
ミトコンドリア標的化抗腫瘍剤
ミトコンドリア標的化抗腫瘍剤、ならびに望ましくない細胞増殖に関連する障害の処置のためにそれらを作製および使用する方法が記載される。一つの局面において、本発明は、式A-Bを有する組成物、または薬学的に許容されるその塩を提供し、式中、Aは分子シャペロン阻害剤であり、Bはミトコンドリア浸透性部分であり、AとBとは任意で連結部分によって連結している。

Notice: Undefined index: DEJ in /mnt/www/gzt_disp.php on line 298
【特許請求の範囲】
【請求項1】
下記式を含む組成物、または薬学的に許容されるその塩:
A-B
式中、Aは分子シャペロン阻害剤であり、Bはミトコンドリア浸透性部分であり、AとBとは任意で連結部分によって連結しており、ここで、Aがシェファーディン(shepherdin)またはその断片である場合、BはアンテナペディアヘリックスIIIホメオドメイン細胞浸透性ペプチド(ANT)またはその断片ではない。
【請求項2】
Aが、アンサマイシンクラスHsp90阻害剤; ゲルダナマイシン類似体Hsp90阻害剤; プリン骨格クラスHsp90阻害剤; レゾルシノールHsp90阻害剤; およびマクロラクトン-Hsp90阻害剤からなる群より選択される小分子である、請求項1記載の組成物。
【請求項3】
AがHsp90のペプチド阻害剤である、請求項1記載の組成物。
【請求項4】
Aが、SEQ ID NO:2(His-Ser-Ser-Gly-Cys)を含むシェファーディンペプチドである、請求項1記載の組成物。
【請求項5】
Aが、Hsp90に結合しかつそれを阻害するSEQ ID NO:1と少なくとも95%同一の配列を含むペプチドである、請求項3記載の組成物。
【請求項6】
Bがミトコンドリア浸透性ペプチドである、請求項1記載の組成物。
【請求項7】
Bが、マイトフュージンペプチド、ミトコンドリア標的化シグナルペプチド、TATペプチド、ANTペプチド、VP22ペプチドまたはPep-1ペプチドからなる群より選択される、請求項6記載の組成物。
【請求項8】
Bが、RNAミトコンドリア浸透性シグナルである、請求項1記載の組成物。
【請求項9】
Bが、グアニジンリッチペプトイド、グアニジンリッチポリカルバメート、β-オリゴアルギニンおよびプロリンリッチデンドリマーからなる群より選択される、請求項1記載の組成物。
【請求項10】
Aが17-アリルアミノ-デメトキシゲルダマイシン(17-AAG)を含む、請求項1記載の組成物。
【請求項11】
Aがラジシコールを含む、請求項1記載の組成物。
【請求項12】
Aがプリン骨格クラスHsp90阻害剤を含む、請求項1記載の組成物。
【請求項13】
Aが、17-ジメチルアミノゲルダナマイシンを含む、請求項1記載の組成物。
【請求項14】
Bが以下である、請求項1記載の組成物:
式中、
R1はH、アルキル、アルケニル、アルキニル、ハロアルキル、アリール、アリールアルキルまたはRaRbRcSiであり;
Ra、RbおよびRcはアルキルまたはアリールより独立して選択され;
nは0、1、2、3、4、5または6である。
【請求項15】
Bが以下である、請求項1記載の組成物:
式中、
