
モノクローナル抗体またはその結合活性断片、ハイブリドーマおよびキット
【課題】 感度、特異性が高く、免疫ブロッティングや免疫沈降に適した、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体を提供することを目的とする。
【解決手段】 16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片であって、16型ヒトパピローマウイルスE6タンパク質と結合し、18型ヒトパピローマウイルスE6タンパク質および/または16型ヒトパピローマウイルスE7タンパク質と実質的に結合しない、モノクローナル抗体またはその結合活性断片、当該抗体を産生するハイブリドーマ、および当該ハイブリドーマを用いることを特徴とする、モノクローナル抗体またはその結合活性断片の調製方法が提供される。
【解決手段】 16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片であって、16型ヒトパピローマウイルスE6タンパク質と結合し、18型ヒトパピローマウイルスE6タンパク質および/または16型ヒトパピローマウイルスE7タンパク質と実質的に結合しない、モノクローナル抗体またはその結合活性断片、当該抗体を産生するハイブリドーマ、および当該ハイブリドーマを用いることを特徴とする、モノクローナル抗体またはその結合活性断片の調製方法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、16型ヒトパピローマウイルス(以下、HPV16とも記す。)のE6タンパク質(以下、HPV16 E6とも記す。)に対するモノクローナル抗体またはその結合活性断片、ハイブリドーマおよびキットに関する。また、本発明は、HPV16 E6に対するモノクローナル抗体またはその結合活性断片の調製方法、および当該抗体を用いる方法に関する。
【背景技術】
【0002】
16型や18型など一群のヒトパピローマウイルス(HPV)は子宮頸がんの90%以上において検出されている。これらのヒトパピローマウイルスは、発がんの原因ウイルスであり、ウイルスのE6とE7遺伝子がその責任遺伝子であることも明らかとなっている。特にHPV16は約半数から検出され、子宮頸がんから最も高頻度に検出される。また、子宮頸がんにおいて発現しているウイルスタンパク質はE6とE7のみであることが報告されている。
【0003】
これまで大腸菌や培養細胞で大量発現させたE6タンパク質と結合する抗体は存在し(非特許文献1:Banks L et al.(1987)、非特許文献2:Banks L et al.(1988)、非特許文献3:Matlashewski G et al)、現在、HPV16および18のE6タンパク質と結合するとされる抗体が、Abcam,Novus Biologicals,GeneTexおよびBiogenesisから市販されている。しかしながら、子宮頸がんで発現しているレベルのE6タンパク質を検出できる抗体は存在しなかった。このため、腫瘍細胞におけるE6タンパク質発現を確認する手段は事実上なく、腫瘍細胞におけるE6タンパク質の検出は極めて困難であった。また、子宮頸がんはHPV感染良性病変である異形成(CIN1,2)から進行するが、高度異形成や上皮内癌(CIN3)でE6,E7タンパク質が高発現すると推定されている。このため、前がん病変からがん化への移行を研究し、または診断する上でもE6,E7タンパク質を検出することが重要だと考えられている。
【0004】
【非特許文献1】Banks L et al., "Identification of human papillomavirus type 18 E6 polypeptide in cells derived from human cervical carcinomas.", J. Gen. Virol., 68, (Pt 5),pp1351-1359 (1987)
【非特許文献2】Banks L, Crawford L., "Analysis of human papillomavirus type 16 polypeptides in transformed primary cells.", Virology, 1988,165(1),pp326-328
【非特許文献3】Matlashewski G, Banks L, Wu-Liao J, Spence P, Pim D, Crawford L., "The expression of human papillomavirus type 18 E6 protein in bacteria and the production of anti-E6 antibodies.", J. Gen. Virol., 1986, 67 (Pt9) ,1909-1916
【非特許文献4】Ed Harlow and David Lane, Antibodies, A Laboratory Manual, 1988, Cold Spring Harbor, Chapter 6
【非特許文献5】Steplewski, Z. et al., Proc. Natl. Acad. Sci. USA,85, 4852, (1988)
【非特許文献6】Kearney JF, Radbruch A, Liesegang B, Rajewsky K., "A new mouse myeloma cell line that has lost immunoglobulin expression but permits the construction of antibody-secreting hybrid cell lines." J. Immunol., 1979, 123(4),1548-1550.
【非特許文献7】S.Sasai et al. :Abst. 3rd International Cell Culture Congress(Sendai) P.46, 1985. Establishment of a medium for mass culture of mammalian cells.
【非特許文献8】Terai, M., Uyama, T., Sugiki, T., Li, X. K., Umezawa, A., and Kiyono, T., "Immortalization of human fetal cells: the life span of umbilical cord blood-derived cells can be prolonged without manipulating p16KNK4a/RB braking pathway.", Mol Biol Cell, 16:1491-1499, 2005.
