
マンガンペルオキシダーゼ変異体及びそれを用いるリグノセルロース系バイオマスの前処理方法
【課題】耐熱性及び耐過酸化水素性に優れたMnP変異体及びそれを用いるリグノセルロース系バイオマスの前処理方法を提供する。
【解決手段】MnP変異体は、特定の配列で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる。リグノセルロース系バイオマスの前処理方法は、リグノセルロース系バイオマスに、前記MnP変異体と、リグニンペルオキシダーゼとを添加する。
【解決手段】MnP変異体は、特定の配列で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる。リグノセルロース系バイオマスの前処理方法は、リグノセルロース系バイオマスに、前記MnP変異体と、リグニンペルオキシダーゼとを添加する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、マンガンペルオキシダーゼ変異体及びそれを用いるリグノセルロース系バイオマスの前処理方法に関する。
【背景技術】
【0002】
近年、自動車用燃料として、ガソリン−エタノール混合燃料を用いることが検討されている。前記エタノールとしては、植物性物質の発酵、蒸留により得たバイオエタノールを用いると、土壌管理を厳密に行うことにより所謂カーボンニュートラル効果を得ることができ、二酸化炭素の排出量を低減して地球温暖化の防止に寄与できるものと考えられている。
【0003】
しかし、前記植物性物質として、例えばサトウキビ、トウモロコシ等の農作物を用いると、該農作物がエタノールの原料として大量に消費されることにより、食料又は飼料としての供給量が減少するという問題がある。そこで、前記植物性物質として、食用または飼料用とならないリグノセルロース系バイオマスを用いてエタノールを製造する技術が検討されている。
【0004】
前記リグノセルロース系バイオマスは、セルロースを含んでおり、該セルロースを酵素糖化によりグルコースに分解し、得られたグルコースを発酵させてバイオエタノールを得ることができる。ところが、前記リグノセルロースは、セルロースの他にヘミセルロース及びリグニンを主な構成成分としており、通常該セルロース及び該ヘミセルロースは、該リグニンと強固に結合することにより保護されている。従って、前記リグノセルロースはそのままでは前記酵素糖化反応が阻害されることとなり、予め前記リグニンを除去しておくことが望ましい。
【0005】
前記リグニンを除去する方法として、微生物または微生物が産生する酵素を用いることが考えられる。前記リグニンを分解する微生物としては白色腐朽菌が知られており、白色腐朽菌はリグニン分解酵素であるマンガンペルオキシダーゼを産生することが知られている(例えば、特許文献1〜3参照)。
【0006】
前記マンガンペルオキシダーゼは、過酸化水素の存在下にMn(II)をMn(III)に酸化する。そして、Mn(III)と有機酸との反応により形成され錯体が、前記リグニンの分解に関与しているものと考えられている(例えば、特許文献4参照)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2002−142772号公報
【特許文献2】特開平9−95885号公報
【特許文献3】特開2006−333758号公報
【特許文献4】特開2009−167542号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
一般に、酵素を工業的に利用する場合には、室温以上の高温で作用させることが好ましい。しかしながら、前記マンガンペルオキシダーゼは、耐熱性が低く、例えば20〜30℃程度の温度で失活するという不都合がある。
【0009】
また、前記マンガンペルオキシダーゼは、過酸化水素存在下でMn(II)をMn(III)に酸化することにより前記リグニンを分解する酵素であるにも拘わらず、過酸化水素に対する耐性が低いという不都合がある。
【0010】
そこで、本発明は、かかる不都合を解消して、耐熱性及び耐過酸化水素性に優れたマンガンペルオキシダーゼ変異体を提供することを目的とする。
【0011】
また、本発明の目的は、前記マンガンペルオキシダーゼ変異体を用いるリグノセルロース系バイオマスの前処理方法を提供することにもある。
【課題を解決するための手段】
【0012】
かかる目的を達成するために、本発明のマンガンペルオキシダーゼ変異体は、野生型マンガンペルオキシダーゼのアミノ酸配列の230〜250番目のアミノ酸をより疎水性の大きなアミノ酸で置換したことを特徴とする。
【0013】
本発明のマンガンペルオキシダーゼ変異体は、野生型マンガンペルオキシダーゼの立体構造において、その内方に位置しているアミノ酸であって、アミノ酸配列の230〜250番目のアミノ酸をより疎水性の大きなアミノ酸で置換している。この結果、本発明のマンガンペルオキシダーゼ変異体は、その立体構造の内方における疎水性相互作用が強化され、該立体構造が安定化するので、優れた耐熱性及び耐過酸化水素性を得ることができると考えられる。
【0014】
本発明のマンガンペルオキシダーゼ変異体は、例えば、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることが好ましい。
【0015】
前記第1のアミノ酸配列は、配列番号3で表されるアミノ酸配列からなる野生型のマンガンペルオキシダーゼにおいて、238番目のM(メチオニン)をI(イソロイシン)にアミノ酸置換したものである。この結果、前記第1のアミノ酸配列又は前記第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体は、優れた耐熱性及び耐過酸化水素性を得ることができる。
【0016】
本発明のマンガンペルオキシダーゼ変異体は、前記第1のアミノ酸配列又は第2のアミノ酸配列からなる組換えタンパク質である。
【0017】
また、本発明は、前記組換えタンパク質をコードすることを特徴とする遺伝子にもある。本発明の遺伝子は、配列番号2で表される塩基配列からなる遺伝子であることが好ましい。
【0018】
本発明のリグニンペルオキシダーゼ変異体は、前記遺伝子を含有する組換えベクターを形質転換体に導入することにより発現させることができる。
【0019】
本発明のリグノセルロース系バイオマスの前処理方法は、リグノセルロース系バイオマスからリグニンを分解して除去するリグノセルロース系バイオマスの前処理方法であって、リグノセルロース系バイオマスに、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体または組換えタンパク質と、リグニンペルオキシダーゼとを添加することを特徴とする。
【0020】
本発明のリグノセルロース系バイオマスの前処理方法では、前記リグノセルロース系バイオマスに、前記リグニンペルオキシダーゼを添加すると、該リグニンペルオキシダーゼが該リグノセルロース系バイオマスのリグニンを分解し、細胞壁を破壊する。
【0021】
一方、前記リグノセルロース系バイオマスに、前記マンガンペルオキシダーゼ変異体を添加すると、該マンガンペルオキシダーゼ変異体は、まず、過酸化水素の存在下にMn(II)をMn(III)に酸化する。生成したMn(III)は、前記のように破壊された細胞壁から、前記リグノセルロース系バイオマスの内部に侵入し、該リグノセルロース系バイオマスの内部の不飽和脂肪酸と錯体を形成する。そして、前記錯体が前記リグノセルロース系バイオマスのリグニンをさらに分解する。
【0022】