Ra、RbおよびRcはアルキルまたはアリールより独立して選択され;
nは1、2または3である。
【請求項16】
Bが(アリール)3P-である、請求項1記載の組成物。
【請求項17】
Bが
である、請求項1記載の組成物。
【請求項18】
Aが以下である、請求項1記載の組成物:
式中、
R2はH、アルキル、アリールまたはアリールアルキルであり; R3はH、アルキルであり; R4はH、アルキル、アルケニル、アリール、アリールアルキル、ORdであり、ここでRdはH、アルキルまたはアリールアルキルである。
【請求項19】
R2がHまたはアルキルであり; R3がH、アルキルであり; R4がHまたはORdであり、ここでRdがH、アルキルである、請求項18記載の組成物。
【請求項20】
R2がHであり; R3がメチルであり; R4がHである、請求項18記載の組成物。
【請求項21】
BがANTまたはそのミトコンドリア浸透性断片を含む、請求項10〜13のいずれか一項記載の組成物。
【請求項22】
AとBとの間に連結部分を含む、請求項1記載の組成物。
【請求項23】
連結部分がペプチドリンカーおよび化学リンカーからなる群より選択される、請求項22記載の組成物。
【請求項24】
リンカー部分が二価でありかつアルキレン、アルケニレン、アルキニレン、シクロアルキレン、アリーレン、ヘテロアリーレンおよびペプチドリンカーからなる群より選択され、ここで該アルキレン、アルケニレンまたはアルキニレンの任意の2個の隣接する炭素-炭素結合が、O、NH、S、PRe、C(O)NRf、アリーレン、ヘテロシクロアルキレンまたはヘテロアリーレンのうち1つまたは複数で置き換えられていてもよく; ここでReおよびRfがアルキルまたはアリールより独立して選択される、請求項1記載の組成物。
【請求項25】
リンカー部分が
である、請求項1記載の組成物。
【請求項26】
リンカー部分がアルキレンである、請求項1記載の組成物。
【請求項27】
リンカー部分が6個の炭素原子を有するアルキレンである、請求項26記載の組成物。
【請求項28】
A-Bが以下である、請求項1記載の組成物、または薬学的に許容されるその塩:
式中、R1はH、アルキル、アルケニル、アルキニル、ハロアルキル、アリール、アリールアルキルまたはRaRbRcSiであり;
R2はH、アルキル、アリールまたはアリールアルキルであり; R3はH、アルキルであり; R4はH、アルキル、アルケニル、アリール、アリールアルキル、ORdであり、ここでRdはH、アルキルまたはアリールアルキルであり;
Ra、RbおよびRcはアルキルまたはアリールより独立して選択され;
nは包含的に1〜10の整数である。
【請求項29】
塩がヘキサフルオロリン酸塩である、請求項28記載の組成物。
【請求項30】
R1がRaRbRcSiであり、ここでRa、RbおよびRcがアルキルまたはアリールより独立して選択され; R2がHであり; R3がH、アルキルであり; R4がHであり; nが1、2、3または4である、請求項26記載の組成物。
【請求項31】
A-Bが
より選択される、請求項1記載の組成物、または薬学的に許容されるその塩。
【請求項32】
塩がヘキサフルオロリン酸塩である、請求項31記載の組成物。
