
マラリアワクチン組成物および細胞媒介免疫性を惹起する成分

広いスペクトルのマラリア寄生体に対して細胞媒介免疫性を惹起し大半のHLA対立遺伝子を網羅するポリエピトープ構築体に基づくマラリアワクチンを提供する。
前記ポリエピトープ構築体中のエピトープは、CD4およびCD8T細胞エピトープ類を含むことが公知のPlasmodium falciparumスポロゾイト周囲タンパク質(CSP)領域に由来しかつCSPの高可変および保存領域両者由来のエピトープ類を含む。
前記ポリエピトープ構築体中のエピトープは、CD4およびCD8T細胞エピトープ類を含むことが公知のPlasmodium falciparumスポロゾイト周囲タンパク質(CSP)領域に由来しかつCSPの高可変および保存領域両者由来のエピトープ類を含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、概括的には、マラリアワクチンに関する。本発明は特に、広いスペクトルのPlasmodium falciparum寄生体種のスポロゾイド周囲タンパク質に対する細胞媒介免疫性を惹起しかつHLA対立遺伝子類の大半により認識されるポリエピトープ構築体に基づくマラリアワクチンを提供する。
【背景技術】
【0002】
マラリアは、アメリカ大陸、アジアおよびアフリカを含む熱帯、亜熱帯で広まっているベクター運搬感染症である。それは、毎年約6億5千万の人々に病気を起こし、百万ないし3百万人を死亡させ、それら人々の大半はサハラ砂漠以南のアフリカにおける幼い子供たちである。
【0003】
本疾患は、属Plasmodiumの原生動物寄生体が原因である。本疾患の最も重篤な病態は、Plasmodium falciparumおよびPlasmodium・vivaxにより惹き起こされるが、他の関連種〔Plasmodium・ovale、Plasmodium・malariae、および時にはPlasmodium・knowlesi〕もまたヒトに感染する。
【0004】
マラリア寄生体は、雌のAnopheles蚊に刺されることにより伝染し、ヒト宿主に対して前記寄生体のスポロゾイト体を運ぶ。前記スポロゾイト類は、血液により肝臓まで運ばれ、そこでそれらは肝細胞中で増殖し、ライフサイクルの次の形態すなわちメロゾイトに変化する。メロゾイト類は、感染肝細胞が破裂すると、最終的には血流中に放出される。次にメロゾイト類は赤血球に感染し、一部は、RBC中で雄および雌の生殖母細胞になる。別の蚊が感染宿主を刺すと、それは、雄および雌の生殖母細胞を摂取し、雌の蚊の内部で融合しスポロゾイトになる。この蚊が次に刺すと、このスポロゾイト類はさらに別の宿主に伝染等する。
【0005】
マラリア感染は、貧血に特徴的な症状(軽度頭痛、息切れ、頻脈等)やさらに発熱、悪寒、悪心、インフルエンザ様疾患のような他の一般的症状、および重篤な場合、昏睡および死亡の原因となる。マラリア伝染は、蚊帳および昆虫忌避剤により蚊に刺されないようにすることによって低下させることができるが、それらはかなり効果的ではあるものの、流通させかつ連続的に適切に使用することが必要であるという欠点を有している。他の対応策には家屋内に殺虫剤を撒くことおよび蚊が産卵している滞留水を排水することによって蚊を制御することが含まれる。残念ながら、殺虫剤の使用は環境上のリスクとなり、ある地方では、全ての滞留水を排水することは実質的に不可能である。
【0006】
Plasmodiumに自然感染した後、ヒト宿主は抗Plasmodium抗体を産生する。しかし、抗体産生だけで前記寄生体を中和する能力(体液性免疫)は、不断の、防御的免疫性としては持続しないし、あるいは十分な記憶特性を有する細胞性免疫も惹起されない。その結果として感染が何度も起こることがあり、疾患治療と予防を複雑化させている。
【0007】
効果的な抗マラリアワクチンを産生するためにいろいろ試みられてきた。Huiらの米国特許第6,660,498号は、ワクチン中で使用するために、バキュロウイルス系を用いて組換えメジャーメロゾイトサーフィスプロテイン(Major Merozoite Surface Protein)1を産生させることを記載している;Knappらの米国特許第5,393,523号はP.falciparumの組換えヒスチジン高含量タンパク質の調製とワクチンにおける用途を記載している;Patarroyoの米国特許第4,957,738号および同第4,735,799号は、後期段階のP.falciparumマラリアに対して抗体を誘発する合成ペプチド化合物混合物を記載している。Asakuraの米国特許第4,643,896号は、新規マラリア関連抗原CRAを開示し、それは、マラリアワクチンとして有用であると記載している。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第6,660,498号
【特許文献2】米国特許第5,393,523号
【特許文献3】米国特許第4,957,738号
【特許文献4】米国特許第4,735,799号
【特許文献5】米国特許第4,643,896号
【発明の開示】
【発明が解決しようとする課題】
【0009】
過去の努力にもかかわらず、マラリアに対して有効なワクチンは残念なことに現在のところ全く得られていない。特に、マラリア寄生体類の全てまたはほとんどの株に対して細胞媒介免疫性を惹起するワクチンは、全く開発されていない。その代わりに、キニーネまたはアルテミシニン誘導体のような予防薬を連続的に服用し、感染リスクを低下させなければならない。これらの予防薬による治療は、しばしば、風土病となっている地方に住んでいる多くの人々にとってあまりにも高価である。さらに、前記寄生体の薬物耐性株が、ますます頻繁に見られるようになっている。
【0010】
前記スポロゾイト周囲タンパク質(本文でCSP)のタンパク質断片をコードするかまたはそれから構築したワクチンはヒトおよび動物モデルの両方で免疫原性およびやや限定された防御能を示している。実際的ではないが最初の成功したマラリアワクチンは、スポロゾイトを運んでいる照射蚊類から構成されていた。細胞免疫応答を惹起するように設計したアジュバント中でデリバリされるサブユニットワクチンは、ヒトにおいてCSPまたはアジュバント単独よりもより高い防御を提供したことは重要である。カルボキシおよびアミノ末端両者の可変領域がさまざまなHLAタイプを有するヒトにより認識されるエピトープ類を含むことが明らかになった。CSPに基づくこれまでのワクチンの大きな欠点は、ある特定のCSP配列のエピトープ類に結合するHLAタイプの広さと細胞および体液性免疫応答の持続性であった。
【0011】
先行技術は、従って、安全で有効なマラリアワクチンをこれまで提供できず、特に、前記寄生体のほとんどの株に対して持続性の細胞媒介免疫応答を効果的に惹起するワクチンを提供してこなかった。
【課題を解決するための手段】
【0012】
本発明は、さまざまなPlasmodium・falciparum株から公知のI型HLAスーパータイプを有するヒト全ての非常に多くが認識するCSPのT細胞エピトープ類を発現するポリエピトープワクチンを設計し開発したことに基づいている。このワクチンは、世界中の感受性ヒト集団においてPlasmodium・falciparumのほとんど全ての株由来のCSPに対して有意な体液性免疫と細胞性免疫を惹起する。
【0013】
本発明は、新規の複数エピトープポリペプチドおよびそれらをコードする遺伝子配列をマラリアワクチンに使用するために提供する。前記ポリペプチド中における複数エピトープ類は、Plasmodium・falciparumスポロゾイト周囲タンパク質(CSP)に由来し、このタンパク質は、マラリアの肝段階において特に重要であることが公知である。エピトープ類をタンパク質の保存領域から選択する通常のワクチン設計に反して、本発明で使用したエピトープ類は、CD4およびCD8T細胞エピトープ類を含むことが公知のCSP高可変領域CD4およびCD8T細胞エピトープ類を含むことが公知のCSP高保存領域から選択した。さらに、選択したエピトープ類は、多くのヒト白血球抗原(HLA)群の全てにわたってヒトでT細胞免疫応答を惹起し、世界中のマラリア寄生体の非常に一般的な株類に典型的である。1態様において、複数エピトープポリペプチドは前記エピトープ間にスペーサーペプチド類を有するように設計することが重要で、それらは、正統的カルボキシ末端(C末)切断のほうに前記ポリペプチドのタンパク質分解を偏らせる。従って、タンパク質分解によりポリペプチドから放出されたペプチド類のアミノ酸配列は、通常は、選択したエピトープ類のアミノ酸配列に対して完全に適合したものとなっており、すなわち、前記スペーサー配列由来のアミノ酸類を含むペプチド類の産生が最小であるかまたは全く起こらない。前記エピトープ類がポリペプチド鎖に沿って直列となっている場合、スペーサーペプチド類を全く添加せず、前記エピトープ下流の“天然の”配列が、正統的カルボキシ末端切断部位を確保する。
【発明の効果】
【0014】
安全で有効なマラリアワクチンを提供する。
【発明を実施するための最良の形態】
【0015】
本発明は、マラリアCSP由来複数のエピトープを含む新規ポリペプチド類(ポリエピトープ類)および前記ポリペプチド類をコードする遺伝子配列を、マラリアワクチンで使用するために開発したことに基づいている。CSP由来エピトープ類は、このタンパク質がマラリアの肝段階で特に重要であることが公知であるので、選択した。P.falciparumのAsembo bay株由来CSPのタンパク質配列を図1に示した(配列番号14)。これまでの抗マラリアワクチンもなんらかの有効性を示したものの、これらのワクチンに対する免疫応答は主に体液性であり、持続期間および株特異性の両者においてかなり不完全である。対照的に、本ワクチンは、細胞媒介免疫性を惹起するように設計した。
【0016】
本発明で使用するためエピトープ類を選択するプロセスは、バイオインフォーマティックスおよびデータ解析の両者を必要とした。旧来のワクチン設計では通常、ある生物の多くの株にまでワクチンの適用性を広げようとして、タンパク質配列の保存領域のみをターゲットとしている。この通常のやり方と対照的に、本発明では、CSPの高可変領域を、選択した保存配列とともに含め、これは、正しいと認められておらず直感に反する手法である。
【0017】
前記ポリペプチドに含めるエピトープ類は、CD8細胞免疫応答を惹起することが公知であるかまたは推定されている。当業者は、CD8応答が細胞内寄生体と戦い撲滅に成功するために必要であること;体液性抗体応答が明らかに不十分であることを認識するであろう。さらに重要なこととして、選択したエピトープ類はヒト白血球抗原(HLA)群中でCD8免疫応答をまとめて惹起し、ヒト集団のほとんど(約98%)を網羅できる。従って、本ワクチンは、マラリア寄生体に暴露されそうな多くのヒトで効果的である。さらに、選択したエピトープ類は、マラリア寄生体の非常に一般的な株に典型的であり、この感染性物質の最も一般的な形態に対して広く保護する。
【0018】
本発明の複数エピトープポリペプチドは通常エピトープ間に“スペーサー”または“リンカー”ペプチド類を有するように設計されていることが重要で、それらは、正統的なカルボキシ末端(C末)切断のほうに前記ポリペプチドのタンパク質分解を偏らせる。“スペーサー”または“リンカー”ペプチド類とは、エピトープ下流の、少なくとも3個のアミノ酸の最小ペプチド配列を意味し、それには、ペプチド切断(タンパク質分解)のための部位が含まれ、前記ペプチド類が読み取り可能に連結されるようにする。すなわち、前記エピトープ類のC末タンパク質分解プロセッシングに干渉しないスペーサーペプチド類を選択し、宿主細胞でこのタンパク質が処理される(タンパク質分解される)時タンパク質分解でポリペプチドから放出されたペプチド類のカルボキシ末端はCSPから放出されるものと同一である。好適には、新しい偶然的カルボキシ末端がスペーサーペプチド内部での切断により全く(あるいはほとんど)産生されない。前記ペプチドエピトープ類のカルボキシ末端は真正である(すなわち、宿主細胞中におけるインビボCSP切断の結果生じるものと同一である)ので、複数エピトープポリペプチド由来ペプチド類は、HLAタンパク質類に結合しCD8細胞免疫応答を惹起すると考えられる。
【0019】
本発明での用途のために選択したエピトープ類を、表1に示した。
【0020】
【表1】

【0021】
この表からわかるとおり、最初の8種のエピトープ類は、CSP可変領域由来である。さらに、前記エピトープ類のうちの7種は、MHCスーパータイプB44に結合することが公知であるかまたは結合すると推定され、第8番目は、スーパータイプA26に結合する。これらのエピトープ類の選択とリンカー配列の詳細な説明は、実施例1と4に示されている。簡単に述べると、P.falciparumのAsembo bay株の配列288−412に対応する可変領域を、公知のT細胞エピトープ類の存在について分析した。アミノ酸22個の配列が上記エピトープ類を高濃度で含むことがわかり、それを用いて他のP.falciparum株由来のCSPタンパク質類配列を含むデータベータから高相同性配列を検索した。検索した配列と比較することで、全ての主要P.falciparum株由来の可変領域T細胞エピトープ類を代表する関連(すなわち高相同性)を有するが同一ではないペプチド配列を選択できた。
【0022】
本発明の実施において、本文に記載のエピトープ類をさまざまな異なる組み合わせで使用できる。例えば、本発明のいくつかの態様において、あるPlasmodiumCSP(通常、P.falciparumCSPであるが必ずしもそうではない)の可変領域から作成されたエピトープ類を構築体中で抗原性配列として単独で使用する。“CSP可変領域”の例は、P.falciparumのAsembo bay株の残基288乃至412(図1で下線を付した対応するCSPタンパク質の一次配列の部分であるが、他のPlasmodium種由来の相同性領域も使用できる。このようなワクチン構築体中において、前記可変領域由来の少なくとも1個のエピトープおよびある場合にはこの可変領域由来の1を超えるエピトープが含まれる。このようなエピトープは、通常は、長さがアミノ酸約5個乃至約15個であろうし、連続アミノ酸の配列を示すであろうし、T細胞エピトープであろう。もし可変領域由来の複数エピトープ類が含まれるならば、それらは隣接し連続する配列としてポリエピトープ構築体中に存在するか、または、それらは本文に述べたようにスペーサーまたはリンカー配列で分離できるか、または、配置を混ぜ合わせることも起こりえ、すなわち、いくつかの配列が隣接し、他が連結配列で分離されている。このタイプの例示的エピトープ類は、配列番号1−8として本文に記載の配列を有するものを含む。一般的に、このようなエピトープ類の長さは、アミノ酸少なくとも約5個から約25個であり、好適には、アミノ酸少なくとも約5個から約20個未満もしくはそれよりはるかに少ないアミノ酸(例 6,7,8,9,10,11,12,13,14,15,16,17,18、または19個のアミノ酸)である。
【0023】
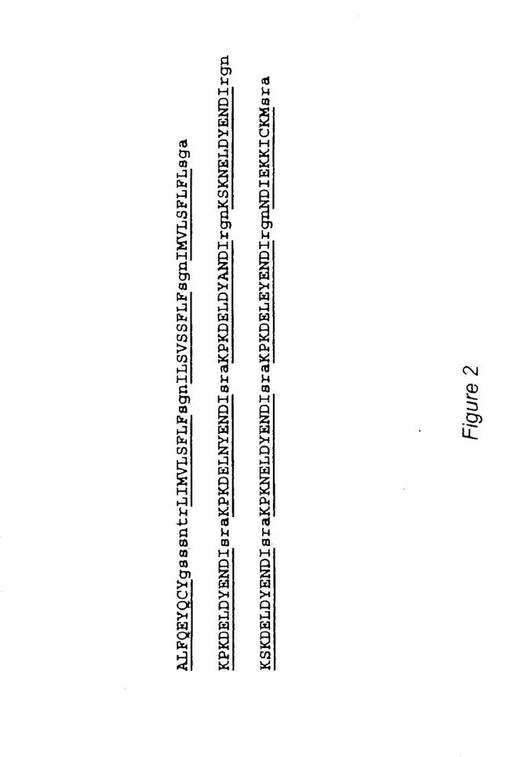
他の態様において、CSP保存領域由来の1個以上のT細胞エピトープ類(本明細書で、“保存CSP”と称することもある)もワクチン構築体中に含まれる。特に、表1の配列番号9−12で示したものなど保存T細胞エピトープ類はスーパータイプA1、A2,A3,A24,A26、B58およびB62への結合能を示し、これらも使用できる。このような保存エピトープ類の選択は、下記の実施例2で詳細に記載されている。簡単に述べると、バイオインフォーマティックスプログラムを用いて、複数(例 少なくとも3個)スーパータイプに結合し異なる(例 少なくとも60種の異なる株全てにわたる)P.falciparum株由来の複数CSP配列全てにわたって非常に高く保存された(例 100%保存)明確なT細胞エピトープ類を、選択した。従って、本発明のいくつかの態様において、可変領域由来の少なくとも1個のエピトープを、少なくとも1個の保存CSP T細胞エピトープと組み合わせてワクチン構築体中に含めることができる。1態様においては、表1のエピトープ類のそれぞれを単独ポリエピトープに含ませる。このようにして一緒に含ませると、本発明のエピトープ類は、ヒト集団の約98%に反応性である。このタイプの例示的ポリエピトープを図2に示した(配列番号13)。
