
ポリジーンを使用する多タンパク質複合体の組換え発現
本発明は、少なくとも3つのポリペプチドをコードするポリジーンをコードする組換えポリヌクレオチドに関するものであり、その中で、ポリジーンを構成する遺伝子の少なくとも1つは、非ウイルス由来であり、ポリジーンを構成する遺伝子によってコードされるポリペプチドの少なくとも2つは各々、該ポリジーンの遺伝子によってコードされる少なくとも1つの他のポリペプチドと少なくとも一過性に相互作用することが可能であり、ポリジーンを構成する遺伝子は各々、少なくとも1つのプロテアーゼ切断部位をコードする配列によって互いに接続される。本発明は、ポリジーンによってコードされるポリタンパク質にも関するものである。本発明のさらなる態様は、組換えポリペプチドを含有するベクター、組換えポリペプチドおよび/またはベクターを含有する宿主細胞ならびに組換えポリペプチドおよび/またはベクターで形質転換された非ヒト遺伝子導入動物である。本発明は、ポリヌクレオチドの生成のための方法、および多タンパク質複合体の製造のための方法にも関するものである。本発明の態様は、遺伝子治療、薬物候補スクリーニング、ワクチン産生および構造的研究のための多タンパク質複合体の結晶化において特に有用である。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ポリジーン(Polygenes)を構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、非ウイルス性由来であり、ポリジーンを構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、該ポリジーンの遺伝子によってコードされる少なくとも1つの他のポリペプチドと少なくとも一過性に相互作用することが各々可能であり、および各ポリジーンを構成する遺伝子が、少なくとも1つのプロテアーゼ切断部位をコードする配列によって、および/または少なくとも1つの自己切断ペプチドをコードする配列によって互いに接続される、単一翻訳領域(ORF)内で各々、少なくとも3つの生物活性のあるポリペプチドを各々コードする少なくとも2つのポリジーンをコードする組換えポリヌクレオチドに関するものである。本発明のさらなる態様は、組換えポリペプチドを含有するベクター、組換えポリヌクレオチドおよび/またはベクターを含有する宿主細胞ならびに組換えポリヌクレオチドおよび/またはベクターで形質転換された非ヒト遺伝子導入動物である。本発明は、ポリヌクレオチドの生成のための、および多タンパク質複合体の製造のための方法にも関するものである。本発明の態様は、遺伝子治療、薬物候補スクリーニング、ワクチン産生および構造研究のための多タンパク質複合体の結晶化において特に有用である。
【0002】
ポストゲノム時代における生物学的研究の尽力に関する強い焦点は、タンパク質の相互作用ネットワーク(相互作用体(interactome))の解明である。同定された多タンパク質(multiprotein)複合体の多くが、ネイティブな細胞中で詳細な分子生物学的分析のための十分量では存在しないため、前記複合体の研究は、大規模な異種性タンパク質生成のための組換え技術に依存している。組換え発現方法は、多タンパク質発現前に、労力および材料の両者において不相応な投資を必要とし、発現後には、改定された発現研究のための多タンパク質構成分を迅速に変化させるための柔軟性を提供しない。
【0003】
多サブユニット複合体を得るために近年使用されている組換え技術が幾多もある。例えば、タンパク質は、可溶性形態または封入体としてのいずれかで、イー・コリィ(E.coli)中での単離物で発現され、精製された後、インビトロで同様に生成されたタンパク質を使用して多タンパク質複合体中に再構成されることが可能である。真核生物細胞(例えば、哺乳動物細胞または酵母細胞)も、一過性発現実験において宿主として使用されることが可能である。この方法論は、効率的なインビトロでの再構成プロトコールの存在に完全に依存する。この戦略が、小さなサブユニットサイズを有するより単純な系については許容可能な結果を生じ得るのに対し、多くの、また、大きなサブユニットを含有するより複雑化した(例えば、より高次の真核動物の、特にヒトの制御複合体の全てに近い)多タンパク質複合体に関しては一般的に適用可能ではない。
【0004】
同時発現は、上述に概略されるインビトロでの再構成の戦略に代わる優れたものとして認識されてきた。幾つもの同時発現系が、原核細胞発現および真核細胞発現両者に関して過去に開発されている。原核細胞系において、同時発現は、選択された遺伝子全てを含有する単一プラスミドを産生することによって、または1つもしくは2つの遺伝子を含有する幾つものプラスミドならびに異なる耐性マーカーおよびレプリコンを同時形質転換することによって達成されることが可能である。
【0005】
真核細胞における同時発現は、バキュロウイルス系を使用することによって具現化されており、初期には、幾つものウイルスによる同時感染によって成功が制限されていたが、後には単一ウイルスから全てのタンパク質を発現させることによってさらに成功し、多くの利点を提唱し、(比較的小さなサブユニットのサイズ、真正のプロセッシングの欠如、真核生物(特にヒト)タンパク質の困難な発現等の)原核細胞系に存在するいくつもの制限を排除した。バキュロウイルス系に関し、単一ウイルスからの発現は、収量を劇的に増大させることが示されている(Berger et al. (2004) Nature Biotech. 22, 1583−1587; Comment (2004), Nature Biotech. 22, vii, New & Views (2004), Nature Biotech. 22, 152, Research Highlights, Nature Methods 2, 7 (2005); Bertolotti−Ciarlet et al. (2003) Vaccine 21 , 3885−3900も参照)のに対し、特に、大規模な産生に関してロジスティックな需要を決定的に低下させる。
【0006】
多タンパク質発現の主要な改良は、WO 2005/085456 A1 (PCT/EP2004/013381;Berger et al. (2004)上述も参照)に開示されている、本発明者によって提供される多重遺伝子発現カセットの作製のためのモジュラー系の提供であった。WO 2005/085456 A1 (PCT/EP2004/013381)によって記載されているMultiBac技術によって、多重遺伝子発現カセットの単純な作製、ならびに発現実験の改変および修正が可能となる(Berger et al. (2004)上述)。
【0007】
しかしながら、特に(真核生物、例えばヒトの遺伝子制御複合体の大部分を構成する)多くの(6、7、8またはそれより多くの)サブユニットを有する多サブユニット複合体の発現の成功およびインビボでの集合に関する障害は、これらのサブユニットの相対的発現レベルが多くの場合、完全には理解されていない機構(例えば、転写および翻訳の効率、タンパク質の安定性、mRNAの安定性および二次構造等)に基づいて典型的に有意に異なるという事実において見出される。結果として、本質的に非平衡の系において、最少量で発現するサブユニットは、複合体の全体的な収量を制限することによって、多サブユニット複合体生成実験の全体的な成功に影響するであろう。したがって、転写/翻訳装置は、過程に組み込まれていない他の成分の過剰量を生成し、したがって細胞内の転写/翻訳資源を「無駄にする」。個々の発現レベルは典型的に、互いに関して何倍も(例えば、最大10倍以上まで)変動し、特に(例えば、多くの真核生物遺伝子制御複合体の場合において、)5、6、8、10以上のサブユニットなど、4個を超える複合体の場合、望ましい多サブユニット複合体の生成の成功に対して抵抗性のある損失を伴う。
【0008】
ピコルナウイルススーパー群のウイルスは、ウイルスプロテアーゼの切断部位によって、または自己切断ペプチドによって互いに接続されるウイルスタンパク質を含むポリタンパク質をコードする単一のまたは2つのORFを含有するセンスの向きでの一本鎖RNA分子からなるゲノムを有する(例えば、Ryan et al. (1997) J. Gener. Virol. 78, 699−723において概説されている)。
【0009】
タンパク質複合体の生成に関して、組換えウイルスを通じてポリタンパク質を発現させる一般的な概念は、TCR(T細胞受容体):CD3複合体の再構成において適用されている(Szymczak et al. (2004) Nature Biotech. 5, 589−594)。著者は、2つの組換えレトロウイルスベクターを使用し、その中で、1つのベクターは、2つのTCRサブユニットをコードする配列を含有したのに対し、他のベクターは、4つのCD3サブユニットを含むポリタンパク質をコードした。サブユニットは、アフトウイルス由来の自己切断2Aペプチド配列によって接続された。このアプローチの1つの不利な点は、完全な複合体を再構成するために、2つの別個のベクターが調製されなければならず、2回の形質移入が必要であることである。
【0010】
ウイルスポリタンパク質のアプローチは、ヘテロ二量体IL−12(Kokuho et al. (1999) Vet. Immunol. Immunopathol. 72, 289−302)ならびにバキュロウイルス多面体由来の核標的指向化シグナルおよび関心対象のタンパク質を含む融合タンパク質(米国特許第5,179,007号)等の小さな構築物のためのバキュロウイルス発現系において適用されている。
【0011】
それゆえ、本発明の基礎となる技術的な課題は、多タンパク質の改良された発現のための新規の系を提供することである。
【0012】
上述の技術的な課題の解決は、特許請求の範囲において定義される態様によって提供される。
【0013】
特に、本発明は、ポリジーンを構成する遺伝子によってコードされるポリペプチドのうちの少なくとも2つが、非ウイルス性由来であり、ポリジーンを構成する遺伝子によってコードされるポリペプチドのうちの少なくとも2つが、該ポリジーンの遺伝子によってコードされる少なくとも1つの他のポリペプチドと少なくとも一過性に相互作用することが各々可能であり、および各ポリジーンを構成する遺伝子が、少なくとも1つのプロテアーゼ切断部位をコードする配列によって、および/または少なくとも1つの自己切断ペプチドをコードする配列によって互いに接続される、単一翻訳領域(ORF)内で各々、少なくとも3つの生物活性のあるポリペプチドを各々コードする少なくとも2つのポリジーンをコードするポリヌクレオチドを提供する。
【0014】
本発明のポリヌクレオチドは、1つ以上の合成ヌクレオチド類似体を含むDNA、RNAまたはポリヌクレオチドであり得る。ポリヌクレオチドは、一本鎖または二本鎖の形態で存在し得る。DNA、特に二本鎖DNAの形態が特に好ましい。本発明のポリヌクレオチドは、化学合成によって生成され得る。本発明の好ましいポリヌクレオチド構築物は、組換え遺伝子技術によって作製される(例えば、Sambrook et al. “Molecular Cloning”, Cold Spring Harbor Laboratory, 1989参照)。
【0015】
本明細書で使用される「ポリジーン(polygenes)」とは、単一ORFにおいて生物活性のある少なくとも3つのポリペプチドをコードする核酸配列である。したがって、ポリジーンを構成する各「遺伝子」とは、ポリペプチド、特にタンパク質または、特異的な、特に構造的、制御的または酵素的機能を有するその断片、変異形、変異体または類似体をコードする核酸配列である。好ましくは、ポリペプチドをコードする「遺伝子」は、構造タンパク質、制御タンパク質もしくは酵素タンパク質またはその断片、変異形、変異体もしくは類似体をコードするcDNAのコード領域を含む。
【0016】
ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「断片」とは、もとのポリペプチドの領域の一部または一領域を意味するものであり、好ましくは、完全なタンパク質の機能の少なくとも1つを保持する断片を意味する。ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「変異形」とは、別の種から派生したもともとのポリペプチドの機能的または非機能的等価物であるポリペプチド、または代替的スプライシングもしくは翻訳後加工から生じるもともとのポリペプチドの機能的もしくは非機能的誘導体を意味する。ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「変異体」とは、自然発生しているタンパク質から、1つ以上のアミノ酸残基の挿入、置換、付加および/または欠失によって派生するポリペプチドを意味する。ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「類似体」とは、非関連アミノ酸配列さえも有し得るが、類似のポリペプチドと同一機能を発揮する、もともとのポリペプチドの機能的等価物を意味する。
【0017】
したがって、核酸レベルにおいて、遺伝子「断片」とは、「断片」が由来するもともとの遺伝子の一部または一領域である。遺伝子「変異形」は、もともとの遺伝子と比較して異なる種において見出される配列を有するかまたは、問題のポリペプチドのスプライシング変異形もしくは翻訳後加工されたバージョンをコードし得る。「変異体」は、1つ以上のヌクレオチドの挿入、置換、付加および/または欠失によって親遺伝子から派生する。遺伝子の「類似体」は、親遺伝子によってコードされるポリペプチドの機能的等価物をコードする。
【0018】
本発明に記載のポリジーン中の遺伝子のうち少なくとも2つは、非ウイルス由来である。「非ウイルス」とは、(機能的タンパク質またはその断片、変異形、類似体もしくは変異体を表す)ぺプチドをコードする核酸配列が、ウイルスのゲノム中に本来見出されないかまたは前記ゲノム由来ではないことを意味する。特に、本発明に記載のポリヌクレオチドのポリジーン中に含まれるヌクレオチド配列は、真核生物および/または原核生物由来である。
【0019】
したがって、本発明によると、多サブユニット集合体の(多タンパク質複合体または代謝経路のメンバーまたは互いに少なくとも一過性に少なくとも潜在的に相互作用するその他のタンパク質等の)サブユニットをコードする遺伝子は、少なくとも2つの翻訳領域(ORF)中に存在する。集合体のサブユニット(ポリペプチド)をコードする配列は、少なくとも2つのポリジーン中に存在し、その中で、各ポリジーンを構成する遺伝子は、プロテアーゼ切断部位をコードする(2つ以上あり得る)配列(すなわち、プロテアーゼの認識部位を含む配列)または少なくとも1つの自己切断ペプチドによって互いに接続されている。
【0020】
本発明の好ましい態様によると、ポリジーンを構成する遺伝子を接続する配列によってコードされる切断部位を切断することのできるプロテアーゼは、本発明のポリヌクレオチドによってコードされる。より好ましくは、プロテアーゼをコードする遺伝子は、ポリジーンの少なくとも1つの一部である。
【0021】
適切なプロテアーゼ切断部位および自己切断ペプチドは、当業者に公知である(例えば、Ryan et al. (1997) J. Gener. Virol. 78, 699−722; Scymczak et al. (2004) Nature Biotech. 5, 589−594参照)。プロテアーゼ切断部位の好ましい例は、ポティウイルスNIaプロテアーゼ(例えば、タバコエッチウイルスプロテアーゼ(tobacco etch virus protease))、ポティウイルスHCプロテアーゼ、ポティウイルスP1(P35)プロテアーゼ、ビオウイルス(byovirus)NIaプロテアーゼ、ビオウイルスRNA−2によりコードされるプロテアーゼ、アフトウイルスLプロテアーゼ、エンテロウイルス2Aプロテアーゼ、ライノウイルス2Aプロテアーゼ、ピコルナ3Cプロテアーゼ、コモウイルス(comovirus)24Kプロテアーゼ、ネポウイルス(nepovirus)24Kプロテアーゼ、RTSV(イネツングロ球状ウイルス(rice tungro spherical virus))3C様プロテアーゼ、PYVF(パースニップ黄色フレックウイルス(Parsnip yellow fleck virus))3C様プロテアーゼ、トロンビン、第Xa因子およびエンテロキナーゼの切断部位である。切断の高い厳密度により、TEV(タバコエッチングウイルス)プロテアーゼの切断部位が特に好ましい。したがって、本発明に記載のポリジーンの遺伝子は好ましくは、Xがいずれかのアミノ酸を表す一般式EXXYXQ(G/S)のアミノ酸配列をコードするヌクレオチド配列を含むヌクレオチドのストレッチによって接続される(TEVによる切断は、QとGとの間またはQとSとの間で生じる)。最も好ましいのは、ENLYFQGおよびENLYFQSをそれぞれコードするリンカーヌクレオチド配列である。
【0022】
好ましい自己切断ペプチド(「シス作用型加水分解性要素」(CHYSEL)とも呼ばれる;deFelipe (2002) Curr. Gene Then 2, 355−378参照)は、ポティウイルスペプチドおよびカルジオウイルス2Aペプチドに由来する。特に好ましい自己切断ペプチドは、FMDV(口蹄疫ウイルス)、ウマ鼻炎Aウイルス、ゾーシーアシグナウイルス(Thosea asigna virus)およびブタテスコウイルス(teschovirus)由来の2Aペプチドより選択される。
【0023】
本発明のポリジーンによりコードされるポリペプチドのうち少なくとも2つは、ポリジーンによってコードされる1つの他のポリペプチドと少なくとも一過性に相互作用することができるかまたは、ポリジーン中に含有される遺伝子によってコードされる別のポリペプチドと少なくとも一過性に相互作用することができると少なくとも思われる。ポリペプチド間で形成される典型的な「相互作用」には、共有結合、水素結合、静電相互作用およびファンデルワールス相互作用が含まれる。「一過性」の相互作用は、二分子、特にタンパク質に対して共通であり、典型的には、酵素とそれらの基質との間、受容体とそれらの(アゴニストまたはアンタゴニスト)リガンドとの間の相互作用、代謝経路のメンバー間の相互作用および制御性の(例えば、遺伝子制御性の)複合体のタンパク質間の相互作用によって表される。
【0024】
