
マラリア原虫由来のCSおよびHBsAg間のハイブリッド蛋白質
【課題】体液性応答だけでなく細胞性免疫応答も産生する、マラリアワクチンにおいて使用する新規な改良された抗原を提供すること。
【解決手段】本発明は、マラリア原虫のCS蛋白質の一部と、B型肝炎ウイルスの表面抗原とからなる新規なハイブリッド蛋白質を提供し、この蛋白質をワクチン処理の目的に使用することが開示されている。
【解決手段】本発明は、マラリア原虫のCS蛋白質の一部と、B型肝炎ウイルスの表面抗原とからなる新規なハイブリッド蛋白質を提供し、この蛋白質をワクチン処理の目的に使用することが開示されている。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はハイブリッド蛋白質、その医薬における使用、特にマラリア感染症の予防における使用、およびこれを含有するワクチンに関する。
【背景技術】
【0002】
マラリアは、世界の主要な衛生問題のうちの一つであり、毎年2〜4百万人の人がこの病気でなくなっている。この病気の最も急性の形態は、原虫寄生体、熱帯熱マラリア原虫(Plasmodium falciparum)によっておこり、マラリアによる死亡のほとんどはこれが原因である。
【0003】
熱帯熱マラリア原虫の生活環は複雑で、完結するためにはヒトおよび蚊の2種の宿主を必要とする。人の感染は感染した蚊の唾液中のスポロゾイトの接種により始まる。スポロゾイトは肝臓に移動し、そこで肝細胞を感染させ、赤血球外細胞内段階を経てメロゾイト段階に分化し、赤血球(RBC)に感染して、無性血液段階の周期的複製を開始する。サイクルは、RBC中の多数のメロゾイトが有性段階ガメトサイトに分化し、これが蚊により摂取され、中部栄養管の一連の段階を経て発達し、スポロゾイトを産生し、これが唾液腺に移動することにより完了する。
【0004】
熱帯熱マラリア原虫のスポロゾイト段階はマラリアワクチンの有効な目標である。スポロゾイトの主要な表面蛋白質は、サーカムスポロゾイト蛋白質(CS蛋白質)として知られる。この7G8株由来の蛋白質はクローンし、発現し、配列決定されている(非特許文献1参照)。7G8株由来の蛋白質は、37回繰り返す4ペプチドAsn−Ala−Asn−Proを含む中央免疫優性繰り返し領域に4個の副繰り返し配列Asn−Val−Asp−Proが散在していることを特徴とする。他の菌株においては、主および副繰り返し配列の数およびその相対的位置が変わる。CS蛋白質の繰り返しのない部分として示される非繰り返しアミノ酸配列からなるNおよびC末端部分がこの中心部分に隣接している。
【0005】
照射したスポロゾイトは、実験的なヒトマラリアに対して著しい保護作用を与える(非特許文献2参照)。しかし、生産が困難なため、照射したスポロゾイトは、ワクチン製造の点から実用的ではない。
サーカムスポロゾイト蛋白質ベースのサブユニットワクチンが提案されている。これらのうち2種のワクチンは臨床試験が行われている。1つは合成ペプチドで、もう1つは組み換え蛋白質である(非特許文献3および非特許文献4参照)。
【0006】
【非特許文献1】ダーム(Dame)ら、サイエンス(Science)225(1984)p593)
【非特許文献2】アメリカン・ジャーナル・オブ・トロピカル・メディスン・アンド・ハイジエン(Am.J.Trop.Med.Hyg.)24:297−402,1975
【非特許文献3】バロウ(Ballou)ら、ランセット(Lancet):i 1277(1987)
【非特許文献4】ヘリントン(Herrington)ら、ネイチャー(Nature)328:257(1987)
【発明の開示】
【発明が解決しようとする課題】
【0007】
これらのワクチンは抗スポロゾイト応答を刺激するのに成功している。しかし、応答の大きさは期待はずれで、応答を全く起こさないワクチンもある。更に、その後の注射に関して抗体レベルの「増加」がなく、in vitro リンパ球増殖検定の結果から、これらの被験者のT細胞のほとんどは、優性繰り返し配列を認識しないことがわかった。それでも、各研究のワクチンは寄生虫血症を発現しなかった。
【課題を解決するための手段】
【0008】
本発明は、体液性応答だけでなく細胞性免疫応答も産生する、マラリアワクチンにおいて使用する新規な改良された抗原を提供する。好ましくは、抗原は免疫優性繰り返し配列に対する中和抗体の産生を誘起する。最も好ましくは、抗原は、CD4+およびCD8+細胞毒性Tリンパ球(CTL)型および遅延型過敏性エフェクターT細胞媒介免疫応答を誘起し、好ましくはTへルパー(TH)記憶細胞も誘起できるものである。
【発明を実施するための最良の形態】
【0009】
従って、本発明は、CS蛋白質の実質的に全てのC末端部分、免疫優性領域の4以上のタンデム繰り返し配列、およびB型肝炎ウイルス由来の表面抗原(HBsAg)を含むハイブリッド蛋白質を提供する。好ましくは、該ハイブリッド蛋白質は、CS蛋白質のC末端部分と実質的に同じである最低160個のアミノ酸を含む配列を含む。CS蛋白質はC末端から最後の12個のアミノ酸を除いたものであってもよい。
【0010】
特に、HBsAgのN末端と直線的リンカーによりフレーム中に融合された熱帯熱マラリア原虫7G8のアミノ酸210〜398に実質的に対応する熱帯熱マラリア原虫のCS蛋白質の一部を含む蛋白質が提供される。該リンカーはHBsAg由来のpreS2の一部を含んでもよい。
【0011】
本発明は、本発明の蛋白質をコードするDNA配列も提供する。
【0012】
特に好ましい具体例は、RTSで表わされるハイブリッド蛋白質である。RTSのアミノ酸配列を第5図に示す。このハイブリッド蛋白質は、
サッカロミセス・セレビシエTDH3遺伝子配列由来のヌクレオチド1059〜1061によりコードされるメチオニン残基[ムスティ・エイ・エム(Musti A.M.)ら、ジン(Gene)1983 25 133−143];
ハイブリッド遺伝子を構築するのに用いられるクローニング法により得られるヌクレオチド配列(1062〜1070)由来の3個のアミノ酸:Met Ala Pro;
熱帯熱マラリア原虫7G8株のサーカムスポロゾイト蛋白質(CSP)のアミノ酸210〜398に相当するヌクレオチド1071〜1637によりコードされる189個のアミノ酸(Dameら、前出);
ハイブリッド遺伝子を構築するために用いられるクローニング法により得られるヌクレオチド1638〜1640によりコードされるアミノ酸(Arg);
ヌクレオチド1641〜1652によりコードされ、B型肝炎ウイルス
(adw血清型)のpreS2蛋白質(9)の4個のカルボキシ末端残基に相当する4個のアミノ酸、Pro Val Thr Asn;
ヌクレオチド1653〜2330によりコードされ、B型肝炎ウイルス(adw血清型)のS蛋白質を特定する226個のアミノ酸
を含む。
【0013】
別の具体例において、熱帯熱マラリア原虫NF54(モレキュラー・アンド・バイオケミカル・パラサイトロジー(Mol.Biochem Parasitol),35:185−190,1989)由来のCSP遺伝子配列を用いて産生され、熱帯熱マラリア原虫NF54由来のCS蛋白質の207〜395領域の実質的に全部を含むRTS*で表わされるハイブリッド蛋白質が提供される。
特に、RTS*は以下のものを含む:
TDH3遺伝子配列由来のヌクレオチド1059〜1061によりコードされるメチオニン;
ハイブリッド遺伝子を構築するのに用いられるクローニング法によりえられるヌクレオチド配列(1062〜1070)由来の3個のアミノ酸:Met Ala Pro;
熱帯熱マラリア原虫NF54株のサーカムスポロゾイト蛋白質(CSP)のアミノ酸207〜395に相当するヌクレオチド1071〜1637によりコードされる189個のアミノ酸鎖(モレキュラー・アンド・バイオケミカル・パラサイトロジー,35:185−190,1989);
ハイブリッド遺伝子を構築するために用いられるクローニング法によりえられるヌクレオチド1638〜1640によりコードされるアミノ酸(Gly);
ヌクレオチド1641〜1652によりコードされ、B型肝炎ウイルス(adw血清型)preS2蛋白質の4個のカルボキシ末端残基に相当する4個のアミノ酸、Pro Val Thr Asn(ネイチャー 280:815−819,1979);
ヌクレオチド1653〜2330によりコードされ、B型肝炎ウイルス(adw血清型)のS蛋白質を特定する226個のアミノ酸鎖(ネイチャー 280:815−819,1979)。
【0014】
RTS*のアミノ酸配列を第9図に示す。
RTS*を含有する発現カセットを構築し、これは以下の様な特性を有する:
エス・セレビシエTDH3遺伝子由来のヌクレオチド1から1058まで伸びるプロモーター配列;
ヌクレオチド1059から始まり、ヌクレオチド2330まで伸びるオープンリーディングフレーム。このオープンリーディングフレームは、そのすぐ後に翻訳終止コドンTAA(ヌクレオチド2331〜2333)が続く。該オープンリーディングフレームは、ハイブリッドRTS*蛋白質を特定するアミノ酸をコードする。
エス・セレビシエARG3遺伝子由来の塩基対2334から3504まで伸びる配列中に含まれる転写終止配列。
【0015】
ヌクレオチド配列を第10図に示す。
【0016】
本発明の蛋白質をコードするDNA配列は、好ましい具体例においては、好ましくは酵母遺伝子由来の転写調節エレメントに隣接し、発現ベクター中に含まれる。
このようなベクターは本発明の別の態様である。好ましいプロモーターは、エス・セレビシエTDH3遺伝子由来のプロモーターである(ムスティら、前出)。
【0017】
本発明はまた、本発明にかかるベクターで形質転換された宿主細胞に関する。宿主細胞は、原核生物であっても真核生物であってもよいが、好ましくは、エス・セレビシエなどの酵母である。このような宿主においては、ハイブリッド蛋白質、例えばRTSは、リポタンパク質粒子として発現される。選択された受容体酵母株は好ましくはすでにそのゲノム中にいくつかの組み込まれたB型肝炎S発現カセットのコピーを有する。得られた株は2種のポリペプチド、SおよびRTS(または他の本発明のハイブリッド蛋白質)を合成し、リポタンパク質粒子の混合物(たとえば、RTS、SまたはRTS*、S)中に自発的にコアセンブルされる。これらの粒子はその表面でハイブリッドのCSP配列を示すのが有利である。これらの混合粒子もまた、本発明に含まれる。有利には、これらの混合粒子中のRTS:SまたはRTS*:Sの比率は1:4である。
【0018】
本発明はまた、適当な希釈剤または担体との混合物中に免疫保護量の本発明の蛋白質または粒子を含むワクチンに関する。
【0019】
本発明のワクチンにおいて、ハイブリッドの水溶液を直接用いてもよい。あるいは、あらかじめ凍結乾燥した、あるいはしていない蛋白質を公知のアジュバントと混合または吸収させてもよく、このようなアジュバントとしてはミョウバン、ムラミルジペプチド、Quil Aなどのサポニンが挙げられるが、これに限定されるわけではない。
【0020】
免疫促進物質も選択的にまたは追加的に含まれる。この免疫促進物質の好ましい具体例は、3脱アシル化モノホスホリルリピッドA(3D−MPL)である。
3脱アシル化モノホスホリルリピッドAはアメリカ合衆国特許第4912094号およびイギリス特許出願第2220211号(リビ(Ribi))から公知であり、米国、モンタナ州、ルビ・イムノケム(Ribi Immunochem)から入手可能である。
【0021】
本発明の蛋白質もこのようなリポソームの微粒子中に封入されてもよい。
【0022】
ワクチン調製物は、ニュー・トレンズ・アンド・デベロプメンツ・イン・バクシンズ(New Trends and Developments in Vaccines,ボーラー(Voller)ら編,ユニバーシティ・パーク・プレス(University Park Press),ボルチモア,メリーランド州,米国,1978)に記載されている。リポソーム中の封入は、例えばフュラートン(Fullerton)(アメリカ合州国特許第4235877号)により記載されている。
【0023】
通常のアジュバントを用いてもよいが、好ましい免疫促進物質は3脱アシル化モノホスホリルリピッドA(3D−MPL)である。
典型的には、3D−MPLを用いた場合、抗原および3D−MPLはミョウバンを加えるかまたは水中油型乳剤中に処方される。3D−MPLはエフェクターT細胞応答の刺激物質であるのでこれを混ぜるのが有利である。
【0024】
従って本発明の好ましい具体例は、本明細書中に記載したように、3D−MPLおよび担体と組み合わせたハイブリッド蛋白質、好ましくはRTSまたはRTS*を含むワクチンを提供する。典型的には、担体は水中油型乳剤またはミョウバンである。
【0025】
最も好ましい具体例においては、ハイブリッド蛋白質は本明細書において記載したような粒子または混合粒子である。
【0026】
各ワクチン用量中の本発明の蛋白質の量は典型的ワクチン中著しい副作用を起こさずに免疫保護応答を誘起できる量に選択される。このような量は、用いた特定の免疫原およびワクチンがアジュバントを含むかどうかにより変わる。一般的に、各用量は1〜1000μg、好ましくは1〜200μg、最も好ましくは10〜100μgの蛋白質を含む。特定のワクチンについて最適な量は、抗体価の観察および他の患者における他の応答を含めた標準的研究により決定できる。初回接種の後、患者は好ましくは約4週間で追加免疫を受け、その後感染の危険がある間は6ヵ月ごとに繰り返し追加免疫を行う。本発明の蛋白質に対する免疫応答はアジュバントおよびまたは免疫促進物質の使用により向上する。