このとき、前記マンガンペルオキシダーゼ変異体は優れた耐過酸化水素性を備えているので、過酸化水素の存在下においても失活することなく、Mn(II)をMn(III)に酸化することができる。また、前記マンガンペルオキシダーゼ変異体は優れた耐熱性を備えているので、室温よりも高い温度、例えば37〜42℃の範囲の温度で作用させることができ、前記リグニンの分解を促進させることができる。
【0023】
また、前記マンガンペルオキシダーゼ変異体によれば、それ自体が前記リグニンの分解を行うことはなく、該マンガンペルオキシダーゼ変異体により酸化されたMn(III)が前記リグニンの分解に関与する。従って、前記マンガンペルオキシダーゼ変異体自体から離間した部位で前記リグニンの分解を行うことができる。
【0024】
尚、本発明のリグノセルロース系バイオマスの前処理方法では、前記マンガンペルオキシダーゼ変異体に代えて、前記組換えタンパク質を用いても、該マンガンペルオキシダーゼ変異体と同一の作用効果を奏することができる。
【0025】
本発明のリグノセルロース系バイオマスの前処理方法では、前記リグノセルロース系バイオマスに、前記リグニンペルオキシダーゼを添加した後、前記マンガンペルオキシダーゼ変異体または組換えタンパク質を添加することが好ましい。このようにすることにより、まず、前記リグニンペルオキシダーゼによりリグニンを分解して前記リグノセルロース系バイオマスの細胞壁を破壊することができ、次いで、破壊された細胞壁からMn(III)が前記リグノセルロース系バイオマスの内部に侵入することができる。
【0026】
従って、前記リグニンペルオキシダーゼと、前記マンガンペルオキシダーゼ変異体または組換えタンパク質との連携を効率よく行うことができる。
【0027】
本発明のリグノセルロース系バイオマスの前処理方法において、前記リグニンペルオキシダーゼは、配列番号3で表されるアミノ酸配列からなる野生型のリグニンペルオキシダーゼにおいて、240番目のH(ヒスチジン)をF(フェニルアラニン)に、241番目のT(トレオニン)をL(ロイシン)に、242番目のI(イソロイシン)をL(ロイシン)に、それぞれアミノ酸置換したリグニンペルオキシダーゼ変異体であることが好ましい。前記野生型のリグニンペルオキシダーゼは、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0028】
前記リグニンペルオキシダーゼ変異体は、優れた耐熱性及び耐過酸化水素性を備えているので、前記マンガンペルオキシダーゼ変異体または組換えタンパク質と同一条件下で作用することができ、該マンガンペルオキシダーゼ変異体または組換えタンパク質との連携をさらに効率よく行うことができる。
【図面の簡単な説明】
【0029】
【図1】野生型のマンガンペルオキシダーゼ及びこれと相同なアミノ酸配列からなるタンパク質48個の系統樹を示す図。
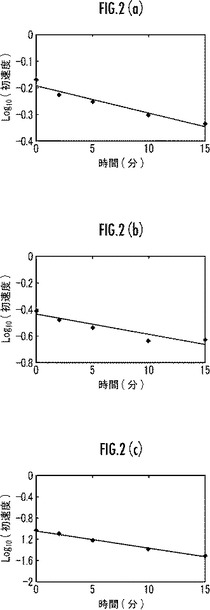
【図2】本発明のマンガンペルオキシダーゼ変異体の37℃における耐熱性を示すグラフ。
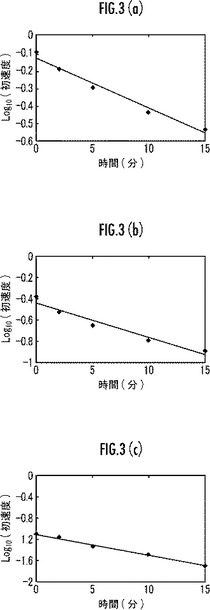
【図3】本発明のマンガンペルオキシダーゼ変異体の42℃における耐熱性を示すグラフ。
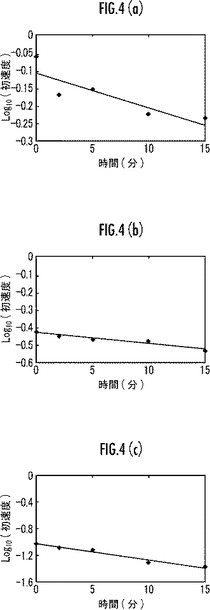
【図4】本発明のマンガンペルオキシダーゼ変異体の耐過酸化水素性を示すグラフ。
【発明を実施するための形態】
【0030】
次に、添付の図面を参照しながら本発明の実施の形態についてさらに詳しく説明する。
【0031】
本実施形態のマンガンペルオキシダーゼ変異体は、白色腐朽菌Phanerochaete chrysosporium UAMH 3641由来の野生型のマンガンペルオキシダーゼの変異体であり、配列番号1で表されるアミノ酸配列からなる。前記野生型のマンガンペルオキシダーゼは、配列番号5で表されるアミノ酸配列からなる。
【0032】
本実施形態のマンガンペルオキシダーゼ変異体の前記配列番号1で表されるアミノ酸配列は、配列番号5で表されるアミノ酸配列において、238番目のM(メチオニン)をI(イソロイシン)にアミノ酸置換したものである。また、本実施形態のマンガンペルオキシダーゼ変異体は、配列番号2で表される塩基配列からなる遺伝子によりコードされ、前記野生型のマンガンペルオキシダーゼは、配列番号6で表される塩基配列からなる遺伝子によりコードされる。
【0033】
前記野生型のマンガンペルオキシダーゼは、耐熱性及び耐過酸化水素性を備えていない。しかし、ウーズ(Woese)らによって示された16srRNAによる系統樹を見ると、80℃以上で至適に生育する生物が該系統樹の根元に多いことが示されている。この事実から、真正細菌、真核生物、古細菌の共通の祖先は超好熱菌と推定され、先祖型のアミノ酸配列を有するか又はそれに近い配列を有するタンパク質は耐熱性を備えているものと考えられる。このような知見は、特開2002−247991号公報に記載されている。
【0034】
そこで、本実施形態では、リグニンを分解する酵素を含み、前記野生型のマンガンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質に関する系統樹を作成し、該系統樹における先祖型のアミノ酸配列について検討した。
【0035】
前記系統樹の作成に当たって、まず、リグニンを分解する酵素を含み、前記野生型のマンガンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質48個をDDBJ内のアミノ酸配列データベースからソフトウェアBLASTPを用いて抽出した。抽出したタンパク質は、リグニンを分解する酵素として、前記野生型のマンガンペルオキシダーゼの他、担子菌由来のマンガンペルオキシダーゼ、子のう菌由来のリグニンペルオキシダーゼ、マンガンペルオキシダーゼとリグニンペルオキシダーゼとのハイブリッド型酵素であるヴァーサタイルペルオキシダーゼ、野生型のリグニンペルオキシダーゼを含んでいる。尚、本明細書では、マンガンペルオキシダーゼを「MnP」、リグニンペルオキシダーゼを「LiP」、ヴァ−サタイルペルオキシダーゼ「VP」と略記することがある。
【0036】
次に、前記野生型MnPと、抽出した48個のタンパク質との合わせて49個のタンパク質について、進化の系統を示す系統樹を作成した。前記系統樹は、ソフトウェアTreefinderとPhyMLとを用いて候補系統樹群を作成し、該候補系統樹群の各候補系統樹について、ソフトウェアPMALのCODEMLを用いて尤度を計算した。そして、最尤度を備える候補系統樹を前記49個のタンパク質の進化の系統を示す系統樹として選択した。選択した系統樹を図1に示す。
【0037】
図1において、符号1は野生型MnP、符号2は野生型LiP、符号3は子のう菌由来LiP、符号4はVPを示す。また、符号5はMnPを示し、符号6はLiPを示す。
【0038】
尚、図1において、野生型MnP1及び野生型LiP2を除く47個のタンパク質は、DDBJ accession numbersにより示している。野生型MnP1及び野生型LiP2を除く47個のタンパク質は、いずれも論文等に発表されており、前記”DDBJ accession numbers”により特定することができる。
【0039】
また、前記野生型LiP2は、配列番号3で表されるアミノ酸配列からなり、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0040】
図1において、分岐点Aに対応するタンパク質は耐熱性を備えており、そのアミノ酸配列の230〜250番目のアミノ酸は、野生型MnP1のアミノ酸配列の230〜250番目のアミノ酸よりも疎水性大きなアミノ酸であると考えられる。
【0041】
そこで、前記野生型MnP1のアミノ酸配列(配列番号5)中、238番目のM(メチオニン)を、Mより疎水性の大きなアミノ酸であるI(イソロイシン)にアミノ酸置換したものを本実施形態の先祖型MnP変異体のアミノ酸配列(配列番号1)とした。
