
ポリアミド原料用カダベリン
【課題】より高反応収率、高精製収率、高濃度、高生産速度および高純度でカダベリンを製造する。
【解決手段】本発明の課題は、高濃度のL−リジン一塩酸塩に、N末端アミノ酸配列に6個のヒスチジンを付与したL−リジン脱炭酸酵素遺伝子を導入した大腸菌の細胞破砕液もしくはL−リジン脱炭酸酵素を細胞表面に局在化させた大腸菌を接触させることにより解決される。更には、L−リジン一塩酸塩にL−リジン脱炭酸酵素を作用させることによりpHを制御する必要がなく、カダベリンを高濃度、高反応収率、高生産速度で生成させ、この反応液をpH13以上にし、極性有機溶媒で抽出し、蒸留することによりカダベリンを採取し、L−リジン一塩酸塩からカダベリンを製造することによって解決される。
【解決手段】本発明の課題は、高濃度のL−リジン一塩酸塩に、N末端アミノ酸配列に6個のヒスチジンを付与したL−リジン脱炭酸酵素遺伝子を導入した大腸菌の細胞破砕液もしくはL−リジン脱炭酸酵素を細胞表面に局在化させた大腸菌を接触させることにより解決される。更には、L−リジン一塩酸塩にL−リジン脱炭酸酵素を作用させることによりpHを制御する必要がなく、カダベリンを高濃度、高反応収率、高生産速度で生成させ、この反応液をpH13以上にし、極性有機溶媒で抽出し、蒸留することによりカダベリンを採取し、L−リジン一塩酸塩からカダベリンを製造することによって解決される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、カダベリンの製造法に関するものである。カダベリンは医薬中間体などの合成原料やポリマー原料として期待され、需要が高まりつつある。
【背景技術】
【0002】
従来、L−リジンを微量のテトラリン過酸化物を含むシクロヘキサノール中で煮沸することによりカダベリンが得られることが知られている(非特許文献1参照。)。しかしながら高温反応であるために大量のエネルギーおよび有機溶媒が必要であるうえに、反応収率が非常に低い(36%)。
【0003】
また、2−シクロヘキセン−1−オンなどのビニルケトン類を触媒としてリジンから合成する方法(非特許文献2、特許文献1参照。)等が知られている。
【0004】
また、この方法によって得られるカダベリンには不純物として、トリ−n−ブチルアミンや2,3,4,5−テトラヒドロピリジンといった塩基性化合物が含有されている。
【0005】
また一般的に、酵素反応進行時にpHが変化する際には、pHを制御するために酸もしくはアルカリが適宜加えられ生成物の濃度が低下し、精製工程において濃縮操作が必要となり効率が悪くなるといった問題点がある。さらには、加えられた酸、アルカリにより酵素が変性失活してしまう。
また、カダベリンは生体内に普遍的に存在する生体アミンであり、その生合成系が解明されつつある(非特許文献3参照。)。また、エシェリシア・コリ(Escherichia coli)由来のL−リジン脱炭酸酵素遺伝子が知られている(非特許文献4参照。)。
【0006】
一方、細胞表面にタンパク質を局在化させる研究は、多くの細菌において行われている。大腸菌について細胞表面にタンパク質を局在化させる研究が活発に行われている(非特許文献5参照)。
【0007】
しかしながら、カダベリンの製造について実際的な製造技術は確立されておらず、効率よく、より温和な条件下で高濃度かつ高純度のカダベリンを製造する方法の開発が望まれている。
【特許文献1】特公平4−10452号公報
【非特許文献1】須山正,金尾清造;アミノ酸の脱炭酸(第4報)薬学雑誌,vol.85(6),P.531−533(1965))
【非特許文献2】Chemistry letters,893(1986)
【非特許文献3】Celia white tabor and Herbert tabor;Microbiological Reviews,vol.49,P.81−99(1985)
【非特許文献4】Shi−yuanmeng and George N. Bennett;Journal of Bacteriology,vol.174,P.2659−2669(1992)
【非特許文献5】George Georgiou,Heather L.Poetschke,Chrisstos Stathopoulos and Joseph A. Francisco;TIBTECH,vol.11,P.6−10(1993)
【発明の開示】
【発明が解決しようとする課題】
【0008】
トリ−n−ブチルアミンおよび2,3,4,5−テトラヒドロピリジンが多く含有されているカダベリンをポリマーの原料とする場合、たとえば、カダベリンを含む原料からポリアミドを加熱重縮合で合成する際には、反応温度が高温であるため、カダベリンに含まれるトリ−n−ブチルアミンや2,3,4,5−テトラヒドロピリジンなどの塩基性化合物がポリアミドの分解反応を起こし得られたポリアミドに耐熱性が不足するといった問題点がある。
【0009】
さらに、トリ−n−ブチルアミンは、水性生物に有毒で、魚類に対する致死量は20−40mg/Lであるといった問題点もある。
【0010】
本発明の課題は、このような不純物の少ないカダベリンの高反応収率、高精製収率、高生産速度、高濃度および高純度での工業的製造方法およびカダベリンの単離方法を提供することである。
【課題を解決するための手段】
【0011】
上記問題点を解決するために、本発明者らは、効率がよく、より温和な条件下で高濃度かつ高純度のカダベリンの製造方法について鋭意研究を行った結果、L−リジン塩水溶液にL−リジン脱炭酸酵素を作用させることによりカダベリンを高反応収率かつ高濃度で得られることを見出した。
【0012】
すなわち、本発明は、「(1)L−リジン塩水溶液に、L−リジン脱炭酸酵素を作用させ、反応液からカダベリンを単離することを特徴とするカダベリンの製造方法。
(2)2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のカダベリン。
(3)トリ−n−ブチルアミンの含有量が0.006wt%以下のカダベリン。
(4)上記(1)の方法で得られたカダベリンまたは上記(2)あるいは(3)記載のカダベリンを原料として含有するポリアミド。」である。
【発明の効果】
【0013】
本発明によれば、高反応収率、高精製収率、高生産速度、高濃度および高純度でのカダベリンの工業的製造方法およびカダベリンの単離方法が提供される。
【発明を実施するための最良の形態】
【0014】
カダベリンとは1,5−ペンタンジアミンのことであり、ポリマー原料、医薬中間体の合成原料などとして有用な化合物である。
【0015】
L−リジン脱炭酸酵素は、L−リジンをカダベリンに転換させる酵素であり、Escherichia coli K12株をはじめとするエシェリシア属微生物のみならず、多くの生物に存在することが知られている。
【0016】
本発明において使用されるL−リジン脱炭酸酵素に特に制限はないが、例えば、バシラス・ハロドゥランス(Bacillus halodurans)、バシラス・サブチリス(Bacillus subtilis)、エシェリシア・コリ(Escherichia coli)、セレノモナス・ルミナンチウム(Selenomonas ruminantium)、ビブリオ・コレラ(Vibrio cholerae)、ビブリオ・パラヘモリティカス(Vibrio parahaemolyticus)、ストレプトマイセス・コエリカーラ(Streptomyces coelicolor)、ストレプトマイセス・ピロサス(Streptomyces pilosus)、エイケネラ・コロデンス(Eikenella corrodens)、イユバクテリウム・アシダミノフィルム(Eubacterium acidaminophilum)、サルモネラ・ティフィムリウム(Salmonella typhimurium)、ハフニア・アルベイ(Hafnia alvei)、ナイセリア・メニンギチデス(Neisseria meningitidis)、テルモプラズマ・アシドフィルム(Thermoplasma acidophilum)、ピロコッカス・アビシ(Pyrococcus abyssi)またはコリネバクテリウム・グルタミカス(Corynebacterium glutamicum)由来のものが好ましく用いられる。さらに好ましくは安全性の認められているエシェリシア・コリ(Escherichia coli)由来のものである。
【0017】
本発明における生成物は、アルカリ性の化合物であるカダベリンであるために、反応が進行するにつれ、反応液のpHはアルカリ側に変化していく。しかし、L−リジン塩水溶液に、L−リジン脱炭酸酵素を作用させれば、反応液のpHを制御する必要がなく効率よくカダベリンを生産できる。用いるL−リジン塩については特に制限はない。好ましくは塩酸塩、硫酸塩、酢酸塩、硝酸塩、炭酸塩である。さらに好ましくはL−リジン一塩酸塩である。
【0018】
L−リジン脱炭酸酵素は本来ビタミンB6と結合し存在するが、L−リジン塩水溶液にビタミンB6を添加することにより、生産速度および反応収率を向上させることができる。ビタミンB6を添加する方法には特に制限はない。反応中に適宜添加しても良い。好ましくはL−リジン脱炭酸酵素が溶液状態であれば、酵素溶液中に添加することが好ましい。反応液中でのビタミンB6の濃度については特に制限はなく。好ましくは反応液に0.05mMのビタミンB6濃度となるように加えればよい。用いるビタミンB6に特に制限はない。好ましくは、ピリドキシン、ピリドキサミン、ピリドキサルおよびピリドキサルリン酸から選ばれる少なくとも1種である。さらに好ましくはピリドキサルリン酸である。
【0019】
また、L−リジン脱炭酸酵素を作用させた後、L−リジン塩が仕込み濃度の20%以下にまでになった反応液からカダベリンを単離することにより、高精製収率でカダベリンが単離できる。20%以下にまでになったことを確認する手段には特に制限はない。好ましくはHPLC法によって確認することができる。さらに好ましくはpHの変化を経時的に測定し、L−リジン塩の濃度と関連づけて反応の終点を確認する方法が採用できる。
【0020】
本発明において使用されるL−リジン脱炭酸酵素を得る方法に特に制限はないが、例えば、L−リジン脱炭酸酵素が細胞内で高発現した組換え細胞(細胞内発現組み換え細胞)などを適当な培地で培養し、増殖した細胞を回収し、当該細胞を破砕すればよい。
【0021】
また、別の方法として、L−リジン脱炭酸酵素が細胞表面で局在化した組み換え細胞(細胞表面発現組み換え細胞)などを適当な培地で培養し、増殖した細胞を回収すればよい。
【0022】
L−リジン脱炭酸酵素が細胞表面で局在化した組み換え細胞とは、少なくともL−リジン脱炭酸酵素が細胞表面に局在化している状態であり、細胞内に同時にL−リジン脱炭酸酵素が存在していても良い。
【0023】
一方、細胞表面にL−リジン脱炭酸酵素を局在化させる具体的な方法としては、例えば、分泌シグナル配列の一部、細胞表面局在タンパク質の一部をコードする遺伝子配列、L−リジン脱炭酸酵素の構造遺伝子配列をこの順で有するDNAを大腸菌に導入すればよい。
【0024】
分泌シグナル配列の一部に制限はない。宿主においてタンパク質を分泌するために必要な配列であればよい。大腸菌においては、例えばリポプロテインの配列の一部であり、具体的には、アミノ酸配列としてMKATKLVLGAVILGSTLLAGCSSNAKIDQ(アミノ酸の一文字表記)と翻訳される遺伝子配列であることが好ましい。
【0025】
細胞表面局在タンパク質の一部をコードする遺伝子配列に特に制限はない。大腸菌においては、例えば外膜結合タンパク質の配列の一部であり、具体的にはOmpA(外膜結合タンパク質)の46番目のアミノ酸から159番目のアミノ酸までの配列の一部であることが好ましい。
【0026】
L−リジン脱炭酸酵素遺伝子、リポプロテイン遺伝子およびOmpA遺伝子をクローニングする方法に特に制限はない。既知の遺伝子情報に基づき、PCR(polymerase chain reaction)法を用いて必要な遺伝領域を増幅取得する方法、既知の遺伝子情報に基づきゲノムライブラリーやcDNAライブラリーより相同性や酵素活性を指標としてクローニングする方法などが挙げられる。本発明においては、これらの遺伝子は、遺伝的多形性などによる変異型も含む。なお、遺伝的多形性とは、遺伝子上の自然突然変異により遺伝子の塩基配列が一部変化しているものをいう。例えばE.coli K12株の染色体DNAよりPCR法を用いてL−リジン脱炭酸酵素をコードする遺伝子であるcadA遺伝子またはldc遺伝子をクローニングする。この際使用する染色体DNAはE.coli由来であればどの菌株由来でもよい。
