
メラノーマ細胞におけるRACK1のアップレギュレーション及びマーカーとしてのその使用
本発明は、哺乳動物のメラニン形成細胞におけるRACK1タンパク質の過剰発現の検出、及びRACK1タンパク質の過剰発現からメラノーマの存在を差し引くことを含む、哺乳動物におけるメラノーマを診断する方法に関する。本発明はまた、メラニン形成細胞におけるRACK1タンパク質の過剰発現、及びRACK1タンパク質の過剰発現から該メラニン形成細胞の腫瘍状態を差し引くことを含む、哺乳動物のメラニン形成細胞の腫瘍状態を決定する方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、癌の分野に含まれ、哺乳動物の癌を診断する方法に関する。
【背景技術】
【0002】
発明の背景
この明細書と実験部分の全体を通じ、さまざまな刊行物を参照する。その刊行物は参考としてこの明細書に組み込まれているものとする。
【0003】
メラノーマは、メラニン形成細胞が変化することによって発達する悪性腫瘍である。メラニン形成細胞は、皮膚、毛髪、目を着色していて、たいていの母斑に高濃度で存在する黒色色素を生成する細胞である。したがってメラノーマの大半は、黒色または茶色である。しかしメラノーマは色素の生成を停止することがある。それが起こるとき、メラノーマはもはや暗い色ではなく、皮膚の色であるピンク、赤、紫である。
【0004】
メラノーマは、皮膚癌の最も深刻な形態である。白色人種におけるその発生率と死亡率は世界的に増加している。メラノーマは最も一般的な皮膚癌ではないが、メラノーマの死者数が最も多い。アメリカ癌学会は、2007年にアメリカ合衆国でメラノーマが新たに約60,000件発生していると見積もっている。
【0005】
誰でもメラノーマになる可能性があるが、いくつかの因子に依存してリスクが大きくなる可能性がある。その因子として特に、太陽への曝露、皮膚にある母斑の数、皮膚のタイプ、遺伝的背景がある。実際、UVA光線とUVB光線の両方とも皮膚癌(メラノーマを含む)を誘発する可能性がある。
【0006】
2つのタイプの母斑の存在、すなわち正常な母斑と、異形成母斑として知られる異型の母斑の存在が、メラノーマのリスクが増大することを示す。遺伝的傾向に関しては、メラノーマと診断された患者の約10%が、メラノーマの履歴がある家族を持つことが知られている。
【0007】
原発性メラノーマは、転移する傾向がある。これは、癌が原発部位から身体の他の場所に広がることを示す。癌細胞は原発腫瘍から離れてリンパ管と血管に侵入し、血流を循環し、身体の別の正常組織内の離れた病巣(転移)で成長する。
【0008】
転移が存在していると、治療への反応が有効でないために予後が厳しい。診断に関しては、色と形が不規則な母斑は、悪性メラノーマまたは前悪性メラノーマが疑われる。視診と皮膚学的検査の後、疑わしい母斑のバイオプシーもつくることができる。そのバイオプシーが悪性である場合には、母斑とその周囲の領域を切除し、広がることを避ける必要がある。
【0009】
原発メラノーマのある患者にとって、臨床と組織病理学的な基準が指標として有用である。しかし正確な診断と予後のためのマーカーがあれば、多数の患者にとって利益があるだろうことが広く認められている。
【0010】
したがってメラノーマを初期に信頼性よく診断することが非常に重要である。
【0011】
メラノーマはウマでもしばしば見られ、成熟したウマにおける最も一般的な皮膚腫瘍である。メラノーマは、アラビア、リピッツァナー、ペルシュロンの血統において非常に一般的に発生することが見られている。なぜならこれらの血統では灰色のウマが多く生まれ、メラノーマはたいてい灰色のウマで見いつかるからである。メラノーマは、一般に、唇、目、耳、唾液腺、肛門、陰茎、膣で見つかる。灰色のウマにおけるメラノーマの大半はゆっくり成長する良性腫瘍であり、他の器官に転移したり広がったりすることは稀である。しかしこの腫瘍は、特に灰色でないウマで他の器官に広がる可能性がある。一般に、この腫瘍は、排尿、排便、交配以外の問題を起こさない。しかしメラノーマは局所的な組織に侵入する可能性があるため、中枢神経系に侵入することによって不自由さや神経の問題を起こす可能性がある。
【0012】
メラノーマはイヌでも観察できる。イヌでは主に皮膚で見られるが、口腔でも見ることができる。メラノーマであると早い段階で診断されると、癌の除去とグレードまたはステージの同定がよりうまくいく可能性がある。良性形態の皮膚腫瘍でさえ、局所的な侵襲性を持つ可能性がある。メラノーマは、身体のあらゆる領域に、その中でも特にリンパ節と肺に転移する(広がる)可能性があるため、イヌにとって将来の見通しは非常に険しく危険である。ネコもメラノーマになる可能性があるが、イヌよりは程度が小さい。
【0013】
したがってメラノーマの診断は、獣医学の観点からも問題がある。
【0014】
メラノーマの診断には経験が必要である。なぜなら初期には、無害の母斑と同じに見えたり、まったく着色していなかったりすることがあるからである。
【0015】
メラノーマであることが確認されて早い段階で治療された場合には、ほぼ100%治癒する。しかしそうでない場合には、この癌が進行して身体の他のいろいろな部分に広がる。そこでは治療が難しく、致命的になる可能性がある。
【0016】
したがってメラノーマを初期に信頼性よく診断することのできる新しいツールが必要とされている。
【0017】
さらに、メラノーマの進行の分子的基礎を理解すると、より有効な治療法を開発する助けになろう。したがってメラニン形成細胞が変化するメカニズムが広く研究されている。
【発明の概要】
【課題を解決するための手段】
【0018】
本発明は、哺乳動物のメラニン形成細胞の腫瘍状態を判断する方法と、哺乳動物でメラノーマを診断する方法に関する。本発明の第1の特徴によれば、この方法は、哺乳動物のメラニン形成細胞におけるRACK-1タンパク質の過剰発現を検出し、過剰発現している場合にはそのメラニン形成細胞が腫瘍であるため、その哺乳動物にメラノーマが存在すると結論するステップを含んでいる。
【0019】
本発明の第2の特徴によれば、この方法は、メラニン形成細胞におけるRACK-1タンパク質の発現レベルを測定し、そのレベルを参照レベルと比較する操作を含んでいる。そのメラニン形成細胞におけるRACK-1タンパク質の発現レベルが参照レベルよりも高い場合には、そのメラニン形成細胞が腫瘍細胞であり、したがってその哺乳動物にメラノーマが存在すると結論される。参照レベルは、正常な非腫瘍メラニン形成細胞、すなわち正常な上皮(皮膚)メラニン形成細胞、または正常な脈絡膜メラニン形成細胞におけるRACK-1タンパク質の発現レベルであることが好ましい。
【0020】
RACK-1タンパク質の発現レベルは、RACK-1のmRNAまたはRACK-1タンパク質のレベルと定義することができる。
【0021】
本発明は、哺乳動物におけるメラノーマの診断方法にも関する。この方法は、その哺乳動物からの生物学的サンプルを、試験管内で、メラニン形成細胞を検出する手段およびRACK-1タンパク質の発現を検出する手段と接触させるステップと、メラニン形成細胞がRACK-1タンパク質の発現と同じ場所に局在しているかどうかを判断するステップを含んでいて、RACK-1タンパク質の発現を示すメラニン形成細胞の存在は、メラノーマであることを示している。
【0022】
サンプルは、バイオプシー(例えばリンパ節、肝臓、肺、心臓、脾臓、目、皮膚のバイオプシー)であることが有利である。
【0023】
本発明の好ましい一実施態様によれば、哺乳動物はヒトである。あるいは哺乳動物は、ブタ、ネコ、イヌ、ウマでもよい。
【0024】
RACK-1タンパク質を特にインサイチュ・イムノアッセイで検出するには、RACK-1に対する抗体を用いることが好ましい。
【0025】
本発明は、哺乳動物でメラノーマを診断するため、RACK-1タンパク質の発現を検出する手段を含むキットにも関する。このようなキットは、メラニン形成細胞を検出する手段をさらに備えていることが有利である。
【0026】
本発明は、腫瘍になったメラニン形成細胞のマーカーとしてRACK-1タンパク質を用いる方法にも関する。
【0027】
本発明は、添付の図面を参照してさらに詳しく説明できる。
【図面の簡単な説明】
【0028】
【図1】図1は、MeLimからのメラノーマの肺転移を示す。図1のAは、さまざまなサイズの多病巣性メラノーマ転移が浸潤した肺の写真である。矢印は、腫瘍組織のいくつかの黒い点を指している。図1のB1〜D1は、MITF抗体を用いて免疫組織化学によって同定したメラニン形成細胞であり、茶色の核顆粒として見ることができる。図1のB2〜D2は、一次抗体なしの非特異的染色が存在していない。図1のB1とB2は正常な皮膚に対応し、C1と図1C2は皮膚メラノーマに対応し、図1のD1とD2はメラノーマの肺への転移に対応する。aは肺胞を表わし、beは気管支表皮を表わし、dは真皮を表わし、eは表皮を表わし、hfは毛包を表わす。矢印は、メラニン形成細胞(黒)とメラノーマ細胞(白)を指す。どの顕微鏡写真のスケール表示棒も100μmである。
【図2】図2は、ブタ組織におけるRACK1 mRNAの発現を示す。脱色した切片に関するRACK1のアンチセンス・プローブ(A)とセンス・プローブ(B)のインサイチュ・ハイブリダイゼーション・オートラジオグラフ。各プローブについて正常な組織(N)を左側に示し(皮膚、肺、肝臓、リンパ節)、腫瘍組織(T)を右側に示してある(皮膚メラノーマ、肺、肝臓、リンパ節から転移したメラノーマ)。アンチセンス・プローブでは、リンパ節を除き、腫瘍では健康な組織または弱っていない組織と比べて信号が強いことに注意されたい。C:アンチセンスRACK1プローブとハイブリダイズさせた肺転移MeLiMメラノーマからの暗視野顕微鏡写真。D:Cからの連続切片におけるヘマトキシリン-エオシン染色であり、C上で腫瘍の着色部位に銀色の顆粒が含まれている。スケール表示棒は、A、Bでは1cm、C、Dでは100μmである。
【図3】図3は、ブタの正常な表皮におけるRACK1の発現を示す。A、C、D:ブタの皮膚に関するRACK1タンパク質(緑)と、MITF(A、C)またはDCT(D)(赤)の二重標識の共焦点顕微鏡分析。対照となるメイシャン・ミニブタからの正常な表皮(A、B)とMeLiM(C、D)。Bは、Aに対応する透過写真である。C:三次元“直交”スライス投射分析が含まれる:中央の大きなパネルは単一の光学スライスを示しており、このスライスには、スライスされたz軸を再構成するためにx軸(緑色の線)とy軸(赤色の線)が規定された。x、zスライス(上)とy、zスライス(右)に関する対応する結果を示してある。青色の線は、z方向の積層体内での中央パネルの画像の位置を示す。核を青色で示してある。ケラチノサイトにはRACK1の細胞質ゾル・スポット信号があり、白い破線で示したメラニン形成細胞にはその信号がないことに注意されたい。点線は、真皮(d)と表皮(e)の分離線を示す。スケール表示棒:5μm。
【図4】図4は、進行段階がさまざまなMeLiMメラノーマにおけるRACK1の細胞分布。RACK1タンパク質(緑)と、MITF(A、C)またはDCT(D)(赤)の二重標識の共焦点顕微鏡分析。A:皮膚メラノーマ。B〜D:リンパ節(B)、肺(C)、心臓(D)へのメラノーマ転移。三次元“直交”スライス投射分析を図3のように示してある。核を青色で示してある。A1:A2に対応する透過写真である。A3:A2挿入図の拡大。A:白い矢印の先端は、MITFに関して陽性の真皮メラノーマ細胞を指しており、これは直交投射による分析である。真皮メラノーマ細胞と表皮ケラチノサイトでRACK1の細胞質ゾル信号が同等であることに注意されたい。RACK1は皮膚メラノーマ細胞と転移メラノーマ細胞で豊富であり、核の周囲に局在している。さらに、転移では、核RACK1が存在している。それを光学スライスと直交投射に黄色い矢印の先端で示してある。A1とA2の点線は、真皮(d)と表皮(e)の分離線を示す。スケール表示棒:5μm。
【図5】図5は、正常なメラニン形成細胞とMeLiMからのメラノーマ細胞におけるβ-カテニンとRHEBの細胞分布を示す。RACK1(赤)とβ-カテニン(A、B)またはRHEB(C、D)(緑)の二重標識の後の正常な皮膚(A1〜A2、C1〜C2)と肺へのメラノーマ転移(B1〜B2、D1〜D2)に関する共焦点顕微鏡写真とそのそれぞれの透過画像を示してある。核対比染色を青色で示してある。矢印は、細胞質ゾルのβ-カテニン(A1)がある正常なメラニン形成細胞、核周辺のRHEB(C1)がある正常なメラニン形成細胞、RACK1なしの正常なメラニン形成細胞を指している。矢印の先端は、RACK1とβ-カテニンに関する核信号のあるメラノーマ細胞(B1)を黄色で、RHEBに関する核信号のあるメラノーマ細胞(D1)を白色で示している。A1の星印は、核β-カテニンのあるケラチノサイトを示している。これはおそらく分裂している細胞である。AとCの点線は、真皮(d)と表皮(e)の分離線を示す。スケール表示棒:5μm。
【図6】図6は、ヒトの皮膚とメラノーマにおけるRACK1を示している。RACK1タンパク質(A、B)、MITF(C、D)に関する二重標識、トプロ3を用いた核対比染色(E、F)の共焦点顕微鏡分析。第1列(A、C、E)は、対照のヒト皮膚。MITF陽性メラニン形成細胞を破線で囲んである。点線は、サンプルの表皮(e)と真皮(d)の境界の分離を示す。第2列(B、D、F)は、リンパ節へのメラノーマの転移である。転移したヒト・メラニン形成細胞の細胞質ではRACK1が高レベルになっているのが見られる。スケール表示棒:10μm。
【図7】図7は、RACK1のレベルがヒトメラノーマでは高いが、母斑ではそうでないことを示している。正常な皮膚(a)、母斑(b)、皮膚メラノーマ(c)、肝臓転移(d)におけるRACK1(第1行)とMITF(第2行)の共焦点顕微鏡写真。SP2ライカ共焦点顕微鏡で単一の光学切片を取得した。RACK1は正常なメラニン形成細胞(a)ではほとんど検出されない。それは母斑(b)の過剰増殖性損傷にも当てはまる。皮膚の切片はさまざまな患者からのものである。矢印の先端は、RACK1が検出されないメラニン形成細胞を指している。矢印は、細胞質RACK1を発現しているメラニン形成細胞を示している。点線は、表皮-真皮の境界を示す。e、表皮;dr、真皮。棒=10μm。
【図8】図8は、ブドウ膜メラノーマにおけるRACK1の過剰発現を示している。この図は、染色体が3モノソミーになったヒト脈絡膜メラノーマでRACK1のレベルが高いことを示している。RACK1(a)とMITF(b)に関する免疫蛍光染色。cでは、DAPI核対比染色。SPEライカ共焦点顕微鏡で単一の光学切片を取得した。棒は10μm。
【図9】図9は、ウマのメラノーマにおけるRACK1の過剰発現を示す。ウマの皮膚悪性メラノーマでRACK1が高レベルになっている。RACK1(a)とMITF(b)に関する免疫蛍光染色。cでは、DAPI核対比染色。アポ・トーム・ツァイス顕微鏡で単一の光学切片を取得した。棒:10μm。
【発明を実施するための形態】
【0029】
本発明に関連して、以下の用語は以下のように定義される。
メラノーマは、メラニン形成細胞の悪性腫瘍である。それは、主に皮膚癌(皮膚メラノーマ)であるが、例えばヒトの眼のメラニン形成細胞(脈絡膜メラノーマ)においても見られるがそれほど頻繁には見られない。
【0030】
転移は、身体の主な部位から他の部位へ癌の広がりである。癌細胞は、一次(又は元の)腫瘍から裂けて、リンパ及び血管に透過し、血流によって循環し、身体の至るところの他の臓器中の離れた場所で沈着し増殖し得る(転移する)。
【0031】
腫瘍は良性又は悪性のいずれかとして分類される。悪性腫瘍は、侵襲及び転移によって拡がることができるが、良性腫瘍は広がらない(そして、局所的に増殖するのみである)。定義としては、用語「癌」は悪性腫瘍にのみ適用される。
【0032】
転移性腫瘍は、癌の最後の段階で非常に一般的である。転移の拡がりは、血液又はリンパを介してあるいはその両方の経路を介して起こり得る。転移の最も一般的な場所は、リンパ節、肝臓、脳及び骨である。
【0033】
癌細胞が広がって新しい腫瘍を形成する時に、続発性又は転移性腫瘍と呼ばれ、その細胞は元の腫瘍の細胞と似ている。このことは、メラノーマが肺に拡がる(転移する)場合に、続発性腫瘍(転移)は、腫瘍性メラニン形成細胞(腫瘍性肺細胞でない)からなることを意味する。肺の疾患は転移性メラノーマである。細胞の腫瘍状態は、その腫瘍又は非-腫瘍状態に関連し、腫瘍細胞は、正常で健常でないが、腫瘍由来である、細胞を意味する。腫瘍性細胞は、良性腫瘍由来でも又は悪性腫瘍由来でもよい。
【0034】
RACK1(又はRACK1)タンパク質は、GeneBankの受入番号NM_006098を有する塩基配列によってコードされるヒトタンパク質(ヒトタンパク質P63244 配列番号69)又はそのアレル変異体、あるいは別の哺乳動物種における任意のオルソログ、例えば、NM_008143によってコードされるネズミ配列、又はNM_214332よってコードされるブタRACK1配列、又はNM_175602によってコードされるウシ配列、又はNM_130734によってコードされるラット配列、又はXM_537934によってコードされるイヌ配列、を意味する。RACK1タンパク質に対応するRNA又はDNAは、以下のタグ:ATTGTAGATG (配列番号1) で検出され得る。
【0035】
本発明は、タンパク質RACK1が、哺乳動物の正常なメラニン形成細胞に関して、腫瘍性メラニン形成細胞において過剰発現する、という発明者らによる発見に依拠する。実際に、正常メラニン形成細胞では、RACK1は、非常に低いレベルで発現し、ほとんど検出されない。しかし、このタンパク質は、腫瘍性メラニン形成細胞、メラノーマ又はメラノーマの転移において顕著に過剰発現することが見出されている。
【0036】
更に、本発明者らは、RACK1の発現が非常に増加し、皮膚性及び脈絡膜のメラノーマにおいて均一に分布しているが、RACK1シグナルは、母斑、すなわち皮膚性良性腫瘍において低くかつ異質である、ことを見出した。
【0037】
本発明はまた、本発明者らの観察によれば、あるメラノーマのみに関連するがすべてのメラノーマには関連しない他の公知のマーカーと比べて、RACK1は、すべての皮膚性及び脈絡膜メラノーマにおいて過剰発現するという、本発明者らによる発見に依拠する。従って、本発明者らのデータは、RACK1がメラニン形成細胞における悪性腫瘍の普遍的なマーカーであることを示す。
【0038】
そのため、第1の態様によれば、本発明は、哺乳動物のメラノーマ細胞の腫瘍状態を決定する方法であって、該メラニン形成細胞におけるRACK1タンパク質の発現又は過剰発現を検出し;及びRACK1タンパク質の発現又は過剰発現を該細胞の腫瘍状態、好ましくは悪性腫瘍状態、と関連づけること、を含む、方法に関する。
【0039】
試験下でのメラニン形成細胞の腫瘍状態は、好ましくは、本発明の方法を実行する前に決定されない。メラニン形成細胞は、好ましくは、樹立された細胞株からのものではない。好ましくは試験下の細胞は、一次細胞である。従って、本発明の方法は、メラニン形成細胞の内の腫瘍性細胞の検出、好ましくは悪性腫瘍性細胞の検出を可能にする。
【0040】
第2の態様によれば、本発明はまた、哺乳動物のメラノーマを診断する方法であって、哺乳動物にメラニン形成細胞中のRACK1タンパク質の発現又は過剰発現を検出し、RACK1タンパク質の発現又は過剰発現をメラノーマの存在と関連づけることを含む方法に関する。
【0041】
該方法は、好ましくは、インビトロ又はエクスビボで行われる、すなわち、RACK1タンパク質の発現又は過剰発現の検出ステップが好ましくはインビトロ又はエクスビボで実行される。あるいは、インビボで行ってもよい。
【0042】
メラニン形成細胞は、メラニンを合成するその能力によって認識される。従って、メラニン合成に関連する任意のタンパク質は、メラニン形成細胞を他の細胞と識別するマーカーとして使用され得る。しかし、ある腫瘍性メラニン形成細胞は、メラニンを産生するその能力を失っていることがあり、メラニン合成経路に関連しない別のマーカーがその場合に使用されることがあることに留意されたい。メラニン形成細胞用の可能性のあるマーカーは、小眼症転写因子MITF、ドパクロムトートメラーゼDCT、チロシナーゼTYR、チロシナーゼ-関連タンパク質1 TRP 1である。
【0043】
RACK1タンパク質の発現は、対照として使用される正常なメラニン形成細胞においてRACK1の発現を超える発現を意味する。本発明によれば、RACK1過剰発現は、例えば入手でき、病原診断のために入手され使用される、パラフィン包埋された少量の一次組織で検出される。更に、本発明のこれらの態様によれば、インサイチュ免疫アッセイと同等又はそれより劣る感度を有する方法が検出に使用されるならば、メラニン形成細胞におけるRACK1発現の存在の1つの検出は、腫瘍性細胞又はメラノーマの存在の指標である。実際に、上記のように、RACK1は正常なメラニン形成細胞で非常に低レベルで発現し、従って、このタンパク質は、標準的方法、例えば検出手段としてのインサイチュ免疫アッセイ、又はインサイチュ免疫アッセイと同等又はそれよりも感度の劣る検出方法によって正常メラニン形成細胞において検出できない。
【0044】
従って、本発明に関連して、インサイチュ免疫アッセイは、該方法を実行するために特に好ましい技術である。実際に、インサイチュ免疫アッセイを用いる、RACK1タンパク質用の陽性細胞の検出は、腫瘍性細胞の存在、よってメラノーマ、の直接的指標である。
【0045】
この態様によれば、本発明の目的は、メラニン形成細胞中のRACK1タンパク質の発現レベルを測定し、そのレベルを参照レベルと比較するステップを含む、哺乳動物のメラニン形成細胞の腫瘍性状態を決定する方法である。
【0046】
前記メラニン形成細胞中の、参照レベルに関してRACK1タンパク質の増加したレベルは、この細胞が腫瘍性細胞(腫瘍状態)、好ましくは悪性腫瘍性細胞である、指標である。
【0047】
試験下のメラニン形成細胞の腫瘍状態は、好ましくは、本発明の方法を実行する前に決定されない。メラニン形成細胞は、このましくは樹立された細胞株由来でない。好ましくは、試験下の細胞は、一次細胞である。
【0048】
第4の態様によれば、本発明は、哺乳動物のメラニン形成細胞中のRACK1タンパク質発現のレベルを測定し、そのレベルを参照レベルと比較するステップを含む、哺乳動物のメラノーマを診断する方法に関する。参照レベルに関して前記メラニン形成細胞のRACK1タンパク質発現の増加したレベルは、メラノーマの存在の指標である。
