAAGAAT−3’(配列番号18)は、Hsd17b4−Rev配列位置2359〜2333に相当する。停止コドンが含まれていた。
Flagタグ付きTysnd1のプライマーは、GenBankアクセッションAK005069の配列に基づいている。
【0026】
<FN>(フォワード)
5’−CCACCATGGACTACAAAGACGATGACGACAAGGGGCGGCAATGGGGAC−3’(配列番号7)は、Kozak配列(CCACCATGG)、FLAGエピトープ(GACTACAAAGACGATGACGACAAG、配列番号8)およびTysnd1配列位置62〜77(GGGCGGCAATGGGGAC、配列番号9)を含む。
<FC1>(フォワード)5’−CCACCATGGGGCGGCAATGGGGAC−3’(配列番号10)は、Kozak配列(CCACCATGG)およびTysnd1配列位置62〜77(GGGCGGCAATGGGGAC、配列番号9)を含む。
<FC2>(リバース)5’−TCAGAGCTTGCTCCGTGGGACCTTGTCGTCATCGTCTTTGTAGTCTTCGGACAGGGGCCGCTGCAG−3’(配列番号11)は、Tysnd1配列位置1724〜1744、FLAGエピトープ(CTTGTCGTCATCGTCTTTGTAGTC、配列番号12)およびヌクレオチド配列位置1745〜1765に相当するTysnd1のC末端からの6個のアミノ酸(ValProArgSerLysLeu、配列番号13)を含む。
【0027】
<細胞およびトランスフェクション>
COS−7(SV40によって形質転換されたサル線維芽細胞)、293FT(SV40の大型T抗原を発現するヒト293細胞の急速成長変異株)およびチャイニーズハムスター卵巣CHO−K1細胞を、10%(v/v)ウシ胎仔血清(Invitrogen)を添加したダルベッコの修飾イーグル培地中で維持し、LipofectAMINE 2000試薬(Invitrogen)をメーカーの取扱説明書に従って使用して一時トランスフェクションした。
【0028】
<緑色蛍光タンパク質(GFP)融合タンパク質の発現ベクターの構築>
全長マウスTysnd1をコードするDNA断片は、プライマーセットB1およびB2を用いたPCRによって、クローニングしたcDNA 1300019N10(GenBankアクセッションAK005069)をテンプレートとして使用して、増幅した。PCR産物は、pGEM−T Easyベクター(Promega)内にサブクローニングした。BamHIによる消化の後、得られた断片は、pEGFP−C1のBamHI部位に挿入した(EGFPは増強GFPである; BD Biosciences Clontech)。得られたベクターは、pEGFP−Tysnd1と名づけた。
【0029】
<エピトープタグ付きラットペルオキシソーム3−オキソアシル補酵素Bチオラーゼ(Acaa1)、ラットアシル補酵素Aオキシダーゼ1(Acox1)、マウスパルミトイルステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)の発現>
以下のベクターを利用した。即ち、pcDNA3.1Zeo中のラットタイプBプレ−3−ケトアシル−CoAチオラーゼ−HA(HA=赤血球凝集素)(九州大学のY.Fujikiより寄贈)、pcDNA3.1/TOPO−HA−Acox1、pcDNA3.1/TOPO−HA−Scp2である。ラットAcox1のN末端へのHAタグ付けは、HAエピトープを含有するフォワードプライマーAX1および停止コドンを含有するリバースプライマーAX2を使用して、PCRをベースとする技法によって行った。同様にHAタグをPCRによって、プライマーS5およびS2ならびにHsd17b4−FwdおよびHsd17b4−Revを使用して、それぞれScp2およびHsd17b4内に導入した。増幅DNA産物は、pcDNA3.1/V5−His−TOPO(Invitrogen)内にクローニングした。プラスミド構築物は全て挿入の方向を点検し、トランスフェクションに使用した。
【0030】
<エピトープタグ付きTysnd1の発現>
培養細胞中でのマウスTysnd1の発現では、2つのベクターを構築した。pcDNA3.1/TOPO−FLAG−Tysnd1は、Tysnd1のN末端にFLAGエピトープを含有する。pcDNA3.1/TOPO−Tysnd1−FLAGは、Tysnd1のC末端にFLAGエピトープを含有する。pcDNA3.1/TOPO−FLAG−Tysnd1を作成するために、FLAGエピトープを含有するフォワードプライマーFN1および停止コドンを含有するリバースプライマーB2を用いて、全長Tysnd1のPCR増幅を実施した。pcDNA3.1/TOPO−Tysnd1−FLAGを構築するために、FLAGエピトープをアミノ酸562と563との間に導入し、これによってTysnd1の未変性PTS1含有C末端を保存した。PCR増幅は、フォワードプライマーFC1およびリバースプライマーFC2を用いて実施した。後者はFLAG配列に加えて、アミノ酸Val−Pro−Arg−Ser−Lys−Leu(位置563〜568)をコードするC末端Tysnd1配列および停止コドンを含有する。構築物は、上述のようにトランスフェクション実験に使用した。
【0031】
<GFP融合タンパク質の局在化>
CHO−K1細胞は、ガラスカバースリップに播種して、発現ベクターpEGFP−Tysnd1およびペルオキシソーム標的化DsRed2レポータータンパク質(BD Biosciences Clontech)をコードしたpDsRed2−Peroxi(Discosoma spからの赤色蛍光タンパク質の変異体2)によって一時トランスフェクションした。EGFP融合タンパク質およびDsRed2−Peroxiタンパク質の生細胞顕微鏡観察は、トランスフェクションの48時間後に、63×対物レンズを用いたレーザー走査共焦点顕微鏡TCS SP2(Leica)で実施した。488nmのアルゴンレーザーを励起に使用した。GFPおよびDsRed2によって放出された蛍光シグナルは、535nmバンドパスフィルタおよび570nmロングパスフィルタをそれぞれ使用して検出した。
【0032】
<免疫ブロッティング分析>
細胞は、上述のようにトランスフェクションした。48時間後、細胞を緩衝液A(Roche Diagnostics GmbHからのComplete(商標)プロテアーゼインヒビターカクテルを追加した、20mMのTris−HCl(pH7.5)、120mMのNaCl、1%のTriton X−100)に溶解させて、12,000×gにて15分間遠心分離にかけた。上澄み画分をSDS−PAGEで分離した。ウェスタンブロット分析を、Hybond−Pポリビニリデンジフルオリド膜(Amersham Biosciences,Inc.)上に電気泳動によって移動させたサンプルを用いて、HA.11モノクローナル抗体(Covance)または抗FLAG(商標)M2抗体(Sigma)および二次抗体、ホースラディッシュペルオキシダーゼに結合したヒツジ抗マウスIgG抗体(Amersham Biosciences,Inc.)を使用して、実施した。抗原−抗体複合体は、ECL plusウェスタンブロッティング検出試薬(Amersham Biosciences,Inc.)を用いて可視化した。タンパク質バンドの見かけの分子量は、予備染色したPrecision Plus Protein(商標)Standards(Bio−Rad)の移動度と比較することによって推定した。
【0033】
<ペルオキシソームの精製>
ペルオキシソームは、以前に記載された3つのプロトコル(Van Veldhoven PP, Baumgart E, Mannaerts GP. Iodixanol (Optiprep), an improved density gradient medium for the iso-osmotic isolation of rat liver peroxisomes. Anal Biochem. 1996 May 15;237(1):17-23; McClelland GB, Khanna S, Gonzalez GF, Butz CE, Brooks GA. Peroxisomal membrane monocarboxylate transporters: evidence for a redox shuttle system? Biochem Biophys Res Commun. 2003 Apr 25;304(1):130-5; Antonenkov VD, Sormunen RT, Hiltunen JK. The behavior of peroxisomes in vitro: mammalian peroxisomes are osmotically sensitive particles. Am J Physiol Cell Physiol. 2004 Dec;287(6):C1623-35)の組合せおよび改良に基づいた方法によって精製した。手短には、雄のウィスターラット(250〜300g)を一晩絶食させた。ジエチルエーテル吸入により動物を殺処分した後、肝臓を門脈を介して灌流した。肝臓を氷冷ホモジナイゼーション緩衝液(0.16Mスクロース、12%(wt/vol)PEG1500、10mM MOPS、pH7.4、1mM EDTA、1mM EGTA、1mM DTT、0.1%(vol/vol)エタノール)中でハサミを用いて細かく刻み、Potter−Elvehjemホモジナイザー(ガラス上のTeflon)でホモジナイゼーションした。核および重ミトコンドリアは、固定角ローター内で3,000×g、4℃にて10分間の遠心分離によってペレット化した。ペレットを再ホモジナイゼーションして、遠心分離を繰り返した。上澄み(ポスト核画分)を合わせて、20,000×gにて20分間遠心分離して、「軽ミトコンドリアペレット」を生成した。このペレットは、ゆるくはめたDounceホモジナイザーを使用して、ホモジナイゼーション緩衝液に再懸濁させた。体積は15ml/開始時の肝臓重量10gに調節して、Optiprep(60%(wt/vol)イオジキサノール、Axis−Shield PoC AS)5体積および希釈媒体(0.16Mのスクロース、12%(wt/vol)のPEG 1500、60mMのMOPS(pH7.4)、6mMのEDTA、6mMのDTT、0.6%(vol/vol)のエタノール)1体積より成る勾配溶液の等体積と混合した。この混合物を近垂直ローター(Beckman Type NVT 65)で180,000×g、4℃にて3.5時間、低加速および減速モードを使用して遠心分離した。遠心分離の後、画分0.5mlを上部から針および注射器によって取り出した。単離媒体を除去するために、画分材料を17,000×gにて20分間沈殿させて、0.1%のTriton X−110および1mMのDTTを含有するPBS緩衝液中に再懸濁させた。
【0034】
<マーカー酵素およびタンパク質含有量>
ペルオキシソーム、ミトコンドリアおよびリソソームのOptiprep勾配画分中での局在を判定するために、カタラーゼ(Johansson LH, Borg LA. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal Biochem. 1988 Oct;174(1):331-6.)、スクシナートデヒドロゲナーゼ(Pennington RJ. Biochemistry of dystrophic muscle. Mitochondrial succinate-tetrazolium reductase and adenosine triphosphatase. Biochem J. 1961 Sep;80:649-54.)およびβ−ガラクトシダーゼ(b−Galアッセイキット、Invitrogen)の活性がそれぞれ測定された。タンパク質含有量は、Bio−Rad DCタンパク質アッセイ試薬キット(Bio−Rad)を使用して測定した。
【0035】
<マウスTysnd1に対するポリクローナル抗体>
ウサギポリクローナル抗体を、Tysnd1のアミノ酸501〜515に相当するポリペプチドSNTRDNNTGATYPHL(配列番号14)と結合したキーホールリンペットヘモシアニンに対して産生し、SCRUM Inc.(東京)によりアフィニティ精製した。ウェスタンブロット分析には、抗体を2μg/mlの濃度で使用した。抗体予備吸収アッセイでは合成ペプチドCSNTRDNNTGATYPHL(配列番号15)をMe2SOに2mg/mlの濃度で溶解させ、次に、ウェスタンブロッティング(40μgペプチド/5μg抗体;約700倍の過剰モル数のペプチド)の前に、一次抗体溶液と混合した。
【0036】
<マウス(ラット)Acaa1に対するポリクローナル抗体>
ウサギポリクローナル抗体を、マウスAcaa1のアミノ酸259〜274に相当するポリペプチドKLKPAFKDGGSTTAGN(配列番号19)と結合したキーホールリンペットヘモシアニンに対して産生し、SCRUM Inc.(東京)によりアフィニティ精製した。ウェスタンブロット分析には、抗体を2μg/mlの濃度で使用した。
【0037】
<低分子干渉RNA(siRNA)>
ヒトTysnd1の発現を下方制御するためのRNA媒介干渉は、Qiagen K.K.から購入した低分子干渉siRNA二本鎖、Hs_Tysnd1_1_HP si RNA (siRNA Tysnd1_1) および Hs_Tysnd1_2_HP si RNA (siRNA Tysnd1_2)を使用して、行われた。siRNA Tysnd1_1は、配列CAG CAG AAA CCT TGC TCT GAA(配列番号20)を標的とし、siRNA Tysnd1_2は、配列CCC GCT GAG CAC TTC CAT GAA(配列番号21)を標的とした。対照siRNA (Qiagen K.K.)は、いかなる既知の哺乳類遺伝子配列に対しても相同性を持たない配列AAT TCT CCG AAC GTG TCA CGT(配列番号22)を標的とした。293FT細胞を、12ウェルプレートで培養した。siRNA(1.2mg)を、0.8mgのpcDNA3.1/TOPO−HA−Acaa1、pcDNA3.1/TOPO−HA−Scp2またはpcDNA3.1/TOPO−HA−Hsd17b4のいずれかとともに同時トランスフェクションした。トランスフェクションを、80%−90%コンフルエントな状態の細胞に、リポフェクトアミン2000(Invitrogen Japan K.K.)を使用して行った。各siRNAサンプルについて、二重のトランスフェクションを行った。トランスフェクション開始後54時間に、細胞を150μlのSDS−PAGEサンプル緩衝液中で溶解した。溶解物を超音波で分解し、95℃で5分間インキュベートし、そしてHA.11モノクローナル抗体(Covance)を使用してウェスタンブロッティングによって分析した。
【0038】
<COS−7細胞からの組み換えTysnd1の精製>
COS−7細胞を、リポフェクトアミン2000トランスフェクション試薬(Invitrogen Japan K.K.)を使用して、cDNA3.1/TOPO−Tysnd1−FLAGでトランスフェクションした。トランスフェクション後48時間に、細胞をPBSで洗い、そして振動下4℃で30分間緩衝液Aを用いて溶解した。細胞溶解物を15,000×gで15分間遠心分離し、細胞残渣の溶解物を取り除いた。この溶解物を、製造業者のプロトコル(Sigma Aldrich Japan K.K.)により、100μlの抗FLAG M2アフィニティービーズを使用して、4℃下で90分間インキュベートした。このビーズを、緩衝液Aで4回洗い、次に緩衝液B(50mM HEPES、pH8.0、115mM NaCl)で4回洗った。このビーズをカラム中に移し、結合していたタンパク質を500μg/mlの3×FLAGペプチドの緩衝液B溶液300μlで溶出した。
【0039】
<組み換えタンパク質のクローニング、発現、および精製>
ラットAcox1、ラットAcaa1、およびマウスScp2のコード配列を、KODポリメラーゼ(TOYOBO CO.LTD)を使用して、PCRで増幅した。結果として得られたAcox1およびScp2のPCR産物を、pTrcHis2−TOPO(Invitrogen Japan K.K.)にクローニングし、C末端にmyc−6xHis タグを有する組み換えタンパク質のための発現ベクターを作った。Acaa1のPCR産物を、pQE−80L発現ベクター(Qiagen K.K.)中にクローニングし、N末端に6xHisのタグを有するAcaa1の発現構築物を作った。Acox1およびScp2の発現ベクターで、Escherichia coli Top10細胞(Invitrogen Japan K.K.)を形質転換した。タンパク質発現の誘導は、製造業者の使用説明書に従い、行った。組換え体Acox1およびScp2を、BD TALON 樹脂(BD Biosciences)を用い、大腸菌の抽出物の可溶性画分から精製した。Acaa1の発現ベクターで、Escherichia coli BL21 Star (DE3)細胞を形質転換した。組換え体Acaa1は、不溶性画分に見られたが、その画分から6Mグアニジン−HClを用いて可溶化された。精製は、BD TALON樹脂を用いて、変性条件下で行われた。精製組換え体Acaa1のリフォールディングを、0.5mM DTTを含有する緩衝液Bでの透析によって行った。組換え体タンパク質は、DC Protein Assay (Bio−Rad Laboratories、Inc)および280nmでの分光光度法を用いて定量した。
【0040】
<インビトロTysnd1プロセッシング活性のアッセイ>
8μlの精製組換え体Tysnd1を、6μlの組換え体Acox1、Scp2、またはAcaa1の50mM Hepes、pH8.0、115mM NaCl、0.2mM DTT溶液と、37℃でインキュベートした。反応を、Laemmliサンプル緩衝液の添加および95℃下での5分間の加熱によって停止した。反応生成物を4%−20%SDS−PAGEによって分離し、Hybond−Pメンブレン上にブロットした。組換え体Acox1およびScp2の開裂生成物を、抗c−Mycモノクローナル抗体(Nacalai Tesque、Inc)を用い、検出した。Acaa1のプロセッシング生成物を、抗Acaa1抗体によるか、または、0.05% Amido Blackを用いたHybond−Pメンブレンの染色によるかして可視化した。
【0041】
<プロセッシングを受けたAcaa1のN末端配列決定>
組換え体Acaa1を、精製Tysnd1と、上記したようにしてインキュベートした。プロセッシングされたAcaa1を、前駆体型から4%−20%SDS−PAGEによって分離し、Hybond−Pメンブレン上にブロットし、そして0.05%Amido Blackを用い染色した。プロセッシングされた41kDaのバンドを、メンブレンから切り取り、そして6サイクルのEdman分解分析にかけた。配列決定分析は、A.Uroshibata(理化学研究所、ゲノム科学総合研究センター、タンパク質基盤研究グループ)によってなされた。
【0042】
<プロテアーゼ阻害活性>
阻害活性のアッセイのために、精製組換え体Tysnd1を、様々なプロテアーゼ阻害剤と30分間25℃で、プレインキュベートした。基質タンパク質の添加の後、インキュベーションを37℃で12時間続行した。以下のプロテアーゼ阻害剤を使用した(最終濃度):ペプスタチンA(1μM)、EGTA(2mM)、EDTA(2mM)、1,10−フェナントロリン(1mM)、1,7−フェナントロリン(1mM)、ベンズアミジン(1mM)、AEBSF(1mM)、アプロチニン(4μg/ml)、ロイペプチン(10μM)、E64(20μM)、NEM(1mM)。プロセッシング生成物を、上記のようにしてアッセイした。
【0043】
<ベザフィブレート処置および細胞下分画>
6頭の7週齢C57BL/6Jオスマウスを、実験に使用した。対照のグループ(n=3)を、標準飼料で飼育し、一方実験グループ(n=3)を、0.5%(v/w)ベザフィブレートを含有する標準飼料で飼育した。細胞下分画では、同じグループに属する3頭の動物の肝臓を混合し、そして上記したようにしてホモジナイゼーションした。ポスト核(PN)画分および軽ミトコンドリア(LM)画分の調製を、「ペルオキシソームの精製」の項で記載したようにして行った。ポストミトコンドリア画分(PM)は、PN画分の20,000×gで20分間の遠心分離の後の上清を示す。
<結果>
(実施例1)
【0044】
<Tysnd1はペルオキシソームに対して標的化される>
Tysnd1はPTS1シグナルを含有するので、このタンパク質がペルオキシソームに対して標的化されているかどうかが調べられた。GFP−Tysnd1融合構築物を、DsRed2−Peroxiをコードするプラスミドと同時トランスフェクションさせた。Tysnd1の全長cDNA配列をGFPのC末端に付加して、融合タンパク質対照のC末端にてPTS1配列を保存した。対照はPTS1シグナルと融合した蛍光タンパク質であり、ペルオキシソームに局在する。GFP蛍光は、多数のスポット様構造物中で観察された(図1A)が、CHO−K1細胞の他の部分では観察されなかった。同じ切片内のGFP−Tysnd1融合タンパク質陽性顆粒は全て、pDsRed2−Peroxiタンパク質(赤色)について陽性であった(図1Bおよび図1C)。この同時局在は、GFP−Tysnd1がペルオキシソームに対して標的化されていたという強力な証拠となる。
(実施例2)
【0045】
<PTS2含有ペルオキシソーム酵素の細胞ベースでの分解>
ペルオキシソームタンパク質の一部は、そのN末端領域に、各前駆体がその最終目的地に到達すると切断されるPTS2ペルオキシソーム標的化配列を含有する(Osumi T, Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T. Amino-terminal presequence of the precursor of peroxisomal 3-ketoacyl-CoA thiolase is a cleavable signal peptide for peroxisomal targeting. Biochem Biophys Res Commun. 1991 Dec 31;181(3):947-54;.Swinkels BW, Gould SJ, Bodnar AG, Rachubinski RA, Subramani S. A novel, cleavable peroxisomal targeting signal at the amino-terminus of the rat 3-ketoacyl-CoA thiolase. EMBO J. 1991 Nov;10(11):3255-62;.Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T, Osumi T. Characterization of the signal peptide at the amino terminus of the rat peroxisomal 3-ketoacyl-CoA thiolase precursor. J Biol Chem. 1994 Feb 25;269(8):6001-10.)。PTS2シグナル配列を切断させるタンパク質分解酵素は、まだ同定されていない。潜在的候補は、既知のペルオキシソームプロテアーゼインスリン分解酵素(Ide)およびペルオキシソーム型Lonプロテアーゼおよび本研究で記載した新規なペルオキシソームプロテアーゼTysnd1である。3つのプロテアーゼうちで、Tysnd1が最も有望な候補である。Ideは大型のタンパク質には作用せず、アミノ酸50個未満の長さのペプチドを代わりに開裂させる一方で(Duckworth, W. C., Bennett, R. G. & Hamel, F. G. (1998) Insulin degradation: progress and potential. Endocr Rev. 1998 Oct;19(5):608-24.;Kurochkin, I. V. (2001) Insulin-degrading enzyme: embarking on amyloid destruction. Trends Biochem Sci. 2001 Jul;26(7):421-5)、ATP依存性Lonプロテアーゼは、変性タンパク質を分解するシャペロンとして機能し得る(Kikuchi M, Hatano N, Yokota S, Shimozawa N, Imanaka T, Taniguchi H. Proteomic analysis of rat liver peroxisome: presence of peroxisome-specific isozyme of Lon protease. J Biol Chem. 2004 Jan 2;279(1):421-8)。
【0046】
PTS2含有シグナル配列の除去でのTysnd1の役割を究明するために、ペルオキシソーム3−オキソアシル補酵素Aチオラーゼ(Acaa1)のプロセッシングが分析された。Acaa1は、残基36個のN末端除去可能配列を持つ大型前駆体として合成される(Osumi T, Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T. Amino-terminal presequence of the precursor of peroxisomal 3-ketoacyl-CoA thiolase is a cleavable signal peptide for peroxisomal targeting. Biochem Biophys Res Commun. 1991 Dec 31;181(3):947-54;.Swinkels BW, Gould SJ, Bodnar AG, Rachubinski RA, Subramani S. A novel, cleavable peroxisomal targeting signal at the amino-terminus of the rat 3-ketoacyl-CoA thiolase. EMBO J. 1991 Nov;10(11):3255-62.;Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T, Osumi T. Characterization of the signal peptide at the amino terminus of the rat peroxisomal 3-ketoacyl-CoA thiolase precursor. J Biol Chem. 1994 Feb 25;269(8):6001-10.)。C末端にHAタグが付いたAcaa1のみがCHO細胞において過剰発現された場合、主に44kDaのプロセッシングされていない前駆体型が検出された。HA−Acaa1は少量のみが成熟41kDa型に変換された(図2、レーン1)。増加する量のpcDNA3.1/TOPO−FLAG−Tysnd1との同時トランスフェクションは、44kDa前駆体の段階的な消失および成熟41kDaタンパク質の並行した増加を引き起こした(図2、レーン2〜4)。同様の結果がCOS−7細胞でも得られた(データは示さず)。結果は、Tysnd1がPTS2を含有ずる先行ペプチドを除去する真のペルオキシソームプロセッシングプロテアーゼであるか、あるいは第二の、まだ同定されていないペルオキシソームプロセッシングプロテアーゼの活性化を通じてAcaa1の成熟を促進することを意味している。