【0027】
L−リジン脱炭酸酵素が細胞表面に局在化していることの確認は、例えばL−リジン脱炭酸酵素を抗原として作製した抗体により、細胞表面発現組み換え細胞を免疫反応後、包埋、薄切りし、電子顕微鏡(免疫電顕法)により観察することで確認できる。
【0028】
本発明では、L−リジン脱炭酸酵素は、L−リジン脱炭酸酵素の細胞内もしくは細胞表面での活性が上昇した組換え細胞から調製されたものが好ましく使用できる。細胞内もしくは細胞表面でL−リジン脱炭酸酵素の活性を上昇させる方法に特に制限はない。具体的には、例えば、L−リジン脱炭酸酵素の酵素量を増加させる方法、もしくは酵素の構造遺伝子自体に変異を導入して、酵素そのものの比活性を上昇させることなどが挙げられる。
【0029】
細胞内もしくは細胞表面の酵素量を増加させる手段としては、遺伝子の転写調節領域の改良、遺伝子のコピー数の増加、蛋白への翻訳の効率化などが挙げられる。
【0030】
転写調節領域の改良とは、遺伝子の転写量を増加させる改変を加えることをいう。例えば、プロモーターに変異を導入することによってプロモーター強化を行い、下流にある遺伝子の転写量を増加させることができる。プロモーターに変異を導入する以外にも、宿主内で強力に発現するプロモーターを導入しても良い。例えば大腸菌においては、lac、tac、trpなどのプロモーターが挙げられる。また、エンハンサーを新たに導入することによって遺伝子の転写量を増加させることができる。染色体DNAのプロモーター等の遺伝子導入については、例えば特開平1−215280号公報に記載されている。
【0031】
遺伝子のコピー数の上昇は、具体的には、遺伝子を多コピー型のベクターに接続して組換えDNAを作製し、該組換えDNAを宿主細胞に保持させることにより達成することができる。ここでベクターとは、プラスミドやファージ等広く用いられているものを含むが、これら以外にも、トランソポゾン(Berg,D.E and Berg.C.M., Bio/Technol.,vol.1,P.417(1983))やMuファージ(特開平2−109985号公報)も含む。遺伝子を相同組換え用プラスミド等を用いた方法で染色体に組み込んでコピー数を上昇させることも可能である。
【0032】
蛋白の翻訳効率を上昇させる方法としては、例えば原核生物においてはSD配列(Shine,J.and Dalgarno,L.,Proc.Natl.Acad.Sci.USA,71,1342−1346 (1974))、真核生物では Kozakのコンセンサス配列(Kozak,M.,Nuc.Acids.Res.,Vol.15,p.8125−8148(1987))を導入、改変することや、使用コドンの最適化(特開昭59−125895)などが挙げられる。
【0033】
L−リシン脱炭酸酵素の細胞内もしくは細胞表面での活性を上昇させる手段としては、L−リジン脱炭酸酵素の構造遺伝子自体に変異を導入して、L−リジン脱炭酸酵素そのものの活性を上昇させることも挙げられる。
【0034】
遺伝子に変異を生じさせるには、部位特異的変異法(Kramer,W. and frita,H.J.,Methods in Enzymology,vol.154,P.350(1987))リコンビナントPCR法(PCR Technology,Stockton Press(1989)、特定の部分のDNAを化学合成する方法、または当該遺伝子をヒドロキシアミン処理する方法や当該遺伝子を保有する菌株を紫外線照射処理、もしくはニトロソグアニジンや亜硝酸などの化学薬剤で処理する方法がある。
【0035】
組換え細胞としては、微生物、動物、植物、または昆虫由来のものが好ましく使用できる。例えば動物を用いる場合、マウス、ラットやそれらの培養細胞などが用いられる。植物を用いる場合、例えばシロイヌナズナ、タバコやそれらの培養細胞が用いられる。また、昆虫を用いる場合、例えばカイコやその培養細胞などが用いられる。また、微生物を用いる場合、例えば、大腸菌などが用いられる。
【0036】
また、L−リジン脱炭酸酵素を複数種組み合わせて使用しても良い。
【0037】
L−リジン脱炭酸酵素を得るために、組換え細胞を培養する方法に特に制限はないが、例えば微生物を培養する場合、使用する培地は、炭素源、窒素源、無機イオンおよび必要に応じその他有機成分を含有する培地が用いられる。例えば、大腸菌の場合しばしばLB培地が用いられる。炭素源としては、グルコース、ラクトース、ガラクトース、フラクトース、アラビノース、マルトース、キシロース、トレハロース、リボースや澱粉の加水分解物などの糖類、グリセロール、マンニトールやソルビトールなどのアルコール類、グルコン酸、フマール酸、クエン酸やコハク酸等の有機酸類を用いることができる。窒素源としては、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム等の無機アンモニウム塩、大豆加水分解物などの有機窒素、アンモニアガス、アンモニア水等を用いることができる。有機微量栄養素としては、各種アミノ酸、ビタミンB1等のビタミン類、RNA等の核酸類などの要求物質または酵母エキス等を適量含有させることが望ましい。それらの他に、必要に応じて、リン酸カルシウム、硫酸カルシウム、鉄イオン、マンガンイオン等が少量添加される。
【0038】
培養条件にも特に制限はなく、例えば大腸菌の場合、好気条件下で16〜72時間程度実施するのが良く、培養温度は30℃〜45℃に、特に好ましくは37℃に、培養pHは5〜8に、特に好ましくはpH7に制御するのがよい。なおpH調整には無機あるいは有機の酸性あるいはアルカリ性物質、さらにアンモニアガス等を使用することができる。
【0039】
増殖した細胞内発現組み換え細胞は、遠心分離等により培養液から回収することができる。回収した細胞内発現組み換え細胞から細胞破砕液を調整するには、通常の方法が用いられる。すなわち、細胞内発現組み換え細胞を超音波処理、ダイノミル、フレンチプレス等の方法にて破砕し、遠心分離により細胞残渣を除去することにより細胞破砕液が得られる。
【0040】
細胞破砕液からL−リジン脱炭酸酵素を精製するには、硫安分画、イオン交換クロマトグラフィー、疎水クロマトグラフィー、アフィニティークロマトグラフィー、ゲル濾過クロマトグラフィー、等電点沈殿、熱処理、pH処理など酵素の精製に通常用いられる手法が適宜組み合わされて用いられる。
【0041】
一般的な工業化される酵素反応は休止細胞を用いるが、本発明においては作用させるL−リジン脱炭酸酵素は、L−リジン脱炭酸酵素活性を有する休止細胞、その細胞破砕液などが使用できるが、細胞破砕液が好ましい。細胞破砕液を用いるとより高生産速度でカダベリンが得られる。
【0042】
さらに、部分的に精製したL−リジン脱炭酸酵素を作用させることによりL−リジンの分解およびカダベリンの分解を抑制することができ、より高収率かつ高純度のカダベリンが得られる。部分的な精製とは、工業的に容易な処理のみ行うことであり例えば熱処理、pH処理等が挙げられる。好ましくは熱処理である。さらに好ましくは単離精製したL−リジン脱炭酸酵素を作用させることである。L−リジン脱炭酸酵素を部分的に精製、または単離精製することで、L−リジンの分解やカダベリンの分解を引き起こす酵素を取り除くことができる。
【0043】
さらに、L−リジン脱炭酸酵素が細胞表面に局在化している細胞を作用させれば、L−リジンの分解およびカダベリンの分解を抑制することができ、より高収率かつ高純度、高生産速度でカダベリンが得られる。さらにはその細胞を回収することで繰り返し酵素反応に利用することができる。しかも酵素の調整が非常に簡便である。
【0044】
一般的に酵素というものは本来あるべきアミノ酸配列で自然界に存在するため、人工的にアミノ酸配列を付与もしくは欠如させると酵素の活性を失うことが多い。しかし本発明において細胞内にL−リジン脱炭酸酵素を発現させる場合、L−リジン脱炭酸酵素のN末端アミノ酸配列に1から30個のアミノ酸配列を付与したL−リジン脱炭酸酵素が好ましく使用できる。ここで、N末端に付与するものは6から10個のヒスチジン残基が好ましい。このように、N末端アミノ酸配列にアミノ酸を付与することで、L−リジン脱炭酸酵素の精製を簡便にできるようにすることができる。N末端アミノ酸配列に1から30個のアミノ酸を付与する方法に特に制限はない。例えばL−リジン脱炭酸酵素遺伝子に付与したいアミノ酸配列に相当する塩基配列を遺伝子工学的手法を用い、組み換えればよい。用いる遺伝子工学的手法に特に制限はないが。好ましくはPCR法やDNA断片同士の連結による方法がある。
【0045】
L−リジン脱炭酸酵素によるL−リジン塩水溶液からカダベリンへの変換は、上記のようにして得られるL−リジン脱炭酸酵素を、L−リジン塩水溶液に接触させることによって行うことができる。
【0046】
用いるL−リジン脱炭酸酵素の量に特に制限はない。L−リジン塩水溶液をカダベリンに変換する反応を触媒するのに十分な量であればよい。好ましくは反応液中の酵素濃度が25から70mg/Lである。好ましくは50mg/Lである。一方、休止細胞を使用する際は、反応液中の細胞濃度が5から15g/Lである。好ましくは10g/Lである。
【0047】
反応温度は、通常、28〜55℃、好ましくは45℃前後である。
【0048】
反応液の状態に特に制限はない。好ましくは静置または攪拌状態である。攪拌状態にする方法には特に制限はない。好ましくは攪拌翼により攪拌する方法である。
【0049】
本発明で使用するL−リジン脱炭酸酵素は、固定化することで繰り返しカダベリン合成反応に使用することができ、酵素の調製に必要なコストを低減させることができる。固定化方法としては、アクリルアミドなどの合成高分子に包括する方法、セファロースやスチレン樹脂を骨格とするイオン交換性担体や疎水性担体に吸着させる方法、または、ガラス担体に共有結合で結合させる方法などが挙げられる。
【0050】
反応時間は、使用する酵素活性、L−リジン塩濃度などの条件によって異なるが、通常、1〜72時間である。また、反応は、L−リジン塩を供給しながら連続的に行ってもよい。
【0051】
このように生成したカダベリンを反応終了後、反応液から採取する方法としては、反応終了液をアルカリでpH12から14にし、極性有機溶媒で抽出すれば良い。
【0052】
用いるアルカリに特に制限はない。好ましくは水酸化ナトリウム、水酸化カリウム、水酸化カルシウムを用いることができる。
【0053】
用いる極性有機溶媒に特に制限はない。好ましくはアニリン、シクロヘキサノン、1−オクタノール、イソブチルアルコール、シクロヘキサノール、クロロホルムを用いることができる。
【0054】
このようにして得られたカダベリンは、トリ−n−ブチルアミンおよび2,3,4,5−テトラヒドロピリジンなどの不純物が少ない。本発明の方法で製造することで、以下の方法で分析した場合に、トリ−n−ブチルアミンの含有量が、0.006wt%以下のカタベリンが得られる。さらに、以下の方法で分析した場合に、2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のものが得られる。さらに、トリ−n−ブチルアミンの含有量が、0.006wt%以下であり、かつ、2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のものが得られる。
【0055】
トリ−n−ブチルアミンおよび2,3,4,5−テトラヒドロピリジンの分析は、GC−MS法によって以下の条件で行う。
【0056】
GC/MS:HP6980/HP5973A
Column:NUKOL 30m×0.24mmI.D. 0.2μm Film
Oven:120℃(一定)
InJ:200℃(Split 10:1)
Flow:He 2.4ml/min (const.Flow)
MS:230℃(SCAN m/z=30〜400)
このようにして得られたカダベリンは、ポリアミドの原料として有用である。
【0057】
ポリアミドの製造方法としては、カダベリンとジカルボン酸の塩、および水の混合物を、加熱して脱水反応を進行させる加熱重縮合法が用いられる。また、加熱重縮合後、固相重合することによって、分子量を上昇させることも可能である。固相重合は、100℃〜融点の温度範囲で、真空中、あるいは不活性ガス中で加熱することにより進行し、加熱重縮合では分子量が不十分なポリアミドを高分子量化することができる。
【0058】
用いるジカルボン酸に特に制限はなく。例えばアジピン酸が好ましい。
【0059】
本発明で得られるカダベリンをポリアミドの原料とすることで、耐熱性の高いポリアミドが得られる。
【実施例】
【0060】