【0049】
本発明の第3及び第4の態様は、RACK1タンパク質発現の発現又は過剰発現の決定が参照レベルとの比較によってなされる、最初の2つの態様に相当する。
【0050】
本発明の方法は、好ましくは、インビトロ又はエクスビボで行われる、すなわち、RACK1タンパク質レベルの決定ステップが、好ましくはインビトロ又はエクスビボで行われる。あるいは、インビボで行ってもよい。
【0051】
本発明の方法では、RACK1タンパク質の発現のためにアッセイされる哺乳動物のメラニン形成細胞は、好ましくは前記哺乳動物の生物試料由来である。比較ステップのために使用される参照レベルは、例えば、非-腫瘍性、すなわち診断される哺乳動物の正常な健常なメラニン形成細胞におけるRACK1タンパク質発現レベルである。正常なメラニン形成細胞は、例えば、正常な表皮メラニン形成細胞又は正常な脈絡膜メラニン形成細胞である。正常なメラニン形成細胞の任意の他の種類は、タンパク質RACK1発現の参照レベルのために使用してもよい。
【0052】
「増加したレベル」とは、レベルが顕著に増加されることである、すなわち、測定されたレベルと参照レベルとの差が統計的観点から有意であり、すなわち測定されたレベルの標準偏差よりも大きい、と理解すべきである。従って、この差は、好ましくは、測定に固有のエラーよりも大きく、そして正常なメラニン形成細胞間で観察された偏差よりも大きい。測定されたレベルと参照レベルとの差は、分析されるメラニン形成細胞に依拠して、及び選択された参照レベルに依拠して顕著に変動し、しかし、腫瘍性細胞のレベルは、検出の限界及び上記のカットオフラインよりも高い。測定されたレベルは、例えば、参照レベルに因って、参照レベルよりも少なくとも5%高くてよく、好ましくは少なくとも10%又は20%高くてよい。本発明の異なった態様では、測定されたレベルは、参照レベルの少なくとも3倍であり、一般的に少なくとも6倍であり、例えば参照レベルよりも少なくとも10倍高い。例えば、参照レベルが非-腫瘍性、すなわち正常な健常なメラニン形成細胞中のRACK1タンパク質発現レベルであるときに、測定されたレベルは、参照レベルの、少なくとも1、5、好ましくは2倍である。
【0053】
本発明はまた、哺乳動物の皮膚断片におけるメラニン形成細胞の腫瘍状態を決定する方法であって、
−該メラニン形成細胞中のRACK1タンパク質の発現レベルを、該哺乳動物の皮膚断片の周囲のケラチノサイトにおけるRACK1タンパク質の発現レベルと比較し;
−発現の等価レベルを該細胞の腫瘍状態と関連づけること、
を含む方法に関する。
本発明はまた、哺乳動物の皮膚メラノーマを診断する方法であって、
-メラニン形成細胞中のRACK1タンパク質の発現レベルを、該哺乳動物の皮膚断片の周囲のケラチノサイトにおけるRACK1タンパク質の発現レベルと比較し;
−発現の等価レベルを該皮膚メラノーマの存在と関連づけること、
を含む方法に関する。
【0054】
メラニン形成細胞及び皮膚断片上のケラチノサイト中のRACK1タンパク質発現のこのような比較は、当業者によって容易になされる。実際に、図3及び4から明らかであるが、一方では、健常なメラニン形成細胞におけるRACK1タンパク質の発現は検出できないが、周囲のケラチノサイトでは検出でき(図3)、他方、RACK1タンパク質の発現レベルは、周囲のケラチノサイトと腫瘍性メラニン形成細胞においては同等である(図4)。本発明によれば、RACK1タンパク質発現レベルは、RACK1タンパク質発現の検出及び/又は上記の発現レベルの推定によって決定又は測定され得る。
【0055】
本発明によれば、その腫瘍状態を定義するために分析されるメラニン形成細胞は、バイオプシー由来でよい。本発明によるバイオプシーは、任意の臓器又は組織由来でよい。バイオプシーは、例えば皮膚又は眼のバイオプシー、又は肝臓、リンパ節、肺、心臓又は脾臓のような内臓由来である。
【0056】
本発明によれば、RACK1タンパク質発現レベルは、診断される細胞中のRACK1タンパク質のレベルの測定によって、あるいは診断される細胞中のRACK1タンパク質に相当するmRNAのレベルの測定によって、決定される。実際に、腫瘍性メラニン形成細胞中のRACK1タンパク質の高レベルは、対応するmRNAの高レベルによるものであり、すなわち、RACK1タンパク質の過剰発現は、腫瘍性細胞中の対応する遺伝子の転写の増加したレベルに主による、ことが、本発明者らによって明らかにされた(実施例の項を参照)。
【0057】
本発明によれば、細胞中のRACK1タンパク質発現レベルは、有利には、インサイチュ免疫アッセイによって決定される。この方法によれば、ペプチド又はタンパク質の発現レベルを特に蛍光によって視覚化及び測定することが可能である。
【0058】
あるいは、RACK1タンパク質に対するmRNAのレベルが決定されるとき、使用される有利な方法は、インサイチュハイブリダイゼーションである。インサイチュハイブリダイゼーション及びインサイチュ免疫アッセイはいずれも、本発明の範囲において当業者に周知である。
【0059】
インサイチュ免疫アッセイについて、RACK1に対する抗体が使用されることになる。かかる抗体は、商業的に入手可能である。インサイチュハイブリダイゼーションに関して、この方法はRACK1 mRNAに特異的にハイブリダイズするプローブの使用を必要とする。好適なプローブの例は、本説明の実験項に記載されている。
【0060】
細胞中のRACK1発現のレベルは、RACK1タンパク質に対応するRNA上でRT-PCRによって決定してもよい。
【0061】
あるいは、RACK1タンパク質の存在又はRACK1タンパク質発現レベルは、免疫学的方法、免疫アッセイ、免疫組織化学アッセイ、共焦点顕微鏡分析、紫外線顕微鏡、電気的検出又はハイブリダイゼーションオートラジオグラフィーを実行することによって、本発明に従って決定される。
【0062】
免疫学的方法は、好ましくは、特定のモノクローナル抗体又はポリクローナル抗体を用いる方法であり、好ましくは、ELISA法、免疫酵素法、免疫蛍光法、放射性-免疫学的方法及び化学-免疫学的方法から選ばれる。最も好ましくは、免疫学的方法は、RACK1タンパク質に対するモノクローナル抗体による、インサイチュ免疫アッセイである。本発明に従うメラノーマは、例えば、皮膚メラノーマ又は脈絡膜メラノーマである。
【0063】
分析されるメラニン形成細胞が、通常メラニン形成細胞が欠けている非-皮膚組織のバイオプシーの一部である時に、本発明の方法による腫瘍性細胞の同定は、メラノーマの転移の指標である。本発明に従って診断される哺乳動物は、好ましくはヒトであり;しかし、本発明はまた、他の哺乳動物、特にペット、好ましくはブタ、ウマ及びイヌにおけるメラノーマの腫瘍性細胞の診断に関する。任意の他の哺乳動物も本発明の範囲内にある。
【0064】
本発明はまた、哺乳動物由来の生物試料と、メラニン形成細胞を検出する手段、及びRACK1タンパク質発現を検出し、メラニン形成細胞がRACK1タンパク質発現と共局在かするか否かを決定する手段とを接触させるステップを含む、哺乳動物におけるメラノーマを診断する方法に関する。RACK1タンパク質発現を示すメラニン形成細胞の存在は、メラノーマの指標である。実際に、健常なメラニン形成細胞では、本発明者らは、RACK1タンパク質発現を検出できなかったが、メラニン形成細胞マーカーの発現を検出した。一方で、腫瘍性メラニン形成細胞では、本発明者らは、RACK1タンパク質及びメラニン形成細胞マーカーの発現を検出した。
【0065】
本方法は、診断される哺乳動物から得た生物試料について、好ましくはインビトロ又はエクスビボで行われ、最も好ましくはインビトロで行われる。従って、本発明の診断方法は、ヒト又は動物の体との任意の相互作用を示すものではなく、後者の存在を必要とする。
【0066】
本方法の実行において使用される生物試料は、好ましくは、メラノーマ又はメラノーマの転移を宿す(host)と疑われた臓器又は組織由来のバイオプシーである。バイオプシーの好ましい例は、リン節、肝臓、肺、心臓、脾臓、眼及び皮膚のバイオプシーである。
【0067】
既に述べたように、RACK1タンパク質発現を検出する手段は、好ましくは、試料中のRACK1タンパク質を検出する手段である。かかる手段は、好ましくは、抗体であり、最も好ましくはモノクローナル抗体である。あるいは、それはポリクローナル抗体でよい。RACK1タンパク質発現を検出する手段は、RACK1タンパク質に相当するmRNAを決定する手段でもよい。
【0068】
前記のRACK1タンパク質発現を検出する手段は、有利には、例えば蛍光タグ又は放射性タグによって標識される。標識は、RACK1を検出するために、及び/又は該手段を定量化するために使用してよい。前記のメラニン形成細胞を検出する手段は、例えば、小眼症転写因子MITFに対する抗体、ドパクロムトートメラーゼDCTに対する抗体、チロシナーゼTYRに対する抗体、チロシナーゼ-関連タンパク質1 TRP 1に対する抗体、又はメラニン合成経路に関連する任意のタンパク質に対する抗体である。
【0069】
有利には、RACK1タンパク質を検出する手段及びメラニン形成細胞を検出する手段はいずれも、抗体であり、該方法は、2つの前記手段(抗体)のための異なった標識(抗体)を有するインサイチュ免疫アッセイによって行われる。
【0070】
本発明の異なった方法によれば、診断される哺乳動物は、好ましくはヒトである。本発明によれば、診断される好ましい哺乳動物は、癌の家族暦を有するヒト、あるいはメラノーマを発症すると疑われるヒト又はかかる疾患を発症する高い危険性が存在すると疑われるヒト、特に、水膨れ又は日焼けによる皮はがれを有する個体(特に、人生の最初の20年で)、色白かつ赤毛の人々、複数の非定型母斑又は形成異常母斑を有するヒト、大きな先天性メラニン形成細胞母斑を有するヒト、及びメラノーマもしくはディスプラスチック・ニーバス症候群(複数の非定型穴)の個人暦又は家族暦を有するヒトである。
【0071】
本発明の別の態様では、診断される哺乳動物は、ブタ、ネコ、イヌ又はウマ、あるいは別のペットである。本発明の診断方法は、診断された哺乳動物で、特に、メラノーマを発症する高い危険性を示す哺乳動物について、より好ましくはメラノーマの素因を示すヒトについて、規則的に繰り返されることが好ましい。本発明の診断法は、少なくとも3ケ月毎に、少なくとも6ケ月毎に、年に1回又は幾分頻繁に繰り返される。転移の前に初期の段階で治療を開始するためには、できるだけ早く、メラノーマを診断することが実際に必須である。
【0072】
本発明の方法は、好ましくは、メラノーマを診断する他の方法と組み合わせて行われる。例えば、本発明の方法は、異なった方法によってなされた最初の診断を確認するために、あるいはメラノーマの疑いを確認するために行われる。あるいは、本発明の方法は、腫瘍性細胞の同定を可能する他の方法と同時に又はその前に行ってよい。
【0073】
本発明の方法は、他の診断方法、例えば解剖病理学的方法と組み合わせて有利に使用される。
【0074】
解剖病理学的方法では、細胞の腫瘍状態は、有糸分裂、空間組織化、及びストローマ及び血管との関連である、3つの異なった基準の決定後に同定され得る。本発明の方法は、これらの基準の少なくとも1つの決定と組み合わせて有利に行われる。
【0075】
本発明はまた、RACK1タンパク質発現を検出するための手段を含む、哺乳動物におけるメラノーマを診断するキットに関する。上記のように、RACK1タンパク質発現を検出するための手段は、好ましくは、RACK1タンパク質を検出するための手段である。前記方法は、例えば抗体、好ましくはモノクローナル抗体であり;それはポリクローナル抗体でもよい。
【0076】
あるいは、前記手段は、RACK1タンパク質に相当するmRNAのレベルを検出するための手段でよい。
【0077】
本発明のキットにおいて使用される手段は、有利には、例えば蛍光又は放射性標識で標識される。
【0078】
上記のキットは、好ましくは、メラニン形成細胞を検出するための手段を更に含む。メラニン形成細胞を検出するための手段は、メラニン形成細胞と非-メラニン形成細胞との識別を可能にする。
【0079】
かかる手段は、例えば、小眼症転写因子MITFに対する抗体、ドパクロムトートメラーゼDCTに対する抗体、チロシナーゼTYRに対する抗体、チロシナーゼ-関連タンパク質1 TRP 1に対する抗体、又はメラニン合成経路に関連する任意のタンパク質に対する抗体である。別の手段も考えられる。
【0080】
好ましくは、前記の更なる手段はまた、有利には、例えば蛍光又は放射性標識によって標識される。前記標識は、有利には、RACK1タンパク質発現を検出する手段に貼り付けられたラベルとは異なる;例えば、異なった波長発光を有する蛍光標識。
【0081】
本発明はまた、哺乳動物、好ましくはヒト又はペットのメラニン形成細胞の、腫瘍状態のマーカー、好ましくは悪性腫瘍状態のマーカーとしてのRACK1タンパク質の使用に関する。
【0082】
本発明の別の目的は、該細胞の腫瘍性状態の指標、好ましくは悪性腫瘍状態の指標としての哺乳動物のメラニン形成細胞中のRACK1タンパク質の発現レベルの使用である。
【0083】
本発明の方法及びキットに関連して上記の特徴及び好ましい態様のすべてもまた、本発明の使用に適用する。
【実施例】
【0084】
メラニン芽種保有リベコフミニブタ(Melanoblastoma-bearing Libechov minipigs)(MeLiM)は、自発的な悪性皮膚メラノーマを提示する。メラニン形成細胞悪性変換をよりよく理解するために、発明者らは、遺伝子発現の連続的解析を、正常皮膚メラニン形成細胞とMeLiMの肺転移由来のメラノーマ細胞とで比較した。メラノーマ進行において、タグ特定により解明された遺伝子はこれまでに報告されていない。この中でもGNB2L1はメラノーマ細胞で過剰発現した。GNB2L1は、活性化Cキナーゼ(PKC)の受容体であるRACK1をコードする。発明者らは、MeLiMのメラノーマに侵された皮膚及び器官においけるRACK1発現を、メラニン形成細胞のマーカーとしてMITFを用いる共焦点顕微鏡法を用いて研究した。RACK1は、正常な表皮メラニン形成細胞では検出されなかった。それとは対照的に、細胞質RACK1は、皮膚腫瘍及び転移の全てのメラノーマ細胞において高度に発現した。PKCβIIはRACK1と共存しなかったが、これはメラノーマ悪性疾患におけるRACK1の役割が、PKCβIIシグナル伝達とは独立していることを示す。RACK1は転移におけるさらなる核である。RHEB及びβ-カテニンも転移の核であるが、RACK1と共存することはない。RACK1の高発現は、ヒト臨床試料の皮膚腫瘍及び転移の全てのメラノーマ細胞において見られたが、RACK1は、健常皮膚のメラニン形成細胞においては検出されなかった。メラノーマ細胞におけるRACK1の発現レベルは、その腫瘍段階に関連するようである。
【0085】
メラノーマはメラニン形成細胞の変換により発症する悪性腫瘍である。白人集団におけるその発生率及び致死率は、全世界的に増加している。転移の存在は、治療に対する非有効な応答のため予後が極めて不良になる。メラノーマ進行の分子的根拠の理解により、より有効な治療の開発を援助することができた。このように、メラニン形成細胞変換のメカニズムは幅広く研究されている。
【0086】
ヒトにおいて、メラノーマは、B-RAFでは66%変異しており、一方INK4a/ARFでは50%欠失している(Chin (1)により概説されている)。この2つの障害の組み合わせは、Ras経路の活性化、網膜芽腫タンパク質のリン酸化、及びTP53分解の増加をもたらす。ただし、細胞変換のその他のメカニズムの存在が示唆されているため、これらの遺伝子が全てメラノーマの主な原因ではない。ヒト及びマウスのメラニン形成細胞腫瘍細胞系列及び試料について、大規模な遺伝子発現プロファイリングが報告されている。RhoC(2)、WNT5a(3)、Notch2(4)、及びGタンパク質及びカルシウム伝達に関連する遺伝子(5)の差次的発現、とりわけ、際立った経路が、細胞接着及び運動性、並びに特定の細胞系列における組織浸潤に携わる。全体的に、メラノーマは、複雑な多重遺伝子疾患であり、そして変換の異なるメカニズムが作用しているはずである。
【0087】
ヒト病理の動物モデルにおいて、皮膚メラノーマに侵されるMeLiM(Melanoblastoma-bearing Libechov Minipig)種は特に興味深い。実際、2月齢のMeLiMは、リンパ節における転移を有する自発的な皮膚メラノーマを提示するが、ヒトに類似する組織学的進化をする内部器官においてはそれほど頻繁ではない(6)。ただし、ヒトにおける大部分の散発性メラノーマを理解するための、これらの遺伝的ブタメラノーマの生物学的解明は、未だ不確かである。MeLiMもであるがヒトにおける疾患の進行に価値有る情報を提供できるか否かを評価するため、発明者らは、MeLiMにおける腫瘍進行に関与する遺伝子を特定し、ヒト皮膚メラノーマにおけるそれらの発現を評価することにした。
【0088】
連続的遺伝子発現解析(serial analysis of gene expression(SAGE))法を選択したのは、マイクロアレイと異なり、本方法は解析する配列に関係なく、完全な細胞発現プロファイルを提供するからである。SAGEライブラリーは、特定細胞型において特異的に発現される遺伝子を解明するための、インシリコ(in silico)での比較が可能である(7)。メラニン形成細胞以外の細胞の寄与を最小化するため、発明者らは、MeLiMの肺メラノーマ転移及び健常なメイシャンミニブタ(Meishan minipig)の皮膚由来のPigMelメラニン形成細胞から培養した一次メラノーマ細胞からSAGEライブリーを構築した(8)。ここでは、悪性と正常メラニン形成細胞との遺伝子発現の比較解析が報告されている。本発明者らは、活性化Cキナーゼ(PKC)の受容体であるRACK1は、MeLiM由来の皮膚腫瘍及び転移の細胞質において発現するが、その発現は正常な皮膚メラニン形成細胞で検出されないことを示す。重要なことには、この腫瘍及び正常のメラニン形成細胞におけるRACK1の差異的発現は、ヒト患者由来の臨床検体でも見られた。これらの結果は、哺乳類におけるメラノーマの進行におけるRACK1の役割を実証するものである。これらは、MeLiM種が、ヒトにおけるメラノーマ進行に関与する複雑なメカニズムを研究するための関連モデルとなることを支持する。
【0089】
材料と方法
ブタ(Sus scrofa domestica)組織
罹患したMeLiMの雄を、National Institute for Agricultural Research(INRA、Jouy-en-Josas, France)で健常なデュロック又はMeLiM種と交配させた(6,9)。動物の処置及び使用は、欧州連合標準に従って、INRA倫理委員会により承認された。フランス(n=13)又はチェコ共和国(n=3)のいずれかで飼育された3月齢以下のMeLiM由来のバイオプシーを使用した。これらには、表在拡散メラノーマ(n=2)及び結節性メラノーマ(n=7)、及びリンパ節(n=13)、肺(n=10)、肝臓(n=1)、心臓(n=2)及び脾臓(n=3)における転移、及び健常皮膚(n=10)が含まれる。健常な有色性メイシャンの背側頸皮膚を、コントロールとして使用した。回収した組織を、4%緩衝化パラホルムアルデヒド(PFA)中で固定化し、パラフィンに包埋した。
【0090】
転移性メラノーマ細胞の単離及びコントロールメラニン形成細胞の培養
若いMeLiM由来の腫瘍肺バイオプシーを、メラノーマ細胞の単離に使用した。ブタメラニン形成細胞細胞の第一代培養物の条件は(8)の記載の通りである。酢酸テトラデカノイル−ホルボールを、培養の第二日目に添加した。48時間後、細胞をリンスし、Dynabead mRNA直接キット結合バッファー(Dynal, Invitrogen Life Technologies, Cergy Pontoise, France)中で溶解させ、そして液体窒素中で凍結させた。コントロールメラニン形成細胞は、非変換PiMeL細胞を37%とした(8)。
【0091】
SAGEライブラリーの構築
メラニンを廃棄するため細胞溶解物を遠心分離し、小型化抽出へのSAGE適用(SAGE)法を用いてライブラリーを作製した(10)。配列決定反応をMWG(Martinsried, Germany)により行った。
【0092】
タグ特定及びプローブのクローニング
SAGEタグを、SAGE 2002 第4.5版ソフトウェアを用いて、配列ファイルから抽出した(7)。統計的有意性を、モンテカルロシミュレーション解析を用いて決定した。P値が0.05以下の場合に有意であるとみなした。タグを、SAGEソフトウェアにより解析した哺乳類Genebankデータベース(http://www.ncbi.nih.gov/Genbank/gbm)、又はAGENAE ESTデータベース(http://www.ncbi.nih.gov/Genbank/gbm)を用いて特定した。ATG開始コドンからのヌクレオチド配列−70〜900bpに対応するブタRACK1部分cDNAを、pCR4TOPOプラスミド(Invitrogen Life Technologies)にサブクローニングした。得られたプラスミドをNotI又はPmeIのいずれかで直線化し、それぞれセンス又はアンチセンスRNAプローブを得た。インビトロにおける転写は(11)に記載の通りに行った。
【0093】
重度有色化試料のインサイチュハイブリダイゼーション
インサイチュハイブリダイゼーションは(12)の記載に切片を漂白する修正を加えて行った。簡単に説明すると、脱パラフィン化した5μmの切片を、0.075%のKMnO4で15分処理し、処理の間及び後に短いリンスを行いながら、5%のシュウ酸中で1分間脱色素化した。切片を4%PFA中で20分固定化し、リンスし、脱水和し、そして空気乾燥した。約15×106 cpm/mlでセンス又はアンチセンス放射標識リボプローブを(11)に記載の通りにハイブリダイズした。スライドを、Biomax MRフィルム(Kodak, France)に3日間感光させ、その後、Kodak NTB2エマルション中に浸漬し4週間感光させた。
【0094】
ヒト組織
クルー機関(Curie Institute)(Paris, France)において、ヒトメラノーマ組織を、リンパ節切除(n=13)、肝切除(n=5)又は表皮切除(n=19)を行った患者から得た。皮膚メラノーマ検体は、2つの表在拡散メラノーマ第IV段階(ブレスロー深さが2.35及び2.5 cm)と3つの再発性皮膚メラノーマからなる。25人の女性及び12人の男性からなる計37人由来の試料を調べた。胸部形成手術から得た正常皮膚を、コントロールとして使用した(n=4)。
