
ポリペプチド

【課題】TRPC6の細胞膜への移行のメカニズムを解明し、当該移行を阻害することによってTRPC6チャネロパチーの予防又は治療に用いることが可能な化合物を提供する。
【解決手段】以下のいずれかの新規なポリペプチドを提供する。(i)特定のアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチド;(ii)前記(i)の部分配列を含むポリペプチドであって、284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、特定のアミノ酸配列の284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド。
【解決手段】以下のいずれかの新規なポリペプチドを提供する。(i)特定のアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチド;(ii)前記(i)の部分配列を含むポリペプチドであって、284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、特定のアミノ酸配列の284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、TRPC6の膜移行を阻害することによってそのチャネル活性を抑制する新規なポリペプチド等に関する。
【背景技術】
【0002】
カルシウム濃度の動的変化は、分泌、収縮、細胞増殖、生存、及び分化を含む様々な細胞応答を引き起こす。Ca2+濃度の増加は、リガンドによる受容体の直接的な活性化、又は細胞内シグナルパスウェイを通じた間接的な活性化の結果、細胞膜上又は小胞体上のCa2+透過性チャネルが開くことによって開始される。
ショウジョウバエのTransient Receptor Potential Canonicalの哺乳類ホモログであるTRPCファミリーは、細胞膜上に存在する6回膜貫通型のイオンチャネルであり、細胞膜におけるCa2+の流入経路として機能するか、膜電位を変えてCa2+流入の駆動力を調節することによって、細胞質の遊離Ca2+濃度の制御に寄与している。哺乳類のTRPCチャネルの7つのサブタイプの中で、TRPC3、6及び7は、配列類似性が高い。これらのタンパク質は、受容体の刺激又はジアシルグリセロール類縁体の投与によって活性化される非選択的なカチオンチャネルである。
【0003】
TRPCチャネルが活性を調節するメカニズムの一つとして、細胞膜へのチャネルの挿入が挙げられる。即ち、TRPCチャネルは、細胞膜と細胞質の間を行き来し、細胞膜に移行することによって活性化される。TRPC3、4及び6は、Gq共役受容体又は受容体チロシンキナーゼの刺激によって細胞膜に移行する(非特許文献1)。プロテインキナーゼC、プロテインキナーゼG、SrcファミリーチロシンキナーゼによってもTRPCチャネルがリン酸化され、これらのチャネルにおける複数のリン酸化部位について報告がされている。
Srcファミリーキナーゼは、すべてのTRPCチャネルに作用する。また、TRPC4及びTRPC6は、上皮成長因子(epidermal growth factor; EGF)の刺激により、Srcファミリーキナーゼによってチロシンのリン酸化を受ける。FynはTRPC6をリン酸化し、ジアシルグリセロール刺激による活性化を増強する(非特許文献2)。しかしながら、リン酸化によってTRPCが活性化されるメカニズムは解明されていなかった。
【0004】
TRPCチャネルの調節の異常が、心疾患、神経変性疾患、呼吸器疾患、腎疾患等の様々な疾患の原因につながることが広く知られている(非特許文献3〜6)。
TRPC6は、血管平滑筋細胞における受容体作動性カチオンチャネルの必須の構成要素であり(非特許文献7)、低酸素性肺高血圧と関係があるとされている(非特許文献8、9)。また、TRPC3及びTRPC6は、カルシニューリンとその下流のエフェクターであるT細胞活性化因子(nuclear factors of activated T-cells, NFAT)を活性化することにより心臓肥大を促進する(非特許文献10〜13)。
【0005】
最近、TRPC6遺伝子の変異が、ヒトの蛋白尿性腎臓疾患である単状糸球体硬化症(focal segmental glomerulosclerosis; FSGS)に関連することが報告された(非特許文献14)。FSGSでは、糸球体上皮細胞(podocyte)と、podocyteの足突起(foot processes)間に形成された細胞間接着装置であるスリット膜とがintegrityを失い、糸球体のバリア機能が損なわれる。
TRPC6は、podocyte内で発現し、スリット膜の重要な構成要素であるNephrinとPodocinに結合して(非特許文献14、15)、腎臓における糸球体のバリア機能に必須の部分を形成している。
マウス腎臓におけるTRPC6の過剰発現は蛋白尿を引き起こすが(非特許文献16)、変異したTRPC6のチャネル活性が、どのように疾患の発症と関連するのかは不明であった。いくつかの変異体(P112Q、R895C、E897K)は、HEK293T細胞で発現させると、アンジオテンシンII受容体仲介性のTRPC6活性化を増大させたが、S270TとN143Sのミスセンス変異体や、57アミノ酸切断変異体(K874X)は、チャネル活性を変化させなかった(非特許文献14、17)。これらのチャネル活性は、下流のNFAT活性化の程度とよく相関した。一方、P112Q変異体はTRPC6の細胞膜での発現を増加させる(非特許文献17)。このことは、TRPC6の細胞膜での発現の変化も疾患の発症に寄与することを示唆している。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Kiselyov K, Patterson RL (2009) Front Biosci 14: 45-58
【非特許文献2】Hisatsune C et al. (2004) J Biol Chem 279(18):18887-18894
【非特許文献3】Abramowitz J, Birnbaumer L (2009) FASEB J 23(2): 297-328
【非特許文献4】Nilius B et al. (2007) Physiol Rev 87(1): 165-217
【非特許文献5】Nilius B et al. (2005) Sci STKE 2005(295): re8
【非特許文献6】Woudenberg-Vrenken TE et al. (2009) Nat Rev Nephrol 5(8): 441-449
【非特許文献7】Inoue R et al. (2001) Circ Res 88(3): 325-332
【非特許文献8】Lin MJ et al. (2004) Circ Res 95(5): 496-505
【非特許文献9】Wang J et al. (2006) Circ Res 98(12): 1528-1537
【非特許文献10】Bush EW et al. (2006) J Biol Chem 281(44): 33487-33496
【非特許文献11】Kuwahara K et al. (2006) J Clin Invest 116(12): 3114-3126
【非特許文献12】Nakayama H et al. (2006) FASEB J 20(10): 1660-1670
【非特許文献13】Onohara N et al. (2006) EMBO J 25(22): 5305-5316
【非特許文献14】Reiser J et al. (2005) Nat Genet 37(7): 739-744
【非特許文献15】Huber TB et al. (2006) Proc Natl Acad Sci U S A 103(46): 17079-17086
【非特許文献16】Moller CC et al. (2007) J Am Soc Nephrol 18(1): 29-36
【非特許文献17】Winn MP et al. (2005) Science 308(5729): 1801-1804
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、TRPC6の細胞膜への移行のメカニズムを解明し、当該移行を阻害することによってTRPC6チャネロパチーの予防又は治療に用いることが可能な医薬を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決するために検討した結果、細胞内のTRPC6は、284位のチロシン(Y284)がリン酸化されるとPLC-γ1と結合し、これにより細胞膜に移行すること;Y284のリン酸化は、TRPC6とNephrinの結合にも必須であること;Nephrinは、Y284がリン酸化されたTRPC6に対してPLC-γ1と競合的に結合し、TRPC6の膜移行を阻害すること;TRPC6との結合部位を含むNephrinの部分ペプチドは、TRPC6とPLC-γ1との結合を阻害する結果、TRPC6の膜移行を抑制してTRPC6のチャネル活性を低下させること、等を見出し、本発明を完成するに至った。
即ち、本発明は、
〔1〕以下のいずれかのポリペプチド:
(i)配列番号:1に記載のアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチド;
(ii)前記(i)の部分配列を含むポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び
(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;
〔2〕前記(ii)に記載のポリペプチドであって、配列番号:3で表される、Nephrinのアミノ酸配列における1216位〜1227位の領域を含む、上記〔1〕に記載のポリペプチド;
〔3〕上記〔1〕又は〔2〕に記載のポリペプチドをコードする核酸;
〔4〕上記〔3〕に記載の核酸を含む発現ベクター;
〔5〕上記〔4〕に記載の発現ベクターを含む形質転換体;
〔6〕配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位に結合する抗体;
〔7〕上記〔1〕若しくは〔2〕に記載のポリペプチド、上記〔3〕に記載の核酸、及び上記〔4〕に記載の発現ベクター、並びに上記〔6〕に記載の抗体からなる群より選択されるいずれか一つを含む医薬組成物;
〔8〕TRPC6チャネロパチーの予防又は治療剤である、上記〔7〕に記載の医薬組成物;
〔9〕前記TRPC6チャネロパチーは、糸球体硬化症、後天性ネフローゼ、特発性肺高血圧症、及び心不全からなる群より選択される、上記〔8〕記載の医薬組成物;
〔10〕候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞へのCa2+イオンの流入を比較する工程と、を含む方法;
〔11〕候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞表面におけるTRPC6の発現を測定する工程と、を含む方法;及び
〔12〕候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物と、配列番号:4で表されるアミノ酸配列を含むポリペプチドとをインキュベートする工程と、
前記候補化合物と前記ポリペプチドとの結合を検出又は測定する工程と、を含む方法、に関する。
【発明の効果】
【0009】
本発明に係るポリペプチドは、TRPC6の膜移行を阻害することによってそのチャネル活性を抑制することができるので、TRPC6のチャネル活性亢進によって発症・進展し得る疾患の予防又は治療に有用である。また、チャネル活性亢進の原因となる分子に特異的に作用すること、元来生体内に存在するタンパク質の一部であることなどから、副作用が少なく、安全性の高い医薬として利用されうる。
【図面の簡単な説明】
【0010】
【図1A】図1は、TRPC6 Y284のリン酸化が、細胞膜への移行に必須であることを示すために行った実験の結果である。図1Aは、EGFによるTRPC6の膜移行の誘導を示す。HA-TRPC6を発現する293T細胞をEGF(200 ng/ml)で、図示された時間刺激した。細胞表面をビオチン化し、ストレプトアビジン−アガロースビーズに結合したタンパク質を、抗α−TRPC6抗体を用いたウエスタンブロッティングで解析した。パネルの右側に、分子量マーカー(kDa)を示す。各ライゼートの一部(2%)をウエスタンブロッティングで解析し、TRPC6の発現レベルを確認した(下パネル)。
【図1B】図1Bは、293T細胞が発現したHA-TRPC6を、Fynの存在下、又は非存在下、α−HA抗体で免疫沈降を行い、α−リン酸化チロシン抗体で検出した結果を示す。
【図1C】図1Cは、HA-TRPC6を発現する293T細胞を、Fynの存在下、又は非存在下で、図1Aの方法に従って、細胞表面のTRPC6をビオチン化して検出した結果を示す。
【図1D】図1Dは、Fynと、チロシンのフェニルアラニン置換を行ったHA-TRPC6の一連の変異体のそれぞれとを、293T細胞で発現させ、図1Aの方法に従って、細胞表面のTRPC6をビオチン化して検出した結果を示す。3から5回の独立した実験の代表的なデータを示す。
【図2A】図2は、PLC-γ1が、TRPC6のリン酸化したY31/Y284に結合し、TRPC6の細胞表面での発現を制御することを示すために行った実験の結果である。図2Aは、リン酸化、又は非リン酸化TRPC6 Y284ペプチドをカップリングゲルに固定し、293T細胞のライゼートとインキュベートした後、結合したタンパク質を、SDS-PAGEと銀染により解析した結果を示す。*で示したバンドを切り出し、トリプシン消化した後、LC-MS/MSによって解析した。
【図2B】図2Bは、Flag-PLC-γ1、Fyn及びHA-TRPC6(野生型、Y31F、Y284F, Y31F/Y284F)を293T細胞で発現させ、図示された抗体を用いて、免疫沈降(IP)及びウエスタンブロット(WB)を行った結果を示す。アスタリスクで示されたバンドは、過剰発現したPLC-γ1との交差反応によるものである。
【図2C】図2Cは、リン酸化若しくは非リン酸化TRPC6 Y31周辺ペプチド(順に配列番号:6、9)、又はリン酸化若しくは非リン酸化Y284周辺のペプチド(順に配列番号:7、10)を用いて、293T細胞のライゼートからのPLC-γ1のpull-downを行った結果を示す。ライゼートの一部(0.5% input)をリカバリーマーカーとして使用した。
【図2D】図2Dは、PLC-γ1の野生型(WT)、SH2点変異(N*、C*)、またはSH2欠失変異(ΔN、ΔC)を用いて、図2Cの方法に従って、ペプチドpull-downアッセイを行った結果を示す。
【図2E】図2Eは、スクランブルsiRNA(コントロール)又はhPLC-γ1 siRNA(Duplex 1又は2)を293T細胞にトランスフェクトし、翌日、HA-TRPC6及びFynをトランスフェクトして、トランスフェクションの1日後、TRPC6の細胞表面での発現を図1Aの方法で測定した結果を示す。
【図2F】図2Fは、Flag-PLC-γ1、Fyn、及びTRPC6(野生型又はΔ98-104)をトランスフェクトした293T細胞を用いて、共免疫沈降を行った結果を示す。
【図2G】図2Gは、98位-104位を欠失したTRPC6(Δ98-104)の細胞表面での発現を表面ビオチン化アッセイで調べた結果である。
【図3A】図3は、生細胞及び腎糸球体におけるTRPC6のチロシンリン酸化に関する実験結果である。図3Aは、図示されたプラスミドをトランスフェクトした293T細胞のライゼートを用い、図示された抗体で免疫沈降を行い、抗α-pY31抗体「又は抗α-pY284抗体で標識した結果を示す。
【図3B】図3Bは、HA-TRPC6を発現する293T細胞をEGFで刺激し/または刺激せずに、抗α-HA抗体による免疫沈降物を、α-リン酸化特異的TRPC6抗体(抗α-pY31抗体又は抗α-pY284抗体)で標識した結果を示す。
【図3C】分離したラット糸球体を、1mM sodium vanadateで30分処理し、又は処理しないで、抗α-pY284抗体でブロットした結果を示す。各サンプルについて、vanadate処理に先立って、10μM PP2で15分、又は5μM SU6656で60分、糸球体を前処理した。
【図4A】図4は、TRPC6 Y284及びNephrin 1216-1227が、両者のチロシンリン酸化依存的相互作用に不可欠であること示す実験結果である。図4Aは、図示されたプラスミドをトランスフェクトした293T細胞のライゼートについて、図示された抗体を用いて免疫沈降及びウエスタンブロッティングを行った結果を示す。
【図4B】図4Bは、FynとHA-TRPC6を共発現する、又はHA-TRPC6のみ発現する293T細胞のライゼートを用い、GST-Nephrin-CD(1102-1252残基)又は予めFynでリン酸化したGST-Nephrin-CDを用いて、in vitroのpull-downアッセイを行った結果を示す。結合したタンパク質を抗α-HA抗体で検出した。GST-Nephrin及びTRPC6のリン酸化が確認された(下のパネル)。
【図4C】図4Cは、Flag-Nephrin、Fyn、及びHA-TRPC6(野生型、又はチロシンをフェニルアラニン置換した一連の変異体)を293T細胞で発現させ、抗α-Flag抗体で行った免疫沈降物と全細胞ライゼートについて、図示された抗体でウエスタンブロッティングを行った結果を示す。
【図4D】図4Dは、Fynと共に、又はFynなしに、野生型HA-TRPC6又はTRPC6のY284F変異体を一過性に発現する293T細胞のライゼートを用いて、GST-Nephrin-CDによってpull-downアッセイを行い、抗α-HA抗体で検出した結果を示す。
【図4E】図4Eは、NephrinまたはリコンビナントNephrin-CDを発現する293T細胞のライゼートについて、ビーズに固定したリン酸化/非リン酸化TRPC6 Y284ペプチド(順に配列番号:7、10)でpull-downし、結合したタンパク質を、α-Nephrin抗体を用いたウエスタンブロッティングにより解析した結果を示す。
【図4F】図4Fは、GST-Nephrin-CDの欠失変異体(1102位-1211位, 1102位-1215位, 1102位-1227位残基を欠失)を用いて、HA-TRPC6及びFynを発現する293T細胞のライゼートからチロシンリン酸化HA-TRPC6をpull-downした結果を示す。CBBはNephrin-CDの染色に用いたクマシーブリリアントブルーを示す。
【図4G】図4Gは、HA-TRPC6、Fyn及びFlag-Nephrin(野生型又は1216位-1227位を欠失したNephrin)を293T細胞にトランスフェクトし、抗α-HA抗体又は抗α-Flag抗体による免疫沈降物について、図示された抗体でウエスタンブロッティングを行った結果を示す。
【図5A】図5は、TRPC6のリン酸化依存的な細胞表面での発現とチャネル活性化を、Nephrinが下方制御することを示す実験結果である。図5Aは、図示されたプラスミドを293T細胞に導入し、共免疫沈降を行った結果を示す。
【図5B】図5Bは、図示されたプラスミドを293T細胞に導入し、表面ビオチン化アッセイを行った結果を示す。
【図5C】図5Cは、図示されたペプチドを293T細胞に導入し、表面ビオチン化アッセイを行った結果を示す。
【図5D】図5Dは、Fura-2を搭載した293T細胞に、図示されたプラスミドを導入し、Ca2+フリーHepes-buffered saline (HBS)、続いてCaCl2を含むHBSで表面かん流し、細胞内カルシウム濃度を測定した結果を示す。細胞内カルシウム濃度([Ca2+]i)は、fura-2蛍光のF340/F380比として、細胞数(n>400)をもとに記録し、平均値をプロットした。
【図5E】図5Eは、図5Dの場合について、ベースラインからのピーク比を各細胞について計算した結果を示す。棒グラフは、5つの独立した実験データの平均と標準誤差を示す。
【図5F】図5Fは、Fura-2を搭載した293T細胞に、図示されたプラスミド及び/又はペプチドを導入し、Ca2+フリーHepes-buffered saline (HBS)、続いてCaCl2を含むHBSで表面かん流し、細胞内カルシウム濃度を測定した結果を示す。細胞内カルシウム濃度([Ca2+]i)は、fura-2蛍光のF340/F380比として、細胞数(n>400)をもとに記録し、平均値をプロットした。
【図5G】図5Gは、図5Fの場合について、ベースラインからのピーク比を各細胞について計算した結果を示す。棒グラフは、5つの独立した実験データの平均と標準誤差を示す。
【図6A】図6は、TRPC6のFSGS関連変異体に対するNephrinの影響を調べた実験結果である。図6Aは、Flag-Nephrin、Fyn、及び各HA-TRPC6変異体を、293T細胞で共発現させ、抗α-Flag抗体による免疫沈降物を、図示された抗体によるウエスタンブロッティングに供した結果を示す。
【図6B】図6Bは、TRPC6-Nephrin結合について、図6Aのdensitometirc tracing(WB:TRPC6)によって定量し、平均値を棒グラフと標準誤差で示した結果を示す(n=3)。数値は、野生型TRPC6で標準化した。
【図6C】図6Cは、TRPC6−PLC-γ1相互作用に対するNephrin発現の影響を、Flag-PLC-γ1、Fyn、及び野生型又は各TRPC6変異体を発現する293T細胞を用いて調べた結果を示す。抗α-Flag抗体による免疫沈降物を、図示された抗体によるウエスタンブロッティングに供した。
【図6D】図6Dは、TRPC6の一連の疾患関連変異体を、Nephrin/Fynとともに、又はNephrin/Fynなしで293T細胞にトランスフェクトし、図1Aと同様に、TRPC6の細胞表面での発現を解析した結果を示す。
【図6E】図6Eは、Nephrin、Fyn、及びTRPC6若しくはTRPC6 N143S変異体を293T細胞で発現させ、[Ca2+]iの変化を図5Dと同様に記録した結果を示す(n=5)。
【図6F】図6Fは、FSGS関連TRPC6変異体のチャネル活性を図6Eと同様に評価した。データは、図5Eと同様に処理した。
【発明を実施するための形態】
【0011】
[ポリペプチド]
本発明に係るポリペプチドは、284位のチロシンがリン酸化されたTRPC6に結合し、TRPC6がPLC-γ1と結合して膜移行することを阻害できるポリペプチドであればどのようなポリペプチドであってもよいが、少なくとも以下の(i)〜(iii)を含む。
(i)配列番号:1で表されるアミノ酸配列を含むポリペプチド;
(ii)配列番号:1で表されるアミノ酸配列の部分配列を含むポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び
(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド。
【0012】
本明細書において、TRPC6の「膜移行」とは、TRPC6が細胞質から細胞膜に移行して、イオンチャネルとして機能できる状態となることを意味する。
【0013】
Nephrinは、約1200アミノ酸からなる膜貫通型タンパク質である。そのC末端側の150アミノ酸が細胞内ドメインであり、この部分にTRPC6との結合部位がある。
(i)のポリペプチドは、配列番号:1で表されるアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチドである。従って、TRPC6との結合部位を含むので、細胞内においてTRPC6に結合することによってTRPC6とPLC-γ1との結合を阻害し、TRPC6の膜移行を阻害しチャネル活性を低下させる。
(i)のポリペプチドは、配列番号:1に記載のアミノ酸配列のみからなるポリペプチドであってもよく、その少なくとも一端に1以上のアミノ酸が結合しているものであってもよい。(i)のポリペプチドの全長は、そのポリペプチドがTRPC6との結合能を有する限り、特に限定されないが、例えば150アミノ酸〜300アミノ酸とすることができる。
