
ペントース糖の発酵
【課題】アルコール発酵ができ、炭素源としてキシロースを利用できる酵母を提供する。
【解決手段】嫌気性菌から得られる真核生物性キシロース異性化酵素をコードする核酸配列で形質転換した宿主細胞であり、発現したときにキシロース異性化酵素をコードする配列は、キシロースをキシルロースに変換する能力を宿主細胞に付与し、それは宿主細胞によりさらに代謝される。従って、宿主細胞は、炭素源としてのキシロースに対して増殖することができる。宿主細胞は酵母または糸状菌のような真核性微生物であることが望ましい。さらにエタノールのような発酵産物の生産プロセスに関し、そのプロセスにおいて、該宿主細胞は増殖及び発酵産物の生産のためにキシロースを利用できる。
【解決手段】嫌気性菌から得られる真核生物性キシロース異性化酵素をコードする核酸配列で形質転換した宿主細胞であり、発現したときにキシロース異性化酵素をコードする配列は、キシロースをキシルロースに変換する能力を宿主細胞に付与し、それは宿主細胞によりさらに代謝される。従って、宿主細胞は、炭素源としてのキシロースに対して増殖することができる。宿主細胞は酵母または糸状菌のような真核性微生物であることが望ましい。さらにエタノールのような発酵産物の生産プロセスに関し、そのプロセスにおいて、該宿主細胞は増殖及び発酵産物の生産のためにキシロースを利用できる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、真核生物性キシロース異性化酵素をコードする核酸配列で形質転換した宿主細胞に関するものである。このキシロース異性化酵素は、宿主細胞で発現し、キシロースをキシルロースに変換する能力を付与する。この宿主細胞は、ペントース含有培地の発酵によるエタノール及びその他の発酵産物の製造プロセスに使用される。さらに本発明は、真核生物性キシロース異性化酵素をコードする核酸配列に関するものである。
【背景技術】
【0002】
この数十年間における古典的化石燃料(石油燃料)の大量消費は、高レベルの汚染の原因となっている。さらに、世界中の石油埋蔵量は無限でないことが認識され、このような認識は、環境問題への関心の高まりと相俟って、CO2排出の60−90%減少を実現できるエタノールのような代替燃料の可能性を研究する新しい動きを生んだ。バイオマス由来のエタノールは、多数の種々の資源から得られるヘキソース糖の発酵により生産することができるが、にも拘らず工業的規模の生産または燃料アルコールに向けられる原料は、ショ糖及びトウモロコシデンプンである。これらの原料の欠点は高価なことである。
【0003】
燃料アルコールの生産拡大には、低コストの供給原料の使用を可能にすることが求められる。現在、植物バイオマスからは唯一リグノセルロース原料がかなり大量に使用可能であり、エタノール生産用の農産物を代替している。リグノセルロース原料の主な発酵可能な糖はグルコース及びキシロースであり、それぞれリグノセルロース中に約40%及び25%含有される。しかし、Saccharomyces cerevisiaeのようなアルコール発酵ができる酵母のほとんどは、炭素源としてキシロースを使用できない。さらに、高収量及び高生産性でキシロースをエタノールに発酵できる微生物は知られていない。リグノセルロース加水分解産物からエタノールを商業的に生産することを可能にするには、これらの性質を持つ微生物が求められるであろう。従って本発明の一つの目的は、アルコール発酵ができ、炭素源としてキシロースを利用できる酵母を提供することである。
【0004】
D−キシロースは、腸内細菌、一部の酵母及び真菌のような種々の微生物によって代謝される。大部分のキシロース利用細菌において、キシロースはキシロース(グルコース)異性化酵素(XI)により直接D−キシルロースに異性化される。しかし、糸状菌及び酵母は、この一ステップの異性化を行うことができず、まずキシロース還元酵素(XR)の作用によりキシロースをキシリトールに還元し、次いでキシリトール脱水素酵素(XDH)によりキシリトールをキシルロースに変換する。最初のステップは補因子としてNAD(P)Hを必要とし、第二ステップはNAD+を必要とする。生成したキシルロースは、その後、キシルロースキナーゼ(XK)によりリン酸化された後にペントースリン酸経路(PPP)に入る。厳密なNADPH依存性のキシロース還元酵素(XR)を持つ微生物では、キシロースからエタノールへの嫌気性発酵は不可能である。キシリトール脱水素酵素(XDH)は厳密にNAD+に依存しているので、酸化還元不均衡(すなわち、NAD+欠乏)を生じるからである。嫌気性条件においてこの酸化還元不均衡を解決するために、微生物はグリセロール及びキシリトールのような副産物を生産する。同様に、キシロースに対するβ−ラクタムの嫌気的な生産も、グルコースに対するβ−ラクタム生産に比較すると生じにくい。これらの低収量の原因は、グルコースを利用する場合に比較して、この経路においてはNADPHの形の還元体を比較的高度に必要とすることにあるように思われる(W.M.van Gulik et al.,Biotechnol Bioeng.Vol.68,No.6,June 20,2000)。
【0005】
Zaldivar et al.(2001,Appl.Microbiol.Biotechnol.56;17-34)に総説されているように、S.cerevisiae及び類似の酵母にキシロース代謝を導入する多くの試みが行われてきた。一つの方法は、少なくとも、キシロース(アルドース)還元酵素及びキシリトール脱水素酵素、すなわちPichia stipitisのXYL1及びXYL2、をコードする遺伝子をS.cerevisiaeに発現させることに関する(US5,866,382;WO95/13362;及びWO97/42307)。この方法は、キシロースに対してS.cerevisiaeの増殖を可能にしたが、主にXR及びXDHの間の酸化還元不均衡の結果として、一般的にエタノール生産性及び/または収量が低く、キシリトールの生産が多いという欠点がある。
【0006】
S.cerevisiae若しくは関連酵母または糸状菌にXIを発現させることにより、酸化還元不均衡及びその結果としてのキシリトール生産と分泌を回避することができるであろう。数種の細菌のキシロース異性化酵素遺伝子がS.cerevisiaeに挿入されたが、S.cerevisiaeにおいて中等温度好性原核生物のXIを発現させても、活性のあるXIを生じなかった(Amore and Hollenverg, 1989, Nucleic Acids Res. 17:7515 ; Amore et al., 1989, Appl. Microbiol. Biotechnol.30:351-357; Chan et al., 1986, Biotechnol. Lett 8:231-234;Chan et al., 1989,Appl.Microbiol.Biotechnol. 31:524-528; Ho et al., 1983, Fed. Proc. Fed. Am. Soc. Exp. Biol. 42 : 2167 ; Hollenberg, 1987, EBC Symposium on Brewer's Yeast, Helsinki(Finland), 24-25 Nov 1986 ; Sarthy et al., 1987, Appl. Envion. Microbiol. 53:1996-2000 ; Ueng et al., 1985, Biotechnol. Lett. 7: 153-158)。しかし、S.cerevisiaeに発現させた好熱性細菌の二種のXIは85℃において1μmol/min/mgの比活性を示した(Bao et al.,1999,Weishengwu-Xuebao 39:49-54;Walfridson et al.,1996,Appl.Environ.Microbiol.61:4184-4190)。しかし、S.cerevisiaeの生理的温度(20−35℃)では、この活性のわずか数%しか維持できず、キシロースから有効にアルコールを発酵するには不十分であった。従って、生理的条件下において十分なXI活性を提供して、炭素源としてキシロースを利用することを可能とするように、酵母で発現することができるXIをコードする核酸に対する要求は依然として存在する。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】US5,866,382
【特許文献2】WO95/13362
【特許文献3】WO97/42307
【非特許文献】
【0008】
【非特許文献1】W.M.van Gulik et al.,Biotechnol Bioeng. Vol.68, No.6, June 20,2000
【非特許文献2】Zaldivar et al., 2001, Appl.Microbiol.Biotechnol.56;17-34)
【非特許文献3】more and Hollenverg, 1989, Nucleic Acids Res. 17:7515
【非特許文献4】Amore et al., 1989, Appl. Microbiol. Biotechnol.30:351-357
【非特許文献5】Chan et al., 1986, Biotechnol. Lett 8:231-234
【非特許文献6】Chan et al., 1989,Appl.Microbiol.Biotechnol. 31:524-528
【非特許文献7】Ho et al., 1983, Fed. Proc. Fed. Am. Soc. Exp. Biol. 42 : 2167
【非特許文献8】Hollenberg, 1987, EBC Symposium on Brewer's Yeast, Helsinki(Finland), 24-25 Nov 1986
【非特許文献9】Sarthy et al., 1987, Appl. Envion. Microbiol. 53:1996-2000
【非特許文献10】Ueng et al., 1985, Biotechnol. Lett. 7: 153-158
【非特許文献11】Bao et al.,1999,Weishengwu-Xuebao 39:49-54;Walfridson et al.,1996,Appl.Environ.Microbiol.61:4184-4190
【発明の概要】
【課題を解決するための手段】
【0009】
[発明の説明]
(定義)
キシロース異性化酵素
本明細書において、酵素「キシロース異性化酵素」(EC5.3.1.5)は、D−キシロースのD−キシルロースへのの直接の異性化、及びその逆の直接の異性化を触媒する酵素として定義される。この酵素はまた、D−キシロースケト異性化酵素としても知られている。一部のキシロース異性化酵素は、D−グルコース及びD−フルクトースの間の変換を触媒することもできるので、時にはグルコース異性化酵素とも呼ばれる。キシロース異性化酵素は補因子としてマグネシウムを必要とする。本発明のキシロース異性化酵素はさらに後述するアミノ酸配列によって定義される。同様に、キシロース異性化酵素はこの酵素をコードするヌクレオチド配列並びに後述されるキシロース異性化酵素をコードする対照ヌクレオチド配列にハイブリダイズするヌクレオチド配列によって定義することができる。
【0010】
本明細書において、キシロース異性化酵素活性の単位(U)は、50mMリン酸緩衝液(pH7.0),10mMキシロース及び10mM MgCl2を含む反応混合物中、37℃で、1分間に1nmolのキシルロースを生産する酵素の量として定義される。生成したキシルロースは、Dische and Borenfreund(1951,J.Biol.Chem.192:583-587)の方法または実施例に記述したHPLCにより測定した。
【0011】
配列の同一性及び類似性
本明細書において配列同一性(sequence identity)とは、配列比較により決定される、二以上のアミノ酸(ポリペプチドまたはタンパク)配列または二以上の核酸(ポリヌクレオチド)配列の関係として定義される。この分野で、「同一性」は、場合によって、アミノ酸配列又は核酸配列の配列間の一致により決定される、アミノ酸または核酸配列間の配列関連性(sequence relatedness)の程度も意味している。二つのアミノ酸配列の「類似性(similarity)」は、あるポリペプチドの他のポリペプチドに対する、アミノ酸配列及び保存的置換アミノ酸を比較して決定される。「同一性」及び「類似性」は、限定はしないが以下に記述されている方法を含む既知方法により容易に計算することができる(Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, New York, 1988; Biocomputing:Informatics and Genome Projects, Smith, D. W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994;Sequence Analysis in Molecular Biology, von Heine, g., Academic Press, 1987; and Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; and Carillo, H., and Lipman, D., SIAM J. Applied Math., 48: 1073(1988)。
【0012】
同一性を決める望ましい方法は、試験する配列間に最大の一致が得られるように設計される。同一性及び類似性を決定する方法は、一般に使用可能なコンピュータプログラムとして作成される。二つの配列間の同一性及び類似性を決定するための望ましいコンピュータプログラム方法には、GCGプログラムパッケージ(Devereux,J., et al., Nucleic Acids Research 12(1):387(1984))、BestFit, BLASTP, BLASTN,及びFASTA(Altschul, S. F. et al., J. Mol. Biol. 215: 403-410 (1990)が含まれる。BLAST XプログラムはNCBI及びその他のサイトから誰でも入手できる(BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda, MD 20894; Altschul, S., et al., J. Mol. Biol. 215: 403-410 (1990)。よく知られたSmith Watermanアルゴリズムも同一性決定に使用することができる。
【0013】
ポリペプチド配列比較のための望ましいパラメーターには以下のアルゴリズムが含まれる:Needleman and Wunsch,J.Mol.Biol.48:443-453 (1970);Comparison matrix:BLOSSUM62 from Hentikoff and Hentikoff,Proc.Natl.Acad.Sci.USA 89:10915-10919(1992);Gap Penalty:12;及びGap Length Penalty:4。これらのパラメーターを使用する有用なプログラムはMadison,WIにあるGenetics Computer Groupの「Ogap」プログラムとして一般に入手することができる。既述のパラメーターはアミノ酸比較(エンドギャップに対するペナルティーなし)のためのデホルトパラメーターである。
【0014】
核酸比較のための望ましいパラメーターは以下のアルゴリズムを含む:アルゴリズム:Needlemann and Wunsch,J.Mol.Biol.48:443-453(1970);Comparison matrix:一致=+10、不一致=0;Gap Penalty:50;Gap Length Penalty:3。Genetics Computer Group,Madison,WIのGapプログラムとして入手できる。上記に示したものは核酸比較のためのデホルトパラメーターである。任意に、アミノ酸類似性の程度を決定する際に、いわゆる「保存的」アミノ酸置換を考慮に入れることもでき、このことは、当業者には明らかである。保存的アミノ酸置換とは類似の側鎖を持つ残基の互換性のことである。例えば、脂肪族側鎖をもつアミノ酸の群はグリシン、アラニン、バリン、ロイシン及びイソロイシン;ヒドロキシ脂肪族側鎖を持つアミノ酸の群はセリン及びトレオニン;アミノ基を含む側鎖を持つアミノ酸の群はアスパラギン及びグルタミン;芳香族側鎖を持つアミノ酸の群はフェニルアラニン、チロシン、及びトリプトファン;塩基性側鎖を持つアミノ酸の群はリシン、アルギニン、及びヒスチジン;硫黄を含む側鎖を持つアミノ酸の群はシステイン及びメチオニンである。望ましい保存的アミノ酸置換群は:バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リシン−アルギニン、アラニン−バリン、及びアスパラギン−グルタミンである。本明細書において開示したアミノ酸配列の置換変異体は、開示配列中の少なくとも一つの残基が除去され、その場所に異なる残基が挿入されたものである。望ましくはアミノ酸の変換は保存的である。天然に存在するアミノ酸の望ましい保存置換は以下のようなものである:アラニンからセリンへ;アルギニンからリシンへ;アスパラギンからグルタミンまたはヒスチジン;アスパラギン酸からグルタミン酸へ;システインからセリンまたはアラニンへ;グルタミンからアスパラギンへ;グルタミン酸からアスパラギン酸へ;ヒスチジンからアスパラギンまたはグルタミンへ;イソロイシンからロイシンまたはバリンへ;ロイシンからイソロイシンまたはバリンへ;リシンからアルギニン;グルタミンまたはグルタミン酸;メチオニンからロイシンまたはイソロイシン;フェニルアラニンからメチオニン、ロイシンまたはチロシンへ;セリンからトレオニンへ;トレオニンからセリンへ;トリプトファンからチロシンへ;チロシンからトリプトファンまたはフェニルアラニンへ;及びバリンからイソロイシンまたはロイシン。
