
ミミズ由来のプロテアーゼ
【課題】ミミズ由来の新規なプロテアーゼの提供。
【解決手段】下記の性質を有するプロテアーゼ:N末端アミノ酸配列:Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln;基質特異性:N-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示す;阻害剤による影響:キモスタチン(Chymostatin)などにより阻害される;最適pHとpH安定性:最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;最適温度と温度安定性:最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない。
【解決手段】下記の性質を有するプロテアーゼ:N末端アミノ酸配列:Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln;基質特異性:N-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示す;阻害剤による影響:キモスタチン(Chymostatin)などにより阻害される;最適pHとpH安定性:最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;最適温度と温度安定性:最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は新規なプロテアーゼに関する。
【背景技術】
【0002】
植物の病気の原因には、細菌、カビ(菌類)などの病原微生物やウイルスなどの他、環境条件等による生理病、遺伝病などが知られている。中でも、カビによる病気が70%以上にもなる。また、植物ウイルスは1000種以上も存在し、植物に感染して農作物の品質の低下や、収量の低下などの被害を及ぼし、その被害額は日本だけでも年間1000億円にものぼると言われている。これらの対策として、農薬の利用や遺伝子組換え、ワクチン療法などが挙げられる。しかし、農薬や遺伝子組み換えは安全性の問題をはらんでおり、ワクチン療法は弱毒ウイルスの開発や接種に労力や注意が必要である(都丸敬一 (2001) 植物のウイルス病物語 始まりからバイオテクノロジーまで. 全国農村教育協会, 106-110)。
【0003】
そこで、天然の抗ウイルス剤を開発することを目的に研究が行われてきた。そして、シマミミズ(Eisenia foetida)から抗植物ウイルス活性を有する物質を既に単離、精製しており、それがセリンプロテアーゼであることを明らかにし、その酵素学的性質や抗植物ウイルス活性についての研究が行われてきた(野田加奈子(2006)大阪府立大学修士論文;山下崇之(2008)大阪府立大学修士論文)。
【0004】
E.foetidaは腐敗した植物などの有機物が豊富な土地に生息し、昔はゴミ捨て場や堆肥舎の近くにも生息し、身近にいるミミズであった。現在は、ゴミ回収の制度が整い堆肥舎がなくなりそれほど身近ではなくなったが、日本だけでなく世界各地でコンポストミミズとして堆肥化によく利用されている。ヨーロッパやシベリア、北アメリカ、イスラエル、チリ、ニュージーランド、オーストラリア、日本など温帯気候下の世界中に広く分布するとされており、熱帯にもところどころにいるようである(渡辺弘之 (2003). ミミズ 嫌われもののはたらきもの. 東海大学出版会)。
【0005】
またE.foetidaを含むミミズの仲間には医薬等への利用が期待される物質が見出されており、E.foetidaの体液は溶血、細胞毒性、血液凝集、抗菌などの作用を示し、この効果を活用した脳血栓の溶血剤や避妊剤などが検討されている(中村好男 (2004). ミミズと土と有機農業. 創森社)。
【0006】
食品の安全性が追求されている今の時代、農作物に対する安全性も重要であり、これからは質とともに安全性も重要視されることが予想される。また、人口増加に伴う食糧危機の点から、農作物の品質の低下や収量の低下は食糧不足に直結する問題である。E.foetidaから安全かつ有効な生物農薬を開発することは、これらの課題の解決に貢献できると期待している。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】都丸敬一 (2001) 植物のウイルス病物語 始まりからバイオテクノロジーまで. 全国農村教育協会, 106-110
【非特許文献2】野田加奈子(2006)大阪府立大学修士論文
【非特許文献3】山下崇之(2008)大阪府立大学修士論文
【非特許文献4】渡辺弘之 (2003) ミミズ 嫌われもののはたらきもの. 東海大学出版会
【非特許文献5】中村好男 (2004). ミミズと土と有機農業. 創森社
【0008】
【非特許文献6】K. Inouye (2006),Protein Expression and Purification 46, 248-255
【非特許文献7】Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109
【非特許文献8】Nobuyoshi Nakajima (1993), Biosci. Biotech. Biochem, 57(10), 1726-1730
【0009】
【非特許文献9】Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109
【非特許文献10】Bradford MM (1976), Anal. Biochem, 72, 248-254
【非特許文献11】深見順一ら(1981)農薬実験法(2)殺菌剤編. ソフトサイエンス社. 110-115
【非特許文献12】脇本哲 (1994). 総説 植物病理学. 養賢堂. 425)
【発明の概要】
【発明が解決しようとする課題】
【0010】
そこで、本発明は、ミミズ(E.foetida)由来の、抗植物ウイルス活性及び抗植物病原細菌活性を有するプロテアーゼ及びその使用、並びにその製造手段を提供するものである。
【課題を解決するための手段】
【0011】
したがって、本発明は、下記の性質:
(1)N末端アミノ酸配列
下記のN末端アミノ酸配列:
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln
を有する;
(2)基質特異性
合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示す;
(3)阻害剤による影響
プロテアーゼ活性が、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N−トシル−L−フェニルアラニンクロメチルケトン(TPCK)、ダイズトリプシンインヒビター(SBTI)により阻害される;
(4)最適pHとpH安定性
最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;および
(5)最適温度と温度安定性
最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない;
を有する、ミミズ(Eisenia foetida)由来のプロテアーゼ、
を提供する。
【0012】
本発明はまた、配列番号:2に記載のアミノ酸配列を含むプロテアーゼ;配列番号:2に記載のアミノ酸配列において、1個〜数個のアミノ酸の置換、欠失及び/又は付加により修飾されているアミノ酸配列からなるプロテアーゼ;配列番号:2に記載のアミノ酸配列に対して少なくとも90%の配列同一性を有するプロテアーゼ;或いは配列番号:1に記載の塩基配列からなる核酸又はその相補鎖に対して高緊縮条件下でハイブリダイズする核酸によりコードされているプロテアーゼ、を提供する。
【0013】
本発明はまた、上記何れかに記載のプロテアーゼをコードする核酸;当該核酸により形質転換された宿主細胞;並びに当該宿主細胞を培養することを含んでなる、上記プロテアーゼの製造方法を提供する。
本発明はまた、上記(1)〜(5)の性質を有するプロテアーゼの製造方法において、当該プロテアーゼをミミズ(Eisenia foetida)から採取することを含んでなる方法を提供する。
【0014】
本発明は更に、前記のプロテアーゼを含んでなる、抗植物病原性ウイルス剤を提供し、当該植物病原性ウイルスは、例えば、キュウリモザイクウイルス(Cucumber mosaic virus;CMV)である。
本発明はまた、前記のプロテアーゼを含んでなる、抗植物病原性細菌剤を提供し、当該植物病原性細菌は、例えば、野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)である。
【図面の簡単な説明】
【0015】
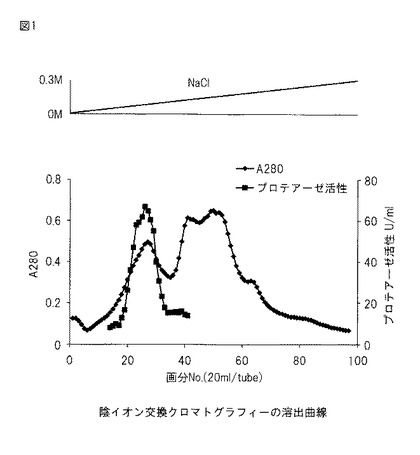
【図1】図1は、本発明のプロテアーゼの精製過程における、陰イオン交換クロマトグラフィーの溶出曲線を示す。
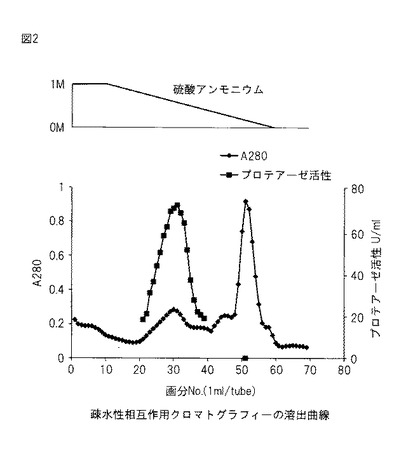
【図2】図2は、本発明のプロテアーゼの精製過程における、疎水性相互作用クロマトグラフィーの溶出曲線を示す。
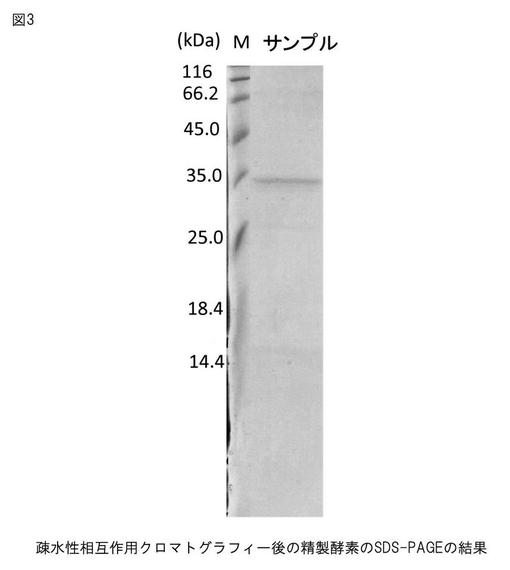
【図3】図3は、図2におけるプロテアーゼ活性画分の、SDS-PAGEによる分析の結果を示す。
【0016】
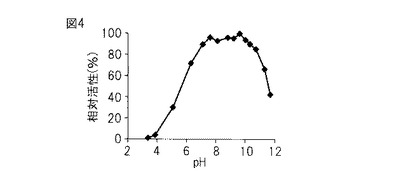
【図4】図4は、本発明のプロテアーゼの最適pHを示すグラフである。
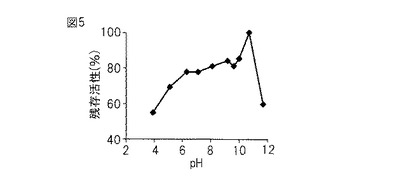
【図5】図5は、本発明のプロテアーゼのpH安定性を示すグラフである。
【図6】図6は、本発明のプロテアーゼの最適温度を示すグラフである。
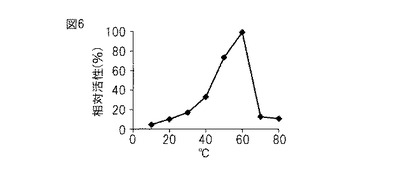
【図7】図7は、本発明のプロテアーゼの温度安定性を示すグラフである。
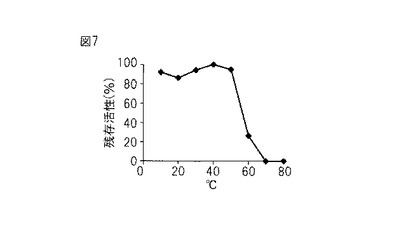