【0014】
上記(ii)のポリペプチドは、Nephrinの細胞内ドメインの一部を含み、且つ配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する限り、どのようなペプチドであってもよい。(ii)のポリペプチドとしては、Nephrin細胞内ドメインの一部の、例えば3アミノ酸〜50アミノ酸の領域を含むポリペプチド、5アミノ酸〜30アミノ酸の領域を含むポリペプチド、8アミノ酸〜20アミノ酸の領域を含むポリペプチド、10アミノ酸〜15アミノ酸の領域を含むポリペプチドが挙げられる。
(ii)のポリペプチドの具体例として、配列番号:5で表されるNephrinのアミノ酸配列における、1216位〜1227位の領域に該当する12アミノ酸(配列番号:3)以下、「Nephrinペプチド」と呼ぶ場合もある)が挙げられる。
【0015】
上記(iii)のポリペプチドは、(i)又は(ii)のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、且つ、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有するポリペプチドである限り、どのようなポリペプチドであってもよい。
本明細書において、用語「アミノ酸」は、その最も広い意味で用いられ、天然のアミノ酸のみならず、非天然アミノ酸も含む。本明細書においては、アミノ酸を慣用的な一文字表記又は三文字表記で示す場合もある。また、TRPC6について「n位のアミノ酸」という場合、配列番号:2で表されるアミノ酸配列のn番目のアミノ酸を示し、Nephrinについて「m位のアミノ酸」という場合、配列番号:5で表されるアミノ酸配列のm番目のアミノ酸を示す。さらに、n位のアミノ酸を別のアミノ酸で置換した場合、「n」の前に置換する前のアミノ酸を、「n」の後ろに置換後のアミノ酸を表す。例えば、TRPC6のY284F変異体は、配列番号:2で表されるアミノ酸配列における284位のチロシン(Y)をフェニルアラニン(F)に置換したものを示す。なお、本明細書において、一文字表記したアミノ酸の前に「p」が付されている場合、当該アミノ酸がリン酸化されていることを示す。例えば、pY284は、284位のチロシンがリン酸化されていることを表す。
【0016】
本明細書において、「1又は数個のアミノ酸が欠失、付加、若しくは置換されているポリペプチド」という場合、欠失等されるアミノ酸の個数は、そのポリペプチドが、Y284がリン酸化されているTRPC6に対する結合能を有する限り特に限定されないが、例えば、1〜9個、1〜5個、又は1〜3個とすることができる。あるいは全体の長さの10%以内、又は5%以内としてもよい。置換又は付加されるアミノ酸は、天然のアミノ酸、非天然アミノ酸、アミノ酸アナログであり得、好ましくは天然アミノ酸である。欠失、付加、置換される場所は、ポリペプチドの末端であっても、中間であってもよく、1ヶ所であっても2ヶ所以上であってもよい。
【0017】
(iii)のポリペプチドの例としては、ヒト以外の動物(例えば、マウス、ラット、ウサギ、ウマ、ヒツジ、イヌ、サル、ブタ等)に由来するNephrinの細胞内ドメイン又はその部分ペプチドが挙げられる。
【0018】
(ii)又は(iii)のポリペプチドが284位のチロシンがリン酸化されたTRPC6と結合能を有するか否かは、タンパク質間の相互作用を検出する公知の方法で確認することができる。例えば、284位のチロシンがリン酸化されたTRPC6、又は284位のチロシンがリン酸化された284位周辺を含むTRPC6の部分ポリペプチド(例えば、配列番号:4で表されるポリペプチド)を固定したプロテインマイクロアレイやアフィニティーカラムを用いる方法、免疫沈降法、表面プラズモン共鳴法等が挙げられる。
【0019】
本発明のポリペプチドは、そのC末端がカルボキシル基又はカルボキシレート基のみでなく、アミドやエステルになっていてもよい。また、本発明のポリペプチドには、ポリペプチドの塩も含まれる。ポリペプチドの塩としては、生理学的に許容される塩基や酸との塩が用いられ、例えば、無機酸(塩酸、臭化水素酸、ヨウ化水素酸、硫酸、リン酸等)の付加塩、有機酸(p-トルエンスルホン酸、メタンスルホン酸、シュウ酸、p-ブロモフェニルスルホン酸、カルボン酸、コハク酸、クエン酸、安息香酸、酢酸等)の付加塩、無機塩基(水酸化アンモニウム又はアルカリ若しくはアルカリ土類金属水酸化物、炭酸塩、重炭酸塩等)、アミノ酸の付加塩等が挙げられる。
【0020】
本発明のポリペプチドは、投与したときに細胞内に導入されるよう、膜透過性ペプチド(cell-penetrating peptide, CPP)と融合させてもよい。CPPは、細胞膜への親和性、細胞内への移行性を有するペプチドの総称であり、HIV-1ウイルスが発現するTrans-activator of transcription protein(TATタンパク質)の11個のアミノ酸からなるドメインであるprotein-transduction domain(PTD)や、ショウジョウバエのAntennapedia、ヘルペスウイルス由来のVP22、オリゴアルギニンなどが知られている。一般に、CPPはアルギニン、リジン、ヒスチジン等の塩基性アミノ酸の含有率が高いものが多い。
【0021】
また、本発明のポリペプチドは、本発明の課題を解決するものである限り、リン酸化、メチル化、アセチル化、アデニリル化、ADPリボシル化、糖鎖付加などの修飾が加えられたものであってもよい。他のペプチドやタンパク質と融合させたものであってもよい。
【0022】
本発明のポリペプチドは、天然物から抽出する方法;液相法、固相法、液相法と固相法を組み合わせたハイブリッド法等の化学合成法;遺伝子組み換え法等、公知のポリペプチドの製造方法によって製造することができる。
天然物から抽出する場合、例えば、ヒト又はその他の動物(例えば、マウス、ラット、ウサギ、ウマ、ヒツジ、イヌ、サル、ブタ等)の組織又は細胞から調製することができる。この場合、組織又は細胞をホモジナイズした後、酸などで抽出し、抽出液を塩析、透析、ゲルろ過、逆相クロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィーなどで精製することによって、精製単離することができる。さらに、必要に応じてプロテアーゼやペプチダーゼで切断してもよい。
【0023】
固相法は、例えば、水酸基を有するレジンの水酸基と、α−アミノ基が保護基で保護された第一のアミノ酸(通常、目的とするペプチドのC末端アミノ酸)のカルボキシ基をエステル化反応させる。エステル化触媒としては、1-メシチレンスルホニル-3-ニトロ-1,2,4-トリアゾール(MSNT)、ジシクロヘキシルカルボジイミド(DCC)、ジイソプロピルカルボジイミド(DIPCDI)等の公知の脱水縮合剤を用いることができる。
次に、第一アミノ酸のα−アミノ基の保護基を脱離させるとともに、主鎖のカルボキシ基以外のすべての官能基が保護された第二のアミノ酸を加え、当該カルボキシ基を活性化させて、第一及び第二のアミノ酸を結合させる。さらに、第二のアミノ酸のα−アミノ基を脱保護し、主鎖のカルボキシ基以外のすべての官能基が保護された第三のアミノ酸を加え、当該カルボキシ基を活性化させて、第二及び第三のアミノ酸を結合させる。これを繰り返して、目的とする長さのペプチドが合成されたら、すべての官能基を脱保護する。
【0024】
固相合成のレジンとしては、Merrifield resin、MBHA resin、Cl-Trt resin、SASRIN resin、Wang resin、Rink amide resin、HMFS resin、Amino-PEGA resin(メルク社)、HMPA-PEGA resin(メルク社)等が挙げられる。これらのレジンは、溶剤(ジメチルホルムアミド(DMF)、2−プロパノール、塩化メチレン等)で洗浄してから用いることができる。
α−アミノ基の保護基としては、ベンジルオキシカルボニル(Cbz又はZ)基、tert−ブトキシカルボニル(Boc)基、フルオレニルメトキシカルボニル(Fmoc)基、ベンジル基、アリル基、アリルオキシカルボニル(Alloc)基等が挙げられる。Cbz基はフッ化水素酸、水素化等によって脱保護でき、Boc基はトリフルオロ酢酸(TFA)により脱保護でき、Fmoc基はピペリジンによる処理で脱保護できる。
α−カルボキシ基の保護は、メチルエステル、エチルエステル、ベンジルエステル、tert−ブチルエステル、シクロヘキシルエステル等を用いることができる。
アミノ酸のその他の官能基として、セリンやトレオニンのヒドロキシ基はベンジル基やtert−ブチル基で保護することができ、チロシンのヒドロキシ基は2−ブロモベンジルオキシカルボニル希やtert−ブチル基で保護する。リジン側鎖のアミノ基、グルタミン酸やアスパラギン酸のカルボキシ基は、α−アミノ基、α−カルボキシ基と同様に保護することができる。
【0025】
カルボキシ基の活性化は、縮合剤を用いて行うことができる。縮合剤としては、例えば、ジシクロヘキシルカルボジイミド(DCC)、ジイソプロピルカルボジイミド(DIPCDI)、1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド(EDCあるいはWSC)、(1H-ベンゾトリアゾール-1-イルオキシ)トリス(ジメチルアミノ)ホスホニウムヘキサフルオロホスファート(BOP)、1-[ビス(ジメチルアミノ)メチル]-1H-ベンゾトリアゾリウム-3-オキシドヘキサフルオロホスファート(HBTU)等が挙げられる。
【0026】
レジンからのペプチド鎖の切断は、TFA、フッ化水素(HF)等の酸で処理することによって行うことができる。
またポリペプチド鎖が長い場合、ある程度固相法によって合成した後、ライゲーション法(例えば、ネイティブケミカルライゲーション法)によって合成したペプチドを連結することも可能である。
【0027】
遺伝子組み換え法によるポリペプチドの製造方法は、後述するように本発明の核酸を用いて行うことができる。
【0028】
[核酸]
本発明の核酸は、本発明のポリペプチドをコードする核酸であり、DNAであってもRNAであってもよい。
本発明の核酸は、本発明のポリペプチドをコードする限り、あらゆる変異体も含むことができる。かかる変異体としては、天然に存在するアレル変異体、天然に存在しない変異体、人為的に改変した変異体を含み、例えば、Nephrinをコードする遺伝子の塩基配列における本発明のポリペプチドに対応する領域において、一又は数個の塩基が欠失、置換、又は付加した核酸が挙げられる。
本発明の核酸の例として、Nephrinペプチドをコードする核酸(配列番号:11)が挙げられる。
【0029】
本発明の核酸は、公知の方法又はそれに準ずる方法で調製することができる。例えば、自動合成装置によって合成することができる。得られたDNAをベクターに挿入するために制限酵素認識部位を加えたり、できたペプチド鎖を酵素などで切り出すためのアミノ酸配列をコードする塩基配列を組み込んでもよい。
また、細胞から全mRNAを抽出し逆転写酵素によりcDNAを調製しても良い。この場合、プライマーとして、ベクターに挿入するために必要な制限酵素認識配列を含むものを使用することができる。得られたcDNA混合物に対し、目的のcDNAを増幅するプライマーを利用してPCR法を行うことにより、目的のcDNAのみが増幅される。このときもプライマーとして、制限酵素認識部位を含むものを用いれば、両端に制限酵素認識配列を有するcDNAを得ることができる。また、逆転写酵素によって得るcDNA混合物に代えて、既存のcDNAライブラリを用いてもよい。
本発明の核酸として、配列番号:11で表される塩基配列を含む核酸が挙げられる。
上述のとおり、本発明のポリペプチドを膜透過性ペプチドと融合させる場合、本発明の核酸は、膜透過性ペプチドをコードする核酸も含む。
【0030】
また、宿主由来のプロテアーゼによる分解を抑制するため、目的のペプチドを他のペプチドとのキメラペプチドとして発現させるキメラタンパク質発現法を用いることもできる。この場合、本発明の核酸としては、目的とするペプチドと、これに結合するペプチドとをコードする核酸が用いられる。
【0031】
[発現ベクター]
本発明の発現ベクターは、本発明の核酸を含む。本発明の核酸はそのまま、又は制限酵素で消化し、又はリンカーを付加して、発現ベクターのプロモータの下流に挿入することができる。ベクターとしては、大腸菌由来プラスミド(pBR322、pBR325、pUC12、pUC13、pUC18、pUC19、pUC118、pBluescript II等)、枯草菌由来プラスミド(pUB110、pTP5、pC1912、pTP4、pE194、pC194等)、酵母由来プラスミド(pSH19、pSH15、YEp、YRp、YIp、YAC等)、バクテリオファージ(λファージ、M13ファージ等)、ウイルス(レトロウイルス、ワクシニアウイルス、アデノウイルス、アデノ随伴ウイルス(AAV)、カリフラワーモザイクウイルス、タバコモザイクウイルス、バキュロウイルス等)、コスミド等が挙げられる。
【0032】
プロモータは、宿主の種類に応じて適宜選択することができる。宿主が動物細胞である場合は、例えば、SV40(simian virus 40)由来プロモータ、CMV(cytomegalovirus)由来プロモータを用いることができる。宿主が大腸菌である場合は、trpプロモータ、T7プロモータ、lacプロモータ等を用いることができる。
発現ベクターには、DNA複製開始点(ori)、選択マーカー(抗生物質抵抗性、栄養要求性等)、エンハンサー、スプライシングシグナル、ポリA付加シグナル、タグ(FLAG、HA、GST、GFPなど)をコードする核酸等を組み込むこともできる。
【0033】
本発明の発現ベクターは、以下に示すとおりin vitroで宿主を形質転換し、本発明のポリペプチドを発現させるために用いてもよく、そのまま対象に投与して、in vivoで本発明のポリペプチドを発現させるために用いることもできる。
【0034】
[形質転換体]
本発明の形質転換体は、本発明の発現ベクターで適当な宿主細胞を形質転換することによって得ることができる。宿主は、ベクターとの関係で適宜選択することができ、例えば、大腸菌、枯草菌、バチルス属菌)、酵母、昆虫又は昆虫細胞、動物細胞等が用いられる。動物細胞として、例えば、HEK293T細胞、CHO細胞、COS細胞、ミエローマ細胞、HeLa細胞、Vero細胞を用いることができる。形質転換は、宿主の種類に応じ、リポフェクション法、リン酸カルシウム法、エレクトロポレーション法、マイクロインジェクション法、パーティクルガン法等、公知の方法に従って行うことができる。
形質転換体を常法に従って培養することにより、目的とするポリペプチドが発現する。培地は、HEK293T細胞の場合、ウシ胎児血清等の血清を添加したDMEM培地を用いることができる。
【0035】
形質転換体の培養物からのポリペプチドの精製は、培養細胞を回収し、適当な緩衝液に懸濁してから超音波処理、凍結融解などの方法により細胞を破壊し、遠心分離やろ過によって粗抽出液を得る。培養液中にポリペプチドが分泌される場合には、上清を回収する。
粗抽出液又は培養上清からの精製も公知の方法又はそれに準ずる方法(例えば、塩析、透析法、限外ろ過法、ゲルろ過法、SDS−PAGE法、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー、逆相高速液体クロマトグラフィー等)で行うことができる。
得られたポリペプチドは、公知の方法又はそれに準ずる方法で遊離体から塩に、又は塩から遊離体に変換してもよい。
【0036】
[抗体]
本明細書において、抗体は、天然に存在するか、または遺伝子組換え技術によって産生される、全長の免疫グロブリン分子(例えば、IgG抗体)、または抗体フラグメントのような免疫グロブリン分子の免疫学的に活性な一部をいう。本明細書において抗体は、キメラ抗体、ヒト化抗体、および完全なヒト抗体を包含する。さらに、いずれのタイプ、クラス、サブクラスも含み、例えば、IgG、IgE、IgM、IgD、IgAなどが含まれる。これらの抗体は、慣用技術を用いて作製することができ、ポリクローナル抗体であっても、モノクローナル抗体であってもよい。
【0037】
本明細書において、抗体フラグメントは、抗体の断片又は抗体の断片に任意の分子を結合させたものであって、対応する全長抗体と同じエピトープを認識するものを意味する。具体的には、VL、VH、CL及びCH1領域からなるFab;2つのFabがヒンジ領域でジスルフィド結合によって連結されているF(ab’)2;VL及びVHからなるFv;VL及びVHを人工のポリペプチドリンカーで連結した一本鎖抗体であるscFv;diabody型、scDb型、tandem scFv型、ロイシンジッパー型などの二重特異性抗体等が挙げられるが、これらに限定されない。
【0038】
本明細書において、キメラ抗体は、異なる種に由来する抗体の断片が連結された抗体を意味する。例えば、可変領域がマウス抗体のアミノ酸配列であり、その他の領域がヒト抗体のアミノ酸配列である抗体が挙げられる。
本明細書において、ヒト化抗体は、非ヒト由来の抗体に特徴的なアミノ酸配列で、ヒト抗体の対応する位置を置換した抗体を意味し、例えば、H鎖及びL鎖のCDRがマウス抗体のアミノ酸配列であり、その他の領域がヒト抗体のアミノ酸配列である抗体が挙げられる。かかる抗体はCDR移植抗体と呼ばれる場合もある。また、ヒト化抗体は、ヒトキメラ抗体を含む場合もある。
【0039】
本発明の一態様において、抗体は、配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位の領域に結合する。当該領域は、TRPC6におけるNephrin及びPLC-γ1との結合領域である。従って、この抗体を細胞内に投与すれば、TRPC6とPLC-γ1との結合が阻害される。
配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位の領域に結合する抗体は、配列番号:4で表されるポリペプチドを抗原として公知の方法により作製することができる。例えば、モノクローナル抗体は、ポリペプチドで免疫した非ヒト哺乳動物から抗体産生細胞を単離し、これを骨髄腫細胞等と融合させてハイブリドーマを作製し、このハイブリドーマが産生した抗体を精製することによって得ることができる。また、ポリクローナル抗体は、ポリペプチドで免疫した動物の血清から得ることができる。
配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位の領域に結合する抗体は、配列番号:4で表されるポリペプチドを抗原として作製した抗体に限定されない。その作製方法に関わらず、ある抗体が配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位に結合する限り、本発明の抗体として用いることができる。
【0040】
[医薬組成物]
一態様において、医薬組成物は、上述したポリペプチド、核酸、又は抗体を有効成分として含み、これらがTRPC6とPLC-γ1との結合を阻害することにより、TRPC6の膜移行、ひいては活性化を抑制する。
上記医薬組成物の投与形態は特に限定されず、経口的投与でも非経口的投与でもよい。非経口投与としては、例えば、筋肉内注射、静脈内注射、皮下注射等の注射投与、経皮投与、経粘膜投与(経鼻、経口腔、経眼、経肺、経膣、経直腸)投与等が上げられる。
上記医薬組成物がポリペプチドを含む場合、代謝及び排泄されやすい性質に鑑みて、各種の修飾を行うことができる。例えば、ポリペプチドにポリエチレングリコール(PEG)や糖鎖を付加して血中滞留時間を長くし、抗原性を低下させることができる。また、ポリ乳酸・グリコール(PLGA)などの生体内分解性の高分子化合、多孔性ヒドロキシアパタイト、リポソーム、表面修飾リポソーム、不飽和脂肪酸で調製したエマルジョン、ナノパーティクル、ナノスフェア等を徐放化基剤として用い、これにポリペプチドを内包させてもよい。経皮投与する場合、弱い電流を皮膚表面に流して角質層を透過させることもできる(イオントフォレシス法)。
【0041】
上記医薬組成物は、有効成分をそのまま用いてもよいし、薬学的に許容できる担体、賦形剤、添加剤等を加えて製剤化してもよい。剤形としては、例えば、液剤(例えば注射剤)、分散剤、懸濁剤、錠剤、丸剤、粉末剤、坐剤、散剤、細粒剤、顆粒剤、カプセル剤、シロップ剤、トローチ剤、吸入剤、軟膏剤、点眼剤、点鼻剤、点耳剤、パップ剤等が挙げられる。
製剤化は、例えば、賦形剤、結合剤、崩壊剤、滑沢剤、溶解剤、溶解補助剤、着色剤、矯味矯臭剤、安定化剤、乳化剤、吸収促進剤、界面活性剤、pH調整剤、防腐剤、抗酸化剤などを適宜使用し、常法により行うことができる。
製剤化に用いられる成分の例としては、精製水、食塩水、リン酸緩衝液、デキストロース、グリセロール、エタノール等薬学的に許容される有機溶剤、動植物油、乳糖、マンニトール、ブドウ糖、ソルビトール、結晶セルロース、ヒドロキシプロピルセルロース、デンプン、コーンスターチ、無水ケイ酸、ケイ酸アルミニウムマグネシウム、コラーゲン、ポリビニルアルコール、ポリビニルピロリドン、カルボキシビニルポリマー、カルボキシメチルセルロースナトリウム、ポリアクリル酸ナトリウム、アルギン酸ナトリウム、水溶性デキストラン、カルボキシメチルスターチナトリウム、ぺクチン、メチルセルロース、エチルセルロース、キサンタンガム、アラビアゴム、トラガント、カゼイン、寒天、ポリエチレングリコール、ジグリセリン、グリセリン、プロピレングリコール、ワセリン、パラフィン、ミリスチン酸オクチルドデシル、ミリスチン酸イソプロピル、高級アルコール、ステアリルアルコール、ステアリン酸、ヒト血清アルブミン、等が挙げられるがこれらに限定されない。
上記医薬組成物が、ポリペプチドを含む場合、ポリペプチド等の経粘膜吸収されにくい難吸収性薬物の吸収を改善する吸収促進剤として、ポリオキシエチレンラウリルエーテル類、ラウリル硫酸ナトリウム、サポニン等の界面活性剤;グリココール酸、デオキシコール酸、タウロコール酸等の胆汁酸塩;EDTA、サリチル酸類等のキレート剤;カプロン酸、カプリン酸、ラウリン酸、オレイン酸、リノール酸、混合ミセル等の脂肪酸類;エナミン誘導体、N-アシルコラーゲンペプチド、N-アシルアミノ酸、シクロデキストリン類、キトサン類、一酸化窒素供与体等を用いることができる。
【0042】
丸剤又は錠剤は、糖衣、胃溶性、腸溶性物質で被覆することもできる。
注射剤は、注射用蒸留水、生理食塩水、プロピレングリコール、ポリエチレングリコール、植物油、アルコール類等を含むことができる。さらに、湿潤剤、乳化剤、分散剤、安定化剤、溶解剤、溶解補助剤、防腐剤等を加えることができる。
【0043】
上記医薬組成物は、TRPC6に関連する疾患の治療又は予防に有効である。TRPC6に関連する疾患としては、TRPC6チャネロパチー(channelopathy)が挙げられる。チャネロパチーとは、イオンチャネルの機能の異常に基づいて発症する疾患の総称である。上記医薬組成物は、TRPC6の機能を抑制するものであるから、TRPC6の発現又は機能が亢進することによって発症するTRPC6チャネロパチーに特に有用である。かかるTRPC6チャネロパチーとしては、例えば、糸球体硬化症、単状糸球体硬化症(FSGS)、後天性ネフローゼ、特発性肺高血圧症、低酸素性肺高血圧症、心臓肥大、心不全、神経変性疾患、呼吸器疾患、腎疾患等が挙げられるがこれらに限定されない。本発明の医薬組成物は、特に遺伝子変異を伴わずにTRPC6の発現が増加、及び/又はTRPC6の機能亢進を生じるTRPC6チャネロパチーに有用である。
哺乳類(例えば、ヒト、マウス、ラット、モルモット、ウサギ、イヌ、ウマ、サル、ブタ等)、特にヒトに投与する場合の投与量は、症状、患者の年齢、性別、体重、感受性差、投与方法、投与間隔、有効成分の種類、製剤の種類によって異なり、特に限定されないが、例えば、30μg〜100g、100μg〜500mg、100μg〜100mgを1回又は数回に分けて投与することができる。注射投与の場合、患者の体重により、1μg/kg〜3000μg/kg、3μg/kg〜1000μg/kgを1回又は数回に分けて投与してもよい。
