
ポックスウイルスゲノムに挿入された相同遺伝子を発現させる組換えポックスウイルス
【課題】本発明は、ある微生物の異なる変異体に由来し50%以上の相同性を有する2つ以上の相同な外来配列を発現させる能力を有する組換えポックスウイルスを提供することを目的とする。
【解決手段】本発明は、ある微生物の異なる変異体に由来し50%以上の相同性を有する2つ以上の相同な外来配列を発現させる能力を有する組換えポックスウイルスに関する。さらに本発明は、そのような組換えポックスウイルスを作製する方法、およびそのような組換えポックスウイルスの医薬またはワクチンとしての使用に関する。また、ヒトを含む生きている動物において、免疫応答に影響を及ぼす方法、好ましくは免疫応答を誘発する方法も提供する。
【解決手段】本発明は、ある微生物の異なる変異体に由来し50%以上の相同性を有する2つ以上の相同な外来配列を発現させる能力を有する組換えポックスウイルスに関する。さらに本発明は、そのような組換えポックスウイルスを作製する方法、およびそのような組換えポックスウイルスの医薬またはワクチンとしての使用に関する。また、ヒトを含む生きている動物において、免疫応答に影響を及ぼす方法、好ましくは免疫応答を誘発する方法も提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、2つ以上の相同な外来遺伝子を発現させる能力を有する組換えポックスウイルスに関する。上記の遺伝子は、ウイルスゲノムにとっては異種であるが、互いに比較した場合には相同である。これらの遺伝子は、特に、ある微生物の近縁変異体またはサブタイプに由来する。さらに本発明は、そのような組換えポックスウイルスを作製する方法、およびそのような組換えポックスウイルスの医薬またはワクチンとしての使用に関する。また、ヒトを含む生きている動物において、免疫応答に影響を及ぼす方法、好ましくは免疫応答を誘発する方法も提供する。
【背景技術】
【0002】
あらゆる生体は、細菌、ウイルス、真菌または寄生虫などの感染性因子または病原性因子の攻撃を絶えず受けている。いわゆる免疫系は、当該生物が、そのような因子によって引き起こされる持続的な感染、疾患または中毒に陥るのを防いでいる。
【0003】
哺乳動物の免疫系は特異的部分と非特異的部分とに分類することができる。ただし両者の間には密接な相互関係がある。非特異的免疫応答は、多種多様な病原性物質または感染性因子に対する即時防御を可能にする。特異的免疫応答は、生物がある物質による攻撃を初めて受けた場合には、誘導期間を経てから生じる。この特異的免疫応答は、主として、抗原特異的な抗体の産生と、マクロファージおよびリンパ球、例えば細胞傷害性T細胞(CTL)の生成とに基づいている。特異的免疫応答は、ある特定の感染から回復した個体はその特定の感染からは保護されるが、他の感染性疾患には依然として罹りやすいという事実の原因である。一般に、同じ感染性因子または極めて似ている感染性因子による2回目の感染では、症状がはるかに軽いか、または症状が全く現れない。このいわゆる免疫は長期間持続し、場合によっては一生涯続くことさえある。その基礎をなしている効果はしばしば免疫記憶と呼ばれ、これを予防接種に利用することができる。
【0004】
予防接種という用語は、ある個体を、無害な部分型または不活化型の感染性因子にばく露して、その個体における免疫応答に影響を及ぼす(好ましくは免疫応答を誘発する)ことにより、その特定感染性因子に対して(一生涯ではないとしても)長期間持続する免疫を生じさせる方法を表す。
【0005】
ヒト痘瘡疾患は天然痘ウイルスによって起こる。天然痘ウイルスはポックスウイルス科に属する。ポックスウイルス科は、脊椎動物細胞および無脊椎動物細胞の細胞質中で複製する複雑なDNAウイルスからなる大きな一科である。
【0006】
ポックスウイルス科は、脊椎動物宿主域および昆虫宿主域に基づいて2つの亜科、すなわちチョルドポックスウイルス亜科とエントモポックスウイルス科とに分類することができる。チョルドポックスウイルス亜科には、とりわけ、オルトポックスウイルス属およびアビポックスウイルス属が含まれている(非特許文献1)。
【0007】
オルトポックスウイルス属には、ヒト痘瘡の原因因子である天然痘ウイルスが含まれると共に、ラクダ痘ウイルス、牛痘ウイルス、羊痘ウイルス、ヤギ痘ウイルス、サル痘ウイルスおよびワクシニアウイルスなどの経済的に重要な他のウイルスも含まれる。この属のメンバーはすべて遺伝的に類縁関係にあり、よく似た形態または宿主域を有する。制限エンドヌクレアーゼ地図も、オルトポックスウイルス属の異なるメンバー間で、90%に達する高い配列一致度を示している(非特許文献2)。
【0008】
ワクシニアウイルス(VV)とは、少なくとも過去100年間にわたって痘瘡の予防接種に使用された因子に与えられた名称である。VVが、長期間にわたる連続継代により、牛痘ウイルスから派生した新種であるのか、今では絶滅したウイルスの生きた代表例である天然痘ウイルスから派生した新種であるのかは分かっていない。あるいは、VVは、遺伝子組み換えの産物なのかもしれない。また、VVの歴史の中で、多くのワクシニア株が生じた。これらのさまざまな株はさまざまな免疫原性を示し、潜在的合併症(その最も重篤なものは種痘後脳炎である)とさまざまな程度に関係づけられている。しかし、これらの株の多くは、痘瘡の予防接種に使用された。例えばNYCBOH株、ウェスタンリザーブ(Western Reserve)株またはワイエス(Wyeth)株は主に米国で使用され、アンカラ(Ankara)株、ベルン(Bern)株、コペンハーゲン(Copenhagen)株、リスター(Lister)株およびMVAは主に欧州での予防接種に使用された。これらさまざまVV株による世界的な予防接種計画の結果、1980年に、WHOはついに、天然痘ウイルスの根絶の成功を宣言した。
【0009】
現在、VVは主に実験株として使用されているが、それ以外に、今でもオルトポックスウイルス属の原型であるとみなされている。これは、VVが最も詳細に特徴づけられたウイルスの一つになった理由でもある(非特許文献3)。VVは、そしてまた最近では他のポックスウイルスも、外来遺伝子を挿入し発現させるために使用されてきた。生きている感染性ポックスウイルスに外来遺伝子を挿入するための基礎技術では、ドナープラスミド中の外来遺伝要素に隣接するポックスDNA配列と、レスキューポックスウイルス(rescuing poxvirus)中に存在する相同な配列との間で、組換えが行われる。遺伝子組換えとは、一般に、2本のDNA鎖間で起こる相同なDNA区間の交換である。ウイルスによっては、RNAがDNAの代わりをする場合もある。相同な核酸区間とは、同じヌクレオチド塩基配列を有する核酸(DNAまたはRNA)の区間である。遺伝子組換えは、感染した宿主細胞内での新しいウイルスゲノムの複製または産生中に自然に起こりうる。したがって、ウイルス遺伝子間の遺伝子組換えは、2種類以上のウイルスまたは他の遺伝子コンストラクトに同時感染した宿主細胞内で起こるウイルス複製サイクル中に起こりうる。第1のゲノムに由来するDNA区間は、第2の同時感染ウイルスのゲノムのうち、そのDNAが第1ウイルスゲノムのDNAと相同である区間を構築する際に、互換的に使用される。
【0010】
挿入されたDNA遺伝子配列を改変感染性ウイルスによってうまく発現させるには、2つの条件が必要である。第1に、改変ウイルスが依然として生存可能であるように、挿入はウイルスの非必須領域になされるべきである。挿入されたDNAを発現させるための第2の条件は、挿入されたDNAに対して適切な関係にあるプロモーターが存在することである。通例、プロモーターは、発現させるべきDNA配列の上流に配置する。
【0011】
例えばB型肝炎ウイルス表面抗原(HBsAg)、インフルエンザウイルス赤血球凝集素(InfHA)または二日熱マラリア原虫(Plasmodium knowlesi)スポロゾイト抗原などを発現させる組換えVVの、感染性疾患予防用の生ワクチンとしての有用性は、実証されており、それに関する総説もある(非特許文献4)。
【0012】
VVのもう一つの利点は、1つのVVゲノム内に複数の外来配列、外来遺伝子または外来抗原を取り込む能力を有することである(非特許文献5)。また、1回の多価ワクチン接種で多くの異種の感染性疾患に対する免疫を誘発することができると報告されている(非特許文献6)。
【0013】
単一のVVによってさまざまな抗原を発現させる例の一つが、Brayらによって記載されている。デングウイルス血清型4の3種類の構造タンパク質、すなわちキャプシド(C)タンパク質、前駆膜(pre−membrane)(prM)タンパク質、エンベロープ(E)タンパク質と、デングウイルス血清型4の2つの非構造タンパク質、すなわちNS1およびNS2aとを発現させることができる組換えVVは、相同なデングウイルス血清型4の攻撃からマウスを保護する能力を有することが示された(非特許文献7)。
【0014】
デングウイルス血清型1(Den−1)からデングウイルス血清型4(Den−4)まで4つの血清型を有するデングウイルスは、ヒトの感染に関して、フラビウイルス属の重要な一メンバーである。デングウイルス感染は、インフルエンザ様の症状から重篤なまたは致死的な病気、すなわちショック症候群(DSS)を伴うデング出血熱(DHF)に及ぶ疾患をもたらす。デングの突発は、媒介動物である蚊が多い熱帯地方および亜熱帯地方の人口密集地域では、依然として公衆衛生上の大きな問題である。
【0015】
デング感染および蚊媒介性フラビウイルスが誘発する他の疾患が、世界の多くの地域で蔓延することを懸念して、デング熱(DF)とデング出血熱(DHF)の両方を防止することができるデングワクチンの開発に向けた努力が盛んに行われるようになり、予防接種を受けた個体を蚊媒介性フラビウイルスの一部または全部によって誘発される感染症から保護するのに役立つワクチンがもたらされている。
【0016】
大半のDF症例が4つの血清型のいずれかによる最初の感染後に顕在化するのに対して、DHF症例は、かなりの部分が、最初に感染したデングウイルスの血清型とは異なる血清型による2回目の感染を受けた患者に起こる。これらの知見から、一定数の症例では、あるデングウイルス血清型に対する抗体を有する個体が適当な間隔を置いて異なるウイルス血清型に逐次的に感染することによってDHFが起こるという仮説がもたらされる。
【0017】
したがって、1つの血清型に対する予防接種は、デングウイルス感染からの完全な保護をもたらすのではなく、同じデングウイルス株による感染からの保護をもたらすにすぎない。さらに重要なことに、1つの血清型に対する予防接種を受けた人では、その人が血清型の異なるデングウイルス株に感染した場合にデング出血熱などの重篤な合併症を発症する危険性が増大している。
【0018】
したがって、4つのデングウイルス血清型のすべてに由来する抗原を有する多価ワクチンが望ましい。
【0019】
これまでは、各VVが異なるウイルスの配列をコードしている一群の組換えVVを混合することによって、多価ワクチンを製造することが提案されていた(非特許文献8)。しかし、そのような多価ワクチンにはいくつかの欠点がある。第1に、いくつかの独立した組換えVVを作製するのは面倒である。個別の製造プロセスだけでなく、品質管理と品質保証にも、著しく時間がかかる。第2に、異なる配列を発現させる組換えウイルスの混合物による感染には、感染事象があまりバランスよく起こらないという危険が常に伴う。主な危険は、多価ワクチンに含まれるすべての異なる組換え体が標的細胞に感染するわけではなく、個々の組換え体による標的細胞の感染しか起こらないというものである。理由の1つとして、組換えウイルスの不均等な分布が考えられる。もう一つの理由として、単一の細胞に感染する際に異なる組換えウイルス間で起こる干渉が考えられる。そのような干渉は重感染現象として知られている。この場合、最終的に感染細胞から発現され、その結果、患者の免疫系に提示されるのは、多価ワクチンの異なる抗原のすべてではなく、一部の抗原だけになる。その結果、多価ワクチンが提示するまたは提示することのできるさまざまな抗原に対する完全な免疫保護は到底もたらされず、一部の抗原に対する免疫保護しか得られないことになる。
【0020】
デングウイルス感染に対するワクチンの場合、多価ワクチンによるアプローチには、デングウイルス2のエンベロープタンパク質に関して報告例(非特許文献9)があるように、異なる配列が異なる量で発現されたり、予測不可能な形で発現されたりすると、その予防接種は患者にとって著しく危険であるという欠点がある。一群の組換えワクシニアウイルスを使った不完全な予防接種では、デングウイルスのすべての血清型に対する免疫保護ではなく、その一部に対する免疫保護しか得られない。残念なことにデング感染の場合、不完全な予防接種は、それによってデングウイルス出血熱などの致死的合併症の危険が増大するため、全く受け入れがたい。
【先行技術文献】
【非特許文献】
【0021】
【非特許文献1】Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第3版,1996年,ISBN:0−7817−0253−4,第83章
【非特許文献2】MackettおよびArchard[1979年]J Gen Virol,45:683−701
【非特許文献3】Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第3版,1996年,ISBN:0−7817−0253−4,第83章および第84章
【非特許文献4】Smith等[1984年]Biotechnology and Genetic Engineering Reviews 2,383−407
【非特許文献5】SmithおよびMoss[1983年]Gene,25(1):21−28
【非特許文献6】Perkusら[1985年]Science,Vol.229,981−984
【非特許文献7】Bray等[1989年]Virology 2853−2856
【非特許文献8】Moss[1990年]Immunology,2,317−327
【非特許文献9】Deubleら[1988年]J. Virol 65:2853
【発明の概要】
【発明が解決しようとする課題】
【0022】
したがって、ある感染性疾患原因微生物の2つ以上の株、クレード、変異体、サブタイプまたは血清型によって起こりうる感染性疾患に対する安定で有効で信頼できるワクチンを提供することが、本発明の目的である。
【0023】
また、すべてのデングウイルス血清型に対する信頼できる予防接種を可能にする、デングウイルス感染に対する安定で有効で信頼できるワクチンを提供することが、本発明のもう一つの目的である。
【課題を解決するための手段】
【0024】
本発明は、ある感染性疾患原因微生物の異なる株、クレード、変異体、サブタイプまたは血清型に由来する相同な遺伝子を1つのポックスウイルスに含めるという着想に基づいている。上述のように、例えばデングウイルスには4つのグループ、サブタイプまたは血清型が存在し、それらはいずれも同じタイプの遺伝子を、例えばキャプシド(C)タンパク質をコードする遺伝子、前駆膜(PrM)タンパク質またはエンベロープ(E)タンパク質をコードする遺伝子などとして含んでいる。しかし、同じタイプの遺伝子の核酸配列は、4つの血清型のすべてで完全に同じわけではなく、完全に相同なわけでもない。例えば、デングウイルス血清型1、2、3および4のPrM遺伝子(PrM1〜4)間で配列を比較したところ(Lasergene 4.05 Magalign,Macintosh)、66.5〜72.9%の配列一致度、すなわち約65〜75%の相同性が明らかになった。感染性疾患原因微生物の異なるサブタイプの遺伝子に見られる相違とばらつきは、それぞれ、1つのサブタイプに対して予防接種しても、同じ微生物の他の変異体による感染に対する保護が自動的にはもたらされないことの理由であると考えられる。そこで、ある感染性疾患原因微生物の異なる株、クレード、変異体、サブタイプまたは血清型に由来する近縁遺伝子または相同遺伝子を含んでいる組換えウイルスを作製することを着想した。しかし、上述したように、相同配列間の相同組換えはウイルスの生活環中に起こり、完全には相同ではないDNA区間の間でさえ起こる。したがって、単一のウイルスゲノムに相同な遺伝子を挿入すると相同組換えが起こり、その結果、挿入した相同遺伝子の欠失が起こると予想された。
【0025】
しかし、少なくとも60%の相同性を有する少なくとも2つの外来遺伝子をゲノム内に含む組換えポックスウイルスを作製したところ、意外にも、前記相同遺伝子はウイルスゲノム中に安定に挿入されたままであることを見い出した。
【0026】
相同な遺伝子(好ましくは少なくとも50%の相同性を有するもの)をウイルスゲノムの異なる挿入部位に挿入した場合でも、それらの遺伝子はウイルスゲノムに安定に挿入されたままである。この場合、前記相同遺伝子間の組換え事象は、ウイルスの増幅やウイルスの生活環にとって重要なウイルス遺伝子の喪失をもたらすと予想された。すなわち、ウイルスの生活環は重大な損傷を受けるだろうと予想された。また、組換え頻度は2つの連鎖遺伝子間の距離に比例するので、異なる挿入部位に位置する2つ以上の相同遺伝子間で起こる組換え事象の頻度は高く、したがって当該遺伝子の欠失及び(又は)著しい干渉が起こるだろうと予想された。したがって、組換え事象が起こらず、相同な遺伝子がウイルスゲノムの異なる挿入部位に安定に挿入されたままであることは、極めて驚くべきことであった。
【0027】
先行技術によれば、日本脳炎ウイルス(JEV)、黄熱ウイルス(YFV)およびデングウイルスなどのフラビウイルスに由来する外来DNAを含む組換えポックスウイルスが知られている(米国特許第5,514,375号)。しかし、前記フラビウイルス由来の各遺伝子は、同じ挿入部位に1回挿入されているだけである。また、適当なコンピュータソフトウェア(Lasergene 4.05 Megaline,Macintosh)で配列比較を行ったところ、ポックスウイルスゲノムに挿入された遺伝子の相同性は、JEV由来の遺伝子の場合は20.2%〜29.6%、YFV由来の遺伝子の場合は29.2%〜45.3%、デングウイルス由来の遺伝子の場合は22.8%〜29.5%であることが明らかになった。
【0028】
変異ワクシニアウイルスアンカラ(Modified Vaccinia Virus Ankara:MVA)の同じ挿入部位(特に欠失部位II)にデングウイルス抗原を挿入することが記載されている国際公開第98/13500号にも、同様のことが言える。
【0029】
米国特許第5,338,683号には、gp13およびgp14ヘルペスウイルス糖タンパク質遺伝子を、単一の組換えポックスウイルスの2つの異なる挿入部位に挿入することが記載されている。しかし両遺伝子は25.2%の相同性しか持っていない。
【0030】
変異ワクシニアウイルスアンカラ(MVA)の同じ挿入部位(欠失部位III)に挿入されたインフルエンザウイルスの赤血球凝集素遺伝子と核タンパク質遺伝子とを配列比較したところ、49.1%の相同性がみとめられた(米国特許第5,676,950号、Sutterら[1994年]Vaccine 12:1032)。
【0031】
米国特許第5,891,442号には、伝染性ファブリキウス嚢病(infectious bursal disease)ウイルスのポリタンパク質VP2、VP3およびVP4のコード配列を有する組換えポックスウイルスが開示されている。これらの遺伝子は41.9%〜50.3%の相同性を持ち、融合されて単一の挿入部位に挿入された。
【0032】
最後に、米国特許第6,217,882号には、52.7%の相同性を有する仮性狂犬病ウイルス抗原gp50およびgp63が同じ部位に挿入されている組換え豚痘ウイルスベクターが記載されている。
【0033】
要約すると、先行技術では、少なくとも50%の相同性を有する相同な遺伝子または配列はすべて、ウイルスゲノム内の同じ挿入部位または単一の挿入部位に挿入されているということができる。
【発明を実施するための最良の形態】
【0034】
本発明によれば、相同な遺伝子または配列は、少なくとも50%の相同性、すなわち50%〜100%の相同性、すなわち少なくとも50%の同一ヌクレオチド塩基を有する。50%未満の相同性を有する遺伝子または配列は異種であるとみなすことができる。本発明では、「相同」または「相同性」という用語を、遺伝子または配列を互いに比較する場合に使用するのに対して、「外来」遺伝子、または「外因性」もしくは「異種」配列という用語は、遺伝子または配列をポックスウイルスゲノムと比較する場合に使用する。すなわち、本発明で使用する場合、これらの用語は、自然界では通常はポックスウイルスに付随して見いだされないDNA配列を表す。したがって本発明は、ウイルスゲノムと比較すると異種であるが、互いには相同である少なくとも2つの遺伝子を含んでいる組換えポックスウイルスに関する。「遺伝子」という用語は、例えばタンパク質、ポリペプチド、ペプチド、抗原などをコードするコード配列を表す。相同な遺伝子から翻訳されるタンパク質、ポリペプチドまたはペプチドは、同じ役割を果たし、同じ機能的特性を示す。相同な遺伝子は通例、異なってはいるが関連のある供給源または生物に由来する。本発明の一実施形態によれば、コード配列における相同性は、好ましくは70%〜80%、より好ましくは80%〜90%、または90%〜100%である。65%〜75%の相同性は最も好ましい。
