
マイコプラズマサブユニットワクチン
本発明は、マイコプラズマ感染を撲滅するためのワクチン、このようなワクチンで使用するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼ、このようなワクチンを製造するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼの使用、このようなワクチンを調製するための方法、並びに前記ワクチンを予防接種された動物と完全細胞ワクチンで予防接種された動物又は野外感染に罹患した動物とを識別するための診断試験に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、とりわけ、マイコプラズマ感染を撲滅するためのワクチン、このようなワクチンにおいて使用するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼ、このようなワクチンを製造するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼの使用、このようなワクチンを調製するための方法、並びに前記ワクチンを予防接種された動物と完全細胞ワクチンで予防接種された動物又は野外感染に罹患した動物を識別するための診断試験に関する。
【背景技術】
【0002】
進化の間に、病原性細菌は、それらの宿主との複雑な相互作用を発達させてきた。これは、しばしば、病原性島、プラスミド、トランスポゾン又はプロファージ上での病原性因子の獲得を伴い、これにより、細菌は、宿主内でコロニーを形成し、生存し、複製することが可能となる。これに対して、最小の自己複製生物であるマイコプラズマ種は、ゲノムを最小サイズへ減少させることによって、グラム陽性細菌から退行性に進化し、その結果、遺伝的資源を節約している。
【0003】
マイコプラズマ種は、地球上で検出される最小の自己複製生物である。それらのゲノムは、マイコプラズマ・ジェニタリウムにおける580キロ塩基対(kb)(Fraser,C.M.et al.,1995,Science270:397−03)からマイコプラズマ・ペネトランスにおける1358kb(Sasaki,Y. et al,2002,Nucleic Acids Res. 30:5293−5300)にわたる。これにより、遺伝的資源の劇的な節約がもたらされ、寄生性の生活様式が必要不可欠となる。病原性マイコプラズマ種は、主に、動物及びヒトに、非定型肺炎、泌尿生殖器感染症及び関節炎を引き起こす(Baseman,J.B.,and J.G. Tully,1997,Emerg.Infect.Dis.3:21−32,Blanchard,A.,and G. F. Browning(eds.).2005.Mycoplasmas:Molecular biology,pathogenicity and strategies for control.Horizon Bioscience,Wymondham,U.K.,Frey,J. 2002.Mycoplasmas of animals,p.73−90.In S. Razin and R. Herrmann(eds.),Molecular biology and pathogenicity of mycoplasmas. Kluwer Academic/Plenum Publishers,New York)。ここでは、ヒト(例えば、M.ジェニタリウム、M.ニューモニエ)、ウシ種(例えば、M.spボバイングループ7)、ブタ(例えば、M.ハイオニューモニエ)又は家禽(例えば、ガリセプチカム)に感染する公知のマイコプラズマ種が存在するという事実のために、本明細書で動物という場合には、これが、とりわけ、ヒト、ウシ種、ブタ及び家禽を含むものと解釈されるべきことが強調される。病原性が、毒素、インベイシン及び細胞溶解素によって概ね決定される他の病原性細菌とは異なり、完全に配列決定された8つの種のゲノム配列分析によって明らかとなったところによれば、病原性マイコプラズマ種は、このような典型的な主要な病原性因子を有していないように見受けられる(Chambaud,I. et al,2001,Nucleic Acids Res.29:2145−2153,Fraser et al.,1995,Science 270:397−403,Himmelreich,R. et al.,1996,Nucleic Acids Res.24:4420−4449,Jaffe,J. D. et al,2004,Genome Res.14:1447−1461,Minion,F.C. et al.,2004,J. Bacteriol.186:7123−7133,Papazisi,L. et al,2003,Microbiology 149:2307−2316,Sasaki,Y. et al.,2002,Nucleic Acids Res.30:5293−5300,Westberg,J. et al.,2004,Genome Res.14:221−227)。
【0004】
PCR法の導入以来、マイコプラズマ感染症の診断は著しく改善され、数個のマイコプラズマ種において、抗原性の可変性が詳しく研究されてきたが、現在のところ、病原性マイコプラズマが、宿主細胞に損傷、炎症及び疾病を引き起こす分子的機序及びエフェクターに関する知識は殆ど得られていない。従って、これらの望ましくない効果を予防することを特に目標とするワクチンの開発は、極めて必要とされている。
【0005】
宿主細胞の損傷、炎症及び疾病を回避又は減弱させる、マイコプラズマ感染を撲滅するためのワクチンを提供することは本発明の1つの目的である。
【0006】
驚くべきことに、新規の主要病原性因子が、マイコプラズマにおける細胞傷害の重要な原因であることが見出された。この病原性因子は、H2O2などの有毒な副産物及びその他の付随する反応性酸素種(ROS)からなるように見受けられた。H2O2/ROSの形成は、グリセロールの代謝に関与する酵素である、酵素マイコプラマのL−α−グリセロリン酸オキシダーゼ(GlpO)の活性と直接関係し得ることを示すことができた。
【0007】
さらに予想外のことに、マイコプラズマ感染によって引き起こされる組織損傷の唯一の原因ではないとしても、H2O2/ROSの形成が中心的な原因であることを示すことができた。これは、今回、初めて、マイコプラズマの主な病原性因子が確定されたことを意味している。
【0008】
マイコプラズマ中のグリセロール代謝経路は、容易に影響又は変化を受けることができず、従って、H2O2/ROSは、マイコプラズマ自体のグリセロール代謝経路の修飾を通じて容易に変化することができない。従って、非マイコプラズマ種での状況とは異なり、生きた弱毒化されたマイコプラズマを開発する経路は、より実効可能性が低いように思われる。
【0009】
しかしながら、驚くべきことに、マイコプラズマのL−α−グリセロリン酸オキシダーゼと反応性を示す抗体は、感染された動物の組織に、ほとんど又は全く損傷を与えない程度まで、H2O2/ROSの産生を抑制できることが、ここに見出された。これは、以下に説明されているように、とりわけ、サブユニットワクチンでの予防接種に道を開く。
【0010】
全ての非マイコプラズマ細菌では、これらの細胞中のカタラーゼが、形成後、H2O2を直ちに分解するので、グリセロリン酸オキシダーゼ活性を有する酵素が細胞内に見出されるという事実に鑑みれば、これは、さらに驚くべきことである。このような場合には、グリセロリン酸オキシダーゼに対して誘導された抗体は、従って、細菌の中に入ることができないので、全く効果を有しない。全ての非マイコプラズマ細菌とは異なり、おそらくは、カタラーゼが欠如している結果、マイコプラズマは、細胞外H2O2の産生をもたらすように、進化の間に、細胞内腔から膜へ、そのマイコプラズマのL−α−グリセロリン酸オキシダーゼを移動させたことがここに見出された。従って、これは、細菌にとって有害でないが、感染された宿主の組織にとっては有害となる。これは、マイコプラズマの場合には、マイコプラズマのL−α−グリセロリン酸オキシダーゼに対する抗体が(他の細菌とは反対に)、細胞外H2O2を放出する酵素の活性を予想外に妨害することを説明し得る。
【0011】
マイコプラズマの酵素である、マイコプラズマのL−α−グリセロリン酸オキシダーゼは、公知の全てのマイコプラズマ種の中に存在する。L−α−グリセロリン酸オキシダーゼに対して行われる全てのさらなる参照は、マイコプラズマのL−α−グリセロリン酸オキシダーゼを表す。以下の表1には、マイコプラズマ種、M.sp.ボバイングループ7(M.sp. bovine group 7)、M.ミコイデスsubsp.カプリ(M.mycoides subsp.Capri)、M.ペネトランス(M.penetrans)、M.ガリセプチカム(M.gallisepticum)、M.モービレ(M.mobile)、M.パルモニス(M.pulmonis)、M.ハイオニューモニエ(M.hyopneumoniae)、M.ニューモニエ(M.pneumoniae)、M.ジェニタリウム(M.genitalium)の株において、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子のEMBL/GenBank受託番号が示されている。さらに、実施例には、マイコプラズマ種、M.ミコイデスS(mall)C(olony)株Afade及びL2の株において、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子のEMBL/Genbank受託番号が示されている。
【0012】
酵素マイコプラズマのL−α−グリセロリン酸オキシダーゼは、全ての公知のマイコプラズマ種の中に存在するという事実のために、マイコプラズマ感染を駆除するための、より具体的には、宿主細胞の損傷、炎症及び疾病を回避又は減弱するための、本タンパク質に対する抗体を産生するためのその使用は、マイコプラズマ種に関わらず、一般的に適用可能であることが明らかとなった。
【0013】
マイコプラズマに対する抗体を産生するための原理は、以下の実施例に説明されている。家畜の多大な喪失を招く重度の感染性疾患である伝染性ウシ胸膜肺炎(CBPP)の発病因子であるマイコプラズマ・ミコイデスsubsp.ミコイデスSCは、マイコプラズマの病原性の分子的基礎を調査するためのモデルとした使用された。M.ミコイデスsubsp.ミコイデスSCは、宿主細胞と接近して生息する、1211kbのゲノムサイズを有する細胞外病原体である(Westberg,J. et al.,2004,Genome Res.14:221−227)。モデルとして、この種を使用することの合理性は、この種の高い病原性及びCBPPの発病因子として明らかに確立されている事実である。さらに、この重いウシの病気は、現在、CBPPの発生に苦しむ国で、家畜の生産に対して著しい社会経済的な重要性を有している。さらに、この伝染病が存在しない国は、再発生する感染の脅威に継続的に曝されている。
【0014】
マイコプラズマ感染を撲滅するために、より具体的には、宿主細胞損傷、炎症及び疾病を回避又は減弱するために、マイコプラズマのL−α−グリセロリン酸オキシダーゼに対する抗体を使用することが可能であることが、ここに確立されたので、次の段階は、インビボ及びインビトロで、抗体を産生するために、マイコプラズマのL−α−グリセロリン酸オキシダーゼを使用することである。マイコプラズマのL−α−グリセロリン酸オキシダーゼのインビボでの使用の例は、ワクチンでの使用である。このようなワクチンは、投与されると、マイコプラズマに対する抗体を誘導する。これは、以下でさらに詳しく考察されている。マイコプラズマのL−α−グリセロリン酸オキシダーゼのインビボ又はインビトロでの使用の例は、ワクチン中で使用するための抗体を産生するための使用である。このようなワクチンも、以下で考察されている。別のワクチンは、マイコプラズマのL−α−グリセロリン酸オキシダーゼ(の免疫原性部分)をコードする遺伝子又はその一部を担持する生組換え担体及びマイコプラズマのL−α−グリセロリン酸オキシダーゼ(の免疫原性部分)をコードする遺伝子又はその一部を含むDNAワクチンを基礎とするワクチンである。
【0015】
様々なマイコプラズマ種において、酵素マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする配列が公知であるので、十分な量で酵素マイコプラズマのL−α−グリセロリン酸オキシダーゼを取得することが、ここに可能となる。これは、例えば、これらのタンパク質をコードする遺伝子を発現するための発現系を使用することによって行うことが可能である。
【0016】
マイコプラズマのL−α−グリセロリン酸オキシダーゼ(の免疫原性部分)をコードする遺伝子又はその一部を発現させるために必要不可欠であるのは、遺伝子がプロモーターの制御下にあるように、遺伝子に機能的に連結された十分なプロモーターである。これは、例えば、標準的な分子生物学の技術を用いて達成することが可能である。(Sambrook,J. and Russell,D.W.,Molecular cloning:a laboratory manual,2001.ISBN 0−87969−577−3)。プロモーターの選択が、タンパク質発現のための宿主細胞として使用される細胞中で遺伝子転写を誘導することが可能な全ての真核、原核又はウイルスプロモータに拡張することは、当業者に自明である。機能的に連結されたプロモーターは、それらが連結された核酸の転写を制御することが可能なプロモーターである。このようなプロモーターは、マイコプラズマプロモーター、例えば、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子のインビボ発現に関与するプロモーター(但し、このプロモーターは、発現のために使用される細胞中で機能的である。)とすることが可能である。プロモーターは、異種のプロモーターとすることも可能である。宿主細胞が細菌である場合には、使用され得る有用な発現調節配列には、Trpプロモーター及びオペレーター(Goeddel,et al.,Nucl.Acids Res.,8,4057,1980);lacプロモーター及びオペレーター(Chang,et al.,Nature,275,615,1978);外膜タンパク質プロモーター(Nakamura,K. and Inouge,M.,EMBO J.,1,771−775,1982);バクテリオファージλプロモーター及びオペレーター(Remaut,E. et al.,Nucl.Acids Res.,11,4677−688,1983);α−アミラーゼ(B.スブチリス)プロモーター及びオペレーター、終結配列並びに選択された宿主細胞と適合性が有るその他の発現増強及び調節配列が含まれる。宿主細胞が酵母である場合、有用な発現調節配列には、例えば、α−接合因子が含まれる。昆虫細胞の場合、バキュロウイルスのポリヘドリン又はp10プロモーターを使用することが可能である(Smith,G.E. et al.,Mol.Cell.Biol.3,2156−65,1983)。宿主細胞が哺乳動物起源である場合、有用な発現調節配列の例には、SV−40プロモーター(Berman,P.W. et al.,Science,222,524−527,1983)又はメタロチオネインプロモーター(Brinster,R.L.,Nature,296,39−42,1982)又は熱ショックプロモーター(Voellmy et al.,Proc.Natl.Acad.Sci.USA,82,4949−53,1985)が含まれる。
【0017】
細菌、酵母、真菌、昆虫及び哺乳動物の細胞発現系は、極めて頻繁に使用される系である。このような系は、本分野において周知であり、一般に、例えば、Invitrogen(www.invitrogen.com)、Novagen(www.merckbiosciences.de)又はClontech Laboratories,Inc.4030 Fabian Way,Palo Alto,California 94303−4607,USAを通じて商業的に入手可能である。これらの発現系に次いで、寄生生物を基礎とした発現系が、極めて魅力的な発現系である。このような系は、例えば、公開番号2714074のフランス特許出願及びUS NTIS公開番号US 08/043109(Hoffman,S. and Rogers,W.:Public.Date 1 December 1993)に記載されている。
【0018】
例えば、ワクチン接種の目的のために、又は抗体を産生させるためにタンパク質が使用される場合には、しかしながら、完全なタンパク質を使用する必要はない。タンパク質に対する免疫応答を誘導することが可能なタンパク質の断片(そのままの状態であるか、又は、例えばKLHなどの担体に連結されている。)、いわゆる、免疫原性断片を使用することも可能である。「免疫原性断片」とは、宿主中で免疫反応を誘導する能力をなお保持する、すなわち、B細胞又はT細胞エピトープを含む完全長タンパク質の断片であると理解される。この時点で、抗原性断片(決定因子)をコードするDNA断片を容易に同定するために、様々な技術が利用可能である。Geysenら(Patent Application WO 84/03564,Patent Application WO 86/06487,US Patent NR.4,833,092,Proc.Natl Acad.Sci.81:3998−4002(1984),J. Imm.Meth.102,259−274(1987))によって記載された方法、いわゆるPEPSCAN法は、タンパク質の免疫学的に重要な領域であるエピトープの検出のための、実行が容易で、迅速な、十分に確立された方法である。本方法は、世界中で使用されており、従って、当業者に周知である。この(経験的な)方法は、B細胞エピトープの検出のために特に適している。また、何れかのタンパク質をコードする遺伝子の配列が与えられれば、現在公知であるエピトープとの配列的及び/又は構造的な一致に基づき、コンピュータアルゴリズムは、免疫学的に重要なエピトープとして、具体的なタンパク質断片を指定することが可能である。これらの領域の決定は、Hopp及びWoods(Proc.Natl.Acad.Sci.78:38248−3828(1981))に従う親水性基準とChou及びFasman(Advances in Enzymology 47:45−148(1987)and US Patent 4,554,101)に従う二次構造要因の組み合わせに基づいている。同様に、Berzofskyの両親媒性基準の補助を得て、コンピュータによって、配列からT細胞エピトープを予測することが可能である(Science 235,1059−1062(1987) and US Patent application NTIS US 07/005,885)。要約された概説は、共通原理に関するShan Lu:Tibtech 9:238−242(1991)、マラリアエピトープに関するGood et al;Science 235:1059−1062(1987)、概説として、Lu;Vaccine 10:3−7(1992),HIV−エピトープに関するBerzowsky;The FASEB Journal 5:2412−2418(1991)に見出される。
【0019】
従って、本発明の第一の実施形態は、マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片と、及び医薬として許容される担体とを含む前記ワクチンに関する。
【0020】
これらの遺伝子の発現産物に基づくこのようなワクチンは、以下に記載されているように、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片を、医薬として許容される担体と混合することによって容易に作製することが可能である。
【0021】
本発明の別の実施形態は、ワクチンにおいて使用するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片に関する。
【0022】
さらに別の実施形態は、マイコプラズマ感染を撲滅するためのワクチンを製造するための、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片の使用に関する。
【0023】
マイコプラズマ感染に対する予防接種のための別の極めて魅力的なアプローチは、医薬として許容される担体と一緒に、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む生組換え担体(LRC)を使用することによるものである。これらのLRCは、さらなる遺伝子情報(この場合には、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片)がクローニングされた微生物又はウイルスである。このようなLRCに感染した動物は、担体の免疫原に対してのみならず、遺伝子コードが、LRC中にさらにクローニングされているタンパク質(例えば、マイコプラズマのL−α−グリセロリン酸オキシダーゼ)の免疫原性部分に対しても免疫原性応答を産生する。細菌のLRCの例として、本分野で公知の弱毒化されたサルモネラ株は、魅力的に使用することが可能である。生組換え担体寄生生物が、とりわけ、Vermeulen,A.N.(Int.Journ.Parasitol.28:1121−1130(1998))によって記載されている。また、LRCウイルスは、核酸を標的細胞中に輸送する手段として使用し得る。生組換え担体ウイルスは、ベクターウイルスとも称される。ベクターとしてしばしば使用されるウイルスは、ワクシニアウイルス(Panicali et al; Proc.Natl.Acad.Sci.USA,79:4927(1982)、ヘルペスウイルス(E.P.A. 0473210A2)及びレトロウイルス(Valerio,D. et al; in Baum,S.J.,Dicke,K.A.,Lotzova,E. and Pluznik,D.H.(Eds.),Experimental Haematology today− 1988. Springer Verlag,New York:pp.92−99(1989))である。
【0024】
