
メリビオースオペロン発現系
本発明は、前記宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオースオペロンのプロモーター領域を含む、宿主において発現可能な新規なベクターに関する。前記核酸配列の発現はメリビオースオペロンの前記プロモーター領域によって制御される。また、原核宿主における核酸配列の制御された異種発現のための前記新規ベクターの使用、ならびに前記宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオース オペロンのプロモーター領域を含む、宿主において発現可能な単離および精製された核酸配列が開示される。前記核酸配列の発現は、メリビオースオペロンの前記プロモーター領域によって制御される。また、前記ベクターまたは前記単離および精製された核酸配列で形質転換された原核宿主、ならびに前記ベクターを使用して宿主においてポリペプチドを産生する方法が開示される。
【発明の詳細な説明】
【発明の分野】
【0001】
本発明は、原核生物の宿主における、例えば組換えタンパク質のようなポリペプチドをコードする核酸の異種発現に適したベクターに関する。より具体的には、本発明は、前記宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオースオペロンのプロモーター領域を含む宿主において発現可能な新しいベクターに関する。前記核酸配列の発現は、メリビオースオペロンの前記プロモーター領域によって制御される。本発明は、さらに例えばポリペプチドをコードする核酸の異種発現に適したこれらのベクターの使用に関する。
【発明の背景】
【0002】
原核生物の系における例えば組換えタンパク質のようなポリペプチドをコードする核酸の異種発現のための多くの系が記載されてきた。しかしながら、原核生物の宿主系におけるほとんどの異種遺伝子発現系は、もっぱら限られた一連の細菌プロモーターに依存してきた。最も幅広く使用されている原核生物のプロモーターには、ラクトース[lac] (Yanisch-Perron et al., 1985, Gene 33, 103-109)、およびトリプトファン[trp] (Goeddel et al., 1980, Nature (London) 287, 411-416) プロモーター、ならびにこれら二つから誘導されたハイブリッドプロモーター [tac および trc] (Brosius, 1984,Gene 27 :161-172 ; Amann and Brosius, 1985, Gene 40,183-190) が含まれる。他の誘導プロモーター系、例えばアラビノースによる誘導のaraBプロモーター(WO 86 04356)、L-ラムノースによる誘導のラムノース プロモーター rhaSB (WO 03068956) またはL-ラムノースによる誘導のラムノース プロモーター rhaBAD (WO 2004/050877)が、タンパク質の異種発現のために十分に記載されてきた。しかしながら、異種遺伝子発現について使用された多くの既知の原核プロモーターが、宿主細胞に対する異種生成物の毒性、生成物の発現の低い割合または非機能的凝集塊(封入体)の形成といった欠点を有する。
【0003】
タンパク質の異種発現において使用されるさらなる誘導性のプロモーター系、例えば、Belyaeva et al., 2000, Mol. Microbiol. 36(1), 211-222 によって記載されたような、メリビオースによって誘導性のメリビオースオペロンが存在する。異種発現について使用されるこれらの誘導性のプロモーター系の大半は、非常に低い誘導率および所望の相同的生成物の低発現に至るという欠点を有する。さらに、これらのプロモーター系は非常に厳密には制御されておらず、従って非誘導性状態における背景活性を誘導し、厳密な発現制御を可能にはしない。例えば、Belyaevaらは、大腸菌のβ-ガラクトシダーゼの発現のために大腸菌のlacZ遺伝子に融合したミリビオースオペロンの異なる断片を使用した。しかしながら、最も高いβ-ガラクトシダーゼ活性を産生した断片(KK43, JK19)は、最も高い背景活性を強く産生した。
【0004】
従って、上述した不利益のない核酸配列の異種発現に適した向上した原核発現系を提供する必要がある。
【発明の概要】
【0005】
前述の記載から明らかなように、これらのおよび他の目的は、所望の異種生成物の高レベルの発現にとって有用である原核プロモーター領域を含む新たなベクターを提供することによって達成される。驚くべきことに、メリビオースオペロンのプロモーター領域が異種生成物の大量の厳密に制御された発現を可能にすることを見出した。第一の側面において、本発明の目的は、前記宿主に対して異種である核酸配列を含む転写単位と機能的に連結したメリビオースオペロンのプロモーター領域を含む宿主において発現可能な新たなベクターを提供することである。前記核酸配列の発現がメリビオースオペロンの前記プロモーター領域によって制御される。また、原核宿主における核酸配列の制御された異種発現に適した前記新規ベクターの使用; 前記宿主に対して異種である核酸配列を含む転写単位と機能的に連結されたメリビオースオペロンのプロモーター領域を含む宿主において発現可能な単離および精製された核酸配列であって、前記核酸配列の発現がメリビオースオペロンの前記プロモーター領域によって制御される核酸配列; 前記ベクターまたは前記単離および精製された核酸配列で形質転換された原核宿主; および前記ベクターを使用する宿主におけるポリペプチドを生成するための方法が提供される。
【0006】
他の対象および利点は、以下の例証的図面を参照して進める以下の詳細な説明および添付の請求項の内容から当業者によって明らかになるであろう。
【発明の詳細な説明】
【0007】
本明細書中で使用されたとき、以下の定義は、本発明の理解を容易にするために提供される。
【0008】
「宿主において発現可能なベクター」または「発現ベクター」は、宿主細胞において特定の核酸配列の転写を可能にする一連の特異的なポリ核酸要素で組換え的または合成的に生成されたポリ核酸構築物である。典型的には、このベクターは、プロモーターに作動可能に連結された、特定の被転写核酸配列を含む転写単位を含む。宿主において発現可能なベクターは、例えば自律的または自己複製的プラスミド、コスミド、ファージ、ウイルスまたはレトロウイルスとすることができる。
【0009】
用語「宿主」、「宿主細胞」および「組換え宿主細胞」は、本明細書中では可換的に使用され、本発明の一以上のベクターまたは単離および精製された核酸配列が導入された原核細胞を指す。これらの用語が特定の対象細胞だけではなく、該細胞の子孫または潜在的子孫をも指すことが理解される。なぜなら、一定の修飾が変異または環境的影響により発生するためである。例えば、事実、子孫は親細胞とは同一ではないが、本明細書中で使用されたときに該用語の範囲内に含まれる。
【0010】
用語「含む(comprise)」は、一般的には含む(include)の意味において使用される。すなわち、一以上の特徴または成分の存在を許容する。
【0011】
「プロモーター」とは、本明細書中に使用されたとき、転写単位の発現を制御する核酸配列をいう。「プロモーター領域」は、細胞内のRNAポリメラーゼを結合することができ、かつ下流(3’方向)のコード配列の転写を開始することができる制御領域である。プロモーター領域内には、転写開始部位(ヌクレアーゼS1でマッピングすることによって保存的に定義される)、ならびに推定上の-35領域およびプリブノウボックスのようなRNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が見出されるであろう。
【0012】
「メリビオースオペロン」とは、Hanatani et al., 1984, J Biol. Chem, 259(3), 1807-12に記載された大腸菌のメリビオースオペロンをいう。メリビオースオペロンは、二つの相違するプロモーターを含む積極的に制御された異化オペロンである。第一のプロモーター(melR プロモーター)は、第二のプロモーター(melAB プロモーター)のメリビオース依存性刺激に不可欠であるmelR 遺伝子の発現を担う。この第二のプロモーターからのメリビオース誘導性の転写は、α-ガラクトシダーゼをコードするmelA遺伝子およびメリビオースパーミアーゼをコードするmelB遺伝子の同時転写を開始する。メリビオースオペロンは、二つの異化制御タンパク質結合部位、位置-81,5のCRP2および位置-195,5のCRP1(melAB プロモーターの転写開始部位の上流)ならびに位置-42,5 (サイト2’)、-62,5 (サイト2)、-100,5 (サイト1)、-120,5 (サイト1’)および-238,5 (サイトR)の5つのMelR 結合部位(melAB プロモーターの転写開始部位の上流)を含む。「メリビオースオペロンのプロモーター領域」は、melAB プロモーター、melR プロモーター、melR遺伝子およびCRP結合部位およびMelR結合部位を含む、melA 遺伝子およびmelB 遺伝子の発現を制御するプロモーター領域を意味する。本明細書で参照された「melAB プロモーター」は、主として転写開始部位、推定上-35領域、プリブノウボックス、CRP結合部位およびMelR結合部位からなる。本明細書中で言及された「CRP1結合部位が欠損したmelAB プロモーター」は、主として転写開始部位、推定上-35領域、プリブノウボックス、CRP結合部位およびサイトRを除いたMelR結合部位からなる。「CRP1結合部位が欠損したmelAB プロモーター」は、抑制されまたは遮断されたCRP1結合部位を含んでいてもよい。これはトランスポゾン支援変異誘発またはノックアウト変異のような既知の技術によって行われ得る。好ましくは、「CRP1結合部位が欠損したmelABプロモーター」は、CRP1結合部位を含まない。
【0013】
メリビオース (6-O-[α]-D-ガラクトピラノシル-D-グルコース)は、酵母での発酵によってラフィノースから得ることができる二糖である。
【0014】
「CRP」は、「異化産物制御タンパク質」を意味する。「CRP」は、しばしば同義的意味をもつ「サイクリックAMP受容タンパク質」として当該技術分野において呼ばれる。CRPは、メリビオースオペロンのような異化オペロンの活性化を媒介するサイクリックAMP (cAMP)によって調節される制御タンパク質である。
【0015】
「エンハンサー」は、転写単位の同定、転写単位に関係した配列の位置、または配列の配向とは無関係に、転写単位の転写を増強するよう作用する核酸配列である。本発明のベクターには任意的にはエンハンサーが含まれる。
【0016】
「転写単位」は、本明細書中において使用されたとき、通常は単一のRNA分子に転写される核酸配列である。転写単位は、機能的に関連したポリペプチド分子をコードする一つの遺伝子(モノシストロニック)または二つの遺伝子(ジシストロニック)またはそれ以上の遺伝子(ポリシストロニック)を含んでもよい。
【0017】
核酸配列は、他の核酸配列と機能的関係に配置されるときに「機能的に連結される」。例えば、シグナル配列についてのDNAは、それがタンパク質の分泌に関与するプレタンパク質として発現される場合、タンパク質についてのDNAと機能的に連結され;プロモーターは、それが配列の転写に影響を与える場合、コード配列に機能的に連結され;またはリボソーム結合部位のような翻訳開始領域は、それがポリペプチドの転写を容易にするように配置される場合、例えばポリペプチドをコードする核酸配列と機能的に連結される。連結は、保存的制限部位でのライゲーションによって達成され得る。このような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーが、従来手法に従って使用される。
【0018】
「核酸」または「核酸配列」または「単離および精製された核酸または核酸配列」は、本明細書中において言及されたとき、DNA、RNA、またはDNA/RNAハイブリッドのいずれであってもよい。核酸または核酸配列がベクター上に配置される場合、それは通常はDNAである。本明細書中で言及されるDNAは、例えば二本鎖DNA、単鎖DNA、一方または両方の鎖が二以上の断片で構成された二本鎖DNA、一方または両方の鎖が非中断性ホスホジエステルバックボーンを有する二本鎖DNA、一以上の単鎖部位および一以上の二本鎖部位を含むDNA、DNA鎖が完全に相補的である二本鎖DNA、DNA鎖が部分的にのみ相補的である二本鎖DNA、環状DNA、共有結合性閉鎖DNA、直鎖状DNA、共有結合性架橋DNA、cDNA、化学合成DNA、半合成DNA、生合成DNA、天然単離DNA、酵素消化DNA、剪断DNA、標識DNA、例えば放射性標識DNAおよび蛍光色素標識DNA、一以上の非天然発生化学種の核酸を含むDNAを含む、任意のポリデオキシヌクレオチド配列とすることができる。DNA配列は、標準的な化学技術、例えば、リン酸トリエステル方法または自動化合成方法およびPCR方法によって合成することができる。精製および単離されたDNA配列はまた、酵素的技術によって生成することができる。
【0019】
本明細書中で言及されるRNAは、例えば単鎖RNA、cRNA、二本鎖RNA、一方または両方の鎖が二以上の断片で構成される二本鎖RNA、一方または両方の鎖が非中断性ホスホジエステルバックボーンを有する二本鎖RNA、一以上の単鎖部位および一以上の二本鎖部位を含むRNA、RNA鎖が完全に相補的である二本鎖RNA、RNA鎖が部分的にのみ相補的である二本鎖RNA、共有結合性架橋RNA、酵素消化RNA、剪断RNA、mRNA、化学合成RNA、半合成RNA、生合成RNA、天然単離RNA、標識RNA、例えば放射性標識RNAおよび蛍光色素標識RNA、一以上の非天然発生化学種の核酸を含むRNAとすることができる。
【0020】
「バリエーション」または「配列のバリエーション」は、保存的核酸の置換による基準配列とは異なる核酸配列を意味する。このとき、一以上の核酸が同じ特徴をもつ他の核酸によって置換される。縮重した配列、欠失および挿入を含む配列が基準配列と同じ機能(機能的に等価)を示す限り、これらの修飾された配列は変異に含まれる。
【0021】
本明細書中で使用されるとき、用語「ポリペプチド」、「ペプチド」、「タンパク質」、「ポリペプチド」および「ペプチド」は可換的に使用され、α-アミノ基と隣接残基のカルボキシ基との間のペプチド結合によって他のアミノ酸残基と連結された一連のアミノ酸残基を指す。
【0022】
用語「単離および精製された核酸配列」とは、核酸配列が本発明に従った状態にあることをいう。該核酸配列は、それらが天然に関与する材料、例えば他の核酸から遊離または実質的に遊離している。該核酸は、自然環境またはそれらが調製される環境(例えば細胞培養)において見出される。調製は、組換え技術によってインビトロまたはインビボにおいて行われる。
【0023】
用語「形質転換」、「形質転換された」または「核酸を宿主細胞に導入すること」は、ベクターのような細胞外核酸が材料を伴ってまたは伴わないで宿主細胞に入る任意のプロセスを意味する。用語「形質転換された細胞(cell transformed)」または「形質転換された細胞 (transformed cell)」は、細胞外核酸が導入され、かつ細胞外核酸を内包する細胞またはその子孫を意味する。核酸は、該核酸が染色体複合体または染色体外要素として複製可能であるように細胞に導入され得る。例えば発現ベクターでの適切な宿主細胞の形質転換は、周知の方法、例えばマイクロインジェクション、エレクトロポーレーション、粒子照射によって、あるいは化学的方法、例えばリン酸カルシウム媒介性形質転換によって達成され得る(例えば、Maniatis et al. 1982, Molecular Cloning, A laboratory Manual, Cold Spring Harbor Laboratory or in Ausubel et al. 1994, Current protocols in molecular biology, John Wiley and Sons.に記載)。
【0024】
「異種核酸配列」または「宿主に対して異種の核酸配列」は、例えば、宿主に対して外来であるポリペプチドのような発現産物(「異種発現」または「異種産物」)、すなわち、宿主とは異なる供与体から発生する核酸配列、あるいは宿主に対して外来であるポリペプチドのような発現産物をコードする化学的に合成された核酸配列をコードする核酸配列を意味する。宿主が特定の原核生物種である場合、異種核酸配列は、好ましくは生物の異なる属またはファミリー、より好ましくは異なる目または綱、特に異なる門および異なるドメイン(領域)から発生する。
【0025】
宿主とは異なる供与体から発生する異種核酸配列は、それが宿主細胞に導入される前、このような修飾された配列が基準配列と同じ機能(機能的に等価)を示す限りにおいて、単鎖核酸または異種核酸配列の一部の変異、挿入、削除または置換によって修飾され得る。本明細書中で参照された異種核酸配列には、生物の異なるドメイン(領域)、例えば、ファージディスプレイライブラリにおいて使用されるヒト抗体のような真核生物(真核起源の)から発生する核酸配列が包含される。核酸配列または核酸配列の一部は、原核生物の宿主の「コドン使用頻度」に応じて修飾される。
【0026】
「シグナル配列」または「シグナルペプチド配列」とは、非細胞質内部位(例えば分泌)へ細胞によって通常輸送されるかまたは膜成分になるタンパク質のNH2-末端に存在する短いアミノ酸配列(すなわち、シグナルペプチド)をコードする核酸配列をいう。シグナルペプチドは、細胞質内から非細胞質内部位へのタンパク質の輸送を方向づける。
【0027】
「翻訳開始領域」は、翻訳開始を促進し、かつシャインダルガーノ配列のようなリボソーム結合部位として機能するシグナル領域である。
【0028】
「転写終結領域」とは、RNAポリメラーゼが転写を終結する配列をいう。転写終結領域は通常転写単位の一部であり、mRNAの安定性を増強する。
【0029】
「抗体」とは、抗原よる刺激後に免疫系のB細胞によって生産される血漿タンパク質のクラスをいう。哺乳動物(例えばヒト)抗体には、IgG、M、A、EまたはDクラスの免疫グロブリンがある。用語「抗体」が本発明の目的のために使用されるとき、これに限定されないが、ポリクローナル抗体、モノクローナル抗体、抗イディオタイプ抗体および自己免疫疾患、糖尿病、多発性硬化症およびリウマチ関節炎において存在する自己抗体、ならびにキメラ抗体が含まれる。基礎的な抗体の構造単位は、四量体構成であることが知られている。各四量体は、ポリペプチド鎖の二つの同一の対で構成されており、各対は、一本の「軽鎖(約25kDa)」および一本の「重鎖(約50-70kDa)」で構成されている。各鎖のアミノ末端部位は、抗原認識の重要な役割を担う約100〜110(あるいはそれ以上)のアミノ酸で構成された可変領域を含む。各鎖のカルボキシ末端部位は、エフェクター機能についての重要な役割を担う定常領域を含む。
【0030】
軽鎖はカッパまたはラムダで分類される。重鎖はガンマ、ミュー、アルファ、デルタ、またはイプシロンとして分類され、抗体のアイソタイプをIgG、IgM、IgA、IgDおよびIgEとして定義する。軽鎖および重鎖では、可変および定常領域は、約12またはそれ以上のアミノ酸で構成された「J」領域によって連結される。重鎖はまた約10以上のアミノ酸の「D」領域を含む。
【0031】
用語「抗体」は、抗体全体およびその結合断片を意味するのに使用される。結合断片は、単鎖断片、Fv断片およびFab断片を含む。用語「Fab断片」は、無処理の抗体のパパイン開裂から生じる結合断片を意味するものとして当該技術分野において使用されるときがある。用語「Fab’およびF(ab’)2」は、ペプシン開裂によって生成された無処理の抗体の結合断片をいうものとして当該技術分野において使用されるときがある。本発明の内容において、Fabは一般的に、抗原の特異的結合に十分な少なくとも実質的に完全な軽鎖および重鎖可変領域と、軽鎖および重鎖の会合を維持するのに十分な軽鎖および重鎖定常領域の一部を有する無処理の抗体の二重鎖結合断片をいうときに使用される。このようなFabの例は、例えばSkerra et al., 1988, Science 240 (4855), 1038-41に記載されている。Fab断片、例えばIgGイディオタイプのFab断片は、無処理の免疫グロブリンの二つの重鎖の間に二つの鎖間ジスルフィド結合を形成する少なくとも一つの二システイン残基を含んでいてもいなくともよい。通常、Fab断片は、全長または実質的に全長の軽鎖を、可変領域および定常領域の少なくともCH1ドメインを含む重鎖と複合させることによって形成される。
