
マイコバクテリア感染症に対するワクチン
【課題】結核又は癩病のようなマイコバクテリア感染症に対するワクチンとして、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原または抗原として有効なそれらの断片をコードするコード配列を含む、ネイキッド核酸構築物の提供。
【解決手段】マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原または抗原として有効なそれらの断片をコードするコード配列であって、哺乳動物宿主細胞中でそのコード配列を発現できるプロモーターに機能可能なように結合されたコード配列を含むネイキッド核酸構築物の、マイコバクテリア感染症に対するワクチンとして使用するための薬剤の製造における使用。
【解決手段】マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原または抗原として有効なそれらの断片をコードするコード配列であって、哺乳動物宿主細胞中でそのコード配列を発現できるプロモーターに機能可能なように結合されたコード配列を含むネイキッド核酸構築物の、マイコバクテリア感染症に対するワクチンとして使用するための薬剤の製造における使用。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、結核、癩病のようなマイコバクテリア感染症に対するワクチンに関する。
【背景技術】
【0002】
結核あるいは癩病に対する防御的な細胞媒介免疫反応がどのようにして獲得され、あるいは発現されるのかということは、古典的な免疫学の中心的な位置にあったにもかかわらず、驚くほどわずかのことしか判っていない。なぜカルメット-ゲラン(Calmette-Guerin) 桿菌(BCG)の生ワクチンを投与することが、あるヒトの集団に対してのみ非常に防御的なのか、あるいは、生菌BCGとは異なり、死菌BCG又は抗原成分では大量に注射してもアジュバントとともに注射しても動物にほんの僅かしか防御を与えないのは何故なのかということは判っていない。
【非特許文献1】Mehra et al (1966): Proc. Natl. Acad. Sci. USA; 83, 7014-7017
【非特許文献2】Thole et al, Infection and Immunity (1990) 58, 80-87
【非特許文献3】Strohmaier et al, J. Gen. Virol., 1982, 59, 205-306
【非特許文献4】Spence et al, J. Gen. Virol., 1989, 70, 2843-51; Smith et al, Gene, 1984, 29, 263-9
【非特許文献5】Geysen et al, Proc. Natl. Acad. Sci, 1984, 81, 3998-4002
【非特許文献6】Hopp and Woods, Ploc, Natl. Acad. Sci., 1981, 78, 3824-8; Berzofsky, Science, 1985, 229, 932-40
【非特許文献7】Ascadi et al, (1991): The New Biologist; 3, 71-81
【非特許文献8】Sambrook et al, (Molecular Cloning: A Laboratory Manual, 1989
【非特許文献9】Invitrogen; Seed and Aruffo, Proc. Natl. Acad. Sci. USA (1987) 84, 3365-3369
【非特許文献10】Gautier et al, Nucl. Acids Res. (1989) 17, 8389
【非特許文献11】Silva et al (1992); Microb. Pathogen. 12, 27-38
【非特許文献12】Invitrogen; Brosius, DNA 8, 759-777, 1989; Brosius, Methods in Enzymology 216, 469-483, 1992
【非特許文献13】Invitrogen, Wang et al, Cell 67, 797-805, 1991
【非特許文献14】Spies and DeMars, Nature 351, 323-324, 1991
【非特許文献15】Seykora et al, Proc. Natl. Acad. Sci. USA 88, 2505-2509, 1991
【非特許文献16】Attaya et al, Nature 355, 647-648, 1992
【発明の開示】
【課題を解決するための手段】
【0003】
癩菌(Mycobacterium leprae)の65 kDa熱ショックタンパク質(MLhsp65) 抗原(Mehra et al (1966): Proc. Natl. Acad. Sci. USA; 83, 7014-7017)に基づいて代替的なワクチンを開発しようとする試みにおいて、我々はこの抗原をコードする発現ベクターを骨髄細胞に安定にトランスフェクトした。このトランスフェクトされた骨髄細胞をマウスに注射すると、マウスは結核の原因物質である結核菌(Mycobacterium tuberculosis)に対して抵抗性を示すことが判った。さらに我々は、MLhsp65 又は癩菌36kDのプロリンリッチ抗原(Thole et al, Infection and Immunity (1990) 58, 80-87)をコードするDNA をマウスにそのまま注射した。これらのマウスも結核菌による感染に対して抵抗性であることが判った。
【0004】
これらの知見は広い用途を有している。従って本発明は、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原又は抗原として有効なそれらの断片をコードするコード配列であって、そのコード配列を哺乳動物宿主細胞中で発現させることができるプロモーターに機能可能なように結合されたコード配列を含むネイキッド(naked) 核酸構築物の、マイコバクテリア感染症に対するワクチンとして使用するための医薬の製造における使用を提供する。
【0005】
本発明はまた、
−マイコバクテリア感染症に対し、ワクチンとしての使用するためのそのようなネイキッド核酸構築物、
−そのようなネイキッド核酸構築物及び許容できる担体又は希釈剤を含むワクチン組成物、
−そのようなネイキッド核酸構築物の有効量を宿主に投与することを含む、マイコバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法、
−マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原又は抗原として有効なそれらの断片をコードするコード配列であって、そのコード配列を骨髄細胞中で発現させることができるプロモーターに機能可能なように結合されたコード配列を含む核酸構築物によりトランスフェクトされた骨髄細胞、
−そのようなトランスフェクトされた骨髄細胞の有効量を宿主に投与することを含む、マイコバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法、及び
−コード配列が、マイコバクテリアプロリンリッチ抗原又は抗原として有効なその断片をコードする、上記のネイキッド核酸構築物も提供するものである。
【0006】
前記のネイキッド核酸構築物は、マイコバクテリアストレスタンパク質もしくはマイコバクテリアプロリンリッチ抗原又は抗原として有効なそれらの断片をコードするコード配列であって、そのコード配列を哺乳動物宿主細胞中で発現させることができるプロモーターに機能可能なように結合されたコード配列を含む。少なくとも1つの別のマイコバクテリアタンパク質又はその断片をコードする核酸であって、機能可能なようにプロモーターに結合されたものがその構築物に含まれていてもよい。典型的には、このようにコードされた別のマイコバクテリアタンパク質またはその断片は、抗原タンパク質又はその抗原性断片である。その別のマイコバクテリアタンパク質またはその断片は、別のマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはその抗原断片であってもよい。
【0007】
前記のネイキッド核酸構築物は、典型的には無細胞でウイルスフリーであり、典型的には単離形態にあるものであり、精製されていてもよい。構築物はDNA であることが好ましいが、RNA 又は修飾核酸であってもよい。このような核酸は、そのバックボーンに修飾を含むことができ、構築物が環状でなく直鎖状である場合に、5'又は3'のいずれか、あるいは両方の末端における付加であり得る。これは、核酸が宿主細胞内、例えば構築物の能力を高める筋肉細胞に取り込まれたときの核酸の寿命を延長するのに役立ち得る。核酸分子に対する公知の修飾としては、メチルホスホネート及びホスホロチオエートバックボーンとすること、及びアクリジンまたはポリリシン鎖を分子の3'及び/または5'末端に付加することが挙げられる。
【0008】
本発明の核酸構築物によりコードされるマイコバクテリアストレスタンパク質は、一般的には、それが由来するマイコバクテリアが環境的なストレス下に置かれたときににその発現が増加するものである。典型的には、マイコバクテリアストレスタンパク質は熱ショックタンパク質であり、例えばそれが由来する細菌が高温、例えば42℃以上にさらされたときに発現が実質的に増加するタンパク質である。
【0009】
マイコバクテリアストレスタンパク質は、典型的には、結核菌、癩菌、ウシ結核菌(Mycobacterium bovis)、トリ結核菌(Mycobacterium avium)又はマイコバクテリウム・バッカエ(Mycobacterium vaccae)から得られる。適当なタンパク質としては、結核菌、癩菌、ウシ結核菌、トリ結核菌又はマイコバクテリウム・バッカエの70、65及び10 kDa熱ショックタンパク質が挙げられる。これらのうち、結核菌、癩菌及びウシ結核菌の65 kDa熱ショックタンパク質が好ましく、癩菌の熱ショックタンパク質が特に好ましい。
【0010】
マイコバクテリアプロリンリッチ抗原は、結核菌、癩菌、ウシ結核菌、トリ結核菌又はマイコバクテリウム・バッカエのプロリンリッチ抗原とすることができる。適当なプロリンリッチ抗原は、癩菌の36 kDaプロリンリッチ抗原である。
【0011】
マイコバクテリアストレスタンパク質またはプロリンリッチ抗原の抗原性断片は、好ましくは最小で5、6、7、8、9、10、15、20、30、40又は50アミノ酸を含む。断片は10、20、30、40又は50アミノ酸長までのものであってもよい。あるいは、20又は10までのアミノ酸残基がストレスタンパク質またはプロリンリッチ抗原のアミノ及び/またはカルボキシ末端から削除されてもよい。