【0015】
核酸配列のハイブリッド形成
本発明のキシロース異性化酵素またはキシルロースキナーゼをコードする核酸配列は、中等度のまたは望ましくは厳密なハイブリッド形成条件下に、SEQ ID NO.2またはSEQ ID NO.4のヌクレオチド配列それぞれとハイブリッド形成する能力によって規定することもできる。厳密なハイブリッド形成条件とは、少なくとも約25、望ましくは約50ヌクレオチド、75または100及び最も望ましくは約200またはそれ以上のヌクレオチドの核酸配列を、約65℃の温度において、1Mの塩、望ましくは6xSSCまたは同等のイオン強度を持つその他の溶液中でハイブリッド形成させることができ、そして0.1Mの塩、またはそれ以下、望ましくは0.2xSSCまたは同等のイオン強度を持つその他の溶液中で65℃において洗浄する条件とここでは定義する。望ましくは、ハイブリッド形成は終夜、すなわち少なくとも10時間行い、望ましくは、洗浄は洗浄液を少なくとも2回交換して少なくとも1時間行う。この条件により、通常約90%またはそれ以上の配列同一性を持つ配列との特異的ハイブリッド形成が可能であろう。
【0016】
中等度の条件とは、少なくとも50ヌクレオチド、望ましくは約200またはそれ以上のヌクレオチドの配列を、約1M塩を含む溶液、望ましくは6xSSCまたは同等のイオン強度のその他の溶液の中で約45℃の温度においてハイブリッド形成させ、そして約1M塩を含む溶液、望ましくは6xSSCまたは相当するイオン強度のその他の溶液の中で室温において洗浄する条件とここでは定義する。望ましくは、ハイブリッド形成は終夜、すなわち少なくとも10時間行い、望ましくは、洗浄は洗浄液を少なくとも2回交換して少なくとも1時間行う。この条件により、通常約50%までの配列同一性を持つ配列との特異的ハイブリッド形成が可能であろう。当業者は、50%及び90%の間の同一性をもつ配列を特異的に同定するためにこれらのハイブリッド形成条件を変更できるであろう。
【0017】
作動的連結
本明細書で使用される「作動的に連結した」とは、機能的に関連するポリヌクレオチド配列の連結のことである。他の核酸配列との機能的な関連を持って配置されている場合に、核酸は「作動的に連結している」。例えば、プロモーターまたはエンハンサーは、それがコード配列の転写に影響するならば、コード配列と作動的に連結している。作動的に連結したとは、典型的には、連結したDNA配列同士が隣接しており、二個のタンパクコード領域を結合する必要がある場合には、連続し、読み枠内にあることを意味する。
【0018】
プロモーター
本明細書で使用される「プロモーター」とは、転写の方向の点で遺伝子の転写開始部位の上流に存在する1またはそれ以上の遺伝子の転写を調節する機能を持つ核酸フラグメントであって、DNA依存RNAポリメラーゼ、転写開始部位、及び限定はしないが、転写因子結合部位、リプレッサー及び活性化タンパク結合部位を含むその他のDNA配列、並びにプロモーターの転写の量を直接的若しくは間接的に調節することが当業者には既知のその他のヌクレオチドの配列によって構造的に同定される。「構成的」プロモーターはほとんどの環境及び発生条件において活性であるプロモーターである。「誘導的」プロモーターは環境及び発生が調節されたときに活性となるプロモーターである。
【図面の簡単な説明】
【0019】
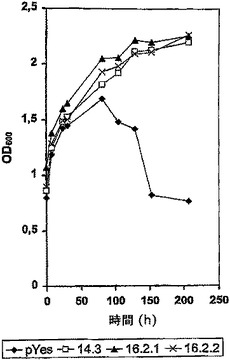
【図1】炭素源として25mMガラクトース及び100mMキシロースを含む培地で増殖した形質転換S.cerevisiaeの増殖曲線。形質転換体pYesは挿入のない酵母発現ベクターを含む。形質転換体14.3,16.2.1及び16.2.2はPiromycessp.E2キシロース異性化酵素コード配列を含むpYESベクターで形質転換されている。
【発明を実施するための形態】
【0020】
[発明の詳細な説明]
本発明の最初の態様は、キシロースからキシルロースへ異性化する能力を持つ形質転換宿主細胞に関するものである。キシロースからキシルロースへ異性化する能力は、キシロース異性化酵素をコードする核酸配列を含む核酸構築物で宿主細胞を形質転換することにより付与される。キシロースからキシルロースへ異性化する形質転換宿主細胞の能力は、キシロースからキシルロースへの直接の異性化である。これは、それぞれキシロース還元酵素およびキシリトール脱水素酵素に触媒されてキシリトール中間体を経由してキシロースからキシルロースへ変換する2ステップ反応に対して、キシロース異性化酵素により触媒される1ステップ反応によりキシロースがキシルロースへ異性化したことを意味すると理解される。
【0021】
核酸配列は、形質転換宿主細胞中で活性形態で発現するキシロース異性化酵素をコードすることが望ましい。従って、宿主細胞中で核酸配列が発現することにより、25℃でタンパクmg当り少なくとも10Uのキシロース異性化酵素比活性、望ましくは25℃で少なくとも20,25,30,50,100,200または300U/mg、の比活性を持つキシロース異性化酵素が生産される。本明細書において、形質転換宿主細胞中で発現したキシロース異性化酵素の比活性は、宿主細胞の細胞分解物、例えば、酵母細胞分解物のタンパクmg当りのキシロース異性化酵素活性の量と定義される。キシロース異性化酵素活性の測定、タンパク量、及び細胞分解物の調製については実施例1に記述されている。また、比活性は、実施例4に示すように測定することもできる。従って、宿主細胞中にヌクレオチド配列を発現させることにより、30℃で少なくとも50U/mgタンパクのキシロース異性化酵素活性、望ましくは30℃で少なくとも100,200,500または750U/mgの比活性を持つキシロース異性化酵素が生成される。
【0022】
望ましくは、宿主細胞中にヌクレオチド配列を発現することにより、キシロースに対するKmが50,40,30または25未満のキシロース異性化酵素が得られ、より望ましくは、キシロースに対するKmが約20mMまたはそれ未満である。
【0023】
キシロース異性化酵素をコードするヌクレオチド配列は、
(a)SEQ ID NO.1のアミノ酸配列と少なくとも40,45,49,50,53,55,60,70,80,90,95,97,98,または99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド配列;
(b)SEQ ID NO.2のヌクレオチド配列と少なくとも40,50,55,56,57,60,70,80,90,95,97,98,または99%の配列同一性を有するヌクレオチド配列を含むヌクレオチド配列;
(c)その相補鎖が、(a)または(b)の核酸分子配列にハイブリダイズするヌクレオチド配列;
(d)遺伝子コードの縮重による、(c)の核酸分子の配列と異なる配列からなるヌクレオチド配列:
からなる群から選択することができる。
【0024】
ヌクレオチド配列は、真核生物性キシロース異性化酵素、すなわち真核生物中に天然に存在するキシロース異性化酵素と同じアミノ酸配列を持つキシロース異性化酵素をコードしていることが望ましい。中等温度好性原核性キシロース異性化酵素に比べ、真核生物性キシロース異性化酵素の発現は、酵母のような真核性宿主細胞においてキシロース異性化酵素が活性形態で発現する可能性を高める。ヌクレオチド配列が植物キシロース異性化酵素(例えば、Hordeum vulgare由来)または菌(fungal)キシロース異性化酵素(例えば、Basidiomycets由来)をコードするのがより望ましい。但し、真核生物性宿主細胞、特に酵母において酵素的に活性形態で発現する可能性をさらに増加させるためには、ヌクレオチド配列が嫌気性菌のキシロース異性化酵素をコードすることが最も望ましい。Neocallimastix,Caecomyces,Piromyces,Orpinomyces,またはRuminomyces科に属する嫌気性菌のキシロース異性化酵素をコードするヌクレオチド配列が最も望ましい。
【0025】
キシロース異性化酵素をコードするヌクレオチド配列で形質転換するための宿主細胞は、細胞の中へ能動的にまたは受動的にキシロースを輸送できる宿主であることが望ましい。宿主細胞は、活性な解糖経路、ペントースリン酸経路を含有していることが望ましく、キシロースから異性化されたキシルロースがピルビン酸に代謝されるようにキシルロースキナーゼを含有していることが望ましい。さらに宿主はピルビン酸を目的とする発酵産物、例えばエタノール、エチレンまたは乳酸に変換するための酵素を含有することが望ましい。望ましい宿主細胞は、天然においてアルコール発酵、望ましくは嫌気性アルコール発酵ができる宿主細胞である。さらに宿主細胞はエタノール及び乳酸、酢酸、ギ酸のような有機酸、及びフルフラール及びヒドロキシメチルフルフラールのような糖分解産物に対して高度な耐性を持っていることが望ましい。宿主細胞のこれらの特性または活性は天然に宿主細胞中に存在することもあるが、遺伝子操作により導入または修飾することができる。適する宿主細胞は細菌または真菌のような微生物であるが、宿主として最も適当なのは酵母または糸状菌である。
【0026】
本明細書において酵母は、真核性微生物として定義され、主として単細胞の形で増殖するEumycotina亜門の全種(Alexopoulos,C.J.,1962,In:Introductory Mycology,John Wiley & Sons,Inc.,New York)を含む。酵母は単細胞葉状体の発芽によっても、また生物体の分裂によっても増殖することができる。宿主細胞として望ましい酵母は、Saccharomyces,Kluyveromyces,Candida,Pichia,Schizosaccharomyces,Hansenula,Kloeckera,Schwanniomyces,及びYarrowia種に属する。酵母は、嫌気性発酵、より望ましくは嫌気性アルコール発酵ができるものが望ましい。
【0027】
本明細書において糸状菌は、Eumycotina亜門の全ての糸状形態を含む真核性微生物として定義される。これらの菌は、キチン、セルロース、及びその他の複合多糖からなる栄養菌糸体によって特徴付けられる。本発明の糸状菌は、形態的、生理的、及び遺伝的に、酵母とは区別される。糸状菌による栄養増殖は菌糸の延長によるものであり、ほとんどの糸状菌の炭素異化は、絶対的好気性による。宿主細胞として望ましい糸状菌はAspergillus,Trichoderma,Humicola,Acremonium,Fusarium,及びPenicillium種に属するものである。
【0028】
数年にわたって、農産糖からバイオ−エタノールを生産するために、種々の生物体の導入が提案されてきた。しかしながら、実用的な主なバイオ−エタノール生産プロセスの全てにおいて、エタノール生産体としてSaccharomyces種の酵母を使用し続けてきた。これは工業的プロセスに対するSaccharomyces種の多くの魅力的な特徴、すなわち、高濃度の酸、エタノール及び浸透圧に対する耐性、嫌気的増殖能力、及び当然ではあるが高いアルコール発酵能力に基づくものである。宿主細胞として望ましい酵母の種は、S.cerevisiae,S.bulderi,S.barnetti,S.exiguus,S.uvarum,S.diastaticus,K.lactis,K.marxianus,K.fragilisを含む。
【0029】
宿主細胞は後に定義する核酸構築物で形質転換され、核酸構築物の一つのコピーを含むものでもよいが、複数のコピーを含むものが望ましい。この核酸構築物はエピソームとして維持され、ARS配列のような自律的複製のための配列を含む。適するエピソーム性核酸構築物は、例えば、酵母2μまたはpKD1(Fleer et al.,1991, Biotechnology 9: 968-975)プラスミドに基づくことができる。しかし、核酸構築物は、宿主細胞のゲノム中に1以上のコピーが組み込まれていることが望ましい。宿主細胞ゲノムへの組み込みは、変則的な組換えによりランダムに生じることもありうるが、菌分子遺伝学の分野でよく知られている相同組換えにより核酸構築物を宿主細胞ゲノムへ組み込むことが望ましい(例えば、WO90/14423,EP−A−0 481 008,EP−A−0 635 574及びUS6,265,186参照)。
【0030】
本発明による望ましい形質転換宿主細胞において、核酸構築物は、宿主細胞に、炭素源として、望ましくは唯一の炭素源としてのキシロースに対して、望ましくは嫌気的条件下で増殖する能力を付与する。それにより形質転換宿主は、好ましくはキシリトールを本質的に生産せず、例えば、当該宿主によって生産されるキシリトールは、検出限界以下、またはモルベースで消費した炭素の5,2,1%未満である。その形質転換宿主細胞は、唯一の炭素源としてのキシロースに対して少なくとも0.01,0.02,0.05,0.1または0.2h−1の速度で増殖する能力を有する。このように本発明の形質転換宿主細胞は、既に定義した比活性レベルでキシロース異性化酵素を発現する。
【0031】
宿主細胞は、(a)宿主細胞中へのキシロース輸送の増加;(b)キシルロースキナーゼ活性の増加;(c)ペントースリン酸経路の流量の増加;(d)カタボライト抑制に対する感受性の減少;(e)エタノール、浸透圧または有機酸に対する耐性の増加;及び(f)副生物生産の減少、からなる群から選択された特徴の一つまたはそれ以上を生じる遺伝子修飾をさらに含むことができる。副生物は目的とする発酵産物以外の炭素含有分子を意味すると理解され、例えば、キシリトール、グリセロール及び/または酢酸が含まれる。そのような遺伝子修飾は、古典的突然変異誘起及びスクリーニング及び/または目的変異体の選別により導入することができる。その他に、遺伝子修飾は、外来性遺伝子の過剰発現及び/または異種遺伝子の発現及び/または内在性遺伝子の不活化を含むことができる。これらの遺伝子は、ヘキソースまたはペントーストランスポーター;S.cerevisiae(XKSI Deng and Ho, 1990, Appl. Biochem. Biotechnol. 24-25: 193-199)またはPiromyces(xylB,すなわちSEQ ID NO.4)のキシルロースキナーゼのようなキシルロースキナーゼ;トランスアルドラーゼ(TAL1)またはトランスケトラーゼ(TKL1)のようなペントースリン酸経路の酵素(例えば、Meinander et al.,1995,Pharmacol.Toxicol.Suppl,2:45参照)、解糖酵素、アルコール脱水素酵素のようなアルコール代謝酵素、をコードする遺伝子から選択されることが望ましい。不活化される望ましい内在性遺伝子としては、例えば、S.cerevisiae HXK2遺伝子(Diderich et al., 2001, Appl. Environ. Microbiol. 67: 1587-1593参照)のようなヘキソースキナーゼ;S.cerevisiae MIG1またはMIG2遺伝子;S.cerevisiae GRE3遺伝子(Traff et al.,2001,Appl.Environm.Microbiol.67:5668-5674)のような(非特異的)アルドース還元酵素遺伝子;S.cerevisiaeグリセロール−リン酸脱水素酵素1及び/または2遺伝子のようなグリセロール代謝に関係する酵素の遺伝子;またはその他の宿主の種の遺伝子の(ハイブリッド形成)相同体、が含まれる。宿主細胞のキシロース代謝に関するその他の望ましい修飾についてはZaldivar et al.(2001、前出)に総説されている。
【0032】
その他の態様において、本発明は、エタノール以外の発酵産物を生産するための形質転換宿主細胞に関係している。その非エタノール発酵産物は原則として酵母または糸状菌のような真核性微生物によって生産することができる大量のまたは精製した化学品である。そのような発酵産物としては、例えば、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質及びセファロスポリンがある。
