
ホスホエノールピルビン酸カルボキシキナーゼおよび/またはピルビン酸正リン酸ジキナーゼをコードする構築物を含む形質転換植物

本発明は、ホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性および/またはピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする、少なくとも1つのコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物を提供する。本構築物は、形質転換植物において、葉の老化中に窒素の転流を引き起こす能力を持っていて、窒素を葉から植物の他の領域まで輸送することができる。本発明は、そのような構築物によって形質転換された植物細胞および植物、形質転換植物を生産する方法、ならびに、老化した植物における窒素の転流率および成長率を上げる方法を提供する。本発明は、遺伝子構築物で形質転換されている収穫された植物の葉(例えば、タバコの葉)、およびそのような収穫された植物の葉を含む喫煙物品も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、形質転換植物の生産に使用される遺伝子構築物に関する。本構築物は、葉の老化中に窒素の転流(remobilisation)を引き起こす能力を持っていて、窒素を葉から植物の他の領域まで輸送することができる。本発明は、そのような構築物で形質転換された植物細胞、および形質転換植物そのものに及ぶ。本発明は、形質転換植物の生産方法、および老化した植物中の窒素転流率を上昇させる方法にも関する。また、本発明は、遺伝子構築物で形質転換されている収穫された植物の葉(例えば、タバコの葉)、およびそのような収穫された植物の葉を含む喫煙物品にも関する。
【背景技術】
【0002】
葉の老化は、葉の細胞が細胞死の前に異なる代謝および構造の変化を受ける植物の発育の一段階である。生理学的および遺伝学的研究では、老化は高度に調節されたプロセスであることが示されている。葉の老化の進行は、葉緑体の分解によって生じるクロロフィルの喪失およびその後の黄変を見かけ上の特徴とする。このような発育段階の特徴である葉のクロロフィル濃度の低下は、例えば、溶媒抽出および分光光度測定によって、あるいは、クロロフィル成分計によって測定することができる。好ましくは一定の条件で育てられた同じ植物について記録された初期の葉のクロロフィル濃度と比較した葉のクロロフィル濃度の低下は、老化を示す。
【0003】
分子的研究では、老化は遺伝子発現における変化と関連していることが示されている。光合成に関与するタンパク質をコードするmRNAのレベルは老化中に減少するが、老化に関与していると思われているタンパク質をコードする遺伝子のmRNAレベルは増加する。老化は、老化関連遺伝子(SAG)として知られている遺伝子によって調節される高度に組織化されたプロセスである。葉の老化は、タンパク質、核酸および膜の劣化、ならびに、この劣化によってその後に生じる発育中の種子、葉または貯蔵器官などの植物の他の領域への栄養分の輸送を伴う。植物の老化の問題の1つは、老化した葉に存在する多くの有用なミネラルおよび栄養分が葉の中に残留し、葉が枯れるにつれて実質的に失われることである。例えば、老化した葉に存在する窒素(アミノ酸のアミン基の形態であってもよい)は、枯れている葉から取り出さなければ無駄になる。
【0004】
従って、特に植物が老化した場合に植物中の窒素の転流を増加させることは、作物生産において重要な有用性を有する可能性がある。最初に、葉から転流される窒素は、より若い葉ならびに発育中の種子に輸送することができる。従って、老化した葉からの窒素の退出効率を上げることによって、種子および植物のより若い部分への窒素の供給を増加させ、それにより、作物収穫率および窒素使用効率を上げることができるかもしれない。これは、世界人口が増加している際には明らかに有益な目的であるが、作物収穫率は需要を満たす程十分に上昇していない。1つの標的となり得る作物は、栄養組織からの窒素の転流が乏しいために低い窒素効率を有するセイヨウアブラナ(学名:Brassica napus)(菜種)である。穀物タンパク質含有量を増加させるという潜在的な利点が大きいという理由から、別の標的作物は小麦である。穀物タンパク質含有量は、小麦の栄養価に影響を与えるだけでなく、穀物の使用および、従って市場価格も決定する。例えば、穀物タンパク質含有量の増加によりパンの体積が増加する。また、タバコの葉の中の残余窒素はニトロソアミンの生成に寄与することが知られているため、窒素の転流を増加させる能力は、タバコ産業において非常に有用になり得る。
【0005】
ホスホエノールピルビン酸カルボキシキナーゼ(PEPCKまたはPCK)[EC4.1.1.49]およびピルビン酸正リン酸ジキナーゼ(PPDK)[EC2.7.9.1]という酵素が知られている。PPDKは原核生物および真核生物の両方に存在し、細菌とより高度な植物との間で配列および三次構造が保存されている(Pocalyko et al., 1990, Biochemistry, 29, 10757-10765)。この酵素は、ピルビン酸+Pi+ATP=PEP+PPi+AMPの反応において、ピルビン酸の可逆的リン酸化を触媒してホスホエノールピルビン酸(PEP)を生成する(Carroll et al., 1990, Federation of European Biochemical Societies, 274, 178-180; Hatch & Slack, 1968, Biochemical Journal, 106, 141-146)。C3およびC4植物の両方において、PPDK遺伝子は、同じ遺伝子から産生される2種類の転写産物を有する変わった構造を有する。長い方の転写産物は葉緑体のタンパク質をコードし、第一エクソンが葉縁体輸送ペプチドをコードするが、短い方の転写産物は、長い方の転写産物の第一イントロン内で別のプロモーターから転写されるため、葉縁体輸送ペプチドをコードする第一エクソンが存在しない。この短い方の転写産物は、PPDKの細胞質基質のイソ型を生成する。この遺伝子構造は、トウモロコシ、イネ、C3およびC4フラベリア属およびシロイヌナズナ(学名:Arabidopsis thaliana)で報告されている。
【0006】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)は、オキサロ酢酸+ATP=PEP+ADP+CO2の反応において、ADP、二酸化炭素およびホスホエノールピルビン酸の可逆的反応を触媒して、ATPおよびオキサロ酢酸を生成する。PCKは広範囲な植物組織の細胞の細胞質基質に存在することが知られている。これらの植物組織としては、発育中の種子、毛状突起および根が挙げられる。植物中のPCKは窒素化合物の代謝が増加した組織内に出現する。
【0007】
本発明者らは、PCKおよび/またはPPDK酵素をコードする遺伝子がプロモーターの制御下で単独あるいは一緒に配置されている複数の遺伝子構築物を作製し、存在すれば、これらの遺伝子の過剰発現が老化した葉の中の窒素の転流にどのような効果を与えたかについて調べた。
【発明の概要】
【0008】
本発明の第1の局面によれば、少なくとも1つのコード配列に作動可能に連結された老化特異的プロモーターを含み、かつホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性および/またはピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする遺伝子構築物が提供される。
【0009】
本発明の第2の局面によれば、少なくとも1つのコード配列に作動可能に連結されたプロモーターを含み、かつホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性およびピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする遺伝子構築物が提供される。
【0010】
本発明者らは、PCKおよびPPDKの2種類の酵素が、老化中の葉からの窒素の転流の間に、様々なアミノ酸の相互転換に関与している可能性があると考えた。本発明者らは仮定に縛られることを望んでないが、PCKおよびPPDKがどのように窒素の転流に影響を与え得るかについて示す推測上の生化学的経路が図18に示されている。従って、本発明者らは、老化中に独立してあるいは同時に植物中のこれらの2種類の酵素の過剰発現を刺激することが窒素の転流に関わっている可能性があると考えた。
【0011】
研究の結果として、本発明者らは、驚くべきことにPCKおよび/またはPPDKをコードする本発明に係る構築物によって、老化した葉からの窒素転流率を上昇させることを見い出した。本発明者らは、窒素はアミノ酸の形態で老化した葉から植物の種子などの植物のより若い部分に移動するのではないかと仮定する。さらに、本発明者らは、これらの酵素が老化した葉の中で過剰発現した際に、栄養植物の成長量の増加(作物収穫率の上昇に該当する)も観察した。従って、本発明者らは、本発明に係る構築物が形質転換植物の生産に有用であり、それにより、老化した葉からの窒素転流率および/または成長率の上昇を示すことができるのではないかと考える。
【0012】
第1または第2の局面の遺伝子構築物中のプロモーターは、RNAポリメラーゼを誘発してPCKおよび/またはPPDK活性を有する少なくとも1種のポリペプチドをコードする少なくとも1つのコード領域に結合しかつその転写を開始することができるものであってもよい。
【0013】
第2の局面の構築物中に存在するプロモーターは、構成性、非構成性または組織特異的であってもよい。好適なプロモーターの例としては、カリフラワーモザイクウイルス35Sプロモーター(完全もしくは切断型)、ルビスコプロモーター、エンドウマメプラストシアニンプロモーター、ノパリンシンターゼプロモーター、クロロフィルr/b結合プロモーター、高分子量グルテニンプロモーター、α,β−グリアジンプロモーター、ホルデインプロモーターまたはパタチンプロモーターが挙げられる。
【0014】
第2の局面の構築物に存在するプロモーターは、老化特異的プロモーターであってもよい。
【0015】
「老化特異的プロモーター(SAG)」は、老化関連遺伝子の発現の制御に関連するプロモーターとすることができる。従って、このプロモーターは、実質的に老化している組織の中でのみ、プロモーターが作動可能に連結されたコード配列(すなわち、遺伝子)の発現を制限することができる。従って、老化特異的プロモーターは、実質的に植物組織が老化している場合にのみ3’タンパク質コード領域の発現が生じるように発生学的に調節される方法で、植物組織中で遺伝子発現を優先的に促進することができるプロモーターとすることができる。当然のことながら、老化は古い葉などの植物の古い部分で生じ、種子などの植物の若い部分では生じない傾向がある。
【0016】
多数の老化関連遺伝子を発現することで知られている植物の一例は、シロイヌナズナである。従って、第1または第2の局面に係る構築物中のプロモーターは、シロイヌナズナの老化関連遺伝子から単離したものであってもよい。Gepsteinら(The Plant Journal, 2003, 36, 629-642)は、モデルとしてシロイヌナズナを用いて、SAGおよびそのプロモーターについて詳しい研究を行なった。当該遺伝子構築物は、本明細書に開示されている任意のSAG由来のプロモーターを含んでいてもよい。例えば、好適なプロモーターは、SAG12、SAG13、SAG101、SAG21およびSAG18からなる群またはその機能的な変異体もしくは機能的な断片から選択されてもよい。
【0017】
好ましいプロモーターは、SAG12およびSAG13プロモーターである。一実施形態では、当該プロモーターは、当業者に知られているSAG12プロモーターまたはその機能的な変異体もしくは断片である(Gan & Amasino, 1997, Plant Physiology, 113: 313-319)。SAG12プロモーターをコードするDNA配列は、本明細書では以下のとおり配列番号16と称する:
配列番号16
TCGAGACCCGATTGTTATTTTTAGACTGAGACAAAAAAGTAGAATCGTTGATTGTTAAAATTTAAAATTAGTTTCATTACGTTTCGATAAAAAAATGATTAGTTTATCATAGCTTAATTATAGCATTGATTTCTAAATTTGTTTTTTGACCACCCTTTTTTCTCTCTTTGGTGTTTTCTTAACATTAGAAGAACCCATAACAATGTACGTTCAAATTAATTAAAAACAATATTTCCAAGTTTTATATACGAAACTTGTTTTTTTTAATGAAAACAGTTGAATAGTTGATTATGAATTAGTTAGATCAATACTCAATATATGATCAATGATGTATATATATGAACTCAGTTGTTATACAAGAAATGAAAATGCTATTTAAATACAGATCATGAAGTGTTAAAAAGTGTCAGAATATGACATGAAGCGTTTTGTCCTACCGGGTATTCGAGTTATAGGTTTGGATCTCTCAAGAATATTTTGGGCCATACTAGTTATATTTGGGCTTAAGCGTTTTGCAAAGAGACGAGGAAGAAAGATTGGGTCAAGTTAACAAAACAGAGACACTCGTATTAGTTGGTACTTTGGTAGCAAGTCGATTTATTTGCCAGTAAAAACTTGGTACACAACTGACAACTCGTATCGTTATTAGTTTGTACTTGGTACCTTTGGTTCAAGAAAAAGTTGATATAGTTAAATCAGTTGTGTTCATGAGGTGATTGTGATTTAATTTGTTGACTAGGGCGATTCCTTCACATCACAATAACAAAGTTTTATAGATTTTTTTTTTATAACATTTTTGCCACGCTTCGTAAAGTTTGGTATTTACACCGCATTTTTCCCTGTACAAGAATTCATATATTATTTATTTATATACTCCAGTTGACAATTATAAGTTTATAACGTTTTTACAATTATTTAAATACCATGTGAAGATCCAAGAATATGTCTTACTTCTTCTTTGTGTAAGAAAACTAACTATATCACTATAATAAAATAATTCTAATCATTATATTTGTAAATATGCAGTTATTTGTCAATTTTGAATTTAGTATTTTAGACGTTATCACTTCAGCCAAATATGATTTGGATTTAAGTCCAAAATGCAATTTCGTACGTATCCCTCTTGTCGTCTAATGATTATTTCAATATTTCTTATATTATCCCTAACTACAGAGCTACATTTATATTGTATTCTAATGACAGGGAAACCTTCATAGAGATTCAGATAGATGAAATTGGTGGGAAACATCATTGAACAGGAAACTTTTAGCAAATCATATCGATTTATCTACAAAAGAATACGTAGCGTAATGAAGTCCACTTGTTGTGAATGACTATGATTTGATCAAATTAGTTAATTTTGTCGAATCATTTTTCTTTTTGATTTGATTAAGCTTTTAACTTGCACGAATGGTTCTCTTGTGAATAAACAGAATCTTTGAATTCAAACTATTTGATTAGTGAAAAGACAAAAGAAGATTCCTTGTTTTTATGTGATTAGTGATTTTGATGCATGAAAGGTACCTACGTACTACAAGAAAAATAAACATGTACGTAACTACGTATCAGCATGTAAAAGTATTTTTTTCCAAATAATTTATACTCATGATAGATTTTTTTTTTTTGAAATGTCAATTAAAAATGCTTTCTTAAATATTAATTTTAATTAATTAAATAAGGAAATATATTTATGCAAAACATCATCAACACATATCCAACTTCGAAAATCTCTATAGTACACAAGTAGAGAAATTAAATTTTACTAGATACAAACTTCCTAATCATCAAATATAAATGTTTACAAAACTAATTAAACCCACCACTAAAATTAACTAAAAATCCGAGCAAAGTGAGTGAACAAGACTTGATTTCAGGTTGATGTAGGACTAAAATGACTACGTATCAAACATCAACGATCATTTAGTTATGTATGAATGAATGTAGTCATTACTTGTAAAACAAAAATGCTTTGATTTGGATCAATCACTTCATGTGAACATTAGCAATTACATCAACCTTATTTTCACTATAAAACCCCATCTCAGTACCCTTCTGAAGTAATCAAATTAAGAGCAAAAGTCATTTAACTTAGG
【0018】
従って、本発明の構築物中のプロモーターは、実質的に配列番号16に示すヌクレオチド配列またはその機能的な変異体もしくは機能的な断片を含んでいてもよい。SAG12プロモーター配列は、米国特許第5,689,042号に記載されているようなシロイヌナズナから得られたものであってもよい。このプロモーター配列は、図3に示すように本発明に係る遺伝子構築物のそれぞれにおいて見られる。プロモーターがSAG12である実施形態では、当然のことながら、当該プロモーターは、配列番号16の1〜2093の各塩基を含んでいてもよい。ただし、プロモーターの機能的な変異体もしくは機能的な断片が本発明の遺伝子構築物に使用されていてもよい。
【0019】
「プロモーターの機能的な変異体もしくは機能的な断片」は、そこに作動可能に連結された任意のコード領域の発現を開始させるのに機能的に十分であるプロモーターの誘導体または部分とすることができる。例えば、プロモーターがSAG12に基づいている実施形態では、当業者であれば、配列番号16が修飾されていても、SAG12プロモーターの部分のみが必要とされてもよく、それによって、本構築物における遺伝子発現がなお開始されることを理解しているであろう。
【0020】
プロモーターの機能的な変異体および機能的な断片は、転写酵素が推定プロモーター領域に結合し、次いで、コード領域の転写を引き起こしてPCKおよび/またはPPDK活性を有するポリペプチドが発現するか否かを評価することによって容易に同定することができる。あるいは、そのような機能的な変異体および断片は、コード領域に結合されている場合には、プロモーター上で突然変異生成を行ない、遺伝子発現が生じ得るか否かを評価することによって調べてもよい。
【0021】
第1の局面の遺伝子構築物は、老化中にPCK活性および/またはPPDK活性を示す少なくとも1種のポリペプチドの発現を生じさせることができる。従って、遺伝子構築物は、(i)ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片および/または(ii)ピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片をコードする少なくとも1つのコード配列を含んでいてもよい。実施例に記載されているように、本発明者らは、PCKおよび/またはPPDK活性を有するポリペプチドに基づいた様々な遺伝子構築物を開発し、これらを図3に示す。
【0022】
第1の局面に係る遺伝子構築物の第1の実施形態では、プロモーターは、PCK活性を示すポリペプチドをコードするコード配列の発現を誘発してもよい。これは、本明細書では「PCK構築物」と称し、図3に示す。従って、第1の実施形態では、遺伝子構築物は、老化特異的プロモーター、およびホスホエノールピルビン酸カルボキシキナーゼ(PCK)をコードするコード配列あるいはその機能的な変異体もしくは断片を含んでいてもよい。本遺伝子構築物は、PPDK活性を有するポリペプチドをコードしないものであってもよい。
【0023】
第1の局面に係る構築物の第2の実施形態では、プロモーターは、PPDK活性を示すポリペプチドをコードするコード配列の発現を誘発してもよい。これは、本明細書では「PPDK構築物」と称し、図3に示す。第2の実施形態では、本遺伝子構築物は、老化特異的プロモーター、およびピルビン酸正リン酸ジキナーゼ(PPDK)をコードするコード配列またはその機能的な変異体もしくは断片を含んでいてもよい。遺伝子構築物は、PCK活性を有するポリペプチドをコードしないものであってもよい。
【0024】
第1の局面に係る構築物の第3の実施形態では、プロモーターは、PCK活性およびPPDK活性を示すポリペプチドをコードする単一のコード配列の発現を誘発してもよい。これを「PCK/PPDK構築物1」と称す。第3の実施形態では、遺伝子構築物は、老化特異的プロモーターならびに(i)ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片および(ii)ピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片をコードするコード配列を含んでいてもよい。第3の実施形態の構築物は、二重活性(すなわち、PCKおよびPPDKの両方の酵素活性)を示す単一の転写産物をコードしてもよい。PCKコード領域は、PPDKコード領域の3'末端に配置されていてもよい。ただし、好ましくは、PCKコード領域は、PPDKコード領域の5'末端に配置されている。
【0025】
第1の局面に係る構築物の第4の実施形態では、プロモーターは、(i)PCK活性を示す第1のポリペプチドをコードする第1のコード配列および(ii)PPDK活性を示す第2のポリペプチドをコードする第2のコード配列の発現を誘発してもよい。これをPCK/PPDK構築物2と称す。よって、第4の実施形態では、本遺伝子構築物は、少なくとも1つの老化特異的プロモーターならびに(i)PCKをコードする第1のコード配列またはその機能的な変異体もしくは断片および(ii)PPDKをコードする第2のコード配列またはその機能的な変異体もしくは断片を含んでいてもよい(すなわち、各酵素に対して1種類の2種類の転写産物がコードされる)。
【0026】
実施例6に記載されているように、本発明者らは、(例えば、「PCK構築物」または「PPDK構築物」のいずれか一方による形質転換によって)宿主細胞中のPCKまたはPPDKのいずれか一方を過剰発現させることで、老化した葉の中の窒素の転流が増加することを見い出した。さらに、本発明者らは、PCKおよびPPDKの単一の構築物によって栄養成長が増加することを見い出した。
【0027】
実施例8に記載されているように、本発明者らは、(例えば、「PCK構築物」または「PPDK構築物」の両方による形質転換によって)宿主細胞中のPCKまたはPPDKを同時に過剰発現させることは、老化した葉の中の窒素の転流を誘発する際に驚くほど有効であることを見い出した。従って、窒素は、例えば輸送アミノ酸として老化した葉から輸送されてもよい。好適な輸送アミノ酸はグルタミンおよび/またはアスパラギンであってもよい。さらに、老化中にPCKおよびPPDKを同時に過剰発現させることによって成長率を上げ、それにより、栄養成長を増加させてもよい。従って、第1の局面の構築物は、PCKおよびPPDKまたはそれらの機能的な変異体もしくは断片をコードするコード配列を含んでいてもよい。
【0028】
第2の局面の構築物がPCKおよびPPDKをコードするコード配列またはそれらの機能的な変異体もしくは断片を含むことはいうまでもない。この2種類の酵素は二重活性を有する単一のポリペプチドとして、あるいは一方がPCK活性を有し、他方がPPDK活性を有する2種類のポリペプチドとしてコードされてもよい。
【0029】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片およびピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片はそれぞれ、植物などの任意の好適な供給源に由来するものであってもよい。各酵素のコード配列は、好適な植物源、例えばシロイヌナズナに由来するものであってもよい。従って、PCK活性を有するポリペプチドをコードするコード配列は、シロイヌナズナに由来するものであってもよい。さらに、PPDK活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ属、トウモロコシ属、フラベリア属またはクレオメ属に由来するものであってもよい。PPDK活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ、トウモロコシ、フラベリア・トリネルビア(Flaveria trinervia)、フラベリア・ビデンティス(Flaveria bidentis)、フラベリア・ブロウニー(Flaveria brownie)またはフウチョウソウに由来するものであってもよい。
【0030】
シロイヌナズナにはPCKをコードする3種類の遺伝子が存在すると考えられている。シロイヌナズナのホスホエノールピルビン酸カルボキシキナーゼ(PCK)の一実施形態をコードするゲノムDNA(gDNA)配列(イントロンおよびエクソンを含む)は、本明細書では以下のとおり配列番号17として示す:
配列番号17
ATGTCGGCCGGTAACGGAAATGCTACTAACGGTGACGGAGGGTTTAGTTTCCCTAAAGGACCGGTGATGCCGAAGATAACGACCGGAGCAGCAAAGAGAGGTAGCGGAGTCTGCCACGACGATAGTGGTCCGACGGTGAATGCCACAACCATCGATGAGCTTCATTCGTTACAGAAGAAACGTTCTGCTCCTACCACACCGATCAACCAAAACGCCGCCGCTGCTTTTGCCGCCGTCTCCGAGGAGGAGCGTCAGAAGATTCAGCTTCAATCTATCAGGTCCTTATAATAACTTCACATATACAGATTATTCATACGTTACTTTTGTTTATAACATACTTTATATCGAATTAAGGAAGATTATTGCGTTTTCGTGTCCGATCATTTTCATGGAAAAAGTGTCTTTTAGCTAAATATATGGTGTAGTATTAAATATTTCTGACGTGATATACACTAAACTTGAAAATTTTCAATTACTATTTCTTCCTTTAATTCGGCAATATAATTTGTTTTTGTTTATTTTTGGATTAGACATTTATGGACAAGTTAATGCGCTATTGTGACTATTACCAGAAAATAATACTTTAATGTACATGACACGTGTTTAAAACGACACGTGGAAACTAATTTTGATTAATTGTGAAACAGTGCATCGTTAGCATCGTTAACGAGAGAGTCAGGACCAAAGGTGGTGAGAGGAGATCCGGCGGAGAAGAAGACCGATGGTTCAACTACTCCGGCGTACGCTCACGGCCAACATCATTCTATCTTTTCTCCGGCTACTGGTGCTGTCAGTGATAGCTCCTTGAAGTTTACTCACGTCCTCTACAATCTTTCGCCTGCAGGTCAACAAATAAACCTAGAATCCGAATCTGAATATTGATAAATGTTTCTGCAACGAGTTTGATAGATTTGGTTTGTGATTTTGTTGTTTGTAGAGCTTTATGAGCAAGCTATTAAGTATGAGAAAGGTTCGTTTATCACTTCTAATGGAGCTTTGGCGACGCTTTCTGGTGCTAAGACTGGTCGTGCTCCCAGAGATAAGCGTGTTGTTAGAGATGCTACTACTGAGGATGAGCTTTGGTGGGGAAAGTGAGTATTCCTAATCTCGATTTTGATTGATGGAGTTTTTGGGTTTATGCTCTGTTTTCGTTTATTGATTTTGGAGTTTGATTTTGATTTTAGGGGTTCGCCGAATATCGAAATGGATGAACATACTTTCATGGTGAACAGAGAAAGAGCTGTTGATTACTTGAATTCCTTGGAAAAGGTATTAAATTTTGAAAACTTTAATCAATGTTGTTGAGTGTAGAACTTTTGATCTAAGTTTATGAAATTTCTGTTGTTGTTGGGGTTTTTAGGTCTTTGTCAATGACCAATACTTAAACTGGGATCCAGAGAACAGAATCAAAGTCAGGATTGTCTCAGCTAGAGCTTACCATTCATTGTTTATGCACAACATGTAAGTAAAATCATTATTGACTCCTTGTATGTCAATCCATTATTGTGGGTGAAAGAAAACAACAAATTAGTAACTGGGGAGGGTGTCAGGTGTATCCGACCAACTCAGGAGGAGCTTGAGAGCTTTGGTACTCCGGATTTTACTATATACAATGCTGGGCAGTTTCCATGTAATCGTTACACTCATTACATGACTTCGTCCACTAGCGTAGACCTTAATCTGGCTAGGAGGGAAATGGTTATACTTGGTACTCAGTATGCTGGGGAAATGAAGAAGGGTCTTTTCAGTGTGATGCATTACCTTATGCCTAAGCGTCGTATTCTCTCCCTTCATTCTGGATGCAATATGGGAAAAGATGGAGATGTTGCTCTCTTCTTTGGACTTTCAGGTATAGTAGAGACAGTACCAACTATGGTGTTGGGTGATGATGGAAGGAACGATAAATCAAATGATACAATACAATTACTGCTGAACTGACTTGAGAACTGCTTGCCTCTTTGTTGAGTTTAGCGGGTGAATTGAGATTGATGATTGTGTTTTTTGTTTTCTATGAATGATGATTTTAGGTACCGGGAAGACAACGCTGTCTACTGATCACAACAGGTATCTTATTGGAGATGATGAGCATTGTTGGACTGAGACTGGTGTTTCGAACATTGAGGGTGGGTGCTATGCTAAGTGTGTTGATCTTTCGAGGGAGAAGGAGCCTGATATCTGGAACGCTATCAAGTTTGGAACAGGTAGAAAGACAGTACGTTGGAATTGTTTTTGAGAAAAAAACATAAAGCAGTGATATAACAATAAGATTCTGATCTTGTTGCAGTTTTGGAAAATGTTGTGTTTGATGAGCACACCAGAGAAGTGGATTACTCTGATAAATCTGTTACAGGTAAAACAATTGTTATTTCTTTCATTCTCTTCGTCCTCACAATTAACAGAATGATCATTTTCGATTCTCTTTGGTTGCAGAGAACACACGTGCTGCCTACCCAATTGAGTTCATTCCAAATGCGAAAATACCTTGTGTTGGTCCACACCCGACAAATGTGATACTTCTGGCTTGTGATGCCTTTGGTGTTCTCCCACCTGTGAGCAAGCTGAATCTGGCACAAACCATGTACCACTTCATCAGTGGTTACACTGCTCTGGTAAGGCCAAAGTAAAAGTCTTTATTTTGCACATCGTCTTCATAAATTTCAAAAGCATAACCAAAGATGTGCAACATATATAGGTTGCTGGCACAGAGGATGGTATCAAGGAGCCAACAGCAACATTCTCAGCTTGCTTTGGTGCAGCTTTCATAATGTTGCATCCCACAAAGTATGCAGCTATGTTAGCTGAGAAGATGAAGTCACAAGGTGCTACTGGTTGGCTCGTCAACACTGGTTGGTCTGGTGGCAGGTATATATGTCCTTCTATGGAAATCGATACAACAAAACGCTGCCTTGTAACACATGTTTGTAGGCTATTAACATGATCTGTAATGTTTTATTTCCTGCAGTTATGGTGTTGGAAACAGAATCAAGCTGGCATACACTAGAAAGATCATCGATGCAATCCATTCGGGCAGTCTCTTGAAGGCAAACTACAAGAAAACCGAAATCTTTGGATTTGAAATCCCAACTGAGATCGAAGGGATACCTTCAGAGATCTTGGACCCCGTCAACTCCGTAAGTTTCTGCAAATCTGTATAATGTAATTGCTTAAGTGATGATGAACAATTTTTTGTTGATTTGGGTTTAATGAAAATGCAGTGGTCTGATAAGAAGGCACACAAAGATACTCTGGTGAAACTGGGAGGTCTGTTCAAGAAGAACTTCGAGGTTTTTGCTAACCATAAGATTGGTGTGATGGTAAGCTTACGGAGGAGATTCTCGCTGCTGGTCCTATCTTTTAG
【0031】
シロイヌナズナのホスホエノールピルビン酸カルボキシキナーゼ(PCK)をコードする相補的DNA(cDNA)配列(エクソンのみ)は、本明細書では以下のとおり配列番号18として示す:
配列番号18
ATGTCGGCCGGTAACGGAAATGCTACTAACGGTGACGGAGGGTTTAGTTTCCCTAAAGGACCGGTGATGCCGAAGATAACGACCGGAGCAGCAAAGAGAGGTAGCGGAGTCTGCCACGACGATAGTGGTCCGACGGTGAATGCCACAACCATCGATGAGCTTCATTCGTTACAGAAGAAACGTTCTGCTCCTACCACACCGATCAACCAAAACGCCGCCGCTGCTTTTGCCGCCGTCTCCGAGGAGGAGCGTCAGAAGATTCAGCTTCAATCTATCAGTGCATCGTTAGCATCGTTAACGAGAGAGTCAGGACCAAAGGTGGTGAGAGGAGATCCGGCGGAGAAGAAGACCGATGGTTCAACTACTCCGGCGTACGCTCACGGCCAACATCATTCTATCTTTTCTCCGGCTACTGGTGCTGTCAGTGATAGCTCCTTGAAGTTTACTCACGTCCTCTACAATCTTTCGCCTGCAGAGCTTTATGAGCAAGCTATTAAGTATGAGAAAGGTTCGTTTATCACTTCTAATGGAGCTTTGGCGACGCTTTCTGGTGCTAAGACTGGTCGTGCTCCCAGAGATAAGCGTGTTGTTAGAGATGCTACTACTGAGGATGAGCTTTGGTGGGGAAAGGGTTCGCCGAATATCGAAATGGATGAACATACTTTCATGGTGAACAGAGAAAGAGCTGTTGATTACTTGAATTCCTTGGAAAAGGTCTTTGTCAATGACCAATACTTAAACTGGGATCCAGAGAACAGAATCAAAGTCAGGATTGTCTCAGCTAGAGCTTACCATTCATTGTTTATGCACAACATGTGTATCCGACCAACTCAGGAGGAGCTTGAGAGCTTTGGTACTCCGGATTTTACTATATACAATGCTGGGCAGTTTCCATGTAATCGTTACACTCATTACATGACTTCGTCCACTAGCGTAGACCTTAATCTGGCTAGGAGGGAAATGGTTATACTTGGTACTCAGTATGCTGGGGAAATGAAGAAGGGTCTTTTCAGTGTGATGCATTACCTTATGCCTAAGCGTCGTATTCTCTCCCTTCATTCTGGATGCAATATGGGAAAAGATGGAGATGTTGCTCTCTTCTTTGGACTTTCAGGTACCGGGAAGACAACGCTGTCTACTGATCACAACAGGTATCTTATTGGAGATGATGAGCATTGTTGGACTGAGACTGGTGTTTCGAACATTGAGGGTGGGTGCTATGCTAAGTGTGTTGATCTTTCGAGGGAGAAGGAGCCTGATATCTGGAACGCTATCAAGTTTGGAACAGTTTTGGAAAATGTTGTGTTTGATGAGCACACCAGAGAAGTGGATTACTCTGATAAATCTGTTACAGAGAACACACGTGCTGCCTACCCAATTGAGTTCATTCCAAATGCGAAAATACCTTGTGTTGGTCCACACCCGACAAATGTGATACTTCTGGCTTGTGATGCCTTTGGTGTTCTCCCACCTGTGAGCAAGCTGAATCTGGCACAAACCATGTACCACTTCATCAGTGGTTACACTGCTCTGGTTGCTGGCACAGAGGATGGTATCAAGGAGCCAACAGCAACATTCTCAGCTTGCTTTGGTGCAGCTTTCATAATGTTGCATCCCACAAAGTATGCAGCTATGTTAGCTGAGAAGATGAAGTCACAAGGTGCTACTGGTTGGCTCGTCAACACTGGTTGGTCTGGTGGCAGTTATGGTGTTGGAAACAGAATCAAGCTGGCATACACTAGAAAGATCATCGATGCAATCCATTCGGGCAGTCTCTTGAAGGCAAACTACAAGAAAACCGAAATCTTTGGATTTGAAATCCCAACTGAGATCGAAGGGATACCTTCAGAGATCTTGGACCCCGTCAACTCCTGGTCTGATAAGAAGGCACACAAAGATACTCTGGTGAAACTGGGAGGTCTGTTCAAGAAGAACTTCGAGGTTTTTGCTAACCATAAGATTGGTGTGATGGTAAGCTTA CGGAGGAGATTCTCGCTGCTGGTCCTATCTTTTAG
【0032】
従って、PCK活性を有するポリペプチドをコードするコード配列は、実質的に配列番号17または配列番号18に示す核酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0033】
シロイヌナズナのPCKのポリペプチド配列は、本明細書では以下のとおり配列番号19として示す:
配列番号19
MSAGNGNATNGDGGFSFPKGPVMPKITTGAAKRGSGVCHDDSGPTVNATTIDELHSLQKKRSAPTTPINQNAAAAFAAVSEEERQKIQLQSISASLASLTRESGPKVVRGDPAEKKTDGSTTPAYAHGQHHSIFSPATGAVSDSSLKFTHVLYNLSPAELYEQAIKYEKGSFITSNGALATLSGAKTGRAPRDKRVVRDATTEDELWWGKGSPNIEMDEHTFMVNRERAVDYLNSLEKVFVNDQYLNWDPENRIKVRIVSARAYHSLFMHNMCIRPTQEELESFGTPDFTIYNAGQFPCNRYTHYMTSSTSVDLNLARREMVILGTQYAGEMKKGLFSVMHYLMPKRRILSLHSGCNMGKDGDVALFFGLSGTGKTTLSTDHNRYLIGDDEHCWTETGVSNIEGGCYAKCVDLSREKEPDIWNAIKFGTVLENVVFDEHTREVDYSDKSVTENTRAAYPIEFIPNAKIPCVGPHPTNVILLACDAFGVLPPVSKLNLAQTMYHFISGYTALVAGTEDGIKEPTATFSACFGAAFIMLHPTKYAAMLAEKMKSQGATGWLVNTGWSGGSYGVGNRIKLAYTRKIIDAIHSGSLLKANYKKTEIFGFEIPTEIEGIPSEILDPVNSWSDKKAHKDTLVKLGGLFKKNFEVFANHKIGVMVSLRRRFSLLVLSF
【0034】
従って、PCK活性を有するポリペプチドは、実質的に配列番号19に示すアミノ酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0035】
シロイヌナズナはPPDKの少なくとも2種類の型、すなわち葉緑体型および細胞質基質型を有し、どちらも遺伝子の5'末端に僅かなスプライシングの変形を有する同じ遺伝子によってコードされ、それにより2種類の型が生じると考えられている。シロイヌナズナのピルビン酸正リン酸ジキナーゼ(PPDK)の両型をコードするgDNA配列(イントロンおよびエクソンを含む)は、本明細書では以下のとおり配列番号20として示す:
配列番号20