【0012】
マイコバクテリアストレスタンパク質またはプロリンリッチ抗原の抗原部位は、標準的な方法を用いて同定することができる。そのような方法は、タンパク質分解酵素又は化学物質を用いてポリペプチドそのものを断片化し、その後各断片の抗体と結合する能力、又は動物もしくは適当なin vitroモデル系中に接種されたときに免疫反応を起こす能力を試験することを含むものとすることができる(Strohmaier et al, J. Gen. Virol., 1982, 59, 205-306)。
【0013】
あるいは、マイコバクテリアストレスタンパク質またはプロリンリッチ抗原をコードするDNA を制限酵素消化または周知の技術により断片化し、発現系に導入して断片を生成してもよい。このような断片は、通常は細菌起源のポリペプチドであるポリペプチドに融合することができる。得られる断片を、これまでに記載されているようにして調べる(Spence et al, J. Gen. Virol., 1989, 70, 2843-51; Smith et al, Gene, 1984, 29, 263-9)。
【0014】
もう1つの方法は、完全長ポリペプチドの全配列をカバーする短いペプチド断片(3〜20アミノ酸長、慣用的には6アミノ酸長)であって、各ペプチドが隣接ペプチドとオーバーラップするものを化学的に合成することである。このようなオーバーラップは1〜10アミノ酸であってよいが、nをペプチドの長さとしてn-1アミノ酸であることが理想的である(Geysen et al, Proc. Natl. Acad. Sci, 1984, 81, 3998-4002)。その後各ペプチドを、通常はペプチドを最初に何らかの担体分子に結合させて免疫反応の誘導を容易にすることを除いてこれまでに記載されたようにして調べる。
【0015】
また、特定の性質、例えば免疫学的に重要な部位と関連していると考えられている親水性について配列を分析するような、予測的な方法がある(Hopp and Woods, Ploc, Natl. Acad. Sci., 1981, 78, 3824-8; Berzofsky, Science, 1985, 229, 932-40)。これらの予測については、その後これまでに記載されたような組換え体ポリペプチドあるいはペプチド法を使用して試験することができる。
【0016】
マイコバクテリアショックタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸配列は、典型的には複製可能な発現ベクターに含まれるものとする。そのような発現ベクターは複製開始起点を含み、該ベクターは細菌宿主細胞のような宿主細胞中で複製されることができ、また核酸配列の発現のためのプロモーター、及び任意にプロモーターのレギュレーターを含んでいる。このようなベクターは、1以上の選択可能マーカー遺伝子、例えば細菌形質転換体を同定するためにはアンピシリン耐性遺伝子、あるいは哺乳動物細胞形質転換体を同定するためにはネオマイシン耐性遺伝子を含んでもよい。この核酸構築物は、任意にプロモーターのためのエンハンサーを含んでもよい。この構築物はまた、機能性タンパク質をコードする核酸に3'において機能可能なように結合したポリアデニル化シグナルを含んでもよい。この構築物はまた、マイコバクテリアストレスタンパク質またはその断片をコードする配列に3'においてターミネーターを含んでもよい。この構築物はまた、マイコバクテリアストレスタンパク質またはその断片をコードする配列に3'において1以上のイントロン又はその他のコード配列を含んでもよい。そのような1以上のイントロンは、構築物が投与される宿主生物からのもの、あるいは別の真核細胞生物からのものとすることができる。
【0017】
上記の核酸構築物において、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはその抗原性断片をコードする核酸配列は、その配列を発現することができるプロモーターに機能可能なように結合されている。「機能可能なように結合された」とは、プロモーターと、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸配列とが、コード配列がプロモーターの制御下に発現し得るような関係にある配置のことをいうものである。従って、例えばプロモーターとコード配列との間に5'非コード配列のようなエレメントがあってもよい。このようなエレメントは、プロモーター配列を得た生物に本来あるものであってもよく、またマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片を得た生物に本来あるものであってもよい。あるいは、前記の1以上のエレメントは、プロモーター配列を得た生物に本来ないもので、またマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片を得た生物にも本来ないものであってもよい。そのような配列は、プロモーターによるコード配列の正確な制御を増強し、あるいは阻害しない場合に構築物に含めることができる。
【0018】
発現ベクターは任意の種類のものであってよい。例えば、ベクターは直鎖状の形態であってもよく、環状の形態であってもよい。共有結合閉環状(CCC) プラスミドDNA は直接筋肉細胞に取り込まれ得るが、そのDNA は細胞のゲノムDNA に組み込まれないことが判っているので、構築物をプラスミドベクターに導入することが好ましい(Ascadi et al, (1991): The New Biologist; 3, 71-81)。当業者であれば、例えばSambrook et al, (Molecular Cloning: A Laboratory Manual, 1989)により記載されたような遺伝子工学技術により修飾されるような広範に入手可能なベクターを原料として、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸配列を含む適当なベクターを製造できるであろう。出発材料として2つの適当なベクターがあり、プラスミドpCDM8 (Invitrogen; Seed and Aruffo, Proc. Natl. Acad. Sci. USA (1987) 84, 3365-3369)及び pHMG (Gautier et al, Nucl. Acids Res. (1989) 17, 8389)である。
【0019】
マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする配列の発現を生起できる任意のプロモーターを、その配列に機能可能なように結合することができる。特に適するプロモーターは、哺乳動物細胞中で発現を生起できるものである。例えば、サイトメガロウイルス(CMV)即時早期遺伝子プロモーターのような、哺乳動物細胞中で発現されるウイルス遺伝子からのプロモーターが適している。また、例えばハウスキーピング遺伝子のプロモーターのような、多くの又は全ての哺乳動物で発現される哺乳動物遺伝子からのプロモーターも適している。例えば、p-ヒドロキシメチル-CoA- レダクターゼ(HMG) プロモーター(Gautier et al (1989))は特に適している。また、マクロファージや樹状細胞のような抗原提示哺乳動物細胞で選択的に発現される遺伝子のプロモーターやその他の調節エレメントも適している。
【0020】
前記核酸構築物は遺伝子治療に適している。特に、例えば結核菌、癩菌及びウシ結核菌により起こるマイコバクテリア感染症に対し、ネイキッドDNA による哺乳動物宿主のワクチン接種に有用である。従って、任意のマイコバクテリア種からの核酸を含む構築物を調製することができる。いくつかのマイコバクテリアストレスタンパク質にはある程度の保存が存在することにより、特定の種からの核酸配列を使用してその種による感染に対してワクチンを接種することは必ずしも必要でない。例えば、ウシ結核菌の生BCG(カルメット-ゲラン桿菌)細胞は、長い間ヒト結核菌に対してヒトにワクチン接種するのに使用されてきた。
【0021】
従って本発明においては、結核菌、癩菌またはウシ結核菌のストレスタンパク質またはプロリンリッチ抗原をコードする核酸構築物を使用して、マイコバクテリアのこれらの3つの種のいずれかによる感染症に対してワクチンを接種することができる。例えば本発明者らにより、癩菌65 kDa hspまたは癩菌36 kDaプロリンリッチ抗原をコードする構築物が、マウスにおいて結核菌に対して有効なワクチンとして作用することが示された。
【0022】
本発明の核酸構築物を用いて、一定の範囲の哺乳動物種にマイコバクテリア感染症に対してワクチン接種することができる。しかし、結核菌に対するヒトのワクチン接種が特に望ましい。ウシ結核菌に対する家畜、シカのワクチン接種も望ましい。また、アナグマは家畜に細菌を伝染させ得るので、ウシ結核菌に対してアナグマにワクチン接種することも望ましい。
【0023】
本発明のネイキッド核酸構築物は、ヒトを含む哺乳動物に任意の適当な経路により投与することができる。適当な経路としては、経口、並びに皮下、筋肉内、静脈内及び皮内等の非経口の経路が挙げられる。
【0024】
好ましい投与経路は、経口投与及び注射であり、典型的には筋肉内又は皮内注射である。ヒト又は動物患者の骨格筋又は皮膚内へのワクチン組成物の注射が特に好ましい。本発明のワクチン組成物の別の投与形態は、バイオリスティック又は「パーティクル・ガン」法である。
【0025】
本発明のネイキッド核酸構築物はそれのみで患者に投与することができ、また、リポソーム又はその他のデリバリー分子とともに投与することができる。有効投与量は、デリバリー分子を使用するか否か、投与経路、ワクチン接種される動物の大きさ等の多くの因子による。典型的な投与量は、投与あたり0.1 〜1000μgの核酸構築物、例えば投与あたり50〜75μg及び100 〜500 μgのような1 〜500 μg、50〜500 μgの範囲である。
【0026】
投与スケジュールは、例えば投与経路、受容体の種、受容体の症状等により変化するが、複数日、複数週、複数月にわたる単回投与又は複数回投与が考えられる。単回投与は、典型的には0.1 〜1000μg、例えば100 〜500 μgの核酸を含むものとし、複数回投与にはそれぞれ0.1 〜1000μg 、例えば100 〜500 μgの核酸を含むものとし、好ましくは筋肉内または皮内注射に適した形態とする。また、本発明の構築物を含む単回又は複数回の核酸ペレット、例えば100 〜500 μgのDNA を含むペレットを受容体の筋肉内または皮内に移植することができる。構築物をバイオリスティック法(biolistic method)により投与する場合は、この経路の効率が高いため、投与量は一般に上記範囲の下限付近となる。そのような投与量としては、例えば構築物の0.1 〜1 μgのような0.1 〜10μgの範囲である。
