
ボルデテラ属菌産生壊死毒素のトキソイドおよび該トキソイドを含む混合ワクチン
【課題】壊死毒素の調製方法、並びにボルデテラ属菌が産生する壊死毒素の改良されたトキソイド及びそのパスツレラ属菌が産生する壊死毒素のトキソイドとの混合トキソイドを提供する。
【解決手段】ボルデテラ・ブロンキセプティカ(気管支敗血症菌)が産生する壊死毒素の含有液を、直接的硫酸化により硫酸基が導入されたクロマト担体ゲルにヘパリン、スルフォプロピル基もしくは分子内にスルフォン基を有するブルー色素が結合されたゲルに接触せしめ、当該壊死毒素を吸着させた後、吸着した壊死毒素をゲルから溶出することによって得られる部分精製壊死毒素、あるいは当該部分精製壊死毒素を他の精製方法と組み合わせてさらに高純度に精製したボルデテラ・ブロンキセプティカ壊死毒素のトキソイド、およびパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする混合ワクチン。
【解決手段】ボルデテラ・ブロンキセプティカ(気管支敗血症菌)が産生する壊死毒素の含有液を、直接的硫酸化により硫酸基が導入されたクロマト担体ゲルにヘパリン、スルフォプロピル基もしくは分子内にスルフォン基を有するブルー色素が結合されたゲルに接触せしめ、当該壊死毒素を吸着させた後、吸着した壊死毒素をゲルから溶出することによって得られる部分精製壊死毒素、あるいは当該部分精製壊死毒素を他の精製方法と組み合わせてさらに高純度に精製したボルデテラ・ブロンキセプティカ壊死毒素のトキソイド、およびパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする混合ワクチン。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、豚の伝染性疾患に対する予防薬に関する。さらに詳細には、改良されたボルデテラ属菌が産生する壊死毒素のトキソイドおよびそのパスツレラ属菌が産生する壊死毒素のトキソイドとの混合トキソイド並びに該トキソイドの調製方法に関する。
【背景技術】
【0002】
萎縮性鼻炎(以下、ARと略称することがある)は豚の慢性呼吸器病で、鼻甲介萎縮、発育遅延、鼻出血、鼻曲がり及び他の呼吸器病の誘発等を主徴とする伝播力の極めて強い疾病である。本病の起因菌としては、毒素原性気管支敗血症菌(以下、Bbと略称することがある)及び毒素原性パスツレラ・マルトシーダ(以下、Pmと略称することがある)が認められており、上部気道粘膜に感染する。Bbは定着因子である赤血球凝集素や線毛を保有しているため鼻粘膜に対する定着性が強いが、それを保有しないPmは単独で定着することが困難である。そのため、Pmの感染が成立するためには予め鼻粘膜を障害する因子の存在が必要である。その最も代表的な因子がBbであり、鼻粘膜障害に関与する病原性因子として、気管細胞毒素や壊死毒素が知られている。実験感染では、子ブタに対しBb、続いて数日後にPmを鼻内感染させることにより野外農場で観察されるような重篤なARを再現することができる。
【0003】
ボルデテラ属菌には、百日咳菌、パラ百日咳菌、気管支敗血症菌、ボルデテラ・アビウム等があり、これらは種々の生物学的活性物質を産生する。Bb壊死毒素(以下、BbTと略称することがある)はこれらの生物学的活性物質の1つであり、いずれのボルデテラ属菌もBbTを産生する。BbTは出血壊死作用、鼻甲介萎縮作用、発育遅延作用、致死作用、血管平滑筋収縮作用、局所の血行障害、脾臓萎縮作用および抗体産生阻害作用などの生物学的活性を有し、ボルデテラ属菌による発症において重要な役割を演じていることが明らかになった。さらに、BbTはBbによる子豚の肺炎病巣形成にも重要な役割を演じていることが明らかにされた(非特許文献1)。
【0004】
一方、パスツレラ・マルトシーダの一部あるいは非定型的パスツレラの一部の菌株は、Pm壊死毒素(以下、PmTと略称することがある)を産生する。PmTは出血壊死作用、鼻甲介萎縮作用、発育遅延作用、致死作用、局所の血行障害および脾臓萎縮作用などの生物学的活性を有し、毒素原性Pmによる発症において重要な役割を演じていることが明らかにされている。また、PmTは毒素原性Pmによる豚の肺炎病巣形成において密接に関係していることが明らかになった(非特許文献2)。BbTおよびPmTはいずれも培地中に産生されず菌体内に蓄積されるが、長期間培養による溶菌に伴って菌体内から培養上清に遊離する。生体内においては溶菌に伴って菌体内から遊離した壊死毒素が作用する。
【0005】
従来知られている細菌由来の壊死毒素含有液、とりわけボルデテラ菌菌体抽出液からの壊死毒素の部分精製方法としてはイオン交換クロマトグラフィーによる方法がある(非特許文献3)。しかしながら、この方法は再現性に乏しい欠点がある。また、同様に壊死毒素の部分精製例として、透析、ショ糖密度勾配超遠心分離及びゲルろ過クロマトグラフィーに基づく方法がある(非特許文献4)が、この方法は再現性に乏しく、さらに、工程が多いにもかかわらず精製度があまり高くないという欠点がある。
【0006】
他の例として、核酸除去剤処理及び陰イオン交換クロマトグラフィー及びハイドロキシアパタイト吸着クロマトグラフィーによる方法がある(非特許文献5)。しかしながら、この方法は核酸除去剤のような高価な試薬を必要とし、工程が多い欠点がある。また、核酸除去剤、透析及び陽イオン交換クロマトグラフィーによる方法もある(非特許文献6)が、この方法は再現性が高く比較的高純度の壊死毒素が得られるものの、前記方法と同様、高価な試薬を必要とする欠点がある。
【0007】
さらに別の例として、塩析及び色素リガンドアフィニティークロマトグラフィーによる方法がある(非特許文献7)が、この方法は再現性は高いものの精製度があまり高くないという欠点がある。
【0008】
従来報告されたAR不活化ワクチンにはBb単味、Pm単味並びにBb、Pm混合不活化ワクチンの3種類があり研究あるいは実用化されている。
【0009】
Bb不活化ワクチンとしては、ホルマリンやグルタールアルデヒド等で不活化された死菌にアルミニウムゲルや油性アジュバントを加えたものがある(非特許文献8、非特許文献9および非特許文献10)。また、Bbの外膜に存在しアデニールサイクラーゼ活性を有する68kDaの蛋白に萎縮性鼻炎の防御効果のあることが明らかにされている(非特許文献11)。さらに、Bb菌の産生する線維状赤血球凝集素を主成分とするコンポーネントワクチンが存在する(非特許文献12および非特許文献13)。抗毒素を与える目的で壊死毒素を加えたワクチンは効果を得ていないという報告もある(非特許文献14および非特許文献15)。
【0010】
一方、Pmの不活化単味ワクチンとしては、PmTトキソイド(非特許文献16および非特許文献17)が研究あるいは実用化されている。
【0011】
さらに、Bb及びPmの混合不活化ワクチンとしては、(1)ARの予防のみを目的とするためBbとPmD型のPmT産生株の死菌を混合したもの、これらにPmT非産生株を加えたもの、あるいはBbとPmTトキソイドを混合したもの(非特許文献18、非特許文献19、非特許文献20および非特許文献21)、(2)ARと肺炎予防を目的とするため、BbとPmD型のPmT産生株、PmA型の死菌を混合したもの(非特許文献22)、(3)ARと肺炎に加えて他の疾病の予防も目的とするためBb、PmA型株、豚丹毒菌の死菌にPmTトキソイドを加えたものが実用化されている(非特許文献23)。
【0012】
ところで、Roop II,R.M.らはBbTがBb感染による豚の鼻甲介萎縮や肺炎病巣形成に重要な役割を演じていることから、BbTトキソイドの必要性を指摘している(非特許文献1)。しかし、BbTとPmTトキソイドとの混合化に関しては全く言及していない。また、Chanter,N.らはBbT及びPmTの共同作用による豚鼻粘膜障害がBb及びPmの増殖に好ましい環境を形成することから、Bbの重要な成分を含有するPmTトキソイドがより効果的であることを提案している(非特許文献24)。
【0013】
しかし、これまでのところ、単にBbT活性を有する画分からなるワクチンではBbT中和抗体を上昇させることに成功していない(非特許文献14、非特許文献15)。
【0014】
一方で、中井らは高度に精製したBbTトキソイドを用いてブタ及びマウスに中和抗体の上昇することを明らかにした(非特許文献25)。しかし、彼らも当該BbTトキソイドとPmTトキソイドとの混合化に関しては全く触れていない。
【非特許文献1】Roop II,R.M.ら、Infection and Immunity,55,p.217〜222(1987)
【非特許文献2】Iwamatsu, S.およびSawada, T.、Japanese Journal of Veterinary Science, 50, p.1200〜1206(1988)
【非特許文献3】Kume,K.ら,Infection and Immunity, 52(4), p.370〜377(1986)
【非特許文献4】Endoh,M.ら,Microbiology and Immunology, 30,(7), p.659〜673(1986)
【非特許文献5】Horiguchi,Y.ら、Microbial Pathogenesis, 6, p.361〜368(1989)
【非特許文献6】Horiguchi,Y.ら、FEMS Microbiology Letters, 66, p.39〜44(1990)
【非特許文献7】Zhang Y. L.およびSekura R. D.、Infection and Immunity, 59(10), ,p.3754〜3759(1991)
【非特許文献8】Goodnow, R. A.、Veterinary Medicine Small Animal Clinician, 72, p.1210〜1212(1977)
【非特許文献9】Nakase,Y.ら、Proceedings of International Pig Veterinary Society Congress, 8,(1976)
【非特許文献10】Giles, C. J.およびSmith, I. M.、Veterinary Bulletin (London), 53, p.327〜338(1983)
【非特許文献11】Montaraz J. A.ら、Infection and Immunity, 47, p.744〜755(1985)
【非特許文献12】Hansen, G. R.ら、In Atrophic rhinitis in pigs (Peterson, K. B.およびNielsen, N.C.編)Communication of the European Communities,Luxembourg, p.89〜97(1983)
【非特許文献13】Ohgitani, T.ら、Vaccine, 9, p.653〜658(1991)
【非特許文献14】Soderlind, O.およびBergstrom, G.,Proceedings of International Pig Veterinary Science Congress, p.175(1984)
【非特許文献15】Marel, C. M. von der.ら、Proceedings of International Pig Veterinary Science Congress, p.170(1984)
【非特許文献16】Pedersen, K. B.およびBarfod, K.、Nordish Veterina Medicine, 31, p.293〜302(1982)
【非特許文献17】Foged, N. T.ら、ヴェテリナリー レコード:Veterinary Record, 125, p.7〜11(1989)
【非特許文献18】Barfod, K.およびPedersen,K. B.、Nordish Veterina Medicine, 36, p.337〜345(1984)
【非特許文献19】Baalsrud, K. J.、Acta Veterina Scandinavia., 28, p.305〜311(1987)
【非特許文献20】de Jong, M. F.ら、Veterinary Quarterly, 9, p.49〜59(1987)
【非特許文献21】Kobisch, M.およびPennings, A.、Veterinary Record, 124, p.57〜61(1989)
【非特許文献22】Ostle, A. G.ら、Veterinary Medicine, p.772〜775(1986)
【非特許文献23】Jayappa, H.ら、International Pig Veterinary Society Congress, p.44(1988)
【非特許文献24】Chanter, N.ら、Research Veterinary Science, 47, p.48〜53(1989)
【非特許文献25】第111回日本獣医学会講演要旨集,p.183(1991)
【発明の開示】
【発明が解決しようとする課題】
【0015】
上述の状況下、壊死毒素の生体防御における役割についてはなお不明な点があり、これらを解明していく上での壊死毒素を大量に調製する方法の開発、ならびに疾病予防のための壊死毒素のトキソイドの開発が望まれている。しかし、壊死毒素は熱に対し不安定で精製が進むにつれて自己凝集しやすくなったり、脂質、酸性蛋白質および疎水性物質等の菌体由来物質と複合体を形成するなどの性質のため精製が困難といった問題点があった。
また、前記BbTとPmTは生物学的活性は類似しているものの、免疫学的に交差しないことが明らかにされており、このようなことから、獣医療上、養豚産業上安全かつ有効なBbT・PmT混合トキソイドの開発が切望されている。
【課題を解決するための手段】
【0016】