以下に実施例を示し、本発明を更に具体的に説明するが、本発明はこれら実施例の記載に限定されるものではない。
【0061】
[L−リジン濃度およびカダベリン濃度のHPLCによる分析方法]
使用カラム:CAPCELL PAK C18(資生堂)
移動相:0.1%(w/w)H3PO4:アセトニトリル=4.5:5.5
検出:UV 360nm
サンプル前処理:分析サンプル25μlに内標として0.03M 1,4−ジアミノブタンを25μl、0.075M 炭酸水素ナトリウムを150μlおよび0.2M 2,4−ジニトロフルオロベンゼンのエタノール溶液を添加混合し37℃で1時間保温する。上記反応溶液50μlを1mlアセトニトリルに溶解後、10,000rpmで5分間遠心した後の10μlをHPLC分析した。
【0062】
参考例1(L−リジン脱炭酸酵素の調整)
(1)L−リジン脱炭酸酵素遺伝子のクローニングおよび細胞内発現ベクターの作製
L−リジン脱炭酸酵素を用いてL−リジン塩からカダベリンに変換させるために、E.coliのL−リジン脱炭酸酵素遺伝子(cadA)のクローニングを行った。
【0063】
データベース(GenBank)に登録されているL−リジン脱炭酸酵素遺伝子(cadA)(Accession No.M76411)の塩基配列を参考にオリゴヌクレオチドプライマー5−atgaacgttattgcaatattg−3’(配列番号:1)、5’−gctgatgggtgagatagaatg−3’(配列番号:2)を合成した。E.coli K12株(ATCC10798)から常法に従い調整したゲノムDNAの溶液を増幅鋳型として0.2mlのミクロ遠心チューブに0.2μlづつ取り、各プライマーを20pmol、20mM トリス塩酸緩衝液(pH8.0)、2.5mM KCl、100μg/mlゼラチン、50μM各dNTP、2単位 LATaqDNAポリメラーゼ(宝酒造製)となるように各試薬を加え、全量を50μlとした。DNAの変性条件を94℃、30秒、プライマーのアニーリング条件を55℃、30秒、DNAプライマーの伸長反応条件を72℃、3分の各条件でBioRad社のサーマルサイクラーを用い、30サイクル反応させた(ポリメラーゼ連鎖反応:以後、PCR法と記す)。尚、本実施例におけるPCR法は特に断らない限り、本条件にて行った。このPCR法により得られた産物を1%アガロースにて電気泳動し、cadA遺伝子を含む約2.1kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpT7−cadAと命名した。
【0064】
このpT7−cadAを増幅鋳型として、オリゴヌクレオチド5’−cgccatgggccatcatcatcatcatcatcatcatatgaacgttattgcaatattg−3’(配列番号:3)、5’−cgcggatccgctgatgggtgagatagaatg−3’(配列番号:4)をプライマーセットとしたPCR法を行った。ここで得られる産物は、オリゴヌクレオチド(配列番号:3)由来の塩基配列と、(配列番号:4)由来の塩基配列が、それぞれcadA遺伝子を含むDNA断片の5’末端、および3’末端に付加されている。この産物を1%アガロースにて電気泳動し、約2.1kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpT7−cadA1と命名した。
【0065】
このpT7−cadA1をNcoI、及びBamHIで消化し、得られた2.1kbのNcoI−BamHI断片を、予めNcoI、及びBamHIで消化しておいたpTV118N(宝酒造製)のNcoI/BamHI間隙に常法に従ったライゲーション反応により挿入し、得られたプラスミドをpLDC1と命名した(図1)。このプラスミドを導入した大腸菌を培養することで、L−リジン脱炭酸酵素のN末端アミノ酸配列に6個のヒスジチン残基が付加された分子量約80kDaの組み換えL−リジン脱炭酸酵素を生産することができる。
【0066】
(2)組み換えL−リジン脱炭酸酵素の産生
pLDC1でE.coli JM109株をアンピシリン耐性に形質転換し、得られた形質転換体をJM109/pLDC1株と命名した。
【0067】
次にJM109/pLDC1株で組み換えL−リジン脱炭酸酵素を産生させた。まず、JM109/pLDC1株をそれぞれ50μg/mlのアンピシリンナトリウムを含んだ滅菌LB培地(Sambrook、J.et.al、2001、 Molecular Cloning 3rd.edition、Cold Spring Harbor Lab.Press)(LB−amp培地)5mlに1白金耳植菌し、37℃で24時間振とうして前培養を行った。
【0068】
この前培養液をLB−amp培地50mlに全量植菌し、37℃、振幅30cmで、180rpmの条件下で3時間培養した後に1mM IPTG(isopropyl−1−thio−β−D−galactoside)添加し、更に4時間培養した。対照実験として、JM109株をpTV118Nで形質転換した形質転換体(以後、JM109/pTV118N株とする)を用い同様の培養を行った。こうして得られた菌体を集め、5mlのTBS緩衝液(宝酒造製)に再懸濁後、超音波破砕および遠心分離により細胞破砕液を調製した。これらのL−リジン脱炭酸酵素活性の測定を単位時間当たりに生成するカダベリンの濃度を測定することにより行った。その結果、対照実験であるJM109/pTV118N株由来の細胞破砕液に対して、JM109/pLDC1株においてはL−リジン脱炭酸酵素活性は約100倍に上昇していた。また、この細胞破砕液をSDS−PAGEで分画し、Penta−His Antibody抗体(QIAGEN社製)でウエスタンブロッティングを行った結果、JM109/pLDC1株由来の細胞破砕液のみから、分子量約80kDaの組み換えL−リジン脱炭酸酵素を検出した。
【0069】
(3)組み換えL−リジン脱炭酸酵素の精製
この組み換えL−リジン脱炭酸酵素は、N末端アミノ酸配列に6個のヒスチジン残基があることから、ニッケルイオンとの相互作用を利用した精製を行った。まず、10mlのキレーティング セファロース ファースト フロー(Chilating Sepharose Fast Flow)担体(アマシャム バイオサイエンス社製)を充填したカラムシステムを構築した。このカラムに50mlの50mM 硫酸ニッケル水溶液、50mlのTBS緩衝液の順で流した後、(2)と同様の方法で得られたJM109/pLDC1株の500mL培養液由来の50ml細胞破砕液を流した。その後、100mlの5mM イミダゾールを含むTBS緩衝液、100mlの50mM イミダゾールを含むTBS緩衝液をこの順序で流した。更に50mlの600mM イミダゾールを含むTBS緩衝液を流した。カラムに流した各々の緩衝液のL−リジン脱炭酸酵素活性を(2)と同様の方法で測定したところ、600mM イミダゾールを含むTBS緩衝液のみに活性があった。また、カラムに流した各々の緩衝液をSDS−PAGEし、クマシーブリリアントブルーで染色したところ、600mM イミダゾールを含むTBS緩衝液から、約80kDaの単一バンドを検出した。また、カラムに流した各々の緩衝液を(2)と同様の方法でウエスタンブロッティングを行ったところ、600mM イミダゾールを含むTBS緩衝液のみに約80kDaタンパク質を検出し、この精製タンパク質はL−リジン脱炭酸酵素活性を有する組み換えL−リジン脱炭酸酵素であることを確認した。
【0070】
(4)L−リジン脱炭酸酵素の細胞表面発現ベクターの作製
まず、リポプロテインの配列の一部および外膜結合タンパク質(OmpA)の46番目のアミノ酸から159番目のアミノ酸までの配列を一つのカセットとしてクローニングした。
【0071】
データベース(GenBank)に登録されている外膜結合タンパク質(ompA)(Accession No.NC_000913)の塩基配列を参考にオリゴヌクレオチドプライマー5−ataaagcttatgaaagctactaaactgg−3’(配列番号:5)、5’−atagtcgacgttgtccggacgagtgccg−3’(配列番号:6)、5’−ataaagcttatgaaagctactaaactggtactgggcgcggtaatcctgggttctactctgctggcaggttgctccagcaacgctaaaatcgatcagaacccgta−3’(配列番号:7)を合成した。E.coli K12株(ATCC10798)から常法に従い調整したゲノムDNAの溶液を増幅鋳型として(配列番号:5)(配列番号:6)(配列番号:7)をプライマーセットとしたPCR法を行った。このPCR法により得られた産物を2%アガロースにて電気泳動し、ompA遺伝子を含む約0.4kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM7と命名した。
【0072】
このpTM7をHindIII、及びSalIで消化し、得られた約0.4kbのHindIII SalI断片を、予めHindIII、及びSalIで消化しておいたpUC19(宝酒造製)のHindIII/SalI間隙に常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM14と命名した。
【0073】
次に、E.coli K12株(ATCC10798)のL−リジン脱炭酸酵素遺伝子(cadA)のクローニングを行った。
【0074】
データベース(GenBank)に登録されているL−リジン脱炭酸酵素遺伝子(cadA)(Accession No.M76411)の塩基配列を参考にオリゴヌクレオチドプライマー5−tatgtcgacatgaacgttattgcaatat −3’(配列番号:8)、5’−ggagagctcttattttttgctttcttcttt−3’(配列番号:9)を合成した。E.coli K12株(ATCC10798)から常法に従い調整したゲノムDNAの溶液を増幅鋳型として(配列番号:8)(配列番号:9)をプライマーセットとしたPCR法を行った。
【0075】
このPCR法により得られた産物を1%アガロースにて電気泳動し、cadA遺伝子を含む約2.1kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM15と命名した。
【0076】
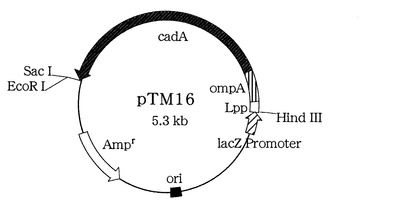
これら調整したpTM15をSalI、及びSacIで消化し、得られた約2.1kbのSalI SacI断片を、予めSalI、及びSacIで消化しておいたpTM14のSalI/SacI間隙に常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM16(図2)と命名した。
【0077】
このプラスミドを導入した大腸菌を培養することで、L−リジン脱炭酸酵素が細胞表面に局在化され、その大腸菌を回収するだけでL−リジン脱炭酸酵素を調整することができるようになる。
【0078】
(5)L−リジン脱炭酸酵素が細胞表面に局在化した細胞の調整
pTM16でE.coli JM109株をアンピシリン耐性に形質転換し、得られた形質転換体をJM109/pTM16株と命名した。
【0079】
次にJM109/pTM16株で組み換えL−リジン脱炭酸酵素を産生させた。まず、JM109/pTM16株をそれぞれ50μg/mlのアンピシリンナトリウムを含んだ滅菌LB培地(LB−amp培地)5mlに1白金耳植菌し、37℃で24時間振とうして前培養を行った。
【0080】
この前培養液をLB−amp培地50mlに全量植菌し、37℃、振幅30cmで、180rpmの条件下で3時間培養した後に1mM IPTGを添加し、更に4時間培養した。こうして得られた菌体を遠心回収し、10mlのTBS緩衝液(宝酒造製)で3回洗浄後、遠心し菌体を回収した。
【0081】
実施例1,2 比較例1(L−リジンの種類の効果)
1000mlのフラスコに、表1に示すL−リジンまたはL−リジン塩を終濃度1.35Mになるように各種L−リジンを加え調整した水溶液500mlを加えた。その後、終濃度50mg/Lになるように参考例1(3)で得た精製した組み換えL−リジン脱炭酸酵素を加え作用させた。45℃で48時間反応した後の反応液中に含まれるカダベリンの濃度を測定した。その結果を表1に示す。
【0082】
【表1】

【0083】
原料として、L−リジン塩水溶液を用いると、より高濃度かつ高反応収率でカダベリンが得られた。
【0084】
実施例3〜7(ビタミンB6の添加の効果)