本発明のポリジーンを構成するヌクレオチド配列によってコードされるポリペプチドは、同一または異別であり得る。したがって、本発明の構築物中に存在する各ポリジーンは、関心対象のタンパク質をコードする各ヌクレオチド配列の1つ以上のコピーを含有し得る。この様式で、例えば、特に、タンパク質が通常、異なるレベルで発現するかおよび/または異なる化学量論における巨大分子集合体中に存在する場合における望ましいタンパク質の最適な発現のために機能する構築物を提供することが可能である。それゆえ、ポリペプチドが、通常使用される系においてほとんど発現しない場合、相当するコード配列の2つ以上のコピーが、本発明の構築物の1つ以上のポリジーン中へ組み込まれ得る。同一のアプローチは、ポリペプチドが、望ましい複合体中の二量体、三量体または多量体として存在する場合、使用され得る。この様式で、本発明の構築物は、当業者が発現させるかおよび/または精製することを望むいずれかの複合体または他の巨大分子集合体の必要条件(発現レベル、化学量論等)に従って、個々に集合され得る。
【0025】
ポリジーンを構成する遺伝子が、多タンパク質複合体のメンバーをコードする遺伝子および代謝経路のメンバーをコードする遺伝子からなる群より選択されることはさらに好ましい。好ましい多タンパク質複合体は、転写因子複合体等の遺伝子制御タンパク質複合体、核内および細胞内輸送に関与する複合体等の輸送複合体、タンパク質折りたたみ複合体、受容体/リガンド複合体、細胞−細胞間認識複合体、アポトーシスに関与する複合体、細胞周期制御に関与する複合体等である。代謝経路のメンバーは例えば、(解糖、糖新生、クエン酸回路、グリコーゲン生合成、ガラクトース経路、カルビン回路等の)炭水化物代謝、(トリアシルグリセロール代謝、脂肪酸の活性化、脂肪酸(偶数鎖/奇数鎖)のβ酸化、α酸化経路、脂肪酸生合成、コレステロール生合成等の)脂質代謝、グルタミン酸反応、クレブス−ヘンゼライト(Henseliet)尿素回路、シキミ酸経路、PheおよびTyr生合成、Trp生合成等の)アミノ酸代謝、(酸化的リン酸化、ATP合成、光合成、メタン代謝等の)エネルギー代謝、核酸代謝(プリンおよびピリミジンの生合成および分解、DNA複製等)のメンバーである。多タンパク質複合体のメンバーおよび代謝経路のメンバーは例えば、その開示内容が参照により本明細書に組み入れられる、http://www.biocarta.com/genes/index.aspおよびG. Michal (ed.) Biochemical Pathways, 1. edition, John Wiley & Sons, Hoboken, NJ, USA, 1999から採取され得る。
【0026】
本発明に記載の各ポリジーンは、少なくとも3つの遺伝子、すなわち生物活性のあるポリペプチドをコードする配列を含有する。より好ましいのは、4、5、6以上またはさらにより多くのタンパク質をコードするポリジーンである。上述のように、ポリペプチドと接続するプロテアーゼ切断部位を切断することのできるプロテアーゼは、ポリジーンの少なくとも1つによってコードされることが好ましい。
【0027】
好ましい態様によると、本発明のポリヌクレオチドは、ポリジーンの1つへ各々作用可能に連結され、したがってポリジーンの発現を調節することのできる少なくとも2つのプロモーター配列を含有する。本発明の構築物中の適切なプロモーターは、polh、p10およびpXIV最晩期バキュロウイルスプロモーター、vp39バキュロウイルス後期プロモーター、vp39polhバキュロウイルス後期/最晩期プロモーター、Pcap/polh、pcna、etl、p35、da26バキュロウイルス初期プロモーター;CMV、SV40、UbC、EF−1α、RSVLTR、MT、PDS47、Ac5、PGALおよびPADHからなる群より選択され得る。プロモーター配列は、全てのポリジーンに関して同一であり得るかまたは、異なるプロモーターが異なるポリジーンに関して選択され得る。
【0028】
好ましくは、本発明のポリジーンを含有する各ORFは、SV40、HSVtkまたはBGH(ウシ成長ホルモン)等の転写終結配列によって隣接される。
【0029】
本発明に記載のポリヌクレオチドは、エンハンサー配列または抑制因子配列等のさらなる制御配列を含有し得る。
【0030】
本発明に記載のポリヌクレオチドが、ベクターまたは宿主細胞中への組み込みのための少なくとも1つの部位を含有することはさらに好ましい。このような組み込み部位によって、(ウイルス等の)ベクターおよび宿主細胞(例えば、真核生物宿主細胞)中へのそれぞれへの簡便なゲノム組み込みまたは一過性組み込みが可能となる。ゲノム組み込みのための部位がより好ましい。
【0031】
特に好ましい組み込み部位は、ウイルス中へのポリヌクレオチドの組み込みと適合性のあるものである。より好ましくは、組み込み部位は、アデノウイルス、アデノ随伴ウイルス(AAV)、自律性パルボウイルス、単純ヘルペスウイルス(HSV)、レトロウイルス、ラジノウイルス、エプスタイン・バーウイルス、レンチウイルス、セムリキ森林ウイルスおよびバキュロウイルスからなる群より選択されるウイルス中へのポリヌクレオチドの組み込みに適合する。
【0032】
さらに好ましい態様において、組み込み部位は、(ヒト細胞、例えばHeLa、Huh7、HEK293、HepG2、KATO−III、IMR32、MT−2、膵臓β細胞、ケラチノサイト、骨髄線維芽細胞、CHP212、初代神経細胞、W12、SK−N−MC、Saos−2、WI38、初代肝細胞、FLC3、143TK−、DLD−1、臍帯静脈細胞、胚性肺線維芽細胞、初代包皮線維芽細胞、骨肉腫細胞、MRC5、MG63細胞等の)哺乳動物細胞、(CPL、FS−13、PK−15等の)ブタ細胞、(MDB、BT等の)ウシ細胞、(FLL−YFT等の)ヒツジ細胞、線虫細胞、(S.セレビシエ、S.ポンベ、C.アルビカンス、P.パストリス等の)酵母細胞、および(S.フルギペルダ(frugiperda)、例えばSf9、Sf21、Express Sf+、High Five H5細胞、ショウジョウバエ(D.melanogaster)、例えばS2シュナイダー細胞等の)昆虫細胞からなる群より好ましく選択され得る真核生物宿主細胞中へのポリヌクレオチドの組み込みに適合する。
【0033】
特に好ましい組み込み部位は、Tn7のトランスポゾン要素、λインテグラーゼ特異的結合部位およびSSR(部位特異的リコンビナーゼ)より選択され、好ましくはcre−lox特異的(LoxP)部位またはFLPリコンビナーゼ特異的組み込み(FRT)部位である。
【0034】
本発明の好ましい態様において、ポリヌクレオチドは、他の毒性物質に対する耐性に基づいて望ましい特性を有する宿主細胞を選択するための1つ以上の耐性マーカーをさらに含む。適切な耐性マーカーの例は、アンピシリン、クロラムフェニコール、ゲンタマイシン、スペクチノマイシンおよび/またはカナマイシンに対する耐性を供与するものである。
【0035】
原核生物宿主細胞中への組み込みのため、本発明のポリヌクレオチドは好ましくは、原核生物宿主中でpir遺伝子に依存した増殖をなすための条件的R6Kγ複製開始点を含む。
【0036】
本発明のポリヌクレオチドの特に好ましい態様は、WO2005/085456A1(PCT/EP2004/013381)に開示される構築物中への発現カセットとしてのポリジーンの挿入によって結果的に生じる。
【0037】
それゆえ、本発明のポリヌクレオチドが、
(a)MCS1に関してプロモーターP1および転写終結(terminator)配列T1によって隣接され、MCS2に関してプロモーターP2および転写終結配列T2によって隣接される多重クローニング部位MCS1またはMCS2を各々含む、頭−頭(head−to−head)、頭−尾(head−to−tail)のまたは尾−尾(tail−to−tail)の配置にある少なくとも2つの発現カセットT1−MCS1−P1およびP2−MCS1−T2
(b)少なくとも2つの制限部位AおよびBを含むプロモーターP1とP2との間にある少なくとも1つの増殖モジュールM
(c)発現カセットの1つと各々隣接する少なくとも2つの制限部位XおよびYを含み、ここで、
(i)制限部位AおよびX並びにBおよびYは互換性があるが、しかし
(ii)AYおよびBXの連結産物が、制限部位A、B、XおよびYに特異的な制限酵素a、b、xまたはyによって酵素的に切断可能ではなく、および
(iii)制限部位AおよびB並びに制限部位XおよびYは互換性がなく、ここで、各ポリジーンは発現カセットの1つ中へ挿入される次の式I
X−T1−MCS1−P1−[A−B]−P2−MCS2−T2−Y (I)
に記載の機能的配置を含むことが好ましい。
【0038】
式(I)の配置を有する構築物のさらに好ましい態様に関して、WO2005/085456 A1(PCT/EP2004/013381)に明確に参照されている。特に、増殖モジュールM中の制限部位AおよびBは、制限部位BstZ17I、SpeI、ClaIおよびNruIまたはそれらのイソ制限酵素(isoschizomer)によって切断される制限部位からなる群より選択される。イソ制限酵素は、同一の切断部位を有する制限酵素である。さらに、制限部位XおよびYの好ましい例は、PmeIおよびAvrIIからなる群より選択される制限部位または、そのイソ制限酵素によって切断される制限部位である。
【0039】
上述の式(I)中に含有される発現カセットのうちの1つ中へ挿入される各ポリジーンを有する特に好ましいポリヌクレオチドは、次の特徴を含む。
(a)プロモーターP1およびP2は、polhおよびp10からなる群より選択され;
(b)転写終結配列は、SV40およびHSVtkからなる群より選択され;
(c)増殖モジュールMにおける制限部位AおよびBは、制限部位BstZ171I、SpeI、ClaIおよびNruIからなる群より選択され;
(d)制限部位XおよびYは、制限部位PmeIおよびAvrIIからなる群より選択され、および
(e)ウイルス組み込みのための部位は、cre−loxおよびTn7からなる群より選択される。式(I)に記載の上述の配置を有するポリヌクレオチドの生成に関して、WO2005/085456 A1(PCT/EP2004/013381)に明確に参照されている。
【0040】
ポリジーンを各々含有する2つ以上のORF内に生物活性のあるいくつものポリペプチドをコードする本発明のポリヌクレオチドの提供は、多サブユニットタンパク質複合体のメンバーをコードする遺伝子のクローニングおよび発現に関し、主要な改良を提供する。一方で、結合されるべきコード配列の膨大なサイズおよび数のため、単一ORF中への全てのサブユニット遺伝子の集合はしばしば、不可能であるかまたは非常に困難である。もう一方で、個別の発現カセット中に各々存在する多サブユニット複合体のいくつものまたは全てのメンバーの効率的な集合はしばしば、非常に非効率的であるようになり、その理由は、全体的な複合体収量が、最少に発現するサブユニットによって決定されるからである。本発明によると、多タンパク質集合体のサブユニットは、少なくとも3つの(好ましくは非ウイルス由来の)ポリペプチドを各々コードする(各々単一のORFを表す)少なくとも2つのポリジーンによってコードされ、結果的に、構築物およびその構成体(特にポリジーンの集合体)の扱いやすいと(特に本発明のポリヌクレオチドが適切なベクター中に存在する場合の)発現効率性との間に最適な妥協を生じる。
【0041】
それゆえ、本発明のさらなる態様は、上述のポリヌクレオチドを含有するベクターである。ベクターは、プラスミド、発現ベクターおよびトランスファーベクターからなる群より選択され得る。より好ましくは、本発明のベクターは、真核生物の遺伝子導入、一過性またはウイルスベクター仲介性遺伝子導入に有用である。
【0042】
特に好ましいベクターは、アデノウイルス、アデノ随伴ウイルス(AAV)、自律性パルボウイルス、単純ヘルペスウイルス(HSV)、レトロウイルス、ラジノウイルス、エプスタイン・バーウイルス、レンチウイルス、セムリキ森林ウイルスおよびバキュロウイルスより選択されるウイルス等の真核生物発現ベクターである。本発明の最も好ましいベクターは、バキュロウイルス発現ベクターである。本発明の好ましいバキュロウイルスは、遺伝子v−cathおよびchiAが機能的に破壊される態様であり、その理由は、このことが、感染およびタンパク質発現の間、細胞内区画の維持の改良に至るからである。v−cath遺伝子は、キチナーゼをコードするウイルスDNAであるchiA上に並置された遺伝子に依存する過程による細胞死の際に活性化されるウイルスプロテアーゼV−CATHをコードする。両遺伝子は好ましくは、V−CATH活性を除去し、chiA遺伝子産物からの干渉なしでキチンアフィニティクロマトグラフィーを利用する選択肢を獲得するために破壊される。機能的に活性のあるv−cathおよびchiA遺伝子を欠失するバキュロウイルス系によって生じる発現産物の品質は、ウイルス依存性タンパク質分解活性および細胞溶解の低下のため、有意に改良される。
【0043】
好ましくは、本発明に記載のベクターは、SSRのための部位を含み、好ましくはcre−lox部位特異的組換えのためのLoxPを含む。より好ましくは、cre−lox部位は、バキュロウイルス遺伝子v−cathおよびchiAのうちの1つまたは両者の機能を破壊するため、それらの中に配置される。
【0044】
本発明のベクターは好ましくは、正確に集合されたベクターでうまく形質移入された宿主の選択のための1つ以上のマーカー遺伝子を含有する。適切なマーカー遺伝子の例は、ルシフェラーゼ、β−Gal、CAT、GFP、BFP、YFP、CFPおよびそれらの変異形等の蛍光タンパク質をコードする遺伝子、およびlacZα遺伝子である。マーカー遺伝子は、上述の例の機能的に等価の変異形、変異体、断片または類似体あるいは、当業者に公知の他の適切なマーカーであり得る。変異形、変異体または類似体は好ましくは、該変異形、変異体または類似体の由来するマーカーと比較して、アミノ酸レベルで少なくとも75%、より好ましくは85%、特に好ましくは90%、特に少なくとも95%の相同性を示す。
【0045】
別の好ましい態様において、本発明のベクターは、トランスポゾン要素、好ましくはTn7結合部位を含む。より好ましくは、このようなトランスポゾン要素、例えばTn7結合部位は、マーカー遺伝子内に配置され、それにより転位による組み込みの成功が、機能的マーカー遺伝子によって提供される表現型を検査することによって評価されることが可能となる。
【0046】
本発明の好ましいトランスファー(transfer)ベクターは、WO2005/085456 A1(PCT/EP2004/13381;そこにそれぞれ開示される配列番号1および2ならびに図1および2参照)に開示されるpFBDMまたはpUCDMに基づいている。本発明のさらに好ましいトランスファーベクターは、上述のpFBDMベクターおよびpUCDMベクターのそれぞれの誘導体に基づいている。
【0047】
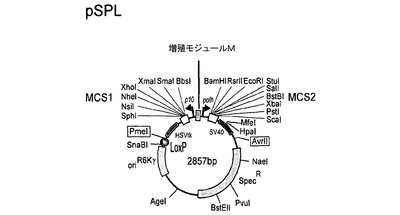
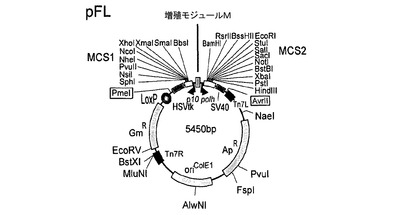
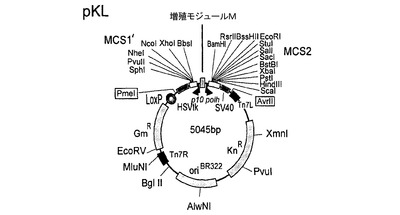
pFBDMおよびpUCDMの特に好ましい誘導体の例は、トランスファーベクターpSPL(図3)、pFL(図4)、pKL(図5)およびpKDM(図6)である。pUCDMのように、pSPLは、条件的な複製開始点(R6Kγ)を含有する。(pFBDMのように)pFLは、高コピー数の複製開始点(ColE1)を含有する。pKDMおよびpKLは、pBR322由来の低コピーの複製開始点を有する。pFBDMとの類似性において、pFL、pKLおよびpKDMは、トランスポゾン要素(Tn7R、Tn7L)を含有する。ベクターpSPL、pFLおよびpKLは、(pUCDMがそうであるように)二重発現カセットに隣接するLoxPの不完全な逆方向反復を有する。ベクターは全て、多遺伝子カセット1を生じるために、上述の増殖モジュール(M)を含有する。pFLおよびpKL(ならびに誘導体)は、受容ベクターであり、pUCDMおよびpSPL(ならびに誘導体)は、Cre仲介性プラスミド融合体における供与ベクターである。
【0048】
本発明の構築物を生じるためのトランスファーベクターに関する上述の好ましい例の重要な特徴は、次の表1に要約されている。
【0049】
【表1】
【0050】
それゆえ、本発明のポリジーンは、制限酵素切断および連結によるかまたは(例えば、BD In−Fusion酵素を使用する)組換えを介してのいずれかで、多重クローニング部位(MCS1およびMCS2)におけるpFBDM、pUCDM、pSPL、pFL、pKLまたはpKDM等のベクター中へと挿入される。バキュロウイルストランスファーベクターpFBDM、pUCDM、pSPL、pFL、pKLおよびpKDMは、改良されたタンパク質生成のために操作された修飾されたレシピエントバキュロウイルスDNAを含み、単純で迅速な方法により、この目的のために仕立てられたイー・コリ細胞中のバキュロウイルスDNAへの遺伝子の組み込みを2つのアクセス部位(attTn7およびLoxP)を介して可能にする。
【0051】
さらなる態様によると、本発明は、本発明のポリヌクレオチドおよび/またはベクターを含有する宿主細胞を提供する。
【0052】
好ましい宿主細胞の例は、ヒト、げっ歯類、CPL、FS−13およびPK−15等のブタの細胞、MDBおよびBT等のウシの細胞、FLL−YFT等のヒツジの細胞等の哺乳動物細胞、線虫(C.elegans)細胞、S.セレビシエ(S.cerevisiae)、S.ポンベ(S.pombe)、P.パストリス(P.pastoris)およびC.アルビカンス(C.albicans)等の酵母細胞、S.フルギペルダ由来の細胞等の昆虫細胞、好ましくはSf9、Sf21、Express Sf+またはHigh Five h5細胞、S2シュナイダー細胞等のショウジョウバエ由来の細胞、およびイー・コリ(E.coli)等の細菌、好ましくはTop10、Dh5α、DH10α、HB101、TG1、BW23473およびBW23474の系である。
【0053】
好ましいヒト細胞は、HeLa、Huh7、HEK293、HepG2、KATO−III、IMR32、MT−2、膵臓β細胞、ケラチノサイト、骨髄線維芽細胞、CHP212、初代神経細胞、W12、SK−N−MC、Saos−2、WI38、初代肝細胞、FLC3、143TK−、DLD−1、臍帯静脈細胞、胚性肺線維芽細胞、初代包皮線維芽細胞、骨肉種細胞、MRC5およびMG63細胞より選択される。