本発明者らは、壊死毒素の効率的な精製方法を見い出すべく種々検討を重ねた結果、(1)壊死毒素含有溶液例えばボルデテラ属菌菌体抽出物を、セルロース硫酸エステルゲル、ヘパリン固定化吸着ゲルもしくは硫酸化アルキル基固定化吸着ゲル等で例示される直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いてクロマトグラフィーを行なうことにより内毒素含有量の少ない部分精製壊死毒素が簡単にしかも比較的高い回収率で得られること、(2)上記直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いて部分精製した壊死毒素をさらに透析、陽イオン交換クロマトグラフィー及びゲルろ過クロマトグラフィーを実施することにより高純度の壊死毒素が簡単に、しかも比較的高回収率で得られること、(3)直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いて部分精製した壊死毒素が未精製のボルデテラ属菌菌体抽出物中壊死毒素に比べ免疫応答が大幅に改善されることの知見を得た。
【0017】
さらに、本発明者らは、ボルデテラ属菌菌体抽出物を直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルに接触せしめ、壊死毒素を吸着させ、夾雑物質と分離し、ゲルから溶出することにより得られた壊死毒素画分を化学的に減毒したものが豚において安全かつ萎縮性鼻炎の予防に有効であることを確認し本発明を完成するに至った。
【0018】
豚に対してPmの感染が成立するためには予め豚鼻粘膜を障害する因子としてのBbの前感染が必要である。BbT抗毒素による免疫をブタに行なった場合、Bb及びPmの攻撃によりPmの鼻粘膜定着並びに鼻甲介萎縮を軽減できる。しかし、個体間で効果に大きな差が認められることから、AR不活化ワクチンの成分としてBbTトキソイドのみでは不十分であることが判明した。
【0019】
本発明者らは、上述の知見を基に混合トキソイドを開発すべく鋭意検討を重ねた結果、ボルデテラ属菌菌体抽出物をセルロース硫酸エステルゲル等を用いて比活性37,000MND/mg蛋白以上に精製した毒素に由来するトキソイドと毒素原性Pm産生PmTに由来するトキソイドを混合することにより得られる混合トキソイドが、豚において安全かつ萎縮性鼻炎の予防に有効であることを確認し、本発明の第二の態様である混合トキソイドを供するに至った。
【0020】
なお、本発明において使用される最少壊死毒単位(MND)とは、リポポリサッカライド(大腸菌、血清型0111:B4由来など)1μg/ml存在下でモルモット皮内に注射され1日後に直径5mm以上の壊死斑を形成する最少毒素量を1MNDと定義される量に基づくものとする。
【発明を実施するための最良の形態】
【0021】
本発明は、壊死毒素含有液をセルロース硫酸エステル等の直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルに接触せしめ、壊死毒素を吸着させた後、ゲルから溶出することを特徴とする壊死毒素の部分精製方法を提供する。また、前記壊死毒素の部分精製方法によって得られたBbTに由来するBbTのトキソイド、並びに該BbTトキソイドと毒素原性Pm産生PmTに由来するトキソイドを混合することにより得られる、豚において安全かつ萎縮性鼻炎の予防に有効な混合トキソイドを提供するものである。
【0022】
本発明において、出発材料である壊死毒素含有液とは、Bb、百日咳菌、パラ百日咳菌及びボルデテラ アビウムの菌体抽出液を含む。または、遺伝子組換え技術によりBbTを発現させた細胞抽出液も本発明の出発材料として含まれる。
【0023】
本発明において好ましい抽出液は、Bbの菌体抽出液であり、Bbをボルデ・ジャング培地、マッコンキー培地などの寒天培地あるいはペプトン培地(Horiguchi, Y.ら,マイクロバイアル パソジェネーシス:Microb. Pathogen., 6,p.361〜368(1989))、コーエン・ウィラー培地、ステナー・ショルテ培地などの液体培地にて、常法により静置培養または振盪培養もしくは通気撹拌培養する。寒天培養菌は、コーンラージ棒等でセルロース硫酸エステルゲルの開始緩衝液に浮遊する。液体培養菌は、遠心分離により菌体を回収し、セルロース硫酸エステルゲルの開始緩衝液に浮遊する。
【0024】
この菌体浮遊液について、Kume, K.らの方法(インフェクション アンド イムニティー:Infect. Immun. 52(4), p.370〜377(1986))、Endoh, M.らの方法(マイクロバイオロジー アンド イムノロジー:Microbial. Immunol., 30(7), p.659〜673(1986))、Horiguchi, Y.らの方法(マイクロバイアル パソジェネーシス:Microbial Pathogen., 6, p.361〜368(1989)あるいはFEMSマイクロバイオロジー レターズ:FEMS Microbiol. Letters, 66, p.39〜44(1990))、Zhang Y. L.およびSekura R. D.の方法(インフェクション アンド イムニティー:Infect. Immun., 59(10), p.3754〜3759(1991))によって、またはセルロース硫酸エステルゲル等の直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いた吸着クロマトグラフィーにより精製を行なうことができるが、本発明においては、直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いた吸着クロマトグラフィーによる精製を実施することを最適な態様とする。
【0025】
この菌体浮遊液を超音波処理、酵素処理、プレス処理、凍結融解の繰り返し等を行なうことにより菌体を破壊し、遠心分離あるいは膜ろ過により菌体残渣を除去する。この菌体抽出物の形であるいは核酸除去剤、塩析、超遠心分離等の前段部分精製処理後に本発明の方法に供される。本発明の方法によれば、前段部分精製処理をあえて行なう必要はなく、菌体抽出液をそのまま硫酸基を有する分子をリガンドとする吸着ゲルあるいは直接硫酸化された吸着ゲルを用いた吸着クロマトグラフィーに付すことができ、工程がきわめて簡便となる。
【0026】
本発明で用いられる直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルについては以下のものが例示される。
【0027】
直接的硫酸化により硫酸基が導入されたクロマト担体ゲルとは、セルロースなどのクロマト担体ゲルが、ピリジンなどの有機溶媒の存在下、クロルスルホン酸、無水硫酸などの硫酸化剤の作用により例えばエステル結合を介してクロマト担体ゲルそのものに直接硫酸基が導入されたものを意味し、結晶セルロースまたは結晶領域および非結晶領域セルロースなどからなる球状セルロースに硫酸エステルが導入されたセルロース硫酸エステルのゲルはその代表である。この場合、得られたセルロース硫酸エステルは原料の形状を保持し、水性溶媒に不溶性であり、物理的安定性に優れ、クロマトグラフィー用ゲルとして好適である。これらの原料セルロース類はすでに市販されており、例えばアビセル(旭化成工業社製)、セルロファインGC−15、同GH−25、同GC−100、同GC−200(チッソ社製)などがある。原料セルロースを前記の方法で硫酸化することもできるし、そのなかのいくつかは硫酸化セルロースとして例えば「硫酸化−セルロファイン」の販売名(チッソ社製)で市販されている。
【0028】
硫酸基を有する分子が共有結合されているクロマト担体ゲルとは、架橋セルロース、架橋デキストランもしくは架橋アガロースゲルなどの天然高分子重合体または親水性ビニルポリマーもしくはスチレンジビニルベンゼン共重合体などの人工高分子重合体からなるクロマト担体ゲルに、高度に硫酸化されたグリコサミノグリカンであるヘパリン、硫酸化アルキル基の一種であるスルフォプロピル基またはスルフォン基を有するブルー色素などがCNBr法などの方法で共有結合的に結合されているものを意味し、すでに数多くのものが市販されている。ヘパリン結合クロマト担体ゲルとしては、「ヘパリンセファロースCL−6B」(ファルマシア社)、「アフィゲルヘパリンゲル」(バイオラッド社)、「ヘパリンセルロファイン」(生化学工業社)など、ブルー色素結合クロマト担体ゲルとしては「アフィゲルブルーゲル」(バイオラッド社)、「AF−ブルートヨパール」(東ソー社)、「ブルーセルロファイン」(生化学工業社)など、さらにスルフォプロピル基結合クロマト担体ゲルとしては「SP−セファデックス」(ファルマシア社)、「SP−トヨパール650M」(東ソー社)などが例示される。
【0029】
本発明において、硫酸基を有する分子をリガンドとする吸着ゲルあるいは直接硫酸化された吸着ゲルを用いて壊死毒素含有液とりわけボルデテラ属菌菌体抽出液中の壊死毒素を精製採取するにあたっては、以下の方法で実施する。ここでは、好適な実施態様であるセルロース硫酸エステルゲルを用いた例について概説するが、他のゲルの場合も同様の条件下で実施することができる。
【0030】
セルロース硫酸エステルゲルを、予め、例えば0〜0.2M塩化ナトリウム添加0.02Mリン酸緩衝液等の中性付近のpH値(pH7〜9)であり、適当な比電導度、すなわちゲルクロマトグラフィーにおける洗浄、溶出の際の一定の基準となる比電導度(その範囲はゲルの種類によっても若干異なるが、通常、20mS/cm以下、一般的には3〜20mS/cm、より一般的には5〜20mS/cmの範囲にある)またはそれ以下の比電導度を有する緩衝液にて平衡化した後に、壊死毒素の吸着操作に供する。セルロース硫酸エステルゲルへの壊死毒素の吸着、ゲルの洗浄、壊死毒素の溶出等一連の精製操作は、バッチ法およびカラム法等の工業的に通常よく用いられる操作方法で行なう。バッチ法で行なう場合は、ボルデテラ属菌菌体抽出液中にセルロース硫酸エステルを投入し、pH7.0〜9.0程度の範囲において0〜30℃程度の温度にて10〜60分程度緩く撹拌して壊死毒素を吸着させる。この際、ボルデテラ属菌菌体抽出液の比電導度が上記所定の範囲、一般的に3〜20mS/cmの範囲、またはそれ以下となるように、適宜濃縮または希釈して吸着操作に付す。
【0031】
吸着終了後、抽出物・ゲル混合液をろ過器上に充填し、吸引ろ過してゲルとろ液を分離する。分離したゲルを、上記所定の範囲の比電導度(一般的には3〜20mS/cm程度)またはそれ以下で、pHが5〜10程度である適当な緩衝液、例えば、0.1M塩化ナトリウム添加0.02Mリン酸緩衝液等を注ぎ吸引して洗浄する。この後、pHが5〜10程度で、比電導度が上記所定の範囲の比電導度以上で、一般的には130mS/cm以下のもの(好ましい範囲は、例えば22〜46mS/cm程度)である適当な緩衝液、0.2〜0.5M塩化ナトリウム添加リン酸緩衝液等を注ぎ、吸着している壊死毒素を溶出する。
【0032】
カラム法にて本発明方法を実施する場合は原材料、洗浄用緩衝液、溶出用緩衝液の条件はバッチ法の場合と同様でよく、これらの通液速度は、10ml/cm2/hr〜500ml/cm2/hr程度に調整して行なうことが推奨される。
【0033】
本発明の精製方法によれば、壊死毒素含有液中の壊死毒素に対する特異的吸着能に優れ、壊死毒素の精製度は数10倍〜110倍に達し、しかも壊死毒素の回収率は40%〜100%と良好である。発熱やショック等副作用の原因物質で壊死毒素とともに菌体破砕抽出液中に大量に含まれる内毒素はセルロース硫酸エステルゲルに吸着しないため、1/30〜1/800量に低減される。得られた精製壊死毒素の比活性が4×105〜3×106MND/mg蛋白と高く、SDS-ポリアクリルアミド電気泳動分析において主要バンドを形成する。
【0034】
上述のとおり、本発明の方法によれば、出発材料の壊死毒素含有液から所望の壊死毒素を良好な収率、純度で採取することができ、その操作も極めて簡便で、また、その精製用クロマトグラフィー吸着体は安価に調製あるいは入手でき、しかも繰り返し使用しても劣化が全くなく経済的にも極めて優れている。従って、本発明の方法は壊死毒素の工業的製造法として極めて優れた方法である。また、本発明の方法は従来の技術である陽イオン交換クロマトグラフィー法、ゲルろ過クロマトグラフィー法等と組み合わせて純度をさらに高めることも可能であり、その際は従来法で得られる結果をはるかに凌ぐ結果を得ることができる。
【0035】
本発明の方法により精製された壊死毒素は、化学薬品で処理することにより、免疫原性を保持しながら毒素活性を消失させトキソイド化される。トキソイド化に際しては一般的に用いられている方法、例えばホルマリンによる方法(Nakai,T.ら,インフェクション アンド イムニティー: Infect.Immun.,46,p.429〜434(1984))やグルタールアルデヒドによる方法(Endoh,M.ら,マイクロバイオロジー アンド イムノロジー:Microbiol.Immunol.,30,p.659〜673(1986))などが適用され得る。さらに、当該トキソイドを体内に投与して液性免疫の惹起を期する場合はアジュバントを加える。加えるアジュバントとしては、アルミニウムゲル、油性アジュバント、サポニンなどが使用できる。当該トキソイドは未精製の壊死毒素画分に比べ免疫原性が高く、かつ安全性にも優れており、このことは後述の実施例のデータにより支持される。
【0036】
本発明の方法によって得られる部分精製壊死毒素は、純度が高く内毒素のほとんどが除去されているため、動物用ワクチンの調製に有用であり、さらに従来の精製方法と組み合わせて純度を高めると各種試薬、医薬品の調製、さらには人体用百日咳ワクチンの調製への展開も可能である。
【0037】
ところで、Bbの自然感染例ではほとんどの場合、BbT中和抗体の出現を認めないが菌凝集抗体は高率に上昇し本病の血清学的診断法として応用されている。本発明者らの調査においてもAR発生農場である15県48農場327頭のBbT中和抗体を検査したところ、1農場(2.1%)の2頭(0.6%)が陽性であるにすぎなかった。従来のBb死菌ワクチンは菌凝集抗体を上昇させるため、診断の妨げとなっていた。しかし、本発明の精製法と従来の精製法を組み合わせることによってBbTの精製度を高めると、中和抗体を付与するものの菌凝集抗体を上昇させないトキソイドの調製が可能となる。このように、ワクチン抗体と感染抗体を血清学的に判別可能にならしめるワクチンやトキソイドは「マーカーワクチン」と呼ばれる。例えば、特定病原体フリー(以下、SPFと略称することがある)農場において、本発明のトキソイドの使用によりARの予防を行ないながら、万一Bbの感染が認められた場合でも菌凝集抗体を測定することにより感染豚を摘発することができる。
