
ポリペプチドの収量を改良する方法
本発明は蛋白質収量を改良するための方法に関する。本方法は、真核生物宿主における1つまたは複数の蛋白質特性について、最適範囲内に入るか最適値にもっと近づくように、一組の関連蛋白質特性の値を修飾するステップを含む。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
[発明の分野]
本発明は、ポリペプチドの収量を改良する方法に関する。特に、本発明は、ポリペプチドの主鎖を修飾するステップによりポリペプチドの収量を改良する方法に関する。
【0002】
[発明の背景]
ゲノムおよびメタゲノムシークエンシングにおける最近の急速な進歩は、潜在的に非常に興味深い多様な蛋白質を表す多くの遺伝子をもたらしている。これらの遺伝子をかなりの水準で発現させる問題は、これらの遺伝子によりコードされる蛋白質の機能性の調査を妨げ、結果として、経済的な実現可能な方法での、このような蛋白質の利用の可能性を阻む。多くの場合、発見された遺伝子は大規模生産にあまり適していないか、または現在の遺伝子工学ツールにかなり利用し難しい生物に由来するため、遺伝子導入系および十分に開発された遺伝子工学ツールが利用可能な、十分に確立された産生宿主を使用することが極めて望ましい。特に糸状菌および酵母等の真核生物は、蛋白質の産生における、特に細胞外蛋白質の産生における、細胞工場として広く使用される。長期にわたり伝統的に使用されているため、これらの種の幾つかは一般に安全である(GRAS)と見なされ、ヒト用製品の製造に非常に興味深いものとなっている。しかし、かなりの改良にもかかわらず、異種遺伝子で得られる産生レベルは大抵、相同遺伝子でみられるものよりはるかに低い。多くの場合、蛋白質の発現は皆無である。
【0003】
蛋白質産生のレベルを上げるために様々な方法がある。これらの中には、強力プロモーターの利用、コピー数の増加、最適コザック(Kozak)配列、mRNA安定要素、最適化コドン使用(国際公開第2008/000632号パンフレット)および遺伝子などがある。しかしながら、これらの戦略は一般に、蛋白質が検出可能なレベルで産生されることを保証するものではない。今日まで、異種蛋白質を産生するための最も成功したアプローチは、それらを、効率的に分泌される相同蛋白質との翻訳融合として発現させることである。それにも拘わらず、産生レベルはやはりそれでもなお著しく後れを取り、多くの場合、発現レベルは問題になるほど低い。一般に、発酵における低発現は、回収における低収量につながる。たとえ発現が最適化されても、最終的な成熟蛋白質生成物は、下流処理での大きな損失のため、やはり非常に低い生成収量に終わるかもしれない。これは、発現された蛋白質がバイオマスと結合したままであるとき、該当するかもしれない。これは高損失に終わるか、あるいは、蛋白質を可溶化するために、高価な、そして時には望ましくない洗剤の使用を必要とする。
【図面の簡単な説明】
【0004】
【図1】図1は、クリベロミセス・ラクテス(K.lactis)発現ベクターpKLPGE−WT(実施例1に記載の構築物)のプラスミドマップを表す。図1は、他のpKLPGE−発現プラスミドの代表的なマップも提供する。PGEコード化遺伝子に関連したLAC4プロモーターおよびamdS選択マーカーカセットを示す。大腸菌(E.coli)DNAは、形質転換前に、制限酵素SacIIで消化することにより、除去できる。
【図2】図2は、発現ベクターpANPGE−3(実施例1に記載の構築物)のプラスミドマップを表す。図2は、他のpANPGE−発現プラスミド類の代表的なマップも提供する。加えて、glaAプロモーターおよび本発明の方法によって変異体PGE酵素をコード化する切断型GlaAの配列およびPGEコード化配列が示される。大腸菌(E.coli)DNAは、アスペルギルス・ニガー(A.niger)菌株の形質転換前に、制限酵素NotIで消化することにより、除去できる。
【図3】図3は、発現ベクターpGBFINZDU−WT(実施例1に記載の構築物)のプラスミドマップを表す。図3は、他のpGBFINZDU−プラスミド、pGBFINZTB−プラスミドおよびpGBFINZTC−プラスミドの代表的なマップも提供する。amdS選択マーカーカセットに関連したglaA隣接領域を示す。加えて、glaAプロモーターの配列ならびに本発明の方法によって変異体酵素をコード化するZDU配列、ZTB配列およびZTC配列が示される。大腸菌(E.coli)DNAは、アスペルギルス・ニガー(A.niger)菌株の形質転換前に、制限酵素NotIで消化することにより、除去できる。
【図4】アスペルギルス・ニガー(A.niger)WT 6およびPGE変異形質転換体pANPGE12#16(A)およびpANPGE13#30(B)のSDS−PAGEおよびウエスタンブロット解析。培養の2日(D2)および3日(D3)の上澄を分析した。14kDaおよび97kDaにおける水平線は、SDS−PAGEおよびウエスタンブロットのアラインメント用である。左側のマーカーの大きさは、SDS−PAGE染色マーカーに相当し、また右側のマーカーはウエスタンブロットマーカーに相当する。
【図5】図5は、全てglaAプロモーターのコントロール下での、異なるZDU構築物を発現する、アスペルギルス・ニガー(A.niger)菌株の、3日間発酵後の培養ブロス中のキチナーゼ活性を表す。シグナル配列、N−末端および蛋白質デザインが修飾されている、変異体SDU構築物を発現するアスペルギルス・ニガー(A.niger)菌株の培養ブロス中のキチナーゼ活性を表す。異なる構築物に関する詳細は、表6に示す。相対的キチナーゼ活性は、OD590測定値として表す。表示した全ての形質転換体群について、3つの形質転換体を単離し、独立に培養した。
【図6】図6は、全てglaAプロモーターのコントロール下での、変異体ZDU構築物を発現する、アスペルギルス・ニガー(A.niger)WT 6菌株およびZDU菌株の、4日間発酵後の、培養ブロスのSDS−PAGEの解析を表す。異なる構築物および発現したZDU蛋白質に関する詳細は、表6に表す。表示した全ての形質転換体群について、3つの形質転換体を単離し、独立に培養した。
【図7】図7は、全てglaAプロモーターのコントロール下での、変異体ZTB構築物を発現するアスペルギルス・ニガー(A.niger)WT 6菌株およびZTB−菌株の、4日間発酵後の、培養ブロスのSDS−PAGE解析を表す。異なる構築物および発現したZTB蛋白質に関する詳細を、表7に表す。表示した全ての形質転換体群について、3つの形質転換体を単離し、独立に培養した。
【図8】図8は、全てglaAプロモーターのコントロール下での、変異体ZTC構築物を発現する、アスペルギルス・ニガー(A.niger)WT 6菌株およびZTC−菌株の、5日間発酵後の、培養ブロスのSDS−PAGE解析を表す。異なる構築物および発現したZTC蛋白質に関する詳細を、表8に表す。表示したZTC−WT形質転換体群について3つの形質転換体を、他の2つの菌株型については2つの菌株を、単離して独立に培養した。
【図9】図9は、局所的な蛋白質特性を表す。
【0005】
[配列番号の説明]
配列番号1:cDNAコドン対最適化(CPO)前胃エステラーゼ(PGE);プロセシングした、すなわちシグナル配列コード化部分がない
配列番号2:蛋白質子ウシ前胃エステラーゼ(PGE)、シグナル配列を含む
配列番号3:DNA PGE蛋白質特性最適化(PFO)変異体KL8、グリコシル化部位1つ追加
配列番号4:蛋白質PGE PFO変異体KL8、グリコシル化部位1つ追加
配列番号5:DNA PGE PFO変異体KL9、グリコシル化部位5つ追加
配列番号6:蛋白質PGE PFO変異体KL9、グリコシル化部位5つ追加
配列番号7:DNA PGE PFO変異体KL11、6.96から7.74へのplシフト
配列番号8:蛋白質 PGE PFO変異体KL11、6.96から7.74へのplシフト
配列番号9:DNA PGE PFO変異体KL12、6.96から6.7へのplシフト
配列番号10:蛋白質PGE PFO変異体KL12、6.96から6.7へのplシフト
配列番号11:DNA PGE、天然のシグナル配列がα−MAT因子シグナルプレ(プロ−)配列に融合したPGE変異体
配列番号12:DNA PGE AN3、Kex部位(KR)とのCPO遺伝子tAG融合物
配列番号13:DNA PGE変異体AN12、6.96から4.6へのplシフト
配列番号14:蛋白質PGE変異体AN12、6.96から4.6へのplシフト
配列番号15:DNA PGE変異体AN13、6.96から4.88へのplシフト
配列番号16:蛋白質PGE変異体AN13、6.96から4.88へのplシフト
配列番号17:DNAキチナーゼ(ZDU)野生型
配列番号18:蛋白質キチナーゼ(ZDU)野生型
配列番号19:DNAキチナーゼ変異体ZDU−6
配列番号20:蛋白質キチナーゼ変異体ZDU−6
配列番号21:DNAキチナーゼ変異体ZDU−7
配列番号22:蛋白質キチナーゼ変異体ZDU−7
配列番号23:DNA β−グルコシダーゼ野生型ZTB−WT
配列番号24:蛋白質β−グルコシダーゼ野生型ZTB−WT
配列番号25:DNA β−グルコシダーゼ変異体ZTB−4
配列番号26:蛋白質β−グルコシダーゼ変異体ZTB−4
配列番号27:DNAエンドグルカナーゼ野生型ZTC−WT
配列番号28:蛋白質エンドグルカナーゼ野生型ZTC−WT
配列番号29:DNAエンドグルカナーゼ変異体ZTC−5
配列番号30:蛋白質エンドグルカナーゼ変異体ZTC−5
【0006】
[詳細な説明]
本発明は、真核生物宿主における1つまたは複数の蛋白質特性について最適範囲内に入るか、または最適値により近づくように、ポリペプチドのアミノ酸主鎖における一組の関連する蛋白質特性値を修飾するステップにより、真核生物宿主細胞によって対象ポリペプチドの分泌を改良する方法に関する。
【0007】
1つの利点は、以前には分泌されないか、または商業的応用は魅力のないほど少量が分泌されるに過ぎなかったが、今では、分泌が改良されたために産業的プロセスに利用可能である、興味深い機能を有する蛋白質である。もう一つの利点は、設計されたポリペプチドがバイオマスから既に分離されているため、下流処理およびポリペプチド類の回収がより容易になることである。
【0008】
本明細書の文脈では、蛋白質特性は、蛋白質アミノ酸配列およびDNA配列から計算により導くことができる性質である。
【0009】
ポリペプチドの修飾は、本明細書では、ポリペプチドのアミノ酸配列の変化をもたらすあらゆる事象と定義される。修飾は、1つまたは複数の修飾と理解される。修飾は、ポリペプチド主鎖における1つまたは複数のアミノ酸の導入(挿入)、置換または除去(削除)によって遂行することが可能である。
【0010】
本明細書の文脈では、用語「分泌」は、細胞外培地、一般に成長培地または生産培地中での、ポリペプチドの出現を指す。分泌されるポリペプチドは、バイオマスを含まない。分泌のレベルは、活性アッセイ(活性単位)、比活性(蛋白質重量当たりの単位)、定量的PAGE解析、定量的質量分析法および抗体法を含む、当該技術分野で周知の方法で測定することが可能である。
【0011】
「ポリペプチドの分泌の改良」という表現は、細胞の、細胞外培地中に分泌されるポリペプチド量の増加を指す。改良は、たとえば細胞内ポリペプチド等の、通常は分泌されないポリペプチドが、今では分泌されるようになることにより反映され得る。改良はまた、たとえばシグナル配列を含むため分泌されると予想されていたのに分泌されなかったポリペプチドが、今では分泌されるようになったということでもある。改良は、もちろん、常に同一宿主遺伝的背景および同一培養または発酵条件に関して測定される。こうした場合、改良された分泌は、たとえば、改良前には可視バンドが皆無であったポリアクリルアミドゲルに蛋白質バンドが出現することから、明白なこともある。
【0012】
あるいは、改良は、非常に少量で分泌されるポリペプチドが、上昇した分泌レベルを示すことにより反映されることもある。
【0013】
一実施形態では、分泌されたポリペプチドの量は、細胞外培地中のポリペプチドの活性を測定することにより決定される。改良前の状況に比べて、細胞外培地中の活性は、少なくとも5%、少なくとも10%、少なくとも15%または少なくとも20%、上昇している可能性がある。好ましくは、活性は少なくとも25%、少なくとも30%、少なくとも35%または少なくとも40%、上昇している。より好ましい実施形態では、活性は、少なくとも45%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも100%、少なくとも200%、少なくとも500%または少なくとも1000%、上昇している。活性は、細胞外培地中で、無活性から若干の活性に上昇していてもよい。
【0014】
任意の真核細胞を、本発明の方法で使用し得る。好ましくは、真核細胞は、哺乳類細胞、昆虫細胞、植物細胞、真菌細胞、または藻類細胞である。好ましい哺乳類細胞としては、たとえばチャイニーズハムスター卵巣(CHO)細胞、COS細胞、293細胞、PerC6細胞、およびハイブリドーマなどが挙げられる。好ましい昆虫細胞としては、たとえばSf9細胞およびSf21細胞およびそれらの派生物などがある。より好ましくは、真核細胞は真菌細胞、すなわち、カンジダ(Candida)、ハンゼヌラ(Hansenula)、クリベロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、シゾサッカロミセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)菌株等の、酵母細胞である。より好ましくは、クリベロミセス・ラクテス(Kluyveromyces lactis)、サッカロミセス・セレビシア(S.cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ヤロウイア・リポリティカ(Yarrowia lipolytica)およびピキア・パストリス(Pichia pastoris)、または糸状菌細胞由来である。最も好ましくは、真核細胞は、糸状菌細胞である。
【0015】
「糸状菌」は、亜門真菌類(Eumycota)および卵菌(Oomycota)の全ての線維状型を含む(Hawksworthらにより、Ainsworth and Bisby’s Dictionary of The Fungi,8th edition,1995,CAB International,University Press,Cambridge,UKで定義されている通り)。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複合多糖類からなる菌糸壁(mycelial wall)を特徴とする。栄養生長は菌糸伸長により、また炭素異化は偏性好気性である。糸状菌菌株としては、アクレモニウム(Acremonium)、アガリクス(Agaricus)、アスペルギルス(Aspergillus)、アウレオバシジウム(Aureobasidium)、クリソスポリウム(Chrysosporium)、コプリナス(Coprinus)、クリプトコッカス(Cryptococcus)、フィリバシジウム(Filibasidium)、フザリウム(Fusarium)、フミコラ(Humicola)、マグナポルテ(Magnaporthe)、ムコール(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカリマスティクス(Neocallimastix)、ニューロスポラ(Neurospora)、ペシロマイセス(Paecilomyces)、ペニシリウム(Penicillium)、ピロミセス(Piromyces)、パネロカエテ(Panerochaete)、プリュロタス(Pleurotus)、シゾフィラム(Schizophyllum)、タラロミセス(Talaromyces)、サーモアスカス(Thermoascus)、シーラビア(Thielavia)、トリポクラジウム(Tolypocladium)、およびトリコデルマ(Trichoderma)などの菌株が挙げられるが、その限りではない。
【0016】
好ましい糸状菌細胞は、アスペルギルス(Aspergillus)属、クリソスポリウム(Chrysosporium)属、ペニシリウム(Penicillium)属、タラロミセス(Talaromyces)属またはトリコデルマ(Trichoderma)属の種属に属し、最も好ましくはアスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ソーヤ(Aspergillus sojae)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、タラロミセス・エメルソニ(Talaromyces emersonii)、アスペルギルス・オリゼー(Aspergillus oryzae)、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)、トリコデルマ・リーセイ(Trichoderma reesei)またはペニシリウム・クリソゲナム(Penicillium chrysogenum)の種属に属する。本発明による宿主細胞がアスペルギルス(Aspergillus)宿主細胞であるとき、その宿主細胞は、好ましくはCBS 513.88またはその派生物である。
【0017】
糸状菌の幾つかの菌株は、たとえば、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection(ATCC))、ドイツ微生物細胞培養コレクション(Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSM))、オランダ微生物株保存センター(Centraalbureau Voor Schimmelcultures(CBS))、および農業研究サービス特許カルチャーコレクション、北部領域研究センター(Agricultural Research Service Patent Culture Collection,Northern Regional Research Center(NRRL))アスペルギルス・ニガー(Aspergillus niger)CBS 513.88、アスペルギルス・オリザエ(Aspergillus oryzae)ATCC 20423、IFO 4177、ATCC 1011、ATCC 9576、ATCC 14488−14491、ATCC 11601、ATCC 12892、ペニシリウム・クリソゲナム(P.chrysogenum)CBS 455.95、ペニシリウム・シトリナム(Penicillium citrinum)ATCC 38065、ペニシリウム・クリソゲナム(Penicillium chrysogenum)P2、タラロミセス・エメルソニ(Talaromyces emersonii)CBS 124.902、アクレモニウム・クリソゲナム(Acremonium chrysogenum)ATCC36225またはATCC 48272、トリコデルマ・リーセイ(Trichoderma reesei)ATCC 26921またはATCC 56765またはATCC 26921、アスペルギルス・ソーヤ(Aspergillus sojae)ATCC11906、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)ATCC44006およびそれらの派生物等、多数のカルチャー・コレクションで公的に、容易に入手できる。
【0018】
本発明の一実施形態では、アスペルギルス・ニガー(A.niger)またはクリベロミセス・ラクテス(K.lactis)が使用される。
【0019】
一実施形態において、真核細胞は、組換え技術でポリペプチドが産生される宿主細胞である。宿主細胞を形質転換または形質移入する好適な方法は、Sambrook,et al.(Molecular Cloning:A Laboratory Manual,2nd ed.Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY,1989)、Davis et al.,Basic Methods in Molecular Biology(1986)および他の実験マニュアルを参照されたい。したがって、本発明はまた、ポリペプチドの分泌を改良するための本発明による方法を対象ポリペプチドに適用し、組換え技術により、本発明に従って修飾されたポリペプチドを産生するステップによって対象ポリペプチドを産生する方法にも関する。本発明はまた、前記組換技術により産生されるポリペプチドにも関する。本発明はまた、ポリペプチドの分泌を改良するための本発明による方法で得られるポリペプチドにも関し;好ましくは前記ポリペプチドは、ポリペプチドの分泌を改良するための本発明による方法で得られる。
【0020】
本発明の方法に従って分泌が改良される対象ポリペプチドは、対象生物活性を有する任意のポリペプチドであってもよい。ポリペプチドは、コラーゲンまたはゼラチン、あるいはそれらの変異体または混成物であってもよい。ポリペプチドは、抗体またはその一部、抗原、凝固因子、酵素、ホルモンまたはホルモン変異体、受容体またはその一部、調節蛋白質、構造蛋白質、レポーター、あるいは血清アルブミン、たとえばウシ血清アルブミンおよびヒト血清アルブミン等、またはトランスフェリン、たとえばラクトフェリン等の、輸送蛋白質、分泌過程に関与する蛋白質、折り畳み過程に関与する蛋白質、シャペロン、ペプチドアミノ酸輸送体、グリコシル化因子、転写因子、合成ペプチドまたはオリゴペプチド、その天然型が細胞内蛋白質であり、かつシグナルペプチドへの融合およびその天然型で既に分泌されているポリペプチドへの融合等の、当該技術分野で周知の方法により分泌される蛋白質であってもよい。このような細胞内蛋白質は、プロテアーゼ、セラミダーゼ類、エポキシド加水分解酵素、アミノペプチダーゼ、アシラーゼ類、アルドラーゼ、ヒドロキシラーゼ、アミノペプチダーゼ、リパーゼ等の酵素であってもよい。ポリペプチドは、その天然型で細胞外に分泌される酵素であってもよい。このような酵素は、酸化還元酵素、トランスフェラーゼ、加水分解酵素、リアーゼ、イソメラーゼ、リガーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、デオキシリボヌクレアーゼ、デキストラナーゼ、エステラーゼの群に属してもよい。酵素は、カルボヒドラーゼ、たとえばエンドグルカナーゼ類、β−グルカナーゼ類、セロビオヒドロラーゼ類またはβ−グルコシダーゼ類等のセルラーゼ類、ヘミセルラーゼ類、あるいはペクチン分解酵素、たとえばキシラナーゼ類、キシロシダーゼ類、マンナナーゼ類、ガラクタナーゼ類、ガラクトシダーゼ類、ペクチンメチルエステラーゼ類、ペクチンリアーゼ類、ペクチン酸リアーゼ類、エンドポリガラクツロナーゼ類、エキソポリガラクツロナーゼ類、ラムノガラクツロナーゼ類、アラバナーゼ類、アラビノフラノシダーゼ類、アラビノキシラン加水分解酵素、ガラクツロナーゼ類、リアーゼ類、またはアミロース分解酵素等;加水分解酵素、イソメラーゼ、またはリガーゼ、フィターゼ等のホスファターゼ類、リパーゼ等のエステラーゼ類、蛋白分解酵素、オキシダーゼ類、トランスフェラーゼ類、またはイソメラーゼ類等の酸化還元酵素であってもよい。酵素は、フィターゼであってもよい。酵素は、アミノペプチダーゼ、アスパラギナーゼ、アミラーゼ、カルボヒドラーゼ、カルボキシペプチダーゼ、エンド−プロテアーゼ、メタロ−プロテアーゼ、セリン−プロテアーゼカタラーゼ、キチナーゼ、クチナーゼ、シクロデキストリングリコシルトランスフェラーゼ、デオキシリボヌクレアーゼ、エステラーゼ、α−ガラクトシダーゼ、β−ガラクトシダーゼ、グルコアミラーゼ、α−グルコシダーゼ、β−グルコシダーゼ、ハロパーオキシダーゼ、蛋白質デアミナーゼ、インベルターゼ、ラッカーゼ、リパーゼ、マンノシダーゼ、ミュータナーゼ、オキシダーゼ、ペクチン分解酵素、ペルオキシダーゼ、ホスホリパーゼ、ポリフェノールオキシダーゼ、リボヌクレアーゼ、トランスグルタミナーゼ、またはグルコースオキシダーゼ、ヘキソースオキシダーゼ、モノオキシゲナーゼであってもよい。分泌が改良されるポリペプチドは、宿主細胞と相同であっても異種であってもよい。相同ポリペプチドの好適な例は、アスペルギルス・ニガー(Aspergillus niger)にクローニングされて産生される、アスペルギルス・ニガー(Aspergillus niger)蛋白質である。異種発現の好適な例としては、糸状菌または酵母にクローニングされて産生される、たとえば大腸菌(E.coli)または桿菌(Bacillus)由来の細菌ポリペプチド、または、糸状菌または酵母にクローニングされて産生される、たとえばウシまたはヤギ由来の哺乳類蛋白質、または酵母にクローニングされて産生される糸状菌ポリペプチド、あるいは、別の真菌にクローニングされて産生される、糸状菌蛋白質などがある。好ましくは、ポリペプチド類をコード化する核酸は、たとえばコドン対最適化により、関連宿主細胞における発現用に最適化される。コドン対最適化は、ポリペプチドをコード化するヌクレオチド配列の改良された発現および/またはコード化されたポリペプチドの改良された産生を得るために、ポリペプチドをコード化するヌクレオチド配列を、それらのコドン使用、特に使用されるコドン対に関して、修飾しておく方法である。コドン対は、コード配列における、一組の、2つの後続するトリプレット(コドン)と定義される。コドン対最適化は、好ましくは国際公開第2008/000632号パンフレットに記載の通りに実施される。
【0021】
好ましくは、修飾されたポリペプチドの特異性は、分泌の改良前と実質的に同じである。これは、たとえば基質特異性または結合特異性が実質的に保持されることを意味する。本明細書の文脈では、用語「実質的に保持される」は、特異性の60%超、65%超、70%超または75%超が保持されることを意味する。好ましくは、特異性の80%超、85%または90%が保持される。最も好ましくは、特異性の95%超、96%、97%、98%または99%が保持される。
【0022】
本発明の方法によれば、改良された分泌の指標である、細胞外培地中の活性レベルは上昇する。しかし、修飾されたポリペプチドの比活性は、低下しない限り、上昇しなくてもよい。したがって、比活性は、好ましくは分泌の改良前と実質的に同じか、それより高い。好ましい実施形態では、比活性は、改良前と実質的に同じである。本明細書の文脈では、「実施的に同レベルの活性」という表現は、親ポリペプチドの活性レベルと、15%未満、好ましくは12%未満または10%未満、より好ましくは8%未満、6%未満または4%未満異なる活性レベルを指す。
【0023】
本明細書の文脈では、用語「ポリペプチド」および蛋白質」は、互換的に使用される。本発明の方法により、あらゆるタイプのポリペプチドが、その分泌を改良することが可能である。好ましい実施形態では、ポリペプチドは、本明細書に前述したリストの1つである。
【0024】
本発明の方法によれば、真核生物宿主における1つまたは複数の蛋白質特性について最適範囲内に入るか、または最適値にもっと近づくように、アミノ酸主鎖における一組の関連する蛋白質特性値が修飾される。
【0025】
修飾されたポリペプチドとリファレンスポリペプチドとの間の蛋白質特性の変化量は、相対的改良(RI)および正規化相対的改良(RIN)という2つの方法で定義することができる。
【0026】
蛋白質特性のRIは、最適値からの蛋白質特性の絶対偏差(D)という観点で定義される:
RI=(DREF−DPFO)/DREF,
ここで、D=|FPOI−F0PT|、FPOIは、対象蛋白質特性値であって、
リファレンスまたはPFOのいずれかであり,FOPTは最適特性値である。
RINは、どの特性が実質的に重要であるかを理解するために、正規化偏差(DN)の観点で定義される。DNは、特性値の上界(UB)および下界(LB)を考慮に入れる(表1参照)。
RIN=DN,REF−DN,PFO,
ここで、FPOI>FOPTであれば、DN=(FPOI−FOPT)/(UB−FOPT)
FPOI<FOPTであれば、DN=(FPOI−FOPT)/(LB−FOPT)
【0027】
本発明の方法によれば、修飾はポリペプチド主鎖に行われる。本明細書の文脈では、用語「主鎖」は、アミノ酸がペプチド結合を介して結合されて、共有結合した一連のアミノ酸を形成するとき、形成される正規構造を指す。本発明では、好ましくは、成熟ポリペプチドの主鎖が修飾される。本発明に関連して、「成熟ポリペプチド」は本明細書では、翻訳および何らかの翻訳後修飾、たとえばN−末端プロセシング、C−末端切断、グリコシル化、リン酸化等々の後に、その最終的な機能形であるポリペプチドとして定義される。修飾前のポリペプチドは、親ポリペプチドまたはリファレンスポリペプチドまたは野生型ポリペプチドと呼ばれ、それに由来する修飾されたポリペプチドと区別される。用語「親ポリペプチド」、「野生型ポリペプチド」および「リファレンス−ポリペプチド」は、本明細書では互換的に使用される。ポリペプチドがキメラポリペプチドであるとき、すなわち効率的に分泌されるポリペプチドとの、好ましくは宿主細胞に由来するポリペプチドとの、翻訳融合物であるとき、キメラポリペプチド全体を本発明に従って修飾することが可能である。キメラポリペプチドが、対象ポリペプチドに融合したリーダーポリペプチドとして効率的に分泌されるポリペプチドを含むとき、対象ポリペプチドは、好ましく修飾される。
【0028】
