
ミトコンドリアDNA可変領域塩基配列による水産魚介類の標識および識別のための方法
【課題】
本発明の課題は、水産魚介類、特に、特定の産地に由来する水産魚介類、一つまたは複数の優良形質を有する優良水産種苗、または特定の品種の水産魚介類を、標識・識別するための簡易かつ正確な方法を提供することである。
【解決手段】
したがって、本発明は、ミトコンドリアDNA D−ループの塩基配列を用いて、一個体または個体群の水産魚介類もしくはその一部またはそれらの加工品の生産履歴を追跡する方法、およびかかる方法に用いられるマーカー遺伝子を決定する方法を提供する。
本発明の課題は、水産魚介類、特に、特定の産地に由来する水産魚介類、一つまたは複数の優良形質を有する優良水産種苗、または特定の品種の水産魚介類を、標識・識別するための簡易かつ正確な方法を提供することである。
【解決手段】
したがって、本発明は、ミトコンドリアDNA D−ループの塩基配列を用いて、一個体または個体群の水産魚介類もしくはその一部またはそれらの加工品の生産履歴を追跡する方法、およびかかる方法に用いられるマーカー遺伝子を決定する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、天然および栽培漁業における水産魚介類の遺伝子識別および標識方法に関する。より具体的には、本発明は、魚介類ミトコンドリアDNAのDループの塩基配列の相違に基づき、魚介類の品種、産地、系統(種苗)を識別および標識する方法、および養殖事業(栽培漁業)において育種された種苗の稚魚の識別および標識のための方法、ならびにかかる方法に用いるマーカー遺伝子の決定および検出の方法に関する。
【背景技術】
【0002】
日本においては海面養殖漁業および栽培漁業の技術開発が従来から進展しているが、特に近年、クロマグロの完全養殖が成功したことを筆頭として、マダイ、ヒラメ、シラスウナギなどにおいて、卵から繁殖可能な成魚を生産し、さらに成魚から次世代を再生産する、天然資源に依存しない完全養殖の技術が確立しつつある。世界的な食糧不足による食糧価格の高騰、日本の食糧自給率の低下、輸入食糧に関する安全性の問題などが火急の懸案である今日、より安全、安価かつ高品質の食糧を提供し得る供給源として、養殖および栽培漁業に対する関心が高まってきている。
【0003】
養殖および栽培漁業においては、優良系統、例えばウイルスや細菌などの病原に対する耐性の高い系統、成長速度が高い系統、繁殖能力の高い系統などを選択して繁殖させることにより、より高品質の人工種苗の育成が行われる。人工種苗の育成、特に食用とされる魚類を継代で完全養殖する試みには莫大な資本が必要であり、その資本回収は容易ではない。また、水産種苗においては、植物種苗のような遺伝子操作による不稔技術の導入は技術的・倫理的に困難であるため、育種・確立した優良種苗(稚魚)を販売・譲渡する場合、譲受人による増殖(繁殖)と販売が容易であり、種苗育成者の権利の保護が非常に困難であることが問題となっている。また、植物種苗については種苗法により登録された種苗(品種)育成者の権利が保護されるが、動物、特に水産魚介類の種苗に関しては、育成者の権利は法的にも実質的な保護がなされていない。このため、養殖・栽培漁業の分野において優良種苗の育成者および放流事業者の権利を保護し、水産業を活性化するために、水産種苗の登録制度の確立と種苗および成魚の標識および同定方法の開発が望まれる。
【0004】
また、栽培漁業、特に放流事業の加入量変動調査のために稚魚を標識する方法の開発も望まれている。例えば、近年、ミナミマグロに次いでタイセイヨウクロマグロも絶滅危惧種に指定され、漁獲制限がなされている。このような種について、人工的に育成された稚魚を放流して自然集団を回復させることが可能であると考えられる。一部において、遺伝子的に均一な種苗の大規模な放流が生物学的多様性を低下させ、ひいては水産資源の衰退を導くことが懸念されているが、むしろ、新たな種苗の放流により他系統との交配が進み、生態系における遺伝子の多様性を高めるので、漁場の回復に寄与すると考えられる。種苗の放流による漁場の回復のためには、遺伝子的組成の異なる新たな種苗の放流、および放流の評価を的確に行う加入量変動調査が必要である。加入量変動調査はまた、自然界における水産資源量の変動を把握するためにも重要であり、漁場における特定の種、品種、系統および種苗の正確な加入量変動調査は、放流事業が生態系に与える影響を把握し、かつ持続可能な漁獲計画のために必要である。
【0005】
放流稚魚を追跡調査するための従来の標識として、例えばサケ稚魚のあぶらびれの切除(北海道札幌市豊平川さけ科学館)、ヒラメ稚魚の無眼側の黒化および焼印(宮津栽培漁業センター)、マダラやトラフグにおける稚魚耳石のALC(アリザリンコンプレクソン)色素標識および腹鰭抜去標識(能登栽培漁業センターおよび南伊豆栽培漁業センター)、さらに近年ではICタグと通信衛星を利用する電子標識などが実施されている。いずれも稚魚に対して物理的に標識を行うものであり、鰭の切除や焼印などの手段は、種苗を傷つける危険があり、また市場における商品価値を損なう恐れがある。ALC色素標識は、種苗を傷つけずに標識を行うので放流後の成長や生存能に影響を与えないという利点がある一方、調査のためには魚を収集・解体して耳石を採取し、蛍光顕微鏡による確認を行わねばならない。また、これらの方法による標識は、放流する全個体に対して標識をする必要があるのみならず、標識個体一代に限られ、F1以降の世代にわたって識別可能なものではない。さらに、マグロやトラフグのような高級魚の場合、検査のために魚を購入することは、費用が莫大となるため実質的に不可能であり、また、クロマグロなどの場合、高価であるのみならず魚体が大きいのでサンプル採取や標識自体が困難であるなど、調査の実施には多くの問題が残されている。
【0006】
水産業における別の懸案事項は、市場に流通する際の水産物の表示に関する問題である。平成11年のJAS法改正により、一般消費者向けに販売される生鮮水産物について、名称、原産地(採捕・生産された水域名または輸出国名)、解凍・養殖の表示が義務づけられている。しかしながら、流通の現場においてこれらの表示の真贋を判定することは実質的に困難であるため、販売業者による水産物の産地(漁場・港)の偽装(国産ウナギとの虚偽表示により外国産ウナギを販売するなど)、種または品種の偽装(例えばクロマグロとの虚偽表示によりキハダマグロを販売するなど)が問題となっている。品種や漁場によって食品としての品質や本来の価格は異なるが、偽装の疑いがある場合の検証は容易ではなく、表示に依存せずに生産履歴を正確かつ容易に追跡できるシステムも存在しないため、食品の表示に対する信頼が失われつつある。したがって、水産物の表示をより厳格にし、ひいては産地消費者がより安心して水産物を選択できるように、流通過程において水産物の生産履歴を厳正に表示し、必要に応じて迅速かつ正確に判定できる方法の開発もまた望まれている。
【0007】
DNA塩基配列の相違に基づいて生物種または亜種内の識別を行う手法は、分子遺伝学、系統進化学などの分野において非常に有用であり、特にヒトにおいては医療、法医学などの分野においても利用されている。魚類においては、例えば、特定の遺伝子マーカーを用いて白化しやすいヒラメを判別する方法が開発されている(特許文献1)。しかしながら、特定の形質をもたらす遺伝子多型をマーカーとする方法では、形質毎にマーカーとなる遺伝子型を同定する必要があり、上記のような養殖および栽培漁業の分野における問題を解決するためには有効ではない。
【0008】
種苗の遺伝子標識を行うために、MHCマイクロサテライトなどのゲノム多型マーカーを用いることも試みられたが、遺伝子組成の限られた養殖の受精槽においてすらMHCの配列の多様性は極めて高く、個体間においても大きく異なるため、MHCハプロタイプにより種苗を標識し同定することは困難であった(本発明者らによる実験:データは示さず)。
【0009】
ミトコンドリアは、核ゲノムとは独立して独自の10数種類の遺伝子群を含む約15,000〜17,000塩基対(ヒトでは16,569塩基対)の環状二本鎖DNAを有する。ミトコンドリアDNA(mtDNA)は母性遺伝により子孫に伝えられるので、遺伝学の分野において利用される。また、mtDNAは、塩基置換速度が核ゲノムと比較して5〜10倍早く、同一種内における変異が多いので、ハプロタイプ識別のために利用されている。例えば、シロザケのmtDNAコントロール領域(制御領域、D−ループ)の多型をハイブリダイゼーションにより検出し、ハプロタイプを判定する方法が開発されている(特許文献2)。
【0010】
Dループは、mtDNA中の非翻訳高度可変領域であり、同一種内においても多く変異が発生する。非特許文献1には、リュウキュウアユについて、自然集団とこれを起源として異なる環境に導入された導入集団の遺伝的組成を比較する目的で、mtDNA Dループのマイクロサテライトを制限酵素切断片長多型(RFLP)分析によりハプロタイプ分析を行い、起源となったリュウキュウアユ自然集団と導入集団との間で遺伝的組成が類似していることを確認したことが記載されている。また、非特許文献2は、キンメダイ属3種のミトコンドリア制御領域(D−ループ)の塩基配列を比較したところ15.5%の塩基置換率であったこと、および、異なる漁場から採集されたキンメダイの全てがそれぞれ独自のハプロタイプを示し、共通の配列を示すものは見られなかったことを記載している。
【0011】
非特許文献3は、ミトコンドリア制御領域中の超可変領域の配列を用いて、クロマグロおよびメカジキの地理的亜種間の遺伝的相関関係を調査した結果、大西洋と地中海とで捕獲されたクロマグロの遺伝子構成に分離は見られなかったことを記載している。一方、非特許文献4は、大西洋クロマグロ当歳魚の集団毎の遺伝的構成をマイクロサテライト遺伝子座とミトコンドリア制御領域を用いて調査した結果、ミトコンドリア制御領域の配列類似性と地理的配置との間に有意な関連性を見出したことを記載している。また、非特許文献5は、ミトコンドリア制御領域5’末端のハプロタイプにより分けられる2つのクレイド(clade)を用いた個体群統計学により、メバチマグロの地理的な遺伝子の流出入を調査することを記載している。
【0012】
これらの文献に開示される方法は、個体のハプロタイプを判定し、特定の地域におけるハプロタイプ頻度を調べることにより、個体群間の関係を調査するためのものである。例えば特許文献2から理解されるように、ある特定のハプロタイプは、複数の個体において観察され、また複数の異なる地域にわたって分布する。したがって、これらの方法によっては、特定の個体または個体群についてハプロタイプまたはハプロタイプ組成を判定することはできるが、その個体が属する母集団(系統または産地)を判定することはできず、またその個体または個体群を遺伝子により標識することも不可能であった。また、例えば非特許文献2から理解されるように、D−ループの塩基配列は同種の個体間であってもハプロタイプの差異が大きいため、種や系統の識別に用いることは不可能であった。
【0013】
非特許文献6は、クロマグロの養殖に関して、養殖系統を継代して育種を行う場合の遺伝的多様性のモニタリングのために、生け簀内で天然親魚約20個体から得られた孵化仔魚について、母系の推定のためにミトコンドリアDNAのDループ前半領域のシークエン20分析を、父系の推定のためにマイクロサテライトマーカー分析を行ったことを記載している。各ロット(同日に回収された受精卵)内で3〜8種類のミトコンドリアDNAハプロタイプが観察され、そのハプロタイプ頻度はロット間で大きく異なり、マイクロサテライトマーカー分析においても、ロット間の遺伝的構成が異なることが明らかとなり、生け簀内の魚群の産卵に関与する親魚が産卵日によって異なることが示唆された。しかしながら、特定の種苗を標識し、放流後および流通の過程においてその履歴を管理・追跡する方法は、未だ見出されていない。
【0014】
このように、水産魚介類の特定の個体あるいは個体群について、その種、品種、系統または産地を簡易かつ正確に識別し、大量の個体を標識することは、従来のいずれの方法によっても行うことができず、かかる方法の開発が望まれていた。
【先行技術文献】
【特許文献】
【0015】
【特許文献1】特開2007-312645号公報
【特許文献2】特開2003-180397号公報
【非特許文献】
【0016】
【非特許文献1】池田他、「マイクロサテライトDNAおよびミトコンドリアDNA DループのPCR−RFLP分析によるリュウキュウアユ導入集団の遺伝的評価」、水産育種、2001年、第31巻、pp.33−37
【非特許文献2】秋本他、「ミトコンドリア制御領域の塩基配列分析による日本周辺漁場におけるキンメダイの集団遺伝構造の解析」、神水研研報第8号、2003年、pp.89-97
【非特許文献3】Carlsson et al., "Microsatellite and mitochondrial DNA analyses of Atlantic bluefin tuna (Thunnus thynnus thynnus) population structure in the Mediterranean Sea", Molecular Ecology, 2004, vol.13, pp.3345-3356
【非特許文献4】Martinez et al., "Genetic diversity and historial demography of Atlantic bigeye tuna (Thunnus obesus)", Molecular Phylogenetics and Evolution, 2006, vol.39, pp.404-416
【非特許文献5】Alvarado Bremer et al., "Comparative phylogeography of Atlantic bluefin tuna and swordfish: the combined population expansion on the regional phylogenies of two highly migratetory pelagic fishes", Molecular Phylogenetics and Evolution, 2005, vol.36, pp.169-187
【非特許文献6】森島他、「養殖クロマグロの遺伝的管理に向けた研究」、平成19年度日本水産学会近畿支部後期例会プログラム、109-D、平成19年12月1日、日本水産学会(http://www.suisan-kinki.org/reikai/h19reikai/h19_kouki/9.pdf)
【発明の概要】
【発明が解決しようとする課題】
【0017】
したがって、本発明は、上記の問題点に鑑み、水産魚介類、特に、特定の産地に由来する水産魚介類、一つまたは複数の優良形質を有する優良水産種苗、または特定の種または品種の水産魚介類の1個体〜大量の個体を、標識・識別するための簡易かつ正確な方法を提供することを目的とする。
【課題を解決するための手段】
【0018】
本発明者らは、魚類の行動様式、繁殖形態に着目した。例えばクロマグロの産卵においては、一個体の雌親魚が一回に約200万〜1,000万個の卵を産卵、孵化する。その受精には複数の雄親魚が関与していると考えられる。また、養殖の場合は特に、一つの養殖槽において一日に産卵する雌親個体の数は少なく、受精卵は約1日で孵化して水面に浮揚し、産卵日毎に回収することが可能である。本発明者らは、これと併せてミトコンドリアDNA(mtDNA)が母系特異的であることに着目した。ミトコンドリアDNAは一般的にサイズが小さく、組み換えが起こらないため個体間での保存性が高いが、Dループの短い領域に変異が集中している。これらのことに焦点を当て鋭意研究を進めた結果、水産魚介類のプロフィールの同定、例えば生産履歴、種、品種、系統または産地などの識別が可能であることを見出し、さらに研究を重ね、発明を完成するに到った。
【0019】
すなわち、本発明は、水産魚介類またはその加工品のミトコンドリアDNA Dループの塩基配列を決定することにより、該水産魚介類またはその加工品の種苗または生産履歴を同定する方法であって、
養殖場において継代した雌親個体から採取された第1の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第1の塩基配列)を決定し、
水産魚介類またはその加工品から採取された第2の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第2の塩基配列)を決定し、