1000mlのフラスコに、L−リジン一塩酸塩を終濃度1.35Mになるように調整した水溶液500mlを加えた。次に表2に示す各種ビタミンB6を終濃度0.05mMになるように添加した。最後に終濃度50mg/Lになるように参考例1(3)で得た精製した組み換えL−リジン脱炭酸酵素を加え作用させた。45℃で48時間反応した後の反応液中に含まれるカダベリンの濃度を測定した。その結果を表2に示す。
【0085】
【表2】
【0086】
この結果、ビタミンB6をL−リジン塩水溶液に添加することにより、より高収率でカダベリンを得られた。
【0087】
実施例8〜11(反応停止の条件の効果)
1000mlのフラスコに、終濃度1.35MになるようにL−リジン一塩酸塩を加え調整した水溶液500mlを加えた。次にピリドキサルリン酸一水和物を終濃度0.05mMになるように添加した。最後に、終濃度50mg/Lになるように参考例1(3)で得た精製した組み換えL−リジン脱炭酸酵素を加え、45℃において作用させた。残存L−リジン濃度をHPLCにより常に測定し、表3に示す残存L−リジン濃度に達した際に反応溶液を80℃に加熱し反応を停止した。これら反応液に水酸化ナトリウムを添加し、反応液のpHを13以上にした。次にクロロホルムを反応液と等量加え、クロロホルム相にカダベリンを抽出した。最後にこのクロロホルム相を、減圧蒸留(30mmHg、80℃)することによりカダベリンを単離した。この単離操作における精製収率を表3にまとめる。
【0088】
【表3】
【0089】
この結果、反応停止条件としてL−リジン塩濃度が仕込み濃度の20%以下になった反応液からカダベリンを単離することにより、高精製収率でカダベリンが得られた。
【0090】
実施例12〜16(得られたL−リジン脱炭酸酵素の精製状態の効果)
参考例1に示す方法により得られたJM109/pLDC1株を培養し表4に示すような精製処理をおこなった。また、L−リジン脱炭酸酵素が細胞表面に局在化したJM109/pTM16株の培養も行った。比較例として参考例1に示す方法により得られたJM109/pTV118N株の培養も行った。
【0091】
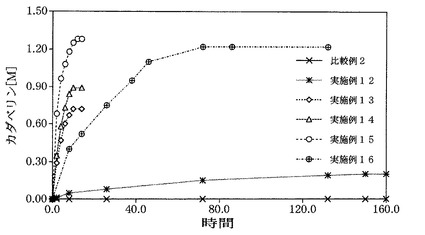
1000mlのフラスコに、終濃度1.35MになるようにL−リジン一塩酸塩を加え調整した水溶液500mlを加えた。次にピリドキサルリン酸一水和物を終濃度0.05mMになるように添加した。最後に、JM109/pTV118N菌体(比較例2)、JM109/pLDC1菌体(実施例12)およびJM109/pTM16菌体(実施例16)は270mlの培養液から得られた菌体(L−リジン脱炭酸酵素濃度を終濃度50mg/L添加することに相当する菌体量)を、また表4に示す各精製状態の組み換えL−リジン脱炭酸酵素(実施例13,14,15)は終濃度50mg/Lになるように加え、45℃において作用させた。経時的にカダベリンの濃度を測定した結果を図3に示す。また、反応が進行しなくなった時間におけるカダベリン収率およびカダベリンの生産速度を求め、表5および図3に示す。
【0092】
【表4】
【0093】
【表5】
【0094】
この結果、組み換えL−リジン脱炭酸酵素を精製することで、高生産速度、高収率でカダベリンが得られた。また、L−リジン脱炭酸酵素を細胞表面に局在化させることで高生産速度、高収率でカダベリンが得られた。
【0095】
参考例2(既存の化学合成法によるカダベリンの調製)
L−リジン一塩酸塩20g(Fluka社製)シクロヘキサノール100ml(シグマアルドリッチジャパン製)に懸濁し、次いで28%ナトリウムメトキシド/メタノール溶液(シグマアルドリッチジャパン製)21.2ml、2−シクロヘキセン−1−オン1ml(シグマアルドリッチジャパン製)を加え、155℃で3時間加熱撹拌した。反応終了後、反応混合物に塩化水素4g(シグマアルドリッチジャパン製)を含むイソプロパノール溶液20ml(シグマアルドリッチジャパン製)を加え、析出した生成物を回収し、乾燥することによりカダベリン二塩酸塩を得た(特公平4−10452の実施例3記載の方法)。この水溶液に、水酸化ナトリウム水溶液を添加することによってカダベリン二塩酸塩をカダベリンに変換し、クロロホルムで抽出して、減圧蒸留(30mmHg、80℃)することにより、カダベリンを得た。
【0096】
実施例17〜21 比較例3(不純物の同定)
実施例12〜16で得られた各反応液を実施例8〜11に示した精製方法によりカダベリンを単離した。不純物の分析を、下記に示す条件でGC−MS法により行い、参考例2で調製したカダベリン(比較例3)の不純物分析と比較した結果を表6に示す。
GC/MS:HP6980/HP5973A
Column:NUKOL 30m×0.24mmI.D. 0.2μm Film
Oven:120℃(一定)
InJ:200℃(Split 10:1)
Flow:He 2.4ml/min (const.Flow)
MS:230℃(SCAN m/z=30〜400)
【0097】
【表6】
【0098】
この結果、L−リジン脱炭酸酵素を用い生成したカダベリンを精製することにより、水性生物に有毒で、また酸と接触すると反応してしまうトリ−n−ブチルアミンの含有量が0.006wt%以下かつ2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下の高純度カダベリンが得られた。
【0099】
参考例3(カダベリンとアジピン酸の塩の調製)
実施例15のカダベリン10.3gを、水25g中に溶解した水溶液を、40℃のウォーターバスに浸して撹拌しているところに、アジピン酸(カーク製)を約1gずつ、中和点付近では約0.2gずつ添加していき、アジピン酸添加量に対する水溶液のpH変化を調べ、中和点を求めると、pH8.66であった。pHが8.66になるように、カダベリンとアジピン酸の等モル塩の50wt%水溶液を調製した。
【0100】
参考例4(カダベリンとアジピン酸の塩の調整)
参考例2のカダベリン10.3gを、水25g中に溶解した水溶液を、40℃のウォーターバスに浸して撹拌しているところに、アジピン酸(カーク製)を約1gずつ、中和点付近では約0.2gずつ添加していき、アジピン酸添加量に対する水溶液のpH変化を調べ、中和点を求めると、pH8.72であった。pHが8.72になるようにカダベリンとアジピン酸の等モル塩の50wt%水溶液を調製した。
【0101】
実施例22(ポリアミドの合成)
参考例3で調製したカダベリンとアジピン酸の等モル塩の50wt%水溶液50.0gを試験管に仕込み、オートクレーブに入れて、密閉し、窒素置換した。ジャケット温度を265℃に設定し、加熱を開始した。缶内圧力が17.5kg/cm2に到達した後、缶内圧力を17.5kg/cm2で3時間保持した。その後、ジャケット温度を275℃に設定し、2時間かけて缶内圧力を常圧に放圧した。その後、缶内温度が245℃に到達した時点で、加熱を停止した。室温に放冷後、試験管をオートクレーブから取り出し、ポリアミドを得た。
【0102】
比較例4
参考例4で調製したカダベリンとアジピン酸の等モル塩の50wt%水溶液50.0gを用いる以外は、実施例22と全く同様の方法でポリアミドを得た。
【0103】
実施例23(ポリアミドの評価)
実施例22で得られたポリアミドおよび比較例4で得られたポリアミドを下記に示す方法で比較評価した。その結果を表7に示す。
【0104】
[ポリアミド中の2,3,4,5−テトラヒドロピリジン、およびトリ−n−ブチルアミンの定量(GC−MS)]
ポリアミド約15gを精秤して、メタノールでソックスレー抽出し、その抽出液を、下記条件でGC−MS分析して、ポリアミド中に含まれる2,3,4,5−テトラヒドロピリジン、およびトリ−n−ブチルアミンを定量した。
装置:ヒューレットパッカード製 HP5890質量検出器
カラム:5%−ジフェニル−95%−ジメチルポリシロキサン
カラム温度:Initial 100℃
Final 250℃
昇温速度:10℃/min
注入口温度:230℃
検出器温度:280℃
キャリアガス:ヘリウム
注入口圧力:50kg/cm2
試料注入量:1μl。
【0105】
[DSC(示差走査熱量測定)]
セイコー電子工業製ロボットDSCRDC220を用い、窒素雰囲気下、ポリアミドを約5mgを採取し、次の条件で測定した。融点+25℃に昇温して3分間保持し、ポリアミドを完全に融解させた後、20℃/分の降温速度で、30℃まで降温し、3分間保持した後、30℃から融点+25℃まで20℃/分の昇温速度で昇温したときに観測される吸熱ピークの温度、および熱量を求めた。
【0106】
[相対粘度(ηr)]
98%硫酸中、0.01g/ml濃度、25℃でオストワルド式粘度計を用いて測定を行った。
【0107】
[溶融滞留試験]
試験管にポリアミド約5gを仕込み、窒素雰囲気下、融点+20℃の温度のシリコンバスに浸漬し、ポリアミドが完全に溶融してから30分間放置した後、ポリアミドを回収して相対粘度測定を行った。
【0108】
【表7】
【0109】
その結果、本発明の方法によって得られた高純度カダベリンを使用することにより、相対粘度(ηr)の保持率の高い(耐熱性の高い)ポリアミドが得られた。
【図面の簡単な説明】
【0110】
【図1】L−リジン脱炭酸酵素細胞内発現ベクターpLDC1のフィジカルマップを示す図である。
【図2】L−リジン脱炭酸酵素細胞表面発現ベクターpTM16のフィジカルマップを示す図である。
【図3】実施例12〜16および比較例2における時間とカダベリン生産量の関係を示す図である。
【技術分野】
【0001】
本発明は、カダベリンの製造法に関するものである。カダベリンは医薬中間体などの合成原料やポリマー原料として期待され、需要が高まりつつある。
【背景技術】
【0002】
従来、L−リジンを微量のテトラリン過酸化物を含むシクロヘキサノール中で煮沸することによりカダベリンが得られることが知られている(非特許文献1参照。)。しかしながら高温反応であるために大量のエネルギーおよび有機溶媒が必要であるうえに、反応収率が非常に低い(36%)。
【0003】
また、2−シクロヘキセン−1−オンなどのビニルケトン類を触媒としてリジンから合成する方法(非特許文献2、特許文献1参照。)等が知られている。
【0004】
また、この方法によって得られるカダベリンには不純物として、トリ−n−ブチルアミンや2,3,4,5−テトラヒドロピリジンといった塩基性化合物が含有されている。
【0005】
また一般的に、酵素反応進行時にpHが変化する際には、pHを制御するために酸もしくはアルカリが適宜加えられ生成物の濃度が低下し、精製工程において濃縮操作が必要となり効率が悪くなるといった問題点がある。さらには、加えられた酸、アルカリにより酵素が変性失活してしまう。
また、カダベリンは生体内に普遍的に存在する生体アミンであり、その生合成系が解明されつつある(非特許文献3参照。)。また、エシェリシア・コリ(Escherichia coli)由来のL−リジン脱炭酸酵素遺伝子が知られている(非特許文献4参照。)。
【0006】
一方、細胞表面にタンパク質を局在化させる研究は、多くの細菌において行われている。大腸菌について細胞表面にタンパク質を局在化させる研究が活発に行われている(非特許文献5参照)。
【0007】
しかしながら、カダベリンの製造について実際的な製造技術は確立されておらず、効率よく、より温和な条件下で高濃度かつ高純度のカダベリンを製造する方法の開発が望まれている。
【特許文献1】特公平4−10452号公報
【非特許文献1】須山正,金尾清造;アミノ酸の脱炭酸(第4報)薬学雑誌,vol.85(6),P.531−533(1965))
【非特許文献2】Chemistry letters,893(1986)
【非特許文献3】Celia white tabor and Herbert tabor;Microbiological Reviews,vol.49,P.81−99(1985)
【非特許文献4】Shi−yuanmeng and George N. Bennett;Journal of Bacteriology,vol.174,P.2659−2669(1992)
【非特許文献5】George Georgiou,Heather L.Poetschke,Chrisstos Stathopoulos and Joseph A. Francisco;TIBTECH,vol.11,P.6−10(1993)
【発明の開示】
【発明が解決しようとする課題】
【0008】
トリ−n−ブチルアミンおよび2,3,4,5−テトラヒドロピリジンが多く含有されているカダベリンをポリマーの原料とする場合、たとえば、カダベリンを含む原料からポリアミドを加熱重縮合で合成する際には、反応温度が高温であるため、カダベリンに含まれるトリ−n−ブチルアミンや2,3,4,5−テトラヒドロピリジンなどの塩基性化合物がポリアミドの分解反応を起こし得られたポリアミドに耐熱性が不足するといった問題点がある。