【0044】
[スクリーニング方法]
本発明のスクリーニング方法は、候補化合物から、TRPC6の活性化を抑制する化合物を選択するための方法である。
当該スクリーニング方法は、その一態様において、候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、候補化合物の存在下と非存在下における前記細胞へのCa2+イオンの流入をモニターする工程と、を含む。
TRPC6及びPLC-γ1を発現する細胞としては、例えば元来双方を発現するpodocyteを用いることもできるし、上述した本発明に係る発現ベクターでPLC-γ1を発現する細胞を形質転換することによって得たものを用いることもできる。PLC-γ1は、全身の様々な細胞で発現している。PLC-γ1を発現しており、本スクリーニング方法に好適に利用できる細胞として、例えばHEK293T細胞が挙げられる。細胞の培地は、使用する細胞によって適宜選択することができる。HEK293T細胞の場合は、10%FBSを加えたDMEM培地を使用することができる。
候補化合物としては、例えば低分子化合物、高分子化合物、ペプチド、タンパク質、核酸が挙げられるがこれらに限定されない。候補化合物は、例えば培地に添加することができる。また候補化合物がペプチドやRNAである場合、これらをコードする核酸を含む発現ベクターで上記細胞を形質転換し、細胞内で発現させてもよい。
【0045】
TRPC6の284位のチロシンをリン酸化するキナーゼとしては、例えば、プロテインキナーゼC、プロテインキナーゼG、非受容体型チロシンキナーゼであるSrcファミリー(Src、Fyn、Yes、Blk、Fgr、Hck、Lck、Lyn)、等を用いることができ、中でもFynが好ましい。
TRPC6の284位のチロシンをリン酸化するキナーゼは、タンパク質としてスクリーニング系に加えてもよいし、当該キナーゼをコードする核酸を含む発現ベクターによりTRPC6及びPLC-γ1を発現する細胞を形質転換し、細胞内でキナーゼを発現させてもよい。キナーゼを加えてインキュベートする工程に先立って、細胞をEGFで刺激しておくことも好ましい。
細胞へのCa2+の流入をモニターする工程も、公知の方法又はそれに準ずる方法で行うことができる。例えば、Fura 2、Fluo 4、Rhod 2などのアセトキシメチルエステル(AM)を細胞内に取り込ませて細胞内のCa2+濃度を測定する方法が挙げられる。Fura-2/AMは、Ca2+の濃度上昇に伴い、335nm〜340nmで励起すると500nmの蛍光強度が増大し、370nm〜380nmで励起すると蛍光強度が減少するので、2波長励起による蛍光強度の比を取ることによって、バックグラウンド蛍光等に左右されず細胞内のカルシウムイオン濃度を性格に測定することができる。
候補化合物の存在下で、候補化合物の非存在下に比較して、細胞内へのCa2+流入が抑制される場合、当該候補化合物はTRPC6の膜移行を阻害して、TRPC6の活性化を抑制する化合物であると特定することができる。こうして特定された化合物は、TRPC6の発現の亢進を通じてCa2+の細胞内への流入が増加することに基づくTRPC6チャネロパチーの治療又は予防薬として有用である。
【0046】
本発明に係るスクリーニング方法は、別の態様において、候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、候補化合物の存在下と非存在下における前記細胞表面におけるTRPC6の発現を測定する工程と、を含む。候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程は上述のとおり行うことができる。
細胞表面におけるTRPC6の発現を測定する工程は、例えば、細胞表面をビオチン化し、ビオチン化されたタンパク質をストレプトアビジンで回収し、当該タンパク質の中にTRPC6が含まれるかどうかを確認することによって行うことができる。回収されたタンパク質にTRPC6が含まれるかどうかの確認は、例えばウエスタンブロッティングにより行うことができる。
候補化合物の存在下で、候補化合物の非存在下に比較して、細胞表面でのTRPC6の発現が低下する場合、当該候補化合物はTRPC6の膜移行を阻害して、TRPC6の活性化を抑制する化合物であると特定することができる。こうして特定された化合物は、TRPC6の発現の亢進を通じてCa2+の細胞内への流入が増加することに基づくTRPC6チャネロパチーの治療又は予防薬として有用である。
【0047】
本発明に係るスクリーニング方法は、さらに別の態様において、候補化合物と配列番号:4で表されるアミノ酸配列を含むポリペプチドとをインキュベートする工程と、前記候補化合物と前記ポリペプチドとの結合を検出する工程と、を含む。
配列番号:4で表されるアミノ酸配列を含むポリペプチドは、例えば、配列番号:4で表されるアミノ酸配列を含む20アミノ酸以下のポリペプチドとすることができ、配列番号:4で表されるアミノ酸配列のN末端及び/又はC末端に任意のアミノ酸が結合したものとすることができる。
候補化合物と、ポリペプチドとの結合は、例えばポリペプチドを固相担体に固定し、標識した候補化合物を加えてインキュベートした後洗浄して、固相担体に捕捉された候補化合物を同定することによって検出又は測定することができる。
候補化合物がポリペプチドやタンパク質である場合、例えば、放射性物質(25I、131I、35S、3H等)、蛍光物質(フルオレセインイソチオシアネート、ローダミン、ダンシルクロリド、フィコエリスリン、テトラメチルローダミンイソチオシアネート、近赤外蛍光材料等)、発光物質(ルシフェラーゼ、ルシフェリン、エクオリン等)、ナノ粒子(金コロイド、量子ドット)、酵素(アルカリホスファターゼ、ペルオキシダーゼ等)で標識することができる。また候補化合物をビオチンで標識し、標識したアビジン又はストレプトアビジンで検出又は測定してもよい。候補化合物とポリペプチドとの結合は、その他の方法、例えば免疫沈降法、表面プラズモン共鳴法等の公知の方法でも検出又は測定することができる。
候補化合物が配列番号:4で表されるアミノ酸配列からなるポリペプチドと結合する場合、当該候補化合物は、284位がリン酸化されたTRPC6とPLC-γ1との結合を拮抗的に阻害しうる。その結果、TRPC6の膜移行を阻害して、TRPC6の活性化を抑制しうる。こうして特定された化合物は、TRPC6の発現の亢進を通じてCa2+の細胞内への流入が増加することに基づくTRPC6チャネロパチーの治療又は予防薬として有用である。
【0048】
本発明のスクリーニング用キットは、本発明のスクリーニング方法に必要な試薬、細胞、培地、使用説明書等を含む。
本発明のスクリーニング用キットは、例えば、TRPC6及びPLC-γ1を発現する293T細胞;TRPC6、PLC-γ1、及びFynを発現する293T細胞;DMEM培地;Fura 2/AM;配列番号:4で表されるアミノ酸配列を含むポリペプチド;及び細胞培養のためのカバースリップ、EGF等の中から適宜選択したものを備えていてもよい。
【0049】
[治療方法]
本発明は、TRPC6とPLC-γ1との結合を阻害する工程を含む、TRPC6チャネロパチーの治療又は予防方法も含む。TRPC6とPLC-γ1との結合を阻害することにより、TRPC6の膜移行を防ぎ、チャネル活性を低下させて、細胞内へのCa2+の過剰な流入を抑制することができる。従って、本発明の治療又は予防方法は、Ca2+の過剰な流入を原因とする疾患に有効である。
TRPC6とPLC-γ1との結合を阻害する工程は、例えば、本発明の医薬組成物を投与することによって行うことができるが、これに限定されない。
【実施例】
【0050】
以下、本発明を実施例に基づいて具体的に説明するが、本発明はこれらに何ら限定されるものではない。
実施例では、以下に示す材料と方法を用いた。
【0051】
抗体と試薬
ウサギ抗α−リン酸化TRPC6抗体(抗α-pY31抗体、抗α-pY284抗体)は、合成した13アミノ酸のオリゴペプチドCNESQDpYLLMDEL(pY31;配列番号:6)及びCLASPApYLSLSSE (pY284;配列番号:7)を抗原として、それぞれ作製した。各オリゴペプチドの最初のシステイン残基は、TRPC6の配列とは異なる。
抗血清は、抗原をSulfoLink(Pierce, Rockford, IL)に固定してアフィニティー精製し、リン酸化されていないペプチドで吸収した。抗α-Flag抗体(M2; Sigma, St. Louis, MO)、抗α-HA抗体(3F10; Roche Diagnostics, Indianapolis, IN)、抗α-リン酸化チロシン抗体(4G10; Upstate , Lake Placid, NY)、ウサギ抗α-TRPC6抗体(Alamone labs, Jerusalem, Israel)、ウサギ抗α-PLC−γ1抗体(Santa Cruz Biotechnology, Inc., Santa Cruz, CA)は、それぞれ購入した。
ウサギ抗α-Nephrin抗体、及び抗α-phospho-Nephrin(pY1204)抗体は、Haritaらの方法(Harita Y et al. (2009) J Biol Chem)に従って調製した。ウエスタンブロッティングは、これらの抗体を1/2000に希釈して行った。
なお、図中、「WB:HA」「WB:Flag」等の表記は、それぞれ抗α−HA抗体、抗α−Flag抗体でウエスタンブロッティングを行ったことを示し、「IP HA」「IP Flag」等の表記は、それぞれ抗α−HA抗体、抗α−Flag抗体で免疫沈降を行ったことを示す。
PP2(Merck KGaA, Darmstadt, Germany)、SU6656(Merck KGaA)、及びEGF(BD Biosciences, Bedford, MA)は購入した。
【0052】
細胞培養とRNAiのトランスフェクション
ヒトHEK293T細胞は、ATCC(Manassas, VA)より購入した。細胞は、10%ウシ胎仔血清を含むDMEM(Dulbecco's modified Eagle's medium)で維持した。
トランスフェクションは、Lipofectamine 2000 reagent (Invitrogen, Carlsbad,
CA)を使用して行った。
PLC−γ1 small interfering RNA (siRNA) Duplex1/2、及びコントロール用のsiCONTROL nontargeting siRNAは、Invitrogenから購入した。Duplex1/2の塩基配列を以下に示す。
Duplex1 (LotNo.87031E10)
CCCUGCGCUGUAAUGAGUUUGAGAU sense (配列番号:12)
AUCUCAAACUCAUUACAGCGCAGGG antisense (配列番号:13)
Duplex2 (LotNo.87031E11)
CCUUGUUGACCUCAUCAGCUACUAU sense (配列番号:14)
AUAGUAGCUGAUGAGGUCAACAAGG antisense (配列番号:15)
これらのsiRNAは、293T細胞にLipofectamine RNAiMAX (Invitrogen)を用いてトランスフェクトした。
【0053】
真核細胞発現ベクター
全長ラットNephrin、全長ラットNephrin-Flag、Y1204F Nephrin、Y1204F Nephrin-Flagをコードする哺乳動物発現プラスミドは、Haritaらの方法(Harita Y et al. (2008) J Biol Chem 283(14): 9177-9186、及びHarita et al, 2009)に従って準備した。
Fynの発現ベクター(Tezuka T et al. (1999) Proc Natl Acad Sci USA 96(2): 435-440)は、東京大学のDr. T. Tezukaより提供を受け、マウスTRPC6-HA(非特許文献2)は、理化学研究所脳科学脳科学総合研究センターのDr. C. Hisatsuneより提供を受け、ヒトFlag-PLC-γ1(Bae SS et al. (2000) FASEB J 14(9):1083-1092)は、Pohang大学(韓国)のDr. B. G. Suhより提供を受けた。PLC-γ1のSH2変異体は、Baeら及びPlattnerらの方法(Bae YS et al. (1998) J Biol Chem 273(8): 4465-4469; Plattner R et al. (2003) Nat Cell Biol 5(4): 309-319)に従って作製した。
マウスTRPC6変異体(Y31F、Y50F、Y85F、Y107F、Y206F、Y208F、Y284F、Y895F、P112Q、N143S、S270T、K874X、R895C、E897K、及び98-104欠失変異体)とNephrin-Flag欠失変異体(1216位-1227位を欠失)をコードする哺乳動物発現プラスミドは、PCRを用いた標準的な方法で作製した。
すべてのコンストラクトについて、DNA配列解析を行い、配列を確認した。PCRで用いたオリゴヌクレオチドは、下表に記載のとおりである。
【0054】
【表1】

【0055】
【表2】
【0056】
【表3】
【0057】
【表4】
【0058】
【表5】
【0059】
【表6】
【0060】
細胞表面のビオチン化
TRPC6の細胞表面における発現は、Winnらの方法に従って解析した(非特許文献17)。それぞれ図中に示されたプラスミドを発現する293T細胞を、2mg/mlのSulfo-NHS-SS-Biotin(Pierce)で表面をビオチン化した(30分、4℃)。ビオチン化されたタンパク質をストレプトアビジンビーズ(Pierce)で回収し、ウエスタンブロットにより解析した。
【0061】
免疫沈降
細胞をLysis buffer(20 mM Tris-HCl(pH 7.4), 150 mM NaCl, 1% Nonidet P-40, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 50 mM NaF, 10g/ml antipain, 10μg/ml leupeptin, 10g/ml aprotinin, 1 mM sodium vanadate)で、氷上に15分置いて溶解した。ライゼートを遠心処理し、M2 α−Flag抗体、又は3F10 α−HA抗体を結合させたビーズと4℃で1時間インキュベートした。TBS-1% NP40でビーズを洗浄し、結合したタンパク質を100mMグリシン−HCl(pH 2.6)で溶出させ、ウエスタンブロッティングに供した。
【0062】
融合タンパク質の細菌発現
GSTタグをつけたNephrinの細胞内ドメイン(Nephrin-CD:アミノ酸1102-1252;配列番号:1)は、Haritaらの方法に従って準備した(Harita et al, 2009)。GST-Nephrin-CD欠失変異体(1102位-1152位、1102位-1205位、1102位-1211位、1102位-1215位、1102位-1227位、1102位-1238位、1102位-1245位をそれぞれ欠失)は、PCRを用いた標準的な方法で作製した。細菌のペレットは、20 mM Tris-HCl (pH 7.5)、150 mM NaCl、1% Nonidet P-40、2 mM dithiothreitol、1mM phenylmethylsulfonyl fluoride、10μg/ml antipain、10μg/ml leupeptinを含む溶液で再懸濁し、超音波処理を行った。不溶性物質は遠心処理により除去した。GST融合タンパク質は、グルタチオン−セファロースカラム(GE Healthcare, Buckinghamshire, UK)により精製し、遊離グルタチオンとともに溶出させた。GST-Nephrin-CD欠失変異体を作製するために用いたオリゴヌクレオチドは、上記表6に示す。
【0063】
Pull-downアッセイ
グルタチオン−セファロースビーズに固定したGST-Nephrin-CDは、Haritaらの方法(Harita et al, 2008; Harita et al, 2009)に従い、Fynによってリン酸化した。
HA-TRPC6を発現する293T細胞のライゼートを、ビーズに固定したリン酸化又は非リン酸化GST-Nephrin-CDと共に4℃で一晩インキュベートした。
ビーズを洗浄バッファー(50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1% Nonidet
P-40)でよく洗浄した。Nephrinと結合したタンパク質を、遊離グルタチオンとともに溶出させ、ウエスタンブロッティングとCBB染色により解析した。ペプチドを用いたPull-downアッセイの場合は、ペプチドをSulfo Link Coupling Gel (Pierce)に固定して行った。
【0064】
質量分析法によるタンパク質の同定
目的のタンパク質を銀染したゲルから切り出し、1pmolのAPI(Achromobacter protease I;40μlのdigestion buffer (10 mM Tris, pH 8.5)中)とともに37℃で一晩静置して消化した。
ペプチドを精製し、高速液体クロマトグラフィー/タンデム質量分析装置(LC/MS; Q-STAR Elite, Applied Biosystems, Carlsbad, CA)を用いたペプチドマスフィンガープリンティングに供した。ペプチドイオンは、ProteinPilotTM (ver. 3.0)ソフトウエアにより解析した。
【0065】
ラット腎糸球体の分離
糸球体は、graded sieving techniqueを用いてラット腎皮質から分離し、タンパク質抽出のために保存した。すべての実験は、東京大学医科学研究所のAnimal Centerのガイドラインに従って行った。Wistarラットは、Charles River Laboratories Japan, Inc (Atsugi, Japan)より購入した。
【0066】
細胞内カルシウム濃度([Ca2+]i)の測定
カバースリップ上で培養した293T細胞を、4μM Fura-2/AM(Dojindo, Kumamoto,
Japan)で満たし、Hepes-buffered saline(HBS; 115 mM NaCl, 5.4 mM KCl, 20 mM HEPES, 1 mM MgCl2, 2 mM CaCl2, 10 mM glucose, pH 7.4)に浸漬させ、30分静置した。
続いて細胞を、室温にて、Ca2+-free HBS(115 mM NaCl, 5.4mM KCl, 20 mM HEPES, 2 mM MgCl2, 10 mM glucose, 0.05 mM EGTA, pH 7.4)及びHBSでかん流した。
Fura-2を搭載した細胞の蛍光は、Olympus IX70顕微鏡で、20×対物レンズ(N.A. 0.75)を用いて観察した。Fura-2の蛍光は、波長340nmと380nmで10秒ごとに交互に励起した。冷却CCDカメラ(ORCA-ER, Hamamatsu Photonics, Hamamatsu, Japan)により像を記録した。
蛍光強度の比(F340/F380)の経時的変化は、各視野のすべての細胞(400〜500)をもとに計算し、平均した。画像収集とデータ解析は、マッキントッシュコンピュータのwritten by T.IのTRPC6I Workbenchソフトウエアによって行った。
【0067】
ペプチドの送達
TRPC6結合領域(1216-1227: CWPEVQCEDPRGI(配列番号:3))を含むNephrinペプチド、又はコントロールペプチド(Nephrin C-terminus; amino acids 1241-1252: CSSLPFELRGHLV(配列番号:8))は、Transport Protein Delivery Reagent (Takara, Tokyo, Japan)を用いて293T細胞に送達させた。
【0068】
統計
数値は、平均±SEMで示す。二つのグループの[Ca2+]iの比較は、Mann-Whitney U-testにより評価した。ウエスタンブロッティングのバンド強度の比較には、Unpaired Student's t-testを用いた。P<0.05を有意とした。
【0069】
実施例1:TRPC6の膜移行におけるTyr284のリン酸化の必要性
TRPCチャネルは細胞膜に移行することによって活性化されると考えられていたことから、表面ビオチン化アッセイによって、TRPC6のリン酸化が膜移行に及ぼす影響を調べた。HAタグで標識したTRPC6を発現する293T細胞をEGFで刺激し、図示される時間が経過した後にビオチン化した。ビオチン化されたTRPC6は、basal conditionでは検出されなかった(図1A)。TRPC6は刺激後1分で細胞膜に現れ、3分後に上限に達し、その状態が60分継続した。構成的に活性なFynの共発現により、チロシンのリン酸化と、細胞表面に局在するTRPC6が増加した(図1B、C)。
TRPC6は、その細胞内ドメインに23のチロシン残基を有する。そこで、どのチロシン残基のリン酸化がTRPC6の膜移行に必要なのか調べるため、Y31、Y50、Y85、Y107、Y206、Y208、Y284、及びY895の8つのチロシン残基のそれぞれをフェニルアラニンに置換して、TRPC6の細胞内局在に与える影響を解析した。Y107、Y206、Y208、及びY284は、TRPC3におけるリン酸化部位として報告されているY49、Y148、Y150、及びY226にそれぞれ対応するものである。Y31、Y50、Y85及びY895は、モチーフサーチプログラムのScan Site (http://scansite.mit.edu)によってSH2含有タンパク質との結合部位と予測された場所である。また、これらの8つのチロシンは、すべて複数の種(ヒト、ラット、マウス、イヌ)において保存されている。
この結果、284位のチロシンをフェニルアラニンに置換したY284F変異は、Fynによって誘導されるTRPC6の細胞表面での発現を著しく減少させた(図1D)。このことから、TRPC6の膜移行には、284位のチロシンのリン酸化が重要であることが示唆された。
【0070】
実施例2:PLC-γ1とリン酸化されたTRPC6との結合がTRPC6の膜移行に及ぼす影響
実施例1の結果から、リン酸化されたTRPC6のY284周辺は、TRPC6の輸送に必要なタンパク質の結合領域になるものと仮定した。TRPC6の輸送に必要なタンパク質を特定するために、リン酸化された、又はリン酸化されていないY284周辺の13アミノ酸のTRPC6ペプチド(配列番号:7)を用いたin vitroの結合アッセイを行った。
両ペプチドをカップリングゲルに固定して、293T細胞ライゼートとインキュベートし、結合したタンパク質をSDS-PAGEに供し、銀染した。図2Aに示すとおり、見かけの分子量が140,000のタンパク質がリン酸化したTRPC6 Y284ペプチドに特異的に結合することが確認された(*で示す)。このバンドのトリプシン消化物を質量分析法で解析したところ、当該タンパク質はPLC-γ1と同定された。
また、共免疫沈降により、TRPC6は、Fyn依存性にPLC-γ1に結合することを確認した(図2B)。Y31Fの変異は、PLC-γ1への結合を部分的に無効とし、Y31とY284がいずれも変異したTRPC6は、PLC-γ1と相互作用しなかった。他のTRPC6変異体は、野生型TRPC6と同等にPLC-γ1に結合した(data not shown)。従って、TRPC6とPLC-γ1の相互作用には、Y284のリン酸化のみでなく、Y31のリン酸化も必要とされることがわかった。
PLC-γ1は、分子の中心に隣り合った2つのSH2ドメイン(N-SH2、C-SH2)を有する。pY31及びpY284ペプチドは、PLC-γ1 N-SH2に選択的に結合する。N-SH2の点変異又は欠失は、結合を強く阻害した。