当業者に周知の通り、成熟中のプロセシングエラーのため、成熟ポリペプチドのN−末端は、成熟ポリペプチドのC−末端と同様、異種であり得る可能性がある。特に、このようなプロセシングエラーは、ポリペプチドの過剰発現で起こり得る。加えて、エキソプロテアーゼ活性は、異質性を生じさせるかもしれない。異質性が起こる程度は、使用される宿主および発酵プリトコールによっても異なる。このようなN−末端およびC−末端プロセシング人工産物は、予想される成熟ポリペプチドと比較して、短いポリペプチド類または長いポリペプチド類をもたらすかもしれない。
【0029】
本発明の一実施形態では、本方法は、
(i)真核生物宿主における、1つまたは複数の蛋白質特性の最適範囲および最適値を決定するステップ、および
(ii)真核生物宿主における、一組の関連した蛋白質特性を決定するステップ(これらの関連した特性の1つまたは複数が、ポリペプチドのアミノ酸主鎖において修飾されるのであれば、その特性は、真核生物宿主によるポリペプチドの分泌を改良するであろう)、および
(iii)(i)で決定されたように、最適範囲内に入るかまたは最適値により近づくように、関連した蛋白質特性値を修飾するステップ(ここで、(i)および(ii)は任意の順序で実施し得る)、
を含む。
【0030】
任意の方法を使用して、一組の関連した特性を決定し得る。一実施形態では、ポリペプチドの分泌を改良するための、関連した一組の特性は下記の通りに決定される:
(i)特定の真核生物宿主における適量の蛋白質の分泌レベルならびにこれらの蛋白質のアミノ酸配列およびDNA配列を含む、データセットSの収集または作成。データセットSは、分泌蛋白質(S+)を含んでもよい。好ましくは、データセットSはまた、非分泌蛋白質(S−)も含む。たとえば、全ての予測される分泌蛋白質をアスペルギルス・ニガー(A.niger)で、発現させることができる(Tsang et al.,2009,Fungal Genetics and Biology,46:S153−160)。分泌される蛋白質は、セットS+に属するが、分泌されない蛋白質は、セットS−に属する。任意の方法を使用して、分泌レベルを測定し得る。あるいは、セットS−は、真核生物宿主における文献で周知の非分泌蛋白質を含んでもよい。Sにおける蛋白質は、真核生物宿主と相同であっても異種であってもよい。
(ii)データセットSにおける全ての蛋白質についての、蛋白質特性(F)の計算。Fは、これらの蛋白質のDNA配列およびアミノ酸配列の両者から誘導することが可能である;
(iii)適切に規定された分類子性能基準に従って、S+とS−とを区別するために、統計分類子の最高の成果を与える、ii)で計算した蛋白質特性のサブセット(Fs)を選択するための統計的分類方法の使用。Fsは、DNA配列(Fs_DNA)およびアミノ酸配列(Fs_AA)の両者から誘導され得る;
【0031】
Fs_AAにおける蛋白質特性は、対応する真核生物宿主における蛋白質分泌を改良するための修飾についての関連する特徴である。
【0032】
好ましくは、成熟ポリペプチドの主鎖は、本発明の方法に従って修飾されるため、蛋白質特性は、一組の成熟蛋白質から計算されることが好ましい。
【0033】
当該技術分野で周知の、標準的統計的分類方法、たとえば、線形判別分類器(Linear Discriminant Classifier(LDC))、二次判別分類器(Quadratic Discriminant Classifier(QDC))、最近傍平均分類器(Nearest Mean Classifier(NMC))、1−/k−最近傍分類器(Nearest Neighbour classifiers)、サポートベクターマシーンおよび決定木等々を使用することができる(Webb,Statistical Pattern Recognition,2nd ed,John Wiley & sons)。このような方法を適用するとき、データセットSは、学習データセットと確認データセットに分けることができ、また当該技術分野で周知の確認方式(たとえば、10分割交差確認)を使用することが可能である。
【0034】
当該技術分野で周知の任意の分類器性能測定、たとえば、特異性、感度、確度、精度および受信者動作特性(ROC)曲線下面積を使用し得る。
【0035】
任意の好適な方法を使用して、蛋白質特性の最適範囲または最適値を決定し得る。
【0036】
一実施形態では、真核生物宿主の蛋白質特性の最適範囲または最適値は、以下の通りに決定される:
i)特定の真核生物宿主における適量の蛋白質の分泌レベルならびにこれらの蛋白質のアミノ酸配列およびDNA配列を含む、データセットSの収集または作成。データセットSは、分泌蛋白質(S+)を含んでもよい。好ましくは、データセットSはまた、非分泌蛋白質(S−)も含む。たとえば、全ての予測される分泌白質をアスペルギルス・ニガー(A.niger)で、発現させることができる(Tsang et al.,2009,Fungal Genetics and Biology,46:S153−160)。分泌される蛋白質は、セットS+に属するが、分泌されない蛋白質は、セットS−に属する。任意の方法を使用して、分泌レベルを測定し得る。あるいは、セットS−は、真核生物宿主における文献で周知の非分泌蛋白質を含んでもよい。Sにおける蛋白質は、真核生物宿主と相同であっても異種であってもよい。
ii)データセットSにおける全ての蛋白質についての、蛋白質特性(F)の計算。Fは、これらの蛋白質のDNA配列およびアミノ酸配列から誘導され得る;
iii)対応する真核生物宿主の各特性についての最適値(F_opt)の決定:最適値は、S+から算出される各蛋白質特性の中心値を算出することによって得てもよい。任意の中心値、たとえば、幾何平均、調和平均、算術平均、トリム平均、最頻値および中央値を使用し得る。算出される中心値は、対応する真核生物宿主に関する特性の最適値である。あるいは、特性値の分布が、選択される確率分布によって十分に説明されるように、S+から算出される各蛋白質特性に確率分布を合わせる。任意の確率分布を使用し得、たとえば正規分布、指数分布、または対数正規分布を使用し得る。確率分布の平均が、対応する真核生物宿主の特性の最適値である。
iv)対応する真核生物宿主について各特性の最適範囲の決定:分泌される蛋白質のみを含むS+セットを考慮すると、蛋白質特性の最適範囲の下界は、S+から算出される蛋白質特性の、0.3−、0.2−、0.15または好ましくは0.10−および0.05−変位値に相当する値と定義される。ここで、値0.3、0.2、0.15、等々は、累積確率を指す。特定の累積確率に相当する変位値は、任意の統計的方法で、たとえば、Statistical Toolbox,Matlab R2007a(The Mathworks Inc)の分位関数を使用して、算出し得る。蛋白質特性の最適範囲の上界は、S+から算出される蛋白質特性の、0.7−、0.8−、0.85または好ましくは0.90−および0.95−変位値に相当する値と定義される。あるいは、分泌蛋白質および非分泌蛋白質の両方を含む全データセットSを考慮すると、蛋白質特性の最適範囲の下界は、それより下の値ではSにおける蛋白質の70%、80%、85%、好ましくは90%または95%が分泌されない低い蛋白質特性の値として定義してもよい;蛋白質特性の最適範囲の上界は、それより上の値ではSにおける蛋白質の70%、80%、85%、好ましくは90%または95%が分泌されない蛋白質特性値として定義される。
【0037】
関連特性および最適範囲および最適値のセットは、宿主細胞ごとに異なるであろう。アスペルギルス・ニガー(A.niger)の場合、蛋白質分泌を増加するために修飾されるべき関連蛋白質特性(Fs_AA)としては、塩基性アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、微小アミノ酸頻度、小アミノ酸頻度、帯電アミノ酸頻度、正味荷電(pH7.2における)、等電点、アスパラギン、アルギニン、イソロイシン、システイン、ヒスチジン、グルタミン、バリン、リシン、グリシン、トレオニンおよびロイシンの頻度、ターン(Garnierにより算出)、EPESTFINDにより算出されるPESTモチーフ、plの局所特性(LF)値、特にLF1およびLF6、GravyスコアのLF値、特にLF2およびLF4、アロマスコアのLF値、特にLF3、LF4およびLF6、原子組成w.r.t.イオウ(S)および局在化特性(たとえば、MultiLoc 局在化予測ツールで予測される)などが挙げられるが、その限りではない。
【0038】
正味荷電は、陽子の電荷と同じ単位を有する。長さ当たりの正味電荷/正味正電荷/正味負電荷/全電荷は、陽子の電荷と同じ単位を有するが、ポリペプチドの長さに正規化される。
【0039】
ポリペプチドの正味荷電は、本明細書では、全てのアミノ酸が溶媒に完全に露出しており、隣接したペプチド類は、所与のアミノ酸のpKに影響を及ぼさず、また構成アミノ酸は、N末端およびC末端と同様、未修飾であると仮定して、推定される。様々なプログラムを使用して、たとえば、Bioinformatics Toolbox of Matlab(バージョンR2008b)の「等電」機能を使用して、またはhttp://emboss.sourceforge.net/で入手可能な、EMBOSS Explorerの「蛋白質統計学(pepstats)」機能を使用して、特定のpHにおけるポリペプチドの正味電荷を算出することができる(初期値pH=7.2による)。
【0040】
長さ当たりの正味荷電は、本明細書では、ポリペプチドの長さで割ったポリペプチドの正味荷電と定義される。
【0041】
長さ当たりの正味正電荷は、本明細書では、pH7.2における、ポリペプチドのN−末端ならびに全てのリシン残基、アルギニン残基およびヒスチジン残基の部分電荷を合計することによって算出される、ポリペプチドの正味正電荷と定義される。長さ当たりの正味正電荷は、ポリペプチドの正味正電荷をポリペプチドの長さで割ることにより決定される。
【0042】
長さ当たりの正味負電荷は、本明細書では、pH7.2における、ポリペプチドのC−末端ならびに全てのアスパラギン酸残基、グルタミン酸残基、システイン残基およびチロシン残基の部分電荷を合計することによって算出される、ポリペプチドの正味負電荷と定義される。長さ当たりの正味負電荷は、ポリペプチドの正味負電荷をポリペプチドの長さで割ることにより決定される。
【0043】
長さ当たりの全電荷は、本明細書では、ポリペプチドの正味陽性電荷(正の数)をポリペプチドの正味負電荷(負の数)だけ減算することにより算出される、ポリペプチドの全電荷と定義される。長さ当たりの全電荷は、ポリペプチドの全電荷をポリペプチドの長さで割ることにより決定される。
【0044】
グレイビー・スコア(gravy score)は、本明細書では、KyteおよびDoolittle(1982)により定義された通り、ポリペプチドの疎水性親水性指数と定義される。各アミノ酸は、4.6と−4.6の間の疎水性スコアを有する。4.6は、最も疎水性の蛋白質に割り当てられ、また−4.6は最も親水性の蛋白質に割り当てられる。ポリペプチドのGRAVYスコアは、好ましくは、Kyte and Doolittle(1982).Kyte,J.and Doolittle,R.1982蛋白質の疎水性親水性特徴を表示する簡単な方法(A simple method for displaying the hydropathic character of a protein.)J.Mol.Biol.,157:105−132.に従って決定される。
【0045】
ポリペプチドのアロマスコアは、本明細書では、ポリペプチド中の3つの芳香族アミノ酸、Phe、TyrおよびTrpの頻度を合計することにより算出される。
【0046】
脂肪族指数は、本明細書では、脂肪族側鎖によって占められる相対体積と定義される。ポリペプチドの脂肪族指数(AI)は、Ikaiの式(1980):AI=f_Ala+af_Val+b(f_Ile+f_Leu)に従って算出される。アミノ酸類アラニン、バリン、イソロイシンおよびロイシンは、脂肪族側鎖を有する。
ここで、aはバリン側鎖の相対体積であり(a=2.9)、bはロイシンおよびイソロイシン側鎖の相対体積である(b=3.9)。f_Ala、f_Val、f_Ileおよびf_Leuは、それぞれ、ポリペプチド中の、アラニン、バリン、イソロイシンおよびロイシンの頻度である。Ikai,A.J.1980球状蛋白質の熱安定性および脂肪族指数(Thermostability and aliphatic index of globular proteins.J.Biochem.,88:1895−1898。)
【0047】
GRAVYおよび脂肪族について、ExPASyサーバー上の蛋白質同定および分析ツール(Protein Identification and Analysis Tools on the ExPASy Server;Gasteiger E.,Hoogland C.,Gattiker A.,Duvaud S.,Wilkins M.R.,Appel R.D.,Bairoch A.;(In)John M.Walker(ed):The Proteomics Protocols Handbook,Humana Press(2005).pp.571−607.も参照することができるであろう。
【0048】
理化学的特性に基づくアミノ酸の種類:
酸性:D,E
脂肪族:A,I,L,V
芳香族:F,W,Y
塩基性:H,K,R
帯電:D,E,H,K,R
非極性:A,C,F,G,I,L,M,P,V,W,Y
極性:D,E,H,K,N,Q,R,S,T
小:A,C,D,G,N,P,S,T,V
微小:A,C,T,S,G
【0049】
配列中の単一要素の組成に基づく特性は、要素iの頻度fiから算出される。頻度と割合は、本明細書では、互換的に使用される。頻度は、配列中の要素総数で割った、要素iが配列中に出現する回数と定義される。配列中の単一要素、たとえばアミノ酸を、多重要素たとえば、微小、酸性に結合することができる。
【0050】
ポリペプチド内のアミノ酸残基の表面接触性は、当該技術分野で周知の任意の方法で決定し得る。
【0051】
ポリペプチドが実験的に解明される構造を有する場合、溶媒接触表面積(ASA)は、Å2で与えられ、またその面積は、蛋白質表面上を、水分子大の球を回転させることによって算出することができる[1]。次いで、ASAを相対表面積(RSA)に変換するが、これは、グリシン[2]またはアラニン[3]と並んだトリペプチドの中央の残基の最大可能露出と比較した、ポリペプチド鎖中の所与のアミノ酸残基のASAとして算出される。閾値アルファより大きいRSA(RSA>=アルファ、0<=アルファ<=1)を有する残基は、露出していると言われ、閾値ベータ未満のRSA(RSA<=アルファ、0<=ベータ<=1)を有する残基は、埋没していると言われる。好ましくは、アルファ>=0.25、より好ましくはアルファ=0.25である。好ましくはベータ<=0.25、より好ましくはベータ=0.25である。
【0052】
ポリペプチドの構造を入手できなければ、表面接触性は、ポリペプチドのアミノ酸配列からも予測することができる。ポリペプチドのアミノ酸配列から表面接触性を予測するために、様々な方法を文献で入手することができ、たとえば[3]、[4]、[5]および[6]に記載されている。好ましくは、RSAは、[4]に記載されているいわゆるNetSurfP法を使用して予測され、これはhttp://www.cbs.dtu.dk/services/NetSurfP/でオンラインアクセスすることができる。このアプリケーションで、表面接触性は、成熟蛋白質のアミノ酸配列から予測される。露出した残基および埋没した残基の定義は、前述と同じである。
[1]Connolly M:分析的分子表面計算(Analytical molecular surface calculation.)。Journal of Applied Crystallography 1983,16(5):548−558.
[2]Chothia C:蛋白質における接触可能表面および埋没表面の性質(The nature of the accessible and buried surfaces in proteins.)。J Mol Biol 1976,105(1):1−12.
[3]Ahmad S,Gromiha MM,Sarai A:アミノ酸配列からの溶媒接触可能性の実数値予測(Real value prediction of solvent accessibility from amino acid sequence.)。Proteins 2003,50(4):629−635.
[4]Bent Petersen et al:溶媒接触性予測に適用される信頼性スコアの包括的割り当て方法(A generic method for assignment of reliability scores applied to solvent accessibility predictions.)。BMC Structural Biology 2009,9:51.
[5]Dor O,Zhou Y:Real−SPINE:蛋白質構造特性の実数値予測のための神経回路網の集積システム(an integrated system of neural networks for real−value prediction of protein structural properties.)。Proteins 2007,68(1):76−81.
[6]Faraggi E,Xue B,Zhou Y:二層神経回路網を介した指導学習による蛋白質の残基溶媒接触可能性および実数値主鎖ねじれ角の予測確度を改良すること(Improving the prediction accuracy of residue solvent accessibility and real−value backbone torsion angles of proteins by guided−learning through a two−layer neural network.)。Proteins 2009,74(4):847−856.
【0053】
アスペルギルス・ニガー(A.niger)の最適値および最適範囲を表1に示す。
【0054】
【表1】

【0055】
【表2】
【0056】
表1において、全蛋白質配列から計算される全ての特性は、全蛋白質の長さに基づく。成熟蛋白質配列、露出残基および埋没残基から計算される全ての特性は、成熟蛋白質の長さに基づく。
【0057】
好ましくは、最適値および範囲特性は、表2から選択される;これらの特性は、一次特性と言われ、その他の特性、すなわち、表2にない表1の特性は、二次特性である。
【0058】
【表3】
【0059】
クリベロミセス・ラクテス(K.lactis)について、好ましい一次特性を表3に示す。
【0060】
【表4】
【0061】
別の実施形態では、ポリペプチドの分泌は、下記のステップで改良される:
i)ポリペプチドの蛋白質特性を計算するステップ、
ii)ポリペプチドの1つまたは複数の蛋白質特性が、最適範囲外であるか、または真核生物宿主の最適値から実質的に逸脱しているかどうかを決定するステップ(ここで、実質的逸脱は、最適値から20%、30%、40%または50%超の差と定義される)、
iii)ポリペプチドの1つまたは複数のFs_AAの値が、最適範囲内に入るか、または適量だけ、最適値に向けて移行するように、好ましくはポリペプチドの蛋白質特性と、蛋白質特性の最適値との差が、10%、15%、20%、または30%超、減少するように、ポリペプチドのアミノ酸配列を合理的に変えるステップ。
【0062】
好ましくは、2つ、3つ、4つまたは5つの蛋白質特性が一緒に修飾され、さらに好ましくは、10超、15または20の蛋白質特性が一緒に組み合わせて修飾される。最も好ましくは、25超または30の蛋白質特性が一緒に修飾される。
【0063】
好ましくは、最適範囲は、表1から選ばれ、さらに好ましくは最適範囲は、表2から選ばれる。あるいは、最適範囲は表3から選ばれる。
【0064】
上記ステップiii)で、ポリペプチドのアミノ酸配列は、当該技術分野で周知の任意の方法で合理的に変えし得る。たとえば、これは、:
(i)相同配列を検索するステップ;
(ii)相同配列を、対象配列と配列比較するステップ;
(iii)蛋白質機能的特性に重要なアミノ酸類を同定するステップ;
(iv)機能的特性を保持しながら、所望のアミノ酸配列特性を導入する;
(vi)所与の宿主に最適なコドンを使用して、最終的な修飾された配列を遺伝子に翻訳するステップ;
(vii)宿主における、再設計されたポリペプチドのクローニングおよび発現
によって達成するステップが可能である。
【0065】
好ましくは、アミノ酸主鎖のアミノ酸類の少なくとも5%が修飾され、より好ましくは、アミノ酸主鎖のアミノ酸類の少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%が修飾される。
【0066】
好ましくは、アミノ酸主鎖の少なくとも5つのアミノ酸類が修飾され、より好ましくはアミノ酸主鎖の少なくとも10のアミノ酸類、さらにより好ましくは少なくとも15のアミノ酸類、さらにより好ましくは少なくとも20のアミノ酸類、さらにより好ましくは少なくとも25のアミノ酸類、さらにより好ましくは少なくとも30のアミノ酸類が修飾される。
【0067】
好ましくは、本発明によれば、一次特性は改良されるが、二次特性は特定の境界内に保たれる。したがって、総体的な最適性スコアFは、全n個の一次特性および全m個の二次特性のDN値に基づいて規定される:
【数1】
【0068】
ηは、0と1の間で0および1を含む(0≦η≦1)の重み係数である。好ましくはη<0.5、より好ましくはη≦0.4、最も好ましくはη=0.3である。ρは、1と5の間で1および5を含み(1≦p≦5)、好ましくはρ=2である(Fは、このとき、ユークリッド距離を表す)。好ましくは、η=0.3およびρ=2である。好ましくは、野生型リファレンス蛋白質に関して、少なくとも5%のF−スコアの改良が達成され、より好ましくは少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%およびさらにより好ましくは少なくとも30%の改良が達成される。
【0069】
好ましくは、少なくとも2、3、4、または5つの特性が修飾され、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が修飾される。好ましくは、少なくとも2、3、4、または5つの特性が改良され、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が改良されるが、好ましくは、10未満、さらにより好ましくは5未満、さらにより好ましくは4未満、の特性が悪化する。好ましくは、その特性は一次特性である。
【0070】
相同配列は、好ましくは、適切な配列データベースのBLAST検索を実施することにより、検索される。相同配列は、対象配列と好ましくは少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、60%、70%、80%、90%、95、%、96%、97%、98%または99%の同一性を有する。最も好ましくは、相同配列は、対象配列と好ましくは約50%の同一性を有する。当業者は、幾つかの異なるコンピュータープログラムを利用して、2つの配列を並べて2つの配列間の相同性を決定できることを知るであろう(D.Sankoff and J.B.Kruskal,(ed.)中のKruskal,J.B.(1983)配列比較の概説(An overview of sequence comparison)、タイムワープ、文字列編集および巨大分子:配列比較の理論と実際(Time warps,string edits and macromolecules:the theory and practice of sequence comparison),pp.1−44AddisonWesley)。当該技術分野で周知の任意の方法を配列比較に使用し得る。2つのアミノ酸配列間または2つのヌクレオチド配列間のパーセント同一性は、たとえば2配列のアラインメントのためのNeedleman and Wunschアルゴリズムを使用して、決定することが可能である(Needleman,S.B. and Wunsch,C.D.(1970)J.Mol.Biol.48,443−453)。
【0071】
対象必須機能特性に重要なアミノ酸類を同定する方法は、当該技術分野で周知である。好適なツールは、対象蛋白質の3D構造または3Dモデルの使用、対象蛋白質または相同蛋白質の突然変異誘発試験、機能的に中性の置換と機能的置換を確立するためのサイト飽和ライブラリーの使用を含む。
【0072】
アミノ酸配列特性を導入するとき、置換は好ましくは、所与の位置に、所要のアミノ酸配列特徴とより多く一致するアミノ酸が相同配列に見られるアミノ酸群から選択される方法で選択される。最新のモデリング技法を使用して、天然の相同体には見られない許容可能な置換を同定することが可能である。所与の折り畳みを採用する新しい配列の生成を許容するモデリング技法に好ましいリファレンスは以下の通りである:
Kuhlman B,Dantas G,Ireton GC,Varani G,Stoddard BL,Baker D(2003)。原子レベルの確度を有する新規な球状蛋白質折り畳みの設計(Design of a novel globular protein fold with atomic−level accuracy)Science 302,1364−8。
Baker D(2006)。巨大分子構造および相互作用の予測および設計(Prediction and design of macromolecular structures and interactions.)。Philos.Trans.R.Soc.Lond.,B,Biol.Sci.361,459−63デノボ蛋白質設計:完全自動化配列選択に向けて(De Novo protein design:towards fully automated sequence selection’) Journal of Molecular Biology,273巻4号,1997年11月7日,789−796ページ Bassil I.Dahiyat,Catherine A.Sarisky,Stephen L.Mayo
【0073】
最新の計算方法は、所与の蛋白質折り畳みを採用することが可能な多数の潜在的配列の生成を許容する。最適な配列を取り出すために使用されるスコアリング機能に特性最適化を導入することによって、所与の産生宿主に最適な配列が、計算による方法で選択され得る。
【0074】
本発明の方法に従って修飾することが可能な蛋白質特性としては、組成特性、生理特性および構造特性などがある。このような特性の好適な例は、アミノ酸の数、分子量、等電点、特定のpHにおける正味荷電、GRAVYスコア、脂肪族指数、不安定指数、組成的特性、C、H、N、O、S原子に関する原子組成、アミノ酸頻度、ジペプチド頻度、トリペプチド頻度、酸性アミノ酸頻度、脂肪族アミノ酸頻度、芳香族アミノ酸頻度、塩基性アミノ酸頻度、グリコシル化パターンおよび帯電アミノ酸頻度ならびに表1に記載の特性である。修飾される特性の組合せも、本発明に包含される。好ましくは2、3、4または5つの蛋白質特性が、一緒に修飾される。より好ましくは、10超、15または20の蛋白質特性が一緒に修飾される。最も好ましくは、25超または30の蛋白質特性が一緒に修飾される。
【0075】
本発明の一実施形態では、1つまたは複数のグリコシル化部位が導入され、他の蛋白質特性も修飾される。本発明の別の実施形態では、帯電アミノ酸頻度が修飾され、他の蛋白質特性も修飾される。本発明の別の実施形態では、極性アミノ酸頻度が修飾され、他の蛋白質特性も修飾される。
【0076】
アミノ酸全体またはDNA配列から計算される蛋白質特性は、蛋白質全体の平均値であり、これから、局所的蛋白質特性が明らかにならない可能性がある。たとえば、ある蛋白質は平均すると親水性であるが、それでも大きな内部疎水性領域を含むこともあり得る。局所的蛋白質特性は、たとえば、Benita et al.(Benita et al.,2006.Molecular and Cellular Proteomics,5:1567−1580)により略述されている方法を用いて、アミノ酸またはDNA配列から計算することができる。
【0077】
ある蛋白質特性の局所特性を計算するために、適当な数のアミノ酸またはヌクレオチドのスライディングウィンドウで、蛋白質特性を局所的に計算することが可能である。次いで、図9に図示する通り、得られた値を、蛋白質のアミノ酸またはDNA配列の長さに沿った曲線としてプロットする。
【0078】
【表5】
【0079】
たとえば、LF1は図1における暗灰色の面積に相当し、LF2は図1における明灰色の面積に相当する。その面積は、台形法を使用して算出することができる(Benita et al.,2006.Molecular and Cellular Proteomics,5:1567−1580)。局所的特性を計算するために、好適な上限および下限、ならびにスライディングウィンドウの大きさを選択することができる。スライディングウィンドウは、任意のサイズのものであってもよい。たとえば、21アミノ酸または塩基対のスライディングウィンドウサイズを使用することができる。上限および下限の値は、曲線における極値ピークを反映するように選択することができる。たとえば、より高い上限は、低いものより多い極値ピークを考慮に入れる。好ましくは、上限および下限は、データセットS+およびS−のFischer基準を最大化するように選択される。Fischer基準(JF)は以下の通りに定義される:
【数2】
ここでμs−およびμs+は、それぞれS−およびS+のセットから計算される局所的特性値の平均値を表し、σ2s−およびσ2s+は、それぞれS−およびS+のセットから計算される局所的特性値の分散を表す。
【0080】