【0042】
本実施形態のMnP変異体(先祖型MnP変異体)は、前記のようにして決定した先祖型MnP変異体のアミノ酸配列に基づいて、次のようにして作成することができる。
【0043】
まず、野生型MnP1の遺伝子がコードされたプラスミドpET21a(+)に、QuikChange(登録商標)Lightning Site-Directed Mutagenesis Kit(STRATAGENE)を用いて、238番目のMをIにアミノ酸置換した祖先型変異を導入し、ベクターとする。目的の変異導入の成否はシークエンシングで確認することができる。
【0044】
次に、前記祖先型変異が導入されたベクターを用いて大腸菌Escherichia coli BL21(DE3)株を形質転換し、TB培地を用い37℃の温度で大量培養する。得られた菌体を超音波で破砕し、30000rpmで20分間遠心して、沈殿を分離する。
【0045】
次に、分離された沈殿を、20mM−Tris-HCl(pH8.5)、2mM−EDTA、2mM−DTT、1%−非イオン性界面活性剤(TritonX−100(商品名))で3回洗浄し、8M−尿素、50mM−Tris-HCl(pH8.5)、2mM−EDTA、1mM−DTTに再懸濁し、4℃の温度に一晩保持して封入体を可溶化する。
【0046】
次に、前記封入体が可溶化された溶液を4倍に希釈することにより尿素の濃度を2Mとし、4℃の温度の暗所に一晩保持してリフォールディングして、本実施形態のMnP変異体(M238I)を得る。このとき、5μM−ヘミン、0.7mM−酸化グルタチオン、10mM−CaCl2(濃度はそれぞれ終濃度である)を添加する。
【0047】
次に、リフォールディングされた本実施形態のMnP変異体(M238I)を、陰イオン交換カラム(DEAE-Sepharose(商品名)、HiTrrapQ HP(商品名)、ResourceQ(商品名))、で分離し、精製する。
【0048】
次に、本実施形態のMnP変異体(M238I)の耐熱性及び耐過酸化水素性を評価した(実施例)。
【0049】
前記耐熱性の評価は、まず、本実施形態のMnP変異体(M238I)48nMを、10mM−酢酸緩衝液(pH6.0)及び1mM−CaCl2からなる溶液中、37℃及び42℃の温度でそれぞれインキュベートした。次に、インキュベート開始後、0分後、2分後、5分後、10分後、15分後に、インキュベートされた本実施形態のMnP変異体(M238I)8nMを、50mM−マロン酸緩衝液(pH4.5)、1.2mM−MnSO4、0.1mM−H2O2を含む溶液に加えて、測定液を調製した。
【0050】
前記測定液中で、本実施形態のMnP変異体(M238I)はMn(III)を生成し、Mn(III)がマロン酸と錯体を形成する。そこで、次に、270nmの波長の光線を用いて、前記測定液の吸光度を測定した。測定中の失活を防ぐために、測定温度は25℃とした。270nmの波長によるモル吸光係数ε270は、ε270=8500である。
【0051】
各試料から得られた初速により、それぞれのインキュベート時間における比活性(μモル/mg分)を算出した。インキュベート開始後0分後及び15分後の比活性について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0052】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフについて、インキュベート温度37℃の場合を図2(a)に、42℃の場合を図3(a)にそれぞれ示す。
【0053】
また、図2(a)及び図3(a)のグラフの傾きから失活速度を求めた。前記失活速度について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0054】
また、前記耐過酸化水素性の評価は、本実施形態のMnP変異体(M238I)48nMを、pH6.0の0.1mM−H2O2溶液中、25℃の温度でインキュベートした以外は、前記耐熱性の評価と全く同一にして、測定液の吸光度を測定することにより行った。
【0055】
各試料から得られた初速により、それぞれのインキュベート時間における比活性(μモル/mg分)を算出した。インキュベート開始後0分後及び15分後の比活性の値を表3に示す。
【0056】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフを図4(a)に示す。また、図4(a)のグラフの傾きから失活速度を求めた。前記失活速度を表3に示す。
【0057】
次に、本実施形態のMnP変異体(M238I)に代えて野生型のMnP1を用いた以外は、本実施形態のMnP変異体(M238I)の場合と全く同一にして野生型のMnP1の耐熱性及び耐過酸化水素性を評価した(比較例1)。
【0058】
前記耐熱性については、インキュベート開始後0分後及び15分後の比活性について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0059】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフについて、インキュベート温度37℃の場合を図2(b)に、42℃の場合を図3(b)にそれぞれ示す。
【0060】
また、図2(b)及び図3(b)のグラフの傾きから失活速度を求めた。前記失活速度について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0061】
また、前記耐過酸化水素性については、インキュベート開始後0分後及び15分後の比活性の値を表3に示す。
【0062】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフを図4(b)に示す。また、図4(b)のグラフの傾きから失活速度を求めた。前記失活速度を表3に示す。
【0063】
次に、前記野生型MnP1のアミノ酸配列(配列番号4)中、238番目のM(メチオニン)を、F(フェニルアラニン)にアミノ酸置換したアミノ酸配列(配列番号7)を備えるMnP変異体(M238F)を作成した。前記MnP変異体(M238F)は、配列番号8で表される塩基配列からなる遺伝子によりコードされる。前記MnP変異体(M238F)は、野生型MnP1の遺伝子がコードされたプラスミドpET21a(+)に、238番目のMをFにアミノ酸置換した変異を導入してベクターとする以外は、本実施形態のMnP変異体(M238I)と全く同一にして作成することができる。
【0064】
次に、本実施形態のMnP変異体(M238I)に代えて、MnP変異体(M238F)を用いた以外は、本実施形態のMnP変異体(M238I)の場合と全く同一にしてMnP変異体(M238F)の耐熱性及び耐過酸化水素性を評価した(比較例2)。
【0065】
前記耐熱性については、インキュベート開始後0分後及び15分後の比活性について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0066】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフについて、インキュベート温度37℃の場合を図2(c)に、42℃の場合を図3(c)にそれぞれ示す。
【0067】
また、図2(c)及び図3(c)のグラフの傾きから失活速度を求めた。前記失活速度について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0068】
前記過酸化水素性については、インキュベート開始後0分後及び15分後の比活性の値を表3に示す。
【0069】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフを図4(c)に示す。また、図4(c)のグラフの傾きから失活速度を求めた。前記失活速度を表3に示す。
