
ポリケタイド合成酵素遺伝子をレポーター遺伝子として利用する方法
【課題】糸状菌において使用でき、検出感度に優れた新規なレポーター遺伝子を提供する。
【解決手段】ポリケタイド合成酵素遺伝子をレポーター遺伝子として有する糸状菌を使用し、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定する。機能既知又は機能未知の転写制御領域と、当該転写制御領域の制御下に発現するポリケタイド合成酵素遺伝子とを含み、発現制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含む核酸構築物。
【解決手段】ポリケタイド合成酵素遺伝子をレポーター遺伝子として有する糸状菌を使用し、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定する。機能既知又は機能未知の転写制御領域と、当該転写制御領域の制御下に発現するポリケタイド合成酵素遺伝子とを含み、発現制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含む核酸構築物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、糸状菌において機能するレポーター遺伝子及び当該レポーター遺伝子を利用した方法、特に当該レポーター遺伝子を利用した方法に関する。
【背景技術】
【0002】
レポーター遺伝子とは、宿主細胞に導入され、発現の有無を可視化できる遺伝子である。すなわち、レポーター遺伝子が発現した宿主細胞では、当該レポーター遺伝子の産物に起因する発光や蛍光が観察される。
【0003】
したがって、レポーター遺伝子を所定の遺伝子と組み換えた組換え体では、レポーター遺伝子の産物に起因する発光や蛍光を観察することで、当該所定の遺伝子の発現パターンを解析することができる。例えば、ある化合物を組換え体に曝露する等の特定の刺激を組換え体に与えたときに発光や蛍光が観察されたとすると、上記所定の遺伝子は、この刺激により発現誘導されるといった知見を得ることができる。あるいは、薬剤応答性プロモーターの下流にレポーター遺伝子を連結した組換え体を使用することで、当該薬剤応答性プロモーターに作用する化合物を新たにスクリーニングすることができる。あるいは、機能未知のプロモーターの下流にレポーター遺伝子を連結した組換え体を使用することで、発現調節作用を有する公知物質に応答性を示す新規なプロモーターをスクリーニングすることができる。
【0004】
一方、創農薬研究プロセスの中で最も重要な過程は、その新薬候補化合物であるリード化合物を見つけ出すことから始まる。しかしながら膨大な種類の化合物で構成される化合物ライブラリーからリード化合物を探索するためには、自動化されたロボットなどを用い、大量の化合物の中から、短時間で目的とするターゲットを選別するハイスループットスクリーニング(HTSスクリーニング)を行う事が望まれている。細胞レベルでのハイスループットスクリーニングにおいて特定の遺伝子発現を評価する場合には、レポーターアッセイがよく用いられている。レポーター遺伝子としては、オワンクラゲ由来のGFP(非特許文献1、非特許文献2)、ホタル由来のルシフェラーゼ、大腸菌由来のGUS(非特許文献3、非特許文献4)などが一般的に使用されている。
【0005】
また、糸状菌の一般的な遺伝子研究分野において、これらのレポーター遺伝子はGFPをはじめとして頻繁に使用されており細胞内での良好な蛍光、発光、発色が確認されている。しかし実際には、糸状菌は、菌糸を絡み合わせながら生育していくという特徴を有しているため、レポーター遺伝子に起因する発光や蛍光の検出感度が十分でないといった問題がある。特に、ハイスループットを目的として、複数のウェルを有するプレートを用いた系では、各ウェルに張り込まれた培地において糸状菌の均一な生育が見られず、レポーター遺伝子に起因する発光や蛍光の検出感度が著しく低下するといった問題がある。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC (1994) Green fluorescent protein as a marker for gene expression. Science 263: 802-805
【非特許文献2】Tsien RY (1998) The green fluorescent protein. Ann. Rev. Biochem 67: 509-544
【非特許文献3】Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6: 3901-3907
【非特許文献4】Jefferson RA, Burgess SM, Hirsh D (1986) β-glucuronidase from Escherichia coli as a gene-fusion marker. Proc. Natl. Acad. Sci. USA 83: 8447-8451
【発明の概要】
【発明が解決しようとする課題】
【0007】
そこで、本発明は、糸状菌において使用でき、検出感度に優れた新規なレポーター遺伝子を提供することを目的とし、当該レポーター遺伝子の発現を検出することを含む方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らは、上述した目的を達成するため鋭意検討した結果、糸状菌を培養した培地中に蓄積されるフラビオリンを検出することで、ポリケタイド合成酵素遺伝子の発現を可視化できることから、当該ポリケタイド合成酵素遺伝子を糸状菌における新規なレポーター遺伝子として使用できることを見いだし、本発明を完成するに至った。
【0009】
本発明は以下を包含する。
【0010】
(1) ポリケタイド合成酵素遺伝子をレポーター遺伝子として有する糸状菌を使用し、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定する方法。
(2) 上記ポリケタイド合成酵素遺伝子は、配列番号2に示すアミノ酸配列を含むタンパク質をコードする、又は配列番号2に示すアミノ酸配列に対して90%以上の類似性を有するアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードすることを特徴とする(1)記載の方法。
(3) 上記ポリケタイド合成酵素遺伝子を、所定の因子に対して転写活性を変動する転写制御領域の制御下に連結してレポーター遺伝子とし、上記糸状菌に対して刺激を与えた後における培地中のフラビオリン量が有意に変動する場合、当該刺激が上記転写制御領域に対して上記所定の因子と同等に作用すると判断することを特徴とする(1)記載の方法。
(4) 上記刺激は、上記糸状菌に対する供試化合物への曝露であることを特徴とする(3)記載の方法。
(5) 上記転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする(3)記載の方法。
(6) 上記転写制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする(3)記載の方法。
(7) 上記転写制御領域として植物病害抵抗性付与剤誘導型プロモーターを使用して植物病害抵抗性付与剤の候補物質をスクリーニングする、上記転写制御領域として電子伝達系阻害剤誘導型プロモーターを使用して電子伝達系阻害剤の候補物質をスクリーニングする、又は上記転写制御領域としてエルゴステロール生合成阻害剤誘導型プロモーターを使用してエルゴステロール生合成阻害剤の候補物質をスクリーニングすることを特徴とする(3)記載の方法。
(8) 上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結してレポーター遺伝子とし、機能既知である発現制御領域の転写活性を変動させる所定の因子を上記糸状菌に作用させたときに、培地中のフラビオリン量が有意に変動する場合、上記機能未知の発現制御領域が上記機能既知の発現制御領域と同等の応答性を示すと判断することを特徴とする(1)記載の方法。
(9) 上記所定の因子は、特定のプロモーターに対して転写活性を促進又は抑制する化合物であることを特徴とする(8)記載の方法。
(10) 上記所定の因子は、植物病害抵抗性付与能を有する化合物、電子伝達系阻害能を有する化合物及びエルゴステロール生合成阻害能を有する化合物からなる群より選ばれる1つの化合物であることを特徴とする(8)記載の方法。
【0011】
(11) 上記機能未知の転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする(8)記載の方法。
(12) 上記所定の因子として植物病害抵抗性付与剤を使用して植物病害抵抗性付与剤誘導型プロモーターをスクリーニングする、上記所定の因子として電子伝達系阻害剤を使用して電子伝達系阻害剤誘導型プロモーターをスクリーニングする、又は上記所定の因子としてエルゴステロール生合成阻害剤を使用してエルゴステロール生合成阻害剤誘導型プロモーターをスクリーニングすることを特徴とする(8)記載の方法。
(13) 上記ポリケタイド合成酵素遺伝子で解析対象の遺伝子を組み換え、培地に生産されるフラビオリンを指標として解析対象の遺伝子の発現パターンを解析することを特徴とする(1)記載の方法。
(14) 導入対象の目的遺伝子と、恒常発現型プロモーターに連結された上記ポリケタイド合成酵素遺伝子とを上記糸状菌に導入し、培地中のフラビオリン量が有意に大である場合、上記目的遺伝子が発現していると判断することを特徴とする(1)記載の方法。
(15) 複数のウェルを有するプレートを使用して当該各ウェルに上記糸状菌を培養し、上記フラビオリンに由来する赤色発色を検出する検出手段を使用して複数のウェルを一括して観察することを特徴とする(1)乃至(14)いずれか一つに記載の方法。
(16) 機能既知又は機能未知の転写制御領域と、当該転写制御領域の制御下に発現するポリケタイド合成酵素遺伝子とを含む核酸構築物。
(17) 上記機能既知の発現制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする(16)記載の核酸構築物。
(18) (16)又は(17)記載の核酸構築物を含む発現ベクター。
(19) (16)又は(17)記載の核酸構築物で形質転換された組み換え糸状菌。
(20) いもち病菌の組換え体であることを特徴とする(19)記載の組み換え糸状菌。
【発明の効果】
【0012】
本発明によれば、糸状菌に対して使用することができ、非常に簡便且つ高感度に発現を可視化できる新規なレポーター遺伝子を提供することができる。したがって、本発明に係るレポーター遺伝子を使用することによって、一般的にレポーター遺伝子の発現を可視化する工程を含む様々な手法を糸状菌に適用することができる。
【図面の簡単な説明】
【0013】
【図1】糸状菌のDHN−メラニン合成経路及び関連酵素を示す概略図である。
【図2】内在ALB1プロモーターALB1Pを既知プロモーターへ置き換えるプラスミドベクターpCHALB1の概略構成図である。
【図3】pCHRDH8PALB1を利用した相同組み換えによるALB1PのRDH8Pへの変換を示す概略図である。
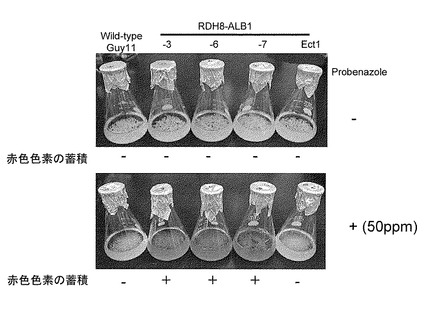
【図4】プロベナゾールでの誘導の有無によるRDH8P-ALB1菌株のYG液体培地上での形態を示す特性図である。
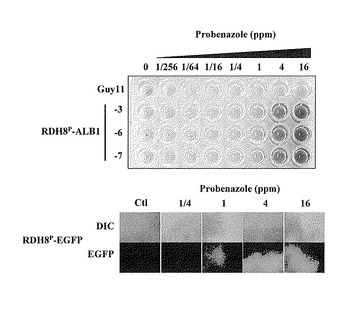
【図5】RDH8Pプロベナゾール応答株を用いた赤色色素フラビオリンの蓄積による遺伝子発現の可視化例とRDH8Pプロベナゾール応答型EGFP株との比較を示す特性図である。
【図6】AOXP電子伝達系阻害剤応答株を用いた赤色色素フラビオリンの蓄積による遺伝子発現の可視化例を示す特性図である。
【図7】CYP51Pエルゴステロール生合成阻害剤応答株を用いた赤色色素フラビオリンの蓄積による遺伝子発現の可視化例を示す特性図である。
【発明を実施するための形態】
【0014】
以下、本発明を図面を参照して詳細に説明する。
レポーター遺伝子
糸状菌において機能する新規レポーター遺伝子は、図1に示すDHN−メラニン合成経路において、アセチルCoA及びマロニルCoAからテトラヒドロキシナフタレン(1,3,6,8−T4HN)の生合成に関与するポリケタイド合成酵素をコードする遺伝子である。ポリケタイド合成酵素遺伝子が発現すると、糸状菌においてテトラヒドロキシナフタレン(1,3,6,8−T4HN)が合成される。このテトラヒドロキシナフタレン(1,3,6,8−T4HN)は、自然酸化により赤色色素であるフラビオリンに変換される。
【0015】
なお、図1に示すように、DHN−メラニン合成経路において、テトラヒドロキシナフタレン(1,3,6,8−T4HN)は、THN還元酵素による還元及びシタロン脱水酵素による脱水によりジヒドロキシナフタレン(1,8−DHN)となる。そして、ジヒドロキシナフタレン(1,8−DHN)が酸化酵素により重合し、黒色のメラニンが生成される。従って、イネいもち病菌などの糸状菌は、通常、黒色となる。例えば、DHN−メラニン合成経路上のTHN還元酵素やシタロン脱水酵素の阻害剤の存在下では、メラニン生合成が阻害され、1,3,6,8−T4HNが蓄積し、蓄積した1,3,6,8−T4HNの自然酸化によりフラビオリンが合成されることになる。したがって、例えば、THN還元酵素阻害剤であるトリシクラゾールの存在下において、イネいもち病菌は赤色を呈する。
【0016】
ただし、イネいもち病菌において、DHN−メラニン合成経路に関与する遺伝子群は、感染器官である付着器形成時に特に強く発現が誘導され、栄養増殖期では培養開始から少なくとも3日間経過しなければ発現が誘導されない。このため、野生型の糸状菌を通常の栄養増殖させている場合、培養開始から約3日までであれば、内在するメラニン生合成に関与する遺伝子群が発現することによる1,3,6,8−T4HNの蓄積及びフラビオリンの合成は生じない。このため、ポリケタイド合成酵素遺伝子を糸状菌におけるレポーター遺伝子として使用することができ、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定(可視化)することができる。
【0017】
ここで、ポリケタイド合成酵素遺伝子としては、特に限定されないが、特に糸状菌に由来することが好ましい。また、ポリケタイド合成酵素遺伝子としては、詳細を後述する、宿主となる糸状菌に由来することが好ましい。さらに、ポリケタイド合成酵素遺伝子としては、一般的に植物病原糸状菌として知られる糸状菌に由来することがより好ましい。なお、植物病原糸状菌におけるDHN−メラニン合成経路及びメラニン合成に関与する遺伝子群については、例えば、Richard J. Howard et al., 1996, Annual Review of Microbiology Vol. 50: 491-512及びKim Langfelder et al., 2003, Fungal Genet. Biol. 38:143-158を参照することができる。すなわち、使用する宿主に応じて、適宜、上述したような文献を参照することで、好ましいポリケタイド合成酵素遺伝子を準備して使用することができる。
【0018】
一例としては、イネいもち病菌に由来するポリケタイド合成酵素遺伝子を挙げることができる。イネいもち病菌に由来するポリケタイド合成酵素遺伝子のコーディング領域の塩基配列を配列番号1に示し、イネいもち病菌に由来するポリケタイド合成酵素遺伝子によりコードされるポリケタイド合成酵素のアミノ酸配列を配列番号2に示す。
【0019】
また、ポリケタイド合成酵素遺伝子としては、配列番号2に示すアミノ酸配列からなるタンパク質をコードする遺伝子に限定されず、例えば配列番号2に示すアミノ酸配列に対して例えば90%以上、好ましくは95%以上、より好ましくは98%以上の類似性を有するアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードする遺伝子であっても良い。ここで、類似性とは、例えばblastアルゴリズムを実装したコンピュータプログラムを用いてデフォルトの設定で求められる値を意味する。
【0020】
