
ミエリン塩基性タンパク質プロモーターの制御下でルシフェラーゼを発現する動物モデル(MBP−LUCI)、及び生物発光インビボ画像化のためのモデルの使用
CNSにおける脱髄及び再髄鞘化事象を転写レベルでインビボで可視化及び定量するための、ミエリン塩基性タンパク質−ルシフェラーゼバイオイメージング非侵襲的モデルが提供される。ルシフェラーゼを発現するトランスジェニック動物を、ホタルルシフェラーゼレポーターにカップリングされたミエリン塩基性タンパク質(MBP)プロモーターを用いて生成した。MBP−luciバイオイメージングモデルは、髄鞘形成の状態及び再髄鞘化を調節する試験化合物の有効性をモニタリングするための手段を提供する。バイオイメージングの利点は、長期間の研究において被験体がそれ自身のコントロールとして役立ち得るということである。同じ被験体が、脱髄及び再髄鞘化プロセスの間、少なくとも10週の期間にわたって連続して追跡され得る。このモデルは、個々の動物画像化応答の正規化を可能にし、そしてかなり分散が減少した良質のデータを提供する。さらに、動物のコホートを異なる時点で屠殺する必要がないので、化合物有効性研究に必要な数を減らすことが可能である。
【発明の詳細な説明】
【技術分野】
【0001】
説明
本発明は、例えば生きた動物のバイオイメージング技術を用いて脱髄/再髄鞘化事象を転写レベルで実時間で可視化及び定量するために使用される、製造されたミエリン塩基性タンパク質−ルシフェラーゼ(MBP−luci)バイオイメージングモデルに関する。
【背景技術】
【0002】
薬物開発では減少率(attrition)が高く、5つの化合物のうち1つだけが開発を切り抜けて承認に至ると説明される(非特許文献1)。さらに、劇的に増加した投資にもかかわらず、新規薬物の導入率は過去30年にわたって比較的一定のままであり、1年あたりに新しい薬物クラスで進展があって最終的に市場に出るのは2つか3つだけである(非特許文献2)。
【0003】
薬物開発の初期段階で適用される分子画像化及び機能的画像化は、生物活性の証拠を提供して的確な薬物効果を裏付けることができる。従って、分子画像化技術における投資は薬物開発を増進すると期待される(非特許文献3)。より従来型の読み取り情報に勝る分子画像化技術の利点は、インビボでの生物学的プロセスを調べるために十分な空間的及び時間的分解能でインタクトな生物において行うことができることである。さらに、これらの分子画像化技術により、同じ動物の異なる時点での繰り返しの非侵襲的で均一でかつ比較的自動化された研究が可能となり、それ故、長期的な研究の統計的検出力を増加させ、そして必要な動物の数及び費用を減少させる。MBP−luciモデルは、インビボでの初期薬物候補スクリーニングの処理能力の増大、さらには生物学的アッセイの感度の増大の両方を後押しする。初期リード化合物はしばしば、販売薬製品としては準最適であるが、インビボでの特異的かつ有意な活性の検出は、特定のCNS疾患の処置を目的として、許容しうるレベルの活性を達成し、かつインビボでの毒性を最少にするための化学構造の最適化をもたらし得る。
【0004】
分子画像化
分子画像化は、生きた生物において細胞及び細胞内レベルでの特定の分子プロセスの評価における画像化技術を利用し統合するための、様々な分野(例えば、細胞分子生物学、化学、医学、薬理学、物理学、生物情報科学及び工学)からのアプローチの収束を指す(非特許文献4)。
【0005】
遺伝子工学の出現は、例えば新薬発見パイプラインを含む応用科学に大きな変化を引き起こした。同じように、動物画像化手順の開発及び開拓は前臨床研究のための新しい手段を提供しつつある(非特許文献5)。動物モデルは伝統的に、実時間で生理学的事象の定量における困難さのために扱いにくい。長年にわたって、新しい画像化方法がこの困難さを克服するために開発されてきた(例えば、MRI、CT、PET)。より最近では、ルシフェラーゼ(ホタルの発光する酵素)のインビボ発現に基づく生物発光(bioluminescense)画像化が標的遺伝子活性の非侵襲性検出のために使用されている。
【0006】
動物工学と分子画像化技術を組み合わせることにより、生きている動物において特定の分子プロセスに関する動的な研究を行うことが可能となった。このアプローチは、前臨床プロトコルに影響を与える可能性があり、従って医学の全ての局面を広く変化させ得る(非特許文献6)。
【0007】
分子画像化:生物発光
インビボ生物発光画像化(BLI)は細胞又は組織からの発光の検出に基づく高感度のツールである。レポーター遺伝子技術の有用性は、インビボ画像化法により生きている動物における特定の細胞及び生物学的プロセスを分析することを可能にする。生物発光(生きている生物による可視光の酵素的生成)は多くの非哺乳動物種において天然に存在する現象である(非特許文献7)。ルシフェラーゼは基質の酸化を触媒して光子を放出する酵素である(非特許文献8)。北アメリカホタル(North American firefly)からの生物発光は最も広く応用される。ホタルルシフェラーゼ遺伝子(luc)発現は酵素ルシフェラーゼを産生し、これが基質D−ルシフェリンを非反応性オキシルシフェリンに変換し、562nmにおける緑色発光を生じる。哺乳動物組織は自然には生物発光しないので、画像を非常に小さいバックグラウンドシグナルで生成することができるので、インビボBLIは非常に魅力的である。
【0008】
BLIには、光レポーターを駆動する選択された遺伝子プロモーターの制御下の生物発光レポーター遺伝子からなる発現カセットを用いて細胞又は組織を遺伝子操作することが必要とされる(図1)。光生成を誘導するために、基質(例えばルシフェリン)を、icv、血管内、腹腔内又は皮下注射により投与する。
【0009】
ルシフェラーゼ/ルシフェリンにより放射される光は、数ミリメートルから数センチメートルの深さの組織を透過することができるが;組織深さ1センチメートルごとに約10倍光子強度が減少する(非特許文献7)。インビボでの生物発光を検出するためには高感度の光検出機器を使用しなければならない。検出器は単位面積あたりに放出される光子の数を測定する。400と1000nmとの間の波長における低レベルの光は、シリコンウエハーに衝突する光子を電子に変換する電荷結合素子カメラを用いて検出することができる(非特許文献9)。ソフトウエアは電子シグナルを2次元画像に変換することができる。ソフトウエアはまた、放射光の強度(検出器に衝突する放射された光子の数)を定量し、そしてこれらの数値を疑似カラー図に変換することもできる。実際のデータは光子カウントで測定されるが、疑似カラー図は迅速な視覚的解釈を可能にする。目的の領域内の定量的測定は、より微細な差異のために必要かもしれない。冷却CCDカメラの使用は熱雑音を低減させ、そして遮光箱(light−tight box)はルシフェラーゼが生成した光を最適に可視化し、定量することを可能にする(非特許文献10)。
【0010】
発光シグナルの解剖学的位置についての自筆文書(autograph)又はX線写真のような別の種類の画像に重ねられたルシフェラーゼ画像を有することは有用である(図2)。ソフトウエアは視覚化及び解釈のために画像を重ねる。
【0011】
脱髄疾患、オリゴデンドロサイト及びミエリン塩基性タンパク質
ミエリン塩基性タンパク質(MBP)は正常なミエリン圧縮及び機能に必要とされる。ミエリンオリゴデンドロサイト糖タンパク質(MOG)及びプロテオリピドタンパク質(PLP)と共に、これらのタンパク質はミエリン構造タンパク質ファミリーのメンバーを構成し、そしてミエリン産生細胞:中枢神経系(CNS)におけるオリゴデンドロサイト及び末梢神経系(PNS)におけるシュワン細胞により合成される。発達するCNSにおいて、MBPの発現は髄鞘形成の期間と一致する。従って、MBPはオリゴデンドロサイト成熟のマーカーとして当該分野で受け入れられている。MBPの高レベル発現(他のタンパク質と共に)は、ミエリン生成と同時に発生し、ミエリン形成の間中継続し、そして軸索破壊の際に終わる(非特許文献11)。
【0012】
ミエリンの完全性を冒す疾患は、冒されたニューロンにおける軸索シグナルの伝導障害を生じ、そしてどのニューロンが含まれるかによって、感覚、運動、認識又は他の機能の障害を生じ得る。脱髄疾患という用語は、疾患の原因よりその効果を示し、そして遺伝的特質、感染性因子、自己免疫反応及び未知の要因により引き起こされ得る。これらは全て、回復性治療の必要性をしばしば伴う大きな満たされていない医療的必要性を有するヒトの状態のままである。
【0013】
中枢神経系の脱髄疾患としては以下が挙げられる:
・多発性硬化症(突発性炎症性脱髄疾患と呼ばれる似た疾患も一緒に)
・横断脊髄炎
・ドヴィック病
・進行性多病巣性白質脳障害
・視神経炎
・白質萎縮症
・ペリツェーウス−メルツバッヒャー病
・ミエリン機能障害/損失は、脊髄損傷、アルツハイマー病及びパーキンソン病、並びに統合失調症とも関連する。
【0014】
末梢神経系の脱髄疾患としては以下が挙げられる:
・ギラン・バレー症候群及びその慢性カウンターパート、慢性炎症性、脱髄性多発ニューロパシー
・末梢神経障害(毒素誘導、糖尿病性、抗MAGなど)
・シャルコー・マリー・ツース病。
【0015】
脱髄のインビボモデル
脱髄疾患と闘うための前臨床候補の開発における共通の段階は、脱髄作用の一部又は全てを再現し、かつ内因性再髄鞘化の能力を有する動物モデルにおいて治療剤候補の作用のアッセイを行うことである。化学的に誘導される脱髄を動物モデルにおいて達成することができ、そして6〜8週齢の若年成体雄性マウスは0.2%クプリゾン(ビス−シクロヘキサノンオキサルジヒドラゾン)の4〜6週の食餌により生じる脱髄を受けやすい(非特許文献12)。クプリゾン処理を用いて、脱髄は脳全体にわたって発生するが、高度に濃縮された白質領域である脳梁内で最も容易に検出される(非特許文献13)。脱髄はクプリゾン食餌開始後3週以内に明らかとなる。食餌を通常の餌に代えることにより、4〜6週以内にほとんど完全な再髄鞘化が可能となる(非特許文献14)。アミロイド前駆体タンパク質又はビールショースキー銀含浸を使用する軸索損傷についての免疫組織化学染色は、8週齢のマウス(Merkler)における軸索損傷をあまり示さない。しかし、軸索離断は6〜7月齢のマウスにおいて有意に増加し、高齢の動物において見られる再髄鞘化の減退に部分的に寄与する(非特許文献15)。分化の際にミエリンと置き換わるオリゴデンドロサイト前駆細胞、並びに小グリア細胞及びマクロファージの両方は、クプリゾン食餌の約4週目にピークに達し、そしてこれらは効率的な再髄鞘化に必要とされる(非特許文献16)。
【0016】
ミエリン含有量の変化を定量するために、脳全体又は小領域を採取し、ドットブロット又はウェスタン分析により評価することができる。あるいは、ミエリンの損失及び修復の小領域を追跡するためのより高解像度の明敏な(長期に対して)アプローチは定量的立体解析学を含む。例えば、コンピューター支援立体解析ツールボックス(computer assisted stereological toolbox)(CAST)システムを使用して、脳梁におけるミエリンルクソールファストブルー(ミエリン染料)の変化を評価することができる。トルイジンブルー及び電子顕微鏡観察は、密集ミエリンが以前の無鞘軸索(naked axons)を正確に鞘で包んでいることを確実にするために必要な最終工程である。長期的な評価に関して、T2強調磁気共鳴画像化法は、クプリゾン食餌を与えられたマウスの脳梁におけるミエリン変化を定量的に評価するための手段を提供する(sanofi−aventis Neurology,2008 非公開の結果)。
【0017】
実時間でインビボで髄鞘形成状態を追跡するMBP−luciバイオイメージングモデル
2003年の文献(非特許文献17)は、MBPプロモーターが、髄鞘形成及び再髄鞘化のタイミングを調節するために重要である4つの広く分布した保存されているモジュール(M1、M2、M3及びM4、0.1から0.4kbの範囲)である、調節エレメントを含有することを報告した(図1、3及び4を参照のこと)。Lac Zをレポーター遺伝子として使用して、近位のモジュールM1及びM2がCNS発達の間にオリゴデンドロサイトにおいて比較的低レベルの発現を駆動し、一方で上流のM3領域は、発達の間中及び成体CNSにおいてオリゴデンドロサイトにおける高レベルの発現を駆動するということが実証された。さらに、このM3領域は、脱髄傷害後の再髄鞘化の間の発現に必要とされる。M4もまたシュワン細胞の髄鞘形成におけるMBP発現を駆動する。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】DiMasi,JA,et al,J Health Econ 22,151−185
【非特許文献2】Lindsay MA,Nature Rev Drug Discovery,2,831−838
【非特許文献3】Rudin M,et al,Progress in Drug Res,2005,vol 62,185−255
【非特許文献4】Massoud T.F.,Genes Dev.17:545−580
【非特許文献5】Maggie A,Ciana P.Nat.Rev.Drug Discvy.4,249−255
【非特許文献6】Maggie A.Trends Pharmacolo.Sci.25,337
【非特許文献7】Contag,CH,Mol.Microbiol.18:593−603
【非特許文献8】Greer LFIII,Luminescence 17:43−74
【非特許文献9】Spibey CP et al Electrophoresis 22:829−836
【非特許文献10】Contag CH,Annu.Rev.Biomed.Eng.4:235−260
【非特許文献11】Gupta et al.Brain Res.464:133−141
【非特許文献12】Ludwin SK ,1978,Lab Invest 39:597−612
【非特許文献13】Merkler et al,2005,NMR Biomed 18:395−403
【非特許文献14】Matsushima et al,2001 ,Brain Pathol 1 1 :107−1 16
【非特許文献15】Irvine KA,et al,2006,J Neuroimmunol 175:69−76
【非特許文献16】Franco RJM,2002,Nat Rev 3:705−714
【非特許文献17】Farhadi et al,The Journal of Neuroscience,November 12,2003,23(32):10214−10223
【発明の概要】
【発明が解決しようとする課題】
【0019】
インビボでのミエリンの変化をアッセイするために動物モデルを使用する現在の方法は時間がかかり、典型的には4〜8週に及び、そして多数の動物を必要とする。ルシフェラーゼ遺伝子を発現する4つの特定の調節領域全てを含むMBP DNAプロモーターは、脳、脊髄、及び/又は末梢神経系におけるミエリン塩基性タンパク質(MBP)転写活性の変化を実時間で追跡するために開発されたマウスMBP−luciトランスジェニックバイオイメージングモデルのための基礎を構成する。この齧歯動物トランスジェニックモデルは、ヒト脱髄疾患の処置のための開発候補を選択するように意図された原理の研究のインビトロ及びインビボでの裏付けのために使用され得、そして現在のモデルに勝る多数の改善を提供し得る:
・ 生物発光により神経系髄鞘形成の変化を非侵襲的に追跡する能力は、死後の組織化学的分析及び組織発現分析に伴うかなりの資源及び労働力の要求を減らす。
・ 長期的な研究は、髄鞘形成の変化に対するモデルの感受性を大いに増加させる。以前のアッセイは各時点で異なるグループの動物を使用しなければならず、そしてこのことが研究の間に収集できるデーター点の数を制限する。別々のグループの動物の必要性のためにさらなる変数が導入される。
・ バイオイメージングデータは迅速に、典型的には同日に処理され、それにより、研究設計を最適化し、そして不必要な資源投資を減らすための調整(すなわち、有意性に達するための研究の長さの延長又は薬物効果が無いことに起因する早期の終了)が可能となる。
・ 動物グループ及び対応する処置は、全て0時点でアッセイすることができ、これにより、処置群あたりに必要とされる動物の数を同時に減らしながら、研究の統計学的有意性が最適化される。研究データは初期値に対して正規化され得、特定の治療の効果に対するより強い結果を支持しながら固有の生物学的変動を大幅に減少する。
【課題を解決するための手段】
【0020】
発明の要旨
本発明は、脱髄(例えば、神経保護、ミエリン保護)及び再髄鞘化(例えば修復)の程度に対する治療実体の効果を評価するための新しいインビボモデルを提供した。本発明のバイオイメージング動物及び化合物スクリーニング方法は、生きている動物における関連性のある実時間高解像度データの生成により、 医薬化合物及び他の治療剤のスクリーニング及び/又は原理の検証の裏付けを可能にする。さらに、可能性のある毒性、PK/PD、及び治療域に関連する評価を得ることができ、それにより霊長類及びヒトにおける投薬の周りの(around)より正確な予測を可能にする。
【0021】
本発明は、生きているトランスジェニックモデル生物を使用する髄鞘形成事象の研究のための改善されたツール及び方法を提供する。本発明は、各被験体が長期間の研究においてそれ自身のベースラインコントロールとして役立ち得るので、他のモデルと比較して必要とされる被験体の全体の数を減らす。従って生物間の変動性が低減し、統計学的結果における信頼性を改善する。本発明の種々の局面において、様々な利点が達成される。様々な用途のいくつかは以下の考察において特徴付けされる。
【0022】
MBP発現との相関により、本発明は、生きているモデル動物において脱髄及び/又は再髄鞘化をモニタリングする方法を提供する。本方法は、MBPプロモーターにより駆動されるルシフェラーゼ遺伝子を発現するモデル生物を生成するように前駆細胞を遺伝子導入で改変することを含み得る。ルシフェラーゼ遺伝子は、発現されると、ルシフェリンのような基質の存在下で光を生じる(生物発光)ように機能することができる。生物からの生物発光をモニタリングすることにより、髄鞘形成及び脱髄の活性を評価することができる。画像化装置及び分析は、生物発光及び髄鞘形成事象を生物の身体の特定の部分に関連付けることを迅速に可能にする。
【0023】
中枢神経系及び末梢神経系の事象を研究するために、これらの神経組織においてミエリンを使用する脊椎動物は、特に適したモデル生物である。哺乳動物は、CNS及びPNSの両方において神経伝導を補助するためにミエリンを使用する生物である。齧歯動物(例えばラット、マウス)、ウサギ、ヒツジ、霊長類、モルモットなどのような一般的な哺乳動物モデルは、本発明の実施における使用に適したモデル生物である。
【0024】
生物のサイズは、それ自体は限定因子ではない。しかし、装置サイズは特定の生物の形状又はサイズに対して最適化され得る。
【0025】
単一のバイオイメージング事象は所望の情報を提供し得る。しかし、本発明の利点は、複数の時点にわたって同じ動物で測定を繰り返して複数の測定を比較する能力を提供する。それにより単一の生物はそれ自体のコントロール又はベースラインとして役立つ。
【0026】
その結果として、単一の生物における1つ又はそれ以上の脱髄又は再髄鞘化間隔にわたるバイオイメージングが可能である。さらに、脱髄又は再髄鞘化間隔による画像化シグナル正規化は、ここで繰り返される間隔を通した画像化は生物におけるMBP転写物のレベルの変化を1つ又はそれ以上の事象にわたって検出するために有効であり、よりロバストなデータを生じる。
【0027】
生物発光シグナルは身体組織により減衰される。従って、身体表面の近位にある神経組織はより強いシグナルを生じる。検出器におけるシグナルを、例えばより高濃度を使用するか又は酵素活性若しくは発現レベルを高めてより多くのルシフェラーゼを反応させることにより、より強いシグナル出力を生じることにより増加させることができる。データ収集はまた、より高感度の検出器を使用して改善され得る。より高感度の検出器は、有意なシグナルを生成させてノイズを最少にするためにしばしば冷却装置を必要とする。データが収集される時間もまたシグナルの量を増加させるために役立つ。
【0028】
本発明のモデル生物は、MBPプロモーターにより駆動されるルシフェラーゼ遺伝子を含むトランスジェニック動物であり得る。この動物は哺乳動物、例えば動物モデルとして使用されるいずれかの哺乳動物であり得る。ラット及びマウスは一般的な動物モデルである。MBPプロモーターの制御下のルシフェラーゼ遺伝子は、プロモーターが活性化されるかオンにされる場合にモデルの特定の標的組織において発現される。
【0029】
MBPプロモーターはM1〜M3を含有していても、M1〜M4を含有していてもよい。
【0030】
シグナル改善は、その毛髪が少ない減衰に寄与するモデルの選択のような簡単な操作によりなされ得る。例えば、その毛髪がC57/B6マウスの毛髪よりも少ない減衰であるモデルは、C57/B6マウスのシグナルを上回るシグナルを生じるだろう。
【0031】
本発明はまた、バイオイメージングのためのモデル生物を作製するための方法を提供する。作製は、例えばマウス系統から開始して、そしてトランスジェニック動物を作製し、次いで生後髄鞘形成のピークにおけるインビボマウス全身画像化が(例えばCNSの)特定の画像化を示す1つ又はそれ以上の系統を選択し;次いで、エクスビボ画像化により所望の領域、例えば主に脳の緻密な(compact)白質領域におけるルシフェラーゼ導入遺伝子の発現が確認される1つ又はそれ以上の系統を選択することにより達成され得る。次いで、ルシフェラーゼ画像強度が脱髄及び再髄鞘化の変化と高度に相関している1つ又はそれ以上の系を選択することができる。改善されたデータのために、適切な操作の間、明確な組織学的脱髄を示す1つ又はそれ以上の系を選択し得る。
【0032】
クプリゾン脱髄モデルは本発明での使用に適している。バイオイメージングのタイミングは、モデル生物の発達特性に依存し、例えば、生後髄鞘形成のピークが一般的に生後約3〜5週齢のG1マウスであることを考慮するようにタイミングを選択してもよい。
【0033】
本発明は、生物におけるミエリンの事象に影響を及ぼし得る化学的化合物、生物学及び他の治療実体をスクリーニングするために有用である。化合物は、遺伝子発現又は細胞内若しくは細胞間のシグナル伝達事象を調節してミエリン事象に影響を及ぼし得る。これらは単にいくつかの具体例であり、特許請求の範囲において特徴付けされる本発明の範囲全部とみなされるべきではない。
【0034】
MBPプロモーターは、記載されるように(例えばFarhadi、上記)、レポーターlac Z遺伝子を使用して十分に特徴付けされているが、MBP−lac Zモデルの使用はβ−ガラクトシダーゼ(lacZレポート遺伝子産物)の検出のための組織採取及び固定化の必要性のために制限される。このプロセスは、生きている動物における検出とは両立できない組織化学的技術を必要とする。この問題を回避するため、ルシフェラーゼ又はGFPレポーターがMBPプロモーターの大部分により選択的に制御されるトランスジェニックバイオイメージングモデルを開発するために、生物発光又は蛍光系の使用が提案されてきた。
【0035】
この新規なバイオイメージングモデルは、生きている動物(例えばマウス)における実時間での脱髄及び/又は再髄鞘化の可視化及び定量のために設計される。このようなモニタリングは、自動化バイオイメージング技術と併せて使用され得る。このモデルは、 標的選択及び化合物進行(progression)に関して重要な方針決定をするために相対的及び定量的に、例えば標的の検証のため(例えばバイオイメージングモデルマウスが所望の特質を有するノックアウトマウス及びトランスジェニックマウスと交配される場合)、そしてまた化合物の検証(例えば、クプリゾン又は実験的自己免疫性脳脊髄炎[EAE]のようなモデルにおける化合物の有効性の測定)のために有用なツールである。
【0036】
