
マイトジェン活性化プロテインキナーゼ調節剤
本発明は、MAPK経路を調節する方法を記載する。さらに本発明は、MAPK媒介疾患の治療のためのMAPK経路中間体の調節のためのMycobacterium wの使用を記載する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、Mycobacterium w(Mw)および/またはその成分を使用したMAPキナーゼの目的別(tailored)調節に関する。
本発明はさらに、MAPキナーゼの一過性または持続性の調節のための手段に関する。
【背景技術】
【0002】
マイトジェン活性化プロテインキナーゼ(MAPK)カスケードは、種々の細胞型が共有する主要なシグナル伝達系である。これらはセリン/スレオニンキナーゼである。これらは、活性化(リン酸化)に際して核に移行する。これらは、重要な細胞周期及び分化特異的タンパク質の発現を調節するものを含む転写因子を含む、多くの異なるタンパク質をリン酸化・活性化する。調節される遺伝子はアポトーシス、炎症、細胞増殖および分化に関与する。
これらのタンパク質は、形態形成、細胞死、ストレス応答、免疫応答、細胞増殖、アポトーシス、パラアポトーシス、細胞生存などを含む種々の細胞応答および生物学的活性を媒介する。
【0003】
MAPKカスケードの活性化は未熟な細胞に限定されず、このカスケードは好中球などの最終的に分化した細胞においても活性化され、このことは、このMAPKカスケードが、最終的に分化した成熟細胞のいくつかの機能においても重要な役割を果たすことを示唆する。
【0004】
2つの異なる細胞におけるMAPキナーゼの活性化は、類似または異なる細胞応答を導くことができる。ERKカスケードは、レセプターチロシンキナーゼ、造血成長因子レセプター、またはいくつかのヘテロトリマーGタンパク質共役レセプターからのシグナルに応答して活性化され、細胞の増殖または分化を促進するシグナルを媒介するようである。
p38およびJNkを含むストレス活性化プロテインキナーゼ(SAPK)は、ヒートショック、高浸透圧、UV照射、タンパク質合成阻害剤または炎症性サイトカインに応答して活性化され、ストレスに対する細胞応答に関与するようである。
【0005】
異なるMAPKサブタイプのカスケードの活性化は、使用される細胞の型および刺激に依存する。各MAPKサブタイプの機能的役割は、細胞の型に従って異なり得る。
Belmontらは、JNKの活性を調節する(必要に応じて阻害または増強する)治療剤の有効用量を、JNK媒介性障害の治療を必要とする対象に投与することによる、JNK媒介性障害を治療する手段を記載している。JNKシグナル伝達経路を刺激する因子は、組織においてプログラムされた細胞死(アポトーシス)を誘導することを含む、多数の方法で使用できる。例えば、UV損傷した細胞の排除は、癌を予防するために使用できる。
【0006】
さらに、MAPK調節剤は、黒色腫の管理において有用である(Clin Cancer Res.2006年4月1日:2371s-2375s)。MAPK調節剤は、パクリタキソールとの相乗作用を有し得る(MoI Pharmacol.2001年8月;60(2):290-301)。MAPK調節剤は、プログラムされた細胞死と関連している(J.Biol Chem.2000年12月15日;275(50):38953-6)。
MAPK調節剤は、抗菌剤などの生物学的治療ならびに化学療法と相乗的に作用する(Oncogene 2003:22、2034-2044)。MAPK調節剤は、化学療法剤に対して抵抗性の細胞の再感受性において有用である(Brit.J.Cancer 2001、85:1175-1184)。MAPK調節剤は、タキソール、シスプラチンなどの癌治療剤の化学療法効果と関連する(Oncogene 2001;20、147-155;Onco gene 2000、19;5142-5152)。
【0007】
ERKの活性化は、細胞の増殖および生存と主に関連しており、全般的に見て、SAPKの活性化は、アポトーシスの誘導に関連している。多数の細胞型を使用して、JNKの持続性の活性化は細胞死を誘導し、ドミナントネガティブ(DN)阻害剤によるJNK活性化の遮断が、一連のアポトーシス刺激による死を防止することが示された。細胞外調節性キナーゼ(MAPK/ERK)のレベルの上昇は、いくつかの癌細胞において活発にかつ頻繁に見出される。p38 MAPK不活化によるIL-2産生のアップレギュレーションは、ERK1/2活性の増加によって媒介される。
【0008】
ERK2は広く分布するプロテインキナーゼであり、上流のMAPキナーゼキナーゼであるMEK1によりThrl83およびTyrl85の両方がリン酸化される場合に、最大の活性に達する(Andersonら、1990、Nature 343、651;Crewsら、1992、Science 258、478)。活性化されると、ERK2は、プロテインキナーゼRsk90(Bjorbaekら、1995、J.Biol.Chem.270、18848)およびMAPK AP2(Rouseら、1994、Cell 78、1027)、ならびにATF2(Raingeaudら、1996、MoI.Cell Biol.16、1247)、Elk-1(Raingeaudら、1996)、c-Fos(Chenら、1993 Proc.Natl.Acad.Sci.USA 90、10952)およびc-Myc(Oliverら、1995、Proc.Soc.Exp.Biol.Med.210、162)などの転写因子を含む多数の調節タンパク質をリン酸化する。ERK2は、Ras/Raf依存的経路の下流の標的でもあり(Moodieら、1993、Science 260、1658)、これらの潜在的に発癌性のタンパク質からのシグナルの中継を助け得る。ERK2は、乳癌細胞の負の増殖制御において役割を果たすことが示されており(FreyおよびMulder、1997、Cancer Res.57、628)、ヒト乳癌におけるERK2の過剰発現が報告されている(Sivaramanら、1997、J Clin.Invest.99、1478)。活性化されたERK2は、エンドセリン刺激された気道平滑筋細胞の増殖にも関与しており、このことは、喘息におけるこのキナーゼの役割を示唆している(Whelchelら、1997、Am.J.Respir.Cell MoI.Biol.16、589)。
【0009】
JNKは、他のMAPKと共に、癌、トロンビンが誘導する血小板凝集、免疫不全障害、自己免疫疾患、細胞死、アレルギー、骨粗しょう症および心疾患に対する細胞応答の媒介において役割を有することに関与している。
JNKは種々の形態の心臓ストレスに対する肥大応答を媒介していることが示されているので、心筋梗塞またはうっ血性心不全などの心血管疾患におけるJNKの役割も報告されている[Circ.Res.83:167-78(1998);Circulation 97:1731-7(1998);J.Biol.Chem.272:28050-6(1997);Circ.Res.79:162-73(1996);Circ.Res.78:947-53(1996);J.Clin.Invest.97:508-14(1996)]。
【0010】
JNKカスケードは、IL-2プロモーターの活性化を含むT細胞活性化においても役割を果たすことが実証されている。したがって、JNKの阻害剤は、病的な免疫応答を変更することにおいて治療的価値を有し得る[J.Immunol.162:3176-87(1999);Eur.J.Immunol.28:3867-77(1998);J.Exp.Med.186:941-53(1997);Eur.J.Immunol.26:989-94(1996)]。
種々の癌におけるJNK活性化の役割も確立されており、癌におけるJNK阻害剤の潜在的な使用が示唆されている。例えば、構成的に活性化されたJNKは、HTLV-1媒介性の腫瘍形成に関連している[Oncogene 13:135-42(1996)]。さらに、p210 BCR-ABL形質転換細胞におけるc-jun遺伝子の調節はJNKの活性に対応し、このことは、慢性骨髄性白血病(CML)の治療におけるJNK阻害剤の役割を示唆している[Blood 92:2450-60(1998)]。
【0011】
JNK、特にJNK3のシグナル伝達は、アルツハイマー病、パーキンソン病、ALS(筋萎縮性側索硬化症)、癲癇およびけいれん、ハンチントン病、外傷性脳損傷、ならびに虚血および出血性卒中などの、アポトーシスが駆動する神経変性疾患の領域に関与している。
JNKは、いくつかの化学療法薬および癌遺伝子産物(例えば、Bcr-AbI、Her-2/Neu、Srcおよび発癌性Ras)により強力に活性化されるが、通常はERKに対する作用を有さない。したがって、SAPKを活性化し、同時にERKをダウンレギュレートする多機能の役割を果たし得る薬剤を設計/産生する必要性が長い間存在していた。
【0012】
変更されたMAPKシグナル伝達経路は、疾患をいくつか挙げると、神経変性障害、自己免疫疾患、腫瘍の発達および進行、化学療法に対する抵抗性、虚血傷害後の損傷などの、種々の疾患状態に関連する。
これらの疾患状態は、MAPKの調節で効果的に治療できる。照射が誘導する癌細胞死ならびに化学療法が誘導する癌の死は、MAKシグナル伝達経路の調節(例えばSAPKのアップレギュレーション)と関連する。
【0013】
SAPKの一過性のアップレギュレーションもまた、自己免疫を誘導することなく免疫応答を誘導するのに有用である。SAPKの一過性のアップレギュレーションは、正常細胞に顕著に影響を与えることなく、癌細胞の死を誘導するのにも有用である。SAPKの一過性のアップレギュレーションは、抗原提示細胞の活性化ならびに成熟T細胞および樹状細胞において役割を果たすことが知られている。Arrighiらは、APC成熟におけるホスホ-p38キナーゼの役割を記載している(The Journal of Immunology、2001、166:3837-3845)。Mercedes Rinconら(Free Radical Biology and Medicine、28巻、9号、2000年5月1日、1328-1337ページ)に記載されたように、SAPKのアップレギュレーションは、T細胞応答において重要な役割を果たす。
