
マウスROSA26遺伝子座からのDNA標的配列を切断するメガヌクレアーゼ変異型及びその使用
2つのI-CreI単量体のうちの一方が、I-CreIの26位〜40位及び44位〜77位にそれぞれ位置するLAGLIDADGコアドメインの2つの機能的サブドメイン内に1つずつ存在する少なくとも2つの置換を有し、マウスROSA26遺伝子座からのDNA標的を切断できるI-CreI変異型。該変異型及びその派生生成物の、興味対象の異種タンパク質を発現するトランスジェニックマウス及び組換えマウス株化細胞を工学的に作製するための使用。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、マウスROSA26遺伝子座からのDNA標的配列を切断するメガヌクレアーゼ変異型(variant)、該変異型をコードするベクター、該ベクターにより改変された細胞、動物若しくは植物、並びに該メガヌクレアーゼ変異型及び派生生成物の、マウスゲノム工学(組換えタンパク質生成、トランスジェニックマウス及び組換えマウス株化細胞の構築)のための使用に関する。
【背景技術】
【0002】
マウスROSA26遺伝子座は、Friedrich及びSorianoにより1991年に、レトロウイルスを感染させた胚性幹(ES)細胞を用いるジーントラップ実験により発見された(Friedrich, G.及びP. Soriano, Genes & Development, 1991, 5, 1513〜1523)。非必須部位であるROSA26遺伝子座のイントロン1で挿入が生じるROSA26マウスジーントラップ系統は、新生児(Friedrich及びSoriano, 1991, 上記)及び造血細胞(Zambrowiczら, Proc. Natl. Acad. Sci. USA, 1997, 94, 3789〜3794)において、胚の発達の間にレポーター遺伝子の遍在的な発現を示す。マウスの第6染色体に位置するROSA26遺伝子座は、3つの転写産物を生成する(図1)。2つの転写産物は、同一の5'末端を共有する共通のプロモーターから発生する(エキソン1及び開始エキソン2)が、どちらも意味のあるORFを含有しない。3つ目のものは、リバース鎖から発生する(Zambrowiczら, 1997, 上記)。マウスROSA26プロモーターの制御下の導入遺伝子は、胚及び成体マウスで遍在的な発現を示す(Soriano, P., Nature Genetics, 1999, 21, 70〜71)。ROSA26遺伝子座をマウスES細胞でターゲティングすることは、トランスジェニックマウスモデルを構築するために広く用いられている(Kisseberthら, Developmental Biology, 1999, 214, 128〜138; Mao X.ら, Proc. Natl. Acad. Sci. USA, 1999, 96, 5037〜5042; Soriano, 1999, 上記; Awatramaniら, Nature Genetics, 2001, 29, 257〜259; Mao X.ら, Blood, 2001, 97, 324〜326; Possematoら., Genesis, 2002, 32, 184〜186; Mao, J.ら, Nucleic Acids Res., 2005, 33, e155; Yuら, Proc. Natl. Acad. Sci. USA, 2005, 102, 8615〜8620; 国際PCT出願WO 99/53017、WO 02/098217、WO 03/020743、WO 2004/063381及びWO 2005/116070))。
【0003】
しかし、マウス細胞における相同組換えの効率は非常に低い(頻度:10-6〜10-9)。
この効率は、標的遺伝子座でのDNA二本鎖破断(double-strand break;DSB)により増強できる。このようなDSBは、定義によれば大きい配列を認識する配列特異的エンドヌクレアーゼであるメガヌクレアーゼにより創出できる(Thierry, A.及びB. Dujon, Nucleic Acids Res., 1992, 20, 5625〜5631)。これらのタンパク質は、生細胞においてユニーク部位を切断し、そのことにより、切断部位の近傍で遺伝子ターゲティングを、1000倍以上増強させる(Puchtaら, Nucleic Acids Res., 1993, 21, 5034〜5040 ; Rouetら, Mol. Cell. Biol., 1994, 14, 8096〜8106 ; Choulikaら, Mol. Cell. Biol., 1995, 15, 1968〜1973; Puchtaら, Proc. Natl. Acad. Sci. U.S.A., 1996, 93, 5055〜5060 ; Sargentら, Mol. Cell. Biol., 1997, 17, 267〜277; Cohen-Tannoudjiら, Mol. Cell. Biol., 1998, 18, 1444〜1448 ; Donohoら, Mol. Cell. Biol., 1998, 18, 4070〜4078; Elliottら, Mol. Cell. Biol., 1998, 18, 93〜101)。
【0004】
しかし、「ホーミングエンドヌクレアーゼ」ともよばれる数百の天然メガヌクレアーゼが同定されているが(Chevalier, B.S.及びB.L. Stoddard, Nucleic Acids Res., 2001, 29, 3757〜3774)、切断され得る配列のレパートリーは、ゲノムの複雑さに取り組むには制限されすぎであり、選択された遺伝子には通常、切断可能部位はない。理論的には、選択された特異性を有する人工配列特異的エンドヌクレアーゼの作製が、この制限を緩和できるはずである。よって、仕立てられた特異性を有するメガヌクレアーゼの作製について、熱心な研究がなされている。
【0005】
最近、ジンクフィンガーと、クラスIIS制限エンドヌクレアーゼであるFokIの触媒ドメインとの融合体を用いて、機能的配列特異的エンドヌクレアーゼが作製された(Smithら, Nucleic Acids Res., 1999, 27, 674〜681; Bibikovaら, Mol. Cell. Biol., 2001, 21, 289〜297 ; Bibikovaら, Genetics, 2002, 161, 1169〜1175 ; Bibikovaら, Science, 2003, 300, 764; Porteus, M.H.及びD. Baltimore, Science, 2003, 300, 763〜; Alwinら, Mol. Ther., 2005, 12, 610〜617; Urnovら, Nature, 2005, 435, 646〜651; Porteus, M.H., Mol. Ther., 2006, 13, 438〜446; 国際PCT出願WO 2007/014275)。このようなヌクレアーゼは、最近、リンパ系列からのヒト細胞におけるILR2G遺伝子を工学的に改変するために用いることができた(Urnovら, Nature, 2005, 435, 646〜651)。
【0006】
Cys2-His2型のジンクフィンガータンパク質(ZFP)の結合特異性は、これらが単純(フィンガーあたり実質的に4残基により駆動される特異性)でモジュール性の(modular)系を示すので、操作するのが容易である(Paboら, Annu. Rev. Biochem., 2001, 70, 313〜340; Jamiesonら, Nat. Rev. Drug Discov., 2003, 2, 361〜368)。Pabo (Rebar, E.J.及びC.O. Pabo, Science, 1994, 263, 671〜673 ; Kim, J.S.及びC.O. Pabo, Proc. Natl. Acad. Sci. U S A, 1998, 95, 2812〜2817)、Klug (Choo, Y.及びA. Klug, Proc. Natl. Acad. Sci. USA, 1994, 91, 11163〜11167; Isalan M.及びA. Klug, Nat. Biotechnol., 2001, 19, 656〜660)及びBarbas (Choo, Y.及びA. Klug, Proc. Natl. Acad. Sci. USA, 1994, 91, 11163〜11167; Isalan M.及びA. Klug, Nat. Biotechnol., 2001, 19, 656〜660)の実験室からの研究は、ほとんどのG/ANNG/ANNG/ANN配列に結合できる新規な人工ZFPの大きいレパートリーをもたらした。
【0007】
にもかかわらず、ZFPは、特に、治療用途のような非常に高レベルの特異性を必要とする用途において制限がある。最近、融合体でのFokIヌクレアーゼ活性が、FokIのいくつかのDNA結合欠陥性変異体の存在下を含んで、1つの認識部位、又はDNAループにより種々の距離で分けられた2つの部位のいずれかと作用することが見出されている(Cattoら, Nucleic Acids Res., 2006, 34, 1711〜1720)。よって、特異性は、哺乳動物細胞及びショウジョウバエ(Drosophila)における毒性により示されるように、非常に退化している(Bibikovaら, Genetics, 2002, 161, 1169〜1175; Bibikovaら, Science, 2003, 300, 764〜)。
【0008】
野生では、メガヌクレアーゼは、ホーミングエンドヌクレアーゼにより実質的に代表される。ホーミングエンドヌクレアーゼ(HE)は、数百のタンパク質ファミリーを含む天然のメガヌクレアーゼの広範なファミリーである(Chevalier, B. S.及びB.L. Stoddard, Nucleic Acids Res., 2001, 29, 3757〜3774)。これらのタンパク質は、「ホーミング」とよばれるプロセスにより伝播される可動性遺伝要素によりコードされる。エンドヌクレアーゼは、可動性要素が存在しない同族の対立遺伝子を切断することにより、受容側遺伝子座に可動性DNAを重複させる相同組換え事象を刺激する。効率及び特異性の点でのそれらの例外的な切断特性に鑑みて、これらは、新規で特異性が高いエンドヌクレアーゼを導くために理想的な足場であり得る。
【0009】
HEは、4つの主要なファミリーに属する。触媒中心に含まれる保存されたペプチドモチーフにちなんで命名されたLAGLIDADGファミリーは、最も普及しており、最もよく特徴付けられている群である。現在、7つの構造が入手可能である。このファミリーからのほとんどのタンパク質が単量体であり、2つのLAGLIDADGモチーフを示すが、いくつかは1つのモチーフのみを有するものの、二量体を形成してパリンドローム又は偽パリンドローム標的配列を切断する。
【0010】
ファミリーのメンバーのうち、LAGLIDADGペプチドが唯一の保存領域であるが、これらのタンパク質は、非常によく似た構造を有する(図2)。触媒コアは、I-CreI(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316)及びI-MsoI (Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)のようなホモ二量体について完全な2回回転対称の、そしてI-SceI (Moureら, J. Mol. Biol., 2003, 334, 685〜69)、I-DmoI (Silvaら, J. Mol. Biol., 1999, 286, 1123〜1136)又はI-AniI (Bolducら, Genes Dev., 2003, 17, 2875〜2888)のような単量体について偽対称を有する2つのDNA結合ドメインで挟まれる。両方の単量体又は両方のドメイン(単量体タンパク質について)は、二価カチオンの周りに組織された触媒コアに貢献する。触媒コアのすぐ上に、2つのLAGLIDADGペプチドが、二量体形成界面において必須の役割を演じる。DNA結合は、DNA主溝の上に位置する2つの典型的なサドル形ββαββ折り畳みに依存する。他のドメインは、例えばPI-PfuI (Ichiyanagiら, J. Mol. Biol., 2000, 300, 889〜901)及びPI-SceI (Moureら, Nat. Struct. Biol., 2002, 9, 764〜770)のようなインテインにおいて見出すことができ、このタンパク質スプライシングドメインはDNA結合にも関与する。
【0011】
I-DmoIドメインのN-末端をI-CreI単量体と融合させることによる機能的キメラメガヌクレアーゼの作製(Chevalierら, Mol. Cell., 2002, 10, 895〜905; Epinatら, Nucleic Acids Res, 2003, 31, 2952〜62; 国際PCT出願WO 03/078619及びWO 2004/031346)は、LAGLIDADGタンパク質の可塑性を証明する。
そのうえ、異なるグループが、I-CreI (Seligmanら, Genetics, 1997, 147, 1653〜1664; Sussmanら, J. Mol. Biol., 2004, 342, 31〜41; 国際PCT出願WO 2006/097784、WO 2006/097853及びWO 2007/049156; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458; Rosenら, Nucleic Acids Res., 2006, 34, 4791〜4800; Smithら, Nucleic Acids Res., Epub 2006年11月27日)、I-SceI (Doyonら, J. Am. Chem. Soc., 2006, 128, 2477〜2484)、PI-SceI (Gimbleら, J. Mol. Biol., 2003, 334, 993〜1008)及びI-MsoI (Ashworthら, Nature, 2006, 441, 656〜659)の特異性を局所的に変更するために合理的アプローチを用いている。
【0012】
さらに、局所的に特異性が変更された数百のI-CreI誘導体が、半合理的(semi-rational)アプローチとハイスループットスクリーニングとを組み合わせることにより工学的に作製された:
- I-CreIの残基Q44、R68及びR70又はQ44、R68、D75及びI77を変異させ、DNA標的(5NNN DNA標的)の±3〜5位のヌクレオチドに対する特異性が変更された変異型のコレクションを、スクリーニングにより同定した(国際PCT出願WO 2006/097784及びWO 2006/097853; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458; Smithら, Nucleic Acids Res., Epub 2006年11月27日)。
【0013】
- I-CreIの残基K28、N30及びQ38、N30、Y33及びQ38又はK28、Y33、Q38及びS40を変異させ、DNA標的(10NNN DNA標的)の±8〜10位のヌクレオチドに対する特異性が変更された変異型のコレクションを、スクリーニングにより同定した(Smithら, Nucleic Acids Res., Epub 2006年11月27日; 国際PCT出願WO 2007/049156)。
I-CreIの残基28〜40及び44〜77は、ホーミングエンドヌクレアーゼハーフサイト(half-site)の別個の部分に結合できる、2つの分離可能な機能的サブドメインを形成することが示された(Smithら Nucleic Acids Res., Epub 2006年11月27日; 国際PCT出願WO 2007/049095)。
【0014】
I-CreIの2つのサブドメインからの変異を同じ単量体内で組み合わせることにより、それぞれのサブドメインが結合する±3〜5位及び±8〜10位のヌクレオチドを含むパリンドロームの組み合わせたDNA標的配列を切断できる新規なキメラ分子(ホモ二量体)の設計が可能になった(Smithら, Nucleic Acids Res., Epub 2006年11月27日; 国際PCT出願WO 2007/049156)。
【0015】
2つの異なる変異型を組み合わせ、それぞれの変異型DNA標的配列の異なる半分の融合により得られるキメラ標的を切断できる機能的へテロ二量体エンドヌクレアーゼに組み立てた(Arnouldら, 上記; 国際PCT出願WO 2006/097854)。興味深いことに、新規なタンパク質は、正確な折り畳み及び安定性と、高い活性と、狭い特異性とを維持した。
【0016】
2つの前の工程の組み合わせにより、4つの異なるサブドメインを含む、より大きいコンビナトリアルアプローチが可能になる。異なるサブドメインは、図3に示すように、別々に改変でき、組み合わせて、選択された特異性を有する完全に再設計されたメガヌクレアーゼ変異型(ヘテロ二量体又は単鎖分子)を得ることができる。第1工程において、新規なメガヌクレアーゼの対を、切断したい標的に由来するパリンドローム標的を切断する新しい分子(「ハーフメガヌクレアーゼ」)に組み合わせる。次いで、このような「ハーフメガヌクレアーゼ」の組み合わせは、興味対象の標的を切断するヘテロ二量体種をもたらすことができる。4組の変異を、モデル標的配列又はヒトRAG1遺伝子からの配列を切断するヘテロ二量体エンドヌクレアーゼに組み立てることが、Smithら(Nucleic Acids Res., Epub 2006年11月27日)に記載されている。
【0017】
しかし、この報告で試験された標的は、パリンドロームI-CreI部位の元来の配列(C1221; 図5)と、±2位及び±1位が同一であった。塩基対±1及び±2は、タンパク質といずれの接触も示さないが、これらの位置は、特に塩基対±1については、含まれる情報がないわけではなく(Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)、さらなる基質特異性の供給源であり得ることが示されている(Argastら, J. Mol. Biol., 1998, 280, 345〜353; Juricaら, Mol. Cell., 1998, 2, 469〜476; Chevalier, B.S.及びB.L. Stoddard, Nucleic Acids Res., 2001, 29, 3757〜3774)。ランダム突然変異を誘発した切断可能なI-CreI標的のインビトロ選択(Argastら, 上記)は、タンパク質結合及び切断活性に対するこれらの4つの塩基対の重要性を明らかにした。活性部位で見出される配向された(ordered)水分子のネットワークが、DNA標的の位置決めに重要であることが示唆された(Chevalierら, Biochemistry, 2004, 43, 14015〜14026)。さらに、I-CreI結合の際にこの領域に出現する広範囲なコンホメーションの変更は、4つの中央のヌクレオチドが、おそらくは、配列依存性のコンホメーションの選好により、基質特異性に貢献できるであろうことを示唆する(Chevalierら, 2003, 上記)。
【発明の概要】
【発明が解決しようとする課題】
【0018】
よって、4つの中央のヌクレオチドgtacを有するパリンドローム配列を切断するホモ二量体として10NNN及び5NNN DNA標的について同定された変異体が、4つの中央のヌクレオチドの変更を含む標的を切断する新しいエンドヌクレアーゼの設計を可能にするかは明らかでなかった。
本発明者らは、I-CreI変異型により切断され得るマウスROSA26遺伝子座における一連のDNA標的を同定した(図17)。図3に記載するコンビナトリアルアプローチを用いて、I-CreIタンパク質のDNA結合ドメインを全体的に再設計し、そのことにより、完全に工学的に改変された特異性を有する新規なメガヌクレアーゼを工学的に作製して、I-CreI C1221 22 bpパリンドローム部位とは、4つの中央のヌクレオチドのうちの1つ(+1位)を含む13ヌクレオチドが異なる(図5)、マウスROSA26遺伝子座からのDNA標的(rosa1)を切断した。
【0019】
組み合わせた変異型を、それぞれヌクレオチド10NNN及び5NNNに対して最初は同定し、工学的に作製されたメガヌクレアーゼの活性に対する標的の4つの中央のヌクレオチドの強い影響が観察されたが、特異性が大きく変更された機能的メガヌクレアーゼを選択した。さらに、工学的に作製されたタンパク質の活性は、連続する2回のランダム突然変異誘発及びスクリーニングにより、I-CreIタンパク質の活性と比較して、著しく改良できた。
【0020】
ROSA26遺伝子座にて2本鎖破断を生じる能力は、該遺伝子座での相同組換えを著しく増強する手段を提供する。つまり、ROSA26遺伝子座を標的にするメガヌクレアーゼは、マウス細胞における効率的な遺伝子挿入を可能にする(図4)。この遺伝子座にて遺伝子を効率的に挿入する(ノックイン)能力は、再現可能な発現レベルを可能にするとともに、挿入を作製するための予測可能なタイムラインを可能にするという利点を有する。可能性のある応用は、マウス細胞における組換えタンパク質の生成、及び例えばタンパク質生成、遺伝子機能研究、薬剤スクリーニングのため、又は疾患モデルとして用いることができるトランスジェニックマウス及び組換えマウス株化細胞の工学的作製を含む。
【課題を解決するための手段】
【0021】
本発明は、2つのI-CreI単量体のうちの少なくとも一方が、I-CreIの26位〜40位及び44位〜77位にそれぞれ位置するLAGLIDADGコアドメインの2つの機能的サブドメイン内に1つずつ存在する少なくとも2つの置換を有し、マウスROSA26遺伝子座からのDNA標的を切断できるI-CreI変異型に関する。
【0022】
本発明による変異型の切断活性は、国際PCT出願WO 2004/067736; Epinatら, Nucleic Acids Res., 2003, 31, 2952〜2962; Chamesら, Nucleic Acids Res., 2005, 33, e178;及びArnouldら, J. Mol. Biol., 2006, 355, 443〜458に記載されるもののような、いずれの公知のインビトロ又はインビボ切断アッセイにより測定できる。例えば、本発明の変異型の切断活性は、酵母又は哺乳動物細胞における直列反復組換えアッセイ(direct repeat recombination assay)により、レポーターベクターを用いて測定できる。レポーターベクターは、酵母又は哺乳動物発現ベクターにクローニングされた、レポーター遺伝子の2つの短縮された非機能的コピー(直列反復)と、ゲノムDNA標的配列とを、介在配列内に含む。変異型の発現は、ゲノムDNA標的配列を切断できる機能的エンドヌクレアーゼをもたらす。この切断は、直列反復間の相同組換えを誘発し、その発現は、適切なアッセイにより監視できる。
【図面の簡単な説明】
【0023】
【図1】マウスROSA26遺伝子座を表す(アクセッション番号EMBL CQ880114;配列番号3)。
【図2】DNA標的に結合したI-CreIホーミングエンドヌクレアーゼの3次元構造を表す。
【図3】I-CreI及び他のLAGLIDADGホーミングエンドヌクレアーゼの特異性を工学的に改変するための2ステップアプローチを示す。
【図4】マウスROSA26遺伝子座を切断するメガヌクレアーゼの使用のためのストラテジーを表す。
【図5】rosa1標的配列及び誘導体を表す。
【図6】pCLS1055ベクターマップを表す。
【図7】pCLS0542ベクターマップを表す。
【図8】I-CreI変異体によるrosa1.3 DNA標的の切断を示す。
【図9】I-CreIコンビナトリアル変異体によるrosa1.4標的の切断を示す。
【図10】pCLS1107ベクターマップを表す。
【図11】ヘテロ二量体I-CreIコンビナトリアル変異体によるrosa1.2及びrosa1標的の切断を示す。
【図12】rosa1標的の切断を示す。
【図13】rosa1標的の切断を示す。
【図14】pCLS1058ベクターマップを表す。
【図15】pCLS1069ベクターマップを表す。
【図16】CHO細胞における染色体外モデルでのI-CreI洗練変異体によるrosa1標的の切断を示す。
【図17】マウスROSA26で見出されるメガヌクレアーゼ標的配列と、該DNA標的のそれぞれを切断できる対応するI-CreI変異型を表す。
【図18】pCLS1675ベクターマップを表す。
【図19】pCLS1761ベクターマップを表す。
【図20】pCLS1762ベクターマップを表す。
【図21】IRES-Hygroマトリクス(pCLS1675)を用いたノックイン(KI)事象のPCR分析を示す。
【発明を実施するための形態】
【0024】
定義
- アミノ酸とは、天然又は合成アミノ酸の鏡像異性体及び立体異性体を含む天然又は合成アミノ酸のことをいう。
ポリペプチド配列中のアミノ酸残基は、本明細書において、1文字コードに従って表し、例えばQはGln又はグルタミン残基を意味し、RはArg又はアルギニン残基を意味し、DはAsp又はアスパラギン酸残基を意味する。
- 酸性アミノ酸とは、アスパラギン酸(D)及びグルタミン酸(E)のことをいう。
- 塩基性アミノ酸とは、リジン(K)、アルギニン(R)及びヒスチジン(H)のことをいう。
- 小さいアミノ酸とは、グリシン(G)及びアラニン(A)のことをいう。
- 芳香族アミノ酸とは、フェニルアラニン(F)、トリプトファン(W)及びチロシン(Y)のことをいう。
【0025】
- ヌクレオチドは、次のように表す:1文字コードは、ヌクレオシドの塩基を表すために用いる:aはアデニンであり、tはチミンであり、cはシトシンであり、gはグアニンである。縮重ヌクレオチドについて、rはg又はa (プリンヌクレオチド)を表し、kはg又はtを表し、sはg又はcを表し、wはa又はtを表し、mはa又はcを表し、yはt又はc (ピリミジンヌクレオチド)を表し、dはg、a又はtを表し、vはg、a又はcを表し、bはg、t又はcを表し、hはa、t又はcを表し、nはg、a、t又はcを表す。
【0026】
- 「メガヌクレアーゼ」により、12〜45 bpの2本鎖DNA標的配列を有するエンドヌクレアーゼを意図する。該メガヌクレアーゼは、各ドメインが単量体上にある二量体酵素、又は単一ポリペプチド上に2つのドメインを含む単量体酵素のいずれかである。
- 「メガヌクレアーゼドメイン」により、メガヌクレアーゼのDNA標的の一方の半分と相互作用し、かつDNA標的の他方の半分と相互作用する同じメガヌクレアーゼの他方のドメインと会合して、該DNA標的を切断できる機能的メガヌクレアーゼを形成できる領域を意図する。
【0027】
- 「メガヌクレアーゼ変異型」又は「変異型」により、野生型メガヌクレアーゼ(天然のメガヌクレアーゼ)のアミノ酸配列における少なくとも1つの残基の、別のアミノ酸での置き換えにより得られるメガヌクレアーゼを意図する。
- 「機能的変異型」により、DNA標的配列、好ましくは、親のメガヌクレアーゼにより切断されない新しい標的を切断できる変異型を意図する。例えば、このような変異型は、DNA標的配列に接触するか、又は直接若しくは間接的に該DNA標的と相互作用する位置にてアミノ酸変動を有する。
- 「新規な特異性を有するメガヌクレアーゼ変異型」により、親のメガヌクレアーゼのものとは異なる切断標的のパターンを有する変異型を意図する。等価で、同様に用いられる用語「新規な特異性」「改変された特異性」「新規な切断特異性」「新規な基質特異性」は、DNA標的配列のヌクレオチドに対する変異型の特異性のことである。
【0028】
- 「I-CreI」により、配列表内の配列番号1の配列に相当する配列SWISSPROT P05725又は配列表内の配列番号133の配列に相当するpdbアクセッションコード1g9yの配列を有する野生型I-CreIを意図する。
- 「ドメイン」又は「コアドメイン」により、約100アミノ酸残基の配列に相当する、LAGLIDADGファミリーのホーミングエンドヌクレアーゼの特徴的なα1β1β2α2β3β4α3折り畳みである「LAGLIDADGホーミングエンドヌクレアーゼコアドメイン」を意図する。該ドメインは、DNA標的の一方の半分と相互作用する逆平行ベータシートに折り畳まれた4つのベータ鎖(β1β2β3β4)を含む。このドメインは、DNA標的の他方の半分と相互作用する別のLAGLIDADGホーミングエンドヌクレアーゼコアドメインと会合して、該DNA標的を切断できる機能エンドヌクレアーゼを形成できる。例えば、二量体ホーミングエンドヌクレアーゼI-CreI (163アミノ酸)の場合、LAGLIDADGホーミングエンドヌクレアーゼコアドメインは、残基6〜94に相当する。
【0029】
- 「単鎖メガヌクレアーゼ」により、ペプチドスペーサーで連結された2つのLAGLIDADGホーミングエンドヌクレアーゼドメイン又はコアドメインを含むメガヌクレアーゼを意図する。単鎖メガヌクレアーゼは、それぞれの親のメガヌクレアーゼ標的配列の1つの異なる半分を含むキメラDNA標的配列を切断できる。
【0030】
- 「サブドメイン」により、ホーミングエンドヌクレアーゼDNA標的ハーフサイト(half-site)の別個の部分と相互作用するLAGLIDADGホーミングエンドヌクレアーゼコアドメインの領域を意図する。2つの異なるサブドメインは独立して挙動し、一方のサブドメインでの変異は、他方のサブドメインの結合及び切断特性を変更しない。よって、2つのサブドメインは、ホーミングエンドヌクレアーゼDNA標的ハーフサイトの別個の部分に結合する。
- 「ベータヘアピン」により、ループ又はターンにより接続されたLAGLIDADGホーミングエンドヌクレアーゼコアドメインの逆平行ベータシートの2つの連続するベータ鎖(β1β2又はβ3β4)を意図する。
【0031】
- 「I-CreI部位」により、I-CreIにより切断される22〜24 bpの2本鎖DNA配列を意図する。I-CreI部位は、野生型(天然)非パリンドロームI-CreIホーミング部位と、C1221ともよばれる配列5'- t-12c-11a-10a-9a-8a-7c-6g-5t-4c-3g-2t-1a+1c+2g+3a+4c+5g+6t+7t+8t+9t+10g+11a+12 (配列番号2; 図5)のような派生パリンドローム配列を含む。
- 「DNA標的」、「DNA標的配列」、「標的配列」、「標的部位」、「標的」、「部位」、「興味対象の部位」、「認識部位」、「認識配列」、「ホーミング認識部位」、「ホーミング部位」、「切断部位」により、I-CreIのようなLAGLIDADGホーミングエンドヌクレアーゼ、又は変異型、又はI-CreIに由来する単鎖キメラメガヌクレアーゼにより認識されかつ切断される20〜24 bpの2本鎖パリンドローム、部分的パリンドローム(偽パリンドローム)又は非パリンドロームのポリヌクレオチド配列を意図する。これらの用語は、そこでメガヌクレアーゼにより2本鎖破断(切断)が誘導される独特のDNAの場所、好ましくはゲノムの場所のことをいう。DNA標的は、C1221について上で示したように、2本鎖ポリヌクレオチドの一方の鎖の5'から3'の配列により定義される。DNA標的の切断は、センス鎖及びアンチセンス鎖のそれぞれについて、+2位及び-2位のヌクレオチドで生じる。そうでないと記載しない限り、I-Cre Iメガヌクレアーゼ変異型によるDNA標的の切断が生じる位置は、DNA標的のセンス鎖上の切断部位に相当する。
【0032】
- 「DNA標的ハーフサイト」、「ハーフ切断部位」又は「ハーフサイト」により、それぞれのLAGLIDADGホーミングエンドヌクレアーゼコアドメインが結合するDNA標的の部分を意図する。
- 「キメラDNA標的」又は「ハイブリッドDNA標的」により、2つの親のメガヌクレアーゼ標的配列の異なる半分の融合を意図する。さらに、該標的の少なくとも一方の半分は、少なくとも2つの別個のサブドメインが結合するヌクレオチドの組み合わせ(組み合わせたDNA標的)を含み得る。
【0033】
- 「マウスROSA26遺伝子座」により、マウス第6染色体に位置し、EMBLアクセッション番号CQ880114 (配列番号3; 13139 bp)に相当する配列を有する遺伝子座を意図する。ROSA26は、3つの転写産物を生成する(図1):2つの転写産物は、同一の5'末端を共有する共通プロモーターから生じるが(エキソン1及びエキソン2開始)、いずれも有効なORFを含有しない。3つ目の転写産物は、リバース鎖から生じる。
- 「マウスROSA26遺伝子座からのDNA標的配列」、「ゲノムDNA標的配列」、「ゲノムDNA切断部位」、「ゲノムDNA標的」又は「ゲノム標的」により、メガヌクレアーゼ変異型により認識され切断されるマウスROSA26遺伝子座の20〜24 bpの配列を意図する。
【0034】
- 「ベクター」により、それが連結された別の核酸を輸送できる核酸分子を意図する。
- 「相同な」により、配列同士の間の相同組換えを導くのに充分な別の配列との同一性を有する、より具体的には少なくとも95%の同一性、好ましくは97%の同一性、より好ましくは99%を有する配列を意図する。
- 「同一性」は、2つの核酸分子又はポリペプチド間の配列同一性のことをいう。同一性は、比較の目的のために整列させ得るそれぞれの配列中の位置の比較により決定できる。比較される配列中の位置が同じ塩基により占められる場合、その分子同士は、その位置において同一である。核酸又はアミノ酸配列間の類似性又は同一性の程度は、核酸配列により共有される位置での同一又は一致するヌクレオチドの数の関数である。種々のアラインメントアルゴリズム及び/又はプログラムを用いて、2つの配列間の同一性を計算することができ、GCG配列解析パッケージ(University of Wisconsin, Madison, Wis.)の一部分として利用可能であり、例えばデフォルト設定で用い得るFASTA又はBLASTを含む。
【0035】
- 「個体」は、哺乳動物、及びその他の脊椎動物(例えば鳥類、魚類及び爬虫類)を含む。用語「哺乳動物」及び「哺乳類」は、本明細書で用いる場合、その子に授乳し、生存する子を出産する(真獣類(eutharian)又は胎盤哺乳類(placental mammals))又は産卵する(後獣類(metatharian)又は無胎盤哺乳類(nonplacental mammals))単孔類、有袋類及び有胎盤類(placental)を含むいずれの脊椎動物のことをいう。哺乳動物の種の例は、ヒト、及びその他の霊長類(例えばサル、チンパンジー)、げっ歯類(例えばラット、マウス、モルモット)、並びにその他、例えばウシ、ブタ及びウマを含む。
- 「変異」により、ポリヌクレオチド(cDNA、遺伝子)又はポリペプチド配列内の1つ又は複数のヌクレオチド/アミノ酸の置換、欠失、挿入を意図する。上記の変異は、遺伝子のコード配列又はその調節配列に影響し得る。これは、ゲノム配列の構造又はコードされたmRNAの構造/安定性にも影響し得る。
【0036】
本発明による変異型は、ホモ二量体又はヘテロ二量体であり得る。好ましくは、ヘテロ二量体の両方の単量体が、26位〜40位及び/又は44位〜77位で変異される。より好ましくは、両方の単量体が、I-CreIの26位〜40位及び44位〜77位の両方に異なる置換を有する。
上記の変異型の好ましい実施形態において、I-CreIの44位〜77位に位置するサブドメイン内の上記の置換は、44位、68位、70位、75位及び/又は77位にある。
【0037】
上記の変異型の別の好ましい実施形態において、I-CreIの26位〜40位に位置するサブドメイン内の上記の置換は、26位、28位、30位、32位、33位、38位及び/又は40位にある。
上記の変異型の別の好ましい実施形態において、上記の置換は、元のアミノ酸の、A、D、E、G、H、K、N、P、Q、R、S、T、Y、C、V、L及びWからなる群より選択されるアミノ酸での置き換えである。
【0038】
上記の変異型の別の好ましい実施形態において、これは、DNA標的配列と接触するか又はDNA主鎖(backbone)若しくはヌクレオチド塩基と直接又は水分子を介して相互作用する他のアミノ酸残基の位置にて、1つ又は複数の変異を含む。これらの残基は、当該技術において公知である(Juricaら, Molecular Cell., 1998, 2, 469〜476; Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)。
特に、付加的な置換は、例えば最終C-末端ループ中の(137位〜143位; Prietoら, Nucleic Acids Res., Epub 2007年4月22日)、リン酸主鎖と接触する位置にて導入され得る。好ましくは、上記の残基は、上記のDNA切断部位の結合及び切断に関与する。より好ましくは、上記の残基は、I-CreIの138位、139位、142位又は143位にある。それぞれの変異が138位と139位の残基の対及び142位と143位の残基の対から選択される残基の異なる対の中にあることを条件として、2つの残基を1つの変異型内で変異させ得る。導入される変異は、最終C-末端ループのアミノ酸の、I-CreI部位のリン酸主鎖との相互作用を改変する。好ましくは、138位又は139位の残基を疎水性アミノ酸により置換して、DNA切断部位のリン酸主鎖との水素結合の形成を回避する。例えば、138位の残基がアラニンにより置換されるか、又は139位の残基がメチオニンにより置換される。142位又は143位の残基を、有利には、小さいアミノ酸、例えばグリシンにより置換して、これらのアミノ酸残基の側鎖のサイズを低減させる。より好ましくは、最終C-末端ループ中の上記の置換は、I-CreI部位の±1〜2位、±6〜7位及び/又は±11〜12位のヌクレオチドに対する変異型の特異性を改変する。
【0039】
上記の変異型の別の好ましい実施形態において、これは、マウスROSA26遺伝子座からのDNA標的配列に対する変異型の結合及び/又は切断特性を改良する1つ又は複数の付加的な変異を含む。
変異される付加的な残基は、I-CreI配列全体、特にI-CreIのC-末端半分(80位〜163位)にあり得る。例えば、変異型は、19位、24位、79位、105位、107位、151位、153位、158位での1つ又は複数の付加的な置換を含む。上記の置換は、有利には、G19S、I24V、S79G、V105A、K107R、V151A、D153G及びK158Eからなる群より選択される。
【0040】
本発明の変異型は、野生型I-CreI (配列番号1又は133)、又は配列番号133と少なくとも85%の同一性、好ましくは少なくとも90%の同一性、より好ましくは少なくとも95%の同一性を有するI-CreI足場タンパク質、例えばI-CreI配列の2位のアラニンの挿入、置換D75N及びC-末端でのAADの挿入(164位〜166位)を有する配列番号4の足場(167アミノ酸)から導き得る。
【0041】
さらに、本発明の変異型は、配列のNH2末端及び/又はCOOH末端に挿入された1つ又は複数の残基を含み得る。例えば、タグ(エピトープ(HA-タグ(YPYDVPDYA;配列番号135)又はS-タグ(KETAAAKFERQHMDS;配列番号136)又はポリヒスチジン配列)を、NH2末端及び/又はCOOH末端に導入する。このタグは、上記の変異型の検出及び/又は精製のために有用である。NH2末端にタグを導入する場合、タグの配列は、変異型の最初のアミノ酸を置き換えるか(少なくとも最初のメチオニン、そして結局は変異型の2番目のアミノ酸;メチオニンで開始するタグ)、又は最初のアミノ酸(メチオニン)と2番目のアミノ酸の間若しくは変異型の最初のアミノ酸と3番目のアミノ酸との間に挿入され得る(メチオニンのないタグ)。
【0042】
変異型は、核局在化シグナル(NLS)も含み得る。上記のNLSは、該変異型を細胞の核内に輸送するために有用である。NLSの例は、KKKRK (配列番号134)である。NLSは、変異型の最初のメチオニンの直後又はN-末端タグの直後に挿入できる。
【0043】
本発明による変異型は、パリンドローム又は偽パリンドロームDNA標的配列を切断できるホモ二量体であり得る。
或いは、上記の変異型は、I-CreIの26位〜40位及び/又は44位〜77位に異なる置換を有する第1及び第2単量体の会合により得られるヘテロ二量体であって、マウスROSA26遺伝子座からの非パリンドロームDNA標的を切断できるヘテロ二量体である。