【0054】
本発明に記載のポリヌクレオチドおよび/またはベクターを含む宿主細胞は、単離された細胞であり得るかまたは、組織もしくは臓器中に存在し得る。
【0055】
本発明のさらなる態様は、本発明の少なくとも1つのポリヌクレオチド配列および/またはベクターで形質転換されている非ヒト遺伝子導入動物に関する。好ましい遺伝子導入動物は、げっ歯類、ブタ、ウシおよび線虫の種である。
【0056】
本発明の遺伝子導入動物は、多タンパク質複合体の役割の解明のために、またはインビボでの生物活性に関して化合物をスクリーニングするために特に有用である。
【0057】
本発明のさらなる態様は、
(a)少なくとも2つのポリジーンを構成する遺伝子のコード領域を提供、好ましくは増幅する工程;
(b)少なくとも1つのプロテアーゼ切断部位および/または少なくとも1つの自己切断ペプチドをコードする配列を該コード領域に提供する工程;
(c)単一ORFが結果として各ポリジーンをもたらすように工程(a)および(b)から結果的に生じる断片を集合させる工程;および
(d)少なくとも2つのポリジーンを単一ポリヌクレオチドに結合する工程
を含む、上述に定義されるポリヌクレオチドの生成のための方法である。
【0058】
本発明の別の局面は、
(a)(好ましくは、本発明のポリヌクレオチドの生成のための上述の方法によって)上述に定義される単一ORF内に少なくとも3つの遺伝子を各々含む少なくとも2つのポリジーンを発生させる工程;および
(b)ポリジーンをプラスミドベクターまたはウイルスベクター中へクローニングする工程;を含み、各ポリジーンの遺伝子のうちの少なくとも1つが非ウイルス由来であり、遺伝子によってコードされるポリペプチドのうちの少なくとも2つが遺伝子によってコードされる1つの他のポリペプチドと少なくとも一過性に相互作用することのできる、本発明に記載のベクターの生成のための方法である。
【0059】
好ましくは、ポリジーン中へ集合する遺伝子のうちの1つは、ポリジーンによってコードされるポリペプチドを接続するプロテアーゼ切断部位を切断することのできるプロテアーゼをコードする遺伝子である。好ましいプロテアーゼは、上述に定義されるとおりである。
【0060】
本発明のポリジーンおよびベクターの構築は、当業者に一般的に公知である多様な分子生物学的技術を通じて実施されることが可能である(例えば、Ausubel et al. (eds.) Current Protocols in Molecular Biology, John Wiley & Sons, Hoboken, NJ, USA, 2003参照)。ポリジーンの生成は、例えば、(例えば、対応するcDNAテンプレートを使用して)特定のポリペプチドをコードするヌクレオチド配列のPCR増幅によって、好ましくは、少なくとも1つのプロテアーゼ切断部位および/または少なくとも1つの自己切断ペプチドをコードする配列を提供するプライマー(5’または3’のいずれか)の使用によって実施され得る。好ましくは、プライマーは、適切な制限酵素の認識配列をさらに含有する。好ましくは、各プライマーは、他のプライマーの制限部位とは異なる制限部位を含有し、それにより結果的に生じる増幅産物を別の増幅産物および/または同一制限部位を含有する線形化されたベクターと指向性の連結をすることが可能である。指向性の連結による構築物の生成のための別の好ましい態様によると、プライマーは、RsrIIまたはBstEII等の自己連結可能ではないオーバーハングを生じる制限酵素の認識配列を含有し得る。プライマー自体が制限部位を含有しない場合、望ましい制限部位を含むアダプターとともに増幅産物を提供することも可能である。もちろん、(増幅のほかに)いずれかの供給源を使用して、望ましいポリペプチドをコードする領域を提供することも可能である。例えば、必要とされる配列は、それ自体既に存在し得るかまたは、配列が適切な制限酵素によって切り出され得る対応するベクター中に既に存在し得る。適切な制限部位等を含有しない構築物は、必要とされる要素を含有する適切なアダプターの連結によって、適切な配列(制限部位、プロテアーゼ切断部位/自己切断可能なペプチド、リンカー等)とともに提供され得る。次に、増幅産物または(制限部位を含有する)その他の適切な構築物が適切な制限酵素で切断される。もちろん、使用されるいずれかのプライマーの配列は、好ましくは適切なベクター中へのポリジーンの最終的な集合後、単一ORFが各ポリジーンを結果として生じるよう、好ましく選択される。
【0061】
本発明の好ましい態様によると、次に、増幅産物または他の適切な配列は、適切なベクター中へ、例えば、上述に参照されるMultiBacベクター系のMCS1またはMCS2で連続してまたは同時に連結され得る。例えば、(WO2005/085456 A1(PCT/EP2004/013381)に記載されるように)1つのポリジーンは、pFDBM中へ導入され得、別のポリジーンは、pUCDM中へ連結され得る。次に、その結果得られる構築物は、関心対象の多タンパク質複合体の発現および生成のために対応する昆虫細胞の感染のために用意されるバキュロウイルス発現ベクターを生じるcre−lox部位特異的組換え(pUCDM誘導体)およびTn7特異的転位(pFBDM誘導体)によって、対応するバクミド(bacmid)の生成のために使用される。
【0062】
ポリジーンを生じ、これらを適切なベクター中へ、制限/連結によって導入するほかに、適切なリコンビナーゼを使用する相同的組換えによってこのような構築物を構築させることも可能である。リコンビナーゼに基づくクローニング技術の適切な例は、BD Biosciences Clontech, Heidelberg, Germany (see Clontechniques, October 2002, p. 10)から市販されているIn−Fusion(登録商標)系およびGene Bridges GmbH, Dresden, Germany (see WO−A−99/29837; http://www.genebridges.com)から市販されているRed(登録商標)/ET(登録商標)組換え系である。
【0063】
In−Fusion(登録商標)系は、対応する相同性領域を有する(適切なベクター等の)線形構築物中へ融合されるべきDNA分子中に15bpの相同性領域を必要とする。したがって、本発明のポリジーンを含有するベクターは、15bpの適切な相同的配列とともに、(プロテアーゼ切断部位および/または自己切断ペプチドをコードするリンカー配列とともに)構成しているコード領域を提供することによって、連続してまたは同時に集合され得る。もし望まれれば、15bpの相同的配列は、構成しているコード領域が望ましい順序で集合されるよう選択され得る。もちろん、適切な相同性領域は、望ましい配列を含有するプライマーを使用する、対応する断片のPCR増幅によって導入され得る。
【0064】
Red(登録商標)/ET(登録商標)組換え系は、40〜60塩基対の相同性配列が必要とされるIn−Fusion(登録商標)系とは異なるが、断片が挿入されるべき構築物は、線形である必要がない。組換えは、宿主中で、好ましくは二重リコンビナーゼ系「ET」(RecE/RecTまたはRedα/Redβ)を発現するイー・コリ系でインビボで実施される。したがって、ポリジーンを構成する断片は、適切なベクターとともに適切な宿主中へ、好ましくはイー・コリ細胞中へ直接形質転換され得る。この様式で、各ポリジーンは、(好ましくは、多タンパク質複合体の各メンバーポリペプチド+少なくとも1つのプロテアーゼ切断部位配列/自己切断ペプチド配列に関するコード領域を含む)断片ごとに連続的に、または全ての断片の同時形質転換のいずれかによって適切なベクター中へ集合され得る。
【0065】
本発明の特に好ましいポリヌクレオチド構築物は、少なくとも同様の長さのポリジーンを含有する。なぜならば、このような構築物から対応するポリタンパク質(polyproteins)を発現させると、かなりの発現レベルを結果的に生じることを本発明者が発見したからである。本発明によると、「同様な長さのポリジーン」とは、ポリジーンのヌクレオチド配列の長さが、50%以下まで、より好ましくは30%以下まで、特に20%以下までまたはさらにより少なく互いに異なることを意味する。
【0066】
さらに、本発明は、
(a)ポリジーンの発現を可能にする条件下で適切な培地中で本発明に記載の宿主細胞を培養する工程;および
(b)ポリジーンによってコードされる発現産物を培地および/または宿主細胞から回収する工程
を含む、インビトロでの多タンパク質複合体の生成のための方法を提供する。
【0067】
本発明は、
(a)上述に定義される単一ORF内で少なくとも3つの遺伝子を各々含む少なくとも2つのポリジーンを発生させる工程;および
(b)ポリジーンを動物中へ形質転換し、それによりポリジーンが該動物中で発現される工程
を含む、インビボでの多タンパク質複合体の生成のための方法にも関する。
【0068】
好ましくは、工程(b)に従ったポリジーンによる動物の形質転換は、ベクター、特にウイルスベクター、より好ましくはバキュロウイルスベクターによって生じる。バキュロウイルスは、哺乳動物手中へのポリジーンの送達に特に有用な媒体である。上述のインビボでの方法は好ましくは、哺乳動物、線虫または昆虫において実施される。適切な動物種の特に好ましい例は、本明細書で上述に定義される。
【0069】
本発明の態様は、タンパク質の多サブユニット集合体に対して向けられるワクチンの調製にも有用である。多サブユニットの複合体はしばしば、個々のタンパク質と比較して異なるエピトープを呈する。それゆえ、本発明に従って生成される多タンパク質複合体は、より適切な様式で自然発生する関連エピトープを呈し、したがって、抗体産生のためのより良好な抗原標的を提供する。
【0070】
近年、重症急性呼吸器症候群(SARS)コロナウイルス由来の4つのタンパク質からなるビリオン様粒子(VLP)が、組換えバキュロウイルス発現ベクターを使用して作製された(Mortola et al. (2004), FEBS Lett. 576, 174−178参照)。ワクチンの調製のためのこのような感染性粒子の効果的な発現は、本発明に従ったポリジーン発現系を使用して大いに促進されるであろう。特に、SARS−VLPの例よりも実質的に多くのポリペプチドを含有する多サブユニット集合体の高収量の発現は、本発明の発現ツールによって利用可能とされる。
【0071】
それゆえ、本発明はさらに、
(a)本発明の少なくとも1つのポリヌクレオチドおよび/またはベクターを哺乳動物へ投与し、それにより本発明のポリジーンが哺乳動物内で発現する工程;
(b)アジュバントを哺乳動物へ場合により投与する工程;および
(c)ポリジーンによりコードされるポリペプチドの少なくとも1つに特異的な抗体および/または前記抗体を産生する脾臓細胞を場合により単離する工程
を含む、ワクチンの生成のための方法に関するものである
【0072】
本発明は、多タンパク質集合体の組換え生成のための簡便かつ単純なアプローチを提供する。これらの多サブユニット集合体は、タンパク質複合体相互作用または他サブユニット集合体を構成するタンパク質の修飾に関して検査され得る。本発明に従って生成される多サブユニット集合体は、医学的価値のある生物学的に有意な活性を発揮し得る候補化合物(有機小分子、核酸、ペプチド、ポリペプチド等)との相互作用に関してもアッセイされ得る。
【0073】
それゆえ、本発明は、タンパク質複合体相互作用またはタンパク質修飾をアッセイするための方法にも関するものである。
【0074】
好ましい態様によると、本発明は、
(a)少なくとも2つのポリジーンを含有する本発明に従った宿主細胞を提供する工程;
(b)ポリジーンの発現を可能にする条件下で宿主細胞を維持する工程;および
(c)ポリジーンによりコードされるポリペプチド間の相互作用または前記ポリペプチドの修飾を検出する工程
を含む、タンパク質複合体の相互作用またはインビトロでの多タンパク質複合体の修飾をスクリーニングするための方法を提供する。
【0075】
本発明の別の好ましい態様は、(i)多タンパク質複合体と相互作用することおよび/または(ii)多タンパク質複合体内でのタンパク質の修飾かおよび/または(iii)多タンパク質複合体内でのもしくは多タンパク質複合体間の相互作用を阻害するかおよび/または多タンパク質複合体内でタンパク質の修飾を阻害することの可能な候補化合物のインビトロでのスクリーニングのための方法であり、
(a)少なくとも2つのポリジーンを含有する本発明に従った宿主細胞を提供する工程;
(b)ポリジーンの発現を可能にする条件下で宿主細胞を維持する工程;
(c)候補化合物を宿主細胞と接触させる工程;
(d)発現産物と候補化合物との相互作用および/または発現産物間の相互作用および/または発現産物の修飾および/または発現産物間の相互作用の阻害を検出する工程
を含む。
【0076】
本発明のポリヌクレオチドおよび/またはベクターは、タンパク質−タンパク質、タンパク質−(多)タンパク質複合体または多タンパク質複合体−多タンパク質複合体の相互作用またはインビボでの多タンパク質複合体の修飾(リン酸化、グリコシル化等)のスクリーニングにも適している。
【0077】
したがって、本発明のさらに好ましい態様は、(i)多タンパク質複合体と相互作用すること、および/または(ii)多タンパク質複合体内でのタンパク質の修飾および/または(iii)多タンパク質複合体内または多タンパク質複合体間の相互作用を阻害することおよび/または多タンパク質複合体内でタンパク質の修飾を阻害することのできる候補化合物のインビボでのスクリーニングのための方法であり、
(a)上述に定義される少なくとも2つのポリジーンを含有する本発明の少なくとも1つのポリヌクレオチドおよび/またはベクターを含む動物を提供し、それによりポリジーンが動物中で発現する工程;
(b)候補化合物を動物へ投与する工程;および
(c)発現産物と候補化合物との相互作用および/または発現産物間の相互作用および/または発現産物の修飾および/または発現産物間の相互作用の阻害を検出する工程
を含む。
【0078】
本発明の多タンパク質発現ツールは、医学的にも使用される。特に、生物活性のある多タンパク質複合体ならびにタンパク質の医学的に有利な組み合わせ、例えば抗体混合物は、場合によりインターロイキンおよび/またはアジュバントと組み合わせて本発明のポリヌクレオチドおよび/または遺伝子送達ベクターによって、動物またはヒトへ投与されることが可能である。
【0079】
したがって、本発明はさらに、遺伝子治療のためのポリジーントランスファー媒体を含む医薬の調製のための、上述のポリヌクレオチドおよび/またはベクターおよび/または宿主細胞の使用に関する。
【0080】
治療適用のための遺伝子送達系を開発するために、多大な尽力がなされている。遺伝子治療は、強い熱意の焦点であったが、過去には非難もあった。今日まで、ヒト疾病に関するインビボおよびエクスビボでの安全で適用可能な臨床的戦略を達成するための過程において、臨床治験における遺伝子治療を評価する上での主要な進展があった(Worgall S. (2004) Peadiatr. Nephrol.参照)。全般的に、遺伝子治療はいまや、遺伝子の補正のための、および治療用遺伝子産物の持続的発現または一過性発現を引き起こす獲得された疾患のための非常に見込みのある手段として有効である(Worgall S.、上述)。ウイルスをベースとした組換えベクター、特に哺乳動物宿主中で複製形質転換受容性ではないものは、哺乳動物細胞遺伝子送達のための強力なツールとして近年出現し、ヒト、霊長類、げっ歯類、ウシ、ブタおよびヒツジの細胞を含む哺乳動物細胞系の全範囲へうまく適用されている(Kost and Condreay (2002) Trends Biotech. 20, 173−180に概説)。エクスビボおよびインビボの両者で複雑な遺伝子トランスファー/治療効果を得るため、単一遺伝子治療によるものよりもむしろ組み合わさった遺伝子治療によるより強力な結果を達成するために、多シストロン性のウイルスベクターに関する需要が高まってきた(de Felipe (2002), Curr. Gene Ther. 2, 355−378; Planelles (2003) Meth. MoI. Biol. 229, 273−284)。例えば、補体の系による不活性化を遮断するためにインビボで投与されるべき担体ウイルス中へのアクセサリー遺伝子の組み込みの要求も、ヒト崩壊促進因子タンパク質ドメイン融合を運搬したバキュロウイルスgp64外被タンパク質を有する偽型バキュロウイルスを使用することによって示されており(Hueser et al. (2001) Nat. Biotech. 19, 451−455)、治療目的のためにトランスファーされるべき多重遺伝子に加え、ウイルス産生レベルに関する組換え修飾を提供する必要性を実証している。
【0081】
したがって、本発明の組換えバキュロウイルスは、遺伝子治療製剤を調製するために好ましい。より好ましくは、本発明の製剤のために使用されるベクターは、
(i)1つ以上の治療用ポリペプチドおよび
(ii)1つ以上のバキュロウイルスタンパク質
をコードする上述に定義される少なくとも2つのポリジーンを含むバキュロウイルスである。
【0082】
好ましい態様において、(ii)に従ったタンパク質は、偽型バキュロウイルスから発現されるヒト化バキュロウイルスタンパク質、好ましくは、ヒト化バキュロウイルス外被タンパク質gp64、例えば崩壊促進因子等のヒトタンパク質と融合されたgp64である。
【0083】
さらに、本発明は、
(a)本発明に従ったポリヌクレオチドを含むポリジーントランスファー媒体を提供する工程;および
(b)遺伝子疾患を罹患している患者へポリジーントランスファー媒体を投与する工程
を含む、インビボでの遺伝子治療法に関するものである。
【0084】
本発明はさらに、
(a)遺伝子疾患を罹患している患者の細胞を回収する工程;
(b)本発明に従ったポリヌクレオチドを含むポリジーントランスファー媒体で、回収された細胞を形質転換する工程;および
(c)形質転換された細胞を患者へ投与する工程
を含む。
【0085】
本発明に従って生成される多タンパク質複合体は、生物物理的研究、特に結晶学的、電子顕微鏡的および/またはNMRの技術を使用する構造研究、特にタンパク質−タンパク質相互作用のための、および薬物開発のためのタンパク質化学的研究において有利に使用され得る。
【0086】
したがって、本発明は、多タンパク質複合体の結晶化のための、本発明のポリヌクレオチドおよび/またはベクターおよび/または宿主細胞の使用に関する。