【0032】
さらに、軽鎖上のC末端システインは、本発明に従って重鎖および軽鎖間の鎖間ジスルフィド結合を削除するためにセリンまたは他のアミノ酸と置換され得る。さらに、典型的には遺伝子工学によって、例えば異なる生物種に属する免疫グロブリン遺伝子区分(例えば、可変領域をコードする区分および定常領域をコードする区分)から構築される抗体であるキメラ抗体が包含される。例えば、マウスモノクローナル抗体由来の遺伝子の可変(V)区分は、ヒト定常(C)区分、例えばIgG1およびIgG4と連結され得る。典型的なキメラ抗体は、マウス抗体由来のVまたは抗原結合ドメインおよびヒト抗体由来のCまたはエフェクタードメインからなるハイブリッドタンパク質である。キメラ抗体は、抗体の可変領域を提供するマウスまたは他の非ヒト抗体と同一または類似の結合特異性および親和性を有する。
【0033】
用語「ヒト抗体」には、天然または人工の加工された親和性変異を含むヒト生殖系列免疫グロブリン配列から誘導された可変および定常領域(存在すれば)を有する抗体が含まれる。本発明のヒト抗体は、ヒト生殖系列免疫グロブリン配列によってコードされないアミノ酸残基を含むことができる(例えば、インビトロでのランダムまたは部位特異的変異誘発またはインビボでの体細胞変異によって誘導された変異)。しかしながら、用語「ヒト抗体」には、他の哺乳動物種(例えばマウス)の生殖系列から誘導されたCDR配列が、ヒトのフレームワーク配列上にドラフトされた抗体(すなわち、ヒト化された抗体)が含まれる。このような「ヒト抗体」の機能的変異体、例えば切断された態様または特定のプロリンまたはシステイン残基が当該技術分野において周知の遺伝子工学手段によって加工された加工性変異体が、上記用語に包含される。このような例は、例えばWO98/02462に見出されるであろう。しかしながら、該用語は、任意のグリコシル化またはペプチドバックボーンの他の化学的修飾とは無関係に、該抗体のアミノ酸配列のみと関係する。
【0034】
一側面において、本発明は、前記宿主と異種である核酸配列を含む転写単位と機能的に連結したメリビオースオペロンのプロモーター領域を含む宿主において発現可能なベクターを提供する。ここで、前記核酸配列の発現は、メリビオースオペロンの前記プロモーター領域によって制御される。
【0035】
本発明によるベクターは、自律的または自己複製的プラスミド、コスミド、ファージ、ウイルスまたはレトロウイルスが好ましい。広い範囲の宿主/ベクターの組み合わせが、本発明の核酸配列を発現するのに使用され得る。有用な発現ベクターは、例えば、染色体、非染色体および/または合成核酸配列のセグメントからなる。適切なベクターには、例えば大腸菌に特異的なベクターのような特異的宿主範囲をもつベクター、およびグラム陰性細菌に有用なベクターのような広い宿主範囲をもつベクターが含まれる。「低コピー数」、「中コピー数」および「高コピー数」のプラスミドが使用され得る。
【0036】
大腸菌内の発現に有用なベクターには、pQE70、pQE60およびpQE-9 (QIAGEN,Inc. ); pBluescript Vektoren、Phagescript Vektoren、pNH8A、pNH16a、pNH18A、pNH46A (Stratagene Cloning Systems, Inc.); ptrc99a、pKK223-3、pKK233-3、pDR540、pRIT5 (Pharmacia Bio-tech, Inc. ); pLG338、pACYC184、pBR322、pUC18、pUCl9、pKC30、pRep4、pACYC177、pACYC184、pRSF1010およびpBW22 [Wilms et al., 2001, Biotechnology and Bioengineering, 73 (2) 95-103] またはこれらの誘導体、例えばプラスミドpBLL15またはプラスミドpAKL15Eがある。さらに有用なプラスミドは当業者に周知であり、例えば、“Cloning Vectors” (Eds. Pouwels P. H. et al. Elsevier, Amsterdam-New York-Oxford, 1985)に記載されている。
【0037】
本発明の好ましいベクターは、自律的または自己複製的プラスミドである。より好ましくは、特異的宿主範囲をもつベクター、例えば大腸菌に特異的なベクターである。最も好ましくは、pBR322、pUC18、pACYC177、pACYC184、pRSF1010およびpBW22またはこれらの誘導体、例えばプラスミドpBLL15またはプラスミドpAKL15E、特に好ましくはpBW22またはこれらの誘導体、例えばプラスミドpBLL15またはプラスミドpAKL15E、より特に好ましくはpBLL15またはpAKL15E、最も特に好ましくはpAKL15Eである。
【0038】
好ましい実施態様において、本発明において使用されるメリビオースオペロンのプロモーター領域はmelABプロモーターである。本発明のより好ましい実施態様において、melABプロモーターは、CRP1結合部位が欠損している。特に好ましい実施態様において、CRP1結合部位が欠損したmelABプロモーターは、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる。通常、プロモーター領域のMelR結合部位は修飾されない。本発明において使用される、メリビオースオペロンのプロモーター領域、melABプロモーター、CRP1結合部位が欠損したmelABプロモーター、ならびに配列番号1からなるCRP1結合部位が欠損したmelABプロモーター、これに相補的な配列およびこれらの変異配列は、通常は大腸菌のメリビオースオペロンまたは他の原核生物、特に腸内細菌のファミリーに属する生物体の機能的に等価なプロモーター領域に由来する。好ましくは、メリビオースオペロンのプロモーター領域、melABプロモーター、CRP1結合部位が欠損したmelABプロモーター、ならびに配列番号1からなるCRP1結合部位が欠損したmelABプロモーター、これに相補的な配列およびこれらの変異配列は、大腸菌のメリビオースオペロン由来である。他の原核生物の機能的に等価なプロモーター領域は、メリビオースによって誘導可能であるプロモーター領域、すなわち、メリビオースの欠如下よりも存在下においてより高い発現活性を示すプロモーター領域を包含する。
【0039】
本発明による転写単位は、通常さらに前記転写単位の翻訳の開始点の翻訳開始領域上流を含み、前記転写開始領域は、配列AGGAGATATACAT(配列番号2)からなる。転写開始領域は核酸配列と機能的に連結される。配列AGGAGATATACAT(配列番号2)は通常は上流に位置し、ATG、GTGまたはTTGになり得る転写単位の翻訳の開始点に直接的に隣接する。
【0040】
通常、前記転写単位はさらに、本発明の異種核酸配列と機能的に連結したシグナル配列を含む。この場合において、ジシストロニックまたはポリシストロニックな転写単位が使用され、シストロンの各々に機能的に連結された異なるまたは同一のシグナル配列が適用され得る。好ましくは、異なるシグナル配列がこのような場合において使用される。使用されたシグナル配列は、原核生物または真核生物のシグナル配列とすることができる。通常、原核生物のシグナル配列が適用される。使用され得および大腸菌にとって特に有用な真核シグナル配列は、例えばヒトセルロプラスミンシグナル配列、ヒト好中球デフェンシン1,2,3前駆体シグナル配列またはWO 03068956に記載されたChlamydomonas reinhardtiiアリルスルファターゼのシグナルペプチドである。通常適用される原核生物のシグナル配列は、糖、アミノ酸、ビタミンおよびイオンのための周辺質結合タンパク質のシグナルペプチド、例えばPelB (エルウィニア属chrysantemi、ペクチン酸塩リアーゼ前駆体)、PelB (エルウィニア属carotovora、ペクチン酸塩リアーゼ前駆体)、PelB (ザントモナス・キャンペストリス、ペクチン酸塩リアーゼ前駆体)、LamB (大腸菌、マルトポリン前駆体)、MalE (大腸菌、マルトース結合タンパク質前駆体)、Bla (大腸菌、β-ラクタマーゼ)、OppA (大腸菌、周辺質糖ペプチド結合タンパク質)、TreA (大腸菌、周辺質トレハロース前駆体)、MppA (大腸菌、周辺質ムレインペプチド結合タンパク質前駆体)、BglX (大腸菌、周辺質ベータ-グルコシダーゼ前駆体)、ArgT (大腸菌、リシン-アルギニン-オルニチン結合周辺質タンパク質前駆体)、MalS (大腸菌、α-アミラーゼ前駆体)、HisJ (大腸菌、ヒスチジン-結合周辺質タンパク質前駆体)、XylF (大腸菌、D-キシロース-結合周辺質タンパク質前駆体)、FecB (大腸菌、ジシトラート(dicitrate)結合周辺質タンパク質前駆体)、OmpA (大腸菌、外膜タンパク質A前駆体) および PhoA (大腸菌、アルカリ性ホスファターゼ前駆体)である。
【0041】
好ましい実施態様において、シグナルペプチドは、大腸菌シグナルペプチドLamB (マルトポリン前駆体)、MalE (マルトース結合タンパク質前駆体)、Bla (β-ラクタマーゼ)、OppA (周辺質オリゴペプチド結合タンパク質)、TreA (周辺質トレハロース前駆体)、MppA (周辺質ムレインペプチド結合タンパク質前駆体)、BglX (周辺質β-グルコシダーゼ前駆体)、ArgT (リシン-アルギニン-オルニチン結合周辺質タンパク質前駆体)、MalS (α-アミラーゼ前駆体)、HisJ (ヒスチジン-結合周辺質タンパク質前駆体)、XylF (D-キシロース-結合周辺質タンパク質前駆体)、FecB (ジシトラート結合周辺質タンパク質前駆体)、OmpA (外膜タンパク質A前駆体)およびPhoA (アルカリ性ホスファターゼ前駆体)から選択される。これらは大腸菌の異種発現について特に有用である。より好ましくは、大腸菌シグナルペプチドLamB (マルトポリン前駆体)、MalE (マルトース結合タンパク質前駆体)、Bla (β-ラクタマーゼ)、TreA (周辺質トレハロース前駆体)、ArgT (リシン-アルギニン-オルニチン結合周辺質タンパク質前駆体)、FecB (ジシトラート結合周辺質タンパク質前駆体)である。最も特に好ましくは、大腸菌シグナルペプチドLamB (マルトポリン前駆体)およびMalE (マルトース結合タンパク質前駆体)である。
【0042】
本発明の発現ベクターにおいて使用されるシグナル配列は、商業的または化学的に合成されることによって得られうる。例えば、シグナル配列は、Van Devanter et. al., Nucleic Acids Res. 12:6159-6168 (1984)に記載されたように自動化合成装置を使用する、Beaucage & Caruthers, Tetrahedron Letts. 22:1859-1862 (1981)に記載された固相ホスホラミダイトトリエステルによって合成され得る。オリゴヌクレオチドの精製は、Pearson & Reanier, J. Chrom. 255:137-149 (1983)に記載されたように天然アクリルアミドゲル電気泳動法またはイオン交換HPLCによって行われ得る。
【0043】
通常、前記転写単位にはさらに、rrnB、RNA I、T7Te、rrnB T1、trp a L126、trp a、tR2、T3Te、P14、tonB t、およびtrp a L153から選択された転写終結領域が含まれる。好ましくは、rrnB 転写終結配列が使用される。
【0044】
本発明による異種核酸配列は、宿主に対して外来種である発現産物をコードする。宿主が大腸菌のような原核生物種である場合、対象の核酸配列は、より好ましくはガンマプロテオバクテリアのような他のクラスに由来、例えばBurkholderia spに由来し、特に異なる門、例えば古細菌に由来し、最も特に真核生物、例えば哺乳動物、特にヒトに由来する。しかしながら、異種核酸配列は、宿主の「コドン使用頻度」に基づいて修飾され得る。本発明による異種配列は、通常は対象の遺伝子である。対象の遺伝子は、好ましくは異種ポリペプチド、例えば構造的、制御的または治療学的タンパク質、または構造のN-もしくはC-末端融合、他のタンパク質(「タグ」)をもつ制御もしくは治療学的タンパク質、例えば緑色蛍光タンパク質または他の融合タンパク質をコードする。異種核酸配列は、RNAの形態において使用され得る転写産物、例えばアンチセンスRNAをコードしてもよい。
【0045】
タンパク質は、宿主細胞および/または細胞外培地の細胞質または周辺質空間内に存在する不溶性凝集塊または可溶性タンパク質として生産され得る。好ましくは、タンパク質は、宿主細胞および/または細胞外培地の周辺質空間内に存在する可溶性タンパク質として生産される。タンパク質の例には、ホルモン、例えば成長ホルモン、成長因子、例えば上皮性成長因子、鎮痛性物質様エンケファリン、酵素様キモトリプシン、抗体、ホルモンに対する受容体が含まれる。また、視覚化マーカー、例えば緑色蛍光タンパク質として通常使用されるタンパク質が含まれる。
【0046】
対象の他のタンパク質は、成長因子受容体(例えば、FGFR、PDGFR、EFG、NGFR、およびVEGF) およびこれらのリガンドである。他のタンパク質は、Gタンパク質受容体であり、物質K受容体、アンジオテンシン受容体、[アルファ]-および[ベータ]-アドレナリン受容体、セロトニン受容体、およびPAF受容体(例えば、Gilman, Ann. Rev. Biochem. 56, 625-649 (1987)を参照)。他のタンパク質は、イオンチャネル(例えば、カルシウム、ナトリウム、カリウムチャネル)、ムスカリン受容体、アセチルコリン受容体、GABA受容体、グルタミン酸受容体、およびドーパミン受容体(Harpold, 米国特許番号5,401,629 および5,436,128を参照)を含む。対象の他のタンパク質は、インテグリンのような付着性タンパク質、および免疫グロブリンスーパーファミリー膜(Springer, Nature 346, 425-433 (1990). Osborn, Cell 62, 3 (1990); Hynes, Cell 69, 11 (1992)を参照)である。他のタンパク質には、サイトカイン、例えばインターロイキンIL-1〜IL-13、腫瘍ネクローシス因子[アルファ]および[ベータ]、インターフェロン[アルファ]、[ベータ]、および[ガンマ]、腫瘍成長因子Beta(TGF-[ベータ]、インターフェロン[アルファ]、[ベータ]、および[ガンマ]、腫瘍成長因子ベータ(TGF-[ベータ])、コロニー刺激因子(CSF)および顆粒球単球コロニー刺激因子(GM-CSF)がある(Human Cytokines: Handbook for Basic & Clinical Research. Aggrawal et al. eds., Blackwell Scientific, Boston, Mass. 1991を参照)。対象の他のタンパク質は、細胞内および細胞間メッセンジャー、例えばアデニル、シクラーゼ、グアニルシクラーゼ、およびホスホリパーゼCである。薬剤はまた、他のタンパク質である。対象の異種タンパク質は、ヒト、哺乳動物または原核生物起源とすることができる。他のタンパク質は、抗原、例えば微生物病原体(ウイルスおよび細菌)および腫瘍由来の糖タンパク質および炭水化物である。
【0047】
他のタンパク質は、キモシン、プロテアーゼ、ポリメラーゼ、デヒドロゲナーゼ、ヌクレアーゼ、グルカナーゼ、オキシダーゼ、α-アミラーゼ、オキシドレダクターゼ、リパーゼ、アミダーゼ、ニトリルヒドラターゼ、エステラーゼまたはニトリラーゼのような酵素である。
【0048】
好ましくは、本発明による異種核酸配列は、ポリペプチド、より好ましくは抗体および最も好ましくはFab断片をコードする。特に、ヒト抗体またはヒト化抗体、より特にはヒトFab断片が核酸配列によってコードされる。核酸配列によってコードされたヒトFab断片は、好ましくはヒト抗体断片または他の哺乳動物種からの少なくとも一つのCDRを移植されたヒト抗体断片である。
【0049】
一以上の好ましい実施態様において、ヒトFab断片は、Knappik et al., 2000, J. Mol. Biol. 296 (1), 57-86によって記載されたCDRにおいて人工的にランダム化された人工のコンセンサスフレームワークベースのヒト抗体ファージライブラリから得られ得る完全なヒトHuCAL-Fabである。
【0050】
他のより好ましい任意の実施態様において、任意的にキメラであってCDRを移植されたヒトFab断片は、非HuCAL Fabである。これは前述のHuCAL Fabの定義とは対照的である。つまり、完全なヒトFab断片の場合において、好ましくはそれはHuCALコンセンサス配列のフレームワークを共有しないが、その非CDR配列部位は、それぞれの可変および定常領域の軽鎖および重鎖の生殖系列コード配列に対して、少なくとも70%、より好ましくは85%、最も好ましくは95%のアミノ酸配列における同一性を示すことを意味し、並びに追加的およびより好ましくは、そのCDRはゲノム親和性変異事象を含むリンパ系細胞の天然に生じるゲノム配列から直接的に得られることを意味する。
【0051】
Fab断片は、好ましくはIgG抗体から誘導され、無処理の免疫グロブリンにおける二つの重鎖間にある二つの鎖間ジスルフィド結合を形成するシステイン残基を含まない。特に、抗体または好ましくはFab断片の重鎖および軽鎖は、ジシストロニック転写単位によってコードされる。各鎖はシグナル配列および転写単位の翻訳の開始点の上流にある同一の(identical)翻訳開始領域に機能的に連結される。好ましくは、転写開始領域は、配列AGGAGATATACAT (配列番号2)からなる。
【0052】
本発明において、シグナル配列および均質核酸配列が発現ベクター内で配置される順序および距離を変えることができる。好ましい実施態様において、シグナル配列は、例えば対象のポリペプチドをコードする核酸配列に対して5’(上流)にある。例えば、シグナルペプチド配列および例えば対象のポリペプチドをコードする核酸配列は、ゼロ〜約1000アミノ酸まで分離させることができる。好ましい実施態様において、例えば対象のポリペプチドをコードするシグナルペプチド配列および核酸配列は、互いに直接的に隣接、すなわちゼロ核酸で分離される。
【0053】
好ましくは、本発明のベクターのプロモーター領域および機能的に連結された転写単位は、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる。
【0054】
より好ましくは、プロモーター領域および本発明のベクターの機能的に連結された転写単位は、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる。
【0055】
また、原核生物の宿主における核酸配列の制御された異種発現のための本発明によるベクターの使用が、本発明によって包含される。発現は、原核生物の宿主に利用可能なメリビオースの量によって制御され得る。通常、培養された原核宿主の培養液中におけるメリビオースの量は、0.01〜100g/L、好ましくは0.1〜10g/L、より好ましくは1〜5g/Lである。
【0056】
好ましくは、異種核酸配列は、ポリペプチド、より好ましくは抗体および最も好ましくはFab断片をコードする。一方、抗体またはFab断片の重鎖および軽鎖は等しい量において発現し、その結果として高濃度の機能的抗体またはFab断片を導く。特に、ヒト抗体またはヒト化抗体、より特にはヒトFab抗体、最も特にはヒトFab断片は、異種核酸配列によってコードされる。
【0057】
高濃度の機能的抗体またはFab断片を得るために、等しい量の重鎖および軽鎖が発現することが重要である。両鎖の一方が他の鎖と比較して過剰生産される場合、望ましくない非還元性の高分子量免疫反応凝集塊が構築され得る。驚くべきことに、本発明のベクターで高力価の機能的抗体が得られ、非常に低量の過剰生産された軽鎖または重鎖または高分子重量の免疫反応性凝集塊のみが構築されることを見出した。通常、抗体またはFab断片の発現量の20%未満、好ましくは10%未満が、過剰生産された軽鎖または重鎖または高分子重量免疫反応性凝集塊として発現される。過剰生産された重鎖および軽鎖の量ならびに高分子量の免疫反応性凝集塊の量は、SDS-PAGEまたはウェスタンブロットを使用して培養された宿主細胞のリゾチーム抽出物のような抗体またはFab断片を発現する宿主の抽出物を分析することによって測定され得る。
【0058】