さらに、ポリケタイド合成酵素遺伝子としては、配列番号2に示すアミノ酸配列に対して1個又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードする遺伝子であっても良い。ここで、複数個のアミノ酸とは、2個から230個、好ましくは2個から150個、より好ましくは2個から100個、さらに好ましくは2個から50個、特に好ましくは2個から20個のアミノ酸を意味する。
【0021】
さらにまた、ポリケタイド合成酵素遺伝子としては、配列番号1に示す塩基配列を有する遺伝子の少なくとも一部に対して、ストリンジェントな条件下でハイブリダイズするポリヌクレオチドを含み、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードするものであってもよい。ここで、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、45℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム)でのハイブリダイゼーション、その後の50〜65℃、0.2〜1×SSC、0.1%SDSでの洗浄が挙げられ、或いはそのような条件として、65〜70℃、1×SSCでのハイブリダイゼーション、その後の65〜70℃、0.3×SSCでの洗浄を挙げることができる。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0022】
なお、配列番号2とは異なるアミノ酸配列をコードするポリケタイド合成酵素遺伝子、配列番号1とは異なる塩基配列を有するポリケタイド合成酵素遺伝子は、当該技術分野で公知の手法によってイネいもち病菌に由来するポリケタイド合成酵素遺伝子を改変することによって得ることができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA Bio社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA Bio社製)を用いて変異が導入される。また、変異導入方法としては、EMS(エチルメタンスルホン酸)、5-ブロモウラシル、2-アミノプリン、ヒドロキシルアミン、N-メチル-N’-ニトロ-Nニトロソグアニジン、その他の発ガン性化合物に代表されるような化学的変異剤を使用する方法でも良いし、X線、アルファ線、ベータ線、ガンマ線、イオンビームに代表されるような放射線処理や紫外線処理による方法でも良い。
【0023】
宿主
上述したポリケタイド合成酵素遺伝子をレポーター遺伝子として使用できる糸状菌としては、特に限定されず使用することができる。また、糸状菌としては、菌糸を形成する真菌であれば特に限定されず使用することができる。上述したレポーター遺伝子として糸状菌由来のポリケタイド合成酵素遺伝子を使用する場合、宿主である糸状菌の菌体内において良好に発現することができる。特に、ポリケタイド合成酵素遺伝子の由来と宿主とが同属の糸状菌であることが好ましく、同種の糸状菌であることがより好ましく、同株の糸状菌であることが更に好ましい。
【0024】
さらに、糸状菌としては、例えば植物病原性糸状菌を使用することが好ましい。植物病原性糸状菌としては、特に限定されないが、例えば、Botrytis属(=Botryotinia属)、Rosellinia属、Aspergillus属(=Eurotium属など)、Phytophthora属、Pythium属、Sclerotinia属、Colletotrichum属(=Glomerella属)、Rhizoctonia属(=Thanatephorus属)、Helicobasidium属、Fusarium属(=Gibberella属、Haematonectoria属、Nectoria属、Calonectoria属)、Magnaporthe属(=Pyricularia属)、Alternaria属、Puccinia属(=Aecidium属)、Uromyces属、Microsphaera属、Blumeria属、Erysiphe属(=Golovinomyces属)、Oidium属(=Sawadae属)、Sphaerotheca属(=Podosphaera属)、Phyllactinia属(=Ovulariopsis属)、Uncinula属(=Golovinomyces属、Erysiphe属)、Podosphaera属(=Sphaerotheca属)、Pythium属、Peronospora属、Ustilago属、Cochliobolus属(=Curvularia属、Bipolaris属、Helminthosporium属)、Cladosporium属(=Mycosphaerella属、Venturia属)、Mycosphaerella属(=Ascochyta属、Cercospora属、Phyllosticta属、Septoria属)、Phomopsis属(=Diaporthe属)、Phoma属(=Leptosphaeria属、Mycosphaerella属)、Venturia属(=Fusicladium属、Spilocaea属)、Claviceps属(=Ustilaginoidea属、Sphaecelia属)、Calonectria属(=Cylindrocladium属)、Verticillium属(=Cosmospora属、Hypocereales属など)、Penicillium属(=Eupenicillium属、Talaromyces属)、Elsinoe属(=Sphaceloma属)、Didymella属(=Ascochyta属、Phoma属など)若しくはRhizopus属に属する菌類を挙げることができる。
【0025】
より具体的に、イネのいもち病菌(Magnaporthe grisea シノニム:Pyricularia oryzae)、ごま葉枯病菌(Cochliobolus miyabeanus)、紋枯病菌(Rhizoctonia solani)、馬鹿苗病菌(Gibberella fujikuroi)、各種作物の苗立枯病菌(Fusarium spp.、Pythium spp.、Rhizopus spp.、Trichoderma sp.)、灰色かび病菌(Botrytis cinerea)、菌核病菌(Sclerotinia sclerotiorum)、ウリ類の炭そ病菌(Colletotrichum lagenarium)、シバのブラウンパッチ病菌(Rhizoctonia solani)等を挙げることができる。
【0026】
レポーター遺伝子の利用形態
上述したポリケタイド合成酵素遺伝子は、上述した宿主においてレポーター遺伝子として機能する。したがって、上述したポリケタイド合成酵素遺伝子は、従来公知のレポーター遺伝子を使用する各種遺伝子工学的手法、分子生物学的手法或いは細胞工学的手法においてレポーター遺伝子として使用することができる。ここでレポーター遺伝子を使用する各種方法は、何ら限定されず、如何なる目的、規模の実験方法及び工業的方法をも含む意味である。
【0027】
通常、ある遺伝子をレポーター遺伝子として使用するには、当該遺伝子の発現を検出、好ましくは可視化できなければならない。ポリケタイド合成酵素遺伝子を外的に導入していない糸状菌では培養開始から約3日間を経過しなければ培地中にフラビオリンを合成しない。しかしながら、ポリケタイド合成酵素遺伝子を外的に導入して発現を誘導すると、比較的早期に培地中にフラビオリンを生産することとなる。したがって、外的に導入したポリケタイド合成酵素遺伝子に起因する、培地中のフラビオリンを検出することで当該遺伝子の発現を検出することができ、ポリケタイド合成酵素遺伝子を糸状菌における有効なレポーター遺伝子として使用することができる。
【0028】
培地中に生産されたフラビオリンを検出するには、特に限定されないが、フラビオリンの赤色を検出すればよい。フラビオリンの赤色は、目視によって検出してもよいし、赤色を光学的に検出できる装置を用いて検出しても良い。このような装置としては、例えば、分光光度計、分光蛍光光度計等を使用することができる。
【0029】
また、フラビオリンによる赤色の濃淡(強度)から、ポリケタイド合成酵素遺伝子の発現の強弱を判定することもできる。すなわち、培地中の赤色が濃い場合にはポリケタイド合成酵素遺伝子が高発現していると判断し、培地中の赤色が薄い場合にはポリケタイド合成酵素遺伝子の発現量が低いと判断することができる。
【0030】
特に、フラビオリンは、レポーター遺伝子を導入した糸状菌を培養した培地中に生産されるため、GFPやGUS等の公知のレポーター遺伝子と比較して、より高感度に検出することができる。GFPやGUS等の公知のレポーター遺伝子を使用した場合、細胞内において蛍光や発色を検出しなければならないため、菌糸を絡み合わせながら生育する糸状菌では非常に検出感度が低くなる。これに対して、上述したポリケタイド合成酵素遺伝子をレポーター遺伝子として使用する場合には、培地中のフラビオリンを検出するため、糸状菌を宿主としても非常に優れた検出感度を達成することができる。
【0031】
したがって、上述したポリケタイド合成酵素遺伝子をレポーター遺伝子とし、宿主として糸状菌を使用する場合には、例えば、複数のウェルを有するプレートを用いて、大量のサンプルを処理することができる。すなわち、レポーター遺伝子としてポリケタイド合成酵素遺伝子を導入した糸状菌を各ウェルに張り込んだ培地にて培養し、プレートリーダーのような大量測定機器により各ウェルにおけるフラビオリンの赤色色素を一括して又は順次に検出する。これにより、レポーター遺伝子の発現をハイスループットで測定することができる。
【0032】
より具体的に、レポーター遺伝子の利用形態としては、特定の遺伝子の発現をレポーター遺伝子により解析する手法が挙げられる。ここで、特定の遺伝子とは、なんら限定されるものではなく、宿主となる糸状菌に内在する各種の遺伝子を挙げることができる。この利用形態では、先ず、上述したポリケタイド合成酵素遺伝子を当該特定の遺伝子と置換するように導入した組み換え糸状菌を準備する。得られた組み換え糸状菌を通常の培地にて培養し、培地に生産されたフラビオリンを検出することでレポーター遺伝子の発現を測定する。例えば、培養条件を異にして組み換え糸状菌を培養し、一方の条件下においてフラビオリンを検出し、他方の条件下においてフラビオリンを検出しなかった場合、上記特定の遺伝子は一方の条件下において特異的に発現されるものと理解することができる。ここで、培養条件とは、温度条件や、培地組成条件、pH条件、化合物の存在・非存在条件等を挙げることができるが、これらに限定されるものではない。
【0033】
また、組み換え糸状菌を培養しながら経時的にフラビオリンの生産量を測定することで、上記特定の遺伝子の発現パターンを解析することができる。この系においては、培養経過に伴って培養条件を種々に変更することができる。例えば、培養開始から所定期間が経過した後に、所定の化合物を培養系に添加したり、温度条件を変更したりすることができる。
【0034】
ところで、レポーター遺伝子の利用形態としては、上記ポリケタイド合成酵素遺伝子を、所定の因子に対して転写活性を変動する転写制御領域の制御下に連結した組み換え糸状菌を用いて、当該転写制御領域に影響を与えうる新規な因子をスクリーニングすることができる。すなわち、この組み換え糸状菌に対して新規な因子を作用させたときに培地中にフラビオリンが生産されていたとすれば、この新規な因子は上記発現制御領域に対して作用し、下流の遺伝子を発現誘導することが判明する。
【0035】
ここで、転写制御領域とは、例えばプロモーターや、その他のシス配列(シス調節配列)を全て包含する意味である。転写制御領域には、プロモーター領域、エンハンサー領域、サイレンサー領域といった転写に関与する因子(転写因子や、アクチベータータンパク質及びサイレンサータンパク質)が結合する領域を意味する。また、所定の因子としては、例えば、化合物、温度、pHといった一般的に遺伝子の転写を制御する種々の因子を挙げることができる。
【0036】
したがって、「所定の因子に対して転写活性を変動する転写制御領域」としては、所定の化合物に応答性を示すプロモーター、所定の温度条件で発現誘導するプロモーター等を挙げることができる。具体的に、生理活性物質をスクリーニングする際には、転写制御領域として、従来公知の生理活性物質応答性プロモーターを使用する。生理活性物質応答性プロモーターとしては、特に制限はなく、スクリーニングしたい生理活性物質に応答するプロモーターから適宜、選択すれば良い。生理活性物質として、農薬、医薬、動物薬等の候補化合物をスクリーニングする場合には、植物病原菌、人畜の病原菌のように防除すべき微生物に有効な作用を有する生理活性物質に応答するプロモーターから選ぶのが良い。また、生理活性物質応答性プロモーターとしては、レポーター遺伝子の発現による代謝産物の測定結果を明確にするために、目的の生理活性物質による活性化がはっきりしていてノイズの少ないプロモーターを使用することが好ましい。
【0037】
さらに具体的に、「所定の因子に対して転写活性を変動する転写制御領域」としては、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を挙げることができる。このようなプロモーターとしては、例えば、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターを挙げることができる。
【0038】
例えば、植物病害抵抗性付与剤誘導型プロモーターの下流にポリケタイド合成酵素遺伝子を連結した組み換え糸状菌に対して供試化合物を作用させたとき、培地中にフラビオリンが検出されたとすれば、当該供試化合物は、植物病害抵抗性付与剤誘導型プロモーターを活性化する作用を有していることが判る。この結果から、当該供試化合物は、植物病害抵抗性付与剤の候補として特定することができる。なお、植物病害抵抗性付与剤とは、植物に対して作用して病原体の感染に対する抵抗性を付与するようなシグナル伝達(サリチル酸経路など)を活性化する物質である。植物病害抵抗性付与剤としては、特に限定されないが、プロベナゾールを挙げることができる。
【0039】
また、例えば、電子伝達系阻害剤誘導型プロモーターの下流にポリケタイド合成酵素遺伝子を連結した組み換え糸状菌に対して供試化合物を作用させたとき、培地中にフラビオリンが検出されたとすれば、当該供試化合物は、電子伝達系阻害剤誘導型プロモーターを活性化する作用を有していることが判る。この結果から、当該供試化合物は、電子伝達系阻害剤の候補として特定することができる。なお、電子伝達系阻害剤とは、糸状菌における電子伝達系を阻害することで糸状菌を衰弱又は死滅若しくは糸状菌の感染力を低下させることが知られている物質である。電子伝達系阻害剤としては、特に限定されないが、複合体1阻害剤であるジフルメトリム、テブフェンピラド、複合体3阻害剤であるクレソキシムメチル、ピリベンカルブ、F1Fo ATPase阻害剤であるオリゴマイシンを挙げることができる。また、電子伝達系阻害剤誘導型プロモーターとしては、特に限定されないがAOXプロモーター(AOXP)を使用することができる。AOXプロモーターは、ミトコンドリアにおける呼吸において、電子伝達系複合体2以外の電子伝達系阻害剤に特異的に応答することが知られている。(Yukioka H, Inagaki S, Tanaka R, Katoh K, Miki N, Mizutani A, Masuko M. (1998) Transcriptional activation of the alternative oxidase gene of the fungus Magnaporthe grisea by a respiratory-inhibiting fungicide and hydrogen peroxide. Biochim Biophys Acta. 1442:161-169)
【0040】