【0027】
本発明のネイキッド核酸構築物をそれのみで投与することも可能であるが、医薬配合物とすることが好ましい。本発明の配合物は、少なくとも1種の活性成分、即ち本発明の核酸構築物を、1種以上の許容される担体及び任意にその他の治療成分とともに含む。1種以上の担体は、配合物の他の成分と適合し、その受容体の心身に有害でないという意味において「許容される」ものでなければならない。リポソームを使用することができ、適当なリポソームとしては、例えば正に荷電する脂質(N-[1-(2,3-ジオレイルオキシ) プロピル]-N,N,N-トリエチルアンモニウム(DOTMA))を含むもの、ジオレオイルホスファチジルエタノールアミン(DOPE)を含むもの、3β-[N-(N',N'-ジメチルアミノエタン)-カルバモイル]コレステロール(Dc-Chol)を含むもの等がある。
【0028】
非経口投与に適した配合物としては、水性及び非水性の殺菌注射溶液があり、これらは酸化防止剤、バッファー、細菌増殖阻害剤、殺菌性抗生物質、及び配合物を意図される受容体の血液と等張にする溶質等を含んでもよい。また、水性及び非水性の殺菌懸濁物があり、これらは懸濁剤及び増粘剤を含んでもよい。さらにリポソーム、又は前記化合物を血液成分もしくは1以上の器官に向かわせるように設計されたその他の微粒子系がある。配合物は単位投与形態、又は複数投与容器、例えばシールしたアンプル又はバイアルの形態とすることができ、使用の直前に注射剤とするために、例えば水のような殺菌液体担体を添加するだけでよい凍結乾燥状態で保存することもできる。注射溶液及び懸濁液は、これまでに開示されているような種類の殺菌粉末、顆粒及び錠剤から即座に調製することができる。
【0029】
本発明の配合物は、上記に具体的に挙げた成分の他に、対象としている配合物の種類に認められた当分野で慣用のその他の物質を含んでもよいことが理解されるべきである。可能な配合物の中では、殺菌された発熱物質を含まない水性及び非水性の溶液が好ましい。本発明の核酸構築物がリポソームに含まれた配合物も好ましい。
【0030】
結核菌、癩菌及びウシ結核菌によるマイコバクテリア感染症に対して有効なワクチンは、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸構築物であってそのコード配列を骨髄細胞内で発現することができるプロモーターに機能可能なように結合されている核酸構築物をトランスフェクトした骨髄細胞から調製することもできる。
【0031】
骨髄細胞は任意の適当な方法によりトランスフェクトすることができる。例えば、核酸構築物を感染性のウイルス粒子、例えばレトロウイルス粒子中にパッケージすることができる。これはSilva et al (1992); Microb. Pathogen. 12, 27-38)により記載された方法により行うことができる。前記構築物は、エレクトロポレーション、リン酸カルシウム沈殿、バイオリスティック法、あるいはネイキッド核酸構築物を溶液中で骨髄細胞と接触させることにより導入することもできる。
【0032】
骨髄細胞に感染させる前記核酸構築物においては、核酸はDNA であってもRNA
であってもよいが、好ましくはDNA である。
【0033】
骨髄細胞にトランスフェクトする核酸構築物は、適当な種類の任意のものとすることができる。典型的には、構築物は発現ベクターの形態であり、例えばレトロウイルス又は本明細書で定義したようなネイキッドDNA 発現ベクターである。例えば構築物は、広く入手可能なpZipNeo ベクターから、又は例えばpCDM8 もしくはpHMGのような本明細書に記載したようなプラスミドベクターから得たレトロウイルスシャトルベクターの形態のものとすることができる。
【0034】
骨髄細胞にトランスフェクトする構築物は、少なくとも1つの別のマイコバクテリアタンパク質又はその断片をコードするコード配列であって、そのコード配列を哺乳動物細胞中で発現させることができるプロモーターに機能可能なように結合したコード配列を含んでいてもよい。典型的には、このようにコードされた別のマイコバクテリアタンパク質又はその断片は、抗原性タンパク質又はその抗原性断片である。この別のマイコバクテリアタンパク質又はその断片は、別のマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはその抗原性断片であってもよい。
【0035】
骨髄細胞にトランスフェクトされる構築物は、任意の適当なプロモーターを含むことができる。特に適するプロモーターは、哺乳動物細胞中で発現を生起できるものである。例えば、サイトメガロウイルス(CMV) 即時早期遺伝子プロモーターのような哺乳動物細胞中で発現されるウイルス遺伝子からのプロモーターが適している。また、例えばハウスキーピング遺伝子のプロモーターのような、多くの又は全ての哺乳動物で発現される哺乳動物遺伝子からのプロモーターも適している。例えば、p-ヒドロキシメチル-CoA-レダクターゼ(HMG) プロモーター(Gautier et al (1989))は特に適している。
【0036】
前記構築物をトランスフェクトした骨髄細胞は任意の適当な方法により投与することができ、例えば非経口的な注射、好ましくは静脈内注射により投与することができる。
【0037】
前記核酸構築物をトランスフェクトした骨髄細胞の任意の有効な量を受容体に投与することができる。典型的には、約1 x 104 から約1 x 108 、例えば約105〜107、例えば1 x 106の骨髄細胞が投与される。
【0038】
受容体に投与されるトランスフェクトされた骨髄細胞は、受容体の免疫系に適合するものであれば任意のものとすることができる。典型的には、細胞又は組織の移植についてと同様に、投与された骨髄細胞の主要組織移植抗原は受容体細胞の主要組織移植抗原と適合する。投与される骨髄細胞は受容個体から得たものであってもよい。
【0039】
前記のトランスフェクトされた骨髄細胞はそれ単独で受容体に投与してもよく、任意の適当な配合物中のものとして投与してもよい。好ましい配合物は受容体の血液と等張の溶液である。
【0040】
以下の実施例により本発明を具体的に説明する。添付の図面は以下の通りである。
【実施例】
【0041】
実施例1
骨髄細胞中で発現された癩菌hsp65(MLhsp65)遺伝子による結核に対する防御
骨髄細胞をBalb/cマウスの大腿骨及び脛骨から取り出した。これらの細胞を、psi-CRE 細胞中の感染性レトロウイルス粒子中にパッケージしたpZIPhsp65 DNAとともにin vitroで培養した(Silva et al (1992); Microb. Pathogen. 12, 27-38)。pZIPhsp65はpZIPML65とも指称され、MLhsp65 遺伝子を含み、Silva et al(1992)により記載されたものである。感染の高い効率及びウイルスDNA の安定な組み込みを確保するため、供与体マウスを最初に5-フルオロウラシルで処置した。高力価のウイルス(5 x 106/ml)と、細胞のウイルス感染を促進するポリブレン(4μg/ml) とをin vitroで使用した。その後、ネオマイシンを培養培地に含めて(0.5 mg/ml) 2日間感染細胞を選択した。
【0042】
受容体マウスは7〜8週齢のBalb/cであり、γ−照射(9.5 Gy)することによりその骨髄細胞を破壊した。マウスに直ちに1 x 106 骨髄細胞を静脈内注射した。これらは、MLhsp65 核酸を含むウイルスを感染させた細胞(BMC-65 細胞) と正常骨髄細胞との混合物 (比1:2)か、正常細胞のみ、あるいはマイコバクテリア遺伝子を持たないベクターを感染させた細胞(BMC-NeO)であった。
【0043】
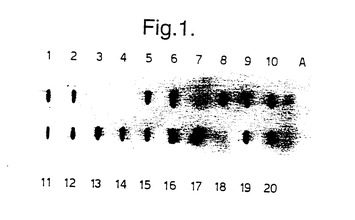
受容体動物中におけるマイコバクテリア遺伝子の発現は、DNA とタンパク質を脾臓細胞及び血液試料からそれぞれ抽出することにより移植後15日に試験した。図1は、DNA サザンブロットハイブリダイゼーションにより、16〜20のマウスからの脾臓細胞がMLhsp65 遺伝子を含んでいたことを示している。抽出されたDNAをニトロセルロース上にブロットし、MLhsp65 遺伝子を含む癩菌3.6 Kb EcoRI DNA断片(Silva et al (1992))を使用してハイブリダイゼーションにより検出した。
【0044】
マウスの別の群について、MLhsp6S-特異的モノクローナル抗体(CL44)を使用してウェスタンブロット検出を行い、20のうち14がその末梢血細胞中にMLhsp65 タンパク質を保有していることが判明した(図2)。組換え体DNA 由来タンパク質(rMLhsp65, 20ng)及び正常マウスの血液細胞から抽出されたタンパク質(細胞溶解物) を陽性及び陰性対照として使用した。結合抗体は、アルカリホスファターゼ抱合ヤギ抗マウスIgにより検出した。
【0045】
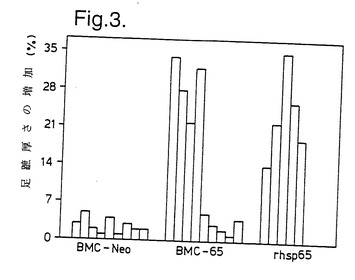
骨髄移植の30日後、別のマウスを遅延型過敏症について調べた(DTH; 5 μg rMLhsp65の注射48時間後の足蹠腫脹)。結果を図3に示す。rMLhsp65を投与された5匹のマウスの全てがDTH 反応を示し、BMC-65を投与された9匹のうち4匹が反応し、BMC-Neo を投与された9匹は反応しなかった。
【0046】
移植36日後に、米国のATCCから入手したよく知られた病原性感染株である結核菌H37Rv の生存可能な5 x 106 をマウスに静脈内感染させた。その3週間後にマウスを屠殺し、肝臓中の生存細菌の数を7H11アガー上のコロニー形成単位としてカウントした。別の対照マウスの群を、フロイント不完全アジュバント(IFA; 25μg)中のrMLhsp65により0日と7日において免疫化し、その後14日に追加免疫した (アジュバントなしの15μg)。
【0047】
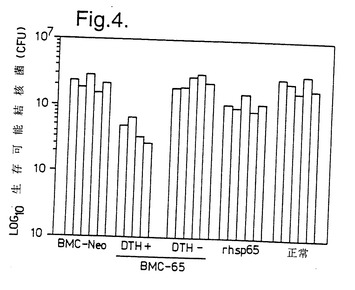
図4は、BMC-65移植後にDTH 反応を示したマウスは結核菌による感染に対しても抵抗性であったが、反応を示さなかったものは抵抗性ではなかったことを示している。rMLhsp65に反応するマウスではより低い程度の保護が見られた。
実施例2
ネイキッド癩菌hsp65 DNA 及び癩菌36 kDaプロリンリッチ抗原DNAの筋肉への直接の注射による結核に対する防御