【0070】
【表1】

【0071】
【表2】
【0072】
【表3】
【0073】
表1〜2及び図2〜3に示すように、本実施例のMnP変異体は、野生型MnP(比較例1)又は238番目のMをFにアミノ酸置換したMnP変異体(比較例2)よりも優れた耐熱性を備えていることが明らかである。
【0074】
また、表3及び図4に示すように、本実施例のMnP変異体は、野生型MnP(比較例1)又は238番目のMをFにアミノ酸置換したMnP変異体(比較例2)よりも優れた耐過酸化水素性を備えていることが明らかである。
【0075】
次に、本実施形態のリグノセルロース系バイオマスの前処理方法について説明する。
【0076】
本実施形態のリグノセルロース系バイオマスの前処理方法は、リグノセルロース系バイオマスを基質として、該基質を糖化酵素により糖化し、得られた糖化溶液を発酵させてバイオエタノールを製造する際に、前処理として該基質のリグニンを除去する方法である。前記リグノセルロース系バイオマスとしては、木材、稲藁、麦藁、バガス、竹、パルプ及びこれらから生じる廃棄物例えば古紙等を挙げることができ、例えば、稲藁を用いることができる。
【0077】
本実施形態のリグノセルロース系バイオマスの前処理方法では、前記リグノセルロース系バイオマスとして稲藁を用いる場合には、まず、カッターミル等により約3mmの長さに粉砕された稲藁に水を添加して、例えば5質量%の濃度の混合液を調製する。
【0078】
次に、前記混合液に、その全量に対し0.1〜0.5μMの範囲のリグニンペルオキシダーゼ(LiP)を添加する。前記LiPは、配列番号3で表されるアミノ酸配列からなる野生型LiPにおいて、240番目のH(ヒスチジン)をF(フェニルアラニン)に、241番目のT(トレオニン)をL(ロイシン)に、242番目のI(イソロイシン)をL(ロイシン)に、それぞれアミノ酸置換したLiP変異体であることが好ましい。前記野生型LiPは、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0079】
前記LiP変異体は、耐熱性及び耐過酸化水素性を備えている。そこで、前記LiP変異体が添加された混合液を、0.1〜0.5mMの過酸化水素の存在下、25〜30℃の範囲の温度に、1〜120時間の範囲の時間保持することにより、前記稲藁のリグニンを分解し、細胞壁を破壊することができる。
【0080】
次に、前記混合液に、その全量に対し0.1〜0.5μMの範囲の本実施形態のMnP変異体(M238I、先祖型MnP変異体)を添加する。本実施形態のMnP変異体は、前述のように耐熱性及び耐過酸化水素性を備えている。
【0081】
そこで、本実施形態のMnP変異体が添加された混合液を、0.1〜0.5mMの過酸化水素の存在下、37〜42℃の範囲の温度に、1〜120時間の範囲の時間保持することにより、前記稲藁に含まれるMn(II)をMn(III)に酸化することができる。生成したMn(III)は、前記LiP変異体により破壊された細胞壁から、前記稲藁の内部に侵入し、該稲藁に含まれる不飽和脂肪酸と錯体を形成する。そして、前記錯体が前記稲藁のリグニンをさらに分解する。
【0082】
また、本実施形態のMnP変異体によれば、それ自体が前記リグニンの分解を行うことはなく、該MnP変異体により酸化されたMn(III)が前記リグニンの分解に関与する。従って、前記MnP変異体自体から離間した部位で前記リグニンの分解を行うことができる。
【0083】
本実施形態のリグノセルロース系バイオマスの前処理方法により処理された前記混合液は、前記リグニンの分解後、さらに糖化酵素を添加することにより、前記稲藁に含まれるセルロース及びヘムセルロースを糖化して、糖化溶液とすることができる。また、前記糖化溶液は、バイオエタノールの原料として用いることができる。
【0084】
尚、本発明のリグノセルロース系バイオマスの前処理方法では、前記MnP変異体に代えて、配列番号1で表される第1のアミノ酸配列からなる組換えタンパク質を用いてもよく、前記MnP変異体と同一の作用効果を奏することができる。
【0085】
また、本発明のリグノセルロース系バイオマスの前処理方法では、前記LiP変異体に代えて、野生型LiP等の他のリグニンペルオキシダーゼを用いてもよい。ただし、この場合には、前記LiP変異体よりも耐熱性又は耐過酸化水素性が低く、本実施形態のMnP変異体と同一条件で用いることが難しくなることがある。
【0086】
また、本実施形態のリグノセルロース系バイオマスの前処理方法では、前記混合液に前記LiP変異体を添加した後、本実施形態のMnP変異体を添加するようにしているが、前記LiP変異体と本実施形態のMnP変異体とを同時に添加するようにしてもよい。
【符号の説明】
【0087】
1…野生型MnP、 2…野生型LiP、 3…子のう菌由来LiP、 4…VP、 5…MnP、 6…LiP、 A…分岐点。
【技術分野】
【0001】
本発明は、マンガンペルオキシダーゼ変異体及びそれを用いるリグノセルロース系バイオマスの前処理方法に関する。
【背景技術】
【0002】
近年、自動車用燃料として、ガソリン−エタノール混合燃料を用いることが検討されている。前記エタノールとしては、植物性物質の発酵、蒸留により得たバイオエタノールを用いると、土壌管理を厳密に行うことにより所謂カーボンニュートラル効果を得ることができ、二酸化炭素の排出量を低減して地球温暖化の防止に寄与できるものと考えられている。
【0003】
しかし、前記植物性物質として、例えばサトウキビ、トウモロコシ等の農作物を用いると、該農作物がエタノールの原料として大量に消費されることにより、食料又は飼料としての供給量が減少するという問題がある。そこで、前記植物性物質として、食用または飼料用とならないリグノセルロース系バイオマスを用いてエタノールを製造する技術が検討されている。
【0004】
前記リグノセルロース系バイオマスは、セルロースを含んでおり、該セルロースを酵素糖化によりグルコースに分解し、得られたグルコースを発酵させてバイオエタノールを得ることができる。ところが、前記リグノセルロースは、セルロースの他にヘミセルロース及びリグニンを主な構成成分としており、通常該セルロース及び該ヘミセルロースは、該リグニンと強固に結合することにより保護されている。従って、前記リグノセルロースはそのままでは前記酵素糖化反応が阻害されることとなり、予め前記リグニンを除去しておくことが望ましい。
【0005】
前記リグニンを除去する方法として、微生物または微生物が産生する酵素を用いることが考えられる。前記リグニンを分解する微生物としては白色腐朽菌が知られており、白色腐朽菌はリグニン分解酵素であるマンガンペルオキシダーゼを産生することが知られている(例えば、特許文献1〜3参照)。
【0006】
前記マンガンペルオキシダーゼは、過酸化水素の存在下にMn(II)をMn(III)に酸化する。そして、Mn(III)と有機酸との反応により形成され錯体が、前記リグニンの分解に関与しているものと考えられている(例えば、特許文献4参照)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2002−142772号公報
【特許文献2】特開平9−95885号公報
【特許文献3】特開2006−333758号公報
【特許文献4】特開2009−167542号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
一般に、酵素を工業的に利用する場合には、室温以上の高温で作用させることが好ましい。しかしながら、前記マンガンペルオキシダーゼは、耐熱性が低く、例えば20〜30℃程度の温度で失活するという不都合がある。