【図8】図8は、本発明のプロテアーゼの、キュウリモザイクウイルス(Cucumber mosaic virus;CMV)に対する阻害を示すグラフである。
【0017】
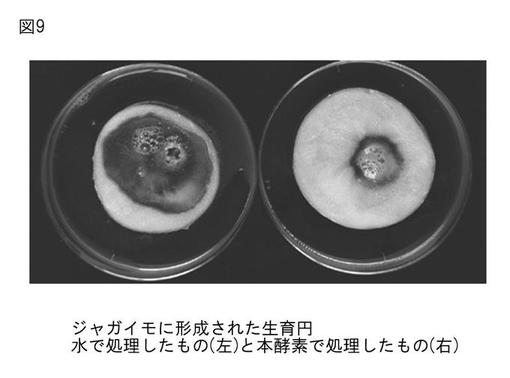
【図9】図9は、本発明のプロテアーゼの、野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)に対する阻害を示す写真である。
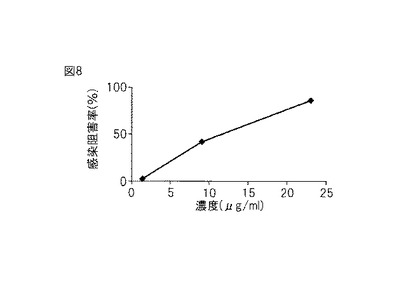
【図10】図10は、本発明のプロテアーゼをコードするDNAのクローニングに使用したクローニングベクターの構造を示す図である。
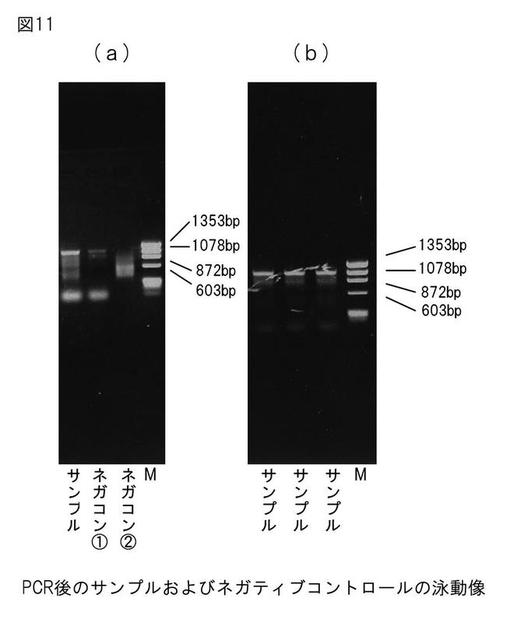
【図11】図11は、PCR増幅産物中の、本発明のプロテアーゼをコードするDNAの検出を示す電気泳動図である。
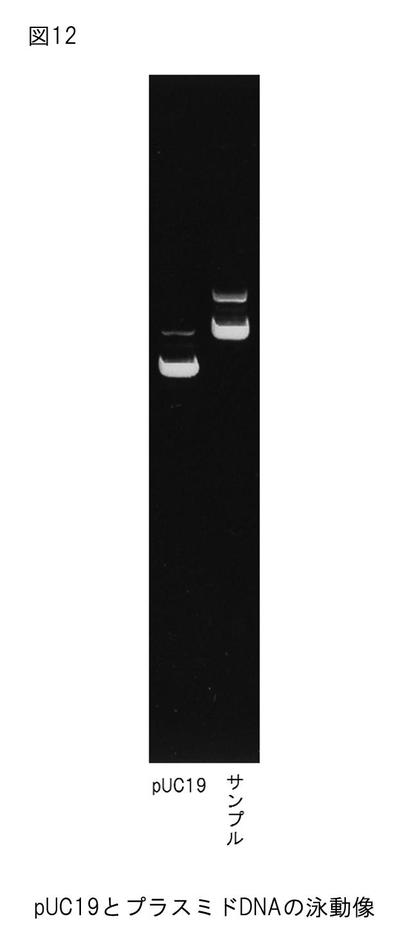
【図12】図12は、本発明のプロテアーゼをコードするDNAを含むプラスミドDNAの電気泳動図である。
【図13】図13は、本発明のプロテアーゼをコードするDNAの塩基配列を及び当該プロテアーゼのアミノ酸配列を示す。
【発明を実施するための形態】
【0018】
本願発明は、下記の性質:
(1)N末端アミノ酸配列
下記のN末端アミノ酸配列:
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln (配列番号:3)
を有する;
(2)基質特異性
合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示す;
(3)阻害剤による影響
プロテアーゼ活性が、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N−トシル−L−フェニルアラニンクロメチルケトン(TPCK)、ダイズトリプシンインヒビター(SBTI)により阻害される;
(4)最適pHとpH安定性
最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;および
(5)最適温度と温度安定性
最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない;
を有する、ミミズ(Eisenia foetida)由来のプロテアーゼ、を提供する。
【0019】
上記のプロテアーゼは、配列番号:2に示すアミノ酸配列を有する。従って本発明はまた、配列番号:2に記載のアミノ酸配列を含むプロテアーゼを提供する。
【0020】
ある酵素につき、そのアミノ酸配列を部分的に改変しても、生来の酵素の活性が維持されることが当業者によりよく知られている。従って本発明はまた、配列番号:2に記載のアミノ酸配列において、1個〜数個のアミノ酸の置換、欠失及び/又は付加により修飾されているアミノ酸配列からなるプロテアーゼ;配列番号:2に記載のアミノ酸配列に対して少なくとも90%の配列同一性を有するプロテアーゼ;或いは配列番号:1に記載の塩基配列からなる核酸又はその相補鎖に対して高緊縮条件下でハイブリダイズする核酸によりコードされているプロテアーゼ、を提供する。
【0021】
酵素の採取方法
本発明のプロテアーゼをミミズから採取するには、例えば、まずは、凍結乾燥したシマミミズから粗酵素液を抽出し、これを陰イオン交換クロマトグラフィー、続いて疎水性相互作用クロマトグラフィーに供する。また、A280およびカゼインを基質としたプロテアーゼ活性を測定しこれを分画の指標とする。最後に、SDS-PAGEを行い純度の確認を行う。
【0022】
プロテアーゼ活性測定方法
サンプル 0.5 mlと1.33% (w/v) カゼイン(0.1 M グリシン-NaOH緩衝液, pH 9.5 に溶解) 1.5 mlを混合し、対照にはサンプル0.5 mlのみを用意し、それぞれ37℃、60分間インキュベートする。これに、0.11 M TCA、0.22 M 酢酸ナトリウム、0.33M 酢酸2 mlを加えよく撹拌し反応を停止する。対照にはその後サンプル 0.5 mlを加る。そして、この反応停止液をNo.2のろ紙を用いてろ過し、ろ液のA275を測定する。ここで、1 U =1分間にA275を0.0074上昇させるのに必要な酵素量(A275 of 0.0074 = 1 μg tyrosine)と定義する(K. Inouye (2006),Protein Expression and Purification 46, 248-255)。
【実施例】
【0023】
次に、本発明を、実施例により更の具体的に説明する。
実施例1. シマミミズからのプロテアーゼの精製
(1)粗酵素液の抽出
凍結乾燥したシマミミズ20 gに50 mM Tris-HCl 緩衝液(pH 7.0)220 mlを加え、30分間氷上で撹拌し、遠心分離(27,700×g、4℃、10 分)した。その上清に終濃度が2%となるようにストレプトマイシン硫酸塩を加えて、30分間氷上で撹拌し、遠心分離(27,700×g、4℃、10 分)して核酸を除去した。次に、この上清を50 mM Tris-HCl 緩衝液(pH 7.0)で3倍に希釈し、氷上で90分間電気透析を行い、グラスフィルターでろ過したものを粗酵素液とした。
【0024】
(2)DEAE-TOYOPEARL 650Mによる陰イオン交換クロマトグラフィー
担体:DEAE-TOYOPEARL 650M
カラム:直径2.5 cm×27 cm
初期緩衝液:50 mM Tris-HCl (pH 7.0)
溶出緩衝液:50 mM Tris-HCl + 0.3 M NaCl (pH 7.0)
カラムを実験台に水平になるように立て、担体を流し込み、500 mlの蒸留水を流して洗い、その後、500 mlの初期緩衝液で平衡化した。次に、粗酵素液をアプライし溶出液のA280が0.1に下がるまで初期緩衝液を流して洗浄を行った。そして、初期緩衝液 1000 mlと溶出緩衝液 1000 mlを架橋して溶出を開始した。試験管1本あたり20 ml、100本の画分に分画し、各画分のA280とプロテアーゼ活性を測定した。
【0025】
結果
陰イオン交換クロマトグラフィーの溶出曲線を図1に示す。約0.07 M NaClでプロテアーゼ活性のピークが得られた。画分No.23〜29を回収し、等量の2M硫酸アンモニウムと混合して、次の疎水性相互作用クロマトグラフィーのサンプルとした。
【0026】
(3)HiTrap Phenyl HPによる疎水性相互作用クロマトグラフィー
カラム:HiTrap Phenyl HP 1ml
初期緩衝液:20 mM Tris-HCl + 1M硫酸アンモニウム (pH 7.0) (吸引ろ過)
溶出緩衝液:20 mM Tris-HCl (pH 7.0) (吸引ろ過)
カラムを実験台に水平になるように立て、適量のMilliQで洗浄し、さらに5mlの溶出緩衝液を流速1 ml/minで流し洗浄した。次に、5 ml〜10 mlの初期緩衝液で平衡化した。
【0027】
陰イオン交換後の活性画分を等量の2M硫酸アンモニウムと混合し、カラムに適用した。初期緩衝液と溶出緩衝液を架橋し、0〜10本までは1M、10〜60本までは1〜0M、60〜70本までは0Mの硫酸アンモニウムで溶出するようにグラジエントの設定を行い、溶出した。エッペンチューブ1本あたり1ml、流速1ml/分で70本の画分に分画し、各画分のA280とプロテアーゼ活性を測定した。
【0028】
結果
上記疎水性相互作用クロマトグラフィーの溶出曲線を図2に示す。約0.6 M硫酸アンモニウムでプロテアーゼ活性のピークが得られた。プロテアーゼ活性の高かった画分No.24〜34をSDS-PAGEのサンプルとした。
【0029】
(4)SDS-PAGE
ランニングゲルとストッキングゲルを、表1のとおりに試薬を混合して作製し、電気泳動層にセットした。疎水性相互作用クロマトグラフィー後の活性画分から10μLとり、10μLの2×サンプル緩衝液と混合して3分間煮沸した。これをゲルに適用し、20 mA、300Vで90分間泳動した。マーカーは次のものを使用した。β−ガラクトシダーゼ(分子量:116,000)、ウシ血清アルブミン(分子量:66,200)、オバルミン(分子量:45,000)、ラクターゼ脱水素酵素(分子量:35,000)、REase Bsp98I(分子量:25,000)、β−ラクトグロブリン(分子量:18,400)、リゾチーム(分子量:14,400)。泳動後のゲルはCBB溶液で染色した。
【0030】
【表1】

【0031】
SDS-PAGEの結果を図3に示す。
また、精製の経過を下記の表2に示す。
【0032】
【表2】
【0033】
考察
図3を見ると、メインバンドの他にもうっすらとバンドが確認できる。図2の疎水性相互作用クロマトグラフィーの溶出曲線を見てもわかるように、画分No.24〜34までは比較的きれいなピークが確認でき多種類のタンパクが混在しているとは考えにくい。したがってメインバンドのほかにうっすらと確認できたバンドは、主に目的タンパクの自己分解産物である可能性が高い。
【0034】
実施例2. 酵素学的性質
(1)N末端アミノ酸配列の解析
精製後の酵素をSDS-PAGEに供した後、PVDF膜に転写した。転写は、300 V、108 mA(1 cm2あたり2 mA)の条件で60分間程度行った。転写後、PVDF膜をMilliQで洗浄し、CBB-R250溶液で染色した後、MilliQで調製した70%メタノールで脱色しMilliQで軽く洗浄してキムタオルの上で乾燥させてから目的のバンドを切り出した。そしてN末端からのアミノ酸配列を決定した。
【0035】
結果
N末端アミノ酸配列を解析した結果、以下の配列が得られた。
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln (配列番号:3)
BLASTを用いて相同性検索を行った結果、相同性を示すタンパク質は見られなかった。また、赤ミミズL.rubellus由来のフィブリン分解酵素とも相同性が見られなかった(Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109;Nobuyoshi Nakajima (1993), Biosci. Biotech. Biochem, 57(10), 1726-1730; Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109)。N末端配列の比較は、下記の表3のとおりである。