上記の変異型により切断されるDNA標的配列は、マウスROSA26遺伝子座のエキソン又はイントロン内にあり得る。
【0044】
上記の変異型の別の好ましい実施形態において、上記のDNA標的は、マウスROSA26遺伝子座の全てをカバーする配列番号5〜30の配列(図17)からなる群より選択される。
【0045】
【表1】

【0046】
より好ましくは、変異型の単量体は、第1及び第2単量体のそれぞれについて少なくとも以下の置換を有する:
- N30H、Y33S、Q44E、R68C、R70S及びD75N (第1単量体)と、N30D、Y33R、Q38T、Q44K、R68E、R70S及びI77R (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号5を切断する(図1及び17;表I)、
- S32N、Y33G、Q44K、R70E及びD75N (第1単量体)と、S32T、Q38W、Q44K、R68E、R70S及びI77R (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号6を切断する(図1及び17;表I)、
- Y33R、Q38N、S40Q、Q44N、R70S、D75R及びI77D (第1単量体)と、N30H、Y33S、Q44A、R70S、D75Q及びI77E (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号7を切断する(図1及び17;表I)、
- K28S、Q38R、S40K、Q44D、R68Y、R70S、D75S及びI77R (第1単量体)と、Y33C、Q38A、R68A、R70K及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号8を切断する(図1及び17;表I)、
【0047】
- Y33C、Q44T、R70S及びD75Y (第1単量体)と、S32D、Q38C、Q44D、R68Y、R70S、D75S及びI77R (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号9を切断する(図1及び17;表I)、
- S32T、Y33C、R68T、R70N及びD75N (第1単量体)と、S32T、Q38W、Q44K、R70E及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号10を切断する(図1及び17;表I)、
- R70S、D75R及びI77Y (第1単量体)と、Y33R、Q38A、S40Q、Q44A、R70S及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号11を切断する(図1及び17;表I)、
- K28S、Q38R、S40K、Q44T、R68N、R70N及びD75N (第1単量体)と、Y33H、Q38S、Q44K、R68Y、R70S、D75Q及びI77N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号12を切断する(図1及び17;表I)、
【0048】
- K28A、Y33S、Q38R、S40K、Q44N、R68Y、R70S、D75R、I77V (第1単量体)と、S32T、Y33C、Q44A、R70S及びD75N (第2単量体);この単量体は、第1イントロン内に位置するROSA26標的配列番号13を切断する(図1及び17;表I)、
- S32D、Y33H、Q44K、R68E、R70S及びI77R (第1単量体)と、S32D、Y33H、Q44D、R68N、R70S及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号14を切断する(図1及び17;表I)、
- N30R、S32D、R68S、R70K及びD75N (第1単量体)と、D75N (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号16を切断する(図1及び17;表I)、
- K28R、Y33A、Q38Y、S40Q、R68Y、R70S、D75R及びI77Q (第1単量体)と、Y33R、Q38A、S40Q、R70S及びI77K (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号17を切断する(図1及び17;表I)、
- K28R、N30D、D75E及びI77R (第1単量体)と、S32D、Q38C、Q44A、R70S、D75R及びI77Y (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号18を切断する(図1及び17;表I)、
- Y33R、Q38A、S40Q、R70S及びD75N (第1単量体)と、R70S、D75Y及びI77R (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号19を切断する(図1及び17;表I)、
【0049】
- Y33R、Q38A、S40Q、Q44N、R68Y、R70S、D75R及びI77V (第1単量体)と、N30R、S32D、Q44T、R68H、R70H及びD75N (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号20を切断する(図1及び17;表I)、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q及びI77N (第1単量体)と、S32A、Y33C、R68Y、R70S、D75R及びI77Q (第2単量体);この変異型は、第2イントロン内に位置するROSA26標的配列番号21を切断する(図1及び17;表I)、
- K28A、Y33S、Q38R、S40K、R68N、R70S、D75N及びI77R (第1単量体)と、S32N、Y33G、Q44A、R68A、R70K及びD75N (第2単量体);この変異型は、第2イントロン内に位置するROSA26標的配列番号22を切断する(図1及び17;表I)、
- N30H、Y33S、Q44Y、R70S及びI77V (第1単量体)と、Y33R、S40Q、Q44A、R70S及びD75N (第2単量体);この変異型は、第2イントロン内に位置するROSA26標的配列番号23を切断する(図1及び17;表I)、
- S32D、Y33H、R68T、R70N及びD75N (第1単量体)と、N30R、S32D、Q44T、R68N、R70N及びD75N (第2単量体);この変異型は、第2イントロンとアンチセンス転写産物内に位置するROSA26標的配列番号24を切断する(図1及び17;表I)、
- S32R、Y33D、Q44A、R70S及びD75N (第1単量体)と、Y33S、Q38R、S40H、R68H、R70H及びD75N (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号25を切断する(図1及び17;表I)、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q及びI77N (第1単量体)と、K28R、Y33A、Q38Y、S40Q、Q44T、R68H、R70H及びD75N (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号26を切断する(図1及び17;表I)、
【0050】
- K28Q、Q38R、S40K、Q44A、R70S、D75E及びI77R (第1単量体)と、N30R、S32D、Q44K、R68Y、R70S、D75Q及びI77N (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号27を切断する(図1及び17;表I)、
- Y33T、Q38A、R68H、R70H及びD75N (第1単量体)と、N30H、Y33S、R70S及びI77K (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号28を切断する(図1及び17;表I)、
- Y33T、S40T、R68A、R70K及びD75N (第1単量体)と、K28R、Y33A、Q38Y、S40Q、R70S及びI77K (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号29を切断する(図1及び17;表I)、並びに
- S32D、Y33H、Q44N、R70S、D75R及びI77D (第1単量体)と、S32D、Q38C、R70S及びI77K (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号30を切断する(図1及び17;表I)。
【0051】
表IのROSA26 DNA標的(配列番号5〜14及び16〜30のヌクレオチド配列)を切断する上記の変異型の例は、配列番号82〜106のアミノ酸配列のいずれかの第1単量体と、配列番号107〜116、4、117〜130のアミノ酸配列のいずれかの第2単量体とをそれぞれ有する変異型を含む(図17)。
【0052】
さらに、以下の変異型は、第2エキソン内に位置するrosa1と命名されるROSA26 DNA標的を切断できる(図1及び17;表I):
- I24V、Q44Y、R70S及びD75N;I24V、Q44Y、R68Y、R70S、D75Y及びI77R;I24V、Q44Y、R70S、D75N及びI77V;I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択される第1単量体と、K28E、Y33R、Q38R、S40R、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40R、Q44A、R70N及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40K、Q44V、R70A及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R70G及びD75Nからなる群より選択される第2単量体とを有する40個の変異型;これらの変異型の例は、表V (第1単量体:m2、m6、m8、m12、m13、m14、m16又はm17 (配列番号39、43、45、49、50、51、53及び54);配列番号60、61、63、65及び66のいずれかの第2単量体)に示す。
- I24V、Q44Y、R70S、D75Y及びI77Tを含む第1単量体と、K28E、Y33R、Q38R、S40R、Q44A、R68S、R70Q及びD75N (第2単量体)とを有する変異型;この変異型の例は、表V (第1単量体m17 (配列番号54)と第2単量体 配列番号62)に示す。
【0053】
- I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択される第1単量体と、K28E、Y33R、Q38R、S40K、Q44A、R70S及びD75N、並びにK28E、Y33R、Q38R、S40K、Q44A、R68T、R70N及びD75Nからなる群より選択される第2単量体とを有する10個の変異型;これらの変異型の例は、表V (第1単量体;m12、m13、m14、m16又はm17 (配列番号49、50、51、53及び54);配列番号64及び67のいずれかの第2単量体)に示す。
- 配列番号72 (MO_1;表VI及びVII)又は配列番号73 (MO_2;表VI及びVII)の配列からなる第1単量体と、配列番号74〜77 (mO_1〜mO_4;表VII)の配列のいずれかからなる第2単量体とを有する変異型;これらの8個の変異型は、rosa1標的についての変異型の切断活性を増大させる付加的な置換を有する。
【0054】
本発明は、上記で定義される配列と少なくとも85%の同一性、好ましくは少なくとも90%の同一性、より好ましくは少なくとも95% (96%、97%、98%、99 %)の同一性を有するI-CreI変異型であって、マウスROSA26遺伝子座からのDNA標的を切断できる変異型を含む。
例えば、本発明は、MO_1及びmO_2から、NLS、タグ又はそれらの両方の挿入により導かれるI-CreI変異型であって、配列番号140〜145の配列からなる群より選択される変異型を含む。
【0055】
ヘテロ二量体変異型は、有利には、2つのI-CreI単量体間に分子間相互作用を作製する第1及び第2単量体の残基に対応する興味対象の少なくとも1対の変異を有する偏性(obligate)ヘテロ二量体変異型であり、ここで、上記の対の第1変異は第1単量体中にあり、上記の対の第2変異は第2単量体中にあり、上記の変異の対は各単量体からの機能的ホモ二量体の形成を妨げ、マウスROSA26遺伝子座からのゲノムDNA標的を切断できる機能的ヘテロ二量体の形成を可能にする。
【0056】
偏性ヘテロ二量体を形成するために、単量体は、第1及び第2単量体のそれぞれについて、以下の変異の対の少なくとも1つを有することが有利である:
a) 8位のグルタミン酸の塩基性アミノ酸、好ましくはアルギニンでの置換(第1単量体)、及び7位のリジンの酸性アミノ酸、好ましくはグルタミン酸での置換(第2単量体);第1単量体は、7位及び96位のリジン残基の少なくとも1つのアルギニンによる置換をさらに含み得る、
b) 61位のグルタミン酸の塩基性アミノ酸、好ましくはアルギニンでの置換(第1単量体)、及び96位のリジンの酸性アミノ酸、好ましくはグルタミン酸での置換(第2単量体);第1単量体は、7位及び96位のリジン残基の少なくとも1つのアルギニンによる置換をさらに含み得る、
c) 97位のロイシンの芳香族アミノ酸、好ましくはフェニルアラニンでの置換(第1単量体)、及び54位のフェニルアラニンの小さいアミノ酸、好ましくはグリシンでの置換(第2単量体);第1単量体は、54位のフェニルアラニンのトリプトファンによる置換をさらに含むことができ、第2単量体は、58位のロイシン又は57位のリジンのメチオニンによる置換をさらに含み得る、
d) 137位のアスパラギン酸の塩基性アミノ酸、好ましくはアルギニンでの置換(第1単量体)、及び51位のアルギニンの酸性アミノ酸、好ましくはグルタミン酸での置換(第2単量体)。
【0057】
例えば、第1単量体は変異D137Rを、第2単量体は変異R51Dを有し得る。偏性ヘテロ二量体メガヌクレアーゼは、有利には、上記のa)、b)、c)又はd)で定義される変異の少なくとも2対を含む。変異の対のうちの1つは、有利にはc)又はd)で定義されるとおりである。好ましくは、一方の単量体は、7位及び96位のリジン残基の酸性アミノ酸(アスパラギン酸(D)又はグルタミン酸(E))、好ましくはアスパラギン酸(K7E及びK96E)による置換を含み、他方の単量体は、8位及び61位のグルタミン酸残基の塩基性アミノ酸(アルギニン(R)又はリジン(K);例えばE8K及びE61R)による置換を含む。より好ましくは、偏性ヘテロ二量体メガヌクレアーゼは、上記のa)、b)及びc)で定義される変異の3対を含む。偏性ヘテロ二量体メガヌクレアーゼは、有利には、(i) E8R、E8K又はE8H、E61R、E61K又はE61H及びL97F、L97W又はL97Y; (ii) K7R、E8R、E61R、K96R及びL97F、或いは(iii) K7R、E8R、F54W、E61R、K96R及びL97Fと、(iv) K7E又はK7D、F54G又はF54A及びK96D又はK96E; (v) K7E、F54G、L58M及びK96E、或いは(vi) K7E、F54G、K57M及びK96Eの変異を少なくとも有する第2単量体(B)とからなる。例えば、第1単量体は、変異K7R、E8R又はE8K、E61R、K96R及びL97F、或いはK7R、E8R又はE8K、F54W、E61R、K96R及びL97Fを有し、第2単量体は、変異K7E、F54G、L58M及びK96E又はK7E、F54G、K57M及びK96Eを有し得る。ヘテロ二量体の例は、配列番号147及び配列番号148である。偏性ヘテロ二量体は、上記で定義される少なくとも1つのNLS及び/又は1つのタグを有し得る。上記のNLS及び/又はタグは、第1及び/又は第2単量体にあり得る。
【0058】
本発明の主題は、上記で定義されるI-CreI変異型に由来する単鎖キメラメガヌクレアーゼ(融合タンパク質)でもある。単鎖メガヌクレアーゼは、2つのI-CreI単量体、2つのI-CreIコアドメイン(I-CreIの6位〜94位)、又はこれら両方の組み合わせを含み得る。好ましくは、2つの単量体/コアドメイン又はこれら両方の組み合わせは、ペプチドリンカーにより連結される。ペプチドリンカーの例は、配列番号149である。単鎖キメラメガヌクレアーゼの例は、配列番号146である。単鎖キメラメガヌクレアーゼは、上記で定義される少なくとも1つのNLS及び/又は1つのタグをさらに含み得る。上記のNLS及び/又はタグは、第1及び/又は第2単量体にあり得る。
【0059】
本発明の主題は、上記で定義される変異型又は単鎖キメラメガヌクレアーゼをコードするポリヌクレオチドフラグメントでもある。上記のポリヌクレオチドは、ホモ二量体若しくはヘテロ二量体の変異型の1つの単量体、又は単鎖キメラメガヌクレアーゼの2つのドメイン/単量体をコードし得る。
【0060】
本発明の主題は、本発明による変異型又は単鎖メガヌクレアーゼの発現のための組換えベクターでもある。組換えベクターは、上記で定義される変異型又は単鎖メガヌクレアーゼをコードする少なくとも1つのポリヌクレオチドフラグメントを含む。好ましい実施形態において、上記のベクターは、それぞれがヘテロ二量体変異型の単量体の一方をコードする2つの異なるポリヌクレオチドフラグメントを含む。
【0061】
本発明において用い得るベクターは、限定されないが、ウイルスベクター、プラスミド、RNAベクター、或いは染色体、非染色体、半合成又は合成の核酸からなり得る線状若しくは環状のDNA又はRNA分子を含む。好ましいベクターは、自律複製できるもの(エピソームベクター)及び/又は連結された核酸の発現を可能にするもの(発現ベクター)である。多数の適切なベクターが当業者に知られ、商業的に入手可能である。
【0062】
ウイルスベクターは、レトロウイルス、アデノウイルス、パルボウイルス(例えばアデノ随伴ウイルス)、コロナウイルス、マイナス鎖RNAウイルス、例えばオルトミクソウイルス(例えばインフルエンザウイルス)、ラブドウイルス(例えば狂犬病及び水疱性口内炎ウイルス)、パラミクソウイルス(例えば麻疹及びセンダイ)、プラス鎖RNAウイルス、例えばピコルナウイルス及びアルファウイルス、並びにアデノウイルス、ヘルペスウイルス(例えば単純ヘルペスウイルス1及び2型、エプスタイン-バーウイルス、サイトメガロウイルス)及びポックスウイルス(例えばワクシニア、鶏痘及びカナリア痘)を含む二本鎖DNAウイルスを含む。その他のウイルスは、例えば、ノーウォークウイルス、トガウイルス、フラビウイルス、レオウイルス、パポバウイルス、ヘパドナウイルス及び肝炎ウイルスを含む。レトロウイルスの例は、トリ白血病肉腫、哺乳類C型、B型ウイルス、D型ウイルス、HTLV-BLV群、レンチウイルス、スプマウイルスを含む(Coffin, J. M., Retroviridae: The viruses and their replication, In Fundamental Virology, 第3版, B. N. Fieldsら編, Lippincott-Raven Publishers, Philadelphia, 1996)。
好ましいベクターは、レンチウイルスベクター、特に自己不活化レンチウイルスベクター(self inactivacting lentiviral vectors)を含む。
【0063】
ベクターは、選択マーカー、例えば真核細胞培養についてネオマイシンホスホトランスフェラーゼ、ヒスチジノールデヒドロゲナーゼ、ジヒドロ葉酸レダクターゼ、ハイグロマイシンホスホトランスフェラーゼ、単純ヘルペスウイルスチミジンキナーゼ、アデノシンデアミナーゼ、グルタミンシンセターゼ及びヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ;エス・セレビシエ(S. cerevisiae)についてTRP1;大腸菌(E. coli)においてテトラサイクリン、リファンピシン又はアンピシリン耐性を含み得る。
【0064】
好ましくは、上記のベクターは、本発明の変異型/単鎖メガヌクレアーゼをコードする配列が、適切な転写及び翻訳制御要素の制御下に位置して、該メガヌクレアーゼの生成又は合成を許容する発現ベクターである。よって、上記のポリヌクレオチドは、発現カセットに含まれる。より具体的には、該ベクターは、複製起点、該コードポリヌクレオチドに機能可能に連結されたプロモーター、リボソーム結合部位、RNAスプライシング部位(ゲノムDNAを用いる場合)、ポリアデニル化部位、及び転写終結部位を含む。これは、エンハンサーも含み得る。プロモーターの選択は、ポリペプチドが発現される細胞に依存する。好ましくは、変異型がヘテロ二量体である場合、各単量体をコードする2つのポリヌクレオチドは、両方のポリヌクレオチドの発現を同時に駆動し得る1つのベクターに含まれる。適切なプロモーターは、組織特異的及び/又は誘導性プロモーターを含む。誘導性プロモーターの例は、重金属のレベルの増加により誘導される真核メタロチオネインプロモーター、イソプロピル-β-D-チオガラクト-ピラノシド(IPTG)に応答して誘導される原核lacZプロモーター、及び温度の増加により誘導される真核熱ショックプロモーターである。組織特異的プロモーターの例は、骨格筋クレアチンキナーゼ、前立腺特異的抗原(PSA)、α-抗トリプシンプロテアーゼ、ヒトサーファクタント(SP)タンパク質A及びB、β-カゼイン及び酸性ホエータンパク質遺伝子である。
【0065】
上記のベクターの別の有利な実施形態によると、これは、上記で定義されるゲノムDNA切断部位を取り囲む領域と相同性を有する配列を含むターゲティング構築物を含む。
或いは、I-CreI変異型/単鎖メガヌクレアーゼをコードするベクターと、ターゲティング構築物を含むベクターとは、異なるベクターである。
より好ましくは、上記のターゲティングDNA構築物は:
a) 上記で定義されるゲノムDNA切断部位を取り囲む領域と相同性を有する配列と、
b) a)に記載の配列で挟まれた、導入される配列と
を含む。
【0066】
マウスROSA26遺伝子座での遺伝子ノックインのために、導入される配列は、外因性遺伝子発現カセット又はその一部分と、結局はHPRT遺伝子のような選択マーカーとを含む。
或いは、導入される配列は、マウスROSA26遺伝子座を、マウスROSA26遺伝子座の特定の配列を改変するため、マウスROSA26遺伝子座若しくはその一部分を減弱又は活性化するため、マウスROSA26遺伝子座の興味対象部位に変異を導入するため、又はマウスROSA26遺伝子座若しくはその一部分を不活性化又は欠失させるために用いられる配列を含む、ある特定の様式で変更させるために用いられる任意のその他の配列であり得る。
【0067】
好ましくは、切断部位を修復するために、少なくとも50 bp、好ましくは100 bpを超える、より好ましくは200 bpを超える相同配列が用いられる。実際に、共有されるDNA相同性は、破断の部位の上流及び下流に接する領域に位置し、導入されるDNA配列は、2つの腕の間に位置するべきである。
よって、ターゲティング構築物は、好ましくは200 pb〜6000 pb、より好ましくは1000 pb〜2000 pbである。
【0068】
配列の挿入のために、DNA相同性は、破断の部位のすぐ上流及び下流の領域に一般的に位置する(破断に隣接する配列;最小修復マトリクス)。しかし、挿入が、切断部位に接する配列の欠失を伴う場合、共有されるDNA相同性は、欠失の領域の上流及び下流の領域に位置する。
例えば、上記で定義される変異型と、それぞれの変異型により作製される切断のそれぞれを修復するための最小マトリクスとにより切断されるマウスROSA26 DNA標的を、図17に示す。
【0069】
本発明の主題は、上記で定義される少なくとも1つのメガヌクレアーゼ(変異型又は単鎖派生キメラメガヌクレアーゼ)及び/又は上記で定義される該メガヌクレアーゼをコードする少なくとも1つの発現ベクターを含むことを特徴とする組成物でもある。
上記の組成物の好ましい実施形態において、これは、上記で定義されるターゲティングDNA構築物を含む。
好ましくは、上記のターゲティングDNA構築物は、組換えベクターに含まれるか、又は本発明によるメガヌクレアーゼをコードするポリヌクレオチドを含む発現ベクターに含まれる。
【0070】
本発明の主題は、さらに、上記で定義されるメガヌクレアーゼ、好ましくは発現ベクターに含まれる1つ又は2つのポリヌクレオチドの、非治療目的でのマウスROSA26遺伝子座でのゲノム工学のための使用である。
上記の使用の有利な実施形態によると、これは、ゲノムDNA標的配列を含むマウスROSA26遺伝子座の興味対象部位内に2本鎖破断を誘発することにより、DNA組換え事象、DNA欠失又は細胞死を誘発するためである。
【0071】
本発明によると、上記の2本鎖破断は、ROSA26遺伝子座内の特定の配列を改変するため、内因性ROSA26遺伝子座を減弱又は活性化するため、ROSA26遺伝子座の興味対象部位に変異を導入するため、外因性遺伝子又はその一部分を導入するため、内因性ROSA26遺伝子座若しくはその一部分を不活性化又は欠失するため、染色体腕を転座させるため、或いはDNAを修復されないままにして分解させるためである。
【0072】
上記の使用の別の有利な実施形態によると、上記の変異型、ポリヌクレオチド、ベクターは、上記で定義されるターゲティングDNA構築物と会合する。
本発明によるメガヌクレアーゼの使用の好ましい実施形態において、これは、少なくとも以下の工程を含む:1) 上記のメガヌクレアーゼの少なくとも1つの認識及び切断部位を含むマウスROSA26 遺伝子座の興味対象部位に、該切断部位を該メガヌクレアーゼと接触させることにより二本鎖破断を導入し、2)標的遺伝子座と相同性を有する配列で挟まれた導入される配列を含むターゲティングDNA構築物を提供する。上記のメガヌクレアーゼは、細胞に直接、又は該メガヌクレアーゼをコードするポリヌクレオチド配列を含み、用いられる細胞におけるその発現に適切な発現ベクターを介して提供され得る。このストラテジーは、DNA配列を標的部位に導入して、例えば、タンパク質生産、遺伝子機能研究、薬剤の開発(薬剤のスクリーニング)のために、又は疾患モデルとして用いることができるノックイントランスジェニックマウス又は組換えマウス株化細胞を作製するために用いられる。
【0073】
本発明の主題は、少なくとも以下の工程を含む、興味対象の生成物を発現するトランスジェニックマウスを作製するための方法でもある:
(a) マウス多能性前駆細胞又はマウス胚に、上記で定義されるメガヌクレアーゼを導入して、該メガヌクレアーゼのDNA認識及び切断部位を含むROSA26遺伝子座の興味対象部位に二本鎖切断を誘発し、同時に又は連続的に、
(b) 工程(a)のマウス前駆細胞又は胚に、切断部位を取り囲む領域と相同性を有する配列で挟まれた興味対象の生成物をコードする配列を少なくとも含むターゲティングDNAを導入して、ターゲティングDNAと染色体DNAとの間の相同組換えにより興味対象配列が挿入されたゲノム改変マウス前駆細胞又は胚を作製し、
(c) 工程(b)のゲノム改変マウス前駆細胞又は胚を、キメラマウスに発達させ、
(d) 工程(c)のキメラマウスからトランスジェニックマウスを誘導する。
【0074】
好ましくは、工程(c)は、工程(b)で作製されたゲノム改変前駆細胞を、キメラマウスを作製するように、胚盤胞へ導入することを含む。
上記の方法の好ましい実施形態によると、これは、任意の手段により、トランスジェニックマウスから興味対象の生成物を回収するさらなる工程(e)を含む。
【0075】
本発明の主題は、少なくとも以下の工程を含む、興味対象の生成物を発現する組換えマウス細胞を作製するための方法でもある:
(a) マウス細胞に、上記で定義されるメガヌクレアーゼを導入して、該メガヌクレアーゼのDNA認識及び切断部位を含むROSA26遺伝子座の興味対象部位に二本鎖切断を誘発し、同時に又は連続的に、
(b) 工程(a)の細胞に、切断部位を取り囲む領域と相同性を有する配列で挟まれた興味対象の生成物をコードする配列を少なくとも含むターゲティングDNAを導入して、ターゲティングDNAと染色体DNAとの間の相同組換えにより興味対象配列が挿入された組換えマウス細胞を作製し、
(c) 工程(b)の組換えマウス細胞を、任意の適切な手段により単離する。
【0076】
上記の方法の好ましい実施形態によると、これは、任意の手段により、組換えマウス細胞から興味対象の生成物を回収するさらなる工程(d)を含む。
ターゲティングDNAは、興味対象部位に上記のターゲティングDNAを導入するために適切な条件下で細胞内に導入される。
ある好ましい実施形態において、ターゲティングDNA構築物はベクターに挿入される。
【0077】
改変される細胞は、興味対象のいずれの細胞であってもよい。トランスジェニックマウスを作製するために、細胞は、当該技術で公知の胚性幹(ES)細胞のような多能性前駆細胞である。組換えマウス株化細胞を作製するために、細胞は、有利には、NSO、SP2/0 (BALB/c骨髄腫; ECACC # 85110503及び# 85072401)、又はL (ATCC # CRL-2648)細胞であり得る。上記のメガヌクレアーゼは、細胞に直接、又は該メガヌクレアーゼをコードするポリヌクレオチド配列を含み、用いられる細胞におけるその発現に適切な発現ベクターを介して提供され得る。
【0078】
興味対象の生成物を発現するトランスジェニック動物/組換え株化細胞を作製するために、ターゲティングDNAは、上記で定義されるように、マウスROSA26遺伝子座中のメガヌクレアーゼ部位の上流及び下流の配列で挟まれた、興味対象の生成物(タンパク質又はRNA)をコードする配列と、結局は選択マーカー遺伝子とを含むことにより、興味対象の外因性配列が、ROSA26遺伝子座のメガヌクレアーゼ部位にて相同組換えにより統合されたゲノム改変細胞(動物前駆細胞又は胚/動物又はヒトの細胞)が作製される。
興味対象の配列は、興味対象のあるタンパク質/ペプチドをコードする任意の遺伝子であり得、限定されないが、レポーター遺伝子、受容体、シグナル伝達分子、転写因子、医薬的に活性なタンパク質及びペプチド、疾患原因遺伝子生成物及び毒素を含む。配列は、例えばsiRNAを含む興味対象のRNA分子もコードし得る。
【0079】
外因性配列の発現は、上記で定義される、内因性ROSA26プロモーター又は構成性若しくは誘導性の異種プロモーター、好ましくは遍在性若しくは組織特異的なプロモーターのいずれかにより駆動され得る。さらに、興味対象の配列の発現は、条件付きであり得る。発現は、部位特異的リコンビナーゼ(Cre、FLPなど)により誘発され得る。
つまり、興味対象配列は、上記の興味対象遺伝子に機能可能に連結された異種プロモーターと、限定されないが、(選択)マーカー遺伝子、リコンビナーゼ認識部位、ポリアデニル化シグナル、スプライス受容配列、イントロン、タンパク質検出のためのタグ及びエンハンサーを含む1つ又は複数の機能的配列とを含み得る適切なカセットに挿入される。
【0080】
或いは、適切なカセットは、上記の興味対象遺伝子に機能可能に連結された配列内リボソーム進入部位(IRES)と、IRES-Hygroマトリクス(pCLS1675)を有する事象、限定されないが、(選択)マーカー遺伝子、リコンビナーゼ認識部位、ポリアデニル化シグナル、スプライス受容配列、イントロン、タンパク質検出のためのタグ及びエンハンサーを含む1つ又は複数の機能的配列とを含み得る。
【0081】
メガヌクレアーゼは、ポリペプチド、又は該ポリペプチドをコードするポリヌクレオチド構築物のいずれかとして用い得る。これは、マウス細胞に、特定の種類の細胞に適する当該技術において公知の任意の簡便な手段により、単独で又は少なくとも適切なビヒクル若しくは担体及び/又はターゲティングDNAのいずれかとともに、導入される。
【0082】
本発明による使用の有利な実施形態によると、メガヌクレアーゼ(ポリペプチド)は、以下のものと会合される(is associated with):
- リポソーム、ポリエチレンイミン(PEI);このような場合、上記の会合物は投与され、よって、標的体細胞に導入される。
- 膜移送ペプチド(membrane translocating peptides) (Bonetta, The Scientist, 2002, 16, 38; Fordら, Gene Ther., 2001, 8, 1〜4 ; Wadia及びDowdy, Curr. Opin. Biotechnol., 2002, 13, 52〜56);このような場合、変異型/単鎖メガヌクレアーゼの配列は、膜移送ペプチドの配列と融合される(融合タンパク質)。
【0083】
本発明による使用の別の有利な実施形態によると、メガヌクレアーゼ(メガヌクレアーゼをコードするポリヌクレオチド)及び/又はターゲティングDNAは、ベクターに挿入される。ターゲティングDNA及び/又はメガヌクレアーゼをコードする核酸を含むベクターは、種々の方法により細胞に導入できる(例えば注入、直接摂取、発射衝撃、リポソーム、エレクトロポレーション)。メガヌクレアーゼは、発現ベクターを用いて、細胞内で安定的又は一過的に発現させ得る。真核細胞における発現の方法は、当該技術において公知である(Current Protocols in Human Genetics: 12章 「Vectors For Gene Therapy」及び13章「Delivery Systems for Gene Therapy」を参照)。所望により、組換えタンパク質中に、核局在化シグナルを組み込んで、核内でそれが発現されることを確実にすることが好ましい。
一旦細胞内に入ると、メガヌクレアーゼと存在するならばターゲティングDNA及び/又はメガヌクレアーゼをコードする核酸を含むベクターは、細胞質から核内の作用部位まで、細胞により移入又は移送される。
【0084】
本発明による使用のある実施形態において、メガヌクレアーゼは、実質的に非免疫原性であり、すなわち、有害免疫応答をほとんど又は全く生じない。この種の有害免疫応答を緩和又は消去するための種々の方法を本発明に従って用いることができる。ある好ましい実施形態において、メガヌクレアーゼは、N-ホルミルメチオニンを実質的に含まない。望ましくない免疫応答を回避する別の様式は、メガヌクレアーゼを、ポリエチレングリコール(「PEG」)又はポリプロピレングリコール(「PPG」) (好ましくは、500〜20,000ダルトンの平均分子量(MW)のもの)とコンジュゲートさせることである。例えばDavisら(US 4,179,337)により記載されるPEG又はPPGとのコンジュゲート形成は、抗ウイルス活性を有する、非免疫原性で、生理活性で、水溶性のエンドヌクレアーゼコンジュゲートを提供できる。ポリエチレン-ポリプロピレングリコールコポリマーを用いる同様の方法は、Saiferら(US 5,006,333)に記載されている。
【0085】
本発明は、上記で定義されるポリヌクレオチド又はベクター、好ましくは発現ベクターで改変された原核又は真核宿主細胞にも関する。
本発明は、細胞の全て又は一部分が上記で定義されるポリヌクレオチド又はベクターで改変されたことを特徴とする非ヒトトランスジェニック動物又はトランスジェニック植物にも関する。
本明細書で用いる場合、細胞とは、原核細胞、例えば細菌細胞、又は真核細胞、例えば動物、植物若しくは酵母細胞のことである。
【0086】
本発明の主題は、上記で定義される少なくとも1つのメガヌクレアーゼ変異型の、別のメガヌクレアーゼを作製するための足場としての使用でもある。例えば、新規な第3世代のメガヌクレアーゼを作製する目的で、3回目の突然変異誘発及び選択/スクリーニングを上記の変異型に対して行うことができる。
【0087】
本発明によるメガヌクレアーゼの異なる使用及び上記のメガヌクレアーゼを用いる方法は、I-CreI変異型、該変異型に由来する単鎖キメラメガヌクレアーゼ、上記で定義される、該変異型若しくは単鎖キメラメガヌクレアーゼをコードするポリヌクレオチド、ベクター、細胞、トランスジェニック植物又は非ヒトトランスジェニック哺乳動物の使用を含む。
【0088】
本発明によるI-CreI変異型は、少なくとも以下の工程を含む、マウスROSA26遺伝子座からのゲノムDNA標的配列を切断できるI-CreI変異型を工学的に作製するための方法により得ることができる:
(a) I-CreIの26位〜40位に位置するLAGLIDADGコアドメインの第1機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第1シリーズを構築し、
(b) I-CreIの44位〜77位に位置するLAGLIDADGコアドメインの第2機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第2シリーズを構築し、
(c) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の-10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(d) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の-5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
【0089】
(e) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(f) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の+3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(g) 工程(c)及び工程(d)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) -10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットと同一であり、(ii) +8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) -5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットと同一であり、(iv) +3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
【0090】
(h) 工程(e)及び工程(f)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) +3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットと同一であり、(ii) -5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(iv) -10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位のヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
(i) 工程(g)及び(h)で得られた変異型を組み合わせて、ヘテロ二量体を形成し、
(j) マウスROSA26遺伝子座からの上記のゲノムDNA標的を切断できる工程(i)からのヘテロ二量体を選択及び/又はスクリーニングする。
【0091】
上記の工程(c)、(d)、(e)又は(f)の1つは省略できる。例えば、工程(c)を省略する場合、工程(d)は、-10位〜-8位及び-5位〜-3位の両方のヌクレオチドトリプレットが上記のゲノム標的の-10位〜-8位及び-5位〜-3位に存在するヌクレオチドトリプレットでそれぞれ置き換えられ、+3位〜+5位及び+8位〜+10位のヌクレオチドトリプレットが、上記のゲノム標的の-5位〜-3位及び-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列でそれぞれ置き換えられた変異I-CreI部位を用いて行われる。
【0092】
工程(a)、(b)、(g)、(h)及び(i)は、上記で定義される、DNA標的配列と接触するか又は該DNA標的と直接若しくは間接的に相互作用する他の位置、変異体の結合及び/又は切断特性を改良する位置、又は機能的ホモ二量体の形成を妨げる位置での、付加的な変異の導入をさらに含み得る。これは、国際PCT出願WO 2004/067736に記載されるようなコンビナトリアルライブラリーを作製することにより行い得る。