【非特許文献9】Kiyono T, Hiraiwa A, Fujita M, Hayashi Y, Akiyama T, Ishibashi M., "Binding of high-risk human papillomavirus E6 oncoproteins to the human homologue of the Drosophila discs large tumor suppressor protein.", Proc. Natl. Acad. Sci. U S A., 1997, 94(21), pp11612-11616
【特許文献1】特開昭60-145088号公報
【発明の開示】
【発明が解決しようとする課題】
【0005】
本発明は、感度、特異性が高く、免疫ブロッティングや免疫沈降に適した、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体を提供することを目的とする。
【課題を解決するための手段】
【0006】
本発明の一の側面によると、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片であって、16型ヒトパピローマウイルスE6タンパク質と結合し、18型ヒトパピローマウイルスE6タンパク質および/または16型ヒトパピローマウイルスE7タンパク質と実質的に結合しない、モノクローナル抗体またはその結合活性断片が提供される。また、本発明の他の側面によると、上記抗体を産生するハイブリドーマが提供される。また、本発明の他の側面によると、上記ハイブリドーマを用いることを特徴とする、モノクローナル抗体またはその結合活性断片の調製方法が提供される。
【0007】
また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を含む、16型ヒトパピローマウイルスE6タンパク質を検出または定量するためのキットが提供される。また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を用いることを特徴とする、16型ヒトパピローマウイルスE6タンパク質を検出または定量する方法が提供される。
【0008】
また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を含む、子宮頸がんの素因、前がん病変および/またはがん病変を診断するためのキットが提供される。また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を用いることを特徴とする、子宮頸がんの素因、前がん病変および/またはがん病変を診断する方法が提供される。
【発明の効果】
【0009】
以下に詳細に説明するように、本発明によると、感度、特異性が高く、免疫ブロッティングや免疫沈降に適した、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体が提供される。
【発明を実施するための最良の形態】
【0010】
以下に、本発明の実施の形態を説明する。もっとも、本発明は、以下に説明する実施の形態によって限定されるものではない。
【0011】
上記したように、本発明の一の側面によると、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片が提供される。ヒトパピローマウイルスは、子宮頸がん等の発がんの原因ウイルスであり、現在、100種類以上が確認されている。ヒトパピローマウイルスは、その感染部位により、上皮型(HPV1,5,8,14,20,21,25,47型等)、粘膜型(HPV6,11,16,18,31,33,35,39,41,45,51,52,56,58,59,68,70型等)に分類され、また、その発癌性により、低リスク群(6,11,41,42,43,44型等)、高リスク群(16,18,31,33,35,39,45,51,52,56,58,59,68,70型等)に分類される。
【0012】
HPVと子宮頸癌との関係は完全には明らかにされていないが、HPVのE6とE7遺伝子がその責任遺伝子であることが明らかとなっている。すなわち、発がん過程においてE6タンパク質がp53の不活化の他テロメラーゼの活性化や細胞トランスフォーメションを引き起こすなど、HPVのE6,E7タンパク質が癌タンパク質の性質を備え、細胞の不死化に関与していることが報告されている。具体的には、HPV16 E6およびE7ならびにHPV18 E6は、それぞれ配列番号1〜3に示すアミノ酸配列を有する。
【0013】
本発明者は、従来存在する抗体よりも感度および特異性に優れ、また、免疫ブロッティングや免疫沈降などの免疫化学的手法に適した、HPV16 E6タンパク質に対するモノクローナル抗体を分泌するハイブリドーマを作製した。
【0014】
従来市販の抗体C1P1は、HPV18 E6に対する抗体として作成されたものであるが(非特許文献1: Banks L et al. (1987)参照)、HPV18および16双方のE6タンパク質を認識するものとして市販されている。一方で、本発明にかかるモノクローナル抗体は非常に特異性が高く、HPV16 E6とは結合するが、HPV18 E6および/またはHPV16 E7とは実質的に結合しない。