また、siRNAによって内因性PLC-γ1をノックダウンすると、細胞膜上のビオチン化TRPC6が著しく減少した。このことは、Fynに誘導されるTRPC6の膜移行にPLC-γ1が不可欠であることを示す。
PLC-γ1はTRPC3のN末端アミノ酸(40位〜46位)と結合し、プレクストリン相同ドメイン(PHドメイン)を形成することが知られている。このPHドメインが、ホスファチジルイノシトール2リン酸(PIP2)に結合し、TRPC3の細胞表面での発現を誘導する。そこで、TRPC6における対応するN末端アミノ酸(98位〜104位)の機能を調べるため、98位〜104位を欠失したTRPC6変異体を用いて共免疫沈降とビオチン化アッセイを行った。当該変異体は、野生型TRPC6と同様に、PLC-γ1への結合能を有し(図2F)、Fyn依存的に細胞膜に移行した(図2G)ことから、98位〜104位の残基ではなく、Y31とY284がPLC-γ1との相互作用及び膜移行に不可欠であることがわかった。
【0071】
実施例3:糸球体におけるSrcファミリーキナーゼによるTRPC6 Y31及びY284のリン酸化
pY31及びpY284を特異的に認識する抗TRPC6抗体を作製した(図3)。各抗体の特異性はTRPC6のY31F及びY284F変異体を用いて確認した(図3A)。EGFもこれらの部位のリン酸化を誘導した(図3B)。単離したラット腎糸球体を、非特異的プロテインホスファターゼ阻害剤vanadateで処理すると、内因性TRPC6のY284のリン酸化は有意に亢進した(図3C)。このリン酸化は、vanadate処理に先立って、Srcファミリーキナーゼの特異的阻害剤PP2又はSU6656で前処理することにより、抑制された。このことは、Y31及びY284のリン酸化がSrcファミリーキナーゼによるものであることを示唆する。
【0072】
実施例4:チロシンをリン酸化されたTRPC6とNephrinとの相互作用
TRPC6はスリット膜の構成要素であるので、他のスリット膜の構成要素の結合が、リン酸化依存的な膜移行に影響を及ぼすか調べた。まず、TRPC6とFynに加え、Nephrin、Neph1、Neph2、又はPodocinを発現する293T細胞を用いた共免疫沈降を行った。その結果、Nephrinは、Fynと共発現させた場合のみTRPC6に結合することを見出した(図4A)。他のスリット膜の構成要素とTRPC6との相互作用は検出されなかった。
NephrinもSrcファミリーキナーゼによってチロシンをリン酸化されることが知られているので、どのリン酸化がTRPC6とNephrinの相互作用に必要なのか調べた。リン酸化されるとNephrinのY1204はPLC-γ1に結合することが知られている(Harita et al., 2009)。しかしながら、このリン酸化部位は、TRPC6との結合には関与しておらず、Y1204の変異体もTRPC6に結合した(図4A)。
次に、GST−Nephrin細胞内ドメイン(Nephrin-CD, 1102位〜1252位;配列番号:1)を、予めin vitroでリン酸化して、又はリン酸化せずに、pull-downアッセイを行った。Fynを共発現させた293T細胞のTRPC6は、in vitroにおいて、Nephrin-CDがFynにリン酸化されているか否かに関わらずGST-Nephrin-CDに結合したが、Fynを共発現させない細胞のTRPC6はGST-Nephrin-CDに結合しなかった(図4B)。従って、TRPC6とNephrinの相互作用にはTRPC6のリン酸化が必要である一方、この相互作用にNephrinのリン酸化は不要であることがわかった。
TRPC6のY31及びY284のリン酸化は、PLC-γ1との相互作用に必要である(図2B)。同様に、チロシンをフェニルアラニンに置換したTRPC6の一連の変異体と、Nephrinとの結合を評価し、TRPC6とNephrinの相互作用に不可欠なチロシンを決定した(図4C)。Y284F変異体はほぼ完全にNephrinと相互作用しなかったが、Y31Fを含む他の変異体は野生型と同様に相互作用したことから、両者の相互作用に不可欠なのはY284のリン酸化であることがわかった。
Y284リン酸化の重要性をさらに確認するため、GST-Nephrin-CDを使ったpull-downアッセイを行った(図4D)。全長TRPC6ほどではなかったが、Y284を囲むTRPC6ペプチド(配列番号:7)もリン酸化されると、293T細胞からNephrinとGST-Nephrin-CDをpull-downし(図4E)、NephrinとTRPC6が直接結合することを示した。
Nephrin細胞質ドメインにおけるTRPC6との結合に必要な領域を特定するため、GST-Nephrin-CDの一連の欠失変異体を構築した。これらの欠失変異体を用いたGST-pull-downアッセイの結果、NephrinとTRPC6の相互作用には、1216位〜1227位を含む残基が不可欠であり(図4F)、この領域を欠失するとNephrinはTRPC6と結合できないことがわかった(図4G)。
【0073】
実施例5:Nephrinによる、リン酸化依存的なTRPC6の細胞表面での発現とチャネル活性化の抑制
PLC-γ1及びNephrinの細胞質ドメインは、いずれもTRPC6の同一のチロシン残基(Y284)に結合するので、これらが競合的に結合する可能性を検討した。図5Aに示されるとおり、Nephrinを共発現させると、TRPC6とPLC-γ1の相互作用は完全に阻害された。
Nephrinは、リン酸化されたY1204がPLC-γ1との結合部位となるが、Y1204は、NephrinのTRPC6結合領域(1216位〜1227位)にも非常に近い。そのため、NephrinによるTRPC6−PLC-γ1複合体形成の阻害は、リン酸化されたNephrin Y1204にPLC-γ1が結合することによる可能性も考えられた。しかしながら、PLC-γ1に結合しないNephrin Y1204F変異体も、TRPC6−PLC-γ1相互作用を阻害した(図5A)。このことから、Nephrinは、NephrinとPLC-γ1との相互作用とは関係なく、TRPC6とPLC-γ1との相互作用を阻害することがわかった。
PLC-γ1はTRPC6の膜移行において重要な役割を果たす(図2E)ので、Nephrinは、TRPC6−PLC-γ1の相互作用を阻害することによって、TRPC6の輸送を阻害する可能性がある。そこで、表面ビオチン化アッセイを行ったところ、Nephrinを共発現させると、TRPC6のFyn誘導性の細胞表面での発現が抑制された(図5B)。TRPC6に結合しないNephrinの欠失変異体(Δ1216-1227)による抑制は、かなり低かった。
次に、Nephrinペプチド(1216-1227残基)を、TRPC6を発現する293T細胞に導入し、Fyn誘導性のTRPC6膜移行を評価した(図5C)。Nephrinペプチドの導入により、Fyn依存性のTRPC6の膜移行が遮断された。一方、コントロールペプチド(NephrinのC末端における同じ長さのペプチド;配列番号:8)を導入しても、TRPC6の膜移行は遮断されなかった。PLC-γ1は、Fynと共発現させた細胞のビオチン化サンプル中で検出された(図5C)。Nephrinペプチドは、PLC-γ1の細胞膜への移行も阻害した。
さらに、TRPC6とFynを発現する293T細胞へのCa2+の流入をモニターしたところ、NephrinはTRPC6チャネル活性を抑制した(図5D、E)。Fynは、TRPC6を発現する細胞におけるCa2+流入を増大させたが、Nephrinを共発現させるとFyn依存性のCa2+流入の増大は有意に抑制された(p<0.001, Mann Whitney test)。この結果は、TRPC6の膜表面発現の阻害効果と一致する。Y284F変異体のFyn誘導性の増加は、同条件ではわずかであった。
また、Nephrinペプチドも、TRPC6チャネル活性に対して、Nephrinと同様に阻害効果を示した(p<0.001, 図5F、G)。この結果は、NephrinペプチドがTRPC6の膜移行を抑制する結果と一致する。
実施例6:TRPC6のFSGS変異体に対するNephrinの阻害効果の低下
TRPC6の変異がFSGSの原因であると特定されたことから、複数のTRPC6チャネル変異体の活性が培養細胞を用いて測定されてきた。しかしながら、それらの結果は、チャネル活性とFSGSの発症の関係を明らかに示すものではなかった。いくつかのTRPC6変異体(P112Q、R895C、E897K)は、チャネル活性を亢進させ、他の変異(N143S、S270T、K874X)は亢進させない。これらの見かけ上矛盾した結果から、podocyte特異的なTRPC6の調節が存在するとの仮説を立てた。
この仮説に基づいて、疾患を引き起こすTRPC6の変異(以下、「疾患関連変異」)であるP112Q、N143S、S270T、K874X、R895C、及びE897Kのそれぞれと、Nephrinとの相互作用を共免疫沈降により解析した(図6A、B)。K874X以外の5つの変異が、TRPC6とNephrinとのリン酸化依存的な相互作用を減少させた。
次に、TRPC6−PLC-γ1相互作用と、TRPC6変異体の細胞表面での発現に対するNephrinの阻害効果を解析した。興味深いことに、Nephrinは、野生型TRPC6の結果と対照的に、TRPC6の疾患関連変異体とPLC-γ1との複合体形成を抑制しなかった。すべてのTRPC6疾患関連変異体について、Nephrinの非存在下における野生型TRPC6と同様に、Fyn誘導性の膜移行が観察された(data not shown)。野生型TRPC6の結果と対照的に、Nephrinは、TRPC6の疾患関連変異体の細胞表面での発現を阻害しなかった(図6D)。
最後に、TRPC6の疾患関連変異体のFyn依存的チャネル活性化にNephrinが及ぼす影響を調べた(図6E、F)。膜局在の結果と一致して、Nephrinは、野生型TRPC6のFyn誘導性のチャネル活性化を抑制したが、疾患関連変異体は、Nephrinに感受性を示さなかった。この結果は、患者のpodocyteにおけるTRPC6の細胞表面での過剰発現と、TRPC6の機能亢進による過剰なCa2+流入を説明し得る。
【配列表フリーテキスト】
【0074】
配列番号:1は、Nephrin細胞内ドメインのアミノ酸配列を示す。
配列番号:2は、全長TRPC6のアミノ酸配列を示す。
配列番号:3は、Nephrinペプチドのアミノ酸配列を示す。
配列番号:4は、TRPC6のpY284周辺ポリペプチドのアミノ酸配列を示す。
配列番号:5は、全長Nephrinのアミノ酸配列を示す。
配列番号:6は、TRPC6のpY31周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:7は、TRPC6のpY284周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:8は、NephrinのC末端1241位−1252位からなるペプチドのアミノ酸配列を示す。
配列番号:9は、TRPC6のY31周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:10は、TRPC6のY284周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:11は、Nephrinペプチドをコードする核酸の塩基配列を示す。
配列番号:12は、siRNA (Duplex1)のセンス鎖の塩基配列を示す。
配列番号:13は、siRNA (Duplex1)のアンチセンス鎖の塩基配列を示す。
配列番号:14は、siRNA (Duplex2)のセンス鎖の塩基配列を示す。
配列番号:15は、siRNA (Duplex1)のアンチセンス鎖の塩基配列を示す。
配列番号:16は、HA-TRPC6のY31F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:17は、HA-TRPC6のY31F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:18は、HA-TRPC6のY50F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:19は、HA-TRPC6のY50F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:20は、HA-TRPC6のY85F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:21は、HA-TRPC6のY85F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:22は、HA-TRPC6のY107F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:23は、HA-TRPC6のY107F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:24は、HA-TRPC6のY206F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:25は、HA-TRPC6のY206F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:26は、HA-TRPC6のY208F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:27は、HA-TRPC6のY208F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:28は、HA-TRPC6のY284F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:29は、HA-TRPC6のY284F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:30は、HA-TRPC6のY895F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:31は、HA-TRPC6のY895F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:32は、HA-TRPC6のFSGS変異体(P112Q変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:33は、HA-TRPC6のFSGS変異体(P112Q変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:34は、HA-TRPC6のFSGS変異体(N143S変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:35は、HA-TRPC6のFSGS変異体(N143S変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:36は、HA-TRPC6のFSGS変異体(S270T変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:37は、HA-TRPC6のFSGS変異体(S270T変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:38は、HA-TRPC6のFSGS変異体(K874X変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:39は、HA-TRPC6のFSGS変異体(K874X変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:40は、HA-TRPC6のFSGS変異体(R895C変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:41は、HA-TRPC6のFSGS変異体(R895C変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:42は、HA-TRPC6のFSGS変異体(E897K変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:43は、HA-TRPC6のFSGS変異体(E897K変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:44は、HA-TRPC6の98位-104位欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:45は、HA-TRPC6の98位-104位欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:46は、Flag-Nephrinの1216位-1227位欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:47は、Flag-Nephrinの1216位-1227位欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:48は、Flag-PLC-γ1のN*(R586K)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:49は、Flag-PLC-γ1のN*(R586K)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:50は、Flag-PLC-γ1のC*(R694K)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:51は、Flag-PLC-γ1のC*(R694K)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:52は、Flag-PLC-γ1のDel(N-SH2)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:53は、Flag-PLC-γ1のDel(N-SH2)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:54は、Flag-PLC-γ1のDel(C-SH2)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:55は、Flag-PLC-γ1のDel(C-SH2)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:56は、GST-Nephrin-CDの1102-1211欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:57は、GST-Nephrin-CDの1102-1211欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:58は、GST-Nephrin-CDの1102-1215欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:59は、GST-Nephrin-CDの1102-1215欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:60は、GST-Nephrin-CDの1102-1227欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:61は、GST-Nephrin-CDの1102-1227欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
【技術分野】
【0001】
本発明は、TRPC6の膜移行を阻害することによってそのチャネル活性を抑制する新規なポリペプチド等に関する。
【背景技術】
【0002】
カルシウム濃度の動的変化は、分泌、収縮、細胞増殖、生存、及び分化を含む様々な細胞応答を引き起こす。Ca2+濃度の増加は、リガンドによる受容体の直接的な活性化、又は細胞内シグナルパスウェイを通じた間接的な活性化の結果、細胞膜上又は小胞体上のCa2+透過性チャネルが開くことによって開始される。
ショウジョウバエのTransient Receptor Potential Canonicalの哺乳類ホモログであるTRPCファミリーは、細胞膜上に存在する6回膜貫通型のイオンチャネルであり、細胞膜におけるCa2+の流入経路として機能するか、膜電位を変えてCa2+流入の駆動力を調節することによって、細胞質の遊離Ca2+濃度の制御に寄与している。哺乳類のTRPCチャネルの7つのサブタイプの中で、TRPC3、6及び7は、配列類似性が高い。これらのタンパク質は、受容体の刺激又はジアシルグリセロール類縁体の投与によって活性化される非選択的なカチオンチャネルである。
【0003】
TRPCチャネルが活性を調節するメカニズムの一つとして、細胞膜へのチャネルの挿入が挙げられる。即ち、TRPCチャネルは、細胞膜と細胞質の間を行き来し、細胞膜に移行することによって活性化される。TRPC3、4及び6は、Gq共役受容体又は受容体チロシンキナーゼの刺激によって細胞膜に移行する(非特許文献1)。プロテインキナーゼC、プロテインキナーゼG、SrcファミリーチロシンキナーゼによってもTRPCチャネルがリン酸化され、これらのチャネルにおける複数のリン酸化部位について報告がされている。
Srcファミリーキナーゼは、すべてのTRPCチャネルに作用する。また、TRPC4及びTRPC6は、上皮成長因子(epidermal growth factor; EGF)の刺激により、Srcファミリーキナーゼによってチロシンのリン酸化を受ける。FynはTRPC6をリン酸化し、ジアシルグリセロール刺激による活性化を増強する(非特許文献2)。しかしながら、リン酸化によってTRPCが活性化されるメカニズムは解明されていなかった。
【0004】
TRPCチャネルの調節の異常が、心疾患、神経変性疾患、呼吸器疾患、腎疾患等の様々な疾患の原因につながることが広く知られている(非特許文献3〜6)。
TRPC6は、血管平滑筋細胞における受容体作動性カチオンチャネルの必須の構成要素であり(非特許文献7)、低酸素性肺高血圧と関係があるとされている(非特許文献8、9)。また、TRPC3及びTRPC6は、カルシニューリンとその下流のエフェクターであるT細胞活性化因子(nuclear factors of activated T-cells, NFAT)を活性化することにより心臓肥大を促進する(非特許文献10〜13)。
【0005】
最近、TRPC6遺伝子の変異が、ヒトの蛋白尿性腎臓疾患である単状糸球体硬化症(focal segmental glomerulosclerosis; FSGS)に関連することが報告された(非特許文献14)。FSGSでは、糸球体上皮細胞(podocyte)と、podocyteの足突起(foot processes)間に形成された細胞間接着装置であるスリット膜とがintegrityを失い、糸球体のバリア機能が損なわれる。
TRPC6は、podocyte内で発現し、スリット膜の重要な構成要素であるNephrinとPodocinに結合して(非特許文献14、15)、腎臓における糸球体のバリア機能に必須の部分を形成している。
マウス腎臓におけるTRPC6の過剰発現は蛋白尿を引き起こすが(非特許文献16)、変異したTRPC6のチャネル活性が、どのように疾患の発症と関連するのかは不明であった。いくつかの変異体(P112Q、R895C、E897K)は、HEK293T細胞で発現させると、アンジオテンシンII受容体仲介性のTRPC6活性化を増大させたが、S270TとN143Sのミスセンス変異体や、57アミノ酸切断変異体(K874X)は、チャネル活性を変化させなかった(非特許文献14、17)。これらのチャネル活性は、下流のNFAT活性化の程度とよく相関した。一方、P112Q変異体はTRPC6の細胞膜での発現を増加させる(非特許文献17)。このことは、TRPC6の細胞膜での発現の変化も疾患の発症に寄与することを示唆している。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Kiselyov K, Patterson RL (2009) Front Biosci 14: 45-58