【0024】
さらに、実施例3に詳細に示したように、さらなる抗原性配列を同定した。簡単に述べると、バイオインフォーマティックス手段を用いて、CSP配列内部で、12スーパータイプ全てに対する9,10、11量体T細胞エピトープ類を高濃度で有する配列クラスターを同定した。最大数のエピトープ類を有するクラスターであるクラスター1および4は、i)Asembo bay株のアミノ酸1−29(配列番号37)とii)Asembo bay株のアミノ酸385−411(配列番号38)をそれぞれ、包含している。これらの配列のいずれかまたは両者とも、本発明のポリエピトープ構築体中に含ませることができる。1態様においては、クラスター1および4は両者ともにポリエピトープ中に存在し、単独の連続配列(例 配列番号39;実施例3参照)として存在する。本文に記載の他のエピトープ類についてと同様に、これらの配列の単一コピーを構築体中で用いることができるし、または、これとは別に、例えば約2乃至約10以上のコピー(例えば、2,3,4,5,6,7,8,9、10,11,12またはそれ以上のコピー)のように多コピーを用いることもできる。さらに、配列番号37−39の配列の1個以上のコピーを、表1のエピトープ類の1個以上すなわち上記で述べた実施例1のCSP可変領域由来のエピトープ類および/または上記で述べた実施例2のCSP由来の1個以上の保存エピトープ類も含む構築体中で用いることができる。
【0025】
本発明の1態様においては、表1のエピトープ12種のそれぞれ少なくとも1個のコピーが、前記ポリエピトープ中に存在する。前記エピトープ12種のそれぞれ少なくとも1個のコピーを含むポリエピトープ例(配列番号13)を図2に示したが、それぞれのエピトープは大文字で記しかつアンダーラインを付した。当業者は、このタイプの構築体中において、前記ポリエピトープ内部のエピトープ類の他の立体構造も可能であることがわかるであろう。例えば、前記エピトープを異なる順序で並べることもできるし、1種以上のエピトープ類の多コピーが存在することもできる(例 1個乃至約10コピー以上が存在できる)し、および/またはクラスター1(配列番号37)及びクラスター4(配列番号38)またはその組み合わせ(配列番号39)のような追加の免疫原性配列も含ませることができる。一般的に、前記ポリエピトープ中のエピトープ類は、単一ポリペプチド鎖として単一プロモータから発現することもあり、もし必要ならば、(適宜)スペーサーまたはリンカー配列によって分離されている。しかし、いくつかの態様において、エピトープまたはエピトープ類グループは、別々のプロモータからすなわち別々のペプチド類またはポリペプチド類として発現できる。いくつかの態様において、(例 全エピトープ類およびエピトープ間のスペーサー配列を含む)ポリエピトープ類は、より大きなポリペプチド内部に含まれることもあり、すなわち、前記ポリエピトープがエピトープでもスペーサー配列でもないアミノ酸配列に隣接されることもある。このような隣接配列はいかなるタイプであってもよく、特定の機能を有することができ、例えば、所望の位置にポリペプチドの転座を向ける配列(例 シグナルペプチド類);問題の分子への結合を促進する配列;特定立体構造の採用を促進する配列;ポリペプチド局在化、同定または単離に有用な配列であり得る。
【0026】
いくつかの態様において、本発明のポリエピトープの各エピトープは、直接隣接しているかまたはスペーサーまたはリンカーペプチドにより隣接エピトープ類から分離されている。このスペーサーペプチド類の機能は、ポリエピトープの正確なカルボキシ末端切断を促進し(タンパク質分解)正統的エピトープ類を放出することである(すなわち、CSPタンパク質のインビボタンパク質分解により産生されたものと同じペプチド配列)。本文に示した例示的ポリエピトープ類中のスペーサー配列に加えて、当業者は、本発明の実施において使用できる他のスペーサーペプチド類も存在することを理解するであろう。当業者はまた、タンパク質分解部位の予測および/または優先的切断部位を含むポリペプチド類設計を可能とするデータベースおよび解析プログラムを知悉している。十分量の正確に処理されたエピトープ類を放出してこのエピトープ類に対する防御性免疫応答を惹起するため生成したポリペプチドが切断される限りにおいて、いかなるスペーサーおよびリンカー配列も用いることができる。
【0027】
1態様において、本発明は、Plasmodium falciparumのAsembo bay株のスポロゾイト周囲タンパク質(CSP)可変領域由来の少なくとも1個のエピトープを含む組換え抗原性ポリペプチドを提供し、そのアミノ酸配列を図1に示し、配列番号14として示した。その一次アミノ酸配列に基づき、このCSPタンパク質は分子量約44,361を有する。本発明によれば、CSP可変領域由来のエピトープを少なくとも1個含む組換え抗原性ポリペプチドは、CSP分子量の約30%以下の分子量、例えば、約13,500kDa以下、例えば約1000kDaから約12,000kDaを有する。
【0028】
例えば図2に示した例としてのポリエピトープのような本発明のポリエピトープに関して、当業者はその配列を改変でき、それでもなお免疫すべき宿主内部に適切なエピトープ類を提供する結果になることがわかるであろう。例えば、適切な免疫応答が惹起される限りにおいて、他のスペーサーペプチドを用いて保存アミノ酸を置換することも可能であろう。
【0029】
本発明は、本文に記載のポリエピトープ類およびそれらをコードする(例 DNA、RNA等)核酸配列を包含し、それらは、通常、発現制御配列に読み取り可能に結合されている。このような核酸配列は、例えば、弱毒化マイコバクテリウムまたは他の菌株、さまざまなウイルスベクター(例 弱毒化アデノウイルスベクター)、プラスミド類、または当業者が思い浮かぶ他の適切なベクター類のようなデリバリビーヒクルまたはベクター中に存在するDNA配列であり得る。免疫すべき宿主にベクターを投与した結果宿主内部でかつこのポリエピトープの正確なタンパク質分解プロセッシングを可能とする条件下で本発明のポリエピトープ産生が起こる限り、いかなるベクターも使用できる。これとは別に、本発明のいくつかの態様において、前記ポリエピトープは、適切な組成物で直接(すなわち、ポリペプチドとして)投与する。
【0030】
本発明は、マラリアに対する免疫応答惹起および/またはマラリアに対する個体のワクチン接種で使用するための組成物を提供する。前記組成物は、本文で記載の1種以上の実質的に精製したポリエピトープ類またはこのポリエピトープ類をコードする核酸配列および薬理学的に適切な担体を含む。ワクチンとしての用途のためのこの種の組成物の調製は、当業者に公知である。典型的には、このような組成物は、液体溶液または懸濁液のいずれかとして調製されるが、しかし、錠剤、丸剤、粉剤等のような固体形状もまた考慮される。液体中溶液または懸濁液に適した投与前固体形状もまた、調製できる。前記調製物もまた、乳化できる。前記活性成分を、製薬上許容できかつ活性成分類に適合する賦形剤と混合することもできる。適切な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノール等であるかまたはそれらの組み合わせである。さらに、前記組成物は、湿潤化または乳化剤、pH緩衝剤等のような助剤を少量含むことができる。さらに、前記組成物は、他のアジュバント類も含むことができる。もしこの組成物の経口形態を投与したいならば、さまざまな増粘剤、着香料、希釈剤、乳化剤、分散助剤または結合剤等を添加できる。本発明の組成物は、投与に適した形状で前記組成物を提供するため、追加のいかなる成分類を含むことができる。製剤中における最終的なポリエピトープまたはコードする核酸の量は、変えることができる。しかし、一般的に、製剤中の量は、約1−99%であろう。
【0031】
本発明はまた、マラリアに対する免疫応答を惹起する方法およびマラリアに対する哺乳類ワクチン接種方法を提供する。前記方法では、一般的に、適切なワクチン接種者を同定すること、薬理学的に許容できる担体中で本明細書に記載したポリエピトープまたはコードする核酸を含む組成物を前記接種者に投与することを含む。本発明のワクチン調製物は、当業者に周知の多くの適切な手段のいかなるものによっても投与でき、注射によって、経口的に、経鼻的に、抗原を含む食物製品摂取等を含むがそれらに限定されない。特定の態様においては、投与の様式は、皮下または筋肉内である。
【0032】
“免疫応答を惹起する”とは、本発明のワクチン調製物の投与が特異的抗体(1乃至1×106、好適には1×103の範囲の力価、より好適には約1×103乃至約1×106の範囲、および最も好適には1×106を超える力価)合成および/または細胞増殖の原因となり、それらは、IFN−γ産生を評価する細胞アッセイによりまたは3Hチミジン取り込み等により測定する。好適な態様において、免疫応答は防御的免疫応答であり、すなわち、免疫応答が、ワクチン接種を受けた個人がさらにマラリア寄生体に攻撃されるのを防御するが、これはいつもそうであるとは限らず、部分防御を提供する免疫応答がそれでもなお非常に有益であることもある。前記方法は、本発明の構築体を含む組成物を薬理学的に許容でき/適合性の担体中で投与することを必要とする。
【0033】
本発明のポリエピトープ中への取り込みのための適切なエピトープ類の選択と分析は、当業者が容易に入手可能なさまざまなデータベースおよびバイオインフォーマティックス解析手段を用いて、実施できる。例(例 細胞障害性Tリンパ球エピトープ推定および分析)としては、NetCTL,NetMHC,EpiJen、MAPPP、MHC−pathway、WAAP、Immune Epitope DatabaseおよびAnalysis Resource(IEDB)等が挙げられるが、それらに限定されない。
【0034】
(実施例)
上述の非限定的実施例は、本発明をさらに説明するために役立つ。
【実施例1】
【0035】
可変エピトープ類の選択:CSP可変領域の解析
【0036】
Plasmodium falciparum3D7由来のCSPタンパク質の可変領域288乃至412(図3中配列番号15)の分析を、IEDBデータベースによればT細胞エピトープ類である配列中で類似性および差を決定するために、行った。同定したT細胞エピトープ類を図3中で配列番号16−21として示した。この分析結果は、アミノ酸22個の配列(配列番号15の残基368−389を図3中でCSP配列内部に四角で囲んで示したが、それは、配列番号38を有する)が、この可変領域で同定したT細胞エピトープ類のかなりの部分を含んでいることを示しており、それを用いてNCBIデータベースを検索して前記アミノ酸ペプチド22個の配列(KPKDELDYANDIEKKICKMEKC 配列番号38)に高い同一性を示すアミノ酸配列を含むタンパク質を同定した。同一性の高いこれらの配列を、さらにIEDBにより解析した。結果を表2に示した。この表からわかるように、類似エピトープ類が、さまざまなP.falciparum単離物から配列10種中で同定され、それらは互いに高い同一性を示し、T細胞エピトープ類であると推定されたかまたは公知であった。
【0037】
【表2】
【0038】
これらの結果に基づき、前記アミノ酸22個の配列のアミノ末端部分を示すアミノ酸12個の配列7種および前記アミノ酸22個のカルボキシ末端近くの一部を示すアミノ酸9個の配列1種を選択して、本発明のポリエピトープへの導入を行った。これらの配列を上記表1に示した(配列番号1−8)。CTLエピトープ類の予測のための前記8種の配列の解析は、NetCTLバイオインフォーマティックスプログラムを用いて、実施した。このプログラムは、T細胞エピトープ類およびひとつのエピトープが結合するHLAスーパータイプ(類)を同定する。この解析は、これらの配列がB44およびA26MHCスーパータイプのHLAバインダーを含むことを明らかにした(表1参照)。
【実施例2】
【0039】
9量体の保存エピトープ類の選択
【0040】
Plasmodium falciparumのAsembo bay株由来CSP配列を、NetCTLバイオインフォーマティックスプログラムを用いて解析し、長さがアミノ酸9個のT細胞エピトープを同定した。結果を表3に示した。
【0041】
【表3】
【0042】
この表からわかるように、総計49種の重要エピトープ(組み合わせスコア>0.75)が同定された。これらの内、36種のみが明確なエピトープを示していた。24種のエピトープは、唯一のスーパータイプにしか結合しないと予測され、8種は2種のスーパータイプに結合すると予測され、4種は3種のスーパータイプに結合すると予測された。3種のスーパータイプ類に結合すると予測される4種のエピトープ類(下記の表4中配列番号9,10,11および12)を、Epitope Conservancy Analysis手段、Immune Epitope Database(IEDB)解析リソースを用いてさらに検討するため、選択した。IEDB手段を用いて、National Center for Biotechnology Information(NCBI)データベース由来の互いに異なる280種のCSP配列中エピトープ類の保存解析を実行した。結果を表4に示した。この表からわかるように、4種のエピトープ類全てが、69を超えるPF株由来CSP配列中で100%保存されている。
【0043】
【表4】
【0044】
配列番号9,10、11および12によって示される保存配列は、従って、ポリエピトープ中に導入するために選択した。
【実施例3】
【0045】
9−11量体エピトープの選択
【0046】
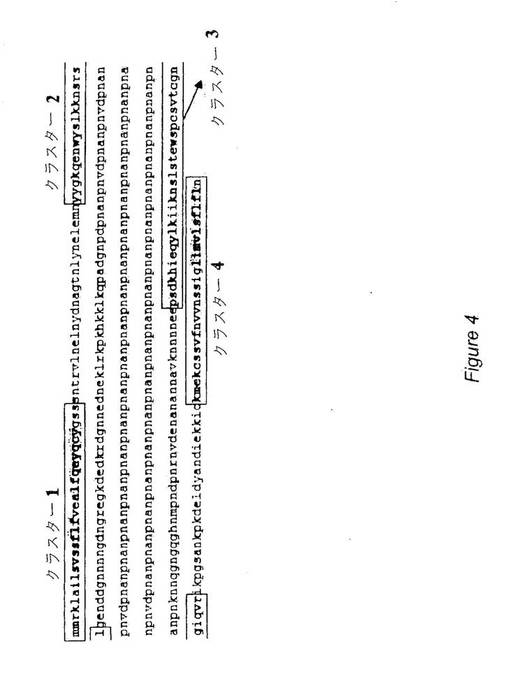
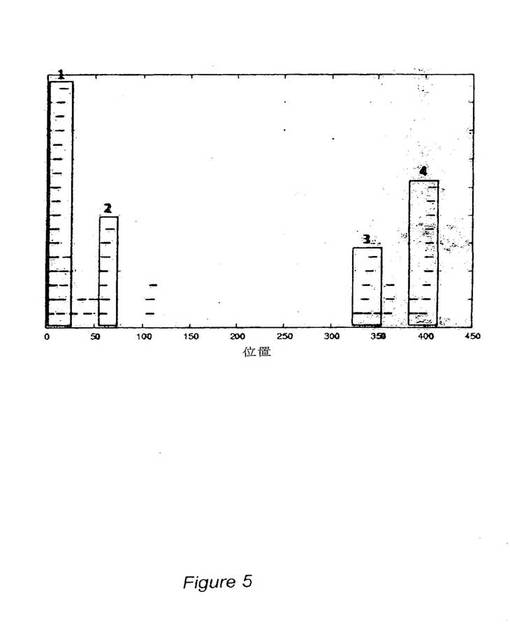
全ての可能なCTL結合エピトープを示すためおよびより長いエピトープ(すなわち、実施例2の9量体類よりも長い)を導入するために、広範囲のNetMHC3.0解析手段を用いて、Plasmodium falciparumのAsembo bay株由来CSP配列中の全てのスーパータイプ12種について、9,10,および11量体T細胞エピトープの同定に使用した。図1は、このCSP配列(配列番号14)を示す。前記12種のスーパータイプのいずれか一つを認識する推定上のT細胞エピトープ(9,10および11量体)の全てをグラフにマッピングし、CSPポリペプチドに沿ったそれらの分布を調べた。この種の解析は、4種の明確な“ホットスポット”(図4および5中の四角で囲んだ配列を参照)を明らかにし、CTLバインダーが最も密に分布していたのは、2種のクラスターであるクラスター1および4中であった。クラスター1は、アミノ酸1−29を包含し、クラスター4は、アミノ酸385−411を包含する。前記2種のクラスターは、上記実施例2で述べた9量体ペプチドについてのこれまでの戦略で選択した4種の“保存”エピトープを全て含んでいる。表5に示したように、総計で、4種の“ホットスポット”が広範囲の対立遺伝子を網羅している。
【0047】
【表5】
【0048】
従って、9,10および11量体CTl結合エピトープを含む好適な構築体は、クラスター1および4のひとつまたは両者のアミノ酸配列を含むであろう。クラスター1は、アミノ酸配列1−29(MMRKLAILSVSSFLFVEALFQEYQCYGSS、配列番号36)を含み、クラスター4は、アミノ酸配列385−412(KMEKCSSVFNVVNSSIGLIMVLSFLFLN,配列番号37)を含む。これら2種の配列を組み合わせると、結果として下記の配列:〔MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN〕(配列番号38)になるであろう。この組み合わせたペプチドは、多数のスーパータイプを示すCTL結合エピトープ類を高密度の集団として含む。本発明のポリエピトープは、先にも述べたように配列番号36,37または38の1種以上のコピーを含むことができる。図6は、配列番号38を含む例示的ポリエピトープ(配列番号39)を図示している。
【実施例4】
【0049】
前記9量体に基づくポリエピトープのスペーサー配列の選択
【0050】
ポリエピトープ構築のためのスペーサー配列の選択は、合理的な特別プロセスを適用することにより実行する。このプロセスにおいて、選択したエピトープ類の配列順序および最適切断に有利であろう短く特異的なアミノ酸配列(スペーサー類)の挿入の両者を、下記のように検討した。