【0027】
本発明の蛋白質は好ましくは酵母、特にサッカロミセス属に属する酵母中に発現される。
【0028】
本発明の別の態様は、本発明のハイブリッド蛋白質の調製法を提供することであり、該方法は、該蛋白質をコードするDNA配列を適当な宿主、好ましくは酵母中で発現し、生成物を回収することからなる。
【0029】
本発明の蛋白質をサッカロミセス株中で発現することが特に好ましい。RTSを例えばこのような菌株中で発現した場合、これは自発的にマルチマーリポタンパク質粒子中で集合する。
これらの粒子は免疫原性が高く、強い体液性応答ならびに免疫学的記憶を誘起し、またCTLおよびDTH型のエフェクターT細胞を誘起することができる。
【0030】
本発明の更に別の態様は、既に記載したように有効量のワクチンを投与することによるマラリア原虫感染症にかかっている患者の治療法である。
【実施例1】
【0031】
1.RIT4383株のRTS、Sの構築
SおよびRTSポリペプチドの両方を含有する粒子の産生に用いるエス・セレビシエRIT4383株は、各蛋白質に対して別の発現カセットを有する。S遺伝子発現カセットは定住Tyレトロトランスポンソンズと相同な直線状組み込みベクターを用いてゲノム中の最低2個の部位で5〜6個のコピー中に組み込まれる。RTS遺伝子発現カセットは、S遺伝子カセットの組み込みに用いたものと同様の直線状組み込みベクターを用いてゲノム中の1または2個の部位で2〜3個のコピー中に組み込まれる。両タイプのカセットからの発現は、酵母TDH3遺伝子由来のプロモーターにより行われる。
【0032】
1.1発現カセットおよび組み込みベクター(pRIT13034)の構築
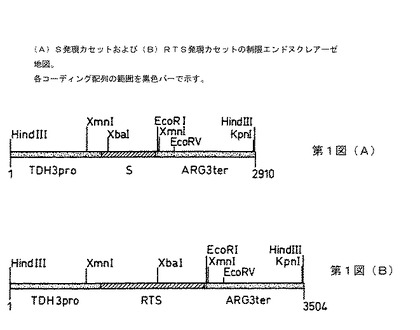
S遺伝子発現カセット(第1A図)はRIT4376株(1)中に見られるものと同じであり、1058bpのTDH3プロモーターフラグメント、681bpのS遺伝子配列、6bpのEcoR1リンカーを含むスペーサーおよびArg3転写ターミネーターを有する1165bpのフラグメントからなる。Sコーディング配列は、adw血清型のウイルスの完全ゲノムクローン(pRIT10616)からのサブクローニングにより得られる。
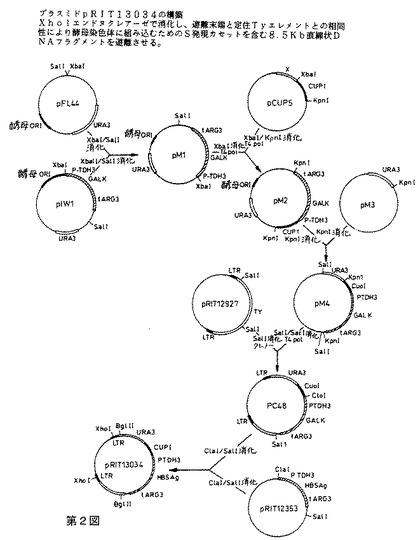
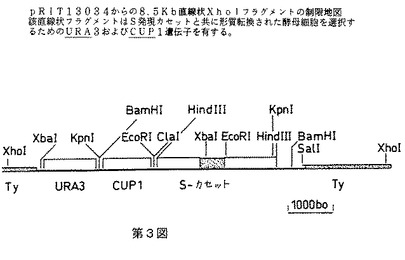
S発現カセットの酵母ゲノム中への組み込みに用いられるTyベクター、pRIT13034の構造を第2図に示す。このタイプの酵母ゲノム中の発現カセットの組み込み用ベクターの構造および使用は、ジェイコブズ(Jacobs)ら(2)に記載されている。pRIT13034はS遺伝子カセット、URA3遺伝子およびpUC9上にクローンされたTyエレメントのコピー中に挿入されたCUP1遺伝子を含有する。URA3遺伝子およびCUP1遺伝子は両方とも形質転換された酵母コロニーを、形質転換されていないコロニーから区別する選択マーカーを提供する。pUC9は、ビエイラ&メッシング(Vieira & Messing)(3)に記載されており、URA3フラグメントはpFL44由来であり[エフ.ラクロート(F.Lacroute),CNRS,ストラスブール]、CUP1フラグメントはpCUP5由来であり(ティー・ブット(T.Butt,)SKF Labs,フィラデルフィア)、 Tyエレメントはジュニアウス(Juniaux)ら(4)からのものであり、S遺伝子カセットはpRIT12353由来である。pRIT13034のXhoIエンドヌクレアーゼを用いた消化により、第3図に示す8500bpの直線状フラグメントが遊離し、これを定住Tyエレメントと遊離末端の相同な組み換えによりゲノム中に組み込むことができる。
【実施例2】
【0033】
Y1295株の構築
受容体株EJ cup1D−3d(trp1、leu2、ura3、cup1、gal1D、MATα)をpRIT13034由来の直線状ベクターフラグメントの形質転換による一次導入に用いた。この株は1個の破砕された遺伝子座cup1、cup1を含む。
直線状XhoIフラグメントで形質転換した後、Ura+コロニーを単離し、銅耐性に関してスクリーンする。サザンブロッティング分析で測定すると、より耐性の強い形質転換体が2〜5個のベクターのコピーを組み込んでいた。2個の形質転換体コロニーのコピー、推定3〜4個のコピーを有するMS9および4〜5個の組み込み直線ベクターのコピーを有するMS23が保持されている。MS23株を次にEJ cup1D−7b株(trp1、ura3、cup1、gal1D、MATα)と交雑させ、ハプロイド有子嚢胞子を回収してMS23−2aを得る。
この菌株を次にMS9と戻し交雑すると得られたLeu−、Trp−ハプロイド分離体(MS54−2c)は5〜6個の組み込まれた発現カセットを含む。サザンブロッティングによると、MS54−2Cは1遺伝子座に4〜5個の組み込みベクターのタンデムコピーを有し、更に別の遺伝子座に組み込まれた1個のコピーを有する。発現カセットの5〜6個のコピー全てはカセットフラグメントを遊離するための全酵母細胞DNAのHindIIIエンドヌクレアーゼでの消化を受けず、S遺伝子特異性プローブを用いたサザンブロッティングにより予想される3kbのバンドが得られる。この菌株の自発的Trp+復帰変異体、MS54−2c−Tが得られる。実験室受入番号Y1295。
【実施例3】
【0034】
RTS発現カセットの構築
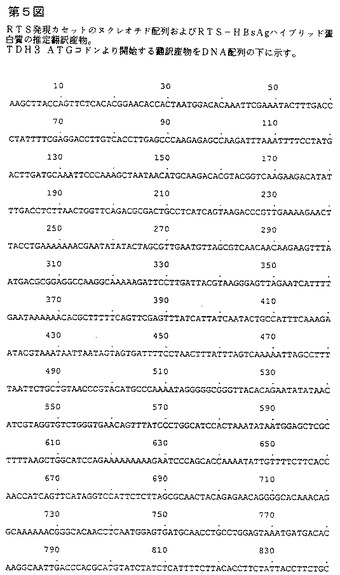
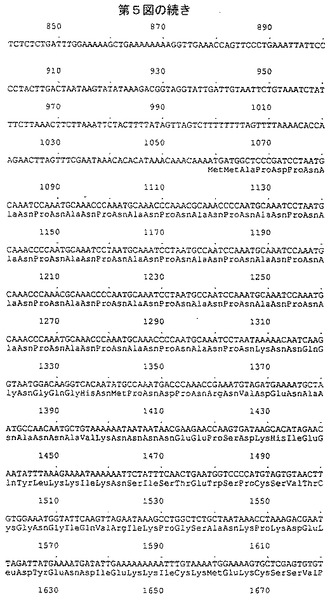
RTSハイブリッド蛋白質の発現カセットを、多段階のクローニング法により構築し、イー・コリ酵母シャトルベクターYep13(6)中にクローンして、プラスミドYep13RTS(第4図)を得る。カセットの構造を第1B図に示す。全ヌクレオチド配列を直接配列決定(コーディング配列および非翻訳調節配列の一部に関して)または関連する文献を参考にして(プロモーターおよびターミネーター配列の一部に関して)決定した。DNA配列を第5図に示す。これは以下の要素を含有する:
エス・セレビシエTDH3遺伝子由来のヌクレオチド1から1058まで伸びるプロモーター配列;
ヌクレオチド1059で始まり、ヌクレオチド2330まで伸びるオープンリーディングフレーム。このオープンリーディングフレームのすぐ後には翻訳終止コドンTAA(ヌクレオチド2331から2333)が続く。オープンリーディングフレームはハイブリッドRTS蛋白質を特定するアミノ酸をコードする;
エス・セレビシエARG3遺伝子(クラビール(Crabeel)ら EMBO J.1983 2:205−212)由来の、2334から3504塩基対まで伸びる配列中に含まれる転写停止配列。
ヌクレオチド1059から2330によりコードされるハイブリッドRTS蛋白質のアミノ酸配列を第5図に示し、これは以下の要素を含む:
TDH3遺伝子配列由来の、ヌクレオチド1059から1061によりコードされるメチオニン残基;
ハイブリッド遺伝子を構築するのに用いられるクローニング法により産生されるヌクレオチド配列(1062〜1070)由来の3個のアミノ酸MetAla Pro;
熱帯熱マラリア原虫7G8株(8)のサーカムスポロゾイト蛋白質(CSP)のアミノ酸210〜398に相当するヌクレオチド1071〜1637によりコードされる189個のアミノ酸鎖;
ハイブリッド遺伝子を構築するのに用いられるクローニング技法により産生されるヌクレオチド1638〜1640によりコードされるアミノ酸(Arg);
ヌクレオチド1641から1652によりコードされ、B型肝炎ウイルス(adw血清型)preS2蛋白質(9)の4個のカルボキシ末端残基に相当する4個のアミノ酸Pro Val Thr Asn;
ヌクレオチド1653から2330によりコードされ、B型肝炎ウイルス(adw血清型)を特定する226個のアミノ酸鎖。
【実施例4】
【0035】
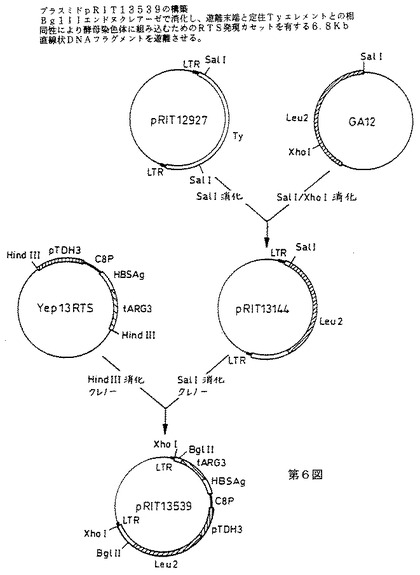
RTSカセット組み込みベクターpRIT13539の構築
RTSカセット組み込みベクターpRIT13539の構造を第6図に示す。
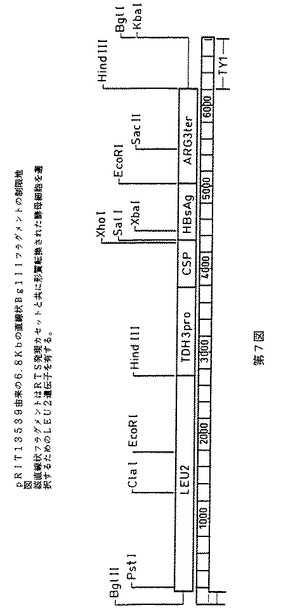
RTS発現カセットをTyベースの組み込みベクターpRIT13144上に挿入する。このベクターは、LEU2遺伝子がベクターpCV9(7)から単離されたSalI−XhoIフラグメントとしてTyエレメントのSalI部位中に挿入されているpRIT12927(2)の誘導体である。従って、RTS発現カセットをpRIT13144中に挿入した後、得られたプラスミドpRIT13539は発現カセットに加えて、選択マーカーとしてLEU2遺伝子を含有する(第5図)。pRIT13539をBglIIエンドヌクレアーゼで消化すると、第7図に示す6800bpの直線状フラグメントが遊離され、これは定住Tyエレメントとの遊離末端の相同組換によりゲノム中に組み込むことができる。
【実施例5】
【0036】
実施例5a Y1295株の形質転換およびRIT4383株(Y1530)の産生
SおよびRTS蛋白質の両方を発現する株を得るために、Y1295を、Leu+コロニーに関する選択を有する6800bpの直線状BglIIフラグメントを用いて形質転換する。種々の割合でゲノム中に存在する両発現カセットのセットを含む数個の組み込み体が得られる。約1:4の割合でRTSおよびS蛋白質を発現する選択された形質転換体の一つは公式名称はRIT4383(実験室受入番号Y1530)である。
【0037】
実施例5b Y1295株の形質転換およびY1631株の産生
(RTSと)類似の構造を、熱帯熱マラリア原虫NF54株(モレキュラー・アンド・バイオケミカル・パラサイトロジー(Mol.Biochem.Parasitol.)35:185−190,1989)由来のCSP遺伝子配列を用いて産生する。得られた融合蛋白質は、熱帯熱マラリア原虫7G8株由来のCSPを用いてえられる構造物と区別するためにRTS*と表わす。
発現カセットの配列を第9図に示す。これは以下のエレメントを含む:
エス・セレビシエTDH3遺伝子由来のヌクレオチド1から1058まで伸びるプロモーター配列;
ヌクレオチド1059で始まり、ヌクレオチド2330まで伸びるオープンリーディングフレーム。このオープンリーディングフレームのすぐ後には翻訳終止コドンTAA(ヌクレオチド2331から2333)が続く。オープンリーディングフレームはハイブリッドRTS*蛋白質を特定するアミノ酸をコードする;
エス・セレビシエARG3遺伝子由来の、2334から3504塩基対まで伸びる配列中に含まれる転写終止配列。
RTS*融合蛋白質をコードするこの発現カセットを、RTS構造物に関して既に記載したのと同じ方法を用いて形質転換し、酵母Y1295株のゲノム中に形質転換し、組み込む。SおよびRTS*蛋白質の両方を発現する形質転換されたクローンが得られる。1つのクローンを、約4S:1RTS*の割合で2個の蛋白質を発現するように選択する。クローンの実験室受入番号はY1631である。
【実施例6】
【0038】
RIT4383株の予備特質化
6.1イムノブロッティング分析
細胞を含まないRIT4383から調製した抽出物を種々の抗体を用いたイムノブロットにより分析した:
S蛋白質に対する単クローン性抗体(Mab HBS1);
RTS蛋白質(Mab167)の繰り返し部分に対する単クローン性抗体;
RTS蛋白質(ウサギ血清No.20)の繰り返しのない配列に対するウサギ血清。
酵母RIT4383株において、2個の発現された生成物を単クローン性抗体HBS1により認識する。即ち、S蛋白質に対応する24KD蛋白質および46KDのRTSハイブリッド蛋白質である。RTSハイブリッド蛋白質も、CSPの繰り返し、および非繰り返しエピトープに対する抗体により検出される。これらの結果から明らかに、RIT4383株が同時に2個のSおよびRTS抗原を、RTS/Sが約1:4の割合で発現することがわかる。
【0039】
6.2CsCl密度勾配遠心
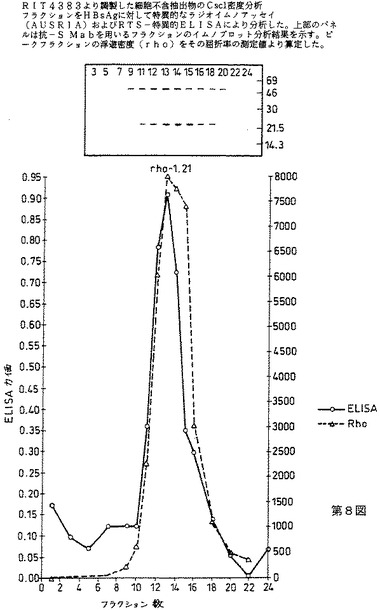
RIT4383株中の粒子の形成を、CsCl密度勾配遠心により分析する。粗抽出物(総蛋白質の±15mg)を、10mlの1.5M CsCl勾配上分析する(40000rpm、+8℃、ベックマン50Tiローター中68時間)。フラクション(0.5ml)を集め、HBsAg(AUSRIA)に対して特異的な放射免疫検定法、RTS特異的ELISAおよび抗HBsAg単クローン性抗体を用いたイムノブロットにより分析する。
第8図に示すように、ELISA、RIAおよびウェスタンブロットピークは、rho=1.21の浮遊密度に対応する勾配の同一フラクション(no.13)で現われ、このことは、SおよびRTSモノマーの両方を含む混合粒子がこの株中に形成されたことを示す。
【実施例7】
【0040】
シードロットの調製
マスターシードロットの製造法
Y1530株(RIT4383)をまず20mlの滅菌YNB(Difco)(デキストロース(0.1%)および1.8%(w/v)寒天(Difco)で補足)をいれたペトリ皿中で30℃で48時間成育させる。表面成長株を滅菌YNBブロス(Difco)(デキストロース(1%)およびグリセロール(10%)を追加)中に懸濁する。この懸濁液を無菌条件下で2mlのポリプロピレンの栓をした滅菌試験管中に分配し(試験管1本につき1ml)、−70℃で貯蔵する。