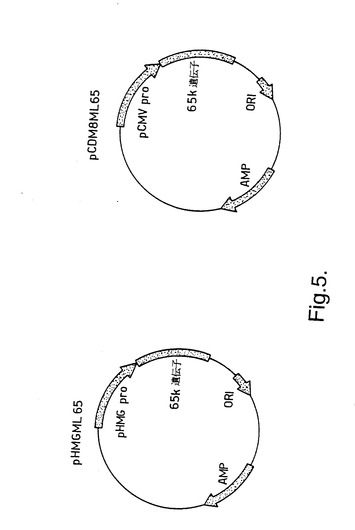
MLhsp65 をコードする遺伝子を標準的な方法により2種の真核生物発現ベクターpCDM8 (Invitrogen; Seed and Aruffo, Proc. Natl. Acad. Sci. USA (1987)84, 3365-3369)及びpHMG (Gautier et al, Nucl. Acids Res. (1989) 17, 8389)にクローン化し、pCDM8ML65 及びpHMGML65を形成した(図5)。即ち、MLhsp65遺伝子を含むDNA 配列を制限エンドヌクレアーゼを用いて大腸菌クローニングベクターpUC8から切り出した(Silva et al (1992))。そしてこの配列をpCDM8 及びpHMGに結合した。pCDM8 及びpHMGは、宿主細胞ゲノムへの組み込みに依存せず、広範なマウス細胞型中で機能しやすい強力なプロモーターを有する発現ベクターである。
【0048】
上記の構築物を標準的な方法により大腸菌中で増殖させた全調製物から精製した。正常Balb/c、CBA/B1O または異系交配させたParkesアルビノマウスに、一方の構築物の50〜75μg を左大腿四頭筋に、他方の構築物の50〜75μgを右大腿四頭筋に注射した。3〜4か月の間に4〜5対の注射をするまで2〜6週の間隔で注射を繰り返した。最後の注射の2週間後、マウスに1 x 106の生存結核菌を腹腔内感染させた。その6週間後マウスを屠殺し、体内器官中の生存細菌数を7H11アガー上でのコロニー形成単位としてカウントした。
【0049】
別の群のマウスに、MLhsp65の代わりに36kDプロリンリッチ抗原の癩菌遺伝子(Thole et al, Infection and Immunity (1990) 58, 80-87)を含むベクターからなるネイキッドDNAを同様に注射した。
【0050】
36 kDプロリンリッチ抗原を含む2種のネイキッドDNA構築物を実際に調製した。1kb EcoR1 断片をpTHL1007 (Thole et al (1990)) から切り出した。この断片を、クローニングベクターpSL301のEcoRIポリリンカー(Invitrogen; Brosius, DNA 8, 759-777, 1989; Brosius, Methods in Enzymology 216, 469-483, 1992)中にクローニングした。
【0051】
0.9 kbのBamH1 断片を得られた構築物から切り出し、pcDNA1/Neo (Invitrogen, Wang et al, Cell 67, 797-805, 1991; Spies and DeMars, Nature 351, 323-324, 1991; Seykora et al, Proc. Natl. Acad. Sci. USA 88, 2505-2509, 1991; Attaya et al, Nature 355, 647-648, 1992)のBamH1 部位にクローン化した。得られた構築物は、マウスに注射した癩菌の36 kD プロリンリッチ抗原の遺伝子を含む2種の構築物の1つであった。他方の構築物は、やはり0.9 kb BamH1断片を切り出し、そしてそれをpHMGのBamH1 部位にクローン化することにより得た。
【0052】
また、さらに別の群に以下のもの、
−生理食塩水のみを筋肉内に、
−IFA 中のrMLhsp65タンパク質を実施例1と同様に、
−ウシ結核菌BCG (1 x 106細胞、0日に皮内に)、あるいは
−挿入遺伝子を含まない空のベクターを筋肉内に注射した。
【0053】
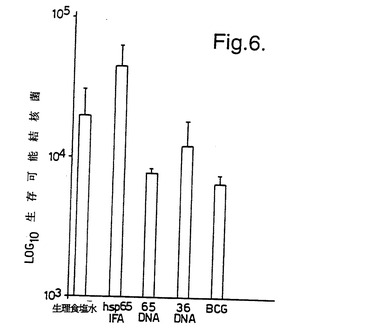
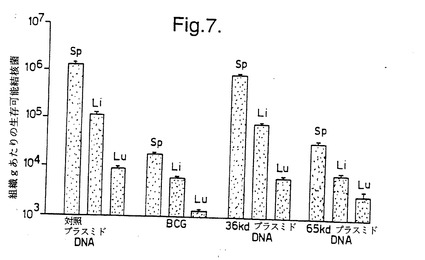
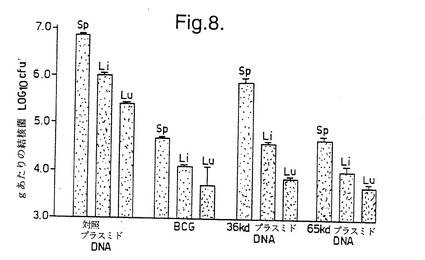
結果を図6〜8に示す。図6は、Balb/cマウスがBCG またはMLhsp65 DNA(65DNA) により有意に防御されたことを示している。図6に示した結果の場合、Balb/cマウスの肝臓中の生存細菌の数をカウントした。図7及び8は、CBA/B1O 及びParkesアルビノマウスの両方が、BCG またはMLhsp65 遺伝子もしくは36 kD プロリンリッチ抗原遺伝子を含むDNA により実質的に防御されたが、空のベクター(対照プラスミドDNA)によっては防御されなかったことを示している。
実施例3
ネイキッドhsp65 DNA の皮膚への直接の注射による結核に対する防御
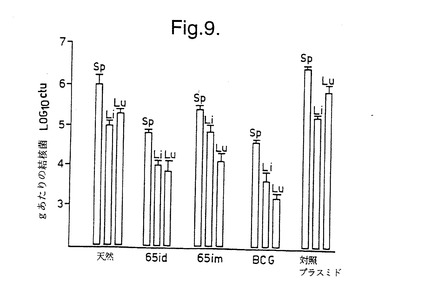
pCDM8ML65 DNA を上記のようにして製造した。Balb/cマウスに、3週間の間隔で3回、50〜75 gのネイキッドDNAを尾底部に皮内注射した。別の群のマウスには、同じ回数で100 〜150 μgの同じDNA を筋肉内に注射するか、50〜75μgのpCDMB(ベクターのみ) を皮内注射した。さらに別の対照群のマウスには、生BCGを上記のように皮内に投与するか、処理しなかった(天然)。最後のDNA注射の8週間後に、マウスに1 x 106の生存可能結核菌M37Rv を腹腔内感染させた。その4週間後にマウスを屠殺し、体内器官中の生存細菌数を7H11アガー上でのコロニー形成単位としてカウントした。結果を図9に示す。この図においては、皮内経路でのpCDM8ML65 DNAにより有効な防御が与えられたことが示されている。
【図面の簡単な説明】
【0054】
【図1】図1は、MLhsp65核酸を含むレトロウイルスを感染させた骨髄細胞と正常骨髄細胞との混合物(比1:2)を注射したBalb/cマウスの脾臓細胞から抽出されたDNAのDNAサザンブロットハイブリダイゼーションの結果を示す。
【図2】図2は、MLhsp65核酸を含むレトロウイルスを感染させた骨髄細胞と正常骨髄細胞との混合物(比1:2)を注射したBalb/cマウスの血液試料から抽出されたタンパク質のウェスタンブロットプロービングの結果を示す。
【図3】図3は、MLhsp65遺伝子を含まないベクターを感染させた骨髄細胞(BMC-Neo)、MLhsp65核酸を含むレトロウイルスを感染させた骨髄細胞(BMC-65)及び組換え体MLhsp65(rhsp65)を注射したマウスについての遅延型過敏症(DTH)試験の結果を示す。
【図4】図4は、図3について挙げたマウスとrMLhsp65により免疫化した別のマウスの群に結核菌の毒性株を感染させた結果を示す。
【図5】図5は、pCDM8ML65 及びpHMGML65のプラスミドマップを示す。
【図6】図6は、生理食塩水(生理食塩水)、rMLhsp65及び不完全フロイントアジュバント(hsp65IFA)、pCDM8ML65及びpHMGML65(65 DNA)、癩菌36 kDプロリンリッチ抗原遺伝子をそれぞれ含むpCDM8及びpHMG(36 DNA)、並びにウシ結核菌BCGを筋肉内に注射したBalb/cマウスの肝臓中の生存可能な結核菌の数を示す。
【図7】図7は、pCDM8及びpHMG (対照プラスミドDNA)、ウシ結核菌BCG、癩菌36 kDプロリンリッチ抗原遺伝子をそれぞれ含むpCDM8及びpHMG(36 kDプラスミドDNA)、並びにpCDM8ML65 及びpHMGML65(65 kDプラスミドDNA)を注射した、異系交配したParkesアルビノマウスの脾臓(Sp)、肝臓(Li)及び肺(Lu)中の生存可能な結核菌の数を示す。
【図8】図8は、pCDM8 及びpHMG (対照プラスミドDNA)、ウシ結核菌BCG、癩菌36 kDプロリンリッチ抗原遺伝子をそれぞれ含むpCDM8及びpHMG(36 kDプラスミドDNA)、並びにpCDM8ML65及びpHMGML65(65 kD プラスミドDNA)を注射したCBA/B1Oマウスの脾臓(Sp)、肝臓(Li)及び肺(Lu)中の生存可能な結核菌の数を示す。
【図9】図9は、非処置Balb/cマウス(天然)、あるいは皮内にpCDM8ML65(65id)、筋肉内にpCDM8ML65(65im)、皮内にウシ結核菌BCGまたは皮内にpCDM8(対照プラスミド)を投与したBalb/cマウスの脾臓(Sp)、肝臓(Li)及び肺(Lu)中の生存可能な結核菌の数を示す。
【技術分野】
【0001】
本発明は、結核、癩病のようなマイコバクテリア感染症に対するワクチンに関する。
【背景技術】
【0002】
結核あるいは癩病に対する防御的な細胞媒介免疫反応がどのようにして獲得され、あるいは発現されるのかということは、古典的な免疫学の中心的な位置にあったにもかかわらず、驚くほどわずかのことしか判っていない。なぜカルメット-ゲラン(Calmette-Guerin) 桿菌(BCG)の生ワクチンを投与することが、あるヒトの集団に対してのみ非常に防御的なのか、あるいは、生菌BCGとは異なり、死菌BCG又は抗原成分では大量に注射してもアジュバントとともに注射しても動物にほんの僅かしか防御を与えないのは何故なのかということは判っていない。
【非特許文献1】Mehra et al (1966): Proc. Natl. Acad. Sci. USA; 83, 7014-7017
【非特許文献2】Thole et al, Infection and Immunity (1990) 58, 80-87
【非特許文献3】Strohmaier et al, J. Gen. Virol., 1982, 59, 205-306
【非特許文献4】Spence et al, J. Gen. Virol., 1989, 70, 2843-51; Smith et al, Gene, 1984, 29, 263-9
【非特許文献5】Geysen et al, Proc. Natl. Acad. Sci, 1984, 81, 3998-4002
【非特許文献6】Hopp and Woods, Ploc, Natl. Acad. Sci., 1981, 78, 3824-8; Berzofsky, Science, 1985, 229, 932-40
【非特許文献7】Ascadi et al, (1991): The New Biologist; 3, 71-81
【非特許文献8】Sambrook et al, (Molecular Cloning: A Laboratory Manual, 1989
【非特許文献9】Invitrogen; Seed and Aruffo, Proc. Natl. Acad. Sci. USA (1987) 84, 3365-3369
【非特許文献10】Gautier et al, Nucl. Acids Res. (1989) 17, 8389
