
ポリペプチド及び組換え微生物
【課題】ポリペプチド及び組換え微生物を提供する。
【解決手段】下記(A)のアミノ酸配列が、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列のN末端側に結合されてなるポリペプチド、及び宿主微生物に当該ポリペプチドのアミノ酸配列をコードする塩基配列を導入した組換え微生物。
(A)配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列
【解決手段】下記(A)のアミノ酸配列が、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列のN末端側に結合されてなるポリペプチド、及び宿主微生物に当該ポリペプチドのアミノ酸配列をコードする塩基配列を導入した組換え微生物。
(A)配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、目的タンパク質の生産に有用なポリペプチド、当該ポリペプチドをコードする塩基配列を導入した組換え微生物、及び当該組換え微生物を用いた目的タンパク質の製造方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、食品、医薬、洗剤、化粧品等の原料、各種化成品原料に至るまで、幅広い分野で行われている。有用タンパク質やポリペプチドの工業的生産では、精製・単離の簡便さから、体内で生産した目的タンパク質等を菌体外へ分泌する能力を持った微生物がよく用いられる。また、より高い生産性を求めて、菌体外分泌・輸送能を有するシグナル配列を目的タンパク質に付加することも行われている。このようなシグナル配列は、通常、分泌タンパク質の成熟領域の上流に見られる。シグナル配列が連結された目的タンパク質は、細胞内で融合タンパク質として合成された後、細胞外へ分泌され、シグナル配列が切断されて成熟タンパクとなる。そのため、培養液から容易に目的タンパク質を精製・回収することができる。
【0003】
このような利点から、種々の微生物でシグナル配列の同定や機能解析が行われている。枯草菌では、アミラーゼをコードする遺伝子をクローニングしてシグナル配列及びプロ配列の切断点を決定し、当該アミラーゼのシグナル配列を目的タンパク質等と融合して、菌体外分泌により目的物質を生産することが試みられている(特許文献1〜3、非特許文献1、2)。例えば、非特許文献3、4、7、8では目的生産物として大腸菌由来βラクタマーゼを、非特許文献5、6ではマウス由来インターフェロンβを、特許文献4ではインターロイキン2を、非特許文献9ではヒト肝炎ウイルス由来PreS2を、それぞれ融合し、菌体外分泌による物質生産性を検討している。また、上記枯草菌アミラーゼのシグナル配列を人為的に改変して、分泌効率を向上させることも行われている(特許文献5)。
【0004】
また、分泌タンパク質の前駆体は、シグナル配列の他にプロ配列を有していることがある。プロ配列は、通常、シグナル配列と同様に成熟タンパク質領域の上流に存在し、細胞内でのタンパク質合成時には付加されているが、細胞外に分泌後には酵素により切断される。プロ配列は、アミノ酸配列の長いタイプ(通常は60〜200アミノ酸程度)と短いタイプ(通常は〜60アミノ酸程度)とに大別され、長いタイプは自身の構造のコンフォメーションを助ける働きをすることが知られているが、短いタイプの機能については未だ不明点が多い。
近年、ラクトコックス・ラクティスを宿主として、スタフィロコッカス・アウレウスのヌクレアーゼ(Nuc)を生産する際に、スタフィロコッカス・アウレウスのヌクレアーゼ(Nuc)のもつ17残基のプロ配列が、分泌性向上に有効であることが報告されている(非特許文献10)。また、スタフィロコッカス・アウレウスのヌクレアーゼ生産において、9残基の合成アミノ酸配列を付加することにより分泌性が向上することも報告されている。(非特許文献10〜13)。また、枯草菌を宿主として、ヒトインターフェロンを生産する際に、枯草菌のアミラーゼAmyEの有するプロ配列の付加が分泌量の向上に寄与することが報告されている(非特許文献14)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開昭59−135892号公報
【特許文献2】特開昭60−188070号公報
【特許文献3】特開昭61−35795号公報
【特許文献4】特開昭62−3796号公報
【特許文献5】特開平1−124385号公報
【非特許文献】
【0006】
【非特許文献1】Yamazaki H, Ohmura K, Nakayama A, Takeichi Y, Otozai K, Yamasaki M, Tamura G, Yamane K (1983) Alpha-amylase genes (amyR2 and amyE+) from an alpha-amylase-hyperproducing Bacillus subtilis strain: molecular cloning and nucleotide sequences. J Bacteriol 156:327-337.
【非特許文献2】Takase T, Mizuno H, Yamane K (1988) NH2-terminal processing of Bacillus subtilis α-amylase. J Biol Chem 263:11548-11553
【非特許文献3】Ohmura K, Shiroza T, Nakamura K, Nakayama A, Yamane K, Yoda K, Yamasaki M, Tamura G.;A Bacillus subtilis secretion vector system derived from the B. subtilis alpha-amylase promoter and signal sequence region, and secretion of Escherichia coli beta-lactamase by the vector system.,J Biochem. 1984 Jan;95(1):87-93.
【非特許文献4】Ohmura K, Nakamura K, Yamazaki H, Shiroza T, Yamane K, Jigami Y, Tanaka H, Yoda K, Yamasaki M, Tamura G., Length and structural effect of signal peptides derived from Bacillus subtilis alpha-amylase on secretion of Escherichia coli beta-lactamase in B. subtilis cells., Nucleic Acids Res. 1984 Jul 11;12(13):5307-19.
【非特許文献5】Shiroza T, Nakazawa K, Tashiro N, Yamane K, Yanagi K, Yamasaki M, Tamura G, Saito H, Kawade Y, Taniguchi T., Synthesis and secretion of biologically active mouse interferon-beta using a Bacillus subtilis alpha-amylase secretion vector., Gene. 1985;34(1):1-8.
【非特許文献6】Nakazawa K, Sasamoto H, Shiraki Y, Harada S, Yanagi K, Yamane K.(1991) Extracellular production of mouse interferon beta by the Bacillus subtilis alpha-amylase secretion vectors: antiviral activity and deduced NH2-terminal amino acid sequences of the secreted proteins. Intervirology. 32(4):216-227.
【非特許文献7】Nakamura K, Itoh Y, Yamane K.(1988) Enhanced secretion of beta-lactamase on structural modification of the Bacillus subtilis alpha-amylase signal peptide. J Biochem. 104(2):265-269.
【非特許文献8】Nakamura K, Fujita Y, Itoh Y, Yamane K. (1989) Modification of length, hydrophobic properties and electric charge of Bacillus subtilis alpha-amylase signal peptide and their different effects on the production of secretory proteins in B. subtilis and Escherichia coli cells. Mol Gen Genet. 216(1):1-9.
【非特許文献9】Honda K, Fujieda H, Ogawa K, Imai M, Yamamoto H, Ikeda T, Yamane K. (1993) Extracellular production of human hepatitis B virus preS2 antigen as hybrid proteins with Bacillus subtilis alpha-amylases in high-salt-concentration media. Appl Microbiol Biotechnol. 40(2-3):341-347.
【非特許文献10】Le Loir Y, Gruss A, Ehrlich SD, Langella P. (1998) A nine-residue synthetic propeptide enhances secretion efficiency of heterologous proteins in Lactococcus lactis.J Bacteriol. 180(7):1895-1903.
【非特許文献11】Le Loir Y, Nouaille S, Commissaire J, Bretigny L, Gruss A, Langella P.(2001) Signal peptide and propeptide optimization for heterologous protein secretion in Lactococcus lactis. Appl Environ Microbiol. 67(9):4119-4127.
【非特許文献12】Avall-Jaaskelainen S, Palva A. (2006) Secretion of biologically active porcine interleukin-2 by Lactococcus lactis. Vet Microbiol. 115(1-3):278-283.
【非特許文献13】Hazebrouck S, Pothelune L, Azevedo V, Corthier G, Wal JM, Langella P. (2007) Efficient production and secretion of bovine beta-lactoglobulin by Lactobacillus casei. Microb Cell Fact. 6:12.
【非特許文献14】Kakeshita H, Kageyama Y, Ara K, Ozaki K, Nakamura K. (2011) Propeptide of Bacillus subtilis amylase enhances extracellular production of human interferon-α in Bacillus subtilis. Appl Microbiol Biotechnol. 89(5):1509-1517.
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、優れた分泌能を有するポリペプチドを提供することを課題とする。また、本発明は当該ポリペプチドをコードする塩基配列を導入した組換え微生物を提供することを課題とする。さらに、本発明は当該組換え微生物を用いたタンパク質又はポリペプチドの製造方法を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者等は上記課題に鑑み、優れた分泌能を有するシグナル配列(シグナルペプチド)について鋭意検討を行った。その結果、配列番号1に示すアミノ酸配列中の少なくとも1位〜44位までの領域を含むアミノ酸配列が、シグナルペプチドとして機能し優れた細胞外分泌能を有すること、このアミノ酸配列の下流に目的タンパク質を結合することで、当該目的タンパク質の分泌生産性が向上することを見出した。本発明はこの知見に基づいて完成されたものである。
【0009】
すなわち、本発明の上記課題は、下記の手段により解決された。
本発明は、下記(A)のアミノ酸配列が、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列のN末端側に結合されてなるポリペプチドに関する。
(A)配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列
また、本発明は、当該ポリペプチドのアミノ酸配列をコードする塩基配列に関する。
また、本発明は、宿主微生物に、当該ポリペプチドのアミノ酸配列をコードする塩基配列を導入した組換え微生物に関する。
さらに、本発明は当該微生物を用いた目的タンパク質又はポリペプチドの製造方法、及び当該微生物を用いた目的タンパク質又はポリペプチドの生産性向上方法に関する。
なお、本発明において、上記(A)のアミノ酸配列を「ペプチド付加プレプロ配列」とも称する。
【発明の効果】
【0010】
本発明のポリペプチドは、N末端側に細胞外への分泌能に優れたアミノ酸配列領域(ペプチド付加プレプロ配列)を有し、目的タンパク質又はポリペプチドを効率よく細胞外に分泌させることができる。本発明のポリペプチドをコードする塩基配列を導入した微生物では、当該ペプチド付加プレプロ配列の働きによって目的タンパク質又はポリペプチドの分泌生産性が向上する。当該微生物を用いた目的タンパク質又はポリペプチドの製造方法は、培養物から容易に目的タンパク質等を回収することができ、製造工程の簡便化と高い生産性とが得られる。しかも、当該製造方法により生産された組換えタンパク質又はポリペプチドは、野生型と同等の活性を維持でき、各種用途に好適に用いることができる。