【0009】
また、前記マンガンペルオキシダーゼは、過酸化水素存在下でMn(II)をMn(III)に酸化することにより前記リグニンを分解する酵素であるにも拘わらず、過酸化水素に対する耐性が低いという不都合がある。
【0010】
そこで、本発明は、かかる不都合を解消して、耐熱性及び耐過酸化水素性に優れたマンガンペルオキシダーゼ変異体を提供することを目的とする。
【0011】
また、本発明の目的は、前記マンガンペルオキシダーゼ変異体を用いるリグノセルロース系バイオマスの前処理方法を提供することにもある。
【課題を解決するための手段】
【0012】
かかる目的を達成するために、本発明のマンガンペルオキシダーゼ変異体は、野生型マンガンペルオキシダーゼのアミノ酸配列の230〜250番目のアミノ酸をより疎水性の大きなアミノ酸で置換したことを特徴とする。
【0013】
本発明のマンガンペルオキシダーゼ変異体は、野生型マンガンペルオキシダーゼの立体構造において、その内方に位置しているアミノ酸であって、アミノ酸配列の230〜250番目のアミノ酸をより疎水性の大きなアミノ酸で置換している。この結果、本発明のマンガンペルオキシダーゼ変異体は、その立体構造の内方における疎水性相互作用が強化され、該立体構造が安定化するので、優れた耐熱性及び耐過酸化水素性を得ることができると考えられる。
【0014】
本発明のマンガンペルオキシダーゼ変異体は、例えば、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることが好ましい。
【0015】
前記第1のアミノ酸配列は、配列番号3で表されるアミノ酸配列からなる野生型のマンガンペルオキシダーゼにおいて、238番目のM(メチオニン)をI(イソロイシン)にアミノ酸置換したものである。この結果、前記第1のアミノ酸配列又は前記第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体は、優れた耐熱性及び耐過酸化水素性を得ることができる。
【0016】
本発明のマンガンペルオキシダーゼ変異体は、前記第1のアミノ酸配列又は第2のアミノ酸配列からなる組換えタンパク質である。
【0017】
また、本発明は、前記組換えタンパク質をコードすることを特徴とする遺伝子にもある。本発明の遺伝子は、配列番号2で表される塩基配列からなる遺伝子であることが好ましい。
【0018】
本発明のリグニンペルオキシダーゼ変異体は、前記遺伝子を含有する組換えベクターを形質転換体に導入することにより発現させることができる。
【0019】
本発明のリグノセルロース系バイオマスの前処理方法は、リグノセルロース系バイオマスからリグニンを分解して除去するリグノセルロース系バイオマスの前処理方法であって、リグノセルロース系バイオマスに、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体または組換えタンパク質と、リグニンペルオキシダーゼとを添加することを特徴とする。
【0020】
本発明のリグノセルロース系バイオマスの前処理方法では、前記リグノセルロース系バイオマスに、前記リグニンペルオキシダーゼを添加すると、該リグニンペルオキシダーゼが該リグノセルロース系バイオマスのリグニンを分解し、細胞壁を破壊する。
【0021】
一方、前記リグノセルロース系バイオマスに、前記マンガンペルオキシダーゼ変異体を添加すると、該マンガンペルオキシダーゼ変異体は、まず、過酸化水素の存在下にMn(II)をMn(III)に酸化する。生成したMn(III)は、前記のように破壊された細胞壁から、前記リグノセルロース系バイオマスの内部に侵入し、該リグノセルロース系バイオマスの内部の不飽和脂肪酸と錯体を形成する。そして、前記錯体が前記リグノセルロース系バイオマスのリグニンをさらに分解する。
【0022】
このとき、前記マンガンペルオキシダーゼ変異体は優れた耐過酸化水素性を備えているので、過酸化水素の存在下においても失活することなく、Mn(II)をMn(III)に酸化することができる。また、前記マンガンペルオキシダーゼ変異体は優れた耐熱性を備えているので、室温よりも高い温度、例えば37〜42℃の範囲の温度で作用させることができ、前記リグニンの分解を促進させることができる。
【0023】
また、前記マンガンペルオキシダーゼ変異体によれば、それ自体が前記リグニンの分解を行うことはなく、該マンガンペルオキシダーゼ変異体により酸化されたMn(III)が前記リグニンの分解に関与する。従って、前記マンガンペルオキシダーゼ変異体自体から離間した部位で前記リグニンの分解を行うことができる。
【0024】
尚、本発明のリグノセルロース系バイオマスの前処理方法では、前記マンガンペルオキシダーゼ変異体に代えて、前記組換えタンパク質を用いても、該マンガンペルオキシダーゼ変異体と同一の作用効果を奏することができる。
【0025】
本発明のリグノセルロース系バイオマスの前処理方法では、前記リグノセルロース系バイオマスに、前記リグニンペルオキシダーゼを添加した後、前記マンガンペルオキシダーゼ変異体または組換えタンパク質を添加することが好ましい。このようにすることにより、まず、前記リグニンペルオキシダーゼによりリグニンを分解して前記リグノセルロース系バイオマスの細胞壁を破壊することができ、次いで、破壊された細胞壁からMn(III)が前記リグノセルロース系バイオマスの内部に侵入することができる。
【0026】
従って、前記リグニンペルオキシダーゼと、前記マンガンペルオキシダーゼ変異体または組換えタンパク質との連携を効率よく行うことができる。
【0027】
本発明のリグノセルロース系バイオマスの前処理方法において、前記リグニンペルオキシダーゼは、配列番号3で表されるアミノ酸配列からなる野生型のリグニンペルオキシダーゼにおいて、240番目のH(ヒスチジン)をF(フェニルアラニン)に、241番目のT(トレオニン)をL(ロイシン)に、242番目のI(イソロイシン)をL(ロイシン)に、それぞれアミノ酸置換したリグニンペルオキシダーゼ変異体であることが好ましい。前記野生型のリグニンペルオキシダーゼは、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0028】
前記リグニンペルオキシダーゼ変異体は、優れた耐熱性及び耐過酸化水素性を備えているので、前記マンガンペルオキシダーゼ変異体または組換えタンパク質と同一条件下で作用することができ、該マンガンペルオキシダーゼ変異体または組換えタンパク質との連携をさらに効率よく行うことができる。
【図面の簡単な説明】
【0029】
【図1】野生型のマンガンペルオキシダーゼ及びこれと相同なアミノ酸配列からなるタンパク質48個の系統樹を示す図。
【図2】本発明のマンガンペルオキシダーゼ変異体の37℃における耐熱性を示すグラフ。
【図3】本発明のマンガンペルオキシダーゼ変異体の42℃における耐熱性を示すグラフ。
【図4】本発明のマンガンペルオキシダーゼ変異体の耐過酸化水素性を示すグラフ。
【発明を実施するための形態】
【0030】
次に、添付の図面を参照しながら本発明の実施の形態についてさらに詳しく説明する。
【0031】
本実施形態のマンガンペルオキシダーゼ変異体は、白色腐朽菌Phanerochaete chrysosporium UAMH 3641由来の野生型のマンガンペルオキシダーゼの変異体であり、配列番号1で表されるアミノ酸配列からなる。前記野生型のマンガンペルオキシダーゼは、配列番号5で表されるアミノ酸配列からなる。
【0032】
本実施形態のマンガンペルオキシダーゼ変異体の前記配列番号1で表されるアミノ酸配列は、配列番号5で表されるアミノ酸配列において、238番目のM(メチオニン)をI(イソロイシン)にアミノ酸置換したものである。また、本実施形態のマンガンペルオキシダーゼ変異体は、配列番号2で表される塩基配列からなる遺伝子によりコードされ、前記野生型のマンガンペルオキシダーゼは、配列番号6で表される塩基配列からなる遺伝子によりコードされる。