【0038】
本発明のもう一つの態様は、前記の精製方法に従って調製されたBbTトキソイドとパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする混合トキソイドを提供することである。
【0039】
本発明において用いられるPmTに関して、出発材料であるPm菌体抽出液とは、毒素原性Pmあるいは非定型的パスツレラ(Kamp,E.M.I.E.et al.,ヴェテリナリー レコード:Vet.Rec.,126,p.434〜437(1990))の菌体抽出液を包含する。または、遺伝子組換え技術によりPmTを発現させた細胞抽出液も本発明の出発材料として含まれる。本発明において好ましい抽出液は、毒素原性Pmの菌体抽出液であり、本菌をデキストロース・スターチ培地、血液寒天培地、YPC培地(Namioka,S.and Murata,M.,コーネル ヴェテリナリアン:Cornell Vet.,51,p.498〜507(1961))などの寒天培地あるいはハートインフュージョン培地、トリプチケース・ソイ・ブロス、Luria-Bertani培地、肉水培地(de Jong,M.F.and Akkermans,J.P.W.M.,ヴェテリナリー クオテリー:Vet.Quart.,8,p.294〜214(1986))などの液体培地で、常法により静置培養または振盪培養もしくは通気撹拌培養する。寒天培養菌は、コーンラージ棒等でトリス緩衝液等に浮遊する。液体培養菌は、遠心分離により菌体を回収し、次の精製工程に適した適当な緩衝液に浮遊する。この菌体浮遊液を超音波処理、酵素処理、プレス処理、凍結融解の繰り返し等を行なうことにより菌体を破壊し、遠心分離あるいは膜ろ過により菌体残渣を除去する。
【0040】
この抽出物をイオン交換クロマトグラフィー、PmTを認識する抗体で作製した親和性クロマトグラフィーなどにより精製を行なう。あるいは毒素原性Pmを液体培地で24〜65時間培養した上清を精製の出発材料に、あるいはそのまま減毒工程に進むことも可能である。
【0041】
上述のように調製した毒素画分をホルマリン、グルタールアルデヒドなどによって処理し減毒を実施する。減毒化終了後、透析などによって緩衝液をリン酸緩衝食塩水に置換し、アジュバントを加える。アジュバントとしては、アルミニウムゲル、油性アジュバント、サポニンなどが使用できる。混合トキソイドは、各々抗原濃度を単味の場合の2倍に調整しアジュバントを加えたものを等量混合することにより製造される。
【0042】
本発明により得られる混合トキソイドは安全性にも優れており、従来のワクチンとは異なってBbT及びPmTを中和する抗体を同時に付与することにより、両毒素による鼻甲介萎縮や発育遅延から豚を予防することができる。
【0043】
本発明により得られるBbTトキソイド並びに混合トキソイドは、妊娠豚に対し分娩の1週間以上前までに2〜6週間隔で2回筋肉内注射されることにより、初乳による移行抗体で産子を予防する。次回の分娩からは分娩前に本トキソイドを1回接種するだけで十分な免疫が成立する。重度のAR感染農場では、7日齢以上の子豚に対し本トキソイドを2〜6週間隔で2回筋肉内注射することにより直接予防することも有効である。
【0044】
実施例
以下、本発明を調製例並びに実施例を挙げより詳細に説明するが、本発明はなんらこれらに限定されるものではない。
調製例1
0℃以下の温度にてピリジン600mlにクロルスルホン酸117gを滴下し、混合する。滴下終了後、混液を加熱し、65〜70℃に昇温する。この中にセルロファインGC-15(チッソ社製)80gを加え、撹拌下65〜70℃にて3時間反応させる。反応終了後、冷却し、10%水酸化ナトリウム水溶液を加えて徐々に中和する。ゲルをろ過分離し、0.01Mリン酸緩衝食塩水で充分に洗浄してセルロース硫酸エステルゲルを得る。
【0045】
調製例2
0℃以下の温度にてピリジン600mlにクロルスルホン酸117gを滴下し、混合する。滴下終了後、混液を加熱し、65〜70℃に昇温する。この中に結晶セルロースであるクロマトグラフィー用アビセル(旭化成工業社製)80gを加え、撹拌下65〜70℃にて4時間保持する。反応終了後、冷却し、10%水酸化ナトリウム水溶液を加えて中和する。ゲルをろ過分離し、0.01Mリン酸緩衝食塩水で充分に洗浄して結晶セルロース硫酸エステルゲルを得る。
【0046】
調製例3
0〜5℃の温度にてピリジン500mlにクロルスルホン酸82gを滴下し、混合する。滴下終了後、混液を加熱し、65〜70℃に昇温する。この中にセルロファインGC-25(チッソ社製)80gを加え、撹拌下65〜70℃にて4時間反応させる。反応終了後、冷却し、10%水酸化ナトリウム水溶液を徐々に加えて中和する。ゲルをろ過分離し、0.01Mリン酸緩衝食塩水(pH7.2)で充分に洗浄してセルロース硫酸エステルゲルを得る。
【0047】
実施例1
(カラムからのグラジェント溶離法)
前記調製例1と同様にして調製したセルロファインGC-15の硫酸化エステル化物をカラム(50mm(内径)×200mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0,比電導度約3ms/cm)を通液して平衡化した。このカラムにBb菌S611株菌体破砕抽出液200mlを通液した。通液後、上記緩衝液にて洗浄し夾雑物質を洗い出した。ついで、0〜0.5M塩化ナトリウム添加リン酸緩衝液(pH8.0、比電導度約3〜46ms/cm)でグラジェント溶出し、総蛋白量9.1mgの壊死毒素を含む画分を得た。壊死毒素の回収率は39.6%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は32.1倍に達した。
【0048】
実施例2
(カラムからのステップワイズ溶離法)
前記調製例1と同様にして調製したセルロファインGC-15の硫酸化エステル化物をカラム(252mm(内径)×210mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0、比電導度約3ms/cm)を通液して平衡化した。このカラムに総蛋白量29〜35gのBbS611株菌体破砕抽出液を通液した。通液後、0〜0.15M塩化ナトリウム添加0.02Mリン酸緩衝液(pH7.8〜8.0、比電導度約3〜17ms/cm)にて洗浄し、夾雑物質を洗い出した。ついで、0.3〜0.5M塩化ナトリウム添加リン酸緩衝液(pH7.5〜7.7、比電導度約30〜46ms/cm)で溶出し、総蛋白量0.2〜1.1gの壊死毒素を含む画分を得た。表1に示したとおり壊死毒素の回収率は76〜100%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は23〜114倍に達し、内毒素は0.1〜3%に低減された。
【0049】
【表1】

【0050】
実施例3
(ヘパリンセファロースカラムからのグラジェント溶離法)
ヘパリンセファロースCL-6B(ファルマシア社製)をカラム(25mm(内径)×140mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0、比電導度約3mS/cm)を通液して平衡化した。このカラムに総蛋白量420mgのBbS611株菌体破砕抽出液を通液した。通液後、上記緩衝液にて洗浄し夾雑物を洗い出した。ついで、0〜0.5M塩化ナトリウム添加リン酸緩衝液(pH8.0、比電導度約3〜46mS/cm)でグラジェント溶出し、総蛋白量24mgの壊死毒素を含む画分を得た。壊死毒素の回収率は40.8%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は7.0倍に達した。分析結果を表2に記した。
【0051】
実施例4
(SP-トヨパール650Mカラムからのグラジェント溶離法)
SP-トヨパール650Mカラム(東ソー社製)をカラム(50mm(内径)×140mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0、比電導度約3mS/cm)を通液して平衡化した。このカラムに総蛋白量631mgのBbS611株菌体破砕抽出液を通液した。通液後、上記緩衝液にて洗浄し夾雑物を洗い出した。ついで、0〜0.5M塩化ナトリウム添加リン酸緩衝液(pH8.0、比電導度約3〜46mS/cm)でグラジェント溶出し、総蛋白量2mgの壊死毒素を含む画分を得た。壊死毒素の回収率は11.5%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は35.5倍に達した。分析結果を表2に記した。
【0052】
【表2】
【0053】
実施例5
(高純度精製)
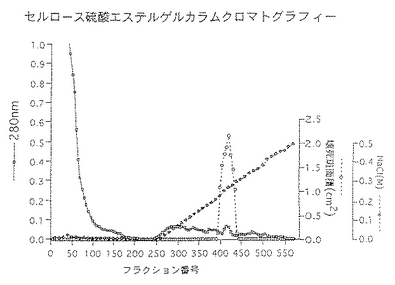
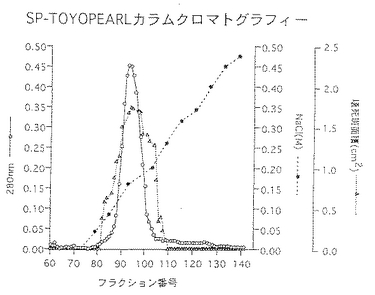
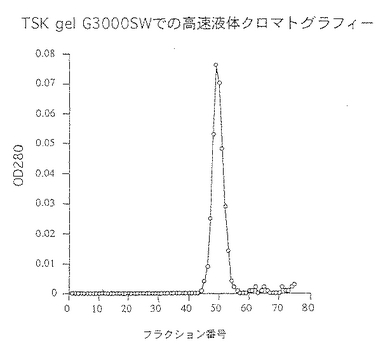
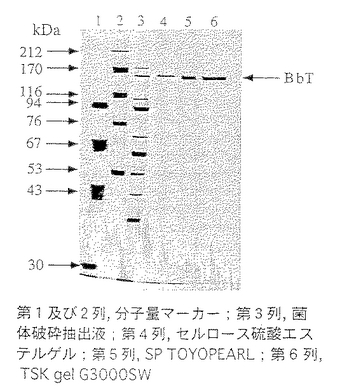
前記実施例1と同様にして調製したセルロース硫酸エステルゲル精製画分を33mMクエン酸・66mMリン酸2ナトリウム・1M尿素(pH5.0)で透析した。同緩衝液で平衡化したSP-Toyopearl 650M(東ソー社製:10mm(内径)×80mm)に総蛋白量9.1mgの透析内容物を通液し、同緩衝液にて洗浄し夾雑物を洗い出した。ついで0〜0.5M塩化ナトリウム添加の上記緩衝液でグラジェント溶出し、総蛋白量3.1mgの壊死毒素を含む画分(SP画分)を得た。50mMリン酸緩衝液(pH7.1)・0.3M硫酸ナトリウム・1M尿素で平衡化したTSK-gel G3000SW (21mm(内径)×600mm)にSP画分を通液し、上記緩衝液にて高速液体クロマトグラフィーを行ない総蛋白量1.3mgの壊死毒素を含む画分(HPLC画分)を得た。これらのクロマトグラムを図1〜図3に示した。また、菌体破砕抽出液〜壊死毒素画分の分析結果を表3及び図4に、精製毒素の性状を表4に記した。
【0054】
【表3】
【0055】
【表4】
【0056】
実施例6
(減毒化)
前記実施例1〜5と同様にして得られた壊死毒素画分にホルマリンを加え37〜40℃で3〜14日間反応させて減毒化を行なった。壊死毒素画分に3/100量の10%ホルマリンを加え、翌日、同量を、翌々日には2/100量を加えた。減毒化終了後、リン酸緩衝食塩液で透析した。
【0057】
実施例7
(アジュバント添加)
前記実施例6と同様にして得られた減毒化壊死毒素に水酸化アルミニウムゲル及び保存剤(チメロサールあるいはホルマリン)を加えた。
【0058】
実施例8
(マウス力価試験)
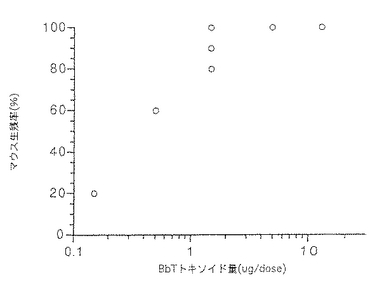
前記実施例7と同様にして得られたトキソイドを5週齢ddYマウスの腹腔内に0.5mlずつ2週間間隔で2回注射した。最終免疫の2週後に約50(25〜100)LD50の壊死毒素で腹腔内攻撃し生死を7日間観察した。結果を図5に示した。トキソイド量とマウス生残率に正の相関関係が認められた。
【0059】
実施例9
(育成ブタでの安全性試験)
前記実施例2と同様にして得られた画分を前記実施例6のとおりに減毒化、前記実施例7のとおりに水酸化アルミニウムゲルを加えたトキソイド2〜50μg/2ml/ドースを調製し、10週齢SPFブタに3週間隔で2回筋肉内注射した。注射による臨床症状や体温の異常はいずれのブタにも認められなかった。
【0060】
実施例10
(妊娠ブタでの有効性試験)
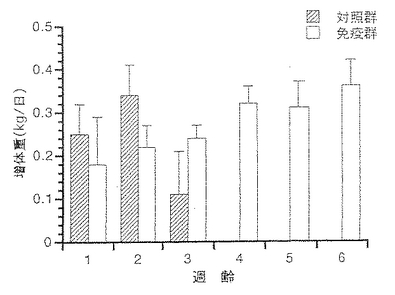
前記実施例5と同様にして得られたHPLC画分を、前記実施例6のとおりに減毒化、前記実施例7のとおりに水酸化アルミニウムゲルを加えたトキソイド(25μg/ml)を妊娠ブタに分娩6及び3週前の2回、2mlずつ筋肉内注射した。対照ブタにはプラセボワクチンを注射した。初乳を十分に摂取させた産子について、3日齢時にBbS611株2億個を全頭の鼻内に、さらに半数の子豚について2及び3週齢時の2回、壊死毒素2600MNDを筋肉内注射により攻撃した。壊死毒素中和抗体価の成績を表5に記した。分娩時の母ブタ血中BbT中和抗体価は32倍、初乳中は256倍、産子の移行抗体価は128倍であった。この移行抗体は6週間以上陽性であった。プラセボ注射ブタ及びその産子は抗体陰性であった。攻撃試験の成績を表6に記した。鼻甲介萎縮は対照群の12頭中9頭に+〜+++のスコアーで認められたが、免疫群には認められなかった。鼻内からの菌回収には両群の差は認められなかった。増体重は、図6に示したとおりBb生菌に加え壊死毒素による攻撃を実施した群間で差が認められた。2週齢時に実施した1回目の壊死毒素攻撃では、対照群の増体率は免疫群の約1/3に低下し、2回目攻撃(3週齢時)により対照群の全頭が死亡した。
【0061】
【表5】
【0062】
【表6】
【0063】
実施例11
(育成ブタでの有効性試験)
前記実施例2と同様にして得られた画分を前記実施例6のとおりに減毒化、前記実施例7のとおりに水酸化アルミニウムゲルを加えたトキソイド2〜50μg/2ml/ドースを調製し、10週齢SPFブタに3週間隔で2回筋肉内注射した。最終免疫から4週後に2億個のBb生菌で鼻内攻撃し、攻撃後3週目までの壊死毒素中和抗体価及を検査した。表7に示したとおり対照ブタは攻撃後もBbT中和抗体陰性であった。免疫群は全頭がBbT中和抗体が陽転した。攻撃により2及び6μg/2ml/ドースのトキソイド注射ブタは急激な中和抗体の上昇が認められたが、20及び50μg/2ml/ドース以上では上昇しなかった。
【0064】