【0035】
本発明の組換えポックスウイルスは、関連する遺伝情報を、1つの単一感染ユニットまたは1つのウイルス粒子だけに含んでいるので、不均等な感染および異なる相同配列の不均衡な発現が起こる危険性はない。したがって、近縁の遺伝子、さらには最近縁の遺伝子、またはほとんど同一な配列を含み、1つの感染細胞中でそれらを発現させる能力を有する本発明の組換えポックスウイルスは、多価ワクチンの作製にとってとりわけ有利である。
【0036】
この利点は、例えばデングウイルスのような、あるウイルスのいくつかの近縁株または血清型が引き起こしうる疾患に対するワクチンの開発にとっては、とりわけ興味深い。異なるデングウイルス血清型の相同遺伝子を含んでいる組換えポックスウイルスを、実施例に記載する。
【0037】
本発明の相同な遺伝子または配列は、例えばベクターウイルスを除く任意のウイルス、任意の細菌、任意の真菌または寄生虫など、任意の微生物に由来することができる。相同な遺伝子または配列は、好ましくは、ある感染性微生物または病原性微生物に由来し、最も好ましくはその微生物の異なる株もしくはクレード、変異体、サブタイプまたは血清型に由来する。
【0038】
「株」または「クレード」という用語は、当業者には周知の技術用語であり、微生物の分類法を表す。この分類体系では、今までに特徴づけられたすべての微生物を、科、属、種、株という階層的序列に分類する(Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第4版,2001)。科のメンバーに関する判定基準が系統学的関係であるのに対して、属は共通する特徴を有する全てのメンバーを含み、種は複製系統を構成して特定の生態的地位を占める多形質群と定義される。「株」または「クレード」という用語は、基本的形態やゲノム構造およびゲノム構成などの共通する特徴を有するが、宿主域、組織向性、地理的分布、弱毒化または病原性などの生物学的特性が異なっている微生物、すなわちウイルスを表す。「変異体」または「血清型」という用語は、軽微なゲノム変異に起因して個別の感染スペクトルまたは抗原特性を示す同じ株(サブタイプともいう)のメンバーを、さらに区別するものである。
【0039】
本発明の別の実施形態では、相同な遺伝子または配列がウイルスから選択され、好ましくはフラビウイルス属に属するウイルス(例えば、好ましくはデングウイルス、ウエストナイルウイルスまたは日本脳炎ウイルスであるが、これらに限らない)、レトロウイルス属に属するウイルス(例えば、好ましくはヒト免疫不全ウイルス(HIV)であるが、これに限らない)、エンテロウイルス属に属するウイルス(例えば、好ましくは手足口病ウイルス、EV71であるが、これらに限らない)、ロタウイルス属に属するウイルス、またはオルトミクソウイルス属に属するウイルス(例えば、好ましくはインフルエンザウイルスであるが、これに限らない)から選択される。フラビウイルスに由来する相同な遺伝子は最も好ましい。
【0040】
別の好ましい実施形態では、相同な遺伝子がデングウイルス遺伝子から選択され、好ましくはC、NS1及び(又は)NS2、あるいは好ましくはE、より好ましくはPrMである。このウイルスの異なる血清型に由来する相同な遺伝子は最も好ましく、それらの遺伝子は、4つのデングウイルス血清型の1つ、2つ、3つまたは全部に由来することができる。
【0041】
さらに別の実施形態では、相同な遺伝子が異なるHIV株またはHIVクレードから選択される。相同な遺伝子は、好ましくは、gag/polコード配列から選択され、より好ましくはenvコード配列から選択され、さらに好ましくは構造及び(又は)調節HIVコード配列の組合せから選択される。
【0042】
本発明に適したベクターウイルスは、選択した宿主細胞(例えば鳥類宿主細胞)中で容易に培養することができるが、ヒトまたはヒト細胞中では、著しく複製欠損性であるか、または実際に複製しないポックスウイルス群から選択される。
【0043】
いくつかの好ましい実施形態では、本発明のポックスウイルスは、カナリア痘ウイルス(Plotkinら[1995年]Dev Biol Stand. vol 84:165−170、Taylorら[1995年]Vaccine,Vol 13.No.6:539−549)、鶏痘ウイルス(Afonsoら[2000年],J Virol,3815−3831、Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第4版,2001,第85章,2916頁)、ペンギン痘ウイルス(Stannardら[1998年]J Gen Virol,79,1637−1649)またはそれらの誘導体を含む群から選択される。これらのウイルスはアビポックスウイルス属に属するので、鳥類細胞中で容易に培養し、増幅することができる。しかし、哺乳類細胞またはヒト細胞では複製欠損性であるため、感染性子孫ウイルスは基本的に産生されないか、ほとんど産生されない。
【0044】
本発明の別の実施形態では、2つ以上の相同な遺伝子を含む組換えポックスウイルスの作製に、ワクシニアウイルス、好ましくは弱毒化ワクシニアウイルスを使用する。
【0045】
ワクシニアウイルス(VV)は短い相同配列の相同組換えを起こすことによって、相同な配列を欠失させうることが知られているが(Howleyら[1996年],Gene 172:233−237)、本発明者らは相同な配列または相同な遺伝子がゲノム中に安定に挿入されている組換えワクシニアウイルスを提供する。これは極めて意外な発見である。というのも、Howelyらによれば、300塩基対(bp)までの短い配列でも、ワクシニアウイルスにおけるゲノムの再編成および相同な配列の欠失を誘発するには十分だったからである。したがって当業者であれば、さらに長い配列ではさらに高い確率で組換え事象が誘発されると予想するだろう。しかし、本発明によれば、完全な相同遺伝子を含む配列でさえ、ワクシニアウイルスのゲノム中に安定に挿入することができる。
【0046】
ワクシニアウイルスの一例は(これに限るわけではないが)、著しく弱毒化され宿主域が制限されたワクシニア株、すなわち変異ワクシニアアンカラ(MVA)である(Sutter,G.ら[1994年]Vaccine 12:1032−40)。MVAは、ワクシニアウイルスのアンカラ株(CVA)をニワトリ胚線維芽細胞で約570代にわたって連続継代することによって作出された(概要についてはMayr,A.ら[1975年]Infection 3,6−14を参照されたい)。長期間にわたるこれらの継代の結果、CVAはそのゲノム配列のうちの約31キロ塩基を欠失させた。得られたウイルス株MVAは、宿主細胞が著しく制限されているとの記載がある(Meyer,H.ら,J. Gen. Virol. 72,1031−1038[1991年])。典型的なMVA株は、ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(European Collection of Animal Cell Cultures)に受託番号ECACC V00120707として寄託されているMVA575である。
【0047】
別の実施形態として、本発明ではMVA−Vero株またはその誘導体を使用することができる。MVA−Vero株はヨーロピアン・コレクション・オブ -アニマル・セル・カルチャーズに、受託番号ECACC V99101431およびECACC 01021411として寄託されている。MVA−Veroの安全性は、国際特許出願第PCT/EP01/02703号に記載の生物学的、化学的および物理的特徴に反映されている。通常のMVAと比較すると、MVA−Veroはゲノム欠失を1つ余分に持っている。
【0048】
本発明ウイルスの「誘導体」という用語は、親ウイルスと同じ特徴を示すがそのゲノムの1つ以上の部分に相違を示す子孫ウイルスを表す。
【0049】
本発明のさらに別の実施形態ではMVA−BNを使用する。MVA−BNはヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズに受託番号ECACC V00083008として寄託されている。MVA−BNウイルスは変異ワクシニアアンカラに由来する著しく弱毒化されたウイルスであることが明らかにされているので、MVA−BNまたはその誘導体を使用することにより、特に安全なウイルスワクチンが作製される。したがって、最も好ましい実施形態では、本発明に従って2つ以上の相同な遺伝子を含むMVA−BNまたはその誘導体を、ウイルスベクターとして使用する。「MVA−BNの誘導体」という用語は、MVA−BNと比較して同じ機能的特徴を有するウイルスを表す。MVA−BNの特徴、あるMVAがMVA−BNおよびその誘導体であるかどうかを調べることを可能にする生物学的アッセイの説明、ならびにMVA−BNまたはその誘導体の作製を可能にする方法は、国際公開第02/42480号(参照により本明細書に組み込まれる)に記載されている。MVA−BNまたはその誘導体の機能的特徴を調べる簡単な方法の一つは、ヒトHaCat細胞におけるその弱毒化および複製の欠如である。
【0050】
本発明の組換えポックスウイルスでは、好ましくは、ポックスウイルス転写制御要素によって、より好ましくはMVA、カナリア痘、鶏痘またはペンギン痘転写制御要素によって、最も好ましくはワクシニアウイルスプロモーターによって、外因性配列の発現を制御する。本発明のポックスウイルス転写制御要素には、他にも、ポックスウイルス系で機能するあらゆる転写制御要素が含まれる。
【0051】
本発明における外因性配列の挿入は、好ましくは、ウイルスゲノムの非必須領域に対して行われる。非必須領域は、例えば、ポックスウイルスの生活環にとって必須でないポックスウイルス遺伝子の遺伝子座またはオープンリーディングフレーム(ORF)である。2つのORFに挟まれた区間を表す遺伝子間領域も、本発明では非必須領域と見なされる。本発明の別の実施形態では、MVAゲノムの天然の欠失部位(参照により本明細書に組み込まれる第PCT/EP96/02926号に開示されているもの)に、外因性配列を挿入する。
【0052】
挿入されるDNAの向きは、本発明の組換えウイルスの機能性または安定性には影響しない。
【0053】
本発明の組換えポックスウイルスは増殖が著しく制限され、その結果、著しく弱毒化されているので、ヒトを含む広範な哺乳動物の処置にとって、さらには免疫不全状態のヒトの処置にとっても、理想的な候補である。したがって本発明は、ヒトを含む動物の生体内で免疫応答を誘発するための医薬組成物およびワクチンも提供する。
【0054】
本医薬組成物は、一般に、1つ以上の薬学的に許容し得るそして/または承認された担体、添加剤、抗生物質、保存剤、アジュバント、希釈剤及び(又は)安定剤を含みうる。そのような補助物質としては、水、食塩水、グリセロール、エタノール、湿潤剤または乳化剤、pH緩衝物質などを挙げることができる。適切な担体は通例、大きくてゆっくり代謝される分子、例えばタンパク質、多糖、ポリ乳酸、ポリグリコール酸、アミノ酸ポリマー、アミノ酸コポリマー、脂質集合体などである。
【0055】
ワクチンを製造するには、本発明の組換えポックスウイルスを生理学的に許容できる形態に変換する。これは、天然痘の予防接種に用いられるポックスウイルスワクチンの製造に関する経験(Stickl,H.ら[1974年]Dtsch. med. Wschr. 99,2386−2392に記載されているもの)に基づいて行うことができる。例えば、精製ウイルスは、5×108 TCID50/mlの力価で、約10mMのトリス(Tris)および140mMのNaCl(pH7.4)中に製剤化して、−80℃で保存される。ワクチン注射剤を製造するには、例えば2%ペプトンおよび1%ヒトアルブミンの存在下で、リン酸緩衝食塩水(PBS)100ml中のウイルス粒子102〜108個を、アンプル(好ましくはガラスアンプル)内で凍結乾燥する。もう一つの選択肢として、ワクチン注射剤は、製剤中のウイルスを段階的に凍結乾燥することによっても製造できる。この製剤は、さらに、例えばマンニトール、デキストラン、糖、グリシン、ラクトースもしくはポリビニルピロリドンなどの添加剤、または、インビボ投与に適した酸化防止剤もしくは不活性ガス、安定剤もしくは組換えタンパク質(例えばヒト血清アルブミン)などの他の助剤も含むことができる。次に、ガラスアンプルを密封して、4℃〜室温で数ヶ月間保存することができる。ただし、必要がない限り、アンプルは−20℃未満の温度で保存することが好ましい。
【0056】
予防接種または治療を行うには、上記の凍結乾燥物を0.1〜0.5mlの水性溶液、好ましくは生理食塩水またはトリス緩衝液に溶解し、それを全身的または局所的に、すなわち非経口投与、皮下投与、筋肉内投与、乱切法、または当業者に知られる他の任意の投与経路によって、投与することができる。当業者であれば、投与様式、投与量および投与回数を、既知の方法で最適化することができる。ただし、最も一般的には、1回目の予防接種注射の約1ヶ月〜6週間後に2回目の注射を行うことによって、患者を予防接種する。
【0057】
本発明の組換えウイルスは、標的細胞への外因性コード配列の導入に使用される。標的細胞への外因性コード配列の導入は、タンパク質、ポリペプチド、ペプチド、抗原およびエピトープをそれぞれインビトロで製造するために使用することができる。また、細胞に相同配列または異種配列を導入する方法は、インビトロ治療およびインビボ治療にも応用することができる。インビトロ治療の場合は、免疫応答を誘発するために、前もって(エクスビボで)本発明の組換えポックスウイルスに感染させておいた単離細胞を、動物の生体に投与する。インビボ治療の場合は、免疫応答を誘発するために、本発明の組換えポックスウイルスを動物の生体に直接投与する。この場合、接種部位を取り巻く細胞は、本発明のウイルスまたはその組換え体に、インビボで直接感染する。感染後、それらの細胞は、外因性コード配列にコードされているタンパク質、ポリペプチド、ペプチドまたは抗原を合成し、次いでそれらを、またはそれらの一部を、細胞表面に提示する。免疫系の専門化した細胞は、そのような外来のタンパク質、ポリペプチド、ペプチド、抗原およびエピトープの提示を認識して、特異的免疫応答を開始する。
【0058】
組換えポックスウイルスを取得する方法またはポックスウイルスゲノムに外因性コード配列を挿入する方法は、当業者にはよく知られている。また、それらの方法は実施例でも説明するし、以下の文献から演繹し、または完遂することもできる。
【0059】
-J.Sambrook,E.F.FritschおよびT.Maniatis著「Molecular Cloning, A laboratory Manual」,第2版,Cold Spring Harbor Laboratory Press,1989年。この文献には、例えばDNAのクローニング、DNAおよびRNAの単離、ウェスタンブロット解析、RT−PCRおよびPCR増幅技術などの標準的な分子生物学技術に関する技術とノウハウが記載されている。
【0060】
-Brian WJ MahyおよびHillar O Kangro編「Virology Methods Manual」,Academic Press,1996年。この文献には、ウイルスの取り扱いと操作に関する技術が記載されている。
【0061】
-AJ DavisonおよびRM Elliott編「Molecular Virology: A Practical Approach」(The Practical Approach Series),IRL Press at Oxford University Press,オックスフォード,1993年の第9章「Expression of genes by Vaccinia virus vectors(ワクシニアウイルスベクターによる遺伝子の発現)」。
【0062】
-「Current Protocols in Molecular Biology」,出版社:John Wiley and Son Inc. 1998年の第16章、第IV節「Expression of proteins in mammalian cells using Vaccinia viral vector(ワクシニアウイルスベクターを使った哺乳動物細胞におけるタンパク質の発現)」。この文献には、MVAの取り扱い、操作および遺伝子工学に関する技術とノウハウが記載されている。
【0063】
本発明の組換えポックスウイルスの作製には、さまざまな方法を応用することができる。ウイルスに挿入すべきDNA配列は、ポックスウイルスの一DNA区間に相同なDNAが挿入されている大腸菌プラスミドコンストラクトに、入れることができる。別途、挿入すべきDNA配列をプロモーターに連結する。このプロモーター−遺伝子連鎖は、非必須遺伝子座を含むポックスDNAの一領域に隣接するDNA配列に相同なDNAがそのプロモーター−遺伝子連鎖の両隣に隣接するように、プラスミドコンストラクト中に配置される。得られたプラスミドコンストラクトを、大腸菌内での増殖によって増幅し、単離する。挿入すべきDNA遺伝子配列を含む単離されたプラスミドを、例えばニワトリ胚線維芽細胞(CEF)などの細胞培養に、ポックスウイルスと共にトランスフェクトする。それぞれプラスミド中およびウイルスゲノム中にある相同なポックスDNA間で相同組換えが起こることにより、外来DNA配列が存在するように改変されたポックスウイルスが得られる。
【0064】
より好ましい実施形態では、適当な細胞培養の細胞、例えばCEF細胞にポックスウイルスを感染させる。次に、その感染細胞に、外来遺伝子(好ましくはポックスウイルス発現制御要素の転写制御を受けるもの)を含む第1プラスミドベクターをトランスフェクトする。上で説明したように、このプラスミドベクターは、ポックスウイルスゲノムの選択した部分への外因性配列の挿入を指示する能力を有する配列も含んでいる。場合により、このプラスミドベクターは、ポックスウイルスプロモーターに作動可能に連結されたマーカー遺伝子及び(又は)選択遺伝子を含んでいるカセットも含有する。好適なマーカー遺伝子または選択遺伝子は、例えば緑色蛍光タンパク質、β−ガラクトシダーゼ、ネオマイシン、ホスホリボシルトランスフェラーゼまたは他のマーカーをコードする遺伝子である。選択カセットまたはマーカーカセットを使用することにより、産生した組換えポックスウイルスの同定と単離が簡単になる。しかし、組換えポックスウイルスは、PCR技術によって同定することもできる。次に、新たな細胞を、上述のようにして得られた組換えポックスウイルスに感染させ、第1ベクターに含まれる遺伝子と相同な遺伝子を含む第2ベクターをトランスフェクトする。この遺伝子がポックスウイルスゲノムの異なる挿入部位に含まれることになっている場合、第2ベクターは、ポックスウイルスのゲノムへの相同遺伝子の組込みを指示する配列も相違する。相同組換えが起こった後に、2つの相同な遺伝子を含む組換えウイルスを単離することができる。組換えウイルス中に3つ以上の相同な遺伝子を導入するには、先の工程で単離された組換えウイルスを感染に使用し、さらにもう一つの相同遺伝子を含むもう一つのベクターをトランスフェクションに使用することによって、感染工程とトランスフェクション工程とを繰り返す。
【0065】
もう一つの選択肢として、上述の感染工程とトランスフェクション工程とは互いに交換することができる。すなわち、適当な細胞を、まず、外来遺伝子を含むプラスミドベクターでトランスフェクトしてから、ポックスウイルスに感染させることもできる。
【0066】
さらにもう一つの選択肢として、各相同遺伝子を異なるウイルスに導入し、ある細胞を、得られた組換えウイルスのすべてに同時感染させ、すべての相同遺伝子を含む組換え体を選別することもできる。
【0067】
さらに本発明は、ポックスウイルスゲノムへの発現可能な相同遺伝子の組込みを指示する2つ以上のプラスミドベクターコンストラクトを含むキットも提供する。そのようなプラスミドベクターは、適当なクローニング部位の他に、ポックスウイルスゲノム中の選択した部分への外因性配列の挿入を指示する能力を有する配列も含んでいる。これらのベクターは、場合により、選択遺伝子カセットまたはマーカー遺伝子カセットも含む。本キットはさらに、1個または数個の相同遺伝子と、場合により選択遺伝子またはマーカー遺伝子とが、上記のベクターコンストラクトによって挿入されている点で組換え型であるウイルスを選択するための手段と説明書も含む。
【0068】