さらに、例えば、エルゴステロール生合成阻害剤誘導型プロモーターの下流にポリケタイド合成酵素遺伝子を連結した組み換え糸状菌に対して供試化合物を作用させたとき、培地中にフラビオリンが検出されたとすれば、当該供試化合物は、エルゴステロール生合成阻害剤誘導型プロモーターを活性化する作用を有していることが判る。この結果から、当該供試化合物は、エルゴステロール生合成阻害剤の候補として特定することができる。なお、エルゴステロール生合成阻害剤とは、糸状菌におけるエルゴステロール生合成経路を阻害することで糸状菌を衰弱又は死滅若しくは糸状菌の感染力を低下させることが知られている物質である。エルゴステロール生合成阻害剤としては、特に限定されないが、オキスポコナゾール、イプコナゾール、トリホリン、プロチオコナゾールを挙げることができる。また、エルゴステロール生合成阻害剤誘導型プロモーターとしては、特に限定されないがCYP51プロモーター(CYP51P)を使用することができる。CYP51プロモーターは、シトクロムP450のひとつで、ステロール生合成経路のラノステロール構造の14-脱メチル化酵素として知られているCYP51遺伝子のプロモーターである。(Ma Z, Proffer TJ, Jacobs JL, Sundin GW. (2006) Overexpression of the 14alpha-demethylase target gene (CYP51) mediates fungicide resistance in Blumeriella jaapii. Appl Environ Microbiol. 72:2581-2585)
【0041】
ところで、レポーター遺伝子の利用形態としては、上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結した組み換え糸状菌を用いて、当該転写制御領域の特性を評価することができる。言い換えれば、上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結した組み換え糸状菌を用いて、公知の因子に対して応答性を示す新規な発現制御領域をスクリーニングすることができる。ここで、機能未知の発現制御領域とは、例えば糸状菌のゲノムに存在する遺伝子のコーディング領域の上流に存在し、当該遺伝子の発現を制御しているものの、どのようなシグナルに依存して発現誘導若しくは発現抑制しているのか不明な領域を意味する。機能未知の発現制御領域としては、コーディング領域の上流に存在する例えば2000塩基、好ましくは1000塩基、より好ましくは500塩基を単離して使用することができる。
【0042】
より具体的に、公知の因子としては、例えば、上述したような化合物、温度、pHといった一般的に遺伝子の転写を制御する種々の因子を挙げることができる。更に具体的には、上述したような、植物病害抵抗性付与能を有する化合物、電子伝達系阻害能を有する化合物或いはエルゴステロール生合成阻害能を有する化合物を使用することによって、新規な植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター或いはエルゴステロール生合成阻害剤誘導型プロモーターをスクリーニングすることができる。
【実施例】
【0043】
以下、実施例により本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0044】
〔実施例1〕
いもち病菌DHN-メラニン生合成に関わるポリケタイド合成酵素遺伝子の発現制御
DHN-メラニン生合成に関わるポリケタイド合成酵素ALB1遺伝子の発現を誘導し、その派生物である赤色色素フラビオリンを合成させることにより、その赤色色素の発色がハイスループットとして利用可能なレポーターであるか否かを調査することを目的に、ALB1遺伝子の発現を既知のプロモーターの制御下に置く実験を試みた。本来ならALB1遺伝子全長の開始コドンより上流領域に既知プロモーターを配置するような構造のプラスミドベクターを作成し、それを宿主へ導入することが望ましいが、いもち病菌ALB1遺伝子の推定全長が6771bpと巨大なため、内在のALB1遺伝子の上流プロモーター領域ALB1Pを他のプロモーターへ置き換える方法をとることにした。ポリケタイド合成酵素遺伝子ALB1の開始コドンより上流のプロモーター領域を相同組み換えによって欠失させ、そこに既知の様々なプロモーター断片を挿入するためのベクターの作成方法および実際に各種プロモーターを挿入したベクターでいもち病菌を形質転換した場合の形質転換体の形状について以下に示す。
【0045】
1)相同組み換えに用いるALB1遺伝子の5’領域およびALB1遺伝子断片のクローニング
Broad Institute のマグナポルテ・グリセア(Magnaporthe grisea)ゲノムデータベース(http://www.broad.mit.edu/annotation/genome/magnaporthe_grisea/MultiHome.html)からALB1遺伝子とその5’領域の塩基配列情報を取得し、その配列情報を参考にALB1の5’領域1440bpを増幅するプライマーペアとして(5‘-CCGAATTCAAGAATGCTTGGACCACACC-3’(配列番号3)及び5‘-ATGGATCCGTACATTTCCGGTTGGATGG-3’(配列番号4))を、またALB1遺伝子の開始コドンから1174bpの断片を増幅するプライマーペアとして(5‘-AAAGATCTGACCATGGCGGACAAGCTGGCTTT-3’(配列番号5)及び5’-GCACTAGTGGATGATGAGGCACGACTTT-3’(配列番号6))のそれぞれのオリゴヌクレオチドを設計した。なおプライマー内に示した下線部分は制限酵素の配列であり、GAATTCはEcoRI、GGATCCはBamHI、AGATCTはBglII、ACTAGTはSpeIサイトを示す。
【0046】
これらのプライマーを用いてマグナポルテ・グリセアGuy11株のゲノムDNAをテンプレートとしてPCR反応を行った。増幅反応は、94℃で3分間鋳型DNAを変性し、94℃で30秒間、55℃で30秒間、72℃で2分間保持するサイクルを35サイクルおこなった後、72℃で5分間完全伸長させ、4℃で保持した。PCR用装置は、PCR Thermal Cycler PERSONAL (タカラバイオ社製)を用いた。このPCRによる増幅断片をアガロース電気泳動にて確認を行ったところ、ALB1の5'領域およびALB1遺伝子断片ともに、予想される1440塩基対、1174塩基対付近の大きさの遺伝子増幅が見られた。この増幅断片をアガロースゲル電気泳動に供したゲル中よりGENECLEAN IIIキット(Qbiogene社製)を用いてDNAを抽出し、これを挿入DNA断片とした。この挿入DNA断片をpGEM-T Easy Vector System(Promega社製)を用いてpGEM-T Easy Vectorに連結させ、連結DNA溶液を得た。連結DNA溶液10μlを氷中でよく冷却した後、氷上解凍したコンピテントセルJM109を100μl加え穏やかに撹拌し、氷中で20分間、続いて42℃で45秒ヒートショック処理を行った。これに、400μlのLB液体培地を加え、37℃で1時間培養後、100μg/mlのアンピシリンを添加したLB平板培地にまき、37℃で一晩培養した。目的のプラスミドDNAを用いて形質転換した大腸菌の単一のコロニーは3mlの100μg/mlのアンピシリンを添加したLB液体培地に植菌し、37℃で一晩振盪培養した。1.5mlの培養液を1.5mlのエッペンドルフチューブに移して15,000×gで1分間遠心分離し、沈殿を100μlの氷冷したTEG(25mM Tris-HCl、10mM EDTA、50mM Glucose、pH8.0)で懸濁し、これに200μlの0.2N NaOH-1%SDSを加えて穏やかに撹拌した後、150μlの3M NaOAc(pH5.2)を加えて混合した。これを15,000×g、4℃で5分間遠心分離し上清を回収し、450μlのフェノール・クロロホルム・イソアミルアルコール(25:24:1)を加えて激しく撹拌した後、15,000×g、室温で5分間遠心分離し上層を回収した。この溶液に-20℃で氷冷した900μlのエタノールを加えて、-20℃で10分間放置後、4℃にて15,000×gで5分間遠心分離した。沈殿を500μlの70%エタノールでリンスしたのち、乾燥させ、最後にRNase(100μg/ml)を含むTE(10mM Tris-HCl、1mM EDTA、pH8.0)50μlに溶解した。得られたプラスミドをEcoRIで切断後、アガロース電気泳動により挿入断片の存在を確認し、このプラスミド中の挿入DNA断片を添付のプロトコールに従い一般的なユニバーサルプライマーSP6(5-GATTTAGGTGACACTATAG-3(配列番号7))およびT7(5-TAATACGACTCACTATAGGG-3(配列番号8))を用い、ABI PRISMTM 377 DNA sequencing system (PE Biosystem社製)にて解析した。その結果、ALB1の5'領域は1440塩基対、ALB1遺伝子は1174塩基対からなることが明らかとなった。また、本ベクターをpGEM-5’ALBおよびpGEM-ALB1174と命名する。
【0047】
2)内在ALB1プロモーターALB1Pを既知プロモーターへ置き換えるプラスミドベクターの作成
pCAMBIA-0380バイナリーベクター(CAMBIA社より購入)を骨格にし、薬剤選択マーカーとしてハイグロマイシンB耐性遺伝子Hphを融合することにより作成した糸状菌形質転換ベクターpCH11(図2)のEcoRI、BamHIサイトにpGEM-5’ALBベクターのEcoRI、BamHI断片を挿入後、続いてBglII、SpeIサイトにpGEM-ALB1174のBglII、SpeI断片を挿入することによりALB1プロモーターALB1Pを欠損すると予想されるプラスミドベクターを作成した。本ベクターをpCHALB1と命名した。本ベクターpCHALB1(図2)は、配置してあるALB1遺伝子開始コドンより上流のHindIIIもしくはBglIIサイトへ既知プロモーターを挿入することにより内在ALB1プロモーターALB1Pを既知プロモーターへ置き換えるプラスミドベクターとして利用できる。
【0048】
〔実施例2〕
pCHALB1へのプロベナゾール誘導型プロモーターRDH8Pの挿入
ALB1遺伝子の上流のALB1P領域を他のプロモーター配列で置き換える例として、RDH8Pプロモーターを用いた場合を以下に示す。RDH8遺伝子は、いもち病菌には殺菌効果が無く、宿主である植物にいもち病菌に対する抵抗性を付与する化合物であるプロベナゾールの存在下で特異的に発現量が亢進する遺伝子として発見された。また、RDH8遺伝子のプロモーター領域を、既知のレポーター遺伝子であるEGFPに連結した解析結果から、RDH8遺伝子のプロモーター領域(RDH8Pプロモーター)は、プロベナゾールの有無によって遺伝子発現を調節することができる発現誘導型プロモーターとして利用できることが明らかとなっている。
【0049】
プロベナゾール誘導型プロモーターRDH8Pの配列は、プライマー5‘-ATAAGGTGCGCGCAAAGGT-3’(配列番号9)及び5‘-CGCAAGCTTAGTTTGGATAGTCGTATGCA-3’ (配列番号10)を用い、いもち病菌ゲノムDNAを鋳型としたPCRによって増幅させ、前述の方法によりpGEM-T Easyベクターに連結した。本ベクターをHindIII処理することにより得たRDH8Pプロモーター配列を同じくHindIII処理したpCHALB1ベクターに連結することにより、RDH8PによりALB1遺伝子を制御可能なプラスミドベクターを作成した。なお、RDH8Pプロモーター配列のベクター内への挿入方向は、PCRおよびシークエンスにより確認し、本ベクターをpCHRDH8PALB1と命名した。
【0050】
(1) pCHRDH8PALB1プラスミドを用いたマグナポルテ・グリセアの形質転換
マグナポルテ・グリセアの形質転換はアグロバクテリウム法(Gento Tsuji, Satoshi Fujii, Naoki Fujihara, Chika Hirose, Seiji Tsuge, Tomonori Shiraishi and Yasuyuki Kubo(2003):Agrobacterium tumefaciens-mediated transformation for random insertional mutagenesis in Colletotrichum lagenarium. Journal of General Plant Pathology 69:230-239)を改良した方法を用いた。予想される相同組み換えによるALB1PのRDH8Pへの変換は図3に示した。得られたRDH8P-ALB1菌株へのベクター挿入の確認は、PCR、シークエンス及びサザン解析によって行った。
【0051】
(2) RDH8P-ALB1菌株の形状
RDH8Pプロモーターは化合物プロベナゾール処理により、条件的に下流遺伝子の発現を顕著に誘導することができる。今回得られたRDH8P-ALB1菌株にプロベナゾールを処理したときに特異的にフラビオリンの赤色色素の蓄積が観察されるか否かを調査するため、YG液体培地でのプロベナゾール添加の有無によるRDH8P-ALB1菌株の色素合成能の観察を行った。
【0052】
プロベナゾールを50ppm添加もしくは無添加のYG液体培地(イーストエクストラクト5g 、グルコース10g/L) 50mlに野生株Guy11および3つのRDH8P-ALB1菌株の胞子を約5×105個移植し、暗黒下、26℃、140 strokes/minの条件で24時間振とう培養を行った。24時間後のサンプルにプロベナゾールを50ppm添加し、再度24時間同条件で振とう培養を行った。また、プロベナゾール無添加のサンプルを対照として用いた。プロベナゾール添加後24時間の写真を図4に示す。その結果、プロベナゾール無添加の全てのサンプルにおいて乳白色の菌糸塊の生育が観察され、培地の色もYG液体培地の色である黄色のまま目立った色素の蓄積は観察されなかった。一方、プロベナゾールを50ppm添加したサンプルでは、対照の野生株Guy11はプロベナゾール無添加区と同様の乳白色の菌糸塊と培地色であったが、3つのRDH8P-ALB1菌株の場合、菌糸塊はほとんど通常と同様の乳白色をしていたのに対し、培地の色は顕著に強い赤色を呈していた。
【0053】
これらの結果から、RDH8P-ALB1菌株ではRDH8Pプロモーター制御下で、プロベナゾールの有無によりRDH8Pプロモーター下流のALB1遺伝子の発現が条件的に調節されており、ALB1遺伝子の発現によりALB1遺伝子の合成産物である1,3,6,8-T4HNが蓄積され、結果として赤色色素フラビオリンの蓄積が観察されたものと考えられた。また、Guy11野生株においては、生育した菌糸のメラニン化が観察されなかった。このことから、菌糸生育に要した48時間という時間内では、いもち病菌のメラニン合成に関与する遺伝子群はほとんど発現しないことを確認できた。
【0054】
このように、ALB1遺伝子の発現制御をフラビオリンの蓄積により可視化できるため、ALB1遺伝子を糸状菌の新しいレポーター遺伝子として利用できる可能性が示された。また、仮に、内在するALB1遺伝子に加えて、RDH8Pプロモーター等の他のプロモーターに置き換えたALB1遺伝子を外的に導入したとしても、外的に導入したALB1遺伝子の発現制御をフラビオリンの蓄積により可視化できることが示された。
【0055】
(3) 96-wellマイクロタイタープレートを用いたハイスループットスクリーニングへの利用
1.プロベナゾールを用いた例
ALB1遺伝子の発現制御による色素蓄積をレポーターに用い、ハイスループットスクリーニングに利用しようとする場合、大量の数のサンプルをアッセイできることが求められる。そこで前述のアッセイのような50mlものハイスケールではなく数100μlというロースケールでのアッセイにも応用可能かを調べるために、RDH8P-ALB1菌株を利用して、96-wellのマイクロタイタープレートを用いたアッセイを行った。YG液体培地に移植したRDH8P-ALB1の3菌株と比較対照として用いた親菌株であるGuy11株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し、26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、プロベナゾールを0、1/256、1/64、1/16、1/4、1、4、16ppmの濃度になるように添加し、再度26℃で約6時間、1000rpmの条件で振とう培養を行った。6時間後のプレートの写真を図5に示す。その結果、RDH8P-ALB1の3菌株ともにプロベナゾール4ppmと16ppmの処理カラムでのみ、薬剤処理後6時間と短い時間でありながら、かなり明瞭な赤色色素の蓄積が確認できた。一方で、Guy11株からは赤色色素の蓄積は全く観察できなかった。この4ppmと16ppmの濃度はRDH8P-ALB1と同様にRDH8Pプロモーターの下流に緑色蛍光色素であるEGFPをレポーター遺伝子として連結した場合の蛍光が観察された濃度と一致しており、この赤色色素を用いたレポーター検出が十分に高感度であることを示している。