【0051】
第一の反復において、全ての関連ペプチドを、最適タンパク質分解に有利なアミノ酸配列を含む短いスペーサーで集合させ、正統的選択ペプチド配列を作成する。前記ポリエピトープの一次配列(図7、配列番号40)をNetCTL解析に供し、スペーサー添加により作成されたペプチド総数を決定した。一次配列についてのこの解析結果により相対的に多数の新規エピトープを確認し、それらは、正統的選択ペプチドよりも優位であった(例 表6にも示したように、実施例において、B7スーパータイプを認識する新規エピトープ10種を作成)。次に、スペーサー領域でアミノ酸配列を他の特異的アミノ酸で置き換えることによって、前記プロセスを再度繰り返すことが必要であった。3−4回反復し、生成したポリエピトープの予測解析によって明らかになったように、このような潜在的な競合性新規エピトープの数を半分に減らした(表7)。最適化ポリエピトープの最終バージョンのアミノ酸配列を、図2に示した(配列番号13)。
【0052】
【表6】
【0053】
【表7】
【0054】
前記ポリエピトープは、8種のHLAスーパータイプに結合可能なエピトープ類を含む。本発明のポリエピトープ中42種のエピトープ類により提供される集団適用範囲(coverage)の徹底した解析を、前記8種のMHCスーパータイプにより示されるHLA対立遺伝子に対して、集団適用範囲アルゴリズム(IEDB)により実施した。この解析の結果、平均集団適用範囲は、97.74%であった。下記の具体的集団クラス中における適用範囲は、オーストラリアで97.45%、欧州で99.67%、北アフリカで99.18%、北東アジアで99.43%、南東アジアで99.81%、サハラ砂漠以南アフリカで98.81%と高い。南アメリカ(ベネズエラおよび一部のブラジル人集団)については、適用範囲が79.34%であり、他のブラジル人集団およびキューバについては、適用範囲は98.79%である。
【0055】
同様の操作を、配列番号38(クラスター1+クラスター4)のような他の配列をポリエピトープ中に含めた時に予測タンパク質分解を最適化しかつ新規の偽エピトープ類の産生を最小とするために、用いることができる。しかし、天然配列が存在しかつ切断部位として一般的に使用されるので、新規エピトープ類の産生は配列番号38の場合たいした問題ではない。
【実施例5】
【0056】
ポリエピトープ製造のための構築体設計と調製
【0057】
下記のセクションで述べるrBCG構築のため、制限エンドヌクレアーゼ(本文で“REs”)(New England Biolabs Beverly、MA)、T4DNAリガーゼ(New England Biolabs Beverly、MA)およびTaqポリメラーゼ(Invitrogen,Carlsbad,CA)を製造業者の指示に従い、用いた;プラスミドDNAは、小規模(Qiagen MiniprepRキット、Santa Clara、CA)または大規模(Qiagen MaxiprepRキット、Santa Clara、CA)プラスミドDNA精製キットを製造業者の指示(Qiagen、Santa Clara、CA)に従い用いて、調製した;ヌクレアーゼ非含有分子生物学等級のMilli−Q水、Tris−HCl(pH7.5)、EDTA pH8.0、1M MgCl−2、100%(v/v)エタノール、超純粋アガロース、およびアガロースゲル電気泳動緩衝液は、Invitrogen,Carlsbad、CAから購入した。RE消化、PCRs、DNA連結反応、およびアガロースゲル電気泳動は、周知の操作(Sambrookら、Molecular Cloning;A Laboratory Manual.1,2,3;1989)(Strausら、Proc Natl Acad Sci USA.Mar;87(5):1889−93;1990)に従い実施した。下記のセクションに述べた各組換えプラスミドのDNA配列を証明するためのヌクレオチド配列決定は、Applied Biosystems自動シークエンサーモデル373Aを用いる従来の自動化DNA配列決定技術により、達成した。
【0058】
PCRプライマーは、Sigma(St.Louis、MO)などの商業的供給業者から購入するかまたはApplied Biosystems DNAシンセサイザー(モデル373A)を用いて合成した。PCRプライマーは濃度150−250μMで使用し、PCR反応のためのアニーリング温度は、クローンマネジャーソフトウェアバージョン4.1(Scientific and Educational Software,Inc.、Durham,NC)を用いて決定した。PCRは、BioRadサーモサイクラー(BioRad,Hercules,CA)中で実施した。増幅用PCRプライマーは、Clone ManagerR(クローンマネジャー)ソフトウェアバージョン4.1(Scientific and Educational Software,Inc.、Durham,NC)を用いて、設計した。その後RE消化物およびPCRを、標準的操作を用いるアガロースゲル電気泳動(Strausら、同上1990;およびSambrookら、同上1989)により解析した。陽性クローンは、適切なREパターンおよび/またはPCRパターンを示すものとして定義する。この操作で同定したプラスミド類を、上記のように、標準的DNA配列決定操作を用いて評価した。
【0059】
DH5aおよびStable2RなどのEscherichia coli株は、Invitrogen(Carlsbad,CA)から購入し、前記組換えプラスミドの最初の宿主として役立った。組換えプラスミドを、上述のとおり(Strausら、同上1990)100−200Ο、15−25μFおよび1.0−2.5kVに設定したGene Pulser(BioRad Laboratories,Hercules,CA)などの高電圧電気パルス装置を用いるエレクトロポレーションによって、E.coli株中に導入した。最適なエレクトロポレーション条件は、megDNA/菌当たりの最大形質転換率が結果として得られる設定を決定することで、確認した。
【0060】
E.coli株は典型的にトリプトン大豆寒天(Difco,Detroit,MI)上でまたはトリプトン大豆培地(Difco,Detroit,MI)中で増殖するが、これは、製造業者の指示に従い作製した。他に断りがなければ、全ての菌は、軽く攪拌しながら5%(v/v)CO2中37℃で増殖させた。適切である時には、前記培地に、抗生物質(Sigma,St.Louis,MO)を添加した。菌株は、典型的に、1ml当たり約109コロニー形成単位(本文で“cfu”と記載)で30%(v/v)グリセロール(Sigma,St.Louis、MO)含有(Difco)中に懸濁させ−80℃で保存した。
【0061】
マイコバクテリウム株は、Middlebrook 7H9またはSaulton Synthetic Mediumのような液体培地中で、好適には37℃で培養した。前記株は、静止または攪拌培養物として維持できる。さらに、BCGの成長速度は、オレイン酸(0.06%v/v;Research Diagnostics Cat.No.01257)およびTyloxapol(0.05%v/v;Research Diagnostics Cat.No.70400)などの界面活性剤の添加により、増強できる。BCG培養物の純度は、リン酸緩衝生理食塩水(本文でPBSと称する)中で系列希釈(例 Neat−10−8から10倍ずつの段階)したBCG培養物のアリコット100mlをMiddlebrook 7H10のような固体培地25−30ml含有3.5インチプレート上に均等に伸ばすことで、評価できる。さらに、培養物純度は、チオグリコール酸培地(Science Lab,カタログ番号1891)および大豆−カゼイン培地(BD,カタログ番号211768)などの市販の培地を用いてさらに評価できる。
【0062】
BCG株AFV−102を発現するPfoA染色体からポリエピトープを発現させるため、前記スペーサー配列を含むポリエピトープをコードするヌクレオチド配列(配列番号13)を化学的に合成し、Mycobacterium bovisのhsp60プロモータおよびMycobacterium tuberculosis抗原85Bのシグナルペプチドに読み取り可能に連結させた。この配列を、BCG AFV−102へのエレクトロポレーションのためにプラスミドベクターpJFINTへ連結させ、AFV−102の染色体への組み込みを実行させた。このベクターは、E.coli colE1複製開始点、転写ターミネータrrnBT1、pEX18gmのT2およびrnhAにより分断された3個の複数クローニング部位、バクレリオファージL5のattPファージ組み込み領域およびバクテリオファージL5のインテグラーゼ遺伝子(GenBank #Z18946)を含んでいる。L5配列のすぐ上流には、Tn10由来カナマイシン耐性対立遺伝子aphA(GenBank#AAM97345)から構成される選択マーカーカセットおよびsacB遺伝子(GenBank#NP_391325)を含ませた。このマーカーカセットは、トランスポゾンTn1000からのγ△リゾルバーゼ結合部位の直接繰り返し単位が隣接していた。このプラスミドはマイコバクテリウム種内で複製ができず、L5 attP配列が、マイコバクテリウム染色体のattB領域との高頻度組換えを可能とし、前記染色体中へのプラスミド配列の組み込みを促進する。次に、γ△リゾルバーゼの導入と10%ショ糖含有固体培地上でのマーカーを含まない株の選択によって、前記マーカーカセットは、インテグラント染色体から除去できる。
【0063】
第2のヌクレオチド配列を化学的に合成し、体液性および細胞性応答を惹起するように設計した前記4個および7個のアミノ酸繰り返し単位の両者を含む改変CSPをコードする。このCSPコード配列は、GPIアンカー部位下流の免疫抑制配列であるシグナルペプチド配列を欠いており、CSPタンパク質の、より少ない数(18)のNANP繰り返し単位をコードする。この配列はまた、hsp60プロモータおよびAg85Bシグナルペプチドに読み取り可能に連結させた。この発現カセットを次に、ポリエピトープ発現配列も含むpJFINTに連結させた。
【0064】
生成したプラスミドを、E.coliStable2中で増殖させ、カナマイシン耐性クローンのプラスミド配列を確認した。このプラスミドを、100mlのE.coli培養物から単離し、BCG Danish 1331のpfo発現誘導体中にエレクトロポレーションした。エレクトロポレーション後、細胞を、10%(v/v)OADCおよび0.05%(v/v)Tyloxapol添加7H9培地中で一晩培養し、50μg/mlカナマイシン含有7H10寒天上に塗布した。プラスミドはマイコバクテリウム複製開始点をコードしないので、全ての試験したコロニー中のカナマイシン耐性は、BCGゲノムのattB部位へのプラスミド組み込みによって付与された。各コロニーをPCR分析のために取り出し、抗原発現分析のため10%(v/v)OADCおよび0.05%(v/v)Tyloxapol添加7H9培地中に接種した。カナマイシン耐性コロニーのPCR特性解析は、AERAS−CSPと命名した組換えBCGの染色体中全プラスミド配列の存在を明らかにした。AERAS−CSP培養物を7H9で洗浄し、これを用いてタンパク質非含有7H9Tyloxapol培養物を接種した。AERAS−CSP培養物上清を遠心分離で採取し、Plasmodium falciparum3D7(ATCC/MR4)のCSPタンパク質に対するウサギポリクローナル抗血清によりイムノブロッティングした。イムノブロッティングは、大きな改変CSPとポリエピトープの両者の存在を明らかにした。
【0065】
このワクチン構築を完成させヒトでの使用に適したものとするため、組み込まれたプラスミドのマーカーカセットを、次に除去した。エレクトロコンピタントAERAS−CSP細胞は、Tn1000のγ△リゾルバーゼをコードするプラスミドpYUB870hyg、sacB対立遺伝子およびハイグロマイシン耐性遺伝子(GenBank #ABD64366)によりエレクトロポレーションした。カナマイシンおよびハイグロマイシンの両者に耐性の形質転換体を7H10培地で選択し、10%(v/v)OADCおよび0.05%(v/v)Tyloxapolを有するが抗生物質を全く含まない7H9液体培地に接種した。7日間増殖させた後、これらの液体培養物の希釈物を10%ショ糖含有7H10に塗布し、aphA−sacBマーカーを切り出しかつpYUB870hygプラスミドを希釈とsacB対立遺伝子に対する選択により欠失した組換え体を選択した。
【0066】
ショ糖耐性形質転換体を、組み込み抗原カセット類のPCR分析のために取り出し、前のようにイムノブロット分析用7H9液体培地に接種した。PCR分析は、この抗原発現カセット類が前記染色体中にまだ存在していることおよびハイグロマイシン耐性マーカーおよびsacB遺伝子が切り出されたことを明らかにした。CSP抗血清による上清と細胞ペレットのイムノブロッティングは、前記カセットの除去がCSPポリエピトープまたは改変CSPの発現を行わないことを明らかにした。
【実施例6】
【0067】
上述のCSPポリエピトープを発現するrBCG AERAS−CSPによるヒトMHC対立遺伝子保有トランスジェニックマウスのワクチン接種により惹起された免疫性
【0068】
MHC I対立遺伝子欠失を有しかつポリエピトープ中でコードされたペプチド類に対応する前記8種の主要ヒトスーパータイプHLA対立遺伝子類A1,A2,A3,A24,A26,B44,B58およびB62のひとつを発現するトランスジェニックSJL/Jマウスを、Jackson Laboratories(Bar Harbor、Maine)から購入する。マウスを、32匹の4群に分け、各群に8HLAタイプのそれぞれについて4匹を含めるようにする。1−4群は、下記のようにワクチン接種する:1群には、皮下でPBS100μlを投与し、2群には、親BCG AERAS−401を皮下で5×105cfuを投与し、3群には、rBCG AERAS−CSPを皮下で5×105cfu投与し、4群には、皮下でAERAS−CSPを5×105cfu投与し、8週後、CMVプロモータ制御下でCSPポリエピトープをコードするDNAワクチンを筋肉内に100μg、投与する。
【0069】
ワクチン接種10週後マウス全部を屠殺し、各HLAタイプの各群から脾臓を採取しプールする。単細胞懸濁液は、この脾臓を70μmのセルストレーナー(BD Biosciences,San Jose,CA)で加圧ろ過することにより、調製する。脾臓細胞を、完全RPMI培地(R10;10%(v/v)ウシ胎児血清(FBS)含有RPMI−1640(Hyclone、Logan,UT))、55μM2−メルカプトエタノール、10mM HEPES,2mM L−グルタミンおよびペニシリン−ストレプトマイシン(Invitrogen、Carlsbad,CA)に再懸濁し、520×gで5分間、4℃で遠心分離する。脾臓1個につき室温で2分間、1mlのACK溶解緩衝液(BioWhittacker、Walkersville、MD)により赤血球を溶解させた後、細胞を洗浄しR10中に再懸濁させる。計数する前に70μmのセルストレーナーにより再度単細胞懸濁液をろ過し、細胞濃度を15×106細胞/mlに調整する。ポリエピトープ中に含ませたものに適合する配列を有するそれぞれのCSP由来ペプチドを、合成した(SynPep Corporation Dublin,CA)。前記ペプチドをR10培地で最終濃度それぞれ1μg/mlおよび5μg/mlに希釈する。サイトカイン発現分析のため、脾臓細胞を3重に2.5×105細胞/ウェルで96ウェル細胞培養プレートに塗布し、抗原とともにまたは抗原なしで72時間培養した。3日間刺激した後上清を採取し、IFN−γを、OptEIATMELISAキット(BD Biosciences,San Jose,CA)を用いるELISAによって製造業者の指示に従い決定する。
【0070】