【0014】
SAPKのダウンレギュレーションは、神経変性疾患における損傷を予防するのに有用である。これは、自己免疫疾患の管理においても有用であり、虚血傷害後の損傷を最小化する。
したがって、SAPKのダウンレギュレーションとERKのアップレギュレーションまたは少なくとも変化を起こさないこととの組み合わせ効果は、上記種々の疾患状態における組織への損傷を予防するのに有用であろう。
【0015】
米国特許第6,994,981号は、パラアポトーシスの調節剤および関連の方法を記載している。EP1208748、WO2004/089929およびWO2006/117567は、MAPK阻害剤に基づく先行技術の特許である。
米国特許第6,852,740号B2は、p38キナーゼ阻害剤としてのピラゾール誘導体を記載している。WO95/31451は、MAPK、特にp38を阻害するピラゾール組成物を記載している。しかし、これらの阻害剤のin vivoでの有効性は、なお調査中である。
【0016】
ストレス活性化経路とマイトジェン活性化ERK経路との間には、負のクロストーク関係が存在する。JNKアクチベータ(例えば、TNF-αおよびセラミド)の生物学的機能のいくつかは、ERK/MAPK経路を介して作用する成長および生存因子に対する細胞の応答を遮断するそれらの能力に寄与している可能性があり、したがって、これらの結果のうち1つを制御することは、シグナル伝達のさらなる歪みを生じ、これらの疾患状態を悪化させる。
ほとんどのMAPKについて、媒介される疾患状態は、SAPKとERKとの間のクロストークを調節することによってMAPK調節に対して作用する薬剤を必要とする。
【0017】
したがって、このような疾患状態の治療について、MAPK経路のレベルの1種または複数の中間体または/およびそれらの比率は一貫しない。特に、1種を超える中間体のレベルが誤って調節される疾患状態の場合、MAPK調節剤が必要であろう。
【0018】
これらの合成MAPK阻害剤の毒性副作用は、下痢、発疹、疲労、手足症候群、脱毛症、悪心、手足皮膚反応、または一般に錯感覚を伴う手掌および足裏上の痛みのある対称的な紅斑性および浮腫性の領域を特徴とする先端紅斑である。時に、指の側方または爪周囲の領域が影響を受けることがある。角質増殖および剥離が一般に生じる。口内炎、脱毛症、掻痒症および爪下綿状出血も観察され、これらの出血は、爪の下の黒または赤のまっすぐな線を特徴とする。これらは、血栓性または塞栓性の機構に起源すると思われる。最初は細菌性心内膜炎の典型的な徴候と思われたが、これらは続いて、高地で、または動脈カテーテルを使用した場合、異なる状況でも存在することが報告された(例えば、抗リン脂質抗体症候群、重症関節リウマチ、閉塞性血栓血管炎、僧帽弁狭窄症)(The Oncologist、12巻、12号、1443-1455、2007年12月)。
【0019】
BCGおよびCpGはSAPKをアップレギュレートでき、同時にERKのアップレギュレーションも観察される。ERKのアップレギュレーションは、副作用を導く炎症性サイトカインであるTNFおよびIL-10のアップレギュレーションを導く。
【0020】
したがって、異常なレベルのSAPKおよびERKに関連する種々の疾患状態を治療するのに有用な、非毒性のMAPK調節剤を開発する必要性が長い間認められてきた。
【発明の概要】
【発明が解決しようとする課題】
【0021】
本発明において、MAPK/ERK中間体レベルの調節を介してシグナル伝達経路を調節するための新規アプローチがとられ、ここで、MAPK/ERK経路は、Mwの用量および細胞の状態に依存してMycobacterium wによって調節/妨害され、したがって低い毒性を有する。また、Mycobacterium w製剤は、百万人を超えるヒト患者の治療のために、種々の適応症に対して既に使用されており、安全性が証明されている。
【課題を解決するための手段】
【0022】
本発明の目的は、マイトジェン活性化プロテインキナーゼ(MAPK)シグナル伝達経路の調節剤を提供することである。
本発明の別の目的は、Mycobacterium wおよび/またはその成分を使用する、マイトジェン活性化プロテインキナーゼの調節を提供することである。
【0023】
本発明の別の目的は、マイトジェン活性化プロテインキナーゼを調節する方法を提供することである。
本発明のなお別の目的は、SAPKがダウンレギュレートされる一方で、ERKレベルは変化を示さない、マイトジェン活性化プロテインキナーゼの調節剤を提供することである。
【0024】
本発明のなお別の目的は、SAPKがアップレギュレートされる一方で、ERKレベルはダウンレギュレートされる、マイトジェン活性化プロテインキナーゼの調節剤を提供することである。
本発明のなお別の目的は、SAPKがアップレギュレートされる一方で、ERKレベルは変化を示さない、マイトジェン活性化プロテインキナーゼの調節剤を提供することである。
本発明のなお別の目的は、哺乳動物において、非経口、腸内および局所経路を介して投与されるMAPK調節剤を提供することである。
【図面の簡単な説明】
【0025】
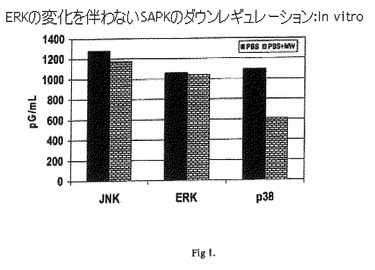
【図1】ERKの変化を伴わないSAPKのダウンレギュレーション:In vitro。
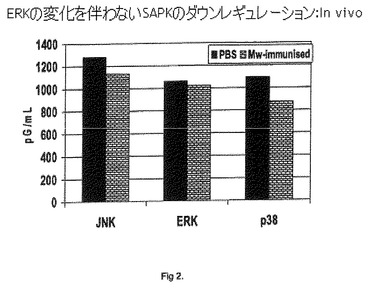
【図2】ERKの変化を伴わないSAPKのダウンレギュレーション:In vivo。
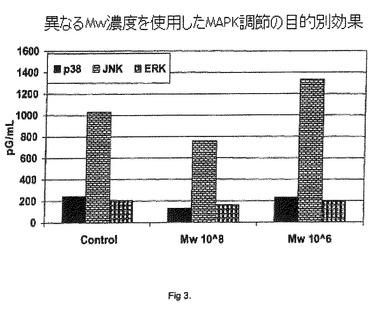
【図3】異なるMw濃度を使用したMAPK調節の目的別効果。
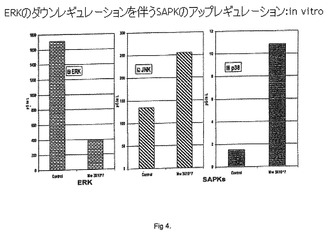
【図4】ERKのダウンレギュレーションを伴うSAPKのアップレギュレーション:in vitro。
【図5】異なるMw濃度を使用したMAPK調節の目的別効果。
【図6】MwによるMAPK調節の寿命。
【図7】ERKレベルの変化を伴わないSAPKの一過性のアップレギュレーション。
【発明を実施するための形態】
【0026】
本発明によれば、MAPK調節剤は以下のプロセスによって調製される。本発明は、医薬組成物、調製方法、HPLC特徴、その安全性および忍容性を含み、使用方法および治療結果は、以下の実施例に記載される。以下は、本発明の例示的な実施例であり、本発明の範囲はこれらの実施例により限定されるべきではない。
【0027】
Mycobacterium wを調製するプロセス:
A. Mycobacterium wの培養
i)培地の調製
Mycobacterium wは、LJ培地などの固体培地またはミドルブルック培地もしくはソートン液体培地などの液体培地で培養する。
より良好な収率のために、ミドルブルック培地を富化する。この培地は好ましくは、グルコース、バクト-トリプトンおよびBSAの添加によって富化できる。これらは好ましくは、20:30:2の比率で使用する。
富化培地を、ミドルブルック培地に添加する。これは、好ましくは15:1〜25:1の比率で、より好ましくは20:1の比率で実施する。
【0028】
ii)バイオリアクタ操作:
a)容器の準備:
容器の内側接触部分(ジョイント、メカニカルシール、O-リング/ガスケット溝など)は、汚染を回避するために適切に洗浄すべきである。この容器を0.1N NaOHで満たし、そのまま24時間放置して、発熱物質および他の汚染物質を除去する。次いで、この容器を最初に酸性化水で洗浄し、次いで通常の水で洗浄する。最後に、この容器を蒸留水ですすぎ(3回)、その後、培地を調製する。
b)バイオリアクタの滅菌:
9Lの蒸留水を含むバイオリアクタを、生蒸気で滅菌する(間接的)。同様に、ミドルブルック培地とともにバイオリアクタを、再度滅菌する。他の添加ビン、入口/出口のエアフィルタなどを、121℃で15分間オートクレーブする(2回)。使用前に、これらを50℃のオーブンで乾燥する。
c)環境パラメータ
i.温度:37±0.5℃
ii.pH:最初6.7〜6.8。
【0029】
B. 回収および濃縮:
これは典型的には、無菌条件下で培養した後、6日目の終わりに行う。細胞の濃縮(ペレット化)は、遠心分離によって実施する。
【0030】
C. 細胞の洗浄:
こうして得たペレットを、通常の生理食塩水で最低3回洗浄する。これは、好ましくは等張の任意の他の流体で洗浄することができる。
【0031】
D. 医薬として許容可能な担体の添加
(発熱物質を含まない通常の生理食塩水をペレットに添加する。)最も好ましくは、発熱物質を含まない等張の任意の流体を医薬的担体として使用できる。担体は、最終形態で所望の活性濃度となるような量で添加する。
【0032】
E. 防腐剤の添加:
細胞/ペレットを汚染なしで維持するために、防腐剤を添加する。好ましくは、0.01%w/vの濃度を有するチメロサールを使用する。
【0033】
F. 最終滅菌:
加熱または電離放射線または滅菌濾過の適用などの種々の物理的方法により、細胞/ペレットを滅菌する。加熱は、乾式加熱または湿式加熱の形態で行うことができる。加熱は、煮沸または低温殺菌の形態とすることもできる。電離放射線は、紫外線またはガンマ線またはマイクロ波または任意の他の形態で行うことができる。
【0034】
G. 品質制御:
i.材料を、純度、無菌性について評価する。
ii.生物を、グラム染色後に抗酸性についてチェックする。
iii.生化学的試験:生物を、以下の生化学的試験に供する。
a)ウレアーゼ
b)Tween 80加水分解
c)ナイアシン試験
d)硝酸還元試験
【0035】
生物は、ウレアーゼ、tween 80加水分解およびナイアシン試験において陰性結果を生じる。生物は、硝酸還元試験では陽性である。
iv.不活化試験:これは、生存生物を見出すために、LJ培地上で産物を培養することにより行う。
v.病原性および/または病原体による汚染
培養した生物を、balb/cマウスに感染させる。死亡するマウスは出ないはずであり、全てが健康のままで体重が増加するはずである。処置の8週間後までに動物を屠殺した時点で、肝臓、肺、脾臓または任意の他の臓器中には、肉眼的病変も顕微鏡的病変も見出されないはずである。