【図面の簡単な説明】
【0011】
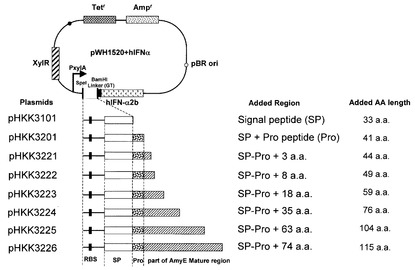
【図1】インターフェロン発現ベクターpHKK3101,pHKK3201、pHKK3221,pHKK3222,pHKK3223,pHKK3224,pHKK3225,及びpHKK3226の構成、並びに導入された枯草菌アミラーゼAmyEシグナル配列、プロ配列、及びAmyE成熟体のN末端アミノ酸配列、及びインターフェロンαの成熟体領域の融合タンパク質の構成を示した図である。併せて、各ベクターの有するAmyE成熟体のN末端アミノ酸配列の長さと、AmyE由来の導入アミノ酸配列の全長を示す。
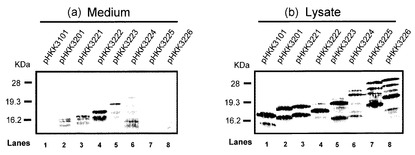
【図2】各発現ベクターをDpr8株に導入して培養し、培養物の(a)Medium(培養液上清)、(b)Lysate(ライゼート)のそれぞれについてウエスタンブロッティングを行い、インターフェロンαを検出した図である。(a)及び(b)の各レーンは、レーン1:pHKK3101、レーン2:pHKK3201、レーン3:pHKK3221、レーン4:pHKK3222、レーン5:pHKK3223、レーン6:pHKK3224、レーン7:pHKK3225、レーン8:pHKK3226導入株をそれぞれ示す。
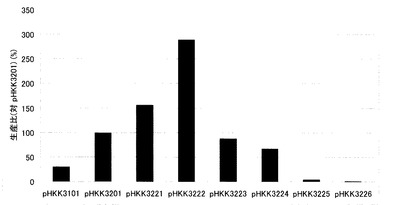
【図3】図2(a)のウエスタンブロッティングで検出されたインターフェロンαのバンドについて、バンド強度からインターフェロンαの菌体外分泌量を数値化し、グラフで示したものである。pHKK3201導入株のインターフェロンα分泌量を100として、各導入株のインターフェロンα分泌量を相対値で表した。
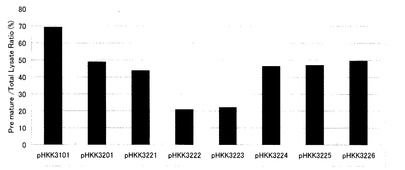
【図4】図2(b)のウエスタンブロッティングで検出された前駆体型インターフェロンと成熟型インターフェロンのバンドについて、それぞれのバンド強度から菌体内蓄積量を数値化し、菌体内の総インターフェロン生産量(前駆体型インターフェロン蓄積量と成熟型インターフェロン蓄積量の和)に対する前駆体型インターフェロン蓄積量の比率を求め、グラフ化したものである。
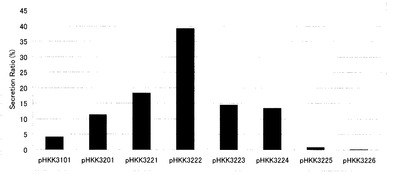
【図5】図3及び4の数値化データを基に、分泌効率(%)=インターフェロン菌体外分泌量/インターフェロン総生産量(菌体外分泌量+ライゼート中の生産量)×100として算出し、グラフ化したものである。
【発明を実施するための形態】
【0012】
本発明のポリペプチドは、配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列(ペプチド付加プレプロ配列)と、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列とを含んでなる。ペプチド付加プレプロ配列は、目的タンパク質又はポリペプチドのアミノ酸配列の上流、すなわちN末端(アミノ末端)側に結合し、目的タンパク質又はポリペプチドを細胞外へ輸送・分泌するシグナルペプチドとして機能する。
【0013】
[ペプチド付加プレプロ配列]
本発明のポリペプチドが有するペプチド付加プレプロ配列は、配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列である。配列番号1に示すアミノ酸配列は、枯草菌のα−アミラーゼAmyE由来のシグナル配列、プロ配列、及び成熟領域のN末端側の一部分からなるアミノ酸配列である。AmyEは、枯草菌体内でシグナル配列、プロ配列及び成熟領域が連結した前駆体として合成され、菌体外に分泌時にシグナル配列が、分泌後にプロ配列がそれぞれ切断されて成熟酵素となる。配列番号1において、1位〜33位のアミノ酸配列がAmyEのシグナル配列に、34位〜41位のアミノ酸配列がAmyEのプロ配列に、42位〜59位のアミノ酸配列がAmyEの成熟タンパク質領域の上流18アミノ酸に相当する。
本発明においては、AmyEのシグナル配列及びプロ配列をまとめてプレプロ配列と呼び、当該配列にAmyEの成熟領域の1部分が付加されているのでペプチド付加プレプロ配列と称した。
AmyEのシグナル配列のアミノ酸配列を配列番号2に、プロ配列のアミノ酸配列を配列番号3に、成熟タンパク質領域のアミノ酸配列を配列番号4にそれぞれ示す。また、AmyEのシグナル配列、プロ配列、及び成熟タンパク質領域のアミノ酸配列をコードする塩基配列の一例を、配列番号5に示す。
【0014】
本発明で用いるペプチド付加プレプロ配列は、少なくとも配列番号1において1位〜44位までのアミノ酸配列を含む。すなわち、AmyEのシグナル配列及びプロ配列をこの順で有し、さらに、プロ配列の後ろに成熟タンパク質領域のN末端側から少なくとも3アミノ酸以上最大で18アミノ酸までが付加されてなることを特徴とする。プロ配列の下流に成熟タンパク質のN末端側の3〜18アミノ酸を付加することで、後述の実施例で実証されているように、分泌効率が向上する。
【0015】
AmyEのシグナル配列及びプロ配列については、これを付加した融合タンパク質において菌体外への分泌性が向上することも報告されている(前記非特許文献14)。また、枯草菌α−アミラーゼのシグナル配列と成熟領域の間に人為的にアミノ酸配列を付加して生産性の効果を検証した報告がある(Itoh Y, Kanoh K, Nakamura K, Takase K, Yamane K. (1990) Artificial insertion of peptides between signal peptide and mature protein: effect on secretion and processing of hybrid thermostable alpha-amylases in Bacillus subtilis and Escherichia coli cells. J Gen Microbiol. 136(8):1551-1558.)。しかしながら、本発明者らが検討したところ、付加されているアミノ酸配列は任意に選択されたものであり、AmyEの成熟タンパク質領域のアミノ酸配列とは全く異なるものであった。
また、α−アミラーゼの成熟体領域に目的タンパク質を融合した融合タンパク質の生産性を評価した報告もある(Honda K, Fujieda H, Ogawa K, Imai M, Yamamoto H, Ikeda T, Yamane K. (1993) Extracellular production of human hepatitis B virus preS2 antigen as hybrid proteins with Bacillus subtilis alpha-amylases in high-salt-concentration media. Appl Microbiol Biotechnol. 40(2-3):341-347.)。しかしながら、付加している成熟体領域が長く本発明のペプチド付加プレプロ配列とは異なり、GSTをキャリアとして使用したタンパク質生産と同じ効果、すなわち発現量の増加や可溶化しやすくする効果を目的として付加されていると考えられ、その効果は本発明とは異なるものである。また、タンパク質生産が特殊な培養・分泌条件下で行われている点においても本発明の製造方法とは異なる。
【0016】
分泌生産性向上の観点から、ペプチド付加プレプロ配列として好ましくは、AmyEのシグナル配列及びプロ配列の下流にAmyEの成熟領域のN末端側3〜10アミノ酸を付加したアミノ酸配列(配列番号1において1位〜44位から1位〜51位までの各アミノ酸配列)であり、より好ましくはプロ配列の後ろにN末端側8〜10アミノ酸を付加したアミノ酸配列(配列番号1の1位〜49位から1位〜51位までの各アミノ酸配列)であり、さらに好ましくはプロ配列の後ろにN末端側8又は10アミノ酸を付加したアミノ酸配列(配列番号1の1位〜49位のアミノ酸配列又は1位〜51位までのアミノ酸配列)であり、特に好ましくはプロ配列の後ろにN末端側8アミノ酸を付加したアミノ酸配列(配列番号1の1位〜49位のアミノ酸配列)である。
【0017】
[目的タンパク質又はポリペプチドのアミノ酸配列]
本発明のポリペプチドは、前述のペプチド付加プレプロ配列のC末端側に、目的タンパク質又はポリペプチドのアミノ酸配列が結合してなる。本発明において、当該目的タンパク質又はポリペプチドとは、枯草菌のAmyEタンパク質以外のタンパク質又はポリペプチドである。前述のように、AmyEとは枯草菌のα−アミラーゼであり、その成熟タンパク質領域のアミノ酸配列は配列番号4で示される。
目的タンパク質又はポリペプチド(以下、目的タンパク質等ともいう)としては、枯草菌のタンパク質AmyE以外であれば特に制限されず、例えば、インターロイキン、上皮成長因子、ケモカイン、エリスロポエチン、腫瘍壊死因子、アンジオスタチン、エンドスタチン、インシュリン、インターフェロン等が挙げられる。なかでも、本発明では目的タンパク質等としてインターフェロンを用いることが好ましい。
【0018】
インターフェロン(Interferon、IFNとも略記する)は、多くの動物細胞にみられるタンパク質で、I型、II型、III型の3つのタイプに大別される。本発明の目的タンパク質等として用いられるのは、I型インターフェロンが好ましい。I型インターフェロンとしては、IFN−α(IFN−α1,2,4,5,6,7,8,10,13,14,16,17,21の13種類)、IFN−β(IFN−β1)、IFN−ω(IFN−ω1)、IFN−ε(IFN−ε1)、IFN−κ(IFN−κ)が挙げられる。I型インターフェロンのなかでも、ヒトI型インターフェロンがより好ましく、ヒトI型インターフェロン−α(hIFN−α2a、hIFN−α2bなど)及びβ(hIFN−β1、hIFN−β2bなど)がさらに好ましく、ヒトI型インターフェロン−αが特に好ましい。
上記インターフェロンのアミノ酸配列情報、及びインターフェロン遺伝子の塩基配列情報は、DDBJ、EMBL、GenBank等の公共の塩基配列データバンク等から得ることができる。一例として、ヒトインターフェロン−α2bの成熟領域のアミノ酸配列を配列番号6に、当該アミノ酸配列をコードする塩基配列の一例を、配列番号17にそれぞれ示す。
【0019】
本発明のポリペプチドに含まれる目的タンパク質等のアミノ酸配列は、当該タンパク質等の成熟領域のみでもよく、当該タンパク質の機能に影響を与えない範囲で他の配列を有していてもよく、例えば、当該アミノ酸配列の合成・調製の際や宿主微生物への導入の際などに人為的に付加した配列(例えば、リンカー配列、アダプター配列など)を適宜有していてもよい。また、成熟領域のN末端側又はC末端側にヒスチジンタグ、FLAGタグ、HAタグやmycタグといったアフィニティタグ、グルタチオン−S−トランスフェラーゼ(GST)、緑色蛍光タンパク質(GFP)、マルトース結合タンパク質(MBP)といったキャリアタンパク質の配列等を有するものであってもよい。
【0020】
[ポリペプチド]
本発明のポリペプチドは、上述したペプチド付加プレプロ配列が、目的タンパク質等のアミノ酸配列のN末端側に結合されたものである。本発明のポリペプチドにおいては、ペプチド付加プレプロ配列と目的タンパク質等とが細胞内で融合タンパク質として発現するように連結されていればよい。ペプチド付加プレプロ配列と目的タンパク質等の成熟領域のアミノ酸配列とが直接結合している場合はもちろん、前述のように、ペプチド付加プレプロ配列と目的タンパク質等のアミノ酸配列とがリンカーを介して結合していてもよいし、ペプチド付加プレプロ配列と目的タンパク質等の成熟領域との間に、制限酵素に由来するアミノ酸、各種のタグや小DNA断片に由来するアミノ酸等の配列を有していてもよい。
【0021】
本発明のポリペプチドは、配列番号1に示すアミノ酸配列情報、及び目的タンパク質のアミノ酸配列情報に基づいて、通常の遺伝子工学的手法により作製できる。具体的には、化学合成による方法、PCR反応による方法、該当配列を含むDNA断片やプラスミドより得る方法などが挙げられる。
【0022】
本発明のポリペプチドは、目的タンパク質等のアミノ酸配列のN末端側にペプチド付加プレプロ配列が結合した構造により、これを発現した微生物では菌体内でペプチド付加プレプロ配列が付加した融合タンパク質が合成され、分泌シグナルの働きにより菌体外に分泌される。ペプチド付加プレプロ配列は、優れた分泌能を有しており、目的タンパク質等を効率よく菌体外に分泌することができる。そのため、ペプチド付加プレプロ配列を結合することにより、菌体内での目的タンパク質前駆体の蓄積が減少する。さらに、菌体外に分泌されたタンパク質は、野生型と同等の生理活性を維持することができる。
【0023】
[ポリペプチドをコードする塩基配列]
本発明の塩基配列は、上述した本発明のポリペプチドのアミノ酸配列をコードするもので、ペプチド付加プレプロ配列をコードする塩基配列の下流に、目的タンパク質等をコードする塩基配列が結合した塩基配列である。上述した本発明のポリペプチドと同様、細胞内で融合タンパク質として発現するよう配列番号1のペプチド付加プレプロ配列をコードする塩基配列と目的タンパク質等コードする塩基配列とが連結されていればよく、リンカー配列等を含むものであってもよい。
ペプチド付加プレプロ配列をコードする塩基配列としては、例えば、配列番号5の塩基配列において1位〜132位から1位〜177位までの領域の各塩基配列が挙げられる。目的タンパク質等をコードする塩基配列として、例えば、配列番号17(hIFN−α2b)の塩基配列が挙げられる。
本発明の塩基配列は、配列番号1のアミノ酸配列をコードする塩基配列、目的タンパク質等をコードする塩基配列の情報をもとに、通常の手法により調製することができる。例えば、塩基配列情報をもとに適当なプライマーを設計し、枯草菌の染色体DNAや目的タンパク質等の遺伝子を有する生物の染色体DNAをそれぞれ鋳型として、PCR反応により合成することができる。また、塩基配列情報をもとに化学合成することもできる。
【0024】
[組換え微生物]
本発明の組換え微生物は、上述した本発明のポリペプチドのアミノ酸配列をコードする塩基配列(遺伝子)を、宿主微生物に導入したものである。