【0056】
2.各種作用点別化合物を利用した例
YG液体培地に移植したRDH8P-ALB1菌株或いはGuy11野生株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、プロベナゾール、及びプロベナゾールとは作用機構が異なる農薬化合物4種を0.016、0.064、0.25、1、4および16ppmの濃度添加し、再度26℃で6時間、1000rpmの条件で振とう培養を行った。6時間後、培地における赤色色素を目視によって評価した結果を表1(Guy11野生株)及び表2(RDH8P-ALB1菌株)に示した。
【0057】
なお、使用した農薬化合物の作用機構は、FRAC FUNGICIDE RESISTANCE ACTION COMMITTEE発行のFRAC Code List 2010(http://www.frac.info/frac/publication/anhang/FRAC_Code_List_2010.pdf)に記載されている。
【0058】
【表1】

【0059】
【表2】
【0060】
表1及び2において、比較対照区は、化合物を添加していない系である。また、赤色色素の蓄積は、赤色の濃淡に基づいて数値化した。また、表中、赤色の蓄積がない場合「−」とし、赤色の蓄積が薄い場合、中間の場合、濃い場合をそれぞれ「+」、「++」及び「+++」とした。
【0061】
その結果、プロベナゾール4ppmと16ppmの処理カラムでのみ薬剤処理後6時間と短い時間からかなり明瞭な赤色色素の蓄積が確認できた。一方で、それ以外の化合物の処理カラムからは赤色色素の蓄積は観察できなかった。また、Guy11野生株では同濃度のプロベナゾールを添加しても赤色の蓄積は観察されなかった。このことから、ALB1遺伝子を発現制御することにより蓄積する赤色色素を利用したレポーターアッセイシステムが化合物スクリーニングに応用可能であると示された。
【0062】
〔実施例3〕
pCHALB1への電子伝達系阻害化合物誘導型プロモーターAOXPの挿入
ALB1遺伝子の上流のALB1P領域を既知のプロモーター配列で置き換える例として、電子伝達系阻害剤のスクリーニングに用いることのできる、電子伝達系阻害剤存在下で特異的に遺伝子の発現を誘導するできるプロモーターAOXPを用いた場合を以下に示す。
【0063】
電子伝達系化合物誘導プロモーターAOXPの配列はBroad Institute のマグナポルテ・グリセア(Magnaporthe grisea)ゲノムデータベース(http://www.broad.mit.edu/annotation/genome/magnaporthe_grisea/MultiHome.html)からAOX 遺伝子の5’領域の塩基配列情報を取得し、その配列情報を参考に一組のオリゴヌクレオチドを設計した(5‘-CCAAGCTTCCTTCCTCACTA-3’(配列番号11)、5‘-CCAAGCTTAAGACGGTACGTTCAGTGT-3’(配列番号12))。マグナポルテ・グリセアGuy11株のゲノムDNAをテンプレートとしてPCR反応を行った。その後、前述の方法によりpGEM-T Easyベクターに連結した。本ベクターをHindIII処理することにより得たAOXPプロモーター配列を、同じくHindIII処理したpCHALB1ベクターに連結することにより、AOXPによりALB1遺伝子を制御可能なプラスミドベクターを作成した。なお、AOXPプロモーター配列のベクター内への挿入方向は、PCRおよびシークエンスにより確認し、本ベクターをpCHAOXPALB1と命名し、本ベクターを用い前述の実施例2のアグロバクテリウム法で形質転換を行った。得られた菌株をAOXP-ALB1菌株と命名し、本菌へのベクター挿入の確認は、PCR、シークエンス及びサザン解析によって行った。
【0064】
(1) 96-wellマイクロタイタープレートを用いたハイスループットスクリーニングへの利用
1.電子伝達系阻害剤を使用した例
AOXP-ALB1菌株を利用して、96-wellのマイクロタイタープレートを用いアッセイを行った。YG液体培地に移植したAOXP-ALB1菌株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し、26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、電子伝達系阻害剤であるジフルメトリム、ピリベンカルブ及びオリゴマイシンを0、1/256、1/64、1/16、1/4、1、4、16ppmの濃度になるように添加し、再度26℃で約12時間、1000rpmの条件で振とう培養を行った。12時間後のプレートの写真を図6に示す。その結果、ジフルメトリムおよびピリベンカルブ処理カラムでは、低濃度の1/256ppmの処理カラムから赤色色素の蓄積が確認でき、1/4ppm及び1ppmに赤色色素のピークを示していた。またオリゴマイシンの場合は、本剤のEC50値が0.006ppmと低いためか高濃度処理では生育阻害が強いため検出できなかったが、低濃度の1/64ppm処理カラムで明瞭な赤色色素の蓄積を確認することができた。
【0065】
2.各種作用点別化合物を利用した例
YG液体培地に移植したAOXP-ALB1菌株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、ジフルメトリム及びピリベンカルブの電子伝達系阻害剤、及びそれらとは作用機構が異なる農薬化合物3種を0.016、0.064、0.25、1、4および16ppmの濃度添加し、再度26℃で12時間、1000rpmの条件で振とう培養を行った。12時間後、培地における赤色色素を目視によって評価した結果を表3に示した。
【0066】
【表3】
【0067】
なお、表3において、比較対照区は、化合物を添加していない系である。また、赤色色素の蓄積は、赤色の濃淡に基づいて数値化した。また、表中、赤色の蓄積がない場合「−」とし、赤色の蓄積が薄い場合、中間の場合、濃い場合をそれぞれ「+」、「++」及び「+++」とした。
【0068】
その結果、予想どおりジフルメトリム及びピリベンカルブといった電子伝達系阻害剤の処理カラムでかなり明瞭な赤色色素の蓄積が確認できた。一方で、それ以外の化合物の処理カラムからは赤色色素の蓄積はほとんど観察できなかった。このことから、ALB1遺伝子を発現制御することによる蓄積する赤色色素を利用したレポーターアッセイシステムが化合物スクリーニングに応用可能であると考えられ、今回のように実際に電子伝達系阻害剤のスクリーニングに利用可能であることが確認できた。
【0069】
〔実施例4〕
pCHALB1へのエルゴステロール生合成阻害化合物誘導型プロモーターCYP51Pの挿入
ALB1遺伝子の上流のALB1P領域を既知のプロモーター配列で置き換える例として、エルゴステロール合成阻害剤のなかでも、Cytochrome P450ステロール14αデメチラーゼ阻害剤のスクリーニングに用いることのできる、ステロール14αデメチラーゼ阻害剤の存在下で特異的に遺伝子の発現を誘導するできるプロモーターCYP51Pを用いた場合を以下に示す。
【0070】
ステロール14αデメチラーゼ阻害剤誘導プロモーターCYP51Pの配列はBroad Instituteのマグナポルテ・グリセア(Magnaporthe grisea)ゲノムデータベース(http://www.broad.mit.edu/annotation/genome/magnaporthe_grisea/MultiHome.html)からCytochrome P450ステロール14αジメチラーゼ(MGG_04628)の5’領域の塩基配列情報を取得し、その配列情報を参考に一組のオリゴヌクレオチドを設計した(5‘-CCAAGCTTATGTGAGCCATCTTCGGATT-3’(配列番号13)、5‘-CCAAGCTTGACGATCGGGTAAGGAGACT-3’(配列番号14))。マグナポルテ・グリセアGuy11株のゲノムDNAをテンプレートとしてPCR反応を行った。その後、前述の方法によりpGEM-T Easyベクターに連結した。本ベクターをHindIII処理することにより得たCYP51Pプロモーター配列を、同じくHindIII処理したpCHALB1ベクターに連結することにより、CYP51PによりALB1遺伝子を制御可能なプラスミドベクターを作成した。なお、CYP51Pプロモーター配列のベクター内への挿入方向は、PCRおよびシークエンスにより確認し、本ベクターをpCH CYP51PALB1と命名し、本ベクターを用い前述の実施例2のアグロバクテリウム法で形質転換を行った。得られた菌株をCYP51P-ALB1菌株と命名し、本菌へのベクター挿入の確認は、PCR、シークエンス及びサザン解析によって行った。
【0071】
(1) 96-wellマイクロタイタープレートを用いたハイスループットスクリーニングへの利用
1.エルゴステロール生合成阻害剤を使用した例
CYP51P-ALB1菌株を利用して、96-wellのマイクロタイタープレートを用いアッセイを行った。YG液体培地に移植したCYP51P-ALB1菌株の胞子懸濁液約5×105 / mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し、26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、ステロール14αデメチラーゼ阻害剤であるオキスポコナゾール、イプコナゾール、トリホリンおよびプロチオコナゾールを0、1/256、1/64、1/16、1/4、1、4、16ppmの濃度になるように添加し、再度26℃で約12時間、1000rpmの条件で振とう培養を行った。12時間後のプレートの写真を図7に示す。その結果、オキスポコナゾール、イプコナゾール、トリホリンおよびプロチオコナゾールともに1ppm〜16ppmの濃度では明瞭な赤色色素の蓄積が確認できた。
【0072】
2.各種作用点別化合物を利用した例
YG液体培地に移植したCYP51P-ALB1菌株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、オキスポコナゾール及びトリホリンのステロール14αデメチラーゼ阻害剤、及びそれらとは作用機構が異なる農薬化合物3種を0.016、0.064、0.25、1、4および16ppmの濃度添加し、再度26℃で12時間、1000rpmの条件で振とう培養を行った。12時間後、培地における赤色色素を目視によって評価した結果を表4に示した。
【0073】
【表4】
【0074】
なお、表4において、比較対照区は、化合物を添加していない系である。また、赤色色素の蓄積は、赤色の濃淡に基づいて数値化した。また、表中、赤色の蓄積がない場合「−」とし、赤色の蓄積が薄い場合、中間の場合、濃い場合をそれぞれ「+」、「++」及び「+++」とした。
【0075】
その結果、予想どおりオキスポコナゾール及びトリホリンのステロール14αデメチラーゼ阻害剤の処理カラムでかなり明瞭な赤色色素の蓄積が確認できた。一方で、それ以外の化合物の処理カラムからは赤色色素の蓄積はほとんど観察できなかった。このことから、ALB1遺伝子を発現制御することによる蓄積する赤色色素を利用したレポーターアッセイシステムが電子伝達系阻害剤とともにエルゴステロール合成阻害剤でも利用できたことから、汎用性があることが確認できた。
【技術分野】
【0001】
本発明は、糸状菌において機能するレポーター遺伝子及び当該レポーター遺伝子を利用した方法、特に当該レポーター遺伝子を利用した方法に関する。
【背景技術】
【0002】
レポーター遺伝子とは、宿主細胞に導入され、発現の有無を可視化できる遺伝子である。すなわち、レポーター遺伝子が発現した宿主細胞では、当該レポーター遺伝子の産物に起因する発光や蛍光が観察される。
【0003】
したがって、レポーター遺伝子を所定の遺伝子と組み換えた組換え体では、レポーター遺伝子の産物に起因する発光や蛍光を観察することで、当該所定の遺伝子の発現パターンを解析することができる。例えば、ある化合物を組換え体に曝露する等の特定の刺激を組換え体に与えたときに発光や蛍光が観察されたとすると、上記所定の遺伝子は、この刺激により発現誘導されるといった知見を得ることができる。あるいは、薬剤応答性プロモーターの下流にレポーター遺伝子を連結した組換え体を使用することで、当該薬剤応答性プロモーターに作用する化合物を新たにスクリーニングすることができる。あるいは、機能未知のプロモーターの下流にレポーター遺伝子を連結した組換え体を使用することで、発現調節作用を有する公知物質に応答性を示す新規なプロモーターをスクリーニングすることができる。
【0004】
一方、創農薬研究プロセスの中で最も重要な過程は、その新薬候補化合物であるリード化合物を見つけ出すことから始まる。しかしながら膨大な種類の化合物で構成される化合物ライブラリーからリード化合物を探索するためには、自動化されたロボットなどを用い、大量の化合物の中から、短時間で目的とするターゲットを選別するハイスループットスクリーニング(HTSスクリーニング)を行う事が望まれている。細胞レベルでのハイスループットスクリーニングにおいて特定の遺伝子発現を評価する場合には、レポーターアッセイがよく用いられている。レポーター遺伝子としては、オワンクラゲ由来のGFP(非特許文献1、非特許文献2)、ホタル由来のルシフェラーゼ、大腸菌由来のGUS(非特許文献3、非特許文献4)などが一般的に使用されている。
【0005】
また、糸状菌の一般的な遺伝子研究分野において、これらのレポーター遺伝子はGFPをはじめとして頻繁に使用されており細胞内での良好な蛍光、発光、発色が確認されている。しかし実際には、糸状菌は、菌糸を絡み合わせながら生育していくという特徴を有しているため、レポーター遺伝子に起因する発光や蛍光の検出感度が十分でないといった問題がある。特に、ハイスループットを目的として、複数のウェルを有するプレートを用いた系では、各ウェルに張り込まれた培地において糸状菌の均一な生育が見られず、レポーター遺伝子に起因する発光や蛍光の検出感度が著しく低下するといった問題がある。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC (1994) Green fluorescent protein as a marker for gene expression. Science 263: 802-805
【非特許文献2】Tsien RY (1998) The green fluorescent protein. Ann. Rev. Biochem 67: 509-544
【非特許文献3】Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6: 3901-3907
【非特許文献4】Jefferson RA, Burgess SM, Hirsh D (1986) β-glucuronidase from Escherichia coli as a gene-fusion marker. Proc. Natl. Acad. Sci. USA 83: 8447-8451
【発明の概要】
【発明が解決しようとする課題】
【0007】
そこで、本発明は、糸状菌において使用でき、検出感度に優れた新規なレポーター遺伝子を提供することを目的とし、当該レポーター遺伝子の発現を検出することを含む方法を提供することを目的とする。
【課題を解決するための手段】
【0008】
本発明者らは、上述した目的を達成するため鋭意検討した結果、糸状菌を培養した培地中に蓄積されるフラビオリンを検出することで、ポリケタイド合成酵素遺伝子の発現を可視化できることから、当該ポリケタイド合成酵素遺伝子を糸状菌における新規なレポーター遺伝子として使用できることを見いだし、本発明を完成するに至った。
【0009】
本発明は以下を包含する。
【0010】
(1) ポリケタイド合成酵素遺伝子をレポーター遺伝子として有する糸状菌を使用し、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定する方法。
(2) 上記ポリケタイド合成酵素遺伝子は、配列番号2に示すアミノ酸配列を含むタンパク質をコードする、又は配列番号2に示すアミノ酸配列に対して90%以上の類似性を有するアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードすることを特徴とする(1)記載の方法。