【0041】
ワーキングシードロットの製造法
マスターシード試験管の1本をすばやく解凍し、その内容物を白金耳で前記のように調製したペトリ皿上に取り出す。30℃で63時間培養した後、表面成長物の一部を、400mlの滅菌YNBブロス(Difco)(デキストロース(2%)を追加)をいれた2Lコニカルフラスコに移す。培養物を30℃で24時間培養した後、無菌条件下で遠心する(6300xgで15分)。ペレットを無菌YNBブロス(Difco)(デキストロース(1%)およびグリセロール(10%)を追加)中に再懸濁する。これを無菌条件下で2mlのガラス栓をした滅菌試験管中に分配し(試験管1本につき0.5ml)、−70℃で貯蔵する。
【実施例8】
【0042】
発酵
接種物の調製
(a)固体培地上の成育
バイアル1つ分のワーキングシードロットを素早く解凍し、デキストロース(1%)および1.8%(w/v)寒天(Difco)を追加した滅菌YNBブロス(Difco)を含有するペトリ皿上に分散させる。ペトリ皿を300℃で48時間培養する。
(b)接種物の成育
ペトリ皿の表面成育物をデキストロース(2%)を追加した滅菌YNBブロス(Difco)中に懸濁し、2個のコニカルフラスコ(2L、フラスコ1個あたり400mlの液体)中に等分する。フラスコをロータリーシェーカー上で30℃で24時間培養する。
【0043】
発酵
5Lの脱イオン水((NH4)2SO4(40g)を追加)を入れた発酵槽(総容量20L)をそのまま清浄なあらかじめフィルターを通した蒸気を用いて2バールgの圧力で121℃で30分間滅菌する。室温に冷却した後、発酵槽中の液体の体積を4Lに調節し、フィルター滅菌したHB4溶液1Lを添加する。発酵は、2個のコニカルフラスコからの接種物(800ml)を添加することにより開始する。
発酵は、フィードバッチ技法を用いて行い、これにより培養物を以下の組成の溶液と共に連続して供給する:
5LのHB4溶液;
4Lのデキストロース80%(121℃で滅菌)
培養密度は30℃およびpH5(NH4OHを添加することにより維持する)での好気的成育により増加する。溶解酸素を、気流および撹拌速度を調節して約50%飽和に維持する。供給物の添加速度は成育速度が最大になり、副生成物のエタノールの形成が最小になるようにあらかじめ決定しておく。
40〜90時間後に発酵を停止する。発酵の終わりで、全培養物体積は10〜18Lであり、乾燥細胞重量は30および100g/Lの間である。培養物を迅速に15〜25℃に冷却し、酵母細胞を遠心分離によりブロスから回収する。濃縮した酵母細胞を、再遠心前にリン酸緩衝液(50mM)で1度洗浄し、続いてポリエチレンの袋中で−70℃で貯蔵する。
【0044】
【表1】

【実施例9】
【0045】
RTS/Sの抽出および精製
抽出法
9.1細胞懸濁液の精製
凍結濃縮酵母細胞(−70℃)を一夜−30℃で解凍し、続いて細胞の入ったポリエチレンの袋を水中(10〜20℃)につけることにより5〜15℃で完全に解凍する。酵母細胞懸濁液を以下の成分を含有する燐酸緩衝溶液(pH8.0)を用いて調製する:エチレンジアミン四酢酸(EDTA)、p−メチルスルホニルフルオリド(PMSF)、イソプロパノールおよびtween20。
【0046】
9.2細胞破砕
細胞を鉛を含まないガラスビーズ(直径0.49〜0.70mm)を含むビーズミル中で破砕する。−20℃の冷却液を循環させて粉砕室を冷却し、粉砕室出口でのホモジネートの温度が15℃を越えないようにする。液体流速は6L/時間、撹拌速度は3000rpmである。このプロセスを2回行う。得られたホモジネートのpHは6.7〜7.5である。
【0047】
9.3ポリエチレングリコール清澄化
ポリエチレングリコール400(PEG400)を破砕した細胞懸濁液に添加して、最終濃度を10%にし(10℃未満で30分、pH6.1)、部分的に清澄化された上清を遠心分離により回収する(J21Bベックマン遠心分離機、JA10ローター、17000gで1時間)。
【0048】
9.4メタノール清澄化
メタノールをpH7でPEGで清澄化した抗原に添加して、5容積のPEGで清澄化した抗原に対してメタノール1容積の割合にする。17000gで20分間遠心分離により清澄化された抗原をえる(J21Bベックマン遠心分離機、JA10ローター)。
【0049】
9.5コロイドシリカ上への吸着/脱着
粗抗原を一夜1.5%(w/v)コロイドシリカ(アエロジル380、デグサ)上に4℃で吸着させる。
ペレットを連続して遠心分離し、NaCl0.9%(w/v)中に再懸濁することにより洗浄(3回)後、1%TWEEN20を含有する10mMピロリン酸緩衝液pH9.5を用いて抗原を脱着させる。
脱着緩衝液体積はメタノールで清澄化した抗原溶液の1/8に相当する。脱着された抗原溶液をL8.70ベックマン超遠心分離ローターR19で、50000gで60分間超遠心分離により回収する。
【0050】
9.6ジアフィルトレーション
精製工程の前に、脱着された抗原を5容積の6Mの尿素、500mMのNaCl、20mMのTRIS−HClでpH8.5で超遠心分離することにより洗浄し、蛋白質および脂質汚染物の大部分を除去する。
つぎに緩衝液を同じシステム(Ultrasette、300kDの公称カットオフのポリスルホン膜を備えたFILTRON)で5容積の10mMのTRIS−HClでpH8.1と交換する。
【0051】
9.7DEAE−TSK650(M)上イオン交換クロマトグラフィー
清澄化した溶液を10mM TRIS緩衝液(pH8.1)で平衡化したアニオン交換カラム(DEAE−TSK650(M))にかける。連続して2容積の10mM TRIS−HCl緩衝液(pH8.1)および40mMNaClを追加した3容積の10mM TRIS−HCl緩衝液(pH8.1)で洗浄した後、1容積未満の150mM NaClを含有する10mM TRIS−HCl緩衝液(pH8.1)を用いて抗原を脱着する。抗原を含むフラクションをプールする。
【0052】
9.8Butyl−TSK650(M)上疎水性相互作用クロマトグラフィー
最終濃度が650mM NaClになるまでNaClを添加した後、抗原溶液を、20mM TRIS−HCl緩衝液、600mM NaCl(pH8.1)で平衡化したButyl-TSK650(M)カラムにかける。抗原の大部分は通過するが、不純物はゲルと結合する。
【0053】
9.9限外濾過による濃縮
HICプールを300kDaの公称カットオフのポリスルホン膜を備えたUltrasetteシステム(FILTRON)により濃縮する。
【0054】
9.10CsCl勾配中での超遠心
CsClをButyl−TSKプールに添加して、最終濃度を1.5Mにする。
50.2Tiベックマンローター中270000gで65時間後、抗原を含有するフラクションを集める。
【0055】
9.11SEPHACRYL S300(HR型)上でのサイズ排除クロマトグラフィー
緩衝液を交換し、低分子量の汚染物質を除去するために、抗原溶液をSEPHACRYL S300 HRカラムにかける。溶出緩衝液は、150mM NaClを含有する10mM燐酸緩衝液(pH7.4)である。
滅菌濾過
150および400μg Lowry/ml間に希釈し、pHを6.8に調節した後、精製した抗原を0.22μm滅菌フィルターを通して濾過することにより滅菌する。得られた溶液は、精製されたRTS/S粒子を含有する。
【実施例10】
【0056】
RTS/Sの免疫学的特性化
10.1抗原性
RTS/S粒子の抗原性を試験するために、異なるエピトープに対する単クローン性抗体を結合させてELISAを何回も行った。
用いた単クローン性抗体(MoAbs)は以下のとおりである:
−MoAb R10: ・CSP領域の繰り返し配列(NANP)に対して特異的
・IgMアイソタイプ
−MoAb RL117: ・CSP領域の非繰り返し配列に対して特異的
−MoAb RF1: ・HBsAgのS配列に対して特異的
・IgGアイソタイプ
Sp2/OAg骨髄種細胞をCSP配列の繰り返しおよび非繰り返し領域の両方を含有する完全に純粋でないRTS様粒子で免疫したBalb/Cマウスの脾細胞と融合することによりMoAb R10およびRL117を室中で調製する。
3バッチのRTS/Sワクチンを分析する:バッチナンバーは24M31、24M32および24M34である。
【0057】
10.2単クローン性抗体R10(抗−繰り返し)との反応
抗−繰り返しMoAb R10を「サンドウィッチ」ELISAに用いる。試験するサンプルをあらかじめMoAb R10をコートしたマイクロタイタープレート中で培養する。ペルオキシダーゼと結合した同じMoAbを次に添加する。37℃で1時間培養し、洗浄した後、オルトフェニレンジアミンH2O2を添加することにより発色する。490nmで吸光度を測定し、抗原濃度に対してプロットする。
結果
3バッチ:24M31、24M32および24M34はMoAb R10と陽性に、適合して反応するので、RTS/S粒子上に繰り返しエピトープが存在することが確認される。最大結合の50%に達するのに必要な抗原量はバッチ24M31、24M32および24M34についてそれぞれ51.2、38.2および60.6ngである。
【0058】
10.3単クローン性抗体RL117(抗−繰り返し)との反応
MoAb RL117のRTS、S粒子との反応性を「サンドウィッチ」ELISA試験で分析する。MoAb RL117はペルオキシダーゼと結合し、RTS/S粒子の検出に用いられ、繰り返し領域に関して特異的なMoAb R10は粒子の捕獲に用いられる。
簡単に言うと、試験するサンプルをあらかじめMoAb R10をコートしたマイクロタイタープレート中で培養する。ペルオキシダーゼと結合したMoAb RL117を次に添加し、37℃で1時間培養し、洗浄した後、オルトフェニレンジアミンH2O2を添加することにより発色する。490nmで吸光度を測定し、抗原濃度に対してプロットする。
結果
3バッチ:24M31、24M32および24M34はMoAb RL117およびR10と陽性に、適合して反応するので、同一のRTS/S粒子上に繰り返しおよび非繰り返しエピトープが存在することが確認される。最大結合の50%に達するのに必要な抗原量はバッチ24M31、24M32および24M34についてそれぞれ169.2、117.6および181.1ngである。
【0059】
10.4単クローン性抗体RF1(抗S)との反応
RTS/S粒子中のSエピトープの存在をペルオキシダーゼと結合したMoAb RF1を検出に用いて「サンドイッチ」ELISA試験により示す。R10MoAbはRTS、S粒子を捕獲するために、マイクロタイタープレート上に直接用いる。
要約すると、試験するサンプルをあらかじめMoAb R10をコートしたマイクロタイタープレート中で培養する。ペルオキシダーゼと結合したMoAb RF1を次に添加する。37℃で1時間培養し、洗浄した後、オルトフェニレンジアミンH2O2を添加することにより発色する。490nmで吸光度を測定し、抗原濃度に対してプロットする。
結果
3バッチ:24M31、24M32および24M34はMoAb RF1およびR10と陽性に反応するので、同じRTS/S粒子上に繰り返しおよびSエピトープが存在することが確認される。最大結合の50%に達するのに必要な抗原量はバッチ24M31、24M32および24M34についてそれぞれ52.3、55.2および106.2ngである。
【実施例11】
【0060】
RTS/S粒子のin vivoでの免疫原性
11.1免疫原性の研究
(RTS、S)粒子の免疫原性に関する試験を、マウスおよびセルコピテカス・イーチオプスサルにおいておこなった。
マウスにおいて、抗CSP抗体をR32tet32抗原を抗繰り返し抗体の検出に用いてELISA試験により分析する。R32tet32はプラスミドpBR322のテトラサイクリン耐性遺伝子の32個のアミノ酸部分と融合した32個のNANP(主)繰り返し配列からなる。組み換え抗原はエシェリヒア.コリ中で産生する。CS蛋白質の非繰り返し配列に対する抗体(Abs)をRLF抗原を用いたELISA試験により測定する。RLF抗原は、インフルエンザウイルスのNS1蛋白質の初めの81個のアミノ酸と融合した繰り返し配列を含まない完全CS蛋白質非翻訳領域からなる。RLF抗原をイー.コリ中で産生する。マウス血清を1:100から初めて連続的に希釈し、力価をELISA試験(1)において光学密度1.0に対応する希釈度の逆数で表わす。抗CSP Absの測定は、各血清に関して行い、力価の等比中項(GMT)を計算する。
抗担体応答を分析するために、抗HBs抗体力価も測定する(プールした血清のみ)。
マウスの試験において、RTS、S粒子の免疫原性を評価するために、HBsAgの免疫原性について通常用いられるBalb/Cマウス(H−2dハプロタイプ)も用いる。免疫化の腹腔内(i.p.)および皮下(s.c.)経路を比較し、免疫促進物質3−脱アシル化モノホスホリルリピッドA(3D−MPL)の免疫応答に関する効果も試験する。
(RTS、S)ワクチンの免疫原性も同様にセルコピテカス・イーチオプスサルにおいて試験する。
選択された個々のまたはプールした血清もそのプラスモジウム.ファルシパラムスポロゾイトによる人肝癌細胞系(HepG2)のin vitroの破壊を抑制する能力に関して試験する(ISI検定(2))。