【0009】
さらに、トリ−n−ブチルアミンは、水性生物に有毒で、魚類に対する致死量は20−40mg/Lであるといった問題点もある。
【0010】
本発明の課題は、このような不純物の少ないカダベリンの高反応収率、高精製収率、高生産速度、高濃度および高純度での工業的製造方法およびカダベリンの単離方法を提供することである。
【課題を解決するための手段】
【0011】
上記問題点を解決するために、本発明者らは、効率がよく、より温和な条件下で高濃度かつ高純度のカダベリンの製造方法について鋭意研究を行った結果、L−リジン塩水溶液にL−リジン脱炭酸酵素を作用させることによりカダベリンを高反応収率かつ高濃度で得られることを見出した。
【0012】
すなわち、本発明は、「(1)L−リジン塩水溶液に、L−リジン脱炭酸酵素を作用させ、反応液からカダベリンを単離することを特徴とするカダベリンの製造方法。
(2)2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のカダベリン。
(3)トリ−n−ブチルアミンの含有量が0.006wt%以下のカダベリン。
(4)上記(1)の方法で得られたカダベリンまたは上記(2)あるいは(3)記載のカダベリンを原料として含有するポリアミド。」である。
【発明の効果】
【0013】
本発明によれば、高反応収率、高精製収率、高生産速度、高濃度および高純度でのカダベリンの工業的製造方法およびカダベリンの単離方法が提供される。
【発明を実施するための最良の形態】
【0014】
カダベリンとは1,5−ペンタンジアミンのことであり、ポリマー原料、医薬中間体の合成原料などとして有用な化合物である。
【0015】
L−リジン脱炭酸酵素は、L−リジンをカダベリンに転換させる酵素であり、Escherichia coli K12株をはじめとするエシェリシア属微生物のみならず、多くの生物に存在することが知られている。
【0016】
本発明において使用されるL−リジン脱炭酸酵素に特に制限はないが、例えば、バシラス・ハロドゥランス(Bacillus halodurans)、バシラス・サブチリス(Bacillus subtilis)、エシェリシア・コリ(Escherichia coli)、セレノモナス・ルミナンチウム(Selenomonas ruminantium)、ビブリオ・コレラ(Vibrio cholerae)、ビブリオ・パラヘモリティカス(Vibrio parahaemolyticus)、ストレプトマイセス・コエリカーラ(Streptomyces coelicolor)、ストレプトマイセス・ピロサス(Streptomyces pilosus)、エイケネラ・コロデンス(Eikenella corrodens)、イユバクテリウム・アシダミノフィルム(Eubacterium acidaminophilum)、サルモネラ・ティフィムリウム(Salmonella typhimurium)、ハフニア・アルベイ(Hafnia alvei)、ナイセリア・メニンギチデス(Neisseria meningitidis)、テルモプラズマ・アシドフィルム(Thermoplasma acidophilum)、ピロコッカス・アビシ(Pyrococcus abyssi)またはコリネバクテリウム・グルタミカス(Corynebacterium glutamicum)由来のものが好ましく用いられる。さらに好ましくは安全性の認められているエシェリシア・コリ(Escherichia coli)由来のものである。
【0017】
本発明における生成物は、アルカリ性の化合物であるカダベリンであるために、反応が進行するにつれ、反応液のpHはアルカリ側に変化していく。しかし、L−リジン塩水溶液に、L−リジン脱炭酸酵素を作用させれば、反応液のpHを制御する必要がなく効率よくカダベリンを生産できる。用いるL−リジン塩については特に制限はない。好ましくは塩酸塩、硫酸塩、酢酸塩、硝酸塩、炭酸塩である。さらに好ましくはL−リジン一塩酸塩である。
【0018】
L−リジン脱炭酸酵素は本来ビタミンB6と結合し存在するが、L−リジン塩水溶液にビタミンB6を添加することにより、生産速度および反応収率を向上させることができる。ビタミンB6を添加する方法には特に制限はない。反応中に適宜添加しても良い。好ましくはL−リジン脱炭酸酵素が溶液状態であれば、酵素溶液中に添加することが好ましい。反応液中でのビタミンB6の濃度については特に制限はなく。好ましくは反応液に0.05mMのビタミンB6濃度となるように加えればよい。用いるビタミンB6に特に制限はない。好ましくは、ピリドキシン、ピリドキサミン、ピリドキサルおよびピリドキサルリン酸から選ばれる少なくとも1種である。さらに好ましくはピリドキサルリン酸である。
【0019】
また、L−リジン脱炭酸酵素を作用させた後、L−リジン塩が仕込み濃度の20%以下にまでになった反応液からカダベリンを単離することにより、高精製収率でカダベリンが単離できる。20%以下にまでになったことを確認する手段には特に制限はない。好ましくはHPLC法によって確認することができる。さらに好ましくはpHの変化を経時的に測定し、L−リジン塩の濃度と関連づけて反応の終点を確認する方法が採用できる。
【0020】
本発明において使用されるL−リジン脱炭酸酵素を得る方法に特に制限はないが、例えば、L−リジン脱炭酸酵素が細胞内で高発現した組換え細胞(細胞内発現組み換え細胞)などを適当な培地で培養し、増殖した細胞を回収し、当該細胞を破砕すればよい。
【0021】
また、別の方法として、L−リジン脱炭酸酵素が細胞表面で局在化した組み換え細胞(細胞表面発現組み換え細胞)などを適当な培地で培養し、増殖した細胞を回収すればよい。
【0022】
L−リジン脱炭酸酵素が細胞表面で局在化した組み換え細胞とは、少なくともL−リジン脱炭酸酵素が細胞表面に局在化している状態であり、細胞内に同時にL−リジン脱炭酸酵素が存在していても良い。
【0023】
一方、細胞表面にL−リジン脱炭酸酵素を局在化させる具体的な方法としては、例えば、分泌シグナル配列の一部、細胞表面局在タンパク質の一部をコードする遺伝子配列、L−リジン脱炭酸酵素の構造遺伝子配列をこの順で有するDNAを大腸菌に導入すればよい。
【0024】
分泌シグナル配列の一部に制限はない。宿主においてタンパク質を分泌するために必要な配列であればよい。大腸菌においては、例えばリポプロテインの配列の一部であり、具体的には、アミノ酸配列としてMKATKLVLGAVILGSTLLAGCSSNAKIDQ(アミノ酸の一文字表記)と翻訳される遺伝子配列であることが好ましい。
【0025】
細胞表面局在タンパク質の一部をコードする遺伝子配列に特に制限はない。大腸菌においては、例えば外膜結合タンパク質の配列の一部であり、具体的にはOmpA(外膜結合タンパク質)の46番目のアミノ酸から159番目のアミノ酸までの配列の一部であることが好ましい。
【0026】
L−リジン脱炭酸酵素遺伝子、リポプロテイン遺伝子およびOmpA遺伝子をクローニングする方法に特に制限はない。既知の遺伝子情報に基づき、PCR(polymerase chain reaction)法を用いて必要な遺伝領域を増幅取得する方法、既知の遺伝子情報に基づきゲノムライブラリーやcDNAライブラリーより相同性や酵素活性を指標としてクローニングする方法などが挙げられる。本発明においては、これらの遺伝子は、遺伝的多形性などによる変異型も含む。なお、遺伝的多形性とは、遺伝子上の自然突然変異により遺伝子の塩基配列が一部変化しているものをいう。例えばE.coli K12株の染色体DNAよりPCR法を用いてL−リジン脱炭酸酵素をコードする遺伝子であるcadA遺伝子またはldc遺伝子をクローニングする。この際使用する染色体DNAはE.coli由来であればどの菌株由来でもよい。
【0027】
L−リジン脱炭酸酵素が細胞表面に局在化していることの確認は、例えばL−リジン脱炭酸酵素を抗原として作製した抗体により、細胞表面発現組み換え細胞を免疫反応後、包埋、薄切りし、電子顕微鏡(免疫電顕法)により観察することで確認できる。
【0028】
本発明では、L−リジン脱炭酸酵素は、L−リジン脱炭酸酵素の細胞内もしくは細胞表面での活性が上昇した組換え細胞から調製されたものが好ましく使用できる。細胞内もしくは細胞表面でL−リジン脱炭酸酵素の活性を上昇させる方法に特に制限はない。具体的には、例えば、L−リジン脱炭酸酵素の酵素量を増加させる方法、もしくは酵素の構造遺伝子自体に変異を導入して、酵素そのものの比活性を上昇させることなどが挙げられる。
【0029】
細胞内もしくは細胞表面の酵素量を増加させる手段としては、遺伝子の転写調節領域の改良、遺伝子のコピー数の増加、蛋白への翻訳の効率化などが挙げられる。
【0030】
転写調節領域の改良とは、遺伝子の転写量を増加させる改変を加えることをいう。例えば、プロモーターに変異を導入することによってプロモーター強化を行い、下流にある遺伝子の転写量を増加させることができる。プロモーターに変異を導入する以外にも、宿主内で強力に発現するプロモーターを導入しても良い。例えば大腸菌においては、lac、tac、trpなどのプロモーターが挙げられる。また、エンハンサーを新たに導入することによって遺伝子の転写量を増加させることができる。染色体DNAのプロモーター等の遺伝子導入については、例えば特開平1−215280号公報に記載されている。
【0031】
遺伝子のコピー数の上昇は、具体的には、遺伝子を多コピー型のベクターに接続して組換えDNAを作製し、該組換えDNAを宿主細胞に保持させることにより達成することができる。ここでベクターとは、プラスミドやファージ等広く用いられているものを含むが、これら以外にも、トランソポゾン(Berg,D.E and Berg.C.M., Bio/Technol.,vol.1,P.417(1983))やMuファージ(特開平2−109985号公報)も含む。遺伝子を相同組換え用プラスミド等を用いた方法で染色体に組み込んでコピー数を上昇させることも可能である。
【0032】
蛋白の翻訳効率を上昇させる方法としては、例えば原核生物においてはSD配列(Shine,J.and Dalgarno,L.,Proc.Natl.Acad.Sci.USA,71,1342−1346 (1974))、真核生物では Kozakのコンセンサス配列(Kozak,M.,Nuc.Acids.Res.,Vol.15,p.8125−8148(1987))を導入、改変することや、使用コドンの最適化(特開昭59−125895)などが挙げられる。
【0033】
L−リシン脱炭酸酵素の細胞内もしくは細胞表面での活性を上昇させる手段としては、L−リジン脱炭酸酵素の構造遺伝子自体に変異を導入して、L−リジン脱炭酸酵素そのものの活性を上昇させることも挙げられる。
【0034】
遺伝子に変異を生じさせるには、部位特異的変異法(Kramer,W. and frita,H.J.,Methods in Enzymology,vol.154,P.350(1987))リコンビナントPCR法(PCR Technology,Stockton Press(1989)、特定の部分のDNAを化学合成する方法、または当該遺伝子をヒドロキシアミン処理する方法や当該遺伝子を保有する菌株を紫外線照射処理、もしくはニトロソグアニジンや亜硝酸などの化学薬剤で処理する方法がある。
【0035】
組換え細胞としては、微生物、動物、植物、または昆虫由来のものが好ましく使用できる。例えば動物を用いる場合、マウス、ラットやそれらの培養細胞などが用いられる。植物を用いる場合、例えばシロイヌナズナ、タバコやそれらの培養細胞が用いられる。また、昆虫を用いる場合、例えばカイコやその培養細胞などが用いられる。また、微生物を用いる場合、例えば、大腸菌などが用いられる。
【0036】
また、L−リジン脱炭酸酵素を複数種組み合わせて使用しても良い。
【0037】
L−リジン脱炭酸酵素を得るために、組換え細胞を培養する方法に特に制限はないが、例えば微生物を培養する場合、使用する培地は、炭素源、窒素源、無機イオンおよび必要に応じその他有機成分を含有する培地が用いられる。例えば、大腸菌の場合しばしばLB培地が用いられる。炭素源としては、グルコース、ラクトース、ガラクトース、フラクトース、アラビノース、マルトース、キシロース、トレハロース、リボースや澱粉の加水分解物などの糖類、グリセロール、マンニトールやソルビトールなどのアルコール類、グルコン酸、フマール酸、クエン酸やコハク酸等の有機酸類を用いることができる。窒素源としては、硫酸アンモニウム、塩化アンモニウム、リン酸アンモニウム等の無機アンモニウム塩、大豆加水分解物などの有機窒素、アンモニアガス、アンモニア水等を用いることができる。有機微量栄養素としては、各種アミノ酸、ビタミンB1等のビタミン類、RNA等の核酸類などの要求物質または酵母エキス等を適量含有させることが望ましい。それらの他に、必要に応じて、リン酸カルシウム、硫酸カルシウム、鉄イオン、マンガンイオン等が少量添加される。