【0047】
したがってTysnd1は、他のPTS2シグナル含有タンパク質全てのプロセッシングに関与し得る。PTS2タンパク質のペルオキシソーム内プロセッシングの生理学的重要性は、まだ理解されていない。ミトコンドリアの場合、前駆体タンパク質の大多数がミトコンドリア標的化のための切断可能アミノ末端延長配列を含有する。ミトコンドリアプロセッシングプロテアーゼによるシグナル配列の除去は、タンパク質折畳みおよびオルガネラ内でのさらなる仕分けのために必要であると考えられる(Gakh O, Cavadini P, Isaya G. Mitochondrial processing peptidases. Biochim Biophys Acta. 2002 Sep 2;1592(1):63-77.)。ミトコンドリアタンパク質とは異なり、ペルオキシソームタンパク質は、事前のアンフォールディングなしに首尾よく移動される(Subramani S, Koller A, Snyder WB. Import of peroxisomal matrix and membrane proteins. Annu Rev Biochem. 2000;69:399-418)。その上、ペルオキシソームマトリクスは単一コンパートメントより成るようであるが、ミトコンドリアスペースは、数個の明確なサブコンパートメントに分割される。近年、Nairらは、PTS2レセプターPex7pがPTS1レセプターPex5と同様に(Dammai V, Subramani S. The human peroxisomal targeting signal receptor, Pex5p, is translocated into the peroxisomal matrix and recycled to the cytosol. Cell. 2001 Apr 20;105(2):187-96)、輸送の「拡張シャトル」(“extended shuttle”)モードに従うことを証明した(Nair DM, Purdue PE, Lazarow PB. Pex7p translocates in and out of peroxisomes in Saccharomyces cerevisiae. J Cell Biol. 2004 Nov 22;167(4):599-604.)。Pex7pは、PTS2タンパク質移入の進行の間にペルオキシソームに入り、サイトゾル中へ再出現して、さらに数回のタンパク質移入を実行する。PTS2タンパク質がシグナル配列を維持した場合、これらのタンパク質はPex7pと共にペルオキシソームに入り、ペルオキシソームから出て、したがってオルガネラへのその蓄積が防止しされ得る。
(実施例3)
【0048】
<PTS1含有ペルオキシソーム酵素のプロセッシング>
PTS2タンパク質に加えて、数個のPTS1ペルオキシソーム酵素もペルオキシソームに移動されたときにプロセッシングを受ける。これらはペルオキシソーム脂肪酸β酸化に関与する鍵酵素である、アシル補酵素Aオキシダーゼ1(Acox1)、ヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)およびステロールキャリアタンパク質2(Scp2)を含む。それゆえ、Tysnd1がラットAcox1、マウスScp2、およびマウスHsd17b4タンパク質を切断できるかどうかが調査された。Acox1は、ポリペプチド成分A(75kDa)、B(53kDa)およびC(22kDa)より成る。それはA2、ABC、およびB2C2を含むヘテロオリゴマーとしても存在する(Tsukamoto T, Yokota S, Fujiki Y. Isolation and characterization of Chinese hamster ovary cell mutants defective in assembly of peroxisomes. J Cell Biol. 1990 Mar;110(3):651-60;Miyazawa S, Hayashi H, Hijikata M, Ishii N, Furuta S, Kagamiyama H, Osumi T, Hashimoto T. Complete nucleotide sequence of cDNA and predicted amino acid sequence of rat acyl-CoA oxidase. J Biol Chem. 1987 Jun 15;262(17):8131-7)。BおよびC断片は、ペルオキシソームへの移動時にAポリペプチドのタンパク質分解変換によって生成される(Miyazawa S, Osumi T, Hashimoto T, Ohno K, Miura S, Fujiki Y. Peroxisome targeting signal of rat liver acyl-coenzyme A oxidase resides at the carboxy terminus. Mol Cell Biol. 1989 Jan;9(1):83-91;Wirtz KW. Phospholipid transfer proteins revisited. Biochem J. 1997 Jun 1;324 (Pt 2):353-60)。分子量59kDaを有するタンパク質、Scp2は、ペルオキシソームにおいて46kDaおよび13kDa断片に変換される(Seedorf U, Ellinghaus P, Roch Nofer J. Sterol carrier protein-2. Biochim Biophys Acta. 2000 Jun 26;1486(1):45-54;Otera H, Nishimura M, Setoguchi K, Mori T, Fujiki Y. Biogenesis of nonspecific lipid transfer protein and sterol carrier protein x: studies using peroxisome assembly-defective pex cell mutants. J Biol Chem. 2001 Jan 26;276(4):2858-64)。ペルオキシソームでは、Scp2はアシルCoAエステルのβ酸化に関与する。その上、Scp2は、α酸化依存性基質の可溶化共タンパク質(solubilizing coprotein)として作用する(Mukherji M, Kershaw NJ, Schofield CJ, Wierzbicki AS, Lloyd MD. Utilization of sterol carrier protein-2 by phytanoyl-CoA 2-hydroxylase in the peroxisomal alpha oxidation of phytanic acid. Chem Biol. 2002 May;9(5):597-605.)。
【0049】
Hsd17b4は、二機能タンパク質であり、2−メチル分岐鎖脂肪酸、C27胆汁酸中間体、超長鎖脂肪酸のペルオキシソームでのβ酸化、およびポリ不飽和脂肪酸の合成に関与する。最近、Hsd17b4活性が、ロイコトリエンの不活性化(Ferdinandusse S, Denis S, Van Roermund CW, Wanders RJ, Dacremont G. Identification of the peroxisomal beta-oxidation enzymes involved in the degradation of long-chain dicarboxylic acids. J Lipid Res. 2004 Jun;45(6):1104-11)および精巣のセルトリ細胞における脂質ホメオスタシスを維持することによって雄性生殖機能(Huyghe S, Schmalbruch H, De Gendt K, Verhoeven G, Guillou F, Van Veldhoven PP, Baes M. Peroxisomal multifunctional protein 2 is essential for lipid homeostasis in Sertoli cells and male fertility in mice. Endocrinology. 2006 Feb 16; [Epub ahead of print as doi:10.1210/en.2005-1571])に必要であることが示された。具体的には、Hsd17b4は、ペルオキシソーム脂肪酸β酸化の第2および第3ステップの、エノイルCoAヒドラターゼ反応および3−ヒドロキシアシル−CoAデヒドロゲナーゼ反応を触媒する(Jiang LL, Kurosawa T, Sato M, Suzuki Y, Hashimoto T. Physiological role of D-3-hydroxyacyl-CoA dehydratase/D-3-hydroxyacyl-CoA dehydrogenase bifunctional protein. J Biochem (Tokyo). 1997 Mar;121(3):506-13)。Hsd17b4(79kDa)はペルオキシソームに到達した後、一箇所で切断されて、35kDaおよび44kDaのサイズを有する別々の酵素成分を生じる。
【0050】
ラットAcox1、マウスScp2およびマウスHsd17b4のタンパク質分解変換に対するTysnd1の効果を評価するために、タンパク質の発現構築物をCOS−7細胞にトランスフェクションした。Acox1、Scp2、およびHsd17b4はそのN末端をHAエピトープでタグ付けして、PTS1シグナルを保存した。ウェスタンブロットでは、Scp2は主として、未プロセッシング酵素型に相当する59kDaタンパク質バンドとして現れた(図3、レーン2)。約48kDaの非常に微弱なバンドは、プロセッシング型に相当する(図3、レーン2)。Tysnd1との同時トランスフェクションは、未プロセッシング59kDa型の著しいレベル低下を、48kDaタンパク質の付随する増加と共に引き起こした(図3、レーン3)。13kDa C末端断片はHAタグを欠いているため、検出されなかった。HA−Acox1を用いてトランスフェクションした細胞からの抽出物の分析は、主要な未プロセッシング75kDa型および48kDaの見かけのサイズを有する酵素のN末端断片の存在を明らかにした(図3、レーン4)。Tysnd1の同時発現は、プロセッシング済48kDaタンパク質の著しい蓄積を引き起こした(図3、レーン5)。同様にHA−Hsd17b4は、Tysnd1と同時発現した場合、プロセッシングを受けN末端の35kDa断片を与えた(図4)。これらのデータは、Tysnd1がペルオキシソーム鍵酵素の細胞プロセッシングに関与していることを明らかに立証するものである。
【0051】
Scp2のタンパク質分解切断は、それぞれ異なる活性を有する2つの断片を生成する。46kDa断片は、分岐鎖脂肪酸チオラーゼとして酵素活性がある(Seedorf U, Brysch P, Engel T, Schrage K, Assmann G. Sterol carrier protein X is peroxisomal 3-oxoacyl coenzyme A thiolase with intrinsic sterol carrier and lipid transfer activity. J Biol Chem. 1994 Aug 19;269(33):21277-83;Antonenkov VD, Van Veldhoven PP, Waelkens E, Mannaerts GP. Substrate specificities of 3-oxoacyl-CoA thiolase A and sterol carrier protein 2/3-oxoacyl-CoA thiolase purified from normal rat liver peroxisomes. Sterol carrier protein 2/3-oxoacyl-CoA thiolase is involved in the metabolism of 2-methyl-branched fatty acids and bile acid intermediates. J Biol Chem. 1997 Oct 10;272(41):26023-31)。非特異的脂質転移タンパク質(nsLTP)としても知られる13kDa断片は、基質、例えば脂肪アシル−CoA誘導体のAcox1への転移において機能し得る(Wouters FS, Bastiaens PI, Wirtz KW, Jovin TM. FRET microscopy demonstrates molecular association of non-specific lipid transfer protein (nsL-TP) with fatty acid oxidation enzymes in peroxisomes. EMBO J. 1998 Dec 15;17(24):7179-89.)。タンパク質(59kDa)の無傷の完全長型は、両方の機能をなお実施できる。したがってプロセッシングの生理学的重要性は明白ではない。Acox1の場合、ペルオキシソーム内プロセッシングの役割は、生成した断片が結合したままであるため、なお一層明らかでない。最近、Woutersら(Wouters FS, Bastiaens PI, Wirtz KW, Jovin TM. FRET microscopy demonstrates molecular association of non-specific lipid transfer protein (nsL-TP) with fatty acid oxidation enzymes in peroxisomes. EMBO J. 1998 Dec 15;17(24):7179-89.)は、nsLTPがAcox1、Acaa1、およびHsd17b4と結合されていることを証明した。これらの結果は、β酸化のペルオキシソーム酵素が機能性複合体内に組織化されていることを示唆する(Wouters FS, Bastiaens PI, Wirtz KW, Jovin TM. FRET microscopy demonstrates molecular association of non-specific lipid transfer protein (nsL-TP) with fatty acid oxidation enzymes in peroxisomes. EMBO J. 1998 Dec 15;17(24):7179-89.)。複合体形成によって、酵素の局所濃度が一時的に増加するため、脂質中間体の酵素間での効率的転移が可能となると考えられる。ペルオキシソーム内プロセッシングは、酵素のコンホメーション変化をもたらし得、これは複合体の形成を向上させると考えられる。Tysnd1は、このプロセスを調節する役割を果たし、したがって向上したペルオキシソーム脂肪酸β酸化を促進し得る。
【0052】
Tysnd1の発現パターンに類似した、55の組織からサンプリングしたマウス遺伝子の同時発現パターン(Mouse Gene Prediction Database; Search Results for 1300019N10Rik (XM_125636.1) and top 100 correlated genes; http://mgpd.med.utoronto.ca/profile.php?nameofgene=XM_125636.1)は、示唆された調節の役割を間接的に支持する。Tysnd1(XM_125636.1)、Acaa1(XM_135249.1)およびScp2(XM_135267.1)転写物は、プロテアーゼ−基質関係から予想されるように、大動脈、褐色脂肪、小腸、肝臓、前立腺および副腎において中〜高レベルで同時発現される。アクセッション番号は、Refseq遺伝子モデルアクセッションであり、参考文献(Mouse Gene Prediction Database; Search Results for 1300019N10Rik (XM_125636.1) and top 100 correlated genes; http://mgpd.med.utoronto.ca/profile.php?nameofgene=XM_125636.1)から引用した。
(実施例4)
【0053】
<Tysnd1の翻訳後プロセッシング>
他のペルオキシソームタンパク質と同様に、Tysnd1はサイトゾル内で合成される。ペルオキシソームへの移入が完了するまで潜在的なTysnd1基質がプロセッシングされないようにするために、タンパク質がペルオキシソームに到達するまでTysnd1のタンパク質分解活性は作動させるべきではない。異なるプロテアーゼが、タンパク質分解活性をサイレンシングさせるのに異なる方法を利用する。大半のプロテアーゼは、不活性前駆体として合成され、したがってタンパク質分解経路の鍵となる事象は、チモーゲンの活性酵素への変換である。
【0054】
推定上のTysnd1プロセッシングを調査するために、タンパク質のN末端またはC末端のどちらかにFLAGエピトープを含有するこの酵素の真核発現ベクターを用いてCOS−7細胞がトランスフェクションされた。C末端へのFLAGエピトープは、アミノ酸562と563との間に導入されて、PTS1シグナルを含有するTysnd1の未変性C末端を保存することに注目すべきである。Tysnd1のN末端でFLAGエピトープをコードするプラスミドを用いたトランスフェクションは、細胞溶解物のウェスタンブロットによって判定されるように、59kDAタンパク質バンドの出現をもたらした。分子量は、cDNA配列の翻訳CDSの計算重量と一致している(図5A、レーン3)。しかしながら約10kDaの追加のバンドも検出された(図5A、レーン3)。この小型タンパク質種は、Tysnd1のN末端の約90個のアミノ酸残基に結合されたFLAGエピトープに相当する(図5B)。他方Tysnd1のC末端でFLAGエピトープをコードするプラスミドを用いて細胞にトランスフェクションした場合、抗体は49kDaの追加のバンドを検出した(図5A、レーン2)。サイズの相違は、10kDa断片がN末端タグ付きTysnd1から分離されることを示し、したがってC末端タグ付きTysnd1タンパク質によって得られた結果を確証している。
【0055】
他のプロテアーゼと同様に、59kDa Tysnd1タンパク質は、触媒的に活性な49kDa型を生成することが予想される不活性前駆体型を示す。9kDa N末端断片の除去は、残基187〜282および311〜536における2つのプロテアーゼ関連ドメインに影響を及ぼさない。結果として、N末端セグメントは、ペルオキシソームに到達するまでTysnd1の活性を抑制する活性化ペプチドであり得る。
【0056】
不活性前駆体の活性Tysnd1への変換は、自己触媒的に、または別の同定されていないプロテアーゼによって、媒介されている可能性がある。活性化のタイミングおよび程度は最終的に、上述のようにペルオキシソーム脂肪酸β酸化の速度を調節し得る。
(実施例5)
【0057】
<マウスTysnd1に対する抗体の特徴づけ>
成熟Tysnd1タンパク質のC末端に近い領域(アミノ酸501〜515)に対するウサギポリクローナル抗体が作成された。抗体の特異性を、E.coli抽出物から精製された組換えマウスTysnd1および一時トランスフェクションされたCOS−7細胞中のN−FLAG−Tysnd1−C産物も認識する能力について試験した。組換えTysnd1のウェスタンブロット分析は、59kDaの予測サイズに相当するシグナルを明らかにした(図6、レーン1)。モックトランスフェクションCOS−7細胞の溶解物では、バンドは検出できなかった(図6、レーン2)。N末端Flagタグ付きTysnd1を発現する細胞は、2つの顕著なバンドを示した(図6、レーン3)。59kDAバンド断片は、アミノ末端領域を含むTysnd1に相当する。49kDA断片は、アミノ末端領域なしのTysnd1タンパク質に相当する(図5を参照)。抗Tysnd1抗体の特異性を試験するために、免疫化に使用された合成ペプチドを用いた競合アッセイが実施された。シグナルは、抗体の予備吸収後に完全に消滅し、高い特異性が立証された(図6、レーン4〜6)。Tysnd1に対する抗体を産生するために使用したペプチドの配列(アミノ酸501〜515)は、ラットTysnd1およびヒトTysnd1で共有され、抗体がこれらの種の組織におけるタンパク質発現を研究するためにも有用であり得ることが示唆される(以下を参照)。
(実施例6)
【0058】
<内在性Tysnd1の局在化およびサイズ>
共焦点顕微鏡法実験はTysnd1のペルオキシソーム局在化を示したが、証拠は人為的条件に基づいている。例えばTysnd1は培養細胞中のTysnd1−GFP産物として過剰発現された。したがってTysnd1のペルオキシソーム局在化を独立して確認することが選択された。ラット肝臓ホモジネートが分画され、抗Tysnd1抗体を使用した免疫ブロッティングによって細胞下画分が分析された。対照マーカー酵素の活性を調べた結果(図7A)は、ペルオキシソームが自己発生性Optiprep勾配の底部付近に位置することを示した(画分18〜20)。ペルオキシソームは上部画分(画分1〜4)に見出された他のオルガネラ(すなわちミトコンドリアおよびリソソーム)からうまく分離され、このサンプルの主なタンパク質含有量を構成する(図7B)。抗Tysnd1抗体を用いた画分のウェスタンブロット分析では、カタラーゼも検出された勾配の領域である、画分18〜20のみで免疫反応性物質が示された(図7C)。これらの結果は、内在性Tysnd1のペルオキシソーム局在化を強力に支持する。ウェスタンブロットによって検出された2つの顕著なバンドは、49kDaおよび27kDaの分子量を有するタンパク質型に相当する(図7C)。興味深いことにCOS−7トランスフェクション細胞では、検出された主なTysnd1種は、無傷のもの(59kDa)および部分的にプロセッシングされたもの(49kDa)に相当するが、27kDa型はほとんど検出できなかった(図5)。これは、Tysnd1のプロセッシング経路を飽和させ得るpcDNA3.1ベクターによって発生された過剰発現の結果であるらしい。
結論として本発明者のデータは、Tysnd1が最終的に少なくとも2つのプロテアーゼ型の形成をもたらす一連のタンパク質分解事象を受け得ることを示唆している。第一の型は、両方のプロテアーゼ関連ドメイン(49kDa)を含有する。第二の型(27kDa)は、C末端プロテアーゼ様ドメイン(308−531)のみを含有する。哺乳類組織における単一翻訳産物からの別々のプロテアーゼドメインの生成は、前例が無いわけではない。最近Calらは、ヒトのポリセラーゼ1(polyserase−I)(Cal S, Quesada V, Garabaya C, Lopez-Otin C. Polyserase-I, a human polyprotease with the ability to generate independent serine protease domains from a single translation product. Proc Natl Acad Sci U S A. 2003 Aug 5;100(16):9185-90)およびポリセラーゼ2(polyserase−2)(Cal S, Quesada V, Llamazares M, Diaz-Perales A, Garabaya C, Lopez-Otin C. Human polyserase-2, a novel enzyme with three tandem serine protease domains in a single polypeptide chain. J Biol Chem. 2005 Jan 21;280(3):1953-61)が、一連のタンパク質分解プロセッシング事象を受けて、3個の独立なセリンプロテアーゼユニットを発生させることを証明した。Tysnd1プロセッシングの複雑な特性は、ペルオキシソームにおける複数の生物学的プロセスの調節を可能にする細胞制御機構を反映しているように思われる。例えばTysnd1の2つの型の、基質特異性およびタンパク質分解効率は異なり得る。
(実施例7)
【0059】
<Tysnd1は、293FT細胞における、Acox1、Scp2、およびHsd17b4のプロセッシングに関与する主要な酵素である>
Tysnd1は、同時発現研究において数個のペルオキシソーム酵素の切断を促進することが示されたが、この過程に他のプロテアーゼも関与し得る可能性が残っている。そこで低分子干渉RNA(siRNA)が利用され、ペルオキシソーム酵素の細胞プロセッシングにおけるTysnd1の相対的貢献度が調査された。siRNAを使う戦略は、細胞における特定のタンパク質の選択的ノックダウンを達成する、強力で新しい手段を提供する(Sandy P, Ventura A, Jacks T. Mammalian RNAi: a practical guide. Biotechniques. 2005 Aug;39(2):215-24.)。試験された多数のマウスおよびヒト細胞系の中で、293FT細胞のみが、トランスフェクション研究において、完全長HA−Acox1、完全長HA−Scp2、および完全長HA−Hsd17b4から生成された検出可能な量の断片を示した。観察されたプロセッシング生成物は、293FT細胞に存在する内在性のペルオキシソームプロセッシングプロテアーゼによって、生成された。本明細書において、ヒトのTYSND1特異的siRNAのTysnd1_1およびTysnd1_2が、Acox1、Scp2、およびHsd17b4のN末端HAタグ含有断片の生成を本質的に阻害することが証明された(図8)。対照的に、対照の非サイレンシングsiRNAは、効果がなかった(図8)。siRNAが標的mRNAを完全にはノックダウンし得ないこと、およびTysnd1が長い半減寿命を有している可能性があることを考慮すると、観察された効果によって、Tysnd1が、Acox1、Scp2、Hsd17b4、およびおそらく他のペルオキシソーム酵素のプロセッシングにおいて、唯一のとまでは行かなくとも主要な役割を果たしていることが、立証される。
(実施例8)
【0060】
<インビトロの研究が、Tysnd1によって直接ペルオキシソーム酵素がプロセッシングされることを立証する>