【0061】
11.2マウスにおける免疫原性
実験1: Al(OH)3上に吸着した臨床的(RTS、S)バッチの免疫原性
方法
免疫化:
10匹のBalb/Cマウスに、あらかじめAl(OH)3上に吸着させた各1μgの3種の(RTS、S)バッチ(24M/31、24M/32および24M/34)を1ヶ月間隔で2回腹腔内注射する。対照マウスに、HBsAg(Engerix−B、バッチENG611B4)を注射する。30日目および45日目に、マウスを採血し、抗体価を各血清について測定する。
血清学的方法:
抗R32tet32および抗RLF価を、それぞれコーティング抗原としてR32tet32およびRLFを用いたELISAにより測定する。マイクロプレートを試験する血清サンプル(1:100から初めて2倍連続希釈12回)とともに培養する。マウスAbsをビオチニル化抗マウスIg、続いてストレプタビジン:ビオチニル化ホースラディッシュペルオキシダーゼコンプレックスおよびオルトフェニレンジアミン/H2O2と反応させる。光学密度を490nmで測定する。力価を光学密度1.0(最大結合の50%)に対応する希釈度の逆数として表わす。各グループのマウスに関して、力価の等比中項(GMT)を計算する。抗HBs抗体価をHollingerの式(3)にしたがって計算し、mIU/mlで表わす。
結果
抗CSP応答:
強い抗R32tet32および抗RLF応答が試験した各(RTS,S)バッチに関して観察された。バッチ間の明らかな違いはない。著しい追加抗原刺激の効果は2回目の投与の後観察される。HbsAg(「Engerix−B」)で免疫したマウスをこの実験において負の対照として用いる。
抗HBs応答:
(RTS、S)バッチはHBsAgキャリアー蛋白質に対する抗体を誘起する。試験はプールした血清のみに関して行った。
【0062】
実験2:Balb/Cマウスにおける(RTS、S)粒子の免疫原性に対する免疫促進物質3D−MPLの効果
本発明者らはマウスにおける(RTS、S)ワクチンの免疫原性に対する免疫促進物質3D−MPLの効果を分析した。腹腔内(i.p.)および皮下(s.c.)経路の両方の免疫化を試験した。
A.ip経路による免疫化
方法
免疫化:
10匹のBalb/CマウスのグループにAl(OH)3単独、またはAl(OH)3+3D−MPL(50μg/用量)上に吸着させた1μgの(RTS、S)(バッチ24M/34)を1ヶ月間隔で2回腹腔内注射する。対照マウスに、150mMNaClを注射する。30日目および45日目に、マウスを採血し、抗体価を各血清について測定する。
血清学的方法:
血清学的方法は前記の始めの実験においてと同様である。
結果
抗CSP応答:
(RTS,S)ワクチンバッチは強い抗R32tet32および抗RLF応答を両処方物において示す。全ての場合において、二回目の免疫化の後に著しい追加抗原刺激の効果が観察された。3D−MPLを含有する処方に関して得られた力価は全ての場合において、アルミニウムのみの処方物より高く、第一抗R32tet32応答(p=0.02)において統計的に著しい増加が観察された。150mM NaClを注射したマウスのグループをこの実験における負の対照として用いた。
抗HB応答:
腹腔内経路により注射された(RTS、S)ワクチン両処方物は強い抗HBs応答を誘起する。著しい追加抗原投与の効果はいずれかの処方物を用いた2回目の免疫化の後に観察される。
【0063】
B.s.c.経路による免疫化
方法
免疫化:
10匹のBalb/Cマウスの群にAl(OH)3のみ、またはAl(OH)3+3D−MPL(50μg/用量)上に吸着させた1μgの(RTS、S)(バッチ24M/34)を1ヶ月間隔で2回皮下注射する。対照マウスに、150mM NaClを注射する。30日目および45日目に、マウスを採血し、抗体価を各血清について測定する。
血清学的方法は前記の始めの実験においてと同様である。
結果
抗CSP応答:
(RTS,S)ワクチンバッチはR32tet32およびRLFに対して正の応答を両処方物において誘起する。全ての場合において、二回目の免疫化の後に著しい追加抗原刺激の効果が観察された。3D−MPL処方に関して抗R32tet32および抗RLF応答(それぞれp=0.18およびp=2.9)の両方に関して統計的に著しい高い力価が45日目に観察される。しかし、一般には、力価は腹腔内経路で観察されたものよりも低い。150mM NaClを注射したマウスのグループをこの実験における負の対照として用いた。
抗HBs応答:
腹腔内経路により注射された(RTS、S)ワクチンの両処方物は良好な抗HBs応答を誘起する。著しい追加抗原刺激の効果はいずれかの処方物を用いた2回目の免疫化の後に観察される。抗CSP応答に関して観察されたように、抗HBs応答は腹腔内経路に比べてこの経路に免疫化によるものは低い。
【0064】
11.3セルコピテカス・イーチオプスにおける免疫原性
Al(OH)3上に吸着した臨床的バッチ24M/32をもちいたセルコピテカス・イーチオプスサルにおける免疫原性を評価した。
方法
免疫化:
5匹のサルに、Al(OH)3(0.5mgAl+++)上に吸着させた20μgの(RTS、S)粒子を0、28および84日目に筋肉内注射する。動物を0、14、28、42、56、66および98日目に採血した。R32tet32、RLFおよびHBs抗原に対する抗体を測定した。
血清学的方法:
抗R32tet32および抗RLF抗体価を、マイクロプレート上にコートしたR32tet32およびRLF抗体をそれぞれ用いたELISAにより測定する。プレートをつぎに試験する血清サンプル(1:10から初めて2倍連続希釈12回)とともに培養する。サル抗体をビオチニル化抗ヒトIg、続いてストレプタビジン:ビオチニル化ホースラディッシュペルオキシダーゼコンプレックスおよびオルトフェニレンジアミン/H2O2により検出する。光学密度を490nmで測定する。力価を光学密度1.0(最大結合の50%)に対応する希釈度の逆数として表わす。各グループに関して、力価の等比中項(GMT)を計算する。抗HBs抗体価をHollingerの式(Hollingerら、1982)にしたがって計算し、mIU/mlで表わす。
結果
抗CSP応答:
(RTS,S)ワクチンはR32tet32およびRLF抗原の両方に対して5匹のサル全てにおいて正の応答を誘起した。著しい追加抗原刺激の効果は2回目の免疫化の14日後(42日目)に観察される。次に抗体価が3回目の免疫化までゆっくり減少するのが観察された。力価はサルJo352の抗RLFの場合以外は3回目の免疫化の14日後(98日)に再び増加した。
3回目の免疫化の後(98日)得られた抗R32tet32力価はしかし2回目の免疫化後(42日)に観察されたものよりも高くない。
抗RLF応答の場合(サルJo353を除く)、3回目の免疫化の後(98日)、2回目の免疫化後(42日)のレベルにくらべて力価の増加が観察された。
抗HB応答:
全てのサルは免疫化の2回目(42日)および3回目(98日)の後著しい追加抗原刺激の効果に関する抗HBs応答を示した。
【0065】
(RTS、S)粒子に対して得られた抗体の生物学的活性
(RTS、S)ワクチンにより誘起された抗体の生物学的機能の測定値として、プールしたマウスおよび個々のさる血清をスポロゾイト侵入阻止(Inhibition of Sporozoite Invation(ISI))検定により試験した(Hollingdaleら、1984)。この検定は熱帯熱マラリア原虫スポロゾイトによる人肝癌細胞系の
in vitroでの侵入を阻止する抗CSP抗体の能力を測定するものでる。
結果
この実験の結果を第1表および第2表に示す。ISI測定値を免疫前の対照血清の活性を抑制0%とした相対的な抑制(%)で表わす。参考のために、試験した血清の抗R32tet32およびRLF抗体価も含める。第1表は試験した全てのマウス血清が非常に強いISI活性を有することを示す。第2表は全ての5匹のサルは対応する免疫血清に比べて98日目に非常に強いISI活性を有することを示す。
結論
(RTS、S)粒子は、マウスおよびサルの両方においてCSPの繰り返しおよび非繰り返しエピトープに対しておよび、HBsAgキャリアーのS蛋白質に対して高い抗体応答を誘起する。
マウスにおける一次抗体応答は3D−MPLの存在により向上する。
腹腔内注射の後得られた抗体価は、皮下経路による免疫価の後得られたものよりも高い。
2種の動物において誘記された抗体は、熱帯熱マラリア原虫スポロゾイトによる培養された人肝癌細胞の侵入を効果的に予防した。
【0066】
【表2】
【0067】
【表3】
【0068】
引例
(1) ハンフォード・エヌ、カベゾン.ティー、コロー・ビー(Hanford N, Cabezon T, Colau B)ら、「コンストラクション・アンド・キャラクタライゼーション・オブ・ア・サッカロミセス・セレビシエ・ストレイン(RIT4376)エクスプレッシング.ヘパティティスBサーフェイス・アンチゲン」(”Construction and Characterization of a Saccharomyces Cerevisiae Strain(RIT4376)Expressing Hepatitis BSurface Antigen”), Postgrad Med J 63,Supp.2:65−70,1987
(2) ジェイコブズ・イー、ルトガーズ・ティー、ボート・ピー(Jacobs E, Rutgers T, Voet P)ら、「シマルタネアス・シンセシス・アンド・アセンブリ・オブ・ベリアス.ヘパティティスBサーフェイス.プロテインズ・イン・サッカロミセス・セレビシエ」(”Simultaneous Synthesis and Assembly of Various Hepatitis B Surface Proteins in Saccharomyces cerevisiae”), Gene 80:279−291,1989
(3) ビーラ・ジェイおよびメシング・ジェイ(Vieira J and Messing J)、「pUCプラスミズ、アンM13mp−デライブド・システム・フォア・インサーション・ミュータゲネシス.アンド.シーケンシング・ウィズ・シンセティック.ユニバーサル・プライマーズ」(”The pUC plasmids,an M13mp7−Derived System for Insertion Mutagenesis and Sequencing with Synthetic Universal Primers”),Gene 19:259−268,1982
(4) ジュニアウクス(Juniaux)Embo J 1:1125−1131,1982.
(5) ヒンネン・エイ、ヒックス・ジェイビイおよびフィンク・ジイアール(Hinnen A, Hicks JBand Fink GR)、「トランスフォーメーション・オブ・イースト」(”Transformation of Yeast”), Proc Natl Acad Sci USA 75:1929−1933,1980.
(6) ブローチ・ジェイアール、ストラザン・ジェイエヌおよびヒックス・ジェイビイ(Broach JR, Strathern JN, and Hicks JB)、「トランスフォーメーション・イン・イースト・デベロプメント・オブ・ア・ハイブリッド・クローニング・ベクター・アンド・アイソレーション・オブ・ザ・CAN1ジーン」(”Transformation in Yeast Development of a Hybrid Cloning Vector and Isolation of the CAN1 Gene”), Gene 8:121−133, 1979
(7) ツァン・エイチ(Zhang H)ら、「ダブル・ストランデッド・SDNAシーケンシング・アズ・ア・チョイス・フォア・DNAシーケンシング」、(”Double Stranded SDNA Sequencing as a Choice for DNA Sequencing”) Nucleic Acids Research 16:1220,1988
(8) ダーム・ジェイビイ、ウィリアムズ・ジェイエル、マッカッチャン.ティーエフ(Dame JB, Williams JL, McCutchan TF)ら、「ストラクチャー・オブ・ザ・ジーン・エンコーディング・ジ・イムノドミナント・サーフェイス・アンチゲン・オン・ザ・スポロゾイツ・オブ・ザ・ヒューマン・マラリア・パラサイト・プラスモジウム・ファルシパラン」(”Structure of the Gene Encoding the Immunodominant Surface
Antigen on the Sporozoites of the Human Malaria Plasmodium falciparum”) Scinence 225:593−599,1984.
(9) バレンツェラ・ピー、グレイ・ピー、キロガ・エム(Valenzuela P, Gray P, Quiroga M)ら、「ヌクレオタイド・シーケンシズ・オブ・ザ・ジーン・コーディング・フォア・ザ・メイジャー・プロテイン・オブ・ヘパティティスBバイラス・サーフェイス・アンチゲン」(”Nucleotide Sequences of the Gene Coding for the Major Protein of Hepatitis B Virus Surface Antigen”) Nature 280:815−819,1979.