上で定義した局所的特徴は、任意の蛋白質特性、たとえばGravyスコア、アロマスコアおよび等電点について算出し得る。
【0081】
首尾よく分泌された蛋白質の配列から誘導できる特性に加えて、特に、標的蛋白質の溶媒接触表面の親水性が上昇すると、細胞から分泌された可溶性蛋白質の増量に大きく成功することが確認された。より詳細には、発現が増加するのみならず、有意に多い蛋白質が、バイオマスまたは他の不溶性物質に付着していない可溶形で、ブロス中に蓄積した。改良された表面親水性を有する蛋白質が与えられれば、いずれも有意により高い分泌で回収することが可能であろう。バイオマスを(濾過または遠心分離によって)除去すると、産生された蛋白質の大部分は濾液中または上澄中にそのまま残る。
【0082】
親水性の上昇は、
・非極性アミノ酸類を、より極性のアミノ酸類に代えること
・より低極性のアミノ酸類を、より極性のアミノ酸類に代えること
・極性アミノ酸類を帯電アミノ酸類に代えること
によって行うことができる。
【0083】
より極性のアミノ酸類または帯電アミノ酸類の数を増加させることにより親水性を上昇させることは、アミノ酸組成を変えることになり、したがって分泌を増加するために適合させることができる組成特性として考えることができる。
【0084】
非極性アミノ酸類は、A、V、L、I、C、M、F群から選択される。アミノ酸類G、P、Y、Wは、極性状況下では非極性として考えることができ、また非極性状況下では極性として考えることができる。より極性の残基は、S、T、N、Q、D、E、H、R、K群から選択される。帯電した残基は、D、E、H、R、K群から選択される。酸性または負に帯電した残基は、E、Dから選択される。塩基性または正に帯電した残基は、H、K、Rから選択される。極性用比較尺度を使用すると:[A、V、L、I、M、F、C]<[G、P、Y、W]<[S、T]<[N、Q、H]<[D、E、K、R]。
【0085】
極めて疎水性の表面領域は、産生宿主における高い産生ストレス、宿主における蛋白質の蓄積、および分泌妨害または無分泌をもたらす、望ましくない凝集またはバイオマスへの望ましくない固着を招く傾向があることが分かっている。総体的親水性を上昇させる置換は、特にこれらの残基が溶媒接触残基(=蛋白質表面残基)を含むとき、分泌改良に非常に有効であることが認められている。より詳細には、接触表面領域において、非極性残基をより極性の残基で置換するとき、極性残基の割合は、組成特性分析の上方境界で定められる極性残基の割合を上回る可能性さえあることが認められている。標的蛋白質の配列特性と宿主要件との非適合性は、標的蛋白質の親水性を上昇させることによって、より詳細には、正と負の電荷が表面全体に一様に分布して、正または負の電荷のホットスポットを防ぐように分布させた追加的電荷を導入することにより、補償することが可能である。
【0086】
特定のアミノ酸配列が与えられれば、幾つかの予測ツールは、どのアミノ酸が表面上にありそうかを予測するために使用できるが、溶媒接触非極性パッチまたは疎水性パッチを予測することが必要なとき、これらのツールの性能は全く貧弱である。したがって、蛋白質接触表面の親水性を調節するためには、3D構造または3D構造モデルが必要である。蛋白質の3D構造は、X線結晶学によって、およびNMRによって、決定することができる。加えて、比較モデリングまたは鋳型に基づくモデリングを使用して、相同蛋白質の3D構造に基づく所与の配列のための信頼できる3Dモデルを構築することが可能である(http://en.wikipedia.org/wiki/Homology modeling)。比較モデリングのための様々なサーバーおよびソフトウェアパッケージは:http://en.wikipedia.org/wiki/Protein structure prediction softwareでみつかる。
【0087】
蛋白質構造予測およびモデリングに関する最近の総説については、Yang Zhang,Current Opinion in Structural Biology 2008,18:342−348を参照されたい。
【0088】
3D構造または3Dモデルの原子座標が与えられれば、当該技術分野で周知の方法により、接触表面を算出することができる。よく知られた方法は、Frederic Richardsにより開発されたローリングボールアルゴリズムによる算出である(1977,「面積、体積、充填および蛋白質構造(Areas,volumes,packing and protein structure.)」、AnnuRev Biophys Bioeng,6:151−176)。http://en.wikipedia.org/wiki/Accessible surface areaも参照されたい。
【0089】
接触表面を決定するためには、置換が多量体(たとえば二量体、三量体、四量体等々)における個々のポリペプチド(モノマー)間の相互作用を妨害するのを回避するために、最終的な成熟蛋白質の四次構造を考えるべきである。
【0090】
表面変調は、下記を含む:
・適切な分泌および回収を妨げかねない、潜在的な粘着性のパッチを生じさせる溶媒から、非極性残基が接触される領域をスポットすること。
・機能的役割を果たす領域、たとえば活性部位全般を、そしてより具体的には基質、共基質およびコファクター用の結合ポケットを除外すること。
・非極性残基を、帯電残基も含む、より極性の残基で置換すること。
・極性残基を、より極性の残基または帯電残基で置換すること。
・高い負電荷を有する領域または高い正電荷を有する領域を回避するために、帯電残基を再分布させること。
・疎水性表面パッチを置換する代わりに、非極性領域の近くにグリコシル化を導入することにより、このような領域を遮蔽することも可能である。
【0091】
一次構造の場合、上昇した親水性は、たとえば下表のように修飾の前後の極性残基の数を比較することによって表される。
【0092】
【表6】
【0093】
接触表面を考えるとき、様々な極性アミノ酸の寄与は、総接触表面に対する、ある特定のアミノ酸またはある特定のアミノ酸群により形成される接触表面の割合として表現することができる。たとえば、帯電残基の総接触表面を算出して、総接触表面積と比較することができる。全ての極性残基を考えることにより、極性接触表面を算出することができる。蛋白質表面の親水性は、非極性表面を犠牲にして極性表面の割合が増加するとき、上昇すると言われる。
【0094】
原則として、グリコシル化を導入し、そのグリコシル化によって遮蔽される領域を推定することもできる。電荷の分布は、視覚的検査を含む任意の利用可能な方法で行い得る。
一実施形態では、改良された分泌のために修飾すべき特性は、表面電荷(再)分布、表面極性−非極性分布、配列モチーフ、たとえばグリコシル化、またはこれらの組合せである。1つの特性、たとえばアミノ酸の修飾は、多くの場合、別の特性の修飾、たとえばC、H、N、O、S原子に関して原子組成の修飾を、もたらすことを当業者は理解するであろう。
【0095】
本発明による方法は、蛋白質産生レベルを高めるための最新の技法と、またはこれらの技法の1つまたは複数の組み合わせと、都合よく組み合わせられることを理解されたい。これらは、強力プロモーター類、コピー数の増加、最適Kozak配列、mRNA安定要素および最適化コドン使用の適用を含むが、その限りではない(国際公開2008/000632号パンフレット)。
【0096】
[実施例]
[菌株]
アスペルギルス・ニガー(A.niger)菌株:WT 1:このアスペルギルス・ニガー(A.niger)菌株を、野生型菌株として使用した。この菌株は、寄託番号CBS 513.88.で、CBS研究所(CBS Institute)に寄託される。
WT 2:このアスペルギルス・ニガー(A.niger)菌株は、グルコアミラーゼをコード化する遺伝子(glaA)の欠失を含むWT 1菌株である。WT 2は、欧州特許出願公開第0 635 574 B1号明細書に記載の「MARKER−GENE FREE」アプローチを使用して構築した。この特許では、CBS 513.88のゲノム中のglaA特異的DNA配列の削除方法が詳細に説明されている。この手順は、最終的に外来DNA配列を全く持たない、MARKER−GENE FREE ΔglaA組換えアスペルギルス・ニガー(A.niger)CBS513.88菌株をもたらした。
WT 3:WT 2における主要な細胞外アスパラギン酸プロテアーゼPepAをコード化しているpepA遺伝子を中断するために、van den Homberghらにより記述されている通り、WT 2のゲノム中のpepA特異的DNA配列を削除した(van den Hombergh JP,Sollewijn Gelpke MD,van de Vondervoort PJ,Buxton FP,Visser J.(1997)−アスペルギルス・ニガー(Aspergillus niger)における3つの酸プロテアーゼの破壊−プロテアーゼスペクトル、細胞内蛋白質分解、および標的蛋白質の分解に及ぼす影響−Eur J Biochem.247(2):605−13)。この手順は、WT 2菌株バックグラウンドに不活性化されたpepA遺伝子を有する、MARKER−GENE FREE WT 3菌株をもたらした。
WT 4:WT 3中のhdfA遺伝子を削除するために、先に、国際公開第05/095624号パンフレットに詳細に記載されている方法を使用して、アスペルギルス・ニガー(Aspergillus niger)WT 4(ΔglaA,ΔpepA,ΔhdfA)を作成した。
WT 5:このアスペルギルス・ニガー(A.niger)菌株は、シュウ酸欠損アスペルギルス・ニガー(A.niger)菌株をもたらす欠失を含むWT 4菌株である。WT 5は、欧州特許出願公開第1157100号明細書および米国特許第6,936,438号明細書に記載の方法を使用することによって構築され、その中では、オキサロ酢酸加水分解酵素をコード化するoahA遺伝子の欠失によってシュウ酸欠損菌株が得られ、菌株WT 5は、WT 4菌株バックグラウンドに不活性化されたoahA遺伝子を有する代表的な菌株として選択された。
WT 6:このアスペルギルス・ニガー(A.niger)菌株は、後続する3ステップで、アルファ−アミラーゼ類をコード化する3遺伝子(amyB、amyBIおよびamyBII)の欠失を含む、WT 5菌株である。欠失ベクターの構築およびこれらの3遺伝子のゲノム欠失は、国際公開第2005095624号パンフレットに詳述されている。国際公開第2005095624号パンフレットに記載のベクターpDEL−AMYA、pDEL−AMYBIおよびpDEL−AMYBIIは、欧州特許出願公開第0 635 574 B1号明細書に記載の「MARKER−GENE FREE」アプローチに従って使用された。上述の手順は、最終的に外来DNA配列を全く持たない、シュウ酸欠損、MARKER−GENE FREE ΔglaA、ΔpepA、ΔhdfA、ΔamyA、ΔamyBIおよびΔamyBIIアミラーゼ−陰性組換えアスペルギルス・ニガー(A.niger)CBS 513.88菌株である、WT 6をもたらした。このように、菌株WT 6は低いアミラーゼバックグラウンドを有し、WT 1と比較して、より効率的な配列のターゲティングに関して高いHR/NHR比を有し、また細胞外蛋白質発現および検出用に最適化されている。
【0097】
クリベロミセス・ラクテス(K.lactis)菌株:クリベロミセス・ラクテス(K.lactis)におけるPGEおよびその変異体の発現を評価するために、2つの菌株を使用した。GG799(New England Biolabs)、および本明細書ではWT 7とも呼ばれ、米国特許第6,265,186 B1号明細書により詳細に記載されている、クリベロミセス・ラクテス(K.lactis)CBS 685.97の派生物。菌株クリベロミセス・ラクテス(K.lactis)WT 7は、突然変異誘発(古典的菌株改良)および遺伝子操作によって、CBS 685.97から誘導した。
【0098】
[キチナーゼ活性アッセイ]
反応混合物は、キチン−アズール(Sigma)3mg、0.1M Na−クエン酸−リン酸バッファー0.5ml、pH5.0の分析試料(培養液)0.1mlを含んでいた。この反応混合物を振盪しながら37℃で24時間、インキュベートし、12000rpmで10分間遠心分離してOD590を測定した。
【0099】
[pNP−β−グルコピラノシドを基質として使用するβ−グルコシダーゼ活性]
50mM酢酸ナトリウムバッファー pH=4.5中、3mM pNP−β−グルコピラノシド(Sigma N7006)原液を調製した。アッセイ:基質原液(3mM)250μl+希釈酵素試料250μLを、40℃でインキュベートした。t=0、10、20および30分に、培養標本100μlと1M炭酸ナトリウム100μlとを混合することによって反応を停止させた。吸光度は、MTP読み取り装置を使用して405nmで測定した。活性は、放出されたpNP μmol/ml/分で表す。
【0100】
[セロビオースを基質として使用する、β−グルコシダーゼ活性]
50mM酢酸ナトリウムバッファー pH=4.5中、最終濃度10mMのセロビオース(SigmaC7252)原液を調製した。アッセイ用に、基質原液(10mM)2000μl+希釈酵素試料100μLを混合し、40℃でインキュベートした。t=0、10、20および30分に、培養標本100μl+50mM水酸化ナトリウム100μlを混合することによって反応を停止させた。試料を限外濾過に付し、ED40パルスアンペロメトリー検出器を具備するDionex DX−500で実施されるパルスアンペロメトリー検出器を用いた高性能陰イオン交換クロマトグラフィ(HPAEC−PAD)を使用して分析した。活性は、放出されたグルコースμmol/ml/分で表す。
【0101】
[AZO−CM−セルロースを使用するエンドグルカナーゼ活性]
本アッセイは、Megazyme手順書S−ACMC 04/07(Megazyme International Ireland Ltd,http://secure.megazyme.com/downloads/en/data/S−ACMC.pdf)に従って実施する。活性は、100mM酢酸ナトリウムバッファー pH4.6中、2%AZO−CM−Celluloseを用いて40℃で測定した。アッセイ用に、基質原液(2%)250μL+希釈酵素溶液250μLを混合した。30分後、沈殿剤溶液1250μLを加えた。沈殿剤溶液(エタノール76%中、300mM酢酸ナトリウムバッファー pH=5と20mM Zn−酢酸)を加えることによって反応を停止させた。1000xgで10分間遠心分離した後、分光光度計を使用して、上澄の590nmにおける吸光度を測定した。活性は、放出された色素μmol/ml/分で表す。
【0102】
[トリブチリンプレートアッセイ]
トリブチリン(C4)を基質として、ローダミンBリパーゼプレートスクリーニングアッセイを行った。ローダミンBプレートアッセイは、試料中の、リパーゼ活性存在のスクリーニングによく使用され、文献(G.Kouker,K.E.Jaeger,Appl.and Environ.Microbiol,1987,211−213)に記載のアッセイから改変された。使用した全ての化学薬品は、分析グレードであった。アラビアゴムエマルジョンは、NaCl 17.9gおよびKH2PO4 0.41gをH2O 400mlに溶解し、最後にグリセロール(87%)540mlを加えることによって作った。アラビアゴム6(6.0)gを徐々に加え、溶解した後、H2Oを加えることにより全量1000mlとした。
ローダミンB溶液は、ローダミンBを20mg/mlの濃度でエタノールに溶解することによって調製した。4%アガロース溶液は、加熱により、アガロース4gをバッファー液(0.1M酢酸塩 pH=5.5)100mlに溶解することによって調製した。リパーゼ活性のスクリーニングに使用した基質は、トリブチリンであった。
プレートアッセイ手順:基質1mlおよびアラビアゴムエマルジョン1.5mlを、バッファー液5mlと混合し、振幅20μmで2×60秒間Soniprepを使用して、または任意選択的に、緑色に設定したUltraturaxで2分間、超音波処理した。この溶液に、熱いアガロース溶液7.5mlをローダミンB 150μlと一緒に加えた。最終溶液をペトリ皿プレートに注入した。プレートは、使用するまで冷蔵庫内に保存した。使用直前に、レプリケーターを使用して、直径3mmの穴を作った。リパーゼ活性を確認する溶液10μlを穴にピペッティングし、その後、プレートを37℃で18〜24時間インキュベートした。穴の周囲の蛍光ハロは、リパーゼ活性を示す。
【0103】
[pNP−ブチレートアッセイ]
前胃エステラーゼ(PGE)活性は、内部酵素標準に対して、最終濃度1mMのパラ−ニトロフェニルブチレートを基質として使用し、37℃で測定した。基質溶液は、アセトニトリル中50mMパラ−ニトロフェニルブチレート原液を作り、それを、0.2%BSAおよび2%Triton X−100を含有する0.1Mリン酸ナトリウムバッファー pH6.7で5倍希釈することによって調製した。0.2%BSAを含有する0.1Mリン酸ナトリウムバッファーpH6.7 120μl、基質溶液15μlを加えた。37℃に予熱した後、適切な希釈度の試料(0.2%BSAを含有する0.1Mリン酸ナトリウムバッファーpH6.7で希釈)15μlを加え、その後、37℃で5分間のインキュベーションの間ずっと、405nmにおける吸光度の上昇を分光光度法で測定した。
試料応答は、ブランクバックグラウンド(試料の代わりに、0.2%BSAを含有する0.1Mリン酸ナトリウムバッファーpH6.7 15μlのインキュベーション)の補正をし、ブランク補正後、一般に0.05〜0.5dAbsの範囲であった。
内部標準を、pH6.0および30℃で実施される、トリブチリンを用いた滴定分析で較正した。PGE試料液(milliQ水で調製)5mlを、予熱したトリブチリン/アラビアゴムエマルジョン(それぞれ、水中93g/Lおよび57g/L)30mLに加えた。放出された遊離脂肪酸を、0.02N NaOHで滴定することによって5分間にわたり測定した。
【0104】
[SDS−PAGE電気泳動]
試料前処理:試料30μlを、水35μlおよびNuPAGE(商標)LDS試料バッファー(4×)Invitrogen 25μlおよびNuPAGE(商標)試料還元剤(Sample Reducing agent)(10×)Invitrogen 10μlに加えた。試料を、サーモミキサー内、70℃で10分間加熱した。SDS−PAGEは、供給者の指示書(Invitrogen:Gel:4〜12%Bis−Trisゲル、バッファー:MES SDS泳動バッファー、実行時間:35分)に従って、二重に実施した。2つのゲルのうちの1つをブロッティングに使用し、試料液10μlおよびマーカーM12(Invitrogen)1μlをゲル上に適用した(NuPAGE(商標) Bis Tris,Invitrogen)。
ゲルは、外部バッファー室内の20倍希釈したMES−SDSバッファー600ml、および内部バッファー室内の酸化防止剤(NuPAGETM Invitrogen)0.5mlを含有する20倍希釈したMES−SDSバッファー200mlを備えたXCELL Surelockを使用して、200Vで動かした。ラン実行後、ゲルを50%メタノール/7%酢酸(50ml)で1時間固定し、脱塩水で2回すすぎ、シプロルビー(Sypro Ruby)(50ml,Invitrogen)で一晩染色した。
画像は、ゲルを脱塩水で10分間洗浄した後、Typhoon 9200(610 BP 30,Green(532nm),PMT 600V,100μm)を使用して作成した。
【0105】
[ウエスタンブロッティング]
[PGEポリクローナル抗体]
PGEポリクローナル抗体は、迅速28日プログラムおよび抗原としての2つの合成PGEペプチドを使用して、Eurogentec(ベルギー)に注文した。PGE抗体は、市販のPiccantase C(DFS)酵素製剤と比較して確認した(データ表示せず)。
ウエスタンブロッティングは、分析S2300の方法に従って実施した。
膜:NC 0.45μm
実行時間:25Vで90分
バッファー:メタノールを含むトランスファーバッファー
【0106】
膜に移動させた後、以下のステップを実施した:
膜を、スキムミルク(PBST中1%のスキムミルク;10mM PBS+0.05%TWEEN20)20ml中で2時間ブロックする。
抗体1:SY0716,ウサギ;抗体40μlを、PBST 20mlに溶解する)室温で一晩(1:500)。
膜を、PBS−Tですすぎ、次にPBSTバッファーで3×20分間洗浄する。
抗体2:ECL Plexヤギ抗ウサギIgG Cy3(GE Healthcare);ECL Plex10μlを、PBST 25mlに溶解する、暗所に保管)1時間。(1:2500)
膜を4回すすぎ、次にPBSTで2×10分間洗浄する
PBSで2×10分間洗浄する
画像は、Typhoon9200(670BP30,緑色(532nm),PMT 450V,100μm)を使用して、膜から作成した。
【0107】
[分子生物学技法]
本明細書の実施例では、当業者に周知の分子生物学技法(see:Sambrook & Russell,Molecular Cloning:A Laboratory Manual,3rd Ed.,CSHL Press,Cold Spring Harbor,NY,2001)を使用して、幾つかの遺伝子を過剰発現させ、その他を、下記の通りに下方制御した。
【0108】
記述され、使用された全ての遺伝子置換ベクターは、周知の原理に従って設計され、またルーチンのクローニング手順に従って構築された。本質的に、これらのベクター類は、宿命的なゲノム遺伝子座における相同組換えを目標とするため、それぞれのORF配列の、約1〜2kbの隣接領域を含む。加えて、これらのベクター類は、形質転換のために、アスペルギルス・ニジュランス(A.nidulans)双方向amdS選択マーカーを、直接反復の中間に含む。本明細書の全実施例で遺伝子欠失に利用された方法は線状DNAを使用するが、これは、二重交差によってゲノムのフランキング配列の相同遺伝子座に組み込まれ、したがって削除されるべき遺伝子がamdS遺伝子に代わる。形質転換後、この直接反復によって、(第2の)相同組換え事象による選択マーカーの除去が可能になる。amdSマーカーの除去は、フルオロアセトアミド培地上にプレーティングすることにより行うことができ、無マーカー遺伝子菌株を選択する結果となる。欧州特許第0 635 574号明細書に「MARKER−GENE FREE」アプローチとしても記述されている、形質転換および後続の対抗選択という、この戦略を使用すれば、amdSマーカーを菌株修飾プログラムで無制限に使用することができる。遺伝子破壊のための基本手順は、国際公開第2006040312号パンフレットの図6に描かれている。欠失ベクター類の一般設計は、以前に欧州特許第635574B号明細書および国際公開第98/46772号パンフレットに記述されており、また欠失ベクターを構築するための一般的クローニングベクターpGBDELの使用および対抗選択手順は、国際公開第06/040312号パンフレットに記述されている。
発現ベクター類、具体的には遺伝子過剰発現用のpGBFIN−発現ベクター類の一般設計、形質転換、マーカー類の使用および選択培地の例は、国際公開第199846772号パンフレット、国際公開第199932617号パンフレット、国際公開第2001121779号パンフレット、国際公開第2005095624号パンフレット、欧州特許第635574B号明細書および国際公開第2005100573号パンフレットにある。
【0109】
[振盪フラスコ発酵]
アスペルギルス・ニガー(A.niger)菌株を、国際公開第99/32617号パンフレットの実施例:「アスペルギルス・ニガー(Aspergillus niger)振盪フラスコ発酵」セクションに記載の通りに、CSL前培養培地20ml中で前培養した(100mlフラスコ、バッフル)。34℃および170rpmで18〜24時間成長させた後、この培養10mlを、発酵培地(FM)に移す。FM中での発酵は、概ね国際公開第99/32617号パンフレットに記載の通り、発酵ブロス100mlが入ったバッフル付き500mlフラスコ内、34℃および170rpmで指示された日数、実施する。
CSL培地は(1リットル当たりの量で):コーン・スティープ・ソリッド(Corn Steep Solids)(Roquette)100g、NaH2PO4*H20 1g、MgSO4*7H2O 0.5g、グルコース*H2O 10gおよびバシルドン(Basildon)(消泡剤)0.25gで構成されていた。成分を脱塩水に溶解し、NaOHまたはH2SO4でpHをpH5.8に調整した;バッフル付き100mlフラスコおよびフォームボールを発酵培地20mlで満たし、120℃で20分間、滅菌した。
発酵培地(FM)は(1リットル当たりの量で)、マルトース*H2O 150g、ソイトン(Soytone)(ペプトン)60g、NaH2PO4*H2O 1g、MgSO4*7H2O 15g、トゥイン(Tween)80 0.08g、バシルドン(Basildon)(消泡剤)0.02g、MES 20g、L−アルギニン 1gで構成されていた。成分を脱塩水に溶解し、NaOHまたはH2SO4でpHをpH6.2に調整した;バッフル付き500mlフラスコおよびフォームボールを発酵ブロス100mlで満たし、120℃で20分間、滅菌した。
クリベロミセス・ラクテス(K.lactis)振盪フラスコ発酵の場合、クリベロミセス・ラクテス(K.lactis)PGE形質転換体の単一コロニーを、1リットル当たり酵母エキス10g、バクトペプトン20g、グルコース40gおよび100mM MES pH6.7を含有する、YEP(4%)−D/MES培地100ml(フラスコ)に接種した。発酵は、280rpmの振盪インキュベーター内、30℃で実施した。上澄を2日および3日に採取し、下記の通りにさらに分析した。
【0110】
[実施例1]
[本発明の方法による、野生型酵素および酵素変異体用のクリベロミセス・ラクテス(K.lactis)発現ベクターおよびアスペルギルス・ニガー(A.niger)発現ベクターの構築]
この実施例では、本発明の酵素の変異体用に、多数の発現ベクターを構築した。クリベロミセス(Kluyveromyces)における発現のための全ての変異体を、pKLAC2発現ベクター(New England Biolabs)に酷似したpKLPGE−ベクターでクローニングした。全てのpKLPGE−ベクター類の一般設計図を図1に示す。アスペルギルス(Aspergillus)における発現用の全ての変異体は、pGBFIN−5発現ベクターまたはpGBTOP−発現ベクターでクローニングした。構築、一般設計図およびこれらのベクター類の使用は、国際公開第199932617号パンフレットに詳述されている。
【0111】
[クリベロミセス・ラクテス(K.lactis)構築物]
ウシ前胃エステラーゼ(PGE)は、産業上興味深い酵素であり、またその全長cDNA配列は、Timmermansら(1994,Gene 147:259−262)により公開されている。クリベロミセス・ラクテス(Kluyveromyces lactis)におけるPGEの発現の場合、このcDNA配列は、最適化されたコドン対(配列番号1)であり、合成的に調製されている(たとえば、DNA2.0,USA,GeneArt、Sloning,Germany)。クリベロミセス・ラクテス(K.lactis)α−因子プレ(プロ−)シグナル配列およびKREAEA Kexプレ(プロ−)−配列プロセシング部位との融合を含む発現構築物を作成した。HindIIIおよびNotI制限部位を介して、合成遺伝子をクリベロミセス・ラクテス(K.lactis)発現ベクターでクローニングし、amdS選択マーカーも含む、pKLPGE−WT(図1)を産生した。加えて、本発明の方法に従って、改良された蛋白質特性を有する幾つかのPGE変異体を設計した。これらの突然変異体は、グリコシル化部位数および/または疎水性に関して、コドン対最適化野生型PGE酵素(配列番号2)と異なっていた。PGE突然変異体酵素コード化遺伝子も、上記の通り、やはり最適化コドン対であり、また合成的に調製されてもいる。XhoIおよびNotIクローニング部位を使用して本質的に前述の通りに、変異体を、クリベロミセス・ラクテス(K.lactis)発現ベクターにクローニングした。PGE構築物のための全ての関連ヌクレオチドおよび蛋白質の詳細を表4に示す。
【0112】
【表7】
【0113】
[アスペルギルス・ニガー(A.niger)構築物]
アスペルギルス・ニガー(A.niger)におけるウシ前胃エステラーゼPGEの発現の場合、cDNA配列は、最適化コドン対(配列番号12)であり、合成的に調製された(たとえば、DNA2.0,USA,GeneArt,Stoning,Germany)。コドン対最適化PGEコード化遺伝子は、切断型グルコアミラーゼキャリア蛋白質(tAG)への融合物として合成的に調製された。図2のpANPGE−3に示す通り、この融合フラグメントを、pGBTOP−アスペルギルス・ニガー(A.niger)発現ベクターに挿入した。
【0114】