宿主微生物としては、グラム陽性菌、グラム陰性菌のいずれを用いてもよいが、タンパク質を菌体外に分泌生産させる能力を有する点からグラム陽性菌が好ましい。グラム陽性菌の中でもバチルス(Bacillus)属細菌がより好ましく、具体的には、枯草菌(Bacillus subtilis)、Bacillus amyloliquefaciens、Bacillus licheniformis、Bacillus stearothermophilus、Bacillus thuringiensis、Bacillus subtilis nattoなどが好ましく用いられる。特に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点から、枯草菌を宿主とすることが好ましい。
【0025】
宿主に導入する塩基配列は、上述した本発明のポリペプチドのアミノ酸配列をコードする塩基配列であり、その調製方法も前述のとおりである。
本発明の組換え微生物には、本発明のポリペプチド配列をコードする塩基配列の他に、発現制御のために有用な塩基配列が適宜導入されていてもよい。例えば、転写、翻訳に関わる制御領域として、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位(SD(Shine-Dalgarno)配列等)及び開始コドンを含む翻訳開始領域、ターミネーター等を、本発明のポリペプチドをコードする塩基配列と適正な形で結合し導入してもよい。また、選択マーカー等を導入してもよい。
【0026】
宿主微生物への塩基配列の導入は、通常の組換えDNA技術を用いて行うことができる。例えば、ポリペプチド配列をコードする塩基配列をベクターに組み込んで、一般的な形質転換法を用いて宿主微生物細胞に取り込ませて導入してもよいし、塩基配列に宿主微生物ゲノムの相同領域を結合したDNA断片を用いて、相同組換えにより宿主微生物ゲノムに直接組み込んでもよい。
【0027】
ベクターを用いる場合、プラスミド等の染色体外で自立増殖・複製するベクターであってもよいし、染色体内に組み込まれるベクターであってもよい。例えば、導入する宿主の種類に応じたプロモーターやターミネーター等の発現調節領域を有する発現ベクターであって、複製開始点や選択マーカー等を有するベクターを用いることができる。具体的には、pHY300、pWH1520、pUB110、pC194、pT127、pE194等のベクターを挙げることができる。
プロモーターやターミネーター、選択マーカー等の種類も特に限定されず、通常使用されるものを適宜選択して用いることができる。プロモーターとしては、例えば、tacプロモーター、lacプロモーター、spacプロモーター、xylプロモーター、T7プロモーター等が用いられる。選択マーカーとしては、クロラムフェニコール、テトラサイクリン、ネオマイシン、カナマイシン、エリスロマイシン等の薬剤耐性遺伝子が挙げられる。
目的塩基配列のベクターへの組み込みは、制限酵素処理やライゲーション等の通常の手法により行うことができる。
【0028】
次いで、構築した組換えベクターを用いて宿主を形質転換し、本発明のポリペプチドをコードする塩基配列が導入された組換え微生物を作製する。形質転換方法は特に限定されず、カルシウムイオンを用いる方法、一般的なコンピテントセル形質転換方法(J.Bacterial.93,1925(1967))、プロトプラスト形質転換法(Mol.Gen.Genet.168,111(1979))、エレクトロポレーション法(FEMS Microbiol.Lett.55,135(1990))、LP形質転換方法(T.Akamatsu及びJ.Sekiguchi,Archives of Microbiology,1987,146,p.353-357;T.Akamatsu及びH.Taguchi,Bioscience,Biotechnology,and Biochemistry,2001,65,4,p.823-829)等通常の方法を用いることができる。
目的の塩基配列が導入されたことの確認は、選択マーカー等を利用することで行うことができる。また、ゲノムを鋳型としたPCR法等によって、目的DNA断片の導入を確認することもできる。
【0029】
[目的タンパク質等の製造]
得られた組換え微生物を用いて、目的タンパク質等を製造する。例えば、組換え微生物を適切な培地を用いて適切な条件下にて培養して、目的タンパク質を産生させ、培養物から目的タンパク質を精製・回収すればよい。本発明では、菌体内で合成された目的タンパク質等が効率よく菌体外に分泌されるので、培養液から目的タンパク質等を回収することができ、精製・単離作業を容易に行うことができる。
【0030】
組換え微生物の培養条件は、宿主の種類に応じて適宜好ましい培養条件を採用することができる。また、用いるベクターやプロモーターに応じて、組換えタンパク質の発現誘導を適宜行ってもよい。例えば、宿主として枯草菌を用いた場合は、2×L培地(実施例参照)を用い、30℃前後で培養を行えばよい。
培養物から目的タンパク質等を精製・単離する方法は特に限定されず、例えば、カラム精製、硫安沈殿、組換えタンパクにタグを付加した場合はタグを用いた精製、透析、フィルター精製等の方法で行うことができる。
【実施例】
【0031】
以下、本発明を実施例に基づきさらに詳細に説明するが、本発明はこれに限定されるものではない。
下記の手法によって、本発明のペプチド付加プレプロ配列及び目的タンパク質をコードする塩基配列を導入した組換え微生物を構築し、目的タンパク質の発現を行った。目的タンパク質として、ヒトインターフェロンα(hIFN‐α)を用いた。
【0032】
[使用菌株、プラスミドベクター及び培地]
(1)使用菌株
大腸菌(エシェリヒア・コリ)C600株(タカラバイオ)
大腸菌(エシェリヒア・コリ)JM109株(タカラバイオ)
枯草菌168株
枯草菌Dpr8株(8種類の分泌型プロテアーゼを欠損する枯草菌変異株(Δepr/ΔwprA/Δmpr/ΔnprB/Δbpr/ΔnprE/Δvpr/ΔaprE);Kodamaら, "Effect of Bacillus subtilis spo0A mutation on cell wall lytic enzymes and extracellular proteases, and prevention of cell lysis", J. Biosci. Bioeng., 2007, 103, p.13-21参照)
(2)プラスミド
pWH1520;耐性マーカー(アンピシリン、テトラサイクリン)、MoBiTec社製
pHKK3100;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列)
pHKK3101;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列、及びhIFN‐α2bの成熟体領域の配列)
pHKK3200;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列及びプロ配列)
pHKK3201;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列及びプロ配列、及びhIFN‐α2bの成熟体領域の配列)
(3)培地
L培地;バクトトリプトン(Difco)1%、イーストエクスラクト(Difco)0.5%、NaCl(和光)0.5%
2×L培地;バクトトリプトン(Difco)2%、イーストエクスラクト(Difco)1%、NaCl(和光)1%
【0033】
1.異種タンパク質hIFN−α発現用ベクターの構築
枯草菌アミラーゼAmyEのシグナル配列(配列番号2)及びプロ配列(配列番号3)をコードするDNA配列を組み込んだ基本ベクターpHKK3200を、特開2008-271956号公報の実施例2の記載に従って構築した。枯草菌168株の染色体を鋳型に、プライマーamyESF−1(ggccACTAGTcttcaaaaaatcaaa(配列番号7)、大文字がSpeI)及びamyESR−2(ggccGGTACCctcattcgatttgttcgc(配列番号8)、大文字がKpnI)を用いてPCR法で増幅したamyE遺伝子のSD配列、シグナル配列及びプロ配列を、pWH1520のマルチクローニングサイトのSpeI−KpnIサイトに導入し、pHKK3200を構築した(Kakeshita et al. 2011 Appl Microbiol Biotechnol 89:1509−1517参照)。
次いで、pORF5−hIFNA2b(インビボジェン製、hIFN−α2b)を鋳型に、プライマーifnaF(ggccGGTACCtgtgatctgcctcaaacc(配列番号9)、大文字がKpnI)及びifnaR(ggccGGATCCttttcattccttacttct(配列番号10)、大文字がBamHI)を用いてPCR法で増幅したインターフェロンα遺伝子の成熟体領域(配列番号17)を、pHKK3200のKpnI−BamHIサイトに導入し、インターフェロンα発現用ベクターpHKK3201を構築した(図1)。
同様の手法で、枯草菌アミラーゼAmyEのシグナル配列をコードするDNA配列を組み込んだ基本ベクターpHKK3100を構築した。pHKK3100に、上記と同様の方法でインターフェロンα遺伝子の成熟体領域を組み込み、インターフェロンα発現用ベクターpHKK3101を作成した(図1)(Kakeshita et al. 2011 Appl Microbiol Biotechnol 89:1509-1517参照)。
【0034】
次に、枯草菌アミラーゼAmyEのシグナル配列、プロ配列、及び成熟領域のN末端側のアミノ酸配列をコードするDNA配列を組み込んだインターフェロンα発現用ベクターpHKK3221〜pHKK3226を構築した。枯草菌168株の染色体を鋳型に、プライマーamyESF−1(ggccACTAGTcttcaaaaaatcaaa(配列番号7)、大文字がSpeI)と、amyES2−5R(ggccGGTACCtgctgtaagctcattcgatttg(配列番号11)、大文字がKpnI)、amyES3R(gaagaatGGTACCgcttttgatcgacgg(配列番号12)、大文字がKpnI)、amyES4R(ggccGGTACCcgaccaattccatgcatgaa(配列番号13)、大文字がKpnI)、amyES5R(ggccGGTACCtgtatatcctgcatcatgaa(配列番号14)、大文字がKpnI)、amyES6R(ggccGGTACCctgatacagccagtaccag(配列番号15)、大文字がKpnI)、又はamyES7R(aaattcttgttcGGTACCtaagtaacgg(配列番号16)、大文字がKpnI)を用いてPCR法で増幅した各PCR産物を、制限酵素SpeI及びKpnIで切断した後、上記で構築したpHKK3201のSpeI−KpnIサイトに導入し、pHKK3221(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側3アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3222(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側8アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3223(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側18アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3224(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側35アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3225(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側63アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、及びpHKK3226(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側74アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)をそれぞれ構築した(図1)。各プラスミドベクターは、AmyE成熟領域をコードする配列とIFNα成熟体領域をコードする配列との間にリンカー配列(Gly-Thr配列)を有する。
なお、上記ベクター構築時の宿主には、大腸菌(エシェリヒア・コリ)C600株及びJM109株(ともにタカラバイオ)を使用した。プラスミド構築時の大腸菌の培養及び枯草菌の前培養時には、L培地を用いた。
【0035】
2.異種タンパク質発現ベクター導入株の作製
上記1.で作成した各インターフェロンα発現ベクターを、形質転換法によって枯草菌変異株Dpr8株を形質転換して、インターフェロンα発現ベクター導入株を得た。形質転換は、SPI培地及びSPII培地を用いてコンピテント法により行い、15μg/mLテトラサイクリンを含むLB寒天培地に生育した菌株を形質転換体として選択した。
【0036】
3.分泌タンパク質の生産性評価
得られた各インターフェロンα発現ベクター導入株について、インターフェロンの生産性をウエスタンブロッティング解析、ELISA解析、及び生理活性測定により評価した。
導入株の培養には、2×L培地を用いた。抗生物質は、テトラサイクリン15μg/mLの濃度で適時使用した。pWH1520のもつキシロースプロモーターの誘導のため、インターフェロンα導入株には、キシロース終濃度0.6%を添加した。培養は、特開2008-271956号公報の実施例2に記載の方法に従い、前培養液を2%植菌し、30℃にて細胞増殖期中期(OD660=0.3)まで培養後、キシロースを最終濃度0.6%添加し、培養した。培養開始後24時間後で集菌し、培養液を得た。
【0037】
(1)ウエスタンブロッティング
得られた培養液から、下記「Medium」及び「Lysate」をそれぞれ調製し、ウエスタンブロッティング用サンプルとして用いた。
「Medium」:培養液を遠心して集菌し、得られた上清液をmediumとした。SDS-PAGEにて展開する際には、等量のサンプルバッファを加えて加熱処理してサンプルとした。
「Lysate」:培養液を遠心して集菌したもの(ペレットの状態)をリゾチーム処理後、バッファ(Tris-HCL)に懸濁し、等量のサンプルバッファを加えて加熱処理した。その後、遠心して得られた上清をサンプルとした。
【0038】