【0087】
本発明のさらなる態様は、
(a)ポリジーンを構成するコード配列のPCR増幅のためのプライマー;
(b)プラスミドベクターまたはウイルスベクター;および
(c)場合により、プラスミドまたはベクターの増殖に適した宿主細胞
を含む多タンパク質複合体の調製のためのキットである。
【0088】
プライマーは、各ポリジーンのための単一ORFを生成するための要求と適合するよう簡便に設計され、プラスミドベクターまたはウイルスベクター中へ連結のための制限部位を(連続的または同時に)含有し得るかおよび/またはプライマーは、(例えば、上述のIn−Fusion(登録商標)系またはRed(登録商標)/ET(登録商標)系を使用する)相同的組換えによってプラスミドベクターまたはウイルスベクター中へポリジーンを集合させるかおよび/または挿入するための配列を含有し得る。
【0089】
本発明は、次の制限のない実施例によってさらに説明される。
【0090】
実施例1:ポリジーンの生成および発現ベクター中への連結
ポリジーンを発生させる原理は、ヒトTATAボックス結合タンパク質(hTBP)中心(hTBPc、位置159で切断された全長タンパク質のc末端断片)を使用することによってここに示される。RsrII制限部位を有するオーバーハングを含有する遺伝子の5’末端にアニールし、アミノ酸スペーサーおよびタバコエッチングウイルス(TEV)切断部位をさらにコードするセンスプライマーを使用するポリメラーゼ連鎖反応(PCR)によって、hTBPcをコードする遺伝子を増幅した。アンチセンスプライマーは、遺伝子の3’末端へアニールし、RsrII制限部位を含有した。RsrIIは、自己連結しない3ヌクレオチドの非対称性オーバーハングを生じる制限酵素であり、制限産物は非対称性であり、連結は方向性産物を生じる。PCR産物をRsrIIで消化し、精製した。PCR産物のDNA(配列番号1)および推定アミノ酸配列(配列番号2)を図1に示す。
【0091】
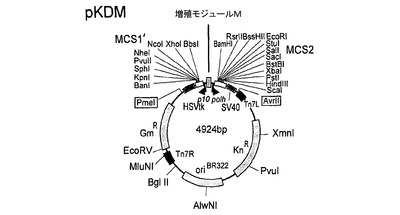
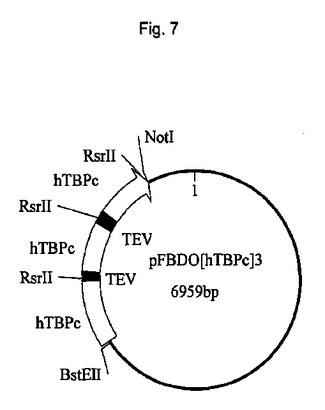
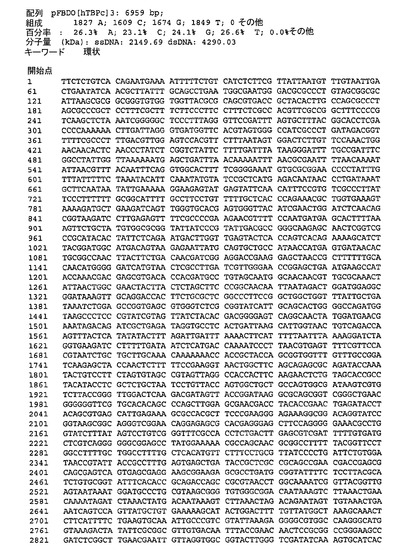
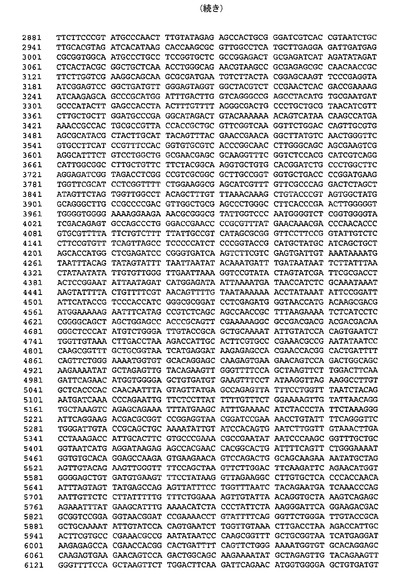
連結は、図2に示されるhTBPcのコンカタマーを生じた。適切なベクター中へのインビトロでの連結反応混合物のサブクローニングは、TEVプロテアーゼ切断部位によって分離される単一ポリタンパク質中で1、2、3および5のhTBPタンパク質をコードするポリジーンを含有する発現構築物を生じた。結果として生じる発現ベクターのうちの1つ(pFBDO[hTBPc]3)の模式的呈示およびヌクレオチド配列(配列番号3)を図3および図4にそれぞれ示す。
【0092】
実施例2:ヒト一般的転写因子のサブユニットをコードするポリジーンを含有するバキュロウイルストランスファーベクターの生成
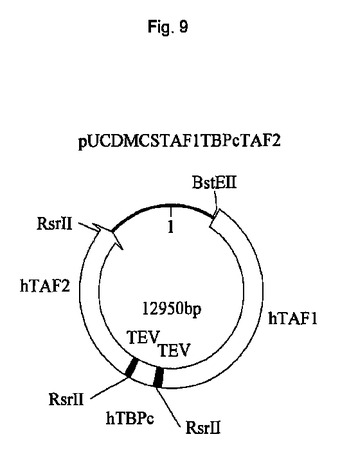
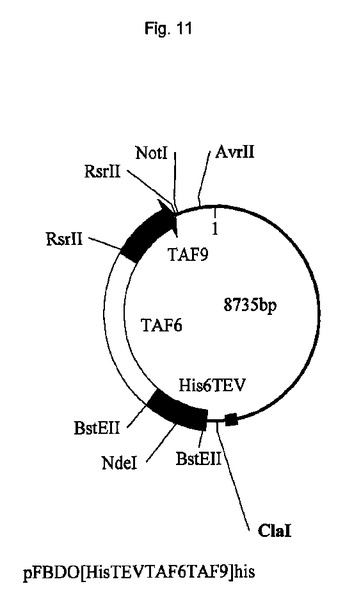
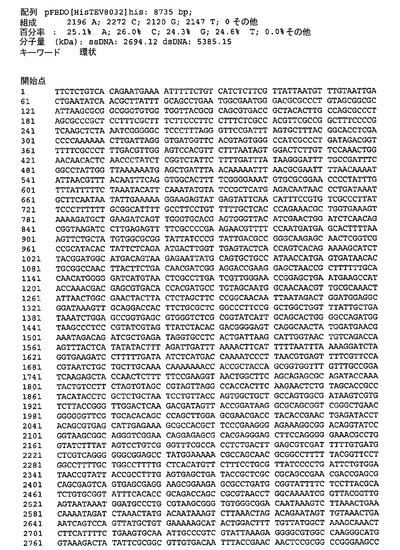
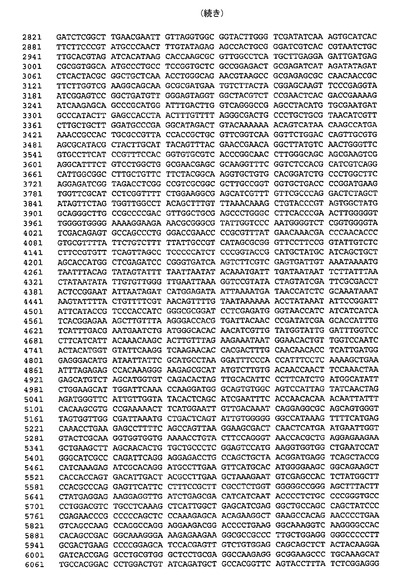
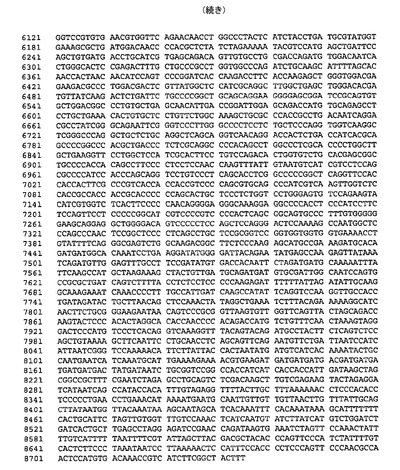
遺伝子が、バキュロウイルス発現のためのトランスファーベクターpUCDM(WO2005/085456 A1(PCT/EP2004/013381)参照)中へ挿入されたhTBPcに加え、ヒトTBP関連因子hTAF1およびhTAF2を含むポリタンパク質をコードする、アミノ酸スペーサーおよびTEVプロテアーゼ部位をコードする配列によって分離されている、ポリジーンを生成した。結果として生じる構築物pUCDMCSTAF1TBPcTAF2の模式的呈示を図9に示す。構築物のヌクレオチド配列を図10(配列番号4)に示す。バキュロウイルス発現のためのトランスファーベクターpFBDM(WO2005/085456 A1(PCT/EP2004/013381)参照)中へ挿入されたTEVプロテアーゼ及びヒトTBP関連因子hTAF6およびhTAF9を含むポリタンパク質をコードするポリジーンを含有するさらなる構築物を生成する。遺伝子は、アミノ酸スペーサーおよびTEVプロテアーゼ部位をコードする配列によって分離されている。結果として生じる構築物pFDDO[HisTEVTAF6TAF9]hisの模式的呈示を図11に示す。この構築物のヌクレオチド配列を図12(配列番号5)に示す。
【0093】
実施例3:バクミド構築物の調製、昆虫細胞の感染およびタンパク質発現
上述の2つのポリジーンを含むバクミド構築物の構築のため、構築物pUCDMCSTAF1TBPcTAF2(pUCDM誘導体)およびpFDDO[HisTEVTAF6TAF9]his(pFBDM誘導体)を各々、WO2005/085456 A1(PCT/EP2004/013381)の実施例5(pUCDMCSTAF1TBPcTAF2用;Cre−lox部位特異的組換え)および実施例6(pFDDO[HisTEVTAF6TATAF9]his用;Tn7転位)に記載されているDH10MultiBacCre細胞中へ導入した。望む場合、一工程転位/cre−lox部位特異的組換えは、同様にWO2005/085456 A1(PCT/EP2004/013381)において記載されているDH10MultiBacCre細胞で実施され得る。確立されたプロトコールに従って、バクミド調製、昆虫細胞の感染およびタンパク質発現を実施した(例えば、O’Reilly et al. (1994) “Baculovirus expression vectors. A laboratory manual“ Oxford University Press, New York − Oxford; ”Bac−to− BacTM Baculovirus Expression Systems Manual” Invitrogen, Life Technologies, Inc., 2000参照)。
【0094】
次の配列リストは、本明細書の一部であり、その中で、配列は次のとおりである。
【0095】
配列番号1は、図1に示されるヒトTATAボックス結合タンパク質(hTPB)中心(hTBPc、位置159で切断された全長タンパク質のc末端断片)をコードするPCR産物のヌクレオチド配列である。
【0096】
配列番号2は、図1に示されるヒトTATAボックス結合タンパク質中心(hTBPc)のアミノ酸配列である。
【0097】
配列番号3は、図8に示されるpFBDO[hTBPc]3のヌクレオチド配列である。
【0098】
配列番号4は、図10に示されるpUCDMCSTAF1TBPcTAF2のヌクレオチド配列である。
【0099】
配列番号5は、図12に示されるpFBDO[HisTEVTAF6TAF9]hisのヌクレオチド配列である。
【図面の簡単な説明】
【0100】
【図1】図1は、ヒトTATAボックス結合タンパク質(hTPB)中心(hTBPc、位置159で切断された全長のタンパク質のc末端断片)をコードするPCR産物のヌクレオチド配列(配列番号1)および推定アミノ酸配列(配列番号2)を示す。(プライマー配列中に存在する)RsrII制限部位の位置が示されている。
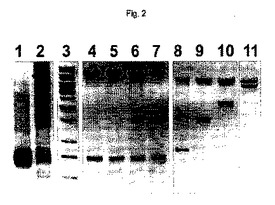
【図2】図2は、hTBC遺伝子セグメントのインビトロでの連結および混合物のサブクローニングのアガロースゲル電気泳動分析の写真を示す。PCRで増幅されたhTPBc遺伝子をRsrIIで消化し、精製した(レーン1)。リガーゼとのインキュベーションは、各1つのORF中で連結された1、2、3およびそれより多くの遺伝子を含有するコンカテマーのラダーを生じる(レーン2、レーン3はMBI DNAマーカー1kbラダーである)。1つのポリジーンを含有するこのように生じた発現構築物の混合物のサブクローニングは、RsrIIを使用して制限消化することによって遊離されることが可能である連結されたhTBPc遺伝子の異なる数を有する(レーン4〜7)。挿入されたポリジーンの外側の消化は、各場合において単一のORFを生じた1(レーン8)、2(レーン9)、3(レーン10)および5(レーン11)のhTBPc遺伝子を証明する。
【図3】図3は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpSPLの模式的呈示を示す。
【図4】図4は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpFLの模式的呈示を示す
【図5】図5は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpKLの模式的呈示を示す。
【図6】図6は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpKDMの模式的呈示を示す。
【図7】図7は、トランスファーベクター構築物pFBDO[hTBPc]3の模式的呈示を示す。
【図8】図8は、pFBDO[hTBPc]3(配列番号3)のヌクレオチド配列を示す。
【図9】図9は、トランスファーベクター構築物pUCDMCSTAF1TBPcTAF2の模式的呈示を示す。
【図10】図10は、pUCDMCSTAF1TBPcTAF2(配列番号4)のヌクレオチド配列を示す。
【図11】図11は、トランスファーベクター構築物pFBDO[HisTEVTAF6TAF9]hisの模式的呈示を示す。
【図12】図12は、pFBDO[HisTEVTAF6TAF9]his(配列番号5)のヌクレオチド配列を示す。
【技術分野】
【0001】
本発明は、ポリジーン(Polygenes)を構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、非ウイルス性由来であり、ポリジーンを構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、該ポリジーンの遺伝子によってコードされる少なくとも1つの他のポリペプチドと少なくとも一過性に相互作用することが各々可能であり、および各ポリジーンを構成する遺伝子が、少なくとも1つのプロテアーゼ切断部位をコードする配列によって、および/または少なくとも1つの自己切断ペプチドをコードする配列によって互いに接続される、単一翻訳領域(ORF)内で各々、少なくとも3つの生物活性のあるポリペプチドを各々コードする少なくとも2つのポリジーンをコードする組換えポリヌクレオチドに関するものである。本発明のさらなる態様は、組換えポリペプチドを含有するベクター、組換えポリヌクレオチドおよび/またはベクターを含有する宿主細胞ならびに組換えポリヌクレオチドおよび/またはベクターで形質転換された非ヒト遺伝子導入動物である。本発明は、ポリヌクレオチドの生成のための、および多タンパク質複合体の製造のための方法にも関するものである。本発明の態様は、遺伝子治療、薬物候補スクリーニング、ワクチン産生および構造研究のための多タンパク質複合体の結晶化において特に有用である。
【0002】
ポストゲノム時代における生物学的研究の尽力に関する強い焦点は、タンパク質の相互作用ネットワーク(相互作用体(interactome))の解明である。同定された多タンパク質(multiprotein)複合体の多くが、ネイティブな細胞中で詳細な分子生物学的分析のための十分量では存在しないため、前記複合体の研究は、大規模な異種性タンパク質生成のための組換え技術に依存している。組換え発現方法は、多タンパク質発現前に、労力および材料の両者において不相応な投資を必要とし、発現後には、改定された発現研究のための多タンパク質構成分を迅速に変化させるための柔軟性を提供しない。
【0003】
多サブユニット複合体を得るために近年使用されている組換え技術が幾多もある。例えば、タンパク質は、可溶性形態または封入体としてのいずれかで、イー・コリィ(E.coli)中での単離物で発現され、精製された後、インビトロで同様に生成されたタンパク質を使用して多タンパク質複合体中に再構成されることが可能である。真核生物細胞(例えば、哺乳動物細胞または酵母細胞)も、一過性発現実験において宿主として使用されることが可能である。この方法論は、効率的なインビトロでの再構成プロトコールの存在に完全に依存する。この戦略が、小さなサブユニットサイズを有するより単純な系については許容可能な結果を生じ得るのに対し、多くの、また、大きなサブユニットを含有するより複雑化した(例えば、より高次の真核動物の、特にヒトの制御複合体の全てに近い)多タンパク質複合体に関しては一般的に適用可能ではない。
【0004】
同時発現は、上述に概略されるインビトロでの再構成の戦略に代わる優れたものとして認識されてきた。幾つもの同時発現系が、原核細胞発現および真核細胞発現両者に関して過去に開発されている。原核細胞系において、同時発現は、選択された遺伝子全てを含有する単一プラスミドを産生することによって、または1つもしくは2つの遺伝子を含有する幾つものプラスミドならびに異なる耐性マーカーおよびレプリコンを同時形質転換することによって達成されることが可能である。
【0005】
真核細胞における同時発現は、バキュロウイルス系を使用することによって具現化されており、初期には、幾つものウイルスによる同時感染によって成功が制限されていたが、後には単一ウイルスから全てのタンパク質を発現させることによってさらに成功し、多くの利点を提唱し、(比較的小さなサブユニットのサイズ、真正のプロセッシングの欠如、真核生物(特にヒト)タンパク質の困難な発現等の)原核細胞系に存在するいくつもの制限を排除した。バキュロウイルス系に関し、単一ウイルスからの発現は、収量を劇的に増大させることが示されている(Berger et al. (2004) Nature Biotech. 22, 1583−1587; Comment (2004), Nature Biotech. 22, vii, New & Views (2004), Nature Biotech. 22, 152, Research Highlights, Nature Methods 2, 7 (2005); Bertolotti−Ciarlet et al. (2003) Vaccine 21 , 3885−3900も参照)のに対し、特に、大規模な産生に関してロジスティックな需要を決定的に低下させる。
【0006】
多タンパク質発現の主要な改良は、WO 2005/085456 A1 (PCT/EP2004/013381;Berger et al. (2004)上述も参照)に開示されている、本発明者によって提供される多重遺伝子発現カセットの作製のためのモジュラー系の提供であった。WO 2005/085456 A1 (PCT/EP2004/013381)によって記載されているMultiBac技術によって、多重遺伝子発現カセットの単純な作製、ならびに発現実験の改変および修正が可能となる(Berger et al. (2004)上述)。
【0007】
しかしながら、特に(真核生物、例えばヒトの遺伝子制御複合体の大部分を構成する)多くの(6、7、8またはそれより多くの)サブユニットを有する多サブユニット複合体の発現の成功およびインビボでの集合に関する障害は、これらのサブユニットの相対的発現レベルが多くの場合、完全には理解されていない機構(例えば、転写および翻訳の効率、タンパク質の安定性、mRNAの安定性および二次構造等)に基づいて典型的に有意に異なるという事実において見出される。結果として、本質的に非平衡の系において、最少量で発現するサブユニットは、複合体の全体的な収量を制限することによって、多サブユニット複合体生成実験の全体的な成功に影響するであろう。したがって、転写/翻訳装置は、過程に組み込まれていない他の成分の過剰量を生成し、したがって細胞内の転写/翻訳資源を「無駄にする」。個々の発現レベルは典型的に、互いに関して何倍も(例えば、最大10倍以上まで)変動し、特に(例えば、多くの真核生物遺伝子制御複合体の場合において、)5、6、8、10以上のサブユニットなど、4個を超える複合体の場合、望ましい多サブユニット複合体の生成の成功に対して抵抗性のある損失を伴う。
【0008】
ピコルナウイルススーパー群のウイルスは、ウイルスプロテアーゼの切断部位によって、または自己切断ペプチドによって互いに接続されるウイルスタンパク質を含むポリタンパク質をコードする単一のまたは2つのORFを含有するセンスの向きでの一本鎖RNA分子からなるゲノムを有する(例えば、Ryan et al. (1997) J. Gener. Virol. 78, 699−723において概説されている)。
【0009】
タンパク質複合体の生成に関して、組換えウイルスを通じてポリタンパク質を発現させる一般的な概念は、TCR(T細胞受容体):CD3複合体の再構成において適用されている(Szymczak et al. (2004) Nature Biotech. 5, 589−594)。著者は、2つの組換えレトロウイルスベクターを使用し、その中で、1つのベクターは、2つのTCRサブユニットをコードする配列を含有したのに対し、他のベクターは、4つのCD3サブユニットを含むポリタンパク質をコードした。サブユニットは、アフトウイルス由来の自己切断2Aペプチド配列によって接続された。このアプローチの1つの不利な点は、完全な複合体を再構成するために、2つの別個のベクターが調製されなければならず、2回の形質移入が必要であることである。
【0010】
ウイルスポリタンパク質のアプローチは、ヘテロ二量体IL−12(Kokuho et al. (1999) Vet. Immunol. Immunopathol. 72, 289−302)ならびにバキュロウイルス多面体由来の核標的指向化シグナルおよび関心対象のタンパク質を含む融合タンパク質(米国特許第5,179,007号)等の小さな構築物のためのバキュロウイルス発現系において適用されている。
【0011】
それゆえ、本発明の基礎となる技術的な課題は、多タンパク質の改良された発現のための新規の系を提供することである。
【0012】
上述の技術的な課題の解決は、特許請求の範囲において定義される態様によって提供される。
【0013】