【0093】
本発明のI-CreI変異型を工学的に作製する方法は、興味対象の遺伝子からのDNA標的に対する変異体の結合及び/又は切断特性を改良するために、変異体全体、又は変異体の一部分、特に変異型のC-末端半分(80位〜163位)に対するランダム変異の導入を含むのが有利である。突然変異誘発は、公知で商業的に利用可能な標準の突然変異誘発の方法に従って、変異型のプールに対してランダム突然変異誘発ライブラリーを作製することにより行うことができる。好ましくは、突然変異誘発は、工程(i)で形成されるか又は工程(j)で得られるヘテロ二量体の一方の単量体の全体の配列に対して、有利には、単量体のプールに対して、好ましくは工程(i)又は(j)のへテロ二量体の両方の単量体に対して行われる。
【0094】
好ましくは、Arnouldら, J. Mol. Biol., Epub 2007年5月10日の図4に記載される方法に従って、2回の選択/スクリーニングを行う。1回目において、ヘテロ二量体の一方の単量体(図4の単量体Y)を、他方の単量体(図4の単量体X)と同時発現させてヘテロ二量体を形成し、改良単量体Y+を、興味対象遺伝子からの標的に対して選択する。2回目において、他方の単量体(単量体X)に突然変異を誘発し、改良単量体Y+と同時発現させてヘテロ二量体を形成し、興味対象遺伝子からの標的に対して選択して、改良された活性を有するメガヌクレアーゼ(X+ Y+)を得る。
【0095】
工程(g)及び(h)における変異の(分子内)組み合わせは、公知のオーバーラップPCR法に従って、2つのサブドメインのそれぞれを含むオーバーラップフラグメントを増幅することにより行ってよい。
工程(i)における変異型の(分子間)組み合わせは、工程(g)からの1つの変異型を、工程(h)からの1つの変異型と同時発現させて、ヘテロ二量体の形成を可能にすることにより行われる。例えば、宿主細胞を、該変異型をコードする1つ又は2つの組換え発現ベクターで改変できる。次いで、国際PCT出願WO 2006/097854及びArnouldら, J. Mol. Biol., 2006, 355, 443〜458に以前に記載されたようにして、細胞を、変異型の発現を可能にする条件下で培養することにより、宿主細胞内でヘテロ二量体が形成される。
【0096】
工程(c)、(d)、(e)、(f)及び/又は(j)における選択及び/又はスクリーニングは、国際PCT出願WO 2004/067736、Arnouldら, J. Mol. Biol., 2006, 355, 443〜458, Epinatら, Nucleic Acids Res., 2003, 31, 2952〜2962及びChamesら, Nucleic Acids Res., 2005, 33, e178に記載されるような、インビトロ又はインビボでの切断アッセイを用いて行うことができる。
上記の方法の別の有利な実施形態によると、工程(c)、(d)、(e)、(f)及び/又は(j)は、インビボで、変異型により作製された変異DNA標的配列内の2本鎖破断が、陽性選択マーカー若しくはレポーター遺伝子の活性化、又は陰性選択マーカー若しくはレポーター遺伝子の不活性化を、該DNA 2本鎖破断の組換え媒介修復により導く条件下で行われる。
【0097】
本発明の主題は、本発明によるマウスROSA26遺伝子座からのDNA標的を切断できる変異型を工学的に作製するために有用な、I-CreIの26位〜40位及び/又は44位〜77位にて変異を有するI-CreI変異型でもある。特に、本発明は、変異型m1〜m18 (表II、配列番号38〜55)、Q44V、R70A及びD75Nを含む変異型(配列番号131;表III)、並びにK28E、Y33R、Q38R、S40R及びD75Nを含む変異型(配列番号132;表III)を含む、上記で定義されるI-CreI変異型を工学的に作製するための方法の工程(c)〜(f)で定義されるI-CreI変異型を含む。本発明は、配列番号60〜67の配列の変異型(表IIIの組み合わせ変異型)を含む、上記で定義されるI-CreI変異型を工学的に作製するための方法の工程(g)及び(h)で定義されるI-CreI変異型も含む。
【0098】
興味対象遺伝子からのDNA標的を切断できる単鎖キメラメガヌクレアーゼは、当該技術において公知の方法により、本発明による変異型から導かれる(Epinatら, Nucleic Acids Res., 2003, 31, 2952〜62; Chevalierら, Mol. Cell., 2002, 10, 895〜905; Steuerら, Chembiochem., 2004, 5, 206〜13; 国際PCT出願WO 03/078619及びWO 2004/031346)。このような方法はいずれも、本発明で定義される変異型に由来する単鎖キメラメガヌクレアーゼを構築するために用い得る。
【0099】
本発明で定義される変異型をコードするポリヌクレオチド配列は、当業者に知られる任意の方法により調製できる。例えば、これらは、cDNA鋳型から、特異的プライマーを用いるポリメラーゼ連鎖反応により増幅される。好ましくは、上記のcDNAのコドンは、所望の発現系における上記のタンパク質の発現に好ましいように選択される。
上記のポリヌクレオチドを含む組換えベクターは、公知の組換えDNA及び遺伝子工学の技術により得て、宿主細胞に導入できる。
【0100】
本発明で定義されるI-CreI変異型又は単鎖誘導体は、上記で定義されるポリペプチドを発現させることにより生成される。好ましくは、該ポリペプチドは、ポリペプチドの発現又は同時発現に適する条件下で、1つの発現ベクター又は2つの発現ベクター(変異型のみの場合)により改変された宿主細胞又はトランスジェニック動物/植物で、発現又は同時発現(変異型のみの場合)され、変異型又は単鎖誘導体は、宿主細胞培養又はトランスジェニック動物/植物から回収される。
【0101】
本発明の実行は、そうでないと記載しない限り、当該技術の範囲内である細胞生物学、細胞培養、分子生物学、トランスジェニック生物学、微生物学、組換えDNA及び免疫学の通常の技術を用いる。このような技術は、文献に充分に説明されている。例えばCurrent Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, 第3版, (Sambrookら, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); Oligonucleotide Synthesis (M. J. Gait編, 1984); Mullisら、米国特許第4,683,195号; Nucleic Acid Hybridization (B. D. Harries及びS. J. Higgins編 1984); Transcription And Translation (B. D. Hames及びS. J. Higgins編 1984); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); Methods In ENZYMOLOGY (J. Abelson及びM. Simon編, Academic Press, Inc., New York)のシリーズ, 特に第154巻及び第155巻(Wuら編)並びに第185巻「Gene Expression Technology」(D. Goeddel編); Gene Transfer Vectors For Mammalian Cells (J. H. Miller及びM. P. Calos編, 1987, Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (Mayer及びWalker編, Academic Press, London, 1987); Handbook Of Experimental Immunology, 第I〜IV巻(D. M. Weir及びC. C. Blackwell編, 1986); 並びにManipulating the Mouse Embryo, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1986)を参照されたい。
【0102】
上記の特徴に加えて、本発明は、本発明によるI-CreIメガヌクレアーゼ変異型及びそれらの使用を説明する実施例並びに添付の図面に言及する以下の記載から明らかになるその他の特徴をさらに含む。添付の図面において:
- 図1は、マウスROSA26遺伝子座を表す(アクセッション番号EMBL CQ880114;配列番号3)。エキソンを箱で囲む(エキソン1:2490位〜2599位;転写産物1からのエキソン2:8228位〜9248位;11845位から開始し、11505位で終了するアンチセンス転写産物とほぼ重複する転写産物2からのエキソン3。今までのところ同定された3つの転写産物を、標的rosa1 (配列番号15)の配列及び位置とともに示す。
【0103】
- 図2は、DNA標的に結合したI-CreIホーミングエンドヌクレアーゼの3次元構造を表す。触媒コアは、DNA主溝の上にサドル形相互作用界面を形成する2つのαββαββα折り畳みで囲まれる。
- 図3は、I-CreI及び他のLAGLIDADGホーミングエンドヌクレアーゼの特異性を工学的に改変するための2ステップアプローチを示す。I-CreI誘導体の大きい集合を、元の足場の半合理的突然変異誘発と、局所的に特異性が変更された機能的変異型のスクリーニングとにより作製する。次いで、コンビナトリアルアプローチを用いて、これらの変異体を、完全に再設計された特異性を有するメガヌクレアーゼに組み立てる。ホモ二量体タンパク質(「ハーフメガヌクレアーゼ」)を、同じαββαββα折り畳み内の2組の変異の組み合わせにより創出し、このような2つの「ハーフメガヌクレアーゼ」の同時発現が、興味対象の標的を切断するヘテロ二量体の種(「カスタムメガヌクレアーゼ」)をもたらし得る。
【0104】
- 図4は、マウスROSA26遺伝子座を切断するメガヌクレアーゼの使用のためのストラテジーを表す。メガヌクレアーゼ誘発相同組換えを用いる遺伝子挿入は、興味対象遺伝子をマウスROSA26遺伝子座にノックインする。イントロン及びエキソンの配列は、相同領域として用い得る。
- 図5は、rosa1標的配列及び誘導体を表す。10GGG_P、5GAT_P及び5TAT_Pは、以前に得られたI-CreI変異体により切断されることが見出された近縁の誘導体である。これらは、C1221 (I-CreI足場タンパク質により切断されるパリンドローム配列)とは、箱で囲んだモチーフが異なる。C1221、10GGG_P、5GAT_P及び5TAT_Pは、最初は24 bp配列で記載されたが、構造データは、22 bpのみがタンパク質/DNA相互作用に関連することを示唆する。しかし、±12位を括弧内に示す。rosa1は、8304位にてマウスROSA26遺伝子座に位置するDNA配列である。rosa1.2標的において、標的の中ほどのGTTC配列は、C1221で見出される塩基であるGTACで置き換えられる。rosa1.3は、rosa1.2の左部分に由来するパリンドローム配列であり、rosa1.4は、rosa1.2の右部分に由来するパリンドローム配列である。図に示すように、10GGG_P、5GAT_P及び5TAT_Pからの箱で囲んだモチーフは、rosa1シリーズの標的で見出される。
【0105】
- 図6は、pCLS1055ベクターマップを表す。
- 図7は、pCLS0542ベクターマップを表す。
- 図8は、I-CreI変異体によるrosa1.3 DNA標的の切断を示す。1次スクリーニングで見出された63個の陽性を、1枚の96ウェルプレート内で再配置し、2次スクリーニングで確認した(4重フォーマットで)。実施例2で選択した22個の変異体を丸で囲む。
- 図9は、I-CreIコンビナトリアル変異体によるrosa1.4標的の切断を示す。1次スクリーニングで見出された69個の陽性を、1枚の96ウェルプレート内で再配置し、2次スクリーニングで確認した(4重フォーマットで)。実施例3で選択した15個の変異体を丸で囲む。
- 図10は、pCLS1107ベクターマップを表す。
【0106】
- 図11は、ヘテロ二量体I-CreIコンビナトリアル変異体によるrosa1.2及びrosa1標的の切断を示す。A. I-CreI変異体の組み合わせのrosa1.2標的を用いたスクリーニングの例。B. I-CreI変異体の同じ組み合わせのrosa1標的を用いたスクリーニング。B5、B6、D5、D6、F5、F6、H5及びH6:pCLS1107空のプラスミドDNAで形質転換された、rosa1.3を切断するI-CreI変異体を発現する酵母株。
- 図12は、rosa1標的の切断を示す。rosa1.4を切断する一連のI-CreI変異体に無作為に突然変異を誘発し、rosa1.3を切断する変異体と同時発現させた。切断を、rosa1標的を用いて試験する。それぞれの4ドットクラスタにおいて、右の2つのドットは、2重での、rosa1を切断する元来のヘテロ二量体の1つに相当し、2つの左のドットは、変異されていないrosa1.3を切断するものと同時発現させた、同じ変異させたrosa1.4を切断するものに相当する(変異体m13、表IV及びVに記載される)。rosa1の改良された切断を示す2つの最適化された変異体を丸で囲み、これらは変異体m13とMO_1 (C10)、又はm13とMO_2 (E2)の同時発現に相当する。MO_1及びMO_2は、表VIにさらに記載される。
【0107】
- 図13は、rosa1標的の切断を示す。rosa1.3を切断する一連のI-CreI変異体に無作為に突然変異を誘発し、rosa1.4を切断する洗練された変異体と同時発現させた。切断は、rosa1標的を用いて試験する。rosa1の効率的な切断を示す変異体を丸で囲む。フィルタにおいて:
- B11は、ヘテロ二量体S19、V24、Y44、R68、S70、N75、V77 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当し;
- C9は、ヘテロ二量体S19、V24、Y44、R68、S70、Q75、I77 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当し;
- C11及びE8は、ヘテロ二量体 V24、Y44、S68、S70、R75、I77、A105 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当し、
- E6は、ヘテロ二量体V24、Y44、S68、S70、R75、I77、G79 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当する。
- H10は陰性対照であり、H11及びH12は異なる強度の陽性対照である。rosa1.3標的を切断する変異体の改良の前後におけるrosa1標的に対するヘテロ二量体の活性を比較するために、各クラスタにおいて、右の2つの点は、実施例5に記載されるヘテロ二量体の1つであり、左の2つの点は、実施例6に記載される変異を有するヘテロ二量体である。
【0108】
- 図14は、pCLS1058ベクターマップを表す。
- 図15は、pCLS1069ベクターマップを表す。
- 図16は、CHO細胞における染色体外モデルでのI-CreI洗練変異体によるrosa1標的の切断を示す。2回のトランスフェクション実験からの値を示す。I-CreI N75及びI-SceIによる同じ実験でのI-CreI及びI-SceI標的の切断は、陽性対照として示す。
- 図17は、マウスROSA26で見出されるメガヌクレアーゼ標的配列と、該DNA標的のそれぞれを切断できる対応するI-CreI変異型を表す。DNA標的の配列(第1列)を、その位置(第2列)とともに示す。標的部位での切断を修復するための最小修復マトリクスを、その最初のヌクレオチド(始点、第5列)と、最後のヌクレオチド(終点、第6列)で示す。各変異型の配列は、記載する位置での残基により規定する。例えば、図17の最初のヘテロ二量体変異型は、K、H、S、S、Q、S、E、C、S、N及びIを28位、30位、32位、33位、38位、40位、44位、68位、70位、75位及び77位にそれぞれ有する第1単量体と、K、D、S、R、T、S、K、E、S、D、Rを28位、30位、32位、33位、38位、40位、44位、68位、70位、75位及び77位にそれぞれ有する第2単量体とからなる。位置は、I-CreI配列SWISSPROT P05725 (配列番号1)を参照にして記載する。I-CreIは、K、N、S、Y、Q、S、Q、R、R、D及びIを、28位、30位、32位、33位、38位、40位、44位、68位、70位、75位及び77位にそれぞれ有する。
【0109】
- 図18は、pCLS1675ベクターマップを表す。
- 図19は、pCLS1761ベクターマップを表す。
- 図20は、pCLS1762ベクターマップを表す。
- 図21は、IRES-Hygroマトリクス(pCLS1675)を用いたノックイン(KI)事象のPCR分析を示す。ROSA26遺伝子座について野生型のクローン及びハイグロマイシンCDSの無作為挿入を有するクローンは、PCRにおいて陰性である。ROSA26遺伝子座でのKI事象を有するクローンは、PCRにおいて陽性である。KI事象及び無作為挿入を有するクローンも、PCRにおいて陽性である。
【実施例】
【0110】
実施例1:マウスROSA26遺伝子座を切断する新規なメガヌクレアーゼを工学的に作製するためのストラテジー
Smithら, Nucleic Acids Res., 2006に記載され、図3に説明されるコンビナトリアルアプローチを用いて、I-CreIのDNA結合ドメインを工学的に作製し、rosa1 (図5)と命名され、マウスROSA26遺伝子座(アクセッション番号CQ880114;配列番号3)のエキソン2の8304位に位置する22 bp (非パリンドローム)の配列を切断した。rosa1配列を切断するメガヌクレアーゼは、遺伝子をマウスROSA26遺伝子座(図4)にノックインするために用いることができた。用途は、以下の分野である。マウス細胞での組換えタンパク質の生成、例えば薬剤スクリーニングの目的での組換え株化細胞の工学的作製、並びに例えば動物モデルとしての使用のためのトランスジェニックマウスの工学的作製。
【0111】
rosa1配列は、部分的に、国際PCT出願WO 2006/097784、WO 2006/097853及びWO 2007/049156; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458並びにSmithら, Nucleic Acids Res., Epub 2006年11月27日に記載されるようにして得られた以前に同定されたメガヌクレアーゼにより切断される10GGG_P、5GAT_P及び5TAT_P標的のパッチワークである(図5)。よって、rosa1は、これらの3つの標的を切断するI-CreI誘導体で見出される変異を組み合わせたメガヌクレアーゼにより切断され得る。
【0112】
10GGG_P、5GAT_P及び5TAT_P配列は、I-CreIにより切断されるパリンドローム配列であるC1221の24 bp誘導体である(国際PCT出願WO 2006/097784、WO 2006/097853及びWO 2007/049156; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458並びにSmithら, Nucleic Acids Res., Epub 2006年11月27日)。しかし、そのDNA標的に結合したI-CreIの構造は、それらの標的の外側の塩基対(-12位及び12位)が、結合及び切断に対して影響しないことを示唆し(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316; Chevalier B.S.及びStoddard B.L., Nucleic Acids Res., 2001, 29, 3757〜3754; Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)、この研究においては、-11位〜11位までを考慮した。よって、標的のrosa1シリーズを、24 bpの代わりに22 bpの配列として規定した。
【0113】
Rosa1は、C1221とは、4 bp中央領域の1塩基対が異なる。その標的に結合したI-CreIタンパク質の構造によると、4つの中央塩基対(-2位〜2位)とI-CreIタンパク質との間に接触はない(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316; Chevalier B.S.及びStoddard B.L., Nucleic Acids Res., 2001, 29, 3757〜3754; Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)。つまり、これらの位置での塩基は、結合効率に影響を与えるとは考えられない。しかし、これらは、この領域の端の2つのニックに起因する切断に影響し得る。よって、-2位〜2位のGTTC配列を、まず、C1221からのGTAC配列で置換して、標的rosa1.2を得た(図5)。
次いで、2つのパリンドローム標的rosa1.3及びrosa1.4を、rosa1.2から導いた(図5)。rosa1.3及びrosa1.4はパリンドロームであるので、これらは、ホモ二量体タンパク質により切断されるはずである。
【0114】
よって、ホモ二量体としてrosa1.3及びrosa1.4配列を切断できるタンパク質をまず設計し(実施例2及び3)、次いで、同時発現させてrosa1を切断するヘテロ二量体を得た(実施例4)。rosa1.2及びrosa1標的を切断するヘテロ二量体を同定できた。rosa1標的についての切断活性を改良するために、rosa1.3及びrosa1.4を切断する選択した一連の変異体を選択して洗練した。選択した変異体にランダム突然変異を誘発し、新規なヘテロ二量体を形成するために用いて、これらをrosa1標的に対してスクリーニングした(実施例5及び6)。最後に、酵母及びCHO細胞において高い切断活性を示すrosa1標的を切断するヘテロ二量体が同定できた。
【0115】
実施例2:rosa1.3を切断するメガヌクレアーゼの作製
この実施例は、I-CreI変異体が、パリンドローム形のrosa1標的の左部分に由来するrosa1.3 DNA標的配列を切断できることを示す(図5)。
この実施例に記載される標的配列は、22 bpのパリンドローム配列である。よって、これらを、最初の11ヌクレオチドのみで、後に接尾辞_Pを付して記載する。例えば、標的rosa1.3は、caacatgatgt_P; 配列番号35とも記載される。
【0116】
rosa1.3標的は、5GAT_Pと、±1位、±2位、±3位、±4位、±5位、±7位、±9位、±10位及び±11位が同様であり、2つの配列は、±6位及び±8位のみが異なる。±6位は、結合及び切断活性に対してほとんど影響しないであろうと仮定した。5GAT_P (caaaacgatgt_P; 配列番号32)を切断できる変異体は、Arnouldら, J Mol Biol. 2006; 355, 443〜458並びに国際PCT出願WO 2006/097784及びWO 2006/097853に記載されるように、I-CreI N75に対する24位、44位、68位、70位、75位及び77位での突然変異誘発により以前に得られた。この実施例において、5GAT_P標的を切断する変異体がrosa1.3標的も切断できるかを調べた。
【0117】
1) 材料及び方法
特異性が変更された変異型をスクリーニングするために用いられるメガヌクレアーゼ変異型を作製する方法及び哺乳動物細胞又は酵母細胞における切断誘発組換えに基づくアッセイは、国際PCT出願WO 2004/067736; Epinatら, Nucleic Acids Res., 2003, 31, 2952〜2962; Chamesら, Nucleic Acids Res., 2005, 33, e178及びArnouldら, J. Mol. Biol., 2006, 355, 443〜458に記載される。これらのアッセイは、機能的LacZレポーター遺伝子をもたらし、これは標準的な方法によりモニターできる。
【0118】
a) 標的ベクターの構築
標的は、以下のようにしてクローニングした。ゲートウェイクローニング配列と接する標的配列に相当するオリゴヌクレオチドをProligoから得た:
5' tggcatacaagtttcaacatgatgtacatcatgttgacaatcgtctgtca 3'(配列番号37)。一本鎖オリゴヌクレオチドのPCR増幅により作製した二本鎖標的DNAを、Gatewayプロトコル(INVITROGEN)を用いて酵母レポーターベクター(pCLS1055, 図6)にクローニングした。酵母レポーターベクターで、エス・セレビシエFYBL2-7B株(MAT a, ura3Δ851, trp1Δ63, leu2Δ1, lys2Δ202)を形質転換した。
【0119】
b) I-CreI変異体
5GAT_Pを切断するI-CreI変異体を、I-CreIの24位、44位、68位、70位、75位及び77位が変異されたライブラリーにおいて、Arnouldら, J. Mol. Biol., 2006, 355, 443〜458、並びに国際PCT出願WO 2006/097784及びWO 2006/097853に以前に記載されたように同定した。これらをDNAベクター(pCLS0542, 図7)にクローニングし、酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae) FYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)で発現させる。
【0120】
c) メガヌクレアーゼ発現クローンの交配と酵母でのスクリーニング
スクリーニングを、以前に記載されたようにして行った(Arnouldら, J. Mol. Biol. 2006, 355, 443〜458)。交配は、コロニーグリッダー(QpixII, Genetix)を用いて行った。変異体を、YPDプレートを覆うナイロンフィルタ上に、低格子密度(low gridding density) (約4スポット/cm2)を用いてグリッドした。2回目のグリッド手順を同じフィルタに対して行って、それぞれの標的について異なるレポーター保持酵母株からなる第2層をスポットした。メンブレンを固体寒天YPDリッチ培地上に置き、30℃にて一晩インキュベートして交配を可能にした。次いで、フィルタを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として有する合成培地上に移し、37℃にて5日間インキュベートして、発現及び標的ベクターを有する二倍体を選択した。5日後に、フィルタを、0.5 Mリン酸ナトリウム緩衝液、pH 7.0中の0.02 % X-Gal、0.1% SDS、6%ジメチルホルムアミド(DMF)、7mM β-メルカプトエタノール、1%アガロースを含む固体アガロース培地上に置き、37℃にてインキュベートして、β-ガラクトシダーゼ活性をモニターした。結果をスキャンにより分析し、適切なソフトウェアを用いて定量を行った。
【0121】
d) 変異体の配列決定
変異体を発現するプラスミドを回収するために、酵母DNAを標準的なプロトコルを用いて抽出し、大腸菌を形質転換するために用いた。変異体ORFの配列決定を、次いで、プラスミドに対してMILLEGEN SAにより行った。或いは、ORFを酵母DNAからPCRにより増幅し(Akadaら, Biotechniques, 2000, 28, 668〜670)、配列決定を、PCR生成物に対して直接、MILLEGEN SAにより行った。
【0122】
2) 結果
I-CreIの24位、44位、68位、70位、75位及び77位が変異されたライブラリーにおいて以前に同定された5GAT_P標的を切断するI-CreI変異体を、rosa1.3 DNA標的(caacatgatgt _P; 配列番号35)に対する切断についてスクリーニングした。合計で63個の陽性クローンが見出され、96ウェルプレートで再配置し、2次スクリーニングにより確認した(図8)。これらの陽性クローンのうち、22個(図8において丸で囲む)を選択した。これらの22個の陽性クローンを配列決定した。これらは、rosa1.3標的を切断する18個の異なる新規なエンドヌクレアーゼに相当することがわかった(m1〜m18と命名;配列番号38〜55;表II)。
【0123】
【表2】
【0124】
実施例3:rosa1.4を切断するメガヌクレアーゼの作製
この実施例は、I-CreI変異体が、パリンドローム形のrosa1標的の右部分に由来するrosa1.4 DNA標的配列を切断できることを示す(図5)。この実施例に記載される全ての標的配列は、22 bpのパリンドローム配列である。よって、これらを、最初の11ヌクレオチドのみで、後に接尾辞_Pを付して記載する。例えば、rosa1.4は、tgggattatgt_P (配列番号10)とよばれる。
【0125】
rosa1.4標的は、5TAT_Pと±1位、±2位、±3位、±4位、±5位及び±7位が、かつ10GGG_Pと±1位、±2位、±7位、±8位、±9位及び±10位が同様である。±6位及び±11位は、結合及び切断活性に対してほとんど影響しないであろうと仮定した。5TAT_Pを切断できる変異体は、Arnouldら, J Mol. Biol., 2006; 355, 443〜458、並びに国際PCT出願WO 2006/097784及びWO 2006/097853に記載されるように、I-CreI N75に対する44位、68位、70位での突然変異誘発により以前に得られた。10GGG_P標的を切断できる変異体は、Smithら, Nucleic Acids Res., Epub 2006年11月27日及び国際PCT出願WO 2007/049156に記載されるように、I-CreI N75に対する28位、30位、33位、38位、40位及び70位での突然変異誘発により得られた。
【0126】
両方の組のタンパク質を70位にて変異させる。しかし、2つの分離可能な機能的サブドメインが存在することが仮定された。このことは、この位置が、標的の±8〜10の塩基に対する特異性にほとんど影響しないことを意味する。
よって、組み合わせた変異体がrosa1.4標的を切断できるかを確認するために、5TAT_P (caaaactatgt_P; 配列番号6)を切断するタンパク質からの44位、68位及び70位での変異を、10GGG_P (cgggacgtcgt_P; 配列番号4)を切断するタンパク質からの28、30、33、38及び40変異と組み合わせた。
【0127】
1) 材料及び方法
実験手順は、実施例2及び以下に記載されるとおりである:
コンビナトリアル変異体の構築
10GGG_P又は5TAT_Pを切断するI-CreI変異体は、10GGG_P又は5TAT_P標的についてそれぞれSmithら, Nucleic Acids Res. Epub 2006年11月27日; 国際PCT出願WO 2007/049156、並びにArnouldら, J. Mol. Biol., 2006, 355, 443〜458; 国際PCT出願WO 2006/097784及びWO 2006/097853で同定された。両方のシリーズからの変異を含むI-CreI由来コード配列を作製するために、I-CreIコード配列の5'末端(aa 1位〜43位)又は3'末端(39位〜167位)を増幅する別々のオーバーラップPCR反応を行った。5'及び3'末端の両方について、PCR増幅を、ベクター(pCLS0542, 図7)に特異的なプライマーGal10F 5'-gcaactttagtgctgacacatacagg-3' (配列番号56)又はGal10R 5'-acaaccttgattggagacttgacc-3' (配列番号57)と、アミノ酸39〜43についてのI-CreIコード配列に特異的なプライマーassF 5'-ctannnttgaccttt-3' (配列番号58)又はassR 5'-aaaggtcaannntag-3' (配列番号59) (ここで、nnnは残基40をコードする)を用いて行う。同じプライマー及び残基40について同じコード配列を用いて行った増幅反応により得られたPCRフラグメントをプールした。次いで、プライマーGal10FとassR又はassFとGal10Rを用いた反応により得られたPCRフラグメントのそれぞれのプールを、等モル比で混合した。最後に、2つのオーバーラップPCRフラグメントのそれぞれの最終プールおよそ25ngと、NcoI及びEagIでの消化により線状にした75 ngのベクターDNA (pCLS0542)とを用いて、サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコル(Gietz, R. D.及びR.A. Woods, Methods Enzymol. 2002, 350, 87〜96)を用いて形質転換した。両方の群の変異を含有するインタクトなコード配列を、酵母におけるインビボ相同組換えにより作製する。
【0128】
2) 結果
I-CreIコンビナトリアル変異体を、I-CreI N75足場上で44位、68位及び70位での変異を、28、30、33、38及び40変異と結び付けることにより構築して、複雑さ(complexity)が2208のライブラリーを得た。コンビナトリアル変異体の例を、表IIIに示す。このライブラリーで酵母を形質転換し、3456個のクローン(多様性の1.5倍)を、rosa1.4 DNA標的(tgggattatgt_P; 配列番号36)に対する切断についてスクリーニングした。合計で69個の陽性クローンが見出され、96ウェルプレートで再配置し、2次スクリーニングにより確認した(図9)。これらの陽性のうち、15個のクローン(図9において丸で囲む)を選択した。配列決定の後に、これらの15クローンは、rosa1.4 DNA標的を切断する8つの異なる新規なエンドヌクレアーゼに相当することがわかった(配列番号60〜67;表III)。
【0129】
【表3】
【0130】
実施例4:rosa1を切断するメガヌクレアーゼの作製
パリンドロームrosa1由来標的(rosa1.3及びrosa1.4)のそれぞれを切断できるI-CreI変異体を、実施例2及び3で同定した。このような変異体の対(一方はrosa1.3を切断し、他方はrosa1.4を切断する)を、酵母において同時発現させた。同時発現の際に、3つの活性分子種、すなわち2つのホモ二量体と1つのヘテロ二量体とが存在するはずである。形成されるはずのヘテロ二量体が非パリンドロームのrosa1及びrosa1.2 DNA標的を切断するかをアッセイした。
【0131】
1) 材料及び方法
a) カナマイシン耐性ベクターでの変異体のクローニング
酵母において2つのI-CreI変異体を同時発現させるために、rosa1.4配列を切断する変異体を、カナマイシン耐性遺伝子をマーカーとして有する酵母発現ベクター(pCLS1107, 図10)にサブクローニングした。変異体を、PCR反応により、pCLS0542及びpCLS1107に共通のプライマー:Gal10F 5'-gcaactttagtgctgacacatacagg-3' (配列番号56)及びGal10R 5'-acaaccttgattggagacttgacc-3' (配列番号57)を用いて増幅した。およそ25 ngのPCRフラグメントとDraIII及びNgoMIVでの消化により線状にした25 ngのベクターDNA (pCLS1107)とを用いて酵母サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコルを用いて形質転換する。I-CreI変異体についてのインタクトなコード配列を、酵母でのインビボ相同組換えにより作製する。pCLS1107ベクターにサブクローニングされたrosa1.4標的を切断する変異体を含有するそれぞれの酵母株を、次いで、rosa1.4標的を発現する酵母と交配させてこれを確認した。プラスミドを発現する変異体を回収するために、酵母DNAを標準的なプロトコルを用いて抽出し、大腸菌を形質転換するのに用い、大腸菌DNAを調製した。
【0132】
b) 変異体同時発現
pCLS0542発現ベクター中のrosa1.3標的を切断する変異体を発現する酵母株を、pCLS1107発現ベクター中のrosa1.4標的を切断する変異体をコードするDNAを用いて形質転換した。形質転換体は、-L Glu + G418培地上で選択した。
【0133】
c) メガヌクレアーゼ同時発現クローンの交配及び酵母でのスクリーニング
交配は、コロニーグリッダー(QpixII, Genetix)を用いて行った。変異体を、YPDプレートを覆うナイロンフィルタ上に、低格子密度(約4スポット/cm2)を用いてグリッドした。2回目のグリッド手順を同じフィルタに対して行って、それぞれの標的について異なるレポーター保持酵母株からなる第2層をスポットした。メンブレンを固体寒天YPDリッチ培地上に置き、30℃にて一晩インキュベートして交配を可能にした。次いで、フィルタを、ロイシン及びトリプトファンを欠き、G418を加え、ガラクトース(1%)を炭素源として有する合成培地上に移し、37℃にて5日間インキュベートして、発現及び標的ベクターを有する二倍体を選択した。5日後に、フィルタを、0.5 Mリン酸ナトリウム緩衝液、pH 7.0中の0.02 % X-Gal、0.1% SDS、6%ジメチルホルムアミド(DMF)、7mM β-メルカプトエタノール、1%アガロースを含む固体アガロース培地上に置き、37℃にてインキュベートして、β-ガラクトシダーゼ活性をモニターした。結果をスキャンにより分析し、適切なソフトウェアを用いて定量を行った。
【0134】
結果
rosa1.3標的を切断する変異体(表IIに記載されるm1〜m18)と、rosa1.4標的を切断する8つの変異体(表IIIに記載される)との同時発現は、全ての場合においてrosa1.2標的の効率的な切断をもたらした(スクリーニングの例を図11Aに示す)。全ての試験した組み合わせを、表IVにまとめる。これらの組み合わせのほとんどは、rosa1.2配列とは+1位の1 bpのみが異なるrosa1天然標的も切断できる(図5)。図11Bに示すように、rosa1天然標的に対して観察されたシグナルは、rosa1.2標的に対して観察されたものと比較して弱い。rosa1 DNA標的を切断する組み合わせを、表Vに示す。
【0135】
【表4】
【0136】
【表5】
【0137】
実施例5 rosa1.4を切断するタンパク質のランダム突然変異誘発及びrosa1.3を切断するタンパク質との組み立てによる、rosa1を切断するメガヌクレアーゼの洗練
非パリンドロームのrosa1.2及びrosa1標的を切断できるI-CreI変異体を、パリンドロームのrosa1.3及びrosa1.4標的を切断する変異体の組み立てにより同定した。しかし、組み合わせは、rosa1.2を効率的に切断できたが、rosa1.2とは1位の1 bpのみが異なるrosa1を弱く切断した。rosa1に対して観察されたシグナルは充分でない。
【0138】
よって、rosa1を切断するタンパク質の組み合わせをランダムに突然変異誘発し、rosa1を効率的に切断する変異体をスクリーニングした。標的に結合したI-CreIタンパク質の構造によると、4つの中央の塩基対(-2位〜2位)とI-CreIタンパク質との間に接触はない(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316; Chevalier, B. S.及びStoddard, B.L. Nucleic Acids Res., 2001, 29, 3757〜3774; Chevalierら, J. Mol. Biol. 2003, 329, 253〜269)。よって、突然変異を誘発する位置の組を合理的に選択することは困難であり、突然変異誘発を、タンパク質のC-末端部分(83個の最後のアミノ酸)又はタンパク質全体に対して行った。ランダム突然変異誘発により、高い複雑さのライブラリーが得られ、試験される変異型ライブラリーの複雑さは、rosa1を切断するヘテロ二量体の2つの成分の一方のみに突然変異を誘発することにより制限される。