【0033】
本発明の核酸構築物による宿主細胞の形質転換及び宿主細胞、望ましくは酵母の前記のようなその他の遺伝子修飾は当業者によく知られている方法により行われる。その方法は例えば標準的教科書から知ることができる、例えば、Sambrook and Russel(2001)“Molecular Cloning: A Laboratory Manual(3rd edition)”,Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, または F. Ausubel et al, eds.,“Current protocols in molecular biology”,Green Publishing and Wiley Interscience, New York (1987)。菌宿主細胞の形質転換及び遺伝子修飾の方法は、例えば、EP−A−0 635 574,WO98/46772,WO99/60102及びWO00/37671から知ることができる。
【0034】
その他の態様において、本発明は、前記で定義したキシロース異性化酵素をコードし、前記の宿主細胞の形質転換に使用されるヌクレオチド配列からなる核酸構築物に関する。核酸構築物において、キシロース異性化酵素をコードするヌクレオチド配列は、後述するように宿主細胞中でヌクレオチド配列の転写を調節し開始するためのプロモーターに作動的に連結していることが望ましい。このプロモーターは、宿主細胞にキシロースをキシルロースに異性化する能力を付与するために、宿主細胞中でキシロース異性化酵素を十分に発現できることが望ましい。プロモーターは、前記のような宿主細胞中で、特異的キシロース異性化酵素を生じさせるものが望ましい。本発明の核酸構築物中において有用なプロモーターは、構成的及び誘導的な天然プロモーター並びに人工的プロモーターである。さらに本発明に使用するための望ましいプロモーターは、カタボライト(グルコース)抑制に感受性がなくそして/または誘導のためのキシロースを必要としないものが望ましいであろう。このような特徴を持つプロモーターは、広く入手可能であり、同業者には知られている。そのようなプロモーターの適当な例は、例えば、酵母のリン酸フルクトキナーゼ(PPK)、トリオースリン酸異性化酵素(TPI)、グリセロアルデヒド−3−リン酸脱水素酵素(GPD,TDH3またはGAPDH)、ピルビン酸キナーゼ(PYK)、ホスホグリンセリン酸キナーゼ(PGK)プロモーターのような解糖遺伝子の酵母プロモーターである。そのようなプロモーターに関する詳細は(WO93/03159)に見ることができる。その他の有用なプロモーターは、リボソームタンパクコード遺伝子プロモーター、ラクターゼ遺伝子プロモーター(LAC4)、アルコール脱水素酵素プロモーター(ADH1,ADH4,など)、及びエノラーゼプロモーター(ENO)である。その他の、構成的及び誘導的プロモーター及びエンハンサーまたは上流活性化配列は当業者に知られているであろう。本発明の核酸構築物に使用されるプロモーターは、必要に応じて、修飾して、その調節特徴を変更することができる。キシロース異性化酵素を発現させるために核酸構築物に使用されるプロモーターは、キシロース異性化酵素を発現させる宿主細胞と同種であることが望ましい。
【0035】
核酸構築物において、キシロース異性化酵素をコードするヌクレオチド配列の3’−末端は、転写ターミネーター配列に作動的に連結していることが望ましい。このターミネーター配列は、選択した宿主細胞、例えば選択した酵母種において作動し得ることが望ましい。いずれの場合にも、ターミネーターの選択は重要ではなく、ターミネーターは酵母ではない真核生物の遺伝子の場合にしばしば作動するが、酵母遺伝子由来のものを用いることができる。転写終結配列はさらにポリアデニル化シグナルを含むことが望ましい。
【0036】
任意に、核酸構築物中に選択マーカーを入れることができる。本明細書で使用する用語「マーカー」とは、マーカーを含有する宿主細胞を、選別、またはスクリーニングすることができる特徴または表現型をコードする遺伝子のことである。マーカー遺伝子は、抗生物質耐性遺伝子とすることができ、それによって形質転換されていない細胞の中から形質転換された細胞を選別するために適当な抗生物質を使用することができる。適当な抗生物質耐性マーカーの例としては、例えば、ジヒドロ葉酸還元酵素、ヒグロマイシン−B−ホスホトランスフェラーゼ、3’−O−ホスホトランスフェラーゼII(カナマイシン、ネオマイシン及びG418耐性)が含まれる。抗生物質耐性マーカーは、倍数体宿主細胞の形質転換には最も便利であるが、しかし栄養要求性マーカー(URA3,TRP1,LEU2)またはS.pombe TPI遺伝子(Russell P R,1985,Gene40:125−130に記載)のような非抗生物質耐性マーカーが使用されることが望ましい。望ましい態様において、核酸構築物により形質転換された宿主細胞はマーカー遺伝子を含まない。マーカー遺伝子を含まない組換え微生物宿主細胞を作る方法はEP−A−0 635 574に開示されており、A.nidulans amdS(アセトアミダーゼ)遺伝子または酵母URA3及びLYS2遺伝子のような二方向性マーカーの使用に基づいている。そのほかには、形質転換細胞をスクリーニングできるように、緑色蛍光タンパク、lacZ,ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、ベータ−グルクロニダーゼのようなスクリーニングに役立つマーカーを本発明の核酸構築物の中に組み込むことができる。
【0037】
さらに任意に本発明の核酸構築物中に存在することができる配列としては、これらに限定はしないが、一つまたはそれ以上のリーダー配列、エンハンサー、組み込み因子、並びに/或いはレポーター遺伝子、イントロン配列、セントロメア、テロメア及び/またはマトリックス接着(MAR)配列がある。本発明の核酸構築物はさらに、ARS配列のような自己複製のための配列を含むことができる。適するエピソーム核酸構築物は、例えば、酵母2μまたはpKD1(Fleer et al., 1991, Biotechnology 9: 968-975)プラスミドを基にすることができる。その他に核酸構築物は、望ましくは相同組換えによる、組み込みのための配列を含むことができる。従ってその配列は、宿主細胞のゲノム中の組み込みの標的部位に対して相同的な配列である。本発明の核酸構築物は、本来既知方法により提供することができ、その方法には核酸/核酸配列を制限及び連結するような技術を含み、その参考文献は例えば、Sambrook and Russel (2001) “Molecular Cloning: A Laboratory Manual (3rd edition), Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, またはF. Ausubel et al., eds.,”Current protocols in molecular biology”,Green Publishing and Wiley Interscience, New York (1987)のような標準的教科書に示されている。
【0038】
そのほかの態様において、本発明はキシロース異性化酵素をコードするヌクレオチドを含む核酸分子に関する。この核酸分子は、
(a)SEQ ID NO.1のアミノ酸配列と少なくとも50,53,54,55,60,70,80,90,95,97,98,または99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードする核酸分子;
(b)SEQ ID NO.2のヌクレオチド配列と少なくとも50,56,57,58,60,70,80,90,95,97,98,または99%の配列同一性を有するヌクレオチド配列を含む核酸分子;
(c)その相補鎖が、(a)または(b)の核酸分子配列にハイブリダイズする核酸分子;及び
(d)遺伝子コードの縮重により(c)の核酸分子配列と異なる配列からなる核酸分子;
からなる群から選択されることが望ましい。
【0039】
また、(a)の核酸分子は、SEQ ID NO.1のアミノ酸配列と少なくとも67,68,69,70,80,90,95,97,98,または99%の配列類似性を有するアミノ酸配列を含むポリペプチドをコードすることができる。(c)の核酸分子は望ましくは前記に定義した中等度の条件、より望ましくは厳密な条件の下にハイブリッド形成する。核酸分子は真核生物由来であることが望ましく、菌のような真核微生物由来であることがより望ましく、前記の嫌気性菌のような嫌気性菌由来であることが最も望ましい。
【0040】
本発明の更に他の態様は、キシルロースキナーゼ、望ましくはD−キシルロースキナーゼをコードするヌクレオチド配列を含む核酸分子に関するものである。本明細書において、D−キシルロースキナーゼ(EC2.7.1.17;D−キシルロキナーゼとも呼ばれる)は、D−キシルロースのキシルロース−5−リン酸への変換を触媒する酵素である。この核酸分子は、
(a)SEQ ID NO.3のアミノ酸配列と少なくとも45,47,48,49,50,55,60,70,80,90,95,97,98,または99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードする核酸分子;
(b)SEQ ID NO.4のヌクレオチド配列と少なくとも30,37,38,39,40,50,60,70,80,90,95,97,98,または99%の配列同一性を有するヌクレオチド配列からなる核酸分子;
(c)その相補鎖が、(a)または(b)の核酸分子配列にハイブリダイズする核酸分子;及び
(d)遺伝子コードの縮重による、(c)の核酸分子配列と異なる配列からなる核酸分子:
からなる群から選択されることが望ましい。
【0041】
また、(a)の核酸分子は、SEQ ID NO.3のアミノ酸配列と少なくとも64,65,66,70,80,90,95,97,98,または99%の配列類似性を有するアミノ酸配列を含むポリペプチドをコードすることができる。(c)の核酸分子は望ましくは前記で定義した中等度の条件、より望ましくは厳密な条件の下にハイブリッド形成する。核酸分子は、真核生物由来であることが望ましく、菌のような真核微生物由来であることがより望ましく、前記の嫌気性菌のような嫌気性菌由来であることが最も望ましい。
【0042】
そのほかの態様において、本発明は、本発明の形質転換宿主細胞がキシロースのようなキシロース資源を含む炭素源の発酵のために使用される、発酵プロセスに関するものである。発酵培地中の炭素源は、キシロース資源に加えてグルコース資源も含むことができる。キシロースまたはグルコースの資源としては、キシロースまたはグルコースそのものでもよく、例えば、リグノセルロース、キシラン、セルロース、デンプンなどのようなキシロースまたはグルコース単位からなる炭水化物のオリゴマーまたはポリマーでもよい。そのような炭水化物からキシロースまたはグルコース単位を遊離させるために(キシラナーゼ、グルカナーゼ、アミラーゼなどのような)、適当な炭水化物分解酵素を、発酵培地に加えてもよく、形質転換宿主細胞に生産させてもよい。後者の場合には、形質転換宿主細胞に遺伝子操作を施し、その炭水化物分解酵素を生産させ、分泌させることができる。望ましいプロセスにおいて、形質転換宿主細胞は、キシロース及びグルコースを共に、望ましくは同時に発酵し、その場合にはグルコース抑制に非感受性の形質転換宿主細胞を使用してジオキシ増殖を阻害することが望ましい。炭素源としてのキシロース(及びグルコース)に加えて、発酵培地はさらに形質転換宿主細胞の増殖に必要な適当な成分を含むであろう。酵母などの微生物を増殖するための発酵培地の組成は、当業者にはよく知られている。
【0043】
発酵プロセスは、エタノール、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、ペニシリンG若しくはペニシリンV及びそれらの発酵誘導体のようなβ−ラクタム並びにセファロスポリンなどの発酵産物を生産するためのプロセスである。発酵プロセスは、好気的または嫌気的な発酵プロセスとすることができる。本明細書において嫌気的発酵プロセスとは、発酵プロセスが無酸素の状態でまたは実質的に酸素が消費されないで(例えば、5mmol/L/h未満)で行われ、有機分子が、電子供与体及び電子受容体のいずれとしても働く発酵プロセスと定義される。酸素が存在しないところでは、糖分解及びバイオマス形成において生成したNADHは、酸化的リン酸化により酸化することはできない。この問題を解決するために多くの微生物は電子及び水素受容体としてピルビン酸またはその誘導体の一つを利用することによって、NAD+を再生する。従って、望ましい嫌気的発酵プロセスにおいて、ピルビン酸は、電子(及び水素)受容体として使用され、エタノール、乳酸、1,3−プロパン−ジオール、エチレン、酢酸またはコハク酸のような発酵産物へと還元される。
【0044】
発酵プロセスは、形質転換宿主細胞にとって最適の温度で行うことが望ましい。従って、ほとんどの酵母または菌の宿主細胞に対して、発酵プロセスは38℃未満の温度で実施される。酵母または糸状菌の宿主細胞に対して、発酵プロセスは35,33,30または28℃未満、且つ20,22,または25℃を超える温度で実施されることが望ましい。
【0045】
望ましいプロセスは、(a)前記に定義した形質転換宿主細胞と共にキシロース資源を含む培地を発酵することによって、宿主細胞にキシロースをエタノールに発酵させ;任意に、(b)エタノールを回収するステップを含むエタノールの生産プロセスである。発酵培地は、グルコース資源を含むことができ、それもエタノールに発酵される。このプロセスにおいて、エタノールの容積生産性は、少なくとも0.5,1.0,1.5,2.0,2.5,3.0,5.0または10.0gエタノール/リットル/時間であることが望ましい。このプロセスにおけるキシロース及び/またはグルコースに対するエタノール収率は、少なくとも50,60,70,90,95または98%であることが望ましい。本明細書においてエタノール収率は、理論的収量(グルコース及びキシロースについて0.51gエタノール/gグルコースまたはキシロース)のパーセンテージとして定義する。
【0046】
その他の態様において、本発明は、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、β−ラクタム抗生物質およびセファロスポリンからなる群から選択される発酵産物の生産プロセスに関する。このプロセスは、(a)前記に定義した形質転換宿主細胞と共にキシロース資源を含む培地を発酵することにより、宿主細胞に、キシロールを発酵産物に発酵させ、任意に、(b)発酵産物を回収する、ステップを含むことが望ましい。望ましいプロセスにおいて、培地はグルコース資源も含む。
【実施例】
【0047】
[実施例1]Piromycesキシラナーゼ異性化酵素及びキシルロースキナーゼcDNAのクローニング
(生物及び増殖条件)
インド象の糞から単離した嫌気性菌Piromyces sp.E2(ATCC76762)を、N2/CO2(80%/20%)中、39℃で、種々の炭素源を添加したM2培地で嫌気的に増殖した(24)。使用した炭素源は、アビセル(微結晶セルロース、タイプPH105、Serva,ドイツ)、フルクトースまたはキシロース(全て0.5%、w/v)であった。増殖が止まった後(水素の発生により判断した)、細胞を遠心分離(15,000xg,4℃,15分間)またはナイロンガーゼ(30μm孔径)による濾過により回収した。
【0048】
(無細胞抽出物の調製)
菌細胞を脱イオン水で洗い、培地成分を除去した。細胞を液体窒素中で凍結し、次いで、乳鉢中で、硝子ビーズ(0.10−0.11mm径)ですりつぶして、無細胞抽出物を調製した。Tris/HCl緩衝液(100mM,pH7.0)を粉末に加え(1:1,w/v)そして15分間解凍した後、懸濁液を遠心分離した(18,000xg,4℃,15分間)。透明な上清を細胞内酵素の原料として使用した。
【0049】
(酵素検定)
キシロース異性化酵素活性は、50mMリン酸緩衝液(pH7.0),10mMキシロース,10mM MgCl2及び適当量の無細胞抽出物を含む反応混合物中37℃で検定した。生成したキシルロースの量は、システイン−カルバゾール法(9)により測定した。キシルロースキナーゼ及びキシロース還元酵素活性はWitteveen et al.(28)による記述にしたがって検定した。活性の1単位は、検定条件の下に、毎分1 nmolのキシルロースを生産する酵素量と定義される。生成したキシルロースは、Dische and Borenfreund(Dische and Borenfreund, 1951, J. Biol. Chem. 192: 583-587)の方法によるか、または80℃のBiorad HPX−87Nカラムを使用し、溶出液として0.01M Na2HPO4を使用して0.6ml/minで溶出するHPLCにより測定した。キシロース及びキシルロースは、内部温度60℃で屈折計により測定した。