【0036】
H. Mycobacterium w成分の調製:
Mw成分は以下の方法によって調製できる。
I.細胞破壊
II.溶媒抽出
III.酵素抽出
【0037】
細胞破壊は、超音波処理または高圧フラクショノメータの使用または浸透圧成分の適用によって実施できる。破壊した細胞を、生理食塩水で洗浄し、遠心分離によって再ペレット化した。
溶媒抽出は、クロロホルム、エタノール、メタノール、アセトン、フェノール、ハロゲン化炭化水素、イソプロピル-アルコール、酢酸、尿素、ヘキサンおよび/または芳香族化合物などの任意の有機溶媒の、個々にまたはそれらの任意の組合せにより、実施できる。
【0038】
酵素抽出は、細胞壁/細胞膜を消化できるタンパク質分解酵素によって実施できる。リゾチーム、リティカーゼおよびプロナーゼが好ましい酵素である。Mwの細胞成分が、Mwの代わりに使用できる。Mwの細胞成分の添加は、製品の有効性の改善をもたらす。
培養した生物を、Balb/cマウスに感染させる。死亡するマウスはでないはずであり、全てが健康なままで体重が増加するはずである。治療の8週間後までに動物を屠殺した時点で、肝臓、肺、脾臓または任意の他の臓器中には、肉眼的病変も顕微鏡的病変も存在しないはずである。
【0039】
こうして調製したMycobacterium wを、そのMAPK調節活性について評価した。以下の実施例は、Mycobacterium wおよび/またはその成分によるMAPKの調節を示す。
以下の実施例は本発明を実証するが、本発明の範囲を限定するものではない。
【実施例】
【0040】
A. NFS60細胞株:サンプル調製
回収後保存した細胞1×107個の細胞ペレットを、以下のように処理する:細胞を、溶解緩衝液#6(PBS(pH7.2-7.4)中、1mM EDTA、0.5% Triton X-100、6M 尿素、10μg/mL ロイペプチン、10μg/mL ペプスタチン、100μM PMSF、3μg/mL アプロチニン、2mM ピロリン酸ナトリウム、1mM 活性化バナジン酸ナトリウム)中に可溶化する。
溶解物を短時間ボルテックスし、10分間氷上に置く。次いで、plastocraft中で10,000rpmで10分間遠心分離する。上清を新たなチューブに移す。6倍希釈の溶解物を、IC Diluent #8(PBS(pH7.2-7.4)中、1mM EDTA、0.5% Triton X-100)を用いて調製し、IC Diluent #3でさらに希釈する。IC Diluent #3中のサンプルを、R&D Systems製のキットを使用したELISAによる分析のために使用する。
【0041】
B. ELISAプレートの調製:
ホスホ-P38(R&D Systems、DuoSet IC カタログ番号DYC869-2)
ホスホ-JNK(R&D Systems、DuoSet IC カタログ番号DYC1018-2)
ホスホ-JNK(R&D Systems、DuoSet IC カタログ番号DYC1387-2)
捕捉抗体を、製造業者の指示に従って作業濃度まで希釈した。96ウェルのマイクロプレートを、1ウェル当たり100μLの希釈したそれぞれの捕捉抗体で即座に被覆した。プレートを密封し、室温で一晩インキュベートした。
【0042】
プレートを吸引し、洗浄緩衝液で洗浄し、このプロセスを2回繰り返して合計3回洗浄した。最後の洗浄後、プレートを裏返し、きれいなブロッティングペーパーで吸い取ることによって、残留した洗浄緩衝液を除去した。各ウェルに300μLのブロック緩衝液を添加することによってプレートをブロッキングした。次いで、プレートを室温で2時間インキュベートし、その後プレートを再度3回洗浄し、吸引し、吸引乾燥した。これで、サンプル添加のためのプレートの準備ができたことになる。各MAPKのためのストリップを、さらなる使用のために、2〜8℃でデシケータ中で保存した。
【0043】
C. 脾細胞の調製:
脾細胞を正常マウスから回収した。マウスを頚椎脱臼によって屠殺した。即座に腹腔を開き、脾臓を単離した。各脾臓を個別にさらに処理した。
各脾臓を、10mlのダルベッコリン酸塩緩衝化生理食塩水(DPBS)で洗浄し、汚染している血液および他の不純物を除去した。脾臓を、無菌シリンジピストンを使用して刻んだ。10mlのRPMI-1640完全培地(10%FBSおよび1%ペニシリン-ストレプトマイシン抗生物質)を添加し、40μmのナイロンセルストレイナーを介して刻んだ脾臓を篩にかけることによって、内容物を50mlのファルコンチューブに移した。細胞を、Hereaeus Multifuge-3 SR中で1500rpmで5分間遠心分離してペレット化した。
【0044】
上清を廃棄し、ペレット中に存在するRBCを、5mlの溶解緩衝液(0.017MのTris-HCI(pH=7.6)中0.144MのNH4CI)で10分間細胞を再懸濁することによって溶解した。反応を、40mlのDPBSの添加によって停止させた。細胞を、Hereaeus Multifuge-3 SR中で1500rpmで5分間遠心分離してペレット化した。得られた細胞の各セットのペレットを、1mlのRPMI-1640完全培地中に再懸濁した。その後、細胞数を、トリパンブルーで細胞を染色することによって、Neubarrチャンバを使用して決定した。細胞懸濁物を、RPMI-1640完全培地で適切に希釈して、107細胞/mlの細胞密度にした。
【0045】
各脾臓から、1mlのRPMI-1640中に細胞を1×107個含む懸濁物5mLを調製した。1mlの各細胞懸濁物のそれぞれを、マイクロタイタープレート中に4連で播種し、そのうち2つを108個のMycobacterium w細胞で刺激した。プレートを6%CO2で37℃で36時間インキュベートした。
48時間後、細胞を回収した。完全にピペッティングした後、細胞をエッペンドルフチューブに移した。細胞を、10,000rpmで10分間の遠心分離によってペレット化した。上清を別のチューブに移した。上清および細胞ペレットの両方を標識し、さらなる分析まで-70℃で保存した。
【0046】
実施例1
ERKの変化を伴わないSAPKのダウンレギュレーション:
Mwのin vitro効果:
脾細胞を、ナイーブBalb/Cマウスから単離し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106個/mLの脾細胞の2セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108のMycobacterium w細胞と共にインキュベートした。
【0047】
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
ホスホ-JNKの結果を、図1中に示されるようにプロットした。108個のMycobacterium wによる脾細胞のin vitro刺激が、48時間にわたりホスホ-JNKレベルの有意な変化を示さないことが観察される。
【0048】
ホスホ-p38の結果を、図1中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激は、108個のMycobacterium w細胞と共に48時間経過した後、ホスホ-p38 MAPKをダウンレギュレートすることが観察される。したがって、Mycobacterium wは、ホスホ-p38レベルをダウンレギュレートする。
ホスホ-ERKの結果を、図1中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激は、対照と比較して、48時間にわたりホスホ-ERKレベルの有意な変化を示さないことが観察される。
【0049】
Mwのin vivo効果:
Balb/Cマウスの群1では0.1mLのPBSで免疫し、群2では0.1mLのMycobacterium w(108細胞)を皮内に投与した後、このBalb/Cマウスから、その7日後に、脾細胞を単離した。細胞を、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
【0050】
結果を図2に示す。Mwで免疫したマウスは、ERKレベルの変化を伴わずに、SAPKのダウンレギュレーションを示すことが観察された。
【0051】
異なるMw濃度を使用した目的別効果:
脾細胞を、ナイーブなBalb/Cマウスから単離し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106個/mLの脾細胞の3セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108個のMycobacterium w細胞と共にインキュベートし、セット3は106個のMycobacterium w細胞と共にインキュベートした。
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
【0052】
ホスホ-JNKの結果を、図3中に示されるようにプロットした。106個のMycobacterium wによる脾細胞のin vitro刺激が、48時間にわたりホスホ-JNKレベルのダウンレギュレーションを示さないことが観察され、その一方で、108個のMycobacterium wで刺激した場合、48時間目の終わりにホスホ-JNKレベルのダウンレギュレーションを示すことが観察される。1998年に、Science 282:2092-2095でDong,Chenらが、およびImmunity 9:575-585でYang,D.Dが、ホスホ-JNKのアップレギュレーションが、T細胞の分化、増殖および活性化に必要であることを記載している。
【0053】
ホスホ-p38の結果を、図3中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激が、108個のMycobacterium w細胞との48時間の後に、ホスホ-p38 MAPKをダウンレギュレートすることが観察されたが、106個のMycobacterium w細胞では観察されなかった。Arrighiらは、APC成熟におけるホスホ-p38キナーゼの役割を記載している(The Journal of Immunology、2001、166:3837-3845)。