CSPポリエピトープペプチド特異は、フローサイトメトリによりIFN−γについて細胞内サイトカイン染色(ICS)により特性解析する。簡単に述べると、上記のように調製した脾臓細胞を陰性対照としてジメチルスルホキシド(DMSO)(Sigma,St.Louis、MO)により刺激し、前記ポリエピトープ対応ペプチドプール(SynPep Corporation Dublin、CA)は、1μg/mlのaCD28およびaCD49d mAbs(BD Biosciences,San Jose,CA)含有R10培地で前希釈する。ホルボール−12−ミリステート−アセテート(0.1μg/ml)およびイオノマイシン(4μg/ml)(PMA/I)、(Sigma,St. Louise,MO))で処理した細胞は、陽性対照として使用した。細胞刺激のため、上記溶液100μlプラス各細胞懸濁液100μlを96ウェルの丸底培養プレート中に添加し、37℃で1時間、5%CO2で培養した。最終懸濁液中のペプチド濃度は1.2μM/ペプチドである。10μg/ml Golgi−Plug(BD Biosciences、San Jose、CA)添加後、プレートをさらに4−5時間インキュベーションする。インキュベーション後、プレートを細胞内サイトカイン染色のために処理する。細胞をPBSで洗浄し、1μlのFcR Block(BD Biosciences,San Jose、US)を含むPBF(PBS+0.5%FBS)50μl中に再懸濁させ、氷上で10分間インキュベーションする。細胞をその後、力価検定済みaCD8−PC5(BD Biosciences,San Jose、CA)抗体で染色する。Cytofix/Cytoperm緩衝液(BD Biosciences,San Jose、CA)を用いて、細胞の透過性を上げる。細胞内サイトカイン染色のため、aIFN−γ Alexa Fluor−488を用いる。インキュベーションおよび洗浄段階の後、細胞ペレットを、1%ホルムアルデヒド(Sigma,MO)を用いて固定した。サンプルを、CyFlow ML(Partec,Muenster,Germany)フローサイトメーターを用いて各サンプルから105標的細胞イベントを採取することによって、分析する。全てのデータ分析は、FlowJoソフトウェア(TreeStar Inc.、USA)により行い、刺激後のIFN−γ陽性CD8+T細胞の百分率は、DMSO単独で刺激した細胞の百分率に対して、計算した。
【0071】
親株のマウスはこれらのエピトープ類全てを認識しかつ提示するために必要なMHC分子類を欠失しているので、3群および4群のマウス由来脾臓細胞によるIFN−γ産生が1群および2群で検出されたバックグラウンドレベルを上回ることは、これらの動物が適切に処理され、AERAS−CSPによりコードされたポリエピトープを認識することおよび所望の免疫応答がワクチン接種により惹起されたことを示唆するであろう。
【実施例7】
【0072】
上述のCSPポリエピトープを発現するrBCG AERAS−CSPによるワクチン接種に対する免疫応答の評価
【0073】
実施例6の場合と同様に、前記ポリエピトープ中でコードされたペプチドに対応する前記8種の主要ヒトスーパータイプHLA対立遺伝子A1,A2,A3,A24,A26,B44,B58およびB62のひとつを発現するトランスジェニックSJL/Jマウスを、Jackson Laboratories(Bar Harbor、Maine)から購入する。マウスを、32匹ずつの4群に分け、各群に8HLAタイプのそれぞれについて4匹の動物を含めるようにする。1−4群は、下記のようにワクチン接種する:1群には、皮下でPBS100μlを投与し、2群には、親BCG AERAS−401を皮下で5×105cfuを投与し、3群には、rBCG AERAS−CSPを皮下で5×105cfu投与し、4群には、皮下でrBCG AERAS−CSPを5×105cfu投与し、8週後、CMVプロモータ制御下でCSPポリエピトープをコードするDNAワクチンを筋肉内に100μg投与する。
【0074】
ワクチン接種2週後マウス全部に少なくとも10回、Plasmodium falciparumCSP発現ハイブリッドPlasmodium bergheiスポロゾイドに感染したAnopheles duremi蚊に刺させることでチャレンジする。ハイブリッドスポロゾイトの構築およびワクチン奏効性研究におけるそれらの用途は、当業者に公知である。rBCG AERAS−CSPを投与した3群および4群のトランスジェニック動物ではマラリアが予防され、一方、1群および2群の動物は、貧血、臓器損傷およびP.bergheiマラリア関連脳病理を発症するであろう。
【実施例8】
【0075】
上述のCSPポリエピトープを発現するrBCG AERAS−CSPをワクチン接種した霊長類の防御
【0076】
アカゲザル10匹に対して、生理食塩水100μlを皮内に(4)、rBCG AERAS−401 5×105cfuを皮内(4)に、rBCG AERAS−CSP 5×105cfuを皮内(8)にワクチン接種する。ワクチン接種2週後、実験的に、動物全てに対して少なくとも100回、Plasmodium falciparumCSタンパク質発現ハイブリッドPlasmodium knowlesiマラリア寄生体類に感染したAnopheles hackeri蚊に刺させてチャレンジする。ハイブリッドスポロゾイトの構築およびワクチン奏効性研究におけるそれらの用途は、当業者に公知である。rBCG AERAS−CSPを投与した全動物はマラリアが予防され、一方、前記ポリエピトープワクチンを投与されなかった1群および2群の動物は、マラリアを発症し、防御されないであろう。
【0077】
本発明を好適な実施の態様について上に述べてきたが、当業者は、付属の請求の範囲の真意と範囲を逸脱することなく変形を加えて実施できることを理解するであろう。従って、本発明は、上述の実施の態様に限定されず、本文に示した明細書の真意と範囲に含まれるあらゆる変形やその均等物を含む。
【産業上の利用可能性】
【0078】
毎年世界で6億人以上が罹患し数百万人が死亡するマラリア感染症の征圧に利用できる。
【図面の簡単な説明】
【0079】
【図1】P.falciparumのAsembo Bay株由来CSPのタンパク質配列(配列番号14)。
【図2】例示的最終反復ポリエピトープのアミノ酸配列(配列番号13)を大略図示したもの。
【図3】CSPタンパク質の可変領域中T細胞エピトープ類。
【図4】前記12種のスーパータイプのいずれかを認識する推定9、10および11量体ペプチド類をCSPポリペプチドに沿ってマッピングしたもの。
【図5】“ホットスポット”クラスター類1−4のグラフ表示。
【図6】クラスター1およびクラスター4の両者の配列を含む例示的ポリエピトープのアミノ酸配列(配列番号39)。
【図7】例示的第一反復ポリエピトープのアミノ酸配列(配列番号40)。
【図8】図7に示した例示的第一反復ポリエピトープをコードする核酸配列(配列番号41)。
【技術分野】
【0001】
本発明は、概括的には、マラリアワクチンに関する。本発明は特に、広いスペクトルのPlasmodium falciparum寄生体種のスポロゾイド周囲タンパク質に対する細胞媒介免疫性を惹起しかつHLA対立遺伝子類の大半により認識されるポリエピトープ構築体に基づくマラリアワクチンを提供する。
【背景技術】
【0002】
マラリアは、アメリカ大陸、アジアおよびアフリカを含む熱帯、亜熱帯で広まっているベクター運搬感染症である。それは、毎年約6億5千万の人々に病気を起こし、百万ないし3百万人を死亡させ、それら人々の大半はサハラ砂漠以南のアフリカにおける幼い子供たちである。
【0003】
本疾患は、属Plasmodiumの原生動物寄生体が原因である。本疾患の最も重篤な病態は、Plasmodium falciparumおよびPlasmodium・vivaxにより惹き起こされるが、他の関連種〔Plasmodium・ovale、Plasmodium・malariae、および時にはPlasmodium・knowlesi〕もまたヒトに感染する。
【0004】
マラリア寄生体は、雌のAnopheles蚊に刺されることにより伝染し、ヒト宿主に対して前記寄生体のスポロゾイト体を運ぶ。前記スポロゾイト類は、血液により肝臓まで運ばれ、そこでそれらは肝細胞中で増殖し、ライフサイクルの次の形態すなわちメロゾイトに変化する。メロゾイト類は、感染肝細胞が破裂すると、最終的には血流中に放出される。次にメロゾイト類は赤血球に感染し、一部は、RBC中で雄および雌の生殖母細胞になる。別の蚊が感染宿主を刺すと、それは、雄および雌の生殖母細胞を摂取し、雌の蚊の内部で融合しスポロゾイトになる。この蚊が次に刺すと、このスポロゾイト類はさらに別の宿主に伝染等する。
【0005】
マラリア感染は、貧血に特徴的な症状(軽度頭痛、息切れ、頻脈等)やさらに発熱、悪寒、悪心、インフルエンザ様疾患のような他の一般的症状、および重篤な場合、昏睡および死亡の原因となる。マラリア伝染は、蚊帳および昆虫忌避剤により蚊に刺されないようにすることによって低下させることができるが、それらはかなり効果的ではあるものの、流通させかつ連続的に適切に使用することが必要であるという欠点を有している。他の対応策には家屋内に殺虫剤を撒くことおよび蚊が産卵している滞留水を排水することによって蚊を制御することが含まれる。残念ながら、殺虫剤の使用は環境上のリスクとなり、ある地方では、全ての滞留水を排水することは実質的に不可能である。
【0006】
Plasmodiumに自然感染した後、ヒト宿主は抗Plasmodium抗体を産生する。しかし、抗体産生だけで前記寄生体を中和する能力(体液性免疫)は、不断の、防御的免疫性としては持続しないし、あるいは十分な記憶特性を有する細胞性免疫も惹起されない。その結果として感染が何度も起こることがあり、疾患治療と予防を複雑化させている。
【0007】
効果的な抗マラリアワクチンを産生するためにいろいろ試みられてきた。Huiらの米国特許第6,660,498号は、ワクチン中で使用するために、バキュロウイルス系を用いて組換えメジャーメロゾイトサーフィスプロテイン(Major Merozoite Surface Protein)1を産生させることを記載している;Knappらの米国特許第5,393,523号はP.falciparumの組換えヒスチジン高含量タンパク質の調製とワクチンにおける用途を記載している;Patarroyoの米国特許第4,957,738号および同第4,735,799号は、後期段階のP.falciparumマラリアに対して抗体を誘発する合成ペプチド化合物混合物を記載している。Asakuraの米国特許第4,643,896号は、新規マラリア関連抗原CRAを開示し、それは、マラリアワクチンとして有用であると記載している。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】米国特許第6,660,498号
【特許文献2】米国特許第5,393,523号
【特許文献3】米国特許第4,957,738号
【特許文献4】米国特許第4,735,799号
【特許文献5】米国特許第4,643,896号
【発明の開示】
【発明が解決しようとする課題】
【0009】
過去の努力にもかかわらず、マラリアに対して有効なワクチンは残念なことに現在のところ全く得られていない。特に、マラリア寄生体類の全てまたはほとんどの株に対して細胞媒介免疫性を惹起するワクチンは、全く開発されていない。その代わりに、キニーネまたはアルテミシニン誘導体のような予防薬を連続的に服用し、感染リスクを低下させなければならない。これらの予防薬による治療は、しばしば、風土病となっている地方に住んでいる多くの人々にとってあまりにも高価である。さらに、前記寄生体の薬物耐性株が、ますます頻繁に見られるようになっている。
【0010】
前記スポロゾイト周囲タンパク質(本文でCSP)のタンパク質断片をコードするかまたはそれから構築したワクチンはヒトおよび動物モデルの両方で免疫原性およびやや限定された防御能を示している。実際的ではないが最初の成功したマラリアワクチンは、スポロゾイトを運んでいる照射蚊類から構成されていた。細胞免疫応答を惹起するように設計したアジュバント中でデリバリされるサブユニットワクチンは、ヒトにおいてCSPまたはアジュバント単独よりもより高い防御を提供したことは重要である。カルボキシおよびアミノ末端両者の可変領域がさまざまなHLAタイプを有するヒトにより認識されるエピトープ類を含むことが明らかになった。CSPに基づくこれまでのワクチンの大きな欠点は、ある特定のCSP配列のエピトープ類に結合するHLAタイプの広さと細胞および体液性免疫応答の持続性であった。
【0011】
先行技術は、従って、安全で有効なマラリアワクチンをこれまで提供できず、特に、前記寄生体のほとんどの株に対して持続性の細胞媒介免疫応答を効果的に惹起するワクチンを提供してこなかった。
【課題を解決するための手段】
【0012】
本発明は、さまざまなPlasmodium・falciparum株から公知のI型HLAスーパータイプを有するヒト全ての非常に多くが認識するCSPのT細胞エピトープ類を発現するポリエピトープワクチンを設計し開発したことに基づいている。このワクチンは、世界中の感受性ヒト集団においてPlasmodium・falciparumのほとんど全ての株由来のCSPに対して有意な体液性免疫と細胞性免疫を惹起する。
【0013】
本発明は、新規の複数エピトープポリペプチドおよびそれらをコードする遺伝子配列をマラリアワクチンに使用するために提供する。前記ポリペプチド中における複数エピトープ類は、Plasmodium・falciparumスポロゾイト周囲タンパク質(CSP)に由来し、このタンパク質は、マラリアの肝段階において特に重要であることが公知である。エピトープ類をタンパク質の保存領域から選択する通常のワクチン設計に反して、本発明で使用したエピトープ類は、CD4およびCD8T細胞エピトープ類を含むことが公知のCSP高可変領域CD4およびCD8T細胞エピトープ類を含むことが公知のCSP高保存領域から選択した。さらに、選択したエピトープ類は、多くのヒト白血球抗原(HLA)群の全てにわたってヒトでT細胞免疫応答を惹起し、世界中のマラリア寄生体の非常に一般的な株類に典型的である。1態様において、複数エピトープポリペプチドは前記エピトープ間にスペーサーペプチド類を有するように設計することが重要で、それらは、正統的カルボキシ末端(C末)切断のほうに前記ポリペプチドのタンパク質分解を偏らせる。従って、タンパク質分解によりポリペプチドから放出されたペプチド類のアミノ酸配列は、通常は、選択したエピトープ類のアミノ酸配列に対して完全に適合したものとなっており、すなわち、前記スペーサー配列由来のアミノ酸類を含むペプチド類の産生が最小であるかまたは全く起こらない。前記エピトープ類がポリペプチド鎖に沿って直列となっている場合、スペーサーペプチド類を全く添加せず、前記エピトープ下流の“天然の”配列が、正統的カルボキシ末端切断部位を確保する。
【発明の効果】
【0014】
安全で有効なマラリアワクチンを提供する。
【発明を実施するための最良の形態】
【0015】
本発明は、マラリアCSP由来複数のエピトープを含む新規ポリペプチド類(ポリエピトープ類)および前記ポリペプチド類をコードする遺伝子配列を、マラリアワクチンで使用するために開発したことに基づいている。CSP由来エピトープ類は、このタンパク質がマラリアの肝段階で特に重要であることが公知であるので、選択した。P.falciparumのAsembo bay株由来CSPのタンパク質配列を図1に示した(配列番号14)。これまでの抗マラリアワクチンもなんらかの有効性を示したものの、これらのワクチンに対する免疫応答は主に体液性であり、持続期間および株特異性の両者においてかなり不完全である。対照的に、本ワクチンは、細胞媒介免疫性を惹起するように設計した。
【0016】
本発明で使用するためエピトープ類を選択するプロセスは、バイオインフォーマティックスおよびデータ解析の両者を必要とした。旧来のワクチン設計では通常、ある生物の多くの株にまでワクチンの適用性を広げようとして、タンパク質配列の保存領域のみをターゲットとしている。この通常のやり方と対照的に、本発明では、CSPの高可変領域を、選択した保存配列とともに含め、これは、正しいと認められておらず直感に反する手法である。
【0017】