【0050】
比活性は、タンパク1mg当りの単位として示す。タンパクは、ウシγ−グロブリンを標準としてBio−Radタンパク試薬(Bio-Rad Laboratories,Richmond,CA,米国)で測定した。
【0051】
(Piromyces sp.E2cDNAライブラリーのランダム配列)
既に記述されている(2)ベクターラムダZAPII中に構築したcDNAライブラリーを使用した。このライブラリーの一部をExAssistヘルパーファージ(Stratagene,La Jolla,CA,米国)による大量切除(mass excission)によりpBluescript SK−クローンに変換した。無作為に取り出したクローンをM13逆プライマーを使用して配列解析し、5’部分の配列を得た。不完全cDNAを使用してプローブを合成し、ライブラリーを再スクリーニングするために使用した。全長配列を得るためにpUC18にサブクローニングを生成さた。配列分析は、dRhodamineターミネーターサイクルシークエンス即時反応DNAシークエンスキット(Perkin-Elmer Applied Biosystems)を使用してABIプリズム310自動化シークエンサーで実施した。
【0052】
(結果)
嫌気性菌Piromyces sp.E2のcDNAライブラリーから無作為に選択したクローンを配列解析し、その結果キシロース異性化酵素及びD−キシルロキナーゼ遺伝子にそれぞれ高い相同性を示す二つのクローン(pH97及びpAK44)を得た。これらのクローンを詳細に解析した。
【0053】
クローンpH97は完全なORFを含有していなかったので、クローンpH97の配列データに基づいて設計したプローブを使用してcDNAライブラリーを再度スクリーニングした。この結果1669bpの挿入配列を有するクローンpR3を得た。キシロース異性化酵素に高い類似性のある437アミノ酸のタンパクをコードするORFを同定することができた。5’非翻訳領域はわずか4bpを含むのみであったが、推定開始メチオニン残基は、既知キシロース異性化酵素配列の整列によく一致した。3’非翻訳領域は351bpの長さであり、嫌気性菌で典型的である、高いAT含有率を有していた。ORFは、基質との相互作用(触媒トリアド(catalytic triad)His102,Asp105,Asp340及びLys235)及びマグネシウムとの結合(Glu232)に重要であることが示されているアミノ酸を含有していた(14,26)。さらに、キシロース異性化酵素のために開発した(20)2個の符号パターン(残基185−194、及び230−237)が存在した。このPiromyces sp.E2キシロース異性化酵素(XylA)はHaemophilus influenzaの酵素(52%同一性、68%類似性)及びHordeum vulgare(49%同一性、67%類似性)と高い相同性を示す。cDNA配列から推定されるポリペプチドは49,395Daの分子量に相当し、5.2の計算値pIを有す。
【0054】
二番目のクローンpAK44は、2041bpの挿入を有し、53,158Daの分子量及び5.0のpIを持つ494アミノ酸のタンパクをコードする完全ORFを含有した。最初のメチオニンに先行して111bpの5’非翻訳領域が存在したが、3’非翻訳領域は445 bpからなっていた。両領域はATが多い。BLAST及びFASTA調査によりキシルロキナーゼとの高い類似性が明らかにされた。Rodriguez−Pefia et al.(22)により定義された2個のリン酸共通領域が部分整列に示されるように位置6−23及び254−270に認められた。さらにPrositeデーターベースに記述されているこの糖キナーゼファミリーの記号が同定された(131−145及び351−372)。Piromyces sp.E2キシルロキナーゼ(XylB)はHaemophilus influenzaのXylBと高い相同性を示した(46%同一性、64%類似性)。
【0055】
[実施例2]酵母発現ベクターの構築
Piromyces sp.E2のキシロース異性化酵素のSaccharomyces cerevisiae中の発現
Piromyces sp.E2のcDNAをpfuポリメラーゼ(Stratgene)を使用するPCR反応に使用した。プライマーはキシロース異性化酵素の5’及び3’末端の配列を使用して設計し、SfiI及びXbaI制限部位を含めた。PCR産物をpPICZαベクター(Invitrogen,Carlsbad,CA,米国)中にクローニングした。キシロース異性化酵素を取り出すために、pPICZαベクターをEcoRI及びXbaIで消化した。消化産物をpYes2ベクター(Invitrogen)に連結した。キシロース異性化酵素を含むpYes2プラスミドをSaccharomyces cerevisiae(株BJ1991,Beth Johns,UvAから供与)に導入した。この株の遺伝型は:matα,leu2,trp1,ura3−251,prb1−1122及びpep4−3、である。形質転換細胞をSCプレート(0.67% YNB培地+0.05% L−Leu+0.05% L−Trp+2%グルコース+2%アガロース)に接種した。
【0056】
形質転換Saccharomyces cerevisiae細胞をグルコース培地で25℃で72時間増殖した(グルコースの代わりにラフィノースを使用することができる)。細胞を回収し、グルコースの代わりにガラクトースを加えたSC培地中に再懸濁した。8時間の誘導後、細胞を回収し、硝子ビーズ(0.10−0.11mm径)及び「破壊緩衝液」(50mMリン酸緩衝液+5%グリセロール+プロテアーゼ阻害剤)を使用して分解した。分解の後混合物を遠心分離(18,000xg,4℃,15分間)した。透明上清を使用して前記方法(実施例1)によりキシロース異性化酵素活性を測定した。10U/mgタンパクの活性が37℃において測定された。
【0057】
[実施例3]キシロース上での形質転換酵母株の増殖
(培地組成)
Saccharomyces cerevisiae株を下記組成のSC−培地上で増殖した:0.67%(w/v)酵母窒素塩基;0.01%(w/v)L−トリプトファン;0.01%(w/v)L−ロイシン及びグルコース、ガラクトースまたはキシロースまたはこれら基質の組合せのいずれか(下記参照)。寒天培地用に培地に2%(w/v)細菌用寒天を加えた。
【0058】
(増殖実験)
挿入のないpYes2で形質転換されたSaccharomyces cerevisiae株BJ1991(遺伝型:matα,leu2,trp1,ura3−251,prb1−1122,pep4−3)及びPiromyces sp.E2キシロース異性化酵素遺伝子を持つpYes2を含む形質転換細胞(16.2.1;16.2.2及び14.3)を、炭素源として10mMグルコースを含むSC−寒天プレート上で増殖した。コロニーが見えるようになったとき、一つのコロニーを使用して、炭素源として100mMキシロース及び25mMガラクトースを含むSVC液体培地に接種した。LKB Ultrospec K分光光度計を使用して600nmの光学密度の増加を測定することにより増殖を監視した。
【0059】
(結果)
増殖実験の結果を図1にまとめた。挿入のないpYes2で形質転換したBJ1991株の培養は80時間までOD600の増加を示した。この後徐々に減少が観察された。これは増殖の末期にしばしば観察される酵母細胞の凝集によるものである。3種の形質転換細胞は、80時間後も増殖を止めず、少なくとも150時間までさらに増加を示した。
【0060】
[実施例4]Saccharomyces cerevisiaeにおいてPiromyces sp.E2キシロース異性化酵素を構成的に発現するための新規で改良された酵母発現ベクターの構築
キシロース異性化酵素をコードするPiromyces sp.E2遺伝子を含むpPICZαベクターを、VentR DNAポリメラーゼ(New England Biolabs)によるPCRの鋳型として使用した。プライマーはキシロース異性化酵素をコードする遺伝子の5’及び3’配列を使用して設計し、EcoRI及びSpeI部位を含めた。さらに、プライマーはpPICZα構築中に認められるXbaI部位を除去し、その代わりに終結コドン(TAA)を入れて設計した。最終産物は、pPICZα構築に認められる追加のアミノ酸(his及びc−Mycタグ)のない、元のオープンリードフレームを復元するように設計した。PCR産物をEcoRI及びSpeIで切り出した。最終産物をpYES2(Invitrogen)由来のベクター中にクローニングした。このベクター中において、キシロース異性化酵素の構成的発現を確実にし、それにより培地にガラクトースを加える必要をなくするために、pYES2中にあるGAL1プロモーターをTPI1プロモーターに置換した。TPI1プロモーターはプラスミドpYX012(R&D systems)の修飾型からクローニングした。このプロモーターをNheI−EcoRIフラグメントとして切り出した。
【0061】
このTPI1プロモーター及びキシロース異性化酵素をコードする遺伝子のPCR産物を共に、SpeI及びXbaIで切り出したpYES2に連結した。このプラスミドを使用してSaccharomyces cerevisiae株CEN.PK113−5D(Peter Kotter,Frankfurtより供与)を形質転換した。この株の遺伝型は:MatA ura3−52である。形質転換細胞は、炭素源として2%グルコースを加えたミネラル培地プレート上で選別した(Verduyn et al.:Effect of benzoic acid on metabolic fluxes in yeasts; a continuous-culture study on the regulation of respiration and alcoholic fermentation.(1992) Yeast 8(7): 501-17)。形質転換していない細胞は、このプレート上では増殖できない。
【0062】
形質転換細胞を、炭素限定恒成分培養において、グルコース/キシロース混合物に対して増殖した。この条件下で増殖した形質転換細胞は、Dersters−Hildersson et al.(Kinetic characterization of D-xylose isomerases by enzymatic assays using D-sorbitol dehydrogenase. Enz. Microb. Techol. 9 (1987) 145-148)により開発された特異的酵素検定により、高いキシロース異性化活性(30℃において800単位/mg)を示す。形質転換S.cerevisiae株の無細胞抽出物中のキシロース異性化酵素のインビトロ活性は、2価カチオン(Mg2+またはCo2+)に依存し、キシロースに対して約20mMの比較的低いKm値が認められた。
【0063】

【技術分野】
【0001】
本発明は、真核生物性キシロース異性化酵素をコードする核酸配列で形質転換した宿主細胞に関するものである。このキシロース異性化酵素は、宿主細胞で発現し、キシロースをキシルロースに変換する能力を付与する。この宿主細胞は、ペントース含有培地の発酵によるエタノール及びその他の発酵産物の製造プロセスに使用される。さらに本発明は、真核生物性キシロース異性化酵素をコードする核酸配列に関するものである。
【背景技術】
【0002】
この数十年間における古典的化石燃料(石油燃料)の大量消費は、高レベルの汚染の原因となっている。さらに、世界中の石油埋蔵量は無限でないことが認識され、このような認識は、環境問題への関心の高まりと相俟って、CO2排出の60−90%減少を実現できるエタノールのような代替燃料の可能性を研究する新しい動きを生んだ。バイオマス由来のエタノールは、多数の種々の資源から得られるヘキソース糖の発酵により生産することができるが、にも拘らず工業的規模の生産または燃料アルコールに向けられる原料は、ショ糖及びトウモロコシデンプンである。これらの原料の欠点は高価なことである。
【0003】
燃料アルコールの生産拡大には、低コストの供給原料の使用を可能にすることが求められる。現在、植物バイオマスからは唯一リグノセルロース原料がかなり大量に使用可能であり、エタノール生産用の農産物を代替している。リグノセルロース原料の主な発酵可能な糖はグルコース及びキシロースであり、それぞれリグノセルロース中に約40%及び25%含有される。しかし、Saccharomyces cerevisiaeのようなアルコール発酵ができる酵母のほとんどは、炭素源としてキシロースを使用できない。さらに、高収量及び高生産性でキシロースをエタノールに発酵できる微生物は知られていない。リグノセルロース加水分解産物からエタノールを商業的に生産することを可能にするには、これらの性質を持つ微生物が求められるであろう。従って本発明の一つの目的は、アルコール発酵ができ、炭素源としてキシロースを利用できる酵母を提供することである。
【0004】
D−キシロースは、腸内細菌、一部の酵母及び真菌のような種々の微生物によって代謝される。大部分のキシロース利用細菌において、キシロースはキシロース(グルコース)異性化酵素(XI)により直接D−キシルロースに異性化される。しかし、糸状菌及び酵母は、この一ステップの異性化を行うことができず、まずキシロース還元酵素(XR)の作用によりキシロースをキシリトールに還元し、次いでキシリトール脱水素酵素(XDH)によりキシリトールをキシルロースに変換する。最初のステップは補因子としてNAD(P)Hを必要とし、第二ステップはNAD+を必要とする。生成したキシルロースは、その後、キシルロースキナーゼ(XK)によりリン酸化された後にペントースリン酸経路(PPP)に入る。厳密なNADPH依存性のキシロース還元酵素(XR)を持つ微生物では、キシロースからエタノールへの嫌気性発酵は不可能である。キシリトール脱水素酵素(XDH)は厳密にNAD+に依存しているので、酸化還元不均衡(すなわち、NAD+欠乏)を生じるからである。嫌気性条件においてこの酸化還元不均衡を解決するために、微生物はグリセロール及びキシリトールのような副産物を生産する。同様に、キシロースに対するβ−ラクタムの嫌気的な生産も、グルコースに対するβ−ラクタム生産に比較すると生じにくい。これらの低収量の原因は、グルコースを利用する場合に比較して、この経路においてはNADPHの形の還元体を比較的高度に必要とすることにあるように思われる(W.M.van Gulik et al.,Biotechnol Bioeng.Vol.68,No.6,June 20,2000)。
【0005】
Zaldivar et al.(2001,Appl.Microbiol.Biotechnol.56;17-34)に総説されているように、S.cerevisiae及び類似の酵母にキシロース代謝を導入する多くの試みが行われてきた。一つの方法は、少なくとも、キシロース(アルドース)還元酵素及びキシリトール脱水素酵素、すなわちPichia stipitisのXYL1及びXYL2、をコードする遺伝子をS.cerevisiaeに発現させることに関する(US5,866,382;WO95/13362;及びWO97/42307)。この方法は、キシロースに対してS.cerevisiaeの増殖を可能にしたが、主にXR及びXDHの間の酸化還元不均衡の結果として、一般的にエタノール生産性及び/または収量が低く、キシリトールの生産が多いという欠点がある。
【0006】