【非特許文献2】Hisatsune C et al. (2004) J Biol Chem 279(18):18887-18894
【非特許文献3】Abramowitz J, Birnbaumer L (2009) FASEB J 23(2): 297-328
【非特許文献4】Nilius B et al. (2007) Physiol Rev 87(1): 165-217
【非特許文献5】Nilius B et al. (2005) Sci STKE 2005(295): re8
【非特許文献6】Woudenberg-Vrenken TE et al. (2009) Nat Rev Nephrol 5(8): 441-449
【非特許文献7】Inoue R et al. (2001) Circ Res 88(3): 325-332
【非特許文献8】Lin MJ et al. (2004) Circ Res 95(5): 496-505
【非特許文献9】Wang J et al. (2006) Circ Res 98(12): 1528-1537
【非特許文献10】Bush EW et al. (2006) J Biol Chem 281(44): 33487-33496
【非特許文献11】Kuwahara K et al. (2006) J Clin Invest 116(12): 3114-3126
【非特許文献12】Nakayama H et al. (2006) FASEB J 20(10): 1660-1670
【非特許文献13】Onohara N et al. (2006) EMBO J 25(22): 5305-5316
【非特許文献14】Reiser J et al. (2005) Nat Genet 37(7): 739-744
【非特許文献15】Huber TB et al. (2006) Proc Natl Acad Sci U S A 103(46): 17079-17086
【非特許文献16】Moller CC et al. (2007) J Am Soc Nephrol 18(1): 29-36
【非特許文献17】Winn MP et al. (2005) Science 308(5729): 1801-1804
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、TRPC6の細胞膜への移行のメカニズムを解明し、当該移行を阻害することによってTRPC6チャネロパチーの予防又は治療に用いることが可能な医薬を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決するために検討した結果、細胞内のTRPC6は、284位のチロシン(Y284)がリン酸化されるとPLC-γ1と結合し、これにより細胞膜に移行すること;Y284のリン酸化は、TRPC6とNephrinの結合にも必須であること;Nephrinは、Y284がリン酸化されたTRPC6に対してPLC-γ1と競合的に結合し、TRPC6の膜移行を阻害すること;TRPC6との結合部位を含むNephrinの部分ペプチドは、TRPC6とPLC-γ1との結合を阻害する結果、TRPC6の膜移行を抑制してTRPC6のチャネル活性を低下させること、等を見出し、本発明を完成するに至った。
即ち、本発明は、
〔1〕以下のいずれかのポリペプチド:
(i)配列番号:1に記載のアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチド;
(ii)前記(i)の部分配列を含むポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び
(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;
〔2〕前記(ii)に記載のポリペプチドであって、配列番号:3で表される、Nephrinのアミノ酸配列における1216位〜1227位の領域を含む、上記〔1〕に記載のポリペプチド;
〔3〕上記〔1〕又は〔2〕に記載のポリペプチドをコードする核酸;
〔4〕上記〔3〕に記載の核酸を含む発現ベクター;
〔5〕上記〔4〕に記載の発現ベクターを含む形質転換体;
〔6〕配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位に結合する抗体;
〔7〕上記〔1〕若しくは〔2〕に記載のポリペプチド、上記〔3〕に記載の核酸、及び上記〔4〕に記載の発現ベクター、並びに上記〔6〕に記載の抗体からなる群より選択されるいずれか一つを含む医薬組成物;
〔8〕TRPC6チャネロパチーの予防又は治療剤である、上記〔7〕に記載の医薬組成物;
〔9〕前記TRPC6チャネロパチーは、糸球体硬化症、後天性ネフローゼ、特発性肺高血圧症、及び心不全からなる群より選択される、上記〔8〕記載の医薬組成物;
〔10〕候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞へのCa2+イオンの流入を比較する工程と、を含む方法;
〔11〕候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞表面におけるTRPC6の発現を測定する工程と、を含む方法;及び
〔12〕候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物と、配列番号:4で表されるアミノ酸配列を含むポリペプチドとをインキュベートする工程と、
前記候補化合物と前記ポリペプチドとの結合を検出又は測定する工程と、を含む方法、に関する。
【発明の効果】
【0009】
本発明に係るポリペプチドは、TRPC6の膜移行を阻害することによってそのチャネル活性を抑制することができるので、TRPC6のチャネル活性亢進によって発症・進展し得る疾患の予防又は治療に有用である。また、チャネル活性亢進の原因となる分子に特異的に作用すること、元来生体内に存在するタンパク質の一部であることなどから、副作用が少なく、安全性の高い医薬として利用されうる。
【図面の簡単な説明】
【0010】
【図1A】図1は、TRPC6 Y284のリン酸化が、細胞膜への移行に必須であることを示すために行った実験の結果である。図1Aは、EGFによるTRPC6の膜移行の誘導を示す。HA-TRPC6を発現する293T細胞をEGF(200 ng/ml)で、図示された時間刺激した。細胞表面をビオチン化し、ストレプトアビジン−アガロースビーズに結合したタンパク質を、抗α−TRPC6抗体を用いたウエスタンブロッティングで解析した。パネルの右側に、分子量マーカー(kDa)を示す。各ライゼートの一部(2%)をウエスタンブロッティングで解析し、TRPC6の発現レベルを確認した(下パネル)。
【図1B】図1Bは、293T細胞が発現したHA-TRPC6を、Fynの存在下、又は非存在下、α−HA抗体で免疫沈降を行い、α−リン酸化チロシン抗体で検出した結果を示す。
【図1C】図1Cは、HA-TRPC6を発現する293T細胞を、Fynの存在下、又は非存在下で、図1Aの方法に従って、細胞表面のTRPC6をビオチン化して検出した結果を示す。
【図1D】図1Dは、Fynと、チロシンのフェニルアラニン置換を行ったHA-TRPC6の一連の変異体のそれぞれとを、293T細胞で発現させ、図1Aの方法に従って、細胞表面のTRPC6をビオチン化して検出した結果を示す。3から5回の独立した実験の代表的なデータを示す。
【図2A】図2は、PLC-γ1が、TRPC6のリン酸化したY31/Y284に結合し、TRPC6の細胞表面での発現を制御することを示すために行った実験の結果である。図2Aは、リン酸化、又は非リン酸化TRPC6 Y284ペプチドをカップリングゲルに固定し、293T細胞のライゼートとインキュベートした後、結合したタンパク質を、SDS-PAGEと銀染により解析した結果を示す。*で示したバンドを切り出し、トリプシン消化した後、LC-MS/MSによって解析した。
【図2B】図2Bは、Flag-PLC-γ1、Fyn及びHA-TRPC6(野生型、Y31F、Y284F, Y31F/Y284F)を293T細胞で発現させ、図示された抗体を用いて、免疫沈降(IP)及びウエスタンブロット(WB)を行った結果を示す。アスタリスクで示されたバンドは、過剰発現したPLC-γ1との交差反応によるものである。
【図2C】図2Cは、リン酸化若しくは非リン酸化TRPC6 Y31周辺ペプチド(順に配列番号:6、9)、又はリン酸化若しくは非リン酸化Y284周辺のペプチド(順に配列番号:7、10)を用いて、293T細胞のライゼートからのPLC-γ1のpull-downを行った結果を示す。ライゼートの一部(0.5% input)をリカバリーマーカーとして使用した。
【図2D】図2Dは、PLC-γ1の野生型(WT)、SH2点変異(N*、C*)、またはSH2欠失変異(ΔN、ΔC)を用いて、図2Cの方法に従って、ペプチドpull-downアッセイを行った結果を示す。
【図2E】図2Eは、スクランブルsiRNA(コントロール)又はhPLC-γ1 siRNA(Duplex 1又は2)を293T細胞にトランスフェクトし、翌日、HA-TRPC6及びFynをトランスフェクトして、トランスフェクションの1日後、TRPC6の細胞表面での発現を図1Aの方法で測定した結果を示す。
【図2F】図2Fは、Flag-PLC-γ1、Fyn、及びTRPC6(野生型又はΔ98-104)をトランスフェクトした293T細胞を用いて、共免疫沈降を行った結果を示す。
【図2G】図2Gは、98位-104位を欠失したTRPC6(Δ98-104)の細胞表面での発現を表面ビオチン化アッセイで調べた結果である。
【図3A】図3は、生細胞及び腎糸球体におけるTRPC6のチロシンリン酸化に関する実験結果である。図3Aは、図示されたプラスミドをトランスフェクトした293T細胞のライゼートを用い、図示された抗体で免疫沈降を行い、抗α-pY31抗体「又は抗α-pY284抗体で標識した結果を示す。
【図3B】図3Bは、HA-TRPC6を発現する293T細胞をEGFで刺激し/または刺激せずに、抗α-HA抗体による免疫沈降物を、α-リン酸化特異的TRPC6抗体(抗α-pY31抗体又は抗α-pY284抗体)で標識した結果を示す。
【図3C】分離したラット糸球体を、1mM sodium vanadateで30分処理し、又は処理しないで、抗α-pY284抗体でブロットした結果を示す。各サンプルについて、vanadate処理に先立って、10μM PP2で15分、又は5μM SU6656で60分、糸球体を前処理した。
【図4A】図4は、TRPC6 Y284及びNephrin 1216-1227が、両者のチロシンリン酸化依存的相互作用に不可欠であること示す実験結果である。図4Aは、図示されたプラスミドをトランスフェクトした293T細胞のライゼートについて、図示された抗体を用いて免疫沈降及びウエスタンブロッティングを行った結果を示す。
【図4B】図4Bは、FynとHA-TRPC6を共発現する、又はHA-TRPC6のみ発現する293T細胞のライゼートを用い、GST-Nephrin-CD(1102-1252残基)又は予めFynでリン酸化したGST-Nephrin-CDを用いて、in vitroのpull-downアッセイを行った結果を示す。結合したタンパク質を抗α-HA抗体で検出した。GST-Nephrin及びTRPC6のリン酸化が確認された(下のパネル)。
【図4C】図4Cは、Flag-Nephrin、Fyn、及びHA-TRPC6(野生型、又はチロシンをフェニルアラニン置換した一連の変異体)を293T細胞で発現させ、抗α-Flag抗体で行った免疫沈降物と全細胞ライゼートについて、図示された抗体でウエスタンブロッティングを行った結果を示す。
【図4D】図4Dは、Fynと共に、又はFynなしに、野生型HA-TRPC6又はTRPC6のY284F変異体を一過性に発現する293T細胞のライゼートを用いて、GST-Nephrin-CDによってpull-downアッセイを行い、抗α-HA抗体で検出した結果を示す。
【図4E】図4Eは、NephrinまたはリコンビナントNephrin-CDを発現する293T細胞のライゼートについて、ビーズに固定したリン酸化/非リン酸化TRPC6 Y284ペプチド(順に配列番号:7、10)でpull-downし、結合したタンパク質を、α-Nephrin抗体を用いたウエスタンブロッティングにより解析した結果を示す。
【図4F】図4Fは、GST-Nephrin-CDの欠失変異体(1102位-1211位, 1102位-1215位, 1102位-1227位残基を欠失)を用いて、HA-TRPC6及びFynを発現する293T細胞のライゼートからチロシンリン酸化HA-TRPC6をpull-downした結果を示す。CBBはNephrin-CDの染色に用いたクマシーブリリアントブルーを示す。
【図4G】図4Gは、HA-TRPC6、Fyn及びFlag-Nephrin(野生型又は1216位-1227位を欠失したNephrin)を293T細胞にトランスフェクトし、抗α-HA抗体又は抗α-Flag抗体による免疫沈降物について、図示された抗体でウエスタンブロッティングを行った結果を示す。
【図5A】図5は、TRPC6のリン酸化依存的な細胞表面での発現とチャネル活性化を、Nephrinが下方制御することを示す実験結果である。図5Aは、図示されたプラスミドを293T細胞に導入し、共免疫沈降を行った結果を示す。
【図5B】図5Bは、図示されたプラスミドを293T細胞に導入し、表面ビオチン化アッセイを行った結果を示す。
【図5C】図5Cは、図示されたペプチドを293T細胞に導入し、表面ビオチン化アッセイを行った結果を示す。
【図5D】図5Dは、Fura-2を搭載した293T細胞に、図示されたプラスミドを導入し、Ca2+フリーHepes-buffered saline (HBS)、続いてCaCl2を含むHBSで表面かん流し、細胞内カルシウム濃度を測定した結果を示す。細胞内カルシウム濃度([Ca2+]i)は、fura-2蛍光のF340/F380比として、細胞数(n>400)をもとに記録し、平均値をプロットした。
【図5E】図5Eは、図5Dの場合について、ベースラインからのピーク比を各細胞について計算した結果を示す。棒グラフは、5つの独立した実験データの平均と標準誤差を示す。
【図5F】図5Fは、Fura-2を搭載した293T細胞に、図示されたプラスミド及び/又はペプチドを導入し、Ca2+フリーHepes-buffered saline (HBS)、続いてCaCl2を含むHBSで表面かん流し、細胞内カルシウム濃度を測定した結果を示す。細胞内カルシウム濃度([Ca2+]i)は、fura-2蛍光のF340/F380比として、細胞数(n>400)をもとに記録し、平均値をプロットした。
【図5G】図5Gは、図5Fの場合について、ベースラインからのピーク比を各細胞について計算した結果を示す。棒グラフは、5つの独立した実験データの平均と標準誤差を示す。
【図6A】図6は、TRPC6のFSGS関連変異体に対するNephrinの影響を調べた実験結果である。図6Aは、Flag-Nephrin、Fyn、及び各HA-TRPC6変異体を、293T細胞で共発現させ、抗α-Flag抗体による免疫沈降物を、図示された抗体によるウエスタンブロッティングに供した結果を示す。
【図6B】図6Bは、TRPC6-Nephrin結合について、図6Aのdensitometirc tracing(WB:TRPC6)によって定量し、平均値を棒グラフと標準誤差で示した結果を示す(n=3)。数値は、野生型TRPC6で標準化した。
【図6C】図6Cは、TRPC6−PLC-γ1相互作用に対するNephrin発現の影響を、Flag-PLC-γ1、Fyn、及び野生型又は各TRPC6変異体を発現する293T細胞を用いて調べた結果を示す。抗α-Flag抗体による免疫沈降物を、図示された抗体によるウエスタンブロッティングに供した。
【図6D】図6Dは、TRPC6の一連の疾患関連変異体を、Nephrin/Fynとともに、又はNephrin/Fynなしで293T細胞にトランスフェクトし、図1Aと同様に、TRPC6の細胞表面での発現を解析した結果を示す。
【図6E】図6Eは、Nephrin、Fyn、及びTRPC6若しくはTRPC6 N143S変異体を293T細胞で発現させ、[Ca2+]iの変化を図5Dと同様に記録した結果を示す(n=5)。
【図6F】図6Fは、FSGS関連TRPC6変異体のチャネル活性を図6Eと同様に評価した。データは、図5Eと同様に処理した。
【発明を実施するための形態】
【0011】
[ポリペプチド]
本発明に係るポリペプチドは、284位のチロシンがリン酸化されたTRPC6に結合し、TRPC6がPLC-γ1と結合して膜移行することを阻害できるポリペプチドであればどのようなポリペプチドであってもよいが、少なくとも以下の(i)〜(iii)を含む。
(i)配列番号:1で表されるアミノ酸配列を含むポリペプチド;
(ii)配列番号:1で表されるアミノ酸配列の部分配列を含むポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び
(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド。
【0012】
本明細書において、TRPC6の「膜移行」とは、TRPC6が細胞質から細胞膜に移行して、イオンチャネルとして機能できる状態となることを意味する。
【0013】
Nephrinは、約1200アミノ酸からなる膜貫通型タンパク質である。そのC末端側の150アミノ酸が細胞内ドメインであり、この部分にTRPC6との結合部位がある。
(i)のポリペプチドは、配列番号:1で表されるアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチドである。従って、TRPC6との結合部位を含むので、細胞内においてTRPC6に結合することによってTRPC6とPLC-γ1との結合を阻害し、TRPC6の膜移行を阻害しチャネル活性を低下させる。
(i)のポリペプチドは、配列番号:1に記載のアミノ酸配列のみからなるポリペプチドであってもよく、その少なくとも一端に1以上のアミノ酸が結合しているものであってもよい。(i)のポリペプチドの全長は、そのポリペプチドがTRPC6との結合能を有する限り、特に限定されないが、例えば150アミノ酸〜300アミノ酸とすることができる。
【0014】
上記(ii)のポリペプチドは、Nephrinの細胞内ドメインの一部を含み、且つ配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する限り、どのようなペプチドであってもよい。(ii)のポリペプチドとしては、Nephrin細胞内ドメインの一部の、例えば3アミノ酸〜50アミノ酸の領域を含むポリペプチド、5アミノ酸〜30アミノ酸の領域を含むポリペプチド、8アミノ酸〜20アミノ酸の領域を含むポリペプチド、10アミノ酸〜15アミノ酸の領域を含むポリペプチドが挙げられる。
(ii)のポリペプチドの具体例として、配列番号:5で表されるNephrinのアミノ酸配列における、1216位〜1227位の領域に該当する12アミノ酸(配列番号:3)以下、「Nephrinペプチド」と呼ぶ場合もある)が挙げられる。
【0015】
上記(iii)のポリペプチドは、(i)又は(ii)のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、且つ、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有するポリペプチドである限り、どのようなポリペプチドであってもよい。
本明細書において、用語「アミノ酸」は、その最も広い意味で用いられ、天然のアミノ酸のみならず、非天然アミノ酸も含む。本明細書においては、アミノ酸を慣用的な一文字表記又は三文字表記で示す場合もある。また、TRPC6について「n位のアミノ酸」という場合、配列番号:2で表されるアミノ酸配列のn番目のアミノ酸を示し、Nephrinについて「m位のアミノ酸」という場合、配列番号:5で表されるアミノ酸配列のm番目のアミノ酸を示す。さらに、n位のアミノ酸を別のアミノ酸で置換した場合、「n」の前に置換する前のアミノ酸を、「n」の後ろに置換後のアミノ酸を表す。例えば、TRPC6のY284F変異体は、配列番号:2で表されるアミノ酸配列における284位のチロシン(Y)をフェニルアラニン(F)に置換したものを示す。なお、本明細書において、一文字表記したアミノ酸の前に「p」が付されている場合、当該アミノ酸がリン酸化されていることを示す。例えば、pY284は、284位のチロシンがリン酸化されていることを表す。
【0016】
本明細書において、「1又は数個のアミノ酸が欠失、付加、若しくは置換されているポリペプチド」という場合、欠失等されるアミノ酸の個数は、そのポリペプチドが、Y284がリン酸化されているTRPC6に対する結合能を有する限り特に限定されないが、例えば、1〜9個、1〜5個、又は1〜3個とすることができる。あるいは全体の長さの10%以内、又は5%以内としてもよい。置換又は付加されるアミノ酸は、天然のアミノ酸、非天然アミノ酸、アミノ酸アナログであり得、好ましくは天然アミノ酸である。欠失、付加、置換される場所は、ポリペプチドの末端であっても、中間であってもよく、1ヶ所であっても2ヶ所以上であってもよい。
【0017】
(iii)のポリペプチドの例としては、ヒト以外の動物(例えば、マウス、ラット、ウサギ、ウマ、ヒツジ、イヌ、サル、ブタ等)に由来するNephrinの細胞内ドメイン又はその部分ペプチドが挙げられる。
【0018】
(ii)又は(iii)のポリペプチドが284位のチロシンがリン酸化されたTRPC6と結合能を有するか否かは、タンパク質間の相互作用を検出する公知の方法で確認することができる。例えば、284位のチロシンがリン酸化されたTRPC6、又は284位のチロシンがリン酸化された284位周辺を含むTRPC6の部分ポリペプチド(例えば、配列番号:4で表されるポリペプチド)を固定したプロテインマイクロアレイやアフィニティーカラムを用いる方法、免疫沈降法、表面プラズモン共鳴法等が挙げられる。
【0019】
本発明のポリペプチドは、そのC末端がカルボキシル基又はカルボキシレート基のみでなく、アミドやエステルになっていてもよい。また、本発明のポリペプチドには、ポリペプチドの塩も含まれる。ポリペプチドの塩としては、生理学的に許容される塩基や酸との塩が用いられ、例えば、無機酸(塩酸、臭化水素酸、ヨウ化水素酸、硫酸、リン酸等)の付加塩、有機酸(p-トルエンスルホン酸、メタンスルホン酸、シュウ酸、p-ブロモフェニルスルホン酸、カルボン酸、コハク酸、クエン酸、安息香酸、酢酸等)の付加塩、無機塩基(水酸化アンモニウム又はアルカリ若しくはアルカリ土類金属水酸化物、炭酸塩、重炭酸塩等)、アミノ酸の付加塩等が挙げられる。
【0020】
本発明のポリペプチドは、投与したときに細胞内に導入されるよう、膜透過性ペプチド(cell-penetrating peptide, CPP)と融合させてもよい。CPPは、細胞膜への親和性、細胞内への移行性を有するペプチドの総称であり、HIV-1ウイルスが発現するTrans-activator of transcription protein(TATタンパク質)の11個のアミノ酸からなるドメインであるprotein-transduction domain(PTD)や、ショウジョウバエのAntennapedia、ヘルペスウイルス由来のVP22、オリゴアルギニンなどが知られている。一般に、CPPはアルギニン、リジン、ヒスチジン等の塩基性アミノ酸の含有率が高いものが多い。
【0021】