また、他の側面において、本発明は、前記宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオースオペロンのプロモーター領域を含む宿主において発現可能な単離および精製された核酸配列を提供する。前記核酸配列の発現は、メリビオースオペロンの前記プロモーター領域によって制御される。melABプロモーターは、好ましいプロモーター領域である。より好ましくはCRP1結合部位が欠損したmelABプロモーターである。最も好ましくは、単離および精製された核酸配列は、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる。特に、単離および精製された核酸配列は、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる。最も特に、単離および精製された核酸配列は、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる。本発明の単離および精製された核酸配列は、当業者に周知の標準的PCRプロトコルおよび方法によって単離され得る。前記精製および単離されたDNA配列はさらに、当業者に周知の一以上の制御配列、例えば核酸配列によってコードされた生成物の発現のために通常使用されるエンハンサーを含むことができる。
【0059】
本発明のベクターまたは単離および精製された核酸配列で効率良くかつ安定的に形質転換された宿主細胞を選択するために、選択可能なマーカー(例えば、抗体に対する耐性)をコードする遺伝子は、対象の核酸配列に沿って宿主細胞に導入され得る。選択可能なマーカーをコードする遺伝子はベクターまたは単離および精製された核酸配列上に配置され得、あるいは別々のベクター上にセパレートフォームで共導入され得る。抗生物質、例えばハイグロマイシン、アンピシリンおよびテトラサイクリンに対する耐性を与えるものを含む様々が選択可能なマーカーが使用され得る。抗生物質の量は、選択的条件を創出するために所望の量に適合され得る。通常、一つの選択可能なマーカーが使用される。同様に、蛍光タンパク質のようなレポーター遺伝子は、形質転換の効率を決定するために、対象の核酸配列に沿って宿主細胞に導入され得る。
【0060】
本発明の他の側面は、本発明のベクターで形質転換された原核生物の宿主を提供することである。本発明の特定の実施態様において、原核生物の宿主は、このような宿主細胞における分泌されたヘテロ二量体タンパク質、例えばFabを発現するジシストロニックな発現カセットにおいて二つの異なるコドン領域を含むプラスミドpBLL15またはプラスミドpAKL15E、好ましくはプラスミドpAKL15Eで形質転換される。好ましくは、このようなヘテロ二量体タンパク質はFabである。本発明の他の実施態様において、原核生物の宿主は、本発明の単離および精製された核酸配列で形質転換される。
【0061】
多様な原核生物の宿主細胞は、通常は本発明の異種核酸配列を発現するのに有用である。これらの宿主には、グラム陰性細胞、例えば大腸菌およびシュードモナス属またはグラム陽性細胞、例えばバチルス属およびストレプトマイセス属が含まれる。好ましくは、宿主細胞はグラム陰性細胞、より好ましくは大腸菌細胞である。使用され得る大腸菌は、例えば株TG1、W3110、DH1、XL1-BlueおよびOrigamiである。これらはDSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany)を介して得ることができる。最も好ましくは、W3110が使用される。宿主細胞にはメリビオースを代謝しないものがある。大腸菌のようにメリビオースを取り込みかつ代謝することができる宿主細胞が修飾され得、メリビオースの取り込みおよび/または代謝に関連した一以上の機能が欠損する。メリビオースの取り込みおよび/または代謝に関連した一以上の機能の欠損は、例えばα-ガラクトシダーゼをコードするmelA遺伝子のようなメリビオースの取り込みおよび/または代謝と関連したタンパク質をコードする遺伝子の発現を抑制または遮断することによって達成され得る。これは、変異誘発またはノックアウト変異を担うトランスポゾンのような既知の技術によって行われ得る。通常、原核生物の宿主は、選択されたシグナル配列に対応し、例えば大腸菌のシグナル配列が使用される場合、宿主細胞は腸内細菌と同じファミリーの一員であり、より好ましくは宿主細胞は大腸菌株である。
【0062】
さらに本発明は、宿主細胞においてポリペプチドを生成するための方法であって、以下の工程を含む方法を提供する。
【0063】
a)ベクターを構築する工程と、
b)原核生物の宿主を前記ベクターで形質転換する工程と、
c)適切な条件下で細胞培養系において前記ポリペプチドを発現させる工程と、
d)前記ポリペプチドを細胞培養系から回収する工程。
【0064】
使用されるベクター、および原核宿主の構築および形質転換は上記に定義されたとおりである。ベクターによって包含された異種核酸配列は、ポリペプチドをコードする。好ましくは生成されたポリペプチドは、抗体および最も好ましくはFab断片である。抗体またはFab断片の重鎖および軽鎖は、細胞培養系で等量において発現され、高濃度の機能的抗体またはFab断片を誘導する。
【0065】
回分培養または流加培養のような連続的または非連続的培養は、培養管、撹拌フラスコまたは細菌発酵槽において適用され得る。通常、宿主細胞は、「栄養酵母培養液」またはKortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたグリセロール含有培養液またはKulla et al., 1983, Arch. Microbiol, 135, 1によって記載された栄養塩培養液のような複合培養液、当該技術分野において既知の従来培養液中において培養される。前記ポリペプチドの発現を実行するために好ましい培養液は、グリセロール含有培養液、より好ましくはKortz et al., 1995, J. Biotechnol. 39, 59-65によって記載された培養液である。
【0066】
培養液は、例えば、緩衝液、塩、ビタミンまたはアミノ酸のような追加成分を添加することによって適切なものとして修飾され得る。異なる培養液または培養液の組み合わせが細胞の培養中に使用され得る。好ましくは、基礎培地として使用される培地は、メリビオースプロモーター領域の厳格な制御を達成するために、メリビオースを含むべきではない。メリビオースは通常、培養液は培養系に依存しながらOD600に達した後に加えられる。通常、培養された原核生物の宿主の培養液中のメリビオースの量は、0.01〜100 g/l、好ましくは0.1〜10 g/l、より好ましくは1〜5 g/lである。回分培養では、通常OD600が0.4以上である。適切なpH範囲は、例えば6〜8、好ましくは7〜7.5であり、適切な培養温度は、10〜40、好ましくは20〜37℃である。細胞は、最大量の発現生成物が蓄積されるまでインキュベートされ、好ましくは1時間〜20日、より好ましくは5時間〜3日間にわたってインキュベートされる。発現生成物の量は、使用される培養系に依存する。撹拌フラスコ培養系では、通常、0.5 g/l 培養液の量の発現生成物が、本発明のベクターで形質転換された宿主で産生されうる。回分および/または流加培養モードにおける発酵槽培養系を使用すると、通常、0.5 g/l 発酵ブロスの量の発現生成物、好ましくは1 g/l以上、より好ましくは1.3 g/l以上の量の発現生成物が得られうる。
【0067】
宿主細胞における発現後、発現産物、例えば対象のポリペプチドは、宿主細胞の培養液から回収され得る。対象のポリペプチドが免疫グロブリン鎖であるとき、重鎖および軽鎖は通常は各々、宿主細胞において発現され、細胞の周辺質に分泌される。発現ベクターのシグナル配列によってコードされるシグナルペプチドは、免疫グロブリン鎖から加工されれる。その後、成熟した重鎖および軽鎖が構築され、完全な抗体またはFab断片が形成される。発現産物の最大の収量を得るために、通常は培養の終わりに細胞が収穫され、かつリゾチーム処理、超音波処理またはフレンチプレスによって溶解される。従って、ポリペプチドは通常、宿主細胞の粗溶解物として最初に得られる。これらはその後、ディファレンシャル沈殿法、モレキュラーシーブ・クロマトグラフィー、イオン交換クロマトグラフィー、等電点電気泳動、ゲル電気泳動、アフィニティー、および免疫アフィニティー・クロマトグラフィーを含む、当業者に既知の標準的なタンパク質精製手順によって精製され得る。これらは十分に知られており、日常的に実施される方法、例えばAusubel et al., supra., and Wu et al. (eds.), Academic Press Inc., N.Y.; Immunochemical Methods In Cell And Molecular Biologyに記載されている。例えば、組換え的に生成された免疫グロブリンまたはFab断片の精製について、それらは、発現した免疫グロブリンが特別に結合し得る標的分子が結合した樹脂を含むカラムを通過させることによって免疫アフィニィティー クロマトグラフィーで精製され得る。
【0068】
本発明はまた、例えば原核生物の宿主においてポリペプチドをコードする核酸の細胞内異種発現のための方法および手段に関する。特に、本発明は原核生物の宿主における異種ポリペプチドの細胞内発現のためのベクターに関する。該ベクターは、前記宿主に対して異種である核酸配列を含む転写単位に機能的に結合されたメリビオースオペロンのプロモーター領域を含む原核生物の宿主内において発現可能である。本発明のベクターのこの実施態様において、核酸配列はベクターおよび異種核酸によってコードされるポリペプチドの発現とともに宿主原核細胞を形質転換させる原核シグナル配列と結合しないので、該ポリペプチドは細胞質から非細胞質まで輸送されないであろう。代わりに、ポリペプチドは封入体の形態における細胞質内または可溶性形態内において発現するであろう。従って、発現したポリペプチドは、細胞、特に細胞抽出物から周知の手順によって単離および精製され得る。本発明はまた、原核宿主細胞内の異種核酸配列の制御された細胞内発現のための前記ベクターの使用;前記ベクターで形質転換された原核宿主または原核宿主細胞;前記ベクターを使用して原核宿主内において異種ポリペプチドの細胞内産生のための方法;およびプロモーター領域を含む異種ポリペプチドの細胞内産生のためのベクター、異種ポリペプチドをコードする異種核酸配列および配列AGGAGATATACATからなる翻訳開始領域を提供する。
【0069】
細胞内発現のために使用され得るベクターの好ましい実施態様において、メリビオースオペロンのプロモーター領域は、好ましくはCRP1結合部位が欠損したmelABプロモーターである。特に好ましくは、CRP1結合部位が欠損したmelABプロモーターは、配列番号1に示される配列、これに相補的な配列およびこれらの変異配列からなる。本発明の他の好ましい実施態様において、ベクターの転写単位にはさらに、前記転写単位の翻訳の開始点の上流にある転写開始領域が含まれる。転写開始領域は配列AGGAGATATACAT(配列番号2)からなる。さらに好ましい実施態様において、細胞内発現のために使用されるベクターには、rrnB転写終結配列のような転写終結領域が含まれる。本発明によれば、異種核酸配列は抗体、抗体断片などのポリペプチドをコードする。
【0070】
当業者は、明細書に記載された発明が特別に記載されたもの以外のバリエーションおよび修飾を含むことを認識するであろう。本発明には本発明の精神または要素的特徴から離れることのない全てのバリエーションおよび修飾が含まれることを理解すべきである。本発明にはまた、個別的または集団的に、この明細書中に言及または示された全ての工程、特徴、組成物および化合物が含まれ、かつ前記工程または特徴の任意かつ全ての組み合わせまたは任意の二以上の組み合わせが含まれる。本発明の開示は、全ての側面において拘束的なものではなく例証として考慮されるべきであり、本発明の範囲は、添付されたクレームによって示される。等価の意味および範囲内に入る全ての変化は、本発明の範囲内に包含されることを意味する。様々な文献がこの明細書を通して引用されているが、引用文献の各々は参照によってその全体が本明細書中に組み込まれる。
【0071】
前述の記述は、以下の例を参照してより完全に理解されるであろう。このような例は、しかしながら、本発明を実施する方法の例証であり、本発明の範囲を制限することを意味するものではない。
【0072】
例
例1
大腸菌W3110ゲノムを、正に制御されたオペロンについてスキャンした。KEGGデータベース上の利用可能なゲノムデータに基づいて(Kyoto Encyclopedia of Genes and Genomes, http://www.genome.ad.jp/kegg/kegg2.html)、正に制御された異化プロモーターを、発現プラスミドにおけるその使用のために同定かつ分析した。プロモーターは厳密に制御されるべきであり、かつ安価かつ無毒で導入されるべきである(つまり、産業的にも有用な化合物であるべきである)。以下の異なる正に制御された異化プロモーターを選択した。
【0073】
正に制御されたプロモーターでの発現プラスミドの構築
−prpプロモーター(誘導性プロピオネート)
−gutAプロモーター(誘導性グルシトール)
−melAB2プロモーター(誘導性メリビオース)
【0074】
プロモーター要素を含む正確なDNA断片を、相当する制御結合部位上の利用可能な情報に基づいて選択した。大腸菌の染色体DNAを「Pitcher et al., 1989, Letters in Applied Microbiology 8, 151-156」に記載された方法によって単離した。プロモーター断片を、以下のプライマーを使用してPCRによって株W3110の染色体DNAから増幅した。ClaIおよびAflIIの制限部位に下線を記した。該断片の配列は以下のとおりである:
【表A】

断片をアガロースゲル電気泳動によって分離し、Qiagen (Hilden, Germany)のgelextraction kit QiaexIIによって単離した。単離された断片をClaIおよびAflIIで切断し、ClaI/AflII-切断pBW22にライゲートした(Wilms et al., 2001, Biotechnology and Bioengineering, 73 (2), 95-103)。配列番号1(pBLL7)からなるmelAB2プロモーターを含む得られたプラスミドを図1に示す。prpプロモーター(pBLL5)およびgutAプロモーター(pBLL6)を含む得られたプラスミドは、ライゲートされたプロモーター領域を除けば同一である。挿入されたプロモーター断片の配列は、シークエンシングによって確認した(Microsynth GmbH, Balgach, Switzerland)。
【0075】
例2
Fab断片発現プラスミドの構築
IPTG-誘導性lacプロモーターに対する代替として(プラスミドpMx9-HuCAL-Fab-H, Knappik et al., 1985, Gene 33, 103-119)、異なる正に制御された発現系をFab-H抗体断片を産生するその能力について分析した。Fab-H断片を、プライマーFab-5(5’-aaa cat atg aaa aag aca gct atc-3’)およびFab-3 (5’-aaa aag ctt tta tca gct ttt cgg ttc-3’)を使用してPCRによってプラスミドpMx9-HuCAL-Fab-Hで増幅した。PCR断片をNdeIおよびHindIIIで切断、かつNdeI/HindIII-切断pBW22に挿入し、L-ラムノース誘導性rhaBADプロモーターを含むプラスミドpBW22-Fab-H を創出した (Volff et al., 1996, Mol. Microbiol. 21, 1037-1047)。同じPCR断片を誘導性プロモーターをもつ異なる発現プラスミドに挿入した。Fab-HおよびmelAB2プロモーター(配列番号3)を含む得られた(推定上の)発現プラスミドpBLL15を図2に示す。prpプロモーター(pBLL13)およびgutAプロモーター(pBLL14)を含む等価なプラスミドが得られた。プラスミドpBW22-Fab-HのFab-H 挿入の配列はシークエンシングによって確認した。
【0076】
例3
Fab断片の発現
株W3110 (DSM 5911, Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany)を、異なる発現プラスミドで形質転換した。プラスミドを、異なる形質転換体から得られたクローンから単離し、制限分析を介してチェックした。プラスミドpBLL14を除いて、全てのプラスミドは、予期された制限パターンを有していた。再度単離されたプラスミドpBLL14は、組換えが原因と考えられる変化したサイズおよび制限パターンを示した。従って、株W3110 (pBLL14)については以下のアッセイにおける試験を行わなかった。残りの株については、活性が倍加されたFab-H抗体断片を分泌する能力について試験を行った。この生産性試験は例4において記載されたように行った。以下の誘導物質を0.2%の濃度において加えた。
【0077】
pBW22-Fab-H L(+)-ラムノース一水和物
pBLL13 プロピオン酸ナトリウム
pBLL15 D(+)-メリビオース一水和物
D(+)-ラフィノース一水和物
D(+)-ガラクトース
【0078】
ドットブロット試験の結果を図3において示す。
【0079】
L-ラムノースおよびメリビオース誘導株W3110(pBW22-Fab-H)およびW3110 (pBLL15)は、有望なドットブロット結果を示した:経時的にシグナルを増加させ、かつほとんどバックグラウンド活性が観察されなかった。活性が倍加された抗体断片の部分は、ELISAを介して定量化された。その結果を以下の表1に要約する。
【表1】
【0080】
表1:異なる発現プラスミドをもつW3110誘導体のELISAの結果。誘導後の時間が示されている。22または25時間後の非誘導の培養物を、非誘導の対照として測定し、株W3110 (pMx9-HuCAL-Fab-H) および TG1F’- (pMx9-HuCAL-Fab-H) の結果を、参照として使用する。Fab-H濃度は、mg/100 OD/Lで与えられる(n.d. 測定されなかった)。
【0081】
全ての株は、4〜6の間のOD600まで対応する誘導物質の存在または欠損下において、あらゆる増殖阻害もなく良好に増殖した。発現プラスミドpBW22-Fab-HおよびpBLL15は、終夜の誘導後に最も高い抗体断片力価を誘導した。メリビオース誘導株W3110 (pBLL15)は、L-ラムノース(pBW22-Fab-H)誘導系と比較して活性抗体断片の形態において遅延性の増加を示した。
【0082】
L-ラムノース誘導性株W3110 (pBW22-Fab-H)を、振盪フラスコ内の呼吸活性のオンライン決定のための新規な測定系である「呼吸活性モニタリングシステム」 (RAMOS, ACBiotec, Julich, Germany) において試験した。通常の実験用振盪フラスコと比較して、抗体力価(ELISAを介して測定)は二倍になった(導入の23時間後に703.64 mg/L/100 OD600)。RAMOS装置を使用する最適化された成長は、活性抗体断片の生産を支持する。
【0083】
例4
振盪フラスコ内におけるメリビオースの誘導
プラスミドpBLL15をもつ大腸菌W3110を、活性が倍加されたFab-H抗体断片を産生するその能力について試験した。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、37 °C] を、20 mlの新鮮グリセロール培地中で稀釈した(1:50)(Kortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたように稀釈)。一方、ビタミン溶液を、Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように使用し、かつ30℃でインキュベートした。培養液が約0.4のOD600に達したとき、メリビオース(0.2%)を加えた。サンプル(1 ml)を異なる時間間隔で採取し、遠心してペレットを−20℃で貯蔵した。凍結した細胞を上述したリゾチーム処理によって溶解させ、上清をドットブロットおよびELISAアッセイにおいて分析した。機能的Fab-H抗体断片の504.28 mg/L/100 OD600 を得た。
【0084】
例5
高分子量凝集塊の出現
高分子量凝集塊が産生されるか否かを見るために、最も高い抗体力価(表1)を示した株W3110 (pBLL15)の抽出物のウェスタンブロットを、抗ヒトFab-H + AP抱合体を使用して行った。培養を例4に記載されたとおりに行った。メリビオースで誘導後9、12および23時間してサンプルを採取した。抗ヒトFab-H + AP結合体を使用した株W3110 (pBLL15)のリゾチーム抽出物のウェスタンブロットを図4に示す。より低濃度の高分子量凝集塊は、より高い力価の機能的抗体断片に対応する。発現系の選択は、抗体断片が形成され:機能的または凝集塊になることに影響を与えるようである。