第1の塩基配列と第2の塩基配列とを比較し、前記水産魚介類またはその加工品が前記雌親個体の後代であることを同定することによって、前記水産魚介類またはその加工品の種苗または生産履歴を同定することを含む、前記方法に関する。
【0020】
また、本発明は、マイクロサテライトにより核型を決定することをさらに含む、前記方法に関する。
【0021】
本発明はさらに、ミトコンドリアDNA Dループの種特異的塩基配列が1から2以上のギャップの挿入により整列化できるか否かの判定をさらに含む、前記方法に関する。
【0022】
本発明はまた、ミトコンドリアDNAのDループ全長の塩基配列を、ミトコンドリアプロリンtRNA特異的プライマーおよびミトコンドリアフェニルアラニンtRNA特異的プライマーを用いて増幅し、両鎖シークエンシングにより決定することをさらに含む、前記方法に関する。
【0023】
また、本発明は、第1の塩基配列と第2の塩基配列との比較が、母系特異的塩基配列が完全に一致するか否かの比較である、前記方法に関する。
さらに、本発明は、水産魚介類またはその加工品が、食品である、前記方法に関する。
【0024】
本明細書において使用される場合、品種とは、同種内の小分類であって、他の集団とは形態、生理、遺伝的に区別し得るような特徴を有する集団を指し、分類学的に亜種として分けられる場合もある。例えば、クロマグロは、一般に、大西洋、地中海および黒海産(タイセイヨウクロマグロ)と、太平洋産(タイヘイヨウクロマグロ)との2つの亜種に分類され、さらに学説では、大西洋産、地中海産および黒海産は別の遺伝学的特徴を有する群に分けられるとされていたが、本明細書中以下に記載する本発明者らの解析により、タイセイヨウクロマグロとタイヘイヨウクロマグロとは別品種として分類される。
【0025】
本明細書において用いる場合、種苗(人工種苗)とは、優れた(または生け簀養殖もしくは放流に適した)形質を有する個体を人為的に選抜して交配し、その次代を系統として養成し、さらに個体または系統の選抜を行うことにより、優良形質を有する系統を繁殖のために確立したものを指し、受精卵、稚魚、仔魚および成魚を含み、養殖のためのものであっても、海川に放流されるためのものであってもよい。
【0026】
本明細書において用いられる場合、生産履歴とは、水産魚介類の個体もしくはその一部またはそれらの加工品が流通の過程にある場合において、生産地(養殖、採取もしくは加工などの場)、生産方法(養殖、採取または加工の方法もしくは条件など)、保存や流通の方法もしくは条件、または加工品である場合原料となる水産魚介類の種、品種もしくは系統などの情報を意味する。また、産地とは、対象となる水産魚介類が由来する水域または施設を意味し、産卵場、餌場、回遊場、漁場、水揚場、孵化および養殖場などを含む。
【0027】
本明細書において用いる場合、母系特異的配列とは、検体となる個体のミトコンドリアDNAのDループ塩基配列と、比較対照となる個体のミトコンドリアDNAのDループ塩基配列とを整列化(アラインメント)して相同性を比較した場合、同一の母系に由来する個体間では完全に一致するが、異母個体との間では、0〜2個程度のギャップの挿入で整列化でき、高度の相同性を示すが完全には一致しない領域の塩基配列を指す。種特異的配列とは、複数の異なる種のDループ塩基配列が数個以上のギャップの挿入で整列化できる場合において、異種間で相同性が高い塩基配列を指す。さらに、本発明者らは、母系特異的配列中に、同一種間でも異なる母系の間で、特に多くの変異が集中する短い領域が存在することを見出した。この領域を本明細書において母系変異集中領域と称する。同様に、異種間でギャップにより整列化できるが多くのギャップが集中する短い領域が存在することを見出し、この領域を異種変異集中領域と称する。
【発明の効果】
【0028】
本発明により、水産魚介類の種苗、産地および品種を簡便かつ迅速に識別することが可能となり、したがって、該水産魚介類の生産履歴および品種などの表示を厳正に管理することができ、種苗や食品としての水産魚介類を、より安全かつ公正に提供することが可能となる。特に輸入やインターネット販売などにより流通される、従来では生産履歴の追跡が困難であった水産物の生産履歴を管理することが可能となり、偽装表示による水産物販売を防止・摘発することができるようになる。また、本発明により、養殖魚介類の優良種苗、例えば、GTL、ウイルス耐性などのの優良種苗を遺伝子識別することが可能となり、種苗のブランド化および育種者の権利の保護がより容易となる。また、本発明により、母系特異的遺伝子配列により標識された種苗を大量生産することができ、これと同時に母系特異的配列のデータベースを構築することにより、放流事業における加入量変動調査が可能となるのみならず、生態系において特定の系統の個体数の増減をモニタリングすることが容易となり、水産魚介類の遺伝的多様性の維持に有用な資料を提供することができる。
【図面の簡単な説明】
【0029】
【図1−1】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−2】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−3】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−4】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−5】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−6】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−7】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−8】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−9】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−10】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−11】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【0030】
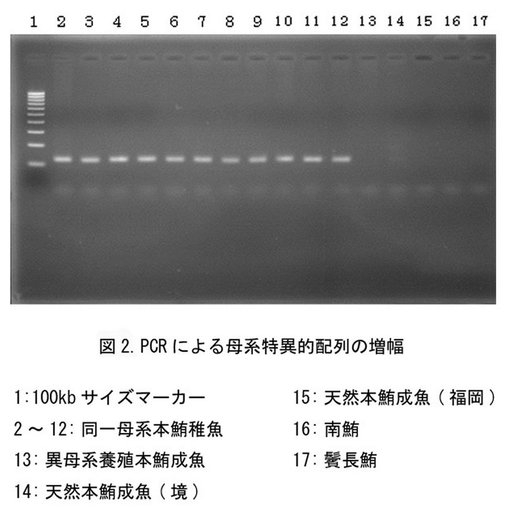
【図2】PCRにより増幅されたマグロ属における母系特異的DNAの電気泳動の結果を示す。
【図3−1】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【図3−2】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【図3−3】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【図3−4】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【0031】
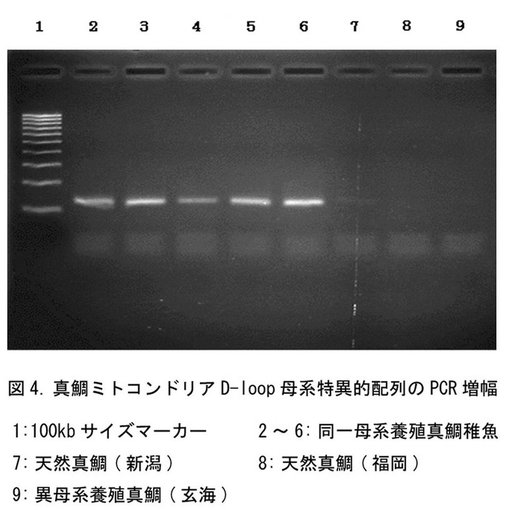
【図4】PCRにより増幅されたマダイにおける母系特異的DNAの電気泳動の結果を示す。
【図5−1】マグロ属のミトコンドリアDNA Dループ塩基配列の比較を示す図である。
【図5−2】マグロ属のミトコンドリアDNA Dループ塩基配列の比較を示す図である。
【図5−3】マグロ属のミトコンドリアDNA Dループ塩基配列の比較を示す図である。
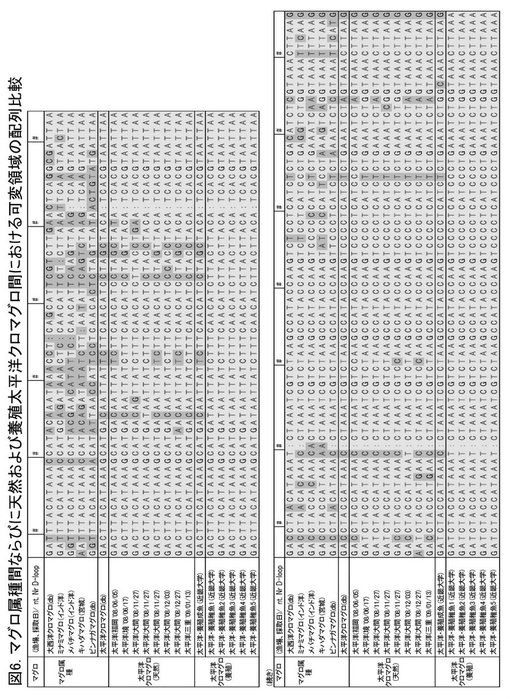
【図6】天然および養殖のマグロのミトコンドリアDNA D−ループ配列の比較を示す図である。
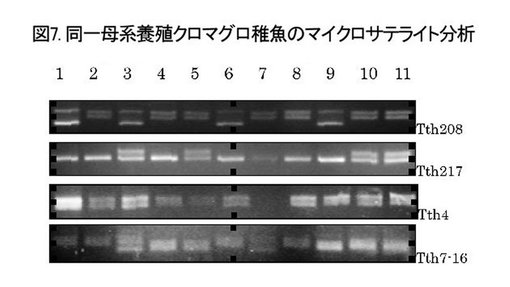
【図7】同一の母系に属する養殖クロマグロ稚魚の4種のマイクロサテライト分析の結果を示すゲル電気泳動の写真である。
【発明を実施するための形態】
【0032】
本発明は、一態様において、水産魚介類またはその加工品のプロフィールを同定する方法を提供する。該方法において、予め採取された水産魚介類のミトコンドリアDNA Dループの塩基配列を決定し、該決定された塩基配列と、同定されるべき検体としての水産魚介類またはその加工品のミトコンドリアDNA Dループの塩基配列とを比較することにより、検体のプロフィールを同定する。プロフィールとは、好ましくは種、品種、系統、生産履歴および/または産地であるが、これらに限定されない。特定の種、品種系統の種苗に特異的な遺伝子マーカーを見つけだすことは容易ではないが、本発明の方法においては、一個体の雌親から多数の次世代の個体が生産される水産魚介類の特徴を利用して、異なる種、系統または母系の間で可変性が特に高いミトコンドリアDループ領域の塩基配列の相違を、種、系統などのマーカーとして用いる。ミトコンドリアDNAは少量のサンプルから得られるコピー数が多く、核DNAが劣化するような条件下においても安定性が高いので、魚類のプロフィールの情報源として最も好適である。
【0033】
一態様において、本発明の方法は、水産魚介類もしくはその一部またはそれらの加工品またはこれらの雌親個体から、ミトコンドリアDNAを含む試料を、生産履歴の初期において予め採取して保存することを含む。生産履歴の初期とは、例えば、採集(漁獲)、生産、卸売、流通、販売などの場を意味し、例えば天然または養殖の魚介類の場合、水揚げした漁業または養殖業の従事者が、鱗、鰭、少量の肉片など該魚介類のミトコンドリアDNAを含むサンプル(試料)を採取し、凍結保存することができる。また、受精卵から養殖した仔魚(稚魚)を販売する場合、同一の雌親が産卵した(同腹の)1以上の受精卵、仔魚もしくは稚魚または雌親の鱗などを採取して凍結保存することができる。本発明の方法により同一母系の稚魚全個体が同一のマーカー(母系特異的配列)により標識されるので、1個体の同腹稚魚のサンプルを保存することにより、放流または販売後の全個体を追跡調査することが可能となる。あるいは、例えばマグロのように解体して流通・販売する場合、卸売場、市場または販売店などにおいて、各個体のサンプルを採取して凍結保存することができる。このようにして、個体の一部をサンプルとして予め採取して保存することにより、特定の個体の全てまたは一部(切り身、加工品など)を販売した後においても、該個体に由来する販売品に含まれるミトコンドリアDNAと、予め採取された同一個体のサンプルに含まれるミトコンドリアDNAとを比較することができる。
【0034】
本発明の方法は、好ましい態様において、前記水産魚介類またはその加工品、または種苗の場合は雌親個体から採取された試料(第1の試料、標準試料)に含まれるミトコンドリアDNA制御領域(D−ループ)の塩基配列中の、母系特異的または種特異的な塩基配列を決定することを含む。例えば、種特異的塩基配列を決定するために、第一に、目的とする種についての指標とする配列を得ることが必要である。具体的には、GeneBankやMitoFish(魚介類ミトコンドリアの公開データベース(http://mitofish.ori.u-tokyo.ac.jp/))などの公共のデータベース(以下dbと略)から、同種または近縁種のmtDNAの塩基配列を検索して用いてもよい。養殖稚魚の標識のための母系特異的配列を得るためには、雌親、または該雌親が産卵した少なくとも1の受精卵、仔魚もしくは稚魚からミトコンドリアDNAを得、公知の分子生物学的手法を用いてDループの塩基配列を決定すればよい。このD−ループ塩基配列とデータベースや他の個体のD−ループ塩基配列とを比較することにより、母系変異集中領域を決定し、これを母系特異的配列として用いることもできる。同一の雌親が産卵した受精卵およびそれに由来する仔魚もしくは稚魚は、全個体が同一の母系特異的配列を有し、この母系特異的配列は他の雌親に由来する個体とは異なる。したがって、1の受精卵、仔魚もしくは稚魚の母系特異的配列が、同一母系集団中の全個体の母系特異的配列となる。
【0035】
母系特異的配列または種特異的配列を決定するために、特定の種、品種、系統または産地に由来する水産魚介類のmtDNA Dループ配列と、他の種、品種、系統または産地に由来する個体の対応する配列とを比較し、識別可能なパーセンテージで塩基の置換が生じる領域を決定する。
【0036】
この領域は、異なる母系の個体間において、例えば800〜1600塩基長の領域中であれば、5〜10個(1%)、好ましくは11〜15個(1.5%)、より好ましくは16〜30個(3%)の塩基の置換を有する領域である。同一母系の個体間においては、この領域の塩基配列が完全に一致することが好ましいが、他の母系との識別が可能である限りにおいて、1または2程度の塩基の置換を含んでもよい。より正確な識別のために、同一の母系の個体間における塩基の置換は、0.2%程度以下、好ましくは約0.1%程度以下、さらに好ましくは0%(完全一致)であることが好ましい。
母系特異的配列の典型的な位置は、Dループのプロリン側末端の塩基を1番目の塩基として、約100〜600番目、好ましくは約150〜450番目の間の塩基配列中に存在する。例えばクロマグロにおいては母系特異的配列は約150〜350番目、マダイにおいては約250〜450番目の間の塩基配列中に存在する。
【0037】
また、種を識別するための領域は、複数の異なる種間で1から2以上のギャップの挿入によりDループ配列が整列化できる場合において、異種間で相同性を示す塩基配列である。異種間での相同性は、90〜97%、好ましくは90〜95%、より好ましくは90〜93%である。複数の配列を比較する多重整列化解析(マルチアラインメント)には、例えばMEGA4(登録商標、Molecular Evolutionary Genetic Analysis Software Version 4.0、Tamura et al., Mol. Biol. Evol., 2007, 24(8):1596-1599)、SEQUENCHER(登録商標)(Gene Codes、米国)などのシークエンス解析のための公共または市販のソフトウェアを用いることができる。
種特異的配列の典型的な位置は、母系特異的配列と部分的に重複する場合もある。例えばクロマグロにおいては種特異的配列は約150〜700番目、特に約150〜250番目および約600〜700番目に見出すことができる。
【0038】
次いで、検体となる水産魚介類から採取された試料(第2の試料、検体)に含まれるミトコンドリアのDループDNAを、上記の母系特異的配列または種特異的配列と比較することにより、その種、品種、系統、生産履歴および産地の識別を行なうことができる。例えば、上記の母系または異種特異的塩基配列に基づいて設計されたプライマーまたはプローブを用いて、公知の手段により、検体のmtDNA中に母系特異的塩基配列または異種特異的塩基配列と同一の配列が含まれるか否かを容易に検出することができる。かかる目的のための公知の手段として、PCR法、RCA法、ダイレクトシークエンス法、TaqMan法、DNAアレイ法、FISH法などが挙げられるがこれらに限定されない。
【0039】
本発明の方法において、さらに、ミトコンドリアDNA Dループの母系または種特異的配列に加えて、他のハプロタイプの比較を組み合わせることにより、識別をより厳密に行ってもよい。通常のハプロタイプ分析では個体間における相違が大きいので、種や種苗の同定には用いることができない。例えば、非特許文献2に示されるとおり、同腹仔のミトコンドリアDNAにおいてすら、マイクロサテライトのハプロタイプは異なる。しかしながら、限られた個体数で繁殖を行う養殖においては、F1におけるハプロタイプの数が比較的少なく、例えば、同じ雌親から生じた11個体の稚魚において、4種のマイクロサテライトにより識別されるハプロタイプは4種類であった(参考例1)。このことを利用して、D−ループによる識別を補強することができる。例えば、放流する人工種苗(稚魚)の標識として、D−ループ母系または種特異的配列のみならず、マイクロサテライトなどにより核型のハプロタイプも決定しておくことにより、万が一、人工種苗と同一の母系に属する個体が天然に存在する場合であっても、採集(漁獲)されたサンプルが天然のものであるか人工種苗であるかを確実に判断することができる。
【0040】