【0033】
前記野生型のマンガンペルオキシダーゼは、耐熱性及び耐過酸化水素性を備えていない。しかし、ウーズ(Woese)らによって示された16srRNAによる系統樹を見ると、80℃以上で至適に生育する生物が該系統樹の根元に多いことが示されている。この事実から、真正細菌、真核生物、古細菌の共通の祖先は超好熱菌と推定され、先祖型のアミノ酸配列を有するか又はそれに近い配列を有するタンパク質は耐熱性を備えているものと考えられる。このような知見は、特開2002−247991号公報に記載されている。
【0034】
そこで、本実施形態では、リグニンを分解する酵素を含み、前記野生型のマンガンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質に関する系統樹を作成し、該系統樹における先祖型のアミノ酸配列について検討した。
【0035】
前記系統樹の作成に当たって、まず、リグニンを分解する酵素を含み、前記野生型のマンガンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質48個をDDBJ内のアミノ酸配列データベースからソフトウェアBLASTPを用いて抽出した。抽出したタンパク質は、リグニンを分解する酵素として、前記野生型のマンガンペルオキシダーゼの他、担子菌由来のマンガンペルオキシダーゼ、子のう菌由来のリグニンペルオキシダーゼ、マンガンペルオキシダーゼとリグニンペルオキシダーゼとのハイブリッド型酵素であるヴァーサタイルペルオキシダーゼ、野生型のリグニンペルオキシダーゼを含んでいる。尚、本明細書では、マンガンペルオキシダーゼを「MnP」、リグニンペルオキシダーゼを「LiP」、ヴァ−サタイルペルオキシダーゼ「VP」と略記することがある。
【0036】
次に、前記野生型MnPと、抽出した48個のタンパク質との合わせて49個のタンパク質について、進化の系統を示す系統樹を作成した。前記系統樹は、ソフトウェアTreefinderとPhyMLとを用いて候補系統樹群を作成し、該候補系統樹群の各候補系統樹について、ソフトウェアPMALのCODEMLを用いて尤度を計算した。そして、最尤度を備える候補系統樹を前記49個のタンパク質の進化の系統を示す系統樹として選択した。選択した系統樹を図1に示す。
【0037】
図1において、符号1は野生型MnP、符号2は野生型LiP、符号3は子のう菌由来LiP、符号4はVPを示す。また、符号5はMnPを示し、符号6はLiPを示す。
【0038】
尚、図1において、野生型MnP1及び野生型LiP2を除く47個のタンパク質は、DDBJ accession numbersにより示している。野生型MnP1及び野生型LiP2を除く47個のタンパク質は、いずれも論文等に発表されており、前記”DDBJ accession numbers”により特定することができる。
【0039】
また、前記野生型LiP2は、配列番号3で表されるアミノ酸配列からなり、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0040】
図1において、分岐点Aに対応するタンパク質は耐熱性を備えており、そのアミノ酸配列の230〜250番目のアミノ酸は、野生型MnP1のアミノ酸配列の230〜250番目のアミノ酸よりも疎水性大きなアミノ酸であると考えられる。
【0041】
そこで、前記野生型MnP1のアミノ酸配列(配列番号5)中、238番目のM(メチオニン)を、Mより疎水性の大きなアミノ酸であるI(イソロイシン)にアミノ酸置換したものを本実施形態の先祖型MnP変異体のアミノ酸配列(配列番号1)とした。
【0042】
本実施形態のMnP変異体(先祖型MnP変異体)は、前記のようにして決定した先祖型MnP変異体のアミノ酸配列に基づいて、次のようにして作成することができる。
【0043】
まず、野生型MnP1の遺伝子がコードされたプラスミドpET21a(+)に、QuikChange(登録商標)Lightning Site-Directed Mutagenesis Kit(STRATAGENE)を用いて、238番目のMをIにアミノ酸置換した祖先型変異を導入し、ベクターとする。目的の変異導入の成否はシークエンシングで確認することができる。
【0044】
次に、前記祖先型変異が導入されたベクターを用いて大腸菌Escherichia coli BL21(DE3)株を形質転換し、TB培地を用い37℃の温度で大量培養する。得られた菌体を超音波で破砕し、30000rpmで20分間遠心して、沈殿を分離する。
【0045】
次に、分離された沈殿を、20mM−Tris-HCl(pH8.5)、2mM−EDTA、2mM−DTT、1%−非イオン性界面活性剤(TritonX−100(商品名))で3回洗浄し、8M−尿素、50mM−Tris-HCl(pH8.5)、2mM−EDTA、1mM−DTTに再懸濁し、4℃の温度に一晩保持して封入体を可溶化する。
【0046】
次に、前記封入体が可溶化された溶液を4倍に希釈することにより尿素の濃度を2Mとし、4℃の温度の暗所に一晩保持してリフォールディングして、本実施形態のMnP変異体(M238I)を得る。このとき、5μM−ヘミン、0.7mM−酸化グルタチオン、10mM−CaCl2(濃度はそれぞれ終濃度である)を添加する。
【0047】
次に、リフォールディングされた本実施形態のMnP変異体(M238I)を、陰イオン交換カラム(DEAE-Sepharose(商品名)、HiTrrapQ HP(商品名)、ResourceQ(商品名))、で分離し、精製する。
【0048】
次に、本実施形態のMnP変異体(M238I)の耐熱性及び耐過酸化水素性を評価した(実施例)。
【0049】
前記耐熱性の評価は、まず、本実施形態のMnP変異体(M238I)48nMを、10mM−酢酸緩衝液(pH6.0)及び1mM−CaCl2からなる溶液中、37℃及び42℃の温度でそれぞれインキュベートした。次に、インキュベート開始後、0分後、2分後、5分後、10分後、15分後に、インキュベートされた本実施形態のMnP変異体(M238I)8nMを、50mM−マロン酸緩衝液(pH4.5)、1.2mM−MnSO4、0.1mM−H2O2を含む溶液に加えて、測定液を調製した。
【0050】
前記測定液中で、本実施形態のMnP変異体(M238I)はMn(III)を生成し、Mn(III)がマロン酸と錯体を形成する。そこで、次に、270nmの波長の光線を用いて、前記測定液の吸光度を測定した。測定中の失活を防ぐために、測定温度は25℃とした。270nmの波長によるモル吸光係数ε270は、ε270=8500である。
【0051】
各試料から得られた初速により、それぞれのインキュベート時間における比活性(μモル/mg分)を算出した。インキュベート開始後0分後及び15分後の比活性について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0052】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフについて、インキュベート温度37℃の場合を図2(a)に、42℃の場合を図3(a)にそれぞれ示す。
【0053】
また、図2(a)及び図3(a)のグラフの傾きから失活速度を求めた。前記失活速度について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0054】
また、前記耐過酸化水素性の評価は、本実施形態のMnP変異体(M238I)48nMを、pH6.0の0.1mM−H2O2溶液中、25℃の温度でインキュベートした以外は、前記耐熱性の評価と全く同一にして、測定液の吸光度を測定することにより行った。