【0085】
例6
シグナルペプチドの影響
大腸菌のゲノムデータベースを使用して、Fab-H断片VL3-CLおよびVH-CHと組み合わせて使用され得る有用なシグナルペプチドを探索した。糖、アミノ酸、ビタミンおよびイオンについて周辺質結合タンパク質からのシグナル配列を選択した。これらの周辺質タンパク質は、他の周辺質タンパク質よりもより広範な研究がなされた比較的均質な群を表わす。これらは一般的には大量に存在するので、これらのシグナル配列は内膜を超えて周辺質へ効果的に輸送されなければならない。全ての可能なシグナルペプチドFab組み合わせを、以下の表2において示したようなSignalPウェブサーバー(http://www.cbs.dtu.dk/services/SignalP-2.0/#submission)を使用して、その配列ペプチドおよび切断サイト可能性について調べた。
【0086】
【表2】
【0087】
【0088】
【0089】
以下の6つの組み合わせを選択した:
- LamB-VL3-CL (マルトポリン前駆体)
- MalE-VH-CH (マルトース結合タンパク質前駆体)
- Bla-VL3-CL (β-ラクタマーゼ)
- TreA-VH-CH (周辺質トレハラーゼ前駆体)
- ArgT-VL3-CL (リシン-アルギニン-オルニチン-結合周辺質タンパク質前駆体)
- FecB-VH-CH (鉄(III)ジシトラート-結合周辺質タンパク質前駆体)
【0090】
シグナルペプチド(SP)とVL3-CLおよびVH-CHとの融合体を生成するための遺伝子融合がオーバーラッピングPCRプライマーで行われ、以下の増幅の表3において要約される。
【表3】
【0091】
シグナルペプチド配列とVL3-CLおよびVH-CH配列との融合は、他の文献 (Horton, R.M., Hunt, H.D., Ho, S.N., Pullen, J.K. and Pease, L.R. (1989) Engineering hybrid genes without the use of restriction enzymes: gene splicing by overlap extension. Gene 77, 61-68) に記載されたように行った。SP-VL3-CL遺伝子を、制限酵素NdeIおよびPstIで切断し、NdeI/PstI切断pBW22 および pBLL7にライゲートした。
【0092】
得られたプラスミドをPstIおよびHindIIIで切断し、PstI/HindIII切断SP-VH-CH遺伝子にライゲートした。bla-VL3-CLおよびfecB-VH-CH遺伝子の統合は可能ではなく、lamB-VL3-CL および malE-VH-CH遺伝子を含むFab-H発現プラスミドのみが試験に供された。L-ラムノース誘導プロモーター(pAKL14)を含むlamB-VL3-CL/malE-VH-CH発現プラスミドを得た。AflII/HindIII断片としてプラスミドpAKL15(例7)から単離されたlamB-VL3-CL/malE-VH-CH遺伝子を、AflII/HindIII-切断pBLL7にライゲートし、pAKL15Eを得た。図5は、メリビオース誘導性プロモーターおよびlamB-VL3-CL/malE-VH-CH (配列番号4)を含む発現プラスミドpAKL15Eを示す。
【0093】
例7
Fab発現上の翻訳開始領域の影響
プラスミドpAKL14およびプラスミドpAKL15EのFab-H遺伝子は、開始コドンの同じDNA配列5’(翻訳開始領域)を含む。最初のプラスミドpMx9-HuCAL-Fab-Hにおいて、両Fab-H遺伝子の翻訳開始領域は異なる。プラスミドpMx9-HuCAL-Fab-HおよびpAKL14/pAKL15Eの翻訳開始領域配列の比較は、以下の表4において示される。
【表4】
【0094】
株W3110(pAKL14)の生産能力を、例4に記載したように振盪フラスコ内において試験した。株はL-ラムノースの存在または欠損下において十分に増殖した。これはFab-Hの生産が細胞の生存能力に影響を与えなかったことを意味する。
【0095】
新しいシグナルペプチド構築物(修飾された翻訳開始シグナルと組み合わされた)は再度、抗体断片力価を328.62 mg/L/100 OD600(プラスミドpMx9-HuCAL-Fab-H由来のMOR 遺伝子構築物を含むプラスミドpBW22-Fab-H)から596.14 mg/L/100 OD600 (プラスミドpAKL14)まで増加し、かつ878.86 mg/L/100 OD600 (プラスミドpAKL15E) まで増加した。pAKL14におけるlamB-VL3-CLおよびmalE-VH-CH遺伝子のシークエンシングは、二つの連続したPCR反応による融合遺伝子の構築が原因であると考えられる三つの塩基の変化を明らかにした。塩基の変化は、以下のアミノ酸の変化を誘導した(誤りのアミノ酸を強調する)。
【表B】
Fab-Hの軽鎖は二つの誤り(D50N, K63N)を含み、かつ重鎖は一つのアミノ酸変化(F156L)を含む。最初のFab-H配列を回復するために、プラスミドpAKL14 (138 bp SexAI/BamHIおよび310 bp BssHII/HindIII断片)由来の二つの断片は、プラスミドpBW22-Fab-H (未変化のFab-H遺伝子配列を含む)の均質な断片に対して変化した。得られたプラスミドpAKL15は、正しいFab-H配列を含む。三つのアミノ酸の変化は、pIが変化しなかったので全体としてFab-Hの性質に影響を与えなかった。従って、機能的Fab-H抗体断片を産生する株W3110 (pAKL15)の能力は、株W3110 (pAKL14)に類似していると考えられるので分析を行わなかった。
【0096】
Fab-H抗体断片の生産能力は、異なる最適化戦略を使用することによって増加され得る。以下の表5はその改良について要約する:
【表5】
【0097】
高Fab-H抗体力価を生産した株をSDS-PAGE(図6)を介して分析した。最も高い機能的Fab-H濃度が、軽鎖および重鎖の平衡量を産生する株において測定された(レーン4および5)。W3110 (pBW22-Fab-H) (レーン3)のようなFab-H断片を含むL-ラムノース誘導株が、軽鎖を激しく過剰産生する。
【0098】
例8
振盪フラスコにおけるメリビオースの誘導
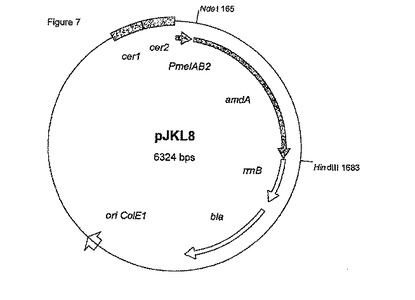
プラスミドpAKL15Eを含む大腸菌W3110は、活性が倍加されたFab-H抗体断片を産生するその能力について試験された。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、37℃] を、20 mlの新鮮グリセロール培地中で稀釈した(1:50)(Kortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたように稀釈)。一方、ビタミン溶液を、Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように使用し、かつ30℃でインキュベートした。培養液が約0.4のOD600に達したとき、メリビオース(0.2%)を加えた。サンプル(1 ml)を異なる時間間隔で採取し、遠心してペレットを-20℃で貯蔵した。凍結された細胞を上述したリゾチーム処理に基づいて溶解し、上清をSDS-PAGEおよびELISAアッセイにおいて分析した。変化したシグナルペプチド(lamB-VL3-CL/malE-VH-CH)でFab-H遺伝子を含むメリビオース誘導株は、最大のFab-H抗体力価を示した(表5)。Fab-Hの軽鎖および重鎖は等量において産生された(図8)。
【0099】
例9
振盪フラスコ内における株KIE153 (Burkholderia sp. DSM9925) からのアミダーゼのメリビオース誘導
プラスミドpJKL8を含む大腸菌W3110は、ラセミピペラジン-2-カルボキサミドを(R)- ピペラジン-2-カルボン酸に転換するその能力について試験を受けた。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、30 °C] を、20 mlの新鮮グリセロール培地中で稀釈した(1:50)(Kortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたように稀釈)。一方、ビタミン溶液を、Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように使用し、かつ30℃でインキュベートした。培養液が約0.8のOD600に達したとき、メリビオース(0.2%)を加えた。19時間の誘導時間後、細胞を-20℃で収穫および貯蔵した。アミダーゼ活性についてEichhorn et al. 1997, Tetrahedron Asymmetry, 8(15), 2533-36に記載したように試験を行った。得られた細胞は、約1 g/ h/ OD600の転換速度で (R)-ピペラジン-2-カルボキサミドを(R)-ピペラジン-2-カルボン酸にエナンチオ選択的に転換した。
【0100】
例10
振盪フラスコ内の単鎖抗体(scFv, S1)のメリビオース誘導
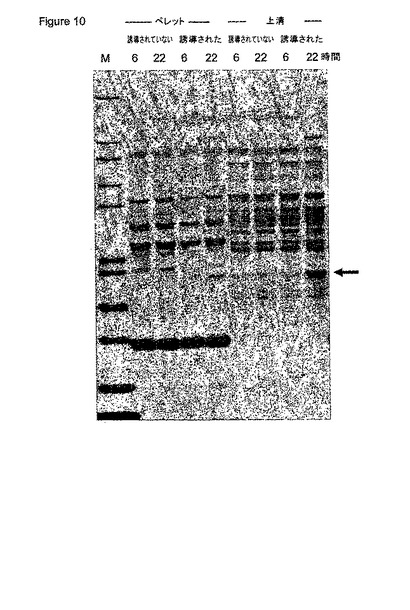
scFv遺伝子を、プライマー5-S (5’-aaa cat atg aaa tac cta ttg cct acg gc-3’) および 3-S1 (5’-aaa aag ctt act acg agg aga cgg-3’)を使用してPCRを介して単離した。対応するS1タンパク質は、大腸菌の周辺質に対するタンパク質の輸送に原因のあるPelBシグナル配列を含む。PCR断片は、NdeI および HindIII で切断され、NdeI/HindIII-切断 pBLL7に挿入され、メリビオース誘導melAB2プロモーター(図9)を含むプラスミドpBLL7-pelB1-S1を作製した。プラスミドpBLL7-S1のS1挿入の配列は、シークエンシングによって確認された。株W3110 (DSM 5911, Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany) をプラスミドpBLL7-pelB1-S1で形質転換した。プラスミドは、異なるクローンから単離され、かつ制限分析によって検証された。大腸菌W3110 (pBLL7-pelB-S1)は、可溶性S1を産生するその能力について試験を受けた。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、30 °C] を、20 mlの新鮮グリセロール培地中で稀釈し(1:50) [Kortz et al., 1995, J. Biotechnol. 39, 59-65に記載されたように稀釈、但しビタミン溶液を除く(Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように)]、30℃でインキュベートした。培養液が約0.4のOD600に達したときにメリビオース(0.2%)を加えた。サンプル(1 mL)を異なる時間間隔で採取し、遠心してペレットを-20℃で貯蔵した。凍結細胞を上述したリゾチーム処理に基づいて溶解し、上清および不溶性タンパク質ペレットをSDS-PAGE(図10)およびBioanalyzerを介して分析した。S1タンパク質(1.7 g/Lx mg/L/100 OD600)の大部分は、可溶性タンパク質画分において産生された。
【図面の簡単な説明】
【0101】
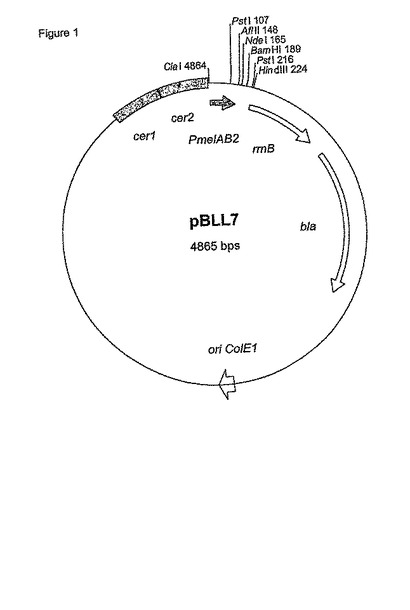
【図1】図1は、メリビオース誘導性melAB2プロモーター(PmelAB2)および転写終結領域(rrnB)を含むプラスミドpBLL7を示す。
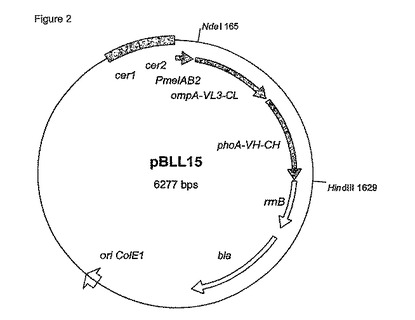
【図2】図2は、メリビオース誘導性プロモーター(PmelAB2)、Fab-H断片の軽鎖および重鎖に機能的に連結したシグナル配列の配列コード(ompA-VL3-CL)および(phoA-VH-CH)、ならびに転写終結領域(rrnB)を含むプラスミドpBLL15を示す。
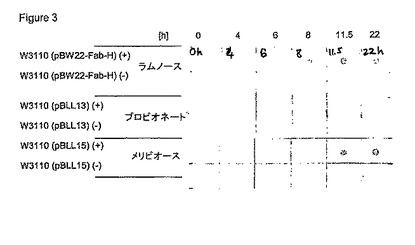
【図3】図3は、異なる発現プラスミドを含む非誘導性(-)および誘導性(+)W3110株のリゾチーム(lysozyme)抽出物のドットブロット結果(Fabを検出するための抗ヒト軽鎖、アルカリ性ペルオキシダーゼを抱合)を示す。時間の間隔を示す。
【図4】図4は、アルカリ性ペルオキシダーゼを抱合したFabを検出するための抗ヒト軽鎖を使用する株W3110(pBLL15)のリゾチーム抽出物のウェスタンブロットを示す。
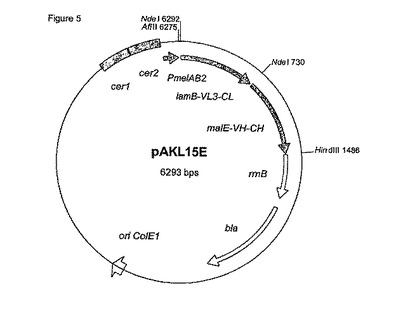
【図5】図5は、メリビオース誘導性プロモーター(PmelAB2)および変化したシグナル配列を含むFab-H遺伝子を含むプラスミドpAKL15Eを示す。
【図6】図6は、高いFab-H抗体濃度を示す異なるW3110株のライソゾーム抽出物のSDS-PAGEを示す。シグナル配列を含まない軽鎖および重鎖を生成する株がネガティブ参照として使用される(レーン1:マーカー; レーン2:W3110 (pMx9-HuCAL-Fab-H); レーン3:W3110 (pBW22-Fab-H); レーン4:W3110 (pBLL15), レーン5:W3110 (pAKL14); レーン6:スタンダード(2 μg))。
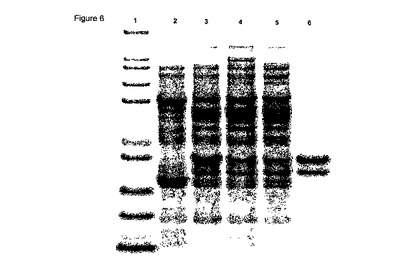
【図7】図7は、メリビオース誘導性プロモーター(PmelAB2)および株KIE153 (Burkholderia sp. DSM9925)由来のアミダーゼをコードする配列、および転写終結領域(rrnB)を含むプラスミドpJKL8を示す。
【図8】図8は、誘導性メリビオースの存在または不存在における株W3110 (pAKL15E)のライソゾーム抽出物のSDS-PAGEを示す。軽鎖および重鎖の位置が示される(レーン1:マーカー; レーン2:W3110 (pAKL15E)、誘導されない; レーン3:W3110 (pAKL15E)、誘導された)。
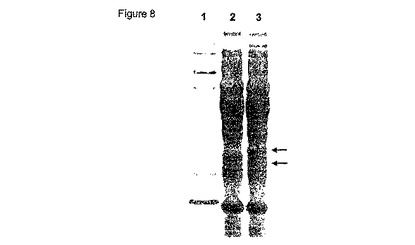
【図9】図9は、メリビオース誘導性melAB2プロモーターおよび単鎖抗体(scFv, S1)をコードする配列を含むプラスミドpBLL7-pelB-S1を示す。S1をコードする配列は、PelBシグナルペプチドをコードする配列によって先行される。
【図10】図10は、非誘導性(-)および誘導性(+)株W3110 (pBLL7-pelB-S1)の未精製抽出物のSDS-PAGEを示す。サンプルは示されたような異なる時間間隔で採取された。リゾチーム処理が可溶性および不溶性タンパク質画分が分析された。矢印はscFvタンパク質を示す。 M = Mark12、インビトロゲンの標準分子量。
【発明の分野】
【0001】
本発明は、原核生物の宿主における、例えば組換えタンパク質のようなポリペプチドをコードする核酸の異種発現に適したベクターに関する。より具体的には、本発明は、前記宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオースオペロンのプロモーター領域を含む宿主において発現可能な新しいベクターに関する。前記核酸配列の発現は、メリビオースオペロンの前記プロモーター領域によって制御される。本発明は、さらに例えばポリペプチドをコードする核酸の異種発現に適したこれらのベクターの使用に関する。
【発明の背景】
【0002】
原核生物の系における例えば組換えタンパク質のようなポリペプチドをコードする核酸の異種発現のための多くの系が記載されてきた。しかしながら、原核生物の宿主系におけるほとんどの異種遺伝子発現系は、もっぱら限られた一連の細菌プロモーターに依存してきた。最も幅広く使用されている原核生物のプロモーターには、ラクトース[lac] (Yanisch-Perron et al., 1985, Gene 33, 103-109)、およびトリプトファン[trp] (Goeddel et al., 1980, Nature (London) 287, 411-416) プロモーター、ならびにこれら二つから誘導されたハイブリッドプロモーター [tac および trc] (Brosius, 1984,Gene 27 :161-172 ; Amann and Brosius, 1985, Gene 40,183-190) が含まれる。他の誘導プロモーター系、例えばアラビノースによる誘導のaraBプロモーター(WO 86 04356)、L-ラムノースによる誘導のラムノース プロモーター rhaSB (WO 03068956) またはL-ラムノースによる誘導のラムノース プロモーター rhaBAD (WO 2004/050877)が、タンパク質の異種発現のために十分に記載されてきた。しかしながら、異種遺伝子発現について使用された多くの既知の原核プロモーターが、宿主細胞に対する異種生成物の毒性、生成物の発現の低い割合または非機能的凝集塊(封入体)の形成といった欠点を有する。
【0003】
タンパク質の異種発現において使用されるさらなる誘導性のプロモーター系、例えば、Belyaeva et al., 2000, Mol. Microbiol. 36(1), 211-222 によって記載されたような、メリビオースによって誘導性のメリビオースオペロンが存在する。異種発現について使用されるこれらの誘導性のプロモーター系の大半は、非常に低い誘導率および所望の相同的生成物の低発現に至るという欠点を有する。さらに、これらのプロモーター系は非常に厳密には制御されておらず、従って非誘導性状態における背景活性を誘導し、厳密な発現制御を可能にはしない。例えば、Belyaevaらは、大腸菌のβ-ガラクトシダーゼの発現のために大腸菌のlacZ遺伝子に融合したミリビオースオペロンの異なる断片を使用した。しかしながら、最も高いβ-ガラクトシダーゼ活性を産生した断片(KK43, JK19)は、最も高い背景活性を強く産生した。
【0004】
従って、上述した不利益のない核酸配列の異種発現に適した向上した原核発現系を提供する必要がある。
【発明の概要】
【0005】
前述の記載から明らかなように、これらのおよび他の目的は、所望の異種生成物の高レベルの発現にとって有用である原核プロモーター領域を含む新たなベクターを提供することによって達成される。驚くべきことに、メリビオースオペロンのプロモーター領域が異種生成物の大量の厳密に制御された発現を可能にすることを見出した。第一の側面において、本発明の目的は、前記宿主に対して異種である核酸配列を含む転写単位と機能的に連結したメリビオースオペロンのプロモーター領域を含む宿主において発現可能な新たなベクターを提供することである。前記核酸配列の発現がメリビオースオペロンの前記プロモーター領域によって制御される。また、原核宿主における核酸配列の制御された異種発現に適した前記新規ベクターの使用; 前記宿主に対して異種である核酸配列を含む転写単位と機能的に連結されたメリビオースオペロンのプロモーター領域を含む宿主において発現可能な単離および精製された核酸配列であって、前記核酸配列の発現がメリビオースオペロンの前記プロモーター領域によって制御される核酸配列; 前記ベクターまたは前記単離および精製された核酸配列で形質転換された原核宿主; および前記ベクターを使用する宿主におけるポリペプチドを生成するための方法が提供される。
【0006】
他の対象および利点は、以下の例証的図面を参照して進める以下の詳細な説明および添付の請求項の内容から当業者によって明らかになるであろう。
【発明の詳細な説明】
【0007】
本明細書中で使用されたとき、以下の定義は、本発明の理解を容易にするために提供される。
【0008】
「宿主において発現可能なベクター」または「発現ベクター」は、宿主細胞において特定の核酸配列の転写を可能にする一連の特異的なポリ核酸要素で組換え的または合成的に生成されたポリ核酸構築物である。典型的には、このベクターは、プロモーターに作動可能に連結された、特定の被転写核酸配列を含む転写単位を含む。宿主において発現可能なベクターは、例えば自律的または自己複製的プラスミド、コスミド、ファージ、ウイルスまたはレトロウイルスとすることができる。
【0009】