前記ポリペプチドに含めるエピトープ類は、CD8細胞免疫応答を惹起することが公知であるかまたは推定されている。当業者は、CD8応答が細胞内寄生体と戦い撲滅に成功するために必要であること;体液性抗体応答が明らかに不十分であることを認識するであろう。さらに重要なこととして、選択したエピトープ類はヒト白血球抗原(HLA)群中でCD8免疫応答をまとめて惹起し、ヒト集団のほとんど(約98%)を網羅できる。従って、本ワクチンは、マラリア寄生体に暴露されそうな多くのヒトで効果的である。さらに、選択したエピトープ類は、マラリア寄生体の非常に一般的な株に典型的であり、この感染性物質の最も一般的な形態に対して広く保護する。
【0018】
本発明の複数エピトープポリペプチドは通常エピトープ間に“スペーサー”または“リンカー”ペプチド類を有するように設計されていることが重要で、それらは、正統的なカルボキシ末端(C末)切断のほうに前記ポリペプチドのタンパク質分解を偏らせる。“スペーサー”または“リンカー”ペプチド類とは、エピトープ下流の、少なくとも3個のアミノ酸の最小ペプチド配列を意味し、それには、ペプチド切断(タンパク質分解)のための部位が含まれ、前記ペプチド類が読み取り可能に連結されるようにする。すなわち、前記エピトープ類のC末タンパク質分解プロセッシングに干渉しないスペーサーペプチド類を選択し、宿主細胞でこのタンパク質が処理される(タンパク質分解される)時タンパク質分解でポリペプチドから放出されたペプチド類のカルボキシ末端はCSPから放出されるものと同一である。好適には、新しい偶然的カルボキシ末端がスペーサーペプチド内部での切断により全く(あるいはほとんど)産生されない。前記ペプチドエピトープ類のカルボキシ末端は真正である(すなわち、宿主細胞中におけるインビボCSP切断の結果生じるものと同一である)ので、複数エピトープポリペプチド由来ペプチド類は、HLAタンパク質類に結合しCD8細胞免疫応答を惹起すると考えられる。
【0019】
本発明での用途のために選択したエピトープ類を、表1に示した。
【0020】
【表1】
【0021】
この表からわかるとおり、最初の8種のエピトープ類は、CSP可変領域由来である。さらに、前記エピトープ類のうちの7種は、MHCスーパータイプB44に結合することが公知であるかまたは結合すると推定され、第8番目は、スーパータイプA26に結合する。これらのエピトープ類の選択とリンカー配列の詳細な説明は、実施例1と4に示されている。簡単に述べると、P.falciparumのAsembo bay株の配列288−412に対応する可変領域を、公知のT細胞エピトープ類の存在について分析した。アミノ酸22個の配列が上記エピトープ類を高濃度で含むことがわかり、それを用いて他のP.falciparum株由来のCSPタンパク質類配列を含むデータベータから高相同性配列を検索した。検索した配列と比較することで、全ての主要P.falciparum株由来の可変領域T細胞エピトープ類を代表する関連(すなわち高相同性)を有するが同一ではないペプチド配列を選択できた。
【0022】
本発明の実施において、本文に記載のエピトープ類をさまざまな異なる組み合わせで使用できる。例えば、本発明のいくつかの態様において、あるPlasmodiumCSP(通常、P.falciparumCSPであるが必ずしもそうではない)の可変領域から作成されたエピトープ類を構築体中で抗原性配列として単独で使用する。“CSP可変領域”の例は、P.falciparumのAsembo bay株の残基288乃至412(図1で下線を付した対応するCSPタンパク質の一次配列の部分であるが、他のPlasmodium種由来の相同性領域も使用できる。このようなワクチン構築体中において、前記可変領域由来の少なくとも1個のエピトープおよびある場合にはこの可変領域由来の1を超えるエピトープが含まれる。このようなエピトープは、通常は、長さがアミノ酸約5個乃至約15個であろうし、連続アミノ酸の配列を示すであろうし、T細胞エピトープであろう。もし可変領域由来の複数エピトープ類が含まれるならば、それらは隣接し連続する配列としてポリエピトープ構築体中に存在するか、または、それらは本文に述べたようにスペーサーまたはリンカー配列で分離できるか、または、配置を混ぜ合わせることも起こりえ、すなわち、いくつかの配列が隣接し、他が連結配列で分離されている。このタイプの例示的エピトープ類は、配列番号1−8として本文に記載の配列を有するものを含む。一般的に、このようなエピトープ類の長さは、アミノ酸少なくとも約5個から約25個であり、好適には、アミノ酸少なくとも約5個から約20個未満もしくはそれよりはるかに少ないアミノ酸(例 6,7,8,9,10,11,12,13,14,15,16,17,18、または19個のアミノ酸)である。
【0023】
他の態様において、CSP保存領域由来の1個以上のT細胞エピトープ類(本明細書で、“保存CSP”と称することもある)もワクチン構築体中に含まれる。特に、表1の配列番号9−12で示したものなど保存T細胞エピトープ類はスーパータイプA1、A2,A3,A24,A26、B58およびB62への結合能を示し、これらも使用できる。このような保存エピトープ類の選択は、下記の実施例2で詳細に記載されている。簡単に述べると、バイオインフォーマティックスプログラムを用いて、複数(例 少なくとも3個)スーパータイプに結合し異なる(例 少なくとも60種の異なる株全てにわたる)P.falciparum株由来の複数CSP配列全てにわたって非常に高く保存された(例 100%保存)明確なT細胞エピトープ類を、選択した。従って、本発明のいくつかの態様において、可変領域由来の少なくとも1個のエピトープを、少なくとも1個の保存CSP T細胞エピトープと組み合わせてワクチン構築体中に含めることができる。1態様においては、表1のエピトープ類のそれぞれを単独ポリエピトープに含ませる。このようにして一緒に含ませると、本発明のエピトープ類は、ヒト集団の約98%に反応性である。このタイプの例示的ポリエピトープを図2に示した(配列番号13)。
【0024】
さらに、実施例3に詳細に示したように、さらなる抗原性配列を同定した。簡単に述べると、バイオインフォーマティックス手段を用いて、CSP配列内部で、12スーパータイプ全てに対する9,10、11量体T細胞エピトープ類を高濃度で有する配列クラスターを同定した。最大数のエピトープ類を有するクラスターであるクラスター1および4は、i)Asembo bay株のアミノ酸1−29(配列番号37)とii)Asembo bay株のアミノ酸385−411(配列番号38)をそれぞれ、包含している。これらの配列のいずれかまたは両者とも、本発明のポリエピトープ構築体中に含ませることができる。1態様においては、クラスター1および4は両者ともにポリエピトープ中に存在し、単独の連続配列(例 配列番号39;実施例3参照)として存在する。本文に記載の他のエピトープ類についてと同様に、これらの配列の単一コピーを構築体中で用いることができるし、または、これとは別に、例えば約2乃至約10以上のコピー(例えば、2,3,4,5,6,7,8,9、10,11,12またはそれ以上のコピー)のように多コピーを用いることもできる。さらに、配列番号37−39の配列の1個以上のコピーを、表1のエピトープ類の1個以上すなわち上記で述べた実施例1のCSP可変領域由来のエピトープ類および/または上記で述べた実施例2のCSP由来の1個以上の保存エピトープ類も含む構築体中で用いることができる。
【0025】
本発明の1態様においては、表1のエピトープ12種のそれぞれ少なくとも1個のコピーが、前記ポリエピトープ中に存在する。前記エピトープ12種のそれぞれ少なくとも1個のコピーを含むポリエピトープ例(配列番号13)を図2に示したが、それぞれのエピトープは大文字で記しかつアンダーラインを付した。当業者は、このタイプの構築体中において、前記ポリエピトープ内部のエピトープ類の他の立体構造も可能であることがわかるであろう。例えば、前記エピトープを異なる順序で並べることもできるし、1種以上のエピトープ類の多コピーが存在することもできる(例 1個乃至約10コピー以上が存在できる)し、および/またはクラスター1(配列番号37)及びクラスター4(配列番号38)またはその組み合わせ(配列番号39)のような追加の免疫原性配列も含ませることができる。一般的に、前記ポリエピトープ中のエピトープ類は、単一ポリペプチド鎖として単一プロモータから発現することもあり、もし必要ならば、(適宜)スペーサーまたはリンカー配列によって分離されている。しかし、いくつかの態様において、エピトープまたはエピトープ類グループは、別々のプロモータからすなわち別々のペプチド類またはポリペプチド類として発現できる。いくつかの態様において、(例 全エピトープ類およびエピトープ間のスペーサー配列を含む)ポリエピトープ類は、より大きなポリペプチド内部に含まれることもあり、すなわち、前記ポリエピトープがエピトープでもスペーサー配列でもないアミノ酸配列に隣接されることもある。このような隣接配列はいかなるタイプであってもよく、特定の機能を有することができ、例えば、所望の位置にポリペプチドの転座を向ける配列(例 シグナルペプチド類);問題の分子への結合を促進する配列;特定立体構造の採用を促進する配列;ポリペプチド局在化、同定または単離に有用な配列であり得る。
【0026】
いくつかの態様において、本発明のポリエピトープの各エピトープは、直接隣接しているかまたはスペーサーまたはリンカーペプチドにより隣接エピトープ類から分離されている。このスペーサーペプチド類の機能は、ポリエピトープの正確なカルボキシ末端切断を促進し(タンパク質分解)正統的エピトープ類を放出することである(すなわち、CSPタンパク質のインビボタンパク質分解により産生されたものと同じペプチド配列)。本文に示した例示的ポリエピトープ類中のスペーサー配列に加えて、当業者は、本発明の実施において使用できる他のスペーサーペプチド類も存在することを理解するであろう。当業者はまた、タンパク質分解部位の予測および/または優先的切断部位を含むポリペプチド類設計を可能とするデータベースおよび解析プログラムを知悉している。十分量の正確に処理されたエピトープ類を放出してこのエピトープ類に対する防御性免疫応答を惹起するため生成したポリペプチドが切断される限りにおいて、いかなるスペーサーおよびリンカー配列も用いることができる。
【0027】
1態様において、本発明は、Plasmodium falciparumのAsembo bay株のスポロゾイト周囲タンパク質(CSP)可変領域由来の少なくとも1個のエピトープを含む組換え抗原性ポリペプチドを提供し、そのアミノ酸配列を図1に示し、配列番号14として示した。その一次アミノ酸配列に基づき、このCSPタンパク質は分子量約44,361を有する。本発明によれば、CSP可変領域由来のエピトープを少なくとも1個含む組換え抗原性ポリペプチドは、CSP分子量の約30%以下の分子量、例えば、約13,500kDa以下、例えば約1000kDaから約12,000kDaを有する。
【0028】
例えば図2に示した例としてのポリエピトープのような本発明のポリエピトープに関して、当業者はその配列を改変でき、それでもなお免疫すべき宿主内部に適切なエピトープ類を提供する結果になることがわかるであろう。例えば、適切な免疫応答が惹起される限りにおいて、他のスペーサーペプチドを用いて保存アミノ酸を置換することも可能であろう。
【0029】
本発明は、本文に記載のポリエピトープ類およびそれらをコードする(例 DNA、RNA等)核酸配列を包含し、それらは、通常、発現制御配列に読み取り可能に結合されている。このような核酸配列は、例えば、弱毒化マイコバクテリウムまたは他の菌株、さまざまなウイルスベクター(例 弱毒化アデノウイルスベクター)、プラスミド類、または当業者が思い浮かぶ他の適切なベクター類のようなデリバリビーヒクルまたはベクター中に存在するDNA配列であり得る。免疫すべき宿主にベクターを投与した結果宿主内部でかつこのポリエピトープの正確なタンパク質分解プロセッシングを可能とする条件下で本発明のポリエピトープ産生が起こる限り、いかなるベクターも使用できる。これとは別に、本発明のいくつかの態様において、前記ポリエピトープは、適切な組成物で直接(すなわち、ポリペプチドとして)投与する。
【0030】
本発明は、マラリアに対する免疫応答惹起および/またはマラリアに対する個体のワクチン接種で使用するための組成物を提供する。前記組成物は、本文で記載の1種以上の実質的に精製したポリエピトープ類またはこのポリエピトープ類をコードする核酸配列および薬理学的に適切な担体を含む。ワクチンとしての用途のためのこの種の組成物の調製は、当業者に公知である。典型的には、このような組成物は、液体溶液または懸濁液のいずれかとして調製されるが、しかし、錠剤、丸剤、粉剤等のような固体形状もまた考慮される。液体中溶液または懸濁液に適した投与前固体形状もまた、調製できる。前記調製物もまた、乳化できる。前記活性成分を、製薬上許容できかつ活性成分類に適合する賦形剤と混合することもできる。適切な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノール等であるかまたはそれらの組み合わせである。さらに、前記組成物は、湿潤化または乳化剤、pH緩衝剤等のような助剤を少量含むことができる。さらに、前記組成物は、他のアジュバント類も含むことができる。もしこの組成物の経口形態を投与したいならば、さまざまな増粘剤、着香料、希釈剤、乳化剤、分散助剤または結合剤等を添加できる。本発明の組成物は、投与に適した形状で前記組成物を提供するため、追加のいかなる成分類を含むことができる。製剤中における最終的なポリエピトープまたはコードする核酸の量は、変えることができる。しかし、一般的に、製剤中の量は、約1−99%であろう。
【0031】
本発明はまた、マラリアに対する免疫応答を惹起する方法およびマラリアに対する哺乳類ワクチン接種方法を提供する。前記方法では、一般的に、適切なワクチン接種者を同定すること、薬理学的に許容できる担体中で本明細書に記載したポリエピトープまたはコードする核酸を含む組成物を前記接種者に投与することを含む。本発明のワクチン調製物は、当業者に周知の多くの適切な手段のいかなるものによっても投与でき、注射によって、経口的に、経鼻的に、抗原を含む食物製品摂取等を含むがそれらに限定されない。特定の態様においては、投与の様式は、皮下または筋肉内である。
【0032】
“免疫応答を惹起する”とは、本発明のワクチン調製物の投与が特異的抗体(1乃至1×106、好適には1×103の範囲の力価、より好適には約1×103乃至約1×106の範囲、および最も好適には1×106を超える力価)合成および/または細胞増殖の原因となり、それらは、IFN−γ産生を評価する細胞アッセイによりまたは3Hチミジン取り込み等により測定する。好適な態様において、免疫応答は防御的免疫応答であり、すなわち、免疫応答が、ワクチン接種を受けた個人がさらにマラリア寄生体に攻撃されるのを防御するが、これはいつもそうであるとは限らず、部分防御を提供する免疫応答がそれでもなお非常に有益であることもある。前記方法は、本発明の構築体を含む組成物を薬理学的に許容でき/適合性の担体中で投与することを必要とする。
【0033】
本発明のポリエピトープ中への取り込みのための適切なエピトープ類の選択と分析は、当業者が容易に入手可能なさまざまなデータベースおよびバイオインフォーマティックス解析手段を用いて、実施できる。例(例 細胞障害性Tリンパ球エピトープ推定および分析)としては、NetCTL,NetMHC,EpiJen、MAPPP、MHC−pathway、WAAP、Immune Epitope DatabaseおよびAnalysis Resource(IEDB)等が挙げられるが、それらに限定されない。
【0034】
(実施例)
上述の非限定的実施例は、本発明をさらに説明するために役立つ。
【実施例1】
【0035】
可変エピトープ類の選択:CSP可変領域の解析
【0036】
Plasmodium falciparum3D7由来のCSPタンパク質の可変領域288乃至412(図3中配列番号15)の分析を、IEDBデータベースによればT細胞エピトープ類である配列中で類似性および差を決定するために、行った。同定したT細胞エピトープ類を図3中で配列番号16−21として示した。この分析結果は、アミノ酸22個の配列(配列番号15の残基368−389を図3中でCSP配列内部に四角で囲んで示したが、それは、配列番号38を有する)が、この可変領域で同定したT細胞エピトープ類のかなりの部分を含んでいることを示しており、それを用いてNCBIデータベースを検索して前記アミノ酸ペプチド22個の配列(KPKDELDYANDIEKKICKMEKC 配列番号38)に高い同一性を示すアミノ酸配列を含むタンパク質を同定した。同一性の高いこれらの配列を、さらにIEDBにより解析した。結果を表2に示した。この表からわかるように、類似エピトープ類が、さまざまなP.falciparum単離物から配列10種中で同定され、それらは互いに高い同一性を示し、T細胞エピトープ類であると推定されたかまたは公知であった。