特に、本発明は、ポリジーンを構成する遺伝子によってコードされるポリペプチドのうちの少なくとも2つが、非ウイルス性由来であり、ポリジーンを構成する遺伝子によってコードされるポリペプチドのうちの少なくとも2つが、該ポリジーンの遺伝子によってコードされる少なくとも1つの他のポリペプチドと少なくとも一過性に相互作用することが各々可能であり、および各ポリジーンを構成する遺伝子が、少なくとも1つのプロテアーゼ切断部位をコードする配列によって、および/または少なくとも1つの自己切断ペプチドをコードする配列によって互いに接続される、単一翻訳領域(ORF)内で各々、少なくとも3つの生物活性のあるポリペプチドを各々コードする少なくとも2つのポリジーンをコードするポリヌクレオチドを提供する。
【0014】
本発明のポリヌクレオチドは、1つ以上の合成ヌクレオチド類似体を含むDNA、RNAまたはポリヌクレオチドであり得る。ポリヌクレオチドは、一本鎖または二本鎖の形態で存在し得る。DNA、特に二本鎖DNAの形態が特に好ましい。本発明のポリヌクレオチドは、化学合成によって生成され得る。本発明の好ましいポリヌクレオチド構築物は、組換え遺伝子技術によって作製される(例えば、Sambrook et al. “Molecular Cloning”, Cold Spring Harbor Laboratory, 1989参照)。
【0015】
本明細書で使用される「ポリジーン(polygenes)」とは、単一ORFにおいて生物活性のある少なくとも3つのポリペプチドをコードする核酸配列である。したがって、ポリジーンを構成する各「遺伝子」とは、ポリペプチド、特にタンパク質または、特異的な、特に構造的、制御的または酵素的機能を有するその断片、変異形、変異体または類似体をコードする核酸配列である。好ましくは、ポリペプチドをコードする「遺伝子」は、構造タンパク質、制御タンパク質もしくは酵素タンパク質またはその断片、変異形、変異体もしくは類似体をコードするcDNAのコード領域を含む。
【0016】
ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「断片」とは、もとのポリペプチドの領域の一部または一領域を意味するものであり、好ましくは、完全なタンパク質の機能の少なくとも1つを保持する断片を意味する。ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「変異形」とは、別の種から派生したもともとのポリペプチドの機能的または非機能的等価物であるポリペプチド、または代替的スプライシングもしくは翻訳後加工から生じるもともとのポリペプチドの機能的もしくは非機能的誘導体を意味する。ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「変異体」とは、自然発生しているタンパク質から、1つ以上のアミノ酸残基の挿入、置換、付加および/または欠失によって派生するポリペプチドを意味する。ポリジーン中に含有される遺伝子によってコードされるポリペプチドの「類似体」とは、非関連アミノ酸配列さえも有し得るが、類似のポリペプチドと同一機能を発揮する、もともとのポリペプチドの機能的等価物を意味する。
【0017】
したがって、核酸レベルにおいて、遺伝子「断片」とは、「断片」が由来するもともとの遺伝子の一部または一領域である。遺伝子「変異形」は、もともとの遺伝子と比較して異なる種において見出される配列を有するかまたは、問題のポリペプチドのスプライシング変異形もしくは翻訳後加工されたバージョンをコードし得る。「変異体」は、1つ以上のヌクレオチドの挿入、置換、付加および/または欠失によって親遺伝子から派生する。遺伝子の「類似体」は、親遺伝子によってコードされるポリペプチドの機能的等価物をコードする。
【0018】
本発明に記載のポリジーン中の遺伝子のうち少なくとも2つは、非ウイルス由来である。「非ウイルス」とは、(機能的タンパク質またはその断片、変異形、類似体もしくは変異体を表す)ぺプチドをコードする核酸配列が、ウイルスのゲノム中に本来見出されないかまたは前記ゲノム由来ではないことを意味する。特に、本発明に記載のポリヌクレオチドのポリジーン中に含まれるヌクレオチド配列は、真核生物および/または原核生物由来である。
【0019】
したがって、本発明によると、多サブユニット集合体の(多タンパク質複合体または代謝経路のメンバーまたは互いに少なくとも一過性に少なくとも潜在的に相互作用するその他のタンパク質等の)サブユニットをコードする遺伝子は、少なくとも2つの翻訳領域(ORF)中に存在する。集合体のサブユニット(ポリペプチド)をコードする配列は、少なくとも2つのポリジーン中に存在し、その中で、各ポリジーンを構成する遺伝子は、プロテアーゼ切断部位をコードする(2つ以上あり得る)配列(すなわち、プロテアーゼの認識部位を含む配列)または少なくとも1つの自己切断ペプチドによって互いに接続されている。
【0020】
本発明の好ましい態様によると、ポリジーンを構成する遺伝子を接続する配列によってコードされる切断部位を切断することのできるプロテアーゼは、本発明のポリヌクレオチドによってコードされる。より好ましくは、プロテアーゼをコードする遺伝子は、ポリジーンの少なくとも1つの一部である。
【0021】
適切なプロテアーゼ切断部位および自己切断ペプチドは、当業者に公知である(例えば、Ryan et al. (1997) J. Gener. Virol. 78, 699−722; Scymczak et al. (2004) Nature Biotech. 5, 589−594参照)。プロテアーゼ切断部位の好ましい例は、ポティウイルスNIaプロテアーゼ(例えば、タバコエッチウイルスプロテアーゼ(tobacco etch virus protease))、ポティウイルスHCプロテアーゼ、ポティウイルスP1(P35)プロテアーゼ、ビオウイルス(byovirus)NIaプロテアーゼ、ビオウイルスRNA−2によりコードされるプロテアーゼ、アフトウイルスLプロテアーゼ、エンテロウイルス2Aプロテアーゼ、ライノウイルス2Aプロテアーゼ、ピコルナ3Cプロテアーゼ、コモウイルス(comovirus)24Kプロテアーゼ、ネポウイルス(nepovirus)24Kプロテアーゼ、RTSV(イネツングロ球状ウイルス(rice tungro spherical virus))3C様プロテアーゼ、PYVF(パースニップ黄色フレックウイルス(Parsnip yellow fleck virus))3C様プロテアーゼ、トロンビン、第Xa因子およびエンテロキナーゼの切断部位である。切断の高い厳密度により、TEV(タバコエッチングウイルス)プロテアーゼの切断部位が特に好ましい。したがって、本発明に記載のポリジーンの遺伝子は好ましくは、Xがいずれかのアミノ酸を表す一般式EXXYXQ(G/S)のアミノ酸配列をコードするヌクレオチド配列を含むヌクレオチドのストレッチによって接続される(TEVによる切断は、QとGとの間またはQとSとの間で生じる)。最も好ましいのは、ENLYFQGおよびENLYFQSをそれぞれコードするリンカーヌクレオチド配列である。
【0022】
好ましい自己切断ペプチド(「シス作用型加水分解性要素」(CHYSEL)とも呼ばれる;deFelipe (2002) Curr. Gene Then 2, 355−378参照)は、ポティウイルスペプチドおよびカルジオウイルス2Aペプチドに由来する。特に好ましい自己切断ペプチドは、FMDV(口蹄疫ウイルス)、ウマ鼻炎Aウイルス、ゾーシーアシグナウイルス(Thosea asigna virus)およびブタテスコウイルス(teschovirus)由来の2Aペプチドより選択される。
【0023】
本発明のポリジーンによりコードされるポリペプチドのうち少なくとも2つは、ポリジーンによってコードされる1つの他のポリペプチドと少なくとも一過性に相互作用することができるかまたは、ポリジーン中に含有される遺伝子によってコードされる別のポリペプチドと少なくとも一過性に相互作用することができると少なくとも思われる。ポリペプチド間で形成される典型的な「相互作用」には、共有結合、水素結合、静電相互作用およびファンデルワールス相互作用が含まれる。「一過性」の相互作用は、二分子、特にタンパク質に対して共通であり、典型的には、酵素とそれらの基質との間、受容体とそれらの(アゴニストまたはアンタゴニスト)リガンドとの間の相互作用、代謝経路のメンバー間の相互作用および制御性の(例えば、遺伝子制御性の)複合体のタンパク質間の相互作用によって表される。
【0024】
本発明のポリジーンを構成するヌクレオチド配列によってコードされるポリペプチドは、同一または異別であり得る。したがって、本発明の構築物中に存在する各ポリジーンは、関心対象のタンパク質をコードする各ヌクレオチド配列の1つ以上のコピーを含有し得る。この様式で、例えば、特に、タンパク質が通常、異なるレベルで発現するかおよび/または異なる化学量論における巨大分子集合体中に存在する場合における望ましいタンパク質の最適な発現のために機能する構築物を提供することが可能である。それゆえ、ポリペプチドが、通常使用される系においてほとんど発現しない場合、相当するコード配列の2つ以上のコピーが、本発明の構築物の1つ以上のポリジーン中へ組み込まれ得る。同一のアプローチは、ポリペプチドが、望ましい複合体中の二量体、三量体または多量体として存在する場合、使用され得る。この様式で、本発明の構築物は、当業者が発現させるかおよび/または精製することを望むいずれかの複合体または他の巨大分子集合体の必要条件(発現レベル、化学量論等)に従って、個々に集合され得る。
【0025】
ポリジーンを構成する遺伝子が、多タンパク質複合体のメンバーをコードする遺伝子および代謝経路のメンバーをコードする遺伝子からなる群より選択されることはさらに好ましい。好ましい多タンパク質複合体は、転写因子複合体等の遺伝子制御タンパク質複合体、核内および細胞内輸送に関与する複合体等の輸送複合体、タンパク質折りたたみ複合体、受容体/リガンド複合体、細胞−細胞間認識複合体、アポトーシスに関与する複合体、細胞周期制御に関与する複合体等である。代謝経路のメンバーは例えば、(解糖、糖新生、クエン酸回路、グリコーゲン生合成、ガラクトース経路、カルビン回路等の)炭水化物代謝、(トリアシルグリセロール代謝、脂肪酸の活性化、脂肪酸(偶数鎖/奇数鎖)のβ酸化、α酸化経路、脂肪酸生合成、コレステロール生合成等の)脂質代謝、グルタミン酸反応、クレブス−ヘンゼライト(Henseliet)尿素回路、シキミ酸経路、PheおよびTyr生合成、Trp生合成等の)アミノ酸代謝、(酸化的リン酸化、ATP合成、光合成、メタン代謝等の)エネルギー代謝、核酸代謝(プリンおよびピリミジンの生合成および分解、DNA複製等)のメンバーである。多タンパク質複合体のメンバーおよび代謝経路のメンバーは例えば、その開示内容が参照により本明細書に組み入れられる、http://www.biocarta.com/genes/index.aspおよびG. Michal (ed.) Biochemical Pathways, 1. edition, John Wiley & Sons, Hoboken, NJ, USA, 1999から採取され得る。
【0026】
本発明に記載の各ポリジーンは、少なくとも3つの遺伝子、すなわち生物活性のあるポリペプチドをコードする配列を含有する。より好ましいのは、4、5、6以上またはさらにより多くのタンパク質をコードするポリジーンである。上述のように、ポリペプチドと接続するプロテアーゼ切断部位を切断することのできるプロテアーゼは、ポリジーンの少なくとも1つによってコードされることが好ましい。
【0027】
好ましい態様によると、本発明のポリヌクレオチドは、ポリジーンの1つへ各々作用可能に連結され、したがってポリジーンの発現を調節することのできる少なくとも2つのプロモーター配列を含有する。本発明の構築物中の適切なプロモーターは、polh、p10およびpXIV最晩期バキュロウイルスプロモーター、vp39バキュロウイルス後期プロモーター、vp39polhバキュロウイルス後期/最晩期プロモーター、Pcap/polh、pcna、etl、p35、da26バキュロウイルス初期プロモーター;CMV、SV40、UbC、EF−1α、RSVLTR、MT、PDS47、Ac5、PGALおよびPADHからなる群より選択され得る。プロモーター配列は、全てのポリジーンに関して同一であり得るかまたは、異なるプロモーターが異なるポリジーンに関して選択され得る。
【0028】
好ましくは、本発明のポリジーンを含有する各ORFは、SV40、HSVtkまたはBGH(ウシ成長ホルモン)等の転写終結配列によって隣接される。
【0029】
本発明に記載のポリヌクレオチドは、エンハンサー配列または抑制因子配列等のさらなる制御配列を含有し得る。
【0030】
本発明に記載のポリヌクレオチドが、ベクターまたは宿主細胞中への組み込みのための少なくとも1つの部位を含有することはさらに好ましい。このような組み込み部位によって、(ウイルス等の)ベクターおよび宿主細胞(例えば、真核生物宿主細胞)中へのそれぞれへの簡便なゲノム組み込みまたは一過性組み込みが可能となる。ゲノム組み込みのための部位がより好ましい。
【0031】
特に好ましい組み込み部位は、ウイルス中へのポリヌクレオチドの組み込みと適合性のあるものである。より好ましくは、組み込み部位は、アデノウイルス、アデノ随伴ウイルス(AAV)、自律性パルボウイルス、単純ヘルペスウイルス(HSV)、レトロウイルス、ラジノウイルス、エプスタイン・バーウイルス、レンチウイルス、セムリキ森林ウイルスおよびバキュロウイルスからなる群より選択されるウイルス中へのポリヌクレオチドの組み込みに適合する。
【0032】
さらに好ましい態様において、組み込み部位は、(ヒト細胞、例えばHeLa、Huh7、HEK293、HepG2、KATO−III、IMR32、MT−2、膵臓β細胞、ケラチノサイト、骨髄線維芽細胞、CHP212、初代神経細胞、W12、SK−N−MC、Saos−2、WI38、初代肝細胞、FLC3、143TK−、DLD−1、臍帯静脈細胞、胚性肺線維芽細胞、初代包皮線維芽細胞、骨肉腫細胞、MRC5、MG63細胞等の)哺乳動物細胞、(CPL、FS−13、PK−15等の)ブタ細胞、(MDB、BT等の)ウシ細胞、(FLL−YFT等の)ヒツジ細胞、線虫細胞、(S.セレビシエ、S.ポンベ、C.アルビカンス、P.パストリス等の)酵母細胞、および(S.フルギペルダ(frugiperda)、例えばSf9、Sf21、Express Sf+、High Five H5細胞、ショウジョウバエ(D.melanogaster)、例えばS2シュナイダー細胞等の)昆虫細胞からなる群より好ましく選択され得る真核生物宿主細胞中へのポリヌクレオチドの組み込みに適合する。
【0033】
特に好ましい組み込み部位は、Tn7のトランスポゾン要素、λインテグラーゼ特異的結合部位およびSSR(部位特異的リコンビナーゼ)より選択され、好ましくはcre−lox特異的(LoxP)部位またはFLPリコンビナーゼ特異的組み込み(FRT)部位である。
【0034】
本発明の好ましい態様において、ポリヌクレオチドは、他の毒性物質に対する耐性に基づいて望ましい特性を有する宿主細胞を選択するための1つ以上の耐性マーカーをさらに含む。適切な耐性マーカーの例は、アンピシリン、クロラムフェニコール、ゲンタマイシン、スペクチノマイシンおよび/またはカナマイシンに対する耐性を供与するものである。
【0035】
原核生物宿主細胞中への組み込みのため、本発明のポリヌクレオチドは好ましくは、原核生物宿主中でpir遺伝子に依存した増殖をなすための条件的R6Kγ複製開始点を含む。
【0036】
本発明のポリヌクレオチドの特に好ましい態様は、WO2005/085456A1(PCT/EP2004/013381)に開示される構築物中への発現カセットとしてのポリジーンの挿入によって結果的に生じる。
【0037】
それゆえ、本発明のポリヌクレオチドが、
(a)MCS1に関してプロモーターP1および転写終結(terminator)配列T1によって隣接され、MCS2に関してプロモーターP2および転写終結配列T2によって隣接される多重クローニング部位MCS1またはMCS2を各々含む、頭−頭(head−to−head)、頭−尾(head−to−tail)のまたは尾−尾(tail−to−tail)の配置にある少なくとも2つの発現カセットT1−MCS1−P1およびP2−MCS1−T2
(b)少なくとも2つの制限部位AおよびBを含むプロモーターP1とP2との間にある少なくとも1つの増殖モジュールM
(c)発現カセットの1つと各々隣接する少なくとも2つの制限部位XおよびYを含み、ここで、
(i)制限部位AおよびX並びにBおよびYは互換性があるが、しかし
(ii)AYおよびBXの連結産物が、制限部位A、B、XおよびYに特異的な制限酵素a、b、xまたはyによって酵素的に切断可能ではなく、および
(iii)制限部位AおよびB並びに制限部位XおよびYは互換性がなく、ここで、各ポリジーンは発現カセットの1つ中へ挿入される次の式I
X−T1−MCS1−P1−[A−B]−P2−MCS2−T2−Y (I)
に記載の機能的配置を含むことが好ましい。
【0038】
式(I)の配置を有する構築物のさらに好ましい態様に関して、WO2005/085456 A1(PCT/EP2004/013381)に明確に参照されている。特に、増殖モジュールM中の制限部位AおよびBは、制限部位BstZ17I、SpeI、ClaIおよびNruIまたはそれらのイソ制限酵素(isoschizomer)によって切断される制限部位からなる群より選択される。イソ制限酵素は、同一の切断部位を有する制限酵素である。さらに、制限部位XおよびYの好ましい例は、PmeIおよびAvrIIからなる群より選択される制限部位または、そのイソ制限酵素によって切断される制限部位である。
【0039】
上述の式(I)中に含有される発現カセットのうちの1つ中へ挿入される各ポリジーンを有する特に好ましいポリヌクレオチドは、次の特徴を含む。
(a)プロモーターP1およびP2は、polhおよびp10からなる群より選択され;
(b)転写終結配列は、SV40およびHSVtkからなる群より選択され;
(c)増殖モジュールMにおける制限部位AおよびBは、制限部位BstZ171I、SpeI、ClaIおよびNruIからなる群より選択され;
(d)制限部位XおよびYは、制限部位PmeIおよびAvrIIからなる群より選択され、および
(e)ウイルス組み込みのための部位は、cre−loxおよびTn7からなる群より選択される。式(I)に記載の上述の配置を有するポリヌクレオチドの生成に関して、WO2005/085456 A1(PCT/EP2004/013381)に明確に参照されている。