本分野で周知の、インビボ相同組換えの技術は、本発明の挿入された核酸の発現を宿主細胞中で誘導することが可能な組換え核酸を、選択した細菌、寄生生物又はウイルスのゲノム中に導入するために使用することが可能である。
【0025】
従って、本発明のさらに別の実施形態は、マイコプラズマのL−α−グリセロリン酸オキシダーゼを又はその免疫原性断片コードする生組換え担体と、及び医薬として許容される担体とを含む、マイコプラズマ感染を撲滅するためのワクチンに関する。
【0026】
機能的に連結されたプロモーターの調節下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む宿主細胞を、本酵素の生産のために使用できることが明らかである。酵素をワクチンとして使用する前に、まず、宿主細胞から酵素を抽出する必要は存在しない。すなわち、宿主細胞は、そのまま使用することも可能である。宿主細胞が、酵素を発現するLRCを含む場合も、同じことが当てはまる。その例は、ウイルス又は細菌のLRCを含む真核細胞である。従って、同じく、本発明の別の実施形態は、機能的に連結されたプロモーターの調節下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む宿主細胞に関する。この形態は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む生組換え担体を含有する宿主細胞にも関する。宿主細胞は、pBR322として細菌を基礎としたプラスミド又はpGEXとして細菌の発現ベクター又はバクテリオファージと組み合わせた、細菌、例えば、エシェリヒア・コリ、バシラス・スブチリス及びラクトバシラス種起源の細胞であり得る。宿主細胞は、真核細胞起源の宿主細胞、例えば、酵母特異的ベクター分子と組み合わせた酵母細胞、又はベクター若しくは組換えバキュロウイスルと組み合わせた、昆虫細胞のようなより高等な真核細胞(Luckow et al; Bio−technology 6:47−55(1988))、例えば、Ti−プラスミドを基礎としたベクター若しくは植物ウイルスベクター(Barton,K.A. et al; Cell 32:1033(1983))と組み合わせた植物細胞、同じく、適切なベクター若しくは組換えウイルスを有するHela細胞、チャイニーズハムスター卵巣細胞(CHO)又はクランデルネコ腎臓細胞のような哺乳動物細胞でもあり得る。例えば、腸上皮、又は例えば呼吸器上皮に感染するサルモネラ担体又はウイルス担体を基礎とする、酵素又はその免疫原性断片を発現することが可能な上記生組換え担体を基礎とする本発明のワクチンは、マイコプラズマの感染の天然経路をよりよく模倣する点で、サブユニットワクチンを上回る利点を有する。さらに、免疫化のために、組換え担体の少量が必要であるに過ぎないので、それらの自己増殖は有利である。
【0027】
予防接種の効率的な別の方法は、当該抗原をコードするDNAを用いた直接の予防接種である。タンパク質をコードするDNAを用いた直接の予防接種は、多くの異なるタンパク質において成功を収めている。(例えば、Donnelly et al.,The Immunologist 2:20−26(1993)に概説されているとおり。)。予防接種のこの方法も、マイコプラズマ感染に対して哺乳動物を予防接種するために極めて魅力的である。従って、本発明の本実施形態のさらに別の形態は、マイコプラズマ感染を撲滅するためのワクチンであり、機能的に連結されたプロモーターの制御下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子又はL−α−グリセロリン酸オキシダーゼの免疫原性断片をコードする前記遺伝子の一部と、及び医薬として許容される担体とを含む前記ワクチンに関する。
【0028】
DNAワクチンは、例えば、無針注射装置を用いて、皮内適用を通じて容易に投与することが可能である。投与のこの方法は、予防接種すべき動物の細胞中にDNAを直接送達する。1と100μgの間の範囲にあるμgのDNAの量は、極めて優れた結果を与える。
【0029】
上記ワクチンは全て、活性な予防接種に貢献する。すなわち、宿主の免疫系は、これらのタンパク質に対する抗体を作製するために、酵素又はその免疫原性断片によって引き起こされる。あるいは、このような抗体は、以下に記載されているように、例えば、ウサギの中で作製することが可能であり、又は抗体産生細胞株から得ることが可能である。次いで、このような抗体は、宿主動物に投与することが可能である。予防接種のこの方法(受動予防接種)は、動物が既に感染しており、天然の免疫応答を惹起させる時間が存在しない場合に選択される予防接種である。これは、免疫無防備状態の動物を予防接種するための好ましい方法でもある。マイコプラズマに対する投与された抗体は、これらの事例において、細菌に直接結合することが可能である。これは、マイコプラズマの増殖を直ちに減少させ、又は停止させるという利点を有する。従って、本発明の本実施形態の1つの別の形態は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片に対する抗体と、及び医薬として許容される担体とを含むワクチンに関する。
【0030】
本実施形態のさらに別の形態において、本発明のワクチンは、同一の宿主に対して病原性である別の生物又はウイルスに由来する1つ又はそれ以上の抗原、このような抗原に対する抗体又はこのような抗原をコードする遺伝情報をさらに含む。
【0031】
好ましい組み合わせワクチンが、マイコプラズマのL−α−グリセロリン酸オキシダーゼに次いで、マイコプラズマ完全細胞調製物を含むワクチンであることは言うまでもない。このような組み合わせワクチンは、マイコプラズマのL−α−グリセロリン酸オキシダーゼの有害な効果のみならず、他のマイコプラズマ関連タンパク質に対しても保護を誘導する。
【0032】
ブタ病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、仮性狂犬病ウイルス(Pseudorabies virus)、ブタインフルエンザウイルス(Porcine influenza virus)、ブタパルボウイルス(Porcine parvo virus)、伝染性胃腸炎ウイルス(Transmissible gastroenteritis virus)、ロタウイルス(Rotavirus)、別のマイコプラズマ属種、特に、マイコプラズマ・ハイオニューモニエ(Mycoplasma hyopneumoniae)、ブラキスピラ・ハイオディセンテリー(Brachyspira hyodysenteriae)、エシェリヒア・コリ(Escherichia coli)、レプトスピラ属種(Leptospira spp)、エリシペロスリクス・リューシオパシエ(Erysipelothrix rhusiopathiae)、ボルデテラ・ブロンキセプティカ(Bordetella bronchiseptica)、ブラキスピラ・ハイオディセンテリー(Brachyspira hyodysenteriae)、シゲラ種(Shigella sp.)、サルモネラ・コレラエスイス(Salmonella choleraesuis)、サルモネラ・チフィムリウム(Salmonella typhimurium)、サルモネラ・エンテリティディス(Salmonella enteritidis)、ヘモフィラス・パラスイス(Haemophilus parasuis)、ラウソニア(Lauwsonia)、パスチュレラ・マルトシダ(Pasteurella multocida)、ストレプトコッカス・スイス(Streptococcus suis)、アクチノバシラス・プリューロニューモニエ(Actinobacillus pleuropneumoniae)、スタフィロコッカス・ハイカス(Staphylococcus hyicus)及びクロストリジウム・ペルフリンゲンス(Clostridium perfringens)の群から選択される。
【0033】
ウシ病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、ウシヘルペスウイルス(Bovine Herpesvirus)、ウシウイルス性下痢症ウイルス(Bovine Viral Diarrhoea virus)、パラインフルエンザ3型ウイルス(Parainfluenza type 3 virus)、ウシパラミクソウイルス(Bovine Paramyxovirus)、口蹄疫ウイルス(Foot and Mouth Disease virus)、ウシ呼吸器多核体ウイルス(Bovine Respiratory Syncytial Virus)、ブタサーコウイルス(porcine circovirus)、ブタ生殖器呼吸器症候群ウイルス(porcine respiratory reproductive syndrome virus)、別のマイコプラズマ属種、パスツレラ・ヘモリティカ(Pasteurella haemolytica)、スタフィロコッカス・オーレウス(Staphylococcus aureus)、エシェリヒア・コリ(Escherichia coli)、レプトスピラ属種(Leptospira spp.)、スタフィロコッカス・ウベリス(Staphylococcus uberis)、タイレリア・パルバ(Theileria parva)、タイレリア・アニュラータ(Theileria annulata)、バベシア・ボビス(Babesia bovis)、バベシア・ビゲミナ(Babesia bigemina)、バベシア・メジャー(Babesia major)、トリパノソーマ種(Trypanosoma species)、アナプラズマ・マージナーレ(Anaplasma marginale)、アナプラズマ・セントラーレ(Anaplasma centrale)及びネオスポラ・カニナム(Neospora caninum)の群から選択される。
【0034】
家禽病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、鶏痘ウイルス(Fowlpox virus)、伝染性気管支炎ウイルス(Infectious Bronchitis virus)、伝染性ファブリキウス嚢病(グンボロ)(Infectious Bursal Disease (Gumboro))、マレック病ウイルス(Marek’s Disease Virus)、ニワトリ貧血因子(Chicken Anaemia agent)、トリレオウイルス(Avian Reovirus)、トルコ鼻気管炎ウイルス(Turkey Rhinotracheitis virus)、ニワトリポックスウイルス(Chicken Poxvirus)、トリ脳脊髄炎ウイルス(Avian Encephalomyelitis virus)、アヒルペストウイルス(Duck Plague virus)、ニューキャッスル病ウイルス(Newcastle Disease virus)、産卵低下症候群ウイルス(Egg Drop syndrome virus)、伝染性喉頭気管ウイルス(Infectious Laryngotracheitis virus)、シチメンチョウのヘルペスウイルス(Herpes Virus of Turkeys)、別のマイコプラズマ属種、とりわけ、マイコプラズマ・ガリセプチカム(Mycoplasma gallisepticum)又はマイコプラズマ・シノビアエ(Mycoplasma synoviae)、ヘモフィルス・パラガリナルム(コリーザ)(Haemophilus paragallinarum (Coryza))、オルニソバクテリウム・ライノトラキア(Ornithobacterium rhinotracheale)、クロストリジウム・ペルフリンゲンス(Clostridium perfringens)、サルモネラ(Salmonella)、カンピロバクター種(Campylobacter species)、E.コリ(E. coli)及びエイメリア(Eimeria)種の群から選択される。
【0035】
ヒト病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、インフルエンザウイルス(influenza virus)、麻疹ウイルス(measles virus)、ムンプスパラミクソウイルス(mumps paramyxovirus)、クロストリジウム・ジフセリエ(Clostridium diphteriae)、クロストリジウム・テタニ(Clostridium tetani)、ボルデテラ・パーチュシス(Bordetella pertussis)、別のマイコプラズマ属種及びポックスウイルス(pox virus)の群から選択される。
【0036】
好ましくは、上記マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片は、以下のマイコプラズマのいずれか:M.ボビス(M.bovis)、M.sp.ボバイングループ7(M.sp .bovine group 7)、M.ミコイデスsubsp.カプリ(M.mycoides subsp. Capri)、M.ペネトランス(M.penetrans)、M.ガリセプチカム(M.gallisepticum)、M.シノビエ(M.synoviae)、M.モービル(M.mobile)、M.パルモニス(M.pulmonis)、M.ハイオニューモニエ(M.hyopneumoniae)、M.ニューモニエ(M.pneumoniae)、M.ジェニタリウム(M.genitalium)、M.ミコイデスS(mall)C(olony)株Afade(M.mycoides S(mall) C(olony) strain Afade)又はM.ミコイデスS(mall)C(olony)株L2(M.mycoides S(mall) C(olony) strain L2)、特に、M.ボビス、M.ハイオニューモニエ、M.ガリセプチカム又はM.シノビエ、より具体的には、M.ボビス又はM.ハイオニューモニエによってコードされるマイコプラズマのL−α−グリセロリン酸オキシダーゼである。
【0037】
本発明の全てのワクチンは、医薬として許容される担体を含む。医薬として許容される担体は、例えば、無菌水又は無菌生理的塩溶液とすることができる。より複雑な形態において、担体は、例えば緩衝液であり得る。
【0038】
ワクチンを調製するための方法は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片、上記生組換え担体、上記遺伝子若しくはその一部、上記宿主細胞又はマイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片に対する抗体と、及び医薬として許容される担体とを混合することを含む。従って、本発明の別の実施形態は、マイコプラズマ感染を撲滅するためのワクチンの調製方法であり、上記マイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片、上記生組換え担体、上記遺伝子若しくはその一部、上記宿主細胞又は上記マイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片に対する抗体と、及び医薬として許容される担体とを混合することを含む、前記方法に関する。
【0039】
本発明のワクチンは、好ましい提示において、アジュバントも含有し得る。一般に、アジュバントは、非特異的様式で、宿主の免疫応答を強化する物質を含む。様々な多数のアジュバントが、本分野において公知である。アジュバントの例は、フロイントの完全及び不完全アジュバント、ビタミンE、非イオン性ブロックポリマー、ムラミルジペプチド、QuillA(R)、鉱物油、例えばBayol(R)又はMarkol(R)、植物油及びCarbopol(R)(ホモポリマー)又はDiluvac(R)Forteである。ワクチンは、いわゆる「ビヒクル」も含み得る。ビヒクルとは、それに対する共有結合なしに、ポリペプチドが付着する化合物である。しばしば使用されるビヒクル化合物は、例えば、水酸化アルミニウム、リン酸アルミニウム又は酸化アルミニウム、シリカ、カオリン及びベントナイトである。
【0040】
抗原がビヒクル中に部分的に埋め込まれている、このようなビヒクルの特殊な形態は、いわゆるISCOMである(EP 109.942,EP 180.564,EP 242.380)。さらに、ワクチンは、1つ又はそれ以上の適切な界面活性化合物又は乳化剤、例えばSpan又はTweenを含み得る。しばしば、例えば、分解し易いポリペプチドを分解から保護して、ワクチンの保存寿命を延長させるために、又は凍結乾燥効率を改善するために、ワクチンは安定化剤と混合される。有用な安定化剤は、とりわけ、SPGA(Bovarnik et al; J. Bacteriology 59:509(1950)、炭水化物、例えば、ソルビトール、マニトール、トレハロース、デンプン、スクロース、デキストラン又はグルコース、アルブミン又はカゼイン又はこれらの分解産物などのタンパク質及びアルカリ金属リン酸塩などの緩衝液である。さらに、ワクチンは、生理的に許容される希釈剤中に懸濁され得る。アジュバント化し、ビヒクル化合物又は希釈剤を添加し、ポリペプチドを乳化し、又は安定化させる別の方法も、本発明において具体化されることは言うまでもない。
【0041】
本発明のワクチンは、タンパク質の1及び100μgの間の量で、極めて適切に投与することが可能であるが、より少量を使用することが原理的に可能である。免疫学的には極めて適切であるが、100μgを超える用量は、商業的な理由で、より魅力的でない。
【0042】
上記LRCウイルス及び細菌などの、弱毒化された生組換え担体に基づくワクチンは、感染中に自身を増幅するので、ずっと少ない用量で投与することが可能である。従って、極めて適切な量は、それぞれ、細菌及びウイルスに対して、103と109CFU/PFUの間の範囲である。
【0043】
多くの投与方法を使用することが可能である。経口適用は、労力を要しないので、投与の極めて魅力的な方法である。経口投与の好ましい方法は、胃の高度に酸性環境を通過した後にのみ崩壊するカプセル中にワクチンを包装することであり、これは、本分野において公知であり、頻繁に使用されている。また、ワクチンは、胃のpHを一時的に増大させるために、本分野で公知の化合物と混合することが可能である。例えば、ワクチンの筋肉内適用による、全身適用も適切である。この経路が採用される場合には、全身適用のために本分野で公知の標準的な手法が非常に適している。
【0044】
マイコプラズマのL−α−グリセロリン酸オキシダーゼに基づくワクチンも、マーカーワクチンとして極めて適切である。マーカーワクチンは、例えば、野生型感染によって誘導される抗体パネルとは異なる、特徴的な抗体パネルに基づいて、予防接種された動物と野外感染した動物との識別を可能とするワクチンである。精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼに基づくワクチンは、本タンパク質に対する抗体を誘導するのみであるが、生きた野生型、生きた弱毒化又は不活化された完全なマイコプラズマに基づくワクチン及び野外感染は、細菌のタンパク質の全て又はほとんどに対して抗体を誘導する。これは、明らかに、極めて異なる抗体パネルを与える。精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼを含むウェル及び別のマイコプラズマのタンパク質を含むウェルを有する単純なELISA検査は、動物から得た血清を試験し、動物が、本発明のワクチンで予防接種されたか、又はマイコプラズマ野外感染に罹患しているかどうかを識別するのに十分である。精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼを含むワクチンを予防接種された動物は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ以外の他のマイコプラズマのタンパク質に対する抗体を有しない。しかしながら、マイコプラズマでの野外感染に遭遇した動物は、全ての免疫原性マイコプラズマタンパク質に対する抗体を有しており、従って、他の非マイコプラズマのL−α−グリセロリン酸オキシダーゼタンパク質に対する抗体も有する。
【0045】
従って、本発明の別の実施形態は、一方で本発明のワクチンを用いた予防接種と、他方で完全細胞ワクチンを用いた予防接種又は野外感染とを識別するための診断検査に関し、このような検査は、精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片を含み、別個に、別の非L−α−グリセロリン酸オキシダーゼタンパク質を含む。
【0046】
上に特定されているように発現された本発明のポリペプチド又はその免疫原性断片は、ポリクローナル、単一特異的又はモノクローナル(又はその誘導体)であり得る抗体を産生するために使用することが可能である。ポリクローナル抗体が望まれる場合には、ポリクローナル血清を作製及び加工するための技術が本分野において周知である(例えば、Mayer and Walter,eds.Immunochemical Methods in Cell and Molecular Biology,Academic Press,London,1987)。本発明のポリペプチド(又はそのバリアント若しくは断片)に対して反応性の本発明のモノクローナル抗体は、本分野でも公知の技術によって、近交系マウスを免疫することによって調製することが可能である(Kohler and Milstein,Nature,256,495−97,1975)。
【0047】