壊死毒素には抗体産生抑制作用のあることが報告されている(Sekiya,K.,マイクロバイオロジー アンド イムノロジー: Microbiol.Immunol.,27,p.905〜915(1983))。攻撃後、対照ブタに中和抗体が上昇しない理由は、攻撃菌の産生するBbTの作用によるものと考えれらえる。従って、免疫群における急激な中和抗体の上昇はBbTの抗体産生抑制作用を防御した結果であることから、育成ブタでは2μgのBbTトキソイドでも有効であることが示唆された。
【0065】
【表7】
【0066】
調製例4
(菌体破砕抽出液:BbTトキソイド1)
BbS611株をペプトン培地で37℃、24時間通気撹拌培養した。菌体を遠心濃縮後、菌体破砕処理し菌体残渣を遠心あるいは膜ろ過(ポアー径0.45μm)で除去した溶液(粗BbT画分)にホルマリンを終濃度0.8%に加えて37〜40℃で7日間減毒した。このトキソイドをリン酸緩衝食塩水に対して透析した。透析内容物に水酸化アルミニウムゲルを1mg(アルミニウム量換算)/mlに加えてBbTトキソイド1とした。
【0067】
調製例5
(BbTトキソイド2)
前記調製例4と同様にして得られた粗BbT画分をセルロース硫酸エステルゲルカラムクロマトグラフィーにより精製した。20mMリン酸緩衝液(pH8.0)で平衡化したセルロース硫酸エステルゲルに粗BbTを通液し、同緩衝液で洗浄した。BbTは1.5MNaCl,1M尿素添加20mMリン酸緩衝液(pH8.0)でセルロース硫酸エステルゲルカラム(7.8cm(内径)×45cm)から溶出した。この溶出画分(CS1画分)を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド2とした。
【0068】
調製例6
(BbTトキソイド3)
前記調製例5と同様にして得られたCS1画分をさらにTSKゲルHW65(2.64cm(内径)×69cm)及びHW75(2.64cm(内径)×64cm)連結カラムで精製した。0.3M硫酸ナトリウム、1M尿素加0.05Mリン酸緩衝液(pH7.2)で平衡化させたカラムにCS1画分を通液し、同緩衝液で分画後、壊死毒活性を有する画分を混合した。この画分を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド3とした。
【0069】
調製例7
(BbTトキソイド4)
前記調製例4と同様にして得られた粗BbT画分をセルロース硫酸エステルゲルクロマトグラフィーにより精製した。0.1MNaCl加20mMリン酸緩衝液(pH8.0)で平衡化したセルロース硫酸エステルゲルに粗BbTを通液し、同緩衝液で洗浄した。BbTは0.5MNaCl加20mMリン酸緩衝液(pH8.0)でセルロース硫酸エステルゲルから溶出した。この画分(CS2画分)を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド4とした。
【0070】
調製例8
(BbTトキソイド5)
前記調製例7と同様にして得られたCS2画分をさらにAffi-gel blueカラム(BioRad社製、100-200メッシュ、5.0cm(内径)×17cm)で精製した。50mM トリス緩衝液(pH 7.5)で平衡化させたAffi-gel blue カラムに同緩衝液に対して透析したCS2画分を通液し、1M リン酸2ナトリウム加50mMトリス緩衝液(pH 7.5)で洗浄した。さらに50mMトリス緩衝液(pH 7.5)で洗浄後、2M塩化マグネシウム、1M尿素加50Mmトリス緩衝液(pH 7.5)でBbTを溶出した。この画分を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド5とした。
【0071】
調製例9
(BbT抗毒素)
前記実施例1及び5と同様にして得られた高度精製BbT画分を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド6とした。さらに、27週齢SPFブタに対して当該トキソイドを3か月間で1mlずつ8回筋肉内接種してBbT抗毒素(中和価4,096倍、菌凝集抗体価20倍以下)を得た。
【0072】
調製例10
(PmTトキソイド)
毒素原性PmD型s70株をハートインフュージョンブロスで37℃、24時間通気撹拌培養した。菌体を遠心濃縮後、菌体破砕処理し菌体残渣を遠心あるいは膜ろ過(ポアー径0.45μm)で除去した溶液(粗PmTと略)を陰イオン交換クロマトグラフィーにより精製した。0〜0.3MNaCl加中性リン酸緩衝液またはトリス緩衝液で平衡化した陰イオン交換体に粗PmTを通液し、同緩衝液で洗浄した。PmTは0.4〜0.5MNaClを含む同緩衝液でイオン交換体から溶出した。この部分精製PmT画分にホルマリンを0.4%に加え、37℃で14日間以上減毒した。このトキソイドをリン酸緩衝食塩水に対して透析した。透析内容物に水酸化アルミニウムゲルを1mg Al/mlに加えてPmTトキソイドとした。
【0073】
実施例12
(各種精製方法により得られたBbTトキソイドのモルモットでの免疫原性)
前記調製例4〜8と同様にして得られた3,000 MND/ml(減毒前活性)のBbTトキソイドをモルモット(Hartley,雌,5週齢)の大腿部筋肉内に0.5ml注射した。注射後28日目に採血し、BbT中和抗体価を検査した。中和抗体測定法は、非働化血清をリン酸緩衝食塩水で2倍階段希釈した。これに等量のBbT(力価 4MND/0.1ml)を加え、氷冷下で一夜反応後、モルモットの皮下に0.1mlずつ注射した。判定は注射後1日目に実施し、中和価は壊死反応を阻止した血清の最大希釈倍数(血清希釈倍率)で示した。
【0074】
表8に示したとおり、比活性が1.4×104 MND/mg蛋白のBbTトキソイド1及び2を注射したモルモットではすべて中和抗体が陰性であった。比活性が3.7×104 MND/mg 蛋白のBbTトキソイド3を注射したモルモットでは3頭中2頭が中和抗体が陽性で、7.8×104MND/mg 蛋白以上では全頭陽転した。
【0075】
BbTは精製の過程で不溶化しやすく、脂質、酸性蛋白など菌体由来の物質と複合体を形成しやすい性質を有している(Wordlaw,A.C.and Parton,R.,Parmacol.Ther.,19,p.1〜53(1983))。そのため、これらの菌体由来物質を低減することにより、より十分な減毒化が可能になるか、あるいはまた毒素活性部位のマスキングを減少させ抗原提示効率が向上することによりBbTトキソイドの免疫原性が高まるものと考えられる。
【0076】
【表8】
【0077】
実施例13
(BbT中和抗体による防御ライン)
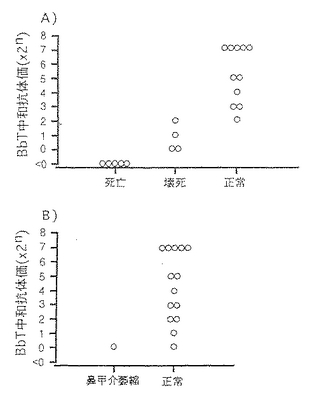
前記調製例10で得られたBbT抗毒素 0.2,1.0,2.0及び20mlを2日齢SPFブタの腹腔内に注射して受身免疫を賦与した。3日齢時にBbS611株108CFUで鼻内攻撃し、さらに7日齢時に粗BbT(1,940MND/頭)で筋肉内攻撃した。図7に示したとおり、BbTの致死作用に対してはBbT中和抗体価1倍以上で防御が認められ、壊死作用に対しては4倍以上で防御が可能であった。
【0078】
実施例14
(PmT中和抗体による防御ライン)
前記調製例10と同様にして得られた粗PmTのトキソイドを等量のフロイント不完全アジュバントで乳化した。8頭のSPF妊娠ブタのうち6頭を免疫群、2頭を対照群とした。免疫群は妊娠期間中に3mlのトキソイド(840 MND/ml)を2〜5回筋肉内注射した。分娩後、初乳を産子に十分飲ませ2日齢または9日齢時に20〜160MNDの粗PmTで筋肉内攻撃した。子ブタは攻撃後14日間観察し鼻甲介病変を観察した。表9に示したとおり、PmTの鼻甲介萎縮作用に対してはPmT中和抗体価2倍以上で防御が認められ、致死作用に対しては2倍以下でも防御が可能であった。
【0079】
【表9】
【0080】
実施例15
(BbT抗毒素受身免疫ブタのBb及びPm攻撃試験)
前記調製例10で得られたBbT抗毒素20mlを2日齢SPFブタの腹腔内に注射して受身免疫を賦与した。3日齢及び7日齢時にそれぞれBbS611株108CFU及びPmD型s70株108CFUで鼻内攻撃し、6週齢で剖検した。表10に示すように、BbT抗毒素でBbの鼻粘膜から大量の菌が回収されたが、Pmの回収は対照に比べると大幅に少なかった。また、対照群の鼻甲介病変は重度(+++)〜極度(++++)であったが、免疫群は一頭が正常(−)で残りが重度及び極度と個体間における差が大きかった。
【0081】
【表10】
【0082】
壊死毒素は菌体内に産生され、溶菌に伴い菌体外に放出される。従って、BbT抗毒素によってBbの鼻粘膜定着を阻止することはできない。Pmが定着するためには予めBbが粘膜に障害を与えていなくてはならない。Bbの産生する鼻粘膜障害因子としては、BbT(Elias,B.ら,ジャパニーズ ジャーナル ヴェテリナリー サイエンス: Jpn.J.Vet.Sci.,52,p.677〜688(1990))及び気管細胞毒素(Cookson,B.T.and Goldman,W.E.,ジャーナル オブ セルラー バイオケミストリー: J.Cell.Biochem.,11B(Suppl.)p.124;Dugal,F.らカナディアン ジャーナル ヴェテリナリー リサーチ:Can.J.Vet.Res.,56,p.260-264(1992))が重要である。気管細胞毒素はPmの定着促進に重要な気管線毛運動停止作用や線毛上皮細胞崩壊作用及び粘液分泌亢進作用を有するため、BbT抗毒素による免疫ではPm定着を部分的にしか阻止することはできない。
【0083】
これらの事実からPmの鼻粘膜への定着を阻止するためには、少なくともBbT及び気管細胞毒に対する免疫を成立させる必要がある。しかし、気管細胞毒トキソイドは精製方法の確立、免疫原性、収量など実用化までには多くの問題を抱えている。そのため、萎縮性鼻炎の主症状を予防し経済的損失を抑制するためのAR不活化ワクチンとしては少なくとも壊死毒混合トキソイドを含有する必要がある。
【0084】
実施例16
(トキソイド混合例)
前記実施例2及び6と同様にして得られた減毒化BbTに水酸化アルミニウムゲルを加え攪拌した。BbT濃度を8,700〜220,000MND/ml(減毒前毒素量BbT蛋白で2〜50μg/mlに相当)に、アルミニウム濃度を0.5〜2.5mg/mlになるよう調製し、保存剤を加えてBbT原液とした。
【0085】
前記調製例11と同様にして得られた水酸化アルミニウムゲル添加減毒化PmTを3,400MND/ml以上(減毒前毒素量)に調製し保存剤を加えてPmT原液とした。保存剤としては、0.01%チメロサールあるいは0.1〜0.4%ホルマリンを使用した。BbT原液及びPmT原液を等量ずつ加えて混合トキソイドを調製した。
【0086】
この混合トキソイドを9週齢SPFブタの頚部筋肉内に3週間隔で2回、2mlずつ接種した。経時的に血清を採取し、BbT中和抗体価並びにPmT中和抗体価を測定した。PmT中和抗体価は牛胎仔肺細胞(Rutter,J.M. and Luther,P.D.,ヴェテリナリー レコード:Vet.Rec.,114,p.393〜396(1984))を用いた中和試験により求めた。ブタでの中和抗体価を表11に、マウスでの力価試験成績を表12に示した。BbT及びPmTの各単味トキソイドと混合トキソイドはブタ及びマウスに注射したとき免疫原性の差が認められず、防御に十分な中和抗体応答が認められ、また安全性も十分であった。
【0087】
【表11】
【0088】
【表12】
【図面の簡単な説明】
【0089】
【図1】図1は本発明による壊死毒素精製法の過程のクロマトグラムを示す。
【0090】
【図2】図2は本発明による壊死毒素精製法の過程のクロマトグラムを示す。
【0091】
【図3】図3は本発明による壊死毒素精製法の過程のクロマトグラムを示す。
【0092】
【図4】図4は本発明による壊死毒素精製法を用いて得られた壊死毒素画分の分析結果を示す。
【0093】
【図5】図5は本発明によって得られるトキソイドのマウス力価試験の結果を示す。
【0094】
【図6】図6は本発明によって得られるトキソイドの妊娠ブタでの有効性試験の結果を示す。
【0095】
【図7】図7はBbT中和抗体による防御ラインの検討結果を示す。
【技術分野】
【0001】
本発明は、豚の伝染性疾患に対する予防薬に関する。さらに詳細には、改良されたボルデテラ属菌が産生する壊死毒素のトキソイドおよびそのパスツレラ属菌が産生する壊死毒素のトキソイドとの混合トキソイド並びに該トキソイドの調製方法に関する。
【背景技術】
【0002】
萎縮性鼻炎(以下、ARと略称することがある)は豚の慢性呼吸器病で、鼻甲介萎縮、発育遅延、鼻出血、鼻曲がり及び他の呼吸器病の誘発等を主徴とする伝播力の極めて強い疾病である。本病の起因菌としては、毒素原性気管支敗血症菌(以下、Bbと略称することがある)及び毒素原性パスツレラ・マルトシーダ(以下、Pmと略称することがある)が認められており、上部気道粘膜に感染する。Bbは定着因子である赤血球凝集素や線毛を保有しているため鼻粘膜に対する定着性が強いが、それを保有しないPmは単独で定着することが困難である。そのため、Pmの感染が成立するためには予め鼻粘膜を障害する因子の存在が必要である。その最も代表的な因子がBbであり、鼻粘膜障害に関与する病原性因子として、気管細胞毒素や壊死毒素が知られている。実験感染では、子ブタに対しBb、続いて数日後にPmを鼻内感染させることにより野外農場で観察されるような重篤なARを再現することができる。
【0003】
ボルデテラ属菌には、百日咳菌、パラ百日咳菌、気管支敗血症菌、ボルデテラ・アビウム等があり、これらは種々の生物学的活性物質を産生する。Bb壊死毒素(以下、BbTと略称することがある)はこれらの生物学的活性物質の1つであり、いずれのボルデテラ属菌もBbTを産生する。BbTは出血壊死作用、鼻甲介萎縮作用、発育遅延作用、致死作用、血管平滑筋収縮作用、局所の血行障害、脾臓萎縮作用および抗体産生阻害作用などの生物学的活性を有し、ボルデテラ属菌による発症において重要な役割を演じていることが明らかになった。さらに、BbTはBbによる子豚の肺炎病巣形成にも重要な役割を演じていることが明らかにされた(非特許文献1)。