仮想キチナーゼ(ZDU,EC3.2.1.14,Uniprot A5AB48)をコード化する野生型アスペルギルス・ニガー(A.niger)遺伝子An08g09030は、アスペルギルス・ニガー(Aniger)ゲノム配列で同定された(EMBL:AM269948−AM270415;Pel et al,「汎用細胞工場アスペルギルス・ニガー(Aspergillus niger)CBS513.88のゲノムシークエンシングおよび分析(Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88.)」。Nat Biotechnol.2007Feb;25(2):221−231)。野生型キチナーゼZDUのcDNA配列は、配列番号17として同定することができ、推定される野生型キチナーゼZDU蛋白質配列は、配列番号18として同定することができる。An08g09030のコード配列は最適化コドン対であり(国際公開第2008000632号パンフレットに詳述されている通り)、またグルコアミラーゼglaAプロモーターの翻訳開始配列は、作成された全ての発現構築物で5’−CACCGTCAAA ATG−3’に修飾されていた(やはり国際公開第2006/077258号パンフレットに詳述されている通り)。加えて、最適翻訳停止配列が使用され、したがって、全ての発現構築物で、野生型5’−TGA−3’翻訳停止配列は5’−TAAA−3に置き換えられた(国際公開第2006/077258号パンフレットに詳述されている通り)。最適化キチナーゼZDU構築物は、PacI−AscIフラグメントとして完全合成し、サブクローニングして配列を検証した。合成されたフラグメント両端のPacI−AscI制限部位を使用して、PacI−AscI消化pGBFIN−5発現ベクターの大きいベクターフラグメントでのクローニングを可能にし、pGBFINZDU−WT発現ベクターを作成した(図3)。
加えて、またZDUキチナーゼの場合と同様の方法で、タラロミセス・エメルソニ(Talaromyces emersonii)β−グルコシダーゼ(ZTB,EC 3.2.1.21,Uniprot Q8X214)およびファネロカエテ・クリソスポリウム(Phanerochaete chrysosporium)エンドグルカナーゼ(ZTC,EC 3.2.1.4,Uniprot Q66NB6)は、最適化コドン対であり(国際公開第2008000632号パンフレットに詳述されている通り)、また全ての適切な制御要素をPacI−AscIフラグメントとしてpGBFIN−5にクローニングし、それぞれpGBFINZTB−WTおよびpGBFINZTC−WTを作成した。
【0115】
本発明の方法による蛋白質特性最適化(PFO)を、ウシ前胃エステラーゼ、アスペルギルス・ニガー(A.niger)キチナーゼ蛋白質配列、タラロミセス・エメルソニ(T.emersonii)β−グルコシダーゼおよびファネロカエテ・クリソスポリウム(P.chrysosporium)エンドグルカナーゼ蛋白質配列に適用した。ウシ前胃エステラーゼの設計された変異体を含むコード配列をEcoRI−SnaBIフラグメントとして完全合成し、配列を検証した。合成フラグメントをpGBTOP−ベクターでクローニングし、pANPGE−発現構築物を作成した。アスペルギルス・ニガー(A.niger)PGE構築物のための全ての関連ヌクレオチドおよび蛋白質の詳細を表5に示す。
【0116】
【表8】
【0117】
キチナーゼ、β−グルコシダーゼおよびエンドグルカナーゼの設計された変異体を含むコード配列をPacI−AscIフラグメントとして完全合成し、サブクローニングして配列を検証した。合成されたフラグメント両端のPacI−AscI制限部位を使用して、PacI−AscI消化pGBFIN−5発現ベクターの大きいベクターフラグメントでのクローニングを可能にし、変異体pGBFIN−発現ベクターを作成した。変異体発現構築物は、下記の通りに命名され、pGBFINZDU−構築物の特徴ならびにそれぞれのヌクレオチド配列および蛋白質配列ののリファレンスは、表6から推定することができ、pGBFINZTB−構築物のそれらは表7から、またpGBFINZTC−構築物のそれらは表8から推定される。
【0118】
【表9】
【0119】
【表10】
【0120】
【表11】
【0121】
[実施例2]
[クリベロミセス・ラクテス(K.lactis)野生型の発現および分泌分析ならびに蛋白質特性最適化PGE]
菌株クリベロミセス・ラクテス(K.lactis)GG799WT7を、amdS選択マーカーも含むクリベロミセス・ラクテス(K.lactis)pKLPGE−構築物(表4)で形質転換した。各形質転換について、20コロニーを、アセトアミドを含有する選択培地で純化した。コロニーの一部を使用して、PCR反応用のDNA鋳型を作成し、各菌株におけるPGE構築物のコピー数を決定した。構築物当たり3つの、PCRスクリーニングで陽性の形質転換体を、酵素基質としてトリブチリンを含有するプレートアッセイでさらにスクリーニングした。野生型PGE酵素の場合、トリブチリンプレートアッセイを使用して、明白な活性ハロを検出することはできなかった。PGE産生用のSDS−PAGEを用いた上澄の分析もまた、陽性結果を示さなかった。意外なことに、最適化蛋白質特性を有するPGE突然変異体5つのうち4つでは、トリブチリンプレートアッセイを使用して、明白な活性ハロを確認することができた。
野生型および突然変異体PGEの多数の形質転換体を、振盪フラスコ内で成長させ、pNP−ブチレートを基質として使用して、ブロスおよび上澄のリパーゼ活性を試験した。PGE突然変異体の様々な活性アッセイの概要を表9に示す。
【0122】
【表12】
【0123】
クリベロミセス・ラクテス(K.lactis)pKLPGE−WT(PGECPO)形質転換体(様々なコピー数)では、0.2U/mlという最大活性が得られた。PGEの蛋白質特性最適化によって、すなわちpKLPGE−12で発現されるとき、このPGE突然変異体の50×を超える活性上昇が確認された。PGE−9、PGE−11およびPGE−12変異体の多数の突然変異体を大規模ベースで発酵させ、改良された分泌を確認した(データ表示せず)。この実施例では、グリコシル化部位数の改変および表面に露出している疎水性酵素部分の極性の変更により(PGEモデリングに基づいて決定される)、我々はクリベロミセス・ラクテス(K.lactis)におけるPGE酵素発現および分泌を劇的に改良できることが分かった。さらに、かなりの量の活性が、上澄にも存在していた。
【0124】
[実施例3]
[アスペルギルス・ニガー(A.niger)における野生型および蛋白質特性最適化PGEの発現および分泌分析]
アスペルギルス・ニガー(A.niger)WT 6を、アスペルギルス・ニジュランス(A.nidulans)amdS選択マーカーおよび変異体pANPGE−プラスミドを担持するpGBAAS構築物で共形質転換した(表5)。全て、国際公開第99/32617号パンフレットに記載の通り、各形質転換について、20コロニーを、アセトアミドを含有する選択培地で純化し、続いてスポアプレートを調製した。真の共形質転換体であった、たとえば、PGEカセットおよびamdSカセットの両者を含んでいた、アスペルギルス・ニガー(A.niger)形質転換体を選択するために、PCR確認する(表示せず)。結果から、選択された形質転換体20のうち少なくとも50%が、PGE発現構築物の1つまたは複数のコピーを含んでいた。形質転換体を含むこれらのPGEを継続した。PGE contransformantsのスポアを回収し、FM培地で振盪フラスコ発酵を実施した。2日目に、上澄試料を採取し、トリブチリンプレートアッセイを使用してリパーゼ活性についてスクリーニングした。
アスペルギルス・ニガー(A.niger)pANPGE−3形質転換体から回収された試料中に、非常に僅かな活性ハロを検出することができた(データ表示せず)。pANPGE−12およびpANPGE−13形質転換体では、大きい活性ハロを検出することができた(データ表示せず)。各構築物pANPGE−3、pANPGE−12およびpANPGE−13について、トリブチリンプレートアッセイで最大のハロを示した形質転換体(1〜3)のリパーゼ活性を、基質としてpNP−ブチレートを使用して試験した。PGE突然変異体の様々な活性アッセイの概要を表10に示す。
【0125】
【表13】
【0126】
WT 6および選択された形質転換体pANPGE−12#16およびpANPGE−13#30の上澄試料を、SDS−PAGEゲル(Invitrogen)およびPGEポリクローナル抗体を使用したウエスタンブロットで、さらに分析した(図4参照)。pANPGE−12のアスペルギルス・ニガー(A.niger)PGEPFO変異体では、SDS−PAGEゲルで、成熟PGEに相当するバンドを検出することができた。PGEポリクローナル抗体PGEを使用して、両形質転換体の上澄中に、交差ハイブリダイズバンドを検出することができた。
最高分子量のバンド(約55kDa)はおそらく成熟PGE突然変異体に相当し、また、より低分子量の交差ハイブリダイズバンドは、蛋白質分解の結果であろう。
蛋白質特性最適化の法則に従って、表面に露出している酵素部分の極性(PGEモデリングに基づいて決定される)を変えることにより、我々はアスペルギルス・ニガー(A.niger)におけるPGE酵素発現を劇的に改良できるという結論に達する。さらに、高い酵素活性も、上澄中で認められた。
【0127】
[実施例4]
[アスペルギルス・ニガー(A.niger)における野生型およびPFO最適化真菌酵素の発現]
実施例1(super)で調製された、pGBFINZDU−発現構築物、pGBFINZTB−発現構築物およびpGBFINZTC−発現構築物を、下記の通り、アスペルギルス・ニガー(A.niger)を使用した形質転換により導入した。WT 6で、異なるpGBFINZDU−ベクター、pGBFINZTB−ベクターおよびpGBFINZTC−ベクター(それぞれ、表6、7および8)を導入するために、国際公開第1998/46772号パンフレットおよび国際公開第1999/32617号パンフレットに記載の通りに、形質転換および後続する形質転換体の選択を実施した。簡単に記載すると、全てのpGBFIN−構築物の線状DNAを単離して、アスペルギルス・ニガー(A.niger)WT 6の形質転換に使用した。形質転換体をアセトアミド培地で選択し、標準手順に従って、コロニーを純化した。コロニーのglaA遺伝子座における組み込みおよびコピー数を、PCRを使用して診断した。類似した推定コピー数(仮想単一コピー)を有する各pGBFINZDU−構築物、pGBFINZTB−構築物およびpGBFINZTC−構築物について、独立した3つの形質転換体を選択し、形質転換プラスミドの数を使用して、たとえば、それぞれ、ZDU−WT−1、ZDU−WT−2、ZDU−WT−3、ZDU−6−1、ZDU−6−2、ZDU−6−3等々と命名した。
上述の通り、34℃および170rpmのインキュベーターシェーカーで、500mlバッフル付き振盪フラスコを使用し、選択されたZDU−菌株、ZTB−菌株およびZTC−菌株ならびにアスペルギルス・ニガー(A.niger)WT 6を使用して、FM培地100mlで、振盪フラスコ実験を実施した。発酵の3日、4日および5日の後、試料を採取し、電気泳動およびキチナーゼ活性により細胞外蛋白質の産生量を測定した。
【0128】
異なる構築物を含むアスペルギルス・ニガー(A.niger)ZDU−5形質転換体の各形質転換体により発現されたキチナーゼの産生を、培養上澄で測定した。3日に測定されたキチナーゼ活性レベルを図5に示す。加えて、4日にサンプリングした培養上澄をSDSゲル電気泳動法および染色で分析した(図6)。これらの結果から、最適化された蛋白質特性は、蛋白質分泌にプラスの影響を与え、そして検出可能な、したがって蛋白質発現レベル上昇およびキチナーゼ酵素活性レベル上昇をもたらすことが明白である。結果を表6にまとめた。
【0129】
異なる構築物を含むアスペルギルス・ニガー(A.niger)ZTB−形質転換体の各形質転換体により発現されたβ−グルコシダーゼの産生を、培養上澄で測定した。4日目にサンプリングした培養上澄をSDSゲル電気泳動法および染色で分析した(図7)。これらの結果から、最適化された蛋白質特性は、蛋白質分泌にプラスの影響を与え、そして検出可能な、したがって、β−グルコシダーゼ酵素のための蛋白質発現レベル上昇をもたらすことが明白である。加えて、3日目にサンプリングした上澄中の活性を、基質としてpNP−β−グルコピラノシドを使用し、pH=4.5および40℃で測定した。蛋白質特性最適化に付しておいたβ−グルコシダーゼの上澄は、コドン最適化遺伝子によりコード化された親β−グルコシダーゼと比較して、最高20倍の活性上昇を示した。空宿主で測定されるバックグラウンドβ−グルコシダーゼ活性は、コドン最適化遺伝子によりコード化された親ベータ−グルコシダーゼの2分の1〜4分の1の低さであった。活性はまた、セロビオースを基質として使用し、pH=4.5および40℃でも測定した。測定された活性上昇は、コドン最適化遺伝子によりコード化された親β−グルコシダーゼと比較して、少なくとも30倍であった(空宿主菌株は、コドン最適化遺伝子によりコード化された親β−グルコシダーゼの3分の1〜10分の1の低さであった)。結果を表7にまとめた。
【0130】
異なる構築物を含むアスペルギルス・ニガー(A.niger)ZTC−形質転換体の各形質転換体により発現されたエンドグルカナーゼの産生を、培養上澄で測定した。4日目にサンプリングした培養上澄をSDSゲル電気泳動法および染色で分析した(図8)。これらの結果から、最適化された蛋白質特性は、蛋白質分泌にプラスの影響を与え、そして検出可能な、したがってエンドグルカナーゼ酵素のための蛋白質発現レベル上昇をもたらすことが明白である。3日目にサンプリングした上澄中のエンドグルカナーゼ活性を、AZO−CM−セルロースを基質として使用し、pH=4.5および40℃で測定した。蛋白質特性最適化に付しておいたエンドグルカナーゼの上澄は、同一宿主で発現されるコドン最適化遺伝子と比較して、350倍を超える活性上昇を示した。空菌株におけるバックグラウンド活性は非常に低いため(SDS−PAGEで検出不能)、活性上昇がそのように高い数字で表されることに留意すべきである。コドン最適化遺伝子によりコード化されたエンドグルカナーゼについては、測定された活性は、ほぼ空宿主菌株で観測されるバックグラウンド活性であった。結果を表8にまとめた。
【0131】
明らかに、これらの実施例は、対象蛋白質および酵素の改良された分泌および産生のために、蛋白質特性最適化のための本発明の方法を、いかに使用できるかを示すものである。さらに、これらの結果は、発現構築物および宿主は、既に幾つかの他の最適化、たとえば強力プロモーター、改良された翻訳開始配列、改良された翻訳停止配列、最適化コドンおよびコドン対使用、および/または蛋白質発現のための改良された宿主等を有するものの、宿主における蛋白質発現を改良するために、本発明の方法を広く利用できることを示している。
【発明の詳細な説明】
【0001】
[発明の分野]
本発明は、ポリペプチドの収量を改良する方法に関する。特に、本発明は、ポリペプチドの主鎖を修飾するステップによりポリペプチドの収量を改良する方法に関する。
【0002】
[発明の背景]
ゲノムおよびメタゲノムシークエンシングにおける最近の急速な進歩は、潜在的に非常に興味深い多様な蛋白質を表す多くの遺伝子をもたらしている。これらの遺伝子をかなりの水準で発現させる問題は、これらの遺伝子によりコードされる蛋白質の機能性の調査を妨げ、結果として、経済的な実現可能な方法での、このような蛋白質の利用の可能性を阻む。多くの場合、発見された遺伝子は大規模生産にあまり適していないか、または現在の遺伝子工学ツールにかなり利用し難しい生物に由来するため、遺伝子導入系および十分に開発された遺伝子工学ツールが利用可能な、十分に確立された産生宿主を使用することが極めて望ましい。特に糸状菌および酵母等の真核生物は、蛋白質の産生における、特に細胞外蛋白質の産生における、細胞工場として広く使用される。長期にわたり伝統的に使用されているため、これらの種の幾つかは一般に安全である(GRAS)と見なされ、ヒト用製品の製造に非常に興味深いものとなっている。しかし、かなりの改良にもかかわらず、異種遺伝子で得られる産生レベルは大抵、相同遺伝子でみられるものよりはるかに低い。多くの場合、蛋白質の発現は皆無である。
【0003】
蛋白質産生のレベルを上げるために様々な方法がある。これらの中には、強力プロモーターの利用、コピー数の増加、最適コザック(Kozak)配列、mRNA安定要素、最適化コドン使用(国際公開第2008/000632号パンフレット)および遺伝子などがある。しかしながら、これらの戦略は一般に、蛋白質が検出可能なレベルで産生されることを保証するものではない。今日まで、異種蛋白質を産生するための最も成功したアプローチは、それらを、効率的に分泌される相同蛋白質との翻訳融合として発現させることである。それにも拘わらず、産生レベルはやはりそれでもなお著しく後れを取り、多くの場合、発現レベルは問題になるほど低い。一般に、発酵における低発現は、回収における低収量につながる。たとえ発現が最適化されても、最終的な成熟蛋白質生成物は、下流処理での大きな損失のため、やはり非常に低い生成収量に終わるかもしれない。これは、発現された蛋白質がバイオマスと結合したままであるとき、該当するかもしれない。これは高損失に終わるか、あるいは、蛋白質を可溶化するために、高価な、そして時には望ましくない洗剤の使用を必要とする。
【図面の簡単な説明】
【0004】
【図1】図1は、クリベロミセス・ラクテス(K.lactis)発現ベクターpKLPGE−WT(実施例1に記載の構築物)のプラスミドマップを表す。図1は、他のpKLPGE−発現プラスミドの代表的なマップも提供する。PGEコード化遺伝子に関連したLAC4プロモーターおよびamdS選択マーカーカセットを示す。大腸菌(E.coli)DNAは、形質転換前に、制限酵素SacIIで消化することにより、除去できる。
【図2】図2は、発現ベクターpANPGE−3(実施例1に記載の構築物)のプラスミドマップを表す。図2は、他のpANPGE−発現プラスミド類の代表的なマップも提供する。加えて、glaAプロモーターおよび本発明の方法によって変異体PGE酵素をコード化する切断型GlaAの配列およびPGEコード化配列が示される。大腸菌(E.coli)DNAは、アスペルギルス・ニガー(A.niger)菌株の形質転換前に、制限酵素NotIで消化することにより、除去できる。
【図3】図3は、発現ベクターpGBFINZDU−WT(実施例1に記載の構築物)のプラスミドマップを表す。図3は、他のpGBFINZDU−プラスミド、pGBFINZTB−プラスミドおよびpGBFINZTC−プラスミドの代表的なマップも提供する。amdS選択マーカーカセットに関連したglaA隣接領域を示す。加えて、glaAプロモーターの配列ならびに本発明の方法によって変異体酵素をコード化するZDU配列、ZTB配列およびZTC配列が示される。大腸菌(E.coli)DNAは、アスペルギルス・ニガー(A.niger)菌株の形質転換前に、制限酵素NotIで消化することにより、除去できる。
【図4】アスペルギルス・ニガー(A.niger)WT 6およびPGE変異形質転換体pANPGE12#16(A)およびpANPGE13#30(B)のSDS−PAGEおよびウエスタンブロット解析。培養の2日(D2)および3日(D3)の上澄を分析した。14kDaおよび97kDaにおける水平線は、SDS−PAGEおよびウエスタンブロットのアラインメント用である。左側のマーカーの大きさは、SDS−PAGE染色マーカーに相当し、また右側のマーカーはウエスタンブロットマーカーに相当する。
【図5】図5は、全てglaAプロモーターのコントロール下での、異なるZDU構築物を発現する、アスペルギルス・ニガー(A.niger)菌株の、3日間発酵後の培養ブロス中のキチナーゼ活性を表す。シグナル配列、N−末端および蛋白質デザインが修飾されている、変異体SDU構築物を発現するアスペルギルス・ニガー(A.niger)菌株の培養ブロス中のキチナーゼ活性を表す。異なる構築物に関する詳細は、表6に示す。相対的キチナーゼ活性は、OD590測定値として表す。表示した全ての形質転換体群について、3つの形質転換体を単離し、独立に培養した。
【図6】図6は、全てglaAプロモーターのコントロール下での、変異体ZDU構築物を発現する、アスペルギルス・ニガー(A.niger)WT 6菌株およびZDU菌株の、4日間発酵後の、培養ブロスのSDS−PAGEの解析を表す。異なる構築物および発現したZDU蛋白質に関する詳細は、表6に表す。表示した全ての形質転換体群について、3つの形質転換体を単離し、独立に培養した。
【図7】図7は、全てglaAプロモーターのコントロール下での、変異体ZTB構築物を発現するアスペルギルス・ニガー(A.niger)WT 6菌株およびZTB−菌株の、4日間発酵後の、培養ブロスのSDS−PAGE解析を表す。異なる構築物および発現したZTB蛋白質に関する詳細を、表7に表す。表示した全ての形質転換体群について、3つの形質転換体を単離し、独立に培養した。
【図8】図8は、全てglaAプロモーターのコントロール下での、変異体ZTC構築物を発現する、アスペルギルス・ニガー(A.niger)WT 6菌株およびZTC−菌株の、5日間発酵後の、培養ブロスのSDS−PAGE解析を表す。異なる構築物および発現したZTC蛋白質に関する詳細を、表8に表す。表示したZTC−WT形質転換体群について3つの形質転換体を、他の2つの菌株型については2つの菌株を、単離して独立に培養した。
【図9】図9は、局所的な蛋白質特性を表す。
【0005】
[配列番号の説明]
配列番号1:cDNAコドン対最適化(CPO)前胃エステラーゼ(PGE);プロセシングした、すなわちシグナル配列コード化部分がない
配列番号2:蛋白質子ウシ前胃エステラーゼ(PGE)、シグナル配列を含む
配列番号3:DNA PGE蛋白質特性最適化(PFO)変異体KL8、グリコシル化部位1つ追加
配列番号4:蛋白質PGE PFO変異体KL8、グリコシル化部位1つ追加
配列番号5:DNA PGE PFO変異体KL9、グリコシル化部位5つ追加
配列番号6:蛋白質PGE PFO変異体KL9、グリコシル化部位5つ追加
配列番号7:DNA PGE PFO変異体KL11、6.96から7.74へのplシフト
配列番号8:蛋白質 PGE PFO変異体KL11、6.96から7.74へのplシフト
配列番号9:DNA PGE PFO変異体KL12、6.96から6.7へのplシフト
配列番号10:蛋白質PGE PFO変異体KL12、6.96から6.7へのplシフト
配列番号11:DNA PGE、天然のシグナル配列がα−MAT因子シグナルプレ(プロ−)配列に融合したPGE変異体
配列番号12:DNA PGE AN3、Kex部位(KR)とのCPO遺伝子tAG融合物
配列番号13:DNA PGE変異体AN12、6.96から4.6へのplシフト
配列番号14:蛋白質PGE変異体AN12、6.96から4.6へのplシフト
配列番号15:DNA PGE変異体AN13、6.96から4.88へのplシフト
配列番号16:蛋白質PGE変異体AN13、6.96から4.88へのplシフト
配列番号17:DNAキチナーゼ(ZDU)野生型
配列番号18:蛋白質キチナーゼ(ZDU)野生型
配列番号19:DNAキチナーゼ変異体ZDU−6
配列番号20:蛋白質キチナーゼ変異体ZDU−6
配列番号21:DNAキチナーゼ変異体ZDU−7
配列番号22:蛋白質キチナーゼ変異体ZDU−7
配列番号23:DNA β−グルコシダーゼ野生型ZTB−WT
配列番号24:蛋白質β−グルコシダーゼ野生型ZTB−WT
配列番号25:DNA β−グルコシダーゼ変異体ZTB−4
配列番号26:蛋白質β−グルコシダーゼ変異体ZTB−4
配列番号27:DNAエンドグルカナーゼ野生型ZTC−WT
配列番号28:蛋白質エンドグルカナーゼ野生型ZTC−WT
配列番号29:DNAエンドグルカナーゼ変異体ZTC−5
配列番号30:蛋白質エンドグルカナーゼ変異体ZTC−5
【0006】
[詳細な説明]
本発明は、真核生物宿主における1つまたは複数の蛋白質特性について最適範囲内に入るか、または最適値により近づくように、ポリペプチドのアミノ酸主鎖における一組の関連する蛋白質特性値を修飾するステップにより、真核生物宿主細胞によって対象ポリペプチドの分泌を改良する方法に関する。
【0007】
1つの利点は、以前には分泌されないか、または商業的応用は魅力のないほど少量が分泌されるに過ぎなかったが、今では、分泌が改良されたために産業的プロセスに利用可能である、興味深い機能を有する蛋白質である。もう一つの利点は、設計されたポリペプチドがバイオマスから既に分離されているため、下流処理およびポリペプチド類の回収がより容易になることである。
【0008】
本明細書の文脈では、蛋白質特性は、蛋白質アミノ酸配列およびDNA配列から計算により導くことができる性質である。
【0009】
ポリペプチドの修飾は、本明細書では、ポリペプチドのアミノ酸配列の変化をもたらすあらゆる事象と定義される。修飾は、1つまたは複数の修飾と理解される。修飾は、ポリペプチド主鎖における1つまたは複数のアミノ酸の導入(挿入)、置換または除去(削除)によって遂行することが可能である。
【0010】
本明細書の文脈では、用語「分泌」は、細胞外培地、一般に成長培地または生産培地中での、ポリペプチドの出現を指す。分泌されるポリペプチドは、バイオマスを含まない。分泌のレベルは、活性アッセイ(活性単位)、比活性(蛋白質重量当たりの単位)、定量的PAGE解析、定量的質量分析法および抗体法を含む、当該技術分野で周知の方法で測定することが可能である。
【0011】
「ポリペプチドの分泌の改良」という表現は、細胞の、細胞外培地中に分泌されるポリペプチド量の増加を指す。改良は、たとえば細胞内ポリペプチド等の、通常は分泌されないポリペプチドが、今では分泌されるようになることにより反映され得る。改良はまた、たとえばシグナル配列を含むため分泌されると予想されていたのに分泌されなかったポリペプチドが、今では分泌されるようになったということでもある。改良は、もちろん、常に同一宿主遺伝的背景および同一培養または発酵条件に関して測定される。こうした場合、改良された分泌は、たとえば、改良前には可視バンドが皆無であったポリアクリルアミドゲルに蛋白質バンドが出現することから、明白なこともある。
【0012】
あるいは、改良は、非常に少量で分泌されるポリペプチドが、上昇した分泌レベルを示すことにより反映されることもある。
【0013】
一実施形態では、分泌されたポリペプチドの量は、細胞外培地中のポリペプチドの活性を測定することにより決定される。改良前の状況に比べて、細胞外培地中の活性は、少なくとも5%、少なくとも10%、少なくとも15%または少なくとも20%、上昇している可能性がある。好ましくは、活性は少なくとも25%、少なくとも30%、少なくとも35%または少なくとも40%、上昇している。より好ましい実施形態では、活性は、少なくとも45%、少なくとも50%、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも100%、少なくとも200%、少なくとも500%または少なくとも1000%、上昇している。活性は、細胞外培地中で、無活性から若干の活性に上昇していてもよい。
【0014】
任意の真核細胞を、本発明の方法で使用し得る。好ましくは、真核細胞は、哺乳類細胞、昆虫細胞、植物細胞、真菌細胞、または藻類細胞である。好ましい哺乳類細胞としては、たとえばチャイニーズハムスター卵巣(CHO)細胞、COS細胞、293細胞、PerC6細胞、およびハイブリドーマなどが挙げられる。好ましい昆虫細胞としては、たとえばSf9細胞およびSf21細胞およびそれらの派生物などがある。より好ましくは、真核細胞は真菌細胞、すなわち、カンジダ(Candida)、ハンゼヌラ(Hansenula)、クリベロミセス(Kluyveromyces)、ピキア(Pichia)、サッカロミセス(Saccharomyces)、シゾサッカロミセス(Schizosaccharomyces)、またはヤロウイア(Yarrowia)菌株等の、酵母細胞である。より好ましくは、クリベロミセス・ラクテス(Kluyveromyces lactis)、サッカロミセス・セレビシア(S.cerevisiae)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、ヤロウイア・リポリティカ(Yarrowia lipolytica)およびピキア・パストリス(Pichia pastoris)、または糸状菌細胞由来である。最も好ましくは、真核細胞は、糸状菌細胞である。
【0015】
「糸状菌」は、亜門真菌類(Eumycota)および卵菌(Oomycota)の全ての線維状型を含む(Hawksworthらにより、Ainsworth and Bisby’s Dictionary of The Fungi,8th edition,1995,CAB International,University Press,Cambridge,UKで定義されている通り)。糸状菌は、キチン、セルロース、グルカン、キトサン、マンナン、および他の複合多糖類からなる菌糸壁(mycelial wall)を特徴とする。栄養生長は菌糸伸長により、また炭素異化は偏性好気性である。糸状菌菌株としては、アクレモニウム(Acremonium)、アガリクス(Agaricus)、アスペルギルス(Aspergillus)、アウレオバシジウム(Aureobasidium)、クリソスポリウム(Chrysosporium)、コプリナス(Coprinus)、クリプトコッカス(Cryptococcus)、フィリバシジウム(Filibasidium)、フザリウム(Fusarium)、フミコラ(Humicola)、マグナポルテ(Magnaporthe)、ムコール(Mucor)、ミセリオフトラ(Myceliophthora)、ネオカリマスティクス(Neocallimastix)、ニューロスポラ(Neurospora)、ペシロマイセス(Paecilomyces)、ペニシリウム(Penicillium)、ピロミセス(Piromyces)、パネロカエテ(Panerochaete)、プリュロタス(Pleurotus)、シゾフィラム(Schizophyllum)、タラロミセス(Talaromyces)、サーモアスカス(Thermoascus)、シーラビア(Thielavia)、トリポクラジウム(Tolypocladium)、およびトリコデルマ(Trichoderma)などの菌株が挙げられるが、その限りではない。