Tysnd1が、数個のペルオキシソーム酵素のプロセッシングに関与していることが、細胞ベースのアッセイで示された。Tysnd1は、その標的を直接にか、または別の真のペルオキシソームプロセッシングプロテアーゼを活性化することによって間接的に、プロセッシングし得る。Tysnd1が直接にペルオキシソーム酵素をプロセッシングするか評価するために、組換え体Tysnd1−FLAGタンパク質を、COS−7細胞で発現させ、そしてアフィニティークロマトグラフィーによって精製した。Tysnd1−FLAGと組換え体Acox1、Scp2、およびAcaa1とをインキュベートすると、無傷のペルオキシソームにおいて生成されたものと同じサイズの開裂生成物が形成された(図9)。Acox1は、切断されて28kDaのC末端断片を生成し、Scp2は、プロセッシングされて16kDaのC末端断片を生成した(図9)。Tysnd1とAcaa1とのインキュベーションは、3kDaのPTS2含有N末端ペプチドの除去をもたらして、成熟Acaa1を生成させた(図9)。N末端配列決定分析によって、Tysnd1はAcaa1を、Cys26残基とSer27残基の間で開裂し、インビボで見られるAcaa1の成熟型を生成させることが確証された。これらの実験は、Tysnd1が真のペルオキシソームのプロセッシングプロテアーゼであることを立証する。
【0061】
Tysnd1によるペルオキシソーム酵素のプロセッシングは、高度に特異的である。数個のペルオキシソーム酵素が切断部位周囲の配列としてAla−[AlaVal]−Proモチーフを共有しているようであるが(Biermann J, van den Bosch H. In vitro processing of the human alkyl-dihydroxyacetonephosphate synthase precursor. Arch Biochem Biophys. 1999 Aug 1;368(1):139-46)、これは一般的法則ではない。例えばScp2は、約アミノ酸25個分離れた2つの位置で切断される。これらの部位の一方は、切断部位周囲にAla−[AlaVal]−Proモチーフを含有する。Acox1において、モチーフはプロセッシング部位周囲には見られなかった。Tysnd1基質切断機構は、単純なペプチド結合認識を包含しないが、むしろ二次および三次構造の認識に基づいていることが示唆される。
(実施例9)
【0062】
<Tysnd1はシステインエンドペプチダーゼである>
Tysnd1のプロテアーゼクラスを決定するために、様々なプロテアーゼインヒビターの存在下で、酵素プロセッシング活性を評価した。この実験のために、2個の基質が選択された。それはAcaa1およびAcox1である。異なるペルオキシソーム標的クラス(PTS1およびPTS2)を代表するのに加えて、Acaa1およびAcox1は、プロセッシング部位を囲むアミノ酸配列が異なる。Tysnd1のプロセッシング活性は、システインプロテアーゼインヒビターN−エチルマレイミド(NEM)(Sigma−Aldrich Japan K.K.)を用いたとき、完全に無くなった(図10)。Tysnd1は、スルフヒドリル活性試薬NEMに対して感受性であるが、2個の他のシステインプロテイナーゼインヒビターであるロイペプチンおよびt−エポキシスクシニル−1−ロイシルアミド−(4−グアニジノ)ブタン(E64)(株式会社ペプチド研究所(大阪府))に対しては抵抗性である。ロイペプチンは、いくつかのリソソームのセリンプロテアーゼおよびシステインプロテアーゼを阻害することが知られ、一方E64はパパインプロテアーゼファミリーの特異的阻害剤である(Salvesen GS, Nagase H. In Proteolytic Enzymes (ed. Beynon R and Bond JS), p. 110. Oxford University Press)。同様の阻害パターンが、ペルオキシソームプロテアーゼIDEについて観察された(IDEはNEMによって阻害されるがロイペプチンおよびE64によっては阻害されない)。金属キレート剤、とりわけ1,10−フェナントロリン(Sigma−Aldrich Japan K.K.)によってTysnd1は部分阻害されるが、これと関連する非キレート化化合物の1,7−フェナントロリン(Wako Pure Chemical Industries,Ltd.)によっては部分阻害されないことは(図10)、Znイオンが、Tysnd1のコンホメーションまたはその基質との複合体形成を安定化させるのに関与している可能性を示す。
(実施例10)
【0063】
<低脂血症薬ベザフィブレートによるTysnd1の誘導>
PPAR−α−アクチベーターであるベザフィブレートは、部分的には、Acaa1、Acox1、およびHsd17b4(これらはTysnd1によってプロセッシングされる)のβ酸化の原因となるペルオキシソーム酵素を誘導することによって、肝臓での脂肪酸のβ酸化を増加させることが知られている(Beier K, Volkl A, Hashimoto T, Fahimi HD. Selective induction of peroxisomal enzymes by the hypolipidemic drug bezafibrate. Detection of modulations by automatic image analysis in conjunction with immunoelectron microscopy and immunoblotting. Eur J Cell Biol. 1988 Aug;46(3):383-93)。それゆえ、Tysnd1もまたベザフィブレートによって誘導されるかどうか試験された。以前の報告に一致して、肝臓のAcox1、Hsd17b4、およびAcaa1のmRNAの誘導が観察された(データは示さず)。肝臓の細胞下分画を行い、Tysnd1タンパク質の含有量が評価された。ベザフィブレート処理は、ペルオキシソームに富む軽ミトコンドリア画分において、49kDa型のTysnd1の著しい増加をもたらした(図11)。対照的に、(ポスト核画分、ポストミトコンドリア画分では、)カタラーゼの総量の僅かな増加がみられ、軽ミトコンドリア画分のカタラーゼの含有量は増加しなかった(図11)。これらのデータは、代謝要求に応じるために、特に脂肪酸のβ酸化が促進される条件で、Tysnd1の発現がその基質タンパク質の発現と同時制御される可能性を示唆する。
(実施例11)
【0064】
<疾患に対するTysnd1のPTS1およびPTS2タンパク質プロセッシング活性の意味>
Acox1、Acaa1、Scp2、Hsd17b4、ならびにおそらく他のPTS2およびPTS1含有ペルオキシソームタンパク質のTysnd1によるプロセッシングは、代謝性疾患例えば食事性肥満、脂肪肝疾患または高コレステロール血症における脂肪酸β酸化、ケトン生成およびコレステロール合成の病原化に関して、潜在的な治療上または診断上の意味を有し得る。Tysnd1と脂肪代謝の関連および微小血管内皮細胞におけるその差次的発現(Keen HL, Ryan MJ, Beyer A, Mathur S, Scheetz TE, Gackle BD, Faraci FM, Casavant TL, Sigmund CD. Gene expression profiling of potential PPARgamma target genes in mouse aorta. Physiol Genomics. 2004 Jun 17;18(1):33-42、この参考文献の表6)も、心臓血管疾患の病原論におけるTysnd1機能不全の役割を意味し得る。心虚血の間、ミトコンドリア脂肪酸酸化の高い速度は、グルコース酸化を阻害する(Dyck JR, Cheng JF, Stanley WC, Barr R, Chandler MP, Brown S, Wallace D, Arrhenius T, Harmon C, Yang G, Nadzan AM, Lopaschuk GD. Malonyl coenzyme a decarboxylase inhibition protects the ischemic heart by inhibiting fatty acid oxidation and stimulating glucose oxidation. Circ Res. 2004 May 14;94(9):e78-84)。ラノラジンおよび長鎖3−ケトアシル補酵素Aチオラーゼインヒビターのトリメタジジンは、脂肪酸酸化を阻害し、したがってグルコース酸化および心臓効率を向上させることが示されている(Lopaschuk GD. Inhibiting fatty acid oxidation as a novel therapeutic approach to treating, ischaemic heart disease. Cardiovasc J S Afr. 2004 Jul;15(4 Suppl 1):S1)。心虚血へのペルオキシソームチオラーゼの寄与は知られているところでは調査されていないが、除外することはできない。Tysnd1の阻害は、チオラーゼの有効性を低下させ、それにより脂肪酸酸化を低下させると同時に心臓効率を向上させる。
【0065】
マウスへの高脂肪食の長期的影響に関する研究は、Acox1、Acaa1の肝臓における発現が著しくアップレギュレートされることを立証している(Murase, T., Mizuno, T., Omachi, T., Onizawa, K., Komine, Y., Kondo, H., Hase, T., Tokimitsu, I., 2001. Dietary diacylglycerol suppresses high fat and high sucrose diet-induced body fat accumulation in C57BL/6J mice. J. Lipid Res. 42, 372–378;Kim S, Sohn I, Ahn JI, Lee KH, Lee YS, Lee YS. Hepatic gene expression profiles in a long-term high-fat diet-induced obesity mouse model. Gene. 2004 Sep 29;340(1):99-109)。したがって成熟Tysnd1の特異的阻害または活性化は、原因となっている肝臓脂質代謝の病態および男性不妊症を制御する新たな戦略を開拓し得る(Huyghe S, Schmalbruch H, De Gendt K, Verhoeven G, Guillou F, Van Veldhoven PP, Baes M. Peroxisomal multifunctional protein 2 is essential for lipid homeostasis in Sertoli cells and male fertility in mice. Endocrinology. 2006 Feb 16; [Epub ahead of print as doi:10.1210/en.2005-1571])。マウスの成熟Scp2の欠損は、フィタン酸の蓄積および成人レフサム病様症候群の発達に関与している(Mukherji M, Kershaw NJ, Schofield CJ, Wierzbicki AS, Lloyd MD. Utilization of sterol carrier protein-2 by phytanoyl-CoA 2-hydroxylase in the peroxisomal alpha oxidation of phytanic acid. Chem Biol. 2002 May;9(5):597-605)。フィタン酸は、ペルオキシソームα−酸化経路でフィタノイル−CoA 2−ヒドロキシラーゼによって分解される。フィタノイル−CoA 2−ヒドロキシラーゼによるフィタノイル−CoAの効率的な2−ヒドロキシル化は成熟Scp2を必要とし、Tysnd1は成熟Scp2の利用可能性を調節する重要な役割を果たし得ている。したがってTysnd1の機能的不全は、レフサム様疾患症状をもたらし得る。
【0066】
Raychaudhuryらによる研究(Raychaudhury B, Banerjee S, Datta SC. Peroxisomal function is altered during leishmania infection. Med Sci Monit. 2003 Apr;9(4):BR125-9)は、リーシュマニア感染が、ペルオキシソーム損傷につながるペルオキシソーム酵素の複数の機能不全を引き起こすことを示した。注目すべきは、本発明者らがリーシュマニア感染時のペルオキシソームタンパク質分解活性の低下も検出したことである。感染時の活性低下は、成熟Tysnd1プロテアーゼ活性またはTysnd1前駆体の成熟Tysnd1への変換を阻害するリーシュマニアタンパク質によって引き起こされている可能性がある。
【図面の簡単な説明】
【0067】
【図1】融合タンパク質GFP−Tysnd1がペルオキシソームに局在することを示す。CHO−K1細胞を、融合されたGFP−Tysnd1およびDsRed2−Peroxiを発現するプラスミドと共に同時トランスフェクションした。生細胞は、蛍光顕微鏡法によって分析し、GFP(A)、DsRed(B)チャネル、および併合シグナル(C)の代表的な画像を示す。
【図2】ラットAcaa1前駆体の成熟Acaa1(mAcaa1)へのプロセッシングに対するTysnd1発現の効果を示す。CHO−K1細胞は、Acaa1−HAおよびTysnd1発現プラスミドの示された組合せを用いて一時トランスフェクションした。プラスミドの総量は、空cDNA3.1の追加により、4μgで一定に維持した。
【図3】Tysnd1の同時発現時のScp2およびAcox1のタンパク質分解変換を示す。COS−7細胞を、FLAG−Tysnd1およびHAマウスScp2またはHAラットAcox1の示された組合せでトランスフェクションした。プラスミドの総量は、pcDNA3.1を使用して4μgに調節した。トランスフェクションの48時間後に細胞溶解物を調製し、12.5%SDS−PAGEによって分離して、抗HA抗体を用いてウェスタンブロットで分析した。矢印は、Scp2およびAcox1前駆体およびその成熟形(mScp2およびmAcox1)の位置を示す。
【図4】Tysnd1の同時発現時のマウスペルオキシソームヒドロキシステロイド(17−β)デヒドロゲナーゼ 4(Hsd17b4)のタンパク質分解変換を示す。COS−7細胞を、FLAG−Tysnd1およびHA−マウスHsd17b4の示された組合せでトランスフェクションした。プラスミドの総量は、pcDNA3.1を使用して4μgに調節した。トランスフェクションの48時間後に細胞溶解物を調製し、12.5%SDS−PAGEゲルによって分離して、抗HA抗体を用いてウェスタンブロットで分析した。矢印は、Hsd17b4前駆体(79kDa)およびHsd17b4成熟形(35kDa)の位置を示す。
【図5】4μgDNAのpcDNA3.1によって(mock、レーン1)、およびC末端にFlagタグをつけたTysnd1によって(レーン3および6)トランスフェクションしたCOS−7細胞におけるTysnd1切断の分析を示す。これらは4%−20%勾配のSDS−PAGEによって分離し、そして抗Tysnd1抗体を使用したウェスタンブロッティングでアッセイした。レーン4−6で使用した抗体は、合成ペプチドCSNTRDNNTGATYPHLを予備吸収させた。
【図6】ポリクローナル抗体のTysnd1との特異的反応性を示す。精製した組換えTysnd1(レーン1および4)、空ベクターpcDNA3.1を用いてトランスフェクションしたCOS−7細胞からの溶解物(レーン2および5)、およびN末端Flagタグ付きTysnd1のベクターを用いてトランスフェクションしたCOS−7細胞からの溶解物(レーン3および6)を勾配4〜20%SDS−PAGEによって分離し、抗Tysnd1抗体を用いてウェスタンブロッティングによってアッセイした。レーン4〜6で使用した抗体は、合成ペプチドCSNTRDNNTGATYPHLを予備吸収させた。
【図7】Optiprep密度勾配での細胞下分画の結果を示す。軽ミトコンドリア画分は、自己形成Optiprep勾配により分画した。画分は、管の上から開始して採取した。カタラーゼ(縞模様の棒)、コハク酸デヒドロゲナーゼ(黒棒)およびβ−ガラクトシダーゼ(中空棒)を、それぞれペルオキシソーム、ミトコンドリアおよびリソソームのマーカー酵素として測定した(図7A)。結果を、全勾配活性のパーセンテージとして与える。図7Bは、タンパク質含有量を示す。等体積の各画分からのタンパク質を、12.5%SDS−PAGEによって分離し、マウスTysnd1に対する抗体を用いて免疫ブロットした(図7C)。
【図8】Tysnd1特異的低分子干渉RNA(siRNA)による、293FT細胞系におけるAcox1、Scp2、およびHsd17b4のプロセッシングの阻害を示す。293FT細胞を、ヒトTYSND1に特異的なsiRNA(Tysnd1_1およびTysnd1_2)または非サイレンシングsiRNA(対照)とともに、Acox1−HA、Scp2−HA、またはHsd17b4−HA発現プラスミドにトランスフェクションさせた。細胞溶解物を、抗HA抗体を用いたウェスタンブロッティングによって分析した。矢印は、Acox1、Scp2、およびHsd17b4の前駆体およびこれらの成熟形(mAox1、mScp2、およびmHsd17b4)の位置を示す。βアクチンのウェスタンブロット分析を、siRNA特異性およびサンプル添加の対照として用いた。
【図9】組み換えTysnd1によるAcox1、Scp2、およびAcaa1のインビトロの切断を示す。組み換えAcox1、Scp2、およびAcaa1を、「材料と方法」で説明されるようにして、E.coli抽出物から精製した。マウスTysnd1を、Tysnd1−Flag発現プラスミドを用い一時トランスフェクションしたCOS−7細胞の抽出物から精製した。Acox1およびScp2のプロセッシングを、抗cMyc抗体を用いたウェスタンブロティングによって分析した。Acaa1切断を、12.5%SDS−PAGEゲルからタンパク質を移した後の、Hybond−Pメンブレンの0.05%Amido Black染色によって検出した。
【図10】Tysnd1によるAcaa1(A)およびAcox1(B)のプロセッシングへの各種プロテアーゼインヒビターの影響を示す。組み換えHis6−Acaa1およびAcox1−myc−His6を、単独で0時間(レーン1)もしくは単独で12時間(レーン2)またはCOS−7細胞溶解物から精製したTysnd1−FLAGとともに12時間(A、レーン3〜14;B、レーン3〜17)インキュベートした。His6−Acaa1のプロセッシングを、抗Acaa1抗体を用いたウェスタンブロットによって評価した。Acox1−myc−His6の切断を、抗cMyc抗体を用いて検出した。
【図11】マウス肝臓におけるTysnd1の含有量へのベザフィブレート処置の効果を示す。マウスを、0.5%(v/w)のベザフィブレートを含有する標準飼料で、3週間飼育した。肝臓細胞下画分を、「材料および方法」でラット肝臓について記載されているようにして調製した。ポスト核画分(PN)、軽ミトコンドリア画分(LM)、およびポストミトコンドリア画分(PM)の100mgのタンパク質を、抗Tysnd1抗体または抗カタラーゼ抗体を用いたウェスタンブロットによって分析した。C、対照マウス;B、ベザフィブレート処置マウスサンプル。
【技術分野】
【0001】
本発明は、ペルオキシソームを標的とするポリペプチドをコードする核酸配列に関し、該ポリペプチドはプロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングする。特定の実施形態において、本発明は、ペルオキシソームプロテアーゼTysnd1が、ペルオキシソームβ酸化およびα酸化に関与するPTS1およびPTS2シグナルを含有する酵素Acaa1、Acox1、Scp2およびHsd17b4を、プロセッシング可能であるという証拠を提供する。本発明はさらに、ペルオキシソームプロテアーゼをコードするこれらの哺乳類核酸のいずれか1つが関与する障害の診断、処置、および予防のための治療、診断および調査方法に関する。
【背景技術】
【0002】
ペルオキシソームは、これまで研究されて来た全ての真核生物に存在するオルガネラである。ペルオキシソームはDNAおよびタンパク質合成能力を欠いているため、全てのペルオキシソームタンパク質は、細胞質ゾルコンパートメント内で合成され、翻訳後にペルオキシソームに仕分けられる(非特許文献1および2)。2つの異なるペルオキシソームシグナル標的化配列(PTS)およびその変異体、C末端PTS1およびN末端PTS2が明らかにされている。ほぼ全てのペルオキシソーム酵素が、その後[STAGQCN]−[KRH]−[LIVMAFY]に拡張されるPTS1シグナル[SA]−K−Lを有し(非特許文献3)、このPTS1シグナルはサイトゾル溶解性レセプターPex5pによって認識される。数個のペルオキシソームタンパク質のみがN末端に位置するPTS2モチーフ[RK]−[LVI]−[X5]−[HQ]−[LAF]を介して標的化される(非特許文献4−6)。PTS1およびPTS2シグナルの両方を欠いている少数のペルオキシソームマトリクスタンパク質は、十分に明らかにされていない内部PTSによってオルガネラに対して標的化されている(非特許文献7)。
【0003】
ペルオキシソームの機能は極めて多様であり、細胞タイプおよび外部刺激に依存している。ヒトにおいては、ペルオキシソームは、多様な同化および異化経路(例えばコレステロール生合成、脂肪酸酸化、プリン代謝、過酸化水素解毒、胆汁酸合成、プラスマロゲン合成、アミノ酸代謝)(非特許文献8および9)、ヒト免疫不全ウイルスおよびロタウイルスの感染性(非特許文献10)および代謝状態とは独立したある種の発生プロセス(非特許文献11)に関与している。酵母においては、ペルオキシソームは、普通でない炭素源、例えばオレイン酸、一級アミン、プリン、D−アミノ酸、およびメタノールの代謝には不可欠である(非特許文献12および13)。植物においては、ペルオキシソームは光呼吸に関与し(非特許文献14)、トリパノソーマにおいては、ペルオキシソームは解糖に関与し(非特許文献15および16)、真菌類においては、それらは二次代謝産物、例えばβ−ラクタムペニシリンの合成に関与している(非特許文献17)。
【0004】
質量分析法(MS)は、配列データベースの急速な発展と相まって、モデル生物におけるペルオキシソームタンパク質組成の特徴づけを世界的に著しく向上させてきた。MSに基づく方法はラット肝臓から、34個の既知の、そして5個の推定のペルオキシソームタンパク質の同定を可能にした(非特許文献18(Kikuchi et al 2004))。複数の遺伝子が、ペルオキシソーム生合成とも呼ばれる、細胞内でのペルオキシソームの生成に関与している。ペルオキシンをコードするいわゆるPEX遺伝子が、機能性ペルオキシソームを欠いた酵母変異株の機能相補性によってクローニングされている(非特許文献7)。配列データベースのスクリーニングによって13個のヒトPEX相同体が同定され、そのうち11個は、ペルオキシソーム障害を有する患者の細胞系にてペルオキシソーム生合成を回復することが証明された(非特許文献19)。ペルオキシソーム生合成の欠陥は、複数の遺伝性ヒト障害、例えば代謝性疾患の中でもレフサム病(非特許文献20)、X染色体連鎖副腎白質ジストロフィー(X−ALD)、メバロン酸尿症(非特許文献21)の原因となっている。
【0005】
ペルオキシソームタンパク質の移入には、著しいタンパク質の修飾が関与していないらしい(非特許文献22)。折畳みポリペプチドおよびタンパク質ダイマーは、ペルオキシソームマトリクス中に取り込ませることができる(非特許文献23および24)。アルコールオキシダーゼモノマーは、酵素的に活性なオクタマーのアセンブリ前に、マトリクス中に取り込まれ(非特許文献25および26)、アラニン:グリオキシル酸アミノトランスフェラーゼ1は、ダイマーとしてでもモノマーとしてでも、同じ効率で取り込ませることができる(非特許文献24)。2つのペルオキシソーム移入モデルが提案されている。1つのモデルでは、ペルオキシソーム移入レセプターがサイトゾルとペルオキシソームとの間で往復することを提案している(非特許文献27−29)。もう一方のモデルでは、ペルオキシソームレセプターがタンパク質をペルオキシソームに引き込むことが示唆されている(非特許文献30−31)。
【0006】
しかしながら、そのプロセッシング、活性化、分解および関連する疾患に関するタンパク質の運命については、タンパク質がいったんペルオキシソーム内に入るとほとんど不明である。これまで2つのプロテアーゼ、すなわちインスリン分解酵素(IDE)(非特許文献32)およびLonプロテアーゼのペルオキシソーム特異的な型(非特許文献18)が、ペルオキシソーム中で実験によって検出されている。IDEは、酸化ペルオキシソームリゾチームの分解で役割を果たし得、一方新規なLONプロテアーゼはペルオキシソーム生合成に関与する可能性がある。
【0007】
Kurochkinら(非特許文献33−35)は、GenBank 139.0のげっ歯類および霊長類mRNA配列の130629個の推定翻訳物を探索した中から、29個の新規なペルオキシソームPTS1標的化タンパク質候補をコンピュータにより同定した。現在Tysnd1またはトリプシンドメイン含有1(GenBankおよびGenPeptアクセッションAK005069およびBAB23793)と呼ばれるそれらの候補の内の一つ1300019N10Rik、ならびにラット(XM_345106およびXP_345107)およびヒト(NM_173555およびNP_775826)でのそのオルソログは、シロイヌナズナに由来するプロテアーゼ関連タンパク質にわずかに似ている。それは、2つのプロテアーゼ関連ドメインであるグルタミルエンドペプチダーゼI(IPR008256)ならびにトリプシン様セリンおよびシステインプロテアーゼ(IPR009003)も含有する。プロテアーゼのグルタミルエンドペプチダーゼIファミリーのメンバーは、セリンタイプのペプチダーゼ活性を保有している。その触媒活動においてセリンを利用するタンパク質分解酵素は遍在的であり、ウイルス、細菌および真核生物に見られる。ペルオキシソームは、それぞれが独自の特異性を備えた、広範囲に渡るプロテアーゼを採用して(recruit)、オルガネラ中のタンパク質の効率的な分解を達成する可能性が高い。
【0008】
Tysnd1は、マウス染色体10に位置する。そのヒトオルソログは、染色体10に対するシンテニック位置に位置する。Tysnd1は脂肪組織、大動脈、肝臓、腎臓および肺で発現される(表1を参照)。同時発現遺伝子は、ペルオキシソーム標的化遺伝子産物をコードすることが知られているPeci、Pex6、Pex16を含む。他の同時発現遺伝子(例えばFsp27およびCas1)は、脂質代謝に関連している。公開されている遺伝子発現リソース(GNF U74A、GNF Atlas 2、マウス遺伝子予測データベース)に由来するデータは、Tysnd1がペルオキシソーム調節脂質代謝に関与することを示唆している。
【0009】
【表1】
【0010】
【非特許文献1】Subramani S, Koller A, Snyder WB. Import of peroxisomal matrix and membrane proteins. Annu Rev Biochem. 2000;69:399-418.