【0038】
培養条件にも特に制限はなく、例えば大腸菌の場合、好気条件下で16〜72時間程度実施するのが良く、培養温度は30℃〜45℃に、特に好ましくは37℃に、培養pHは5〜8に、特に好ましくはpH7に制御するのがよい。なおpH調整には無機あるいは有機の酸性あるいはアルカリ性物質、さらにアンモニアガス等を使用することができる。
【0039】
増殖した細胞内発現組み換え細胞は、遠心分離等により培養液から回収することができる。回収した細胞内発現組み換え細胞から細胞破砕液を調整するには、通常の方法が用いられる。すなわち、細胞内発現組み換え細胞を超音波処理、ダイノミル、フレンチプレス等の方法にて破砕し、遠心分離により細胞残渣を除去することにより細胞破砕液が得られる。
【0040】
細胞破砕液からL−リジン脱炭酸酵素を精製するには、硫安分画、イオン交換クロマトグラフィー、疎水クロマトグラフィー、アフィニティークロマトグラフィー、ゲル濾過クロマトグラフィー、等電点沈殿、熱処理、pH処理など酵素の精製に通常用いられる手法が適宜組み合わされて用いられる。
【0041】
一般的な工業化される酵素反応は休止細胞を用いるが、本発明においては作用させるL−リジン脱炭酸酵素は、L−リジン脱炭酸酵素活性を有する休止細胞、その細胞破砕液などが使用できるが、細胞破砕液が好ましい。細胞破砕液を用いるとより高生産速度でカダベリンが得られる。
【0042】
さらに、部分的に精製したL−リジン脱炭酸酵素を作用させることによりL−リジンの分解およびカダベリンの分解を抑制することができ、より高収率かつ高純度のカダベリンが得られる。部分的な精製とは、工業的に容易な処理のみ行うことであり例えば熱処理、pH処理等が挙げられる。好ましくは熱処理である。さらに好ましくは単離精製したL−リジン脱炭酸酵素を作用させることである。L−リジン脱炭酸酵素を部分的に精製、または単離精製することで、L−リジンの分解やカダベリンの分解を引き起こす酵素を取り除くことができる。
【0043】
さらに、L−リジン脱炭酸酵素が細胞表面に局在化している細胞を作用させれば、L−リジンの分解およびカダベリンの分解を抑制することができ、より高収率かつ高純度、高生産速度でカダベリンが得られる。さらにはその細胞を回収することで繰り返し酵素反応に利用することができる。しかも酵素の調整が非常に簡便である。
【0044】
一般的に酵素というものは本来あるべきアミノ酸配列で自然界に存在するため、人工的にアミノ酸配列を付与もしくは欠如させると酵素の活性を失うことが多い。しかし本発明において細胞内にL−リジン脱炭酸酵素を発現させる場合、L−リジン脱炭酸酵素のN末端アミノ酸配列に1から30個のアミノ酸配列を付与したL−リジン脱炭酸酵素が好ましく使用できる。ここで、N末端に付与するものは6から10個のヒスチジン残基が好ましい。このように、N末端アミノ酸配列にアミノ酸を付与することで、L−リジン脱炭酸酵素の精製を簡便にできるようにすることができる。N末端アミノ酸配列に1から30個のアミノ酸を付与する方法に特に制限はない。例えばL−リジン脱炭酸酵素遺伝子に付与したいアミノ酸配列に相当する塩基配列を遺伝子工学的手法を用い、組み換えればよい。用いる遺伝子工学的手法に特に制限はないが。好ましくはPCR法やDNA断片同士の連結による方法がある。
【0045】
L−リジン脱炭酸酵素によるL−リジン塩水溶液からカダベリンへの変換は、上記のようにして得られるL−リジン脱炭酸酵素を、L−リジン塩水溶液に接触させることによって行うことができる。
【0046】
用いるL−リジン脱炭酸酵素の量に特に制限はない。L−リジン塩水溶液をカダベリンに変換する反応を触媒するのに十分な量であればよい。好ましくは反応液中の酵素濃度が25から70mg/Lである。好ましくは50mg/Lである。一方、休止細胞を使用する際は、反応液中の細胞濃度が5から15g/Lである。好ましくは10g/Lである。
【0047】
反応温度は、通常、28〜55℃、好ましくは45℃前後である。
【0048】
反応液の状態に特に制限はない。好ましくは静置または攪拌状態である。攪拌状態にする方法には特に制限はない。好ましくは攪拌翼により攪拌する方法である。
【0049】
本発明で使用するL−リジン脱炭酸酵素は、固定化することで繰り返しカダベリン合成反応に使用することができ、酵素の調製に必要なコストを低減させることができる。固定化方法としては、アクリルアミドなどの合成高分子に包括する方法、セファロースやスチレン樹脂を骨格とするイオン交換性担体や疎水性担体に吸着させる方法、または、ガラス担体に共有結合で結合させる方法などが挙げられる。
【0050】
反応時間は、使用する酵素活性、L−リジン塩濃度などの条件によって異なるが、通常、1〜72時間である。また、反応は、L−リジン塩を供給しながら連続的に行ってもよい。
【0051】
このように生成したカダベリンを反応終了後、反応液から採取する方法としては、反応終了液をアルカリでpH12から14にし、極性有機溶媒で抽出すれば良い。
【0052】
用いるアルカリに特に制限はない。好ましくは水酸化ナトリウム、水酸化カリウム、水酸化カルシウムを用いることができる。
【0053】
用いる極性有機溶媒に特に制限はない。好ましくはアニリン、シクロヘキサノン、1−オクタノール、イソブチルアルコール、シクロヘキサノール、クロロホルムを用いることができる。
【0054】
このようにして得られたカダベリンは、トリ−n−ブチルアミンおよび2,3,4,5−テトラヒドロピリジンなどの不純物が少ない。本発明の方法で製造することで、以下の方法で分析した場合に、トリ−n−ブチルアミンの含有量が、0.006wt%以下のカタベリンが得られる。さらに、以下の方法で分析した場合に、2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のものが得られる。さらに、トリ−n−ブチルアミンの含有量が、0.006wt%以下であり、かつ、2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のものが得られる。
【0055】
トリ−n−ブチルアミンおよび2,3,4,5−テトラヒドロピリジンの分析は、GC−MS法によって以下の条件で行う。
【0056】
GC/MS:HP6980/HP5973A
Column:NUKOL 30m×0.24mmI.D. 0.2μm Film
Oven:120℃(一定)
InJ:200℃(Split 10:1)
Flow:He 2.4ml/min (const.Flow)
MS:230℃(SCAN m/z=30〜400)
このようにして得られたカダベリンは、ポリアミドの原料として有用である。
【0057】
ポリアミドの製造方法としては、カダベリンとジカルボン酸の塩、および水の混合物を、加熱して脱水反応を進行させる加熱重縮合法が用いられる。また、加熱重縮合後、固相重合することによって、分子量を上昇させることも可能である。固相重合は、100℃〜融点の温度範囲で、真空中、あるいは不活性ガス中で加熱することにより進行し、加熱重縮合では分子量が不十分なポリアミドを高分子量化することができる。
【0058】
用いるジカルボン酸に特に制限はなく。例えばアジピン酸が好ましい。
【0059】
本発明で得られるカダベリンをポリアミドの原料とすることで、耐熱性の高いポリアミドが得られる。
【実施例】
【0060】
以下に実施例を示し、本発明を更に具体的に説明するが、本発明はこれら実施例の記載に限定されるものではない。
【0061】
[L−リジン濃度およびカダベリン濃度のHPLCによる分析方法]
使用カラム:CAPCELL PAK C18(資生堂)
移動相:0.1%(w/w)H3PO4:アセトニトリル=4.5:5.5
検出:UV 360nm
サンプル前処理:分析サンプル25μlに内標として0.03M 1,4−ジアミノブタンを25μl、0.075M 炭酸水素ナトリウムを150μlおよび0.2M 2,4−ジニトロフルオロベンゼンのエタノール溶液を添加混合し37℃で1時間保温する。上記反応溶液50μlを1mlアセトニトリルに溶解後、10,000rpmで5分間遠心した後の10μlをHPLC分析した。
【0062】
参考例1(L−リジン脱炭酸酵素の調整)
(1)L−リジン脱炭酸酵素遺伝子のクローニングおよび細胞内発現ベクターの作製
L−リジン脱炭酸酵素を用いてL−リジン塩からカダベリンに変換させるために、E.coliのL−リジン脱炭酸酵素遺伝子(cadA)のクローニングを行った。
【0063】
データベース(GenBank)に登録されているL−リジン脱炭酸酵素遺伝子(cadA)(Accession No.M76411)の塩基配列を参考にオリゴヌクレオチドプライマー5−atgaacgttattgcaatattg−3’(配列番号:1)、5’−gctgatgggtgagatagaatg−3’(配列番号:2)を合成した。E.coli K12株(ATCC10798)から常法に従い調整したゲノムDNAの溶液を増幅鋳型として0.2mlのミクロ遠心チューブに0.2μlづつ取り、各プライマーを20pmol、20mM トリス塩酸緩衝液(pH8.0)、2.5mM KCl、100μg/mlゼラチン、50μM各dNTP、2単位 LATaqDNAポリメラーゼ(宝酒造製)となるように各試薬を加え、全量を50μlとした。DNAの変性条件を94℃、30秒、プライマーのアニーリング条件を55℃、30秒、DNAプライマーの伸長反応条件を72℃、3分の各条件でBioRad社のサーマルサイクラーを用い、30サイクル反応させた(ポリメラーゼ連鎖反応:以後、PCR法と記す)。尚、本実施例におけるPCR法は特に断らない限り、本条件にて行った。このPCR法により得られた産物を1%アガロースにて電気泳動し、cadA遺伝子を含む約2.1kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpT7−cadAと命名した。
【0064】
このpT7−cadAを増幅鋳型として、オリゴヌクレオチド5’−cgccatgggccatcatcatcatcatcatcatcatatgaacgttattgcaatattg−3’(配列番号:3)、5’−cgcggatccgctgatgggtgagatagaatg−3’(配列番号:4)をプライマーセットとしたPCR法を行った。ここで得られる産物は、オリゴヌクレオチド(配列番号:3)由来の塩基配列と、(配列番号:4)由来の塩基配列が、それぞれcadA遺伝子を含むDNA断片の5’末端、および3’末端に付加されている。この産物を1%アガロースにて電気泳動し、約2.1kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpT7−cadA1と命名した。
【0065】
このpT7−cadA1をNcoI、及びBamHIで消化し、得られた2.1kbのNcoI−BamHI断片を、予めNcoI、及びBamHIで消化しておいたpTV118N(宝酒造製)のNcoI/BamHI間隙に常法に従ったライゲーション反応により挿入し、得られたプラスミドをpLDC1と命名した(図1)。このプラスミドを導入した大腸菌を培養することで、L−リジン脱炭酸酵素のN末端アミノ酸配列に6個のヒスジチン残基が付加された分子量約80kDaの組み換えL−リジン脱炭酸酵素を生産することができる。
【0066】
(2)組み換えL−リジン脱炭酸酵素の産生
pLDC1でE.coli JM109株をアンピシリン耐性に形質転換し、得られた形質転換体をJM109/pLDC1株と命名した。
【0067】
次にJM109/pLDC1株で組み換えL−リジン脱炭酸酵素を産生させた。まず、JM109/pLDC1株をそれぞれ50μg/mlのアンピシリンナトリウムを含んだ滅菌LB培地(Sambrook、J.et.al、2001、 Molecular Cloning 3rd.edition、Cold Spring Harbor Lab.Press)(LB−amp培地)5mlに1白金耳植菌し、37℃で24時間振とうして前培養を行った。
【0068】