ウエスタンブロッティングは、セミドライシステム(バイオラッド)を用いて行った。SDS−PAGEによるタンパク質分離後、タンパク質をイモビロンPVDF膜(ミリポア)に転写した。タンパク質の検出は、イムノスターLD検出システム(商品名、和光純薬工業株式会社製)を用いた。抗インターフェロンα抗体は、(PeproTech EC LTD)のものを、HRP標識二次抗体は、GEヘルスケアのものを使用して検出した。
インターフェロンの分泌量及び細胞内蓄積量の測定は、ウエスタンブロッティング法により検出したバンドの濃淡を、ImageJ(National Institutes of Health, USA)により数値化することで行った。
【0039】
(1−1)インターフェロン分泌量の測定
各導入株のMediumサンプルを用いて、菌体外に分泌されたインターフェロンをウエスタンブロッティング法により検出した(図2(a))。
次いで、得られたインターフェロンのバンドの濃淡を、ImageJにより数値化し、各導入株のインターフェロン分泌量とした(図3)。なお、コントロールにはシグナル配列とプロ配列を持つpHKK3201導入株を用い、pHKK3201導入株での分泌量を100として、各導入株における分泌量を相対値(%)で示した。
【0040】
図3から明らかなように、シグナル配列とプロ配列に成熟領域の3アミノ酸残基を付加したpHKK3221導入株、及び8アミノ酸残基を付加したpHKK3222導入株では、コントロールのpHKK3201導入株に比べインターフェロン分泌量(生産量)が大きく上昇した。また、シグナル配列とプロ配列に18アミノ酸残基を付加したpHKK3223導入株でも、コントロールpHKK3201導入株と同程度の生産性があった。特に、8アミノ酸残基を付加したpHKK3222導入株では、インターフェロンの生産量が最大となり、シグナル配列のみのpHKK3101導入株と比べて9.5倍、シグナル配列とプロ配列を有するpHKK3201導入株と比べて2.9倍の生産性向上が確認された。
また、35アミノ酸残基を付加したpHKK3224導入株は、シグナル配列のみのpHKK3101導入株と比べて生産性は向上しているものの、シグナル配列とプロ配列を有するpHKK3201導入株に比べると多少生産性が低下した。63アミノ酸残基以上を付加した導入株では、インターフェロンの分泌はほとんど確認されなかった。
【0041】
(1−2)菌体内のインターフェロン蓄積量の測定
各導入株のLysateサンプルを用いて、菌体内のインターフェロンをウエスタンブロッティング法により検出した(図2(b))。
その結果、各サンプルにおいて、菌体内に前駆体型インターフェロンと成熟型インターフェロンの両者が確認された。なお、前駆体型インターフェロンとは、シグナル配列が切断されていない状態であり、成熟型インターフェロンとは、付加したアミノ酸配列からシグナル配列が切断された状態である。
菌体外へ分泌されるタンパク質は、シグナル配列(シグナルペプチド)をアミノ末端側に持ち、これは菌体外へ分泌される際にシグナルペプチダーゼよって切断される。細胞内への前駆体の蓄積は、シグナルペプチドが切断されておらず、菌体外への正常な分泌ができていないことを示している。宿主自体が持つ分泌タンパク質が菌体内に蓄積する現象は、分泌輸送経路構成因子の欠損や変異による機能不全があった場合に見られるもので、通常は観察されない。異種タンパク質発現においても、菌体内に前駆体タンパク質が蓄積していることは、分泌効率が悪いことを示している。異種タンパク質の細胞内蓄積は宿主への負荷となるため、効率的な異種タンパク質生産という点からは、蓄積が少ない状態が好ましい。
【0042】
次いで、得られた前駆体型インターフェロンと成熟型インターフェロンのバンドについて、濃淡をImageJにより数値化し、菌体内の前駆体型インターフェロン蓄積量及び成熟型インターフェロン蓄積量を算出した。これらをもとに、菌体内の総インターフェロン生産量(前駆体型インターフェロン蓄積量と成熟型インターフェロン蓄積量の和)に対する前駆体型インターフェロン蓄積量の比率を求め、前駆体型インターフェロン蓄積率(%)とした(図4)。
【0043】
図4から明らかなように、シグナル配列とプロ配列に3アミノ酸残基を付加したpHKK3221導入株、8アミノ酸残基を付加したpHKK3222導入株、18アミノ酸残基を付加したpHKK3223導入株では、シグナル配列とプロ配列を有するpHKK3201導入株に比べて、前駆体型インターフェロンの蓄積が減少していた。前駆体の蓄積減少は、特にpHKK3222導入株及びpHKK3223導入株で顕著であった。
この結果から、菌体内での前駆体の蓄積解消が、インターフェロン生産量及び分泌効率の向上につながっている可能性が高いと考えられる。
【0044】
(1−3)インターフェロン分泌効率
前記(1−1)で得られた培養液中に分泌されたインターフェロン量、及び(1−2)で得られたライゼート中のインターフェロン総量(前駆体蓄積量+成熟体蓄積量)から、各導入株における分泌効率を下記式により求めた。結果を図5に示す。
分泌効率(%)=培地中に分泌されたインターフェロン量/インターフェロン総生産量 インターフェロン総生産量=培地中に分泌されたインターフェロン量+ライゼート中のインターフェロン総生産量(前駆体+成熟体)
【0045】
図5から明らかなように、シグナル配列とプロ配列を有するpHKK3201導入株に比べて、pHKK3221導入株、pHKK3222導入株、pHKK3223導入株では分泌効率が向上していた。特に、シグナル配列とプロ配列に8アミノ酸残基を付加したpHKK3222導入株では分泌効率が約4倍と飛躍的に向上した。
【0046】
(2)ELISA法による菌体外分泌されたインターフェロン量の測定
上記(1)ウエスタンブロッティングに用いた「Medium」サンプルと同様の調製手順で、pHKK3201導入株及びpHKK3222導入株の培養液を遠心して集菌し、得られた上清液をELISA用サンプルとした。測定には、IFN-α Human ELISAキット(R&Dsystems,Inc)を使用した。測定実験は、鎌倉テクノサイエンスに委託して行われた。
ELISA法により算出された培養液上清中に含まれるインターフェロンの総量を、表1の「インターフェロン総量」欄に示した。
【0047】
(3)インターフェロン活性測定
前記(1−1)でインターフェロン生産量が最大であったpHKK3222を用いて、シグナル配列とプロ配列に8アミノ酸残基が付加された組換えインターフェロンの生理活性を測定した。測定実験は、鎌倉テクノサイエンスに委託して行った。
上記(2)のELISA法と同様の手順で、pHKK3201導入株及びpHKK3222導入株の培養上清液を調製して、測定試料とした。なお、バックグラウンドとして、インターフェロンをコードする遺伝子を有しないプラスミドを導入した株を用いて測定し、100IU/mL以下であることを確認している。
生理活性測定は、FL細胞に対するSindbisウイルスの感染を指標として計測した。標準品には、インターフェロンA注射用(hIFNα2b、シェリング・プラウ株式会社(現 MSD株式会社)製)を使用した。単位は、国際単位(IU)を使用した。
インターフェロンの生理活性は、インターフェロンに接触させたFL細胞にシンドビスウイルスを感染させることによる抗ウイルス作用を指標とした生理活性測定法を用いた。無血清培地A(5%ウシ胎児血清、10mM HEPES緩衝液)で4×105cells/mLの細胞液を調製し、マイクロプレートに10μL/well播種した。炭酸ガスインキュベーター(5%CO2、37℃)内で20〜28時間培養後、2倍段階希釈した標準品または測定試料5μL/wellを添加した。炭酸ガスインキュベーター内でさらに20〜28時間培養した後、マイクロプレート内の培養液を除去し、培地B 100μL/wellで1回洗浄後除去した。各試料に、無血清培地B(0.5%BSA、20mM HEPES緩衝液)で適宜希釈したウイルス液100μL/wellを添加し、マイクロプレートをインキュベーター内(37℃)で18〜28時間培養した。マイクロプレート内のウイルス液を廃棄し、染色固定液(0.05w/v%クリスタルバイオレット、10v/v%エタノールを含む中性ホルマリン液)に1〜2時間浸し、水洗後、乾燥した。
得られたサンプルを、マイクロプレートリーダーを用いて吸光度(波長595nm)を測定した。Cell Controlの吸光度を100%、Virus Controlの吸光度を0%とし、各wellの吸光度%を算出した。吸光度50%の希釈倍数を力価とし、標準品(インターフェロンA注射用300(IFN−α2b)、市販品)の力価により測定試料の力価を補正し、試料のIFN力価を相対的に算出した。
上記測定により算出された培養上清液サンプル中の総インターフェロン活性を、表1の「総インターフェロン活性」欄に示した。
【0048】
次いで、この培養上清中の総インターフェロン活性と、上記(2)ELISA法により算出された培養液上清中に含まれるインターフェロン総量とから、分泌された組換えインターフェロンの比活性を算出した。比活性=総インターフェロン活性/インターフェロン総量 として算出し、結果は表1の「比活性」欄に示した。当該比活性は、同タンパク質量における組換えインターフェロンタンパク質の活性能を示している。
【0049】
【表1】

【0050】
表1から明らかなように、「比活性」の値はpHKK3201導入株を用いた系とpHKK3222導入株を用いた系とで同程度であった。このことから、pHKK3222導入株(シグナル配列+プロ配列+8アミノ酸残基を付加)により菌体外に産出された組換えインターフェロンは、pHKK3201導入株(シグナル配列+プロ配列を付加)により菌体外に産出された組換えインターフェロンと同等の生理活性を有することが確認された。なお、本発明者らは以前に、シグナル配列+インターフェロン導入株(本発明のpHKK3101株に相当、菌体外分泌時に当該シグナル配列は切断済み)及びシグナル配列+プロ配列+インターフェロン導入株(本発明のpHKK3201導入株に相当)を用いて、上記(2)及び(3)と同様のインターフェロン活性測定を行っており、両導入株により菌体外に産出された組換えインターフェロンが、野生型と同等の生理活性を有することを既に確認済みである(前記非特許文献14参照)。これらの結果から、pHKK3222導入株(シグナル配列+プロ配列+8アミノ酸残基を付加)により菌体外に産出された組換えインターフェロンは、野生型と同等の生理活性を保持しており、成熟領域の8アミノ酸残基を付加したことによる機能的な悪影響はないことが確認された。
【技術分野】
【0001】
本発明は、目的タンパク質の生産に有用なポリペプチド、当該ポリペプチドをコードする塩基配列を導入した組換え微生物、及び当該組換え微生物を用いた目的タンパク質の製造方法に関する。
【背景技術】
【0002】
微生物による有用物質の工業的生産は、食品、医薬、洗剤、化粧品等の原料、各種化成品原料に至るまで、幅広い分野で行われている。有用タンパク質やポリペプチドの工業的生産では、精製・単離の簡便さから、体内で生産した目的タンパク質等を菌体外へ分泌する能力を持った微生物がよく用いられる。また、より高い生産性を求めて、菌体外分泌・輸送能を有するシグナル配列を目的タンパク質に付加することも行われている。このようなシグナル配列は、通常、分泌タンパク質の成熟領域の上流に見られる。シグナル配列が連結された目的タンパク質は、細胞内で融合タンパク質として合成された後、細胞外へ分泌され、シグナル配列が切断されて成熟タンパクとなる。そのため、培養液から容易に目的タンパク質を精製・回収することができる。
【0003】
このような利点から、種々の微生物でシグナル配列の同定や機能解析が行われている。枯草菌では、アミラーゼをコードする遺伝子をクローニングしてシグナル配列及びプロ配列の切断点を決定し、当該アミラーゼのシグナル配列を目的タンパク質等と融合して、菌体外分泌により目的物質を生産することが試みられている(特許文献1〜3、非特許文献1、2)。例えば、非特許文献3、4、7、8では目的生産物として大腸菌由来βラクタマーゼを、非特許文献5、6ではマウス由来インターフェロンβを、特許文献4ではインターロイキン2を、非特許文献9ではヒト肝炎ウイルス由来PreS2を、それぞれ融合し、菌体外分泌による物質生産性を検討している。また、上記枯草菌アミラーゼのシグナル配列を人為的に改変して、分泌効率を向上させることも行われている(特許文献5)。
【0004】
また、分泌タンパク質の前駆体は、シグナル配列の他にプロ配列を有していることがある。プロ配列は、通常、シグナル配列と同様に成熟タンパク質領域の上流に存在し、細胞内でのタンパク質合成時には付加されているが、細胞外に分泌後には酵素により切断される。プロ配列は、アミノ酸配列の長いタイプ(通常は60〜200アミノ酸程度)と短いタイプ(通常は〜60アミノ酸程度)とに大別され、長いタイプは自身の構造のコンフォメーションを助ける働きをすることが知られているが、短いタイプの機能については未だ不明点が多い。
近年、ラクトコックス・ラクティスを宿主として、スタフィロコッカス・アウレウスのヌクレアーゼ(Nuc)を生産する際に、スタフィロコッカス・アウレウスのヌクレアーゼ(Nuc)のもつ17残基のプロ配列が、分泌性向上に有効であることが報告されている(非特許文献10)。また、スタフィロコッカス・アウレウスのヌクレアーゼ生産において、9残基の合成アミノ酸配列を付加することにより分泌性が向上することも報告されている。(非特許文献10〜13)。また、枯草菌を宿主として、ヒトインターフェロンを生産する際に、枯草菌のアミラーゼAmyEの有するプロ配列の付加が分泌量の向上に寄与することが報告されている(非特許文献14)。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開昭59−135892号公報
【特許文献2】特開昭60−188070号公報
【特許文献3】特開昭61−35795号公報
【特許文献4】特開昭62−3796号公報
【特許文献5】特開平1−124385号公報
【非特許文献】
【0006】
【非特許文献1】Yamazaki H, Ohmura K, Nakayama A, Takeichi Y, Otozai K, Yamasaki M, Tamura G, Yamane K (1983) Alpha-amylase genes (amyR2 and amyE+) from an alpha-amylase-hyperproducing Bacillus subtilis strain: molecular cloning and nucleotide sequences. J Bacteriol 156:327-337.