【0015】
また、好ましくは、本発明にかかるモノクローナル抗体は、感度が非常に高く、通常の免疫ブロッティングにより30pgレベルのHPV16 E6を検出できる。一方で、市販の抗体C1P1のHPV16 E6に対する感度はおおよそ1μgである。子宮頸がんでは、約5pg/μg総タンパク質のHPV16 E6が発現しているが、本発明にかかるモノクローナル抗体によると、従来はできなかった子宮頸がんで発現しているレベルのE6タンパク質を容易に検出できる。
【0016】
上記モノクローナル抗体を産生するハイブリドーマの作製にあたっては、通常のハイブリドーマ作製方法を用いることができる(非特許文献4:Ed Harlow et al.等参照)。一般に、抗原として天然または合成のHPV18 E6またはその断片で動物を免疫し、得られた抗体産生細胞を骨髄腫細胞などと融合させる方法が用いられる。
【0017】
免疫動物として、例えば哺乳動物(例、マウス,ラット,モルモット,ネコ,イヌ,サル,ウサギ,ヒツジ,ヤギ,ハムスターなど),鳥類(例、ニワトリ,ガチョウ,アヒルなど)などを用いることができる。特に、マウス,ラット,ウサギなどが好ましい。免疫動物に誘導される抗体産生細胞としては、抗体を産生する細胞であれば脾臓,リンパ節,末梢血リンパ球などいずれ由来のものでもよいが、特に脾臓細胞が好適に用いられる。免疫方法は、例えばマウスを免疫する場合、皮下、腹腔内、静脈内、筋肉内、皮内等のいずれのルートからでも可能であるが、主として皮下、腹腔内、静脈内に(とりわけ皮下)投与することが好ましい。また、免疫間隔、免疫量等は適宜設定することができるが、例えば2週間隔で約2〜6回免疫し、最終免疫後、約1〜5回、好ましくは約2〜4日後に脾臓細胞を摘出する方法がよく用いられる。免疫量は1回にペプチド量として、マウス当り約0.1μg以上、好ましくは約10μg〜300μg用いることが望ましい。また、脾臓を摘出する前に、部分採血を行い、血中の抗体価の上昇を確認した上で、脾臓細胞を用いる融合実験を行うことが望ましい。
【0018】
得られた抗体産生細胞を増殖性の細胞と融合してハイブリドーマとすることにより、安定的に抗体を産生することが可能になる。融合相手の細胞としては特に骨髄腫細胞(例えば、NS−1,P3−X63−AgUI,X45,SP2,X68−Ag8)が好適に用いられる。特にMOPC21(BALB/cマウス)由来の骨髄腫細胞、X68−Ag8およびその改良株が好ましく用いられる。これら改良株あるいは他の骨髄腫由来の細胞株は8−アザグアニン耐性であり、ヒポキサンチン・グアニンホスホリボシルトランスフェラーゼを欠くため、HAT培地(ヒポキサンチン−アミノプテリン−チミジン含有培地)では成育できず、融合後のハイブリドーマの選択に有利である。
【0019】
細胞融合は公知の方法に従って実施することができる。融合剤として、例えばポリエチレングリコール(PEG),センダイウイルスなどを用いることができる。特にPEGが好適に用いられる。PEGは、平均分子量1000〜6000のものが好ましく、特に好ましくはPEG4000が用いられる。PEGは好ましくは10〜80%さらに好ましくは40〜50%の濃度範囲で用いる。細胞融合は以下のように行うことができる。すなわち、抗体産生細胞と骨髄腫細胞をRPMI1640等の培地で洗い、これらを通常2:1〜10:1の比率で混合し、室温で700×g,5分間遠心して細胞ペレットを得る。細胞ペレットを37℃の恒温槽で温めながらほぐし、予め温めておいたPEG溶液を徐々に加えながら良く混ぜる。通常、PEG量は108個細胞当たり1mL加えられる。ついで、予め温めておいた培地を徐々に滴下、混和し、PEG濃度を下げる。通常は10分位の時間をかけ1〜30mLの培地を加える。室温で遠心して細胞を集め、10%ウシ胎児血清(以下、FCSと略記することがある)を含む培地で脾細胞として1〜2×106/mLの細胞濃度で96穴プレートに100μLずつ分注する。一夜放置後、HATおよび10%FCSを含む培地(HAT培地)100μLを加える。この操作を省略するため細胞融合後、細胞をHAT培地に懸濁してまいても良い。2〜3週間の間に数回の培地交換を行う。培地交換の方法としては通常プレートの各穴から培地100〜200μLを除き、新鮮なHAT培地100〜200μLを加える。この間、ハイブリドーマの出現が認められれば、出来るだけ早くスクリーニングを行い、目的とする抗体を産生しているハイブリドーマの成育しているウェルを検出する。得られたハイブリドーマは直ちにクローニングを行う。
【0020】
このような抗体産生ハイブリドーマのスクリーニングには、ELISA法など公知の種々の方法が使用できる。抗体活性陽性のハイブリドーマは直ちにクローニングに供されるが、通常これは限界希釈法などで容易に実施される。クローン化されたハイブリドーマの培養上清については、上記の方法でその抗体価を測定し、安定的に力価の高い抗体を産生するハイブリドーマを選択することができる。さらに、交差することが望まれないタンパク質(HPV18 E6等)への結合活性に基づいて抗体のスクリーニングをさらに行うことができる。
【0021】