【図面の簡単な説明】
【0069】
【図1】第1図(A)S発現カセットおよび(B)RTS発現カセットの制限エンドヌクレアーゼ地図を示す。各コーディング配列の範囲を黒色バーで示す。
【図2】プラスミドpRIT13034の構築を示す第2図である。Xholエンドヌクレアーゼで消化し、遊離末端と定住Tyエレメントとの相同性により酵母染色体に組み込むためのS発現カセットを含む8.5kb直線状DNAフラグメントを遊離させる。
【図3】pRIT13034由来の8.5kb直線状Xholフラグメントの制限地図を示す第3図である。該直線状フラグメントは、S発現カセットと共に形質転換された酵母細胞を選択するためのURA3およびCUP1遺伝子を有する。
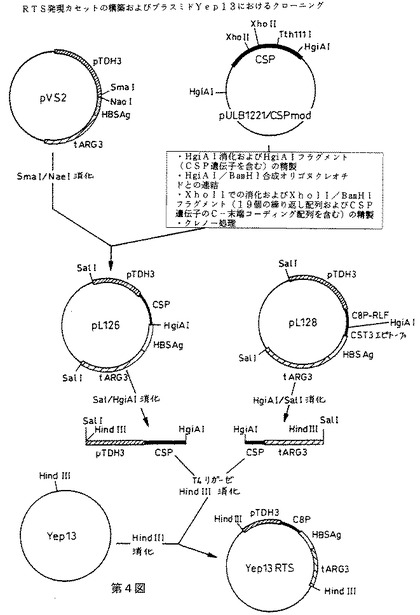
【図4】RTS発現カセットの構築およびプラスミドYep13におけるクローニングを示す第4図である。
【図5−1】RTS発現カセットのヌクレオチド配列およびRTS−HBsAgハイブリッドタンパク質の推定翻訳産物を示す第5図である。TDH3 ATGコドンより開始する翻訳産物をDNA配列の下に示す。
【図5−2】第5図の続きを示す。
【図5−3】第5図の続きを示す。
【図5−4】第5図の続きを示す。
【図5−5】第5図の続きを示す。
【図6】プラスミドpRIT13539の構築を示す第6図である。BglIIエンドヌクレアーゼで消化し、遊離末端と定住Tyエレメントとの相同性により酵母染色体に組み込むためのRTS発現カセットを有する6.8kb直線状DNAフラングメントを遊離させる。
【図7】pRIT13539由来の6.8kb直線状BglIIフラグメントの制限地図を示す第7図である。該直線状フラグメントは、RTS発現カセットと共に形質転換された酵母細胞を選択するためのLEU2遺伝子を有する。
【図8】RIT4383より調製した細胞不含抽出物のCsCl密度分析を示す第8図である。フラクションをHBsAgに対して特異的なラジオイムノアッセイ(AUSRIA)およびRTS−特異的ELISAにより分析した。上部のパネルは、抗−S Mabを用いるフラクションのイムノブロット分析結果を示す。ピークフラクションの浮遊密度(rho)をその屈折率の測定値により算定した。
【図9−1】RTS*発現カセットのヌクレオチド配列およびハイブリッドタンパク質の推定翻訳産物を示す第9図である。
【図9−2】第9図の続きを示す。
【図9−3】第9図の続きを示す。
【図9−4】第9図の続きを示す。
【図9−5】第9図の続きを示す。
【図9−6】第9図の続きを示す。
【図9−7】第9図の続きを示す。
【技術分野】
【0001】
本発明はハイブリッド蛋白質、その医薬における使用、特にマラリア感染症の予防における使用、およびこれを含有するワクチンに関する。
【背景技術】
【0002】
マラリアは、世界の主要な衛生問題のうちの一つであり、毎年2〜4百万人の人がこの病気でなくなっている。この病気の最も急性の形態は、原虫寄生体、熱帯熱マラリア原虫(Plasmodium falciparum)によっておこり、マラリアによる死亡のほとんどはこれが原因である。
【0003】
熱帯熱マラリア原虫の生活環は複雑で、完結するためにはヒトおよび蚊の2種の宿主を必要とする。人の感染は感染した蚊の唾液中のスポロゾイトの接種により始まる。スポロゾイトは肝臓に移動し、そこで肝細胞を感染させ、赤血球外細胞内段階を経てメロゾイト段階に分化し、赤血球(RBC)に感染して、無性血液段階の周期的複製を開始する。サイクルは、RBC中の多数のメロゾイトが有性段階ガメトサイトに分化し、これが蚊により摂取され、中部栄養管の一連の段階を経て発達し、スポロゾイトを産生し、これが唾液腺に移動することにより完了する。
【0004】
熱帯熱マラリア原虫のスポロゾイト段階はマラリアワクチンの有効な目標である。スポロゾイトの主要な表面蛋白質は、サーカムスポロゾイト蛋白質(CS蛋白質)として知られる。この7G8株由来の蛋白質はクローンし、発現し、配列決定されている(非特許文献1参照)。7G8株由来の蛋白質は、37回繰り返す4ペプチドAsn−Ala−Asn−Proを含む中央免疫優性繰り返し領域に4個の副繰り返し配列Asn−Val−Asp−Proが散在していることを特徴とする。他の菌株においては、主および副繰り返し配列の数およびその相対的位置が変わる。CS蛋白質の繰り返しのない部分として示される非繰り返しアミノ酸配列からなるNおよびC末端部分がこの中心部分に隣接している。
【0005】
照射したスポロゾイトは、実験的なヒトマラリアに対して著しい保護作用を与える(非特許文献2参照)。しかし、生産が困難なため、照射したスポロゾイトは、ワクチン製造の点から実用的ではない。
サーカムスポロゾイト蛋白質ベースのサブユニットワクチンが提案されている。これらのうち2種のワクチンは臨床試験が行われている。1つは合成ペプチドで、もう1つは組み換え蛋白質である(非特許文献3および非特許文献4参照)。
【0006】
【非特許文献1】ダーム(Dame)ら、サイエンス(Science)225(1984)p593)
【非特許文献2】アメリカン・ジャーナル・オブ・トロピカル・メディスン・アンド・ハイジエン(Am.J.Trop.Med.Hyg.)24:297−402,1975
【非特許文献3】バロウ(Ballou)ら、ランセット(Lancet):i 1277(1987)
【非特許文献4】ヘリントン(Herrington)ら、ネイチャー(Nature)328:257(1987)
【発明の開示】
【発明が解決しようとする課題】
【0007】
これらのワクチンは抗スポロゾイト応答を刺激するのに成功している。しかし、応答の大きさは期待はずれで、応答を全く起こさないワクチンもある。更に、その後の注射に関して抗体レベルの「増加」がなく、in vitro リンパ球増殖検定の結果から、これらの被験者のT細胞のほとんどは、優性繰り返し配列を認識しないことがわかった。それでも、各研究のワクチンは寄生虫血症を発現しなかった。
【課題を解決するための手段】
【0008】
本発明は、体液性応答だけでなく細胞性免疫応答も産生する、マラリアワクチンにおいて使用する新規な改良された抗原を提供する。好ましくは、抗原は免疫優性繰り返し配列に対する中和抗体の産生を誘起する。最も好ましくは、抗原は、CD4+およびCD8+細胞毒性Tリンパ球(CTL)型および遅延型過敏性エフェクターT細胞媒介免疫応答を誘起し、好ましくはTへルパー(TH)記憶細胞も誘起できるものである。
【発明を実施するための最良の形態】
【0009】
従って、本発明は、CS蛋白質の実質的に全てのC末端部分、免疫優性領域の4以上のタンデム繰り返し配列、およびB型肝炎ウイルス由来の表面抗原(HBsAg)を含むハイブリッド蛋白質を提供する。好ましくは、該ハイブリッド蛋白質は、CS蛋白質のC末端部分と実質的に同じである最低160個のアミノ酸を含む配列を含む。CS蛋白質はC末端から最後の12個のアミノ酸を除いたものであってもよい。
【0010】
特に、HBsAgのN末端と直線的リンカーによりフレーム中に融合された熱帯熱マラリア原虫7G8のアミノ酸210〜398に実質的に対応する熱帯熱マラリア原虫のCS蛋白質の一部を含む蛋白質が提供される。該リンカーはHBsAg由来のpreS2の一部を含んでもよい。
【0011】
本発明は、本発明の蛋白質をコードするDNA配列も提供する。
【0012】
特に好ましい具体例は、RTSで表わされるハイブリッド蛋白質である。RTSのアミノ酸配列を第5図に示す。このハイブリッド蛋白質は、
サッカロミセス・セレビシエTDH3遺伝子配列由来のヌクレオチド1059〜1061によりコードされるメチオニン残基[ムスティ・エイ・エム(Musti A.M.)ら、ジン(Gene)1983 25 133−143];
ハイブリッド遺伝子を構築するのに用いられるクローニング法により得られるヌクレオチド配列(1062〜1070)由来の3個のアミノ酸:Met Ala Pro;
熱帯熱マラリア原虫7G8株のサーカムスポロゾイト蛋白質(CSP)のアミノ酸210〜398に相当するヌクレオチド1071〜1637によりコードされる189個のアミノ酸(Dameら、前出);
ハイブリッド遺伝子を構築するために用いられるクローニング法により得られるヌクレオチド1638〜1640によりコードされるアミノ酸(Arg);
ヌクレオチド1641〜1652によりコードされ、B型肝炎ウイルス
(adw血清型)のpreS2蛋白質(9)の4個のカルボキシ末端残基に相当する4個のアミノ酸、Pro Val Thr Asn;
ヌクレオチド1653〜2330によりコードされ、B型肝炎ウイルス(adw血清型)のS蛋白質を特定する226個のアミノ酸
を含む。
【0013】
別の具体例において、熱帯熱マラリア原虫NF54(モレキュラー・アンド・バイオケミカル・パラサイトロジー(Mol.Biochem Parasitol),35:185−190,1989)由来のCSP遺伝子配列を用いて産生され、熱帯熱マラリア原虫NF54由来のCS蛋白質の207〜395領域の実質的に全部を含むRTS*で表わされるハイブリッド蛋白質が提供される。
特に、RTS*は以下のものを含む:
TDH3遺伝子配列由来のヌクレオチド1059〜1061によりコードされるメチオニン;
ハイブリッド遺伝子を構築するのに用いられるクローニング法によりえられるヌクレオチド配列(1062〜1070)由来の3個のアミノ酸:Met Ala Pro;
熱帯熱マラリア原虫NF54株のサーカムスポロゾイト蛋白質(CSP)のアミノ酸207〜395に相当するヌクレオチド1071〜1637によりコードされる189個のアミノ酸鎖(モレキュラー・アンド・バイオケミカル・パラサイトロジー,35:185−190,1989);
ハイブリッド遺伝子を構築するために用いられるクローニング法によりえられるヌクレオチド1638〜1640によりコードされるアミノ酸(Gly);
ヌクレオチド1641〜1652によりコードされ、B型肝炎ウイルス(adw血清型)preS2蛋白質の4個のカルボキシ末端残基に相当する4個のアミノ酸、Pro Val Thr Asn(ネイチャー 280:815−819,1979);
ヌクレオチド1653〜2330によりコードされ、B型肝炎ウイルス(adw血清型)のS蛋白質を特定する226個のアミノ酸鎖(ネイチャー 280:815−819,1979)。
【0014】
RTS*のアミノ酸配列を第9図に示す。
RTS*を含有する発現カセットを構築し、これは以下の様な特性を有する:
エス・セレビシエTDH3遺伝子由来のヌクレオチド1から1058まで伸びるプロモーター配列;
ヌクレオチド1059から始まり、ヌクレオチド2330まで伸びるオープンリーディングフレーム。このオープンリーディングフレームは、そのすぐ後に翻訳終止コドンTAA(ヌクレオチド2331〜2333)が続く。該オープンリーディングフレームは、ハイブリッドRTS*蛋白質を特定するアミノ酸をコードする。
エス・セレビシエARG3遺伝子由来の塩基対2334から3504まで伸びる配列中に含まれる転写終止配列。
【0015】
ヌクレオチド配列を第10図に示す。
【0016】
本発明の蛋白質をコードするDNA配列は、好ましい具体例においては、好ましくは酵母遺伝子由来の転写調節エレメントに隣接し、発現ベクター中に含まれる。
このようなベクターは本発明の別の態様である。好ましいプロモーターは、エス・セレビシエTDH3遺伝子由来のプロモーターである(ムスティら、前出)。
【0017】
本発明はまた、本発明にかかるベクターで形質転換された宿主細胞に関する。宿主細胞は、原核生物であっても真核生物であってもよいが、好ましくは、エス・セレビシエなどの酵母である。このような宿主においては、ハイブリッド蛋白質、例えばRTSは、リポタンパク質粒子として発現される。選択された受容体酵母株は好ましくはすでにそのゲノム中にいくつかの組み込まれたB型肝炎S発現カセットのコピーを有する。得られた株は2種のポリペプチド、SおよびRTS(または他の本発明のハイブリッド蛋白質)を合成し、リポタンパク質粒子の混合物(たとえば、RTS、SまたはRTS*、S)中に自発的にコアセンブルされる。これらの粒子はその表面でハイブリッドのCSP配列を示すのが有利である。これらの混合粒子もまた、本発明に含まれる。有利には、これらの混合粒子中のRTS:SまたはRTS*:Sの比率は1:4である。
【0018】
本発明はまた、適当な希釈剤または担体との混合物中に免疫保護量の本発明の蛋白質または粒子を含むワクチンに関する。
【0019】
本発明のワクチンにおいて、ハイブリッドの水溶液を直接用いてもよい。あるいは、あらかじめ凍結乾燥した、あるいはしていない蛋白質を公知のアジュバントと混合または吸収させてもよく、このようなアジュバントとしてはミョウバン、ムラミルジペプチド、Quil Aなどのサポニンが挙げられるが、これに限定されるわけではない。
【0020】
免疫促進物質も選択的にまたは追加的に含まれる。この免疫促進物質の好ましい具体例は、3脱アシル化モノホスホリルリピッドA(3D−MPL)である。
3脱アシル化モノホスホリルリピッドAはアメリカ合衆国特許第4912094号およびイギリス特許出願第2220211号(リビ(Ribi))から公知であり、米国、モンタナ州、ルビ・イムノケム(Ribi Immunochem)から入手可能である。
【0021】
本発明の蛋白質もこのようなリポソームの微粒子中に封入されてもよい。
【0022】
ワクチン調製物は、ニュー・トレンズ・アンド・デベロプメンツ・イン・バクシンズ(New Trends and Developments in Vaccines,ボーラー(Voller)ら編,ユニバーシティ・パーク・プレス(University Park Press),ボルチモア,メリーランド州,米国,1978)に記載されている。リポソーム中の封入は、例えばフュラートン(Fullerton)(アメリカ合州国特許第4235877号)により記載されている。
【0023】
通常のアジュバントを用いてもよいが、好ましい免疫促進物質は3脱アシル化モノホスホリルリピッドA(3D−MPL)である。
典型的には、3D−MPLを用いた場合、抗原および3D−MPLはミョウバンを加えるかまたは水中油型乳剤中に処方される。3D−MPLはエフェクターT細胞応答の刺激物質であるのでこれを混ぜるのが有利である。
【0024】
従って本発明の好ましい具体例は、本明細書中に記載したように、3D−MPLおよび担体と組み合わせたハイブリッド蛋白質、好ましくはRTSまたはRTS*を含むワクチンを提供する。典型的には、担体は水中油型乳剤またはミョウバンである。
【0025】
最も好ましい具体例においては、ハイブリッド蛋白質は本明細書において記載したような粒子または混合粒子である。
【0026】
各ワクチン用量中の本発明の蛋白質の量は典型的ワクチン中著しい副作用を起こさずに免疫保護応答を誘起できる量に選択される。このような量は、用いた特定の免疫原およびワクチンがアジュバントを含むかどうかにより変わる。一般的に、各用量は1〜1000μg、好ましくは1〜200μg、最も好ましくは10〜100μgの蛋白質を含む。特定のワクチンについて最適な量は、抗体価の観察および他の患者における他の応答を含めた標準的研究により決定できる。初回接種の後、患者は好ましくは約4週間で追加免疫を受け、その後感染の危険がある間は6ヵ月ごとに繰り返し追加免疫を行う。本発明の蛋白質に対する免疫応答はアジュバントおよびまたは免疫促進物質の使用により向上する。
【0027】
本発明の蛋白質は好ましくは酵母、特にサッカロミセス属に属する酵母中に発現される。
【0028】
本発明の別の態様は、本発明のハイブリッド蛋白質の調製法を提供することであり、該方法は、該蛋白質をコードするDNA配列を適当な宿主、好ましくは酵母中で発現し、生成物を回収することからなる。
【0029】
本発明の蛋白質をサッカロミセス株中で発現することが特に好ましい。RTSを例えばこのような菌株中で発現した場合、これは自発的にマルチマーリポタンパク質粒子中で集合する。
これらの粒子は免疫原性が高く、強い体液性応答ならびに免疫学的記憶を誘起し、またCTLおよびDTH型のエフェクターT細胞を誘起することができる。
【0030】
本発明の更に別の態様は、既に記載したように有効量のワクチンを投与することによるマラリア原虫感染症にかかっている患者の治療法である。
【実施例1】
【0031】
1.RIT4383株のRTS、Sの構築
SおよびRTSポリペプチドの両方を含有する粒子の産生に用いるエス・セレビシエRIT4383株は、各蛋白質に対して別の発現カセットを有する。S遺伝子発現カセットは定住Tyレトロトランスポンソンズと相同な直線状組み込みベクターを用いてゲノム中の最低2個の部位で5〜6個のコピー中に組み込まれる。RTS遺伝子発現カセットは、S遺伝子カセットの組み込みに用いたものと同様の直線状組み込みベクターを用いてゲノム中の1または2個の部位で2〜3個のコピー中に組み込まれる。両タイプのカセットからの発現は、酵母TDH3遺伝子由来のプロモーターにより行われる。
【0032】
1.1発現カセットおよび組み込みベクター(pRIT13034)の構築