【非特許文献11】Silva et al (1992); Microb. Pathogen. 12, 27-38
【非特許文献12】Invitrogen; Brosius, DNA 8, 759-777, 1989; Brosius, Methods in Enzymology 216, 469-483, 1992
【非特許文献13】Invitrogen, Wang et al, Cell 67, 797-805, 1991
【非特許文献14】Spies and DeMars, Nature 351, 323-324, 1991
【非特許文献15】Seykora et al, Proc. Natl. Acad. Sci. USA 88, 2505-2509, 1991
【非特許文献16】Attaya et al, Nature 355, 647-648, 1992
【発明の開示】
【課題を解決するための手段】
【0003】
癩菌(Mycobacterium leprae)の65 kDa熱ショックタンパク質(MLhsp65) 抗原(Mehra et al (1966): Proc. Natl. Acad. Sci. USA; 83, 7014-7017)に基づいて代替的なワクチンを開発しようとする試みにおいて、我々はこの抗原をコードする発現ベクターを骨髄細胞に安定にトランスフェクトした。このトランスフェクトされた骨髄細胞をマウスに注射すると、マウスは結核の原因物質である結核菌(Mycobacterium tuberculosis)に対して抵抗性を示すことが判った。さらに我々は、MLhsp65 又は癩菌36kDのプロリンリッチ抗原(Thole et al, Infection and Immunity (1990) 58, 80-87)をコードするDNA をマウスにそのまま注射した。これらのマウスも結核菌による感染に対して抵抗性であることが判った。
【0004】
これらの知見は広い用途を有している。従って本発明は、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原又は抗原として有効なそれらの断片をコードするコード配列であって、そのコード配列を哺乳動物宿主細胞中で発現させることができるプロモーターに機能可能なように結合されたコード配列を含むネイキッド(naked) 核酸構築物の、マイコバクテリア感染症に対するワクチンとして使用するための医薬の製造における使用を提供する。
【0005】
本発明はまた、
−マイコバクテリア感染症に対し、ワクチンとしての使用するためのそのようなネイキッド核酸構築物、
−そのようなネイキッド核酸構築物及び許容できる担体又は希釈剤を含むワクチン組成物、
−そのようなネイキッド核酸構築物の有効量を宿主に投与することを含む、マイコバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法、
−マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原又は抗原として有効なそれらの断片をコードするコード配列であって、そのコード配列を骨髄細胞中で発現させることができるプロモーターに機能可能なように結合されたコード配列を含む核酸構築物によりトランスフェクトされた骨髄細胞、
−そのようなトランスフェクトされた骨髄細胞の有効量を宿主に投与することを含む、マイコバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法、及び
−コード配列が、マイコバクテリアプロリンリッチ抗原又は抗原として有効なその断片をコードする、上記のネイキッド核酸構築物も提供するものである。
【0006】
前記のネイキッド核酸構築物は、マイコバクテリアストレスタンパク質もしくはマイコバクテリアプロリンリッチ抗原又は抗原として有効なそれらの断片をコードするコード配列であって、そのコード配列を哺乳動物宿主細胞中で発現させることができるプロモーターに機能可能なように結合されたコード配列を含む。少なくとも1つの別のマイコバクテリアタンパク質又はその断片をコードする核酸であって、機能可能なようにプロモーターに結合されたものがその構築物に含まれていてもよい。典型的には、このようにコードされた別のマイコバクテリアタンパク質またはその断片は、抗原タンパク質又はその抗原性断片である。その別のマイコバクテリアタンパク質またはその断片は、別のマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはその抗原断片であってもよい。
【0007】
前記のネイキッド核酸構築物は、典型的には無細胞でウイルスフリーであり、典型的には単離形態にあるものであり、精製されていてもよい。構築物はDNA であることが好ましいが、RNA 又は修飾核酸であってもよい。このような核酸は、そのバックボーンに修飾を含むことができ、構築物が環状でなく直鎖状である場合に、5'又は3'のいずれか、あるいは両方の末端における付加であり得る。これは、核酸が宿主細胞内、例えば構築物の能力を高める筋肉細胞に取り込まれたときの核酸の寿命を延長するのに役立ち得る。核酸分子に対する公知の修飾としては、メチルホスホネート及びホスホロチオエートバックボーンとすること、及びアクリジンまたはポリリシン鎖を分子の3'及び/または5'末端に付加することが挙げられる。
【0008】
本発明の核酸構築物によりコードされるマイコバクテリアストレスタンパク質は、一般的には、それが由来するマイコバクテリアが環境的なストレス下に置かれたときににその発現が増加するものである。典型的には、マイコバクテリアストレスタンパク質は熱ショックタンパク質であり、例えばそれが由来する細菌が高温、例えば42℃以上にさらされたときに発現が実質的に増加するタンパク質である。
【0009】
マイコバクテリアストレスタンパク質は、典型的には、結核菌、癩菌、ウシ結核菌(Mycobacterium bovis)、トリ結核菌(Mycobacterium avium)又はマイコバクテリウム・バッカエ(Mycobacterium vaccae)から得られる。適当なタンパク質としては、結核菌、癩菌、ウシ結核菌、トリ結核菌又はマイコバクテリウム・バッカエの70、65及び10 kDa熱ショックタンパク質が挙げられる。これらのうち、結核菌、癩菌及びウシ結核菌の65 kDa熱ショックタンパク質が好ましく、癩菌の熱ショックタンパク質が特に好ましい。
【0010】
マイコバクテリアプロリンリッチ抗原は、結核菌、癩菌、ウシ結核菌、トリ結核菌又はマイコバクテリウム・バッカエのプロリンリッチ抗原とすることができる。適当なプロリンリッチ抗原は、癩菌の36 kDaプロリンリッチ抗原である。
【0011】
マイコバクテリアストレスタンパク質またはプロリンリッチ抗原の抗原性断片は、好ましくは最小で5、6、7、8、9、10、15、20、30、40又は50アミノ酸を含む。断片は10、20、30、40又は50アミノ酸長までのものであってもよい。あるいは、20又は10までのアミノ酸残基がストレスタンパク質またはプロリンリッチ抗原のアミノ及び/またはカルボキシ末端から削除されてもよい。
【0012】
マイコバクテリアストレスタンパク質またはプロリンリッチ抗原の抗原部位は、標準的な方法を用いて同定することができる。そのような方法は、タンパク質分解酵素又は化学物質を用いてポリペプチドそのものを断片化し、その後各断片の抗体と結合する能力、又は動物もしくは適当なin vitroモデル系中に接種されたときに免疫反応を起こす能力を試験することを含むものとすることができる(Strohmaier et al, J. Gen. Virol., 1982, 59, 205-306)。
【0013】
あるいは、マイコバクテリアストレスタンパク質またはプロリンリッチ抗原をコードするDNA を制限酵素消化または周知の技術により断片化し、発現系に導入して断片を生成してもよい。このような断片は、通常は細菌起源のポリペプチドであるポリペプチドに融合することができる。得られる断片を、これまでに記載されているようにして調べる(Spence et al, J. Gen. Virol., 1989, 70, 2843-51; Smith et al, Gene, 1984, 29, 263-9)。
【0014】
もう1つの方法は、完全長ポリペプチドの全配列をカバーする短いペプチド断片(3〜20アミノ酸長、慣用的には6アミノ酸長)であって、各ペプチドが隣接ペプチドとオーバーラップするものを化学的に合成することである。このようなオーバーラップは1〜10アミノ酸であってよいが、nをペプチドの長さとしてn-1アミノ酸であることが理想的である(Geysen et al, Proc. Natl. Acad. Sci, 1984, 81, 3998-4002)。その後各ペプチドを、通常はペプチドを最初に何らかの担体分子に結合させて免疫反応の誘導を容易にすることを除いてこれまでに記載されたようにして調べる。
【0015】
また、特定の性質、例えば免疫学的に重要な部位と関連していると考えられている親水性について配列を分析するような、予測的な方法がある(Hopp and Woods, Ploc, Natl. Acad. Sci., 1981, 78, 3824-8; Berzofsky, Science, 1985, 229, 932-40)。これらの予測については、その後これまでに記載されたような組換え体ポリペプチドあるいはペプチド法を使用して試験することができる。
【0016】
マイコバクテリアショックタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸配列は、典型的には複製可能な発現ベクターに含まれるものとする。そのような発現ベクターは複製開始起点を含み、該ベクターは細菌宿主細胞のような宿主細胞中で複製されることができ、また核酸配列の発現のためのプロモーター、及び任意にプロモーターのレギュレーターを含んでいる。このようなベクターは、1以上の選択可能マーカー遺伝子、例えば細菌形質転換体を同定するためにはアンピシリン耐性遺伝子、あるいは哺乳動物細胞形質転換体を同定するためにはネオマイシン耐性遺伝子を含んでもよい。この核酸構築物は、任意にプロモーターのためのエンハンサーを含んでもよい。この構築物はまた、機能性タンパク質をコードする核酸に3'において機能可能なように結合したポリアデニル化シグナルを含んでもよい。この構築物はまた、マイコバクテリアストレスタンパク質またはその断片をコードする配列に3'においてターミネーターを含んでもよい。この構築物はまた、マイコバクテリアストレスタンパク質またはその断片をコードする配列に3'において1以上のイントロン又はその他のコード配列を含んでもよい。そのような1以上のイントロンは、構築物が投与される宿主生物からのもの、あるいは別の真核細胞生物からのものとすることができる。