【0004】
一方、パスツレラ・マルトシーダの一部あるいは非定型的パスツレラの一部の菌株は、Pm壊死毒素(以下、PmTと略称することがある)を産生する。PmTは出血壊死作用、鼻甲介萎縮作用、発育遅延作用、致死作用、局所の血行障害および脾臓萎縮作用などの生物学的活性を有し、毒素原性Pmによる発症において重要な役割を演じていることが明らかにされている。また、PmTは毒素原性Pmによる豚の肺炎病巣形成において密接に関係していることが明らかになった(非特許文献2)。BbTおよびPmTはいずれも培地中に産生されず菌体内に蓄積されるが、長期間培養による溶菌に伴って菌体内から培養上清に遊離する。生体内においては溶菌に伴って菌体内から遊離した壊死毒素が作用する。
【0005】
従来知られている細菌由来の壊死毒素含有液、とりわけボルデテラ菌菌体抽出液からの壊死毒素の部分精製方法としてはイオン交換クロマトグラフィーによる方法がある(非特許文献3)。しかしながら、この方法は再現性に乏しい欠点がある。また、同様に壊死毒素の部分精製例として、透析、ショ糖密度勾配超遠心分離及びゲルろ過クロマトグラフィーに基づく方法がある(非特許文献4)が、この方法は再現性に乏しく、さらに、工程が多いにもかかわらず精製度があまり高くないという欠点がある。
【0006】
他の例として、核酸除去剤処理及び陰イオン交換クロマトグラフィー及びハイドロキシアパタイト吸着クロマトグラフィーによる方法がある(非特許文献5)。しかしながら、この方法は核酸除去剤のような高価な試薬を必要とし、工程が多い欠点がある。また、核酸除去剤、透析及び陽イオン交換クロマトグラフィーによる方法もある(非特許文献6)が、この方法は再現性が高く比較的高純度の壊死毒素が得られるものの、前記方法と同様、高価な試薬を必要とする欠点がある。
【0007】
さらに別の例として、塩析及び色素リガンドアフィニティークロマトグラフィーによる方法がある(非特許文献7)が、この方法は再現性は高いものの精製度があまり高くないという欠点がある。
【0008】
従来報告されたAR不活化ワクチンにはBb単味、Pm単味並びにBb、Pm混合不活化ワクチンの3種類があり研究あるいは実用化されている。
【0009】
Bb不活化ワクチンとしては、ホルマリンやグルタールアルデヒド等で不活化された死菌にアルミニウムゲルや油性アジュバントを加えたものがある(非特許文献8、非特許文献9および非特許文献10)。また、Bbの外膜に存在しアデニールサイクラーゼ活性を有する68kDaの蛋白に萎縮性鼻炎の防御効果のあることが明らかにされている(非特許文献11)。さらに、Bb菌の産生する線維状赤血球凝集素を主成分とするコンポーネントワクチンが存在する(非特許文献12および非特許文献13)。抗毒素を与える目的で壊死毒素を加えたワクチンは効果を得ていないという報告もある(非特許文献14および非特許文献15)。
【0010】
一方、Pmの不活化単味ワクチンとしては、PmTトキソイド(非特許文献16および非特許文献17)が研究あるいは実用化されている。
【0011】
さらに、Bb及びPmの混合不活化ワクチンとしては、(1)ARの予防のみを目的とするためBbとPmD型のPmT産生株の死菌を混合したもの、これらにPmT非産生株を加えたもの、あるいはBbとPmTトキソイドを混合したもの(非特許文献18、非特許文献19、非特許文献20および非特許文献21)、(2)ARと肺炎予防を目的とするため、BbとPmD型のPmT産生株、PmA型の死菌を混合したもの(非特許文献22)、(3)ARと肺炎に加えて他の疾病の予防も目的とするためBb、PmA型株、豚丹毒菌の死菌にPmTトキソイドを加えたものが実用化されている(非特許文献23)。
【0012】
ところで、Roop II,R.M.らはBbTがBb感染による豚の鼻甲介萎縮や肺炎病巣形成に重要な役割を演じていることから、BbTトキソイドの必要性を指摘している(非特許文献1)。しかし、BbTとPmTトキソイドとの混合化に関しては全く言及していない。また、Chanter,N.らはBbT及びPmTの共同作用による豚鼻粘膜障害がBb及びPmの増殖に好ましい環境を形成することから、Bbの重要な成分を含有するPmTトキソイドがより効果的であることを提案している(非特許文献24)。
【0013】
しかし、これまでのところ、単にBbT活性を有する画分からなるワクチンではBbT中和抗体を上昇させることに成功していない(非特許文献14、非特許文献15)。
【0014】
一方で、中井らは高度に精製したBbTトキソイドを用いてブタ及びマウスに中和抗体の上昇することを明らかにした(非特許文献25)。しかし、彼らも当該BbTトキソイドとPmTトキソイドとの混合化に関しては全く触れていない。
【非特許文献1】Roop II,R.M.ら、Infection and Immunity,55,p.217〜222(1987)
【非特許文献2】Iwamatsu, S.およびSawada, T.、Japanese Journal of Veterinary Science, 50, p.1200〜1206(1988)
【非特許文献3】Kume,K.ら,Infection and Immunity, 52(4), p.370〜377(1986)
【非特許文献4】Endoh,M.ら,Microbiology and Immunology, 30,(7), p.659〜673(1986)
【非特許文献5】Horiguchi,Y.ら、Microbial Pathogenesis, 6, p.361〜368(1989)
【非特許文献6】Horiguchi,Y.ら、FEMS Microbiology Letters, 66, p.39〜44(1990)
【非特許文献7】Zhang Y. L.およびSekura R. D.、Infection and Immunity, 59(10), ,p.3754〜3759(1991)
【非特許文献8】Goodnow, R. A.、Veterinary Medicine Small Animal Clinician, 72, p.1210〜1212(1977)
【非特許文献9】Nakase,Y.ら、Proceedings of International Pig Veterinary Society Congress, 8,(1976)
【非特許文献10】Giles, C. J.およびSmith, I. M.、Veterinary Bulletin (London), 53, p.327〜338(1983)
【非特許文献11】Montaraz J. A.ら、Infection and Immunity, 47, p.744〜755(1985)
【非特許文献12】Hansen, G. R.ら、In Atrophic rhinitis in pigs (Peterson, K. B.およびNielsen, N.C.編)Communication of the European Communities,Luxembourg, p.89〜97(1983)
【非特許文献13】Ohgitani, T.ら、Vaccine, 9, p.653〜658(1991)
【非特許文献14】Soderlind, O.およびBergstrom, G.,Proceedings of International Pig Veterinary Science Congress, p.175(1984)
【非特許文献15】Marel, C. M. von der.ら、Proceedings of International Pig Veterinary Science Congress, p.170(1984)
【非特許文献16】Pedersen, K. B.およびBarfod, K.、Nordish Veterina Medicine, 31, p.293〜302(1982)
【非特許文献17】Foged, N. T.ら、ヴェテリナリー レコード:Veterinary Record, 125, p.7〜11(1989)
【非特許文献18】Barfod, K.およびPedersen,K. B.、Nordish Veterina Medicine, 36, p.337〜345(1984)
【非特許文献19】Baalsrud, K. J.、Acta Veterina Scandinavia., 28, p.305〜311(1987)
【非特許文献20】de Jong, M. F.ら、Veterinary Quarterly, 9, p.49〜59(1987)
【非特許文献21】Kobisch, M.およびPennings, A.、Veterinary Record, 124, p.57〜61(1989)
【非特許文献22】Ostle, A. G.ら、Veterinary Medicine, p.772〜775(1986)
【非特許文献23】Jayappa, H.ら、International Pig Veterinary Society Congress, p.44(1988)
【非特許文献24】Chanter, N.ら、Research Veterinary Science, 47, p.48〜53(1989)
【非特許文献25】第111回日本獣医学会講演要旨集,p.183(1991)
【発明の開示】
【発明が解決しようとする課題】
【0015】
上述の状況下、壊死毒素の生体防御における役割についてはなお不明な点があり、これらを解明していく上での壊死毒素を大量に調製する方法の開発、ならびに疾病予防のための壊死毒素のトキソイドの開発が望まれている。しかし、壊死毒素は熱に対し不安定で精製が進むにつれて自己凝集しやすくなったり、脂質、酸性蛋白質および疎水性物質等の菌体由来物質と複合体を形成するなどの性質のため精製が困難といった問題点があった。
また、前記BbTとPmTは生物学的活性は類似しているものの、免疫学的に交差しないことが明らかにされており、このようなことから、獣医療上、養豚産業上安全かつ有効なBbT・PmT混合トキソイドの開発が切望されている。
【課題を解決するための手段】
【0016】
本発明者らは、壊死毒素の効率的な精製方法を見い出すべく種々検討を重ねた結果、(1)壊死毒素含有溶液例えばボルデテラ属菌菌体抽出物を、セルロース硫酸エステルゲル、ヘパリン固定化吸着ゲルもしくは硫酸化アルキル基固定化吸着ゲル等で例示される直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いてクロマトグラフィーを行なうことにより内毒素含有量の少ない部分精製壊死毒素が簡単にしかも比較的高い回収率で得られること、(2)上記直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いて部分精製した壊死毒素をさらに透析、陽イオン交換クロマトグラフィー及びゲルろ過クロマトグラフィーを実施することにより高純度の壊死毒素が簡単に、しかも比較的高回収率で得られること、(3)直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いて部分精製した壊死毒素が未精製のボルデテラ属菌菌体抽出物中壊死毒素に比べ免疫応答が大幅に改善されることの知見を得た。
【0017】
さらに、本発明者らは、ボルデテラ属菌菌体抽出物を直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルに接触せしめ、壊死毒素を吸着させ、夾雑物質と分離し、ゲルから溶出することにより得られた壊死毒素画分を化学的に減毒したものが豚において安全かつ萎縮性鼻炎の予防に有効であることを確認し本発明を完成するに至った。
【0018】
豚に対してPmの感染が成立するためには予め豚鼻粘膜を障害する因子としてのBbの前感染が必要である。BbT抗毒素による免疫をブタに行なった場合、Bb及びPmの攻撃によりPmの鼻粘膜定着並びに鼻甲介萎縮を軽減できる。しかし、個体間で効果に大きな差が認められることから、AR不活化ワクチンの成分としてBbTトキソイドのみでは不十分であることが判明した。
【0019】
本発明者らは、上述の知見を基に混合トキソイドを開発すべく鋭意検討を重ねた結果、ボルデテラ属菌菌体抽出物をセルロース硫酸エステルゲル等を用いて比活性37,000MND/mg蛋白以上に精製した毒素に由来するトキソイドと毒素原性Pm産生PmTに由来するトキソイドを混合することにより得られる混合トキソイドが、豚において安全かつ萎縮性鼻炎の予防に有効であることを確認し、本発明の第二の態様である混合トキソイドを供するに至った。
【0020】
なお、本発明において使用される最少壊死毒単位(MND)とは、リポポリサッカライド(大腸菌、血清型0111:B4由来など)1μg/ml存在下でモルモット皮内に注射され1日後に直径5mm以上の壊死斑を形成する最少毒素量を1MNDと定義される量に基づくものとする。
【発明を実施するための最良の形態】
【0021】
本発明は、壊死毒素含有液をセルロース硫酸エステル等の直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルに接触せしめ、壊死毒素を吸着させた後、ゲルから溶出することを特徴とする壊死毒素の部分精製方法を提供する。また、前記壊死毒素の部分精製方法によって得られたBbTに由来するBbTのトキソイド、並びに該BbTトキソイドと毒素原性Pm産生PmTに由来するトキソイドを混合することにより得られる、豚において安全かつ萎縮性鼻炎の予防に有効な混合トキソイドを提供するものである。
【0022】
本発明において、出発材料である壊死毒素含有液とは、Bb、百日咳菌、パラ百日咳菌及びボルデテラ アビウムの菌体抽出液を含む。または、遺伝子組換え技術によりBbTを発現させた細胞抽出液も本発明の出発材料として含まれる。
【0023】