また、本発明のポリペプチドは、本発明の課題を解決するものである限り、リン酸化、メチル化、アセチル化、アデニリル化、ADPリボシル化、糖鎖付加などの修飾が加えられたものであってもよい。他のペプチドやタンパク質と融合させたものであってもよい。
【0022】
本発明のポリペプチドは、天然物から抽出する方法;液相法、固相法、液相法と固相法を組み合わせたハイブリッド法等の化学合成法;遺伝子組み換え法等、公知のポリペプチドの製造方法によって製造することができる。
天然物から抽出する場合、例えば、ヒト又はその他の動物(例えば、マウス、ラット、ウサギ、ウマ、ヒツジ、イヌ、サル、ブタ等)の組織又は細胞から調製することができる。この場合、組織又は細胞をホモジナイズした後、酸などで抽出し、抽出液を塩析、透析、ゲルろ過、逆相クロマトグラフィー、イオン交換クロマトグラフィー、アフィニティークロマトグラフィーなどで精製することによって、精製単離することができる。さらに、必要に応じてプロテアーゼやペプチダーゼで切断してもよい。
【0023】
固相法は、例えば、水酸基を有するレジンの水酸基と、α−アミノ基が保護基で保護された第一のアミノ酸(通常、目的とするペプチドのC末端アミノ酸)のカルボキシ基をエステル化反応させる。エステル化触媒としては、1-メシチレンスルホニル-3-ニトロ-1,2,4-トリアゾール(MSNT)、ジシクロヘキシルカルボジイミド(DCC)、ジイソプロピルカルボジイミド(DIPCDI)等の公知の脱水縮合剤を用いることができる。
次に、第一アミノ酸のα−アミノ基の保護基を脱離させるとともに、主鎖のカルボキシ基以外のすべての官能基が保護された第二のアミノ酸を加え、当該カルボキシ基を活性化させて、第一及び第二のアミノ酸を結合させる。さらに、第二のアミノ酸のα−アミノ基を脱保護し、主鎖のカルボキシ基以外のすべての官能基が保護された第三のアミノ酸を加え、当該カルボキシ基を活性化させて、第二及び第三のアミノ酸を結合させる。これを繰り返して、目的とする長さのペプチドが合成されたら、すべての官能基を脱保護する。
【0024】
固相合成のレジンとしては、Merrifield resin、MBHA resin、Cl-Trt resin、SASRIN resin、Wang resin、Rink amide resin、HMFS resin、Amino-PEGA resin(メルク社)、HMPA-PEGA resin(メルク社)等が挙げられる。これらのレジンは、溶剤(ジメチルホルムアミド(DMF)、2−プロパノール、塩化メチレン等)で洗浄してから用いることができる。
α−アミノ基の保護基としては、ベンジルオキシカルボニル(Cbz又はZ)基、tert−ブトキシカルボニル(Boc)基、フルオレニルメトキシカルボニル(Fmoc)基、ベンジル基、アリル基、アリルオキシカルボニル(Alloc)基等が挙げられる。Cbz基はフッ化水素酸、水素化等によって脱保護でき、Boc基はトリフルオロ酢酸(TFA)により脱保護でき、Fmoc基はピペリジンによる処理で脱保護できる。
α−カルボキシ基の保護は、メチルエステル、エチルエステル、ベンジルエステル、tert−ブチルエステル、シクロヘキシルエステル等を用いることができる。
アミノ酸のその他の官能基として、セリンやトレオニンのヒドロキシ基はベンジル基やtert−ブチル基で保護することができ、チロシンのヒドロキシ基は2−ブロモベンジルオキシカルボニル希やtert−ブチル基で保護する。リジン側鎖のアミノ基、グルタミン酸やアスパラギン酸のカルボキシ基は、α−アミノ基、α−カルボキシ基と同様に保護することができる。
【0025】
カルボキシ基の活性化は、縮合剤を用いて行うことができる。縮合剤としては、例えば、ジシクロヘキシルカルボジイミド(DCC)、ジイソプロピルカルボジイミド(DIPCDI)、1-エチル-3-(3-ジメチルアミノプロピル)カルボジイミド(EDCあるいはWSC)、(1H-ベンゾトリアゾール-1-イルオキシ)トリス(ジメチルアミノ)ホスホニウムヘキサフルオロホスファート(BOP)、1-[ビス(ジメチルアミノ)メチル]-1H-ベンゾトリアゾリウム-3-オキシドヘキサフルオロホスファート(HBTU)等が挙げられる。
【0026】
レジンからのペプチド鎖の切断は、TFA、フッ化水素(HF)等の酸で処理することによって行うことができる。
またポリペプチド鎖が長い場合、ある程度固相法によって合成した後、ライゲーション法(例えば、ネイティブケミカルライゲーション法)によって合成したペプチドを連結することも可能である。
【0027】
遺伝子組み換え法によるポリペプチドの製造方法は、後述するように本発明の核酸を用いて行うことができる。
【0028】
[核酸]
本発明の核酸は、本発明のポリペプチドをコードする核酸であり、DNAであってもRNAであってもよい。
本発明の核酸は、本発明のポリペプチドをコードする限り、あらゆる変異体も含むことができる。かかる変異体としては、天然に存在するアレル変異体、天然に存在しない変異体、人為的に改変した変異体を含み、例えば、Nephrinをコードする遺伝子の塩基配列における本発明のポリペプチドに対応する領域において、一又は数個の塩基が欠失、置換、又は付加した核酸が挙げられる。
本発明の核酸の例として、Nephrinペプチドをコードする核酸(配列番号:11)が挙げられる。
【0029】
本発明の核酸は、公知の方法又はそれに準ずる方法で調製することができる。例えば、自動合成装置によって合成することができる。得られたDNAをベクターに挿入するために制限酵素認識部位を加えたり、できたペプチド鎖を酵素などで切り出すためのアミノ酸配列をコードする塩基配列を組み込んでもよい。
また、細胞から全mRNAを抽出し逆転写酵素によりcDNAを調製しても良い。この場合、プライマーとして、ベクターに挿入するために必要な制限酵素認識配列を含むものを使用することができる。得られたcDNA混合物に対し、目的のcDNAを増幅するプライマーを利用してPCR法を行うことにより、目的のcDNAのみが増幅される。このときもプライマーとして、制限酵素認識部位を含むものを用いれば、両端に制限酵素認識配列を有するcDNAを得ることができる。また、逆転写酵素によって得るcDNA混合物に代えて、既存のcDNAライブラリを用いてもよい。
本発明の核酸として、配列番号:11で表される塩基配列を含む核酸が挙げられる。
上述のとおり、本発明のポリペプチドを膜透過性ペプチドと融合させる場合、本発明の核酸は、膜透過性ペプチドをコードする核酸も含む。
【0030】
また、宿主由来のプロテアーゼによる分解を抑制するため、目的のペプチドを他のペプチドとのキメラペプチドとして発現させるキメラタンパク質発現法を用いることもできる。この場合、本発明の核酸としては、目的とするペプチドと、これに結合するペプチドとをコードする核酸が用いられる。
【0031】
[発現ベクター]
本発明の発現ベクターは、本発明の核酸を含む。本発明の核酸はそのまま、又は制限酵素で消化し、又はリンカーを付加して、発現ベクターのプロモータの下流に挿入することができる。ベクターとしては、大腸菌由来プラスミド(pBR322、pBR325、pUC12、pUC13、pUC18、pUC19、pUC118、pBluescript II等)、枯草菌由来プラスミド(pUB110、pTP5、pC1912、pTP4、pE194、pC194等)、酵母由来プラスミド(pSH19、pSH15、YEp、YRp、YIp、YAC等)、バクテリオファージ(λファージ、M13ファージ等)、ウイルス(レトロウイルス、ワクシニアウイルス、アデノウイルス、アデノ随伴ウイルス(AAV)、カリフラワーモザイクウイルス、タバコモザイクウイルス、バキュロウイルス等)、コスミド等が挙げられる。
【0032】
プロモータは、宿主の種類に応じて適宜選択することができる。宿主が動物細胞である場合は、例えば、SV40(simian virus 40)由来プロモータ、CMV(cytomegalovirus)由来プロモータを用いることができる。宿主が大腸菌である場合は、trpプロモータ、T7プロモータ、lacプロモータ等を用いることができる。
発現ベクターには、DNA複製開始点(ori)、選択マーカー(抗生物質抵抗性、栄養要求性等)、エンハンサー、スプライシングシグナル、ポリA付加シグナル、タグ(FLAG、HA、GST、GFPなど)をコードする核酸等を組み込むこともできる。
【0033】
本発明の発現ベクターは、以下に示すとおりin vitroで宿主を形質転換し、本発明のポリペプチドを発現させるために用いてもよく、そのまま対象に投与して、in vivoで本発明のポリペプチドを発現させるために用いることもできる。
【0034】
[形質転換体]
本発明の形質転換体は、本発明の発現ベクターで適当な宿主細胞を形質転換することによって得ることができる。宿主は、ベクターとの関係で適宜選択することができ、例えば、大腸菌、枯草菌、バチルス属菌)、酵母、昆虫又は昆虫細胞、動物細胞等が用いられる。動物細胞として、例えば、HEK293T細胞、CHO細胞、COS細胞、ミエローマ細胞、HeLa細胞、Vero細胞を用いることができる。形質転換は、宿主の種類に応じ、リポフェクション法、リン酸カルシウム法、エレクトロポレーション法、マイクロインジェクション法、パーティクルガン法等、公知の方法に従って行うことができる。
形質転換体を常法に従って培養することにより、目的とするポリペプチドが発現する。培地は、HEK293T細胞の場合、ウシ胎児血清等の血清を添加したDMEM培地を用いることができる。
【0035】
形質転換体の培養物からのポリペプチドの精製は、培養細胞を回収し、適当な緩衝液に懸濁してから超音波処理、凍結融解などの方法により細胞を破壊し、遠心分離やろ過によって粗抽出液を得る。培養液中にポリペプチドが分泌される場合には、上清を回収する。
粗抽出液又は培養上清からの精製も公知の方法又はそれに準ずる方法(例えば、塩析、透析法、限外ろ過法、ゲルろ過法、SDS−PAGE法、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー、逆相高速液体クロマトグラフィー等)で行うことができる。
得られたポリペプチドは、公知の方法又はそれに準ずる方法で遊離体から塩に、又は塩から遊離体に変換してもよい。
【0036】
[抗体]
本明細書において、抗体は、天然に存在するか、または遺伝子組換え技術によって産生される、全長の免疫グロブリン分子(例えば、IgG抗体)、または抗体フラグメントのような免疫グロブリン分子の免疫学的に活性な一部をいう。本明細書において抗体は、キメラ抗体、ヒト化抗体、および完全なヒト抗体を包含する。さらに、いずれのタイプ、クラス、サブクラスも含み、例えば、IgG、IgE、IgM、IgD、IgAなどが含まれる。これらの抗体は、慣用技術を用いて作製することができ、ポリクローナル抗体であっても、モノクローナル抗体であってもよい。
【0037】
本明細書において、抗体フラグメントは、抗体の断片又は抗体の断片に任意の分子を結合させたものであって、対応する全長抗体と同じエピトープを認識するものを意味する。具体的には、VL、VH、CL及びCH1領域からなるFab;2つのFabがヒンジ領域でジスルフィド結合によって連結されているF(ab’)2;VL及びVHからなるFv;VL及びVHを人工のポリペプチドリンカーで連結した一本鎖抗体であるscFv;diabody型、scDb型、tandem scFv型、ロイシンジッパー型などの二重特異性抗体等が挙げられるが、これらに限定されない。
【0038】
本明細書において、キメラ抗体は、異なる種に由来する抗体の断片が連結された抗体を意味する。例えば、可変領域がマウス抗体のアミノ酸配列であり、その他の領域がヒト抗体のアミノ酸配列である抗体が挙げられる。
本明細書において、ヒト化抗体は、非ヒト由来の抗体に特徴的なアミノ酸配列で、ヒト抗体の対応する位置を置換した抗体を意味し、例えば、H鎖及びL鎖のCDRがマウス抗体のアミノ酸配列であり、その他の領域がヒト抗体のアミノ酸配列である抗体が挙げられる。かかる抗体はCDR移植抗体と呼ばれる場合もある。また、ヒト化抗体は、ヒトキメラ抗体を含む場合もある。
【0039】
本発明の一態様において、抗体は、配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位の領域に結合する。当該領域は、TRPC6におけるNephrin及びPLC-γ1との結合領域である。従って、この抗体を細胞内に投与すれば、TRPC6とPLC-γ1との結合が阻害される。
配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位の領域に結合する抗体は、配列番号:4で表されるポリペプチドを抗原として公知の方法により作製することができる。例えば、モノクローナル抗体は、ポリペプチドで免疫した非ヒト哺乳動物から抗体産生細胞を単離し、これを骨髄腫細胞等と融合させてハイブリドーマを作製し、このハイブリドーマが産生した抗体を精製することによって得ることができる。また、ポリクローナル抗体は、ポリペプチドで免疫した動物の血清から得ることができる。
配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位の領域に結合する抗体は、配列番号:4で表されるポリペプチドを抗原として作製した抗体に限定されない。その作製方法に関わらず、ある抗体が配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位に結合する限り、本発明の抗体として用いることができる。
【0040】
[医薬組成物]
一態様において、医薬組成物は、上述したポリペプチド、核酸、又は抗体を有効成分として含み、これらがTRPC6とPLC-γ1との結合を阻害することにより、TRPC6の膜移行、ひいては活性化を抑制する。
上記医薬組成物の投与形態は特に限定されず、経口的投与でも非経口的投与でもよい。非経口投与としては、例えば、筋肉内注射、静脈内注射、皮下注射等の注射投与、経皮投与、経粘膜投与(経鼻、経口腔、経眼、経肺、経膣、経直腸)投与等が上げられる。
上記医薬組成物がポリペプチドを含む場合、代謝及び排泄されやすい性質に鑑みて、各種の修飾を行うことができる。例えば、ポリペプチドにポリエチレングリコール(PEG)や糖鎖を付加して血中滞留時間を長くし、抗原性を低下させることができる。また、ポリ乳酸・グリコール(PLGA)などの生体内分解性の高分子化合、多孔性ヒドロキシアパタイト、リポソーム、表面修飾リポソーム、不飽和脂肪酸で調製したエマルジョン、ナノパーティクル、ナノスフェア等を徐放化基剤として用い、これにポリペプチドを内包させてもよい。経皮投与する場合、弱い電流を皮膚表面に流して角質層を透過させることもできる(イオントフォレシス法)。
【0041】
上記医薬組成物は、有効成分をそのまま用いてもよいし、薬学的に許容できる担体、賦形剤、添加剤等を加えて製剤化してもよい。剤形としては、例えば、液剤(例えば注射剤)、分散剤、懸濁剤、錠剤、丸剤、粉末剤、坐剤、散剤、細粒剤、顆粒剤、カプセル剤、シロップ剤、トローチ剤、吸入剤、軟膏剤、点眼剤、点鼻剤、点耳剤、パップ剤等が挙げられる。
製剤化は、例えば、賦形剤、結合剤、崩壊剤、滑沢剤、溶解剤、溶解補助剤、着色剤、矯味矯臭剤、安定化剤、乳化剤、吸収促進剤、界面活性剤、pH調整剤、防腐剤、抗酸化剤などを適宜使用し、常法により行うことができる。
製剤化に用いられる成分の例としては、精製水、食塩水、リン酸緩衝液、デキストロース、グリセロール、エタノール等薬学的に許容される有機溶剤、動植物油、乳糖、マンニトール、ブドウ糖、ソルビトール、結晶セルロース、ヒドロキシプロピルセルロース、デンプン、コーンスターチ、無水ケイ酸、ケイ酸アルミニウムマグネシウム、コラーゲン、ポリビニルアルコール、ポリビニルピロリドン、カルボキシビニルポリマー、カルボキシメチルセルロースナトリウム、ポリアクリル酸ナトリウム、アルギン酸ナトリウム、水溶性デキストラン、カルボキシメチルスターチナトリウム、ぺクチン、メチルセルロース、エチルセルロース、キサンタンガム、アラビアゴム、トラガント、カゼイン、寒天、ポリエチレングリコール、ジグリセリン、グリセリン、プロピレングリコール、ワセリン、パラフィン、ミリスチン酸オクチルドデシル、ミリスチン酸イソプロピル、高級アルコール、ステアリルアルコール、ステアリン酸、ヒト血清アルブミン、等が挙げられるがこれらに限定されない。
上記医薬組成物が、ポリペプチドを含む場合、ポリペプチド等の経粘膜吸収されにくい難吸収性薬物の吸収を改善する吸収促進剤として、ポリオキシエチレンラウリルエーテル類、ラウリル硫酸ナトリウム、サポニン等の界面活性剤;グリココール酸、デオキシコール酸、タウロコール酸等の胆汁酸塩;EDTA、サリチル酸類等のキレート剤;カプロン酸、カプリン酸、ラウリン酸、オレイン酸、リノール酸、混合ミセル等の脂肪酸類;エナミン誘導体、N-アシルコラーゲンペプチド、N-アシルアミノ酸、シクロデキストリン類、キトサン類、一酸化窒素供与体等を用いることができる。
【0042】
丸剤又は錠剤は、糖衣、胃溶性、腸溶性物質で被覆することもできる。
注射剤は、注射用蒸留水、生理食塩水、プロピレングリコール、ポリエチレングリコール、植物油、アルコール類等を含むことができる。さらに、湿潤剤、乳化剤、分散剤、安定化剤、溶解剤、溶解補助剤、防腐剤等を加えることができる。
【0043】
上記医薬組成物は、TRPC6に関連する疾患の治療又は予防に有効である。TRPC6に関連する疾患としては、TRPC6チャネロパチー(channelopathy)が挙げられる。チャネロパチーとは、イオンチャネルの機能の異常に基づいて発症する疾患の総称である。上記医薬組成物は、TRPC6の機能を抑制するものであるから、TRPC6の発現又は機能が亢進することによって発症するTRPC6チャネロパチーに特に有用である。かかるTRPC6チャネロパチーとしては、例えば、糸球体硬化症、単状糸球体硬化症(FSGS)、後天性ネフローゼ、特発性肺高血圧症、低酸素性肺高血圧症、心臓肥大、心不全、神経変性疾患、呼吸器疾患、腎疾患等が挙げられるがこれらに限定されない。本発明の医薬組成物は、特に遺伝子変異を伴わずにTRPC6の発現が増加、及び/又はTRPC6の機能亢進を生じるTRPC6チャネロパチーに有用である。
哺乳類(例えば、ヒト、マウス、ラット、モルモット、ウサギ、イヌ、ウマ、サル、ブタ等)、特にヒトに投与する場合の投与量は、症状、患者の年齢、性別、体重、感受性差、投与方法、投与間隔、有効成分の種類、製剤の種類によって異なり、特に限定されないが、例えば、30μg〜100g、100μg〜500mg、100μg〜100mgを1回又は数回に分けて投与することができる。注射投与の場合、患者の体重により、1μg/kg〜3000μg/kg、3μg/kg〜1000μg/kgを1回又は数回に分けて投与してもよい。
【0044】
[スクリーニング方法]
本発明のスクリーニング方法は、候補化合物から、TRPC6の活性化を抑制する化合物を選択するための方法である。
当該スクリーニング方法は、その一態様において、候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、候補化合物の存在下と非存在下における前記細胞へのCa2+イオンの流入をモニターする工程と、を含む。
TRPC6及びPLC-γ1を発現する細胞としては、例えば元来双方を発現するpodocyteを用いることもできるし、上述した本発明に係る発現ベクターでPLC-γ1を発現する細胞を形質転換することによって得たものを用いることもできる。PLC-γ1は、全身の様々な細胞で発現している。PLC-γ1を発現しており、本スクリーニング方法に好適に利用できる細胞として、例えばHEK293T細胞が挙げられる。細胞の培地は、使用する細胞によって適宜選択することができる。HEK293T細胞の場合は、10%FBSを加えたDMEM培地を使用することができる。
候補化合物としては、例えば低分子化合物、高分子化合物、ペプチド、タンパク質、核酸が挙げられるがこれらに限定されない。候補化合物は、例えば培地に添加することができる。また候補化合物がペプチドやRNAである場合、これらをコードする核酸を含む発現ベクターで上記細胞を形質転換し、細胞内で発現させてもよい。
【0045】
TRPC6の284位のチロシンをリン酸化するキナーゼとしては、例えば、プロテインキナーゼC、プロテインキナーゼG、非受容体型チロシンキナーゼであるSrcファミリー(Src、Fyn、Yes、Blk、Fgr、Hck、Lck、Lyn)、等を用いることができ、中でもFynが好ましい。
TRPC6の284位のチロシンをリン酸化するキナーゼは、タンパク質としてスクリーニング系に加えてもよいし、当該キナーゼをコードする核酸を含む発現ベクターによりTRPC6及びPLC-γ1を発現する細胞を形質転換し、細胞内でキナーゼを発現させてもよい。キナーゼを加えてインキュベートする工程に先立って、細胞をEGFで刺激しておくことも好ましい。
細胞へのCa2+の流入をモニターする工程も、公知の方法又はそれに準ずる方法で行うことができる。例えば、Fura 2、Fluo 4、Rhod 2などのアセトキシメチルエステル(AM)を細胞内に取り込ませて細胞内のCa2+濃度を測定する方法が挙げられる。Fura-2/AMは、Ca2+の濃度上昇に伴い、335nm〜340nmで励起すると500nmの蛍光強度が増大し、370nm〜380nmで励起すると蛍光強度が減少するので、2波長励起による蛍光強度の比を取ることによって、バックグラウンド蛍光等に左右されず細胞内のカルシウムイオン濃度を性格に測定することができる。
候補化合物の存在下で、候補化合物の非存在下に比較して、細胞内へのCa2+流入が抑制される場合、当該候補化合物はTRPC6の膜移行を阻害して、TRPC6の活性化を抑制する化合物であると特定することができる。こうして特定された化合物は、TRPC6の発現の亢進を通じてCa2+の細胞内への流入が増加することに基づくTRPC6チャネロパチーの治療又は予防薬として有用である。
【0046】
本発明に係るスクリーニング方法は、別の態様において、候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、候補化合物の存在下と非存在下における前記細胞表面におけるTRPC6の発現を測定する工程と、を含む。候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程は上述のとおり行うことができる。
細胞表面におけるTRPC6の発現を測定する工程は、例えば、細胞表面をビオチン化し、ビオチン化されたタンパク質をストレプトアビジンで回収し、当該タンパク質の中にTRPC6が含まれるかどうかを確認することによって行うことができる。回収されたタンパク質にTRPC6が含まれるかどうかの確認は、例えばウエスタンブロッティングにより行うことができる。
候補化合物の存在下で、候補化合物の非存在下に比較して、細胞表面でのTRPC6の発現が低下する場合、当該候補化合物はTRPC6の膜移行を阻害して、TRPC6の活性化を抑制する化合物であると特定することができる。こうして特定された化合物は、TRPC6の発現の亢進を通じてCa2+の細胞内への流入が増加することに基づくTRPC6チャネロパチーの治療又は予防薬として有用である。
【0047】
本発明に係るスクリーニング方法は、さらに別の態様において、候補化合物と配列番号:4で表されるアミノ酸配列を含むポリペプチドとをインキュベートする工程と、前記候補化合物と前記ポリペプチドとの結合を検出する工程と、を含む。
配列番号:4で表されるアミノ酸配列を含むポリペプチドは、例えば、配列番号:4で表されるアミノ酸配列を含む20アミノ酸以下のポリペプチドとすることができ、配列番号:4で表されるアミノ酸配列のN末端及び/又はC末端に任意のアミノ酸が結合したものとすることができる。
候補化合物と、ポリペプチドとの結合は、例えばポリペプチドを固相担体に固定し、標識した候補化合物を加えてインキュベートした後洗浄して、固相担体に捕捉された候補化合物を同定することによって検出又は測定することができる。