【0017】
上記の核酸構築物において、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはその抗原性断片をコードする核酸配列は、その配列を発現することができるプロモーターに機能可能なように結合されている。「機能可能なように結合された」とは、プロモーターと、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸配列とが、コード配列がプロモーターの制御下に発現し得るような関係にある配置のことをいうものである。従って、例えばプロモーターとコード配列との間に5'非コード配列のようなエレメントがあってもよい。このようなエレメントは、プロモーター配列を得た生物に本来あるものであってもよく、またマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片を得た生物に本来あるものであってもよい。あるいは、前記の1以上のエレメントは、プロモーター配列を得た生物に本来ないもので、またマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片を得た生物にも本来ないものであってもよい。そのような配列は、プロモーターによるコード配列の正確な制御を増強し、あるいは阻害しない場合に構築物に含めることができる。
【0018】
発現ベクターは任意の種類のものであってよい。例えば、ベクターは直鎖状の形態であってもよく、環状の形態であってもよい。共有結合閉環状(CCC) プラスミドDNA は直接筋肉細胞に取り込まれ得るが、そのDNA は細胞のゲノムDNA に組み込まれないことが判っているので、構築物をプラスミドベクターに導入することが好ましい(Ascadi et al, (1991): The New Biologist; 3, 71-81)。当業者であれば、例えばSambrook et al, (Molecular Cloning: A Laboratory Manual, 1989)により記載されたような遺伝子工学技術により修飾されるような広範に入手可能なベクターを原料として、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸配列を含む適当なベクターを製造できるであろう。出発材料として2つの適当なベクターがあり、プラスミドpCDM8 (Invitrogen; Seed and Aruffo, Proc. Natl. Acad. Sci. USA (1987) 84, 3365-3369)及び pHMG (Gautier et al, Nucl. Acids Res. (1989) 17, 8389)である。
【0019】
マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする配列の発現を生起できる任意のプロモーターを、その配列に機能可能なように結合することができる。特に適するプロモーターは、哺乳動物細胞中で発現を生起できるものである。例えば、サイトメガロウイルス(CMV)即時早期遺伝子プロモーターのような、哺乳動物細胞中で発現されるウイルス遺伝子からのプロモーターが適している。また、例えばハウスキーピング遺伝子のプロモーターのような、多くの又は全ての哺乳動物で発現される哺乳動物遺伝子からのプロモーターも適している。例えば、p-ヒドロキシメチル-CoA- レダクターゼ(HMG) プロモーター(Gautier et al (1989))は特に適している。また、マクロファージや樹状細胞のような抗原提示哺乳動物細胞で選択的に発現される遺伝子のプロモーターやその他の調節エレメントも適している。
【0020】
前記核酸構築物は遺伝子治療に適している。特に、例えば結核菌、癩菌及びウシ結核菌により起こるマイコバクテリア感染症に対し、ネイキッドDNA による哺乳動物宿主のワクチン接種に有用である。従って、任意のマイコバクテリア種からの核酸を含む構築物を調製することができる。いくつかのマイコバクテリアストレスタンパク質にはある程度の保存が存在することにより、特定の種からの核酸配列を使用してその種による感染に対してワクチンを接種することは必ずしも必要でない。例えば、ウシ結核菌の生BCG(カルメット-ゲラン桿菌)細胞は、長い間ヒト結核菌に対してヒトにワクチン接種するのに使用されてきた。
【0021】
従って本発明においては、結核菌、癩菌またはウシ結核菌のストレスタンパク質またはプロリンリッチ抗原をコードする核酸構築物を使用して、マイコバクテリアのこれらの3つの種のいずれかによる感染症に対してワクチンを接種することができる。例えば本発明者らにより、癩菌65 kDa hspまたは癩菌36 kDaプロリンリッチ抗原をコードする構築物が、マウスにおいて結核菌に対して有効なワクチンとして作用することが示された。
【0022】
本発明の核酸構築物を用いて、一定の範囲の哺乳動物種にマイコバクテリア感染症に対してワクチン接種することができる。しかし、結核菌に対するヒトのワクチン接種が特に望ましい。ウシ結核菌に対する家畜、シカのワクチン接種も望ましい。また、アナグマは家畜に細菌を伝染させ得るので、ウシ結核菌に対してアナグマにワクチン接種することも望ましい。
【0023】
本発明のネイキッド核酸構築物は、ヒトを含む哺乳動物に任意の適当な経路により投与することができる。適当な経路としては、経口、並びに皮下、筋肉内、静脈内及び皮内等の非経口の経路が挙げられる。
【0024】
好ましい投与経路は、経口投与及び注射であり、典型的には筋肉内又は皮内注射である。ヒト又は動物患者の骨格筋又は皮膚内へのワクチン組成物の注射が特に好ましい。本発明のワクチン組成物の別の投与形態は、バイオリスティック又は「パーティクル・ガン」法である。
【0025】
本発明のネイキッド核酸構築物はそれのみで患者に投与することができ、また、リポソーム又はその他のデリバリー分子とともに投与することができる。有効投与量は、デリバリー分子を使用するか否か、投与経路、ワクチン接種される動物の大きさ等の多くの因子による。典型的な投与量は、投与あたり0.1 〜1000μgの核酸構築物、例えば投与あたり50〜75μg及び100 〜500 μgのような1 〜500 μg、50〜500 μgの範囲である。
【0026】
投与スケジュールは、例えば投与経路、受容体の種、受容体の症状等により変化するが、複数日、複数週、複数月にわたる単回投与又は複数回投与が考えられる。単回投与は、典型的には0.1 〜1000μg、例えば100 〜500 μgの核酸を含むものとし、複数回投与にはそれぞれ0.1 〜1000μg 、例えば100 〜500 μgの核酸を含むものとし、好ましくは筋肉内または皮内注射に適した形態とする。また、本発明の構築物を含む単回又は複数回の核酸ペレット、例えば100 〜500 μgのDNA を含むペレットを受容体の筋肉内または皮内に移植することができる。構築物をバイオリスティック法(biolistic method)により投与する場合は、この経路の効率が高いため、投与量は一般に上記範囲の下限付近となる。そのような投与量としては、例えば構築物の0.1 〜1 μgのような0.1 〜10μgの範囲である。
【0027】
本発明のネイキッド核酸構築物をそれのみで投与することも可能であるが、医薬配合物とすることが好ましい。本発明の配合物は、少なくとも1種の活性成分、即ち本発明の核酸構築物を、1種以上の許容される担体及び任意にその他の治療成分とともに含む。1種以上の担体は、配合物の他の成分と適合し、その受容体の心身に有害でないという意味において「許容される」ものでなければならない。リポソームを使用することができ、適当なリポソームとしては、例えば正に荷電する脂質(N-[1-(2,3-ジオレイルオキシ) プロピル]-N,N,N-トリエチルアンモニウム(DOTMA))を含むもの、ジオレオイルホスファチジルエタノールアミン(DOPE)を含むもの、3β-[N-(N',N'-ジメチルアミノエタン)-カルバモイル]コレステロール(Dc-Chol)を含むもの等がある。
【0028】
非経口投与に適した配合物としては、水性及び非水性の殺菌注射溶液があり、これらは酸化防止剤、バッファー、細菌増殖阻害剤、殺菌性抗生物質、及び配合物を意図される受容体の血液と等張にする溶質等を含んでもよい。また、水性及び非水性の殺菌懸濁物があり、これらは懸濁剤及び増粘剤を含んでもよい。さらにリポソーム、又は前記化合物を血液成分もしくは1以上の器官に向かわせるように設計されたその他の微粒子系がある。配合物は単位投与形態、又は複数投与容器、例えばシールしたアンプル又はバイアルの形態とすることができ、使用の直前に注射剤とするために、例えば水のような殺菌液体担体を添加するだけでよい凍結乾燥状態で保存することもできる。注射溶液及び懸濁液は、これまでに開示されているような種類の殺菌粉末、顆粒及び錠剤から即座に調製することができる。
【0029】
本発明の配合物は、上記に具体的に挙げた成分の他に、対象としている配合物の種類に認められた当分野で慣用のその他の物質を含んでもよいことが理解されるべきである。可能な配合物の中では、殺菌された発熱物質を含まない水性及び非水性の溶液が好ましい。本発明の核酸構築物がリポソームに含まれた配合物も好ましい。
【0030】
結核菌、癩菌及びウシ結核菌によるマイコバクテリア感染症に対して有効なワクチンは、マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはそれらの断片をコードする核酸構築物であってそのコード配列を骨髄細胞内で発現することができるプロモーターに機能可能なように結合されている核酸構築物をトランスフェクトした骨髄細胞から調製することもできる。
【0031】
骨髄細胞は任意の適当な方法によりトランスフェクトすることができる。例えば、核酸構築物を感染性のウイルス粒子、例えばレトロウイルス粒子中にパッケージすることができる。これはSilva et al (1992); Microb. Pathogen. 12, 27-38)により記載された方法により行うことができる。前記構築物は、エレクトロポレーション、リン酸カルシウム沈殿、バイオリスティック法、あるいはネイキッド核酸構築物を溶液中で骨髄細胞と接触させることにより導入することもできる。
【0032】
骨髄細胞に感染させる前記核酸構築物においては、核酸はDNA であってもRNA
であってもよいが、好ましくはDNA である。
【0033】