S.cerevisiae若しくは関連酵母または糸状菌にXIを発現させることにより、酸化還元不均衡及びその結果としてのキシリトール生産と分泌を回避することができるであろう。数種の細菌のキシロース異性化酵素遺伝子がS.cerevisiaeに挿入されたが、S.cerevisiaeにおいて中等温度好性原核生物のXIを発現させても、活性のあるXIを生じなかった(Amore and Hollenverg, 1989, Nucleic Acids Res. 17:7515 ; Amore et al., 1989, Appl. Microbiol. Biotechnol.30:351-357; Chan et al., 1986, Biotechnol. Lett 8:231-234;Chan et al., 1989,Appl.Microbiol.Biotechnol. 31:524-528; Ho et al., 1983, Fed. Proc. Fed. Am. Soc. Exp. Biol. 42 : 2167 ; Hollenberg, 1987, EBC Symposium on Brewer's Yeast, Helsinki(Finland), 24-25 Nov 1986 ; Sarthy et al., 1987, Appl. Envion. Microbiol. 53:1996-2000 ; Ueng et al., 1985, Biotechnol. Lett. 7: 153-158)。しかし、S.cerevisiaeに発現させた好熱性細菌の二種のXIは85℃において1μmol/min/mgの比活性を示した(Bao et al.,1999,Weishengwu-Xuebao 39:49-54;Walfridson et al.,1996,Appl.Environ.Microbiol.61:4184-4190)。しかし、S.cerevisiaeの生理的温度(20−35℃)では、この活性のわずか数%しか維持できず、キシロースから有効にアルコールを発酵するには不十分であった。従って、生理的条件下において十分なXI活性を提供して、炭素源としてキシロースを利用することを可能とするように、酵母で発現することができるXIをコードする核酸に対する要求は依然として存在する。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】US5,866,382
【特許文献2】WO95/13362
【特許文献3】WO97/42307
【非特許文献】
【0008】
【非特許文献1】W.M.van Gulik et al.,Biotechnol Bioeng. Vol.68, No.6, June 20,2000
【非特許文献2】Zaldivar et al., 2001, Appl.Microbiol.Biotechnol.56;17-34)
【非特許文献3】more and Hollenverg, 1989, Nucleic Acids Res. 17:7515
【非特許文献4】Amore et al., 1989, Appl. Microbiol. Biotechnol.30:351-357
【非特許文献5】Chan et al., 1986, Biotechnol. Lett 8:231-234
【非特許文献6】Chan et al., 1989,Appl.Microbiol.Biotechnol. 31:524-528
【非特許文献7】Ho et al., 1983, Fed. Proc. Fed. Am. Soc. Exp. Biol. 42 : 2167
【非特許文献8】Hollenberg, 1987, EBC Symposium on Brewer's Yeast, Helsinki(Finland), 24-25 Nov 1986
【非特許文献9】Sarthy et al., 1987, Appl. Envion. Microbiol. 53:1996-2000
【非特許文献10】Ueng et al., 1985, Biotechnol. Lett. 7: 153-158
【非特許文献11】Bao et al.,1999,Weishengwu-Xuebao 39:49-54;Walfridson et al.,1996,Appl.Environ.Microbiol.61:4184-4190
【発明の概要】
【課題を解決するための手段】
【0009】
[発明の説明]
(定義)
キシロース異性化酵素
本明細書において、酵素「キシロース異性化酵素」(EC5.3.1.5)は、D−キシロースのD−キシルロースへのの直接の異性化、及びその逆の直接の異性化を触媒する酵素として定義される。この酵素はまた、D−キシロースケト異性化酵素としても知られている。一部のキシロース異性化酵素は、D−グルコース及びD−フルクトースの間の変換を触媒することもできるので、時にはグルコース異性化酵素とも呼ばれる。キシロース異性化酵素は補因子としてマグネシウムを必要とする。本発明のキシロース異性化酵素はさらに後述するアミノ酸配列によって定義される。同様に、キシロース異性化酵素はこの酵素をコードするヌクレオチド配列並びに後述されるキシロース異性化酵素をコードする対照ヌクレオチド配列にハイブリダイズするヌクレオチド配列によって定義することができる。
【0010】
本明細書において、キシロース異性化酵素活性の単位(U)は、50mMリン酸緩衝液(pH7.0),10mMキシロース及び10mM MgCl2を含む反応混合物中、37℃で、1分間に1nmolのキシルロースを生産する酵素の量として定義される。生成したキシルロースは、Dische and Borenfreund(1951,J.Biol.Chem.192:583-587)の方法または実施例に記述したHPLCにより測定した。
【0011】
配列の同一性及び類似性
本明細書において配列同一性(sequence identity)とは、配列比較により決定される、二以上のアミノ酸(ポリペプチドまたはタンパク)配列または二以上の核酸(ポリヌクレオチド)配列の関係として定義される。この分野で、「同一性」は、場合によって、アミノ酸配列又は核酸配列の配列間の一致により決定される、アミノ酸または核酸配列間の配列関連性(sequence relatedness)の程度も意味している。二つのアミノ酸配列の「類似性(similarity)」は、あるポリペプチドの他のポリペプチドに対する、アミノ酸配列及び保存的置換アミノ酸を比較して決定される。「同一性」及び「類似性」は、限定はしないが以下に記述されている方法を含む既知方法により容易に計算することができる(Computational Molecular Biology, Lesk, A. M., ed., Oxford University Press, New York, 1988; Biocomputing:Informatics and Genome Projects, Smith, D. W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey, 1994;Sequence Analysis in Molecular Biology, von Heine, g., Academic Press, 1987; and Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991; and Carillo, H., and Lipman, D., SIAM J. Applied Math., 48: 1073(1988)。
【0012】
同一性を決める望ましい方法は、試験する配列間に最大の一致が得られるように設計される。同一性及び類似性を決定する方法は、一般に使用可能なコンピュータプログラムとして作成される。二つの配列間の同一性及び類似性を決定するための望ましいコンピュータプログラム方法には、GCGプログラムパッケージ(Devereux,J., et al., Nucleic Acids Research 12(1):387(1984))、BestFit, BLASTP, BLASTN,及びFASTA(Altschul, S. F. et al., J. Mol. Biol. 215: 403-410 (1990)が含まれる。BLAST XプログラムはNCBI及びその他のサイトから誰でも入手できる(BLAST Manual, Altschul, S., et al., NCBI NLM NIH Bethesda, MD 20894; Altschul, S., et al., J. Mol. Biol. 215: 403-410 (1990)。よく知られたSmith Watermanアルゴリズムも同一性決定に使用することができる。
【0013】
ポリペプチド配列比較のための望ましいパラメーターには以下のアルゴリズムが含まれる:Needleman and Wunsch,J.Mol.Biol.48:443-453 (1970);Comparison matrix:BLOSSUM62 from Hentikoff and Hentikoff,Proc.Natl.Acad.Sci.USA 89:10915-10919(1992);Gap Penalty:12;及びGap Length Penalty:4。これらのパラメーターを使用する有用なプログラムはMadison,WIにあるGenetics Computer Groupの「Ogap」プログラムとして一般に入手することができる。既述のパラメーターはアミノ酸比較(エンドギャップに対するペナルティーなし)のためのデホルトパラメーターである。
【0014】
核酸比較のための望ましいパラメーターは以下のアルゴリズムを含む:アルゴリズム:Needlemann and Wunsch,J.Mol.Biol.48:443-453(1970);Comparison matrix:一致=+10、不一致=0;Gap Penalty:50;Gap Length Penalty:3。Genetics Computer Group,Madison,WIのGapプログラムとして入手できる。上記に示したものは核酸比較のためのデホルトパラメーターである。任意に、アミノ酸類似性の程度を決定する際に、いわゆる「保存的」アミノ酸置換を考慮に入れることもでき、このことは、当業者には明らかである。保存的アミノ酸置換とは類似の側鎖を持つ残基の互換性のことである。例えば、脂肪族側鎖をもつアミノ酸の群はグリシン、アラニン、バリン、ロイシン及びイソロイシン;ヒドロキシ脂肪族側鎖を持つアミノ酸の群はセリン及びトレオニン;アミノ基を含む側鎖を持つアミノ酸の群はアスパラギン及びグルタミン;芳香族側鎖を持つアミノ酸の群はフェニルアラニン、チロシン、及びトリプトファン;塩基性側鎖を持つアミノ酸の群はリシン、アルギニン、及びヒスチジン;硫黄を含む側鎖を持つアミノ酸の群はシステイン及びメチオニンである。望ましい保存的アミノ酸置換群は:バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リシン−アルギニン、アラニン−バリン、及びアスパラギン−グルタミンである。本明細書において開示したアミノ酸配列の置換変異体は、開示配列中の少なくとも一つの残基が除去され、その場所に異なる残基が挿入されたものである。望ましくはアミノ酸の変換は保存的である。天然に存在するアミノ酸の望ましい保存置換は以下のようなものである:アラニンからセリンへ;アルギニンからリシンへ;アスパラギンからグルタミンまたはヒスチジン;アスパラギン酸からグルタミン酸へ;システインからセリンまたはアラニンへ;グルタミンからアスパラギンへ;グルタミン酸からアスパラギン酸へ;ヒスチジンからアスパラギンまたはグルタミンへ;イソロイシンからロイシンまたはバリンへ;ロイシンからイソロイシンまたはバリンへ;リシンからアルギニン;グルタミンまたはグルタミン酸;メチオニンからロイシンまたはイソロイシン;フェニルアラニンからメチオニン、ロイシンまたはチロシンへ;セリンからトレオニンへ;トレオニンからセリンへ;トリプトファンからチロシンへ;チロシンからトリプトファンまたはフェニルアラニンへ;及びバリンからイソロイシンまたはロイシン。
【0015】
核酸配列のハイブリッド形成
本発明のキシロース異性化酵素またはキシルロースキナーゼをコードする核酸配列は、中等度のまたは望ましくは厳密なハイブリッド形成条件下に、SEQ ID NO.2またはSEQ ID NO.4のヌクレオチド配列それぞれとハイブリッド形成する能力によって規定することもできる。厳密なハイブリッド形成条件とは、少なくとも約25、望ましくは約50ヌクレオチド、75または100及び最も望ましくは約200またはそれ以上のヌクレオチドの核酸配列を、約65℃の温度において、1Mの塩、望ましくは6xSSCまたは同等のイオン強度を持つその他の溶液中でハイブリッド形成させることができ、そして0.1Mの塩、またはそれ以下、望ましくは0.2xSSCまたは同等のイオン強度を持つその他の溶液中で65℃において洗浄する条件とここでは定義する。望ましくは、ハイブリッド形成は終夜、すなわち少なくとも10時間行い、望ましくは、洗浄は洗浄液を少なくとも2回交換して少なくとも1時間行う。この条件により、通常約90%またはそれ以上の配列同一性を持つ配列との特異的ハイブリッド形成が可能であろう。
【0016】
中等度の条件とは、少なくとも50ヌクレオチド、望ましくは約200またはそれ以上のヌクレオチドの配列を、約1M塩を含む溶液、望ましくは6xSSCまたは同等のイオン強度のその他の溶液の中で約45℃の温度においてハイブリッド形成させ、そして約1M塩を含む溶液、望ましくは6xSSCまたは相当するイオン強度のその他の溶液の中で室温において洗浄する条件とここでは定義する。望ましくは、ハイブリッド形成は終夜、すなわち少なくとも10時間行い、望ましくは、洗浄は洗浄液を少なくとも2回交換して少なくとも1時間行う。この条件により、通常約50%までの配列同一性を持つ配列との特異的ハイブリッド形成が可能であろう。当業者は、50%及び90%の間の同一性をもつ配列を特異的に同定するためにこれらのハイブリッド形成条件を変更できるであろう。
【0017】
作動的連結
本明細書で使用される「作動的に連結した」とは、機能的に関連するポリヌクレオチド配列の連結のことである。他の核酸配列との機能的な関連を持って配置されている場合に、核酸は「作動的に連結している」。例えば、プロモーターまたはエンハンサーは、それがコード配列の転写に影響するならば、コード配列と作動的に連結している。作動的に連結したとは、典型的には、連結したDNA配列同士が隣接しており、二個のタンパクコード領域を結合する必要がある場合には、連続し、読み枠内にあることを意味する。
【0018】
プロモーター
本明細書で使用される「プロモーター」とは、転写の方向の点で遺伝子の転写開始部位の上流に存在する1またはそれ以上の遺伝子の転写を調節する機能を持つ核酸フラグメントであって、DNA依存RNAポリメラーゼ、転写開始部位、及び限定はしないが、転写因子結合部位、リプレッサー及び活性化タンパク結合部位を含むその他のDNA配列、並びにプロモーターの転写の量を直接的若しくは間接的に調節することが当業者には既知のその他のヌクレオチドの配列によって構造的に同定される。「構成的」プロモーターはほとんどの環境及び発生条件において活性であるプロモーターである。「誘導的」プロモーターは環境及び発生が調節されたときに活性となるプロモーターである。
【図面の簡単な説明】
【0019】
【図1】炭素源として25mMガラクトース及び100mMキシロースを含む培地で増殖した形質転換S.cerevisiaeの増殖曲線。形質転換体pYesは挿入のない酵母発現ベクターを含む。形質転換体14.3,16.2.1及び16.2.2はPiromycessp.E2キシロース異性化酵素コード配列を含むpYESベクターで形質転換されている。
【発明を実施するための形態】
【0020】
[発明の詳細な説明]
本発明の最初の態様は、キシロースからキシルロースへ異性化する能力を持つ形質転換宿主細胞に関するものである。キシロースからキシルロースへ異性化する能力は、キシロース異性化酵素をコードする核酸配列を含む核酸構築物で宿主細胞を形質転換することにより付与される。キシロースからキシルロースへ異性化する形質転換宿主細胞の能力は、キシロースからキシルロースへの直接の異性化である。これは、それぞれキシロース還元酵素およびキシリトール脱水素酵素に触媒されてキシリトール中間体を経由してキシロースからキシルロースへ変換する2ステップ反応に対して、キシロース異性化酵素により触媒される1ステップ反応によりキシロースがキシルロースへ異性化したことを意味すると理解される。
【0021】
核酸配列は、形質転換宿主細胞中で活性形態で発現するキシロース異性化酵素をコードすることが望ましい。従って、宿主細胞中で核酸配列が発現することにより、25℃でタンパクmg当り少なくとも10Uのキシロース異性化酵素比活性、望ましくは25℃で少なくとも20,25,30,50,100,200または300U/mg、の比活性を持つキシロース異性化酵素が生産される。本明細書において、形質転換宿主細胞中で発現したキシロース異性化酵素の比活性は、宿主細胞の細胞分解物、例えば、酵母細胞分解物のタンパクmg当りのキシロース異性化酵素活性の量と定義される。キシロース異性化酵素活性の測定、タンパク量、及び細胞分解物の調製については実施例1に記述されている。また、比活性は、実施例4に示すように測定することもできる。従って、宿主細胞中にヌクレオチド配列を発現させることにより、30℃で少なくとも50U/mgタンパクのキシロース異性化酵素活性、望ましくは30℃で少なくとも100,200,500または750U/mgの比活性を持つキシロース異性化酵素が生成される。
【0022】
望ましくは、宿主細胞中にヌクレオチド配列を発現することにより、キシロースに対するKmが50,40,30または25未満のキシロース異性化酵素が得られ、より望ましくは、キシロースに対するKmが約20mMまたはそれ未満である。
【0023】
キシロース異性化酵素をコードするヌクレオチド配列は、