本発明において好ましい抽出液は、Bbの菌体抽出液であり、Bbをボルデ・ジャング培地、マッコンキー培地などの寒天培地あるいはペプトン培地(Horiguchi, Y.ら,マイクロバイアル パソジェネーシス:Microb. Pathogen., 6,p.361〜368(1989))、コーエン・ウィラー培地、ステナー・ショルテ培地などの液体培地にて、常法により静置培養または振盪培養もしくは通気撹拌培養する。寒天培養菌は、コーンラージ棒等でセルロース硫酸エステルゲルの開始緩衝液に浮遊する。液体培養菌は、遠心分離により菌体を回収し、セルロース硫酸エステルゲルの開始緩衝液に浮遊する。
【0024】
この菌体浮遊液について、Kume, K.らの方法(インフェクション アンド イムニティー:Infect. Immun. 52(4), p.370〜377(1986))、Endoh, M.らの方法(マイクロバイオロジー アンド イムノロジー:Microbial. Immunol., 30(7), p.659〜673(1986))、Horiguchi, Y.らの方法(マイクロバイアル パソジェネーシス:Microbial Pathogen., 6, p.361〜368(1989)あるいはFEMSマイクロバイオロジー レターズ:FEMS Microbiol. Letters, 66, p.39〜44(1990))、Zhang Y. L.およびSekura R. D.の方法(インフェクション アンド イムニティー:Infect. Immun., 59(10), p.3754〜3759(1991))によって、またはセルロース硫酸エステルゲル等の直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いた吸着クロマトグラフィーにより精製を行なうことができるが、本発明においては、直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを用いた吸着クロマトグラフィーによる精製を実施することを最適な態様とする。
【0025】
この菌体浮遊液を超音波処理、酵素処理、プレス処理、凍結融解の繰り返し等を行なうことにより菌体を破壊し、遠心分離あるいは膜ろ過により菌体残渣を除去する。この菌体抽出物の形であるいは核酸除去剤、塩析、超遠心分離等の前段部分精製処理後に本発明の方法に供される。本発明の方法によれば、前段部分精製処理をあえて行なう必要はなく、菌体抽出液をそのまま硫酸基を有する分子をリガンドとする吸着ゲルあるいは直接硫酸化された吸着ゲルを用いた吸着クロマトグラフィーに付すことができ、工程がきわめて簡便となる。
【0026】
本発明で用いられる直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルについては以下のものが例示される。
【0027】
直接的硫酸化により硫酸基が導入されたクロマト担体ゲルとは、セルロースなどのクロマト担体ゲルが、ピリジンなどの有機溶媒の存在下、クロルスルホン酸、無水硫酸などの硫酸化剤の作用により例えばエステル結合を介してクロマト担体ゲルそのものに直接硫酸基が導入されたものを意味し、結晶セルロースまたは結晶領域および非結晶領域セルロースなどからなる球状セルロースに硫酸エステルが導入されたセルロース硫酸エステルのゲルはその代表である。この場合、得られたセルロース硫酸エステルは原料の形状を保持し、水性溶媒に不溶性であり、物理的安定性に優れ、クロマトグラフィー用ゲルとして好適である。これらの原料セルロース類はすでに市販されており、例えばアビセル(旭化成工業社製)、セルロファインGC−15、同GH−25、同GC−100、同GC−200(チッソ社製)などがある。原料セルロースを前記の方法で硫酸化することもできるし、そのなかのいくつかは硫酸化セルロースとして例えば「硫酸化−セルロファイン」の販売名(チッソ社製)で市販されている。
【0028】
硫酸基を有する分子が共有結合されているクロマト担体ゲルとは、架橋セルロース、架橋デキストランもしくは架橋アガロースゲルなどの天然高分子重合体または親水性ビニルポリマーもしくはスチレンジビニルベンゼン共重合体などの人工高分子重合体からなるクロマト担体ゲルに、高度に硫酸化されたグリコサミノグリカンであるヘパリン、硫酸化アルキル基の一種であるスルフォプロピル基またはスルフォン基を有するブルー色素などがCNBr法などの方法で共有結合的に結合されているものを意味し、すでに数多くのものが市販されている。ヘパリン結合クロマト担体ゲルとしては、「ヘパリンセファロースCL−6B」(ファルマシア社)、「アフィゲルヘパリンゲル」(バイオラッド社)、「ヘパリンセルロファイン」(生化学工業社)など、ブルー色素結合クロマト担体ゲルとしては「アフィゲルブルーゲル」(バイオラッド社)、「AF−ブルートヨパール」(東ソー社)、「ブルーセルロファイン」(生化学工業社)など、さらにスルフォプロピル基結合クロマト担体ゲルとしては「SP−セファデックス」(ファルマシア社)、「SP−トヨパール650M」(東ソー社)などが例示される。
【0029】
本発明において、硫酸基を有する分子をリガンドとする吸着ゲルあるいは直接硫酸化された吸着ゲルを用いて壊死毒素含有液とりわけボルデテラ属菌菌体抽出液中の壊死毒素を精製採取するにあたっては、以下の方法で実施する。ここでは、好適な実施態様であるセルロース硫酸エステルゲルを用いた例について概説するが、他のゲルの場合も同様の条件下で実施することができる。
【0030】
セルロース硫酸エステルゲルを、予め、例えば0〜0.2M塩化ナトリウム添加0.02Mリン酸緩衝液等の中性付近のpH値(pH7〜9)であり、適当な比電導度、すなわちゲルクロマトグラフィーにおける洗浄、溶出の際の一定の基準となる比電導度(その範囲はゲルの種類によっても若干異なるが、通常、20mS/cm以下、一般的には3〜20mS/cm、より一般的には5〜20mS/cmの範囲にある)またはそれ以下の比電導度を有する緩衝液にて平衡化した後に、壊死毒素の吸着操作に供する。セルロース硫酸エステルゲルへの壊死毒素の吸着、ゲルの洗浄、壊死毒素の溶出等一連の精製操作は、バッチ法およびカラム法等の工業的に通常よく用いられる操作方法で行なう。バッチ法で行なう場合は、ボルデテラ属菌菌体抽出液中にセルロース硫酸エステルを投入し、pH7.0〜9.0程度の範囲において0〜30℃程度の温度にて10〜60分程度緩く撹拌して壊死毒素を吸着させる。この際、ボルデテラ属菌菌体抽出液の比電導度が上記所定の範囲、一般的に3〜20mS/cmの範囲、またはそれ以下となるように、適宜濃縮または希釈して吸着操作に付す。
【0031】
吸着終了後、抽出物・ゲル混合液をろ過器上に充填し、吸引ろ過してゲルとろ液を分離する。分離したゲルを、上記所定の範囲の比電導度(一般的には3〜20mS/cm程度)またはそれ以下で、pHが5〜10程度である適当な緩衝液、例えば、0.1M塩化ナトリウム添加0.02Mリン酸緩衝液等を注ぎ吸引して洗浄する。この後、pHが5〜10程度で、比電導度が上記所定の範囲の比電導度以上で、一般的には130mS/cm以下のもの(好ましい範囲は、例えば22〜46mS/cm程度)である適当な緩衝液、0.2〜0.5M塩化ナトリウム添加リン酸緩衝液等を注ぎ、吸着している壊死毒素を溶出する。
【0032】
カラム法にて本発明方法を実施する場合は原材料、洗浄用緩衝液、溶出用緩衝液の条件はバッチ法の場合と同様でよく、これらの通液速度は、10ml/cm2/hr〜500ml/cm2/hr程度に調整して行なうことが推奨される。
【0033】
本発明の精製方法によれば、壊死毒素含有液中の壊死毒素に対する特異的吸着能に優れ、壊死毒素の精製度は数10倍〜110倍に達し、しかも壊死毒素の回収率は40%〜100%と良好である。発熱やショック等副作用の原因物質で壊死毒素とともに菌体破砕抽出液中に大量に含まれる内毒素はセルロース硫酸エステルゲルに吸着しないため、1/30〜1/800量に低減される。得られた精製壊死毒素の比活性が4×105〜3×106MND/mg蛋白と高く、SDS-ポリアクリルアミド電気泳動分析において主要バンドを形成する。
【0034】
上述のとおり、本発明の方法によれば、出発材料の壊死毒素含有液から所望の壊死毒素を良好な収率、純度で採取することができ、その操作も極めて簡便で、また、その精製用クロマトグラフィー吸着体は安価に調製あるいは入手でき、しかも繰り返し使用しても劣化が全くなく経済的にも極めて優れている。従って、本発明の方法は壊死毒素の工業的製造法として極めて優れた方法である。また、本発明の方法は従来の技術である陽イオン交換クロマトグラフィー法、ゲルろ過クロマトグラフィー法等と組み合わせて純度をさらに高めることも可能であり、その際は従来法で得られる結果をはるかに凌ぐ結果を得ることができる。
【0035】
本発明の方法により精製された壊死毒素は、化学薬品で処理することにより、免疫原性を保持しながら毒素活性を消失させトキソイド化される。トキソイド化に際しては一般的に用いられている方法、例えばホルマリンによる方法(Nakai,T.ら,インフェクション アンド イムニティー: Infect.Immun.,46,p.429〜434(1984))やグルタールアルデヒドによる方法(Endoh,M.ら,マイクロバイオロジー アンド イムノロジー:Microbiol.Immunol.,30,p.659〜673(1986))などが適用され得る。さらに、当該トキソイドを体内に投与して液性免疫の惹起を期する場合はアジュバントを加える。加えるアジュバントとしては、アルミニウムゲル、油性アジュバント、サポニンなどが使用できる。当該トキソイドは未精製の壊死毒素画分に比べ免疫原性が高く、かつ安全性にも優れており、このことは後述の実施例のデータにより支持される。
【0036】
本発明の方法によって得られる部分精製壊死毒素は、純度が高く内毒素のほとんどが除去されているため、動物用ワクチンの調製に有用であり、さらに従来の精製方法と組み合わせて純度を高めると各種試薬、医薬品の調製、さらには人体用百日咳ワクチンの調製への展開も可能である。
【0037】
ところで、Bbの自然感染例ではほとんどの場合、BbT中和抗体の出現を認めないが菌凝集抗体は高率に上昇し本病の血清学的診断法として応用されている。本発明者らの調査においてもAR発生農場である15県48農場327頭のBbT中和抗体を検査したところ、1農場(2.1%)の2頭(0.6%)が陽性であるにすぎなかった。従来のBb死菌ワクチンは菌凝集抗体を上昇させるため、診断の妨げとなっていた。しかし、本発明の精製法と従来の精製法を組み合わせることによってBbTの精製度を高めると、中和抗体を付与するものの菌凝集抗体を上昇させないトキソイドの調製が可能となる。このように、ワクチン抗体と感染抗体を血清学的に判別可能にならしめるワクチンやトキソイドは「マーカーワクチン」と呼ばれる。例えば、特定病原体フリー(以下、SPFと略称することがある)農場において、本発明のトキソイドの使用によりARの予防を行ないながら、万一Bbの感染が認められた場合でも菌凝集抗体を測定することにより感染豚を摘発することができる。
【0038】
本発明のもう一つの態様は、前記の精製方法に従って調製されたBbTトキソイドとパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする混合トキソイドを提供することである。
【0039】
本発明において用いられるPmTに関して、出発材料であるPm菌体抽出液とは、毒素原性Pmあるいは非定型的パスツレラ(Kamp,E.M.I.E.et al.,ヴェテリナリー レコード:Vet.Rec.,126,p.434〜437(1990))の菌体抽出液を包含する。または、遺伝子組換え技術によりPmTを発現させた細胞抽出液も本発明の出発材料として含まれる。本発明において好ましい抽出液は、毒素原性Pmの菌体抽出液であり、本菌をデキストロース・スターチ培地、血液寒天培地、YPC培地(Namioka,S.and Murata,M.,コーネル ヴェテリナリアン:Cornell Vet.,51,p.498〜507(1961))などの寒天培地あるいはハートインフュージョン培地、トリプチケース・ソイ・ブロス、Luria-Bertani培地、肉水培地(de Jong,M.F.and Akkermans,J.P.W.M.,ヴェテリナリー クオテリー:Vet.Quart.,8,p.294〜214(1986))などの液体培地で、常法により静置培養または振盪培養もしくは通気撹拌培養する。寒天培養菌は、コーンラージ棒等でトリス緩衝液等に浮遊する。液体培養菌は、遠心分離により菌体を回収し、次の精製工程に適した適当な緩衝液に浮遊する。この菌体浮遊液を超音波処理、酵素処理、プレス処理、凍結融解の繰り返し等を行なうことにより菌体を破壊し、遠心分離あるいは膜ろ過により菌体残渣を除去する。
【0040】
この抽出物をイオン交換クロマトグラフィー、PmTを認識する抗体で作製した親和性クロマトグラフィーなどにより精製を行なう。あるいは毒素原性Pmを液体培地で24〜65時間培養した上清を精製の出発材料に、あるいはそのまま減毒工程に進むことも可能である。
【0041】
上述のように調製した毒素画分をホルマリン、グルタールアルデヒドなどによって処理し減毒を実施する。減毒化終了後、透析などによって緩衝液をリン酸緩衝食塩水に置換し、アジュバントを加える。アジュバントとしては、アルミニウムゲル、油性アジュバント、サポニンなどが使用できる。混合トキソイドは、各々抗原濃度を単味の場合の2倍に調整しアジュバントを加えたものを等量混合することにより製造される。
【0042】
本発明により得られる混合トキソイドは安全性にも優れており、従来のワクチンとは異なってBbT及びPmTを中和する抗体を同時に付与することにより、両毒素による鼻甲介萎縮や発育遅延から豚を予防することができる。
【0043】
本発明により得られるBbTトキソイド並びに混合トキソイドは、妊娠豚に対し分娩の1週間以上前までに2〜6週間隔で2回筋肉内注射されることにより、初乳による移行抗体で産子を予防する。次回の分娩からは分娩前に本トキソイドを1回接種するだけで十分な免疫が成立する。重度のAR感染農場では、7日齢以上の子豚に対し本トキソイドを2〜6週間隔で2回筋肉内注射することにより直接予防することも有効である。
【0044】
実施例