【0036】
【表3】
【0037】
(2)基質特異性の測定
2.5 mMの各合成基質溶液(70%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)100 μLと、酵素100 μLを混合し、37℃で30分間反応させ、405 nmにおける吸光度を測定した。コントロールとしては、酵素の代わりに同量の0.1 M グリシン-NaOH 緩衝液(pH 9.5)を用いた。測定値が一番大きかったものを100として、各pHでの測定値から相対活性値を算出した。なお、使用した合成基質は次の5つである。
【0038】
(1)N-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド(キモトリプシン用基質)
(2)N-グルタリル-L-Phe-p-ニトロアニリド(キモトリプシン用基質)
(3)N-α-ベンゾイル-L-Arg-p-ニトロアニリド(トリプシン用基質)
(4)N-サクシニル-Ala-Ala-Pro-Leu-p-ニトロアニリド(エラスターゼ用基質)
(5)N-サクシニル-Ala-Ala-Ala-p-ニトロアニリド(エラスターゼ用基質)
【0039】
結果
表4に示すとおり、本酵素は合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu-p-ニトロアニリドに対して高い活性を示した。
【0040】
【表4】
【0041】
(3)阻害剤による影響
酵素60μLと、終濃度が以下に示す濃度になるように各プロテアーゼ阻害剤60μLを混合し、37℃で30分間プレインキュベートした。コントロールとしては、プロテアーゼ阻害剤の代わりに同量の水を用いた。プレインキュベート後、5 mMのN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド溶液(4%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)60μLを加え、37℃で20分間反応させ、405 nmにおける吸光度を測定した。コントロールの測定値を阻害率0%として、各阻害剤の阻害率を算出した。
【0042】
(1)0.01 mg/ml ダイズトリプシンインヒビター(SBTI)
(2)1 mM フェニルメチルスルホニルフルオリド(PMSF)
(3)0.1 mg/ml キモスタチン(Chymostatin)
(4)1mM N-トシル-L-フェニルアラニンクロロメチルケトン(TPCK)
(5)0.01 mg/ml ストレプトマイセス ズブチリシンインヒビター(SSI)
(6)1mM 2,2'-ビピリジル
(7)1mM 1,10-フェナンスロリン(Phenanthoroline)
(8)1mM EDTA
(9)0.1 mM E-64
注:(1)〜(5)はセリンプロテアーゼ阻害剤であり、(6)〜(8)はメタロプロテアーゼ阻害剤であり、(9)はシステインプロテアーゼ阻害剤である。
【0043】
結果
表5に示すとおり、本酵素のプロテアーゼ活性は、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N-トシル-L-フェニルアラニンクロロメチルケトン(TPCK)及びダイズトリプシンインヒビター(SBTI)のセリンプロテアーゼ阻害剤に強く阻害されたが、他の阻害剤にはそれほど影響を受けなかった。
【0044】
【表5】
【0045】
(4)最適pH
2%アゾカゼイン100 μL、pH 3.4〜11.7の広域緩衝液(0.2 Mホウ酸、0.05 Mクエン酸、0.1 Mリン酸三ナトリウム十二水和物)300 μL、酵素100 μLをエッペンチューブに混合し、37℃で90分間反応させた。コントロールとしては、2%アゾカゼイン100 μL、pH 3.4〜11.7の広域緩衝液(0.2 Mホウ酸、0.05 Mクエン酸、0.1 Mリン酸三ナトリウム十二水和物)400 μLを混合したものを用いた。反応後、10%TCA溶液500 μLを加えて完全に反応を止め、遠心分離(17,400×g、室温、10 分)して、その上清の366 nmにおける吸光度を測定した。測定値が一番大きかったものを100として、各pHでの測定値から相対活性値を算出した。
【0046】
結果
図4に示すとおり、最適pHは9.6付近にある。
【0047】
(5)pH安定性
pH 3.9〜11.7の広域緩衝液90 μL、酵素10 μLをエッペンチューブに混合し、37℃で60分間プレインキュベートした。コントロールとしてはpH 3.9〜11.7の広域緩衝液100 μLを用いた。プレインキュベート後、1%アゾカゼイン(0.5 M グリシン-NaOH 緩衝液, pH 9.5に溶解)400 μLを加え、37℃で90分間反応させた。反応後、10%TCA 溶液500 μLを加えて完全に反応を止め、遠心分離(17,400×g、室温、10 分)して、その上清の366 nmにおける吸光度を測定した。測定値が一番大きかったものを100として、各pHでの測定値から相対活性値を算出した。
【0048】
結果
図5に示すとおり、pH 6〜11で80%以上の安定性があった。
【0049】
(6)最適温度
2.5 mMのN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド溶液(4%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)80 μLを10〜80℃の各温度で5分間プレインキュベートし、酵素80 μLを加え、各温度で20分間反応させ、405 nmにおける吸光度を測定した。コントロールとしては、酵素の代わりに同量の0.1 M グリシン-NaOH 緩衝液(pH 9.5)を加えたものを用いた。測定値が一番大きかったものを100として、各温度での測定値から相対活性値を算出した。
【0050】
結果
図6に示すとおり、最適温度は約60℃である。
【0051】
(7)温度安定性
酵素80 μLを10〜80℃の各温度で30分間プレインキュベートした。コントロールとしては、酵素の代わりに同量の0.1 M グリシン-NaOH 緩衝液(pH 9.5)を用いた。プレインキュベート後、2.5 mMのN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド溶液(4%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)80 μLを加えて、37℃で30分間反応させ、405 nmにおける吸光度を測定した。測定値が一番大きかったものを100として、各温度での測定値から相対活性値を算出した。
結果
図7に示すとおり、加熱によるプロテアーゼ活性の低下は50℃まで見られなかった。
【0052】
実施例3. プロテアーゼの農業分野への利用
本実施例では、本酵素の応用利用を目的とした研究を行った。まず、農作物に甚大な被害をもたらせているキュウリモザイクウイルス(Cucumber mosaic virus;CMV)に対する感染阻害効果を本酵素が有するのか検定した。検定方法は混合接種法で行った。この方法は、感染阻害作用の有無を確認するには最適な方法である。
【0053】
また、農作物の被害は植物ウイルスだけではなく植物病原菌によっても引き起こされている。そこで、抗細菌活性も有するのか検定した結果、植物病原菌の一つである野菜類軟腐病(Erwinia carotovora subsp.carotovora)に対して抗細菌活性を示した。
【0054】
(1)酵素濃度の決定
酵素の濃度は、BSAを標準タンパク質としてブラッドフォードの方法(Bradford MM (1976), Anal. Biochem, 72, 248-254)によって決定した。
【0055】
(2)植物ウイルスに対する感染阻害作用の検定
検定植物:ジュウロクササゲ(Vigna unguiculate cv. kurodane-sanjaku)
検定ウイルス:キュウリモザイクウイルス(Cucumber mosaic virus;CMV)のpepo系統
バーミキュライトを満たした鉢にササゲの種子をまき温室内に置き、発芽した後、土を入れたセルトレイに苗を移植し育種した。実験には、初生葉が最大に展開したものを用い、1試料あたり5〜10個体を用いた。
【0056】
まず、0.1Mリン酸緩衝液(pH 7.0)(1 Mリン酸緩衝液は1M KH2PO4と1M K2HPO4を2:3で混合し、pH 7.0に調整したもの)でCMV溶液を100 μg/mlに調製した。試料は20 mM Tris-HCl 緩衝液m(pH 7.0)で各濃度に希釈した。これらを表6のとおりエッペンチューブに加え、サンプル溶液とコントロール溶液を調製し、それぞれをボルテックスでよく混合して氷中に30分間置いた。
【0057】
【表6】
【0058】
ササゲへの接種は混合接種法で行った。最大に展開したササゲの初生葉に、サンプル葉とコントロール葉を区別するためにマーカー用インクで印をし、研磨剤としてカーボランダムをうっすらと満遍なくふりかけた。調製したサンプル溶液とコントロール溶液を氷中に置いて30分後に、一方の葉にサンプル溶液を、他方の葉にはコントロール溶液を綿棒で優しく接種した。接種後、すぐに流水でカーボランダムを洗い流した。ウイルスに感染すると葉に病班を形成するので、接種して二日後にこの病班の数を測定し感染阻害率を算出した(深見順一ら(1981)農薬実験法(2)殺菌剤編. ソフトサイエンス社. 110-115)。
【0059】
【数1】
【0060】
結果
CMVに対する感染阻害効果を検定した結果、本酵素は、約23 μg/mlで平均86%の感染阻害率を示し、約9 μg/mlで平均42%の感染阻害率を示した。
【0061】
(3)野菜類軟腐病に対する生育阻害作用の検定
検定植物:男爵イモ(Solanum tuberosum L.)