ホスホ-ERKの結果を、図3中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激が、対照と比較して、48時間にわたりホスホ-ERKレベルの有意な変化を示さないことが観察される。
【0054】
実施例2
ERKのダウンレギュレーションを伴うSAPKのアップレギュレーション:
Mwのin vitro効果:
NFS60細胞を、10%FBS、1%抗生物質および10nG/mLのIL-3を含むDMEM培地中で培養した。細胞を、細胞1×105個の濃度でマイクロタイターウェル中にプレートした。ウェルの数を2セットに分割した。セット1を対照としてPBSと共にインキュベートし、セット2を3×107個のMycobacterium w細胞と共にインキュベートした。
【0055】
培養1、2、4、8および24時間の時点で、細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
対照と比較したMycobacterium wで刺激された細胞中のホスホ-JNKレベルは、4時間目まで有意な変化を示さず、8時間目の終わりから24時間目の時点で2倍になる(図4)。
【0056】
対照と比較したMycobacterium wで刺激された細胞中のホスホ-p38レベルは、示差的な用量依存的効果を示す。3×107個の用量のMycobacterium w細胞では、ホスホ-p38レベルは急上昇し、4〜8時間目にわたり高いままであり、次いで低下し始め、24時間目の終わりには対照よりも低い(図4)。刺激のための3×107個の濃度のMycobacterium w細胞でNFS-60細胞が殺傷されるのは驚くべきことであった。
【0057】
対照におけるホスホ-ERKレベルは、8時間目まで有意な変化を示さず、24時間目の終わりには2倍になる。これは、細胞数の増加に起因する可能性がある。図4中に示されるように、Mycobacterium wで刺激された細胞は、最初の1時間目の時点でそれ自体ホスホ-ERKレベルの増加を示して2倍になり、8時間目の時点で、1時間目の半分まで低下し、24時間目の時点で、そのレベルは、その時点での対照の1/4とほぼ同じになる。この低下は、生存細胞数の減少と関連した。
したがって、ホスホ-ERKレベルの同時の低下を伴ったSAPK(ホスホ-p38レベルおよびホスホ-JNKレベル)の上昇は、NFS60細胞の死と関連する。
【0058】
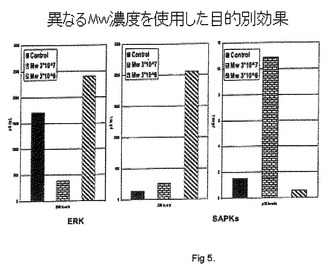
異なるMw濃度を使用した目的別効果:
NFS60細胞を、10%FBS、1%抗生物質および10nG/mLのIL-3を含むDMEM培地中で培養した。細胞を、1×105細胞の濃度でマイクロタイターウェル中にプレートした。ウェルの数を5セットに分割した。セット1は対照としてPBSと共にインキュベートし、残りは各々、3×107、1×107、7×106および3×106個のMycobacterium w細胞と共にそれぞれインキュベートした。
【0059】
培養1、2、4、8および24時間の時点で、細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
3×107個のMycobacterium wで刺激された細胞中のホスホ-JNKレベルは、対照と比較して、4時間目まで有意な変化を示さず、8時間目の終わりから24時間目で2倍になるが(図5)、一方、より低い濃度では、レベルの上昇は遅延し試験期間中には効果はない(図5)。
【0060】
対照と比較したMycobacterium wで刺激された細胞中のホスホ-p38レベルは、示差的な用量依存的効果を示す。3×107個の用量のMycobacterium w細胞では、ホスホ-p38レベルは急上昇し、4〜8時間にわたり高いままであり、次いで低下し始め、24時間目の終わりでは対照よりも低い。他の全てのMycobacterium w濃度で、ホスホ-p38レベルは、対照と同様にダウンレギュレートされる(図5)。
【0061】
対照におけるホスホ-ERKレベルは、8時間目まで有意な変化を示さず、24時間目の終わりには2倍になる。これは、細胞数の増加に起因する可能性がある。図5中に示されるように、Mycobacterium wで刺激された細胞は、8時間目の時点で1時間目の半分までの低下を示すが、24時間目の時点では、そのレベルは、その時点での対照の1/4とほぼ同じになる。この低下は、生存細胞数の減少と関連した。
【0062】
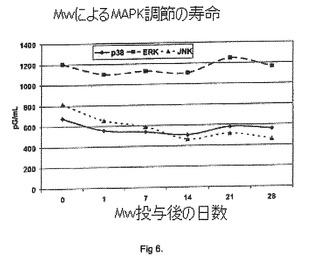
実施例3
MAPK調節の寿命:
Balb/Cマウスの群1では1mLのPBSで免疫し、群2〜6では1mLのMycobacterium w(109細胞)を静脈内に投与し、これらのBalb/Cマウスから、脾細胞を単離した。群1および2を1日目に屠殺し、群3を7日目に、群4を14日目に、群5を21日目に、群6を28日目に屠殺し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106/mLの脾細胞の3セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108個のMycobacterium w細胞と共にインキュベートし、セット3は106個のMycobacterium w細胞と共にインキュベートした。
【0063】
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
図6中に示されるように、ホスホ-JNKレベルは、免疫の24時間後から28日目(試験期間)までダウンレギュレーションを示す。
【0064】
図6中に示されるように、ホスホ-p38レベルも、免疫の24時間後から21日目までダウンレギュレーションを示した。
図6中に示されるように、ホスホ-ERKレベルは、免疫の24時間後から14日目までダウンレギュレーションを示し、その後再び正常レベルとなる。
【0065】
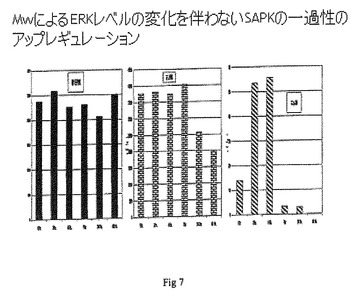
実施例4
ERKの変化を伴わないSAPKの一過性のアップレギュレーション:
脾細胞を、ナイーブなBalb/Cマウスから単離し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106個/mLの脾細胞の3セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108のMycobacterium w細胞と共にインキュベートし、セット3は106個のMycobacterium w細胞と共にインキュベートした。
【0066】
培養1、2、4、8および24時間の時点で、細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
ELISAにより測定したホスホ-JNKレベルは、106個のMycobacterium wによるホスホロ(phosphor)ホスホ-JNKのアップレギュレーションを示す。図7中に示されるように、このアップレギュレーションは、1〜8時間目だけの一過的なものであるが、24時間目の時点で、このレベルはダウンレギュレートされる。
【0067】
ELISAにより測定したホスホ-p38レベルは、108個でのホスホロ(phosphor)ホスホ-p38のアップレギュレーションを示す。図7中に示されるように、このアップレギュレーションは、1〜4時間目だけの一過的なものであるが、8〜24時間目では、このレベルはダウンレギュレートされる。
SAPKのアップレギュレーションは、Mercedes Rinconら(Free Radical Biology and Medicine、28巻、9号、2000年5月1日、1328-1337ページ)に記載されるように、T細胞応答において重要な役割を果たす。これらの知見は、より低いレベルと同様に用量依存的であり、図7中に示されるように、細胞106個のMycobacterium wは、この特性を示さず、一方でERKでは、このような調節は観察されなかった。
【技術分野】
【0001】
本発明は、Mycobacterium w(Mw)および/またはその成分を使用したMAPキナーゼの目的別(tailored)調節に関する。
本発明はさらに、MAPキナーゼの一過性または持続性の調節のための手段に関する。
【背景技術】
【0002】
マイトジェン活性化プロテインキナーゼ(MAPK)カスケードは、種々の細胞型が共有する主要なシグナル伝達系である。これらはセリン/スレオニンキナーゼである。これらは、活性化(リン酸化)に際して核に移行する。これらは、重要な細胞周期及び分化特異的タンパク質の発現を調節するものを含む転写因子を含む、多くの異なるタンパク質をリン酸化・活性化する。調節される遺伝子はアポトーシス、炎症、細胞増殖および分化に関与する。
これらのタンパク質は、形態形成、細胞死、ストレス応答、免疫応答、細胞増殖、アポトーシス、パラアポトーシス、細胞生存などを含む種々の細胞応答および生物学的活性を媒介する。
【0003】
MAPKカスケードの活性化は未熟な細胞に限定されず、このカスケードは好中球などの最終的に分化した細胞においても活性化され、このことは、このMAPKカスケードが、最終的に分化した成熟細胞のいくつかの機能においても重要な役割を果たすことを示唆する。
【0004】
2つの異なる細胞におけるMAPキナーゼの活性化は、類似または異なる細胞応答を導くことができる。ERKカスケードは、レセプターチロシンキナーゼ、造血成長因子レセプター、またはいくつかのヘテロトリマーGタンパク質共役レセプターからのシグナルに応答して活性化され、細胞の増殖または分化を促進するシグナルを媒介するようである。
p38およびJNkを含むストレス活性化プロテインキナーゼ(SAPK)は、ヒートショック、高浸透圧、UV照射、タンパク質合成阻害剤または炎症性サイトカインに応答して活性化され、ストレスに対する細胞応答に関与するようである。
【0005】
異なるMAPKサブタイプのカスケードの活性化は、使用される細胞の型および刺激に依存する。各MAPKサブタイプの機能的役割は、細胞の型に従って異なり得る。