本発明のバイオイメージングモデル(MBP−luci TG)は開発されて脱髄/再髄鞘化事象をインビボで定量するために使用された。本バイオイメージングモデルの利点は、長期の研究が可能となり、それによって各生物がそれ自身のコントロールとして役立つことができるということである。それ故、特定の時点における動物の犠牲が避けられる。
【0037】
個々のマウスは脱髄及び再髄鞘化プロセスを通して継続的に追跡され得る。
【0038】
バイオイメージング方法論は、生きているマウスにおいて生物学的応答を追跡するために必要とする時間及び資源がより少ない。
【図面の簡単な説明】
【0039】
【図1】ミエリン発現における実時間変化を追跡するために使用されるMBPルシフェラーゼトランスジェニックモデルの図式的要約を示す。
【図2】内在性MBPプロモーターが、発達の間及び成人期中にオリゴデンドロサイトにおけるMBPの発現を異なって調節する4つのエレメントを有するということを示す。(HF Farhadi:J.Neurosc.2003,23 (32),10214−10223)。M1/M2は両方とも生後初期段階の転写物を調節する。M3は成熟の全体にわたる発現の転写物調節に関与する。M4は、MBP転写物のシュワン細胞発現に寄与する。MBP−luci導入遺伝子構造は2つの形態のMBPプロモーターを有する。系121はM4エレメントを含み、そして脳、脊髄及び末梢神経系においてルシフェラーゼを発現すると予測される。系171はより短いMBPプロモーターからなり(例えばM4エレメントを欠く)、そしてバイオイメージング分析により確認されたように、主に脳で発現する。
【図3】ルシフェラーゼ発現ベクターにおける5K bpの5’DNAを有するMBPプロモーターを含むMBP−luci導入遺伝子発現カセットを示す。
【図4】ルシフェラーゼ発現ベクターにおける10K bpの5’DNAを有するMBPプロモーターを含むMBP−luci導入遺伝子発現カセットを示す。
【図5】MBP−Luci導入遺伝子系のスクリーニングツリーの例を示す。
【図6】MBP−luci創始者の第一レベルのスクリーニングからのインビボルシフェラーゼ画像を示す。ルシフェラーゼ強度は105(+++)から103(+)(光子/cm2/秒)の範囲に及ぶ。
【図7】年齢の増加に伴う脳におけるルシフェラーゼ画像の減少を示す、7週(A)及び10ヶ月(B)でのバイオイメージングモデル生物を示し、これは生後初期の発達後に観察される髄鞘形成の現象と相関する。
【図8】MBP−luciマウスのインビボでのバイオイメージングを示す:頭蓋(ROI:対象の領域)において測定された2.7x103光子/sに相当するバックグラウンド生物発光を有する、左側のネガティブ導入遺伝子野生型マウス(WT)。ヘテロ接合MBP−luciマウスは右に示され、そしてROI生物発光は1.327x105光子/sに相当する。ホモ接合MBP−luciマウスは中央に示され、ROI値3.2924 x105 光子/sを有し、これはほぼ予測通りであり、ヘテロ接合の値の2倍である。
【図9】CNS特異的時間的ルシフェラーゼ発現を示す。示されるようにMBP−luciトランスジェニックマウスからの頭蓋生物発光は、4週(A及び*)から8(B及び**)まで減少し、Taqman分析(C)により決定された内在性MBP転写物レベルに対応している。
【図10】MBP−luciマウスにおけるルシフェラーゼのCNS特異的及び小領域発現を示す。MBP−luciトランスジェニックマウス脳の2つの矢状(sagital)切断(A及びB)後(followed)のスライスのバイオイメージングは、脳における脳梁小領域がその最も高い生物発光シグナルを含み、そしてクプリゾン病変モデルにおける最も高い脱髄及び再髄鞘化と相関するということを明らかにした。
【図11】MBP−luciモデルにおけるバイオイメージング及びクプリゾン病変食餌において起こることが知られている他の生物学的応答を示す。ここで クプリゾン食餌は4週間与えられ、そしてグラフの左に示されるバイオイメージング及び細胞変化を生じる。示される(矢印)各週におけるバイオイメージングは、内在性MBP発現レベルと平行して変化すると予測される(Matsushima GK and Morell P,Brain Pathology 11,1−10,2001)。
【図12】野生型C57 BL/6Jマウスにおける連続的なクプリゾン食餌処置に応じた内在性MBP定常状態mRNAの変化を示す。8週齢のマウスをそれらの食餌でクプリゾンに12週まで曝露させた(黒三角)。第二のグループにおいて(白三角)、クプリゾン餌を6週の曝露後に取り除き、そしてマウスをさらに6週間回復させた。mRNAレベルについてのデータは、ノーザンブロットによる単一測定であり、そして3つのコントロールの平均と比較してプロットされている(Jurevics H,et al,Journal of Neurochemistry,2002,82,126−136)。
【図13】MBP−Luciバイオイメージングモデルに対する証明されたクプリゾン食餌の影響を示す。画像化シグナルは、内在性MBP遺伝子の発現パターンを明確に再現し得る。クプリゾン餌を与えた間(0週から4週)、2〜3倍の画像化シグナル減少があり、そしてクプリゾン餌の除去後(4週から7週)の3〜4倍の画像化シグナル増加もあった。
【図14】MBP−luciモデルにおける脳梁(corpus collosum)のルクソールファストブルー(LFB)染色を示す。マウスをクプリゾン又は通常餌のいずれかで4週間処置し、次いで両方とも通常食餌に戻した。6週目に、クプリゾン処置されたMBP−luciの脳梁(corpus collosum)(B)における明瞭な脱髄が、通常食餌処置グループ(A)と比較してLFB染色を使用して組織化学的に検出され得る。髄鞘形成における構造的変化の組織化学的アッセイは、処置されたMBP−luciモデルにおけるバイオイメージングシグナルにおける変化と相関する。
【図15】ペルオキシソーム増殖因子活性化受容体デルタ(PPARδ)アゴニストツール化合物(本明細書以下では’517)が、自発的再髄鞘化の期間の間に系171 hetマウスにおけるルシフェラーゼの画像化シグナルを増強することを示す。8週齢MBP−luciマウス(系171ヘテロ接合性、B6C3H系統)を0.2%クプリゾン含有餌で4週間飼育し、次いで通常餌にして再髄鞘化させた。次いでマウスにビヒクル(0.6%カルボキシメチルセルロールナトリウム塩及び0.5%Tween80)又は30mg/kg PPARδアゴニストツール化合物’517のいずれかを8日間、1日に2回経口投与し、そして示された時点で画像化した。データを0週ベースラインシグナルに対して正規化した。ツール化合物’517(n=12)は、ビヒクルグループ(n=12)と比較してルシフェラーゼシグナルの30〜100%の相対的な増加を生じ、これはオリゴデンドロサイト前駆細胞分化の刺激に対する化合物の効果に起因するものと考えられている(例えば、インビトロでの知見と一致する)。
【図16】PPARδアゴニストツール化合物’517は、再髄鞘化期の間のルクソールファストブルー(LFB)ミエリン染色を改善する(系171ヘテロ接合、B6C3H系統)。ホルマリン固定パラフィン埋包脳からの傍矢状組織切断を、脳梁におけるミエリンの定量的評価のためにLFBで染色した。各時点についての染色した切片をスコア付けして0(完全な髄鞘形成)から5(完全な脱髄)までのスケールで等級分けした。スコア付けシステムは以下のとおりであった:0=正常ミエリン、脱髄無し、1=最小限の局在化した脱髄、2=軽度から中程度、局在化した脱髄、3=中程度、局所的に極度の脱髄、4=重症、局所的に極度の脱髄、5=重症、広範な脱髄。クプリゾン4週後のマウスからのLFB染色した脳切片の組織学的評価により、系171ヘテロ接合マウス(n=5)における脳梁の中程度から重症の脱髄が確認された。4週から5週の再髄鞘化期の間のvでの処置は、LFBにより7週の時点で決定した場合に、ツール化合物’517グループ(n=5)をビヒクルコントロールグループ(n=3)と比較して、ミエリンの測定可能な増加を生じた。小さいnの数にもかかわらず、組織学的データはインビボルシフェラーゼバイオイメージングデータを支持しており、このことは、ビヒクルコントロールと比較した場合の、’517化合物で処置されたマウスにおける増加した再髄鞘化を示している。
【図17】エストロゲン受容体ベータ(ERβ)アゴニスト(本明細書以下では’5a)30mg/kg及びポジティブコントロールクエチアピン10mg/kgは、クプリゾンモデルにおける脱髄期の間、保護的である。MBP−luc系171ヘテロ接合 B6C3Hマウスにクプリゾン食餌を4週間与え、そして’5a(10mg/kg又は30mg/kg)又はポジティブコントロールクエチアピン(QTP)を経口投与した。マウスを0週(ベースライン)、3週及び4週目に画像化し、データを0週ベースラインに対して正規化した。QTPグループはビヒクルコントロールに対して3週及び4週の両方の時点において10mg/kgで画像化シグナルの有意な増加を示した。QTP処置グループについての結果は、Yanbo Zhangらにより公開されたデータ(表題「Quetiapine alleviates the cuprizone−induced white matter pathology in brain of C57BL/6 mouse」(Schizophrenia Research,2008,Dec 106,182−91))と一致する。30mg/kgでの化合物’5aは、ビヒクルと比較してクプリゾン食餌での3週目(傾向)及び4週目(有意)に増強されたシグナルを示した。10 mg/kgでの’5aは3週目及び4週目の両方において有意な効果を有していなかった。結果は、MBP−luci画像化モデルが、クプリゾンモデルにおける用量依存性変化を検出するために十分感受性であることを示唆する。
【図18】’5a(30mg/kg)及びQTP(10mg/kg)のグループは、ビヒクルコントロールグループと比較して3週目及び4週目に有意に高い導入遺伝子活性を有する。画像化モデルデータは、QTP及び’5aの両方がクプリゾン誘導脳脱髄及びミエリン崩壊を弱めるということを支持する。
【図19】ホモ接合マウスとヘテロ接合マウス(B6C3H系171)との間のクプリゾンモデル画像化シグナル比較を示す。N3世代ヘテロ接合マウスを交配させてMBP−luci対立遺伝子についてのホモ接合マウスを作製した。ホモ接合マウスは、クプリゾン処置の間にヘテロ接合マウスよりも2倍大きなバイオイメージングシグナル域を示した。ホモ接合体におけるレポーター遺伝子の2つのコピーは、脱髄期(例えばクプリゾン食餌における4週後)の間2倍より大きなシグナル減少及び再髄鞘化期(例えばクプリゾン食事の除去及び通常食へ戻した1週後)の間に2倍のシグナル増加を示した。 ヘテロ接合系171(B6C3H系統)がクプリゾンモデルにおいて機能し、そして化合物効果を検出し得るということをデータが実証したが、このモデルは、バイオイメージングシグナル強度を増加させるために交配してホモ接合性にすることによりさらに改善され得る。これはまた、マウスコロニーがホモ接合体として維持され得るので、モデル産生を効率化し、そして遺伝子型解析の費用を減少させる。全体的に、より大きな画像化域は、薬理的化合物プロファイリング研究における化合物の効果の検出を増強し得る。
【図20】LFBで染色されたホルマリン固定されたパラフィン包埋マウス脳からの傍矢状組織切片における脳梁の顕微鏡写真(括弧内の濃い色の長手方向の構造−201)は、ミエリン状態について評価された白質の領域を示す。この領域を、脳梁のルクソールファストブルー(LFB)染色の定量的デジタル画像化分析のために使用した。
【図21】3つの異なるMBP−luci系(系171 B6C3Hヘテロ接合系統、系121 C57BL/6ヘテロ接合系統及び系171B6C3Hホモ接合系統)についての画像化域及び組織学領域(histology window)の比較。マウスに0.2%クプリゾンを含む食餌を4週間又は5週間与えた。画像化データを0週ベースライン測定に対して正規化した。各研究の終わりに、マウス脳を採取し、そして連続パラフィン切片をミエリンについてルクソールファストブルー(LFB)で染色した。平均定性的LFBスコア(0〜5)を表に示す。系171 B6C3Hホモ接合マウスは、最も大きな画像化シグナル減少を示し、そしてまた定性的な組織学により評価した場合に最も重症の脱髄を示した。系171 B6C3Hヘテロ接合マウスは、最も小さな画像化域を示し、そしてまた4週目に最も少ない組織学的脱髄を示した。
【図22】クプリゾンモデルにおける系171ホモ接合マウスの使用を、クエチアピン(QTP)に対する処置応答の実証によりさらに検証した。マウスにクプリゾン食餌を5週間与え、そして同時に、毎日QTP(10mg/kg)を経口投与した。マウスを0週目(ベースライン)、3週目、及び5週目に画像化した。データを0週ベースライン測定に対して正規化した。QTP(10mg/kg)は、ビヒクルコントロールと比較して、3週目及び4週目の両方の時点で画像化シグナルの有意な増加を生じた。結果は系171ヘテロ接合マウスで得られたものと一致する(図17及び18)。データは、系171ホモ接合マウスが、クプリゾンモデルにおけるミエリン発現及び完全性の維持に対する化合物の効果を評価するために使用され得るということを示す。
【図23】系121マウスからのエクスビボ脊髄画像化。発光の重ね合わせは、導入遺伝子発現が脳及び脊髄の白質領域に局在化していたということを説明する。このことは、動物全身バイオイメージング実験から至った結論をさらに支持する。
【図24】C57Bl/6マウス;野生型、系171ヘテロ接合及び系171ホモ接合マウスにおける脳梁のルクソールファストブルー(LFB)染色の定量的デジタル画像解析。脳梁内のポジティブLFB染色を有する領域のパーセントの定量を、染色されたスライドのスキャンされたデジタル画像での脳梁の手書きで輪郭線を描いた領域に対してAperio(R)カラーデコンボリューションアルゴリズムを使用して計算した。動物1体について1つの断面を評価した。グループあたりの動物の数は9と10の間で変動した。LFB染色に対してポジティブな領域のパーセントを動物ごとに、そしてグループに関して計算した。グループ間の統計学的有意性を、対応t検定により評価した。171ホモ接合マウス及び野生C57 BL/6マウスの両方が、4週のクプリゾン食餌後に重症の脱髄を示す(ポジティブ染色%は40〜60%の間である)。対照的に、系171ヘテロ接合マウスは、軽度の脱髄しか示さない(ポジティブ染色%は65〜80%の間である)。従って、系171ホモ接合マウスを、クプリゾン誘導脱髄モデルにおける使用に好ましい系として同定した。
【発明を実施するための形態】
【0040】
発明の説明
以下の5つの基準は、バイオイメージングモデルについての選択を順次最適化するために首尾よく適用された:
特定の実施態様は、5K又は10Kのベクターのいずれかを用いて以下のプロセスにしたがって遺伝子導入操作されたマウスの系から作製され得る。以下に記載される結果は、選択プロセスが35のトランスジェニック系で開始するようにしたものからである。図5はこのプロセスを図表でまとめる。
【0041】
生成されたトランスジェニック系から、生後髄鞘形成のピーク(例えば3〜5週G1マウス)におけるインビボでの動物(例えばマウス)全身画像化がCNS特異的画像化を示す系を選択する。選択された35の系から6つを次の選択段階に進めた。
【0042】
次に、エクスビボ画像化により主に脳の白質領域でのルシフェラーゼ導入遺伝子発現が確認された系を選択した。工程1からの6つの系のうち5つを次の段階に進めた。
【0043】
次いで、ルシフェラーゼ画像強度が、(この例では)クプリゾン脱髄モデルにおいて誘導される脱髄及び再髄鞘化の変化と高度に相関する系を選択した。工程2からの5つの系のうち3つを次の段階に進めた。
【0044】
次に、上記クプリゾンモデルにおいて明らかな組織学的脱髄を示した系を選択した。工程3からの3つのうち2つを好ましい系として選択した。
【0045】
構想の決定的な証拠として、本発明者らは、A003398711(PPARδ選択的アゴニスト)の有効性がバイオイメージングモデルにおいて最適に検出可能である1つの系を選択した。
【0046】
系171(B6C3系統、ヘテロ接合)と指定された模範的な系を、上記の5つの基準を使用して選択し、そして好ましいモデルとして使用した。
【0047】
このプロセスのより詳細な説明は以下の実施例に記載される。
【0048】
定義
別に指示がなければ本明細書で使用される用語は、一般的な口語的用法と異なり得る科学用語で通常使用される意味を有する。
【0049】
遺伝子は、転写される領域だけでなく転写されない領域を含むように広く解釈されるべきである。
【0050】
化合物は、化学的化合物、例えば有機化学実体、生物学的化合物、例えば抗体及び抗原認識フラグメント並びに構築物、核酸、例えばRNAiなどを含むように広く解釈される。
【0051】
ホタルルシフェラーゼを発現するトランスジェニックマウスを作製した。これらの動物において、レポーター遺伝子ルシフェラーゼを、MBPプロモーターに連結し、それ故、MBP発現がオンになった場合に例えばCNSの白質(有髄)領域の細胞においてルシフェラーゼの発現を駆動する。基質ルシフェリンの全身注射(IV、IP、SC)は、生きているマウスの頭部から検出可能で定量可能な光シグナルを生じる。MBP−luci系を選択するためにクプリゾン脱髄モデルを適用し、そしてこれらの動物にルシフェリンを繰り返し注射することにより、脱髄及び再髄鞘化を非侵襲性生物発光画像化により長期的に、例えば2ヶ月間にわたって連続的にモニタリング及び定量することができる。このモデルは、クプリゾン誘導脱髄及びPPARδ化合物誘導再髄鞘化を首尾よく定量的にモニタリングした。
【0052】
検出器技術における進歩は、感度及び画質におけるかなりの改善をもたらした。現在では光子は、光子がシリコンウェハに衝突するにつれて光子を電子に変換する専門化された電荷結合素子(CDD)カメラにより検出される。CCDカメラは入射光子の強度を電荷パターンにコード化し、次いでこれが処理されて画像を生じる。ノイズは、CDカメラを過冷却し、そしてカメラを遮光箱中に備え付けることにより低減される。これらのカメラは一般的には画像取得及び解析の間コンピューターにより制御される。かなり小さく、従って実験作業台上に適応され得る第二世代のカメラシステムは、技術を日常的な実験にとって実行可能で実用的なものにした。Xenogene Companyはバイオイメージング技術を商業化した。
【0053】
利用可能な画像化様式のうち、生物発光又は蛍光に基づく光学技術は最も入手し易く、かつ容易に操作されるとして現れた。生物発光画像化(BLI)は、組織からのバックグラウンド発光(ノイズ)が極めて低いので、優れた感度により特徴づけられる。今まで、BLIは様々な動物モデルにおいて細胞運動、腫瘍進行、遺伝子発現、及びウイルス感染のような生物学的プロセスをモニタリングするために首尾よく使用されてきた。
【0054】
ホタルルシフェラーゼは活性のためにATPのような細胞内補因子を必要とする。これにより、この酵素を発現するように遺伝子操作された細胞にその使用が限定される。結果として、多くの有用な画像化適用、例えば循環する因子の分布、細胞外抗原発現の検出、及び内在性細胞の標識は、ホタルルシフェラーゼ画像化を受け入れにくい。ホタルルシフェラーゼのさらなる欠点は、固定された細胞及び組織サンプルにおいてそれを検出するための代替の基質がないことである。このことは、インビボ画像化を顕微分析と相関させることを困難にした。
【0055】
内部器官から放射された光の検出感度は、ルシフェラーゼ発現のレベル、標識された細胞の体内での深さ(光子が組織を通って移動しなければならない距離t)、及び検出システムの感度を含むいくつかの要因に依存する。
【0056】
非侵襲性動物全身画像化を使用するルシフェラーゼレポーター発現カセットの発現のモニタリングは記載されている(Contag,C.,米国特許第5,650,135号、1997年7月22日、(参照により本明細書に加入される);Contag,P.,et al,Nature Medicine 4(2):245−247,1998;Contag,C.,et al,OSA TOPS on Biomedical Optical Spectroscopy and Diagnostics 3:220−224,1996; Contag,C.H.,,Photochemistry and Photobiology 66(4):523−531,1997;Contag,C.H.,al,Molecular Microbiology 18(4):593−603,1995)。このような画像化は、典型的には少なくとも1つの光検出器装置要素、例えば電荷結合素子(CCD)カメラを使用する。
【0057】
MBP遺伝子の制御エレメント
ミエリン塩基性タンパク質(MBP)は、主に神経系において発現されるポリペプチドのファミリーであり、髄鞘形成において重要な役割を果たす。例えば分化オリゴデンドロサイトにおけるMBPの発現は、主に転写レベルで調節される。Journal of Neuroscienceにおいて、FarhadiらはMBP遺伝子の発現を時間的に制御する新しい調節性組み合わせエレメント(combinatorial element)を記載した。
【0058】
Farhadiらは、グリアが髄鞘形成の開始の間及び後の様々な段階でMBPの発現を制御するために調節配列の異なる組み合わせを使用するということを示した。
【0059】
ミエリン塩基性タンパク質(MBP)は、正常なミエリン圧縮に必要であり、そしてMSのような実験的及びヒト脱髄疾患の両方に関与している。
【0060】
ミエリン生物学の理解をさらに進歩させるため、そして生きた動物においてミエリン増強化合物を試験するために。本発明は、本発明のMBP−luciモデルを生成するために、MBPプロモーターの性質及び最も鋭敏なルシフェラーゼレポーター技術を使用した。今や本モデルは生きている動物におけるミエリン遺伝子転写応答の高感度のインビボ測定を可能にする。
【0061】
MBP−luciレポートカセットの構築
129SvEv BACライブラリー(Cell&Molecular Technologies)を、MBPプロモーターM3領域に位置するプローブを用いてスクリーニングした。このプローブは507bpであり、プライマー対(5’−actccttaccacacttcttgcagg−3’ 5’−tctattgggtgatgtgtgccatc−3)を用いて生成された。(配列番号1及び2) MBP BACをサザン解析により同じプローブを用いて確認した(EcoRIを使用して消化された7.6Kのフラグメント及び/又はBamHIを用いて消化された13.8Kのフラグメント)。
【0062】
高忠実度PCRによりプライマーセット(MBP−L−SP2 5−gggggatccacctgggacgtagcttttgctg及びMBP−AP1 5−ggggtttaaactccggaagctgctgtggg)(配列番号3及び4)を用いて増幅された、M1からM4まで(10K)を含む「長い」MBPプロモーターを、Invitrogenのxl−topoベクター中にクローンして、中間ベクターを生成した(Topo MBP10kベクター)。次いで、MBP 10kプロモーター(BamH1及びPmeIフラグメント)をpGL3 hygro neoベクター中に挿入した(BglII及びPmeI部位)。最終的な10KベクターをpGL3−hygro−long MBP−luciと呼ぶ(例えば図4を参照のこと)。
【0063】
高忠実度PCRによりプライマーセット(MBP−S−SP2 5−gggggatccatccctggatgcctcagaagag及びMBP−AP1 5−ggggtttaaactccggaagctgctgtggg)(配列番号5及び6)を用いて増幅された、M1〜M3(5k)を含む「短い」MBPプロモーターをInvitrogenのp2.1−topoベクター中にクローンして中間ベクターを生成した(Topo MBP5kベクター)。次いでMBP 5kプロモーター(BamH1及びPmeIフラグメント)をpGL3 hygro neoベクター(BglII及びPmeI部位)中に挿入した。最終的な5KベクターをpGL3−hygro−short MBP−luciと呼ぶ。(例えば図3を参照のこと)。