以下、本発明を調製例並びに実施例を挙げより詳細に説明するが、本発明はなんらこれらに限定されるものではない。
調製例1
0℃以下の温度にてピリジン600mlにクロルスルホン酸117gを滴下し、混合する。滴下終了後、混液を加熱し、65〜70℃に昇温する。この中にセルロファインGC-15(チッソ社製)80gを加え、撹拌下65〜70℃にて3時間反応させる。反応終了後、冷却し、10%水酸化ナトリウム水溶液を加えて徐々に中和する。ゲルをろ過分離し、0.01Mリン酸緩衝食塩水で充分に洗浄してセルロース硫酸エステルゲルを得る。
【0045】
調製例2
0℃以下の温度にてピリジン600mlにクロルスルホン酸117gを滴下し、混合する。滴下終了後、混液を加熱し、65〜70℃に昇温する。この中に結晶セルロースであるクロマトグラフィー用アビセル(旭化成工業社製)80gを加え、撹拌下65〜70℃にて4時間保持する。反応終了後、冷却し、10%水酸化ナトリウム水溶液を加えて中和する。ゲルをろ過分離し、0.01Mリン酸緩衝食塩水で充分に洗浄して結晶セルロース硫酸エステルゲルを得る。
【0046】
調製例3
0〜5℃の温度にてピリジン500mlにクロルスルホン酸82gを滴下し、混合する。滴下終了後、混液を加熱し、65〜70℃に昇温する。この中にセルロファインGC-25(チッソ社製)80gを加え、撹拌下65〜70℃にて4時間反応させる。反応終了後、冷却し、10%水酸化ナトリウム水溶液を徐々に加えて中和する。ゲルをろ過分離し、0.01Mリン酸緩衝食塩水(pH7.2)で充分に洗浄してセルロース硫酸エステルゲルを得る。
【0047】
実施例1
(カラムからのグラジェント溶離法)
前記調製例1と同様にして調製したセルロファインGC-15の硫酸化エステル化物をカラム(50mm(内径)×200mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0,比電導度約3ms/cm)を通液して平衡化した。このカラムにBb菌S611株菌体破砕抽出液200mlを通液した。通液後、上記緩衝液にて洗浄し夾雑物質を洗い出した。ついで、0〜0.5M塩化ナトリウム添加リン酸緩衝液(pH8.0、比電導度約3〜46ms/cm)でグラジェント溶出し、総蛋白量9.1mgの壊死毒素を含む画分を得た。壊死毒素の回収率は39.6%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は32.1倍に達した。
【0048】
実施例2
(カラムからのステップワイズ溶離法)
前記調製例1と同様にして調製したセルロファインGC-15の硫酸化エステル化物をカラム(252mm(内径)×210mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0、比電導度約3ms/cm)を通液して平衡化した。このカラムに総蛋白量29〜35gのBbS611株菌体破砕抽出液を通液した。通液後、0〜0.15M塩化ナトリウム添加0.02Mリン酸緩衝液(pH7.8〜8.0、比電導度約3〜17ms/cm)にて洗浄し、夾雑物質を洗い出した。ついで、0.3〜0.5M塩化ナトリウム添加リン酸緩衝液(pH7.5〜7.7、比電導度約30〜46ms/cm)で溶出し、総蛋白量0.2〜1.1gの壊死毒素を含む画分を得た。表1に示したとおり壊死毒素の回収率は76〜100%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は23〜114倍に達し、内毒素は0.1〜3%に低減された。
【0049】
【表1】
【0050】
実施例3
(ヘパリンセファロースカラムからのグラジェント溶離法)
ヘパリンセファロースCL-6B(ファルマシア社製)をカラム(25mm(内径)×140mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0、比電導度約3mS/cm)を通液して平衡化した。このカラムに総蛋白量420mgのBbS611株菌体破砕抽出液を通液した。通液後、上記緩衝液にて洗浄し夾雑物を洗い出した。ついで、0〜0.5M塩化ナトリウム添加リン酸緩衝液(pH8.0、比電導度約3〜46mS/cm)でグラジェント溶出し、総蛋白量24mgの壊死毒素を含む画分を得た。壊死毒素の回収率は40.8%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は7.0倍に達した。分析結果を表2に記した。
【0051】
実施例4
(SP-トヨパール650Mカラムからのグラジェント溶離法)
SP-トヨパール650Mカラム(東ソー社製)をカラム(50mm(内径)×140mm)に充填し、これに0.02Mリン酸緩衝液(pH8.0、比電導度約3mS/cm)を通液して平衡化した。このカラムに総蛋白量631mgのBbS611株菌体破砕抽出液を通液した。通液後、上記緩衝液にて洗浄し夾雑物を洗い出した。ついで、0〜0.5M塩化ナトリウム添加リン酸緩衝液(pH8.0、比電導度約3〜46mS/cm)でグラジェント溶出し、総蛋白量2mgの壊死毒素を含む画分を得た。壊死毒素の回収率は11.5%で、精製度(精製壊死毒素画分の比活性/菌体破砕抽出液の比活性)は35.5倍に達した。分析結果を表2に記した。
【0052】
【表2】
【0053】
実施例5
(高純度精製)
前記実施例1と同様にして調製したセルロース硫酸エステルゲル精製画分を33mMクエン酸・66mMリン酸2ナトリウム・1M尿素(pH5.0)で透析した。同緩衝液で平衡化したSP-Toyopearl 650M(東ソー社製:10mm(内径)×80mm)に総蛋白量9.1mgの透析内容物を通液し、同緩衝液にて洗浄し夾雑物を洗い出した。ついで0〜0.5M塩化ナトリウム添加の上記緩衝液でグラジェント溶出し、総蛋白量3.1mgの壊死毒素を含む画分(SP画分)を得た。50mMリン酸緩衝液(pH7.1)・0.3M硫酸ナトリウム・1M尿素で平衡化したTSK-gel G3000SW (21mm(内径)×600mm)にSP画分を通液し、上記緩衝液にて高速液体クロマトグラフィーを行ない総蛋白量1.3mgの壊死毒素を含む画分(HPLC画分)を得た。これらのクロマトグラムを図1〜図3に示した。また、菌体破砕抽出液〜壊死毒素画分の分析結果を表3及び図4に、精製毒素の性状を表4に記した。
【0054】
【表3】
【0055】
【表4】
【0056】
実施例6
(減毒化)
前記実施例1〜5と同様にして得られた壊死毒素画分にホルマリンを加え37〜40℃で3〜14日間反応させて減毒化を行なった。壊死毒素画分に3/100量の10%ホルマリンを加え、翌日、同量を、翌々日には2/100量を加えた。減毒化終了後、リン酸緩衝食塩液で透析した。
【0057】
実施例7
(アジュバント添加)
前記実施例6と同様にして得られた減毒化壊死毒素に水酸化アルミニウムゲル及び保存剤(チメロサールあるいはホルマリン)を加えた。
【0058】
実施例8
(マウス力価試験)
前記実施例7と同様にして得られたトキソイドを5週齢ddYマウスの腹腔内に0.5mlずつ2週間間隔で2回注射した。最終免疫の2週後に約50(25〜100)LD50の壊死毒素で腹腔内攻撃し生死を7日間観察した。結果を図5に示した。トキソイド量とマウス生残率に正の相関関係が認められた。
【0059】
実施例9
(育成ブタでの安全性試験)
前記実施例2と同様にして得られた画分を前記実施例6のとおりに減毒化、前記実施例7のとおりに水酸化アルミニウムゲルを加えたトキソイド2〜50μg/2ml/ドースを調製し、10週齢SPFブタに3週間隔で2回筋肉内注射した。注射による臨床症状や体温の異常はいずれのブタにも認められなかった。
【0060】
実施例10
(妊娠ブタでの有効性試験)
前記実施例5と同様にして得られたHPLC画分を、前記実施例6のとおりに減毒化、前記実施例7のとおりに水酸化アルミニウムゲルを加えたトキソイド(25μg/ml)を妊娠ブタに分娩6及び3週前の2回、2mlずつ筋肉内注射した。対照ブタにはプラセボワクチンを注射した。初乳を十分に摂取させた産子について、3日齢時にBbS611株2億個を全頭の鼻内に、さらに半数の子豚について2及び3週齢時の2回、壊死毒素2600MNDを筋肉内注射により攻撃した。壊死毒素中和抗体価の成績を表5に記した。分娩時の母ブタ血中BbT中和抗体価は32倍、初乳中は256倍、産子の移行抗体価は128倍であった。この移行抗体は6週間以上陽性であった。プラセボ注射ブタ及びその産子は抗体陰性であった。攻撃試験の成績を表6に記した。鼻甲介萎縮は対照群の12頭中9頭に+〜+++のスコアーで認められたが、免疫群には認められなかった。鼻内からの菌回収には両群の差は認められなかった。増体重は、図6に示したとおりBb生菌に加え壊死毒素による攻撃を実施した群間で差が認められた。2週齢時に実施した1回目の壊死毒素攻撃では、対照群の増体率は免疫群の約1/3に低下し、2回目攻撃(3週齢時)により対照群の全頭が死亡した。
【0061】
【表5】
【0062】
【表6】
【0063】
実施例11
(育成ブタでの有効性試験)
前記実施例2と同様にして得られた画分を前記実施例6のとおりに減毒化、前記実施例7のとおりに水酸化アルミニウムゲルを加えたトキソイド2〜50μg/2ml/ドースを調製し、10週齢SPFブタに3週間隔で2回筋肉内注射した。最終免疫から4週後に2億個のBb生菌で鼻内攻撃し、攻撃後3週目までの壊死毒素中和抗体価及を検査した。表7に示したとおり対照ブタは攻撃後もBbT中和抗体陰性であった。免疫群は全頭がBbT中和抗体が陽転した。攻撃により2及び6μg/2ml/ドースのトキソイド注射ブタは急激な中和抗体の上昇が認められたが、20及び50μg/2ml/ドース以上では上昇しなかった。
【0064】
壊死毒素には抗体産生抑制作用のあることが報告されている(Sekiya,K.,マイクロバイオロジー アンド イムノロジー: Microbiol.Immunol.,27,p.905〜915(1983))。攻撃後、対照ブタに中和抗体が上昇しない理由は、攻撃菌の産生するBbTの作用によるものと考えれらえる。従って、免疫群における急激な中和抗体の上昇はBbTの抗体産生抑制作用を防御した結果であることから、育成ブタでは2μgのBbTトキソイドでも有効であることが示唆された。
【0065】
【表7】
【0066】
調製例4
(菌体破砕抽出液:BbTトキソイド1)
BbS611株をペプトン培地で37℃、24時間通気撹拌培養した。菌体を遠心濃縮後、菌体破砕処理し菌体残渣を遠心あるいは膜ろ過(ポアー径0.45μm)で除去した溶液(粗BbT画分)にホルマリンを終濃度0.8%に加えて37〜40℃で7日間減毒した。このトキソイドをリン酸緩衝食塩水に対して透析した。透析内容物に水酸化アルミニウムゲルを1mg(アルミニウム量換算)/mlに加えてBbTトキソイド1とした。
【0067】
調製例5
(BbTトキソイド2)
前記調製例4と同様にして得られた粗BbT画分をセルロース硫酸エステルゲルカラムクロマトグラフィーにより精製した。20mMリン酸緩衝液(pH8.0)で平衡化したセルロース硫酸エステルゲルに粗BbTを通液し、同緩衝液で洗浄した。BbTは1.5MNaCl,1M尿素添加20mMリン酸緩衝液(pH8.0)でセルロース硫酸エステルゲルカラム(7.8cm(内径)×45cm)から溶出した。この溶出画分(CS1画分)を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド2とした。
【0068】
調製例6
(BbTトキソイド3)
前記調製例5と同様にして得られたCS1画分をさらにTSKゲルHW65(2.64cm(内径)×69cm)及びHW75(2.64cm(内径)×64cm)連結カラムで精製した。0.3M硫酸ナトリウム、1M尿素加0.05Mリン酸緩衝液(pH7.2)で平衡化させたカラムにCS1画分を通液し、同緩衝液で分画後、壊死毒活性を有する画分を混合した。この画分を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド3とした。
【0069】
調製例7
(BbTトキソイド4)
前記調製例4と同様にして得られた粗BbT画分をセルロース硫酸エステルゲルクロマトグラフィーにより精製した。0.1MNaCl加20mMリン酸緩衝液(pH8.0)で平衡化したセルロース硫酸エステルゲルに粗BbTを通液し、同緩衝液で洗浄した。BbTは0.5MNaCl加20mMリン酸緩衝液(pH8.0)でセルロース硫酸エステルゲルから溶出した。この画分(CS2画分)を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド4とした。
【0070】
調製例8
(BbTトキソイド5)
前記調製例7と同様にして得られたCS2画分をさらにAffi-gel blueカラム(BioRad社製、100-200メッシュ、5.0cm(内径)×17cm)で精製した。50mM トリス緩衝液(pH 7.5)で平衡化させたAffi-gel blue カラムに同緩衝液に対して透析したCS2画分を通液し、1M リン酸2ナトリウム加50mMトリス緩衝液(pH 7.5)で洗浄した。さらに50mMトリス緩衝液(pH 7.5)で洗浄後、2M塩化マグネシウム、1M尿素加50Mmトリス緩衝液(pH 7.5)でBbTを溶出した。この画分を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド5とした。
【0071】
調製例9
(BbT抗毒素)
前記実施例1及び5と同様にして得られた高度精製BbT画分を前記調製例4と同様に減毒処理を行ない、透析後、水酸化アルミニウムゲルを加えてBbTトキソイド6とした。さらに、27週齢SPFブタに対して当該トキソイドを3か月間で1mlずつ8回筋肉内接種してBbT抗毒素(中和価4,096倍、菌凝集抗体価20倍以下)を得た。
【0072】
調製例10
(PmTトキソイド)