この前培養液をLB−amp培地50mlに全量植菌し、37℃、振幅30cmで、180rpmの条件下で3時間培養した後に1mM IPTG(isopropyl−1−thio−β−D−galactoside)添加し、更に4時間培養した。対照実験として、JM109株をpTV118Nで形質転換した形質転換体(以後、JM109/pTV118N株とする)を用い同様の培養を行った。こうして得られた菌体を集め、5mlのTBS緩衝液(宝酒造製)に再懸濁後、超音波破砕および遠心分離により細胞破砕液を調製した。これらのL−リジン脱炭酸酵素活性の測定を単位時間当たりに生成するカダベリンの濃度を測定することにより行った。その結果、対照実験であるJM109/pTV118N株由来の細胞破砕液に対して、JM109/pLDC1株においてはL−リジン脱炭酸酵素活性は約100倍に上昇していた。また、この細胞破砕液をSDS−PAGEで分画し、Penta−His Antibody抗体(QIAGEN社製)でウエスタンブロッティングを行った結果、JM109/pLDC1株由来の細胞破砕液のみから、分子量約80kDaの組み換えL−リジン脱炭酸酵素を検出した。
【0069】
(3)組み換えL−リジン脱炭酸酵素の精製
この組み換えL−リジン脱炭酸酵素は、N末端アミノ酸配列に6個のヒスチジン残基があることから、ニッケルイオンとの相互作用を利用した精製を行った。まず、10mlのキレーティング セファロース ファースト フロー(Chilating Sepharose Fast Flow)担体(アマシャム バイオサイエンス社製)を充填したカラムシステムを構築した。このカラムに50mlの50mM 硫酸ニッケル水溶液、50mlのTBS緩衝液の順で流した後、(2)と同様の方法で得られたJM109/pLDC1株の500mL培養液由来の50ml細胞破砕液を流した。その後、100mlの5mM イミダゾールを含むTBS緩衝液、100mlの50mM イミダゾールを含むTBS緩衝液をこの順序で流した。更に50mlの600mM イミダゾールを含むTBS緩衝液を流した。カラムに流した各々の緩衝液のL−リジン脱炭酸酵素活性を(2)と同様の方法で測定したところ、600mM イミダゾールを含むTBS緩衝液のみに活性があった。また、カラムに流した各々の緩衝液をSDS−PAGEし、クマシーブリリアントブルーで染色したところ、600mM イミダゾールを含むTBS緩衝液から、約80kDaの単一バンドを検出した。また、カラムに流した各々の緩衝液を(2)と同様の方法でウエスタンブロッティングを行ったところ、600mM イミダゾールを含むTBS緩衝液のみに約80kDaタンパク質を検出し、この精製タンパク質はL−リジン脱炭酸酵素活性を有する組み換えL−リジン脱炭酸酵素であることを確認した。
【0070】
(4)L−リジン脱炭酸酵素の細胞表面発現ベクターの作製
まず、リポプロテインの配列の一部および外膜結合タンパク質(OmpA)の46番目のアミノ酸から159番目のアミノ酸までの配列を一つのカセットとしてクローニングした。
【0071】
データベース(GenBank)に登録されている外膜結合タンパク質(ompA)(Accession No.NC_000913)の塩基配列を参考にオリゴヌクレオチドプライマー5−ataaagcttatgaaagctactaaactgg−3’(配列番号:5)、5’−atagtcgacgttgtccggacgagtgccg−3’(配列番号:6)、5’−ataaagcttatgaaagctactaaactggtactgggcgcggtaatcctgggttctactctgctggcaggttgctccagcaacgctaaaatcgatcagaacccgta−3’(配列番号:7)を合成した。E.coli K12株(ATCC10798)から常法に従い調整したゲノムDNAの溶液を増幅鋳型として(配列番号:5)(配列番号:6)(配列番号:7)をプライマーセットとしたPCR法を行った。このPCR法により得られた産物を2%アガロースにて電気泳動し、ompA遺伝子を含む約0.4kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM7と命名した。
【0072】
このpTM7をHindIII、及びSalIで消化し、得られた約0.4kbのHindIII SalI断片を、予めHindIII、及びSalIで消化しておいたpUC19(宝酒造製)のHindIII/SalI間隙に常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM14と命名した。
【0073】
次に、E.coli K12株(ATCC10798)のL−リジン脱炭酸酵素遺伝子(cadA)のクローニングを行った。
【0074】
データベース(GenBank)に登録されているL−リジン脱炭酸酵素遺伝子(cadA)(Accession No.M76411)の塩基配列を参考にオリゴヌクレオチドプライマー5−tatgtcgacatgaacgttattgcaatat −3’(配列番号:8)、5’−ggagagctcttattttttgctttcttcttt−3’(配列番号:9)を合成した。E.coli K12株(ATCC10798)から常法に従い調整したゲノムDNAの溶液を増幅鋳型として(配列番号:8)(配列番号:9)をプライマーセットとしたPCR法を行った。
【0075】
このPCR法により得られた産物を1%アガロースにて電気泳動し、cadA遺伝子を含む約2.1kbのDNA断片を常法に従い調整した。この断片を、プラスミドベクターpT7blue(Novagen社製)のEcoRV部位の3’−末端にT塩基が付加された間隙に、常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM15と命名した。
【0076】
これら調整したpTM15をSalI、及びSacIで消化し、得られた約2.1kbのSalI SacI断片を、予めSalI、及びSacIで消化しておいたpTM14のSalI/SacI間隙に常法に従ったライゲーション反応により挿入し、得られたプラスミドをpTM16(図2)と命名した。
【0077】
このプラスミドを導入した大腸菌を培養することで、L−リジン脱炭酸酵素が細胞表面に局在化され、その大腸菌を回収するだけでL−リジン脱炭酸酵素を調整することができるようになる。
【0078】
(5)L−リジン脱炭酸酵素が細胞表面に局在化した細胞の調整
pTM16でE.coli JM109株をアンピシリン耐性に形質転換し、得られた形質転換体をJM109/pTM16株と命名した。
【0079】
次にJM109/pTM16株で組み換えL−リジン脱炭酸酵素を産生させた。まず、JM109/pTM16株をそれぞれ50μg/mlのアンピシリンナトリウムを含んだ滅菌LB培地(LB−amp培地)5mlに1白金耳植菌し、37℃で24時間振とうして前培養を行った。
【0080】
この前培養液をLB−amp培地50mlに全量植菌し、37℃、振幅30cmで、180rpmの条件下で3時間培養した後に1mM IPTGを添加し、更に4時間培養した。こうして得られた菌体を遠心回収し、10mlのTBS緩衝液(宝酒造製)で3回洗浄後、遠心し菌体を回収した。
【0081】
実施例1,2 比較例1(L−リジンの種類の効果)
1000mlのフラスコに、表1に示すL−リジンまたはL−リジン塩を終濃度1.35Mになるように各種L−リジンを加え調整した水溶液500mlを加えた。その後、終濃度50mg/Lになるように参考例1(3)で得た精製した組み換えL−リジン脱炭酸酵素を加え作用させた。45℃で48時間反応した後の反応液中に含まれるカダベリンの濃度を測定した。その結果を表1に示す。
【0082】
【表1】
【0083】
原料として、L−リジン塩水溶液を用いると、より高濃度かつ高反応収率でカダベリンが得られた。
【0084】
実施例3〜7(ビタミンB6の添加の効果)
1000mlのフラスコに、L−リジン一塩酸塩を終濃度1.35Mになるように調整した水溶液500mlを加えた。次に表2に示す各種ビタミンB6を終濃度0.05mMになるように添加した。最後に終濃度50mg/Lになるように参考例1(3)で得た精製した組み換えL−リジン脱炭酸酵素を加え作用させた。45℃で48時間反応した後の反応液中に含まれるカダベリンの濃度を測定した。その結果を表2に示す。
【0085】
【表2】
【0086】
この結果、ビタミンB6をL−リジン塩水溶液に添加することにより、より高収率でカダベリンを得られた。
【0087】
実施例8〜11(反応停止の条件の効果)
1000mlのフラスコに、終濃度1.35MになるようにL−リジン一塩酸塩を加え調整した水溶液500mlを加えた。次にピリドキサルリン酸一水和物を終濃度0.05mMになるように添加した。最後に、終濃度50mg/Lになるように参考例1(3)で得た精製した組み換えL−リジン脱炭酸酵素を加え、45℃において作用させた。残存L−リジン濃度をHPLCにより常に測定し、表3に示す残存L−リジン濃度に達した際に反応溶液を80℃に加熱し反応を停止した。これら反応液に水酸化ナトリウムを添加し、反応液のpHを13以上にした。次にクロロホルムを反応液と等量加え、クロロホルム相にカダベリンを抽出した。最後にこのクロロホルム相を、減圧蒸留(30mmHg、80℃)することによりカダベリンを単離した。この単離操作における精製収率を表3にまとめる。
【0088】
【表3】
【0089】
この結果、反応停止条件としてL−リジン塩濃度が仕込み濃度の20%以下になった反応液からカダベリンを単離することにより、高精製収率でカダベリンが得られた。
【0090】
実施例12〜16(得られたL−リジン脱炭酸酵素の精製状態の効果)
参考例1に示す方法により得られたJM109/pLDC1株を培養し表4に示すような精製処理をおこなった。また、L−リジン脱炭酸酵素が細胞表面に局在化したJM109/pTM16株の培養も行った。比較例として参考例1に示す方法により得られたJM109/pTV118N株の培養も行った。
【0091】
1000mlのフラスコに、終濃度1.35MになるようにL−リジン一塩酸塩を加え調整した水溶液500mlを加えた。次にピリドキサルリン酸一水和物を終濃度0.05mMになるように添加した。最後に、JM109/pTV118N菌体(比較例2)、JM109/pLDC1菌体(実施例12)およびJM109/pTM16菌体(実施例16)は270mlの培養液から得られた菌体(L−リジン脱炭酸酵素濃度を終濃度50mg/L添加することに相当する菌体量)を、また表4に示す各精製状態の組み換えL−リジン脱炭酸酵素(実施例13,14,15)は終濃度50mg/Lになるように加え、45℃において作用させた。経時的にカダベリンの濃度を測定した結果を図3に示す。また、反応が進行しなくなった時間におけるカダベリン収率およびカダベリンの生産速度を求め、表5および図3に示す。
【0092】
【表4】
【0093】
【表5】
【0094】
この結果、組み換えL−リジン脱炭酸酵素を精製することで、高生産速度、高収率でカダベリンが得られた。また、L−リジン脱炭酸酵素を細胞表面に局在化させることで高生産速度、高収率でカダベリンが得られた。
【0095】
参考例2(既存の化学合成法によるカダベリンの調製)
L−リジン一塩酸塩20g(Fluka社製)シクロヘキサノール100ml(シグマアルドリッチジャパン製)に懸濁し、次いで28%ナトリウムメトキシド/メタノール溶液(シグマアルドリッチジャパン製)21.2ml、2−シクロヘキセン−1−オン1ml(シグマアルドリッチジャパン製)を加え、155℃で3時間加熱撹拌した。反応終了後、反応混合物に塩化水素4g(シグマアルドリッチジャパン製)を含むイソプロパノール溶液20ml(シグマアルドリッチジャパン製)を加え、析出した生成物を回収し、乾燥することによりカダベリン二塩酸塩を得た(特公平4−10452の実施例3記載の方法)。この水溶液に、水酸化ナトリウム水溶液を添加することによってカダベリン二塩酸塩をカダベリンに変換し、クロロホルムで抽出して、減圧蒸留(30mmHg、80℃)することにより、カダベリンを得た。
【0096】
実施例17〜21 比較例3(不純物の同定)
実施例12〜16で得られた各反応液を実施例8〜11に示した精製方法によりカダベリンを単離した。不純物の分析を、下記に示す条件でGC−MS法により行い、参考例2で調製したカダベリン(比較例3)の不純物分析と比較した結果を表6に示す。
GC/MS:HP6980/HP5973A
Column:NUKOL 30m×0.24mmI.D. 0.2μm Film
Oven:120℃(一定)
InJ:200℃(Split 10:1)
Flow:He 2.4ml/min (const.Flow)
MS:230℃(SCAN m/z=30〜400)
【0097】