本発明の方法は、好ましい態様において、前記母系特異的塩基配列、母系変異集中領域または異種特異的塩基配列に基づくプライマーまたはプローブの作成を含む。プライマーは、Dループ全長の増幅のためのプライマー、または、母系特異的塩基配列、母系変異集中領域もしくは種特異的塩基配列の特異的増幅のためのプライマーを含む。プローブは、母系特異的塩基配列、母系変異集中領域または種特異的塩基配列に特異的に結合する核酸、例えばオリゴヌクレオチドを含む。目的とする母系特異的または種特異的塩基配列の一部に対して少なくとも部分的に相補的となるように設計したプライマーまたはプローブを用いて、定法により、検体中に目的の配列を含むmtDNAが含まれるか否かを容易に決定することができる。具体的には、例えば、栽培漁業における加入量変動調査において、検体となる魚の個体のmtDNAを鋳型として、調査対象の種苗(母系)に特異的な配列に基づいて設計されたプライマーを用いてPCRやダイレクトシークエンスを行うことにより、該個体が目的の種苗であるか否かを容易に判定することができる。あるいは、産地または系統などによりデータベース化した複数〜多数の母系特異的配列と、検体の配列とを、DNAアレイ法などにより比較し、検体がデータベース中に記載される産地または系統に該当するか否かを容易に判定することができる。
【0041】
別の態様において、前記水産魚介類もしくはその一部またはそれらの加工品に、これらが前記母系特異的または異種特異的塩基配列により標識されていることを記載する文書またはラベルを添付してもよい。この文書またはラベルはまた、それらが添付されている水産魚介類は、その生産履歴の初期において予めサンプルが採取され保存されているか、および/または既に母系または種特異的配列が決定されており、必要に応じて(例えば該水産魚介類の購入者または使用者の希望により)、該水産魚介類に含まれるミトコンドリアDNAの塩基配列を母系または種特異的配列と比較することが可能であることを示すことができる。かかる文書またはラベルを添付された水産魚介類やその加工品は、生産者から離れて流通した後においても生産履歴を追跡することが可能であり、また、遺伝子標識により検査を行うため産地や品種の偽装が困難となることから、より安全性の高い食品を提供することが可能となる。
【0042】
一態様において、同一の種、品種、系統、生産履歴および/または産地に属する複数の雌親に由来する複数の母系特異的または種特異的配列を、産地、品種または系統によりプールしてデータベース化し、検体の同定に役立てることもできる。例えば、養殖場において恒常的に雌個体の母系特異的配列を取得してデータベース化することにより、継代養殖され流通されているブリやマグロなどの水産魚介類が由来する養殖場および出荷時期などを追跡することができる。または、特定の産地について同様のデータベースを構築し、産地表示の真偽を判定するために役立てることもできる。具体的には、目的の水域におけるサンプリングにより得られる対照試料を基に、1または2以上の母系が存在する集団として、産地を識別する。例えば、特定の水域における定着性が強く母系の数が比較的少ない集団であれば、該集団における母系特異的配列をプールしたデータベースを作成することにより、該データベースに含まれる母系特異的配列により産地を特定することができる。
【0043】
別の態様において、本発明は、上記の方法において、検体となる水産魚介類もしくはその一部またはそれらの加工品から、ミトコンドリアDNAを含む試料を採取して保存することを含む。本発明の方法によって生産履歴を追跡することができる水産魚介類もしくはその一部またはそれらの加工品は、生きている魚介類であっても、または生鮮、加熱調理済み、乾燥もしくは冷凍の魚介類であっても、それらの鱗、鰭、鰓、尾、皮膚、骨、体液、筋肉または内臓組織などであってもよい。一般に、ミトコンドリアは一つの細胞内に多数(数百〜数千個)存在するため、少量のサンプルから充分なmtDNAを得ることができ、また核DNAと比較して構造的に安定であるため、核DNAが劣化する条件下においても抽出できる。したがって、本発明の方法により、食品もしくは動物飼料またはそれらの原材料として加工された水産魚介類の生産履歴を調査することが可能となる。特に、魚類の種や亜種の判定は、流通の経路の下流、すなわち、対象となる魚介類が既に販売、加工、調理された状況において主として行われることが予測されるが、本発明の方法に従ってミトコンドリアDNAを利用することにより、加工・調理・冷凍・解凍などの処理の後であっても、少量のサンプルが得られれば、魚種を判定することができる。より正確な生産履歴調査のために、組織試料は、非加熱の試料の場合は約0.2g以上(鱗の場合数枚)、加熱された試料の場合は約1g〜5gであることが好ましい。採取した試料は、サンプルチューブに密封し、−20℃〜−80℃で凍結保存する。ミトコンドリアDNAの抽出・精製は、定法にしたがってアルカリ法などにより行い、TEに溶解する。長期保存する場合は、アルコールで沈殿させて−20℃〜−80℃で保存するか、またはポリマー、ビーズもしくは紙などの担体を用いてDNAを室温で長期保存する方法も開発されている(例えば「DNAブック」:Kawai他、Genome Research,2003,13:1488-1495を参照)。
【0044】
本発明の一態様において、養殖または栽培漁業において育成された種苗の受精卵、仔魚または稚魚の生産履歴を追跡する方法を提供する。養殖又は栽培漁業においては、親魚の産卵期の観察により、同腹の(雌親が同じ)受精卵および仔魚(稚魚)の群を得ることが可能である。本発明の方法により、同時期に浮遊した個体群を、同一の母系特異的配列により遺伝子標識された種苗とみなすことができる。したがって、継代養殖や優良株との交配によって育成した優良種苗を、雌親によって分けた群として識別することにより、各々の群の雌親のmtDNAのDループ母系特異的塩基配列により標識された母系群とすることができる。例えばマグロなどの場合、一個体の雌魚から生産された数百万〜数千万個体の種苗の全てに、魚体を傷つけることも遺伝子組み替えなどを行うこともなく、均一な標識を行うことができ、放流後の追跡調査、特に母系別の加入量変動調査を容易に行うことができる。また、種苗の近親交配による遺伝的障害を避ける指標を提供することもできる。また、他者が不当に種苗を複製した場合にこれを摘発することが技術的に可能となるので、種苗を育成する業者の保護に役立てることができる。
【0045】
本発明の方法は、水産魚介類の種類によって限定されるものではないが、特に、広域に流通し、品種、産地または種苗により安全性、栄養価、価格などが変化する水産魚介類の生産履歴の追跡に、特に有用である。具体的には、スズキ目サバ亜目サバ科マグロ属(例えば、クロマグロ、ミナミマグロ、キハダマグロ、メバチマグロ、ビンナガマグロなど)、同サワラ属(サワラなど)および同カツオ属(カツオなど)、スズキ目スズキ亜目アジ科ブリ属(ブリ、カンパチ、ヒラマサなど)および同マアジ属(マアジなど)、スズキ目スズキ亜目スズキ科(スズキなど)、スズキ目スズキ亜目タイ科マダイ亜科(マダイなど)、フグ目フグ科(トラフグ、マフグなど)、カレイ目カレイ亜目(ヒラメ、カレイ)、サケ目サケ科サケ属(ベニザケ、シロザケ、ギンザケ、カラフトマス、マスノスケなど)、ウナギ目ウナギ亜目(ウナギ、アナゴ、ハモ)、貝類(ハマグリ、アサリ、ホタテ、シジミ、およびサザエ)などが挙げられる。
【0046】
一態様において、本発明の方法は、ミトコンドリアDNAのDループ全長の塩基配列を、ミトコンドリアプロリンtRNA特異的プライマーおよびミトコンドリアフェニルアラニンtRNA特異的プライマーを用いて両鎖シークエンシングにより決定することをさらに含む。ミトコンドリアDNAに含まれる遺伝子の配置や塩基配列は、水産魚介類の近縁種の間で、Dループを除いて、保存性が高い。Dループは0.8〜1.5kb程度の塩基長を有する高度可変領域であるが、その3’末端側において隣接するプロリンtRNA遺伝子および5’末端側において隣接するフェニルアラニンtRNA遺伝子の塩基配列は保存性が高い。したがって、Dループを挟んでプロリンtRNAおよびフェニルアラニンtRNAの配列に対して各々プライマーを設計することにより、Dループの全長をPCRで増幅することができる。プロリンtRNAおよびフェニルラニンtRNAのプライマーによりPCRでDループDNAを増幅した後、酵素などによりPCR産物中のプライマーを消化および未反応dNTPを不活性化し、次いで、増幅に使用した2種のPCRプライマーの各々を用いて、それぞれの末端からシークエンスを行なう。配列の信頼性の観点から、両鎖シークエンスを行うことが好ましい。シークエンスの精度をあげるために相補鎖が一致した場合のみDループの塩基配列とする。両鎖が重複しない領域は得られたDループ内側から再度シークエンスする。
【実施例】
【0047】
以下に例を示して本発明をより詳細に説明するが、本発明の範囲は、これらの例によって何ら限定されるものではない。また、本発明の方法を適宜改変することは、当業者の通常の技術の範囲内である。
【0048】
材料および方法
1.ミトコンドリアDNAの抽出・精製法
試薬の組成:
(1)P1溶液(再懸濁試薬:250ml、2〜8℃保存)
0.5M EDTA 5.0ml
1M Tris−HCl(pH 8.0) 12.5ml
滅菌水で250mlに増量し、オートクレーブ滅菌する。
(2)P2溶液(溶菌試薬:250ml、室温保存)
SDS 2.5g
NaOH 2.0g
滅菌水で250mlに増量する。
(3)P3溶液(除蛋白沈殿試薬:500ml、4〜8℃保存)
酢酸カリウム 147.3g
酢酸 60.0ml
滅菌水で500mlに増量し、pH 5.5に調整し、オートクレーブ滅菌する。
【0049】
ミトコンドリアDNAの抽出:
(1)水産魚介類の組織約0.2g(鱗の場合数枚)の試料を、サンプルチューブに移し、P1溶液を0.4ml添加し、ペスツル棒でサンプルを潰す。
(2)P2溶液を0.4ml添加し、ペスツル棒で軽く混合する。
(3)P3溶液を0.4ml添加し、チューブの底を指で弾いて攪拌する。
(4)室温で5分間、20,000×gで遠心分離し、上澄を別のチューブに移す。
(5)フェノール・クロロホルム溶液を0.4ml添加し、ボルテックスミキサーにかける。
(6)再び遠心分離を行ない、上澄を回収する。
(7)5、6の操作を繰り返す。
(8)得られた上澄の0.6倍容積のイソプロパノールを加える。
(9)20,000×gで30分間遠心分離し、上澄を捨て、70%エタノールで沈殿を洗浄する。
(10)再び遠心分離した後、上澄を捨て、65℃で5分間静置して乾燥させる。
(11)0.1MのTE 30μlに溶解させ、ミトコンドリアDNA溶液とする。
【0050】
2.DループDNAの増幅
例えば以下の手順に従ってミトコンドリアのDNAのDループ塩基配列を増幅する:
(1)魚介類ミトコンドリアのDNAの塩基配列は、MitoFish(http://mitofish.ori.u-tokyo.ac.jp/)、GeneBank(http://www.ncbi.nlm.nih.gov/Genbank/)などの公開データベース(以下dbと略)から得られる、目的の種と同種、あるいは近縁種のmtDNAの塩基配列中、Dループの両側にあるプロリンtRNAおよびフェニルラニンtRNAの遺伝子配列からPCRプライマーを作製する。なお、dbに含まれていない新規の試料の場合は、近縁種のプライマーで代替えする(成功例:ブリ(実施例3を参照))か、あるいはミトコンドリアDNAのショットガンシークエンスなどにより配列を得る。
【0051】
(2)以下の組成でPCR反応用カクテルを作成し、試料のミトコンドリアDNAを0.5μl添加する:
10×PCRバッファー:2μl、25mM MgCl2:2μl、2mM dNTPs:2μl、5U/μl Taqポリメラーゼ:0.1μl、16.7pmol/μlプライマー(フォワード):1μl、16.7pmol/μlプライマー(リバース):1μl、精製水:11.3μl。
【0052】
(3)熱変性95℃ 10分、DNA増幅(95℃ 1分、65℃ 30秒、72℃ 1分)×34サイクル、72℃ 5分の反応条件で、PCRを行う。
(4)PCR反応液7.1μlを384ウェルPCRプレートに分注し、Exo SAP IT(USB Co.)を2.9μl添加する。
(5)37℃で15分間反応後、80℃で15分間熱変性する。
【0053】
3.DループDNA両鎖シークエンス法
PCRでDNAを増幅した後、Exo SAP IT(USB Co.)などにより、PCR産物中のプライマーを消化および未反応dNTPを不活性化し、次いで、増幅に使用した2種のPCRプライマーの各々を用いて、それぞれの末端から、例えば以下の手順でシークエンスを行なう。
【0054】
(1)Exo SAP ITの反応液を2分割し(各5μl)、384ウェルPCRプレートの空のウェルに移す。
(2)シークエンス用カクテルをABI社マニュアルに従って以下の組成で作製する:
フォワード(またはリバース)プライマー:16.7pmol/μlを1μl、5×シークエンスバッファー:2μl、Bigdye:2μl。
(3)ヒートシールで密封し、熱変性96℃ 1分、伸長(96℃ 10秒、50℃ 5秒、60℃ 4分)×30サイクルの条件でサイクルシークエンス反応を行う。
(4)ヒートシールをはがし、125mMのEDTAを2.5μl、エタノールを25μl添加し、1時間、室温で静置する。
(5)4℃、3,273×gで30分間遠心分離する。
(6)PCRプレートを逆さまにしてエタノールを除去する。
(7)冷却した70%エタノールを35μl添加し、同条件で遠心分離する。
(8)エタノールを除去し、65℃に5分間置いて、乾燥させる。
(9)ホルムアミドを5μl添加し、熱変性する。
(10)ABI373型シークエンサーでシークエンスする。
【0055】
4.多重整列化解析
以下に、「SEQUENCHER(登録商標)」を用いる場合の解析手順を示す。
(1)SEQUENCHER(登録商標)にシークエンスのデータをインポートする。
(2)末端のクオリティーの低い配列を除去する。
(3)データベース、または過去のシークエンスデータをシークエンチャーにインポートする。
(4)例えば以下の条件設定において、複数の配列のコンティグを作成し、相同性に基づいて、母系特異的配列、母系変異集中領域および同属異種特異的配列を決定する:
Assembly Algorithm :Dirty Dataをチェック
Optimize Gap placement :Use Re Aligner,Prefer 3’ Gap Placementをチェック
Minimum Match Percentage:80
Minimum Overlap :20。
【0056】
実施例1:クロマグロ母系特異的塩基配列の抽出と種苗の遺伝子標識
方法:
1)「MitoFish」(http://mitofish.ori.u-tokyo.ac.jp)から、本まぐろ(太平洋クロマグロ:Thunnus orientalis)のミトコンドリアDNAの塩基配列(GeneBank登録番号AB185022:配列番号1、以下dbと記載)を検索し、プロリンtRNAとプェニルアラニンtRNAの塩基配列からDループ増幅のためのPCRプライマーを以下の通り作製した。
本まぐろのDループDNA増幅のためのPCR共通プライマー配列
フォワードプライマー(プロリン側):
5’-TACCCCTAACTCCCAAAGCTAGG-3’(配列番号2)
リバースプライマー(フェニルアラニン側):
5’-GCTTTCTAGGGCCCATCTTAAC-3’(配列番号3)
【0057】
2)境港と福岡で水揚げされた天然本まぐろ(太平洋クロマグロ)成魚と養殖本まぐろ(太平洋クロマグロ)成魚、各1尾ずつ、および同時に採取した浮遊受精卵を孵化した本まぐろ稚魚11尾からミトコンドリアを抽出し、PCRの鋳型DNAとした。
3)上記の方法でDループDNAを増幅した後に塩基配列決定を行ない、データベースの本まぐろ(db)のDループの塩基配列(配列番号1)と比較した。
【0058】
結果:
多重整列化の結果を図1に示す(なお、各々のDループ塩基配列は、境港:配列番号20、福岡:配列番号21、養殖成魚:配列番号22、養殖稚魚1〜11:配列番号23〜33とする)。図中、下部の配列はデータベース(db)の太平洋クロマグロのミトコンドリアDループの塩基配列におけるnt(ヌクレオチド)1からnt865までであり、その下部の黒点は、該db太平洋クロマグロと比較した他のマグロとの間で塩基の相違があることを示す。上記プライマーを用いて決定した太平洋クロマグロ14個体のDループDNAは、dbと同じ865塩基であり、各塩基配列はギャップを加えることなく多重整列化することができた。天然および養殖の各成魚間において、個体間の配列の塩基相同性は高く、わずかに6〜22の異なった塩基相違点が太平洋クロマグロ(db)の前半nt175〜586間の29部位に存在していた(表1)。
【0059】
【表1】