用語「宿主」、「宿主細胞」および「組換え宿主細胞」は、本明細書中では可換的に使用され、本発明の一以上のベクターまたは単離および精製された核酸配列が導入された原核細胞を指す。これらの用語が特定の対象細胞だけではなく、該細胞の子孫または潜在的子孫をも指すことが理解される。なぜなら、一定の修飾が変異または環境的影響により発生するためである。例えば、事実、子孫は親細胞とは同一ではないが、本明細書中で使用されたときに該用語の範囲内に含まれる。
【0010】
用語「含む(comprise)」は、一般的には含む(include)の意味において使用される。すなわち、一以上の特徴または成分の存在を許容する。
【0011】
「プロモーター」とは、本明細書中に使用されたとき、転写単位の発現を制御する核酸配列をいう。「プロモーター領域」は、細胞内のRNAポリメラーゼを結合することができ、かつ下流(3’方向)のコード配列の転写を開始することができる制御領域である。プロモーター領域内には、転写開始部位(ヌクレアーゼS1でマッピングすることによって保存的に定義される)、ならびに推定上の-35領域およびプリブノウボックスのようなRNAポリメラーゼの結合を担うタンパク質結合ドメイン(コンセンサス配列)が見出されるであろう。
【0012】
「メリビオースオペロン」とは、Hanatani et al., 1984, J Biol. Chem, 259(3), 1807-12に記載された大腸菌のメリビオースオペロンをいう。メリビオースオペロンは、二つの相違するプロモーターを含む積極的に制御された異化オペロンである。第一のプロモーター(melR プロモーター)は、第二のプロモーター(melAB プロモーター)のメリビオース依存性刺激に不可欠であるmelR 遺伝子の発現を担う。この第二のプロモーターからのメリビオース誘導性の転写は、α-ガラクトシダーゼをコードするmelA遺伝子およびメリビオースパーミアーゼをコードするmelB遺伝子の同時転写を開始する。メリビオースオペロンは、二つの異化制御タンパク質結合部位、位置-81,5のCRP2および位置-195,5のCRP1(melAB プロモーターの転写開始部位の上流)ならびに位置-42,5 (サイト2’)、-62,5 (サイト2)、-100,5 (サイト1)、-120,5 (サイト1’)および-238,5 (サイトR)の5つのMelR 結合部位(melAB プロモーターの転写開始部位の上流)を含む。「メリビオースオペロンのプロモーター領域」は、melAB プロモーター、melR プロモーター、melR遺伝子およびCRP結合部位およびMelR結合部位を含む、melA 遺伝子およびmelB 遺伝子の発現を制御するプロモーター領域を意味する。本明細書で参照された「melAB プロモーター」は、主として転写開始部位、推定上-35領域、プリブノウボックス、CRP結合部位およびMelR結合部位からなる。本明細書中で言及された「CRP1結合部位が欠損したmelAB プロモーター」は、主として転写開始部位、推定上-35領域、プリブノウボックス、CRP結合部位およびサイトRを除いたMelR結合部位からなる。「CRP1結合部位が欠損したmelAB プロモーター」は、抑制されまたは遮断されたCRP1結合部位を含んでいてもよい。これはトランスポゾン支援変異誘発またはノックアウト変異のような既知の技術によって行われ得る。好ましくは、「CRP1結合部位が欠損したmelABプロモーター」は、CRP1結合部位を含まない。
【0013】
メリビオース (6-O-[α]-D-ガラクトピラノシル-D-グルコース)は、酵母での発酵によってラフィノースから得ることができる二糖である。
【0014】
「CRP」は、「異化産物制御タンパク質」を意味する。「CRP」は、しばしば同義的意味をもつ「サイクリックAMP受容タンパク質」として当該技術分野において呼ばれる。CRPは、メリビオースオペロンのような異化オペロンの活性化を媒介するサイクリックAMP (cAMP)によって調節される制御タンパク質である。
【0015】
「エンハンサー」は、転写単位の同定、転写単位に関係した配列の位置、または配列の配向とは無関係に、転写単位の転写を増強するよう作用する核酸配列である。本発明のベクターには任意的にはエンハンサーが含まれる。
【0016】
「転写単位」は、本明細書中において使用されたとき、通常は単一のRNA分子に転写される核酸配列である。転写単位は、機能的に関連したポリペプチド分子をコードする一つの遺伝子(モノシストロニック)または二つの遺伝子(ジシストロニック)またはそれ以上の遺伝子(ポリシストロニック)を含んでもよい。
【0017】
核酸配列は、他の核酸配列と機能的関係に配置されるときに「機能的に連結される」。例えば、シグナル配列についてのDNAは、それがタンパク質の分泌に関与するプレタンパク質として発現される場合、タンパク質についてのDNAと機能的に連結され;プロモーターは、それが配列の転写に影響を与える場合、コード配列に機能的に連結され;またはリボソーム結合部位のような翻訳開始領域は、それがポリペプチドの転写を容易にするように配置される場合、例えばポリペプチドをコードする核酸配列と機能的に連結される。連結は、保存的制限部位でのライゲーションによって達成され得る。このような部位が存在しない場合、合成オリゴヌクレオチドアダプターまたはリンカーが、従来手法に従って使用される。
【0018】
「核酸」または「核酸配列」または「単離および精製された核酸または核酸配列」は、本明細書中において言及されたとき、DNA、RNA、またはDNA/RNAハイブリッドのいずれであってもよい。核酸または核酸配列がベクター上に配置される場合、それは通常はDNAである。本明細書中で言及されるDNAは、例えば二本鎖DNA、単鎖DNA、一方または両方の鎖が二以上の断片で構成された二本鎖DNA、一方または両方の鎖が非中断性ホスホジエステルバックボーンを有する二本鎖DNA、一以上の単鎖部位および一以上の二本鎖部位を含むDNA、DNA鎖が完全に相補的である二本鎖DNA、DNA鎖が部分的にのみ相補的である二本鎖DNA、環状DNA、共有結合性閉鎖DNA、直鎖状DNA、共有結合性架橋DNA、cDNA、化学合成DNA、半合成DNA、生合成DNA、天然単離DNA、酵素消化DNA、剪断DNA、標識DNA、例えば放射性標識DNAおよび蛍光色素標識DNA、一以上の非天然発生化学種の核酸を含むDNAを含む、任意のポリデオキシヌクレオチド配列とすることができる。DNA配列は、標準的な化学技術、例えば、リン酸トリエステル方法または自動化合成方法およびPCR方法によって合成することができる。精製および単離されたDNA配列はまた、酵素的技術によって生成することができる。
【0019】
本明細書中で言及されるRNAは、例えば単鎖RNA、cRNA、二本鎖RNA、一方または両方の鎖が二以上の断片で構成される二本鎖RNA、一方または両方の鎖が非中断性ホスホジエステルバックボーンを有する二本鎖RNA、一以上の単鎖部位および一以上の二本鎖部位を含むRNA、RNA鎖が完全に相補的である二本鎖RNA、RNA鎖が部分的にのみ相補的である二本鎖RNA、共有結合性架橋RNA、酵素消化RNA、剪断RNA、mRNA、化学合成RNA、半合成RNA、生合成RNA、天然単離RNA、標識RNA、例えば放射性標識RNAおよび蛍光色素標識RNA、一以上の非天然発生化学種の核酸を含むRNAとすることができる。
【0020】
「バリエーション」または「配列のバリエーション」は、保存的核酸の置換による基準配列とは異なる核酸配列を意味する。このとき、一以上の核酸が同じ特徴をもつ他の核酸によって置換される。縮重した配列、欠失および挿入を含む配列が基準配列と同じ機能(機能的に等価)を示す限り、これらの修飾された配列は変異に含まれる。
【0021】
本明細書中で使用されるとき、用語「ポリペプチド」、「ペプチド」、「タンパク質」、「ポリペプチド」および「ペプチド」は可換的に使用され、α-アミノ基と隣接残基のカルボキシ基との間のペプチド結合によって他のアミノ酸残基と連結された一連のアミノ酸残基を指す。
【0022】
用語「単離および精製された核酸配列」とは、核酸配列が本発明に従った状態にあることをいう。該核酸配列は、それらが天然に関与する材料、例えば他の核酸から遊離または実質的に遊離している。該核酸は、自然環境またはそれらが調製される環境(例えば細胞培養)において見出される。調製は、組換え技術によってインビトロまたはインビボにおいて行われる。
【0023】
用語「形質転換」、「形質転換された」または「核酸を宿主細胞に導入すること」は、ベクターのような細胞外核酸が材料を伴ってまたは伴わないで宿主細胞に入る任意のプロセスを意味する。用語「形質転換された細胞(cell transformed)」または「形質転換された細胞 (transformed cell)」は、細胞外核酸が導入され、かつ細胞外核酸を内包する細胞またはその子孫を意味する。核酸は、該核酸が染色体複合体または染色体外要素として複製可能であるように細胞に導入され得る。例えば発現ベクターでの適切な宿主細胞の形質転換は、周知の方法、例えばマイクロインジェクション、エレクトロポーレーション、粒子照射によって、あるいは化学的方法、例えばリン酸カルシウム媒介性形質転換によって達成され得る(例えば、Maniatis et al. 1982, Molecular Cloning, A laboratory Manual, Cold Spring Harbor Laboratory or in Ausubel et al. 1994, Current protocols in molecular biology, John Wiley and Sons.に記載)。
【0024】
「異種核酸配列」または「宿主に対して異種の核酸配列」は、例えば、宿主に対して外来であるポリペプチドのような発現産物(「異種発現」または「異種産物」)、すなわち、宿主とは異なる供与体から発生する核酸配列、あるいは宿主に対して外来であるポリペプチドのような発現産物をコードする化学的に合成された核酸配列をコードする核酸配列を意味する。宿主が特定の原核生物種である場合、異種核酸配列は、好ましくは生物の異なる属またはファミリー、より好ましくは異なる目または綱、特に異なる門および異なるドメイン(領域)から発生する。
【0025】
宿主とは異なる供与体から発生する異種核酸配列は、それが宿主細胞に導入される前、このような修飾された配列が基準配列と同じ機能(機能的に等価)を示す限りにおいて、単鎖核酸または異種核酸配列の一部の変異、挿入、削除または置換によって修飾され得る。本明細書中で参照された異種核酸配列には、生物の異なるドメイン(領域)、例えば、ファージディスプレイライブラリにおいて使用されるヒト抗体のような真核生物(真核起源の)から発生する核酸配列が包含される。核酸配列または核酸配列の一部は、原核生物の宿主の「コドン使用頻度」に応じて修飾される。
【0026】
「シグナル配列」または「シグナルペプチド配列」とは、非細胞質内部位(例えば分泌)へ細胞によって通常輸送されるかまたは膜成分になるタンパク質のNH2-末端に存在する短いアミノ酸配列(すなわち、シグナルペプチド)をコードする核酸配列をいう。シグナルペプチドは、細胞質内から非細胞質内部位へのタンパク質の輸送を方向づける。
【0027】
「翻訳開始領域」は、翻訳開始を促進し、かつシャインダルガーノ配列のようなリボソーム結合部位として機能するシグナル領域である。
【0028】
「転写終結領域」とは、RNAポリメラーゼが転写を終結する配列をいう。転写終結領域は通常転写単位の一部であり、mRNAの安定性を増強する。
【0029】
「抗体」とは、抗原よる刺激後に免疫系のB細胞によって生産される血漿タンパク質のクラスをいう。哺乳動物(例えばヒト)抗体には、IgG、M、A、EまたはDクラスの免疫グロブリンがある。用語「抗体」が本発明の目的のために使用されるとき、これに限定されないが、ポリクローナル抗体、モノクローナル抗体、抗イディオタイプ抗体および自己免疫疾患、糖尿病、多発性硬化症およびリウマチ関節炎において存在する自己抗体、ならびにキメラ抗体が含まれる。基礎的な抗体の構造単位は、四量体構成であることが知られている。各四量体は、ポリペプチド鎖の二つの同一の対で構成されており、各対は、一本の「軽鎖(約25kDa)」および一本の「重鎖(約50-70kDa)」で構成されている。各鎖のアミノ末端部位は、抗原認識の重要な役割を担う約100〜110(あるいはそれ以上)のアミノ酸で構成された可変領域を含む。各鎖のカルボキシ末端部位は、エフェクター機能についての重要な役割を担う定常領域を含む。
【0030】
軽鎖はカッパまたはラムダで分類される。重鎖はガンマ、ミュー、アルファ、デルタ、またはイプシロンとして分類され、抗体のアイソタイプをIgG、IgM、IgA、IgDおよびIgEとして定義する。軽鎖および重鎖では、可変および定常領域は、約12またはそれ以上のアミノ酸で構成された「J」領域によって連結される。重鎖はまた約10以上のアミノ酸の「D」領域を含む。
【0031】
用語「抗体」は、抗体全体およびその結合断片を意味するのに使用される。結合断片は、単鎖断片、Fv断片およびFab断片を含む。用語「Fab断片」は、無処理の抗体のパパイン開裂から生じる結合断片を意味するものとして当該技術分野において使用されるときがある。用語「Fab’およびF(ab’)2」は、ペプシン開裂によって生成された無処理の抗体の結合断片をいうものとして当該技術分野において使用されるときがある。本発明の内容において、Fabは一般的に、抗原の特異的結合に十分な少なくとも実質的に完全な軽鎖および重鎖可変領域と、軽鎖および重鎖の会合を維持するのに十分な軽鎖および重鎖定常領域の一部を有する無処理の抗体の二重鎖結合断片をいうときに使用される。このようなFabの例は、例えばSkerra et al., 1988, Science 240 (4855), 1038-41に記載されている。Fab断片、例えばIgGイディオタイプのFab断片は、無処理の免疫グロブリンの二つの重鎖の間に二つの鎖間ジスルフィド結合を形成する少なくとも一つの二システイン残基を含んでいてもいなくともよい。通常、Fab断片は、全長または実質的に全長の軽鎖を、可変領域および定常領域の少なくともCH1ドメインを含む重鎖と複合させることによって形成される。
【0032】
さらに、軽鎖上のC末端システインは、本発明に従って重鎖および軽鎖間の鎖間ジスルフィド結合を削除するためにセリンまたは他のアミノ酸と置換され得る。さらに、典型的には遺伝子工学によって、例えば異なる生物種に属する免疫グロブリン遺伝子区分(例えば、可変領域をコードする区分および定常領域をコードする区分)から構築される抗体であるキメラ抗体が包含される。例えば、マウスモノクローナル抗体由来の遺伝子の可変(V)区分は、ヒト定常(C)区分、例えばIgG1およびIgG4と連結され得る。典型的なキメラ抗体は、マウス抗体由来のVまたは抗原結合ドメインおよびヒト抗体由来のCまたはエフェクタードメインからなるハイブリッドタンパク質である。キメラ抗体は、抗体の可変領域を提供するマウスまたは他の非ヒト抗体と同一または類似の結合特異性および親和性を有する。
【0033】
用語「ヒト抗体」には、天然または人工の加工された親和性変異を含むヒト生殖系列免疫グロブリン配列から誘導された可変および定常領域(存在すれば)を有する抗体が含まれる。本発明のヒト抗体は、ヒト生殖系列免疫グロブリン配列によってコードされないアミノ酸残基を含むことができる(例えば、インビトロでのランダムまたは部位特異的変異誘発またはインビボでの体細胞変異によって誘導された変異)。しかしながら、用語「ヒト抗体」には、他の哺乳動物種(例えばマウス)の生殖系列から誘導されたCDR配列が、ヒトのフレームワーク配列上にドラフトされた抗体(すなわち、ヒト化された抗体)が含まれる。このような「ヒト抗体」の機能的変異体、例えば切断された態様または特定のプロリンまたはシステイン残基が当該技術分野において周知の遺伝子工学手段によって加工された加工性変異体が、上記用語に包含される。このような例は、例えばWO98/02462に見出されるであろう。しかしながら、該用語は、任意のグリコシル化またはペプチドバックボーンの他の化学的修飾とは無関係に、該抗体のアミノ酸配列のみと関係する。
【0034】
一側面において、本発明は、前記宿主と異種である核酸配列を含む転写単位と機能的に連結したメリビオースオペロンのプロモーター領域を含む宿主において発現可能なベクターを提供する。ここで、前記核酸配列の発現は、メリビオースオペロンの前記プロモーター領域によって制御される。
【0035】
本発明によるベクターは、自律的または自己複製的プラスミド、コスミド、ファージ、ウイルスまたはレトロウイルスが好ましい。広い範囲の宿主/ベクターの組み合わせが、本発明の核酸配列を発現するのに使用され得る。有用な発現ベクターは、例えば、染色体、非染色体および/または合成核酸配列のセグメントからなる。適切なベクターには、例えば大腸菌に特異的なベクターのような特異的宿主範囲をもつベクター、およびグラム陰性細菌に有用なベクターのような広い宿主範囲をもつベクターが含まれる。「低コピー数」、「中コピー数」および「高コピー数」のプラスミドが使用され得る。
【0036】
大腸菌内の発現に有用なベクターには、pQE70、pQE60およびpQE-9 (QIAGEN,Inc. ); pBluescript Vektoren、Phagescript Vektoren、pNH8A、pNH16a、pNH18A、pNH46A (Stratagene Cloning Systems, Inc.); ptrc99a、pKK223-3、pKK233-3、pDR540、pRIT5 (Pharmacia Bio-tech, Inc. ); pLG338、pACYC184、pBR322、pUC18、pUCl9、pKC30、pRep4、pACYC177、pACYC184、pRSF1010およびpBW22 [Wilms et al., 2001, Biotechnology and Bioengineering, 73 (2) 95-103] またはこれらの誘導体、例えばプラスミドpBLL15またはプラスミドpAKL15Eがある。さらに有用なプラスミドは当業者に周知であり、例えば、“Cloning Vectors” (Eds. Pouwels P. H. et al. Elsevier, Amsterdam-New York-Oxford, 1985)に記載されている。
【0037】
本発明の好ましいベクターは、自律的または自己複製的プラスミドである。より好ましくは、特異的宿主範囲をもつベクター、例えば大腸菌に特異的なベクターである。最も好ましくは、pBR322、pUC18、pACYC177、pACYC184、pRSF1010およびpBW22またはこれらの誘導体、例えばプラスミドpBLL15またはプラスミドpAKL15E、特に好ましくはpBW22またはこれらの誘導体、例えばプラスミドpBLL15またはプラスミドpAKL15E、より特に好ましくはpBLL15またはpAKL15E、最も特に好ましくはpAKL15Eである。
【0038】
好ましい実施態様において、本発明において使用されるメリビオースオペロンのプロモーター領域はmelABプロモーターである。本発明のより好ましい実施態様において、melABプロモーターは、CRP1結合部位が欠損している。特に好ましい実施態様において、CRP1結合部位が欠損したmelABプロモーターは、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる。通常、プロモーター領域のMelR結合部位は修飾されない。本発明において使用される、メリビオースオペロンのプロモーター領域、melABプロモーター、CRP1結合部位が欠損したmelABプロモーター、ならびに配列番号1からなるCRP1結合部位が欠損したmelABプロモーター、これに相補的な配列およびこれらの変異配列は、通常は大腸菌のメリビオースオペロンまたは他の原核生物、特に腸内細菌のファミリーに属する生物体の機能的に等価なプロモーター領域に由来する。好ましくは、メリビオースオペロンのプロモーター領域、melABプロモーター、CRP1結合部位が欠損したmelABプロモーター、ならびに配列番号1からなるCRP1結合部位が欠損したmelABプロモーター、これに相補的な配列およびこれらの変異配列は、大腸菌のメリビオースオペロン由来である。他の原核生物の機能的に等価なプロモーター領域は、メリビオースによって誘導可能であるプロモーター領域、すなわち、メリビオースの欠如下よりも存在下においてより高い発現活性を示すプロモーター領域を包含する。
【0039】
本発明による転写単位は、通常さらに前記転写単位の翻訳の開始点の翻訳開始領域上流を含み、前記転写開始領域は、配列AGGAGATATACAT(配列番号2)からなる。転写開始領域は核酸配列と機能的に連結される。配列AGGAGATATACAT(配列番号2)は通常は上流に位置し、ATG、GTGまたはTTGになり得る転写単位の翻訳の開始点に直接的に隣接する。