【0064】
両方のpGL3−hygo−MBPプラスミドからのDNA配列はM1、M2、M3及びM4の読み取りを確かにした。さらに、これらのプラスミドの293T細胞への一過性トランスフェクションは、検出可能なルシフェラーゼ活性を生じた。
【0065】
動物の取り扱い及びトランスジェニックマウスの生成
全ての動物の作業は連邦政府のガイドラインにしたがって行った。マウスの3つの異なる系統(FVB、B6C3及びC57 BL/6)を使用した。画像化をイソフルラン(Baxter,Deerfield,IL)を用いた吸入麻酔下で行い;完全に回復するまでマウスを観察した。
【0066】
トランスジェニックマウスを以下のようにして生成した:pGL3−hygro−MBP10k−luci又はpGL3−hygro−MBP5k−luciプラスミドのいずれかをNot I及びBamH1酵素で消化した。次いで、MBPプロモーター、ルシフェラーゼ及びポリアデニル化シグナルを含むフラグメントをゲル精製した。トランスジェニックマウスをFVB、B6C3又はC57BL/6の胚への標準的な前核注入(pronuclear injection)により生成した。手短には、前核マイクロ注入の間に、MBP−luci遺伝子カセットDNAを受精直後のマウス卵子に直接導入する。細い針を使用して、DNAを精液由来の大きな雄性前核に注入する。DNAはしばしば、1回又は2回の細胞分裂が起こった後、ゲノム中のランダムな位置に多くの縦列に配置されたコピーとして一体化する傾向がある。従って、得られたマウスは部分的にトランスジェニックである。トランスジェニック細胞が生殖系に寄与する場合、いくつかのトランスジェニック卵子又は精液が産生され、そしてマウスの次の世代は完全にトランスジェニックとなる。
【0067】
トランスジェニック創始者及びそれらのTg+G1子孫を、ホタルルシフェラーゼ遺伝子に特異的なプライマー(PCRプライマー:5’gaaatgtccgttcggttggcagaagc−3’、及び5’ccaaaaccgtgatggaatggaacaaca−3’)(配列番号7及び8)を使用して、尾部生検DNAのポリメラーゼ連鎖反応(PCR)により同定した。
【0068】
25のポジティブ創始者の子孫を、インビボ画像化システム(IVIS 100; Xenogen,Alameda,CA)を使用して画像化し、そして6つのトランスジェニック系を脳画像化シグナルを用いて同定した(2つのFVB系及び4つのB6C3HF1系)。脳画像化ポジティブC57 BL/6創始者はこの実行において生成されなかったので、1つのFVB系をC57BL/6マウスと戻し交配してC57 BL6系統を達成した。続いてB6C3系171マウスを、異種交配により生殖させてホモ接合トランスジェニック交配対を達成した。B6C3F 1系171もC57アルビノ系と戻し交配した。
【0069】
表1
MBP−luci系をスクリーニング選択するためにインビボ生物発光画像化を使用した。G1マウスをイソフルランで麻酔し、そして250mg/kgの用量のルシフェリンを尾部静脈を通して又は皮下注射した。ルシフェリン注射の8分後にマウスを画像化した。6つの系を脳画像化シグナルを用いて同定した(図5、2つのFVB系統:58及び121、並びに4つのB6C3系統:12、23、85及び171)。系58を除いて、他の5つの系は脳の白質領域においてエクスビボルシフェラーゼ画像化シグナルを示した。
【0070】
表1は、尾部生検PCR遺伝子型により生後まもなく同定された35のトランスジェニックDNAポジティブ創始者マウスからのデータを示す。15のDNAポジティブ創始者系はMBP−10k luciを生じ、そして20のDNAポジティブ創始者系はMBP−5k luciを生じた。この出願全体を通して、導入遺伝子及びトランスジェニックマウスはMBP−luciと略される。
【0071】
【表1】

【0072】
図7及び9は、CNS小領域及び年齢関連性髄鞘形成とよく相関しているMBP−luci生物発光を示す。
【0073】
エクスビボ画像化及びルミノメーターアッセイ
単離された器官のエクスビボルシフェラーゼ画像化を、ルシフェリンの皮下(SC)注射(250mg/kg)の10分後にCO2により動物を安楽死させた直後に行った。切開した器官をプラスチックシートで覆った黒い紙上に置き、そしてIVISにより画像化した;強い生物発光シグナルが、切開後20〜30分以内に検出可能なままであった。画像解析及び生物発光定量をLiving Imageソフトウェア(Xenogen Corp.)を使用して行った。
【0074】
組織サンプルを阻害剤を含む溶解緩衝液中に入れた(Passive Lysis Buffer [Promega]及びComplete Mini Protease Inhibitor Cocktail [Roche,Indianapolis,IN])。組織を組織ホモジナイザーを使用してホモジナイズした。組織を少しの間超音波処理してさらにホモジナイズした。組織ホモジネートを遠心分離し、そして清澄化した溶解物をルミノメーターアッセイに使用した。ルミノメーターアッセイのために、ルシフェラーゼアッセイ基質(Luciferase Assay System,Promega)を製造者により示されるとおりに調製した。組織ホモジネート(20μl)及び基質(100μl)を混合し、そして測定をルミノメーターで行った。バックグラウンド発光読み取りを得て、バックグラウンド読み取りを発光データから差し引いた。タンパク質濃度をBCAタンパク質アッセイキット(Pierce,Rockford,IL)を使用して製造者のプロことルにしたがって決定した。それぞれのタンパク質溶解物についての発光をタンパク質1マイクログラムあたりの任意単位の光として計算した。
【0075】
クプリゾン誘導脱髄及び組織学的検証
4週間にわたってクプリゾンをマウスに投与することにより、脳梁の広範な脱髄が生じた。クプリゾン誘導脱髄は重大な小神経膠細胞症及びマクロファージ動員を伴うが(Bakker and Ludwin,J Neurol Sci 78:125−37,1987; Hiremath et al.,J Neuroimmunol 92:38−49,1998; McMahon et al.,J Neuroimmunol 130:32−45,2002)、最小のT細胞応答を有する(Matsushima and Morell,Brain Pathol 11:107−16,2001)。このモデルにおけるミエリン損傷部位の一貫して予測可能な性質は、脳梁髄鞘形成における容易に定量可能な変化を生じる。これらの変化は、オリゴデンドロサイト前駆細胞による新規の髄鞘形成から生じるかもしれないが、免疫調節性の機構による末端脱髄の防止(Pluchino et al.,Nature 436:266−71,2005)は現実味のある代替の説明かもしれない。
【0076】
上で概説したように、MBP−ルシフェラーゼトランスジェニック(MBP−luci Tg)マウスの多数の系統を、クプリゾン誘導脱髄/再髄鞘化事象のインビボ評価のために評価した。ミエリン塩基性タンパク質(MBP)プロモーター駆動ルシフェラーゼ(luci)の発現は、MBPタンパク質を発現しているトランスジェニック(Tg)哺乳動物の脳におけるミエリンのインビボバイオイメージング定量を可能にした。例えばこのモデルは、それらの食餌中に0.2%クプリゾンを与えた野生型C57/BL6マウスを使用し得る。以前のモデルは、様々な化合物処置の後にミエリンの評価のために多数の時点で最終的に屠殺することを必要とした。多数の動物が必要とされるので、動物間変動性は、有意性を達成するためにさらなる被験体(より大きなn)を必要とする因子であった。
【0077】
MBP 5k−luci系171(B6C3)マウスは、4週間の食餌中0.2%のクプリゾンで、ルクソールファストブルー(LFB)組織化学的染色により評価した場合、脳の脳梁における顕著で有意な脱髄を示した。この脱髄は、バイオイメージングインビボルシフェラーゼシグナルの低下とさらに関連付けられた。
【0078】
しかし、MBP 10−Luci系121マウスのFVB系統は、比較できる脱髄を示さなかった。有意な脱髄を生じ得る食餌中の様々な量のクプリゾン及びクプリゾン処置の様々な期間の見込みがある投与計画を同定しようとして、さらなる研究をFVBマウスにおいて行った。結果は脳梁におけるLFB評価により中程度の量の脱髄しか示さなかった。従って、系171を優先的に開発した。
【0079】
クプリゾンモデルに対する異なる系統の影響:
トランスジェニックマウスは、その高い繁殖力に起因してしばしばFVB/NJ (FVB)系統を使用して作製されてきた。FVB系統マウスはまた、それらの白くて比較的光吸収性でない毛皮の色に起因してトランスジェニックバイオイメージングモデルのために広く使用される。剃毛のような毛髪の除去もまた、FVB又は他の系統において毛髪又は毛皮による吸収又は散乱に起因するシグナル損失を低減するために使用され得る。
【0080】
クプリゾンモデルについて系統間の差異が観察されたので、系統の選択は結果に影響を及ぼし得る。特定の目的のために最適な系統の選択は、例えば選択されたアッセイ及び装置に依存性の所定の最適化と考えられる。しかし、トランスジェニック哺乳動物の作製は限定因子とはみなされない;むしろ、例えば、髄鞘形成に影響する条件に対する特定の系統及びトランスジェニック系の感受性は、データ品質を改善するための選択基準として使用される。選択された髄鞘形成/脱髄事象に依存して、系統又は遺伝的バックグラウンドの選択は結果に影響を及ぼし得る。特定の脱髄モデルは特定の系統においてより良好に機能し得ると考えられる。モデル及び系統のこのような選択は、アッセイ開発の一部として日常的と考えられるだろう。脱髄及び再髄鞘化を研究するためにクプリゾン食餌は優れたモデルであり、系統差異で外見上観察されるこのモデルにおける強い遺伝要因が存在する。
【0081】
FVB系統:
FVB系統からのマウスを一部分においてそれらの白色毛皮色のために選択した。FVBは大部分のトランスジェニック実験及びその後の遺伝子解析に適したシステムを提供する。例えば、同系交配FVB系統は、活発な生殖能及び一貫して多い同腹子を特徴とする。これは大きな集団を生成する際に費用及び労力を減らす。さらに、FVB受精卵は大きく突出した前核を含み、これはDNAの微量注入を容易にする。さらに、FVB系統はアルビノ毛皮色を有し、それによりバイオイメージングの第一候補となっている。これらの特徴はFVB系統をトランスジェニックバイオイメージングモデルを用いた研究のための使用に有利にする。しかし、他の系統を、それらが所望の特徴を示す場合は使用することができる。
【0082】
FVB系統マウスは体重減少という点で0.2%クプリゾンに対して非常に感受性である。1週あたり2〜3回の通常餌/トランスゲル(transgel)栄養補助剤が重篤な体重減少及び毒性を避けるために必要とされる。さらに、発明者らの経験により、FVB系統マウスが様々なクプリゾン食餌投与計画からの最少の組織学的脱髄を示すということが示された。
【0083】
従って、その後の検証及び応用を容易にするために、MBP 10k−luci系121(FVB系統)をC57 Bl/6と戻し交配した。
【0084】
B6C3/Tac系統:
B6C3ハイブリッド系統は、C57 BL/6Ntac雌性マウスをTaconic USの市販のコロニーからのC3H/HeNTac雄性マウスと異種交配させることにより発生させることができる。これは黒色又はアグーチの毛皮色を有する。B6C3は、C57BL/6及びC3Hが異なる遺伝子座においてヘテロ接合性であり、そしてそれらが同じ遺伝子座においてホモ接合性である。
【0085】
B6C3/Tacマウスは明白な組織学的脱髄を示した。具体的には、バイオイメージングモデル系171 MBP 5k−Luciホモ接合マウスにける脱髄は、野生型C57BL6と比較して少しだけ規模が小さく、軽度の変動性であった。系171 MBP 5k−Luciヘテロ接合マウスにおける脱髄は、C57BL6及び系171 MBP 5k−Luciホモ接合マウスと比較して、かなり重症度が低く、より局在化しており、そしてより変動性であった。これらの結果は、MBP−luciモデルが、髄鞘形成に影響を及ぼす事象に対する個体、系統又は種の感受性を決定する際に有用であり得る。さらに、MBP−luciの構成は黒色毛皮の哺乳動物においても有用性を失わない。
【0086】
BALB/cJ系統:
BALB/cJマウスにおける皮質脱髄に対するクプリゾンの効果も調べた。これらのマウスにおいて、皮質脱髄は部分的のみであった。
【0087】
さらに、皮質小グリア細胞蓄積はBALB/cJマウスにおいて顕著に増加したが、一方で小グリア細胞はC57BL/6マウスの皮質には存在しなかった。従って系統差は異なる研究目的を支持するために有用であり得る。
【0088】
C57 BL/6J Jax系統:
C57BL/6遺伝的背景の動物は多くのクプリゾンモデル研究に適しており、そしていくつかの研究所で過去30年にわたって使用されてきた。8週齢のC57BL/6マウスに食餌中0.2%のクプリゾンを与えた場合、成熟オリゴデンドログリア(olidgodendroglia)は特異的に侵襲され(膨大な量のミエリンの支持の代謝要求を満たすことができない)、そしてアポトーシスを受ける。この事象には小グリア細胞の動員及びミエリンのファゴサイトーシスが直ぐ後に続く。形態学的研究と合わせたミエリン遺伝子発現の研究は、連続した代謝負荷に曝されても、オリゴデンドログリア前駆細胞が増殖し、そして脱髄領域に侵入するということを示した。クプリゾン負荷が終了した場合、ほとんど完全な再髄鞘化がせいぜい数週間以内に起こる。可溶性因子による異なる細胞型間の細胞間情報伝達が推測され得る。本発明の方法及びモデルは、そこで細胞間情報伝達事象を研究するため、例えば、推定因子が髄鞘形成のための動員に関与するか否かを決定するため、動員を容易にする化合物をスクリーニングするため、及び動員を阻害する化合物をスクリーニングするために有用であり得る。
【0089】
さらに、MBP−luciモデルの再現性は、脱髄及び又は再髄鞘化のプロセスを加速するか又は抑制し得る操作(例えば、共通の遺伝的背景の利用可能なノックアウト若しくはトランスジェニック、又は妨害するRNA若しくは薬理的処置)の試験を可能にし得るということを示す。
【0090】
MBP−luciモデルの改善
系171(B6C3H系統、ヘテロ接合体)は髄鞘形成/脱髄研究において役立つことが示されたが、このモデルは、(1) バイオイメージングシグナル強度を増大させ、そしてモデル製造及び遺伝子型解析費用を減らすために、繁殖させてホモ接合性(homozygocity)とすることにより;(2) 白色毛皮で画像化シグナル減衰を減らし、そして複数回のNair剃毛後の皮膚反応を低減するために、繁殖させてC57系統のようなアルビノ系統とすることにより;(3) 社内開発CNSクプリゾンモデル系統と整合させるために系121をC57 BL/6系統と繁殖させることによりさらに改善され得る。
【0091】
本発明者らは今や、系171ホモ接合が系171ヘテロ接合系の2倍を超えるバイオイメージング域を示す(細胞あたりレポーター遺伝子カセットの2つのコピー)をいうことを実証した。他の実験もまた、アルビノC57がクプリゾンモデルに対して応答することを実証した。
【0092】
画像化システム及びデータ解析
生物発光を、IVIS画像化システム(Xenogen Corp.,Alameda,CA)を使用して非侵襲的に測定した。画像はルシフェリンの腹腔内(i.p.)注射(250mg/kg−1;Xenogene Corp.)の10分後に、文章中に他の記載がなければ、60秒収集、ビニング10で撮影した。画像収集の間、マウスを約2%イソフルラン(Abbott Laboratories Ltd.,Kent,United Kingdom)の吸入により継続的に鎮静させた。
【0093】
画像化システム説明:
本発明を証明するデータ収集のためにIVIS(R)Imaging System 100 (Xenogene)を使用した。Xenogenの高感度のIVIS(R)Imaging System 100シリーズは、例えば10〜25cmの調整可能な矩形視野を提供し、5匹のマウス又は2匹の大きなラットの画像化、さらには1つの標準的なマイクロタイタープレートも可能である。このシステムは25mm(1.0インチ)平方の背面薄肉化(back−thinned)裏面入射型CCD(電荷結合素子)カメラを特徴とし、これは電子的バックグラウンドを最小にして感度を最大にするために閉鎖循環冷却システム(液体窒素を用いない)により約−90〜−120℃、例えば−105℃に低温冷却される。CCDカメラは、特にスペクトルの赤色領域における高効率光子検出用に設計されている。これは非常に少ない数の光子を検出でき、さらに伝統的なカメラのように動作し;その広いシグナル範囲での画像の表示はXenogenのLiving Image(R)ソフトウェアの機能である。異なる帯域幅をとり出すための6ポジションフィルタホイールがある。このスペクトル情報は発生源の細胞の深さ及び分布についてより多く明らかにすることができる。CCDは冷却され、そして電子的読み出しは、実時間インビボ画像を作成するために集積されたデータが非常に低いノイズを有するように最適化される。
【0094】
遮光性画像化チャンバー
高い遮光性の低バックグラウンド画像化チャンバーは、IVIS(R)Imaging System 100シリーズを標準的な実験室の照明環境で使用することを可能にする。画像化チャンバー内のサンプル棚は視野を調節するために上下に移動する。研究者は動物全体を見ることも、追加の詳細のために一部に焦点を合わせることもできる。棚は麻酔された例えばマウス又はラットの満足な状態を高めるために加熱される。このシステムは、ウェブサイトページに示される、Xenogen−the XGI−8 Gas Anesthesia Systemからの加熱されたサンプル棚、ガス麻酔接続、及び全体ガス麻酔オプションのような動物操作の特徴を含む。より大きな画像化チャンバーはより大きな被験体又はより多数の被験体の使用を可能にし得る。
【0095】
インビボ生物発光アッセイのためのルシフェリンの製造:
以下の材料を実施例で使用した:
ホタルD−ルシフェリンカリウム塩 1.0g/バイアル(例えばXenogen XR−1001又はBiosynth L−8220)。
Mg2+及びCa2+を含まないDPBS
ボトルトップフィルター0.2um。
【0096】
以下の手順を画像化に使用した:
ルシフェリンのDPBS中25mg/mlのストック溶液を調製し、そして0.2umフィルターを通して滅菌ろ過した。5mlのアリコートを−20℃で保存した。注射用量は10ul/体重1gであった。各マウスを対象に250mgルシフェリン/体重1kgを投与した(例えば20gのマウスについては、200ulを注射してルシフェリン2.0mgを送達する)。ルシフェリンを画像化の数分前に皮下(SC)、又は腹腔内(IP)又は静脈内(IV)注射した。ルシフェリン動態研究を、ピークシグナル域を決定するために各動物モデルについて場合により行った。
【0097】
3.6 MBP−luci画像化法:
上記のように、マウスに250mg/kg D−ルシフェリンをSC、IP又はIVにより注射した。5分後(静脈内)又は8分後(腹腔内又はSC)に、マウスをIVIS 100(Xenogen)を使用して16分間(サイズ8で8つの画像について60秒の画像化及び60秒の間隔)画像化した。生物発光を定量するために、興味のある同じ円形の領域を各マウスの頭部領域を取り囲むように位置決めし、そして画像化シグナルを、LIVINGIMAGEソフトウエア(バージョン2.5、Xenogen)を使用して平均放射輝度(光子/秒/cm2/ステラジアン(steridian))として定量した。興味のある頭部領域を、一定面積に保ち、そして全て実験内で位置決めした。データを各動物について処置開始時の生物発光に対して正規化した。
【0098】
3.7 統計学的解析
統計学的解析のために、EverStat V5及びSigma Stat統計ソフトウェアパッケージを使用した。グループにおける画像化の平均を平均値とみなし、そして全てのグループについてのSEを計算した。
2つのグループの平均値を比較する場合、ウィルコクソン対比較(paired)検定又はウィルコクソン非対比較(unpaired)検定を行った。両側P値<0.05を統計学的に有意とみなした。
【実施例】
【0099】
トランスジェニックマウス生成
「長い」プロモーターはM1、M2、M3、M4を含む約10KBであり、そして「短い」プロモーターはM1、M2及びM3を含む約5KBであった。これらを高忠実度PCR法を用いてMBP遺伝子を含有するマウス細菌人工染色体(BAC)からクローン化した。次いで各プロモーターフラグメントをベクター中に、例えばpGL3−hygroベクターのポリリンク(poly link)部位に(into the into the)クローン化した(図1及び図2)。
【0100】
プラスミドをNot I及びBamH1で制限してMBP−luciトランスジェニック発現カセットを放出させ(図3)、これらを、標準的な前核微量注入技術を使用してFVB/Tac系統及びB6C3/Tac系統でトランスジェニックマウスを生成させるために使用した。
【0101】
トランスジェニック(Tg)動物生成のための一般的な方策は当該分野で周知であり、例えばPinkert,C.A.(ed.) 1994.Transgenic animal technology:A laboratory handbook.Academic Press,Inc.,San Diego,Calif.;Monastersky G.M.and Robl,J.M.(ed.)(1995) Strategies in transgenic animal science.ASM Press.Washington D.C.に記載される。
【0102】
脱髄/再髄鞘化事象と相関したMBP−Luci導入遺伝子シグナル
白質領域のルシフェラーゼ発現が観察されたMBP10K−luciトランスジェニック系121(FVB系統)を、立証実験のためのクプリゾンモデルにおいて使用した。図11に示されるように、第一のクプリゾン研究を、ルシフェラーゼ画像化を1週目、2週目及び4週目に繰り返し(0.2%クプリゾン食餌)、続いて通常のクプリゾン非含有食餌に戻して5週目、6週目及び7週目に画像化した(RE 1週から3週)。図に示されるように、この系121からのルシフェラーゼ発現は、クプリゾン誘導脱髄及び再髄鞘化の時間経過と明らかに相関していた。このことは、公開されている内在性MBP mRNA研究(Jurevics et al.,Journal of Neurochemistry,2002,82,126−136)と一致する。FVB系統マウスは1つの系統以外であり、そして体重減少により見られるように0.2%クプリゾンに対して感受性であることが知られている。通常、1週当たり2〜3回の通常餌/トランスゲル栄養補助剤が、この系統における重篤な体重減少及び毒性を回避するために必要とされる。
【0103】
クプリゾンモデル検証:
周知でありかつ広く使用される脱髄/再髄鞘化モデルの1つはマウスにおけるクプリゾンモデルである。このモデルは、例えば連続した4〜6週の間、粉末状齧歯動物試料(lab chow)で投与されるクプリゾン、銅キレート剤(典型的には約0.2%質量/質量; ビスシクロヘキサノンオキサルジヒドラゾン、CAS番号370−81−0、Sigma C9012)の食餌消費を含む(例えば:Matsushima及びMorell,2001を参照のこと)。クプリゾンは成熟オリゴデンドロサイトに対して選択的に毒性であることが示されている。クプリゾン食餌の通常食餌へのその後の切り替えにより、回復をもたらす環境が生じ、その結果、クプリゾン食餌停止の4週〜6週後に、マウスは脳梁において高い再髄鞘化を示す。従って、クプリゾンモデルは、その中で脱髄及び再髄鞘化の局面を研究するための完全なインビボパラダイムを提供する(図11及び12)。
【0104】