【0060】
塩基相違点はnt175〜200に7部位、およびnt273〜305に5部位に相違点が集中していたためにその間の配列を母系特異的塩基配列と規定した。一方、養殖稚魚11尾間では、配列は完全に一致しているために、同時に浮遊した受精卵は同一雌親由来であり、雌親が異なる他の成魚との相違点は特定部位に集中していることが明らかとなった。またこの結果から、同一の雌親に由来する受精卵や稚魚は、そのうちの1個体のミトコンドリアD−ループの塩基配列を決定することにより、全個体を同じ母系集団として容易に標識できることが示された。
【0061】
応用例1:クロマグロ母系特異的塩基配列の特定と簡易検出法
母系特異的塩基配列、特に母系変異集中領域の塩基配列を増幅するPCR用プライマーを作製することにより、検体から簡易かつ迅速な方法により母系特異的シグナルを得、その母系性から検体の生産履歴を追跡することができる。以下に例を示す。
【0062】
方法:
以下のプライマー対を作製し、天然および養殖の太平洋クロマグロ成魚3尾および稚魚11尾、ならびにミナミマグロおよびビンナガマグロ1尾ずつから抽出したミトコンドリアDNAを鋳型として、タッチダウンPCRにより増幅を行なった。
母系特異的PCRフォワードプライマー(図1、同一母系太平洋クロマグロnt.175〜200:母系特異的変異点7ヶ所):
5’-CTTTCAACATCTTACTTACATCACG-3’(配列番号4)
母系特異的PCRリバースプライマー(図1、同一母系太平洋クロマグロnt.273〜305:母系特異的変異点5カ所):
5’-TTCTTACTGCGTTTAAGTTTACCAG-3(配列番号5)
【0063】
結果:
図2に、PCR増幅産物のアガロース電気泳動の結果を示す。母系が異なる天然太平洋クロマグロおよび養殖太平洋クロマグロからはシグナルは検出されなかったが、同一母系の養殖稚魚11尾から135塩基対の母系特異的DNAが増幅された。したがって、同一母系の養殖稚魚(種苗)が、母系特異的塩基配列によって遺伝子標識されており、該母系特異的塩基配列に特異的に設計されたプライマーを用いたPCRにより容易に検出し得ることを確認した。本明細書において示した系統または産地以外の種苗においても、同様に母系特異的なプライマーを設計し、検体のリアルタイムPCRを行なえば、シグナルの有無によって簡便かつ迅速に母系識別を行うことができる。
【0064】
実施例2:マダイ母系特異的塩基配列と種苗の遺伝子標識
方法:
1)「MitoFish」からマダイ(Pagrus major)のミトコンドリアDNA塩基配列(GeneBank登録番号AP002949:配列番号6)を入手し、プロリンtRNAおよびフェニルアラニンtRNAの塩基配列から以下のPCRプライマーを設計して、マダイDループ増幅と塩基配列決定に使用した:
フォワードプライマー(クロマグロ(db)フェニルアラニンtRNA塩基配列から代用):
5’-GCTTTCTAGGGCCCATCTTAAC-3’(配列番号7)
リバースプライマーI(マダイdbプロリンtRNAから設計):
5’-TTTAACCCCCACCATTGGCTCCC-3’(配列番号8)
リバースプライマーII(マダイDループ内塩基配列より再設計):
5’-AGAGGGGGGTAACAATGAGATC-3’(配列番号9)
【0065】
2)マダイのプロリンtRNAおよびフェニルアラニンtRNAの塩基配列はいずれもクロマグロの対応する配列に一致していたが、リバース側において1塩基が異なっていたため、マダイdbのフェニルアラニンtRNA塩基配列を利用した(リバースプライマーI)。塩基配列に関しては、最初に読めたDループ3’末端側の塩基配列から第2のリバースプライマーIIを作製し、完全に両鎖の塩基配列を読み取った。
【0066】
3)PCRの鋳型DNAには、福岡玄海の養殖成魚と山口、福岡の天然成魚各1尾ずつ、および同日浮遊しに採取された受精卵を孵化成長させた養殖種苗10尾からミトコンドリアを抽出し、PCR鋳型DNAとした。上記の方法でDループDNA増幅および塩基配列決定を行ない、データベースのマダイの塩基配列(以下dbと記載)を基準に多重整列化を行なった(図3)。
【0067】
結果:
マダイのDループはいずれも約1054塩基対からなり、太平洋クロマグロ(db)Dループ(869塩基対)より189塩基対長い。4箇所にギャップを挿入することにより多重整列化が可能となった(図3)。特に養殖稚魚10尾間ではDループの配列は全て同一であった。同時に浮遊し採取された受精卵から生じる稚魚のミトコンドリアDループは同じ塩基配列を有することから、これらの稚魚が同一雌親に由来することが明らかとなり、また、同一の雌親に由来する受精卵や稚魚は、そのうちの1個体のミトコンドリアD−ループの塩基配列を決定することにより、全個体を同じ母系集団として容易に標識できることが示された。一方、産地の異なる各個体間では、約1054塩基対中、9〜24塩基の相違点が見られた(表2)。これらの変異のうち、マダイdbのDループnt.273〜311間に9カ所、nt370〜416間に9箇所が集中していたことから、この間を母系特異的領域とした。
【0068】
【表2】
【0069】
応用例2:マダイ母系特異的塩基配列の特定と簡易検出法
方法:
実施例2において決定した母系特異的領域の塩基配列に基づいて以下のプライマーを設計し、天然および養殖のマダイ異母系成魚3尾および同一母系稚魚5尾から抽出したミトコンドリアDNAを鋳型として、タッチダウンPCRにより増幅を行なった:
マダイ母系特異的PCRフォワードプライマー(図3、マダイ稚魚nt.273〜308:母系特異的変異点8ヶ所):
5’-AGAAATAGCAAGTGTTCAAGTATTTATCCCCAAAAC-3’(配列番号10)
マダイ母系特異的PCRリバースプライマー(図3、マダイ稚魚nt.402〜375:母系特異的変異点5カ所):
5’-GTGTTGGTACTTGGTAGACATTTGTCG -3‘(配列番号11)
【0070】
結果:
図4に、PCR増幅産物のアガロース電気泳動の結果を示す。母系が異なる天然および養殖成魚からはシグナルは検出されなかったが、同一母系の養殖稚魚5尾から129塩基対の母系特異的DNAが増幅された。したがって、同一母系の養殖稚魚(種苗)が、母系特異的塩基配列によって遺伝子標識されており、該母系特異的塩基配列に特異的に設計されたプライマーを用いたPCRにより容易に検出し得ることを確認した。
【0071】
実施例3:ブリのDループ解析例とクロマグロ、マダイとの比較
マダイ、ブリ、クロマグロは養殖魚売上げの90%を占め、いずれもスズキ目に属するがマダイはスズキ亜目タイ科,マグロはサバ亜目サバ科、ブリはスズキ亜目アジ科に属する。スズキ目はその他カツオ、マアジなど重要な回遊水産魚を数多く含む。これらの遺伝的類縁性を知るためにDループを比較した。ただし、ブリのミトコンドリアDNA塩基配列はデータベースに登録されていないため、クロマグロのtRNAプライマーで代用した。
【0072】
方法:
鳥取、福岡で水揚げされた天然ブリのミトコンドリアDNAを鋳型として、実施例1において使用したクロマグロのDループ両端プライマー(配列番号2および3)を用いて、約900塩基対のDNAを増幅した。両端プライマーの各々を用いて両鎖シークエンスを行なったところ、フェニルアラニン側では一度で全長を読み取ることができなかったため、第一のプライマーにより得た部分配列から次のプライマーを設計し、全長の塩基配列を決定した。
ブリ逆方向プライマー: 5’-GGTTTAACGCGCAAAAGCCGAG-3’(配列番号12)
【0073】
結果:
鳥取ブリ(配列番号13)と福岡ブリ(配列番号14)とのDループ塩基配列を整列化させるために、配列中1カ所にギャップを必要とした。699塩基中の690塩基配列が相同であり、相違塩基は8塩基であった。さらに試料を増やすことにより、ブリのミトコンドリアDNA Dループの塩基配列で品種(亜種)識別ができる。
なお、同じスズキ目間では、マダイ、マグロおよびブリの間では、随所に短い(最長33個)相同塩基はあるが、ギャップ挿入数が多く、多重整列化ができない状態であった。
【0074】
実施例4:マグロの種、品種および産地の識別と生産履歴管理(トレーサビリテイー)
MitoFishデータベースから入手した本まぐろ(太平洋クロマグロ:Thunnus orientalis)のミトコンドリアDNAの塩基配列(GeneBank登録番号AB185022:配列番号1)を標準材料として、以下の一般に市販されているマグロ属の種および亜種との間でDループの塩基配列の比較解析を行った:
【0075】
大西洋クロマグロ(Thunnus thynnus):ミトコンドリアDNA塩基配列(GeneBank登録番号AY302574:配列番号15)
ビンナガマグロ(Thunnus alalunga):ミトコンドリアDNA塩基配列(GeneBank登録番号AB101291:配列番号16)
メバチマグロ(インド洋産:Thunnus obesus:配列番号17)
キハダマグロ(宮城沖産:Thunnus albacares:配列番号18)
ミナミマグロ(オーストラリア産:Thunnus maccoyii:配列番号19)
【0076】
結果:
上記マグロ6種のDループ塩基配列は869塩基前後であったが、1種あたり10部位前後のギャップを挿入することにより、多重整列化を行うことができた。ただし、ギャップ挿入は22部位あり、特に母系特異的配列前半のnt172〜232に9部位が集中していた。各品種間では869塩基中39〜77塩基の違いがあったが(表3)、その他の領域では相同性を保っていた。特に、nt173〜215にギャップ12部位を含む35部位とnt628〜672にギャップ3部位を含む20部位に相違塩基が集中していた(図5−1〜3)。いずれも部分的に太平洋クロマグロ母系特異的配列と重複するが、ギャップの位置および変異の数が前記母系特異的配列とは明確に異なるので、種識別領域(同属異種識別領域)として特定することができる。
【0077】
【表3】
【0078】
常法に従い、残る相同領域から遺伝的類似性を求めた結果、大西洋クロマグロ/太平洋クロマグロ、ミナミマグロ/太平洋クロマグロ、大西洋クロマグロ/ミナミマグロ間の相違塩基数とギャップ挿入数はそれぞれ69(+8)、77(+5)、49(+2)であった。いずれも「トロ」の材料とされるこれらの種が、遺伝的には互いに遠位にあることが明らかとなった。
【0079】
この結果により、ミトコンドリアDループの塩基配列の相違により、マグロ属およびその種間の識別が可能であり、食品表示の偽装の防止および摘発などに有用であることが示唆される。ただし、同じ太平洋クロマグロ間でも最大22個の相違点があるが(表1)、マグロ属の種間識別はギャップ数を加味した相違塩基数で判断できる。
【0080】
実施例5:マグロの産地識別
方法:
図1および図3に示すように、マグロおよびマダイのDループ塩基配列を母系により識別することが可能となったが、母系は産卵場所や魚群,あるいは棲息海域で規定される可能性があるので、産地識別に利用できる可能性を調べた。棲息海域が広い回遊魚であるマグロのDループ塩基配列が産地によってどの程度異なるか、メバチマグロ、キハダマグロにおいて比較した。
【0081】
結果:
下記の産地が異なる個体のDループ塩基配列の比較では、ギャップを加えることなしに配列を整列化できた。以下に、同種のマグロでの産地による塩基配列の変異数を示す(括弧内の数値=相違塩基部位数/比較塩基部位数):
メバチマグロ :パラオ/インド洋(18/569)
大西洋/パラオ(28/569)
大西洋/インド洋(35/569)
キハダマグロ :インド洋/宮城沖(12/756)
太平洋クロマグロ:(6〜22/869)
【0082】
同種間の多重整列化ではギャップの挿入は必要なかったが、産地が異なる場合、太平洋クロマグロの母系特異的塩基配列に相当する領域に相違が見られた。すなわち、この領域の多型を用いて産地を識別し得ることが示唆される。
【0083】
本発明の方法により、産地の異なるマダイ、マグロのDループの塩基配列の相違で母系を識別することができるが、逆に塩基配列の相違から該当する産地を推定する場合、商品の生産履歴管理と同様に、比較対照となる試料を必要とする。好ましくは、産地または棲息海域に存在する魚群のDループ塩基配列データベースを作成し、データベース中の全配列と比較する。データベース作成と並行して産地識別を行ってもよい。
【0084】
実施例6:天然および養殖のマグロのD−ループ配列の比較
天然のマグロ集団と養殖稚魚との遺伝的相違を調べるために、築地市場に集積される天然のクロマグロと、近畿大学水産研究所から供与されたクロマグロ受精卵および稚魚のD−ループ塩基配列を、上記の方法により決定して比較した。
結果
漁場や水揚場の異なる535尾の太平洋クロマグロのうち、養殖稚魚と同じDループ配列を有する個体は存在しなかった(図6)。この結果から、本発明の方法を実際に用いて、例えば、漁獲された集団中の人工種苗を同定することができ、放流事業における加入量変動調査や、漁獲高に対する放流事業の貢献度を判定することや、育成・販売した種苗が他者により複製された場合に追跡を行うことなどが可能であることの裏付けがなされた。
【0085】
参考例1:マイクロサテライトによるハプロタイプ分析
同一母系の養殖クロマグロ稚魚11尾について、公知のマイクロサテライトマーカーTth208、Tth217、Tth4およびTth7-16(Clark et al., 2004, Mol. Ecol. Notes, 4:70-73)を用いてハプロタイプ分析を行った。少なくとも4パターンのハプロタイプが観察された(図7)ことから、少なくとも3個体の雄親と雌親(1個体)が産卵時に存在したことが示唆される。一方、天然のクロマグロについて同様の分析を行ったところ、分析を行った全個体間でマイクロサテライトパターンが異なり、同一のハプロタイプを示す個体は存在しなかった(データは示さず)。これらの結果から、遺伝的多様性が小さい養殖魚の場合は産卵時に存在する親魚の個体数が少ないので、受精卵全てのマイクロサテライトによるハプロタイプを決定することが可能であり、したがって、mtDNA Dループによる母系識別にマイクロサテライト分析による雄系識別を組み合わせることにより、より精度が高い系統標識が可能になることが示唆される。
【産業上の利用可能性】
【0086】
本発明は、水産資源保護として有用な魚介類のトレーサビリティーの提供を可能とするものであり、このトレーサビリティの提供により、食品としての魚介類の生産履歴または養殖もしくは栽培漁業のための稚魚の追跡(調査)を可能にすることにより、魚介類の品質および産地を保証し、生産履歴表示の偽装を摘発、防止し、また優良種苗の遺伝子標識による識別および養殖・育種者の権利の確立に寄与するものである。本発明はさらに、放流事業に用いる大量の稚魚の標識と洋上での追跡調査および加入量変動調査を可能にすることにより、水産資源の遺伝的多様性の維持および排他的経済区域内での栽培漁業の発展に寄与するものである。
【技術分野】
【0001】
本発明は、天然および栽培漁業における水産魚介類の遺伝子識別および標識方法に関する。より具体的には、本発明は、魚介類ミトコンドリアDNAのDループの塩基配列の相違に基づき、魚介類の品種、産地、系統(種苗)を識別および標識する方法、および養殖事業(栽培漁業)において育種された種苗の稚魚の識別および標識のための方法、ならびにかかる方法に用いるマーカー遺伝子の決定および検出の方法に関する。
【背景技術】
【0002】
日本においては海面養殖漁業および栽培漁業の技術開発が従来から進展しているが、特に近年、クロマグロの完全養殖が成功したことを筆頭として、マダイ、ヒラメ、シラスウナギなどにおいて、卵から繁殖可能な成魚を生産し、さらに成魚から次世代を再生産する、天然資源に依存しない完全養殖の技術が確立しつつある。世界的な食糧不足による食糧価格の高騰、日本の食糧自給率の低下、輸入食糧に関する安全性の問題などが火急の懸案である今日、より安全、安価かつ高品質の食糧を提供し得る供給源として、養殖および栽培漁業に対する関心が高まってきている。
【0003】
養殖および栽培漁業においては、優良系統、例えばウイルスや細菌などの病原に対する耐性の高い系統、成長速度が高い系統、繁殖能力の高い系統などを選択して繁殖させることにより、より高品質の人工種苗の育成が行われる。人工種苗の育成、特に食用とされる魚類を継代で完全養殖する試みには莫大な資本が必要であり、その資本回収は容易ではない。また、水産種苗においては、植物種苗のような遺伝子操作による不稔技術の導入は技術的・倫理的に困難であるため、育種・確立した優良種苗(稚魚)を販売・譲渡する場合、譲受人による増殖(繁殖)と販売が容易であり、種苗育成者の権利の保護が非常に困難であることが問題となっている。また、植物種苗については種苗法により登録された種苗(品種)育成者の権利が保護されるが、動物、特に水産魚介類の種苗に関しては、育成者の権利は法的にも実質的な保護がなされていない。このため、養殖・栽培漁業の分野において優良種苗の育成者および放流事業者の権利を保護し、水産業を活性化するために、水産種苗の登録制度の確立と種苗および成魚の標識および同定方法の開発が望まれる。
【0004】
また、栽培漁業、特に放流事業の加入量変動調査のために稚魚を標識する方法の開発も望まれている。例えば、近年、ミナミマグロに次いでタイセイヨウクロマグロも絶滅危惧種に指定され、漁獲制限がなされている。このような種について、人工的に育成された稚魚を放流して自然集団を回復させることが可能であると考えられる。一部において、遺伝子的に均一な種苗の大規模な放流が生物学的多様性を低下させ、ひいては水産資源の衰退を導くことが懸念されているが、むしろ、新たな種苗の放流により他系統との交配が進み、生態系における遺伝子の多様性を高めるので、漁場の回復に寄与すると考えられる。種苗の放流による漁場の回復のためには、遺伝子的組成の異なる新たな種苗の放流、および放流の評価を的確に行う加入量変動調査が必要である。加入量変動調査はまた、自然界における水産資源量の変動を把握するためにも重要であり、漁場における特定の種、品種、系統および種苗の正確な加入量変動調査は、放流事業が生態系に与える影響を把握し、かつ持続可能な漁獲計画のために必要である。
【0005】
放流稚魚を追跡調査するための従来の標識として、例えばサケ稚魚のあぶらびれの切除(北海道札幌市豊平川さけ科学館)、ヒラメ稚魚の無眼側の黒化および焼印(宮津栽培漁業センター)、マダラやトラフグにおける稚魚耳石のALC(アリザリンコンプレクソン)色素標識および腹鰭抜去標識(能登栽培漁業センターおよび南伊豆栽培漁業センター)、さらに近年ではICタグと通信衛星を利用する電子標識などが実施されている。いずれも稚魚に対して物理的に標識を行うものであり、鰭の切除や焼印などの手段は、種苗を傷つける危険があり、また市場における商品価値を損なう恐れがある。ALC色素標識は、種苗を傷つけずに標識を行うので放流後の成長や生存能に影響を与えないという利点がある一方、調査のためには魚を収集・解体して耳石を採取し、蛍光顕微鏡による確認を行わねばならない。また、これらの方法による標識は、放流する全個体に対して標識をする必要があるのみならず、標識個体一代に限られ、F1以降の世代にわたって識別可能なものではない。さらに、マグロやトラフグのような高級魚の場合、検査のために魚を購入することは、費用が莫大となるため実質的に不可能であり、また、クロマグロなどの場合、高価であるのみならず魚体が大きいのでサンプル採取や標識自体が困難であるなど、調査の実施には多くの問題が残されている。
【0006】
水産業における別の懸案事項は、市場に流通する際の水産物の表示に関する問題である。平成11年のJAS法改正により、一般消費者向けに販売される生鮮水産物について、名称、原産地(採捕・生産された水域名または輸出国名)、解凍・養殖の表示が義務づけられている。しかしながら、流通の現場においてこれらの表示の真贋を判定することは実質的に困難であるため、販売業者による水産物の産地(漁場・港)の偽装(国産ウナギとの虚偽表示により外国産ウナギを販売するなど)、種または品種の偽装(例えばクロマグロとの虚偽表示によりキハダマグロを販売するなど)が問題となっている。品種や漁場によって食品としての品質や本来の価格は異なるが、偽装の疑いがある場合の検証は容易ではなく、表示に依存せずに生産履歴を正確かつ容易に追跡できるシステムも存在しないため、食品の表示に対する信頼が失われつつある。したがって、水産物の表示をより厳格にし、ひいては産地消費者がより安心して水産物を選択できるように、流通過程において水産物の生産履歴を厳正に表示し、必要に応じて迅速かつ正確に判定できる方法の開発もまた望まれている。