【0040】
通常、前記転写単位はさらに、本発明の異種核酸配列と機能的に連結したシグナル配列を含む。この場合において、ジシストロニックまたはポリシストロニックな転写単位が使用され、シストロンの各々に機能的に連結された異なるまたは同一のシグナル配列が適用され得る。好ましくは、異なるシグナル配列がこのような場合において使用される。使用されたシグナル配列は、原核生物または真核生物のシグナル配列とすることができる。通常、原核生物のシグナル配列が適用される。使用され得および大腸菌にとって特に有用な真核シグナル配列は、例えばヒトセルロプラスミンシグナル配列、ヒト好中球デフェンシン1,2,3前駆体シグナル配列またはWO 03068956に記載されたChlamydomonas reinhardtiiアリルスルファターゼのシグナルペプチドである。通常適用される原核生物のシグナル配列は、糖、アミノ酸、ビタミンおよびイオンのための周辺質結合タンパク質のシグナルペプチド、例えばPelB (エルウィニア属chrysantemi、ペクチン酸塩リアーゼ前駆体)、PelB (エルウィニア属carotovora、ペクチン酸塩リアーゼ前駆体)、PelB (ザントモナス・キャンペストリス、ペクチン酸塩リアーゼ前駆体)、LamB (大腸菌、マルトポリン前駆体)、MalE (大腸菌、マルトース結合タンパク質前駆体)、Bla (大腸菌、β-ラクタマーゼ)、OppA (大腸菌、周辺質糖ペプチド結合タンパク質)、TreA (大腸菌、周辺質トレハロース前駆体)、MppA (大腸菌、周辺質ムレインペプチド結合タンパク質前駆体)、BglX (大腸菌、周辺質ベータ-グルコシダーゼ前駆体)、ArgT (大腸菌、リシン-アルギニン-オルニチン結合周辺質タンパク質前駆体)、MalS (大腸菌、α-アミラーゼ前駆体)、HisJ (大腸菌、ヒスチジン-結合周辺質タンパク質前駆体)、XylF (大腸菌、D-キシロース-結合周辺質タンパク質前駆体)、FecB (大腸菌、ジシトラート(dicitrate)結合周辺質タンパク質前駆体)、OmpA (大腸菌、外膜タンパク質A前駆体) および PhoA (大腸菌、アルカリ性ホスファターゼ前駆体)である。
【0041】
好ましい実施態様において、シグナルペプチドは、大腸菌シグナルペプチドLamB (マルトポリン前駆体)、MalE (マルトース結合タンパク質前駆体)、Bla (β-ラクタマーゼ)、OppA (周辺質オリゴペプチド結合タンパク質)、TreA (周辺質トレハロース前駆体)、MppA (周辺質ムレインペプチド結合タンパク質前駆体)、BglX (周辺質β-グルコシダーゼ前駆体)、ArgT (リシン-アルギニン-オルニチン結合周辺質タンパク質前駆体)、MalS (α-アミラーゼ前駆体)、HisJ (ヒスチジン-結合周辺質タンパク質前駆体)、XylF (D-キシロース-結合周辺質タンパク質前駆体)、FecB (ジシトラート結合周辺質タンパク質前駆体)、OmpA (外膜タンパク質A前駆体)およびPhoA (アルカリ性ホスファターゼ前駆体)から選択される。これらは大腸菌の異種発現について特に有用である。より好ましくは、大腸菌シグナルペプチドLamB (マルトポリン前駆体)、MalE (マルトース結合タンパク質前駆体)、Bla (β-ラクタマーゼ)、TreA (周辺質トレハロース前駆体)、ArgT (リシン-アルギニン-オルニチン結合周辺質タンパク質前駆体)、FecB (ジシトラート結合周辺質タンパク質前駆体)である。最も特に好ましくは、大腸菌シグナルペプチドLamB (マルトポリン前駆体)およびMalE (マルトース結合タンパク質前駆体)である。
【0042】
本発明の発現ベクターにおいて使用されるシグナル配列は、商業的または化学的に合成されることによって得られうる。例えば、シグナル配列は、Van Devanter et. al., Nucleic Acids Res. 12:6159-6168 (1984)に記載されたように自動化合成装置を使用する、Beaucage & Caruthers, Tetrahedron Letts. 22:1859-1862 (1981)に記載された固相ホスホラミダイトトリエステルによって合成され得る。オリゴヌクレオチドの精製は、Pearson & Reanier, J. Chrom. 255:137-149 (1983)に記載されたように天然アクリルアミドゲル電気泳動法またはイオン交換HPLCによって行われ得る。
【0043】
通常、前記転写単位にはさらに、rrnB、RNA I、T7Te、rrnB T1、trp a L126、trp a、tR2、T3Te、P14、tonB t、およびtrp a L153から選択された転写終結領域が含まれる。好ましくは、rrnB 転写終結配列が使用される。
【0044】
本発明による異種核酸配列は、宿主に対して外来種である発現産物をコードする。宿主が大腸菌のような原核生物種である場合、対象の核酸配列は、より好ましくはガンマプロテオバクテリアのような他のクラスに由来、例えばBurkholderia spに由来し、特に異なる門、例えば古細菌に由来し、最も特に真核生物、例えば哺乳動物、特にヒトに由来する。しかしながら、異種核酸配列は、宿主の「コドン使用頻度」に基づいて修飾され得る。本発明による異種配列は、通常は対象の遺伝子である。対象の遺伝子は、好ましくは異種ポリペプチド、例えば構造的、制御的または治療学的タンパク質、または構造のN-もしくはC-末端融合、他のタンパク質(「タグ」)をもつ制御もしくは治療学的タンパク質、例えば緑色蛍光タンパク質または他の融合タンパク質をコードする。異種核酸配列は、RNAの形態において使用され得る転写産物、例えばアンチセンスRNAをコードしてもよい。
【0045】
タンパク質は、宿主細胞および/または細胞外培地の細胞質または周辺質空間内に存在する不溶性凝集塊または可溶性タンパク質として生産され得る。好ましくは、タンパク質は、宿主細胞および/または細胞外培地の周辺質空間内に存在する可溶性タンパク質として生産される。タンパク質の例には、ホルモン、例えば成長ホルモン、成長因子、例えば上皮性成長因子、鎮痛性物質様エンケファリン、酵素様キモトリプシン、抗体、ホルモンに対する受容体が含まれる。また、視覚化マーカー、例えば緑色蛍光タンパク質として通常使用されるタンパク質が含まれる。
【0046】
対象の他のタンパク質は、成長因子受容体(例えば、FGFR、PDGFR、EFG、NGFR、およびVEGF) およびこれらのリガンドである。他のタンパク質は、Gタンパク質受容体であり、物質K受容体、アンジオテンシン受容体、[アルファ]-および[ベータ]-アドレナリン受容体、セロトニン受容体、およびPAF受容体(例えば、Gilman, Ann. Rev. Biochem. 56, 625-649 (1987)を参照)。他のタンパク質は、イオンチャネル(例えば、カルシウム、ナトリウム、カリウムチャネル)、ムスカリン受容体、アセチルコリン受容体、GABA受容体、グルタミン酸受容体、およびドーパミン受容体(Harpold, 米国特許番号5,401,629 および5,436,128を参照)を含む。対象の他のタンパク質は、インテグリンのような付着性タンパク質、および免疫グロブリンスーパーファミリー膜(Springer, Nature 346, 425-433 (1990). Osborn, Cell 62, 3 (1990); Hynes, Cell 69, 11 (1992)を参照)である。他のタンパク質には、サイトカイン、例えばインターロイキンIL-1〜IL-13、腫瘍ネクローシス因子[アルファ]および[ベータ]、インターフェロン[アルファ]、[ベータ]、および[ガンマ]、腫瘍成長因子Beta(TGF-[ベータ]、インターフェロン[アルファ]、[ベータ]、および[ガンマ]、腫瘍成長因子ベータ(TGF-[ベータ])、コロニー刺激因子(CSF)および顆粒球単球コロニー刺激因子(GM-CSF)がある(Human Cytokines: Handbook for Basic & Clinical Research. Aggrawal et al. eds., Blackwell Scientific, Boston, Mass. 1991を参照)。対象の他のタンパク質は、細胞内および細胞間メッセンジャー、例えばアデニル、シクラーゼ、グアニルシクラーゼ、およびホスホリパーゼCである。薬剤はまた、他のタンパク質である。対象の異種タンパク質は、ヒト、哺乳動物または原核生物起源とすることができる。他のタンパク質は、抗原、例えば微生物病原体(ウイルスおよび細菌)および腫瘍由来の糖タンパク質および炭水化物である。
【0047】
他のタンパク質は、キモシン、プロテアーゼ、ポリメラーゼ、デヒドロゲナーゼ、ヌクレアーゼ、グルカナーゼ、オキシダーゼ、α-アミラーゼ、オキシドレダクターゼ、リパーゼ、アミダーゼ、ニトリルヒドラターゼ、エステラーゼまたはニトリラーゼのような酵素である。
【0048】
好ましくは、本発明による異種核酸配列は、ポリペプチド、より好ましくは抗体および最も好ましくはFab断片をコードする。特に、ヒト抗体またはヒト化抗体、より特にはヒトFab断片が核酸配列によってコードされる。核酸配列によってコードされたヒトFab断片は、好ましくはヒト抗体断片または他の哺乳動物種からの少なくとも一つのCDRを移植されたヒト抗体断片である。
【0049】
一以上の好ましい実施態様において、ヒトFab断片は、Knappik et al., 2000, J. Mol. Biol. 296 (1), 57-86によって記載されたCDRにおいて人工的にランダム化された人工のコンセンサスフレームワークベースのヒト抗体ファージライブラリから得られ得る完全なヒトHuCAL-Fabである。
【0050】
他のより好ましい任意の実施態様において、任意的にキメラであってCDRを移植されたヒトFab断片は、非HuCAL Fabである。これは前述のHuCAL Fabの定義とは対照的である。つまり、完全なヒトFab断片の場合において、好ましくはそれはHuCALコンセンサス配列のフレームワークを共有しないが、その非CDR配列部位は、それぞれの可変および定常領域の軽鎖および重鎖の生殖系列コード配列に対して、少なくとも70%、より好ましくは85%、最も好ましくは95%のアミノ酸配列における同一性を示すことを意味し、並びに追加的およびより好ましくは、そのCDRはゲノム親和性変異事象を含むリンパ系細胞の天然に生じるゲノム配列から直接的に得られることを意味する。
【0051】
Fab断片は、好ましくはIgG抗体から誘導され、無処理の免疫グロブリンにおける二つの重鎖間にある二つの鎖間ジスルフィド結合を形成するシステイン残基を含まない。特に、抗体または好ましくはFab断片の重鎖および軽鎖は、ジシストロニック転写単位によってコードされる。各鎖はシグナル配列および転写単位の翻訳の開始点の上流にある同一の(identical)翻訳開始領域に機能的に連結される。好ましくは、転写開始領域は、配列AGGAGATATACAT (配列番号2)からなる。
【0052】
本発明において、シグナル配列および均質核酸配列が発現ベクター内で配置される順序および距離を変えることができる。好ましい実施態様において、シグナル配列は、例えば対象のポリペプチドをコードする核酸配列に対して5’(上流)にある。例えば、シグナルペプチド配列および例えば対象のポリペプチドをコードする核酸配列は、ゼロ〜約1000アミノ酸まで分離させることができる。好ましい実施態様において、例えば対象のポリペプチドをコードするシグナルペプチド配列および核酸配列は、互いに直接的に隣接、すなわちゼロ核酸で分離される。
【0053】
好ましくは、本発明のベクターのプロモーター領域および機能的に連結された転写単位は、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる。
【0054】
より好ましくは、プロモーター領域および本発明のベクターの機能的に連結された転写単位は、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる。
【0055】
また、原核生物の宿主における核酸配列の制御された異種発現のための本発明によるベクターの使用が、本発明によって包含される。発現は、原核生物の宿主に利用可能なメリビオースの量によって制御され得る。通常、培養された原核宿主の培養液中におけるメリビオースの量は、0.01〜100g/L、好ましくは0.1〜10g/L、より好ましくは1〜5g/Lである。
【0056】
好ましくは、異種核酸配列は、ポリペプチド、より好ましくは抗体および最も好ましくはFab断片をコードする。一方、抗体またはFab断片の重鎖および軽鎖は等しい量において発現し、その結果として高濃度の機能的抗体またはFab断片を導く。特に、ヒト抗体またはヒト化抗体、より特にはヒトFab抗体、最も特にはヒトFab断片は、異種核酸配列によってコードされる。
【0057】
高濃度の機能的抗体またはFab断片を得るために、等しい量の重鎖および軽鎖が発現することが重要である。両鎖の一方が他の鎖と比較して過剰生産される場合、望ましくない非還元性の高分子量免疫反応凝集塊が構築され得る。驚くべきことに、本発明のベクターで高力価の機能的抗体が得られ、非常に低量の過剰生産された軽鎖または重鎖または高分子重量の免疫反応性凝集塊のみが構築されることを見出した。通常、抗体またはFab断片の発現量の20%未満、好ましくは10%未満が、過剰生産された軽鎖または重鎖または高分子重量免疫反応性凝集塊として発現される。過剰生産された重鎖および軽鎖の量ならびに高分子量の免疫反応性凝集塊の量は、SDS-PAGEまたはウェスタンブロットを使用して培養された宿主細胞のリゾチーム抽出物のような抗体またはFab断片を発現する宿主の抽出物を分析することによって測定され得る。
【0058】
また、他の側面において、本発明は、前記宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオースオペロンのプロモーター領域を含む宿主において発現可能な単離および精製された核酸配列を提供する。前記核酸配列の発現は、メリビオースオペロンの前記プロモーター領域によって制御される。melABプロモーターは、好ましいプロモーター領域である。より好ましくはCRP1結合部位が欠損したmelABプロモーターである。最も好ましくは、単離および精製された核酸配列は、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる。特に、単離および精製された核酸配列は、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる。最も特に、単離および精製された核酸配列は、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる。本発明の単離および精製された核酸配列は、当業者に周知の標準的PCRプロトコルおよび方法によって単離され得る。前記精製および単離されたDNA配列はさらに、当業者に周知の一以上の制御配列、例えば核酸配列によってコードされた生成物の発現のために通常使用されるエンハンサーを含むことができる。
【0059】
本発明のベクターまたは単離および精製された核酸配列で効率良くかつ安定的に形質転換された宿主細胞を選択するために、選択可能なマーカー(例えば、抗体に対する耐性)をコードする遺伝子は、対象の核酸配列に沿って宿主細胞に導入され得る。選択可能なマーカーをコードする遺伝子はベクターまたは単離および精製された核酸配列上に配置され得、あるいは別々のベクター上にセパレートフォームで共導入され得る。抗生物質、例えばハイグロマイシン、アンピシリンおよびテトラサイクリンに対する耐性を与えるものを含む様々が選択可能なマーカーが使用され得る。抗生物質の量は、選択的条件を創出するために所望の量に適合され得る。通常、一つの選択可能なマーカーが使用される。同様に、蛍光タンパク質のようなレポーター遺伝子は、形質転換の効率を決定するために、対象の核酸配列に沿って宿主細胞に導入され得る。
【0060】
本発明の他の側面は、本発明のベクターで形質転換された原核生物の宿主を提供することである。本発明の特定の実施態様において、原核生物の宿主は、このような宿主細胞における分泌されたヘテロ二量体タンパク質、例えばFabを発現するジシストロニックな発現カセットにおいて二つの異なるコドン領域を含むプラスミドpBLL15またはプラスミドpAKL15E、好ましくはプラスミドpAKL15Eで形質転換される。好ましくは、このようなヘテロ二量体タンパク質はFabである。本発明の他の実施態様において、原核生物の宿主は、本発明の単離および精製された核酸配列で形質転換される。
【0061】
多様な原核生物の宿主細胞は、通常は本発明の異種核酸配列を発現するのに有用である。これらの宿主には、グラム陰性細胞、例えば大腸菌およびシュードモナス属またはグラム陽性細胞、例えばバチルス属およびストレプトマイセス属が含まれる。好ましくは、宿主細胞はグラム陰性細胞、より好ましくは大腸菌細胞である。使用され得る大腸菌は、例えば株TG1、W3110、DH1、XL1-BlueおよびOrigamiである。これらはDSMZ (Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany)を介して得ることができる。最も好ましくは、W3110が使用される。宿主細胞にはメリビオースを代謝しないものがある。大腸菌のようにメリビオースを取り込みかつ代謝することができる宿主細胞が修飾され得、メリビオースの取り込みおよび/または代謝に関連した一以上の機能が欠損する。メリビオースの取り込みおよび/または代謝に関連した一以上の機能の欠損は、例えばα-ガラクトシダーゼをコードするmelA遺伝子のようなメリビオースの取り込みおよび/または代謝と関連したタンパク質をコードする遺伝子の発現を抑制または遮断することによって達成され得る。これは、変異誘発またはノックアウト変異を担うトランスポゾンのような既知の技術によって行われ得る。通常、原核生物の宿主は、選択されたシグナル配列に対応し、例えば大腸菌のシグナル配列が使用される場合、宿主細胞は腸内細菌と同じファミリーの一員であり、より好ましくは宿主細胞は大腸菌株である。
【0062】
さらに本発明は、宿主細胞においてポリペプチドを生成するための方法であって、以下の工程を含む方法を提供する。
【0063】
a)ベクターを構築する工程と、
b)原核生物の宿主を前記ベクターで形質転換する工程と、
c)適切な条件下で細胞培養系において前記ポリペプチドを発現させる工程と、
d)前記ポリペプチドを細胞培養系から回収する工程。
【0064】
使用されるベクター、および原核宿主の構築および形質転換は上記に定義されたとおりである。ベクターによって包含された異種核酸配列は、ポリペプチドをコードする。好ましくは生成されたポリペプチドは、抗体および最も好ましくはFab断片である。抗体またはFab断片の重鎖および軽鎖は、細胞培養系で等量において発現され、高濃度の機能的抗体またはFab断片を誘導する。
【0065】
回分培養または流加培養のような連続的または非連続的培養は、培養管、撹拌フラスコまたは細菌発酵槽において適用され得る。通常、宿主細胞は、「栄養酵母培養液」またはKortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたグリセロール含有培養液またはKulla et al., 1983, Arch. Microbiol, 135, 1によって記載された栄養塩培養液のような複合培養液、当該技術分野において既知の従来培養液中において培養される。前記ポリペプチドの発現を実行するために好ましい培養液は、グリセロール含有培養液、より好ましくはKortz et al., 1995, J. Biotechnol. 39, 59-65によって記載された培養液である。
【0066】
培養液は、例えば、緩衝液、塩、ビタミンまたはアミノ酸のような追加成分を添加することによって適切なものとして修飾され得る。異なる培養液または培養液の組み合わせが細胞の培養中に使用され得る。好ましくは、基礎培地として使用される培地は、メリビオースプロモーター領域の厳格な制御を達成するために、メリビオースを含むべきではない。メリビオースは通常、培養液は培養系に依存しながらOD600に達した後に加えられる。通常、培養された原核生物の宿主の培養液中のメリビオースの量は、0.01〜100 g/l、好ましくは0.1〜10 g/l、より好ましくは1〜5 g/lである。回分培養では、通常OD600が0.4以上である。適切なpH範囲は、例えば6〜8、好ましくは7〜7.5であり、適切な培養温度は、10〜40、好ましくは20〜37℃である。細胞は、最大量の発現生成物が蓄積されるまでインキュベートされ、好ましくは1時間〜20日、より好ましくは5時間〜3日間にわたってインキュベートされる。発現生成物の量は、使用される培養系に依存する。撹拌フラスコ培養系では、通常、0.5 g/l 培養液の量の発現生成物が、本発明のベクターで形質転換された宿主で産生されうる。回分および/または流加培養モードにおける発酵槽培養系を使用すると、通常、0.5 g/l 発酵ブロスの量の発現生成物、好ましくは1 g/l以上、より好ましくは1.3 g/l以上の量の発現生成物が得られうる。