さらなる証拠として、MBP 5k−luci系171(B6C3系統)を試験した。この系統もまた、図13に示されるように、クプリゾン誘導脱髄及び再髄鞘化事象に対して相関関係にある画像化応答を示した。7匹のTg+マウスを0.2%クプリゾンで6週間処置し、そして3匹のTg+マウスを通常食餌で6週間処置した。全ての7匹のマウスが0.2%クプリゾン食餌に耐え、そして15〜25%の間の平均体重減少を有していた。クプリゾン処置と共に、有意な画像化シグナル低下があった(脱髄)。例えば、0週目から4週目で43%のシグナル低下、そして0週目から6週目で74%のシグナル低下があった。
【0105】
MBP−luciマウスクプリゾンモデル組織学的検証
組織学的検証については、目的はバイオイメージングモデルにおいて、クプリゾン処置の間のレポーター遺伝子の応答が、クプリゾン処置マウスの脳梁における構造的に検出可能な脱髄と相関することを確認することであった。これらの病的状況を、ルクソールファストブルー(LFB)染色により可視化した(図20及び24を参照のこと)。
【0106】
具体的には、最初の試験において0.2%クプリゾン食餌を用いたMBP 10k−luci系121(FVB系統)について、LFB染色によりごくわずかな脱髄しか検出されなかった。これらのFVB系統マウスについて明瞭な組織学的脱髄を生成させるために、様々なクプリゾン食餌投与計画にしたがって重篤な体重減少を回避するよう試みた。0.2%、0.175%及び0.15%のクプリゾン用量グループ(6週の研究)は、重篤な体重減少及び毒性を避けるために1週あたり3〜4回の通常餌/トランスゲル栄養補助剤を必要とした。また、クプリゾン濃度を曝露時間が延びるにつれてさらに低くした。クプリゾン濃度(0.14%、0.12%及び0.1%)及び処置期間(7週及び9週)を用いた研究を、週に1回までの通常餌/トランスゲル栄養補助剤を用いて試験した。しかし、全てのデータは、FVB系統マウス(8週齢、体重28.5g±3g)が多様なクプリゾン食餌投与計画からのより少ない組織学的脱髄を有するということを示した。この研究モデルの予備的開発のための特に好ましい実施態様としてはFVB系を選択しなかった。FVB系統は画像化及びクプリゾン毒性に対する感受性については良好であるが、体重減少は、別の系統を使用することによりこれらの予備研究において容易に回避できる交絡変数を導入し得る。
【0107】
別の系統では、MBP 5k−luci系171(B6C3)マウス(8週齢、体重25g±3g)が明瞭な組織学的脱髄を示した。
【0108】
マウス脳組織を、6週の0.2%クプリゾン処置の終わりに集めた。7匹のクプリゾン処置されたマウスは全て、脳梁領域において明瞭な脱髄を有し、そして3匹のコントロールマウスは全て、脳梁領域において正常な髄鞘形成を示した。MBP 5K−luci系171からのこれらのデータは、画像化シグナルがクプリゾン誘導脱髄期を追跡するものであるというさらなる証拠を提供する。
【0109】
さらなる定量的LFB分析(図14)は、MBP−Luci B6C3系171ホモ接合マウス(8週齢、体重21g±3g)が、B6C3H系171ヘテロ接合マウス(8週齢、体重25.5g±3g)と比較してより一貫性のある、わずかにより重症な脱髄を4週目に示すということを実証した。楕円中のより濃い領域は染色されたミエリンを示す。
【0110】
さらに、0.2%クプリゾンを与えられたC57BI/6系統野生型雄性マウス(8週齢、体重20g±3g)は最も重症で一貫した脱髄を示した。C57 BL/6系統は、予備研究において記載されるクプリゾンモデル及びPPARδ試験化合物効果についてのポジティブコントロールとして役立った。
【0111】
1.MBP−luciマウスは、PPARδ選択的アゴニストツール化合物のCNS再髄鞘化に対するポジティブな効果を裏付ける:
MBP−ルシフェラーゼマウスを、このインビボバイオイメージングモデルがペルオキシソーム増殖因子活性化受容体δ(PPARδ)アゴニストツール化合物(‘571)のCNS再髄鞘化に対する効果を検出するために使用され得るか否かを評価するために使用した。
【0112】
ペルオキシソーム増殖因子活性化受容体(PPAR)は、標的遺伝子の発現を調節する転写因子として機能する核受容体スーパーファミリーに属する。他の転写因子とは異なり、核受容体の活性は、生物学的膜に容易に浸透する対応するリガンド−親水性小分子に結合することにより調節され得る。核受容体ファミリーについての複雑な細胞シグナル経路にもかかわらず、核受容体を薬物標的として使用する長い成功の歴史がある。PPARは細胞分化、発達及び代謝の調節において不可欠な役割を果たす。PPARは、今までに同定された別々の遺伝子によりコードされる3つの密接に関連したアイソフォームを有する:一般的にPPARα、PPARγ及びPPARδとして知られ、PPARβとしても知られる;J.Berger and D.E.Miller,Annu.Rev.Med.,2002,53,409−435)。それぞれの受容体サブタイプはシグニチャー(signature)DNA結合ドメイン(DBD)及びリガンド結合ドメイン(LBD)を有し、両方ともリガンド活性化遺伝子発現に必要である。PPARはレチノイドX受容体(RXR)とヘテロ二量体として結合する。
【0113】
PPARδはCNSにおいてかなり発現されるように思われる;しかしそこでのその機能の多くはまだ不確定なままである。しかし、1つだけ興味がもたれるのは、PPARδがCNSの主要な脂質産生細胞である齧歯動物のオリゴデンドロサイトにおいて発現されたという発見である(J.Granneman,et al.,J.Neurosci.Res.,1998,51,563−573)。さらに、PPARδ選択的アゴニストがマウス飼育(cultures)においてオリゴデンドログリアミエリン遺伝子発現及びミエリン鞘径を有意に増加させるということが見出されたということもわかった(I.Saluja et al.,Glia,2001,33,194−204)。PPARδノックアウトマウスは、より小さな脳全体のサイズ及び白質における減少したレベルの髄鞘形成を有する(Mol cell Biology 200 20:5119)。さらに、PPARδアゴニストは、多発性硬化症の実験的自己免疫性脳脊髄炎(EAE)モデルにおいて保護的効果を及ぼす(Polak et al.,J Neuroimmunology 2005 168:65−75)。
【0114】
共同研究者らは、選択的PPARδアゴニストが神経組織において機能的役割を果たし、そしてオリゴデンドロサイト前駆細胞分化を刺激するということを以前に実証した。経口で生物が利用可能な脳浸透性のPPARδ選択的アゴニストであるSAR117145は、齧歯動物及びヒトのオリゴデンドロサイト前駆細胞分化をインビトロで濃度依存性の様式で刺激した;PPARδ選択的アンタゴニストでブロックされ得る効果。
【0115】
ラットオリゴデンドロサイトにおいて、ミエリン塩基性タンパク質の増加した発現に下流PPAR標的Angptl4の増加したmRNA発現が先行し、そしてこの上方調節はPPARδのレンチウイルスshRNAノックダウンでブロックされた。マウスに2%クプリゾンの食餌を4種間与えた急性脱髄のマウスクプリゾンモデルにおいて、SAR117145はCNS再髄鞘化を増強し、Angptl4 mRNA発現を増加させ、そして脳梁を横切る軸索伝導を改善した。Angptl4のCNS活性化には腓腹筋の増加した発現が付随し、これが潜在的な代替マーカーとして役立ち得ると示唆される。これらのデータは、PPARδアゴニストがCNS再髄鞘化を増強し、そして軸索機能を改善し得ることを実証し、そして脱髄障害の処置のための内在性修復プロセスの刺激におけるそれらの潜在的用途を示唆する(米国特許出願第20070149580号、USE OF PEROXISOME PROLIFERATOR ACTIVATED RECEPTOR DELTA AGONISTS FOR THE TREATMENT OF MS AND OTHER DEMYELINATING DISEASES)。
【0116】
PPARδアゴニストツール化合物(’517;図15)を、B6C3H系171ヘテロ接合マウスで試験した。8週齢のマウスを0.2%クプリゾンを含む食餌で4週間飼育し、次いで再髄鞘化のために通常餌を与えた。次いでマウスに、週に2回、ビヒクル(0.6%カルボキシメチルセルロースナトリウム塩及び0.5%Tween80)又は30mg/kg PPARδアゴニストツール化合物’517のいずれかを8日間経口投与し、そしてグラフに示された時点で画像化した。データを0週ベースラインシグナルに対して正規化した。ツール化合物’517のグループ(n=15)ではビヒクルグループ(n=13)よりも30〜100%の相対的なルシフェラーゼシグナル増加があり、これはオリゴデンドロサイト前駆細胞分化の増強に起因すると本発明者らは考えている。
【0117】
’517の効果を、同じ研究においてルクソールファストブルー(LFB)染色によりさらに組織学的に確認した(図16)。ホルマリン固定パラフィン包埋脳からの傍矢状組織断面を、脳梁におけるミエリンの定性的評価のためにLFBで染色した。各時点について染色した断面をスコア付けし、そして0(完全な髄鞘形成)から5(完全な脱髄)までのスケールで等級付けした。スコア付けシステムは以下のとおりであった:0=正常ミエリン、脱髄無し、1=最小限の局在化した脱髄、2=軽度から中程度、局在化した脱髄、3=中程度、局所的に極度の脱髄、4=重症、局所的に極度の脱髄、5=重症、広範な脱髄。クプリゾンで4週後のマウスからのLFB染色した脳断面の組織学的評価は、系171ヘテロ接合マウス(n=5)における脳梁の中程度から重症の脱髄を裏付けた。再髄鞘化期の間の4週から5週目の’517での処置は、ツール化合物’517グループ(N=5)をビヒクルコントロールグループ(N=3)と比較して、7週目の時点でLFBにより決定した場合に測定可能な増加を生じた。少ないnの数にもかかわらず、組織学的データはインビボルシフェラーゼバイオイメージングデータを支持しており、ビヒクルコントロールと比較した場合に’517化合物で処置されたマウスにおける増加した再髄鞘化を示した。
【0118】
系171ヘテロ接合マウスを用いた第二のPPARδ 517研究は同様の結果を示した(データは示しておらず、そしてまた’517処置を3週延長した)。第一の研究(図15)と一致して、第二の研究もまた、’517がオリゴデンドロサイト前駆細胞分化を再髄鞘化プロセスの間の初期の回復期に加速したということを示唆する(M Lindner and S Heine,et al,Neuropathology and Applied Neurobiology,34,105−114,2008)。
【0119】
2.MBP−luciマウスはERβアゴニスト、’5a又はポジティブコントロール化合物QTPのミエリン発現に対する保護効果を検出する
エストロゲン受容体(ER)は、特異的応答エレメントによりDNAに対して直接的に作用し、そして遺伝子発現を調節する、ステロイド核受容体のファミリーに属する。ERの2つのサブタイプ、ERα及びERβが存在する。ERαは子宮、前立腺、卵巣、骨、乳房及び脳において高度に発現され、一方ERβは、結腸、前立腺、卵巣、骨髄及び脳に存在する。ERβの選択的標的化は、ERαの副作用を避けるための魅力的な治療アプローチである。ERサブタイプ選択的化合物が同定されている。
【0120】
例えば、ERβ(ERαではない)アゴニストは、オリゴデンドロサイト及び神経芽細胞腫細胞のアポトーシスをインビトロで保護することが示されている。さらに、ERβ又はER−アゴニストはEAEを寛解させ、そして神経保護効果を有する(ミエリン及び軸索の保存)。以前のPPARδアゴニストツール化合物’517に基づいて、MBP−luci系を、ERβアゴニスト、’5aをクプリゾンモデルにおいて概略を描く(profile)ために使用した。さらに、本発明者らは、AstraZenecaの統合失調症薬クエチアピン(10mg/kg、PO、qd)を、クプリゾン誘導脱髄に対するその保護効果(Schizophrenia Research,2008 Dec 106,182−91)に基づいてポジティブコントロールとして含めた。2つの独立した研究をB6CH系171ヘテロ接合マウスを用いて行った。
【0121】
第一の研究(図17)は、ERβアゴニスト‘5a及びポジティブコントロールクエチアピン(QTP)が、クプリゾンモデルにおいて脱髄期の間保護的であることを示した。MBP−luc系171ヘテロ接合B6C3Hマウスに、クプリゾンの食餌を4週間与え、そして’5a(10mg/kg、N=12又は30mg/kg N=16)又はQTP(10mg/kg、N=14)を経口投薬した。マウスを0週目(ベースライン)、3週目、及び4週目に画像化し、データを0週目のベースラインに対して正規化した。以前の研究(図15)と同様に、ビヒクルグループは49%(3週)及び45%(4週)のバイオイメージングシグナル減少を生じた。QTPグループは、10mg/kgで3週及び4週の両方の時点についてビヒクルコントロールに対して画像化シグナルにおいて有意な増加を示した。QTP処置グループについての結果は、Zhangらにより公開されたデータ(Schizophrenia Research,2008,106:182−91)と一致する。化合物’5aは30mg/kgで4週目にビヒクルに勝る有意な増加を示したが、10mg/kgでは3週でも4週でも有意な差異を示さなかった。これらの結果は、MBP−luci画像化モデルをクプリゾンモデルにおける用量依存性保護効果を評価するために使用することができるということを示唆する。
【0122】
さらなる研究(図18)を、ビヒクルについてのより大きなコホート及び’5a化合物30mg/kg処置グループを用いたこと以外は、第一の研究の結果(図17)を確認するために設計した。第一の研究の結果と一致して、マウスをQTPで10mg/kgで処置することにより(N=17)、ビヒクルで処置されたコントロール(N=25)と3週目(25%対47%の減少、p=0.028)及び4週目(6%の増加対25%の減少)に比較した場合にクプリゾン誘導シグナル減少の統計学的に有意な阻害をもたらした。さらに、30mg/kgでの’5a(N=27)は、ビヒクルコントロールグループと3週目(31%対47%の減少、p=0.0079)及び4週目(6%減少対25%減少、p=0.0015)に比較して、有意に高い導入遺伝子活性を生じた。
【0123】
これらの二つの研究は、30mg/kgでの‘5aはCNSのバイオイメージングシグナルにおけるクプリゾン食餌に誘導される減少を有意に防止するが、10mg/kgのポジティブコントロールQTPと同じ程度ではない(これらの実験において使用された条件下で)。以前の研究がCNS髄鞘形成の程度とMBP−luciバイオイメージングシグナルとの間の直接的な関係を示しているので、これらの現在の結果は、’5a及びQTPの両方がクプリゾン餌投与の間のCNSにおける脱髄を防止するということを示唆する。画像化モデルデータは、QTP及び’5aの両方がクプリゾン誘導脳脱髄及びミエリン破壊を軽減するということを支持する。
【0124】
3.クプリゾンモデルにおけるMBP−luci系の比較
様々な系統バックグラウンドの6つのトランスジェニック系を、異なるバイオイメージング適用のために生成した。
【0125】
クプリゾンモデルにおけるB6C3H系171ホモ接合マウス及びヘテロ接合画像化シグナル(図19)を比較した。ホモ接合体におけるレポーター遺伝子の2つのコピーは、脱髄期の間に2倍より大きいシグナル減少を示し、そして再髄鞘化期の間に2倍のシグナル増加を示した。ヘテロ接合系171(B6C3H系統)がクプリゾンモデルにおいて機能し、化合物効果を検出し得るということが実証されるが、バイオイメージングシグナル強度を増大させるためにホモ接合性に繁殖させることによりこのモデルをさらに改善し得るということが予測される。マウスコロニーはホモ接合体として維持され得るので、これはまた、モデル製造を効率化して遺伝子型同定費用を減少させるだろう。さらに、画像化域が大きくなれば、モデルは化合物誘導変化の検出に対してより高感度になる。
【0126】
図20は、クプリゾンモデルにおける3つの異なる系からの組織学的LFBデータの比較を示す。定量的LFBデータにより、ホモ接合マウスが最も重症なクプリゾン誘導脱髄(典型的に使用される野生型C57 BL/6マウス系統に見られるものと同様の重症度)を示したという、系171 B6C3Hを用いたバイオイメージング結果が裏付けられた。系171ヘテロ接合マウス系はより低い重症度の脱髄を示したが、これらの系171ヘテロ接合マウスは最も少ない量のクプリゾン誘導脱髄を示した。
【0127】
図21において、最大のバイオイメージングシグナル減少を有するMBP−luci系が、組織学的に評価された場合に最も高い脱髄も有していたことを実証する。3つの異なるMBP−luci系(系171 B6C3Hヘテロ接合(het)系統、系121 C57BL/6ヘテロ接合系統及び系171B6C3Hホモ接合系統)をバイオイメージング及びルクソールファストブルー(LFB;ミエリン染色)組織学により比較した。マウスを4週間又は5週間、0.2%クプリゾンを含有する食餌で飼育した。画像化データを0週のベースライン測定に対して正規化した。各研究の終わりに、マウス脳を採取し、そして連続的パラフィン切片をLFBを用いてミエリンについて染色した。平均定性的LFBスコア(0〜5)を図21の表に示した。系171 B6C3Hホモ接合マウスは、最大の画像化シグナル減少を示し、そしてまた定性的組織学的評価により評価した場合に最も重症な脱髄を実証した。系171 B6C3Hヘテロ接合マウスは、最も小さな画像化域、及び4週目に最も少ない組織学的脱髄も示した。系171 B6C3Hホモ接合マウスは、クプリゾン食餌前のそれらのベースライン画像化を参照して、クプリゾン食餌の間に最も大きなバイオイメージングシグナル減少を有していた。例えば、クプリゾン食餌で3週後に、171 B6C3Hホモ接合マウスは72%のシグナル減少(0週を参照して3週、p<0.05)を有していたが、系171 B6C3Hヘテロ接合マウスは45%のバイオイメージングシグナル減少(0週を参照して、p<0.05)を有していた。系121 C57 BL/6ヘテロ接合マウスは、ルシフェラーゼシグナルにおける最少の減少を有していた(0週に対して3週で33%の減少、p<0.05)。
【0128】
MBP−luciモデルの感受性及び応答性を、図22に示される171 B6C3Hホモ接合マウスのQTP(10mg/kg)を用いた処置によりさらに確認した。171 B6C3Hヘテロ接合マウス(図17及び18)を使用した結果と一致して、QTP(10mg/kg)はバイオイメージングシグナル減少を有意に防止した。これらの結果に基づいて、系171 B6C3Hホモ接合マウスをクプリゾン誘導脱髄バイオイメージングモデルのための最適な系として同定した。
【0129】
4.MBP−luciモデルのさらなる適用
MBP−luciマウスは脳及び脊髄の両方ルシフェラーゼ発現を有する。図23に示されるように、発光シグナルは主に脳及び脊髄の白質領域からのものであった。クプリゾンモデル適用について首尾よく実証された脳からのルシフェラーゼ画像化に加えて、脊髄からのルシフェラーゼ画像化シグナルは、MSの実験的アレルギー性脳脊髄炎(EAE)において使用され得、又は脊髄モデルとして応用され得る。
【技術分野】
【0001】
説明
本発明は、例えば生きた動物のバイオイメージング技術を用いて脱髄/再髄鞘化事象を転写レベルで実時間で可視化及び定量するために使用される、製造されたミエリン塩基性タンパク質−ルシフェラーゼ(MBP−luci)バイオイメージングモデルに関する。
【背景技術】
【0002】
薬物開発では減少率(attrition)が高く、5つの化合物のうち1つだけが開発を切り抜けて承認に至ると説明される(非特許文献1)。さらに、劇的に増加した投資にもかかわらず、新規薬物の導入率は過去30年にわたって比較的一定のままであり、1年あたりに新しい薬物クラスで進展があって最終的に市場に出るのは2つか3つだけである(非特許文献2)。
【0003】
薬物開発の初期段階で適用される分子画像化及び機能的画像化は、生物活性の証拠を提供して的確な薬物効果を裏付けることができる。従って、分子画像化技術における投資は薬物開発を増進すると期待される(非特許文献3)。より従来型の読み取り情報に勝る分子画像化技術の利点は、インビボでの生物学的プロセスを調べるために十分な空間的及び時間的分解能でインタクトな生物において行うことができることである。さらに、これらの分子画像化技術により、同じ動物の異なる時点での繰り返しの非侵襲的で均一でかつ比較的自動化された研究が可能となり、それ故、長期的な研究の統計的検出力を増加させ、そして必要な動物の数及び費用を減少させる。MBP−luciモデルは、インビボでの初期薬物候補スクリーニングの処理能力の増大、さらには生物学的アッセイの感度の増大の両方を後押しする。初期リード化合物はしばしば、販売薬製品としては準最適であるが、インビボでの特異的かつ有意な活性の検出は、特定のCNS疾患の処置を目的として、許容しうるレベルの活性を達成し、かつインビボでの毒性を最少にするための化学構造の最適化をもたらし得る。
【0004】
分子画像化
分子画像化は、生きた生物において細胞及び細胞内レベルでの特定の分子プロセスの評価における画像化技術を利用し統合するための、様々な分野(例えば、細胞分子生物学、化学、医学、薬理学、物理学、生物情報科学及び工学)からのアプローチの収束を指す(非特許文献4)。
【0005】
遺伝子工学の出現は、例えば新薬発見パイプラインを含む応用科学に大きな変化を引き起こした。同じように、動物画像化手順の開発及び開拓は前臨床研究のための新しい手段を提供しつつある(非特許文献5)。動物モデルは伝統的に、実時間で生理学的事象の定量における困難さのために扱いにくい。長年にわたって、新しい画像化方法がこの困難さを克服するために開発されてきた(例えば、MRI、CT、PET)。より最近では、ルシフェラーゼ(ホタルの発光する酵素)のインビボ発現に基づく生物発光(bioluminescense)画像化が標的遺伝子活性の非侵襲性検出のために使用されている。
【0006】
動物工学と分子画像化技術を組み合わせることにより、生きている動物において特定の分子プロセスに関する動的な研究を行うことが可能となった。このアプローチは、前臨床プロトコルに影響を与える可能性があり、従って医学の全ての局面を広く変化させ得る(非特許文献6)。
【0007】
分子画像化:生物発光
インビボ生物発光画像化(BLI)は細胞又は組織からの発光の検出に基づく高感度のツールである。レポーター遺伝子技術の有用性は、インビボ画像化法により生きている動物における特定の細胞及び生物学的プロセスを分析することを可能にする。生物発光(生きている生物による可視光の酵素的生成)は多くの非哺乳動物種において天然に存在する現象である(非特許文献7)。ルシフェラーゼは基質の酸化を触媒して光子を放出する酵素である(非特許文献8)。北アメリカホタル(North American firefly)からの生物発光は最も広く応用される。ホタルルシフェラーゼ遺伝子(luc)発現は酵素ルシフェラーゼを産生し、これが基質D−ルシフェリンを非反応性オキシルシフェリンに変換し、562nmにおける緑色発光を生じる。哺乳動物組織は自然には生物発光しないので、画像を非常に小さいバックグラウンドシグナルで生成することができるので、インビボBLIは非常に魅力的である。
【0008】
BLIには、光レポーターを駆動する選択された遺伝子プロモーターの制御下の生物発光レポーター遺伝子からなる発現カセットを用いて細胞又は組織を遺伝子操作することが必要とされる(図1)。光生成を誘導するために、基質(例えばルシフェリン)を、icv、血管内、腹腔内又は皮下注射により投与する。
【0009】