【0007】
DNA塩基配列の相違に基づいて生物種または亜種内の識別を行う手法は、分子遺伝学、系統進化学などの分野において非常に有用であり、特にヒトにおいては医療、法医学などの分野においても利用されている。魚類においては、例えば、特定の遺伝子マーカーを用いて白化しやすいヒラメを判別する方法が開発されている(特許文献1)。しかしながら、特定の形質をもたらす遺伝子多型をマーカーとする方法では、形質毎にマーカーとなる遺伝子型を同定する必要があり、上記のような養殖および栽培漁業の分野における問題を解決するためには有効ではない。
【0008】
種苗の遺伝子標識を行うために、MHCマイクロサテライトなどのゲノム多型マーカーを用いることも試みられたが、遺伝子組成の限られた養殖の受精槽においてすらMHCの配列の多様性は極めて高く、個体間においても大きく異なるため、MHCハプロタイプにより種苗を標識し同定することは困難であった(本発明者らによる実験:データは示さず)。
【0009】
ミトコンドリアは、核ゲノムとは独立して独自の10数種類の遺伝子群を含む約15,000〜17,000塩基対(ヒトでは16,569塩基対)の環状二本鎖DNAを有する。ミトコンドリアDNA(mtDNA)は母性遺伝により子孫に伝えられるので、遺伝学の分野において利用される。また、mtDNAは、塩基置換速度が核ゲノムと比較して5〜10倍早く、同一種内における変異が多いので、ハプロタイプ識別のために利用されている。例えば、シロザケのmtDNAコントロール領域(制御領域、D−ループ)の多型をハイブリダイゼーションにより検出し、ハプロタイプを判定する方法が開発されている(特許文献2)。
【0010】
Dループは、mtDNA中の非翻訳高度可変領域であり、同一種内においても多く変異が発生する。非特許文献1には、リュウキュウアユについて、自然集団とこれを起源として異なる環境に導入された導入集団の遺伝的組成を比較する目的で、mtDNA Dループのマイクロサテライトを制限酵素切断片長多型(RFLP)分析によりハプロタイプ分析を行い、起源となったリュウキュウアユ自然集団と導入集団との間で遺伝的組成が類似していることを確認したことが記載されている。また、非特許文献2は、キンメダイ属3種のミトコンドリア制御領域(D−ループ)の塩基配列を比較したところ15.5%の塩基置換率であったこと、および、異なる漁場から採集されたキンメダイの全てがそれぞれ独自のハプロタイプを示し、共通の配列を示すものは見られなかったことを記載している。
【0011】
非特許文献3は、ミトコンドリア制御領域中の超可変領域の配列を用いて、クロマグロおよびメカジキの地理的亜種間の遺伝的相関関係を調査した結果、大西洋と地中海とで捕獲されたクロマグロの遺伝子構成に分離は見られなかったことを記載している。一方、非特許文献4は、大西洋クロマグロ当歳魚の集団毎の遺伝的構成をマイクロサテライト遺伝子座とミトコンドリア制御領域を用いて調査した結果、ミトコンドリア制御領域の配列類似性と地理的配置との間に有意な関連性を見出したことを記載している。また、非特許文献5は、ミトコンドリア制御領域5’末端のハプロタイプにより分けられる2つのクレイド(clade)を用いた個体群統計学により、メバチマグロの地理的な遺伝子の流出入を調査することを記載している。
【0012】
これらの文献に開示される方法は、個体のハプロタイプを判定し、特定の地域におけるハプロタイプ頻度を調べることにより、個体群間の関係を調査するためのものである。例えば特許文献2から理解されるように、ある特定のハプロタイプは、複数の個体において観察され、また複数の異なる地域にわたって分布する。したがって、これらの方法によっては、特定の個体または個体群についてハプロタイプまたはハプロタイプ組成を判定することはできるが、その個体が属する母集団(系統または産地)を判定することはできず、またその個体または個体群を遺伝子により標識することも不可能であった。また、例えば非特許文献2から理解されるように、D−ループの塩基配列は同種の個体間であってもハプロタイプの差異が大きいため、種や系統の識別に用いることは不可能であった。
【0013】
非特許文献6は、クロマグロの養殖に関して、養殖系統を継代して育種を行う場合の遺伝的多様性のモニタリングのために、生け簀内で天然親魚約20個体から得られた孵化仔魚について、母系の推定のためにミトコンドリアDNAのDループ前半領域のシークエン20分析を、父系の推定のためにマイクロサテライトマーカー分析を行ったことを記載している。各ロット(同日に回収された受精卵)内で3〜8種類のミトコンドリアDNAハプロタイプが観察され、そのハプロタイプ頻度はロット間で大きく異なり、マイクロサテライトマーカー分析においても、ロット間の遺伝的構成が異なることが明らかとなり、生け簀内の魚群の産卵に関与する親魚が産卵日によって異なることが示唆された。しかしながら、特定の種苗を標識し、放流後および流通の過程においてその履歴を管理・追跡する方法は、未だ見出されていない。
【0014】
このように、水産魚介類の特定の個体あるいは個体群について、その種、品種、系統または産地を簡易かつ正確に識別し、大量の個体を標識することは、従来のいずれの方法によっても行うことができず、かかる方法の開発が望まれていた。
【先行技術文献】
【特許文献】
【0015】
【特許文献1】特開2007-312645号公報
【特許文献2】特開2003-180397号公報
【非特許文献】
【0016】
【非特許文献1】池田他、「マイクロサテライトDNAおよびミトコンドリアDNA DループのPCR−RFLP分析によるリュウキュウアユ導入集団の遺伝的評価」、水産育種、2001年、第31巻、pp.33−37
【非特許文献2】秋本他、「ミトコンドリア制御領域の塩基配列分析による日本周辺漁場におけるキンメダイの集団遺伝構造の解析」、神水研研報第8号、2003年、pp.89-97
【非特許文献3】Carlsson et al., "Microsatellite and mitochondrial DNA analyses of Atlantic bluefin tuna (Thunnus thynnus thynnus) population structure in the Mediterranean Sea", Molecular Ecology, 2004, vol.13, pp.3345-3356
【非特許文献4】Martinez et al., "Genetic diversity and historial demography of Atlantic bigeye tuna (Thunnus obesus)", Molecular Phylogenetics and Evolution, 2006, vol.39, pp.404-416
【非特許文献5】Alvarado Bremer et al., "Comparative phylogeography of Atlantic bluefin tuna and swordfish: the combined population expansion on the regional phylogenies of two highly migratetory pelagic fishes", Molecular Phylogenetics and Evolution, 2005, vol.36, pp.169-187
【非特許文献6】森島他、「養殖クロマグロの遺伝的管理に向けた研究」、平成19年度日本水産学会近畿支部後期例会プログラム、109-D、平成19年12月1日、日本水産学会(http://www.suisan-kinki.org/reikai/h19reikai/h19_kouki/9.pdf)
【発明の概要】
【発明が解決しようとする課題】
【0017】
したがって、本発明は、上記の問題点に鑑み、水産魚介類、特に、特定の産地に由来する水産魚介類、一つまたは複数の優良形質を有する優良水産種苗、または特定の種または品種の水産魚介類の1個体〜大量の個体を、標識・識別するための簡易かつ正確な方法を提供することを目的とする。
【課題を解決するための手段】
【0018】
本発明者らは、魚類の行動様式、繁殖形態に着目した。例えばクロマグロの産卵においては、一個体の雌親魚が一回に約200万〜1,000万個の卵を産卵、孵化する。その受精には複数の雄親魚が関与していると考えられる。また、養殖の場合は特に、一つの養殖槽において一日に産卵する雌親個体の数は少なく、受精卵は約1日で孵化して水面に浮揚し、産卵日毎に回収することが可能である。本発明者らは、これと併せてミトコンドリアDNA(mtDNA)が母系特異的であることに着目した。ミトコンドリアDNAは一般的にサイズが小さく、組み換えが起こらないため個体間での保存性が高いが、Dループの短い領域に変異が集中している。これらのことに焦点を当て鋭意研究を進めた結果、水産魚介類のプロフィールの同定、例えば生産履歴、種、品種、系統または産地などの識別が可能であることを見出し、さらに研究を重ね、発明を完成するに到った。
【0019】
すなわち、本発明は、水産魚介類またはその加工品のミトコンドリアDNA Dループの塩基配列を決定することにより、該水産魚介類またはその加工品の種苗または生産履歴を同定する方法であって、
養殖場において継代した雌親個体から採取された第1の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第1の塩基配列)を決定し、
水産魚介類またはその加工品から採取された第2の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第2の塩基配列)を決定し、
第1の塩基配列と第2の塩基配列とを比較し、前記水産魚介類またはその加工品が前記雌親個体の後代であることを同定することによって、前記水産魚介類またはその加工品の種苗または生産履歴を同定することを含む、前記方法に関する。
【0020】
また、本発明は、マイクロサテライトにより核型を決定することをさらに含む、前記方法に関する。
【0021】
本発明はさらに、ミトコンドリアDNA Dループの種特異的塩基配列が1から2以上のギャップの挿入により整列化できるか否かの判定をさらに含む、前記方法に関する。
【0022】
本発明はまた、ミトコンドリアDNAのDループ全長の塩基配列を、ミトコンドリアプロリンtRNA特異的プライマーおよびミトコンドリアフェニルアラニンtRNA特異的プライマーを用いて増幅し、両鎖シークエンシングにより決定することをさらに含む、前記方法に関する。
【0023】
また、本発明は、第1の塩基配列と第2の塩基配列との比較が、母系特異的塩基配列が完全に一致するか否かの比較である、前記方法に関する。
さらに、本発明は、水産魚介類またはその加工品が、食品である、前記方法に関する。
【0024】
本明細書において使用される場合、品種とは、同種内の小分類であって、他の集団とは形態、生理、遺伝的に区別し得るような特徴を有する集団を指し、分類学的に亜種として分けられる場合もある。例えば、クロマグロは、一般に、大西洋、地中海および黒海産(タイセイヨウクロマグロ)と、太平洋産(タイヘイヨウクロマグロ)との2つの亜種に分類され、さらに学説では、大西洋産、地中海産および黒海産は別の遺伝学的特徴を有する群に分けられるとされていたが、本明細書中以下に記載する本発明者らの解析により、タイセイヨウクロマグロとタイヘイヨウクロマグロとは別品種として分類される。
【0025】
本明細書において用いる場合、種苗(人工種苗)とは、優れた(または生け簀養殖もしくは放流に適した)形質を有する個体を人為的に選抜して交配し、その次代を系統として養成し、さらに個体または系統の選抜を行うことにより、優良形質を有する系統を繁殖のために確立したものを指し、受精卵、稚魚、仔魚および成魚を含み、養殖のためのものであっても、海川に放流されるためのものであってもよい。
【0026】
本明細書において用いられる場合、生産履歴とは、水産魚介類の個体もしくはその一部またはそれらの加工品が流通の過程にある場合において、生産地(養殖、採取もしくは加工などの場)、生産方法(養殖、採取または加工の方法もしくは条件など)、保存や流通の方法もしくは条件、または加工品である場合原料となる水産魚介類の種、品種もしくは系統などの情報を意味する。また、産地とは、対象となる水産魚介類が由来する水域または施設を意味し、産卵場、餌場、回遊場、漁場、水揚場、孵化および養殖場などを含む。
【0027】
本明細書において用いる場合、母系特異的配列とは、検体となる個体のミトコンドリアDNAのDループ塩基配列と、比較対照となる個体のミトコンドリアDNAのDループ塩基配列とを整列化(アラインメント)して相同性を比較した場合、同一の母系に由来する個体間では完全に一致するが、異母個体との間では、0〜2個程度のギャップの挿入で整列化でき、高度の相同性を示すが完全には一致しない領域の塩基配列を指す。種特異的配列とは、複数の異なる種のDループ塩基配列が数個以上のギャップの挿入で整列化できる場合において、異種間で相同性が高い塩基配列を指す。さらに、本発明者らは、母系特異的配列中に、同一種間でも異なる母系の間で、特に多くの変異が集中する短い領域が存在することを見出した。この領域を本明細書において母系変異集中領域と称する。同様に、異種間でギャップにより整列化できるが多くのギャップが集中する短い領域が存在することを見出し、この領域を異種変異集中領域と称する。
【発明の効果】
【0028】
本発明により、水産魚介類の種苗、産地および品種を簡便かつ迅速に識別することが可能となり、したがって、該水産魚介類の生産履歴および品種などの表示を厳正に管理することができ、種苗や食品としての水産魚介類を、より安全かつ公正に提供することが可能となる。特に輸入やインターネット販売などにより流通される、従来では生産履歴の追跡が困難であった水産物の生産履歴を管理することが可能となり、偽装表示による水産物販売を防止・摘発することができるようになる。また、本発明により、養殖魚介類の優良種苗、例えば、GTL、ウイルス耐性などのの優良種苗を遺伝子識別することが可能となり、種苗のブランド化および育種者の権利の保護がより容易となる。また、本発明により、母系特異的遺伝子配列により標識された種苗を大量生産することができ、これと同時に母系特異的配列のデータベースを構築することにより、放流事業における加入量変動調査が可能となるのみならず、生態系において特定の系統の個体数の増減をモニタリングすることが容易となり、水産魚介類の遺伝的多様性の維持に有用な資料を提供することができる。
【図面の簡単な説明】
【0029】
【図1−1】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−2】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−3】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−4】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−5】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−6】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−7】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−8】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−9】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−10】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【図1−11】マグロ属のミトコンドリアのDループ塩基配列の比較を示す図である。
【0030】
【図2】PCRにより増幅されたマグロ属における母系特異的DNAの電気泳動の結果を示す。
【図3−1】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【図3−2】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【図3−3】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【図3−4】天然および養殖マダイのミトコンドリアDNA Dループ塩基配列の多様性を示す図である。
【0031】
【図4】PCRにより増幅されたマダイにおける母系特異的DNAの電気泳動の結果を示す。
【図5−1】マグロ属のミトコンドリアDNA Dループ塩基配列の比較を示す図である。
【図5−2】マグロ属のミトコンドリアDNA Dループ塩基配列の比較を示す図である。
【図5−3】マグロ属のミトコンドリアDNA Dループ塩基配列の比較を示す図である。
【図6】天然および養殖のマグロのミトコンドリアDNA D−ループ配列の比較を示す図である。
【図7】同一の母系に属する養殖クロマグロ稚魚の4種のマイクロサテライト分析の結果を示すゲル電気泳動の写真である。
【発明を実施するための形態】
【0032】
本発明は、一態様において、水産魚介類またはその加工品のプロフィールを同定する方法を提供する。該方法において、予め採取された水産魚介類のミトコンドリアDNA Dループの塩基配列を決定し、該決定された塩基配列と、同定されるべき検体としての水産魚介類またはその加工品のミトコンドリアDNA Dループの塩基配列とを比較することにより、検体のプロフィールを同定する。プロフィールとは、好ましくは種、品種、系統、生産履歴および/または産地であるが、これらに限定されない。特定の種、品種系統の種苗に特異的な遺伝子マーカーを見つけだすことは容易ではないが、本発明の方法においては、一個体の雌親から多数の次世代の個体が生産される水産魚介類の特徴を利用して、異なる種、系統または母系の間で可変性が特に高いミトコンドリアDループ領域の塩基配列の相違を、種、系統などのマーカーとして用いる。ミトコンドリアDNAは少量のサンプルから得られるコピー数が多く、核DNAが劣化するような条件下においても安定性が高いので、魚類のプロフィールの情報源として最も好適である。
【0033】
一態様において、本発明の方法は、水産魚介類もしくはその一部またはそれらの加工品またはこれらの雌親個体から、ミトコンドリアDNAを含む試料を、生産履歴の初期において予め採取して保存することを含む。生産履歴の初期とは、例えば、採集(漁獲)、生産、卸売、流通、販売などの場を意味し、例えば天然または養殖の魚介類の場合、水揚げした漁業または養殖業の従事者が、鱗、鰭、少量の肉片など該魚介類のミトコンドリアDNAを含むサンプル(試料)を採取し、凍結保存することができる。また、受精卵から養殖した仔魚(稚魚)を販売する場合、同一の雌親が産卵した(同腹の)1以上の受精卵、仔魚もしくは稚魚または雌親の鱗などを採取して凍結保存することができる。本発明の方法により同一母系の稚魚全個体が同一のマーカー(母系特異的配列)により標識されるので、1個体の同腹稚魚のサンプルを保存することにより、放流または販売後の全個体を追跡調査することが可能となる。あるいは、例えばマグロのように解体して流通・販売する場合、卸売場、市場または販売店などにおいて、各個体のサンプルを採取して凍結保存することができる。このようにして、個体の一部をサンプルとして予め採取して保存することにより、特定の個体の全てまたは一部(切り身、加工品など)を販売した後においても、該個体に由来する販売品に含まれるミトコンドリアDNAと、予め採取された同一個体のサンプルに含まれるミトコンドリアDNAとを比較することができる。
【0034】