(a)SEQ ID NO.1のアミノ酸配列と少なくとも40,45,49,50,53,55,60,70,80,90,95,97,98,または99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードするヌクレオチド配列;
(b)SEQ ID NO.2のヌクレオチド配列と少なくとも40,50,55,56,57,60,70,80,90,95,97,98,または99%の配列同一性を有するヌクレオチド配列を含むヌクレオチド配列;
(c)その相補鎖が、(a)または(b)の核酸分子配列にハイブリダイズするヌクレオチド配列;
(d)遺伝子コードの縮重による、(c)の核酸分子の配列と異なる配列からなるヌクレオチド配列:
からなる群から選択することができる。
【0024】
ヌクレオチド配列は、真核生物性キシロース異性化酵素、すなわち真核生物中に天然に存在するキシロース異性化酵素と同じアミノ酸配列を持つキシロース異性化酵素をコードしていることが望ましい。中等温度好性原核性キシロース異性化酵素に比べ、真核生物性キシロース異性化酵素の発現は、酵母のような真核性宿主細胞においてキシロース異性化酵素が活性形態で発現する可能性を高める。ヌクレオチド配列が植物キシロース異性化酵素(例えば、Hordeum vulgare由来)または菌(fungal)キシロース異性化酵素(例えば、Basidiomycets由来)をコードするのがより望ましい。但し、真核生物性宿主細胞、特に酵母において酵素的に活性形態で発現する可能性をさらに増加させるためには、ヌクレオチド配列が嫌気性菌のキシロース異性化酵素をコードすることが最も望ましい。Neocallimastix,Caecomyces,Piromyces,Orpinomyces,またはRuminomyces科に属する嫌気性菌のキシロース異性化酵素をコードするヌクレオチド配列が最も望ましい。
【0025】
キシロース異性化酵素をコードするヌクレオチド配列で形質転換するための宿主細胞は、細胞の中へ能動的にまたは受動的にキシロースを輸送できる宿主であることが望ましい。宿主細胞は、活性な解糖経路、ペントースリン酸経路を含有していることが望ましく、キシロースから異性化されたキシルロースがピルビン酸に代謝されるようにキシルロースキナーゼを含有していることが望ましい。さらに宿主はピルビン酸を目的とする発酵産物、例えばエタノール、エチレンまたは乳酸に変換するための酵素を含有することが望ましい。望ましい宿主細胞は、天然においてアルコール発酵、望ましくは嫌気性アルコール発酵ができる宿主細胞である。さらに宿主細胞はエタノール及び乳酸、酢酸、ギ酸のような有機酸、及びフルフラール及びヒドロキシメチルフルフラールのような糖分解産物に対して高度な耐性を持っていることが望ましい。宿主細胞のこれらの特性または活性は天然に宿主細胞中に存在することもあるが、遺伝子操作により導入または修飾することができる。適する宿主細胞は細菌または真菌のような微生物であるが、宿主として最も適当なのは酵母または糸状菌である。
【0026】
本明細書において酵母は、真核性微生物として定義され、主として単細胞の形で増殖するEumycotina亜門の全種(Alexopoulos,C.J.,1962,In:Introductory Mycology,John Wiley & Sons,Inc.,New York)を含む。酵母は単細胞葉状体の発芽によっても、また生物体の分裂によっても増殖することができる。宿主細胞として望ましい酵母は、Saccharomyces,Kluyveromyces,Candida,Pichia,Schizosaccharomyces,Hansenula,Kloeckera,Schwanniomyces,及びYarrowia種に属する。酵母は、嫌気性発酵、より望ましくは嫌気性アルコール発酵ができるものが望ましい。
【0027】
本明細書において糸状菌は、Eumycotina亜門の全ての糸状形態を含む真核性微生物として定義される。これらの菌は、キチン、セルロース、及びその他の複合多糖からなる栄養菌糸体によって特徴付けられる。本発明の糸状菌は、形態的、生理的、及び遺伝的に、酵母とは区別される。糸状菌による栄養増殖は菌糸の延長によるものであり、ほとんどの糸状菌の炭素異化は、絶対的好気性による。宿主細胞として望ましい糸状菌はAspergillus,Trichoderma,Humicola,Acremonium,Fusarium,及びPenicillium種に属するものである。
【0028】
数年にわたって、農産糖からバイオ−エタノールを生産するために、種々の生物体の導入が提案されてきた。しかしながら、実用的な主なバイオ−エタノール生産プロセスの全てにおいて、エタノール生産体としてSaccharomyces種の酵母を使用し続けてきた。これは工業的プロセスに対するSaccharomyces種の多くの魅力的な特徴、すなわち、高濃度の酸、エタノール及び浸透圧に対する耐性、嫌気的増殖能力、及び当然ではあるが高いアルコール発酵能力に基づくものである。宿主細胞として望ましい酵母の種は、S.cerevisiae,S.bulderi,S.barnetti,S.exiguus,S.uvarum,S.diastaticus,K.lactis,K.marxianus,K.fragilisを含む。
【0029】
宿主細胞は後に定義する核酸構築物で形質転換され、核酸構築物の一つのコピーを含むものでもよいが、複数のコピーを含むものが望ましい。この核酸構築物はエピソームとして維持され、ARS配列のような自律的複製のための配列を含む。適するエピソーム性核酸構築物は、例えば、酵母2μまたはpKD1(Fleer et al.,1991, Biotechnology 9: 968-975)プラスミドに基づくことができる。しかし、核酸構築物は、宿主細胞のゲノム中に1以上のコピーが組み込まれていることが望ましい。宿主細胞ゲノムへの組み込みは、変則的な組換えによりランダムに生じることもありうるが、菌分子遺伝学の分野でよく知られている相同組換えにより核酸構築物を宿主細胞ゲノムへ組み込むことが望ましい(例えば、WO90/14423,EP−A−0 481 008,EP−A−0 635 574及びUS6,265,186参照)。
【0030】
本発明による望ましい形質転換宿主細胞において、核酸構築物は、宿主細胞に、炭素源として、望ましくは唯一の炭素源としてのキシロースに対して、望ましくは嫌気的条件下で増殖する能力を付与する。それにより形質転換宿主は、好ましくはキシリトールを本質的に生産せず、例えば、当該宿主によって生産されるキシリトールは、検出限界以下、またはモルベースで消費した炭素の5,2,1%未満である。その形質転換宿主細胞は、唯一の炭素源としてのキシロースに対して少なくとも0.01,0.02,0.05,0.1または0.2h−1の速度で増殖する能力を有する。このように本発明の形質転換宿主細胞は、既に定義した比活性レベルでキシロース異性化酵素を発現する。
【0031】
宿主細胞は、(a)宿主細胞中へのキシロース輸送の増加;(b)キシルロースキナーゼ活性の増加;(c)ペントースリン酸経路の流量の増加;(d)カタボライト抑制に対する感受性の減少;(e)エタノール、浸透圧または有機酸に対する耐性の増加;及び(f)副生物生産の減少、からなる群から選択された特徴の一つまたはそれ以上を生じる遺伝子修飾をさらに含むことができる。副生物は目的とする発酵産物以外の炭素含有分子を意味すると理解され、例えば、キシリトール、グリセロール及び/または酢酸が含まれる。そのような遺伝子修飾は、古典的突然変異誘起及びスクリーニング及び/または目的変異体の選別により導入することができる。その他に、遺伝子修飾は、外来性遺伝子の過剰発現及び/または異種遺伝子の発現及び/または内在性遺伝子の不活化を含むことができる。これらの遺伝子は、ヘキソースまたはペントーストランスポーター;S.cerevisiae(XKSI Deng and Ho, 1990, Appl. Biochem. Biotechnol. 24-25: 193-199)またはPiromyces(xylB,すなわちSEQ ID NO.4)のキシルロースキナーゼのようなキシルロースキナーゼ;トランスアルドラーゼ(TAL1)またはトランスケトラーゼ(TKL1)のようなペントースリン酸経路の酵素(例えば、Meinander et al.,1995,Pharmacol.Toxicol.Suppl,2:45参照)、解糖酵素、アルコール脱水素酵素のようなアルコール代謝酵素、をコードする遺伝子から選択されることが望ましい。不活化される望ましい内在性遺伝子としては、例えば、S.cerevisiae HXK2遺伝子(Diderich et al., 2001, Appl. Environ. Microbiol. 67: 1587-1593参照)のようなヘキソースキナーゼ;S.cerevisiae MIG1またはMIG2遺伝子;S.cerevisiae GRE3遺伝子(Traff et al.,2001,Appl.Environm.Microbiol.67:5668-5674)のような(非特異的)アルドース還元酵素遺伝子;S.cerevisiaeグリセロール−リン酸脱水素酵素1及び/または2遺伝子のようなグリセロール代謝に関係する酵素の遺伝子;またはその他の宿主の種の遺伝子の(ハイブリッド形成)相同体、が含まれる。宿主細胞のキシロース代謝に関するその他の望ましい修飾についてはZaldivar et al.(2001、前出)に総説されている。
【0032】
その他の態様において、本発明は、エタノール以外の発酵産物を生産するための形質転換宿主細胞に関係している。その非エタノール発酵産物は原則として酵母または糸状菌のような真核性微生物によって生産することができる大量のまたは精製した化学品である。そのような発酵産物としては、例えば、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質及びセファロスポリンがある。
【0033】
本発明の核酸構築物による宿主細胞の形質転換及び宿主細胞、望ましくは酵母の前記のようなその他の遺伝子修飾は当業者によく知られている方法により行われる。その方法は例えば標準的教科書から知ることができる、例えば、Sambrook and Russel(2001)“Molecular Cloning: A Laboratory Manual(3rd edition)”,Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, または F. Ausubel et al, eds.,“Current protocols in molecular biology”,Green Publishing and Wiley Interscience, New York (1987)。菌宿主細胞の形質転換及び遺伝子修飾の方法は、例えば、EP−A−0 635 574,WO98/46772,WO99/60102及びWO00/37671から知ることができる。
【0034】
その他の態様において、本発明は、前記で定義したキシロース異性化酵素をコードし、前記の宿主細胞の形質転換に使用されるヌクレオチド配列からなる核酸構築物に関する。核酸構築物において、キシロース異性化酵素をコードするヌクレオチド配列は、後述するように宿主細胞中でヌクレオチド配列の転写を調節し開始するためのプロモーターに作動的に連結していることが望ましい。このプロモーターは、宿主細胞にキシロースをキシルロースに異性化する能力を付与するために、宿主細胞中でキシロース異性化酵素を十分に発現できることが望ましい。プロモーターは、前記のような宿主細胞中で、特異的キシロース異性化酵素を生じさせるものが望ましい。本発明の核酸構築物中において有用なプロモーターは、構成的及び誘導的な天然プロモーター並びに人工的プロモーターである。さらに本発明に使用するための望ましいプロモーターは、カタボライト(グルコース)抑制に感受性がなくそして/または誘導のためのキシロースを必要としないものが望ましいであろう。このような特徴を持つプロモーターは、広く入手可能であり、同業者には知られている。そのようなプロモーターの適当な例は、例えば、酵母のリン酸フルクトキナーゼ(PPK)、トリオースリン酸異性化酵素(TPI)、グリセロアルデヒド−3−リン酸脱水素酵素(GPD,TDH3またはGAPDH)、ピルビン酸キナーゼ(PYK)、ホスホグリンセリン酸キナーゼ(PGK)プロモーターのような解糖遺伝子の酵母プロモーターである。そのようなプロモーターに関する詳細は(WO93/03159)に見ることができる。その他の有用なプロモーターは、リボソームタンパクコード遺伝子プロモーター、ラクターゼ遺伝子プロモーター(LAC4)、アルコール脱水素酵素プロモーター(ADH1,ADH4,など)、及びエノラーゼプロモーター(ENO)である。その他の、構成的及び誘導的プロモーター及びエンハンサーまたは上流活性化配列は当業者に知られているであろう。本発明の核酸構築物に使用されるプロモーターは、必要に応じて、修飾して、その調節特徴を変更することができる。キシロース異性化酵素を発現させるために核酸構築物に使用されるプロモーターは、キシロース異性化酵素を発現させる宿主細胞と同種であることが望ましい。
【0035】
核酸構築物において、キシロース異性化酵素をコードするヌクレオチド配列の3’−末端は、転写ターミネーター配列に作動的に連結していることが望ましい。このターミネーター配列は、選択した宿主細胞、例えば選択した酵母種において作動し得ることが望ましい。いずれの場合にも、ターミネーターの選択は重要ではなく、ターミネーターは酵母ではない真核生物の遺伝子の場合にしばしば作動するが、酵母遺伝子由来のものを用いることができる。転写終結配列はさらにポリアデニル化シグナルを含むことが望ましい。
【0036】
任意に、核酸構築物中に選択マーカーを入れることができる。本明細書で使用する用語「マーカー」とは、マーカーを含有する宿主細胞を、選別、またはスクリーニングすることができる特徴または表現型をコードする遺伝子のことである。マーカー遺伝子は、抗生物質耐性遺伝子とすることができ、それによって形質転換されていない細胞の中から形質転換された細胞を選別するために適当な抗生物質を使用することができる。適当な抗生物質耐性マーカーの例としては、例えば、ジヒドロ葉酸還元酵素、ヒグロマイシン−B−ホスホトランスフェラーゼ、3’−O−ホスホトランスフェラーゼII(カナマイシン、ネオマイシン及びG418耐性)が含まれる。抗生物質耐性マーカーは、倍数体宿主細胞の形質転換には最も便利であるが、しかし栄養要求性マーカー(URA3,TRP1,LEU2)またはS.pombe TPI遺伝子(Russell P R,1985,Gene40:125−130に記載)のような非抗生物質耐性マーカーが使用されることが望ましい。望ましい態様において、核酸構築物により形質転換された宿主細胞はマーカー遺伝子を含まない。マーカー遺伝子を含まない組換え微生物宿主細胞を作る方法はEP−A−0 635 574に開示されており、A.nidulans amdS(アセトアミダーゼ)遺伝子または酵母URA3及びLYS2遺伝子のような二方向性マーカーの使用に基づいている。そのほかには、形質転換細胞をスクリーニングできるように、緑色蛍光タンパク、lacZ,ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ、ベータ−グルクロニダーゼのようなスクリーニングに役立つマーカーを本発明の核酸構築物の中に組み込むことができる。
【0037】
さらに任意に本発明の核酸構築物中に存在することができる配列としては、これらに限定はしないが、一つまたはそれ以上のリーダー配列、エンハンサー、組み込み因子、並びに/或いはレポーター遺伝子、イントロン配列、セントロメア、テロメア及び/またはマトリックス接着(MAR)配列がある。本発明の核酸構築物はさらに、ARS配列のような自己複製のための配列を含むことができる。適するエピソーム核酸構築物は、例えば、酵母2μまたはpKD1(Fleer et al., 1991, Biotechnology 9: 968-975)プラスミドを基にすることができる。その他に核酸構築物は、望ましくは相同組換えによる、組み込みのための配列を含むことができる。従ってその配列は、宿主細胞のゲノム中の組み込みの標的部位に対して相同的な配列である。本発明の核酸構築物は、本来既知方法により提供することができ、その方法には核酸/核酸配列を制限及び連結するような技術を含み、その参考文献は例えば、Sambrook and Russel (2001) “Molecular Cloning: A Laboratory Manual (3rd edition), Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, またはF. Ausubel et al., eds.,”Current protocols in molecular biology”,Green Publishing and Wiley Interscience, New York (1987)のような標準的教科書に示されている。