検定菌:野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)
傷やかさぶたがない直径9〜10 cm程度の男爵イモを用意し、爪で傷つけないように水道水で土や分泌物等を洗い流した。(以下の操作は全てクリンベンチ内で行った)ビーカーに70%エタノールを満たし、洗ったジャガイモを20秒程度入れ、乾かした。その後、手袋をして切り口に触れないように1 cm間隔でなるべく平行になるように輪切りにし、シャーレに入れた。このジャガイモの中央にサンプル(27 μg/ml)を50 μL滴下し、さらに滴下したサンプルの中央に点接種で植菌した。コントロールは滅菌水を用いた。シャーレ内の乾燥を防ぐために、キムワイプを滅菌水で湿らせてシャーレ内に置いてからパラフィルムをシャーレのふちに巻いて、34℃で3日インキュベートした。菌が生育すると、生育円が観察されるのでこの生育円の大きさで生育阻害作用の有無を判断した。
【0062】
結果
野菜類軟腐病菌の生育円の様子を図9に示す。滅菌水で処理したものに比べて、本酵素(27 μg/ml)で処理したものは生育円が小さいのが確認できた。
【0063】
(4)考察
本酵素もCMVに対して感染阻害効果を示し、約23 μg/mlで86%、約9μg/mlで42%の感染阻害率を示した。以前の研究では、同じセリンプロテアーゼであるα-キモトリプシンやズブチリシンのIC50はそれぞれ396 μ/ml、74.3 μg/mlであった(山下ら, 大阪府立大学修士論文)。これらの値と比べてもE.foetida由来のプロテアーゼは低濃度で効果を示しており、CMVに対する感染阻害効果はかなり特異的であるといえる。
【0064】
野菜類軟腐病はハクサイ、ダイコン、ニンジン、ジャガイモなど多種類の作物にみられる世界中に分布する重要病害である(脇本哲 (1994). 総説 植物病理学. 養賢堂. 425)。この病原菌は、Erwinia carotovora subsp.carotovoraであり、植物の傷口や昆虫の食痕、気孔、水孔などから組織内に侵入し、増殖に伴って生産するペクチナーゼによって宿主細胞を遊離させ組織を軟腐させる。腐敗した組織は悪臭を放つ。防除法としては、抵抗性遺伝子の導入や薬剤散布がなされている(都丸敬一 (2001). 全国農村教育協会. 3, 106-110)。
【0065】
この病原菌に対して生育阻害効果があるのか検定したところ、生育円が小さく抑えられているのが観察できた(図9)。この結果より、本酵素は抗植物病原細菌活性も有する可能性が示唆された。
E.foetida由来のプロテアーゼは有望な抗植物ウイルス剤の候補であり、植物病原菌の生育阻害作用もあることが示唆されたので、スペクトルの広い生物農薬として利用できる可能性があると考えられる。
【0066】
実施例4. プロテアーゼのcDNAクローニング
(1)mRNAの抽出
以下の操作はRNase freeの条件下で行った。
凍結乾燥後に破砕したシマミミズ100 mgを1.5 ml容エッペンチューブに入れ、1mlのISOGENを加えて完全に溶解させた。これを遠心分離(12,100×g、4℃、15 分)し、上清のみを別のエッペンチューブに移して5分間室温で放置した。これに0.2 mlのクロロホルムを加えボルテックスでよく混合して3分間室温で放置し、遠心分離(12,100×g、4℃、15 分)した。遠心分離後、3相に分かれたので水層(上層)のみを回収し、これに0.4 mlのイソプロパノールを加え10分間室温で放置してから遠心分離(12,100×g、4℃、10 分)した。遠心分離後、上清を捨て、沈殿に1 mlの75%エタノールを加えて軽くボルテックスし遠心分離(12,100×g、4℃、5分)した。遠心分離後、上清を完全に捨てて、沈殿に20μLのDEPC処理水を加え完全に溶解させてmRNA溶液とした。(mRNA溶液は1μg/mlに調製した。)mRNA溶液は−80℃で保存した。
【0067】
(2)逆転写PCRによるcDNAの合成
以下の操作はRNase freeの条件下で行った。
Oligo(dT)17-APプライマー(100 μM)1μL、dNTP Mix(2mM)5μL、mRNA溶液1μL、DEPC処理水5μLをPCR用のチューブに混合し、65℃で5分間置いた後すぐに氷中に3分間置いた。これに、5×First-Standard Buffer 4μL、0.1 M DTT 2μL、RNase インヒビター1μLを加え、42℃で2分間置いた。その後、Super Script II RT(逆転写酵素)1μLを加えて、(42℃, 50 分)→(70℃, 15 分)→(4℃,∞)の条件で逆転写PCRを開始し、cDNAを得た。cDNAは−20℃で保存した。
(Oligo(dT)17-APプライマー)
GGCCACGCGTCGACTAGTACTTTTTTTTTTTTTTTTT (配列番号:4)
【0068】
(3)PCRによる目的DNAの増幅
本酵素のN末端アミノ酸配列から設計したプライマーをセンスプライマー、Oligo(dT)17-APプライマーのアダプタープライマーをアンチセンスプライマーとして用いて、PCRを行った。表7のとおりサンプルおよびネガティブコントロール(ネガ・コン)をそれぞれ全量20 μL調製し、{(98℃, 10 秒)→(58℃, 30 秒)→(72℃, 1分)}×40サイクル→(4℃, ∞)の条件でPCRを行った。PCR後、0.001%エチジウムブロマイドを含む1%アガロースゲルで電気泳動を行い、目的DNAの増幅の確認をした。
(センスプライマー) GGIGARATHATHCCICAYAAYGCITA (配列番号:5)
(アンチセンスプライマー) GGCCACGCGTCGACTAGTAC (配列番号:6)
【0069】
【表7】
【0070】
(4)ライゲーション反応と形質転換
アガロースゲル電気泳動後、目的のバンドをカッターで切りだし、ゲル抽出を行った。ゲル抽出は、QIAquick Gel Extraction Kit (50)(QIAGEN社)を使用し、付属のプロトコールに従った。
【0071】
次に、pCR 2.1-TOPOベクター(invitrogen社)(図10)を用いてライゲーション反応および形質転換を行った。まず、ゲル抽出液1 μL、Salt Solution 0.5 μL、TOPOベクター0.5 μL、滅菌MilliQ μLをチューブにとり(計3μL)、穏やかに混和し室温に5分間置いてライゲーション反応させた。このうち2 μLを大腸菌(One shot E.coli)25 μLに添加し穏やかに撹拌して氷上で20分間静置し、42℃で30秒間ヒートショックを与えて形質転換した。その後、125 μLのS.O.C(室温)を加え37℃で1時間振とうしながらインキュベートした。インキュベート後、IPTG、X-Galを含むLB+アンピシリンプレートにスプレッドし、37℃で一晩インキュベートした。
【0072】
(5)プラスミドDNAの抽出と遺伝子の解析
生じたコロニーのうち、白もしくはうすい水色のコロニーをピックアップし、LB+アンピシリンプレートにレプリカを作成して、さらにコロニーPCRを行いインサートを確認した。レプリカは37℃で一晩インキュベートした。レプリカのコロニーが生えると、そのうちコロニーPCRでインサートが確認できたもののコロニーを1白金耳ピックアップし、5 mlのLB+アンピシリン液体培地に植菌し37℃で20時間、振盪しながらインキュベートした。
【0073】
20時間のインキュベート後、エッペンチューブに培養液を入れ遠心分離して上清を捨て菌体を回収した。この菌体から、QIAprep Spin Miniprep Kit (50)(QIAGEN社)を使用し、付属のプロトコールに従ってプラスミドDNAを抽出した。
抽出したプラスミドDNAを濃度既知のpUC19とともに泳動し、その発光度合いを比較してプラスミドDNAの濃度を決定した。そして、プラスミドDNA (0.25 μg/μL)2μL、滅菌MilliQ 17 μL、プライマー1種(M13-20プライマー、M13-Revプライマーの両方を用意)1μLをチューブに入れ、遺伝子の解析を行った。
【0074】
コロニーPCRは次のとおりに行った。Green Taq 10 μL、M13-20プライマー(100 μM)0.1 μL、M13-Revプライマー(100 μM)0.1 μL、滅菌MilliQ 9.8 μLをチューブにとり、これにコロニーをつついた爪楊枝の先をつけて、上記(3)と同じ条件でPCRおよびアガロースゲル電気泳動を行った。
(M13-20プライマー)GTAAAACGACGGCCAGT (配列番号:7)
(M13-Revプライマー)CAGGAAACAGCTATGAC (配列番号:8)
【0075】
結果
PCRの結果を図11に示す。マーカーとしてX174-HaeIIを用いた。図11の(a)中のネガティブコントロール(1)及び(2)の泳動像から判断し、図11の(b)中の1078 bpの位置のバンドを切り出してゲル抽出を行った。
抽出したプラスミドDNAの泳動結果を図12に示す。マーカーとしてX174-HaeIIを用いた。pUC19(0.25 μg/μL)の発光度合いと比較して抽出したプラスミドDNAの濃度を決定し、遺伝子の解析を行った。得られた塩基配列と終止コドンまでのアミノ酸配列を図13(配列番号1及び配列番号2)に示す。
【技術分野】
【0001】
本発明は新規なプロテアーゼに関する。
【背景技術】
【0002】
植物の病気の原因には、細菌、カビ(菌類)などの病原微生物やウイルスなどの他、環境条件等による生理病、遺伝病などが知られている。中でも、カビによる病気が70%以上にもなる。また、植物ウイルスは1000種以上も存在し、植物に感染して農作物の品質の低下や、収量の低下などの被害を及ぼし、その被害額は日本だけでも年間1000億円にものぼると言われている。これらの対策として、農薬の利用や遺伝子組換え、ワクチン療法などが挙げられる。しかし、農薬や遺伝子組み換えは安全性の問題をはらんでおり、ワクチン療法は弱毒ウイルスの開発や接種に労力や注意が必要である(都丸敬一 (2001) 植物のウイルス病物語 始まりからバイオテクノロジーまで. 全国農村教育協会, 106-110)。
【0003】
そこで、天然の抗ウイルス剤を開発することを目的に研究が行われてきた。そして、シマミミズ(Eisenia foetida)から抗植物ウイルス活性を有する物質を既に単離、精製しており、それがセリンプロテアーゼであることを明らかにし、その酵素学的性質や抗植物ウイルス活性についての研究が行われてきた(野田加奈子(2006)大阪府立大学修士論文;山下崇之(2008)大阪府立大学修士論文)。
【0004】