さらに別の実施形態として、本発明は、本発明の組換えポックスウイルスに由来するDNA配列もしくはその一部、または本発明の組換えポックスウイルスに相同なDNA配列もしくはその一部を包含する。そのような配列は、本発明の相同遺伝子の1つの少なくとも断片を含む外因性配列の少なくとも一部と、本発明のポックスウイルスゲノム配列の少なくとも断片とを含み、前記ポックスウイルスゲノム配列は、好ましくは、前記外因性配列に隣接している。
【0069】
そのようなDNA配列は、本ウイルスまたはその誘導体を同定しまたは単離するために使用することができ、例えばPCRプライマーやハイブリダイゼーションプローブを作製するためにそれらを使用したり、アレイ技術にそれらを使用したりすることができる。
【図面の簡単な説明】
【0070】
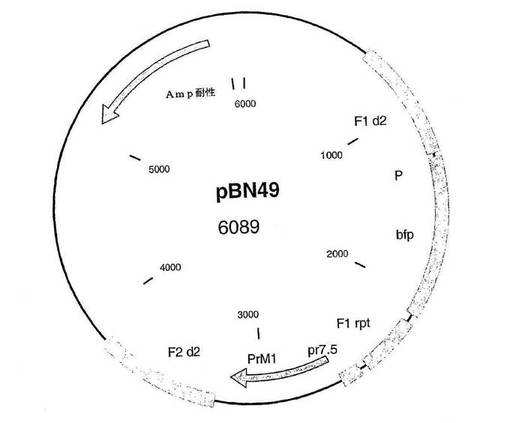
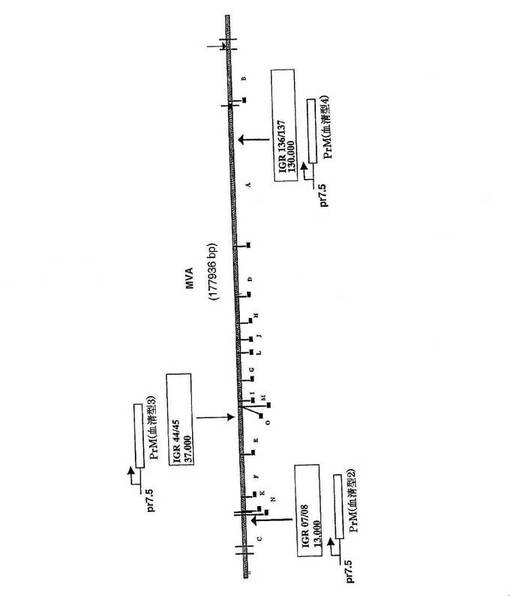
【図1】実施例1に記載のMVAゲノム中の4つのPrM(=前駆膜)遺伝子(血清型1〜4)の挿入部位を表す概略図である。
【図2】挿入プラスミドベクターコンストラクトを表す。
【図3】挿入プラスミドベクターコンストラクトを表す。
【図4】挿入プラスミドベクターコンストラクトを表す。
【図5】挿入プラスミドベクターコンストラクトを表す。
【図6】挿入プラスミドベクターコンストラクトを表す。
【図7】挿入プラスミドベクターコンストラクトを表す。
【図8】挿入プラスミドベクターコンストラクトを表す。
【図9】挿入プラスミドベクターコンストラクトを表す。
【図10】4種類の挿入ベクター(pBN49、pBN50、pBN40、pBN39)のベクタークローニング戦略のPCR検証を示す。
【図11】4つの相同なデングウイルスPrM遺伝子を含む組換えポックスウイルスのPCR検証(実施例1)を示す。
【図12】挿入プラスミドベクターコンストラクトを表す。
【図13】挿入プラスミドベクターコンストラクトを表す。
【図14】挿入プラスミドベクターコンストラクトを表す。
【図15】挿入プラスミドベクターコンストラクトを表す。
【図16】挿入プラスミドベクターコンストラクトを表す。
【図17】挿入プラスミドベクターコンストラクトを表す。
【図18】実施例2に記載のMVAゲノム中の3つのPrM遺伝子(血清型2〜4)の挿入部位を表す概略図である。
【図19】3つの相同なデングウイルスPrM遺伝子が遺伝子間領域に挿入されている組換えポックスウイルスのPCR検証(実施例2)を示す。
【0071】
図面の簡単な説明
図1:図1は、実施例1に記載のMVAゲノム中の4つのPrM(=前駆膜)遺伝子(血清型1〜4)の挿入部位を表す概略図である。
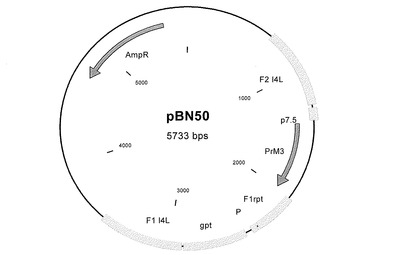
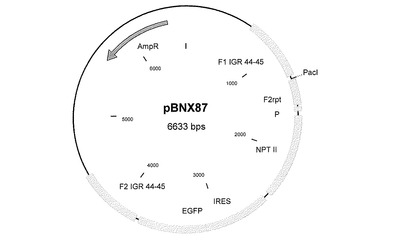
図2〜9及び図12〜17:図2〜9および図12〜17は、挿入プラスミドベクターコンストラクトを表す図であり、ベクター名、そのサイズ、および以下に示すような興味ある配列の位置が記載されている:AmpR=アンピシリン耐性遺伝子、bfp=青色蛍光タンパク質遺伝子、dA=欠失部位A、dE=欠失部位E、d2=欠失部位2、Ecogpt=大腸菌グアノシンホスホリボシルトランスフェラーゼ遺伝子、EGFP=強化緑色蛍光タンパク質遺伝子、F1=隣接配列1、F2=隣接配列2、I4L=遺伝子間領域I4L、IGR=遺伝子間領域、NPT II=ネオマイシン耐性遺伝子、P=ポックスウイルスプロモーター、pr7.5=ワクシニアプロモーター7.5、PrM=デングウイルスの前駆膜遺伝子、数字はそれが4つの血清型のうちのどれに由来するかを示している、rpt=隣接配列の反復である。
【0072】
図10:図10は、4種類の挿入ベクター(pBN49、pBN50、pBN40、pBN39)のベクタークローニング戦略のPCR検証を示す。各プラスミドを4種類のPCRプライマーの組合せで試験した。各組合せは、それぞれ、別個の一挿入部位に組み込まれた別個の一PrM配列に特異的である。
【0073】
図11:図11は、4つの相同なデングウイルスPrM遺伝子を含む組換えポックスウイルスのPCR検証(実施例1)を示す。ゲルの上側には、組換えウイルスのさまざまなPCR結果を示し、下側には、表記の対照プラスミドの同じPCR反応の結果を示す。相同な配列を含むプラスミドをpBN39、pBN49またはpBN50と呼ぶ。PrMは、挿入されたデングウイルス前駆膜遺伝子を意味し、数字はそれが4つの血清型のどれに由来するかを示している。dA=欠失部位A、dE=欠失部位E、d2=欠失部位2、I4L=遺伝子間領域I4Lは、外因性DNAの挿入部位を表す。
【0074】
図18:図18は、実施例2に記載のMVAゲノム中の3つのPrM遺伝子(血清型2〜4)の挿入部位を表す概略図である。
【0075】
図19:図19は、3つの相同なデングウイルスPrM遺伝子が遺伝子間領域に挿入されている組換えポックスウイルスのPCR検証(実施例2)を示す。上図は、PrM2に特異的なPCR反応の結果を表し、中図はPrM3に特異的なPCR反応の結果を表し、下図はPrM4に特異的なPCR反応の結果を表す。レーン8は、対照プラスミドの同じPCR反応を表す。レーン2は空ベクター対照MVAを表す。PrMは、挿入されたデングウイルス前駆膜遺伝子を意味し、数字はそれが4つの血清型のどれに由来するかを示している。M=分子量マーカー。
【0076】
以下に実施例を挙げて本発明をさらに詳しく説明する。しかしながら当業者は、これらの実施例が、決して本発明によって提供される技術の適用範囲を限定するものではないことを十分に理解している。
【実施例1】
【0077】
挿入ベクター
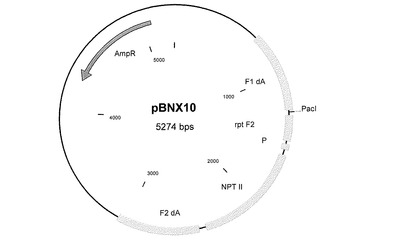
欠失部位A用の挿入ベクター
ゲノム位置7608〜7609に相当するMVAゲノムのいわゆる欠失部位Aまたは欠失部位1に外因性配列を挿入するために、欠失部位Aに隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適当なコンピュータソフトウェア(DNAsis、日立ソフトウェアエンジニアリング、米国サンブルーノ)を使って適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーターの転写制御を受けるNPT II遺伝子(ネオマイシン耐性)などの選択遺伝子カセットを導入する。また、欠失部位Aに挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図2に示す(pBNX10)。
【0078】
欠失部位E用の挿入ベクター
ゲノム位置170480〜170481に相当するMVAゲノムのいわゆる欠失部位Eまたは欠失部位4に外因性配列を挿入するために、欠失部位Eに隣り合う約600bpの隣接配列を含むベクターを構築した。このベクターは上述したように設計し、構築される。隣接配列の間に、ポックスウイルスプロモーターの転写制御を受けるEGFP遺伝子(緑色蛍光タンパク質、Clonetech)を配置する。また、欠失部位Aに挿入しようとする追加の遺伝子または配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図3に示す(pBNX32)。
【0079】
欠失部位2用の挿入ベクター
ゲノム位置20718〜20719に相当するMVAゲノムのいわゆる欠失部位2に外因性配列を挿入するために、欠失部位2に隣り合う約600bpの隣接配列を含むベクターを構築した。このベクターは上述したように設計し、構築される。隣接配列の間に、ポックスウイルスプロモーターの転写制御を受けるhbfp遺伝子(humanized blue fluorescing protein:ヒト化青色蛍光タンパク質、Pavalkis GNら)を配置する。また、欠失部位2に挿入しようとする追加の遺伝子または配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図4に示す(pBNX36)。
【0080】
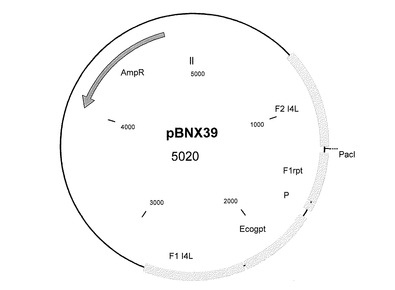
遺伝子間領域I4L用の挿入ベクター
ゲノム位置56760に相当するORF I3LとI4Lの間の遺伝子間領域に外因性配列を挿入するために、I4L遺伝子座の遺伝子間領域に隣り合う約600bpの隣接配列を含むベクターを構築した。このベクターは上述したように設計し、構築される。隣接配列の間に、ポックスウイルスプロモーターの転写制御を受けるEcogpt遺伝子(またはgpt、これは大腸菌から単離されるホスホリボシルトランスフェラーゼ遺伝子を意味する)を配置する。また、I4L ORF後の遺伝子間領域に挿入しようとする追加の遺伝子または配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図5に示す(pBNX39)。
【0081】
数個の相同な遺伝子がゲノム内に組み込まれている組換えポックスウイルスの構築
挿入ベクター
デングウイルスの4つの血清型に由来する4つのPrM遺伝子をMVAゲノムに挿入するために、4つの独立した組換えベクターを使用した。
【0082】
これらのベクターは、上に詳述したように、相同組換えにより狙って挿入するために、MVAゲノムに相同な配列を有する。また、各ベクターは選択遺伝子カセットまたはレポーター遺伝子カセットも持っている。
【0083】
4つのデングウイルス血清型のPrM配列を、オリゴアニーリングとPCR増幅とによって、合成的に作製した。それらのPrM配列をポックスウイルスプロモーター要素の下流にクローニングして、発現カセットを形成させた。次にこの発現カセットを、適切な挿入ベクターコンストラクトのクローニング部位にクローニングした。
【0084】
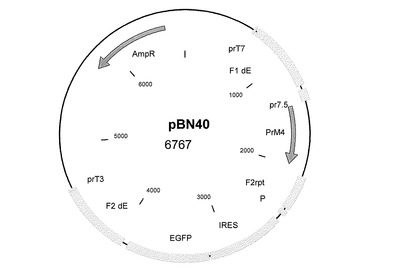
結果として、欠失部位A用の挿入ベクターコンストラクトは、デングウイルス血清型2のPrM遺伝子を含有した(図6−pBN39)。欠失部位2用の挿入ベクターコンストラクトは、デングウイルス血清型1のPrM遺伝子を含有した(図7−pBN49)。遺伝子間領域I4L用の挿入ベクターコンストラクトは、デングウイルス血清型3のPrM遺伝子を含有した(図8−pBN50)。欠失部位E用の挿入ベクターコンストラクトは、デングウイルス血清型4のPrM遺伝子を含有した(図9−pBN40)。
【0085】
挿入ベクターのPCR検証
クローニング戦略を検証するために、PCRアッセイを行った。これらのPCRアッセイの場合、選択したプライマー対は、挿入部位に関して特異的な隣接配列に特異的に結合するプライマーと、高度に相同なデングウイルスPrM遺伝子の1つに特異的に結合する第2プライマーとの組合せである。
【0086】
デングウイルス血清型2のPrM遺伝子を含有する欠失部位A用の挿入ベクターは、プライマーoBN93(CGCGGATCCATGCTGAACATCTTGAACAGGAGACGCAGA.SEQ ID NO:1)およびoBN477(CATGATAAGAGATTGTATCAG.SEQ ID NO:2)を使って選別した。
【0087】
デングウイルス血清型1のPrM遺伝子を含有する欠失部位2用の挿入ベクターは、プライマーoBN194(ATGTTGAACATAATGAACAGGAGGAAAAGATCTGTGACCATGCTCCTCATGCTGCTGCCCACAGCCCTGGCGTTCCATCT.SEQ ID NO:3)およびoBN476(GATTTTGCTATTCAGTGGACTGGATG.SEQ ID NO:4)を使って選別した。
【0088】
デングウイルス血清型3のPrM遺伝子を含有する遺伝子間領域I4L用の挿入ベクターは、プライマーoBN255(CCTTAATCGAATTCTCATGTCATGGATGGGGTAACCAGCATTAATAGT.SEQ ID NO:5)およびoBN479(GCTCCCATTCAATTCACATTGG.SEQ ID NO:6)を使って選別した。
【0089】
デングウイルス血清型4のPrM遺伝子を含有する欠失部位E用の挿入ベクターは、プライマーoBN210(ATCCCATTCCTGAATGTGGTGTTAAAGCTACTGAGCGCTTCTCTCGTCTCCGTTCTCCGCTCTGGGTGCATGTCCCATAC.SEQ ID NO:7)およびoBN478(GTACATGGATGATATAGATATG.SEQ ID NO:8)を使って選別した。
【0090】
PCR実験は、10×PCR緩衝液、MgCl2緩衝液およびTaq DNAポリメラーゼを含むTaq DNAポリメラーゼキット(Roche、カタログ番号201205)またはこれと等価な試薬類を使って、サーマルサイクラーGeneAmp9700(Perkin Elmer)で行う。一般に、PCR反応は、45μlの主混合物、試料DNAおよび必要に応じてddH2Oを含む50μlの総反応液量を使って調製した。主混合物は、30.75μlのddH2O、5μlの10×緩衝液、1μlのdNTP混合物(各10mM)、2.5μlの各プライマー(5pmol/ml)、3μlのMgCl2(25mM)および0.25μlのTaqポリメラーゼ(5U/μl)を使って調製すべきである。
【0091】
増幅は以下のプログラムを使って行った。
1)変性:4分 94℃
2)30サイクル:
変性:30秒 94℃
アニーリング:30秒 55℃
伸長:1〜3分 72℃
3)伸長:7分 72℃
4)保存 4℃。
【0092】
挿入される遺伝子のサイズに基づいて、伸長時間は少なくとも1分/kbにするべきである。
【0093】
図10に示すPCR結果は、上記の単独挿入物に使用したプライマーの組合せの特異性を実証している。
【0094】
プライマーの組合せoBN194/oBN476は、欠失部位2およびインサートとしてのPrM1に特異的である。プラスミドpBN49の予想PCR断片は678bpのサイズを有する(ゲルの上側、レーン3に示す)。
【0095】
プライマーの組合せoBN255/oBN479は、遺伝子間領域I4LおよびインサートとしてのPrM3に特異的である。プラスミドpBN50の予想PCR断片は825bpのサイズを有する(ゲルの上側、レーン9に示す)。
【0096】
プライマーの組合せoBN210/oBN478は、欠失部位EおよびインサートとしてのPrM4に特異的である。プラスミドpBN40の予想PCR断片は607bpのサイズを有する(ゲルの下側、レーン5に示す)。
【0097】
プライマーの組合せoBN93/oBN477は、欠失部位AおよびインサートとしてのPrM2に特異的である。プラスミドpBN39の予想PCR断片は636bpのサイズを有する(ゲルの下側、レーン11に示す)。
【0098】
相同組換えによる組換えMVAの作製
組換えMVAによって外来遺伝子を発現させるには、これらの遺伝子を、相同組換えと呼ばれる過程によって、ウイルスゲノム中に挿入する必要がある。そのために、興味ある外来遺伝子を、上述のように、挿入ベクター中にクローニングしておいた。細胞をMVA−BNに感染させた後に、このベクターをトランスフェクトする必要がある。感染とトランスフェクションとを受けた細胞の細胞質では、組換えが起こるだろう。やはり挿入ベクターに含まれている選択及び(又は)レポーターカセットを利用して、組換えウイルスを含む細胞を同定し、単離する。
【0099】
相同組換え
相同組換えを行うために、DMEM(ダルベッコ変法イーグル培地,Gibco BRL)+10%ウシ胎仔血清(FCS)またはVP−SFM(Gibco BRL)+4mmol/l L−グルタミン(無血清製造法の場合)を使って、BHK(ベビーハムスター腎)細胞またはCEF(初代ニワトリ胚線維芽細胞)を6穴プレートに播種する。
【0100】
細胞はまだ増殖期にある必要があるので、トランスフェクションの日に60〜80%コンフルエントに達するべきである。感染に用いる感染多重度(moi)を決定するには細胞数を知っておく必要があるので、播種前に細胞数を数えた。
【0101】
感染を行うために、500μlの希釈液が0.01のmoiを与える適当なウイルス量を含むように、MVAストックをDMEM/FCSまたはVP−SFM/L−グルタミンに希釈する。細胞は、播種後に1回分裂すると仮定する。細胞から培地を取り除き、細胞を500μlの希釈ウイルスに、室温で振とうしながら1時間感染させる。接種物を除去し、細胞をDMEM/VP−SFMで洗浄する。感染細胞を、それぞれ1.6mlのDMEM/FCSおよびVP−SFM/L−グルタミン中に入れておき、その間に、トランスフェクション反応(キアゲン・エフェクテン・キット(Qiagen Effectene Kit))の準備をする。
【0102】
トランスフェクションには、「エフェクテン(Effectene)」トランスフェクションキット(Qiagen)を使用する。線状化した挿入ベクター1〜5μg(多重トランスフェクションの場合は総量)と緩衝液ECとのトランスフェクション混合物を、最終体積が150μlになるように調製する。DNA 1μgにつき8.0μlのエンハンサー(Enhancer)を加え、ボルテックスし、室温で5分間インキュベートする。次に、DNA 1μgにつき25μlのエフェクテンを、原液のチューブをボルテックスしてから加え、その溶液をボルテックスによって十分に混合し、室温で10分間インキュベートする。それぞれ600μlのDMEM/FCSおよびVP−SFM/L−グルタミンを加え、混合した後、そのトランスフェクション混合物全体を、既に培地で覆われている細胞に加える。培養皿を穏やかに振とうしてトランスフェクション反応液を混合する。37℃、5%CO2で、一晩インキュベートする。翌日、培地を除去し、新鮮なDMEM/FCSまたはVP−SFM/L−グルタミンと置き換える。インキュベーションを3日目まで続ける。
【0103】
収集するために、細胞を培地中にこすり取り、得られた細胞懸濁液を適当なチューブに移して、短期保存の場合は−20℃で凍結し、長期保存の場合は−80℃で凍結する。
【0104】
MVAへのPrM4の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型4のPrM遺伝子と、レポーター遺伝子としてEGPF遺伝子とを含んでいる挿入ベクターpBN40をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子EGFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスはプラーク精製によって精製する必要がある。
【0105】
プラーク精製を行うには、感染細胞(蛍光細胞または染色細胞)をピペットチップで単離し、200μlのPBSまたは培地に再懸濁し、吸引する。次に、約106個の細胞が入っている新しい培養皿を、再懸濁したプラーク100μlに感染させる。48時間後に、細胞を300μlのPBSにとりだす。懸濁液からDNAを抽出し、PCR分析によって選別する。予想されるバンドを示すクローンを選択し、新しい6穴プレートをさまざまな量のこのウイルスに感染させる。ウェルに1%アガロースを重層することにより、ウイルスのさらなる伝播を避ける。48時間後に、組換えウイルスクローンを含む感染細胞を単離する。
【0106】
この操作を、PCR分析で野生型MVA−BNを検出することができなくなるまで繰り返す。
【0107】
4回のプラーク精製後に、予想される挿入物を選択的に増幅するプライマー対(上述のoBN210およびoBN478)と、対照として挿入部位である欠失部位Eを特異的に認識するプライマー対(oBN453:GTTGAAGGATTCACTTCCGTGGA,SEQ ID NO:9およびoBN454:GCATTCACAGATTCTATTGTGAGTC,SEQ ID NO:10)とを使って、PCRアッセイにより、組換えウイルスMVA−PrM4を同定した。
【0108】
MVA−PrM4へのPrM2の挿入
上記のプロトコールに従って細胞をMVA−PrM4に感染させ、さらに、デングウイルス血清型2のPrM遺伝子と、選択遺伝子としてNPT II(ネオマイシン耐性遺伝子)とを含んでいる挿入ベクターpBN39をトランスフェクトした。抗生物質耐性遺伝子を発現させる組換えMVAを精製するには、プラーク精製前に選択条件下でウイルス増幅を3回行うことが推奨される。ネオマイシンホスホトランスフェラーゼ選択を行うために、G418を培地に加える。G418はネオマイシンの誘導体であり、リボソームの作用を妨害することにより、タンパク質生合成を阻害する。NPT遺伝子活性はG418をリン酸化によって失活させる。
【0109】
ネオマイシン選択下で16回のプラーク精製後に、予想される挿入物を選択的に増幅するプライマー対(上記のoBN93およびoBN477)と、対照として挿入部位である欠失部位Aを特異的に認識するプライマー対(上記のoBN477およびoBN452:GTTTCATCAGAAATGACTCCATGAAA,SEQ ID NO:11)とを使って、PCRアッセイにより、組換えウイルスMVA−PrM4/PrM2を同定した。さらに、欠失部位EへのPrM4の挿入も、プライマー対oBN210−oBN478およびoBN453−oBN454を使って確認する。