組織を、4%PFA(n=38)又はブアン固定液(n=2)のいずれかにおいて固定し、パラフィンに包埋させた。
【0095】
抗体(免疫標識用の希釈物)
使用したマウスモノクローナル抗体は、抗−MITF(1:50)(クローン C5+D5、Zymed, Clinisciences, Montrouge, France)、抗-RACK1(1:150)及び抗-β-カテニン(1:100)(Transduction Laboratories, BD Biosciences, Le Pont de Claix, France)であり、及びポリクローナルPEP-8抗-DCT(1:1000)、抗-RHEB(1:100)及び抗-PKCβII(1:200)(Santa Cruz Biotechnology, Tebu-bio, Le Perray, France)であった。交差反応及び当該抗体のブタ組織に対する特異性を、ウェスタンブロットを用いて確認した。
ウマ試料のアッセイ用に、MITF抗体(C5+D5、Zymed)を1:100で希釈し、分子グレードのBSA0.1 mg/mlを、ステップ毎に正常なヤギ血清に添加した。
【0096】
免疫染色及び共焦点顕微鏡法
ブタ切片はクエン酸バッファー(pH6)中で、ヒト切片はTris-EDTA(pH9)、0.05% Tween20中で、マイクロ波照射脱パラフィン化切片により、抗原の回収を行った。免疫組織化学については、一次抗体をアビジン−ビオチン複合体(ABC Elite, Vector, Biovalley, France)と反応させた。二重の免疫蛍光法については、4℃で一晩アプライした抗体を、抗マウスアイソタイプ又は抗-ウサギ抗体に曝し、一方はAlexa Fluor 555で標識し、他方はビオチンと結合させて、Alexa Fluor 488標識化ストレプトアビジン(Molecular Probes, Invitrogen Life Technologies, France)に曝した。核対比染色は、Topro 3(Molecular probes)を用いて行った。切片を、LicaレーザーTCS SP2スキャニング共焦点顕微鏡で、0.7 μm厚の光学的切片を作製して観察した。一次抗体不存在のコントロールは非特異的標識を示さない。共焦点画像は、コンピュータプログラム Leica Lite又はZeiss LSM Image Browserで処理した。示された全画像は、個々の切片のz系、加えて指示がある場合は、積層の直行画像(orthogonal projections)である。最終的な図は、Adobe Photoshop (Adobe Systems, USA)で編集した。
【0097】
結果
MeLiM由来の転移メラノーマの特徴付け
リンパ節、肝臓、心臓及び肺においてメラノーマ転移を発症した若いMeLiMを、図1Aに示した。組織切片において、メラノーマ細胞の特定を、メラニン形成細胞において特異的な核シグナルを発生する小眼球症転写因子MITFに対する抗体を用いて行った。免疫組織化学により、MITFで標識したコントロール皮膚表皮の基底層において、正常なメラニン形成細胞が示される(図1B1)。皮膚メラノーマ及び転移における腫瘍細胞も、MITFで標識された(図1C1〜D1)。非特異標識は、本抗体では検出されなかった(図1B2〜D2)。すなわち、MITFは、当該ブタにおけるメラノーマ進行の研究に有用な、メラニン形成細胞系列の感受性マーカーである。
【0098】
転移性メラノーマ細胞とメラニン形成細胞の比較発現解析
肺転移由来のメラノーマ細胞を単離するため、腫瘍の一次培養を、ブタメラニン形成細胞増殖のために最適化した条件で行った(8)。48時間後、培養における接着細胞は、主にメラニン形成細胞であった。2500万個の転移性メラノーマ細胞(MMC)及びPigMel正常メラニン形成細胞(NM)からSAGEライブラリーを構築した。NM及びMMCライブリーから、それぞれ5,466個及び6,131個の異なるタグ(転写物)に対応する、総数11,300個及び11,700個のタグを配列決定した。70個のタグは、2つのライブラリーにおいて差異的に表された(P<0.05)。45個(77%)は発現した配列タグに対応し、その37個(53%)は、既知のcDNAに対応した。タグの大半は、高レベルの遺伝子発現に対応した。特定された1遺伝子は、RNAプロセシング及びタンパク質合成(20%)、転写(7%)、又はシグナル伝達(4%)に関与する。遺伝子が示すその機能的クラスにより任意に配列させたタグのリストは、NMと比較して、MMCを増加及び減少させ、それをそれぞれ表1及び2に示す。GNB2L1のmRNAは、MMC及びNMのカウントが、それぞれ31及び13である豊富なタグに対応した。
タグ配列:ATTGTAGATG(配列番号1)
【0099】
本発明者らは、メラニン形成細胞及びメラノーマにおけるGNB2L1発現をより詳細に研究することを選択したのは、それが、活性化Cキナーゼの受容体であるRACK1(NM-214332、ヒトRACK1 P63244 配列番号69と同一である)をコードし、そのmRNAがヒト癌性細胞においてアップレギュレーションされることがわかっている(13)ためである。
【0100】
【表1】

【0101】
【表2】
【0102】
【表3】
【0103】
【表4】
【0104】
【表5】
【0105】
メラノーマにおけるRACK1 mRNAの過剰発現
RACK1 mRAの分布パターンを特定するため、本発明者らは、正常な皮膚メラノーマ、及び肺、肝臓及びリンパ節由来の転移性メラノーマ試料のブタ切片において、インサイチュハイブリダイゼーションを行った。重度有色化腫瘍領域のバックグラウンドを回避するため、メラニンを酸化し同時にmRNAを維持する漂白処理を追加した。アンチセンス及びセンスプローブで得られるフィルムオートラジオグラフィーから、ヒトリンパ節で報告される(13)ような、シグナルの強いリンパ節以外にも、健常組織において、RACK1 mRNAのわずかなシグナルが示された(図2A)。これとは対照的に、皮膚メラノーマ、肺及び肝転移試料の腫瘍部位では強いシグナルが観察され、非腫瘍部位は、より低いシグナルが示された(図2A)。センスプローブのオートラジオグラフィーシグナルは、ほとんどなかった(図2B)。エマルションオートグラフィー上の暗視野照明は、肺メラノーマの腫瘍領域上の銀粒子を強調する(図2C)。これらの結果により、SAGEデータにより予測されるようなメラノーマにおけるRACK1 mRNAの過剰発現が確認された。
【0106】
皮膚由来のメラニン形成細胞における、及びブタにおけるメラノーマ進行の際の、RACK1タンパク質局在化
RACK1 mRNAの過剰発現が機能的有意性を有するか否かを調べるため、本発明者らは、メラニン形成細胞のマーカーとしてMITFを用いる二重免疫染色を用い、共焦点顕微鏡によりRACK1細胞分布を解析した。本発明者らは、最初に、コントロールメイシャン及び健常なMeLiMの皮膚、すなわち、正常なブタ表皮におけるRACL1の発現を、RACK1タンパク質(蛍光で緑にラベルした)の共焦点顕微鏡解析、及びブタ皮膚におけるMITF又はDCTの二重標識により解析した。本発明者らは、ケラチノサイトにおけるRACK1細胞質の斑点状シグナル、及びメラニン形成細胞におけるその不存在を観察した(図3)。
【0107】
RACK1タンパク質は、表皮において発現し、ケラチノサイトにおける細胞質に広く局在化した(図3A〜D)。メイシャン及びMeLiM皮膚のMITF陽性メラニン形成細胞において、RACK1発現は検出されなかった(図3A、C)。これと一致して、メラノソームに限定される酵素である、ドパクロムトートメラーゼ(DCT)を試験する場合、RACK1及びDCTの二重標識は、正常皮膚において明らかに重複しなかった(図3D)。次に、本発明者らは、腫瘍組織、特に異なる進行段階でのMeLiMメラノーマにおけるRACK1の細胞分布を、RACK1タンパク質の共焦点顕微鏡解析(緑)、及びMITFの二重標識(赤)により解析した(図4)。解析した組織は、リンパ節、肺及び心臓における皮膚メラノーマ及びメラノーマ転移である。これは、真皮メラノーマ細胞及び表皮ケラチノサイトで、同程度のRACK1細胞質シグナルであることに注意すべきである。RACK1は、核周囲に局在化する皮膚及び転移性のメラノーマ細胞に豊富である。さらに転移において、核RACK1が存在する。
【0108】
皮膚メラノーマにおいて、有色化MITF陽性細胞の巣は、RACK1タンパク質を表皮ケラチノサイトと同程度に強く発現した(図4A)。RACK1は、細胞質に局在化し、その大半は核周囲領域であった(図4のA3)。リンパ節、肺及び心臓メラノーマ転移において、RACK1タンパク質は、MITF−陽性細胞で豊富であった。RACK1の準細胞性分布は、細胞質性であった。ただし、核斑点に対するさらなる標識が、MMCの15%で観察された(図4B〜D、黄色矢印)。
すなわち、腫瘍細胞におけるRACK1過剰発現は、MMC(転移性メラノーマ細胞)におけるさらなる核局在化を伴い、異なる進行段階のメラノーマ細胞において細胞質性として観察される。
【0109】
ブタメラノーマ細胞におけるRACK1の推定パートナー
RACK1作用の分子的メカニズムを探索するために、発明者らは、メラノーマ細胞中のRACK1タンパク質のパートナーを探した。RACK1は、PKCβIIの細胞内受容体として同定された(14)。発明者らは、メラノーマ進行中のPKCβII発現を研究した。PKCβII免疫蛍光は、NMにおいて弱い細胞質シグナルを示した(正常メラニン形成細胞はなかった)。重複蛍光はRACK1で検出できないが、MMCは更なる別個の核ドットを示した。比較のために、RACK1シグナルと重複する強いPKCβIIシグナルを組織リンパ球で検出した。このことは、メラニン形成細胞が豊富なPKCβIIを発現しないことを示唆している。
【0110】
β-カテニン変異は、メラノーマにおいて有る役割を果たすと考えられ(15)、β-カテインは細胞接着において重要である。そのため、発明者らは、共焦点顕微鏡、及びRACK1(赤色)とβ-カテニン(緑色)の二重標識後の正常皮膚及び肺でのメラノーマ転移の各々の送信画像によって、正常メラニン形成細胞及びMeLiM由来のメラノーマ細胞中のβ-カテニンを試験した。β-カテニンは、偏在的に発現され、正常ブタ表皮の細胞膜に局在化される(図5A)。
【0111】
MMCでは、膜及び細胞質β-カテニンシグナルが核標識と共に通常視覚化された。しかし、β-カテニン及びRACK1シグナルは、核内には局在化しなかった(図5B)。
【0112】
現在のSAGE試験においてMMC中に発現された別のタグは、小GTPaseのRasスーパーファミリーのマンバーである、脳内に豊富にあるRasホモログをコードするRHEBとして同定された。RHEBは、成長因子、栄養素及びPI3Kを連結するアミノ酸及びラパマイシン(TOR)シグナルの標的に応答して、細胞成長の制御において重要な役割を果たす(16)。従って、発明者らは、MeLiM腫瘍におけるRHEB局在化を試験した。RHEBシグナルは、NMにおいて核周囲であるが、MMC上では拡散し、核のように見えた(図5C〜D)。メラニン形成細胞及び腫瘍細胞においてPACK1シグナルについて何の局在化は見られなかった。
【0113】
これら結果は、β-カテニン、RHEB及びPKCβIIが、メラノーマ進行中のPACK1の主な機能性パートナーではないことを示唆している。
【0114】
ヒト皮膚及びメラノーマ試料中のPACK1タンパク質局在化
最後に、発明者らは、ヒトメラニン形成細胞及びメラノーマ細胞のマーカーとしてMITFを用いて、PACK1の過剰発現がヒトメラノーマ試料においても起こるか否かについて試験した(17)。正常皮膚では、PACK1は、ケラチノサイトに対してメラニン形成細胞において、MITF二重標識(図6C)又はTopro 3での核対比染色によって(図6E)、PACK1タンパク質の共焦点顕微鏡によってわずかに検出された(図6A)。皮膚メラノーマ及びリンパ節及び肝臓でのメラノーマの転移において、MITF-陽性細胞は、細胞形状の基礎となるPACK1の強力な顆粒細胞質パターンを示した。RACK1過剰発現は、試験した20試料の各々で一貫して観察された(図6B、D及びF)。これらのメラノーマ試料中には核PACK1標識は見られなかった。
【0115】
ヒトメラノーマ及び母斑におけるPACK1発現の比較
ヒトメラノーマにおけるPACK1の発現を更に調べるために、発明者らは、一連の試料中のその存在を確認した。メラニン形成マーカーとしてMITFを用いて、4つの正常皮膚バイオプシー由来のホルマリン固定組織、14つの母斑及び5つの皮膚マラノーマを分析した(17)。結果を表3に纏める。
【0116】
【表6】
【0117】
正常皮膚では、PACK1はメラニン形成細胞中に無視できる量で存在したが、隣接するケラチノサイトは、強力な細胞質性シグナルを示した(図7A)。母斑では、PACK1シグナルは、所定の母斑内で及び2つのパターンを示す母斑間で異質であった:14つの母斑の内7つにおいて、PACK1シグナルは、検出されず(矢印の頭、図7B)、また僅かでも検出されなかた(矢印、図7B)。表皮又は真皮メラノーマにおいて検出される時に、シグナルは必ず膜性であった。
【0118】
纏めると、これらの結果は、RACK1 mRNA及びタンパク質が、ブタメラノーマにおいて見出されたように、ヒトメラノーマ中でアップレギュレートされることを示している。このことは、メラノーマ腫瘍状態とPACK1の高レベルとの関係を示している。
【0119】
脈絡膜メラノーマ中のPACK1過剰発現
脈絡膜メラノーマ(又はブドウ膜メラノーマ)は、ヒトの悪性眼腫瘍の最も一般的な形態である。染色体3モノソミー、腫瘍中の珍しい染色体異常が、ブドウ膜メラノーマの約半分で検出された。染色体3モノソミーは腫瘍悪性度と関連する(38)。
【0120】
発明者らは、染色体3モノソミー(N=6)を有する又は無しのブドウ膜メラノーマ上の除核生成物から作製された組織スライドを分析した。該分析は、上記と本質的に同一のプロトコルによって実行した。RACK1は、染色体3モノソミーを有する(図8で説明される)及び有さない、ブドウ膜メラノーマの2つの種類の全ての腫瘍性メラニン形成細胞で高レベルで発現される。
【0121】
ウマメラノーマ中のPACK1発現
発明者らは、PACK1が、ウマの健常皮膚の正常メラニン形成細胞において検出可能なレベルで発現されないことを観察した。
【0122】
発明者らはまた、悪性腫瘍由来の6試料中のPACK1発現を試験した。ウマ皮膚悪性メラノーマにおけるPACK1発現を示す図9に見られるように、それらは、PACK1発現にすべて陽性であった。このことはブタ及びヒトで既に得られた結果を確認するものである。
【0123】
これらのデータは、ヒト試料の分析について述べたのと本質的に同一のプロトコルを用いて得た。
【0124】
皮膚メラノーマ中のPACK1過剰発現の一般的性質
すべてのメラノーマが一次クローン事象の同一のセットを示さず、そのいくつかはより頻繁に起こることは、良く知られている。一次クローン事象に関する知見に関する最近のレビューでは、Bennett(39)は、ヒト皮膚メラノーマにおける最も効力を有する遺伝子を報告している(この文献の表1参照)。これらのデータから推論されるように、すべての皮膚メラノーマ中では影響を与える遺伝子はない。このことは、メラノーマ由来の一次腫瘍の根底にあるメカニズムは外因性であることを示唆している。例えば、CDKN2A遺伝子は、78%の皮膚メラノーマがクローン的に改変されており;B-RAF遺伝子は47%の皮膚メラノーマは改変されている。
【0125】
これに対して、本発明者らは、彼らが試験した皮膚及びブドウ膜メラノーマ由来のすべての試料におけるPACK1の過剰発現を検出した。このことは、メラニン形成細胞中の一次腫瘍を誘発するメカニズムに関係なく、PACK1がメラノーマの普遍的なマーカーであり得ることを示唆している。遺伝子的に外因性の腫瘍中でのPACK1のこの系統的な過剰発現は、任意のメラノーマの腫瘍進行の更なるステップとしてのPACK1の正の調節によって説明され得る。
【0126】
従って、ヒト及び他の哺乳動物のためのメラノーマ診断において非常に有用である、PACK1は皮膚及びブドウ膜メラノーマの任意のメラニン形成細胞中で過剰発現される単一のマーカーであると考えられる。
【0127】
考察
皮膚悪性メラノーマについて数少ない実験動物モデルが存在する。ネズミモデルは、それぞれ癌原遺伝子及び腫瘍抑制遺伝子において、機能亢進及び機能性突然変異の喪失との組合せを必要とする(1)。高い浸透率で皮膚メラノーマを誘導するために、新生マウスでは重度の日焼けが必要とされることがある(18)。対照的に、MeLiMブタモデルは、ヒトメラノーマに類似する病理組織学的特徴を有する自発的な皮膚メラノーマを示す(6)。ここで、発明者らは、MeLiMの遺伝性疾患が、ブタ及びヒトにおけるメラノーマの発症に関連する調節複合体を特定するために有用であるという証拠を提供する。実際に、発明者らは、細胞内受容体RACK1がブタメラノーマ及びヒトメラノーマで過剰発現されることを見出した。このことは、MeLiMモデル予測がヒトメラノーマにおいて実証される最初のことである;これは、同様なメカニズムがブタ及びヒトにおけるメラニン形成細胞の悪性形質転換においては働いていることを示している。
【0128】
SAGE分析では、RACK1タグは、正常な皮膚メラニン形成細胞由来のライブラリーよりも転移性メラノーマ細胞の一次培養物から確立されたライブラリーにおいてより多かった。一環して、RACK1 mRNA及びタンパク質は、正常表皮メラニン形成細胞において僅かに検出されたが、皮膚性及び転移性メラノーマの腫瘍性細胞においては高レベルで見出された。このことは、これまで試験したすべてのブタ(n=38)及びヒト(n=20)試料に当てはまる。
【0129】
RACK1は、他のタンパク質に対して安定又は可逆的のいずれかの結合能力を与える7つの内部Trp-Asp 40(WD40)反復を含む(19)。RACK1と相互作用する60つのタンパク質のほとんどは、これまで記載されている(ヒトタンパク質参照データBベース)及びRACK1は様々な細胞コンパートメントにおいてソレラパートナーに結合することが分かった(19)。従って、RACK1は、細胞の適応プロセスにおいて中心的な役割を果たすと考えられている。
【0130】
RACK1は、活性化されたPKCβIIのための標的タンパク質として最初に同定された(14)。一方、PKCβは、ヒトの培養されたメラニン形成細胞において発現される(20)。PKCβ及びRACK1が複合化され、メラノソーム上に固定される時に、メラニン形成の重要な酵素である(21)PKCβホスホリレートチロシナーゼは、正常なメラニン形成細胞では、PKCβがメラニン形成を刺激することによって主に働くことを示唆している。一方、間接的な観察は、メラノーマ発症においてPKCβIIの可能な役割について議論する。第1に、PKCβIIは、肥満細胞においてAKTを直接リン酸化することが分かり(22)、そして、AKTは、細胞生存及び悪性メラノーマの発症を促進することが知られている(23)。第2に、WNT5Aは、ヒトメラノーマ細胞の顕著な侵襲的挙動の強力なマーカーとして同定され、WNT5aは、PKCを活性化するその能力によってこの効果を仲介すると考えられる(3)。これらの観察に基づいて、発明者らは、ブタ及びヒトメラノーマにおけるPKCβII発現を分析した。PKCβIIは、正常な皮膚メラニン形成細胞においてはほとんど検出されず、正常な皮膚メラニン形成細胞と比べて悪性メラニン形成細胞においてアップレギュレートされなかった。更に、PKCβ発現は、メラノーマ細胞株の90%において喪失される(24)。しかし、腫瘍性メラニン形成細胞におけるRACK1の過剰発現の効果がPKCβIIに依拠することはないであろう。しかし、RACK1は他のPKCアイソフォームと相互作用することができた。
【0131】
発明者らは、ブタの転移性メラノーマ細胞の15%の核においてRACK1を検出した。RACK1の核転位は、既に観察されている。すなわち、げっ歯類のアルコールへの急性曝露の後に、RACK1は、PKCβIIから脱共役し、海馬ニューロンの核に転位した(25)。核では、RACK1はBDNFを含む数個の遺伝子の誘導を介在する(26)。BDN増加Fは、エタノール取り込みを負に制御するドーパミンD3受容体の亢進された発現をもたらす(27)。メラノーマ転移が、一次性皮膚悪性メラノーマよりBDNF及びその受容体をより頻繁に発現することが報告されているので(28)、発明者らは、BDNFがMeLiM転移において発現されるか否かを試験した。しかし、BDNF発現はMeLiM転移のメラノーマ細胞では検出されなかった。このことは、核RACK1が、メラニン形成細胞におけるBDNF非依存的細胞反応を介在することを示唆している。
【0132】
ヒト癌におけるインビボでのRACK1のアップレギュレーションを検出した研究はほとんどなく(13,29)、メラニン形成細胞との関係ではまったくない。RACK1 mRNAは、5つの非小細胞肺癌で強力に発現されていることが見出され;大きな血管の内皮は、これらの腫瘍におけるRACK1発現の主な部位として同定された(13)。 RACK1 mRNAはまた、結腸直腸癌の11の症例において高度に発現され、非癌部位よりも癌細胞において強い発現を示した(29)。しかし、腫瘍性メラニン形成細胞におけるRACK1の過剰発現の開示はなかった。
【0133】
ここで、発明者らは、RACK1 mRNA及びその対応するタンパク質が皮膚及び転移性メラノーマにおいて系統的に過剰発現されたことを示す;RACK1レベルは、形質転換されたメラニン形成細胞それ自体において上昇した。
【0134】