E.foetidaは腐敗した植物などの有機物が豊富な土地に生息し、昔はゴミ捨て場や堆肥舎の近くにも生息し、身近にいるミミズであった。現在は、ゴミ回収の制度が整い堆肥舎がなくなりそれほど身近ではなくなったが、日本だけでなく世界各地でコンポストミミズとして堆肥化によく利用されている。ヨーロッパやシベリア、北アメリカ、イスラエル、チリ、ニュージーランド、オーストラリア、日本など温帯気候下の世界中に広く分布するとされており、熱帯にもところどころにいるようである(渡辺弘之 (2003). ミミズ 嫌われもののはたらきもの. 東海大学出版会)。
【0005】
またE.foetidaを含むミミズの仲間には医薬等への利用が期待される物質が見出されており、E.foetidaの体液は溶血、細胞毒性、血液凝集、抗菌などの作用を示し、この効果を活用した脳血栓の溶血剤や避妊剤などが検討されている(中村好男 (2004). ミミズと土と有機農業. 創森社)。
【0006】
食品の安全性が追求されている今の時代、農作物に対する安全性も重要であり、これからは質とともに安全性も重要視されることが予想される。また、人口増加に伴う食糧危機の点から、農作物の品質の低下や収量の低下は食糧不足に直結する問題である。E.foetidaから安全かつ有効な生物農薬を開発することは、これらの課題の解決に貢献できると期待している。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】都丸敬一 (2001) 植物のウイルス病物語 始まりからバイオテクノロジーまで. 全国農村教育協会, 106-110
【非特許文献2】野田加奈子(2006)大阪府立大学修士論文
【非特許文献3】山下崇之(2008)大阪府立大学修士論文
【非特許文献4】渡辺弘之 (2003) ミミズ 嫌われもののはたらきもの. 東海大学出版会
【非特許文献5】中村好男 (2004). ミミズと土と有機農業. 創森社
【0008】
【非特許文献6】K. Inouye (2006),Protein Expression and Purification 46, 248-255
【非特許文献7】Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109
【非特許文献8】Nobuyoshi Nakajima (1993), Biosci. Biotech. Biochem, 57(10), 1726-1730
【0009】
【非特許文献9】Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109
【非特許文献10】Bradford MM (1976), Anal. Biochem, 72, 248-254
【非特許文献11】深見順一ら(1981)農薬実験法(2)殺菌剤編. ソフトサイエンス社. 110-115
【非特許文献12】脇本哲 (1994). 総説 植物病理学. 養賢堂. 425)
【発明の概要】
【発明が解決しようとする課題】
【0010】
そこで、本発明は、ミミズ(E.foetida)由来の、抗植物ウイルス活性及び抗植物病原細菌活性を有するプロテアーゼ及びその使用、並びにその製造手段を提供するものである。
【課題を解決するための手段】
【0011】
したがって、本発明は、下記の性質:
(1)N末端アミノ酸配列
下記のN末端アミノ酸配列:
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln
を有する;
(2)基質特異性
合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示す;
(3)阻害剤による影響
プロテアーゼ活性が、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N−トシル−L−フェニルアラニンクロメチルケトン(TPCK)、ダイズトリプシンインヒビター(SBTI)により阻害される;
(4)最適pHとpH安定性
最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;および
(5)最適温度と温度安定性
最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない;
を有する、ミミズ(Eisenia foetida)由来のプロテアーゼ、
を提供する。
【0012】
本発明はまた、配列番号:2に記載のアミノ酸配列を含むプロテアーゼ;配列番号:2に記載のアミノ酸配列において、1個〜数個のアミノ酸の置換、欠失及び/又は付加により修飾されているアミノ酸配列からなるプロテアーゼ;配列番号:2に記載のアミノ酸配列に対して少なくとも90%の配列同一性を有するプロテアーゼ;或いは配列番号:1に記載の塩基配列からなる核酸又はその相補鎖に対して高緊縮条件下でハイブリダイズする核酸によりコードされているプロテアーゼ、を提供する。
【0013】
本発明はまた、上記何れかに記載のプロテアーゼをコードする核酸;当該核酸により形質転換された宿主細胞;並びに当該宿主細胞を培養することを含んでなる、上記プロテアーゼの製造方法を提供する。
本発明はまた、上記(1)〜(5)の性質を有するプロテアーゼの製造方法において、当該プロテアーゼをミミズ(Eisenia foetida)から採取することを含んでなる方法を提供する。
【0014】
本発明は更に、前記のプロテアーゼを含んでなる、抗植物病原性ウイルス剤を提供し、当該植物病原性ウイルスは、例えば、キュウリモザイクウイルス(Cucumber mosaic virus;CMV)である。
本発明はまた、前記のプロテアーゼを含んでなる、抗植物病原性細菌剤を提供し、当該植物病原性細菌は、例えば、野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)である。
【図面の簡単な説明】
【0015】
【図1】図1は、本発明のプロテアーゼの精製過程における、陰イオン交換クロマトグラフィーの溶出曲線を示す。
【図2】図2は、本発明のプロテアーゼの精製過程における、疎水性相互作用クロマトグラフィーの溶出曲線を示す。
【図3】図3は、図2におけるプロテアーゼ活性画分の、SDS-PAGEによる分析の結果を示す。
【0016】
【図4】図4は、本発明のプロテアーゼの最適pHを示すグラフである。
【図5】図5は、本発明のプロテアーゼのpH安定性を示すグラフである。
【図6】図6は、本発明のプロテアーゼの最適温度を示すグラフである。
【図7】図7は、本発明のプロテアーゼの温度安定性を示すグラフである。
【図8】図8は、本発明のプロテアーゼの、キュウリモザイクウイルス(Cucumber mosaic virus;CMV)に対する阻害を示すグラフである。
【0017】
【図9】図9は、本発明のプロテアーゼの、野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)に対する阻害を示す写真である。
【図10】図10は、本発明のプロテアーゼをコードするDNAのクローニングに使用したクローニングベクターの構造を示す図である。
【図11】図11は、PCR増幅産物中の、本発明のプロテアーゼをコードするDNAの検出を示す電気泳動図である。
【図12】図12は、本発明のプロテアーゼをコードするDNAを含むプラスミドDNAの電気泳動図である。
【図13】図13は、本発明のプロテアーゼをコードするDNAの塩基配列を及び当該プロテアーゼのアミノ酸配列を示す。
【発明を実施するための形態】
【0018】
本願発明は、下記の性質:
(1)N末端アミノ酸配列
下記のN末端アミノ酸配列:
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln (配列番号:3)
を有する;
(2)基質特異性
合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示す;
(3)阻害剤による影響
プロテアーゼ活性が、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N−トシル−L−フェニルアラニンクロメチルケトン(TPCK)、ダイズトリプシンインヒビター(SBTI)により阻害される;
(4)最適pHとpH安定性
最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;および
(5)最適温度と温度安定性
最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない;
を有する、ミミズ(Eisenia foetida)由来のプロテアーゼ、を提供する。
【0019】
上記のプロテアーゼは、配列番号:2に示すアミノ酸配列を有する。従って本発明はまた、配列番号:2に記載のアミノ酸配列を含むプロテアーゼを提供する。
【0020】
ある酵素につき、そのアミノ酸配列を部分的に改変しても、生来の酵素の活性が維持されることが当業者によりよく知られている。従って本発明はまた、配列番号:2に記載のアミノ酸配列において、1個〜数個のアミノ酸の置換、欠失及び/又は付加により修飾されているアミノ酸配列からなるプロテアーゼ;配列番号:2に記載のアミノ酸配列に対して少なくとも90%の配列同一性を有するプロテアーゼ;或いは配列番号:1に記載の塩基配列からなる核酸又はその相補鎖に対して高緊縮条件下でハイブリダイズする核酸によりコードされているプロテアーゼ、を提供する。
【0021】
酵素の採取方法
本発明のプロテアーゼをミミズから採取するには、例えば、まずは、凍結乾燥したシマミミズから粗酵素液を抽出し、これを陰イオン交換クロマトグラフィー、続いて疎水性相互作用クロマトグラフィーに供する。また、A280およびカゼインを基質としたプロテアーゼ活性を測定しこれを分画の指標とする。最後に、SDS-PAGEを行い純度の確認を行う。
【0022】
プロテアーゼ活性測定方法
サンプル 0.5 mlと1.33% (w/v) カゼイン(0.1 M グリシン-NaOH緩衝液, pH 9.5 に溶解) 1.5 mlを混合し、対照にはサンプル0.5 mlのみを用意し、それぞれ37℃、60分間インキュベートする。これに、0.11 M TCA、0.22 M 酢酸ナトリウム、0.33M 酢酸2 mlを加えよく撹拌し反応を停止する。対照にはその後サンプル 0.5 mlを加る。そして、この反応停止液をNo.2のろ紙を用いてろ過し、ろ液のA275を測定する。ここで、1 U =1分間にA275を0.0074上昇させるのに必要な酵素量(A275 of 0.0074 = 1 μg tyrosine)と定義する(K. Inouye (2006),Protein Expression and Purification 46, 248-255)。