【非特許文献2】Takase T, Mizuno H, Yamane K (1988) NH2-terminal processing of Bacillus subtilis α-amylase. J Biol Chem 263:11548-11553
【非特許文献3】Ohmura K, Shiroza T, Nakamura K, Nakayama A, Yamane K, Yoda K, Yamasaki M, Tamura G.;A Bacillus subtilis secretion vector system derived from the B. subtilis alpha-amylase promoter and signal sequence region, and secretion of Escherichia coli beta-lactamase by the vector system.,J Biochem. 1984 Jan;95(1):87-93.
【非特許文献4】Ohmura K, Nakamura K, Yamazaki H, Shiroza T, Yamane K, Jigami Y, Tanaka H, Yoda K, Yamasaki M, Tamura G., Length and structural effect of signal peptides derived from Bacillus subtilis alpha-amylase on secretion of Escherichia coli beta-lactamase in B. subtilis cells., Nucleic Acids Res. 1984 Jul 11;12(13):5307-19.
【非特許文献5】Shiroza T, Nakazawa K, Tashiro N, Yamane K, Yanagi K, Yamasaki M, Tamura G, Saito H, Kawade Y, Taniguchi T., Synthesis and secretion of biologically active mouse interferon-beta using a Bacillus subtilis alpha-amylase secretion vector., Gene. 1985;34(1):1-8.
【非特許文献6】Nakazawa K, Sasamoto H, Shiraki Y, Harada S, Yanagi K, Yamane K.(1991) Extracellular production of mouse interferon beta by the Bacillus subtilis alpha-amylase secretion vectors: antiviral activity and deduced NH2-terminal amino acid sequences of the secreted proteins. Intervirology. 32(4):216-227.
【非特許文献7】Nakamura K, Itoh Y, Yamane K.(1988) Enhanced secretion of beta-lactamase on structural modification of the Bacillus subtilis alpha-amylase signal peptide. J Biochem. 104(2):265-269.
【非特許文献8】Nakamura K, Fujita Y, Itoh Y, Yamane K. (1989) Modification of length, hydrophobic properties and electric charge of Bacillus subtilis alpha-amylase signal peptide and their different effects on the production of secretory proteins in B. subtilis and Escherichia coli cells. Mol Gen Genet. 216(1):1-9.
【非特許文献9】Honda K, Fujieda H, Ogawa K, Imai M, Yamamoto H, Ikeda T, Yamane K. (1993) Extracellular production of human hepatitis B virus preS2 antigen as hybrid proteins with Bacillus subtilis alpha-amylases in high-salt-concentration media. Appl Microbiol Biotechnol. 40(2-3):341-347.
【非特許文献10】Le Loir Y, Gruss A, Ehrlich SD, Langella P. (1998) A nine-residue synthetic propeptide enhances secretion efficiency of heterologous proteins in Lactococcus lactis.J Bacteriol. 180(7):1895-1903.
【非特許文献11】Le Loir Y, Nouaille S, Commissaire J, Bretigny L, Gruss A, Langella P.(2001) Signal peptide and propeptide optimization for heterologous protein secretion in Lactococcus lactis. Appl Environ Microbiol. 67(9):4119-4127.
【非特許文献12】Avall-Jaaskelainen S, Palva A. (2006) Secretion of biologically active porcine interleukin-2 by Lactococcus lactis. Vet Microbiol. 115(1-3):278-283.
【非特許文献13】Hazebrouck S, Pothelune L, Azevedo V, Corthier G, Wal JM, Langella P. (2007) Efficient production and secretion of bovine beta-lactoglobulin by Lactobacillus casei. Microb Cell Fact. 6:12.
【非特許文献14】Kakeshita H, Kageyama Y, Ara K, Ozaki K, Nakamura K. (2011) Propeptide of Bacillus subtilis amylase enhances extracellular production of human interferon-α in Bacillus subtilis. Appl Microbiol Biotechnol. 89(5):1509-1517.
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、優れた分泌能を有するポリペプチドを提供することを課題とする。また、本発明は当該ポリペプチドをコードする塩基配列を導入した組換え微生物を提供することを課題とする。さらに、本発明は当該組換え微生物を用いたタンパク質又はポリペプチドの製造方法を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者等は上記課題に鑑み、優れた分泌能を有するシグナル配列(シグナルペプチド)について鋭意検討を行った。その結果、配列番号1に示すアミノ酸配列中の少なくとも1位〜44位までの領域を含むアミノ酸配列が、シグナルペプチドとして機能し優れた細胞外分泌能を有すること、このアミノ酸配列の下流に目的タンパク質を結合することで、当該目的タンパク質の分泌生産性が向上することを見出した。本発明はこの知見に基づいて完成されたものである。
【0009】
すなわち、本発明の上記課題は、下記の手段により解決された。
本発明は、下記(A)のアミノ酸配列が、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列のN末端側に結合されてなるポリペプチドに関する。
(A)配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列
また、本発明は、当該ポリペプチドのアミノ酸配列をコードする塩基配列に関する。
また、本発明は、宿主微生物に、当該ポリペプチドのアミノ酸配列をコードする塩基配列を導入した組換え微生物に関する。
さらに、本発明は当該微生物を用いた目的タンパク質又はポリペプチドの製造方法、及び当該微生物を用いた目的タンパク質又はポリペプチドの生産性向上方法に関する。
なお、本発明において、上記(A)のアミノ酸配列を「ペプチド付加プレプロ配列」とも称する。
【発明の効果】
【0010】
本発明のポリペプチドは、N末端側に細胞外への分泌能に優れたアミノ酸配列領域(ペプチド付加プレプロ配列)を有し、目的タンパク質又はポリペプチドを効率よく細胞外に分泌させることができる。本発明のポリペプチドをコードする塩基配列を導入した微生物では、当該ペプチド付加プレプロ配列の働きによって目的タンパク質又はポリペプチドの分泌生産性が向上する。当該微生物を用いた目的タンパク質又はポリペプチドの製造方法は、培養物から容易に目的タンパク質等を回収することができ、製造工程の簡便化と高い生産性とが得られる。しかも、当該製造方法により生産された組換えタンパク質又はポリペプチドは、野生型と同等の活性を維持でき、各種用途に好適に用いることができる。
【図面の簡単な説明】
【0011】
【図1】インターフェロン発現ベクターpHKK3101,pHKK3201、pHKK3221,pHKK3222,pHKK3223,pHKK3224,pHKK3225,及びpHKK3226の構成、並びに導入された枯草菌アミラーゼAmyEシグナル配列、プロ配列、及びAmyE成熟体のN末端アミノ酸配列、及びインターフェロンαの成熟体領域の融合タンパク質の構成を示した図である。併せて、各ベクターの有するAmyE成熟体のN末端アミノ酸配列の長さと、AmyE由来の導入アミノ酸配列の全長を示す。
【図2】各発現ベクターをDpr8株に導入して培養し、培養物の(a)Medium(培養液上清)、(b)Lysate(ライゼート)のそれぞれについてウエスタンブロッティングを行い、インターフェロンαを検出した図である。(a)及び(b)の各レーンは、レーン1:pHKK3101、レーン2:pHKK3201、レーン3:pHKK3221、レーン4:pHKK3222、レーン5:pHKK3223、レーン6:pHKK3224、レーン7:pHKK3225、レーン8:pHKK3226導入株をそれぞれ示す。
【図3】図2(a)のウエスタンブロッティングで検出されたインターフェロンαのバンドについて、バンド強度からインターフェロンαの菌体外分泌量を数値化し、グラフで示したものである。pHKK3201導入株のインターフェロンα分泌量を100として、各導入株のインターフェロンα分泌量を相対値で表した。
【図4】図2(b)のウエスタンブロッティングで検出された前駆体型インターフェロンと成熟型インターフェロンのバンドについて、それぞれのバンド強度から菌体内蓄積量を数値化し、菌体内の総インターフェロン生産量(前駆体型インターフェロン蓄積量と成熟型インターフェロン蓄積量の和)に対する前駆体型インターフェロン蓄積量の比率を求め、グラフ化したものである。
【図5】図3及び4の数値化データを基に、分泌効率(%)=インターフェロン菌体外分泌量/インターフェロン総生産量(菌体外分泌量+ライゼート中の生産量)×100として算出し、グラフ化したものである。
【発明を実施するための形態】
【0012】
本発明のポリペプチドは、配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列(ペプチド付加プレプロ配列)と、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列とを含んでなる。ペプチド付加プレプロ配列は、目的タンパク質又はポリペプチドのアミノ酸配列の上流、すなわちN末端(アミノ末端)側に結合し、目的タンパク質又はポリペプチドを細胞外へ輸送・分泌するシグナルペプチドとして機能する。
【0013】
[ペプチド付加プレプロ配列]
本発明のポリペプチドが有するペプチド付加プレプロ配列は、配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列である。配列番号1に示すアミノ酸配列は、枯草菌のα−アミラーゼAmyE由来のシグナル配列、プロ配列、及び成熟領域のN末端側の一部分からなるアミノ酸配列である。AmyEは、枯草菌体内でシグナル配列、プロ配列及び成熟領域が連結した前駆体として合成され、菌体外に分泌時にシグナル配列が、分泌後にプロ配列がそれぞれ切断されて成熟酵素となる。配列番号1において、1位〜33位のアミノ酸配列がAmyEのシグナル配列に、34位〜41位のアミノ酸配列がAmyEのプロ配列に、42位〜59位のアミノ酸配列がAmyEの成熟タンパク質領域の上流18アミノ酸に相当する。