【非特許文献2】Purdue PE, Lazarow PB. Peroxisome biogenesis. Annu Rev Cell Dev Biol. 2001;17:701-52.
【非特許文献3】Olivier LM, Kovacs W, Masuda K, Keller GA, Krisans SK. Identification of peroxisomal targeting signals in cholesterol biosynthetic enzymes. AA-CoA thiolase, hmg-coa synthase, MPPD, and FPP synthase. J Lipid Res. 2000 Dec;41(12):1921-35.
【非特許文献4】Osumi T, Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T. Amino-terminal presequence of the precursor of peroxisomal 3-ketoacyl-CoA thiolase is a cleavable signal peptide for peroxisomal targeting. Biochem Biophys Res Commun. 1991 Dec 31;181(3):947-54.
【非特許文献5】Swinkels BW, Gould SJ, Bodnar AG, Rachubinski RA, Subramani S. A novel, cleavable peroxisomal targeting signal at the amino-terminus of the rat 3-ketoacyl-CoA thiolase. EMBO J. 1991 Nov;10(11):3255-62.
【非特許文献6】Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T, Osumi T. Characterization of the signal peptide at the amino terminus of the rat peroxisomal 3-ketoacyl-CoA thiolase precursor. J Biol Chem. 1994 Feb 25;269(8):6001-10.
【非特許文献7】Hettema EH, Distel B, Tabak HF. Import of proteins into peroxisomes. Biochim Biophys Acta. 1999 Aug 12;1451(1):17-34.
【非特許文献8】Titorenko VI, Rachubinski RA. The life cycle of the peroxisome. Nat Rev Mol Cell Biol. 2001 May;2(5):357-68.
【非特許文献9】Titorenko VI, Rachubinski RA. The peroxisome: orchestrating important developmental decisions from inside the cell. J Cell Biol. 2004 Mar 1;164(5):641-5.
【非特許文献10】Cohen GB, Rangan VS, Chen BK, Smith S, Baltimore D. The human thioesterase II protein binds to a site on HIV-1 Nef critical for CD4 down-regulation. J Biol Chem. 2000 Jul 28;275(30):23097-105.
【非特許文献11】Gavva NR, Wen SC, Daftari P, Moniwa M, Yang WM,Yang-Feng LP, Seto E, Davie JR, Shen CK. NAPP2, a peroxisomal membrane protein, is also a transcriptional corepressor. Genomics. 2002 Mar;79(3):423-31.
【非特許文献12】Van der Klei IJ, Veenhuis M. Yeast peroxisomes: function and biogenesis of a versatile cell organelle. Trends Microbiol. 1997 Dec;5(12):502-9.
【非特許文献13】Veenhuis M, Kiel JA, Van Der Klei IJ. Peroxisome assembly in yeast. Microsc Res Tech. 2003 Jun 1;61(2):139-50.
【非特許文献14】Olsen LJ. The surprising complexity of peroxisome biogenesis. Plant Mol Biol. 1998 Sep;38(1-2):163-89.
【非特許文献15】Hannaert V, Michels PA. Structure, function, and biogenesis of glycosomes in kinetoplastida. J Bioenerg Biomembr. 1994 Apr;26(2):205-12.
【非特許文献16】Moyersoen J, Choe J, Fan E, Hol WG, Michels PA. Biogenesis of peroxisomes and glycosomes: trypanosomatid glycosome assembly is a promising new drug target. FEMS Microbiol Rev. 2004 Nov;28(5):603-43.
【非特許文献17】Van de Kamp M, Driessen AJ, Konings WN. Compartmentalization and transport in beta-lactam antibiotic biosynthesis by filamentous fungi. Antonie Van Leeuwenhoek. 1999 Jan-Feb;75(1-2):41-78.
【非特許文献18】Kikuchi M, Hatano N, Yokota S, Shimozawa N, Imanaka T, Taniguchi H. Proteomic analysis of rat liver peroxisome: presence of peroxisome-specific isozyme of Lon protease. J Biol Chem. 2004 Jan 2;279(1):421-8.
【非特許文献19】Gould SJ, Valle D. Peroxisome biogenesis disorders: genetics and cell biology. Trends Genet. 2000 Aug;16(8):340-5.
【非特許文献20】Jansen GA, Waterham HR, Wanders RJ. Molecular basis of Refsum disease: sequence variations in phytanoyl-CoA hydroxylase (PHYH) and the PTS2 receptor (PEX7). Hum Mutat. 2004 Mar;23(3):209-18.
【非特許文献21】Sacksteder KA, Gould SJ. The genetics of peroxisome biogenesis. Annu Rev Genet. 2000;34:623-652.
【非特許文献22】Titorenko VI, Smith JJ, Szilard RK, Rachubinski RA. Pex20p of the yeast Yarrowia lipolytica is required for the oligomerization of thiolase in the cytosol and for its targeting to the peroxisome. J Cell Biol. 1998 Jul 27;142(2):403-20.
【非特許文献23】Glover JR, Andrews DW, Rachubinski RA. Saccharomyces cerevisiae peroxisomal thiolase is imported as a dimer. Proc Natl Acad Sci U. S. A. 1994 Oct 25;91(22):10541-5.
【非特許文献24】Leiper JM, Oatey PB, Danpure CJ. Inhibition of alanine:glyoxylate aminotransferase 1 dimerization is a prerequisite for its peroxisome-to-mitochondrion mistargeting in primary hyperoxaluria type 1. J Cell Biol. 1996 Nov;135(4):939-51.
【非特許文献25】Bellion E, Goodman JM. Proton ionophores prevent assembly of a peroxisomal protein. Cell. 1987 Jan 16;48(1):165-73.
【非特許文献26】Waterham HR, Russell KA, Vries Y, Cregg JM. Peroxisomal targeting, import, and assembly of alcohol oxidase in Pichia pastoris. J Cell Biol. 1997 Dec 15;139(6):1419-31.
【非特許文献27】Marzioch M, Erdmann R, Veenhuis M, Kunau WH. PAS7 encodes a novel yeast member of the WD-40 protein family essential for import of 3-oxoacyl-CoA thiolase, a PTS2-containing protein, into peroxisomes. EMBO J. 1994 Oct 17;13(20):4908-18.
【非特許文献28】Albertini M, Rehling P, Erdmann R, Girzalsky W, Kiel JA, Veenhuis M, Kunau WH. Pex14p, a peroxisomal membrane protein binding both receptors of the two PTS-dependent import pathways. Cell. 1997 Apr 4;89(1):83-92.
【非特許文献29】Elgersma Y, Elgersma-Hooisma M, Wenzel T, McCafferyJM, Farquhar MG, Subramani S. A mobile PTS2 receptor for peroxisomal protein import in Pichia pastoris. J Cell Biol. 1998 Feb 23;140(4):807-20.
【非特許文献30】Szilard RK, Titorenko VI, Veenhuis M, Rachubinski RA.Pay32p of the yeast Yarrowia lipolytica is an intraperoxisomal component of the matrix protein translocation machinery. J Cell Biol. 1995 Dec;131(6 Pt 1):1453-69.
【非特許文献31】Zhang JW, Lazarow PB. PEB1 (PAS7) in Saccharomyces cerevisiae encodes a hydrophilic, intra-peroxisomal protein that is a member of the WD repeat family and is essential for the import of thiolase into peroxisomes. J Cell Biol. 1995 Apr;129(1):65-80.
【非特許文献32】Morita M, Kurochkin IV, Motojima K, Goto S, TakanoT, Okamura S, Sato R, Yokota S, Imanaka T. Insulin-degrading enzyme exists inside of rat liver peroxisomes and degrades oxidized proteins. Cell Struct Funct. 2000 Oct;25(5):309-15.
【非特許文献33】Kurochkin IV, RIKEN GER Group, GSL Members, Konagaya A, Nagashima T, Schönbach C. Identification of Potential Peroxisomal Proteins in Mouse. Genome Res. 2003 Jun; 13(6b):1560. [http://www.genome.org/cgi/content/full/13/6b/1560]
【非特許文献34】Kurochkin IV, Schönbach C. Konagaya A. In Silico Prediction of Peroxisomal Proteins in Mouse. Genome Informatics 2003 Dec; 14:539-40 [http://www.jsbi.org/journal/GIW03/GIW03P068.pdf]
【非特許文献35】Kurochkin IV, Nagashima T, Konagaya A, Schönbach C.Sequence-based Discovery of the Human and Rodent Peroxisomal Proteome. Appl Bioinformatics; in press June 2005.
【非特許文献36】UCSC Genome browser; mouse Tysnd1 http://genome.ucsc.edu/cgi-bin/hgTracks?db=mm5&position=chr10:61457382-61464634&hgsid=39784308
【非特許文献37】UCSC Genome browser; human TYSND1 http://genome.ucsc.edu/cgi-bin/hgTracks?db=hg17&position=chr10:71568974-71575956&hgsid=39784308
【非特許文献38】Gene Entrez mouse Tysnd1 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene&cmd=Retrieve&dopt=Graphics&list_uids=71767
【非特許文献39】Gene Entrez human TYSND1 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene&cmd=Retrieve&dopt=Graphics&list_uids=219743
【非特許文献40】Swissprot/TrEMBL mouse Tysnd1 and human TYSND1 http://kr.expasy.org/cgi-bin/sprot-search-de?TYSND1
【非特許文献41】InterPro domain: Peptidase, trypsin-like serine and cysteine proteases http://www.ebi.ac.uk/interpro/ISpy?mode=single&ac=[SwissProt/TrEMBL accession number]
【非特許文献42】UCSC Genome browser; gene expression for mouse Tysnd1 and neighbors http://genome.ucsc.edu/cgi-bin/hgNear?hgsid=39784308&org=Mouse&db=mm5&near_search=AK005069&submit=Go%21&near.order=expGnfAtlas2&near.count=50
【非特許文献43】Mouse Gene Prediction Database; Search Results for 1300019N10Rik (XM_125636.1) and top 100 correlated genes; http://mgpd.med.utoronto.ca/profile.php?nameofgene=XM_125636.1
【非特許文献44】UCSC Genome browser; gene expression for human TYSND1 and neighbors http://genome.ucsc.edu/cgi-bin/hgNear?hgsid=73232388&org=Human&db=hg18&near_search=TYSND1&submit=Go%21&near_order=expGnfAtlas2&near.count=50
【非特許文献45】Array Prospector gene expression for human TYSND1 (SwissProt/TrEBML accession Q96AR5) http://string.embl-heidelberg.de:8080/prophecies_cgi/new_prophecies/new_prophecies.pl?genename=Q96AR5&species=9606&colorcode=1&maxgenes=10&submit=SUBMIT
【発明の開示】
【発明が解決しようとする課題】
【0011】
発明の概要
【0012】
Tysnd1またはトリプシンドメイン含有1は最初に、仮説上のタンパク質として報告された(Kawai J, et al., RIKEN Genome Exploration Research Group Phase II Team and the FANTOM Consortium. Functional annotation of a full-length mouse cDNA collection. Nature. 2001 Feb 8;409(6821):685-90; Okazaki Y et al., FANTOM Consortium; RIKEN Genome Exploration Research Group Phase, I & II Team. Analysis of the mouse transcriptome based on functional annotation of 60,770 full-length cDNAs. Nature. 2002 Dec 5;420(6915):563-73.)。公開データベースで利用可能なタンパク質レベルに対する機能情報(Gene Entrez(Gene Entrez human TYSND1 http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene&cmd=Retrieve&dopt=Graphics&list_uids=219743)、SwissProt/TrEMBL(Swissprot/TrEMBL mouse Tysnd1 and human TYSND1 http://kr.expasy.org/cgi-bin/sprot-search-de?TYSND1))はコンピュータにより推測され、タンパク質モチーフおよび配列相同性検索に基づいている。本明細書において初めて、Tysnd1タンパク質機能のインビトロにおける実証について記載される。その機能としては、1)ペルオキシソームへの局在化、2)ペルオキシソームプロテアーゼ機能、3)PTS2含有ペルオキシソーム酵素のタンパク質分解開裂、4)PTS1含有ペルオキシソーム酵素のタンパク質分解開裂、5)前駆型Tysnd1の成熟Tysnd1への翻訳後のプロセッシングが挙げられる。機能2)〜4)は、Tysnd1の基質として、ラット3−ケトアシル−CoAチオラーゼB、ペルオキシソームの前駆体(RGD1562373_predicted)、ラットアシル補酵素Aオキシダーゼ1(Acox1)、マウスステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−b)デヒドロゲナーゼ4(Hsd17b4)を用いて証明された。
【0013】
1つの態様において、本発明は、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む、細胞におけるペルオキシソームの生成および機能に関連する障害を処置するための医薬組成物を提供する。
さらなる態様において、本発明は、細胞内におけるペルオキシソームの生成および/または機能に関連する疾患または症候群に診断された被検体(例えば、ヒト患者)を処置する方法であり、該被検体にプロテアーゼ活性を有しそしてペルオキシソーム酵素をプロセッシングするペプチドを投与する工程を含む方法を提供する。
【課題を解決するための手段】
【0014】
ポリペプチドは好ましくは、Tysnd1によってコードされる。ペルオキシソーム酵素は、3−オキソアシル補酵素Bチオラーゼ(以降、Acaa1と呼ぶ)、アシル補酵素Aオキシダーゼ1(Acox1)、パルミトイルステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ 4(Hsd17b4)からなる群より選択される少なくとも1つであってよい。疾患は、代謝性疾患の中でも、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、メバロン酸尿症、および男性不妊症からなる群より選択されてもよい。
【0015】
別の態様において、本発明は、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む、細胞におけるペルオキシソームの生成および機能に関連する障害を診断する医薬組成物を提供する。
【0016】
ポリペプチドは好ましくは、Tysnd1によってコードされる。ペルオキシソーム酵素は、3−オキソアシル補酵素Bチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、パルミトイルステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ 4(Hsd17b4)からなる群より選択される少なくとも1つであってよい。疾患は、代謝性疾患の中でもレフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、メバロン酸尿症および男性不妊症からなる群より選択されてもよい。
発明の詳細な説明
【発明を実施するための最良の形態】
【0017】
定義