【0110】
MVAへのPrM1の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型1のPrM遺伝子と、レポーター遺伝子としてhbfp(ヒト化青色蛍光タンパク質の遺伝子)とを含んでいる挿入ベクターpBN49をトランスフェクトした。合成されたhbfpタンパク質は、3日目に、組換えウイルスに感染した細胞中に検出することができる。得られた組換えウイルスをプラーク精製によって精製した。10回のプラーク精製後に、予想される挿入物を選択的に増幅するプライマー対(上述のoBN194およびoBN476)と、対照として挿入部位である欠失部位2を特異的に認識するプライマー対(oBN54:CGGGGTACCCGACGAACAAGGAACTGTAGCAGAGGCATC,SEQ ID NO:12およびoBN56:AACTGCAGTTGTTCGTATGTCATAAATTCTTTAATTAT,SEQ ID NO:13)とを使って、PCRアッセイにより、組換えウイルスMVA−PrM1を同定した。
【0111】
MVAへのPrM3の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型3のPrM遺伝子と、レポーター遺伝子としてEcogpt遺伝子(Ecogpt、略してgptは、ホスホリボシルトランスフェラーゼ遺伝子を意味する)とを含んでいる挿入ベクターpBN50をトランスフェクトした。得られた組換えウイルスを、ミコフェノール酸、キサンチンおよびヒポキサンチンの添加により、ホスホリボシルトランスフェラーゼ代謝選択下で、プラーク精製を3回行うことによって精製した。ミコフェノール酸(MPA)はイノシン一リン酸デヒドロゲナーゼを阻害し、大半の細胞株でプリン合成の遮断およびウイルス複製の阻害をもたらす。この遮断は、構成的プロモーターからEcogptを発現させると共に、基質キサンチンおよびヒポキサンチンを与えることによって、克服することができる。
【0112】
予想される挿入物を選択的に増幅するプライマー対(上述のoBN255およびoBN479)と、対照として挿入部位I4Lを特異的に認識するプライマー対(oBN499:CAACTCTCTTCTTGATTACC,SEQ ID NO:14およびoBN500:CGATCAAAGTCAATCTATG,SEQ ID NO:15)とを使って、PCRアッセイにより、生成した組換えウイルスMVA−PrM3を同定した。
【0113】
MVA−PrM1とMVA−PrM3の同時感染
上記のプロトコールに従って、細胞を、等量のMVA−PrM1とMVA−PrM3に感染させた。青色蛍光クローンをホスホリボシルトランスフェラーゼ代謝選択下で3回プラーク精製した後、上記のプライマー対(上記のoBN255およびoBN479、oBN499およびoBN500、oBN194およびoBN476、oBN54およびoBN56)を使ったPCRによって、組換えウイルスを分析した。得られた組換えウイルスをMVA−PrM1/PrM3と名付けた。
【0114】
MVA−PrM1/PrM3とMVA−PrM2/PrM4の同時感染
上記のプロトコールに従って、細胞を、等量のMVA−PrM1/PrM3とMVA−PrM2/PrM4に感染させた。ホスホリボシルトランスフェラーゼ代謝選択およびネオマイシン選択下で、プラーク精製を行った。緑色および青色の蛍光を惹起する組換えウイルスを単離し、上記プライマー対(上記のoBN255およびoBN479、oBN499およびoBN500、oBN194およびoBN476、oBN54およびoBN56、oBN93およびoBN477、oBN477およびoBN452、oBN210およびoBN478、oBN453およびoBN454))を使ったPCRによって分析した。
【0115】
4つのPrM遺伝子をすべて含んでいる組換えウイルス(クローン20)のPCR分析を図11に示す。ゲルの上側は、この組換えウイルスのさまざまなPCR結果を示し、下側は、対照プラスミド(表示のもの)に関する同じPCR反応の結果を示す。レーン1、10および11は1kbおよび100bpの分子マーカーを表す。
【0116】
プライマーの組合せoBN210/oBN478は、欠失部位EおよびインサートとしてのPrM4に特異的である。組換えウイルスおよびプラスミドpBN40の予想PCR断片は607bpのサイズを有する(レーン2に示す)。
【0117】
プライマーの組合せoBN453/oBN454は、欠失部位Eに特異的である。組換えウイルスの予想PCR断片は2.7kbであり、野生型ウイルスの予想バンドは2.3kbである(レーン3に示す)。ゲルの上側でも、野生型ウイルスに特異的なバンドを同定することができる。これは、野生型ウイルスがまだ完全にはこの組換えウイルス調製物から除去されていないことを意味する。さらなるプラーク精製が必要である。
【0118】
プライマーの組合せoBN93/oBN477は、欠失部位AおよびインサートとしてのPrM2に特異的である。組換えウイルスとプラスミドpBN39の予想PCR断片は636bpのサイズを有する(レーン4に示す)。
【0119】
プライマーの組合せoBN477/oBN452は、欠失部位Aに特異的である。組換えウイルスの予想PCR断片は4.1kbであり、野生型ウイルスの予想バンドは2.7kbである(レーン5に示す)。野生型ウイルスに特異的なバンドをゲルの上側で同定することができる。
【0120】
プライマーの組合せoBN255/oBN479は、遺伝子間領域I4LおよびインサートとしてのPrM3に特異的である。組換えウイルスとプラスミドpBN50の予想PCR断片は825bpのサイズを有する(レーン6に示す)。
【0121】
プライマーの組合せoBN499/oBN500は、I4Lの遺伝子間領域に特異的である。組換えウイルスの予想PCR断片は1.0kbであり、野生型ウイルスの予想バンドは0.3kbである(レーン7に示す)。
【0122】
プライマーの組合せoBN194/oBN476は、欠失部位2およびインサートとしてのPrM1に特異的である。組換えウイルスとプラスミドpBN49の予想PCR断片は678bpのサイズを有する(レーン8に示す)。
【0123】
プライマーの組合せoBN54/oBN56は、欠失部位2に特異的である。組換えウイルスの予想PCR断片は1.6kbであり、野生型ウイルスの予想バンドは0.9kbである(レーン9に示す)。野生型ウイルスに特異的なバンドをゲルの上側で同定することができる。
【0124】
もう一つの選択肢として、4種類のウイルスを作製し、細胞を4種類のウイルスのすべてに同時感染させ、組換え体を選別することもできる。
【0125】
さらなる選択マーカーまたは耐性マーカーを含む組換えベクターを使って、改良を行うこともできる。
【実施例2】
【0126】
挿入ベクター
遺伝子間領域136−137(IGR 136−137)用の組換えベクター
MVAゲノムのゲノム位置129,940に相当するいわゆる遺伝子間領域(IGR)136−137に外因性配列を挿入するために、この挿入部位に隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーター(P)の転写制御を受けるNPT II遺伝子(ネオマイシン耐性)などの選択遺伝子カセットを導入する。また、IGR 136−137に挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位(PacI)も存在する。そのような本発明のベクターコンストラクトの一つを図12に示す(pBNX67)。
【0127】
遺伝子間領域07−08(IGR 07−08)用の組換えベクター
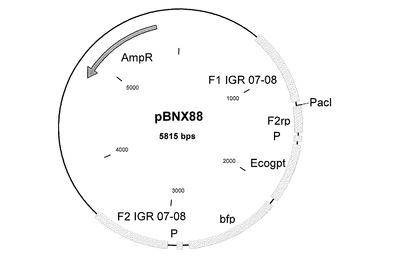
MVAゲノムのゲノム位置12,800に相当する遺伝子間領域(IGR)07−08に外因性配列を挿入するために、この挿入部位に隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーター(P)の転写制御を受けるEcogpt遺伝子(グアニンホスホリボシルトランスフェラーゼ)などの選択遺伝子カセットを導入する。また、IGR 07−08に挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位(PacI)も存在する。そのような本発明のベクターコンストラクトの一つを図13に示す(pBNX88)。
【0128】
遺伝子間領域44−45(IGR 44−45)用の組換えベクター
MVAゲノムのゲノム位置37,330に相当する遺伝子間領域(IGR)44−45に外因性配列を挿入するために、この挿入部位に隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーター(P)の転写制御を受けるNPT II遺伝子(ネオマイシン耐性)などの選択遺伝子カセットを導入する。また、IGR 44−45に挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位(PacI)も存在する。そのような本発明のベクターコンストラクトの一つを図14に示す(pBNX87)。
【0129】
数個の相同な遺伝子がゲノム内に組み込まれている組換えポックスウイルスの構築
挿入ベクター
デングウイルスの血清型2、3および4に由来する3つのPrM遺伝子をMVAゲノムに挿入するために、3つの独立した組換えベクターを使用した。
【0130】
これらのベクターは、上に詳述したように、相同組換えによる挿入をターゲティングするために、MVAゲノムに相同な配列を有する。また、各ベクターは選択およびレポーター遺伝子カセットも持っている。
【0131】
3つのデングウイルス血清型のPrM配列を、実施例1で説明したように、合成的に作製した。
【0132】
結果として、IGR136−137用の挿入ベクターコンストラクトは、デングウイルス血清型4のPrM遺伝子を含有した(図15−pBN27)。IGR 07−08用の挿入ベクターコンストラクトは、デングウイルス血清型2のPrM遺伝子を含有し(図16−pBN34)、IGR 44−45用の挿入ベクターコンストラクトは、デングウイルス血清型3のPrM遺伝子を含有した(図17−pBN47)。
【0133】
相同組換えによる組換えMVAの作製
相同組換えによる組換えMVAの作製を、実施例1で説明したように行った。MVAゲノムにおけるPrM4、PrM3およびPrM2の挿入部位を、図18に示す。
【0134】
MVAへのPrM4の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型4のPrM遺伝子と、レポーター遺伝子としてEGFP遺伝子とを含んでいる挿入ベクターpBN27をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子EGFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスは実施例1で説明したようにプラーク精製によって精製する必要がある。4回のプラーク精製後に、挿入部位IGR136−137を選択的に増幅するプライマー対(oBN1008:gataccgatcacgttcta,SEQ ID NO:16およびoBN1009:ggatatgattatgtagag,SEQ ID NO:17)を使って、PCRアッセイにより、組換えウイルスMVA−PrM4を同定した。
【0135】
MVAへのPrM2の挿入
上記のプロトコールに従って細胞をMVA−PrM4に感染させ、さらに、デングウイルス血清型2のPrM遺伝子と、レポーター遺伝子としてBFP遺伝子とを含んでいる挿入ベクターpBN34をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子BFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスは実施例1で説明したようにプラーク精製によって精製する必要がある。6回のプラーク精製後に、組換えウイルスMVA−PrM4+PrM2をさらに継代し、増幅させて、粗製ストックを調製した。挿入部位IGR07−08を選択的に増幅するプライマー対(oBN 903:ctggataaatacgaggacgtg,SEQ ID NO:18およびoBN904:gacaattatccgacgcaccg,SEQ ID NO:19)を使って、PCRアッセイにより、組換え体を同定した。
【0136】
MVAへのPrM3の挿入。
【0137】
上記のプロトコールに従って細胞をMVA−PrM2+4に感染させ、さらに、デングウイルス血清型3のPrM遺伝子と、レポーター遺伝子としてEGFP遺伝子とを含んでいる挿入ベクターpBN47をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子EGFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスは実施例1で説明したようにプラーク精製によって精製する必要がある。3回のプラーク精製後に、挿入部位IGR44−45を選択的に増幅するプライマー対(oBN904:cgttagacaacacaccgacgatgg,SEQ ID NO:20およびoBN905:cggatgaaaaatttttggaag,SEQ ID NO:21)を使って、PCRアッセイにより、組換えウイルスMVA−PrM4+3+2を同定した。
【0138】
3つのデングウイルスPrM遺伝子を含む組換えウイルスのPCR分析を図19に示す。PCR実験は実施例1で説明したように行う。プライマーの組合せoBN1008およびoBN1009は、PrM4挿入物を含むIGR136−137に特異的である(図19、下図)。組換えウイルスの予想PCR断片は、プラスミド陽性対照(レーン8)と同様に、1kbのサイズを有する(レーン4、5および6に示す)。PrM4を持たない空ベクター対照は、190bpの予想断片を示す(レーン2)。レーンMは分子量マーカーを表し、レーン1、3および7は空である。プライマーの組合せoBN902およびoBN903は、PrM2挿入物を含むIGR07−08に特異的である(図19、上図)。組換えウイルスの予想PCR断片は、プラスミド陽性対照(レーン8)と同様に、960bpのサイズを有する(レーン4〜6に示す)。PrM2を持たない空ベクター対照は、190bpの予想断片を示す(レーン2)。プライマーの組合せoBN904およびoBN905は、PrM3挿入物を含むIGR44−45に特異的である(図19、中図)。組換えウイルスの予想PCR断片は、プラスミド陽性対照(レーン8)と同様に、932bpのサイズを有する(レーン4〜6に示す)。PrM2を持たない空ベクター対照は、185bpの予想断片を示す(レーン2)。
【技術分野】
【0001】
本発明は、2つ以上の相同な外来遺伝子を発現させる能力を有する組換えポックスウイルスに関する。上記の遺伝子は、ウイルスゲノムにとっては異種であるが、互いに比較した場合には相同である。これらの遺伝子は、特に、ある微生物の近縁変異体またはサブタイプに由来する。さらに本発明は、そのような組換えポックスウイルスを作製する方法、およびそのような組換えポックスウイルスの医薬またはワクチンとしての使用に関する。また、ヒトを含む生きている動物において、免疫応答に影響を及ぼす方法、好ましくは免疫応答を誘発する方法も提供する。
【背景技術】
【0002】
あらゆる生体は、細菌、ウイルス、真菌または寄生虫などの感染性因子または病原性因子の攻撃を絶えず受けている。いわゆる免疫系は、当該生物が、そのような因子によって引き起こされる持続的な感染、疾患または中毒に陥るのを防いでいる。
【0003】
哺乳動物の免疫系は特異的部分と非特異的部分とに分類することができる。ただし両者の間には密接な相互関係がある。非特異的免疫応答は、多種多様な病原性物質または感染性因子に対する即時防御を可能にする。特異的免疫応答は、生物がある物質による攻撃を初めて受けた場合には、誘導期間を経てから生じる。この特異的免疫応答は、主として、抗原特異的な抗体の産生と、マクロファージおよびリンパ球、例えば細胞傷害性T細胞(CTL)の生成とに基づいている。特異的免疫応答は、ある特定の感染から回復した個体はその特定の感染からは保護されるが、他の感染性疾患には依然として罹りやすいという事実の原因である。一般に、同じ感染性因子または極めて似ている感染性因子による2回目の感染では、症状がはるかに軽いか、または症状が全く現れない。このいわゆる免疫は長期間持続し、場合によっては一生涯続くことさえある。その基礎をなしている効果はしばしば免疫記憶と呼ばれ、これを予防接種に利用することができる。
【0004】
予防接種という用語は、ある個体を、無害な部分型または不活化型の感染性因子にばく露して、その個体における免疫応答に影響を及ぼす(好ましくは免疫応答を誘発する)ことにより、その特定感染性因子に対して(一生涯ではないとしても)長期間持続する免疫を生じさせる方法を表す。
【0005】
ヒト痘瘡疾患は天然痘ウイルスによって起こる。天然痘ウイルスはポックスウイルス科に属する。ポックスウイルス科は、脊椎動物細胞および無脊椎動物細胞の細胞質中で複製する複雑なDNAウイルスからなる大きな一科である。
【0006】
ポックスウイルス科は、脊椎動物宿主域および昆虫宿主域に基づいて2つの亜科、すなわちチョルドポックスウイルス亜科とエントモポックスウイルス科とに分類することができる。チョルドポックスウイルス亜科には、とりわけ、オルトポックスウイルス属およびアビポックスウイルス属が含まれている(非特許文献1)。
【0007】
オルトポックスウイルス属には、ヒト痘瘡の原因因子である天然痘ウイルスが含まれると共に、ラクダ痘ウイルス、牛痘ウイルス、羊痘ウイルス、ヤギ痘ウイルス、サル痘ウイルスおよびワクシニアウイルスなどの経済的に重要な他のウイルスも含まれる。この属のメンバーはすべて遺伝的に類縁関係にあり、よく似た形態または宿主域を有する。制限エンドヌクレアーゼ地図も、オルトポックスウイルス属の異なるメンバー間で、90%に達する高い配列一致度を示している(非特許文献2)。
【0008】
ワクシニアウイルス(VV)とは、少なくとも過去100年間にわたって痘瘡の予防接種に使用された因子に与えられた名称である。VVが、長期間にわたる連続継代により、牛痘ウイルスから派生した新種であるのか、今では絶滅したウイルスの生きた代表例である天然痘ウイルスから派生した新種であるのかは分かっていない。あるいは、VVは、遺伝子組み換えの産物なのかもしれない。また、VVの歴史の中で、多くのワクシニア株が生じた。これらのさまざまな株はさまざまな免疫原性を示し、潜在的合併症(その最も重篤なものは種痘後脳炎である)とさまざまな程度に関係づけられている。しかし、これらの株の多くは、痘瘡の予防接種に使用された。例えばNYCBOH株、ウェスタンリザーブ(Western Reserve)株またはワイエス(Wyeth)株は主に米国で使用され、アンカラ(Ankara)株、ベルン(Bern)株、コペンハーゲン(Copenhagen)株、リスター(Lister)株およびMVAは主に欧州での予防接種に使用された。これらさまざまVV株による世界的な予防接種計画の結果、1980年に、WHOはついに、天然痘ウイルスの根絶の成功を宣言した。
【0009】
現在、VVは主に実験株として使用されているが、それ以外に、今でもオルトポックスウイルス属の原型であるとみなされている。これは、VVが最も詳細に特徴づけられたウイルスの一つになった理由でもある(非特許文献3)。VVは、そしてまた最近では他のポックスウイルスも、外来遺伝子を挿入し発現させるために使用されてきた。生きている感染性ポックスウイルスに外来遺伝子を挿入するための基礎技術では、ドナープラスミド中の外来遺伝要素に隣接するポックスDNA配列と、レスキューポックスウイルス(rescuing poxvirus)中に存在する相同な配列との間で、組換えが行われる。遺伝子組換えとは、一般に、2本のDNA鎖間で起こる相同なDNA区間の交換である。ウイルスによっては、RNAがDNAの代わりをする場合もある。相同な核酸区間とは、同じヌクレオチド塩基配列を有する核酸(DNAまたはRNA)の区間である。遺伝子組換えは、感染した宿主細胞内での新しいウイルスゲノムの複製または産生中に自然に起こりうる。したがって、ウイルス遺伝子間の遺伝子組換えは、2種類以上のウイルスまたは他の遺伝子コンストラクトに同時感染した宿主細胞内で起こるウイルス複製サイクル中に起こりうる。第1のゲノムに由来するDNA区間は、第2の同時感染ウイルスのゲノムのうち、そのDNAが第1ウイルスゲノムのDNAと相同である区間を構築する際に、互換的に使用される。
【0010】
挿入されたDNA遺伝子配列を改変感染性ウイルスによってうまく発現させるには、2つの条件が必要である。第1に、改変ウイルスが依然として生存可能であるように、挿入はウイルスの非必須領域になされるべきである。挿入されたDNAを発現させるための第2の条件は、挿入されたDNAに対して適切な関係にあるプロモーターが存在することである。通例、プロモーターは、発現させるべきDNA配列の上流に配置する。