Belmontらは、JNKの活性を調節する(必要に応じて阻害または増強する)治療剤の有効用量を、JNK媒介性障害の治療を必要とする対象に投与することによる、JNK媒介性障害を治療する手段を記載している。JNKシグナル伝達経路を刺激する因子は、組織においてプログラムされた細胞死(アポトーシス)を誘導することを含む、多数の方法で使用できる。例えば、UV損傷した細胞の排除は、癌を予防するために使用できる。
【0006】
さらに、MAPK調節剤は、黒色腫の管理において有用である(Clin Cancer Res.2006年4月1日:2371s-2375s)。MAPK調節剤は、パクリタキソールとの相乗作用を有し得る(MoI Pharmacol.2001年8月;60(2):290-301)。MAPK調節剤は、プログラムされた細胞死と関連している(J.Biol Chem.2000年12月15日;275(50):38953-6)。
MAPK調節剤は、抗菌剤などの生物学的治療ならびに化学療法と相乗的に作用する(Oncogene 2003:22、2034-2044)。MAPK調節剤は、化学療法剤に対して抵抗性の細胞の再感受性において有用である(Brit.J.Cancer 2001、85:1175-1184)。MAPK調節剤は、タキソール、シスプラチンなどの癌治療剤の化学療法効果と関連する(Oncogene 2001;20、147-155;Onco gene 2000、19;5142-5152)。
【0007】
ERKの活性化は、細胞の増殖および生存と主に関連しており、全般的に見て、SAPKの活性化は、アポトーシスの誘導に関連している。多数の細胞型を使用して、JNKの持続性の活性化は細胞死を誘導し、ドミナントネガティブ(DN)阻害剤によるJNK活性化の遮断が、一連のアポトーシス刺激による死を防止することが示された。細胞外調節性キナーゼ(MAPK/ERK)のレベルの上昇は、いくつかの癌細胞において活発にかつ頻繁に見出される。p38 MAPK不活化によるIL-2産生のアップレギュレーションは、ERK1/2活性の増加によって媒介される。
【0008】
ERK2は広く分布するプロテインキナーゼであり、上流のMAPキナーゼキナーゼであるMEK1によりThrl83およびTyrl85の両方がリン酸化される場合に、最大の活性に達する(Andersonら、1990、Nature 343、651;Crewsら、1992、Science 258、478)。活性化されると、ERK2は、プロテインキナーゼRsk90(Bjorbaekら、1995、J.Biol.Chem.270、18848)およびMAPK AP2(Rouseら、1994、Cell 78、1027)、ならびにATF2(Raingeaudら、1996、MoI.Cell Biol.16、1247)、Elk-1(Raingeaudら、1996)、c-Fos(Chenら、1993 Proc.Natl.Acad.Sci.USA 90、10952)およびc-Myc(Oliverら、1995、Proc.Soc.Exp.Biol.Med.210、162)などの転写因子を含む多数の調節タンパク質をリン酸化する。ERK2は、Ras/Raf依存的経路の下流の標的でもあり(Moodieら、1993、Science 260、1658)、これらの潜在的に発癌性のタンパク質からのシグナルの中継を助け得る。ERK2は、乳癌細胞の負の増殖制御において役割を果たすことが示されており(FreyおよびMulder、1997、Cancer Res.57、628)、ヒト乳癌におけるERK2の過剰発現が報告されている(Sivaramanら、1997、J Clin.Invest.99、1478)。活性化されたERK2は、エンドセリン刺激された気道平滑筋細胞の増殖にも関与しており、このことは、喘息におけるこのキナーゼの役割を示唆している(Whelchelら、1997、Am.J.Respir.Cell MoI.Biol.16、589)。
【0009】
JNKは、他のMAPKと共に、癌、トロンビンが誘導する血小板凝集、免疫不全障害、自己免疫疾患、細胞死、アレルギー、骨粗しょう症および心疾患に対する細胞応答の媒介において役割を有することに関与している。
JNKは種々の形態の心臓ストレスに対する肥大応答を媒介していることが示されているので、心筋梗塞またはうっ血性心不全などの心血管疾患におけるJNKの役割も報告されている[Circ.Res.83:167-78(1998);Circulation 97:1731-7(1998);J.Biol.Chem.272:28050-6(1997);Circ.Res.79:162-73(1996);Circ.Res.78:947-53(1996);J.Clin.Invest.97:508-14(1996)]。
【0010】
JNKカスケードは、IL-2プロモーターの活性化を含むT細胞活性化においても役割を果たすことが実証されている。したがって、JNKの阻害剤は、病的な免疫応答を変更することにおいて治療的価値を有し得る[J.Immunol.162:3176-87(1999);Eur.J.Immunol.28:3867-77(1998);J.Exp.Med.186:941-53(1997);Eur.J.Immunol.26:989-94(1996)]。
種々の癌におけるJNK活性化の役割も確立されており、癌におけるJNK阻害剤の潜在的な使用が示唆されている。例えば、構成的に活性化されたJNKは、HTLV-1媒介性の腫瘍形成に関連している[Oncogene 13:135-42(1996)]。さらに、p210 BCR-ABL形質転換細胞におけるc-jun遺伝子の調節はJNKの活性に対応し、このことは、慢性骨髄性白血病(CML)の治療におけるJNK阻害剤の役割を示唆している[Blood 92:2450-60(1998)]。
【0011】
JNK、特にJNK3のシグナル伝達は、アルツハイマー病、パーキンソン病、ALS(筋萎縮性側索硬化症)、癲癇およびけいれん、ハンチントン病、外傷性脳損傷、ならびに虚血および出血性卒中などの、アポトーシスが駆動する神経変性疾患の領域に関与している。
JNKは、いくつかの化学療法薬および癌遺伝子産物(例えば、Bcr-AbI、Her-2/Neu、Srcおよび発癌性Ras)により強力に活性化されるが、通常はERKに対する作用を有さない。したがって、SAPKを活性化し、同時にERKをダウンレギュレートする多機能の役割を果たし得る薬剤を設計/産生する必要性が長い間存在していた。
【0012】
変更されたMAPKシグナル伝達経路は、疾患をいくつか挙げると、神経変性障害、自己免疫疾患、腫瘍の発達および進行、化学療法に対する抵抗性、虚血傷害後の損傷などの、種々の疾患状態に関連する。
これらの疾患状態は、MAPKの調節で効果的に治療できる。照射が誘導する癌細胞死ならびに化学療法が誘導する癌の死は、MAKシグナル伝達経路の調節(例えばSAPKのアップレギュレーション)と関連する。
【0013】
SAPKの一過性のアップレギュレーションもまた、自己免疫を誘導することなく免疫応答を誘導するのに有用である。SAPKの一過性のアップレギュレーションは、正常細胞に顕著に影響を与えることなく、癌細胞の死を誘導するのにも有用である。SAPKの一過性のアップレギュレーションは、抗原提示細胞の活性化ならびに成熟T細胞および樹状細胞において役割を果たすことが知られている。Arrighiらは、APC成熟におけるホスホ-p38キナーゼの役割を記載している(The Journal of Immunology、2001、166:3837-3845)。Mercedes Rinconら(Free Radical Biology and Medicine、28巻、9号、2000年5月1日、1328-1337ページ)に記載されたように、SAPKのアップレギュレーションは、T細胞応答において重要な役割を果たす。
【0014】
SAPKのダウンレギュレーションは、神経変性疾患における損傷を予防するのに有用である。これは、自己免疫疾患の管理においても有用であり、虚血傷害後の損傷を最小化する。
したがって、SAPKのダウンレギュレーションとERKのアップレギュレーションまたは少なくとも変化を起こさないこととの組み合わせ効果は、上記種々の疾患状態における組織への損傷を予防するのに有用であろう。
【0015】
米国特許第6,994,981号は、パラアポトーシスの調節剤および関連の方法を記載している。EP1208748、WO2004/089929およびWO2006/117567は、MAPK阻害剤に基づく先行技術の特許である。
米国特許第6,852,740号B2は、p38キナーゼ阻害剤としてのピラゾール誘導体を記載している。WO95/31451は、MAPK、特にp38を阻害するピラゾール組成物を記載している。しかし、これらの阻害剤のin vivoでの有効性は、なお調査中である。
【0016】
ストレス活性化経路とマイトジェン活性化ERK経路との間には、負のクロストーク関係が存在する。JNKアクチベータ(例えば、TNF-αおよびセラミド)の生物学的機能のいくつかは、ERK/MAPK経路を介して作用する成長および生存因子に対する細胞の応答を遮断するそれらの能力に寄与している可能性があり、したがって、これらの結果のうち1つを制御することは、シグナル伝達のさらなる歪みを生じ、これらの疾患状態を悪化させる。
ほとんどのMAPKについて、媒介される疾患状態は、SAPKとERKとの間のクロストークを調節することによってMAPK調節に対して作用する薬剤を必要とする。
【0017】
したがって、このような疾患状態の治療について、MAPK経路のレベルの1種または複数の中間体または/およびそれらの比率は一貫しない。特に、1種を超える中間体のレベルが誤って調節される疾患状態の場合、MAPK調節剤が必要であろう。
【0018】