本発明の実施は、別途指示しない限り、当分野の技術範囲内である、分子生物学の従来技法を利用する。そのような技法は、文献に十分説明されている。
【0018】
以下の定義が、以下に文書化した説明で使用される具体的な用語について与えられている。
【0019】
「ペルオキシソーム」という用語は、本明細書で使用されるとき、真核細胞に見出される酵素を保持した、膜結合の小胞を指す。ペルオキシソームは時に、マイクロボディと呼ばれる。ペルオキシソームは、そのような酵素を残りの細胞部分から隔離する役割を果たし、細胞代謝を組織化する。
【0020】
ポリペプチドを「プロセッシングする」という用語は、本明細書で使用されるとき、成熟タンパク質またはペプチドを形成するための、細胞における合成されたポリペプチドの開裂(特に加水分解開裂)を指す。ポリペプチドは、他の基の付加などによってアミノ酸残基を修飾されるか、その活性な三次元コンホーメーションに折畳まれるか、あるいは分解のために標的化されてもよい。
【0021】
「Tysnd1」という用語は、第10染色体(マウス)に位置するゲノムDNA配列およびその遺伝子産物を指す。そのヒトオルソログは第10染色体に対するシンテニック位置に位置する。Tysnd1の特色付けは上の表1に示されている。
本発明はここで、本発明の実施形態が示されている添付図を参照して以下でより十分に説明される。
【0022】
[実施形態]
<材料および方法>
<発現ベクターの構築に使用するプライマー>
プライマーB1およびB2は、GenBankアクセッションAK005069のTysnd1ヌクレオチド配列に基づいている。B1およびB2は、構築プラスミドベクター、pEGFP−Tysnd1に使用した。
<B1>(フォワード)5’−GGATCCATGGGGCGGCAATGGGGAC−3’(配列番号1)は、BamHI部位(GGATCC)およびTysnd1配列位置59〜77を含有する。
<B2>(リバース)5’−GGATCCTCAGAGCTTGCTCCGTGGGAC−3’(配列番号2)は、BamHI部位(GGATCC)およびTysnd1配列位置1745〜1765を含有する。
プライマーAX1およびAX2は、GenBankアクセッションBC085743のラットAcox1ヌクレオチド配列に基づいている。
【0023】
<AX1>(フォワード)
(化1)
5’-ACCATGGGCTACCCTTACGACGTGCCTGACTACGCCAACCCCGACCTGCGCAAGGAGC-3’ (配列番号3)
は、ATG開始コドンを包囲する合成Kozak配列(太字)を含有する。HA−tagペプチドTyrAspValProAspTyrAlaは、下線の配列によってコードされる。プライマーの位置37〜58は、BC085743のAcox1配列位置94〜115に相当する。
<AX2>(リバース)5’−TCAAAGCTTGGACTGCAGGGGCTTC−3’(配列番号4)は、Acox1配列位置2052〜2076を含有する。停止コドンが含まれていた。
プライマーS5およびS2は、GenBankアクセッションBC034613のマウスScp2ヌクレオチド配列に基づいている。
【0024】
<S5>(フォワード)
(化2)
5’-ACCATGGGCTACCCTTACGACGTGCCTGACTACGCCCCTTCTGTCGCTTTGAA-3’
ATCTCC (配列番号5)
は、ATG開始コドンを包囲する合成Kozak配列(太字)を含有する。HA−tagペプチドTyrAspValProAspTyrAlaは、下線の配列によってコードされる。プライマーの位置37〜59は、Scp2配列位置25〜57に相当する。
<S2>(リバース)5’−CTCCTCACAGCTTAGCTTTGC−3’(配列番号6)は、Scp2配列位置1649〜1669に相当する。停止コドンが含まれていた。
プライマーHsd17b4−FwdおよびHsd17b4−Revは、GenBankアクセッションAK004866のマウスHsd17b4ヌクレオチド配列に基づいている。
【0025】
<Hsd17b4−Fwd>(フォワード)
5’−ACCATGGGCTACCCTTACGACGTGCCTGACTACGCCGCTTCGCCGCTGAGGTTCGAC−3’(配列番号17)
は、ATG開始コドンを包囲する合成Kozak配列(ACCATGG)を含有する。HA−tagペプチドTyrAspValProAspTyrAlaは、下線の配列によってコードされる。プライマーの位置37〜57は、Hsd17b4配列位置155〜175に相当する。
<Hsd17b4−Rev>(リバース)
5’−TCAGAGCTTGGCATAGTCTTTAAGAAT−3’(配列番号18)は、Hsd17b4−Rev配列位置2359〜2333に相当する。停止コドンが含まれていた。
Flagタグ付きTysnd1のプライマーは、GenBankアクセッションAK005069の配列に基づいている。
【0026】
<FN>(フォワード)
5’−CCACCATGGACTACAAAGACGATGACGACAAGGGGCGGCAATGGGGAC−3’(配列番号7)は、Kozak配列(CCACCATGG)、FLAGエピトープ(GACTACAAAGACGATGACGACAAG、配列番号8)およびTysnd1配列位置62〜77(GGGCGGCAATGGGGAC、配列番号9)を含む。
<FC1>(フォワード)5’−CCACCATGGGGCGGCAATGGGGAC−3’(配列番号10)は、Kozak配列(CCACCATGG)およびTysnd1配列位置62〜77(GGGCGGCAATGGGGAC、配列番号9)を含む。
<FC2>(リバース)5’−TCAGAGCTTGCTCCGTGGGACCTTGTCGTCATCGTCTTTGTAGTCTTCGGACAGGGGCCGCTGCAG−3’(配列番号11)は、Tysnd1配列位置1724〜1744、FLAGエピトープ(CTTGTCGTCATCGTCTTTGTAGTC、配列番号12)およびヌクレオチド配列位置1745〜1765に相当するTysnd1のC末端からの6個のアミノ酸(ValProArgSerLysLeu、配列番号13)を含む。
【0027】
<細胞およびトランスフェクション>
COS−7(SV40によって形質転換されたサル線維芽細胞)、293FT(SV40の大型T抗原を発現するヒト293細胞の急速成長変異株)およびチャイニーズハムスター卵巣CHO−K1細胞を、10%(v/v)ウシ胎仔血清(Invitrogen)を添加したダルベッコの修飾イーグル培地中で維持し、LipofectAMINE 2000試薬(Invitrogen)をメーカーの取扱説明書に従って使用して一時トランスフェクションした。
【0028】
<緑色蛍光タンパク質(GFP)融合タンパク質の発現ベクターの構築>
全長マウスTysnd1をコードするDNA断片は、プライマーセットB1およびB2を用いたPCRによって、クローニングしたcDNA 1300019N10(GenBankアクセッションAK005069)をテンプレートとして使用して、増幅した。PCR産物は、pGEM−T Easyベクター(Promega)内にサブクローニングした。BamHIによる消化の後、得られた断片は、pEGFP−C1のBamHI部位に挿入した(EGFPは増強GFPである; BD Biosciences Clontech)。得られたベクターは、pEGFP−Tysnd1と名づけた。
【0029】
<エピトープタグ付きラットペルオキシソーム3−オキソアシル補酵素Bチオラーゼ(Acaa1)、ラットアシル補酵素Aオキシダーゼ1(Acox1)、マウスパルミトイルステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)の発現>
以下のベクターを利用した。即ち、pcDNA3.1Zeo中のラットタイプBプレ−3−ケトアシル−CoAチオラーゼ−HA(HA=赤血球凝集素)(九州大学のY.Fujikiより寄贈)、pcDNA3.1/TOPO−HA−Acox1、pcDNA3.1/TOPO−HA−Scp2である。ラットAcox1のN末端へのHAタグ付けは、HAエピトープを含有するフォワードプライマーAX1および停止コドンを含有するリバースプライマーAX2を使用して、PCRをベースとする技法によって行った。同様にHAタグをPCRによって、プライマーS5およびS2ならびにHsd17b4−FwdおよびHsd17b4−Revを使用して、それぞれScp2およびHsd17b4内に導入した。増幅DNA産物は、pcDNA3.1/V5−His−TOPO(Invitrogen)内にクローニングした。プラスミド構築物は全て挿入の方向を点検し、トランスフェクションに使用した。
【0030】
<エピトープタグ付きTysnd1の発現>
培養細胞中でのマウスTysnd1の発現では、2つのベクターを構築した。pcDNA3.1/TOPO−FLAG−Tysnd1は、Tysnd1のN末端にFLAGエピトープを含有する。pcDNA3.1/TOPO−Tysnd1−FLAGは、Tysnd1のC末端にFLAGエピトープを含有する。pcDNA3.1/TOPO−FLAG−Tysnd1を作成するために、FLAGエピトープを含有するフォワードプライマーFN1および停止コドンを含有するリバースプライマーB2を用いて、全長Tysnd1のPCR増幅を実施した。pcDNA3.1/TOPO−Tysnd1−FLAGを構築するために、FLAGエピトープをアミノ酸562と563との間に導入し、これによってTysnd1の未変性PTS1含有C末端を保存した。PCR増幅は、フォワードプライマーFC1およびリバースプライマーFC2を用いて実施した。後者はFLAG配列に加えて、アミノ酸Val−Pro−Arg−Ser−Lys−Leu(位置563〜568)をコードするC末端Tysnd1配列および停止コドンを含有する。構築物は、上述のようにトランスフェクション実験に使用した。
【0031】
<GFP融合タンパク質の局在化>
CHO−K1細胞は、ガラスカバースリップに播種して、発現ベクターpEGFP−Tysnd1およびペルオキシソーム標的化DsRed2レポータータンパク質(BD Biosciences Clontech)をコードしたpDsRed2−Peroxi(Discosoma spからの赤色蛍光タンパク質の変異体2)によって一時トランスフェクションした。EGFP融合タンパク質およびDsRed2−Peroxiタンパク質の生細胞顕微鏡観察は、トランスフェクションの48時間後に、63×対物レンズを用いたレーザー走査共焦点顕微鏡TCS SP2(Leica)で実施した。488nmのアルゴンレーザーを励起に使用した。GFPおよびDsRed2によって放出された蛍光シグナルは、535nmバンドパスフィルタおよび570nmロングパスフィルタをそれぞれ使用して検出した。
【0032】
<免疫ブロッティング分析>
細胞は、上述のようにトランスフェクションした。48時間後、細胞を緩衝液A(Roche Diagnostics GmbHからのComplete(商標)プロテアーゼインヒビターカクテルを追加した、20mMのTris−HCl(pH7.5)、120mMのNaCl、1%のTriton X−100)に溶解させて、12,000×gにて15分間遠心分離にかけた。上澄み画分をSDS−PAGEで分離した。ウェスタンブロット分析を、Hybond−Pポリビニリデンジフルオリド膜(Amersham Biosciences,Inc.)上に電気泳動によって移動させたサンプルを用いて、HA.11モノクローナル抗体(Covance)または抗FLAG(商標)M2抗体(Sigma)および二次抗体、ホースラディッシュペルオキシダーゼに結合したヒツジ抗マウスIgG抗体(Amersham Biosciences,Inc.)を使用して、実施した。抗原−抗体複合体は、ECL plusウェスタンブロッティング検出試薬(Amersham Biosciences,Inc.)を用いて可視化した。タンパク質バンドの見かけの分子量は、予備染色したPrecision Plus Protein(商標)Standards(Bio−Rad)の移動度と比較することによって推定した。
【0033】
<ペルオキシソームの精製>
ペルオキシソームは、以前に記載された3つのプロトコル(Van Veldhoven PP, Baumgart E, Mannaerts GP. Iodixanol (Optiprep), an improved density gradient medium for the iso-osmotic isolation of rat liver peroxisomes. Anal Biochem. 1996 May 15;237(1):17-23; McClelland GB, Khanna S, Gonzalez GF, Butz CE, Brooks GA. Peroxisomal membrane monocarboxylate transporters: evidence for a redox shuttle system? Biochem Biophys Res Commun. 2003 Apr 25;304(1):130-5; Antonenkov VD, Sormunen RT, Hiltunen JK. The behavior of peroxisomes in vitro: mammalian peroxisomes are osmotically sensitive particles. Am J Physiol Cell Physiol. 2004 Dec;287(6):C1623-35)の組合せおよび改良に基づいた方法によって精製した。手短には、雄のウィスターラット(250〜300g)を一晩絶食させた。ジエチルエーテル吸入により動物を殺処分した後、肝臓を門脈を介して灌流した。肝臓を氷冷ホモジナイゼーション緩衝液(0.16Mスクロース、12%(wt/vol)PEG1500、10mM MOPS、pH7.4、1mM EDTA、1mM EGTA、1mM DTT、0.1%(vol/vol)エタノール)中でハサミを用いて細かく刻み、Potter−Elvehjemホモジナイザー(ガラス上のTeflon)でホモジナイゼーションした。核および重ミトコンドリアは、固定角ローター内で3,000×g、4℃にて10分間の遠心分離によってペレット化した。ペレットを再ホモジナイゼーションして、遠心分離を繰り返した。上澄み(ポスト核画分)を合わせて、20,000×gにて20分間遠心分離して、「軽ミトコンドリアペレット」を生成した。このペレットは、ゆるくはめたDounceホモジナイザーを使用して、ホモジナイゼーション緩衝液に再懸濁させた。体積は15ml/開始時の肝臓重量10gに調節して、Optiprep(60%(wt/vol)イオジキサノール、Axis−Shield PoC AS)5体積および希釈媒体(0.16Mのスクロース、12%(wt/vol)のPEG 1500、60mMのMOPS(pH7.4)、6mMのEDTA、6mMのDTT、0.6%(vol/vol)のエタノール)1体積より成る勾配溶液の等体積と混合した。この混合物を近垂直ローター(Beckman Type NVT 65)で180,000×g、4℃にて3.5時間、低加速および減速モードを使用して遠心分離した。遠心分離の後、画分0.5mlを上部から針および注射器によって取り出した。単離媒体を除去するために、画分材料を17,000×gにて20分間沈殿させて、0.1%のTriton X−110および1mMのDTTを含有するPBS緩衝液中に再懸濁させた。
【0034】
<マーカー酵素およびタンパク質含有量>
ペルオキシソーム、ミトコンドリアおよびリソソームのOptiprep勾配画分中での局在を判定するために、カタラーゼ(Johansson LH, Borg LA. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal Biochem. 1988 Oct;174(1):331-6.)、スクシナートデヒドロゲナーゼ(Pennington RJ. Biochemistry of dystrophic muscle. Mitochondrial succinate-tetrazolium reductase and adenosine triphosphatase. Biochem J. 1961 Sep;80:649-54.)およびβ−ガラクトシダーゼ(b−Galアッセイキット、Invitrogen)の活性がそれぞれ測定された。タンパク質含有量は、Bio−Rad DCタンパク質アッセイ試薬キット(Bio−Rad)を使用して測定した。
【0035】
<マウスTysnd1に対するポリクローナル抗体>
ウサギポリクローナル抗体を、Tysnd1のアミノ酸501〜515に相当するポリペプチドSNTRDNNTGATYPHL(配列番号14)と結合したキーホールリンペットヘモシアニンに対して産生し、SCRUM Inc.(東京)によりアフィニティ精製した。ウェスタンブロット分析には、抗体を2μg/mlの濃度で使用した。抗体予備吸収アッセイでは合成ペプチドCSNTRDNNTGATYPHL(配列番号15)をMe2SOに2mg/mlの濃度で溶解させ、次に、ウェスタンブロッティング(40μgペプチド/5μg抗体;約700倍の過剰モル数のペプチド)の前に、一次抗体溶液と混合した。
【0036】
<マウス(ラット)Acaa1に対するポリクローナル抗体>
ウサギポリクローナル抗体を、マウスAcaa1のアミノ酸259〜274に相当するポリペプチドKLKPAFKDGGSTTAGN(配列番号19)と結合したキーホールリンペットヘモシアニンに対して産生し、SCRUM Inc.(東京)によりアフィニティ精製した。ウェスタンブロット分析には、抗体を2μg/mlの濃度で使用した。
【0037】
<低分子干渉RNA(siRNA)>
ヒトTysnd1の発現を下方制御するためのRNA媒介干渉は、Qiagen K.K.から購入した低分子干渉siRNA二本鎖、Hs_Tysnd1_1_HP si RNA (siRNA Tysnd1_1) および Hs_Tysnd1_2_HP si RNA (siRNA Tysnd1_2)を使用して、行われた。siRNA Tysnd1_1は、配列CAG CAG AAA CCT TGC TCT GAA(配列番号20)を標的とし、siRNA Tysnd1_2は、配列CCC GCT GAG CAC TTC CAT GAA(配列番号21)を標的とした。対照siRNA (Qiagen K.K.)は、いかなる既知の哺乳類遺伝子配列に対しても相同性を持たない配列AAT TCT CCG AAC GTG TCA CGT(配列番号22)を標的とした。293FT細胞を、12ウェルプレートで培養した。siRNA(1.2mg)を、0.8mgのpcDNA3.1/TOPO−HA−Acaa1、pcDNA3.1/TOPO−HA−Scp2またはpcDNA3.1/TOPO−HA−Hsd17b4のいずれかとともに同時トランスフェクションした。トランスフェクションを、80%−90%コンフルエントな状態の細胞に、リポフェクトアミン2000(Invitrogen Japan K.K.)を使用して行った。各siRNAサンプルについて、二重のトランスフェクションを行った。トランスフェクション開始後54時間に、細胞を150μlのSDS−PAGEサンプル緩衝液中で溶解した。溶解物を超音波で分解し、95℃で5分間インキュベートし、そしてHA.11モノクローナル抗体(Covance)を使用してウェスタンブロッティングによって分析した。
【0038】
<COS−7細胞からの組み換えTysnd1の精製>
COS−7細胞を、リポフェクトアミン2000トランスフェクション試薬(Invitrogen Japan K.K.)を使用して、cDNA3.1/TOPO−Tysnd1−FLAGでトランスフェクションした。トランスフェクション後48時間に、細胞をPBSで洗い、そして振動下4℃で30分間緩衝液Aを用いて溶解した。細胞溶解物を15,000×gで15分間遠心分離し、細胞残渣の溶解物を取り除いた。この溶解物を、製造業者のプロトコル(Sigma Aldrich Japan K.K.)により、100μlの抗FLAG M2アフィニティービーズを使用して、4℃下で90分間インキュベートした。このビーズを、緩衝液Aで4回洗い、次に緩衝液B(50mM HEPES、pH8.0、115mM NaCl)で4回洗った。このビーズをカラム中に移し、結合していたタンパク質を500μg/mlの3×FLAGペプチドの緩衝液B溶液300μlで溶出した。
【0039】
<組み換えタンパク質のクローニング、発現、および精製>
ラットAcox1、ラットAcaa1、およびマウスScp2のコード配列を、KODポリメラーゼ(TOYOBO CO.LTD)を使用して、PCRで増幅した。結果として得られたAcox1およびScp2のPCR産物を、pTrcHis2−TOPO(Invitrogen Japan K.K.)にクローニングし、C末端にmyc−6xHis タグを有する組み換えタンパク質のための発現ベクターを作った。Acaa1のPCR産物を、pQE−80L発現ベクター(Qiagen K.K.)中にクローニングし、N末端に6xHisのタグを有するAcaa1の発現構築物を作った。Acox1およびScp2の発現ベクターで、Escherichia coli Top10細胞(Invitrogen Japan K.K.)を形質転換した。タンパク質発現の誘導は、製造業者の使用説明書に従い、行った。組換え体Acox1およびScp2を、BD TALON 樹脂(BD Biosciences)を用い、大腸菌の抽出物の可溶性画分から精製した。Acaa1の発現ベクターで、Escherichia coli BL21 Star (DE3)細胞を形質転換した。組換え体Acaa1は、不溶性画分に見られたが、その画分から6Mグアニジン−HClを用いて可溶化された。精製は、BD TALON樹脂を用いて、変性条件下で行われた。精製組換え体Acaa1のリフォールディングを、0.5mM DTTを含有する緩衝液Bでの透析によって行った。組換え体タンパク質は、DC Protein Assay (Bio−Rad Laboratories、Inc)および280nmでの分光光度法を用いて定量した。
【0040】
<インビトロTysnd1プロセッシング活性のアッセイ>
8μlの精製組換え体Tysnd1を、6μlの組換え体Acox1、Scp2、またはAcaa1の50mM Hepes、pH8.0、115mM NaCl、0.2mM DTT溶液と、37℃でインキュベートした。反応を、Laemmliサンプル緩衝液の添加および95℃下での5分間の加熱によって停止した。反応生成物を4%−20%SDS−PAGEによって分離し、Hybond−Pメンブレン上にブロットした。組換え体Acox1およびScp2の開裂生成物を、抗c−Mycモノクローナル抗体(Nacalai Tesque、Inc)を用い、検出した。Acaa1のプロセッシング生成物を、抗Acaa1抗体によるか、または、0.05% Amido Blackを用いたHybond−Pメンブレンの染色によるかして可視化した。
【0041】
<プロセッシングを受けたAcaa1のN末端配列決定>
組換え体Acaa1を、精製Tysnd1と、上記したようにしてインキュベートした。プロセッシングされたAcaa1を、前駆体型から4%−20%SDS−PAGEによって分離し、Hybond−Pメンブレン上にブロットし、そして0.05%Amido Blackを用い染色した。プロセッシングされた41kDaのバンドを、メンブレンから切り取り、そして6サイクルのEdman分解分析にかけた。配列決定分析は、A.Uroshibata(理化学研究所、ゲノム科学総合研究センター、タンパク質基盤研究グループ)によってなされた。