【0011】
例えばB型肝炎ウイルス表面抗原(HBsAg)、インフルエンザウイルス赤血球凝集素(InfHA)または二日熱マラリア原虫(Plasmodium knowlesi)スポロゾイト抗原などを発現させる組換えVVの、感染性疾患予防用の生ワクチンとしての有用性は、実証されており、それに関する総説もある(非特許文献4)。
【0012】
VVのもう一つの利点は、1つのVVゲノム内に複数の外来配列、外来遺伝子または外来抗原を取り込む能力を有することである(非特許文献5)。また、1回の多価ワクチン接種で多くの異種の感染性疾患に対する免疫を誘発することができると報告されている(非特許文献6)。
【0013】
単一のVVによってさまざまな抗原を発現させる例の一つが、Brayらによって記載されている。デングウイルス血清型4の3種類の構造タンパク質、すなわちキャプシド(C)タンパク質、前駆膜(pre−membrane)(prM)タンパク質、エンベロープ(E)タンパク質と、デングウイルス血清型4の2つの非構造タンパク質、すなわちNS1およびNS2aとを発現させることができる組換えVVは、相同なデングウイルス血清型4の攻撃からマウスを保護する能力を有することが示された(非特許文献7)。
【0014】
デングウイルス血清型1(Den−1)からデングウイルス血清型4(Den−4)まで4つの血清型を有するデングウイルスは、ヒトの感染に関して、フラビウイルス属の重要な一メンバーである。デングウイルス感染は、インフルエンザ様の症状から重篤なまたは致死的な病気、すなわちショック症候群(DSS)を伴うデング出血熱(DHF)に及ぶ疾患をもたらす。デングの突発は、媒介動物である蚊が多い熱帯地方および亜熱帯地方の人口密集地域では、依然として公衆衛生上の大きな問題である。
【0015】
デング感染および蚊媒介性フラビウイルスが誘発する他の疾患が、世界の多くの地域で蔓延することを懸念して、デング熱(DF)とデング出血熱(DHF)の両方を防止することができるデングワクチンの開発に向けた努力が盛んに行われるようになり、予防接種を受けた個体を蚊媒介性フラビウイルスの一部または全部によって誘発される感染症から保護するのに役立つワクチンがもたらされている。
【0016】
大半のDF症例が4つの血清型のいずれかによる最初の感染後に顕在化するのに対して、DHF症例は、かなりの部分が、最初に感染したデングウイルスの血清型とは異なる血清型による2回目の感染を受けた患者に起こる。これらの知見から、一定数の症例では、あるデングウイルス血清型に対する抗体を有する個体が適当な間隔を置いて異なるウイルス血清型に逐次的に感染することによってDHFが起こるという仮説がもたらされる。
【0017】
したがって、1つの血清型に対する予防接種は、デングウイルス感染からの完全な保護をもたらすのではなく、同じデングウイルス株による感染からの保護をもたらすにすぎない。さらに重要なことに、1つの血清型に対する予防接種を受けた人では、その人が血清型の異なるデングウイルス株に感染した場合にデング出血熱などの重篤な合併症を発症する危険性が増大している。
【0018】
したがって、4つのデングウイルス血清型のすべてに由来する抗原を有する多価ワクチンが望ましい。
【0019】
これまでは、各VVが異なるウイルスの配列をコードしている一群の組換えVVを混合することによって、多価ワクチンを製造することが提案されていた(非特許文献8)。しかし、そのような多価ワクチンにはいくつかの欠点がある。第1に、いくつかの独立した組換えVVを作製するのは面倒である。個別の製造プロセスだけでなく、品質管理と品質保証にも、著しく時間がかかる。第2に、異なる配列を発現させる組換えウイルスの混合物による感染には、感染事象があまりバランスよく起こらないという危険が常に伴う。主な危険は、多価ワクチンに含まれるすべての異なる組換え体が標的細胞に感染するわけではなく、個々の組換え体による標的細胞の感染しか起こらないというものである。理由の1つとして、組換えウイルスの不均等な分布が考えられる。もう一つの理由として、単一の細胞に感染する際に異なる組換えウイルス間で起こる干渉が考えられる。そのような干渉は重感染現象として知られている。この場合、最終的に感染細胞から発現され、その結果、患者の免疫系に提示されるのは、多価ワクチンの異なる抗原のすべてではなく、一部の抗原だけになる。その結果、多価ワクチンが提示するまたは提示することのできるさまざまな抗原に対する完全な免疫保護は到底もたらされず、一部の抗原に対する免疫保護しか得られないことになる。
【0020】
デングウイルス感染に対するワクチンの場合、多価ワクチンによるアプローチには、デングウイルス2のエンベロープタンパク質に関して報告例(非特許文献9)があるように、異なる配列が異なる量で発現されたり、予測不可能な形で発現されたりすると、その予防接種は患者にとって著しく危険であるという欠点がある。一群の組換えワクシニアウイルスを使った不完全な予防接種では、デングウイルスのすべての血清型に対する免疫保護ではなく、その一部に対する免疫保護しか得られない。残念なことにデング感染の場合、不完全な予防接種は、それによってデングウイルス出血熱などの致死的合併症の危険が増大するため、全く受け入れがたい。
【先行技術文献】
【非特許文献】
【0021】
【非特許文献1】Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第3版,1996年,ISBN:0−7817−0253−4,第83章
【非特許文献2】MackettおよびArchard[1979年]J Gen Virol,45:683−701
【非特許文献3】Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第3版,1996年,ISBN:0−7817−0253−4,第83章および第84章
【非特許文献4】Smith等[1984年]Biotechnology and Genetic Engineering Reviews 2,383−407
【非特許文献5】SmithおよびMoss[1983年]Gene,25(1):21−28
【非特許文献6】Perkusら[1985年]Science,Vol.229,981−984
【非特許文献7】Bray等[1989年]Virology 2853−2856
【非特許文献8】Moss[1990年]Immunology,2,317−327
【非特許文献9】Deubleら[1988年]J. Virol 65:2853
【発明の概要】
【発明が解決しようとする課題】
【0022】
したがって、ある感染性疾患原因微生物の2つ以上の株、クレード、変異体、サブタイプまたは血清型によって起こりうる感染性疾患に対する安定で有効で信頼できるワクチンを提供することが、本発明の目的である。
【0023】
また、すべてのデングウイルス血清型に対する信頼できる予防接種を可能にする、デングウイルス感染に対する安定で有効で信頼できるワクチンを提供することが、本発明のもう一つの目的である。
【課題を解決するための手段】
【0024】
本発明は、ある感染性疾患原因微生物の異なる株、クレード、変異体、サブタイプまたは血清型に由来する相同な遺伝子を1つのポックスウイルスに含めるという着想に基づいている。上述のように、例えばデングウイルスには4つのグループ、サブタイプまたは血清型が存在し、それらはいずれも同じタイプの遺伝子を、例えばキャプシド(C)タンパク質をコードする遺伝子、前駆膜(PrM)タンパク質またはエンベロープ(E)タンパク質をコードする遺伝子などとして含んでいる。しかし、同じタイプの遺伝子の核酸配列は、4つの血清型のすべてで完全に同じわけではなく、完全に相同なわけでもない。例えば、デングウイルス血清型1、2、3および4のPrM遺伝子(PrM1〜4)間で配列を比較したところ(Lasergene 4.05 Magalign,Macintosh)、66.5〜72.9%の配列一致度、すなわち約65〜75%の相同性が明らかになった。感染性疾患原因微生物の異なるサブタイプの遺伝子に見られる相違とばらつきは、それぞれ、1つのサブタイプに対して予防接種しても、同じ微生物の他の変異体による感染に対する保護が自動的にはもたらされないことの理由であると考えられる。そこで、ある感染性疾患原因微生物の異なる株、クレード、変異体、サブタイプまたは血清型に由来する近縁遺伝子または相同遺伝子を含んでいる組換えウイルスを作製することを着想した。しかし、上述したように、相同配列間の相同組換えはウイルスの生活環中に起こり、完全には相同ではないDNA区間の間でさえ起こる。したがって、単一のウイルスゲノムに相同な遺伝子を挿入すると相同組換えが起こり、その結果、挿入した相同遺伝子の欠失が起こると予想された。
【0025】
しかし、少なくとも60%の相同性を有する少なくとも2つの外来遺伝子をゲノム内に含む組換えポックスウイルスを作製したところ、意外にも、前記相同遺伝子はウイルスゲノム中に安定に挿入されたままであることを見い出した。
【0026】
相同な遺伝子(好ましくは少なくとも50%の相同性を有するもの)をウイルスゲノムの異なる挿入部位に挿入した場合でも、それらの遺伝子はウイルスゲノムに安定に挿入されたままである。この場合、前記相同遺伝子間の組換え事象は、ウイルスの増幅やウイルスの生活環にとって重要なウイルス遺伝子の喪失をもたらすと予想された。すなわち、ウイルスの生活環は重大な損傷を受けるだろうと予想された。また、組換え頻度は2つの連鎖遺伝子間の距離に比例するので、異なる挿入部位に位置する2つ以上の相同遺伝子間で起こる組換え事象の頻度は高く、したがって当該遺伝子の欠失及び(又は)著しい干渉が起こるだろうと予想された。したがって、組換え事象が起こらず、相同な遺伝子がウイルスゲノムの異なる挿入部位に安定に挿入されたままであることは、極めて驚くべきことであった。
【0027】
先行技術によれば、日本脳炎ウイルス(JEV)、黄熱ウイルス(YFV)およびデングウイルスなどのフラビウイルスに由来する外来DNAを含む組換えポックスウイルスが知られている(米国特許第5,514,375号)。しかし、前記フラビウイルス由来の各遺伝子は、同じ挿入部位に1回挿入されているだけである。また、適当なコンピュータソフトウェア(Lasergene 4.05 Megaline,Macintosh)で配列比較を行ったところ、ポックスウイルスゲノムに挿入された遺伝子の相同性は、JEV由来の遺伝子の場合は20.2%〜29.6%、YFV由来の遺伝子の場合は29.2%〜45.3%、デングウイルス由来の遺伝子の場合は22.8%〜29.5%であることが明らかになった。
【0028】
変異ワクシニアウイルスアンカラ(Modified Vaccinia Virus Ankara:MVA)の同じ挿入部位(特に欠失部位II)にデングウイルス抗原を挿入することが記載されている国際公開第98/13500号にも、同様のことが言える。
【0029】
米国特許第5,338,683号には、gp13およびgp14ヘルペスウイルス糖タンパク質遺伝子を、単一の組換えポックスウイルスの2つの異なる挿入部位に挿入することが記載されている。しかし両遺伝子は25.2%の相同性しか持っていない。
【0030】
変異ワクシニアウイルスアンカラ(MVA)の同じ挿入部位(欠失部位III)に挿入されたインフルエンザウイルスの赤血球凝集素遺伝子と核タンパク質遺伝子とを配列比較したところ、49.1%の相同性がみとめられた(米国特許第5,676,950号、Sutterら[1994年]Vaccine 12:1032)。
【0031】
米国特許第5,891,442号には、伝染性ファブリキウス嚢病(infectious bursal disease)ウイルスのポリタンパク質VP2、VP3およびVP4のコード配列を有する組換えポックスウイルスが開示されている。これらの遺伝子は41.9%〜50.3%の相同性を持ち、融合されて単一の挿入部位に挿入された。
【0032】
最後に、米国特許第6,217,882号には、52.7%の相同性を有する仮性狂犬病ウイルス抗原gp50およびgp63が同じ部位に挿入されている組換え豚痘ウイルスベクターが記載されている。
【0033】
要約すると、先行技術では、少なくとも50%の相同性を有する相同な遺伝子または配列はすべて、ウイルスゲノム内の同じ挿入部位または単一の挿入部位に挿入されているということができる。
【発明を実施するための最良の形態】
【0034】
本発明によれば、相同な遺伝子または配列は、少なくとも50%の相同性、すなわち50%〜100%の相同性、すなわち少なくとも50%の同一ヌクレオチド塩基を有する。50%未満の相同性を有する遺伝子または配列は異種であるとみなすことができる。本発明では、「相同」または「相同性」という用語を、遺伝子または配列を互いに比較する場合に使用するのに対して、「外来」遺伝子、または「外因性」もしくは「異種」配列という用語は、遺伝子または配列をポックスウイルスゲノムと比較する場合に使用する。すなわち、本発明で使用する場合、これらの用語は、自然界では通常はポックスウイルスに付随して見いだされないDNA配列を表す。したがって本発明は、ウイルスゲノムと比較すると異種であるが、互いには相同である少なくとも2つの遺伝子を含んでいる組換えポックスウイルスに関する。「遺伝子」という用語は、例えばタンパク質、ポリペプチド、ペプチド、抗原などをコードするコード配列を表す。相同な遺伝子から翻訳されるタンパク質、ポリペプチドまたはペプチドは、同じ役割を果たし、同じ機能的特性を示す。相同な遺伝子は通例、異なってはいるが関連のある供給源または生物に由来する。本発明の一実施形態によれば、コード配列における相同性は、好ましくは70%〜80%、より好ましくは80%〜90%、または90%〜100%である。65%〜75%の相同性は最も好ましい。
【0035】
本発明の組換えポックスウイルスは、関連する遺伝情報を、1つの単一感染ユニットまたは1つのウイルス粒子だけに含んでいるので、不均等な感染および異なる相同配列の不均衡な発現が起こる危険性はない。したがって、近縁の遺伝子、さらには最近縁の遺伝子、またはほとんど同一な配列を含み、1つの感染細胞中でそれらを発現させる能力を有する本発明の組換えポックスウイルスは、多価ワクチンの作製にとってとりわけ有利である。
【0036】
この利点は、例えばデングウイルスのような、あるウイルスのいくつかの近縁株または血清型が引き起こしうる疾患に対するワクチンの開発にとっては、とりわけ興味深い。異なるデングウイルス血清型の相同遺伝子を含んでいる組換えポックスウイルスを、実施例に記載する。
【0037】
本発明の相同な遺伝子または配列は、例えばベクターウイルスを除く任意のウイルス、任意の細菌、任意の真菌または寄生虫など、任意の微生物に由来することができる。相同な遺伝子または配列は、好ましくは、ある感染性微生物または病原性微生物に由来し、最も好ましくはその微生物の異なる株もしくはクレード、変異体、サブタイプまたは血清型に由来する。
【0038】
「株」または「クレード」という用語は、当業者には周知の技術用語であり、微生物の分類法を表す。この分類体系では、今までに特徴づけられたすべての微生物を、科、属、種、株という階層的序列に分類する(Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第4版,2001)。科のメンバーに関する判定基準が系統学的関係であるのに対して、属は共通する特徴を有する全てのメンバーを含み、種は複製系統を構成して特定の生態的地位を占める多形質群と定義される。「株」または「クレード」という用語は、基本的形態やゲノム構造およびゲノム構成などの共通する特徴を有するが、宿主域、組織向性、地理的分布、弱毒化または病原性などの生物学的特性が異なっている微生物、すなわちウイルスを表す。「変異体」または「血清型」という用語は、軽微なゲノム変異に起因して個別の感染スペクトルまたは抗原特性を示す同じ株(サブタイプともいう)のメンバーを、さらに区別するものである。
【0039】
本発明の別の実施形態では、相同な遺伝子または配列がウイルスから選択され、好ましくはフラビウイルス属に属するウイルス(例えば、好ましくはデングウイルス、ウエストナイルウイルスまたは日本脳炎ウイルスであるが、これらに限らない)、レトロウイルス属に属するウイルス(例えば、好ましくはヒト免疫不全ウイルス(HIV)であるが、これに限らない)、エンテロウイルス属に属するウイルス(例えば、好ましくは手足口病ウイルス、EV71であるが、これらに限らない)、ロタウイルス属に属するウイルス、またはオルトミクソウイルス属に属するウイルス(例えば、好ましくはインフルエンザウイルスであるが、これに限らない)から選択される。フラビウイルスに由来する相同な遺伝子は最も好ましい。
【0040】
別の好ましい実施形態では、相同な遺伝子がデングウイルス遺伝子から選択され、好ましくはC、NS1及び(又は)NS2、あるいは好ましくはE、より好ましくはPrMである。このウイルスの異なる血清型に由来する相同な遺伝子は最も好ましく、それらの遺伝子は、4つのデングウイルス血清型の1つ、2つ、3つまたは全部に由来することができる。
【0041】
さらに別の実施形態では、相同な遺伝子が異なるHIV株またはHIVクレードから選択される。相同な遺伝子は、好ましくは、gag/polコード配列から選択され、より好ましくはenvコード配列から選択され、さらに好ましくは構造及び(又は)調節HIVコード配列の組合せから選択される。
【0042】
本発明に適したベクターウイルスは、選択した宿主細胞(例えば鳥類宿主細胞)中で容易に培養することができるが、ヒトまたはヒト細胞中では、著しく複製欠損性であるか、または実際に複製しないポックスウイルス群から選択される。
【0043】
いくつかの好ましい実施形態では、本発明のポックスウイルスは、カナリア痘ウイルス(Plotkinら[1995年]Dev Biol Stand. vol 84:165−170、Taylorら[1995年]Vaccine,Vol 13.No.6:539−549)、鶏痘ウイルス(Afonsoら[2000年],J Virol,3815−3831、Fields B.N.編「Fields Virology」,Lippincott−Raven Publishers,第4版,2001,第85章,2916頁)、ペンギン痘ウイルス(Stannardら[1998年]J Gen Virol,79,1637−1649)またはそれらの誘導体を含む群から選択される。これらのウイルスはアビポックスウイルス属に属するので、鳥類細胞中で容易に培養し、増幅することができる。しかし、哺乳類細胞またはヒト細胞では複製欠損性であるため、感染性子孫ウイルスは基本的に産生されないか、ほとんど産生されない。
【0044】
本発明の別の実施形態では、2つ以上の相同な遺伝子を含む組換えポックスウイルスの作製に、ワクシニアウイルス、好ましくは弱毒化ワクシニアウイルスを使用する。
【0045】
ワクシニアウイルス(VV)は短い相同配列の相同組換えを起こすことによって、相同な配列を欠失させうることが知られているが(Howleyら[1996年],Gene 172:233−237)、本発明者らは相同な配列または相同な遺伝子がゲノム中に安定に挿入されている組換えワクシニアウイルスを提供する。これは極めて意外な発見である。というのも、Howelyらによれば、300塩基対(bp)までの短い配列でも、ワクシニアウイルスにおけるゲノムの再編成および相同な配列の欠失を誘発するには十分だったからである。したがって当業者であれば、さらに長い配列ではさらに高い確率で組換え事象が誘発されると予想するだろう。しかし、本発明によれば、完全な相同遺伝子を含む配列でさえ、ワクシニアウイルスのゲノム中に安定に挿入することができる。
【0046】
ワクシニアウイルスの一例は(これに限るわけではないが)、著しく弱毒化され宿主域が制限されたワクシニア株、すなわち変異ワクシニアアンカラ(MVA)である(Sutter,G.ら[1994年]Vaccine 12:1032−40)。MVAは、ワクシニアウイルスのアンカラ株(CVA)をニワトリ胚線維芽細胞で約570代にわたって連続継代することによって作出された(概要についてはMayr,A.ら[1975年]Infection 3,6−14を参照されたい)。長期間にわたるこれらの継代の結果、CVAはそのゲノム配列のうちの約31キロ塩基を欠失させた。得られたウイルス株MVAは、宿主細胞が著しく制限されているとの記載がある(Meyer,H.ら,J. Gen. Virol. 72,1031−1038[1991年])。典型的なMVA株は、ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(European Collection of Animal Cell Cultures)に受託番号ECACC V00120707として寄託されているMVA575である。