上記モノクローナル抗体を産生するハイブリドーマとして、ハイブリドーマC−26,N−20G8,N−46A4またはN−47C5(FERM AP−20921,FERM AP−20922,FERM AP−20923またはFERM AP−20924)が、独立行政法人産業技術総合研究所特許生物寄託センターに寄託されている。
【0022】
本発明にかかるモノクローナル抗体の生成および蓄積は、本発明にかかるハイブリドーマを通常液体培地、好ましくは無血清培地中、またはヒト以外の温血動物(通常はマウス)の腹腔内で培養し実施することができる。特に高純度の抗体が必要な場合、無血清培地を使用することが望ましい。具体的には、液体培地として、例えば、抗生物質(ゲンタマイシン等)およびIL−6を添加したGIT培地(日本製薬株式会社)などを用いることができる。培養は通常約3〜60日間、好ましくは約3〜10日間、約30〜38℃、好ましくは約37℃で実施される。また、マウス腹腔内への移植については、1匹当り約2×105〜5×107個,好ましくは約1〜5×106個の抗体産生ハイブリドーマを腹腔内に移入し、約10〜35日間の飼育で十分量の抗体含有腹水液3〜10mlが得られる。
【0023】
液体培地あるいは腹水液中の抗体の精製は、公知の生化学的手法を組み合わせることにより実施できる。例えば、抗体含有液を遠心分離後、硫酸アンモニウムや硫酸ナトリウムを用いて上清液を塩析する。得られたタンパク質沈殿物を適当な緩衝液に溶解し透析後カラムクロマトグラフィー(DEAEイオン交換カラム,ヒドロキシアパタイトカラム,ゲルろ過カラム,プロテインAカラム,プロテインGカラムなど),イムノアフィニティークロマトグラフィーなどに供し、目的とする抗体を分離し、精製することができる。以上のような分離および精製操作により、例えば500mLの液体培地からタンパク質重量比で90%以上の純度の抗体を約5〜30mg得ることができる。また50mLの腹水液からは同様の抗体が10〜200mg得られる。特に、プロテインAあるいはGカラムを用いたクロマトグラフィーにゲルろ過カラム担体、DEAEイオン交換カラム担体、ヒドロキシアパタイトなどを用いたクロマトグラフィーを組み合わせることによりさらに高純度の抗体を得ることができる。
【0024】
以上のようにして得られた抗体は、タンパク質分解酵素(パパイン,ペプシンなど)処理ならびに還元剤処理などにより、結合能を保持したままFab,Fab’あるいはF(ab’)2断片を得ることができ、本発明にかかる抗体と同様の目的で用いることができる。また、抗体の抗原認識部位を含む可変領域をコードするDNAを取得し、これに遺伝子操作技術(非特許文献5:Steplewski Z et al.)を用いてヒトIgGの定常領域コードする遺伝子を結合させ、マウス−ヒトキメラ抗体を作製することもできる。
【0025】
上記したように、本発明にかかるモノクローナル抗体には、より高い感度および特異性で、16型ヒトパピローマウイルスE6タンパク質を検出または定量することができ、ひいては、子宮頸がんの素因、前がん病変および/またはがん病変の診断に好適に適用することができる。被検試料として、血漿、血清、尿、脳脊髄液、腹水、胸水、羊水等の体液や、痰、便などを用いることができる。これらの試料は、そのまま、または各種緩衝液で希釈または抽出後濃縮し、免疫学的測定の試料とすることができる。また、本発明にかかるモノクローナル抗体によると、これらのためのキットを提供することができる。キットは、免疫化学的測定に用いられるその他の試薬をさらに含むことができる。
【実施例】
【0026】
以下に、本発明の実施例を、添付図面を参照しながら説明する。もっとも、本発明は、以下に説明する実施例によって限定されるものではない。
【0027】
[実験例1:免疫感作]
〔材料〕
・抗原:抗体作成のために、HPV16 E6タンパク質のアミノ末端16アミノ酸残基またはカルボキシ末端12アミノ酸残基からなる合成ペプチド1、2を免疫原として使用した(図1参照)。
ペプチド1:MFQDPQERPRKLPQLC(配列番号4)
ペプチド2:CRSSRTRRETQL(配列番号5)
・ミエローマ細胞株:ハイブリドーマ作成のために、マウス骨髄腫細胞株(P3-X63-Ag8.653)を使用した(非特許文献6:Kearney JF et al.参照)。
・培地:ハイブリドーマの培養のために、ゲンタマイシンを終濃度0.1mg/mlで、rHuman IL-6(Invitrogen Diaclone #IM-14)を終濃度1ng/mlで含む無血清培地GIT(日本製薬株式会社)を使用した(非特許文献7:S.Sasai et al.、特許文献1:特開昭60-145088号参照)。
【0028】
〔プロトコール〕
HPV16 E6に対する抗体を産生するハイブリドーマを、非特許文献4(Ed Harlow et al.)に記載の手順に準じて作製した。前記免疫原をフロイントコンプリートアジュバンド(FCA)とともに投与することで、マウスを免疫感作した。このマウスから採取した脾臓細胞とマウス骨髄腫細胞株(P3-X63-Ag8.653)とをPEGを用いて細胞融合させることで、ハイブリドーマを作製した。得られたハイブリドーマを、抗原として用いた上記ペプチド、および全長E6タンパク質を用いてスクリーニングおよびクローニングした。ペプチド1を抗原としたものからはハイブリドーマN−20G8,N−46A4およびN−47C5が、ペプチド2を抗原としたものからはハイブリドーマC−26が得られた。