【実施例】
【0023】
次に、本発明を、実施例により更の具体的に説明する。
実施例1. シマミミズからのプロテアーゼの精製
(1)粗酵素液の抽出
凍結乾燥したシマミミズ20 gに50 mM Tris-HCl 緩衝液(pH 7.0)220 mlを加え、30分間氷上で撹拌し、遠心分離(27,700×g、4℃、10 分)した。その上清に終濃度が2%となるようにストレプトマイシン硫酸塩を加えて、30分間氷上で撹拌し、遠心分離(27,700×g、4℃、10 分)して核酸を除去した。次に、この上清を50 mM Tris-HCl 緩衝液(pH 7.0)で3倍に希釈し、氷上で90分間電気透析を行い、グラスフィルターでろ過したものを粗酵素液とした。
【0024】
(2)DEAE-TOYOPEARL 650Mによる陰イオン交換クロマトグラフィー
担体:DEAE-TOYOPEARL 650M
カラム:直径2.5 cm×27 cm
初期緩衝液:50 mM Tris-HCl (pH 7.0)
溶出緩衝液:50 mM Tris-HCl + 0.3 M NaCl (pH 7.0)
カラムを実験台に水平になるように立て、担体を流し込み、500 mlの蒸留水を流して洗い、その後、500 mlの初期緩衝液で平衡化した。次に、粗酵素液をアプライし溶出液のA280が0.1に下がるまで初期緩衝液を流して洗浄を行った。そして、初期緩衝液 1000 mlと溶出緩衝液 1000 mlを架橋して溶出を開始した。試験管1本あたり20 ml、100本の画分に分画し、各画分のA280とプロテアーゼ活性を測定した。
【0025】
結果
陰イオン交換クロマトグラフィーの溶出曲線を図1に示す。約0.07 M NaClでプロテアーゼ活性のピークが得られた。画分No.23〜29を回収し、等量の2M硫酸アンモニウムと混合して、次の疎水性相互作用クロマトグラフィーのサンプルとした。
【0026】
(3)HiTrap Phenyl HPによる疎水性相互作用クロマトグラフィー
カラム:HiTrap Phenyl HP 1ml
初期緩衝液:20 mM Tris-HCl + 1M硫酸アンモニウム (pH 7.0) (吸引ろ過)
溶出緩衝液:20 mM Tris-HCl (pH 7.0) (吸引ろ過)
カラムを実験台に水平になるように立て、適量のMilliQで洗浄し、さらに5mlの溶出緩衝液を流速1 ml/minで流し洗浄した。次に、5 ml〜10 mlの初期緩衝液で平衡化した。
【0027】
陰イオン交換後の活性画分を等量の2M硫酸アンモニウムと混合し、カラムに適用した。初期緩衝液と溶出緩衝液を架橋し、0〜10本までは1M、10〜60本までは1〜0M、60〜70本までは0Mの硫酸アンモニウムで溶出するようにグラジエントの設定を行い、溶出した。エッペンチューブ1本あたり1ml、流速1ml/分で70本の画分に分画し、各画分のA280とプロテアーゼ活性を測定した。
【0028】
結果
上記疎水性相互作用クロマトグラフィーの溶出曲線を図2に示す。約0.6 M硫酸アンモニウムでプロテアーゼ活性のピークが得られた。プロテアーゼ活性の高かった画分No.24〜34をSDS-PAGEのサンプルとした。
【0029】
(4)SDS-PAGE
ランニングゲルとストッキングゲルを、表1のとおりに試薬を混合して作製し、電気泳動層にセットした。疎水性相互作用クロマトグラフィー後の活性画分から10μLとり、10μLの2×サンプル緩衝液と混合して3分間煮沸した。これをゲルに適用し、20 mA、300Vで90分間泳動した。マーカーは次のものを使用した。β−ガラクトシダーゼ(分子量:116,000)、ウシ血清アルブミン(分子量:66,200)、オバルミン(分子量:45,000)、ラクターゼ脱水素酵素(分子量:35,000)、REase Bsp98I(分子量:25,000)、β−ラクトグロブリン(分子量:18,400)、リゾチーム(分子量:14,400)。泳動後のゲルはCBB溶液で染色した。
【0030】
【表1】
【0031】
SDS-PAGEの結果を図3に示す。
また、精製の経過を下記の表2に示す。
【0032】
【表2】
【0033】
考察
図3を見ると、メインバンドの他にもうっすらとバンドが確認できる。図2の疎水性相互作用クロマトグラフィーの溶出曲線を見てもわかるように、画分No.24〜34までは比較的きれいなピークが確認でき多種類のタンパクが混在しているとは考えにくい。したがってメインバンドのほかにうっすらと確認できたバンドは、主に目的タンパクの自己分解産物である可能性が高い。
【0034】
実施例2. 酵素学的性質
(1)N末端アミノ酸配列の解析
精製後の酵素をSDS-PAGEに供した後、PVDF膜に転写した。転写は、300 V、108 mA(1 cm2あたり2 mA)の条件で60分間程度行った。転写後、PVDF膜をMilliQで洗浄し、CBB-R250溶液で染色した後、MilliQで調製した70%メタノールで脱色しMilliQで軽く洗浄してキムタオルの上で乾燥させてから目的のバンドを切り出した。そしてN末端からのアミノ酸配列を決定した。
【0035】
結果
N末端アミノ酸配列を解析した結果、以下の配列が得られた。
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln (配列番号:3)
BLASTを用いて相同性検索を行った結果、相同性を示すタンパク質は見られなかった。また、赤ミミズL.rubellus由来のフィブリン分解酵素とも相同性が見られなかった(Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109;Nobuyoshi Nakajima (1993), Biosci. Biotech. Biochem, 57(10), 1726-1730; Feng Wang, Chao Wang (2003), Biotechnol. Lett, 25, 1105-1109)。N末端配列の比較は、下記の表3のとおりである。
【0036】
【表3】
【0037】
(2)基質特異性の測定
2.5 mMの各合成基質溶液(70%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)100 μLと、酵素100 μLを混合し、37℃で30分間反応させ、405 nmにおける吸光度を測定した。コントロールとしては、酵素の代わりに同量の0.1 M グリシン-NaOH 緩衝液(pH 9.5)を用いた。測定値が一番大きかったものを100として、各pHでの測定値から相対活性値を算出した。なお、使用した合成基質は次の5つである。
【0038】
(1)N-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド(キモトリプシン用基質)
(2)N-グルタリル-L-Phe-p-ニトロアニリド(キモトリプシン用基質)
(3)N-α-ベンゾイル-L-Arg-p-ニトロアニリド(トリプシン用基質)
(4)N-サクシニル-Ala-Ala-Pro-Leu-p-ニトロアニリド(エラスターゼ用基質)
(5)N-サクシニル-Ala-Ala-Ala-p-ニトロアニリド(エラスターゼ用基質)
【0039】
結果
表4に示すとおり、本酵素は合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu-p-ニトロアニリドに対して高い活性を示した。
【0040】
【表4】
【0041】
(3)阻害剤による影響
酵素60μLと、終濃度が以下に示す濃度になるように各プロテアーゼ阻害剤60μLを混合し、37℃で30分間プレインキュベートした。コントロールとしては、プロテアーゼ阻害剤の代わりに同量の水を用いた。プレインキュベート後、5 mMのN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド溶液(4%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)60μLを加え、37℃で20分間反応させ、405 nmにおける吸光度を測定した。コントロールの測定値を阻害率0%として、各阻害剤の阻害率を算出した。
【0042】
(1)0.01 mg/ml ダイズトリプシンインヒビター(SBTI)
(2)1 mM フェニルメチルスルホニルフルオリド(PMSF)
(3)0.1 mg/ml キモスタチン(Chymostatin)
(4)1mM N-トシル-L-フェニルアラニンクロロメチルケトン(TPCK)
(5)0.01 mg/ml ストレプトマイセス ズブチリシンインヒビター(SSI)
(6)1mM 2,2'-ビピリジル
(7)1mM 1,10-フェナンスロリン(Phenanthoroline)
(8)1mM EDTA
(9)0.1 mM E-64
注:(1)〜(5)はセリンプロテアーゼ阻害剤であり、(6)〜(8)はメタロプロテアーゼ阻害剤であり、(9)はシステインプロテアーゼ阻害剤である。
【0043】
結果
表5に示すとおり、本酵素のプロテアーゼ活性は、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N-トシル-L-フェニルアラニンクロロメチルケトン(TPCK)及びダイズトリプシンインヒビター(SBTI)のセリンプロテアーゼ阻害剤に強く阻害されたが、他の阻害剤にはそれほど影響を受けなかった。
【0044】
【表5】
【0045】
(4)最適pH
2%アゾカゼイン100 μL、pH 3.4〜11.7の広域緩衝液(0.2 Mホウ酸、0.05 Mクエン酸、0.1 Mリン酸三ナトリウム十二水和物)300 μL、酵素100 μLをエッペンチューブに混合し、37℃で90分間反応させた。コントロールとしては、2%アゾカゼイン100 μL、pH 3.4〜11.7の広域緩衝液(0.2 Mホウ酸、0.05 Mクエン酸、0.1 Mリン酸三ナトリウム十二水和物)400 μLを混合したものを用いた。反応後、10%TCA溶液500 μLを加えて完全に反応を止め、遠心分離(17,400×g、室温、10 分)して、その上清の366 nmにおける吸光度を測定した。測定値が一番大きかったものを100として、各pHでの測定値から相対活性値を算出した。
【0046】
結果
図4に示すとおり、最適pHは9.6付近にある。
【0047】
(5)pH安定性