本発明の抗体の大規模な作製方法も、本分野において公知である。このような方法は、ファージディスプレイのために、繊維状ファージ中に、本発明のタンパク質をコードする遺伝情報(の断片)をクローニングすることに基づいている。このような技術は、とりわけ、http://aximt1.imt.uni−marburg.de/〜rek/aepphage.htmlの「filamentous phage display」の「Antibody Engineering Page」並びにCortese,R. et al.,(1994) in Trends Biotechn.12:262−267.,Clackson,T. & Wells,J.A.(1994) in Trends Biotechn.12:173−183,Marks,J.D. et al.,(1992) in J. Biol.Chem.267:16007−16010,Winter,G. et al.,(1994) in Annu.Rev.Immunol.12:433−455及びLittle,M.et al.,(1994) Biotechn.Adv.12:539−555による概説論文に記載されている。続いて、camelid重鎖抗体を発現するcamelid発現ライブラリーをスクリーニングするために、前記ファージを使用する。(Muyldermans,S. and Lauwereys,M.,Journ. Molec. Recogn. 12:131−140(1999) and Ghahroudi,M.A. et al.,FEBS Letters 414:512−526(1997)).所望の抗体を発現するライブラリーから得られた細胞は複製され、続いて、抗体の大規模発現のために使用することが可能である。
【0048】
実施例
【実施例1】
【0049】
株、細胞、増殖条件及びDNA抽出。
【0050】
別段の記載がなければ、M.ミコイデスsubsp.ミコイデスSC株Afade(Farcha Laboratory,N’Djamena,Chadで、1968年に単離された高度に病原性の野外株)を、病原性研究のために使用した。この株は、天然及び実験条件下で、CBPPを引き起こす。別段の記載がある場合には、活性なグリセロール取り込み系GtsABCを欠く、より病原性が低い株であるM.ミコイデスsubsp.ミコイデスSC株L2を使用した。さらに、標準株PGI並びにアフリカ及び欧州の発生に由来する10個の他の株も、遺伝子分析のために、本研究で使用した。マイコプラズマの培養物は、108から109コロニー形成単位/mL(cfu/mL)の密度になるようにマイコプラズマブロス培地中で、又は固体マイコプラズマ寒天培地(Axcell Biotechnologies,St. Genis l’Argentiere,France)上で増殖させた。生きたM.ミコイデスsubsp.ミコイデスSCの増殖及び取り扱いは、BL3封じ込め安全標準を満たす生物学的安全性研究室の中で行った。DNA抽出は、以前に記載したとおり行った(Cheng,X.,J. et al.,1995,Microbiology 141:3221−3228)。遺伝子操作及びサブクローニングについては、エシェリヒア・コリ株DH5α[F−φ80dlacZデルタM15デルタ(lacZYA−argF)U169deoRrecA1endA1hsdR17(rK−、mK+)phoAsupE44λ−thi−1gyrA96relA1]及びBL21(DE3)[F’dcm ompThsdS(rB−mB−)galλ(DE3)]を使用した。組換えポリヒスチジンN及びC末端融合タンパク質の発現のために、pETHIS−1発現ベクター(Schaller,A.et al.,1999,Microbiology145:2105−2116)を使用した。
【0051】
ECaNEp細胞は、地元の食肉処理場から得られた胎児から調製し、24ウェルマイクロタイタープレート中の、7%ウシ胎児血清(FCS)及びペニシリン(100IU/mL)が補充されたEME−Earle培地中に維持し、加湿された5%CO2雰囲気中、37℃で、2×105細胞/ウェルの集密度で使用した。FCS及び細胞培地は、Seromed(Biochrom,Munich,Germany)から購入した。細胞は、PCRを使用してマイコプラズマによる汚染について、又は免疫染色を用いてBVDウイルスによる汚染について、定型的にスクリーニングした。
【0052】
PCR増幅、サザンブロット及びDNA配列分析。
【0053】
glpO特異的DNAプローブは、標準的なPCR条件を使用し(Cheng,X.,J. et al.,1995,Microbiology 141:3221−3228)、オリゴヌクレオチドプライマーglpO EcoRI N
【0054】
【化1】

及びglpO NotI C
【0055】
【化2】
を用いて、ジゴキシゲニン−11−dUTP(Dig)(Roche Diagnostics,Rotkreuz,Switzerland)の存在下でPCRによって構築した。マイコプラズマから得たゲノムDNAの抽出及びサザンブロット分析は、以前に記載されたとおりに行った(Pilo,P.et al.,2003,Vet.Microbiol.92:37−48)。
【0056】
DNA配列決定は、DNA Sequenator AB3100遺伝子分析装置及びTaq色素デオキシターミネータサイクル配列決定キット(Applied Biosystems,Norwalk,Conn.)を用い、オリゴヌクレオチドプライマーglpO EcoRI N及びglpO NotI Cを用いて、並びにglpO内部プライマーを用いるプライマーウォーキングによって行った。DNA及び推測されたアミノ酸配列は、PC/GeneプログラムPROSITE(Bairoch,A.,P. Bucher,and K. Hofmann. 1995,Nucleic Acids Res.24:189−196)を用いて分析した。GenBank及びEMBLデータベース中の配列との配列比較は、BLAST(Altschul,S.F. et al.,1997,Nucleic Acids Res.25:3389−3402)を用いて行った。タンパク質の分析は、プログラムMotif Scan(http://hits.isb−sib.ch/cgi−bin/hits motifscan)及びTMpred(http://www.ch.embnet.org/software/TMPRED form.html)及び「toppred」ソフトウェア(von Heijne,G. 1992,J. MoI. Biol. 225:487−94)を用いることによって実施した。
【実施例2】
【0057】
組換えGlpOのクローニング、部位指定突然変異誘発及び発現。
【0058】
それぞれ、EcoRI及びNotIに対する制限部位を含有するプライマーglpO EcoRI N及びd glpO NotI Cを用いて、まず、M.ミコイデスsubsp.ミコイデスSC株AfadeのglpO遺伝子を増幅した。さらに、プライマーglpO NotI Cは、変異されたTGGTrpコドンを含んだ。(適切なヌクレオチド置換を有する)プライマー対glpO mut1L
【0059】
【化3】
を用いるオーバーラップ伸長PCR法(Braman,J.,C. Papworth,and A. Greener. 1996,Methods Mol.Biol.57:31−44)を用いて、glpO遺伝子中の他の2つのマイコプラズマ特異的TGATrpコドンを、TGGTrpコドンと置換した。EcoRI及びNotI切断部位を隣接させることによって、PCR産物をpETHIS−1中にクローニングした。他のタンパク質に対して以前に記載されたように、発現及びポリヒスチジン尾部が付着された融合タンパク質のNi2+キレートクロマトグラフィーを用いた精製のために、DNA配列決定によって構築物を分析し、E.コリBL21(DE3)中に導入した(Schaller,A.,et al.,1999,Microbiology 145:2105−2116)。
【実施例3】
【0060】
血清、ポリクローナル抗体、イムノグロブリン精製及びFab調製。
【0061】
アフリカンM.ミコイデスsubsp.ミコイデスSC株Afadeを用いた、管理された実験的感染から得たウシ血清は、Abdo及び共同研究者によって詳しく記載されている(Abdo,E.−M.et al.,1998,Vet.Microbiol.59:109−122)。組換えGlpOに対して誘導されたポリクローナル単一特異的血清は、アジュバント10(Gerbu Biotechnik GmbH,Gaiberg,Germany)500μLと混合された、PBS緩衝液pH8.0(50mMNa2HPO4/NaH2PO4 pH8.0,140mMNaCl)500μL中の精製された、組換えポリヒスチジン尾部が付加されたタンパク質GlpO160μgでウサギを皮下免疫化した後、2及び4週目に、タンパク質40及び20μgで強化免疫することによって取得した。最後の強化免疫から10日後に、ウサギから採血した。血液試料から抗血清を調製し、−20℃で保存した。
【0062】
製造業者によって指示されたとおり、HiTrap Protein Gキット(Amersham Pharmacia Biotech,Uppsala,Sweden)を用いて、ウサギ抗GlpO血清及び同じウサギの前血清から得られた免疫グロブリンG(IgG)画分を精製した。製造業者の指示書に従い、ImmunoPure Fab Preparationキット(Pierce,Rockford,Ill.)を用いて、Fab断片の調製を行った。PBS緩衝液pH7.4に対して、Fab断片を一晩透析し、次いで、フィルター滅菌した。Bradfordの方法(Bradford,M.M.1976,Anal.Biochem.72:248−254.)を用いて、タンパク質濃度を測定した。リポタンパク質LppCに対して誘導された単一特異的ウサギ血清は、以前に記載されている(Pilo P. et al.,2003,Vet.Res.34:761−775.)。
【実施例4】
【0063】
イムノブロット分析、TritonX−114分配及び増殖阻害試験
マイコプラズマから得た総抗原は、以前に記載したとおり調製した(Fleury,B. et al,2001,J. Clin.Microbiol.39:2814−2822)。1:2000希釈のウシ血清及び1:1000希釈のウサギ単一特異的血清抗GlpOを用いて、イムノブロッティングを行った。
【0064】
TritonX−114分配法(Bordier,C. 1981,J.Biol.Chem.256:1604−1607)により、固定相培養から得られたM.ミコイデスsubsp.ミコイデスSC総抗原を疎水性及び親水性画分へ分割した。TritonX−114界面活性剤相及び水相から得た試料を、GlpOに対して誘導された単一特異的ポリクローナル抗体及びM.ミコイデスsubsp.ミコイデスSCに対して誘導されたウシ血清を用いたイムノブロッティングによって分析した。
【0065】
希釈されていない補体除去された(decomplemented)単一特異的ウサギ抗GlpO血清、精製されたIgG又は抗GlpOIgGのFab断片5及び10μLを、マイコプラズマ含有寒天培地上に滴加し、37℃で4日間プレートを温置することによって、増殖阻害試験を行った。M.ミコイデスsubsp.ミコイデスSCに対する血清を陽性対照として使用し、LppQに対する単一特異的ウサギ血清を陰性対照として使用した。以前に記載されているように、光学顕微鏡下で増殖阻害の観察を実施した(Papazisi,L. et al,2003,Microbiology 149:2307−2316,Poveda,J. B.,and R. Nicholas,1998,Methods Mol.Biol.104:105−111)。
【実施例5】
【0066】
走査型電子顕微鏡(SEM)及び免疫金標識
電子顕微鏡のために、予めポリ−L−リジンで被覆された、金又は白金がスパッタリングされたカバーガラス上のマイコプラズマブロス培地中で、M.ミコイデスsubsp.ミコイデスSCを37℃で5日間培養した。37℃のPBS緩衝液pH7.4で、細胞を3回洗浄し、室温で30分間、PBS中の4%パラホルムアルデヒド中に固定した。PBSで洗浄した後、0.2Mグリシン及び1%BSAが補充されたPBS緩衝液中、室温で15分間、カバーガラスをブロッキングし、その後、4℃で一晩、1%BSAが補充されたPBS中、1:100又は1:50希釈されたウサギ血清抗GlpOから得られたIgGとともにカバーガラスを温置した。次いで、PBSで10分間、試料を洗浄し、室温で90分間、PBS中に1:50希釈された、15nmのコロイド状金が抱合されたヤギ抗ウサギIgG(British Biocell International,Cardiff,UK)で標識した。0.1Mカコジル酸塩緩衝液pH7.4でカバーガラスを洗浄し、標準的なプロトコールに従って、SEMのために加工した。要約すれば、0.13Mカコジル酸塩緩衝液pH7.4中の0.11%ルテニウムレッドを加えた1.33%OsO4中で15分間、試料をオスミウム酸染色し、0.1Mカコジル酸塩緩衝液で洗浄し、増加するエタノール系列を通じて脱水し、ヘキサメチルジシラザンの蒸発によって乾燥させた(Sigma,Buchs,Switzerland)。
【0067】
5kVの加速電圧、8mmの作動距離及び50,000から100,000倍までの拡大で、二次電子シグナル及び対応する後方散乱された電子シグナルを、高解像度電解放出走査型電子顕微鏡DSM982Gemini(Zeiss,Oberkochen,Germany)中で調べた。
【0068】
対照実験は、一次抗体の省略、並びに、それぞれウサギ抗カルシトニン抗体の使用(Anawa Biomedical Services and Products,Zurich Switzerland)及びウサギ前免疫血清の使用を含んだ。
【実施例6】
【0069】
H2O2産生の定量及び阻害アッセイ
H2O2産生を測定するために、約5×108cfu/mLの密度になるまで、37℃で3日間、マイコプラズマ培養培地中でM.ミコイデスsubsp.ミコイデスSCの株を増殖させた。4℃で10分間、8000×gで培養物を遠心し、温置培地(67.6mM HEPES pH7.3,140mM NaCl,7mM MgCl2)中で1回洗浄し、109cfu/mLの密度で、予め加温された37℃の温置培地中に再懸濁し、1mLの分取試料中に分けて、37℃で1時間温置した。H2O2産生を誘導するために、100μMの最終濃度(ウシ血清中での生理的濃度)で、マイコプラズマ懸濁液にグリセロールを添加した。グリセロールの添加から0、1、2、5、10、20及び120分後に、以前に記載されたように(Vilei,E.M.,and J. Frey,2001,Clin.Diagn.Lab.Immunol.8:85−92)、過酸化物試験(Merck KgaA,Darmstadt,Germany)を用いて、H2O2の産生を測定した。マイコプラズマのL−α−グリセロリン酸オキシダーゼGlpOを阻害するために、0.26μg/mLから2.6μg/mLまでの濃度での温置に続いて、PBS緩衝液で2回洗浄することによって、GlpOに対して誘導されたIgGの精製されたFab断片でマイコプラズマを前処理した。H2O2産生の誘導後に、M.ミコイデスsubsp.ミコイデスSC細胞の生存性を評価するために、アッセイの終了時に、反応アッセイの分取試料をマイコプラズマ寒天プレート上に播種し、37℃で増殖させた。
【実施例7】
【0070】
細胞毒性活性の評価
周密状態に達するまで、胚性ウシ鼻上皮細胞(ECaNEp細胞)(Schweizer,M.,and E. Peterhans,1999,J. Gen. Virol. 80:1147−1155)を、24ウェルプレート中で増殖させた。アッセイの前に、培地を除去し、補充物を含まないMEM−Earle培地200μLによって、又は100Mグリセロールを補充したMEM−Earle培地によって置き換えた。次いで、50マイコプラズマ/細胞の感染効率(MOI)で、ECaNEp細胞を感染させた。GlpO活性を遮断するために、0.26μg/mLの抗GlpOFab断片でM.ミコイデスsubsp.ミコイデスSCを前処理した。0.75%クリスタルバイオレット、0.25%NaCl、1.75%ホルムアルデヒド及び50%エタノールでの固定及び染色後に、生きたECaNEp細胞を計数し、感染後の様々な時点で、位相差顕微鏡下で写真を撮影した。LppCはグリセロール代謝と相関しないので、M.ミコイデスsubsp.ミコイデスSCの膜リポタンパク質LppCに対して誘導されたポリクローナルIgGから得られる精製されたFab断片を対照として使用した。ECaNEp細胞との接触前に、M.ミコイデスsubsp.ミコイデスSCの増殖中に産生されたH2O2が、ウシ細胞に対して有害であるかどうかを測定するために、0.22μmフィルター(Millipore,Bedford,Mass.)を通して、グリセロールの存在下で増殖されたM.ミコイデスsubsp.ミコイデスSCの培養物の上清をろ過し、ECaNEp細胞に添加した。ECaNEp細胞の生存性は、トリパンブルー排除によって評価した。
【実施例8】
【0071】
ECaNEp細胞中のH2O2及びその他のROSによって引き起こされた酸化的ストレスの検出
ECaNEp細胞中の細胞内ROSによる酸化的ストレスを評価するために、5(及び6)−クロロメチルー2’,7’−ジクロロジヒドロフルオレセインジアセタート−アセチルエステル(CM−H2DCFDA; Molecular Probes,Eugene,Oreg.)の酸化を使用した。この色素は細胞の中に入り、ROSによる細胞内酸化後に蛍光シグナルを産生する。以下の修飾を用いて、細胞毒性活性の評価に使用したものと同じ条件を用いた。対照として、H2O2及びROSによる酸化を防ぐために、ECaNEp細胞を含有するウェルの半分を、30mMN−アセチル−L−システイン(NAC)で処理した。次いで、10μMCM−H2DCFDAとともに、ECaNEp細胞を1時間温置し、MEM−Earle培地で1回洗浄した。次いで、グリセロールの存在下又は不存在下において、500マイコプラズマ/細胞のMOIで細胞を感染させた。GlpO活性を遮断するために、0.26μg/mLの抗GlpOFab断片でM.ミコイデスsubsp.ミコイデスSCを前処理した。対照として、150μMから4.4mMにわたるH2O2溶液で、細胞を20分間処理した。Nikon Eclipse TE300顕微鏡を用いた蛍光顕微鏡法によって、マイコプラズマでの感染から20分後に、細胞内H2O2及びROSをモニターした。CM−H2DCFDAを含む全ての工程(この化学物質の取り扱いを含む。)が、暗所で行われたことに注目されたい。
【0072】
ヌクレオチド配列受託番号。
【0073】
M.ミコイデスsubsp.ミコイデスSC株Afade及びL2から得られたglpOのヌクレオチド配列に対するEMBL/GenBank受託番号は、それぞれ、AJ581566及びAJ581564である。マイコプラズマsp.ボバイングループ7株PG50及びM.ミコイデスsubsp.カプリ株PG3から得られたglpO遺伝子の配列は、それぞれ、AJ581565及びAJ581567の受託番号で寄託された。
【実施例9】
【0074】
glpO遺伝子の遺伝的及び機能的分析
M.ミコイデスsubsp.ミコイデスSCから得られたglpO遺伝子は、378アミノ酸ポリペプチドのGlpOをコードしており、42.7kDaの予想分子量及び8.14のpIを有する。本遺伝子は、3つのTGATrpコドンを含有する。本タンパク質は、アミノ酸位置8から36に推定FAD結合部位を有し、表面露出されているように見受けられる。TMpredは、アミノ酸6から25及び140から157にわたる2つの顕著な膜貫通領域を同定した。高度に病原性の株Afade中において、glpO遺伝子の後には、遺伝子glpK(推定グリセロールリン酸キナーゼ)及びglpF(推定グリセロール促進物質因子)が続く。標準株PG1についても、同じ遺伝子配置が観察される。glpOに対する遺伝子プローブを用いたゲノムDNAのサザンブロット分析は、M.ミコイデスsubsp.ミコイデスSC株Afade、L2、標準株PGI並びに分析されたアフリカ及び欧州での発症から得られた10個の他の株中に、glpOが存在することを示した(データは図示せず。)。単一特異的抗GlpOIgGを用いた総抗原のイムノブロット分析は、別個の45kDaタンパク質バンドを明らかにし、検査されたM.ミコイデスsubsp.ミコイデスSCの全ての株中に、glpO遺伝子の発現を確認した(データは図示せず。)。M.ミコイデスsubsp.ミコイデスSC株Afade及び株L2から得られたGlpOのアミノ酸配列は、標準株PGI(48)のGlpOのものと同一であることが明らかとなり、様々なマイコプラズマ種のグリセロール−3−リン酸脱水素酵素と類似性を示した(表1)。
【0075】
ゲノム配列分析は、マイコプラズマがカタラーゼ及びジスムターゼ活性を欠如することを予測する。細胞質のGlpO活性は、細胞内H2O2毒性を誘導し、次いで、細胞内H2O2毒性がマイコプラズマ自体に対して有害であるので、本発明者らは、本酵素が表面膜に存在しているに違いないという仮説を立てた。「toppred」ソフトウェアを用いた、(表1に列記されている)様々な病原性マイコプラズマ種由来のGlpO又は相同的酵素の配列分析によって膜貫通構造が明らかとなり、これらの酵素のドメインが表面に位置すると予想された。M.ミコイデスsubsp.ミコイデスSCの細胞表面にGlpOが存在することの直接的な証拠は、単一特異的ウサギ抗GlpO血清から得られるIgGを用いた免疫金標識後の走査型電子顕微鏡(SEM)によって得られた(図1a−d)。マイコプラズマの膜中にGlpOが存在することを確認するために、M.ミコイデスsubsp.ミコイデスSCの総タンパク質を、TritonX−114相分配に供した。抗GlpO血清は、TritonX−114相中の45kDaタンパク質(GlpO)(これは、水相には存在しない。)と強く反応し、GlpOがM.ミコイデスsubsp.ミコイデスSCの内在性膜タンパク質であることを示している(図1e)。この株で実験的に感染されたウシから得たウシ血清を用いた、M.ミコイデスsubsp.ミコイデスSCのTritonX−114抽出物のイムノブロット分析は、GlpOとともにゲル上を同時に移動するGlpO特異的バンドを45kDaの場所に明らかにし(データは示さず。)、ウシのCBPP感染中に、GlpOが血清転換に関与することを示している。
【0076】