【0016】
好ましい糸状菌細胞は、アスペルギルス(Aspergillus)属、クリソスポリウム(Chrysosporium)属、ペニシリウム(Penicillium)属、タラロミセス(Talaromyces)属またはトリコデルマ(Trichoderma)属の種属に属し、最も好ましくはアスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・フォエティダス(Aspergillus foetidus)、アスペルギルス・ソーヤ(Aspergillus sojae)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、タラロミセス・エメルソニ(Talaromyces emersonii)、アスペルギルス・オリゼー(Aspergillus oryzae)、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)、トリコデルマ・リーセイ(Trichoderma reesei)またはペニシリウム・クリソゲナム(Penicillium chrysogenum)の種属に属する。本発明による宿主細胞がアスペルギルス(Aspergillus)宿主細胞であるとき、その宿主細胞は、好ましくはCBS 513.88またはその派生物である。
【0017】
糸状菌の幾つかの菌株は、たとえば、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection(ATCC))、ドイツ微生物細胞培養コレクション(Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(DSM))、オランダ微生物株保存センター(Centraalbureau Voor Schimmelcultures(CBS))、および農業研究サービス特許カルチャーコレクション、北部領域研究センター(Agricultural Research Service Patent Culture Collection,Northern Regional Research Center(NRRL))アスペルギルス・ニガー(Aspergillus niger)CBS 513.88、アスペルギルス・オリザエ(Aspergillus oryzae)ATCC 20423、IFO 4177、ATCC 1011、ATCC 9576、ATCC 14488−14491、ATCC 11601、ATCC 12892、ペニシリウム・クリソゲナム(P.chrysogenum)CBS 455.95、ペニシリウム・シトリナム(Penicillium citrinum)ATCC 38065、ペニシリウム・クリソゲナム(Penicillium chrysogenum)P2、タラロミセス・エメルソニ(Talaromyces emersonii)CBS 124.902、アクレモニウム・クリソゲナム(Acremonium chrysogenum)ATCC36225またはATCC 48272、トリコデルマ・リーセイ(Trichoderma reesei)ATCC 26921またはATCC 56765またはATCC 26921、アスペルギルス・ソーヤ(Aspergillus sojae)ATCC11906、クリソスポリウム・ラクノウェンス(Chrysosporium lucknowense)ATCC44006およびそれらの派生物等、多数のカルチャー・コレクションで公的に、容易に入手できる。
【0018】
本発明の一実施形態では、アスペルギルス・ニガー(A.niger)またはクリベロミセス・ラクテス(K.lactis)が使用される。
【0019】
一実施形態において、真核細胞は、組換え技術でポリペプチドが産生される宿主細胞である。宿主細胞を形質転換または形質移入する好適な方法は、Sambrook,et al.(Molecular Cloning:A Laboratory Manual,2nd ed.Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,NY,1989)、Davis et al.,Basic Methods in Molecular Biology(1986)および他の実験マニュアルを参照されたい。したがって、本発明はまた、ポリペプチドの分泌を改良するための本発明による方法を対象ポリペプチドに適用し、組換え技術により、本発明に従って修飾されたポリペプチドを産生するステップによって対象ポリペプチドを産生する方法にも関する。本発明はまた、前記組換技術により産生されるポリペプチドにも関する。本発明はまた、ポリペプチドの分泌を改良するための本発明による方法で得られるポリペプチドにも関し;好ましくは前記ポリペプチドは、ポリペプチドの分泌を改良するための本発明による方法で得られる。
【0020】
本発明の方法に従って分泌が改良される対象ポリペプチドは、対象生物活性を有する任意のポリペプチドであってもよい。ポリペプチドは、コラーゲンまたはゼラチン、あるいはそれらの変異体または混成物であってもよい。ポリペプチドは、抗体またはその一部、抗原、凝固因子、酵素、ホルモンまたはホルモン変異体、受容体またはその一部、調節蛋白質、構造蛋白質、レポーター、あるいは血清アルブミン、たとえばウシ血清アルブミンおよびヒト血清アルブミン等、またはトランスフェリン、たとえばラクトフェリン等の、輸送蛋白質、分泌過程に関与する蛋白質、折り畳み過程に関与する蛋白質、シャペロン、ペプチドアミノ酸輸送体、グリコシル化因子、転写因子、合成ペプチドまたはオリゴペプチド、その天然型が細胞内蛋白質であり、かつシグナルペプチドへの融合およびその天然型で既に分泌されているポリペプチドへの融合等の、当該技術分野で周知の方法により分泌される蛋白質であってもよい。このような細胞内蛋白質は、プロテアーゼ、セラミダーゼ類、エポキシド加水分解酵素、アミノペプチダーゼ、アシラーゼ類、アルドラーゼ、ヒドロキシラーゼ、アミノペプチダーゼ、リパーゼ等の酵素であってもよい。ポリペプチドは、その天然型で細胞外に分泌される酵素であってもよい。このような酵素は、酸化還元酵素、トランスフェラーゼ、加水分解酵素、リアーゼ、イソメラーゼ、リガーゼ、カタラーゼ、セルラーゼ、キチナーゼ、クチナーゼ、デオキシリボヌクレアーゼ、デキストラナーゼ、エステラーゼの群に属してもよい。酵素は、カルボヒドラーゼ、たとえばエンドグルカナーゼ類、β−グルカナーゼ類、セロビオヒドロラーゼ類またはβ−グルコシダーゼ類等のセルラーゼ類、ヘミセルラーゼ類、あるいはペクチン分解酵素、たとえばキシラナーゼ類、キシロシダーゼ類、マンナナーゼ類、ガラクタナーゼ類、ガラクトシダーゼ類、ペクチンメチルエステラーゼ類、ペクチンリアーゼ類、ペクチン酸リアーゼ類、エンドポリガラクツロナーゼ類、エキソポリガラクツロナーゼ類、ラムノガラクツロナーゼ類、アラバナーゼ類、アラビノフラノシダーゼ類、アラビノキシラン加水分解酵素、ガラクツロナーゼ類、リアーゼ類、またはアミロース分解酵素等;加水分解酵素、イソメラーゼ、またはリガーゼ、フィターゼ等のホスファターゼ類、リパーゼ等のエステラーゼ類、蛋白分解酵素、オキシダーゼ類、トランスフェラーゼ類、またはイソメラーゼ類等の酸化還元酵素であってもよい。酵素は、フィターゼであってもよい。酵素は、アミノペプチダーゼ、アスパラギナーゼ、アミラーゼ、カルボヒドラーゼ、カルボキシペプチダーゼ、エンド−プロテアーゼ、メタロ−プロテアーゼ、セリン−プロテアーゼカタラーゼ、キチナーゼ、クチナーゼ、シクロデキストリングリコシルトランスフェラーゼ、デオキシリボヌクレアーゼ、エステラーゼ、α−ガラクトシダーゼ、β−ガラクトシダーゼ、グルコアミラーゼ、α−グルコシダーゼ、β−グルコシダーゼ、ハロパーオキシダーゼ、蛋白質デアミナーゼ、インベルターゼ、ラッカーゼ、リパーゼ、マンノシダーゼ、ミュータナーゼ、オキシダーゼ、ペクチン分解酵素、ペルオキシダーゼ、ホスホリパーゼ、ポリフェノールオキシダーゼ、リボヌクレアーゼ、トランスグルタミナーゼ、またはグルコースオキシダーゼ、ヘキソースオキシダーゼ、モノオキシゲナーゼであってもよい。分泌が改良されるポリペプチドは、宿主細胞と相同であっても異種であってもよい。相同ポリペプチドの好適な例は、アスペルギルス・ニガー(Aspergillus niger)にクローニングされて産生される、アスペルギルス・ニガー(Aspergillus niger)蛋白質である。異種発現の好適な例としては、糸状菌または酵母にクローニングされて産生される、たとえば大腸菌(E.coli)または桿菌(Bacillus)由来の細菌ポリペプチド、または、糸状菌または酵母にクローニングされて産生される、たとえばウシまたはヤギ由来の哺乳類蛋白質、または酵母にクローニングされて産生される糸状菌ポリペプチド、あるいは、別の真菌にクローニングされて産生される、糸状菌蛋白質などがある。好ましくは、ポリペプチド類をコード化する核酸は、たとえばコドン対最適化により、関連宿主細胞における発現用に最適化される。コドン対最適化は、ポリペプチドをコード化するヌクレオチド配列の改良された発現および/またはコード化されたポリペプチドの改良された産生を得るために、ポリペプチドをコード化するヌクレオチド配列を、それらのコドン使用、特に使用されるコドン対に関して、修飾しておく方法である。コドン対は、コード配列における、一組の、2つの後続するトリプレット(コドン)と定義される。コドン対最適化は、好ましくは国際公開第2008/000632号パンフレットに記載の通りに実施される。
【0021】
好ましくは、修飾されたポリペプチドの特異性は、分泌の改良前と実質的に同じである。これは、たとえば基質特異性または結合特異性が実質的に保持されることを意味する。本明細書の文脈では、用語「実質的に保持される」は、特異性の60%超、65%超、70%超または75%超が保持されることを意味する。好ましくは、特異性の80%超、85%または90%が保持される。最も好ましくは、特異性の95%超、96%、97%、98%または99%が保持される。
【0022】
本発明の方法によれば、改良された分泌の指標である、細胞外培地中の活性レベルは上昇する。しかし、修飾されたポリペプチドの比活性は、低下しない限り、上昇しなくてもよい。したがって、比活性は、好ましくは分泌の改良前と実質的に同じか、それより高い。好ましい実施形態では、比活性は、改良前と実質的に同じである。本明細書の文脈では、「実施的に同レベルの活性」という表現は、親ポリペプチドの活性レベルと、15%未満、好ましくは12%未満または10%未満、より好ましくは8%未満、6%未満または4%未満異なる活性レベルを指す。
【0023】
本明細書の文脈では、用語「ポリペプチド」および蛋白質」は、互換的に使用される。本発明の方法により、あらゆるタイプのポリペプチドが、その分泌を改良することが可能である。好ましい実施形態では、ポリペプチドは、本明細書に前述したリストの1つである。
【0024】
本発明の方法によれば、真核生物宿主における1つまたは複数の蛋白質特性について最適範囲内に入るか、または最適値にもっと近づくように、アミノ酸主鎖における一組の関連する蛋白質特性値が修飾される。
【0025】
修飾されたポリペプチドとリファレンスポリペプチドとの間の蛋白質特性の変化量は、相対的改良(RI)および正規化相対的改良(RIN)という2つの方法で定義することができる。
【0026】
蛋白質特性のRIは、最適値からの蛋白質特性の絶対偏差(D)という観点で定義される:
RI=(DREF−DPFO)/DREF,
ここで、D=|FPOI−F0PT|、FPOIは、対象蛋白質特性値であって、
リファレンスまたはPFOのいずれかであり,FOPTは最適特性値である。
RINは、どの特性が実質的に重要であるかを理解するために、正規化偏差(DN)の観点で定義される。DNは、特性値の上界(UB)および下界(LB)を考慮に入れる(表1参照)。
RIN=DN,REF−DN,PFO,
ここで、FPOI>FOPTであれば、DN=(FPOI−FOPT)/(UB−FOPT)
FPOI<FOPTであれば、DN=(FPOI−FOPT)/(LB−FOPT)
【0027】
本発明の方法によれば、修飾はポリペプチド主鎖に行われる。本明細書の文脈では、用語「主鎖」は、アミノ酸がペプチド結合を介して結合されて、共有結合した一連のアミノ酸を形成するとき、形成される正規構造を指す。本発明では、好ましくは、成熟ポリペプチドの主鎖が修飾される。本発明に関連して、「成熟ポリペプチド」は本明細書では、翻訳および何らかの翻訳後修飾、たとえばN−末端プロセシング、C−末端切断、グリコシル化、リン酸化等々の後に、その最終的な機能形であるポリペプチドとして定義される。修飾前のポリペプチドは、親ポリペプチドまたはリファレンスポリペプチドまたは野生型ポリペプチドと呼ばれ、それに由来する修飾されたポリペプチドと区別される。用語「親ポリペプチド」、「野生型ポリペプチド」および「リファレンス−ポリペプチド」は、本明細書では互換的に使用される。ポリペプチドがキメラポリペプチドであるとき、すなわち効率的に分泌されるポリペプチドとの、好ましくは宿主細胞に由来するポリペプチドとの、翻訳融合物であるとき、キメラポリペプチド全体を本発明に従って修飾することが可能である。キメラポリペプチドが、対象ポリペプチドに融合したリーダーポリペプチドとして効率的に分泌されるポリペプチドを含むとき、対象ポリペプチドは、好ましく修飾される。
【0028】
当業者に周知の通り、成熟中のプロセシングエラーのため、成熟ポリペプチドのN−末端は、成熟ポリペプチドのC−末端と同様、異種であり得る可能性がある。特に、このようなプロセシングエラーは、ポリペプチドの過剰発現で起こり得る。加えて、エキソプロテアーゼ活性は、異質性を生じさせるかもしれない。異質性が起こる程度は、使用される宿主および発酵プリトコールによっても異なる。このようなN−末端およびC−末端プロセシング人工産物は、予想される成熟ポリペプチドと比較して、短いポリペプチド類または長いポリペプチド類をもたらすかもしれない。
【0029】
本発明の一実施形態では、本方法は、
(i)真核生物宿主における、1つまたは複数の蛋白質特性の最適範囲および最適値を決定するステップ、および
(ii)真核生物宿主における、一組の関連した蛋白質特性を決定するステップ(これらの関連した特性の1つまたは複数が、ポリペプチドのアミノ酸主鎖において修飾されるのであれば、その特性は、真核生物宿主によるポリペプチドの分泌を改良するであろう)、および
(iii)(i)で決定されたように、最適範囲内に入るかまたは最適値により近づくように、関連した蛋白質特性値を修飾するステップ(ここで、(i)および(ii)は任意の順序で実施し得る)、
を含む。
【0030】
任意の方法を使用して、一組の関連した特性を決定し得る。一実施形態では、ポリペプチドの分泌を改良するための、関連した一組の特性は下記の通りに決定される:
(i)特定の真核生物宿主における適量の蛋白質の分泌レベルならびにこれらの蛋白質のアミノ酸配列およびDNA配列を含む、データセットSの収集または作成。データセットSは、分泌蛋白質(S+)を含んでもよい。好ましくは、データセットSはまた、非分泌蛋白質(S−)も含む。たとえば、全ての予測される分泌蛋白質をアスペルギルス・ニガー(A.niger)で、発現させることができる(Tsang et al.,2009,Fungal Genetics and Biology,46:S153−160)。分泌される蛋白質は、セットS+に属するが、分泌されない蛋白質は、セットS−に属する。任意の方法を使用して、分泌レベルを測定し得る。あるいは、セットS−は、真核生物宿主における文献で周知の非分泌蛋白質を含んでもよい。Sにおける蛋白質は、真核生物宿主と相同であっても異種であってもよい。
(ii)データセットSにおける全ての蛋白質についての、蛋白質特性(F)の計算。Fは、これらの蛋白質のDNA配列およびアミノ酸配列の両者から誘導することが可能である;
(iii)適切に規定された分類子性能基準に従って、S+とS−とを区別するために、統計分類子の最高の成果を与える、ii)で計算した蛋白質特性のサブセット(Fs)を選択するための統計的分類方法の使用。Fsは、DNA配列(Fs_DNA)およびアミノ酸配列(Fs_AA)の両者から誘導され得る;
【0031】
Fs_AAにおける蛋白質特性は、対応する真核生物宿主における蛋白質分泌を改良するための修飾についての関連する特徴である。
【0032】
好ましくは、成熟ポリペプチドの主鎖は、本発明の方法に従って修飾されるため、蛋白質特性は、一組の成熟蛋白質から計算されることが好ましい。
【0033】
当該技術分野で周知の、標準的統計的分類方法、たとえば、線形判別分類器(Linear Discriminant Classifier(LDC))、二次判別分類器(Quadratic Discriminant Classifier(QDC))、最近傍平均分類器(Nearest Mean Classifier(NMC))、1−/k−最近傍分類器(Nearest Neighbour classifiers)、サポートベクターマシーンおよび決定木等々を使用することができる(Webb,Statistical Pattern Recognition,2nd ed,John Wiley & sons)。このような方法を適用するとき、データセットSは、学習データセットと確認データセットに分けることができ、また当該技術分野で周知の確認方式(たとえば、10分割交差確認)を使用することが可能である。
【0034】
当該技術分野で周知の任意の分類器性能測定、たとえば、特異性、感度、確度、精度および受信者動作特性(ROC)曲線下面積を使用し得る。
【0035】
任意の好適な方法を使用して、蛋白質特性の最適範囲または最適値を決定し得る。
【0036】
一実施形態では、真核生物宿主の蛋白質特性の最適範囲または最適値は、以下の通りに決定される:
i)特定の真核生物宿主における適量の蛋白質の分泌レベルならびにこれらの蛋白質のアミノ酸配列およびDNA配列を含む、データセットSの収集または作成。データセットSは、分泌蛋白質(S+)を含んでもよい。好ましくは、データセットSはまた、非分泌蛋白質(S−)も含む。たとえば、全ての予測される分泌白質をアスペルギルス・ニガー(A.niger)で、発現させることができる(Tsang et al.,2009,Fungal Genetics and Biology,46:S153−160)。分泌される蛋白質は、セットS+に属するが、分泌されない蛋白質は、セットS−に属する。任意の方法を使用して、分泌レベルを測定し得る。あるいは、セットS−は、真核生物宿主における文献で周知の非分泌蛋白質を含んでもよい。Sにおける蛋白質は、真核生物宿主と相同であっても異種であってもよい。
ii)データセットSにおける全ての蛋白質についての、蛋白質特性(F)の計算。Fは、これらの蛋白質のDNA配列およびアミノ酸配列から誘導され得る;
iii)対応する真核生物宿主の各特性についての最適値(F_opt)の決定:最適値は、S+から算出される各蛋白質特性の中心値を算出することによって得てもよい。任意の中心値、たとえば、幾何平均、調和平均、算術平均、トリム平均、最頻値および中央値を使用し得る。算出される中心値は、対応する真核生物宿主に関する特性の最適値である。あるいは、特性値の分布が、選択される確率分布によって十分に説明されるように、S+から算出される各蛋白質特性に確率分布を合わせる。任意の確率分布を使用し得、たとえば正規分布、指数分布、または対数正規分布を使用し得る。確率分布の平均が、対応する真核生物宿主の特性の最適値である。
iv)対応する真核生物宿主について各特性の最適範囲の決定:分泌される蛋白質のみを含むS+セットを考慮すると、蛋白質特性の最適範囲の下界は、S+から算出される蛋白質特性の、0.3−、0.2−、0.15または好ましくは0.10−および0.05−変位値に相当する値と定義される。ここで、値0.3、0.2、0.15、等々は、累積確率を指す。特定の累積確率に相当する変位値は、任意の統計的方法で、たとえば、Statistical Toolbox,Matlab R2007a(The Mathworks Inc)の分位関数を使用して、算出し得る。蛋白質特性の最適範囲の上界は、S+から算出される蛋白質特性の、0.7−、0.8−、0.85または好ましくは0.90−および0.95−変位値に相当する値と定義される。あるいは、分泌蛋白質および非分泌蛋白質の両方を含む全データセットSを考慮すると、蛋白質特性の最適範囲の下界は、それより下の値ではSにおける蛋白質の70%、80%、85%、好ましくは90%または95%が分泌されない低い蛋白質特性の値として定義してもよい;蛋白質特性の最適範囲の上界は、それより上の値ではSにおける蛋白質の70%、80%、85%、好ましくは90%または95%が分泌されない蛋白質特性値として定義される。
【0037】
関連特性および最適範囲および最適値のセットは、宿主細胞ごとに異なるであろう。アスペルギルス・ニガー(A.niger)の場合、蛋白質分泌を増加するために修飾されるべき関連蛋白質特性(Fs_AA)としては、塩基性アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、微小アミノ酸頻度、小アミノ酸頻度、帯電アミノ酸頻度、正味荷電(pH7.2における)、等電点、アスパラギン、アルギニン、イソロイシン、システイン、ヒスチジン、グルタミン、バリン、リシン、グリシン、トレオニンおよびロイシンの頻度、ターン(Garnierにより算出)、EPESTFINDにより算出されるPESTモチーフ、plの局所特性(LF)値、特にLF1およびLF6、GravyスコアのLF値、特にLF2およびLF4、アロマスコアのLF値、特にLF3、LF4およびLF6、原子組成w.r.t.イオウ(S)および局在化特性(たとえば、MultiLoc 局在化予測ツールで予測される)などが挙げられるが、その限りではない。
【0038】
正味荷電は、陽子の電荷と同じ単位を有する。長さ当たりの正味電荷/正味正電荷/正味負電荷/全電荷は、陽子の電荷と同じ単位を有するが、ポリペプチドの長さに正規化される。
【0039】
ポリペプチドの正味荷電は、本明細書では、全てのアミノ酸が溶媒に完全に露出しており、隣接したペプチド類は、所与のアミノ酸のpKに影響を及ぼさず、また構成アミノ酸は、N末端およびC末端と同様、未修飾であると仮定して、推定される。様々なプログラムを使用して、たとえば、Bioinformatics Toolbox of Matlab(バージョンR2008b)の「等電」機能を使用して、またはhttp://emboss.sourceforge.net/で入手可能な、EMBOSS Explorerの「蛋白質統計学(pepstats)」機能を使用して、特定のpHにおけるポリペプチドの正味電荷を算出することができる(初期値pH=7.2による)。
【0040】
長さ当たりの正味荷電は、本明細書では、ポリペプチドの長さで割ったポリペプチドの正味荷電と定義される。
【0041】
長さ当たりの正味正電荷は、本明細書では、pH7.2における、ポリペプチドのN−末端ならびに全てのリシン残基、アルギニン残基およびヒスチジン残基の部分電荷を合計することによって算出される、ポリペプチドの正味正電荷と定義される。長さ当たりの正味正電荷は、ポリペプチドの正味正電荷をポリペプチドの長さで割ることにより決定される。
【0042】
長さ当たりの正味負電荷は、本明細書では、pH7.2における、ポリペプチドのC−末端ならびに全てのアスパラギン酸残基、グルタミン酸残基、システイン残基およびチロシン残基の部分電荷を合計することによって算出される、ポリペプチドの正味負電荷と定義される。長さ当たりの正味負電荷は、ポリペプチドの正味負電荷をポリペプチドの長さで割ることにより決定される。
【0043】
長さ当たりの全電荷は、本明細書では、ポリペプチドの正味陽性電荷(正の数)をポリペプチドの正味負電荷(負の数)だけ減算することにより算出される、ポリペプチドの全電荷と定義される。長さ当たりの全電荷は、ポリペプチドの全電荷をポリペプチドの長さで割ることにより決定される。
【0044】
グレイビー・スコア(gravy score)は、本明細書では、KyteおよびDoolittle(1982)により定義された通り、ポリペプチドの疎水性親水性指数と定義される。各アミノ酸は、4.6と−4.6の間の疎水性スコアを有する。4.6は、最も疎水性の蛋白質に割り当てられ、また−4.6は最も親水性の蛋白質に割り当てられる。ポリペプチドのGRAVYスコアは、好ましくは、Kyte and Doolittle(1982).Kyte,J.and Doolittle,R.1982蛋白質の疎水性親水性特徴を表示する簡単な方法(A simple method for displaying the hydropathic character of a protein.)J.Mol.Biol.,157:105−132.に従って決定される。
【0045】
ポリペプチドのアロマスコアは、本明細書では、ポリペプチド中の3つの芳香族アミノ酸、Phe、TyrおよびTrpの頻度を合計することにより算出される。
【0046】
脂肪族指数は、本明細書では、脂肪族側鎖によって占められる相対体積と定義される。ポリペプチドの脂肪族指数(AI)は、Ikaiの式(1980):AI=f_Ala+af_Val+b(f_Ile+f_Leu)に従って算出される。アミノ酸類アラニン、バリン、イソロイシンおよびロイシンは、脂肪族側鎖を有する。
ここで、aはバリン側鎖の相対体積であり(a=2.9)、bはロイシンおよびイソロイシン側鎖の相対体積である(b=3.9)。f_Ala、f_Val、f_Ileおよびf_Leuは、それぞれ、ポリペプチド中の、アラニン、バリン、イソロイシンおよびロイシンの頻度である。Ikai,A.J.1980球状蛋白質の熱安定性および脂肪族指数(Thermostability and aliphatic index of globular proteins.J.Biochem.,88:1895−1898。)
【0047】
GRAVYおよび脂肪族について、ExPASyサーバー上の蛋白質同定および分析ツール(Protein Identification and Analysis Tools on the ExPASy Server;Gasteiger E.,Hoogland C.,Gattiker A.,Duvaud S.,Wilkins M.R.,Appel R.D.,Bairoch A.;(In)John M.Walker(ed):The Proteomics Protocols Handbook,Humana Press(2005).pp.571−607.も参照することができるであろう。
【0048】
理化学的特性に基づくアミノ酸の種類:
酸性:D,E
脂肪族:A,I,L,V
芳香族:F,W,Y
塩基性:H,K,R
帯電:D,E,H,K,R
非極性:A,C,F,G,I,L,M,P,V,W,Y
極性:D,E,H,K,N,Q,R,S,T
小:A,C,D,G,N,P,S,T,V
微小:A,C,T,S,G
【0049】
配列中の単一要素の組成に基づく特性は、要素iの頻度fiから算出される。頻度と割合は、本明細書では、互換的に使用される。頻度は、配列中の要素総数で割った、要素iが配列中に出現する回数と定義される。配列中の単一要素、たとえばアミノ酸を、多重要素たとえば、微小、酸性に結合することができる。
【0050】
ポリペプチド内のアミノ酸残基の表面接触性は、当該技術分野で周知の任意の方法で決定し得る。
【0051】
ポリペプチドが実験的に解明される構造を有する場合、溶媒接触表面積(ASA)は、Å2で与えられ、またその面積は、蛋白質表面上を、水分子大の球を回転させることによって算出することができる[1]。次いで、ASAを相対表面積(RSA)に変換するが、これは、グリシン[2]またはアラニン[3]と並んだトリペプチドの中央の残基の最大可能露出と比較した、ポリペプチド鎖中の所与のアミノ酸残基のASAとして算出される。閾値アルファより大きいRSA(RSA>=アルファ、0<=アルファ<=1)を有する残基は、露出していると言われ、閾値ベータ未満のRSA(RSA<=アルファ、0<=ベータ<=1)を有する残基は、埋没していると言われる。好ましくは、アルファ>=0.25、より好ましくはアルファ=0.25である。好ましくはベータ<=0.25、より好ましくはベータ=0.25である。
【0052】
ポリペプチドの構造を入手できなければ、表面接触性は、ポリペプチドのアミノ酸配列からも予測することができる。ポリペプチドのアミノ酸配列から表面接触性を予測するために、様々な方法を文献で入手することができ、たとえば[3]、[4]、[5]および[6]に記載されている。好ましくは、RSAは、[4]に記載されているいわゆるNetSurfP法を使用して予測され、これはhttp://www.cbs.dtu.dk/services/NetSurfP/でオンラインアクセスすることができる。このアプリケーションで、表面接触性は、成熟蛋白質のアミノ酸配列から予測される。露出した残基および埋没した残基の定義は、前述と同じである。
[1]Connolly M:分析的分子表面計算(Analytical molecular surface calculation.)。Journal of Applied Crystallography 1983,16(5):548−558.