ATGACAAGTATGATCGTGAAGACAACGCCGGAGCTCTTCAAAGGAAATGGAGTGTTCCGTACGGATCATCTCGGAGAAAACCGAATGGTTAGTCGATCAAACCGGCTAGGTGATGGATCAAACCGTTTCCCTAGAACCGGTACAATCCATTGCCAACGGTTAAGCATAGCAAAGACCGGTTTGCATCGTGAGACGAAGGCTCGAGCCATACTTAGCCCTGTGTCCGATCCGGCCGCTTCCATAGCCCAAAAGGTAAGCCTTTCCATTTCAATCATTCTGGTGTATTTTCACCATAAAATTTTATACACTTTTTTATTACGTTTTGTTTTATGATTCTGACGTGAGATTCTTGAGAGAAACTATCACCGATCATTGGGTCGAACCATCTAGCAGCTCAATTATTATCGGTTATAACCCTACCGGTTATAGAATACAAAACAGGTTACGCCATTGTGACATTTGCTTTGTGATCTTGTGAGACGATTAATTATTTGATGTTGATTGGTTTCGTTACTCTTGTTTAAACAATCGAACGGTTCAAACTAATACACACATGTGATGTGAGATCATTTCGGTAGTAATACCAAATAGCGTCTGGCCTAAATTATGAAAGTACTATTTTGAATTAAATTATTGTGGAAACATGAACTTATTTAAATTCAAGTATTTTCGAAATTTGTAATAAAAAAAAACTTTTCCTCTAGATTCATTAGCCCTACTTTTCGTAGAAACAACTTTAATGTATTCAAAGACCACTTTGCTGCTTAAGTCAGACTCTTGTGCCACTTGGTAGATCCACCAATGCCACGTTTTGTTATTGTGCCAAAGAATACGTGAATATGTCCAAACGGCAATCAAATTCTTGGCGTAAAACACAAAAATTATGATACTAGTTTAAATCCACAATTCACCTTCACCATAAAGAATTCATGTATTAGAGATGGTATGACAAGAACTGGTTGAATTTGATGACATTTGTTTGCTATTGTTTTGGTTAAGTAAAAGTTTTGTTAAAAAGGAAAATAGCATCGGTAGTGGCAGATAGCAAGTGTGTGAGTGAGATCAGATATGGTTGACACATCTATGACGAGTCATCGCAACGAAACTTCTTTAATTTTGGTCAATTATATTACAATTTAGCATTTCGAGGTTGGAATTTTGGAATGATCTCTTGATAAGATAATAATGTATTTTTGATGACGTATCCATCAAAACTATAAATGATTTATATTAAATATGAAATTTCGACTGTATACAAGTTTTTATATTATAAAATTATTCGATGTACATATGATCATAATAACTTTACTATATATATAGATACGTATATGTGTTCTTAAACTTGCACAAACATTTCTGCAATCTAAACCTCAATCAAAACAAACAAACAAAAAACCATGATGCAGCGAGTATTCACCTTTGGAAAAGGAAGAAGCGAAGGCAACAAGGGCATGAAGTCCTTGGTATGTTACCAATACCATCATCATGATCATATCAATTCATTAATAATTTAGTGTTTGCTATTTTCAAGAACCATTTATCAAAAATGTTAATTGTTGTTGTGTATGAAGTTGGGAGGGAAAGGAGCCAACCTGGCGGAGATGGCTAGCATAGGCTTGTCGGTGCCGCCGGGGCTAACCATATCGACGGAGGCTTGTCAGCAGTATCAGATCGCCGGCAAAAAGCTTCCAGAAGGTTTATGGGAAGAGATCTTAGAAGGTCTTAGCTTCATCGAACGTGACATTGGAGCTTCCCTCGCTGATCCCTCCAAGCCACTCCTCCTCTCTGTTCGCTCCGGCGCCGCCGTAAGTTAATTATAACTTTTTTTCTTGACTATTTTTATTTTAAGGATTTTTTCTAATGTTAAATTTCTGTTTTTTTTTCTTTCTATGTTTTCTTTAATCTTTTGAAGATTTTTTGACGCAGATTTTGACTTGTTAGATTTCTTTTATTGAAGTTGAGATCCAAATATTTTTTTGGTTATTTTGCCATTTGGCCGTTTTTGGAAGAGTTTAAAATGTACTAGATAGAAAATGAATAAGTTTTGTGGCTATTGAAAGACCTAATGATTTTGGTATTCAAACTATAACGTAGAAAATGAAGATCTTTCGTTTATCTATTTTTAAAACAGAACTACATTGACTTGTCTTTGATCGATATTTTGCATTGTAGATCTCAATGCCTGGTATGATGGACACTGTACTTAACCTTGGCTTGAACGACCAAGTCGTCGTTGGTCTGGCCGCAAAAAGCGGAGAGCGTTTTGCTTACGATTCGTTCCGGCGTTTTCTTGATATGTTTGGTGATGTTGTAAGTCCTCTGTTTTTCAATACTATTTCAGGTAACTTGCATGACAAGAAAATTCTTTGACCTACCTTATAATTGTTTTCTTGATCAATAAAAGGTGATGGGAATTCCACACGCCAAGTTTGAAGAGAAGTTAGAGAGAATGAAGGAGAGGAAAGGAGTTAAAAATGACACTGACTTAAGCGCGGCTGATCTCAAGGAATTGGTTGAGCAGTACAAGAGTGTTTACTTAGAGGCCAAGGGTCAAGAGTTTCCTTCAGGTTTGTTTTGATTCCTACTTGAGGTCAAGTGATAAAAATTAGTTATTAGTTACAAATGTTTAAACGGGGTTAATTGCAGATCCAAAGAAGCAATTGGAGCTAGCGATTGAAGCGGTATTCGATTCTTGGGATAGCCCGAGAGCGAACAAGTACAGAAGTATTAACCAGATAACTGGATTGAAAGGAACCGCGGTTAACATTCAGTGTATGGTGTTTGGAAACATGGGGGACACTTCAGGGACTGGTGTTCTCTTCACTAGGAACCCTAGCACAGGAGAGAAGAAGCTTTATGGCGAGTTTCTAGTTAATGCTCAGGTTTGGCATCTATCACAATGTGTGAATCTCATATCAACAAGTAAGCCCATACTCATTAAATATTGGTTTTGGGACAGGGAGAGGATGTGGTTGCAGGGATAAGAACACCAGAAGATTTGGATACAATGAAGAGATTTATGCCTGAGGCTTACGCTGAACTTGTTGAGAACTGCAACATCTTAGAAAGACATTACAAAGACATGATGGTTGATACACATAAACAATACTTCAATTAGTCCTCATCAACAATTCTTTAGTAATTTAAACAAAATCTCAAATGTGTATTGCAGGATATTGAATTCACAGTACAAGAAGAGAGATTGTGGATGCTGCAATGCAGAGCGGGTAAGCGAACGGGTAAAGGCGCCGTGAAGATAGCAGTTGATATGGTAGGTGAAGGGCTTGTTGAGAAATCTTCTGCTATCAAAATGGTGGAGCCTCAACATCTTGATCAACTACTTCACCCACAGGTACAAACTCAAATATTCATCTTCTTCTTTTTTCATAGTCATAAACTTGATGTTGAAACCAAAATTCGAAACTTACTGGTAATGATTGGTTCACTTGAACAAGAACTAATGGGTTTAAGACGTTTAGGGTTTAGGAGTAAAAGCAGAGATGATTGTCTGACACGTAACCGATGAATAGGGTTTGGAAATTTTGATTCAGAGGTCAATGAAGGTTTTTTTTTTTTTTTTTTATTGATGGATTAGTTTCATGATCCATCGGGGTATCGTGAAAAAGTGGTGGCCAAAGGCTTACCTGCGTCACCAGGAGCGGCGGTTGGACAGGTTGTGTTCACGGCGGAGGAAGCCGAAGCTTGGCATTCTCAGGGTAAAACTGTGATTCTGGTTCGAACTGAGACAAGCCCTGACGATGTGGGAGGTATGCACGCAGCGGAAGGTATATTGACGGCTAGAGGAGGAATGACGTCACACGCGGCTGTTGTTGCTCGCGGTTGGGGAAAATGTTGCATTGCTGGTTGTTCCGAGATTCGTGTCGACGAGAACCACAAGGTTTTTGGATTCGATTTTAGAAACTTGTCATATAAGTTAGGGGAAGATTGTTTCTAAAGTTAGGGTTTAAAAATTTTCAGGTTCTATTGATTGGAGATTTGACGATTAATGAAGGCGAATGGATCTCAATGAACGGATCAACCGGTGAGGTTATATTAGGGAAACAAGCATTGGCTCCTCCGGCTTTAAGTCCAGATTTGGAGACTTTCATGTCCTGGGCTGATGCAATCAGACGTCTCAAGGTGTTTATGAGTTTCTGTTCCTTTAACTTGTTTGATATTTTTAAACTTTCTAACTCAAATGTTCGATGACCGATAAGGTTATGGCGAATGCGGATACACCTGAAGACGCCATTGCAGCTAGGAAAAACGGAGCTCAAGGAATCGGGCTTTGTAGGACAGAGCATATGGTAACTCCTCCTCTGTACTTGATTTCATGTTTTTGATGATTTAGATTGTTTGTATCCAAATGTTTAATGTTGTCTTTGGTTTGGTTTTTAAGTTCTTTGGAGCAGATAGGATTAAAGCAGTGAGAAAGATGATAATGGCGGTAACAACAGAGCAAAGGAAAGCTTCTCTCGACATCTTGCTTCCTTACCAACGTTCGGATTTCGAAGGGATCTTCCGTGCTATGGATGGTAAATGTTTTGAGTCGTCTCTCTAAAATGTATCACAACTTAAAACATGCCTAAACCTTTTTATTTTTCTAGGTTTACCGGTAACAATCCGTTTGTTAGACCCTCCGCTTCACGAGTTTCTCCCGGAAGGCGACTTGGACAACATTGTACATGAGCTAGCTGAAGAAACTGGTGTGAAAGAAGATGAAGTCTTGTCACGGATAGAGAAACTCTCTGAAGTGAATCCAATGCTTGGTTTCCGCGGTTGCAGGTTTCTTACTCTCTTTGTTTCTCTCTGTCTCTTTGCACCTGAAGAACAATCTGATGATCGGTAAACTTGTACGTTATAGGCTCGGAATATCGTATCCAGAGCTAACGGAGATGCAAGCGCGTGCAATTTTTGAAGCTGCAGCGTCAATGCAGGACCAAGGTGTTACTGTCATTCCTGAGATTATGGTTCCACTTGTAGGAACTCCTCAGGAATTGGGTCACCAAGTTGATGTAATTCGTAAAGTTGCAAAGAAAGTATTTGCTGAGAAGGGTCATACCGTGAGCTACAAGGTTGGGACAATGATTGAGATCCCTCGAGCCGCGCTCATTGCAGATGAGGTAAATGTAACAAGACACAAAATGTGTTTTAGGCACTTGAAACCATGTTGCTATTTGCTAAGTAGGAACCTTTTTCTTTTGACAGATTGCGAAAGAGGCGGAGTTTTTCTCGTTCGGGACAAACGACTTGACGCAGATGACGTTTGGATACAGTAGAGACGATGTCGGCAAGTTTCTACCGATTTACCTCGCCAAAGGAATCTTACAGCACGACCCTTTTGAGGTATAATGACTACCATTTCGTTTGCTCTCTATCCATAGGATAAAATCTTGATAGCCATTTTTTTGTGTTTGGACCAGGTTCTTGATCAGCAAGGTGTAGGGCAATTGATCAAGATGGCGACAGAAAAAGGACGAGCAGCTAGGCCTAGCCTCAAGGTTGGGATATGTGGAGAACATGGAGGAGATCCATCTTCTGTGGGATTCTTTGCTGAAGCAGGACTTGACTATGTCTCTTGTTCTCCTTTCAGGTAATTGATTAATTTCCAAACCAATAAACACTTTTTTTACAACACTATTGTATAACTCAGATTGATGTAATTTTGGGATTTCTGTTGTTGTTGTTGTTGTTGTTGTTGTTGCAGGGTTCCAATTGCAAGGCTTGCAGCTGCTCAAGTAGTTGTTGCATGA
【0036】
シロイヌナズナのピルビン酸正リン酸ジキナーゼ(PPDK)の細胞質基質型をコードするcDNA配列は、本明細書では以下のとおり配列番号21として示す:
配列番号21
ATGATGCAGCGAGTATTCACCTTTGGAAAAGGAAGAAGCGAAGGCAACAAGGGCATGAAGTCCTTGTTGGGAGGGAAAGGAGCCAACCTGGCGGAGATGGCTAGCATAGGCTTGTCGGTGCCGCCGGGGCTAACCATATCGACGGAGGCTTGTCAGCAGTATCAGATCGCCGGCAAAAAGCTTCCAGAAGGTTTATGGGAAGAGATCTTAGAAGGTCTTAGCTTCATCGAACGTGACATTGGAGCTTCCCTCGCTGATCCCTCCAAGCCACTCCTCCTCTCTGTTCGCTCCGGCGCCGCCATCTCAATGCCTGGTATGATGGACACTGTACTTAACCTTGGCTTGAACGACCAAGTCGTCGTTGGTCTGGCCGCAAAAAGCGGAGAGCGTTTTGCTTACGATTCGTTCCGGCGTTTTCTTGATATGTTTGGTGATGTTGTGATGGGAATTCCACACGCCAAGTTTGAAGAGAAGTTAGAGAGAATGAAGGAGAGGAAAGGAGTTAAAAATGACACTGACTTAAGCGCGGCTGATCTCAAGGAATTGGTTGAGCAGTACAAGAGTGTTTACTTAGAGGCCAAGGGTCAAGAGTTTCCTTCAGATCCAAAGAAGCAATTGGAGCTAGCGATTGAAGCGGTATTCGATTCTTGGGATAGCCCGAGAGCGAACAAGTACAGAAGTATTAACCAGATAACTGGATTGAAAGGAACCGCGGTTAACATTCAGTGTATGGTGTTTGGAAACATGGGGGACACTTCAGGGACTGGTGTTCTCTTCACTAGGAACCCTAGCACAGGAGAGAAGAAGCTTTATGGCGAGTTTCTAGTTAATGCTCAGGTTTGGCATCTATCACAATGTGTGAATCTCATATCAACAAGGATAAGAACACCAGAAGATTTGGATACAATGAAGAGATTTATGCCTGAGGCTTACGCTGAACTTGTTGAGAACTGCAACATCTTAGAAAGACATTACAAAGACATGATGGATATTGAATTCACAGTACAAGAAGAGAGATTGTGGATGCTGCAATGCAGAGCGGGTAAGCGAACGGGTAAAGGCGCCGTGAAGATAGCAGTTGATATGGTAGGTGAAGGGCTTGTTGAGAAATCTTCTGCTATCAAAATGGTGGAGCCTCAACATCTTGATCAACTACTTCACCCACAGTTTCATGATCCATCGGGGTATCGTGAAAAAGTGGTGGCCAAAGGCTTACCTGCGTCACCAGGAGCGGCGGTTGGACAGGTTGTGTTCACGGCGGAGGAAGCCGAAGCTTGGCATTCTCAGGGTAAAACTGTGATTCTGGTTCGAACTGAGACAAGCCCTGACGATGTGGGAGGTATGCACGCAGCGGAAGGTATATTGACGGCTAGAGGAGGAATGACGTCACACGCGGCTGTTGTTGCTCGCGGTTGGGGAAAATGTTGCATTGCTGGTTGTTCCGAGATTCGTGTCGACGAGAACCACAAGGTTCTATTGATTGGAGATTTGACGATTAATGAAGGCGAATGGATCTCAATGAACGGATCAACCGGTGAGGTTATATTAGGGAAACAAGCATTGGCTCCTCCGGCTTTAAGTCCAGATTTGGAGACTTTCATGTCCTGGGCTGATGCAATCAGACGTCTCAAGGTTATGGCGAATGCGGATACACCTGAAGACGCCATTGCAGCTAGGAAAAACGGAGCTCAAGGAATCGGGCTTTGTAGGACAGAGCATATGATTGTTTGTATCCAAATGTTTAATGTTGTCTTTGGTTTGGTTTTTAAGTTCTTTGGAGCAGATAGGATTAAAGCAGTGAGAAAGATGATAATGGCGGTAACAACAGAGCAAAGGAAAGCTTCTCTCGACATCTTGCTTCCTTACCAACGTTCGGATTTCGAAGGGATCTTCCGTGCTATGGATGGTTTACCGGTAACAATCCGTTTGTTAGACCCTCCGCTTCACGAGTTTCTCCCGGAAGGCGACTTGGACAACATTGTACATGAGCTAGCTGAAGAAACTGGTGTGAAAGAAGATGAAGTCTTGTCACGGATAGAGAAACTCTCTGAAGTGAATCCAATGCTTGGTTTCCGCGGTTGCAGGCTCGGAATATCGTATCCAGAGCTAACGGAGATGCAAGCGCGTGCAATTTTTGAAGCTGCAGCGTCAATGCAGGACCAAGGTGTTACTGTCATTCCTGAGATTATGGTTCCACTTGTAGGAACTCCTCAGGAATTGGGTCACCAAGTTGATGTAATTCGTAAAGTTGCAAAGAAAGTATTTGCTGAGAAGGGTCATACCGTGAGCTACAAGGTTGGGACAATGATTGAGATCCCTCGAGCCGCGCTCATTGCAGATGAGATTGCGAAAGAGGCGGAGTTTTTCTCGTTCGGGACAAACGACTTGACGCAGATGACGTTTGGATACAGTAGAGACGATGTCGGCAAGTTTCTACCGATTTACCTCGCCAAAGGAATCTTACAGCACGACCCTTTTGAGGTTCTTGATCAGCAAGGTGTAGGGCAATTGATCAAGATGGCGACAGAAAAAGGACGAGCAGCTAGGCCTAGCCTCAAGGTTGGGATATGTGGAGAACATGGAGGAGATCCATCTTCTGTGGGATTCTTTGCTGAAGCAGGACTTGACTATGTCTCTTGTTCTCCTTTCAGGGTTCCAATTGCAAGGCTTGCAGCTGCTCAAGTAGTTGTTGCATGA
【0037】
シロイヌナズナのピルビン酸正リン酸ジキナーゼ(PPDK)の葉緑体型をコードするcDNA配列は、本明細書では以下のとおり配列番号22として示す:
配列番号22
ATGACAAGTATGATCGTGAAGACAACGCCGGAGCTCTTCAAAGGAAATGGAGTGTTCCGTACGGATCATCTCGGAGAAAACCGAATGGTTAGTCGATCAAACCGGCTAGGTGATGGATCAAACCGTTTCCCTAGAACCGGTACAATCCATTGCCAACGGTTAAGCATAGCAAAGACCGGTTTGCATCGTGAGACGAAGGCTCGAGCCATACTTAGCCCTGTGTCCGATCCGGCCGCTTCCATAGCCCAAAAGCGAGTATTCACCTTTGGAAAAGGAAGAAGCGAAGGCAACAAGGGCATGAAGTCCTTGTTGGGAGGGAAAGGAGCCAACCTGGCGGAGATGGCTAGCATAGGCTTGTCGGTGCCGCCGGGGCTAACCATATCGACGGAGGCTTGTCAGCAGTATCAGATCGCCGGCAAAAAGCTTCCAGAAGGTTTATGGGAAGAGATCTTAGAAGGTCTTAGCTTCATCGAACGTGACATTGGAGCTTCCCTCGCTGATCCCTCCAAGCCACTCCTCCTCTCTGTTCGCTCCGGCGCCGCCATCTCAATGCCTGGTATGATGGACACTGTACTTAACCTTGGCTTGAACGACCAAGTCGTCGTTGGTCTGGCCGCAAAAAGCGGAGAGCGTTTTGCTTACGATTCGTTCCGGCGTTTTCTTGATATGTTTGGTGATGTTGTGATGGGAATTCCACACGCCAAGTTTGAAGAGAAGTTAGAGAGAATGAAGGAGAGGAAAGGAGTTAAAAATGACACTGACTTAAGCGCGGCTGATCTCAAGGAATTGGTTGAGCAGTACAAGAGTGTTTACTTAGAGGCCAAGGGTCAAGAGTTTCCTTCAGATCCAAAGAAGCAATTGGAGCTAGCGATTGAAGCGGTATTCGATTCTTGGGATAGCCCGAGAGCGAACAAGTACAGAAGTATTAACCAGATAACTGGATTGAAAGGAACCGCGGTTAACATTCAGTGTATGGTGTTTGGAAACATGGGGGACACTTCAGGGACTGGTGTTCTCTTCACTAGGAACCCTAGCACAGGAGAGAAGAAGCTTTATGGCGAGTTTCTAGTTAATGCTCAGGTTTGGCATCTATCACAATGTGTGAATCTCATATCAACAAGGATAAGAACACCAGAAGATTTGGATACAATGAAGAGATTTATGCCTGAGGCTTACGCTGAACTTGTTGAGAACTGCAACATCTTAGAAAGACATTACAAAGACATGATGGATATTGAATTCACAGTACAAGAAGAGAGATTGTGGATGCTGCAATGCAGAGCGGGTAAGCGAACGGGTAAAGGCGCCGTGAAGATAGCAGTTGATATGGTAGGTGAAGGGCTTGTTGAGAAATCTTCTGCTATCAAAATGGTGGAGCCTCAACATCTTGATCAACTACTTCACCCACAGTTTCATGATCCATCGGGGTATCGTGAAAAAGTGGTGGCCAAAGGCTTACCTGCGTCACCAGGAGCGGCGGTTGGACAGGTTGTGTTCACGGCGGAGGAAGCCGAAGCTTGGCATTCTCAGGGTAAAACTGTGATTCTGGTTCGAACTGAGACAAGCCCTGACGATGTGGGAGGTATGCACGCAGCGGAAGGTATATTGACGGCTAGAGGAGGAATGACGTCACACGCGGCTGTTGTTGCTCGCGGTTGGGGAAAATGTTGCATTGCTGGTTGTTCCGAGATTCGTGTCGACGAGAACCACAAGGTTCTATTGATTGGAGATTTGACGATTAATGAAGGCGAATGGATCTCAATGAACGGATCAACCGGTGAGGTTATATTAGGGAAACAAGCATTGGCTCCTCCGGCTTTAAGTCCAGATTTGGAGACTTTCATGTCCTGGGCTGATGCAATCAGACGTCTCAAGGTTATGGCGAATGCGGATACACCTGAAGACGCCATTGCAGCTAGGAAAAACGGAGCTCAAGGAATCGGGCTTTGTAGGACAGAGCATATGATTGTTTGTATCCAAATGTTTAATGTTGTCTTTGGTTTGGTTTTTAAGTTCTTTGGAGCAGATAGGATTAAAGCAGTGAGAAAGATGATAATGGCGGTAACAACAGAGCAAAGGAAAGCTTCTCTCGACATCTTGCTTCCTTACCAACGTTCGGATTTCGAAGGGATCTTCCGTGCTATGGATGGTTTACCGGTAACAATCCGTTTGTTAGACCCTCCGCTTCACGAGTTTCTCCCGGAAGGCGACTTGGACAACATTGTACATGAGCTAGCTGAAGAAACTGGTGTGAAAGAAGATGAAGTCTTGTCACGGATAGAGAAACTCTCTGAAGTGAATCCAATGCTTGGTTTCCGCGGTTGCAGGCTCGGAATATCGTATCCAGAGCTAACGGAGATGCAAGCGCGTGCAATTTTTGAAGCTGCAGCGTCAATGCAGGACCAAGGTGTTACTGTCATTCCTGAGATTATGGTTCCACTTGTAGGAACTCCTCAGGAATTGGGTCACCAAGTTGATGTAATTCGTAAAGTTGCAAAGAAAGTATTTGCTGAGAAGGGTCATACCGTGAGCTACAAGGTTGGGACAATGATTGAGATCCCTCGAGCCGCGCTCATTGCAGATGAGATTGCGAAAGAGGCGGAGTTTTTCTCGTTCGGGACAAACGACTTGACGCAGATGACGTTTGGATACAGTAGAGACGATGTCGGCAAGTTTCTACCGATTTACCTCGCCAAAGGAATCTTACAGCACGACCCTTTTGAGGTTCTTGATCAGCAAGGTGTAGGGCAATTGATCAAGATGGCGACAGAAAAAGGACGAGCAGCTAGGCCTAGCCTCAAGGTTGGGATATGTGGAGAACATGGAGGAGATCCATCTTCTGTGGGATTCTTTGCTGAAGCAGGACTTGACTATGTCTCTTGTTCTCCTTTCAGGGTTCCAATTGCAAGGCTTGCAGCTGCTCAAGTAGTTGTTGCATGA
【0038】
従って、PPDK活性を有するポリペプチドをコードするコード配列は、実質的に配列番号20、配列番号21または配列番号22に示す核酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0039】
シロイヌナズナPPDKの細胞質基質型のポリペプチド配列は、本明細書では以下のとおり配列番号23として示す:
配列番号23
MMQRVFTFGKGRSEGNKGMKSLLGGKGANLAEMASIGLSVPPGLTISTEACQQYQIAGKKLPEGLWEEILEGLSFIERDIGASLADPSKPLLLSVRSGAAISMPGMMDTVLNLGLNDQVVVGLAAKSGERFAYDSFRRFLDMFGDVVMGIPHAKFEEKLERMKERKGVKNDTDLSAADLKELVEQYKSVYLEAKGQEFPSDPKKQLELAIEAVFDSWDSPRANKYRSINQITGLKGTAVNIQCMVFGNMGDTSGTGVLFTRNPSTGEKKLYGEFLVNAQVWHLSQCVNLISTRIRTPEDLDTMKRFMPEAYAELVENCNILERHYKDMMDIEFTVQEERLWMLQCRAGKRTGKGAVKIAVDMVGEGLVEKSSAIKMVEPQHLDQLLHPQFHDPSGYREKVVAKGLPASPGAAVGQVVFTAEEAEAWHSQGKTVILVRTETSPDDVGGMHAAEGILTARGGMTSHAAVVARGWGKCCIAGCSEIRVDENHKVLLIGDLTINEGEWISMNGSTGEVILGKQALAPPALSPDLETFMSWADAIRRLKVMANADTPEDAIAARKNGAQGIGLCRTEHMIVCIQMFNVVFGLVFKFFGADRIKAVRKMIMAVTTEQRKASLDILLPYQRSDFEGIFRAMDGLPVTIRLLDPPLHEFLPEGDLDNIVHELAEETGVKEDEVLSRIEKLSEVNPMLGFRGCRLGISYPELTEMQARAIFEAAASMQDQGVTVIPEIMVPLVGTPQELGHQVDVIRKVAKKVFAEKGHTVSYKVGTMIEIPRAALIADEIAKEAEFFSFGTNDLTQMTFGYSRDDVGKFLPIYLAKGILQHDPFEVLDQQGVGQLIKMATEKGRAARPSLKVGICGEHGGDPSSVGFFAEAGLDYVSCSPFRVPIARLAAAQVVVA
【0040】
シロイヌナズナのPPDKの葉緑体型のポリペプチド配列は、本明細書では以下のとおり配列番号24として示す:
配列番号24
MTSMIVKTTPELFKGNGVFRTDHLGENRMVSRSNRLGDGSNRFPRTGTIHCQRLSIAKTGLHRETKARAILSPVSDPAASIAQKRVFTFGKGRSEGNKGMKSLLGGKGANLAEMASIGLSVPPGLTISTEACQQYQIAGKKLPEGLWEEILEGLSFIERDIGASLADPSKPLLLSVRSGAAISMPGMMDTVLNLGLNDQVVVGLAAKSGERFAYDSFRRFLDMFGDVVMGIPHAKFEEKLERMKERKGVKNDTDLSAADLKELVEQYKSVYLEAKGQEFPSDPKKQLELAIEAVFDSWDSPRANKYRSINQITGLKGTAVNIQCMVFGNMGDTSGTGVLFTRNPSTGEKKLYGEFLVNAQVWHLSQCVNLISTRIRTPEDLDTMKRFMPEAYAELVENCNILERHYKDMMDIEFTVQEERLWMLQCRAGKRTGKGAVKIAVDMVGEGLVEKSSAIKMVEPQHLDQLLHPQFHDPSGYREKVVAKGLPASPGAAVGQVVFTAEEAEAWHSQGKTVILVRTETSPDDVGGMHAAEGILTARGGMTSHAAVVARGWGKCCIAGCSEIRVDENHKVLLIGDLTINEGEWISMNGSTGEVILGKQALAPPALSPDLETFMSWADAIRRLKVMANADTPEDAIAARKNGAQGIGLCRTEHMIVCIQMFNVVFGLVFKFFGADRIKAVRKMIMAVTTEQRKASLDILLPYQRSDFEGIFRAMDGLPVTIRLLDPPLHEFLPEGDLDNIVHELAEETGVKEDEVLSRIEKLSEVNPMLGFRGCRLGISYPELTEMQARAIFEAAASMQDQGVTVIPEIMVPLVGTPQELGHQVDVIRKVAKKVFAEKGHTVSYKVGTMIEIPRAALIADEIAKEAEFFSFGTNDLTQMTFGYSRDDVGKFLPIYLAKGILQHDPFEVLDQQGVGQLIKMATEKGRAARPSLKVGICGEHGGDPSSVGFFAEAGLDYVSCSPFRVPIARLAAAQVVVA
【0041】
従って、PPDK活性を有するポリペプチドは、実質的に配列番号23または配列番号24に示すアミノ酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。最も高いPPDK発現量が、ほぼ例外なくPPDKをコードするgDNAを内部に含む細胞株のみで認められたため、イントロンがPPDKの発現にプラスの影響を与えたのではないかと思われる。従って、一実施形態では、本構築物は、PPDKおよび/またはPCKのいずれか一方をコードする遺伝子のcDNAを含んでいてもよい。
【0042】
本発明の遺伝子構築物は、発現カセットの形態であってもよく、宿主細胞における少なくとも1つのコード配列の発現に適したものであればよい。本発明の遺伝子構築物は、それがベクターに組み込まれることなく宿主細胞に導入されてもよい。例えば、核酸分子であり得る遺伝子構築物は、リポソームまたはウイルス粒子内に組み込まれてもよい。あるいは、精製された核酸分子(例えば、ヒストンを含まないDNA、すなわち裸のDNA)が、例えば、直接的なエンドサイトーシス取り込みなどの好適な手段によって宿主細胞に直接挿入されてもよい。本遺伝子構築物は、形質移入、感染、微量注入、細胞融合、原形質融合または衝撃照射(ballistic bombardment)によって宿主対象(例えば、植物)の細胞に直接導入されていてもよい。あるいは、本発明の遺伝子構築物は、パーティクルガン法を用いて宿主細胞に直接導入されていてもよい。あるいは、当該遺伝子構築物は、好適な宿主細胞における発現のために組換え型ベクターの内部に含まれていてもよい。
【0043】
従って、第3の局面では、第1または第2の局面に係る遺伝子構築物を含む組換え型ベクターが提供される。組換え型ベクターは、プラスミド、コスミドまたはファージであってもよい。そのような組換え型ベクターは、宿主細胞を本発明の遺伝子構築物で形質転換し、かつその中の発現カセットを複製するのに非常に有用である。当業者であれば、本発明の遺伝子構築物は、発現のために多くの種類の骨格ベクターと組み合わせ得ることを理解しているであろう。骨格ベクターは、例えば、1つのベクターが大腸菌およびアグロバクテリウム・ツメファシエンスの両方で複製することができるバイナリーベクターであってもよい。例えば、好適なベクターは、pBIN19などのpBINプラスミドであってもよい。しかし、好ましい骨格ベクターはBNP1380000001であり、これは、pBINPLUS(F. A. van Engelen et al. Transgenic Research (1995) 4, 288-290)に基づいており、かつSAG12プロモーターを内部に含む。
【0044】
組換え型ベクターは、プロモーター(例えば、老化関連プロモーター)に加えて、様々な他の機能的な成分、および少なくとも1つの(PCKおよび/またはPPDKをコードする)コード配列を含んでいてもよい。例えば、組換え型ベクターは、宿主細胞中の細胞質基質内で自己複製するように設計されていてもよい。この場合、組換え型ベクター内に、DNA複製を誘発または調節する成分が必要とされる場合がある。あるいは、組換え型ベクターは、宿主細胞のゲノムに組み込まれるように設計されていてもよい。この場合、(例えば相同組換えによる)標的組換えを支持するDNA配列が想定される。
【0045】
また、組換え型ベクターは、クローン化プロセスにおける選択可能なマーカーとして、すなわち、形質移入または形質転換されている細胞の選択を可能にするため、および異種のDNAが組み込まれているベクターを内部に含む細胞の選択を可能にするために使用できる遺伝子のDNAコードを含んでいてもよい。当該ベクターは、コード配列の発現の調節に関与するDNA、または発現されるポリペプチドをある種の宿主細胞、例えば葉緑体に導くためのDNAを含んでいてもよい。従って、第3の局面のベクターは、選択可能なマーカー遺伝子(例えば、抗生物質耐性遺伝子)、ポリペプチド終結シグナルおよびタンパク質標的配列(例えば、葉緑体輸送ペプチド)からなる群から選択される少なくとも1つのさらなる成分を含んでいてもよい。
【0046】
好適なマーカー遺伝子の例としては、抗生物質耐性遺伝子、例えば、カナマイシン、ジェネテシン(G418)およびハイグロマイシン(npt−II、hyg−B)に対して耐性を示す遺伝子;除草剤耐性遺伝子、例えば、ホスフィノトリシンおよびスルホンアミド系除草剤に対して耐性を示す遺伝子(それぞれbarおよびsuI:欧州特許出願公開第242246号、欧州特許出願公開第0249637号);およびスクリーニング可能なマーカー、例えば、β−グルクロニダーゼ(英国特許第2197653号)、ルシフェラーゼおよび緑色蛍光タンパク質(GFP)が挙げられる。当該マーカー遺伝子は、第2のプロモーター(老化関連プロモーターでなくてもよい)によって制御されてもよく、細胞における発現を可能にし、種子の中に存在していても存在していなくてもよく、それにより、植物の発育の任意の段階でマーカーを含有する細胞または組織の選択を可能にする。好適な第2のプロモーターは、アグロバクテリウムのノパリン合成酵素遺伝子のプロモーターおよび35Sカリフラワーモザイクウイルス(CaMV)転写産物をコードする遺伝子に由来するプロモーターである。ただし、任意の他の好適な第2のプロモーターを使用してもよい。
【0047】
本発明の遺伝子構築物の様々な実施形態は図1に示すクローン化手順を用いて作製してもよく、それについて以下のようにまとめることができる。PCKおよびPPDKをコードする遺伝子のgDNAおよびcDNA型は、好適なプライマーを用いるPCRによってgDNAまたはcDNA鋳型から増幅してもよい。PCR産物は、アガロースゲル電気泳動法を用いて調べてもよい。次いで、PCR産物を、クローン化目的のための好適なベクター(例えば、pCR 4Blunt-TOPOベクター(Invitrogen社))に結合してもよい。PCR産物を内部に含むベクターを大腸菌などの好適な宿主中で増殖させてもよい。次いで、大腸菌コロニーを、好適なプライマーを用いるPCRによってスクリーニングしてもよく、正しい制限酵素消化パターンを示すプラスミド中の挿入断片を好適なプライマーを用いて配列決定してもよい。
【0048】
TOPOcDNA(PCKまたはPPDK)またはTOPOgDNA(PCKまたはPPDK)を保有する大腸菌のコロニーは、好適な量の各プラスミドを産生するために培養してもよく、次いでそれを精製してもよい。次いで、プラスミドを消化して、PPDKまたはPCKをコードするDNA断片を遊離させた後、これを、pBNPプラスミドなどの好適なプロモーター(例えば、SAGプロモーター)を内部に含むベクターにクローン化してもよい。得られたPPDK構築物をBNP−PPDKcDNAおよびBNP−PPDKgDNAと命名し、得られたPCK構築物をpALBNP1(cDNA)およびpALBNP2(gDNA)と命名した。第3の局面に係るベクターの実施形態は、実質的に図3に示すものであってもよい。
【0049】
第4の局面では、同じ条件で培養された野生型植物中のPCKおよび/またはPPDKの対応する濃度よりも試験植物の葉の中のPCKおよび/またはPPDKの濃度を上げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために試験植物における植物の代謝を変えることを含む方法が提供される。
【0050】
第5の局面では、同じ条件で培養された野生型植物中の窒素の対応する濃度よりも試験植物の葉の中の窒素濃度を下げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために試験植物における植物の代謝を変えることを含む方法が提供される。
【0051】
第6の局面では、同じ条件で培養された野生型植物の対応する成長率と比較して試験植物の成長率を上げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために試験植物における植物の代謝を変えることを含む方法が提供される。
【0052】
植物の葉の中の窒素濃度および植物の成長率の測定方法は実施例に記載されている。第4、第5または第6の局面の方法は、第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで試験植物の細胞を形質転換することを含んでもよい。本遺伝子構築物またはベクターは、任意の好適な手段によって宿主細胞に導入してもよい。
【0053】
第7の局面では、第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係る組換え型ベクターを含む細胞が提供される。
【0054】
当該細胞は植物細胞であってもよい。本発明者らが、宿主細胞中にPCKおよび/またはPPDKを過剰発現させることが老化した葉の中の窒素の転流を誘発する際に驚くほど有効であることを観察したように、第7の局面の細胞は、PCKおよび/またはPPDKが過剰発現されるように、第1または第2の局面の1種または複数の構築物あるいは第3の局面の1種または複数のベクターを含んでいてもよい。
【0055】
例えば、宿主細胞は、第1の局面の遺伝子構築物の第1の実施形態(すなわちPCK構築物)のみで形質転換されていてもよい。あるいは、宿主細胞は、第2の局面の遺伝子構築物の第2の実施形態(すなわちPPDK構築物)のみで形質転換されていてもよい。別の実施形態では、宿主細胞は、PCKおよびPPDKが宿主細胞において発現されるように、第1の局面の遺伝子構築物の第1および第2の実施形態で形質転換されていてもよい。あるいは、宿主細胞は、PCKおよびPPDKが宿主細胞において発現されるように、第1の局面の構築物の第3または第4の実施形態(すなわちPCK/PPDK構築物1または2)で形質転換されていてもよい。また、宿主細胞が、PCKおよびPPDKの両方をコードする第2の局面の構築物で形質転換されていてもよいことも想定される。
【0056】
当該細胞は、公知の技術を用いて本発明に係る遺伝子構築物またはベクターで形質転換されていてもよい。遺伝子構築物を宿主細胞に導入するのに適した手段は、例えば、欧州特許出願公開第0116718号および第0270822号に記載されているように当該技術分野で知られている手段による、アグロバクテリウムが保持するディスアーム型Tiプラスミドベクターの使用を含んでもよい。さらなる方法は、植物原形質を形質転換することであってもよく、この方法では、最初に細胞壁を除去し、核酸を導入し、次いで細胞壁を再編成する。次いで、形質転換された細胞を植物に成長させてもよい。
【0057】
第8の局面では、第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターを含む形質転換植物が提供される。
【0058】
第8の局面に係る形質転換植物としては、アブラナ属などのアブラナ科が挙げられる。当該植物は、セイヨウアブラナ(菜種)であってもよい。
【0059】
第8の局面に係る形質転換植物のさらなる例としては、コムギ連(Triticeae)などのイネ科が挙げられる。当該植物は、コムギ属(Triticum spp.)(小麦)であってもよい。小麦中の種子タンパク質含有量を増加させることによって、パンなどの小麦を含む食品の体積を増加させてもよい。
【0060】
第8の局面に係る好適な形質転換植物のさらなる例としては、植物のナス科が挙げられ、ナス科の例としては、例えば、チョウセンアサガオ、ナス、マンドレイク、ベラドンナ(deadly nightshade (belladonna))、トウガラシ(パプリカ、カラトウガラシ)、ジャガイモおよびタバコが挙げられる。ナス科の好適な属の一例はニコチアナである。ニコチアナの好適な種は、タバコ植物、または単にタバコと称される場合がある。植物を第1または第2の局面の遺伝子構築物あるいは第3の局面のベクターで形質転換する様々な方法が知られており、本発明で使用することができる。
【0061】
例えば、タバコは、以下のように形質転換してもよい。タバコ(学名:Nicotiana tabacum)を、基本的にHorsch et al. (Science 227: 1229-1231, 1985)に記載されているように葉片共培養法を用いて形質転換する。最も若い2枚の広がった葉を7週齢のタバコ植物から採取してもよく、8%のドメストス(商標)中で10分間表面滅菌し、滅菌蒸留水で6回洗浄してもよい。葉片をNo.6コルク穿孔器を用いて切断し、約2分間かけて(本発明に係る)適当なバイナリーベクターを含有するアグロバクテリウム懸濁液に入れてもよい。この葉片を、2枚の無菌濾紙の間に穏やかにブロットしてもよい。10枚の葉片をLS3%スクロース+2μMのBAP+0.2μMのNAAを含むプレート上に置き、次いで、これを成長室(growth room)で2日間培養してもよい。葉片は、クラフォラン500g/lおよびのカナマイシン100g/lを添加したLS+3%スクロース+2μMのBAP+0.2μMのNAAを含むプレートに移してもよい。この葉片は、2週間後に上記培地を含む新しいプレートに移してもよい。さらに2週間後、この葉片は、クラフォラン500mg/lおよびカナマイシン100mg/lを添加したLS+3%スクロース+0.5μMのBAPを含むプレートに移してもよい。この葉片は、2週間ごとに新しい培地に移してもよい。苗条が現れたら、それらを切除し、クラフォラン500mg/lを添加したLS+3%スクロースを入れた瓶に移してもよい。瓶の中の苗条は、約4週間後、LS+3%スクロース+クラフォラン250mg/lに移してもよい。さらに3〜4週間後、この植物を、LS+3%スクロース(抗生物質なし)に移して、根づかせてもよい。植物が根づいたら、温室の土壌に移してもよい。
【0062】
第9の局面では、第8の局面に係る形質転換植物から得られた植物繁殖用製品が提供される。
【0063】
「植物繁殖用製品」は、そこからさらなる植物を生産することができる任意の植物であってもよい。好適には、この植物繁殖用製品は種子であってもよい。
【0064】
本発明の第10の局面では、同じ条件で培養された対応する野生型植物よりも高い割合で窒素を転流させる形質転換植物の生産方法であって、
i)第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで植物細胞を形質転換する工程と、
ii)形質転換細胞から植物を再生させる工程と、
を含む方法が提供される。
【0065】
第11の局面では、同じ条件で培養された対応する野生型植物よりも高い成長率を有する形質転換植物の生産方法であって、
i)第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで植物細胞を形質転換する工程と、
ii)形質転換細胞から植物を再生させる工程と、
を含む方法が提供される。
【0066】
好ましくかつ有利なことに、本発明に係る方法は、生産される植物の健康または適応度を損なうことはない。好ましくは、本方法は、第1または第2の局面の遺伝子構築物あるいは第3の局面のベクターを用いて、試験植物および好ましくはその葉を形質転換することを含む。本発明者らは、宿主細胞中でPCKおよび/またはPPDKを過剰発現させることが老化した葉の中の窒素の転流を誘発する際に有効であることを観察した。従って、第10および第11の局面の方法は、PCKおよびPPDKの両方が過剰発現されるように、本発明の1種または複数の構築物で試験植物を形質転換することを含むことが好ましい。例えば、試験植物は、本発明の第1の局面の遺伝子構築物の第1の実施形態(すなわちPCK構築物)およびさらに第1の局面の構築物の第2の実施形態(すなわちPPDK構築物)で形質転換してもよい。従って、これらの2種類の構築物の形質転換によって両方の酵素の過剰発現が生じる。あるいは、試験植物は、PCKおよびPPDKをそれぞれコードする本発明の第1の局面の構築物の第3または第4の実施形態で形質転換してもよい。あるいは、試験植物は、PCKおよびPPDKの両方をコードする本発明の第2の局面の構築物で形質転換してもよい。
【0067】
本発明者らは、PCKおよびPPDKをコードする1種以上の構築物で形質転換された試験植物の植物の葉が老化の開始時に窒素の転流の増加を示し、従って、窒素濃度がその葉において低下するすことを観察した。さらに、本発明者らは栄養成長率の上昇を観察した。本発明者らは、仮定に縛られてはいないが、葉の中の窒素濃度の低下によって成長率の上昇、従って作物収穫率の上昇が誘発され得ると考える。
【0068】
本発明の第12の局面では、同じ条件で栽培された野生型植物から採取された、収穫された葉の中の対応する窒素濃度よりも低濃度の窒素を含有する収穫された葉が提供され、ここで、この葉は、第8の局面に係る形質転換植物から収穫されたもの、あるいは第10または第11の局面に係る方法によって生産されたものである。
【0069】
本発明の第13の局面では、老化した葉の中の窒素濃度を低下させることができる変異型タバコ植物から得られる窒素が減少したタバコを含む喫煙物品が提供される。
【0070】
窒素が減少したタバコには、同じ条件で栽培された野生型植物中の対応する濃度よりも窒素濃度の低いタバコが含まれる。そのような喫煙物品は、変異型タバコ植物から得られたタバコを含んでもよく、このタバコは、本発明の第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで形質転換されたものであってもよい。
【0071】
本明細書で使用する「喫煙物品」という用語は、タバコ、タバコ誘導体、膨張タバコ、再生タバコまたはタバコ代用品および、また不燃性(heat-not-burn)製品をベースとするか否かに関わらず、喫煙に適した製品、例えば、手巻きタバコ、紙巻タバコ、葉巻およびシガリロ(小型葉巻)を含むことができる。
【0072】
当然のことながら、本発明は、実質的に本明細書で参照されている配列のいずれかのアミノ酸または核酸配列(その機能的な変異体もしくは機能的な断片を含む)を含む、任意の核酸、ペプチド、その変異体、誘導体または類似体に及ぶ。「実質的にアミノ酸/ポリヌクレオチド/ポリペチド配列」、「機能的な変異体」および「機能的な断片」という用語は、本明細書において参照される配列のいずれか1つを含むアミノ酸/ポリヌクレオチド/ポリペチド配列との少なくとも40%の配列同一性、例えば、(PCK酵素の一実施形態をコードする)配列番号17として同定される遺伝子との40%の同一性、配列番号19(すなわちPCK酵素の一実施形態)として同定されたポリペチドとの40%の同一性、(PPDK酵素の一実施形態をコードする)配列番号21として同定された遺伝子との40%の同一性、または配列番号23(すなわちPPDK酵素の一実施形態)として同定されたポリペチドとの40%の同一性を有する配列とすることができる。
【0073】
また、参照されている配列のいずれかに対して65%超、より好ましくは70%超、さらにより好ましくは75%超、なおより好ましくは80%超の配列の同一性を有するアミノ酸/ポリヌクレオチド/ポリペプチド配列も想定される。好ましくは、アミノ酸/ポリヌクレオチド/ポリペプチド配列は、本明細書で参照されている配列のいずれかとの少なくとも85%の同一性、より好ましくは少なくとも90%の同一性、さらにより好ましくは少なくとも92%の同一性、なおより好ましくは少なくとも95%の同一性、なおより好ましくは少なくとも97%の同一性、なおより好ましくは少なくとも98%の同一性、最も好ましくは少なくとも99%の同一性を有する。