(3) 上記ポリケタイド合成酵素遺伝子を、所定の因子に対して転写活性を変動する転写制御領域の制御下に連結してレポーター遺伝子とし、上記糸状菌に対して刺激を与えた後における培地中のフラビオリン量が有意に変動する場合、当該刺激が上記転写制御領域に対して上記所定の因子と同等に作用すると判断することを特徴とする(1)記載の方法。
(4) 上記刺激は、上記糸状菌に対する供試化合物への曝露であることを特徴とする(3)記載の方法。
(5) 上記転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする(3)記載の方法。
(6) 上記転写制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする(3)記載の方法。
(7) 上記転写制御領域として植物病害抵抗性付与剤誘導型プロモーターを使用して植物病害抵抗性付与剤の候補物質をスクリーニングする、上記転写制御領域として電子伝達系阻害剤誘導型プロモーターを使用して電子伝達系阻害剤の候補物質をスクリーニングする、又は上記転写制御領域としてエルゴステロール生合成阻害剤誘導型プロモーターを使用してエルゴステロール生合成阻害剤の候補物質をスクリーニングすることを特徴とする(3)記載の方法。
(8) 上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結してレポーター遺伝子とし、機能既知である発現制御領域の転写活性を変動させる所定の因子を上記糸状菌に作用させたときに、培地中のフラビオリン量が有意に変動する場合、上記機能未知の発現制御領域が上記機能既知の発現制御領域と同等の応答性を示すと判断することを特徴とする(1)記載の方法。
(9) 上記所定の因子は、特定のプロモーターに対して転写活性を促進又は抑制する化合物であることを特徴とする(8)記載の方法。
(10) 上記所定の因子は、植物病害抵抗性付与能を有する化合物、電子伝達系阻害能を有する化合物及びエルゴステロール生合成阻害能を有する化合物からなる群より選ばれる1つの化合物であることを特徴とする(8)記載の方法。
【0011】
(11) 上記機能未知の転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする(8)記載の方法。
(12) 上記所定の因子として植物病害抵抗性付与剤を使用して植物病害抵抗性付与剤誘導型プロモーターをスクリーニングする、上記所定の因子として電子伝達系阻害剤を使用して電子伝達系阻害剤誘導型プロモーターをスクリーニングする、又は上記所定の因子としてエルゴステロール生合成阻害剤を使用してエルゴステロール生合成阻害剤誘導型プロモーターをスクリーニングすることを特徴とする(8)記載の方法。
(13) 上記ポリケタイド合成酵素遺伝子で解析対象の遺伝子を組み換え、培地に生産されるフラビオリンを指標として解析対象の遺伝子の発現パターンを解析することを特徴とする(1)記載の方法。
(14) 導入対象の目的遺伝子と、恒常発現型プロモーターに連結された上記ポリケタイド合成酵素遺伝子とを上記糸状菌に導入し、培地中のフラビオリン量が有意に大である場合、上記目的遺伝子が発現していると判断することを特徴とする(1)記載の方法。
(15) 複数のウェルを有するプレートを使用して当該各ウェルに上記糸状菌を培養し、上記フラビオリンに由来する赤色発色を検出する検出手段を使用して複数のウェルを一括して観察することを特徴とする(1)乃至(14)いずれか一つに記載の方法。
(16) 機能既知又は機能未知の転写制御領域と、当該転写制御領域の制御下に発現するポリケタイド合成酵素遺伝子とを含む核酸構築物。
(17) 上記機能既知の発現制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする(16)記載の核酸構築物。
(18) (16)又は(17)記載の核酸構築物を含む発現ベクター。
(19) (16)又は(17)記載の核酸構築物で形質転換された組み換え糸状菌。
(20) いもち病菌の組換え体であることを特徴とする(19)記載の組み換え糸状菌。
【発明の効果】
【0012】
本発明によれば、糸状菌に対して使用することができ、非常に簡便且つ高感度に発現を可視化できる新規なレポーター遺伝子を提供することができる。したがって、本発明に係るレポーター遺伝子を使用することによって、一般的にレポーター遺伝子の発現を可視化する工程を含む様々な手法を糸状菌に適用することができる。
【図面の簡単な説明】
【0013】
【図1】糸状菌のDHN−メラニン合成経路及び関連酵素を示す概略図である。
【図2】内在ALB1プロモーターALB1Pを既知プロモーターへ置き換えるプラスミドベクターpCHALB1の概略構成図である。
【図3】pCHRDH8PALB1を利用した相同組み換えによるALB1PのRDH8Pへの変換を示す概略図である。
【図4】プロベナゾールでの誘導の有無によるRDH8P-ALB1菌株のYG液体培地上での形態を示す特性図である。
【図5】RDH8Pプロベナゾール応答株を用いた赤色色素フラビオリンの蓄積による遺伝子発現の可視化例とRDH8Pプロベナゾール応答型EGFP株との比較を示す特性図である。
【図6】AOXP電子伝達系阻害剤応答株を用いた赤色色素フラビオリンの蓄積による遺伝子発現の可視化例を示す特性図である。
【図7】CYP51Pエルゴステロール生合成阻害剤応答株を用いた赤色色素フラビオリンの蓄積による遺伝子発現の可視化例を示す特性図である。
【発明を実施するための形態】
【0014】
以下、本発明を図面を参照して詳細に説明する。
レポーター遺伝子
糸状菌において機能する新規レポーター遺伝子は、図1に示すDHN−メラニン合成経路において、アセチルCoA及びマロニルCoAからテトラヒドロキシナフタレン(1,3,6,8−T4HN)の生合成に関与するポリケタイド合成酵素をコードする遺伝子である。ポリケタイド合成酵素遺伝子が発現すると、糸状菌においてテトラヒドロキシナフタレン(1,3,6,8−T4HN)が合成される。このテトラヒドロキシナフタレン(1,3,6,8−T4HN)は、自然酸化により赤色色素であるフラビオリンに変換される。
【0015】
なお、図1に示すように、DHN−メラニン合成経路において、テトラヒドロキシナフタレン(1,3,6,8−T4HN)は、THN還元酵素による還元及びシタロン脱水酵素による脱水によりジヒドロキシナフタレン(1,8−DHN)となる。そして、ジヒドロキシナフタレン(1,8−DHN)が酸化酵素により重合し、黒色のメラニンが生成される。従って、イネいもち病菌などの糸状菌は、通常、黒色となる。例えば、DHN−メラニン合成経路上のTHN還元酵素やシタロン脱水酵素の阻害剤の存在下では、メラニン生合成が阻害され、1,3,6,8−T4HNが蓄積し、蓄積した1,3,6,8−T4HNの自然酸化によりフラビオリンが合成されることになる。したがって、例えば、THN還元酵素阻害剤であるトリシクラゾールの存在下において、イネいもち病菌は赤色を呈する。
【0016】
ただし、イネいもち病菌において、DHN−メラニン合成経路に関与する遺伝子群は、感染器官である付着器形成時に特に強く発現が誘導され、栄養増殖期では培養開始から少なくとも3日間経過しなければ発現が誘導されない。このため、野生型の糸状菌を通常の栄養増殖させている場合、培養開始から約3日までであれば、内在するメラニン生合成に関与する遺伝子群が発現することによる1,3,6,8−T4HNの蓄積及びフラビオリンの合成は生じない。このため、ポリケタイド合成酵素遺伝子を糸状菌におけるレポーター遺伝子として使用することができ、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定(可視化)することができる。
【0017】
ここで、ポリケタイド合成酵素遺伝子としては、特に限定されないが、特に糸状菌に由来することが好ましい。また、ポリケタイド合成酵素遺伝子としては、詳細を後述する、宿主となる糸状菌に由来することが好ましい。さらに、ポリケタイド合成酵素遺伝子としては、一般的に植物病原糸状菌として知られる糸状菌に由来することがより好ましい。なお、植物病原糸状菌におけるDHN−メラニン合成経路及びメラニン合成に関与する遺伝子群については、例えば、Richard J. Howard et al., 1996, Annual Review of Microbiology Vol. 50: 491-512及びKim Langfelder et al., 2003, Fungal Genet. Biol. 38:143-158を参照することができる。すなわち、使用する宿主に応じて、適宜、上述したような文献を参照することで、好ましいポリケタイド合成酵素遺伝子を準備して使用することができる。
【0018】
一例としては、イネいもち病菌に由来するポリケタイド合成酵素遺伝子を挙げることができる。イネいもち病菌に由来するポリケタイド合成酵素遺伝子のコーディング領域の塩基配列を配列番号1に示し、イネいもち病菌に由来するポリケタイド合成酵素遺伝子によりコードされるポリケタイド合成酵素のアミノ酸配列を配列番号2に示す。
【0019】
また、ポリケタイド合成酵素遺伝子としては、配列番号2に示すアミノ酸配列からなるタンパク質をコードする遺伝子に限定されず、例えば配列番号2に示すアミノ酸配列に対して例えば90%以上、好ましくは95%以上、より好ましくは98%以上の類似性を有するアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードする遺伝子であっても良い。ここで、類似性とは、例えばblastアルゴリズムを実装したコンピュータプログラムを用いてデフォルトの設定で求められる値を意味する。
【0020】
さらに、ポリケタイド合成酵素遺伝子としては、配列番号2に示すアミノ酸配列に対して1個又は複数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードする遺伝子であっても良い。ここで、複数個のアミノ酸とは、2個から230個、好ましくは2個から150個、より好ましくは2個から100個、さらに好ましくは2個から50個、特に好ましくは2個から20個のアミノ酸を意味する。
【0021】
さらにまた、ポリケタイド合成酵素遺伝子としては、配列番号1に示す塩基配列を有する遺伝子の少なくとも一部に対して、ストリンジェントな条件下でハイブリダイズするポリヌクレオチドを含み、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードするものであってもよい。ここで、ストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、45℃、6×SSC(塩化ナトリウム/クエン酸ナトリウム)でのハイブリダイゼーション、その後の50〜65℃、0.2〜1×SSC、0.1%SDSでの洗浄が挙げられ、或いはそのような条件として、65〜70℃、1×SSCでのハイブリダイゼーション、その後の65〜70℃、0.3×SSCでの洗浄を挙げることができる。ハイブリダイゼーションは、J. Sambrook et al. Molecular Cloning, A Laboratory Manual,2nd Ed., Cold Spring Harbor Laboratory(1989)に記載されている方法等、従来公知の方法で行うことができる。
【0022】
なお、配列番号2とは異なるアミノ酸配列をコードするポリケタイド合成酵素遺伝子、配列番号1とは異なる塩基配列を有するポリケタイド合成酵素遺伝子は、当該技術分野で公知の手法によってイネいもち病菌に由来するポリケタイド合成酵素遺伝子を改変することによって得ることができる。塩基配列に変異を導入するには、Kunkel法またはGapped duplex法等の公知手法又はこれに準ずる方法により行うことができ、例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutant-KやMutant-G(何れも商品名、TAKARA Bio社製))等を用いて、あるいはLA PCR in vitro Mutagenesisシリーズキット(商品名、TAKARA Bio社製)を用いて変異が導入される。また、変異導入方法としては、EMS(エチルメタンスルホン酸)、5-ブロモウラシル、2-アミノプリン、ヒドロキシルアミン、N-メチル-N’-ニトロ-Nニトロソグアニジン、その他の発ガン性化合物に代表されるような化学的変異剤を使用する方法でも良いし、X線、アルファ線、ベータ線、ガンマ線、イオンビームに代表されるような放射線処理や紫外線処理による方法でも良い。
【0023】
宿主
上述したポリケタイド合成酵素遺伝子をレポーター遺伝子として使用できる糸状菌としては、特に限定されず使用することができる。また、糸状菌としては、菌糸を形成する真菌であれば特に限定されず使用することができる。上述したレポーター遺伝子として糸状菌由来のポリケタイド合成酵素遺伝子を使用する場合、宿主である糸状菌の菌体内において良好に発現することができる。特に、ポリケタイド合成酵素遺伝子の由来と宿主とが同属の糸状菌であることが好ましく、同種の糸状菌であることがより好ましく、同株の糸状菌であることが更に好ましい。
【0024】
さらに、糸状菌としては、例えば植物病原性糸状菌を使用することが好ましい。植物病原性糸状菌としては、特に限定されないが、例えば、Botrytis属(=Botryotinia属)、Rosellinia属、Aspergillus属(=Eurotium属など)、Phytophthora属、Pythium属、Sclerotinia属、Colletotrichum属(=Glomerella属)、Rhizoctonia属(=Thanatephorus属)、Helicobasidium属、Fusarium属(=Gibberella属、Haematonectoria属、Nectoria属、Calonectoria属)、Magnaporthe属(=Pyricularia属)、Alternaria属、Puccinia属(=Aecidium属)、Uromyces属、Microsphaera属、Blumeria属、Erysiphe属(=Golovinomyces属)、Oidium属(=Sawadae属)、Sphaerotheca属(=Podosphaera属)、Phyllactinia属(=Ovulariopsis属)、Uncinula属(=Golovinomyces属、Erysiphe属)、Podosphaera属(=Sphaerotheca属)、Pythium属、Peronospora属、Ustilago属、Cochliobolus属(=Curvularia属、Bipolaris属、Helminthosporium属)、Cladosporium属(=Mycosphaerella属、Venturia属)、Mycosphaerella属(=Ascochyta属、Cercospora属、Phyllosticta属、Septoria属)、Phomopsis属(=Diaporthe属)、Phoma属(=Leptosphaeria属、Mycosphaerella属)、Venturia属(=Fusicladium属、Spilocaea属)、Claviceps属(=Ustilaginoidea属、Sphaecelia属)、Calonectria属(=Cylindrocladium属)、Verticillium属(=Cosmospora属、Hypocereales属など)、Penicillium属(=Eupenicillium属、Talaromyces属)、Elsinoe属(=Sphaceloma属)、Didymella属(=Ascochyta属、Phoma属など)若しくはRhizopus属に属する菌類を挙げることができる。
【0025】
より具体的に、イネのいもち病菌(Magnaporthe grisea シノニム:Pyricularia oryzae)、ごま葉枯病菌(Cochliobolus miyabeanus)、紋枯病菌(Rhizoctonia solani)、馬鹿苗病菌(Gibberella fujikuroi)、各種作物の苗立枯病菌(Fusarium spp.、Pythium spp.、Rhizopus spp.、Trichoderma sp.)、灰色かび病菌(Botrytis cinerea)、菌核病菌(Sclerotinia sclerotiorum)、ウリ類の炭そ病菌(Colletotrichum lagenarium)、シバのブラウンパッチ病菌(Rhizoctonia solani)等を挙げることができる。