マイコプラズマのL−α−グリセロリン酸オキシダーゼとしてのGlpOの機能を確認するために、本発明者らは、単一特異的抗GlpOIgGのFab断片の存在下又は不存在下で、M.ミコイデスsubsp.ミコイデスSCの純粋培養物にグリセロールを添加した後、H2O2の産生を測定した。図1fに示されているように、中指数関数期の増殖相にある培養物に100μMグリセロールを添加することによって、直ちに、増殖培地中へH2O2が放出されて、10分後に150μMに達し、この濃度は、最長2時間保持された(図1f)。0.26μg/mLの最小濃度の、単一特異的ポリクローナル抗GlpO抗体又は抗GlpOIgGのFab断片で、マイコプラズマ懸濁液を前処理すると、H2O2の放出を特異的に遮断することができたが、対照抗体として使用された抗LppCIgGのFab断片によっては遮断できなかった(図1f)。LppCは、グリセロール代謝とは関係していない、M.ミコイデスsubsp.ミコイデスSCの表面リポタンパク質である。グリセロールの補充後に、M.ミコイデスsubsp.ミコイデスSCの培養物にカタラーゼを添加することによって、1μMの濃度(これは、本アッセイの検出レベルである。)を下回るまで、培地中のH2O2レベルが低下した。抗GlpO抗体又は抗GlpOFab断片の添加なし又は添加ありで、M.ミコイデスsubsp.ミコイデスSCの世代時間は3.3時間であった。さらに、抗GlpO抗体は、血清滴下増殖阻害試験において効果を有しなかった(データは示さず。)。100μMグリセロールの存在下で、2時間増殖され、150μMH2O2を産生した純粋培養物は、3.2時間の世代時間を示したのに対して、グリセロールが添加されなかった対照培養物は、3.3時間の世代時間を有した。このため、マイコプラズマの生存性は、グリセロールによって誘導されたH2O2の産生によって影響を受けなかった。
【実施例10】
【0077】
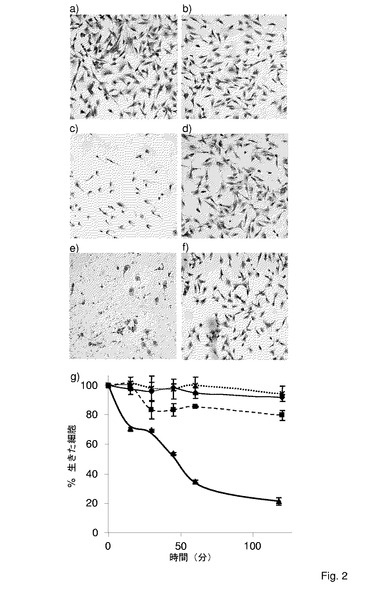
ウシ上皮細胞に対するM.ミコイデスsubsp.ミコイデスSCの細胞毒性
グリセロール代謝から生じるH2O2及び関連のROSの産生が、M.ミコイデスsubsp.ミコイデスSCの病原性に寄与しているかどうかを評価するために、本発明者らは、様々な条件下で、胚性仔ウシ鼻上皮(ECaNEp)細胞に対する細胞毒性を分析した(図2)。図2bに示されているように、50マイコプラズマ/細胞の感染効率でのM.ミコイデスsubsp.ミコイデスSC株AfadeによるECaNEp細胞の感染は、培地中のグリセロールの不存在下で、感染から1時間後に弱い細胞毒性効果をもたらすに過ぎなかった。しかしながら、グリセロールの生理的濃度の存在下で、マイコプラズマでECaNEp細胞の感染を行った場合には、多くのECaNEp細胞が表面から剥離され、続いて、完全な溶解を起こした(図2c)。H2O2産生を付随的に遮断する0.26μg/mLの濃度の抗GlpOFab断片でマイコプラズマを前処理すると、この細胞毒性効果を遮断することが可能であった。これらの条件下で、ECaNEp細胞は、感染から1時間後に形態的変化を示さなかった(図2d)。グリセロールを含有する培地中で増殖されたM.ミコイデスsubsp.ミコイデスSCの細胞毒性は、対照として使用された抗LppCFab断片によって阻害されなかった(図2e)。活性なグリセロール輸送体GtsABCに対する遺伝子を欠如し、H2O2のごく少量を産生するより病原性が低い株(L2)を用いると、グリセロールの存在下でECaNEp細胞を感染させたときに、弱い細胞毒性効果が観察されるのみであった(図2f)。対照実験において、グリセロール、抗GlpOFab若しくは抗体LppCFab断片のみ又は組み合わせを添加しても、ECaNEp細胞に影響を及ぼさなかった(データは図示せず。)。細胞毒性実験の速度論(図2g)は、M.ミコイデスsubsp.ミコイデスSCで感染されたECaNEp細胞へのグリセロールの添加後に、細胞毒性の誘導が急速に起こり、2時間後には、75%の死亡率に達することを示している。より高い感染効率(500マイコプラズマ/細胞)では、グリセロールの添加から30分後に、細胞の完全な死亡率に達した。抗GlpOFabでのマイコプラズマの前処理は、観察期間全体にわたって、細胞の死亡を完全に阻害した(図2g)。添加されたグリセロールの不存在下で、M.ミコイデスsubsp.ミコイデスSCでのECaNEp細胞の感染は、弱い細胞毒性効果を示すに過ぎず(図2g)、これも抗GlpOFabによって阻害することが可能であり、従って、細胞又は培地中へのグリセロールの残留量によるものであり得る。興味深いことに、約150μMH2O2を含有するグリセロールの存在下で増殖されたM.ミコイデスsubsp.ミコイデスSC培養物のろ過された上清又は細胞培養物への150μMH2O2の添加は、1時間の曝露後に、ECaNEp細胞に対して目に見える細胞毒性効果を有しなかった。外来H2O2のECaNEp細胞培養物への添加によって誘導された細胞毒性は、まず、4.4mMの濃度で達成され、細胞の多くが、曝露の1時間後に死滅した。この濃度は、グリセロールの存在下での、M.ミコイデスsubsp.ミコイデスSCの増殖培地中で測定された濃度より30倍高い。
【0078】
M.ミコイデスsubsp.ミコイデスSCでの感染又はグリセロールの添加後に、細胞内H2O2及び他のROSによって引き起こされたECaNEp細胞中で酸化的ストレスをモニターするために、本発明者らは、CM−H2DCFDAでECaNEp細胞を前処理し、蛍光顕微鏡によって、この化合物の細胞内酸化を検出した(図3)。CM−H2DCFDAの酸化によるエステル基の切断が、高度に蛍光性のジクロロフルオレセイン(DCF)誘導体の形成を細胞中にもたらすが、酸化されていないCM−H2DCFDAは非蛍光である。図3cからd中に示されているように、M.ミコイデスsubsp.ミコイデスSCでのECaNEp細胞の感染は、グリセロールの添加から20分後に、ECaNEp細胞中に蛍光の強い誘導をもたらし、細胞内H2O2又はROSの存在を反映している。これに対して、グリセロールの添加なしの感染されたECaNEp細胞中には、蛍光は検出されなかった(図3a−b)。ECaNEp細胞の感染前に、抗GlpO抗体でマイコプラズマを処理すると、CM−H2DCFDAの細胞内酸化は起こらなかった(図3e−f)。さらに、CM−H2DCFDAの細胞内酸化は、細胞培地にグリセロールを添加する前に、抗酸化剤NACでECaNEp細胞を処理することによって遮断された(図3g−h)。グリセロールの添加から10分後に、M.ミコイデスsubsp.ミコイデスSCによって放出された濃度に対応する濃度である150μMの外来H2O2を添加すると、ECaNEp細胞の細胞内酸化は起こらなかった。細胞毒性の誘導の場合と同様、ECaNEp細胞中での細胞内CM−H2DCFDAの酸化を達成するために、4.4mM外来H2O2の濃度が必要であった(図3i−j)。
【0079】
結論
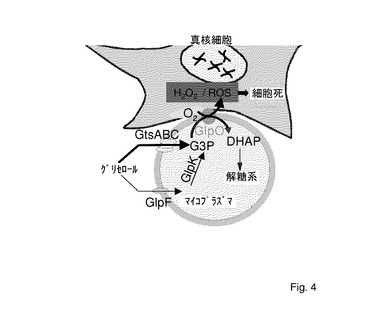
本発明において、M.ミコイデスsubsp.ミコイデスSCのGlpOは、ECaNEp細胞に対する細胞毒性において中心的な役割を果たす膜タンパク質として同定された。グリセロールの生理的濃度の存在下で、109cfu/mlの密度のM.ミコイデスsubsp.ミコイデスSCは、最大150μMまで、培地中にH2O2の比較的大量を放出する。H2O2のこの量は、グルコースの存在下で増殖されたマイコプラズマ(ちょうど50%、H2O2産生を刺激すると報告された。)によって産生される量と比べると、約20倍である。グリセロールの生理的濃度の存在下で、ECaNEp細胞をM.ミコイデスsubsp.ミコイデスSCに曝露すると、H2O2が、まず、それらのサイトゾル中に検出され、続いて、細胞死が起こった。本発明者らは、マイコプラズマによって培地中へ放出されたH2O2濃度と、CM−H2DCFCA酸化及び細胞死を引き起こすために必要とされる濃度との間に、顕著な齟齬を認めた。従って、150μMH2O2を含有するろ過されたマイコプラズマの増殖培地又はECaNEp細胞への150μM外来H2O2の添加は、サイトゾル中での検出可能なCM−H2DCFDA酸化をもたらさず、1時間以内の細胞死ももたらさなかった。このことは、毒性化合物を宿主細胞に首尾よく誘導するためには、マイコプラズマと宿主細胞間の密接な接触が必要であることを示している。この文脈において、M.ミコイデスsubsp.ミコイデスSC、及び、さらには、多くの他の病原性マイコプラズマは、それらの宿主細胞に強固に付着するが、貫通はしないことに注目すべきである。さらに、ECaNEp細胞に対して、グリセロールによって誘導される細胞毒性効果を得るためには、グリセロールの活発な取り込みが必要である。グリセロールの活発な取り込みに必要とされるGtsABC輸送体を欠如し、著しくより低い速度で、1/10未満のH2O2を産生するM.ミコイデスsubsp.ミコイデスSCの欧州株L2は、同一の実験条件下で、ECaNEp細胞に対して、弱い細胞毒性効果を有するに過ぎなかった。株L2によるグリセロールの残りの取り込みは、グリセロール促進物質因子と推定されるGlpFによって媒介される、より低効率の輸送体経路を用いて行われると考えられる。
【0080】
我々の結果に基づいて、本発明者らは、M.ミコイデスsubsp.ミコイデスSCによる真核細胞への細胞損傷を引き起こすための以下のモデルを提案する(図4)。間質液中に存在するグリセロールは、高活性のABCグリセロール輸送体(GtsA、GtsB及びGtsC)を介して活発に取り込まれ、続いて、グリセロール−3−リン酸へリン酸化される。次いで、これは、GlpOによって、O2の存在下でDHAPへと酸化され、これは解糖系に入り、H2O2の一分子を産生する。マイコプラズマの宿主細胞膜との緊密な接触によって促進されて、H2O2及び付随するROSは宿主細胞に入る。宿主細胞内では、H2O2及びROSは、細胞傷害の強力な媒介物質及び炎症性プロセスの誘導物質として作用する。これらは、組織細胞を直接障害し、又は宿主細胞遺伝子発現(例えば、NF−κBの活性化を介して、又はFenton反応を介して、炎症促進性遺伝子)を誘導することによって、宿主を損傷すると予想される(Crichton,R. R. et al.2002,J. Inorg.Biochem.91:9−18)。興味深いことに、マイコプラズマは、貪食細胞中で呼吸性バーストを誘導することが以前に示されており、宿主によって生成されたROSが、組織損傷に対してさらに寄与し得ることを示唆する。
【0081】
H2O2及びROSは、NF−κBを活性化することも可能であり、このようにすることによって、真核生物宿主中で、多様な免疫及び炎症促進性遺伝子の発現を誘導し、これは、マイコプラズマによって引き起こされる気道感染において特に重要な可能性がある機序である。
【0082】
マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はマイコプラズマのL−α−グリセロリン酸オキシダーゼに対する抗体に基づくワクチンは、H2O2及びROSの産生を首尾よく抑制し、このようにすることによって、本ワクチンは、宿主細胞の損傷、炎症及び疾病を予防する。
【0083】
【表1】
【図面の簡単な説明】
【0084】
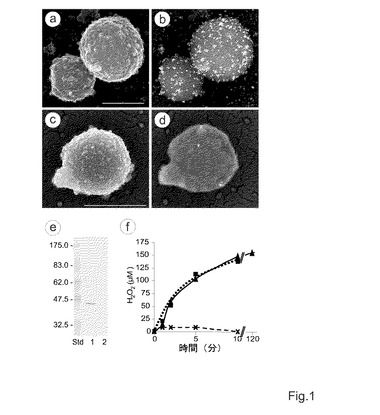
【図1】マイコプラズマのL−α−グリセロリン酸オキシダーゼの位置及び活性。a−d、それぞれ、抗GlpOから得られたIgG(a,b)及び免疫前血清(c、d)とともに温置されたM.ミコイデスsubsp.ミコイデスSC株Afadeの免疫金標識を示す走査型電子顕微鏡写真。二次電子顕微鏡写真は、細胞表面(a,c)を示しており、後方散乱された電子顕微鏡写真は、15nmのコロイド状金が抱合された二次抗体(b,d)を明らかにする。スケールバー、500nm。e、GlpO抗血清を用いた、M.ミコイデスsubsp.ミコイデスSC株AfadeのTritonX−114分画された総抗原のイムノブロット分析。レーン1、界面活性剤相、タンパク質20μg;レーン2、水相、タンパク質20μg;Std、分子量標準、f、100μMグリセロールの添加後における、M.ミコイデスsubsp.ミコイデスSC株Afadeの過酸化水素産生。示されているデータは、5つの独立した測定の平均値である。各測定値の標準偏差は、平均値の5%以下であった。三角及び実線、処理されていないM.ミコイデスsubsp.ミコイデスSC;十字及び破線、抗GlpOIgG由来のFab断片で前処理されたM.ミコイデスsubsp.ミコイデスSC;四角及び点線、抗LppC由来のFab断片で前処理されたM.ミコイデスsubsp.ミコイデスSC。
【図2】ECaNEp細胞に対するM.ミコイデスsubsp.ミコイデスSCの細胞毒性(a−f)及び細胞生存性アッセイ(g)。a、ECaNEp細胞(対照);b、M.ミコイデスsubsp.ミコイデスSC株Afadeでの感染から1時間後のECaNEp;c、グリセロール(100μM)の存在下における、株Afadeでの感染から1時間後のECaNEp;d、抗GlpOIgG由来のFab断片で前処理された株Afadeでの、グリセロール(100μM)の存在下における感染から1時間後のECaNEp;e、抗LppCIgG由来のFab断片で前処理された株Afadeでの、グリセロール(100μM)の存在下における感染から1時間後のECaNEp;f、グリセロール(100μM)の存在下における、M.ミコイデスsubsp.ミコイデスSC株L2(活性なグリセロール取り込み系を欠く。)で感染されたECaNEp細胞。全ての実験で、感染効率は、50マイコプラズマ/細胞であった。g、50マイコプラズマ/細胞の感染効率で、M.ミコイデスsubsp.ミコイデスSC株Afadeに感染させた後の様々な時点における生きたECaNEp細胞。三角及び太い実線、100μMグリセロールの存在下での、株Afadeによる感染;十字及び点線、抗GlpOFab断片(0.26μg/mL)で前処理された株AdadeSCによる、100μMグリセロールの存在下での感染;四角及び破線、グリセロールなしの株Afade感染;ダイヤモンド及び細い実線、ECaNEp細胞のみ(対照)。
【図3】500マイコプラズマ/細胞の感染効率で、M.ミコイデスsubsp.ミコイデスSC株Afadeに感染されたECaNEp細胞又はH2O2で処理されたECaNEp細胞中の細胞内H2O2及びROSの検出。ECaNEp細胞の位相差顕微鏡写真(a、c、e、g及びi)及び蛍光顕微鏡写真(b、d、f、h及びj);グリセロールの不存在下における、M.ミコイデスsubsp.ミコイデスSC株Afadeでの感染から20分後(a,b);グリセロールが補充された培地中で、株Afadeに感染(c、d);グリセロールが補充された培地中において、抗GlpOIgG由来のFab断片で前処理された株Adadeで感染(e、f);N−アセチル−L−システインで前処理され、次いで、グリセロールを加えた培地中、株Adadeで感染(g、h);4.4mMH2O2が補充された培地中で、20分間温置(i、j)。
【図4】M.ミコイデスsubsp.ミコイデスSCによって宿主細胞炎症を誘発するためのモデル。GtsABC:活性なグリセロール輸送体及びリン酸化系;GlpF:グリセロール促進物質因子;GlpK:グリセロールキナーゼ;G3P:グリセロール−3−リン酸;DHAP:ジヒドロキシアセトンリン酸;GlpO:マイコプラズマのL−α−グリセロリン酸オキシダーゼ。太い矢印は、M.ミコイデスsubsp.ミコイデスSCの主要な病原性経路を示す。
【技術分野】
【0001】
本発明は、とりわけ、マイコプラズマ感染を撲滅するためのワクチン、このようなワクチンにおいて使用するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼ、このようなワクチンを製造するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼの使用、このようなワクチンを調製するための方法、並びに前記ワクチンを予防接種された動物と完全細胞ワクチンで予防接種された動物又は野外感染に罹患した動物を識別するための診断試験に関する。
【背景技術】
【0002】
進化の間に、病原性細菌は、それらの宿主との複雑な相互作用を発達させてきた。これは、しばしば、病原性島、プラスミド、トランスポゾン又はプロファージ上での病原性因子の獲得を伴い、これにより、細菌は、宿主内でコロニーを形成し、生存し、複製することが可能となる。これに対して、最小の自己複製生物であるマイコプラズマ種は、ゲノムを最小サイズへ減少させることによって、グラム陽性細菌から退行性に進化し、その結果、遺伝的資源を節約している。
【0003】
マイコプラズマ種は、地球上で検出される最小の自己複製生物である。それらのゲノムは、マイコプラズマ・ジェニタリウムにおける580キロ塩基対(kb)(Fraser,C.M.et al.,1995,Science270:397−03)からマイコプラズマ・ペネトランスにおける1358kb(Sasaki,Y. et al,2002,Nucleic Acids Res. 30:5293−5300)にわたる。これにより、遺伝的資源の劇的な節約がもたらされ、寄生性の生活様式が必要不可欠となる。病原性マイコプラズマ種は、主に、動物及びヒトに、非定型肺炎、泌尿生殖器感染症及び関節炎を引き起こす(Baseman,J.B.,and J.G. Tully,1997,Emerg.Infect.Dis.3:21−32,Blanchard,A.,and G. F. Browning(eds.).2005.Mycoplasmas:Molecular biology,pathogenicity and strategies for control.Horizon Bioscience,Wymondham,U.K.,Frey,J. 2002.Mycoplasmas of animals,p.73−90.In S. Razin and R. Herrmann(eds.),Molecular biology and pathogenicity of mycoplasmas. Kluwer Academic/Plenum Publishers,New York)。ここでは、ヒト(例えば、M.ジェニタリウム、M.ニューモニエ)、ウシ種(例えば、M.spボバイングループ7)、ブタ(例えば、M.ハイオニューモニエ)又は家禽(例えば、ガリセプチカム)に感染する公知のマイコプラズマ種が存在するという事実のために、本明細書で動物という場合には、これが、とりわけ、ヒト、ウシ種、ブタ及び家禽を含むものと解釈されるべきことが強調される。病原性が、毒素、インベイシン及び細胞溶解素によって概ね決定される他の病原性細菌とは異なり、完全に配列決定された8つの種のゲノム配列分析によって明らかとなったところによれば、病原性マイコプラズマ種は、このような典型的な主要な病原性因子を有していないように見受けられる(Chambaud,I. et al,2001,Nucleic Acids Res.29:2145−2153,Fraser et al.,1995,Science 270:397−403,Himmelreich,R. et al.,1996,Nucleic Acids Res.24:4420−4449,Jaffe,J. D. et al,2004,Genome Res.14:1447−1461,Minion,F.C. et al.,2004,J. Bacteriol.186:7123−7133,Papazisi,L. et al,2003,Microbiology 149:2307−2316,Sasaki,Y. et al.,2002,Nucleic Acids Res.30:5293−5300,Westberg,J. et al.,2004,Genome Res.14:221−227)。
【0004】
PCR法の導入以来、マイコプラズマ感染症の診断は著しく改善され、数個のマイコプラズマ種において、抗原性の可変性が詳しく研究されてきたが、現在のところ、病原性マイコプラズマが、宿主細胞に損傷、炎症及び疾病を引き起こす分子的機序及びエフェクターに関する知識は殆ど得られていない。従って、これらの望ましくない効果を予防することを特に目標とするワクチンの開発は、極めて必要とされている。
【0005】
宿主細胞の損傷、炎症及び疾病を回避又は減弱させる、マイコプラズマ感染を撲滅するためのワクチンを提供することは本発明の1つの目的である。
【0006】
驚くべきことに、新規の主要病原性因子が、マイコプラズマにおける細胞傷害の重要な原因であることが見出された。この病原性因子は、H2O2などの有毒な副産物及びその他の付随する反応性酸素種(ROS)からなるように見受けられた。H2O2/ROSの形成は、グリセロールの代謝に関与する酵素である、酵素マイコプラマのL−α−グリセロリン酸オキシダーゼ(GlpO)の活性と直接関係し得ることを示すことができた。
【0007】
さらに予想外のことに、マイコプラズマ感染によって引き起こされる組織損傷の唯一の原因ではないとしても、H2O2/ROSの形成が中心的な原因であることを示すことができた。これは、今回、初めて、マイコプラズマの主な病原性因子が確定されたことを意味している。
【0008】
マイコプラズマ中のグリセロール代謝経路は、容易に影響又は変化を受けることができず、従って、H2O2/ROSは、マイコプラズマ自体のグリセロール代謝経路の修飾を通じて容易に変化することができない。従って、非マイコプラズマ種での状況とは異なり、生きた弱毒化されたマイコプラズマを開発する経路は、より実効可能性が低いように思われる。
【0009】
しかしながら、驚くべきことに、マイコプラズマのL−α−グリセロリン酸オキシダーゼと反応性を示す抗体は、感染された動物の組織に、ほとんど又は全く損傷を与えない程度まで、H2O2/ROSの産生を抑制できることが、ここに見出された。これは、以下に説明されているように、とりわけ、サブユニットワクチンでの予防接種に道を開く。
【0010】
全ての非マイコプラズマ細菌では、これらの細胞中のカタラーゼが、形成後、H2O2を直ちに分解するので、グリセロリン酸オキシダーゼ活性を有する酵素が細胞内に見出されるという事実に鑑みれば、これは、さらに驚くべきことである。このような場合には、グリセロリン酸オキシダーゼに対して誘導された抗体は、従って、細菌の中に入ることができないので、全く効果を有しない。全ての非マイコプラズマ細菌とは異なり、おそらくは、カタラーゼが欠如している結果、マイコプラズマは、細胞外H2O2の産生をもたらすように、進化の間に、細胞内腔から膜へ、そのマイコプラズマのL−α−グリセロリン酸オキシダーゼを移動させたことがここに見出された。従って、これは、細菌にとって有害でないが、感染された宿主の組織にとっては有害となる。これは、マイコプラズマの場合には、マイコプラズマのL−α−グリセロリン酸オキシダーゼに対する抗体が(他の細菌とは反対に)、細胞外H2O2を放出する酵素の活性を予想外に妨害することを説明し得る。