S遺伝子発現カセット(第1A図)はRIT4376株(1)中に見られるものと同じであり、1058bpのTDH3プロモーターフラグメント、681bpのS遺伝子配列、6bpのEcoR1リンカーを含むスペーサーおよびArg3転写ターミネーターを有する1165bpのフラグメントからなる。Sコーディング配列は、adw血清型のウイルスの完全ゲノムクローン(pRIT10616)からのサブクローニングにより得られる。
S発現カセットの酵母ゲノム中への組み込みに用いられるTyベクター、pRIT13034の構造を第2図に示す。このタイプの酵母ゲノム中の発現カセットの組み込み用ベクターの構造および使用は、ジェイコブズ(Jacobs)ら(2)に記載されている。pRIT13034はS遺伝子カセット、URA3遺伝子およびpUC9上にクローンされたTyエレメントのコピー中に挿入されたCUP1遺伝子を含有する。URA3遺伝子およびCUP1遺伝子は両方とも形質転換された酵母コロニーを、形質転換されていないコロニーから区別する選択マーカーを提供する。pUC9は、ビエイラ&メッシング(Vieira & Messing)(3)に記載されており、URA3フラグメントはpFL44由来であり[エフ.ラクロート(F.Lacroute),CNRS,ストラスブール]、CUP1フラグメントはpCUP5由来であり(ティー・ブット(T.Butt,)SKF Labs,フィラデルフィア)、 Tyエレメントはジュニアウス(Juniaux)ら(4)からのものであり、S遺伝子カセットはpRIT12353由来である。pRIT13034のXhoIエンドヌクレアーゼを用いた消化により、第3図に示す8500bpの直線状フラグメントが遊離し、これを定住Tyエレメントと遊離末端の相同な組み換えによりゲノム中に組み込むことができる。
【実施例2】
【0033】
Y1295株の構築
受容体株EJ cup1D−3d(trp1、leu2、ura3、cup1、gal1D、MATα)をpRIT13034由来の直線状ベクターフラグメントの形質転換による一次導入に用いた。この株は1個の破砕された遺伝子座cup1、cup1を含む。
直線状XhoIフラグメントで形質転換した後、Ura+コロニーを単離し、銅耐性に関してスクリーンする。サザンブロッティング分析で測定すると、より耐性の強い形質転換体が2〜5個のベクターのコピーを組み込んでいた。2個の形質転換体コロニーのコピー、推定3〜4個のコピーを有するMS9および4〜5個の組み込み直線ベクターのコピーを有するMS23が保持されている。MS23株を次にEJ cup1D−7b株(trp1、ura3、cup1、gal1D、MATα)と交雑させ、ハプロイド有子嚢胞子を回収してMS23−2aを得る。
この菌株を次にMS9と戻し交雑すると得られたLeu−、Trp−ハプロイド分離体(MS54−2c)は5〜6個の組み込まれた発現カセットを含む。サザンブロッティングによると、MS54−2Cは1遺伝子座に4〜5個の組み込みベクターのタンデムコピーを有し、更に別の遺伝子座に組み込まれた1個のコピーを有する。発現カセットの5〜6個のコピー全てはカセットフラグメントを遊離するための全酵母細胞DNAのHindIIIエンドヌクレアーゼでの消化を受けず、S遺伝子特異性プローブを用いたサザンブロッティングにより予想される3kbのバンドが得られる。この菌株の自発的Trp+復帰変異体、MS54−2c−Tが得られる。実験室受入番号Y1295。
【実施例3】
【0034】
RTS発現カセットの構築
RTSハイブリッド蛋白質の発現カセットを、多段階のクローニング法により構築し、イー・コリ酵母シャトルベクターYep13(6)中にクローンして、プラスミドYep13RTS(第4図)を得る。カセットの構造を第1B図に示す。全ヌクレオチド配列を直接配列決定(コーディング配列および非翻訳調節配列の一部に関して)または関連する文献を参考にして(プロモーターおよびターミネーター配列の一部に関して)決定した。DNA配列を第5図に示す。これは以下の要素を含有する:
エス・セレビシエTDH3遺伝子由来のヌクレオチド1から1058まで伸びるプロモーター配列;
ヌクレオチド1059で始まり、ヌクレオチド2330まで伸びるオープンリーディングフレーム。このオープンリーディングフレームのすぐ後には翻訳終止コドンTAA(ヌクレオチド2331から2333)が続く。オープンリーディングフレームはハイブリッドRTS蛋白質を特定するアミノ酸をコードする;
エス・セレビシエARG3遺伝子(クラビール(Crabeel)ら EMBO J.1983 2:205−212)由来の、2334から3504塩基対まで伸びる配列中に含まれる転写停止配列。
ヌクレオチド1059から2330によりコードされるハイブリッドRTS蛋白質のアミノ酸配列を第5図に示し、これは以下の要素を含む:
TDH3遺伝子配列由来の、ヌクレオチド1059から1061によりコードされるメチオニン残基;
ハイブリッド遺伝子を構築するのに用いられるクローニング法により産生されるヌクレオチド配列(1062〜1070)由来の3個のアミノ酸MetAla Pro;
熱帯熱マラリア原虫7G8株(8)のサーカムスポロゾイト蛋白質(CSP)のアミノ酸210〜398に相当するヌクレオチド1071〜1637によりコードされる189個のアミノ酸鎖;
ハイブリッド遺伝子を構築するのに用いられるクローニング技法により産生されるヌクレオチド1638〜1640によりコードされるアミノ酸(Arg);
ヌクレオチド1641から1652によりコードされ、B型肝炎ウイルス(adw血清型)preS2蛋白質(9)の4個のカルボキシ末端残基に相当する4個のアミノ酸Pro Val Thr Asn;
ヌクレオチド1653から2330によりコードされ、B型肝炎ウイルス(adw血清型)を特定する226個のアミノ酸鎖。
【実施例4】
【0035】
RTSカセット組み込みベクターpRIT13539の構築
RTSカセット組み込みベクターpRIT13539の構造を第6図に示す。
RTS発現カセットをTyベースの組み込みベクターpRIT13144上に挿入する。このベクターは、LEU2遺伝子がベクターpCV9(7)から単離されたSalI−XhoIフラグメントとしてTyエレメントのSalI部位中に挿入されているpRIT12927(2)の誘導体である。従って、RTS発現カセットをpRIT13144中に挿入した後、得られたプラスミドpRIT13539は発現カセットに加えて、選択マーカーとしてLEU2遺伝子を含有する(第5図)。pRIT13539をBglIIエンドヌクレアーゼで消化すると、第7図に示す6800bpの直線状フラグメントが遊離され、これは定住Tyエレメントとの遊離末端の相同組換によりゲノム中に組み込むことができる。
【実施例5】
【0036】
実施例5a Y1295株の形質転換およびRIT4383株(Y1530)の産生
SおよびRTS蛋白質の両方を発現する株を得るために、Y1295を、Leu+コロニーに関する選択を有する6800bpの直線状BglIIフラグメントを用いて形質転換する。種々の割合でゲノム中に存在する両発現カセットのセットを含む数個の組み込み体が得られる。約1:4の割合でRTSおよびS蛋白質を発現する選択された形質転換体の一つは公式名称はRIT4383(実験室受入番号Y1530)である。
【0037】
実施例5b Y1295株の形質転換およびY1631株の産生
(RTSと)類似の構造を、熱帯熱マラリア原虫NF54株(モレキュラー・アンド・バイオケミカル・パラサイトロジー(Mol.Biochem.Parasitol.)35:185−190,1989)由来のCSP遺伝子配列を用いて産生する。得られた融合蛋白質は、熱帯熱マラリア原虫7G8株由来のCSPを用いてえられる構造物と区別するためにRTS*と表わす。
発現カセットの配列を第9図に示す。これは以下のエレメントを含む:
エス・セレビシエTDH3遺伝子由来のヌクレオチド1から1058まで伸びるプロモーター配列;
ヌクレオチド1059で始まり、ヌクレオチド2330まで伸びるオープンリーディングフレーム。このオープンリーディングフレームのすぐ後には翻訳終止コドンTAA(ヌクレオチド2331から2333)が続く。オープンリーディングフレームはハイブリッドRTS*蛋白質を特定するアミノ酸をコードする;
エス・セレビシエARG3遺伝子由来の、2334から3504塩基対まで伸びる配列中に含まれる転写終止配列。
RTS*融合蛋白質をコードするこの発現カセットを、RTS構造物に関して既に記載したのと同じ方法を用いて形質転換し、酵母Y1295株のゲノム中に形質転換し、組み込む。SおよびRTS*蛋白質の両方を発現する形質転換されたクローンが得られる。1つのクローンを、約4S:1RTS*の割合で2個の蛋白質を発現するように選択する。クローンの実験室受入番号はY1631である。
【実施例6】
【0038】
RIT4383株の予備特質化
6.1イムノブロッティング分析
細胞を含まないRIT4383から調製した抽出物を種々の抗体を用いたイムノブロットにより分析した:
S蛋白質に対する単クローン性抗体(Mab HBS1);
RTS蛋白質(Mab167)の繰り返し部分に対する単クローン性抗体;
RTS蛋白質(ウサギ血清No.20)の繰り返しのない配列に対するウサギ血清。
酵母RIT4383株において、2個の発現された生成物を単クローン性抗体HBS1により認識する。即ち、S蛋白質に対応する24KD蛋白質および46KDのRTSハイブリッド蛋白質である。RTSハイブリッド蛋白質も、CSPの繰り返し、および非繰り返しエピトープに対する抗体により検出される。これらの結果から明らかに、RIT4383株が同時に2個のSおよびRTS抗原を、RTS/Sが約1:4の割合で発現することがわかる。
【0039】
6.2CsCl密度勾配遠心
RIT4383株中の粒子の形成を、CsCl密度勾配遠心により分析する。粗抽出物(総蛋白質の±15mg)を、10mlの1.5M CsCl勾配上分析する(40000rpm、+8℃、ベックマン50Tiローター中68時間)。フラクション(0.5ml)を集め、HBsAg(AUSRIA)に対して特異的な放射免疫検定法、RTS特異的ELISAおよび抗HBsAg単クローン性抗体を用いたイムノブロットにより分析する。
第8図に示すように、ELISA、RIAおよびウェスタンブロットピークは、rho=1.21の浮遊密度に対応する勾配の同一フラクション(no.13)で現われ、このことは、SおよびRTSモノマーの両方を含む混合粒子がこの株中に形成されたことを示す。
【実施例7】
【0040】
シードロットの調製
マスターシードロットの製造法
Y1530株(RIT4383)をまず20mlの滅菌YNB(Difco)(デキストロース(0.1%)および1.8%(w/v)寒天(Difco)で補足)をいれたペトリ皿中で30℃で48時間成育させる。表面成長株を滅菌YNBブロス(Difco)(デキストロース(1%)およびグリセロール(10%)を追加)中に懸濁する。この懸濁液を無菌条件下で2mlのポリプロピレンの栓をした滅菌試験管中に分配し(試験管1本につき1ml)、−70℃で貯蔵する。
【0041】
ワーキングシードロットの製造法
マスターシード試験管の1本をすばやく解凍し、その内容物を白金耳で前記のように調製したペトリ皿上に取り出す。30℃で63時間培養した後、表面成長物の一部を、400mlの滅菌YNBブロス(Difco)(デキストロース(2%)を追加)をいれた2Lコニカルフラスコに移す。培養物を30℃で24時間培養した後、無菌条件下で遠心する(6300xgで15分)。ペレットを無菌YNBブロス(Difco)(デキストロース(1%)およびグリセロール(10%)を追加)中に再懸濁する。これを無菌条件下で2mlのガラス栓をした滅菌試験管中に分配し(試験管1本につき0.5ml)、−70℃で貯蔵する。
【実施例8】
【0042】
発酵
接種物の調製
(a)固体培地上の成育
バイアル1つ分のワーキングシードロットを素早く解凍し、デキストロース(1%)および1.8%(w/v)寒天(Difco)を追加した滅菌YNBブロス(Difco)を含有するペトリ皿上に分散させる。ペトリ皿を300℃で48時間培養する。
(b)接種物の成育
ペトリ皿の表面成育物をデキストロース(2%)を追加した滅菌YNBブロス(Difco)中に懸濁し、2個のコニカルフラスコ(2L、フラスコ1個あたり400mlの液体)中に等分する。フラスコをロータリーシェーカー上で30℃で24時間培養する。
【0043】
発酵
5Lの脱イオン水((NH4)2SO4(40g)を追加)を入れた発酵槽(総容量20L)をそのまま清浄なあらかじめフィルターを通した蒸気を用いて2バールgの圧力で121℃で30分間滅菌する。室温に冷却した後、発酵槽中の液体の体積を4Lに調節し、フィルター滅菌したHB4溶液1Lを添加する。発酵は、2個のコニカルフラスコからの接種物(800ml)を添加することにより開始する。
発酵は、フィードバッチ技法を用いて行い、これにより培養物を以下の組成の溶液と共に連続して供給する:
5LのHB4溶液;
4Lのデキストロース80%(121℃で滅菌)
培養密度は30℃およびpH5(NH4OHを添加することにより維持する)での好気的成育により増加する。溶解酸素を、気流および撹拌速度を調節して約50%飽和に維持する。供給物の添加速度は成育速度が最大になり、副生成物のエタノールの形成が最小になるようにあらかじめ決定しておく。
40〜90時間後に発酵を停止する。発酵の終わりで、全培養物体積は10〜18Lであり、乾燥細胞重量は30および100g/Lの間である。培養物を迅速に15〜25℃に冷却し、酵母細胞を遠心分離によりブロスから回収する。濃縮した酵母細胞を、再遠心前にリン酸緩衝液(50mM)で1度洗浄し、続いてポリエチレンの袋中で−70℃で貯蔵する。
【0044】
【表1】
【実施例9】
【0045】
RTS/Sの抽出および精製
抽出法
9.1細胞懸濁液の精製
凍結濃縮酵母細胞(−70℃)を一夜−30℃で解凍し、続いて細胞の入ったポリエチレンの袋を水中(10〜20℃)につけることにより5〜15℃で完全に解凍する。酵母細胞懸濁液を以下の成分を含有する燐酸緩衝溶液(pH8.0)を用いて調製する:エチレンジアミン四酢酸(EDTA)、p−メチルスルホニルフルオリド(PMSF)、イソプロパノールおよびtween20。
【0046】
9.2細胞破砕
細胞を鉛を含まないガラスビーズ(直径0.49〜0.70mm)を含むビーズミル中で破砕する。−20℃の冷却液を循環させて粉砕室を冷却し、粉砕室出口でのホモジネートの温度が15℃を越えないようにする。液体流速は6L/時間、撹拌速度は3000rpmである。このプロセスを2回行う。得られたホモジネートのpHは6.7〜7.5である。
【0047】
9.3ポリエチレングリコール清澄化
ポリエチレングリコール400(PEG400)を破砕した細胞懸濁液に添加して、最終濃度を10%にし(10℃未満で30分、pH6.1)、部分的に清澄化された上清を遠心分離により回収する(J21Bベックマン遠心分離機、JA10ローター、17000gで1時間)。
【0048】
9.4メタノール清澄化
メタノールをpH7でPEGで清澄化した抗原に添加して、5容積のPEGで清澄化した抗原に対してメタノール1容積の割合にする。17000gで20分間遠心分離により清澄化された抗原をえる(J21Bベックマン遠心分離機、JA10ローター)。
【0049】
9.5コロイドシリカ上への吸着/脱着
粗抗原を一夜1.5%(w/v)コロイドシリカ(アエロジル380、デグサ)上に4℃で吸着させる。
ペレットを連続して遠心分離し、NaCl0.9%(w/v)中に再懸濁することにより洗浄(3回)後、1%TWEEN20を含有する10mMピロリン酸緩衝液pH9.5を用いて抗原を脱着させる。
脱着緩衝液体積はメタノールで清澄化した抗原溶液の1/8に相当する。脱着された抗原溶液をL8.70ベックマン超遠心分離ローターR19で、50000gで60分間超遠心分離により回収する。
【0050】
9.6ジアフィルトレーション
精製工程の前に、脱着された抗原を5容積の6Mの尿素、500mMのNaCl、20mMのTRIS−HClでpH8.5で超遠心分離することにより洗浄し、蛋白質および脂質汚染物の大部分を除去する。
つぎに緩衝液を同じシステム(Ultrasette、300kDの公称カットオフのポリスルホン膜を備えたFILTRON)で5容積の10mMのTRIS−HClでpH8.1と交換する。
【0051】
9.7DEAE−TSK650(M)上イオン交換クロマトグラフィー
清澄化した溶液を10mM TRIS緩衝液(pH8.1)で平衡化したアニオン交換カラム(DEAE−TSK650(M))にかける。連続して2容積の10mM TRIS−HCl緩衝液(pH8.1)および40mMNaClを追加した3容積の10mM TRIS−HCl緩衝液(pH8.1)で洗浄した後、1容積未満の150mM NaClを含有する10mM TRIS−HCl緩衝液(pH8.1)を用いて抗原を脱着する。抗原を含むフラクションをプールする。
【0052】
9.8Butyl−TSK650(M)上疎水性相互作用クロマトグラフィー
最終濃度が650mM NaClになるまでNaClを添加した後、抗原溶液を、20mM TRIS−HCl緩衝液、600mM NaCl(pH8.1)で平衡化したButyl-TSK650(M)カラムにかける。抗原の大部分は通過するが、不純物はゲルと結合する。
【0053】
9.9限外濾過による濃縮
HICプールを300kDaの公称カットオフのポリスルホン膜を備えたUltrasetteシステム(FILTRON)により濃縮する。