【0040】
ポリジーンを各々含有する2つ以上のORF内に生物活性のあるいくつものポリペプチドをコードする本発明のポリヌクレオチドの提供は、多サブユニットタンパク質複合体のメンバーをコードする遺伝子のクローニングおよび発現に関し、主要な改良を提供する。一方で、結合されるべきコード配列の膨大なサイズおよび数のため、単一ORF中への全てのサブユニット遺伝子の集合はしばしば、不可能であるかまたは非常に困難である。もう一方で、個別の発現カセット中に各々存在する多サブユニット複合体のいくつものまたは全てのメンバーの効率的な集合はしばしば、非常に非効率的であるようになり、その理由は、全体的な複合体収量が、最少に発現するサブユニットによって決定されるからである。本発明によると、多タンパク質集合体のサブユニットは、少なくとも3つの(好ましくは非ウイルス由来の)ポリペプチドを各々コードする(各々単一のORFを表す)少なくとも2つのポリジーンによってコードされ、結果的に、構築物およびその構成体(特にポリジーンの集合体)の扱いやすいと(特に本発明のポリヌクレオチドが適切なベクター中に存在する場合の)発現効率性との間に最適な妥協を生じる。
【0041】
それゆえ、本発明のさらなる態様は、上述のポリヌクレオチドを含有するベクターである。ベクターは、プラスミド、発現ベクターおよびトランスファーベクターからなる群より選択され得る。より好ましくは、本発明のベクターは、真核生物の遺伝子導入、一過性またはウイルスベクター仲介性遺伝子導入に有用である。
【0042】
特に好ましいベクターは、アデノウイルス、アデノ随伴ウイルス(AAV)、自律性パルボウイルス、単純ヘルペスウイルス(HSV)、レトロウイルス、ラジノウイルス、エプスタイン・バーウイルス、レンチウイルス、セムリキ森林ウイルスおよびバキュロウイルスより選択されるウイルス等の真核生物発現ベクターである。本発明の最も好ましいベクターは、バキュロウイルス発現ベクターである。本発明の好ましいバキュロウイルスは、遺伝子v−cathおよびchiAが機能的に破壊される態様であり、その理由は、このことが、感染およびタンパク質発現の間、細胞内区画の維持の改良に至るからである。v−cath遺伝子は、キチナーゼをコードするウイルスDNAであるchiA上に並置された遺伝子に依存する過程による細胞死の際に活性化されるウイルスプロテアーゼV−CATHをコードする。両遺伝子は好ましくは、V−CATH活性を除去し、chiA遺伝子産物からの干渉なしでキチンアフィニティクロマトグラフィーを利用する選択肢を獲得するために破壊される。機能的に活性のあるv−cathおよびchiA遺伝子を欠失するバキュロウイルス系によって生じる発現産物の品質は、ウイルス依存性タンパク質分解活性および細胞溶解の低下のため、有意に改良される。
【0043】
好ましくは、本発明に記載のベクターは、SSRのための部位を含み、好ましくはcre−lox部位特異的組換えのためのLoxPを含む。より好ましくは、cre−lox部位は、バキュロウイルス遺伝子v−cathおよびchiAのうちの1つまたは両者の機能を破壊するため、それらの中に配置される。
【0044】
本発明のベクターは好ましくは、正確に集合されたベクターでうまく形質移入された宿主の選択のための1つ以上のマーカー遺伝子を含有する。適切なマーカー遺伝子の例は、ルシフェラーゼ、β−Gal、CAT、GFP、BFP、YFP、CFPおよびそれらの変異形等の蛍光タンパク質をコードする遺伝子、およびlacZα遺伝子である。マーカー遺伝子は、上述の例の機能的に等価の変異形、変異体、断片または類似体あるいは、当業者に公知の他の適切なマーカーであり得る。変異形、変異体または類似体は好ましくは、該変異形、変異体または類似体の由来するマーカーと比較して、アミノ酸レベルで少なくとも75%、より好ましくは85%、特に好ましくは90%、特に少なくとも95%の相同性を示す。
【0045】
別の好ましい態様において、本発明のベクターは、トランスポゾン要素、好ましくはTn7結合部位を含む。より好ましくは、このようなトランスポゾン要素、例えばTn7結合部位は、マーカー遺伝子内に配置され、それにより転位による組み込みの成功が、機能的マーカー遺伝子によって提供される表現型を検査することによって評価されることが可能となる。
【0046】
本発明の好ましいトランスファー(transfer)ベクターは、WO2005/085456 A1(PCT/EP2004/13381;そこにそれぞれ開示される配列番号1および2ならびに図1および2参照)に開示されるpFBDMまたはpUCDMに基づいている。本発明のさらに好ましいトランスファーベクターは、上述のpFBDMベクターおよびpUCDMベクターのそれぞれの誘導体に基づいている。
【0047】
pFBDMおよびpUCDMの特に好ましい誘導体の例は、トランスファーベクターpSPL(図3)、pFL(図4)、pKL(図5)およびpKDM(図6)である。pUCDMのように、pSPLは、条件的な複製開始点(R6Kγ)を含有する。(pFBDMのように)pFLは、高コピー数の複製開始点(ColE1)を含有する。pKDMおよびpKLは、pBR322由来の低コピーの複製開始点を有する。pFBDMとの類似性において、pFL、pKLおよびpKDMは、トランスポゾン要素(Tn7R、Tn7L)を含有する。ベクターpSPL、pFLおよびpKLは、(pUCDMがそうであるように)二重発現カセットに隣接するLoxPの不完全な逆方向反復を有する。ベクターは全て、多遺伝子カセット1を生じるために、上述の増殖モジュール(M)を含有する。pFLおよびpKL(ならびに誘導体)は、受容ベクターであり、pUCDMおよびpSPL(ならびに誘導体)は、Cre仲介性プラスミド融合体における供与ベクターである。
【0048】
本発明の構築物を生じるためのトランスファーベクターに関する上述の好ましい例の重要な特徴は、次の表1に要約されている。
【0049】
【表1】
【0050】
それゆえ、本発明のポリジーンは、制限酵素切断および連結によるかまたは(例えば、BD In−Fusion酵素を使用する)組換えを介してのいずれかで、多重クローニング部位(MCS1およびMCS2)におけるpFBDM、pUCDM、pSPL、pFL、pKLまたはpKDM等のベクター中へと挿入される。バキュロウイルストランスファーベクターpFBDM、pUCDM、pSPL、pFL、pKLおよびpKDMは、改良されたタンパク質生成のために操作された修飾されたレシピエントバキュロウイルスDNAを含み、単純で迅速な方法により、この目的のために仕立てられたイー・コリ細胞中のバキュロウイルスDNAへの遺伝子の組み込みを2つのアクセス部位(attTn7およびLoxP)を介して可能にする。
【0051】
さらなる態様によると、本発明は、本発明のポリヌクレオチドおよび/またはベクターを含有する宿主細胞を提供する。
【0052】
好ましい宿主細胞の例は、ヒト、げっ歯類、CPL、FS−13およびPK−15等のブタの細胞、MDBおよびBT等のウシの細胞、FLL−YFT等のヒツジの細胞等の哺乳動物細胞、線虫(C.elegans)細胞、S.セレビシエ(S.cerevisiae)、S.ポンベ(S.pombe)、P.パストリス(P.pastoris)およびC.アルビカンス(C.albicans)等の酵母細胞、S.フルギペルダ由来の細胞等の昆虫細胞、好ましくはSf9、Sf21、Express Sf+またはHigh Five h5細胞、S2シュナイダー細胞等のショウジョウバエ由来の細胞、およびイー・コリ(E.coli)等の細菌、好ましくはTop10、Dh5α、DH10α、HB101、TG1、BW23473およびBW23474の系である。
【0053】
好ましいヒト細胞は、HeLa、Huh7、HEK293、HepG2、KATO−III、IMR32、MT−2、膵臓β細胞、ケラチノサイト、骨髄線維芽細胞、CHP212、初代神経細胞、W12、SK−N−MC、Saos−2、WI38、初代肝細胞、FLC3、143TK−、DLD−1、臍帯静脈細胞、胚性肺線維芽細胞、初代包皮線維芽細胞、骨肉種細胞、MRC5およびMG63細胞より選択される。
【0054】
本発明に記載のポリヌクレオチドおよび/またはベクターを含む宿主細胞は、単離された細胞であり得るかまたは、組織もしくは臓器中に存在し得る。
【0055】
本発明のさらなる態様は、本発明の少なくとも1つのポリヌクレオチド配列および/またはベクターで形質転換されている非ヒト遺伝子導入動物に関する。好ましい遺伝子導入動物は、げっ歯類、ブタ、ウシおよび線虫の種である。
【0056】
本発明の遺伝子導入動物は、多タンパク質複合体の役割の解明のために、またはインビボでの生物活性に関して化合物をスクリーニングするために特に有用である。
【0057】
本発明のさらなる態様は、
(a)少なくとも2つのポリジーンを構成する遺伝子のコード領域を提供、好ましくは増幅する工程;
(b)少なくとも1つのプロテアーゼ切断部位および/または少なくとも1つの自己切断ペプチドをコードする配列を該コード領域に提供する工程;
(c)単一ORFが結果として各ポリジーンをもたらすように工程(a)および(b)から結果的に生じる断片を集合させる工程;および
(d)少なくとも2つのポリジーンを単一ポリヌクレオチドに結合する工程
を含む、上述に定義されるポリヌクレオチドの生成のための方法である。
【0058】
本発明の別の局面は、
(a)(好ましくは、本発明のポリヌクレオチドの生成のための上述の方法によって)上述に定義される単一ORF内に少なくとも3つの遺伝子を各々含む少なくとも2つのポリジーンを発生させる工程;および
(b)ポリジーンをプラスミドベクターまたはウイルスベクター中へクローニングする工程;を含み、各ポリジーンの遺伝子のうちの少なくとも1つが非ウイルス由来であり、遺伝子によってコードされるポリペプチドのうちの少なくとも2つが遺伝子によってコードされる1つの他のポリペプチドと少なくとも一過性に相互作用することのできる、本発明に記載のベクターの生成のための方法である。
【0059】
好ましくは、ポリジーン中へ集合する遺伝子のうちの1つは、ポリジーンによってコードされるポリペプチドを接続するプロテアーゼ切断部位を切断することのできるプロテアーゼをコードする遺伝子である。好ましいプロテアーゼは、上述に定義されるとおりである。
【0060】
本発明のポリジーンおよびベクターの構築は、当業者に一般的に公知である多様な分子生物学的技術を通じて実施されることが可能である(例えば、Ausubel et al. (eds.) Current Protocols in Molecular Biology, John Wiley & Sons, Hoboken, NJ, USA, 2003参照)。ポリジーンの生成は、例えば、(例えば、対応するcDNAテンプレートを使用して)特定のポリペプチドをコードするヌクレオチド配列のPCR増幅によって、好ましくは、少なくとも1つのプロテアーゼ切断部位および/または少なくとも1つの自己切断ペプチドをコードする配列を提供するプライマー(5’または3’のいずれか)の使用によって実施され得る。好ましくは、プライマーは、適切な制限酵素の認識配列をさらに含有する。好ましくは、各プライマーは、他のプライマーの制限部位とは異なる制限部位を含有し、それにより結果的に生じる増幅産物を別の増幅産物および/または同一制限部位を含有する線形化されたベクターと指向性の連結をすることが可能である。指向性の連結による構築物の生成のための別の好ましい態様によると、プライマーは、RsrIIまたはBstEII等の自己連結可能ではないオーバーハングを生じる制限酵素の認識配列を含有し得る。プライマー自体が制限部位を含有しない場合、望ましい制限部位を含むアダプターとともに増幅産物を提供することも可能である。もちろん、(増幅のほかに)いずれかの供給源を使用して、望ましいポリペプチドをコードする領域を提供することも可能である。例えば、必要とされる配列は、それ自体既に存在し得るかまたは、配列が適切な制限酵素によって切り出され得る対応するベクター中に既に存在し得る。適切な制限部位等を含有しない構築物は、必要とされる要素を含有する適切なアダプターの連結によって、適切な配列(制限部位、プロテアーゼ切断部位/自己切断可能なペプチド、リンカー等)とともに提供され得る。次に、増幅産物または(制限部位を含有する)その他の適切な構築物が適切な制限酵素で切断される。もちろん、使用されるいずれかのプライマーの配列は、好ましくは適切なベクター中へのポリジーンの最終的な集合後、単一ORFが各ポリジーンを結果として生じるよう、好ましく選択される。
【0061】
本発明の好ましい態様によると、次に、増幅産物または他の適切な配列は、適切なベクター中へ、例えば、上述に参照されるMultiBacベクター系のMCS1またはMCS2で連続してまたは同時に連結され得る。例えば、(WO2005/085456 A1(PCT/EP2004/013381)に記載されるように)1つのポリジーンは、pFDBM中へ導入され得、別のポリジーンは、pUCDM中へ連結され得る。次に、その結果得られる構築物は、関心対象の多タンパク質複合体の発現および生成のために対応する昆虫細胞の感染のために用意されるバキュロウイルス発現ベクターを生じるcre−lox部位特異的組換え(pUCDM誘導体)およびTn7特異的転位(pFBDM誘導体)によって、対応するバクミド(bacmid)の生成のために使用される。
【0062】
ポリジーンを生じ、これらを適切なベクター中へ、制限/連結によって導入するほかに、適切なリコンビナーゼを使用する相同的組換えによってこのような構築物を構築させることも可能である。リコンビナーゼに基づくクローニング技術の適切な例は、BD Biosciences Clontech, Heidelberg, Germany (see Clontechniques, October 2002, p. 10)から市販されているIn−Fusion(登録商標)系およびGene Bridges GmbH, Dresden, Germany (see WO−A−99/29837; http://www.genebridges.com)から市販されているRed(登録商標)/ET(登録商標)組換え系である。
【0063】
In−Fusion(登録商標)系は、対応する相同性領域を有する(適切なベクター等の)線形構築物中へ融合されるべきDNA分子中に15bpの相同性領域を必要とする。したがって、本発明のポリジーンを含有するベクターは、15bpの適切な相同的配列とともに、(プロテアーゼ切断部位および/または自己切断ペプチドをコードするリンカー配列とともに)構成しているコード領域を提供することによって、連続してまたは同時に集合され得る。もし望まれれば、15bpの相同的配列は、構成しているコード領域が望ましい順序で集合されるよう選択され得る。もちろん、適切な相同性領域は、望ましい配列を含有するプライマーを使用する、対応する断片のPCR増幅によって導入され得る。
【0064】
Red(登録商標)/ET(登録商標)組換え系は、40〜60塩基対の相同性配列が必要とされるIn−Fusion(登録商標)系とは異なるが、断片が挿入されるべき構築物は、線形である必要がない。組換えは、宿主中で、好ましくは二重リコンビナーゼ系「ET」(RecE/RecTまたはRedα/Redβ)を発現するイー・コリ系でインビボで実施される。したがって、ポリジーンを構成する断片は、適切なベクターとともに適切な宿主中へ、好ましくはイー・コリ細胞中へ直接形質転換され得る。この様式で、各ポリジーンは、(好ましくは、多タンパク質複合体の各メンバーポリペプチド+少なくとも1つのプロテアーゼ切断部位配列/自己切断ペプチド配列に関するコード領域を含む)断片ごとに連続的に、または全ての断片の同時形質転換のいずれかによって適切なベクター中へ集合され得る。
【0065】
本発明の特に好ましいポリヌクレオチド構築物は、少なくとも同様の長さのポリジーンを含有する。なぜならば、このような構築物から対応するポリタンパク質(polyproteins)を発現させると、かなりの発現レベルを結果的に生じることを本発明者が発見したからである。本発明によると、「同様な長さのポリジーン」とは、ポリジーンのヌクレオチド配列の長さが、50%以下まで、より好ましくは30%以下まで、特に20%以下までまたはさらにより少なく互いに異なることを意味する。
【0066】
さらに、本発明は、
(a)ポリジーンの発現を可能にする条件下で適切な培地中で本発明に記載の宿主細胞を培養する工程;および
(b)ポリジーンによってコードされる発現産物を培地および/または宿主細胞から回収する工程
を含む、インビトロでの多タンパク質複合体の生成のための方法を提供する。
【0067】
本発明は、
(a)上述に定義される単一ORF内で少なくとも3つの遺伝子を各々含む少なくとも2つのポリジーンを発生させる工程;および
(b)ポリジーンを動物中へ形質転換し、それによりポリジーンが該動物中で発現される工程
を含む、インビボでの多タンパク質複合体の生成のための方法にも関する。
【0068】
好ましくは、工程(b)に従ったポリジーンによる動物の形質転換は、ベクター、特にウイルスベクター、より好ましくはバキュロウイルスベクターによって生じる。バキュロウイルスは、哺乳動物手中へのポリジーンの送達に特に有用な媒体である。上述のインビボでの方法は好ましくは、哺乳動物、線虫または昆虫において実施される。適切な動物種の特に好ましい例は、本明細書で上述に定義される。
【0069】
本発明の態様は、タンパク質の多サブユニット集合体に対して向けられるワクチンの調製にも有用である。多サブユニットの複合体はしばしば、個々のタンパク質と比較して異なるエピトープを呈する。それゆえ、本発明に従って生成される多タンパク質複合体は、より適切な様式で自然発生する関連エピトープを呈し、したがって、抗体産生のためのより良好な抗原標的を提供する。
【0070】
近年、重症急性呼吸器症候群(SARS)コロナウイルス由来の4つのタンパク質からなるビリオン様粒子(VLP)が、組換えバキュロウイルス発現ベクターを使用して作製された(Mortola et al. (2004), FEBS Lett. 576, 174−178参照)。ワクチンの調製のためのこのような感染性粒子の効果的な発現は、本発明に従ったポリジーン発現系を使用して大いに促進されるであろう。特に、SARS−VLPの例よりも実質的に多くのポリペプチドを含有する多サブユニット集合体の高収量の発現は、本発明の発現ツールによって利用可能とされる。
【0071】