【0042】
<プロテアーゼ阻害活性>
阻害活性のアッセイのために、精製組換え体Tysnd1を、様々なプロテアーゼ阻害剤と30分間25℃で、プレインキュベートした。基質タンパク質の添加の後、インキュベーションを37℃で12時間続行した。以下のプロテアーゼ阻害剤を使用した(最終濃度):ペプスタチンA(1μM)、EGTA(2mM)、EDTA(2mM)、1,10−フェナントロリン(1mM)、1,7−フェナントロリン(1mM)、ベンズアミジン(1mM)、AEBSF(1mM)、アプロチニン(4μg/ml)、ロイペプチン(10μM)、E64(20μM)、NEM(1mM)。プロセッシング生成物を、上記のようにしてアッセイした。
【0043】
<ベザフィブレート処置および細胞下分画>
6頭の7週齢C57BL/6Jオスマウスを、実験に使用した。対照のグループ(n=3)を、標準飼料で飼育し、一方実験グループ(n=3)を、0.5%(v/w)ベザフィブレートを含有する標準飼料で飼育した。細胞下分画では、同じグループに属する3頭の動物の肝臓を混合し、そして上記したようにしてホモジナイゼーションした。ポスト核(PN)画分および軽ミトコンドリア(LM)画分の調製を、「ペルオキシソームの精製」の項で記載したようにして行った。ポストミトコンドリア画分(PM)は、PN画分の20,000×gで20分間の遠心分離の後の上清を示す。
<結果>
(実施例1)
【0044】
<Tysnd1はペルオキシソームに対して標的化される>
Tysnd1はPTS1シグナルを含有するので、このタンパク質がペルオキシソームに対して標的化されているかどうかが調べられた。GFP−Tysnd1融合構築物を、DsRed2−Peroxiをコードするプラスミドと同時トランスフェクションさせた。Tysnd1の全長cDNA配列をGFPのC末端に付加して、融合タンパク質対照のC末端にてPTS1配列を保存した。対照はPTS1シグナルと融合した蛍光タンパク質であり、ペルオキシソームに局在する。GFP蛍光は、多数のスポット様構造物中で観察された(図1A)が、CHO−K1細胞の他の部分では観察されなかった。同じ切片内のGFP−Tysnd1融合タンパク質陽性顆粒は全て、pDsRed2−Peroxiタンパク質(赤色)について陽性であった(図1Bおよび図1C)。この同時局在は、GFP−Tysnd1がペルオキシソームに対して標的化されていたという強力な証拠となる。
(実施例2)
【0045】
<PTS2含有ペルオキシソーム酵素の細胞ベースでの分解>
ペルオキシソームタンパク質の一部は、そのN末端領域に、各前駆体がその最終目的地に到達すると切断されるPTS2ペルオキシソーム標的化配列を含有する(Osumi T, Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T. Amino-terminal presequence of the precursor of peroxisomal 3-ketoacyl-CoA thiolase is a cleavable signal peptide for peroxisomal targeting. Biochem Biophys Res Commun. 1991 Dec 31;181(3):947-54;.Swinkels BW, Gould SJ, Bodnar AG, Rachubinski RA, Subramani S. A novel, cleavable peroxisomal targeting signal at the amino-terminus of the rat 3-ketoacyl-CoA thiolase. EMBO J. 1991 Nov;10(11):3255-62;.Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T, Osumi T. Characterization of the signal peptide at the amino terminus of the rat peroxisomal 3-ketoacyl-CoA thiolase precursor. J Biol Chem. 1994 Feb 25;269(8):6001-10.)。PTS2シグナル配列を切断させるタンパク質分解酵素は、まだ同定されていない。潜在的候補は、既知のペルオキシソームプロテアーゼインスリン分解酵素(Ide)およびペルオキシソーム型Lonプロテアーゼおよび本研究で記載した新規なペルオキシソームプロテアーゼTysnd1である。3つのプロテアーゼうちで、Tysnd1が最も有望な候補である。Ideは大型のタンパク質には作用せず、アミノ酸50個未満の長さのペプチドを代わりに開裂させる一方で(Duckworth, W. C., Bennett, R. G. & Hamel, F. G. (1998) Insulin degradation: progress and potential. Endocr Rev. 1998 Oct;19(5):608-24.;Kurochkin, I. V. (2001) Insulin-degrading enzyme: embarking on amyloid destruction. Trends Biochem Sci. 2001 Jul;26(7):421-5)、ATP依存性Lonプロテアーゼは、変性タンパク質を分解するシャペロンとして機能し得る(Kikuchi M, Hatano N, Yokota S, Shimozawa N, Imanaka T, Taniguchi H. Proteomic analysis of rat liver peroxisome: presence of peroxisome-specific isozyme of Lon protease. J Biol Chem. 2004 Jan 2;279(1):421-8)。
【0046】
PTS2含有シグナル配列の除去でのTysnd1の役割を究明するために、ペルオキシソーム3−オキソアシル補酵素Aチオラーゼ(Acaa1)のプロセッシングが分析された。Acaa1は、残基36個のN末端除去可能配列を持つ大型前駆体として合成される(Osumi T, Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T. Amino-terminal presequence of the precursor of peroxisomal 3-ketoacyl-CoA thiolase is a cleavable signal peptide for peroxisomal targeting. Biochem Biophys Res Commun. 1991 Dec 31;181(3):947-54;.Swinkels BW, Gould SJ, Bodnar AG, Rachubinski RA, Subramani S. A novel, cleavable peroxisomal targeting signal at the amino-terminus of the rat 3-ketoacyl-CoA thiolase. EMBO J. 1991 Nov;10(11):3255-62.;Tsukamoto T, Hata S, Yokota S, Miura S, Fujiki Y, Hijikata M, Miyazawa S, Hashimoto T, Osumi T. Characterization of the signal peptide at the amino terminus of the rat peroxisomal 3-ketoacyl-CoA thiolase precursor. J Biol Chem. 1994 Feb 25;269(8):6001-10.)。C末端にHAタグが付いたAcaa1のみがCHO細胞において過剰発現された場合、主に44kDaのプロセッシングされていない前駆体型が検出された。HA−Acaa1は少量のみが成熟41kDa型に変換された(図2、レーン1)。増加する量のpcDNA3.1/TOPO−FLAG−Tysnd1との同時トランスフェクションは、44kDa前駆体の段階的な消失および成熟41kDaタンパク質の並行した増加を引き起こした(図2、レーン2〜4)。同様の結果がCOS−7細胞でも得られた(データは示さず)。結果は、Tysnd1がPTS2を含有ずる先行ペプチドを除去する真のペルオキシソームプロセッシングプロテアーゼであるか、あるいは第二の、まだ同定されていないペルオキシソームプロセッシングプロテアーゼの活性化を通じてAcaa1の成熟を促進することを意味している。
【0047】
したがってTysnd1は、他のPTS2シグナル含有タンパク質全てのプロセッシングに関与し得る。PTS2タンパク質のペルオキシソーム内プロセッシングの生理学的重要性は、まだ理解されていない。ミトコンドリアの場合、前駆体タンパク質の大多数がミトコンドリア標的化のための切断可能アミノ末端延長配列を含有する。ミトコンドリアプロセッシングプロテアーゼによるシグナル配列の除去は、タンパク質折畳みおよびオルガネラ内でのさらなる仕分けのために必要であると考えられる(Gakh O, Cavadini P, Isaya G. Mitochondrial processing peptidases. Biochim Biophys Acta. 2002 Sep 2;1592(1):63-77.)。ミトコンドリアタンパク質とは異なり、ペルオキシソームタンパク質は、事前のアンフォールディングなしに首尾よく移動される(Subramani S, Koller A, Snyder WB. Import of peroxisomal matrix and membrane proteins. Annu Rev Biochem. 2000;69:399-418)。その上、ペルオキシソームマトリクスは単一コンパートメントより成るようであるが、ミトコンドリアスペースは、数個の明確なサブコンパートメントに分割される。近年、Nairらは、PTS2レセプターPex7pがPTS1レセプターPex5と同様に(Dammai V, Subramani S. The human peroxisomal targeting signal receptor, Pex5p, is translocated into the peroxisomal matrix and recycled to the cytosol. Cell. 2001 Apr 20;105(2):187-96)、輸送の「拡張シャトル」(“extended shuttle”)モードに従うことを証明した(Nair DM, Purdue PE, Lazarow PB. Pex7p translocates in and out of peroxisomes in Saccharomyces cerevisiae. J Cell Biol. 2004 Nov 22;167(4):599-604.)。Pex7pは、PTS2タンパク質移入の進行の間にペルオキシソームに入り、サイトゾル中へ再出現して、さらに数回のタンパク質移入を実行する。PTS2タンパク質がシグナル配列を維持した場合、これらのタンパク質はPex7pと共にペルオキシソームに入り、ペルオキシソームから出て、したがってオルガネラへのその蓄積が防止しされ得る。
(実施例3)
【0048】
<PTS1含有ペルオキシソーム酵素のプロセッシング>
PTS2タンパク質に加えて、数個のPTS1ペルオキシソーム酵素もペルオキシソームに移動されたときにプロセッシングを受ける。これらはペルオキシソーム脂肪酸β酸化に関与する鍵酵素である、アシル補酵素Aオキシダーゼ1(Acox1)、ヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)およびステロールキャリアタンパク質2(Scp2)を含む。それゆえ、Tysnd1がラットAcox1、マウスScp2、およびマウスHsd17b4タンパク質を切断できるかどうかが調査された。Acox1は、ポリペプチド成分A(75kDa)、B(53kDa)およびC(22kDa)より成る。それはA2、ABC、およびB2C2を含むヘテロオリゴマーとしても存在する(Tsukamoto T, Yokota S, Fujiki Y. Isolation and characterization of Chinese hamster ovary cell mutants defective in assembly of peroxisomes. J Cell Biol. 1990 Mar;110(3):651-60;Miyazawa S, Hayashi H, Hijikata M, Ishii N, Furuta S, Kagamiyama H, Osumi T, Hashimoto T. Complete nucleotide sequence of cDNA and predicted amino acid sequence of rat acyl-CoA oxidase. J Biol Chem. 1987 Jun 15;262(17):8131-7)。BおよびC断片は、ペルオキシソームへの移動時にAポリペプチドのタンパク質分解変換によって生成される(Miyazawa S, Osumi T, Hashimoto T, Ohno K, Miura S, Fujiki Y. Peroxisome targeting signal of rat liver acyl-coenzyme A oxidase resides at the carboxy terminus. Mol Cell Biol. 1989 Jan;9(1):83-91;Wirtz KW. Phospholipid transfer proteins revisited. Biochem J. 1997 Jun 1;324 (Pt 2):353-60)。分子量59kDaを有するタンパク質、Scp2は、ペルオキシソームにおいて46kDaおよび13kDa断片に変換される(Seedorf U, Ellinghaus P, Roch Nofer J. Sterol carrier protein-2. Biochim Biophys Acta. 2000 Jun 26;1486(1):45-54;Otera H, Nishimura M, Setoguchi K, Mori T, Fujiki Y. Biogenesis of nonspecific lipid transfer protein and sterol carrier protein x: studies using peroxisome assembly-defective pex cell mutants. J Biol Chem. 2001 Jan 26;276(4):2858-64)。ペルオキシソームでは、Scp2はアシルCoAエステルのβ酸化に関与する。その上、Scp2は、α酸化依存性基質の可溶化共タンパク質(solubilizing coprotein)として作用する(Mukherji M, Kershaw NJ, Schofield CJ, Wierzbicki AS, Lloyd MD. Utilization of sterol carrier protein-2 by phytanoyl-CoA 2-hydroxylase in the peroxisomal alpha oxidation of phytanic acid. Chem Biol. 2002 May;9(5):597-605.)。
【0049】
Hsd17b4は、二機能タンパク質であり、2−メチル分岐鎖脂肪酸、C27胆汁酸中間体、超長鎖脂肪酸のペルオキシソームでのβ酸化、およびポリ不飽和脂肪酸の合成に関与する。最近、Hsd17b4活性が、ロイコトリエンの不活性化(Ferdinandusse S, Denis S, Van Roermund CW, Wanders RJ, Dacremont G. Identification of the peroxisomal beta-oxidation enzymes involved in the degradation of long-chain dicarboxylic acids. J Lipid Res. 2004 Jun;45(6):1104-11)および精巣のセルトリ細胞における脂質ホメオスタシスを維持することによって雄性生殖機能(Huyghe S, Schmalbruch H, De Gendt K, Verhoeven G, Guillou F, Van Veldhoven PP, Baes M. Peroxisomal multifunctional protein 2 is essential for lipid homeostasis in Sertoli cells and male fertility in mice. Endocrinology. 2006 Feb 16; [Epub ahead of print as doi:10.1210/en.2005-1571])に必要であることが示された。具体的には、Hsd17b4は、ペルオキシソーム脂肪酸β酸化の第2および第3ステップの、エノイルCoAヒドラターゼ反応および3−ヒドロキシアシル−CoAデヒドロゲナーゼ反応を触媒する(Jiang LL, Kurosawa T, Sato M, Suzuki Y, Hashimoto T. Physiological role of D-3-hydroxyacyl-CoA dehydratase/D-3-hydroxyacyl-CoA dehydrogenase bifunctional protein. J Biochem (Tokyo). 1997 Mar;121(3):506-13)。Hsd17b4(79kDa)はペルオキシソームに到達した後、一箇所で切断されて、35kDaおよび44kDaのサイズを有する別々の酵素成分を生じる。
【0050】
ラットAcox1、マウスScp2およびマウスHsd17b4のタンパク質分解変換に対するTysnd1の効果を評価するために、タンパク質の発現構築物をCOS−7細胞にトランスフェクションした。Acox1、Scp2、およびHsd17b4はそのN末端をHAエピトープでタグ付けして、PTS1シグナルを保存した。ウェスタンブロットでは、Scp2は主として、未プロセッシング酵素型に相当する59kDaタンパク質バンドとして現れた(図3、レーン2)。約48kDaの非常に微弱なバンドは、プロセッシング型に相当する(図3、レーン2)。Tysnd1との同時トランスフェクションは、未プロセッシング59kDa型の著しいレベル低下を、48kDaタンパク質の付随する増加と共に引き起こした(図3、レーン3)。13kDa C末端断片はHAタグを欠いているため、検出されなかった。HA−Acox1を用いてトランスフェクションした細胞からの抽出物の分析は、主要な未プロセッシング75kDa型および48kDaの見かけのサイズを有する酵素のN末端断片の存在を明らかにした(図3、レーン4)。Tysnd1の同時発現は、プロセッシング済48kDaタンパク質の著しい蓄積を引き起こした(図3、レーン5)。同様にHA−Hsd17b4は、Tysnd1と同時発現した場合、プロセッシングを受けN末端の35kDa断片を与えた(図4)。これらのデータは、Tysnd1がペルオキシソーム鍵酵素の細胞プロセッシングに関与していることを明らかに立証するものである。
【0051】
Scp2のタンパク質分解切断は、それぞれ異なる活性を有する2つの断片を生成する。46kDa断片は、分岐鎖脂肪酸チオラーゼとして酵素活性がある(Seedorf U, Brysch P, Engel T, Schrage K, Assmann G. Sterol carrier protein X is peroxisomal 3-oxoacyl coenzyme A thiolase with intrinsic sterol carrier and lipid transfer activity. J Biol Chem. 1994 Aug 19;269(33):21277-83;Antonenkov VD, Van Veldhoven PP, Waelkens E, Mannaerts GP. Substrate specificities of 3-oxoacyl-CoA thiolase A and sterol carrier protein 2/3-oxoacyl-CoA thiolase purified from normal rat liver peroxisomes. Sterol carrier protein 2/3-oxoacyl-CoA thiolase is involved in the metabolism of 2-methyl-branched fatty acids and bile acid intermediates. J Biol Chem. 1997 Oct 10;272(41):26023-31)。非特異的脂質転移タンパク質(nsLTP)としても知られる13kDa断片は、基質、例えば脂肪アシル−CoA誘導体のAcox1への転移において機能し得る(Wouters FS, Bastiaens PI, Wirtz KW, Jovin TM. FRET microscopy demonstrates molecular association of non-specific lipid transfer protein (nsL-TP) with fatty acid oxidation enzymes in peroxisomes. EMBO J. 1998 Dec 15;17(24):7179-89.)。タンパク質(59kDa)の無傷の完全長型は、両方の機能をなお実施できる。したがってプロセッシングの生理学的重要性は明白ではない。Acox1の場合、ペルオキシソーム内プロセッシングの役割は、生成した断片が結合したままであるため、なお一層明らかでない。最近、Woutersら(Wouters FS, Bastiaens PI, Wirtz KW, Jovin TM. FRET microscopy demonstrates molecular association of non-specific lipid transfer protein (nsL-TP) with fatty acid oxidation enzymes in peroxisomes. EMBO J. 1998 Dec 15;17(24):7179-89.)は、nsLTPがAcox1、Acaa1、およびHsd17b4と結合されていることを証明した。これらの結果は、β酸化のペルオキシソーム酵素が機能性複合体内に組織化されていることを示唆する(Wouters FS, Bastiaens PI, Wirtz KW, Jovin TM. FRET microscopy demonstrates molecular association of non-specific lipid transfer protein (nsL-TP) with fatty acid oxidation enzymes in peroxisomes. EMBO J. 1998 Dec 15;17(24):7179-89.)。複合体形成によって、酵素の局所濃度が一時的に増加するため、脂質中間体の酵素間での効率的転移が可能となると考えられる。ペルオキシソーム内プロセッシングは、酵素のコンホメーション変化をもたらし得、これは複合体の形成を向上させると考えられる。Tysnd1は、このプロセスを調節する役割を果たし、したがって向上したペルオキシソーム脂肪酸β酸化を促進し得る。