【0074】
当業者には、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算方法が分かっている。2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合を計算するために、最初に2つの配列のアラインメントを作成した後、配列の同一性値を計算しなければならない。2つの配列の同一性の割合は、(i)配列を整列させるために使用される方法、例えば、ClustalW、BLAST、FASTA、Smith−Waterman法(異なるプログラムで実行される)、あるいは3D比較による構造アラインメント、および(ii)アラインメント法(例えば、ローカルまたはグローバルアラインメント)によって使用されるパラメータ、使用される対のスコア行列(例えば、BLOSUM62、PAM250、Gonnetなど)、およびギャップペナルティ(例えば、関数および定数)によって異なる値を取る場合もある。
【0075】
アラインメント作成後の、2つの配列間の同一性の割合に関する計算方法は数多くある。例えば、(i)最も短い配列の長さ、(ii)アラインメントの長さ、(iii)配列の平均長、(iv)ギャップのない位置の数、または(iv)オーバーハングを除く等価位置の数によって同一部分の数を割ってもよい。さらに、当然のことながら、同一性の割合は長さにも強く依存する。従って、一対の配列が短い程、偶然に生じることを予測し得る配列の同一性が高くなる。
【0076】
従って、当然のことながら、タンパク質またはDNA配列の正確なアラインメントは複雑なプロセスである。有名なマルチプルアライメントプログラムであるClustalW(Thompson et al., 1994, Nucleic Acids Research, 22, 4673-4680; Thompson et al., 1997, Nucleic Acids Research, 24, 4876-4882)は、本発明に従ってタンパク質またはDNAのマルチプルアラインメントを生成する好ましい方法である。ClustalWの好適なパラメータは、以下のとおりであってもよい:DNAアラインメントについては:ギャップ開始ペナルティー(Gap Open Penalty)=15.0、ギャップ伸長ペナルティー(Gap Extension Penalty)=6.66、および行列=同一性。タンパク質アラインメントについては:ギャップ開始ペナルティー=10.0、ギャップ伸長ペナルティー=0.2、および行列=Gonnet。DNAおよびタンパク質アラインメントについては:ENDGAP=−1およびGAPDIST=4。当業者であれば、最適な配列アラインメントのためにこれらのパラメータおよび他のパラメータを変える必要があるかも知れないことに気付いているであろう。
【0077】
好ましくは、次に、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算は、(N/T)*100(式中、Nは、同一の残基を共有する配列の位置の数であり、Tは、ギャップを含めるがオーバーハングを除いて比較される位置の合計数である)としてそのようなアラインメントから計算する。従って、2つの配列間の同一性の割合の最も好ましい計算方法は、(i)好適な組み合わせのパラメータ(例えば、上記のようなパラメータ)を用いるClustalWプログラムを用いて配列アラインメントを作成すること、および(ii)NおよびTの値を式:配列の同一性=(N/T)*100に挿入することを含む。
【0078】
同様の配列の他の同定方法が当該技術分野で知られている。例えば、実質的に類似したヌクレオチド配列は、厳密な条件で、配列番号16、17、18、20、21、22に示されている配列またはそれらの相補体とハイブリッド形成する配列によってコードされる。厳密な条件とは、ヌクレオチドが約45℃で3×塩化ナトリウム/クエン酸ナトリウム(SSC)中でフィルター結合DNAまたはRNAとハイブリッド形成した後、約20〜65℃で0.2×SSC/0.1%SDS中で少なくとも1回洗浄されることを意味する。あるいは、実質的に同様のポリペプチドは、配列番号19、23または24に示されている配列とは、少なくとも1個、ただし5、10、20、50または100個未満のアミノ酸が異なっていてもよい。
【0079】
遺伝子コードの縮重により、任意の核酸配列を、それによってコードされるタンパク質の配列に実質的に影響を与えることなく変化または変更させてその機能的な変異体を得ることができることは明らかである。好適なヌクレオチド変異体は、配列内の同じアミノ酸をコードする異なるコドンの置換によって静的な変化(silent change)を生じさせることにより変化した配列を有する変異体である。他の好適な変異体は、相同ヌクレオチド配列を有するが、置換されたアミノ酸と同様の生物物理学的特性の側鎖を有するアミノ酸をコードする異なるコドンの置換によって保存的変化を生じさせることにより変化した配列の全てもしくは一部を含む変異体である。例えば、小型で非極性の疎水性アミノ酸としては、グリシン、アラニン、ロイシン、イソロイシン、バリン、プロリンおよびメチオニンが挙げられる。大型で非極性の疎水性アミノ酸としては、フェニルアラニン、トリプトファンおよびチロシンが挙げられる。中立極性のアミノ酸としては、セリン、トレオニン、システイン、アスパラギンおよびグルタミンが挙げられる。正電荷を持つ(塩基性)アミノ酸としては、リジン、アルギニンおよびヒスチジンが挙げられる。負の電荷を持つ(酸性)アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。従って、当然のことながら、アミノ酸は同様の生物物理学的特性を有するアミノ酸で置換することができ、当業者には、これらのアミノ酸をコードするヌクレオチド配列が分かっている。
【0080】
本明細書(添付の特許請求の範囲、要約書および図面のいずれも含む)に記載されている全ての特徴、および/またはそのように開示されている任意の方法またはプロセスの全ての工程は、そのような特徴および/または工程の少なくとも一部が相互に排他的である組み合わせを除く任意の組み合わせで、上記の局面のいずれかと組み合わせてもよい。
【0081】
本発明のさらなる理解のため、および本発明の実施形態の実施方法を示すために、ここで、例として添付の図を参照する。
【図面の簡単な説明】
【0082】
【図1】遺伝子構築物のBNP−PPDKcDNAおよびBNP−PPDKgDNAのクローン化に使用される手順を示す。(a)シロイヌナズナPPDKの細胞質基質のイソ型のcDNAおよびgDNA型のPCR増幅、(b)クローニングベクターpCR 4Blunt-TOPOへの挿入、(c)AvrIIおよびBamHIによる、PPDKを放出するためのクローニングベクター(左側)および標的SAG12含有ベクターpBNP(右側)の制限エンドヌクレアーゼ消化、(d)PPDKのpBNPへの連結反応および、AvrIIおよびBamHIによる構築物の制限エンドヌクレアーゼ消化によって産生されたDNA断片を示すアガロースゲル、および(e)アグロバクテリウム媒介性形質転換によってシロイヌナズナ生態型Columbia 0に導入される構築物。
【図2a】本発明に係る発現ベクターの作製に使用されるプラスミドpCR4 BLUNT-TOPOを示す。
【図2b】PPDKをコードするcDNAまたはgDNAがAvrIIおよびBamHI消化を用いてpCR4 BLUNT-TOPOに挿入されたことをまとめた表である。
【図3a】SAG12プロモーターを含有するプラスミドpBNPを示す。
【図3b】PPDK挿入断片(cDNAまたはgDNA)がAvrIIおよびBamHI消化を用いてpBNPに導入され、かつPCK挿入断片(cDNAまたはgDNA)がXbaIおよびSacI消化を用いてpBNPに導入され、その結果、本発明の実施形態に係るベクターが得られたことをまとめた表である。
【図4】ウェスタンブロットによる形質転換SAG12−PPDKシロイヌナズナ細胞株の選択を示す。SAG12−PPDKcDNA細胞株を上側に示し、SAG12−PPDKgDNA細胞株を下側に示す。
【図5】野生型、ΔPPDKおよび5種類の独立したSAG12−PPDKgDNA系統における5週目以降のPPDK発現量の定量化を示す。
【図6】(b)K326および(c)バーレー21タバコ系統におけるSAG12−PPDK(cDNAおよびgDNA)の選択を示す。
【図7】K326タバコの熟した葉におけるPPDKの過剰発現を示す。
【図8】時間に対する、シロイヌナズナの様々な細胞株、すなわち、野生型、ΔPPDKおよびSAG12−PPDKgDNAのロゼットの写真を示す。
【図9】9週目のシロイヌナズナの生殖組織の質量を示す。
【図10】播種後3〜9週目の野生型およびSAG12−PPDgDNA植物の総植物生質量(すなわち、ロゼット+生殖組織)を示す。
【図11】7週目における野生型、ΔPPDKおよびSAG12−PPDKgDNA植物の葉の窒素含有量を示す。
【図12】野生型、ΔPPDKおよびSAG12−PPDK植物の個々の種子の窒素含有量を示す。
【図13】野生型、ΔPPDKおよびSAG12−PPDKgDNAシロイヌナズナ細胞株の葉の総遊離アミノ酸含有量を示す。
【図14】様々なタバコの細胞株の種子の質量を示し、ゼロコピーは野生型であり、G4(gDNA)、G10、G8およびC10(cDNA)はSAG12−PPDK系統である。
【図15】様々なタバコの細胞株の種子の窒素含有量を示し、ゼロコピーは野生型であり、G4(gDNA)、G10、G8およびC10(cDNA)はSAG12−PPDK系統である。
【図16】PCK/PPDK二重挿入断片のためのウェスタンブロット法の結果である。全ての系統は、SAG12プロモーターの活性と相関する特に7週目に、より高濃度のPCKを示す。
【図17】形質転換した植物のPCKおよびPPDKの両発現によって、対照7と比較してロゼット質量が増加することを示す。
【図18】窒素の転流が、輸送アミノ酸であるアスパラギンおよびグルタミンのPCKおよびPPDK依存的産生によって生じ得ることを示す仮定上の生化学的経路を示す。
【発明を実施するための形態】
【実施例】
【0083】
実施例1−SAG12プロモーターシロイヌナズナPPDK植物形質転換構築物の作製
一連のSAG12−PPDKおよびPCK発現ベクターを、図3に示すように作製し、次いで、形質転換した植物においてPPDKおよびPCKの発現を増加させるそれらの能力を分析した。一実施形態では、本構築物は、PCKまたはその機能的な変異体もしくは断片をコードし、かつPPDKをコードしないものであってもよい。別の実施形態では、本構築物は、PPDKまたはその機能的な変異体もしくは断片をコードし、かつPCKをコードしないものであってもよい。ただし、さらに別の局面では、本構築物は、PCKまたはその機能的な変異体もしくは断片およびPPDKまたはその機能的な変異体もしくは断片をコードするものであってもよい。後述するように、最初にシロイヌナズナのPPDK遺伝子のgDNAを単離する。
【0084】
gDNAの単離
シロイヌナズナ生態型Columbia 0からのgDNAを、推奨されている手順に従ってDNeasy植物ミニキット(DNeasy Plant Mini Kit)(Qiagen社)を用いて葉から抽出した。gDNAを、表1にまとめたプライマー配列を用いる下記のようなPCR反応のための鋳型として使用した。
【0085】
【表1】

【0086】
RNAの単離およびcDNAの合成
全RNAを7日齢のシロイヌナズナの子葉から抽出した。リボヌクレアーゼを含まない機器を用いて氷上でRNAの抽出を行い、ピロ炭酸ジエチル(DEPC、Sigma−Aldrich社)で処理した水を用いて溶液を調製した。水1リットルにつきDEPC1mlを添加し、混合物を換気フード内で一晩撹拌した後、加圧滅菌した。TriPure単離試薬(Roche社)を用いてRNAを抽出した。乳鉢と乳棒を用いて組織200mgを液体窒素中で粉砕し、TriPure単離試薬1mlを添加し、推奨されている手順を続けた。RNAペレットは、60℃まで10分間予熱した無傷のRNA溶液(RNA Secure)(Ambion社)20μlに再懸濁した。試料を、13,000rpmで、4℃で5分間遠心分離し、上澄みを清潔な1.5mlのマイクロ遠心分離管に移し、汚染性の破片を除去した。
【0087】
RNAの量および純度は、Eppendorf Biophotometer分光光度計を用いて260および280nmで読み取ることによって、分光測光法で測定した(Maniatis et al., 1982, Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory)。また、RNAは、0.5×TBEに溶かした1.5%(w/v)のアガロース(Melford Laboratories社)を用いるアガロースゲル電気泳動を用いて調べた。試料を1×RNA試料緩衝液に懸濁し、泳動用緩衝液として0.5×TBEを用いて80Vの電気泳動に供した。4×作業濃度のRNA試料緩衝液は、0.002%(w/v)の臭化エチジウム(Sigma−Aldrich社)、2×トリス−ホウ酸−EDTA緩衝液(2×TBE)(2×TBEは、180mMのトリス−HCl、180mMのホウ酸および8mMのEDTA(pH8.3)を含有していた)、13%のフィコール400(Sigma−Aldrich社)、0.01%のブロムフェノールブルーおよび7Mの尿素(Ficher Scientific社))を含有していた。
【0088】
cDNAを合成するためのRNAの逆転写は、25μlの最終体積に対して、2μgのRNA、1μgのOligo(dT)(15)プライマー(Roche社)、1×モロニーマウス白血病ウイルス(MMLV)緩衝液(Promega社)、0.4mMのdNTP(Bioline社)、40単位の組み換え型RNasinリボヌクレアーゼ阻害剤(Promega社)、200単位のMMLV逆転写酵素(Promega社)およびヌクレアーゼ非含有水を用いて行った。この試料を42℃で1時間培養した。
【0089】
シロイヌナズナPPDKの増幅
PPDKの細胞質基質のイソ型をコードする遺伝子のgDNAおよびcDNA型を、表1に示すAvrII制限部位を含む順方向プライマー(AtCytFWD−AvrII、配列番号1)およびBamHI制限部位を含む逆方向プライマー(AtCytREV−BamHI、配列番号2)を用いるPCRによって、gDNAまたはcDNAの鋳型から増幅した。PCR反応混合物は、1×HF緩衝液(NEB社)、2mMの塩化マグネシウム(MgCl2、NEB社)、0.5mMのdNTPs(Bioline社)、100ngの鋳型(cDNAまたはgDNA)、0.5μMの各プライマーおよび1単位のPhusion High-Fidelity DNAポリメラーゼ(NEB社)を含有していた。熱循環は、98℃で30秒間の最初の変性工程、次いで、98℃で10秒間、55℃で30秒間および72℃で2分間の30サイクル、72℃で5分間の最終的な伸長工程によって、Techneサーマルサイクラー(Techne Thermal Cycler)を用いて行った。
【0090】
シロイヌナズナPPDKのcDNAおよびgDNAを単離および調製した後、それらを使用して、図1にまとめた手順を用いて様々な構築物を作製した。老化中にPPDKを過剰発現させるために、シロイヌナズナのPPDKの細胞質基質のイソ型のcDNAおよびgDNA型を、pBNPベクター中の老化誘発SAG12プロモーターに融合した。
(a)AvrIIおよびBamHI制限部位をそれぞれ含み、かつ2.4および4.3kb産物を産生するプライマーを用いる、シロイヌナズナのPPDKの細胞質基質のイソ型のcDNAおよびgDNA型のPCR増幅。
(b)クローニングベクターpCR 4Blunt-TOPOへの挿入。PPDK挿入断片全体を配列決定し、予想した配列と同一であることが分かった。
(c)AvrIIおよびBamHIによるクローニングベクターおよび目的ベクターpBNPの制限エンドヌクレアーゼ消化。
(d)BNPへの連結反応、および構築物のAvrIIおよびBamHIによる制限エンドヌクレアーゼ消化によって産生されたDNA断片を示すアガロースゲル。予想されたバンドの大きさは、cDNA構築物では14.8kbおよび2.4kb、gDNA構築物では14.8kbおよび4.4kbであった。
(e)アグロバクテリウム媒介性の形質転換によってシロイヌナズナ生態型Columbia 0に導入された構築物。構築物は、連結反応部位全体にわたって配列決定した。遺伝子nptIIはネオマイシンホスホトランスフェラーゼをコードし、植物中にカナマイシン耐性を付与する。LBおよびRB:T−DNAの左右それぞれの境界;nos Pro:ノパリンシンターゼプロモーター;nos t:ノパリンシンターゼターミネーター;SAG12 Pro:シロイヌナズナSAG12遺伝子のプロモーター。
【0091】
図1を参照しながら、構築物の作製について以下に詳細に説明する。
【0092】
pCR 4Blunt-TOPOベクターへのクローン化
PCR産物を、0.5×TBEに溶かした1%(w/v)のアガロース、0.5μgml−1の臭化エチジウム(Sigma−Aldrich社)を使用するアガロースゲル電気泳動を用いて調べた。試料を、1×DNA試料緩衝液(6×作業濃度のDNA試料緩衝液は、50mMのトリス(ヒドロキシメチル)アミノメタン(トリス−HCl、Melford Laboratories社)、60%のグリセリンおよび0.25%(w/v)のブロモフェノールブルー(Sigma−Aldrich社)を含有していた)に懸濁し、泳動用緩衝液として0.5×TBEを用いる80Vの電気泳動に供した。
【0093】
2.6kbのPPDKcDNAバンドおよび4.4kbのPPDKgDNAバンドを、推奨されている手順に従ってQIAQuick PCR精製キット(QIAQuick PCR Purification Kit)(Qiagen社)を用いて精製し、30μlの分子生物学等級水(BDH Laboratory Supplies社)を用いて溶出した。PCR産物は、推奨されている手順に従ってpCR 4Blunt-TOPOベクター(Invitrogen社)に連結させた(平滑末端)。クローン化反応物(2μl)を、推奨されている手順に従って50μlのSub-cloning Efficiency DH5α大腸菌(Invitrogen社)に形質転換した。形質転換した大腸菌細胞は、ルリア−ベルターニ(LB)寒天上で37℃で一晩増殖させた。ルリア−ベルターニ培地(LB培地)は、10gl−1のバクト−トリプトン(Bacto-Tryptone)(BD)、5gl−1のバクト酵母抽出物(Bacto-Yeast Extract)(Oxoid社)および85mMの塩化ナトリウム(Fisher Scientific社)を含んでいた。このpHは、高圧蒸気滅菌前に10Mの水酸化ナトリウムで7.0に調整した。1.5%(w/v)の寒天(BD)を50μgml−1のカナマイシン(Melford Laboratories社)を含むLB培地に添加してLB寒天を調製した。
【0094】
大腸菌のコロニーは、1×NH4緩衝液(Bioline社)、2.5mMのMgCl2(Bioline社)、0.5mMのdNTPs(Bioline社)、0.3μMの各プライマー(AtCytFWD−AvrIIおよびAtCytREV−BamHI)、すなわち配列番号1および2、ならびに0.5単位のBioTaq NAポリメラーゼ(Bioline社)を用いるPCRによってスクリーニングした。熱循環は、95℃で5分間の最初の変性工程、次いで、95℃で30秒間、55℃で30秒間および72℃で4分30秒間の30サイクル、72℃で5分間の最終的な伸長工程によって行った。PCR産物を、上記のように1%(w/v)のアガロースゲル電気泳動を用いて調べた。所望の挿入断片を含むコロニーを、50μgml−1のカナマイシンを含む5mlのLB培地において37℃の振盪培養器内で一晩増殖させた。プラスミドDNAを、推奨されている手順に従ってQIAPrepスピンミニプレップキット(QIAPrep Spin Miniprep Kit)(Qiagen社)を用いて抽出した。DNAは、1×BamHI緩衝液中の10単位のBamHI(NEB1)によって37℃で1時間消化した。消化は、上記のように1%(w/v)のアガロースゲル電気泳動を用いて調べた。正しい制限酵素消化パターンを示すプラスミ中の挿入断片を、3730DNA分析機(Applied Biosystems社)を用いて、表1に示すプライマーAtCytFWD−AvrII(配列番号1)、AtPPDKSeqF1(配列番号3)、AtPPDKSeqF2(配列番号4)、AtPPDKSeqR1(配列番号5)、AtPPDKSeqR2(配列番号6)およびAtCytREV−BamHI(配列番号2)を用いて配列決定した。配列は、BioEdit(Ibis Biosciences社)で分析した。
【0095】
これらの構築物をTOPO−PPDKcDNAおよびTOPO−PPDKgDNAと命名し、図2aおよび図2bに示す。
【0096】
pBNP構築物のクローン化
PPDKを、老化誘発プロモーターSAG12の制御下でBNP1380000001バイナリーベクターに連結させた。SAG12を含む骨格プラスミド(BNP1380000001)は、pBINPLUS(F. A. van Engelen et al. Transgenic Research (1995) 4, 288-290)に基づくものであり、図3aに示す。最初に、TOPO−cDNAまたはTOPO−gDNAを保有する大腸菌のコロニーを使用して、50μgml−1のカナマイシンを含む25mlのLB培地に植菌した。これらの培養物を37℃の振盪培養器内で一晩培養し、推奨されている手順に従ってプラスミドミディキット(Plasmid Midi Kit)(Qiagen社)を用いてプラスミドDNAを抽出した。プラスミドミディキットを用いて、50μgml−1のカナマイシンを含む100mlの培養物からpBNP1380000001ベクターを精製した。
【0097】
次いで、これらのプラスミドを、制限酵素AvrIIおよびBamHIによって消化した。消化反応物は、37℃で一晩培養し、2μgのDNA(BNP社)または4μgのDNA(TOPO−PPDKcDNAおよびTOPO−PPDKgDNA)のいずれか一方、1×緩衝液2、10単位のAvrIIおよび10単位のBamHIを含んでいた。試料は、0.8%(w/v)のアガロース、0.5×TBEおよび25μMのクリスタルバイオレット(Hopkin and Williams社)を用いるクリスタルバイオレットアガロースゲル電気泳動(Rand, 1996, Crystal violet can be used to visualize DNA bands during gel electrophoresis and to improve cloning efficiency(クリスタルバイオレットは、ゲル電気泳動中にDNAバンドを視覚化し、かつクローン化効率を向上させるために使用することができる). Elsevier Trends Journals Technical Tips Online.)によって分離し、クリスタルバイオレット試料緩衝液(250μMのクリスタルバイオレット(Hopkin and Williams社)および30%(w/v)のスクロース(Fisher Scientific社)および泳動用緩衝液としての25μMのクリスタルバイオレットを含有する0.5×TBEを用いて、約2時間50Vの電気泳動に供した。推奨されている手順に従ってQIAQuickゲル抽出キット(QIAQuick Gel Extraction Kit)(Qiagen社)を用いるゲルバンド抽出を使用して、14.4kbのBNP、2.6kbのPPDKcDNAおよび4.4kbのPPDKgDNA断片を抽出した。断片は、臭化エチジウムを用いる1%のアガロースゲル電気泳動を用いてHyperLadder I(Bioline社)に対して調査および定量化した。
【0098】
ゲル抽出したgDNA断片は、gDNA断片(4.4kb)およびTOPO骨格(3.9kb)と同様の大きさであるため、ホスファターゼで処理してあらゆる汚染性TOPO骨格との連結反応を防止した。エビアルカリホスファターゼ(SAP、1単位、Roche社)を、1×脱リン酸化緩衝液(Roche社)に入れた1μgのゲル抽出したgDNA断片に添加し、この反応物を37℃で30分間培養した後、65℃で10分間失活させた。連結反応は、10:1のモル比で消化したBNPを有するcDNAまたはgDNA断片のいずれか一方、1×連結反応緩衝液(NEB社)および1単位のT4DNAリガーゼ(NEB社)を用いて行った。連結反応物を16℃で一晩培養し、2μlを使用して、推奨されている手順に従ってLibrary−Efficiency(登録商標)DH5α大腸菌(Invitrogen社)を形質転換した。
【0099】
形質転換した大腸菌を、50μgml−1のカナマイシンを含むLB寒天上に移し、37℃で一晩培養した。大腸菌のコロニーは、1×NH4緩衝液、2.5mMのMgCl2、0.5mMのdNTP、0.3μMの表1に示す各プライマー(AtPPDKexon15FWD(配列番号7)およびBNPnostREV(配列番号8)、および0.5単位のBioTaq DNAポリメラーゼを用いるPCRによってスクリーニングした。熱循環は、95℃で5分間の最初の変性工程、次いで、95℃で30秒間、55℃で30秒間および72℃で3分間の30サイクル、72℃で5分間の最終的な伸長工程によって行った。PCR産物は、上記のように1%(w/v)のアガロースゲル電気泳動を用いて調べた。所望の挿入断片を含むコロニーを、50μgml−1のカナマイシンを含む5mlのLB培地において37℃の振盪培養器内で一晩増殖させた。プラスミドDNAは、推奨されている手順に従ってQIAPrepスピンミニプレップキット(QIAPrep Spin Miniprep Kit)を用いて抽出した。DNAは、上記のように1%(w/v)のアガロースゲル電気泳動に供する前に37℃で1時間、1×BamHI緩衝液中で10単位のBamHIおよび10単位のStuIで消化した。正しい制限酵素消化パターンを示す、cDNAおよびgDNA連結反応によってそれぞれ得られた1つのコロニーを選択し、上述のように、表1に示すプライマーBNP−SAG12FWD(配列番号9)を用いて配列決定した。
【0100】
これらの構築物をBNP−PPDKcDNAおよびBNP−PPDKgDNAと命名し、図3aおよび図3bに示す。
【0101】
実施例2−BNP−PPDKcDNAおよびBNP−PPDKgDNAによるシロイヌナズナの形質転換
アグロバクテリウムチュメファシエンスのGV3101−R株を、電気穿孔によって、図3に示すプラスミドBNP−PPDKcDNAおよびBNP−PPDKgDNAで形質転換した。それらの調製については、実施例1に記載されている。
【0102】
電気的形質転換受容性(electrocompetent)アグロバクテリウムを、25mgl−1のリファンピシン(Sigma−Aldrich社)を含むLB培地の培養物から産生し、30℃および、600nmにおいて0.4〜0.6の光学濃度で増殖させ、Eppendorf Biophotometer分光光度計を用いて測定した。500mlの培養物を4000gで15分間遠心分離し、上澄みを捨て、細胞を500mlの10%冷グリセリン(Fisher Scientific社)に再懸濁した。遠心分離および再懸濁を250ml、次いで10ml、最後に2mlのグリセリンを用いて繰り返した。細胞は、50μlの分割量に等分し、液体窒素中で急速冷凍し、−80℃で保存した。電気穿孔は、BioRad遺伝子導入装置(BioRad Gene Pulser)を用いて行った。プラスミドDNA(200ng)を、上記のように予備冷却した電気穿孔法用キュベット(Gene Pulser Cuvette、BioRad社)に入れた50μlのアグロバクテリウム細胞に添加し、この細胞を氷上で5分間培養した。このキュベットに対して、2.5mVのパルス、400オームの抵抗および25μFの静電容量を用いた電気穿孔を行った。1mlのSOC培地(Super Optimal Broth, Catabolite Repression)は、20g/lのバクト−トリプトン、5g/lのバクト酵母抽出物、85mMの塩化ナトリウムおよび250mMの塩化カリウムを含んでいた。10Mの水酸化ナトリウムを使用して、高圧蒸気滅菌前にpHを7.0に調整した。使用前に、最終濃度が10mMになるように無菌塩化マグネシウムを添加し、最終濃度が20mMになるように無菌グルコース(Fisher Scientific社)を添加した。この細胞を30℃の振盪培養器内で2時間培養した後、50μgml−1のカナマイシンおよび50μgml−1のリファンピシンを含むLB寒天上に移した。
【0103】
30℃で2日間の培養後に、コロニーをPCR、次いで制限酵素消化によってスクリーニングした。PCRのスクリーニングで陽性であり、かつ期待した制限消化パターンを示すコロニーを、50μlml−1のカナマイシンおよび50μlml−1のリファンピシンを含む5mlのLB培地に植菌し、30℃の振盪培養器内で一晩培養した。翌日、この培養物600μlを使用して上記のように500mlのLB培地に植菌し、花浸漬(floral dipping)に使用する前に、30℃の振盪培養器内で30時間培養した。
【0104】
シロイヌナズナ植物は4週齢で花浸漬のために使用した。全ての構築物を、野生型の生態型Columbia 0に形質転換した。上記のように調製したアグロバクテリウム培養物を5,000gで、4℃で15分間遠心分離し、細胞ペレットは、250mlの無菌の5%スクロース(Fisher Scientific社)溶液(w/v)および0.05%Silwett L-77(OSi Specialties社)に再懸濁した。植物は、穏やかに撹拌しながら細胞懸濁液に約10秒間浸漬した後、ポリエチレン製ラップで蓋をし、24時間直射日光を避けた。この後、植物を標準的な規制飼育に戻し、種子を収穫した。
【0105】
形質転換シロイヌナズナの選択
形質転換した植物からの種子は、Harrison et al. (2006), Plant Methods, 2, 19に従って、50μgml−1のカナマイシン(Dufecha Biochemie社)に関して選択した。抗生物質抵抗性を示すT1植物は結実するまで成長し、自己授粉した。この種子を上記のように選択し、3:1の耐性:非耐性比を示すT2系統を、導入遺伝子の単一コピーを保有するものとして選択した。これらの植物は再び自己授粉し、上記のように選択した。100%の耐性を示す子孫(T3)を産生したT2系統をホモ接合性として選択し、全ての実験に使用した。メンデル遺伝学を使用して、導入遺伝子に対してホモ接合性であり、かつ単一の導入遺伝子コピーを含むT2世代植物を選択した。SAG12−PPDKcDNAおよびSAG12−PPDKgDNAで形質転換した各植物に対して、5種類の独立した単一の挿入断片のホモ接合性系統を選択した。
【0106】
実施例3−BNP−PPDKcDNAおよびBNP−PPDKgDNAによるタバコの形質転換
形質転換受容性アグロバクテリウムチュメファシエンス株LBA4404を、実施例2に記載されているように電気穿孔法によって、プラスミドBNP、BNP−PPDKcDNAおよびBNPPPDKgDNAで形質転換した。電気穿孔後、電気穿孔した細胞に1mlのLB培地を添加し、次いで、その細胞を、50μgml−1カナマイシンおよび100μgml−1スペクチノマイシン(Sigma−Aldrich社)を含むLB寒天上に移す前に、28℃の振盪培養器内で2時間培養した。28℃で2日間の培養後、1つのコロニーを使用して、50μgml−1カナマイシンおよび100μgml−1スペクチノマイシンを含む50mlのLB培地に植菌した。この培養物を、28℃の振盪培養器内で3日間培養した。プラスミドDNAを抽出し、制限酵素消化によって分析し、50μlの培養物を使用して、50μgml−1のカナマイシンおよび100μgml−1のスペクチノマイシンを含む50mlのLB培地に植菌した。この培養物を、28℃の振盪培養器内で一晩培養した。
【0107】
タバコ栽培品種バーレー21およびK326を種子から栽培し、最も若い葉を8週齢の植物から切除し、8%(v/v)ドメストス濃厚漂白剤(Domestos社)で10分間滅菌した後、無菌蒸留水ですすいだ。No.6コルク穿孔器を使用して葉片に穴を開け、次いで、これを25mlのアグロバクテリウム培養物に2分間入れた。次いで、葉片を、2.2μMの6−ベンジルアミノプリン(BAP)および0.27μMのα−ナフタレン酢酸(NAA)を含むMS培地上に下側を下に向けて置いた。これらを、22℃の成長室に2日間置いた。次いで、葉片を、500μgml−1のクラフォラン(Roussel Laboratories社)(共培養されていない対照)または500μgml−1のクラフォランおよび100μgml−1のカナマイシンを含む上記のような選択的なMS培地に移した。葉片は、6週間にわたって14日毎に、上記のような新しい選択的なMS培地に移した。次いで、カルスおよび苗条塊(shoot clump)を葉片から除去し、0.5μMのBAP、500μgml−1のクラフォランおよび100μgml−1のカナマイシンを含むLS培地に入れた。2週間後、苗条を0.5μMのBAP、500μgml−1のクラフォランを含みかつカナマイシンを含まないLS培地を入れた150mlの瓶に移した。
【0108】
それから3週間後、優性の苗条を、250μgml−1のクラフォランを含みかつBAPまたはカナマイシンを含まないLS培地に移した。さらに3週間後、苗条を、抗生物質またはBAPを含まないをLS培地に移すことによって、さらに浄化した。十分な根が生えたら、植物を温室内の土壌に移した。
【0109】
形質転換タバコの選択
定量的PCR(Q−PCR)を使用して、T0およびT1植物中の導入遺伝子のコピー数を定量化した。可能であれば、単一の挿入断片T0およびホモ接合性T1植物を分析のために選択した。導入遺伝子の検出のために、表1に示すようなプライマーBNP−1271F(配列番号10)およびBNP−1334R(配列番号11)を使用し、Vic/TAMARA標識プローブBNP1291TV(配列番号12)を使用して、nptII導入遺伝子にアニールした。内部定量化のために、プライマーNtCyc−184F(配列番号13)およびNtCyc−316R(配列番号14)を使用し、FAM/TAMARA標識プローブNtCyc−267T(配列番号15)を使用して、サイクロフィリンをコードする内因性遺伝子にアニールした。定量化は、比較Ct法によるものであった(ΔΔCt method, Bubner & Baldwin (2004), Plant Cell Reports, 23, 263-271)。反応混合物は、1×ユニバーサルマスターミックス(Universal Master Mix)(ABI社)、0.9μMの各プライマー、0.2μMの各プローブ(BNPおよびNtCycプライマーおよびプローブに対して別の反応を用いる)および推奨されている手順に従ってDNeasy植物ミニキット(Qiagen社)を用いて葉の組織から抽出した約500ngのgDNA鋳型を含んでいた。熱循環は、95℃で10分間の最初の変性工程、次いで、95℃で15秒間、60℃で1分間の40サイクルによって、7900HT FastリアルタイムPCRシステム(7900HT Fast Real-Time PCR System)(Applied Biosystems社)で行った。データは、付属のSDS2.2ソフトウェア(Applied Biosystems社)を用いて分析した。
【0110】
形質転換したT0世代のK326およびバーレー21タバコの組織培養における再生後、定量的PCR(Q−PCR)を使用して、導入遺伝子の単一コピーを有する植物を選択した。単一の挿入断片の植物は、導入遺伝子サイレンシングの可能性を減少させるように選択した。Q−PCRは、ネオマイシンホスホトランスフェラーゼをコードするnptII導入遺伝子に相補的なオリゴヌクレオチドを用いて行った。この遺伝子は、構築物pBNP、BNP−PPDKcDNAおよびBNP−PPDKgDNA中の植物ゲノム内に運ばれたT−DNAの左右の境界の間に存在していた。pBNPのみによって形質転換した植物は、組織培養による再生が空のベクターによって形質転換された植物とPPDKコード配列を含むベクターによって形質転換された植物との間で一致することを確認するための空のベクター対照として機能した。形質転換が成功したことを確認するための再生およびQ−PCR検査後、これらの植菌を破棄した。
【0111】
各構築物(SAG12−PPDKcDNAおよびSAG12−PPDKgDNA)ならびに各栽培品種(K326およびバーレー21)のために、6種類の植物を選択した。単一の挿入断片の植物のみを選択することができなかったため、6種類の単一の挿入断片の植物が入手できなかった場合は、可能な限り最も少ないコピー数を有する植物を選択した。これらの植物は自己授粉し、その種子を収穫した。
【0112】
選択したT0植物ごとに14株の子孫を育て、Q−PCRを繰り返してホモ接合性植物を選択した。ホモ接合性植物の使用によって、将来の世代における導入遺伝子の安定性を確保しなければならない。T0親ごとに、導入遺伝子を保有している4株の子孫を選択した。しかし、全てのT0親に対して、ホモ接合性の子孫を必ずしも選択することができるわけではなかった。親のコピー数が3つ以上であった場合は、より少ないコピー数を有する植物を選択して、導入遺伝子サイレンシングの可能性を減少させかつ、また必要であれば、T2世代からのホモ接合体の選択を単純化した。このようにして、各構築物および各栽培品種のために、5種類の独立した系統から、全てのさらなる実験のためにより少ない導入遺伝子コピー数を有する4種類の生物学的複製(同胞)を選択した。
【0113】
実施例4−形質転換したシロイヌナズナ植物中のPPDKタンパク質の検出および定量化
タンパク質の抽出
シロイヌナズナの葉の組織(100mg)を、マイクロ乳棒を用いて1.5mlの微小遠心管中で液体窒素冷却して粉砕し、400μlの抽出緩衝液(リン酸カリウム緩衝液(+プロテアーゼ阻害剤反応混液(PIC、Sigma社)))を添加した。タンパク質の濃度は、推奨された手順に従ってBio−Radタンパク質アッセイ試薬(Bio-Rad Protein Assay Reagent)(Bio−Rad社)を用いてブラッドフォードアッセイ(Jones et al., 1989, Journal of Chemical Ecology, 15, 979-992)で測定した。
【0114】
ポリアクリルアミドゲル電気泳動
タンパク質は、10%(v/v)アクリルアミド(37.5:1=アクリル:ビスアクリル、Severn Biotech社)、50%(v/v)免疫ブロット分解緩衝液(セクション2.10を参照)、0.05%(w/v)ペルオキソ二硫酸アンモニウム(APS、AnalaR社)、および0.05%(v/v)N,N,N’,N’−テトラメチルエチレンジアミン(TEMED、Severn Biotech社)を含む分解ゲルにおけるポリアクリルアミドゲル電気泳動(PAGE社)によって分離した。濃縮用ゲルは、上記のような5%アクリルアミド、50%(v/v)免疫ブロット濃縮用緩衝液、0.06%(w/v)APSおよび0.1%(v/v)TEMEDを含んでいた。電気泳動は、1×免疫ブロット試料緩衝液(66mMのトリス−HCl、10%(v/v)グリセリン、0.7mMのSDS、0.7Mのβ−メルカプトエタノール(BDH Laboratory Supplies社)および0.05%(w/v)ブロモフェノールブルー)および免疫ブロット泳動用緩衝液、(25mMのトリス−HCl、0.29mMのグリシン(Fisher Scientific社)および3.5mMのSDS)を用いて抽出した20μgの総タンパク質に対して70mAの電流で1時間30分間行った。
【0115】
二組のゲルを同時に泳動した。一方を免疫ブロット分析(以下に記載する)のために使用し、他方を推奨された手順に従ってGelCode青色安全タンパク質染色(GelCode Blue Safe Protein Stain)(Thermo Scientific社)を用いて染色した。ゲル乾燥膜(Gel Drying Film)(Promega社)を使用して染色したゲルを乾燥した。
【0116】
免疫ブロット分析
免疫ブロット分析用のゲルからのタンパク質を、Protean極厚ブロット紙(Protean Extra-Thick Blot Paper)(Bio−Rad社)、および免疫ブロット用転写緩衝液(48mMのトリス−HCl、39mMのグリシン、1.3mMのSDSおよび20%(v/v)メタノール(Fisher Scientific社))を用いて、セミドライブロッター(Semi-Dry Blotter)(Bio−Rad社)中でProtan BA83硝酸セルロース膜(Schleicher and Schuell社)に15Vで1時間転写した。タンパク質の転写を確認するために、ポンソー染色(0.5%(w/v)ポンソーS(Fluka社)および1%(w/v)氷酢酸(Fisher Scientific社))をブロットした後の膜に塗布し、次いで、その膜を蒸留水ですすいだ。
【0117】
PPDK免疫ブロット分析用のブロッキング緩衝液は、1%(w/v)乾燥脱脂乳粉末(Marvel社)および0.1%(v/v)ポリオキシエチレンモノラウリン酸ソルビタン(TWEEN20、Sigma−Aldrich社)のリン酸緩衝食塩水(PBS:1.5mMの正トリン酸二水素カリウム(KH2PO4、AnalaR社)、8.1mMの正リン酸二水素ナトリウム、2.7mMの塩化カリウムおよび137mMの塩化ナトリウム)を用いて毎日更新した。塩酸を使用してpHを7.4に調整した。膜は、振盪機上の室温のブロッキング緩衝液中で1時間培養した。一次ハイブリッド形成は、ウサギ抗PPDK抗体(Chris Chastain、ミネソタ州立大学)の1:10,000の希釈液を用いて行い、次いで、ブロッキング緩衝液でそれぞれ5分間の洗浄を3回行った。二次ハイブリッド形成は、ロバ抗ラビットビオチン化全抗体(GE Healthcare社)の1:1000の希釈液を用いて行い、次いで、上記のように3回洗浄した。三次ハイブリッド形成は、ストレプトアビジンビオチン化ワサビペルオキシダーゼ複合体(GE Healthcare社)の1:1000の希釈液を用いて行い、次いで、各0.1%(v/v)TWEEN20を含むPBS中で5分間の洗浄を3回行った。
【0118】
次いで、膜を蒸留水中で3回すすいだ。検出は、推奨されている手順に従って、Western Lightning化学発光試薬(Western Lightning Chemiluminescence Reagent)(増感ルミノール(Enhanced Luminol)、Perkin Elmer社)を用いて行った。
【0119】
図4を参照すると、ウェスタンブロットによる形質転換SAG12−PPDKシロイヌナズナ細胞株の選択が示されている。タンパク質を、8週齢のシロイヌナズナ野生型、ΔPPDK、SAG12−PPDKcDNAおよびSAG12−PPDKgDNA植物の葉の組織から抽出し、より高濃度のPPDKを発現している株を選択するために免疫ブロット分析に供した。組み換え型トウモロコシPPDK(30μg)を陽性対照として使用した。PPDKは、野生型またはPPDK変異型植物(ΔPPDK)において検出されなかった。SAG12−PPDKgDNA植物は、SAG12−PPDKcDNAよりも高濃度のPPDKを発現していた。その後の全ての分析は、5種類のSAG12−PPDKgDNA系統に対して行った。
【0120】
PPDKは、野生型およびΔPPDK植物中では検出することができず、いくつかのSAG12−PPDKcDNA植物中では低濃度で検出することができ、全てのSAG12−PPDKgDNA系統で検出することができた。これらの系統のうち3つにおいて、野生型またはSAG12−PPDKcDNA系統に比べ非常により高い量のPPDKが検出された。従って、イントロンの存在は、PPDKの発現レベルに対してプラスの効果があると思われ、全てのさらなる実験での使用のためにSAG12−PPDKgDNA系統を選択した。
【0121】
SAG12−PPDKgDNA植物におけるPPDKの累積の開始および程度を測定するために、PPDKの免疫ブロットを、野生型、ΔPPDKおよびSAG12−PPDKgDNA植物に対して行った。
【0122】
図5を参照すると、野生型、ΔPPDKおよび5週目以降の5種類の独立したSAG12−PPDKgDNA系統におけるPPDK発現量の定量化が示されている。PPDK発現量は、5〜9週目の各時点の野生型の割合として計算した。データは、これらの5つの時点の平均として示されている。エラーバーは標準誤差(SEM)を示す。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は、分散分析を用いて試験した(F=4.775、df=6、p=0.001)。野生型と有意差が認められる系統は、G12.3(p=0.005)およびG16.1(p=0.004)であった。
【0123】