骨髄細胞にトランスフェクトする核酸構築物は、適当な種類の任意のものとすることができる。典型的には、構築物は発現ベクターの形態であり、例えばレトロウイルス又は本明細書で定義したようなネイキッドDNA 発現ベクターである。例えば構築物は、広く入手可能なpZipNeo ベクターから、又は例えばpCDM8 もしくはpHMGのような本明細書に記載したようなプラスミドベクターから得たレトロウイルスシャトルベクターの形態のものとすることができる。
【0034】
骨髄細胞にトランスフェクトする構築物は、少なくとも1つの別のマイコバクテリアタンパク質又はその断片をコードするコード配列であって、そのコード配列を哺乳動物細胞中で発現させることができるプロモーターに機能可能なように結合したコード配列を含んでいてもよい。典型的には、このようにコードされた別のマイコバクテリアタンパク質又はその断片は、抗原性タンパク質又はその抗原性断片である。この別のマイコバクテリアタンパク質又はその断片は、別のマイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原またはその抗原性断片であってもよい。
【0035】
骨髄細胞にトランスフェクトされる構築物は、任意の適当なプロモーターを含むことができる。特に適するプロモーターは、哺乳動物細胞中で発現を生起できるものである。例えば、サイトメガロウイルス(CMV) 即時早期遺伝子プロモーターのような哺乳動物細胞中で発現されるウイルス遺伝子からのプロモーターが適している。また、例えばハウスキーピング遺伝子のプロモーターのような、多くの又は全ての哺乳動物で発現される哺乳動物遺伝子からのプロモーターも適している。例えば、p-ヒドロキシメチル-CoA-レダクターゼ(HMG) プロモーター(Gautier et al (1989))は特に適している。
【0036】
前記構築物をトランスフェクトした骨髄細胞は任意の適当な方法により投与することができ、例えば非経口的な注射、好ましくは静脈内注射により投与することができる。
【0037】
前記核酸構築物をトランスフェクトした骨髄細胞の任意の有効な量を受容体に投与することができる。典型的には、約1 x 104 から約1 x 108 、例えば約105〜107、例えば1 x 106の骨髄細胞が投与される。
【0038】
受容体に投与されるトランスフェクトされた骨髄細胞は、受容体の免疫系に適合するものであれば任意のものとすることができる。典型的には、細胞又は組織の移植についてと同様に、投与された骨髄細胞の主要組織移植抗原は受容体細胞の主要組織移植抗原と適合する。投与される骨髄細胞は受容個体から得たものであってもよい。
【0039】
前記のトランスフェクトされた骨髄細胞はそれ単独で受容体に投与してもよく、任意の適当な配合物中のものとして投与してもよい。好ましい配合物は受容体の血液と等張の溶液である。
【0040】
以下の実施例により本発明を具体的に説明する。添付の図面は以下の通りである。
【実施例】
【0041】
実施例1
骨髄細胞中で発現された癩菌hsp65(MLhsp65)遺伝子による結核に対する防御
骨髄細胞をBalb/cマウスの大腿骨及び脛骨から取り出した。これらの細胞を、psi-CRE 細胞中の感染性レトロウイルス粒子中にパッケージしたpZIPhsp65 DNAとともにin vitroで培養した(Silva et al (1992); Microb. Pathogen. 12, 27-38)。pZIPhsp65はpZIPML65とも指称され、MLhsp65 遺伝子を含み、Silva et al(1992)により記載されたものである。感染の高い効率及びウイルスDNA の安定な組み込みを確保するため、供与体マウスを最初に5-フルオロウラシルで処置した。高力価のウイルス(5 x 106/ml)と、細胞のウイルス感染を促進するポリブレン(4μg/ml) とをin vitroで使用した。その後、ネオマイシンを培養培地に含めて(0.5 mg/ml) 2日間感染細胞を選択した。
【0042】
受容体マウスは7〜8週齢のBalb/cであり、γ−照射(9.5 Gy)することによりその骨髄細胞を破壊した。マウスに直ちに1 x 106 骨髄細胞を静脈内注射した。これらは、MLhsp65 核酸を含むウイルスを感染させた細胞(BMC-65 細胞) と正常骨髄細胞との混合物 (比1:2)か、正常細胞のみ、あるいはマイコバクテリア遺伝子を持たないベクターを感染させた細胞(BMC-NeO)であった。
【0043】
受容体動物中におけるマイコバクテリア遺伝子の発現は、DNA とタンパク質を脾臓細胞及び血液試料からそれぞれ抽出することにより移植後15日に試験した。図1は、DNA サザンブロットハイブリダイゼーションにより、16〜20のマウスからの脾臓細胞がMLhsp65 遺伝子を含んでいたことを示している。抽出されたDNAをニトロセルロース上にブロットし、MLhsp65 遺伝子を含む癩菌3.6 Kb EcoRI DNA断片(Silva et al (1992))を使用してハイブリダイゼーションにより検出した。
【0044】
マウスの別の群について、MLhsp6S-特異的モノクローナル抗体(CL44)を使用してウェスタンブロット検出を行い、20のうち14がその末梢血細胞中にMLhsp65 タンパク質を保有していることが判明した(図2)。組換え体DNA 由来タンパク質(rMLhsp65, 20ng)及び正常マウスの血液細胞から抽出されたタンパク質(細胞溶解物) を陽性及び陰性対照として使用した。結合抗体は、アルカリホスファターゼ抱合ヤギ抗マウスIgにより検出した。
【0045】
骨髄移植の30日後、別のマウスを遅延型過敏症について調べた(DTH; 5 μg rMLhsp65の注射48時間後の足蹠腫脹)。結果を図3に示す。rMLhsp65を投与された5匹のマウスの全てがDTH 反応を示し、BMC-65を投与された9匹のうち4匹が反応し、BMC-Neo を投与された9匹は反応しなかった。
【0046】
移植36日後に、米国のATCCから入手したよく知られた病原性感染株である結核菌H37Rv の生存可能な5 x 106 をマウスに静脈内感染させた。その3週間後にマウスを屠殺し、肝臓中の生存細菌の数を7H11アガー上のコロニー形成単位としてカウントした。別の対照マウスの群を、フロイント不完全アジュバント(IFA; 25μg)中のrMLhsp65により0日と7日において免疫化し、その後14日に追加免疫した (アジュバントなしの15μg)。
【0047】
図4は、BMC-65移植後にDTH 反応を示したマウスは結核菌による感染に対しても抵抗性であったが、反応を示さなかったものは抵抗性ではなかったことを示している。rMLhsp65に反応するマウスではより低い程度の保護が見られた。
実施例2
ネイキッド癩菌hsp65 DNA 及び癩菌36 kDaプロリンリッチ抗原DNAの筋肉への直接の注射による結核に対する防御
MLhsp65 をコードする遺伝子を標準的な方法により2種の真核生物発現ベクターpCDM8 (Invitrogen; Seed and Aruffo, Proc. Natl. Acad. Sci. USA (1987)84, 3365-3369)及びpHMG (Gautier et al, Nucl. Acids Res. (1989) 17, 8389)にクローン化し、pCDM8ML65 及びpHMGML65を形成した(図5)。即ち、MLhsp65遺伝子を含むDNA 配列を制限エンドヌクレアーゼを用いて大腸菌クローニングベクターpUC8から切り出した(Silva et al (1992))。そしてこの配列をpCDM8 及びpHMGに結合した。pCDM8 及びpHMGは、宿主細胞ゲノムへの組み込みに依存せず、広範なマウス細胞型中で機能しやすい強力なプロモーターを有する発現ベクターである。
【0048】
上記の構築物を標準的な方法により大腸菌中で増殖させた全調製物から精製した。正常Balb/c、CBA/B1O または異系交配させたParkesアルビノマウスに、一方の構築物の50〜75μg を左大腿四頭筋に、他方の構築物の50〜75μgを右大腿四頭筋に注射した。3〜4か月の間に4〜5対の注射をするまで2〜6週の間隔で注射を繰り返した。最後の注射の2週間後、マウスに1 x 106の生存結核菌を腹腔内感染させた。その6週間後マウスを屠殺し、体内器官中の生存細菌数を7H11アガー上でのコロニー形成単位としてカウントした。
【0049】
別の群のマウスに、MLhsp65の代わりに36kDプロリンリッチ抗原の癩菌遺伝子(Thole et al, Infection and Immunity (1990) 58, 80-87)を含むベクターからなるネイキッドDNAを同様に注射した。
【0050】
36 kDプロリンリッチ抗原を含む2種のネイキッドDNA構築物を実際に調製した。1kb EcoR1 断片をpTHL1007 (Thole et al (1990)) から切り出した。この断片を、クローニングベクターpSL301のEcoRIポリリンカー(Invitrogen; Brosius, DNA 8, 759-777, 1989; Brosius, Methods in Enzymology 216, 469-483, 1992)中にクローニングした。
【0051】
0.9 kbのBamH1 断片を得られた構築物から切り出し、pcDNA1/Neo (Invitrogen, Wang et al, Cell 67, 797-805, 1991; Spies and DeMars, Nature 351, 323-324, 1991; Seykora et al, Proc. Natl. Acad. Sci. USA 88, 2505-2509, 1991; Attaya et al, Nature 355, 647-648, 1992)のBamH1 部位にクローン化した。得られた構築物は、マウスに注射した癩菌の36 kD プロリンリッチ抗原の遺伝子を含む2種の構築物の1つであった。他方の構築物は、やはり0.9 kb BamH1断片を切り出し、そしてそれをpHMGのBamH1 部位にクローン化することにより得た。
【0052】
また、さらに別の群に以下のもの、
−生理食塩水のみを筋肉内に、
−IFA 中のrMLhsp65タンパク質を実施例1と同様に、
−ウシ結核菌BCG (1 x 106細胞、0日に皮内に)、あるいは
−挿入遺伝子を含まない空のベクターを筋肉内に注射した。
【0053】
結果を図6〜8に示す。図6は、Balb/cマウスがBCG またはMLhsp65 DNA(65DNA) により有意に防御されたことを示している。図6に示した結果の場合、Balb/cマウスの肝臓中の生存細菌の数をカウントした。図7及び8は、CBA/B1O 及びParkesアルビノマウスの両方が、BCG またはMLhsp65 遺伝子もしくは36 kD プロリンリッチ抗原遺伝子を含むDNA により実質的に防御されたが、空のベクター(対照プラスミドDNA)によっては防御されなかったことを示している。