【0052】
Tysnd1の発現パターンに類似した、55の組織からサンプリングしたマウス遺伝子の同時発現パターン(Mouse Gene Prediction Database; Search Results for 1300019N10Rik (XM_125636.1) and top 100 correlated genes; http://mgpd.med.utoronto.ca/profile.php?nameofgene=XM_125636.1)は、示唆された調節の役割を間接的に支持する。Tysnd1(XM_125636.1)、Acaa1(XM_135249.1)およびScp2(XM_135267.1)転写物は、プロテアーゼ−基質関係から予想されるように、大動脈、褐色脂肪、小腸、肝臓、前立腺および副腎において中〜高レベルで同時発現される。アクセッション番号は、Refseq遺伝子モデルアクセッションであり、参考文献(Mouse Gene Prediction Database; Search Results for 1300019N10Rik (XM_125636.1) and top 100 correlated genes; http://mgpd.med.utoronto.ca/profile.php?nameofgene=XM_125636.1)から引用した。
(実施例4)
【0053】
<Tysnd1の翻訳後プロセッシング>
他のペルオキシソームタンパク質と同様に、Tysnd1はサイトゾル内で合成される。ペルオキシソームへの移入が完了するまで潜在的なTysnd1基質がプロセッシングされないようにするために、タンパク質がペルオキシソームに到達するまでTysnd1のタンパク質分解活性は作動させるべきではない。異なるプロテアーゼが、タンパク質分解活性をサイレンシングさせるのに異なる方法を利用する。大半のプロテアーゼは、不活性前駆体として合成され、したがってタンパク質分解経路の鍵となる事象は、チモーゲンの活性酵素への変換である。
【0054】
推定上のTysnd1プロセッシングを調査するために、タンパク質のN末端またはC末端のどちらかにFLAGエピトープを含有するこの酵素の真核発現ベクターを用いてCOS−7細胞がトランスフェクションされた。C末端へのFLAGエピトープは、アミノ酸562と563との間に導入されて、PTS1シグナルを含有するTysnd1の未変性C末端を保存することに注目すべきである。Tysnd1のN末端でFLAGエピトープをコードするプラスミドを用いたトランスフェクションは、細胞溶解物のウェスタンブロットによって判定されるように、59kDAタンパク質バンドの出現をもたらした。分子量は、cDNA配列の翻訳CDSの計算重量と一致している(図5A、レーン3)。しかしながら約10kDaの追加のバンドも検出された(図5A、レーン3)。この小型タンパク質種は、Tysnd1のN末端の約90個のアミノ酸残基に結合されたFLAGエピトープに相当する(図5B)。他方Tysnd1のC末端でFLAGエピトープをコードするプラスミドを用いて細胞にトランスフェクションした場合、抗体は49kDaの追加のバンドを検出した(図5A、レーン2)。サイズの相違は、10kDa断片がN末端タグ付きTysnd1から分離されることを示し、したがってC末端タグ付きTysnd1タンパク質によって得られた結果を確証している。
【0055】
他のプロテアーゼと同様に、59kDa Tysnd1タンパク質は、触媒的に活性な49kDa型を生成することが予想される不活性前駆体型を示す。9kDa N末端断片の除去は、残基187〜282および311〜536における2つのプロテアーゼ関連ドメインに影響を及ぼさない。結果として、N末端セグメントは、ペルオキシソームに到達するまでTysnd1の活性を抑制する活性化ペプチドであり得る。
【0056】
不活性前駆体の活性Tysnd1への変換は、自己触媒的に、または別の同定されていないプロテアーゼによって、媒介されている可能性がある。活性化のタイミングおよび程度は最終的に、上述のようにペルオキシソーム脂肪酸β酸化の速度を調節し得る。
(実施例5)
【0057】
<マウスTysnd1に対する抗体の特徴づけ>
成熟Tysnd1タンパク質のC末端に近い領域(アミノ酸501〜515)に対するウサギポリクローナル抗体が作成された。抗体の特異性を、E.coli抽出物から精製された組換えマウスTysnd1および一時トランスフェクションされたCOS−7細胞中のN−FLAG−Tysnd1−C産物も認識する能力について試験した。組換えTysnd1のウェスタンブロット分析は、59kDaの予測サイズに相当するシグナルを明らかにした(図6、レーン1)。モックトランスフェクションCOS−7細胞の溶解物では、バンドは検出できなかった(図6、レーン2)。N末端Flagタグ付きTysnd1を発現する細胞は、2つの顕著なバンドを示した(図6、レーン3)。59kDAバンド断片は、アミノ末端領域を含むTysnd1に相当する。49kDA断片は、アミノ末端領域なしのTysnd1タンパク質に相当する(図5を参照)。抗Tysnd1抗体の特異性を試験するために、免疫化に使用された合成ペプチドを用いた競合アッセイが実施された。シグナルは、抗体の予備吸収後に完全に消滅し、高い特異性が立証された(図6、レーン4〜6)。Tysnd1に対する抗体を産生するために使用したペプチドの配列(アミノ酸501〜515)は、ラットTysnd1およびヒトTysnd1で共有され、抗体がこれらの種の組織におけるタンパク質発現を研究するためにも有用であり得ることが示唆される(以下を参照)。
(実施例6)
【0058】
<内在性Tysnd1の局在化およびサイズ>
共焦点顕微鏡法実験はTysnd1のペルオキシソーム局在化を示したが、証拠は人為的条件に基づいている。例えばTysnd1は培養細胞中のTysnd1−GFP産物として過剰発現された。したがってTysnd1のペルオキシソーム局在化を独立して確認することが選択された。ラット肝臓ホモジネートが分画され、抗Tysnd1抗体を使用した免疫ブロッティングによって細胞下画分が分析された。対照マーカー酵素の活性を調べた結果(図7A)は、ペルオキシソームが自己発生性Optiprep勾配の底部付近に位置することを示した(画分18〜20)。ペルオキシソームは上部画分(画分1〜4)に見出された他のオルガネラ(すなわちミトコンドリアおよびリソソーム)からうまく分離され、このサンプルの主なタンパク質含有量を構成する(図7B)。抗Tysnd1抗体を用いた画分のウェスタンブロット分析では、カタラーゼも検出された勾配の領域である、画分18〜20のみで免疫反応性物質が示された(図7C)。これらの結果は、内在性Tysnd1のペルオキシソーム局在化を強力に支持する。ウェスタンブロットによって検出された2つの顕著なバンドは、49kDaおよび27kDaの分子量を有するタンパク質型に相当する(図7C)。興味深いことにCOS−7トランスフェクション細胞では、検出された主なTysnd1種は、無傷のもの(59kDa)および部分的にプロセッシングされたもの(49kDa)に相当するが、27kDa型はほとんど検出できなかった(図5)。これは、Tysnd1のプロセッシング経路を飽和させ得るpcDNA3.1ベクターによって発生された過剰発現の結果であるらしい。
結論として本発明者のデータは、Tysnd1が最終的に少なくとも2つのプロテアーゼ型の形成をもたらす一連のタンパク質分解事象を受け得ることを示唆している。第一の型は、両方のプロテアーゼ関連ドメイン(49kDa)を含有する。第二の型(27kDa)は、C末端プロテアーゼ様ドメイン(308−531)のみを含有する。哺乳類組織における単一翻訳産物からの別々のプロテアーゼドメインの生成は、前例が無いわけではない。最近Calらは、ヒトのポリセラーゼ1(polyserase−I)(Cal S, Quesada V, Garabaya C, Lopez-Otin C. Polyserase-I, a human polyprotease with the ability to generate independent serine protease domains from a single translation product. Proc Natl Acad Sci U S A. 2003 Aug 5;100(16):9185-90)およびポリセラーゼ2(polyserase−2)(Cal S, Quesada V, Llamazares M, Diaz-Perales A, Garabaya C, Lopez-Otin C. Human polyserase-2, a novel enzyme with three tandem serine protease domains in a single polypeptide chain. J Biol Chem. 2005 Jan 21;280(3):1953-61)が、一連のタンパク質分解プロセッシング事象を受けて、3個の独立なセリンプロテアーゼユニットを発生させることを証明した。Tysnd1プロセッシングの複雑な特性は、ペルオキシソームにおける複数の生物学的プロセスの調節を可能にする細胞制御機構を反映しているように思われる。例えばTysnd1の2つの型の、基質特異性およびタンパク質分解効率は異なり得る。
(実施例7)
【0059】
<Tysnd1は、293FT細胞における、Acox1、Scp2、およびHsd17b4のプロセッシングに関与する主要な酵素である>
Tysnd1は、同時発現研究において数個のペルオキシソーム酵素の切断を促進することが示されたが、この過程に他のプロテアーゼも関与し得る可能性が残っている。そこで低分子干渉RNA(siRNA)が利用され、ペルオキシソーム酵素の細胞プロセッシングにおけるTysnd1の相対的貢献度が調査された。siRNAを使う戦略は、細胞における特定のタンパク質の選択的ノックダウンを達成する、強力で新しい手段を提供する(Sandy P, Ventura A, Jacks T. Mammalian RNAi: a practical guide. Biotechniques. 2005 Aug;39(2):215-24.)。試験された多数のマウスおよびヒト細胞系の中で、293FT細胞のみが、トランスフェクション研究において、完全長HA−Acox1、完全長HA−Scp2、および完全長HA−Hsd17b4から生成された検出可能な量の断片を示した。観察されたプロセッシング生成物は、293FT細胞に存在する内在性のペルオキシソームプロセッシングプロテアーゼによって、生成された。本明細書において、ヒトのTYSND1特異的siRNAのTysnd1_1およびTysnd1_2が、Acox1、Scp2、およびHsd17b4のN末端HAタグ含有断片の生成を本質的に阻害することが証明された(図8)。対照的に、対照の非サイレンシングsiRNAは、効果がなかった(図8)。siRNAが標的mRNAを完全にはノックダウンし得ないこと、およびTysnd1が長い半減寿命を有している可能性があることを考慮すると、観察された効果によって、Tysnd1が、Acox1、Scp2、Hsd17b4、およびおそらく他のペルオキシソーム酵素のプロセッシングにおいて、唯一のとまでは行かなくとも主要な役割を果たしていることが、立証される。
(実施例8)
【0060】
<インビトロの研究が、Tysnd1によって直接ペルオキシソーム酵素がプロセッシングされることを立証する>
Tysnd1が、数個のペルオキシソーム酵素のプロセッシングに関与していることが、細胞ベースのアッセイで示された。Tysnd1は、その標的を直接にか、または別の真のペルオキシソームプロセッシングプロテアーゼを活性化することによって間接的に、プロセッシングし得る。Tysnd1が直接にペルオキシソーム酵素をプロセッシングするか評価するために、組換え体Tysnd1−FLAGタンパク質を、COS−7細胞で発現させ、そしてアフィニティークロマトグラフィーによって精製した。Tysnd1−FLAGと組換え体Acox1、Scp2、およびAcaa1とをインキュベートすると、無傷のペルオキシソームにおいて生成されたものと同じサイズの開裂生成物が形成された(図9)。Acox1は、切断されて28kDaのC末端断片を生成し、Scp2は、プロセッシングされて16kDaのC末端断片を生成した(図9)。Tysnd1とAcaa1とのインキュベーションは、3kDaのPTS2含有N末端ペプチドの除去をもたらして、成熟Acaa1を生成させた(図9)。N末端配列決定分析によって、Tysnd1はAcaa1を、Cys26残基とSer27残基の間で開裂し、インビボで見られるAcaa1の成熟型を生成させることが確証された。これらの実験は、Tysnd1が真のペルオキシソームのプロセッシングプロテアーゼであることを立証する。
【0061】
Tysnd1によるペルオキシソーム酵素のプロセッシングは、高度に特異的である。数個のペルオキシソーム酵素が切断部位周囲の配列としてAla−[AlaVal]−Proモチーフを共有しているようであるが(Biermann J, van den Bosch H. In vitro processing of the human alkyl-dihydroxyacetonephosphate synthase precursor. Arch Biochem Biophys. 1999 Aug 1;368(1):139-46)、これは一般的法則ではない。例えばScp2は、約アミノ酸25個分離れた2つの位置で切断される。これらの部位の一方は、切断部位周囲にAla−[AlaVal]−Proモチーフを含有する。Acox1において、モチーフはプロセッシング部位周囲には見られなかった。Tysnd1基質切断機構は、単純なペプチド結合認識を包含しないが、むしろ二次および三次構造の認識に基づいていることが示唆される。
(実施例9)
【0062】
<Tysnd1はシステインエンドペプチダーゼである>
Tysnd1のプロテアーゼクラスを決定するために、様々なプロテアーゼインヒビターの存在下で、酵素プロセッシング活性を評価した。この実験のために、2個の基質が選択された。それはAcaa1およびAcox1である。異なるペルオキシソーム標的クラス(PTS1およびPTS2)を代表するのに加えて、Acaa1およびAcox1は、プロセッシング部位を囲むアミノ酸配列が異なる。Tysnd1のプロセッシング活性は、システインプロテアーゼインヒビターN−エチルマレイミド(NEM)(Sigma−Aldrich Japan K.K.)を用いたとき、完全に無くなった(図10)。Tysnd1は、スルフヒドリル活性試薬NEMに対して感受性であるが、2個の他のシステインプロテイナーゼインヒビターであるロイペプチンおよびt−エポキシスクシニル−1−ロイシルアミド−(4−グアニジノ)ブタン(E64)(株式会社ペプチド研究所(大阪府))に対しては抵抗性である。ロイペプチンは、いくつかのリソソームのセリンプロテアーゼおよびシステインプロテアーゼを阻害することが知られ、一方E64はパパインプロテアーゼファミリーの特異的阻害剤である(Salvesen GS, Nagase H. In Proteolytic Enzymes (ed. Beynon R and Bond JS), p. 110. Oxford University Press)。同様の阻害パターンが、ペルオキシソームプロテアーゼIDEについて観察された(IDEはNEMによって阻害されるがロイペプチンおよびE64によっては阻害されない)。金属キレート剤、とりわけ1,10−フェナントロリン(Sigma−Aldrich Japan K.K.)によってTysnd1は部分阻害されるが、これと関連する非キレート化化合物の1,7−フェナントロリン(Wako Pure Chemical Industries,Ltd.)によっては部分阻害されないことは(図10)、Znイオンが、Tysnd1のコンホメーションまたはその基質との複合体形成を安定化させるのに関与している可能性を示す。
(実施例10)
【0063】
<低脂血症薬ベザフィブレートによるTysnd1の誘導>
PPAR−α−アクチベーターであるベザフィブレートは、部分的には、Acaa1、Acox1、およびHsd17b4(これらはTysnd1によってプロセッシングされる)のβ酸化の原因となるペルオキシソーム酵素を誘導することによって、肝臓での脂肪酸のβ酸化を増加させることが知られている(Beier K, Volkl A, Hashimoto T, Fahimi HD. Selective induction of peroxisomal enzymes by the hypolipidemic drug bezafibrate. Detection of modulations by automatic image analysis in conjunction with immunoelectron microscopy and immunoblotting. Eur J Cell Biol. 1988 Aug;46(3):383-93)。それゆえ、Tysnd1もまたベザフィブレートによって誘導されるかどうか試験された。以前の報告に一致して、肝臓のAcox1、Hsd17b4、およびAcaa1のmRNAの誘導が観察された(データは示さず)。肝臓の細胞下分画を行い、Tysnd1タンパク質の含有量が評価された。ベザフィブレート処理は、ペルオキシソームに富む軽ミトコンドリア画分において、49kDa型のTysnd1の著しい増加をもたらした(図11)。対照的に、(ポスト核画分、ポストミトコンドリア画分では、)カタラーゼの総量の僅かな増加がみられ、軽ミトコンドリア画分のカタラーゼの含有量は増加しなかった(図11)。これらのデータは、代謝要求に応じるために、特に脂肪酸のβ酸化が促進される条件で、Tysnd1の発現がその基質タンパク質の発現と同時制御される可能性を示唆する。
(実施例11)
【0064】
<疾患に対するTysnd1のPTS1およびPTS2タンパク質プロセッシング活性の意味>
Acox1、Acaa1、Scp2、Hsd17b4、ならびにおそらく他のPTS2およびPTS1含有ペルオキシソームタンパク質のTysnd1によるプロセッシングは、代謝性疾患例えば食事性肥満、脂肪肝疾患または高コレステロール血症における脂肪酸β酸化、ケトン生成およびコレステロール合成の病原化に関して、潜在的な治療上または診断上の意味を有し得る。Tysnd1と脂肪代謝の関連および微小血管内皮細胞におけるその差次的発現(Keen HL, Ryan MJ, Beyer A, Mathur S, Scheetz TE, Gackle BD, Faraci FM, Casavant TL, Sigmund CD. Gene expression profiling of potential PPARgamma target genes in mouse aorta. Physiol Genomics. 2004 Jun 17;18(1):33-42、この参考文献の表6)も、心臓血管疾患の病原論におけるTysnd1機能不全の役割を意味し得る。心虚血の間、ミトコンドリア脂肪酸酸化の高い速度は、グルコース酸化を阻害する(Dyck JR, Cheng JF, Stanley WC, Barr R, Chandler MP, Brown S, Wallace D, Arrhenius T, Harmon C, Yang G, Nadzan AM, Lopaschuk GD. Malonyl coenzyme a decarboxylase inhibition protects the ischemic heart by inhibiting fatty acid oxidation and stimulating glucose oxidation. Circ Res. 2004 May 14;94(9):e78-84)。ラノラジンおよび長鎖3−ケトアシル補酵素Aチオラーゼインヒビターのトリメタジジンは、脂肪酸酸化を阻害し、したがってグルコース酸化および心臓効率を向上させることが示されている(Lopaschuk GD. Inhibiting fatty acid oxidation as a novel therapeutic approach to treating, ischaemic heart disease. Cardiovasc J S Afr. 2004 Jul;15(4 Suppl 1):S1)。心虚血へのペルオキシソームチオラーゼの寄与は知られているところでは調査されていないが、除外することはできない。Tysnd1の阻害は、チオラーゼの有効性を低下させ、それにより脂肪酸酸化を低下させると同時に心臓効率を向上させる。
【0065】
マウスへの高脂肪食の長期的影響に関する研究は、Acox1、Acaa1の肝臓における発現が著しくアップレギュレートされることを立証している(Murase, T., Mizuno, T., Omachi, T., Onizawa, K., Komine, Y., Kondo, H., Hase, T., Tokimitsu, I., 2001. Dietary diacylglycerol suppresses high fat and high sucrose diet-induced body fat accumulation in C57BL/6J mice. J. Lipid Res. 42, 372–378;Kim S, Sohn I, Ahn JI, Lee KH, Lee YS, Lee YS. Hepatic gene expression profiles in a long-term high-fat diet-induced obesity mouse model. Gene. 2004 Sep 29;340(1):99-109)。したがって成熟Tysnd1の特異的阻害または活性化は、原因となっている肝臓脂質代謝の病態および男性不妊症を制御する新たな戦略を開拓し得る(Huyghe S, Schmalbruch H, De Gendt K, Verhoeven G, Guillou F, Van Veldhoven PP, Baes M. Peroxisomal multifunctional protein 2 is essential for lipid homeostasis in Sertoli cells and male fertility in mice. Endocrinology. 2006 Feb 16; [Epub ahead of print as doi:10.1210/en.2005-1571])。マウスの成熟Scp2の欠損は、フィタン酸の蓄積および成人レフサム病様症候群の発達に関与している(Mukherji M, Kershaw NJ, Schofield CJ, Wierzbicki AS, Lloyd MD. Utilization of sterol carrier protein-2 by phytanoyl-CoA 2-hydroxylase in the peroxisomal alpha oxidation of phytanic acid. Chem Biol. 2002 May;9(5):597-605)。フィタン酸は、ペルオキシソームα−酸化経路でフィタノイル−CoA 2−ヒドロキシラーゼによって分解される。フィタノイル−CoA 2−ヒドロキシラーゼによるフィタノイル−CoAの効率的な2−ヒドロキシル化は成熟Scp2を必要とし、Tysnd1は成熟Scp2の利用可能性を調節する重要な役割を果たし得ている。したがってTysnd1の機能的不全は、レフサム様疾患症状をもたらし得る。