タンパク質は、播種後3週目から9週目までの週間隔で収穫された葉の組織から抽出した。PPDKは、5週目以降の野生型植物中で低量検出され、これにより、転写産物の観察された増加によってタンパク質の発現量が増加することが確認された。SAG12−PPDKgDNA植物では、PPDKの累積の開始も5週目に観察されたが、PPDK発現量は野生型よりも多い。PPDKタンパク質は、ΔPPDK植物ではいずれの段階においても検出されなかった。
【0124】
葉のタンパク質抽出物中のPPDKの定量化を可能にする方法も最適化した。異なる量の組み換え型トウモロコシPPDKタンパク質をSDS−PAGEおよび免疫ブロット分析に供した。バンド強度は、AlphaEase画像化ソフトウェアおよび作成された標準曲線を用いて計算した。回帰直線はSigmaPlotソフトウェアを用いて計算し、次いで、これを使用して植物の葉の試料中のPPDK量を計算した。免疫ブロットの検出レベルが変化する可能性があるため、異なる免疫ブロットの間の真の比較を可能にするために、全ての免疫ブロットに対して少なくとも3種類の標準物質を含める必要があった。異なる抽出物中のPPDK発現量の定量化のためのこの技術を使用して、異なる形質転換系統を選択および比較した。
【0125】
PPDKタンパク質発現量は、各時点における各系統の少なくとも4個の生物学的複製について、組み換え型トウモロコシPPDKタンパク質の公知の標準物質を参照することによって計算した。分析から、SAG12−PPDKgDNA中のPPDK発現量が5週目以降の野生型における発現量よりも高いことが分かった。PPDK発現量は、各SAG12−PPDKgDNA系統について5週目以降の各時点の野生型の割合、および各SAG12−PPDKgDNA系統の老化期間におけるPPDKの累積を定量化するためのこれらの値からの平均として計算した。図5に示すように、これによって、SAG12−PPDKgDNA系統をPPDK発現量の増加に従って順序付けることができた。
【0126】
形質転換植物中に高い量で蓄積したタンパク質が失活する場合があるため、タンパク質発現量の増加が必ずしも酵素活性の増加を暗示するというわけではない。PPDKは暗所で可逆的にリン酸化され、その結果、PPDKは失活する。抗体は、PPDKのリン酸化された型(リン酸化PPDK)に抗して増加し、PPDKの失活型の特異的検出を可能にするが、リン酸化状態に関係なく以前に使用された抗体(抗PPDK)はPPDKを検出する(Chastain et al., 2002, Plant Physiology, 128, 1368-1378)。抗リン酸化PPDK抗体を使用して、免疫ブロット時には抗PPDK抗体によって最初に検出され、次いで剥離され、かつ再検出される播種後3週目から9週目までに経時的に失活したPPDKを検出した。
【0127】
驚くべきことに、失活した(リン酸化された)PPDKは、複数の免疫ブロットにおいて野生型植物中に超低濃度でのみ、かつ老化の後期段階の間にのみ検出することができた。SAG12−PPDKgDNA植物では、失活した時点に応じて、PPDKは、より高濃度で検出された。しかし、PPDK発現量の増加開始直後の時点では、総PPDKの高い発現量にも関わらず、失活したPPDKは、ほとんどあるいは全く検出されなかった。これらの結果は、一部のPPDKの失活がSAG12−PPDKgDNA植物中に生じる可能性があるが、酵素的に活性なPPDKの発現量の増加は、少なくとも老化の初期段階の間にSAG12−PPDK植物において存在することを示唆している。
【0128】
実施例5−形質転換したタバコ(N.tabacum)植物中のPPDKタンパク質の検出および定量化
タバコの葉の組織(100mg)を、マイクロ乳棒を用いて1.5mlの微小遠心管の中で液体窒素下で粉砕し、400μlの抽出緩衝液(オーバーコート緩衝液(Overcoat Buffer)(PIC社))を添加した。タンパク質の濃度は、推奨されている手順に従って、Bio−Radタンパク質アッセイ試薬(Bio−Rad社)を用いてブラッドフォードアッセイ(Jones et al., 1989)で測定した。タンパク質は、実施例4に記載されているようにポリアクリルアミドゲル電気泳動(PAGE社)によって分離した。実施例4に記載されている免疫ブロットによってタンパク質を分析し、かつPPDKタンパク質を定量化した。
【0129】
PPDKに対する免疫ブロットは、分離および暗所での30℃の培養によって老化が誘発されたK326およびバーレー21のT1世代タバコの葉から抽出した葉のタンパク質に対して行った。老化は、植物が成熟期に達する前に最も高いPPDK発現量を有する形質転換系統を特定し、かつ他の系統を破棄できるように誘発した。K326植物では、PPDKは、3種類のSAG12−PPDKgDNA系統(G4、G8およびG10)および1種類のSAG12−PPDKcDNA系統(C10)において高い発現量で検出された。バーレー21植物では、PPDKは、4種類のSAG12−PPDKgDNA系統(G3、G7、G15およびG23)に大量に存在したが、K326系統に認められる程に高い発現量では存在していなかった。最も高いPPDK発現量を有するK326およびバーレー21のそれぞれの4種類の系統をさらなる全ての分析のために使用した。ほぼ例外なくSAG12−PPDKgDNA系統のみに最も高いPPDK発現量が観察されたため、イントロンがPPDK発現に対してプラスの効果を有していると思われる。
【0130】
図6を参照すると、以下のように、SAG12−PPDK−K326およびバーレー21タバコ系統の選択が示されている:
(b)形質転換K326系統を選択するためのPPDK免疫ブロット。老化は、分離および暗所での30℃で3日間の培養によって6週齢の植物の緑葉において誘発された。タンパク質を抽出し、免疫ブロット分析に供して、より高濃度のPPDKを発現している系統を選択した。組み換え型トウモロコシPPDK(50μg)を陽性対照として使用した。各親のT0植物からの1つの子孫に関する1つの代表的な免疫ブロットが示されている。SAG12−PPDKgDNA親系統G4、G8およびG10から、および、SAG12−PPDKcDNA系統C10からの植物をその後の全ての分析に使用した。
(c)セクション(b)におけるK326に関して、形質転換バーレー21系統を選択するためのPPDK免疫ブロット。SAG12−PPDKgDNA親系統G3、G7、G15およびG23からの植物をその後の全ての分析のために使用した。
【0131】
PPDKに対する免疫ブロットを、野生型、ゼロコピー(陰性の分離体)、SAG12−PPDKgDNA−K326、SAG12−PPDKcDNA−K326およびSAG12−PPDKgDNA−バーレー21タバコに対して行って、PPDKの発現の開始を測定し、かつ形質転換した植物中のPPDKの過剰発現の程度を定量化した。タンパク質は、K326野生型および形質転換した植物の葉1(最も古い)〜3ヵ月齢の植物の8(若い)から抽出し、PPDKに対する免疫ブロットを行った。
【0132】
図7を参照すると、K326タバコの熟した葉におけるPPDKの過剰発現が示されている。PPDK発現量は免疫ブロットから計算した。データは、ゼロコピー植物の10個の生物学的複製および各SAG12−PPDK系統の4個の生物学的複製の平均として示されている。エラーバーは標準誤差を示す。SAG12−PPDK系統は、PPDK発現量の上昇で配置されている。遺伝子型間の差は、分散分析を用いて試験した(F=6.995、df=4、p=0.001)。ゼロコピー植物と有意差が認められる系統は、G10(p=0.006)、G8(p=0.003)およびC10(p=0.000)であった。
【0133】
PPDK発現量は、野生型植物のより古い葉の中でより高く、これは、PPDKが、タバコならびにシロイヌナズナにおける老化中に自然に上方制御されることを示している。形質転換したK326植物では、PPDK発現量は同様に、より古い葉においてより高く、野生型植物において非常により高かった。また、最も高いPPDK発現量を有する形質転換した系統のより若い葉においてPPDK発現量はより高かった。免疫ブロットを、ゼロコピー植物(陰性の分離体)および形質転換した植物の「熟した」葉から抽出したタンパク質に対して行った。熟した葉は、収穫可能な段階にあり、かつ老化の後期段階にある葉である。PPDK発現量を定量化した。PPDK発現量は、ゼロコピー植物と比較して、全ての4種類の独立した形質転換した系統の熟した葉においてより高かった。SAG12プロモーターを用いる老化中のPPDKの過剰発現は、K326タバコでは成功していた。
【0134】
実施例6−形質転換したシロイヌナズナ植物の表現型分析
シロイヌナズナ植物の成長分析
図8を参照すると、シロイヌナズナにおけるPPDKgDNAの過剰発現によって、より大きなロゼットを有する植物が生産されることが明らかに示されている。播種後3、5、7および9週目に野生型、ΔPPDKおよびSAG12−PPDKgDNA植物を撮影した。SAG12−PPDKgDNA植物は野生型よりも大きかった。PPDK植物と野生型植物との間に識別可能な差はなかった。
【0135】
ロゼットおよび生殖組織の生質量は、0.1mgの単位まで、メトラー・トレドAB104−S(Mettler Toledo AB104-S)天秤を用いて精密に測定した。ロゼット組織の乾燥質量は、Edwards Super Modulyo凍結乾燥器(Edwards Super Modulyo Freeze Dryer)を用いて一晩凍結乾燥した試料から、0.1mgの単位まで精密に測定した。
【0136】
図9を参照すると、9週目の生殖組織の質量が示されている。より高い生殖組織質量を有するより高いPPDK発現量を有する系統について、PPDKに対する用量反応が観察された。総生殖組織(茎、茎生葉、花および長角果)を計量した。値は3個の生物学的複製の平均である。エラーバーは標準誤差を示す。形質転換SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は分散分析を用いて試験した(F=8.062、df=6、p=0.001)。全てのSAG12−PPDKgDNA系統は、野生型とは有意に異なっていた(G9.4:p=0.031、G5.4:p=0.000、G19.2:p=0.000、G12.3:p=0.001、G16.1:p=0.001)。傾向は絶対ではないが、より高いPPDK発現量を有する系統はより高い生殖組織質量を有する傾向にあった。総植物生質量の割合としての生殖組織も計算したが、分散分析を用いて試験した場合、野生型、ΔPPDKまたはSAG12−PPDKgDNA植物では有意差は認められなかった(F=0.544、df=6、p=0.767、データは示さない)。
【0137】
しかし、総植物質量の割合としての生殖組織の比率は変化せず、これは、植物は全体としてより大きく、かつ栄養組織と生殖組織との資源分配は変化しないことを示唆していた。ΔPPDK植物における総植物生質量および生殖組織質量も測定したが、野生型とは有意差は認められなかった。ロゼットの乾燥質量も測定したところ、総植物生質量および生殖組織質量に関しては、SAG12−PPDKgDNA植物が野生型よりも有意に高い質量を有することが分かった。
【0138】
図10を参照すると、播種後3週目から9週目までの野生型およびSAG12−PPDgDNA植物の総植物生質量(ロゼット+生殖組織)が示されている。データは、野生型では3個の生物学的複製、SAG12−PPDKgDNAでは15個の生物学的複製(5種類の独立した系統ごとに3個の植物)の平均として示されている。エラーバーは標準誤差を示す。野生型およびSAG12−PPDKgDNAの総植物質量は、各時点でスチューデントのt−検定を用いて比較した。ロゼット質量は、9週目では、SAG12−PPDKgDNA植物の方が有意に高かった(p=0.032)。
【0139】
5週目では、SAG12プロモーターによるPPDK過剰発現の開始前に、優位差は認められなかった。全ての5種類の独立したSAG12−PPDKgDNA系統のロゼットの乾燥質量は増加した。質量が大きくPPDK含有量の高い系統については、PPDK発現量に対する用量反応が生殖組織質量に関して観察された。ロゼットの乾燥質量をΔPPDK植物についても測定し、野生型植物とは有意差は認められなかった。
【0140】
シロイヌナズナの葉の表面積の測定
ロゼットの表面積は、単一の成葉の試料(通常は葉10枚)を採取し、上記のように葉およびロゼット全体の質量を測定し、かつルーラーに並べてそれらの葉を撮影して測定した。葉の表面積は、ImageJソフトウェア(アメリカ国立衛生研究所)を用いて測定し、ここから、ロゼット全体の表面積を計算した。
【0141】
ロゼットの表面積は、PPDK過剰発現の開始後にSAG12−PPDKgDNA植物で増加した。6週目および7週目にSAG12−PPDKgDNA植物の表面積は有意により大きかったが、8週目および9週目では、表面積は野生型とは有意差は認められなかった。表面積のこの大きい減少は、野生型と比較してSAG12−PPDKgDNA植物中の栄養分の転流が増加したことに起因していると思われた。ΔPPDK植物の表面積は野生型植物とは有意差は認められなかった。
【0142】
クロロフィル濃度の測定
クロロフィル含有量は、CCM−200手持ち式クロロフィル計(Opti-Sciences社)を用いて、相対的な単位で測定した。
【0143】
シロイヌナズナの蛍光測定
FV/FM比を、Hansatech FMS2蛍光光度計を用いて測定した。葉は、測定前に一晩、製造業者によって提供されたリーフクリップ(leaf clip)を用いて暗順応させた。老化の開始が野生型植物に対してSAG12−PPDKgDNAまたはΔPPDK植物において変化したか否かを判定するために、FV/FM比を測定した。FV/FMは、光化学系IIの量子効率の推定値であり、光阻害が生じると減少する。クロロフィル含有量の減少が検出される前に、光合成の減少が老化の初期に生じるため、FV/FMは老化の開始の有用な基準である。FV/FMは、播種後(葉がFV/FMを測定するのに十分な程大きくなる)4週目から9週目までに経時的に測定した。野生型、SAG12−PPDKgDNAおよびΔPPDK植物のFV/FMは、7週目に最大となり、その後減少し、これは、全ての3種類の遺伝子型において老化の開始のタイミングが同じであったことを示していた。しかし、SAG12−PPDKgDNA植物のFV/FMは、8週目に野生型よりも有意に高く、これは、老化の後期段階における長期にわたる光合成作用を示していた。
【0144】
窒素含有量
窒素分析用の組織を、Edwards Super Modulyo凍結乾燥器を用いて凍結乾燥した。シロイヌナズナの葉の組織(25mg)またはタバコの葉の組織(100mg)を、窒素非含有秤量紙(Elementar Analysensysteme社)に包装した。推奨されている設定を用いて、Rapid N III窒素分析装置(Rapid N III Nitrogen Analyzer)(Elementar Analysensysteme社)を使用して乾燥重量の割合として窒素含有量を測定した。アスパラギン酸(Sigma−Aldrich社)を標準物質として使用した。葉の窒素含有量は、シロイヌナズナの野生型、SAG12−PPDKgDNAおよびΔPPDK植物において、播種後3週目から9週間目までに経時的に測定した。
【0145】
図11を参照すると、7週目における野生型、ΔPPDKおよびSAG12−PPDKgDNA植物の葉の窒素含有量が示されている。データは、野生型およびΔPPDKでは8個の生物学的複製、各SAG12−PPDKgDNA系統では4個の生物学的複製の平均として示している。エラーバーは標準誤差である。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は分散分析を用いて試験した(F=6.047、df=6、p=0.000)。ΔPPDK植物および全てのSAG12−PPDKgDNA系統は、野生型とは有意に異なっていた(ΔPPDK:p=0.004、G9.4:p=0.000、G5.4:p=0.000、G19.2:p=0.001、G12.3:p=0.007、G16.1:p=0.006)。
【0146】
葉の窒素は、7週目以降は、野生型と比較して全ての5種類の独立したSAG12−PPDKgDNA系統において有意により低く、これは、老化中にPPDK発現量が増加することによって窒素の転流効率を高めることができるという仮定を支持するものであった。PPDKの発現および活性は、SAG12−PPDKgDNA植物において6週目にピークに達し、これは、PPDK発現量の増加と測定可能な葉の窒素の減少との間に時間の遅れが生じたことを示唆しており、この時間の遅れは、タンパク質アミノ酸を輸送アミノ酸(アスパラギンおよびグルタミン)に変換し、かつそれらを葉から輸送するのに要する時間に起因していると思われた。
【0147】
シロイヌナズナ種子の分析
個々の種子の質量および窒素含有量を、野生型、SAG12−PPDKgDNAおよびΔPPDKシロイヌナズナ植物の種子について測定した。
【0148】
図12を参照すると、野生型、ΔPPDKおよびSAG12−PPDK植物の個々の種子の窒素含有量が示されている。SAG12−PPDKgDNA植物の種子窒素含有量において有意な増加が認められた。データは、野生型およびΔPPDKでは8個の生物学的複製、各SAG12−PPDKgDNA系統では4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は分散分析を用いて試験した(F=6.704、df=6、p=0.000)。野生型と有意差が認められる系統は、G19.2(p=0.000)、G12.3(p=0.005)およびG16.1(p=0.002)であった。より高いPPDK発現量を有する系統は、より高い種子窒素質量を有する傾向がある。従って、個々の種子質量は、野生型に対して全ての5種類の独立したSAG12−PPDKgDNA系統で増加し、より高い種子質量を有するより高いPPDK発現量を有する植物については、PPDK発現量に対する用量反応が観察された。各植物から収穫された全種子を計量し、野生型とSAG12−PPDKgDNA植物との間に有意差はなく、SAG12−PPDK植物において種子の大きさがそのように増加したことによって総種子収穫量は損なわれなかった。
【0149】
シロイヌナズナの葉の組織の遊離アミノ酸含有量
葉の組織(100mg)を、マイクロ乳棒(Eppendorf社)を用いて、1.5mlの微小遠心管中で液体窒素冷却して粉砕し、300μlの無菌脱イオン水を添加した。試料は、13,000rpmで、4℃で5分間遠心分離にかけた。タンパク質の濃度は、推奨されている手順に従って、Bio−Radタンパク質アッセイ試薬(Bio−Rad社)を用いるブラッドフォードアッセイ(Jones et al., 1989)で測定した。推奨されている手順に係って、EZfaastアミノ酸試料試験キット(EZfaast Amino Acid Sample Testing Kit)(Phenomenex社)を用いるアミノ酸分析のために試料を調製した。10mMのギ酸アンモニウム(BDH laboratory supplies社)の50%無菌蒸留水と50%超高純度メタノール(Romil社)溶液に試料を再懸濁した。
【0150】
次いで、試料を、EZfaast250×3.0mmAAA−MSカラム(Phenomenex社)ならびに移動相として10mMのギ酸アンモニウム/50%(v/v)無菌蒸留水および50%(v/v)超高純度メタノールを用いるQ Trap LC/MS/MS(Applied Biosystems社/MDS SCIEX社)による液体クロマトグラフィー−質量分析(LC−MS)にかけた。質量分析計は、EZfaastキット(EZfaast Kit)(Phemonenex社)で推奨されているような条件を用いて正イオンモードで使用した。結果は、付属のAnalystソフトウェア(Applied Biosystems社/MDS SCIEX社)で分析した。
【0151】
総遊離アミノ酸含有量は、野生型、SAG12−PPDKgDNAおよびΔPPDKシロイヌナズナ植物の葉において、播種後3週目から9週目までに経時的に測定した。
【0152】
図13を参照すると、PPDKを過剰発現しているシロイヌナズナの葉の中で総遊離アミノ酸含有量が増加することが示されている。データは、野生型およびΔPPDKでは6個の生物学的複製、各SAG12−PPDKgDNA系統では4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。全ての5種類のSAG12−PPDKgDNA系統が、野生型よりも高い総アミノ酸含有量を有するが、ばらつきが高く、かつ分散分析を用いて試験した場合、有意差は認められなかった(F=1.314、df=6、p=0.289)。
【0153】
SAG12−PPDKgDNA植物では、葉の窒素含有量が野生型よりも有意に低くなるときと同じでかつSAG12−PPDKgDNA植物の葉の中のPPDK発現量が最大になったときから1週間後である7週目の時点で、総遊離アミノ酸含有量は、野生型植物よりも有意に高かった。全ての5種類の独立したSAG12−PPDKgDNA系統で増加が生じたが、ばらつきは高く、かつ個々の系統と野生型との間に有意差は認められなかった。この増加は、この時点で、アミノ酸の排出が生じ得るよりも大きな割合でアミノ酸の産生が生じるため、アミノ酸は葉の老化中に蓄積することを示唆している。総遊離アミノ酸は、野生型と比較してΔPPDK植物に有意差は認められなかった。
【0154】
輸送アミノ酸(グルタミンおよびアスパラギン)も測定し、総遊離アミノ酸の割合として表した。SAG12−PPDKgDNA植物では、輸送アミノ酸含有量は、7週目および8週目において野生型植物よりも有意に高かった。この増加は、全ての5種類の独立したSAG12−PPDKgDNA系統において生じたが、ばらつきは高く、かつ個々の系統と野生型の差は有意ではなかった。従って、SAG12−PPDKgDNA植物における総遊離アミノ酸含有量の増加に加えて、輸送アミノ酸の含有量は、全体の割合として増加する。これも、グルタミンおよびアスパラギンの産生がこれらの時点において葉の排出量を上回ることを示唆し、老化中のPPDK発現量の増加によってアミノ酸の相互転換効率が上昇し、その結果、輸送アミノ酸が産生されるという仮定を支持している。ΔPPDK植物では、野生型に対する有意差は観察されなかった。
【0155】
実施例7−形質転換したタバコ植物の表現型分析
タバコの葉の窒素含有量の分析
葉の窒素含有量を、陰性の分離体およびSAG12−PPDK−K326タバコの熟した葉について測定した。熟した葉は、タバコの生産のために収穫の準備ができいている葉である。4種類の独立したSAG12−PPDK系統のいくつかでは、SAG12−PPDK植物の葉の窒素含有量は陰性の分離体植物よりも低かった。
【0156】
熟した葉は、タバコの生産のために収穫される段階にあるため、葉の窒素含有量を測定するために使用した。しかし、熟した葉における葉の窒素含有量にほとんど差がなかったという事実は、必ずしも窒素の転流が増加しなかったことを暗示しているわけではない。老化しているSAG12−PPDKgDNAシロイヌナズナ植物の時間的経過において、葉の窒素含有量は野生型よりも有意に低かったが、老化の後半までにその差は非常により小さくなった。従って、タバコの葉の窒素含有量における差は老化の初期に生じ、葉の窒素含有量を測定するときまでに差が減少するという可能性がある。
【0157】
タバコの葉のアミノ酸含有量の分析
分離および暗所での30℃で3日間の培養によって老化が誘発されたK326タバコの成葉についてアミノ酸含有量を測定した。これによって、老化の誘発の前後で同じ葉を比較することができた。葉の分離および暗所での培養による老化の誘発は、重複が最大である年齢に関連する老化による遺伝子発現パターンを示す。
【0158】
K326タバコでは、陰性の分離体植物およびSAG12−PPDK植物における老化の誘発後に総アミノ酸含有量が増加した。この増加は、老化の誘発前の総アミノ酸含有量の割合として計算した。SAG12−PPDK系統C10については、この増加は、陰性の分離体植物よりも有意に小さかったが、他のSAG12−PPDK系統はどれも陰性の分離体植物に対して有意差を示さなかった。従って、老化中のPPDKの過剰発現は、暗所で誘発された老化の誘発後の総アミノ酸含有量にほとんど影響を与えることはないと思われた。老化の誘発後に、K326タバコについて、輸送アミノ酸(グルタミンおよびアスパラギン)含有量も増加したが、この増加は、陰性の分離体およびSAG12−PPDK系統において生じ、老化の誘発後の増加の程度において遺伝子型間に有意差はなかった。
【0159】
タバコの種子の分析
全ての4種類の独立した形質転換系統について、個々の種子の質量をK326タバコについて測定し、陰性の分離体植物と比較して、SAG12−PPDKgDNAおよびSAG12−PPDKcDNA植物の方が有意に高かった。莢あたりの種子の質量も測定し、SAG12−PPDKgDNAおよびSAG12−PPDKcDNA植物の方が有意に高かった。
【0160】
図14を参照すると、種子の大きさがSAG12−PPDK−K326タバコで増加することが示されている。K326ゼロコピー(SAG12−PPDK挿入断片の陰性の分離体である植物)およびSAG12−PPDK植物の個々の種子の質量は、約1000個の種子を撮影、計数および計量して計算した。データは、ゼロコピー植物では10個の生物学的複製、各SAG12−PPDK系統では4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDK系統は、熟した葉におけるPPDK発現量の昇順で配置されている。種子の質量はSAG12−PPDK植物の方が高い。遺伝子型間の差は分散分析を用いて試験した(F=4.870、df=4、p=0.006)。ゼロコピー植物との有意差が認められる系統は、G8(p=0.005)およびC10(p=0.002)であった。
【0161】
種子の窒素含有量の割合は、全ての4種類の独立したSAG12−PPDKgDNAおよびSAG12−PPDKcDNA系統の方が高かったが、有意差は認められなかった。しかし、種子あたりの窒素の質量は、SAG12−PPDKgDNAおよびSAG12−PPDKcDNA植物の方が有意に高かった。
【0162】
図15を参照すると、種子の窒素含有量がSAG12−PPDK−K326タバコで増加することが明らかに示されている。K326ゼロコピーおよびSAG12−PPDK植物の個々の種子中の窒素の質量は、種子の質量および窒素含有量のデータから計算した。データは、ゼロコピー植物では10個の生物学的複製、各SAG12−PPDKでは4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDK系統は、熟した葉におけるPPDK発現量の昇順で配置されている。個々の種子中の窒素の質量は、SAG12−PPDK植物の方が高い。遺伝子型間の差は分散分析を用いて試験した(F=7.807、df=4、p=0.001)。ゼロコピー植物との有意差が認められる系統は、G8(p=0.000)およびC10(p=0.000)であった。
【0163】
これは、種子への窒素の供給がSAG12−PPDKgDNAおよびSAG12−PPDKcDNA−K326タバコ植物で増加したことを示唆している。K326植物では、PPDKに対する用量反応が観察された。熟した葉のPPDK発現量がより高い植物は、種子の個々の種子の質量がより大きく、莢あたりの種子の質量がより高く、かつ個々の種子中の窒素の質量が増加していた。PPDK含有量の増加が種子への窒素の供給の増加と関係があるように思われるため、この結果は窒素の転流におけるPPDKの役割を強く支持するものである。
【0164】
要約すると、SAG12−PPDK−K326タバコでは、個々の種子の質量(図14)および種子あたりの窒素の質量(図15)はどちらも増加し、この結果は、PPDKの過剰発現によって窒素の転流が増加したことを示唆していた。従って、シロイヌナズナSAG12−PPDKgDNA植物に関しては、老化している葉の輸送アミノ酸含有量の増加が期待される。ただし、シロイヌナズナでは、自然に老化した葉についてアミノ酸含有量を測定したが、タバコでは、葉の分離および暗所での培養によって老化を誘発した。暗所で誘発された老化と年齢に関連する老化とでは生じるプロセスが異なるため、このタバコの葉で生じているプロセスは、シロイヌナズナの葉におけるプロセスに類似している可能性は低かった。
【0165】
実施例8−PCKおよびPPDKを過剰発現しているシロイヌナズナ植物の世代
実施例1および2に記載されているように、PCKのコード領域およびゲノムクローンを、シロイヌナズナに形質転換されたBNP1380000001バイナリーベクター中の老化関連遺伝子12(SAG12)プロモーターに融合させることによって、老化中にシロイヌナズナPCK(At4g37870.1)の過剰な発現が達成された。
【0166】
最初にAtPCKコード配列をシロイヌナズナcDNAから単離し、PPDKに関して実施例1に記載されているように、PCRを用いてゲノム配列をシロイヌナズナgDNAから単離した。ただし、PCRプライマーは、シロイヌナズナAtPCK遺伝子の開始および終止コドンを含むように設計されており、図3aに示すその後のBNP1380000001ベクターへの遺伝子の連結反応を容易にするために、表1に示すように、順方向プライマー(AtPCK−XbaI−FOR)(配列番号25)内にXbaI制限部位、そして逆方向プライマー(AtPCK−SacI−REV)(配列番号26)内にSacI制限部位も含む。
【0167】
実施例1に従って、各PCR反応に使用されるcDNAおよびgDNA鋳型を調製した。PCR反応混合物は、1×HF緩衝液(NEB社)、2mMの塩化マグネシウム(NEB社)、0.5mMのdNTP(Bioline社)、100ngの鋳型(cDNAまたはgDNA)、0.5μMの各プライマーおよび1単位のPhusion High-FidelityDNAポリメラーゼ(NEB社)を含んでいた。熱循環は、98℃で30秒間の最初の変性工程、次いで、98℃で10秒間、60℃で30秒間の35サイクル、および72℃でコード領域に対して2分30秒間、ゲノムクローンに対して4分30秒間の伸長時間によって、Techneサーマルサイクラーを用いて行った。最後の工程では、72℃で10分間の最終的な伸長を行った。
【0168】
これらのPCR産物によって、コード配列(cDNA)に対する2kbのバンドおよびPCKのゲノム配列に対する3.5kbのバンドが得られ、これらをPEG沈殿させた。実施例1に記載されているPPDK同様に、増幅したDNAを、推奨されている手順に従って、pCR4 Blunt-TOPOベクター(Invitrogen社)に連結させた(平滑末端)。次いで、このプラスミドを、Library Efficiency DH5α大腸菌細胞に形質転換した。カナマイシン(50μgml−1)を選択的な抗生物質として使用した。陽性のコロニーを、コード領域およびゲノムクローンのためのプライマーAtPCK−XbaI−FOR(配列番号25)およびAtPCK−SacI−REV(配列番号26)を用いるコロニーPCRによって選択した。陽性のコロニーを、50μgml−1カナマイシンを含む5mlのLB培地において、37℃の振盪培養器内で一晩増殖させた。
【0169】
プラスミドDNAは、QIAprepスピンミニプレップキット(Qiagen社)を用いて単離し、挿入断片の大きさを連続的な酵素制限消化によって分析した。これは、10単位のXbaIおよび1×XbaI緩衝液によって1μgのDNAを37℃で2時間消化し、次いで10単位SacIおよび1×SacI緩衝液を37℃で一晩添加することによって行った。正しい挿入断片を含むプラスミドDNAは、AtPCK−XbaI−FOR/AtPCK−SacI−REVを用いて配列決定した。配列はBioEditを用いて分析し、増幅したAtPCKコード配列およびゲノム配列はBLASTXを用いて確認した。
【0170】
AtPCKコード領域およびゲノムクローンを図3aに示すpBNPベクターに連結させるために、2種類のプラスミドをそれぞれ表す大腸菌コロニーを使用して、50μgml−1のカナマイシンを含む25mlのLB培地に植菌した。培養物は37℃で一晩振盪させ続け、プラスミドDNAは、QIAfilterプラスミドミディキット(QIAfilter plasmid midi kit)(Qiagen社)を用いて精製した。pBNPベクターは、同一の様式だが、50μgml−1のカナマイシンを含む100mlの培養物から精製した。上述したものと同じ連続的な酵素消化を、全ての精製したプラスミドDNAに対して行った。コード領域およびゲノムクローンを含むプラスミドのために3μgのDNAを消化し、pBNPベクターのために1μgのDNAを消化した。試料はクリスタルバイオレットゲル電気泳動によって分離し、その産物はQiaquickゲル抽出キット(Qiagen社)を用いて精製した。14.4kbのpBNPベクター産物をアルカリホスファターゼで処理して自己連結反応を防止し、コード領域およびゲノムクローン挿入断片は、XbaI/SacI消化を用いてpBNPベクターに連結させた。Library efficiency DH5α大腸菌細胞は2μlの連結反応物で形質転換し、陽性のコロニーを、AtPCK−XbaI−FORおよびAtPCK−SacI−REVを用いるPCRによってスクリーニングした。
【0171】
プラスミドDNAを、QIAprepスピンミニプレップキット(Qiagen社)を用いて、所望の挿入断片を含むコロニーから抽出し、続いて、DNAを10単位のXbaI、15単位のSacIおよび1×XbaI緩衝液によって37℃で2時間消化した。期待される正しい制限酵素消化パターンを示したpBNPベクター中のコード配列およびにゲノム配列挿入断片それぞれの2つの別々のコロニーをBNP−SAG12FWDプライマー(配列番号9)を用いて選択および配列決定した。この配列はBioEditプログラムを用いて分析した。図3bに示すように、作製した構築物はpALBNP1(コード配列)およびpALBNP2(ゲノム配列)と命名した。
【0172】
5種類のホモ接合性単一挿入断片系統を作製し、PCK特異的抗体を用いて、実施例3に記載されているような免疫ブロットによってPCKの過剰発現を確認した。ポリクローナル抗血清は、PCKアミノ酸配列に対して設計された合成ペプチドを用いてウサギで産生した。この配列は、(i)DEHCWTETGVSNIEG(配列番号27)および(ii)CVDLSREKEPDIWNA(配列番号28)であり、それらは、化学的に合成した後、ウサギに混注される前に、キーホールリンペットヘモシアニンに連結したものであった。抗体を用いて得られた結果を図16に示す。図16は、形質転換した植物中のPCKタンパク質濃度が上昇したことを示している。
【0173】
SAG12−PCK−PPDK細胞株は、単一のSAG12−PCK形質転換体(3)1および(19)4を、強いSAG12−PPDK過剰発現系統と交差させることによって作製した。図17に示すように、PCKおよびPPDKの両方の濃度が上昇した植物の分析によって、ロゼットの質量が増加していることが分かった。
【0174】
最後に、本発明者らは仮定に縛られることを望んでいないが、図18は、PCKおよびPPDKがどのように窒素転流に影響を与え得るかについて示す推測上の生化学的経路を示す。
【0175】
PCK/PPDK二重構築物
本発明者らは、図3に示すPCKおよびPPDK単一構築物が、植物に形質転換されると、これらの酵素が老化した葉で過剰発現した場合に、老化した葉からの窒素の転流率の上昇、さらに栄養植物成長量の増加(作物収穫率の上昇に該当する)を引き起こすことを観察した。従って、本発明者らは、PCKおよびPPDKをコードする遺伝子がどちらもSAG12プロモーターの制御下でpBNP130000001に挿入されている2種類の二重構築物を作製することに決めた。
【0176】
第1の二重構築物は、XbaI/SacI消化を用いて、BNP−PPDKgDNAのPPDKをコードするgDNA断片の下流(すなわち3'末端)にgDNAをコードするPCKを連結させることによって作製した。従って、プラスミド内のSAG12プロモーターは、PPDK遺伝子およびPCK遺伝子の両方の発現に関与した。
【0177】
第2の二重構築物は、AvrII/BamHI消化を用いて、pALBNP2のPCKをコードするgDNA断片のすぐ下流にPPDKをコードするgDNAを連結させることによって作製した。この場合も、SAG12はPPDK遺伝子およびPCK遺伝子の両方の発現に関与した。
【技術分野】
【0001】
本発明は、形質転換植物の生産に使用される遺伝子構築物に関する。本構築物は、葉の老化中に窒素の転流(remobilisation)を引き起こす能力を持っていて、窒素を葉から植物の他の領域まで輸送することができる。本発明は、そのような構築物で形質転換された植物細胞、および形質転換植物そのものに及ぶ。本発明は、形質転換植物の生産方法、および老化した植物中の窒素転流率を上昇させる方法にも関する。また、本発明は、遺伝子構築物で形質転換されている収穫された植物の葉(例えば、タバコの葉)、およびそのような収穫された植物の葉を含む喫煙物品にも関する。
【背景技術】
【0002】
葉の老化は、葉の細胞が細胞死の前に異なる代謝および構造の変化を受ける植物の発育の一段階である。生理学的および遺伝学的研究では、老化は高度に調節されたプロセスであることが示されている。葉の老化の進行は、葉緑体の分解によって生じるクロロフィルの喪失およびその後の黄変を見かけ上の特徴とする。このような発育段階の特徴である葉のクロロフィル濃度の低下は、例えば、溶媒抽出および分光光度測定によって、あるいは、クロロフィル成分計によって測定することができる。好ましくは一定の条件で育てられた同じ植物について記録された初期の葉のクロロフィル濃度と比較した葉のクロロフィル濃度の低下は、老化を示す。
【0003】
分子的研究では、老化は遺伝子発現における変化と関連していることが示されている。光合成に関与するタンパク質をコードするmRNAのレベルは老化中に減少するが、老化に関与していると思われているタンパク質をコードする遺伝子のmRNAレベルは増加する。老化は、老化関連遺伝子(SAG)として知られている遺伝子によって調節される高度に組織化されたプロセスである。葉の老化は、タンパク質、核酸および膜の劣化、ならびに、この劣化によってその後に生じる発育中の種子、葉または貯蔵器官などの植物の他の領域への栄養分の輸送を伴う。植物の老化の問題の1つは、老化した葉に存在する多くの有用なミネラルおよび栄養分が葉の中に残留し、葉が枯れるにつれて実質的に失われることである。例えば、老化した葉に存在する窒素(アミノ酸のアミン基の形態であってもよい)は、枯れている葉から取り出さなければ無駄になる。
【0004】
従って、特に植物が老化した場合に植物中の窒素の転流を増加させることは、作物生産において重要な有用性を有する可能性がある。最初に、葉から転流される窒素は、より若い葉ならびに発育中の種子に輸送することができる。従って、老化した葉からの窒素の退出効率を上げることによって、種子および植物のより若い部分への窒素の供給を増加させ、それにより、作物収穫率および窒素使用効率を上げることができるかもしれない。これは、世界人口が増加している際には明らかに有益な目的であるが、作物収穫率は需要を満たす程十分に上昇していない。1つの標的となり得る作物は、栄養組織からの窒素の転流が乏しいために低い窒素効率を有するセイヨウアブラナ(学名:Brassica napus)(菜種)である。穀物タンパク質含有量を増加させるという潜在的な利点が大きいという理由から、別の標的作物は小麦である。穀物タンパク質含有量は、小麦の栄養価に影響を与えるだけでなく、穀物の使用および、従って市場価格も決定する。例えば、穀物タンパク質含有量の増加によりパンの体積が増加する。また、タバコの葉の中の残余窒素はニトロソアミンの生成に寄与することが知られているため、窒素の転流を増加させる能力は、タバコ産業において非常に有用になり得る。
【0005】
ホスホエノールピルビン酸カルボキシキナーゼ(PEPCKまたはPCK)[EC4.1.1.49]およびピルビン酸正リン酸ジキナーゼ(PPDK)[EC2.7.9.1]という酵素が知られている。PPDKは原核生物および真核生物の両方に存在し、細菌とより高度な植物との間で配列および三次構造が保存されている(Pocalyko et al., 1990, Biochemistry, 29, 10757-10765)。この酵素は、ピルビン酸+Pi+ATP=PEP+PPi+AMPの反応において、ピルビン酸の可逆的リン酸化を触媒してホスホエノールピルビン酸(PEP)を生成する(Carroll et al., 1990, Federation of European Biochemical Societies, 274, 178-180; Hatch & Slack, 1968, Biochemical Journal, 106, 141-146)。C3およびC4植物の両方において、PPDK遺伝子は、同じ遺伝子から産生される2種類の転写産物を有する変わった構造を有する。長い方の転写産物は葉緑体のタンパク質をコードし、第一エクソンが葉縁体輸送ペプチドをコードするが、短い方の転写産物は、長い方の転写産物の第一イントロン内で別のプロモーターから転写されるため、葉縁体輸送ペプチドをコードする第一エクソンが存在しない。この短い方の転写産物は、PPDKの細胞質基質のイソ型を生成する。この遺伝子構造は、トウモロコシ、イネ、C3およびC4フラベリア属およびシロイヌナズナ(学名:Arabidopsis thaliana)で報告されている。
【0006】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)は、オキサロ酢酸+ATP=PEP+ADP+CO2の反応において、ADP、二酸化炭素およびホスホエノールピルビン酸の可逆的反応を触媒して、ATPおよびオキサロ酢酸を生成する。PCKは広範囲な植物組織の細胞の細胞質基質に存在することが知られている。これらの植物組織としては、発育中の種子、毛状突起および根が挙げられる。植物中のPCKは窒素化合物の代謝が増加した組織内に出現する。
【0007】
本発明者らは、PCKおよび/またはPPDK酵素をコードする遺伝子がプロモーターの制御下で単独あるいは一緒に配置されている複数の遺伝子構築物を作製し、存在すれば、これらの遺伝子の過剰発現が老化した葉の中の窒素の転流にどのような効果を与えたかについて調べた。
【発明の概要】
【0008】
本発明の第1の局面によれば、少なくとも1つのコード配列に作動可能に連結された老化特異的プロモーターを含み、かつホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性および/またはピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする遺伝子構築物が提供される。
【0009】
本発明の第2の局面によれば、少なくとも1つのコード配列に作動可能に連結されたプロモーターを含み、かつホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性およびピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする遺伝子構築物が提供される。
【0010】
本発明者らは、PCKおよびPPDKの2種類の酵素が、老化中の葉からの窒素の転流の間に、様々なアミノ酸の相互転換に関与している可能性があると考えた。本発明者らは仮定に縛られることを望んでないが、PCKおよびPPDKがどのように窒素の転流に影響を与え得るかについて示す推測上の生化学的経路が図18に示されている。従って、本発明者らは、老化中に独立してあるいは同時に植物中のこれらの2種類の酵素の過剰発現を刺激することが窒素の転流に関わっている可能性があると考えた。
【0011】
研究の結果として、本発明者らは、驚くべきことにPCKおよび/またはPPDKをコードする本発明に係る構築物によって、老化した葉からの窒素転流率を上昇させることを見い出した。本発明者らは、窒素はアミノ酸の形態で老化した葉から植物の種子などの植物のより若い部分に移動するのではないかと仮定する。さらに、本発明者らは、これらの酵素が老化した葉の中で過剰発現した際に、栄養植物の成長量の増加(作物収穫率の上昇に該当する)も観察した。従って、本発明者らは、本発明に係る構築物が形質転換植物の生産に有用であり、それにより、老化した葉からの窒素転流率および/または成長率の上昇を示すことができるのではないかと考える。
【0012】
第1または第2の局面の遺伝子構築物中のプロモーターは、RNAポリメラーゼを誘発してPCKおよび/またはPPDK活性を有する少なくとも1種のポリペプチドをコードする少なくとも1つのコード領域に結合しかつその転写を開始することができるものであってもよい。
【0013】
第2の局面の構築物中に存在するプロモーターは、構成性、非構成性または組織特異的であってもよい。好適なプロモーターの例としては、カリフラワーモザイクウイルス35Sプロモーター(完全もしくは切断型)、ルビスコプロモーター、エンドウマメプラストシアニンプロモーター、ノパリンシンターゼプロモーター、クロロフィルr/b結合プロモーター、高分子量グルテニンプロモーター、α,β−グリアジンプロモーター、ホルデインプロモーターまたはパタチンプロモーターが挙げられる。