ルシフェラーゼ/ルシフェリンにより放射される光は、数ミリメートルから数センチメートルの深さの組織を透過することができるが;組織深さ1センチメートルごとに約10倍光子強度が減少する(非特許文献7)。インビボでの生物発光を検出するためには高感度の光検出機器を使用しなければならない。検出器は単位面積あたりに放出される光子の数を測定する。400と1000nmとの間の波長における低レベルの光は、シリコンウエハーに衝突する光子を電子に変換する電荷結合素子カメラを用いて検出することができる(非特許文献9)。ソフトウエアは電子シグナルを2次元画像に変換することができる。ソフトウエアはまた、放射光の強度(検出器に衝突する放射された光子の数)を定量し、そしてこれらの数値を疑似カラー図に変換することもできる。実際のデータは光子カウントで測定されるが、疑似カラー図は迅速な視覚的解釈を可能にする。目的の領域内の定量的測定は、より微細な差異のために必要かもしれない。冷却CCDカメラの使用は熱雑音を低減させ、そして遮光箱(light−tight box)はルシフェラーゼが生成した光を最適に可視化し、定量することを可能にする(非特許文献10)。
【0010】
発光シグナルの解剖学的位置についての自筆文書(autograph)又はX線写真のような別の種類の画像に重ねられたルシフェラーゼ画像を有することは有用である(図2)。ソフトウエアは視覚化及び解釈のために画像を重ねる。
【0011】
脱髄疾患、オリゴデンドロサイト及びミエリン塩基性タンパク質
ミエリン塩基性タンパク質(MBP)は正常なミエリン圧縮及び機能に必要とされる。ミエリンオリゴデンドロサイト糖タンパク質(MOG)及びプロテオリピドタンパク質(PLP)と共に、これらのタンパク質はミエリン構造タンパク質ファミリーのメンバーを構成し、そしてミエリン産生細胞:中枢神経系(CNS)におけるオリゴデンドロサイト及び末梢神経系(PNS)におけるシュワン細胞により合成される。発達するCNSにおいて、MBPの発現は髄鞘形成の期間と一致する。従って、MBPはオリゴデンドロサイト成熟のマーカーとして当該分野で受け入れられている。MBPの高レベル発現(他のタンパク質と共に)は、ミエリン生成と同時に発生し、ミエリン形成の間中継続し、そして軸索破壊の際に終わる(非特許文献11)。
【0012】
ミエリンの完全性を冒す疾患は、冒されたニューロンにおける軸索シグナルの伝導障害を生じ、そしてどのニューロンが含まれるかによって、感覚、運動、認識又は他の機能の障害を生じ得る。脱髄疾患という用語は、疾患の原因よりその効果を示し、そして遺伝的特質、感染性因子、自己免疫反応及び未知の要因により引き起こされ得る。これらは全て、回復性治療の必要性をしばしば伴う大きな満たされていない医療的必要性を有するヒトの状態のままである。
【0013】
中枢神経系の脱髄疾患としては以下が挙げられる:
・多発性硬化症(突発性炎症性脱髄疾患と呼ばれる似た疾患も一緒に)
・横断脊髄炎
・ドヴィック病
・進行性多病巣性白質脳障害
・視神経炎
・白質萎縮症
・ペリツェーウス−メルツバッヒャー病
・ミエリン機能障害/損失は、脊髄損傷、アルツハイマー病及びパーキンソン病、並びに統合失調症とも関連する。
【0014】
末梢神経系の脱髄疾患としては以下が挙げられる:
・ギラン・バレー症候群及びその慢性カウンターパート、慢性炎症性、脱髄性多発ニューロパシー
・末梢神経障害(毒素誘導、糖尿病性、抗MAGなど)
・シャルコー・マリー・ツース病。
【0015】
脱髄のインビボモデル
脱髄疾患と闘うための前臨床候補の開発における共通の段階は、脱髄作用の一部又は全てを再現し、かつ内因性再髄鞘化の能力を有する動物モデルにおいて治療剤候補の作用のアッセイを行うことである。化学的に誘導される脱髄を動物モデルにおいて達成することができ、そして6〜8週齢の若年成体雄性マウスは0.2%クプリゾン(ビス−シクロヘキサノンオキサルジヒドラゾン)の4〜6週の食餌により生じる脱髄を受けやすい(非特許文献12)。クプリゾン処理を用いて、脱髄は脳全体にわたって発生するが、高度に濃縮された白質領域である脳梁内で最も容易に検出される(非特許文献13)。脱髄はクプリゾン食餌開始後3週以内に明らかとなる。食餌を通常の餌に代えることにより、4〜6週以内にほとんど完全な再髄鞘化が可能となる(非特許文献14)。アミロイド前駆体タンパク質又はビールショースキー銀含浸を使用する軸索損傷についての免疫組織化学染色は、8週齢のマウス(Merkler)における軸索損傷をあまり示さない。しかし、軸索離断は6〜7月齢のマウスにおいて有意に増加し、高齢の動物において見られる再髄鞘化の減退に部分的に寄与する(非特許文献15)。分化の際にミエリンと置き換わるオリゴデンドロサイト前駆細胞、並びに小グリア細胞及びマクロファージの両方は、クプリゾン食餌の約4週目にピークに達し、そしてこれらは効率的な再髄鞘化に必要とされる(非特許文献16)。
【0016】
ミエリン含有量の変化を定量するために、脳全体又は小領域を採取し、ドットブロット又はウェスタン分析により評価することができる。あるいは、ミエリンの損失及び修復の小領域を追跡するためのより高解像度の明敏な(長期に対して)アプローチは定量的立体解析学を含む。例えば、コンピューター支援立体解析ツールボックス(computer assisted stereological toolbox)(CAST)システムを使用して、脳梁におけるミエリンルクソールファストブルー(ミエリン染料)の変化を評価することができる。トルイジンブルー及び電子顕微鏡観察は、密集ミエリンが以前の無鞘軸索(naked axons)を正確に鞘で包んでいることを確実にするために必要な最終工程である。長期的な評価に関して、T2強調磁気共鳴画像化法は、クプリゾン食餌を与えられたマウスの脳梁におけるミエリン変化を定量的に評価するための手段を提供する(sanofi−aventis Neurology,2008 非公開の結果)。
【0017】
実時間でインビボで髄鞘形成状態を追跡するMBP−luciバイオイメージングモデル
2003年の文献(非特許文献17)は、MBPプロモーターが、髄鞘形成及び再髄鞘化のタイミングを調節するために重要である4つの広く分布した保存されているモジュール(M1、M2、M3及びM4、0.1から0.4kbの範囲)である、調節エレメントを含有することを報告した(図1、3及び4を参照のこと)。Lac Zをレポーター遺伝子として使用して、近位のモジュールM1及びM2がCNS発達の間にオリゴデンドロサイトにおいて比較的低レベルの発現を駆動し、一方で上流のM3領域は、発達の間中及び成体CNSにおいてオリゴデンドロサイトにおける高レベルの発現を駆動するということが実証された。さらに、このM3領域は、脱髄傷害後の再髄鞘化の間の発現に必要とされる。M4もまたシュワン細胞の髄鞘形成におけるMBP発現を駆動する。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】DiMasi,JA,et al,J Health Econ 22,151−185
【非特許文献2】Lindsay MA,Nature Rev Drug Discovery,2,831−838
【非特許文献3】Rudin M,et al,Progress in Drug Res,2005,vol 62,185−255
【非特許文献4】Massoud T.F.,Genes Dev.17:545−580
【非特許文献5】Maggie A,Ciana P.Nat.Rev.Drug Discvy.4,249−255
【非特許文献6】Maggie A.Trends Pharmacolo.Sci.25,337
【非特許文献7】Contag,CH,Mol.Microbiol.18:593−603
【非特許文献8】Greer LFIII,Luminescence 17:43−74
【非特許文献9】Spibey CP et al Electrophoresis 22:829−836
【非特許文献10】Contag CH,Annu.Rev.Biomed.Eng.4:235−260
【非特許文献11】Gupta et al.Brain Res.464:133−141
【非特許文献12】Ludwin SK ,1978,Lab Invest 39:597−612
【非特許文献13】Merkler et al,2005,NMR Biomed 18:395−403
【非特許文献14】Matsushima et al,2001 ,Brain Pathol 1 1 :107−1 16
【非特許文献15】Irvine KA,et al,2006,J Neuroimmunol 175:69−76
【非特許文献16】Franco RJM,2002,Nat Rev 3:705−714
【非特許文献17】Farhadi et al,The Journal of Neuroscience,November 12,2003,23(32):10214−10223
【発明の概要】
【発明が解決しようとする課題】
【0019】
インビボでのミエリンの変化をアッセイするために動物モデルを使用する現在の方法は時間がかかり、典型的には4〜8週に及び、そして多数の動物を必要とする。ルシフェラーゼ遺伝子を発現する4つの特定の調節領域全てを含むMBP DNAプロモーターは、脳、脊髄、及び/又は末梢神経系におけるミエリン塩基性タンパク質(MBP)転写活性の変化を実時間で追跡するために開発されたマウスMBP−luciトランスジェニックバイオイメージングモデルのための基礎を構成する。この齧歯動物トランスジェニックモデルは、ヒト脱髄疾患の処置のための開発候補を選択するように意図された原理の研究のインビトロ及びインビボでの裏付けのために使用され得、そして現在のモデルに勝る多数の改善を提供し得る:
・ 生物発光により神経系髄鞘形成の変化を非侵襲的に追跡する能力は、死後の組織化学的分析及び組織発現分析に伴うかなりの資源及び労働力の要求を減らす。
・ 長期的な研究は、髄鞘形成の変化に対するモデルの感受性を大いに増加させる。以前のアッセイは各時点で異なるグループの動物を使用しなければならず、そしてこのことが研究の間に収集できるデーター点の数を制限する。別々のグループの動物の必要性のためにさらなる変数が導入される。
・ バイオイメージングデータは迅速に、典型的には同日に処理され、それにより、研究設計を最適化し、そして不必要な資源投資を減らすための調整(すなわち、有意性に達するための研究の長さの延長又は薬物効果が無いことに起因する早期の終了)が可能となる。
・ 動物グループ及び対応する処置は、全て0時点でアッセイすることができ、これにより、処置群あたりに必要とされる動物の数を同時に減らしながら、研究の統計学的有意性が最適化される。研究データは初期値に対して正規化され得、特定の治療の効果に対するより強い結果を支持しながら固有の生物学的変動を大幅に減少する。
【課題を解決するための手段】
【0020】
発明の要旨
本発明は、脱髄(例えば、神経保護、ミエリン保護)及び再髄鞘化(例えば修復)の程度に対する治療実体の効果を評価するための新しいインビボモデルを提供した。本発明のバイオイメージング動物及び化合物スクリーニング方法は、生きている動物における関連性のある実時間高解像度データの生成により、 医薬化合物及び他の治療剤のスクリーニング及び/又は原理の検証の裏付けを可能にする。さらに、可能性のある毒性、PK/PD、及び治療域に関連する評価を得ることができ、それにより霊長類及びヒトにおける投薬の周りの(around)より正確な予測を可能にする。
【0021】
本発明は、生きているトランスジェニックモデル生物を使用する髄鞘形成事象の研究のための改善されたツール及び方法を提供する。本発明は、各被験体が長期間の研究においてそれ自身のベースラインコントロールとして役立ち得るので、他のモデルと比較して必要とされる被験体の全体の数を減らす。従って生物間の変動性が低減し、統計学的結果における信頼性を改善する。本発明の種々の局面において、様々な利点が達成される。様々な用途のいくつかは以下の考察において特徴付けされる。
【0022】
MBP発現との相関により、本発明は、生きているモデル動物において脱髄及び/又は再髄鞘化をモニタリングする方法を提供する。本方法は、MBPプロモーターにより駆動されるルシフェラーゼ遺伝子を発現するモデル生物を生成するように前駆細胞を遺伝子導入で改変することを含み得る。ルシフェラーゼ遺伝子は、発現されると、ルシフェリンのような基質の存在下で光を生じる(生物発光)ように機能することができる。生物からの生物発光をモニタリングすることにより、髄鞘形成及び脱髄の活性を評価することができる。画像化装置及び分析は、生物発光及び髄鞘形成事象を生物の身体の特定の部分に関連付けることを迅速に可能にする。
【0023】
中枢神経系及び末梢神経系の事象を研究するために、これらの神経組織においてミエリンを使用する脊椎動物は、特に適したモデル生物である。哺乳動物は、CNS及びPNSの両方において神経伝導を補助するためにミエリンを使用する生物である。齧歯動物(例えばラット、マウス)、ウサギ、ヒツジ、霊長類、モルモットなどのような一般的な哺乳動物モデルは、本発明の実施における使用に適したモデル生物である。
【0024】
生物のサイズは、それ自体は限定因子ではない。しかし、装置サイズは特定の生物の形状又はサイズに対して最適化され得る。
【0025】
単一のバイオイメージング事象は所望の情報を提供し得る。しかし、本発明の利点は、複数の時点にわたって同じ動物で測定を繰り返して複数の測定を比較する能力を提供する。それにより単一の生物はそれ自体のコントロール又はベースラインとして役立つ。
【0026】
その結果として、単一の生物における1つ又はそれ以上の脱髄又は再髄鞘化間隔にわたるバイオイメージングが可能である。さらに、脱髄又は再髄鞘化間隔による画像化シグナル正規化は、ここで繰り返される間隔を通した画像化は生物におけるMBP転写物のレベルの変化を1つ又はそれ以上の事象にわたって検出するために有効であり、よりロバストなデータを生じる。
【0027】
生物発光シグナルは身体組織により減衰される。従って、身体表面の近位にある神経組織はより強いシグナルを生じる。検出器におけるシグナルを、例えばより高濃度を使用するか又は酵素活性若しくは発現レベルを高めてより多くのルシフェラーゼを反応させることにより、より強いシグナル出力を生じることにより増加させることができる。データ収集はまた、より高感度の検出器を使用して改善され得る。より高感度の検出器は、有意なシグナルを生成させてノイズを最少にするためにしばしば冷却装置を必要とする。データが収集される時間もまたシグナルの量を増加させるために役立つ。
【0028】
本発明のモデル生物は、MBPプロモーターにより駆動されるルシフェラーゼ遺伝子を含むトランスジェニック動物であり得る。この動物は哺乳動物、例えば動物モデルとして使用されるいずれかの哺乳動物であり得る。ラット及びマウスは一般的な動物モデルである。MBPプロモーターの制御下のルシフェラーゼ遺伝子は、プロモーターが活性化されるかオンにされる場合にモデルの特定の標的組織において発現される。
【0029】
MBPプロモーターはM1〜M3を含有していても、M1〜M4を含有していてもよい。
【0030】
シグナル改善は、その毛髪が少ない減衰に寄与するモデルの選択のような簡単な操作によりなされ得る。例えば、その毛髪がC57/B6マウスの毛髪よりも少ない減衰であるモデルは、C57/B6マウスのシグナルを上回るシグナルを生じるだろう。
【0031】
本発明はまた、バイオイメージングのためのモデル生物を作製するための方法を提供する。作製は、例えばマウス系統から開始して、そしてトランスジェニック動物を作製し、次いで生後髄鞘形成のピークにおけるインビボマウス全身画像化が(例えばCNSの)特定の画像化を示す1つ又はそれ以上の系統を選択し;次いで、エクスビボ画像化により所望の領域、例えば主に脳の緻密な(compact)白質領域におけるルシフェラーゼ導入遺伝子の発現が確認される1つ又はそれ以上の系統を選択することにより達成され得る。次いで、ルシフェラーゼ画像強度が脱髄及び再髄鞘化の変化と高度に相関している1つ又はそれ以上の系を選択することができる。改善されたデータのために、適切な操作の間、明確な組織学的脱髄を示す1つ又はそれ以上の系を選択し得る。
【0032】
クプリゾン脱髄モデルは本発明での使用に適している。バイオイメージングのタイミングは、モデル生物の発達特性に依存し、例えば、生後髄鞘形成のピークが一般的に生後約3〜5週齢のG1マウスであることを考慮するようにタイミングを選択してもよい。
【0033】
本発明は、生物におけるミエリンの事象に影響を及ぼし得る化学的化合物、生物学及び他の治療実体をスクリーニングするために有用である。化合物は、遺伝子発現又は細胞内若しくは細胞間のシグナル伝達事象を調節してミエリン事象に影響を及ぼし得る。これらは単にいくつかの具体例であり、特許請求の範囲において特徴付けされる本発明の範囲全部とみなされるべきではない。
【0034】
MBPプロモーターは、記載されるように(例えばFarhadi、上記)、レポーターlac Z遺伝子を使用して十分に特徴付けされているが、MBP−lac Zモデルの使用はβ−ガラクトシダーゼ(lacZレポート遺伝子産物)の検出のための組織採取及び固定化の必要性のために制限される。このプロセスは、生きている動物における検出とは両立できない組織化学的技術を必要とする。この問題を回避するため、ルシフェラーゼ又はGFPレポーターがMBPプロモーターの大部分により選択的に制御されるトランスジェニックバイオイメージングモデルを開発するために、生物発光又は蛍光系の使用が提案されてきた。
【0035】
この新規なバイオイメージングモデルは、生きている動物(例えばマウス)における実時間での脱髄及び/又は再髄鞘化の可視化及び定量のために設計される。このようなモニタリングは、自動化バイオイメージング技術と併せて使用され得る。このモデルは、 標的選択及び化合物進行(progression)に関して重要な方針決定をするために相対的及び定量的に、例えば標的の検証のため(例えばバイオイメージングモデルマウスが所望の特質を有するノックアウトマウス及びトランスジェニックマウスと交配される場合)、そしてまた化合物の検証(例えば、クプリゾン又は実験的自己免疫性脳脊髄炎[EAE]のようなモデルにおける化合物の有効性の測定)のために有用なツールである。
【0036】
本発明のバイオイメージングモデル(MBP−luci TG)は開発されて脱髄/再髄鞘化事象をインビボで定量するために使用された。本バイオイメージングモデルの利点は、長期の研究が可能となり、それによって各生物がそれ自身のコントロールとして役立つことができるということである。それ故、特定の時点における動物の犠牲が避けられる。
【0037】
個々のマウスは脱髄及び再髄鞘化プロセスを通して継続的に追跡され得る。
【0038】
バイオイメージング方法論は、生きているマウスにおいて生物学的応答を追跡するために必要とする時間及び資源がより少ない。
【図面の簡単な説明】
【0039】
【図1】ミエリン発現における実時間変化を追跡するために使用されるMBPルシフェラーゼトランスジェニックモデルの図式的要約を示す。
【図2】内在性MBPプロモーターが、発達の間及び成人期中にオリゴデンドロサイトにおけるMBPの発現を異なって調節する4つのエレメントを有するということを示す。(HF Farhadi:J.Neurosc.2003,23 (32),10214−10223)。M1/M2は両方とも生後初期段階の転写物を調節する。M3は成熟の全体にわたる発現の転写物調節に関与する。M4は、MBP転写物のシュワン細胞発現に寄与する。MBP−luci導入遺伝子構造は2つの形態のMBPプロモーターを有する。系121はM4エレメントを含み、そして脳、脊髄及び末梢神経系においてルシフェラーゼを発現すると予測される。系171はより短いMBPプロモーターからなり(例えばM4エレメントを欠く)、そしてバイオイメージング分析により確認されたように、主に脳で発現する。
【図3】ルシフェラーゼ発現ベクターにおける5K bpの5’DNAを有するMBPプロモーターを含むMBP−luci導入遺伝子発現カセットを示す。
【図4】ルシフェラーゼ発現ベクターにおける10K bpの5’DNAを有するMBPプロモーターを含むMBP−luci導入遺伝子発現カセットを示す。
【図5】MBP−Luci導入遺伝子系のスクリーニングツリーの例を示す。
【図6】MBP−luci創始者の第一レベルのスクリーニングからのインビボルシフェラーゼ画像を示す。ルシフェラーゼ強度は105(+++)から103(+)(光子/cm2/秒)の範囲に及ぶ。
【図7】年齢の増加に伴う脳におけるルシフェラーゼ画像の減少を示す、7週(A)及び10ヶ月(B)でのバイオイメージングモデル生物を示し、これは生後初期の発達後に観察される髄鞘形成の現象と相関する。
【図8】MBP−luciマウスのインビボでのバイオイメージングを示す:頭蓋(ROI:対象の領域)において測定された2.7x103光子/sに相当するバックグラウンド生物発光を有する、左側のネガティブ導入遺伝子野生型マウス(WT)。ヘテロ接合MBP−luciマウスは右に示され、そしてROI生物発光は1.327x105光子/sに相当する。ホモ接合MBP−luciマウスは中央に示され、ROI値3.2924 x105 光子/sを有し、これはほぼ予測通りであり、ヘテロ接合の値の2倍である。
【図9】CNS特異的時間的ルシフェラーゼ発現を示す。示されるようにMBP−luciトランスジェニックマウスからの頭蓋生物発光は、4週(A及び*)から8(B及び**)まで減少し、Taqman分析(C)により決定された内在性MBP転写物レベルに対応している。
【図10】MBP−luciマウスにおけるルシフェラーゼのCNS特異的及び小領域発現を示す。MBP−luciトランスジェニックマウス脳の2つの矢状(sagital)切断(A及びB)後(followed)のスライスのバイオイメージングは、脳における脳梁小領域がその最も高い生物発光シグナルを含み、そしてクプリゾン病変モデルにおける最も高い脱髄及び再髄鞘化と相関するということを明らかにした。
【図11】MBP−luciモデルにおけるバイオイメージング及びクプリゾン病変食餌において起こることが知られている他の生物学的応答を示す。ここで クプリゾン食餌は4週間与えられ、そしてグラフの左に示されるバイオイメージング及び細胞変化を生じる。示される(矢印)各週におけるバイオイメージングは、内在性MBP発現レベルと平行して変化すると予測される(Matsushima GK and Morell P,Brain Pathology 11,1−10,2001)。
【図12】野生型C57 BL/6Jマウスにおける連続的なクプリゾン食餌処置に応じた内在性MBP定常状態mRNAの変化を示す。8週齢のマウスをそれらの食餌でクプリゾンに12週まで曝露させた(黒三角)。第二のグループにおいて(白三角)、クプリゾン餌を6週の曝露後に取り除き、そしてマウスをさらに6週間回復させた。mRNAレベルについてのデータは、ノーザンブロットによる単一測定であり、そして3つのコントロールの平均と比較してプロットされている(Jurevics H,et al,Journal of Neurochemistry,2002,82,126−136)。
【図13】MBP−Luciバイオイメージングモデルに対する証明されたクプリゾン食餌の影響を示す。画像化シグナルは、内在性MBP遺伝子の発現パターンを明確に再現し得る。クプリゾン餌を与えた間(0週から4週)、2〜3倍の画像化シグナル減少があり、そしてクプリゾン餌の除去後(4週から7週)の3〜4倍の画像化シグナル増加もあった。
【図14】MBP−luciモデルにおける脳梁(corpus collosum)のルクソールファストブルー(LFB)染色を示す。マウスをクプリゾン又は通常餌のいずれかで4週間処置し、次いで両方とも通常食餌に戻した。6週目に、クプリゾン処置されたMBP−luciの脳梁(corpus collosum)(B)における明瞭な脱髄が、通常食餌処置グループ(A)と比較してLFB染色を使用して組織化学的に検出され得る。髄鞘形成における構造的変化の組織化学的アッセイは、処置されたMBP−luciモデルにおけるバイオイメージングシグナルにおける変化と相関する。