つまり、rosa1.4を切断するタンパク質に突然変異を誘発し、rosa1.3を切断するタンパク質と同時発現させたときにこれらがrosa1を効率的に切断できるかを試験した。
【0139】
1) 材料及び方法
a) ランダム突然変異誘発
ランダム突然変異誘発ライブラリーを、選択された変異体のプールに対して、PCRにより、Mn2+又はdNTPの誘導体、例えば8-オキソ-dGTP及びdPTPを用いて、JBS dNTP-Mutageneisキット中のJENA BIOSCIENCE GmbHからのプロトコルに記載されるようにして2ステップPCRプロセスで創出した。タンパク質全体に対するランダム突然変異誘発のために、用いたプライマーは:preATGCreFor 5'-gcataaattactatacttctatagacacgcaaacacaaatacacagcggcctt gccacc-3'; 配列番号68)及びICreIpostRev (5'-ggctcgaggagctcgtctagaggatcgctcgag ttatcagtcggccgc -3'; 配列番号69)である。タンパク質のC-末端部分に対する突然変異誘発のために、用いたプライマーは、dNTP誘導体を用いてAA78a83For (5'-ttaagcgaaatcaagccg-3'; 配列番号70)及びICreIpostRevである。タンパク質の残りは、高忠実度のtaqポリメラーゼを用い、dNTP誘導体を用いずに、プライマーpreATGCreFor及びAA78a83Rev (5'-cggcttgatttcgcttaa-3'; 配列番号71)を用いて増幅する。
【0140】
変異体のプールを、PCR反応により、pCLS0542 (図7)及びpCLS1107 (図10)に共通するこれらのプライマーを用いて増幅した。およそ75 ngのPCRフラグメントとDraIII及びNgoMIVでの消化により線状にした75 ngのベクターDNA (pCLS1107)を用いて、酵母サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコルを用いて形質転換する。I-CreI変異体についてのインタクトなコード配列のライブラリーを、酵母でのインビボ相同組換えにより作製する。陽性をもたらすクローンを、実施例2に記載されるようにして配列決定により確認した。
【0141】
b) rosa1標的を含有する酵母株でのロイシン発現ベクター中の変異体のクローニング
rosa1標的を酵母レポーターベクター(pCLS1055、図6)中に含有する酵母FYBL2-7B株(MAT a, ura3Δ851, trp1Δ63, leu2Δ1, lys2Δ202)を、LEU2遺伝子をマーカーとして有するpCLS0542ベクター中のrosa1.3標的を切断する変異体で、高効率LiAc形質転換プロトコルを用いて形質転換する。得られた酵母株を、実施例4に記載される交配アッセイのための標的として用いる。
【0142】
2) 結果
rosa1.4を切断する4つの変異体(ERRR / AHQ、ERRR / ARN、ERRK / AHQ及びERRK / VRA、表IVによる)をプールし、全てのタンパク質又はタンパク質のC-末端部分に対してランダム突然変異を誘発し、酵母を形質転換した。4464個の形質転換されたクローンを、次いで、実施例2に記載されるものから選択された、(i) レポータープラスミド内にrosa1標的を含有し、(ii) rosa1.3標的を切断する変異型を発現する酵母株と交配させた。I-CreI V24 Y44 S68 S70 R75 I77 (すなわちVYSSRI)変異体、又はI-CreI V24 Y44 R68 S70 Q75 I77 (すなわちVYRSQI)変異体、又はI-CreI V24 Y44 R68 S70 Y75 T77 (すなわちVYRSYT)変異体(表IIを参照)を発現する3つのこのような株を用いた。2つのクローンは、このような酵母株と交配させたときに、同じ酵母株と突然変異誘発する前の変異体と比較してrosa1標的のよりよい切断を引き起こすことがわかった。結論として、VYSSRI、VYRSQI又はVYRSYTとヘテロ二量体を形成するときにrosa1を効率的に切断できる2つのタンパク質(表VI)が同定された(図12)。
【0143】
【表6】
【0144】
実施例6:rosa1.3を切断するタンパク質のランダム突然変異誘発及びrosa1.4を切断する洗練されたタンパク質との組み立てによる、rosa1を切断するメガヌクレアーゼの洗練
rosa1標的を切断できるI-CreI変異体を、rosa1.3を切断する変異体及びrosa1.4を切断する洗練された変異体の組み立てにより同定した。メガヌクレアーゼの活性を増大させるために、rosa1を切断するヘテロ二量体の第2成分に突然変異を誘発した。この実施例において、rosa1.3を切断する変異体に突然変異を誘発し、その後、実施例5で同定されたrosa1.4を切断する洗練された変異体との組み合わせでrosa1をより効率的に切断する変異体をスクリーニングした。
【0145】
1) 材料及び方法
a) ランダム突然変異誘発:
ランダム突然変異誘発ライブラリーを、選択された変異体のプールに対して、PCRにより、Mn2+又はdNTPの誘導体、例えば8-オキソ-dGTP及びdPTPを用いて、JBS dNTP-Mutageneisキット中のJENA BIOSCIENCE GmbHからのプロトコルに記載されるようにして2ステップPCRプロセスで創出した。タンパク質全体に対するランダム突然変異誘発のために、用いたプライマーは:preATGCreFor 5'-gcataaattactatacttctatagacacgcaaacacaaatacacagcggcctt gccacc-3'; 配列番号68)及びICreIpostRev (5'-ggctcgaggagctcgtctagaggatcgctcgag ttatcagtcggccgc -3'; 配列番号69)である。タンパク質のC-末端部分に対する突然変異誘発のために、用いたプライマーは、dNTP誘導体を用いてAA78a83For (5'-ttaagcgaaatcaagccg-3'; 配列番号70)及びICreIpostRevであった。タンパク質の残りは、高忠実度のtaqポリメラーゼを用い、dNTP誘導体を用いずに、プライマーpreATGCreFor及びAA78a83Rev (5'-cggcttgatttcgcttaa-3'; 配列番号71)を用いて増幅する。
【0146】
変異体のプールを、PCR反応により、pCLS0542 (図7)及びpCLS1107 (図10)に共通するこれらのプライマーを用いて増幅した。およそ75 ngのPCRフラグメントとNcoI及びEagIでの消化により線状にした75 ngのベクターDNA (pCLS0542)を用いて、酵母サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコルを用いて形質転換する。I-CreI変異体についてのインタクトなコード配列のライブラリーを、酵母でのインビボ相同組換えにより作製する。陽性をもたらすクローンを、実施例2に記載されるようにして配列決定により確認した。
【0147】
b) rosa1標的を含有する酵母株でのカナマイシン発現ベクター中の変異体のクローニング
rosa1標的を酵母レポーターベクター(pCLS1055、図6)中に含有する酵母FYBL2-7B株(MAT a, ura3Δ851, trp1Δ63, leu2Δ1, lys2Δ202)を、pCLS1107ベクター中のrosa1.4標的を切断するMO_1及びMO_2洗練変異体で、高効率LiAc形質転換プロトコルを用いて形質転換する。変異体-標的酵母を、実施例4に記載される交配アッセイのための標的として用いる。
【0148】
2) 結果
rosa1.3を切断する4つの変異体の2つのプール(プール1: VYRSNI、VYYSYR、VYRSNV及びVYNSRIと、プール2: VYYSYR、VYSSRI、VYRSQI及びVYRSYT、表Vによる)に、全てのタンパク質又はタンパク質のC-末端部分に対してランダム突然変異を誘発し、酵母を形質転換した。8928個の形質転換されたクローンを、(i) レポータープラスミド内にrosa1標的を含有し、(ii) rosa1.4標的を切断する変異型を発現する酵母株と交配させた。I-CreI E28 R33 R38 R40 A44 H68 Q70 N75 A105 R107 (すなわちMO_1)変異体、I-CreI E28 R33 R38 K40 A44 H68 Q70 N75 A105 R107 A151 G153 E158 (すなわちMO_2)変異体のいずれかを発現する2つのこのような株を用いた。5つのクローンは、このような酵母株と交配させたときに、同じ酵母株と突然変異誘発する前の変異体と比較してrosa1標的のよりよい切断を引き起こすことがわかった(図13)。配列決定の後に、これらは、4つのタンパク質に相当することがわかった。結論として、MO_1又はMO_2とヘテロ二量体を形成したときにrosa1を効率的に切断できる4つのタンパク質が同定された(表VII)。
【0149】
【表7】
【0150】
実施例7:CHO細胞における染色体外モデルでのrosa1標的切断の確認
実施例6において、酵母においてrosa1標的を効率的に切断できるI-CreI洗練変異体を同定した。この実施例では、CHO細胞においてrosa1標的を切断する変異体の2つの組み合わせの能力を、哺乳動物細胞における染色体外アッセイを用いて試験した。
【0151】
1) 材料及び方法
a) CHOスクリーニングのためのベクターでのrosa1標的のクローニング
標的は、以下のようにしてクローニングした。ゲートウェイクローニング配列と接する標的配列に相当するオリゴヌクレオチドをProligoから得た:
5' tggcatacaagtttcaacatgatgtacatcatgttgacaatcgtctgtca 3'(配列番号37)。一本鎖オリゴヌクレオチドのPCR増幅により作製した二本鎖標的DNAを、Gatewayプロトコル(INVITROGEN)を用いてCHOレポーターベクター(pCLS1058、図14)にクローニングした。
【0152】
b) メガヌクレアーゼの再クローニング
実施例6で同定したI-CreI N75、I-SceI及びI-CreI変異体のORFをPCRにより増幅し、配列決定した(MILLEGEN)。次いで、ORFをGatewayプロトコル(INVITROGEN)を用いて再クローニングした。ORFを、ProligoからのプライマーB1F: 5' ggggacaagtttgtacaaaaaagcaggcttcgaaggagatagaaccatggccaataccaaatataacaaagagttcc 3' (配列番号78)及びB2R: 5' ggggaccactttgtacaagaaagctgggtttagtcggccgccggggaggatttcttcttctcgc 3' (配列番号79)を用いる酵母DNAのPCRにより増幅した。PCR生成物を、InvitrogenからのCHOゲートウェイ発現ベクターpCDNA6.2にクローニングした(pCLS1069、図15)。得られたクローンを、実施例2に記載されるようにして配列決定により確認した。
【0153】
c) 哺乳動物細胞での染色体外アッセイ
CHO細胞を、Polyfectトランスフェクション試薬を、供給業者(QIAGEN)のプロトコルに従って用いてトランスフェクションした。トランスフェクションの72時間後に、培養培地を回収し、150μlの溶解/視覚化バッファーを、β-ガラクトシダーゼ液体アッセイのために加えた(典型的には、1リットルのバッファーは、100 mlの溶解バッファー(Tris-HCl 10 mM pH 7.5、NaCl 150 mM、Triton X100 0.1%、BSA 0.1 mg/ml、プロテアーゼ阻害剤)、10 mlのMg 100×バッファー(MgCl2 100 mM、β-メルカプトエタノール35%)、110 ml ONPG 8 mg/ml及び780 mlのリン酸ナトリウム0.1M pH 7.5を含有する)。37℃でのインキュベーションの後に、光学密度を420 nmで測定した。全体のプロセスを、自動化されたBioCel (登録商標)プラットフォーム(VELOCITY11)で行う。
【0154】
2) 結果
図16に示す2回の実験の結果は、I-CreI変異体の2つの組み合わせ(mO_2 / MO_1及びmO_2 / MO_2)が、I-CreI標的(tcaaaacgtcgtgagacagtttgg、配列番号80)に対するI-CreI N75、又はI-SceI標的(tagggataacagggtaat、配列番号81)に対するI-SceIの活性と同様の活性で、CHO細胞においてrosa1標的を切断できることを示す。
【0155】
実施例8:マウス細胞におけるROSA26遺伝子座でのゲノム工学
酵母及び哺乳動物細胞(CHO K1細胞)での染色体外アッセイにおいてrosa1標的を効率的に切断できるI-CreI洗練変異体を、実施例6及び7で同定した。マウスL細胞においてROSA26遺伝子座にて相同組換えを誘発する2つのI-CreI洗練変異体の1つの組み合わせの能力を、この実施例において試験した。
【0156】
1) 材料及び方法
a) ノックイン(KI)マトリクス
2つのマウスROSA26ホモロジーアーム、CQ880114配列(配列表の配列番号3に相当)の6283〜8317のHG ROSA26及び8313〜10319のHD ROSA26の間にクローニングされたハイグロマイシン耐性遺伝子コード配列(CDS)を含む2つのノックインマトリクスを構築した。得られたプラスミドは、pCLS1679及びpCLS1675であった(図18のプラスミドマップ)。pCLS1679において、SV40ポリAに機能可能に連結されたハイグロマイシン耐性遺伝子のコード配列(hygro CDS)を、pBR322ベクター(PROMEGA)に、HG ROSA26とHD ROSA26との間でクローニングした。pCLS1675は、pCLS1679とは、hygro CDSのすぐ上流の配列内リボソーム進入部位(IRES; 配列番号139)の挿入が異なる。
【0157】
b) メガヌクレアーゼのクローニング
I-CreI洗練変異体mO_2及びMO_1のORFは、実施例6 (表VII)に記載される。変異体発現を、ヒト伸長因子1アルファ(EF1α)プロモーター又はサイトメガロウイルス最初期(CMV)プロモーターの制御下の2つの発現ベクターにおいて作製した(pCLS1069、図15)。変異体を、pCLS1069に、CMVプロモーターの下で実施例7に記載されるようにしてクローニングした。得られたプラスミドを、配列決定により確認した(MILLEGEN)。pCLS1761 (図19)及びpCLS1762 (図20)において、mO_2及びMO_1 I-CreI変異体はそれぞれ、EF1αプロモーターの制御下である。
【0158】
c) マウスL細胞でのノックイン実験
マウスL細胞(ATCC # CRL-2648)を、10%胎児ウシ血清、ペニシリン、ストレプトマイシン及びファンギゾンを補った完全DMEM培地(DMEM Glutamax, GIBCO)で培養する。細胞を、リポフェクタミン試薬(INVITROGEN)を製造業者により推奨される手順に従って用いて、トランスフェクションする。トランスフェクションの2日後に、選択を、完全培地中の0.6 mg/mlのハイグロマイシンを用いて行う。選択の2週間後に、耐性クローンを、ClonePixロボット(GENETIX)を用いて採取する。クローンを、0.6 mg/mlのハイグロマイシンを補った完全培地中で、96ウェルプレートにおいて1週間増殖させる。ゲノムDNAを、96ウェルプレートで培養される耐性クローンから、ZR96キット(ZYMO RESEARCH)を用いて抽出する。
【0159】
c) ノックイン事象のPCR分析
ノックイン事象を、HG ROSA26ホモロジーアームの上流のマウスROSA26配列、及びハイグロマイシンCDSにそれぞれ位置するプライマー対KI_GHG_S5 (5' tagtatacagaaactgttgcatcgc 3'; 配列番号137)及びHygSeqRev (5'cgtctgctgctccatacaag 3'; 配列番号138)を用いる、ゲノムDNAに対するPCR分析により検出して、KI特異的PCR増幅を得る(図21)。
【0160】
2) 結果
この実施例で用いたROSA26メガヌクレアーゼは、実施例6 (表VII)に記載され、かつヒト伸長因子1アルファ(EF1α)プロモーター(pCLS1761及びpCLS1762)又はサイトメガロウイルス最初期(CMV)プロモーター(pCLS1069、図15)の制御下で2つの発現ベクターにクローニングされたmO_2及びMO_1である。マウスL細胞に、3つのベクターを同時トランスフェクションさせた:mO_2及びMO_1 ROSA26メガヌクレアーゼを発現する2つのプラスミドと、KIマトリクス。メガヌクレアーゼは、それぞれEF1αプロモーターを有するpCLS1761及びpCLS1762にクローニングされ、KIマトリクスはpCLS1675にクローニングされた。
【0161】
合計で2600000個のマウスL細胞に、2μgのKIマトリクスベクター及び5μg又は10μgのそれぞれのメガヌクレアーゼ発現ベクターを同時トランスフェクションさせた。自発的KI頻度の対照として、同数の細胞に、2μgのKIマトリクスベクターのみをトランスフェクションさせた。トランスフェクション効率(40%)を、蛍光マーカー発現プラスミドを用いるFACS分析により決定した。耐性クローンの頻度は、ハイグロマイシン耐性クローンの総数を計数し、トランスフェクション効率で校正することにより決定した。それぞれ2605個、1197個及び1902個のハイグロマイシン耐性クローンを得た(表VIII)。92個又は184個のクローンを、条件あたり採取し、材料及び方法に記載されるようにしてPCRにより分析した。結果を表VIIIに示す。
【0162】
【表8】
【0163】
ROSA26メガヌクレアーゼとKIマトリクスの同時トランスフェクションは、L細胞においてマウスROSA26遺伝子座にて相同組換えを4.9×10-4の最大頻度で誘発した。KIマトリクス単独でのトランスフェクションを用いて、自発的相同組換えは観察されなかった。この実施例は、ROSA26メガヌクレアーゼが、マウスL細胞においてマウスROSA26遺伝子座にて相同組換えを誘発する能力を示す。
【0164】
実施例9:mO_2及びMO_1に由来するメガヌクレアーゼ
メガヌクレアーゼ構築物は、mO_2 (配列番号75)及びMO_1 (配列番号72)から、Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, 第3版, (Sambrookら, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press)に充分に説明される分子生物学及び組換えDNAの通常の技術を用いて工学的に作製した。
【0165】
NLS (KKKRK; 配列番号134)を、MO_1及びmO_2の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、それぞれ配列番号140及び141を有する。
タグ(TagHA; YPYDVPDYA; 配列番号135)を、mO_2の最初(M1)及び3番目(N3)のアミノ酸の間に挿入した。得られた変異型は、配列番号142である。
タグ(STag; KETAAAKFERQHMDS; 配列番号136)を、MO_1の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、配列番号143である。
タグ(TagHA; YPYDVPDYA; 配列番号135)及びNLS (KKKRK; 配列番号134)を、mO_2の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、配列番号144である。
タグ(STag; KETAAAKFERQHMDS; 配列番号136)及びNLS (KKKRK; 配列番号134)を、MO_1の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、配列番号145である。
【0166】
mO_2単量体(配列番号75の3位〜164位)からリンカー(GGSDKYNQALSKYNQALSKYNQALSGGGGS; 配列番号149)により分けられたMO_1単量体 (配列番号72の1位〜166位) を含む単鎖メガヌクレアーゼを構築した。得られた単鎖メガヌクレアーゼは、配列番号146である。
mO_2/MO_1に由来する偏性ヘテロ二量体を、mO_2単量体にE8K及びE61R変異を、そしてMO_1単量体にK7E及びK96E変異を導入することにより工学的に作製した。得られたヘテロ二量体は、配列番号147及び配列番号148からなる。
【技術分野】
【0001】
本発明は、マウスROSA26遺伝子座からのDNA標的配列を切断するメガヌクレアーゼ変異型(variant)、該変異型をコードするベクター、該ベクターにより改変された細胞、動物若しくは植物、並びに該メガヌクレアーゼ変異型及び派生生成物の、マウスゲノム工学(組換えタンパク質生成、トランスジェニックマウス及び組換えマウス株化細胞の構築)のための使用に関する。
【背景技術】
【0002】
マウスROSA26遺伝子座は、Friedrich及びSorianoにより1991年に、レトロウイルスを感染させた胚性幹(ES)細胞を用いるジーントラップ実験により発見された(Friedrich, G.及びP. Soriano, Genes & Development, 1991, 5, 1513〜1523)。非必須部位であるROSA26遺伝子座のイントロン1で挿入が生じるROSA26マウスジーントラップ系統は、新生児(Friedrich及びSoriano, 1991, 上記)及び造血細胞(Zambrowiczら, Proc. Natl. Acad. Sci. USA, 1997, 94, 3789〜3794)において、胚の発達の間にレポーター遺伝子の遍在的な発現を示す。マウスの第6染色体に位置するROSA26遺伝子座は、3つの転写産物を生成する(図1)。2つの転写産物は、同一の5'末端を共有する共通のプロモーターから発生する(エキソン1及び開始エキソン2)が、どちらも意味のあるORFを含有しない。3つ目のものは、リバース鎖から発生する(Zambrowiczら, 1997, 上記)。マウスROSA26プロモーターの制御下の導入遺伝子は、胚及び成体マウスで遍在的な発現を示す(Soriano, P., Nature Genetics, 1999, 21, 70〜71)。ROSA26遺伝子座をマウスES細胞でターゲティングすることは、トランスジェニックマウスモデルを構築するために広く用いられている(Kisseberthら, Developmental Biology, 1999, 214, 128〜138; Mao X.ら, Proc. Natl. Acad. Sci. USA, 1999, 96, 5037〜5042; Soriano, 1999, 上記; Awatramaniら, Nature Genetics, 2001, 29, 257〜259; Mao X.ら, Blood, 2001, 97, 324〜326; Possematoら., Genesis, 2002, 32, 184〜186; Mao, J.ら, Nucleic Acids Res., 2005, 33, e155; Yuら, Proc. Natl. Acad. Sci. USA, 2005, 102, 8615〜8620; 国際PCT出願WO 99/53017、WO 02/098217、WO 03/020743、WO 2004/063381及びWO 2005/116070))。
【0003】
しかし、マウス細胞における相同組換えの効率は非常に低い(頻度:10-6〜10-9)。
この効率は、標的遺伝子座でのDNA二本鎖破断(double-strand break;DSB)により増強できる。このようなDSBは、定義によれば大きい配列を認識する配列特異的エンドヌクレアーゼであるメガヌクレアーゼにより創出できる(Thierry, A.及びB. Dujon, Nucleic Acids Res., 1992, 20, 5625〜5631)。これらのタンパク質は、生細胞においてユニーク部位を切断し、そのことにより、切断部位の近傍で遺伝子ターゲティングを、1000倍以上増強させる(Puchtaら, Nucleic Acids Res., 1993, 21, 5034〜5040 ; Rouetら, Mol. Cell. Biol., 1994, 14, 8096〜8106 ; Choulikaら, Mol. Cell. Biol., 1995, 15, 1968〜1973; Puchtaら, Proc. Natl. Acad. Sci. U.S.A., 1996, 93, 5055〜5060 ; Sargentら, Mol. Cell. Biol., 1997, 17, 267〜277; Cohen-Tannoudjiら, Mol. Cell. Biol., 1998, 18, 1444〜1448 ; Donohoら, Mol. Cell. Biol., 1998, 18, 4070〜4078; Elliottら, Mol. Cell. Biol., 1998, 18, 93〜101)。
【0004】
しかし、「ホーミングエンドヌクレアーゼ」ともよばれる数百の天然メガヌクレアーゼが同定されているが(Chevalier, B.S.及びB.L. Stoddard, Nucleic Acids Res., 2001, 29, 3757〜3774)、切断され得る配列のレパートリーは、ゲノムの複雑さに取り組むには制限されすぎであり、選択された遺伝子には通常、切断可能部位はない。理論的には、選択された特異性を有する人工配列特異的エンドヌクレアーゼの作製が、この制限を緩和できるはずである。よって、仕立てられた特異性を有するメガヌクレアーゼの作製について、熱心な研究がなされている。
【0005】
最近、ジンクフィンガーと、クラスIIS制限エンドヌクレアーゼであるFokIの触媒ドメインとの融合体を用いて、機能的配列特異的エンドヌクレアーゼが作製された(Smithら, Nucleic Acids Res., 1999, 27, 674〜681; Bibikovaら, Mol. Cell. Biol., 2001, 21, 289〜297 ; Bibikovaら, Genetics, 2002, 161, 1169〜1175 ; Bibikovaら, Science, 2003, 300, 764; Porteus, M.H.及びD. Baltimore, Science, 2003, 300, 763〜; Alwinら, Mol. Ther., 2005, 12, 610〜617; Urnovら, Nature, 2005, 435, 646〜651; Porteus, M.H., Mol. Ther., 2006, 13, 438〜446; 国際PCT出願WO 2007/014275)。このようなヌクレアーゼは、最近、リンパ系列からのヒト細胞におけるILR2G遺伝子を工学的に改変するために用いることができた(Urnovら, Nature, 2005, 435, 646〜651)。
【0006】
Cys2-His2型のジンクフィンガータンパク質(ZFP)の結合特異性は、これらが単純(フィンガーあたり実質的に4残基により駆動される特異性)でモジュール性の(modular)系を示すので、操作するのが容易である(Paboら, Annu. Rev. Biochem., 2001, 70, 313〜340; Jamiesonら, Nat. Rev. Drug Discov., 2003, 2, 361〜368)。Pabo (Rebar, E.J.及びC.O. Pabo, Science, 1994, 263, 671〜673 ; Kim, J.S.及びC.O. Pabo, Proc. Natl. Acad. Sci. U S A, 1998, 95, 2812〜2817)、Klug (Choo, Y.及びA. Klug, Proc. Natl. Acad. Sci. USA, 1994, 91, 11163〜11167; Isalan M.及びA. Klug, Nat. Biotechnol., 2001, 19, 656〜660)及びBarbas (Choo, Y.及びA. Klug, Proc. Natl. Acad. Sci. USA, 1994, 91, 11163〜11167; Isalan M.及びA. Klug, Nat. Biotechnol., 2001, 19, 656〜660)の実験室からの研究は、ほとんどのG/ANNG/ANNG/ANN配列に結合できる新規な人工ZFPの大きいレパートリーをもたらした。
【0007】
にもかかわらず、ZFPは、特に、治療用途のような非常に高レベルの特異性を必要とする用途において制限がある。最近、融合体でのFokIヌクレアーゼ活性が、FokIのいくつかのDNA結合欠陥性変異体の存在下を含んで、1つの認識部位、又はDNAループにより種々の距離で分けられた2つの部位のいずれかと作用することが見出されている(Cattoら, Nucleic Acids Res., 2006, 34, 1711〜1720)。よって、特異性は、哺乳動物細胞及びショウジョウバエ(Drosophila)における毒性により示されるように、非常に退化している(Bibikovaら, Genetics, 2002, 161, 1169〜1175; Bibikovaら, Science, 2003, 300, 764〜)。
【0008】
野生では、メガヌクレアーゼは、ホーミングエンドヌクレアーゼにより実質的に代表される。ホーミングエンドヌクレアーゼ(HE)は、数百のタンパク質ファミリーを含む天然のメガヌクレアーゼの広範なファミリーである(Chevalier, B. S.及びB.L. Stoddard, Nucleic Acids Res., 2001, 29, 3757〜3774)。これらのタンパク質は、「ホーミング」とよばれるプロセスにより伝播される可動性遺伝要素によりコードされる。エンドヌクレアーゼは、可動性要素が存在しない同族の対立遺伝子を切断することにより、受容側遺伝子座に可動性DNAを重複させる相同組換え事象を刺激する。効率及び特異性の点でのそれらの例外的な切断特性に鑑みて、これらは、新規で特異性が高いエンドヌクレアーゼを導くために理想的な足場であり得る。
【0009】
HEは、4つの主要なファミリーに属する。触媒中心に含まれる保存されたペプチドモチーフにちなんで命名されたLAGLIDADGファミリーは、最も普及しており、最もよく特徴付けられている群である。現在、7つの構造が入手可能である。このファミリーからのほとんどのタンパク質が単量体であり、2つのLAGLIDADGモチーフを示すが、いくつかは1つのモチーフのみを有するものの、二量体を形成してパリンドローム又は偽パリンドローム標的配列を切断する。
【0010】
ファミリーのメンバーのうち、LAGLIDADGペプチドが唯一の保存領域であるが、これらのタンパク質は、非常によく似た構造を有する(図2)。触媒コアは、I-CreI(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316)及びI-MsoI (Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)のようなホモ二量体について完全な2回回転対称の、そしてI-SceI (Moureら, J. Mol. Biol., 2003, 334, 685〜69)、I-DmoI (Silvaら, J. Mol. Biol., 1999, 286, 1123〜1136)又はI-AniI (Bolducら, Genes Dev., 2003, 17, 2875〜2888)のような単量体について偽対称を有する2つのDNA結合ドメインで挟まれる。両方の単量体又は両方のドメイン(単量体タンパク質について)は、二価カチオンの周りに組織された触媒コアに貢献する。触媒コアのすぐ上に、2つのLAGLIDADGペプチドが、二量体形成界面において必須の役割を演じる。DNA結合は、DNA主溝の上に位置する2つの典型的なサドル形ββαββ折り畳みに依存する。他のドメインは、例えばPI-PfuI (Ichiyanagiら, J. Mol. Biol., 2000, 300, 889〜901)及びPI-SceI (Moureら, Nat. Struct. Biol., 2002, 9, 764〜770)のようなインテインにおいて見出すことができ、このタンパク質スプライシングドメインはDNA結合にも関与する。
【0011】
I-DmoIドメインのN-末端をI-CreI単量体と融合させることによる機能的キメラメガヌクレアーゼの作製(Chevalierら, Mol. Cell., 2002, 10, 895〜905; Epinatら, Nucleic Acids Res, 2003, 31, 2952〜62; 国際PCT出願WO 03/078619及びWO 2004/031346)は、LAGLIDADGタンパク質の可塑性を証明する。
そのうえ、異なるグループが、I-CreI (Seligmanら, Genetics, 1997, 147, 1653〜1664; Sussmanら, J. Mol. Biol., 2004, 342, 31〜41; 国際PCT出願WO 2006/097784、WO 2006/097853及びWO 2007/049156; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458; Rosenら, Nucleic Acids Res., 2006, 34, 4791〜4800; Smithら, Nucleic Acids Res., Epub 2006年11月27日)、I-SceI (Doyonら, J. Am. Chem. Soc., 2006, 128, 2477〜2484)、PI-SceI (Gimbleら, J. Mol. Biol., 2003, 334, 993〜1008)及びI-MsoI (Ashworthら, Nature, 2006, 441, 656〜659)の特異性を局所的に変更するために合理的アプローチを用いている。
【0012】
さらに、局所的に特異性が変更された数百のI-CreI誘導体が、半合理的(semi-rational)アプローチとハイスループットスクリーニングとを組み合わせることにより工学的に作製された:
- I-CreIの残基Q44、R68及びR70又はQ44、R68、D75及びI77を変異させ、DNA標的(5NNN DNA標的)の±3〜5位のヌクレオチドに対する特異性が変更された変異型のコレクションを、スクリーニングにより同定した(国際PCT出願WO 2006/097784及びWO 2006/097853; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458; Smithら, Nucleic Acids Res., Epub 2006年11月27日)。
【0013】
- I-CreIの残基K28、N30及びQ38、N30、Y33及びQ38又はK28、Y33、Q38及びS40を変異させ、DNA標的(10NNN DNA標的)の±8〜10位のヌクレオチドに対する特異性が変更された変異型のコレクションを、スクリーニングにより同定した(Smithら, Nucleic Acids Res., Epub 2006年11月27日; 国際PCT出願WO 2007/049156)。
I-CreIの残基28〜40及び44〜77は、ホーミングエンドヌクレアーゼハーフサイト(half-site)の別個の部分に結合できる、2つの分離可能な機能的サブドメインを形成することが示された(Smithら Nucleic Acids Res., Epub 2006年11月27日; 国際PCT出願WO 2007/049095)。
【0014】
I-CreIの2つのサブドメインからの変異を同じ単量体内で組み合わせることにより、それぞれのサブドメインが結合する±3〜5位及び±8〜10位のヌクレオチドを含むパリンドロームの組み合わせたDNA標的配列を切断できる新規なキメラ分子(ホモ二量体)の設計が可能になった(Smithら, Nucleic Acids Res., Epub 2006年11月27日; 国際PCT出願WO 2007/049156)。
【0015】
2つの異なる変異型を組み合わせ、それぞれの変異型DNA標的配列の異なる半分の融合により得られるキメラ標的を切断できる機能的へテロ二量体エンドヌクレアーゼに組み立てた(Arnouldら, 上記; 国際PCT出願WO 2006/097854)。興味深いことに、新規なタンパク質は、正確な折り畳み及び安定性と、高い活性と、狭い特異性とを維持した。
【0016】
2つの前の工程の組み合わせにより、4つの異なるサブドメインを含む、より大きいコンビナトリアルアプローチが可能になる。異なるサブドメインは、図3に示すように、別々に改変でき、組み合わせて、選択された特異性を有する完全に再設計されたメガヌクレアーゼ変異型(ヘテロ二量体又は単鎖分子)を得ることができる。第1工程において、新規なメガヌクレアーゼの対を、切断したい標的に由来するパリンドローム標的を切断する新しい分子(「ハーフメガヌクレアーゼ」)に組み合わせる。次いで、このような「ハーフメガヌクレアーゼ」の組み合わせは、興味対象の標的を切断するヘテロ二量体種をもたらすことができる。4組の変異を、モデル標的配列又はヒトRAG1遺伝子からの配列を切断するヘテロ二量体エンドヌクレアーゼに組み立てることが、Smithら(Nucleic Acids Res., Epub 2006年11月27日)に記載されている。
【0017】
しかし、この報告で試験された標的は、パリンドロームI-CreI部位の元来の配列(C1221; 図5)と、±2位及び±1位が同一であった。塩基対±1及び±2は、タンパク質といずれの接触も示さないが、これらの位置は、特に塩基対±1については、含まれる情報がないわけではなく(Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)、さらなる基質特異性の供給源であり得ることが示されている(Argastら, J. Mol. Biol., 1998, 280, 345〜353; Juricaら, Mol. Cell., 1998, 2, 469〜476; Chevalier, B.S.及びB.L. Stoddard, Nucleic Acids Res., 2001, 29, 3757〜3774)。ランダム突然変異を誘発した切断可能なI-CreI標的のインビトロ選択(Argastら, 上記)は、タンパク質結合及び切断活性に対するこれらの4つの塩基対の重要性を明らかにした。活性部位で見出される配向された(ordered)水分子のネットワークが、DNA標的の位置決めに重要であることが示唆された(Chevalierら, Biochemistry, 2004, 43, 14015〜14026)。さらに、I-CreI結合の際にこの領域に出現する広範囲なコンホメーションの変更は、4つの中央のヌクレオチドが、おそらくは、配列依存性のコンホメーションの選好により、基質特異性に貢献できるであろうことを示唆する(Chevalierら, 2003, 上記)。
【発明の概要】
【発明が解決しようとする課題】
【0018】
よって、4つの中央のヌクレオチドgtacを有するパリンドローム配列を切断するホモ二量体として10NNN及び5NNN DNA標的について同定された変異体が、4つの中央のヌクレオチドの変更を含む標的を切断する新しいエンドヌクレアーゼの設計を可能にするかは明らかでなかった。
本発明者らは、I-CreI変異型により切断され得るマウスROSA26遺伝子座における一連のDNA標的を同定した(図17)。図3に記載するコンビナトリアルアプローチを用いて、I-CreIタンパク質のDNA結合ドメインを全体的に再設計し、そのことにより、完全に工学的に改変された特異性を有する新規なメガヌクレアーゼを工学的に作製して、I-CreI C1221 22 bpパリンドローム部位とは、4つの中央のヌクレオチドのうちの1つ(+1位)を含む13ヌクレオチドが異なる(図5)、マウスROSA26遺伝子座からのDNA標的(rosa1)を切断した。
【0019】