それゆえ、本発明はさらに、
(a)本発明の少なくとも1つのポリヌクレオチドおよび/またはベクターを哺乳動物へ投与し、それにより本発明のポリジーンが哺乳動物内で発現する工程;
(b)アジュバントを哺乳動物へ場合により投与する工程;および
(c)ポリジーンによりコードされるポリペプチドの少なくとも1つに特異的な抗体および/または前記抗体を産生する脾臓細胞を場合により単離する工程
を含む、ワクチンの生成のための方法に関するものである
【0072】
本発明は、多タンパク質集合体の組換え生成のための簡便かつ単純なアプローチを提供する。これらの多サブユニット集合体は、タンパク質複合体相互作用または他サブユニット集合体を構成するタンパク質の修飾に関して検査され得る。本発明に従って生成される多サブユニット集合体は、医学的価値のある生物学的に有意な活性を発揮し得る候補化合物(有機小分子、核酸、ペプチド、ポリペプチド等)との相互作用に関してもアッセイされ得る。
【0073】
それゆえ、本発明は、タンパク質複合体相互作用またはタンパク質修飾をアッセイするための方法にも関するものである。
【0074】
好ましい態様によると、本発明は、
(a)少なくとも2つのポリジーンを含有する本発明に従った宿主細胞を提供する工程;
(b)ポリジーンの発現を可能にする条件下で宿主細胞を維持する工程;および
(c)ポリジーンによりコードされるポリペプチド間の相互作用または前記ポリペプチドの修飾を検出する工程
を含む、タンパク質複合体の相互作用またはインビトロでの多タンパク質複合体の修飾をスクリーニングするための方法を提供する。
【0075】
本発明の別の好ましい態様は、(i)多タンパク質複合体と相互作用することおよび/または(ii)多タンパク質複合体内でのタンパク質の修飾かおよび/または(iii)多タンパク質複合体内でのもしくは多タンパク質複合体間の相互作用を阻害するかおよび/または多タンパク質複合体内でタンパク質の修飾を阻害することの可能な候補化合物のインビトロでのスクリーニングのための方法であり、
(a)少なくとも2つのポリジーンを含有する本発明に従った宿主細胞を提供する工程;
(b)ポリジーンの発現を可能にする条件下で宿主細胞を維持する工程;
(c)候補化合物を宿主細胞と接触させる工程;
(d)発現産物と候補化合物との相互作用および/または発現産物間の相互作用および/または発現産物の修飾および/または発現産物間の相互作用の阻害を検出する工程
を含む。
【0076】
本発明のポリヌクレオチドおよび/またはベクターは、タンパク質−タンパク質、タンパク質−(多)タンパク質複合体または多タンパク質複合体−多タンパク質複合体の相互作用またはインビボでの多タンパク質複合体の修飾(リン酸化、グリコシル化等)のスクリーニングにも適している。
【0077】
したがって、本発明のさらに好ましい態様は、(i)多タンパク質複合体と相互作用すること、および/または(ii)多タンパク質複合体内でのタンパク質の修飾および/または(iii)多タンパク質複合体内または多タンパク質複合体間の相互作用を阻害することおよび/または多タンパク質複合体内でタンパク質の修飾を阻害することのできる候補化合物のインビボでのスクリーニングのための方法であり、
(a)上述に定義される少なくとも2つのポリジーンを含有する本発明の少なくとも1つのポリヌクレオチドおよび/またはベクターを含む動物を提供し、それによりポリジーンが動物中で発現する工程;
(b)候補化合物を動物へ投与する工程;および
(c)発現産物と候補化合物との相互作用および/または発現産物間の相互作用および/または発現産物の修飾および/または発現産物間の相互作用の阻害を検出する工程
を含む。
【0078】
本発明の多タンパク質発現ツールは、医学的にも使用される。特に、生物活性のある多タンパク質複合体ならびにタンパク質の医学的に有利な組み合わせ、例えば抗体混合物は、場合によりインターロイキンおよび/またはアジュバントと組み合わせて本発明のポリヌクレオチドおよび/または遺伝子送達ベクターによって、動物またはヒトへ投与されることが可能である。
【0079】
したがって、本発明はさらに、遺伝子治療のためのポリジーントランスファー媒体を含む医薬の調製のための、上述のポリヌクレオチドおよび/またはベクターおよび/または宿主細胞の使用に関する。
【0080】
治療適用のための遺伝子送達系を開発するために、多大な尽力がなされている。遺伝子治療は、強い熱意の焦点であったが、過去には非難もあった。今日まで、ヒト疾病に関するインビボおよびエクスビボでの安全で適用可能な臨床的戦略を達成するための過程において、臨床治験における遺伝子治療を評価する上での主要な進展があった(Worgall S. (2004) Peadiatr. Nephrol.参照)。全般的に、遺伝子治療はいまや、遺伝子の補正のための、および治療用遺伝子産物の持続的発現または一過性発現を引き起こす獲得された疾患のための非常に見込みのある手段として有効である(Worgall S.、上述)。ウイルスをベースとした組換えベクター、特に哺乳動物宿主中で複製形質転換受容性ではないものは、哺乳動物細胞遺伝子送達のための強力なツールとして近年出現し、ヒト、霊長類、げっ歯類、ウシ、ブタおよびヒツジの細胞を含む哺乳動物細胞系の全範囲へうまく適用されている(Kost and Condreay (2002) Trends Biotech. 20, 173−180に概説)。エクスビボおよびインビボの両者で複雑な遺伝子トランスファー/治療効果を得るため、単一遺伝子治療によるものよりもむしろ組み合わさった遺伝子治療によるより強力な結果を達成するために、多シストロン性のウイルスベクターに関する需要が高まってきた(de Felipe (2002), Curr. Gene Ther. 2, 355−378; Planelles (2003) Meth. MoI. Biol. 229, 273−284)。例えば、補体の系による不活性化を遮断するためにインビボで投与されるべき担体ウイルス中へのアクセサリー遺伝子の組み込みの要求も、ヒト崩壊促進因子タンパク質ドメイン融合を運搬したバキュロウイルスgp64外被タンパク質を有する偽型バキュロウイルスを使用することによって示されており(Hueser et al. (2001) Nat. Biotech. 19, 451−455)、治療目的のためにトランスファーされるべき多重遺伝子に加え、ウイルス産生レベルに関する組換え修飾を提供する必要性を実証している。
【0081】
したがって、本発明の組換えバキュロウイルスは、遺伝子治療製剤を調製するために好ましい。より好ましくは、本発明の製剤のために使用されるベクターは、
(i)1つ以上の治療用ポリペプチドおよび
(ii)1つ以上のバキュロウイルスタンパク質
をコードする上述に定義される少なくとも2つのポリジーンを含むバキュロウイルスである。
【0082】
好ましい態様において、(ii)に従ったタンパク質は、偽型バキュロウイルスから発現されるヒト化バキュロウイルスタンパク質、好ましくは、ヒト化バキュロウイルス外被タンパク質gp64、例えば崩壊促進因子等のヒトタンパク質と融合されたgp64である。
【0083】
さらに、本発明は、
(a)本発明に従ったポリヌクレオチドを含むポリジーントランスファー媒体を提供する工程;および
(b)遺伝子疾患を罹患している患者へポリジーントランスファー媒体を投与する工程
を含む、インビボでの遺伝子治療法に関するものである。
【0084】
本発明はさらに、
(a)遺伝子疾患を罹患している患者の細胞を回収する工程;
(b)本発明に従ったポリヌクレオチドを含むポリジーントランスファー媒体で、回収された細胞を形質転換する工程;および
(c)形質転換された細胞を患者へ投与する工程
を含む。
【0085】
本発明に従って生成される多タンパク質複合体は、生物物理的研究、特に結晶学的、電子顕微鏡的および/またはNMRの技術を使用する構造研究、特にタンパク質−タンパク質相互作用のための、および薬物開発のためのタンパク質化学的研究において有利に使用され得る。
【0086】
したがって、本発明は、多タンパク質複合体の結晶化のための、本発明のポリヌクレオチドおよび/またはベクターおよび/または宿主細胞の使用に関する。
【0087】
本発明のさらなる態様は、
(a)ポリジーンを構成するコード配列のPCR増幅のためのプライマー;
(b)プラスミドベクターまたはウイルスベクター;および
(c)場合により、プラスミドまたはベクターの増殖に適した宿主細胞
を含む多タンパク質複合体の調製のためのキットである。
【0088】
プライマーは、各ポリジーンのための単一ORFを生成するための要求と適合するよう簡便に設計され、プラスミドベクターまたはウイルスベクター中へ連結のための制限部位を(連続的または同時に)含有し得るかおよび/またはプライマーは、(例えば、上述のIn−Fusion(登録商標)系またはRed(登録商標)/ET(登録商標)系を使用する)相同的組換えによってプラスミドベクターまたはウイルスベクター中へポリジーンを集合させるかおよび/または挿入するための配列を含有し得る。
【0089】
本発明は、次の制限のない実施例によってさらに説明される。
【0090】
実施例1:ポリジーンの生成および発現ベクター中への連結
ポリジーンを発生させる原理は、ヒトTATAボックス結合タンパク質(hTBP)中心(hTBPc、位置159で切断された全長タンパク質のc末端断片)を使用することによってここに示される。RsrII制限部位を有するオーバーハングを含有する遺伝子の5’末端にアニールし、アミノ酸スペーサーおよびタバコエッチングウイルス(TEV)切断部位をさらにコードするセンスプライマーを使用するポリメラーゼ連鎖反応(PCR)によって、hTBPcをコードする遺伝子を増幅した。アンチセンスプライマーは、遺伝子の3’末端へアニールし、RsrII制限部位を含有した。RsrIIは、自己連結しない3ヌクレオチドの非対称性オーバーハングを生じる制限酵素であり、制限産物は非対称性であり、連結は方向性産物を生じる。PCR産物をRsrIIで消化し、精製した。PCR産物のDNA(配列番号1)および推定アミノ酸配列(配列番号2)を図1に示す。
【0091】
連結は、図2に示されるhTBPcのコンカタマーを生じた。適切なベクター中へのインビトロでの連結反応混合物のサブクローニングは、TEVプロテアーゼ切断部位によって分離される単一ポリタンパク質中で1、2、3および5のhTBPタンパク質をコードするポリジーンを含有する発現構築物を生じた。結果として生じる発現ベクターのうちの1つ(pFBDO[hTBPc]3)の模式的呈示およびヌクレオチド配列(配列番号3)を図3および図4にそれぞれ示す。
【0092】
実施例2:ヒト一般的転写因子のサブユニットをコードするポリジーンを含有するバキュロウイルストランスファーベクターの生成
遺伝子が、バキュロウイルス発現のためのトランスファーベクターpUCDM(WO2005/085456 A1(PCT/EP2004/013381)参照)中へ挿入されたhTBPcに加え、ヒトTBP関連因子hTAF1およびhTAF2を含むポリタンパク質をコードする、アミノ酸スペーサーおよびTEVプロテアーゼ部位をコードする配列によって分離されている、ポリジーンを生成した。結果として生じる構築物pUCDMCSTAF1TBPcTAF2の模式的呈示を図9に示す。構築物のヌクレオチド配列を図10(配列番号4)に示す。バキュロウイルス発現のためのトランスファーベクターpFBDM(WO2005/085456 A1(PCT/EP2004/013381)参照)中へ挿入されたTEVプロテアーゼ及びヒトTBP関連因子hTAF6およびhTAF9を含むポリタンパク質をコードするポリジーンを含有するさらなる構築物を生成する。遺伝子は、アミノ酸スペーサーおよびTEVプロテアーゼ部位をコードする配列によって分離されている。結果として生じる構築物pFDDO[HisTEVTAF6TAF9]hisの模式的呈示を図11に示す。この構築物のヌクレオチド配列を図12(配列番号5)に示す。
【0093】
実施例3:バクミド構築物の調製、昆虫細胞の感染およびタンパク質発現
上述の2つのポリジーンを含むバクミド構築物の構築のため、構築物pUCDMCSTAF1TBPcTAF2(pUCDM誘導体)およびpFDDO[HisTEVTAF6TAF9]his(pFBDM誘導体)を各々、WO2005/085456 A1(PCT/EP2004/013381)の実施例5(pUCDMCSTAF1TBPcTAF2用;Cre−lox部位特異的組換え)および実施例6(pFDDO[HisTEVTAF6TATAF9]his用;Tn7転位)に記載されているDH10MultiBacCre細胞中へ導入した。望む場合、一工程転位/cre−lox部位特異的組換えは、同様にWO2005/085456 A1(PCT/EP2004/013381)において記載されているDH10MultiBacCre細胞で実施され得る。確立されたプロトコールに従って、バクミド調製、昆虫細胞の感染およびタンパク質発現を実施した(例えば、O’Reilly et al. (1994) “Baculovirus expression vectors. A laboratory manual“ Oxford University Press, New York − Oxford; ”Bac−to− BacTM Baculovirus Expression Systems Manual” Invitrogen, Life Technologies, Inc., 2000参照)。
【0094】
次の配列リストは、本明細書の一部であり、その中で、配列は次のとおりである。
【0095】
配列番号1は、図1に示されるヒトTATAボックス結合タンパク質(hTPB)中心(hTBPc、位置159で切断された全長タンパク質のc末端断片)をコードするPCR産物のヌクレオチド配列である。
【0096】
配列番号2は、図1に示されるヒトTATAボックス結合タンパク質中心(hTBPc)のアミノ酸配列である。
【0097】
配列番号3は、図8に示されるpFBDO[hTBPc]3のヌクレオチド配列である。
【0098】
配列番号4は、図10に示されるpUCDMCSTAF1TBPcTAF2のヌクレオチド配列である。
【0099】
配列番号5は、図12に示されるpFBDO[HisTEVTAF6TAF9]hisのヌクレオチド配列である。
【図面の簡単な説明】
【0100】
【図1】図1は、ヒトTATAボックス結合タンパク質(hTPB)中心(hTBPc、位置159で切断された全長のタンパク質のc末端断片)をコードするPCR産物のヌクレオチド配列(配列番号1)および推定アミノ酸配列(配列番号2)を示す。(プライマー配列中に存在する)RsrII制限部位の位置が示されている。
【図2】図2は、hTBC遺伝子セグメントのインビトロでの連結および混合物のサブクローニングのアガロースゲル電気泳動分析の写真を示す。PCRで増幅されたhTPBc遺伝子をRsrIIで消化し、精製した(レーン1)。リガーゼとのインキュベーションは、各1つのORF中で連結された1、2、3およびそれより多くの遺伝子を含有するコンカテマーのラダーを生じる(レーン2、レーン3はMBI DNAマーカー1kbラダーである)。1つのポリジーンを含有するこのように生じた発現構築物の混合物のサブクローニングは、RsrIIを使用して制限消化することによって遊離されることが可能である連結されたhTBPc遺伝子の異なる数を有する(レーン4〜7)。挿入されたポリジーンの外側の消化は、各場合において単一のORFを生じた1(レーン8)、2(レーン9)、3(レーン10)および5(レーン11)のhTBPc遺伝子を証明する。
【図3】図3は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpSPLの模式的呈示を示す。
【図4】図4は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpFLの模式的呈示を示す
【図5】図5は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpKLの模式的呈示を示す。
【図6】図6は、本発明の好ましいトランスファーベクター構築物の基礎となる基本的なトランスファーベクターpKDMの模式的呈示を示す。
【図7】図7は、トランスファーベクター構築物pFBDO[hTBPc]3の模式的呈示を示す。
【図8】図8は、pFBDO[hTBPc]3(配列番号3)のヌクレオチド配列を示す。
【図9】図9は、トランスファーベクター構築物pUCDMCSTAF1TBPcTAF2の模式的呈示を示す。
【図10】図10は、pUCDMCSTAF1TBPcTAF2(配列番号4)のヌクレオチド配列を示す。
【図11】図11は、トランスファーベクター構築物pFBDO[HisTEVTAF6TAF9]hisの模式的呈示を示す。
【図12】図12は、pFBDO[HisTEVTAF6TAF9]his(配列番号5)のヌクレオチド配列を示す。
【特許請求の範囲】
【請求項1】
単一翻訳領域(ORF)内で各々、少なくとも3つの生物活性のあるポリペプチドを各々コードする少なくとも2つのポリジーンをコードするポリヌクレオチドであって、前記ポリジーンを構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、非ウイルス性由来であり、前記ポリジーンを構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、該ポリジーンの遺伝子によってコードされる少なくとも1つの他のポリペプチドと少なくとも一過性に相互作用することが各々可能であり、および各ポリジーンを構成する遺伝子が、少なくとも1つのプロテアーゼ切断部位をコードする配列によって、および/または少なくとも1つの自己切断ペプチドをコードする配列によって互いに接続されている、前記ポリヌクレオチド。
【請求項2】
各ポリジーンが、前記ポリジーンの発現を調節することが可能であるプロモーター配列へ操作可能に連結される、請求項1に記載のポリヌクレオチド。
【請求項3】
各ORFが、転写終結配列によって隣接される、請求項1または2に記載のポリヌクレオチド。
【請求項4】