RACK1過剰発現が悪性の皮膚メラノーマの進行に役立つかもしれないとの提案は、培養された細胞株において行われたいくつかの実験によって支持されている。実際に、増加及び減少しているRACK1レベルは、細胞形質転換に関連した複数の細胞機能を明らかにした。RACK1の過剰発現は、3T3繊維芽細胞の成長を阻害することがわかった(30,31)。しかし、RACK1タンパク質レベルは培養された細胞において活性に調節されることを示唆するが、RACK1翻訳のモルホリノオリゴヌクレオチドによる遮断もまた、3T3繊維芽細胞の成長を減少させた(31)。興味深いことに、RACK1の過剰発現は、MCF-7乳癌細胞の増殖率を亢進し、このことは、繊維芽細胞及び腫瘍性細胞におけるRACK1の識別効果を示す(32)。従って、RACK1は、PC-12褐色細胞腫細胞において血清枯渇によって(33)、あるいはW7.2 T細胞においていくつかのアポトーシス刺激によって(34)誘導されたアポトーシスを阻害した。より具体的には、RACK1を過剰発現するMeWoヒトメラノーマ細胞は、UV-誘導アポトーシスに対する増加した抵抗性を示すことがわかった(35)。この効果は、PKCによるJNKの活性化を促進するためにアダプタータンパク質としてRACK1によって介在される(35)。加えて、正常な非-運動性MCF7癌細胞におけるRACK1過剰発現は、創傷治癒アッセイにおいてその転移能力を増加し、Transwellアッセイにおいてその運動性を誘導することが報告された。従って、siRNAを用いるRACK1発現の抑制は、前立腺癌DU 145細胞において反対の効果を有し、ranswellアッセイにおいてその転移を阻害した(36)。
【0135】
RACK1アップレギュレーションによる細胞転移の促進は、IGF-1Rとタンパク質ホスファターゼ2a(PP2A)又はβ1インテグリンのいずれかとの関連に因ることが最近提案された。実際に、RACK1を過剰発現する細胞では、RACK1はPP2Aを放出し、β1インテグリンと会合し、よって、インテグリンシグナル化を亢進し、増加した細胞転移能力を与える。
【0136】
RACK1の過剰発現はまた、いくつかのヒト卵巣癌株のインシュリン-及びIGF-1-促進された足場-非依存的成長を亢進することができる(37)。再度、この効果は、インシュリン受容体(IR)IGF-1R及びSTAT3との関連でアダプタータンパク質として働くRACK1によって介在される(37)。
【0137】
興味深いことに、MeWoメラニン形成細胞におけるRACK1の過剰発現は、STAT3のインシュリン-誘導活性化を亢進することがわかった、このことは、該経路の成分がこれらのメラノーマ-誘導細胞(37)に存在することを示唆する。最後に、RACK1用のsiRNAを用いるMeWoメラニン形成細胞におけるRACK1発現の阻害は、皮下注射後にヌードマウスの腫瘍成長を顕著に減少させた(35)。
【0138】
これらの観察の各々は、RACK1過剰発現が、MeLiMブタモデル及びヒト患者における悪性皮膚メラノーマの進行において原因となる事象であるという見解を支持する。
【0139】
結論として、RACK1は、細胞増殖、アポトーシスに対する抵抗性、転移及び拡散能力に関連するいくつかのシグナル経路の集中点として現れ、その調整解除は、メラニン形成細胞結合において腫瘍性形質転換をもたらす可能性がある。
【0140】
参考文献
1. Chin L. The genetics of malignant melanoma: lessons from mouse and man. Nat Rev Cancer 2003; 3: 559-70.
2. Clark EA, Golub TR1 Lander ES, Hynes RO. Genomic analysis of metastasis reveals an essential role for RhoC. Nature 2000; 406: 532-5.
3. Weeraratna AT, Jiang Y, Hostetter G, et al. Wnt5a signaling directly affects cell motility and invasion of metastatic melanoma. Cancer Cell 2002; 1: 279-88.
4. Hoek K, Rimm DL, Williams KR, et al. Expression profiling reveals novel pathways in the transformation of melanocytes to melanomas. Cancer Res 2004; 64: 5270-82.
5. Weeraratna AT, Becker D, Carr KM, et al. Generation and analysis of melanoma SAGE libraries: SAGE advice on the melanoma transcriptome. Oncogene 2004; 23: 2264-74.
6. Vincent-Naulleau S, Le Chalony C, Leplat JJ, et al. Clinical and histopathological characterization of cutaneous melanomas in the melanoblastoma-bearing Libechov minipig model. Pigment Cell Res 2004; 17:24-35.
7. Velculescu VE, Zhang L, Vogelstein B, Kinzler KW. Serial analysis of gene expression. Science 1995; 270: 484-7.
8. JuIe S, Bosse P, Egidy G, Panthier JJ. Establishment and characterization of a normal melanocyte cell line derived from pig skin. Pigment Cell Res 2003; 16: 407-10.
9. Le Chalony C, Renard C, Vincent-Naulleau S, et al. CDKN2A region polymorphism and genetic susceptibility to melanoma in the MeLiM swine model of familial melanoma, lnt J Cancer 2003; 103: 631-5.
10. Virion B, Cheval L, Buhler JM, et al. Serial microanalysis of renal transcriptomes. Proc Natl Acad Sci U S A 1999; 96: 15286-91.
11. Egidy G, Juillerat-Jeanneret L, Jeannin JF, et al. Modulation of human colon tumorstromal interactions by the endothelin system. Am J Pathol 2000; 157: 1863-74.
12. Johren O, Sanvitto GL, Egidy G, Saavedra JM. Angiotensin II AT1A receptor mRNA expression is induced by estrogen-progesterone in dopaminergic neurons of the female rat arcuate nucleus. J Neurosci 1997; 17: 8283-92.
13. Berns H, Humar R, Hengerer B, Kiefer FN, Battegay EJ. RACK1 is up-regulated in angiogenesis and human carcinomas. Faseb J 2000; 14: 2549-58.
14. Ron D, Jiang Z, Yao L, et al. Coordinated movement of RACK1 with activated betallPKC. J Biol Chem 1999; 274: 27039-46.
15. Rubinfeld B, Robbins P, El-Gamil M, et al. Stabilization of beta-catenin by genetic defects in melanoma cell lines. Science 1997; 275: 1790-2.
16. Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell 2006; 124: 471-84.
17. King R, Weilbaecher KN, McGiII G, et al. Microphthalmia transcription factor. A sensitive and specific melanocyte marker for MelanomaDiagnosis. Am J Pathol 1999; 155: 731-8.
19. McCahill A, Warwicker J, Bolger GB, Houslay MD, Yarwood SJ. The RACK1 scaffold protein: a dynamic cog in cell response mechanisms. Mol Pharmacol 2002; 62: 1261-73.
20. Oka M, Ogita K, Ando H, Kikkawa U, lchihashi M. Differential down-regulation of protein kinase C subspecies in normal human melanocytes: possible involvement of the zeta subspecies in growth regulation. J Invest Dermatol 1995; 105: 567-71.
21. Park HY, Wu H, Killoran CE, Gilchrest BA. The receptor for activated C-kinase-1 (RACK1) anchors activated PKC-beta on melanosomes. J Cell Sci 2004; 117: 3659-68.
22. Kawakami Y, Nishimoto H, Kitaura J, et al. Protein kinase C betall regulates Akt phosphorylation on Ser-473 in a cell type- and stimulus-specific fashion. J Biol Chem 2004; 279: 47720-5.
23. Stahl JM, Sharma A, Cheung M, et al. Deregulated Akt3 activity promotes development of malignant melanoma. Cancer Res 2004; 64: 7002-10.
24. Gilhooly EM, Morse-Gaudio M, Bianchi L, et al. Loss of expression of protein kinase C beta is a common phenomenon in human malignant melanoma: a result of transformation or differentiation? Melanoma Res 2001; 11: 355-69.
25. Ron D, Vagts AJ, Dohrman DP, et al. Uncoupling of betallPKC from its targeting protein RACK1 in response to ethanol in cultured cells and mouse brain. Faseb J 2000; 14: 2303-14.
26. McGough NN, He DY, Logrip ML, et al. RACK1 and brain-derived neurotrophic factor: a homeostatic pathway that regulates alcohol addiction. J Neurosci 2004; 24: 10542-52.
27. Jeanblanc J, He DY, McGough NN, et al. The dopamine D3 receptor is part of a homeostatic pathway regulating ethanol consumption. J Neurosci 2006; 26: 1457-64.
28. lnnominato PF, Libbrecht L, van den Oord JJ. Expression of neurotrophins and their receptors in pigment cell lesions of the skin. J Pathol 2001; 194: 95-100.
29. Saito A, Fujii G, Sato Y, et al. Detection of genes expressed in primary colon cancers by in situ hybridisation: overexpression of RACK1. Mol Pathol 2002; 55: 34-9.
30. Chang BY, Conroy KB, Machleder EM, Cartwright CA. RACK1, a receptor for activated C kinase and a homolog of the beta subunit of G proteins, inhibits activity of src tyrosine kinases and growth of NIH 3T3 cells. Mol Cell Biol 1998; 18: 3245-56.
31. Hermanto U, Zong CS, Li W, Wang LH. RACK1, an insulin-like growth factor I (IGF-I) receptor-interacting protein, modulates IGF-1-dependent integrin signaling and promotes cell spreading and contact with extracellular matrix. Mol Cell Biol 2002; 22: 2345- 65.
32. Kiely PA, Sant A, O'Connor R. RACK1 is an insulin-like growth factor 1 (IGF-1) receptor- interacting protein that can regulate IGF-1-mediated Akt activation and protection from cell death. J Biol Chem 2002; 277: 22581-9.
33. Choi DS, Young H, McMahon T, Wang D, Messing RO. The mouse RACK1 gene is regulated by nuclear factor-kappa B and contributes to cell survival. Mol Pharmacol 2003; 64: 1541-8.
34. Mourtada-Maarabouni M, Kirkham L, Farzaneh F, Williams GT. Functional expression cloning reveals a central role for the receptor for activated protein kinase C 1 (RACK1) in T cell apoptosis. J Leukoc Biol 2005; 78: 503-14.
35. Lopez-Bergami P, Habelhah H, Bhoumik A, et al. RACK1 mediates activation of JNK by protein kinase C [corrected]. Mol Cell 2005; 19:3 09-20.
36. Kiely PA, O'Gorman D, Luong K, Ron D, O'Connor R. Insulin-like growth factor I controls a mutually exclusive association of RACK1 with protein phosphatase 2A and betai integrin to promote cell migration. Mol Cell Biol 2006; 26: 4041-51.
37. Zhang W, Zong CS, Hermanto U, et al. RACK1 Recruits STAT3 Specifically to Insulin and Insulin-Like Growth Factor 1 Receptors for Activation, Which Is Important for Regulating Anchorage-Independent Growth. Mol Cell Biol 2006; 26: 413-24.
38. White VA, et al. Correlation of cytogenetic abnormalities with the outcome of patients with uveal melanoma. Cancer 1998; 83(2): 354-9.
39. Bennett D. C. How to make a melanoma:what do we know of the primary clonal events? Pigment Cell Melanoma Res. 2008; 21: 27-38.
【技術分野】
【0001】
発明の分野
本発明は、癌の分野に含まれ、哺乳動物の癌を診断する方法に関する。
【背景技術】
【0002】
発明の背景
この明細書と実験部分の全体を通じ、さまざまな刊行物を参照する。その刊行物は参考としてこの明細書に組み込まれているものとする。
【0003】
メラノーマは、メラニン形成細胞が変化することによって発達する悪性腫瘍である。メラニン形成細胞は、皮膚、毛髪、目を着色していて、たいていの母斑に高濃度で存在する黒色色素を生成する細胞である。したがってメラノーマの大半は、黒色または茶色である。しかしメラノーマは色素の生成を停止することがある。それが起こるとき、メラノーマはもはや暗い色ではなく、皮膚の色であるピンク、赤、紫である。
【0004】
メラノーマは、皮膚癌の最も深刻な形態である。白色人種におけるその発生率と死亡率は世界的に増加している。メラノーマは最も一般的な皮膚癌ではないが、メラノーマの死者数が最も多い。アメリカ癌学会は、2007年にアメリカ合衆国でメラノーマが新たに約60,000件発生していると見積もっている。
【0005】
誰でもメラノーマになる可能性があるが、いくつかの因子に依存してリスクが大きくなる可能性がある。その因子として特に、太陽への曝露、皮膚にある母斑の数、皮膚のタイプ、遺伝的背景がある。実際、UVA光線とUVB光線の両方とも皮膚癌(メラノーマを含む)を誘発する可能性がある。
【0006】
2つのタイプの母斑の存在、すなわち正常な母斑と、異形成母斑として知られる異型の母斑の存在が、メラノーマのリスクが増大することを示す。遺伝的傾向に関しては、メラノーマと診断された患者の約10%が、メラノーマの履歴がある家族を持つことが知られている。
【0007】
原発性メラノーマは、転移する傾向がある。これは、癌が原発部位から身体の他の場所に広がることを示す。癌細胞は原発腫瘍から離れてリンパ管と血管に侵入し、血流を循環し、身体の別の正常組織内の離れた病巣(転移)で成長する。
【0008】
転移が存在していると、治療への反応が有効でないために予後が厳しい。診断に関しては、色と形が不規則な母斑は、悪性メラノーマまたは前悪性メラノーマが疑われる。視診と皮膚学的検査の後、疑わしい母斑のバイオプシーもつくることができる。そのバイオプシーが悪性である場合には、母斑とその周囲の領域を切除し、広がることを避ける必要がある。
【0009】
原発メラノーマのある患者にとって、臨床と組織病理学的な基準が指標として有用である。しかし正確な診断と予後のためのマーカーがあれば、多数の患者にとって利益があるだろうことが広く認められている。
【0010】
したがってメラノーマを初期に信頼性よく診断することが非常に重要である。
【0011】
メラノーマはウマでもしばしば見られ、成熟したウマにおける最も一般的な皮膚腫瘍である。メラノーマは、アラビア、リピッツァナー、ペルシュロンの血統において非常に一般的に発生することが見られている。なぜならこれらの血統では灰色のウマが多く生まれ、メラノーマはたいてい灰色のウマで見いつかるからである。メラノーマは、一般に、唇、目、耳、唾液腺、肛門、陰茎、膣で見つかる。灰色のウマにおけるメラノーマの大半はゆっくり成長する良性腫瘍であり、他の器官に転移したり広がったりすることは稀である。しかしこの腫瘍は、特に灰色でないウマで他の器官に広がる可能性がある。一般に、この腫瘍は、排尿、排便、交配以外の問題を起こさない。しかしメラノーマは局所的な組織に侵入する可能性があるため、中枢神経系に侵入することによって不自由さや神経の問題を起こす可能性がある。
【0012】
メラノーマはイヌでも観察できる。イヌでは主に皮膚で見られるが、口腔でも見ることができる。メラノーマであると早い段階で診断されると、癌の除去とグレードまたはステージの同定がよりうまくいく可能性がある。良性形態の皮膚腫瘍でさえ、局所的な侵襲性を持つ可能性がある。メラノーマは、身体のあらゆる領域に、その中でも特にリンパ節と肺に転移する(広がる)可能性があるため、イヌにとって将来の見通しは非常に険しく危険である。ネコもメラノーマになる可能性があるが、イヌよりは程度が小さい。
【0013】
したがってメラノーマの診断は、獣医学の観点からも問題がある。
【0014】
メラノーマの診断には経験が必要である。なぜなら初期には、無害の母斑と同じに見えたり、まったく着色していなかったりすることがあるからである。
【0015】
メラノーマであることが確認されて早い段階で治療された場合には、ほぼ100%治癒する。しかしそうでない場合には、この癌が進行して身体の他のいろいろな部分に広がる。そこでは治療が難しく、致命的になる可能性がある。
【0016】
したがってメラノーマを初期に信頼性よく診断することのできる新しいツールが必要とされている。
【0017】