【0038】
そのほかの態様において、本発明はキシロース異性化酵素をコードするヌクレオチドを含む核酸分子に関する。この核酸分子は、
(a)SEQ ID NO.1のアミノ酸配列と少なくとも50,53,54,55,60,70,80,90,95,97,98,または99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードする核酸分子;
(b)SEQ ID NO.2のヌクレオチド配列と少なくとも50,56,57,58,60,70,80,90,95,97,98,または99%の配列同一性を有するヌクレオチド配列を含む核酸分子;
(c)その相補鎖が、(a)または(b)の核酸分子配列にハイブリダイズする核酸分子;及び
(d)遺伝子コードの縮重により(c)の核酸分子配列と異なる配列からなる核酸分子;
からなる群から選択されることが望ましい。
【0039】
また、(a)の核酸分子は、SEQ ID NO.1のアミノ酸配列と少なくとも67,68,69,70,80,90,95,97,98,または99%の配列類似性を有するアミノ酸配列を含むポリペプチドをコードすることができる。(c)の核酸分子は望ましくは前記に定義した中等度の条件、より望ましくは厳密な条件の下にハイブリッド形成する。核酸分子は真核生物由来であることが望ましく、菌のような真核微生物由来であることがより望ましく、前記の嫌気性菌のような嫌気性菌由来であることが最も望ましい。
【0040】
本発明の更に他の態様は、キシルロースキナーゼ、望ましくはD−キシルロースキナーゼをコードするヌクレオチド配列を含む核酸分子に関するものである。本明細書において、D−キシルロースキナーゼ(EC2.7.1.17;D−キシルロキナーゼとも呼ばれる)は、D−キシルロースのキシルロース−5−リン酸への変換を触媒する酵素である。この核酸分子は、
(a)SEQ ID NO.3のアミノ酸配列と少なくとも45,47,48,49,50,55,60,70,80,90,95,97,98,または99%の配列同一性を有するアミノ酸配列を含むポリペプチドをコードする核酸分子;
(b)SEQ ID NO.4のヌクレオチド配列と少なくとも30,37,38,39,40,50,60,70,80,90,95,97,98,または99%の配列同一性を有するヌクレオチド配列からなる核酸分子;
(c)その相補鎖が、(a)または(b)の核酸分子配列にハイブリダイズする核酸分子;及び
(d)遺伝子コードの縮重による、(c)の核酸分子配列と異なる配列からなる核酸分子:
からなる群から選択されることが望ましい。
【0041】
また、(a)の核酸分子は、SEQ ID NO.3のアミノ酸配列と少なくとも64,65,66,70,80,90,95,97,98,または99%の配列類似性を有するアミノ酸配列を含むポリペプチドをコードすることができる。(c)の核酸分子は望ましくは前記で定義した中等度の条件、より望ましくは厳密な条件の下にハイブリッド形成する。核酸分子は、真核生物由来であることが望ましく、菌のような真核微生物由来であることがより望ましく、前記の嫌気性菌のような嫌気性菌由来であることが最も望ましい。
【0042】
そのほかの態様において、本発明は、本発明の形質転換宿主細胞がキシロースのようなキシロース資源を含む炭素源の発酵のために使用される、発酵プロセスに関するものである。発酵培地中の炭素源は、キシロース資源に加えてグルコース資源も含むことができる。キシロースまたはグルコースの資源としては、キシロースまたはグルコースそのものでもよく、例えば、リグノセルロース、キシラン、セルロース、デンプンなどのようなキシロースまたはグルコース単位からなる炭水化物のオリゴマーまたはポリマーでもよい。そのような炭水化物からキシロースまたはグルコース単位を遊離させるために(キシラナーゼ、グルカナーゼ、アミラーゼなどのような)、適当な炭水化物分解酵素を、発酵培地に加えてもよく、形質転換宿主細胞に生産させてもよい。後者の場合には、形質転換宿主細胞に遺伝子操作を施し、その炭水化物分解酵素を生産させ、分泌させることができる。望ましいプロセスにおいて、形質転換宿主細胞は、キシロース及びグルコースを共に、望ましくは同時に発酵し、その場合にはグルコース抑制に非感受性の形質転換宿主細胞を使用してジオキシ増殖を阻害することが望ましい。炭素源としてのキシロース(及びグルコース)に加えて、発酵培地はさらに形質転換宿主細胞の増殖に必要な適当な成分を含むであろう。酵母などの微生物を増殖するための発酵培地の組成は、当業者にはよく知られている。
【0043】
発酵プロセスは、エタノール、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、ペニシリンG若しくはペニシリンV及びそれらの発酵誘導体のようなβ−ラクタム並びにセファロスポリンなどの発酵産物を生産するためのプロセスである。発酵プロセスは、好気的または嫌気的な発酵プロセスとすることができる。本明細書において嫌気的発酵プロセスとは、発酵プロセスが無酸素の状態でまたは実質的に酸素が消費されないで(例えば、5mmol/L/h未満)で行われ、有機分子が、電子供与体及び電子受容体のいずれとしても働く発酵プロセスと定義される。酸素が存在しないところでは、糖分解及びバイオマス形成において生成したNADHは、酸化的リン酸化により酸化することはできない。この問題を解決するために多くの微生物は電子及び水素受容体としてピルビン酸またはその誘導体の一つを利用することによって、NAD+を再生する。従って、望ましい嫌気的発酵プロセスにおいて、ピルビン酸は、電子(及び水素)受容体として使用され、エタノール、乳酸、1,3−プロパン−ジオール、エチレン、酢酸またはコハク酸のような発酵産物へと還元される。
【0044】
発酵プロセスは、形質転換宿主細胞にとって最適の温度で行うことが望ましい。従って、ほとんどの酵母または菌の宿主細胞に対して、発酵プロセスは38℃未満の温度で実施される。酵母または糸状菌の宿主細胞に対して、発酵プロセスは35,33,30または28℃未満、且つ20,22,または25℃を超える温度で実施されることが望ましい。
【0045】
望ましいプロセスは、(a)前記に定義した形質転換宿主細胞と共にキシロース資源を含む培地を発酵することによって、宿主細胞にキシロースをエタノールに発酵させ;任意に、(b)エタノールを回収するステップを含むエタノールの生産プロセスである。発酵培地は、グルコース資源を含むことができ、それもエタノールに発酵される。このプロセスにおいて、エタノールの容積生産性は、少なくとも0.5,1.0,1.5,2.0,2.5,3.0,5.0または10.0gエタノール/リットル/時間であることが望ましい。このプロセスにおけるキシロース及び/またはグルコースに対するエタノール収率は、少なくとも50,60,70,90,95または98%であることが望ましい。本明細書においてエタノール収率は、理論的収量(グルコース及びキシロースについて0.51gエタノール/gグルコースまたはキシロース)のパーセンテージとして定義する。
【0046】
その他の態様において、本発明は、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、β−ラクタム抗生物質およびセファロスポリンからなる群から選択される発酵産物の生産プロセスに関する。このプロセスは、(a)前記に定義した形質転換宿主細胞と共にキシロース資源を含む培地を発酵することにより、宿主細胞に、キシロールを発酵産物に発酵させ、任意に、(b)発酵産物を回収する、ステップを含むことが望ましい。望ましいプロセスにおいて、培地はグルコース資源も含む。
【実施例】
【0047】
[実施例1]Piromycesキシラナーゼ異性化酵素及びキシルロースキナーゼcDNAのクローニング
(生物及び増殖条件)
インド象の糞から単離した嫌気性菌Piromyces sp.E2(ATCC76762)を、N2/CO2(80%/20%)中、39℃で、種々の炭素源を添加したM2培地で嫌気的に増殖した(24)。使用した炭素源は、アビセル(微結晶セルロース、タイプPH105、Serva,ドイツ)、フルクトースまたはキシロース(全て0.5%、w/v)であった。増殖が止まった後(水素の発生により判断した)、細胞を遠心分離(15,000xg,4℃,15分間)またはナイロンガーゼ(30μm孔径)による濾過により回収した。
【0048】
(無細胞抽出物の調製)
菌細胞を脱イオン水で洗い、培地成分を除去した。細胞を液体窒素中で凍結し、次いで、乳鉢中で、硝子ビーズ(0.10−0.11mm径)ですりつぶして、無細胞抽出物を調製した。Tris/HCl緩衝液(100mM,pH7.0)を粉末に加え(1:1,w/v)そして15分間解凍した後、懸濁液を遠心分離した(18,000xg,4℃,15分間)。透明な上清を細胞内酵素の原料として使用した。
【0049】
(酵素検定)
キシロース異性化酵素活性は、50mMリン酸緩衝液(pH7.0),10mMキシロース,10mM MgCl2及び適当量の無細胞抽出物を含む反応混合物中37℃で検定した。生成したキシルロースの量は、システイン−カルバゾール法(9)により測定した。キシルロースキナーゼ及びキシロース還元酵素活性はWitteveen et al.(28)による記述にしたがって検定した。活性の1単位は、検定条件の下に、毎分1 nmolのキシルロースを生産する酵素量と定義される。生成したキシルロースは、Dische and Borenfreund(Dische and Borenfreund, 1951, J. Biol. Chem. 192: 583-587)の方法によるか、または80℃のBiorad HPX−87Nカラムを使用し、溶出液として0.01M Na2HPO4を使用して0.6ml/minで溶出するHPLCにより測定した。キシロース及びキシルロースは、内部温度60℃で屈折計により測定した。
【0050】
比活性は、タンパク1mg当りの単位として示す。タンパクは、ウシγ−グロブリンを標準としてBio−Radタンパク試薬(Bio-Rad Laboratories,Richmond,CA,米国)で測定した。
【0051】
(Piromyces sp.E2cDNAライブラリーのランダム配列)
既に記述されている(2)ベクターラムダZAPII中に構築したcDNAライブラリーを使用した。このライブラリーの一部をExAssistヘルパーファージ(Stratagene,La Jolla,CA,米国)による大量切除(mass excission)によりpBluescript SK−クローンに変換した。無作為に取り出したクローンをM13逆プライマーを使用して配列解析し、5’部分の配列を得た。不完全cDNAを使用してプローブを合成し、ライブラリーを再スクリーニングするために使用した。全長配列を得るためにpUC18にサブクローニングを生成さた。配列分析は、dRhodamineターミネーターサイクルシークエンス即時反応DNAシークエンスキット(Perkin-Elmer Applied Biosystems)を使用してABIプリズム310自動化シークエンサーで実施した。
【0052】
(結果)
嫌気性菌Piromyces sp.E2のcDNAライブラリーから無作為に選択したクローンを配列解析し、その結果キシロース異性化酵素及びD−キシルロキナーゼ遺伝子にそれぞれ高い相同性を示す二つのクローン(pH97及びpAK44)を得た。これらのクローンを詳細に解析した。
【0053】
クローンpH97は完全なORFを含有していなかったので、クローンpH97の配列データに基づいて設計したプローブを使用してcDNAライブラリーを再度スクリーニングした。この結果1669bpの挿入配列を有するクローンpR3を得た。キシロース異性化酵素に高い類似性のある437アミノ酸のタンパクをコードするORFを同定することができた。5’非翻訳領域はわずか4bpを含むのみであったが、推定開始メチオニン残基は、既知キシロース異性化酵素配列の整列によく一致した。3’非翻訳領域は351bpの長さであり、嫌気性菌で典型的である、高いAT含有率を有していた。ORFは、基質との相互作用(触媒トリアド(catalytic triad)His102,Asp105,Asp340及びLys235)及びマグネシウムとの結合(Glu232)に重要であることが示されているアミノ酸を含有していた(14,26)。さらに、キシロース異性化酵素のために開発した(20)2個の符号パターン(残基185−194、及び230−237)が存在した。このPiromyces sp.E2キシロース異性化酵素(XylA)はHaemophilus influenzaの酵素(52%同一性、68%類似性)及びHordeum vulgare(49%同一性、67%類似性)と高い相同性を示す。cDNA配列から推定されるポリペプチドは49,395Daの分子量に相当し、5.2の計算値pIを有す。
【0054】
二番目のクローンpAK44は、2041bpの挿入を有し、53,158Daの分子量及び5.0のpIを持つ494アミノ酸のタンパクをコードする完全ORFを含有した。最初のメチオニンに先行して111bpの5’非翻訳領域が存在したが、3’非翻訳領域は445 bpからなっていた。両領域はATが多い。BLAST及びFASTA調査によりキシルロキナーゼとの高い類似性が明らかにされた。Rodriguez−Pefia et al.(22)により定義された2個のリン酸共通領域が部分整列に示されるように位置6−23及び254−270に認められた。さらにPrositeデーターベースに記述されているこの糖キナーゼファミリーの記号が同定された(131−145及び351−372)。Piromyces sp.E2キシルロキナーゼ(XylB)はHaemophilus influenzaのXylBと高い相同性を示した(46%同一性、64%類似性)。
【0055】
[実施例2]酵母発現ベクターの構築
Piromyces sp.E2のキシロース異性化酵素のSaccharomyces cerevisiae中の発現
Piromyces sp.E2のcDNAをpfuポリメラーゼ(Stratgene)を使用するPCR反応に使用した。プライマーはキシロース異性化酵素の5’及び3’末端の配列を使用して設計し、SfiI及びXbaI制限部位を含めた。PCR産物をpPICZαベクター(Invitrogen,Carlsbad,CA,米国)中にクローニングした。キシロース異性化酵素を取り出すために、pPICZαベクターをEcoRI及びXbaIで消化した。消化産物をpYes2ベクター(Invitrogen)に連結した。キシロース異性化酵素を含むpYes2プラスミドをSaccharomyces cerevisiae(株BJ1991,Beth Johns,UvAから供与)に導入した。この株の遺伝型は:matα,leu2,trp1,ura3−251,prb1−1122及びpep4−3、である。形質転換細胞をSCプレート(0.67% YNB培地+0.05% L−Leu+0.05% L−Trp+2%グルコース+2%アガロース)に接種した。
【0056】
形質転換Saccharomyces cerevisiae細胞をグルコース培地で25℃で72時間増殖した(グルコースの代わりにラフィノースを使用することができる)。細胞を回収し、グルコースの代わりにガラクトースを加えたSC培地中に再懸濁した。8時間の誘導後、細胞を回収し、硝子ビーズ(0.10−0.11mm径)及び「破壊緩衝液」(50mMリン酸緩衝液+5%グリセロール+プロテアーゼ阻害剤)を使用して分解した。分解の後混合物を遠心分離(18,000xg,4℃,15分間)した。透明上清を使用して前記方法(実施例1)によりキシロース異性化酵素活性を測定した。10U/mgタンパクの活性が37℃において測定された。
【0057】
[実施例3]キシロース上での形質転換酵母株の増殖
(培地組成)
Saccharomyces cerevisiae株を下記組成のSC−培地上で増殖した:0.67%(w/v)酵母窒素塩基;0.01%(w/v)L−トリプトファン;0.01%(w/v)L−ロイシン及びグルコース、ガラクトースまたはキシロースまたはこれら基質の組合せのいずれか(下記参照)。寒天培地用に培地に2%(w/v)細菌用寒天を加えた。
【0058】
(増殖実験)
挿入のないpYes2で形質転換されたSaccharomyces cerevisiae株BJ1991(遺伝型:matα,leu2,trp1,ura3−251,prb1−1122,pep4−3)及びPiromyces sp.E2キシロース異性化酵素遺伝子を持つpYes2を含む形質転換細胞(16.2.1;16.2.2及び14.3)を、炭素源として10mMグルコースを含むSC−寒天プレート上で増殖した。コロニーが見えるようになったとき、一つのコロニーを使用して、炭素源として100mMキシロース及び25mMガラクトースを含むSVC液体培地に接種した。LKB Ultrospec K分光光度計を使用して600nmの光学密度の増加を測定することにより増殖を監視した。