候補化合物がポリペプチドやタンパク質である場合、例えば、放射性物質(25I、131I、35S、3H等)、蛍光物質(フルオレセインイソチオシアネート、ローダミン、ダンシルクロリド、フィコエリスリン、テトラメチルローダミンイソチオシアネート、近赤外蛍光材料等)、発光物質(ルシフェラーゼ、ルシフェリン、エクオリン等)、ナノ粒子(金コロイド、量子ドット)、酵素(アルカリホスファターゼ、ペルオキシダーゼ等)で標識することができる。また候補化合物をビオチンで標識し、標識したアビジン又はストレプトアビジンで検出又は測定してもよい。候補化合物とポリペプチドとの結合は、その他の方法、例えば免疫沈降法、表面プラズモン共鳴法等の公知の方法でも検出又は測定することができる。
候補化合物が配列番号:4で表されるアミノ酸配列からなるポリペプチドと結合する場合、当該候補化合物は、284位がリン酸化されたTRPC6とPLC-γ1との結合を拮抗的に阻害しうる。その結果、TRPC6の膜移行を阻害して、TRPC6の活性化を抑制しうる。こうして特定された化合物は、TRPC6の発現の亢進を通じてCa2+の細胞内への流入が増加することに基づくTRPC6チャネロパチーの治療又は予防薬として有用である。
【0048】
本発明のスクリーニング用キットは、本発明のスクリーニング方法に必要な試薬、細胞、培地、使用説明書等を含む。
本発明のスクリーニング用キットは、例えば、TRPC6及びPLC-γ1を発現する293T細胞;TRPC6、PLC-γ1、及びFynを発現する293T細胞;DMEM培地;Fura 2/AM;配列番号:4で表されるアミノ酸配列を含むポリペプチド;及び細胞培養のためのカバースリップ、EGF等の中から適宜選択したものを備えていてもよい。
【0049】
[治療方法]
本発明は、TRPC6とPLC-γ1との結合を阻害する工程を含む、TRPC6チャネロパチーの治療又は予防方法も含む。TRPC6とPLC-γ1との結合を阻害することにより、TRPC6の膜移行を防ぎ、チャネル活性を低下させて、細胞内へのCa2+の過剰な流入を抑制することができる。従って、本発明の治療又は予防方法は、Ca2+の過剰な流入を原因とする疾患に有効である。
TRPC6とPLC-γ1との結合を阻害する工程は、例えば、本発明の医薬組成物を投与することによって行うことができるが、これに限定されない。
【実施例】
【0050】
以下、本発明を実施例に基づいて具体的に説明するが、本発明はこれらに何ら限定されるものではない。
実施例では、以下に示す材料と方法を用いた。
【0051】
抗体と試薬
ウサギ抗α−リン酸化TRPC6抗体(抗α-pY31抗体、抗α-pY284抗体)は、合成した13アミノ酸のオリゴペプチドCNESQDpYLLMDEL(pY31;配列番号:6)及びCLASPApYLSLSSE (pY284;配列番号:7)を抗原として、それぞれ作製した。各オリゴペプチドの最初のシステイン残基は、TRPC6の配列とは異なる。
抗血清は、抗原をSulfoLink(Pierce, Rockford, IL)に固定してアフィニティー精製し、リン酸化されていないペプチドで吸収した。抗α-Flag抗体(M2; Sigma, St. Louis, MO)、抗α-HA抗体(3F10; Roche Diagnostics, Indianapolis, IN)、抗α-リン酸化チロシン抗体(4G10; Upstate , Lake Placid, NY)、ウサギ抗α-TRPC6抗体(Alamone labs, Jerusalem, Israel)、ウサギ抗α-PLC−γ1抗体(Santa Cruz Biotechnology, Inc., Santa Cruz, CA)は、それぞれ購入した。
ウサギ抗α-Nephrin抗体、及び抗α-phospho-Nephrin(pY1204)抗体は、Haritaらの方法(Harita Y et al. (2009) J Biol Chem)に従って調製した。ウエスタンブロッティングは、これらの抗体を1/2000に希釈して行った。
なお、図中、「WB:HA」「WB:Flag」等の表記は、それぞれ抗α−HA抗体、抗α−Flag抗体でウエスタンブロッティングを行ったことを示し、「IP HA」「IP Flag」等の表記は、それぞれ抗α−HA抗体、抗α−Flag抗体で免疫沈降を行ったことを示す。
PP2(Merck KGaA, Darmstadt, Germany)、SU6656(Merck KGaA)、及びEGF(BD Biosciences, Bedford, MA)は購入した。
【0052】
細胞培養とRNAiのトランスフェクション
ヒトHEK293T細胞は、ATCC(Manassas, VA)より購入した。細胞は、10%ウシ胎仔血清を含むDMEM(Dulbecco's modified Eagle's medium)で維持した。
トランスフェクションは、Lipofectamine 2000 reagent (Invitrogen, Carlsbad,
CA)を使用して行った。
PLC−γ1 small interfering RNA (siRNA) Duplex1/2、及びコントロール用のsiCONTROL nontargeting siRNAは、Invitrogenから購入した。Duplex1/2の塩基配列を以下に示す。
Duplex1 (LotNo.87031E10)
CCCUGCGCUGUAAUGAGUUUGAGAU sense (配列番号:12)
AUCUCAAACUCAUUACAGCGCAGGG antisense (配列番号:13)
Duplex2 (LotNo.87031E11)
CCUUGUUGACCUCAUCAGCUACUAU sense (配列番号:14)
AUAGUAGCUGAUGAGGUCAACAAGG antisense (配列番号:15)
これらのsiRNAは、293T細胞にLipofectamine RNAiMAX (Invitrogen)を用いてトランスフェクトした。
【0053】
真核細胞発現ベクター
全長ラットNephrin、全長ラットNephrin-Flag、Y1204F Nephrin、Y1204F Nephrin-Flagをコードする哺乳動物発現プラスミドは、Haritaらの方法(Harita Y et al. (2008) J Biol Chem 283(14): 9177-9186、及びHarita et al, 2009)に従って準備した。
Fynの発現ベクター(Tezuka T et al. (1999) Proc Natl Acad Sci USA 96(2): 435-440)は、東京大学のDr. T. Tezukaより提供を受け、マウスTRPC6-HA(非特許文献2)は、理化学研究所脳科学脳科学総合研究センターのDr. C. Hisatsuneより提供を受け、ヒトFlag-PLC-γ1(Bae SS et al. (2000) FASEB J 14(9):1083-1092)は、Pohang大学(韓国)のDr. B. G. Suhより提供を受けた。PLC-γ1のSH2変異体は、Baeら及びPlattnerらの方法(Bae YS et al. (1998) J Biol Chem 273(8): 4465-4469; Plattner R et al. (2003) Nat Cell Biol 5(4): 309-319)に従って作製した。
マウスTRPC6変異体(Y31F、Y50F、Y85F、Y107F、Y206F、Y208F、Y284F、Y895F、P112Q、N143S、S270T、K874X、R895C、E897K、及び98-104欠失変異体)とNephrin-Flag欠失変異体(1216位-1227位を欠失)をコードする哺乳動物発現プラスミドは、PCRを用いた標準的な方法で作製した。
すべてのコンストラクトについて、DNA配列解析を行い、配列を確認した。PCRで用いたオリゴヌクレオチドは、下表に記載のとおりである。
【0054】
【表1】
【0055】
【表2】
【0056】
【表3】
【0057】
【表4】
【0058】
【表5】
【0059】
【表6】
【0060】
細胞表面のビオチン化
TRPC6の細胞表面における発現は、Winnらの方法に従って解析した(非特許文献17)。それぞれ図中に示されたプラスミドを発現する293T細胞を、2mg/mlのSulfo-NHS-SS-Biotin(Pierce)で表面をビオチン化した(30分、4℃)。ビオチン化されたタンパク質をストレプトアビジンビーズ(Pierce)で回収し、ウエスタンブロットにより解析した。
【0061】
免疫沈降
細胞をLysis buffer(20 mM Tris-HCl(pH 7.4), 150 mM NaCl, 1% Nonidet P-40, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 50 mM NaF, 10g/ml antipain, 10μg/ml leupeptin, 10g/ml aprotinin, 1 mM sodium vanadate)で、氷上に15分置いて溶解した。ライゼートを遠心処理し、M2 α−Flag抗体、又は3F10 α−HA抗体を結合させたビーズと4℃で1時間インキュベートした。TBS-1% NP40でビーズを洗浄し、結合したタンパク質を100mMグリシン−HCl(pH 2.6)で溶出させ、ウエスタンブロッティングに供した。
【0062】
融合タンパク質の細菌発現
GSTタグをつけたNephrinの細胞内ドメイン(Nephrin-CD:アミノ酸1102-1252;配列番号:1)は、Haritaらの方法に従って準備した(Harita et al, 2009)。GST-Nephrin-CD欠失変異体(1102位-1152位、1102位-1205位、1102位-1211位、1102位-1215位、1102位-1227位、1102位-1238位、1102位-1245位をそれぞれ欠失)は、PCRを用いた標準的な方法で作製した。細菌のペレットは、20 mM Tris-HCl (pH 7.5)、150 mM NaCl、1% Nonidet P-40、2 mM dithiothreitol、1mM phenylmethylsulfonyl fluoride、10μg/ml antipain、10μg/ml leupeptinを含む溶液で再懸濁し、超音波処理を行った。不溶性物質は遠心処理により除去した。GST融合タンパク質は、グルタチオン−セファロースカラム(GE Healthcare, Buckinghamshire, UK)により精製し、遊離グルタチオンとともに溶出させた。GST-Nephrin-CD欠失変異体を作製するために用いたオリゴヌクレオチドは、上記表6に示す。
【0063】
Pull-downアッセイ
グルタチオン−セファロースビーズに固定したGST-Nephrin-CDは、Haritaらの方法(Harita et al, 2008; Harita et al, 2009)に従い、Fynによってリン酸化した。
HA-TRPC6を発現する293T細胞のライゼートを、ビーズに固定したリン酸化又は非リン酸化GST-Nephrin-CDと共に4℃で一晩インキュベートした。
ビーズを洗浄バッファー(50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1% Nonidet
P-40)でよく洗浄した。Nephrinと結合したタンパク質を、遊離グルタチオンとともに溶出させ、ウエスタンブロッティングとCBB染色により解析した。ペプチドを用いたPull-downアッセイの場合は、ペプチドをSulfo Link Coupling Gel (Pierce)に固定して行った。
【0064】
質量分析法によるタンパク質の同定
目的のタンパク質を銀染したゲルから切り出し、1pmolのAPI(Achromobacter protease I;40μlのdigestion buffer (10 mM Tris, pH 8.5)中)とともに37℃で一晩静置して消化した。
ペプチドを精製し、高速液体クロマトグラフィー/タンデム質量分析装置(LC/MS; Q-STAR Elite, Applied Biosystems, Carlsbad, CA)を用いたペプチドマスフィンガープリンティングに供した。ペプチドイオンは、ProteinPilotTM (ver. 3.0)ソフトウエアにより解析した。
【0065】
ラット腎糸球体の分離
糸球体は、graded sieving techniqueを用いてラット腎皮質から分離し、タンパク質抽出のために保存した。すべての実験は、東京大学医科学研究所のAnimal Centerのガイドラインに従って行った。Wistarラットは、Charles River Laboratories Japan, Inc (Atsugi, Japan)より購入した。
【0066】
細胞内カルシウム濃度([Ca2+]i)の測定
カバースリップ上で培養した293T細胞を、4μM Fura-2/AM(Dojindo, Kumamoto,
Japan)で満たし、Hepes-buffered saline(HBS; 115 mM NaCl, 5.4 mM KCl, 20 mM HEPES, 1 mM MgCl2, 2 mM CaCl2, 10 mM glucose, pH 7.4)に浸漬させ、30分静置した。
続いて細胞を、室温にて、Ca2+-free HBS(115 mM NaCl, 5.4mM KCl, 20 mM HEPES, 2 mM MgCl2, 10 mM glucose, 0.05 mM EGTA, pH 7.4)及びHBSでかん流した。
Fura-2を搭載した細胞の蛍光は、Olympus IX70顕微鏡で、20×対物レンズ(N.A. 0.75)を用いて観察した。Fura-2の蛍光は、波長340nmと380nmで10秒ごとに交互に励起した。冷却CCDカメラ(ORCA-ER, Hamamatsu Photonics, Hamamatsu, Japan)により像を記録した。
蛍光強度の比(F340/F380)の経時的変化は、各視野のすべての細胞(400〜500)をもとに計算し、平均した。画像収集とデータ解析は、マッキントッシュコンピュータのwritten by T.IのTRPC6I Workbenchソフトウエアによって行った。
【0067】
ペプチドの送達
TRPC6結合領域(1216-1227: CWPEVQCEDPRGI(配列番号:3))を含むNephrinペプチド、又はコントロールペプチド(Nephrin C-terminus; amino acids 1241-1252: CSSLPFELRGHLV(配列番号:8))は、Transport Protein Delivery Reagent (Takara, Tokyo, Japan)を用いて293T細胞に送達させた。
【0068】
統計
数値は、平均±SEMで示す。二つのグループの[Ca2+]iの比較は、Mann-Whitney U-testにより評価した。ウエスタンブロッティングのバンド強度の比較には、Unpaired Student's t-testを用いた。P<0.05を有意とした。
【0069】
実施例1:TRPC6の膜移行におけるTyr284のリン酸化の必要性
TRPCチャネルは細胞膜に移行することによって活性化されると考えられていたことから、表面ビオチン化アッセイによって、TRPC6のリン酸化が膜移行に及ぼす影響を調べた。HAタグで標識したTRPC6を発現する293T細胞をEGFで刺激し、図示される時間が経過した後にビオチン化した。ビオチン化されたTRPC6は、basal conditionでは検出されなかった(図1A)。TRPC6は刺激後1分で細胞膜に現れ、3分後に上限に達し、その状態が60分継続した。構成的に活性なFynの共発現により、チロシンのリン酸化と、細胞表面に局在するTRPC6が増加した(図1B、C)。
TRPC6は、その細胞内ドメインに23のチロシン残基を有する。そこで、どのチロシン残基のリン酸化がTRPC6の膜移行に必要なのか調べるため、Y31、Y50、Y85、Y107、Y206、Y208、Y284、及びY895の8つのチロシン残基のそれぞれをフェニルアラニンに置換して、TRPC6の細胞内局在に与える影響を解析した。Y107、Y206、Y208、及びY284は、TRPC3におけるリン酸化部位として報告されているY49、Y148、Y150、及びY226にそれぞれ対応するものである。Y31、Y50、Y85及びY895は、モチーフサーチプログラムのScan Site (http://scansite.mit.edu)によってSH2含有タンパク質との結合部位と予測された場所である。また、これらの8つのチロシンは、すべて複数の種(ヒト、ラット、マウス、イヌ)において保存されている。
この結果、284位のチロシンをフェニルアラニンに置換したY284F変異は、Fynによって誘導されるTRPC6の細胞表面での発現を著しく減少させた(図1D)。このことから、TRPC6の膜移行には、284位のチロシンのリン酸化が重要であることが示唆された。
【0070】
実施例2:PLC-γ1とリン酸化されたTRPC6との結合がTRPC6の膜移行に及ぼす影響
実施例1の結果から、リン酸化されたTRPC6のY284周辺は、TRPC6の輸送に必要なタンパク質の結合領域になるものと仮定した。TRPC6の輸送に必要なタンパク質を特定するために、リン酸化された、又はリン酸化されていないY284周辺の13アミノ酸のTRPC6ペプチド(配列番号:7)を用いたin vitroの結合アッセイを行った。
両ペプチドをカップリングゲルに固定して、293T細胞ライゼートとインキュベートし、結合したタンパク質をSDS-PAGEに供し、銀染した。図2Aに示すとおり、見かけの分子量が140,000のタンパク質がリン酸化したTRPC6 Y284ペプチドに特異的に結合することが確認された(*で示す)。このバンドのトリプシン消化物を質量分析法で解析したところ、当該タンパク質はPLC-γ1と同定された。
また、共免疫沈降により、TRPC6は、Fyn依存性にPLC-γ1に結合することを確認した(図2B)。Y31Fの変異は、PLC-γ1への結合を部分的に無効とし、Y31とY284がいずれも変異したTRPC6は、PLC-γ1と相互作用しなかった。他のTRPC6変異体は、野生型TRPC6と同等にPLC-γ1に結合した(data not shown)。従って、TRPC6とPLC-γ1の相互作用には、Y284のリン酸化のみでなく、Y31のリン酸化も必要とされることがわかった。
PLC-γ1は、分子の中心に隣り合った2つのSH2ドメイン(N-SH2、C-SH2)を有する。pY31及びpY284ペプチドは、PLC-γ1 N-SH2に選択的に結合する。N-SH2の点変異又は欠失は、結合を強く阻害した。
また、siRNAによって内因性PLC-γ1をノックダウンすると、細胞膜上のビオチン化TRPC6が著しく減少した。このことは、Fynに誘導されるTRPC6の膜移行にPLC-γ1が不可欠であることを示す。
PLC-γ1はTRPC3のN末端アミノ酸(40位〜46位)と結合し、プレクストリン相同ドメイン(PHドメイン)を形成することが知られている。このPHドメインが、ホスファチジルイノシトール2リン酸(PIP2)に結合し、TRPC3の細胞表面での発現を誘導する。そこで、TRPC6における対応するN末端アミノ酸(98位〜104位)の機能を調べるため、98位〜104位を欠失したTRPC6変異体を用いて共免疫沈降とビオチン化アッセイを行った。当該変異体は、野生型TRPC6と同様に、PLC-γ1への結合能を有し(図2F)、Fyn依存的に細胞膜に移行した(図2G)ことから、98位〜104位の残基ではなく、Y31とY284がPLC-γ1との相互作用及び膜移行に不可欠であることがわかった。
【0071】
実施例3:糸球体におけるSrcファミリーキナーゼによるTRPC6 Y31及びY284のリン酸化
pY31及びpY284を特異的に認識する抗TRPC6抗体を作製した(図3)。各抗体の特異性はTRPC6のY31F及びY284F変異体を用いて確認した(図3A)。EGFもこれらの部位のリン酸化を誘導した(図3B)。単離したラット腎糸球体を、非特異的プロテインホスファターゼ阻害剤vanadateで処理すると、内因性TRPC6のY284のリン酸化は有意に亢進した(図3C)。このリン酸化は、vanadate処理に先立って、Srcファミリーキナーゼの特異的阻害剤PP2又はSU6656で前処理することにより、抑制された。このことは、Y31及びY284のリン酸化がSrcファミリーキナーゼによるものであることを示唆する。
【0072】
実施例4:チロシンをリン酸化されたTRPC6とNephrinとの相互作用
TRPC6はスリット膜の構成要素であるので、他のスリット膜の構成要素の結合が、リン酸化依存的な膜移行に影響を及ぼすか調べた。まず、TRPC6とFynに加え、Nephrin、Neph1、Neph2、又はPodocinを発現する293T細胞を用いた共免疫沈降を行った。その結果、Nephrinは、Fynと共発現させた場合のみTRPC6に結合することを見出した(図4A)。他のスリット膜の構成要素とTRPC6との相互作用は検出されなかった。
NephrinもSrcファミリーキナーゼによってチロシンをリン酸化されることが知られているので、どのリン酸化がTRPC6とNephrinの相互作用に必要なのか調べた。リン酸化されるとNephrinのY1204はPLC-γ1に結合することが知られている(Harita et al., 2009)。しかしながら、このリン酸化部位は、TRPC6との結合には関与しておらず、Y1204の変異体もTRPC6に結合した(図4A)。
次に、GST−Nephrin細胞内ドメイン(Nephrin-CD, 1102位〜1252位;配列番号:1)を、予めin vitroでリン酸化して、又はリン酸化せずに、pull-downアッセイを行った。Fynを共発現させた293T細胞のTRPC6は、in vitroにおいて、Nephrin-CDがFynにリン酸化されているか否かに関わらずGST-Nephrin-CDに結合したが、Fynを共発現させない細胞のTRPC6はGST-Nephrin-CDに結合しなかった(図4B)。従って、TRPC6とNephrinの相互作用にはTRPC6のリン酸化が必要である一方、この相互作用にNephrinのリン酸化は不要であることがわかった。
TRPC6のY31及びY284のリン酸化は、PLC-γ1との相互作用に必要である(図2B)。同様に、チロシンをフェニルアラニンに置換したTRPC6の一連の変異体と、Nephrinとの結合を評価し、TRPC6とNephrinの相互作用に不可欠なチロシンを決定した(図4C)。Y284F変異体はほぼ完全にNephrinと相互作用しなかったが、Y31Fを含む他の変異体は野生型と同様に相互作用したことから、両者の相互作用に不可欠なのはY284のリン酸化であることがわかった。
Y284リン酸化の重要性をさらに確認するため、GST-Nephrin-CDを使ったpull-downアッセイを行った(図4D)。全長TRPC6ほどではなかったが、Y284を囲むTRPC6ペプチド(配列番号:7)もリン酸化されると、293T細胞からNephrinとGST-Nephrin-CDをpull-downし(図4E)、NephrinとTRPC6が直接結合することを示した。
Nephrin細胞質ドメインにおけるTRPC6との結合に必要な領域を特定するため、GST-Nephrin-CDの一連の欠失変異体を構築した。これらの欠失変異体を用いたGST-pull-downアッセイの結果、NephrinとTRPC6の相互作用には、1216位〜1227位を含む残基が不可欠であり(図4F)、この領域を欠失するとNephrinはTRPC6と結合できないことがわかった(図4G)。
【0073】
実施例5:Nephrinによる、リン酸化依存的なTRPC6の細胞表面での発現とチャネル活性化の抑制
PLC-γ1及びNephrinの細胞質ドメインは、いずれもTRPC6の同一のチロシン残基(Y284)に結合するので、これらが競合的に結合する可能性を検討した。図5Aに示されるとおり、Nephrinを共発現させると、TRPC6とPLC-γ1の相互作用は完全に阻害された。