本発明においては、AmyEのシグナル配列及びプロ配列をまとめてプレプロ配列と呼び、当該配列にAmyEの成熟領域の1部分が付加されているのでペプチド付加プレプロ配列と称した。
AmyEのシグナル配列のアミノ酸配列を配列番号2に、プロ配列のアミノ酸配列を配列番号3に、成熟タンパク質領域のアミノ酸配列を配列番号4にそれぞれ示す。また、AmyEのシグナル配列、プロ配列、及び成熟タンパク質領域のアミノ酸配列をコードする塩基配列の一例を、配列番号5に示す。
【0014】
本発明で用いるペプチド付加プレプロ配列は、少なくとも配列番号1において1位〜44位までのアミノ酸配列を含む。すなわち、AmyEのシグナル配列及びプロ配列をこの順で有し、さらに、プロ配列の後ろに成熟タンパク質領域のN末端側から少なくとも3アミノ酸以上最大で18アミノ酸までが付加されてなることを特徴とする。プロ配列の下流に成熟タンパク質のN末端側の3〜18アミノ酸を付加することで、後述の実施例で実証されているように、分泌効率が向上する。
【0015】
AmyEのシグナル配列及びプロ配列については、これを付加した融合タンパク質において菌体外への分泌性が向上することも報告されている(前記非特許文献14)。また、枯草菌α−アミラーゼのシグナル配列と成熟領域の間に人為的にアミノ酸配列を付加して生産性の効果を検証した報告がある(Itoh Y, Kanoh K, Nakamura K, Takase K, Yamane K. (1990) Artificial insertion of peptides between signal peptide and mature protein: effect on secretion and processing of hybrid thermostable alpha-amylases in Bacillus subtilis and Escherichia coli cells. J Gen Microbiol. 136(8):1551-1558.)。しかしながら、本発明者らが検討したところ、付加されているアミノ酸配列は任意に選択されたものであり、AmyEの成熟タンパク質領域のアミノ酸配列とは全く異なるものであった。
また、α−アミラーゼの成熟体領域に目的タンパク質を融合した融合タンパク質の生産性を評価した報告もある(Honda K, Fujieda H, Ogawa K, Imai M, Yamamoto H, Ikeda T, Yamane K. (1993) Extracellular production of human hepatitis B virus preS2 antigen as hybrid proteins with Bacillus subtilis alpha-amylases in high-salt-concentration media. Appl Microbiol Biotechnol. 40(2-3):341-347.)。しかしながら、付加している成熟体領域が長く本発明のペプチド付加プレプロ配列とは異なり、GSTをキャリアとして使用したタンパク質生産と同じ効果、すなわち発現量の増加や可溶化しやすくする効果を目的として付加されていると考えられ、その効果は本発明とは異なるものである。また、タンパク質生産が特殊な培養・分泌条件下で行われている点においても本発明の製造方法とは異なる。
【0016】
分泌生産性向上の観点から、ペプチド付加プレプロ配列として好ましくは、AmyEのシグナル配列及びプロ配列の下流にAmyEの成熟領域のN末端側3〜10アミノ酸を付加したアミノ酸配列(配列番号1において1位〜44位から1位〜51位までの各アミノ酸配列)であり、より好ましくはプロ配列の後ろにN末端側8〜10アミノ酸を付加したアミノ酸配列(配列番号1の1位〜49位から1位〜51位までの各アミノ酸配列)であり、さらに好ましくはプロ配列の後ろにN末端側8又は10アミノ酸を付加したアミノ酸配列(配列番号1の1位〜49位のアミノ酸配列又は1位〜51位までのアミノ酸配列)であり、特に好ましくはプロ配列の後ろにN末端側8アミノ酸を付加したアミノ酸配列(配列番号1の1位〜49位のアミノ酸配列)である。
【0017】
[目的タンパク質又はポリペプチドのアミノ酸配列]
本発明のポリペプチドは、前述のペプチド付加プレプロ配列のC末端側に、目的タンパク質又はポリペプチドのアミノ酸配列が結合してなる。本発明において、当該目的タンパク質又はポリペプチドとは、枯草菌のAmyEタンパク質以外のタンパク質又はポリペプチドである。前述のように、AmyEとは枯草菌のα−アミラーゼであり、その成熟タンパク質領域のアミノ酸配列は配列番号4で示される。
目的タンパク質又はポリペプチド(以下、目的タンパク質等ともいう)としては、枯草菌のタンパク質AmyE以外であれば特に制限されず、例えば、インターロイキン、上皮成長因子、ケモカイン、エリスロポエチン、腫瘍壊死因子、アンジオスタチン、エンドスタチン、インシュリン、インターフェロン等が挙げられる。なかでも、本発明では目的タンパク質等としてインターフェロンを用いることが好ましい。
【0018】
インターフェロン(Interferon、IFNとも略記する)は、多くの動物細胞にみられるタンパク質で、I型、II型、III型の3つのタイプに大別される。本発明の目的タンパク質等として用いられるのは、I型インターフェロンが好ましい。I型インターフェロンとしては、IFN−α(IFN−α1,2,4,5,6,7,8,10,13,14,16,17,21の13種類)、IFN−β(IFN−β1)、IFN−ω(IFN−ω1)、IFN−ε(IFN−ε1)、IFN−κ(IFN−κ)が挙げられる。I型インターフェロンのなかでも、ヒトI型インターフェロンがより好ましく、ヒトI型インターフェロン−α(hIFN−α2a、hIFN−α2bなど)及びβ(hIFN−β1、hIFN−β2bなど)がさらに好ましく、ヒトI型インターフェロン−αが特に好ましい。
上記インターフェロンのアミノ酸配列情報、及びインターフェロン遺伝子の塩基配列情報は、DDBJ、EMBL、GenBank等の公共の塩基配列データバンク等から得ることができる。一例として、ヒトインターフェロン−α2bの成熟領域のアミノ酸配列を配列番号6に、当該アミノ酸配列をコードする塩基配列の一例を、配列番号17にそれぞれ示す。
【0019】
本発明のポリペプチドに含まれる目的タンパク質等のアミノ酸配列は、当該タンパク質等の成熟領域のみでもよく、当該タンパク質の機能に影響を与えない範囲で他の配列を有していてもよく、例えば、当該アミノ酸配列の合成・調製の際や宿主微生物への導入の際などに人為的に付加した配列(例えば、リンカー配列、アダプター配列など)を適宜有していてもよい。また、成熟領域のN末端側又はC末端側にヒスチジンタグ、FLAGタグ、HAタグやmycタグといったアフィニティタグ、グルタチオン−S−トランスフェラーゼ(GST)、緑色蛍光タンパク質(GFP)、マルトース結合タンパク質(MBP)といったキャリアタンパク質の配列等を有するものであってもよい。
【0020】
[ポリペプチド]
本発明のポリペプチドは、上述したペプチド付加プレプロ配列が、目的タンパク質等のアミノ酸配列のN末端側に結合されたものである。本発明のポリペプチドにおいては、ペプチド付加プレプロ配列と目的タンパク質等とが細胞内で融合タンパク質として発現するように連結されていればよい。ペプチド付加プレプロ配列と目的タンパク質等の成熟領域のアミノ酸配列とが直接結合している場合はもちろん、前述のように、ペプチド付加プレプロ配列と目的タンパク質等のアミノ酸配列とがリンカーを介して結合していてもよいし、ペプチド付加プレプロ配列と目的タンパク質等の成熟領域との間に、制限酵素に由来するアミノ酸、各種のタグや小DNA断片に由来するアミノ酸等の配列を有していてもよい。
【0021】
本発明のポリペプチドは、配列番号1に示すアミノ酸配列情報、及び目的タンパク質のアミノ酸配列情報に基づいて、通常の遺伝子工学的手法により作製できる。具体的には、化学合成による方法、PCR反応による方法、該当配列を含むDNA断片やプラスミドより得る方法などが挙げられる。
【0022】
本発明のポリペプチドは、目的タンパク質等のアミノ酸配列のN末端側にペプチド付加プレプロ配列が結合した構造により、これを発現した微生物では菌体内でペプチド付加プレプロ配列が付加した融合タンパク質が合成され、分泌シグナルの働きにより菌体外に分泌される。ペプチド付加プレプロ配列は、優れた分泌能を有しており、目的タンパク質等を効率よく菌体外に分泌することができる。そのため、ペプチド付加プレプロ配列を結合することにより、菌体内での目的タンパク質前駆体の蓄積が減少する。さらに、菌体外に分泌されたタンパク質は、野生型と同等の生理活性を維持することができる。
【0023】
[ポリペプチドをコードする塩基配列]
本発明の塩基配列は、上述した本発明のポリペプチドのアミノ酸配列をコードするもので、ペプチド付加プレプロ配列をコードする塩基配列の下流に、目的タンパク質等をコードする塩基配列が結合した塩基配列である。上述した本発明のポリペプチドと同様、細胞内で融合タンパク質として発現するよう配列番号1のペプチド付加プレプロ配列をコードする塩基配列と目的タンパク質等コードする塩基配列とが連結されていればよく、リンカー配列等を含むものであってもよい。
ペプチド付加プレプロ配列をコードする塩基配列としては、例えば、配列番号5の塩基配列において1位〜132位から1位〜177位までの領域の各塩基配列が挙げられる。目的タンパク質等をコードする塩基配列として、例えば、配列番号17(hIFN−α2b)の塩基配列が挙げられる。
本発明の塩基配列は、配列番号1のアミノ酸配列をコードする塩基配列、目的タンパク質等をコードする塩基配列の情報をもとに、通常の手法により調製することができる。例えば、塩基配列情報をもとに適当なプライマーを設計し、枯草菌の染色体DNAや目的タンパク質等の遺伝子を有する生物の染色体DNAをそれぞれ鋳型として、PCR反応により合成することができる。また、塩基配列情報をもとに化学合成することもできる。
【0024】
[組換え微生物]
本発明の組換え微生物は、上述した本発明のポリペプチドのアミノ酸配列をコードする塩基配列(遺伝子)を、宿主微生物に導入したものである。
宿主微生物としては、グラム陽性菌、グラム陰性菌のいずれを用いてもよいが、タンパク質を菌体外に分泌生産させる能力を有する点からグラム陽性菌が好ましい。グラム陽性菌の中でもバチルス(Bacillus)属細菌がより好ましく、具体的には、枯草菌(Bacillus subtilis)、Bacillus amyloliquefaciens、Bacillus licheniformis、Bacillus stearothermophilus、Bacillus thuringiensis、Bacillus subtilis nattoなどが好ましく用いられる。特に、全ゲノム情報が明らかにされ、遺伝子工学、ゲノム工学技術が確立されている点から、枯草菌を宿主とすることが好ましい。
【0025】
宿主に導入する塩基配列は、上述した本発明のポリペプチドのアミノ酸配列をコードする塩基配列であり、その調製方法も前述のとおりである。
本発明の組換え微生物には、本発明のポリペプチド配列をコードする塩基配列の他に、発現制御のために有用な塩基配列が適宜導入されていてもよい。例えば、転写、翻訳に関わる制御領域として、プロモーター及び転写開始点を含む転写開始制御領域、リボソーム結合部位(SD(Shine-Dalgarno)配列等)及び開始コドンを含む翻訳開始領域、ターミネーター等を、本発明のポリペプチドをコードする塩基配列と適正な形で結合し導入してもよい。また、選択マーカー等を導入してもよい。
【0026】
宿主微生物への塩基配列の導入は、通常の組換えDNA技術を用いて行うことができる。例えば、ポリペプチド配列をコードする塩基配列をベクターに組み込んで、一般的な形質転換法を用いて宿主微生物細胞に取り込ませて導入してもよいし、塩基配列に宿主微生物ゲノムの相同領域を結合したDNA断片を用いて、相同組換えにより宿主微生物ゲノムに直接組み込んでもよい。
【0027】
ベクターを用いる場合、プラスミド等の染色体外で自立増殖・複製するベクターであってもよいし、染色体内に組み込まれるベクターであってもよい。例えば、導入する宿主の種類に応じたプロモーターやターミネーター等の発現調節領域を有する発現ベクターであって、複製開始点や選択マーカー等を有するベクターを用いることができる。具体的には、pHY300、pWH1520、pUB110、pC194、pT127、pE194等のベクターを挙げることができる。