【0059】
(結果)
増殖実験の結果を図1にまとめた。挿入のないpYes2で形質転換したBJ1991株の培養は80時間までOD600の増加を示した。この後徐々に減少が観察された。これは増殖の末期にしばしば観察される酵母細胞の凝集によるものである。3種の形質転換細胞は、80時間後も増殖を止めず、少なくとも150時間までさらに増加を示した。
【0060】
[実施例4]Saccharomyces cerevisiaeにおいてPiromyces sp.E2キシロース異性化酵素を構成的に発現するための新規で改良された酵母発現ベクターの構築
キシロース異性化酵素をコードするPiromyces sp.E2遺伝子を含むpPICZαベクターを、VentR DNAポリメラーゼ(New England Biolabs)によるPCRの鋳型として使用した。プライマーはキシロース異性化酵素をコードする遺伝子の5’及び3’配列を使用して設計し、EcoRI及びSpeI部位を含めた。さらに、プライマーはpPICZα構築中に認められるXbaI部位を除去し、その代わりに終結コドン(TAA)を入れて設計した。最終産物は、pPICZα構築に認められる追加のアミノ酸(his及びc−Mycタグ)のない、元のオープンリードフレームを復元するように設計した。PCR産物をEcoRI及びSpeIで切り出した。最終産物をpYES2(Invitrogen)由来のベクター中にクローニングした。このベクター中において、キシロース異性化酵素の構成的発現を確実にし、それにより培地にガラクトースを加える必要をなくするために、pYES2中にあるGAL1プロモーターをTPI1プロモーターに置換した。TPI1プロモーターはプラスミドpYX012(R&D systems)の修飾型からクローニングした。このプロモーターをNheI−EcoRIフラグメントとして切り出した。
【0061】
このTPI1プロモーター及びキシロース異性化酵素をコードする遺伝子のPCR産物を共に、SpeI及びXbaIで切り出したpYES2に連結した。このプラスミドを使用してSaccharomyces cerevisiae株CEN.PK113−5D(Peter Kotter,Frankfurtより供与)を形質転換した。この株の遺伝型は:MatA ura3−52である。形質転換細胞は、炭素源として2%グルコースを加えたミネラル培地プレート上で選別した(Verduyn et al.:Effect of benzoic acid on metabolic fluxes in yeasts; a continuous-culture study on the regulation of respiration and alcoholic fermentation.(1992) Yeast 8(7): 501-17)。形質転換していない細胞は、このプレート上では増殖できない。
【0062】
形質転換細胞を、炭素限定恒成分培養において、グルコース/キシロース混合物に対して増殖した。この条件下で増殖した形質転換細胞は、Dersters−Hildersson et al.(Kinetic characterization of D-xylose isomerases by enzymatic assays using D-sorbitol dehydrogenase. Enz. Microb. Techol. 9 (1987) 145-148)により開発された特異的酵素検定により、高いキシロース異性化活性(30℃において800単位/mg)を示す。形質転換S.cerevisiae株の無細胞抽出物中のキシロース異性化酵素のインビトロ活性は、2価カチオン(Mg2+またはCo2+)に依存し、キシロースに対して約20mMの比較的低いKm値が認められた。
【0063】
【特許請求の範囲】
【請求項1】
SEQ ID NO.1のアミノ酸配列と少なくとも70%配列同一性(sequence identity)を有するアミノ酸配列(配列同一性は、Needleman and Wunsch,J.Mol.Biol.48:443-453 (1970)に記載のアルゴリズムを使用し、Hentikoff and Hentikoff,Proc.Natl.Acad.Sci.USA 89:10915-10919(1992)に記載のBLOSSUM62 Comparison matrixを使用し、Gap Penalty:12及びGap Length Penalty:4で同定した)を含むキシロース異性化酵素をコードするヌクレオチド配列を含む核酸構築物で形質転換した真菌宿主細胞であって、
該形質転換で、該核酸構築物により、キシロースを炭素源として利用する能力を付与された、真菌宿主細胞。
【請求項2】
前記アミノ酸配列は、
(i)SEQ ID NO.1のアミノ酸配列の185−194の位置のアミノ酸配列、
(ii)SEQ ID NO.1のアミノ酸配列の230−237の位置のアミノ酸配列、
(iii)SEQ ID NO.1のアミノ酸配列の102の位置のヒスチジン、105の位置のアスパラギン酸、340の位置のアスパラギン酸、及び235の位置のリジン、並びに
(iv)SEQ ID NO.1のアミノ酸配列の233の位置のグルタミン
を含む、請求項1に記載の真菌宿主細胞。
【請求項3】
前記宿主細胞が、Saccharomyces,Kluyveromyces,Candida,Pichia,Schizosaccharmyces,Hansenula,Kloeckera,Schwanniomyces,及びYarrowia属の何れか一つに属する酵母である、請求項1又は2に記載の形質転換宿主細胞。
【請求項4】
前記酵母が、S.cerevisiae,S.bulderi,S.barnetti,S.exiguus,S.uvarum,S.diastaticus,K.lactis,K.marxianus,及びK.fragilis種の何れか一つに属する、請求項3に記載の形質転換宿主細胞。
【請求項5】
前記宿主細胞が、Aspergillus,Trichoderma,Humicola,Acremonium,Fusarium,及びPenicillium属の何れか一つに属する糸状菌である、請求項1又は2に記載の形質転換宿主細胞。
【請求項6】
キシロースをキシルロースに異性化する能力を宿主細胞に付与するために、キシロース異性化酵素をコードする前記ヌクレオチド配列を、前記宿主細胞中で該キシロース異性化酵素を充分に発現させるプロモーターに作動的に連結している、請求項1から5の何れか1項に記載の形質転換宿主細胞。
【請求項7】
前記宿主細胞中で、該プロモーターがカタボライト抑制に非感受性である、請求項6に記載の形質転換宿主細胞。
【請求項8】
前記宿主細胞が
(a)該宿主細胞中へのキシロース輸送の増加;
(b)キシルロースキナーゼ活性の増加;
(c)ペントースリン酸経路の流量の増加;
(d)カタボライト抑制に対する感受性の減少;
(e)エタノール、浸透圧または有機酸に対する耐性の増加;及び
(f)副産物生産の減少、
からなる群から選択された特徴を生じる遺伝子修飾を含む、請求項1から7の何れか1項に記載の形質転換宿主細胞。
【請求項9】
前記遺伝子修飾が、内在性遺伝子の過剰発現、異種遺伝子の発現、またはそれらの組合せからなり、それらによって、該遺伝子が;ヘキソースまたはペントーストランスポーター、キシルロースキナーゼ;ペントースリン酸経路系酵素、解糖系酵素、及びエタノール代謝酵素をコードする遺伝子からなる群から選択される、請求項8に記載の形質転換宿主細胞。
【請求項10】
前記遺伝子修飾が内在性遺伝子の不活化からなり、それによって、該遺伝子がヘキソースキナーゼ遺伝子、Saccharomyces MIG1及びMIG2遺伝子、並びにそれらの遺伝子の塩基配列に対する相補配列を有するポリヌクレオチドとストリジェントな条件でハイブリダイズし、ヘキソキナーゼ、MIG1又はMIG2活性を有するタンパク質をコードする遺伝子ホモログからなる群から選択される、請求項8に記載の形質転換宿主細胞。
【請求項11】
前記宿主細胞が、該宿主細胞に、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質及びセファロスポリンを生産する能力を付与する一つまたはそれ以上の酵素を発現している、請求項1から10の何れか1項に記載の形質転換宿主細胞。
【請求項12】
前記宿主細胞が、アルコール脱水素酵素活性を減少させる遺伝子修飾を含む、請求項11に記載の形質転換宿主細胞。
【請求項13】
(a)請求項1から10の何れか一項に記載の形質転換真菌宿主細胞であって、アルコール発酵能力を有する細胞と共にキシロース資源を含有する培地を発酵することによって、該宿主細胞によってキシロースをエタノールに発酵させ、任意に
(b)エタノールを回収する、
ステップを含む、エタノールの生産プロセス。
【請求項14】
前記培地がグルコース資源も含む、請求項13に記載のプロセス。
【請求項15】
エタノールの容積生産性が、少なくとも0.5g/リットル/時間である、請求項13又は14に記載のプロセス。
【請求項16】
エタノール収率が少なくとも50%である、請求項13から15の何れか1項に記載のプロセス。
【請求項17】
(a)請求項11又は12に記載の形質転換真菌宿主細胞と共にキシロース資源を含む培地を発酵することによって、該真菌宿主細胞によりキシロースを発酵産物に発酵させ、任意に
(b)発酵産物を回収する、
ステップを含む、乳酸、酢酸、コハク酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質及びセファロスポリンからなる群から選択される発酵産物を生産するプロセス。
【請求項18】
該培地がグルコース資源も含む、請求項17に記載のプロセス。
【請求項1】
SEQ ID NO.1のアミノ酸配列と少なくとも70%配列同一性(sequence identity)を有するアミノ酸配列(配列同一性は、Needleman and Wunsch,J.Mol.Biol.48:443-453 (1970)に記載のアルゴリズムを使用し、Hentikoff and Hentikoff,Proc.Natl.Acad.Sci.USA 89:10915-10919(1992)に記載のBLOSSUM62 Comparison matrixを使用し、Gap Penalty:12及びGap Length Penalty:4で同定した)を含むキシロース異性化酵素をコードするヌクレオチド配列を含む核酸構築物で形質転換した真菌宿主細胞であって、
該形質転換で、該核酸構築物により、キシロースを炭素源として利用する能力を付与された、真菌宿主細胞。
【請求項2】
前記アミノ酸配列は、
(i)SEQ ID NO.1のアミノ酸配列の185−194の位置のアミノ酸配列、
(ii)SEQ ID NO.1のアミノ酸配列の230−237の位置のアミノ酸配列、
(iii)SEQ ID NO.1のアミノ酸配列の102の位置のヒスチジン、105の位置のアスパラギン酸、340の位置のアスパラギン酸、及び235の位置のリジン、並びに
(iv)SEQ ID NO.1のアミノ酸配列の233の位置のグルタミン
を含む、請求項1に記載の真菌宿主細胞。
【請求項3】
前記宿主細胞が、Saccharomyces,Kluyveromyces,Candida,Pichia,Schizosaccharmyces,Hansenula,Kloeckera,Schwanniomyces,及びYarrowia属の何れか一つに属する酵母である、請求項1又は2に記載の形質転換宿主細胞。
【請求項4】
前記酵母が、S.cerevisiae,S.bulderi,S.barnetti,S.exiguus,S.uvarum,S.diastaticus,K.lactis,K.marxianus,及びK.fragilis種の何れか一つに属する、請求項3に記載の形質転換宿主細胞。
【請求項5】
前記宿主細胞が、Aspergillus,Trichoderma,Humicola,Acremonium,Fusarium,及びPenicillium属の何れか一つに属する糸状菌である、請求項1又は2に記載の形質転換宿主細胞。
【請求項6】
キシロースをキシルロースに異性化する能力を宿主細胞に付与するために、キシロース異性化酵素をコードする前記ヌクレオチド配列を、前記宿主細胞中で該キシロース異性化酵素を充分に発現させるプロモーターに作動的に連結している、請求項1から5の何れか1項に記載の形質転換宿主細胞。
【請求項7】
前記宿主細胞中で、該プロモーターがカタボライト抑制に非感受性である、請求項6に記載の形質転換宿主細胞。
【請求項8】
前記宿主細胞が
(a)該宿主細胞中へのキシロース輸送の増加;
(b)キシルロースキナーゼ活性の増加;
(c)ペントースリン酸経路の流量の増加;
(d)カタボライト抑制に対する感受性の減少;
(e)エタノール、浸透圧または有機酸に対する耐性の増加;及び
(f)副産物生産の減少、
からなる群から選択された特徴を生じる遺伝子修飾を含む、請求項1から7の何れか1項に記載の形質転換宿主細胞。
【請求項9】
前記遺伝子修飾が、内在性遺伝子の過剰発現、異種遺伝子の発現、またはそれらの組合せからなり、それらによって、該遺伝子が;ヘキソースまたはペントーストランスポーター、キシルロースキナーゼ;ペントースリン酸経路系酵素、解糖系酵素、及びエタノール代謝酵素をコードする遺伝子からなる群から選択される、請求項8に記載の形質転換宿主細胞。
【請求項10】
前記遺伝子修飾が内在性遺伝子の不活化からなり、それによって、該遺伝子がヘキソースキナーゼ遺伝子、Saccharomyces MIG1及びMIG2遺伝子、並びにそれらの遺伝子の塩基配列に対する相補配列を有するポリヌクレオチドとストリジェントな条件でハイブリダイズし、ヘキソキナーゼ、MIG1又はMIG2活性を有するタンパク質をコードする遺伝子ホモログからなる群から選択される、請求項8に記載の形質転換宿主細胞。
【請求項11】
前記宿主細胞が、該宿主細胞に、乳酸、酢酸、コハク酸、アミノ酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質及びセファロスポリンを生産する能力を付与する一つまたはそれ以上の酵素を発現している、請求項1から10の何れか1項に記載の形質転換宿主細胞。
【請求項12】
前記宿主細胞が、アルコール脱水素酵素活性を減少させる遺伝子修飾を含む、請求項11に記載の形質転換宿主細胞。
【請求項13】
(a)請求項1から10の何れか一項に記載の形質転換真菌宿主細胞であって、アルコール発酵能力を有する細胞と共にキシロース資源を含有する培地を発酵することによって、該宿主細胞によってキシロースをエタノールに発酵させ、任意に
(b)エタノールを回収する、
ステップを含む、エタノールの生産プロセス。
【請求項14】
前記培地がグルコース資源も含む、請求項13に記載のプロセス。
【請求項15】
エタノールの容積生産性が、少なくとも0.5g/リットル/時間である、請求項13又は14に記載のプロセス。
【請求項16】
エタノール収率が少なくとも50%である、請求項13から15の何れか1項に記載のプロセス。
【請求項17】
(a)請求項11又は12に記載の形質転換真菌宿主細胞と共にキシロース資源を含む培地を発酵することによって、該真菌宿主細胞によりキシロースを発酵産物に発酵させ、任意に
(b)発酵産物を回収する、
ステップを含む、乳酸、酢酸、コハク酸、1,3−プロパン−ジオール、エチレン、グリセロール、β−ラクタム抗生物質及びセファロスポリンからなる群から選択される発酵産物を生産するプロセス。
【請求項18】
該培地がグルコース資源も含む、請求項17に記載のプロセス。
【図1】

【公開番号】特開2012−231794(P2012−231794A)
【公開日】平成24年11月29日(2012.11.29)
【国際特許分類】
【出願番号】特願2012−167585(P2012−167585)
【出願日】平成24年7月27日(2012.7.27)
【分割の表示】特願2009−112827(P2009−112827)の分割
【原出願日】平成15年1月23日(2003.1.23)
【出願人】(511122374)シー5 イースト カンパニー ビー.ブイ. (2)
【Fターム(参考)】
【公開日】平成24年11月29日(2012.11.29)
【国際特許分類】
【出願日】平成24年7月27日(2012.7.27)
【分割の表示】特願2009−112827(P2009−112827)の分割
【原出願日】平成15年1月23日(2003.1.23)
【出願人】(511122374)シー5 イースト カンパニー ビー.ブイ. (2)
【Fターム(参考)】
[ Back to top ]