【0055】
各試料から得られた初速により、それぞれのインキュベート時間における比活性(μモル/mg分)を算出した。インキュベート開始後0分後及び15分後の比活性の値を表3に示す。
【0056】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフを図4(a)に示す。また、図4(a)のグラフの傾きから失活速度を求めた。前記失活速度を表3に示す。
【0057】
次に、本実施形態のMnP変異体(M238I)に代えて野生型のMnP1を用いた以外は、本実施形態のMnP変異体(M238I)の場合と全く同一にして野生型のMnP1の耐熱性及び耐過酸化水素性を評価した(比較例1)。
【0058】
前記耐熱性については、インキュベート開始後0分後及び15分後の比活性について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0059】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフについて、インキュベート温度37℃の場合を図2(b)に、42℃の場合を図3(b)にそれぞれ示す。
【0060】
また、図2(b)及び図3(b)のグラフの傾きから失活速度を求めた。前記失活速度について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0061】
また、前記耐過酸化水素性については、インキュベート開始後0分後及び15分後の比活性の値を表3に示す。
【0062】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフを図4(b)に示す。また、図4(b)のグラフの傾きから失活速度を求めた。前記失活速度を表3に示す。
【0063】
次に、前記野生型MnP1のアミノ酸配列(配列番号4)中、238番目のM(メチオニン)を、F(フェニルアラニン)にアミノ酸置換したアミノ酸配列(配列番号7)を備えるMnP変異体(M238F)を作成した。前記MnP変異体(M238F)は、配列番号8で表される塩基配列からなる遺伝子によりコードされる。前記MnP変異体(M238F)は、野生型MnP1の遺伝子がコードされたプラスミドpET21a(+)に、238番目のMをFにアミノ酸置換した変異を導入してベクターとする以外は、本実施形態のMnP変異体(M238I)と全く同一にして作成することができる。
【0064】
次に、本実施形態のMnP変異体(M238I)に代えて、MnP変異体(M238F)を用いた以外は、本実施形態のMnP変異体(M238I)の場合と全く同一にしてMnP変異体(M238F)の耐熱性及び耐過酸化水素性を評価した(比較例2)。
【0065】
前記耐熱性については、インキュベート開始後0分後及び15分後の比活性について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0066】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフについて、インキュベート温度37℃の場合を図2(c)に、42℃の場合を図3(c)にそれぞれ示す。
【0067】
また、図2(c)及び図3(c)のグラフの傾きから失活速度を求めた。前記失活速度について、インキュベート温度37℃のときの値を表1に、42℃のときの値を表2にそれぞれ示す。
【0068】
前記過酸化水素性については、インキュベート開始後0分後及び15分後の比活性の値を表3に示す。
【0069】
また、初速の常用対数を縦軸とし、時間を横軸としたグラフを図4(c)に示す。また、図4(c)のグラフの傾きから失活速度を求めた。前記失活速度を表3に示す。
【0070】
【表1】
【0071】
【表2】
【0072】
【表3】
【0073】
表1〜2及び図2〜3に示すように、本実施例のMnP変異体は、野生型MnP(比較例1)又は238番目のMをFにアミノ酸置換したMnP変異体(比較例2)よりも優れた耐熱性を備えていることが明らかである。
【0074】
また、表3及び図4に示すように、本実施例のMnP変異体は、野生型MnP(比較例1)又は238番目のMをFにアミノ酸置換したMnP変異体(比較例2)よりも優れた耐過酸化水素性を備えていることが明らかである。
【0075】
次に、本実施形態のリグノセルロース系バイオマスの前処理方法について説明する。
【0076】
本実施形態のリグノセルロース系バイオマスの前処理方法は、リグノセルロース系バイオマスを基質として、該基質を糖化酵素により糖化し、得られた糖化溶液を発酵させてバイオエタノールを製造する際に、前処理として該基質のリグニンを除去する方法である。前記リグノセルロース系バイオマスとしては、木材、稲藁、麦藁、バガス、竹、パルプ及びこれらから生じる廃棄物例えば古紙等を挙げることができ、例えば、稲藁を用いることができる。
【0077】
本実施形態のリグノセルロース系バイオマスの前処理方法では、前記リグノセルロース系バイオマスとして稲藁を用いる場合には、まず、カッターミル等により約3mmの長さに粉砕された稲藁に水を添加して、例えば5質量%の濃度の混合液を調製する。
【0078】
次に、前記混合液に、その全量に対し0.1〜0.5μMの範囲のリグニンペルオキシダーゼ(LiP)を添加する。前記LiPは、配列番号3で表されるアミノ酸配列からなる野生型LiPにおいて、240番目のH(ヒスチジン)をF(フェニルアラニン)に、241番目のT(トレオニン)をL(ロイシン)に、242番目のI(イソロイシン)をL(ロイシン)に、それぞれアミノ酸置換したLiP変異体であることが好ましい。前記野生型LiPは、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0079】
前記LiP変異体は、耐熱性及び耐過酸化水素性を備えている。そこで、前記LiP変異体が添加された混合液を、0.1〜0.5mMの過酸化水素の存在下、25〜30℃の範囲の温度に、1〜120時間の範囲の時間保持することにより、前記稲藁のリグニンを分解し、細胞壁を破壊することができる。
【0080】
次に、前記混合液に、その全量に対し0.1〜0.5μMの範囲の本実施形態のMnP変異体(M238I、先祖型MnP変異体)を添加する。本実施形態のMnP変異体は、前述のように耐熱性及び耐過酸化水素性を備えている。
【0081】
そこで、本実施形態のMnP変異体が添加された混合液を、0.1〜0.5mMの過酸化水素の存在下、37〜42℃の範囲の温度に、1〜120時間の範囲の時間保持することにより、前記稲藁に含まれるMn(II)をMn(III)に酸化することができる。生成したMn(III)は、前記LiP変異体により破壊された細胞壁から、前記稲藁の内部に侵入し、該稲藁に含まれる不飽和脂肪酸と錯体を形成する。そして、前記錯体が前記稲藁のリグニンをさらに分解する。
【0082】
また、本実施形態のMnP変異体によれば、それ自体が前記リグニンの分解を行うことはなく、該MnP変異体により酸化されたMn(III)が前記リグニンの分解に関与する。従って、前記MnP変異体自体から離間した部位で前記リグニンの分解を行うことができる。
【0083】
本実施形態のリグノセルロース系バイオマスの前処理方法により処理された前記混合液は、前記リグニンの分解後、さらに糖化酵素を添加することにより、前記稲藁に含まれるセルロース及びヘムセルロースを糖化して、糖化溶液とすることができる。また、前記糖化溶液は、バイオエタノールの原料として用いることができる。
【0084】
尚、本発明のリグノセルロース系バイオマスの前処理方法では、前記MnP変異体に代えて、配列番号1で表される第1のアミノ酸配列からなる組換えタンパク質を用いてもよく、前記MnP変異体と同一の作用効果を奏することができる。