【0029】
[実験例2:抗体の特徴づけ]
〔実験例2−1:免疫ブロッティング〕
得られたハイブリドーマにより産生されるモノクローナル抗体の検出感度および特異性を試験した。プラスミドまたはレトロウイルスベクターを用いてHPV16 E6を発現させた細胞、および子宮頸がん細胞株を用いた。免疫ブロッティングは、非特許文献8(Terai et al.)に記載された方法に準じて行った。5%(v/v)プロテアーゼインヒビターカクテル(ナカライテスク株式会社)を添加したリシスバッファー(0.5% NP4O,1mM DTT,50mM NaCl,25mM Tris-HCl,pH8.0,0.02% NaN3)を用いて全細胞タンパク質を抽出した。細胞タンパク質をSDS-PAGEにより分離し、セミドライトランスファーシステム(アトー株式会社)によりJmmobilon-Pメンブレン(Millipore,Bedford,MA)にブロットした。上記ハイブリドーマ細胞株の調整培地、および市販の抗E6モノクローナル抗体(C1P5, Carbiochem, Ab-1)を、それぞれ1/2および1/100の希釈率でプローブとして用いた。二次抗体として、ホースラディッシュペルオキシダーゼで標識した抗マウス免疫グロブリンを用いた。ローディングコントロールとして、1/1000の希釈率の抗βアクチンヤギポリクローナル抗体(Santa Cruz, sc-1616)および抗ヤギ免疫グロブリンでβアクチンをプローブした。また、ローディングコントロールとして、抗HA抗体(クローン16B12)でHAタグをプローブした。Lumi-light plusウェスタンブロッティング基質(Roche)により視覚化したタンパク質を、LAS3000 CCDイメージングシステム(富士写真フイルム株式会社)により検出および定量化した。
【0030】
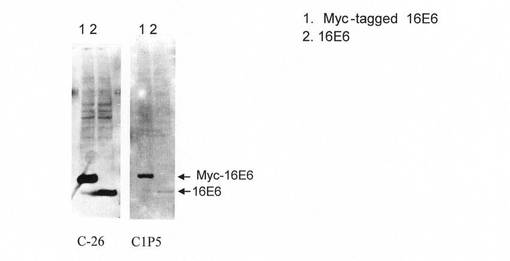
図2に示すように、ハイブリドーマ細胞株C−26から産生された抗体は、プラスミドを用いて発現させたE6タンパク質を、最もよく使われている市販抗体(C1P5)よりも約10倍の感度で検出できた。しかしながら、C−26から産生された抗体は、レトロウイルスベクターを用いて発現させたE6タンパク質や子宮頸がん細胞株で発現するE6タンパク質を検出することはできなかった(図示せず)。
【0031】
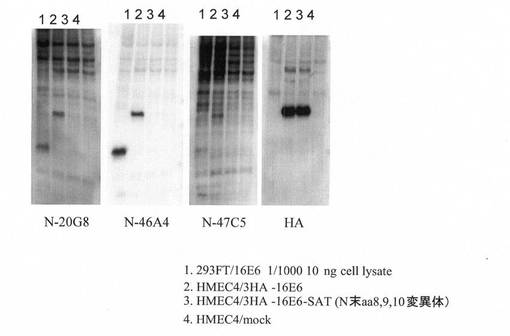
また、図3に示すように、図2においてC1P5(市販抗体)がかろうじて検出できた量の1/1000量(10ng)のE6タンパク質をN−46A4等は余裕を持って検出できた。その他のクローンも同様にE6タンパク質を高い感度で検出可能であった。しかし、得られた抗体はN末端に変異を導入したE6タンパク質(N末端から8番目(Arg)、9番目(Pro)、10番目(Arg)のアミノ酸残基をそれぞれSer,Ala,Thrに置換したもの)は認識しなかった。ここから、これらの抗体はE6タンパク質のN末端を特異的に認識していることが確認される。
【0032】
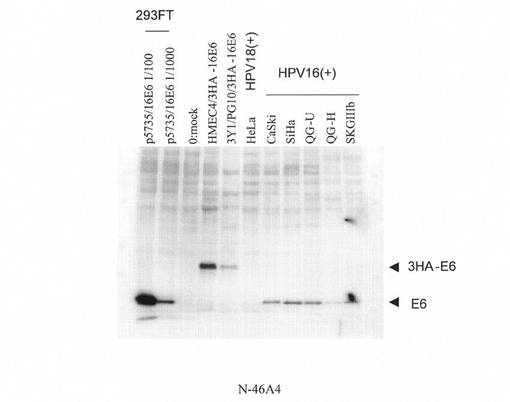
また、図4に示すように、ハイブリドーマN−46A4から産生された抗体は、HPV16陽性である全ての子宮頸がん細胞株(CaSki,SiHa,QG−U,Qg−H,SKGIIIb)で発現するE6タンパク質を10μgの細胞タンパク質を用いて検出できた。しかし、ハイブリドーマN−46A4から産生された抗体は、HPV18陽性のHeLa細胞のE6は認識しなかった。
【0033】
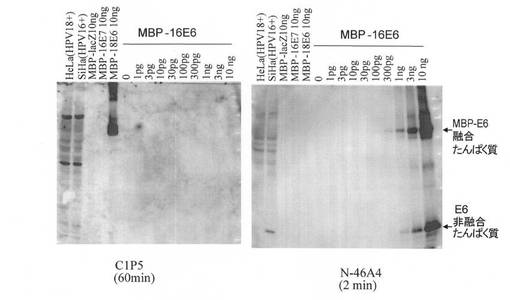
同様に、大腸菌で発現させ、すでに定量されたMBP融合タンパク質を用いて、免疫ブロッティング法により抗体の感度を定量した。図5に示すように、ハイブリドーマN−46A4から産生された抗体は、MBP−16E6 100pg(E6タンパク質の重量に換算すると約30pg)を2分の露出で検出できた。しかしながら、ハイブリドーマN−46A4から産生された抗体は、その100倍量(10ng)のMBP−18E6、MBP−16E7およびMBP−lacZタンパク質とは2分の露出でも全く反応しなかった。市販の抗体C1P5は、10ngのMBP−18E6を1分の露出で検出できたが(図示せず)、10ngのMBP−16E6を60分の露出によっても全く検出できなかった。
【0034】
なお、大腸菌での合成に用いたMBP−E6の遺伝子にはE6の開始コドンが残っており、非融合のE6タンパク質も一部産生されたと思われる。クマシーブリリアントブルー(CBB)染色の結果から(図示せず)、非融合16E6タンパク質の量はMBP−16E6の約10%程度であった。図5の結果と併せると、上記抗体N−46A4による非融合16E6タンパク質の検出感度は約30pgであり、融合タンパク質から推定される感度と大きな開きはなかった。
【0035】
このように、上記抗体は約30pgのHPV16 E6タンパク質を検出できる一方で、その100倍量のHPV18 E6およびHPV16 E7であっても免疫ブロッティングで検出しない。すなわち、HPV18 E6およびHPV16 E7と比べて100倍以上の感度で、上記抗体はHPV16 E6を検出できることがわかる。