【0054】
9.10CsCl勾配中での超遠心
CsClをButyl−TSKプールに添加して、最終濃度を1.5Mにする。
50.2Tiベックマンローター中270000gで65時間後、抗原を含有するフラクションを集める。
【0055】
9.11SEPHACRYL S300(HR型)上でのサイズ排除クロマトグラフィー
緩衝液を交換し、低分子量の汚染物質を除去するために、抗原溶液をSEPHACRYL S300 HRカラムにかける。溶出緩衝液は、150mM NaClを含有する10mM燐酸緩衝液(pH7.4)である。
滅菌濾過
150および400μg Lowry/ml間に希釈し、pHを6.8に調節した後、精製した抗原を0.22μm滅菌フィルターを通して濾過することにより滅菌する。得られた溶液は、精製されたRTS/S粒子を含有する。
【実施例10】
【0056】
RTS/Sの免疫学的特性化
10.1抗原性
RTS/S粒子の抗原性を試験するために、異なるエピトープに対する単クローン性抗体を結合させてELISAを何回も行った。
用いた単クローン性抗体(MoAbs)は以下のとおりである:
−MoAb R10: ・CSP領域の繰り返し配列(NANP)に対して特異的
・IgMアイソタイプ
−MoAb RL117: ・CSP領域の非繰り返し配列に対して特異的
−MoAb RF1: ・HBsAgのS配列に対して特異的
・IgGアイソタイプ
Sp2/OAg骨髄種細胞をCSP配列の繰り返しおよび非繰り返し領域の両方を含有する完全に純粋でないRTS様粒子で免疫したBalb/Cマウスの脾細胞と融合することによりMoAb R10およびRL117を室中で調製する。
3バッチのRTS/Sワクチンを分析する:バッチナンバーは24M31、24M32および24M34である。
【0057】
10.2単クローン性抗体R10(抗−繰り返し)との反応
抗−繰り返しMoAb R10を「サンドウィッチ」ELISAに用いる。試験するサンプルをあらかじめMoAb R10をコートしたマイクロタイタープレート中で培養する。ペルオキシダーゼと結合した同じMoAbを次に添加する。37℃で1時間培養し、洗浄した後、オルトフェニレンジアミンH2O2を添加することにより発色する。490nmで吸光度を測定し、抗原濃度に対してプロットする。
結果
3バッチ:24M31、24M32および24M34はMoAb R10と陽性に、適合して反応するので、RTS/S粒子上に繰り返しエピトープが存在することが確認される。最大結合の50%に達するのに必要な抗原量はバッチ24M31、24M32および24M34についてそれぞれ51.2、38.2および60.6ngである。
【0058】
10.3単クローン性抗体RL117(抗−繰り返し)との反応
MoAb RL117のRTS、S粒子との反応性を「サンドウィッチ」ELISA試験で分析する。MoAb RL117はペルオキシダーゼと結合し、RTS/S粒子の検出に用いられ、繰り返し領域に関して特異的なMoAb R10は粒子の捕獲に用いられる。
簡単に言うと、試験するサンプルをあらかじめMoAb R10をコートしたマイクロタイタープレート中で培養する。ペルオキシダーゼと結合したMoAb RL117を次に添加し、37℃で1時間培養し、洗浄した後、オルトフェニレンジアミンH2O2を添加することにより発色する。490nmで吸光度を測定し、抗原濃度に対してプロットする。
結果
3バッチ:24M31、24M32および24M34はMoAb RL117およびR10と陽性に、適合して反応するので、同一のRTS/S粒子上に繰り返しおよび非繰り返しエピトープが存在することが確認される。最大結合の50%に達するのに必要な抗原量はバッチ24M31、24M32および24M34についてそれぞれ169.2、117.6および181.1ngである。
【0059】
10.4単クローン性抗体RF1(抗S)との反応
RTS/S粒子中のSエピトープの存在をペルオキシダーゼと結合したMoAb RF1を検出に用いて「サンドイッチ」ELISA試験により示す。R10MoAbはRTS、S粒子を捕獲するために、マイクロタイタープレート上に直接用いる。
要約すると、試験するサンプルをあらかじめMoAb R10をコートしたマイクロタイタープレート中で培養する。ペルオキシダーゼと結合したMoAb RF1を次に添加する。37℃で1時間培養し、洗浄した後、オルトフェニレンジアミンH2O2を添加することにより発色する。490nmで吸光度を測定し、抗原濃度に対してプロットする。
結果
3バッチ:24M31、24M32および24M34はMoAb RF1およびR10と陽性に反応するので、同じRTS/S粒子上に繰り返しおよびSエピトープが存在することが確認される。最大結合の50%に達するのに必要な抗原量はバッチ24M31、24M32および24M34についてそれぞれ52.3、55.2および106.2ngである。
【実施例11】
【0060】
RTS/S粒子のin vivoでの免疫原性
11.1免疫原性の研究
(RTS、S)粒子の免疫原性に関する試験を、マウスおよびセルコピテカス・イーチオプスサルにおいておこなった。
マウスにおいて、抗CSP抗体をR32tet32抗原を抗繰り返し抗体の検出に用いてELISA試験により分析する。R32tet32はプラスミドpBR322のテトラサイクリン耐性遺伝子の32個のアミノ酸部分と融合した32個のNANP(主)繰り返し配列からなる。組み換え抗原はエシェリヒア.コリ中で産生する。CS蛋白質の非繰り返し配列に対する抗体(Abs)をRLF抗原を用いたELISA試験により測定する。RLF抗原は、インフルエンザウイルスのNS1蛋白質の初めの81個のアミノ酸と融合した繰り返し配列を含まない完全CS蛋白質非翻訳領域からなる。RLF抗原をイー.コリ中で産生する。マウス血清を1:100から初めて連続的に希釈し、力価をELISA試験(1)において光学密度1.0に対応する希釈度の逆数で表わす。抗CSP Absの測定は、各血清に関して行い、力価の等比中項(GMT)を計算する。
抗担体応答を分析するために、抗HBs抗体力価も測定する(プールした血清のみ)。
マウスの試験において、RTS、S粒子の免疫原性を評価するために、HBsAgの免疫原性について通常用いられるBalb/Cマウス(H−2dハプロタイプ)も用いる。免疫化の腹腔内(i.p.)および皮下(s.c.)経路を比較し、免疫促進物質3−脱アシル化モノホスホリルリピッドA(3D−MPL)の免疫応答に関する効果も試験する。
(RTS、S)ワクチンの免疫原性も同様にセルコピテカス・イーチオプスサルにおいて試験する。
選択された個々のまたはプールした血清もそのプラスモジウム.ファルシパラムスポロゾイトによる人肝癌細胞系(HepG2)のin vitroの破壊を抑制する能力に関して試験する(ISI検定(2))。
【0061】
11.2マウスにおける免疫原性
実験1: Al(OH)3上に吸着した臨床的(RTS、S)バッチの免疫原性
方法
免疫化:
10匹のBalb/Cマウスに、あらかじめAl(OH)3上に吸着させた各1μgの3種の(RTS、S)バッチ(24M/31、24M/32および24M/34)を1ヶ月間隔で2回腹腔内注射する。対照マウスに、HBsAg(Engerix−B、バッチENG611B4)を注射する。30日目および45日目に、マウスを採血し、抗体価を各血清について測定する。
血清学的方法:
抗R32tet32および抗RLF価を、それぞれコーティング抗原としてR32tet32およびRLFを用いたELISAにより測定する。マイクロプレートを試験する血清サンプル(1:100から初めて2倍連続希釈12回)とともに培養する。マウスAbsをビオチニル化抗マウスIg、続いてストレプタビジン:ビオチニル化ホースラディッシュペルオキシダーゼコンプレックスおよびオルトフェニレンジアミン/H2O2と反応させる。光学密度を490nmで測定する。力価を光学密度1.0(最大結合の50%)に対応する希釈度の逆数として表わす。各グループのマウスに関して、力価の等比中項(GMT)を計算する。抗HBs抗体価をHollingerの式(3)にしたがって計算し、mIU/mlで表わす。
結果
抗CSP応答:
強い抗R32tet32および抗RLF応答が試験した各(RTS,S)バッチに関して観察された。バッチ間の明らかな違いはない。著しい追加抗原刺激の効果は2回目の投与の後観察される。HbsAg(「Engerix−B」)で免疫したマウスをこの実験において負の対照として用いる。
抗HBs応答:
(RTS、S)バッチはHBsAgキャリアー蛋白質に対する抗体を誘起する。試験はプールした血清のみに関して行った。
【0062】
実験2:Balb/Cマウスにおける(RTS、S)粒子の免疫原性に対する免疫促進物質3D−MPLの効果
本発明者らはマウスにおける(RTS、S)ワクチンの免疫原性に対する免疫促進物質3D−MPLの効果を分析した。腹腔内(i.p.)および皮下(s.c.)経路の両方の免疫化を試験した。
A.ip経路による免疫化
方法
免疫化:
10匹のBalb/CマウスのグループにAl(OH)3単独、またはAl(OH)3+3D−MPL(50μg/用量)上に吸着させた1μgの(RTS、S)(バッチ24M/34)を1ヶ月間隔で2回腹腔内注射する。対照マウスに、150mMNaClを注射する。30日目および45日目に、マウスを採血し、抗体価を各血清について測定する。
血清学的方法:
血清学的方法は前記の始めの実験においてと同様である。
結果
抗CSP応答:
(RTS,S)ワクチンバッチは強い抗R32tet32および抗RLF応答を両処方物において示す。全ての場合において、二回目の免疫化の後に著しい追加抗原刺激の効果が観察された。3D−MPLを含有する処方に関して得られた力価は全ての場合において、アルミニウムのみの処方物より高く、第一抗R32tet32応答(p=0.02)において統計的に著しい増加が観察された。150mM NaClを注射したマウスのグループをこの実験における負の対照として用いた。
抗HB応答:
腹腔内経路により注射された(RTS、S)ワクチン両処方物は強い抗HBs応答を誘起する。著しい追加抗原投与の効果はいずれかの処方物を用いた2回目の免疫化の後に観察される。
【0063】
B.s.c.経路による免疫化
方法
免疫化:
10匹のBalb/Cマウスの群にAl(OH)3のみ、またはAl(OH)3+3D−MPL(50μg/用量)上に吸着させた1μgの(RTS、S)(バッチ24M/34)を1ヶ月間隔で2回皮下注射する。対照マウスに、150mM NaClを注射する。30日目および45日目に、マウスを採血し、抗体価を各血清について測定する。
血清学的方法は前記の始めの実験においてと同様である。
結果
抗CSP応答:
(RTS,S)ワクチンバッチはR32tet32およびRLFに対して正の応答を両処方物において誘起する。全ての場合において、二回目の免疫化の後に著しい追加抗原刺激の効果が観察された。3D−MPL処方に関して抗R32tet32および抗RLF応答(それぞれp=0.18およびp=2.9)の両方に関して統計的に著しい高い力価が45日目に観察される。しかし、一般には、力価は腹腔内経路で観察されたものよりも低い。150mM NaClを注射したマウスのグループをこの実験における負の対照として用いた。
抗HBs応答:
腹腔内経路により注射された(RTS、S)ワクチンの両処方物は良好な抗HBs応答を誘起する。著しい追加抗原刺激の効果はいずれかの処方物を用いた2回目の免疫化の後に観察される。抗CSP応答に関して観察されたように、抗HBs応答は腹腔内経路に比べてこの経路に免疫化によるものは低い。
【0064】
11.3セルコピテカス・イーチオプスにおける免疫原性
Al(OH)3上に吸着した臨床的バッチ24M/32をもちいたセルコピテカス・イーチオプスサルにおける免疫原性を評価した。
方法
免疫化:
5匹のサルに、Al(OH)3(0.5mgAl+++)上に吸着させた20μgの(RTS、S)粒子を0、28および84日目に筋肉内注射する。動物を0、14、28、42、56、66および98日目に採血した。R32tet32、RLFおよびHBs抗原に対する抗体を測定した。
血清学的方法:
抗R32tet32および抗RLF抗体価を、マイクロプレート上にコートしたR32tet32およびRLF抗体をそれぞれ用いたELISAにより測定する。プレートをつぎに試験する血清サンプル(1:10から初めて2倍連続希釈12回)とともに培養する。サル抗体をビオチニル化抗ヒトIg、続いてストレプタビジン:ビオチニル化ホースラディッシュペルオキシダーゼコンプレックスおよびオルトフェニレンジアミン/H2O2により検出する。光学密度を490nmで測定する。力価を光学密度1.0(最大結合の50%)に対応する希釈度の逆数として表わす。各グループに関して、力価の等比中項(GMT)を計算する。抗HBs抗体価をHollingerの式(Hollingerら、1982)にしたがって計算し、mIU/mlで表わす。
結果
抗CSP応答:
(RTS,S)ワクチンはR32tet32およびRLF抗原の両方に対して5匹のサル全てにおいて正の応答を誘起した。著しい追加抗原刺激の効果は2回目の免疫化の14日後(42日目)に観察される。次に抗体価が3回目の免疫化までゆっくり減少するのが観察された。力価はサルJo352の抗RLFの場合以外は3回目の免疫化の14日後(98日)に再び増加した。
3回目の免疫化の後(98日)得られた抗R32tet32力価はしかし2回目の免疫化後(42日)に観察されたものよりも高くない。
抗RLF応答の場合(サルJo353を除く)、3回目の免疫化の後(98日)、2回目の免疫化後(42日)のレベルにくらべて力価の増加が観察された。
抗HB応答:
全てのサルは免疫化の2回目(42日)および3回目(98日)の後著しい追加抗原刺激の効果に関する抗HBs応答を示した。
【0065】
(RTS、S)粒子に対して得られた抗体の生物学的活性
(RTS、S)ワクチンにより誘起された抗体の生物学的機能の測定値として、プールしたマウスおよび個々のさる血清をスポロゾイト侵入阻止(Inhibition of Sporozoite Invation(ISI))検定により試験した(Hollingdaleら、1984)。この検定は熱帯熱マラリア原虫スポロゾイトによる人肝癌細胞系の
in vitroでの侵入を阻止する抗CSP抗体の能力を測定するものでる。
結果
この実験の結果を第1表および第2表に示す。ISI測定値を免疫前の対照血清の活性を抑制0%とした相対的な抑制(%)で表わす。参考のために、試験した血清の抗R32tet32およびRLF抗体価も含める。第1表は試験した全てのマウス血清が非常に強いISI活性を有することを示す。第2表は全ての5匹のサルは対応する免疫血清に比べて98日目に非常に強いISI活性を有することを示す。
結論
(RTS、S)粒子は、マウスおよびサルの両方においてCSPの繰り返しおよび非繰り返しエピトープに対しておよび、HBsAgキャリアーのS蛋白質に対して高い抗体応答を誘起する。
マウスにおける一次抗体応答は3D−MPLの存在により向上する。
腹腔内注射の後得られた抗体価は、皮下経路による免疫価の後得られたものよりも高い。
2種の動物において誘記された抗体は、熱帯熱マラリア原虫スポロゾイトによる培養された人肝癌細胞の侵入を効果的に予防した。
【0066】
【表2】
【0067】
【表3】
【0068】
引例
(1) ハンフォード・エヌ、カベゾン.ティー、コロー・ビー(Hanford N, Cabezon T, Colau B)ら、「コンストラクション・アンド・キャラクタライゼーション・オブ・ア・サッカロミセス・セレビシエ・ストレイン(RIT4376)エクスプレッシング.ヘパティティスBサーフェイス・アンチゲン」(”Construction and Characterization of a Saccharomyces Cerevisiae Strain(RIT4376)Expressing Hepatitis BSurface Antigen”), Postgrad Med J 63,Supp.2:65−70,1987
(2) ジェイコブズ・イー、ルトガーズ・ティー、ボート・ピー(Jacobs E, Rutgers T, Voet P)ら、「シマルタネアス・シンセシス・アンド・アセンブリ・オブ・ベリアス.ヘパティティスBサーフェイス.プロテインズ・イン・サッカロミセス・セレビシエ」(”Simultaneous Synthesis and Assembly of Various Hepatitis B Surface Proteins in Saccharomyces cerevisiae”), Gene 80:279−291,1989
(3) ビーラ・ジェイおよびメシング・ジェイ(Vieira J and Messing J)、「pUCプラスミズ、アンM13mp−デライブド・システム・フォア・インサーション・ミュータゲネシス.アンド.シーケンシング・ウィズ・シンセティック.ユニバーサル・プライマーズ」(”The pUC plasmids,an M13mp7−Derived System for Insertion Mutagenesis and Sequencing with Synthetic Universal Primers”),Gene 19:259−268,1982
(4) ジュニアウクス(Juniaux)Embo J 1:1125−1131,1982.