【0037】
【表2】
【0038】
これらの結果に基づき、前記アミノ酸22個の配列のアミノ末端部分を示すアミノ酸12個の配列7種および前記アミノ酸22個のカルボキシ末端近くの一部を示すアミノ酸9個の配列1種を選択して、本発明のポリエピトープへの導入を行った。これらの配列を上記表1に示した(配列番号1−8)。CTLエピトープ類の予測のための前記8種の配列の解析は、NetCTLバイオインフォーマティックスプログラムを用いて、実施した。このプログラムは、T細胞エピトープ類およびひとつのエピトープが結合するHLAスーパータイプ(類)を同定する。この解析は、これらの配列がB44およびA26MHCスーパータイプのHLAバインダーを含むことを明らかにした(表1参照)。
【実施例2】
【0039】
9量体の保存エピトープ類の選択
【0040】
Plasmodium falciparumのAsembo bay株由来CSP配列を、NetCTLバイオインフォーマティックスプログラムを用いて解析し、長さがアミノ酸9個のT細胞エピトープを同定した。結果を表3に示した。
【0041】
【表3】
【0042】
この表からわかるように、総計49種の重要エピトープ(組み合わせスコア>0.75)が同定された。これらの内、36種のみが明確なエピトープを示していた。24種のエピトープは、唯一のスーパータイプにしか結合しないと予測され、8種は2種のスーパータイプに結合すると予測され、4種は3種のスーパータイプに結合すると予測された。3種のスーパータイプ類に結合すると予測される4種のエピトープ類(下記の表4中配列番号9,10,11および12)を、Epitope Conservancy Analysis手段、Immune Epitope Database(IEDB)解析リソースを用いてさらに検討するため、選択した。IEDB手段を用いて、National Center for Biotechnology Information(NCBI)データベース由来の互いに異なる280種のCSP配列中エピトープ類の保存解析を実行した。結果を表4に示した。この表からわかるように、4種のエピトープ類全てが、69を超えるPF株由来CSP配列中で100%保存されている。
【0043】
【表4】
【0044】
配列番号9,10、11および12によって示される保存配列は、従って、ポリエピトープ中に導入するために選択した。
【実施例3】
【0045】
9−11量体エピトープの選択
【0046】
全ての可能なCTL結合エピトープを示すためおよびより長いエピトープ(すなわち、実施例2の9量体類よりも長い)を導入するために、広範囲のNetMHC3.0解析手段を用いて、Plasmodium falciparumのAsembo bay株由来CSP配列中の全てのスーパータイプ12種について、9,10,および11量体T細胞エピトープの同定に使用した。図1は、このCSP配列(配列番号14)を示す。前記12種のスーパータイプのいずれか一つを認識する推定上のT細胞エピトープ(9,10および11量体)の全てをグラフにマッピングし、CSPポリペプチドに沿ったそれらの分布を調べた。この種の解析は、4種の明確な“ホットスポット”(図4および5中の四角で囲んだ配列を参照)を明らかにし、CTLバインダーが最も密に分布していたのは、2種のクラスターであるクラスター1および4中であった。クラスター1は、アミノ酸1−29を包含し、クラスター4は、アミノ酸385−411を包含する。前記2種のクラスターは、上記実施例2で述べた9量体ペプチドについてのこれまでの戦略で選択した4種の“保存”エピトープを全て含んでいる。表5に示したように、総計で、4種の“ホットスポット”が広範囲の対立遺伝子を網羅している。
【0047】
【表5】
【0048】
従って、9,10および11量体CTl結合エピトープを含む好適な構築体は、クラスター1および4のひとつまたは両者のアミノ酸配列を含むであろう。クラスター1は、アミノ酸配列1−29(MMRKLAILSVSSFLFVEALFQEYQCYGSS、配列番号36)を含み、クラスター4は、アミノ酸配列385−412(KMEKCSSVFNVVNSSIGLIMVLSFLFLN,配列番号37)を含む。これら2種の配列を組み合わせると、結果として下記の配列:〔MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN〕(配列番号38)になるであろう。この組み合わせたペプチドは、多数のスーパータイプを示すCTL結合エピトープ類を高密度の集団として含む。本発明のポリエピトープは、先にも述べたように配列番号36,37または38の1種以上のコピーを含むことができる。図6は、配列番号38を含む例示的ポリエピトープ(配列番号39)を図示している。
【実施例4】
【0049】
前記9量体に基づくポリエピトープのスペーサー配列の選択
【0050】
ポリエピトープ構築のためのスペーサー配列の選択は、合理的な特別プロセスを適用することにより実行する。このプロセスにおいて、選択したエピトープ類の配列順序および最適切断に有利であろう短く特異的なアミノ酸配列(スペーサー類)の挿入の両者を、下記のように検討した。
【0051】
第一の反復において、全ての関連ペプチドを、最適タンパク質分解に有利なアミノ酸配列を含む短いスペーサーで集合させ、正統的選択ペプチド配列を作成する。前記ポリエピトープの一次配列(図7、配列番号40)をNetCTL解析に供し、スペーサー添加により作成されたペプチド総数を決定した。一次配列についてのこの解析結果により相対的に多数の新規エピトープを確認し、それらは、正統的選択ペプチドよりも優位であった(例 表6にも示したように、実施例において、B7スーパータイプを認識する新規エピトープ10種を作成)。次に、スペーサー領域でアミノ酸配列を他の特異的アミノ酸で置き換えることによって、前記プロセスを再度繰り返すことが必要であった。3−4回反復し、生成したポリエピトープの予測解析によって明らかになったように、このような潜在的な競合性新規エピトープの数を半分に減らした(表7)。最適化ポリエピトープの最終バージョンのアミノ酸配列を、図2に示した(配列番号13)。
【0052】
【表6】
【0053】
【表7】
【0054】
前記ポリエピトープは、8種のHLAスーパータイプに結合可能なエピトープ類を含む。本発明のポリエピトープ中42種のエピトープ類により提供される集団適用範囲(coverage)の徹底した解析を、前記8種のMHCスーパータイプにより示されるHLA対立遺伝子に対して、集団適用範囲アルゴリズム(IEDB)により実施した。この解析の結果、平均集団適用範囲は、97.74%であった。下記の具体的集団クラス中における適用範囲は、オーストラリアで97.45%、欧州で99.67%、北アフリカで99.18%、北東アジアで99.43%、南東アジアで99.81%、サハラ砂漠以南アフリカで98.81%と高い。南アメリカ(ベネズエラおよび一部のブラジル人集団)については、適用範囲が79.34%であり、他のブラジル人集団およびキューバについては、適用範囲は98.79%である。
【0055】
同様の操作を、配列番号38(クラスター1+クラスター4)のような他の配列をポリエピトープ中に含めた時に予測タンパク質分解を最適化しかつ新規の偽エピトープ類の産生を最小とするために、用いることができる。しかし、天然配列が存在しかつ切断部位として一般的に使用されるので、新規エピトープ類の産生は配列番号38の場合たいした問題ではない。
【実施例5】
【0056】
ポリエピトープ製造のための構築体設計と調製
【0057】
下記のセクションで述べるrBCG構築のため、制限エンドヌクレアーゼ(本文で“REs”)(New England Biolabs Beverly、MA)、T4DNAリガーゼ(New England Biolabs Beverly、MA)およびTaqポリメラーゼ(Invitrogen,Carlsbad,CA)を製造業者の指示に従い、用いた;プラスミドDNAは、小規模(Qiagen MiniprepRキット、Santa Clara、CA)または大規模(Qiagen MaxiprepRキット、Santa Clara、CA)プラスミドDNA精製キットを製造業者の指示(Qiagen、Santa Clara、CA)に従い用いて、調製した;ヌクレアーゼ非含有分子生物学等級のMilli−Q水、Tris−HCl(pH7.5)、EDTA pH8.0、1M MgCl−2、100%(v/v)エタノール、超純粋アガロース、およびアガロースゲル電気泳動緩衝液は、Invitrogen,Carlsbad、CAから購入した。RE消化、PCRs、DNA連結反応、およびアガロースゲル電気泳動は、周知の操作(Sambrookら、Molecular Cloning;A Laboratory Manual.1,2,3;1989)(Strausら、Proc Natl Acad Sci USA.Mar;87(5):1889−93;1990)に従い実施した。下記のセクションに述べた各組換えプラスミドのDNA配列を証明するためのヌクレオチド配列決定は、Applied Biosystems自動シークエンサーモデル373Aを用いる従来の自動化DNA配列決定技術により、達成した。
【0058】
PCRプライマーは、Sigma(St.Louis、MO)などの商業的供給業者から購入するかまたはApplied Biosystems DNAシンセサイザー(モデル373A)を用いて合成した。PCRプライマーは濃度150−250μMで使用し、PCR反応のためのアニーリング温度は、クローンマネジャーソフトウェアバージョン4.1(Scientific and Educational Software,Inc.、Durham,NC)を用いて決定した。PCRは、BioRadサーモサイクラー(BioRad,Hercules,CA)中で実施した。増幅用PCRプライマーは、Clone ManagerR(クローンマネジャー)ソフトウェアバージョン4.1(Scientific and Educational Software,Inc.、Durham,NC)を用いて、設計した。その後RE消化物およびPCRを、標準的操作を用いるアガロースゲル電気泳動(Strausら、同上1990;およびSambrookら、同上1989)により解析した。陽性クローンは、適切なREパターンおよび/またはPCRパターンを示すものとして定義する。この操作で同定したプラスミド類を、上記のように、標準的DNA配列決定操作を用いて評価した。
【0059】
DH5aおよびStable2RなどのEscherichia coli株は、Invitrogen(Carlsbad,CA)から購入し、前記組換えプラスミドの最初の宿主として役立った。組換えプラスミドを、上述のとおり(Strausら、同上1990)100−200Ο、15−25μFおよび1.0−2.5kVに設定したGene Pulser(BioRad Laboratories,Hercules,CA)などの高電圧電気パルス装置を用いるエレクトロポレーションによって、E.coli株中に導入した。最適なエレクトロポレーション条件は、megDNA/菌当たりの最大形質転換率が結果として得られる設定を決定することで、確認した。
【0060】
E.coli株は典型的にトリプトン大豆寒天(Difco,Detroit,MI)上でまたはトリプトン大豆培地(Difco,Detroit,MI)中で増殖するが、これは、製造業者の指示に従い作製した。他に断りがなければ、全ての菌は、軽く攪拌しながら5%(v/v)CO2中37℃で増殖させた。適切である時には、前記培地に、抗生物質(Sigma,St.Louis,MO)を添加した。菌株は、典型的に、1ml当たり約109コロニー形成単位(本文で“cfu”と記載)で30%(v/v)グリセロール(Sigma,St.Louis、MO)含有(Difco)中に懸濁させ−80℃で保存した。
【0061】
マイコバクテリウム株は、Middlebrook 7H9またはSaulton Synthetic Mediumのような液体培地中で、好適には37℃で培養した。前記株は、静止または攪拌培養物として維持できる。さらに、BCGの成長速度は、オレイン酸(0.06%v/v;Research Diagnostics Cat.No.01257)およびTyloxapol(0.05%v/v;Research Diagnostics Cat.No.70400)などの界面活性剤の添加により、増強できる。BCG培養物の純度は、リン酸緩衝生理食塩水(本文でPBSと称する)中で系列希釈(例 Neat−10−8から10倍ずつの段階)したBCG培養物のアリコット100mlをMiddlebrook 7H10のような固体培地25−30ml含有3.5インチプレート上に均等に伸ばすことで、評価できる。さらに、培養物純度は、チオグリコール酸培地(Science Lab,カタログ番号1891)および大豆−カゼイン培地(BD,カタログ番号211768)などの市販の培地を用いてさらに評価できる。
【0062】
BCG株AFV−102を発現するPfoA染色体からポリエピトープを発現させるため、前記スペーサー配列を含むポリエピトープをコードするヌクレオチド配列(配列番号13)を化学的に合成し、Mycobacterium bovisのhsp60プロモータおよびMycobacterium tuberculosis抗原85Bのシグナルペプチドに読み取り可能に連結させた。この配列を、BCG AFV−102へのエレクトロポレーションのためにプラスミドベクターpJFINTへ連結させ、AFV−102の染色体への組み込みを実行させた。このベクターは、E.coli colE1複製開始点、転写ターミネータrrnBT1、pEX18gmのT2およびrnhAにより分断された3個の複数クローニング部位、バクレリオファージL5のattPファージ組み込み領域およびバクテリオファージL5のインテグラーゼ遺伝子(GenBank #Z18946)を含んでいる。L5配列のすぐ上流には、Tn10由来カナマイシン耐性対立遺伝子aphA(GenBank#AAM97345)から構成される選択マーカーカセットおよびsacB遺伝子(GenBank#NP_391325)を含ませた。このマーカーカセットは、トランスポゾンTn1000からのγ△リゾルバーゼ結合部位の直接繰り返し単位が隣接していた。このプラスミドはマイコバクテリウム種内で複製ができず、L5 attP配列が、マイコバクテリウム染色体のattB領域との高頻度組換えを可能とし、前記染色体中へのプラスミド配列の組み込みを促進する。次に、γ△リゾルバーゼの導入と10%ショ糖含有固体培地上でのマーカーを含まない株の選択によって、前記マーカーカセットは、インテグラント染色体から除去できる。
【0063】
第2のヌクレオチド配列を化学的に合成し、体液性および細胞性応答を惹起するように設計した前記4個および7個のアミノ酸繰り返し単位の両者を含む改変CSPをコードする。このCSPコード配列は、GPIアンカー部位下流の免疫抑制配列であるシグナルペプチド配列を欠いており、CSPタンパク質の、より少ない数(18)のNANP繰り返し単位をコードする。この配列はまた、hsp60プロモータおよびAg85Bシグナルペプチドに読み取り可能に連結させた。この発現カセットを次に、ポリエピトープ発現配列も含むpJFINTに連結させた。
【0064】