pH 3.9〜11.7の広域緩衝液90 μL、酵素10 μLをエッペンチューブに混合し、37℃で60分間プレインキュベートした。コントロールとしてはpH 3.9〜11.7の広域緩衝液100 μLを用いた。プレインキュベート後、1%アゾカゼイン(0.5 M グリシン-NaOH 緩衝液, pH 9.5に溶解)400 μLを加え、37℃で90分間反応させた。反応後、10%TCA 溶液500 μLを加えて完全に反応を止め、遠心分離(17,400×g、室温、10 分)して、その上清の366 nmにおける吸光度を測定した。測定値が一番大きかったものを100として、各pHでの測定値から相対活性値を算出した。
【0048】
結果
図5に示すとおり、pH 6〜11で80%以上の安定性があった。
【0049】
(6)最適温度
2.5 mMのN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド溶液(4%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)80 μLを10〜80℃の各温度で5分間プレインキュベートし、酵素80 μLを加え、各温度で20分間反応させ、405 nmにおける吸光度を測定した。コントロールとしては、酵素の代わりに同量の0.1 M グリシン-NaOH 緩衝液(pH 9.5)を加えたものを用いた。測定値が一番大きかったものを100として、各温度での測定値から相対活性値を算出した。
【0050】
結果
図6に示すとおり、最適温度は約60℃である。
【0051】
(7)温度安定性
酵素80 μLを10〜80℃の各温度で30分間プレインキュベートした。コントロールとしては、酵素の代わりに同量の0.1 M グリシン-NaOH 緩衝液(pH 9.5)を用いた。プレインキュベート後、2.5 mMのN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリド溶液(4%DMSOを含む0.1 M グリシン-NaOH 緩衝液, pH 9.5に溶解)80 μLを加えて、37℃で30分間反応させ、405 nmにおける吸光度を測定した。測定値が一番大きかったものを100として、各温度での測定値から相対活性値を算出した。
結果
図7に示すとおり、加熱によるプロテアーゼ活性の低下は50℃まで見られなかった。
【0052】
実施例3. プロテアーゼの農業分野への利用
本実施例では、本酵素の応用利用を目的とした研究を行った。まず、農作物に甚大な被害をもたらせているキュウリモザイクウイルス(Cucumber mosaic virus;CMV)に対する感染阻害効果を本酵素が有するのか検定した。検定方法は混合接種法で行った。この方法は、感染阻害作用の有無を確認するには最適な方法である。
【0053】
また、農作物の被害は植物ウイルスだけではなく植物病原菌によっても引き起こされている。そこで、抗細菌活性も有するのか検定した結果、植物病原菌の一つである野菜類軟腐病(Erwinia carotovora subsp.carotovora)に対して抗細菌活性を示した。
【0054】
(1)酵素濃度の決定
酵素の濃度は、BSAを標準タンパク質としてブラッドフォードの方法(Bradford MM (1976), Anal. Biochem, 72, 248-254)によって決定した。
【0055】
(2)植物ウイルスに対する感染阻害作用の検定
検定植物:ジュウロクササゲ(Vigna unguiculate cv. kurodane-sanjaku)
検定ウイルス:キュウリモザイクウイルス(Cucumber mosaic virus;CMV)のpepo系統
バーミキュライトを満たした鉢にササゲの種子をまき温室内に置き、発芽した後、土を入れたセルトレイに苗を移植し育種した。実験には、初生葉が最大に展開したものを用い、1試料あたり5〜10個体を用いた。
【0056】
まず、0.1Mリン酸緩衝液(pH 7.0)(1 Mリン酸緩衝液は1M KH2PO4と1M K2HPO4を2:3で混合し、pH 7.0に調整したもの)でCMV溶液を100 μg/mlに調製した。試料は20 mM Tris-HCl 緩衝液m(pH 7.0)で各濃度に希釈した。これらを表6のとおりエッペンチューブに加え、サンプル溶液とコントロール溶液を調製し、それぞれをボルテックスでよく混合して氷中に30分間置いた。
【0057】
【表6】
【0058】
ササゲへの接種は混合接種法で行った。最大に展開したササゲの初生葉に、サンプル葉とコントロール葉を区別するためにマーカー用インクで印をし、研磨剤としてカーボランダムをうっすらと満遍なくふりかけた。調製したサンプル溶液とコントロール溶液を氷中に置いて30分後に、一方の葉にサンプル溶液を、他方の葉にはコントロール溶液を綿棒で優しく接種した。接種後、すぐに流水でカーボランダムを洗い流した。ウイルスに感染すると葉に病班を形成するので、接種して二日後にこの病班の数を測定し感染阻害率を算出した(深見順一ら(1981)農薬実験法(2)殺菌剤編. ソフトサイエンス社. 110-115)。
【0059】
【数1】
【0060】
結果
CMVに対する感染阻害効果を検定した結果、本酵素は、約23 μg/mlで平均86%の感染阻害率を示し、約9 μg/mlで平均42%の感染阻害率を示した。
【0061】
(3)野菜類軟腐病に対する生育阻害作用の検定
検定植物:男爵イモ(Solanum tuberosum L.)
検定菌:野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)
傷やかさぶたがない直径9〜10 cm程度の男爵イモを用意し、爪で傷つけないように水道水で土や分泌物等を洗い流した。(以下の操作は全てクリンベンチ内で行った)ビーカーに70%エタノールを満たし、洗ったジャガイモを20秒程度入れ、乾かした。その後、手袋をして切り口に触れないように1 cm間隔でなるべく平行になるように輪切りにし、シャーレに入れた。このジャガイモの中央にサンプル(27 μg/ml)を50 μL滴下し、さらに滴下したサンプルの中央に点接種で植菌した。コントロールは滅菌水を用いた。シャーレ内の乾燥を防ぐために、キムワイプを滅菌水で湿らせてシャーレ内に置いてからパラフィルムをシャーレのふちに巻いて、34℃で3日インキュベートした。菌が生育すると、生育円が観察されるのでこの生育円の大きさで生育阻害作用の有無を判断した。
【0062】
結果
野菜類軟腐病菌の生育円の様子を図9に示す。滅菌水で処理したものに比べて、本酵素(27 μg/ml)で処理したものは生育円が小さいのが確認できた。
【0063】
(4)考察
本酵素もCMVに対して感染阻害効果を示し、約23 μg/mlで86%、約9μg/mlで42%の感染阻害率を示した。以前の研究では、同じセリンプロテアーゼであるα-キモトリプシンやズブチリシンのIC50はそれぞれ396 μ/ml、74.3 μg/mlであった(山下ら, 大阪府立大学修士論文)。これらの値と比べてもE.foetida由来のプロテアーゼは低濃度で効果を示しており、CMVに対する感染阻害効果はかなり特異的であるといえる。
【0064】
野菜類軟腐病はハクサイ、ダイコン、ニンジン、ジャガイモなど多種類の作物にみられる世界中に分布する重要病害である(脇本哲 (1994). 総説 植物病理学. 養賢堂. 425)。この病原菌は、Erwinia carotovora subsp.carotovoraであり、植物の傷口や昆虫の食痕、気孔、水孔などから組織内に侵入し、増殖に伴って生産するペクチナーゼによって宿主細胞を遊離させ組織を軟腐させる。腐敗した組織は悪臭を放つ。防除法としては、抵抗性遺伝子の導入や薬剤散布がなされている(都丸敬一 (2001). 全国農村教育協会. 3, 106-110)。
【0065】
この病原菌に対して生育阻害効果があるのか検定したところ、生育円が小さく抑えられているのが観察できた(図9)。この結果より、本酵素は抗植物病原細菌活性も有する可能性が示唆された。
E.foetida由来のプロテアーゼは有望な抗植物ウイルス剤の候補であり、植物病原菌の生育阻害作用もあることが示唆されたので、スペクトルの広い生物農薬として利用できる可能性があると考えられる。
【0066】
実施例4. プロテアーゼのcDNAクローニング
(1)mRNAの抽出
以下の操作はRNase freeの条件下で行った。
凍結乾燥後に破砕したシマミミズ100 mgを1.5 ml容エッペンチューブに入れ、1mlのISOGENを加えて完全に溶解させた。これを遠心分離(12,100×g、4℃、15 分)し、上清のみを別のエッペンチューブに移して5分間室温で放置した。これに0.2 mlのクロロホルムを加えボルテックスでよく混合して3分間室温で放置し、遠心分離(12,100×g、4℃、15 分)した。遠心分離後、3相に分かれたので水層(上層)のみを回収し、これに0.4 mlのイソプロパノールを加え10分間室温で放置してから遠心分離(12,100×g、4℃、10 分)した。遠心分離後、上清を捨て、沈殿に1 mlの75%エタノールを加えて軽くボルテックスし遠心分離(12,100×g、4℃、5分)した。遠心分離後、上清を完全に捨てて、沈殿に20μLのDEPC処理水を加え完全に溶解させてmRNA溶液とした。(mRNA溶液は1μg/mlに調製した。)mRNA溶液は−80℃で保存した。
【0067】
(2)逆転写PCRによるcDNAの合成
以下の操作はRNase freeの条件下で行った。
Oligo(dT)17-APプライマー(100 μM)1μL、dNTP Mix(2mM)5μL、mRNA溶液1μL、DEPC処理水5μLをPCR用のチューブに混合し、65℃で5分間置いた後すぐに氷中に3分間置いた。これに、5×First-Standard Buffer 4μL、0.1 M DTT 2μL、RNase インヒビター1μLを加え、42℃で2分間置いた。その後、Super Script II RT(逆転写酵素)1μLを加えて、(42℃, 50 分)→(70℃, 15 分)→(4℃,∞)の条件で逆転写PCRを開始し、cDNAを得た。cDNAは−20℃で保存した。
(Oligo(dT)17-APプライマー)
GGCCACGCGTCGACTAGTACTTTTTTTTTTTTTTTTT (配列番号:4)
【0068】
(3)PCRによる目的DNAの増幅
本酵素のN末端アミノ酸配列から設計したプライマーをセンスプライマー、Oligo(dT)17-APプライマーのアダプタープライマーをアンチセンスプライマーとして用いて、PCRを行った。表7のとおりサンプルおよびネガティブコントロール(ネガ・コン)をそれぞれ全量20 μL調製し、{(98℃, 10 秒)→(58℃, 30 秒)→(72℃, 1分)}×40サイクル→(4℃, ∞)の条件でPCRを行った。PCR後、0.001%エチジウムブロマイドを含む1%アガロースゲルで電気泳動を行い、目的DNAの増幅の確認をした。