【請求項33】
A-Bが以下である、請求項1記載の組成物:
式中、qは1、2、3、4、5または6であり; Xは薬学的に許容される対イオンである。
【請求項34】
qが3である、請求項33記載の組成物。
【請求項35】
アリールがフェニルであり、qが3である、請求項33記載の組成物。
【請求項36】
Xがヘキサフルオロリン酸塩である、請求項33記載の組成物。
【請求項37】
A-Bが
である、請求項1記載の組成物。
【請求項38】
がん細胞死を誘発するために十分な量で請求項1〜16のいずれか一項記載のミトコンドリア標的化シャペロン阻害剤を対象に投与する段階を含む、がん細胞死または腫瘍細胞死を誘発するための方法。
【請求項39】
がんまたは腫瘍を有する対象を同定する段階;
該がんまたは腫瘍の細胞が対照細胞に比べてミトコンドリアにおけるシャペロンの増加した濃度を有するか否かを決定する段階;
ミトコンドリアにおけるシャペロンの増加したレベルを対象が有する場合、式
A-B
を含むミトコンドリア標的化シャペロン阻害剤を哺乳動物に投与する段階であって、式中Aがシャペロン阻害剤であり、Bがミトコンドリア浸透性部分であり、AとBとが任意で連結部分によって連結している、段階
を含む、がん細胞死または腫瘍細胞死を誘発するための方法。
【請求項40】
少なくとも1つのシャペロンおよびシクロフィリンDを含む試料を与える段階;
試料と試験薬剤とを接触させる段階; ならびに
試験薬剤の存在および非存在下での試料中のシャペロンおよびシクロフィリンDの結合を検出する段階
を含む、シャペロン活性を阻害するための候補薬剤を同定するための方法であって、
結合を阻害する試験薬剤がシャペロン活性を阻害するための候補薬剤である、方法。
【請求項41】
シャペロンがHsp60、HspA9、Hsp90またはTRAP-1である、請求項40記載の方法。
【請求項1】
下記式を含む組成物、または薬学的に許容されるその塩:
A-B
式中、Aは分子シャペロン阻害剤であり、Bはミトコンドリア浸透性部分であり、AとBとは任意で連結部分によって連結しており、ここで、Aがシェファーディン(shepherdin)またはその断片である場合、BはアンテナペディアヘリックスIIIホメオドメイン細胞浸透性ペプチド(ANT)またはその断片ではない。
【請求項2】
Aが、アンサマイシンクラスHsp90阻害剤; ゲルダナマイシン類似体Hsp90阻害剤; プリン骨格クラスHsp90阻害剤; レゾルシノールHsp90阻害剤; およびマクロラクトン-Hsp90阻害剤からなる群より選択される小分子である、請求項1記載の組成物。
【請求項3】
AがHsp90のペプチド阻害剤である、請求項1記載の組成物。
【請求項4】
Aが、SEQ ID NO:2(His-Ser-Ser-Gly-Cys)を含むシェファーディンペプチドである、請求項1記載の組成物。
【請求項5】
Aが、Hsp90に結合しかつそれを阻害するSEQ ID NO:1と少なくとも95%同一の配列を含むペプチドである、請求項3記載の組成物。
【請求項6】
Bがミトコンドリア浸透性ペプチドである、請求項1記載の組成物。
【請求項7】