生成したプラスミドを、E.coliStable2中で増殖させ、カナマイシン耐性クローンのプラスミド配列を確認した。このプラスミドを、100mlのE.coli培養物から単離し、BCG Danish 1331のpfo発現誘導体中にエレクトロポレーションした。エレクトロポレーション後、細胞を、10%(v/v)OADCおよび0.05%(v/v)Tyloxapol添加7H9培地中で一晩培養し、50μg/mlカナマイシン含有7H10寒天上に塗布した。プラスミドはマイコバクテリウム複製開始点をコードしないので、全ての試験したコロニー中のカナマイシン耐性は、BCGゲノムのattB部位へのプラスミド組み込みによって付与された。各コロニーをPCR分析のために取り出し、抗原発現分析のため10%(v/v)OADCおよび0.05%(v/v)Tyloxapol添加7H9培地中に接種した。カナマイシン耐性コロニーのPCR特性解析は、AERAS−CSPと命名した組換えBCGの染色体中全プラスミド配列の存在を明らかにした。AERAS−CSP培養物を7H9で洗浄し、これを用いてタンパク質非含有7H9Tyloxapol培養物を接種した。AERAS−CSP培養物上清を遠心分離で採取し、Plasmodium falciparum3D7(ATCC/MR4)のCSPタンパク質に対するウサギポリクローナル抗血清によりイムノブロッティングした。イムノブロッティングは、大きな改変CSPとポリエピトープの両者の存在を明らかにした。
【0065】
このワクチン構築を完成させヒトでの使用に適したものとするため、組み込まれたプラスミドのマーカーカセットを、次に除去した。エレクトロコンピタントAERAS−CSP細胞は、Tn1000のγ△リゾルバーゼをコードするプラスミドpYUB870hyg、sacB対立遺伝子およびハイグロマイシン耐性遺伝子(GenBank #ABD64366)によりエレクトロポレーションした。カナマイシンおよびハイグロマイシンの両者に耐性の形質転換体を7H10培地で選択し、10%(v/v)OADCおよび0.05%(v/v)Tyloxapolを有するが抗生物質を全く含まない7H9液体培地に接種した。7日間増殖させた後、これらの液体培養物の希釈物を10%ショ糖含有7H10に塗布し、aphA−sacBマーカーを切り出しかつpYUB870hygプラスミドを希釈とsacB対立遺伝子に対する選択により欠失した組換え体を選択した。
【0066】
ショ糖耐性形質転換体を、組み込み抗原カセット類のPCR分析のために取り出し、前のようにイムノブロット分析用7H9液体培地に接種した。PCR分析は、この抗原発現カセット類が前記染色体中にまだ存在していることおよびハイグロマイシン耐性マーカーおよびsacB遺伝子が切り出されたことを明らかにした。CSP抗血清による上清と細胞ペレットのイムノブロッティングは、前記カセットの除去がCSPポリエピトープまたは改変CSPの発現を行わないことを明らかにした。
【実施例6】
【0067】
上述のCSPポリエピトープを発現するrBCG AERAS−CSPによるヒトMHC対立遺伝子保有トランスジェニックマウスのワクチン接種により惹起された免疫性
【0068】
MHC I対立遺伝子欠失を有しかつポリエピトープ中でコードされたペプチド類に対応する前記8種の主要ヒトスーパータイプHLA対立遺伝子類A1,A2,A3,A24,A26,B44,B58およびB62のひとつを発現するトランスジェニックSJL/Jマウスを、Jackson Laboratories(Bar Harbor、Maine)から購入する。マウスを、32匹の4群に分け、各群に8HLAタイプのそれぞれについて4匹を含めるようにする。1−4群は、下記のようにワクチン接種する:1群には、皮下でPBS100μlを投与し、2群には、親BCG AERAS−401を皮下で5×105cfuを投与し、3群には、rBCG AERAS−CSPを皮下で5×105cfu投与し、4群には、皮下でAERAS−CSPを5×105cfu投与し、8週後、CMVプロモータ制御下でCSPポリエピトープをコードするDNAワクチンを筋肉内に100μg、投与する。
【0069】
ワクチン接種10週後マウス全部を屠殺し、各HLAタイプの各群から脾臓を採取しプールする。単細胞懸濁液は、この脾臓を70μmのセルストレーナー(BD Biosciences,San Jose,CA)で加圧ろ過することにより、調製する。脾臓細胞を、完全RPMI培地(R10;10%(v/v)ウシ胎児血清(FBS)含有RPMI−1640(Hyclone、Logan,UT))、55μM2−メルカプトエタノール、10mM HEPES,2mM L−グルタミンおよびペニシリン−ストレプトマイシン(Invitrogen、Carlsbad,CA)に再懸濁し、520×gで5分間、4℃で遠心分離する。脾臓1個につき室温で2分間、1mlのACK溶解緩衝液(BioWhittacker、Walkersville、MD)により赤血球を溶解させた後、細胞を洗浄しR10中に再懸濁させる。計数する前に70μmのセルストレーナーにより再度単細胞懸濁液をろ過し、細胞濃度を15×106細胞/mlに調整する。ポリエピトープ中に含ませたものに適合する配列を有するそれぞれのCSP由来ペプチドを、合成した(SynPep Corporation Dublin,CA)。前記ペプチドをR10培地で最終濃度それぞれ1μg/mlおよび5μg/mlに希釈する。サイトカイン発現分析のため、脾臓細胞を3重に2.5×105細胞/ウェルで96ウェル細胞培養プレートに塗布し、抗原とともにまたは抗原なしで72時間培養した。3日間刺激した後上清を採取し、IFN−γを、OptEIATMELISAキット(BD Biosciences,San Jose,CA)を用いるELISAによって製造業者の指示に従い決定する。
【0070】
CSPポリエピトープペプチド特異は、フローサイトメトリによりIFN−γについて細胞内サイトカイン染色(ICS)により特性解析する。簡単に述べると、上記のように調製した脾臓細胞を陰性対照としてジメチルスルホキシド(DMSO)(Sigma,St.Louis、MO)により刺激し、前記ポリエピトープ対応ペプチドプール(SynPep Corporation Dublin、CA)は、1μg/mlのaCD28およびaCD49d mAbs(BD Biosciences,San Jose,CA)含有R10培地で前希釈する。ホルボール−12−ミリステート−アセテート(0.1μg/ml)およびイオノマイシン(4μg/ml)(PMA/I)、(Sigma,St. Louise,MO))で処理した細胞は、陽性対照として使用した。細胞刺激のため、上記溶液100μlプラス各細胞懸濁液100μlを96ウェルの丸底培養プレート中に添加し、37℃で1時間、5%CO2で培養した。最終懸濁液中のペプチド濃度は1.2μM/ペプチドである。10μg/ml Golgi−Plug(BD Biosciences、San Jose、CA)添加後、プレートをさらに4−5時間インキュベーションする。インキュベーション後、プレートを細胞内サイトカイン染色のために処理する。細胞をPBSで洗浄し、1μlのFcR Block(BD Biosciences,San Jose、US)を含むPBF(PBS+0.5%FBS)50μl中に再懸濁させ、氷上で10分間インキュベーションする。細胞をその後、力価検定済みaCD8−PC5(BD Biosciences,San Jose、CA)抗体で染色する。Cytofix/Cytoperm緩衝液(BD Biosciences,San Jose、CA)を用いて、細胞の透過性を上げる。細胞内サイトカイン染色のため、aIFN−γ Alexa Fluor−488を用いる。インキュベーションおよび洗浄段階の後、細胞ペレットを、1%ホルムアルデヒド(Sigma,MO)を用いて固定した。サンプルを、CyFlow ML(Partec,Muenster,Germany)フローサイトメーターを用いて各サンプルから105標的細胞イベントを採取することによって、分析する。全てのデータ分析は、FlowJoソフトウェア(TreeStar Inc.、USA)により行い、刺激後のIFN−γ陽性CD8+T細胞の百分率は、DMSO単独で刺激した細胞の百分率に対して、計算した。
【0071】
親株のマウスはこれらのエピトープ類全てを認識しかつ提示するために必要なMHC分子類を欠失しているので、3群および4群のマウス由来脾臓細胞によるIFN−γ産生が1群および2群で検出されたバックグラウンドレベルを上回ることは、これらの動物が適切に処理され、AERAS−CSPによりコードされたポリエピトープを認識することおよび所望の免疫応答がワクチン接種により惹起されたことを示唆するであろう。
【実施例7】
【0072】
上述のCSPポリエピトープを発現するrBCG AERAS−CSPによるワクチン接種に対する免疫応答の評価
【0073】
実施例6の場合と同様に、前記ポリエピトープ中でコードされたペプチドに対応する前記8種の主要ヒトスーパータイプHLA対立遺伝子A1,A2,A3,A24,A26,B44,B58およびB62のひとつを発現するトランスジェニックSJL/Jマウスを、Jackson Laboratories(Bar Harbor、Maine)から購入する。マウスを、32匹ずつの4群に分け、各群に8HLAタイプのそれぞれについて4匹の動物を含めるようにする。1−4群は、下記のようにワクチン接種する:1群には、皮下でPBS100μlを投与し、2群には、親BCG AERAS−401を皮下で5×105cfuを投与し、3群には、rBCG AERAS−CSPを皮下で5×105cfu投与し、4群には、皮下でrBCG AERAS−CSPを5×105cfu投与し、8週後、CMVプロモータ制御下でCSPポリエピトープをコードするDNAワクチンを筋肉内に100μg投与する。
【0074】
ワクチン接種2週後マウス全部に少なくとも10回、Plasmodium falciparumCSP発現ハイブリッドPlasmodium bergheiスポロゾイドに感染したAnopheles duremi蚊に刺させることでチャレンジする。ハイブリッドスポロゾイトの構築およびワクチン奏効性研究におけるそれらの用途は、当業者に公知である。rBCG AERAS−CSPを投与した3群および4群のトランスジェニック動物ではマラリアが予防され、一方、1群および2群の動物は、貧血、臓器損傷およびP.bergheiマラリア関連脳病理を発症するであろう。
【実施例8】
【0075】
上述のCSPポリエピトープを発現するrBCG AERAS−CSPをワクチン接種した霊長類の防御
【0076】
アカゲザル10匹に対して、生理食塩水100μlを皮内に(4)、rBCG AERAS−401 5×105cfuを皮内(4)に、rBCG AERAS−CSP 5×105cfuを皮内(8)にワクチン接種する。ワクチン接種2週後、実験的に、動物全てに対して少なくとも100回、Plasmodium falciparumCSタンパク質発現ハイブリッドPlasmodium knowlesiマラリア寄生体類に感染したAnopheles hackeri蚊に刺させてチャレンジする。ハイブリッドスポロゾイトの構築およびワクチン奏効性研究におけるそれらの用途は、当業者に公知である。rBCG AERAS−CSPを投与した全動物はマラリアが予防され、一方、前記ポリエピトープワクチンを投与されなかった1群および2群の動物は、マラリアを発症し、防御されないであろう。
【0077】
本発明を好適な実施の態様について上に述べてきたが、当業者は、付属の請求の範囲の真意と範囲を逸脱することなく変形を加えて実施できることを理解するであろう。従って、本発明は、上述の実施の態様に限定されず、本文に示した明細書の真意と範囲に含まれるあらゆる変形やその均等物を含む。
【産業上の利用可能性】
【0078】
毎年世界で6億人以上が罹患し数百万人が死亡するマラリア感染症の征圧に利用できる。
【図面の簡単な説明】
【0079】
【図1】P.falciparumのAsembo Bay株由来CSPのタンパク質配列(配列番号14)。
【図2】例示的最終反復ポリエピトープのアミノ酸配列(配列番号13)を大略図示したもの。
【図3】CSPタンパク質の可変領域中T細胞エピトープ類。
【図4】前記12種のスーパータイプのいずれかを認識する推定9、10および11量体ペプチド類をCSPポリペプチドに沿ってマッピングしたもの。
【図5】“ホットスポット”クラスター類1−4のグラフ表示。
【図6】クラスター1およびクラスター4の両者の配列を含む例示的ポリエピトープのアミノ酸配列(配列番号39)。
【図7】例示的第一反復ポリエピトープのアミノ酸配列(配列番号40)。
【図8】図7に示した例示的第一反復ポリエピトープをコードする核酸配列(配列番号41)。
【特許請求の範囲】
【請求項1】
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);
NDIEKKICKM(配列番号8);
ILSVSSFLF(配列番号9);
ALFQEYQCY(配列番号10);
LIMVLSFLF(配列番号11);および
IMVLSFLFL(配列番号12);
から構成される群から選択された2種以上のアミノ酸配列を含む組換え抗原性ポリペプチドであって、
前記2種以上のアミノ酸配列ペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする組換え抗原性ポリペプチド。
【請求項2】
i) KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、および
KPKDELEYEMDI(配列番号7);
から構成される群から選択されたアミノ酸配列を有する3種以上のペプチド;
ii) アミノ酸配列NDIEKKICKM(配列番号8)のペプチド;
iii) アミノ酸配列ILSVSSFLF(配列番号9)のペプチド;
iv) アミノ酸配列ALFQEYQCY(配列番号10)のペプチド;および
v) LIMVLSFLF(配列番号11)およびIMVLSFLFL(配列番号12)から構成される群から選択されるアミノ酸配列を有する1種以上のペプチド;
を含む組換え抗原性ポリペプチドであって、
前記抗原性ポリペプチドにおけるペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする組換え抗原性ポリペプチド。
【請求項3】
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);
NDIEKKICKM(配列番号8);
ILSVSSFLF(配列番号9);
ALFQEYQCY(配列番号10);
LIMVLSFLF(配列番号11);および
IMVLSFLFL(配列番号12);
から構成される群から選択された2種以上のアミノ酸配列を含む抗原性ポリペプチドをコードする核酸であって、
前記2種以上のアミノ酸配列ペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする抗原性ポリペプチドをコードする核酸。
【請求項4】
i) KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、および
KPKDELEYEMDI(配列番号7);
から構成される群から選択されたアミノ酸配列を有する3種以上のペプチド;
ii) アミノ酸配列NDIEKKICKM(配列番号8)のペプチド;
iii) アミノ酸配列ILSVSSFLF(配列番号9)のペプチド;
iv) アミノ酸配列ALFQEYQCY(配列番号10)のペプチド;および
v) LIMVLSFLF(配列番号11)およびIMVLSFLFL(配列番号12)から構成される群から選択されるアミノ酸配列を有する1種以上のペプチド類;
を含む抗原性ポリペプチドをコードする核酸であって、
前記抗原性ポリペプチドにおけるペプチド類が、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする抗原性ポリペプチドをコードする核酸。
【請求項5】
マラリアに対して個体を免疫する方法であって、
前記個体に対して
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);
NDIEKKICKM(配列番号8);
ILSVSSFLF(配列番号9);
ALFQEYQCY(配列番号10);
LIMVLSFLF(配列番号11);および
IMVLSFLFL(配列番号12);
から構成される群から選択された2種以上のアミノ酸配列を含む組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸を投与する過程を含み、
前記2種以上のアミノ酸配列ペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とし、前記組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸が前記個体を免疫するために十分な量で投与されることを特徴とする免疫する方法。