[2]Chothia C:蛋白質における接触可能表面および埋没表面の性質(The nature of the accessible and buried surfaces in proteins.)。J Mol Biol 1976,105(1):1−12.
[3]Ahmad S,Gromiha MM,Sarai A:アミノ酸配列からの溶媒接触可能性の実数値予測(Real value prediction of solvent accessibility from amino acid sequence.)。Proteins 2003,50(4):629−635.
[4]Bent Petersen et al:溶媒接触性予測に適用される信頼性スコアの包括的割り当て方法(A generic method for assignment of reliability scores applied to solvent accessibility predictions.)。BMC Structural Biology 2009,9:51.
[5]Dor O,Zhou Y:Real−SPINE:蛋白質構造特性の実数値予測のための神経回路網の集積システム(an integrated system of neural networks for real−value prediction of protein structural properties.)。Proteins 2007,68(1):76−81.
[6]Faraggi E,Xue B,Zhou Y:二層神経回路網を介した指導学習による蛋白質の残基溶媒接触可能性および実数値主鎖ねじれ角の予測確度を改良すること(Improving the prediction accuracy of residue solvent accessibility and real−value backbone torsion angles of proteins by guided−learning through a two−layer neural network.)。Proteins 2009,74(4):847−856.
【0053】
アスペルギルス・ニガー(A.niger)の最適値および最適範囲を表1に示す。
【0054】
【表1】
【0055】
【表2】
【0056】
表1において、全蛋白質配列から計算される全ての特性は、全蛋白質の長さに基づく。成熟蛋白質配列、露出残基および埋没残基から計算される全ての特性は、成熟蛋白質の長さに基づく。
【0057】
好ましくは、最適値および範囲特性は、表2から選択される;これらの特性は、一次特性と言われ、その他の特性、すなわち、表2にない表1の特性は、二次特性である。
【0058】
【表3】
【0059】
クリベロミセス・ラクテス(K.lactis)について、好ましい一次特性を表3に示す。
【0060】
【表4】
【0061】
別の実施形態では、ポリペプチドの分泌は、下記のステップで改良される:
i)ポリペプチドの蛋白質特性を計算するステップ、
ii)ポリペプチドの1つまたは複数の蛋白質特性が、最適範囲外であるか、または真核生物宿主の最適値から実質的に逸脱しているかどうかを決定するステップ(ここで、実質的逸脱は、最適値から20%、30%、40%または50%超の差と定義される)、
iii)ポリペプチドの1つまたは複数のFs_AAの値が、最適範囲内に入るか、または適量だけ、最適値に向けて移行するように、好ましくはポリペプチドの蛋白質特性と、蛋白質特性の最適値との差が、10%、15%、20%、または30%超、減少するように、ポリペプチドのアミノ酸配列を合理的に変えるステップ。
【0062】
好ましくは、2つ、3つ、4つまたは5つの蛋白質特性が一緒に修飾され、さらに好ましくは、10超、15または20の蛋白質特性が一緒に組み合わせて修飾される。最も好ましくは、25超または30の蛋白質特性が一緒に修飾される。
【0063】
好ましくは、最適範囲は、表1から選ばれ、さらに好ましくは最適範囲は、表2から選ばれる。あるいは、最適範囲は表3から選ばれる。
【0064】
上記ステップiii)で、ポリペプチドのアミノ酸配列は、当該技術分野で周知の任意の方法で合理的に変えし得る。たとえば、これは、:
(i)相同配列を検索するステップ;
(ii)相同配列を、対象配列と配列比較するステップ;
(iii)蛋白質機能的特性に重要なアミノ酸類を同定するステップ;
(iv)機能的特性を保持しながら、所望のアミノ酸配列特性を導入する;
(vi)所与の宿主に最適なコドンを使用して、最終的な修飾された配列を遺伝子に翻訳するステップ;
(vii)宿主における、再設計されたポリペプチドのクローニングおよび発現
によって達成するステップが可能である。
【0065】
好ましくは、アミノ酸主鎖のアミノ酸類の少なくとも5%が修飾され、より好ましくは、アミノ酸主鎖のアミノ酸類の少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%が修飾される。
【0066】
好ましくは、アミノ酸主鎖の少なくとも5つのアミノ酸類が修飾され、より好ましくはアミノ酸主鎖の少なくとも10のアミノ酸類、さらにより好ましくは少なくとも15のアミノ酸類、さらにより好ましくは少なくとも20のアミノ酸類、さらにより好ましくは少なくとも25のアミノ酸類、さらにより好ましくは少なくとも30のアミノ酸類が修飾される。
【0067】
好ましくは、本発明によれば、一次特性は改良されるが、二次特性は特定の境界内に保たれる。したがって、総体的な最適性スコアFは、全n個の一次特性および全m個の二次特性のDN値に基づいて規定される:
【数1】
【0068】
ηは、0と1の間で0および1を含む(0≦η≦1)の重み係数である。好ましくはη<0.5、より好ましくはη≦0.4、最も好ましくはη=0.3である。ρは、1と5の間で1および5を含み(1≦p≦5)、好ましくはρ=2である(Fは、このとき、ユークリッド距離を表す)。好ましくは、η=0.3およびρ=2である。好ましくは、野生型リファレンス蛋白質に関して、少なくとも5%のF−スコアの改良が達成され、より好ましくは少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%およびさらにより好ましくは少なくとも30%の改良が達成される。
【0069】
好ましくは、少なくとも2、3、4、または5つの特性が修飾され、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が修飾される。好ましくは、少なくとも2、3、4、または5つの特性が改良され、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が改良されるが、好ましくは、10未満、さらにより好ましくは5未満、さらにより好ましくは4未満、の特性が悪化する。好ましくは、その特性は一次特性である。
【0070】
相同配列は、好ましくは、適切な配列データベースのBLAST検索を実施することにより、検索される。相同配列は、対象配列と好ましくは少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、60%、70%、80%、90%、95、%、96%、97%、98%または99%の同一性を有する。最も好ましくは、相同配列は、対象配列と好ましくは約50%の同一性を有する。当業者は、幾つかの異なるコンピュータープログラムを利用して、2つの配列を並べて2つの配列間の相同性を決定できることを知るであろう(D.Sankoff and J.B.Kruskal,(ed.)中のKruskal,J.B.(1983)配列比較の概説(An overview of sequence comparison)、タイムワープ、文字列編集および巨大分子:配列比較の理論と実際(Time warps,string edits and macromolecules:the theory and practice of sequence comparison),pp.1−44AddisonWesley)。当該技術分野で周知の任意の方法を配列比較に使用し得る。2つのアミノ酸配列間または2つのヌクレオチド配列間のパーセント同一性は、たとえば2配列のアラインメントのためのNeedleman and Wunschアルゴリズムを使用して、決定することが可能である(Needleman,S.B. and Wunsch,C.D.(1970)J.Mol.Biol.48,443−453)。
【0071】
対象必須機能特性に重要なアミノ酸類を同定する方法は、当該技術分野で周知である。好適なツールは、対象蛋白質の3D構造または3Dモデルの使用、対象蛋白質または相同蛋白質の突然変異誘発試験、機能的に中性の置換と機能的置換を確立するためのサイト飽和ライブラリーの使用を含む。
【0072】
アミノ酸配列特性を導入するとき、置換は好ましくは、所与の位置に、所要のアミノ酸配列特徴とより多く一致するアミノ酸が相同配列に見られるアミノ酸群から選択される方法で選択される。最新のモデリング技法を使用して、天然の相同体には見られない許容可能な置換を同定することが可能である。所与の折り畳みを採用する新しい配列の生成を許容するモデリング技法に好ましいリファレンスは以下の通りである:
Kuhlman B,Dantas G,Ireton GC,Varani G,Stoddard BL,Baker D(2003)。原子レベルの確度を有する新規な球状蛋白質折り畳みの設計(Design of a novel globular protein fold with atomic−level accuracy)Science 302,1364−8。
Baker D(2006)。巨大分子構造および相互作用の予測および設計(Prediction and design of macromolecular structures and interactions.)。Philos.Trans.R.Soc.Lond.,B,Biol.Sci.361,459−63デノボ蛋白質設計:完全自動化配列選択に向けて(De Novo protein design:towards fully automated sequence selection’) Journal of Molecular Biology,273巻4号,1997年11月7日,789−796ページ Bassil I.Dahiyat,Catherine A.Sarisky,Stephen L.Mayo
【0073】
最新の計算方法は、所与の蛋白質折り畳みを採用することが可能な多数の潜在的配列の生成を許容する。最適な配列を取り出すために使用されるスコアリング機能に特性最適化を導入することによって、所与の産生宿主に最適な配列が、計算による方法で選択され得る。
【0074】
本発明の方法に従って修飾することが可能な蛋白質特性としては、組成特性、生理特性および構造特性などがある。このような特性の好適な例は、アミノ酸の数、分子量、等電点、特定のpHにおける正味荷電、GRAVYスコア、脂肪族指数、不安定指数、組成的特性、C、H、N、O、S原子に関する原子組成、アミノ酸頻度、ジペプチド頻度、トリペプチド頻度、酸性アミノ酸頻度、脂肪族アミノ酸頻度、芳香族アミノ酸頻度、塩基性アミノ酸頻度、グリコシル化パターンおよび帯電アミノ酸頻度ならびに表1に記載の特性である。修飾される特性の組合せも、本発明に包含される。好ましくは2、3、4または5つの蛋白質特性が、一緒に修飾される。より好ましくは、10超、15または20の蛋白質特性が一緒に修飾される。最も好ましくは、25超または30の蛋白質特性が一緒に修飾される。
【0075】
本発明の一実施形態では、1つまたは複数のグリコシル化部位が導入され、他の蛋白質特性も修飾される。本発明の別の実施形態では、帯電アミノ酸頻度が修飾され、他の蛋白質特性も修飾される。本発明の別の実施形態では、極性アミノ酸頻度が修飾され、他の蛋白質特性も修飾される。
【0076】
アミノ酸全体またはDNA配列から計算される蛋白質特性は、蛋白質全体の平均値であり、これから、局所的蛋白質特性が明らかにならない可能性がある。たとえば、ある蛋白質は平均すると親水性であるが、それでも大きな内部疎水性領域を含むこともあり得る。局所的蛋白質特性は、たとえば、Benita et al.(Benita et al.,2006.Molecular and Cellular Proteomics,5:1567−1580)により略述されている方法を用いて、アミノ酸またはDNA配列から計算することができる。
【0077】
ある蛋白質特性の局所特性を計算するために、適当な数のアミノ酸またはヌクレオチドのスライディングウィンドウで、蛋白質特性を局所的に計算することが可能である。次いで、図9に図示する通り、得られた値を、蛋白質のアミノ酸またはDNA配列の長さに沿った曲線としてプロットする。
【0078】
【表5】
【0079】
たとえば、LF1は図1における暗灰色の面積に相当し、LF2は図1における明灰色の面積に相当する。その面積は、台形法を使用して算出することができる(Benita et al.,2006.Molecular and Cellular Proteomics,5:1567−1580)。局所的特性を計算するために、好適な上限および下限、ならびにスライディングウィンドウの大きさを選択することができる。スライディングウィンドウは、任意のサイズのものであってもよい。たとえば、21アミノ酸または塩基対のスライディングウィンドウサイズを使用することができる。上限および下限の値は、曲線における極値ピークを反映するように選択することができる。たとえば、より高い上限は、低いものより多い極値ピークを考慮に入れる。好ましくは、上限および下限は、データセットS+およびS−のFischer基準を最大化するように選択される。Fischer基準(JF)は以下の通りに定義される:
【数2】
ここでμs−およびμs+は、それぞれS−およびS+のセットから計算される局所的特性値の平均値を表し、σ2s−およびσ2s+は、それぞれS−およびS+のセットから計算される局所的特性値の分散を表す。
【0080】
上で定義した局所的特徴は、任意の蛋白質特性、たとえばGravyスコア、アロマスコアおよび等電点について算出し得る。
【0081】
首尾よく分泌された蛋白質の配列から誘導できる特性に加えて、特に、標的蛋白質の溶媒接触表面の親水性が上昇すると、細胞から分泌された可溶性蛋白質の増量に大きく成功することが確認された。より詳細には、発現が増加するのみならず、有意に多い蛋白質が、バイオマスまたは他の不溶性物質に付着していない可溶形で、ブロス中に蓄積した。改良された表面親水性を有する蛋白質が与えられれば、いずれも有意により高い分泌で回収することが可能であろう。バイオマスを(濾過または遠心分離によって)除去すると、産生された蛋白質の大部分は濾液中または上澄中にそのまま残る。
【0082】
親水性の上昇は、
・非極性アミノ酸類を、より極性のアミノ酸類に代えること
・より低極性のアミノ酸類を、より極性のアミノ酸類に代えること
・極性アミノ酸類を帯電アミノ酸類に代えること
によって行うことができる。
【0083】
より極性のアミノ酸類または帯電アミノ酸類の数を増加させることにより親水性を上昇させることは、アミノ酸組成を変えることになり、したがって分泌を増加するために適合させることができる組成特性として考えることができる。
【0084】
非極性アミノ酸類は、A、V、L、I、C、M、F群から選択される。アミノ酸類G、P、Y、Wは、極性状況下では非極性として考えることができ、また非極性状況下では極性として考えることができる。より極性の残基は、S、T、N、Q、D、E、H、R、K群から選択される。帯電した残基は、D、E、H、R、K群から選択される。酸性または負に帯電した残基は、E、Dから選択される。塩基性または正に帯電した残基は、H、K、Rから選択される。極性用比較尺度を使用すると:[A、V、L、I、M、F、C]<[G、P、Y、W]<[S、T]<[N、Q、H]<[D、E、K、R]。
【0085】
極めて疎水性の表面領域は、産生宿主における高い産生ストレス、宿主における蛋白質の蓄積、および分泌妨害または無分泌をもたらす、望ましくない凝集またはバイオマスへの望ましくない固着を招く傾向があることが分かっている。総体的親水性を上昇させる置換は、特にこれらの残基が溶媒接触残基(=蛋白質表面残基)を含むとき、分泌改良に非常に有効であることが認められている。より詳細には、接触表面領域において、非極性残基をより極性の残基で置換するとき、極性残基の割合は、組成特性分析の上方境界で定められる極性残基の割合を上回る可能性さえあることが認められている。標的蛋白質の配列特性と宿主要件との非適合性は、標的蛋白質の親水性を上昇させることによって、より詳細には、正と負の電荷が表面全体に一様に分布して、正または負の電荷のホットスポットを防ぐように分布させた追加的電荷を導入することにより、補償することが可能である。
【0086】
特定のアミノ酸配列が与えられれば、幾つかの予測ツールは、どのアミノ酸が表面上にありそうかを予測するために使用できるが、溶媒接触非極性パッチまたは疎水性パッチを予測することが必要なとき、これらのツールの性能は全く貧弱である。したがって、蛋白質接触表面の親水性を調節するためには、3D構造または3D構造モデルが必要である。蛋白質の3D構造は、X線結晶学によって、およびNMRによって、決定することができる。加えて、比較モデリングまたは鋳型に基づくモデリングを使用して、相同蛋白質の3D構造に基づく所与の配列のための信頼できる3Dモデルを構築することが可能である(http://en.wikipedia.org/wiki/Homology modeling)。比較モデリングのための様々なサーバーおよびソフトウェアパッケージは:http://en.wikipedia.org/wiki/Protein structure prediction softwareでみつかる。
【0087】
蛋白質構造予測およびモデリングに関する最近の総説については、Yang Zhang,Current Opinion in Structural Biology 2008,18:342−348を参照されたい。
【0088】
3D構造または3Dモデルの原子座標が与えられれば、当該技術分野で周知の方法により、接触表面を算出することができる。よく知られた方法は、Frederic Richardsにより開発されたローリングボールアルゴリズムによる算出である(1977,「面積、体積、充填および蛋白質構造(Areas,volumes,packing and protein structure.)」、AnnuRev Biophys Bioeng,6:151−176)。http://en.wikipedia.org/wiki/Accessible surface areaも参照されたい。
【0089】
接触表面を決定するためには、置換が多量体(たとえば二量体、三量体、四量体等々)における個々のポリペプチド(モノマー)間の相互作用を妨害するのを回避するために、最終的な成熟蛋白質の四次構造を考えるべきである。
【0090】
表面変調は、下記を含む:
・適切な分泌および回収を妨げかねない、潜在的な粘着性のパッチを生じさせる溶媒から、非極性残基が接触される領域をスポットすること。
・機能的役割を果たす領域、たとえば活性部位全般を、そしてより具体的には基質、共基質およびコファクター用の結合ポケットを除外すること。
・非極性残基を、帯電残基も含む、より極性の残基で置換すること。
・極性残基を、より極性の残基または帯電残基で置換すること。
・高い負電荷を有する領域または高い正電荷を有する領域を回避するために、帯電残基を再分布させること。
・疎水性表面パッチを置換する代わりに、非極性領域の近くにグリコシル化を導入することにより、このような領域を遮蔽することも可能である。
【0091】
一次構造の場合、上昇した親水性は、たとえば下表のように修飾の前後の極性残基の数を比較することによって表される。
【0092】
【表6】
【0093】
接触表面を考えるとき、様々な極性アミノ酸の寄与は、総接触表面に対する、ある特定のアミノ酸またはある特定のアミノ酸群により形成される接触表面の割合として表現することができる。たとえば、帯電残基の総接触表面を算出して、総接触表面積と比較することができる。全ての極性残基を考えることにより、極性接触表面を算出することができる。蛋白質表面の親水性は、非極性表面を犠牲にして極性表面の割合が増加するとき、上昇すると言われる。
【0094】
原則として、グリコシル化を導入し、そのグリコシル化によって遮蔽される領域を推定することもできる。電荷の分布は、視覚的検査を含む任意の利用可能な方法で行い得る。
一実施形態では、改良された分泌のために修飾すべき特性は、表面電荷(再)分布、表面極性−非極性分布、配列モチーフ、たとえばグリコシル化、またはこれらの組合せである。1つの特性、たとえばアミノ酸の修飾は、多くの場合、別の特性の修飾、たとえばC、H、N、O、S原子に関して原子組成の修飾を、もたらすことを当業者は理解するであろう。
【0095】
本発明による方法は、蛋白質産生レベルを高めるための最新の技法と、またはこれらの技法の1つまたは複数の組み合わせと、都合よく組み合わせられることを理解されたい。これらは、強力プロモーター類、コピー数の増加、最適Kozak配列、mRNA安定要素および最適化コドン使用の適用を含むが、その限りではない(国際公開2008/000632号パンフレット)。
【0096】
[実施例]
[菌株]
アスペルギルス・ニガー(A.niger)菌株:WT 1:このアスペルギルス・ニガー(A.niger)菌株を、野生型菌株として使用した。この菌株は、寄託番号CBS 513.88.で、CBS研究所(CBS Institute)に寄託される。
WT 2:このアスペルギルス・ニガー(A.niger)菌株は、グルコアミラーゼをコード化する遺伝子(glaA)の欠失を含むWT 1菌株である。WT 2は、欧州特許出願公開第0 635 574 B1号明細書に記載の「MARKER−GENE FREE」アプローチを使用して構築した。この特許では、CBS 513.88のゲノム中のglaA特異的DNA配列の削除方法が詳細に説明されている。この手順は、最終的に外来DNA配列を全く持たない、MARKER−GENE FREE ΔglaA組換えアスペルギルス・ニガー(A.niger)CBS513.88菌株をもたらした。
WT 3:WT 2における主要な細胞外アスパラギン酸プロテアーゼPepAをコード化しているpepA遺伝子を中断するために、van den Homberghらにより記述されている通り、WT 2のゲノム中のpepA特異的DNA配列を削除した(van den Hombergh JP,Sollewijn Gelpke MD,van de Vondervoort PJ,Buxton FP,Visser J.(1997)−アスペルギルス・ニガー(Aspergillus niger)における3つの酸プロテアーゼの破壊−プロテアーゼスペクトル、細胞内蛋白質分解、および標的蛋白質の分解に及ぼす影響−Eur J Biochem.247(2):605−13)。この手順は、WT 2菌株バックグラウンドに不活性化されたpepA遺伝子を有する、MARKER−GENE FREE WT 3菌株をもたらした。
WT 4:WT 3中のhdfA遺伝子を削除するために、先に、国際公開第05/095624号パンフレットに詳細に記載されている方法を使用して、アスペルギルス・ニガー(Aspergillus niger)WT 4(ΔglaA,ΔpepA,ΔhdfA)を作成した。
WT 5:このアスペルギルス・ニガー(A.niger)菌株は、シュウ酸欠損アスペルギルス・ニガー(A.niger)菌株をもたらす欠失を含むWT 4菌株である。WT 5は、欧州特許出願公開第1157100号明細書および米国特許第6,936,438号明細書に記載の方法を使用することによって構築され、その中では、オキサロ酢酸加水分解酵素をコード化するoahA遺伝子の欠失によってシュウ酸欠損菌株が得られ、菌株WT 5は、WT 4菌株バックグラウンドに不活性化されたoahA遺伝子を有する代表的な菌株として選択された。
WT 6:このアスペルギルス・ニガー(A.niger)菌株は、後続する3ステップで、アルファ−アミラーゼ類をコード化する3遺伝子(amyB、amyBIおよびamyBII)の欠失を含む、WT 5菌株である。欠失ベクターの構築およびこれらの3遺伝子のゲノム欠失は、国際公開第2005095624号パンフレットに詳述されている。国際公開第2005095624号パンフレットに記載のベクターpDEL−AMYA、pDEL−AMYBIおよびpDEL−AMYBIIは、欧州特許出願公開第0 635 574 B1号明細書に記載の「MARKER−GENE FREE」アプローチに従って使用された。上述の手順は、最終的に外来DNA配列を全く持たない、シュウ酸欠損、MARKER−GENE FREE ΔglaA、ΔpepA、ΔhdfA、ΔamyA、ΔamyBIおよびΔamyBIIアミラーゼ−陰性組換えアスペルギルス・ニガー(A.niger)CBS 513.88菌株である、WT 6をもたらした。このように、菌株WT 6は低いアミラーゼバックグラウンドを有し、WT 1と比較して、より効率的な配列のターゲティングに関して高いHR/NHR比を有し、また細胞外蛋白質発現および検出用に最適化されている。
【0097】
クリベロミセス・ラクテス(K.lactis)菌株:クリベロミセス・ラクテス(K.lactis)におけるPGEおよびその変異体の発現を評価するために、2つの菌株を使用した。GG799(New England Biolabs)、および本明細書ではWT 7とも呼ばれ、米国特許第6,265,186 B1号明細書により詳細に記載されている、クリベロミセス・ラクテス(K.lactis)CBS 685.97の派生物。菌株クリベロミセス・ラクテス(K.lactis)WT 7は、突然変異誘発(古典的菌株改良)および遺伝子操作によって、CBS 685.97から誘導した。
【0098】
[キチナーゼ活性アッセイ]
反応混合物は、キチン−アズール(Sigma)3mg、0.1M Na−クエン酸−リン酸バッファー0.5ml、pH5.0の分析試料(培養液)0.1mlを含んでいた。この反応混合物を振盪しながら37℃で24時間、インキュベートし、12000rpmで10分間遠心分離してOD590を測定した。
【0099】
[pNP−β−グルコピラノシドを基質として使用するβ−グルコシダーゼ活性]
50mM酢酸ナトリウムバッファー pH=4.5中、3mM pNP−β−グルコピラノシド(Sigma N7006)原液を調製した。アッセイ:基質原液(3mM)250μl+希釈酵素試料250μLを、40℃でインキュベートした。t=0、10、20および30分に、培養標本100μlと1M炭酸ナトリウム100μlとを混合することによって反応を停止させた。吸光度は、MTP読み取り装置を使用して405nmで測定した。活性は、放出されたpNP μmol/ml/分で表す。
【0100】
[セロビオースを基質として使用する、β−グルコシダーゼ活性]