さらに、メラノーマの進行の分子的基礎を理解すると、より有効な治療法を開発する助けになろう。したがってメラニン形成細胞が変化するメカニズムが広く研究されている。
【発明の概要】
【課題を解決するための手段】
【0018】
本発明は、哺乳動物のメラニン形成細胞の腫瘍状態を判断する方法と、哺乳動物でメラノーマを診断する方法に関する。本発明の第1の特徴によれば、この方法は、哺乳動物のメラニン形成細胞におけるRACK-1タンパク質の過剰発現を検出し、過剰発現している場合にはそのメラニン形成細胞が腫瘍であるため、その哺乳動物にメラノーマが存在すると結論するステップを含んでいる。
【0019】
本発明の第2の特徴によれば、この方法は、メラニン形成細胞におけるRACK-1タンパク質の発現レベルを測定し、そのレベルを参照レベルと比較する操作を含んでいる。そのメラニン形成細胞におけるRACK-1タンパク質の発現レベルが参照レベルよりも高い場合には、そのメラニン形成細胞が腫瘍細胞であり、したがってその哺乳動物にメラノーマが存在すると結論される。参照レベルは、正常な非腫瘍メラニン形成細胞、すなわち正常な上皮(皮膚)メラニン形成細胞、または正常な脈絡膜メラニン形成細胞におけるRACK-1タンパク質の発現レベルであることが好ましい。
【0020】
RACK-1タンパク質の発現レベルは、RACK-1のmRNAまたはRACK-1タンパク質のレベルと定義することができる。
【0021】
本発明は、哺乳動物におけるメラノーマの診断方法にも関する。この方法は、その哺乳動物からの生物学的サンプルを、試験管内で、メラニン形成細胞を検出する手段およびRACK-1タンパク質の発現を検出する手段と接触させるステップと、メラニン形成細胞がRACK-1タンパク質の発現と同じ場所に局在しているかどうかを判断するステップを含んでいて、RACK-1タンパク質の発現を示すメラニン形成細胞の存在は、メラノーマであることを示している。
【0022】
サンプルは、バイオプシー(例えばリンパ節、肝臓、肺、心臓、脾臓、目、皮膚のバイオプシー)であることが有利である。
【0023】
本発明の好ましい一実施態様によれば、哺乳動物はヒトである。あるいは哺乳動物は、ブタ、ネコ、イヌ、ウマでもよい。
【0024】
RACK-1タンパク質を特にインサイチュ・イムノアッセイで検出するには、RACK-1に対する抗体を用いることが好ましい。
【0025】
本発明は、哺乳動物でメラノーマを診断するため、RACK-1タンパク質の発現を検出する手段を含むキットにも関する。このようなキットは、メラニン形成細胞を検出する手段をさらに備えていることが有利である。
【0026】
本発明は、腫瘍になったメラニン形成細胞のマーカーとしてRACK-1タンパク質を用いる方法にも関する。
【0027】
本発明は、添付の図面を参照してさらに詳しく説明できる。
【図面の簡単な説明】
【0028】
【図1】図1は、MeLimからのメラノーマの肺転移を示す。図1のAは、さまざまなサイズの多病巣性メラノーマ転移が浸潤した肺の写真である。矢印は、腫瘍組織のいくつかの黒い点を指している。図1のB1〜D1は、MITF抗体を用いて免疫組織化学によって同定したメラニン形成細胞であり、茶色の核顆粒として見ることができる。図1のB2〜D2は、一次抗体なしの非特異的染色が存在していない。図1のB1とB2は正常な皮膚に対応し、C1と図1C2は皮膚メラノーマに対応し、図1のD1とD2はメラノーマの肺への転移に対応する。aは肺胞を表わし、beは気管支表皮を表わし、dは真皮を表わし、eは表皮を表わし、hfは毛包を表わす。矢印は、メラニン形成細胞(黒)とメラノーマ細胞(白)を指す。どの顕微鏡写真のスケール表示棒も100μmである。
【図2】図2は、ブタ組織におけるRACK1 mRNAの発現を示す。脱色した切片に関するRACK1のアンチセンス・プローブ(A)とセンス・プローブ(B)のインサイチュ・ハイブリダイゼーション・オートラジオグラフ。各プローブについて正常な組織(N)を左側に示し(皮膚、肺、肝臓、リンパ節)、腫瘍組織(T)を右側に示してある(皮膚メラノーマ、肺、肝臓、リンパ節から転移したメラノーマ)。アンチセンス・プローブでは、リンパ節を除き、腫瘍では健康な組織または弱っていない組織と比べて信号が強いことに注意されたい。C:アンチセンスRACK1プローブとハイブリダイズさせた肺転移MeLiMメラノーマからの暗視野顕微鏡写真。D:Cからの連続切片におけるヘマトキシリン-エオシン染色であり、C上で腫瘍の着色部位に銀色の顆粒が含まれている。スケール表示棒は、A、Bでは1cm、C、Dでは100μmである。
【図3】図3は、ブタの正常な表皮におけるRACK1の発現を示す。A、C、D:ブタの皮膚に関するRACK1タンパク質(緑)と、MITF(A、C)またはDCT(D)(赤)の二重標識の共焦点顕微鏡分析。対照となるメイシャン・ミニブタからの正常な表皮(A、B)とMeLiM(C、D)。Bは、Aに対応する透過写真である。C:三次元“直交”スライス投射分析が含まれる:中央の大きなパネルは単一の光学スライスを示しており、このスライスには、スライスされたz軸を再構成するためにx軸(緑色の線)とy軸(赤色の線)が規定された。x、zスライス(上)とy、zスライス(右)に関する対応する結果を示してある。青色の線は、z方向の積層体内での中央パネルの画像の位置を示す。核を青色で示してある。ケラチノサイトにはRACK1の細胞質ゾル・スポット信号があり、白い破線で示したメラニン形成細胞にはその信号がないことに注意されたい。点線は、真皮(d)と表皮(e)の分離線を示す。スケール表示棒:5μm。
【図4】図4は、進行段階がさまざまなMeLiMメラノーマにおけるRACK1の細胞分布。RACK1タンパク質(緑)と、MITF(A、C)またはDCT(D)(赤)の二重標識の共焦点顕微鏡分析。A:皮膚メラノーマ。B〜D:リンパ節(B)、肺(C)、心臓(D)へのメラノーマ転移。三次元“直交”スライス投射分析を図3のように示してある。核を青色で示してある。A1:A2に対応する透過写真である。A3:A2挿入図の拡大。A:白い矢印の先端は、MITFに関して陽性の真皮メラノーマ細胞を指しており、これは直交投射による分析である。真皮メラノーマ細胞と表皮ケラチノサイトでRACK1の細胞質ゾル信号が同等であることに注意されたい。RACK1は皮膚メラノーマ細胞と転移メラノーマ細胞で豊富であり、核の周囲に局在している。さらに、転移では、核RACK1が存在している。それを光学スライスと直交投射に黄色い矢印の先端で示してある。A1とA2の点線は、真皮(d)と表皮(e)の分離線を示す。スケール表示棒:5μm。
【図5】図5は、正常なメラニン形成細胞とMeLiMからのメラノーマ細胞におけるβ-カテニンとRHEBの細胞分布を示す。RACK1(赤)とβ-カテニン(A、B)またはRHEB(C、D)(緑)の二重標識の後の正常な皮膚(A1〜A2、C1〜C2)と肺へのメラノーマ転移(B1〜B2、D1〜D2)に関する共焦点顕微鏡写真とそのそれぞれの透過画像を示してある。核対比染色を青色で示してある。矢印は、細胞質ゾルのβ-カテニン(A1)がある正常なメラニン形成細胞、核周辺のRHEB(C1)がある正常なメラニン形成細胞、RACK1なしの正常なメラニン形成細胞を指している。矢印の先端は、RACK1とβ-カテニンに関する核信号のあるメラノーマ細胞(B1)を黄色で、RHEBに関する核信号のあるメラノーマ細胞(D1)を白色で示している。A1の星印は、核β-カテニンのあるケラチノサイトを示している。これはおそらく分裂している細胞である。AとCの点線は、真皮(d)と表皮(e)の分離線を示す。スケール表示棒:5μm。
【図6】図6は、ヒトの皮膚とメラノーマにおけるRACK1を示している。RACK1タンパク質(A、B)、MITF(C、D)に関する二重標識、トプロ3を用いた核対比染色(E、F)の共焦点顕微鏡分析。第1列(A、C、E)は、対照のヒト皮膚。MITF陽性メラニン形成細胞を破線で囲んである。点線は、サンプルの表皮(e)と真皮(d)の境界の分離を示す。第2列(B、D、F)は、リンパ節へのメラノーマの転移である。転移したヒト・メラニン形成細胞の細胞質ではRACK1が高レベルになっているのが見られる。スケール表示棒:10μm。
【図7】図7は、RACK1のレベルがヒトメラノーマでは高いが、母斑ではそうでないことを示している。正常な皮膚(a)、母斑(b)、皮膚メラノーマ(c)、肝臓転移(d)におけるRACK1(第1行)とMITF(第2行)の共焦点顕微鏡写真。SP2ライカ共焦点顕微鏡で単一の光学切片を取得した。RACK1は正常なメラニン形成細胞(a)ではほとんど検出されない。それは母斑(b)の過剰増殖性損傷にも当てはまる。皮膚の切片はさまざまな患者からのものである。矢印の先端は、RACK1が検出されないメラニン形成細胞を指している。矢印は、細胞質RACK1を発現しているメラニン形成細胞を示している。点線は、表皮-真皮の境界を示す。e、表皮;dr、真皮。棒=10μm。
【図8】図8は、ブドウ膜メラノーマにおけるRACK1の過剰発現を示している。この図は、染色体が3モノソミーになったヒト脈絡膜メラノーマでRACK1のレベルが高いことを示している。RACK1(a)とMITF(b)に関する免疫蛍光染色。cでは、DAPI核対比染色。SPEライカ共焦点顕微鏡で単一の光学切片を取得した。棒は10μm。
【図9】図9は、ウマのメラノーマにおけるRACK1の過剰発現を示す。ウマの皮膚悪性メラノーマでRACK1が高レベルになっている。RACK1(a)とMITF(b)に関する免疫蛍光染色。cでは、DAPI核対比染色。アポ・トーム・ツァイス顕微鏡で単一の光学切片を取得した。棒:10μm。
【発明を実施するための形態】
【0029】
本発明に関連して、以下の用語は以下のように定義される。
メラノーマは、メラニン形成細胞の悪性腫瘍である。それは、主に皮膚癌(皮膚メラノーマ)であるが、例えばヒトの眼のメラニン形成細胞(脈絡膜メラノーマ)においても見られるがそれほど頻繁には見られない。
【0030】
転移は、身体の主な部位から他の部位へ癌の広がりである。癌細胞は、一次(又は元の)腫瘍から裂けて、リンパ及び血管に透過し、血流によって循環し、身体の至るところの他の臓器中の離れた場所で沈着し増殖し得る(転移する)。
【0031】
腫瘍は良性又は悪性のいずれかとして分類される。悪性腫瘍は、侵襲及び転移によって拡がることができるが、良性腫瘍は広がらない(そして、局所的に増殖するのみである)。定義としては、用語「癌」は悪性腫瘍にのみ適用される。
【0032】
転移性腫瘍は、癌の最後の段階で非常に一般的である。転移の拡がりは、血液又はリンパを介してあるいはその両方の経路を介して起こり得る。転移の最も一般的な場所は、リンパ節、肝臓、脳及び骨である。
【0033】
癌細胞が広がって新しい腫瘍を形成する時に、続発性又は転移性腫瘍と呼ばれ、その細胞は元の腫瘍の細胞と似ている。このことは、メラノーマが肺に拡がる(転移する)場合に、続発性腫瘍(転移)は、腫瘍性メラニン形成細胞(腫瘍性肺細胞でない)からなることを意味する。肺の疾患は転移性メラノーマである。細胞の腫瘍状態は、その腫瘍又は非-腫瘍状態に関連し、腫瘍細胞は、正常で健常でないが、腫瘍由来である、細胞を意味する。腫瘍性細胞は、良性腫瘍由来でも又は悪性腫瘍由来でもよい。
【0034】
RACK1(又はRACK1)タンパク質は、GeneBankの受入番号NM_006098を有する塩基配列によってコードされるヒトタンパク質(ヒトタンパク質P63244 配列番号69)又はそのアレル変異体、あるいは別の哺乳動物種における任意のオルソログ、例えば、NM_008143によってコードされるネズミ配列、又はNM_214332よってコードされるブタRACK1配列、又はNM_175602によってコードされるウシ配列、又はNM_130734によってコードされるラット配列、又はXM_537934によってコードされるイヌ配列、を意味する。RACK1タンパク質に対応するRNA又はDNAは、以下のタグ:ATTGTAGATG (配列番号1) で検出され得る。
【0035】
本発明は、タンパク質RACK1が、哺乳動物の正常なメラニン形成細胞に関して、腫瘍性メラニン形成細胞において過剰発現する、という発明者らによる発見に依拠する。実際に、正常メラニン形成細胞では、RACK1は、非常に低いレベルで発現し、ほとんど検出されない。しかし、このタンパク質は、腫瘍性メラニン形成細胞、メラノーマ又はメラノーマの転移において顕著に過剰発現することが見出されている。
【0036】
更に、本発明者らは、RACK1の発現が非常に増加し、皮膚性及び脈絡膜のメラノーマにおいて均一に分布しているが、RACK1シグナルは、母斑、すなわち皮膚性良性腫瘍において低くかつ異質である、ことを見出した。
【0037】
本発明はまた、本発明者らの観察によれば、あるメラノーマのみに関連するがすべてのメラノーマには関連しない他の公知のマーカーと比べて、RACK1は、すべての皮膚性及び脈絡膜メラノーマにおいて過剰発現するという、本発明者らによる発見に依拠する。従って、本発明者らのデータは、RACK1がメラニン形成細胞における悪性腫瘍の普遍的なマーカーであることを示す。
【0038】
そのため、第1の態様によれば、本発明は、哺乳動物のメラノーマ細胞の腫瘍状態を決定する方法であって、該メラニン形成細胞におけるRACK1タンパク質の発現又は過剰発現を検出し;及びRACK1タンパク質の発現又は過剰発現を該細胞の腫瘍状態、好ましくは悪性腫瘍状態、と関連づけること、を含む、方法に関する。
【0039】
試験下でのメラニン形成細胞の腫瘍状態は、好ましくは、本発明の方法を実行する前に決定されない。メラニン形成細胞は、好ましくは、樹立された細胞株からのものではない。好ましくは試験下の細胞は、一次細胞である。従って、本発明の方法は、メラニン形成細胞の内の腫瘍性細胞の検出、好ましくは悪性腫瘍性細胞の検出を可能にする。
【0040】
第2の態様によれば、本発明はまた、哺乳動物のメラノーマを診断する方法であって、哺乳動物にメラニン形成細胞中のRACK1タンパク質の発現又は過剰発現を検出し、RACK1タンパク質の発現又は過剰発現をメラノーマの存在と関連づけることを含む方法に関する。
【0041】
該方法は、好ましくは、インビトロ又はエクスビボで行われる、すなわち、RACK1タンパク質の発現又は過剰発現の検出ステップが好ましくはインビトロ又はエクスビボで実行される。あるいは、インビボで行ってもよい。
【0042】
メラニン形成細胞は、メラニンを合成するその能力によって認識される。従って、メラニン合成に関連する任意のタンパク質は、メラニン形成細胞を他の細胞と識別するマーカーとして使用され得る。しかし、ある腫瘍性メラニン形成細胞は、メラニンを産生するその能力を失っていることがあり、メラニン合成経路に関連しない別のマーカーがその場合に使用されることがあることに留意されたい。メラニン形成細胞用の可能性のあるマーカーは、小眼症転写因子MITF、ドパクロムトートメラーゼDCT、チロシナーゼTYR、チロシナーゼ-関連タンパク質1 TRP 1である。
【0043】
RACK1タンパク質の発現は、対照として使用される正常なメラニン形成細胞においてRACK1の発現を超える発現を意味する。本発明によれば、RACK1過剰発現は、例えば入手でき、病原診断のために入手され使用される、パラフィン包埋された少量の一次組織で検出される。更に、本発明のこれらの態様によれば、インサイチュ免疫アッセイと同等又はそれより劣る感度を有する方法が検出に使用されるならば、メラニン形成細胞におけるRACK1発現の存在の1つの検出は、腫瘍性細胞又はメラノーマの存在の指標である。実際に、上記のように、RACK1は正常なメラニン形成細胞で非常に低レベルで発現し、従って、このタンパク質は、標準的方法、例えば検出手段としてのインサイチュ免疫アッセイ、又はインサイチュ免疫アッセイと同等又はそれよりも感度の劣る検出方法によって正常メラニン形成細胞において検出できない。
【0044】
従って、本発明に関連して、インサイチュ免疫アッセイは、該方法を実行するために特に好ましい技術である。実際に、インサイチュ免疫アッセイを用いる、RACK1タンパク質用の陽性細胞の検出は、腫瘍性細胞の存在、よってメラノーマ、の直接的指標である。
【0045】
この態様によれば、本発明の目的は、メラニン形成細胞中のRACK1タンパク質の発現レベルを測定し、そのレベルを参照レベルと比較するステップを含む、哺乳動物のメラニン形成細胞の腫瘍性状態を決定する方法である。
【0046】
前記メラニン形成細胞中の、参照レベルに関してRACK1タンパク質の増加したレベルは、この細胞が腫瘍性細胞(腫瘍状態)、好ましくは悪性腫瘍性細胞である、指標である。
【0047】
試験下のメラニン形成細胞の腫瘍状態は、好ましくは、本発明の方法を実行する前に決定されない。メラニン形成細胞は、このましくは樹立された細胞株由来でない。好ましくは、試験下の細胞は、一次細胞である。
【0048】
第4の態様によれば、本発明は、哺乳動物のメラニン形成細胞中のRACK1タンパク質発現のレベルを測定し、そのレベルを参照レベルと比較するステップを含む、哺乳動物のメラノーマを診断する方法に関する。参照レベルに関して前記メラニン形成細胞のRACK1タンパク質発現の増加したレベルは、メラノーマの存在の指標である。
【0049】
本発明の第3及び第4の態様は、RACK1タンパク質発現の発現又は過剰発現の決定が参照レベルとの比較によってなされる、最初の2つの態様に相当する。
【0050】
本発明の方法は、好ましくは、インビトロ又はエクスビボで行われる、すなわち、RACK1タンパク質レベルの決定ステップが、好ましくはインビトロ又はエクスビボで行われる。あるいは、インビボで行ってもよい。
【0051】
本発明の方法では、RACK1タンパク質の発現のためにアッセイされる哺乳動物のメラニン形成細胞は、好ましくは前記哺乳動物の生物試料由来である。比較ステップのために使用される参照レベルは、例えば、非-腫瘍性、すなわち診断される哺乳動物の正常な健常なメラニン形成細胞におけるRACK1タンパク質発現レベルである。正常なメラニン形成細胞は、例えば、正常な表皮メラニン形成細胞又は正常な脈絡膜メラニン形成細胞である。正常なメラニン形成細胞の任意の他の種類は、タンパク質RACK1発現の参照レベルのために使用してもよい。
【0052】
「増加したレベル」とは、レベルが顕著に増加されることである、すなわち、測定されたレベルと参照レベルとの差が統計的観点から有意であり、すなわち測定されたレベルの標準偏差よりも大きい、と理解すべきである。従って、この差は、好ましくは、測定に固有のエラーよりも大きく、そして正常なメラニン形成細胞間で観察された偏差よりも大きい。測定されたレベルと参照レベルとの差は、分析されるメラニン形成細胞に依拠して、及び選択された参照レベルに依拠して顕著に変動し、しかし、腫瘍性細胞のレベルは、検出の限界及び上記のカットオフラインよりも高い。測定されたレベルは、例えば、参照レベルに因って、参照レベルよりも少なくとも5%高くてよく、好ましくは少なくとも10%又は20%高くてよい。本発明の異なった態様では、測定されたレベルは、参照レベルの少なくとも3倍であり、一般的に少なくとも6倍であり、例えば参照レベルよりも少なくとも10倍高い。例えば、参照レベルが非-腫瘍性、すなわち正常な健常なメラニン形成細胞中のRACK1タンパク質発現レベルであるときに、測定されたレベルは、参照レベルの、少なくとも1、5、好ましくは2倍である。
【0053】
本発明はまた、哺乳動物の皮膚断片におけるメラニン形成細胞の腫瘍状態を決定する方法であって、
−該メラニン形成細胞中のRACK1タンパク質の発現レベルを、該哺乳動物の皮膚断片の周囲のケラチノサイトにおけるRACK1タンパク質の発現レベルと比較し;
−発現の等価レベルを該細胞の腫瘍状態と関連づけること、
を含む方法に関する。
本発明はまた、哺乳動物の皮膚メラノーマを診断する方法であって、
-メラニン形成細胞中のRACK1タンパク質の発現レベルを、該哺乳動物の皮膚断片の周囲のケラチノサイトにおけるRACK1タンパク質の発現レベルと比較し;
−発現の等価レベルを該皮膚メラノーマの存在と関連づけること、
を含む方法に関する。
【0054】
メラニン形成細胞及び皮膚断片上のケラチノサイト中のRACK1タンパク質発現のこのような比較は、当業者によって容易になされる。実際に、図3及び4から明らかであるが、一方では、健常なメラニン形成細胞におけるRACK1タンパク質の発現は検出できないが、周囲のケラチノサイトでは検出でき(図3)、他方、RACK1タンパク質の発現レベルは、周囲のケラチノサイトと腫瘍性メラニン形成細胞においては同等である(図4)。本発明によれば、RACK1タンパク質発現レベルは、RACK1タンパク質発現の検出及び/又は上記の発現レベルの推定によって決定又は測定され得る。
【0055】
本発明によれば、その腫瘍状態を定義するために分析されるメラニン形成細胞は、バイオプシー由来でよい。本発明によるバイオプシーは、任意の臓器又は組織由来でよい。バイオプシーは、例えば皮膚又は眼のバイオプシー、又は肝臓、リンパ節、肺、心臓又は脾臓のような内臓由来である。
【0056】
本発明によれば、RACK1タンパク質発現レベルは、診断される細胞中のRACK1タンパク質のレベルの測定によって、あるいは診断される細胞中のRACK1タンパク質に相当するmRNAのレベルの測定によって、決定される。実際に、腫瘍性メラニン形成細胞中のRACK1タンパク質の高レベルは、対応するmRNAの高レベルによるものであり、すなわち、RACK1タンパク質の過剰発現は、腫瘍性細胞中の対応する遺伝子の転写の増加したレベルに主による、ことが、本発明者らによって明らかにされた(実施例の項を参照)。
【0057】
本発明によれば、細胞中のRACK1タンパク質発現レベルは、有利には、インサイチュ免疫アッセイによって決定される。この方法によれば、ペプチド又はタンパク質の発現レベルを特に蛍光によって視覚化及び測定することが可能である。
【0058】
あるいは、RACK1タンパク質に対するmRNAのレベルが決定されるとき、使用される有利な方法は、インサイチュハイブリダイゼーションである。インサイチュハイブリダイゼーション及びインサイチュ免疫アッセイはいずれも、本発明の範囲において当業者に周知である。
【0059】
インサイチュ免疫アッセイについて、RACK1に対する抗体が使用されることになる。かかる抗体は、商業的に入手可能である。インサイチュハイブリダイゼーションに関して、この方法はRACK1 mRNAに特異的にハイブリダイズするプローブの使用を必要とする。好適なプローブの例は、本説明の実験項に記載されている。
【0060】