【0085】
また、本発明のリグノセルロース系バイオマスの前処理方法では、前記LiP変異体に代えて、野生型LiP等の他のリグニンペルオキシダーゼを用いてもよい。ただし、この場合には、前記LiP変異体よりも耐熱性又は耐過酸化水素性が低く、本実施形態のMnP変異体と同一条件で用いることが難しくなることがある。
【0086】
また、本実施形態のリグノセルロース系バイオマスの前処理方法では、前記混合液に前記LiP変異体を添加した後、本実施形態のMnP変異体を添加するようにしているが、前記LiP変異体と本実施形態のMnP変異体とを同時に添加するようにしてもよい。
【符号の説明】
【0087】
1…野生型MnP、 2…野生型LiP、 3…子のう菌由来LiP、 4…VP、 5…MnP、 6…LiP、 A…分岐点。
【特許請求の範囲】
【請求項1】
野生型マンガンペルオキシダーゼのアミノ酸配列の230〜250番目のアミノ酸をより疎水性の大きなアミノ酸で置換したことを特徴とするマンガンペルオキシダーゼ変異体。
【請求項2】
請求項1記載のマンガンペルオキシダーゼ変異体であって、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とするマンガンペルオキシダーゼ変異体。
【請求項3】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とする組換えタンパク質。
【請求項4】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードすることを特徴とする遺伝子。
【請求項5】
請求項4記載の遺伝子において、配列番号2で表される塩基配列からなることを特徴とする遺伝子。
【請求項6】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有することを特徴とする組換えベクター。
【請求項7】
請求項6記載の組換えベクターにおいて、配列番号2で表される塩基配列からなる遺伝子を含有することを特徴とする組換えベクター。
【請求項8】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【請求項9】
請求項8記載の形質転換体において、配列番号2で表される塩基配列からなる遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【請求項10】
リグノセルロース系バイオマスからリグニンを分解して除去するリグノセルロース系バイオマスの前処理方法であって、
リグノセルロース系バイオマスに、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体または組換えタンパク質と、リグニンペルオキシダーゼとを添加することを特徴とするリグノセルロース系バイオマスの前処理方法。
【請求項11】
請求項10記載のリグノセルロース系バイオマスの前処理方法において、
前記リグノセルロース系バイオマスに、前記リグニンペルオキシダーゼを添加した後、前記配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体または組換えタンパク質を添加することを特徴とするリグノセルロース系バイオマスの前処理方法。
【請求項12】
請求項10又は請求項11記載のリグノセルロース系バイオマスの前処理方法において、
前記リグニンペルオキシダーゼは、配列番号3で表される第3のアミノ酸配列、又は第3のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第4のアミノ酸配列からなるリグニンペルオキシダーゼ変異体であることを特徴とするリグノセルロース系バイオマスの前処理方法。
【請求項1】
野生型マンガンペルオキシダーゼのアミノ酸配列の230〜250番目のアミノ酸をより疎水性の大きなアミノ酸で置換したことを特徴とするマンガンペルオキシダーゼ変異体。
【請求項2】
請求項1記載のマンガンペルオキシダーゼ変異体であって、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とするマンガンペルオキシダーゼ変異体。
【請求項3】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とする組換えタンパク質。
【請求項4】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードすることを特徴とする遺伝子。
【請求項5】
請求項4記載の遺伝子において、配列番号2で表される塩基配列からなることを特徴とする遺伝子。
【請求項6】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有することを特徴とする組換えベクター。
【請求項7】
請求項6記載の組換えベクターにおいて、配列番号2で表される塩基配列からなる遺伝子を含有することを特徴とする組換えベクター。
【請求項8】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【請求項9】
請求項8記載の形質転換体において、配列番号2で表される塩基配列からなる遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【請求項10】
リグノセルロース系バイオマスからリグニンを分解して除去するリグノセルロース系バイオマスの前処理方法であって、
リグノセルロース系バイオマスに、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体または組換えタンパク質と、リグニンペルオキシダーゼとを添加することを特徴とするリグノセルロース系バイオマスの前処理方法。
【請求項11】
請求項10記載のリグノセルロース系バイオマスの前処理方法において、
前記リグノセルロース系バイオマスに、前記リグニンペルオキシダーゼを添加した後、前記配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなるマンガンペルオキシダーゼ変異体または組換えタンパク質を添加することを特徴とするリグノセルロース系バイオマスの前処理方法。
【請求項12】
請求項10又は請求項11記載のリグノセルロース系バイオマスの前処理方法において、
前記リグニンペルオキシダーゼは、配列番号3で表される第3のアミノ酸配列、又は第3のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第4のアミノ酸配列からなるリグニンペルオキシダーゼ変異体であることを特徴とするリグノセルロース系バイオマスの前処理方法。
【図1】


【図2】
【図3】
【図4】
【図2】
【図3】
【図4】
【公開番号】特開2012−175911(P2012−175911A)
【公開日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願番号】特願2011−39990(P2011−39990)
【出願日】平成23年2月25日(2011.2.25)
【出願人】(592068200)学校法人東京薬科大学 (32)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
【公開日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願日】平成23年2月25日(2011.2.25)
【出願人】(592068200)学校法人東京薬科大学 (32)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
[ Back to top ]