【0011】
マイコプラズマの酵素である、マイコプラズマのL−α−グリセロリン酸オキシダーゼは、公知の全てのマイコプラズマ種の中に存在する。L−α−グリセロリン酸オキシダーゼに対して行われる全てのさらなる参照は、マイコプラズマのL−α−グリセロリン酸オキシダーゼを表す。以下の表1には、マイコプラズマ種、M.sp.ボバイングループ7(M.sp. bovine group 7)、M.ミコイデスsubsp.カプリ(M.mycoides subsp.Capri)、M.ペネトランス(M.penetrans)、M.ガリセプチカム(M.gallisepticum)、M.モービレ(M.mobile)、M.パルモニス(M.pulmonis)、M.ハイオニューモニエ(M.hyopneumoniae)、M.ニューモニエ(M.pneumoniae)、M.ジェニタリウム(M.genitalium)の株において、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子のEMBL/GenBank受託番号が示されている。さらに、実施例には、マイコプラズマ種、M.ミコイデスS(mall)C(olony)株Afade及びL2の株において、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子のEMBL/Genbank受託番号が示されている。
【0012】
酵素マイコプラズマのL−α−グリセロリン酸オキシダーゼは、全ての公知のマイコプラズマ種の中に存在するという事実のために、マイコプラズマ感染を駆除するための、より具体的には、宿主細胞の損傷、炎症及び疾病を回避又は減弱するための、本タンパク質に対する抗体を産生するためのその使用は、マイコプラズマ種に関わらず、一般的に適用可能であることが明らかとなった。
【0013】
マイコプラズマに対する抗体を産生するための原理は、以下の実施例に説明されている。家畜の多大な喪失を招く重度の感染性疾患である伝染性ウシ胸膜肺炎(CBPP)の発病因子であるマイコプラズマ・ミコイデスsubsp.ミコイデスSCは、マイコプラズマの病原性の分子的基礎を調査するためのモデルとした使用された。M.ミコイデスsubsp.ミコイデスSCは、宿主細胞と接近して生息する、1211kbのゲノムサイズを有する細胞外病原体である(Westberg,J. et al.,2004,Genome Res.14:221−227)。モデルとして、この種を使用することの合理性は、この種の高い病原性及びCBPPの発病因子として明らかに確立されている事実である。さらに、この重いウシの病気は、現在、CBPPの発生に苦しむ国で、家畜の生産に対して著しい社会経済的な重要性を有している。さらに、この伝染病が存在しない国は、再発生する感染の脅威に継続的に曝されている。
【0014】
マイコプラズマ感染を撲滅するために、より具体的には、宿主細胞損傷、炎症及び疾病を回避又は減弱するために、マイコプラズマのL−α−グリセロリン酸オキシダーゼに対する抗体を使用することが可能であることが、ここに確立されたので、次の段階は、インビボ及びインビトロで、抗体を産生するために、マイコプラズマのL−α−グリセロリン酸オキシダーゼを使用することである。マイコプラズマのL−α−グリセロリン酸オキシダーゼのインビボでの使用の例は、ワクチンでの使用である。このようなワクチンは、投与されると、マイコプラズマに対する抗体を誘導する。これは、以下でさらに詳しく考察されている。マイコプラズマのL−α−グリセロリン酸オキシダーゼのインビボ又はインビトロでの使用の例は、ワクチン中で使用するための抗体を産生するための使用である。このようなワクチンも、以下で考察されている。別のワクチンは、マイコプラズマのL−α−グリセロリン酸オキシダーゼ(の免疫原性部分)をコードする遺伝子又はその一部を担持する生組換え担体及びマイコプラズマのL−α−グリセロリン酸オキシダーゼ(の免疫原性部分)をコードする遺伝子又はその一部を含むDNAワクチンを基礎とするワクチンである。
【0015】
様々なマイコプラズマ種において、酵素マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする配列が公知であるので、十分な量で酵素マイコプラズマのL−α−グリセロリン酸オキシダーゼを取得することが、ここに可能となる。これは、例えば、これらのタンパク質をコードする遺伝子を発現するための発現系を使用することによって行うことが可能である。
【0016】
マイコプラズマのL−α−グリセロリン酸オキシダーゼ(の免疫原性部分)をコードする遺伝子又はその一部を発現させるために必要不可欠であるのは、遺伝子がプロモーターの制御下にあるように、遺伝子に機能的に連結された十分なプロモーターである。これは、例えば、標準的な分子生物学の技術を用いて達成することが可能である。(Sambrook,J. and Russell,D.W.,Molecular cloning:a laboratory manual,2001.ISBN 0−87969−577−3)。プロモーターの選択が、タンパク質発現のための宿主細胞として使用される細胞中で遺伝子転写を誘導することが可能な全ての真核、原核又はウイルスプロモータに拡張することは、当業者に自明である。機能的に連結されたプロモーターは、それらが連結された核酸の転写を制御することが可能なプロモーターである。このようなプロモーターは、マイコプラズマプロモーター、例えば、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子のインビボ発現に関与するプロモーター(但し、このプロモーターは、発現のために使用される細胞中で機能的である。)とすることが可能である。プロモーターは、異種のプロモーターとすることも可能である。宿主細胞が細菌である場合には、使用され得る有用な発現調節配列には、Trpプロモーター及びオペレーター(Goeddel,et al.,Nucl.Acids Res.,8,4057,1980);lacプロモーター及びオペレーター(Chang,et al.,Nature,275,615,1978);外膜タンパク質プロモーター(Nakamura,K. and Inouge,M.,EMBO J.,1,771−775,1982);バクテリオファージλプロモーター及びオペレーター(Remaut,E. et al.,Nucl.Acids Res.,11,4677−688,1983);α−アミラーゼ(B.スブチリス)プロモーター及びオペレーター、終結配列並びに選択された宿主細胞と適合性が有るその他の発現増強及び調節配列が含まれる。宿主細胞が酵母である場合、有用な発現調節配列には、例えば、α−接合因子が含まれる。昆虫細胞の場合、バキュロウイルスのポリヘドリン又はp10プロモーターを使用することが可能である(Smith,G.E. et al.,Mol.Cell.Biol.3,2156−65,1983)。宿主細胞が哺乳動物起源である場合、有用な発現調節配列の例には、SV−40プロモーター(Berman,P.W. et al.,Science,222,524−527,1983)又はメタロチオネインプロモーター(Brinster,R.L.,Nature,296,39−42,1982)又は熱ショックプロモーター(Voellmy et al.,Proc.Natl.Acad.Sci.USA,82,4949−53,1985)が含まれる。
【0017】
細菌、酵母、真菌、昆虫及び哺乳動物の細胞発現系は、極めて頻繁に使用される系である。このような系は、本分野において周知であり、一般に、例えば、Invitrogen(www.invitrogen.com)、Novagen(www.merckbiosciences.de)又はClontech Laboratories,Inc.4030 Fabian Way,Palo Alto,California 94303−4607,USAを通じて商業的に入手可能である。これらの発現系に次いで、寄生生物を基礎とした発現系が、極めて魅力的な発現系である。このような系は、例えば、公開番号2714074のフランス特許出願及びUS NTIS公開番号US 08/043109(Hoffman,S. and Rogers,W.:Public.Date 1 December 1993)に記載されている。
【0018】
例えば、ワクチン接種の目的のために、又は抗体を産生させるためにタンパク質が使用される場合には、しかしながら、完全なタンパク質を使用する必要はない。タンパク質に対する免疫応答を誘導することが可能なタンパク質の断片(そのままの状態であるか、又は、例えばKLHなどの担体に連結されている。)、いわゆる、免疫原性断片を使用することも可能である。「免疫原性断片」とは、宿主中で免疫反応を誘導する能力をなお保持する、すなわち、B細胞又はT細胞エピトープを含む完全長タンパク質の断片であると理解される。この時点で、抗原性断片(決定因子)をコードするDNA断片を容易に同定するために、様々な技術が利用可能である。Geysenら(Patent Application WO 84/03564,Patent Application WO 86/06487,US Patent NR.4,833,092,Proc.Natl Acad.Sci.81:3998−4002(1984),J. Imm.Meth.102,259−274(1987))によって記載された方法、いわゆるPEPSCAN法は、タンパク質の免疫学的に重要な領域であるエピトープの検出のための、実行が容易で、迅速な、十分に確立された方法である。本方法は、世界中で使用されており、従って、当業者に周知である。この(経験的な)方法は、B細胞エピトープの検出のために特に適している。また、何れかのタンパク質をコードする遺伝子の配列が与えられれば、現在公知であるエピトープとの配列的及び/又は構造的な一致に基づき、コンピュータアルゴリズムは、免疫学的に重要なエピトープとして、具体的なタンパク質断片を指定することが可能である。これらの領域の決定は、Hopp及びWoods(Proc.Natl.Acad.Sci.78:38248−3828(1981))に従う親水性基準とChou及びFasman(Advances in Enzymology 47:45−148(1987)and US Patent 4,554,101)に従う二次構造要因の組み合わせに基づいている。同様に、Berzofskyの両親媒性基準の補助を得て、コンピュータによって、配列からT細胞エピトープを予測することが可能である(Science 235,1059−1062(1987) and US Patent application NTIS US 07/005,885)。要約された概説は、共通原理に関するShan Lu:Tibtech 9:238−242(1991)、マラリアエピトープに関するGood et al;Science 235:1059−1062(1987)、概説として、Lu;Vaccine 10:3−7(1992),HIV−エピトープに関するBerzowsky;The FASEB Journal 5:2412−2418(1991)に見出される。
【0019】
従って、本発明の第一の実施形態は、マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片と、及び医薬として許容される担体とを含む前記ワクチンに関する。
【0020】
これらの遺伝子の発現産物に基づくこのようなワクチンは、以下に記載されているように、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片を、医薬として許容される担体と混合することによって容易に作製することが可能である。
【0021】
本発明の別の実施形態は、ワクチンにおいて使用するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片に関する。
【0022】
さらに別の実施形態は、マイコプラズマ感染を撲滅するためのワクチンを製造するための、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片の使用に関する。
【0023】
マイコプラズマ感染に対する予防接種のための別の極めて魅力的なアプローチは、医薬として許容される担体と一緒に、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む生組換え担体(LRC)を使用することによるものである。これらのLRCは、さらなる遺伝子情報(この場合には、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片)がクローニングされた微生物又はウイルスである。このようなLRCに感染した動物は、担体の免疫原に対してのみならず、遺伝子コードが、LRC中にさらにクローニングされているタンパク質(例えば、マイコプラズマのL−α−グリセロリン酸オキシダーゼ)の免疫原性部分に対しても免疫原性応答を産生する。細菌のLRCの例として、本分野で公知の弱毒化されたサルモネラ株は、魅力的に使用することが可能である。生組換え担体寄生生物が、とりわけ、Vermeulen,A.N.(Int.Journ.Parasitol.28:1121−1130(1998))によって記載されている。また、LRCウイルスは、核酸を標的細胞中に輸送する手段として使用し得る。生組換え担体ウイルスは、ベクターウイルスとも称される。ベクターとしてしばしば使用されるウイルスは、ワクシニアウイルス(Panicali et al; Proc.Natl.Acad.Sci.USA,79:4927(1982)、ヘルペスウイルス(E.P.A. 0473210A2)及びレトロウイルス(Valerio,D. et al; in Baum,S.J.,Dicke,K.A.,Lotzova,E. and Pluznik,D.H.(Eds.),Experimental Haematology today− 1988. Springer Verlag,New York:pp.92−99(1989))である。
【0024】
本分野で周知の、インビボ相同組換えの技術は、本発明の挿入された核酸の発現を宿主細胞中で誘導することが可能な組換え核酸を、選択した細菌、寄生生物又はウイルスのゲノム中に導入するために使用することが可能である。
【0025】
従って、本発明のさらに別の実施形態は、マイコプラズマのL−α−グリセロリン酸オキシダーゼを又はその免疫原性断片コードする生組換え担体と、及び医薬として許容される担体とを含む、マイコプラズマ感染を撲滅するためのワクチンに関する。
【0026】
機能的に連結されたプロモーターの調節下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む宿主細胞を、本酵素の生産のために使用できることが明らかである。酵素をワクチンとして使用する前に、まず、宿主細胞から酵素を抽出する必要は存在しない。すなわち、宿主細胞は、そのまま使用することも可能である。宿主細胞が、酵素を発現するLRCを含む場合も、同じことが当てはまる。その例は、ウイルス又は細菌のLRCを含む真核細胞である。従って、同じく、本発明の別の実施形態は、機能的に連結されたプロモーターの調節下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む宿主細胞に関する。この形態は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする遺伝子又はその断片を含む生組換え担体を含有する宿主細胞にも関する。宿主細胞は、pBR322として細菌を基礎としたプラスミド又はpGEXとして細菌の発現ベクター又はバクテリオファージと組み合わせた、細菌、例えば、エシェリヒア・コリ、バシラス・スブチリス及びラクトバシラス種起源の細胞であり得る。宿主細胞は、真核細胞起源の宿主細胞、例えば、酵母特異的ベクター分子と組み合わせた酵母細胞、又はベクター若しくは組換えバキュロウイスルと組み合わせた、昆虫細胞のようなより高等な真核細胞(Luckow et al; Bio−technology 6:47−55(1988))、例えば、Ti−プラスミドを基礎としたベクター若しくは植物ウイルスベクター(Barton,K.A. et al; Cell 32:1033(1983))と組み合わせた植物細胞、同じく、適切なベクター若しくは組換えウイルスを有するHela細胞、チャイニーズハムスター卵巣細胞(CHO)又はクランデルネコ腎臓細胞のような哺乳動物細胞でもあり得る。例えば、腸上皮、又は例えば呼吸器上皮に感染するサルモネラ担体又はウイルス担体を基礎とする、酵素又はその免疫原性断片を発現することが可能な上記生組換え担体を基礎とする本発明のワクチンは、マイコプラズマの感染の天然経路をよりよく模倣する点で、サブユニットワクチンを上回る利点を有する。さらに、免疫化のために、組換え担体の少量が必要であるに過ぎないので、それらの自己増殖は有利である。
【0027】
予防接種の効率的な別の方法は、当該抗原をコードするDNAを用いた直接の予防接種である。タンパク質をコードするDNAを用いた直接の予防接種は、多くの異なるタンパク質において成功を収めている。(例えば、Donnelly et al.,The Immunologist 2:20−26(1993)に概説されているとおり。)。予防接種のこの方法も、マイコプラズマ感染に対して哺乳動物を予防接種するために極めて魅力的である。従って、本発明の本実施形態のさらに別の形態は、マイコプラズマ感染を撲滅するためのワクチンであり、機能的に連結されたプロモーターの制御下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子又はL−α−グリセロリン酸オキシダーゼの免疫原性断片をコードする前記遺伝子の一部と、及び医薬として許容される担体とを含む前記ワクチンに関する。
【0028】
DNAワクチンは、例えば、無針注射装置を用いて、皮内適用を通じて容易に投与することが可能である。投与のこの方法は、予防接種すべき動物の細胞中にDNAを直接送達する。1と100μgの間の範囲にあるμgのDNAの量は、極めて優れた結果を与える。
【0029】
上記ワクチンは全て、活性な予防接種に貢献する。すなわち、宿主の免疫系は、これらのタンパク質に対する抗体を作製するために、酵素又はその免疫原性断片によって引き起こされる。あるいは、このような抗体は、以下に記載されているように、例えば、ウサギの中で作製することが可能であり、又は抗体産生細胞株から得ることが可能である。次いで、このような抗体は、宿主動物に投与することが可能である。予防接種のこの方法(受動予防接種)は、動物が既に感染しており、天然の免疫応答を惹起させる時間が存在しない場合に選択される予防接種である。これは、免疫無防備状態の動物を予防接種するための好ましい方法でもある。マイコプラズマに対する投与された抗体は、これらの事例において、細菌に直接結合することが可能である。これは、マイコプラズマの増殖を直ちに減少させ、又は停止させるという利点を有する。従って、本発明の本実施形態の1つの別の形態は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片に対する抗体と、及び医薬として許容される担体とを含むワクチンに関する。
【0030】
本実施形態のさらに別の形態において、本発明のワクチンは、同一の宿主に対して病原性である別の生物又はウイルスに由来する1つ又はそれ以上の抗原、このような抗原に対する抗体又はこのような抗原をコードする遺伝情報をさらに含む。
【0031】
好ましい組み合わせワクチンが、マイコプラズマのL−α−グリセロリン酸オキシダーゼに次いで、マイコプラズマ完全細胞調製物を含むワクチンであることは言うまでもない。このような組み合わせワクチンは、マイコプラズマのL−α−グリセロリン酸オキシダーゼの有害な効果のみならず、他のマイコプラズマ関連タンパク質に対しても保護を誘導する。
【0032】
ブタ病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、仮性狂犬病ウイルス(Pseudorabies virus)、ブタインフルエンザウイルス(Porcine influenza virus)、ブタパルボウイルス(Porcine parvo virus)、伝染性胃腸炎ウイルス(Transmissible gastroenteritis virus)、ロタウイルス(Rotavirus)、別のマイコプラズマ属種、特に、マイコプラズマ・ハイオニューモニエ(Mycoplasma hyopneumoniae)、ブラキスピラ・ハイオディセンテリー(Brachyspira hyodysenteriae)、エシェリヒア・コリ(Escherichia coli)、レプトスピラ属種(Leptospira spp)、エリシペロスリクス・リューシオパシエ(Erysipelothrix rhusiopathiae)、ボルデテラ・ブロンキセプティカ(Bordetella bronchiseptica)、ブラキスピラ・ハイオディセンテリー(Brachyspira hyodysenteriae)、シゲラ種(Shigella sp.)、サルモネラ・コレラエスイス(Salmonella choleraesuis)、サルモネラ・チフィムリウム(Salmonella typhimurium)、サルモネラ・エンテリティディス(Salmonella enteritidis)、ヘモフィラス・パラスイス(Haemophilus parasuis)、ラウソニア(Lauwsonia)、パスチュレラ・マルトシダ(Pasteurella multocida)、ストレプトコッカス・スイス(Streptococcus suis)、アクチノバシラス・プリューロニューモニエ(Actinobacillus pleuropneumoniae)、スタフィロコッカス・ハイカス(Staphylococcus hyicus)及びクロストリジウム・ペルフリンゲンス(Clostridium perfringens)の群から選択される。