実施例3
ネイキッドhsp65 DNA の皮膚への直接の注射による結核に対する防御
pCDM8ML65 DNA を上記のようにして製造した。Balb/cマウスに、3週間の間隔で3回、50〜75 gのネイキッドDNAを尾底部に皮内注射した。別の群のマウスには、同じ回数で100 〜150 μgの同じDNA を筋肉内に注射するか、50〜75μgのpCDMB(ベクターのみ) を皮内注射した。さらに別の対照群のマウスには、生BCGを上記のように皮内に投与するか、処理しなかった(天然)。最後のDNA注射の8週間後に、マウスに1 x 106の生存可能結核菌M37Rv を腹腔内感染させた。その4週間後にマウスを屠殺し、体内器官中の生存細菌数を7H11アガー上でのコロニー形成単位としてカウントした。結果を図9に示す。この図においては、皮内経路でのpCDM8ML65 DNAにより有効な防御が与えられたことが示されている。
【図面の簡単な説明】
【0054】
【図1】図1は、MLhsp65核酸を含むレトロウイルスを感染させた骨髄細胞と正常骨髄細胞との混合物(比1:2)を注射したBalb/cマウスの脾臓細胞から抽出されたDNAのDNAサザンブロットハイブリダイゼーションの結果を示す。
【図2】図2は、MLhsp65核酸を含むレトロウイルスを感染させた骨髄細胞と正常骨髄細胞との混合物(比1:2)を注射したBalb/cマウスの血液試料から抽出されたタンパク質のウェスタンブロットプロービングの結果を示す。
【図3】図3は、MLhsp65遺伝子を含まないベクターを感染させた骨髄細胞(BMC-Neo)、MLhsp65核酸を含むレトロウイルスを感染させた骨髄細胞(BMC-65)及び組換え体MLhsp65(rhsp65)を注射したマウスについての遅延型過敏症(DTH)試験の結果を示す。
【図4】図4は、図3について挙げたマウスとrMLhsp65により免疫化した別のマウスの群に結核菌の毒性株を感染させた結果を示す。
【図5】図5は、pCDM8ML65 及びpHMGML65のプラスミドマップを示す。
【図6】図6は、生理食塩水(生理食塩水)、rMLhsp65及び不完全フロイントアジュバント(hsp65IFA)、pCDM8ML65及びpHMGML65(65 DNA)、癩菌36 kDプロリンリッチ抗原遺伝子をそれぞれ含むpCDM8及びpHMG(36 DNA)、並びにウシ結核菌BCGを筋肉内に注射したBalb/cマウスの肝臓中の生存可能な結核菌の数を示す。
【図7】図7は、pCDM8及びpHMG (対照プラスミドDNA)、ウシ結核菌BCG、癩菌36 kDプロリンリッチ抗原遺伝子をそれぞれ含むpCDM8及びpHMG(36 kDプラスミドDNA)、並びにpCDM8ML65 及びpHMGML65(65 kDプラスミドDNA)を注射した、異系交配したParkesアルビノマウスの脾臓(Sp)、肝臓(Li)及び肺(Lu)中の生存可能な結核菌の数を示す。
【図8】図8は、pCDM8 及びpHMG (対照プラスミドDNA)、ウシ結核菌BCG、癩菌36 kDプロリンリッチ抗原遺伝子をそれぞれ含むpCDM8及びpHMG(36 kDプラスミドDNA)、並びにpCDM8ML65及びpHMGML65(65 kD プラスミドDNA)を注射したCBA/B1Oマウスの脾臓(Sp)、肝臓(Li)及び肺(Lu)中の生存可能な結核菌の数を示す。
【図9】図9は、非処置Balb/cマウス(天然)、あるいは皮内にpCDM8ML65(65id)、筋肉内にpCDM8ML65(65im)、皮内にウシ結核菌BCGまたは皮内にpCDM8(対照プラスミド)を投与したBalb/cマウスの脾臓(Sp)、肝臓(Li)及び肺(Lu)中の生存可能な結核菌の数を示す。
【特許請求の範囲】
【請求項1】
マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原または抗
原として有効なそれらの断片をコードするコード配列であって、哺乳動物宿主細胞中でそのコード配列を発現できるプロモーターに機能可能なように結合されたコード配列を含むネイキッド核酸構築物の、マイコバクテリア感染症に対するワクチンとして使用するための薬剤の製造における使用。
【請求項2】
ネイキッド核酸構築物のコード配列が、マイコバクテリア熱ショックタンパ
ク質をコードする請求項1に記載の使用。
【請求項3】
ネイキッド核酸構築物のコード配列が、結核菌、癩菌もしくはウシ結核菌の
70 kDa、65 kDa若しくは10 kDa熱ショックタンパク質またはそれらの抗原として有効な断片をコードする請求項2に記載の使用。
【請求項4】
ネイキッド核酸構築物のコード配列が癩菌の65 kDaタンパク質をコードする
請求項3に記載の使用。
【請求項5】
ネイキッド核酸構築物のコード配列が、結核菌、癩菌またはウシ結核菌のプ
ロリンリッチ抗原をコードする請求項1に記載の使用。
【請求項6】
ネイキッド核酸構築物のコード配列が癩菌の36 kDaプロリンリッチ抗原をコ
ードする請求項5に記載の使用。
【請求項7】
ネイキッド核酸構築物がDNA 構築物である請求項1〜6のいずれかに記載の
使用。
【請求項8】
ネイキッド核酸構築物がプラスミドである請求項1〜7のいずれかに記載の
使用。
【請求項9】
ネイキッド核酸構築物がRNA 構築物である請求項1〜6のいずれかに記載の
使用。
【請求項10】
マイコバクテリア感染症に対するワクチンとして使用するための請求項1〜
9のいずれかに定義されたネイキッド核酸構築物。
【請求項11】
請求項1〜9のいずれかに定義されたネイキッド核酸構築物及び許容される
担体または希釈剤を含むワクチン組成物。
【請求項12】
請求項1〜8のいずれかに定義されたネイキッド核酸構築物の有効量を宿主
に投与することを含む、マイコバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法。
【請求項13】
マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原または抗
原として有効なそれらの断片をコードするコード配列であって、骨髄細胞中でそのコード配列を発現できるプロモーターに機能可能なように結合されたコード配列を含む核酸構築物がトランスフェクトされた骨髄細胞。
【請求項14】
請求項13に定義された骨髄細胞の有効量を宿主に投与することを含む、マイ
コバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法。
【請求項15】
コード配列が、マイコバクテリアプロリンリッチ抗原またはその抗原として
有効な断片をコードする、請求項1またはそれに従属する請求項5〜9のいずれかに定義されたネイキッド核酸構築物。
【請求項1】
マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原または抗
原として有効なそれらの断片をコードするコード配列であって、哺乳動物宿主細胞中でそのコード配列を発現できるプロモーターに機能可能なように結合されたコード配列を含むネイキッド核酸構築物の、マイコバクテリア感染症に対するワクチンとして使用するための薬剤の製造における使用。
【請求項2】
ネイキッド核酸構築物のコード配列が、マイコバクテリア熱ショックタンパ
ク質をコードする請求項1に記載の使用。
【請求項3】
ネイキッド核酸構築物のコード配列が、結核菌、癩菌もしくはウシ結核菌の
70 kDa、65 kDa若しくは10 kDa熱ショックタンパク質またはそれらの抗原として有効な断片をコードする請求項2に記載の使用。
【請求項4】
ネイキッド核酸構築物のコード配列が癩菌の65 kDaタンパク質をコードする
請求項3に記載の使用。
【請求項5】
ネイキッド核酸構築物のコード配列が、結核菌、癩菌またはウシ結核菌のプ
ロリンリッチ抗原をコードする請求項1に記載の使用。
【請求項6】
ネイキッド核酸構築物のコード配列が癩菌の36 kDaプロリンリッチ抗原をコ
ードする請求項5に記載の使用。
【請求項7】
ネイキッド核酸構築物がDNA 構築物である請求項1〜6のいずれかに記載の
使用。
【請求項8】
ネイキッド核酸構築物がプラスミドである請求項1〜7のいずれかに記載の
使用。
【請求項9】
ネイキッド核酸構築物がRNA 構築物である請求項1〜6のいずれかに記載の
使用。
【請求項10】
マイコバクテリア感染症に対するワクチンとして使用するための請求項1〜
9のいずれかに定義されたネイキッド核酸構築物。
【請求項11】
請求項1〜9のいずれかに定義されたネイキッド核酸構築物及び許容される
担体または希釈剤を含むワクチン組成物。
【請求項12】
請求項1〜8のいずれかに定義されたネイキッド核酸構築物の有効量を宿主
に投与することを含む、マイコバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法。
【請求項13】
マイコバクテリアストレスタンパク質もしくはプロリンリッチ抗原または抗
原として有効なそれらの断片をコードするコード配列であって、骨髄細胞中でそのコード配列を発現できるプロモーターに機能可能なように結合されたコード配列を含む核酸構築物がトランスフェクトされた骨髄細胞。
【請求項14】
請求項13に定義された骨髄細胞の有効量を宿主に投与することを含む、マイ
コバクテリア感染症に対して哺乳動物宿主にワクチン接種する方法。
【請求項15】
コード配列が、マイコバクテリアプロリンリッチ抗原またはその抗原として
有効な断片をコードする、請求項1またはそれに従属する請求項5〜9のいずれかに定義されたネイキッド核酸構築物。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2007−332149(P2007−332149A)
【公開日】平成19年12月27日(2007.12.27)
【国際特許分類】
【出願番号】特願2007−190620(P2007−190620)
【出願日】平成19年7月23日(2007.7.23)
【分割の表示】特願平7−529472の分割
【原出願日】平成7年5月18日(1995.5.18)
【出願人】(597166578)メディカル リサーチ カウンシル (60)
【Fターム(参考)】
【公開日】平成19年12月27日(2007.12.27)
【国際特許分類】
【出願日】平成19年7月23日(2007.7.23)
【分割の表示】特願平7−529472の分割
【原出願日】平成7年5月18日(1995.5.18)
【出願人】(597166578)メディカル リサーチ カウンシル (60)
【Fターム(参考)】
[ Back to top ]