50mM酢酸ナトリウムバッファー pH=4.5中、最終濃度10mMのセロビオース(SigmaC7252)原液を調製した。アッセイ用に、基質原液(10mM)2000μl+希釈酵素試料100μLを混合し、40℃でインキュベートした。t=0、10、20および30分に、培養標本100μl+50mM水酸化ナトリウム100μlを混合することによって反応を停止させた。試料を限外濾過に付し、ED40パルスアンペロメトリー検出器を具備するDionex DX−500で実施されるパルスアンペロメトリー検出器を用いた高性能陰イオン交換クロマトグラフィ(HPAEC−PAD)を使用して分析した。活性は、放出されたグルコースμmol/ml/分で表す。
【0101】
[AZO−CM−セルロースを使用するエンドグルカナーゼ活性]
本アッセイは、Megazyme手順書S−ACMC 04/07(Megazyme International Ireland Ltd,http://secure.megazyme.com/downloads/en/data/S−ACMC.pdf)に従って実施する。活性は、100mM酢酸ナトリウムバッファー pH4.6中、2%AZO−CM−Celluloseを用いて40℃で測定した。アッセイ用に、基質原液(2%)250μL+希釈酵素溶液250μLを混合した。30分後、沈殿剤溶液1250μLを加えた。沈殿剤溶液(エタノール76%中、300mM酢酸ナトリウムバッファー pH=5と20mM Zn−酢酸)を加えることによって反応を停止させた。1000xgで10分間遠心分離した後、分光光度計を使用して、上澄の590nmにおける吸光度を測定した。活性は、放出された色素μmol/ml/分で表す。
【0102】
[トリブチリンプレートアッセイ]
トリブチリン(C4)を基質として、ローダミンBリパーゼプレートスクリーニングアッセイを行った。ローダミンBプレートアッセイは、試料中の、リパーゼ活性存在のスクリーニングによく使用され、文献(G.Kouker,K.E.Jaeger,Appl.and Environ.Microbiol,1987,211−213)に記載のアッセイから改変された。使用した全ての化学薬品は、分析グレードであった。アラビアゴムエマルジョンは、NaCl 17.9gおよびKH2PO4 0.41gをH2O 400mlに溶解し、最後にグリセロール(87%)540mlを加えることによって作った。アラビアゴム6(6.0)gを徐々に加え、溶解した後、H2Oを加えることにより全量1000mlとした。
ローダミンB溶液は、ローダミンBを20mg/mlの濃度でエタノールに溶解することによって調製した。4%アガロース溶液は、加熱により、アガロース4gをバッファー液(0.1M酢酸塩 pH=5.5)100mlに溶解することによって調製した。リパーゼ活性のスクリーニングに使用した基質は、トリブチリンであった。
プレートアッセイ手順:基質1mlおよびアラビアゴムエマルジョン1.5mlを、バッファー液5mlと混合し、振幅20μmで2×60秒間Soniprepを使用して、または任意選択的に、緑色に設定したUltraturaxで2分間、超音波処理した。この溶液に、熱いアガロース溶液7.5mlをローダミンB 150μlと一緒に加えた。最終溶液をペトリ皿プレートに注入した。プレートは、使用するまで冷蔵庫内に保存した。使用直前に、レプリケーターを使用して、直径3mmの穴を作った。リパーゼ活性を確認する溶液10μlを穴にピペッティングし、その後、プレートを37℃で18〜24時間インキュベートした。穴の周囲の蛍光ハロは、リパーゼ活性を示す。
【0103】
[pNP−ブチレートアッセイ]
前胃エステラーゼ(PGE)活性は、内部酵素標準に対して、最終濃度1mMのパラ−ニトロフェニルブチレートを基質として使用し、37℃で測定した。基質溶液は、アセトニトリル中50mMパラ−ニトロフェニルブチレート原液を作り、それを、0.2%BSAおよび2%Triton X−100を含有する0.1Mリン酸ナトリウムバッファー pH6.7で5倍希釈することによって調製した。0.2%BSAを含有する0.1Mリン酸ナトリウムバッファーpH6.7 120μl、基質溶液15μlを加えた。37℃に予熱した後、適切な希釈度の試料(0.2%BSAを含有する0.1Mリン酸ナトリウムバッファーpH6.7で希釈)15μlを加え、その後、37℃で5分間のインキュベーションの間ずっと、405nmにおける吸光度の上昇を分光光度法で測定した。
試料応答は、ブランクバックグラウンド(試料の代わりに、0.2%BSAを含有する0.1Mリン酸ナトリウムバッファーpH6.7 15μlのインキュベーション)の補正をし、ブランク補正後、一般に0.05〜0.5dAbsの範囲であった。
内部標準を、pH6.0および30℃で実施される、トリブチリンを用いた滴定分析で較正した。PGE試料液(milliQ水で調製)5mlを、予熱したトリブチリン/アラビアゴムエマルジョン(それぞれ、水中93g/Lおよび57g/L)30mLに加えた。放出された遊離脂肪酸を、0.02N NaOHで滴定することによって5分間にわたり測定した。
【0104】
[SDS−PAGE電気泳動]
試料前処理:試料30μlを、水35μlおよびNuPAGE(商標)LDS試料バッファー(4×)Invitrogen 25μlおよびNuPAGE(商標)試料還元剤(Sample Reducing agent)(10×)Invitrogen 10μlに加えた。試料を、サーモミキサー内、70℃で10分間加熱した。SDS−PAGEは、供給者の指示書(Invitrogen:Gel:4〜12%Bis−Trisゲル、バッファー:MES SDS泳動バッファー、実行時間:35分)に従って、二重に実施した。2つのゲルのうちの1つをブロッティングに使用し、試料液10μlおよびマーカーM12(Invitrogen)1μlをゲル上に適用した(NuPAGE(商標) Bis Tris,Invitrogen)。
ゲルは、外部バッファー室内の20倍希釈したMES−SDSバッファー600ml、および内部バッファー室内の酸化防止剤(NuPAGETM Invitrogen)0.5mlを含有する20倍希釈したMES−SDSバッファー200mlを備えたXCELL Surelockを使用して、200Vで動かした。ラン実行後、ゲルを50%メタノール/7%酢酸(50ml)で1時間固定し、脱塩水で2回すすぎ、シプロルビー(Sypro Ruby)(50ml,Invitrogen)で一晩染色した。
画像は、ゲルを脱塩水で10分間洗浄した後、Typhoon 9200(610 BP 30,Green(532nm),PMT 600V,100μm)を使用して作成した。
【0105】
[ウエスタンブロッティング]
[PGEポリクローナル抗体]
PGEポリクローナル抗体は、迅速28日プログラムおよび抗原としての2つの合成PGEペプチドを使用して、Eurogentec(ベルギー)に注文した。PGE抗体は、市販のPiccantase C(DFS)酵素製剤と比較して確認した(データ表示せず)。
ウエスタンブロッティングは、分析S2300の方法に従って実施した。
膜:NC 0.45μm
実行時間:25Vで90分
バッファー:メタノールを含むトランスファーバッファー
【0106】
膜に移動させた後、以下のステップを実施した:
膜を、スキムミルク(PBST中1%のスキムミルク;10mM PBS+0.05%TWEEN20)20ml中で2時間ブロックする。
抗体1:SY0716,ウサギ;抗体40μlを、PBST 20mlに溶解する)室温で一晩(1:500)。
膜を、PBS−Tですすぎ、次にPBSTバッファーで3×20分間洗浄する。
抗体2:ECL Plexヤギ抗ウサギIgG Cy3(GE Healthcare);ECL Plex10μlを、PBST 25mlに溶解する、暗所に保管)1時間。(1:2500)
膜を4回すすぎ、次にPBSTで2×10分間洗浄する
PBSで2×10分間洗浄する
画像は、Typhoon9200(670BP30,緑色(532nm),PMT 450V,100μm)を使用して、膜から作成した。
【0107】
[分子生物学技法]
本明細書の実施例では、当業者に周知の分子生物学技法(see:Sambrook & Russell,Molecular Cloning:A Laboratory Manual,3rd Ed.,CSHL Press,Cold Spring Harbor,NY,2001)を使用して、幾つかの遺伝子を過剰発現させ、その他を、下記の通りに下方制御した。
【0108】
記述され、使用された全ての遺伝子置換ベクターは、周知の原理に従って設計され、またルーチンのクローニング手順に従って構築された。本質的に、これらのベクター類は、宿命的なゲノム遺伝子座における相同組換えを目標とするため、それぞれのORF配列の、約1〜2kbの隣接領域を含む。加えて、これらのベクター類は、形質転換のために、アスペルギルス・ニジュランス(A.nidulans)双方向amdS選択マーカーを、直接反復の中間に含む。本明細書の全実施例で遺伝子欠失に利用された方法は線状DNAを使用するが、これは、二重交差によってゲノムのフランキング配列の相同遺伝子座に組み込まれ、したがって削除されるべき遺伝子がamdS遺伝子に代わる。形質転換後、この直接反復によって、(第2の)相同組換え事象による選択マーカーの除去が可能になる。amdSマーカーの除去は、フルオロアセトアミド培地上にプレーティングすることにより行うことができ、無マーカー遺伝子菌株を選択する結果となる。欧州特許第0 635 574号明細書に「MARKER−GENE FREE」アプローチとしても記述されている、形質転換および後続の対抗選択という、この戦略を使用すれば、amdSマーカーを菌株修飾プログラムで無制限に使用することができる。遺伝子破壊のための基本手順は、国際公開第2006040312号パンフレットの図6に描かれている。欠失ベクター類の一般設計は、以前に欧州特許第635574B号明細書および国際公開第98/46772号パンフレットに記述されており、また欠失ベクターを構築するための一般的クローニングベクターpGBDELの使用および対抗選択手順は、国際公開第06/040312号パンフレットに記述されている。
発現ベクター類、具体的には遺伝子過剰発現用のpGBFIN−発現ベクター類の一般設計、形質転換、マーカー類の使用および選択培地の例は、国際公開第199846772号パンフレット、国際公開第199932617号パンフレット、国際公開第2001121779号パンフレット、国際公開第2005095624号パンフレット、欧州特許第635574B号明細書および国際公開第2005100573号パンフレットにある。
【0109】
[振盪フラスコ発酵]
アスペルギルス・ニガー(A.niger)菌株を、国際公開第99/32617号パンフレットの実施例:「アスペルギルス・ニガー(Aspergillus niger)振盪フラスコ発酵」セクションに記載の通りに、CSL前培養培地20ml中で前培養した(100mlフラスコ、バッフル)。34℃および170rpmで18〜24時間成長させた後、この培養10mlを、発酵培地(FM)に移す。FM中での発酵は、概ね国際公開第99/32617号パンフレットに記載の通り、発酵ブロス100mlが入ったバッフル付き500mlフラスコ内、34℃および170rpmで指示された日数、実施する。
CSL培地は(1リットル当たりの量で):コーン・スティープ・ソリッド(Corn Steep Solids)(Roquette)100g、NaH2PO4*H20 1g、MgSO4*7H2O 0.5g、グルコース*H2O 10gおよびバシルドン(Basildon)(消泡剤)0.25gで構成されていた。成分を脱塩水に溶解し、NaOHまたはH2SO4でpHをpH5.8に調整した;バッフル付き100mlフラスコおよびフォームボールを発酵培地20mlで満たし、120℃で20分間、滅菌した。
発酵培地(FM)は(1リットル当たりの量で)、マルトース*H2O 150g、ソイトン(Soytone)(ペプトン)60g、NaH2PO4*H2O 1g、MgSO4*7H2O 15g、トゥイン(Tween)80 0.08g、バシルドン(Basildon)(消泡剤)0.02g、MES 20g、L−アルギニン 1gで構成されていた。成分を脱塩水に溶解し、NaOHまたはH2SO4でpHをpH6.2に調整した;バッフル付き500mlフラスコおよびフォームボールを発酵ブロス100mlで満たし、120℃で20分間、滅菌した。
クリベロミセス・ラクテス(K.lactis)振盪フラスコ発酵の場合、クリベロミセス・ラクテス(K.lactis)PGE形質転換体の単一コロニーを、1リットル当たり酵母エキス10g、バクトペプトン20g、グルコース40gおよび100mM MES pH6.7を含有する、YEP(4%)−D/MES培地100ml(フラスコ)に接種した。発酵は、280rpmの振盪インキュベーター内、30℃で実施した。上澄を2日および3日に採取し、下記の通りにさらに分析した。
【0110】
[実施例1]
[本発明の方法による、野生型酵素および酵素変異体用のクリベロミセス・ラクテス(K.lactis)発現ベクターおよびアスペルギルス・ニガー(A.niger)発現ベクターの構築]
この実施例では、本発明の酵素の変異体用に、多数の発現ベクターを構築した。クリベロミセス(Kluyveromyces)における発現のための全ての変異体を、pKLAC2発現ベクター(New England Biolabs)に酷似したpKLPGE−ベクターでクローニングした。全てのpKLPGE−ベクター類の一般設計図を図1に示す。アスペルギルス(Aspergillus)における発現用の全ての変異体は、pGBFIN−5発現ベクターまたはpGBTOP−発現ベクターでクローニングした。構築、一般設計図およびこれらのベクター類の使用は、国際公開第199932617号パンフレットに詳述されている。
【0111】
[クリベロミセス・ラクテス(K.lactis)構築物]
ウシ前胃エステラーゼ(PGE)は、産業上興味深い酵素であり、またその全長cDNA配列は、Timmermansら(1994,Gene 147:259−262)により公開されている。クリベロミセス・ラクテス(Kluyveromyces lactis)におけるPGEの発現の場合、このcDNA配列は、最適化されたコドン対(配列番号1)であり、合成的に調製されている(たとえば、DNA2.0,USA,GeneArt、Sloning,Germany)。クリベロミセス・ラクテス(K.lactis)α−因子プレ(プロ−)シグナル配列およびKREAEA Kexプレ(プロ−)−配列プロセシング部位との融合を含む発現構築物を作成した。HindIIIおよびNotI制限部位を介して、合成遺伝子をクリベロミセス・ラクテス(K.lactis)発現ベクターでクローニングし、amdS選択マーカーも含む、pKLPGE−WT(図1)を産生した。加えて、本発明の方法に従って、改良された蛋白質特性を有する幾つかのPGE変異体を設計した。これらの突然変異体は、グリコシル化部位数および/または疎水性に関して、コドン対最適化野生型PGE酵素(配列番号2)と異なっていた。PGE突然変異体酵素コード化遺伝子も、上記の通り、やはり最適化コドン対であり、また合成的に調製されてもいる。XhoIおよびNotIクローニング部位を使用して本質的に前述の通りに、変異体を、クリベロミセス・ラクテス(K.lactis)発現ベクターにクローニングした。PGE構築物のための全ての関連ヌクレオチドおよび蛋白質の詳細を表4に示す。
【0112】
【表7】
【0113】
[アスペルギルス・ニガー(A.niger)構築物]
アスペルギルス・ニガー(A.niger)におけるウシ前胃エステラーゼPGEの発現の場合、cDNA配列は、最適化コドン対(配列番号12)であり、合成的に調製された(たとえば、DNA2.0,USA,GeneArt,Stoning,Germany)。コドン対最適化PGEコード化遺伝子は、切断型グルコアミラーゼキャリア蛋白質(tAG)への融合物として合成的に調製された。図2のpANPGE−3に示す通り、この融合フラグメントを、pGBTOP−アスペルギルス・ニガー(A.niger)発現ベクターに挿入した。
【0114】
仮想キチナーゼ(ZDU,EC3.2.1.14,Uniprot A5AB48)をコード化する野生型アスペルギルス・ニガー(A.niger)遺伝子An08g09030は、アスペルギルス・ニガー(Aniger)ゲノム配列で同定された(EMBL:AM269948−AM270415;Pel et al,「汎用細胞工場アスペルギルス・ニガー(Aspergillus niger)CBS513.88のゲノムシークエンシングおよび分析(Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS513.88.)」。Nat Biotechnol.2007Feb;25(2):221−231)。野生型キチナーゼZDUのcDNA配列は、配列番号17として同定することができ、推定される野生型キチナーゼZDU蛋白質配列は、配列番号18として同定することができる。An08g09030のコード配列は最適化コドン対であり(国際公開第2008000632号パンフレットに詳述されている通り)、またグルコアミラーゼglaAプロモーターの翻訳開始配列は、作成された全ての発現構築物で5’−CACCGTCAAA ATG−3’に修飾されていた(やはり国際公開第2006/077258号パンフレットに詳述されている通り)。加えて、最適翻訳停止配列が使用され、したがって、全ての発現構築物で、野生型5’−TGA−3’翻訳停止配列は5’−TAAA−3に置き換えられた(国際公開第2006/077258号パンフレットに詳述されている通り)。最適化キチナーゼZDU構築物は、PacI−AscIフラグメントとして完全合成し、サブクローニングして配列を検証した。合成されたフラグメント両端のPacI−AscI制限部位を使用して、PacI−AscI消化pGBFIN−5発現ベクターの大きいベクターフラグメントでのクローニングを可能にし、pGBFINZDU−WT発現ベクターを作成した(図3)。
加えて、またZDUキチナーゼの場合と同様の方法で、タラロミセス・エメルソニ(Talaromyces emersonii)β−グルコシダーゼ(ZTB,EC 3.2.1.21,Uniprot Q8X214)およびファネロカエテ・クリソスポリウム(Phanerochaete chrysosporium)エンドグルカナーゼ(ZTC,EC 3.2.1.4,Uniprot Q66NB6)は、最適化コドン対であり(国際公開第2008000632号パンフレットに詳述されている通り)、また全ての適切な制御要素をPacI−AscIフラグメントとしてpGBFIN−5にクローニングし、それぞれpGBFINZTB−WTおよびpGBFINZTC−WTを作成した。
【0115】
本発明の方法による蛋白質特性最適化(PFO)を、ウシ前胃エステラーゼ、アスペルギルス・ニガー(A.niger)キチナーゼ蛋白質配列、タラロミセス・エメルソニ(T.emersonii)β−グルコシダーゼおよびファネロカエテ・クリソスポリウム(P.chrysosporium)エンドグルカナーゼ蛋白質配列に適用した。ウシ前胃エステラーゼの設計された変異体を含むコード配列をEcoRI−SnaBIフラグメントとして完全合成し、配列を検証した。合成フラグメントをpGBTOP−ベクターでクローニングし、pANPGE−発現構築物を作成した。アスペルギルス・ニガー(A.niger)PGE構築物のための全ての関連ヌクレオチドおよび蛋白質の詳細を表5に示す。
【0116】
【表8】
【0117】
キチナーゼ、β−グルコシダーゼおよびエンドグルカナーゼの設計された変異体を含むコード配列をPacI−AscIフラグメントとして完全合成し、サブクローニングして配列を検証した。合成されたフラグメント両端のPacI−AscI制限部位を使用して、PacI−AscI消化pGBFIN−5発現ベクターの大きいベクターフラグメントでのクローニングを可能にし、変異体pGBFIN−発現ベクターを作成した。変異体発現構築物は、下記の通りに命名され、pGBFINZDU−構築物の特徴ならびにそれぞれのヌクレオチド配列および蛋白質配列ののリファレンスは、表6から推定することができ、pGBFINZTB−構築物のそれらは表7から、またpGBFINZTC−構築物のそれらは表8から推定される。
【0118】
【表9】
【0119】
【表10】
【0120】
【表11】
【0121】
[実施例2]
[クリベロミセス・ラクテス(K.lactis)野生型の発現および分泌分析ならびに蛋白質特性最適化PGE]
菌株クリベロミセス・ラクテス(K.lactis)GG799WT7を、amdS選択マーカーも含むクリベロミセス・ラクテス(K.lactis)pKLPGE−構築物(表4)で形質転換した。各形質転換について、20コロニーを、アセトアミドを含有する選択培地で純化した。コロニーの一部を使用して、PCR反応用のDNA鋳型を作成し、各菌株におけるPGE構築物のコピー数を決定した。構築物当たり3つの、PCRスクリーニングで陽性の形質転換体を、酵素基質としてトリブチリンを含有するプレートアッセイでさらにスクリーニングした。野生型PGE酵素の場合、トリブチリンプレートアッセイを使用して、明白な活性ハロを検出することはできなかった。PGE産生用のSDS−PAGEを用いた上澄の分析もまた、陽性結果を示さなかった。意外なことに、最適化蛋白質特性を有するPGE突然変異体5つのうち4つでは、トリブチリンプレートアッセイを使用して、明白な活性ハロを確認することができた。
野生型および突然変異体PGEの多数の形質転換体を、振盪フラスコ内で成長させ、pNP−ブチレートを基質として使用して、ブロスおよび上澄のリパーゼ活性を試験した。PGE突然変異体の様々な活性アッセイの概要を表9に示す。
【0122】
【表12】
【0123】
クリベロミセス・ラクテス(K.lactis)pKLPGE−WT(PGECPO)形質転換体(様々なコピー数)では、0.2U/mlという最大活性が得られた。PGEの蛋白質特性最適化によって、すなわちpKLPGE−12で発現されるとき、このPGE突然変異体の50×を超える活性上昇が確認された。PGE−9、PGE−11およびPGE−12変異体の多数の突然変異体を大規模ベースで発酵させ、改良された分泌を確認した(データ表示せず)。この実施例では、グリコシル化部位数の改変および表面に露出している疎水性酵素部分の極性の変更により(PGEモデリングに基づいて決定される)、我々はクリベロミセス・ラクテス(K.lactis)におけるPGE酵素発現および分泌を劇的に改良できることが分かった。さらに、かなりの量の活性が、上澄にも存在していた。
【0124】
[実施例3]
[アスペルギルス・ニガー(A.niger)における野生型および蛋白質特性最適化PGEの発現および分泌分析]
アスペルギルス・ニガー(A.niger)WT 6を、アスペルギルス・ニジュランス(A.nidulans)amdS選択マーカーおよび変異体pANPGE−プラスミドを担持するpGBAAS構築物で共形質転換した(表5)。全て、国際公開第99/32617号パンフレットに記載の通り、各形質転換について、20コロニーを、アセトアミドを含有する選択培地で純化し、続いてスポアプレートを調製した。真の共形質転換体であった、たとえば、PGEカセットおよびamdSカセットの両者を含んでいた、アスペルギルス・ニガー(A.niger)形質転換体を選択するために、PCR確認する(表示せず)。結果から、選択された形質転換体20のうち少なくとも50%が、PGE発現構築物の1つまたは複数のコピーを含んでいた。形質転換体を含むこれらのPGEを継続した。PGE contransformantsのスポアを回収し、FM培地で振盪フラスコ発酵を実施した。2日目に、上澄試料を採取し、トリブチリンプレートアッセイを使用してリパーゼ活性についてスクリーニングした。
アスペルギルス・ニガー(A.niger)pANPGE−3形質転換体から回収された試料中に、非常に僅かな活性ハロを検出することができた(データ表示せず)。pANPGE−12およびpANPGE−13形質転換体では、大きい活性ハロを検出することができた(データ表示せず)。各構築物pANPGE−3、pANPGE−12およびpANPGE−13について、トリブチリンプレートアッセイで最大のハロを示した形質転換体(1〜3)のリパーゼ活性を、基質としてpNP−ブチレートを使用して試験した。PGE突然変異体の様々な活性アッセイの概要を表10に示す。
【0125】
【表13】
【0126】
WT 6および選択された形質転換体pANPGE−12#16およびpANPGE−13#30の上澄試料を、SDS−PAGEゲル(Invitrogen)およびPGEポリクローナル抗体を使用したウエスタンブロットで、さらに分析した(図4参照)。pANPGE−12のアスペルギルス・ニガー(A.niger)PGEPFO変異体では、SDS−PAGEゲルで、成熟PGEに相当するバンドを検出することができた。PGEポリクローナル抗体PGEを使用して、両形質転換体の上澄中に、交差ハイブリダイズバンドを検出することができた。
最高分子量のバンド(約55kDa)はおそらく成熟PGE突然変異体に相当し、また、より低分子量の交差ハイブリダイズバンドは、蛋白質分解の結果であろう。
蛋白質特性最適化の法則に従って、表面に露出している酵素部分の極性(PGEモデリングに基づいて決定される)を変えることにより、我々はアスペルギルス・ニガー(A.niger)におけるPGE酵素発現を劇的に改良できるという結論に達する。さらに、高い酵素活性も、上澄中で認められた。
【0127】
[実施例4]
[アスペルギルス・ニガー(A.niger)における野生型およびPFO最適化真菌酵素の発現]
実施例1(super)で調製された、pGBFINZDU−発現構築物、pGBFINZTB−発現構築物およびpGBFINZTC−発現構築物を、下記の通り、アスペルギルス・ニガー(A.niger)を使用した形質転換により導入した。WT 6で、異なるpGBFINZDU−ベクター、pGBFINZTB−ベクターおよびpGBFINZTC−ベクター(それぞれ、表6、7および8)を導入するために、国際公開第1998/46772号パンフレットおよび国際公開第1999/32617号パンフレットに記載の通りに、形質転換および後続する形質転換体の選択を実施した。簡単に記載すると、全てのpGBFIN−構築物の線状DNAを単離して、アスペルギルス・ニガー(A.niger)WT 6の形質転換に使用した。形質転換体をアセトアミド培地で選択し、標準手順に従って、コロニーを純化した。コロニーのglaA遺伝子座における組み込みおよびコピー数を、PCRを使用して診断した。類似した推定コピー数(仮想単一コピー)を有する各pGBFINZDU−構築物、pGBFINZTB−構築物およびpGBFINZTC−構築物について、独立した3つの形質転換体を選択し、形質転換プラスミドの数を使用して、たとえば、それぞれ、ZDU−WT−1、ZDU−WT−2、ZDU−WT−3、ZDU−6−1、ZDU−6−2、ZDU−6−3等々と命名した。