組み合わせた変異型を、それぞれヌクレオチド10NNN及び5NNNに対して最初は同定し、工学的に作製されたメガヌクレアーゼの活性に対する標的の4つの中央のヌクレオチドの強い影響が観察されたが、特異性が大きく変更された機能的メガヌクレアーゼを選択した。さらに、工学的に作製されたタンパク質の活性は、連続する2回のランダム突然変異誘発及びスクリーニングにより、I-CreIタンパク質の活性と比較して、著しく改良できた。
【0020】
ROSA26遺伝子座にて2本鎖破断を生じる能力は、該遺伝子座での相同組換えを著しく増強する手段を提供する。つまり、ROSA26遺伝子座を標的にするメガヌクレアーゼは、マウス細胞における効率的な遺伝子挿入を可能にする(図4)。この遺伝子座にて遺伝子を効率的に挿入する(ノックイン)能力は、再現可能な発現レベルを可能にするとともに、挿入を作製するための予測可能なタイムラインを可能にするという利点を有する。可能性のある応用は、マウス細胞における組換えタンパク質の生成、及び例えばタンパク質生成、遺伝子機能研究、薬剤スクリーニングのため、又は疾患モデルとして用いることができるトランスジェニックマウス及び組換えマウス株化細胞の工学的作製を含む。
【課題を解決するための手段】
【0021】
本発明は、2つのI-CreI単量体のうちの少なくとも一方が、I-CreIの26位〜40位及び44位〜77位にそれぞれ位置するLAGLIDADGコアドメインの2つの機能的サブドメイン内に1つずつ存在する少なくとも2つの置換を有し、マウスROSA26遺伝子座からのDNA標的を切断できるI-CreI変異型に関する。
【0022】
本発明による変異型の切断活性は、国際PCT出願WO 2004/067736; Epinatら, Nucleic Acids Res., 2003, 31, 2952〜2962; Chamesら, Nucleic Acids Res., 2005, 33, e178;及びArnouldら, J. Mol. Biol., 2006, 355, 443〜458に記載されるもののような、いずれの公知のインビトロ又はインビボ切断アッセイにより測定できる。例えば、本発明の変異型の切断活性は、酵母又は哺乳動物細胞における直列反復組換えアッセイ(direct repeat recombination assay)により、レポーターベクターを用いて測定できる。レポーターベクターは、酵母又は哺乳動物発現ベクターにクローニングされた、レポーター遺伝子の2つの短縮された非機能的コピー(直列反復)と、ゲノムDNA標的配列とを、介在配列内に含む。変異型の発現は、ゲノムDNA標的配列を切断できる機能的エンドヌクレアーゼをもたらす。この切断は、直列反復間の相同組換えを誘発し、その発現は、適切なアッセイにより監視できる。
【図面の簡単な説明】
【0023】
【図1】マウスROSA26遺伝子座を表す(アクセッション番号EMBL CQ880114;配列番号3)。
【図2】DNA標的に結合したI-CreIホーミングエンドヌクレアーゼの3次元構造を表す。
【図3】I-CreI及び他のLAGLIDADGホーミングエンドヌクレアーゼの特異性を工学的に改変するための2ステップアプローチを示す。
【図4】マウスROSA26遺伝子座を切断するメガヌクレアーゼの使用のためのストラテジーを表す。
【図5】rosa1標的配列及び誘導体を表す。
【図6】pCLS1055ベクターマップを表す。
【図7】pCLS0542ベクターマップを表す。
【図8】I-CreI変異体によるrosa1.3 DNA標的の切断を示す。
【図9】I-CreIコンビナトリアル変異体によるrosa1.4標的の切断を示す。
【図10】pCLS1107ベクターマップを表す。
【図11】ヘテロ二量体I-CreIコンビナトリアル変異体によるrosa1.2及びrosa1標的の切断を示す。
【図12】rosa1標的の切断を示す。
【図13】rosa1標的の切断を示す。
【図14】pCLS1058ベクターマップを表す。
【図15】pCLS1069ベクターマップを表す。
【図16】CHO細胞における染色体外モデルでのI-CreI洗練変異体によるrosa1標的の切断を示す。
【図17】マウスROSA26で見出されるメガヌクレアーゼ標的配列と、該DNA標的のそれぞれを切断できる対応するI-CreI変異型を表す。
【図18】pCLS1675ベクターマップを表す。
【図19】pCLS1761ベクターマップを表す。
【図20】pCLS1762ベクターマップを表す。
【図21】IRES-Hygroマトリクス(pCLS1675)を用いたノックイン(KI)事象のPCR分析を示す。
【発明を実施するための形態】
【0024】
定義
- アミノ酸とは、天然又は合成アミノ酸の鏡像異性体及び立体異性体を含む天然又は合成アミノ酸のことをいう。
ポリペプチド配列中のアミノ酸残基は、本明細書において、1文字コードに従って表し、例えばQはGln又はグルタミン残基を意味し、RはArg又はアルギニン残基を意味し、DはAsp又はアスパラギン酸残基を意味する。
- 酸性アミノ酸とは、アスパラギン酸(D)及びグルタミン酸(E)のことをいう。
- 塩基性アミノ酸とは、リジン(K)、アルギニン(R)及びヒスチジン(H)のことをいう。
- 小さいアミノ酸とは、グリシン(G)及びアラニン(A)のことをいう。
- 芳香族アミノ酸とは、フェニルアラニン(F)、トリプトファン(W)及びチロシン(Y)のことをいう。
【0025】
- ヌクレオチドは、次のように表す:1文字コードは、ヌクレオシドの塩基を表すために用いる:aはアデニンであり、tはチミンであり、cはシトシンであり、gはグアニンである。縮重ヌクレオチドについて、rはg又はa (プリンヌクレオチド)を表し、kはg又はtを表し、sはg又はcを表し、wはa又はtを表し、mはa又はcを表し、yはt又はc (ピリミジンヌクレオチド)を表し、dはg、a又はtを表し、vはg、a又はcを表し、bはg、t又はcを表し、hはa、t又はcを表し、nはg、a、t又はcを表す。
【0026】
- 「メガヌクレアーゼ」により、12〜45 bpの2本鎖DNA標的配列を有するエンドヌクレアーゼを意図する。該メガヌクレアーゼは、各ドメインが単量体上にある二量体酵素、又は単一ポリペプチド上に2つのドメインを含む単量体酵素のいずれかである。
- 「メガヌクレアーゼドメイン」により、メガヌクレアーゼのDNA標的の一方の半分と相互作用し、かつDNA標的の他方の半分と相互作用する同じメガヌクレアーゼの他方のドメインと会合して、該DNA標的を切断できる機能的メガヌクレアーゼを形成できる領域を意図する。
【0027】
- 「メガヌクレアーゼ変異型」又は「変異型」により、野生型メガヌクレアーゼ(天然のメガヌクレアーゼ)のアミノ酸配列における少なくとも1つの残基の、別のアミノ酸での置き換えにより得られるメガヌクレアーゼを意図する。
- 「機能的変異型」により、DNA標的配列、好ましくは、親のメガヌクレアーゼにより切断されない新しい標的を切断できる変異型を意図する。例えば、このような変異型は、DNA標的配列に接触するか、又は直接若しくは間接的に該DNA標的と相互作用する位置にてアミノ酸変動を有する。
- 「新規な特異性を有するメガヌクレアーゼ変異型」により、親のメガヌクレアーゼのものとは異なる切断標的のパターンを有する変異型を意図する。等価で、同様に用いられる用語「新規な特異性」「改変された特異性」「新規な切断特異性」「新規な基質特異性」は、DNA標的配列のヌクレオチドに対する変異型の特異性のことである。
【0028】
- 「I-CreI」により、配列表内の配列番号1の配列に相当する配列SWISSPROT P05725又は配列表内の配列番号133の配列に相当するpdbアクセッションコード1g9yの配列を有する野生型I-CreIを意図する。
- 「ドメイン」又は「コアドメイン」により、約100アミノ酸残基の配列に相当する、LAGLIDADGファミリーのホーミングエンドヌクレアーゼの特徴的なα1β1β2α2β3β4α3折り畳みである「LAGLIDADGホーミングエンドヌクレアーゼコアドメイン」を意図する。該ドメインは、DNA標的の一方の半分と相互作用する逆平行ベータシートに折り畳まれた4つのベータ鎖(β1β2β3β4)を含む。このドメインは、DNA標的の他方の半分と相互作用する別のLAGLIDADGホーミングエンドヌクレアーゼコアドメインと会合して、該DNA標的を切断できる機能エンドヌクレアーゼを形成できる。例えば、二量体ホーミングエンドヌクレアーゼI-CreI (163アミノ酸)の場合、LAGLIDADGホーミングエンドヌクレアーゼコアドメインは、残基6〜94に相当する。
【0029】
- 「単鎖メガヌクレアーゼ」により、ペプチドスペーサーで連結された2つのLAGLIDADGホーミングエンドヌクレアーゼドメイン又はコアドメインを含むメガヌクレアーゼを意図する。単鎖メガヌクレアーゼは、それぞれの親のメガヌクレアーゼ標的配列の1つの異なる半分を含むキメラDNA標的配列を切断できる。
【0030】
- 「サブドメイン」により、ホーミングエンドヌクレアーゼDNA標的ハーフサイト(half-site)の別個の部分と相互作用するLAGLIDADGホーミングエンドヌクレアーゼコアドメインの領域を意図する。2つの異なるサブドメインは独立して挙動し、一方のサブドメインでの変異は、他方のサブドメインの結合及び切断特性を変更しない。よって、2つのサブドメインは、ホーミングエンドヌクレアーゼDNA標的ハーフサイトの別個の部分に結合する。
- 「ベータヘアピン」により、ループ又はターンにより接続されたLAGLIDADGホーミングエンドヌクレアーゼコアドメインの逆平行ベータシートの2つの連続するベータ鎖(β1β2又はβ3β4)を意図する。
【0031】
- 「I-CreI部位」により、I-CreIにより切断される22〜24 bpの2本鎖DNA配列を意図する。I-CreI部位は、野生型(天然)非パリンドロームI-CreIホーミング部位と、C1221ともよばれる配列5'- t-12c-11a-10a-9a-8a-7c-6g-5t-4c-3g-2t-1a+1c+2g+3a+4c+5g+6t+7t+8t+9t+10g+11a+12 (配列番号2; 図5)のような派生パリンドローム配列を含む。
- 「DNA標的」、「DNA標的配列」、「標的配列」、「標的部位」、「標的」、「部位」、「興味対象の部位」、「認識部位」、「認識配列」、「ホーミング認識部位」、「ホーミング部位」、「切断部位」により、I-CreIのようなLAGLIDADGホーミングエンドヌクレアーゼ、又は変異型、又はI-CreIに由来する単鎖キメラメガヌクレアーゼにより認識されかつ切断される20〜24 bpの2本鎖パリンドローム、部分的パリンドローム(偽パリンドローム)又は非パリンドロームのポリヌクレオチド配列を意図する。これらの用語は、そこでメガヌクレアーゼにより2本鎖破断(切断)が誘導される独特のDNAの場所、好ましくはゲノムの場所のことをいう。DNA標的は、C1221について上で示したように、2本鎖ポリヌクレオチドの一方の鎖の5'から3'の配列により定義される。DNA標的の切断は、センス鎖及びアンチセンス鎖のそれぞれについて、+2位及び-2位のヌクレオチドで生じる。そうでないと記載しない限り、I-Cre Iメガヌクレアーゼ変異型によるDNA標的の切断が生じる位置は、DNA標的のセンス鎖上の切断部位に相当する。
【0032】
- 「DNA標的ハーフサイト」、「ハーフ切断部位」又は「ハーフサイト」により、それぞれのLAGLIDADGホーミングエンドヌクレアーゼコアドメインが結合するDNA標的の部分を意図する。
- 「キメラDNA標的」又は「ハイブリッドDNA標的」により、2つの親のメガヌクレアーゼ標的配列の異なる半分の融合を意図する。さらに、該標的の少なくとも一方の半分は、少なくとも2つの別個のサブドメインが結合するヌクレオチドの組み合わせ(組み合わせたDNA標的)を含み得る。
【0033】
- 「マウスROSA26遺伝子座」により、マウス第6染色体に位置し、EMBLアクセッション番号CQ880114 (配列番号3; 13139 bp)に相当する配列を有する遺伝子座を意図する。ROSA26は、3つの転写産物を生成する(図1):2つの転写産物は、同一の5'末端を共有する共通プロモーターから生じるが(エキソン1及びエキソン2開始)、いずれも有効なORFを含有しない。3つ目の転写産物は、リバース鎖から生じる。
- 「マウスROSA26遺伝子座からのDNA標的配列」、「ゲノムDNA標的配列」、「ゲノムDNA切断部位」、「ゲノムDNA標的」又は「ゲノム標的」により、メガヌクレアーゼ変異型により認識され切断されるマウスROSA26遺伝子座の20〜24 bpの配列を意図する。
【0034】
- 「ベクター」により、それが連結された別の核酸を輸送できる核酸分子を意図する。
- 「相同な」により、配列同士の間の相同組換えを導くのに充分な別の配列との同一性を有する、より具体的には少なくとも95%の同一性、好ましくは97%の同一性、より好ましくは99%を有する配列を意図する。
- 「同一性」は、2つの核酸分子又はポリペプチド間の配列同一性のことをいう。同一性は、比較の目的のために整列させ得るそれぞれの配列中の位置の比較により決定できる。比較される配列中の位置が同じ塩基により占められる場合、その分子同士は、その位置において同一である。核酸又はアミノ酸配列間の類似性又は同一性の程度は、核酸配列により共有される位置での同一又は一致するヌクレオチドの数の関数である。種々のアラインメントアルゴリズム及び/又はプログラムを用いて、2つの配列間の同一性を計算することができ、GCG配列解析パッケージ(University of Wisconsin, Madison, Wis.)の一部分として利用可能であり、例えばデフォルト設定で用い得るFASTA又はBLASTを含む。
【0035】
- 「個体」は、哺乳動物、及びその他の脊椎動物(例えば鳥類、魚類及び爬虫類)を含む。用語「哺乳動物」及び「哺乳類」は、本明細書で用いる場合、その子に授乳し、生存する子を出産する(真獣類(eutharian)又は胎盤哺乳類(placental mammals))又は産卵する(後獣類(metatharian)又は無胎盤哺乳類(nonplacental mammals))単孔類、有袋類及び有胎盤類(placental)を含むいずれの脊椎動物のことをいう。哺乳動物の種の例は、ヒト、及びその他の霊長類(例えばサル、チンパンジー)、げっ歯類(例えばラット、マウス、モルモット)、並びにその他、例えばウシ、ブタ及びウマを含む。
- 「変異」により、ポリヌクレオチド(cDNA、遺伝子)又はポリペプチド配列内の1つ又は複数のヌクレオチド/アミノ酸の置換、欠失、挿入を意図する。上記の変異は、遺伝子のコード配列又はその調節配列に影響し得る。これは、ゲノム配列の構造又はコードされたmRNAの構造/安定性にも影響し得る。
【0036】
本発明による変異型は、ホモ二量体又はヘテロ二量体であり得る。好ましくは、ヘテロ二量体の両方の単量体が、26位〜40位及び/又は44位〜77位で変異される。より好ましくは、両方の単量体が、I-CreIの26位〜40位及び44位〜77位の両方に異なる置換を有する。
上記の変異型の好ましい実施形態において、I-CreIの44位〜77位に位置するサブドメイン内の上記の置換は、44位、68位、70位、75位及び/又は77位にある。
【0037】
上記の変異型の別の好ましい実施形態において、I-CreIの26位〜40位に位置するサブドメイン内の上記の置換は、26位、28位、30位、32位、33位、38位及び/又は40位にある。
上記の変異型の別の好ましい実施形態において、上記の置換は、元のアミノ酸の、A、D、E、G、H、K、N、P、Q、R、S、T、Y、C、V、L及びWからなる群より選択されるアミノ酸での置き換えである。
【0038】
上記の変異型の別の好ましい実施形態において、これは、DNA標的配列と接触するか又はDNA主鎖(backbone)若しくはヌクレオチド塩基と直接又は水分子を介して相互作用する他のアミノ酸残基の位置にて、1つ又は複数の変異を含む。これらの残基は、当該技術において公知である(Juricaら, Molecular Cell., 1998, 2, 469〜476; Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)。
特に、付加的な置換は、例えば最終C-末端ループ中の(137位〜143位; Prietoら, Nucleic Acids Res., Epub 2007年4月22日)、リン酸主鎖と接触する位置にて導入され得る。好ましくは、上記の残基は、上記のDNA切断部位の結合及び切断に関与する。より好ましくは、上記の残基は、I-CreIの138位、139位、142位又は143位にある。それぞれの変異が138位と139位の残基の対及び142位と143位の残基の対から選択される残基の異なる対の中にあることを条件として、2つの残基を1つの変異型内で変異させ得る。導入される変異は、最終C-末端ループのアミノ酸の、I-CreI部位のリン酸主鎖との相互作用を改変する。好ましくは、138位又は139位の残基を疎水性アミノ酸により置換して、DNA切断部位のリン酸主鎖との水素結合の形成を回避する。例えば、138位の残基がアラニンにより置換されるか、又は139位の残基がメチオニンにより置換される。142位又は143位の残基を、有利には、小さいアミノ酸、例えばグリシンにより置換して、これらのアミノ酸残基の側鎖のサイズを低減させる。より好ましくは、最終C-末端ループ中の上記の置換は、I-CreI部位の±1〜2位、±6〜7位及び/又は±11〜12位のヌクレオチドに対する変異型の特異性を改変する。
【0039】
上記の変異型の別の好ましい実施形態において、これは、マウスROSA26遺伝子座からのDNA標的配列に対する変異型の結合及び/又は切断特性を改良する1つ又は複数の付加的な変異を含む。
変異される付加的な残基は、I-CreI配列全体、特にI-CreIのC-末端半分(80位〜163位)にあり得る。例えば、変異型は、19位、24位、79位、105位、107位、151位、153位、158位での1つ又は複数の付加的な置換を含む。上記の置換は、有利には、G19S、I24V、S79G、V105A、K107R、V151A、D153G及びK158Eからなる群より選択される。
【0040】
本発明の変異型は、野生型I-CreI (配列番号1又は133)、又は配列番号133と少なくとも85%の同一性、好ましくは少なくとも90%の同一性、より好ましくは少なくとも95%の同一性を有するI-CreI足場タンパク質、例えばI-CreI配列の2位のアラニンの挿入、置換D75N及びC-末端でのAADの挿入(164位〜166位)を有する配列番号4の足場(167アミノ酸)から導き得る。
【0041】
さらに、本発明の変異型は、配列のNH2末端及び/又はCOOH末端に挿入された1つ又は複数の残基を含み得る。例えば、タグ(エピトープ(HA-タグ(YPYDVPDYA;配列番号135)又はS-タグ(KETAAAKFERQHMDS;配列番号136)又はポリヒスチジン配列)を、NH2末端及び/又はCOOH末端に導入する。このタグは、上記の変異型の検出及び/又は精製のために有用である。NH2末端にタグを導入する場合、タグの配列は、変異型の最初のアミノ酸を置き換えるか(少なくとも最初のメチオニン、そして結局は変異型の2番目のアミノ酸;メチオニンで開始するタグ)、又は最初のアミノ酸(メチオニン)と2番目のアミノ酸の間若しくは変異型の最初のアミノ酸と3番目のアミノ酸との間に挿入され得る(メチオニンのないタグ)。
【0042】
変異型は、核局在化シグナル(NLS)も含み得る。上記のNLSは、該変異型を細胞の核内に輸送するために有用である。NLSの例は、KKKRK (配列番号134)である。NLSは、変異型の最初のメチオニンの直後又はN-末端タグの直後に挿入できる。
【0043】
本発明による変異型は、パリンドローム又は偽パリンドロームDNA標的配列を切断できるホモ二量体であり得る。
或いは、上記の変異型は、I-CreIの26位〜40位及び/又は44位〜77位に異なる置換を有する第1及び第2単量体の会合により得られるヘテロ二量体であって、マウスROSA26遺伝子座からの非パリンドロームDNA標的を切断できるヘテロ二量体である。
上記の変異型により切断されるDNA標的配列は、マウスROSA26遺伝子座のエキソン又はイントロン内にあり得る。
【0044】
上記の変異型の別の好ましい実施形態において、上記のDNA標的は、マウスROSA26遺伝子座の全てをカバーする配列番号5〜30の配列(図17)からなる群より選択される。
【0045】
【表1】
【0046】
より好ましくは、変異型の単量体は、第1及び第2単量体のそれぞれについて少なくとも以下の置換を有する:
- N30H、Y33S、Q44E、R68C、R70S及びD75N (第1単量体)と、N30D、Y33R、Q38T、Q44K、R68E、R70S及びI77R (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号5を切断する(図1及び17;表I)、
- S32N、Y33G、Q44K、R70E及びD75N (第1単量体)と、S32T、Q38W、Q44K、R68E、R70S及びI77R (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号6を切断する(図1及び17;表I)、
- Y33R、Q38N、S40Q、Q44N、R70S、D75R及びI77D (第1単量体)と、N30H、Y33S、Q44A、R70S、D75Q及びI77E (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号7を切断する(図1及び17;表I)、
- K28S、Q38R、S40K、Q44D、R68Y、R70S、D75S及びI77R (第1単量体)と、Y33C、Q38A、R68A、R70K及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号8を切断する(図1及び17;表I)、
【0047】
- Y33C、Q44T、R70S及びD75Y (第1単量体)と、S32D、Q38C、Q44D、R68Y、R70S、D75S及びI77R (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号9を切断する(図1及び17;表I)、
- S32T、Y33C、R68T、R70N及びD75N (第1単量体)と、S32T、Q38W、Q44K、R70E及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号10を切断する(図1及び17;表I)、
- R70S、D75R及びI77Y (第1単量体)と、Y33R、Q38A、S40Q、Q44A、R70S及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号11を切断する(図1及び17;表I)、
- K28S、Q38R、S40K、Q44T、R68N、R70N及びD75N (第1単量体)と、Y33H、Q38S、Q44K、R68Y、R70S、D75Q及びI77N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号12を切断する(図1及び17;表I)、
【0048】
- K28A、Y33S、Q38R、S40K、Q44N、R68Y、R70S、D75R、I77V (第1単量体)と、S32T、Y33C、Q44A、R70S及びD75N (第2単量体);この単量体は、第1イントロン内に位置するROSA26標的配列番号13を切断する(図1及び17;表I)、
- S32D、Y33H、Q44K、R68E、R70S及びI77R (第1単量体)と、S32D、Y33H、Q44D、R68N、R70S及びD75N (第2単量体);この変異型は、第1イントロン内に位置するROSA26標的配列番号14を切断する(図1及び17;表I)、
- N30R、S32D、R68S、R70K及びD75N (第1単量体)と、D75N (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号16を切断する(図1及び17;表I)、
- K28R、Y33A、Q38Y、S40Q、R68Y、R70S、D75R及びI77Q (第1単量体)と、Y33R、Q38A、S40Q、R70S及びI77K (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号17を切断する(図1及び17;表I)、
- K28R、N30D、D75E及びI77R (第1単量体)と、S32D、Q38C、Q44A、R70S、D75R及びI77Y (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号18を切断する(図1及び17;表I)、
- Y33R、Q38A、S40Q、R70S及びD75N (第1単量体)と、R70S、D75Y及びI77R (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号19を切断する(図1及び17;表I)、
【0049】
- Y33R、Q38A、S40Q、Q44N、R68Y、R70S、D75R及びI77V (第1単量体)と、N30R、S32D、Q44T、R68H、R70H及びD75N (第2単量体);この変異型は、第2エキソン内に位置するROSA26標的配列番号20を切断する(図1及び17;表I)、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q及びI77N (第1単量体)と、S32A、Y33C、R68Y、R70S、D75R及びI77Q (第2単量体);この変異型は、第2イントロン内に位置するROSA26標的配列番号21を切断する(図1及び17;表I)、
- K28A、Y33S、Q38R、S40K、R68N、R70S、D75N及びI77R (第1単量体)と、S32N、Y33G、Q44A、R68A、R70K及びD75N (第2単量体);この変異型は、第2イントロン内に位置するROSA26標的配列番号22を切断する(図1及び17;表I)、
- N30H、Y33S、Q44Y、R70S及びI77V (第1単量体)と、Y33R、S40Q、Q44A、R70S及びD75N (第2単量体);この変異型は、第2イントロン内に位置するROSA26標的配列番号23を切断する(図1及び17;表I)、
- S32D、Y33H、R68T、R70N及びD75N (第1単量体)と、N30R、S32D、Q44T、R68N、R70N及びD75N (第2単量体);この変異型は、第2イントロンとアンチセンス転写産物内に位置するROSA26標的配列番号24を切断する(図1及び17;表I)、
- S32R、Y33D、Q44A、R70S及びD75N (第1単量体)と、Y33S、Q38R、S40H、R68H、R70H及びD75N (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号25を切断する(図1及び17;表I)、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q及びI77N (第1単量体)と、K28R、Y33A、Q38Y、S40Q、Q44T、R68H、R70H及びD75N (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号26を切断する(図1及び17;表I)、
【0050】
- K28Q、Q38R、S40K、Q44A、R70S、D75E及びI77R (第1単量体)と、N30R、S32D、Q44K、R68Y、R70S、D75Q及びI77N (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号27を切断する(図1及び17;表I)、
- Y33T、Q38A、R68H、R70H及びD75N (第1単量体)と、N30H、Y33S、R70S及びI77K (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号28を切断する(図1及び17;表I)、
- Y33T、S40T、R68A、R70K及びD75N (第1単量体)と、K28R、Y33A、Q38Y、S40Q、R70S及びI77K (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号29を切断する(図1及び17;表I)、並びに
- S32D、Y33H、Q44N、R70S、D75R及びI77D (第1単量体)と、S32D、Q38C、R70S及びI77K (第2単量体);この変異型は、第3エキソン又はアンチセンス転写産物内に位置するROSA26標的配列番号30を切断する(図1及び17;表I)。
【0051】
表IのROSA26 DNA標的(配列番号5〜14及び16〜30のヌクレオチド配列)を切断する上記の変異型の例は、配列番号82〜106のアミノ酸配列のいずれかの第1単量体と、配列番号107〜116、4、117〜130のアミノ酸配列のいずれかの第2単量体とをそれぞれ有する変異型を含む(図17)。
【0052】
さらに、以下の変異型は、第2エキソン内に位置するrosa1と命名されるROSA26 DNA標的を切断できる(図1及び17;表I):
- I24V、Q44Y、R70S及びD75N;I24V、Q44Y、R68Y、R70S、D75Y及びI77R;I24V、Q44Y、R70S、D75N及びI77V;I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択される第1単量体と、K28E、Y33R、Q38R、S40R、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40R、Q44A、R70N及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40K、Q44V、R70A及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R70G及びD75Nからなる群より選択される第2単量体とを有する40個の変異型;これらの変異型の例は、表V (第1単量体:m2、m6、m8、m12、m13、m14、m16又はm17 (配列番号39、43、45、49、50、51、53及び54);配列番号60、61、63、65及び66のいずれかの第2単量体)に示す。
- I24V、Q44Y、R70S、D75Y及びI77Tを含む第1単量体と、K28E、Y33R、Q38R、S40R、Q44A、R68S、R70Q及びD75N (第2単量体)とを有する変異型;この変異型の例は、表V (第1単量体m17 (配列番号54)と第2単量体 配列番号62)に示す。
【0053】
- I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択される第1単量体と、K28E、Y33R、Q38R、S40K、Q44A、R70S及びD75N、並びにK28E、Y33R、Q38R、S40K、Q44A、R68T、R70N及びD75Nからなる群より選択される第2単量体とを有する10個の変異型;これらの変異型の例は、表V (第1単量体;m12、m13、m14、m16又はm17 (配列番号49、50、51、53及び54);配列番号64及び67のいずれかの第2単量体)に示す。
- 配列番号72 (MO_1;表VI及びVII)又は配列番号73 (MO_2;表VI及びVII)の配列からなる第1単量体と、配列番号74〜77 (mO_1〜mO_4;表VII)の配列のいずれかからなる第2単量体とを有する変異型;これらの8個の変異型は、rosa1標的についての変異型の切断活性を増大させる付加的な置換を有する。
【0054】
本発明は、上記で定義される配列と少なくとも85%の同一性、好ましくは少なくとも90%の同一性、より好ましくは少なくとも95% (96%、97%、98%、99 %)の同一性を有するI-CreI変異型であって、マウスROSA26遺伝子座からのDNA標的を切断できる変異型を含む。
例えば、本発明は、MO_1及びmO_2から、NLS、タグ又はそれらの両方の挿入により導かれるI-CreI変異型であって、配列番号140〜145の配列からなる群より選択される変異型を含む。
【0055】
ヘテロ二量体変異型は、有利には、2つのI-CreI単量体間に分子間相互作用を作製する第1及び第2単量体の残基に対応する興味対象の少なくとも1対の変異を有する偏性(obligate)ヘテロ二量体変異型であり、ここで、上記の対の第1変異は第1単量体中にあり、上記の対の第2変異は第2単量体中にあり、上記の変異の対は各単量体からの機能的ホモ二量体の形成を妨げ、マウスROSA26遺伝子座からのゲノムDNA標的を切断できる機能的ヘテロ二量体の形成を可能にする。
【0056】
偏性ヘテロ二量体を形成するために、単量体は、第1及び第2単量体のそれぞれについて、以下の変異の対の少なくとも1つを有することが有利である:
a) 8位のグルタミン酸の塩基性アミノ酸、好ましくはアルギニンでの置換(第1単量体)、及び7位のリジンの酸性アミノ酸、好ましくはグルタミン酸での置換(第2単量体);第1単量体は、7位及び96位のリジン残基の少なくとも1つのアルギニンによる置換をさらに含み得る、
b) 61位のグルタミン酸の塩基性アミノ酸、好ましくはアルギニンでの置換(第1単量体)、及び96位のリジンの酸性アミノ酸、好ましくはグルタミン酸での置換(第2単量体);第1単量体は、7位及び96位のリジン残基の少なくとも1つのアルギニンによる置換をさらに含み得る、
c) 97位のロイシンの芳香族アミノ酸、好ましくはフェニルアラニンでの置換(第1単量体)、及び54位のフェニルアラニンの小さいアミノ酸、好ましくはグリシンでの置換(第2単量体);第1単量体は、54位のフェニルアラニンのトリプトファンによる置換をさらに含むことができ、第2単量体は、58位のロイシン又は57位のリジンのメチオニンによる置換をさらに含み得る、
d) 137位のアスパラギン酸の塩基性アミノ酸、好ましくはアルギニンでの置換(第1単量体)、及び51位のアルギニンの酸性アミノ酸、好ましくはグルタミン酸での置換(第2単量体)。
【0057】
例えば、第1単量体は変異D137Rを、第2単量体は変異R51Dを有し得る。偏性ヘテロ二量体メガヌクレアーゼは、有利には、上記のa)、b)、c)又はd)で定義される変異の少なくとも2対を含む。変異の対のうちの1つは、有利にはc)又はd)で定義されるとおりである。好ましくは、一方の単量体は、7位及び96位のリジン残基の酸性アミノ酸(アスパラギン酸(D)又はグルタミン酸(E))、好ましくはアスパラギン酸(K7E及びK96E)による置換を含み、他方の単量体は、8位及び61位のグルタミン酸残基の塩基性アミノ酸(アルギニン(R)又はリジン(K);例えばE8K及びE61R)による置換を含む。より好ましくは、偏性ヘテロ二量体メガヌクレアーゼは、上記のa)、b)及びc)で定義される変異の3対を含む。偏性ヘテロ二量体メガヌクレアーゼは、有利には、(i) E8R、E8K又はE8H、E61R、E61K又はE61H及びL97F、L97W又はL97Y; (ii) K7R、E8R、E61R、K96R及びL97F、或いは(iii) K7R、E8R、F54W、E61R、K96R及びL97Fと、(iv) K7E又はK7D、F54G又はF54A及びK96D又はK96E; (v) K7E、F54G、L58M及びK96E、或いは(vi) K7E、F54G、K57M及びK96Eの変異を少なくとも有する第2単量体(B)とからなる。例えば、第1単量体は、変異K7R、E8R又はE8K、E61R、K96R及びL97F、或いはK7R、E8R又はE8K、F54W、E61R、K96R及びL97Fを有し、第2単量体は、変異K7E、F54G、L58M及びK96E又はK7E、F54G、K57M及びK96Eを有し得る。ヘテロ二量体の例は、配列番号147及び配列番号148である。偏性ヘテロ二量体は、上記で定義される少なくとも1つのNLS及び/又は1つのタグを有し得る。上記のNLS及び/又はタグは、第1及び/又は第2単量体にあり得る。
【0058】
本発明の主題は、上記で定義されるI-CreI変異型に由来する単鎖キメラメガヌクレアーゼ(融合タンパク質)でもある。単鎖メガヌクレアーゼは、2つのI-CreI単量体、2つのI-CreIコアドメイン(I-CreIの6位〜94位)、又はこれら両方の組み合わせを含み得る。好ましくは、2つの単量体/コアドメイン又はこれら両方の組み合わせは、ペプチドリンカーにより連結される。ペプチドリンカーの例は、配列番号149である。単鎖キメラメガヌクレアーゼの例は、配列番号146である。単鎖キメラメガヌクレアーゼは、上記で定義される少なくとも1つのNLS及び/又は1つのタグをさらに含み得る。上記のNLS及び/又はタグは、第1及び/又は第2単量体にあり得る。
【0059】
本発明の主題は、上記で定義される変異型又は単鎖キメラメガヌクレアーゼをコードするポリヌクレオチドフラグメントでもある。上記のポリヌクレオチドは、ホモ二量体若しくはヘテロ二量体の変異型の1つの単量体、又は単鎖キメラメガヌクレアーゼの2つのドメイン/単量体をコードし得る。
【0060】
本発明の主題は、本発明による変異型又は単鎖メガヌクレアーゼの発現のための組換えベクターでもある。組換えベクターは、上記で定義される変異型又は単鎖メガヌクレアーゼをコードする少なくとも1つのポリヌクレオチドフラグメントを含む。好ましい実施形態において、上記のベクターは、それぞれがヘテロ二量体変異型の単量体の一方をコードする2つの異なるポリヌクレオチドフラグメントを含む。
【0061】
本発明において用い得るベクターは、限定されないが、ウイルスベクター、プラスミド、RNAベクター、或いは染色体、非染色体、半合成又は合成の核酸からなり得る線状若しくは環状のDNA又はRNA分子を含む。好ましいベクターは、自律複製できるもの(エピソームベクター)及び/又は連結された核酸の発現を可能にするもの(発現ベクター)である。多数の適切なベクターが当業者に知られ、商業的に入手可能である。
【0062】
ウイルスベクターは、レトロウイルス、アデノウイルス、パルボウイルス(例えばアデノ随伴ウイルス)、コロナウイルス、マイナス鎖RNAウイルス、例えばオルトミクソウイルス(例えばインフルエンザウイルス)、ラブドウイルス(例えば狂犬病及び水疱性口内炎ウイルス)、パラミクソウイルス(例えば麻疹及びセンダイ)、プラス鎖RNAウイルス、例えばピコルナウイルス及びアルファウイルス、並びにアデノウイルス、ヘルペスウイルス(例えば単純ヘルペスウイルス1及び2型、エプスタイン-バーウイルス、サイトメガロウイルス)及びポックスウイルス(例えばワクシニア、鶏痘及びカナリア痘)を含む二本鎖DNAウイルスを含む。その他のウイルスは、例えば、ノーウォークウイルス、トガウイルス、フラビウイルス、レオウイルス、パポバウイルス、ヘパドナウイルス及び肝炎ウイルスを含む。レトロウイルスの例は、トリ白血病肉腫、哺乳類C型、B型ウイルス、D型ウイルス、HTLV-BLV群、レンチウイルス、スプマウイルスを含む(Coffin, J. M., Retroviridae: The viruses and their replication, In Fundamental Virology, 第3版, B. N. Fieldsら編, Lippincott-Raven Publishers, Philadelphia, 1996)。
好ましいベクターは、レンチウイルスベクター、特に自己不活化レンチウイルスベクター(self inactivacting lentiviral vectors)を含む。
【0063】
ベクターは、選択マーカー、例えば真核細胞培養についてネオマイシンホスホトランスフェラーゼ、ヒスチジノールデヒドロゲナーゼ、ジヒドロ葉酸レダクターゼ、ハイグロマイシンホスホトランスフェラーゼ、単純ヘルペスウイルスチミジンキナーゼ、アデノシンデアミナーゼ、グルタミンシンセターゼ及びヒポキサンチン-グアニンホスホリボシルトランスフェラーゼ;エス・セレビシエ(S. cerevisiae)についてTRP1;大腸菌(E. coli)においてテトラサイクリン、リファンピシン又はアンピシリン耐性を含み得る。
【0064】