【0067】
宿主細胞における発現後、発現産物、例えば対象のポリペプチドは、宿主細胞の培養液から回収され得る。対象のポリペプチドが免疫グロブリン鎖であるとき、重鎖および軽鎖は通常は各々、宿主細胞において発現され、細胞の周辺質に分泌される。発現ベクターのシグナル配列によってコードされるシグナルペプチドは、免疫グロブリン鎖から加工されれる。その後、成熟した重鎖および軽鎖が構築され、完全な抗体またはFab断片が形成される。発現産物の最大の収量を得るために、通常は培養の終わりに細胞が収穫され、かつリゾチーム処理、超音波処理またはフレンチプレスによって溶解される。従って、ポリペプチドは通常、宿主細胞の粗溶解物として最初に得られる。これらはその後、ディファレンシャル沈殿法、モレキュラーシーブ・クロマトグラフィー、イオン交換クロマトグラフィー、等電点電気泳動、ゲル電気泳動、アフィニティー、および免疫アフィニティー・クロマトグラフィーを含む、当業者に既知の標準的なタンパク質精製手順によって精製され得る。これらは十分に知られており、日常的に実施される方法、例えばAusubel et al., supra., and Wu et al. (eds.), Academic Press Inc., N.Y.; Immunochemical Methods In Cell And Molecular Biologyに記載されている。例えば、組換え的に生成された免疫グロブリンまたはFab断片の精製について、それらは、発現した免疫グロブリンが特別に結合し得る標的分子が結合した樹脂を含むカラムを通過させることによって免疫アフィニィティー クロマトグラフィーで精製され得る。
【0068】
本発明はまた、例えば原核生物の宿主においてポリペプチドをコードする核酸の細胞内異種発現のための方法および手段に関する。特に、本発明は原核生物の宿主における異種ポリペプチドの細胞内発現のためのベクターに関する。該ベクターは、前記宿主に対して異種である核酸配列を含む転写単位に機能的に結合されたメリビオースオペロンのプロモーター領域を含む原核生物の宿主内において発現可能である。本発明のベクターのこの実施態様において、核酸配列はベクターおよび異種核酸によってコードされるポリペプチドの発現とともに宿主原核細胞を形質転換させる原核シグナル配列と結合しないので、該ポリペプチドは細胞質から非細胞質まで輸送されないであろう。代わりに、ポリペプチドは封入体の形態における細胞質内または可溶性形態内において発現するであろう。従って、発現したポリペプチドは、細胞、特に細胞抽出物から周知の手順によって単離および精製され得る。本発明はまた、原核宿主細胞内の異種核酸配列の制御された細胞内発現のための前記ベクターの使用;前記ベクターで形質転換された原核宿主または原核宿主細胞;前記ベクターを使用して原核宿主内において異種ポリペプチドの細胞内産生のための方法;およびプロモーター領域を含む異種ポリペプチドの細胞内産生のためのベクター、異種ポリペプチドをコードする異種核酸配列および配列AGGAGATATACATからなる翻訳開始領域を提供する。
【0069】
細胞内発現のために使用され得るベクターの好ましい実施態様において、メリビオースオペロンのプロモーター領域は、好ましくはCRP1結合部位が欠損したmelABプロモーターである。特に好ましくは、CRP1結合部位が欠損したmelABプロモーターは、配列番号1に示される配列、これに相補的な配列およびこれらの変異配列からなる。本発明の他の好ましい実施態様において、ベクターの転写単位にはさらに、前記転写単位の翻訳の開始点の上流にある転写開始領域が含まれる。転写開始領域は配列AGGAGATATACAT(配列番号2)からなる。さらに好ましい実施態様において、細胞内発現のために使用されるベクターには、rrnB転写終結配列のような転写終結領域が含まれる。本発明によれば、異種核酸配列は抗体、抗体断片などのポリペプチドをコードする。
【0070】
当業者は、明細書に記載された発明が特別に記載されたもの以外のバリエーションおよび修飾を含むことを認識するであろう。本発明には本発明の精神または要素的特徴から離れることのない全てのバリエーションおよび修飾が含まれることを理解すべきである。本発明にはまた、個別的または集団的に、この明細書中に言及または示された全ての工程、特徴、組成物および化合物が含まれ、かつ前記工程または特徴の任意かつ全ての組み合わせまたは任意の二以上の組み合わせが含まれる。本発明の開示は、全ての側面において拘束的なものではなく例証として考慮されるべきであり、本発明の範囲は、添付されたクレームによって示される。等価の意味および範囲内に入る全ての変化は、本発明の範囲内に包含されることを意味する。様々な文献がこの明細書を通して引用されているが、引用文献の各々は参照によってその全体が本明細書中に組み込まれる。
【0071】
前述の記述は、以下の例を参照してより完全に理解されるであろう。このような例は、しかしながら、本発明を実施する方法の例証であり、本発明の範囲を制限することを意味するものではない。
【0072】
例
例1
大腸菌W3110ゲノムを、正に制御されたオペロンについてスキャンした。KEGGデータベース上の利用可能なゲノムデータに基づいて(Kyoto Encyclopedia of Genes and Genomes, http://www.genome.ad.jp/kegg/kegg2.html)、正に制御された異化プロモーターを、発現プラスミドにおけるその使用のために同定かつ分析した。プロモーターは厳密に制御されるべきであり、かつ安価かつ無毒で導入されるべきである(つまり、産業的にも有用な化合物であるべきである)。以下の異なる正に制御された異化プロモーターを選択した。
【0073】
正に制御されたプロモーターでの発現プラスミドの構築
−prpプロモーター(誘導性プロピオネート)
−gutAプロモーター(誘導性グルシトール)
−melAB2プロモーター(誘導性メリビオース)
【0074】
プロモーター要素を含む正確なDNA断片を、相当する制御結合部位上の利用可能な情報に基づいて選択した。大腸菌の染色体DNAを「Pitcher et al., 1989, Letters in Applied Microbiology 8, 151-156」に記載された方法によって単離した。プロモーター断片を、以下のプライマーを使用してPCRによって株W3110の染色体DNAから増幅した。ClaIおよびAflIIの制限部位に下線を記した。該断片の配列は以下のとおりである:
【表A】
断片をアガロースゲル電気泳動によって分離し、Qiagen (Hilden, Germany)のgelextraction kit QiaexIIによって単離した。単離された断片をClaIおよびAflIIで切断し、ClaI/AflII-切断pBW22にライゲートした(Wilms et al., 2001, Biotechnology and Bioengineering, 73 (2), 95-103)。配列番号1(pBLL7)からなるmelAB2プロモーターを含む得られたプラスミドを図1に示す。prpプロモーター(pBLL5)およびgutAプロモーター(pBLL6)を含む得られたプラスミドは、ライゲートされたプロモーター領域を除けば同一である。挿入されたプロモーター断片の配列は、シークエンシングによって確認した(Microsynth GmbH, Balgach, Switzerland)。
【0075】
例2
Fab断片発現プラスミドの構築
IPTG-誘導性lacプロモーターに対する代替として(プラスミドpMx9-HuCAL-Fab-H, Knappik et al., 1985, Gene 33, 103-119)、異なる正に制御された発現系をFab-H抗体断片を産生するその能力について分析した。Fab-H断片を、プライマーFab-5(5’-aaa cat atg aaa aag aca gct atc-3’)およびFab-3 (5’-aaa aag ctt tta tca gct ttt cgg ttc-3’)を使用してPCRによってプラスミドpMx9-HuCAL-Fab-Hで増幅した。PCR断片をNdeIおよびHindIIIで切断、かつNdeI/HindIII-切断pBW22に挿入し、L-ラムノース誘導性rhaBADプロモーターを含むプラスミドpBW22-Fab-H を創出した (Volff et al., 1996, Mol. Microbiol. 21, 1037-1047)。同じPCR断片を誘導性プロモーターをもつ異なる発現プラスミドに挿入した。Fab-HおよびmelAB2プロモーター(配列番号3)を含む得られた(推定上の)発現プラスミドpBLL15を図2に示す。prpプロモーター(pBLL13)およびgutAプロモーター(pBLL14)を含む等価なプラスミドが得られた。プラスミドpBW22-Fab-HのFab-H 挿入の配列はシークエンシングによって確認した。
【0076】
例3
Fab断片の発現
株W3110 (DSM 5911, Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany)を、異なる発現プラスミドで形質転換した。プラスミドを、異なる形質転換体から得られたクローンから単離し、制限分析を介してチェックした。プラスミドpBLL14を除いて、全てのプラスミドは、予期された制限パターンを有していた。再度単離されたプラスミドpBLL14は、組換えが原因と考えられる変化したサイズおよび制限パターンを示した。従って、株W3110 (pBLL14)については以下のアッセイにおける試験を行わなかった。残りの株については、活性が倍加されたFab-H抗体断片を分泌する能力について試験を行った。この生産性試験は例4において記載されたように行った。以下の誘導物質を0.2%の濃度において加えた。
【0077】
pBW22-Fab-H L(+)-ラムノース一水和物
pBLL13 プロピオン酸ナトリウム
pBLL15 D(+)-メリビオース一水和物
D(+)-ラフィノース一水和物
D(+)-ガラクトース
【0078】
ドットブロット試験の結果を図3において示す。
【0079】
L-ラムノースおよびメリビオース誘導株W3110(pBW22-Fab-H)およびW3110 (pBLL15)は、有望なドットブロット結果を示した:経時的にシグナルを増加させ、かつほとんどバックグラウンド活性が観察されなかった。活性が倍加された抗体断片の部分は、ELISAを介して定量化された。その結果を以下の表1に要約する。
【表1】
【0080】
表1:異なる発現プラスミドをもつW3110誘導体のELISAの結果。誘導後の時間が示されている。22または25時間後の非誘導の培養物を、非誘導の対照として測定し、株W3110 (pMx9-HuCAL-Fab-H) および TG1F’- (pMx9-HuCAL-Fab-H) の結果を、参照として使用する。Fab-H濃度は、mg/100 OD/Lで与えられる(n.d. 測定されなかった)。
【0081】
全ての株は、4〜6の間のOD600まで対応する誘導物質の存在または欠損下において、あらゆる増殖阻害もなく良好に増殖した。発現プラスミドpBW22-Fab-HおよびpBLL15は、終夜の誘導後に最も高い抗体断片力価を誘導した。メリビオース誘導株W3110 (pBLL15)は、L-ラムノース(pBW22-Fab-H)誘導系と比較して活性抗体断片の形態において遅延性の増加を示した。
【0082】
L-ラムノース誘導性株W3110 (pBW22-Fab-H)を、振盪フラスコ内の呼吸活性のオンライン決定のための新規な測定系である「呼吸活性モニタリングシステム」 (RAMOS, ACBiotec, Julich, Germany) において試験した。通常の実験用振盪フラスコと比較して、抗体力価(ELISAを介して測定)は二倍になった(導入の23時間後に703.64 mg/L/100 OD600)。RAMOS装置を使用する最適化された成長は、活性抗体断片の生産を支持する。
【0083】
例4
振盪フラスコ内におけるメリビオースの誘導
プラスミドpBLL15をもつ大腸菌W3110を、活性が倍加されたFab-H抗体断片を産生するその能力について試験した。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、37 °C] を、20 mlの新鮮グリセロール培地中で稀釈した(1:50)(Kortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたように稀釈)。一方、ビタミン溶液を、Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように使用し、かつ30℃でインキュベートした。培養液が約0.4のOD600に達したとき、メリビオース(0.2%)を加えた。サンプル(1 ml)を異なる時間間隔で採取し、遠心してペレットを−20℃で貯蔵した。凍結した細胞を上述したリゾチーム処理によって溶解させ、上清をドットブロットおよびELISAアッセイにおいて分析した。機能的Fab-H抗体断片の504.28 mg/L/100 OD600 を得た。
【0084】
例5
高分子量凝集塊の出現
高分子量凝集塊が産生されるか否かを見るために、最も高い抗体力価(表1)を示した株W3110 (pBLL15)の抽出物のウェスタンブロットを、抗ヒトFab-H + AP抱合体を使用して行った。培養を例4に記載されたとおりに行った。メリビオースで誘導後9、12および23時間してサンプルを採取した。抗ヒトFab-H + AP結合体を使用した株W3110 (pBLL15)のリゾチーム抽出物のウェスタンブロットを図4に示す。より低濃度の高分子量凝集塊は、より高い力価の機能的抗体断片に対応する。発現系の選択は、抗体断片が形成され:機能的または凝集塊になることに影響を与えるようである。
【0085】
例6
シグナルペプチドの影響
大腸菌のゲノムデータベースを使用して、Fab-H断片VL3-CLおよびVH-CHと組み合わせて使用され得る有用なシグナルペプチドを探索した。糖、アミノ酸、ビタミンおよびイオンについて周辺質結合タンパク質からのシグナル配列を選択した。これらの周辺質タンパク質は、他の周辺質タンパク質よりもより広範な研究がなされた比較的均質な群を表わす。これらは一般的には大量に存在するので、これらのシグナル配列は内膜を超えて周辺質へ効果的に輸送されなければならない。全ての可能なシグナルペプチドFab組み合わせを、以下の表2において示したようなSignalPウェブサーバー(http://www.cbs.dtu.dk/services/SignalP-2.0/#submission)を使用して、その配列ペプチドおよび切断サイト可能性について調べた。
【0086】
【表2】
【0087】
【0088】
【0089】
以下の6つの組み合わせを選択した:
- LamB-VL3-CL (マルトポリン前駆体)
- MalE-VH-CH (マルトース結合タンパク質前駆体)
- Bla-VL3-CL (β-ラクタマーゼ)
- TreA-VH-CH (周辺質トレハラーゼ前駆体)
- ArgT-VL3-CL (リシン-アルギニン-オルニチン-結合周辺質タンパク質前駆体)
- FecB-VH-CH (鉄(III)ジシトラート-結合周辺質タンパク質前駆体)
【0090】
シグナルペプチド(SP)とVL3-CLおよびVH-CHとの融合体を生成するための遺伝子融合がオーバーラッピングPCRプライマーで行われ、以下の増幅の表3において要約される。
【表3】
【0091】
シグナルペプチド配列とVL3-CLおよびVH-CH配列との融合は、他の文献 (Horton, R.M., Hunt, H.D., Ho, S.N., Pullen, J.K. and Pease, L.R. (1989) Engineering hybrid genes without the use of restriction enzymes: gene splicing by overlap extension. Gene 77, 61-68) に記載されたように行った。SP-VL3-CL遺伝子を、制限酵素NdeIおよびPstIで切断し、NdeI/PstI切断pBW22 および pBLL7にライゲートした。
【0092】
得られたプラスミドをPstIおよびHindIIIで切断し、PstI/HindIII切断SP-VH-CH遺伝子にライゲートした。bla-VL3-CLおよびfecB-VH-CH遺伝子の統合は可能ではなく、lamB-VL3-CL および malE-VH-CH遺伝子を含むFab-H発現プラスミドのみが試験に供された。L-ラムノース誘導プロモーター(pAKL14)を含むlamB-VL3-CL/malE-VH-CH発現プラスミドを得た。AflII/HindIII断片としてプラスミドpAKL15(例7)から単離されたlamB-VL3-CL/malE-VH-CH遺伝子を、AflII/HindIII-切断pBLL7にライゲートし、pAKL15Eを得た。図5は、メリビオース誘導性プロモーターおよびlamB-VL3-CL/malE-VH-CH (配列番号4)を含む発現プラスミドpAKL15Eを示す。
【0093】
例7
Fab発現上の翻訳開始領域の影響
プラスミドpAKL14およびプラスミドpAKL15EのFab-H遺伝子は、開始コドンの同じDNA配列5’(翻訳開始領域)を含む。最初のプラスミドpMx9-HuCAL-Fab-Hにおいて、両Fab-H遺伝子の翻訳開始領域は異なる。プラスミドpMx9-HuCAL-Fab-HおよびpAKL14/pAKL15Eの翻訳開始領域配列の比較は、以下の表4において示される。
【表4】
【0094】
株W3110(pAKL14)の生産能力を、例4に記載したように振盪フラスコ内において試験した。株はL-ラムノースの存在または欠損下において十分に増殖した。これはFab-Hの生産が細胞の生存能力に影響を与えなかったことを意味する。
【0095】
新しいシグナルペプチド構築物(修飾された翻訳開始シグナルと組み合わされた)は再度、抗体断片力価を328.62 mg/L/100 OD600(プラスミドpMx9-HuCAL-Fab-H由来のMOR 遺伝子構築物を含むプラスミドpBW22-Fab-H)から596.14 mg/L/100 OD600 (プラスミドpAKL14)まで増加し、かつ878.86 mg/L/100 OD600 (プラスミドpAKL15E) まで増加した。pAKL14におけるlamB-VL3-CLおよびmalE-VH-CH遺伝子のシークエンシングは、二つの連続したPCR反応による融合遺伝子の構築が原因であると考えられる三つの塩基の変化を明らかにした。塩基の変化は、以下のアミノ酸の変化を誘導した(誤りのアミノ酸を強調する)。
【表B】
Fab-Hの軽鎖は二つの誤り(D50N, K63N)を含み、かつ重鎖は一つのアミノ酸変化(F156L)を含む。最初のFab-H配列を回復するために、プラスミドpAKL14 (138 bp SexAI/BamHIおよび310 bp BssHII/HindIII断片)由来の二つの断片は、プラスミドpBW22-Fab-H (未変化のFab-H遺伝子配列を含む)の均質な断片に対して変化した。得られたプラスミドpAKL15は、正しいFab-H配列を含む。三つのアミノ酸の変化は、pIが変化しなかったので全体としてFab-Hの性質に影響を与えなかった。従って、機能的Fab-H抗体断片を産生する株W3110 (pAKL15)の能力は、株W3110 (pAKL14)に類似していると考えられるので分析を行わなかった。
【0096】
Fab-H抗体断片の生産能力は、異なる最適化戦略を使用することによって増加され得る。以下の表5はその改良について要約する:
【表5】
【0097】
高Fab-H抗体力価を生産した株をSDS-PAGE(図6)を介して分析した。最も高い機能的Fab-H濃度が、軽鎖および重鎖の平衡量を産生する株において測定された(レーン4および5)。W3110 (pBW22-Fab-H) (レーン3)のようなFab-H断片を含むL-ラムノース誘導株が、軽鎖を激しく過剰産生する。
【0098】
例8
振盪フラスコにおけるメリビオースの誘導