【0047】
別の実施形態として、本発明ではMVA−Vero株またはその誘導体を使用することができる。MVA−Vero株はヨーロピアン・コレクション・オブ -アニマル・セル・カルチャーズに、受託番号ECACC V99101431およびECACC 01021411として寄託されている。MVA−Veroの安全性は、国際特許出願第PCT/EP01/02703号に記載の生物学的、化学的および物理的特徴に反映されている。通常のMVAと比較すると、MVA−Veroはゲノム欠失を1つ余分に持っている。
【0048】
本発明ウイルスの「誘導体」という用語は、親ウイルスと同じ特徴を示すがそのゲノムの1つ以上の部分に相違を示す子孫ウイルスを表す。
【0049】
本発明のさらに別の実施形態ではMVA−BNを使用する。MVA−BNはヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズに受託番号ECACC V00083008として寄託されている。MVA−BNウイルスは変異ワクシニアアンカラに由来する著しく弱毒化されたウイルスであることが明らかにされているので、MVA−BNまたはその誘導体を使用することにより、特に安全なウイルスワクチンが作製される。したがって、最も好ましい実施形態では、本発明に従って2つ以上の相同な遺伝子を含むMVA−BNまたはその誘導体を、ウイルスベクターとして使用する。「MVA−BNの誘導体」という用語は、MVA−BNと比較して同じ機能的特徴を有するウイルスを表す。MVA−BNの特徴、あるMVAがMVA−BNおよびその誘導体であるかどうかを調べることを可能にする生物学的アッセイの説明、ならびにMVA−BNまたはその誘導体の作製を可能にする方法は、国際公開第02/42480号(参照により本明細書に組み込まれる)に記載されている。MVA−BNまたはその誘導体の機能的特徴を調べる簡単な方法の一つは、ヒトHaCat細胞におけるその弱毒化および複製の欠如である。
【0050】
本発明の組換えポックスウイルスでは、好ましくは、ポックスウイルス転写制御要素によって、より好ましくはMVA、カナリア痘、鶏痘またはペンギン痘転写制御要素によって、最も好ましくはワクシニアウイルスプロモーターによって、外因性配列の発現を制御する。本発明のポックスウイルス転写制御要素には、他にも、ポックスウイルス系で機能するあらゆる転写制御要素が含まれる。
【0051】
本発明における外因性配列の挿入は、好ましくは、ウイルスゲノムの非必須領域に対して行われる。非必須領域は、例えば、ポックスウイルスの生活環にとって必須でないポックスウイルス遺伝子の遺伝子座またはオープンリーディングフレーム(ORF)である。2つのORFに挟まれた区間を表す遺伝子間領域も、本発明では非必須領域と見なされる。本発明の別の実施形態では、MVAゲノムの天然の欠失部位(参照により本明細書に組み込まれる第PCT/EP96/02926号に開示されているもの)に、外因性配列を挿入する。
【0052】
挿入されるDNAの向きは、本発明の組換えウイルスの機能性または安定性には影響しない。
【0053】
本発明の組換えポックスウイルスは増殖が著しく制限され、その結果、著しく弱毒化されているので、ヒトを含む広範な哺乳動物の処置にとって、さらには免疫不全状態のヒトの処置にとっても、理想的な候補である。したがって本発明は、ヒトを含む動物の生体内で免疫応答を誘発するための医薬組成物およびワクチンも提供する。
【0054】
本医薬組成物は、一般に、1つ以上の薬学的に許容し得るそして/または承認された担体、添加剤、抗生物質、保存剤、アジュバント、希釈剤及び(又は)安定剤を含みうる。そのような補助物質としては、水、食塩水、グリセロール、エタノール、湿潤剤または乳化剤、pH緩衝物質などを挙げることができる。適切な担体は通例、大きくてゆっくり代謝される分子、例えばタンパク質、多糖、ポリ乳酸、ポリグリコール酸、アミノ酸ポリマー、アミノ酸コポリマー、脂質集合体などである。
【0055】
ワクチンを製造するには、本発明の組換えポックスウイルスを生理学的に許容できる形態に変換する。これは、天然痘の予防接種に用いられるポックスウイルスワクチンの製造に関する経験(Stickl,H.ら[1974年]Dtsch. med. Wschr. 99,2386−2392に記載されているもの)に基づいて行うことができる。例えば、精製ウイルスは、5×108 TCID50/mlの力価で、約10mMのトリス(Tris)および140mMのNaCl(pH7.4)中に製剤化して、−80℃で保存される。ワクチン注射剤を製造するには、例えば2%ペプトンおよび1%ヒトアルブミンの存在下で、リン酸緩衝食塩水(PBS)100ml中のウイルス粒子102〜108個を、アンプル(好ましくはガラスアンプル)内で凍結乾燥する。もう一つの選択肢として、ワクチン注射剤は、製剤中のウイルスを段階的に凍結乾燥することによっても製造できる。この製剤は、さらに、例えばマンニトール、デキストラン、糖、グリシン、ラクトースもしくはポリビニルピロリドンなどの添加剤、または、インビボ投与に適した酸化防止剤もしくは不活性ガス、安定剤もしくは組換えタンパク質(例えばヒト血清アルブミン)などの他の助剤も含むことができる。次に、ガラスアンプルを密封して、4℃〜室温で数ヶ月間保存することができる。ただし、必要がない限り、アンプルは−20℃未満の温度で保存することが好ましい。
【0056】
予防接種または治療を行うには、上記の凍結乾燥物を0.1〜0.5mlの水性溶液、好ましくは生理食塩水またはトリス緩衝液に溶解し、それを全身的または局所的に、すなわち非経口投与、皮下投与、筋肉内投与、乱切法、または当業者に知られる他の任意の投与経路によって、投与することができる。当業者であれば、投与様式、投与量および投与回数を、既知の方法で最適化することができる。ただし、最も一般的には、1回目の予防接種注射の約1ヶ月〜6週間後に2回目の注射を行うことによって、患者を予防接種する。
【0057】
本発明の組換えウイルスは、標的細胞への外因性コード配列の導入に使用される。標的細胞への外因性コード配列の導入は、タンパク質、ポリペプチド、ペプチド、抗原およびエピトープをそれぞれインビトロで製造するために使用することができる。また、細胞に相同配列または異種配列を導入する方法は、インビトロ治療およびインビボ治療にも応用することができる。インビトロ治療の場合は、免疫応答を誘発するために、前もって(エクスビボで)本発明の組換えポックスウイルスに感染させておいた単離細胞を、動物の生体に投与する。インビボ治療の場合は、免疫応答を誘発するために、本発明の組換えポックスウイルスを動物の生体に直接投与する。この場合、接種部位を取り巻く細胞は、本発明のウイルスまたはその組換え体に、インビボで直接感染する。感染後、それらの細胞は、外因性コード配列にコードされているタンパク質、ポリペプチド、ペプチドまたは抗原を合成し、次いでそれらを、またはそれらの一部を、細胞表面に提示する。免疫系の専門化した細胞は、そのような外来のタンパク質、ポリペプチド、ペプチド、抗原およびエピトープの提示を認識して、特異的免疫応答を開始する。
【0058】
組換えポックスウイルスを取得する方法またはポックスウイルスゲノムに外因性コード配列を挿入する方法は、当業者にはよく知られている。また、それらの方法は実施例でも説明するし、以下の文献から演繹し、または完遂することもできる。
【0059】
-J.Sambrook,E.F.FritschおよびT.Maniatis著「Molecular Cloning, A laboratory Manual」,第2版,Cold Spring Harbor Laboratory Press,1989年。この文献には、例えばDNAのクローニング、DNAおよびRNAの単離、ウェスタンブロット解析、RT−PCRおよびPCR増幅技術などの標準的な分子生物学技術に関する技術とノウハウが記載されている。
【0060】
-Brian WJ MahyおよびHillar O Kangro編「Virology Methods Manual」,Academic Press,1996年。この文献には、ウイルスの取り扱いと操作に関する技術が記載されている。
【0061】
-AJ DavisonおよびRM Elliott編「Molecular Virology: A Practical Approach」(The Practical Approach Series),IRL Press at Oxford University Press,オックスフォード,1993年の第9章「Expression of genes by Vaccinia virus vectors(ワクシニアウイルスベクターによる遺伝子の発現)」。
【0062】
-「Current Protocols in Molecular Biology」,出版社:John Wiley and Son Inc. 1998年の第16章、第IV節「Expression of proteins in mammalian cells using Vaccinia viral vector(ワクシニアウイルスベクターを使った哺乳動物細胞におけるタンパク質の発現)」。この文献には、MVAの取り扱い、操作および遺伝子工学に関する技術とノウハウが記載されている。
【0063】
本発明の組換えポックスウイルスの作製には、さまざまな方法を応用することができる。ウイルスに挿入すべきDNA配列は、ポックスウイルスの一DNA区間に相同なDNAが挿入されている大腸菌プラスミドコンストラクトに、入れることができる。別途、挿入すべきDNA配列をプロモーターに連結する。このプロモーター−遺伝子連鎖は、非必須遺伝子座を含むポックスDNAの一領域に隣接するDNA配列に相同なDNAがそのプロモーター−遺伝子連鎖の両隣に隣接するように、プラスミドコンストラクト中に配置される。得られたプラスミドコンストラクトを、大腸菌内での増殖によって増幅し、単離する。挿入すべきDNA遺伝子配列を含む単離されたプラスミドを、例えばニワトリ胚線維芽細胞(CEF)などの細胞培養に、ポックスウイルスと共にトランスフェクトする。それぞれプラスミド中およびウイルスゲノム中にある相同なポックスDNA間で相同組換えが起こることにより、外来DNA配列が存在するように改変されたポックスウイルスが得られる。
【0064】
より好ましい実施形態では、適当な細胞培養の細胞、例えばCEF細胞にポックスウイルスを感染させる。次に、その感染細胞に、外来遺伝子(好ましくはポックスウイルス発現制御要素の転写制御を受けるもの)を含む第1プラスミドベクターをトランスフェクトする。上で説明したように、このプラスミドベクターは、ポックスウイルスゲノムの選択した部分への外因性配列の挿入を指示する能力を有する配列も含んでいる。場合により、このプラスミドベクターは、ポックスウイルスプロモーターに作動可能に連結されたマーカー遺伝子及び(又は)選択遺伝子を含んでいるカセットも含有する。好適なマーカー遺伝子または選択遺伝子は、例えば緑色蛍光タンパク質、β−ガラクトシダーゼ、ネオマイシン、ホスホリボシルトランスフェラーゼまたは他のマーカーをコードする遺伝子である。選択カセットまたはマーカーカセットを使用することにより、産生した組換えポックスウイルスの同定と単離が簡単になる。しかし、組換えポックスウイルスは、PCR技術によって同定することもできる。次に、新たな細胞を、上述のようにして得られた組換えポックスウイルスに感染させ、第1ベクターに含まれる遺伝子と相同な遺伝子を含む第2ベクターをトランスフェクトする。この遺伝子がポックスウイルスゲノムの異なる挿入部位に含まれることになっている場合、第2ベクターは、ポックスウイルスのゲノムへの相同遺伝子の組込みを指示する配列も相違する。相同組換えが起こった後に、2つの相同な遺伝子を含む組換えウイルスを単離することができる。組換えウイルス中に3つ以上の相同な遺伝子を導入するには、先の工程で単離された組換えウイルスを感染に使用し、さらにもう一つの相同遺伝子を含むもう一つのベクターをトランスフェクションに使用することによって、感染工程とトランスフェクション工程とを繰り返す。
【0065】
もう一つの選択肢として、上述の感染工程とトランスフェクション工程とは互いに交換することができる。すなわち、適当な細胞を、まず、外来遺伝子を含むプラスミドベクターでトランスフェクトしてから、ポックスウイルスに感染させることもできる。
【0066】
さらにもう一つの選択肢として、各相同遺伝子を異なるウイルスに導入し、ある細胞を、得られた組換えウイルスのすべてに同時感染させ、すべての相同遺伝子を含む組換え体を選別することもできる。
【0067】
さらに本発明は、ポックスウイルスゲノムへの発現可能な相同遺伝子の組込みを指示する2つ以上のプラスミドベクターコンストラクトを含むキットも提供する。そのようなプラスミドベクターは、適当なクローニング部位の他に、ポックスウイルスゲノム中の選択した部分への外因性配列の挿入を指示する能力を有する配列も含んでいる。これらのベクターは、場合により、選択遺伝子カセットまたはマーカー遺伝子カセットも含む。本キットはさらに、1個または数個の相同遺伝子と、場合により選択遺伝子またはマーカー遺伝子とが、上記のベクターコンストラクトによって挿入されている点で組換え型であるウイルスを選択するための手段と説明書も含む。
【0068】
さらに別の実施形態として、本発明は、本発明の組換えポックスウイルスに由来するDNA配列もしくはその一部、または本発明の組換えポックスウイルスに相同なDNA配列もしくはその一部を包含する。そのような配列は、本発明の相同遺伝子の1つの少なくとも断片を含む外因性配列の少なくとも一部と、本発明のポックスウイルスゲノム配列の少なくとも断片とを含み、前記ポックスウイルスゲノム配列は、好ましくは、前記外因性配列に隣接している。
【0069】
そのようなDNA配列は、本ウイルスまたはその誘導体を同定しまたは単離するために使用することができ、例えばPCRプライマーやハイブリダイゼーションプローブを作製するためにそれらを使用したり、アレイ技術にそれらを使用したりすることができる。
【図面の簡単な説明】
【0070】
【図1】実施例1に記載のMVAゲノム中の4つのPrM(=前駆膜)遺伝子(血清型1〜4)の挿入部位を表す概略図である。
【図2】挿入プラスミドベクターコンストラクトを表す。
【図3】挿入プラスミドベクターコンストラクトを表す。
【図4】挿入プラスミドベクターコンストラクトを表す。
【図5】挿入プラスミドベクターコンストラクトを表す。
【図6】挿入プラスミドベクターコンストラクトを表す。
【図7】挿入プラスミドベクターコンストラクトを表す。
【図8】挿入プラスミドベクターコンストラクトを表す。
【図9】挿入プラスミドベクターコンストラクトを表す。
【図10】4種類の挿入ベクター(pBN49、pBN50、pBN40、pBN39)のベクタークローニング戦略のPCR検証を示す。
【図11】4つの相同なデングウイルスPrM遺伝子を含む組換えポックスウイルスのPCR検証(実施例1)を示す。
【図12】挿入プラスミドベクターコンストラクトを表す。
【図13】挿入プラスミドベクターコンストラクトを表す。
【図14】挿入プラスミドベクターコンストラクトを表す。
【図15】挿入プラスミドベクターコンストラクトを表す。
【図16】挿入プラスミドベクターコンストラクトを表す。
【図17】挿入プラスミドベクターコンストラクトを表す。
【図18】実施例2に記載のMVAゲノム中の3つのPrM遺伝子(血清型2〜4)の挿入部位を表す概略図である。
【図19】3つの相同なデングウイルスPrM遺伝子が遺伝子間領域に挿入されている組換えポックスウイルスのPCR検証(実施例2)を示す。
【0071】
図面の簡単な説明
図1:図1は、実施例1に記載のMVAゲノム中の4つのPrM(=前駆膜)遺伝子(血清型1〜4)の挿入部位を表す概略図である。
図2〜9及び図12〜17:図2〜9および図12〜17は、挿入プラスミドベクターコンストラクトを表す図であり、ベクター名、そのサイズ、および以下に示すような興味ある配列の位置が記載されている:AmpR=アンピシリン耐性遺伝子、bfp=青色蛍光タンパク質遺伝子、dA=欠失部位A、dE=欠失部位E、d2=欠失部位2、Ecogpt=大腸菌グアノシンホスホリボシルトランスフェラーゼ遺伝子、EGFP=強化緑色蛍光タンパク質遺伝子、F1=隣接配列1、F2=隣接配列2、I4L=遺伝子間領域I4L、IGR=遺伝子間領域、NPT II=ネオマイシン耐性遺伝子、P=ポックスウイルスプロモーター、pr7.5=ワクシニアプロモーター7.5、PrM=デングウイルスの前駆膜遺伝子、数字はそれが4つの血清型のうちのどれに由来するかを示している、rpt=隣接配列の反復である。
【0072】
図10:図10は、4種類の挿入ベクター(pBN49、pBN50、pBN40、pBN39)のベクタークローニング戦略のPCR検証を示す。各プラスミドを4種類のPCRプライマーの組合せで試験した。各組合せは、それぞれ、別個の一挿入部位に組み込まれた別個の一PrM配列に特異的である。
【0073】
図11:図11は、4つの相同なデングウイルスPrM遺伝子を含む組換えポックスウイルスのPCR検証(実施例1)を示す。ゲルの上側には、組換えウイルスのさまざまなPCR結果を示し、下側には、表記の対照プラスミドの同じPCR反応の結果を示す。相同な配列を含むプラスミドをpBN39、pBN49またはpBN50と呼ぶ。PrMは、挿入されたデングウイルス前駆膜遺伝子を意味し、数字はそれが4つの血清型のどれに由来するかを示している。dA=欠失部位A、dE=欠失部位E、d2=欠失部位2、I4L=遺伝子間領域I4Lは、外因性DNAの挿入部位を表す。
【0074】
図18:図18は、実施例2に記載のMVAゲノム中の3つのPrM遺伝子(血清型2〜4)の挿入部位を表す概略図である。
【0075】
図19:図19は、3つの相同なデングウイルスPrM遺伝子が遺伝子間領域に挿入されている組換えポックスウイルスのPCR検証(実施例2)を示す。上図は、PrM2に特異的なPCR反応の結果を表し、中図はPrM3に特異的なPCR反応の結果を表し、下図はPrM4に特異的なPCR反応の結果を表す。レーン8は、対照プラスミドの同じPCR反応を表す。レーン2は空ベクター対照MVAを表す。PrMは、挿入されたデングウイルス前駆膜遺伝子を意味し、数字はそれが4つの血清型のどれに由来するかを示している。M=分子量マーカー。
【0076】
以下に実施例を挙げて本発明をさらに詳しく説明する。しかしながら当業者は、これらの実施例が、決して本発明によって提供される技術の適用範囲を限定するものではないことを十分に理解している。
【実施例1】
【0077】
挿入ベクター
欠失部位A用の挿入ベクター
ゲノム位置7608〜7609に相当するMVAゲノムのいわゆる欠失部位Aまたは欠失部位1に外因性配列を挿入するために、欠失部位Aに隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適当なコンピュータソフトウェア(DNAsis、日立ソフトウェアエンジニアリング、米国サンブルーノ)を使って適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーターの転写制御を受けるNPT II遺伝子(ネオマイシン耐性)などの選択遺伝子カセットを導入する。また、欠失部位Aに挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図2に示す(pBNX10)。
【0078】
欠失部位E用の挿入ベクター
ゲノム位置170480〜170481に相当するMVAゲノムのいわゆる欠失部位Eまたは欠失部位4に外因性配列を挿入するために、欠失部位Eに隣り合う約600bpの隣接配列を含むベクターを構築した。このベクターは上述したように設計し、構築される。隣接配列の間に、ポックスウイルスプロモーターの転写制御を受けるEGFP遺伝子(緑色蛍光タンパク質、Clonetech)を配置する。また、欠失部位Aに挿入しようとする追加の遺伝子または配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図3に示す(pBNX32)。
【0079】
欠失部位2用の挿入ベクター
ゲノム位置20718〜20719に相当するMVAゲノムのいわゆる欠失部位2に外因性配列を挿入するために、欠失部位2に隣り合う約600bpの隣接配列を含むベクターを構築した。このベクターは上述したように設計し、構築される。隣接配列の間に、ポックスウイルスプロモーターの転写制御を受けるhbfp遺伝子(humanized blue fluorescing protein:ヒト化青色蛍光タンパク質、Pavalkis GNら)を配置する。また、欠失部位2に挿入しようとする追加の遺伝子または配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図4に示す(pBNX36)。