【表6】
【0098】
この結果、L−リジン脱炭酸酵素を用い生成したカダベリンを精製することにより、水性生物に有毒で、また酸と接触すると反応してしまうトリ−n−ブチルアミンの含有量が0.006wt%以下かつ2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下の高純度カダベリンが得られた。
【0099】
参考例3(カダベリンとアジピン酸の塩の調製)
実施例15のカダベリン10.3gを、水25g中に溶解した水溶液を、40℃のウォーターバスに浸して撹拌しているところに、アジピン酸(カーク製)を約1gずつ、中和点付近では約0.2gずつ添加していき、アジピン酸添加量に対する水溶液のpH変化を調べ、中和点を求めると、pH8.66であった。pHが8.66になるように、カダベリンとアジピン酸の等モル塩の50wt%水溶液を調製した。
【0100】
参考例4(カダベリンとアジピン酸の塩の調整)
参考例2のカダベリン10.3gを、水25g中に溶解した水溶液を、40℃のウォーターバスに浸して撹拌しているところに、アジピン酸(カーク製)を約1gずつ、中和点付近では約0.2gずつ添加していき、アジピン酸添加量に対する水溶液のpH変化を調べ、中和点を求めると、pH8.72であった。pHが8.72になるようにカダベリンとアジピン酸の等モル塩の50wt%水溶液を調製した。
【0101】
実施例22(ポリアミドの合成)
参考例3で調製したカダベリンとアジピン酸の等モル塩の50wt%水溶液50.0gを試験管に仕込み、オートクレーブに入れて、密閉し、窒素置換した。ジャケット温度を265℃に設定し、加熱を開始した。缶内圧力が17.5kg/cm2に到達した後、缶内圧力を17.5kg/cm2で3時間保持した。その後、ジャケット温度を275℃に設定し、2時間かけて缶内圧力を常圧に放圧した。その後、缶内温度が245℃に到達した時点で、加熱を停止した。室温に放冷後、試験管をオートクレーブから取り出し、ポリアミドを得た。
【0102】
比較例4
参考例4で調製したカダベリンとアジピン酸の等モル塩の50wt%水溶液50.0gを用いる以外は、実施例22と全く同様の方法でポリアミドを得た。
【0103】
実施例23(ポリアミドの評価)
実施例22で得られたポリアミドおよび比較例4で得られたポリアミドを下記に示す方法で比較評価した。その結果を表7に示す。
【0104】
[ポリアミド中の2,3,4,5−テトラヒドロピリジン、およびトリ−n−ブチルアミンの定量(GC−MS)]
ポリアミド約15gを精秤して、メタノールでソックスレー抽出し、その抽出液を、下記条件でGC−MS分析して、ポリアミド中に含まれる2,3,4,5−テトラヒドロピリジン、およびトリ−n−ブチルアミンを定量した。
装置:ヒューレットパッカード製 HP5890質量検出器
カラム:5%−ジフェニル−95%−ジメチルポリシロキサン
カラム温度:Initial 100℃
Final 250℃
昇温速度:10℃/min
注入口温度:230℃
検出器温度:280℃
キャリアガス:ヘリウム
注入口圧力:50kg/cm2
試料注入量:1μl。
【0105】
[DSC(示差走査熱量測定)]
セイコー電子工業製ロボットDSCRDC220を用い、窒素雰囲気下、ポリアミドを約5mgを採取し、次の条件で測定した。融点+25℃に昇温して3分間保持し、ポリアミドを完全に融解させた後、20℃/分の降温速度で、30℃まで降温し、3分間保持した後、30℃から融点+25℃まで20℃/分の昇温速度で昇温したときに観測される吸熱ピークの温度、および熱量を求めた。
【0106】
[相対粘度(ηr)]
98%硫酸中、0.01g/ml濃度、25℃でオストワルド式粘度計を用いて測定を行った。
【0107】
[溶融滞留試験]
試験管にポリアミド約5gを仕込み、窒素雰囲気下、融点+20℃の温度のシリコンバスに浸漬し、ポリアミドが完全に溶融してから30分間放置した後、ポリアミドを回収して相対粘度測定を行った。
【0108】
【表7】
【0109】
その結果、本発明の方法によって得られた高純度カダベリンを使用することにより、相対粘度(ηr)の保持率の高い(耐熱性の高い)ポリアミドが得られた。
【図面の簡単な説明】
【0110】
【図1】L−リジン脱炭酸酵素細胞内発現ベクターpLDC1のフィジカルマップを示す図である。
【図2】L−リジン脱炭酸酵素細胞表面発現ベクターpTM16のフィジカルマップを示す図である。
【図3】実施例12〜16および比較例2における時間とカダベリン生産量の関係を示す図である。
【特許請求の範囲】
【請求項1】
L−リジン塩水溶液に、L−リジン脱炭酸酵素を作用させ、反応液からカダベリンを単離することを特徴とするカダベリンの製造方法。
【請求項2】
L−リジン塩が、塩酸塩、硫酸塩、酢酸塩、硝酸塩、炭酸塩のいずれかであることを特徴とする請求項1に記載のカダベリンの製造方法。
【請求項3】
L−リジン塩が、一塩酸塩であることを特徴とする請求項1に記載のカダベリンの製造方法。
【請求項4】
ビタミンB6を添加することを特徴とする請求項1から3のいずれかに記載のカダベリンの製造方法。
【請求項5】
ビタミンB6が、ピリドキサルリン酸であることを特徴とする請求項4に記載のカダベリンの製造方法。
【請求項6】
L−リジン脱炭酸酵素を作用させた後、L−リジン塩の濃度が仕込み濃度の20%以下になった反応液からカダベリンを単離することを特徴とする請求項1から5のいずれかに記載のカダベリンの製造方法。
【請求項7】
L−リジン脱炭酸酵素が、エシェリシア(Escherichia)属由来であることを特徴とする請求項1から6のいずれかに記載のカダベリンの製造方法。
【請求項8】
L−リジン脱炭酸酵素が、L−リジン脱炭酸酵素活性を有する細胞の破砕液に含まれるL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項9】
L−リジン脱炭酸酵素が、部分的に精製されたL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項10】
L−リジン脱炭酸酵素が、単離精製されたL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項11】
L−リジン脱炭酸酵素が、細胞表面に局在化しているL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項12】
L−リジン脱炭酸酵素が、N末端アミノ酸配列に1から30個のアミノ酸を付与した組換えL−リジン脱炭酸酵素であることを特徴とする請求項1から11のいずれかに記載のカダベリンの製造方法。
【請求項13】
L−リジン脱炭酸酵素が、N末端アミノ酸配列に6から10個のヒスチジン残基を付与した組換えL−リジン脱炭酸酵素であることを特徴とする請求項12に記載のカダベリンの製造方法。
【請求項14】
反応後の反応液をアルカリでpH12から14にし、極性有機溶媒でカダベリンを抽出することを特徴とする請求項1から13のいずれかに記載のカダベリンの製造方法。
【請求項15】
アルカリが、水酸化ナトリウム、水酸化カリウム、水酸化カルシウムのいずれかであることを特徴とする請求項14に記載のカダベリンの製造方法。
【請求項16】
極性有機溶媒がアニリン、シクロヘキサノン、1−オクタノール、イソブチルアルコール、シクロヘキサノール、クロロホルムのいずれかであることを特徴とする請求項14に記載のカダベリンの製造方法。
【請求項17】
2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のカダベリン。
【請求項18】
トリ−n−ブチルアミンの含有量が0.006wt%以下のカダベリン。
【請求項19】
2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下かつトリ−n−ブチルアミンの含有量が0.006%以下のカダベリン。
【請求項20】
請求項1から16のいずれか1項記載の方法で得られるカダベリンもしくは請求項17から19のいずれか1項記載のカダベリンを原料として含有するポリアミド。
【請求項1】
L−リジン塩水溶液に、L−リジン脱炭酸酵素を作用させ、反応液からカダベリンを単離することを特徴とするカダベリンの製造方法。
【請求項2】
L−リジン塩が、塩酸塩、硫酸塩、酢酸塩、硝酸塩、炭酸塩のいずれかであることを特徴とする請求項1に記載のカダベリンの製造方法。
【請求項3】
L−リジン塩が、一塩酸塩であることを特徴とする請求項1に記載のカダベリンの製造方法。
【請求項4】
ビタミンB6を添加することを特徴とする請求項1から3のいずれかに記載のカダベリンの製造方法。
【請求項5】
ビタミンB6が、ピリドキサルリン酸であることを特徴とする請求項4に記載のカダベリンの製造方法。
【請求項6】
L−リジン脱炭酸酵素を作用させた後、L−リジン塩の濃度が仕込み濃度の20%以下になった反応液からカダベリンを単離することを特徴とする請求項1から5のいずれかに記載のカダベリンの製造方法。
【請求項7】
L−リジン脱炭酸酵素が、エシェリシア(Escherichia)属由来であることを特徴とする請求項1から6のいずれかに記載のカダベリンの製造方法。
【請求項8】
L−リジン脱炭酸酵素が、L−リジン脱炭酸酵素活性を有する細胞の破砕液に含まれるL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項9】
L−リジン脱炭酸酵素が、部分的に精製されたL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項10】
L−リジン脱炭酸酵素が、単離精製されたL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項11】
L−リジン脱炭酸酵素が、細胞表面に局在化しているL−リジン脱炭酸酵素であることを特徴とする請求項1から7のいずれかに記載のカダベリンの製造方法。
【請求項12】
L−リジン脱炭酸酵素が、N末端アミノ酸配列に1から30個のアミノ酸を付与した組換えL−リジン脱炭酸酵素であることを特徴とする請求項1から11のいずれかに記載のカダベリンの製造方法。
【請求項13】
L−リジン脱炭酸酵素が、N末端アミノ酸配列に6から10個のヒスチジン残基を付与した組換えL−リジン脱炭酸酵素であることを特徴とする請求項12に記載のカダベリンの製造方法。
【請求項14】
反応後の反応液をアルカリでpH12から14にし、極性有機溶媒でカダベリンを抽出することを特徴とする請求項1から13のいずれかに記載のカダベリンの製造方法。
【請求項15】
アルカリが、水酸化ナトリウム、水酸化カリウム、水酸化カルシウムのいずれかであることを特徴とする請求項14に記載のカダベリンの製造方法。
【請求項16】
極性有機溶媒がアニリン、シクロヘキサノン、1−オクタノール、イソブチルアルコール、シクロヘキサノール、クロロホルムのいずれかであることを特徴とする請求項14に記載のカダベリンの製造方法。
【請求項17】
2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下のカダベリン。
【請求項18】
トリ−n−ブチルアミンの含有量が0.006wt%以下のカダベリン。
【請求項19】
2,3,4,5−テトラヒドロピリジンの含有量が1.4wt%以下かつトリ−n−ブチルアミンの含有量が0.006%以下のカダベリン。
【請求項20】
請求項1から16のいずれか1項記載の方法で得られるカダベリンもしくは請求項17から19のいずれか1項記載のカダベリンを原料として含有するポリアミド。
【図1】


【図2】
【図3】
【図2】
【図3】
【公開番号】特開2009−28045(P2009−28045A)
【公開日】平成21年2月12日(2009.2.12)
【国際特許分類】
【出願番号】特願2008−195749(P2008−195749)
【出願日】平成20年7月30日(2008.7.30)
【分割の表示】特願2002−257374(P2002−257374)の分割
【原出願日】平成14年9月3日(2002.9.3)
【出願人】(000003159)東レ株式会社 (7,677)
【Fターム(参考)】
【公開日】平成21年2月12日(2009.2.12)
【国際特許分類】
【出願日】平成20年7月30日(2008.7.30)
【分割の表示】特願2002−257374(P2002−257374)の分割
【原出願日】平成14年9月3日(2002.9.3)
【出願人】(000003159)東レ株式会社 (7,677)
【Fターム(参考)】
[ Back to top ]