Bが、マイトフュージンペプチド、ミトコンドリア標的化シグナルペプチド、TATペプチド、ANTペプチド、VP22ペプチドまたはPep-1ペプチドからなる群より選択される、請求項6記載の組成物。
【請求項8】
Bが、RNAミトコンドリア浸透性シグナルである、請求項1記載の組成物。
【請求項9】
Bが、グアニジンリッチペプトイド、グアニジンリッチポリカルバメート、β-オリゴアルギニンおよびプロリンリッチデンドリマーからなる群より選択される、請求項1記載の組成物。
【請求項10】
Aが17-アリルアミノ-デメトキシゲルダマイシン(17-AAG)を含む、請求項1記載の組成物。
【請求項11】
Aがラジシコールを含む、請求項1記載の組成物。
【請求項12】
Aがプリン骨格クラスHsp90阻害剤を含む、請求項1記載の組成物。
【請求項13】
Aが、17-ジメチルアミノゲルダナマイシンを含む、請求項1記載の組成物。
【請求項14】
Bが以下である、請求項1記載の組成物:
式中、
R1はH、アルキル、アルケニル、アルキニル、ハロアルキル、アリール、アリールアルキルまたはRaRbRcSiであり;
Ra、RbおよびRcはアルキルまたはアリールより独立して選択され;
nは0、1、2、3、4、5または6である。
【請求項15】
Bが以下である、請求項1記載の組成物:
式中、
Ra、RbおよびRcはアルキルまたはアリールより独立して選択され;
nは1、2または3である。
【請求項16】
Bが(アリール)3P-である、請求項1記載の組成物。
【請求項17】
Bが
である、請求項1記載の組成物。
【請求項18】
Aが以下である、請求項1記載の組成物:
式中、
R2はH、アルキル、アリールまたはアリールアルキルであり; R3はH、アルキルであり; R4はH、アルキル、アルケニル、アリール、アリールアルキル、ORdであり、ここでRdはH、アルキルまたはアリールアルキルである。
【請求項19】
R2がHまたはアルキルであり; R3がH、アルキルであり; R4がHまたはORdであり、ここでRdがH、アルキルである、請求項18記載の組成物。
【請求項20】
R2がHであり; R3がメチルであり; R4がHである、請求項18記載の組成物。
【請求項21】
BがANTまたはそのミトコンドリア浸透性断片を含む、請求項10〜13のいずれか一項記載の組成物。
【請求項22】
AとBとの間に連結部分を含む、請求項1記載の組成物。
【請求項23】
連結部分がペプチドリンカーおよび化学リンカーからなる群より選択される、請求項22記載の組成物。
【請求項24】
リンカー部分が二価でありかつアルキレン、アルケニレン、アルキニレン、シクロアルキレン、アリーレン、ヘテロアリーレンおよびペプチドリンカーからなる群より選択され、ここで該アルキレン、アルケニレンまたはアルキニレンの任意の2個の隣接する炭素-炭素結合が、O、NH、S、PRe、C(O)NRf、アリーレン、ヘテロシクロアルキレンまたはヘテロアリーレンのうち1つまたは複数で置き換えられていてもよく; ここでReおよびRfがアルキルまたはアリールより独立して選択される、請求項1記載の組成物。
【請求項25】
リンカー部分が
である、請求項1記載の組成物。
【請求項26】
リンカー部分がアルキレンである、請求項1記載の組成物。
【請求項27】