【0014】
第2の局面の構築物に存在するプロモーターは、老化特異的プロモーターであってもよい。
【0015】
「老化特異的プロモーター(SAG)」は、老化関連遺伝子の発現の制御に関連するプロモーターとすることができる。従って、このプロモーターは、実質的に老化している組織の中でのみ、プロモーターが作動可能に連結されたコード配列(すなわち、遺伝子)の発現を制限することができる。従って、老化特異的プロモーターは、実質的に植物組織が老化している場合にのみ3’タンパク質コード領域の発現が生じるように発生学的に調節される方法で、植物組織中で遺伝子発現を優先的に促進することができるプロモーターとすることができる。当然のことながら、老化は古い葉などの植物の古い部分で生じ、種子などの植物の若い部分では生じない傾向がある。
【0016】
多数の老化関連遺伝子を発現することで知られている植物の一例は、シロイヌナズナである。従って、第1または第2の局面に係る構築物中のプロモーターは、シロイヌナズナの老化関連遺伝子から単離したものであってもよい。Gepsteinら(The Plant Journal, 2003, 36, 629-642)は、モデルとしてシロイヌナズナを用いて、SAGおよびそのプロモーターについて詳しい研究を行なった。当該遺伝子構築物は、本明細書に開示されている任意のSAG由来のプロモーターを含んでいてもよい。例えば、好適なプロモーターは、SAG12、SAG13、SAG101、SAG21およびSAG18からなる群またはその機能的な変異体もしくは機能的な断片から選択されてもよい。
【0017】
好ましいプロモーターは、SAG12およびSAG13プロモーターである。一実施形態では、当該プロモーターは、当業者に知られているSAG12プロモーターまたはその機能的な変異体もしくは断片である(Gan & Amasino, 1997, Plant Physiology, 113: 313-319)。SAG12プロモーターをコードするDNA配列は、本明細書では以下のとおり配列番号16と称する:
配列番号16
TCGAGACCCGATTGTTATTTTTAGACTGAGACAAAAAAGTAGAATCGTTGATTGTTAAAATTTAAAATTAGTTTCATTACGTTTCGATAAAAAAATGATTAGTTTATCATAGCTTAATTATAGCATTGATTTCTAAATTTGTTTTTTGACCACCCTTTTTTCTCTCTTTGGTGTTTTCTTAACATTAGAAGAACCCATAACAATGTACGTTCAAATTAATTAAAAACAATATTTCCAAGTTTTATATACGAAACTTGTTTTTTTTAATGAAAACAGTTGAATAGTTGATTATGAATTAGTTAGATCAATACTCAATATATGATCAATGATGTATATATATGAACTCAGTTGTTATACAAGAAATGAAAATGCTATTTAAATACAGATCATGAAGTGTTAAAAAGTGTCAGAATATGACATGAAGCGTTTTGTCCTACCGGGTATTCGAGTTATAGGTTTGGATCTCTCAAGAATATTTTGGGCCATACTAGTTATATTTGGGCTTAAGCGTTTTGCAAAGAGACGAGGAAGAAAGATTGGGTCAAGTTAACAAAACAGAGACACTCGTATTAGTTGGTACTTTGGTAGCAAGTCGATTTATTTGCCAGTAAAAACTTGGTACACAACTGACAACTCGTATCGTTATTAGTTTGTACTTGGTACCTTTGGTTCAAGAAAAAGTTGATATAGTTAAATCAGTTGTGTTCATGAGGTGATTGTGATTTAATTTGTTGACTAGGGCGATTCCTTCACATCACAATAACAAAGTTTTATAGATTTTTTTTTTATAACATTTTTGCCACGCTTCGTAAAGTTTGGTATTTACACCGCATTTTTCCCTGTACAAGAATTCATATATTATTTATTTATATACTCCAGTTGACAATTATAAGTTTATAACGTTTTTACAATTATTTAAATACCATGTGAAGATCCAAGAATATGTCTTACTTCTTCTTTGTGTAAGAAAACTAACTATATCACTATAATAAAATAATTCTAATCATTATATTTGTAAATATGCAGTTATTTGTCAATTTTGAATTTAGTATTTTAGACGTTATCACTTCAGCCAAATATGATTTGGATTTAAGTCCAAAATGCAATTTCGTACGTATCCCTCTTGTCGTCTAATGATTATTTCAATATTTCTTATATTATCCCTAACTACAGAGCTACATTTATATTGTATTCTAATGACAGGGAAACCTTCATAGAGATTCAGATAGATGAAATTGGTGGGAAACATCATTGAACAGGAAACTTTTAGCAAATCATATCGATTTATCTACAAAAGAATACGTAGCGTAATGAAGTCCACTTGTTGTGAATGACTATGATTTGATCAAATTAGTTAATTTTGTCGAATCATTTTTCTTTTTGATTTGATTAAGCTTTTAACTTGCACGAATGGTTCTCTTGTGAATAAACAGAATCTTTGAATTCAAACTATTTGATTAGTGAAAAGACAAAAGAAGATTCCTTGTTTTTATGTGATTAGTGATTTTGATGCATGAAAGGTACCTACGTACTACAAGAAAAATAAACATGTACGTAACTACGTATCAGCATGTAAAAGTATTTTTTTCCAAATAATTTATACTCATGATAGATTTTTTTTTTTTGAAATGTCAATTAAAAATGCTTTCTTAAATATTAATTTTAATTAATTAAATAAGGAAATATATTTATGCAAAACATCATCAACACATATCCAACTTCGAAAATCTCTATAGTACACAAGTAGAGAAATTAAATTTTACTAGATACAAACTTCCTAATCATCAAATATAAATGTTTACAAAACTAATTAAACCCACCACTAAAATTAACTAAAAATCCGAGCAAAGTGAGTGAACAAGACTTGATTTCAGGTTGATGTAGGACTAAAATGACTACGTATCAAACATCAACGATCATTTAGTTATGTATGAATGAATGTAGTCATTACTTGTAAAACAAAAATGCTTTGATTTGGATCAATCACTTCATGTGAACATTAGCAATTACATCAACCTTATTTTCACTATAAAACCCCATCTCAGTACCCTTCTGAAGTAATCAAATTAAGAGCAAAAGTCATTTAACTTAGG
【0018】
従って、本発明の構築物中のプロモーターは、実質的に配列番号16に示すヌクレオチド配列またはその機能的な変異体もしくは機能的な断片を含んでいてもよい。SAG12プロモーター配列は、米国特許第5,689,042号に記載されているようなシロイヌナズナから得られたものであってもよい。このプロモーター配列は、図3に示すように本発明に係る遺伝子構築物のそれぞれにおいて見られる。プロモーターがSAG12である実施形態では、当然のことながら、当該プロモーターは、配列番号16の1〜2093の各塩基を含んでいてもよい。ただし、プロモーターの機能的な変異体もしくは機能的な断片が本発明の遺伝子構築物に使用されていてもよい。
【0019】
「プロモーターの機能的な変異体もしくは機能的な断片」は、そこに作動可能に連結された任意のコード領域の発現を開始させるのに機能的に十分であるプロモーターの誘導体または部分とすることができる。例えば、プロモーターがSAG12に基づいている実施形態では、当業者であれば、配列番号16が修飾されていても、SAG12プロモーターの部分のみが必要とされてもよく、それによって、本構築物における遺伝子発現がなお開始されることを理解しているであろう。
【0020】
プロモーターの機能的な変異体および機能的な断片は、転写酵素が推定プロモーター領域に結合し、次いで、コード領域の転写を引き起こしてPCKおよび/またはPPDK活性を有するポリペプチドが発現するか否かを評価することによって容易に同定することができる。あるいは、そのような機能的な変異体および断片は、コード領域に結合されている場合には、プロモーター上で突然変異生成を行ない、遺伝子発現が生じ得るか否かを評価することによって調べてもよい。
【0021】
第1の局面の遺伝子構築物は、老化中にPCK活性および/またはPPDK活性を示す少なくとも1種のポリペプチドの発現を生じさせることができる。従って、遺伝子構築物は、(i)ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片および/または(ii)ピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片をコードする少なくとも1つのコード配列を含んでいてもよい。実施例に記載されているように、本発明者らは、PCKおよび/またはPPDK活性を有するポリペプチドに基づいた様々な遺伝子構築物を開発し、これらを図3に示す。
【0022】
第1の局面に係る遺伝子構築物の第1の実施形態では、プロモーターは、PCK活性を示すポリペプチドをコードするコード配列の発現を誘発してもよい。これは、本明細書では「PCK構築物」と称し、図3に示す。従って、第1の実施形態では、遺伝子構築物は、老化特異的プロモーター、およびホスホエノールピルビン酸カルボキシキナーゼ(PCK)をコードするコード配列あるいはその機能的な変異体もしくは断片を含んでいてもよい。本遺伝子構築物は、PPDK活性を有するポリペプチドをコードしないものであってもよい。
【0023】
第1の局面に係る構築物の第2の実施形態では、プロモーターは、PPDK活性を示すポリペプチドをコードするコード配列の発現を誘発してもよい。これは、本明細書では「PPDK構築物」と称し、図3に示す。第2の実施形態では、本遺伝子構築物は、老化特異的プロモーター、およびピルビン酸正リン酸ジキナーゼ(PPDK)をコードするコード配列またはその機能的な変異体もしくは断片を含んでいてもよい。遺伝子構築物は、PCK活性を有するポリペプチドをコードしないものであってもよい。
【0024】
第1の局面に係る構築物の第3の実施形態では、プロモーターは、PCK活性およびPPDK活性を示すポリペプチドをコードする単一のコード配列の発現を誘発してもよい。これを「PCK/PPDK構築物1」と称す。第3の実施形態では、遺伝子構築物は、老化特異的プロモーターならびに(i)ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片および(ii)ピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片をコードするコード配列を含んでいてもよい。第3の実施形態の構築物は、二重活性(すなわち、PCKおよびPPDKの両方の酵素活性)を示す単一の転写産物をコードしてもよい。PCKコード領域は、PPDKコード領域の3'末端に配置されていてもよい。ただし、好ましくは、PCKコード領域は、PPDKコード領域の5'末端に配置されている。
【0025】
第1の局面に係る構築物の第4の実施形態では、プロモーターは、(i)PCK活性を示す第1のポリペプチドをコードする第1のコード配列および(ii)PPDK活性を示す第2のポリペプチドをコードする第2のコード配列の発現を誘発してもよい。これをPCK/PPDK構築物2と称す。よって、第4の実施形態では、本遺伝子構築物は、少なくとも1つの老化特異的プロモーターならびに(i)PCKをコードする第1のコード配列またはその機能的な変異体もしくは断片および(ii)PPDKをコードする第2のコード配列またはその機能的な変異体もしくは断片を含んでいてもよい(すなわち、各酵素に対して1種類の2種類の転写産物がコードされる)。
【0026】
実施例6に記載されているように、本発明者らは、(例えば、「PCK構築物」または「PPDK構築物」のいずれか一方による形質転換によって)宿主細胞中のPCKまたはPPDKのいずれか一方を過剰発現させることで、老化した葉の中の窒素の転流が増加することを見い出した。さらに、本発明者らは、PCKおよびPPDKの単一の構築物によって栄養成長が増加することを見い出した。
【0027】
実施例8に記載されているように、本発明者らは、(例えば、「PCK構築物」または「PPDK構築物」の両方による形質転換によって)宿主細胞中のPCKまたはPPDKを同時に過剰発現させることは、老化した葉の中の窒素の転流を誘発する際に驚くほど有効であることを見い出した。従って、窒素は、例えば輸送アミノ酸として老化した葉から輸送されてもよい。好適な輸送アミノ酸はグルタミンおよび/またはアスパラギンであってもよい。さらに、老化中にPCKおよびPPDKを同時に過剰発現させることによって成長率を上げ、それにより、栄養成長を増加させてもよい。従って、第1の局面の構築物は、PCKおよびPPDKまたはそれらの機能的な変異体もしくは断片をコードするコード配列を含んでいてもよい。
【0028】
第2の局面の構築物がPCKおよびPPDKをコードするコード配列またはそれらの機能的な変異体もしくは断片を含むことはいうまでもない。この2種類の酵素は二重活性を有する単一のポリペプチドとして、あるいは一方がPCK活性を有し、他方がPPDK活性を有する2種類のポリペプチドとしてコードされてもよい。
【0029】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片およびピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片はそれぞれ、植物などの任意の好適な供給源に由来するものであってもよい。各酵素のコード配列は、好適な植物源、例えばシロイヌナズナに由来するものであってもよい。従って、PCK活性を有するポリペプチドをコードするコード配列は、シロイヌナズナに由来するものであってもよい。さらに、PPDK活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ属、トウモロコシ属、フラベリア属またはクレオメ属に由来するものであってもよい。PPDK活性を有するポリペプチドをコードするコード配列は、シロイヌナズナ、トウモロコシ、フラベリア・トリネルビア(Flaveria trinervia)、フラベリア・ビデンティス(Flaveria bidentis)、フラベリア・ブロウニー(Flaveria brownie)またはフウチョウソウに由来するものであってもよい。
【0030】
シロイヌナズナにはPCKをコードする3種類の遺伝子が存在すると考えられている。シロイヌナズナのホスホエノールピルビン酸カルボキシキナーゼ(PCK)の一実施形態をコードするゲノムDNA(gDNA)配列(イントロンおよびエクソンを含む)は、本明細書では以下のとおり配列番号17として示す:
配列番号17
ATGTCGGCCGGTAACGGAAATGCTACTAACGGTGACGGAGGGTTTAGTTTCCCTAAAGGACCGGTGATGCCGAAGATAACGACCGGAGCAGCAAAGAGAGGTAGCGGAGTCTGCCACGACGATAGTGGTCCGACGGTGAATGCCACAACCATCGATGAGCTTCATTCGTTACAGAAGAAACGTTCTGCTCCTACCACACCGATCAACCAAAACGCCGCCGCTGCTTTTGCCGCCGTCTCCGAGGAGGAGCGTCAGAAGATTCAGCTTCAATCTATCAGGTCCTTATAATAACTTCACATATACAGATTATTCATACGTTACTTTTGTTTATAACATACTTTATATCGAATTAAGGAAGATTATTGCGTTTTCGTGTCCGATCATTTTCATGGAAAAAGTGTCTTTTAGCTAAATATATGGTGTAGTATTAAATATTTCTGACGTGATATACACTAAACTTGAAAATTTTCAATTACTATTTCTTCCTTTAATTCGGCAATATAATTTGTTTTTGTTTATTTTTGGATTAGACATTTATGGACAAGTTAATGCGCTATTGTGACTATTACCAGAAAATAATACTTTAATGTACATGACACGTGTTTAAAACGACACGTGGAAACTAATTTTGATTAATTGTGAAACAGTGCATCGTTAGCATCGTTAACGAGAGAGTCAGGACCAAAGGTGGTGAGAGGAGATCCGGCGGAGAAGAAGACCGATGGTTCAACTACTCCGGCGTACGCTCACGGCCAACATCATTCTATCTTTTCTCCGGCTACTGGTGCTGTCAGTGATAGCTCCTTGAAGTTTACTCACGTCCTCTACAATCTTTCGCCTGCAGGTCAACAAATAAACCTAGAATCCGAATCTGAATATTGATAAATGTTTCTGCAACGAGTTTGATAGATTTGGTTTGTGATTTTGTTGTTTGTAGAGCTTTATGAGCAAGCTATTAAGTATGAGAAAGGTTCGTTTATCACTTCTAATGGAGCTTTGGCGACGCTTTCTGGTGCTAAGACTGGTCGTGCTCCCAGAGATAAGCGTGTTGTTAGAGATGCTACTACTGAGGATGAGCTTTGGTGGGGAAAGTGAGTATTCCTAATCTCGATTTTGATTGATGGAGTTTTTGGGTTTATGCTCTGTTTTCGTTTATTGATTTTGGAGTTTGATTTTGATTTTAGGGGTTCGCCGAATATCGAAATGGATGAACATACTTTCATGGTGAACAGAGAAAGAGCTGTTGATTACTTGAATTCCTTGGAAAAGGTATTAAATTTTGAAAACTTTAATCAATGTTGTTGAGTGTAGAACTTTTGATCTAAGTTTATGAAATTTCTGTTGTTGTTGGGGTTTTTAGGTCTTTGTCAATGACCAATACTTAAACTGGGATCCAGAGAACAGAATCAAAGTCAGGATTGTCTCAGCTAGAGCTTACCATTCATTGTTTATGCACAACATGTAAGTAAAATCATTATTGACTCCTTGTATGTCAATCCATTATTGTGGGTGAAAGAAAACAACAAATTAGTAACTGGGGAGGGTGTCAGGTGTATCCGACCAACTCAGGAGGAGCTTGAGAGCTTTGGTACTCCGGATTTTACTATATACAATGCTGGGCAGTTTCCATGTAATCGTTACACTCATTACATGACTTCGTCCACTAGCGTAGACCTTAATCTGGCTAGGAGGGAAATGGTTATACTTGGTACTCAGTATGCTGGGGAAATGAAGAAGGGTCTTTTCAGTGTGATGCATTACCTTATGCCTAAGCGTCGTATTCTCTCCCTTCATTCTGGATGCAATATGGGAAAAGATGGAGATGTTGCTCTCTTCTTTGGACTTTCAGGTATAGTAGAGACAGTACCAACTATGGTGTTGGGTGATGATGGAAGGAACGATAAATCAAATGATACAATACAATTACTGCTGAACTGACTTGAGAACTGCTTGCCTCTTTGTTGAGTTTAGCGGGTGAATTGAGATTGATGATTGTGTTTTTTGTTTTCTATGAATGATGATTTTAGGTACCGGGAAGACAACGCTGTCTACTGATCACAACAGGTATCTTATTGGAGATGATGAGCATTGTTGGACTGAGACTGGTGTTTCGAACATTGAGGGTGGGTGCTATGCTAAGTGTGTTGATCTTTCGAGGGAGAAGGAGCCTGATATCTGGAACGCTATCAAGTTTGGAACAGGTAGAAAGACAGTACGTTGGAATTGTTTTTGAGAAAAAAACATAAAGCAGTGATATAACAATAAGATTCTGATCTTGTTGCAGTTTTGGAAAATGTTGTGTTTGATGAGCACACCAGAGAAGTGGATTACTCTGATAAATCTGTTACAGGTAAAACAATTGTTATTTCTTTCATTCTCTTCGTCCTCACAATTAACAGAATGATCATTTTCGATTCTCTTTGGTTGCAGAGAACACACGTGCTGCCTACCCAATTGAGTTCATTCCAAATGCGAAAATACCTTGTGTTGGTCCACACCCGACAAATGTGATACTTCTGGCTTGTGATGCCTTTGGTGTTCTCCCACCTGTGAGCAAGCTGAATCTGGCACAAACCATGTACCACTTCATCAGTGGTTACACTGCTCTGGTAAGGCCAAAGTAAAAGTCTTTATTTTGCACATCGTCTTCATAAATTTCAAAAGCATAACCAAAGATGTGCAACATATATAGGTTGCTGGCACAGAGGATGGTATCAAGGAGCCAACAGCAACATTCTCAGCTTGCTTTGGTGCAGCTTTCATAATGTTGCATCCCACAAAGTATGCAGCTATGTTAGCTGAGAAGATGAAGTCACAAGGTGCTACTGGTTGGCTCGTCAACACTGGTTGGTCTGGTGGCAGGTATATATGTCCTTCTATGGAAATCGATACAACAAAACGCTGCCTTGTAACACATGTTTGTAGGCTATTAACATGATCTGTAATGTTTTATTTCCTGCAGTTATGGTGTTGGAAACAGAATCAAGCTGGCATACACTAGAAAGATCATCGATGCAATCCATTCGGGCAGTCTCTTGAAGGCAAACTACAAGAAAACCGAAATCTTTGGATTTGAAATCCCAACTGAGATCGAAGGGATACCTTCAGAGATCTTGGACCCCGTCAACTCCGTAAGTTTCTGCAAATCTGTATAATGTAATTGCTTAAGTGATGATGAACAATTTTTTGTTGATTTGGGTTTAATGAAAATGCAGTGGTCTGATAAGAAGGCACACAAAGATACTCTGGTGAAACTGGGAGGTCTGTTCAAGAAGAACTTCGAGGTTTTTGCTAACCATAAGATTGGTGTGATGGTAAGCTTACGGAGGAGATTCTCGCTGCTGGTCCTATCTTTTAG
【0031】
シロイヌナズナのホスホエノールピルビン酸カルボキシキナーゼ(PCK)をコードする相補的DNA(cDNA)配列(エクソンのみ)は、本明細書では以下のとおり配列番号18として示す:
配列番号18
ATGTCGGCCGGTAACGGAAATGCTACTAACGGTGACGGAGGGTTTAGTTTCCCTAAAGGACCGGTGATGCCGAAGATAACGACCGGAGCAGCAAAGAGAGGTAGCGGAGTCTGCCACGACGATAGTGGTCCGACGGTGAATGCCACAACCATCGATGAGCTTCATTCGTTACAGAAGAAACGTTCTGCTCCTACCACACCGATCAACCAAAACGCCGCCGCTGCTTTTGCCGCCGTCTCCGAGGAGGAGCGTCAGAAGATTCAGCTTCAATCTATCAGTGCATCGTTAGCATCGTTAACGAGAGAGTCAGGACCAAAGGTGGTGAGAGGAGATCCGGCGGAGAAGAAGACCGATGGTTCAACTACTCCGGCGTACGCTCACGGCCAACATCATTCTATCTTTTCTCCGGCTACTGGTGCTGTCAGTGATAGCTCCTTGAAGTTTACTCACGTCCTCTACAATCTTTCGCCTGCAGAGCTTTATGAGCAAGCTATTAAGTATGAGAAAGGTTCGTTTATCACTTCTAATGGAGCTTTGGCGACGCTTTCTGGTGCTAAGACTGGTCGTGCTCCCAGAGATAAGCGTGTTGTTAGAGATGCTACTACTGAGGATGAGCTTTGGTGGGGAAAGGGTTCGCCGAATATCGAAATGGATGAACATACTTTCATGGTGAACAGAGAAAGAGCTGTTGATTACTTGAATTCCTTGGAAAAGGTCTTTGTCAATGACCAATACTTAAACTGGGATCCAGAGAACAGAATCAAAGTCAGGATTGTCTCAGCTAGAGCTTACCATTCATTGTTTATGCACAACATGTGTATCCGACCAACTCAGGAGGAGCTTGAGAGCTTTGGTACTCCGGATTTTACTATATACAATGCTGGGCAGTTTCCATGTAATCGTTACACTCATTACATGACTTCGTCCACTAGCGTAGACCTTAATCTGGCTAGGAGGGAAATGGTTATACTTGGTACTCAGTATGCTGGGGAAATGAAGAAGGGTCTTTTCAGTGTGATGCATTACCTTATGCCTAAGCGTCGTATTCTCTCCCTTCATTCTGGATGCAATATGGGAAAAGATGGAGATGTTGCTCTCTTCTTTGGACTTTCAGGTACCGGGAAGACAACGCTGTCTACTGATCACAACAGGTATCTTATTGGAGATGATGAGCATTGTTGGACTGAGACTGGTGTTTCGAACATTGAGGGTGGGTGCTATGCTAAGTGTGTTGATCTTTCGAGGGAGAAGGAGCCTGATATCTGGAACGCTATCAAGTTTGGAACAGTTTTGGAAAATGTTGTGTTTGATGAGCACACCAGAGAAGTGGATTACTCTGATAAATCTGTTACAGAGAACACACGTGCTGCCTACCCAATTGAGTTCATTCCAAATGCGAAAATACCTTGTGTTGGTCCACACCCGACAAATGTGATACTTCTGGCTTGTGATGCCTTTGGTGTTCTCCCACCTGTGAGCAAGCTGAATCTGGCACAAACCATGTACCACTTCATCAGTGGTTACACTGCTCTGGTTGCTGGCACAGAGGATGGTATCAAGGAGCCAACAGCAACATTCTCAGCTTGCTTTGGTGCAGCTTTCATAATGTTGCATCCCACAAAGTATGCAGCTATGTTAGCTGAGAAGATGAAGTCACAAGGTGCTACTGGTTGGCTCGTCAACACTGGTTGGTCTGGTGGCAGTTATGGTGTTGGAAACAGAATCAAGCTGGCATACACTAGAAAGATCATCGATGCAATCCATTCGGGCAGTCTCTTGAAGGCAAACTACAAGAAAACCGAAATCTTTGGATTTGAAATCCCAACTGAGATCGAAGGGATACCTTCAGAGATCTTGGACCCCGTCAACTCCTGGTCTGATAAGAAGGCACACAAAGATACTCTGGTGAAACTGGGAGGTCTGTTCAAGAAGAACTTCGAGGTTTTTGCTAACCATAAGATTGGTGTGATGGTAAGCTTA CGGAGGAGATTCTCGCTGCTGGTCCTATCTTTTAG
【0032】
従って、PCK活性を有するポリペプチドをコードするコード配列は、実質的に配列番号17または配列番号18に示す核酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0033】
シロイヌナズナのPCKのポリペプチド配列は、本明細書では以下のとおり配列番号19として示す:
配列番号19
MSAGNGNATNGDGGFSFPKGPVMPKITTGAAKRGSGVCHDDSGPTVNATTIDELHSLQKKRSAPTTPINQNAAAAFAAVSEEERQKIQLQSISASLASLTRESGPKVVRGDPAEKKTDGSTTPAYAHGQHHSIFSPATGAVSDSSLKFTHVLYNLSPAELYEQAIKYEKGSFITSNGALATLSGAKTGRAPRDKRVVRDATTEDELWWGKGSPNIEMDEHTFMVNRERAVDYLNSLEKVFVNDQYLNWDPENRIKVRIVSARAYHSLFMHNMCIRPTQEELESFGTPDFTIYNAGQFPCNRYTHYMTSSTSVDLNLARREMVILGTQYAGEMKKGLFSVMHYLMPKRRILSLHSGCNMGKDGDVALFFGLSGTGKTTLSTDHNRYLIGDDEHCWTETGVSNIEGGCYAKCVDLSREKEPDIWNAIKFGTVLENVVFDEHTREVDYSDKSVTENTRAAYPIEFIPNAKIPCVGPHPTNVILLACDAFGVLPPVSKLNLAQTMYHFISGYTALVAGTEDGIKEPTATFSACFGAAFIMLHPTKYAAMLAEKMKSQGATGWLVNTGWSGGSYGVGNRIKLAYTRKIIDAIHSGSLLKANYKKTEIFGFEIPTEIEGIPSEILDPVNSWSDKKAHKDTLVKLGGLFKKNFEVFANHKIGVMVSLRRRFSLLVLSF
【0034】
従って、PCK活性を有するポリペプチドは、実質的に配列番号19に示すアミノ酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0035】
シロイヌナズナはPPDKの少なくとも2種類の型、すなわち葉緑体型および細胞質基質型を有し、どちらも遺伝子の5'末端に僅かなスプライシングの変形を有する同じ遺伝子によってコードされ、それにより2種類の型が生じると考えられている。シロイヌナズナのピルビン酸正リン酸ジキナーゼ(PPDK)の両型をコードするgDNA配列(イントロンおよびエクソンを含む)は、本明細書では以下のとおり配列番号20として示す:
配列番号20
ATGACAAGTATGATCGTGAAGACAACGCCGGAGCTCTTCAAAGGAAATGGAGTGTTCCGTACGGATCATCTCGGAGAAAACCGAATGGTTAGTCGATCAAACCGGCTAGGTGATGGATCAAACCGTTTCCCTAGAACCGGTACAATCCATTGCCAACGGTTAAGCATAGCAAAGACCGGTTTGCATCGTGAGACGAAGGCTCGAGCCATACTTAGCCCTGTGTCCGATCCGGCCGCTTCCATAGCCCAAAAGGTAAGCCTTTCCATTTCAATCATTCTGGTGTATTTTCACCATAAAATTTTATACACTTTTTTATTACGTTTTGTTTTATGATTCTGACGTGAGATTCTTGAGAGAAACTATCACCGATCATTGGGTCGAACCATCTAGCAGCTCAATTATTATCGGTTATAACCCTACCGGTTATAGAATACAAAACAGGTTACGCCATTGTGACATTTGCTTTGTGATCTTGTGAGACGATTAATTATTTGATGTTGATTGGTTTCGTTACTCTTGTTTAAACAATCGAACGGTTCAAACTAATACACACATGTGATGTGAGATCATTTCGGTAGTAATACCAAATAGCGTCTGGCCTAAATTATGAAAGTACTATTTTGAATTAAATTATTGTGGAAACATGAACTTATTTAAATTCAAGTATTTTCGAAATTTGTAATAAAAAAAAACTTTTCCTCTAGATTCATTAGCCCTACTTTTCGTAGAAACAACTTTAATGTATTCAAAGACCACTTTGCTGCTTAAGTCAGACTCTTGTGCCACTTGGTAGATCCACCAATGCCACGTTTTGTTATTGTGCCAAAGAATACGTGAATATGTCCAAACGGCAATCAAATTCTTGGCGTAAAACACAAAAATTATGATACTAGTTTAAATCCACAATTCACCTTCACCATAAAGAATTCATGTATTAGAGATGGTATGACAAGAACTGGTTGAATTTGATGACATTTGTTTGCTATTGTTTTGGTTAAGTAAAAGTTTTGTTAAAAAGGAAAATAGCATCGGTAGTGGCAGATAGCAAGTGTGTGAGTGAGATCAGATATGGTTGACACATCTATGACGAGTCATCGCAACGAAACTTCTTTAATTTTGGTCAATTATATTACAATTTAGCATTTCGAGGTTGGAATTTTGGAATGATCTCTTGATAAGATAATAATGTATTTTTGATGACGTATCCATCAAAACTATAAATGATTTATATTAAATATGAAATTTCGACTGTATACAAGTTTTTATATTATAAAATTATTCGATGTACATATGATCATAATAACTTTACTATATATATAGATACGTATATGTGTTCTTAAACTTGCACAAACATTTCTGCAATCTAAACCTCAATCAAAACAAACAAACAAAAAACCATGATGCAGCGAGTATTCACCTTTGGAAAAGGAAGAAGCGAAGGCAACAAGGGCATGAAGTCCTTGGTATGTTACCAATACCATCATCATGATCATATCAATTCATTAATAATTTAGTGTTTGCTATTTTCAAGAACCATTTATCAAAAATGTTAATTGTTGTTGTGTATGAAGTTGGGAGGGAAAGGAGCCAACCTGGCGGAGATGGCTAGCATAGGCTTGTCGGTGCCGCCGGGGCTAACCATATCGACGGAGGCTTGTCAGCAGTATCAGATCGCCGGCAAAAAGCTTCCAGAAGGTTTATGGGAAGAGATCTTAGAAGGTCTTAGCTTCATCGAACGTGACATTGGAGCTTCCCTCGCTGATCCCTCCAAGCCACTCCTCCTCTCTGTTCGCTCCGGCGCCGCCGTAAGTTAATTATAACTTTTTTTCTTGACTATTTTTATTTTAAGGATTTTTTCTAATGTTAAATTTCTGTTTTTTTTTCTTTCTATGTTTTCTTTAATCTTTTGAAGATTTTTTGACGCAGATTTTGACTTGTTAGATTTCTTTTATTGAAGTTGAGATCCAAATATTTTTTTGGTTATTTTGCCATTTGGCCGTTTTTGGAAGAGTTTAAAATGTACTAGATAGAAAATGAATAAGTTTTGTGGCTATTGAAAGACCTAATGATTTTGGTATTCAAACTATAACGTAGAAAATGAAGATCTTTCGTTTATCTATTTTTAAAACAGAACTACATTGACTTGTCTTTGATCGATATTTTGCATTGTAGATCTCAATGCCTGGTATGATGGACACTGTACTTAACCTTGGCTTGAACGACCAAGTCGTCGTTGGTCTGGCCGCAAAAAGCGGAGAGCGTTTTGCTTACGATTCGTTCCGGCGTTTTCTTGATATGTTTGGTGATGTTGTAAGTCCTCTGTTTTTCAATACTATTTCAGGTAACTTGCATGACAAGAAAATTCTTTGACCTACCTTATAATTGTTTTCTTGATCAATAAAAGGTGATGGGAATTCCACACGCCAAGTTTGAAGAGAAGTTAGAGAGAATGAAGGAGAGGAAAGGAGTTAAAAATGACACTGACTTAAGCGCGGCTGATCTCAAGGAATTGGTTGAGCAGTACAAGAGTGTTTACTTAGAGGCCAAGGGTCAAGAGTTTCCTTCAGGTTTGTTTTGATTCCTACTTGAGGTCAAGTGATAAAAATTAGTTATTAGTTACAAATGTTTAAACGGGGTTAATTGCAGATCCAAAGAAGCAATTGGAGCTAGCGATTGAAGCGGTATTCGATTCTTGGGATAGCCCGAGAGCGAACAAGTACAGAAGTATTAACCAGATAACTGGATTGAAAGGAACCGCGGTTAACATTCAGTGTATGGTGTTTGGAAACATGGGGGACACTTCAGGGACTGGTGTTCTCTTCACTAGGAACCCTAGCACAGGAGAGAAGAAGCTTTATGGCGAGTTTCTAGTTAATGCTCAGGTTTGGCATCTATCACAATGTGTGAATCTCATATCAACAAGTAAGCCCATACTCATTAAATATTGGTTTTGGGACAGGGAGAGGATGTGGTTGCAGGGATAAGAACACCAGAAGATTTGGATACAATGAAGAGATTTATGCCTGAGGCTTACGCTGAACTTGTTGAGAACTGCAACATCTTAGAAAGACATTACAAAGACATGATGGTTGATACACATAAACAATACTTCAATTAGTCCTCATCAACAATTCTTTAGTAATTTAAACAAAATCTCAAATGTGTATTGCAGGATATTGAATTCACAGTACAAGAAGAGAGATTGTGGATGCTGCAATGCAGAGCGGGTAAGCGAACGGGTAAAGGCGCCGTGAAGATAGCAGTTGATATGGTAGGTGAAGGGCTTGTTGAGAAATCTTCTGCTATCAAAATGGTGGAGCCTCAACATCTTGATCAACTACTTCACCCACAGGTACAAACTCAAATATTCATCTTCTTCTTTTTTCATAGTCATAAACTTGATGTTGAAACCAAAATTCGAAACTTACTGGTAATGATTGGTTCACTTGAACAAGAACTAATGGGTTTAAGACGTTTAGGGTTTAGGAGTAAAAGCAGAGATGATTGTCTGACACGTAACCGATGAATAGGGTTTGGAAATTTTGATTCAGAGGTCAATGAAGGTTTTTTTTTTTTTTTTTTATTGATGGATTAGTTTCATGATCCATCGGGGTATCGTGAAAAAGTGGTGGCCAAAGGCTTACCTGCGTCACCAGGAGCGGCGGTTGGACAGGTTGTGTTCACGGCGGAGGAAGCCGAAGCTTGGCATTCTCAGGGTAAAACTGTGATTCTGGTTCGAACTGAGACAAGCCCTGACGATGTGGGAGGTATGCACGCAGCGGAAGGTATATTGACGGCTAGAGGAGGAATGACGTCACACGCGGCTGTTGTTGCTCGCGGTTGGGGAAAATGTTGCATTGCTGGTTGTTCCGAGATTCGTGTCGACGAGAACCACAAGGTTTTTGGATTCGATTTTAGAAACTTGTCATATAAGTTAGGGGAAGATTGTTTCTAAAGTTAGGGTTTAAAAATTTTCAGGTTCTATTGATTGGAGATTTGACGATTAATGAAGGCGAATGGATCTCAATGAACGGATCAACCGGTGAGGTTATATTAGGGAAACAAGCATTGGCTCCTCCGGCTTTAAGTCCAGATTTGGAGACTTTCATGTCCTGGGCTGATGCAATCAGACGTCTCAAGGTGTTTATGAGTTTCTGTTCCTTTAACTTGTTTGATATTTTTAAACTTTCTAACTCAAATGTTCGATGACCGATAAGGTTATGGCGAATGCGGATACACCTGAAGACGCCATTGCAGCTAGGAAAAACGGAGCTCAAGGAATCGGGCTTTGTAGGACAGAGCATATGGTAACTCCTCCTCTGTACTTGATTTCATGTTTTTGATGATTTAGATTGTTTGTATCCAAATGTTTAATGTTGTCTTTGGTTTGGTTTTTAAGTTCTTTGGAGCAGATAGGATTAAAGCAGTGAGAAAGATGATAATGGCGGTAACAACAGAGCAAAGGAAAGCTTCTCTCGACATCTTGCTTCCTTACCAACGTTCGGATTTCGAAGGGATCTTCCGTGCTATGGATGGTAAATGTTTTGAGTCGTCTCTCTAAAATGTATCACAACTTAAAACATGCCTAAACCTTTTTATTTTTCTAGGTTTACCGGTAACAATCCGTTTGTTAGACCCTCCGCTTCACGAGTTTCTCCCGGAAGGCGACTTGGACAACATTGTACATGAGCTAGCTGAAGAAACTGGTGTGAAAGAAGATGAAGTCTTGTCACGGATAGAGAAACTCTCTGAAGTGAATCCAATGCTTGGTTTCCGCGGTTGCAGGTTTCTTACTCTCTTTGTTTCTCTCTGTCTCTTTGCACCTGAAGAACAATCTGATGATCGGTAAACTTGTACGTTATAGGCTCGGAATATCGTATCCAGAGCTAACGGAGATGCAAGCGCGTGCAATTTTTGAAGCTGCAGCGTCAATGCAGGACCAAGGTGTTACTGTCATTCCTGAGATTATGGTTCCACTTGTAGGAACTCCTCAGGAATTGGGTCACCAAGTTGATGTAATTCGTAAAGTTGCAAAGAAAGTATTTGCTGAGAAGGGTCATACCGTGAGCTACAAGGTTGGGACAATGATTGAGATCCCTCGAGCCGCGCTCATTGCAGATGAGGTAAATGTAACAAGACACAAAATGTGTTTTAGGCACTTGAAACCATGTTGCTATTTGCTAAGTAGGAACCTTTTTCTTTTGACAGATTGCGAAAGAGGCGGAGTTTTTCTCGTTCGGGACAAACGACTTGACGCAGATGACGTTTGGATACAGTAGAGACGATGTCGGCAAGTTTCTACCGATTTACCTCGCCAAAGGAATCTTACAGCACGACCCTTTTGAGGTATAATGACTACCATTTCGTTTGCTCTCTATCCATAGGATAAAATCTTGATAGCCATTTTTTTGTGTTTGGACCAGGTTCTTGATCAGCAAGGTGTAGGGCAATTGATCAAGATGGCGACAGAAAAAGGACGAGCAGCTAGGCCTAGCCTCAAGGTTGGGATATGTGGAGAACATGGAGGAGATCCATCTTCTGTGGGATTCTTTGCTGAAGCAGGACTTGACTATGTCTCTTGTTCTCCTTTCAGGTAATTGATTAATTTCCAAACCAATAAACACTTTTTTTACAACACTATTGTATAACTCAGATTGATGTAATTTTGGGATTTCTGTTGTTGTTGTTGTTGTTGTTGTTGTTGCAGGGTTCCAATTGCAAGGCTTGCAGCTGCTCAAGTAGTTGTTGCATGA
【0036】
シロイヌナズナのピルビン酸正リン酸ジキナーゼ(PPDK)の細胞質基質型をコードするcDNA配列は、本明細書では以下のとおり配列番号21として示す:
配列番号21