好ましくは、上記のベクターは、本発明の変異型/単鎖メガヌクレアーゼをコードする配列が、適切な転写及び翻訳制御要素の制御下に位置して、該メガヌクレアーゼの生成又は合成を許容する発現ベクターである。よって、上記のポリヌクレオチドは、発現カセットに含まれる。より具体的には、該ベクターは、複製起点、該コードポリヌクレオチドに機能可能に連結されたプロモーター、リボソーム結合部位、RNAスプライシング部位(ゲノムDNAを用いる場合)、ポリアデニル化部位、及び転写終結部位を含む。これは、エンハンサーも含み得る。プロモーターの選択は、ポリペプチドが発現される細胞に依存する。好ましくは、変異型がヘテロ二量体である場合、各単量体をコードする2つのポリヌクレオチドは、両方のポリヌクレオチドの発現を同時に駆動し得る1つのベクターに含まれる。適切なプロモーターは、組織特異的及び/又は誘導性プロモーターを含む。誘導性プロモーターの例は、重金属のレベルの増加により誘導される真核メタロチオネインプロモーター、イソプロピル-β-D-チオガラクト-ピラノシド(IPTG)に応答して誘導される原核lacZプロモーター、及び温度の増加により誘導される真核熱ショックプロモーターである。組織特異的プロモーターの例は、骨格筋クレアチンキナーゼ、前立腺特異的抗原(PSA)、α-抗トリプシンプロテアーゼ、ヒトサーファクタント(SP)タンパク質A及びB、β-カゼイン及び酸性ホエータンパク質遺伝子である。
【0065】
上記のベクターの別の有利な実施形態によると、これは、上記で定義されるゲノムDNA切断部位を取り囲む領域と相同性を有する配列を含むターゲティング構築物を含む。
或いは、I-CreI変異型/単鎖メガヌクレアーゼをコードするベクターと、ターゲティング構築物を含むベクターとは、異なるベクターである。
より好ましくは、上記のターゲティングDNA構築物は:
a) 上記で定義されるゲノムDNA切断部位を取り囲む領域と相同性を有する配列と、
b) a)に記載の配列で挟まれた、導入される配列と
を含む。
【0066】
マウスROSA26遺伝子座での遺伝子ノックインのために、導入される配列は、外因性遺伝子発現カセット又はその一部分と、結局はHPRT遺伝子のような選択マーカーとを含む。
或いは、導入される配列は、マウスROSA26遺伝子座を、マウスROSA26遺伝子座の特定の配列を改変するため、マウスROSA26遺伝子座若しくはその一部分を減弱又は活性化するため、マウスROSA26遺伝子座の興味対象部位に変異を導入するため、又はマウスROSA26遺伝子座若しくはその一部分を不活性化又は欠失させるために用いられる配列を含む、ある特定の様式で変更させるために用いられる任意のその他の配列であり得る。
【0067】
好ましくは、切断部位を修復するために、少なくとも50 bp、好ましくは100 bpを超える、より好ましくは200 bpを超える相同配列が用いられる。実際に、共有されるDNA相同性は、破断の部位の上流及び下流に接する領域に位置し、導入されるDNA配列は、2つの腕の間に位置するべきである。
よって、ターゲティング構築物は、好ましくは200 pb〜6000 pb、より好ましくは1000 pb〜2000 pbである。
【0068】
配列の挿入のために、DNA相同性は、破断の部位のすぐ上流及び下流の領域に一般的に位置する(破断に隣接する配列;最小修復マトリクス)。しかし、挿入が、切断部位に接する配列の欠失を伴う場合、共有されるDNA相同性は、欠失の領域の上流及び下流の領域に位置する。
例えば、上記で定義される変異型と、それぞれの変異型により作製される切断のそれぞれを修復するための最小マトリクスとにより切断されるマウスROSA26 DNA標的を、図17に示す。
【0069】
本発明の主題は、上記で定義される少なくとも1つのメガヌクレアーゼ(変異型又は単鎖派生キメラメガヌクレアーゼ)及び/又は上記で定義される該メガヌクレアーゼをコードする少なくとも1つの発現ベクターを含むことを特徴とする組成物でもある。
上記の組成物の好ましい実施形態において、これは、上記で定義されるターゲティングDNA構築物を含む。
好ましくは、上記のターゲティングDNA構築物は、組換えベクターに含まれるか、又は本発明によるメガヌクレアーゼをコードするポリヌクレオチドを含む発現ベクターに含まれる。
【0070】
本発明の主題は、さらに、上記で定義されるメガヌクレアーゼ、好ましくは発現ベクターに含まれる1つ又は2つのポリヌクレオチドの、非治療目的でのマウスROSA26遺伝子座でのゲノム工学のための使用である。
上記の使用の有利な実施形態によると、これは、ゲノムDNA標的配列を含むマウスROSA26遺伝子座の興味対象部位内に2本鎖破断を誘発することにより、DNA組換え事象、DNA欠失又は細胞死を誘発するためである。
【0071】
本発明によると、上記の2本鎖破断は、ROSA26遺伝子座内の特定の配列を改変するため、内因性ROSA26遺伝子座を減弱又は活性化するため、ROSA26遺伝子座の興味対象部位に変異を導入するため、外因性遺伝子又はその一部分を導入するため、内因性ROSA26遺伝子座若しくはその一部分を不活性化又は欠失するため、染色体腕を転座させるため、或いはDNAを修復されないままにして分解させるためである。
【0072】
上記の使用の別の有利な実施形態によると、上記の変異型、ポリヌクレオチド、ベクターは、上記で定義されるターゲティングDNA構築物と会合する。
本発明によるメガヌクレアーゼの使用の好ましい実施形態において、これは、少なくとも以下の工程を含む:1) 上記のメガヌクレアーゼの少なくとも1つの認識及び切断部位を含むマウスROSA26 遺伝子座の興味対象部位に、該切断部位を該メガヌクレアーゼと接触させることにより二本鎖破断を導入し、2)標的遺伝子座と相同性を有する配列で挟まれた導入される配列を含むターゲティングDNA構築物を提供する。上記のメガヌクレアーゼは、細胞に直接、又は該メガヌクレアーゼをコードするポリヌクレオチド配列を含み、用いられる細胞におけるその発現に適切な発現ベクターを介して提供され得る。このストラテジーは、DNA配列を標的部位に導入して、例えば、タンパク質生産、遺伝子機能研究、薬剤の開発(薬剤のスクリーニング)のために、又は疾患モデルとして用いることができるノックイントランスジェニックマウス又は組換えマウス株化細胞を作製するために用いられる。
【0073】
本発明の主題は、少なくとも以下の工程を含む、興味対象の生成物を発現するトランスジェニックマウスを作製するための方法でもある:
(a) マウス多能性前駆細胞又はマウス胚に、上記で定義されるメガヌクレアーゼを導入して、該メガヌクレアーゼのDNA認識及び切断部位を含むROSA26遺伝子座の興味対象部位に二本鎖切断を誘発し、同時に又は連続的に、
(b) 工程(a)のマウス前駆細胞又は胚に、切断部位を取り囲む領域と相同性を有する配列で挟まれた興味対象の生成物をコードする配列を少なくとも含むターゲティングDNAを導入して、ターゲティングDNAと染色体DNAとの間の相同組換えにより興味対象配列が挿入されたゲノム改変マウス前駆細胞又は胚を作製し、
(c) 工程(b)のゲノム改変マウス前駆細胞又は胚を、キメラマウスに発達させ、
(d) 工程(c)のキメラマウスからトランスジェニックマウスを誘導する。
【0074】
好ましくは、工程(c)は、工程(b)で作製されたゲノム改変前駆細胞を、キメラマウスを作製するように、胚盤胞へ導入することを含む。
上記の方法の好ましい実施形態によると、これは、任意の手段により、トランスジェニックマウスから興味対象の生成物を回収するさらなる工程(e)を含む。
【0075】
本発明の主題は、少なくとも以下の工程を含む、興味対象の生成物を発現する組換えマウス細胞を作製するための方法でもある:
(a) マウス細胞に、上記で定義されるメガヌクレアーゼを導入して、該メガヌクレアーゼのDNA認識及び切断部位を含むROSA26遺伝子座の興味対象部位に二本鎖切断を誘発し、同時に又は連続的に、
(b) 工程(a)の細胞に、切断部位を取り囲む領域と相同性を有する配列で挟まれた興味対象の生成物をコードする配列を少なくとも含むターゲティングDNAを導入して、ターゲティングDNAと染色体DNAとの間の相同組換えにより興味対象配列が挿入された組換えマウス細胞を作製し、
(c) 工程(b)の組換えマウス細胞を、任意の適切な手段により単離する。
【0076】
上記の方法の好ましい実施形態によると、これは、任意の手段により、組換えマウス細胞から興味対象の生成物を回収するさらなる工程(d)を含む。
ターゲティングDNAは、興味対象部位に上記のターゲティングDNAを導入するために適切な条件下で細胞内に導入される。
ある好ましい実施形態において、ターゲティングDNA構築物はベクターに挿入される。
【0077】
改変される細胞は、興味対象のいずれの細胞であってもよい。トランスジェニックマウスを作製するために、細胞は、当該技術で公知の胚性幹(ES)細胞のような多能性前駆細胞である。組換えマウス株化細胞を作製するために、細胞は、有利には、NSO、SP2/0 (BALB/c骨髄腫; ECACC # 85110503及び# 85072401)、又はL (ATCC # CRL-2648)細胞であり得る。上記のメガヌクレアーゼは、細胞に直接、又は該メガヌクレアーゼをコードするポリヌクレオチド配列を含み、用いられる細胞におけるその発現に適切な発現ベクターを介して提供され得る。
【0078】
興味対象の生成物を発現するトランスジェニック動物/組換え株化細胞を作製するために、ターゲティングDNAは、上記で定義されるように、マウスROSA26遺伝子座中のメガヌクレアーゼ部位の上流及び下流の配列で挟まれた、興味対象の生成物(タンパク質又はRNA)をコードする配列と、結局は選択マーカー遺伝子とを含むことにより、興味対象の外因性配列が、ROSA26遺伝子座のメガヌクレアーゼ部位にて相同組換えにより統合されたゲノム改変細胞(動物前駆細胞又は胚/動物又はヒトの細胞)が作製される。
興味対象の配列は、興味対象のあるタンパク質/ペプチドをコードする任意の遺伝子であり得、限定されないが、レポーター遺伝子、受容体、シグナル伝達分子、転写因子、医薬的に活性なタンパク質及びペプチド、疾患原因遺伝子生成物及び毒素を含む。配列は、例えばsiRNAを含む興味対象のRNA分子もコードし得る。
【0079】
外因性配列の発現は、上記で定義される、内因性ROSA26プロモーター又は構成性若しくは誘導性の異種プロモーター、好ましくは遍在性若しくは組織特異的なプロモーターのいずれかにより駆動され得る。さらに、興味対象の配列の発現は、条件付きであり得る。発現は、部位特異的リコンビナーゼ(Cre、FLPなど)により誘発され得る。
つまり、興味対象配列は、上記の興味対象遺伝子に機能可能に連結された異種プロモーターと、限定されないが、(選択)マーカー遺伝子、リコンビナーゼ認識部位、ポリアデニル化シグナル、スプライス受容配列、イントロン、タンパク質検出のためのタグ及びエンハンサーを含む1つ又は複数の機能的配列とを含み得る適切なカセットに挿入される。
【0080】
或いは、適切なカセットは、上記の興味対象遺伝子に機能可能に連結された配列内リボソーム進入部位(IRES)と、IRES-Hygroマトリクス(pCLS1675)を有する事象、限定されないが、(選択)マーカー遺伝子、リコンビナーゼ認識部位、ポリアデニル化シグナル、スプライス受容配列、イントロン、タンパク質検出のためのタグ及びエンハンサーを含む1つ又は複数の機能的配列とを含み得る。
【0081】
メガヌクレアーゼは、ポリペプチド、又は該ポリペプチドをコードするポリヌクレオチド構築物のいずれかとして用い得る。これは、マウス細胞に、特定の種類の細胞に適する当該技術において公知の任意の簡便な手段により、単独で又は少なくとも適切なビヒクル若しくは担体及び/又はターゲティングDNAのいずれかとともに、導入される。
【0082】
本発明による使用の有利な実施形態によると、メガヌクレアーゼ(ポリペプチド)は、以下のものと会合される(is associated with):
- リポソーム、ポリエチレンイミン(PEI);このような場合、上記の会合物は投与され、よって、標的体細胞に導入される。
- 膜移送ペプチド(membrane translocating peptides) (Bonetta, The Scientist, 2002, 16, 38; Fordら, Gene Ther., 2001, 8, 1〜4 ; Wadia及びDowdy, Curr. Opin. Biotechnol., 2002, 13, 52〜56);このような場合、変異型/単鎖メガヌクレアーゼの配列は、膜移送ペプチドの配列と融合される(融合タンパク質)。
【0083】
本発明による使用の別の有利な実施形態によると、メガヌクレアーゼ(メガヌクレアーゼをコードするポリヌクレオチド)及び/又はターゲティングDNAは、ベクターに挿入される。ターゲティングDNA及び/又はメガヌクレアーゼをコードする核酸を含むベクターは、種々の方法により細胞に導入できる(例えば注入、直接摂取、発射衝撃、リポソーム、エレクトロポレーション)。メガヌクレアーゼは、発現ベクターを用いて、細胞内で安定的又は一過的に発現させ得る。真核細胞における発現の方法は、当該技術において公知である(Current Protocols in Human Genetics: 12章 「Vectors For Gene Therapy」及び13章「Delivery Systems for Gene Therapy」を参照)。所望により、組換えタンパク質中に、核局在化シグナルを組み込んで、核内でそれが発現されることを確実にすることが好ましい。
一旦細胞内に入ると、メガヌクレアーゼと存在するならばターゲティングDNA及び/又はメガヌクレアーゼをコードする核酸を含むベクターは、細胞質から核内の作用部位まで、細胞により移入又は移送される。
【0084】
本発明による使用のある実施形態において、メガヌクレアーゼは、実質的に非免疫原性であり、すなわち、有害免疫応答をほとんど又は全く生じない。この種の有害免疫応答を緩和又は消去するための種々の方法を本発明に従って用いることができる。ある好ましい実施形態において、メガヌクレアーゼは、N-ホルミルメチオニンを実質的に含まない。望ましくない免疫応答を回避する別の様式は、メガヌクレアーゼを、ポリエチレングリコール(「PEG」)又はポリプロピレングリコール(「PPG」) (好ましくは、500〜20,000ダルトンの平均分子量(MW)のもの)とコンジュゲートさせることである。例えばDavisら(US 4,179,337)により記載されるPEG又はPPGとのコンジュゲート形成は、抗ウイルス活性を有する、非免疫原性で、生理活性で、水溶性のエンドヌクレアーゼコンジュゲートを提供できる。ポリエチレン-ポリプロピレングリコールコポリマーを用いる同様の方法は、Saiferら(US 5,006,333)に記載されている。
【0085】
本発明は、上記で定義されるポリヌクレオチド又はベクター、好ましくは発現ベクターで改変された原核又は真核宿主細胞にも関する。
本発明は、細胞の全て又は一部分が上記で定義されるポリヌクレオチド又はベクターで改変されたことを特徴とする非ヒトトランスジェニック動物又はトランスジェニック植物にも関する。
本明細書で用いる場合、細胞とは、原核細胞、例えば細菌細胞、又は真核細胞、例えば動物、植物若しくは酵母細胞のことである。
【0086】
本発明の主題は、上記で定義される少なくとも1つのメガヌクレアーゼ変異型の、別のメガヌクレアーゼを作製するための足場としての使用でもある。例えば、新規な第3世代のメガヌクレアーゼを作製する目的で、3回目の突然変異誘発及び選択/スクリーニングを上記の変異型に対して行うことができる。
【0087】
本発明によるメガヌクレアーゼの異なる使用及び上記のメガヌクレアーゼを用いる方法は、I-CreI変異型、該変異型に由来する単鎖キメラメガヌクレアーゼ、上記で定義される、該変異型若しくは単鎖キメラメガヌクレアーゼをコードするポリヌクレオチド、ベクター、細胞、トランスジェニック植物又は非ヒトトランスジェニック哺乳動物の使用を含む。
【0088】
本発明によるI-CreI変異型は、少なくとも以下の工程を含む、マウスROSA26遺伝子座からのゲノムDNA標的配列を切断できるI-CreI変異型を工学的に作製するための方法により得ることができる:
(a) I-CreIの26位〜40位に位置するLAGLIDADGコアドメインの第1機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第1シリーズを構築し、
(b) I-CreIの44位〜77位に位置するLAGLIDADGコアドメインの第2機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第2シリーズを構築し、
(c) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の-10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(d) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の-5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
【0089】
(e) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(f) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の+3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(g) 工程(c)及び工程(d)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) -10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットと同一であり、(ii) +8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) -5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットと同一であり、(iv) +3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
【0090】
(h) 工程(e)及び工程(f)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) +3位〜+5位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットと同一であり、(ii) -5位〜-3位のヌクレオチドトリプレットが、ゲノム標的の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(iv) -10位〜-8位のヌクレオチドトリプレットが、ゲノム標的の+8位〜+10位のヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
(i) 工程(g)及び(h)で得られた変異型を組み合わせて、ヘテロ二量体を形成し、
(j) マウスROSA26遺伝子座からの上記のゲノムDNA標的を切断できる工程(i)からのヘテロ二量体を選択及び/又はスクリーニングする。
【0091】
上記の工程(c)、(d)、(e)又は(f)の1つは省略できる。例えば、工程(c)を省略する場合、工程(d)は、-10位〜-8位及び-5位〜-3位の両方のヌクレオチドトリプレットが上記のゲノム標的の-10位〜-8位及び-5位〜-3位に存在するヌクレオチドトリプレットでそれぞれ置き換えられ、+3位〜+5位及び+8位〜+10位のヌクレオチドトリプレットが、上記のゲノム標的の-5位〜-3位及び-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列でそれぞれ置き換えられた変異I-CreI部位を用いて行われる。
【0092】
工程(a)、(b)、(g)、(h)及び(i)は、上記で定義される、DNA標的配列と接触するか又は該DNA標的と直接若しくは間接的に相互作用する他の位置、変異体の結合及び/又は切断特性を改良する位置、又は機能的ホモ二量体の形成を妨げる位置での、付加的な変異の導入をさらに含み得る。これは、国際PCT出願WO 2004/067736に記載されるようなコンビナトリアルライブラリーを作製することにより行い得る。
【0093】
本発明のI-CreI変異型を工学的に作製する方法は、興味対象の遺伝子からのDNA標的に対する変異体の結合及び/又は切断特性を改良するために、変異体全体、又は変異体の一部分、特に変異型のC-末端半分(80位〜163位)に対するランダム変異の導入を含むのが有利である。突然変異誘発は、公知で商業的に利用可能な標準の突然変異誘発の方法に従って、変異型のプールに対してランダム突然変異誘発ライブラリーを作製することにより行うことができる。好ましくは、突然変異誘発は、工程(i)で形成されるか又は工程(j)で得られるヘテロ二量体の一方の単量体の全体の配列に対して、有利には、単量体のプールに対して、好ましくは工程(i)又は(j)のへテロ二量体の両方の単量体に対して行われる。
【0094】
好ましくは、Arnouldら, J. Mol. Biol., Epub 2007年5月10日の図4に記載される方法に従って、2回の選択/スクリーニングを行う。1回目において、ヘテロ二量体の一方の単量体(図4の単量体Y)を、他方の単量体(図4の単量体X)と同時発現させてヘテロ二量体を形成し、改良単量体Y+を、興味対象遺伝子からの標的に対して選択する。2回目において、他方の単量体(単量体X)に突然変異を誘発し、改良単量体Y+と同時発現させてヘテロ二量体を形成し、興味対象遺伝子からの標的に対して選択して、改良された活性を有するメガヌクレアーゼ(X+ Y+)を得る。
【0095】
工程(g)及び(h)における変異の(分子内)組み合わせは、公知のオーバーラップPCR法に従って、2つのサブドメインのそれぞれを含むオーバーラップフラグメントを増幅することにより行ってよい。
工程(i)における変異型の(分子間)組み合わせは、工程(g)からの1つの変異型を、工程(h)からの1つの変異型と同時発現させて、ヘテロ二量体の形成を可能にすることにより行われる。例えば、宿主細胞を、該変異型をコードする1つ又は2つの組換え発現ベクターで改変できる。次いで、国際PCT出願WO 2006/097854及びArnouldら, J. Mol. Biol., 2006, 355, 443〜458に以前に記載されたようにして、細胞を、変異型の発現を可能にする条件下で培養することにより、宿主細胞内でヘテロ二量体が形成される。
【0096】
工程(c)、(d)、(e)、(f)及び/又は(j)における選択及び/又はスクリーニングは、国際PCT出願WO 2004/067736、Arnouldら, J. Mol. Biol., 2006, 355, 443〜458, Epinatら, Nucleic Acids Res., 2003, 31, 2952〜2962及びChamesら, Nucleic Acids Res., 2005, 33, e178に記載されるような、インビトロ又はインビボでの切断アッセイを用いて行うことができる。
上記の方法の別の有利な実施形態によると、工程(c)、(d)、(e)、(f)及び/又は(j)は、インビボで、変異型により作製された変異DNA標的配列内の2本鎖破断が、陽性選択マーカー若しくはレポーター遺伝子の活性化、又は陰性選択マーカー若しくはレポーター遺伝子の不活性化を、該DNA 2本鎖破断の組換え媒介修復により導く条件下で行われる。
【0097】
本発明の主題は、本発明によるマウスROSA26遺伝子座からのDNA標的を切断できる変異型を工学的に作製するために有用な、I-CreIの26位〜40位及び/又は44位〜77位にて変異を有するI-CreI変異型でもある。特に、本発明は、変異型m1〜m18 (表II、配列番号38〜55)、Q44V、R70A及びD75Nを含む変異型(配列番号131;表III)、並びにK28E、Y33R、Q38R、S40R及びD75Nを含む変異型(配列番号132;表III)を含む、上記で定義されるI-CreI変異型を工学的に作製するための方法の工程(c)〜(f)で定義されるI-CreI変異型を含む。本発明は、配列番号60〜67の配列の変異型(表IIIの組み合わせ変異型)を含む、上記で定義されるI-CreI変異型を工学的に作製するための方法の工程(g)及び(h)で定義されるI-CreI変異型も含む。
【0098】
興味対象遺伝子からのDNA標的を切断できる単鎖キメラメガヌクレアーゼは、当該技術において公知の方法により、本発明による変異型から導かれる(Epinatら, Nucleic Acids Res., 2003, 31, 2952〜62; Chevalierら, Mol. Cell., 2002, 10, 895〜905; Steuerら, Chembiochem., 2004, 5, 206〜13; 国際PCT出願WO 03/078619及びWO 2004/031346)。このような方法はいずれも、本発明で定義される変異型に由来する単鎖キメラメガヌクレアーゼを構築するために用い得る。
【0099】
本発明で定義される変異型をコードするポリヌクレオチド配列は、当業者に知られる任意の方法により調製できる。例えば、これらは、cDNA鋳型から、特異的プライマーを用いるポリメラーゼ連鎖反応により増幅される。好ましくは、上記のcDNAのコドンは、所望の発現系における上記のタンパク質の発現に好ましいように選択される。
上記のポリヌクレオチドを含む組換えベクターは、公知の組換えDNA及び遺伝子工学の技術により得て、宿主細胞に導入できる。
【0100】
本発明で定義されるI-CreI変異型又は単鎖誘導体は、上記で定義されるポリペプチドを発現させることにより生成される。好ましくは、該ポリペプチドは、ポリペプチドの発現又は同時発現に適する条件下で、1つの発現ベクター又は2つの発現ベクター(変異型のみの場合)により改変された宿主細胞又はトランスジェニック動物/植物で、発現又は同時発現(変異型のみの場合)され、変異型又は単鎖誘導体は、宿主細胞培養又はトランスジェニック動物/植物から回収される。
【0101】
本発明の実行は、そうでないと記載しない限り、当該技術の範囲内である細胞生物学、細胞培養、分子生物学、トランスジェニック生物学、微生物学、組換えDNA及び免疫学の通常の技術を用いる。このような技術は、文献に充分に説明されている。例えばCurrent Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, 第3版, (Sambrookら, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); Oligonucleotide Synthesis (M. J. Gait編, 1984); Mullisら、米国特許第4,683,195号; Nucleic Acid Hybridization (B. D. Harries及びS. J. Higgins編 1984); Transcription And Translation (B. D. Hames及びS. J. Higgins編 1984); Culture Of Animal Cells (R. I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); Methods In ENZYMOLOGY (J. Abelson及びM. Simon編, Academic Press, Inc., New York)のシリーズ, 特に第154巻及び第155巻(Wuら編)並びに第185巻「Gene Expression Technology」(D. Goeddel編); Gene Transfer Vectors For Mammalian Cells (J. H. Miller及びM. P. Calos編, 1987, Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (Mayer及びWalker編, Academic Press, London, 1987); Handbook Of Experimental Immunology, 第I〜IV巻(D. M. Weir及びC. C. Blackwell編, 1986); 並びにManipulating the Mouse Embryo, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1986)を参照されたい。
【0102】
上記の特徴に加えて、本発明は、本発明によるI-CreIメガヌクレアーゼ変異型及びそれらの使用を説明する実施例並びに添付の図面に言及する以下の記載から明らかになるその他の特徴をさらに含む。添付の図面において:
- 図1は、マウスROSA26遺伝子座を表す(アクセッション番号EMBL CQ880114;配列番号3)。エキソンを箱で囲む(エキソン1:2490位〜2599位;転写産物1からのエキソン2:8228位〜9248位;11845位から開始し、11505位で終了するアンチセンス転写産物とほぼ重複する転写産物2からのエキソン3。今までのところ同定された3つの転写産物を、標的rosa1 (配列番号15)の配列及び位置とともに示す。
【0103】
- 図2は、DNA標的に結合したI-CreIホーミングエンドヌクレアーゼの3次元構造を表す。触媒コアは、DNA主溝の上にサドル形相互作用界面を形成する2つのαββαββα折り畳みで囲まれる。
- 図3は、I-CreI及び他のLAGLIDADGホーミングエンドヌクレアーゼの特異性を工学的に改変するための2ステップアプローチを示す。I-CreI誘導体の大きい集合を、元の足場の半合理的突然変異誘発と、局所的に特異性が変更された機能的変異型のスクリーニングとにより作製する。次いで、コンビナトリアルアプローチを用いて、これらの変異体を、完全に再設計された特異性を有するメガヌクレアーゼに組み立てる。ホモ二量体タンパク質(「ハーフメガヌクレアーゼ」)を、同じαββαββα折り畳み内の2組の変異の組み合わせにより創出し、このような2つの「ハーフメガヌクレアーゼ」の同時発現が、興味対象の標的を切断するヘテロ二量体の種(「カスタムメガヌクレアーゼ」)をもたらし得る。
【0104】
- 図4は、マウスROSA26遺伝子座を切断するメガヌクレアーゼの使用のためのストラテジーを表す。メガヌクレアーゼ誘発相同組換えを用いる遺伝子挿入は、興味対象遺伝子をマウスROSA26遺伝子座にノックインする。イントロン及びエキソンの配列は、相同領域として用い得る。
- 図5は、rosa1標的配列及び誘導体を表す。10GGG_P、5GAT_P及び5TAT_Pは、以前に得られたI-CreI変異体により切断されることが見出された近縁の誘導体である。これらは、C1221 (I-CreI足場タンパク質により切断されるパリンドローム配列)とは、箱で囲んだモチーフが異なる。C1221、10GGG_P、5GAT_P及び5TAT_Pは、最初は24 bp配列で記載されたが、構造データは、22 bpのみがタンパク質/DNA相互作用に関連することを示唆する。しかし、±12位を括弧内に示す。rosa1は、8304位にてマウスROSA26遺伝子座に位置するDNA配列である。rosa1.2標的において、標的の中ほどのGTTC配列は、C1221で見出される塩基であるGTACで置き換えられる。rosa1.3は、rosa1.2の左部分に由来するパリンドローム配列であり、rosa1.4は、rosa1.2の右部分に由来するパリンドローム配列である。図に示すように、10GGG_P、5GAT_P及び5TAT_Pからの箱で囲んだモチーフは、rosa1シリーズの標的で見出される。
【0105】
- 図6は、pCLS1055ベクターマップを表す。
- 図7は、pCLS0542ベクターマップを表す。
- 図8は、I-CreI変異体によるrosa1.3 DNA標的の切断を示す。1次スクリーニングで見出された63個の陽性を、1枚の96ウェルプレート内で再配置し、2次スクリーニングで確認した(4重フォーマットで)。実施例2で選択した22個の変異体を丸で囲む。
- 図9は、I-CreIコンビナトリアル変異体によるrosa1.4標的の切断を示す。1次スクリーニングで見出された69個の陽性を、1枚の96ウェルプレート内で再配置し、2次スクリーニングで確認した(4重フォーマットで)。実施例3で選択した15個の変異体を丸で囲む。
- 図10は、pCLS1107ベクターマップを表す。
【0106】
- 図11は、ヘテロ二量体I-CreIコンビナトリアル変異体によるrosa1.2及びrosa1標的の切断を示す。A. I-CreI変異体の組み合わせのrosa1.2標的を用いたスクリーニングの例。B. I-CreI変異体の同じ組み合わせのrosa1標的を用いたスクリーニング。B5、B6、D5、D6、F5、F6、H5及びH6:pCLS1107空のプラスミドDNAで形質転換された、rosa1.3を切断するI-CreI変異体を発現する酵母株。
- 図12は、rosa1標的の切断を示す。rosa1.4を切断する一連のI-CreI変異体に無作為に突然変異を誘発し、rosa1.3を切断する変異体と同時発現させた。切断を、rosa1標的を用いて試験する。それぞれの4ドットクラスタにおいて、右の2つのドットは、2重での、rosa1を切断する元来のヘテロ二量体の1つに相当し、2つの左のドットは、変異されていないrosa1.3を切断するものと同時発現させた、同じ変異させたrosa1.4を切断するものに相当する(変異体m13、表IV及びVに記載される)。rosa1の改良された切断を示す2つの最適化された変異体を丸で囲み、これらは変異体m13とMO_1 (C10)、又はm13とMO_2 (E2)の同時発現に相当する。MO_1及びMO_2は、表VIにさらに記載される。
【0107】
- 図13は、rosa1標的の切断を示す。rosa1.3を切断する一連のI-CreI変異体に無作為に突然変異を誘発し、rosa1.4を切断する洗練された変異体と同時発現させた。切断は、rosa1標的を用いて試験する。rosa1の効率的な切断を示す変異体を丸で囲む。フィルタにおいて:
- B11は、ヘテロ二量体S19、V24、Y44、R68、S70、N75、V77 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当し;
- C9は、ヘテロ二量体S19、V24、Y44、R68、S70、Q75、I77 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当し;
- C11及びE8は、ヘテロ二量体 V24、Y44、S68、S70、R75、I77、A105 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当し、
- E6は、ヘテロ二量体V24、Y44、S68、S70、R75、I77、G79 + E28、R33、R38、K40、A44、H68、Q70、A105、R107、A151、G153、E158に相当する。
- H10は陰性対照であり、H11及びH12は異なる強度の陽性対照である。rosa1.3標的を切断する変異体の改良の前後におけるrosa1標的に対するヘテロ二量体の活性を比較するために、各クラスタにおいて、右の2つの点は、実施例5に記載されるヘテロ二量体の1つであり、左の2つの点は、実施例6に記載される変異を有するヘテロ二量体である。
【0108】
- 図14は、pCLS1058ベクターマップを表す。
- 図15は、pCLS1069ベクターマップを表す。
- 図16は、CHO細胞における染色体外モデルでのI-CreI洗練変異体によるrosa1標的の切断を示す。2回のトランスフェクション実験からの値を示す。I-CreI N75及びI-SceIによる同じ実験でのI-CreI及びI-SceI標的の切断は、陽性対照として示す。
- 図17は、マウスROSA26で見出されるメガヌクレアーゼ標的配列と、該DNA標的のそれぞれを切断できる対応するI-CreI変異型を表す。DNA標的の配列(第1列)を、その位置(第2列)とともに示す。標的部位での切断を修復するための最小修復マトリクスを、その最初のヌクレオチド(始点、第5列)と、最後のヌクレオチド(終点、第6列)で示す。各変異型の配列は、記載する位置での残基により規定する。例えば、図17の最初のヘテロ二量体変異型は、K、H、S、S、Q、S、E、C、S、N及びIを28位、30位、32位、33位、38位、40位、44位、68位、70位、75位及び77位にそれぞれ有する第1単量体と、K、D、S、R、T、S、K、E、S、D、Rを28位、30位、32位、33位、38位、40位、44位、68位、70位、75位及び77位にそれぞれ有する第2単量体とからなる。位置は、I-CreI配列SWISSPROT P05725 (配列番号1)を参照にして記載する。I-CreIは、K、N、S、Y、Q、S、Q、R、R、D及びIを、28位、30位、32位、33位、38位、40位、44位、68位、70位、75位及び77位にそれぞれ有する。
【0109】
- 図18は、pCLS1675ベクターマップを表す。
- 図19は、pCLS1761ベクターマップを表す。
- 図20は、pCLS1762ベクターマップを表す。
- 図21は、IRES-Hygroマトリクス(pCLS1675)を用いたノックイン(KI)事象のPCR分析を示す。ROSA26遺伝子座について野生型のクローン及びハイグロマイシンCDSの無作為挿入を有するクローンは、PCRにおいて陰性である。ROSA26遺伝子座でのKI事象を有するクローンは、PCRにおいて陽性である。KI事象及び無作為挿入を有するクローンも、PCRにおいて陽性である。
【実施例】
【0110】
実施例1:マウスROSA26遺伝子座を切断する新規なメガヌクレアーゼを工学的に作製するためのストラテジー
Smithら, Nucleic Acids Res., 2006に記載され、図3に説明されるコンビナトリアルアプローチを用いて、I-CreIのDNA結合ドメインを工学的に作製し、rosa1 (図5)と命名され、マウスROSA26遺伝子座(アクセッション番号CQ880114;配列番号3)のエキソン2の8304位に位置する22 bp (非パリンドローム)の配列を切断した。rosa1配列を切断するメガヌクレアーゼは、遺伝子をマウスROSA26遺伝子座(図4)にノックインするために用いることができた。用途は、以下の分野である。マウス細胞での組換えタンパク質の生成、例えば薬剤スクリーニングの目的での組換え株化細胞の工学的作製、並びに例えば動物モデルとしての使用のためのトランスジェニックマウスの工学的作製。
【0111】
rosa1配列は、部分的に、国際PCT出願WO 2006/097784、WO 2006/097853及びWO 2007/049156; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458並びにSmithら, Nucleic Acids Res., Epub 2006年11月27日に記載されるようにして得られた以前に同定されたメガヌクレアーゼにより切断される10GGG_P、5GAT_P及び5TAT_P標的のパッチワークである(図5)。よって、rosa1は、これらの3つの標的を切断するI-CreI誘導体で見出される変異を組み合わせたメガヌクレアーゼにより切断され得る。
【0112】
10GGG_P、5GAT_P及び5TAT_P配列は、I-CreIにより切断されるパリンドローム配列であるC1221の24 bp誘導体である(国際PCT出願WO 2006/097784、WO 2006/097853及びWO 2007/049156; Arnouldら, J. Mol. Biol., 2006, 355, 443〜458並びにSmithら, Nucleic Acids Res., Epub 2006年11月27日)。しかし、そのDNA標的に結合したI-CreIの構造は、それらの標的の外側の塩基対(-12位及び12位)が、結合及び切断に対して影響しないことを示唆し(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316; Chevalier B.S.及びStoddard B.L., Nucleic Acids Res., 2001, 29, 3757〜3754; Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)、この研究においては、-11位〜11位までを考慮した。よって、標的のrosa1シリーズを、24 bpの代わりに22 bpの配列として規定した。