【0080】
遺伝子間領域I4L用の挿入ベクター
ゲノム位置56760に相当するORF I3LとI4Lの間の遺伝子間領域に外因性配列を挿入するために、I4L遺伝子座の遺伝子間領域に隣り合う約600bpの隣接配列を含むベクターを構築した。このベクターは上述したように設計し、構築される。隣接配列の間に、ポックスウイルスプロモーターの転写制御を受けるEcogpt遺伝子(またはgpt、これは大腸菌から単離されるホスホリボシルトランスフェラーゼ遺伝子を意味する)を配置する。また、I4L ORF後の遺伝子間領域に挿入しようとする追加の遺伝子または配列を挿入するためのクローニング部位も存在する。そのような本発明のベクターコンストラクトの一つを図5に示す(pBNX39)。
【0081】
数個の相同な遺伝子がゲノム内に組み込まれている組換えポックスウイルスの構築
挿入ベクター
デングウイルスの4つの血清型に由来する4つのPrM遺伝子をMVAゲノムに挿入するために、4つの独立した組換えベクターを使用した。
【0082】
これらのベクターは、上に詳述したように、相同組換えにより狙って挿入するために、MVAゲノムに相同な配列を有する。また、各ベクターは選択遺伝子カセットまたはレポーター遺伝子カセットも持っている。
【0083】
4つのデングウイルス血清型のPrM配列を、オリゴアニーリングとPCR増幅とによって、合成的に作製した。それらのPrM配列をポックスウイルスプロモーター要素の下流にクローニングして、発現カセットを形成させた。次にこの発現カセットを、適切な挿入ベクターコンストラクトのクローニング部位にクローニングした。
【0084】
結果として、欠失部位A用の挿入ベクターコンストラクトは、デングウイルス血清型2のPrM遺伝子を含有した(図6−pBN39)。欠失部位2用の挿入ベクターコンストラクトは、デングウイルス血清型1のPrM遺伝子を含有した(図7−pBN49)。遺伝子間領域I4L用の挿入ベクターコンストラクトは、デングウイルス血清型3のPrM遺伝子を含有した(図8−pBN50)。欠失部位E用の挿入ベクターコンストラクトは、デングウイルス血清型4のPrM遺伝子を含有した(図9−pBN40)。
【0085】
挿入ベクターのPCR検証
クローニング戦略を検証するために、PCRアッセイを行った。これらのPCRアッセイの場合、選択したプライマー対は、挿入部位に関して特異的な隣接配列に特異的に結合するプライマーと、高度に相同なデングウイルスPrM遺伝子の1つに特異的に結合する第2プライマーとの組合せである。
【0086】
デングウイルス血清型2のPrM遺伝子を含有する欠失部位A用の挿入ベクターは、プライマーoBN93(CGCGGATCCATGCTGAACATCTTGAACAGGAGACGCAGA.SEQ ID NO:1)およびoBN477(CATGATAAGAGATTGTATCAG.SEQ ID NO:2)を使って選別した。
【0087】
デングウイルス血清型1のPrM遺伝子を含有する欠失部位2用の挿入ベクターは、プライマーoBN194(ATGTTGAACATAATGAACAGGAGGAAAAGATCTGTGACCATGCTCCTCATGCTGCTGCCCACAGCCCTGGCGTTCCATCT.SEQ ID NO:3)およびoBN476(GATTTTGCTATTCAGTGGACTGGATG.SEQ ID NO:4)を使って選別した。
【0088】
デングウイルス血清型3のPrM遺伝子を含有する遺伝子間領域I4L用の挿入ベクターは、プライマーoBN255(CCTTAATCGAATTCTCATGTCATGGATGGGGTAACCAGCATTAATAGT.SEQ ID NO:5)およびoBN479(GCTCCCATTCAATTCACATTGG.SEQ ID NO:6)を使って選別した。
【0089】
デングウイルス血清型4のPrM遺伝子を含有する欠失部位E用の挿入ベクターは、プライマーoBN210(ATCCCATTCCTGAATGTGGTGTTAAAGCTACTGAGCGCTTCTCTCGTCTCCGTTCTCCGCTCTGGGTGCATGTCCCATAC.SEQ ID NO:7)およびoBN478(GTACATGGATGATATAGATATG.SEQ ID NO:8)を使って選別した。
【0090】
PCR実験は、10×PCR緩衝液、MgCl2緩衝液およびTaq DNAポリメラーゼを含むTaq DNAポリメラーゼキット(Roche、カタログ番号201205)またはこれと等価な試薬類を使って、サーマルサイクラーGeneAmp9700(Perkin Elmer)で行う。一般に、PCR反応は、45μlの主混合物、試料DNAおよび必要に応じてddH2Oを含む50μlの総反応液量を使って調製した。主混合物は、30.75μlのddH2O、5μlの10×緩衝液、1μlのdNTP混合物(各10mM)、2.5μlの各プライマー(5pmol/ml)、3μlのMgCl2(25mM)および0.25μlのTaqポリメラーゼ(5U/μl)を使って調製すべきである。
【0091】
増幅は以下のプログラムを使って行った。
1)変性:4分 94℃
2)30サイクル:
変性:30秒 94℃
アニーリング:30秒 55℃
伸長:1〜3分 72℃
3)伸長:7分 72℃
4)保存 4℃。
【0092】
挿入される遺伝子のサイズに基づいて、伸長時間は少なくとも1分/kbにするべきである。
【0093】
図10に示すPCR結果は、上記の単独挿入物に使用したプライマーの組合せの特異性を実証している。
【0094】
プライマーの組合せoBN194/oBN476は、欠失部位2およびインサートとしてのPrM1に特異的である。プラスミドpBN49の予想PCR断片は678bpのサイズを有する(ゲルの上側、レーン3に示す)。
【0095】
プライマーの組合せoBN255/oBN479は、遺伝子間領域I4LおよびインサートとしてのPrM3に特異的である。プラスミドpBN50の予想PCR断片は825bpのサイズを有する(ゲルの上側、レーン9に示す)。
【0096】
プライマーの組合せoBN210/oBN478は、欠失部位EおよびインサートとしてのPrM4に特異的である。プラスミドpBN40の予想PCR断片は607bpのサイズを有する(ゲルの下側、レーン5に示す)。
【0097】
プライマーの組合せoBN93/oBN477は、欠失部位AおよびインサートとしてのPrM2に特異的である。プラスミドpBN39の予想PCR断片は636bpのサイズを有する(ゲルの下側、レーン11に示す)。
【0098】
相同組換えによる組換えMVAの作製
組換えMVAによって外来遺伝子を発現させるには、これらの遺伝子を、相同組換えと呼ばれる過程によって、ウイルスゲノム中に挿入する必要がある。そのために、興味ある外来遺伝子を、上述のように、挿入ベクター中にクローニングしておいた。細胞をMVA−BNに感染させた後に、このベクターをトランスフェクトする必要がある。感染とトランスフェクションとを受けた細胞の細胞質では、組換えが起こるだろう。やはり挿入ベクターに含まれている選択及び(又は)レポーターカセットを利用して、組換えウイルスを含む細胞を同定し、単離する。
【0099】
相同組換え
相同組換えを行うために、DMEM(ダルベッコ変法イーグル培地,Gibco BRL)+10%ウシ胎仔血清(FCS)またはVP−SFM(Gibco BRL)+4mmol/l L−グルタミン(無血清製造法の場合)を使って、BHK(ベビーハムスター腎)細胞またはCEF(初代ニワトリ胚線維芽細胞)を6穴プレートに播種する。
【0100】
細胞はまだ増殖期にある必要があるので、トランスフェクションの日に60〜80%コンフルエントに達するべきである。感染に用いる感染多重度(moi)を決定するには細胞数を知っておく必要があるので、播種前に細胞数を数えた。
【0101】
感染を行うために、500μlの希釈液が0.01のmoiを与える適当なウイルス量を含むように、MVAストックをDMEM/FCSまたはVP−SFM/L−グルタミンに希釈する。細胞は、播種後に1回分裂すると仮定する。細胞から培地を取り除き、細胞を500μlの希釈ウイルスに、室温で振とうしながら1時間感染させる。接種物を除去し、細胞をDMEM/VP−SFMで洗浄する。感染細胞を、それぞれ1.6mlのDMEM/FCSおよびVP−SFM/L−グルタミン中に入れておき、その間に、トランスフェクション反応(キアゲン・エフェクテン・キット(Qiagen Effectene Kit))の準備をする。
【0102】
トランスフェクションには、「エフェクテン(Effectene)」トランスフェクションキット(Qiagen)を使用する。線状化した挿入ベクター1〜5μg(多重トランスフェクションの場合は総量)と緩衝液ECとのトランスフェクション混合物を、最終体積が150μlになるように調製する。DNA 1μgにつき8.0μlのエンハンサー(Enhancer)を加え、ボルテックスし、室温で5分間インキュベートする。次に、DNA 1μgにつき25μlのエフェクテンを、原液のチューブをボルテックスしてから加え、その溶液をボルテックスによって十分に混合し、室温で10分間インキュベートする。それぞれ600μlのDMEM/FCSおよびVP−SFM/L−グルタミンを加え、混合した後、そのトランスフェクション混合物全体を、既に培地で覆われている細胞に加える。培養皿を穏やかに振とうしてトランスフェクション反応液を混合する。37℃、5%CO2で、一晩インキュベートする。翌日、培地を除去し、新鮮なDMEM/FCSまたはVP−SFM/L−グルタミンと置き換える。インキュベーションを3日目まで続ける。
【0103】
収集するために、細胞を培地中にこすり取り、得られた細胞懸濁液を適当なチューブに移して、短期保存の場合は−20℃で凍結し、長期保存の場合は−80℃で凍結する。
【0104】
MVAへのPrM4の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型4のPrM遺伝子と、レポーター遺伝子としてEGPF遺伝子とを含んでいる挿入ベクターpBN40をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子EGFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスはプラーク精製によって精製する必要がある。
【0105】
プラーク精製を行うには、感染細胞(蛍光細胞または染色細胞)をピペットチップで単離し、200μlのPBSまたは培地に再懸濁し、吸引する。次に、約106個の細胞が入っている新しい培養皿を、再懸濁したプラーク100μlに感染させる。48時間後に、細胞を300μlのPBSにとりだす。懸濁液からDNAを抽出し、PCR分析によって選別する。予想されるバンドを示すクローンを選択し、新しい6穴プレートをさまざまな量のこのウイルスに感染させる。ウェルに1%アガロースを重層することにより、ウイルスのさらなる伝播を避ける。48時間後に、組換えウイルスクローンを含む感染細胞を単離する。
【0106】
この操作を、PCR分析で野生型MVA−BNを検出することができなくなるまで繰り返す。
【0107】
4回のプラーク精製後に、予想される挿入物を選択的に増幅するプライマー対(上述のoBN210およびoBN478)と、対照として挿入部位である欠失部位Eを特異的に認識するプライマー対(oBN453:GTTGAAGGATTCACTTCCGTGGA,SEQ ID NO:9およびoBN454:GCATTCACAGATTCTATTGTGAGTC,SEQ ID NO:10)とを使って、PCRアッセイにより、組換えウイルスMVA−PrM4を同定した。
【0108】
MVA−PrM4へのPrM2の挿入
上記のプロトコールに従って細胞をMVA−PrM4に感染させ、さらに、デングウイルス血清型2のPrM遺伝子と、選択遺伝子としてNPT II(ネオマイシン耐性遺伝子)とを含んでいる挿入ベクターpBN39をトランスフェクトした。抗生物質耐性遺伝子を発現させる組換えMVAを精製するには、プラーク精製前に選択条件下でウイルス増幅を3回行うことが推奨される。ネオマイシンホスホトランスフェラーゼ選択を行うために、G418を培地に加える。G418はネオマイシンの誘導体であり、リボソームの作用を妨害することにより、タンパク質生合成を阻害する。NPT遺伝子活性はG418をリン酸化によって失活させる。
【0109】
ネオマイシン選択下で16回のプラーク精製後に、予想される挿入物を選択的に増幅するプライマー対(上記のoBN93およびoBN477)と、対照として挿入部位である欠失部位Aを特異的に認識するプライマー対(上記のoBN477およびoBN452:GTTTCATCAGAAATGACTCCATGAAA,SEQ ID NO:11)とを使って、PCRアッセイにより、組換えウイルスMVA−PrM4/PrM2を同定した。さらに、欠失部位EへのPrM4の挿入も、プライマー対oBN210−oBN478およびoBN453−oBN454を使って確認する。
【0110】
MVAへのPrM1の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型1のPrM遺伝子と、レポーター遺伝子としてhbfp(ヒト化青色蛍光タンパク質の遺伝子)とを含んでいる挿入ベクターpBN49をトランスフェクトした。合成されたhbfpタンパク質は、3日目に、組換えウイルスに感染した細胞中に検出することができる。得られた組換えウイルスをプラーク精製によって精製した。10回のプラーク精製後に、予想される挿入物を選択的に増幅するプライマー対(上述のoBN194およびoBN476)と、対照として挿入部位である欠失部位2を特異的に認識するプライマー対(oBN54:CGGGGTACCCGACGAACAAGGAACTGTAGCAGAGGCATC,SEQ ID NO:12およびoBN56:AACTGCAGTTGTTCGTATGTCATAAATTCTTTAATTAT,SEQ ID NO:13)とを使って、PCRアッセイにより、組換えウイルスMVA−PrM1を同定した。
【0111】
MVAへのPrM3の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型3のPrM遺伝子と、レポーター遺伝子としてEcogpt遺伝子(Ecogpt、略してgptは、ホスホリボシルトランスフェラーゼ遺伝子を意味する)とを含んでいる挿入ベクターpBN50をトランスフェクトした。得られた組換えウイルスを、ミコフェノール酸、キサンチンおよびヒポキサンチンの添加により、ホスホリボシルトランスフェラーゼ代謝選択下で、プラーク精製を3回行うことによって精製した。ミコフェノール酸(MPA)はイノシン一リン酸デヒドロゲナーゼを阻害し、大半の細胞株でプリン合成の遮断およびウイルス複製の阻害をもたらす。この遮断は、構成的プロモーターからEcogptを発現させると共に、基質キサンチンおよびヒポキサンチンを与えることによって、克服することができる。
【0112】
予想される挿入物を選択的に増幅するプライマー対(上述のoBN255およびoBN479)と、対照として挿入部位I4Lを特異的に認識するプライマー対(oBN499:CAACTCTCTTCTTGATTACC,SEQ ID NO:14およびoBN500:CGATCAAAGTCAATCTATG,SEQ ID NO:15)とを使って、PCRアッセイにより、生成した組換えウイルスMVA−PrM3を同定した。
【0113】
MVA−PrM1とMVA−PrM3の同時感染
上記のプロトコールに従って、細胞を、等量のMVA−PrM1とMVA−PrM3に感染させた。青色蛍光クローンをホスホリボシルトランスフェラーゼ代謝選択下で3回プラーク精製した後、上記のプライマー対(上記のoBN255およびoBN479、oBN499およびoBN500、oBN194およびoBN476、oBN54およびoBN56)を使ったPCRによって、組換えウイルスを分析した。得られた組換えウイルスをMVA−PrM1/PrM3と名付けた。
【0114】
MVA−PrM1/PrM3とMVA−PrM2/PrM4の同時感染
上記のプロトコールに従って、細胞を、等量のMVA−PrM1/PrM3とMVA−PrM2/PrM4に感染させた。ホスホリボシルトランスフェラーゼ代謝選択およびネオマイシン選択下で、プラーク精製を行った。緑色および青色の蛍光を惹起する組換えウイルスを単離し、上記プライマー対(上記のoBN255およびoBN479、oBN499およびoBN500、oBN194およびoBN476、oBN54およびoBN56、oBN93およびoBN477、oBN477およびoBN452、oBN210およびoBN478、oBN453およびoBN454))を使ったPCRによって分析した。
【0115】
4つのPrM遺伝子をすべて含んでいる組換えウイルス(クローン20)のPCR分析を図11に示す。ゲルの上側は、この組換えウイルスのさまざまなPCR結果を示し、下側は、対照プラスミド(表示のもの)に関する同じPCR反応の結果を示す。レーン1、10および11は1kbおよび100bpの分子マーカーを表す。
【0116】
プライマーの組合せoBN210/oBN478は、欠失部位EおよびインサートとしてのPrM4に特異的である。組換えウイルスおよびプラスミドpBN40の予想PCR断片は607bpのサイズを有する(レーン2に示す)。
【0117】
プライマーの組合せoBN453/oBN454は、欠失部位Eに特異的である。組換えウイルスの予想PCR断片は2.7kbであり、野生型ウイルスの予想バンドは2.3kbである(レーン3に示す)。ゲルの上側でも、野生型ウイルスに特異的なバンドを同定することができる。これは、野生型ウイルスがまだ完全にはこの組換えウイルス調製物から除去されていないことを意味する。さらなるプラーク精製が必要である。
【0118】
プライマーの組合せoBN93/oBN477は、欠失部位AおよびインサートとしてのPrM2に特異的である。組換えウイルスとプラスミドpBN39の予想PCR断片は636bpのサイズを有する(レーン4に示す)。
【0119】
プライマーの組合せoBN477/oBN452は、欠失部位Aに特異的である。組換えウイルスの予想PCR断片は4.1kbであり、野生型ウイルスの予想バンドは2.7kbである(レーン5に示す)。野生型ウイルスに特異的なバンドをゲルの上側で同定することができる。
【0120】
プライマーの組合せoBN255/oBN479は、遺伝子間領域I4LおよびインサートとしてのPrM3に特異的である。組換えウイルスとプラスミドpBN50の予想PCR断片は825bpのサイズを有する(レーン6に示す)。
【0121】
プライマーの組合せoBN499/oBN500は、I4Lの遺伝子間領域に特異的である。組換えウイルスの予想PCR断片は1.0kbであり、野生型ウイルスの予想バンドは0.3kbである(レーン7に示す)。
【0122】
プライマーの組合せoBN194/oBN476は、欠失部位2およびインサートとしてのPrM1に特異的である。組換えウイルスとプラスミドpBN49の予想PCR断片は678bpのサイズを有する(レーン8に示す)。
【0123】
プライマーの組合せoBN54/oBN56は、欠失部位2に特異的である。組換えウイルスの予想PCR断片は1.6kbであり、野生型ウイルスの予想バンドは0.9kbである(レーン9に示す)。野生型ウイルスに特異的なバンドをゲルの上側で同定することができる。
【0124】
もう一つの選択肢として、4種類のウイルスを作製し、細胞を4種類のウイルスのすべてに同時感染させ、組換え体を選別することもできる。
【0125】
さらなる選択マーカーまたは耐性マーカーを含む組換えベクターを使って、改良を行うこともできる。
【実施例2】
【0126】
挿入ベクター
遺伝子間領域136−137(IGR 136−137)用の組換えベクター
MVAゲノムのゲノム位置129,940に相当するいわゆる遺伝子間領域(IGR)136−137に外因性配列を挿入するために、この挿入部位に隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーター(P)の転写制御を受けるNPT II遺伝子(ネオマイシン耐性)などの選択遺伝子カセットを導入する。また、IGR 136−137に挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位(PacI)も存在する。そのような本発明のベクターコンストラクトの一つを図12に示す(pBNX67)。
【0127】
遺伝子間領域07−08(IGR 07−08)用の組換えベクター
MVAゲノムのゲノム位置12,800に相当する遺伝子間領域(IGR)07−08に外因性配列を挿入するために、この挿入部位に隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーター(P)の転写制御を受けるEcogpt遺伝子(グアニンホスホリボシルトランスフェラーゼ)などの選択遺伝子カセットを導入する。また、IGR 07−08に挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位(PacI)も存在する。そのような本発明のベクターコンストラクトの一つを図13に示す(pBNX88)。