本発明の方法は、好ましい態様において、前記水産魚介類またはその加工品、または種苗の場合は雌親個体から採取された試料(第1の試料、標準試料)に含まれるミトコンドリアDNA制御領域(D−ループ)の塩基配列中の、母系特異的または種特異的な塩基配列を決定することを含む。例えば、種特異的塩基配列を決定するために、第一に、目的とする種についての指標とする配列を得ることが必要である。具体的には、GeneBankやMitoFish(魚介類ミトコンドリアの公開データベース(http://mitofish.ori.u-tokyo.ac.jp/))などの公共のデータベース(以下dbと略)から、同種または近縁種のmtDNAの塩基配列を検索して用いてもよい。養殖稚魚の標識のための母系特異的配列を得るためには、雌親、または該雌親が産卵した少なくとも1の受精卵、仔魚もしくは稚魚からミトコンドリアDNAを得、公知の分子生物学的手法を用いてDループの塩基配列を決定すればよい。このD−ループ塩基配列とデータベースや他の個体のD−ループ塩基配列とを比較することにより、母系変異集中領域を決定し、これを母系特異的配列として用いることもできる。同一の雌親が産卵した受精卵およびそれに由来する仔魚もしくは稚魚は、全個体が同一の母系特異的配列を有し、この母系特異的配列は他の雌親に由来する個体とは異なる。したがって、1の受精卵、仔魚もしくは稚魚の母系特異的配列が、同一母系集団中の全個体の母系特異的配列となる。
【0035】
母系特異的配列または種特異的配列を決定するために、特定の種、品種、系統または産地に由来する水産魚介類のmtDNA Dループ配列と、他の種、品種、系統または産地に由来する個体の対応する配列とを比較し、識別可能なパーセンテージで塩基の置換が生じる領域を決定する。
【0036】
この領域は、異なる母系の個体間において、例えば800〜1600塩基長の領域中であれば、5〜10個(1%)、好ましくは11〜15個(1.5%)、より好ましくは16〜30個(3%)の塩基の置換を有する領域である。同一母系の個体間においては、この領域の塩基配列が完全に一致することが好ましいが、他の母系との識別が可能である限りにおいて、1または2程度の塩基の置換を含んでもよい。より正確な識別のために、同一の母系の個体間における塩基の置換は、0.2%程度以下、好ましくは約0.1%程度以下、さらに好ましくは0%(完全一致)であることが好ましい。
母系特異的配列の典型的な位置は、Dループのプロリン側末端の塩基を1番目の塩基として、約100〜600番目、好ましくは約150〜450番目の間の塩基配列中に存在する。例えばクロマグロにおいては母系特異的配列は約150〜350番目、マダイにおいては約250〜450番目の間の塩基配列中に存在する。
【0037】
また、種を識別するための領域は、複数の異なる種間で1から2以上のギャップの挿入によりDループ配列が整列化できる場合において、異種間で相同性を示す塩基配列である。異種間での相同性は、90〜97%、好ましくは90〜95%、より好ましくは90〜93%である。複数の配列を比較する多重整列化解析(マルチアラインメント)には、例えばMEGA4(登録商標、Molecular Evolutionary Genetic Analysis Software Version 4.0、Tamura et al., Mol. Biol. Evol., 2007, 24(8):1596-1599)、SEQUENCHER(登録商標)(Gene Codes、米国)などのシークエンス解析のための公共または市販のソフトウェアを用いることができる。
種特異的配列の典型的な位置は、母系特異的配列と部分的に重複する場合もある。例えばクロマグロにおいては種特異的配列は約150〜700番目、特に約150〜250番目および約600〜700番目に見出すことができる。
【0038】
次いで、検体となる水産魚介類から採取された試料(第2の試料、検体)に含まれるミトコンドリアのDループDNAを、上記の母系特異的配列または種特異的配列と比較することにより、その種、品種、系統、生産履歴および産地の識別を行なうことができる。例えば、上記の母系または異種特異的塩基配列に基づいて設計されたプライマーまたはプローブを用いて、公知の手段により、検体のmtDNA中に母系特異的塩基配列または異種特異的塩基配列と同一の配列が含まれるか否かを容易に検出することができる。かかる目的のための公知の手段として、PCR法、RCA法、ダイレクトシークエンス法、TaqMan法、DNAアレイ法、FISH法などが挙げられるがこれらに限定されない。
【0039】
本発明の方法において、さらに、ミトコンドリアDNA Dループの母系または種特異的配列に加えて、他のハプロタイプの比較を組み合わせることにより、識別をより厳密に行ってもよい。通常のハプロタイプ分析では個体間における相違が大きいので、種や種苗の同定には用いることができない。例えば、非特許文献2に示されるとおり、同腹仔のミトコンドリアDNAにおいてすら、マイクロサテライトのハプロタイプは異なる。しかしながら、限られた個体数で繁殖を行う養殖においては、F1におけるハプロタイプの数が比較的少なく、例えば、同じ雌親から生じた11個体の稚魚において、4種のマイクロサテライトにより識別されるハプロタイプは4種類であった(参考例1)。このことを利用して、D−ループによる識別を補強することができる。例えば、放流する人工種苗(稚魚)の標識として、D−ループ母系または種特異的配列のみならず、マイクロサテライトなどにより核型のハプロタイプも決定しておくことにより、万が一、人工種苗と同一の母系に属する個体が天然に存在する場合であっても、採集(漁獲)されたサンプルが天然のものであるか人工種苗であるかを確実に判断することができる。
【0040】
本発明の方法は、好ましい態様において、前記母系特異的塩基配列、母系変異集中領域または異種特異的塩基配列に基づくプライマーまたはプローブの作成を含む。プライマーは、Dループ全長の増幅のためのプライマー、または、母系特異的塩基配列、母系変異集中領域もしくは種特異的塩基配列の特異的増幅のためのプライマーを含む。プローブは、母系特異的塩基配列、母系変異集中領域または種特異的塩基配列に特異的に結合する核酸、例えばオリゴヌクレオチドを含む。目的とする母系特異的または種特異的塩基配列の一部に対して少なくとも部分的に相補的となるように設計したプライマーまたはプローブを用いて、定法により、検体中に目的の配列を含むmtDNAが含まれるか否かを容易に決定することができる。具体的には、例えば、栽培漁業における加入量変動調査において、検体となる魚の個体のmtDNAを鋳型として、調査対象の種苗(母系)に特異的な配列に基づいて設計されたプライマーを用いてPCRやダイレクトシークエンスを行うことにより、該個体が目的の種苗であるか否かを容易に判定することができる。あるいは、産地または系統などによりデータベース化した複数〜多数の母系特異的配列と、検体の配列とを、DNAアレイ法などにより比較し、検体がデータベース中に記載される産地または系統に該当するか否かを容易に判定することができる。
【0041】
別の態様において、前記水産魚介類もしくはその一部またはそれらの加工品に、これらが前記母系特異的または異種特異的塩基配列により標識されていることを記載する文書またはラベルを添付してもよい。この文書またはラベルはまた、それらが添付されている水産魚介類は、その生産履歴の初期において予めサンプルが採取され保存されているか、および/または既に母系または種特異的配列が決定されており、必要に応じて(例えば該水産魚介類の購入者または使用者の希望により)、該水産魚介類に含まれるミトコンドリアDNAの塩基配列を母系または種特異的配列と比較することが可能であることを示すことができる。かかる文書またはラベルを添付された水産魚介類やその加工品は、生産者から離れて流通した後においても生産履歴を追跡することが可能であり、また、遺伝子標識により検査を行うため産地や品種の偽装が困難となることから、より安全性の高い食品を提供することが可能となる。
【0042】
一態様において、同一の種、品種、系統、生産履歴および/または産地に属する複数の雌親に由来する複数の母系特異的または種特異的配列を、産地、品種または系統によりプールしてデータベース化し、検体の同定に役立てることもできる。例えば、養殖場において恒常的に雌個体の母系特異的配列を取得してデータベース化することにより、継代養殖され流通されているブリやマグロなどの水産魚介類が由来する養殖場および出荷時期などを追跡することができる。または、特定の産地について同様のデータベースを構築し、産地表示の真偽を判定するために役立てることもできる。具体的には、目的の水域におけるサンプリングにより得られる対照試料を基に、1または2以上の母系が存在する集団として、産地を識別する。例えば、特定の水域における定着性が強く母系の数が比較的少ない集団であれば、該集団における母系特異的配列をプールしたデータベースを作成することにより、該データベースに含まれる母系特異的配列により産地を特定することができる。
【0043】
別の態様において、本発明は、上記の方法において、検体となる水産魚介類もしくはその一部またはそれらの加工品から、ミトコンドリアDNAを含む試料を採取して保存することを含む。本発明の方法によって生産履歴を追跡することができる水産魚介類もしくはその一部またはそれらの加工品は、生きている魚介類であっても、または生鮮、加熱調理済み、乾燥もしくは冷凍の魚介類であっても、それらの鱗、鰭、鰓、尾、皮膚、骨、体液、筋肉または内臓組織などであってもよい。一般に、ミトコンドリアは一つの細胞内に多数(数百〜数千個)存在するため、少量のサンプルから充分なmtDNAを得ることができ、また核DNAと比較して構造的に安定であるため、核DNAが劣化する条件下においても抽出できる。したがって、本発明の方法により、食品もしくは動物飼料またはそれらの原材料として加工された水産魚介類の生産履歴を調査することが可能となる。特に、魚類の種や亜種の判定は、流通の経路の下流、すなわち、対象となる魚介類が既に販売、加工、調理された状況において主として行われることが予測されるが、本発明の方法に従ってミトコンドリアDNAを利用することにより、加工・調理・冷凍・解凍などの処理の後であっても、少量のサンプルが得られれば、魚種を判定することができる。より正確な生産履歴調査のために、組織試料は、非加熱の試料の場合は約0.2g以上(鱗の場合数枚)、加熱された試料の場合は約1g〜5gであることが好ましい。採取した試料は、サンプルチューブに密封し、−20℃〜−80℃で凍結保存する。ミトコンドリアDNAの抽出・精製は、定法にしたがってアルカリ法などにより行い、TEに溶解する。長期保存する場合は、アルコールで沈殿させて−20℃〜−80℃で保存するか、またはポリマー、ビーズもしくは紙などの担体を用いてDNAを室温で長期保存する方法も開発されている(例えば「DNAブック」:Kawai他、Genome Research,2003,13:1488-1495を参照)。
【0044】
本発明の一態様において、養殖または栽培漁業において育成された種苗の受精卵、仔魚または稚魚の生産履歴を追跡する方法を提供する。養殖又は栽培漁業においては、親魚の産卵期の観察により、同腹の(雌親が同じ)受精卵および仔魚(稚魚)の群を得ることが可能である。本発明の方法により、同時期に浮遊した個体群を、同一の母系特異的配列により遺伝子標識された種苗とみなすことができる。したがって、継代養殖や優良株との交配によって育成した優良種苗を、雌親によって分けた群として識別することにより、各々の群の雌親のmtDNAのDループ母系特異的塩基配列により標識された母系群とすることができる。例えばマグロなどの場合、一個体の雌魚から生産された数百万〜数千万個体の種苗の全てに、魚体を傷つけることも遺伝子組み替えなどを行うこともなく、均一な標識を行うことができ、放流後の追跡調査、特に母系別の加入量変動調査を容易に行うことができる。また、種苗の近親交配による遺伝的障害を避ける指標を提供することもできる。また、他者が不当に種苗を複製した場合にこれを摘発することが技術的に可能となるので、種苗を育成する業者の保護に役立てることができる。
【0045】
本発明の方法は、水産魚介類の種類によって限定されるものではないが、特に、広域に流通し、品種、産地または種苗により安全性、栄養価、価格などが変化する水産魚介類の生産履歴の追跡に、特に有用である。具体的には、スズキ目サバ亜目サバ科マグロ属(例えば、クロマグロ、ミナミマグロ、キハダマグロ、メバチマグロ、ビンナガマグロなど)、同サワラ属(サワラなど)および同カツオ属(カツオなど)、スズキ目スズキ亜目アジ科ブリ属(ブリ、カンパチ、ヒラマサなど)および同マアジ属(マアジなど)、スズキ目スズキ亜目スズキ科(スズキなど)、スズキ目スズキ亜目タイ科マダイ亜科(マダイなど)、フグ目フグ科(トラフグ、マフグなど)、カレイ目カレイ亜目(ヒラメ、カレイ)、サケ目サケ科サケ属(ベニザケ、シロザケ、ギンザケ、カラフトマス、マスノスケなど)、ウナギ目ウナギ亜目(ウナギ、アナゴ、ハモ)、貝類(ハマグリ、アサリ、ホタテ、シジミ、およびサザエ)などが挙げられる。
【0046】
一態様において、本発明の方法は、ミトコンドリアDNAのDループ全長の塩基配列を、ミトコンドリアプロリンtRNA特異的プライマーおよびミトコンドリアフェニルアラニンtRNA特異的プライマーを用いて両鎖シークエンシングにより決定することをさらに含む。ミトコンドリアDNAに含まれる遺伝子の配置や塩基配列は、水産魚介類の近縁種の間で、Dループを除いて、保存性が高い。Dループは0.8〜1.5kb程度の塩基長を有する高度可変領域であるが、その3’末端側において隣接するプロリンtRNA遺伝子および5’末端側において隣接するフェニルアラニンtRNA遺伝子の塩基配列は保存性が高い。したがって、Dループを挟んでプロリンtRNAおよびフェニルアラニンtRNAの配列に対して各々プライマーを設計することにより、Dループの全長をPCRで増幅することができる。プロリンtRNAおよびフェニルラニンtRNAのプライマーによりPCRでDループDNAを増幅した後、酵素などによりPCR産物中のプライマーを消化および未反応dNTPを不活性化し、次いで、増幅に使用した2種のPCRプライマーの各々を用いて、それぞれの末端からシークエンスを行なう。配列の信頼性の観点から、両鎖シークエンスを行うことが好ましい。シークエンスの精度をあげるために相補鎖が一致した場合のみDループの塩基配列とする。両鎖が重複しない領域は得られたDループ内側から再度シークエンスする。
【実施例】
【0047】
以下に例を示して本発明をより詳細に説明するが、本発明の範囲は、これらの例によって何ら限定されるものではない。また、本発明の方法を適宜改変することは、当業者の通常の技術の範囲内である。
【0048】
材料および方法
1.ミトコンドリアDNAの抽出・精製法
試薬の組成:
(1)P1溶液(再懸濁試薬:250ml、2〜8℃保存)
0.5M EDTA 5.0ml
1M Tris−HCl(pH 8.0) 12.5ml
滅菌水で250mlに増量し、オートクレーブ滅菌する。
(2)P2溶液(溶菌試薬:250ml、室温保存)
SDS 2.5g
NaOH 2.0g
滅菌水で250mlに増量する。
(3)P3溶液(除蛋白沈殿試薬:500ml、4〜8℃保存)
酢酸カリウム 147.3g
酢酸 60.0ml
滅菌水で500mlに増量し、pH 5.5に調整し、オートクレーブ滅菌する。
【0049】
ミトコンドリアDNAの抽出:
(1)水産魚介類の組織約0.2g(鱗の場合数枚)の試料を、サンプルチューブに移し、P1溶液を0.4ml添加し、ペスツル棒でサンプルを潰す。
(2)P2溶液を0.4ml添加し、ペスツル棒で軽く混合する。
(3)P3溶液を0.4ml添加し、チューブの底を指で弾いて攪拌する。
(4)室温で5分間、20,000×gで遠心分離し、上澄を別のチューブに移す。
(5)フェノール・クロロホルム溶液を0.4ml添加し、ボルテックスミキサーにかける。
(6)再び遠心分離を行ない、上澄を回収する。
(7)5、6の操作を繰り返す。
(8)得られた上澄の0.6倍容積のイソプロパノールを加える。
(9)20,000×gで30分間遠心分離し、上澄を捨て、70%エタノールで沈殿を洗浄する。
(10)再び遠心分離した後、上澄を捨て、65℃で5分間静置して乾燥させる。
(11)0.1MのTE 30μlに溶解させ、ミトコンドリアDNA溶液とする。
【0050】
2.DループDNAの増幅
例えば以下の手順に従ってミトコンドリアのDNAのDループ塩基配列を増幅する:
(1)魚介類ミトコンドリアのDNAの塩基配列は、MitoFish(http://mitofish.ori.u-tokyo.ac.jp/)、GeneBank(http://www.ncbi.nlm.nih.gov/Genbank/)などの公開データベース(以下dbと略)から得られる、目的の種と同種、あるいは近縁種のmtDNAの塩基配列中、Dループの両側にあるプロリンtRNAおよびフェニルラニンtRNAの遺伝子配列からPCRプライマーを作製する。なお、dbに含まれていない新規の試料の場合は、近縁種のプライマーで代替えする(成功例:ブリ(実施例3を参照))か、あるいはミトコンドリアDNAのショットガンシークエンスなどにより配列を得る。
【0051】
(2)以下の組成でPCR反応用カクテルを作成し、試料のミトコンドリアDNAを0.5μl添加する:
10×PCRバッファー:2μl、25mM MgCl2:2μl、2mM dNTPs:2μl、5U/μl Taqポリメラーゼ:0.1μl、16.7pmol/μlプライマー(フォワード):1μl、16.7pmol/μlプライマー(リバース):1μl、精製水:11.3μl。
【0052】
(3)熱変性95℃ 10分、DNA増幅(95℃ 1分、65℃ 30秒、72℃ 1分)×34サイクル、72℃ 5分の反応条件で、PCRを行う。
(4)PCR反応液7.1μlを384ウェルPCRプレートに分注し、Exo SAP IT(USB Co.)を2.9μl添加する。
(5)37℃で15分間反応後、80℃で15分間熱変性する。
【0053】
3.DループDNA両鎖シークエンス法
PCRでDNAを増幅した後、Exo SAP IT(USB Co.)などにより、PCR産物中のプライマーを消化および未反応dNTPを不活性化し、次いで、増幅に使用した2種のPCRプライマーの各々を用いて、それぞれの末端から、例えば以下の手順でシークエンスを行なう。
【0054】
(1)Exo SAP ITの反応液を2分割し(各5μl)、384ウェルPCRプレートの空のウェルに移す。
(2)シークエンス用カクテルをABI社マニュアルに従って以下の組成で作製する:
フォワード(またはリバース)プライマー:16.7pmol/μlを1μl、5×シークエンスバッファー:2μl、Bigdye:2μl。
(3)ヒートシールで密封し、熱変性96℃ 1分、伸長(96℃ 10秒、50℃ 5秒、60℃ 4分)×30サイクルの条件でサイクルシークエンス反応を行う。
(4)ヒートシールをはがし、125mMのEDTAを2.5μl、エタノールを25μl添加し、1時間、室温で静置する。
(5)4℃、3,273×gで30分間遠心分離する。
(6)PCRプレートを逆さまにしてエタノールを除去する。
(7)冷却した70%エタノールを35μl添加し、同条件で遠心分離する。
(8)エタノールを除去し、65℃に5分間置いて、乾燥させる。
(9)ホルムアミドを5μl添加し、熱変性する。
(10)ABI373型シークエンサーでシークエンスする。
【0055】
4.多重整列化解析
以下に、「SEQUENCHER(登録商標)」を用いる場合の解析手順を示す。
(1)SEQUENCHER(登録商標)にシークエンスのデータをインポートする。
(2)末端のクオリティーの低い配列を除去する。
(3)データベース、または過去のシークエンスデータをシークエンチャーにインポートする。
(4)例えば以下の条件設定において、複数の配列のコンティグを作成し、相同性に基づいて、母系特異的配列、母系変異集中領域および同属異種特異的配列を決定する:
Assembly Algorithm :Dirty Dataをチェック
Optimize Gap placement :Use Re Aligner,Prefer 3’ Gap Placementをチェック
Minimum Match Percentage:80
Minimum Overlap :20。
【0056】
実施例1:クロマグロ母系特異的塩基配列の抽出と種苗の遺伝子標識
方法:
1)「MitoFish」(http://mitofish.ori.u-tokyo.ac.jp)から、本まぐろ(太平洋クロマグロ:Thunnus orientalis)のミトコンドリアDNAの塩基配列(GeneBank登録番号AB185022:配列番号1、以下dbと記載)を検索し、プロリンtRNAとプェニルアラニンtRNAの塩基配列からDループ増幅のためのPCRプライマーを以下の通り作製した。