【図15】ペルオキシソーム増殖因子活性化受容体デルタ(PPARδ)アゴニストツール化合物(本明細書以下では’517)が、自発的再髄鞘化の期間の間に系171 hetマウスにおけるルシフェラーゼの画像化シグナルを増強することを示す。8週齢MBP−luciマウス(系171ヘテロ接合性、B6C3H系統)を0.2%クプリゾン含有餌で4週間飼育し、次いで通常餌にして再髄鞘化させた。次いでマウスにビヒクル(0.6%カルボキシメチルセルロールナトリウム塩及び0.5%Tween80)又は30mg/kg PPARδアゴニストツール化合物’517のいずれかを8日間、1日に2回経口投与し、そして示された時点で画像化した。データを0週ベースラインシグナルに対して正規化した。ツール化合物’517(n=12)は、ビヒクルグループ(n=12)と比較してルシフェラーゼシグナルの30〜100%の相対的な増加を生じ、これはオリゴデンドロサイト前駆細胞分化の刺激に対する化合物の効果に起因するものと考えられている(例えば、インビトロでの知見と一致する)。
【図16】PPARδアゴニストツール化合物’517は、再髄鞘化期の間のルクソールファストブルー(LFB)ミエリン染色を改善する(系171ヘテロ接合、B6C3H系統)。ホルマリン固定パラフィン埋包脳からの傍矢状組織切断を、脳梁におけるミエリンの定量的評価のためにLFBで染色した。各時点についての染色した切片をスコア付けして0(完全な髄鞘形成)から5(完全な脱髄)までのスケールで等級分けした。スコア付けシステムは以下のとおりであった:0=正常ミエリン、脱髄無し、1=最小限の局在化した脱髄、2=軽度から中程度、局在化した脱髄、3=中程度、局所的に極度の脱髄、4=重症、局所的に極度の脱髄、5=重症、広範な脱髄。クプリゾン4週後のマウスからのLFB染色した脳切片の組織学的評価により、系171ヘテロ接合マウス(n=5)における脳梁の中程度から重症の脱髄が確認された。4週から5週の再髄鞘化期の間のvでの処置は、LFBにより7週の時点で決定した場合に、ツール化合物’517グループ(n=5)をビヒクルコントロールグループ(n=3)と比較して、ミエリンの測定可能な増加を生じた。小さいnの数にもかかわらず、組織学的データはインビボルシフェラーゼバイオイメージングデータを支持しており、このことは、ビヒクルコントロールと比較した場合の、’517化合物で処置されたマウスにおける増加した再髄鞘化を示している。
【図17】エストロゲン受容体ベータ(ERβ)アゴニスト(本明細書以下では’5a)30mg/kg及びポジティブコントロールクエチアピン10mg/kgは、クプリゾンモデルにおける脱髄期の間、保護的である。MBP−luc系171ヘテロ接合 B6C3Hマウスにクプリゾン食餌を4週間与え、そして’5a(10mg/kg又は30mg/kg)又はポジティブコントロールクエチアピン(QTP)を経口投与した。マウスを0週(ベースライン)、3週及び4週目に画像化し、データを0週ベースラインに対して正規化した。QTPグループはビヒクルコントロールに対して3週及び4週の両方の時点において10mg/kgで画像化シグナルの有意な増加を示した。QTP処置グループについての結果は、Yanbo Zhangらにより公開されたデータ(表題「Quetiapine alleviates the cuprizone−induced white matter pathology in brain of C57BL/6 mouse」(Schizophrenia Research,2008,Dec 106,182−91))と一致する。30mg/kgでの化合物’5aは、ビヒクルと比較してクプリゾン食餌での3週目(傾向)及び4週目(有意)に増強されたシグナルを示した。10 mg/kgでの’5aは3週目及び4週目の両方において有意な効果を有していなかった。結果は、MBP−luci画像化モデルが、クプリゾンモデルにおける用量依存性変化を検出するために十分感受性であることを示唆する。
【図18】’5a(30mg/kg)及びQTP(10mg/kg)のグループは、ビヒクルコントロールグループと比較して3週目及び4週目に有意に高い導入遺伝子活性を有する。画像化モデルデータは、QTP及び’5aの両方がクプリゾン誘導脳脱髄及びミエリン崩壊を弱めるということを支持する。
【図19】ホモ接合マウスとヘテロ接合マウス(B6C3H系171)との間のクプリゾンモデル画像化シグナル比較を示す。N3世代ヘテロ接合マウスを交配させてMBP−luci対立遺伝子についてのホモ接合マウスを作製した。ホモ接合マウスは、クプリゾン処置の間にヘテロ接合マウスよりも2倍大きなバイオイメージングシグナル域を示した。ホモ接合体におけるレポーター遺伝子の2つのコピーは、脱髄期(例えばクプリゾン食餌における4週後)の間2倍より大きなシグナル減少及び再髄鞘化期(例えばクプリゾン食事の除去及び通常食へ戻した1週後)の間に2倍のシグナル増加を示した。 ヘテロ接合系171(B6C3H系統)がクプリゾンモデルにおいて機能し、そして化合物効果を検出し得るということをデータが実証したが、このモデルは、バイオイメージングシグナル強度を増加させるために交配してホモ接合性にすることによりさらに改善され得る。これはまた、マウスコロニーがホモ接合体として維持され得るので、モデル産生を効率化し、そして遺伝子型解析の費用を減少させる。全体的に、より大きな画像化域は、薬理的化合物プロファイリング研究における化合物の効果の検出を増強し得る。
【図20】LFBで染色されたホルマリン固定されたパラフィン包埋マウス脳からの傍矢状組織切片における脳梁の顕微鏡写真(括弧内の濃い色の長手方向の構造−201)は、ミエリン状態について評価された白質の領域を示す。この領域を、脳梁のルクソールファストブルー(LFB)染色の定量的デジタル画像化分析のために使用した。
【図21】3つの異なるMBP−luci系(系171 B6C3Hヘテロ接合系統、系121 C57BL/6ヘテロ接合系統及び系171B6C3Hホモ接合系統)についての画像化域及び組織学領域(histology window)の比較。マウスに0.2%クプリゾンを含む食餌を4週間又は5週間与えた。画像化データを0週ベースライン測定に対して正規化した。各研究の終わりに、マウス脳を採取し、そして連続パラフィン切片をミエリンについてルクソールファストブルー(LFB)で染色した。平均定性的LFBスコア(0〜5)を表に示す。系171 B6C3Hホモ接合マウスは、最も大きな画像化シグナル減少を示し、そしてまた定性的な組織学により評価した場合に最も重症の脱髄を示した。系171 B6C3Hヘテロ接合マウスは、最も小さな画像化域を示し、そしてまた4週目に最も少ない組織学的脱髄を示した。
【図22】クプリゾンモデルにおける系171ホモ接合マウスの使用を、クエチアピン(QTP)に対する処置応答の実証によりさらに検証した。マウスにクプリゾン食餌を5週間与え、そして同時に、毎日QTP(10mg/kg)を経口投与した。マウスを0週目(ベースライン)、3週目、及び5週目に画像化した。データを0週ベースライン測定に対して正規化した。QTP(10mg/kg)は、ビヒクルコントロールと比較して、3週目及び4週目の両方の時点で画像化シグナルの有意な増加を生じた。結果は系171ヘテロ接合マウスで得られたものと一致する(図17及び18)。データは、系171ホモ接合マウスが、クプリゾンモデルにおけるミエリン発現及び完全性の維持に対する化合物の効果を評価するために使用され得るということを示す。
【図23】系121マウスからのエクスビボ脊髄画像化。発光の重ね合わせは、導入遺伝子発現が脳及び脊髄の白質領域に局在化していたということを説明する。このことは、動物全身バイオイメージング実験から至った結論をさらに支持する。
【図24】C57Bl/6マウス;野生型、系171ヘテロ接合及び系171ホモ接合マウスにおける脳梁のルクソールファストブルー(LFB)染色の定量的デジタル画像解析。脳梁内のポジティブLFB染色を有する領域のパーセントの定量を、染色されたスライドのスキャンされたデジタル画像での脳梁の手書きで輪郭線を描いた領域に対してAperio(R)カラーデコンボリューションアルゴリズムを使用して計算した。動物1体について1つの断面を評価した。グループあたりの動物の数は9と10の間で変動した。LFB染色に対してポジティブな領域のパーセントを動物ごとに、そしてグループに関して計算した。グループ間の統計学的有意性を、対応t検定により評価した。171ホモ接合マウス及び野生C57 BL/6マウスの両方が、4週のクプリゾン食餌後に重症の脱髄を示す(ポジティブ染色%は40〜60%の間である)。対照的に、系171ヘテロ接合マウスは、軽度の脱髄しか示さない(ポジティブ染色%は65〜80%の間である)。従って、系171ホモ接合マウスを、クプリゾン誘導脱髄モデルにおける使用に好ましい系として同定した。
【発明を実施するための形態】
【0040】
発明の説明
以下の5つの基準は、バイオイメージングモデルについての選択を順次最適化するために首尾よく適用された:
特定の実施態様は、5K又は10Kのベクターのいずれかを用いて以下のプロセスにしたがって遺伝子導入操作されたマウスの系から作製され得る。以下に記載される結果は、選択プロセスが35のトランスジェニック系で開始するようにしたものからである。図5はこのプロセスを図表でまとめる。
【0041】
生成されたトランスジェニック系から、生後髄鞘形成のピーク(例えば3〜5週G1マウス)におけるインビボでの動物(例えばマウス)全身画像化がCNS特異的画像化を示す系を選択する。選択された35の系から6つを次の選択段階に進めた。
【0042】
次に、エクスビボ画像化により主に脳の白質領域でのルシフェラーゼ導入遺伝子発現が確認された系を選択した。工程1からの6つの系のうち5つを次の段階に進めた。
【0043】
次いで、ルシフェラーゼ画像強度が、(この例では)クプリゾン脱髄モデルにおいて誘導される脱髄及び再髄鞘化の変化と高度に相関する系を選択した。工程2からの5つの系のうち3つを次の段階に進めた。
【0044】
次に、上記クプリゾンモデルにおいて明らかな組織学的脱髄を示した系を選択した。工程3からの3つのうち2つを好ましい系として選択した。
【0045】
構想の決定的な証拠として、本発明者らは、A003398711(PPARδ選択的アゴニスト)の有効性がバイオイメージングモデルにおいて最適に検出可能である1つの系を選択した。
【0046】
系171(B6C3系統、ヘテロ接合)と指定された模範的な系を、上記の5つの基準を使用して選択し、そして好ましいモデルとして使用した。
【0047】
このプロセスのより詳細な説明は以下の実施例に記載される。
【0048】
定義
別に指示がなければ本明細書で使用される用語は、一般的な口語的用法と異なり得る科学用語で通常使用される意味を有する。
【0049】
遺伝子は、転写される領域だけでなく転写されない領域を含むように広く解釈されるべきである。
【0050】
化合物は、化学的化合物、例えば有機化学実体、生物学的化合物、例えば抗体及び抗原認識フラグメント並びに構築物、核酸、例えばRNAiなどを含むように広く解釈される。
【0051】
ホタルルシフェラーゼを発現するトランスジェニックマウスを作製した。これらの動物において、レポーター遺伝子ルシフェラーゼを、MBPプロモーターに連結し、それ故、MBP発現がオンになった場合に例えばCNSの白質(有髄)領域の細胞においてルシフェラーゼの発現を駆動する。基質ルシフェリンの全身注射(IV、IP、SC)は、生きているマウスの頭部から検出可能で定量可能な光シグナルを生じる。MBP−luci系を選択するためにクプリゾン脱髄モデルを適用し、そしてこれらの動物にルシフェリンを繰り返し注射することにより、脱髄及び再髄鞘化を非侵襲性生物発光画像化により長期的に、例えば2ヶ月間にわたって連続的にモニタリング及び定量することができる。このモデルは、クプリゾン誘導脱髄及びPPARδ化合物誘導再髄鞘化を首尾よく定量的にモニタリングした。
【0052】
検出器技術における進歩は、感度及び画質におけるかなりの改善をもたらした。現在では光子は、光子がシリコンウェハに衝突するにつれて光子を電子に変換する専門化された電荷結合素子(CDD)カメラにより検出される。CCDカメラは入射光子の強度を電荷パターンにコード化し、次いでこれが処理されて画像を生じる。ノイズは、CDカメラを過冷却し、そしてカメラを遮光箱中に備え付けることにより低減される。これらのカメラは一般的には画像取得及び解析の間コンピューターにより制御される。かなり小さく、従って実験作業台上に適応され得る第二世代のカメラシステムは、技術を日常的な実験にとって実行可能で実用的なものにした。Xenogene Companyはバイオイメージング技術を商業化した。
【0053】
利用可能な画像化様式のうち、生物発光又は蛍光に基づく光学技術は最も入手し易く、かつ容易に操作されるとして現れた。生物発光画像化(BLI)は、組織からのバックグラウンド発光(ノイズ)が極めて低いので、優れた感度により特徴づけられる。今まで、BLIは様々な動物モデルにおいて細胞運動、腫瘍進行、遺伝子発現、及びウイルス感染のような生物学的プロセスをモニタリングするために首尾よく使用されてきた。
【0054】
ホタルルシフェラーゼは活性のためにATPのような細胞内補因子を必要とする。これにより、この酵素を発現するように遺伝子操作された細胞にその使用が限定される。結果として、多くの有用な画像化適用、例えば循環する因子の分布、細胞外抗原発現の検出、及び内在性細胞の標識は、ホタルルシフェラーゼ画像化を受け入れにくい。ホタルルシフェラーゼのさらなる欠点は、固定された細胞及び組織サンプルにおいてそれを検出するための代替の基質がないことである。このことは、インビボ画像化を顕微分析と相関させることを困難にした。
【0055】
内部器官から放射された光の検出感度は、ルシフェラーゼ発現のレベル、標識された細胞の体内での深さ(光子が組織を通って移動しなければならない距離t)、及び検出システムの感度を含むいくつかの要因に依存する。
【0056】
非侵襲性動物全身画像化を使用するルシフェラーゼレポーター発現カセットの発現のモニタリングは記載されている(Contag,C.,米国特許第5,650,135号、1997年7月22日、(参照により本明細書に加入される);Contag,P.,et al,Nature Medicine 4(2):245−247,1998;Contag,C.,et al,OSA TOPS on Biomedical Optical Spectroscopy and Diagnostics 3:220−224,1996; Contag,C.H.,,Photochemistry and Photobiology 66(4):523−531,1997;Contag,C.H.,al,Molecular Microbiology 18(4):593−603,1995)。このような画像化は、典型的には少なくとも1つの光検出器装置要素、例えば電荷結合素子(CCD)カメラを使用する。
【0057】
MBP遺伝子の制御エレメント
ミエリン塩基性タンパク質(MBP)は、主に神経系において発現されるポリペプチドのファミリーであり、髄鞘形成において重要な役割を果たす。例えば分化オリゴデンドロサイトにおけるMBPの発現は、主に転写レベルで調節される。Journal of Neuroscienceにおいて、FarhadiらはMBP遺伝子の発現を時間的に制御する新しい調節性組み合わせエレメント(combinatorial element)を記載した。
【0058】
Farhadiらは、グリアが髄鞘形成の開始の間及び後の様々な段階でMBPの発現を制御するために調節配列の異なる組み合わせを使用するということを示した。
【0059】
ミエリン塩基性タンパク質(MBP)は、正常なミエリン圧縮に必要であり、そしてMSのような実験的及びヒト脱髄疾患の両方に関与している。
【0060】
ミエリン生物学の理解をさらに進歩させるため、そして生きた動物においてミエリン増強化合物を試験するために。本発明は、本発明のMBP−luciモデルを生成するために、MBPプロモーターの性質及び最も鋭敏なルシフェラーゼレポーター技術を使用した。今や本モデルは生きている動物におけるミエリン遺伝子転写応答の高感度のインビボ測定を可能にする。
【0061】
MBP−luciレポートカセットの構築
129SvEv BACライブラリー(Cell&Molecular Technologies)を、MBPプロモーターM3領域に位置するプローブを用いてスクリーニングした。このプローブは507bpであり、プライマー対(5’−actccttaccacacttcttgcagg−3’ 5’−tctattgggtgatgtgtgccatc−3)を用いて生成された。(配列番号1及び2) MBP BACをサザン解析により同じプローブを用いて確認した(EcoRIを使用して消化された7.6Kのフラグメント及び/又はBamHIを用いて消化された13.8Kのフラグメント)。
【0062】
高忠実度PCRによりプライマーセット(MBP−L−SP2 5−gggggatccacctgggacgtagcttttgctg及びMBP−AP1 5−ggggtttaaactccggaagctgctgtggg)(配列番号3及び4)を用いて増幅された、M1からM4まで(10K)を含む「長い」MBPプロモーターを、Invitrogenのxl−topoベクター中にクローンして、中間ベクターを生成した(Topo MBP10kベクター)。次いで、MBP 10kプロモーター(BamH1及びPmeIフラグメント)をpGL3 hygro neoベクター中に挿入した(BglII及びPmeI部位)。最終的な10KベクターをpGL3−hygro−long MBP−luciと呼ぶ(例えば図4を参照のこと)。
【0063】
高忠実度PCRによりプライマーセット(MBP−S−SP2 5−gggggatccatccctggatgcctcagaagag及びMBP−AP1 5−ggggtttaaactccggaagctgctgtggg)(配列番号5及び6)を用いて増幅された、M1〜M3(5k)を含む「短い」MBPプロモーターをInvitrogenのp2.1−topoベクター中にクローンして中間ベクターを生成した(Topo MBP5kベクター)。次いでMBP 5kプロモーター(BamH1及びPmeIフラグメント)をpGL3 hygro neoベクター(BglII及びPmeI部位)中に挿入した。最終的な5KベクターをpGL3−hygro−short MBP−luciと呼ぶ。(例えば図3を参照のこと)。
【0064】
両方のpGL3−hygo−MBPプラスミドからのDNA配列はM1、M2、M3及びM4の読み取りを確かにした。さらに、これらのプラスミドの293T細胞への一過性トランスフェクションは、検出可能なルシフェラーゼ活性を生じた。
【0065】
動物の取り扱い及びトランスジェニックマウスの生成
全ての動物の作業は連邦政府のガイドラインにしたがって行った。マウスの3つの異なる系統(FVB、B6C3及びC57 BL/6)を使用した。画像化をイソフルラン(Baxter,Deerfield,IL)を用いた吸入麻酔下で行い;完全に回復するまでマウスを観察した。
【0066】
トランスジェニックマウスを以下のようにして生成した:pGL3−hygro−MBP10k−luci又はpGL3−hygro−MBP5k−luciプラスミドのいずれかをNot I及びBamH1酵素で消化した。次いで、MBPプロモーター、ルシフェラーゼ及びポリアデニル化シグナルを含むフラグメントをゲル精製した。トランスジェニックマウスをFVB、B6C3又はC57BL/6の胚への標準的な前核注入(pronuclear injection)により生成した。手短には、前核マイクロ注入の間に、MBP−luci遺伝子カセットDNAを受精直後のマウス卵子に直接導入する。細い針を使用して、DNAを精液由来の大きな雄性前核に注入する。DNAはしばしば、1回又は2回の細胞分裂が起こった後、ゲノム中のランダムな位置に多くの縦列に配置されたコピーとして一体化する傾向がある。従って、得られたマウスは部分的にトランスジェニックである。トランスジェニック細胞が生殖系に寄与する場合、いくつかのトランスジェニック卵子又は精液が産生され、そしてマウスの次の世代は完全にトランスジェニックとなる。
【0067】
トランスジェニック創始者及びそれらのTg+G1子孫を、ホタルルシフェラーゼ遺伝子に特異的なプライマー(PCRプライマー:5’gaaatgtccgttcggttggcagaagc−3’、及び5’ccaaaaccgtgatggaatggaacaaca−3’)(配列番号7及び8)を使用して、尾部生検DNAのポリメラーゼ連鎖反応(PCR)により同定した。
【0068】
25のポジティブ創始者の子孫を、インビボ画像化システム(IVIS 100; Xenogen,Alameda,CA)を使用して画像化し、そして6つのトランスジェニック系を脳画像化シグナルを用いて同定した(2つのFVB系及び4つのB6C3HF1系)。脳画像化ポジティブC57 BL/6創始者はこの実行において生成されなかったので、1つのFVB系をC57BL/6マウスと戻し交配してC57 BL6系統を達成した。続いてB6C3系171マウスを、異種交配により生殖させてホモ接合トランスジェニック交配対を達成した。B6C3F 1系171もC57アルビノ系と戻し交配した。
【0069】
表1
MBP−luci系をスクリーニング選択するためにインビボ生物発光画像化を使用した。G1マウスをイソフルランで麻酔し、そして250mg/kgの用量のルシフェリンを尾部静脈を通して又は皮下注射した。ルシフェリン注射の8分後にマウスを画像化した。6つの系を脳画像化シグナルを用いて同定した(図5、2つのFVB系統:58及び121、並びに4つのB6C3系統:12、23、85及び171)。系58を除いて、他の5つの系は脳の白質領域においてエクスビボルシフェラーゼ画像化シグナルを示した。
【0070】
表1は、尾部生検PCR遺伝子型により生後まもなく同定された35のトランスジェニックDNAポジティブ創始者マウスからのデータを示す。15のDNAポジティブ創始者系はMBP−10k luciを生じ、そして20のDNAポジティブ創始者系はMBP−5k luciを生じた。この出願全体を通して、導入遺伝子及びトランスジェニックマウスはMBP−luciと略される。
【0071】
【表1】
【0072】
図7及び9は、CNS小領域及び年齢関連性髄鞘形成とよく相関しているMBP−luci生物発光を示す。
【0073】
エクスビボ画像化及びルミノメーターアッセイ
単離された器官のエクスビボルシフェラーゼ画像化を、ルシフェリンの皮下(SC)注射(250mg/kg)の10分後にCO2により動物を安楽死させた直後に行った。切開した器官をプラスチックシートで覆った黒い紙上に置き、そしてIVISにより画像化した;強い生物発光シグナルが、切開後20〜30分以内に検出可能なままであった。画像解析及び生物発光定量をLiving Imageソフトウェア(Xenogen Corp.)を使用して行った。
【0074】
組織サンプルを阻害剤を含む溶解緩衝液中に入れた(Passive Lysis Buffer [Promega]及びComplete Mini Protease Inhibitor Cocktail [Roche,Indianapolis,IN])。組織を組織ホモジナイザーを使用してホモジナイズした。組織を少しの間超音波処理してさらにホモジナイズした。組織ホモジネートを遠心分離し、そして清澄化した溶解物をルミノメーターアッセイに使用した。ルミノメーターアッセイのために、ルシフェラーゼアッセイ基質(Luciferase Assay System,Promega)を製造者により示されるとおりに調製した。組織ホモジネート(20μl)及び基質(100μl)を混合し、そして測定をルミノメーターで行った。バックグラウンド発光読み取りを得て、バックグラウンド読み取りを発光データから差し引いた。タンパク質濃度をBCAタンパク質アッセイキット(Pierce,Rockford,IL)を使用して製造者のプロことルにしたがって決定した。それぞれのタンパク質溶解物についての発光をタンパク質1マイクログラムあたりの任意単位の光として計算した。
【0075】
クプリゾン誘導脱髄及び組織学的検証