【0128】
遺伝子間領域44−45(IGR 44−45)用の組換えベクター
MVAゲノムのゲノム位置37,330に相当する遺伝子間領域(IGR)44−45に外因性配列を挿入するために、この挿入部位に隣り合う約600bpの隣接配列を含むプラスミドベクターを構築した。MVA−BNゲノムDNAからこの隣接配列を単離するために、適切なPCRプライマーを設計することができる。そのようなプライマーは、制限酵素部位を有する伸長部分を含み、その部分を使って、隣接配列をベクタープラスミドにクローニングすることができる。これらの隣接配列の間に、例えばポックスウイルスプロモーター(P)の転写制御を受けるNPT II遺伝子(ネオマイシン耐性)などの選択遺伝子カセットを導入する。また、IGR 44−45に挿入しようとする追加の遺伝子または外因性配列を挿入するためのクローニング部位(PacI)も存在する。そのような本発明のベクターコンストラクトの一つを図14に示す(pBNX87)。
【0129】
数個の相同な遺伝子がゲノム内に組み込まれている組換えポックスウイルスの構築
挿入ベクター
デングウイルスの血清型2、3および4に由来する3つのPrM遺伝子をMVAゲノムに挿入するために、3つの独立した組換えベクターを使用した。
【0130】
これらのベクターは、上に詳述したように、相同組換えによる挿入をターゲティングするために、MVAゲノムに相同な配列を有する。また、各ベクターは選択およびレポーター遺伝子カセットも持っている。
【0131】
3つのデングウイルス血清型のPrM配列を、実施例1で説明したように、合成的に作製した。
【0132】
結果として、IGR136−137用の挿入ベクターコンストラクトは、デングウイルス血清型4のPrM遺伝子を含有した(図15−pBN27)。IGR 07−08用の挿入ベクターコンストラクトは、デングウイルス血清型2のPrM遺伝子を含有し(図16−pBN34)、IGR 44−45用の挿入ベクターコンストラクトは、デングウイルス血清型3のPrM遺伝子を含有した(図17−pBN47)。
【0133】
相同組換えによる組換えMVAの作製
相同組換えによる組換えMVAの作製を、実施例1で説明したように行った。MVAゲノムにおけるPrM4、PrM3およびPrM2の挿入部位を、図18に示す。
【0134】
MVAへのPrM4の挿入
1回目は、上述のプロトコールに従って細胞をMVA−BNに感染させ、さらに、デングウイルス血清型4のPrM遺伝子と、レポーター遺伝子としてEGFP遺伝子とを含んでいる挿入ベクターpBN27をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子EGFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスは実施例1で説明したようにプラーク精製によって精製する必要がある。4回のプラーク精製後に、挿入部位IGR136−137を選択的に増幅するプライマー対(oBN1008:gataccgatcacgttcta,SEQ ID NO:16およびoBN1009:ggatatgattatgtagag,SEQ ID NO:17)を使って、PCRアッセイにより、組換えウイルスMVA−PrM4を同定した。
【0135】
MVAへのPrM2の挿入
上記のプロトコールに従って細胞をMVA−PrM4に感染させ、さらに、デングウイルス血清型2のPrM遺伝子と、レポーター遺伝子としてBFP遺伝子とを含んでいる挿入ベクターpBN34をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子BFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスは実施例1で説明したようにプラーク精製によって精製する必要がある。6回のプラーク精製後に、組換えウイルスMVA−PrM4+PrM2をさらに継代し、増幅させて、粗製ストックを調製した。挿入部位IGR07−08を選択的に増幅するプライマー対(oBN 903:ctggataaatacgaggacgtg,SEQ ID NO:18およびoBN904:gacaattatccgacgcaccg,SEQ ID NO:19)を使って、PCRアッセイにより、組換え体を同定した。
【0136】
MVAへのPrM3の挿入。
【0137】
上記のプロトコールに従って細胞をMVA−PrM2+4に感染させ、さらに、デングウイルス血清型3のPrM遺伝子と、レポーター遺伝子としてEGFP遺伝子とを含んでいる挿入ベクターpBN47をトランスフェクトした。トランスフェクトしたベクターはレポーター遺伝子EGFPを含んでいるので、組換えウイルスに感染した細胞には、合成されたタンパク質を、遅くとも3日目には検出することができる。得られた組換えウイルスは実施例1で説明したようにプラーク精製によって精製する必要がある。3回のプラーク精製後に、挿入部位IGR44−45を選択的に増幅するプライマー対(oBN904:cgttagacaacacaccgacgatgg,SEQ ID NO:20およびoBN905:cggatgaaaaatttttggaag,SEQ ID NO:21)を使って、PCRアッセイにより、組換えウイルスMVA−PrM4+3+2を同定した。
【0138】
3つのデングウイルスPrM遺伝子を含む組換えウイルスのPCR分析を図19に示す。PCR実験は実施例1で説明したように行う。プライマーの組合せoBN1008およびoBN1009は、PrM4挿入物を含むIGR136−137に特異的である(図19、下図)。組換えウイルスの予想PCR断片は、プラスミド陽性対照(レーン8)と同様に、1kbのサイズを有する(レーン4、5および6に示す)。PrM4を持たない空ベクター対照は、190bpの予想断片を示す(レーン2)。レーンMは分子量マーカーを表し、レーン1、3および7は空である。プライマーの組合せoBN902およびoBN903は、PrM2挿入物を含むIGR07−08に特異的である(図19、上図)。組換えウイルスの予想PCR断片は、プラスミド陽性対照(レーン8)と同様に、960bpのサイズを有する(レーン4〜6に示す)。PrM2を持たない空ベクター対照は、190bpの予想断片を示す(レーン2)。プライマーの組合せoBN904およびoBN905は、PrM3挿入物を含むIGR44−45に特異的である(図19、中図)。組換えウイルスの予想PCR断片は、プラスミド陽性対照(レーン8)と同様に、932bpのサイズを有する(レーン4〜6に示す)。PrM2を持たない空ベクター対照は、185bpの予想断片を示す(レーン2)。
【特許請求の範囲】
【請求項1】
少なくとも2つの相同な外来遺伝子を含み、当該遺伝子のそれぞれがウイルスゲノムの異なる挿入部位に挿入されている、組換えポックスウイルス。
【請求項2】
当該遺伝子が少なくとも50%の相同性を有する、請求項1記載の組換えポックスウイルス。
【請求項3】
少なくとも2つの相同な外来遺伝子を含み、それらの遺伝子が少なくとも60%の相同性を有する、組換えポックスウイルス。
【請求項4】
当該遺伝子が65〜75%の相同性を有する、請求項2又は3記載の組換えポックスウイルス。
【請求項5】
当該遺伝子がフラビウイルスに由来する、請求項1〜4のいずれか1つに記載の組換えポックスウイルス。
【請求項6】
上記フラビウイルスがデングウイルスである、請求項5記載の組換えポックスウイルス。
【請求項7】
当該遺伝子が、上記ウイルスの少なくとも2種類の異なる血清型に由来する少なくとも2つの相同な遺伝子である、請求項5又は6記載の組換えポックスウイルス。
【請求項8】
当該遺伝子が少なくとも2つのPrM遺伝子である、請求項5〜7のいずれか1つに記載の組換えポックスウイルス。
【請求項9】
当該遺伝子が4つのPrM遺伝子である、請求項5〜8のいずれか1つに記載の組換えポックスウイルス。
【請求項10】
上記ポックスウイルスがワクシニアウイルスである、請求項1〜9のいずれか1つに記載の組換えポックスウイルス。
【請求項11】
上記ワクシニアウイルスが変異ワクシニアアンカラ(Modified Vaccinia Ankara)(MVA)ウイルスである、請求項10記載の組換えポックスウイルス。
【請求項12】
上記MVAが、ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(European Collection of Animal Cell Cultures)(ECACC)に番号V00083008として寄託されているMVA−BNである、請求項11記載の組換えポックスウイルス。
【請求項13】
上記ポックスウイルスが、ヒト細胞を含む哺乳動物細胞中で、複製欠損性または複製不能である、請求項1〜12のいずれか1つに記載の組換えポックスウイルス。
【請求項14】
当該遺伝子が、ポックスウイルスゲノムの天然の欠失部位及び/又は遺伝子間領域に挿入されている、請求項1〜13のいずれか1つに記載の組換えポックスウイルス。
【請求項15】
医薬またはワクチンとしての請求項1〜14のいずれか1つに記載の組換えポックスウイルス。
【請求項16】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスを含むワクチン。
【請求項17】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスと、薬学的に許容し得る担体、希釈剤、アジュバント及び/又は添加剤とを含む医薬組成物。
【請求項18】
ヒトを含む生きている動物の免疫応答に影響を与える、好ましくは免疫応答を誘発するための、請求項1から14のいずれか1つに記載の組換えポックスウイルス、請求項16に記載のワクチン、または請求項17記載の調合物。
【請求項19】
医薬を製造するために、請求項1〜14のいずれか1つに記載の組換えポックスウイルスを使用する方法。
【請求項20】
ヒトを含む生きている動物において、免疫応答に影響を与える、好ましくは免疫応答を誘発する方法であって、請求項1〜14のいずれか1つに記載の組換えポックスウイルス、請求項16記載のワクチン、または請求項17記載の調合物の治療上有効な量を、処置すべき動物またはヒトに投与することを含む、上記方法。
【請求項21】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスを含む細胞。
【請求項22】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスを製造する方法であって、
-細胞にポックスウイルスを感染させる工程、
-ポックスウイルスゲノムに対して異種遺伝子及びポックスウイルスゲノム配列(これはポックスウイルスゲノムの挿入部位への前記異種遺伝子の組込みを指示する能力を有する)を含む第1ベクターコンストラクトを、上記感染細胞にトランスフェクトする工程、
-産生した組換えポックスウイルスを同定し、単離し、場合により精製する工程、
-先の工程で得られた組換えポックスウイルスを細胞の感染に使用すると共に、ポックスウイルスゲノムに対して異種でありかつ第1ベクターコンストラクトの遺伝子に対して相同であるもう一つの遺伝子を含む追加のベクターコンストラクトを使用することによって、上記の工程を繰り返す工程、
を含む、上記方法。
【請求項23】
-各コンストラクトがポックスウイルス発現制御要素の転写制御を受ける遺伝子を含む2つ以上のベクターコンストラクトであって、異なるベクターに含まれる遺伝子が相同な遺伝子であり、しかも各遺伝子には、ポックスウイルスゲノムへの当該遺伝子の組込みを指示する能力を有するポックスウイルスDNA配列が隣接している、上記ベクターコンストラクト、および
-前記相同遺伝子がゲノムに組み込まれている組換えポックスウイルスを同定するか、及び/又は選択するための手段、
を含むキット。
【請求項24】
各相同遺伝子には、ポックスウイルスゲノムの異なる挿入部位への各ベクターコンストラクトの前記相同遺伝子の組込みを指示する能力を有するポックスウイルスDNA配列が隣接している、請求項23記載のキット。
【請求項25】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスの組換えポックスウイルスゲノムに由来するかまたは同組換えポックスウイルスゲノムに相同なDNA配列であって、少なくとも2つの相同な遺伝子と、ポックスウイルスゲノムの配列の少なくとも一部とを含むDNA配列。
【請求項26】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスに感染した細胞を検出する方法であって、請求項25記載のDNA配列を前記細胞に投与することを含む、上記方法。
【請求項27】
請求項1〜14記載の組換えポックスウイルスを同定する方法であって、請求項25記載のDNA配列を前記ウイルスに投与することを含む、上記方法。
【請求項1】
少なくとも2つの相同な外来遺伝子を含み、当該遺伝子のそれぞれがウイルスゲノムの異なる挿入部位に挿入されている、組換えポックスウイルス。
【請求項2】
当該遺伝子が少なくとも50%の相同性を有する、請求項1記載の組換えポックスウイルス。
【請求項3】
少なくとも2つの相同な外来遺伝子を含み、それらの遺伝子が少なくとも60%の相同性を有する、組換えポックスウイルス。
【請求項4】
当該遺伝子が65〜75%の相同性を有する、請求項2又は3記載の組換えポックスウイルス。
【請求項5】
当該遺伝子がフラビウイルスに由来する、請求項1〜4のいずれか1つに記載の組換えポックスウイルス。
【請求項6】
上記フラビウイルスがデングウイルスである、請求項5記載の組換えポックスウイルス。
【請求項7】
当該遺伝子が、上記ウイルスの少なくとも2種類の異なる血清型に由来する少なくとも2つの相同な遺伝子である、請求項5又は6記載の組換えポックスウイルス。
【請求項8】
当該遺伝子が少なくとも2つのPrM遺伝子である、請求項5〜7のいずれか1つに記載の組換えポックスウイルス。
【請求項9】
当該遺伝子が4つのPrM遺伝子である、請求項5〜8のいずれか1つに記載の組換えポックスウイルス。
【請求項10】
上記ポックスウイルスがワクシニアウイルスである、請求項1〜9のいずれか1つに記載の組換えポックスウイルス。
【請求項11】
上記ワクシニアウイルスが変異ワクシニアアンカラ(Modified Vaccinia Ankara)(MVA)ウイルスである、請求項10記載の組換えポックスウイルス。
【請求項12】
上記MVAが、ヨーロピアン・コレクション・オブ・アニマル・セル・カルチャーズ(European Collection of Animal Cell Cultures)(ECACC)に番号V00083008として寄託されているMVA−BNである、請求項11記載の組換えポックスウイルス。
【請求項13】
上記ポックスウイルスが、ヒト細胞を含む哺乳動物細胞中で、複製欠損性または複製不能である、請求項1〜12のいずれか1つに記載の組換えポックスウイルス。
【請求項14】
当該遺伝子が、ポックスウイルスゲノムの天然の欠失部位及び/又は遺伝子間領域に挿入されている、請求項1〜13のいずれか1つに記載の組換えポックスウイルス。
【請求項15】
医薬またはワクチンとしての請求項1〜14のいずれか1つに記載の組換えポックスウイルス。
【請求項16】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスを含むワクチン。
【請求項17】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスと、薬学的に許容し得る担体、希釈剤、アジュバント及び/又は添加剤とを含む医薬組成物。
【請求項18】
ヒトを含む生きている動物の免疫応答に影響を与える、好ましくは免疫応答を誘発するための、請求項1から14のいずれか1つに記載の組換えポックスウイルス、請求項16に記載のワクチン、または請求項17記載の調合物。
【請求項19】
医薬を製造するために、請求項1〜14のいずれか1つに記載の組換えポックスウイルスを使用する方法。
【請求項20】
ヒトを含む生きている動物において、免疫応答に影響を与える、好ましくは免疫応答を誘発する方法であって、請求項1〜14のいずれか1つに記載の組換えポックスウイルス、請求項16記載のワクチン、または請求項17記載の調合物の治療上有効な量を、処置すべき動物またはヒトに投与することを含む、上記方法。
【請求項21】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスを含む細胞。
【請求項22】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスを製造する方法であって、
-細胞にポックスウイルスを感染させる工程、
-ポックスウイルスゲノムに対して異種遺伝子及びポックスウイルスゲノム配列(これはポックスウイルスゲノムの挿入部位への前記異種遺伝子の組込みを指示する能力を有する)を含む第1ベクターコンストラクトを、上記感染細胞にトランスフェクトする工程、
-産生した組換えポックスウイルスを同定し、単離し、場合により精製する工程、
-先の工程で得られた組換えポックスウイルスを細胞の感染に使用すると共に、ポックスウイルスゲノムに対して異種でありかつ第1ベクターコンストラクトの遺伝子に対して相同であるもう一つの遺伝子を含む追加のベクターコンストラクトを使用することによって、上記の工程を繰り返す工程、
を含む、上記方法。
【請求項23】
-各コンストラクトがポックスウイルス発現制御要素の転写制御を受ける遺伝子を含む2つ以上のベクターコンストラクトであって、異なるベクターに含まれる遺伝子が相同な遺伝子であり、しかも各遺伝子には、ポックスウイルスゲノムへの当該遺伝子の組込みを指示する能力を有するポックスウイルスDNA配列が隣接している、上記ベクターコンストラクト、および
-前記相同遺伝子がゲノムに組み込まれている組換えポックスウイルスを同定するか、及び/又は選択するための手段、
を含むキット。
【請求項24】
各相同遺伝子には、ポックスウイルスゲノムの異なる挿入部位への各ベクターコンストラクトの前記相同遺伝子の組込みを指示する能力を有するポックスウイルスDNA配列が隣接している、請求項23記載のキット。
【請求項25】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスの組換えポックスウイルスゲノムに由来するかまたは同組換えポックスウイルスゲノムに相同なDNA配列であって、少なくとも2つの相同な遺伝子と、ポックスウイルスゲノムの配列の少なくとも一部とを含むDNA配列。
【請求項26】
請求項1〜14のいずれか1つに記載の組換えポックスウイルスに感染した細胞を検出する方法であって、請求項25記載のDNA配列を前記細胞に投与することを含む、上記方法。
【請求項27】
請求項1〜14記載の組換えポックスウイルスを同定する方法であって、請求項25記載のDNA配列を前記ウイルスに投与することを含む、上記方法。
【図2】


【図3】

【図5】
【図6】

【図8】
【図9】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図19】

【図1】
【図4】

【図7】
【図10】
【図11】

【図18】
【図3】
【図5】
【図6】
【図8】
【図9】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図19】
【図1】
【図4】
【図7】
【図10】
【図11】
【図18】
【公開番号】特開2010−259446(P2010−259446A)
【公開日】平成22年11月18日(2010.11.18)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−176512(P2010−176512)
【出願日】平成22年8月5日(2010.8.5)
【分割の表示】特願2004−506501(P2004−506501)の分割
【原出願日】平成15年5月14日(2003.5.14)
【出願人】(502240076)バヴァリアン・ノルディック・アクティーゼルスカブ (18)
【Fターム(参考)】
【公開日】平成22年11月18日(2010.11.18)
【国際特許分類】
【出願番号】特願2010−176512(P2010−176512)
【出願日】平成22年8月5日(2010.8.5)
【分割の表示】特願2004−506501(P2004−506501)の分割
【原出願日】平成15年5月14日(2003.5.14)
【出願人】(502240076)バヴァリアン・ノルディック・アクティーゼルスカブ (18)
【Fターム(参考)】
[ Back to top ]