【0033】
ウシ病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、ウシヘルペスウイルス(Bovine Herpesvirus)、ウシウイルス性下痢症ウイルス(Bovine Viral Diarrhoea virus)、パラインフルエンザ3型ウイルス(Parainfluenza type 3 virus)、ウシパラミクソウイルス(Bovine Paramyxovirus)、口蹄疫ウイルス(Foot and Mouth Disease virus)、ウシ呼吸器多核体ウイルス(Bovine Respiratory Syncytial Virus)、ブタサーコウイルス(porcine circovirus)、ブタ生殖器呼吸器症候群ウイルス(porcine respiratory reproductive syndrome virus)、別のマイコプラズマ属種、パスツレラ・ヘモリティカ(Pasteurella haemolytica)、スタフィロコッカス・オーレウス(Staphylococcus aureus)、エシェリヒア・コリ(Escherichia coli)、レプトスピラ属種(Leptospira spp.)、スタフィロコッカス・ウベリス(Staphylococcus uberis)、タイレリア・パルバ(Theileria parva)、タイレリア・アニュラータ(Theileria annulata)、バベシア・ボビス(Babesia bovis)、バベシア・ビゲミナ(Babesia bigemina)、バベシア・メジャー(Babesia major)、トリパノソーマ種(Trypanosoma species)、アナプラズマ・マージナーレ(Anaplasma marginale)、アナプラズマ・セントラーレ(Anaplasma centrale)及びネオスポラ・カニナム(Neospora caninum)の群から選択される。
【0034】
家禽病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、鶏痘ウイルス(Fowlpox virus)、伝染性気管支炎ウイルス(Infectious Bronchitis virus)、伝染性ファブリキウス嚢病(グンボロ)(Infectious Bursal Disease (Gumboro))、マレック病ウイルス(Marek’s Disease Virus)、ニワトリ貧血因子(Chicken Anaemia agent)、トリレオウイルス(Avian Reovirus)、トルコ鼻気管炎ウイルス(Turkey Rhinotracheitis virus)、ニワトリポックスウイルス(Chicken Poxvirus)、トリ脳脊髄炎ウイルス(Avian Encephalomyelitis virus)、アヒルペストウイルス(Duck Plague virus)、ニューキャッスル病ウイルス(Newcastle Disease virus)、産卵低下症候群ウイルス(Egg Drop syndrome virus)、伝染性喉頭気管ウイルス(Infectious Laryngotracheitis virus)、シチメンチョウのヘルペスウイルス(Herpes Virus of Turkeys)、別のマイコプラズマ属種、とりわけ、マイコプラズマ・ガリセプチカム(Mycoplasma gallisepticum)又はマイコプラズマ・シノビアエ(Mycoplasma synoviae)、ヘモフィルス・パラガリナルム(コリーザ)(Haemophilus paragallinarum (Coryza))、オルニソバクテリウム・ライノトラキア(Ornithobacterium rhinotracheale)、クロストリジウム・ペルフリンゲンス(Clostridium perfringens)、サルモネラ(Salmonella)、カンピロバクター種(Campylobacter species)、E.コリ(E. coli)及びエイメリア(Eimeria)種の群から選択される。
【0035】
ヒト病原性のマイコプラズマ種に対する保護用のマイコプラズマワクチンが作製される場合には、このような生物及びウイルスは、好ましくは、インフルエンザウイルス(influenza virus)、麻疹ウイルス(measles virus)、ムンプスパラミクソウイルス(mumps paramyxovirus)、クロストリジウム・ジフセリエ(Clostridium diphteriae)、クロストリジウム・テタニ(Clostridium tetani)、ボルデテラ・パーチュシス(Bordetella pertussis)、別のマイコプラズマ属種及びポックスウイルス(pox virus)の群から選択される。
【0036】
好ましくは、上記マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片は、以下のマイコプラズマのいずれか:M.ボビス(M.bovis)、M.sp.ボバイングループ7(M.sp .bovine group 7)、M.ミコイデスsubsp.カプリ(M.mycoides subsp. Capri)、M.ペネトランス(M.penetrans)、M.ガリセプチカム(M.gallisepticum)、M.シノビエ(M.synoviae)、M.モービル(M.mobile)、M.パルモニス(M.pulmonis)、M.ハイオニューモニエ(M.hyopneumoniae)、M.ニューモニエ(M.pneumoniae)、M.ジェニタリウム(M.genitalium)、M.ミコイデスS(mall)C(olony)株Afade(M.mycoides S(mall) C(olony) strain Afade)又はM.ミコイデスS(mall)C(olony)株L2(M.mycoides S(mall) C(olony) strain L2)、特に、M.ボビス、M.ハイオニューモニエ、M.ガリセプチカム又はM.シノビエ、より具体的には、M.ボビス又はM.ハイオニューモニエによってコードされるマイコプラズマのL−α−グリセロリン酸オキシダーゼである。
【0037】
本発明の全てのワクチンは、医薬として許容される担体を含む。医薬として許容される担体は、例えば、無菌水又は無菌生理的塩溶液とすることができる。より複雑な形態において、担体は、例えば緩衝液であり得る。
【0038】
ワクチンを調製するための方法は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片、上記生組換え担体、上記遺伝子若しくはその一部、上記宿主細胞又はマイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片に対する抗体と、及び医薬として許容される担体とを混合することを含む。従って、本発明の別の実施形態は、マイコプラズマ感染を撲滅するためのワクチンの調製方法であり、上記マイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片、上記生組換え担体、上記遺伝子若しくはその一部、上記宿主細胞又は上記マイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片に対する抗体と、及び医薬として許容される担体とを混合することを含む、前記方法に関する。
【0039】
本発明のワクチンは、好ましい提示において、アジュバントも含有し得る。一般に、アジュバントは、非特異的様式で、宿主の免疫応答を強化する物質を含む。様々な多数のアジュバントが、本分野において公知である。アジュバントの例は、フロイントの完全及び不完全アジュバント、ビタミンE、非イオン性ブロックポリマー、ムラミルジペプチド、QuillA(R)、鉱物油、例えばBayol(R)又はMarkol(R)、植物油及びCarbopol(R)(ホモポリマー)又はDiluvac(R)Forteである。ワクチンは、いわゆる「ビヒクル」も含み得る。ビヒクルとは、それに対する共有結合なしに、ポリペプチドが付着する化合物である。しばしば使用されるビヒクル化合物は、例えば、水酸化アルミニウム、リン酸アルミニウム又は酸化アルミニウム、シリカ、カオリン及びベントナイトである。
【0040】
抗原がビヒクル中に部分的に埋め込まれている、このようなビヒクルの特殊な形態は、いわゆるISCOMである(EP 109.942,EP 180.564,EP 242.380)。さらに、ワクチンは、1つ又はそれ以上の適切な界面活性化合物又は乳化剤、例えばSpan又はTweenを含み得る。しばしば、例えば、分解し易いポリペプチドを分解から保護して、ワクチンの保存寿命を延長させるために、又は凍結乾燥効率を改善するために、ワクチンは安定化剤と混合される。有用な安定化剤は、とりわけ、SPGA(Bovarnik et al; J. Bacteriology 59:509(1950)、炭水化物、例えば、ソルビトール、マニトール、トレハロース、デンプン、スクロース、デキストラン又はグルコース、アルブミン又はカゼイン又はこれらの分解産物などのタンパク質及びアルカリ金属リン酸塩などの緩衝液である。さらに、ワクチンは、生理的に許容される希釈剤中に懸濁され得る。アジュバント化し、ビヒクル化合物又は希釈剤を添加し、ポリペプチドを乳化し、又は安定化させる別の方法も、本発明において具体化されることは言うまでもない。
【0041】
本発明のワクチンは、タンパク質の1及び100μgの間の量で、極めて適切に投与することが可能であるが、より少量を使用することが原理的に可能である。免疫学的には極めて適切であるが、100μgを超える用量は、商業的な理由で、より魅力的でない。
【0042】
上記LRCウイルス及び細菌などの、弱毒化された生組換え担体に基づくワクチンは、感染中に自身を増幅するので、ずっと少ない用量で投与することが可能である。従って、極めて適切な量は、それぞれ、細菌及びウイルスに対して、103と109CFU/PFUの間の範囲である。
【0043】
多くの投与方法を使用することが可能である。経口適用は、労力を要しないので、投与の極めて魅力的な方法である。経口投与の好ましい方法は、胃の高度に酸性環境を通過した後にのみ崩壊するカプセル中にワクチンを包装することであり、これは、本分野において公知であり、頻繁に使用されている。また、ワクチンは、胃のpHを一時的に増大させるために、本分野で公知の化合物と混合することが可能である。例えば、ワクチンの筋肉内適用による、全身適用も適切である。この経路が採用される場合には、全身適用のために本分野で公知の標準的な手法が非常に適している。
【0044】
マイコプラズマのL−α−グリセロリン酸オキシダーゼに基づくワクチンも、マーカーワクチンとして極めて適切である。マーカーワクチンは、例えば、野生型感染によって誘導される抗体パネルとは異なる、特徴的な抗体パネルに基づいて、予防接種された動物と野外感染した動物との識別を可能とするワクチンである。精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼに基づくワクチンは、本タンパク質に対する抗体を誘導するのみであるが、生きた野生型、生きた弱毒化又は不活化された完全なマイコプラズマに基づくワクチン及び野外感染は、細菌のタンパク質の全て又はほとんどに対して抗体を誘導する。これは、明らかに、極めて異なる抗体パネルを与える。精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼを含むウェル及び別のマイコプラズマのタンパク質を含むウェルを有する単純なELISA検査は、動物から得た血清を試験し、動物が、本発明のワクチンで予防接種されたか、又はマイコプラズマ野外感染に罹患しているかどうかを識別するのに十分である。精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼを含むワクチンを予防接種された動物は、マイコプラズマのL−α−グリセロリン酸オキシダーゼ以外の他のマイコプラズマのタンパク質に対する抗体を有しない。しかしながら、マイコプラズマでの野外感染に遭遇した動物は、全ての免疫原性マイコプラズマタンパク質に対する抗体を有しており、従って、他の非マイコプラズマのL−α−グリセロリン酸オキシダーゼタンパク質に対する抗体も有する。
【0045】
従って、本発明の別の実施形態は、一方で本発明のワクチンを用いた予防接種と、他方で完全細胞ワクチンを用いた予防接種又は野外感染とを識別するための診断検査に関し、このような検査は、精製されたマイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片を含み、別個に、別の非L−α−グリセロリン酸オキシダーゼタンパク質を含む。
【0046】
上に特定されているように発現された本発明のポリペプチド又はその免疫原性断片は、ポリクローナル、単一特異的又はモノクローナル(又はその誘導体)であり得る抗体を産生するために使用することが可能である。ポリクローナル抗体が望まれる場合には、ポリクローナル血清を作製及び加工するための技術が本分野において周知である(例えば、Mayer and Walter,eds.Immunochemical Methods in Cell and Molecular Biology,Academic Press,London,1987)。本発明のポリペプチド(又はそのバリアント若しくは断片)に対して反応性の本発明のモノクローナル抗体は、本分野でも公知の技術によって、近交系マウスを免疫することによって調製することが可能である(Kohler and Milstein,Nature,256,495−97,1975)。
【0047】
本発明の抗体の大規模な作製方法も、本分野において公知である。このような方法は、ファージディスプレイのために、繊維状ファージ中に、本発明のタンパク質をコードする遺伝情報(の断片)をクローニングすることに基づいている。このような技術は、とりわけ、http://aximt1.imt.uni−marburg.de/〜rek/aepphage.htmlの「filamentous phage display」の「Antibody Engineering Page」並びにCortese,R. et al.,(1994) in Trends Biotechn.12:262−267.,Clackson,T. & Wells,J.A.(1994) in Trends Biotechn.12:173−183,Marks,J.D. et al.,(1992) in J. Biol.Chem.267:16007−16010,Winter,G. et al.,(1994) in Annu.Rev.Immunol.12:433−455及びLittle,M.et al.,(1994) Biotechn.Adv.12:539−555による概説論文に記載されている。続いて、camelid重鎖抗体を発現するcamelid発現ライブラリーをスクリーニングするために、前記ファージを使用する。(Muyldermans,S. and Lauwereys,M.,Journ. Molec. Recogn. 12:131−140(1999) and Ghahroudi,M.A. et al.,FEBS Letters 414:512−526(1997)).所望の抗体を発現するライブラリーから得られた細胞は複製され、続いて、抗体の大規模発現のために使用することが可能である。
【0048】
実施例
【実施例1】
【0049】
株、細胞、増殖条件及びDNA抽出。
【0050】
別段の記載がなければ、M.ミコイデスsubsp.ミコイデスSC株Afade(Farcha Laboratory,N’Djamena,Chadで、1968年に単離された高度に病原性の野外株)を、病原性研究のために使用した。この株は、天然及び実験条件下で、CBPPを引き起こす。別段の記載がある場合には、活性なグリセロール取り込み系GtsABCを欠く、より病原性が低い株であるM.ミコイデスsubsp.ミコイデスSC株L2を使用した。さらに、標準株PGI並びにアフリカ及び欧州の発生に由来する10個の他の株も、遺伝子分析のために、本研究で使用した。マイコプラズマの培養物は、108から109コロニー形成単位/mL(cfu/mL)の密度になるようにマイコプラズマブロス培地中で、又は固体マイコプラズマ寒天培地(Axcell Biotechnologies,St. Genis l’Argentiere,France)上で増殖させた。生きたM.ミコイデスsubsp.ミコイデスSCの増殖及び取り扱いは、BL3封じ込め安全標準を満たす生物学的安全性研究室の中で行った。DNA抽出は、以前に記載したとおり行った(Cheng,X.,J. et al.,1995,Microbiology 141:3221−3228)。遺伝子操作及びサブクローニングについては、エシェリヒア・コリ株DH5α[F−φ80dlacZデルタM15デルタ(lacZYA−argF)U169deoRrecA1endA1hsdR17(rK−、mK+)phoAsupE44λ−thi−1gyrA96relA1]及びBL21(DE3)[F’dcm ompThsdS(rB−mB−)galλ(DE3)]を使用した。組換えポリヒスチジンN及びC末端融合タンパク質の発現のために、pETHIS−1発現ベクター(Schaller,A.et al.,1999,Microbiology145:2105−2116)を使用した。
【0051】
ECaNEp細胞は、地元の食肉処理場から得られた胎児から調製し、24ウェルマイクロタイタープレート中の、7%ウシ胎児血清(FCS)及びペニシリン(100IU/mL)が補充されたEME−Earle培地中に維持し、加湿された5%CO2雰囲気中、37℃で、2×105細胞/ウェルの集密度で使用した。FCS及び細胞培地は、Seromed(Biochrom,Munich,Germany)から購入した。細胞は、PCRを使用してマイコプラズマによる汚染について、又は免疫染色を用いてBVDウイルスによる汚染について、定型的にスクリーニングした。
【0052】
PCR増幅、サザンブロット及びDNA配列分析。
【0053】
glpO特異的DNAプローブは、標準的なPCR条件を使用し(Cheng,X.,J. et al.,1995,Microbiology 141:3221−3228)、オリゴヌクレオチドプライマーglpO EcoRI N
【0054】
【化1】
及びglpO NotI C
【0055】
【化2】
を用いて、ジゴキシゲニン−11−dUTP(Dig)(Roche Diagnostics,Rotkreuz,Switzerland)の存在下でPCRによって構築した。マイコプラズマから得たゲノムDNAの抽出及びサザンブロット分析は、以前に記載されたとおりに行った(Pilo,P.et al.,2003,Vet.Microbiol.92:37−48)。
【0056】
DNA配列決定は、DNA Sequenator AB3100遺伝子分析装置及びTaq色素デオキシターミネータサイクル配列決定キット(Applied Biosystems,Norwalk,Conn.)を用い、オリゴヌクレオチドプライマーglpO EcoRI N及びglpO NotI Cを用いて、並びにglpO内部プライマーを用いるプライマーウォーキングによって行った。DNA及び推測されたアミノ酸配列は、PC/GeneプログラムPROSITE(Bairoch,A.,P. Bucher,and K. Hofmann. 1995,Nucleic Acids Res.24:189−196)を用いて分析した。GenBank及びEMBLデータベース中の配列との配列比較は、BLAST(Altschul,S.F. et al.,1997,Nucleic Acids Res.25:3389−3402)を用いて行った。タンパク質の分析は、プログラムMotif Scan(http://hits.isb−sib.ch/cgi−bin/hits motifscan)及びTMpred(http://www.ch.embnet.org/software/TMPRED form.html)及び「toppred」ソフトウェア(von Heijne,G. 1992,J. MoI. Biol. 225:487−94)を用いることによって実施した。
【実施例2】
【0057】
組換えGlpOのクローニング、部位指定突然変異誘発及び発現。
【0058】
それぞれ、EcoRI及びNotIに対する制限部位を含有するプライマーglpO EcoRI N及びd glpO NotI Cを用いて、まず、M.ミコイデスsubsp.ミコイデスSC株AfadeのglpO遺伝子を増幅した。さらに、プライマーglpO NotI Cは、変異されたTGGTrpコドンを含んだ。(適切なヌクレオチド置換を有する)プライマー対glpO mut1L
【0059】
【化3】
を用いるオーバーラップ伸長PCR法(Braman,J.,C. Papworth,and A. Greener. 1996,Methods Mol.Biol.57:31−44)を用いて、glpO遺伝子中の他の2つのマイコプラズマ特異的TGATrpコドンを、TGGTrpコドンと置換した。EcoRI及びNotI切断部位を隣接させることによって、PCR産物をpETHIS−1中にクローニングした。他のタンパク質に対して以前に記載されたように、発現及びポリヒスチジン尾部が付着された融合タンパク質のNi2+キレートクロマトグラフィーを用いた精製のために、DNA配列決定によって構築物を分析し、E.コリBL21(DE3)中に導入した(Schaller,A.,et al.,1999,Microbiology 145:2105−2116)。
【実施例3】
【0060】
血清、ポリクローナル抗体、イムノグロブリン精製及びFab調製。
【0061】