毒素原性PmD型s70株をハートインフュージョンブロスで37℃、24時間通気撹拌培養した。菌体を遠心濃縮後、菌体破砕処理し菌体残渣を遠心あるいは膜ろ過(ポアー径0.45μm)で除去した溶液(粗PmTと略)を陰イオン交換クロマトグラフィーにより精製した。0〜0.3MNaCl加中性リン酸緩衝液またはトリス緩衝液で平衡化した陰イオン交換体に粗PmTを通液し、同緩衝液で洗浄した。PmTは0.4〜0.5MNaClを含む同緩衝液でイオン交換体から溶出した。この部分精製PmT画分にホルマリンを0.4%に加え、37℃で14日間以上減毒した。このトキソイドをリン酸緩衝食塩水に対して透析した。透析内容物に水酸化アルミニウムゲルを1mg Al/mlに加えてPmTトキソイドとした。
【0073】
実施例12
(各種精製方法により得られたBbTトキソイドのモルモットでの免疫原性)
前記調製例4〜8と同様にして得られた3,000 MND/ml(減毒前活性)のBbTトキソイドをモルモット(Hartley,雌,5週齢)の大腿部筋肉内に0.5ml注射した。注射後28日目に採血し、BbT中和抗体価を検査した。中和抗体測定法は、非働化血清をリン酸緩衝食塩水で2倍階段希釈した。これに等量のBbT(力価 4MND/0.1ml)を加え、氷冷下で一夜反応後、モルモットの皮下に0.1mlずつ注射した。判定は注射後1日目に実施し、中和価は壊死反応を阻止した血清の最大希釈倍数(血清希釈倍率)で示した。
【0074】
表8に示したとおり、比活性が1.4×104 MND/mg蛋白のBbTトキソイド1及び2を注射したモルモットではすべて中和抗体が陰性であった。比活性が3.7×104 MND/mg 蛋白のBbTトキソイド3を注射したモルモットでは3頭中2頭が中和抗体が陽性で、7.8×104MND/mg 蛋白以上では全頭陽転した。
【0075】
BbTは精製の過程で不溶化しやすく、脂質、酸性蛋白など菌体由来の物質と複合体を形成しやすい性質を有している(Wordlaw,A.C.and Parton,R.,Parmacol.Ther.,19,p.1〜53(1983))。そのため、これらの菌体由来物質を低減することにより、より十分な減毒化が可能になるか、あるいはまた毒素活性部位のマスキングを減少させ抗原提示効率が向上することによりBbTトキソイドの免疫原性が高まるものと考えられる。
【0076】
【表8】
【0077】
実施例13
(BbT中和抗体による防御ライン)
前記調製例10で得られたBbT抗毒素 0.2,1.0,2.0及び20mlを2日齢SPFブタの腹腔内に注射して受身免疫を賦与した。3日齢時にBbS611株108CFUで鼻内攻撃し、さらに7日齢時に粗BbT(1,940MND/頭)で筋肉内攻撃した。図7に示したとおり、BbTの致死作用に対してはBbT中和抗体価1倍以上で防御が認められ、壊死作用に対しては4倍以上で防御が可能であった。
【0078】
実施例14
(PmT中和抗体による防御ライン)
前記調製例10と同様にして得られた粗PmTのトキソイドを等量のフロイント不完全アジュバントで乳化した。8頭のSPF妊娠ブタのうち6頭を免疫群、2頭を対照群とした。免疫群は妊娠期間中に3mlのトキソイド(840 MND/ml)を2〜5回筋肉内注射した。分娩後、初乳を産子に十分飲ませ2日齢または9日齢時に20〜160MNDの粗PmTで筋肉内攻撃した。子ブタは攻撃後14日間観察し鼻甲介病変を観察した。表9に示したとおり、PmTの鼻甲介萎縮作用に対してはPmT中和抗体価2倍以上で防御が認められ、致死作用に対しては2倍以下でも防御が可能であった。
【0079】
【表9】
【0080】
実施例15
(BbT抗毒素受身免疫ブタのBb及びPm攻撃試験)
前記調製例10で得られたBbT抗毒素20mlを2日齢SPFブタの腹腔内に注射して受身免疫を賦与した。3日齢及び7日齢時にそれぞれBbS611株108CFU及びPmD型s70株108CFUで鼻内攻撃し、6週齢で剖検した。表10に示すように、BbT抗毒素でBbの鼻粘膜から大量の菌が回収されたが、Pmの回収は対照に比べると大幅に少なかった。また、対照群の鼻甲介病変は重度(+++)〜極度(++++)であったが、免疫群は一頭が正常(−)で残りが重度及び極度と個体間における差が大きかった。
【0081】
【表10】
【0082】
壊死毒素は菌体内に産生され、溶菌に伴い菌体外に放出される。従って、BbT抗毒素によってBbの鼻粘膜定着を阻止することはできない。Pmが定着するためには予めBbが粘膜に障害を与えていなくてはならない。Bbの産生する鼻粘膜障害因子としては、BbT(Elias,B.ら,ジャパニーズ ジャーナル ヴェテリナリー サイエンス: Jpn.J.Vet.Sci.,52,p.677〜688(1990))及び気管細胞毒素(Cookson,B.T.and Goldman,W.E.,ジャーナル オブ セルラー バイオケミストリー: J.Cell.Biochem.,11B(Suppl.)p.124;Dugal,F.らカナディアン ジャーナル ヴェテリナリー リサーチ:Can.J.Vet.Res.,56,p.260-264(1992))が重要である。気管細胞毒素はPmの定着促進に重要な気管線毛運動停止作用や線毛上皮細胞崩壊作用及び粘液分泌亢進作用を有するため、BbT抗毒素による免疫ではPm定着を部分的にしか阻止することはできない。
【0083】
これらの事実からPmの鼻粘膜への定着を阻止するためには、少なくともBbT及び気管細胞毒に対する免疫を成立させる必要がある。しかし、気管細胞毒トキソイドは精製方法の確立、免疫原性、収量など実用化までには多くの問題を抱えている。そのため、萎縮性鼻炎の主症状を予防し経済的損失を抑制するためのAR不活化ワクチンとしては少なくとも壊死毒混合トキソイドを含有する必要がある。
【0084】
実施例16
(トキソイド混合例)
前記実施例2及び6と同様にして得られた減毒化BbTに水酸化アルミニウムゲルを加え攪拌した。BbT濃度を8,700〜220,000MND/ml(減毒前毒素量BbT蛋白で2〜50μg/mlに相当)に、アルミニウム濃度を0.5〜2.5mg/mlになるよう調製し、保存剤を加えてBbT原液とした。
【0085】
前記調製例11と同様にして得られた水酸化アルミニウムゲル添加減毒化PmTを3,400MND/ml以上(減毒前毒素量)に調製し保存剤を加えてPmT原液とした。保存剤としては、0.01%チメロサールあるいは0.1〜0.4%ホルマリンを使用した。BbT原液及びPmT原液を等量ずつ加えて混合トキソイドを調製した。
【0086】
この混合トキソイドを9週齢SPFブタの頚部筋肉内に3週間隔で2回、2mlずつ接種した。経時的に血清を採取し、BbT中和抗体価並びにPmT中和抗体価を測定した。PmT中和抗体価は牛胎仔肺細胞(Rutter,J.M. and Luther,P.D.,ヴェテリナリー レコード:Vet.Rec.,114,p.393〜396(1984))を用いた中和試験により求めた。ブタでの中和抗体価を表11に、マウスでの力価試験成績を表12に示した。BbT及びPmTの各単味トキソイドと混合トキソイドはブタ及びマウスに注射したとき免疫原性の差が認められず、防御に十分な中和抗体応答が認められ、また安全性も十分であった。
【0087】
【表11】
【0088】
【表12】
【図面の簡単な説明】
【0089】
【図1】図1は本発明による壊死毒素精製法の過程のクロマトグラムを示す。
【0090】
【図2】図2は本発明による壊死毒素精製法の過程のクロマトグラムを示す。
【0091】
【図3】図3は本発明による壊死毒素精製法の過程のクロマトグラムを示す。
【0092】
【図4】図4は本発明による壊死毒素精製法を用いて得られた壊死毒素画分の分析結果を示す。
【0093】
【図5】図5は本発明によって得られるトキソイドのマウス力価試験の結果を示す。
【0094】
【図6】図6は本発明によって得られるトキソイドの妊娠ブタでの有効性試験の結果を示す。
【0095】
【図7】図7はBbT中和抗体による防御ラインの検討結果を示す。
【特許請求の範囲】
【請求項1】
ボルデテラ・ブロンキセプティカ(気管支敗血症菌)が産生する壊死毒素の含有液を、直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルまたは天然高分子重合体または人工高分子重合体からなるクロマト担体ゲルにヘパリン、スルフォプロピル基もしくは分子内にスルフォン基を有するブルー色素が単独でもしくは組み合わされて結合されたゲルに接触せしめ、当該壊死毒素を吸着させた後、吸着した壊死毒素をゲルから溶出することによって得られる部分精製壊死毒素、あるいは当該部分精製壊死毒素を他の精製方法と組み合わせてさらに高純度に精製したものを、化学的処理により免疫原性を保持しながら活性を消失させた、37,000最少壊死毒単位(以下、MNDと略称する)/mg蛋白以上の比活性を有するボルデテラ・ブロンキセプティカ壊死毒素のトキソイド。
【請求項2】
該部分精製壊死毒素が、下記の工程:
(1) 直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを、pH7〜9、3〜20mS/cmの範囲にある所定の比電導度以下の緩衝液であらかじめ処理して平衡化したのち、
(2) ボルデテラ・ブロンキセプティカ壊死毒素含有液をpH7〜9、温度0〜30℃、3〜20mS/cmの範囲にある所定の比電導度以下の条件下で直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルに接触させ、
(3) 壊死毒素のゲルからの溶出処理に先だって、吸着ゲルをpH5〜10、3〜20mS/cmの範囲にある所定の比電導度以下の緩衝液で洗浄し、
(4) 壊死毒素のゲルからの溶出を、pH5〜10、3〜20mS/cmの範囲にある所定の比電導度以上〜130mS/cmの緩衝液を用いて行なう
からなる方法によって得られる部分精製壊死毒素である請求項1に記載のボルデテラ・ブロンキセプティカ壊死毒素のトキソイド。
【請求項3】
ボルデテラ属菌が産生する壊死毒素のトキソイドおよびパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする混合ワクチン。
【請求項4】
請求項1または2に記載のボルデテラ・ブロンキセプティカ(気管支敗血症菌)が産生する壊死毒素のトキソイドおよびパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする請求項3に記載の混合ワクチン。
【請求項5】
37,000MND/mg蛋白以上の比活性を有するボルデテラ属菌が産生する壊死毒素のトキソイドを含有する請求項3に記載の混合ワクチン。
【請求項1】
ボルデテラ・ブロンキセプティカ(気管支敗血症菌)が産生する壊死毒素の含有液を、直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルまたは天然高分子重合体または人工高分子重合体からなるクロマト担体ゲルにヘパリン、スルフォプロピル基もしくは分子内にスルフォン基を有するブルー色素が単独でもしくは組み合わされて結合されたゲルに接触せしめ、当該壊死毒素を吸着させた後、吸着した壊死毒素をゲルから溶出することによって得られる部分精製壊死毒素、あるいは当該部分精製壊死毒素を他の精製方法と組み合わせてさらに高純度に精製したものを、化学的処理により免疫原性を保持しながら活性を消失させた、37,000最少壊死毒単位(以下、MNDと略称する)/mg蛋白以上の比活性を有するボルデテラ・ブロンキセプティカ壊死毒素のトキソイド。
【請求項2】
該部分精製壊死毒素が、下記の工程:
(1) 直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルを、pH7〜9、3〜20mS/cmの範囲にある所定の比電導度以下の緩衝液であらかじめ処理して平衡化したのち、
(2) ボルデテラ・ブロンキセプティカ壊死毒素含有液をpH7〜9、温度0〜30℃、3〜20mS/cmの範囲にある所定の比電導度以下の条件下で直接的硫酸化により硫酸基が導入されたクロマト担体ゲルもしくは硫酸基を有する分子が共有結合されているクロマト担体ゲルに接触させ、
(3) 壊死毒素のゲルからの溶出処理に先だって、吸着ゲルをpH5〜10、3〜20mS/cmの範囲にある所定の比電導度以下の緩衝液で洗浄し、
(4) 壊死毒素のゲルからの溶出を、pH5〜10、3〜20mS/cmの範囲にある所定の比電導度以上〜130mS/cmの緩衝液を用いて行なう
からなる方法によって得られる部分精製壊死毒素である請求項1に記載のボルデテラ・ブロンキセプティカ壊死毒素のトキソイド。
【請求項3】
ボルデテラ属菌が産生する壊死毒素のトキソイドおよびパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする混合ワクチン。
【請求項4】
請求項1または2に記載のボルデテラ・ブロンキセプティカ(気管支敗血症菌)が産生する壊死毒素のトキソイドおよびパスツレラ属菌が産生する壊死毒素のトキソイドを主成分とする請求項3に記載の混合ワクチン。
【請求項5】
37,000MND/mg蛋白以上の比活性を有するボルデテラ属菌が産生する壊死毒素のトキソイドを含有する請求項3に記載の混合ワクチン。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2006−342175(P2006−342175A)
【公開日】平成18年12月21日(2006.12.21)
【国際特許分類】
【出願番号】特願2006−214582(P2006−214582)
【出願日】平成18年8月7日(2006.8.7)
【分割の表示】特願平8−501925の分割
【原出願日】平成7年6月7日(1995.6.7)
【出願人】(000173555)財団法人化学及血清療法研究所 (86)
【Fターム(参考)】
【公開日】平成18年12月21日(2006.12.21)
【国際特許分類】
【出願日】平成18年8月7日(2006.8.7)
【分割の表示】特願平8−501925の分割
【原出願日】平成7年6月7日(1995.6.7)
【出願人】(000173555)財団法人化学及血清療法研究所 (86)
【Fターム(参考)】
[ Back to top ]