プラスミドpAKL15Eを含む大腸菌W3110は、活性が倍加されたFab-H抗体断片を産生するその能力について試験された。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、37℃] を、20 mlの新鮮グリセロール培地中で稀釈した(1:50)(Kortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたように稀釈)。一方、ビタミン溶液を、Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように使用し、かつ30℃でインキュベートした。培養液が約0.4のOD600に達したとき、メリビオース(0.2%)を加えた。サンプル(1 ml)を異なる時間間隔で採取し、遠心してペレットを-20℃で貯蔵した。凍結された細胞を上述したリゾチーム処理に基づいて溶解し、上清をSDS-PAGEおよびELISAアッセイにおいて分析した。変化したシグナルペプチド(lamB-VL3-CL/malE-VH-CH)でFab-H遺伝子を含むメリビオース誘導株は、最大のFab-H抗体力価を示した(表5)。Fab-Hの軽鎖および重鎖は等量において産生された(図8)。
【0099】
例9
振盪フラスコ内における株KIE153 (Burkholderia sp. DSM9925) からのアミダーゼのメリビオース誘導
プラスミドpJKL8を含む大腸菌W3110は、ラセミピペラジン-2-カルボキサミドを(R)- ピペラジン-2-カルボン酸に転換するその能力について試験を受けた。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、30 °C] を、20 mlの新鮮グリセロール培地中で稀釈した(1:50)(Kortz et al., 1995, J. Biotechnol. 39, 59-65によって記載されたように稀釈)。一方、ビタミン溶液を、Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように使用し、かつ30℃でインキュベートした。培養液が約0.8のOD600に達したとき、メリビオース(0.2%)を加えた。19時間の誘導時間後、細胞を-20℃で収穫および貯蔵した。アミダーゼ活性についてEichhorn et al. 1997, Tetrahedron Asymmetry, 8(15), 2533-36に記載したように試験を行った。得られた細胞は、約1 g/ h/ OD600の転換速度で (R)-ピペラジン-2-カルボキサミドを(R)-ピペラジン-2-カルボン酸にエナンチオ選択的に転換した。
【0100】
例10
振盪フラスコ内の単鎖抗体(scFv, S1)のメリビオース誘導
scFv遺伝子を、プライマー5-S (5’-aaa cat atg aaa tac cta ttg cct acg gc-3’) および 3-S1 (5’-aaa aag ctt act acg agg aga cgg-3’)を使用してPCRを介して単離した。対応するS1タンパク質は、大腸菌の周辺質に対するタンパク質の輸送に原因のあるPelBシグナル配列を含む。PCR断片は、NdeI および HindIII で切断され、NdeI/HindIII-切断 pBLL7に挿入され、メリビオース誘導melAB2プロモーター(図9)を含むプラスミドpBLL7-pelB1-S1を作製した。プラスミドpBLL7-S1のS1挿入の配列は、シークエンシングによって確認された。株W3110 (DSM 5911, Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Braunschweig, Germany) をプラスミドpBLL7-pelB1-S1で形質転換した。プラスミドは、異なるクローンから単離され、かつ制限分析によって検証された。大腸菌W3110 (pBLL7-pelB-S1)は、可溶性S1を産生するその能力について試験を受けた。終夜培養液 [NYB培地中(10 g/lトリプトン、5 g/l酵母抽出物、5 g/l塩化ナトリウム) 100 μg/mlのアンピシリンを補充、30 °C] を、20 mlの新鮮グリセロール培地中で稀釈し(1:50) [Kortz et al., 1995, J. Biotechnol. 39, 59-65に記載されたように稀釈、但しビタミン溶液を除く(Kulla et al., 1983, Arch. Microbiol, 135, 1によって記載されたように)]、30℃でインキュベートした。培養液が約0.4のOD600に達したときにメリビオース(0.2%)を加えた。サンプル(1 mL)を異なる時間間隔で採取し、遠心してペレットを-20℃で貯蔵した。凍結細胞を上述したリゾチーム処理に基づいて溶解し、上清および不溶性タンパク質ペレットをSDS-PAGE(図10)およびBioanalyzerを介して分析した。S1タンパク質(1.7 g/Lx mg/L/100 OD600)の大部分は、可溶性タンパク質画分において産生された。
【図面の簡単な説明】
【0101】
【図1】図1は、メリビオース誘導性melAB2プロモーター(PmelAB2)および転写終結領域(rrnB)を含むプラスミドpBLL7を示す。
【図2】図2は、メリビオース誘導性プロモーター(PmelAB2)、Fab-H断片の軽鎖および重鎖に機能的に連結したシグナル配列の配列コード(ompA-VL3-CL)および(phoA-VH-CH)、ならびに転写終結領域(rrnB)を含むプラスミドpBLL15を示す。
【図3】図3は、異なる発現プラスミドを含む非誘導性(-)および誘導性(+)W3110株のリゾチーム(lysozyme)抽出物のドットブロット結果(Fabを検出するための抗ヒト軽鎖、アルカリ性ペルオキシダーゼを抱合)を示す。時間の間隔を示す。
【図4】図4は、アルカリ性ペルオキシダーゼを抱合したFabを検出するための抗ヒト軽鎖を使用する株W3110(pBLL15)のリゾチーム抽出物のウェスタンブロットを示す。
【図5】図5は、メリビオース誘導性プロモーター(PmelAB2)および変化したシグナル配列を含むFab-H遺伝子を含むプラスミドpAKL15Eを示す。
【図6】図6は、高いFab-H抗体濃度を示す異なるW3110株のライソゾーム抽出物のSDS-PAGEを示す。シグナル配列を含まない軽鎖および重鎖を生成する株がネガティブ参照として使用される(レーン1:マーカー; レーン2:W3110 (pMx9-HuCAL-Fab-H); レーン3:W3110 (pBW22-Fab-H); レーン4:W3110 (pBLL15), レーン5:W3110 (pAKL14); レーン6:スタンダード(2 μg))。
【図7】図7は、メリビオース誘導性プロモーター(PmelAB2)および株KIE153 (Burkholderia sp. DSM9925)由来のアミダーゼをコードする配列、および転写終結領域(rrnB)を含むプラスミドpJKL8を示す。
【図8】図8は、誘導性メリビオースの存在または不存在における株W3110 (pAKL15E)のライソゾーム抽出物のSDS-PAGEを示す。軽鎖および重鎖の位置が示される(レーン1:マーカー; レーン2:W3110 (pAKL15E)、誘導されない; レーン3:W3110 (pAKL15E)、誘導された)。
【図9】図9は、メリビオース誘導性melAB2プロモーターおよび単鎖抗体(scFv, S1)をコードする配列を含むプラスミドpBLL7-pelB-S1を示す。S1をコードする配列は、PelBシグナルペプチドをコードする配列によって先行される。
【図10】図10は、非誘導性(-)および誘導性(+)株W3110 (pBLL7-pelB-S1)の未精製抽出物のSDS-PAGEを示す。サンプルは示されたような異なる時間間隔で採取された。リゾチーム処理が可溶性および不溶性タンパク質画分が分析された。矢印はscFvタンパク質を示す。 M = Mark12、インビトロゲンの標準分子量。
【特許請求の範囲】
【請求項1】
宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオース オペロンのプロモーター領域を含む、宿主において発現可能なベクターであって、前記核酸配列の発現がメリビオース オペロンの前記プロモーター領域によって制御されるベクター。
【請求項2】
前記プロモーター領域が、melABプロモーターである請求項1に記載のベクター。
【請求項3】
前記melABプロモーターが、CRP1結合部位を欠損している請求項2に記載のベクター。
【請求項4】
前記CRP1結合部位を欠損したmelABプロモーターが、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる請求項3に記載のベクター。
【請求項5】
前記転写単位がさらに、前記転写単位の翻訳の開始点の翻訳開始領域上流を含む請求項1〜4に記載のベクターであって、前記翻訳開始領域が、配列AGGAGATATACAT(配列番号2)からなり、前記翻訳開始領域が、前記核酸配列に機能的に連結されるベクター。
【請求項6】
前記転写単位が、前記核酸配列に機能的に連結したシグナル配列をさらに含む請求1〜5のいずれか一項に記載のベクター。
【請求項7】
前記シグナル配列が、原核シグナル配列である請求項6に記載のベクター。
【請求項8】
前記原核シグナル配列が、糖、アミノ酸、ビタミンおよびイオンのための周辺質結合タンパク質のシグナルペプチドから選択される請求項7のベクター。
【請求項9】
前記転写単位がさらに、rrnB転写終結配列である転写終結領域を含む請求1〜8のいずれか一項に記載のベクター。
【請求項10】
前記核酸配列が、ポリペプチドをコードする請求1〜9のいずれか一項に記載のベクター。
【請求項11】
前記核酸配列が、抗体をコードする請求1〜9のいずれか一項に記載のベクター。
【請求項12】
前記核酸配列が、Fab断片をコードする請求1〜9のいずれか一項に記載のベクター。
【請求項13】
前記Fab断片の重鎖および軽鎖がジシストロニックな転写単位によってコードされ、各鎖が、シグナル配列および転写単位の翻訳の開始点の上流にある同一の翻訳開始領域に機能的に連結された請求項12に記載のベクター。
【請求項14】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる請求項1〜13のいずれか一項に記載のベクター。
【請求項15】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる請求項1〜13のいずれか一項に記載のベクター。
【請求項16】
前記ベクターが、自律的または自己複製的プラスミド、コスミド、ファージ、ウイルスまたはレトロウイルスである請求項1〜15のいずれか一項に記載のベクター。
【請求項17】
原核宿主における核酸配列の制御された異種発現のための、請求項1〜16のいずれか一項に記載のベクターの使用。
【請求項18】
前記核酸配列が、ポリペプチドをコードする請求項17に記載のベクターの使用。
【請求項19】
前記ポリペプチドがFab断片であり、Fab断片の重鎖および軽鎖が等量において発現する、請求項18に記載のベクターの使用。
【請求項20】
宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオース オペロンのプロモーター領域を含む、宿主において発現可能な単離および精製された核酸配列であって、前記核酸配列の発現が、前記メリビオース オペロンのプロモーター領域によって制御される核酸配列。
【請求項21】
前記プロモーター領域が、melABプロモーターである請求項20に記載の単離および精製された核酸配列。
【請求項22】
前記melABプロモーターが、CRP1結合部位を欠損している請求項21に記載の単離および精製された核酸配列。
【請求項23】
前記CRP1結合部位を欠損したmelABプロモーターが、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる、請求項22に記載の単離および精製された核酸配列。
【請求項24】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる、請求項20〜23のいずれか一項に記載の単離および精製された核酸配列。
【請求項25】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる、請求項20〜23のいずれか一項に記載の単離および精製された核酸配列。
【請求項26】
プラスミドpBLL15。
【請求項27】
プラスミドpAKL15E。
【請求項28】
請求項1〜16のいずれか一項に記載のベクターで形質転換された原核宿主。
【請求項29】
請求項20〜25のいずれか一項に記載の単離および精製された核酸配列で形質転換された原核宿主。
【請求項30】
請求項26または27に記載のプラスミドで形質転換された原核宿主。
【請求項31】
宿主においてポリペプチドを産生するための方法であって、以下の工程を含む方法。
a)請求項10〜16のいずれか一項に記載のベクターを構築する工程と、
b)前記ベクターで原核宿主を形質転換する工程と、
c)適切な条件下の細胞培養系において前記ポリペプチドの発現を可能にする工程と、
d)前記ポリペプチドを前記細胞培養系から回収する工程。
【請求項32】
前記産生されたポリペプチドがFab断片であり、Fab断片の重鎖および軽鎖が前記細胞培養系で等量において発現される請求項31に記載の方法。
【請求項33】
前記ポリペプチドの発現が、グリセロール含有培養液中において行われる請求項31または32に記載の方法。
【請求項1】
宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオース オペロンのプロモーター領域を含む、宿主において発現可能なベクターであって、前記核酸配列の発現がメリビオース オペロンの前記プロモーター領域によって制御されるベクター。
【請求項2】
前記プロモーター領域が、melABプロモーターである請求項1に記載のベクター。
【請求項3】
前記melABプロモーターが、CRP1結合部位を欠損している請求項2に記載のベクター。
【請求項4】
前記CRP1結合部位を欠損したmelABプロモーターが、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる請求項3に記載のベクター。
【請求項5】
前記転写単位がさらに、前記転写単位の翻訳の開始点の翻訳開始領域上流を含む請求項1〜4に記載のベクターであって、前記翻訳開始領域が、配列AGGAGATATACAT(配列番号2)からなり、前記翻訳開始領域が、前記核酸配列に機能的に連結されるベクター。
【請求項6】
前記転写単位が、前記核酸配列に機能的に連結したシグナル配列をさらに含む請求1〜5のいずれか一項に記載のベクター。
【請求項7】
前記シグナル配列が、原核シグナル配列である請求項6に記載のベクター。
【請求項8】
前記原核シグナル配列が、糖、アミノ酸、ビタミンおよびイオンのための周辺質結合タンパク質のシグナルペプチドから選択される請求項7のベクター。
【請求項9】
前記転写単位がさらに、rrnB転写終結配列である転写終結領域を含む請求1〜8のいずれか一項に記載のベクター。
【請求項10】
前記核酸配列が、ポリペプチドをコードする請求1〜9のいずれか一項に記載のベクター。
【請求項11】
前記核酸配列が、抗体をコードする請求1〜9のいずれか一項に記載のベクター。
【請求項12】
前記核酸配列が、Fab断片をコードする請求1〜9のいずれか一項に記載のベクター。
【請求項13】
前記Fab断片の重鎖および軽鎖がジシストロニックな転写単位によってコードされ、各鎖が、シグナル配列および転写単位の翻訳の開始点の上流にある同一の翻訳開始領域に機能的に連結された請求項12に記載のベクター。
【請求項14】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる請求項1〜13のいずれか一項に記載のベクター。
【請求項15】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる請求項1〜13のいずれか一項に記載のベクター。
【請求項16】
前記ベクターが、自律的または自己複製的プラスミド、コスミド、ファージ、ウイルスまたはレトロウイルスである請求項1〜15のいずれか一項に記載のベクター。
【請求項17】
原核宿主における核酸配列の制御された異種発現のための、請求項1〜16のいずれか一項に記載のベクターの使用。
【請求項18】
前記核酸配列が、ポリペプチドをコードする請求項17に記載のベクターの使用。
【請求項19】
前記ポリペプチドがFab断片であり、Fab断片の重鎖および軽鎖が等量において発現する、請求項18に記載のベクターの使用。
【請求項20】
宿主に対して異種である核酸配列を含む転写単位に機能的に連結されたメリビオース オペロンのプロモーター領域を含む、宿主において発現可能な単離および精製された核酸配列であって、前記核酸配列の発現が、前記メリビオース オペロンのプロモーター領域によって制御される核酸配列。
【請求項21】
前記プロモーター領域が、melABプロモーターである請求項20に記載の単離および精製された核酸配列。
【請求項22】
前記melABプロモーターが、CRP1結合部位を欠損している請求項21に記載の単離および精製された核酸配列。
【請求項23】
前記CRP1結合部位を欠損したmelABプロモーターが、配列番号1の配列、これに相補的な配列およびこれらの変異配列からなる、請求項22に記載の単離および精製された核酸配列。
【請求項24】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号3の配列、これに相補的な配列およびこれらの変異配列からなる、請求項20〜23のいずれか一項に記載の単離および精製された核酸配列。
【請求項25】
前記プロモーター領域および前記機能的に連結された転写単位が、配列番号4の配列、これに相補的な配列およびこれらの変異配列からなる、請求項20〜23のいずれか一項に記載の単離および精製された核酸配列。
【請求項26】
プラスミドpBLL15。
【請求項27】
プラスミドpAKL15E。
【請求項28】
請求項1〜16のいずれか一項に記載のベクターで形質転換された原核宿主。
【請求項29】
請求項20〜25のいずれか一項に記載の単離および精製された核酸配列で形質転換された原核宿主。
【請求項30】
請求項26または27に記載のプラスミドで形質転換された原核宿主。
【請求項31】
宿主においてポリペプチドを産生するための方法であって、以下の工程を含む方法。
a)請求項10〜16のいずれか一項に記載のベクターを構築する工程と、
b)前記ベクターで原核宿主を形質転換する工程と、
c)適切な条件下の細胞培養系において前記ポリペプチドの発現を可能にする工程と、
d)前記ポリペプチドを前記細胞培養系から回収する工程。
【請求項32】
前記産生されたポリペプチドがFab断片であり、Fab断片の重鎖および軽鎖が前記細胞培養系で等量において発現される請求項31に記載の方法。
【請求項33】
前記ポリペプチドの発現が、グリセロール含有培養液中において行われる請求項31または32に記載の方法。
【図1】

【図2】
【図3】
【図4】

【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2008−522598(P2008−522598A)
【公表日】平成20年7月3日(2008.7.3)
【国際特許分類】
【出願番号】特願2007−544798(P2007−544798)
【出願日】平成17年12月5日(2005.12.5)
【国際出願番号】PCT/EP2005/013012
【国際公開番号】WO2006/061173
【国際公開日】平成18年6月15日(2006.6.15)
【出願人】(398075600)ロンザ ア−ゲ− (58)
【出願人】(594029230)モルフォシス・アクチェンゲゼルシャフト (9)
【氏名又は名称原語表記】MORPHOSYS AG
【Fターム(参考)】
【公表日】平成20年7月3日(2008.7.3)
【国際特許分類】
【出願日】平成17年12月5日(2005.12.5)
【国際出願番号】PCT/EP2005/013012
【国際公開番号】WO2006/061173
【国際公開日】平成18年6月15日(2006.6.15)
【出願人】(398075600)ロンザ ア−ゲ− (58)
【出願人】(594029230)モルフォシス・アクチェンゲゼルシャフト (9)
【氏名又は名称原語表記】MORPHOSYS AG
【Fターム(参考)】
[ Back to top ]