ATGATGCAGCGAGTATTCACCTTTGGAAAAGGAAGAAGCGAAGGCAACAAGGGCATGAAGTCCTTGTTGGGAGGGAAAGGAGCCAACCTGGCGGAGATGGCTAGCATAGGCTTGTCGGTGCCGCCGGGGCTAACCATATCGACGGAGGCTTGTCAGCAGTATCAGATCGCCGGCAAAAAGCTTCCAGAAGGTTTATGGGAAGAGATCTTAGAAGGTCTTAGCTTCATCGAACGTGACATTGGAGCTTCCCTCGCTGATCCCTCCAAGCCACTCCTCCTCTCTGTTCGCTCCGGCGCCGCCATCTCAATGCCTGGTATGATGGACACTGTACTTAACCTTGGCTTGAACGACCAAGTCGTCGTTGGTCTGGCCGCAAAAAGCGGAGAGCGTTTTGCTTACGATTCGTTCCGGCGTTTTCTTGATATGTTTGGTGATGTTGTGATGGGAATTCCACACGCCAAGTTTGAAGAGAAGTTAGAGAGAATGAAGGAGAGGAAAGGAGTTAAAAATGACACTGACTTAAGCGCGGCTGATCTCAAGGAATTGGTTGAGCAGTACAAGAGTGTTTACTTAGAGGCCAAGGGTCAAGAGTTTCCTTCAGATCCAAAGAAGCAATTGGAGCTAGCGATTGAAGCGGTATTCGATTCTTGGGATAGCCCGAGAGCGAACAAGTACAGAAGTATTAACCAGATAACTGGATTGAAAGGAACCGCGGTTAACATTCAGTGTATGGTGTTTGGAAACATGGGGGACACTTCAGGGACTGGTGTTCTCTTCACTAGGAACCCTAGCACAGGAGAGAAGAAGCTTTATGGCGAGTTTCTAGTTAATGCTCAGGTTTGGCATCTATCACAATGTGTGAATCTCATATCAACAAGGATAAGAACACCAGAAGATTTGGATACAATGAAGAGATTTATGCCTGAGGCTTACGCTGAACTTGTTGAGAACTGCAACATCTTAGAAAGACATTACAAAGACATGATGGATATTGAATTCACAGTACAAGAAGAGAGATTGTGGATGCTGCAATGCAGAGCGGGTAAGCGAACGGGTAAAGGCGCCGTGAAGATAGCAGTTGATATGGTAGGTGAAGGGCTTGTTGAGAAATCTTCTGCTATCAAAATGGTGGAGCCTCAACATCTTGATCAACTACTTCACCCACAGTTTCATGATCCATCGGGGTATCGTGAAAAAGTGGTGGCCAAAGGCTTACCTGCGTCACCAGGAGCGGCGGTTGGACAGGTTGTGTTCACGGCGGAGGAAGCCGAAGCTTGGCATTCTCAGGGTAAAACTGTGATTCTGGTTCGAACTGAGACAAGCCCTGACGATGTGGGAGGTATGCACGCAGCGGAAGGTATATTGACGGCTAGAGGAGGAATGACGTCACACGCGGCTGTTGTTGCTCGCGGTTGGGGAAAATGTTGCATTGCTGGTTGTTCCGAGATTCGTGTCGACGAGAACCACAAGGTTCTATTGATTGGAGATTTGACGATTAATGAAGGCGAATGGATCTCAATGAACGGATCAACCGGTGAGGTTATATTAGGGAAACAAGCATTGGCTCCTCCGGCTTTAAGTCCAGATTTGGAGACTTTCATGTCCTGGGCTGATGCAATCAGACGTCTCAAGGTTATGGCGAATGCGGATACACCTGAAGACGCCATTGCAGCTAGGAAAAACGGAGCTCAAGGAATCGGGCTTTGTAGGACAGAGCATATGATTGTTTGTATCCAAATGTTTAATGTTGTCTTTGGTTTGGTTTTTAAGTTCTTTGGAGCAGATAGGATTAAAGCAGTGAGAAAGATGATAATGGCGGTAACAACAGAGCAAAGGAAAGCTTCTCTCGACATCTTGCTTCCTTACCAACGTTCGGATTTCGAAGGGATCTTCCGTGCTATGGATGGTTTACCGGTAACAATCCGTTTGTTAGACCCTCCGCTTCACGAGTTTCTCCCGGAAGGCGACTTGGACAACATTGTACATGAGCTAGCTGAAGAAACTGGTGTGAAAGAAGATGAAGTCTTGTCACGGATAGAGAAACTCTCTGAAGTGAATCCAATGCTTGGTTTCCGCGGTTGCAGGCTCGGAATATCGTATCCAGAGCTAACGGAGATGCAAGCGCGTGCAATTTTTGAAGCTGCAGCGTCAATGCAGGACCAAGGTGTTACTGTCATTCCTGAGATTATGGTTCCACTTGTAGGAACTCCTCAGGAATTGGGTCACCAAGTTGATGTAATTCGTAAAGTTGCAAAGAAAGTATTTGCTGAGAAGGGTCATACCGTGAGCTACAAGGTTGGGACAATGATTGAGATCCCTCGAGCCGCGCTCATTGCAGATGAGATTGCGAAAGAGGCGGAGTTTTTCTCGTTCGGGACAAACGACTTGACGCAGATGACGTTTGGATACAGTAGAGACGATGTCGGCAAGTTTCTACCGATTTACCTCGCCAAAGGAATCTTACAGCACGACCCTTTTGAGGTTCTTGATCAGCAAGGTGTAGGGCAATTGATCAAGATGGCGACAGAAAAAGGACGAGCAGCTAGGCCTAGCCTCAAGGTTGGGATATGTGGAGAACATGGAGGAGATCCATCTTCTGTGGGATTCTTTGCTGAAGCAGGACTTGACTATGTCTCTTGTTCTCCTTTCAGGGTTCCAATTGCAAGGCTTGCAGCTGCTCAAGTAGTTGTTGCATGA
【0037】
シロイヌナズナのピルビン酸正リン酸ジキナーゼ(PPDK)の葉緑体型をコードするcDNA配列は、本明細書では以下のとおり配列番号22として示す:
配列番号22
ATGACAAGTATGATCGTGAAGACAACGCCGGAGCTCTTCAAAGGAAATGGAGTGTTCCGTACGGATCATCTCGGAGAAAACCGAATGGTTAGTCGATCAAACCGGCTAGGTGATGGATCAAACCGTTTCCCTAGAACCGGTACAATCCATTGCCAACGGTTAAGCATAGCAAAGACCGGTTTGCATCGTGAGACGAAGGCTCGAGCCATACTTAGCCCTGTGTCCGATCCGGCCGCTTCCATAGCCCAAAAGCGAGTATTCACCTTTGGAAAAGGAAGAAGCGAAGGCAACAAGGGCATGAAGTCCTTGTTGGGAGGGAAAGGAGCCAACCTGGCGGAGATGGCTAGCATAGGCTTGTCGGTGCCGCCGGGGCTAACCATATCGACGGAGGCTTGTCAGCAGTATCAGATCGCCGGCAAAAAGCTTCCAGAAGGTTTATGGGAAGAGATCTTAGAAGGTCTTAGCTTCATCGAACGTGACATTGGAGCTTCCCTCGCTGATCCCTCCAAGCCACTCCTCCTCTCTGTTCGCTCCGGCGCCGCCATCTCAATGCCTGGTATGATGGACACTGTACTTAACCTTGGCTTGAACGACCAAGTCGTCGTTGGTCTGGCCGCAAAAAGCGGAGAGCGTTTTGCTTACGATTCGTTCCGGCGTTTTCTTGATATGTTTGGTGATGTTGTGATGGGAATTCCACACGCCAAGTTTGAAGAGAAGTTAGAGAGAATGAAGGAGAGGAAAGGAGTTAAAAATGACACTGACTTAAGCGCGGCTGATCTCAAGGAATTGGTTGAGCAGTACAAGAGTGTTTACTTAGAGGCCAAGGGTCAAGAGTTTCCTTCAGATCCAAAGAAGCAATTGGAGCTAGCGATTGAAGCGGTATTCGATTCTTGGGATAGCCCGAGAGCGAACAAGTACAGAAGTATTAACCAGATAACTGGATTGAAAGGAACCGCGGTTAACATTCAGTGTATGGTGTTTGGAAACATGGGGGACACTTCAGGGACTGGTGTTCTCTTCACTAGGAACCCTAGCACAGGAGAGAAGAAGCTTTATGGCGAGTTTCTAGTTAATGCTCAGGTTTGGCATCTATCACAATGTGTGAATCTCATATCAACAAGGATAAGAACACCAGAAGATTTGGATACAATGAAGAGATTTATGCCTGAGGCTTACGCTGAACTTGTTGAGAACTGCAACATCTTAGAAAGACATTACAAAGACATGATGGATATTGAATTCACAGTACAAGAAGAGAGATTGTGGATGCTGCAATGCAGAGCGGGTAAGCGAACGGGTAAAGGCGCCGTGAAGATAGCAGTTGATATGGTAGGTGAAGGGCTTGTTGAGAAATCTTCTGCTATCAAAATGGTGGAGCCTCAACATCTTGATCAACTACTTCACCCACAGTTTCATGATCCATCGGGGTATCGTGAAAAAGTGGTGGCCAAAGGCTTACCTGCGTCACCAGGAGCGGCGGTTGGACAGGTTGTGTTCACGGCGGAGGAAGCCGAAGCTTGGCATTCTCAGGGTAAAACTGTGATTCTGGTTCGAACTGAGACAAGCCCTGACGATGTGGGAGGTATGCACGCAGCGGAAGGTATATTGACGGCTAGAGGAGGAATGACGTCACACGCGGCTGTTGTTGCTCGCGGTTGGGGAAAATGTTGCATTGCTGGTTGTTCCGAGATTCGTGTCGACGAGAACCACAAGGTTCTATTGATTGGAGATTTGACGATTAATGAAGGCGAATGGATCTCAATGAACGGATCAACCGGTGAGGTTATATTAGGGAAACAAGCATTGGCTCCTCCGGCTTTAAGTCCAGATTTGGAGACTTTCATGTCCTGGGCTGATGCAATCAGACGTCTCAAGGTTATGGCGAATGCGGATACACCTGAAGACGCCATTGCAGCTAGGAAAAACGGAGCTCAAGGAATCGGGCTTTGTAGGACAGAGCATATGATTGTTTGTATCCAAATGTTTAATGTTGTCTTTGGTTTGGTTTTTAAGTTCTTTGGAGCAGATAGGATTAAAGCAGTGAGAAAGATGATAATGGCGGTAACAACAGAGCAAAGGAAAGCTTCTCTCGACATCTTGCTTCCTTACCAACGTTCGGATTTCGAAGGGATCTTCCGTGCTATGGATGGTTTACCGGTAACAATCCGTTTGTTAGACCCTCCGCTTCACGAGTTTCTCCCGGAAGGCGACTTGGACAACATTGTACATGAGCTAGCTGAAGAAACTGGTGTGAAAGAAGATGAAGTCTTGTCACGGATAGAGAAACTCTCTGAAGTGAATCCAATGCTTGGTTTCCGCGGTTGCAGGCTCGGAATATCGTATCCAGAGCTAACGGAGATGCAAGCGCGTGCAATTTTTGAAGCTGCAGCGTCAATGCAGGACCAAGGTGTTACTGTCATTCCTGAGATTATGGTTCCACTTGTAGGAACTCCTCAGGAATTGGGTCACCAAGTTGATGTAATTCGTAAAGTTGCAAAGAAAGTATTTGCTGAGAAGGGTCATACCGTGAGCTACAAGGTTGGGACAATGATTGAGATCCCTCGAGCCGCGCTCATTGCAGATGAGATTGCGAAAGAGGCGGAGTTTTTCTCGTTCGGGACAAACGACTTGACGCAGATGACGTTTGGATACAGTAGAGACGATGTCGGCAAGTTTCTACCGATTTACCTCGCCAAAGGAATCTTACAGCACGACCCTTTTGAGGTTCTTGATCAGCAAGGTGTAGGGCAATTGATCAAGATGGCGACAGAAAAAGGACGAGCAGCTAGGCCTAGCCTCAAGGTTGGGATATGTGGAGAACATGGAGGAGATCCATCTTCTGTGGGATTCTTTGCTGAAGCAGGACTTGACTATGTCTCTTGTTCTCCTTTCAGGGTTCCAATTGCAAGGCTTGCAGCTGCTCAAGTAGTTGTTGCATGA
【0038】
従って、PPDK活性を有するポリペプチドをコードするコード配列は、実質的に配列番号20、配列番号21または配列番号22に示す核酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。
【0039】
シロイヌナズナPPDKの細胞質基質型のポリペプチド配列は、本明細書では以下のとおり配列番号23として示す:
配列番号23
MMQRVFTFGKGRSEGNKGMKSLLGGKGANLAEMASIGLSVPPGLTISTEACQQYQIAGKKLPEGLWEEILEGLSFIERDIGASLADPSKPLLLSVRSGAAISMPGMMDTVLNLGLNDQVVVGLAAKSGERFAYDSFRRFLDMFGDVVMGIPHAKFEEKLERMKERKGVKNDTDLSAADLKELVEQYKSVYLEAKGQEFPSDPKKQLELAIEAVFDSWDSPRANKYRSINQITGLKGTAVNIQCMVFGNMGDTSGTGVLFTRNPSTGEKKLYGEFLVNAQVWHLSQCVNLISTRIRTPEDLDTMKRFMPEAYAELVENCNILERHYKDMMDIEFTVQEERLWMLQCRAGKRTGKGAVKIAVDMVGEGLVEKSSAIKMVEPQHLDQLLHPQFHDPSGYREKVVAKGLPASPGAAVGQVVFTAEEAEAWHSQGKTVILVRTETSPDDVGGMHAAEGILTARGGMTSHAAVVARGWGKCCIAGCSEIRVDENHKVLLIGDLTINEGEWISMNGSTGEVILGKQALAPPALSPDLETFMSWADAIRRLKVMANADTPEDAIAARKNGAQGIGLCRTEHMIVCIQMFNVVFGLVFKFFGADRIKAVRKMIMAVTTEQRKASLDILLPYQRSDFEGIFRAMDGLPVTIRLLDPPLHEFLPEGDLDNIVHELAEETGVKEDEVLSRIEKLSEVNPMLGFRGCRLGISYPELTEMQARAIFEAAASMQDQGVTVIPEIMVPLVGTPQELGHQVDVIRKVAKKVFAEKGHTVSYKVGTMIEIPRAALIADEIAKEAEFFSFGTNDLTQMTFGYSRDDVGKFLPIYLAKGILQHDPFEVLDQQGVGQLIKMATEKGRAARPSLKVGICGEHGGDPSSVGFFAEAGLDYVSCSPFRVPIARLAAAQVVVA
【0040】
シロイヌナズナのPPDKの葉緑体型のポリペプチド配列は、本明細書では以下のとおり配列番号24として示す:
配列番号24
MTSMIVKTTPELFKGNGVFRTDHLGENRMVSRSNRLGDGSNRFPRTGTIHCQRLSIAKTGLHRETKARAILSPVSDPAASIAQKRVFTFGKGRSEGNKGMKSLLGGKGANLAEMASIGLSVPPGLTISTEACQQYQIAGKKLPEGLWEEILEGLSFIERDIGASLADPSKPLLLSVRSGAAISMPGMMDTVLNLGLNDQVVVGLAAKSGERFAYDSFRRFLDMFGDVVMGIPHAKFEEKLERMKERKGVKNDTDLSAADLKELVEQYKSVYLEAKGQEFPSDPKKQLELAIEAVFDSWDSPRANKYRSINQITGLKGTAVNIQCMVFGNMGDTSGTGVLFTRNPSTGEKKLYGEFLVNAQVWHLSQCVNLISTRIRTPEDLDTMKRFMPEAYAELVENCNILERHYKDMMDIEFTVQEERLWMLQCRAGKRTGKGAVKIAVDMVGEGLVEKSSAIKMVEPQHLDQLLHPQFHDPSGYREKVVAKGLPASPGAAVGQVVFTAEEAEAWHSQGKTVILVRTETSPDDVGGMHAAEGILTARGGMTSHAAVVARGWGKCCIAGCSEIRVDENHKVLLIGDLTINEGEWISMNGSTGEVILGKQALAPPALSPDLETFMSWADAIRRLKVMANADTPEDAIAARKNGAQGIGLCRTEHMIVCIQMFNVVFGLVFKFFGADRIKAVRKMIMAVTTEQRKASLDILLPYQRSDFEGIFRAMDGLPVTIRLLDPPLHEFLPEGDLDNIVHELAEETGVKEDEVLSRIEKLSEVNPMLGFRGCRLGISYPELTEMQARAIFEAAASMQDQGVTVIPEIMVPLVGTPQELGHQVDVIRKVAKKVFAEKGHTVSYKVGTMIEIPRAALIADEIAKEAEFFSFGTNDLTQMTFGYSRDDVGKFLPIYLAKGILQHDPFEVLDQQGVGQLIKMATEKGRAARPSLKVGICGEHGGDPSSVGFFAEAGLDYVSCSPFRVPIARLAAAQVVVA
【0041】
従って、PPDK活性を有するポリペプチドは、実質的に配列番号23または配列番号24に示すアミノ酸配列またはその機能的な変異体もしくは断片を含んでいてもよい。最も高いPPDK発現量が、ほぼ例外なくPPDKをコードするgDNAを内部に含む細胞株のみで認められたため、イントロンがPPDKの発現にプラスの影響を与えたのではないかと思われる。従って、一実施形態では、本構築物は、PPDKおよび/またはPCKのいずれか一方をコードする遺伝子のcDNAを含んでいてもよい。
【0042】
本発明の遺伝子構築物は、発現カセットの形態であってもよく、宿主細胞における少なくとも1つのコード配列の発現に適したものであればよい。本発明の遺伝子構築物は、それがベクターに組み込まれることなく宿主細胞に導入されてもよい。例えば、核酸分子であり得る遺伝子構築物は、リポソームまたはウイルス粒子内に組み込まれてもよい。あるいは、精製された核酸分子(例えば、ヒストンを含まないDNA、すなわち裸のDNA)が、例えば、直接的なエンドサイトーシス取り込みなどの好適な手段によって宿主細胞に直接挿入されてもよい。本遺伝子構築物は、形質移入、感染、微量注入、細胞融合、原形質融合または衝撃照射(ballistic bombardment)によって宿主対象(例えば、植物)の細胞に直接導入されていてもよい。あるいは、本発明の遺伝子構築物は、パーティクルガン法を用いて宿主細胞に直接導入されていてもよい。あるいは、当該遺伝子構築物は、好適な宿主細胞における発現のために組換え型ベクターの内部に含まれていてもよい。
【0043】
従って、第3の局面では、第1または第2の局面に係る遺伝子構築物を含む組換え型ベクターが提供される。組換え型ベクターは、プラスミド、コスミドまたはファージであってもよい。そのような組換え型ベクターは、宿主細胞を本発明の遺伝子構築物で形質転換し、かつその中の発現カセットを複製するのに非常に有用である。当業者であれば、本発明の遺伝子構築物は、発現のために多くの種類の骨格ベクターと組み合わせ得ることを理解しているであろう。骨格ベクターは、例えば、1つのベクターが大腸菌およびアグロバクテリウム・ツメファシエンスの両方で複製することができるバイナリーベクターであってもよい。例えば、好適なベクターは、pBIN19などのpBINプラスミドであってもよい。しかし、好ましい骨格ベクターはBNP1380000001であり、これは、pBINPLUS(F. A. van Engelen et al. Transgenic Research (1995) 4, 288-290)に基づいており、かつSAG12プロモーターを内部に含む。
【0044】
組換え型ベクターは、プロモーター(例えば、老化関連プロモーター)に加えて、様々な他の機能的な成分、および少なくとも1つの(PCKおよび/またはPPDKをコードする)コード配列を含んでいてもよい。例えば、組換え型ベクターは、宿主細胞中の細胞質基質内で自己複製するように設計されていてもよい。この場合、組換え型ベクター内に、DNA複製を誘発または調節する成分が必要とされる場合がある。あるいは、組換え型ベクターは、宿主細胞のゲノムに組み込まれるように設計されていてもよい。この場合、(例えば相同組換えによる)標的組換えを支持するDNA配列が想定される。
【0045】
また、組換え型ベクターは、クローン化プロセスにおける選択可能なマーカーとして、すなわち、形質移入または形質転換されている細胞の選択を可能にするため、および異種のDNAが組み込まれているベクターを内部に含む細胞の選択を可能にするために使用できる遺伝子のDNAコードを含んでいてもよい。当該ベクターは、コード配列の発現の調節に関与するDNA、または発現されるポリペプチドをある種の宿主細胞、例えば葉緑体に導くためのDNAを含んでいてもよい。従って、第3の局面のベクターは、選択可能なマーカー遺伝子(例えば、抗生物質耐性遺伝子)、ポリペプチド終結シグナルおよびタンパク質標的配列(例えば、葉緑体輸送ペプチド)からなる群から選択される少なくとも1つのさらなる成分を含んでいてもよい。
【0046】
好適なマーカー遺伝子の例としては、抗生物質耐性遺伝子、例えば、カナマイシン、ジェネテシン(G418)およびハイグロマイシン(npt−II、hyg−B)に対して耐性を示す遺伝子;除草剤耐性遺伝子、例えば、ホスフィノトリシンおよびスルホンアミド系除草剤に対して耐性を示す遺伝子(それぞれbarおよびsuI:欧州特許出願公開第242246号、欧州特許出願公開第0249637号);およびスクリーニング可能なマーカー、例えば、β−グルクロニダーゼ(英国特許第2197653号)、ルシフェラーゼおよび緑色蛍光タンパク質(GFP)が挙げられる。当該マーカー遺伝子は、第2のプロモーター(老化関連プロモーターでなくてもよい)によって制御されてもよく、細胞における発現を可能にし、種子の中に存在していても存在していなくてもよく、それにより、植物の発育の任意の段階でマーカーを含有する細胞または組織の選択を可能にする。好適な第2のプロモーターは、アグロバクテリウムのノパリン合成酵素遺伝子のプロモーターおよび35Sカリフラワーモザイクウイルス(CaMV)転写産物をコードする遺伝子に由来するプロモーターである。ただし、任意の他の好適な第2のプロモーターを使用してもよい。
【0047】
本発明の遺伝子構築物の様々な実施形態は図1に示すクローン化手順を用いて作製してもよく、それについて以下のようにまとめることができる。PCKおよびPPDKをコードする遺伝子のgDNAおよびcDNA型は、好適なプライマーを用いるPCRによってgDNAまたはcDNA鋳型から増幅してもよい。PCR産物は、アガロースゲル電気泳動法を用いて調べてもよい。次いで、PCR産物を、クローン化目的のための好適なベクター(例えば、pCR 4Blunt-TOPOベクター(Invitrogen社))に結合してもよい。PCR産物を内部に含むベクターを大腸菌などの好適な宿主中で増殖させてもよい。次いで、大腸菌コロニーを、好適なプライマーを用いるPCRによってスクリーニングしてもよく、正しい制限酵素消化パターンを示すプラスミド中の挿入断片を好適なプライマーを用いて配列決定してもよい。
【0048】
TOPOcDNA(PCKまたはPPDK)またはTOPOgDNA(PCKまたはPPDK)を保有する大腸菌のコロニーは、好適な量の各プラスミドを産生するために培養してもよく、次いでそれを精製してもよい。次いで、プラスミドを消化して、PPDKまたはPCKをコードするDNA断片を遊離させた後、これを、pBNPプラスミドなどの好適なプロモーター(例えば、SAGプロモーター)を内部に含むベクターにクローン化してもよい。得られたPPDK構築物をBNP−PPDKcDNAおよびBNP−PPDKgDNAと命名し、得られたPCK構築物をpALBNP1(cDNA)およびpALBNP2(gDNA)と命名した。第3の局面に係るベクターの実施形態は、実質的に図3に示すものであってもよい。
【0049】
第4の局面では、同じ条件で培養された野生型植物中のPCKおよび/またはPPDKの対応する濃度よりも試験植物の葉の中のPCKおよび/またはPPDKの濃度を上げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために試験植物における植物の代謝を変えることを含む方法が提供される。
【0050】
第5の局面では、同じ条件で培養された野生型植物中の窒素の対応する濃度よりも試験植物の葉の中の窒素濃度を下げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために試験植物における植物の代謝を変えることを含む方法が提供される。
【0051】
第6の局面では、同じ条件で培養された野生型植物の対応する成長率と比較して試験植物の成長率を上げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために試験植物における植物の代謝を変えることを含む方法が提供される。
【0052】
植物の葉の中の窒素濃度および植物の成長率の測定方法は実施例に記載されている。第4、第5または第6の局面の方法は、第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで試験植物の細胞を形質転換することを含んでもよい。本遺伝子構築物またはベクターは、任意の好適な手段によって宿主細胞に導入してもよい。
【0053】
第7の局面では、第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係る組換え型ベクターを含む細胞が提供される。
【0054】
当該細胞は植物細胞であってもよい。本発明者らが、宿主細胞中にPCKおよび/またはPPDKを過剰発現させることが老化した葉の中の窒素の転流を誘発する際に驚くほど有効であることを観察したように、第7の局面の細胞は、PCKおよび/またはPPDKが過剰発現されるように、第1または第2の局面の1種または複数の構築物あるいは第3の局面の1種または複数のベクターを含んでいてもよい。
【0055】
例えば、宿主細胞は、第1の局面の遺伝子構築物の第1の実施形態(すなわちPCK構築物)のみで形質転換されていてもよい。あるいは、宿主細胞は、第2の局面の遺伝子構築物の第2の実施形態(すなわちPPDK構築物)のみで形質転換されていてもよい。別の実施形態では、宿主細胞は、PCKおよびPPDKが宿主細胞において発現されるように、第1の局面の遺伝子構築物の第1および第2の実施形態で形質転換されていてもよい。あるいは、宿主細胞は、PCKおよびPPDKが宿主細胞において発現されるように、第1の局面の構築物の第3または第4の実施形態(すなわちPCK/PPDK構築物1または2)で形質転換されていてもよい。また、宿主細胞が、PCKおよびPPDKの両方をコードする第2の局面の構築物で形質転換されていてもよいことも想定される。
【0056】
当該細胞は、公知の技術を用いて本発明に係る遺伝子構築物またはベクターで形質転換されていてもよい。遺伝子構築物を宿主細胞に導入するのに適した手段は、例えば、欧州特許出願公開第0116718号および第0270822号に記載されているように当該技術分野で知られている手段による、アグロバクテリウムが保持するディスアーム型Tiプラスミドベクターの使用を含んでもよい。さらなる方法は、植物原形質を形質転換することであってもよく、この方法では、最初に細胞壁を除去し、核酸を導入し、次いで細胞壁を再編成する。次いで、形質転換された細胞を植物に成長させてもよい。
【0057】
第8の局面では、第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターを含む形質転換植物が提供される。
【0058】
第8の局面に係る形質転換植物としては、アブラナ属などのアブラナ科が挙げられる。当該植物は、セイヨウアブラナ(菜種)であってもよい。
【0059】
第8の局面に係る形質転換植物のさらなる例としては、コムギ連(Triticeae)などのイネ科が挙げられる。当該植物は、コムギ属(Triticum spp.)(小麦)であってもよい。小麦中の種子タンパク質含有量を増加させることによって、パンなどの小麦を含む食品の体積を増加させてもよい。
【0060】
第8の局面に係る好適な形質転換植物のさらなる例としては、植物のナス科が挙げられ、ナス科の例としては、例えば、チョウセンアサガオ、ナス、マンドレイク、ベラドンナ(deadly nightshade (belladonna))、トウガラシ(パプリカ、カラトウガラシ)、ジャガイモおよびタバコが挙げられる。ナス科の好適な属の一例はニコチアナである。ニコチアナの好適な種は、タバコ植物、または単にタバコと称される場合がある。植物を第1または第2の局面の遺伝子構築物あるいは第3の局面のベクターで形質転換する様々な方法が知られており、本発明で使用することができる。
【0061】
例えば、タバコは、以下のように形質転換してもよい。タバコ(学名:Nicotiana tabacum)を、基本的にHorsch et al. (Science 227: 1229-1231, 1985)に記載されているように葉片共培養法を用いて形質転換する。最も若い2枚の広がった葉を7週齢のタバコ植物から採取してもよく、8%のドメストス(商標)中で10分間表面滅菌し、滅菌蒸留水で6回洗浄してもよい。葉片をNo.6コルク穿孔器を用いて切断し、約2分間かけて(本発明に係る)適当なバイナリーベクターを含有するアグロバクテリウム懸濁液に入れてもよい。この葉片を、2枚の無菌濾紙の間に穏やかにブロットしてもよい。10枚の葉片をLS3%スクロース+2μMのBAP+0.2μMのNAAを含むプレート上に置き、次いで、これを成長室(growth room)で2日間培養してもよい。葉片は、クラフォラン500g/lおよびのカナマイシン100g/lを添加したLS+3%スクロース+2μMのBAP+0.2μMのNAAを含むプレートに移してもよい。この葉片は、2週間後に上記培地を含む新しいプレートに移してもよい。さらに2週間後、この葉片は、クラフォラン500mg/lおよびカナマイシン100mg/lを添加したLS+3%スクロース+0.5μMのBAPを含むプレートに移してもよい。この葉片は、2週間ごとに新しい培地に移してもよい。苗条が現れたら、それらを切除し、クラフォラン500mg/lを添加したLS+3%スクロースを入れた瓶に移してもよい。瓶の中の苗条は、約4週間後、LS+3%スクロース+クラフォラン250mg/lに移してもよい。さらに3〜4週間後、この植物を、LS+3%スクロース(抗生物質なし)に移して、根づかせてもよい。植物が根づいたら、温室の土壌に移してもよい。
【0062】
第9の局面では、第8の局面に係る形質転換植物から得られた植物繁殖用製品が提供される。
【0063】
「植物繁殖用製品」は、そこからさらなる植物を生産することができる任意の植物であってもよい。好適には、この植物繁殖用製品は種子であってもよい。
【0064】
本発明の第10の局面では、同じ条件で培養された対応する野生型植物よりも高い割合で窒素を転流させる形質転換植物の生産方法であって、
i)第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで植物細胞を形質転換する工程と、
ii)形質転換細胞から植物を再生させる工程と、
を含む方法が提供される。
【0065】
第11の局面では、同じ条件で培養された対応する野生型植物よりも高い成長率を有する形質転換植物の生産方法であって、
i)第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで植物細胞を形質転換する工程と、
ii)形質転換細胞から植物を再生させる工程と、
を含む方法が提供される。
【0066】
好ましくかつ有利なことに、本発明に係る方法は、生産される植物の健康または適応度を損なうことはない。好ましくは、本方法は、第1または第2の局面の遺伝子構築物あるいは第3の局面のベクターを用いて、試験植物および好ましくはその葉を形質転換することを含む。本発明者らは、宿主細胞中でPCKおよび/またはPPDKを過剰発現させることが老化した葉の中の窒素の転流を誘発する際に有効であることを観察した。従って、第10および第11の局面の方法は、PCKおよびPPDKの両方が過剰発現されるように、本発明の1種または複数の構築物で試験植物を形質転換することを含むことが好ましい。例えば、試験植物は、本発明の第1の局面の遺伝子構築物の第1の実施形態(すなわちPCK構築物)およびさらに第1の局面の構築物の第2の実施形態(すなわちPPDK構築物)で形質転換してもよい。従って、これらの2種類の構築物の形質転換によって両方の酵素の過剰発現が生じる。あるいは、試験植物は、PCKおよびPPDKをそれぞれコードする本発明の第1の局面の構築物の第3または第4の実施形態で形質転換してもよい。あるいは、試験植物は、PCKおよびPPDKの両方をコードする本発明の第2の局面の構築物で形質転換してもよい。
【0067】
本発明者らは、PCKおよびPPDKをコードする1種以上の構築物で形質転換された試験植物の植物の葉が老化の開始時に窒素の転流の増加を示し、従って、窒素濃度がその葉において低下するすことを観察した。さらに、本発明者らは栄養成長率の上昇を観察した。本発明者らは、仮定に縛られてはいないが、葉の中の窒素濃度の低下によって成長率の上昇、従って作物収穫率の上昇が誘発され得ると考える。
【0068】
本発明の第12の局面では、同じ条件で栽培された野生型植物から採取された、収穫された葉の中の対応する窒素濃度よりも低濃度の窒素を含有する収穫された葉が提供され、ここで、この葉は、第8の局面に係る形質転換植物から収穫されたもの、あるいは第10または第11の局面に係る方法によって生産されたものである。
【0069】
本発明の第13の局面では、老化した葉の中の窒素濃度を低下させることができる変異型タバコ植物から得られる窒素が減少したタバコを含む喫煙物品が提供される。
【0070】
窒素が減少したタバコには、同じ条件で栽培された野生型植物中の対応する濃度よりも窒素濃度の低いタバコが含まれる。そのような喫煙物品は、変異型タバコ植物から得られたタバコを含んでもよく、このタバコは、本発明の第1または第2の局面に係る遺伝子構築物あるいは第3の局面に係るベクターで形質転換されたものであってもよい。
【0071】
本明細書で使用する「喫煙物品」という用語は、タバコ、タバコ誘導体、膨張タバコ、再生タバコまたはタバコ代用品および、また不燃性(heat-not-burn)製品をベースとするか否かに関わらず、喫煙に適した製品、例えば、手巻きタバコ、紙巻タバコ、葉巻およびシガリロ(小型葉巻)を含むことができる。
【0072】
当然のことながら、本発明は、実質的に本明細書で参照されている配列のいずれかのアミノ酸または核酸配列(その機能的な変異体もしくは機能的な断片を含む)を含む、任意の核酸、ペプチド、その変異体、誘導体または類似体に及ぶ。「実質的にアミノ酸/ポリヌクレオチド/ポリペチド配列」、「機能的な変異体」および「機能的な断片」という用語は、本明細書において参照される配列のいずれか1つを含むアミノ酸/ポリヌクレオチド/ポリペチド配列との少なくとも40%の配列同一性、例えば、(PCK酵素の一実施形態をコードする)配列番号17として同定される遺伝子との40%の同一性、配列番号19(すなわちPCK酵素の一実施形態)として同定されたポリペチドとの40%の同一性、(PPDK酵素の一実施形態をコードする)配列番号21として同定された遺伝子との40%の同一性、または配列番号23(すなわちPPDK酵素の一実施形態)として同定されたポリペチドとの40%の同一性を有する配列とすることができる。
【0073】
また、参照されている配列のいずれかに対して65%超、より好ましくは70%超、さらにより好ましくは75%超、なおより好ましくは80%超の配列の同一性を有するアミノ酸/ポリヌクレオチド/ポリペプチド配列も想定される。好ましくは、アミノ酸/ポリヌクレオチド/ポリペプチド配列は、本明細書で参照されている配列のいずれかとの少なくとも85%の同一性、より好ましくは少なくとも90%の同一性、さらにより好ましくは少なくとも92%の同一性、なおより好ましくは少なくとも95%の同一性、なおより好ましくは少なくとも97%の同一性、なおより好ましくは少なくとも98%の同一性、最も好ましくは少なくとも99%の同一性を有する。
【0074】
当業者には、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算方法が分かっている。2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合を計算するために、最初に2つの配列のアラインメントを作成した後、配列の同一性値を計算しなければならない。2つの配列の同一性の割合は、(i)配列を整列させるために使用される方法、例えば、ClustalW、BLAST、FASTA、Smith−Waterman法(異なるプログラムで実行される)、あるいは3D比較による構造アラインメント、および(ii)アラインメント法(例えば、ローカルまたはグローバルアラインメント)によって使用されるパラメータ、使用される対のスコア行列(例えば、BLOSUM62、PAM250、Gonnetなど)、およびギャップペナルティ(例えば、関数および定数)によって異なる値を取る場合もある。
【0075】
アラインメント作成後の、2つの配列間の同一性の割合に関する計算方法は数多くある。例えば、(i)最も短い配列の長さ、(ii)アラインメントの長さ、(iii)配列の平均長、(iv)ギャップのない位置の数、または(iv)オーバーハングを除く等価位置の数によって同一部分の数を割ってもよい。さらに、当然のことながら、同一性の割合は長さにも強く依存する。従って、一対の配列が短い程、偶然に生じることを予測し得る配列の同一性が高くなる。
【0076】
従って、当然のことながら、タンパク質またはDNA配列の正確なアラインメントは複雑なプロセスである。有名なマルチプルアライメントプログラムであるClustalW(Thompson et al., 1994, Nucleic Acids Research, 22, 4673-4680; Thompson et al., 1997, Nucleic Acids Research, 24, 4876-4882)は、本発明に従ってタンパク質またはDNAのマルチプルアラインメントを生成する好ましい方法である。ClustalWの好適なパラメータは、以下のとおりであってもよい:DNAアラインメントについては:ギャップ開始ペナルティー(Gap Open Penalty)=15.0、ギャップ伸長ペナルティー(Gap Extension Penalty)=6.66、および行列=同一性。タンパク質アラインメントについては:ギャップ開始ペナルティー=10.0、ギャップ伸長ペナルティー=0.2、および行列=Gonnet。DNAおよびタンパク質アラインメントについては:ENDGAP=−1およびGAPDIST=4。当業者であれば、最適な配列アラインメントのためにこれらのパラメータおよび他のパラメータを変える必要があるかも知れないことに気付いているであろう。
【0077】
好ましくは、次に、2つのアミノ酸/ポリヌクレオチド/ポリペプチド配列間の同一性の割合の計算は、(N/T)*100(式中、Nは、同一の残基を共有する配列の位置の数であり、Tは、ギャップを含めるがオーバーハングを除いて比較される位置の合計数である)としてそのようなアラインメントから計算する。従って、2つの配列間の同一性の割合の最も好ましい計算方法は、(i)好適な組み合わせのパラメータ(例えば、上記のようなパラメータ)を用いるClustalWプログラムを用いて配列アラインメントを作成すること、および(ii)NおよびTの値を式:配列の同一性=(N/T)*100に挿入することを含む。
【0078】
同様の配列の他の同定方法が当該技術分野で知られている。例えば、実質的に類似したヌクレオチド配列は、厳密な条件で、配列番号16、17、18、20、21、22に示されている配列またはそれらの相補体とハイブリッド形成する配列によってコードされる。厳密な条件とは、ヌクレオチドが約45℃で3×塩化ナトリウム/クエン酸ナトリウム(SSC)中でフィルター結合DNAまたはRNAとハイブリッド形成した後、約20〜65℃で0.2×SSC/0.1%SDS中で少なくとも1回洗浄されることを意味する。あるいは、実質的に同様のポリペプチドは、配列番号19、23または24に示されている配列とは、少なくとも1個、ただし5、10、20、50または100個未満のアミノ酸が異なっていてもよい。
【0079】
遺伝子コードの縮重により、任意の核酸配列を、それによってコードされるタンパク質の配列に実質的に影響を与えることなく変化または変更させてその機能的な変異体を得ることができることは明らかである。好適なヌクレオチド変異体は、配列内の同じアミノ酸をコードする異なるコドンの置換によって静的な変化(silent change)を生じさせることにより変化した配列を有する変異体である。他の好適な変異体は、相同ヌクレオチド配列を有するが、置換されたアミノ酸と同様の生物物理学的特性の側鎖を有するアミノ酸をコードする異なるコドンの置換によって保存的変化を生じさせることにより変化した配列の全てもしくは一部を含む変異体である。例えば、小型で非極性の疎水性アミノ酸としては、グリシン、アラニン、ロイシン、イソロイシン、バリン、プロリンおよびメチオニンが挙げられる。大型で非極性の疎水性アミノ酸としては、フェニルアラニン、トリプトファンおよびチロシンが挙げられる。中立極性のアミノ酸としては、セリン、トレオニン、システイン、アスパラギンおよびグルタミンが挙げられる。正電荷を持つ(塩基性)アミノ酸としては、リジン、アルギニンおよびヒスチジンが挙げられる。負の電荷を持つ(酸性)アミノ酸としては、アスパラギン酸およびグルタミン酸が挙げられる。従って、当然のことながら、アミノ酸は同様の生物物理学的特性を有するアミノ酸で置換することができ、当業者には、これらのアミノ酸をコードするヌクレオチド配列が分かっている。
【0080】
本明細書(添付の特許請求の範囲、要約書および図面のいずれも含む)に記載されている全ての特徴、および/またはそのように開示されている任意の方法またはプロセスの全ての工程は、そのような特徴および/または工程の少なくとも一部が相互に排他的である組み合わせを除く任意の組み合わせで、上記の局面のいずれかと組み合わせてもよい。
【0081】
本発明のさらなる理解のため、および本発明の実施形態の実施方法を示すために、ここで、例として添付の図を参照する。
【図面の簡単な説明】
【0082】
【図1】遺伝子構築物のBNP−PPDKcDNAおよびBNP−PPDKgDNAのクローン化に使用される手順を示す。(a)シロイヌナズナPPDKの細胞質基質のイソ型のcDNAおよびgDNA型のPCR増幅、(b)クローニングベクターpCR 4Blunt-TOPOへの挿入、(c)AvrIIおよびBamHIによる、PPDKを放出するためのクローニングベクター(左側)および標的SAG12含有ベクターpBNP(右側)の制限エンドヌクレアーゼ消化、(d)PPDKのpBNPへの連結反応および、AvrIIおよびBamHIによる構築物の制限エンドヌクレアーゼ消化によって産生されたDNA断片を示すアガロースゲル、および(e)アグロバクテリウム媒介性形質転換によってシロイヌナズナ生態型Columbia 0に導入される構築物。
【図2a】本発明に係る発現ベクターの作製に使用されるプラスミドpCR4 BLUNT-TOPOを示す。
【図2b】PPDKをコードするcDNAまたはgDNAがAvrIIおよびBamHI消化を用いてpCR4 BLUNT-TOPOに挿入されたことをまとめた表である。
【図3a】SAG12プロモーターを含有するプラスミドpBNPを示す。
【図3b】PPDK挿入断片(cDNAまたはgDNA)がAvrIIおよびBamHI消化を用いてpBNPに導入され、かつPCK挿入断片(cDNAまたはgDNA)がXbaIおよびSacI消化を用いてpBNPに導入され、その結果、本発明の実施形態に係るベクターが得られたことをまとめた表である。
【図4】ウェスタンブロットによる形質転換SAG12−PPDKシロイヌナズナ細胞株の選択を示す。SAG12−PPDKcDNA細胞株を上側に示し、SAG12−PPDKgDNA細胞株を下側に示す。
【図5】野生型、ΔPPDKおよび5種類の独立したSAG12−PPDKgDNA系統における5週目以降のPPDK発現量の定量化を示す。
【図6】(b)K326および(c)バーレー21タバコ系統におけるSAG12−PPDK(cDNAおよびgDNA)の選択を示す。
【図7】K326タバコの熟した葉におけるPPDKの過剰発現を示す。
【図8】時間に対する、シロイヌナズナの様々な細胞株、すなわち、野生型、ΔPPDKおよびSAG12−PPDKgDNAのロゼットの写真を示す。
【図9】9週目のシロイヌナズナの生殖組織の質量を示す。
【図10】播種後3〜9週目の野生型およびSAG12−PPDgDNA植物の総植物生質量(すなわち、ロゼット+生殖組織)を示す。
【図11】7週目における野生型、ΔPPDKおよびSAG12−PPDKgDNA植物の葉の窒素含有量を示す。
【図12】野生型、ΔPPDKおよびSAG12−PPDK植物の個々の種子の窒素含有量を示す。
【図13】野生型、ΔPPDKおよびSAG12−PPDKgDNAシロイヌナズナ細胞株の葉の総遊離アミノ酸含有量を示す。
【図14】様々なタバコの細胞株の種子の質量を示し、ゼロコピーは野生型であり、G4(gDNA)、G10、G8およびC10(cDNA)はSAG12−PPDK系統である。
【図15】様々なタバコの細胞株の種子の窒素含有量を示し、ゼロコピーは野生型であり、G4(gDNA)、G10、G8およびC10(cDNA)はSAG12−PPDK系統である。
【図16】PCK/PPDK二重挿入断片のためのウェスタンブロット法の結果である。全ての系統は、SAG12プロモーターの活性と相関する特に7週目に、より高濃度のPCKを示す。
【図17】形質転換した植物のPCKおよびPPDKの両発現によって、対照7と比較してロゼット質量が増加することを示す。
【図18】窒素の転流が、輸送アミノ酸であるアスパラギンおよびグルタミンのPCKおよびPPDK依存的産生によって生じ得ることを示す仮定上の生化学的経路を示す。
【発明を実施するための形態】
【実施例】
【0083】
実施例1−SAG12プロモーターシロイヌナズナPPDK植物形質転換構築物の作製