【0036】
これらの結果を基にSiHa細胞タンパク質20μg中の16E6タンパク質量を半定量すると約100pgとなった。なお、MBP融合タンパク質の定量は、総タンパク質量とMBP−E6の位置のバンドの濃さから判定した。
【0037】
なお、C1P5は18E6とβガラクトシダーゼとの融合タンパク質を抗原として取られた抗体であり、16E6にも反応すると報告されている(実際図2でも検出できている)。また、非特許文献1(Banks L et al. (1987))においては[35S]システイン標識したHeLa細胞で発現しているHPV18 E6タンパク質を免疫沈降で検出できることが示されている。C1P5はHPV18 E6タンパク質に対してはかなり良い抗体であることが分かったが、HPV16 E6タンパク質に対しては免疫ブロッティングでも免疫沈降法でも非常に感度の悪い(ほぼ使い物にならない)抗体であることが分かった。具体的には、C1P5は10ngのMBP−18E6タンパク質を1分以内の露光で楽々検出できた、おそらく1ngも検出できると推定されるのに対し、MBP−16E6は10ngを60分の露光でも全く検出できなかった。図2,3の結果と併せると、C1P5の16E6タンパク質の検出感度は約300ng(MBP−16E6の重量で約1μg)と推定される。
【0038】
〔実験例2−2:免疫沈降〕
得られたハイブリドーマにより産生されるモノクローナル抗体の免疫沈降の効率を試験した。レトロウイルスベクターを用いてHPV16 E6タンパク質を発現させた細胞、および子宮頸がん細胞株を用いた。免疫沈降は、非特許文献9(Kiyono T)に記載された方法に準じて行った。2%(v/v)プロテアーゼインヒビターカクテル(ナカライテスク株式会社)を添加したリシスバッファー(10mM Tris-HCl(pH7.8),1% NP-40,150mM NaCl,1mM EDTA)を用いて全細胞タンパク質を抽出した。得られた抽出液と、上記ハイブリドーマ細胞株の調整培地1mLまたは市販の抗Myc抗体(9E10.2)もしくは抗HA抗体(16B12)との混合物にプロテインGセファロース4B(Pharmacia)を加え4℃で1時間インキュベートした。その後、プロテインGセファロース4Bを洗浄し、SDS-PAGEおよび抗HA抗体(16B12)による免疫ブロッティングにより免疫沈降の効率を確認した。
【0039】
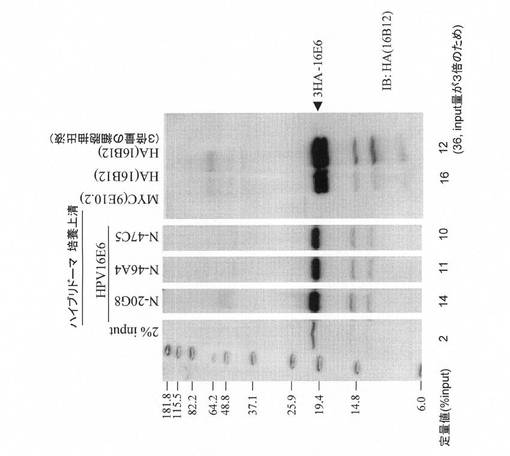
図6に示すように、各抗体はHAタグを3個もつE6タンパク質を効率よく免疫沈降した。抗HA抗体の方が免疫沈降効率が高いが、これはエピトープを3個持つため相乗的に効率が上がっているためと考えられる。各抗体も約10〜14%のE6タンパク質を免疫沈降しており、一般的にその効率は高いと考えられる
【0040】
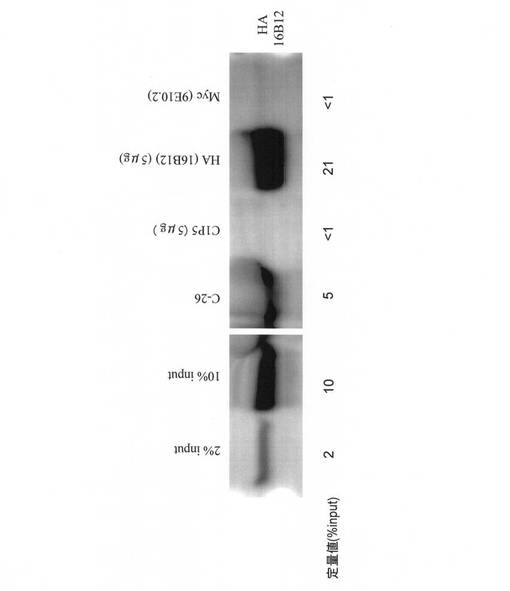
図6とほぼ同じ条件でハイブリドーマC−26から産生された抗体と市販の抗体C1P5の免疫沈降の効率を比較した。図7に示すように、C−26から産生された抗体は、HAタグを3個もつE6タンパク質を約5%免疫沈降した。これに対し、C1P5は抗原をほとんど(1%以下)免疫沈降できなかった。図5の結果と併せると、少なくとも我々の実験条件ではC1P5はHPV18 E6タンパク質に特異的な抗体ではあるが、免疫ブロッティングでも免疫沈降でもHPV16 E6タンパク質の検出感度は著しく低いことが分かる。対照と して市販の抗HA抗体(16B12)を用いた。図6と同様に抗HA抗体の方が免疫沈降効率が高いが、これはエピトープを3個持つため相乗的に効率が上がっているためと考えられる。なお、C−26から産生された抗体は免疫沈降にも使えるが、その効率は、図6のN−20G8,N−46A4,N−47C5から産生された抗体の方が優れていた。
【0041】
表1に以上の結果の概要を示す。
【表1】

【図面の簡単な説明】
【0042】
【図1】HPV16 E6のORFがコードするタンパク質と抗原に用いたペプチドのアミノ酸配列を示す。ペプチド1:MFQDPQERPRKLPQLC(配列番号1),ペプチド2:CRSSRTRRETQL(配列番号2)。
【図2】本発明にかかるモノクローナル抗体(C−26)による免疫ブロッティングの結果を示す。比較として従来市販の抗体C1P5によるものも示す。レーン1:293FT/p2816:pEF-MYC16E6SD(293FT細胞にMYCタグ融合HPV16 E6の発現ベクターを導入したもの。);レーン2:293FT/p5735:pEF6/16E6SD(293FT細胞にHPV16 E6の発現ベクターを導入したもの。)。各々、総細胞タンパク質10μg。露光時間:C−26=60分;C1P5=164分。
【図3】本発明にかかるモノクローナル抗体(N−20G8,N−46A4,N−47C5)による免疫ブロッティングの結果を示す。レーン1:293FT/p5735:pEF6/16E6SD;レーン2:HMEC4/3HA-16E6(HMEC4細胞に3HAタグ融合HPV16 E6発現ベクターを導入したもの。);レーン3:HMEC4/3HA-16E6-SAT(HPV16 E6のN末端変異体を発現する以外はレーン2と同じ);レーン4:HMEC4/mock(HMEC4細胞に発現ベクターを導入していないもの。)。総細胞タンパク質:レーン1 10ng,レーン2〜3 20μg。