Nephrinは、リン酸化されたY1204がPLC-γ1との結合部位となるが、Y1204は、NephrinのTRPC6結合領域(1216位〜1227位)にも非常に近い。そのため、NephrinによるTRPC6−PLC-γ1複合体形成の阻害は、リン酸化されたNephrin Y1204にPLC-γ1が結合することによる可能性も考えられた。しかしながら、PLC-γ1に結合しないNephrin Y1204F変異体も、TRPC6−PLC-γ1相互作用を阻害した(図5A)。このことから、Nephrinは、NephrinとPLC-γ1との相互作用とは関係なく、TRPC6とPLC-γ1との相互作用を阻害することがわかった。
PLC-γ1はTRPC6の膜移行において重要な役割を果たす(図2E)ので、Nephrinは、TRPC6−PLC-γ1の相互作用を阻害することによって、TRPC6の輸送を阻害する可能性がある。そこで、表面ビオチン化アッセイを行ったところ、Nephrinを共発現させると、TRPC6のFyn誘導性の細胞表面での発現が抑制された(図5B)。TRPC6に結合しないNephrinの欠失変異体(Δ1216-1227)による抑制は、かなり低かった。
次に、Nephrinペプチド(1216-1227残基)を、TRPC6を発現する293T細胞に導入し、Fyn誘導性のTRPC6膜移行を評価した(図5C)。Nephrinペプチドの導入により、Fyn依存性のTRPC6の膜移行が遮断された。一方、コントロールペプチド(NephrinのC末端における同じ長さのペプチド;配列番号:8)を導入しても、TRPC6の膜移行は遮断されなかった。PLC-γ1は、Fynと共発現させた細胞のビオチン化サンプル中で検出された(図5C)。Nephrinペプチドは、PLC-γ1の細胞膜への移行も阻害した。
さらに、TRPC6とFynを発現する293T細胞へのCa2+の流入をモニターしたところ、NephrinはTRPC6チャネル活性を抑制した(図5D、E)。Fynは、TRPC6を発現する細胞におけるCa2+流入を増大させたが、Nephrinを共発現させるとFyn依存性のCa2+流入の増大は有意に抑制された(p<0.001, Mann Whitney test)。この結果は、TRPC6の膜表面発現の阻害効果と一致する。Y284F変異体のFyn誘導性の増加は、同条件ではわずかであった。
また、Nephrinペプチドも、TRPC6チャネル活性に対して、Nephrinと同様に阻害効果を示した(p<0.001, 図5F、G)。この結果は、NephrinペプチドがTRPC6の膜移行を抑制する結果と一致する。
実施例6:TRPC6のFSGS変異体に対するNephrinの阻害効果の低下
TRPC6の変異がFSGSの原因であると特定されたことから、複数のTRPC6チャネル変異体の活性が培養細胞を用いて測定されてきた。しかしながら、それらの結果は、チャネル活性とFSGSの発症の関係を明らかに示すものではなかった。いくつかのTRPC6変異体(P112Q、R895C、E897K)は、チャネル活性を亢進させ、他の変異(N143S、S270T、K874X)は亢進させない。これらの見かけ上矛盾した結果から、podocyte特異的なTRPC6の調節が存在するとの仮説を立てた。
この仮説に基づいて、疾患を引き起こすTRPC6の変異(以下、「疾患関連変異」)であるP112Q、N143S、S270T、K874X、R895C、及びE897Kのそれぞれと、Nephrinとの相互作用を共免疫沈降により解析した(図6A、B)。K874X以外の5つの変異が、TRPC6とNephrinとのリン酸化依存的な相互作用を減少させた。
次に、TRPC6−PLC-γ1相互作用と、TRPC6変異体の細胞表面での発現に対するNephrinの阻害効果を解析した。興味深いことに、Nephrinは、野生型TRPC6の結果と対照的に、TRPC6の疾患関連変異体とPLC-γ1との複合体形成を抑制しなかった。すべてのTRPC6疾患関連変異体について、Nephrinの非存在下における野生型TRPC6と同様に、Fyn誘導性の膜移行が観察された(data not shown)。野生型TRPC6の結果と対照的に、Nephrinは、TRPC6の疾患関連変異体の細胞表面での発現を阻害しなかった(図6D)。
最後に、TRPC6の疾患関連変異体のFyn依存的チャネル活性化にNephrinが及ぼす影響を調べた(図6E、F)。膜局在の結果と一致して、Nephrinは、野生型TRPC6のFyn誘導性のチャネル活性化を抑制したが、疾患関連変異体は、Nephrinに感受性を示さなかった。この結果は、患者のpodocyteにおけるTRPC6の細胞表面での過剰発現と、TRPC6の機能亢進による過剰なCa2+流入を説明し得る。
【配列表フリーテキスト】
【0074】
配列番号:1は、Nephrin細胞内ドメインのアミノ酸配列を示す。
配列番号:2は、全長TRPC6のアミノ酸配列を示す。
配列番号:3は、Nephrinペプチドのアミノ酸配列を示す。
配列番号:4は、TRPC6のpY284周辺ポリペプチドのアミノ酸配列を示す。
配列番号:5は、全長Nephrinのアミノ酸配列を示す。
配列番号:6は、TRPC6のpY31周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:7は、TRPC6のpY284周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:8は、NephrinのC末端1241位−1252位からなるペプチドのアミノ酸配列を示す。
配列番号:9は、TRPC6のY31周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:10は、TRPC6のY284周辺ペプチドのN末端にCysを付加したペプチドのアミノ酸配列を示す。
配列番号:11は、Nephrinペプチドをコードする核酸の塩基配列を示す。
配列番号:12は、siRNA (Duplex1)のセンス鎖の塩基配列を示す。
配列番号:13は、siRNA (Duplex1)のアンチセンス鎖の塩基配列を示す。
配列番号:14は、siRNA (Duplex2)のセンス鎖の塩基配列を示す。
配列番号:15は、siRNA (Duplex1)のアンチセンス鎖の塩基配列を示す。
配列番号:16は、HA-TRPC6のY31F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:17は、HA-TRPC6のY31F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:18は、HA-TRPC6のY50F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:19は、HA-TRPC6のY50F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:20は、HA-TRPC6のY85F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:21は、HA-TRPC6のY85F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:22は、HA-TRPC6のY107F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:23は、HA-TRPC6のY107F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:24は、HA-TRPC6のY206F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:25は、HA-TRPC6のY206F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:26は、HA-TRPC6のY208F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:27は、HA-TRPC6のY208F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:28は、HA-TRPC6のY284F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:29は、HA-TRPC6のY284F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:30は、HA-TRPC6のY895F変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:31は、HA-TRPC6のY895F変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:32は、HA-TRPC6のFSGS変異体(P112Q変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:33は、HA-TRPC6のFSGS変異体(P112Q変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:34は、HA-TRPC6のFSGS変異体(N143S変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:35は、HA-TRPC6のFSGS変異体(N143S変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:36は、HA-TRPC6のFSGS変異体(S270T変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:37は、HA-TRPC6のFSGS変異体(S270T変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:38は、HA-TRPC6のFSGS変異体(K874X変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:39は、HA-TRPC6のFSGS変異体(K874X変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:40は、HA-TRPC6のFSGS変異体(R895C変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:41は、HA-TRPC6のFSGS変異体(R895C変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:42は、HA-TRPC6のFSGS変異体(E897K変異体)プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:43は、HA-TRPC6のFSGS変異体(E897K変異体)プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:44は、HA-TRPC6の98位-104位欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:45は、HA-TRPC6の98位-104位欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:46は、Flag-Nephrinの1216位-1227位欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:47は、Flag-Nephrinの1216位-1227位欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:48は、Flag-PLC-γ1のN*(R586K)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:49は、Flag-PLC-γ1のN*(R586K)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:50は、Flag-PLC-γ1のC*(R694K)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:51は、Flag-PLC-γ1のC*(R694K)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:52は、Flag-PLC-γ1のDel(N-SH2)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:53は、Flag-PLC-γ1のDel(N-SH2)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:54は、Flag-PLC-γ1のDel(C-SH2)変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:55は、Flag-PLC-γ1のDel(C-SH2)変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:56は、GST-Nephrin-CDの1102-1211欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:57は、GST-Nephrin-CDの1102-1211欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:58は、GST-Nephrin-CDの1102-1215欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:59は、GST-Nephrin-CDの1102-1215欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
配列番号:60は、GST-Nephrin-CDの1102-1227欠失変異体プラスミドの作製に用いたフォワードプライマーの塩基配列を示す。
配列番号:61は、GST-Nephrin-CDの1102-1227欠失変異体プラスミドの作製に用いたリバースプライマーの塩基配列を示す。
【特許請求の範囲】
【請求項1】
以下のいずれかのポリペプチド:
(i)配列番号:1に記載のアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチド;
(ii)前記(i)の部分配列を含むポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び
(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド。
【請求項2】
前記(ii)に記載のポリペプチドであって、配列番号:3で表される、Nephrinのアミノ酸配列における1216位〜1227位の領域を含む、請求項1に記載のポリペプチド。
【請求項3】
請求項1又は2に記載のポリペプチドをコードする核酸。
【請求項4】
請求項3に記載の核酸を含む発現ベクター。
【請求項5】
請求項4に記載の発現ベクターを含む形質転換体。
【請求項6】
配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位に結合する抗体。
【請求項7】
請求項1若しくは2に記載のポリペプチド、請求項3に記載の核酸、及び請求項4に記載の発現ベクター、並びに請求項6に記載の抗体からなる群より選択されるいずれか一つを含む医薬組成物。
【請求項8】
TRPC6チャネロパチーの予防又は治療剤である、請求項7に記載の医薬組成物。
【請求項9】
前記TRPC6チャネロパチーは、糸球体硬化症、後天性ネフローゼ、特発性肺高血圧症、及び心不全からなる群より選択される、請求項8記載の医薬組成物。
【請求項10】
候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞へのCa2+イオンの流入を比較する工程と、を含む方法。
【請求項11】
候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞表面におけるTRPC6の発現を測定する工程と、を含む方法。
【請求項12】
候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物と、配列番号:4で表されるアミノ酸配列を含むポリペプチドとをインキュベートする工程と、
前記候補化合物と前記ポリペプチドとの結合を検出又は測定する工程と、を含む方法。
【請求項1】
以下のいずれかのポリペプチド:
(i)配列番号:1に記載のアミノ酸配列からなるNephrinの細胞内ドメインを含むポリペプチド;
(ii)前記(i)の部分配列を含むポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド;及び
(iii)前記(i)又は(ii)に記載のポリペプチドのアミノ酸配列において、1又は数個のアミノ酸が欠失、付加若しくは置換されているポリペプチドであって、配列番号:2で表されるアミノ酸配列における284位のチロシンがリン酸化されたTRPC6と結合能を有する、ポリペプチド。
【請求項2】
前記(ii)に記載のポリペプチドであって、配列番号:3で表される、Nephrinのアミノ酸配列における1216位〜1227位の領域を含む、請求項1に記載のポリペプチド。
【請求項3】
請求項1又は2に記載のポリペプチドをコードする核酸。
【請求項4】
請求項3に記載の核酸を含む発現ベクター。
【請求項5】
請求項4に記載の発現ベクターを含む形質転換体。
【請求項6】
配列番号:2で表されるTRPC6のアミノ酸配列における279位〜290位に結合する抗体。
【請求項7】
請求項1若しくは2に記載のポリペプチド、請求項3に記載の核酸、及び請求項4に記載の発現ベクター、並びに請求項6に記載の抗体からなる群より選択されるいずれか一つを含む医薬組成物。
【請求項8】
TRPC6チャネロパチーの予防又は治療剤である、請求項7に記載の医薬組成物。
【請求項9】
前記TRPC6チャネロパチーは、糸球体硬化症、後天性ネフローゼ、特発性肺高血圧症、及び心不全からなる群より選択される、請求項8記載の医薬組成物。
【請求項10】
候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞へのCa2+イオンの流入を比較する工程と、を含む方法。
【請求項11】
候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物の存在下又は非存在下で、TRPC6及びPLC-γ1を発現する細胞を、TRPC6の284位のチロシンをリン酸化するキナーゼとインキュベートする工程と、
候補化合物の存在下と非存在下における前記細胞表面におけるTRPC6の発現を測定する工程と、を含む方法。
【請求項12】
候補化合物から、TRPC6の活性化を抑制する化合物を選択するためのスクリーニング方法であって、
候補化合物と、配列番号:4で表されるアミノ酸配列を含むポリペプチドとをインキュベートする工程と、
前記候補化合物と前記ポリペプチドとの結合を検出又は測定する工程と、を含む方法。
【図1A】

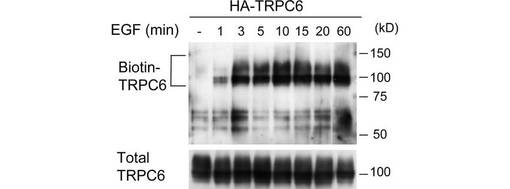
【図1B】
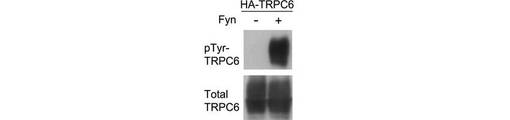
【図1C】
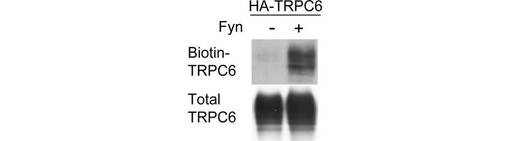
【図1D】
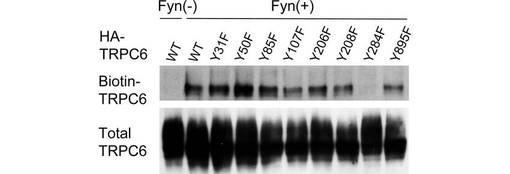
【図2A】
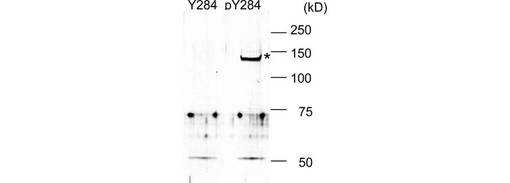
【図2B】
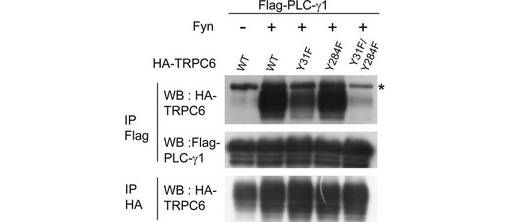
【図2C】

【図2D】

【図2E】
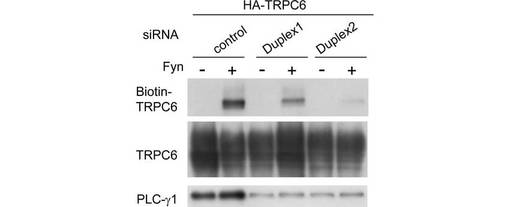
【図2F】
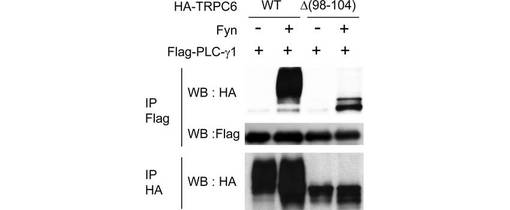
【図2G】
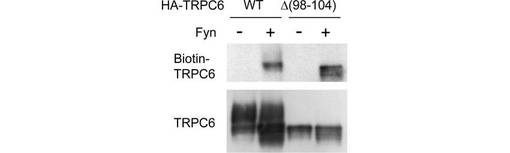
【図3A】
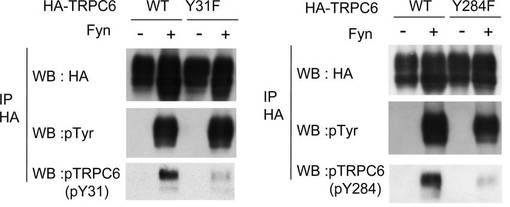
【図3B】
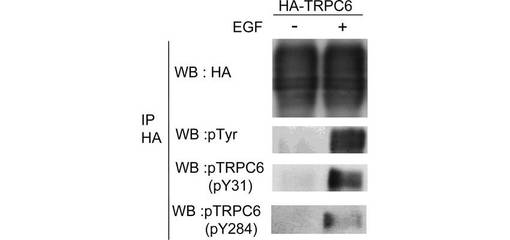
【図3C】
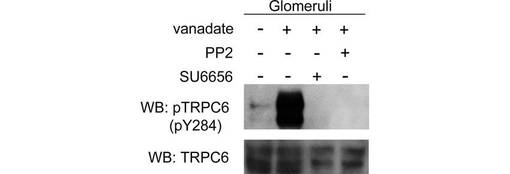
【図4A】
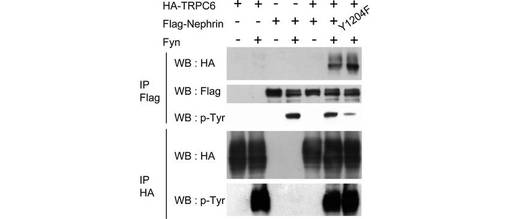
【図4B】
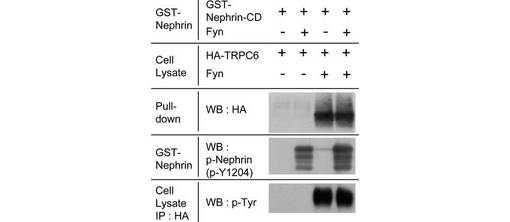
【図4C】
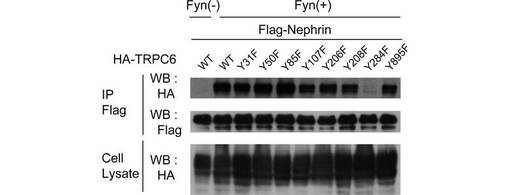
【図4D】
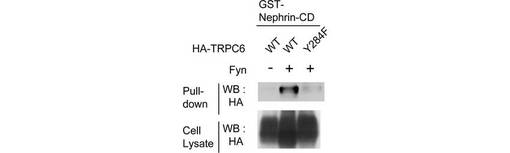
【図4E】
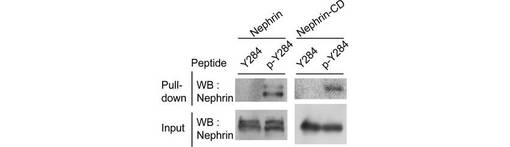
【図4F】
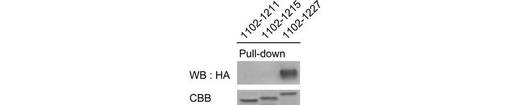
【図4G】
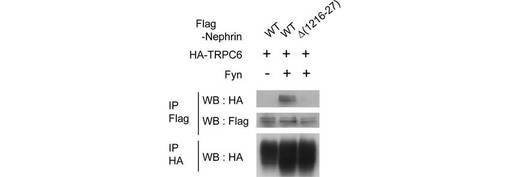
【図5A】
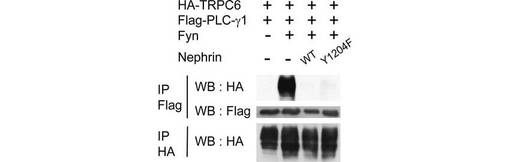
【図5B】
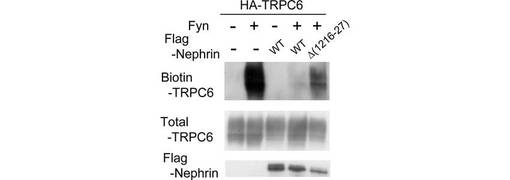
【図5C】
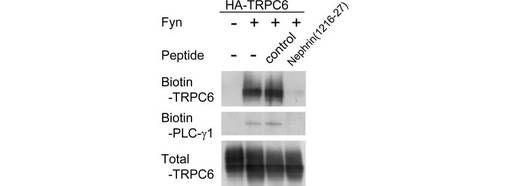
【図5D】
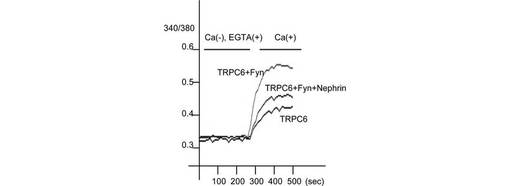
【図5E】
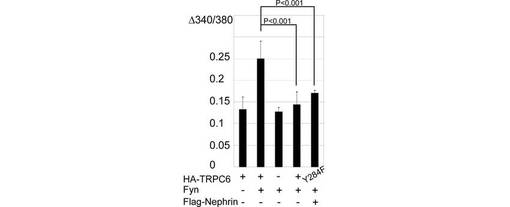
【図5F】
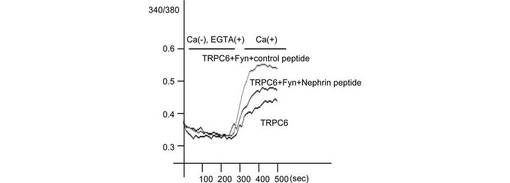
【図5G】
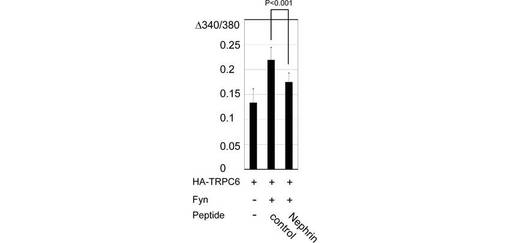
【図6A】
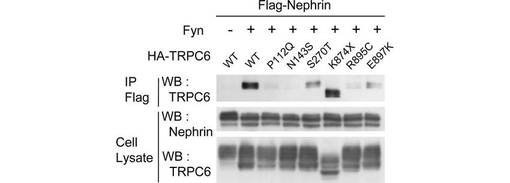
【図6B】
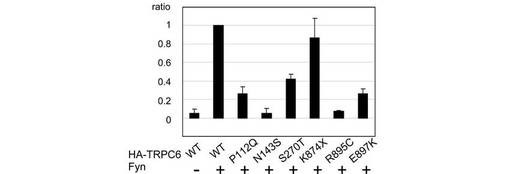
【図6C】
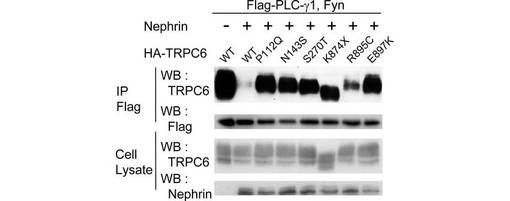
【図6D】
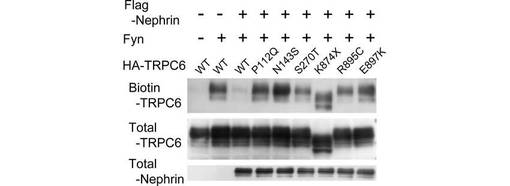
【図6E】
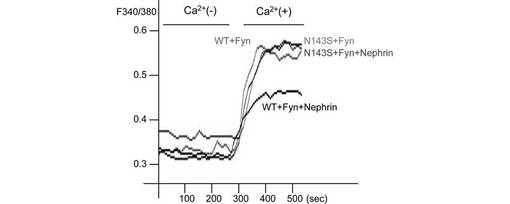
【図6F】
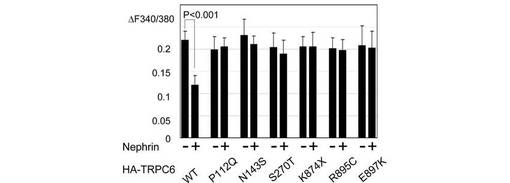
【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図2C】
【図2D】
【図2E】
【図2F】
【図2G】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【公開番号】特開2012−29662(P2012−29662A)
【公開日】平成24年2月16日(2012.2.16)
【国際特許分類】
【出願番号】特願2010−174155(P2010−174155)
【出願日】平成22年8月3日(2010.8.3)
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【出願人】(598041566)学校法人北里研究所 (180)
【出願人】(899000068)学校法人早稲田大学 (602)
【Fターム(参考)】
【公開日】平成24年2月16日(2012.2.16)
【国際特許分類】
【出願日】平成22年8月3日(2010.8.3)
【出願人】(504137912)国立大学法人 東京大学 (1,942)
【出願人】(598041566)学校法人北里研究所 (180)
【出願人】(899000068)学校法人早稲田大学 (602)
【Fターム(参考)】
[ Back to top ]