リンカー部分が6個の炭素原子を有するアルキレンである、請求項26記載の組成物。
【請求項28】
A-Bが以下である、請求項1記載の組成物、または薬学的に許容されるその塩:
式中、R1はH、アルキル、アルケニル、アルキニル、ハロアルキル、アリール、アリールアルキルまたはRaRbRcSiであり;
R2はH、アルキル、アリールまたはアリールアルキルであり; R3はH、アルキルであり; R4はH、アルキル、アルケニル、アリール、アリールアルキル、ORdであり、ここでRdはH、アルキルまたはアリールアルキルであり;
Ra、RbおよびRcはアルキルまたはアリールより独立して選択され;
nは包含的に1〜10の整数である。
【請求項29】
塩がヘキサフルオロリン酸塩である、請求項28記載の組成物。
【請求項30】
R1がRaRbRcSiであり、ここでRa、RbおよびRcがアルキルまたはアリールより独立して選択され; R2がHであり; R3がH、アルキルであり; R4がHであり; nが1、2、3または4である、請求項26記載の組成物。
【請求項31】
A-Bが
より選択される、請求項1記載の組成物、または薬学的に許容されるその塩。
【請求項32】
塩がヘキサフルオロリン酸塩である、請求項31記載の組成物。
【請求項33】
A-Bが以下である、請求項1記載の組成物:
式中、qは1、2、3、4、5または6であり; Xは薬学的に許容される対イオンである。
【請求項34】
qが3である、請求項33記載の組成物。
【請求項35】
アリールがフェニルであり、qが3である、請求項33記載の組成物。
【請求項36】
Xがヘキサフルオロリン酸塩である、請求項33記載の組成物。
【請求項37】
A-Bが
である、請求項1記載の組成物。
【請求項38】
がん細胞死を誘発するために十分な量で請求項1〜16のいずれか一項記載のミトコンドリア標的化シャペロン阻害剤を対象に投与する段階を含む、がん細胞死または腫瘍細胞死を誘発するための方法。
【請求項39】
がんまたは腫瘍を有する対象を同定する段階;
該がんまたは腫瘍の細胞が対照細胞に比べてミトコンドリアにおけるシャペロンの増加した濃度を有するか否かを決定する段階;
ミトコンドリアにおけるシャペロンの増加したレベルを対象が有する場合、式
A-B
を含むミトコンドリア標的化シャペロン阻害剤を哺乳動物に投与する段階であって、式中Aがシャペロン阻害剤であり、Bがミトコンドリア浸透性部分であり、AとBとが任意で連結部分によって連結している、段階
を含む、がん細胞死または腫瘍細胞死を誘発するための方法。
【請求項40】
少なくとも1つのシャペロンおよびシクロフィリンDを含む試料を与える段階;
試料と試験薬剤とを接触させる段階; ならびに
試験薬剤の存在および非存在下での試料中のシャペロンおよびシクロフィリンDの結合を検出する段階
を含む、シャペロン活性を阻害するための候補薬剤を同定するための方法であって、
結合を阻害する試験薬剤がシャペロン活性を阻害するための候補薬剤である、方法。
【請求項41】
シャペロンがHsp60、HspA9、Hsp90またはTRAP-1である、請求項40記載の方法。
【図1】

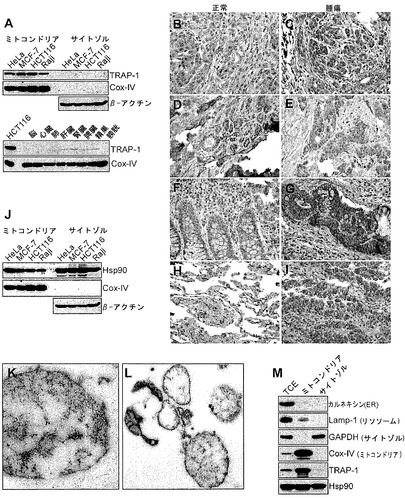
【図2】