【0113】
Rosa1は、C1221とは、4 bp中央領域の1塩基対が異なる。その標的に結合したI-CreIタンパク質の構造によると、4つの中央塩基対(-2位〜2位)とI-CreIタンパク質との間に接触はない(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316; Chevalier B.S.及びStoddard B.L., Nucleic Acids Res., 2001, 29, 3757〜3754; Chevalierら, J. Mol. Biol., 2003, 329, 253〜269)。つまり、これらの位置での塩基は、結合効率に影響を与えるとは考えられない。しかし、これらは、この領域の端の2つのニックに起因する切断に影響し得る。よって、-2位〜2位のGTTC配列を、まず、C1221からのGTAC配列で置換して、標的rosa1.2を得た(図5)。
次いで、2つのパリンドローム標的rosa1.3及びrosa1.4を、rosa1.2から導いた(図5)。rosa1.3及びrosa1.4はパリンドロームであるので、これらは、ホモ二量体タンパク質により切断されるはずである。
【0114】
よって、ホモ二量体としてrosa1.3及びrosa1.4配列を切断できるタンパク質をまず設計し(実施例2及び3)、次いで、同時発現させてrosa1を切断するヘテロ二量体を得た(実施例4)。rosa1.2及びrosa1標的を切断するヘテロ二量体を同定できた。rosa1標的についての切断活性を改良するために、rosa1.3及びrosa1.4を切断する選択した一連の変異体を選択して洗練した。選択した変異体にランダム突然変異を誘発し、新規なヘテロ二量体を形成するために用いて、これらをrosa1標的に対してスクリーニングした(実施例5及び6)。最後に、酵母及びCHO細胞において高い切断活性を示すrosa1標的を切断するヘテロ二量体が同定できた。
【0115】
実施例2:rosa1.3を切断するメガヌクレアーゼの作製
この実施例は、I-CreI変異体が、パリンドローム形のrosa1標的の左部分に由来するrosa1.3 DNA標的配列を切断できることを示す(図5)。
この実施例に記載される標的配列は、22 bpのパリンドローム配列である。よって、これらを、最初の11ヌクレオチドのみで、後に接尾辞_Pを付して記載する。例えば、標的rosa1.3は、caacatgatgt_P; 配列番号35とも記載される。
【0116】
rosa1.3標的は、5GAT_Pと、±1位、±2位、±3位、±4位、±5位、±7位、±9位、±10位及び±11位が同様であり、2つの配列は、±6位及び±8位のみが異なる。±6位は、結合及び切断活性に対してほとんど影響しないであろうと仮定した。5GAT_P (caaaacgatgt_P; 配列番号32)を切断できる変異体は、Arnouldら, J Mol Biol. 2006; 355, 443〜458並びに国際PCT出願WO 2006/097784及びWO 2006/097853に記載されるように、I-CreI N75に対する24位、44位、68位、70位、75位及び77位での突然変異誘発により以前に得られた。この実施例において、5GAT_P標的を切断する変異体がrosa1.3標的も切断できるかを調べた。
【0117】
1) 材料及び方法
特異性が変更された変異型をスクリーニングするために用いられるメガヌクレアーゼ変異型を作製する方法及び哺乳動物細胞又は酵母細胞における切断誘発組換えに基づくアッセイは、国際PCT出願WO 2004/067736; Epinatら, Nucleic Acids Res., 2003, 31, 2952〜2962; Chamesら, Nucleic Acids Res., 2005, 33, e178及びArnouldら, J. Mol. Biol., 2006, 355, 443〜458に記載される。これらのアッセイは、機能的LacZレポーター遺伝子をもたらし、これは標準的な方法によりモニターできる。
【0118】
a) 標的ベクターの構築
標的は、以下のようにしてクローニングした。ゲートウェイクローニング配列と接する標的配列に相当するオリゴヌクレオチドをProligoから得た:
5' tggcatacaagtttcaacatgatgtacatcatgttgacaatcgtctgtca 3'(配列番号37)。一本鎖オリゴヌクレオチドのPCR増幅により作製した二本鎖標的DNAを、Gatewayプロトコル(INVITROGEN)を用いて酵母レポーターベクター(pCLS1055, 図6)にクローニングした。酵母レポーターベクターで、エス・セレビシエFYBL2-7B株(MAT a, ura3Δ851, trp1Δ63, leu2Δ1, lys2Δ202)を形質転換した。
【0119】
b) I-CreI変異体
5GAT_Pを切断するI-CreI変異体を、I-CreIの24位、44位、68位、70位、75位及び77位が変異されたライブラリーにおいて、Arnouldら, J. Mol. Biol., 2006, 355, 443〜458、並びに国際PCT出願WO 2006/097784及びWO 2006/097853に以前に記載されたように同定した。これらをDNAベクター(pCLS0542, 図7)にクローニングし、酵母サッカロミセス・セレビシエ(Saccharomyces cerevisiae) FYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)で発現させる。
【0120】
c) メガヌクレアーゼ発現クローンの交配と酵母でのスクリーニング
スクリーニングを、以前に記載されたようにして行った(Arnouldら, J. Mol. Biol. 2006, 355, 443〜458)。交配は、コロニーグリッダー(QpixII, Genetix)を用いて行った。変異体を、YPDプレートを覆うナイロンフィルタ上に、低格子密度(low gridding density) (約4スポット/cm2)を用いてグリッドした。2回目のグリッド手順を同じフィルタに対して行って、それぞれの標的について異なるレポーター保持酵母株からなる第2層をスポットした。メンブレンを固体寒天YPDリッチ培地上に置き、30℃にて一晩インキュベートして交配を可能にした。次いで、フィルタを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として有する合成培地上に移し、37℃にて5日間インキュベートして、発現及び標的ベクターを有する二倍体を選択した。5日後に、フィルタを、0.5 Mリン酸ナトリウム緩衝液、pH 7.0中の0.02 % X-Gal、0.1% SDS、6%ジメチルホルムアミド(DMF)、7mM β-メルカプトエタノール、1%アガロースを含む固体アガロース培地上に置き、37℃にてインキュベートして、β-ガラクトシダーゼ活性をモニターした。結果をスキャンにより分析し、適切なソフトウェアを用いて定量を行った。
【0121】
d) 変異体の配列決定
変異体を発現するプラスミドを回収するために、酵母DNAを標準的なプロトコルを用いて抽出し、大腸菌を形質転換するために用いた。変異体ORFの配列決定を、次いで、プラスミドに対してMILLEGEN SAにより行った。或いは、ORFを酵母DNAからPCRにより増幅し(Akadaら, Biotechniques, 2000, 28, 668〜670)、配列決定を、PCR生成物に対して直接、MILLEGEN SAにより行った。
【0122】
2) 結果
I-CreIの24位、44位、68位、70位、75位及び77位が変異されたライブラリーにおいて以前に同定された5GAT_P標的を切断するI-CreI変異体を、rosa1.3 DNA標的(caacatgatgt _P; 配列番号35)に対する切断についてスクリーニングした。合計で63個の陽性クローンが見出され、96ウェルプレートで再配置し、2次スクリーニングにより確認した(図8)。これらの陽性クローンのうち、22個(図8において丸で囲む)を選択した。これらの22個の陽性クローンを配列決定した。これらは、rosa1.3標的を切断する18個の異なる新規なエンドヌクレアーゼに相当することがわかった(m1〜m18と命名;配列番号38〜55;表II)。
【0123】
【表2】
【0124】
実施例3:rosa1.4を切断するメガヌクレアーゼの作製
この実施例は、I-CreI変異体が、パリンドローム形のrosa1標的の右部分に由来するrosa1.4 DNA標的配列を切断できることを示す(図5)。この実施例に記載される全ての標的配列は、22 bpのパリンドローム配列である。よって、これらを、最初の11ヌクレオチドのみで、後に接尾辞_Pを付して記載する。例えば、rosa1.4は、tgggattatgt_P (配列番号10)とよばれる。
【0125】
rosa1.4標的は、5TAT_Pと±1位、±2位、±3位、±4位、±5位及び±7位が、かつ10GGG_Pと±1位、±2位、±7位、±8位、±9位及び±10位が同様である。±6位及び±11位は、結合及び切断活性に対してほとんど影響しないであろうと仮定した。5TAT_Pを切断できる変異体は、Arnouldら, J Mol. Biol., 2006; 355, 443〜458、並びに国際PCT出願WO 2006/097784及びWO 2006/097853に記載されるように、I-CreI N75に対する44位、68位、70位での突然変異誘発により以前に得られた。10GGG_P標的を切断できる変異体は、Smithら, Nucleic Acids Res., Epub 2006年11月27日及び国際PCT出願WO 2007/049156に記載されるように、I-CreI N75に対する28位、30位、33位、38位、40位及び70位での突然変異誘発により得られた。
【0126】
両方の組のタンパク質を70位にて変異させる。しかし、2つの分離可能な機能的サブドメインが存在することが仮定された。このことは、この位置が、標的の±8〜10の塩基に対する特異性にほとんど影響しないことを意味する。
よって、組み合わせた変異体がrosa1.4標的を切断できるかを確認するために、5TAT_P (caaaactatgt_P; 配列番号6)を切断するタンパク質からの44位、68位及び70位での変異を、10GGG_P (cgggacgtcgt_P; 配列番号4)を切断するタンパク質からの28、30、33、38及び40変異と組み合わせた。
【0127】
1) 材料及び方法
実験手順は、実施例2及び以下に記載されるとおりである:
コンビナトリアル変異体の構築
10GGG_P又は5TAT_Pを切断するI-CreI変異体は、10GGG_P又は5TAT_P標的についてそれぞれSmithら, Nucleic Acids Res. Epub 2006年11月27日; 国際PCT出願WO 2007/049156、並びにArnouldら, J. Mol. Biol., 2006, 355, 443〜458; 国際PCT出願WO 2006/097784及びWO 2006/097853で同定された。両方のシリーズからの変異を含むI-CreI由来コード配列を作製するために、I-CreIコード配列の5'末端(aa 1位〜43位)又は3'末端(39位〜167位)を増幅する別々のオーバーラップPCR反応を行った。5'及び3'末端の両方について、PCR増幅を、ベクター(pCLS0542, 図7)に特異的なプライマーGal10F 5'-gcaactttagtgctgacacatacagg-3' (配列番号56)又はGal10R 5'-acaaccttgattggagacttgacc-3' (配列番号57)と、アミノ酸39〜43についてのI-CreIコード配列に特異的なプライマーassF 5'-ctannnttgaccttt-3' (配列番号58)又はassR 5'-aaaggtcaannntag-3' (配列番号59) (ここで、nnnは残基40をコードする)を用いて行う。同じプライマー及び残基40について同じコード配列を用いて行った増幅反応により得られたPCRフラグメントをプールした。次いで、プライマーGal10FとassR又はassFとGal10Rを用いた反応により得られたPCRフラグメントのそれぞれのプールを、等モル比で混合した。最後に、2つのオーバーラップPCRフラグメントのそれぞれの最終プールおよそ25ngと、NcoI及びEagIでの消化により線状にした75 ngのベクターDNA (pCLS0542)とを用いて、サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコル(Gietz, R. D.及びR.A. Woods, Methods Enzymol. 2002, 350, 87〜96)を用いて形質転換した。両方の群の変異を含有するインタクトなコード配列を、酵母におけるインビボ相同組換えにより作製する。
【0128】
2) 結果
I-CreIコンビナトリアル変異体を、I-CreI N75足場上で44位、68位及び70位での変異を、28、30、33、38及び40変異と結び付けることにより構築して、複雑さ(complexity)が2208のライブラリーを得た。コンビナトリアル変異体の例を、表IIIに示す。このライブラリーで酵母を形質転換し、3456個のクローン(多様性の1.5倍)を、rosa1.4 DNA標的(tgggattatgt_P; 配列番号36)に対する切断についてスクリーニングした。合計で69個の陽性クローンが見出され、96ウェルプレートで再配置し、2次スクリーニングにより確認した(図9)。これらの陽性のうち、15個のクローン(図9において丸で囲む)を選択した。配列決定の後に、これらの15クローンは、rosa1.4 DNA標的を切断する8つの異なる新規なエンドヌクレアーゼに相当することがわかった(配列番号60〜67;表III)。
【0129】
【表3】
【0130】
実施例4:rosa1を切断するメガヌクレアーゼの作製
パリンドロームrosa1由来標的(rosa1.3及びrosa1.4)のそれぞれを切断できるI-CreI変異体を、実施例2及び3で同定した。このような変異体の対(一方はrosa1.3を切断し、他方はrosa1.4を切断する)を、酵母において同時発現させた。同時発現の際に、3つの活性分子種、すなわち2つのホモ二量体と1つのヘテロ二量体とが存在するはずである。形成されるはずのヘテロ二量体が非パリンドロームのrosa1及びrosa1.2 DNA標的を切断するかをアッセイした。
【0131】
1) 材料及び方法
a) カナマイシン耐性ベクターでの変異体のクローニング
酵母において2つのI-CreI変異体を同時発現させるために、rosa1.4配列を切断する変異体を、カナマイシン耐性遺伝子をマーカーとして有する酵母発現ベクター(pCLS1107, 図10)にサブクローニングした。変異体を、PCR反応により、pCLS0542及びpCLS1107に共通のプライマー:Gal10F 5'-gcaactttagtgctgacacatacagg-3' (配列番号56)及びGal10R 5'-acaaccttgattggagacttgacc-3' (配列番号57)を用いて増幅した。およそ25 ngのPCRフラグメントとDraIII及びNgoMIVでの消化により線状にした25 ngのベクターDNA (pCLS1107)とを用いて酵母サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコルを用いて形質転換する。I-CreI変異体についてのインタクトなコード配列を、酵母でのインビボ相同組換えにより作製する。pCLS1107ベクターにサブクローニングされたrosa1.4標的を切断する変異体を含有するそれぞれの酵母株を、次いで、rosa1.4標的を発現する酵母と交配させてこれを確認した。プラスミドを発現する変異体を回収するために、酵母DNAを標準的なプロトコルを用いて抽出し、大腸菌を形質転換するのに用い、大腸菌DNAを調製した。
【0132】
b) 変異体同時発現
pCLS0542発現ベクター中のrosa1.3標的を切断する変異体を発現する酵母株を、pCLS1107発現ベクター中のrosa1.4標的を切断する変異体をコードするDNAを用いて形質転換した。形質転換体は、-L Glu + G418培地上で選択した。
【0133】
c) メガヌクレアーゼ同時発現クローンの交配及び酵母でのスクリーニング
交配は、コロニーグリッダー(QpixII, Genetix)を用いて行った。変異体を、YPDプレートを覆うナイロンフィルタ上に、低格子密度(約4スポット/cm2)を用いてグリッドした。2回目のグリッド手順を同じフィルタに対して行って、それぞれの標的について異なるレポーター保持酵母株からなる第2層をスポットした。メンブレンを固体寒天YPDリッチ培地上に置き、30℃にて一晩インキュベートして交配を可能にした。次いで、フィルタを、ロイシン及びトリプトファンを欠き、G418を加え、ガラクトース(1%)を炭素源として有する合成培地上に移し、37℃にて5日間インキュベートして、発現及び標的ベクターを有する二倍体を選択した。5日後に、フィルタを、0.5 Mリン酸ナトリウム緩衝液、pH 7.0中の0.02 % X-Gal、0.1% SDS、6%ジメチルホルムアミド(DMF)、7mM β-メルカプトエタノール、1%アガロースを含む固体アガロース培地上に置き、37℃にてインキュベートして、β-ガラクトシダーゼ活性をモニターした。結果をスキャンにより分析し、適切なソフトウェアを用いて定量を行った。
【0134】
結果
rosa1.3標的を切断する変異体(表IIに記載されるm1〜m18)と、rosa1.4標的を切断する8つの変異体(表IIIに記載される)との同時発現は、全ての場合においてrosa1.2標的の効率的な切断をもたらした(スクリーニングの例を図11Aに示す)。全ての試験した組み合わせを、表IVにまとめる。これらの組み合わせのほとんどは、rosa1.2配列とは+1位の1 bpのみが異なるrosa1天然標的も切断できる(図5)。図11Bに示すように、rosa1天然標的に対して観察されたシグナルは、rosa1.2標的に対して観察されたものと比較して弱い。rosa1 DNA標的を切断する組み合わせを、表Vに示す。
【0135】
【表4】
【0136】
【表5】
【0137】
実施例5 rosa1.4を切断するタンパク質のランダム突然変異誘発及びrosa1.3を切断するタンパク質との組み立てによる、rosa1を切断するメガヌクレアーゼの洗練
非パリンドロームのrosa1.2及びrosa1標的を切断できるI-CreI変異体を、パリンドロームのrosa1.3及びrosa1.4標的を切断する変異体の組み立てにより同定した。しかし、組み合わせは、rosa1.2を効率的に切断できたが、rosa1.2とは1位の1 bpのみが異なるrosa1を弱く切断した。rosa1に対して観察されたシグナルは充分でない。
【0138】
よって、rosa1を切断するタンパク質の組み合わせをランダムに突然変異誘発し、rosa1を効率的に切断する変異体をスクリーニングした。標的に結合したI-CreIタンパク質の構造によると、4つの中央の塩基対(-2位〜2位)とI-CreIタンパク質との間に接触はない(Chevalierら, Nat. Struct. Biol., 2001, 8, 312〜316; Chevalier, B. S.及びStoddard, B.L. Nucleic Acids Res., 2001, 29, 3757〜3774; Chevalierら, J. Mol. Biol. 2003, 329, 253〜269)。よって、突然変異を誘発する位置の組を合理的に選択することは困難であり、突然変異誘発を、タンパク質のC-末端部分(83個の最後のアミノ酸)又はタンパク質全体に対して行った。ランダム突然変異誘発により、高い複雑さのライブラリーが得られ、試験される変異型ライブラリーの複雑さは、rosa1を切断するヘテロ二量体の2つの成分の一方のみに突然変異を誘発することにより制限される。
つまり、rosa1.4を切断するタンパク質に突然変異を誘発し、rosa1.3を切断するタンパク質と同時発現させたときにこれらがrosa1を効率的に切断できるかを試験した。
【0139】
1) 材料及び方法
a) ランダム突然変異誘発
ランダム突然変異誘発ライブラリーを、選択された変異体のプールに対して、PCRにより、Mn2+又はdNTPの誘導体、例えば8-オキソ-dGTP及びdPTPを用いて、JBS dNTP-Mutageneisキット中のJENA BIOSCIENCE GmbHからのプロトコルに記載されるようにして2ステップPCRプロセスで創出した。タンパク質全体に対するランダム突然変異誘発のために、用いたプライマーは:preATGCreFor 5'-gcataaattactatacttctatagacacgcaaacacaaatacacagcggcctt gccacc-3'; 配列番号68)及びICreIpostRev (5'-ggctcgaggagctcgtctagaggatcgctcgag ttatcagtcggccgc -3'; 配列番号69)である。タンパク質のC-末端部分に対する突然変異誘発のために、用いたプライマーは、dNTP誘導体を用いてAA78a83For (5'-ttaagcgaaatcaagccg-3'; 配列番号70)及びICreIpostRevである。タンパク質の残りは、高忠実度のtaqポリメラーゼを用い、dNTP誘導体を用いずに、プライマーpreATGCreFor及びAA78a83Rev (5'-cggcttgatttcgcttaa-3'; 配列番号71)を用いて増幅する。
【0140】
変異体のプールを、PCR反応により、pCLS0542 (図7)及びpCLS1107 (図10)に共通するこれらのプライマーを用いて増幅した。およそ75 ngのPCRフラグメントとDraIII及びNgoMIVでの消化により線状にした75 ngのベクターDNA (pCLS1107)を用いて、酵母サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコルを用いて形質転換する。I-CreI変異体についてのインタクトなコード配列のライブラリーを、酵母でのインビボ相同組換えにより作製する。陽性をもたらすクローンを、実施例2に記載されるようにして配列決定により確認した。
【0141】
b) rosa1標的を含有する酵母株でのロイシン発現ベクター中の変異体のクローニング
rosa1標的を酵母レポーターベクター(pCLS1055、図6)中に含有する酵母FYBL2-7B株(MAT a, ura3Δ851, trp1Δ63, leu2Δ1, lys2Δ202)を、LEU2遺伝子をマーカーとして有するpCLS0542ベクター中のrosa1.3標的を切断する変異体で、高効率LiAc形質転換プロトコルを用いて形質転換する。得られた酵母株を、実施例4に記載される交配アッセイのための標的として用いる。
【0142】
2) 結果
rosa1.4を切断する4つの変異体(ERRR / AHQ、ERRR / ARN、ERRK / AHQ及びERRK / VRA、表IVによる)をプールし、全てのタンパク質又はタンパク質のC-末端部分に対してランダム突然変異を誘発し、酵母を形質転換した。4464個の形質転換されたクローンを、次いで、実施例2に記載されるものから選択された、(i) レポータープラスミド内にrosa1標的を含有し、(ii) rosa1.3標的を切断する変異型を発現する酵母株と交配させた。I-CreI V24 Y44 S68 S70 R75 I77 (すなわちVYSSRI)変異体、又はI-CreI V24 Y44 R68 S70 Q75 I77 (すなわちVYRSQI)変異体、又はI-CreI V24 Y44 R68 S70 Y75 T77 (すなわちVYRSYT)変異体(表IIを参照)を発現する3つのこのような株を用いた。2つのクローンは、このような酵母株と交配させたときに、同じ酵母株と突然変異誘発する前の変異体と比較してrosa1標的のよりよい切断を引き起こすことがわかった。結論として、VYSSRI、VYRSQI又はVYRSYTとヘテロ二量体を形成するときにrosa1を効率的に切断できる2つのタンパク質(表VI)が同定された(図12)。
【0143】
【表6】
【0144】
実施例6:rosa1.3を切断するタンパク質のランダム突然変異誘発及びrosa1.4を切断する洗練されたタンパク質との組み立てによる、rosa1を切断するメガヌクレアーゼの洗練
rosa1標的を切断できるI-CreI変異体を、rosa1.3を切断する変異体及びrosa1.4を切断する洗練された変異体の組み立てにより同定した。メガヌクレアーゼの活性を増大させるために、rosa1を切断するヘテロ二量体の第2成分に突然変異を誘発した。この実施例において、rosa1.3を切断する変異体に突然変異を誘発し、その後、実施例5で同定されたrosa1.4を切断する洗練された変異体との組み合わせでrosa1をより効率的に切断する変異体をスクリーニングした。
【0145】
1) 材料及び方法
a) ランダム突然変異誘発:
ランダム突然変異誘発ライブラリーを、選択された変異体のプールに対して、PCRにより、Mn2+又はdNTPの誘導体、例えば8-オキソ-dGTP及びdPTPを用いて、JBS dNTP-Mutageneisキット中のJENA BIOSCIENCE GmbHからのプロトコルに記載されるようにして2ステップPCRプロセスで創出した。タンパク質全体に対するランダム突然変異誘発のために、用いたプライマーは:preATGCreFor 5'-gcataaattactatacttctatagacacgcaaacacaaatacacagcggcctt gccacc-3'; 配列番号68)及びICreIpostRev (5'-ggctcgaggagctcgtctagaggatcgctcgag ttatcagtcggccgc -3'; 配列番号69)である。タンパク質のC-末端部分に対する突然変異誘発のために、用いたプライマーは、dNTP誘導体を用いてAA78a83For (5'-ttaagcgaaatcaagccg-3'; 配列番号70)及びICreIpostRevであった。タンパク質の残りは、高忠実度のtaqポリメラーゼを用い、dNTP誘導体を用いずに、プライマーpreATGCreFor及びAA78a83Rev (5'-cggcttgatttcgcttaa-3'; 配列番号71)を用いて増幅する。
【0146】
変異体のプールを、PCR反応により、pCLS0542 (図7)及びpCLS1107 (図10)に共通するこれらのプライマーを用いて増幅した。およそ75 ngのPCRフラグメントとNcoI及びEagIでの消化により線状にした75 ngのベクターDNA (pCLS0542)を用いて、酵母サッカロミセス・セレビシエFYC2-6A株(MATα, trp1Δ63, leu2Δ1, his3Δ200)を、高効率LiAc形質転換プロトコルを用いて形質転換する。I-CreI変異体についてのインタクトなコード配列のライブラリーを、酵母でのインビボ相同組換えにより作製する。陽性をもたらすクローンを、実施例2に記載されるようにして配列決定により確認した。
【0147】
b) rosa1標的を含有する酵母株でのカナマイシン発現ベクター中の変異体のクローニング
rosa1標的を酵母レポーターベクター(pCLS1055、図6)中に含有する酵母FYBL2-7B株(MAT a, ura3Δ851, trp1Δ63, leu2Δ1, lys2Δ202)を、pCLS1107ベクター中のrosa1.4標的を切断するMO_1及びMO_2洗練変異体で、高効率LiAc形質転換プロトコルを用いて形質転換する。変異体-標的酵母を、実施例4に記載される交配アッセイのための標的として用いる。
【0148】
2) 結果
rosa1.3を切断する4つの変異体の2つのプール(プール1: VYRSNI、VYYSYR、VYRSNV及びVYNSRIと、プール2: VYYSYR、VYSSRI、VYRSQI及びVYRSYT、表Vによる)に、全てのタンパク質又はタンパク質のC-末端部分に対してランダム突然変異を誘発し、酵母を形質転換した。8928個の形質転換されたクローンを、(i) レポータープラスミド内にrosa1標的を含有し、(ii) rosa1.4標的を切断する変異型を発現する酵母株と交配させた。I-CreI E28 R33 R38 R40 A44 H68 Q70 N75 A105 R107 (すなわちMO_1)変異体、I-CreI E28 R33 R38 K40 A44 H68 Q70 N75 A105 R107 A151 G153 E158 (すなわちMO_2)変異体のいずれかを発現する2つのこのような株を用いた。5つのクローンは、このような酵母株と交配させたときに、同じ酵母株と突然変異誘発する前の変異体と比較してrosa1標的のよりよい切断を引き起こすことがわかった(図13)。配列決定の後に、これらは、4つのタンパク質に相当することがわかった。結論として、MO_1又はMO_2とヘテロ二量体を形成したときにrosa1を効率的に切断できる4つのタンパク質が同定された(表VII)。
【0149】
【表7】
【0150】
実施例7:CHO細胞における染色体外モデルでのrosa1標的切断の確認
実施例6において、酵母においてrosa1標的を効率的に切断できるI-CreI洗練変異体を同定した。この実施例では、CHO細胞においてrosa1標的を切断する変異体の2つの組み合わせの能力を、哺乳動物細胞における染色体外アッセイを用いて試験した。
【0151】
1) 材料及び方法
a) CHOスクリーニングのためのベクターでのrosa1標的のクローニング
標的は、以下のようにしてクローニングした。ゲートウェイクローニング配列と接する標的配列に相当するオリゴヌクレオチドをProligoから得た:
5' tggcatacaagtttcaacatgatgtacatcatgttgacaatcgtctgtca 3'(配列番号37)。一本鎖オリゴヌクレオチドのPCR増幅により作製した二本鎖標的DNAを、Gatewayプロトコル(INVITROGEN)を用いてCHOレポーターベクター(pCLS1058、図14)にクローニングした。
【0152】
b) メガヌクレアーゼの再クローニング
実施例6で同定したI-CreI N75、I-SceI及びI-CreI変異体のORFをPCRにより増幅し、配列決定した(MILLEGEN)。次いで、ORFをGatewayプロトコル(INVITROGEN)を用いて再クローニングした。ORFを、ProligoからのプライマーB1F: 5' ggggacaagtttgtacaaaaaagcaggcttcgaaggagatagaaccatggccaataccaaatataacaaagagttcc 3' (配列番号78)及びB2R: 5' ggggaccactttgtacaagaaagctgggtttagtcggccgccggggaggatttcttcttctcgc 3' (配列番号79)を用いる酵母DNAのPCRにより増幅した。PCR生成物を、InvitrogenからのCHOゲートウェイ発現ベクターpCDNA6.2にクローニングした(pCLS1069、図15)。得られたクローンを、実施例2に記載されるようにして配列決定により確認した。
【0153】
c) 哺乳動物細胞での染色体外アッセイ
CHO細胞を、Polyfectトランスフェクション試薬を、供給業者(QIAGEN)のプロトコルに従って用いてトランスフェクションした。トランスフェクションの72時間後に、培養培地を回収し、150μlの溶解/視覚化バッファーを、β-ガラクトシダーゼ液体アッセイのために加えた(典型的には、1リットルのバッファーは、100 mlの溶解バッファー(Tris-HCl 10 mM pH 7.5、NaCl 150 mM、Triton X100 0.1%、BSA 0.1 mg/ml、プロテアーゼ阻害剤)、10 mlのMg 100×バッファー(MgCl2 100 mM、β-メルカプトエタノール35%)、110 ml ONPG 8 mg/ml及び780 mlのリン酸ナトリウム0.1M pH 7.5を含有する)。37℃でのインキュベーションの後に、光学密度を420 nmで測定した。全体のプロセスを、自動化されたBioCel (登録商標)プラットフォーム(VELOCITY11)で行う。
【0154】
2) 結果
図16に示す2回の実験の結果は、I-CreI変異体の2つの組み合わせ(mO_2 / MO_1及びmO_2 / MO_2)が、I-CreI標的(tcaaaacgtcgtgagacagtttgg、配列番号80)に対するI-CreI N75、又はI-SceI標的(tagggataacagggtaat、配列番号81)に対するI-SceIの活性と同様の活性で、CHO細胞においてrosa1標的を切断できることを示す。
【0155】
実施例8:マウス細胞におけるROSA26遺伝子座でのゲノム工学
酵母及び哺乳動物細胞(CHO K1細胞)での染色体外アッセイにおいてrosa1標的を効率的に切断できるI-CreI洗練変異体を、実施例6及び7で同定した。マウスL細胞においてROSA26遺伝子座にて相同組換えを誘発する2つのI-CreI洗練変異体の1つの組み合わせの能力を、この実施例において試験した。
【0156】
1) 材料及び方法
a) ノックイン(KI)マトリクス
2つのマウスROSA26ホモロジーアーム、CQ880114配列(配列表の配列番号3に相当)の6283〜8317のHG ROSA26及び8313〜10319のHD ROSA26の間にクローニングされたハイグロマイシン耐性遺伝子コード配列(CDS)を含む2つのノックインマトリクスを構築した。得られたプラスミドは、pCLS1679及びpCLS1675であった(図18のプラスミドマップ)。pCLS1679において、SV40ポリAに機能可能に連結されたハイグロマイシン耐性遺伝子のコード配列(hygro CDS)を、pBR322ベクター(PROMEGA)に、HG ROSA26とHD ROSA26との間でクローニングした。pCLS1675は、pCLS1679とは、hygro CDSのすぐ上流の配列内リボソーム進入部位(IRES; 配列番号139)の挿入が異なる。
【0157】
b) メガヌクレアーゼのクローニング
I-CreI洗練変異体mO_2及びMO_1のORFは、実施例6 (表VII)に記載される。変異体発現を、ヒト伸長因子1アルファ(EF1α)プロモーター又はサイトメガロウイルス最初期(CMV)プロモーターの制御下の2つの発現ベクターにおいて作製した(pCLS1069、図15)。変異体を、pCLS1069に、CMVプロモーターの下で実施例7に記載されるようにしてクローニングした。得られたプラスミドを、配列決定により確認した(MILLEGEN)。pCLS1761 (図19)及びpCLS1762 (図20)において、mO_2及びMO_1 I-CreI変異体はそれぞれ、EF1αプロモーターの制御下である。
【0158】
c) マウスL細胞でのノックイン実験
マウスL細胞(ATCC # CRL-2648)を、10%胎児ウシ血清、ペニシリン、ストレプトマイシン及びファンギゾンを補った完全DMEM培地(DMEM Glutamax, GIBCO)で培養する。細胞を、リポフェクタミン試薬(INVITROGEN)を製造業者により推奨される手順に従って用いて、トランスフェクションする。トランスフェクションの2日後に、選択を、完全培地中の0.6 mg/mlのハイグロマイシンを用いて行う。選択の2週間後に、耐性クローンを、ClonePixロボット(GENETIX)を用いて採取する。クローンを、0.6 mg/mlのハイグロマイシンを補った完全培地中で、96ウェルプレートにおいて1週間増殖させる。ゲノムDNAを、96ウェルプレートで培養される耐性クローンから、ZR96キット(ZYMO RESEARCH)を用いて抽出する。
【0159】
c) ノックイン事象のPCR分析
ノックイン事象を、HG ROSA26ホモロジーアームの上流のマウスROSA26配列、及びハイグロマイシンCDSにそれぞれ位置するプライマー対KI_GHG_S5 (5' tagtatacagaaactgttgcatcgc 3'; 配列番号137)及びHygSeqRev (5'cgtctgctgctccatacaag 3'; 配列番号138)を用いる、ゲノムDNAに対するPCR分析により検出して、KI特異的PCR増幅を得る(図21)。
【0160】
2) 結果
この実施例で用いたROSA26メガヌクレアーゼは、実施例6 (表VII)に記載され、かつヒト伸長因子1アルファ(EF1α)プロモーター(pCLS1761及びpCLS1762)又はサイトメガロウイルス最初期(CMV)プロモーター(pCLS1069、図15)の制御下で2つの発現ベクターにクローニングされたmO_2及びMO_1である。マウスL細胞に、3つのベクターを同時トランスフェクションさせた:mO_2及びMO_1 ROSA26メガヌクレアーゼを発現する2つのプラスミドと、KIマトリクス。メガヌクレアーゼは、それぞれEF1αプロモーターを有するpCLS1761及びpCLS1762にクローニングされ、KIマトリクスはpCLS1675にクローニングされた。
【0161】
合計で2600000個のマウスL細胞に、2μgのKIマトリクスベクター及び5μg又は10μgのそれぞれのメガヌクレアーゼ発現ベクターを同時トランスフェクションさせた。自発的KI頻度の対照として、同数の細胞に、2μgのKIマトリクスベクターのみをトランスフェクションさせた。トランスフェクション効率(40%)を、蛍光マーカー発現プラスミドを用いるFACS分析により決定した。耐性クローンの頻度は、ハイグロマイシン耐性クローンの総数を計数し、トランスフェクション効率で校正することにより決定した。それぞれ2605個、1197個及び1902個のハイグロマイシン耐性クローンを得た(表VIII)。92個又は184個のクローンを、条件あたり採取し、材料及び方法に記載されるようにしてPCRにより分析した。結果を表VIIIに示す。
【0162】
【表8】
【0163】
ROSA26メガヌクレアーゼとKIマトリクスの同時トランスフェクションは、L細胞においてマウスROSA26遺伝子座にて相同組換えを4.9×10-4の最大頻度で誘発した。KIマトリクス単独でのトランスフェクションを用いて、自発的相同組換えは観察されなかった。この実施例は、ROSA26メガヌクレアーゼが、マウスL細胞においてマウスROSA26遺伝子座にて相同組換えを誘発する能力を示す。
【0164】
実施例9:mO_2及びMO_1に由来するメガヌクレアーゼ
メガヌクレアーゼ構築物は、mO_2 (配列番号75)及びMO_1 (配列番号72)から、Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, 第3版, (Sambrookら, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press)に充分に説明される分子生物学及び組換えDNAの通常の技術を用いて工学的に作製した。
【0165】
NLS (KKKRK; 配列番号134)を、MO_1及びmO_2の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、それぞれ配列番号140及び141を有する。
タグ(TagHA; YPYDVPDYA; 配列番号135)を、mO_2の最初(M1)及び3番目(N3)のアミノ酸の間に挿入した。得られた変異型は、配列番号142である。
タグ(STag; KETAAAKFERQHMDS; 配列番号136)を、MO_1の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、配列番号143である。
タグ(TagHA; YPYDVPDYA; 配列番号135)及びNLS (KKKRK; 配列番号134)を、mO_2の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、配列番号144である。
タグ(STag; KETAAAKFERQHMDS; 配列番号136)及びNLS (KKKRK; 配列番号134)を、MO_1の最初(M1)及び2番目(A2)のアミノ酸の間に挿入した。得られた変異型は、配列番号145である。
【0166】
mO_2単量体(配列番号75の3位〜164位)からリンカー(GGSDKYNQALSKYNQALSKYNQALSGGGGS; 配列番号149)により分けられたMO_1単量体 (配列番号72の1位〜166位) を含む単鎖メガヌクレアーゼを構築した。得られた単鎖メガヌクレアーゼは、配列番号146である。