アフリカンM.ミコイデスsubsp.ミコイデスSC株Afadeを用いた、管理された実験的感染から得たウシ血清は、Abdo及び共同研究者によって詳しく記載されている(Abdo,E.−M.et al.,1998,Vet.Microbiol.59:109−122)。組換えGlpOに対して誘導されたポリクローナル単一特異的血清は、アジュバント10(Gerbu Biotechnik GmbH,Gaiberg,Germany)500μLと混合された、PBS緩衝液pH8.0(50mMNa2HPO4/NaH2PO4 pH8.0,140mMNaCl)500μL中の精製された、組換えポリヒスチジン尾部が付加されたタンパク質GlpO160μgでウサギを皮下免疫化した後、2及び4週目に、タンパク質40及び20μgで強化免疫することによって取得した。最後の強化免疫から10日後に、ウサギから採血した。血液試料から抗血清を調製し、−20℃で保存した。
【0062】
製造業者によって指示されたとおり、HiTrap Protein Gキット(Amersham Pharmacia Biotech,Uppsala,Sweden)を用いて、ウサギ抗GlpO血清及び同じウサギの前血清から得られた免疫グロブリンG(IgG)画分を精製した。製造業者の指示書に従い、ImmunoPure Fab Preparationキット(Pierce,Rockford,Ill.)を用いて、Fab断片の調製を行った。PBS緩衝液pH7.4に対して、Fab断片を一晩透析し、次いで、フィルター滅菌した。Bradfordの方法(Bradford,M.M.1976,Anal.Biochem.72:248−254.)を用いて、タンパク質濃度を測定した。リポタンパク質LppCに対して誘導された単一特異的ウサギ血清は、以前に記載されている(Pilo P. et al.,2003,Vet.Res.34:761−775.)。
【実施例4】
【0063】
イムノブロット分析、TritonX−114分配及び増殖阻害試験
マイコプラズマから得た総抗原は、以前に記載したとおり調製した(Fleury,B. et al,2001,J. Clin.Microbiol.39:2814−2822)。1:2000希釈のウシ血清及び1:1000希釈のウサギ単一特異的血清抗GlpOを用いて、イムノブロッティングを行った。
【0064】
TritonX−114分配法(Bordier,C. 1981,J.Biol.Chem.256:1604−1607)により、固定相培養から得られたM.ミコイデスsubsp.ミコイデスSC総抗原を疎水性及び親水性画分へ分割した。TritonX−114界面活性剤相及び水相から得た試料を、GlpOに対して誘導された単一特異的ポリクローナル抗体及びM.ミコイデスsubsp.ミコイデスSCに対して誘導されたウシ血清を用いたイムノブロッティングによって分析した。
【0065】
希釈されていない補体除去された(decomplemented)単一特異的ウサギ抗GlpO血清、精製されたIgG又は抗GlpOIgGのFab断片5及び10μLを、マイコプラズマ含有寒天培地上に滴加し、37℃で4日間プレートを温置することによって、増殖阻害試験を行った。M.ミコイデスsubsp.ミコイデスSCに対する血清を陽性対照として使用し、LppQに対する単一特異的ウサギ血清を陰性対照として使用した。以前に記載されているように、光学顕微鏡下で増殖阻害の観察を実施した(Papazisi,L. et al,2003,Microbiology 149:2307−2316,Poveda,J. B.,and R. Nicholas,1998,Methods Mol.Biol.104:105−111)。
【実施例5】
【0066】
走査型電子顕微鏡(SEM)及び免疫金標識
電子顕微鏡のために、予めポリ−L−リジンで被覆された、金又は白金がスパッタリングされたカバーガラス上のマイコプラズマブロス培地中で、M.ミコイデスsubsp.ミコイデスSCを37℃で5日間培養した。37℃のPBS緩衝液pH7.4で、細胞を3回洗浄し、室温で30分間、PBS中の4%パラホルムアルデヒド中に固定した。PBSで洗浄した後、0.2Mグリシン及び1%BSAが補充されたPBS緩衝液中、室温で15分間、カバーガラスをブロッキングし、その後、4℃で一晩、1%BSAが補充されたPBS中、1:100又は1:50希釈されたウサギ血清抗GlpOから得られたIgGとともにカバーガラスを温置した。次いで、PBSで10分間、試料を洗浄し、室温で90分間、PBS中に1:50希釈された、15nmのコロイド状金が抱合されたヤギ抗ウサギIgG(British Biocell International,Cardiff,UK)で標識した。0.1Mカコジル酸塩緩衝液pH7.4でカバーガラスを洗浄し、標準的なプロトコールに従って、SEMのために加工した。要約すれば、0.13Mカコジル酸塩緩衝液pH7.4中の0.11%ルテニウムレッドを加えた1.33%OsO4中で15分間、試料をオスミウム酸染色し、0.1Mカコジル酸塩緩衝液で洗浄し、増加するエタノール系列を通じて脱水し、ヘキサメチルジシラザンの蒸発によって乾燥させた(Sigma,Buchs,Switzerland)。
【0067】
5kVの加速電圧、8mmの作動距離及び50,000から100,000倍までの拡大で、二次電子シグナル及び対応する後方散乱された電子シグナルを、高解像度電解放出走査型電子顕微鏡DSM982Gemini(Zeiss,Oberkochen,Germany)中で調べた。
【0068】
対照実験は、一次抗体の省略、並びに、それぞれウサギ抗カルシトニン抗体の使用(Anawa Biomedical Services and Products,Zurich Switzerland)及びウサギ前免疫血清の使用を含んだ。
【実施例6】
【0069】
H2O2産生の定量及び阻害アッセイ
H2O2産生を測定するために、約5×108cfu/mLの密度になるまで、37℃で3日間、マイコプラズマ培養培地中でM.ミコイデスsubsp.ミコイデスSCの株を増殖させた。4℃で10分間、8000×gで培養物を遠心し、温置培地(67.6mM HEPES pH7.3,140mM NaCl,7mM MgCl2)中で1回洗浄し、109cfu/mLの密度で、予め加温された37℃の温置培地中に再懸濁し、1mLの分取試料中に分けて、37℃で1時間温置した。H2O2産生を誘導するために、100μMの最終濃度(ウシ血清中での生理的濃度)で、マイコプラズマ懸濁液にグリセロールを添加した。グリセロールの添加から0、1、2、5、10、20及び120分後に、以前に記載されたように(Vilei,E.M.,and J. Frey,2001,Clin.Diagn.Lab.Immunol.8:85−92)、過酸化物試験(Merck KgaA,Darmstadt,Germany)を用いて、H2O2の産生を測定した。マイコプラズマのL−α−グリセロリン酸オキシダーゼGlpOを阻害するために、0.26μg/mLから2.6μg/mLまでの濃度での温置に続いて、PBS緩衝液で2回洗浄することによって、GlpOに対して誘導されたIgGの精製されたFab断片でマイコプラズマを前処理した。H2O2産生の誘導後に、M.ミコイデスsubsp.ミコイデスSC細胞の生存性を評価するために、アッセイの終了時に、反応アッセイの分取試料をマイコプラズマ寒天プレート上に播種し、37℃で増殖させた。
【実施例7】
【0070】
細胞毒性活性の評価
周密状態に達するまで、胚性ウシ鼻上皮細胞(ECaNEp細胞)(Schweizer,M.,and E. Peterhans,1999,J. Gen. Virol. 80:1147−1155)を、24ウェルプレート中で増殖させた。アッセイの前に、培地を除去し、補充物を含まないMEM−Earle培地200μLによって、又は100Mグリセロールを補充したMEM−Earle培地によって置き換えた。次いで、50マイコプラズマ/細胞の感染効率(MOI)で、ECaNEp細胞を感染させた。GlpO活性を遮断するために、0.26μg/mLの抗GlpOFab断片でM.ミコイデスsubsp.ミコイデスSCを前処理した。0.75%クリスタルバイオレット、0.25%NaCl、1.75%ホルムアルデヒド及び50%エタノールでの固定及び染色後に、生きたECaNEp細胞を計数し、感染後の様々な時点で、位相差顕微鏡下で写真を撮影した。LppCはグリセロール代謝と相関しないので、M.ミコイデスsubsp.ミコイデスSCの膜リポタンパク質LppCに対して誘導されたポリクローナルIgGから得られる精製されたFab断片を対照として使用した。ECaNEp細胞との接触前に、M.ミコイデスsubsp.ミコイデスSCの増殖中に産生されたH2O2が、ウシ細胞に対して有害であるかどうかを測定するために、0.22μmフィルター(Millipore,Bedford,Mass.)を通して、グリセロールの存在下で増殖されたM.ミコイデスsubsp.ミコイデスSCの培養物の上清をろ過し、ECaNEp細胞に添加した。ECaNEp細胞の生存性は、トリパンブルー排除によって評価した。
【実施例8】
【0071】
ECaNEp細胞中のH2O2及びその他のROSによって引き起こされた酸化的ストレスの検出
ECaNEp細胞中の細胞内ROSによる酸化的ストレスを評価するために、5(及び6)−クロロメチルー2’,7’−ジクロロジヒドロフルオレセインジアセタート−アセチルエステル(CM−H2DCFDA; Molecular Probes,Eugene,Oreg.)の酸化を使用した。この色素は細胞の中に入り、ROSによる細胞内酸化後に蛍光シグナルを産生する。以下の修飾を用いて、細胞毒性活性の評価に使用したものと同じ条件を用いた。対照として、H2O2及びROSによる酸化を防ぐために、ECaNEp細胞を含有するウェルの半分を、30mMN−アセチル−L−システイン(NAC)で処理した。次いで、10μMCM−H2DCFDAとともに、ECaNEp細胞を1時間温置し、MEM−Earle培地で1回洗浄した。次いで、グリセロールの存在下又は不存在下において、500マイコプラズマ/細胞のMOIで細胞を感染させた。GlpO活性を遮断するために、0.26μg/mLの抗GlpOFab断片でM.ミコイデスsubsp.ミコイデスSCを前処理した。対照として、150μMから4.4mMにわたるH2O2溶液で、細胞を20分間処理した。Nikon Eclipse TE300顕微鏡を用いた蛍光顕微鏡法によって、マイコプラズマでの感染から20分後に、細胞内H2O2及びROSをモニターした。CM−H2DCFDAを含む全ての工程(この化学物質の取り扱いを含む。)が、暗所で行われたことに注目されたい。
【0072】
ヌクレオチド配列受託番号。
【0073】
M.ミコイデスsubsp.ミコイデスSC株Afade及びL2から得られたglpOのヌクレオチド配列に対するEMBL/GenBank受託番号は、それぞれ、AJ581566及びAJ581564である。マイコプラズマsp.ボバイングループ7株PG50及びM.ミコイデスsubsp.カプリ株PG3から得られたglpO遺伝子の配列は、それぞれ、AJ581565及びAJ581567の受託番号で寄託された。
【実施例9】
【0074】
glpO遺伝子の遺伝的及び機能的分析
M.ミコイデスsubsp.ミコイデスSCから得られたglpO遺伝子は、378アミノ酸ポリペプチドのGlpOをコードしており、42.7kDaの予想分子量及び8.14のpIを有する。本遺伝子は、3つのTGATrpコドンを含有する。本タンパク質は、アミノ酸位置8から36に推定FAD結合部位を有し、表面露出されているように見受けられる。TMpredは、アミノ酸6から25及び140から157にわたる2つの顕著な膜貫通領域を同定した。高度に病原性の株Afade中において、glpO遺伝子の後には、遺伝子glpK(推定グリセロールリン酸キナーゼ)及びglpF(推定グリセロール促進物質因子)が続く。標準株PG1についても、同じ遺伝子配置が観察される。glpOに対する遺伝子プローブを用いたゲノムDNAのサザンブロット分析は、M.ミコイデスsubsp.ミコイデスSC株Afade、L2、標準株PGI並びに分析されたアフリカ及び欧州での発症から得られた10個の他の株中に、glpOが存在することを示した(データは図示せず。)。単一特異的抗GlpOIgGを用いた総抗原のイムノブロット分析は、別個の45kDaタンパク質バンドを明らかにし、検査されたM.ミコイデスsubsp.ミコイデスSCの全ての株中に、glpO遺伝子の発現を確認した(データは図示せず。)。M.ミコイデスsubsp.ミコイデスSC株Afade及び株L2から得られたGlpOのアミノ酸配列は、標準株PGI(48)のGlpOのものと同一であることが明らかとなり、様々なマイコプラズマ種のグリセロール−3−リン酸脱水素酵素と類似性を示した(表1)。
【0075】
ゲノム配列分析は、マイコプラズマがカタラーゼ及びジスムターゼ活性を欠如することを予測する。細胞質のGlpO活性は、細胞内H2O2毒性を誘導し、次いで、細胞内H2O2毒性がマイコプラズマ自体に対して有害であるので、本発明者らは、本酵素が表面膜に存在しているに違いないという仮説を立てた。「toppred」ソフトウェアを用いた、(表1に列記されている)様々な病原性マイコプラズマ種由来のGlpO又は相同的酵素の配列分析によって膜貫通構造が明らかとなり、これらの酵素のドメインが表面に位置すると予想された。M.ミコイデスsubsp.ミコイデスSCの細胞表面にGlpOが存在することの直接的な証拠は、単一特異的ウサギ抗GlpO血清から得られるIgGを用いた免疫金標識後の走査型電子顕微鏡(SEM)によって得られた(図1a−d)。マイコプラズマの膜中にGlpOが存在することを確認するために、M.ミコイデスsubsp.ミコイデスSCの総タンパク質を、TritonX−114相分配に供した。抗GlpO血清は、TritonX−114相中の45kDaタンパク質(GlpO)(これは、水相には存在しない。)と強く反応し、GlpOがM.ミコイデスsubsp.ミコイデスSCの内在性膜タンパク質であることを示している(図1e)。この株で実験的に感染されたウシから得たウシ血清を用いた、M.ミコイデスsubsp.ミコイデスSCのTritonX−114抽出物のイムノブロット分析は、GlpOとともにゲル上を同時に移動するGlpO特異的バンドを45kDaの場所に明らかにし(データは示さず。)、ウシのCBPP感染中に、GlpOが血清転換に関与することを示している。
【0076】
マイコプラズマのL−α−グリセロリン酸オキシダーゼとしてのGlpOの機能を確認するために、本発明者らは、単一特異的抗GlpOIgGのFab断片の存在下又は不存在下で、M.ミコイデスsubsp.ミコイデスSCの純粋培養物にグリセロールを添加した後、H2O2の産生を測定した。図1fに示されているように、中指数関数期の増殖相にある培養物に100μMグリセロールを添加することによって、直ちに、増殖培地中へH2O2が放出されて、10分後に150μMに達し、この濃度は、最長2時間保持された(図1f)。0.26μg/mLの最小濃度の、単一特異的ポリクローナル抗GlpO抗体又は抗GlpOIgGのFab断片で、マイコプラズマ懸濁液を前処理すると、H2O2の放出を特異的に遮断することができたが、対照抗体として使用された抗LppCIgGのFab断片によっては遮断できなかった(図1f)。LppCは、グリセロール代謝とは関係していない、M.ミコイデスsubsp.ミコイデスSCの表面リポタンパク質である。グリセロールの補充後に、M.ミコイデスsubsp.ミコイデスSCの培養物にカタラーゼを添加することによって、1μMの濃度(これは、本アッセイの検出レベルである。)を下回るまで、培地中のH2O2レベルが低下した。抗GlpO抗体又は抗GlpOFab断片の添加なし又は添加ありで、M.ミコイデスsubsp.ミコイデスSCの世代時間は3.3時間であった。さらに、抗GlpO抗体は、血清滴下増殖阻害試験において効果を有しなかった(データは示さず。)。100μMグリセロールの存在下で、2時間増殖され、150μMH2O2を産生した純粋培養物は、3.2時間の世代時間を示したのに対して、グリセロールが添加されなかった対照培養物は、3.3時間の世代時間を有した。このため、マイコプラズマの生存性は、グリセロールによって誘導されたH2O2の産生によって影響を受けなかった。
【実施例10】
【0077】
ウシ上皮細胞に対するM.ミコイデスsubsp.ミコイデスSCの細胞毒性
グリセロール代謝から生じるH2O2及び関連のROSの産生が、M.ミコイデスsubsp.ミコイデスSCの病原性に寄与しているかどうかを評価するために、本発明者らは、様々な条件下で、胚性仔ウシ鼻上皮(ECaNEp)細胞に対する細胞毒性を分析した(図2)。図2bに示されているように、50マイコプラズマ/細胞の感染効率でのM.ミコイデスsubsp.ミコイデスSC株AfadeによるECaNEp細胞の感染は、培地中のグリセロールの不存在下で、感染から1時間後に弱い細胞毒性効果をもたらすに過ぎなかった。しかしながら、グリセロールの生理的濃度の存在下で、マイコプラズマでECaNEp細胞の感染を行った場合には、多くのECaNEp細胞が表面から剥離され、続いて、完全な溶解を起こした(図2c)。H2O2産生を付随的に遮断する0.26μg/mLの濃度の抗GlpOFab断片でマイコプラズマを前処理すると、この細胞毒性効果を遮断することが可能であった。これらの条件下で、ECaNEp細胞は、感染から1時間後に形態的変化を示さなかった(図2d)。グリセロールを含有する培地中で増殖されたM.ミコイデスsubsp.ミコイデスSCの細胞毒性は、対照として使用された抗LppCFab断片によって阻害されなかった(図2e)。活性なグリセロール輸送体GtsABCに対する遺伝子を欠如し、H2O2のごく少量を産生するより病原性が低い株(L2)を用いると、グリセロールの存在下でECaNEp細胞を感染させたときに、弱い細胞毒性効果が観察されるのみであった(図2f)。対照実験において、グリセロール、抗GlpOFab若しくは抗体LppCFab断片のみ又は組み合わせを添加しても、ECaNEp細胞に影響を及ぼさなかった(データは図示せず。)。細胞毒性実験の速度論(図2g)は、M.ミコイデスsubsp.ミコイデスSCで感染されたECaNEp細胞へのグリセロールの添加後に、細胞毒性の誘導が急速に起こり、2時間後には、75%の死亡率に達することを示している。より高い感染効率(500マイコプラズマ/細胞)では、グリセロールの添加から30分後に、細胞の完全な死亡率に達した。抗GlpOFabでのマイコプラズマの前処理は、観察期間全体にわたって、細胞の死亡を完全に阻害した(図2g)。添加されたグリセロールの不存在下で、M.ミコイデスsubsp.ミコイデスSCでのECaNEp細胞の感染は、弱い細胞毒性効果を示すに過ぎず(図2g)、これも抗GlpOFabによって阻害することが可能であり、従って、細胞又は培地中へのグリセロールの残留量によるものであり得る。興味深いことに、約150μMH2O2を含有するグリセロールの存在下で増殖されたM.ミコイデスsubsp.ミコイデスSC培養物のろ過された上清又は細胞培養物への150μMH2O2の添加は、1時間の曝露後に、ECaNEp細胞に対して目に見える細胞毒性効果を有しなかった。外来H2O2のECaNEp細胞培養物への添加によって誘導された細胞毒性は、まず、4.4mMの濃度で達成され、細胞の多くが、曝露の1時間後に死滅した。この濃度は、グリセロールの存在下での、M.ミコイデスsubsp.ミコイデスSCの増殖培地中で測定された濃度より30倍高い。
【0078】
M.ミコイデスsubsp.ミコイデスSCでの感染又はグリセロールの添加後に、細胞内H2O2及び他のROSによって引き起こされたECaNEp細胞中で酸化的ストレスをモニターするために、本発明者らは、CM−H2DCFDAでECaNEp細胞を前処理し、蛍光顕微鏡によって、この化合物の細胞内酸化を検出した(図3)。CM−H2DCFDAの酸化によるエステル基の切断が、高度に蛍光性のジクロロフルオレセイン(DCF)誘導体の形成を細胞中にもたらすが、酸化されていないCM−H2DCFDAは非蛍光である。図3cからd中に示されているように、M.ミコイデスsubsp.ミコイデスSCでのECaNEp細胞の感染は、グリセロールの添加から20分後に、ECaNEp細胞中に蛍光の強い誘導をもたらし、細胞内H2O2又はROSの存在を反映している。これに対して、グリセロールの添加なしの感染されたECaNEp細胞中には、蛍光は検出されなかった(図3a−b)。ECaNEp細胞の感染前に、抗GlpO抗体でマイコプラズマを処理すると、CM−H2DCFDAの細胞内酸化は起こらなかった(図3e−f)。さらに、CM−H2DCFDAの細胞内酸化は、細胞培地にグリセロールを添加する前に、抗酸化剤NACでECaNEp細胞を処理することによって遮断された(図3g−h)。グリセロールの添加から10分後に、M.ミコイデスsubsp.ミコイデスSCによって放出された濃度に対応する濃度である150μMの外来H2O2を添加すると、ECaNEp細胞の細胞内酸化は起こらなかった。細胞毒性の誘導の場合と同様、ECaNEp細胞中での細胞内CM−H2DCFDAの酸化を達成するために、4.4mM外来H2O2の濃度が必要であった(図3i−j)。
【0079】
結論
本発明において、M.ミコイデスsubsp.ミコイデスSCのGlpOは、ECaNEp細胞に対する細胞毒性において中心的な役割を果たす膜タンパク質として同定された。グリセロールの生理的濃度の存在下で、109cfu/mlの密度のM.ミコイデスsubsp.ミコイデスSCは、最大150μMまで、培地中にH2O2の比較的大量を放出する。H2O2のこの量は、グルコースの存在下で増殖されたマイコプラズマ(ちょうど50%、H2O2産生を刺激すると報告された。)によって産生される量と比べると、約20倍である。グリセロールの生理的濃度の存在下で、ECaNEp細胞をM.ミコイデスsubsp.ミコイデスSCに曝露すると、H2O2が、まず、それらのサイトゾル中に検出され、続いて、細胞死が起こった。本発明者らは、マイコプラズマによって培地中へ放出されたH2O2濃度と、CM−H2DCFCA酸化及び細胞死を引き起こすために必要とされる濃度との間に、顕著な齟齬を認めた。従って、150μMH2O2を含有するろ過されたマイコプラズマの増殖培地又はECaNEp細胞への150μM外来H2O2の添加は、サイトゾル中での検出可能なCM−H2DCFDA酸化をもたらさず、1時間以内の細胞死ももたらさなかった。このことは、毒性化合物を宿主細胞に首尾よく誘導するためには、マイコプラズマと宿主細胞間の密接な接触が必要であることを示している。この文脈において、M.ミコイデスsubsp.ミコイデスSC、及び、さらには、多くの他の病原性マイコプラズマは、それらの宿主細胞に強固に付着するが、貫通はしないことに注目すべきである。さらに、ECaNEp細胞に対して、グリセロールによって誘導される細胞毒性効果を得るためには、グリセロールの活発な取り込みが必要である。グリセロールの活発な取り込みに必要とされるGtsABC輸送体を欠如し、著しくより低い速度で、1/10未満のH2O2を産生するM.ミコイデスsubsp.ミコイデスSCの欧州株L2は、同一の実験条件下で、ECaNEp細胞に対して、弱い細胞毒性効果を有するに過ぎなかった。株L2によるグリセロールの残りの取り込みは、グリセロール促進物質因子と推定されるGlpFによって媒介される、より低効率の輸送体経路を用いて行われると考えられる。