(5) ヒンネン・エイ、ヒックス・ジェイビイおよびフィンク・ジイアール(Hinnen A, Hicks JBand Fink GR)、「トランスフォーメーション・オブ・イースト」(”Transformation of Yeast”), Proc Natl Acad Sci USA 75:1929−1933,1980.
(6) ブローチ・ジェイアール、ストラザン・ジェイエヌおよびヒックス・ジェイビイ(Broach JR, Strathern JN, and Hicks JB)、「トランスフォーメーション・イン・イースト・デベロプメント・オブ・ア・ハイブリッド・クローニング・ベクター・アンド・アイソレーション・オブ・ザ・CAN1ジーン」(”Transformation in Yeast Development of a Hybrid Cloning Vector and Isolation of the CAN1 Gene”), Gene 8:121−133, 1979
(7) ツァン・エイチ(Zhang H)ら、「ダブル・ストランデッド・SDNAシーケンシング・アズ・ア・チョイス・フォア・DNAシーケンシング」、(”Double Stranded SDNA Sequencing as a Choice for DNA Sequencing”) Nucleic Acids Research 16:1220,1988
(8) ダーム・ジェイビイ、ウィリアムズ・ジェイエル、マッカッチャン.ティーエフ(Dame JB, Williams JL, McCutchan TF)ら、「ストラクチャー・オブ・ザ・ジーン・エンコーディング・ジ・イムノドミナント・サーフェイス・アンチゲン・オン・ザ・スポロゾイツ・オブ・ザ・ヒューマン・マラリア・パラサイト・プラスモジウム・ファルシパラン」(”Structure of the Gene Encoding the Immunodominant Surface
Antigen on the Sporozoites of the Human Malaria Plasmodium falciparum”) Scinence 225:593−599,1984.
(9) バレンツェラ・ピー、グレイ・ピー、キロガ・エム(Valenzuela P, Gray P, Quiroga M)ら、「ヌクレオタイド・シーケンシズ・オブ・ザ・ジーン・コーディング・フォア・ザ・メイジャー・プロテイン・オブ・ヘパティティスBバイラス・サーフェイス・アンチゲン」(”Nucleotide Sequences of the Gene Coding for the Major Protein of Hepatitis B Virus Surface Antigen”) Nature 280:815−819,1979.
【図面の簡単な説明】
【0069】
【図1】第1図(A)S発現カセットおよび(B)RTS発現カセットの制限エンドヌクレアーゼ地図を示す。各コーディング配列の範囲を黒色バーで示す。
【図2】プラスミドpRIT13034の構築を示す第2図である。Xholエンドヌクレアーゼで消化し、遊離末端と定住Tyエレメントとの相同性により酵母染色体に組み込むためのS発現カセットを含む8.5kb直線状DNAフラグメントを遊離させる。
【図3】pRIT13034由来の8.5kb直線状Xholフラグメントの制限地図を示す第3図である。該直線状フラグメントは、S発現カセットと共に形質転換された酵母細胞を選択するためのURA3およびCUP1遺伝子を有する。
【図4】RTS発現カセットの構築およびプラスミドYep13におけるクローニングを示す第4図である。
【図5−1】RTS発現カセットのヌクレオチド配列およびRTS−HBsAgハイブリッドタンパク質の推定翻訳産物を示す第5図である。TDH3 ATGコドンより開始する翻訳産物をDNA配列の下に示す。
【図5−2】第5図の続きを示す。
【図5−3】第5図の続きを示す。
【図5−4】第5図の続きを示す。
【図5−5】第5図の続きを示す。
【図6】プラスミドpRIT13539の構築を示す第6図である。BglIIエンドヌクレアーゼで消化し、遊離末端と定住Tyエレメントとの相同性により酵母染色体に組み込むためのRTS発現カセットを有する6.8kb直線状DNAフラングメントを遊離させる。
【図7】pRIT13539由来の6.8kb直線状BglIIフラグメントの制限地図を示す第7図である。該直線状フラグメントは、RTS発現カセットと共に形質転換された酵母細胞を選択するためのLEU2遺伝子を有する。
【図8】RIT4383より調製した細胞不含抽出物のCsCl密度分析を示す第8図である。フラクションをHBsAgに対して特異的なラジオイムノアッセイ(AUSRIA)およびRTS−特異的ELISAにより分析した。上部のパネルは、抗−S Mabを用いるフラクションのイムノブロット分析結果を示す。ピークフラクションの浮遊密度(rho)をその屈折率の測定値により算定した。
【図9−1】RTS*発現カセットのヌクレオチド配列およびハイブリッドタンパク質の推定翻訳産物を示す第9図である。
【図9−2】第9図の続きを示す。
【図9−3】第9図の続きを示す。
【図9−4】第9図の続きを示す。
【図9−5】第9図の続きを示す。
【図9−6】第9図の続きを示す。
【図9−7】第9図の続きを示す。
【特許請求の範囲】
【請求項1】
実質的にマラリア原虫のCS蛋白質の全てのC末端部分、CS蛋白質免疫優性領域の4以上のタンデム繰り返し配列およびB型肝炎ウイルス由来の表面抗原(HBsAg)からなることを特徴とするハイブリッド蛋白質。
【請求項2】
CS蛋白質がHBsAgと融合している請求項1に記載のハイブリッド蛋白質。
【請求項3】
請求項1または2に記載のハイブリッド蛋白質をコードするDNA。
【請求項4】
転写調節エレメントと結合している請求項3に記載のDNAを含有するベクター。
【請求項5】
請求項4のベクターで形質転換した宿主。
【請求項6】
宿主がエス・セレビシエである請求項5に記載の宿主。
【請求項7】
加えて、B型肝炎表面抗原をコードする遺伝子で形質転換された請求項6に記載の宿主。
【請求項8】
請求項1または2に記載のハイブリッド蛋白質からなるマルチマーリポ蛋白質粒子。
【請求項9】
請求項1または2に記載のハイブリッド蛋白質およびB型肝炎表面抗原からなる混合マルチマーリポ蛋白質粒子。
【請求項10】
ハイブリッド蛋白質の表面抗原に対する割合が1:4である請求項9に記載の粒子。
【請求項11】
適当な希釈剤または担体との混合物中、請求項1、2および8〜10のいずれか1つに記載の免疫保護量の粒子または蛋白質からなるワクチン。
【請求項12】
加えて、3−脱アシル化モノホスホリルリピッドAからなる請求項11記載のワクチン。
【請求項13】
加えて、ミョウバンからなる請求項12記載のワクチン。
【請求項14】
水中油型エマルジョン中に調製された請求項12記載のワクチン。
【請求項15】
適当な宿主中で請求項1または2に記載の蛋白質をコードするDNAを発現し、生成物を回収することからなる請求項1または2に記載のハイブリッド蛋白質の生成法。
【請求項16】
請求項1または2に記載の蛋白質およびB型肝炎表面抗原をコードするDNAをサッカロミセス属の株中に発現することからなる請求項8〜10に記載の粒子の生成法。
【請求項1】
実質的にマラリア原虫のCS蛋白質の全てのC末端部分、CS蛋白質免疫優性領域の4以上のタンデム繰り返し配列およびB型肝炎ウイルス由来の表面抗原(HBsAg)からなることを特徴とするハイブリッド蛋白質。
【請求項2】
CS蛋白質がHBsAgと融合している請求項1に記載のハイブリッド蛋白質。
【請求項3】
請求項1または2に記載のハイブリッド蛋白質をコードするDNA。
【請求項4】
転写調節エレメントと結合している請求項3に記載のDNAを含有するベクター。
【請求項5】
請求項4のベクターで形質転換した宿主。
【請求項6】
宿主がエス・セレビシエである請求項5に記載の宿主。
【請求項7】
加えて、B型肝炎表面抗原をコードする遺伝子で形質転換された請求項6に記載の宿主。
【請求項8】
請求項1または2に記載のハイブリッド蛋白質からなるマルチマーリポ蛋白質粒子。
【請求項9】
請求項1または2に記載のハイブリッド蛋白質およびB型肝炎表面抗原からなる混合マルチマーリポ蛋白質粒子。
【請求項10】
ハイブリッド蛋白質の表面抗原に対する割合が1:4である請求項9に記載の粒子。
【請求項11】
適当な希釈剤または担体との混合物中、請求項1、2および8〜10のいずれか1つに記載の免疫保護量の粒子または蛋白質からなるワクチン。
【請求項12】
加えて、3−脱アシル化モノホスホリルリピッドAからなる請求項11記載のワクチン。
【請求項13】
加えて、ミョウバンからなる請求項12記載のワクチン。
【請求項14】
水中油型エマルジョン中に調製された請求項12記載のワクチン。
【請求項15】
適当な宿主中で請求項1または2に記載の蛋白質をコードするDNAを発現し、生成物を回収することからなる請求項1または2に記載のハイブリッド蛋白質の生成法。
【請求項16】
請求項1または2に記載の蛋白質およびB型肝炎表面抗原をコードするDNAをサッカロミセス属の株中に発現することからなる請求項8〜10に記載の粒子の生成法。
【図1】

【図2】
【図3】
【図4】
【図5−1】
【図5−2】
【図5−3】

【図5−4】

【図5−5】

【図6】
【図7】
【図8】
【図9−1】

【図9−2】

【図9−3】

【図9−4】

【図9−5】

【図9−6】

【図9−7】

【図2】
【図3】
【図4】
【図5−1】
【図5−2】
【図5−3】
【図5−4】
【図5−5】
【図6】
【図7】
【図8】
【図9−1】
【図9−2】
【図9−3】
【図9−4】
【図9−5】
【図9−6】
【図9−7】
【公開番号】特開2007−209343(P2007−209343A)
【公開日】平成19年8月23日(2007.8.23)
【国際特許分類】
【出願番号】特願2007−66032(P2007−66032)
【出願日】平成19年3月15日(2007.3.15)
【分割の表示】特願平5−508957の分割
【原出願日】平成4年11月11日(1992.11.11)
【出願人】(595136380)グラクソスミスクライン・バイオロジカルス・ソシエテ・アノニム (10)
【氏名又は名称原語表記】GLAXOSMITHKLINE BIOLOGICALS S.A.
【Fターム(参考)】
【公開日】平成19年8月23日(2007.8.23)
【国際特許分類】
【出願日】平成19年3月15日(2007.3.15)
【分割の表示】特願平5−508957の分割
【原出願日】平成4年11月11日(1992.11.11)
【出願人】(595136380)グラクソスミスクライン・バイオロジカルス・ソシエテ・アノニム (10)
【氏名又は名称原語表記】GLAXOSMITHKLINE BIOLOGICALS S.A.
【Fターム(参考)】
[ Back to top ]