プロテアーゼ切断部位が、ポティウイルスNIaプロテアーゼ、ポティウイルスHCプロテアーゼ、ポティウイルスP1(P35)プロテアーゼ、ビオウイルスNIaプロテアーゼ、ビオウイルスRNA−2によりコードされるプロテアーゼ、アフトウイルスLプロテアーゼ、エンテロウイルス2Aプロテアーゼ、ライノウイルス2Aプロテアーゼ、ピコルナ3Cプロテアーゼ、コモウイルス24Kプロテアーゼ、ネポウイルス24Kプロテアーゼ、RTSV(イネツングロ球状ウイルス)3C様プロテアーゼ、PYVF(パースニップ黄色フレックウイルス)3C様プロテアーゼ、トロンビン、第Xa因子およびエンテロキナーゼの切断部位からなる群より選択される、請求項1〜3のいずれか一項に記載のポリヌクレオチド。
【請求項5】
プロテアーゼ切断部位が、TEV(タバコエッチウイルス)プロテアーゼ切断部位である、請求項4に記載のポリヌクレオチド。
【請求項6】
自己切断ペプチドが、アフトウイルス2Aペプチドおよびカルジオウイルス2Aペプチドからなる群より選択される、請求項1〜4のいずれか一項に記載のポリヌクレオチド。
【請求項7】
自己切断ペプチドが、FMDV(口蹄疫ウイルス)2Aペプチドである、請求項6に記載のポリヌクレオチド。
【請求項8】
プロテアーゼ切断部位を切断することのできるプロテアーゼのための遺伝子をさらに含有する、請求項1〜7のいずれか一項に記載のポリヌクレオチド。
【請求項9】
プロテアーゼ遺伝子が、ポリジーンの少なくとも1つの一部である、請求項5に記載のポリヌクレオチド。
【請求項10】
少なくとも1つのポリジーンが、少なくとも4つの遺伝子を含む、請求項1〜9のいずれか一項に記載のポリヌクレオチド。
【請求項11】
各ポリジーンが、少なくとも4つの遺伝子を含む、請求項10に記載のポリヌクレオチド。
【請求項12】
ポリジーンのヌクレオチド配列の長さが、50%未満まで、好ましくは30%未満まで、より好ましくは20%まで互いに異なる、請求項1〜11のいずれか一項に記載のポリヌクレオチド。
【請求項13】
ベクターまたは宿主細胞中へ少なくとも1つの部位を組み込むために前記部位をさらに含む、請求項1〜12のいずれか一項に記載のポリヌクレオチド。
【請求項14】
ポリジーンを構成する遺伝子が、多タンパク質複合体のメンバーをコードする遺伝子と代謝経路のメンバーをコードする遺伝子とからなる群より選択される、請求項1〜13のいずれか一項に記載のポリヌクレオチド。
【請求項15】
(a)MCS1に関してプロモーターP1および転写終結配列T1によって隣接されおよびMCS2に関してプロモーターP2および転写終結配列T2によって隣接される多重クローニング部位MCS1またはMCS2を各々含む、頭−頭、頭−尾または尾−尾の配置にある少なくとも2つの発現カセットT1−MCS1−P1およびP2−MCS1−T2
(b)少なくとも2つの制限部位AおよびBを含むプロモーターP1とP2との間にある少なくとも1つの増殖モジュールM
(c)発現カセットの1つと各々隣接する少なくとも2つの部位XおよびYを含み、ここで
(d)制限部位AおよびX並びにBおよびYは互換性があるが、しかし
(e)AYおよびBXの連結産物は、制限部位A、B、XおよびYにそれぞれ特異的な制限酵素a、b、xまたはyによって酵素的に切断可能ではなく、および
(f)制限部位AおよびB並びにXおよびYは互換性がなく、および、各ポリジーンが発現カセットの1つ中へ挿入される、式I
X−T1−MCS1−P1−[A−B]−P2−MCS2−T2−Y (I)
に記載の機能的配置を含む、請求項1〜14のいずれか一項に記載のポリヌクレオチド。
【請求項16】
増殖モジュールM中の制限部位AおよびBが、制限部位BstZ17I、SpeI、ClaIおよびNruIまたはそれらのイソ制限酵素によって切断される制限部位からなる群より選択される、請求項15に記載のポリヌクレオチド。
【請求項17】
制限部位XおよびYが、制限部位PmeIおよびAvrIIまたはそれらのイソ制限酵素によって切断される制限部位からなる群より選択される、請求項15または16に記載のポリヌクレオチド。
【請求項18】
(a)プロモーターP1およびP2が、polhおよびp10からなる群より選択され;
(b)転写終結配列が、SV40およびHSVtkからなる群より選択され;
(c)増殖モジュールM中の制限部位AおよびBが、制限部位BstZ171I、SpeI、ClaIおよびNruIからなる群より選択され;
(d)制限部位XおよびYが、制限部位PmeIおよびAvrIIからなる群より選択され;および
(e)ウイルス組み込みのための部位が、cre−loxおよびTn7からなる群より選択される、請求項15〜17のいずれか一項に記載のポリヌクレオチド。
【請求項19】
請求項1〜18のいずれか一項に記載のポリヌクレオチドを含むベクター。
【請求項20】
真核生物発現ベクターまたはトランスファーベクターである、請求項19に記載のベクター。
【請求項21】
ウイルスである、請求項19または20に記載のベクター。
【請求項22】
バキュロウイルス発現ベクターである、請求項21に記載のベクター。
【請求項23】
バキュロウイルス遺伝子v−cathおよびchiAが、機能的に破壊されている、請求項22に記載のベクター。
【請求項24】
部位特異的リコンビナーゼ(SSR)のための部位を含む、請求項19〜23のいずれか一項に記載のベクター。
【請求項25】
cre−lox特異的組換えのためのLoxP部位を含む、請求項24に記載のベクター。
【請求項26】
トランスポゾン要素を含む、請求項19〜25のいずれか一項に記載のベクター。
【請求項27】
トランスポゾン要素が、Tn7結合部位である、請求項26に記載のベクター。
【請求項28】
請求項1〜18のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターを含有する宿主細胞。
【請求項29】
細菌、酵母細胞、昆虫細胞、線形動物細胞および哺乳動物細胞からなる群より選択される、請求項28に記載の宿主細胞。
【請求項30】
イー・コリ、S.セレビシエ、P.パストリス、S.ポンベ、C.アルビカンス、Sf9細胞、線虫細胞、ヒト細胞、げっ歯類細胞、ブタ細胞、ウシ細胞およびヒツジ細胞からなる群より選択される、請求項29に記載の宿主細胞。
【請求項31】
請求項1〜28のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターで形質転換された遺伝子導入非ヒト動物。
【請求項32】
げっ歯類、ブタ、ウシおよび線虫より選択される、請求項31に記載の動物。
【請求項33】
(a)ポリジーンを構成する遺伝子のコード領域を増幅させる工程;
(b)該コード領域に、少なくとも1つのプロテアーゼ切断部位および/または少なくとも1つの自己切断ペプチドをコードする配列を提供する工程;
(c)単一ORFが結果的に各ポリジーンをもたらすように、工程(a)および(b)から結果的に生じる断片を集合させる工程;
(d)少なくとも2つのポリジーンを単一ポリヌクレオチドに結合する工程
を含む、請求項1〜18のいずれか一項に記載のポリヌクレオチドの生成のための方法。
【請求項34】
(a)ポリジーンの発現を可能にする条件下で、請求項28〜30のいずれか一項に記載の宿主細胞を適切な培地中で培養する工程;および
(b)ポリジーンによってコードされる発現産物を培地および/または宿主細胞から回収する工程
を含む、インビトロでの多タンパク質複合体の産生のための方法。
【請求項35】
遺伝子治療のためのポリジーントランスファー媒体を含む医薬の調製のための、請求項1〜18のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターおよび/または請求項28〜30のいずれか一項に記載の宿主細胞の使用。
【請求項36】
多タンパク質複合体の結晶化のための、請求項1〜18のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターおよび/または請求項28〜30のいずれか一項に記載の宿主細胞の使用。
【請求項1】
単一翻訳領域(ORF)内で各々、少なくとも3つの生物活性のあるポリペプチドを各々コードする少なくとも2つのポリジーンをコードするポリヌクレオチドであって、前記ポリジーンを構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、非ウイルス性由来であり、前記ポリジーンを構成する遺伝子によってコードされるポリペプチドの少なくとも2つが、該ポリジーンの遺伝子によってコードされる少なくとも1つの他のポリペプチドと少なくとも一過性に相互作用することが各々可能であり、および各ポリジーンを構成する遺伝子が、少なくとも1つのプロテアーゼ切断部位をコードする配列によって、および/または少なくとも1つの自己切断ペプチドをコードする配列によって互いに接続されている、前記ポリヌクレオチド。
【請求項2】
各ポリジーンが、前記ポリジーンの発現を調節することが可能であるプロモーター配列へ操作可能に連結される、請求項1に記載のポリヌクレオチド。
【請求項3】
各ORFが、転写終結配列によって隣接される、請求項1または2に記載のポリヌクレオチド。
【請求項4】
プロテアーゼ切断部位が、ポティウイルスNIaプロテアーゼ、ポティウイルスHCプロテアーゼ、ポティウイルスP1(P35)プロテアーゼ、ビオウイルスNIaプロテアーゼ、ビオウイルスRNA−2によりコードされるプロテアーゼ、アフトウイルスLプロテアーゼ、エンテロウイルス2Aプロテアーゼ、ライノウイルス2Aプロテアーゼ、ピコルナ3Cプロテアーゼ、コモウイルス24Kプロテアーゼ、ネポウイルス24Kプロテアーゼ、RTSV(イネツングロ球状ウイルス)3C様プロテアーゼ、PYVF(パースニップ黄色フレックウイルス)3C様プロテアーゼ、トロンビン、第Xa因子およびエンテロキナーゼの切断部位からなる群より選択される、請求項1〜3のいずれか一項に記載のポリヌクレオチド。
【請求項5】
プロテアーゼ切断部位が、TEV(タバコエッチウイルス)プロテアーゼ切断部位である、請求項4に記載のポリヌクレオチド。
【請求項6】
自己切断ペプチドが、アフトウイルス2Aペプチドおよびカルジオウイルス2Aペプチドからなる群より選択される、請求項1〜4のいずれか一項に記載のポリヌクレオチド。
【請求項7】
自己切断ペプチドが、FMDV(口蹄疫ウイルス)2Aペプチドである、請求項6に記載のポリヌクレオチド。
【請求項8】
プロテアーゼ切断部位を切断することのできるプロテアーゼのための遺伝子をさらに含有する、請求項1〜7のいずれか一項に記載のポリヌクレオチド。
【請求項9】
プロテアーゼ遺伝子が、ポリジーンの少なくとも1つの一部である、請求項5に記載のポリヌクレオチド。
【請求項10】
少なくとも1つのポリジーンが、少なくとも4つの遺伝子を含む、請求項1〜9のいずれか一項に記載のポリヌクレオチド。
【請求項11】
各ポリジーンが、少なくとも4つの遺伝子を含む、請求項10に記載のポリヌクレオチド。
【請求項12】
ポリジーンのヌクレオチド配列の長さが、50%未満まで、好ましくは30%未満まで、より好ましくは20%まで互いに異なる、請求項1〜11のいずれか一項に記載のポリヌクレオチド。
【請求項13】
ベクターまたは宿主細胞中へ少なくとも1つの部位を組み込むために前記部位をさらに含む、請求項1〜12のいずれか一項に記載のポリヌクレオチド。
【請求項14】
ポリジーンを構成する遺伝子が、多タンパク質複合体のメンバーをコードする遺伝子と代謝経路のメンバーをコードする遺伝子とからなる群より選択される、請求項1〜13のいずれか一項に記載のポリヌクレオチド。
【請求項15】
(a)MCS1に関してプロモーターP1および転写終結配列T1によって隣接されおよびMCS2に関してプロモーターP2および転写終結配列T2によって隣接される多重クローニング部位MCS1またはMCS2を各々含む、頭−頭、頭−尾または尾−尾の配置にある少なくとも2つの発現カセットT1−MCS1−P1およびP2−MCS1−T2
(b)少なくとも2つの制限部位AおよびBを含むプロモーターP1とP2との間にある少なくとも1つの増殖モジュールM
(c)発現カセットの1つと各々隣接する少なくとも2つの部位XおよびYを含み、ここで
(d)制限部位AおよびX並びにBおよびYは互換性があるが、しかし
(e)AYおよびBXの連結産物は、制限部位A、B、XおよびYにそれぞれ特異的な制限酵素a、b、xまたはyによって酵素的に切断可能ではなく、および
(f)制限部位AおよびB並びにXおよびYは互換性がなく、および、各ポリジーンが発現カセットの1つ中へ挿入される、式I
X−T1−MCS1−P1−[A−B]−P2−MCS2−T2−Y (I)
に記載の機能的配置を含む、請求項1〜14のいずれか一項に記載のポリヌクレオチド。
【請求項16】
増殖モジュールM中の制限部位AおよびBが、制限部位BstZ17I、SpeI、ClaIおよびNruIまたはそれらのイソ制限酵素によって切断される制限部位からなる群より選択される、請求項15に記載のポリヌクレオチド。
【請求項17】
制限部位XおよびYが、制限部位PmeIおよびAvrIIまたはそれらのイソ制限酵素によって切断される制限部位からなる群より選択される、請求項15または16に記載のポリヌクレオチド。
【請求項18】
(a)プロモーターP1およびP2が、polhおよびp10からなる群より選択され;
(b)転写終結配列が、SV40およびHSVtkからなる群より選択され;
(c)増殖モジュールM中の制限部位AおよびBが、制限部位BstZ171I、SpeI、ClaIおよびNruIからなる群より選択され;
(d)制限部位XおよびYが、制限部位PmeIおよびAvrIIからなる群より選択され;および
(e)ウイルス組み込みのための部位が、cre−loxおよびTn7からなる群より選択される、請求項15〜17のいずれか一項に記載のポリヌクレオチド。
【請求項19】
請求項1〜18のいずれか一項に記載のポリヌクレオチドを含むベクター。
【請求項20】
真核生物発現ベクターまたはトランスファーベクターである、請求項19に記載のベクター。
【請求項21】
ウイルスである、請求項19または20に記載のベクター。
【請求項22】
バキュロウイルス発現ベクターである、請求項21に記載のベクター。
【請求項23】
バキュロウイルス遺伝子v−cathおよびchiAが、機能的に破壊されている、請求項22に記載のベクター。
【請求項24】
部位特異的リコンビナーゼ(SSR)のための部位を含む、請求項19〜23のいずれか一項に記載のベクター。
【請求項25】
cre−lox特異的組換えのためのLoxP部位を含む、請求項24に記載のベクター。
【請求項26】
トランスポゾン要素を含む、請求項19〜25のいずれか一項に記載のベクター。
【請求項27】
トランスポゾン要素が、Tn7結合部位である、請求項26に記載のベクター。
【請求項28】
請求項1〜18のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターを含有する宿主細胞。
【請求項29】
細菌、酵母細胞、昆虫細胞、線形動物細胞および哺乳動物細胞からなる群より選択される、請求項28に記載の宿主細胞。
【請求項30】
イー・コリ、S.セレビシエ、P.パストリス、S.ポンベ、C.アルビカンス、Sf9細胞、線虫細胞、ヒト細胞、げっ歯類細胞、ブタ細胞、ウシ細胞およびヒツジ細胞からなる群より選択される、請求項29に記載の宿主細胞。
【請求項31】
請求項1〜28のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターで形質転換された遺伝子導入非ヒト動物。
【請求項32】
げっ歯類、ブタ、ウシおよび線虫より選択される、請求項31に記載の動物。
【請求項33】
(a)ポリジーンを構成する遺伝子のコード領域を増幅させる工程;
(b)該コード領域に、少なくとも1つのプロテアーゼ切断部位および/または少なくとも1つの自己切断ペプチドをコードする配列を提供する工程;
(c)単一ORFが結果的に各ポリジーンをもたらすように、工程(a)および(b)から結果的に生じる断片を集合させる工程;
(d)少なくとも2つのポリジーンを単一ポリヌクレオチドに結合する工程
を含む、請求項1〜18のいずれか一項に記載のポリヌクレオチドの生成のための方法。
【請求項34】
(a)ポリジーンの発現を可能にする条件下で、請求項28〜30のいずれか一項に記載の宿主細胞を適切な培地中で培養する工程;および
(b)ポリジーンによってコードされる発現産物を培地および/または宿主細胞から回収する工程
を含む、インビトロでの多タンパク質複合体の産生のための方法。
【請求項35】
遺伝子治療のためのポリジーントランスファー媒体を含む医薬の調製のための、請求項1〜18のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターおよび/または請求項28〜30のいずれか一項に記載の宿主細胞の使用。
【請求項36】
多タンパク質複合体の結晶化のための、請求項1〜18のいずれか一項に記載のポリヌクレオチドおよび/または請求項19〜27のいずれか一項に記載のベクターおよび/または請求項28〜30のいずれか一項に記載の宿主細胞の使用。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【図8−3】

【図9】
【図10−1】
【図10−2】
【図10−3】

【図10−4】

【図10−5】

【図11】
【図12−1】
【図12−2】
【図12−3】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【図8−3】
【図9】
【図10−1】
【図10−2】
【図10−3】
【図10−4】
【図10−5】
【図11】
【図12−1】
【図12−2】
【図12−3】
【公表番号】特表2009−514541(P2009−514541A)
【公表日】平成21年4月9日(2009.4.9)
【国際特許分類】
【出願番号】特願2008−539315(P2008−539315)
【出願日】平成18年11月6日(2006.11.6)
【国際出願番号】PCT/EP2006/010608
【国際公開番号】WO2007/054250
【国際公開日】平成19年5月18日(2007.5.18)
【出願人】(508139000)
【氏名又は名称原語表記】ETH Zurich
【Fターム(参考)】
【公表日】平成21年4月9日(2009.4.9)
【国際特許分類】
【出願日】平成18年11月6日(2006.11.6)
【国際出願番号】PCT/EP2006/010608
【国際公開番号】WO2007/054250
【国際公開日】平成19年5月18日(2007.5.18)
【出願人】(508139000)
【氏名又は名称原語表記】ETH Zurich
【Fターム(参考)】
[ Back to top ]