【0026】
レポーター遺伝子の利用形態
上述したポリケタイド合成酵素遺伝子は、上述した宿主においてレポーター遺伝子として機能する。したがって、上述したポリケタイド合成酵素遺伝子は、従来公知のレポーター遺伝子を使用する各種遺伝子工学的手法、分子生物学的手法或いは細胞工学的手法においてレポーター遺伝子として使用することができる。ここでレポーター遺伝子を使用する各種方法は、何ら限定されず、如何なる目的、規模の実験方法及び工業的方法をも含む意味である。
【0027】
通常、ある遺伝子をレポーター遺伝子として使用するには、当該遺伝子の発現を検出、好ましくは可視化できなければならない。ポリケタイド合成酵素遺伝子を外的に導入していない糸状菌では培養開始から約3日間を経過しなければ培地中にフラビオリンを合成しない。しかしながら、ポリケタイド合成酵素遺伝子を外的に導入して発現を誘導すると、比較的早期に培地中にフラビオリンを生産することとなる。したがって、外的に導入したポリケタイド合成酵素遺伝子に起因する、培地中のフラビオリンを検出することで当該遺伝子の発現を検出することができ、ポリケタイド合成酵素遺伝子を糸状菌における有効なレポーター遺伝子として使用することができる。
【0028】
培地中に生産されたフラビオリンを検出するには、特に限定されないが、フラビオリンの赤色を検出すればよい。フラビオリンの赤色は、目視によって検出してもよいし、赤色を光学的に検出できる装置を用いて検出しても良い。このような装置としては、例えば、分光光度計、分光蛍光光度計等を使用することができる。
【0029】
また、フラビオリンによる赤色の濃淡(強度)から、ポリケタイド合成酵素遺伝子の発現の強弱を判定することもできる。すなわち、培地中の赤色が濃い場合にはポリケタイド合成酵素遺伝子が高発現していると判断し、培地中の赤色が薄い場合にはポリケタイド合成酵素遺伝子の発現量が低いと判断することができる。
【0030】
特に、フラビオリンは、レポーター遺伝子を導入した糸状菌を培養した培地中に生産されるため、GFPやGUS等の公知のレポーター遺伝子と比較して、より高感度に検出することができる。GFPやGUS等の公知のレポーター遺伝子を使用した場合、細胞内において蛍光や発色を検出しなければならないため、菌糸を絡み合わせながら生育する糸状菌では非常に検出感度が低くなる。これに対して、上述したポリケタイド合成酵素遺伝子をレポーター遺伝子として使用する場合には、培地中のフラビオリンを検出するため、糸状菌を宿主としても非常に優れた検出感度を達成することができる。
【0031】
したがって、上述したポリケタイド合成酵素遺伝子をレポーター遺伝子とし、宿主として糸状菌を使用する場合には、例えば、複数のウェルを有するプレートを用いて、大量のサンプルを処理することができる。すなわち、レポーター遺伝子としてポリケタイド合成酵素遺伝子を導入した糸状菌を各ウェルに張り込んだ培地にて培養し、プレートリーダーのような大量測定機器により各ウェルにおけるフラビオリンの赤色色素を一括して又は順次に検出する。これにより、レポーター遺伝子の発現をハイスループットで測定することができる。
【0032】
より具体的に、レポーター遺伝子の利用形態としては、特定の遺伝子の発現をレポーター遺伝子により解析する手法が挙げられる。ここで、特定の遺伝子とは、なんら限定されるものではなく、宿主となる糸状菌に内在する各種の遺伝子を挙げることができる。この利用形態では、先ず、上述したポリケタイド合成酵素遺伝子を当該特定の遺伝子と置換するように導入した組み換え糸状菌を準備する。得られた組み換え糸状菌を通常の培地にて培養し、培地に生産されたフラビオリンを検出することでレポーター遺伝子の発現を測定する。例えば、培養条件を異にして組み換え糸状菌を培養し、一方の条件下においてフラビオリンを検出し、他方の条件下においてフラビオリンを検出しなかった場合、上記特定の遺伝子は一方の条件下において特異的に発現されるものと理解することができる。ここで、培養条件とは、温度条件や、培地組成条件、pH条件、化合物の存在・非存在条件等を挙げることができるが、これらに限定されるものではない。
【0033】
また、組み換え糸状菌を培養しながら経時的にフラビオリンの生産量を測定することで、上記特定の遺伝子の発現パターンを解析することができる。この系においては、培養経過に伴って培養条件を種々に変更することができる。例えば、培養開始から所定期間が経過した後に、所定の化合物を培養系に添加したり、温度条件を変更したりすることができる。
【0034】
ところで、レポーター遺伝子の利用形態としては、上記ポリケタイド合成酵素遺伝子を、所定の因子に対して転写活性を変動する転写制御領域の制御下に連結した組み換え糸状菌を用いて、当該転写制御領域に影響を与えうる新規な因子をスクリーニングすることができる。すなわち、この組み換え糸状菌に対して新規な因子を作用させたときに培地中にフラビオリンが生産されていたとすれば、この新規な因子は上記発現制御領域に対して作用し、下流の遺伝子を発現誘導することが判明する。
【0035】
ここで、転写制御領域とは、例えばプロモーターや、その他のシス配列(シス調節配列)を全て包含する意味である。転写制御領域には、プロモーター領域、エンハンサー領域、サイレンサー領域といった転写に関与する因子(転写因子や、アクチベータータンパク質及びサイレンサータンパク質)が結合する領域を意味する。また、所定の因子としては、例えば、化合物、温度、pHといった一般的に遺伝子の転写を制御する種々の因子を挙げることができる。
【0036】
したがって、「所定の因子に対して転写活性を変動する転写制御領域」としては、所定の化合物に応答性を示すプロモーター、所定の温度条件で発現誘導するプロモーター等を挙げることができる。具体的に、生理活性物質をスクリーニングする際には、転写制御領域として、従来公知の生理活性物質応答性プロモーターを使用する。生理活性物質応答性プロモーターとしては、特に制限はなく、スクリーニングしたい生理活性物質に応答するプロモーターから適宜、選択すれば良い。生理活性物質として、農薬、医薬、動物薬等の候補化合物をスクリーニングする場合には、植物病原菌、人畜の病原菌のように防除すべき微生物に有効な作用を有する生理活性物質に応答するプロモーターから選ぶのが良い。また、生理活性物質応答性プロモーターとしては、レポーター遺伝子の発現による代謝産物の測定結果を明確にするために、目的の生理活性物質による活性化がはっきりしていてノイズの少ないプロモーターを使用することが好ましい。
【0037】
さらに具体的に、「所定の因子に対して転写活性を変動する転写制御領域」としては、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を挙げることができる。このようなプロモーターとしては、例えば、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターを挙げることができる。
【0038】
例えば、植物病害抵抗性付与剤誘導型プロモーターの下流にポリケタイド合成酵素遺伝子を連結した組み換え糸状菌に対して供試化合物を作用させたとき、培地中にフラビオリンが検出されたとすれば、当該供試化合物は、植物病害抵抗性付与剤誘導型プロモーターを活性化する作用を有していることが判る。この結果から、当該供試化合物は、植物病害抵抗性付与剤の候補として特定することができる。なお、植物病害抵抗性付与剤とは、植物に対して作用して病原体の感染に対する抵抗性を付与するようなシグナル伝達(サリチル酸経路など)を活性化する物質である。植物病害抵抗性付与剤としては、特に限定されないが、プロベナゾールを挙げることができる。
【0039】
また、例えば、電子伝達系阻害剤誘導型プロモーターの下流にポリケタイド合成酵素遺伝子を連結した組み換え糸状菌に対して供試化合物を作用させたとき、培地中にフラビオリンが検出されたとすれば、当該供試化合物は、電子伝達系阻害剤誘導型プロモーターを活性化する作用を有していることが判る。この結果から、当該供試化合物は、電子伝達系阻害剤の候補として特定することができる。なお、電子伝達系阻害剤とは、糸状菌における電子伝達系を阻害することで糸状菌を衰弱又は死滅若しくは糸状菌の感染力を低下させることが知られている物質である。電子伝達系阻害剤としては、特に限定されないが、複合体1阻害剤であるジフルメトリム、テブフェンピラド、複合体3阻害剤であるクレソキシムメチル、ピリベンカルブ、F1Fo ATPase阻害剤であるオリゴマイシンを挙げることができる。また、電子伝達系阻害剤誘導型プロモーターとしては、特に限定されないがAOXプロモーター(AOXP)を使用することができる。AOXプロモーターは、ミトコンドリアにおける呼吸において、電子伝達系複合体2以外の電子伝達系阻害剤に特異的に応答することが知られている。(Yukioka H, Inagaki S, Tanaka R, Katoh K, Miki N, Mizutani A, Masuko M. (1998) Transcriptional activation of the alternative oxidase gene of the fungus Magnaporthe grisea by a respiratory-inhibiting fungicide and hydrogen peroxide. Biochim Biophys Acta. 1442:161-169)
【0040】
さらに、例えば、エルゴステロール生合成阻害剤誘導型プロモーターの下流にポリケタイド合成酵素遺伝子を連結した組み換え糸状菌に対して供試化合物を作用させたとき、培地中にフラビオリンが検出されたとすれば、当該供試化合物は、エルゴステロール生合成阻害剤誘導型プロモーターを活性化する作用を有していることが判る。この結果から、当該供試化合物は、エルゴステロール生合成阻害剤の候補として特定することができる。なお、エルゴステロール生合成阻害剤とは、糸状菌におけるエルゴステロール生合成経路を阻害することで糸状菌を衰弱又は死滅若しくは糸状菌の感染力を低下させることが知られている物質である。エルゴステロール生合成阻害剤としては、特に限定されないが、オキスポコナゾール、イプコナゾール、トリホリン、プロチオコナゾールを挙げることができる。また、エルゴステロール生合成阻害剤誘導型プロモーターとしては、特に限定されないがCYP51プロモーター(CYP51P)を使用することができる。CYP51プロモーターは、シトクロムP450のひとつで、ステロール生合成経路のラノステロール構造の14-脱メチル化酵素として知られているCYP51遺伝子のプロモーターである。(Ma Z, Proffer TJ, Jacobs JL, Sundin GW. (2006) Overexpression of the 14alpha-demethylase target gene (CYP51) mediates fungicide resistance in Blumeriella jaapii. Appl Environ Microbiol. 72:2581-2585)
【0041】
ところで、レポーター遺伝子の利用形態としては、上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結した組み換え糸状菌を用いて、当該転写制御領域の特性を評価することができる。言い換えれば、上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結した組み換え糸状菌を用いて、公知の因子に対して応答性を示す新規な発現制御領域をスクリーニングすることができる。ここで、機能未知の発現制御領域とは、例えば糸状菌のゲノムに存在する遺伝子のコーディング領域の上流に存在し、当該遺伝子の発現を制御しているものの、どのようなシグナルに依存して発現誘導若しくは発現抑制しているのか不明な領域を意味する。機能未知の発現制御領域としては、コーディング領域の上流に存在する例えば2000塩基、好ましくは1000塩基、より好ましくは500塩基を単離して使用することができる。
【0042】
より具体的に、公知の因子としては、例えば、上述したような化合物、温度、pHといった一般的に遺伝子の転写を制御する種々の因子を挙げることができる。更に具体的には、上述したような、植物病害抵抗性付与能を有する化合物、電子伝達系阻害能を有する化合物或いはエルゴステロール生合成阻害能を有する化合物を使用することによって、新規な植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター或いはエルゴステロール生合成阻害剤誘導型プロモーターをスクリーニングすることができる。
【実施例】
【0043】
以下、実施例により本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0044】
〔実施例1〕
いもち病菌DHN-メラニン生合成に関わるポリケタイド合成酵素遺伝子の発現制御
DHN-メラニン生合成に関わるポリケタイド合成酵素ALB1遺伝子の発現を誘導し、その派生物である赤色色素フラビオリンを合成させることにより、その赤色色素の発色がハイスループットとして利用可能なレポーターであるか否かを調査することを目的に、ALB1遺伝子の発現を既知のプロモーターの制御下に置く実験を試みた。本来ならALB1遺伝子全長の開始コドンより上流領域に既知プロモーターを配置するような構造のプラスミドベクターを作成し、それを宿主へ導入することが望ましいが、いもち病菌ALB1遺伝子の推定全長が6771bpと巨大なため、内在のALB1遺伝子の上流プロモーター領域ALB1Pを他のプロモーターへ置き換える方法をとることにした。ポリケタイド合成酵素遺伝子ALB1の開始コドンより上流のプロモーター領域を相同組み換えによって欠失させ、そこに既知の様々なプロモーター断片を挿入するためのベクターの作成方法および実際に各種プロモーターを挿入したベクターでいもち病菌を形質転換した場合の形質転換体の形状について以下に示す。
【0045】
1)相同組み換えに用いるALB1遺伝子の5’領域およびALB1遺伝子断片のクローニング
Broad Institute のマグナポルテ・グリセア(Magnaporthe grisea)ゲノムデータベース(http://www.broad.mit.edu/annotation/genome/magnaporthe_grisea/MultiHome.html)からALB1遺伝子とその5’領域の塩基配列情報を取得し、その配列情報を参考にALB1の5’領域1440bpを増幅するプライマーペアとして(5‘-CCGAATTCAAGAATGCTTGGACCACACC-3’(配列番号3)及び5‘-ATGGATCCGTACATTTCCGGTTGGATGG-3’(配列番号4))を、またALB1遺伝子の開始コドンから1174bpの断片を増幅するプライマーペアとして(5‘-AAAGATCTGACCATGGCGGACAAGCTGGCTTT-3’(配列番号5)及び5’-GCACTAGTGGATGATGAGGCACGACTTT-3’(配列番号6))のそれぞれのオリゴヌクレオチドを設計した。なおプライマー内に示した下線部分は制限酵素の配列であり、GAATTCはEcoRI、GGATCCはBamHI、AGATCTはBglII、ACTAGTはSpeIサイトを示す。
【0046】