細胞中のRACK1発現のレベルは、RACK1タンパク質に対応するRNA上でRT-PCRによって決定してもよい。
【0061】
あるいは、RACK1タンパク質の存在又はRACK1タンパク質発現レベルは、免疫学的方法、免疫アッセイ、免疫組織化学アッセイ、共焦点顕微鏡分析、紫外線顕微鏡、電気的検出又はハイブリダイゼーションオートラジオグラフィーを実行することによって、本発明に従って決定される。
【0062】
免疫学的方法は、好ましくは、特定のモノクローナル抗体又はポリクローナル抗体を用いる方法であり、好ましくは、ELISA法、免疫酵素法、免疫蛍光法、放射性-免疫学的方法及び化学-免疫学的方法から選ばれる。最も好ましくは、免疫学的方法は、RACK1タンパク質に対するモノクローナル抗体による、インサイチュ免疫アッセイである。本発明に従うメラノーマは、例えば、皮膚メラノーマ又は脈絡膜メラノーマである。
【0063】
分析されるメラニン形成細胞が、通常メラニン形成細胞が欠けている非-皮膚組織のバイオプシーの一部である時に、本発明の方法による腫瘍性細胞の同定は、メラノーマの転移の指標である。本発明に従って診断される哺乳動物は、好ましくはヒトであり;しかし、本発明はまた、他の哺乳動物、特にペット、好ましくはブタ、ウマ及びイヌにおけるメラノーマの腫瘍性細胞の診断に関する。任意の他の哺乳動物も本発明の範囲内にある。
【0064】
本発明はまた、哺乳動物由来の生物試料と、メラニン形成細胞を検出する手段、及びRACK1タンパク質発現を検出し、メラニン形成細胞がRACK1タンパク質発現と共局在かするか否かを決定する手段とを接触させるステップを含む、哺乳動物におけるメラノーマを診断する方法に関する。RACK1タンパク質発現を示すメラニン形成細胞の存在は、メラノーマの指標である。実際に、健常なメラニン形成細胞では、本発明者らは、RACK1タンパク質発現を検出できなかったが、メラニン形成細胞マーカーの発現を検出した。一方で、腫瘍性メラニン形成細胞では、本発明者らは、RACK1タンパク質及びメラニン形成細胞マーカーの発現を検出した。
【0065】
本方法は、診断される哺乳動物から得た生物試料について、好ましくはインビトロ又はエクスビボで行われ、最も好ましくはインビトロで行われる。従って、本発明の診断方法は、ヒト又は動物の体との任意の相互作用を示すものではなく、後者の存在を必要とする。
【0066】
本方法の実行において使用される生物試料は、好ましくは、メラノーマ又はメラノーマの転移を宿す(host)と疑われた臓器又は組織由来のバイオプシーである。バイオプシーの好ましい例は、リン節、肝臓、肺、心臓、脾臓、眼及び皮膚のバイオプシーである。
【0067】
既に述べたように、RACK1タンパク質発現を検出する手段は、好ましくは、試料中のRACK1タンパク質を検出する手段である。かかる手段は、好ましくは、抗体であり、最も好ましくはモノクローナル抗体である。あるいは、それはポリクローナル抗体でよい。RACK1タンパク質発現を検出する手段は、RACK1タンパク質に相当するmRNAを決定する手段でもよい。
【0068】
前記のRACK1タンパク質発現を検出する手段は、有利には、例えば蛍光タグ又は放射性タグによって標識される。標識は、RACK1を検出するために、及び/又は該手段を定量化するために使用してよい。前記のメラニン形成細胞を検出する手段は、例えば、小眼症転写因子MITFに対する抗体、ドパクロムトートメラーゼDCTに対する抗体、チロシナーゼTYRに対する抗体、チロシナーゼ-関連タンパク質1 TRP 1に対する抗体、又はメラニン合成経路に関連する任意のタンパク質に対する抗体である。
【0069】
有利には、RACK1タンパク質を検出する手段及びメラニン形成細胞を検出する手段はいずれも、抗体であり、該方法は、2つの前記手段(抗体)のための異なった標識(抗体)を有するインサイチュ免疫アッセイによって行われる。
【0070】
本発明の異なった方法によれば、診断される哺乳動物は、好ましくはヒトである。本発明によれば、診断される好ましい哺乳動物は、癌の家族暦を有するヒト、あるいはメラノーマを発症すると疑われるヒト又はかかる疾患を発症する高い危険性が存在すると疑われるヒト、特に、水膨れ又は日焼けによる皮はがれを有する個体(特に、人生の最初の20年で)、色白かつ赤毛の人々、複数の非定型母斑又は形成異常母斑を有するヒト、大きな先天性メラニン形成細胞母斑を有するヒト、及びメラノーマもしくはディスプラスチック・ニーバス症候群(複数の非定型穴)の個人暦又は家族暦を有するヒトである。
【0071】
本発明の別の態様では、診断される哺乳動物は、ブタ、ネコ、イヌ又はウマ、あるいは別のペットである。本発明の診断方法は、診断された哺乳動物で、特に、メラノーマを発症する高い危険性を示す哺乳動物について、より好ましくはメラノーマの素因を示すヒトについて、規則的に繰り返されることが好ましい。本発明の診断法は、少なくとも3ケ月毎に、少なくとも6ケ月毎に、年に1回又は幾分頻繁に繰り返される。転移の前に初期の段階で治療を開始するためには、できるだけ早く、メラノーマを診断することが実際に必須である。
【0072】
本発明の方法は、好ましくは、メラノーマを診断する他の方法と組み合わせて行われる。例えば、本発明の方法は、異なった方法によってなされた最初の診断を確認するために、あるいはメラノーマの疑いを確認するために行われる。あるいは、本発明の方法は、腫瘍性細胞の同定を可能する他の方法と同時に又はその前に行ってよい。
【0073】
本発明の方法は、他の診断方法、例えば解剖病理学的方法と組み合わせて有利に使用される。
【0074】
解剖病理学的方法では、細胞の腫瘍状態は、有糸分裂、空間組織化、及びストローマ及び血管との関連である、3つの異なった基準の決定後に同定され得る。本発明の方法は、これらの基準の少なくとも1つの決定と組み合わせて有利に行われる。
【0075】
本発明はまた、RACK1タンパク質発現を検出するための手段を含む、哺乳動物におけるメラノーマを診断するキットに関する。上記のように、RACK1タンパク質発現を検出するための手段は、好ましくは、RACK1タンパク質を検出するための手段である。前記方法は、例えば抗体、好ましくはモノクローナル抗体であり;それはポリクローナル抗体でもよい。
【0076】
あるいは、前記手段は、RACK1タンパク質に相当するmRNAのレベルを検出するための手段でよい。
【0077】
本発明のキットにおいて使用される手段は、有利には、例えば蛍光又は放射性標識で標識される。
【0078】
上記のキットは、好ましくは、メラニン形成細胞を検出するための手段を更に含む。メラニン形成細胞を検出するための手段は、メラニン形成細胞と非-メラニン形成細胞との識別を可能にする。
【0079】
かかる手段は、例えば、小眼症転写因子MITFに対する抗体、ドパクロムトートメラーゼDCTに対する抗体、チロシナーゼTYRに対する抗体、チロシナーゼ-関連タンパク質1 TRP 1に対する抗体、又はメラニン合成経路に関連する任意のタンパク質に対する抗体である。別の手段も考えられる。
【0080】
好ましくは、前記の更なる手段はまた、有利には、例えば蛍光又は放射性標識によって標識される。前記標識は、有利には、RACK1タンパク質発現を検出する手段に貼り付けられたラベルとは異なる;例えば、異なった波長発光を有する蛍光標識。
【0081】
本発明はまた、哺乳動物、好ましくはヒト又はペットのメラニン形成細胞の、腫瘍状態のマーカー、好ましくは悪性腫瘍状態のマーカーとしてのRACK1タンパク質の使用に関する。
【0082】
本発明の別の目的は、該細胞の腫瘍性状態の指標、好ましくは悪性腫瘍状態の指標としての哺乳動物のメラニン形成細胞中のRACK1タンパク質の発現レベルの使用である。
【0083】
本発明の方法及びキットに関連して上記の特徴及び好ましい態様のすべてもまた、本発明の使用に適用する。
【実施例】
【0084】
メラニン芽種保有リベコフミニブタ(Melanoblastoma-bearing Libechov minipigs)(MeLiM)は、自発的な悪性皮膚メラノーマを提示する。メラニン形成細胞悪性変換をよりよく理解するために、発明者らは、遺伝子発現の連続的解析を、正常皮膚メラニン形成細胞とMeLiMの肺転移由来のメラノーマ細胞とで比較した。メラノーマ進行において、タグ特定により解明された遺伝子はこれまでに報告されていない。この中でもGNB2L1はメラノーマ細胞で過剰発現した。GNB2L1は、活性化Cキナーゼ(PKC)の受容体であるRACK1をコードする。発明者らは、MeLiMのメラノーマに侵された皮膚及び器官においけるRACK1発現を、メラニン形成細胞のマーカーとしてMITFを用いる共焦点顕微鏡法を用いて研究した。RACK1は、正常な表皮メラニン形成細胞では検出されなかった。それとは対照的に、細胞質RACK1は、皮膚腫瘍及び転移の全てのメラノーマ細胞において高度に発現した。PKCβIIはRACK1と共存しなかったが、これはメラノーマ悪性疾患におけるRACK1の役割が、PKCβIIシグナル伝達とは独立していることを示す。RACK1は転移におけるさらなる核である。RHEB及びβ-カテニンも転移の核であるが、RACK1と共存することはない。RACK1の高発現は、ヒト臨床試料の皮膚腫瘍及び転移の全てのメラノーマ細胞において見られたが、RACK1は、健常皮膚のメラニン形成細胞においては検出されなかった。メラノーマ細胞におけるRACK1の発現レベルは、その腫瘍段階に関連するようである。
【0085】
メラノーマはメラニン形成細胞の変換により発症する悪性腫瘍である。白人集団におけるその発生率及び致死率は、全世界的に増加している。転移の存在は、治療に対する非有効な応答のため予後が極めて不良になる。メラノーマ進行の分子的根拠の理解により、より有効な治療の開発を援助することができた。このように、メラニン形成細胞変換のメカニズムは幅広く研究されている。
【0086】
ヒトにおいて、メラノーマは、B-RAFでは66%変異しており、一方INK4a/ARFでは50%欠失している(Chin (1)により概説されている)。この2つの障害の組み合わせは、Ras経路の活性化、網膜芽腫タンパク質のリン酸化、及びTP53分解の増加をもたらす。ただし、細胞変換のその他のメカニズムの存在が示唆されているため、これらの遺伝子が全てメラノーマの主な原因ではない。ヒト及びマウスのメラニン形成細胞腫瘍細胞系列及び試料について、大規模な遺伝子発現プロファイリングが報告されている。RhoC(2)、WNT5a(3)、Notch2(4)、及びGタンパク質及びカルシウム伝達に関連する遺伝子(5)の差次的発現、とりわけ、際立った経路が、細胞接着及び運動性、並びに特定の細胞系列における組織浸潤に携わる。全体的に、メラノーマは、複雑な多重遺伝子疾患であり、そして変換の異なるメカニズムが作用しているはずである。
【0087】
ヒト病理の動物モデルにおいて、皮膚メラノーマに侵されるMeLiM(Melanoblastoma-bearing Libechov Minipig)種は特に興味深い。実際、2月齢のMeLiMは、リンパ節における転移を有する自発的な皮膚メラノーマを提示するが、ヒトに類似する組織学的進化をする内部器官においてはそれほど頻繁ではない(6)。ただし、ヒトにおける大部分の散発性メラノーマを理解するための、これらの遺伝的ブタメラノーマの生物学的解明は、未だ不確かである。MeLiMもであるがヒトにおける疾患の進行に価値有る情報を提供できるか否かを評価するため、発明者らは、MeLiMにおける腫瘍進行に関与する遺伝子を特定し、ヒト皮膚メラノーマにおけるそれらの発現を評価することにした。
【0088】
連続的遺伝子発現解析(serial analysis of gene expression(SAGE))法を選択したのは、マイクロアレイと異なり、本方法は解析する配列に関係なく、完全な細胞発現プロファイルを提供するからである。SAGEライブラリーは、特定細胞型において特異的に発現される遺伝子を解明するための、インシリコ(in silico)での比較が可能である(7)。メラニン形成細胞以外の細胞の寄与を最小化するため、発明者らは、MeLiMの肺メラノーマ転移及び健常なメイシャンミニブタ(Meishan minipig)の皮膚由来のPigMelメラニン形成細胞から培養した一次メラノーマ細胞からSAGEライブリーを構築した(8)。ここでは、悪性と正常メラニン形成細胞との遺伝子発現の比較解析が報告されている。本発明者らは、活性化Cキナーゼ(PKC)の受容体であるRACK1は、MeLiM由来の皮膚腫瘍及び転移の細胞質において発現するが、その発現は正常な皮膚メラニン形成細胞で検出されないことを示す。重要なことには、この腫瘍及び正常のメラニン形成細胞におけるRACK1の差異的発現は、ヒト患者由来の臨床検体でも見られた。これらの結果は、哺乳類におけるメラノーマの進行におけるRACK1の役割を実証するものである。これらは、MeLiM種が、ヒトにおけるメラノーマ進行に関与する複雑なメカニズムを研究するための関連モデルとなることを支持する。
【0089】
材料と方法
ブタ(Sus scrofa domestica)組織
罹患したMeLiMの雄を、National Institute for Agricultural Research(INRA、Jouy-en-Josas, France)で健常なデュロック又はMeLiM種と交配させた(6,9)。動物の処置及び使用は、欧州連合標準に従って、INRA倫理委員会により承認された。フランス(n=13)又はチェコ共和国(n=3)のいずれかで飼育された3月齢以下のMeLiM由来のバイオプシーを使用した。これらには、表在拡散メラノーマ(n=2)及び結節性メラノーマ(n=7)、及びリンパ節(n=13)、肺(n=10)、肝臓(n=1)、心臓(n=2)及び脾臓(n=3)における転移、及び健常皮膚(n=10)が含まれる。健常な有色性メイシャンの背側頸皮膚を、コントロールとして使用した。回収した組織を、4%緩衝化パラホルムアルデヒド(PFA)中で固定化し、パラフィンに包埋した。
【0090】
転移性メラノーマ細胞の単離及びコントロールメラニン形成細胞の培養
若いMeLiM由来の腫瘍肺バイオプシーを、メラノーマ細胞の単離に使用した。ブタメラニン形成細胞細胞の第一代培養物の条件は(8)の記載の通りである。酢酸テトラデカノイル−ホルボールを、培養の第二日目に添加した。48時間後、細胞をリンスし、Dynabead mRNA直接キット結合バッファー(Dynal, Invitrogen Life Technologies, Cergy Pontoise, France)中で溶解させ、そして液体窒素中で凍結させた。コントロールメラニン形成細胞は、非変換PiMeL細胞を37%とした(8)。
【0091】
SAGEライブラリーの構築
メラニンを廃棄するため細胞溶解物を遠心分離し、小型化抽出へのSAGE適用(SAGE)法を用いてライブラリーを作製した(10)。配列決定反応をMWG(Martinsried, Germany)により行った。
【0092】
タグ特定及びプローブのクローニング
SAGEタグを、SAGE 2002 第4.5版ソフトウェアを用いて、配列ファイルから抽出した(7)。統計的有意性を、モンテカルロシミュレーション解析を用いて決定した。P値が0.05以下の場合に有意であるとみなした。タグを、SAGEソフトウェアにより解析した哺乳類Genebankデータベース(http://www.ncbi.nih.gov/Genbank/gbm)、又はAGENAE ESTデータベース(http://www.ncbi.nih.gov/Genbank/gbm)を用いて特定した。ATG開始コドンからのヌクレオチド配列−70〜900bpに対応するブタRACK1部分cDNAを、pCR4TOPOプラスミド(Invitrogen Life Technologies)にサブクローニングした。得られたプラスミドをNotI又はPmeIのいずれかで直線化し、それぞれセンス又はアンチセンスRNAプローブを得た。インビトロにおける転写は(11)に記載の通りに行った。
【0093】
重度有色化試料のインサイチュハイブリダイゼーション
インサイチュハイブリダイゼーションは(12)の記載に切片を漂白する修正を加えて行った。簡単に説明すると、脱パラフィン化した5μmの切片を、0.075%のKMnO4で15分処理し、処理の間及び後に短いリンスを行いながら、5%のシュウ酸中で1分間脱色素化した。切片を4%PFA中で20分固定化し、リンスし、脱水和し、そして空気乾燥した。約15×106 cpm/mlでセンス又はアンチセンス放射標識リボプローブを(11)に記載の通りにハイブリダイズした。スライドを、Biomax MRフィルム(Kodak, France)に3日間感光させ、その後、Kodak NTB2エマルション中に浸漬し4週間感光させた。
【0094】
ヒト組織
クルー機関(Curie Institute)(Paris, France)において、ヒトメラノーマ組織を、リンパ節切除(n=13)、肝切除(n=5)又は表皮切除(n=19)を行った患者から得た。皮膚メラノーマ検体は、2つの表在拡散メラノーマ第IV段階(ブレスロー深さが2.35及び2.5 cm)と3つの再発性皮膚メラノーマからなる。25人の女性及び12人の男性からなる計37人由来の試料を調べた。胸部形成手術から得た正常皮膚を、コントロールとして使用した(n=4)。
組織を、4%PFA(n=38)又はブアン固定液(n=2)のいずれかにおいて固定し、パラフィンに包埋させた。
【0095】
抗体(免疫標識用の希釈物)
使用したマウスモノクローナル抗体は、抗−MITF(1:50)(クローン C5+D5、Zymed, Clinisciences, Montrouge, France)、抗-RACK1(1:150)及び抗-β-カテニン(1:100)(Transduction Laboratories, BD Biosciences, Le Pont de Claix, France)であり、及びポリクローナルPEP-8抗-DCT(1:1000)、抗-RHEB(1:100)及び抗-PKCβII(1:200)(Santa Cruz Biotechnology, Tebu-bio, Le Perray, France)であった。交差反応及び当該抗体のブタ組織に対する特異性を、ウェスタンブロットを用いて確認した。
ウマ試料のアッセイ用に、MITF抗体(C5+D5、Zymed)を1:100で希釈し、分子グレードのBSA0.1 mg/mlを、ステップ毎に正常なヤギ血清に添加した。
【0096】
免疫染色及び共焦点顕微鏡法
ブタ切片はクエン酸バッファー(pH6)中で、ヒト切片はTris-EDTA(pH9)、0.05% Tween20中で、マイクロ波照射脱パラフィン化切片により、抗原の回収を行った。免疫組織化学については、一次抗体をアビジン−ビオチン複合体(ABC Elite, Vector, Biovalley, France)と反応させた。二重の免疫蛍光法については、4℃で一晩アプライした抗体を、抗マウスアイソタイプ又は抗-ウサギ抗体に曝し、一方はAlexa Fluor 555で標識し、他方はビオチンと結合させて、Alexa Fluor 488標識化ストレプトアビジン(Molecular Probes, Invitrogen Life Technologies, France)に曝した。核対比染色は、Topro 3(Molecular probes)を用いて行った。切片を、LicaレーザーTCS SP2スキャニング共焦点顕微鏡で、0.7 μm厚の光学的切片を作製して観察した。一次抗体不存在のコントロールは非特異的標識を示さない。共焦点画像は、コンピュータプログラム Leica Lite又はZeiss LSM Image Browserで処理した。示された全画像は、個々の切片のz系、加えて指示がある場合は、積層の直行画像(orthogonal projections)である。最終的な図は、Adobe Photoshop (Adobe Systems, USA)で編集した。
【0097】
結果
MeLiM由来の転移メラノーマの特徴付け
リンパ節、肝臓、心臓及び肺においてメラノーマ転移を発症した若いMeLiMを、図1Aに示した。組織切片において、メラノーマ細胞の特定を、メラニン形成細胞において特異的な核シグナルを発生する小眼球症転写因子MITFに対する抗体を用いて行った。免疫組織化学により、MITFで標識したコントロール皮膚表皮の基底層において、正常なメラニン形成細胞が示される(図1B1)。皮膚メラノーマ及び転移における腫瘍細胞も、MITFで標識された(図1C1〜D1)。非特異標識は、本抗体では検出されなかった(図1B2〜D2)。すなわち、MITFは、当該ブタにおけるメラノーマ進行の研究に有用な、メラニン形成細胞系列の感受性マーカーである。
【0098】
転移性メラノーマ細胞とメラニン形成細胞の比較発現解析
肺転移由来のメラノーマ細胞を単離するため、腫瘍の一次培養を、ブタメラニン形成細胞増殖のために最適化した条件で行った(8)。48時間後、培養における接着細胞は、主にメラニン形成細胞であった。2500万個の転移性メラノーマ細胞(MMC)及びPigMel正常メラニン形成細胞(NM)からSAGEライブラリーを構築した。NM及びMMCライブリーから、それぞれ5,466個及び6,131個の異なるタグ(転写物)に対応する、総数11,300個及び11,700個のタグを配列決定した。70個のタグは、2つのライブラリーにおいて差異的に表された(P<0.05)。45個(77%)は発現した配列タグに対応し、その37個(53%)は、既知のcDNAに対応した。タグの大半は、高レベルの遺伝子発現に対応した。特定された1遺伝子は、RNAプロセシング及びタンパク質合成(20%)、転写(7%)、又はシグナル伝達(4%)に関与する。遺伝子が示すその機能的クラスにより任意に配列させたタグのリストは、NMと比較して、MMCを増加及び減少させ、それをそれぞれ表1及び2に示す。GNB2L1のmRNAは、MMC及びNMのカウントが、それぞれ31及び13である豊富なタグに対応した。
タグ配列:ATTGTAGATG(配列番号1)
【0099】
本発明者らは、メラニン形成細胞及びメラノーマにおけるGNB2L1発現をより詳細に研究することを選択したのは、それが、活性化Cキナーゼの受容体であるRACK1(NM-214332、ヒトRACK1 P63244 配列番号69と同一である)をコードし、そのmRNAがヒト癌性細胞においてアップレギュレーションされることがわかっている(13)ためである。
【0100】
【表1】
【0101】
【表2】
【0102】
【表3】
【0103】
【表4】
【0104】
【表5】