4週間にわたってクプリゾンをマウスに投与することにより、脳梁の広範な脱髄が生じた。クプリゾン誘導脱髄は重大な小神経膠細胞症及びマクロファージ動員を伴うが(Bakker and Ludwin,J Neurol Sci 78:125−37,1987; Hiremath et al.,J Neuroimmunol 92:38−49,1998; McMahon et al.,J Neuroimmunol 130:32−45,2002)、最小のT細胞応答を有する(Matsushima and Morell,Brain Pathol 11:107−16,2001)。このモデルにおけるミエリン損傷部位の一貫して予測可能な性質は、脳梁髄鞘形成における容易に定量可能な変化を生じる。これらの変化は、オリゴデンドロサイト前駆細胞による新規の髄鞘形成から生じるかもしれないが、免疫調節性の機構による末端脱髄の防止(Pluchino et al.,Nature 436:266−71,2005)は現実味のある代替の説明かもしれない。
【0076】
上で概説したように、MBP−ルシフェラーゼトランスジェニック(MBP−luci Tg)マウスの多数の系統を、クプリゾン誘導脱髄/再髄鞘化事象のインビボ評価のために評価した。ミエリン塩基性タンパク質(MBP)プロモーター駆動ルシフェラーゼ(luci)の発現は、MBPタンパク質を発現しているトランスジェニック(Tg)哺乳動物の脳におけるミエリンのインビボバイオイメージング定量を可能にした。例えばこのモデルは、それらの食餌中に0.2%クプリゾンを与えた野生型C57/BL6マウスを使用し得る。以前のモデルは、様々な化合物処置の後にミエリンの評価のために多数の時点で最終的に屠殺することを必要とした。多数の動物が必要とされるので、動物間変動性は、有意性を達成するためにさらなる被験体(より大きなn)を必要とする因子であった。
【0077】
MBP 5k−luci系171(B6C3)マウスは、4週間の食餌中0.2%のクプリゾンで、ルクソールファストブルー(LFB)組織化学的染色により評価した場合、脳の脳梁における顕著で有意な脱髄を示した。この脱髄は、バイオイメージングインビボルシフェラーゼシグナルの低下とさらに関連付けられた。
【0078】
しかし、MBP 10−Luci系121マウスのFVB系統は、比較できる脱髄を示さなかった。有意な脱髄を生じ得る食餌中の様々な量のクプリゾン及びクプリゾン処置の様々な期間の見込みがある投与計画を同定しようとして、さらなる研究をFVBマウスにおいて行った。結果は脳梁におけるLFB評価により中程度の量の脱髄しか示さなかった。従って、系171を優先的に開発した。
【0079】
クプリゾンモデルに対する異なる系統の影響:
トランスジェニックマウスは、その高い繁殖力に起因してしばしばFVB/NJ (FVB)系統を使用して作製されてきた。FVB系統マウスはまた、それらの白くて比較的光吸収性でない毛皮の色に起因してトランスジェニックバイオイメージングモデルのために広く使用される。剃毛のような毛髪の除去もまた、FVB又は他の系統において毛髪又は毛皮による吸収又は散乱に起因するシグナル損失を低減するために使用され得る。
【0080】
クプリゾンモデルについて系統間の差異が観察されたので、系統の選択は結果に影響を及ぼし得る。特定の目的のために最適な系統の選択は、例えば選択されたアッセイ及び装置に依存性の所定の最適化と考えられる。しかし、トランスジェニック哺乳動物の作製は限定因子とはみなされない;むしろ、例えば、髄鞘形成に影響する条件に対する特定の系統及びトランスジェニック系の感受性は、データ品質を改善するための選択基準として使用される。選択された髄鞘形成/脱髄事象に依存して、系統又は遺伝的バックグラウンドの選択は結果に影響を及ぼし得る。特定の脱髄モデルは特定の系統においてより良好に機能し得ると考えられる。モデル及び系統のこのような選択は、アッセイ開発の一部として日常的と考えられるだろう。脱髄及び再髄鞘化を研究するためにクプリゾン食餌は優れたモデルであり、系統差異で外見上観察されるこのモデルにおける強い遺伝要因が存在する。
【0081】
FVB系統:
FVB系統からのマウスを一部分においてそれらの白色毛皮色のために選択した。FVBは大部分のトランスジェニック実験及びその後の遺伝子解析に適したシステムを提供する。例えば、同系交配FVB系統は、活発な生殖能及び一貫して多い同腹子を特徴とする。これは大きな集団を生成する際に費用及び労力を減らす。さらに、FVB受精卵は大きく突出した前核を含み、これはDNAの微量注入を容易にする。さらに、FVB系統はアルビノ毛皮色を有し、それによりバイオイメージングの第一候補となっている。これらの特徴はFVB系統をトランスジェニックバイオイメージングモデルを用いた研究のための使用に有利にする。しかし、他の系統を、それらが所望の特徴を示す場合は使用することができる。
【0082】
FVB系統マウスは体重減少という点で0.2%クプリゾンに対して非常に感受性である。1週あたり2〜3回の通常餌/トランスゲル(transgel)栄養補助剤が重篤な体重減少及び毒性を避けるために必要とされる。さらに、発明者らの経験により、FVB系統マウスが様々なクプリゾン食餌投与計画からの最少の組織学的脱髄を示すということが示された。
【0083】
従って、その後の検証及び応用を容易にするために、MBP 10k−luci系121(FVB系統)をC57 Bl/6と戻し交配した。
【0084】
B6C3/Tac系統:
B6C3ハイブリッド系統は、C57 BL/6Ntac雌性マウスをTaconic USの市販のコロニーからのC3H/HeNTac雄性マウスと異種交配させることにより発生させることができる。これは黒色又はアグーチの毛皮色を有する。B6C3は、C57BL/6及びC3Hが異なる遺伝子座においてヘテロ接合性であり、そしてそれらが同じ遺伝子座においてホモ接合性である。
【0085】
B6C3/Tacマウスは明白な組織学的脱髄を示した。具体的には、バイオイメージングモデル系171 MBP 5k−Luciホモ接合マウスにける脱髄は、野生型C57BL6と比較して少しだけ規模が小さく、軽度の変動性であった。系171 MBP 5k−Luciヘテロ接合マウスにおける脱髄は、C57BL6及び系171 MBP 5k−Luciホモ接合マウスと比較して、かなり重症度が低く、より局在化しており、そしてより変動性であった。これらの結果は、MBP−luciモデルが、髄鞘形成に影響を及ぼす事象に対する個体、系統又は種の感受性を決定する際に有用であり得る。さらに、MBP−luciの構成は黒色毛皮の哺乳動物においても有用性を失わない。
【0086】
BALB/cJ系統:
BALB/cJマウスにおける皮質脱髄に対するクプリゾンの効果も調べた。これらのマウスにおいて、皮質脱髄は部分的のみであった。
【0087】
さらに、皮質小グリア細胞蓄積はBALB/cJマウスにおいて顕著に増加したが、一方で小グリア細胞はC57BL/6マウスの皮質には存在しなかった。従って系統差は異なる研究目的を支持するために有用であり得る。
【0088】
C57 BL/6J Jax系統:
C57BL/6遺伝的背景の動物は多くのクプリゾンモデル研究に適しており、そしていくつかの研究所で過去30年にわたって使用されてきた。8週齢のC57BL/6マウスに食餌中0.2%のクプリゾンを与えた場合、成熟オリゴデンドログリア(olidgodendroglia)は特異的に侵襲され(膨大な量のミエリンの支持の代謝要求を満たすことができない)、そしてアポトーシスを受ける。この事象には小グリア細胞の動員及びミエリンのファゴサイトーシスが直ぐ後に続く。形態学的研究と合わせたミエリン遺伝子発現の研究は、連続した代謝負荷に曝されても、オリゴデンドログリア前駆細胞が増殖し、そして脱髄領域に侵入するということを示した。クプリゾン負荷が終了した場合、ほとんど完全な再髄鞘化がせいぜい数週間以内に起こる。可溶性因子による異なる細胞型間の細胞間情報伝達が推測され得る。本発明の方法及びモデルは、そこで細胞間情報伝達事象を研究するため、例えば、推定因子が髄鞘形成のための動員に関与するか否かを決定するため、動員を容易にする化合物をスクリーニングするため、及び動員を阻害する化合物をスクリーニングするために有用であり得る。
【0089】
さらに、MBP−luciモデルの再現性は、脱髄及び又は再髄鞘化のプロセスを加速するか又は抑制し得る操作(例えば、共通の遺伝的背景の利用可能なノックアウト若しくはトランスジェニック、又は妨害するRNA若しくは薬理的処置)の試験を可能にし得るということを示す。
【0090】
MBP−luciモデルの改善
系171(B6C3H系統、ヘテロ接合体)は髄鞘形成/脱髄研究において役立つことが示されたが、このモデルは、(1) バイオイメージングシグナル強度を増大させ、そしてモデル製造及び遺伝子型解析費用を減らすために、繁殖させてホモ接合性(homozygocity)とすることにより;(2) 白色毛皮で画像化シグナル減衰を減らし、そして複数回のNair剃毛後の皮膚反応を低減するために、繁殖させてC57系統のようなアルビノ系統とすることにより;(3) 社内開発CNSクプリゾンモデル系統と整合させるために系121をC57 BL/6系統と繁殖させることによりさらに改善され得る。
【0091】
本発明者らは今や、系171ホモ接合が系171ヘテロ接合系の2倍を超えるバイオイメージング域を示す(細胞あたりレポーター遺伝子カセットの2つのコピー)をいうことを実証した。他の実験もまた、アルビノC57がクプリゾンモデルに対して応答することを実証した。
【0092】
画像化システム及びデータ解析
生物発光を、IVIS画像化システム(Xenogen Corp.,Alameda,CA)を使用して非侵襲的に測定した。画像はルシフェリンの腹腔内(i.p.)注射(250mg/kg−1;Xenogene Corp.)の10分後に、文章中に他の記載がなければ、60秒収集、ビニング10で撮影した。画像収集の間、マウスを約2%イソフルラン(Abbott Laboratories Ltd.,Kent,United Kingdom)の吸入により継続的に鎮静させた。
【0093】
画像化システム説明:
本発明を証明するデータ収集のためにIVIS(R)Imaging System 100 (Xenogene)を使用した。Xenogenの高感度のIVIS(R)Imaging System 100シリーズは、例えば10〜25cmの調整可能な矩形視野を提供し、5匹のマウス又は2匹の大きなラットの画像化、さらには1つの標準的なマイクロタイタープレートも可能である。このシステムは25mm(1.0インチ)平方の背面薄肉化(back−thinned)裏面入射型CCD(電荷結合素子)カメラを特徴とし、これは電子的バックグラウンドを最小にして感度を最大にするために閉鎖循環冷却システム(液体窒素を用いない)により約−90〜−120℃、例えば−105℃に低温冷却される。CCDカメラは、特にスペクトルの赤色領域における高効率光子検出用に設計されている。これは非常に少ない数の光子を検出でき、さらに伝統的なカメラのように動作し;その広いシグナル範囲での画像の表示はXenogenのLiving Image(R)ソフトウェアの機能である。異なる帯域幅をとり出すための6ポジションフィルタホイールがある。このスペクトル情報は発生源の細胞の深さ及び分布についてより多く明らかにすることができる。CCDは冷却され、そして電子的読み出しは、実時間インビボ画像を作成するために集積されたデータが非常に低いノイズを有するように最適化される。
【0094】
遮光性画像化チャンバー
高い遮光性の低バックグラウンド画像化チャンバーは、IVIS(R)Imaging System 100シリーズを標準的な実験室の照明環境で使用することを可能にする。画像化チャンバー内のサンプル棚は視野を調節するために上下に移動する。研究者は動物全体を見ることも、追加の詳細のために一部に焦点を合わせることもできる。棚は麻酔された例えばマウス又はラットの満足な状態を高めるために加熱される。このシステムは、ウェブサイトページに示される、Xenogen−the XGI−8 Gas Anesthesia Systemからの加熱されたサンプル棚、ガス麻酔接続、及び全体ガス麻酔オプションのような動物操作の特徴を含む。より大きな画像化チャンバーはより大きな被験体又はより多数の被験体の使用を可能にし得る。
【0095】
インビボ生物発光アッセイのためのルシフェリンの製造:
以下の材料を実施例で使用した:
ホタルD−ルシフェリンカリウム塩 1.0g/バイアル(例えばXenogen XR−1001又はBiosynth L−8220)。
Mg2+及びCa2+を含まないDPBS
ボトルトップフィルター0.2um。
【0096】
以下の手順を画像化に使用した:
ルシフェリンのDPBS中25mg/mlのストック溶液を調製し、そして0.2umフィルターを通して滅菌ろ過した。5mlのアリコートを−20℃で保存した。注射用量は10ul/体重1gであった。各マウスを対象に250mgルシフェリン/体重1kgを投与した(例えば20gのマウスについては、200ulを注射してルシフェリン2.0mgを送達する)。ルシフェリンを画像化の数分前に皮下(SC)、又は腹腔内(IP)又は静脈内(IV)注射した。ルシフェリン動態研究を、ピークシグナル域を決定するために各動物モデルについて場合により行った。
【0097】
3.6 MBP−luci画像化法:
上記のように、マウスに250mg/kg D−ルシフェリンをSC、IP又はIVにより注射した。5分後(静脈内)又は8分後(腹腔内又はSC)に、マウスをIVIS 100(Xenogen)を使用して16分間(サイズ8で8つの画像について60秒の画像化及び60秒の間隔)画像化した。生物発光を定量するために、興味のある同じ円形の領域を各マウスの頭部領域を取り囲むように位置決めし、そして画像化シグナルを、LIVINGIMAGEソフトウエア(バージョン2.5、Xenogen)を使用して平均放射輝度(光子/秒/cm2/ステラジアン(steridian))として定量した。興味のある頭部領域を、一定面積に保ち、そして全て実験内で位置決めした。データを各動物について処置開始時の生物発光に対して正規化した。
【0098】
3.7 統計学的解析
統計学的解析のために、EverStat V5及びSigma Stat統計ソフトウェアパッケージを使用した。グループにおける画像化の平均を平均値とみなし、そして全てのグループについてのSEを計算した。
2つのグループの平均値を比較する場合、ウィルコクソン対比較(paired)検定又はウィルコクソン非対比較(unpaired)検定を行った。両側P値<0.05を統計学的に有意とみなした。
【実施例】
【0099】
トランスジェニックマウス生成
「長い」プロモーターはM1、M2、M3、M4を含む約10KBであり、そして「短い」プロモーターはM1、M2及びM3を含む約5KBであった。これらを高忠実度PCR法を用いてMBP遺伝子を含有するマウス細菌人工染色体(BAC)からクローン化した。次いで各プロモーターフラグメントをベクター中に、例えばpGL3−hygroベクターのポリリンク(poly link)部位に(into the into the)クローン化した(図1及び図2)。
【0100】
プラスミドをNot I及びBamH1で制限してMBP−luciトランスジェニック発現カセットを放出させ(図3)、これらを、標準的な前核微量注入技術を使用してFVB/Tac系統及びB6C3/Tac系統でトランスジェニックマウスを生成させるために使用した。
【0101】
トランスジェニック(Tg)動物生成のための一般的な方策は当該分野で周知であり、例えばPinkert,C.A.(ed.) 1994.Transgenic animal technology:A laboratory handbook.Academic Press,Inc.,San Diego,Calif.;Monastersky G.M.and Robl,J.M.(ed.)(1995) Strategies in transgenic animal science.ASM Press.Washington D.C.に記載される。
【0102】
脱髄/再髄鞘化事象と相関したMBP−Luci導入遺伝子シグナル
白質領域のルシフェラーゼ発現が観察されたMBP10K−luciトランスジェニック系121(FVB系統)を、立証実験のためのクプリゾンモデルにおいて使用した。図11に示されるように、第一のクプリゾン研究を、ルシフェラーゼ画像化を1週目、2週目及び4週目に繰り返し(0.2%クプリゾン食餌)、続いて通常のクプリゾン非含有食餌に戻して5週目、6週目及び7週目に画像化した(RE 1週から3週)。図に示されるように、この系121からのルシフェラーゼ発現は、クプリゾン誘導脱髄及び再髄鞘化の時間経過と明らかに相関していた。このことは、公開されている内在性MBP mRNA研究(Jurevics et al.,Journal of Neurochemistry,2002,82,126−136)と一致する。FVB系統マウスは1つの系統以外であり、そして体重減少により見られるように0.2%クプリゾンに対して感受性であることが知られている。通常、1週当たり2〜3回の通常餌/トランスゲル栄養補助剤が、この系統における重篤な体重減少及び毒性を回避するために必要とされる。
【0103】
クプリゾンモデル検証:
周知でありかつ広く使用される脱髄/再髄鞘化モデルの1つはマウスにおけるクプリゾンモデルである。このモデルは、例えば連続した4〜6週の間、粉末状齧歯動物試料(lab chow)で投与されるクプリゾン、銅キレート剤(典型的には約0.2%質量/質量; ビスシクロヘキサノンオキサルジヒドラゾン、CAS番号370−81−0、Sigma C9012)の食餌消費を含む(例えば:Matsushima及びMorell,2001を参照のこと)。クプリゾンは成熟オリゴデンドロサイトに対して選択的に毒性であることが示されている。クプリゾン食餌の通常食餌へのその後の切り替えにより、回復をもたらす環境が生じ、その結果、クプリゾン食餌停止の4週〜6週後に、マウスは脳梁において高い再髄鞘化を示す。従って、クプリゾンモデルは、その中で脱髄及び再髄鞘化の局面を研究するための完全なインビボパラダイムを提供する(図11及び12)。
【0104】
さらなる証拠として、MBP 5k−luci系171(B6C3系統)を試験した。この系統もまた、図13に示されるように、クプリゾン誘導脱髄及び再髄鞘化事象に対して相関関係にある画像化応答を示した。7匹のTg+マウスを0.2%クプリゾンで6週間処置し、そして3匹のTg+マウスを通常食餌で6週間処置した。全ての7匹のマウスが0.2%クプリゾン食餌に耐え、そして15〜25%の間の平均体重減少を有していた。クプリゾン処置と共に、有意な画像化シグナル低下があった(脱髄)。例えば、0週目から4週目で43%のシグナル低下、そして0週目から6週目で74%のシグナル低下があった。
【0105】
MBP−luciマウスクプリゾンモデル組織学的検証
組織学的検証については、目的はバイオイメージングモデルにおいて、クプリゾン処置の間のレポーター遺伝子の応答が、クプリゾン処置マウスの脳梁における構造的に検出可能な脱髄と相関することを確認することであった。これらの病的状況を、ルクソールファストブルー(LFB)染色により可視化した(図20及び24を参照のこと)。
【0106】
具体的には、最初の試験において0.2%クプリゾン食餌を用いたMBP 10k−luci系121(FVB系統)について、LFB染色によりごくわずかな脱髄しか検出されなかった。これらのFVB系統マウスについて明瞭な組織学的脱髄を生成させるために、様々なクプリゾン食餌投与計画にしたがって重篤な体重減少を回避するよう試みた。0.2%、0.175%及び0.15%のクプリゾン用量グループ(6週の研究)は、重篤な体重減少及び毒性を避けるために1週あたり3〜4回の通常餌/トランスゲル栄養補助剤を必要とした。また、クプリゾン濃度を曝露時間が延びるにつれてさらに低くした。クプリゾン濃度(0.14%、0.12%及び0.1%)及び処置期間(7週及び9週)を用いた研究を、週に1回までの通常餌/トランスゲル栄養補助剤を用いて試験した。しかし、全てのデータは、FVB系統マウス(8週齢、体重28.5g±3g)が多様なクプリゾン食餌投与計画からのより少ない組織学的脱髄を有するということを示した。この研究モデルの予備的開発のための特に好ましい実施態様としてはFVB系を選択しなかった。FVB系統は画像化及びクプリゾン毒性に対する感受性については良好であるが、体重減少は、別の系統を使用することによりこれらの予備研究において容易に回避できる交絡変数を導入し得る。
【0107】
別の系統では、MBP 5k−luci系171(B6C3)マウス(8週齢、体重25g±3g)が明瞭な組織学的脱髄を示した。
【0108】
マウス脳組織を、6週の0.2%クプリゾン処置の終わりに集めた。7匹のクプリゾン処置されたマウスは全て、脳梁領域において明瞭な脱髄を有し、そして3匹のコントロールマウスは全て、脳梁領域において正常な髄鞘形成を示した。MBP 5K−luci系171からのこれらのデータは、画像化シグナルがクプリゾン誘導脱髄期を追跡するものであるというさらなる証拠を提供する。
【0109】
さらなる定量的LFB分析(図14)は、MBP−Luci B6C3系171ホモ接合マウス(8週齢、体重21g±3g)が、B6C3H系171ヘテロ接合マウス(8週齢、体重25.5g±3g)と比較してより一貫性のある、わずかにより重症な脱髄を4週目に示すということを実証した。楕円中のより濃い領域は染色されたミエリンを示す。
【0110】
さらに、0.2%クプリゾンを与えられたC57BI/6系統野生型雄性マウス(8週齢、体重20g±3g)は最も重症で一貫した脱髄を示した。C57 BL/6系統は、予備研究において記載されるクプリゾンモデル及びPPARδ試験化合物効果についてのポジティブコントロールとして役立った。
【0111】
1.MBP−luciマウスは、PPARδ選択的アゴニストツール化合物のCNS再髄鞘化に対するポジティブな効果を裏付ける:
MBP−ルシフェラーゼマウスを、このインビボバイオイメージングモデルがペルオキシソーム増殖因子活性化受容体δ(PPARδ)アゴニストツール化合物(‘571)のCNS再髄鞘化に対する効果を検出するために使用され得るか否かを評価するために使用した。
【0112】
ペルオキシソーム増殖因子活性化受容体(PPAR)は、標的遺伝子の発現を調節する転写因子として機能する核受容体スーパーファミリーに属する。他の転写因子とは異なり、核受容体の活性は、生物学的膜に容易に浸透する対応するリガンド−親水性小分子に結合することにより調節され得る。核受容体ファミリーについての複雑な細胞シグナル経路にもかかわらず、核受容体を薬物標的として使用する長い成功の歴史がある。PPARは細胞分化、発達及び代謝の調節において不可欠な役割を果たす。PPARは、今までに同定された別々の遺伝子によりコードされる3つの密接に関連したアイソフォームを有する:一般的にPPARα、PPARγ及びPPARδとして知られ、PPARβとしても知られる;J.Berger and D.E.Miller,Annu.Rev.Med.,2002,53,409−435)。それぞれの受容体サブタイプはシグニチャー(signature)DNA結合ドメイン(DBD)及びリガンド結合ドメイン(LBD)を有し、両方ともリガンド活性化遺伝子発現に必要である。PPARはレチノイドX受容体(RXR)とヘテロ二量体として結合する。
【0113】
PPARδはCNSにおいてかなり発現されるように思われる;しかしそこでのその機能の多くはまだ不確定なままである。しかし、1つだけ興味がもたれるのは、PPARδがCNSの主要な脂質産生細胞である齧歯動物のオリゴデンドロサイトにおいて発現されたという発見である(J.Granneman,et al.,J.Neurosci.Res.,1998,51,563−573)。さらに、PPARδ選択的アゴニストがマウス飼育(cultures)においてオリゴデンドログリアミエリン遺伝子発現及びミエリン鞘径を有意に増加させるということが見出されたということもわかった(I.Saluja et al.,Glia,2001,33,194−204)。PPARδノックアウトマウスは、より小さな脳全体のサイズ及び白質における減少したレベルの髄鞘形成を有する(Mol cell Biology 200 20:5119)。さらに、PPARδアゴニストは、多発性硬化症の実験的自己免疫性脳脊髄炎(EAE)モデルにおいて保護的効果を及ぼす(Polak et al.,J Neuroimmunology 2005 168:65−75)。