これらのプライマーを用いてマグナポルテ・グリセアGuy11株のゲノムDNAをテンプレートとしてPCR反応を行った。増幅反応は、94℃で3分間鋳型DNAを変性し、94℃で30秒間、55℃で30秒間、72℃で2分間保持するサイクルを35サイクルおこなった後、72℃で5分間完全伸長させ、4℃で保持した。PCR用装置は、PCR Thermal Cycler PERSONAL (タカラバイオ社製)を用いた。このPCRによる増幅断片をアガロース電気泳動にて確認を行ったところ、ALB1の5'領域およびALB1遺伝子断片ともに、予想される1440塩基対、1174塩基対付近の大きさの遺伝子増幅が見られた。この増幅断片をアガロースゲル電気泳動に供したゲル中よりGENECLEAN IIIキット(Qbiogene社製)を用いてDNAを抽出し、これを挿入DNA断片とした。この挿入DNA断片をpGEM-T Easy Vector System(Promega社製)を用いてpGEM-T Easy Vectorに連結させ、連結DNA溶液を得た。連結DNA溶液10μlを氷中でよく冷却した後、氷上解凍したコンピテントセルJM109を100μl加え穏やかに撹拌し、氷中で20分間、続いて42℃で45秒ヒートショック処理を行った。これに、400μlのLB液体培地を加え、37℃で1時間培養後、100μg/mlのアンピシリンを添加したLB平板培地にまき、37℃で一晩培養した。目的のプラスミドDNAを用いて形質転換した大腸菌の単一のコロニーは3mlの100μg/mlのアンピシリンを添加したLB液体培地に植菌し、37℃で一晩振盪培養した。1.5mlの培養液を1.5mlのエッペンドルフチューブに移して15,000×gで1分間遠心分離し、沈殿を100μlの氷冷したTEG(25mM Tris-HCl、10mM EDTA、50mM Glucose、pH8.0)で懸濁し、これに200μlの0.2N NaOH-1%SDSを加えて穏やかに撹拌した後、150μlの3M NaOAc(pH5.2)を加えて混合した。これを15,000×g、4℃で5分間遠心分離し上清を回収し、450μlのフェノール・クロロホルム・イソアミルアルコール(25:24:1)を加えて激しく撹拌した後、15,000×g、室温で5分間遠心分離し上層を回収した。この溶液に-20℃で氷冷した900μlのエタノールを加えて、-20℃で10分間放置後、4℃にて15,000×gで5分間遠心分離した。沈殿を500μlの70%エタノールでリンスしたのち、乾燥させ、最後にRNase(100μg/ml)を含むTE(10mM Tris-HCl、1mM EDTA、pH8.0)50μlに溶解した。得られたプラスミドをEcoRIで切断後、アガロース電気泳動により挿入断片の存在を確認し、このプラスミド中の挿入DNA断片を添付のプロトコールに従い一般的なユニバーサルプライマーSP6(5-GATTTAGGTGACACTATAG-3(配列番号7))およびT7(5-TAATACGACTCACTATAGGG-3(配列番号8))を用い、ABI PRISMTM 377 DNA sequencing system (PE Biosystem社製)にて解析した。その結果、ALB1の5'領域は1440塩基対、ALB1遺伝子は1174塩基対からなることが明らかとなった。また、本ベクターをpGEM-5’ALBおよびpGEM-ALB1174と命名する。
【0047】
2)内在ALB1プロモーターALB1Pを既知プロモーターへ置き換えるプラスミドベクターの作成
pCAMBIA-0380バイナリーベクター(CAMBIA社より購入)を骨格にし、薬剤選択マーカーとしてハイグロマイシンB耐性遺伝子Hphを融合することにより作成した糸状菌形質転換ベクターpCH11(図2)のEcoRI、BamHIサイトにpGEM-5’ALBベクターのEcoRI、BamHI断片を挿入後、続いてBglII、SpeIサイトにpGEM-ALB1174のBglII、SpeI断片を挿入することによりALB1プロモーターALB1Pを欠損すると予想されるプラスミドベクターを作成した。本ベクターをpCHALB1と命名した。本ベクターpCHALB1(図2)は、配置してあるALB1遺伝子開始コドンより上流のHindIIIもしくはBglIIサイトへ既知プロモーターを挿入することにより内在ALB1プロモーターALB1Pを既知プロモーターへ置き換えるプラスミドベクターとして利用できる。
【0048】
〔実施例2〕
pCHALB1へのプロベナゾール誘導型プロモーターRDH8Pの挿入
ALB1遺伝子の上流のALB1P領域を他のプロモーター配列で置き換える例として、RDH8Pプロモーターを用いた場合を以下に示す。RDH8遺伝子は、いもち病菌には殺菌効果が無く、宿主である植物にいもち病菌に対する抵抗性を付与する化合物であるプロベナゾールの存在下で特異的に発現量が亢進する遺伝子として発見された。また、RDH8遺伝子のプロモーター領域を、既知のレポーター遺伝子であるEGFPに連結した解析結果から、RDH8遺伝子のプロモーター領域(RDH8Pプロモーター)は、プロベナゾールの有無によって遺伝子発現を調節することができる発現誘導型プロモーターとして利用できることが明らかとなっている。
【0049】
プロベナゾール誘導型プロモーターRDH8Pの配列は、プライマー5‘-ATAAGGTGCGCGCAAAGGT-3’(配列番号9)及び5‘-CGCAAGCTTAGTTTGGATAGTCGTATGCA-3’ (配列番号10)を用い、いもち病菌ゲノムDNAを鋳型としたPCRによって増幅させ、前述の方法によりpGEM-T Easyベクターに連結した。本ベクターをHindIII処理することにより得たRDH8Pプロモーター配列を同じくHindIII処理したpCHALB1ベクターに連結することにより、RDH8PによりALB1遺伝子を制御可能なプラスミドベクターを作成した。なお、RDH8Pプロモーター配列のベクター内への挿入方向は、PCRおよびシークエンスにより確認し、本ベクターをpCHRDH8PALB1と命名した。
【0050】
(1) pCHRDH8PALB1プラスミドを用いたマグナポルテ・グリセアの形質転換
マグナポルテ・グリセアの形質転換はアグロバクテリウム法(Gento Tsuji, Satoshi Fujii, Naoki Fujihara, Chika Hirose, Seiji Tsuge, Tomonori Shiraishi and Yasuyuki Kubo(2003):Agrobacterium tumefaciens-mediated transformation for random insertional mutagenesis in Colletotrichum lagenarium. Journal of General Plant Pathology 69:230-239)を改良した方法を用いた。予想される相同組み換えによるALB1PのRDH8Pへの変換は図3に示した。得られたRDH8P-ALB1菌株へのベクター挿入の確認は、PCR、シークエンス及びサザン解析によって行った。
【0051】
(2) RDH8P-ALB1菌株の形状
RDH8Pプロモーターは化合物プロベナゾール処理により、条件的に下流遺伝子の発現を顕著に誘導することができる。今回得られたRDH8P-ALB1菌株にプロベナゾールを処理したときに特異的にフラビオリンの赤色色素の蓄積が観察されるか否かを調査するため、YG液体培地でのプロベナゾール添加の有無によるRDH8P-ALB1菌株の色素合成能の観察を行った。
【0052】
プロベナゾールを50ppm添加もしくは無添加のYG液体培地(イーストエクストラクト5g 、グルコース10g/L) 50mlに野生株Guy11および3つのRDH8P-ALB1菌株の胞子を約5×105個移植し、暗黒下、26℃、140 strokes/minの条件で24時間振とう培養を行った。24時間後のサンプルにプロベナゾールを50ppm添加し、再度24時間同条件で振とう培養を行った。また、プロベナゾール無添加のサンプルを対照として用いた。プロベナゾール添加後24時間の写真を図4に示す。その結果、プロベナゾール無添加の全てのサンプルにおいて乳白色の菌糸塊の生育が観察され、培地の色もYG液体培地の色である黄色のまま目立った色素の蓄積は観察されなかった。一方、プロベナゾールを50ppm添加したサンプルでは、対照の野生株Guy11はプロベナゾール無添加区と同様の乳白色の菌糸塊と培地色であったが、3つのRDH8P-ALB1菌株の場合、菌糸塊はほとんど通常と同様の乳白色をしていたのに対し、培地の色は顕著に強い赤色を呈していた。
【0053】
これらの結果から、RDH8P-ALB1菌株ではRDH8Pプロモーター制御下で、プロベナゾールの有無によりRDH8Pプロモーター下流のALB1遺伝子の発現が条件的に調節されており、ALB1遺伝子の発現によりALB1遺伝子の合成産物である1,3,6,8-T4HNが蓄積され、結果として赤色色素フラビオリンの蓄積が観察されたものと考えられた。また、Guy11野生株においては、生育した菌糸のメラニン化が観察されなかった。このことから、菌糸生育に要した48時間という時間内では、いもち病菌のメラニン合成に関与する遺伝子群はほとんど発現しないことを確認できた。
【0054】
このように、ALB1遺伝子の発現制御をフラビオリンの蓄積により可視化できるため、ALB1遺伝子を糸状菌の新しいレポーター遺伝子として利用できる可能性が示された。また、仮に、内在するALB1遺伝子に加えて、RDH8Pプロモーター等の他のプロモーターに置き換えたALB1遺伝子を外的に導入したとしても、外的に導入したALB1遺伝子の発現制御をフラビオリンの蓄積により可視化できることが示された。
【0055】
(3) 96-wellマイクロタイタープレートを用いたハイスループットスクリーニングへの利用
1.プロベナゾールを用いた例
ALB1遺伝子の発現制御による色素蓄積をレポーターに用い、ハイスループットスクリーニングに利用しようとする場合、大量の数のサンプルをアッセイできることが求められる。そこで前述のアッセイのような50mlものハイスケールではなく数100μlというロースケールでのアッセイにも応用可能かを調べるために、RDH8P-ALB1菌株を利用して、96-wellのマイクロタイタープレートを用いたアッセイを行った。YG液体培地に移植したRDH8P-ALB1の3菌株と比較対照として用いた親菌株であるGuy11株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し、26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、プロベナゾールを0、1/256、1/64、1/16、1/4、1、4、16ppmの濃度になるように添加し、再度26℃で約6時間、1000rpmの条件で振とう培養を行った。6時間後のプレートの写真を図5に示す。その結果、RDH8P-ALB1の3菌株ともにプロベナゾール4ppmと16ppmの処理カラムでのみ、薬剤処理後6時間と短い時間でありながら、かなり明瞭な赤色色素の蓄積が確認できた。一方で、Guy11株からは赤色色素の蓄積は全く観察できなかった。この4ppmと16ppmの濃度はRDH8P-ALB1と同様にRDH8Pプロモーターの下流に緑色蛍光色素であるEGFPをレポーター遺伝子として連結した場合の蛍光が観察された濃度と一致しており、この赤色色素を用いたレポーター検出が十分に高感度であることを示している。
【0056】
2.各種作用点別化合物を利用した例
YG液体培地に移植したRDH8P-ALB1菌株或いはGuy11野生株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、プロベナゾール、及びプロベナゾールとは作用機構が異なる農薬化合物4種を0.016、0.064、0.25、1、4および16ppmの濃度添加し、再度26℃で6時間、1000rpmの条件で振とう培養を行った。6時間後、培地における赤色色素を目視によって評価した結果を表1(Guy11野生株)及び表2(RDH8P-ALB1菌株)に示した。
【0057】
なお、使用した農薬化合物の作用機構は、FRAC FUNGICIDE RESISTANCE ACTION COMMITTEE発行のFRAC Code List 2010(http://www.frac.info/frac/publication/anhang/FRAC_Code_List_2010.pdf)に記載されている。
【0058】
【表1】
【0059】
【表2】
【0060】
表1及び2において、比較対照区は、化合物を添加していない系である。また、赤色色素の蓄積は、赤色の濃淡に基づいて数値化した。また、表中、赤色の蓄積がない場合「−」とし、赤色の蓄積が薄い場合、中間の場合、濃い場合をそれぞれ「+」、「++」及び「+++」とした。
【0061】
その結果、プロベナゾール4ppmと16ppmの処理カラムでのみ薬剤処理後6時間と短い時間からかなり明瞭な赤色色素の蓄積が確認できた。一方で、それ以外の化合物の処理カラムからは赤色色素の蓄積は観察できなかった。また、Guy11野生株では同濃度のプロベナゾールを添加しても赤色の蓄積は観察されなかった。このことから、ALB1遺伝子を発現制御することにより蓄積する赤色色素を利用したレポーターアッセイシステムが化合物スクリーニングに応用可能であると示された。
【0062】
〔実施例3〕
pCHALB1への電子伝達系阻害化合物誘導型プロモーターAOXPの挿入
ALB1遺伝子の上流のALB1P領域を既知のプロモーター配列で置き換える例として、電子伝達系阻害剤のスクリーニングに用いることのできる、電子伝達系阻害剤存在下で特異的に遺伝子の発現を誘導するできるプロモーターAOXPを用いた場合を以下に示す。
【0063】
電子伝達系化合物誘導プロモーターAOXPの配列はBroad Institute のマグナポルテ・グリセア(Magnaporthe grisea)ゲノムデータベース(http://www.broad.mit.edu/annotation/genome/magnaporthe_grisea/MultiHome.html)からAOX 遺伝子の5’領域の塩基配列情報を取得し、その配列情報を参考に一組のオリゴヌクレオチドを設計した(5‘-CCAAGCTTCCTTCCTCACTA-3’(配列番号11)、5‘-CCAAGCTTAAGACGGTACGTTCAGTGT-3’(配列番号12))。マグナポルテ・グリセアGuy11株のゲノムDNAをテンプレートとしてPCR反応を行った。その後、前述の方法によりpGEM-T Easyベクターに連結した。本ベクターをHindIII処理することにより得たAOXPプロモーター配列を、同じくHindIII処理したpCHALB1ベクターに連結することにより、AOXPによりALB1遺伝子を制御可能なプラスミドベクターを作成した。なお、AOXPプロモーター配列のベクター内への挿入方向は、PCRおよびシークエンスにより確認し、本ベクターをpCHAOXPALB1と命名し、本ベクターを用い前述の実施例2のアグロバクテリウム法で形質転換を行った。得られた菌株をAOXP-ALB1菌株と命名し、本菌へのベクター挿入の確認は、PCR、シークエンス及びサザン解析によって行った。
【0064】
(1) 96-wellマイクロタイタープレートを用いたハイスループットスクリーニングへの利用
1.電子伝達系阻害剤を使用した例
AOXP-ALB1菌株を利用して、96-wellのマイクロタイタープレートを用いアッセイを行った。YG液体培地に移植したAOXP-ALB1菌株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し、26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、電子伝達系阻害剤であるジフルメトリム、ピリベンカルブ及びオリゴマイシンを0、1/256、1/64、1/16、1/4、1、4、16ppmの濃度になるように添加し、再度26℃で約12時間、1000rpmの条件で振とう培養を行った。12時間後のプレートの写真を図6に示す。その結果、ジフルメトリムおよびピリベンカルブ処理カラムでは、低濃度の1/256ppmの処理カラムから赤色色素の蓄積が確認でき、1/4ppm及び1ppmに赤色色素のピークを示していた。またオリゴマイシンの場合は、本剤のEC50値が0.006ppmと低いためか高濃度処理では生育阻害が強いため検出できなかったが、低濃度の1/64ppm処理カラムで明瞭な赤色色素の蓄積を確認することができた。
【0065】
2.各種作用点別化合物を利用した例
YG液体培地に移植したAOXP-ALB1菌株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、ジフルメトリム及びピリベンカルブの電子伝達系阻害剤、及びそれらとは作用機構が異なる農薬化合物3種を0.016、0.064、0.25、1、4および16ppmの濃度添加し、再度26℃で12時間、1000rpmの条件で振とう培養を行った。12時間後、培地における赤色色素を目視によって評価した結果を表3に示した。
【0066】
【表3】
【0067】
なお、表3において、比較対照区は、化合物を添加していない系である。また、赤色色素の蓄積は、赤色の濃淡に基づいて数値化した。また、表中、赤色の蓄積がない場合「−」とし、赤色の蓄積が薄い場合、中間の場合、濃い場合をそれぞれ「+」、「++」及び「+++」とした。