【図4】本発明にかかるモノクローナル抗体(N−46A4)による免疫ブロッティングの結果を示す。293FT/p5735/16E6(293FT細胞にHPV16 E6の発現ベクターを導入したもの。);0:mock(293FT細胞に発現ベクターを導入していないもの。);HMEC4/3HA-16E6(HMEC4細胞に3HA-16E6の発現ベクターを導入したもの。);3Y1/PG10/3HA-16E6(3Y1/PG10細胞に3HA-16E6の発現ベクターを導入したもの。)。総細胞タンパク質:293FT/p5735/16E6 1/100, 1/1000は各々100ng,10ng,その他は10μg。
【図5】本発明にかかるモノクローナル抗体(N−46A4)による免疫ブロッティングの結果を示す。比較として従来市販の抗体C1P5によるものも示す。総細胞タンパク質:HelaおよびSiHa 20μg,その他図示の通り。
【図6】本発明にかかるモノクローナル抗体(N−20G8,N−46A4,N−47C5)による免疫沈降の結果を示す。293FT/3HA-16E6SD(293FT細胞に一過性遺伝子導入により3HA-16E6を高発現させた細胞)の細胞抽出液を用いた。免疫沈降のネガティブコントロールとして抗Myc抗体(9E10.2)を、ポジティブコントロールとして抗HA抗体(16B12)を用いた。定量のために、用いた細胞抽出液の2容量%をSDS-PAGEおよび免疫ブロッティングに供した。
【図7】本発明にかかるモノクローナル抗体(C−26)による免疫沈降の結果を示す。免疫沈降のネガティブコントロールとして抗Myc抗体(9E10.2)を、ポジティブコントロールとして抗HA抗体(16B12)を用いた。定量のために、用いた細胞抽出液の2、10容量%をSDS-PAGEおよび免疫ブロッティングに供した。
【技術分野】
【0001】
本発明は、16型ヒトパピローマウイルス(以下、HPV16とも記す。)のE6タンパク質(以下、HPV16 E6とも記す。)に対するモノクローナル抗体またはその結合活性断片、ハイブリドーマおよびキットに関する。また、本発明は、HPV16 E6に対するモノクローナル抗体またはその結合活性断片の調製方法、および当該抗体を用いる方法に関する。
【背景技術】
【0002】
16型や18型など一群のヒトパピローマウイルス(HPV)は子宮頸がんの90%以上において検出されている。これらのヒトパピローマウイルスは、発がんの原因ウイルスであり、ウイルスのE6とE7遺伝子がその責任遺伝子であることも明らかとなっている。特にHPV16は約半数から検出され、子宮頸がんから最も高頻度に検出される。また、子宮頸がんにおいて発現しているウイルスタンパク質はE6とE7のみであることが報告されている。
【0003】
これまで大腸菌や培養細胞で大量発現させたE6タンパク質と結合する抗体は存在し(非特許文献1:Banks L et al.(1987)、非特許文献2:Banks L et al.(1988)、非特許文献3:Matlashewski G et al)、現在、HPV16および18のE6タンパク質と結合するとされる抗体が、Abcam,Novus Biologicals,GeneTexおよびBiogenesisから市販されている。しかしながら、子宮頸がんで発現しているレベルのE6タンパク質を検出できる抗体は存在しなかった。このため、腫瘍細胞におけるE6タンパク質発現を確認する手段は事実上なく、腫瘍細胞におけるE6タンパク質の検出は極めて困難であった。また、子宮頸がんはHPV感染良性病変である異形成(CIN1,2)から進行するが、高度異形成や上皮内癌(CIN3)でE6,E7タンパク質が高発現すると推定されている。このため、前がん病変からがん化への移行を研究し、または診断する上でもE6,E7タンパク質を検出することが重要だと考えられている。
【0004】
【非特許文献1】Banks L et al., "Identification of human papillomavirus type 18 E6 polypeptide in cells derived from human cervical carcinomas.", J. Gen. Virol., 68, (Pt 5),pp1351-1359 (1987)
【非特許文献2】Banks L, Crawford L., "Analysis of human papillomavirus type 16 polypeptides in transformed primary cells.", Virology, 1988,165(1),pp326-328
【非特許文献3】Matlashewski G, Banks L, Wu-Liao J, Spence P, Pim D, Crawford L., "The expression of human papillomavirus type 18 E6 protein in bacteria and the production of anti-E6 antibodies.", J. Gen. Virol., 1986, 67 (Pt9) ,1909-1916