(センスプライマー) GGIGARATHATHCCICAYAAYGCITA (配列番号:5)
(アンチセンスプライマー) GGCCACGCGTCGACTAGTAC (配列番号:6)
【0069】
【表7】
【0070】
(4)ライゲーション反応と形質転換
アガロースゲル電気泳動後、目的のバンドをカッターで切りだし、ゲル抽出を行った。ゲル抽出は、QIAquick Gel Extraction Kit (50)(QIAGEN社)を使用し、付属のプロトコールに従った。
【0071】
次に、pCR 2.1-TOPOベクター(invitrogen社)(図10)を用いてライゲーション反応および形質転換を行った。まず、ゲル抽出液1 μL、Salt Solution 0.5 μL、TOPOベクター0.5 μL、滅菌MilliQ μLをチューブにとり(計3μL)、穏やかに混和し室温に5分間置いてライゲーション反応させた。このうち2 μLを大腸菌(One shot E.coli)25 μLに添加し穏やかに撹拌して氷上で20分間静置し、42℃で30秒間ヒートショックを与えて形質転換した。その後、125 μLのS.O.C(室温)を加え37℃で1時間振とうしながらインキュベートした。インキュベート後、IPTG、X-Galを含むLB+アンピシリンプレートにスプレッドし、37℃で一晩インキュベートした。
【0072】
(5)プラスミドDNAの抽出と遺伝子の解析
生じたコロニーのうち、白もしくはうすい水色のコロニーをピックアップし、LB+アンピシリンプレートにレプリカを作成して、さらにコロニーPCRを行いインサートを確認した。レプリカは37℃で一晩インキュベートした。レプリカのコロニーが生えると、そのうちコロニーPCRでインサートが確認できたもののコロニーを1白金耳ピックアップし、5 mlのLB+アンピシリン液体培地に植菌し37℃で20時間、振盪しながらインキュベートした。
【0073】
20時間のインキュベート後、エッペンチューブに培養液を入れ遠心分離して上清を捨て菌体を回収した。この菌体から、QIAprep Spin Miniprep Kit (50)(QIAGEN社)を使用し、付属のプロトコールに従ってプラスミドDNAを抽出した。
抽出したプラスミドDNAを濃度既知のpUC19とともに泳動し、その発光度合いを比較してプラスミドDNAの濃度を決定した。そして、プラスミドDNA (0.25 μg/μL)2μL、滅菌MilliQ 17 μL、プライマー1種(M13-20プライマー、M13-Revプライマーの両方を用意)1μLをチューブに入れ、遺伝子の解析を行った。
【0074】
コロニーPCRは次のとおりに行った。Green Taq 10 μL、M13-20プライマー(100 μM)0.1 μL、M13-Revプライマー(100 μM)0.1 μL、滅菌MilliQ 9.8 μLをチューブにとり、これにコロニーをつついた爪楊枝の先をつけて、上記(3)と同じ条件でPCRおよびアガロースゲル電気泳動を行った。
(M13-20プライマー)GTAAAACGACGGCCAGT (配列番号:7)
(M13-Revプライマー)CAGGAAACAGCTATGAC (配列番号:8)
【0075】
結果
PCRの結果を図11に示す。マーカーとしてX174-HaeIIを用いた。図11の(a)中のネガティブコントロール(1)及び(2)の泳動像から判断し、図11の(b)中の1078 bpの位置のバンドを切り出してゲル抽出を行った。
抽出したプラスミドDNAの泳動結果を図12に示す。マーカーとしてX174-HaeIIを用いた。pUC19(0.25 μg/μL)の発光度合いと比較して抽出したプラスミドDNAの濃度を決定し、遺伝子の解析を行った。得られた塩基配列と終止コドンまでのアミノ酸配列を図13(配列番号1及び配列番号2)に示す。
【特許請求の範囲】
【請求項1】
下記の性質:
(1)N末端アミノ酸配列
下記のN末端アミノ酸配列:
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln
を有する;
(2)基質特異性
合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示した;
(3)阻害剤による影響
プロテアーゼ活性が、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N−トシル−L−フェニルアラニンクロメチルケトン(TPCK)、ダイズトリプシンインヒビター(SBTI)により阻害される;
(4)最適pHとpH安定性
最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;および
(5)最適温度と温度安定性
最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない;
を有する、ミミズ(Eisenia foetida)由来のプロテアーゼ。
【請求項2】
配列番号:2に記載のアミノ酸配列を含むプロテアーゼ。
【請求項3】
配列番号:2に記載のアミノ酸配列において、1個〜数個のアミノ酸の置換、欠失及び/又は付加により修飾されているアミノ酸配列からなるプロテアーゼ。
【請求項4】
配列番号:2に記載のアミノ酸配列に対して少なくとも90%の配列同一性を有するプロテアーゼ。
【請求項5】
配列番号:1に記載の塩基配列からなる核酸又はその相補鎖に対して高緊縮条件下でハイブリダイズする核酸によりコードされているプロテアーゼ。
【請求項6】
請求項2〜5の何れか1項に記載のプロテアーゼをコードする核酸。
【請求項7】
請求項6に記載の核酸により形質転換された宿主細胞。
【請求項8】
請求項1〜5のいずれか1項に記載のプロテアーゼの製造方法において、請求項7に記載の宿主細胞を培養することを含んで成る方法。
【請求項9】
請求項1に記載のプロテアーゼの製造方法において、当該プロテアーゼをミミズ(Eisenia foetida)から採取することを含んでなる方法。
【請求項10】
請求項1〜5のいずれか1項に記載のプロテアーゼを含んでなる、抗植物病原性ウイルス剤。
【請求項11】
前記植物病原性ウイルスが、キュウリモザイクウイルス(Cucumber mosaic virus;CMV)である、請求項10に記載の抗植物病原性ウイルス剤。
【請求項12】
請求項1〜5のいずれか1項に記載のプロテアーゼを含んでなる、抗植物病原性細菌剤。
【請求項13】
前記植物病原性細菌が、野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)である、請求項12に記載の抗植物病原性ウイルス剤。
【請求項1】
下記の性質:
(1)N末端アミノ酸配列
下記のN末端アミノ酸配列:
Gly-Glu-Ile-Ile-Pro-His-Asn-Ala-Tyr-Leu-Arg-Tyr-Asp-Asp-Gln
を有する;
(2)基質特異性
合成基質であるN-サクシニル-Ala-Ala-Pro-Phe-p-ニトロアニリドおよびN-サクシニル-Ala-Ala-Pro-Leu- p-ニトロアニリドに対して高い活性を示した;
(3)阻害剤による影響
プロテアーゼ活性が、キモスタチン(Chymostatin)、フェニルメチルスルホニルフルオリド(PMSF)、N−トシル−L−フェニルアラニンクロメチルケトン(TPCK)、ダイズトリプシンインヒビター(SBTI)により阻害される;
(4)最適pHとpH安定性
最適pHは9.6であり、pH 6〜11(37℃で60分間)で80%以上の安定性を有する;および
(5)最適温度と温度安定性
最適温度は60℃であり、加熱(10〜80℃での30分間プレインキュベート)によるプロテアーゼ活性の低下は50℃まで見られない;
を有する、ミミズ(Eisenia foetida)由来のプロテアーゼ。
【請求項2】
配列番号:2に記載のアミノ酸配列を含むプロテアーゼ。
【請求項3】
配列番号:2に記載のアミノ酸配列において、1個〜数個のアミノ酸の置換、欠失及び/又は付加により修飾されているアミノ酸配列からなるプロテアーゼ。
【請求項4】
配列番号:2に記載のアミノ酸配列に対して少なくとも90%の配列同一性を有するプロテアーゼ。
【請求項5】
配列番号:1に記載の塩基配列からなる核酸又はその相補鎖に対して高緊縮条件下でハイブリダイズする核酸によりコードされているプロテアーゼ。
【請求項6】
請求項2〜5の何れか1項に記載のプロテアーゼをコードする核酸。
【請求項7】
請求項6に記載の核酸により形質転換された宿主細胞。
【請求項8】
請求項1〜5のいずれか1項に記載のプロテアーゼの製造方法において、請求項7に記載の宿主細胞を培養することを含んで成る方法。
【請求項9】
請求項1に記載のプロテアーゼの製造方法において、当該プロテアーゼをミミズ(Eisenia foetida)から採取することを含んでなる方法。
【請求項10】
請求項1〜5のいずれか1項に記載のプロテアーゼを含んでなる、抗植物病原性ウイルス剤。
【請求項11】
前記植物病原性ウイルスが、キュウリモザイクウイルス(Cucumber mosaic virus;CMV)である、請求項10に記載の抗植物病原性ウイルス剤。
【請求項12】
請求項1〜5のいずれか1項に記載のプロテアーゼを含んでなる、抗植物病原性細菌剤。
【請求項13】
前記植物病原性細菌が、野菜類軟腐病菌(Erwinia carotovora subsp.carotovora)である、請求項12に記載の抗植物病原性ウイルス剤。
【図1】

【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図10】
【図13】

【図3】
【図9】
【図11】
【図12】
【図2】
【図4】
【図5】
【図6】
【図7】
【図8】
【図10】
【図13】
【図3】
【図9】
【図11】
【図12】
【公開番号】特開2011−177105(P2011−177105A)
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【出願番号】特願2010−44395(P2010−44395)
【出願日】平成22年3月1日(2010.3.1)
【出願人】(505127721)公立大学法人大阪府立大学 (688)
【Fターム(参考)】
【公開日】平成23年9月15日(2011.9.15)
【国際特許分類】
【出願日】平成22年3月1日(2010.3.1)
【出願人】(505127721)公立大学法人大阪府立大学 (688)
【Fターム(参考)】
[ Back to top ]