これらの合成MAPK阻害剤の毒性副作用は、下痢、発疹、疲労、手足症候群、脱毛症、悪心、手足皮膚反応、または一般に錯感覚を伴う手掌および足裏上の痛みのある対称的な紅斑性および浮腫性の領域を特徴とする先端紅斑である。時に、指の側方または爪周囲の領域が影響を受けることがある。角質増殖および剥離が一般に生じる。口内炎、脱毛症、掻痒症および爪下綿状出血も観察され、これらの出血は、爪の下の黒または赤のまっすぐな線を特徴とする。これらは、血栓性または塞栓性の機構に起源すると思われる。最初は細菌性心内膜炎の典型的な徴候と思われたが、これらは続いて、高地で、または動脈カテーテルを使用した場合、異なる状況でも存在することが報告された(例えば、抗リン脂質抗体症候群、重症関節リウマチ、閉塞性血栓血管炎、僧帽弁狭窄症)(The Oncologist、12巻、12号、1443-1455、2007年12月)。
【0019】
BCGおよびCpGはSAPKをアップレギュレートでき、同時にERKのアップレギュレーションも観察される。ERKのアップレギュレーションは、副作用を導く炎症性サイトカインであるTNFおよびIL-10のアップレギュレーションを導く。
【0020】
したがって、異常なレベルのSAPKおよびERKに関連する種々の疾患状態を治療するのに有用な、非毒性のMAPK調節剤を開発する必要性が長い間認められてきた。
【発明の概要】
【発明が解決しようとする課題】
【0021】
本発明において、MAPK/ERK中間体レベルの調節を介してシグナル伝達経路を調節するための新規アプローチがとられ、ここで、MAPK/ERK経路は、Mwの用量および細胞の状態に依存してMycobacterium wによって調節/妨害され、したがって低い毒性を有する。また、Mycobacterium w製剤は、百万人を超えるヒト患者の治療のために、種々の適応症に対して既に使用されており、安全性が証明されている。
【課題を解決するための手段】
【0022】
本発明の目的は、マイトジェン活性化プロテインキナーゼ(MAPK)シグナル伝達経路の調節剤を提供することである。
本発明の別の目的は、Mycobacterium wおよび/またはその成分を使用する、マイトジェン活性化プロテインキナーゼの調節を提供することである。
【0023】
本発明の別の目的は、マイトジェン活性化プロテインキナーゼを調節する方法を提供することである。
本発明のなお別の目的は、SAPKがダウンレギュレートされる一方で、ERKレベルは変化を示さない、マイトジェン活性化プロテインキナーゼの調節剤を提供することである。
【0024】
本発明のなお別の目的は、SAPKがアップレギュレートされる一方で、ERKレベルはダウンレギュレートされる、マイトジェン活性化プロテインキナーゼの調節剤を提供することである。
本発明のなお別の目的は、SAPKがアップレギュレートされる一方で、ERKレベルは変化を示さない、マイトジェン活性化プロテインキナーゼの調節剤を提供することである。
本発明のなお別の目的は、哺乳動物において、非経口、腸内および局所経路を介して投与されるMAPK調節剤を提供することである。
【図面の簡単な説明】
【0025】
【図1】ERKの変化を伴わないSAPKのダウンレギュレーション:In vitro。
【図2】ERKの変化を伴わないSAPKのダウンレギュレーション:In vivo。
【図3】異なるMw濃度を使用したMAPK調節の目的別効果。
【図4】ERKのダウンレギュレーションを伴うSAPKのアップレギュレーション:in vitro。
【図5】異なるMw濃度を使用したMAPK調節の目的別効果。
【図6】MwによるMAPK調節の寿命。
【図7】ERKレベルの変化を伴わないSAPKの一過性のアップレギュレーション。
【発明を実施するための形態】
【0026】
本発明によれば、MAPK調節剤は以下のプロセスによって調製される。本発明は、医薬組成物、調製方法、HPLC特徴、その安全性および忍容性を含み、使用方法および治療結果は、以下の実施例に記載される。以下は、本発明の例示的な実施例であり、本発明の範囲はこれらの実施例により限定されるべきではない。
【0027】
Mycobacterium wを調製するプロセス:
A. Mycobacterium wの培養
i)培地の調製
Mycobacterium wは、LJ培地などの固体培地またはミドルブルック培地もしくはソートン液体培地などの液体培地で培養する。
より良好な収率のために、ミドルブルック培地を富化する。この培地は好ましくは、グルコース、バクト-トリプトンおよびBSAの添加によって富化できる。これらは好ましくは、20:30:2の比率で使用する。
富化培地を、ミドルブルック培地に添加する。これは、好ましくは15:1〜25:1の比率で、より好ましくは20:1の比率で実施する。
【0028】
ii)バイオリアクタ操作:
a)容器の準備:
容器の内側接触部分(ジョイント、メカニカルシール、O-リング/ガスケット溝など)は、汚染を回避するために適切に洗浄すべきである。この容器を0.1N NaOHで満たし、そのまま24時間放置して、発熱物質および他の汚染物質を除去する。次いで、この容器を最初に酸性化水で洗浄し、次いで通常の水で洗浄する。最後に、この容器を蒸留水ですすぎ(3回)、その後、培地を調製する。
b)バイオリアクタの滅菌:
9Lの蒸留水を含むバイオリアクタを、生蒸気で滅菌する(間接的)。同様に、ミドルブルック培地とともにバイオリアクタを、再度滅菌する。他の添加ビン、入口/出口のエアフィルタなどを、121℃で15分間オートクレーブする(2回)。使用前に、これらを50℃のオーブンで乾燥する。
c)環境パラメータ
i.温度:37±0.5℃
ii.pH:最初6.7〜6.8。
【0029】
B. 回収および濃縮:
これは典型的には、無菌条件下で培養した後、6日目の終わりに行う。細胞の濃縮(ペレット化)は、遠心分離によって実施する。
【0030】
C. 細胞の洗浄:
こうして得たペレットを、通常の生理食塩水で最低3回洗浄する。これは、好ましくは等張の任意の他の流体で洗浄することができる。
【0031】
D. 医薬として許容可能な担体の添加
(発熱物質を含まない通常の生理食塩水をペレットに添加する。)最も好ましくは、発熱物質を含まない等張の任意の流体を医薬的担体として使用できる。担体は、最終形態で所望の活性濃度となるような量で添加する。
【0032】
E. 防腐剤の添加:
細胞/ペレットを汚染なしで維持するために、防腐剤を添加する。好ましくは、0.01%w/vの濃度を有するチメロサールを使用する。
【0033】
F. 最終滅菌:
加熱または電離放射線または滅菌濾過の適用などの種々の物理的方法により、細胞/ペレットを滅菌する。加熱は、乾式加熱または湿式加熱の形態で行うことができる。加熱は、煮沸または低温殺菌の形態とすることもできる。電離放射線は、紫外線またはガンマ線またはマイクロ波または任意の他の形態で行うことができる。
【0034】
G. 品質制御:
i.材料を、純度、無菌性について評価する。
ii.生物を、グラム染色後に抗酸性についてチェックする。
iii.生化学的試験:生物を、以下の生化学的試験に供する。
a)ウレアーゼ
b)Tween 80加水分解
c)ナイアシン試験
d)硝酸還元試験
【0035】
生物は、ウレアーゼ、tween 80加水分解およびナイアシン試験において陰性結果を生じる。生物は、硝酸還元試験では陽性である。
iv.不活化試験:これは、生存生物を見出すために、LJ培地上で産物を培養することにより行う。
v.病原性および/または病原体による汚染
培養した生物を、balb/cマウスに感染させる。死亡するマウスは出ないはずであり、全てが健康のままで体重が増加するはずである。処置の8週間後までに動物を屠殺した時点で、肝臓、肺、脾臓または任意の他の臓器中には、肉眼的病変も顕微鏡的病変も見出されないはずである。
【0036】
H. Mycobacterium w成分の調製:
Mw成分は以下の方法によって調製できる。
I.細胞破壊
II.溶媒抽出
III.酵素抽出
【0037】
細胞破壊は、超音波処理または高圧フラクショノメータの使用または浸透圧成分の適用によって実施できる。破壊した細胞を、生理食塩水で洗浄し、遠心分離によって再ペレット化した。
溶媒抽出は、クロロホルム、エタノール、メタノール、アセトン、フェノール、ハロゲン化炭化水素、イソプロピル-アルコール、酢酸、尿素、ヘキサンおよび/または芳香族化合物などの任意の有機溶媒の、個々にまたはそれらの任意の組合せにより、実施できる。
【0038】
酵素抽出は、細胞壁/細胞膜を消化できるタンパク質分解酵素によって実施できる。リゾチーム、リティカーゼおよびプロナーゼが好ましい酵素である。Mwの細胞成分が、Mwの代わりに使用できる。Mwの細胞成分の添加は、製品の有効性の改善をもたらす。
培養した生物を、Balb/cマウスに感染させる。死亡するマウスはでないはずであり、全てが健康なままで体重が増加するはずである。治療の8週間後までに動物を屠殺した時点で、肝臓、肺、脾臓または任意の他の臓器中には、肉眼的病変も顕微鏡的病変も存在しないはずである。
【0039】
こうして調製したMycobacterium wを、そのMAPK調節活性について評価した。以下の実施例は、Mycobacterium wおよび/またはその成分によるMAPKの調節を示す。
以下の実施例は本発明を実証するが、本発明の範囲を限定するものではない。
【実施例】
【0040】
A. NFS60細胞株:サンプル調製
回収後保存した細胞1×107個の細胞ペレットを、以下のように処理する:細胞を、溶解緩衝液#6(PBS(pH7.2-7.4)中、1mM EDTA、0.5% Triton X-100、6M 尿素、10μg/mL ロイペプチン、10μg/mL ペプスタチン、100μM PMSF、3μg/mL アプロチニン、2mM ピロリン酸ナトリウム、1mM 活性化バナジン酸ナトリウム)中に可溶化する。
溶解物を短時間ボルテックスし、10分間氷上に置く。次いで、plastocraft中で10,000rpmで10分間遠心分離する。上清を新たなチューブに移す。6倍希釈の溶解物を、IC Diluent #8(PBS(pH7.2-7.4)中、1mM EDTA、0.5% Triton X-100)を用いて調製し、IC Diluent #3でさらに希釈する。IC Diluent #3中のサンプルを、R&D Systems製のキットを使用したELISAによる分析のために使用する。
【0041】
B. ELISAプレートの調製:
ホスホ-P38(R&D Systems、DuoSet IC カタログ番号DYC869-2)
ホスホ-JNK(R&D Systems、DuoSet IC カタログ番号DYC1018-2)
ホスホ-JNK(R&D Systems、DuoSet IC カタログ番号DYC1387-2)
捕捉抗体を、製造業者の指示に従って作業濃度まで希釈した。96ウェルのマイクロプレートを、1ウェル当たり100μLの希釈したそれぞれの捕捉抗体で即座に被覆した。プレートを密封し、室温で一晩インキュベートした。
【0042】
プレートを吸引し、洗浄緩衝液で洗浄し、このプロセスを2回繰り返して合計3回洗浄した。最後の洗浄後、プレートを裏返し、きれいなブロッティングペーパーで吸い取ることによって、残留した洗浄緩衝液を除去した。各ウェルに300μLのブロック緩衝液を添加することによってプレートをブロッキングした。次いで、プレートを室温で2時間インキュベートし、その後プレートを再度3回洗浄し、吸引し、吸引乾燥した。これで、サンプル添加のためのプレートの準備ができたことになる。各MAPKのためのストリップを、さらなる使用のために、2〜8℃でデシケータ中で保存した。
【0043】
C. 脾細胞の調製:
脾細胞を正常マウスから回収した。マウスを頚椎脱臼によって屠殺した。即座に腹腔を開き、脾臓を単離した。各脾臓を個別にさらに処理した。
各脾臓を、10mlのダルベッコリン酸塩緩衝化生理食塩水(DPBS)で洗浄し、汚染している血液および他の不純物を除去した。脾臓を、無菌シリンジピストンを使用して刻んだ。10mlのRPMI-1640完全培地(10%FBSおよび1%ペニシリン-ストレプトマイシン抗生物質)を添加し、40μmのナイロンセルストレイナーを介して刻んだ脾臓を篩にかけることによって、内容物を50mlのファルコンチューブに移した。細胞を、Hereaeus Multifuge-3 SR中で1500rpmで5分間遠心分離してペレット化した。
【0044】
上清を廃棄し、ペレット中に存在するRBCを、5mlの溶解緩衝液(0.017MのTris-HCI(pH=7.6)中0.144MのNH4CI)で10分間細胞を再懸濁することによって溶解した。反応を、40mlのDPBSの添加によって停止させた。細胞を、Hereaeus Multifuge-3 SR中で1500rpmで5分間遠心分離してペレット化した。得られた細胞の各セットのペレットを、1mlのRPMI-1640完全培地中に再懸濁した。その後、細胞数を、トリパンブルーで細胞を染色することによって、Neubarrチャンバを使用して決定した。細胞懸濁物を、RPMI-1640完全培地で適切に希釈して、107細胞/mlの細胞密度にした。
【0045】
各脾臓から、1mlのRPMI-1640中に細胞を1×107個含む懸濁物5mLを調製した。1mlの各細胞懸濁物のそれぞれを、マイクロタイタープレート中に4連で播種し、そのうち2つを108個のMycobacterium w細胞で刺激した。プレートを6%CO2で37℃で36時間インキュベートした。
48時間後、細胞を回収した。完全にピペッティングした後、細胞をエッペンドルフチューブに移した。細胞を、10,000rpmで10分間の遠心分離によってペレット化した。上清を別のチューブに移した。上清および細胞ペレットの両方を標識し、さらなる分析まで-70℃で保存した。
【0046】
実施例1
ERKの変化を伴わないSAPKのダウンレギュレーション:
Mwのin vitro効果:
脾細胞を、ナイーブBalb/Cマウスから単離し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106個/mLの脾細胞の2セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108のMycobacterium w細胞と共にインキュベートした。
【0047】
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
ホスホ-JNKの結果を、図1中に示されるようにプロットした。108個のMycobacterium wによる脾細胞のin vitro刺激が、48時間にわたりホスホ-JNKレベルの有意な変化を示さないことが観察される。
【0048】
ホスホ-p38の結果を、図1中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激は、108個のMycobacterium w細胞と共に48時間経過した後、ホスホ-p38 MAPKをダウンレギュレートすることが観察される。したがって、Mycobacterium wは、ホスホ-p38レベルをダウンレギュレートする。
ホスホ-ERKの結果を、図1中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激は、対照と比較して、48時間にわたりホスホ-ERKレベルの有意な変化を示さないことが観察される。
【0049】
Mwのin vivo効果:
Balb/Cマウスの群1では0.1mLのPBSで免疫し、群2では0.1mLのMycobacterium w(108細胞)を皮内に投与した後、このBalb/Cマウスから、その7日後に、脾細胞を単離した。細胞を、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
【0050】
結果を図2に示す。Mwで免疫したマウスは、ERKレベルの変化を伴わずに、SAPKのダウンレギュレーションを示すことが観察された。
【0051】
異なるMw濃度を使用した目的別効果:
脾細胞を、ナイーブなBalb/Cマウスから単離し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106個/mLの脾細胞の3セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108個のMycobacterium w細胞と共にインキュベートし、セット3は106個のMycobacterium w細胞と共にインキュベートした。
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
【0052】
ホスホ-JNKの結果を、図3中に示されるようにプロットした。106個のMycobacterium wによる脾細胞のin vitro刺激が、48時間にわたりホスホ-JNKレベルのダウンレギュレーションを示さないことが観察され、その一方で、108個のMycobacterium wで刺激した場合、48時間目の終わりにホスホ-JNKレベルのダウンレギュレーションを示すことが観察される。1998年に、Science 282:2092-2095でDong,Chenらが、およびImmunity 9:575-585でYang,D.Dが、ホスホ-JNKのアップレギュレーションが、T細胞の分化、増殖および活性化に必要であることを記載している。
【0053】
ホスホ-p38の結果を、図3中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激が、108個のMycobacterium w細胞との48時間の後に、ホスホ-p38 MAPKをダウンレギュレートすることが観察されたが、106個のMycobacterium w細胞では観察されなかった。Arrighiらは、APC成熟におけるホスホ-p38キナーゼの役割を記載している(The Journal of Immunology、2001、166:3837-3845)。
ホスホ-ERKの結果を、図3中に示されるようにプロットした。Mycobacterium wによる脾細胞のin vitro刺激が、対照と比較して、48時間にわたりホスホ-ERKレベルの有意な変化を示さないことが観察される。
【0054】
実施例2
ERKのダウンレギュレーションを伴うSAPKのアップレギュレーション:
Mwのin vitro効果:
NFS60細胞を、10%FBS、1%抗生物質および10nG/mLのIL-3を含むDMEM培地中で培養した。細胞を、細胞1×105個の濃度でマイクロタイターウェル中にプレートした。ウェルの数を2セットに分割した。セット1を対照としてPBSと共にインキュベートし、セット2を3×107個のMycobacterium w細胞と共にインキュベートした。
【0055】
培養1、2、4、8および24時間の時点で、細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
対照と比較したMycobacterium wで刺激された細胞中のホスホ-JNKレベルは、4時間目まで有意な変化を示さず、8時間目の終わりから24時間目の時点で2倍になる(図4)。
【0056】
対照と比較したMycobacterium wで刺激された細胞中のホスホ-p38レベルは、示差的な用量依存的効果を示す。3×107個の用量のMycobacterium w細胞では、ホスホ-p38レベルは急上昇し、4〜8時間目にわたり高いままであり、次いで低下し始め、24時間目の終わりには対照よりも低い(図4)。刺激のための3×107個の濃度のMycobacterium w細胞でNFS-60細胞が殺傷されるのは驚くべきことであった。
【0057】
対照におけるホスホ-ERKレベルは、8時間目まで有意な変化を示さず、24時間目の終わりには2倍になる。これは、細胞数の増加に起因する可能性がある。図4中に示されるように、Mycobacterium wで刺激された細胞は、最初の1時間目の時点でそれ自体ホスホ-ERKレベルの増加を示して2倍になり、8時間目の時点で、1時間目の半分まで低下し、24時間目の時点で、そのレベルは、その時点での対照の1/4とほぼ同じになる。この低下は、生存細胞数の減少と関連した。
したがって、ホスホ-ERKレベルの同時の低下を伴ったSAPK(ホスホ-p38レベルおよびホスホ-JNKレベル)の上昇は、NFS60細胞の死と関連する。
【0058】
異なるMw濃度を使用した目的別効果:
NFS60細胞を、10%FBS、1%抗生物質および10nG/mLのIL-3を含むDMEM培地中で培養した。細胞を、1×105細胞の濃度でマイクロタイターウェル中にプレートした。ウェルの数を5セットに分割した。セット1は対照としてPBSと共にインキュベートし、残りは各々、3×107、1×107、7×106および3×106個のMycobacterium w細胞と共にそれぞれインキュベートした。
【0059】
培養1、2、4、8および24時間の時点で、細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
3×107個のMycobacterium wで刺激された細胞中のホスホ-JNKレベルは、対照と比較して、4時間目まで有意な変化を示さず、8時間目の終わりから24時間目で2倍になるが(図5)、一方、より低い濃度では、レベルの上昇は遅延し試験期間中には効果はない(図5)。
【0060】
対照と比較したMycobacterium wで刺激された細胞中のホスホ-p38レベルは、示差的な用量依存的効果を示す。3×107個の用量のMycobacterium w細胞では、ホスホ-p38レベルは急上昇し、4〜8時間にわたり高いままであり、次いで低下し始め、24時間目の終わりでは対照よりも低い。他の全てのMycobacterium w濃度で、ホスホ-p38レベルは、対照と同様にダウンレギュレートされる(図5)。
【0061】
対照におけるホスホ-ERKレベルは、8時間目まで有意な変化を示さず、24時間目の終わりには2倍になる。これは、細胞数の増加に起因する可能性がある。図5中に示されるように、Mycobacterium wで刺激された細胞は、8時間目の時点で1時間目の半分までの低下を示すが、24時間目の時点では、そのレベルは、その時点での対照の1/4とほぼ同じになる。この低下は、生存細胞数の減少と関連した。
【0062】
実施例3
MAPK調節の寿命:
Balb/Cマウスの群1では1mLのPBSで免疫し、群2〜6では1mLのMycobacterium w(109細胞)を静脈内に投与し、これらのBalb/Cマウスから、脾細胞を単離した。群1および2を1日目に屠殺し、群3を7日目に、群4を14日目に、群5を21日目に、群6を28日目に屠殺し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106/mLの脾細胞の3セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108個のMycobacterium w細胞と共にインキュベートし、セット3は106個のMycobacterium w細胞と共にインキュベートした。
【0063】
培養48時間の時点で細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
図6中に示されるように、ホスホ-JNKレベルは、免疫の24時間後から28日目(試験期間)までダウンレギュレーションを示す。
【0064】
図6中に示されるように、ホスホ-p38レベルも、免疫の24時間後から21日目までダウンレギュレーションを示した。
図6中に示されるように、ホスホ-ERKレベルは、免疫の24時間後から14日目までダウンレギュレーションを示し、その後再び正常レベルとなる。
【0065】
実施例4
ERKの変化を伴わないSAPKの一過性のアップレギュレーション:
脾細胞を、ナイーブなBalb/Cマウスから単離し、マイクロタイタープレート中で、10%FBSおよび1%抗生物質を含むRPMI 1640培地中で培養した。細胞を、それぞれ106個/mLの脾細胞の3セットに分割した。セット1はPBSと共にインキュベートし(対照)、セット2は108のMycobacterium w細胞と共にインキュベートし、セット3は106個のMycobacterium w細胞と共にインキュベートした。
【0066】
培養1、2、4、8および24時間の時点で、細胞を回収し、MAPK ELISA(ホスホ-JNKアッセイ、ホスホ-p38アッセイおよびホスホ-ERKアッセイ)を、R&D Systems製の上記市販キットを使用して、製造業者の指示に従って実施した。
ELISAにより測定したホスホ-JNKレベルは、106個のMycobacterium wによるホスホロ(phosphor)ホスホ-JNKのアップレギュレーションを示す。図7中に示されるように、このアップレギュレーションは、1〜8時間目だけの一過的なものであるが、24時間目の時点で、このレベルはダウンレギュレートされる。
【0067】
ELISAにより測定したホスホ-p38レベルは、108個でのホスホロ(phosphor)ホスホ-p38のアップレギュレーションを示す。図7中に示されるように、このアップレギュレーションは、1〜4時間目だけの一過的なものであるが、8〜24時間目では、このレベルはダウンレギュレートされる。
SAPKのアップレギュレーションは、Mercedes Rinconら(Free Radical Biology and Medicine、28巻、9号、2000年5月1日、1328-1337ページ)に記載されるように、T細胞応答において重要な役割を果たす。これらの知見は、より低いレベルと同様に用量依存的であり、図7中に示されるように、細胞106個のMycobacterium wは、この特性を示さず、一方でERKでは、このような調節は観察されなかった。
【特許請求の範囲】
【請求項1】
Mycobacterium w(Mw)および/またはその成分は、哺乳動物および/または哺乳動物細胞におけるマイトジェン活性化プロテインキナーゼの調節剤である。
【請求項2】
Mycobacterium wおよび/またはその成分を投与する工程を含む、マイトジェン活性化プロテインキナーゼの調節を必要とする哺乳動物においてマイトジェン活性化プロテインキナーゼを調節する方法。
【請求項3】
使用されるMycobacterium wおよび/またはその成分の濃度が、1012〜101個のMycobacterium w細胞の範囲である、請求項2に記載のマイトジェン活性化プロテインキナーゼを調節する方法。
【請求項4】
使用されるMycobacterium wおよび/またはその成分の好ましい濃度が、108〜106個のMycobacterium w細胞の範囲である、請求項2に記載のマイトジェン活性化プロテインキナーゼを調節する方法。
【請求項5】
ERKレベルの変化を伴わないSAPKの一過性のアップレギュレーションを含む、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【請求項6】
ERKレベルのダウンレギュレーションを伴うSAPKの一過性のアップレギュレーションを含む、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【請求項7】
ERKレベルの有意な変更を伴わないSAPKのダウンレギュレーションを含む、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【請求項8】
Mycobacterium wおよび/またはその成分の量を変化させることにより調節が決定される、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【請求項1】
Mycobacterium w(Mw)および/またはその成分は、哺乳動物および/または哺乳動物細胞におけるマイトジェン活性化プロテインキナーゼの調節剤である。
【請求項2】
Mycobacterium wおよび/またはその成分を投与する工程を含む、マイトジェン活性化プロテインキナーゼの調節を必要とする哺乳動物においてマイトジェン活性化プロテインキナーゼを調節する方法。
【請求項3】
使用されるMycobacterium wおよび/またはその成分の濃度が、1012〜101個のMycobacterium w細胞の範囲である、請求項2に記載のマイトジェン活性化プロテインキナーゼを調節する方法。
【請求項4】
使用されるMycobacterium wおよび/またはその成分の好ましい濃度が、108〜106個のMycobacterium w細胞の範囲である、請求項2に記載のマイトジェン活性化プロテインキナーゼを調節する方法。
【請求項5】
ERKレベルの変化を伴わないSAPKの一過性のアップレギュレーションを含む、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【請求項6】
ERKレベルのダウンレギュレーションを伴うSAPKの一過性のアップレギュレーションを含む、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【請求項7】
ERKレベルの有意な変更を伴わないSAPKのダウンレギュレーションを含む、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【請求項8】
Mycobacterium wおよび/またはその成分の量を変化させることにより調節が決定される、請求項2に記載のマイトジェン活性化プロテインキナーゼの調節。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2010−531347(P2010−531347A)
【公表日】平成22年9月24日(2010.9.24)
【国際特許分類】
【出願番号】特願2010−514167(P2010−514167)
【出願日】平成20年6月26日(2008.6.26)
【国際出願番号】PCT/IB2008/001675
【国際公開番号】WO2009/001204
【国際公開日】平成20年12月31日(2008.12.31)
【出願人】(507421393)カディラ ファーマシューティカルズ リミテッド (7)
【Fターム(参考)】
【公表日】平成22年9月24日(2010.9.24)
【国際特許分類】
【出願日】平成20年6月26日(2008.6.26)
【国際出願番号】PCT/IB2008/001675
【国際公開番号】WO2009/001204
【国際公開日】平成20年12月31日(2008.12.31)
【出願人】(507421393)カディラ ファーマシューティカルズ リミテッド (7)
【Fターム(参考)】
[ Back to top ]