一連のSAG12−PPDKおよびPCK発現ベクターを、図3に示すように作製し、次いで、形質転換した植物においてPPDKおよびPCKの発現を増加させるそれらの能力を分析した。一実施形態では、本構築物は、PCKまたはその機能的な変異体もしくは断片をコードし、かつPPDKをコードしないものであってもよい。別の実施形態では、本構築物は、PPDKまたはその機能的な変異体もしくは断片をコードし、かつPCKをコードしないものであってもよい。ただし、さらに別の局面では、本構築物は、PCKまたはその機能的な変異体もしくは断片およびPPDKまたはその機能的な変異体もしくは断片をコードするものであってもよい。後述するように、最初にシロイヌナズナのPPDK遺伝子のgDNAを単離する。
【0084】
gDNAの単離
シロイヌナズナ生態型Columbia 0からのgDNAを、推奨されている手順に従ってDNeasy植物ミニキット(DNeasy Plant Mini Kit)(Qiagen社)を用いて葉から抽出した。gDNAを、表1にまとめたプライマー配列を用いる下記のようなPCR反応のための鋳型として使用した。
【0085】
【表1】
【0086】
RNAの単離およびcDNAの合成
全RNAを7日齢のシロイヌナズナの子葉から抽出した。リボヌクレアーゼを含まない機器を用いて氷上でRNAの抽出を行い、ピロ炭酸ジエチル(DEPC、Sigma−Aldrich社)で処理した水を用いて溶液を調製した。水1リットルにつきDEPC1mlを添加し、混合物を換気フード内で一晩撹拌した後、加圧滅菌した。TriPure単離試薬(Roche社)を用いてRNAを抽出した。乳鉢と乳棒を用いて組織200mgを液体窒素中で粉砕し、TriPure単離試薬1mlを添加し、推奨されている手順を続けた。RNAペレットは、60℃まで10分間予熱した無傷のRNA溶液(RNA Secure)(Ambion社)20μlに再懸濁した。試料を、13,000rpmで、4℃で5分間遠心分離し、上澄みを清潔な1.5mlのマイクロ遠心分離管に移し、汚染性の破片を除去した。
【0087】
RNAの量および純度は、Eppendorf Biophotometer分光光度計を用いて260および280nmで読み取ることによって、分光測光法で測定した(Maniatis et al., 1982, Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory)。また、RNAは、0.5×TBEに溶かした1.5%(w/v)のアガロース(Melford Laboratories社)を用いるアガロースゲル電気泳動を用いて調べた。試料を1×RNA試料緩衝液に懸濁し、泳動用緩衝液として0.5×TBEを用いて80Vの電気泳動に供した。4×作業濃度のRNA試料緩衝液は、0.002%(w/v)の臭化エチジウム(Sigma−Aldrich社)、2×トリス−ホウ酸−EDTA緩衝液(2×TBE)(2×TBEは、180mMのトリス−HCl、180mMのホウ酸および8mMのEDTA(pH8.3)を含有していた)、13%のフィコール400(Sigma−Aldrich社)、0.01%のブロムフェノールブルーおよび7Mの尿素(Ficher Scientific社))を含有していた。
【0088】
cDNAを合成するためのRNAの逆転写は、25μlの最終体積に対して、2μgのRNA、1μgのOligo(dT)(15)プライマー(Roche社)、1×モロニーマウス白血病ウイルス(MMLV)緩衝液(Promega社)、0.4mMのdNTP(Bioline社)、40単位の組み換え型RNasinリボヌクレアーゼ阻害剤(Promega社)、200単位のMMLV逆転写酵素(Promega社)およびヌクレアーゼ非含有水を用いて行った。この試料を42℃で1時間培養した。
【0089】
シロイヌナズナPPDKの増幅
PPDKの細胞質基質のイソ型をコードする遺伝子のgDNAおよびcDNA型を、表1に示すAvrII制限部位を含む順方向プライマー(AtCytFWD−AvrII、配列番号1)およびBamHI制限部位を含む逆方向プライマー(AtCytREV−BamHI、配列番号2)を用いるPCRによって、gDNAまたはcDNAの鋳型から増幅した。PCR反応混合物は、1×HF緩衝液(NEB社)、2mMの塩化マグネシウム(MgCl2、NEB社)、0.5mMのdNTPs(Bioline社)、100ngの鋳型(cDNAまたはgDNA)、0.5μMの各プライマーおよび1単位のPhusion High-Fidelity DNAポリメラーゼ(NEB社)を含有していた。熱循環は、98℃で30秒間の最初の変性工程、次いで、98℃で10秒間、55℃で30秒間および72℃で2分間の30サイクル、72℃で5分間の最終的な伸長工程によって、Techneサーマルサイクラー(Techne Thermal Cycler)を用いて行った。
【0090】
シロイヌナズナPPDKのcDNAおよびgDNAを単離および調製した後、それらを使用して、図1にまとめた手順を用いて様々な構築物を作製した。老化中にPPDKを過剰発現させるために、シロイヌナズナのPPDKの細胞質基質のイソ型のcDNAおよびgDNA型を、pBNPベクター中の老化誘発SAG12プロモーターに融合した。
(a)AvrIIおよびBamHI制限部位をそれぞれ含み、かつ2.4および4.3kb産物を産生するプライマーを用いる、シロイヌナズナのPPDKの細胞質基質のイソ型のcDNAおよびgDNA型のPCR増幅。
(b)クローニングベクターpCR 4Blunt-TOPOへの挿入。PPDK挿入断片全体を配列決定し、予想した配列と同一であることが分かった。
(c)AvrIIおよびBamHIによるクローニングベクターおよび目的ベクターpBNPの制限エンドヌクレアーゼ消化。
(d)BNPへの連結反応、および構築物のAvrIIおよびBamHIによる制限エンドヌクレアーゼ消化によって産生されたDNA断片を示すアガロースゲル。予想されたバンドの大きさは、cDNA構築物では14.8kbおよび2.4kb、gDNA構築物では14.8kbおよび4.4kbであった。
(e)アグロバクテリウム媒介性の形質転換によってシロイヌナズナ生態型Columbia 0に導入された構築物。構築物は、連結反応部位全体にわたって配列決定した。遺伝子nptIIはネオマイシンホスホトランスフェラーゼをコードし、植物中にカナマイシン耐性を付与する。LBおよびRB:T−DNAの左右それぞれの境界;nos Pro:ノパリンシンターゼプロモーター;nos t:ノパリンシンターゼターミネーター;SAG12 Pro:シロイヌナズナSAG12遺伝子のプロモーター。
【0091】
図1を参照しながら、構築物の作製について以下に詳細に説明する。
【0092】
pCR 4Blunt-TOPOベクターへのクローン化
PCR産物を、0.5×TBEに溶かした1%(w/v)のアガロース、0.5μgml−1の臭化エチジウム(Sigma−Aldrich社)を使用するアガロースゲル電気泳動を用いて調べた。試料を、1×DNA試料緩衝液(6×作業濃度のDNA試料緩衝液は、50mMのトリス(ヒドロキシメチル)アミノメタン(トリス−HCl、Melford Laboratories社)、60%のグリセリンおよび0.25%(w/v)のブロモフェノールブルー(Sigma−Aldrich社)を含有していた)に懸濁し、泳動用緩衝液として0.5×TBEを用いる80Vの電気泳動に供した。
【0093】
2.6kbのPPDKcDNAバンドおよび4.4kbのPPDKgDNAバンドを、推奨されている手順に従ってQIAQuick PCR精製キット(QIAQuick PCR Purification Kit)(Qiagen社)を用いて精製し、30μlの分子生物学等級水(BDH Laboratory Supplies社)を用いて溶出した。PCR産物は、推奨されている手順に従ってpCR 4Blunt-TOPOベクター(Invitrogen社)に連結させた(平滑末端)。クローン化反応物(2μl)を、推奨されている手順に従って50μlのSub-cloning Efficiency DH5α大腸菌(Invitrogen社)に形質転換した。形質転換した大腸菌細胞は、ルリア−ベルターニ(LB)寒天上で37℃で一晩増殖させた。ルリア−ベルターニ培地(LB培地)は、10gl−1のバクト−トリプトン(Bacto-Tryptone)(BD)、5gl−1のバクト酵母抽出物(Bacto-Yeast Extract)(Oxoid社)および85mMの塩化ナトリウム(Fisher Scientific社)を含んでいた。このpHは、高圧蒸気滅菌前に10Mの水酸化ナトリウムで7.0に調整した。1.5%(w/v)の寒天(BD)を50μgml−1のカナマイシン(Melford Laboratories社)を含むLB培地に添加してLB寒天を調製した。
【0094】
大腸菌のコロニーは、1×NH4緩衝液(Bioline社)、2.5mMのMgCl2(Bioline社)、0.5mMのdNTPs(Bioline社)、0.3μMの各プライマー(AtCytFWD−AvrIIおよびAtCytREV−BamHI)、すなわち配列番号1および2、ならびに0.5単位のBioTaq NAポリメラーゼ(Bioline社)を用いるPCRによってスクリーニングした。熱循環は、95℃で5分間の最初の変性工程、次いで、95℃で30秒間、55℃で30秒間および72℃で4分30秒間の30サイクル、72℃で5分間の最終的な伸長工程によって行った。PCR産物を、上記のように1%(w/v)のアガロースゲル電気泳動を用いて調べた。所望の挿入断片を含むコロニーを、50μgml−1のカナマイシンを含む5mlのLB培地において37℃の振盪培養器内で一晩増殖させた。プラスミドDNAを、推奨されている手順に従ってQIAPrepスピンミニプレップキット(QIAPrep Spin Miniprep Kit)(Qiagen社)を用いて抽出した。DNAは、1×BamHI緩衝液中の10単位のBamHI(NEB1)によって37℃で1時間消化した。消化は、上記のように1%(w/v)のアガロースゲル電気泳動を用いて調べた。正しい制限酵素消化パターンを示すプラスミ中の挿入断片を、3730DNA分析機(Applied Biosystems社)を用いて、表1に示すプライマーAtCytFWD−AvrII(配列番号1)、AtPPDKSeqF1(配列番号3)、AtPPDKSeqF2(配列番号4)、AtPPDKSeqR1(配列番号5)、AtPPDKSeqR2(配列番号6)およびAtCytREV−BamHI(配列番号2)を用いて配列決定した。配列は、BioEdit(Ibis Biosciences社)で分析した。
【0095】
これらの構築物をTOPO−PPDKcDNAおよびTOPO−PPDKgDNAと命名し、図2aおよび図2bに示す。
【0096】
pBNP構築物のクローン化
PPDKを、老化誘発プロモーターSAG12の制御下でBNP1380000001バイナリーベクターに連結させた。SAG12を含む骨格プラスミド(BNP1380000001)は、pBINPLUS(F. A. van Engelen et al. Transgenic Research (1995) 4, 288-290)に基づくものであり、図3aに示す。最初に、TOPO−cDNAまたはTOPO−gDNAを保有する大腸菌のコロニーを使用して、50μgml−1のカナマイシンを含む25mlのLB培地に植菌した。これらの培養物を37℃の振盪培養器内で一晩培養し、推奨されている手順に従ってプラスミドミディキット(Plasmid Midi Kit)(Qiagen社)を用いてプラスミドDNAを抽出した。プラスミドミディキットを用いて、50μgml−1のカナマイシンを含む100mlの培養物からpBNP1380000001ベクターを精製した。
【0097】
次いで、これらのプラスミドを、制限酵素AvrIIおよびBamHIによって消化した。消化反応物は、37℃で一晩培養し、2μgのDNA(BNP社)または4μgのDNA(TOPO−PPDKcDNAおよびTOPO−PPDKgDNA)のいずれか一方、1×緩衝液2、10単位のAvrIIおよび10単位のBamHIを含んでいた。試料は、0.8%(w/v)のアガロース、0.5×TBEおよび25μMのクリスタルバイオレット(Hopkin and Williams社)を用いるクリスタルバイオレットアガロースゲル電気泳動(Rand, 1996, Crystal violet can be used to visualize DNA bands during gel electrophoresis and to improve cloning efficiency(クリスタルバイオレットは、ゲル電気泳動中にDNAバンドを視覚化し、かつクローン化効率を向上させるために使用することができる). Elsevier Trends Journals Technical Tips Online.)によって分離し、クリスタルバイオレット試料緩衝液(250μMのクリスタルバイオレット(Hopkin and Williams社)および30%(w/v)のスクロース(Fisher Scientific社)および泳動用緩衝液としての25μMのクリスタルバイオレットを含有する0.5×TBEを用いて、約2時間50Vの電気泳動に供した。推奨されている手順に従ってQIAQuickゲル抽出キット(QIAQuick Gel Extraction Kit)(Qiagen社)を用いるゲルバンド抽出を使用して、14.4kbのBNP、2.6kbのPPDKcDNAおよび4.4kbのPPDKgDNA断片を抽出した。断片は、臭化エチジウムを用いる1%のアガロースゲル電気泳動を用いてHyperLadder I(Bioline社)に対して調査および定量化した。
【0098】
ゲル抽出したgDNA断片は、gDNA断片(4.4kb)およびTOPO骨格(3.9kb)と同様の大きさであるため、ホスファターゼで処理してあらゆる汚染性TOPO骨格との連結反応を防止した。エビアルカリホスファターゼ(SAP、1単位、Roche社)を、1×脱リン酸化緩衝液(Roche社)に入れた1μgのゲル抽出したgDNA断片に添加し、この反応物を37℃で30分間培養した後、65℃で10分間失活させた。連結反応は、10:1のモル比で消化したBNPを有するcDNAまたはgDNA断片のいずれか一方、1×連結反応緩衝液(NEB社)および1単位のT4DNAリガーゼ(NEB社)を用いて行った。連結反応物を16℃で一晩培養し、2μlを使用して、推奨されている手順に従ってLibrary−Efficiency(登録商標)DH5α大腸菌(Invitrogen社)を形質転換した。
【0099】
形質転換した大腸菌を、50μgml−1のカナマイシンを含むLB寒天上に移し、37℃で一晩培養した。大腸菌のコロニーは、1×NH4緩衝液、2.5mMのMgCl2、0.5mMのdNTP、0.3μMの表1に示す各プライマー(AtPPDKexon15FWD(配列番号7)およびBNPnostREV(配列番号8)、および0.5単位のBioTaq DNAポリメラーゼを用いるPCRによってスクリーニングした。熱循環は、95℃で5分間の最初の変性工程、次いで、95℃で30秒間、55℃で30秒間および72℃で3分間の30サイクル、72℃で5分間の最終的な伸長工程によって行った。PCR産物は、上記のように1%(w/v)のアガロースゲル電気泳動を用いて調べた。所望の挿入断片を含むコロニーを、50μgml−1のカナマイシンを含む5mlのLB培地において37℃の振盪培養器内で一晩増殖させた。プラスミドDNAは、推奨されている手順に従ってQIAPrepスピンミニプレップキット(QIAPrep Spin Miniprep Kit)を用いて抽出した。DNAは、上記のように1%(w/v)のアガロースゲル電気泳動に供する前に37℃で1時間、1×BamHI緩衝液中で10単位のBamHIおよび10単位のStuIで消化した。正しい制限酵素消化パターンを示す、cDNAおよびgDNA連結反応によってそれぞれ得られた1つのコロニーを選択し、上述のように、表1に示すプライマーBNP−SAG12FWD(配列番号9)を用いて配列決定した。
【0100】
これらの構築物をBNP−PPDKcDNAおよびBNP−PPDKgDNAと命名し、図3aおよび図3bに示す。
【0101】
実施例2−BNP−PPDKcDNAおよびBNP−PPDKgDNAによるシロイヌナズナの形質転換
アグロバクテリウムチュメファシエンスのGV3101−R株を、電気穿孔によって、図3に示すプラスミドBNP−PPDKcDNAおよびBNP−PPDKgDNAで形質転換した。それらの調製については、実施例1に記載されている。
【0102】
電気的形質転換受容性(electrocompetent)アグロバクテリウムを、25mgl−1のリファンピシン(Sigma−Aldrich社)を含むLB培地の培養物から産生し、30℃および、600nmにおいて0.4〜0.6の光学濃度で増殖させ、Eppendorf Biophotometer分光光度計を用いて測定した。500mlの培養物を4000gで15分間遠心分離し、上澄みを捨て、細胞を500mlの10%冷グリセリン(Fisher Scientific社)に再懸濁した。遠心分離および再懸濁を250ml、次いで10ml、最後に2mlのグリセリンを用いて繰り返した。細胞は、50μlの分割量に等分し、液体窒素中で急速冷凍し、−80℃で保存した。電気穿孔は、BioRad遺伝子導入装置(BioRad Gene Pulser)を用いて行った。プラスミドDNA(200ng)を、上記のように予備冷却した電気穿孔法用キュベット(Gene Pulser Cuvette、BioRad社)に入れた50μlのアグロバクテリウム細胞に添加し、この細胞を氷上で5分間培養した。このキュベットに対して、2.5mVのパルス、400オームの抵抗および25μFの静電容量を用いた電気穿孔を行った。1mlのSOC培地(Super Optimal Broth, Catabolite Repression)は、20g/lのバクト−トリプトン、5g/lのバクト酵母抽出物、85mMの塩化ナトリウムおよび250mMの塩化カリウムを含んでいた。10Mの水酸化ナトリウムを使用して、高圧蒸気滅菌前にpHを7.0に調整した。使用前に、最終濃度が10mMになるように無菌塩化マグネシウムを添加し、最終濃度が20mMになるように無菌グルコース(Fisher Scientific社)を添加した。この細胞を30℃の振盪培養器内で2時間培養した後、50μgml−1のカナマイシンおよび50μgml−1のリファンピシンを含むLB寒天上に移した。
【0103】
30℃で2日間の培養後に、コロニーをPCR、次いで制限酵素消化によってスクリーニングした。PCRのスクリーニングで陽性であり、かつ期待した制限消化パターンを示すコロニーを、50μlml−1のカナマイシンおよび50μlml−1のリファンピシンを含む5mlのLB培地に植菌し、30℃の振盪培養器内で一晩培養した。翌日、この培養物600μlを使用して上記のように500mlのLB培地に植菌し、花浸漬(floral dipping)に使用する前に、30℃の振盪培養器内で30時間培養した。
【0104】
シロイヌナズナ植物は4週齢で花浸漬のために使用した。全ての構築物を、野生型の生態型Columbia 0に形質転換した。上記のように調製したアグロバクテリウム培養物を5,000gで、4℃で15分間遠心分離し、細胞ペレットは、250mlの無菌の5%スクロース(Fisher Scientific社)溶液(w/v)および0.05%Silwett L-77(OSi Specialties社)に再懸濁した。植物は、穏やかに撹拌しながら細胞懸濁液に約10秒間浸漬した後、ポリエチレン製ラップで蓋をし、24時間直射日光を避けた。この後、植物を標準的な規制飼育に戻し、種子を収穫した。
【0105】
形質転換シロイヌナズナの選択
形質転換した植物からの種子は、Harrison et al. (2006), Plant Methods, 2, 19に従って、50μgml−1のカナマイシン(Dufecha Biochemie社)に関して選択した。抗生物質抵抗性を示すT1植物は結実するまで成長し、自己授粉した。この種子を上記のように選択し、3:1の耐性:非耐性比を示すT2系統を、導入遺伝子の単一コピーを保有するものとして選択した。これらの植物は再び自己授粉し、上記のように選択した。100%の耐性を示す子孫(T3)を産生したT2系統をホモ接合性として選択し、全ての実験に使用した。メンデル遺伝学を使用して、導入遺伝子に対してホモ接合性であり、かつ単一の導入遺伝子コピーを含むT2世代植物を選択した。SAG12−PPDKcDNAおよびSAG12−PPDKgDNAで形質転換した各植物に対して、5種類の独立した単一の挿入断片のホモ接合性系統を選択した。
【0106】
実施例3−BNP−PPDKcDNAおよびBNP−PPDKgDNAによるタバコの形質転換
形質転換受容性アグロバクテリウムチュメファシエンス株LBA4404を、実施例2に記載されているように電気穿孔法によって、プラスミドBNP、BNP−PPDKcDNAおよびBNPPPDKgDNAで形質転換した。電気穿孔後、電気穿孔した細胞に1mlのLB培地を添加し、次いで、その細胞を、50μgml−1カナマイシンおよび100μgml−1スペクチノマイシン(Sigma−Aldrich社)を含むLB寒天上に移す前に、28℃の振盪培養器内で2時間培養した。28℃で2日間の培養後、1つのコロニーを使用して、50μgml−1カナマイシンおよび100μgml−1スペクチノマイシンを含む50mlのLB培地に植菌した。この培養物を、28℃の振盪培養器内で3日間培養した。プラスミドDNAを抽出し、制限酵素消化によって分析し、50μlの培養物を使用して、50μgml−1のカナマイシンおよび100μgml−1のスペクチノマイシンを含む50mlのLB培地に植菌した。この培養物を、28℃の振盪培養器内で一晩培養した。
【0107】
タバコ栽培品種バーレー21およびK326を種子から栽培し、最も若い葉を8週齢の植物から切除し、8%(v/v)ドメストス濃厚漂白剤(Domestos社)で10分間滅菌した後、無菌蒸留水ですすいだ。No.6コルク穿孔器を使用して葉片に穴を開け、次いで、これを25mlのアグロバクテリウム培養物に2分間入れた。次いで、葉片を、2.2μMの6−ベンジルアミノプリン(BAP)および0.27μMのα−ナフタレン酢酸(NAA)を含むMS培地上に下側を下に向けて置いた。これらを、22℃の成長室に2日間置いた。次いで、葉片を、500μgml−1のクラフォラン(Roussel Laboratories社)(共培養されていない対照)または500μgml−1のクラフォランおよび100μgml−1のカナマイシンを含む上記のような選択的なMS培地に移した。葉片は、6週間にわたって14日毎に、上記のような新しい選択的なMS培地に移した。次いで、カルスおよび苗条塊(shoot clump)を葉片から除去し、0.5μMのBAP、500μgml−1のクラフォランおよび100μgml−1のカナマイシンを含むLS培地に入れた。2週間後、苗条を0.5μMのBAP、500μgml−1のクラフォランを含みかつカナマイシンを含まないLS培地を入れた150mlの瓶に移した。
【0108】
それから3週間後、優性の苗条を、250μgml−1のクラフォランを含みかつBAPまたはカナマイシンを含まないLS培地に移した。さらに3週間後、苗条を、抗生物質またはBAPを含まないをLS培地に移すことによって、さらに浄化した。十分な根が生えたら、植物を温室内の土壌に移した。
【0109】
形質転換タバコの選択
定量的PCR(Q−PCR)を使用して、T0およびT1植物中の導入遺伝子のコピー数を定量化した。可能であれば、単一の挿入断片T0およびホモ接合性T1植物を分析のために選択した。導入遺伝子の検出のために、表1に示すようなプライマーBNP−1271F(配列番号10)およびBNP−1334R(配列番号11)を使用し、Vic/TAMARA標識プローブBNP1291TV(配列番号12)を使用して、nptII導入遺伝子にアニールした。内部定量化のために、プライマーNtCyc−184F(配列番号13)およびNtCyc−316R(配列番号14)を使用し、FAM/TAMARA標識プローブNtCyc−267T(配列番号15)を使用して、サイクロフィリンをコードする内因性遺伝子にアニールした。定量化は、比較Ct法によるものであった(ΔΔCt method, Bubner & Baldwin (2004), Plant Cell Reports, 23, 263-271)。反応混合物は、1×ユニバーサルマスターミックス(Universal Master Mix)(ABI社)、0.9μMの各プライマー、0.2μMの各プローブ(BNPおよびNtCycプライマーおよびプローブに対して別の反応を用いる)および推奨されている手順に従ってDNeasy植物ミニキット(Qiagen社)を用いて葉の組織から抽出した約500ngのgDNA鋳型を含んでいた。熱循環は、95℃で10分間の最初の変性工程、次いで、95℃で15秒間、60℃で1分間の40サイクルによって、7900HT FastリアルタイムPCRシステム(7900HT Fast Real-Time PCR System)(Applied Biosystems社)で行った。データは、付属のSDS2.2ソフトウェア(Applied Biosystems社)を用いて分析した。
【0110】
形質転換したT0世代のK326およびバーレー21タバコの組織培養における再生後、定量的PCR(Q−PCR)を使用して、導入遺伝子の単一コピーを有する植物を選択した。単一の挿入断片の植物は、導入遺伝子サイレンシングの可能性を減少させるように選択した。Q−PCRは、ネオマイシンホスホトランスフェラーゼをコードするnptII導入遺伝子に相補的なオリゴヌクレオチドを用いて行った。この遺伝子は、構築物pBNP、BNP−PPDKcDNAおよびBNP−PPDKgDNA中の植物ゲノム内に運ばれたT−DNAの左右の境界の間に存在していた。pBNPのみによって形質転換した植物は、組織培養による再生が空のベクターによって形質転換された植物とPPDKコード配列を含むベクターによって形質転換された植物との間で一致することを確認するための空のベクター対照として機能した。形質転換が成功したことを確認するための再生およびQ−PCR検査後、これらの植菌を破棄した。
【0111】
各構築物(SAG12−PPDKcDNAおよびSAG12−PPDKgDNA)ならびに各栽培品種(K326およびバーレー21)のために、6種類の植物を選択した。単一の挿入断片の植物のみを選択することができなかったため、6種類の単一の挿入断片の植物が入手できなかった場合は、可能な限り最も少ないコピー数を有する植物を選択した。これらの植物は自己授粉し、その種子を収穫した。
【0112】
選択したT0植物ごとに14株の子孫を育て、Q−PCRを繰り返してホモ接合性植物を選択した。ホモ接合性植物の使用によって、将来の世代における導入遺伝子の安定性を確保しなければならない。T0親ごとに、導入遺伝子を保有している4株の子孫を選択した。しかし、全てのT0親に対して、ホモ接合性の子孫を必ずしも選択することができるわけではなかった。親のコピー数が3つ以上であった場合は、より少ないコピー数を有する植物を選択して、導入遺伝子サイレンシングの可能性を減少させかつ、また必要であれば、T2世代からのホモ接合体の選択を単純化した。このようにして、各構築物および各栽培品種のために、5種類の独立した系統から、全てのさらなる実験のためにより少ない導入遺伝子コピー数を有する4種類の生物学的複製(同胞)を選択した。
【0113】
実施例4−形質転換したシロイヌナズナ植物中のPPDKタンパク質の検出および定量化
タンパク質の抽出
シロイヌナズナの葉の組織(100mg)を、マイクロ乳棒を用いて1.5mlの微小遠心管中で液体窒素冷却して粉砕し、400μlの抽出緩衝液(リン酸カリウム緩衝液(+プロテアーゼ阻害剤反応混液(PIC、Sigma社)))を添加した。タンパク質の濃度は、推奨された手順に従ってBio−Radタンパク質アッセイ試薬(Bio-Rad Protein Assay Reagent)(Bio−Rad社)を用いてブラッドフォードアッセイ(Jones et al., 1989, Journal of Chemical Ecology, 15, 979-992)で測定した。
【0114】
ポリアクリルアミドゲル電気泳動
タンパク質は、10%(v/v)アクリルアミド(37.5:1=アクリル:ビスアクリル、Severn Biotech社)、50%(v/v)免疫ブロット分解緩衝液(セクション2.10を参照)、0.05%(w/v)ペルオキソ二硫酸アンモニウム(APS、AnalaR社)、および0.05%(v/v)N,N,N’,N’−テトラメチルエチレンジアミン(TEMED、Severn Biotech社)を含む分解ゲルにおけるポリアクリルアミドゲル電気泳動(PAGE社)によって分離した。濃縮用ゲルは、上記のような5%アクリルアミド、50%(v/v)免疫ブロット濃縮用緩衝液、0.06%(w/v)APSおよび0.1%(v/v)TEMEDを含んでいた。電気泳動は、1×免疫ブロット試料緩衝液(66mMのトリス−HCl、10%(v/v)グリセリン、0.7mMのSDS、0.7Mのβ−メルカプトエタノール(BDH Laboratory Supplies社)および0.05%(w/v)ブロモフェノールブルー)および免疫ブロット泳動用緩衝液、(25mMのトリス−HCl、0.29mMのグリシン(Fisher Scientific社)および3.5mMのSDS)を用いて抽出した20μgの総タンパク質に対して70mAの電流で1時間30分間行った。
【0115】
二組のゲルを同時に泳動した。一方を免疫ブロット分析(以下に記載する)のために使用し、他方を推奨された手順に従ってGelCode青色安全タンパク質染色(GelCode Blue Safe Protein Stain)(Thermo Scientific社)を用いて染色した。ゲル乾燥膜(Gel Drying Film)(Promega社)を使用して染色したゲルを乾燥した。
【0116】
免疫ブロット分析
免疫ブロット分析用のゲルからのタンパク質を、Protean極厚ブロット紙(Protean Extra-Thick Blot Paper)(Bio−Rad社)、および免疫ブロット用転写緩衝液(48mMのトリス−HCl、39mMのグリシン、1.3mMのSDSおよび20%(v/v)メタノール(Fisher Scientific社))を用いて、セミドライブロッター(Semi-Dry Blotter)(Bio−Rad社)中でProtan BA83硝酸セルロース膜(Schleicher and Schuell社)に15Vで1時間転写した。タンパク質の転写を確認するために、ポンソー染色(0.5%(w/v)ポンソーS(Fluka社)および1%(w/v)氷酢酸(Fisher Scientific社))をブロットした後の膜に塗布し、次いで、その膜を蒸留水ですすいだ。
【0117】
PPDK免疫ブロット分析用のブロッキング緩衝液は、1%(w/v)乾燥脱脂乳粉末(Marvel社)および0.1%(v/v)ポリオキシエチレンモノラウリン酸ソルビタン(TWEEN20、Sigma−Aldrich社)のリン酸緩衝食塩水(PBS:1.5mMの正トリン酸二水素カリウム(KH2PO4、AnalaR社)、8.1mMの正リン酸二水素ナトリウム、2.7mMの塩化カリウムおよび137mMの塩化ナトリウム)を用いて毎日更新した。塩酸を使用してpHを7.4に調整した。膜は、振盪機上の室温のブロッキング緩衝液中で1時間培養した。一次ハイブリッド形成は、ウサギ抗PPDK抗体(Chris Chastain、ミネソタ州立大学)の1:10,000の希釈液を用いて行い、次いで、ブロッキング緩衝液でそれぞれ5分間の洗浄を3回行った。二次ハイブリッド形成は、ロバ抗ラビットビオチン化全抗体(GE Healthcare社)の1:1000の希釈液を用いて行い、次いで、上記のように3回洗浄した。三次ハイブリッド形成は、ストレプトアビジンビオチン化ワサビペルオキシダーゼ複合体(GE Healthcare社)の1:1000の希釈液を用いて行い、次いで、各0.1%(v/v)TWEEN20を含むPBS中で5分間の洗浄を3回行った。
【0118】
次いで、膜を蒸留水中で3回すすいだ。検出は、推奨されている手順に従って、Western Lightning化学発光試薬(Western Lightning Chemiluminescence Reagent)(増感ルミノール(Enhanced Luminol)、Perkin Elmer社)を用いて行った。
【0119】
図4を参照すると、ウェスタンブロットによる形質転換SAG12−PPDKシロイヌナズナ細胞株の選択が示されている。タンパク質を、8週齢のシロイヌナズナ野生型、ΔPPDK、SAG12−PPDKcDNAおよびSAG12−PPDKgDNA植物の葉の組織から抽出し、より高濃度のPPDKを発現している株を選択するために免疫ブロット分析に供した。組み換え型トウモロコシPPDK(30μg)を陽性対照として使用した。PPDKは、野生型またはPPDK変異型植物(ΔPPDK)において検出されなかった。SAG12−PPDKgDNA植物は、SAG12−PPDKcDNAよりも高濃度のPPDKを発現していた。その後の全ての分析は、5種類のSAG12−PPDKgDNA系統に対して行った。
【0120】
PPDKは、野生型およびΔPPDK植物中では検出することができず、いくつかのSAG12−PPDKcDNA植物中では低濃度で検出することができ、全てのSAG12−PPDKgDNA系統で検出することができた。これらの系統のうち3つにおいて、野生型またはSAG12−PPDKcDNA系統に比べ非常により高い量のPPDKが検出された。従って、イントロンの存在は、PPDKの発現レベルに対してプラスの効果があると思われ、全てのさらなる実験での使用のためにSAG12−PPDKgDNA系統を選択した。
【0121】
SAG12−PPDKgDNA植物におけるPPDKの累積の開始および程度を測定するために、PPDKの免疫ブロットを、野生型、ΔPPDKおよびSAG12−PPDKgDNA植物に対して行った。
【0122】
図5を参照すると、野生型、ΔPPDKおよび5週目以降の5種類の独立したSAG12−PPDKgDNA系統におけるPPDK発現量の定量化が示されている。PPDK発現量は、5〜9週目の各時点の野生型の割合として計算した。データは、これらの5つの時点の平均として示されている。エラーバーは標準誤差(SEM)を示す。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は、分散分析を用いて試験した(F=4.775、df=6、p=0.001)。野生型と有意差が認められる系統は、G12.3(p=0.005)およびG16.1(p=0.004)であった。
【0123】
タンパク質は、播種後3週目から9週目までの週間隔で収穫された葉の組織から抽出した。PPDKは、5週目以降の野生型植物中で低量検出され、これにより、転写産物の観察された増加によってタンパク質の発現量が増加することが確認された。SAG12−PPDKgDNA植物では、PPDKの累積の開始も5週目に観察されたが、PPDK発現量は野生型よりも多い。PPDKタンパク質は、ΔPPDK植物ではいずれの段階においても検出されなかった。
【0124】
葉のタンパク質抽出物中のPPDKの定量化を可能にする方法も最適化した。異なる量の組み換え型トウモロコシPPDKタンパク質をSDS−PAGEおよび免疫ブロット分析に供した。バンド強度は、AlphaEase画像化ソフトウェアおよび作成された標準曲線を用いて計算した。回帰直線はSigmaPlotソフトウェアを用いて計算し、次いで、これを使用して植物の葉の試料中のPPDK量を計算した。免疫ブロットの検出レベルが変化する可能性があるため、異なる免疫ブロットの間の真の比較を可能にするために、全ての免疫ブロットに対して少なくとも3種類の標準物質を含める必要があった。異なる抽出物中のPPDK発現量の定量化のためのこの技術を使用して、異なる形質転換系統を選択および比較した。
【0125】
PPDKタンパク質発現量は、各時点における各系統の少なくとも4個の生物学的複製について、組み換え型トウモロコシPPDKタンパク質の公知の標準物質を参照することによって計算した。分析から、SAG12−PPDKgDNA中のPPDK発現量が5週目以降の野生型における発現量よりも高いことが分かった。PPDK発現量は、各SAG12−PPDKgDNA系統について5週目以降の各時点の野生型の割合、および各SAG12−PPDKgDNA系統の老化期間におけるPPDKの累積を定量化するためのこれらの値からの平均として計算した。図5に示すように、これによって、SAG12−PPDKgDNA系統をPPDK発現量の増加に従って順序付けることができた。
【0126】
形質転換植物中に高い量で蓄積したタンパク質が失活する場合があるため、タンパク質発現量の増加が必ずしも酵素活性の増加を暗示するというわけではない。PPDKは暗所で可逆的にリン酸化され、その結果、PPDKは失活する。抗体は、PPDKのリン酸化された型(リン酸化PPDK)に抗して増加し、PPDKの失活型の特異的検出を可能にするが、リン酸化状態に関係なく以前に使用された抗体(抗PPDK)はPPDKを検出する(Chastain et al., 2002, Plant Physiology, 128, 1368-1378)。抗リン酸化PPDK抗体を使用して、免疫ブロット時には抗PPDK抗体によって最初に検出され、次いで剥離され、かつ再検出される播種後3週目から9週目までに経時的に失活したPPDKを検出した。
【0127】
驚くべきことに、失活した(リン酸化された)PPDKは、複数の免疫ブロットにおいて野生型植物中に超低濃度でのみ、かつ老化の後期段階の間にのみ検出することができた。SAG12−PPDKgDNA植物では、失活した時点に応じて、PPDKは、より高濃度で検出された。しかし、PPDK発現量の増加開始直後の時点では、総PPDKの高い発現量にも関わらず、失活したPPDKは、ほとんどあるいは全く検出されなかった。これらの結果は、一部のPPDKの失活がSAG12−PPDKgDNA植物中に生じる可能性があるが、酵素的に活性なPPDKの発現量の増加は、少なくとも老化の初期段階の間にSAG12−PPDK植物において存在することを示唆している。
【0128】
実施例5−形質転換したタバコ(N.tabacum)植物中のPPDKタンパク質の検出および定量化
タバコの葉の組織(100mg)を、マイクロ乳棒を用いて1.5mlの微小遠心管の中で液体窒素下で粉砕し、400μlの抽出緩衝液(オーバーコート緩衝液(Overcoat Buffer)(PIC社))を添加した。タンパク質の濃度は、推奨されている手順に従って、Bio−Radタンパク質アッセイ試薬(Bio−Rad社)を用いてブラッドフォードアッセイ(Jones et al., 1989)で測定した。タンパク質は、実施例4に記載されているようにポリアクリルアミドゲル電気泳動(PAGE社)によって分離した。実施例4に記載されている免疫ブロットによってタンパク質を分析し、かつPPDKタンパク質を定量化した。
【0129】
PPDKに対する免疫ブロットは、分離および暗所での30℃の培養によって老化が誘発されたK326およびバーレー21のT1世代タバコの葉から抽出した葉のタンパク質に対して行った。老化は、植物が成熟期に達する前に最も高いPPDK発現量を有する形質転換系統を特定し、かつ他の系統を破棄できるように誘発した。K326植物では、PPDKは、3種類のSAG12−PPDKgDNA系統(G4、G8およびG10)および1種類のSAG12−PPDKcDNA系統(C10)において高い発現量で検出された。バーレー21植物では、PPDKは、4種類のSAG12−PPDKgDNA系統(G3、G7、G15およびG23)に大量に存在したが、K326系統に認められる程に高い発現量では存在していなかった。最も高いPPDK発現量を有するK326およびバーレー21のそれぞれの4種類の系統をさらなる全ての分析のために使用した。ほぼ例外なくSAG12−PPDKgDNA系統のみに最も高いPPDK発現量が観察されたため、イントロンがPPDK発現に対してプラスの効果を有していると思われる。
【0130】
図6を参照すると、以下のように、SAG12−PPDK−K326およびバーレー21タバコ系統の選択が示されている:
(b)形質転換K326系統を選択するためのPPDK免疫ブロット。老化は、分離および暗所での30℃で3日間の培養によって6週齢の植物の緑葉において誘発された。タンパク質を抽出し、免疫ブロット分析に供して、より高濃度のPPDKを発現している系統を選択した。組み換え型トウモロコシPPDK(50μg)を陽性対照として使用した。各親のT0植物からの1つの子孫に関する1つの代表的な免疫ブロットが示されている。SAG12−PPDKgDNA親系統G4、G8およびG10から、および、SAG12−PPDKcDNA系統C10からの植物をその後の全ての分析に使用した。
(c)セクション(b)におけるK326に関して、形質転換バーレー21系統を選択するためのPPDK免疫ブロット。SAG12−PPDKgDNA親系統G3、G7、G15およびG23からの植物をその後の全ての分析のために使用した。
【0131】
PPDKに対する免疫ブロットを、野生型、ゼロコピー(陰性の分離体)、SAG12−PPDKgDNA−K326、SAG12−PPDKcDNA−K326およびSAG12−PPDKgDNA−バーレー21タバコに対して行って、PPDKの発現の開始を測定し、かつ形質転換した植物中のPPDKの過剰発現の程度を定量化した。タンパク質は、K326野生型および形質転換した植物の葉1(最も古い)〜3ヵ月齢の植物の8(若い)から抽出し、PPDKに対する免疫ブロットを行った。
【0132】
図7を参照すると、K326タバコの熟した葉におけるPPDKの過剰発現が示されている。PPDK発現量は免疫ブロットから計算した。データは、ゼロコピー植物の10個の生物学的複製および各SAG12−PPDK系統の4個の生物学的複製の平均として示されている。エラーバーは標準誤差を示す。SAG12−PPDK系統は、PPDK発現量の上昇で配置されている。遺伝子型間の差は、分散分析を用いて試験した(F=6.995、df=4、p=0.001)。ゼロコピー植物と有意差が認められる系統は、G10(p=0.006)、G8(p=0.003)およびC10(p=0.000)であった。
【0133】
PPDK発現量は、野生型植物のより古い葉の中でより高く、これは、PPDKが、タバコならびにシロイヌナズナにおける老化中に自然に上方制御されることを示している。形質転換したK326植物では、PPDK発現量は同様に、より古い葉においてより高く、野生型植物において非常により高かった。また、最も高いPPDK発現量を有する形質転換した系統のより若い葉においてPPDK発現量はより高かった。免疫ブロットを、ゼロコピー植物(陰性の分離体)および形質転換した植物の「熟した」葉から抽出したタンパク質に対して行った。熟した葉は、収穫可能な段階にあり、かつ老化の後期段階にある葉である。PPDK発現量を定量化した。PPDK発現量は、ゼロコピー植物と比較して、全ての4種類の独立した形質転換した系統の熟した葉においてより高かった。SAG12プロモーターを用いる老化中のPPDKの過剰発現は、K326タバコでは成功していた。
【0134】
実施例6−形質転換したシロイヌナズナ植物の表現型分析
シロイヌナズナ植物の成長分析
図8を参照すると、シロイヌナズナにおけるPPDKgDNAの過剰発現によって、より大きなロゼットを有する植物が生産されることが明らかに示されている。播種後3、5、7および9週目に野生型、ΔPPDKおよびSAG12−PPDKgDNA植物を撮影した。SAG12−PPDKgDNA植物は野生型よりも大きかった。PPDK植物と野生型植物との間に識別可能な差はなかった。
【0135】
ロゼットおよび生殖組織の生質量は、0.1mgの単位まで、メトラー・トレドAB104−S(Mettler Toledo AB104-S)天秤を用いて精密に測定した。ロゼット組織の乾燥質量は、Edwards Super Modulyo凍結乾燥器(Edwards Super Modulyo Freeze Dryer)を用いて一晩凍結乾燥した試料から、0.1mgの単位まで精密に測定した。
【0136】
図9を参照すると、9週目の生殖組織の質量が示されている。より高い生殖組織質量を有するより高いPPDK発現量を有する系統について、PPDKに対する用量反応が観察された。総生殖組織(茎、茎生葉、花および長角果)を計量した。値は3個の生物学的複製の平均である。エラーバーは標準誤差を示す。形質転換SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は分散分析を用いて試験した(F=8.062、df=6、p=0.001)。全てのSAG12−PPDKgDNA系統は、野生型とは有意に異なっていた(G9.4:p=0.031、G5.4:p=0.000、G19.2:p=0.000、G12.3:p=0.001、G16.1:p=0.001)。傾向は絶対ではないが、より高いPPDK発現量を有する系統はより高い生殖組織質量を有する傾向にあった。総植物生質量の割合としての生殖組織も計算したが、分散分析を用いて試験した場合、野生型、ΔPPDKまたはSAG12−PPDKgDNA植物では有意差は認められなかった(F=0.544、df=6、p=0.767、データは示さない)。