本まぐろのDループDNA増幅のためのPCR共通プライマー配列
フォワードプライマー(プロリン側):
5’-TACCCCTAACTCCCAAAGCTAGG-3’(配列番号2)
リバースプライマー(フェニルアラニン側):
5’-GCTTTCTAGGGCCCATCTTAAC-3’(配列番号3)
【0057】
2)境港と福岡で水揚げされた天然本まぐろ(太平洋クロマグロ)成魚と養殖本まぐろ(太平洋クロマグロ)成魚、各1尾ずつ、および同時に採取した浮遊受精卵を孵化した本まぐろ稚魚11尾からミトコンドリアを抽出し、PCRの鋳型DNAとした。
3)上記の方法でDループDNAを増幅した後に塩基配列決定を行ない、データベースの本まぐろ(db)のDループの塩基配列(配列番号1)と比較した。
【0058】
結果:
多重整列化の結果を図1に示す(なお、各々のDループ塩基配列は、境港:配列番号20、福岡:配列番号21、養殖成魚:配列番号22、養殖稚魚1〜11:配列番号23〜33とする)。図中、下部の配列はデータベース(db)の太平洋クロマグロのミトコンドリアDループの塩基配列におけるnt(ヌクレオチド)1からnt865までであり、その下部の黒点は、該db太平洋クロマグロと比較した他のマグロとの間で塩基の相違があることを示す。上記プライマーを用いて決定した太平洋クロマグロ14個体のDループDNAは、dbと同じ865塩基であり、各塩基配列はギャップを加えることなく多重整列化することができた。天然および養殖の各成魚間において、個体間の配列の塩基相同性は高く、わずかに6〜22の異なった塩基相違点が太平洋クロマグロ(db)の前半nt175〜586間の29部位に存在していた(表1)。
【0059】
【表1】
【0060】
塩基相違点はnt175〜200に7部位、およびnt273〜305に5部位に相違点が集中していたためにその間の配列を母系特異的塩基配列と規定した。一方、養殖稚魚11尾間では、配列は完全に一致しているために、同時に浮遊した受精卵は同一雌親由来であり、雌親が異なる他の成魚との相違点は特定部位に集中していることが明らかとなった。またこの結果から、同一の雌親に由来する受精卵や稚魚は、そのうちの1個体のミトコンドリアD−ループの塩基配列を決定することにより、全個体を同じ母系集団として容易に標識できることが示された。
【0061】
応用例1:クロマグロ母系特異的塩基配列の特定と簡易検出法
母系特異的塩基配列、特に母系変異集中領域の塩基配列を増幅するPCR用プライマーを作製することにより、検体から簡易かつ迅速な方法により母系特異的シグナルを得、その母系性から検体の生産履歴を追跡することができる。以下に例を示す。
【0062】
方法:
以下のプライマー対を作製し、天然および養殖の太平洋クロマグロ成魚3尾および稚魚11尾、ならびにミナミマグロおよびビンナガマグロ1尾ずつから抽出したミトコンドリアDNAを鋳型として、タッチダウンPCRにより増幅を行なった。
母系特異的PCRフォワードプライマー(図1、同一母系太平洋クロマグロnt.175〜200:母系特異的変異点7ヶ所):
5’-CTTTCAACATCTTACTTACATCACG-3’(配列番号4)
母系特異的PCRリバースプライマー(図1、同一母系太平洋クロマグロnt.273〜305:母系特異的変異点5カ所):
5’-TTCTTACTGCGTTTAAGTTTACCAG-3(配列番号5)
【0063】
結果:
図2に、PCR増幅産物のアガロース電気泳動の結果を示す。母系が異なる天然太平洋クロマグロおよび養殖太平洋クロマグロからはシグナルは検出されなかったが、同一母系の養殖稚魚11尾から135塩基対の母系特異的DNAが増幅された。したがって、同一母系の養殖稚魚(種苗)が、母系特異的塩基配列によって遺伝子標識されており、該母系特異的塩基配列に特異的に設計されたプライマーを用いたPCRにより容易に検出し得ることを確認した。本明細書において示した系統または産地以外の種苗においても、同様に母系特異的なプライマーを設計し、検体のリアルタイムPCRを行なえば、シグナルの有無によって簡便かつ迅速に母系識別を行うことができる。
【0064】
実施例2:マダイ母系特異的塩基配列と種苗の遺伝子標識
方法:
1)「MitoFish」からマダイ(Pagrus major)のミトコンドリアDNA塩基配列(GeneBank登録番号AP002949:配列番号6)を入手し、プロリンtRNAおよびフェニルアラニンtRNAの塩基配列から以下のPCRプライマーを設計して、マダイDループ増幅と塩基配列決定に使用した:
フォワードプライマー(クロマグロ(db)フェニルアラニンtRNA塩基配列から代用):
5’-GCTTTCTAGGGCCCATCTTAAC-3’(配列番号7)
リバースプライマーI(マダイdbプロリンtRNAから設計):
5’-TTTAACCCCCACCATTGGCTCCC-3’(配列番号8)
リバースプライマーII(マダイDループ内塩基配列より再設計):
5’-AGAGGGGGGTAACAATGAGATC-3’(配列番号9)
【0065】
2)マダイのプロリンtRNAおよびフェニルアラニンtRNAの塩基配列はいずれもクロマグロの対応する配列に一致していたが、リバース側において1塩基が異なっていたため、マダイdbのフェニルアラニンtRNA塩基配列を利用した(リバースプライマーI)。塩基配列に関しては、最初に読めたDループ3’末端側の塩基配列から第2のリバースプライマーIIを作製し、完全に両鎖の塩基配列を読み取った。
【0066】
3)PCRの鋳型DNAには、福岡玄海の養殖成魚と山口、福岡の天然成魚各1尾ずつ、および同日浮遊しに採取された受精卵を孵化成長させた養殖種苗10尾からミトコンドリアを抽出し、PCR鋳型DNAとした。上記の方法でDループDNA増幅および塩基配列決定を行ない、データベースのマダイの塩基配列(以下dbと記載)を基準に多重整列化を行なった(図3)。
【0067】
結果:
マダイのDループはいずれも約1054塩基対からなり、太平洋クロマグロ(db)Dループ(869塩基対)より189塩基対長い。4箇所にギャップを挿入することにより多重整列化が可能となった(図3)。特に養殖稚魚10尾間ではDループの配列は全て同一であった。同時に浮遊し採取された受精卵から生じる稚魚のミトコンドリアDループは同じ塩基配列を有することから、これらの稚魚が同一雌親に由来することが明らかとなり、また、同一の雌親に由来する受精卵や稚魚は、そのうちの1個体のミトコンドリアD−ループの塩基配列を決定することにより、全個体を同じ母系集団として容易に標識できることが示された。一方、産地の異なる各個体間では、約1054塩基対中、9〜24塩基の相違点が見られた(表2)。これらの変異のうち、マダイdbのDループnt.273〜311間に9カ所、nt370〜416間に9箇所が集中していたことから、この間を母系特異的領域とした。
【0068】
【表2】
【0069】
応用例2:マダイ母系特異的塩基配列の特定と簡易検出法
方法:
実施例2において決定した母系特異的領域の塩基配列に基づいて以下のプライマーを設計し、天然および養殖のマダイ異母系成魚3尾および同一母系稚魚5尾から抽出したミトコンドリアDNAを鋳型として、タッチダウンPCRにより増幅を行なった:
マダイ母系特異的PCRフォワードプライマー(図3、マダイ稚魚nt.273〜308:母系特異的変異点8ヶ所):
5’-AGAAATAGCAAGTGTTCAAGTATTTATCCCCAAAAC-3’(配列番号10)
マダイ母系特異的PCRリバースプライマー(図3、マダイ稚魚nt.402〜375:母系特異的変異点5カ所):
5’-GTGTTGGTACTTGGTAGACATTTGTCG -3‘(配列番号11)
【0070】
結果:
図4に、PCR増幅産物のアガロース電気泳動の結果を示す。母系が異なる天然および養殖成魚からはシグナルは検出されなかったが、同一母系の養殖稚魚5尾から129塩基対の母系特異的DNAが増幅された。したがって、同一母系の養殖稚魚(種苗)が、母系特異的塩基配列によって遺伝子標識されており、該母系特異的塩基配列に特異的に設計されたプライマーを用いたPCRにより容易に検出し得ることを確認した。
【0071】
実施例3:ブリのDループ解析例とクロマグロ、マダイとの比較
マダイ、ブリ、クロマグロは養殖魚売上げの90%を占め、いずれもスズキ目に属するがマダイはスズキ亜目タイ科,マグロはサバ亜目サバ科、ブリはスズキ亜目アジ科に属する。スズキ目はその他カツオ、マアジなど重要な回遊水産魚を数多く含む。これらの遺伝的類縁性を知るためにDループを比較した。ただし、ブリのミトコンドリアDNA塩基配列はデータベースに登録されていないため、クロマグロのtRNAプライマーで代用した。
【0072】
方法:
鳥取、福岡で水揚げされた天然ブリのミトコンドリアDNAを鋳型として、実施例1において使用したクロマグロのDループ両端プライマー(配列番号2および3)を用いて、約900塩基対のDNAを増幅した。両端プライマーの各々を用いて両鎖シークエンスを行なったところ、フェニルアラニン側では一度で全長を読み取ることができなかったため、第一のプライマーにより得た部分配列から次のプライマーを設計し、全長の塩基配列を決定した。
ブリ逆方向プライマー: 5’-GGTTTAACGCGCAAAAGCCGAG-3’(配列番号12)
【0073】
結果:
鳥取ブリ(配列番号13)と福岡ブリ(配列番号14)とのDループ塩基配列を整列化させるために、配列中1カ所にギャップを必要とした。699塩基中の690塩基配列が相同であり、相違塩基は8塩基であった。さらに試料を増やすことにより、ブリのミトコンドリアDNA Dループの塩基配列で品種(亜種)識別ができる。
なお、同じスズキ目間では、マダイ、マグロおよびブリの間では、随所に短い(最長33個)相同塩基はあるが、ギャップ挿入数が多く、多重整列化ができない状態であった。
【0074】
実施例4:マグロの種、品種および産地の識別と生産履歴管理(トレーサビリテイー)
MitoFishデータベースから入手した本まぐろ(太平洋クロマグロ:Thunnus orientalis)のミトコンドリアDNAの塩基配列(GeneBank登録番号AB185022:配列番号1)を標準材料として、以下の一般に市販されているマグロ属の種および亜種との間でDループの塩基配列の比較解析を行った:
【0075】
大西洋クロマグロ(Thunnus thynnus):ミトコンドリアDNA塩基配列(GeneBank登録番号AY302574:配列番号15)
ビンナガマグロ(Thunnus alalunga):ミトコンドリアDNA塩基配列(GeneBank登録番号AB101291:配列番号16)
メバチマグロ(インド洋産:Thunnus obesus:配列番号17)
キハダマグロ(宮城沖産:Thunnus albacares:配列番号18)
ミナミマグロ(オーストラリア産:Thunnus maccoyii:配列番号19)
【0076】
結果:
上記マグロ6種のDループ塩基配列は869塩基前後であったが、1種あたり10部位前後のギャップを挿入することにより、多重整列化を行うことができた。ただし、ギャップ挿入は22部位あり、特に母系特異的配列前半のnt172〜232に9部位が集中していた。各品種間では869塩基中39〜77塩基の違いがあったが(表3)、その他の領域では相同性を保っていた。特に、nt173〜215にギャップ12部位を含む35部位とnt628〜672にギャップ3部位を含む20部位に相違塩基が集中していた(図5−1〜3)。いずれも部分的に太平洋クロマグロ母系特異的配列と重複するが、ギャップの位置および変異の数が前記母系特異的配列とは明確に異なるので、種識別領域(同属異種識別領域)として特定することができる。
【0077】
【表3】
【0078】
常法に従い、残る相同領域から遺伝的類似性を求めた結果、大西洋クロマグロ/太平洋クロマグロ、ミナミマグロ/太平洋クロマグロ、大西洋クロマグロ/ミナミマグロ間の相違塩基数とギャップ挿入数はそれぞれ69(+8)、77(+5)、49(+2)であった。いずれも「トロ」の材料とされるこれらの種が、遺伝的には互いに遠位にあることが明らかとなった。
【0079】
この結果により、ミトコンドリアDループの塩基配列の相違により、マグロ属およびその種間の識別が可能であり、食品表示の偽装の防止および摘発などに有用であることが示唆される。ただし、同じ太平洋クロマグロ間でも最大22個の相違点があるが(表1)、マグロ属の種間識別はギャップ数を加味した相違塩基数で判断できる。
【0080】
実施例5:マグロの産地識別
方法:
図1および図3に示すように、マグロおよびマダイのDループ塩基配列を母系により識別することが可能となったが、母系は産卵場所や魚群,あるいは棲息海域で規定される可能性があるので、産地識別に利用できる可能性を調べた。棲息海域が広い回遊魚であるマグロのDループ塩基配列が産地によってどの程度異なるか、メバチマグロ、キハダマグロにおいて比較した。
【0081】
結果:
下記の産地が異なる個体のDループ塩基配列の比較では、ギャップを加えることなしに配列を整列化できた。以下に、同種のマグロでの産地による塩基配列の変異数を示す(括弧内の数値=相違塩基部位数/比較塩基部位数):
メバチマグロ :パラオ/インド洋(18/569)
大西洋/パラオ(28/569)
大西洋/インド洋(35/569)
キハダマグロ :インド洋/宮城沖(12/756)
太平洋クロマグロ:(6〜22/869)
【0082】
同種間の多重整列化ではギャップの挿入は必要なかったが、産地が異なる場合、太平洋クロマグロの母系特異的塩基配列に相当する領域に相違が見られた。すなわち、この領域の多型を用いて産地を識別し得ることが示唆される。
【0083】
本発明の方法により、産地の異なるマダイ、マグロのDループの塩基配列の相違で母系を識別することができるが、逆に塩基配列の相違から該当する産地を推定する場合、商品の生産履歴管理と同様に、比較対照となる試料を必要とする。好ましくは、産地または棲息海域に存在する魚群のDループ塩基配列データベースを作成し、データベース中の全配列と比較する。データベース作成と並行して産地識別を行ってもよい。
【0084】
実施例6:天然および養殖のマグロのD−ループ配列の比較
天然のマグロ集団と養殖稚魚との遺伝的相違を調べるために、築地市場に集積される天然のクロマグロと、近畿大学水産研究所から供与されたクロマグロ受精卵および稚魚のD−ループ塩基配列を、上記の方法により決定して比較した。
結果
漁場や水揚場の異なる535尾の太平洋クロマグロのうち、養殖稚魚と同じDループ配列を有する個体は存在しなかった(図6)。この結果から、本発明の方法を実際に用いて、例えば、漁獲された集団中の人工種苗を同定することができ、放流事業における加入量変動調査や、漁獲高に対する放流事業の貢献度を判定することや、育成・販売した種苗が他者により複製された場合に追跡を行うことなどが可能であることの裏付けがなされた。
【0085】
参考例1:マイクロサテライトによるハプロタイプ分析
同一母系の養殖クロマグロ稚魚11尾について、公知のマイクロサテライトマーカーTth208、Tth217、Tth4およびTth7-16(Clark et al., 2004, Mol. Ecol. Notes, 4:70-73)を用いてハプロタイプ分析を行った。少なくとも4パターンのハプロタイプが観察された(図7)ことから、少なくとも3個体の雄親と雌親(1個体)が産卵時に存在したことが示唆される。一方、天然のクロマグロについて同様の分析を行ったところ、分析を行った全個体間でマイクロサテライトパターンが異なり、同一のハプロタイプを示す個体は存在しなかった(データは示さず)。これらの結果から、遺伝的多様性が小さい養殖魚の場合は産卵時に存在する親魚の個体数が少ないので、受精卵全てのマイクロサテライトによるハプロタイプを決定することが可能であり、したがって、mtDNA Dループによる母系識別にマイクロサテライト分析による雄系識別を組み合わせることにより、より精度が高い系統標識が可能になることが示唆される。
【産業上の利用可能性】
【0086】
本発明は、水産資源保護として有用な魚介類のトレーサビリティーの提供を可能とするものであり、このトレーサビリティの提供により、食品としての魚介類の生産履歴または養殖もしくは栽培漁業のための稚魚の追跡(調査)を可能にすることにより、魚介類の品質および産地を保証し、生産履歴表示の偽装を摘発、防止し、また優良種苗の遺伝子標識による識別および養殖・育種者の権利の確立に寄与するものである。本発明はさらに、放流事業に用いる大量の稚魚の標識と洋上での追跡調査および加入量変動調査を可能にすることにより、水産資源の遺伝的多様性の維持および排他的経済区域内での栽培漁業の発展に寄与するものである。
【特許請求の範囲】
【請求項1】
水産魚介類またはその加工品のミトコンドリアDNA Dループの塩基配列を決定することにより、該水産魚介類またはその加工品の種苗または生産履歴を同定する方法であって、
養殖場において継代した雌親個体から採取された第1の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第1の塩基配列)を決定し、
水産魚介類またはその加工品から採取された第2の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第2の塩基配列)を決定し、
第1の塩基配列と第2の塩基配列とを比較し、前記水産魚介類またはその加工品が前記雌親個体の後代であることを同定することによって、前記水産魚介類またはその加工品の種苗または生産履歴を同定することを含む、前記方法。
【請求項2】
マイクロサテライトにより核型を決定することをさらに含む、請求項1に記載の方法。
【請求項3】
ミトコンドリアDNA Dループの種特異的塩基配列が1から2以上のギャップの挿入により整列化できるか否かの判定をさらに含む、請求項1または2に記載の方法。
【請求項4】
ミトコンドリアDNAのDループ全長の塩基配列を、ミトコンドリアプロリンtRNA特異的プライマーおよびミトコンドリアフェニルアラニンtRNA特異的プライマーを用いて増幅し、両鎖シークエンシングにより決定することをさらに含む、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
第1の塩基配列と第2の塩基配列との比較が、母系特異的塩基配列が完全に一致するか否かの比較である、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
水産魚介類またはその加工品が、食品である、請求項1〜5のいずれか一項に記載の方法。
【請求項1】
水産魚介類またはその加工品のミトコンドリアDNA Dループの塩基配列を決定することにより、該水産魚介類またはその加工品の種苗または生産履歴を同定する方法であって、
養殖場において継代した雌親個体から採取された第1の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第1の塩基配列)を決定し、
水産魚介類またはその加工品から採取された第2の試料のミトコンドリアDNA Dループの母系特異的塩基配列(第2の塩基配列)を決定し、
第1の塩基配列と第2の塩基配列とを比較し、前記水産魚介類またはその加工品が前記雌親個体の後代であることを同定することによって、前記水産魚介類またはその加工品の種苗または生産履歴を同定することを含む、前記方法。
【請求項2】
マイクロサテライトにより核型を決定することをさらに含む、請求項1に記載の方法。
【請求項3】
ミトコンドリアDNA Dループの種特異的塩基配列が1から2以上のギャップの挿入により整列化できるか否かの判定をさらに含む、請求項1または2に記載の方法。
【請求項4】
ミトコンドリアDNAのDループ全長の塩基配列を、ミトコンドリアプロリンtRNA特異的プライマーおよびミトコンドリアフェニルアラニンtRNA特異的プライマーを用いて増幅し、両鎖シークエンシングにより決定することをさらに含む、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
第1の塩基配列と第2の塩基配列との比較が、母系特異的塩基配列が完全に一致するか否かの比較である、請求項1〜4のいずれか一項に記載の方法。
【請求項6】
水産魚介類またはその加工品が、食品である、請求項1〜5のいずれか一項に記載の方法。
【図1−1】