【0114】
共同研究者らは、選択的PPARδアゴニストが神経組織において機能的役割を果たし、そしてオリゴデンドロサイト前駆細胞分化を刺激するということを以前に実証した。経口で生物が利用可能な脳浸透性のPPARδ選択的アゴニストであるSAR117145は、齧歯動物及びヒトのオリゴデンドロサイト前駆細胞分化をインビトロで濃度依存性の様式で刺激した;PPARδ選択的アンタゴニストでブロックされ得る効果。
【0115】
ラットオリゴデンドロサイトにおいて、ミエリン塩基性タンパク質の増加した発現に下流PPAR標的Angptl4の増加したmRNA発現が先行し、そしてこの上方調節はPPARδのレンチウイルスshRNAノックダウンでブロックされた。マウスに2%クプリゾンの食餌を4種間与えた急性脱髄のマウスクプリゾンモデルにおいて、SAR117145はCNS再髄鞘化を増強し、Angptl4 mRNA発現を増加させ、そして脳梁を横切る軸索伝導を改善した。Angptl4のCNS活性化には腓腹筋の増加した発現が付随し、これが潜在的な代替マーカーとして役立ち得ると示唆される。これらのデータは、PPARδアゴニストがCNS再髄鞘化を増強し、そして軸索機能を改善し得ることを実証し、そして脱髄障害の処置のための内在性修復プロセスの刺激におけるそれらの潜在的用途を示唆する(米国特許出願第20070149580号、USE OF PEROXISOME PROLIFERATOR ACTIVATED RECEPTOR DELTA AGONISTS FOR THE TREATMENT OF MS AND OTHER DEMYELINATING DISEASES)。
【0116】
PPARδアゴニストツール化合物(’517;図15)を、B6C3H系171ヘテロ接合マウスで試験した。8週齢のマウスを0.2%クプリゾンを含む食餌で4週間飼育し、次いで再髄鞘化のために通常餌を与えた。次いでマウスに、週に2回、ビヒクル(0.6%カルボキシメチルセルロースナトリウム塩及び0.5%Tween80)又は30mg/kg PPARδアゴニストツール化合物’517のいずれかを8日間経口投与し、そしてグラフに示された時点で画像化した。データを0週ベースラインシグナルに対して正規化した。ツール化合物’517のグループ(n=15)ではビヒクルグループ(n=13)よりも30〜100%の相対的なルシフェラーゼシグナル増加があり、これはオリゴデンドロサイト前駆細胞分化の増強に起因すると本発明者らは考えている。
【0117】
’517の効果を、同じ研究においてルクソールファストブルー(LFB)染色によりさらに組織学的に確認した(図16)。ホルマリン固定パラフィン包埋脳からの傍矢状組織断面を、脳梁におけるミエリンの定性的評価のためにLFBで染色した。各時点について染色した断面をスコア付けし、そして0(完全な髄鞘形成)から5(完全な脱髄)までのスケールで等級付けした。スコア付けシステムは以下のとおりであった:0=正常ミエリン、脱髄無し、1=最小限の局在化した脱髄、2=軽度から中程度、局在化した脱髄、3=中程度、局所的に極度の脱髄、4=重症、局所的に極度の脱髄、5=重症、広範な脱髄。クプリゾンで4週後のマウスからのLFB染色した脳断面の組織学的評価は、系171ヘテロ接合マウス(n=5)における脳梁の中程度から重症の脱髄を裏付けた。再髄鞘化期の間の4週から5週目の’517での処置は、ツール化合物’517グループ(N=5)をビヒクルコントロールグループ(N=3)と比較して、7週目の時点でLFBにより決定した場合に測定可能な増加を生じた。少ないnの数にもかかわらず、組織学的データはインビボルシフェラーゼバイオイメージングデータを支持しており、ビヒクルコントロールと比較した場合に’517化合物で処置されたマウスにおける増加した再髄鞘化を示した。
【0118】
系171ヘテロ接合マウスを用いた第二のPPARδ 517研究は同様の結果を示した(データは示しておらず、そしてまた’517処置を3週延長した)。第一の研究(図15)と一致して、第二の研究もまた、’517がオリゴデンドロサイト前駆細胞分化を再髄鞘化プロセスの間の初期の回復期に加速したということを示唆する(M Lindner and S Heine,et al,Neuropathology and Applied Neurobiology,34,105−114,2008)。
【0119】
2.MBP−luciマウスはERβアゴニスト、’5a又はポジティブコントロール化合物QTPのミエリン発現に対する保護効果を検出する
エストロゲン受容体(ER)は、特異的応答エレメントによりDNAに対して直接的に作用し、そして遺伝子発現を調節する、ステロイド核受容体のファミリーに属する。ERの2つのサブタイプ、ERα及びERβが存在する。ERαは子宮、前立腺、卵巣、骨、乳房及び脳において高度に発現され、一方ERβは、結腸、前立腺、卵巣、骨髄及び脳に存在する。ERβの選択的標的化は、ERαの副作用を避けるための魅力的な治療アプローチである。ERサブタイプ選択的化合物が同定されている。
【0120】
例えば、ERβ(ERαではない)アゴニストは、オリゴデンドロサイト及び神経芽細胞腫細胞のアポトーシスをインビトロで保護することが示されている。さらに、ERβ又はER−アゴニストはEAEを寛解させ、そして神経保護効果を有する(ミエリン及び軸索の保存)。以前のPPARδアゴニストツール化合物’517に基づいて、MBP−luci系を、ERβアゴニスト、’5aをクプリゾンモデルにおいて概略を描く(profile)ために使用した。さらに、本発明者らは、AstraZenecaの統合失調症薬クエチアピン(10mg/kg、PO、qd)を、クプリゾン誘導脱髄に対するその保護効果(Schizophrenia Research,2008 Dec 106,182−91)に基づいてポジティブコントロールとして含めた。2つの独立した研究をB6CH系171ヘテロ接合マウスを用いて行った。
【0121】
第一の研究(図17)は、ERβアゴニスト‘5a及びポジティブコントロールクエチアピン(QTP)が、クプリゾンモデルにおいて脱髄期の間保護的であることを示した。MBP−luc系171ヘテロ接合B6C3Hマウスに、クプリゾンの食餌を4週間与え、そして’5a(10mg/kg、N=12又は30mg/kg N=16)又はQTP(10mg/kg、N=14)を経口投薬した。マウスを0週目(ベースライン)、3週目、及び4週目に画像化し、データを0週目のベースラインに対して正規化した。以前の研究(図15)と同様に、ビヒクルグループは49%(3週)及び45%(4週)のバイオイメージングシグナル減少を生じた。QTPグループは、10mg/kgで3週及び4週の両方の時点についてビヒクルコントロールに対して画像化シグナルにおいて有意な増加を示した。QTP処置グループについての結果は、Zhangらにより公開されたデータ(Schizophrenia Research,2008,106:182−91)と一致する。化合物’5aは30mg/kgで4週目にビヒクルに勝る有意な増加を示したが、10mg/kgでは3週でも4週でも有意な差異を示さなかった。これらの結果は、MBP−luci画像化モデルをクプリゾンモデルにおける用量依存性保護効果を評価するために使用することができるということを示唆する。
【0122】
さらなる研究(図18)を、ビヒクルについてのより大きなコホート及び’5a化合物30mg/kg処置グループを用いたこと以外は、第一の研究の結果(図17)を確認するために設計した。第一の研究の結果と一致して、マウスをQTPで10mg/kgで処置することにより(N=17)、ビヒクルで処置されたコントロール(N=25)と3週目(25%対47%の減少、p=0.028)及び4週目(6%の増加対25%の減少)に比較した場合にクプリゾン誘導シグナル減少の統計学的に有意な阻害をもたらした。さらに、30mg/kgでの’5a(N=27)は、ビヒクルコントロールグループと3週目(31%対47%の減少、p=0.0079)及び4週目(6%減少対25%減少、p=0.0015)に比較して、有意に高い導入遺伝子活性を生じた。
【0123】
これらの二つの研究は、30mg/kgでの‘5aはCNSのバイオイメージングシグナルにおけるクプリゾン食餌に誘導される減少を有意に防止するが、10mg/kgのポジティブコントロールQTPと同じ程度ではない(これらの実験において使用された条件下で)。以前の研究がCNS髄鞘形成の程度とMBP−luciバイオイメージングシグナルとの間の直接的な関係を示しているので、これらの現在の結果は、’5a及びQTPの両方がクプリゾン餌投与の間のCNSにおける脱髄を防止するということを示唆する。画像化モデルデータは、QTP及び’5aの両方がクプリゾン誘導脳脱髄及びミエリン破壊を軽減するということを支持する。
【0124】
3.クプリゾンモデルにおけるMBP−luci系の比較
様々な系統バックグラウンドの6つのトランスジェニック系を、異なるバイオイメージング適用のために生成した。
【0125】
クプリゾンモデルにおけるB6C3H系171ホモ接合マウス及びヘテロ接合画像化シグナル(図19)を比較した。ホモ接合体におけるレポーター遺伝子の2つのコピーは、脱髄期の間に2倍より大きいシグナル減少を示し、そして再髄鞘化期の間に2倍のシグナル増加を示した。ヘテロ接合系171(B6C3H系統)がクプリゾンモデルにおいて機能し、化合物効果を検出し得るということが実証されるが、バイオイメージングシグナル強度を増大させるためにホモ接合性に繁殖させることによりこのモデルをさらに改善し得るということが予測される。マウスコロニーはホモ接合体として維持され得るので、これはまた、モデル製造を効率化して遺伝子型同定費用を減少させるだろう。さらに、画像化域が大きくなれば、モデルは化合物誘導変化の検出に対してより高感度になる。
【0126】
図20は、クプリゾンモデルにおける3つの異なる系からの組織学的LFBデータの比較を示す。定量的LFBデータにより、ホモ接合マウスが最も重症なクプリゾン誘導脱髄(典型的に使用される野生型C57 BL/6マウス系統に見られるものと同様の重症度)を示したという、系171 B6C3Hを用いたバイオイメージング結果が裏付けられた。系171ヘテロ接合マウス系はより低い重症度の脱髄を示したが、これらの系171ヘテロ接合マウスは最も少ない量のクプリゾン誘導脱髄を示した。
【0127】
図21において、最大のバイオイメージングシグナル減少を有するMBP−luci系が、組織学的に評価された場合に最も高い脱髄も有していたことを実証する。3つの異なるMBP−luci系(系171 B6C3Hヘテロ接合(het)系統、系121 C57BL/6ヘテロ接合系統及び系171B6C3Hホモ接合系統)をバイオイメージング及びルクソールファストブルー(LFB;ミエリン染色)組織学により比較した。マウスを4週間又は5週間、0.2%クプリゾンを含有する食餌で飼育した。画像化データを0週のベースライン測定に対して正規化した。各研究の終わりに、マウス脳を採取し、そして連続的パラフィン切片をLFBを用いてミエリンについて染色した。平均定性的LFBスコア(0〜5)を図21の表に示した。系171 B6C3Hホモ接合マウスは、最大の画像化シグナル減少を示し、そしてまた定性的組織学的評価により評価した場合に最も重症な脱髄を実証した。系171 B6C3Hヘテロ接合マウスは、最も小さな画像化域、及び4週目に最も少ない組織学的脱髄も示した。系171 B6C3Hホモ接合マウスは、クプリゾン食餌前のそれらのベースライン画像化を参照して、クプリゾン食餌の間に最も大きなバイオイメージングシグナル減少を有していた。例えば、クプリゾン食餌で3週後に、171 B6C3Hホモ接合マウスは72%のシグナル減少(0週を参照して3週、p<0.05)を有していたが、系171 B6C3Hヘテロ接合マウスは45%のバイオイメージングシグナル減少(0週を参照して、p<0.05)を有していた。系121 C57 BL/6ヘテロ接合マウスは、ルシフェラーゼシグナルにおける最少の減少を有していた(0週に対して3週で33%の減少、p<0.05)。
【0128】
MBP−luciモデルの感受性及び応答性を、図22に示される171 B6C3Hホモ接合マウスのQTP(10mg/kg)を用いた処置によりさらに確認した。171 B6C3Hヘテロ接合マウス(図17及び18)を使用した結果と一致して、QTP(10mg/kg)はバイオイメージングシグナル減少を有意に防止した。これらの結果に基づいて、系171 B6C3Hホモ接合マウスをクプリゾン誘導脱髄バイオイメージングモデルのための最適な系として同定した。
【0129】
4.MBP−luciモデルのさらなる適用
MBP−luciマウスは脳及び脊髄の両方ルシフェラーゼ発現を有する。図23に示されるように、発光シグナルは主に脳及び脊髄の白質領域からのものであった。クプリゾンモデル適用について首尾よく実証された脳からのルシフェラーゼ画像化に加えて、脊髄からのルシフェラーゼ画像化シグナルは、MSの実験的アレルギー性脳脊髄炎(EAE)において使用され得、又は脊髄モデルとして応用され得る。
【特許請求の範囲】
【請求項1】
生きている生物において脱髄又は再髄鞘化をモニタリングする方法であって:
ミエリン塩基性タンパク質(MBP)プロモーターにより駆動されるルシフェラーゼ遺伝子を発現するように該生物を遺伝子導入で改変する工程;
該生物からの生物発光をモニタリングする工程;及び
該光の出力を該生物の身体の特定の部分に関連付ける工程
を含む、上記方法。
【請求項2】
モデル生物が哺乳動物である、請求項1に記載の方法。
【請求項3】
哺乳動物がマウスである、請求項2に記載の方法。
【請求項4】
同じ生物において、最初のモニタリングとは別の時点でモニタリングを繰り返す工程;及び
同じ生物における髄鞘形成の変化を観察するために該モニタリング事象の出力を比較する工程
をさらに含む、請求項1に記載の方法。
【請求項5】
脱髄間隔又は再髄鞘化間隔を通してのシグナル正規化を画像化する工程をさらに含み、ここで該間隔を通しての画像化は、該間隔に亘る該生物におけるMBP転写物のレベルの変化を検出するために効果的である、
請求項4に記載の方法。
【請求項6】
ミエリン塩基性タンパク質(MBP)プロモーターにより駆動されるルシフェラーゼ遺伝子を含むトランスジェニック生物。
【請求項7】
前記生物がマウスである、請求項6に記載のトランスジェニック生物。
【請求項8】
ミエリン塩基性タンパク質(MBP)プロモーターの制御下にあるルシフェラーゼ遺伝子を発現するトランスジェニック生物。
【請求項9】
前記モデル生物がマウスである、請求項8に記載のトランスジェニック動物。
【請求項10】
前記MBPプロモーターがM1〜M4を含む、請求項6に記載のトランスジェニック生物。
【請求項11】
前記MBPプロモーターがM1〜M3を含む、請求項6に記載のトランスジェニック生物。
【請求項12】
前記生物が哺乳動物であり、該哺乳動物が白髪又は減少した毛髪を有する、請求項1に記載の方法。
【請求項13】
前記哺乳動物の毛髪が530〜640nmの波長範囲内で吸収する光はC57/B6よりも少ない、請求項12に記載の方法。
【請求項14】
バイオイメージング系統を発生させるための方法であって:
(1) 生後髄鞘形成のピークにおけるインビボマウス全身画像化がCNS特異的画像化を示す系を選択する工程;
(2) エクスビボ画像化により、ルシフェラーゼ導入遺伝子発現が主に脳又は脊髄の白質領域にあることが確認される系を選択する工程;
(3) ルシフェラーゼ画像強度が、脱髄モデルにおいて誘導された脱髄及び再髄鞘化における変化と高度に相関している系を選択する工程;及び
(4) 明瞭な組織学的脱髄を示す系をモデルとして選択する工程
を含む、上記方法。
【請求項15】
前記生後髄鞘形成のピークが約3〜5週のG1マウスである、請求項14に記載の方法。
【請求項16】
モデルがクプリゾン脱髄モデルである、請求項14に記載の方法。
【請求項17】
少なくとも1つの脱髄または再髄鞘化事象に対する化合物の効果をモニタリングする、請求項1に記載の方法。
【請求項18】
遺伝子事象の効果をモニタリングする、請求項1に記載の方法。
【請求項19】
髄鞘形成のモニタリングのための改善された方法であって、該改善がMBP−ルシフェラーゼを発現するトランスジェニック動物を画像化する工程を含む、方法。
【請求項20】
髄鞘形成研究における変動性を低減させる方法であって、MBP制御ルシフェラーゼを動物において少なくとも2つの異なる時点においてモニタリングする工程を含む、上記方法。
【請求項21】
前記生物の身体の特定の部分が、末梢神経系及び中枢神経系からなる群より選択される、請求項1に記載の方法。
【請求項1】
生きている生物において脱髄又は再髄鞘化をモニタリングする方法であって:
ミエリン塩基性タンパク質(MBP)プロモーターにより駆動されるルシフェラーゼ遺伝子を発現するように該生物を遺伝子導入で改変する工程;
該生物からの生物発光をモニタリングする工程;及び
該光の出力を該生物の身体の特定の部分に関連付ける工程
を含む、上記方法。
【請求項2】
モデル生物が哺乳動物である、請求項1に記載の方法。
【請求項3】
哺乳動物がマウスである、請求項2に記載の方法。
【請求項4】
同じ生物において、最初のモニタリングとは別の時点でモニタリングを繰り返す工程;及び
同じ生物における髄鞘形成の変化を観察するために該モニタリング事象の出力を比較する工程
をさらに含む、請求項1に記載の方法。
【請求項5】
脱髄間隔又は再髄鞘化間隔を通してのシグナル正規化を画像化する工程をさらに含み、ここで該間隔を通しての画像化は、該間隔に亘る該生物におけるMBP転写物のレベルの変化を検出するために効果的である、
請求項4に記載の方法。
【請求項6】
ミエリン塩基性タンパク質(MBP)プロモーターにより駆動されるルシフェラーゼ遺伝子を含むトランスジェニック生物。
【請求項7】
前記生物がマウスである、請求項6に記載のトランスジェニック生物。
【請求項8】
ミエリン塩基性タンパク質(MBP)プロモーターの制御下にあるルシフェラーゼ遺伝子を発現するトランスジェニック生物。
【請求項9】
前記モデル生物がマウスである、請求項8に記載のトランスジェニック動物。
【請求項10】
前記MBPプロモーターがM1〜M4を含む、請求項6に記載のトランスジェニック生物。
【請求項11】
前記MBPプロモーターがM1〜M3を含む、請求項6に記載のトランスジェニック生物。
【請求項12】
前記生物が哺乳動物であり、該哺乳動物が白髪又は減少した毛髪を有する、請求項1に記載の方法。
【請求項13】
前記哺乳動物の毛髪が530〜640nmの波長範囲内で吸収する光はC57/B6よりも少ない、請求項12に記載の方法。
【請求項14】
バイオイメージング系統を発生させるための方法であって:
(1) 生後髄鞘形成のピークにおけるインビボマウス全身画像化がCNS特異的画像化を示す系を選択する工程;
(2) エクスビボ画像化により、ルシフェラーゼ導入遺伝子発現が主に脳又は脊髄の白質領域にあることが確認される系を選択する工程;
(3) ルシフェラーゼ画像強度が、脱髄モデルにおいて誘導された脱髄及び再髄鞘化における変化と高度に相関している系を選択する工程;及び
(4) 明瞭な組織学的脱髄を示す系をモデルとして選択する工程
を含む、上記方法。
【請求項15】
前記生後髄鞘形成のピークが約3〜5週のG1マウスである、請求項14に記載の方法。
【請求項16】
モデルがクプリゾン脱髄モデルである、請求項14に記載の方法。
【請求項17】
少なくとも1つの脱髄または再髄鞘化事象に対する化合物の効果をモニタリングする、請求項1に記載の方法。
【請求項18】
遺伝子事象の効果をモニタリングする、請求項1に記載の方法。
【請求項19】
髄鞘形成のモニタリングのための改善された方法であって、該改善がMBP−ルシフェラーゼを発現するトランスジェニック動物を画像化する工程を含む、方法。
【請求項20】
髄鞘形成研究における変動性を低減させる方法であって、MBP制御ルシフェラーゼを動物において少なくとも2つの異なる時点においてモニタリングする工程を含む、上記方法。
【請求項21】
前記生物の身体の特定の部分が、末梢神経系及び中枢神経系からなる群より選択される、請求項1に記載の方法。
【図1】

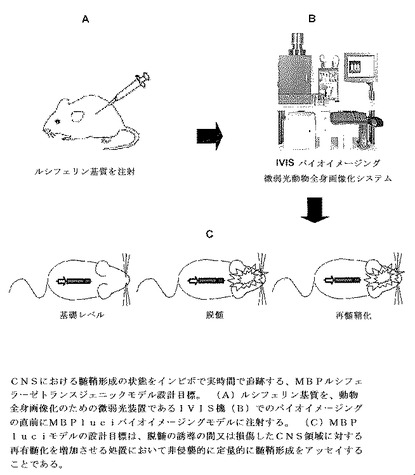
【図2】

【図3】
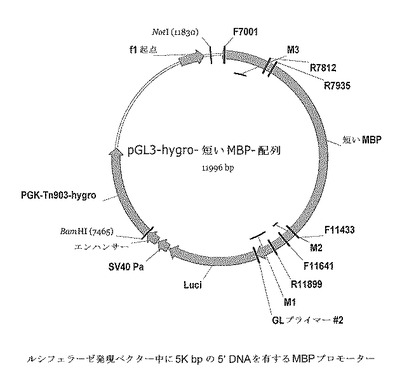
【図4】
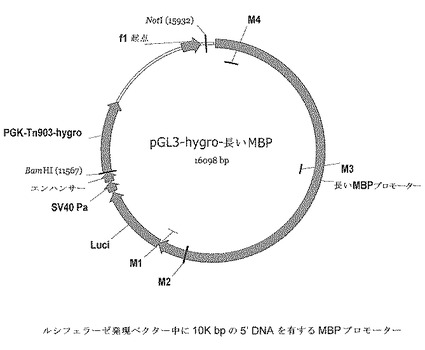
【図5】
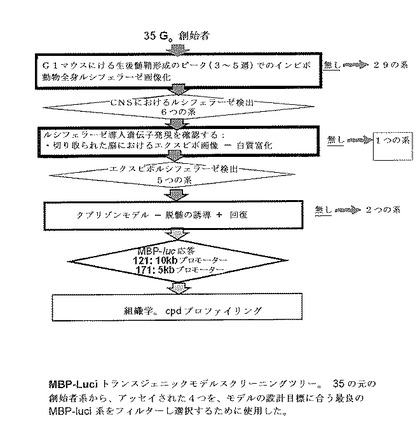
【図11】

【図12】
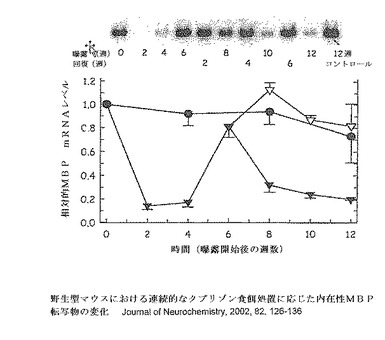
【図15】
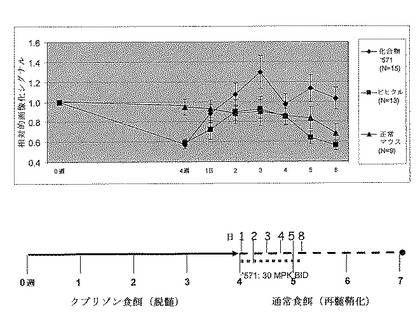
【図16】
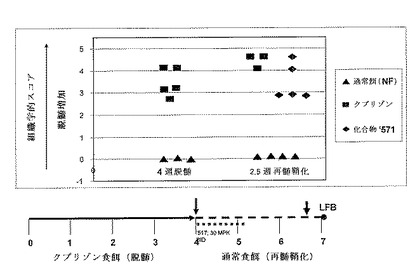
【図17】
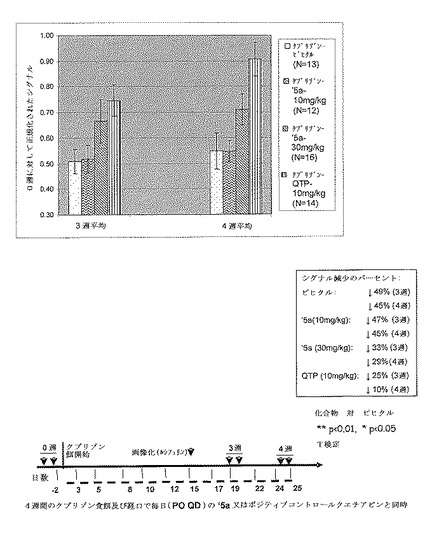
【図18】
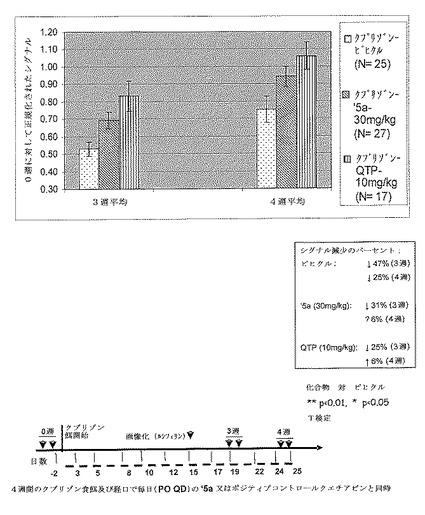
【図19】
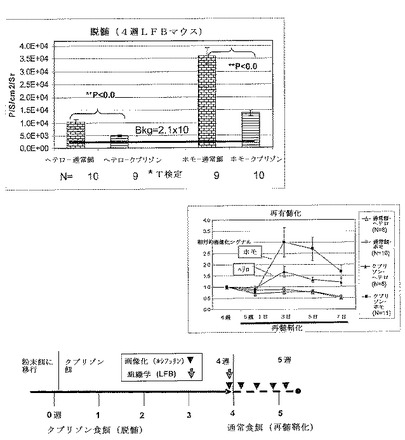
【図21】
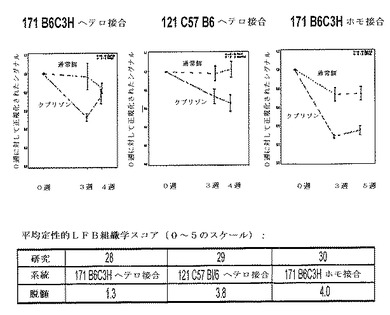
【図22】
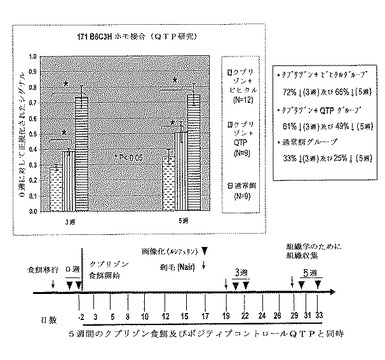
【図24】
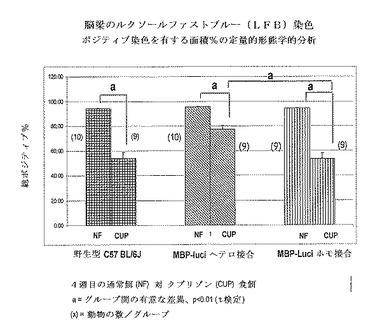
【図6】
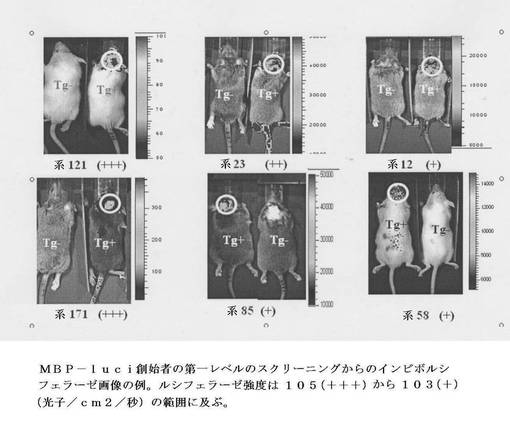
【図7】
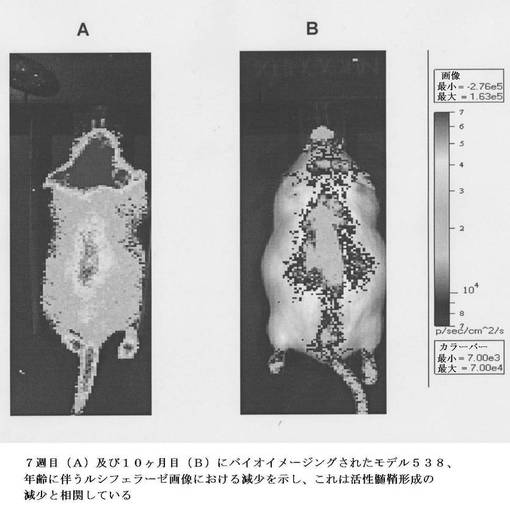
【図8】
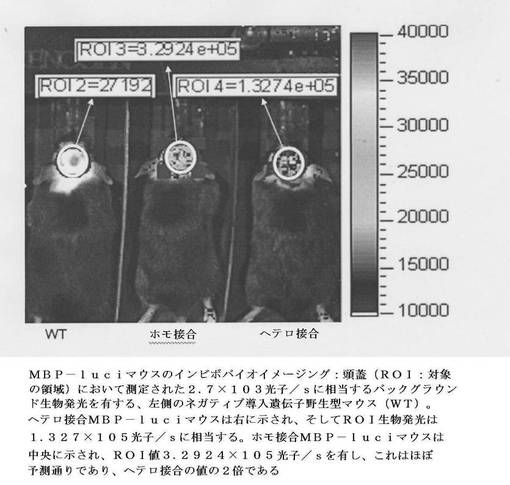
【図9】
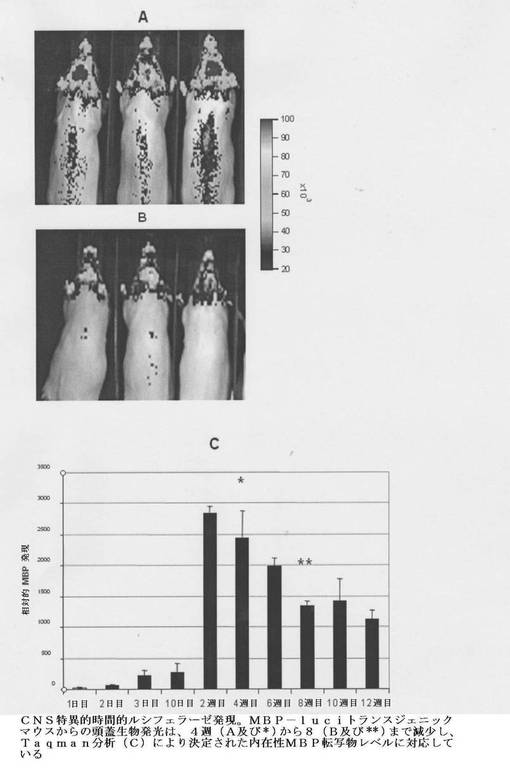
【図10】
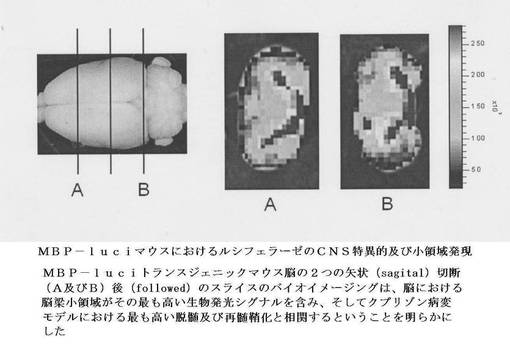
【図13】
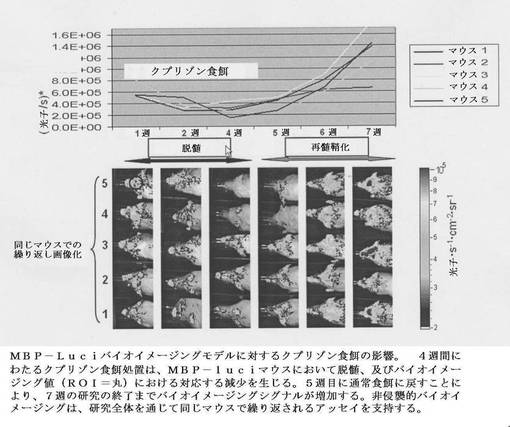
【図14】
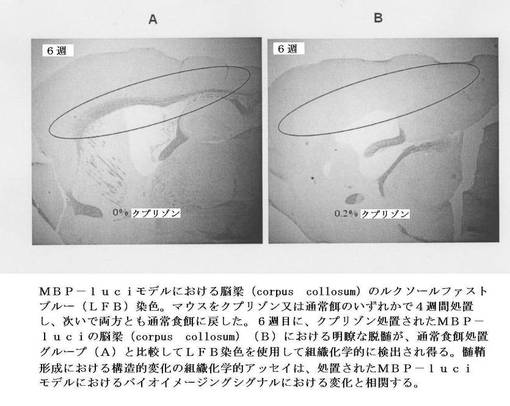
【図20】
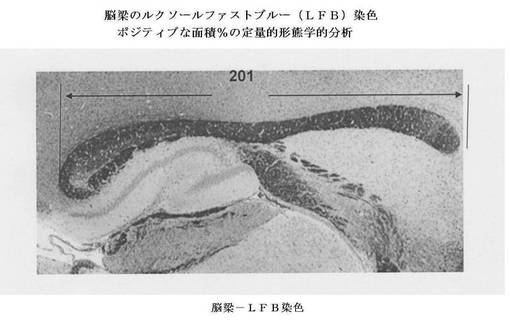
【図23】

【図2】
【図3】
【図4】
【図5】
【図11】
【図12】
【図15】
【図16】
【図17】
【図18】
【図19】
【図21】
【図22】
【図24】
【図6】
【図7】
【図8】
【図9】
【図10】
【図13】
【図14】
【図20】
【図23】
【公表番号】特表2013−514534(P2013−514534A)
【公表日】平成25年4月25日(2013.4.25)
【国際特許分類】
【出願番号】特願2012−544586(P2012−544586)
【出願日】平成22年12月3日(2010.12.3)
【国際出願番号】PCT/US2010/058823
【国際公開番号】WO2011/084281
【国際公開日】平成23年7月14日(2011.7.14)
【出願人】(504456798)サノフイ (433)
【Fターム(参考)】
【公表日】平成25年4月25日(2013.4.25)
【国際特許分類】
【出願日】平成22年12月3日(2010.12.3)
【国際出願番号】PCT/US2010/058823
【国際公開番号】WO2011/084281
【国際公開日】平成23年7月14日(2011.7.14)
【出願人】(504456798)サノフイ (433)
【Fターム(参考)】
[ Back to top ]