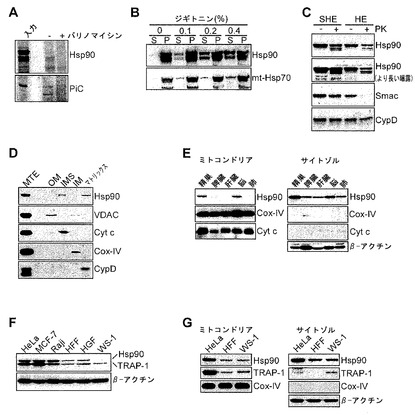
【図3】
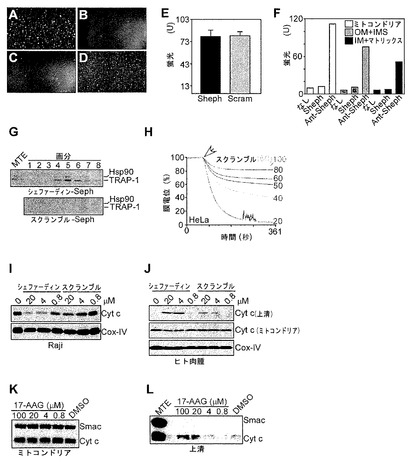
【図4】
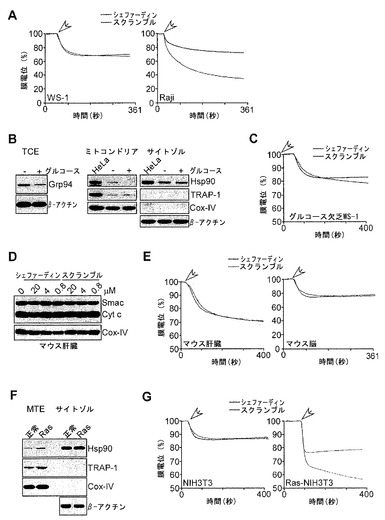
【図5】
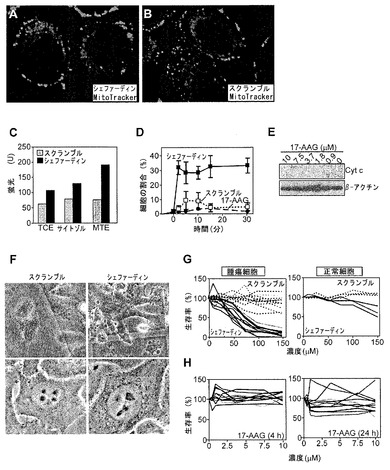
【図6】
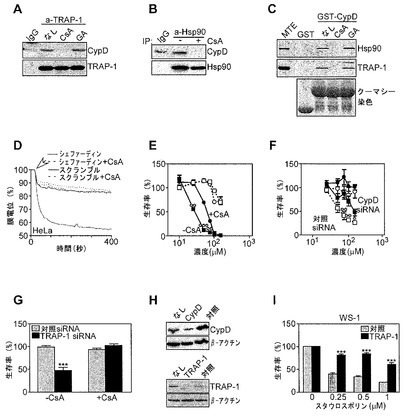
【図7】
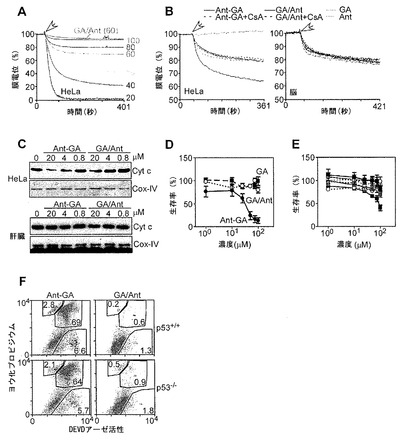
【図8】
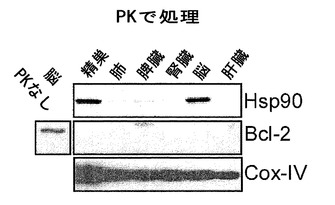
【図9】
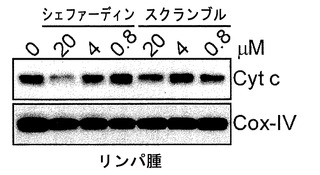
【図10】
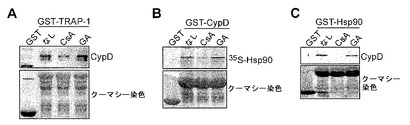
【図11】
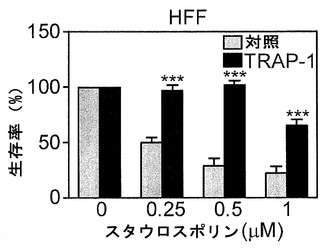
【図12】
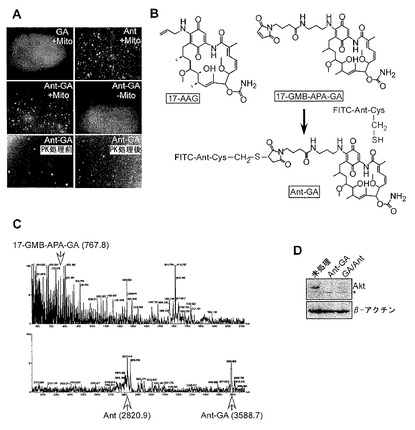
【図13】
【図14−1】

【図14−2】

【図14−3】

【図15】
【図16】
【図17】
【図18】
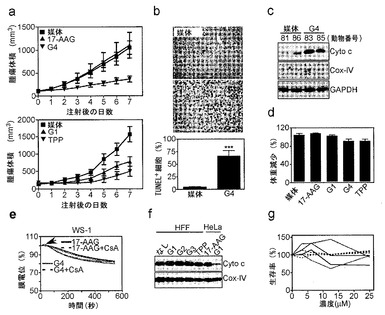
【図19】
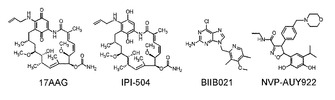
【図20】
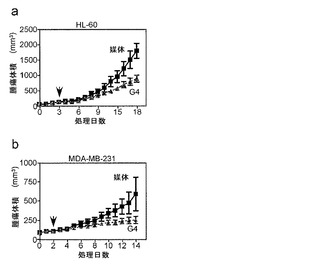
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14−1】
【図14−2】
【図14−3】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【公表番号】特表2011−501731(P2011−501731A)
【公表日】平成23年1月13日(2011.1.13)
【国際特許分類】
【出願番号】特願2010−524247(P2010−524247)
【出願日】平成20年9月10日(2008.9.10)
【国際出願番号】PCT/US2008/075895
【国際公開番号】WO2009/036092
【国際公開日】平成21年3月19日(2009.3.19)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.WINDOWS
【出願人】(505231659)ユニバーシティ オブ マサチューセッツ (23)
【Fターム(参考)】
【公表日】平成23年1月13日(2011.1.13)
【国際特許分類】
【出願日】平成20年9月10日(2008.9.10)
【国際出願番号】PCT/US2008/075895
【国際公開番号】WO2009/036092
【国際公開日】平成21年3月19日(2009.3.19)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.WINDOWS
【出願人】(505231659)ユニバーシティ オブ マサチューセッツ (23)
【Fターム(参考)】
[ Back to top ]