【図1−2】

【図1−3】

【図1−4】

【図1−5】

【図1−6】

【図1−7】

【図1−8】

【図1−9】

【図1−10】

【図1−11】

【図2】
【図3−1】

【図3−2】

【図3−3】

【図3−4】

【図4】
【図5−1】

【図5−2】

【図5−3】

【図6】
【図7】
【図1−2】
【図1−3】
【図1−4】
【図1−5】
【図1−6】
【図1−7】
【図1−8】
【図1−9】
【図1−10】
【図1−11】
【図2】
【図3−1】
【図3−2】
【図3−3】
【図3−4】
【図4】
【図5−1】
【図5−2】
【図5−3】
【図6】
【図7】
【公開番号】特開2012−105682(P2012−105682A)
【公開日】平成24年6月7日(2012.6.7)
【国際特許分類】
【出願番号】特願2012−50265(P2012−50265)
【出願日】平成24年3月7日(2012.3.7)
【分割の表示】特願2009−269544(P2009−269544)の分割
【原出願日】平成21年11月27日(2009.11.27)
【出願人】(508350889)有限会社ジェノテックス (2)
【Fターム(参考)】
【公開日】平成24年6月7日(2012.6.7)
【国際特許分類】
【出願日】平成24年3月7日(2012.3.7)
【分割の表示】特願2009−269544(P2009−269544)の分割
【原出願日】平成21年11月27日(2009.11.27)
【出願人】(508350889)有限会社ジェノテックス (2)
【Fターム(参考)】
[ Back to top ]