【請求項6】
マラリアに対して個体を免疫する方法であって、
前記個体に対して
i) KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、および
KPKDELEYEMDI(配列番号7);
から構成される群から選択されたアミノ酸配列を有する3種以上のペプチド;
ii) アミノ酸配列NDIEKKICKM(配列番号8)のペプチド;
iii) アミノ酸配列ILSVSSFLF(配列番号9)のペプチド;
iv) アミノ酸配列ALFQEYQCY(配列番号10)のペプチド;および
v) LIMVLSFLF(配列番号11)およびIMVLSFLFL(配列番号12)から構成される群から選択されるアミノ酸配列を有する1種以上のペプチド;
を含む組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸を投与する過程を含み、
前記抗原性ポリペプチド中のペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることと、前記組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸が前記個体を免疫するために十分な量で投与されることとを特徴とする免疫する方法。
【請求項7】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーを含む組換え抗原性ポリペプチド。
【請求項8】
前記アミノ酸配列の多コピーが存在するとともに、i)互いに直接隣接して配置されているかまたはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする請求項7記載の組換え抗原性ポリペプチド。
【請求項9】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーを含む抗原性ポリペプチドをコードする核酸。
【請求項10】
前記アミノ酸配列の多コピーが存在するとともに、i)互いに直接隣接して配置されているかまたはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする請求項9記載の核酸。
【請求項11】
マラリアに対して個体を免疫する方法であって、
前記個体に対してアミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーを含む組み換え抗原性ポリペプチドを投与する過程を含むことを特徴とする免疫する方法。
【請求項12】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項1記載の組換え抗原性ポリペプチド。
【請求項13】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項2記載の組換え抗原性ポリペプチド。
【請求項14】
前記核酸が、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらにコードする請求項3記載の核酸。
【請求項15】
前記核酸が、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらにコードする請求項4記載の核酸。
【請求項16】
前記組換え抗原性ポリペプチドが、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項5記載の方法。
【請求項17】
前記組換え抗原性ポリペプチドが、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項6記載の方法。
【請求項18】
組換え抗原性ポリペプチドが分子量13,500kDa以下を有し、Plasmodium falciparumのAsembo bay株のスポロゾイト周囲タンパク質(CSP)可変領域由来のエピトープを少なくとも1個含む組換え抗原性ポリペプチド。
【請求項19】
前記少なくとも1個のCSP可変領域由来のエピトープがT細胞エピトープである請求項18記載の組換え抗原性ポリペプチド。
【請求項20】
前記少なくとも1個のCSP可変領域由来のエピトープが
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);および
NDIEKKICKM(配列番号8)
から構成される群から選択した配列番号で表されるアミノ酸配列を有することを特徴とする請求項18記載の組換え抗原性ポリペプチド。
【請求項21】
前記組換え抗原性ポリペプチドが、1個以上の保存CSPエピトープをさらに含む請求項18記載の組換え抗原性ポリペプチド。
【請求項22】
前記1個以上の保存CSPエピトープが、配列番号9、配列番号10、配列番号11および配列番号12から構成される群から選択した配列番号で表されるアミノ酸配列を有する請求項21記載の組換え抗原性ポリペプチド。
【請求項23】
前記組換え抗原性ポリペプチドが、配列番号36、配列番号37、および配列番号38から構成される群から選択した1個以上のエピトープをさらに含む請求項18記載の組換え抗原性ポリペプチド。
【請求項1】
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);
NDIEKKICKM(配列番号8);
ILSVSSFLF(配列番号9);
ALFQEYQCY(配列番号10);
LIMVLSFLF(配列番号11);および
IMVLSFLFL(配列番号12);
から構成される群から選択された2種以上のアミノ酸配列を含む組換え抗原性ポリペプチドであって、
前記2種以上のアミノ酸配列ペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする組換え抗原性ポリペプチド。
【請求項2】
i) KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、および
KPKDELEYEMDI(配列番号7);
から構成される群から選択されたアミノ酸配列を有する3種以上のペプチド;
ii) アミノ酸配列NDIEKKICKM(配列番号8)のペプチド;
iii) アミノ酸配列ILSVSSFLF(配列番号9)のペプチド;
iv) アミノ酸配列ALFQEYQCY(配列番号10)のペプチド;および
v) LIMVLSFLF(配列番号11)およびIMVLSFLFL(配列番号12)から構成される群から選択されるアミノ酸配列を有する1種以上のペプチド;
を含む組換え抗原性ポリペプチドであって、
前記抗原性ポリペプチドにおけるペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする組換え抗原性ポリペプチド。
【請求項3】
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);
NDIEKKICKM(配列番号8);
ILSVSSFLF(配列番号9);
ALFQEYQCY(配列番号10);
LIMVLSFLF(配列番号11);および
IMVLSFLFL(配列番号12);
から構成される群から選択された2種以上のアミノ酸配列を含む抗原性ポリペプチドをコードする核酸であって、
前記2種以上のアミノ酸配列ペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする抗原性ポリペプチドをコードする核酸。
【請求項4】
i) KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、および
KPKDELEYEMDI(配列番号7);
から構成される群から選択されたアミノ酸配列を有する3種以上のペプチド;
ii) アミノ酸配列NDIEKKICKM(配列番号8)のペプチド;
iii) アミノ酸配列ILSVSSFLF(配列番号9)のペプチド;
iv) アミノ酸配列ALFQEYQCY(配列番号10)のペプチド;および
v) LIMVLSFLF(配列番号11)およびIMVLSFLFL(配列番号12)から構成される群から選択されるアミノ酸配列を有する1種以上のペプチド類;
を含む抗原性ポリペプチドをコードする核酸であって、
前記抗原性ポリペプチドにおけるペプチド類が、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする抗原性ポリペプチドをコードする核酸。
【請求項5】
マラリアに対して個体を免疫する方法であって、
前記個体に対して
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);
NDIEKKICKM(配列番号8);
ILSVSSFLF(配列番号9);
ALFQEYQCY(配列番号10);
LIMVLSFLF(配列番号11);および
IMVLSFLFL(配列番号12);
から構成される群から選択された2種以上のアミノ酸配列を含む組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸を投与する過程を含み、
前記2種以上のアミノ酸配列ペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とし、前記組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸が前記個体を免疫するために十分な量で投与されることを特徴とする免疫する方法。
【請求項6】
マラリアに対して個体を免疫する方法であって、
前記個体に対して
i) KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、および
KPKDELEYEMDI(配列番号7);
から構成される群から選択されたアミノ酸配列を有する3種以上のペプチド;
ii) アミノ酸配列NDIEKKICKM(配列番号8)のペプチド;
iii) アミノ酸配列ILSVSSFLF(配列番号9)のペプチド;
iv) アミノ酸配列ALFQEYQCY(配列番号10)のペプチド;および
v) LIMVLSFLF(配列番号11)およびIMVLSFLFL(配列番号12)から構成される群から選択されるアミノ酸配列を有する1種以上のペプチド;
を含む組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸を投与する過程を含み、
前記抗原性ポリペプチド中のペプチドが、i)互いに直接隣接して配置されているか;またはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることと、前記組換え抗原性ポリペプチドまたは前記組換え抗原性ポリペプチドをコードする核酸が前記個体を免疫するために十分な量で投与されることとを特徴とする免疫する方法。
【請求項7】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーを含む組換え抗原性ポリペプチド。
【請求項8】
前記アミノ酸配列の多コピーが存在するとともに、i)互いに直接隣接して配置されているかまたはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする請求項7記載の組換え抗原性ポリペプチド。
【請求項9】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーを含む抗原性ポリペプチドをコードする核酸。
【請求項10】
前記アミノ酸配列の多コピーが存在するとともに、i)互いに直接隣接して配置されているかまたはii)前記組換え抗原性ポリペプチド内部において正統的カルボキシ末端タンパク質分解切断を可能とするスペーサーペプチドによって互いに分離されていることを特徴とする請求項9記載の核酸。
【請求項11】
マラリアに対して個体を免疫する方法であって、
前記個体に対してアミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーを含む組み換え抗原性ポリペプチドを投与する過程を含むことを特徴とする免疫する方法。
【請求項12】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項1記載の組換え抗原性ポリペプチド。
【請求項13】
アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項2記載の組換え抗原性ポリペプチド。
【請求項14】
前記核酸が、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらにコードする請求項3記載の核酸。
【請求項15】
前記核酸が、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらにコードする請求項4記載の核酸。
【請求項16】
前記組換え抗原性ポリペプチドが、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項5記載の方法。
【請求項17】
前記組換え抗原性ポリペプチドが、アミノ酸配列MMRKLAILSVSSFLFVEALFQEYQCYGSSKMEKCSSVFNVVNSSIGLIMVLSFLFLN(配列番号38)の1個以上のコピーをさらに含む請求項6記載の方法。
【請求項18】
組換え抗原性ポリペプチドが分子量13,500kDa以下を有し、Plasmodium falciparumのAsembo bay株のスポロゾイト周囲タンパク質(CSP)可変領域由来のエピトープを少なくとも1個含む組換え抗原性ポリペプチド。
【請求項19】
前記少なくとも1個のCSP可変領域由来のエピトープがT細胞エピトープである請求項18記載の組換え抗原性ポリペプチド。
【請求項20】
前記少なくとも1個のCSP可変領域由来のエピトープが
KPKDELDYENDI(配列番号1)、
KPKDELNYENDI(配列番号2)、
KPKDELDYANDI(配列番号3)、
KSKNELDYENDI(配列番号4)、
KSKDELDYENDI(配列番号5)、
KPKNELDYEMDI(配列番号6)、
KPKDELEYEMDI(配列番号7);および
NDIEKKICKM(配列番号8)
から構成される群から選択した配列番号で表されるアミノ酸配列を有することを特徴とする請求項18記載の組換え抗原性ポリペプチド。
【請求項21】
前記組換え抗原性ポリペプチドが、1個以上の保存CSPエピトープをさらに含む請求項18記載の組換え抗原性ポリペプチド。
【請求項22】
前記1個以上の保存CSPエピトープが、配列番号9、配列番号10、配列番号11および配列番号12から構成される群から選択した配列番号で表されるアミノ酸配列を有する請求項21記載の組換え抗原性ポリペプチド。
【請求項23】
前記組換え抗原性ポリペプチドが、配列番号36、配列番号37、および配列番号38から構成される群から選択した1個以上のエピトープをさらに含む請求項18記載の組換え抗原性ポリペプチド。
【図1】


【図2】
【図3】

【図4】
【図5】
【図6】

【図7】

【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2011−509672(P2011−509672A)
【公表日】平成23年3月31日(2011.3.31)
【国際特許分類】
【出願番号】特願2010−543174(P2010−543174)
【出願日】平成21年1月12日(2009.1.12)
【国際出願番号】PCT/US2009/030734
【国際公開番号】WO2009/091692
【国際公開日】平成21年7月23日(2009.7.23)
【出願人】(507158972)アエラス グローバル ティービー ワクチン ファウンデーション (2)
【Fターム(参考)】
【公表日】平成23年3月31日(2011.3.31)
【国際特許分類】
【出願日】平成21年1月12日(2009.1.12)
【国際出願番号】PCT/US2009/030734
【国際公開番号】WO2009/091692
【国際公開日】平成21年7月23日(2009.7.23)
【出願人】(507158972)アエラス グローバル ティービー ワクチン ファウンデーション (2)
【Fターム(参考)】
[ Back to top ]