【0105】
メラノーマにおけるRACK1 mRNAの過剰発現
RACK1 mRAの分布パターンを特定するため、本発明者らは、正常な皮膚メラノーマ、及び肺、肝臓及びリンパ節由来の転移性メラノーマ試料のブタ切片において、インサイチュハイブリダイゼーションを行った。重度有色化腫瘍領域のバックグラウンドを回避するため、メラニンを酸化し同時にmRNAを維持する漂白処理を追加した。アンチセンス及びセンスプローブで得られるフィルムオートラジオグラフィーから、ヒトリンパ節で報告される(13)ような、シグナルの強いリンパ節以外にも、健常組織において、RACK1 mRNAのわずかなシグナルが示された(図2A)。これとは対照的に、皮膚メラノーマ、肺及び肝転移試料の腫瘍部位では強いシグナルが観察され、非腫瘍部位は、より低いシグナルが示された(図2A)。センスプローブのオートラジオグラフィーシグナルは、ほとんどなかった(図2B)。エマルションオートグラフィー上の暗視野照明は、肺メラノーマの腫瘍領域上の銀粒子を強調する(図2C)。これらの結果により、SAGEデータにより予測されるようなメラノーマにおけるRACK1 mRNAの過剰発現が確認された。
【0106】
皮膚由来のメラニン形成細胞における、及びブタにおけるメラノーマ進行の際の、RACK1タンパク質局在化
RACK1 mRNAの過剰発現が機能的有意性を有するか否かを調べるため、本発明者らは、メラニン形成細胞のマーカーとしてMITFを用いる二重免疫染色を用い、共焦点顕微鏡によりRACK1細胞分布を解析した。本発明者らは、最初に、コントロールメイシャン及び健常なMeLiMの皮膚、すなわち、正常なブタ表皮におけるRACL1の発現を、RACK1タンパク質(蛍光で緑にラベルした)の共焦点顕微鏡解析、及びブタ皮膚におけるMITF又はDCTの二重標識により解析した。本発明者らは、ケラチノサイトにおけるRACK1細胞質の斑点状シグナル、及びメラニン形成細胞におけるその不存在を観察した(図3)。
【0107】
RACK1タンパク質は、表皮において発現し、ケラチノサイトにおける細胞質に広く局在化した(図3A〜D)。メイシャン及びMeLiM皮膚のMITF陽性メラニン形成細胞において、RACK1発現は検出されなかった(図3A、C)。これと一致して、メラノソームに限定される酵素である、ドパクロムトートメラーゼ(DCT)を試験する場合、RACK1及びDCTの二重標識は、正常皮膚において明らかに重複しなかった(図3D)。次に、本発明者らは、腫瘍組織、特に異なる進行段階でのMeLiMメラノーマにおけるRACK1の細胞分布を、RACK1タンパク質の共焦点顕微鏡解析(緑)、及びMITFの二重標識(赤)により解析した(図4)。解析した組織は、リンパ節、肺及び心臓における皮膚メラノーマ及びメラノーマ転移である。これは、真皮メラノーマ細胞及び表皮ケラチノサイトで、同程度のRACK1細胞質シグナルであることに注意すべきである。RACK1は、核周囲に局在化する皮膚及び転移性のメラノーマ細胞に豊富である。さらに転移において、核RACK1が存在する。
【0108】
皮膚メラノーマにおいて、有色化MITF陽性細胞の巣は、RACK1タンパク質を表皮ケラチノサイトと同程度に強く発現した(図4A)。RACK1は、細胞質に局在化し、その大半は核周囲領域であった(図4のA3)。リンパ節、肺及び心臓メラノーマ転移において、RACK1タンパク質は、MITF−陽性細胞で豊富であった。RACK1の準細胞性分布は、細胞質性であった。ただし、核斑点に対するさらなる標識が、MMCの15%で観察された(図4B〜D、黄色矢印)。
すなわち、腫瘍細胞におけるRACK1過剰発現は、MMC(転移性メラノーマ細胞)におけるさらなる核局在化を伴い、異なる進行段階のメラノーマ細胞において細胞質性として観察される。
【0109】
ブタメラノーマ細胞におけるRACK1の推定パートナー
RACK1作用の分子的メカニズムを探索するために、発明者らは、メラノーマ細胞中のRACK1タンパク質のパートナーを探した。RACK1は、PKCβIIの細胞内受容体として同定された(14)。発明者らは、メラノーマ進行中のPKCβII発現を研究した。PKCβII免疫蛍光は、NMにおいて弱い細胞質シグナルを示した(正常メラニン形成細胞はなかった)。重複蛍光はRACK1で検出できないが、MMCは更なる別個の核ドットを示した。比較のために、RACK1シグナルと重複する強いPKCβIIシグナルを組織リンパ球で検出した。このことは、メラニン形成細胞が豊富なPKCβIIを発現しないことを示唆している。
【0110】
β-カテニン変異は、メラノーマにおいて有る役割を果たすと考えられ(15)、β-カテインは細胞接着において重要である。そのため、発明者らは、共焦点顕微鏡、及びRACK1(赤色)とβ-カテニン(緑色)の二重標識後の正常皮膚及び肺でのメラノーマ転移の各々の送信画像によって、正常メラニン形成細胞及びMeLiM由来のメラノーマ細胞中のβ-カテニンを試験した。β-カテニンは、偏在的に発現され、正常ブタ表皮の細胞膜に局在化される(図5A)。
【0111】
MMCでは、膜及び細胞質β-カテニンシグナルが核標識と共に通常視覚化された。しかし、β-カテニン及びRACK1シグナルは、核内には局在化しなかった(図5B)。
【0112】
現在のSAGE試験においてMMC中に発現された別のタグは、小GTPaseのRasスーパーファミリーのマンバーである、脳内に豊富にあるRasホモログをコードするRHEBとして同定された。RHEBは、成長因子、栄養素及びPI3Kを連結するアミノ酸及びラパマイシン(TOR)シグナルの標的に応答して、細胞成長の制御において重要な役割を果たす(16)。従って、発明者らは、MeLiM腫瘍におけるRHEB局在化を試験した。RHEBシグナルは、NMにおいて核周囲であるが、MMC上では拡散し、核のように見えた(図5C〜D)。メラニン形成細胞及び腫瘍細胞においてPACK1シグナルについて何の局在化は見られなかった。
【0113】
これら結果は、β-カテニン、RHEB及びPKCβIIが、メラノーマ進行中のPACK1の主な機能性パートナーではないことを示唆している。
【0114】
ヒト皮膚及びメラノーマ試料中のPACK1タンパク質局在化
最後に、発明者らは、ヒトメラニン形成細胞及びメラノーマ細胞のマーカーとしてMITFを用いて、PACK1の過剰発現がヒトメラノーマ試料においても起こるか否かについて試験した(17)。正常皮膚では、PACK1は、ケラチノサイトに対してメラニン形成細胞において、MITF二重標識(図6C)又はTopro 3での核対比染色によって(図6E)、PACK1タンパク質の共焦点顕微鏡によってわずかに検出された(図6A)。皮膚メラノーマ及びリンパ節及び肝臓でのメラノーマの転移において、MITF-陽性細胞は、細胞形状の基礎となるPACK1の強力な顆粒細胞質パターンを示した。RACK1過剰発現は、試験した20試料の各々で一貫して観察された(図6B、D及びF)。これらのメラノーマ試料中には核PACK1標識は見られなかった。
【0115】
ヒトメラノーマ及び母斑におけるPACK1発現の比較
ヒトメラノーマにおけるPACK1の発現を更に調べるために、発明者らは、一連の試料中のその存在を確認した。メラニン形成マーカーとしてMITFを用いて、4つの正常皮膚バイオプシー由来のホルマリン固定組織、14つの母斑及び5つの皮膚マラノーマを分析した(17)。結果を表3に纏める。
【0116】
【表6】
【0117】
正常皮膚では、PACK1はメラニン形成細胞中に無視できる量で存在したが、隣接するケラチノサイトは、強力な細胞質性シグナルを示した(図7A)。母斑では、PACK1シグナルは、所定の母斑内で及び2つのパターンを示す母斑間で異質であった:14つの母斑の内7つにおいて、PACK1シグナルは、検出されず(矢印の頭、図7B)、また僅かでも検出されなかた(矢印、図7B)。表皮又は真皮メラノーマにおいて検出される時に、シグナルは必ず膜性であった。
【0118】
纏めると、これらの結果は、RACK1 mRNA及びタンパク質が、ブタメラノーマにおいて見出されたように、ヒトメラノーマ中でアップレギュレートされることを示している。このことは、メラノーマ腫瘍状態とPACK1の高レベルとの関係を示している。
【0119】
脈絡膜メラノーマ中のPACK1過剰発現
脈絡膜メラノーマ(又はブドウ膜メラノーマ)は、ヒトの悪性眼腫瘍の最も一般的な形態である。染色体3モノソミー、腫瘍中の珍しい染色体異常が、ブドウ膜メラノーマの約半分で検出された。染色体3モノソミーは腫瘍悪性度と関連する(38)。
【0120】
発明者らは、染色体3モノソミー(N=6)を有する又は無しのブドウ膜メラノーマ上の除核生成物から作製された組織スライドを分析した。該分析は、上記と本質的に同一のプロトコルによって実行した。RACK1は、染色体3モノソミーを有する(図8で説明される)及び有さない、ブドウ膜メラノーマの2つの種類の全ての腫瘍性メラニン形成細胞で高レベルで発現される。
【0121】
ウマメラノーマ中のPACK1発現
発明者らは、PACK1が、ウマの健常皮膚の正常メラニン形成細胞において検出可能なレベルで発現されないことを観察した。
【0122】
発明者らはまた、悪性腫瘍由来の6試料中のPACK1発現を試験した。ウマ皮膚悪性メラノーマにおけるPACK1発現を示す図9に見られるように、それらは、PACK1発現にすべて陽性であった。このことはブタ及びヒトで既に得られた結果を確認するものである。
【0123】
これらのデータは、ヒト試料の分析について述べたのと本質的に同一のプロトコルを用いて得た。
【0124】
皮膚メラノーマ中のPACK1過剰発現の一般的性質
すべてのメラノーマが一次クローン事象の同一のセットを示さず、そのいくつかはより頻繁に起こることは、良く知られている。一次クローン事象に関する知見に関する最近のレビューでは、Bennett(39)は、ヒト皮膚メラノーマにおける最も効力を有する遺伝子を報告している(この文献の表1参照)。これらのデータから推論されるように、すべての皮膚メラノーマ中では影響を与える遺伝子はない。このことは、メラノーマ由来の一次腫瘍の根底にあるメカニズムは外因性であることを示唆している。例えば、CDKN2A遺伝子は、78%の皮膚メラノーマがクローン的に改変されており;B-RAF遺伝子は47%の皮膚メラノーマは改変されている。
【0125】
これに対して、本発明者らは、彼らが試験した皮膚及びブドウ膜メラノーマ由来のすべての試料におけるPACK1の過剰発現を検出した。このことは、メラニン形成細胞中の一次腫瘍を誘発するメカニズムに関係なく、PACK1がメラノーマの普遍的なマーカーであり得ることを示唆している。遺伝子的に外因性の腫瘍中でのPACK1のこの系統的な過剰発現は、任意のメラノーマの腫瘍進行の更なるステップとしてのPACK1の正の調節によって説明され得る。
【0126】
従って、ヒト及び他の哺乳動物のためのメラノーマ診断において非常に有用である、PACK1は皮膚及びブドウ膜メラノーマの任意のメラニン形成細胞中で過剰発現される単一のマーカーであると考えられる。
【0127】
考察
皮膚悪性メラノーマについて数少ない実験動物モデルが存在する。ネズミモデルは、それぞれ癌原遺伝子及び腫瘍抑制遺伝子において、機能亢進及び機能性突然変異の喪失との組合せを必要とする(1)。高い浸透率で皮膚メラノーマを誘導するために、新生マウスでは重度の日焼けが必要とされることがある(18)。対照的に、MeLiMブタモデルは、ヒトメラノーマに類似する病理組織学的特徴を有する自発的な皮膚メラノーマを示す(6)。ここで、発明者らは、MeLiMの遺伝性疾患が、ブタ及びヒトにおけるメラノーマの発症に関連する調節複合体を特定するために有用であるという証拠を提供する。実際に、発明者らは、細胞内受容体RACK1がブタメラノーマ及びヒトメラノーマで過剰発現されることを見出した。このことは、MeLiMモデル予測がヒトメラノーマにおいて実証される最初のことである;これは、同様なメカニズムがブタ及びヒトにおけるメラニン形成細胞の悪性形質転換においては働いていることを示している。
【0128】
SAGE分析では、RACK1タグは、正常な皮膚メラニン形成細胞由来のライブラリーよりも転移性メラノーマ細胞の一次培養物から確立されたライブラリーにおいてより多かった。一環して、RACK1 mRNA及びタンパク質は、正常表皮メラニン形成細胞において僅かに検出されたが、皮膚性及び転移性メラノーマの腫瘍性細胞においては高レベルで見出された。このことは、これまで試験したすべてのブタ(n=38)及びヒト(n=20)試料に当てはまる。
【0129】
RACK1は、他のタンパク質に対して安定又は可逆的のいずれかの結合能力を与える7つの内部Trp-Asp 40(WD40)反復を含む(19)。RACK1と相互作用する60つのタンパク質のほとんどは、これまで記載されている(ヒトタンパク質参照データBベース)及びRACK1は様々な細胞コンパートメントにおいてソレラパートナーに結合することが分かった(19)。従って、RACK1は、細胞の適応プロセスにおいて中心的な役割を果たすと考えられている。
【0130】
RACK1は、活性化されたPKCβIIのための標的タンパク質として最初に同定された(14)。一方、PKCβは、ヒトの培養されたメラニン形成細胞において発現される(20)。PKCβ及びRACK1が複合化され、メラノソーム上に固定される時に、メラニン形成の重要な酵素である(21)PKCβホスホリレートチロシナーゼは、正常なメラニン形成細胞では、PKCβがメラニン形成を刺激することによって主に働くことを示唆している。一方、間接的な観察は、メラノーマ発症においてPKCβIIの可能な役割について議論する。第1に、PKCβIIは、肥満細胞においてAKTを直接リン酸化することが分かり(22)、そして、AKTは、細胞生存及び悪性メラノーマの発症を促進することが知られている(23)。第2に、WNT5Aは、ヒトメラノーマ細胞の顕著な侵襲的挙動の強力なマーカーとして同定され、WNT5aは、PKCを活性化するその能力によってこの効果を仲介すると考えられる(3)。これらの観察に基づいて、発明者らは、ブタ及びヒトメラノーマにおけるPKCβII発現を分析した。PKCβIIは、正常な皮膚メラニン形成細胞においてはほとんど検出されず、正常な皮膚メラニン形成細胞と比べて悪性メラニン形成細胞においてアップレギュレートされなかった。更に、PKCβ発現は、メラノーマ細胞株の90%において喪失される(24)。しかし、腫瘍性メラニン形成細胞におけるRACK1の過剰発現の効果がPKCβIIに依拠することはないであろう。しかし、RACK1は他のPKCアイソフォームと相互作用することができた。
【0131】
発明者らは、ブタの転移性メラノーマ細胞の15%の核においてRACK1を検出した。RACK1の核転位は、既に観察されている。すなわち、げっ歯類のアルコールへの急性曝露の後に、RACK1は、PKCβIIから脱共役し、海馬ニューロンの核に転位した(25)。核では、RACK1はBDNFを含む数個の遺伝子の誘導を介在する(26)。BDN増加Fは、エタノール取り込みを負に制御するドーパミンD3受容体の亢進された発現をもたらす(27)。メラノーマ転移が、一次性皮膚悪性メラノーマよりBDNF及びその受容体をより頻繁に発現することが報告されているので(28)、発明者らは、BDNFがMeLiM転移において発現されるか否かを試験した。しかし、BDNF発現はMeLiM転移のメラノーマ細胞では検出されなかった。このことは、核RACK1が、メラニン形成細胞におけるBDNF非依存的細胞反応を介在することを示唆している。
【0132】
ヒト癌におけるインビボでのRACK1のアップレギュレーションを検出した研究はほとんどなく(13,29)、メラニン形成細胞との関係ではまったくない。RACK1 mRNAは、5つの非小細胞肺癌で強力に発現されていることが見出され;大きな血管の内皮は、これらの腫瘍におけるRACK1発現の主な部位として同定された(13)。 RACK1 mRNAはまた、結腸直腸癌の11の症例において高度に発現され、非癌部位よりも癌細胞において強い発現を示した(29)。しかし、腫瘍性メラニン形成細胞におけるRACK1の過剰発現の開示はなかった。
【0133】
ここで、発明者らは、RACK1 mRNA及びその対応するタンパク質が皮膚及び転移性メラノーマにおいて系統的に過剰発現されたことを示す;RACK1レベルは、形質転換されたメラニン形成細胞それ自体において上昇した。
【0134】
RACK1過剰発現が悪性の皮膚メラノーマの進行に役立つかもしれないとの提案は、培養された細胞株において行われたいくつかの実験によって支持されている。実際に、増加及び減少しているRACK1レベルは、細胞形質転換に関連した複数の細胞機能を明らかにした。RACK1の過剰発現は、3T3繊維芽細胞の成長を阻害することがわかった(30,31)。しかし、RACK1タンパク質レベルは培養された細胞において活性に調節されることを示唆するが、RACK1翻訳のモルホリノオリゴヌクレオチドによる遮断もまた、3T3繊維芽細胞の成長を減少させた(31)。興味深いことに、RACK1の過剰発現は、MCF-7乳癌細胞の増殖率を亢進し、このことは、繊維芽細胞及び腫瘍性細胞におけるRACK1の識別効果を示す(32)。従って、RACK1は、PC-12褐色細胞腫細胞において血清枯渇によって(33)、あるいはW7.2 T細胞においていくつかのアポトーシス刺激によって(34)誘導されたアポトーシスを阻害した。より具体的には、RACK1を過剰発現するMeWoヒトメラノーマ細胞は、UV-誘導アポトーシスに対する増加した抵抗性を示すことがわかった(35)。この効果は、PKCによるJNKの活性化を促進するためにアダプタータンパク質としてRACK1によって介在される(35)。加えて、正常な非-運動性MCF7癌細胞におけるRACK1過剰発現は、創傷治癒アッセイにおいてその転移能力を増加し、Transwellアッセイにおいてその運動性を誘導することが報告された。従って、siRNAを用いるRACK1発現の抑制は、前立腺癌DU 145細胞において反対の効果を有し、ranswellアッセイにおいてその転移を阻害した(36)。
【0135】
RACK1アップレギュレーションによる細胞転移の促進は、IGF-1Rとタンパク質ホスファターゼ2a(PP2A)又はβ1インテグリンのいずれかとの関連に因ることが最近提案された。実際に、RACK1を過剰発現する細胞では、RACK1はPP2Aを放出し、β1インテグリンと会合し、よって、インテグリンシグナル化を亢進し、増加した細胞転移能力を与える。
【0136】
RACK1の過剰発現はまた、いくつかのヒト卵巣癌株のインシュリン-及びIGF-1-促進された足場-非依存的成長を亢進することができる(37)。再度、この効果は、インシュリン受容体(IR)IGF-1R及びSTAT3との関連でアダプタータンパク質として働くRACK1によって介在される(37)。
【0137】
興味深いことに、MeWoメラニン形成細胞におけるRACK1の過剰発現は、STAT3のインシュリン-誘導活性化を亢進することがわかった、このことは、該経路の成分がこれらのメラノーマ-誘導細胞(37)に存在することを示唆する。最後に、RACK1用のsiRNAを用いるMeWoメラニン形成細胞におけるRACK1発現の阻害は、皮下注射後にヌードマウスの腫瘍成長を顕著に減少させた(35)。
【0138】
これらの観察の各々は、RACK1過剰発現が、MeLiMブタモデル及びヒト患者における悪性皮膚メラノーマの進行において原因となる事象であるという見解を支持する。
【0139】
結論として、RACK1は、細胞増殖、アポトーシスに対する抵抗性、転移及び拡散能力に関連するいくつかのシグナル経路の集中点として現れ、その調整解除は、メラニン形成細胞結合において腫瘍性形質転換をもたらす可能性がある。
【0140】
参考文献
1. Chin L. The genetics of malignant melanoma: lessons from mouse and man. Nat Rev Cancer 2003; 3: 559-70.
2. Clark EA, Golub TR1 Lander ES, Hynes RO. Genomic analysis of metastasis reveals an essential role for RhoC. Nature 2000; 406: 532-5.
3. Weeraratna AT, Jiang Y, Hostetter G, et al. Wnt5a signaling directly affects cell motility and invasion of metastatic melanoma. Cancer Cell 2002; 1: 279-88.
4. Hoek K, Rimm DL, Williams KR, et al. Expression profiling reveals novel pathways in the transformation of melanocytes to melanomas. Cancer Res 2004; 64: 5270-82.
5. Weeraratna AT, Becker D, Carr KM, et al. Generation and analysis of melanoma SAGE libraries: SAGE advice on the melanoma transcriptome. Oncogene 2004; 23: 2264-74.
6. Vincent-Naulleau S, Le Chalony C, Leplat JJ, et al. Clinical and histopathological characterization of cutaneous melanomas in the melanoblastoma-bearing Libechov minipig model. Pigment Cell Res 2004; 17:24-35.
7. Velculescu VE, Zhang L, Vogelstein B, Kinzler KW. Serial analysis of gene expression. Science 1995; 270: 484-7.
8. JuIe S, Bosse P, Egidy G, Panthier JJ. Establishment and characterization of a normal melanocyte cell line derived from pig skin. Pigment Cell Res 2003; 16: 407-10.