【0066】
Raychaudhuryらによる研究(Raychaudhury B, Banerjee S, Datta SC. Peroxisomal function is altered during leishmania infection. Med Sci Monit. 2003 Apr;9(4):BR125-9)は、リーシュマニア感染が、ペルオキシソーム損傷につながるペルオキシソーム酵素の複数の機能不全を引き起こすことを示した。注目すべきは、本発明者らがリーシュマニア感染時のペルオキシソームタンパク質分解活性の低下も検出したことである。感染時の活性低下は、成熟Tysnd1プロテアーゼ活性またはTysnd1前駆体の成熟Tysnd1への変換を阻害するリーシュマニアタンパク質によって引き起こされている可能性がある。
【図面の簡単な説明】
【0067】
【図1】融合タンパク質GFP−Tysnd1がペルオキシソームに局在することを示す。CHO−K1細胞を、融合されたGFP−Tysnd1およびDsRed2−Peroxiを発現するプラスミドと共に同時トランスフェクションした。生細胞は、蛍光顕微鏡法によって分析し、GFP(A)、DsRed(B)チャネル、および併合シグナル(C)の代表的な画像を示す。
【図2】ラットAcaa1前駆体の成熟Acaa1(mAcaa1)へのプロセッシングに対するTysnd1発現の効果を示す。CHO−K1細胞は、Acaa1−HAおよびTysnd1発現プラスミドの示された組合せを用いて一時トランスフェクションした。プラスミドの総量は、空cDNA3.1の追加により、4μgで一定に維持した。
【図3】Tysnd1の同時発現時のScp2およびAcox1のタンパク質分解変換を示す。COS−7細胞を、FLAG−Tysnd1およびHAマウスScp2またはHAラットAcox1の示された組合せでトランスフェクションした。プラスミドの総量は、pcDNA3.1を使用して4μgに調節した。トランスフェクションの48時間後に細胞溶解物を調製し、12.5%SDS−PAGEによって分離して、抗HA抗体を用いてウェスタンブロットで分析した。矢印は、Scp2およびAcox1前駆体およびその成熟形(mScp2およびmAcox1)の位置を示す。
【図4】Tysnd1の同時発現時のマウスペルオキシソームヒドロキシステロイド(17−β)デヒドロゲナーゼ 4(Hsd17b4)のタンパク質分解変換を示す。COS−7細胞を、FLAG−Tysnd1およびHA−マウスHsd17b4の示された組合せでトランスフェクションした。プラスミドの総量は、pcDNA3.1を使用して4μgに調節した。トランスフェクションの48時間後に細胞溶解物を調製し、12.5%SDS−PAGEゲルによって分離して、抗HA抗体を用いてウェスタンブロットで分析した。矢印は、Hsd17b4前駆体(79kDa)およびHsd17b4成熟形(35kDa)の位置を示す。
【図5】4μgDNAのpcDNA3.1によって(mock、レーン1)、およびC末端にFlagタグをつけたTysnd1によって(レーン3および6)トランスフェクションしたCOS−7細胞におけるTysnd1切断の分析を示す。これらは4%−20%勾配のSDS−PAGEによって分離し、そして抗Tysnd1抗体を使用したウェスタンブロッティングでアッセイした。レーン4−6で使用した抗体は、合成ペプチドCSNTRDNNTGATYPHLを予備吸収させた。
【図6】ポリクローナル抗体のTysnd1との特異的反応性を示す。精製した組換えTysnd1(レーン1および4)、空ベクターpcDNA3.1を用いてトランスフェクションしたCOS−7細胞からの溶解物(レーン2および5)、およびN末端Flagタグ付きTysnd1のベクターを用いてトランスフェクションしたCOS−7細胞からの溶解物(レーン3および6)を勾配4〜20%SDS−PAGEによって分離し、抗Tysnd1抗体を用いてウェスタンブロッティングによってアッセイした。レーン4〜6で使用した抗体は、合成ペプチドCSNTRDNNTGATYPHLを予備吸収させた。
【図7】Optiprep密度勾配での細胞下分画の結果を示す。軽ミトコンドリア画分は、自己形成Optiprep勾配により分画した。画分は、管の上から開始して採取した。カタラーゼ(縞模様の棒)、コハク酸デヒドロゲナーゼ(黒棒)およびβ−ガラクトシダーゼ(中空棒)を、それぞれペルオキシソーム、ミトコンドリアおよびリソソームのマーカー酵素として測定した(図7A)。結果を、全勾配活性のパーセンテージとして与える。図7Bは、タンパク質含有量を示す。等体積の各画分からのタンパク質を、12.5%SDS−PAGEによって分離し、マウスTysnd1に対する抗体を用いて免疫ブロットした(図7C)。
【図8】Tysnd1特異的低分子干渉RNA(siRNA)による、293FT細胞系におけるAcox1、Scp2、およびHsd17b4のプロセッシングの阻害を示す。293FT細胞を、ヒトTYSND1に特異的なsiRNA(Tysnd1_1およびTysnd1_2)または非サイレンシングsiRNA(対照)とともに、Acox1−HA、Scp2−HA、またはHsd17b4−HA発現プラスミドにトランスフェクションさせた。細胞溶解物を、抗HA抗体を用いたウェスタンブロッティングによって分析した。矢印は、Acox1、Scp2、およびHsd17b4の前駆体およびこれらの成熟形(mAox1、mScp2、およびmHsd17b4)の位置を示す。βアクチンのウェスタンブロット分析を、siRNA特異性およびサンプル添加の対照として用いた。
【図9】組み換えTysnd1によるAcox1、Scp2、およびAcaa1のインビトロの切断を示す。組み換えAcox1、Scp2、およびAcaa1を、「材料と方法」で説明されるようにして、E.coli抽出物から精製した。マウスTysnd1を、Tysnd1−Flag発現プラスミドを用い一時トランスフェクションしたCOS−7細胞の抽出物から精製した。Acox1およびScp2のプロセッシングを、抗cMyc抗体を用いたウェスタンブロティングによって分析した。Acaa1切断を、12.5%SDS−PAGEゲルからタンパク質を移した後の、Hybond−Pメンブレンの0.05%Amido Black染色によって検出した。
【図10】Tysnd1によるAcaa1(A)およびAcox1(B)のプロセッシングへの各種プロテアーゼインヒビターの影響を示す。組み換えHis6−Acaa1およびAcox1−myc−His6を、単独で0時間(レーン1)もしくは単独で12時間(レーン2)またはCOS−7細胞溶解物から精製したTysnd1−FLAGとともに12時間(A、レーン3〜14;B、レーン3〜17)インキュベートした。His6−Acaa1のプロセッシングを、抗Acaa1抗体を用いたウェスタンブロットによって評価した。Acox1−myc−His6の切断を、抗cMyc抗体を用いて検出した。
【図11】マウス肝臓におけるTysnd1の含有量へのベザフィブレート処置の効果を示す。マウスを、0.5%(v/w)のベザフィブレートを含有する標準飼料で、3週間飼育した。肝臓細胞下画分を、「材料および方法」でラット肝臓について記載されているようにして調製した。ポスト核画分(PN)、軽ミトコンドリア画分(LM)、およびポストミトコンドリア画分(PM)の100mgのタンパク質を、抗Tysnd1抗体または抗カタラーゼ抗体を用いたウェスタンブロットによって分析した。C、対照マウス;B、ベザフィブレート処置マウスサンプル。
【特許請求の範囲】
【請求項1】
細胞におけるペルオキシソームの生成および/または機能に関連する疾患または症候群を診断するための医薬組成物であって、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む医薬組成物。
【請求項2】
ポリペプチドがTysnd1によってコードされたペプチド、その哺乳類またはヒトオルソログ、その成熟型およびプロテアーゼドメインを含有するその断片からなる群より選択される、請求項1に記載の医薬組成物。
【請求項3】
ペルオキシソーム酵素が、哺乳類またはヒトの3−オキソアシル補酵素Aチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、ステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)からなる群より選択される少なくとも1つである、請求項1に記載の医薬組成物。
【請求項4】
疾患または症候群が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項1に記載の医薬組成物。
【請求項5】
ポリヌクレオチドが、保存的アミノ酸置換を含むアミノ酸配列を有する、請求項1に記載の医薬組成物。
【請求項6】
細胞におけるペルオキシソームの生成および/または機能に関連する疾患または症候群を治療するための医薬組成物であって、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む医薬組成物。
【請求項7】
ポリペプチドがTysnd1によってコードされたペプチド、その哺乳類またはヒトオルソログ、その成熟型およびプロテアーゼドメインを含有するその断片からなる群より選択される、請求項6に記載の医薬組成物。
【請求項8】
ペルオキシソーム酵素が、哺乳類またはヒトの3−オキソアシル補酵素Aチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、ステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)からなる群より選択される少なくとも1つである、請求項6に記載の医薬組成物。
【請求項9】
疾患または症候群が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項6に記載の医薬組成物。
【請求項10】
ポリヌクレオチドが、保存的アミノ酸置換を含むアミノ酸配列を有する、請求項6に記載の医薬組成物。
【請求項11】
Tysnd1およびその哺乳類またはヒトオルソログの生物活性のモジュレーター、アクチベーターまたはインヒビターであって、Tysnd1アンチセンス核酸、リボザイム、センスポリヌクレオチド、二重鎖RNA、RNAi、アプタマー、小分子アンタゴニストまたは小分子アゴニストからなる群より選択される、モジュレーター、アクチベーターまたはインヒビター。
【請求項12】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドに結合する抗体。
【請求項13】
前記抗体がモノクローナル抗体である、請求項12に記載の抗体。
【請求項14】
抗体がヒト化抗体である、請求項12に記載の抗体。
【請求項15】
少なくとも1つの容器、請求項1〜10のいずれか一項に記載の医薬組成物および/または請求項11に記載のモジュレーター、アクチベーターまたはインヒビターおよび/または請求項12〜14のいずれか一項に記載の抗体を含むキット。
【請求項16】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドのサンプル中での存在または量を測定するための方法であって、
a)サンプルを提供するステップと、
b)サンプルに請求項12〜14のいずれか一項に記載の抗体を接触させるステップと、
c)前記ポリペプチドに結合した抗体の存在または量を測定するステップと
を含む方法。
【請求項17】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドに結合する薬剤を特定する方法であって、
a)前記ポリペプチドに候補薬剤を接触させるステップと、
b)前記候補薬剤が前記ポリペプチドに結合するかどうかを判定するステップと
を含む方法。
【請求項18】
薬剤がペルオキシソームタンパク質、そのペプチド断片、小分子アンタゴニストまたは小分子アゴニストである、請求項17に記載の方法。
【請求項19】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドの生物活性を調節、アンタゴナイズまたは増進する方法であって、ポリペプチドを発現する細胞または組織サンプルに、ポリペプチドの生物活性を調節、アンタゴナイズまたは増進するために十分な量の前記ポリペプチドに結合する化合物を接触させるステップを含む方法。
【請求項20】
細胞または組織サンプルが哺乳類、ヒトまたは患者からである、請求項19に記載の方法。
【請求項21】
哺乳類、ヒトまたは患者が、代謝経路調節に関連する疾患に罹患している、請求項20に記載の方法。
【請求項22】
疾患が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項21に記載の方法。
【請求項23】
細胞におけるペルオキシソームの生成および/または機能に関連する疾患または症候群と診断された被検体を処置する方法であって、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを該被検体に投与するステップを含む方法。
【請求項24】
ポリペプチドがTysnd1によってコードされたペプチド、その哺乳類またはヒトオルソログ、その成熟型およびプロテアーゼドメインを含有するその断片からなる群より選択される、請求項23に記載の方法。
【請求項25】
ペルオキシソーム酵素が、哺乳類またはヒトの3−オキソアシル補酵素Aチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、ステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)からなる群より選択される少なくとも1つである、請求項23に記載の方法。
【請求項26】
疾患または症候群が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項23に記載の方法。
【請求項27】
ポリヌクレオチドが、保存的アミノ酸置換を含むアミノ酸配列を有する、請求項23に記載の方法。
【請求項1】
細胞におけるペルオキシソームの生成および/または機能に関連する疾患または症候群を診断するための医薬組成物であって、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む医薬組成物。
【請求項2】
ポリペプチドがTysnd1によってコードされたペプチド、その哺乳類またはヒトオルソログ、その成熟型およびプロテアーゼドメインを含有するその断片からなる群より選択される、請求項1に記載の医薬組成物。
【請求項3】
ペルオキシソーム酵素が、哺乳類またはヒトの3−オキソアシル補酵素Aチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、ステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)からなる群より選択される少なくとも1つである、請求項1に記載の医薬組成物。
【請求項4】
疾患または症候群が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項1に記載の医薬組成物。
【請求項5】
ポリヌクレオチドが、保存的アミノ酸置換を含むアミノ酸配列を有する、請求項1に記載の医薬組成物。
【請求項6】
細胞におけるペルオキシソームの生成および/または機能に関連する疾患または症候群を治療するための医薬組成物であって、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを含む医薬組成物。
【請求項7】
ポリペプチドがTysnd1によってコードされたペプチド、その哺乳類またはヒトオルソログ、その成熟型およびプロテアーゼドメインを含有するその断片からなる群より選択される、請求項6に記載の医薬組成物。
【請求項8】
ペルオキシソーム酵素が、哺乳類またはヒトの3−オキソアシル補酵素Aチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、ステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)からなる群より選択される少なくとも1つである、請求項6に記載の医薬組成物。
【請求項9】
疾患または症候群が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項6に記載の医薬組成物。
【請求項10】
ポリヌクレオチドが、保存的アミノ酸置換を含むアミノ酸配列を有する、請求項6に記載の医薬組成物。
【請求項11】
Tysnd1およびその哺乳類またはヒトオルソログの生物活性のモジュレーター、アクチベーターまたはインヒビターであって、Tysnd1アンチセンス核酸、リボザイム、センスポリヌクレオチド、二重鎖RNA、RNAi、アプタマー、小分子アンタゴニストまたは小分子アゴニストからなる群より選択される、モジュレーター、アクチベーターまたはインヒビター。
【請求項12】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドに結合する抗体。
【請求項13】
前記抗体がモノクローナル抗体である、請求項12に記載の抗体。
【請求項14】
抗体がヒト化抗体である、請求項12に記載の抗体。
【請求項15】
少なくとも1つの容器、請求項1〜10のいずれか一項に記載の医薬組成物および/または請求項11に記載のモジュレーター、アクチベーターまたはインヒビターおよび/または請求項12〜14のいずれか一項に記載の抗体を含むキット。
【請求項16】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドのサンプル中での存在または量を測定するための方法であって、
a)サンプルを提供するステップと、
b)サンプルに請求項12〜14のいずれか一項に記載の抗体を接触させるステップと、
c)前記ポリペプチドに結合した抗体の存在または量を測定するステップと
を含む方法。
【請求項17】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドに結合する薬剤を特定する方法であって、
a)前記ポリペプチドに候補薬剤を接触させるステップと、
b)前記候補薬剤が前記ポリペプチドに結合するかどうかを判定するステップと
を含む方法。
【請求項18】
薬剤がペルオキシソームタンパク質、そのペプチド断片、小分子アンタゴニストまたは小分子アゴニストである、請求項17に記載の方法。
【請求項19】
プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドの生物活性を調節、アンタゴナイズまたは増進する方法であって、ポリペプチドを発現する細胞または組織サンプルに、ポリペプチドの生物活性を調節、アンタゴナイズまたは増進するために十分な量の前記ポリペプチドに結合する化合物を接触させるステップを含む方法。
【請求項20】
細胞または組織サンプルが哺乳類、ヒトまたは患者からである、請求項19に記載の方法。
【請求項21】
哺乳類、ヒトまたは患者が、代謝経路調節に関連する疾患に罹患している、請求項20に記載の方法。
【請求項22】
疾患が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項21に記載の方法。
【請求項23】
細胞におけるペルオキシソームの生成および/または機能に関連する疾患または症候群と診断された被検体を処置する方法であって、プロテアーゼ活性を有し、ペルオキシソーム酵素をプロセッシングするポリペプチドを該被検体に投与するステップを含む方法。
【請求項24】
ポリペプチドがTysnd1によってコードされたペプチド、その哺乳類またはヒトオルソログ、その成熟型およびプロテアーゼドメインを含有するその断片からなる群より選択される、請求項23に記載の方法。
【請求項25】
ペルオキシソーム酵素が、哺乳類またはヒトの3−オキソアシル補酵素Aチオラーゼ(Acaa1)、アシル補酵素Aオキシダーゼ1(Acox1)、ステロールキャリアタンパク質2(Scp2)、およびヒドロキシステロイド(17−β)デヒドロゲナーゼ4(Hsd17b4)からなる群より選択される少なくとも1つである、請求項23に記載の方法。
【請求項26】
疾患または症候群が、代謝性疾患の中でも、代謝性症候群、レフサム病、X染色体連鎖副腎白質ジストロフィー(X−ALD)、およびメバロン酸尿症、男性不妊症、心臓血管疾患、ペルオキシソームの機能不全に関連する皮膚、肝臓、肺、腎臓、睾丸、脳または中枢神経系の疾患からなる群より選択される、請求項23に記載の方法。
【請求項27】
ポリヌクレオチドが、保存的アミノ酸置換を含むアミノ酸配列を有する、請求項23に記載の方法。
【図8】

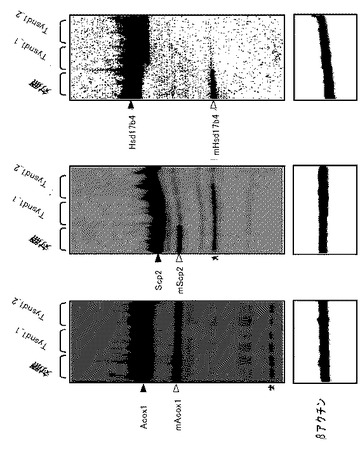
【図1】
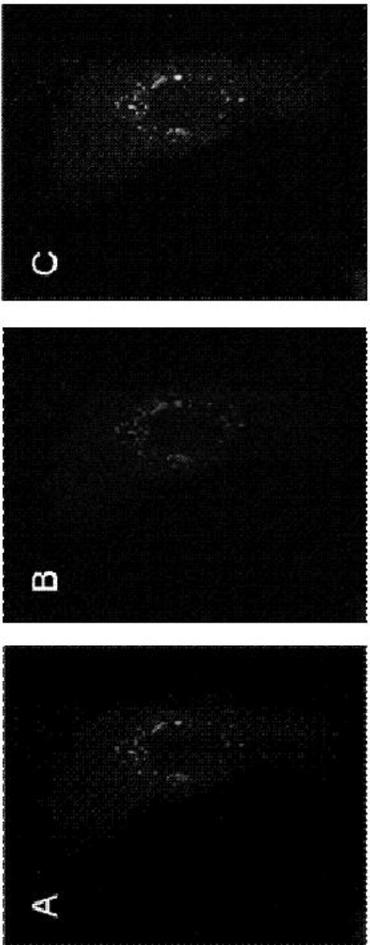
【図2】
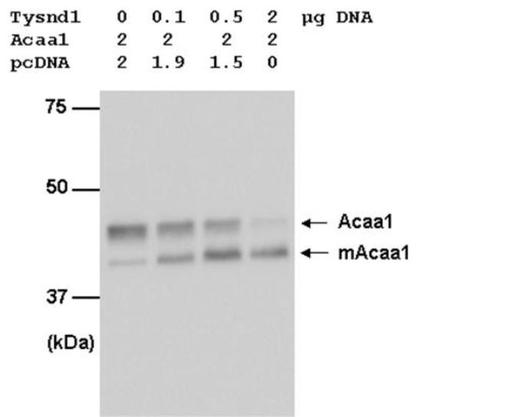
【図3】
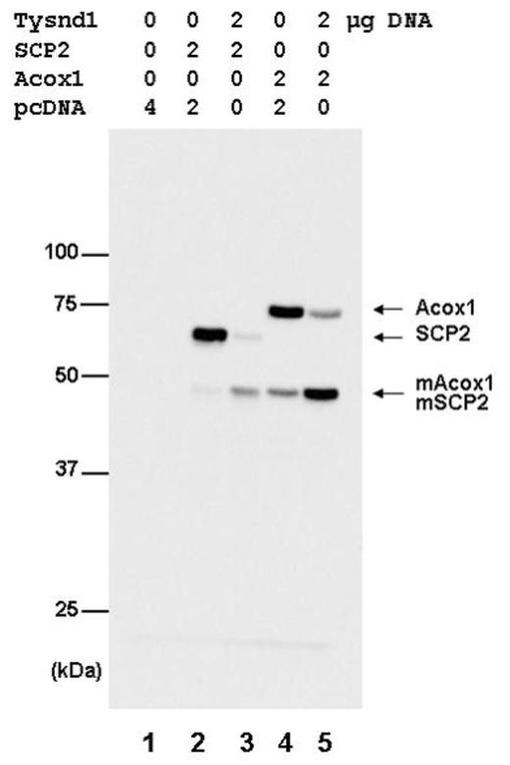
【図4】
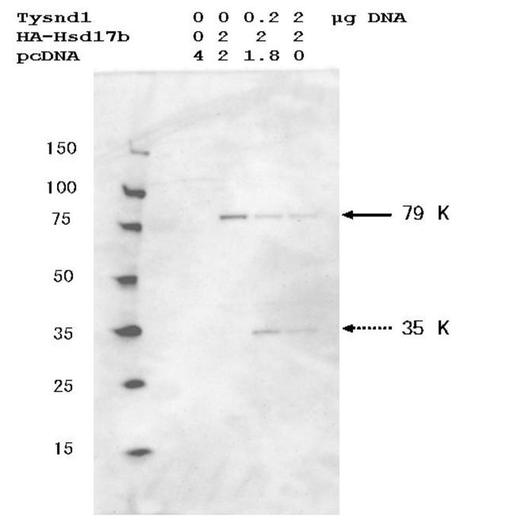
【図5】
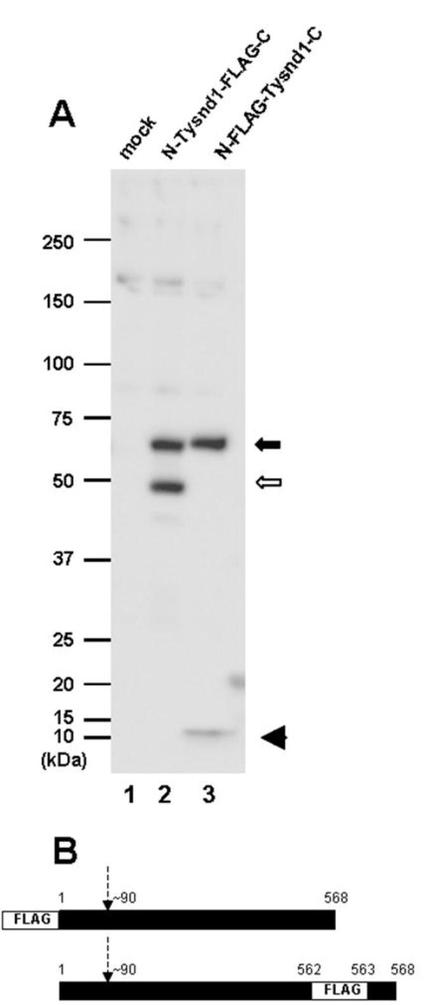
【図6】
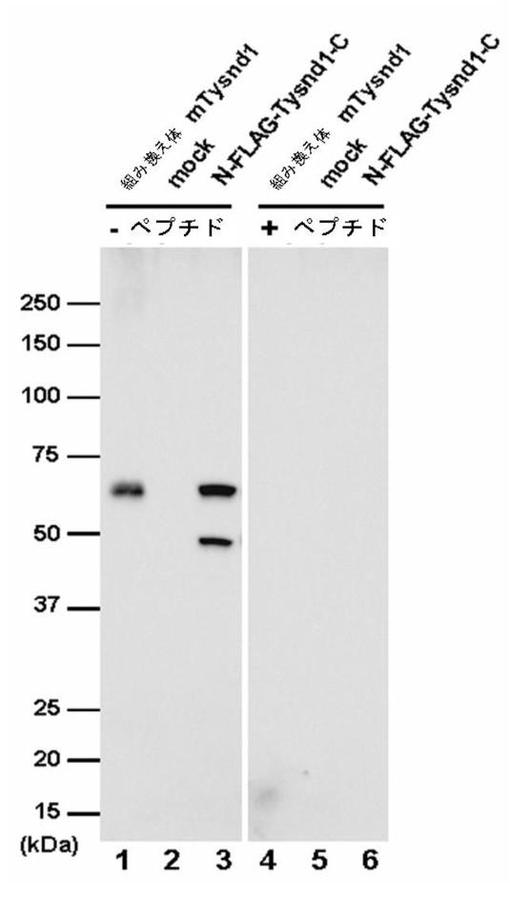
【図7】
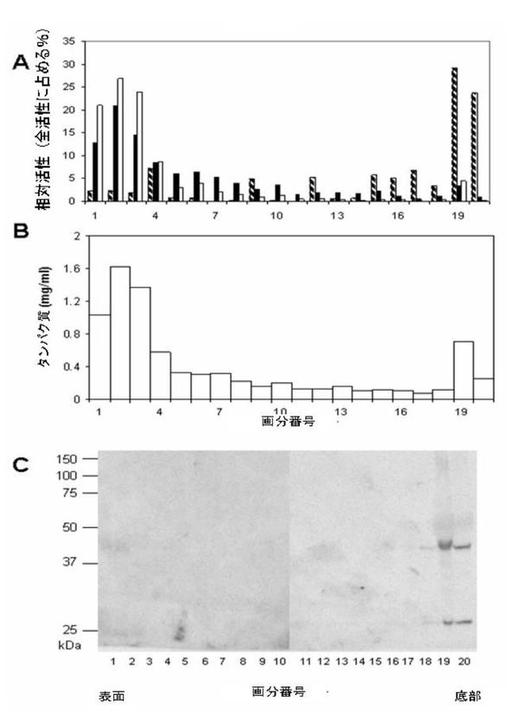
【図9】
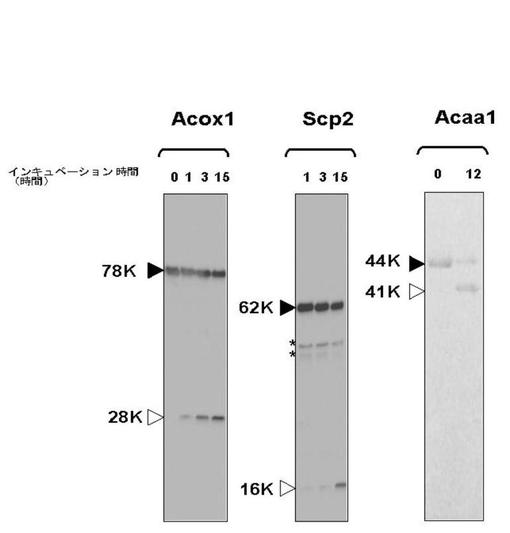
【図10】

【図11】
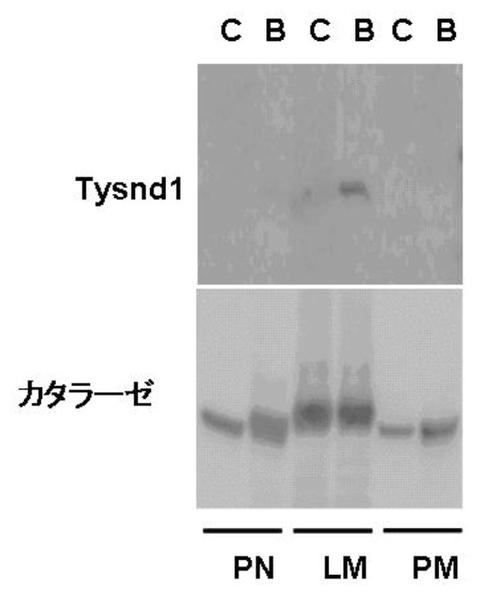
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図9】
【図10】
【図11】
【公表番号】特表2008−543274(P2008−543274A)
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願番号】特願2007−555819(P2007−555819)
【出願日】平成18年6月9日(2006.6.9)
【国際出願番号】PCT/JP2006/312085
【国際公開番号】WO2006/132445
【国際公開日】平成18年12月14日(2006.12.14)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.TEFLON
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【出願人】(504013775)学校法人 埼玉医科大学 (39)
【Fターム(参考)】
【公表日】平成20年12月4日(2008.12.4)
【国際特許分類】
【出願日】平成18年6月9日(2006.6.9)
【国際出願番号】PCT/JP2006/312085
【国際公開番号】WO2006/132445
【国際公開日】平成18年12月14日(2006.12.14)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.TEFLON
【出願人】(503359821)独立行政法人理化学研究所 (1,056)
【出願人】(504013775)学校法人 埼玉医科大学 (39)
【Fターム(参考)】
[ Back to top ]