mO_2/MO_1に由来する偏性ヘテロ二量体を、mO_2単量体にE8K及びE61R変異を、そしてMO_1単量体にK7E及びK96E変異を導入することにより工学的に作製した。得られたヘテロ二量体は、配列番号147及び配列番号148からなる。
【特許請求の範囲】
【請求項1】
2つのI-CreI単量体のうちの一方が、I-CreIの26位〜40位及び44位〜77位にそれぞれ位置するLAGLIDADGコアドメインの2つの機能的サブドメイン内に1つずつ存在する少なくとも2つの置換を有し、マウスROSA26遺伝子座からのDNA標的を切断でき、かつ少なくとも以下の工程:
(a) I-CreIの26位〜40位に位置するLAGLIDADGコアドメインの第1機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第1シリーズを構築し、
(b) I-CreIの44位〜77位に位置するLAGLIDADGコアドメインの第2機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第2シリーズを構築し、
(c) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の-10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(d) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の-5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(e) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(f) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の+3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(g) 工程(c)及び工程(d)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) -10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットと同一であり、(ii) +8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) -5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットと同一であり、(iv) +3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
(h) 工程(e)及び工程(f)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) +3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットと同一であり、(ii) -5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(iv) -10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位のヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
(i) 工程(g)及び/又は(h)で得られた変異型を組み合わせて、ヘテロ二量体を形成し、
(j) マウスROSA26遺伝子座からの上記のDNA標的配列を切断できる工程(i)からのヘテロ二量体を選択及び/又はスクリーニングする
を含む方法により得ることができるI-CreI変異型。
【請求項2】
I-CreIの44位〜77位に位置するサブドメイン内の前記置換が、44位、68位、70位、75位及び/又は77位にある請求項1に記載の変異型。
【請求項3】
I-CreIの26位〜40位に位置するサブドメイン内の前記置換が、26位、28位、30位、32位、33位、38位及び/又は40位にある請求項1に記載の変異型。
【請求項4】
I-CreI部位の±1〜2位、±6〜7位及び/又は±11〜12位のヌクレオチドに対する変異型の特異性を改変する1つ又は複数の置換をI-CreIの137位〜143位に含む請求項1〜3のいずれか1項に記載の変異型。
【請求項5】
マウスROSA26遺伝子座からのDNA標的配列に対する変異型の結合及び/又は切断特性を改良する1つ又は複数の置換をI-CreI配列全体に含む請求項1〜4のいずれか1項に記載の変異型。
【請求項6】
G19S、I24V、S79G、V105A、K107R、V151A、D153G及びK158Eからなる群より選択される1つ又は複数の置換を含む請求項5に記載の変異型。
【請求項7】
前記置換が、元のアミノ酸の、A、D、E、G、H、K、N、P、Q、R、S、T、Y、C、W、L及びVからなる群より選択されるアミノ酸での置き換えである請求項1〜6のいずれか1項に記載の変異型。
【請求項8】
I-CreIの26位〜40位及び/又は44位〜77位に異なる変異を有する第1及び第2単量体の会合により得られ、マウスROSA26遺伝子座からの非パリンドロームDNA標的配列を切断できるヘテロ二量体である請求項1〜7のいずれか1項に記載の変異型。
【請求項9】
前記DNA標的が、配列番号5〜14及び16〜30の配列からなる群より選択される請求項8に記載の変異型。
【請求項10】
第1単量体及び第2単量体が、それぞれ少なくとも以下の置換:
- N30H、Y33S、Q44E、R68C、R70S、D75N及びN30D、Y33R、Q38T、Q44K、R68E、R70S、I77R、
- S32N、Y33G、Q44K、R70E、D75N及びS32T、Q38W、Q44K、R68E、R70S、I77R、
- Y33R、Q38N、S40Q、Q44N、R70S、D75R、I77D及びN30H、Y33S、Q44A、R70S、D75Q、I77E、
- K28S、Q38R、S40K、Q44D、R68Y、R70S、D75S、I77R及びY33C、Q38A、R68A、R70K、D75N、
- Y33C、Q44T、R70S、D75Y及びS32D、Q38C、Q44D、R68Y、R70S、D75S、I77R、
- S32T、Y33C、R68T、R70N、D75N及びS32T、Q38W、Q44K、R70E、D75N、
- R70S、D75R、I77Y及びY33R、Q38A、S40Q、Q44A、R70S、D75N、
- K28S、Q38R、S40K、Q44T、R68N、R70N、D75N及びY33H、Q38S、Q44K、R68Y、R70S、D75Q、I77N、
- K28A、Y33S、Q38R、S40K、Q44N、R68Y、R70S、D75R、I77V及びS32T、Y33C、Q44A、R70S、D75N、
- S32D、Y33H、Q44K、R68E、R70S、I77R及びS32D、Y33H、Q44D、R68N、R70S、D75N、
- N30R、S32D、R68S、R70K、D75N及びD75N、
- K28R、Y33A、Q38Y、S40Q、R68Y、R70S、D75R、I77Q及びY33R、Q38A、S40Q、R70S、I77K、
- K28R、N30D、D75E、I77R及びS32D、Q38C、Q44A、R70S、D75R、I77Y、
- Y33R、Q38A、S40Q、R70S、D75N及びR70S、D75Y、I77R、
- Y33R、Q38A、S40Q、Q44N、R68Y、R70S、D75R、I77V及びN30R、S32D、Q44T、R68H、R70H、D75N、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q、I77N及びS32A、Y33C、R68Y、R70S、D75R、I77Q、
- K28A、Y33S、Q38R、S40K、R68N、R70S、D75N、I77R及びS32N、Y33G、Q44A、R68A、R70K、D75N、
- N30H、Y33S、Q44Y、R70S、I77V及びY33R、S40Q、Q44A、R70S、D75N、
- S32D、Y33H、R68T、R70N、D75N及びN30R、S32D、Q44T、R68N、R70N、D75N、
- S32R、Y33D、Q44A、R70S、D75N及びY33S、Q38R、S40H、R68H、R70H、D75N、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q、I77N及びK28R、Y33A、Q38Y、S40Q、Q44T、R68H、R70H、D75N、
- K28Q、Q38R、S40K、Q44A、R70S、D75E、I77R及びN30R、S32D、Q44K、R68Y、R70S、D75Q、I77N、
- Y33T、Q38A、R68H、R70H、D75N及びN30H、Y33S、R70S、I77K、
- Y33T、S40T、R68A、R70K、D75N及びK28R、Y33A、Q38Y、S40Q、R70S、並びに
- S32D、Y33H、Q44N、R70S、D75R、I77D及びS32D、Q38C、R70S、I77K
を含む請求項9に記載の変異型。
【請求項11】
それぞれ、第1単量体が配列番号82〜106の配列のいずれかのものであり、第2単量体が配列番号107〜116、4、117〜130の配列のいずれかのものである請求項10に記載の変異型。
【請求項12】
前記DNA標的が配列番号15である請求項8に記載の変異型。
【請求項13】
第1単量体が、I24V、Q44Y、R70S及びD75N;I24V、Q44Y、R68Y、R70S、D75Y及びI77R;I24V、Q44Y、R70S、D75N及びI77V;I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択され、第2単量体が、K28E、Y33R、Q38R、S40R、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40R、Q44A、R70N及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40K、Q44V、R70A及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R70G及びD75Nからなる群より選択される請求項12に記載の変異型。
【請求項14】
第1単量体が、配列番号39、43、45、49、50、51、53及び54の配列からなる群より選択され、第2単量体が、配列番号60、61、63、65及び66の配列からなる群より選択される請求項13に記載の変異型。
【請求項15】
第1単量体及び第2単量体がそれぞれ、少なくとも以下の置換:I24V、Q44Y、R70S、D75Y、I77T及びK28E、Y33R、Q38R、S40R、Q44A、R68S、R70Q、D75Nを含む請求項12に記載の変異型。
【請求項16】
第1単量体及び第2単量体がそれぞれ、配列番号54及び配列番号62である請求項15に記載の変異型。
【請求項17】
第1単量体が、I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択され、第2単量体が、K28E、Y33R、Q38R、S40K、Q44A、R70S、D75N及びK28E、Y33R、Q38R、S40K、Q44A、R68T、R70N、D75Nからなる群より選択される請求項12に記載の変異型。
【請求項18】
第1単量体が、配列番号49、50、51、53及び54の配列からなる群より選択され、第2単量体が、配列番号64及び67の配列からなる群より選択される請求項17に記載の変異型。
【請求項19】
第1単量体が、配列番号72及び73の配列からなる群より選択され、第2単量体が、配列番号74〜77の配列からなる群より選択される請求項12に記載の変異型。
【請求項20】
核局在化シグナル及び/又はタグを含む請求項1〜19のいずれか1項に記載の変異型。
【請求項21】
請求項11、14、16、18及び19で定義される配列と少なくとも95%の配列同一性を有する請求項1〜10、12、13、15、17及び20のいずれか1項に記載の変異型。
【請求項22】
偏性ヘテロ二量体であり、第1単量体がD137R変異をさらに含み、第2単量体がR51D変異をさらに含む請求項8〜21のいずれか1項に記載の変異型。
【請求項23】
偏性ヘテロ二量体であり、第1単量体がE8R又はE8K及びE61R変異をさらに含み、第2単量体がK7E及びK96E変異をさらに含む請求項8〜21のいずれか1項に記載の変異型。
【請求項24】
請求項1〜23のいずれか1項に記載の1つの変異型の2つの単量体又はコアドメイン或いはその両方の組み合わせを含む単鎖キメラメガヌクレアーゼ。
【請求項25】
ペプチドリンカーで連結された請求項10、11及び13〜23のいずれか1項で定義される第1及び第2単量体を含む請求項24に記載の単鎖メガヌクレアーゼ。
【請求項26】
請求項1〜23のいずれか1項に記載の変異型又は請求項24若しくは25に記載の単鎖キメラメガヌクレアーゼをコードするポリヌクレオチドフラグメント。
【請求項27】
請求項26に記載の少なくとも1つのポリヌクレオチドフラグメントを含む発現ベクター。
【請求項28】
2つの異なるポリヌクレオチドフラグメントを含み、そのそれぞれが、請求項8〜23のいずれか1項に記載のヘテロ二量体変異型の単量体の1つをコードする請求項27に記載の発現ベクター。
【請求項29】
請求項1、8、9及び12のいずれか1項で定義されるマウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する配列で挟まれたマウスROSA26遺伝子座に導入される配列を含むターゲティング構築物を含むベクター。
【請求項30】
請求項1、8、9及び12のいずれか1項で定義されるマウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する配列で挟まれたマウスROSA26遺伝子座に導入される配列を含むターゲティング構築物を含む請求項27又は28に記載のベクター。
【請求項31】
導入される前記配列が、異種プロモーターに機能可能に連結された興味対象の生成物をコードするDNA配列を含むカセットである請求項29又は30に記載のベクター。
【請求項32】
導入される前記配列が、興味対象の生成物をコードするDNA配列を含むカセットであり、前記カセットのマウスROSA26遺伝子座への導入が、内因性ROSA26プロモーターからの前記生成物の発現を誘発する請求項29又は30に記載のベクター。
【請求項33】
マウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する前記配列が、配列番号3の
3131位〜3330位、3401位〜3600位、4628位〜4827位、5495位〜5694位、5519位〜5718位、5817位〜6016位、5903位〜6102位、6320位〜6519位、7305位〜7504位、7981位〜8180位、8215位〜8414位、8305位〜8504位、8494位〜8693位、8589位〜8788位、8660位〜8859位、8921位〜9120位、9191位〜9390位、9467位〜9666位、10174位〜10373位、11469位〜11668位、12302位〜12501位、12325位〜12524位、12446位〜12465位、12702位〜12901位、12815位〜13014位及び12865位〜13064位
から選択される配列を含むマウスROSA26遺伝子座のフラグメントである請求項29〜32のいずれか1項に記載のベクター。
【請求項34】
マウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する前記配列が、前記切断部位の上流及び下流の配列を含むマウスROSA26遺伝子座のフラグメントであり、そのことにより切断部位に隣接するコード配列の欠失を可能にする請求項29〜32のいずれか1項に記載のベクター。
【請求項35】
請求項1〜23のいずれか1項に記載の少なくとも1つの変異型、請求項24若しくは25に記載の1つの単鎖キメラメガヌクレアーゼ及び/又は請求項27、28及び30〜34のいずれか1項に記載の少なくとも1つの発現ベクターを含む組成物。
【請求項36】
請求項31〜34のいずれか1項で定義される前記変異型のゲノムDNA標的部位を取り囲む領域と相同性を有する配列で挟まれたマウスROSA26遺伝子座に導入される配列を含むターゲティングDNA構築物を含む請求項35に記載の変異型。
【請求項37】
請求項26に記載のポリヌクレオチド又は請求項27〜34のいずれか1項に記載のベクターにより改変された宿主細胞。
【請求項38】
請求項26又は28で定義される1つ又は2つのポリヌクレオチドフラグメントを含む非ヒトトランスジェニック動物。
【請求項39】
請求項26又は28で定義される1つ又は2つのポリヌクレオチドフラグメントを含むトランスジェニック植物。
【請求項40】
請求項1〜23のいずれか1項に記載の少なくとも1つの変異型、請求項24若しくは25に記載の1つの単鎖キメラメガヌクレアーゼ、請求項27〜34のいずれか1項に記載の1つのベクターの、非治療目的でのマウスゲノムの工学的改変のための使用。
【請求項41】
前記変異型、単鎖キメラメガヌクレアーゼ又はベクターが、請求項29〜34のいずれか1項で定義されるターゲティングDNA構築物と会合する請求項40に記載の使用。
【請求項42】
前記ターゲティングDNA構築物が、請求項31〜34のいずれか1項で定義されるマウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する配列で挟まれた興味対象の生成物をコードするDNA配列と結局はマーカー遺伝子とを含むカセットを含む請求項41に記載の使用。
【請求項43】
興味対象の異種タンパク質を発現するトランスジェニックマウス又は組換えマウス株化細胞を作製するための請求項40〜42のいずれか1項に記載の使用。
【請求項1】
2つのI-CreI単量体のうちの一方が、I-CreIの26位〜40位及び44位〜77位にそれぞれ位置するLAGLIDADGコアドメインの2つの機能的サブドメイン内に1つずつ存在する少なくとも2つの置換を有し、マウスROSA26遺伝子座からのDNA標的を切断でき、かつ少なくとも以下の工程:
(a) I-CreIの26位〜40位に位置するLAGLIDADGコアドメインの第1機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第1シリーズを構築し、
(b) I-CreIの44位〜77位に位置するLAGLIDADGコアドメインの第2機能的サブドメイン内に少なくとも1つの置換を有するI-CreI変異型の第2シリーズを構築し、
(c) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の-10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(d) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の-5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットで置き換えられ、(ii) +3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(e) 工程(a)の第1シリーズから、少なくとも(i) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(f) 工程(b)の第2シリーズから、少なくとも(i) I-CreI部位の+3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットで置き換えられ、(ii) -5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列で置き換えられた変異I-CreI部位を切断できる変異型を選択及び/又はスクリーニングし、
(g) 工程(c)及び工程(d)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) -10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットと同一であり、(ii) +8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-10位〜-8位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) -5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットと同一であり、(iv) +3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の-5位〜-3位に存在するヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
(h) 工程(e)及び工程(f)からの2つの変異型の26位〜40位及び44位〜77位の変異を、単一の変異型に組み合わせて、(i) +3位〜+5位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットと同一であり、(ii) -5位〜-3位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+3位〜+5位に存在するヌクレオチドトリプレットの逆相補配列と同一であり、(iii) I-CreI部位の+8位〜+10位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位に存在するヌクレオチドトリプレットで置き換えられ、(iv) -10位〜-8位のヌクレオチドトリプレットが、マウスROSA26遺伝子座からのDNA標的配列の+8位〜+10位のヌクレオチドトリプレットの逆相補配列と同一である配列を切断する新規なホモ二量体I-CreI変異型を得て、
(i) 工程(g)及び/又は(h)で得られた変異型を組み合わせて、ヘテロ二量体を形成し、
(j) マウスROSA26遺伝子座からの上記のDNA標的配列を切断できる工程(i)からのヘテロ二量体を選択及び/又はスクリーニングする
を含む方法により得ることができるI-CreI変異型。
【請求項2】
I-CreIの44位〜77位に位置するサブドメイン内の前記置換が、44位、68位、70位、75位及び/又は77位にある請求項1に記載の変異型。
【請求項3】
I-CreIの26位〜40位に位置するサブドメイン内の前記置換が、26位、28位、30位、32位、33位、38位及び/又は40位にある請求項1に記載の変異型。
【請求項4】
I-CreI部位の±1〜2位、±6〜7位及び/又は±11〜12位のヌクレオチドに対する変異型の特異性を改変する1つ又は複数の置換をI-CreIの137位〜143位に含む請求項1〜3のいずれか1項に記載の変異型。
【請求項5】
マウスROSA26遺伝子座からのDNA標的配列に対する変異型の結合及び/又は切断特性を改良する1つ又は複数の置換をI-CreI配列全体に含む請求項1〜4のいずれか1項に記載の変異型。
【請求項6】
G19S、I24V、S79G、V105A、K107R、V151A、D153G及びK158Eからなる群より選択される1つ又は複数の置換を含む請求項5に記載の変異型。
【請求項7】
前記置換が、元のアミノ酸の、A、D、E、G、H、K、N、P、Q、R、S、T、Y、C、W、L及びVからなる群より選択されるアミノ酸での置き換えである請求項1〜6のいずれか1項に記載の変異型。
【請求項8】
I-CreIの26位〜40位及び/又は44位〜77位に異なる変異を有する第1及び第2単量体の会合により得られ、マウスROSA26遺伝子座からの非パリンドロームDNA標的配列を切断できるヘテロ二量体である請求項1〜7のいずれか1項に記載の変異型。
【請求項9】
前記DNA標的が、配列番号5〜14及び16〜30の配列からなる群より選択される請求項8に記載の変異型。
【請求項10】
第1単量体及び第2単量体が、それぞれ少なくとも以下の置換:
- N30H、Y33S、Q44E、R68C、R70S、D75N及びN30D、Y33R、Q38T、Q44K、R68E、R70S、I77R、
- S32N、Y33G、Q44K、R70E、D75N及びS32T、Q38W、Q44K、R68E、R70S、I77R、
- Y33R、Q38N、S40Q、Q44N、R70S、D75R、I77D及びN30H、Y33S、Q44A、R70S、D75Q、I77E、
- K28S、Q38R、S40K、Q44D、R68Y、R70S、D75S、I77R及びY33C、Q38A、R68A、R70K、D75N、
- Y33C、Q44T、R70S、D75Y及びS32D、Q38C、Q44D、R68Y、R70S、D75S、I77R、
- S32T、Y33C、R68T、R70N、D75N及びS32T、Q38W、Q44K、R70E、D75N、
- R70S、D75R、I77Y及びY33R、Q38A、S40Q、Q44A、R70S、D75N、
- K28S、Q38R、S40K、Q44T、R68N、R70N、D75N及びY33H、Q38S、Q44K、R68Y、R70S、D75Q、I77N、
- K28A、Y33S、Q38R、S40K、Q44N、R68Y、R70S、D75R、I77V及びS32T、Y33C、Q44A、R70S、D75N、
- S32D、Y33H、Q44K、R68E、R70S、I77R及びS32D、Y33H、Q44D、R68N、R70S、D75N、
- N30R、S32D、R68S、R70K、D75N及びD75N、
- K28R、Y33A、Q38Y、S40Q、R68Y、R70S、D75R、I77Q及びY33R、Q38A、S40Q、R70S、I77K、
- K28R、N30D、D75E、I77R及びS32D、Q38C、Q44A、R70S、D75R、I77Y、
- Y33R、Q38A、S40Q、R70S、D75N及びR70S、D75Y、I77R、
- Y33R、Q38A、S40Q、Q44N、R68Y、R70S、D75R、I77V及びN30R、S32D、Q44T、R68H、R70H、D75N、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q、I77N及びS32A、Y33C、R68Y、R70S、D75R、I77Q、
- K28A、Y33S、Q38R、S40K、R68N、R70S、D75N、I77R及びS32N、Y33G、Q44A、R68A、R70K、D75N、
- N30H、Y33S、Q44Y、R70S、I77V及びY33R、S40Q、Q44A、R70S、D75N、
- S32D、Y33H、R68T、R70N、D75N及びN30R、S32D、Q44T、R68N、R70N、D75N、
- S32R、Y33D、Q44A、R70S、D75N及びY33S、Q38R、S40H、R68H、R70H、D75N、
- Y33P、S40Q、Q44K、R68Y、R70S、D75Q、I77N及びK28R、Y33A、Q38Y、S40Q、Q44T、R68H、R70H、D75N、
- K28Q、Q38R、S40K、Q44A、R70S、D75E、I77R及びN30R、S32D、Q44K、R68Y、R70S、D75Q、I77N、
- Y33T、Q38A、R68H、R70H、D75N及びN30H、Y33S、R70S、I77K、
- Y33T、S40T、R68A、R70K、D75N及びK28R、Y33A、Q38Y、S40Q、R70S、並びに
- S32D、Y33H、Q44N、R70S、D75R、I77D及びS32D、Q38C、R70S、I77K
を含む請求項9に記載の変異型。
【請求項11】
それぞれ、第1単量体が配列番号82〜106の配列のいずれかのものであり、第2単量体が配列番号107〜116、4、117〜130の配列のいずれかのものである請求項10に記載の変異型。
【請求項12】
前記DNA標的が配列番号15である請求項8に記載の変異型。
【請求項13】
第1単量体が、I24V、Q44Y、R70S及びD75N;I24V、Q44Y、R68Y、R70S、D75Y及びI77R;I24V、Q44Y、R70S、D75N及びI77V;I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択され、第2単量体が、K28E、Y33R、Q38R、S40R、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40R、Q44A、R70N及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R68H、R70Q及びD75N;K28E、Y33R、Q38R、S40K、Q44V、R70A及びD75N;K28E、Y33R、Q38R、S40K、Q44A、R70G及びD75Nからなる群より選択される請求項12に記載の変異型。
【請求項14】
第1単量体が、配列番号39、43、45、49、50、51、53及び54の配列からなる群より選択され、第2単量体が、配列番号60、61、63、65及び66の配列からなる群より選択される請求項13に記載の変異型。
【請求項15】
第1単量体及び第2単量体がそれぞれ、少なくとも以下の置換:I24V、Q44Y、R70S、D75Y、I77T及びK28E、Y33R、Q38R、S40R、Q44A、R68S、R70Q、D75Nを含む請求項12に記載の変異型。
【請求項16】
第1単量体及び第2単量体がそれぞれ、配列番号54及び配列番号62である請求項15に記載の変異型。
【請求項17】
第1単量体が、I24V、Q44Y、R68N、R70S及びD75R;I24V、Q44Y、R68S、R70S及びD75R;I24V、Q44Y、R70S及びD75Q;I24V、Q44Y、R68Y、R70S、D75R及びI77V;I24V、Q44Y、R70S、D75Y及びI77Tからなる群より選択され、第2単量体が、K28E、Y33R、Q38R、S40K、Q44A、R70S、D75N及びK28E、Y33R、Q38R、S40K、Q44A、R68T、R70N、D75Nからなる群より選択される請求項12に記載の変異型。
【請求項18】
第1単量体が、配列番号49、50、51、53及び54の配列からなる群より選択され、第2単量体が、配列番号64及び67の配列からなる群より選択される請求項17に記載の変異型。
【請求項19】
第1単量体が、配列番号72及び73の配列からなる群より選択され、第2単量体が、配列番号74〜77の配列からなる群より選択される請求項12に記載の変異型。
【請求項20】
核局在化シグナル及び/又はタグを含む請求項1〜19のいずれか1項に記載の変異型。
【請求項21】
請求項11、14、16、18及び19で定義される配列と少なくとも95%の配列同一性を有する請求項1〜10、12、13、15、17及び20のいずれか1項に記載の変異型。
【請求項22】
偏性ヘテロ二量体であり、第1単量体がD137R変異をさらに含み、第2単量体がR51D変異をさらに含む請求項8〜21のいずれか1項に記載の変異型。
【請求項23】
偏性ヘテロ二量体であり、第1単量体がE8R又はE8K及びE61R変異をさらに含み、第2単量体がK7E及びK96E変異をさらに含む請求項8〜21のいずれか1項に記載の変異型。
【請求項24】
請求項1〜23のいずれか1項に記載の1つの変異型の2つの単量体又はコアドメイン或いはその両方の組み合わせを含む単鎖キメラメガヌクレアーゼ。
【請求項25】
ペプチドリンカーで連結された請求項10、11及び13〜23のいずれか1項で定義される第1及び第2単量体を含む請求項24に記載の単鎖メガヌクレアーゼ。
【請求項26】
請求項1〜23のいずれか1項に記載の変異型又は請求項24若しくは25に記載の単鎖キメラメガヌクレアーゼをコードするポリヌクレオチドフラグメント。
【請求項27】
請求項26に記載の少なくとも1つのポリヌクレオチドフラグメントを含む発現ベクター。
【請求項28】
2つの異なるポリヌクレオチドフラグメントを含み、そのそれぞれが、請求項8〜23のいずれか1項に記載のヘテロ二量体変異型の単量体の1つをコードする請求項27に記載の発現ベクター。
【請求項29】
請求項1、8、9及び12のいずれか1項で定義されるマウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する配列で挟まれたマウスROSA26遺伝子座に導入される配列を含むターゲティング構築物を含むベクター。
【請求項30】
請求項1、8、9及び12のいずれか1項で定義されるマウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する配列で挟まれたマウスROSA26遺伝子座に導入される配列を含むターゲティング構築物を含む請求項27又は28に記載のベクター。
【請求項31】
導入される前記配列が、異種プロモーターに機能可能に連結された興味対象の生成物をコードするDNA配列を含むカセットである請求項29又は30に記載のベクター。
【請求項32】
導入される前記配列が、興味対象の生成物をコードするDNA配列を含むカセットであり、前記カセットのマウスROSA26遺伝子座への導入が、内因性ROSA26プロモーターからの前記生成物の発現を誘発する請求項29又は30に記載のベクター。
【請求項33】
マウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する前記配列が、配列番号3の
3131位〜3330位、3401位〜3600位、4628位〜4827位、5495位〜5694位、5519位〜5718位、5817位〜6016位、5903位〜6102位、6320位〜6519位、7305位〜7504位、7981位〜8180位、8215位〜8414位、8305位〜8504位、8494位〜8693位、8589位〜8788位、8660位〜8859位、8921位〜9120位、9191位〜9390位、9467位〜9666位、10174位〜10373位、11469位〜11668位、12302位〜12501位、12325位〜12524位、12446位〜12465位、12702位〜12901位、12815位〜13014位及び12865位〜13064位
から選択される配列を含むマウスROSA26遺伝子座のフラグメントである請求項29〜32のいずれか1項に記載のベクター。
【請求項34】
マウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する前記配列が、前記切断部位の上流及び下流の配列を含むマウスROSA26遺伝子座のフラグメントであり、そのことにより切断部位に隣接するコード配列の欠失を可能にする請求項29〜32のいずれか1項に記載のベクター。
【請求項35】
請求項1〜23のいずれか1項に記載の少なくとも1つの変異型、請求項24若しくは25に記載の1つの単鎖キメラメガヌクレアーゼ及び/又は請求項27、28及び30〜34のいずれか1項に記載の少なくとも1つの発現ベクターを含む組成物。
【請求項36】
請求項31〜34のいずれか1項で定義される前記変異型のゲノムDNA標的部位を取り囲む領域と相同性を有する配列で挟まれたマウスROSA26遺伝子座に導入される配列を含むターゲティングDNA構築物を含む請求項35に記載の変異型。
【請求項37】
請求項26に記載のポリヌクレオチド又は請求項27〜34のいずれか1項に記載のベクターにより改変された宿主細胞。
【請求項38】
請求項26又は28で定義される1つ又は2つのポリヌクレオチドフラグメントを含む非ヒトトランスジェニック動物。
【請求項39】
請求項26又は28で定義される1つ又は2つのポリヌクレオチドフラグメントを含むトランスジェニック植物。
【請求項40】
請求項1〜23のいずれか1項に記載の少なくとも1つの変異型、請求項24若しくは25に記載の1つの単鎖キメラメガヌクレアーゼ、請求項27〜34のいずれか1項に記載の1つのベクターの、非治療目的でのマウスゲノムの工学的改変のための使用。
【請求項41】
前記変異型、単鎖キメラメガヌクレアーゼ又はベクターが、請求項29〜34のいずれか1項で定義されるターゲティングDNA構築物と会合する請求項40に記載の使用。
【請求項42】
前記ターゲティングDNA構築物が、請求項31〜34のいずれか1項で定義されるマウスROSA26遺伝子座に存在するゲノムDNA切断部位を取り囲む領域と相同性を有する配列で挟まれた興味対象の生成物をコードするDNA配列と結局はマーカー遺伝子とを含むカセットを含む請求項41に記載の使用。
【請求項43】
興味対象の異種タンパク質を発現するトランスジェニックマウス又は組換えマウス株化細胞を作製するための請求項40〜42のいずれか1項に記載の使用。
【図1】

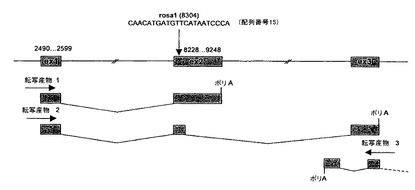
【図2】
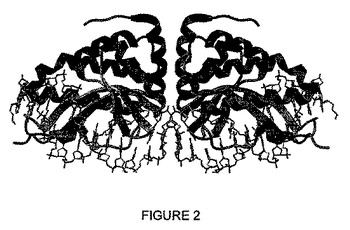
【図3】
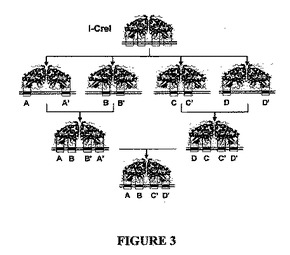
【図4】
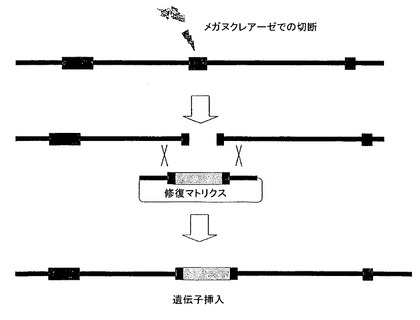
【図5】

【図6】
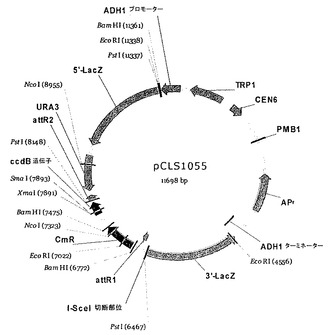
【図7】
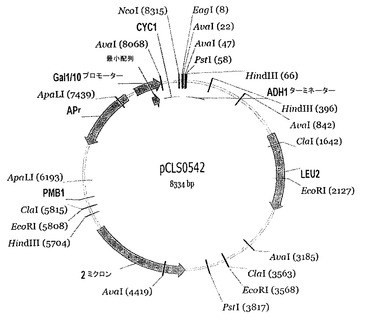
【図8】
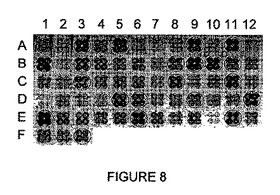
【図9】
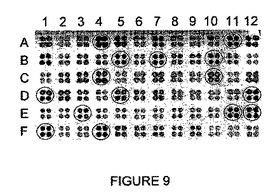
【図10】
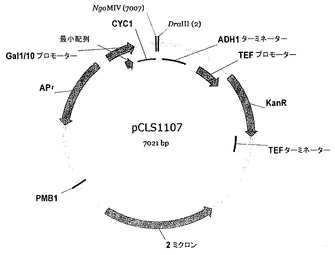
【図11A】

【図11B】

【図12】
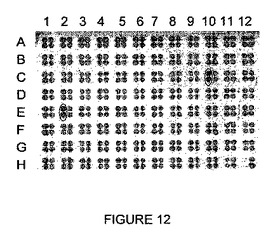
【図13】
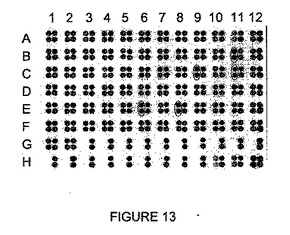
【図14】
【図15】
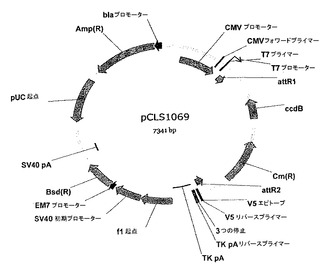
【図16】
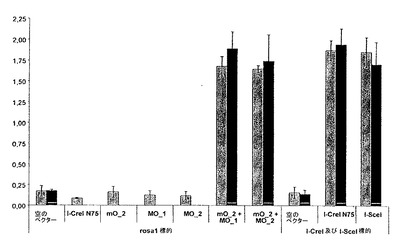
【図17】
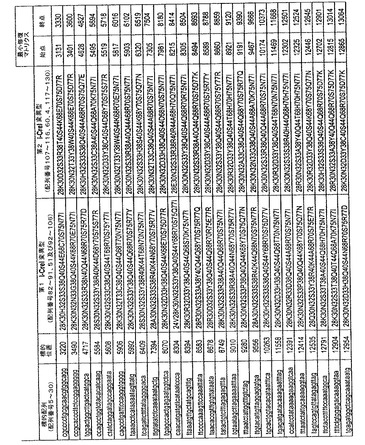
【図18】
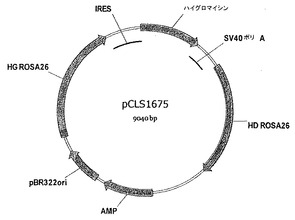
【図19】
【図20】
【図21】
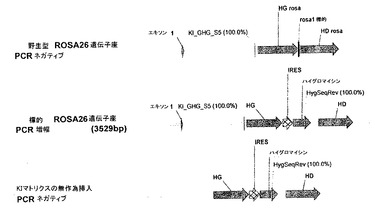
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11A】
【図11B】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【公表番号】特表2010−528648(P2010−528648A)
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願番号】特願2010−510912(P2010−510912)
【出願日】平成20年6月6日(2008.6.6)
【国際出願番号】PCT/IB2008/002500
【国際公開番号】WO2008/152523
【国際公開日】平成20年12月18日(2008.12.18)
【出願人】(508245231)
【氏名又は名称原語表記】CELLECTIS
【住所又は居所原語表記】102 avenue Gaston Roussel,F−93235 Romainville Cedex,France
【Fターム(参考)】
【公表日】平成22年8月26日(2010.8.26)
【国際特許分類】
【出願日】平成20年6月6日(2008.6.6)
【国際出願番号】PCT/IB2008/002500
【国際公開番号】WO2008/152523
【国際公開日】平成20年12月18日(2008.12.18)
【出願人】(508245231)
【氏名又は名称原語表記】CELLECTIS
【住所又は居所原語表記】102 avenue Gaston Roussel,F−93235 Romainville Cedex,France
【Fターム(参考)】
[ Back to top ]