【0080】
我々の結果に基づいて、本発明者らは、M.ミコイデスsubsp.ミコイデスSCによる真核細胞への細胞損傷を引き起こすための以下のモデルを提案する(図4)。間質液中に存在するグリセロールは、高活性のABCグリセロール輸送体(GtsA、GtsB及びGtsC)を介して活発に取り込まれ、続いて、グリセロール−3−リン酸へリン酸化される。次いで、これは、GlpOによって、O2の存在下でDHAPへと酸化され、これは解糖系に入り、H2O2の一分子を産生する。マイコプラズマの宿主細胞膜との緊密な接触によって促進されて、H2O2及び付随するROSは宿主細胞に入る。宿主細胞内では、H2O2及びROSは、細胞傷害の強力な媒介物質及び炎症性プロセスの誘導物質として作用する。これらは、組織細胞を直接障害し、又は宿主細胞遺伝子発現(例えば、NF−κBの活性化を介して、又はFenton反応を介して、炎症促進性遺伝子)を誘導することによって、宿主を損傷すると予想される(Crichton,R. R. et al.2002,J. Inorg.Biochem.91:9−18)。興味深いことに、マイコプラズマは、貪食細胞中で呼吸性バーストを誘導することが以前に示されており、宿主によって生成されたROSが、組織損傷に対してさらに寄与し得ることを示唆する。
【0081】
H2O2及びROSは、NF−κBを活性化することも可能であり、このようにすることによって、真核生物宿主中で、多様な免疫及び炎症促進性遺伝子の発現を誘導し、これは、マイコプラズマによって引き起こされる気道感染において特に重要な可能性がある機序である。
【0082】
マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はマイコプラズマのL−α−グリセロリン酸オキシダーゼに対する抗体に基づくワクチンは、H2O2及びROSの産生を首尾よく抑制し、このようにすることによって、本ワクチンは、宿主細胞の損傷、炎症及び疾病を予防する。
【0083】
【表1】
【図面の簡単な説明】
【0084】
【図1】マイコプラズマのL−α−グリセロリン酸オキシダーゼの位置及び活性。a−d、それぞれ、抗GlpOから得られたIgG(a,b)及び免疫前血清(c、d)とともに温置されたM.ミコイデスsubsp.ミコイデスSC株Afadeの免疫金標識を示す走査型電子顕微鏡写真。二次電子顕微鏡写真は、細胞表面(a,c)を示しており、後方散乱された電子顕微鏡写真は、15nmのコロイド状金が抱合された二次抗体(b,d)を明らかにする。スケールバー、500nm。e、GlpO抗血清を用いた、M.ミコイデスsubsp.ミコイデスSC株AfadeのTritonX−114分画された総抗原のイムノブロット分析。レーン1、界面活性剤相、タンパク質20μg;レーン2、水相、タンパク質20μg;Std、分子量標準、f、100μMグリセロールの添加後における、M.ミコイデスsubsp.ミコイデスSC株Afadeの過酸化水素産生。示されているデータは、5つの独立した測定の平均値である。各測定値の標準偏差は、平均値の5%以下であった。三角及び実線、処理されていないM.ミコイデスsubsp.ミコイデスSC;十字及び破線、抗GlpOIgG由来のFab断片で前処理されたM.ミコイデスsubsp.ミコイデスSC;四角及び点線、抗LppC由来のFab断片で前処理されたM.ミコイデスsubsp.ミコイデスSC。
【図2】ECaNEp細胞に対するM.ミコイデスsubsp.ミコイデスSCの細胞毒性(a−f)及び細胞生存性アッセイ(g)。a、ECaNEp細胞(対照);b、M.ミコイデスsubsp.ミコイデスSC株Afadeでの感染から1時間後のECaNEp;c、グリセロール(100μM)の存在下における、株Afadeでの感染から1時間後のECaNEp;d、抗GlpOIgG由来のFab断片で前処理された株Afadeでの、グリセロール(100μM)の存在下における感染から1時間後のECaNEp;e、抗LppCIgG由来のFab断片で前処理された株Afadeでの、グリセロール(100μM)の存在下における感染から1時間後のECaNEp;f、グリセロール(100μM)の存在下における、M.ミコイデスsubsp.ミコイデスSC株L2(活性なグリセロール取り込み系を欠く。)で感染されたECaNEp細胞。全ての実験で、感染効率は、50マイコプラズマ/細胞であった。g、50マイコプラズマ/細胞の感染効率で、M.ミコイデスsubsp.ミコイデスSC株Afadeに感染させた後の様々な時点における生きたECaNEp細胞。三角及び太い実線、100μMグリセロールの存在下での、株Afadeによる感染;十字及び点線、抗GlpOFab断片(0.26μg/mL)で前処理された株AdadeSCによる、100μMグリセロールの存在下での感染;四角及び破線、グリセロールなしの株Afade感染;ダイヤモンド及び細い実線、ECaNEp細胞のみ(対照)。
【図3】500マイコプラズマ/細胞の感染効率で、M.ミコイデスsubsp.ミコイデスSC株Afadeに感染されたECaNEp細胞又はH2O2で処理されたECaNEp細胞中の細胞内H2O2及びROSの検出。ECaNEp細胞の位相差顕微鏡写真(a、c、e、g及びi)及び蛍光顕微鏡写真(b、d、f、h及びj);グリセロールの不存在下における、M.ミコイデスsubsp.ミコイデスSC株Afadeでの感染から20分後(a,b);グリセロールが補充された培地中で、株Afadeに感染(c、d);グリセロールが補充された培地中において、抗GlpOIgG由来のFab断片で前処理された株Adadeで感染(e、f);N−アセチル−L−システインで前処理され、次いで、グリセロールを加えた培地中、株Adadeで感染(g、h);4.4mMH2O2が補充された培地中で、20分間温置(i、j)。
【図4】M.ミコイデスsubsp.ミコイデスSCによって宿主細胞炎症を誘発するためのモデル。GtsABC:活性なグリセロール輸送体及びリン酸化系;GlpF:グリセロール促進物質因子;GlpK:グリセロールキナーゼ;G3P:グリセロール−3−リン酸;DHAP:ジヒドロキシアセトンリン酸;GlpO:マイコプラズマのL−α−グリセロリン酸オキシダーゼ。太い矢印は、M.ミコイデスsubsp.ミコイデスSCの主要な病原性経路を示す。
【特許請求の範囲】
【請求項1】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項2】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする生組換え担体と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項3】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする生組換え担体を含む宿主細胞と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項4】
マイコプラズマ感染を撲滅するためのワクチンであり、機能的に連結されたプロモーターの制御下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子又はL−α−グリセロリン酸オキシダーゼの免疫原性断片をコードする前記遺伝子の一部と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項5】
マイコプラズマ感染を撲滅するためのワクチンであり、機能的に連結されたプロモーターの制御下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子又はL−α−グリセロリン酸オキシダーゼの免疫原性断片をコードする前記遺伝子の一部を含む宿主細胞と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項6】
同一の宿主に対して病原性である別の微生物又はウイルスに由来する1つ又はそれ以上の抗原、このような抗原に対する抗体又はこのような抗原をコードする遺伝情報をさらに含むことを特徴とする、請求項1から5に記載のワクチン。
【請求項7】
ウイルス又は微生物が、仮性狂犬病ウイルス、ブタインフルエンザウイルス、ブタパルボウイルス、伝染性胃腸炎ウイルス、ロタウイルス、別のマイコプラズマ属種、特に、マイコプラズマ・ハイオニューモニエ、ブラキスピラ・ハイオディセンテリー、エシェリヒア・コリ、レプトスピラ属種、エリシペロスリクス・リューシオパシエ、ボルデテラ・ブロンキセプティカ、ブラキスピラ・ハイオディセンテリー、シゲラ種、サルモネラ・コレラエスイス、サルモネラ・チフィムリウム、サルモネラ・エンテリティディス、ヘモフィラス・パラスイス、ラウソニア、パスチュレラ・マルトシダ、ストレプトコッカス・スイス、アクチノバシラス・プリューロニューモニエ、スタフィロコッカス・ハイカス及びクロストリジウム・ペルフリンゲンスの群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項8】
ウイルス又は微生物が、ウシヘルペスウイルス、ウシウイルス性下痢症ウイルス、パラインフルエンザ3型ウイルス、ウシパラミクソウイルス、口蹄疫ウイルス、ウシ呼吸器多核体ウイルス、ブタサーコウイルス、ブタ生殖器呼吸器症候群ウイルス、別のマイコプラズマ属種、パスツレラ・ヘモリティカ、スタフィロコッカス・オーレウス、エシェリヒア・コリ、レプトスピラ属種、スタフィロコッカス・ウベリス、タイレリア・パルバ、タイレリア・アニュラータ、バベシア・ボビス、バベシア・ビゲミナ、バベシア・メジャー、トリパノソーマ種、アナプラズマ・マージナーレ、アナプラズマ・セントラーレ及びネオスポラ・カニナムからなる群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項9】
ウイルス又は微生物が、鶏痘ウイルス、伝染性気管支炎ウイルス、伝染性ファブリキウス嚢病ウイルス(グンボロ)、マレック病ウイルス、ニワトリ貧血因子、トリレオウイルス、トルコ鼻気管炎ウイルス、ニワトリポックスウイルス、トリ脳脊髄炎ウイルス、アヒルペストウイルス、ニューキャッスル病ウイルス、産卵低下症候群ウイルス、伝染性喉頭気管ウイルス、シチメンチョウのヘルペスウイルス、別のマイコプラズマ属種、とりわけ、マイコプラズマ・ガリセプチカム又はマイコプラズマ・シノビアエ、ヘモフィルス・パラガリナルム(コリーザ)、オルニソバクテリウム・ライノトラキア、クロストリジウム・ペルフリンゲンス、サルモネラ、カンピロバクター種、E.コリ及びエイメリア種からなる群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項10】
ウイルス又は微生物が、インフルエンザウイルス、麻疹ウイルス、ムンプスパラミクソウイルス、クロストリジウム・ジフセリエ、クロストリジウム・テタニ、ボルデテラ・パーチュシス及びポックスウイルスからなる群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項11】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片に対する抗体と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項12】
アジュバントを含むことを特徴とする、請求項1から11に記載のワクチン。
【請求項13】
マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片が、M.ボビス、M.sp.ボバイングループ7、M.ミコイデスsubsp.カプリ、M.ペネトランス、M.ガリセプチカム、M.シノビエ、M.モービレ、M.パルモニス、M.ハイオニューモニエ、M.ニューモニエ、M.ジェニタリウム、M.ミコイデスS(mall)C(olony)株Afade又はM.ミコイデスS(mall)C(olony)株L2によってコードされるマイコプラズマのL−α−グリセロリン酸オキシダーゼであることを特徴とする、請求項1から12に記載のワクチン。
【請求項14】
ワクチンにおいて使用するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片。
【請求項15】
マイコプラズマ感染を撲滅するためのワクチンを製造するための、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片の使用。
【請求項16】
請求項1から14に記載のワクチンを調製するための方法であり、請求項1に記載されているマイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片、請求項2に記載されている生組換え担体、請求項4に記載されている遺伝子若しくはその一部、請求項3及び5に記載の宿主細胞又は請求項11に記載されているマイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片に対する抗体と、及び医薬として許容される担体とを混合することを含む、前記方法。
【請求項17】
一方で請求項1から14に記載のワクチンを用いた予防接種と、他方で完全細胞ワクチン又は野外感染を用いた予防接種とを識別するための診断検査であり、精製されたマイコプラズマのL−α−グリセロリンサノキシダーゼ又はその免疫原性断片を含むことを特徴とする前記検査。
【請求項1】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項2】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする生組換え担体と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項3】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片をコードする生組換え担体を含む宿主細胞と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項4】
マイコプラズマ感染を撲滅するためのワクチンであり、機能的に連結されたプロモーターの制御下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子又はL−α−グリセロリン酸オキシダーゼの免疫原性断片をコードする前記遺伝子の一部と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項5】
マイコプラズマ感染を撲滅するためのワクチンであり、機能的に連結されたプロモーターの制御下にある、マイコプラズマのL−α−グリセロリン酸オキシダーゼをコードする遺伝子又はL−α−グリセロリン酸オキシダーゼの免疫原性断片をコードする前記遺伝子の一部を含む宿主細胞と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項6】
同一の宿主に対して病原性である別の微生物又はウイルスに由来する1つ又はそれ以上の抗原、このような抗原に対する抗体又はこのような抗原をコードする遺伝情報をさらに含むことを特徴とする、請求項1から5に記載のワクチン。
【請求項7】
ウイルス又は微生物が、仮性狂犬病ウイルス、ブタインフルエンザウイルス、ブタパルボウイルス、伝染性胃腸炎ウイルス、ロタウイルス、別のマイコプラズマ属種、特に、マイコプラズマ・ハイオニューモニエ、ブラキスピラ・ハイオディセンテリー、エシェリヒア・コリ、レプトスピラ属種、エリシペロスリクス・リューシオパシエ、ボルデテラ・ブロンキセプティカ、ブラキスピラ・ハイオディセンテリー、シゲラ種、サルモネラ・コレラエスイス、サルモネラ・チフィムリウム、サルモネラ・エンテリティディス、ヘモフィラス・パラスイス、ラウソニア、パスチュレラ・マルトシダ、ストレプトコッカス・スイス、アクチノバシラス・プリューロニューモニエ、スタフィロコッカス・ハイカス及びクロストリジウム・ペルフリンゲンスの群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項8】
ウイルス又は微生物が、ウシヘルペスウイルス、ウシウイルス性下痢症ウイルス、パラインフルエンザ3型ウイルス、ウシパラミクソウイルス、口蹄疫ウイルス、ウシ呼吸器多核体ウイルス、ブタサーコウイルス、ブタ生殖器呼吸器症候群ウイルス、別のマイコプラズマ属種、パスツレラ・ヘモリティカ、スタフィロコッカス・オーレウス、エシェリヒア・コリ、レプトスピラ属種、スタフィロコッカス・ウベリス、タイレリア・パルバ、タイレリア・アニュラータ、バベシア・ボビス、バベシア・ビゲミナ、バベシア・メジャー、トリパノソーマ種、アナプラズマ・マージナーレ、アナプラズマ・セントラーレ及びネオスポラ・カニナムからなる群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項9】
ウイルス又は微生物が、鶏痘ウイルス、伝染性気管支炎ウイルス、伝染性ファブリキウス嚢病ウイルス(グンボロ)、マレック病ウイルス、ニワトリ貧血因子、トリレオウイルス、トルコ鼻気管炎ウイルス、ニワトリポックスウイルス、トリ脳脊髄炎ウイルス、アヒルペストウイルス、ニューキャッスル病ウイルス、産卵低下症候群ウイルス、伝染性喉頭気管ウイルス、シチメンチョウのヘルペスウイルス、別のマイコプラズマ属種、とりわけ、マイコプラズマ・ガリセプチカム又はマイコプラズマ・シノビアエ、ヘモフィルス・パラガリナルム(コリーザ)、オルニソバクテリウム・ライノトラキア、クロストリジウム・ペルフリンゲンス、サルモネラ、カンピロバクター種、E.コリ及びエイメリア種からなる群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項10】
ウイルス又は微生物が、インフルエンザウイルス、麻疹ウイルス、ムンプスパラミクソウイルス、クロストリジウム・ジフセリエ、クロストリジウム・テタニ、ボルデテラ・パーチュシス及びポックスウイルスからなる群から選択されることを特徴とする、請求項6に記載のワクチン。
【請求項11】
マイコプラズマ感染を撲滅するためのワクチンであり、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片に対する抗体と、及び医薬として許容される担体とを含むことを特徴とする前記ワクチン。
【請求項12】
アジュバントを含むことを特徴とする、請求項1から11に記載のワクチン。
【請求項13】
マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片が、M.ボビス、M.sp.ボバイングループ7、M.ミコイデスsubsp.カプリ、M.ペネトランス、M.ガリセプチカム、M.シノビエ、M.モービレ、M.パルモニス、M.ハイオニューモニエ、M.ニューモニエ、M.ジェニタリウム、M.ミコイデスS(mall)C(olony)株Afade又はM.ミコイデスS(mall)C(olony)株L2によってコードされるマイコプラズマのL−α−グリセロリン酸オキシダーゼであることを特徴とする、請求項1から12に記載のワクチン。
【請求項14】
ワクチンにおいて使用するためのマイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片。
【請求項15】
マイコプラズマ感染を撲滅するためのワクチンを製造するための、マイコプラズマのL−α−グリセロリン酸オキシダーゼ又はその免疫原性断片の使用。
【請求項16】
請求項1から14に記載のワクチンを調製するための方法であり、請求項1に記載されているマイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片、請求項2に記載されている生組換え担体、請求項4に記載されている遺伝子若しくはその一部、請求項3及び5に記載の宿主細胞又は請求項11に記載されているマイコプラズマのL−α−グリセロリン酸オキシダーゼ若しくはその免疫原性断片に対する抗体と、及び医薬として許容される担体とを混合することを含む、前記方法。
【請求項17】
一方で請求項1から14に記載のワクチンを用いた予防接種と、他方で完全細胞ワクチン又は野外感染を用いた予防接種とを識別するための診断検査であり、精製されたマイコプラズマのL−α−グリセロリンサノキシダーゼ又はその免疫原性断片を含むことを特徴とする前記検査。
【図1】

【図2】
【図3】

【図4】
【図2】
【図3】
【図4】
【公表番号】特表2009−500007(P2009−500007A)
【公表日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願番号】特願2008−518870(P2008−518870)
【出願日】平成18年7月6日(2006.7.6)
【国際出願番号】PCT/EP2006/063948
【国際公開番号】WO2007/006712
【国際公開日】平成19年1月18日(2007.1.18)
【出願人】(508004074)ユニバーシテイ・オブ・ベルン (2)
【Fターム(参考)】
【公表日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願日】平成18年7月6日(2006.7.6)
【国際出願番号】PCT/EP2006/063948
【国際公開番号】WO2007/006712
【国際公開日】平成19年1月18日(2007.1.18)
【出願人】(508004074)ユニバーシテイ・オブ・ベルン (2)
【Fターム(参考)】
[ Back to top ]