プロモーターやターミネーター、選択マーカー等の種類も特に限定されず、通常使用されるものを適宜選択して用いることができる。プロモーターとしては、例えば、tacプロモーター、lacプロモーター、spacプロモーター、xylプロモーター、T7プロモーター等が用いられる。選択マーカーとしては、クロラムフェニコール、テトラサイクリン、ネオマイシン、カナマイシン、エリスロマイシン等の薬剤耐性遺伝子が挙げられる。
目的塩基配列のベクターへの組み込みは、制限酵素処理やライゲーション等の通常の手法により行うことができる。
【0028】
次いで、構築した組換えベクターを用いて宿主を形質転換し、本発明のポリペプチドをコードする塩基配列が導入された組換え微生物を作製する。形質転換方法は特に限定されず、カルシウムイオンを用いる方法、一般的なコンピテントセル形質転換方法(J.Bacterial.93,1925(1967))、プロトプラスト形質転換法(Mol.Gen.Genet.168,111(1979))、エレクトロポレーション法(FEMS Microbiol.Lett.55,135(1990))、LP形質転換方法(T.Akamatsu及びJ.Sekiguchi,Archives of Microbiology,1987,146,p.353-357;T.Akamatsu及びH.Taguchi,Bioscience,Biotechnology,and Biochemistry,2001,65,4,p.823-829)等通常の方法を用いることができる。
目的の塩基配列が導入されたことの確認は、選択マーカー等を利用することで行うことができる。また、ゲノムを鋳型としたPCR法等によって、目的DNA断片の導入を確認することもできる。
【0029】
[目的タンパク質等の製造]
得られた組換え微生物を用いて、目的タンパク質等を製造する。例えば、組換え微生物を適切な培地を用いて適切な条件下にて培養して、目的タンパク質を産生させ、培養物から目的タンパク質を精製・回収すればよい。本発明では、菌体内で合成された目的タンパク質等が効率よく菌体外に分泌されるので、培養液から目的タンパク質等を回収することができ、精製・単離作業を容易に行うことができる。
【0030】
組換え微生物の培養条件は、宿主の種類に応じて適宜好ましい培養条件を採用することができる。また、用いるベクターやプロモーターに応じて、組換えタンパク質の発現誘導を適宜行ってもよい。例えば、宿主として枯草菌を用いた場合は、2×L培地(実施例参照)を用い、30℃前後で培養を行えばよい。
培養物から目的タンパク質等を精製・単離する方法は特に限定されず、例えば、カラム精製、硫安沈殿、組換えタンパクにタグを付加した場合はタグを用いた精製、透析、フィルター精製等の方法で行うことができる。
【実施例】
【0031】
以下、本発明を実施例に基づきさらに詳細に説明するが、本発明はこれに限定されるものではない。
下記の手法によって、本発明のペプチド付加プレプロ配列及び目的タンパク質をコードする塩基配列を導入した組換え微生物を構築し、目的タンパク質の発現を行った。目的タンパク質として、ヒトインターフェロンα(hIFN‐α)を用いた。
【0032】
[使用菌株、プラスミドベクター及び培地]
(1)使用菌株
大腸菌(エシェリヒア・コリ)C600株(タカラバイオ)
大腸菌(エシェリヒア・コリ)JM109株(タカラバイオ)
枯草菌168株
枯草菌Dpr8株(8種類の分泌型プロテアーゼを欠損する枯草菌変異株(Δepr/ΔwprA/Δmpr/ΔnprB/Δbpr/ΔnprE/Δvpr/ΔaprE);Kodamaら, "Effect of Bacillus subtilis spo0A mutation on cell wall lytic enzymes and extracellular proteases, and prevention of cell lysis", J. Biosci. Bioeng., 2007, 103, p.13-21参照)
(2)プラスミド
pWH1520;耐性マーカー(アンピシリン、テトラサイクリン)、MoBiTec社製
pHKK3100;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列)
pHKK3101;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列、及びhIFN‐α2bの成熟体領域の配列)
pHKK3200;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列及びプロ配列)
pHKK3201;耐性マーカー(アンピシリン、テトラサイクリン)、挿入配列(枯草菌アミラーゼAmyEのシグナル配列及びプロ配列、及びhIFN‐α2bの成熟体領域の配列)
(3)培地
L培地;バクトトリプトン(Difco)1%、イーストエクスラクト(Difco)0.5%、NaCl(和光)0.5%
2×L培地;バクトトリプトン(Difco)2%、イーストエクスラクト(Difco)1%、NaCl(和光)1%
【0033】
1.異種タンパク質hIFN−α発現用ベクターの構築
枯草菌アミラーゼAmyEのシグナル配列(配列番号2)及びプロ配列(配列番号3)をコードするDNA配列を組み込んだ基本ベクターpHKK3200を、特開2008-271956号公報の実施例2の記載に従って構築した。枯草菌168株の染色体を鋳型に、プライマーamyESF−1(ggccACTAGTcttcaaaaaatcaaa(配列番号7)、大文字がSpeI)及びamyESR−2(ggccGGTACCctcattcgatttgttcgc(配列番号8)、大文字がKpnI)を用いてPCR法で増幅したamyE遺伝子のSD配列、シグナル配列及びプロ配列を、pWH1520のマルチクローニングサイトのSpeI−KpnIサイトに導入し、pHKK3200を構築した(Kakeshita et al. 2011 Appl Microbiol Biotechnol 89:1509−1517参照)。
次いで、pORF5−hIFNA2b(インビボジェン製、hIFN−α2b)を鋳型に、プライマーifnaF(ggccGGTACCtgtgatctgcctcaaacc(配列番号9)、大文字がKpnI)及びifnaR(ggccGGATCCttttcattccttacttct(配列番号10)、大文字がBamHI)を用いてPCR法で増幅したインターフェロンα遺伝子の成熟体領域(配列番号17)を、pHKK3200のKpnI−BamHIサイトに導入し、インターフェロンα発現用ベクターpHKK3201を構築した(図1)。
同様の手法で、枯草菌アミラーゼAmyEのシグナル配列をコードするDNA配列を組み込んだ基本ベクターpHKK3100を構築した。pHKK3100に、上記と同様の方法でインターフェロンα遺伝子の成熟体領域を組み込み、インターフェロンα発現用ベクターpHKK3101を作成した(図1)(Kakeshita et al. 2011 Appl Microbiol Biotechnol 89:1509-1517参照)。
【0034】
次に、枯草菌アミラーゼAmyEのシグナル配列、プロ配列、及び成熟領域のN末端側のアミノ酸配列をコードするDNA配列を組み込んだインターフェロンα発現用ベクターpHKK3221〜pHKK3226を構築した。枯草菌168株の染色体を鋳型に、プライマーamyESF−1(ggccACTAGTcttcaaaaaatcaaa(配列番号7)、大文字がSpeI)と、amyES2−5R(ggccGGTACCtgctgtaagctcattcgatttg(配列番号11)、大文字がKpnI)、amyES3R(gaagaatGGTACCgcttttgatcgacgg(配列番号12)、大文字がKpnI)、amyES4R(ggccGGTACCcgaccaattccatgcatgaa(配列番号13)、大文字がKpnI)、amyES5R(ggccGGTACCtgtatatcctgcatcatgaa(配列番号14)、大文字がKpnI)、amyES6R(ggccGGTACCctgatacagccagtaccag(配列番号15)、大文字がKpnI)、又はamyES7R(aaattcttgttcGGTACCtaagtaacgg(配列番号16)、大文字がKpnI)を用いてPCR法で増幅した各PCR産物を、制限酵素SpeI及びKpnIで切断した後、上記で構築したpHKK3201のSpeI−KpnIサイトに導入し、pHKK3221(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側3アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3222(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側8アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3223(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側18アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3224(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側35アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、pHKK3225(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側63アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)、及びpHKK3226(挿入配列;アミラーゼAmyEのシグナル配列、プロ配列及びAmyEの成熟領域のアミノ末端側74アミノ残基と、IFNα成熟体の融合タンパク質をコードするDNA配列)をそれぞれ構築した(図1)。各プラスミドベクターは、AmyE成熟領域をコードする配列とIFNα成熟体領域をコードする配列との間にリンカー配列(Gly-Thr配列)を有する。
なお、上記ベクター構築時の宿主には、大腸菌(エシェリヒア・コリ)C600株及びJM109株(ともにタカラバイオ)を使用した。プラスミド構築時の大腸菌の培養及び枯草菌の前培養時には、L培地を用いた。
【0035】
2.異種タンパク質発現ベクター導入株の作製
上記1.で作成した各インターフェロンα発現ベクターを、形質転換法によって枯草菌変異株Dpr8株を形質転換して、インターフェロンα発現ベクター導入株を得た。形質転換は、SPI培地及びSPII培地を用いてコンピテント法により行い、15μg/mLテトラサイクリンを含むLB寒天培地に生育した菌株を形質転換体として選択した。
【0036】
3.分泌タンパク質の生産性評価
得られた各インターフェロンα発現ベクター導入株について、インターフェロンの生産性をウエスタンブロッティング解析、ELISA解析、及び生理活性測定により評価した。
導入株の培養には、2×L培地を用いた。抗生物質は、テトラサイクリン15μg/mLの濃度で適時使用した。pWH1520のもつキシロースプロモーターの誘導のため、インターフェロンα導入株には、キシロース終濃度0.6%を添加した。培養は、特開2008-271956号公報の実施例2に記載の方法に従い、前培養液を2%植菌し、30℃にて細胞増殖期中期(OD660=0.3)まで培養後、キシロースを最終濃度0.6%添加し、培養した。培養開始後24時間後で集菌し、培養液を得た。
【0037】
(1)ウエスタンブロッティング
得られた培養液から、下記「Medium」及び「Lysate」をそれぞれ調製し、ウエスタンブロッティング用サンプルとして用いた。
「Medium」:培養液を遠心して集菌し、得られた上清液をmediumとした。SDS-PAGEにて展開する際には、等量のサンプルバッファを加えて加熱処理してサンプルとした。
「Lysate」:培養液を遠心して集菌したもの(ペレットの状態)をリゾチーム処理後、バッファ(Tris-HCL)に懸濁し、等量のサンプルバッファを加えて加熱処理した。その後、遠心して得られた上清をサンプルとした。
【0038】
ウエスタンブロッティングは、セミドライシステム(バイオラッド)を用いて行った。SDS−PAGEによるタンパク質分離後、タンパク質をイモビロンPVDF膜(ミリポア)に転写した。タンパク質の検出は、イムノスターLD検出システム(商品名、和光純薬工業株式会社製)を用いた。抗インターフェロンα抗体は、(PeproTech EC LTD)のものを、HRP標識二次抗体は、GEヘルスケアのものを使用して検出した。
インターフェロンの分泌量及び細胞内蓄積量の測定は、ウエスタンブロッティング法により検出したバンドの濃淡を、ImageJ(National Institutes of Health, USA)により数値化することで行った。
【0039】
(1−1)インターフェロン分泌量の測定
各導入株のMediumサンプルを用いて、菌体外に分泌されたインターフェロンをウエスタンブロッティング法により検出した(図2(a))。