【0068】
その結果、予想どおりジフルメトリム及びピリベンカルブといった電子伝達系阻害剤の処理カラムでかなり明瞭な赤色色素の蓄積が確認できた。一方で、それ以外の化合物の処理カラムからは赤色色素の蓄積はほとんど観察できなかった。このことから、ALB1遺伝子を発現制御することによる蓄積する赤色色素を利用したレポーターアッセイシステムが化合物スクリーニングに応用可能であると考えられ、今回のように実際に電子伝達系阻害剤のスクリーニングに利用可能であることが確認できた。
【0069】
〔実施例4〕
pCHALB1へのエルゴステロール生合成阻害化合物誘導型プロモーターCYP51Pの挿入
ALB1遺伝子の上流のALB1P領域を既知のプロモーター配列で置き換える例として、エルゴステロール合成阻害剤のなかでも、Cytochrome P450ステロール14αデメチラーゼ阻害剤のスクリーニングに用いることのできる、ステロール14αデメチラーゼ阻害剤の存在下で特異的に遺伝子の発現を誘導するできるプロモーターCYP51Pを用いた場合を以下に示す。
【0070】
ステロール14αデメチラーゼ阻害剤誘導プロモーターCYP51Pの配列はBroad Instituteのマグナポルテ・グリセア(Magnaporthe grisea)ゲノムデータベース(http://www.broad.mit.edu/annotation/genome/magnaporthe_grisea/MultiHome.html)からCytochrome P450ステロール14αジメチラーゼ(MGG_04628)の5’領域の塩基配列情報を取得し、その配列情報を参考に一組のオリゴヌクレオチドを設計した(5‘-CCAAGCTTATGTGAGCCATCTTCGGATT-3’(配列番号13)、5‘-CCAAGCTTGACGATCGGGTAAGGAGACT-3’(配列番号14))。マグナポルテ・グリセアGuy11株のゲノムDNAをテンプレートとしてPCR反応を行った。その後、前述の方法によりpGEM-T Easyベクターに連結した。本ベクターをHindIII処理することにより得たCYP51Pプロモーター配列を、同じくHindIII処理したpCHALB1ベクターに連結することにより、CYP51PによりALB1遺伝子を制御可能なプラスミドベクターを作成した。なお、CYP51Pプロモーター配列のベクター内への挿入方向は、PCRおよびシークエンスにより確認し、本ベクターをpCH CYP51PALB1と命名し、本ベクターを用い前述の実施例2のアグロバクテリウム法で形質転換を行った。得られた菌株をCYP51P-ALB1菌株と命名し、本菌へのベクター挿入の確認は、PCR、シークエンス及びサザン解析によって行った。
【0071】
(1) 96-wellマイクロタイタープレートを用いたハイスループットスクリーニングへの利用
1.エルゴステロール生合成阻害剤を使用した例
CYP51P-ALB1菌株を利用して、96-wellのマイクロタイタープレートを用いアッセイを行った。YG液体培地に移植したCYP51P-ALB1菌株の胞子懸濁液約5×105 / mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し、26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、ステロール14αデメチラーゼ阻害剤であるオキスポコナゾール、イプコナゾール、トリホリンおよびプロチオコナゾールを0、1/256、1/64、1/16、1/4、1、4、16ppmの濃度になるように添加し、再度26℃で約12時間、1000rpmの条件で振とう培養を行った。12時間後のプレートの写真を図7に示す。その結果、オキスポコナゾール、イプコナゾール、トリホリンおよびプロチオコナゾールともに1ppm〜16ppmの濃度では明瞭な赤色色素の蓄積が確認できた。
【0072】
2.各種作用点別化合物を利用した例
YG液体培地に移植したCYP51P-ALB1菌株の胞子懸濁液約5×105/mlを200μlずつ96-well丸底マイクロタイタープレート(TPP社製)に分注し26℃で30時間、1000rpmの条件で激しく振とう培養を行った。その後、オキスポコナゾール及びトリホリンのステロール14αデメチラーゼ阻害剤、及びそれらとは作用機構が異なる農薬化合物3種を0.016、0.064、0.25、1、4および16ppmの濃度添加し、再度26℃で12時間、1000rpmの条件で振とう培養を行った。12時間後、培地における赤色色素を目視によって評価した結果を表4に示した。
【0073】
【表4】
【0074】
なお、表4において、比較対照区は、化合物を添加していない系である。また、赤色色素の蓄積は、赤色の濃淡に基づいて数値化した。また、表中、赤色の蓄積がない場合「−」とし、赤色の蓄積が薄い場合、中間の場合、濃い場合をそれぞれ「+」、「++」及び「+++」とした。
【0075】
その結果、予想どおりオキスポコナゾール及びトリホリンのステロール14αデメチラーゼ阻害剤の処理カラムでかなり明瞭な赤色色素の蓄積が確認できた。一方で、それ以外の化合物の処理カラムからは赤色色素の蓄積はほとんど観察できなかった。このことから、ALB1遺伝子を発現制御することによる蓄積する赤色色素を利用したレポーターアッセイシステムが電子伝達系阻害剤とともにエルゴステロール合成阻害剤でも利用できたことから、汎用性があることが確認できた。
【特許請求の範囲】
【請求項1】
ポリケタイド合成酵素遺伝子をレポーター遺伝子として有する糸状菌を使用し、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定する方法。
【請求項2】
上記ポリケタイド合成酵素遺伝子は、配列番号2に示すアミノ酸配列を含むタンパク質をコードする、又は配列番号2に示すアミノ酸配列に対して90%以上の類似性を有するアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードすることを特徴とする請求項1記載の方法。
【請求項3】
上記ポリケタイド合成酵素遺伝子を、所定の因子に対して転写活性を変動する転写制御領域の制御下に連結してレポーター遺伝子とし、上記糸状菌に対して刺激を与えた後における培地中のフラビオリン量が有意に変動する場合、当該刺激が上記転写制御領域に対して上記所定の因子と同等に作用すると判断することを特徴とする請求項1記載の方法。
【請求項4】
上記刺激は、上記糸状菌に対する供試化合物への曝露であることを特徴とする請求項3記載の方法。
【請求項5】
上記転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする請求項3記載の方法。
【請求項6】
上記転写制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする請求項3記載の方法。
【請求項7】
上記転写制御領域として植物病害抵抗性付与剤誘導型プロモーターを使用して植物病害抵抗性付与剤の候補物質をスクリーニングする、上記転写制御領域として電子伝達系阻害剤誘導型プロモーターを使用して電子伝達系阻害剤の候補物質をスクリーニングする、又は上記転写制御領域としてエルゴステロール生合成阻害剤誘導型プロモーターを使用してエルゴステロール生合成阻害剤の候補物質をスクリーニングすることを特徴とする請求項3記載の方法。
【請求項8】
上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結してレポーター遺伝子とし、機能既知である発現制御領域の転写活性を変動させる所定の因子を上記糸状菌に作用させたときに、培地中のフラビオリン量が有意に変動する場合、上記機能未知の発現制御領域が上記機能既知の発現制御領域と同等の応答性を示すと判断することを特徴とする請求項1記載の方法。
【請求項9】
上記所定の因子は、特定のプロモーターに対して転写活性を促進又は抑制する化合物であることを特徴とする請求項8記載の方法。
【請求項10】
上記所定の因子は、植物病害抵抗性付与能を有する化合物、電子伝達系阻害能を有する化合物及びエルゴステロール生合成阻害能を有する化合物からなる群より選ばれる1つの化合物であることを特徴とする請求項8記載の方法。
【請求項11】
上記機能未知の転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする請求項8記載の方法。
【請求項12】
上記所定の因子として植物病害抵抗性付与剤を使用して植物病害抵抗性付与剤誘導型プロモーターをスクリーニングする、上記所定の因子として電子伝達系阻害剤を使用して電子伝達系阻害剤誘導型プロモーターをスクリーニングする、又は上記所定の因子としてエルゴステロール生合成阻害剤を使用してエルゴステロール生合成阻害剤誘導型プロモーターをスクリーニングすることを特徴とする請求項8記載の方法。
【請求項13】
上記ポリケタイド合成酵素遺伝子で解析対象の遺伝子を組み換え、培地に生産されるフラビオリンを指標として解析対象の遺伝子の発現パターンを解析することを特徴とする請求項1記載の方法。
【請求項14】
導入対象の目的遺伝子と、恒常発現型プロモーターに連結された上記ポリケタイド合成酵素遺伝子とを上記糸状菌に導入し、培地中のフラビオリン量が有意に大である場合、上記目的遺伝子が発現していると判断することを特徴とする請求項1記載の方法。
【請求項15】
複数のウェルを有するプレートを使用して当該各ウェルに上記糸状菌を培養し、上記フラビオリンに由来する赤色発色を検出する検出手段を使用して複数のウェルを一括して観察することを特徴とする請求項1乃至14いずれか一項記載の方法。
【請求項16】
機能既知又は機能未知の転写制御領域と、当該転写制御領域の制御下に発現するポリケタイド合成酵素遺伝子とを含む核酸構築物。
【請求項17】
上記機能既知の発現制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする請求項16記載の核酸構築物。
【請求項18】
請求項16又は17記載の核酸構築物を含む発現ベクター。
【請求項19】
請求項16又は17記載の核酸構築物で形質転換された組み換え糸状菌。
【請求項20】
いもち病菌の組換え体であることを特徴とする請求項19記載の組み換え糸状菌。
【請求項1】
ポリケタイド合成酵素遺伝子をレポーター遺伝子として有する糸状菌を使用し、当該遺伝子の発現を培地に生産されるフラビオリンを指標として判定する方法。
【請求項2】
上記ポリケタイド合成酵素遺伝子は、配列番号2に示すアミノ酸配列を含むタンパク質をコードする、又は配列番号2に示すアミノ酸配列に対して90%以上の類似性を有するアミノ酸配列を含むタンパク質であり、且つアセチルCoA及びマロニルCoAからテトラヒドロキシナフタレンを合成する活性を有するタンパク質をコードすることを特徴とする請求項1記載の方法。
【請求項3】
上記ポリケタイド合成酵素遺伝子を、所定の因子に対して転写活性を変動する転写制御領域の制御下に連結してレポーター遺伝子とし、上記糸状菌に対して刺激を与えた後における培地中のフラビオリン量が有意に変動する場合、当該刺激が上記転写制御領域に対して上記所定の因子と同等に作用すると判断することを特徴とする請求項1記載の方法。
【請求項4】
上記刺激は、上記糸状菌に対する供試化合物への曝露であることを特徴とする請求項3記載の方法。
【請求項5】
上記転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする請求項3記載の方法。
【請求項6】
上記転写制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする請求項3記載の方法。
【請求項7】
上記転写制御領域として植物病害抵抗性付与剤誘導型プロモーターを使用して植物病害抵抗性付与剤の候補物質をスクリーニングする、上記転写制御領域として電子伝達系阻害剤誘導型プロモーターを使用して電子伝達系阻害剤の候補物質をスクリーニングする、又は上記転写制御領域としてエルゴステロール生合成阻害剤誘導型プロモーターを使用してエルゴステロール生合成阻害剤の候補物質をスクリーニングすることを特徴とする請求項3記載の方法。
【請求項8】
上記ポリケタイド合成酵素遺伝子を、機能未知の発現制御領域の制御下に連結してレポーター遺伝子とし、機能既知である発現制御領域の転写活性を変動させる所定の因子を上記糸状菌に作用させたときに、培地中のフラビオリン量が有意に変動する場合、上記機能未知の発現制御領域が上記機能既知の発現制御領域と同等の応答性を示すと判断することを特徴とする請求項1記載の方法。
【請求項9】
上記所定の因子は、特定のプロモーターに対して転写活性を促進又は抑制する化合物であることを特徴とする請求項8記載の方法。
【請求項10】
上記所定の因子は、植物病害抵抗性付与能を有する化合物、電子伝達系阻害能を有する化合物及びエルゴステロール生合成阻害能を有する化合物からなる群より選ばれる1つの化合物であることを特徴とする請求項8記載の方法。
【請求項11】
上記機能未知の転写制御領域は、所定の化合物の存在下で転写活性を促進又は抑制するプロモーター領域を含むことを特徴とする請求項8記載の方法。
【請求項12】
上記所定の因子として植物病害抵抗性付与剤を使用して植物病害抵抗性付与剤誘導型プロモーターをスクリーニングする、上記所定の因子として電子伝達系阻害剤を使用して電子伝達系阻害剤誘導型プロモーターをスクリーニングする、又は上記所定の因子としてエルゴステロール生合成阻害剤を使用してエルゴステロール生合成阻害剤誘導型プロモーターをスクリーニングすることを特徴とする請求項8記載の方法。
【請求項13】
上記ポリケタイド合成酵素遺伝子で解析対象の遺伝子を組み換え、培地に生産されるフラビオリンを指標として解析対象の遺伝子の発現パターンを解析することを特徴とする請求項1記載の方法。
【請求項14】
導入対象の目的遺伝子と、恒常発現型プロモーターに連結された上記ポリケタイド合成酵素遺伝子とを上記糸状菌に導入し、培地中のフラビオリン量が有意に大である場合、上記目的遺伝子が発現していると判断することを特徴とする請求項1記載の方法。
【請求項15】
複数のウェルを有するプレートを使用して当該各ウェルに上記糸状菌を培養し、上記フラビオリンに由来する赤色発色を検出する検出手段を使用して複数のウェルを一括して観察することを特徴とする請求項1乃至14いずれか一項記載の方法。
【請求項16】
機能既知又は機能未知の転写制御領域と、当該転写制御領域の制御下に発現するポリケタイド合成酵素遺伝子とを含む核酸構築物。
【請求項17】
上記機能既知の発現制御領域は、植物病害抵抗性付与剤誘導型プロモーター、電子伝達系阻害剤誘導型プロモーター及びエルゴステロール生合成阻害剤誘導型プロモーターからなる群から選ばれる1つのプロモーター領域を含むことを特徴とする請求項16記載の核酸構築物。
【請求項18】
請求項16又は17記載の核酸構築物を含む発現ベクター。
【請求項19】
請求項16又は17記載の核酸構築物で形質転換された組み換え糸状菌。
【請求項20】
いもち病菌の組換え体であることを特徴とする請求項19記載の組み換え糸状菌。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】

【図7】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2011−188819(P2011−188819A)
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願番号】特願2010−58032(P2010−58032)
【出願日】平成22年3月15日(2010.3.15)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(000000169)クミアイ化学工業株式会社 (86)
【Fターム(参考)】
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願日】平成22年3月15日(2010.3.15)
【出願人】(501167644)独立行政法人農業生物資源研究所 (200)
【出願人】(000000169)クミアイ化学工業株式会社 (86)
【Fターム(参考)】
[ Back to top ]