上述の通り、34℃および170rpmのインキュベーターシェーカーで、500mlバッフル付き振盪フラスコを使用し、選択されたZDU−菌株、ZTB−菌株およびZTC−菌株ならびにアスペルギルス・ニガー(A.niger)WT 6を使用して、FM培地100mlで、振盪フラスコ実験を実施した。発酵の3日、4日および5日の後、試料を採取し、電気泳動およびキチナーゼ活性により細胞外蛋白質の産生量を測定した。
【0128】
異なる構築物を含むアスペルギルス・ニガー(A.niger)ZDU−5形質転換体の各形質転換体により発現されたキチナーゼの産生を、培養上澄で測定した。3日に測定されたキチナーゼ活性レベルを図5に示す。加えて、4日にサンプリングした培養上澄をSDSゲル電気泳動法および染色で分析した(図6)。これらの結果から、最適化された蛋白質特性は、蛋白質分泌にプラスの影響を与え、そして検出可能な、したがって蛋白質発現レベル上昇およびキチナーゼ酵素活性レベル上昇をもたらすことが明白である。結果を表6にまとめた。
【0129】
異なる構築物を含むアスペルギルス・ニガー(A.niger)ZTB−形質転換体の各形質転換体により発現されたβ−グルコシダーゼの産生を、培養上澄で測定した。4日目にサンプリングした培養上澄をSDSゲル電気泳動法および染色で分析した(図7)。これらの結果から、最適化された蛋白質特性は、蛋白質分泌にプラスの影響を与え、そして検出可能な、したがって、β−グルコシダーゼ酵素のための蛋白質発現レベル上昇をもたらすことが明白である。加えて、3日目にサンプリングした上澄中の活性を、基質としてpNP−β−グルコピラノシドを使用し、pH=4.5および40℃で測定した。蛋白質特性最適化に付しておいたβ−グルコシダーゼの上澄は、コドン最適化遺伝子によりコード化された親β−グルコシダーゼと比較して、最高20倍の活性上昇を示した。空宿主で測定されるバックグラウンドβ−グルコシダーゼ活性は、コドン最適化遺伝子によりコード化された親ベータ−グルコシダーゼの2分の1〜4分の1の低さであった。活性はまた、セロビオースを基質として使用し、pH=4.5および40℃でも測定した。測定された活性上昇は、コドン最適化遺伝子によりコード化された親β−グルコシダーゼと比較して、少なくとも30倍であった(空宿主菌株は、コドン最適化遺伝子によりコード化された親β−グルコシダーゼの3分の1〜10分の1の低さであった)。結果を表7にまとめた。
【0130】
異なる構築物を含むアスペルギルス・ニガー(A.niger)ZTC−形質転換体の各形質転換体により発現されたエンドグルカナーゼの産生を、培養上澄で測定した。4日目にサンプリングした培養上澄をSDSゲル電気泳動法および染色で分析した(図8)。これらの結果から、最適化された蛋白質特性は、蛋白質分泌にプラスの影響を与え、そして検出可能な、したがってエンドグルカナーゼ酵素のための蛋白質発現レベル上昇をもたらすことが明白である。3日目にサンプリングした上澄中のエンドグルカナーゼ活性を、AZO−CM−セルロースを基質として使用し、pH=4.5および40℃で測定した。蛋白質特性最適化に付しておいたエンドグルカナーゼの上澄は、同一宿主で発現されるコドン最適化遺伝子と比較して、350倍を超える活性上昇を示した。空菌株におけるバックグラウンド活性は非常に低いため(SDS−PAGEで検出不能)、活性上昇がそのように高い数字で表されることに留意すべきである。コドン最適化遺伝子によりコード化されたエンドグルカナーゼについては、測定された活性は、ほぼ空宿主菌株で観測されるバックグラウンド活性であった。結果を表8にまとめた。
【0131】
明らかに、これらの実施例は、対象蛋白質および酵素の改良された分泌および産生のために、蛋白質特性最適化のための本発明の方法を、いかに使用できるかを示すものである。さらに、これらの結果は、発現構築物および宿主は、既に幾つかの他の最適化、たとえば強力プロモーター、改良された翻訳開始配列、改良された翻訳停止配列、最適化コドンおよびコドン対使用、および/または蛋白質発現のための改良された宿主等を有するものの、宿主における蛋白質発現を改良するために、本発明の方法を広く利用できることを示している。
【特許請求の範囲】
【請求項1】
真核生物宿主細胞によって対象ポリペプチドの分泌を改良するための方法であって、前記真核生物宿主における1つまたは複数の蛋白質特性について最適範囲内に入るか、または最適値により近づくように、前記ポリペプチドのアミノ酸主鎖における一組の関連する蛋白質特性の値を修飾するステップを含む方法。
【請求項2】
(i)前記真核生物宿主における1つまたは複数の蛋白質特性について最適範囲および最適値を決定するステップ、および
(ii)前記真核生物宿主における一組の関連する蛋白質特性を決定するステップであって、前記ポリペプチドのアミノ酸主鎖におけるこれらの関連特性の1つまたは複数が修飾されれば、前記真核生物宿主による前記ポリペプチドの前記分泌を改良するであろう特性を決定するステップ、および、
(iii)(i)で決定される通り、前記最適範囲内に入るか、または前記最適値により近づくように前記関連する蛋白質特性の値を修飾するステップ、
を含み、
(i)および(ii)は、任意の順序で実施し得る、
請求項1に記載の方法。
【請求項3】
関連した一組の特性が、
a.特定の真核生物宿主における適量の蛋白質の分泌レベルおよびこれらの蛋白質の前記アミノ酸配列およびDNA配列を含む、データセットSを収集または作成するステップ;
b.前記データセットSにおける全ての蛋白質について蛋白質特性(F)を計算するステップ;
c.適切に規定された分類子性能基準に従って、前記データセットSにおける分泌蛋白質S+と非分泌蛋白質S−とを区別するために、統計分類子の最高の成果を与える蛋白質特性のサブセット(Fs)を選択するための統計的分類方法を使用するステップ;
によって決定される、請求項1または2に記載の方法。
【請求項4】
前記蛋白質特性が一組の成熟蛋白質から算出される、請求項3に記載の方法。
【請求項5】
真核生物宿主の蛋白質特性の最適範囲または最適値が、
a.特定の真核生物宿主における適量の蛋白質の分泌レベルおよびこれらの蛋白質の前記アミノ酸配列およびDNA配列を含む、データセットSを収集または作成するステップ;
b.前記データセットSにおける全ての蛋白質について蛋白質特性(F)を計算するステップ;
c.前記特性の値の分布が前記選択される確立分布によって十分に説明されるように、S+から算出される各蛋白質特性に確率分布を合わせるステップにより、前記真核生物宿主の各特性の最適値(F_opt)を決定するステップ;
d.前記真核生物宿主について各特性の最適範囲を決定するステップ;
によって決定される、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
真核生物宿主によるポリペプチドの分泌を改良する方法であって、
i)前記ポリペプチドについて、蛋白質特性を計算するステップ、
ii)前記ポリペプチドの1つまたは複数の蛋白質特性が、最適範囲外であるかまたは前記真核生物宿主の最適値から実質的に逸脱しているかどうかを決定するステップ、
iii)前記ポリペプチドの1つまたは複数のFs_AAの値が、前記最適範囲内に入るか、または、RIまたはRINにより規定される適量だけ、前記最適値に向けて移行するように、前記ポリペプチドのアミノ酸配列を合理的に変えるステップであって、ここで、RIまたはRINで規定される変化は、好ましくは10%超、15%、20%、および最も好ましくは30%超である、ステップ
を含む方法。
【請求項7】
前記ポリペプチドの主鎖が、下記の特徴:アミノ酸の数、分子量、等電点、特定のpHにおける正味荷電、GRAVYスコア、脂肪族指数、不安定指数、組成的特性、C、H、N、O、S原子の原子組成、アミノ酸頻度、ジペプチド頻度、トリペプチド頻度、酸性アミノ酸頻度、脂肪族アミノ酸頻度、芳香族アミノ酸頻度、塩基性アミノ酸頻度、局所的特徴、局在化特性、グリコシル化パターンおよび帯電アミノ酸頻度;
の1つまたは複数に関して修飾される、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記ポリペプチドの主鎖が、下記の特性:塩基性アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、微小アミノ酸頻度、小アミノ酸頻度、帯電アミノ酸頻度、pH7.2における正味電荷、等電点、Asn、Arg、Ile、Cys、His、Gln、Val、Lys、Gly、ThrおよびLeuのそれぞれの頻度、局在化特性、Garnierにより算出されるターン、EPESTFINDにより算出されるPESTモチーフ、plのLF値、GravyスコアのLF値、アロマスコアのLF値、イオウ(S)組成;
の1つまたは複数に関して修飾される、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
前記ポリペプチドの主鎖が:pl、正味電荷、長さ当たりの正味荷電、長さ当たりの正味正電荷、長さ当たりの正味負電荷、長さ当たりの全電荷、グレイビースコア、アロマスコア、脂肪族指数、微小アミノ酸頻度、小アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、帯電アミノ酸頻度、酸性アミノ酸頻度、塩基性アミノ酸頻度、脂肪族アミノ酸頻度、Ala、Arg、Asn、Asp、Cys、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Phe、Pro、Ser、Thr、Trp、TyrおよびValのそれぞれの頻度、からなる群から選択される1つまたは複数の特性に関して修飾される、請求項1〜7のいずれか一項に記載の方法。
【請求項10】
前記ポリペプチドの主鎖が、pl、正味荷電(pH7.2)、長さ当たりの正味荷電(pH7.2)、長さ当たりの正味正電荷(pH7.2)、長さ当たりの全電荷(pH7.2)、脂肪族指数、小アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、帯電アミノ酸頻度、アミノ酸頻度、Arg、Gln、Glu、Lys、PheおよびThrのそれぞれの頻度からなる群から選択される1つまたは複数の特徴に関して修飾される、請求項1〜9のいずれか一項に記載の方法。
【請求項11】
前記ポリペプチドの主鎖が、グリコシル化部位、グレイビースコア、極性アミノ酸頻度、非極性アミノ酸頻度、帯電アミノ酸頻度、酸性アミノ酸頻度、塩基性アミノ酸頻度、Glu、LysおよびThrのそれぞれの頻度からなる群から選択される1つまたは複数の特性に関して修飾される、請求項1〜9のいずれか一項に記載の方法。
【請求項12】
前記アミノ酸主鎖のアミノ酸類の少なくとも5%が修飾される、より好ましくは前記アミノ酸主鎖のアミノ酸類の少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%が修飾される、請求項1〜11のいずれか一項に記載の方法。
【請求項13】
前記アミノ酸主鎖の少なくとも5つのアミノ酸類が修飾される、より好ましくは前記アミノ酸主鎖の少なくとも10のアミノ酸類、さらにより好ましくは少なくとも15のアミノ酸類、さらにより好ましくは少なくとも20のアミノ酸類、さらにより好ましくは少なくとも25のアミノ酸類、さらにより好ましくは少なくとも30のアミノ酸類が修飾される、請求項1〜11のいずれか一項に記載の方法。
【請求項14】
野生型リファレンス蛋白質に関して、少なくとも5%のF−スコアの改良が達成される、より好ましくは少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%、さらにより好ましくは少なくとも30%の改良が達成され、前記F−スコアは式:
【数1】
式中、ηは0と1の間で0および1を含む(0≦η≦1)の重み係数であって、好ましくはη0.5以下、より好ましくはη0.4以下、最も好ましくはη=0.3であり、ρは、1と5の間で1および5を含み(1≦p≦5)、好ましくはρ=2である)
にしたがって算出される、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
少なくとも2、3、4、または5つの特性が修飾される、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が修飾される、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
少なくとも2、3、4、または5つの特性が改良される、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が改良され、好ましくは10未満、さらにより好ましくは5未満、さらにより好ましくは4未満の特性が悪化する、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
前記特性は一次特性である、請求項1〜16のいずれか一項に記載の方法。
【請求項18】
前記ポリペプチドの主鎖が1つまたは複数の他の蛋白質特性に関して修飾される、請求項1または17に記載の方法。
【請求項19】
前記成熟ポリペプチドの主鎖が修飾される、請求項1〜18のいずれか一項に記載の方法。
【請求項20】
前記真核細胞が、酵母細胞または糸状菌細胞である、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
前記ポリペプチドが哺乳類ポリペプチドまたは細菌ポリペプチドである、請求項1〜20のいずれか一項に記載の方法。
【請求項22】
前記修飾されるポリペプチドの特異性が実質的に分泌の改良前と同じに保持される、請求項1〜21のいずれか一項に記載の方法。
【請求項23】
前記修飾されるポリペプチドの比活性が実質的に分泌の改良前と同じに保持される、請求項1〜22のいずれか一項に記載の方法。
【請求項24】
前記分泌の改良が前記活性の上昇によって測定され、また前記細胞外培地中の活性が少なくとも5%上昇する、請求項1〜23のいずれか一項に記載の方法。
【請求項25】
前記ポリペプチドが酵素、膜蛋白質、ホルモンまたは受容体である、請求項1〜24のいずれか一項に記載の方法。
【請求項26】
請求項1〜25のいずれかに記載の方法を、対象ポリペプチドに適用するステップ、および組換え技術により修飾されたポリペプチドを産生するステップを含む、対象ポリペプチドを産生する方法。
【請求項27】
請求項26に記載の方法で得られるポリペプチド。
【請求項28】
請求項26に記載の方法で入手できるポリペプチド。
【請求項29】
請求項1〜25に記載の方法で得られる修飾されたポリペプチド。
【請求項30】
請求項1〜25に記載の方法で入手できる修飾されたポリペプチド。
【請求項1】
真核生物宿主細胞によって対象ポリペプチドの分泌を改良するための方法であって、前記真核生物宿主における1つまたは複数の蛋白質特性について最適範囲内に入るか、または最適値により近づくように、前記ポリペプチドのアミノ酸主鎖における一組の関連する蛋白質特性の値を修飾するステップを含む方法。
【請求項2】
(i)前記真核生物宿主における1つまたは複数の蛋白質特性について最適範囲および最適値を決定するステップ、および
(ii)前記真核生物宿主における一組の関連する蛋白質特性を決定するステップであって、前記ポリペプチドのアミノ酸主鎖におけるこれらの関連特性の1つまたは複数が修飾されれば、前記真核生物宿主による前記ポリペプチドの前記分泌を改良するであろう特性を決定するステップ、および、
(iii)(i)で決定される通り、前記最適範囲内に入るか、または前記最適値により近づくように前記関連する蛋白質特性の値を修飾するステップ、
を含み、
(i)および(ii)は、任意の順序で実施し得る、
請求項1に記載の方法。
【請求項3】
関連した一組の特性が、
a.特定の真核生物宿主における適量の蛋白質の分泌レベルおよびこれらの蛋白質の前記アミノ酸配列およびDNA配列を含む、データセットSを収集または作成するステップ;
b.前記データセットSにおける全ての蛋白質について蛋白質特性(F)を計算するステップ;
c.適切に規定された分類子性能基準に従って、前記データセットSにおける分泌蛋白質S+と非分泌蛋白質S−とを区別するために、統計分類子の最高の成果を与える蛋白質特性のサブセット(Fs)を選択するための統計的分類方法を使用するステップ;
によって決定される、請求項1または2に記載の方法。
【請求項4】
前記蛋白質特性が一組の成熟蛋白質から算出される、請求項3に記載の方法。
【請求項5】
真核生物宿主の蛋白質特性の最適範囲または最適値が、
a.特定の真核生物宿主における適量の蛋白質の分泌レベルおよびこれらの蛋白質の前記アミノ酸配列およびDNA配列を含む、データセットSを収集または作成するステップ;
b.前記データセットSにおける全ての蛋白質について蛋白質特性(F)を計算するステップ;
c.前記特性の値の分布が前記選択される確立分布によって十分に説明されるように、S+から算出される各蛋白質特性に確率分布を合わせるステップにより、前記真核生物宿主の各特性の最適値(F_opt)を決定するステップ;
d.前記真核生物宿主について各特性の最適範囲を決定するステップ;
によって決定される、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
真核生物宿主によるポリペプチドの分泌を改良する方法であって、
i)前記ポリペプチドについて、蛋白質特性を計算するステップ、
ii)前記ポリペプチドの1つまたは複数の蛋白質特性が、最適範囲外であるかまたは前記真核生物宿主の最適値から実質的に逸脱しているかどうかを決定するステップ、
iii)前記ポリペプチドの1つまたは複数のFs_AAの値が、前記最適範囲内に入るか、または、RIまたはRINにより規定される適量だけ、前記最適値に向けて移行するように、前記ポリペプチドのアミノ酸配列を合理的に変えるステップであって、ここで、RIまたはRINで規定される変化は、好ましくは10%超、15%、20%、および最も好ましくは30%超である、ステップ
を含む方法。
【請求項7】
前記ポリペプチドの主鎖が、下記の特徴:アミノ酸の数、分子量、等電点、特定のpHにおける正味荷電、GRAVYスコア、脂肪族指数、不安定指数、組成的特性、C、H、N、O、S原子の原子組成、アミノ酸頻度、ジペプチド頻度、トリペプチド頻度、酸性アミノ酸頻度、脂肪族アミノ酸頻度、芳香族アミノ酸頻度、塩基性アミノ酸頻度、局所的特徴、局在化特性、グリコシル化パターンおよび帯電アミノ酸頻度;
の1つまたは複数に関して修飾される、請求項1〜6のいずれか一項に記載の方法。
【請求項8】
前記ポリペプチドの主鎖が、下記の特性:塩基性アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、微小アミノ酸頻度、小アミノ酸頻度、帯電アミノ酸頻度、pH7.2における正味電荷、等電点、Asn、Arg、Ile、Cys、His、Gln、Val、Lys、Gly、ThrおよびLeuのそれぞれの頻度、局在化特性、Garnierにより算出されるターン、EPESTFINDにより算出されるPESTモチーフ、plのLF値、GravyスコアのLF値、アロマスコアのLF値、イオウ(S)組成;
の1つまたは複数に関して修飾される、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
前記ポリペプチドの主鎖が:pl、正味電荷、長さ当たりの正味荷電、長さ当たりの正味正電荷、長さ当たりの正味負電荷、長さ当たりの全電荷、グレイビースコア、アロマスコア、脂肪族指数、微小アミノ酸頻度、小アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、帯電アミノ酸頻度、酸性アミノ酸頻度、塩基性アミノ酸頻度、脂肪族アミノ酸頻度、Ala、Arg、Asn、Asp、Cys、Gln、Glu、Gly、His、Ile、Leu、Lys、Met、Phe、Pro、Ser、Thr、Trp、TyrおよびValのそれぞれの頻度、からなる群から選択される1つまたは複数の特性に関して修飾される、請求項1〜7のいずれか一項に記載の方法。
【請求項10】
前記ポリペプチドの主鎖が、pl、正味荷電(pH7.2)、長さ当たりの正味荷電(pH7.2)、長さ当たりの正味正電荷(pH7.2)、長さ当たりの全電荷(pH7.2)、脂肪族指数、小アミノ酸頻度、極性アミノ酸頻度、非極性アミノ酸頻度、帯電アミノ酸頻度、アミノ酸頻度、Arg、Gln、Glu、Lys、PheおよびThrのそれぞれの頻度からなる群から選択される1つまたは複数の特徴に関して修飾される、請求項1〜9のいずれか一項に記載の方法。
【請求項11】
前記ポリペプチドの主鎖が、グリコシル化部位、グレイビースコア、極性アミノ酸頻度、非極性アミノ酸頻度、帯電アミノ酸頻度、酸性アミノ酸頻度、塩基性アミノ酸頻度、Glu、LysおよびThrのそれぞれの頻度からなる群から選択される1つまたは複数の特性に関して修飾される、請求項1〜9のいずれか一項に記載の方法。
【請求項12】
前記アミノ酸主鎖のアミノ酸類の少なくとも5%が修飾される、より好ましくは前記アミノ酸主鎖のアミノ酸類の少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%が修飾される、請求項1〜11のいずれか一項に記載の方法。
【請求項13】
前記アミノ酸主鎖の少なくとも5つのアミノ酸類が修飾される、より好ましくは前記アミノ酸主鎖の少なくとも10のアミノ酸類、さらにより好ましくは少なくとも15のアミノ酸類、さらにより好ましくは少なくとも20のアミノ酸類、さらにより好ましくは少なくとも25のアミノ酸類、さらにより好ましくは少なくとも30のアミノ酸類が修飾される、請求項1〜11のいずれか一項に記載の方法。
【請求項14】
野生型リファレンス蛋白質に関して、少なくとも5%のF−スコアの改良が達成される、より好ましくは少なくとも10%、さらにより好ましくは少なくとも15%、さらにより好ましくは少なくとも20%、さらにより好ましくは少なくとも30%の改良が達成され、前記F−スコアは式:
【数1】
式中、ηは0と1の間で0および1を含む(0≦η≦1)の重み係数であって、好ましくはη0.5以下、より好ましくはη0.4以下、最も好ましくはη=0.3であり、ρは、1と5の間で1および5を含み(1≦p≦5)、好ましくはρ=2である)
にしたがって算出される、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
少なくとも2、3、4、または5つの特性が修飾される、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が修飾される、請求項1〜14のいずれか一項に記載の方法。
【請求項16】
少なくとも2、3、4、または5つの特性が改良される、より好ましくは少なくとも10、さらにより好ましくは少なくとも15、さらにより好ましくは少なくとも20、さらにより好ましくは少なくとも25、およびさらにより好ましくは少なくとも30の特性が改良され、好ましくは10未満、さらにより好ましくは5未満、さらにより好ましくは4未満の特性が悪化する、請求項1〜15のいずれか一項に記載の方法。
【請求項17】
前記特性は一次特性である、請求項1〜16のいずれか一項に記載の方法。
【請求項18】
前記ポリペプチドの主鎖が1つまたは複数の他の蛋白質特性に関して修飾される、請求項1または17に記載の方法。
【請求項19】
前記成熟ポリペプチドの主鎖が修飾される、請求項1〜18のいずれか一項に記載の方法。
【請求項20】
前記真核細胞が、酵母細胞または糸状菌細胞である、請求項1〜19のいずれか一項に記載の方法。
【請求項21】
前記ポリペプチドが哺乳類ポリペプチドまたは細菌ポリペプチドである、請求項1〜20のいずれか一項に記載の方法。
【請求項22】
前記修飾されるポリペプチドの特異性が実質的に分泌の改良前と同じに保持される、請求項1〜21のいずれか一項に記載の方法。
【請求項23】
前記修飾されるポリペプチドの比活性が実質的に分泌の改良前と同じに保持される、請求項1〜22のいずれか一項に記載の方法。
【請求項24】
前記分泌の改良が前記活性の上昇によって測定され、また前記細胞外培地中の活性が少なくとも5%上昇する、請求項1〜23のいずれか一項に記載の方法。
【請求項25】
前記ポリペプチドが酵素、膜蛋白質、ホルモンまたは受容体である、請求項1〜24のいずれか一項に記載の方法。
【請求項26】
請求項1〜25のいずれかに記載の方法を、対象ポリペプチドに適用するステップ、および組換え技術により修飾されたポリペプチドを産生するステップを含む、対象ポリペプチドを産生する方法。
【請求項27】
請求項26に記載の方法で得られるポリペプチド。
【請求項28】
請求項26に記載の方法で入手できるポリペプチド。
【請求項29】
請求項1〜25に記載の方法で得られる修飾されたポリペプチド。
【請求項30】
請求項1〜25に記載の方法で入手できる修飾されたポリペプチド。
【図1】

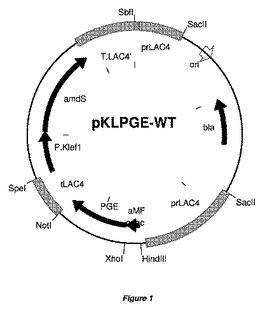
【図2】
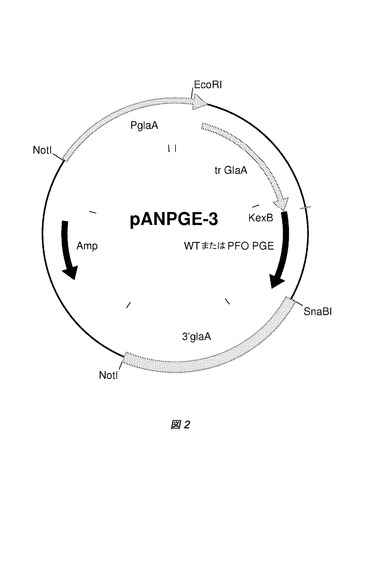
【図3】
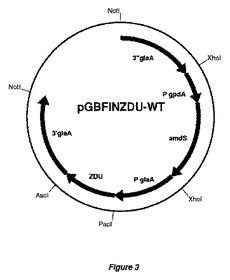
【図4】
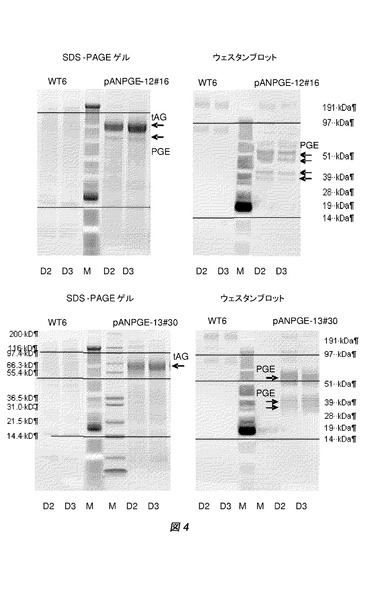
【図5】
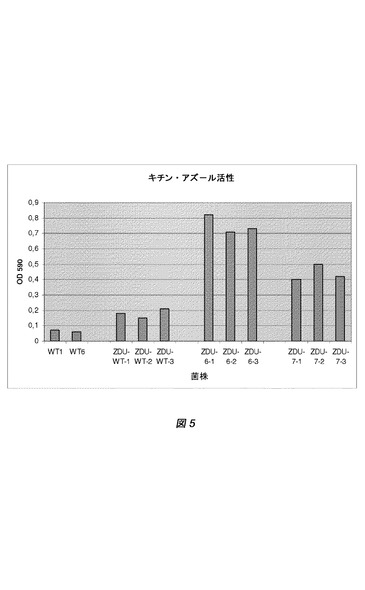
【図6】
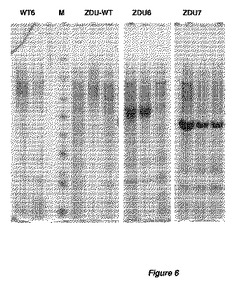
【図7】
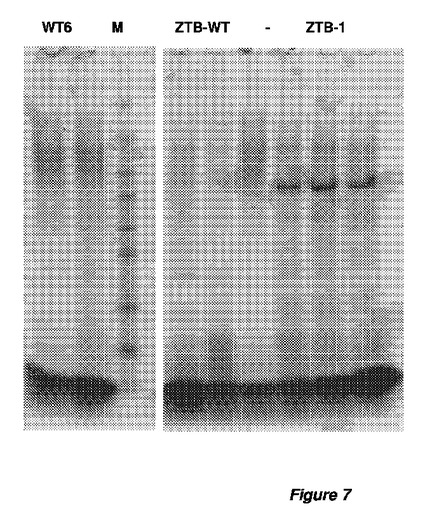
【図8】
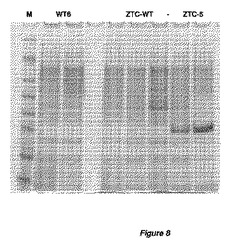
【図9】
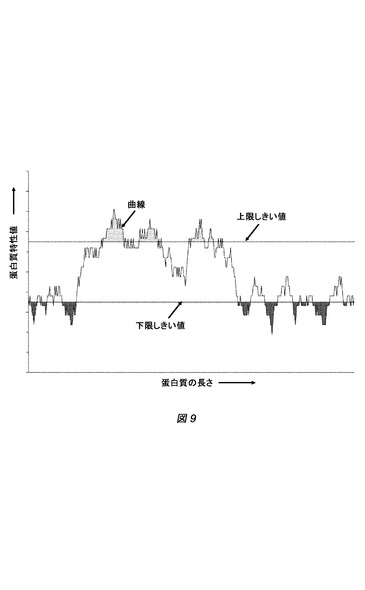
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2012−520064(P2012−520064A)
【公表日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願番号】特願2011−553416(P2011−553416)
【出願日】平成22年3月8日(2010.3.8)
【国際出願番号】PCT/EP2010/052918
【国際公開番号】WO2010/102982
【国際公開日】平成22年9月16日(2010.9.16)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
【公表日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願日】平成22年3月8日(2010.3.8)
【国際出願番号】PCT/EP2010/052918
【国際公開番号】WO2010/102982
【国際公開日】平成22年9月16日(2010.9.16)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【Fターム(参考)】
[ Back to top ]