次いで、得られたインターフェロンのバンドの濃淡を、ImageJにより数値化し、各導入株のインターフェロン分泌量とした(図3)。なお、コントロールにはシグナル配列とプロ配列を持つpHKK3201導入株を用い、pHKK3201導入株での分泌量を100として、各導入株における分泌量を相対値(%)で示した。
【0040】
図3から明らかなように、シグナル配列とプロ配列に成熟領域の3アミノ酸残基を付加したpHKK3221導入株、及び8アミノ酸残基を付加したpHKK3222導入株では、コントロールのpHKK3201導入株に比べインターフェロン分泌量(生産量)が大きく上昇した。また、シグナル配列とプロ配列に18アミノ酸残基を付加したpHKK3223導入株でも、コントロールpHKK3201導入株と同程度の生産性があった。特に、8アミノ酸残基を付加したpHKK3222導入株では、インターフェロンの生産量が最大となり、シグナル配列のみのpHKK3101導入株と比べて9.5倍、シグナル配列とプロ配列を有するpHKK3201導入株と比べて2.9倍の生産性向上が確認された。
また、35アミノ酸残基を付加したpHKK3224導入株は、シグナル配列のみのpHKK3101導入株と比べて生産性は向上しているものの、シグナル配列とプロ配列を有するpHKK3201導入株に比べると多少生産性が低下した。63アミノ酸残基以上を付加した導入株では、インターフェロンの分泌はほとんど確認されなかった。
【0041】
(1−2)菌体内のインターフェロン蓄積量の測定
各導入株のLysateサンプルを用いて、菌体内のインターフェロンをウエスタンブロッティング法により検出した(図2(b))。
その結果、各サンプルにおいて、菌体内に前駆体型インターフェロンと成熟型インターフェロンの両者が確認された。なお、前駆体型インターフェロンとは、シグナル配列が切断されていない状態であり、成熟型インターフェロンとは、付加したアミノ酸配列からシグナル配列が切断された状態である。
菌体外へ分泌されるタンパク質は、シグナル配列(シグナルペプチド)をアミノ末端側に持ち、これは菌体外へ分泌される際にシグナルペプチダーゼよって切断される。細胞内への前駆体の蓄積は、シグナルペプチドが切断されておらず、菌体外への正常な分泌ができていないことを示している。宿主自体が持つ分泌タンパク質が菌体内に蓄積する現象は、分泌輸送経路構成因子の欠損や変異による機能不全があった場合に見られるもので、通常は観察されない。異種タンパク質発現においても、菌体内に前駆体タンパク質が蓄積していることは、分泌効率が悪いことを示している。異種タンパク質の細胞内蓄積は宿主への負荷となるため、効率的な異種タンパク質生産という点からは、蓄積が少ない状態が好ましい。
【0042】
次いで、得られた前駆体型インターフェロンと成熟型インターフェロンのバンドについて、濃淡をImageJにより数値化し、菌体内の前駆体型インターフェロン蓄積量及び成熟型インターフェロン蓄積量を算出した。これらをもとに、菌体内の総インターフェロン生産量(前駆体型インターフェロン蓄積量と成熟型インターフェロン蓄積量の和)に対する前駆体型インターフェロン蓄積量の比率を求め、前駆体型インターフェロン蓄積率(%)とした(図4)。
【0043】
図4から明らかなように、シグナル配列とプロ配列に3アミノ酸残基を付加したpHKK3221導入株、8アミノ酸残基を付加したpHKK3222導入株、18アミノ酸残基を付加したpHKK3223導入株では、シグナル配列とプロ配列を有するpHKK3201導入株に比べて、前駆体型インターフェロンの蓄積が減少していた。前駆体の蓄積減少は、特にpHKK3222導入株及びpHKK3223導入株で顕著であった。
この結果から、菌体内での前駆体の蓄積解消が、インターフェロン生産量及び分泌効率の向上につながっている可能性が高いと考えられる。
【0044】
(1−3)インターフェロン分泌効率
前記(1−1)で得られた培養液中に分泌されたインターフェロン量、及び(1−2)で得られたライゼート中のインターフェロン総量(前駆体蓄積量+成熟体蓄積量)から、各導入株における分泌効率を下記式により求めた。結果を図5に示す。
分泌効率(%)=培地中に分泌されたインターフェロン量/インターフェロン総生産量 インターフェロン総生産量=培地中に分泌されたインターフェロン量+ライゼート中のインターフェロン総生産量(前駆体+成熟体)
【0045】
図5から明らかなように、シグナル配列とプロ配列を有するpHKK3201導入株に比べて、pHKK3221導入株、pHKK3222導入株、pHKK3223導入株では分泌効率が向上していた。特に、シグナル配列とプロ配列に8アミノ酸残基を付加したpHKK3222導入株では分泌効率が約4倍と飛躍的に向上した。
【0046】
(2)ELISA法による菌体外分泌されたインターフェロン量の測定
上記(1)ウエスタンブロッティングに用いた「Medium」サンプルと同様の調製手順で、pHKK3201導入株及びpHKK3222導入株の培養液を遠心して集菌し、得られた上清液をELISA用サンプルとした。測定には、IFN-α Human ELISAキット(R&Dsystems,Inc)を使用した。測定実験は、鎌倉テクノサイエンスに委託して行われた。
ELISA法により算出された培養液上清中に含まれるインターフェロンの総量を、表1の「インターフェロン総量」欄に示した。
【0047】
(3)インターフェロン活性測定
前記(1−1)でインターフェロン生産量が最大であったpHKK3222を用いて、シグナル配列とプロ配列に8アミノ酸残基が付加された組換えインターフェロンの生理活性を測定した。測定実験は、鎌倉テクノサイエンスに委託して行った。
上記(2)のELISA法と同様の手順で、pHKK3201導入株及びpHKK3222導入株の培養上清液を調製して、測定試料とした。なお、バックグラウンドとして、インターフェロンをコードする遺伝子を有しないプラスミドを導入した株を用いて測定し、100IU/mL以下であることを確認している。
生理活性測定は、FL細胞に対するSindbisウイルスの感染を指標として計測した。標準品には、インターフェロンA注射用(hIFNα2b、シェリング・プラウ株式会社(現 MSD株式会社)製)を使用した。単位は、国際単位(IU)を使用した。
インターフェロンの生理活性は、インターフェロンに接触させたFL細胞にシンドビスウイルスを感染させることによる抗ウイルス作用を指標とした生理活性測定法を用いた。無血清培地A(5%ウシ胎児血清、10mM HEPES緩衝液)で4×105cells/mLの細胞液を調製し、マイクロプレートに10μL/well播種した。炭酸ガスインキュベーター(5%CO2、37℃)内で20〜28時間培養後、2倍段階希釈した標準品または測定試料5μL/wellを添加した。炭酸ガスインキュベーター内でさらに20〜28時間培養した後、マイクロプレート内の培養液を除去し、培地B 100μL/wellで1回洗浄後除去した。各試料に、無血清培地B(0.5%BSA、20mM HEPES緩衝液)で適宜希釈したウイルス液100μL/wellを添加し、マイクロプレートをインキュベーター内(37℃)で18〜28時間培養した。マイクロプレート内のウイルス液を廃棄し、染色固定液(0.05w/v%クリスタルバイオレット、10v/v%エタノールを含む中性ホルマリン液)に1〜2時間浸し、水洗後、乾燥した。
得られたサンプルを、マイクロプレートリーダーを用いて吸光度(波長595nm)を測定した。Cell Controlの吸光度を100%、Virus Controlの吸光度を0%とし、各wellの吸光度%を算出した。吸光度50%の希釈倍数を力価とし、標準品(インターフェロンA注射用300(IFN−α2b)、市販品)の力価により測定試料の力価を補正し、試料のIFN力価を相対的に算出した。
上記測定により算出された培養上清液サンプル中の総インターフェロン活性を、表1の「総インターフェロン活性」欄に示した。
【0048】
次いで、この培養上清中の総インターフェロン活性と、上記(2)ELISA法により算出された培養液上清中に含まれるインターフェロン総量とから、分泌された組換えインターフェロンの比活性を算出した。比活性=総インターフェロン活性/インターフェロン総量 として算出し、結果は表1の「比活性」欄に示した。当該比活性は、同タンパク質量における組換えインターフェロンタンパク質の活性能を示している。
【0049】
【表1】
【0050】
表1から明らかなように、「比活性」の値はpHKK3201導入株を用いた系とpHKK3222導入株を用いた系とで同程度であった。このことから、pHKK3222導入株(シグナル配列+プロ配列+8アミノ酸残基を付加)により菌体外に産出された組換えインターフェロンは、pHKK3201導入株(シグナル配列+プロ配列を付加)により菌体外に産出された組換えインターフェロンと同等の生理活性を有することが確認された。なお、本発明者らは以前に、シグナル配列+インターフェロン導入株(本発明のpHKK3101株に相当、菌体外分泌時に当該シグナル配列は切断済み)及びシグナル配列+プロ配列+インターフェロン導入株(本発明のpHKK3201導入株に相当)を用いて、上記(2)及び(3)と同様のインターフェロン活性測定を行っており、両導入株により菌体外に産出された組換えインターフェロンが、野生型と同等の生理活性を有することを既に確認済みである(前記非特許文献14参照)。これらの結果から、pHKK3222導入株(シグナル配列+プロ配列+8アミノ酸残基を付加)により菌体外に産出された組換えインターフェロンは、野生型と同等の生理活性を保持しており、成熟領域の8アミノ酸残基を付加したことによる機能的な悪影響はないことが確認された。
【特許請求の範囲】
【請求項1】
下記(A)のアミノ酸配列が、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列のN末端側に結合されてなるポリペプチド。
(A)配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列
【請求項2】
前記目的タンパク質又はポリペプチドが、インターフェロンである請求項1記載のポリペプチド。
【請求項3】
前記(A)のアミノ酸配列が、配列番号1に示すアミノ酸配列の1位〜49位までのアミノ酸配列である請求項1又は2記載のポリペプチド。
【請求項4】
請求項1〜3のいずれか1項に記載のポリペプチドのアミノ酸配列をコードする塩基配列。
【請求項5】
宿主微生物に、請求項1〜3のいずれか1項に記載のポリペプチドのアミノ酸配列をコードする塩基配列を導入した組換え微生物。
【請求項6】
前記宿主が、バチルス(Bacillus)属に属する微生物である請求項5記載の組換え微生物。
【請求項7】
請求項5又は6記載の組換え微生物を用いた目的タンパク質又はポリペプチドの製造方法。
【請求項8】
請求項5又は6記載の組換え微生物を用いた目的タンパク質又はポリペプチドの生産性向上方法。
【請求項1】
下記(A)のアミノ酸配列が、枯草菌のAmyEタンパク質以外の目的タンパク質又はポリペプチドのアミノ酸配列のN末端側に結合されてなるポリペプチド。
(A)配列番号1に示すアミノ酸配列中の連続した一部分又は全部のアミノ酸配列であって、少なくとも1位〜44位までの領域を含むアミノ酸配列
【請求項2】
前記目的タンパク質又はポリペプチドが、インターフェロンである請求項1記載のポリペプチド。
【請求項3】
前記(A)のアミノ酸配列が、配列番号1に示すアミノ酸配列の1位〜49位までのアミノ酸配列である請求項1又は2記載のポリペプチド。
【請求項4】
請求項1〜3のいずれか1項に記載のポリペプチドのアミノ酸配列をコードする塩基配列。
【請求項5】
宿主微生物に、請求項1〜3のいずれか1項に記載のポリペプチドのアミノ酸配列をコードする塩基配列を導入した組換え微生物。
【請求項6】
前記宿主が、バチルス(Bacillus)属に属する微生物である請求項5記載の組換え微生物。
【請求項7】
請求項5又は6記載の組換え微生物を用いた目的タンパク質又はポリペプチドの製造方法。
【請求項8】
請求項5又は6記載の組換え微生物を用いた目的タンパク質又はポリペプチドの生産性向上方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2013−66434(P2013−66434A)
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2011−207792(P2011−207792)
【出願日】平成23年9月22日(2011.9.22)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
【公開日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成23年9月22日(2011.9.22)
【出願人】(000000918)花王株式会社 (8,290)
【Fターム(参考)】
[ Back to top ]