9. Le Chalony C, Renard C, Vincent-Naulleau S, et al. CDKN2A region polymorphism and genetic susceptibility to melanoma in the MeLiM swine model of familial melanoma, lnt J Cancer 2003; 103: 631-5.
10. Virion B, Cheval L, Buhler JM, et al. Serial microanalysis of renal transcriptomes. Proc Natl Acad Sci U S A 1999; 96: 15286-91.
11. Egidy G, Juillerat-Jeanneret L, Jeannin JF, et al. Modulation of human colon tumorstromal interactions by the endothelin system. Am J Pathol 2000; 157: 1863-74.
12. Johren O, Sanvitto GL, Egidy G, Saavedra JM. Angiotensin II AT1A receptor mRNA expression is induced by estrogen-progesterone in dopaminergic neurons of the female rat arcuate nucleus. J Neurosci 1997; 17: 8283-92.
13. Berns H, Humar R, Hengerer B, Kiefer FN, Battegay EJ. RACK1 is up-regulated in angiogenesis and human carcinomas. Faseb J 2000; 14: 2549-58.
14. Ron D, Jiang Z, Yao L, et al. Coordinated movement of RACK1 with activated betallPKC. J Biol Chem 1999; 274: 27039-46.
15. Rubinfeld B, Robbins P, El-Gamil M, et al. Stabilization of beta-catenin by genetic defects in melanoma cell lines. Science 1997; 275: 1790-2.
16. Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell 2006; 124: 471-84.
17. King R, Weilbaecher KN, McGiII G, et al. Microphthalmia transcription factor. A sensitive and specific melanocyte marker for MelanomaDiagnosis. Am J Pathol 1999; 155: 731-8.
19. McCahill A, Warwicker J, Bolger GB, Houslay MD, Yarwood SJ. The RACK1 scaffold protein: a dynamic cog in cell response mechanisms. Mol Pharmacol 2002; 62: 1261-73.
20. Oka M, Ogita K, Ando H, Kikkawa U, lchihashi M. Differential down-regulation of protein kinase C subspecies in normal human melanocytes: possible involvement of the zeta subspecies in growth regulation. J Invest Dermatol 1995; 105: 567-71.
21. Park HY, Wu H, Killoran CE, Gilchrest BA. The receptor for activated C-kinase-1 (RACK1) anchors activated PKC-beta on melanosomes. J Cell Sci 2004; 117: 3659-68.
22. Kawakami Y, Nishimoto H, Kitaura J, et al. Protein kinase C betall regulates Akt phosphorylation on Ser-473 in a cell type- and stimulus-specific fashion. J Biol Chem 2004; 279: 47720-5.
23. Stahl JM, Sharma A, Cheung M, et al. Deregulated Akt3 activity promotes development of malignant melanoma. Cancer Res 2004; 64: 7002-10.
24. Gilhooly EM, Morse-Gaudio M, Bianchi L, et al. Loss of expression of protein kinase C beta is a common phenomenon in human malignant melanoma: a result of transformation or differentiation? Melanoma Res 2001; 11: 355-69.
25. Ron D, Vagts AJ, Dohrman DP, et al. Uncoupling of betallPKC from its targeting protein RACK1 in response to ethanol in cultured cells and mouse brain. Faseb J 2000; 14: 2303-14.
26. McGough NN, He DY, Logrip ML, et al. RACK1 and brain-derived neurotrophic factor: a homeostatic pathway that regulates alcohol addiction. J Neurosci 2004; 24: 10542-52.
27. Jeanblanc J, He DY, McGough NN, et al. The dopamine D3 receptor is part of a homeostatic pathway regulating ethanol consumption. J Neurosci 2006; 26: 1457-64.
28. lnnominato PF, Libbrecht L, van den Oord JJ. Expression of neurotrophins and their receptors in pigment cell lesions of the skin. J Pathol 2001; 194: 95-100.
29. Saito A, Fujii G, Sato Y, et al. Detection of genes expressed in primary colon cancers by in situ hybridisation: overexpression of RACK1. Mol Pathol 2002; 55: 34-9.
30. Chang BY, Conroy KB, Machleder EM, Cartwright CA. RACK1, a receptor for activated C kinase and a homolog of the beta subunit of G proteins, inhibits activity of src tyrosine kinases and growth of NIH 3T3 cells. Mol Cell Biol 1998; 18: 3245-56.
31. Hermanto U, Zong CS, Li W, Wang LH. RACK1, an insulin-like growth factor I (IGF-I) receptor-interacting protein, modulates IGF-1-dependent integrin signaling and promotes cell spreading and contact with extracellular matrix. Mol Cell Biol 2002; 22: 2345- 65.
32. Kiely PA, Sant A, O'Connor R. RACK1 is an insulin-like growth factor 1 (IGF-1) receptor- interacting protein that can regulate IGF-1-mediated Akt activation and protection from cell death. J Biol Chem 2002; 277: 22581-9.
33. Choi DS, Young H, McMahon T, Wang D, Messing RO. The mouse RACK1 gene is regulated by nuclear factor-kappa B and contributes to cell survival. Mol Pharmacol 2003; 64: 1541-8.
34. Mourtada-Maarabouni M, Kirkham L, Farzaneh F, Williams GT. Functional expression cloning reveals a central role for the receptor for activated protein kinase C 1 (RACK1) in T cell apoptosis. J Leukoc Biol 2005; 78: 503-14.
35. Lopez-Bergami P, Habelhah H, Bhoumik A, et al. RACK1 mediates activation of JNK by protein kinase C [corrected]. Mol Cell 2005; 19:3 09-20.
36. Kiely PA, O'Gorman D, Luong K, Ron D, O'Connor R. Insulin-like growth factor I controls a mutually exclusive association of RACK1 with protein phosphatase 2A and betai integrin to promote cell migration. Mol Cell Biol 2006; 26: 4041-51.
37. Zhang W, Zong CS, Hermanto U, et al. RACK1 Recruits STAT3 Specifically to Insulin and Insulin-Like Growth Factor 1 Receptors for Activation, Which Is Important for Regulating Anchorage-Independent Growth. Mol Cell Biol 2006; 26: 413-24.
38. White VA, et al. Correlation of cytogenetic abnormalities with the outcome of patients with uveal melanoma. Cancer 1998; 83(2): 354-9.
39. Bennett D. C. How to make a melanoma:what do we know of the primary clonal events? Pigment Cell Melanoma Res. 2008; 21: 27-38.
【特許請求の範囲】
【請求項1】
哺乳動物のメラノーマ細胞の腫瘍状態を決定する方法であって、
−該メラニン形成細胞におけるRACK1タンパク質の発現又は過剰発現を検出し;及び
-RACK1タンパク質の発現又は過剰発現を該細胞の腫瘍状態と関連づけること、
を含む、方法。
【請求項2】
哺乳動物のメラニン形成細胞を診断する方法であって、
-該細胞の生物試料からメラニン形成細胞中のRACK1タンパク質の発現又は過剰発現を検出し;及び
-RACK1タンパク質の発現又は過剰発現をメラニン形成細胞の存在と関連づけること、
を含む、方法。
【請求項3】
哺乳動物のメラニン形成細胞の腫瘍状態を決定する方法であって、
-該メラニン形成細胞中のRACK1タンパク質の発現レベルを測定し;及び
-そのレベルを参照レベルと比較すること、参照レベルに関して該メラニン形成細胞中のRACK1タンパク質発現の増加したレベルは、該細胞の腫瘍状態を示す、
を含む、方法。
【請求項4】
哺乳動物のメラニン形成細胞を診断する方法であって、
-該哺乳動物の生物試料からメラニン形成細胞中のRACK1タンパク質の発現レベルを測定し;及び
-そのレベルを参照レベルと比較すること、参照レベルに関して該メラニン形成細胞中のRACK1タンパク質発現の増加したレベルは、メラニン形成細胞の存在を示す、
を含む、方法。
【請求項5】
前記参照レベルが、哺乳動物の非-腫瘍メラニン形成細胞中のRACK1タンパク質発現のレベルでる、請求項3又は4記載の方法。
【請求項6】
前記非-腫瘍メラニン形成細胞が、正常表皮性メラニン形成細胞又は正常脈絡膜メラニン形成細胞である、請求項5記載の方法。
【請求項7】
前記RACK1タンパク質発現レベルが、RACK1タンパク質のレベルによって定義される、請求項3又は4記載の方法。
【請求項8】
前記RACK1タンパク質発現レベルが、インサイチュ免疫アッセイによって測定される、請求項7記載の方法。
【請求項9】
前記RACK1タンパク質発現レベルが、RACK1タンパク質に対応するmRNAレベルによって定義される、請求項3又は4記載の方法。
【請求項10】
前記のRACK1タンパク質に対応するmRNAレベルが、インサイチュハイブリダイゼーションによって測定される、請求項9記載の方法。
【請求項11】
前記メラニン形成細胞がバイオプシー由来である、請求項1〜4のいずれか1項記載の方法。
【請求項12】
前記のRACK1タンパク質の発現もしくは過剰発現又はRACK1タンパク質発現のレベルが、免疫学的方法、免疫アッセイ、免疫組織化学アッセイ、共焦点顕微鏡分析、紫外線顕微鏡、電気化学的検出又はハイブリダイゼーションオートラジオグラフィーを実行することによって測定される、請求項1〜4のいずれか1項記載の方法。
【請求項13】
前記免疫学的方法が、特定のモノクローナル又はポリクローナル抗体を使用する方法、ELISA法、免疫酵素的方法、免疫蛍光法、放射-免疫学的方法又は化学-免疫学的方法である、請求項12記載の方法。
【請求項14】
前記メラニン形成細胞が皮膚メラノーマ又は脈絡膜メラノーマである、請求項2又は4記載の方法。
【請求項15】
哺乳動物におけるメラノーマを診断する方法であって、
-該哺乳動物由来の生物的試料を、メラニン形成細胞を検出する手段及びRACK1タンパク質発現を検出する手段と、インビトロで接触させ;及び
-メラニン形成細胞がRACK1タンパク質発現と共局在化するか否かを決定すること、ここで、RACK1タンパク質発現を示すメラニン形成細胞の存在がメラノーマの指標である、
を含む、方法。
【請求項16】
前記生物試料がバイオプシーである、請求項15記載の方法。
【請求項17】
前記バイオプシーが、リンパ節(lymp node)、肝臓、肺、心臓、脾臓、眼又は皮膚のバイオプシーである、請求項16記載の方法。
【請求項18】
前記哺乳動物がヒトである、請求項1〜4及び15のいずれか1項記載の方法。
【請求項19】
前記哺乳動物が、ブタ、ネコ、イヌ又はウマである、請求項1〜4及び15のいずれか1項記載の方法。
【請求項20】
前記のRACK1タンパク質発現を検出するための手段が、RACK1タンパク質を検出するための手段である、請求項15記載の方法。
【請求項21】
前記のRACK1タンパク質発現を検出するための手段が、抗体、好ましくはモノクローナル抗体である、請求項15記載の方法。
【請求項22】
RACK1タンパク質発現を検出するための手段を含む、哺乳動物におけるメラノーマを診断するためのキット。
【請求項23】
前記手段が、RACK1タンパク質を検出するための手段である、請求項22記載のキット。
【請求項24】
前記手段が、抗体、好ましくはモノクローナル抗体である、請求項22記載のキット。
【請求項25】
メラニン形成細胞を検出するための手段を更に含む、請求項22記載のキット。
【請求項26】
前記のメラニン形成細胞を検出するための手段が、小眼症転写因子MITFに対する抗体、ドパクロムトートメラーゼDCTに対する抗体、チロシナーゼTYRに対する抗体、チロシナーゼ-関連タンパク質1 TRP 1に対する抗体、又はメラニン合成経路に関連する任意のタンパク質に対する抗体である、請求項25記載のキット。
【請求項27】
哺乳動物のメラニン形成細胞の腫瘍状態のためのマーカーとしての、RACK1タンパク質の使用。
【請求項28】
その腫瘍状態の指標としての、哺乳動物のメラニン形成細胞におけるRACK1タンパク質の発現レベルの使用。
【請求項1】
哺乳動物のメラノーマ細胞の腫瘍状態を決定する方法であって、
−該メラニン形成細胞におけるRACK1タンパク質の発現又は過剰発現を検出し;及び
-RACK1タンパク質の発現又は過剰発現を該細胞の腫瘍状態と関連づけること、
を含む、方法。
【請求項2】
哺乳動物のメラニン形成細胞を診断する方法であって、
-該細胞の生物試料からメラニン形成細胞中のRACK1タンパク質の発現又は過剰発現を検出し;及び
-RACK1タンパク質の発現又は過剰発現をメラニン形成細胞の存在と関連づけること、
を含む、方法。
【請求項3】
哺乳動物のメラニン形成細胞の腫瘍状態を決定する方法であって、
-該メラニン形成細胞中のRACK1タンパク質の発現レベルを測定し;及び
-そのレベルを参照レベルと比較すること、参照レベルに関して該メラニン形成細胞中のRACK1タンパク質発現の増加したレベルは、該細胞の腫瘍状態を示す、
を含む、方法。
【請求項4】
哺乳動物のメラニン形成細胞を診断する方法であって、
-該哺乳動物の生物試料からメラニン形成細胞中のRACK1タンパク質の発現レベルを測定し;及び
-そのレベルを参照レベルと比較すること、参照レベルに関して該メラニン形成細胞中のRACK1タンパク質発現の増加したレベルは、メラニン形成細胞の存在を示す、
を含む、方法。
【請求項5】
前記参照レベルが、哺乳動物の非-腫瘍メラニン形成細胞中のRACK1タンパク質発現のレベルでる、請求項3又は4記載の方法。
【請求項6】
前記非-腫瘍メラニン形成細胞が、正常表皮性メラニン形成細胞又は正常脈絡膜メラニン形成細胞である、請求項5記載の方法。
【請求項7】
前記RACK1タンパク質発現レベルが、RACK1タンパク質のレベルによって定義される、請求項3又は4記載の方法。
【請求項8】
前記RACK1タンパク質発現レベルが、インサイチュ免疫アッセイによって測定される、請求項7記載の方法。
【請求項9】
前記RACK1タンパク質発現レベルが、RACK1タンパク質に対応するmRNAレベルによって定義される、請求項3又は4記載の方法。
【請求項10】
前記のRACK1タンパク質に対応するmRNAレベルが、インサイチュハイブリダイゼーションによって測定される、請求項9記載の方法。
【請求項11】
前記メラニン形成細胞がバイオプシー由来である、請求項1〜4のいずれか1項記載の方法。
【請求項12】
前記のRACK1タンパク質の発現もしくは過剰発現又はRACK1タンパク質発現のレベルが、免疫学的方法、免疫アッセイ、免疫組織化学アッセイ、共焦点顕微鏡分析、紫外線顕微鏡、電気化学的検出又はハイブリダイゼーションオートラジオグラフィーを実行することによって測定される、請求項1〜4のいずれか1項記載の方法。
【請求項13】
前記免疫学的方法が、特定のモノクローナル又はポリクローナル抗体を使用する方法、ELISA法、免疫酵素的方法、免疫蛍光法、放射-免疫学的方法又は化学-免疫学的方法である、請求項12記載の方法。
【請求項14】
前記メラニン形成細胞が皮膚メラノーマ又は脈絡膜メラノーマである、請求項2又は4記載の方法。
【請求項15】
哺乳動物におけるメラノーマを診断する方法であって、
-該哺乳動物由来の生物的試料を、メラニン形成細胞を検出する手段及びRACK1タンパク質発現を検出する手段と、インビトロで接触させ;及び
-メラニン形成細胞がRACK1タンパク質発現と共局在化するか否かを決定すること、ここで、RACK1タンパク質発現を示すメラニン形成細胞の存在がメラノーマの指標である、
を含む、方法。
【請求項16】
前記生物試料がバイオプシーである、請求項15記載の方法。
【請求項17】
前記バイオプシーが、リンパ節(lymp node)、肝臓、肺、心臓、脾臓、眼又は皮膚のバイオプシーである、請求項16記載の方法。
【請求項18】
前記哺乳動物がヒトである、請求項1〜4及び15のいずれか1項記載の方法。
【請求項19】
前記哺乳動物が、ブタ、ネコ、イヌ又はウマである、請求項1〜4及び15のいずれか1項記載の方法。
【請求項20】
前記のRACK1タンパク質発現を検出するための手段が、RACK1タンパク質を検出するための手段である、請求項15記載の方法。
【請求項21】
前記のRACK1タンパク質発現を検出するための手段が、抗体、好ましくはモノクローナル抗体である、請求項15記載の方法。
【請求項22】
RACK1タンパク質発現を検出するための手段を含む、哺乳動物におけるメラノーマを診断するためのキット。
【請求項23】
前記手段が、RACK1タンパク質を検出するための手段である、請求項22記載のキット。
【請求項24】
前記手段が、抗体、好ましくはモノクローナル抗体である、請求項22記載のキット。
【請求項25】
メラニン形成細胞を検出するための手段を更に含む、請求項22記載のキット。
【請求項26】
前記のメラニン形成細胞を検出するための手段が、小眼症転写因子MITFに対する抗体、ドパクロムトートメラーゼDCTに対する抗体、チロシナーゼTYRに対する抗体、チロシナーゼ-関連タンパク質1 TRP 1に対する抗体、又はメラニン合成経路に関連する任意のタンパク質に対する抗体である、請求項25記載のキット。
【請求項27】
哺乳動物のメラニン形成細胞の腫瘍状態のためのマーカーとしての、RACK1タンパク質の使用。
【請求項28】
その腫瘍状態の指標としての、哺乳動物のメラニン形成細胞におけるRACK1タンパク質の発現レベルの使用。
【図1】

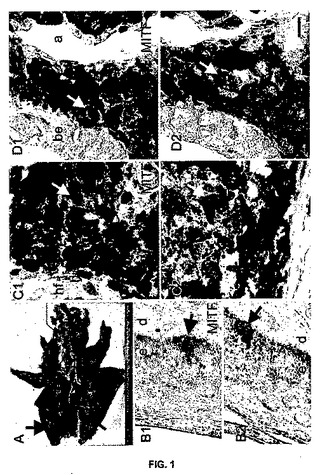
【図2】
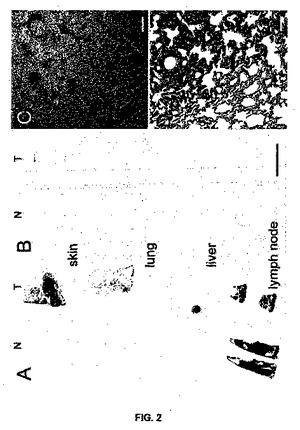
【図3】
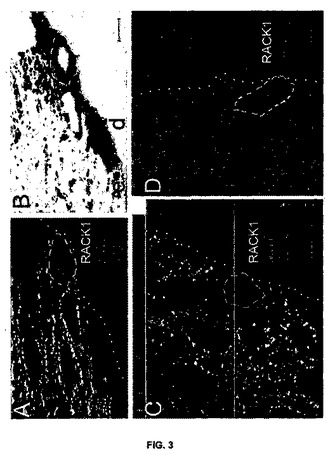
【図4】

【図5】
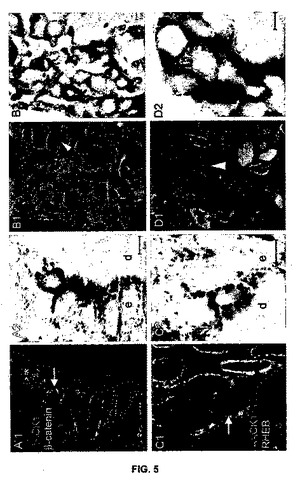
【図6】
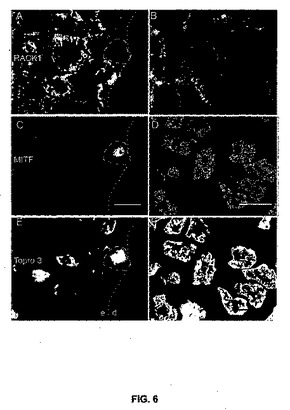
【図7】
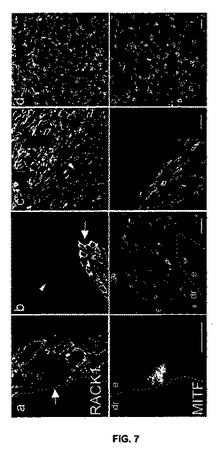
【図8】

【図9】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2010−534840(P2010−534840A)
【公表日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願番号】特願2010−518552(P2010−518552)
【出願日】平成20年7月28日(2008.7.28)
【国際出願番号】PCT/EP2008/006189
【国際公開番号】WO2009/015846
【国際公開日】平成21年2月5日(2009.2.5)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(501089863)サントル ナシオナル ドゥ ラ ルシェルシェサイアンティフィク(セエヌエールエス) (173)
【出願人】(500056471)
【出願人】(510028637)エコール ナシオナル ベテリネール ダルフォール (1)
【出願人】(594184698)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック(イエヌエールア) (3)
【Fターム(参考)】
【公表日】平成22年11月11日(2010.11.11)
【国際特許分類】
【出願日】平成20年7月28日(2008.7.28)
【国際出願番号】PCT/EP2008/006189
【国際公開番号】WO2009/015846
【国際公開日】平成21年2月5日(2009.2.5)
【出願人】(596009674)アンスティテュ・パストゥール (23)
【出願人】(501089863)サントル ナシオナル ドゥ ラ ルシェルシェサイアンティフィク(セエヌエールエス) (173)
【出願人】(500056471)
【出願人】(510028637)エコール ナシオナル ベテリネール ダルフォール (1)
【出願人】(594184698)アンスティテュ ナシオナル ドゥ ラ ルシェルシュ アグロノミック(イエヌエールア) (3)
【Fターム(参考)】
[ Back to top ]