【0137】
しかし、総植物質量の割合としての生殖組織の比率は変化せず、これは、植物は全体としてより大きく、かつ栄養組織と生殖組織との資源分配は変化しないことを示唆していた。ΔPPDK植物における総植物生質量および生殖組織質量も測定したが、野生型とは有意差は認められなかった。ロゼットの乾燥質量も測定したところ、総植物生質量および生殖組織質量に関しては、SAG12−PPDKgDNA植物が野生型よりも有意に高い質量を有することが分かった。
【0138】
図10を参照すると、播種後3週目から9週目までの野生型およびSAG12−PPDgDNA植物の総植物生質量(ロゼット+生殖組織)が示されている。データは、野生型では3個の生物学的複製、SAG12−PPDKgDNAでは15個の生物学的複製(5種類の独立した系統ごとに3個の植物)の平均として示されている。エラーバーは標準誤差を示す。野生型およびSAG12−PPDKgDNAの総植物質量は、各時点でスチューデントのt−検定を用いて比較した。ロゼット質量は、9週目では、SAG12−PPDKgDNA植物の方が有意に高かった(p=0.032)。
【0139】
5週目では、SAG12プロモーターによるPPDK過剰発現の開始前に、優位差は認められなかった。全ての5種類の独立したSAG12−PPDKgDNA系統のロゼットの乾燥質量は増加した。質量が大きくPPDK含有量の高い系統については、PPDK発現量に対する用量反応が生殖組織質量に関して観察された。ロゼットの乾燥質量をΔPPDK植物についても測定し、野生型植物とは有意差は認められなかった。
【0140】
シロイヌナズナの葉の表面積の測定
ロゼットの表面積は、単一の成葉の試料(通常は葉10枚)を採取し、上記のように葉およびロゼット全体の質量を測定し、かつルーラーに並べてそれらの葉を撮影して測定した。葉の表面積は、ImageJソフトウェア(アメリカ国立衛生研究所)を用いて測定し、ここから、ロゼット全体の表面積を計算した。
【0141】
ロゼットの表面積は、PPDK過剰発現の開始後にSAG12−PPDKgDNA植物で増加した。6週目および7週目にSAG12−PPDKgDNA植物の表面積は有意により大きかったが、8週目および9週目では、表面積は野生型とは有意差は認められなかった。表面積のこの大きい減少は、野生型と比較してSAG12−PPDKgDNA植物中の栄養分の転流が増加したことに起因していると思われた。ΔPPDK植物の表面積は野生型植物とは有意差は認められなかった。
【0142】
クロロフィル濃度の測定
クロロフィル含有量は、CCM−200手持ち式クロロフィル計(Opti-Sciences社)を用いて、相対的な単位で測定した。
【0143】
シロイヌナズナの蛍光測定
FV/FM比を、Hansatech FMS2蛍光光度計を用いて測定した。葉は、測定前に一晩、製造業者によって提供されたリーフクリップ(leaf clip)を用いて暗順応させた。老化の開始が野生型植物に対してSAG12−PPDKgDNAまたはΔPPDK植物において変化したか否かを判定するために、FV/FM比を測定した。FV/FMは、光化学系IIの量子効率の推定値であり、光阻害が生じると減少する。クロロフィル含有量の減少が検出される前に、光合成の減少が老化の初期に生じるため、FV/FMは老化の開始の有用な基準である。FV/FMは、播種後(葉がFV/FMを測定するのに十分な程大きくなる)4週目から9週目までに経時的に測定した。野生型、SAG12−PPDKgDNAおよびΔPPDK植物のFV/FMは、7週目に最大となり、その後減少し、これは、全ての3種類の遺伝子型において老化の開始のタイミングが同じであったことを示していた。しかし、SAG12−PPDKgDNA植物のFV/FMは、8週目に野生型よりも有意に高く、これは、老化の後期段階における長期にわたる光合成作用を示していた。
【0144】
窒素含有量
窒素分析用の組織を、Edwards Super Modulyo凍結乾燥器を用いて凍結乾燥した。シロイヌナズナの葉の組織(25mg)またはタバコの葉の組織(100mg)を、窒素非含有秤量紙(Elementar Analysensysteme社)に包装した。推奨されている設定を用いて、Rapid N III窒素分析装置(Rapid N III Nitrogen Analyzer)(Elementar Analysensysteme社)を使用して乾燥重量の割合として窒素含有量を測定した。アスパラギン酸(Sigma−Aldrich社)を標準物質として使用した。葉の窒素含有量は、シロイヌナズナの野生型、SAG12−PPDKgDNAおよびΔPPDK植物において、播種後3週目から9週間目までに経時的に測定した。
【0145】
図11を参照すると、7週目における野生型、ΔPPDKおよびSAG12−PPDKgDNA植物の葉の窒素含有量が示されている。データは、野生型およびΔPPDKでは8個の生物学的複製、各SAG12−PPDKgDNA系統では4個の生物学的複製の平均として示している。エラーバーは標準誤差である。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は分散分析を用いて試験した(F=6.047、df=6、p=0.000)。ΔPPDK植物および全てのSAG12−PPDKgDNA系統は、野生型とは有意に異なっていた(ΔPPDK:p=0.004、G9.4:p=0.000、G5.4:p=0.000、G19.2:p=0.001、G12.3:p=0.007、G16.1:p=0.006)。
【0146】
葉の窒素は、7週目以降は、野生型と比較して全ての5種類の独立したSAG12−PPDKgDNA系統において有意により低く、これは、老化中にPPDK発現量が増加することによって窒素の転流効率を高めることができるという仮定を支持するものであった。PPDKの発現および活性は、SAG12−PPDKgDNA植物において6週目にピークに達し、これは、PPDK発現量の増加と測定可能な葉の窒素の減少との間に時間の遅れが生じたことを示唆しており、この時間の遅れは、タンパク質アミノ酸を輸送アミノ酸(アスパラギンおよびグルタミン)に変換し、かつそれらを葉から輸送するのに要する時間に起因していると思われた。
【0147】
シロイヌナズナ種子の分析
個々の種子の質量および窒素含有量を、野生型、SAG12−PPDKgDNAおよびΔPPDKシロイヌナズナ植物の種子について測定した。
【0148】
図12を参照すると、野生型、ΔPPDKおよびSAG12−PPDK植物の個々の種子の窒素含有量が示されている。SAG12−PPDKgDNA植物の種子窒素含有量において有意な増加が認められた。データは、野生型およびΔPPDKでは8個の生物学的複製、各SAG12−PPDKgDNA系統では4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。遺伝子型間の差は分散分析を用いて試験した(F=6.704、df=6、p=0.000)。野生型と有意差が認められる系統は、G19.2(p=0.000)、G12.3(p=0.005)およびG16.1(p=0.002)であった。より高いPPDK発現量を有する系統は、より高い種子窒素質量を有する傾向がある。従って、個々の種子質量は、野生型に対して全ての5種類の独立したSAG12−PPDKgDNA系統で増加し、より高い種子質量を有するより高いPPDK発現量を有する植物については、PPDK発現量に対する用量反応が観察された。各植物から収穫された全種子を計量し、野生型とSAG12−PPDKgDNA植物との間に有意差はなく、SAG12−PPDK植物において種子の大きさがそのように増加したことによって総種子収穫量は損なわれなかった。
【0149】
シロイヌナズナの葉の組織の遊離アミノ酸含有量
葉の組織(100mg)を、マイクロ乳棒(Eppendorf社)を用いて、1.5mlの微小遠心管中で液体窒素冷却して粉砕し、300μlの無菌脱イオン水を添加した。試料は、13,000rpmで、4℃で5分間遠心分離にかけた。タンパク質の濃度は、推奨されている手順に従って、Bio−Radタンパク質アッセイ試薬(Bio−Rad社)を用いるブラッドフォードアッセイ(Jones et al., 1989)で測定した。推奨されている手順に係って、EZfaastアミノ酸試料試験キット(EZfaast Amino Acid Sample Testing Kit)(Phenomenex社)を用いるアミノ酸分析のために試料を調製した。10mMのギ酸アンモニウム(BDH laboratory supplies社)の50%無菌蒸留水と50%超高純度メタノール(Romil社)溶液に試料を再懸濁した。
【0150】
次いで、試料を、EZfaast250×3.0mmAAA−MSカラム(Phenomenex社)ならびに移動相として10mMのギ酸アンモニウム/50%(v/v)無菌蒸留水および50%(v/v)超高純度メタノールを用いるQ Trap LC/MS/MS(Applied Biosystems社/MDS SCIEX社)による液体クロマトグラフィー−質量分析(LC−MS)にかけた。質量分析計は、EZfaastキット(EZfaast Kit)(Phemonenex社)で推奨されているような条件を用いて正イオンモードで使用した。結果は、付属のAnalystソフトウェア(Applied Biosystems社/MDS SCIEX社)で分析した。
【0151】
総遊離アミノ酸含有量は、野生型、SAG12−PPDKgDNAおよびΔPPDKシロイヌナズナ植物の葉において、播種後3週目から9週目までに経時的に測定した。
【0152】
図13を参照すると、PPDKを過剰発現しているシロイヌナズナの葉の中で総遊離アミノ酸含有量が増加することが示されている。データは、野生型およびΔPPDKでは6個の生物学的複製、各SAG12−PPDKgDNA系統では4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDKgDNA系統は、PPDK発現量の昇順で配置されている。全ての5種類のSAG12−PPDKgDNA系統が、野生型よりも高い総アミノ酸含有量を有するが、ばらつきが高く、かつ分散分析を用いて試験した場合、有意差は認められなかった(F=1.314、df=6、p=0.289)。
【0153】
SAG12−PPDKgDNA植物では、葉の窒素含有量が野生型よりも有意に低くなるときと同じでかつSAG12−PPDKgDNA植物の葉の中のPPDK発現量が最大になったときから1週間後である7週目の時点で、総遊離アミノ酸含有量は、野生型植物よりも有意に高かった。全ての5種類の独立したSAG12−PPDKgDNA系統で増加が生じたが、ばらつきは高く、かつ個々の系統と野生型との間に有意差は認められなかった。この増加は、この時点で、アミノ酸の排出が生じ得るよりも大きな割合でアミノ酸の産生が生じるため、アミノ酸は葉の老化中に蓄積することを示唆している。総遊離アミノ酸は、野生型と比較してΔPPDK植物に有意差は認められなかった。
【0154】
輸送アミノ酸(グルタミンおよびアスパラギン)も測定し、総遊離アミノ酸の割合として表した。SAG12−PPDKgDNA植物では、輸送アミノ酸含有量は、7週目および8週目において野生型植物よりも有意に高かった。この増加は、全ての5種類の独立したSAG12−PPDKgDNA系統において生じたが、ばらつきは高く、かつ個々の系統と野生型の差は有意ではなかった。従って、SAG12−PPDKgDNA植物における総遊離アミノ酸含有量の増加に加えて、輸送アミノ酸の含有量は、全体の割合として増加する。これも、グルタミンおよびアスパラギンの産生がこれらの時点において葉の排出量を上回ることを示唆し、老化中のPPDK発現量の増加によってアミノ酸の相互転換効率が上昇し、その結果、輸送アミノ酸が産生されるという仮定を支持している。ΔPPDK植物では、野生型に対する有意差は観察されなかった。
【0155】
実施例7−形質転換したタバコ植物の表現型分析
タバコの葉の窒素含有量の分析
葉の窒素含有量を、陰性の分離体およびSAG12−PPDK−K326タバコの熟した葉について測定した。熟した葉は、タバコの生産のために収穫の準備ができいている葉である。4種類の独立したSAG12−PPDK系統のいくつかでは、SAG12−PPDK植物の葉の窒素含有量は陰性の分離体植物よりも低かった。
【0156】
熟した葉は、タバコの生産のために収穫される段階にあるため、葉の窒素含有量を測定するために使用した。しかし、熟した葉における葉の窒素含有量にほとんど差がなかったという事実は、必ずしも窒素の転流が増加しなかったことを暗示しているわけではない。老化しているSAG12−PPDKgDNAシロイヌナズナ植物の時間的経過において、葉の窒素含有量は野生型よりも有意に低かったが、老化の後半までにその差は非常により小さくなった。従って、タバコの葉の窒素含有量における差は老化の初期に生じ、葉の窒素含有量を測定するときまでに差が減少するという可能性がある。
【0157】
タバコの葉のアミノ酸含有量の分析
分離および暗所での30℃で3日間の培養によって老化が誘発されたK326タバコの成葉についてアミノ酸含有量を測定した。これによって、老化の誘発の前後で同じ葉を比較することができた。葉の分離および暗所での培養による老化の誘発は、重複が最大である年齢に関連する老化による遺伝子発現パターンを示す。
【0158】
K326タバコでは、陰性の分離体植物およびSAG12−PPDK植物における老化の誘発後に総アミノ酸含有量が増加した。この増加は、老化の誘発前の総アミノ酸含有量の割合として計算した。SAG12−PPDK系統C10については、この増加は、陰性の分離体植物よりも有意に小さかったが、他のSAG12−PPDK系統はどれも陰性の分離体植物に対して有意差を示さなかった。従って、老化中のPPDKの過剰発現は、暗所で誘発された老化の誘発後の総アミノ酸含有量にほとんど影響を与えることはないと思われた。老化の誘発後に、K326タバコについて、輸送アミノ酸(グルタミンおよびアスパラギン)含有量も増加したが、この増加は、陰性の分離体およびSAG12−PPDK系統において生じ、老化の誘発後の増加の程度において遺伝子型間に有意差はなかった。
【0159】
タバコの種子の分析
全ての4種類の独立した形質転換系統について、個々の種子の質量をK326タバコについて測定し、陰性の分離体植物と比較して、SAG12−PPDKgDNAおよびSAG12−PPDKcDNA植物の方が有意に高かった。莢あたりの種子の質量も測定し、SAG12−PPDKgDNAおよびSAG12−PPDKcDNA植物の方が有意に高かった。
【0160】
図14を参照すると、種子の大きさがSAG12−PPDK−K326タバコで増加することが示されている。K326ゼロコピー(SAG12−PPDK挿入断片の陰性の分離体である植物)およびSAG12−PPDK植物の個々の種子の質量は、約1000個の種子を撮影、計数および計量して計算した。データは、ゼロコピー植物では10個の生物学的複製、各SAG12−PPDK系統では4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDK系統は、熟した葉におけるPPDK発現量の昇順で配置されている。種子の質量はSAG12−PPDK植物の方が高い。遺伝子型間の差は分散分析を用いて試験した(F=4.870、df=4、p=0.006)。ゼロコピー植物との有意差が認められる系統は、G8(p=0.005)およびC10(p=0.002)であった。
【0161】
種子の窒素含有量の割合は、全ての4種類の独立したSAG12−PPDKgDNAおよびSAG12−PPDKcDNA系統の方が高かったが、有意差は認められなかった。しかし、種子あたりの窒素の質量は、SAG12−PPDKgDNAおよびSAG12−PPDKcDNA植物の方が有意に高かった。
【0162】
図15を参照すると、種子の窒素含有量がSAG12−PPDK−K326タバコで増加することが明らかに示されている。K326ゼロコピーおよびSAG12−PPDK植物の個々の種子中の窒素の質量は、種子の質量および窒素含有量のデータから計算した。データは、ゼロコピー植物では10個の生物学的複製、各SAG12−PPDKでは4個の生物学的複製の平均として示されている。エラーバーは標準誤差である。SAG12−PPDK系統は、熟した葉におけるPPDK発現量の昇順で配置されている。個々の種子中の窒素の質量は、SAG12−PPDK植物の方が高い。遺伝子型間の差は分散分析を用いて試験した(F=7.807、df=4、p=0.001)。ゼロコピー植物との有意差が認められる系統は、G8(p=0.000)およびC10(p=0.000)であった。
【0163】
これは、種子への窒素の供給がSAG12−PPDKgDNAおよびSAG12−PPDKcDNA−K326タバコ植物で増加したことを示唆している。K326植物では、PPDKに対する用量反応が観察された。熟した葉のPPDK発現量がより高い植物は、種子の個々の種子の質量がより大きく、莢あたりの種子の質量がより高く、かつ個々の種子中の窒素の質量が増加していた。PPDK含有量の増加が種子への窒素の供給の増加と関係があるように思われるため、この結果は窒素の転流におけるPPDKの役割を強く支持するものである。
【0164】
要約すると、SAG12−PPDK−K326タバコでは、個々の種子の質量(図14)および種子あたりの窒素の質量(図15)はどちらも増加し、この結果は、PPDKの過剰発現によって窒素の転流が増加したことを示唆していた。従って、シロイヌナズナSAG12−PPDKgDNA植物に関しては、老化している葉の輸送アミノ酸含有量の増加が期待される。ただし、シロイヌナズナでは、自然に老化した葉についてアミノ酸含有量を測定したが、タバコでは、葉の分離および暗所での培養によって老化を誘発した。暗所で誘発された老化と年齢に関連する老化とでは生じるプロセスが異なるため、このタバコの葉で生じているプロセスは、シロイヌナズナの葉におけるプロセスに類似している可能性は低かった。
【0165】
実施例8−PCKおよびPPDKを過剰発現しているシロイヌナズナ植物の世代
実施例1および2に記載されているように、PCKのコード領域およびゲノムクローンを、シロイヌナズナに形質転換されたBNP1380000001バイナリーベクター中の老化関連遺伝子12(SAG12)プロモーターに融合させることによって、老化中にシロイヌナズナPCK(At4g37870.1)の過剰な発現が達成された。
【0166】
最初にAtPCKコード配列をシロイヌナズナcDNAから単離し、PPDKに関して実施例1に記載されているように、PCRを用いてゲノム配列をシロイヌナズナgDNAから単離した。ただし、PCRプライマーは、シロイヌナズナAtPCK遺伝子の開始および終止コドンを含むように設計されており、図3aに示すその後のBNP1380000001ベクターへの遺伝子の連結反応を容易にするために、表1に示すように、順方向プライマー(AtPCK−XbaI−FOR)(配列番号25)内にXbaI制限部位、そして逆方向プライマー(AtPCK−SacI−REV)(配列番号26)内にSacI制限部位も含む。
【0167】
実施例1に従って、各PCR反応に使用されるcDNAおよびgDNA鋳型を調製した。PCR反応混合物は、1×HF緩衝液(NEB社)、2mMの塩化マグネシウム(NEB社)、0.5mMのdNTP(Bioline社)、100ngの鋳型(cDNAまたはgDNA)、0.5μMの各プライマーおよび1単位のPhusion High-FidelityDNAポリメラーゼ(NEB社)を含んでいた。熱循環は、98℃で30秒間の最初の変性工程、次いで、98℃で10秒間、60℃で30秒間の35サイクル、および72℃でコード領域に対して2分30秒間、ゲノムクローンに対して4分30秒間の伸長時間によって、Techneサーマルサイクラーを用いて行った。最後の工程では、72℃で10分間の最終的な伸長を行った。
【0168】
これらのPCR産物によって、コード配列(cDNA)に対する2kbのバンドおよびPCKのゲノム配列に対する3.5kbのバンドが得られ、これらをPEG沈殿させた。実施例1に記載されているPPDK同様に、増幅したDNAを、推奨されている手順に従って、pCR4 Blunt-TOPOベクター(Invitrogen社)に連結させた(平滑末端)。次いで、このプラスミドを、Library Efficiency DH5α大腸菌細胞に形質転換した。カナマイシン(50μgml−1)を選択的な抗生物質として使用した。陽性のコロニーを、コード領域およびゲノムクローンのためのプライマーAtPCK−XbaI−FOR(配列番号25)およびAtPCK−SacI−REV(配列番号26)を用いるコロニーPCRによって選択した。陽性のコロニーを、50μgml−1カナマイシンを含む5mlのLB培地において、37℃の振盪培養器内で一晩増殖させた。
【0169】
プラスミドDNAは、QIAprepスピンミニプレップキット(Qiagen社)を用いて単離し、挿入断片の大きさを連続的な酵素制限消化によって分析した。これは、10単位のXbaIおよび1×XbaI緩衝液によって1μgのDNAを37℃で2時間消化し、次いで10単位SacIおよび1×SacI緩衝液を37℃で一晩添加することによって行った。正しい挿入断片を含むプラスミドDNAは、AtPCK−XbaI−FOR/AtPCK−SacI−REVを用いて配列決定した。配列はBioEditを用いて分析し、増幅したAtPCKコード配列およびゲノム配列はBLASTXを用いて確認した。
【0170】
AtPCKコード領域およびゲノムクローンを図3aに示すpBNPベクターに連結させるために、2種類のプラスミドをそれぞれ表す大腸菌コロニーを使用して、50μgml−1のカナマイシンを含む25mlのLB培地に植菌した。培養物は37℃で一晩振盪させ続け、プラスミドDNAは、QIAfilterプラスミドミディキット(QIAfilter plasmid midi kit)(Qiagen社)を用いて精製した。pBNPベクターは、同一の様式だが、50μgml−1のカナマイシンを含む100mlの培養物から精製した。上述したものと同じ連続的な酵素消化を、全ての精製したプラスミドDNAに対して行った。コード領域およびゲノムクローンを含むプラスミドのために3μgのDNAを消化し、pBNPベクターのために1μgのDNAを消化した。試料はクリスタルバイオレットゲル電気泳動によって分離し、その産物はQiaquickゲル抽出キット(Qiagen社)を用いて精製した。14.4kbのpBNPベクター産物をアルカリホスファターゼで処理して自己連結反応を防止し、コード領域およびゲノムクローン挿入断片は、XbaI/SacI消化を用いてpBNPベクターに連結させた。Library efficiency DH5α大腸菌細胞は2μlの連結反応物で形質転換し、陽性のコロニーを、AtPCK−XbaI−FORおよびAtPCK−SacI−REVを用いるPCRによってスクリーニングした。
【0171】
プラスミドDNAを、QIAprepスピンミニプレップキット(Qiagen社)を用いて、所望の挿入断片を含むコロニーから抽出し、続いて、DNAを10単位のXbaI、15単位のSacIおよび1×XbaI緩衝液によって37℃で2時間消化した。期待される正しい制限酵素消化パターンを示したpBNPベクター中のコード配列およびにゲノム配列挿入断片それぞれの2つの別々のコロニーをBNP−SAG12FWDプライマー(配列番号9)を用いて選択および配列決定した。この配列はBioEditプログラムを用いて分析した。図3bに示すように、作製した構築物はpALBNP1(コード配列)およびpALBNP2(ゲノム配列)と命名した。
【0172】
5種類のホモ接合性単一挿入断片系統を作製し、PCK特異的抗体を用いて、実施例3に記載されているような免疫ブロットによってPCKの過剰発現を確認した。ポリクローナル抗血清は、PCKアミノ酸配列に対して設計された合成ペプチドを用いてウサギで産生した。この配列は、(i)DEHCWTETGVSNIEG(配列番号27)および(ii)CVDLSREKEPDIWNA(配列番号28)であり、それらは、化学的に合成した後、ウサギに混注される前に、キーホールリンペットヘモシアニンに連結したものであった。抗体を用いて得られた結果を図16に示す。図16は、形質転換した植物中のPCKタンパク質濃度が上昇したことを示している。
【0173】
SAG12−PCK−PPDK細胞株は、単一のSAG12−PCK形質転換体(3)1および(19)4を、強いSAG12−PPDK過剰発現系統と交差させることによって作製した。図17に示すように、PCKおよびPPDKの両方の濃度が上昇した植物の分析によって、ロゼットの質量が増加していることが分かった。
【0174】
最後に、本発明者らは仮定に縛られることを望んでいないが、図18は、PCKおよびPPDKがどのように窒素転流に影響を与え得るかについて示す推測上の生化学的経路を示す。
【0175】
PCK/PPDK二重構築物
本発明者らは、図3に示すPCKおよびPPDK単一構築物が、植物に形質転換されると、これらの酵素が老化した葉で過剰発現した場合に、老化した葉からの窒素の転流率の上昇、さらに栄養植物成長量の増加(作物収穫率の上昇に該当する)を引き起こすことを観察した。従って、本発明者らは、PCKおよびPPDKをコードする遺伝子がどちらもSAG12プロモーターの制御下でpBNP130000001に挿入されている2種類の二重構築物を作製することに決めた。
【0176】
第1の二重構築物は、XbaI/SacI消化を用いて、BNP−PPDKgDNAのPPDKをコードするgDNA断片の下流(すなわち3'末端)にgDNAをコードするPCKを連結させることによって作製した。従って、プラスミド内のSAG12プロモーターは、PPDK遺伝子およびPCK遺伝子の両方の発現に関与した。
【0177】
第2の二重構築物は、AvrII/BamHI消化を用いて、pALBNP2のPCKをコードするgDNA断片のすぐ下流にPPDKをコードするgDNAを連結させることによって作製した。この場合も、SAG12はPPDK遺伝子およびPCK遺伝子の両方の発現に関与した。
【特許請求の範囲】
【請求項1】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性および/またはピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする、少なくとも1つのコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物。
【請求項2】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性およびピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする、少なくとも1つのコード配列に作動可能に連結されたプロモーターを含む遺伝子構築物。
【請求項3】
前記プロモーターが老化特異的プロモーターである、請求項2に記載の遺伝子構築物。
【請求項4】
前記プロモーターがシロイヌナズナ中の老化関連遺伝子から単離されたものである、請求項1乃至3いずれか1項に記載の遺伝子構築物。
【請求項5】
前記プロモーターが、SAG12、SAG13、SAG101、SAG21およびSAG18またはその機能的な変異体もしくは断片からなる群から選択される、請求項1乃至4いずれか1項に記載の遺伝子構築物。
【請求項6】
前記プロモーターがSAG12プロモーターまたはその機能的な変異体もしくは断片である、請求項1乃至5いずれか1項に記載の遺伝子構築物。
【請求項7】
前記プロモーターが、実質的に配列番号16に示されているヌクレオチド配列またはその機能的な変異体もしくは断片を含む、請求項1乃至6いずれか1項に記載の遺伝子構築物。
【請求項8】
前記少なくとも1つのコード配列が、(i)ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片および/または(ii)ピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片をコードする、請求項1に記載の遺伝子構築物。
【請求項9】
PCK活性を有する前記ポリペプチドをコードする前記コード配列がシロイヌナズナに由来する、請求項1乃至8いずれか1項に記載の遺伝子構築物。
【請求項10】
PPDK活性を有する前記ポリペプチドをコードする前記コード配列が、シロイヌナズナ属、トウモロコシ属、フラベリア属またはクレオメ属に由来する、請求項1乃至9いずれか1項に記載の遺伝子構築物。
【請求項11】
PCK活性を有するポリペチドをコードする前記コード配列が、実質的に配列番号17または配列番号18に示されている核酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至10いずれか1項に記載の遺伝子構築物。
【請求項12】
PCK活性を有する前記ポリペチドが、実質的に配列番号19に示されているアミノ酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至11いずれか1項に記載の遺伝子構築物。
【請求項13】
PPDK活性を有するポリペチドをコードする前記コード配列が、実質的に配列番号20、配列番号21または配列番号22に示されている核酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至12いずれか1項に記載の遺伝子構築物。
【請求項14】
PPDK活性を有する前記ポリペチドが、実質的に配列番号23または配列番号24に示されているアミノ酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至13いずれか1項に記載の遺伝子構築物。
【請求項15】
前記構築物がPCKまたはその機能的な変異体もしくは断片をコードしかつPPDKをコードしない、請求項1に記載の遺伝子構築物。
【請求項16】
前記構築物がPPDKまたはその機能的な変異体もしくは断片をコードし、かつPCKをコードしない、請求項1に記載の遺伝子構築物。
【請求項17】
前記構築物がPCKまたはその機能的な変異体もしくは断片およびPPDKまたはその機能的な変異体もしくは断片をコードする、請求項1乃至14いずれか1項に記載の遺伝子構築物。
【請求項18】
請求項1乃至17いずれか1項に記載の遺伝子構築物を含む組み換え型ベクター。
【請求項19】
実質的に図3に示す組み換え型ベクター。
【請求項20】
同じ条件で培養された野生型植物中のPCKおよび/またはPPDKの対応する濃度よりも試験植物の葉の中のPCKおよび/またはPPDKの濃度を上げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために前記試験植物における植物の代謝を変えることを含む方法。
【請求項21】
同じ条件で培養された野生型植物中の対応する窒素濃度よりも試験植物の葉の中の窒素濃度を下げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために前記試験植物における植物の代謝を変えることを含む方法。
【請求項22】
同じ条件で培養された野生型植物の対応する成長率と比較して試験植物の成長率を上げる方法であって、葉の老化の開始後の植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために前記試験植物における植物の代謝を変えることを含む方法。
【請求項23】
請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターによって試験植物の細胞を形質転換することを含む、請求項20乃至22いずれか1項に記載の方法。
【請求項24】
同じ条件で培養された前記野生型植物中のPCKおよびPPDKの対応する濃度よりも前記植物の葉の中のPCKおよびPPDKの濃度を上げることを含む、請求項20乃至23いずれか1項に記載の方法。
【請求項25】
請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載の組み換え型ベクターを含む細胞。
【請求項26】
請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載の組み換え型ベクターを含む形質転換植物。
【請求項27】
前記植物はアブラナ科に由来する、請求項26に記載の形質転換植物。
【請求項28】
前記植物はイネ科に由来する、請求項26に記載の形質転換植物。
【請求項29】
前記植物はナス科に由来する、請求項26に記載の形質転換植物。
【請求項30】
請求項26乃至29いずれか1項に記載の形質転換植物から入手可能な植物繁用製品。
【請求項31】
前記植物繁殖用製品が種子である、請求項30に記載の植物繁殖用製品。
【請求項32】
同じ条件で培養された対応する野生型植物よりも高い割合で窒素を転流させる形質転換植物を生産する方法であって、
i)請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターによって植物細胞を形質転換する工程と、
ii)前記形質転換した細胞から植物を再生させる工程と、
を含む方法。
【請求項33】
同じ条件で培養された対応する野生型植物よりも高い成長率を有する形質転換植物を生産する方法であって、
i)請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターによって植物細胞を形質転換する工程と、
ii)前記形質転換した細胞から植物を再生させる工程と、
を含む方法。
【請求項34】
同じ条件で栽培された野生型植物から採取した、収穫された葉の中の対応する窒素濃度よりも低濃度の窒素を含有する収穫された葉であって、前記葉が、請求項26乃至29いずれか1項に記載の形質転換植物から収穫されたものまたは請求項32または33に記載の方法によって生産されたものである収穫された葉。
【請求項35】
変異型タバコ植物から得られた、窒素が減少したタバコを含む喫煙物品であって、前記変異体が老化した葉の中の窒素濃度を下げることができる喫煙物品。
【請求項36】
前記喫煙物品が、請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターをによって形質転換されている変異型タバコ植物から得られたタバコから製造されている、請求項35に記載の喫煙物品。
【請求項1】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性および/またはピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする、少なくとも1つのコード配列に作動可能に連結された老化特異的プロモーターを含む遺伝子構築物。
【請求項2】
ホスホエノールピルビン酸カルボキシキナーゼ(PCK)活性およびピルビン酸正リン酸ジキナーゼ(PPDK)活性を有する少なくとも1種のポリペプチドをコードする、少なくとも1つのコード配列に作動可能に連結されたプロモーターを含む遺伝子構築物。
【請求項3】
前記プロモーターが老化特異的プロモーターである、請求項2に記載の遺伝子構築物。
【請求項4】
前記プロモーターがシロイヌナズナ中の老化関連遺伝子から単離されたものである、請求項1乃至3いずれか1項に記載の遺伝子構築物。
【請求項5】
前記プロモーターが、SAG12、SAG13、SAG101、SAG21およびSAG18またはその機能的な変異体もしくは断片からなる群から選択される、請求項1乃至4いずれか1項に記載の遺伝子構築物。
【請求項6】
前記プロモーターがSAG12プロモーターまたはその機能的な変異体もしくは断片である、請求項1乃至5いずれか1項に記載の遺伝子構築物。
【請求項7】
前記プロモーターが、実質的に配列番号16に示されているヌクレオチド配列またはその機能的な変異体もしくは断片を含む、請求項1乃至6いずれか1項に記載の遺伝子構築物。
【請求項8】
前記少なくとも1つのコード配列が、(i)ホスホエノールピルビン酸カルボキシキナーゼ(PCK)またはその機能的な変異体もしくは断片および/または(ii)ピルビン酸正リン酸ジキナーゼ(PPDK)またはその機能的な変異体もしくは断片をコードする、請求項1に記載の遺伝子構築物。
【請求項9】
PCK活性を有する前記ポリペプチドをコードする前記コード配列がシロイヌナズナに由来する、請求項1乃至8いずれか1項に記載の遺伝子構築物。
【請求項10】
PPDK活性を有する前記ポリペプチドをコードする前記コード配列が、シロイヌナズナ属、トウモロコシ属、フラベリア属またはクレオメ属に由来する、請求項1乃至9いずれか1項に記載の遺伝子構築物。
【請求項11】
PCK活性を有するポリペチドをコードする前記コード配列が、実質的に配列番号17または配列番号18に示されている核酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至10いずれか1項に記載の遺伝子構築物。
【請求項12】
PCK活性を有する前記ポリペチドが、実質的に配列番号19に示されているアミノ酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至11いずれか1項に記載の遺伝子構築物。
【請求項13】
PPDK活性を有するポリペチドをコードする前記コード配列が、実質的に配列番号20、配列番号21または配列番号22に示されている核酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至12いずれか1項に記載の遺伝子構築物。
【請求項14】
PPDK活性を有する前記ポリペチドが、実質的に配列番号23または配列番号24に示されているアミノ酸配列またはその機能的な変異体もしくは断片を含む、請求項1乃至13いずれか1項に記載の遺伝子構築物。
【請求項15】
前記構築物がPCKまたはその機能的な変異体もしくは断片をコードしかつPPDKをコードしない、請求項1に記載の遺伝子構築物。
【請求項16】
前記構築物がPPDKまたはその機能的な変異体もしくは断片をコードし、かつPCKをコードしない、請求項1に記載の遺伝子構築物。
【請求項17】
前記構築物がPCKまたはその機能的な変異体もしくは断片およびPPDKまたはその機能的な変異体もしくは断片をコードする、請求項1乃至14いずれか1項に記載の遺伝子構築物。
【請求項18】
請求項1乃至17いずれか1項に記載の遺伝子構築物を含む組み換え型ベクター。
【請求項19】
実質的に図3に示す組み換え型ベクター。
【請求項20】
同じ条件で培養された野生型植物中のPCKおよび/またはPPDKの対応する濃度よりも試験植物の葉の中のPCKおよび/またはPPDKの濃度を上げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために前記試験植物における植物の代謝を変えることを含む方法。
【請求項21】
同じ条件で培養された野生型植物中の対応する窒素濃度よりも試験植物の葉の中の窒素濃度を下げる方法であって、葉の老化の開始後に植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために前記試験植物における植物の代謝を変えることを含む方法。
【請求項22】
同じ条件で培養された野生型植物の対応する成長率と比較して試験植物の成長率を上げる方法であって、葉の老化の開始後の植物の葉の中のPCKおよび/またはPPDKの濃度の上昇を達成するために前記試験植物における植物の代謝を変えることを含む方法。
【請求項23】
請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターによって試験植物の細胞を形質転換することを含む、請求項20乃至22いずれか1項に記載の方法。
【請求項24】
同じ条件で培養された前記野生型植物中のPCKおよびPPDKの対応する濃度よりも前記植物の葉の中のPCKおよびPPDKの濃度を上げることを含む、請求項20乃至23いずれか1項に記載の方法。
【請求項25】
請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載の組み換え型ベクターを含む細胞。
【請求項26】
請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載の組み換え型ベクターを含む形質転換植物。
【請求項27】
前記植物はアブラナ科に由来する、請求項26に記載の形質転換植物。
【請求項28】
前記植物はイネ科に由来する、請求項26に記載の形質転換植物。
【請求項29】
前記植物はナス科に由来する、請求項26に記載の形質転換植物。
【請求項30】
請求項26乃至29いずれか1項に記載の形質転換植物から入手可能な植物繁用製品。
【請求項31】
前記植物繁殖用製品が種子である、請求項30に記載の植物繁殖用製品。
【請求項32】
同じ条件で培養された対応する野生型植物よりも高い割合で窒素を転流させる形質転換植物を生産する方法であって、
i)請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターによって植物細胞を形質転換する工程と、
ii)前記形質転換した細胞から植物を再生させる工程と、
を含む方法。
【請求項33】
同じ条件で培養された対応する野生型植物よりも高い成長率を有する形質転換植物を生産する方法であって、
i)請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターによって植物細胞を形質転換する工程と、
ii)前記形質転換した細胞から植物を再生させる工程と、
を含む方法。
【請求項34】
同じ条件で栽培された野生型植物から採取した、収穫された葉の中の対応する窒素濃度よりも低濃度の窒素を含有する収穫された葉であって、前記葉が、請求項26乃至29いずれか1項に記載の形質転換植物から収穫されたものまたは請求項32または33に記載の方法によって生産されたものである収穫された葉。
【請求項35】
変異型タバコ植物から得られた、窒素が減少したタバコを含む喫煙物品であって、前記変異体が老化した葉の中の窒素濃度を下げることができる喫煙物品。
【請求項36】
前記喫煙物品が、請求項1乃至17いずれか1項に記載の遺伝子構築物または請求項18または19に記載のベクターをによって形質転換されている変異型タバコ植物から得られたタバコから製造されている、請求項35に記載の喫煙物品。
【図1】


【図2a】

【図3a】
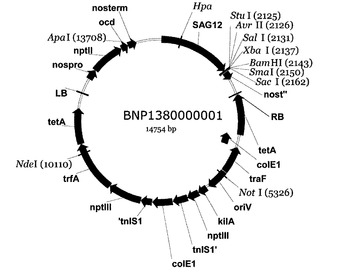
【図3b】
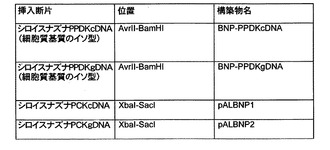
【図4】
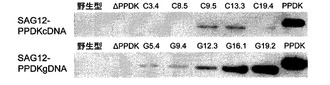
【図5】
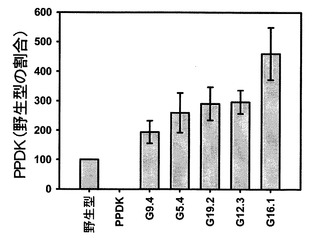
【図6】
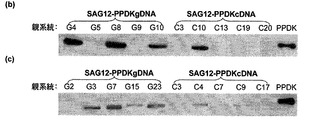
【図7】
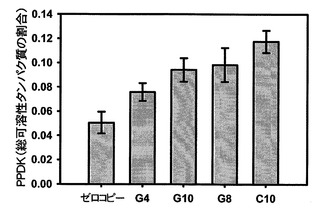
【図8】

【図9】
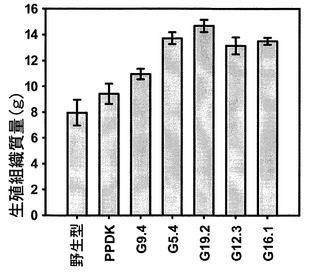
【図10】
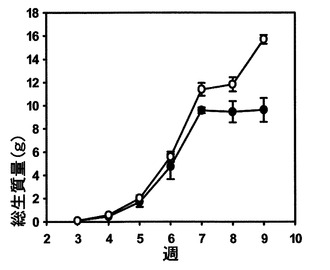
【図11】
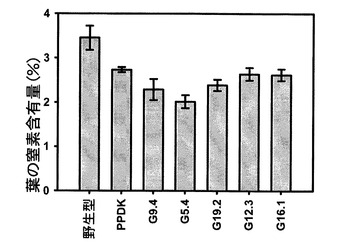
【図12】

【図13】
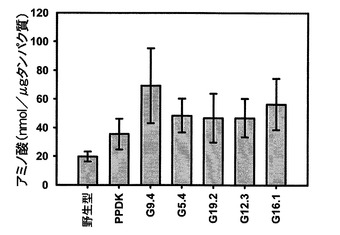
【図14】
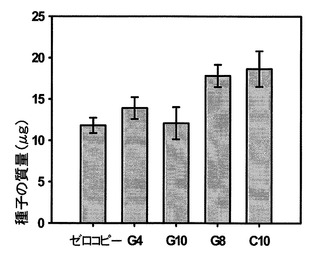
【図15】

【図16】
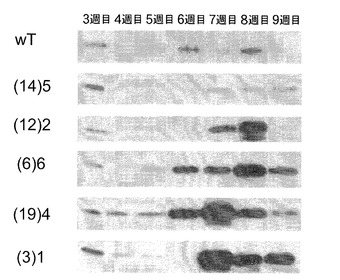
【図17】
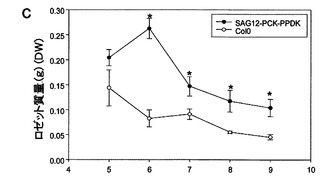
【図18】

【図2b】

【図2a】
【図3a】
【図3b】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図2b】
【公表番号】特表2012−518995(P2012−518995A)
【公表日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2011−551528(P2011−551528)
【出願日】平成22年2月25日(2010.2.25)
【国際出願番号】PCT/GB2010/050321
【国際公開番号】WO2010/097623
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
【公表日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成22年2月25日(2010.2.25)
【国際出願番号】PCT/GB2010/050321
【国際公開番号】WO2010/097623
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(301004857)アドヴァンスト・テクノロジーズ(ケンブリッジ)リミテッド (8)
【Fターム(参考)】
[ Back to top ]