【非特許文献4】Ed Harlow and David Lane, Antibodies, A Laboratory Manual, 1988, Cold Spring Harbor, Chapter 6
【非特許文献5】Steplewski, Z. et al., Proc. Natl. Acad. Sci. USA,85, 4852, (1988)
【非特許文献6】Kearney JF, Radbruch A, Liesegang B, Rajewsky K., "A new mouse myeloma cell line that has lost immunoglobulin expression but permits the construction of antibody-secreting hybrid cell lines." J. Immunol., 1979, 123(4),1548-1550.
【非特許文献7】S.Sasai et al. :Abst. 3rd International Cell Culture Congress(Sendai) P.46, 1985. Establishment of a medium for mass culture of mammalian cells.
【非特許文献8】Terai, M., Uyama, T., Sugiki, T., Li, X. K., Umezawa, A., and Kiyono, T., "Immortalization of human fetal cells: the life span of umbilical cord blood-derived cells can be prolonged without manipulating p16KNK4a/RB braking pathway.", Mol Biol Cell, 16:1491-1499, 2005.
【非特許文献9】Kiyono T, Hiraiwa A, Fujita M, Hayashi Y, Akiyama T, Ishibashi M., "Binding of high-risk human papillomavirus E6 oncoproteins to the human homologue of the Drosophila discs large tumor suppressor protein.", Proc. Natl. Acad. Sci. U S A., 1997, 94(21), pp11612-11616
【特許文献1】特開昭60-145088号公報
【発明の開示】
【発明が解決しようとする課題】
【0005】
本発明は、感度、特異性が高く、免疫ブロッティングや免疫沈降に適した、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体を提供することを目的とする。
【課題を解決するための手段】
【0006】
本発明の一の側面によると、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片であって、16型ヒトパピローマウイルスE6タンパク質と結合し、18型ヒトパピローマウイルスE6タンパク質および/または16型ヒトパピローマウイルスE7タンパク質と実質的に結合しない、モノクローナル抗体またはその結合活性断片が提供される。また、本発明の他の側面によると、上記抗体を産生するハイブリドーマが提供される。また、本発明の他の側面によると、上記ハイブリドーマを用いることを特徴とする、モノクローナル抗体またはその結合活性断片の調製方法が提供される。
【0007】
また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を含む、16型ヒトパピローマウイルスE6タンパク質を検出または定量するためのキットが提供される。また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を用いることを特徴とする、16型ヒトパピローマウイルスE6タンパク質を検出または定量する方法が提供される。
【0008】
また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を含む、子宮頸がんの素因、前がん病変および/またはがん病変を診断するためのキットが提供される。また、本発明の他の側面によると、上記モノクローナル抗体またはその結合活性断片を用いることを特徴とする、子宮頸がんの素因、前がん病変および/またはがん病変を診断する方法が提供される。
【発明の効果】
【0009】
以下に詳細に説明するように、本発明によると、感度、特異性が高く、免疫ブロッティングや免疫沈降に適した、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体が提供される。
【発明を実施するための最良の形態】
【0010】
以下に、本発明の実施の形態を説明する。もっとも、本発明は、以下に説明する実施の形態によって限定されるものではない。
【0011】
上記したように、本発明の一の側面によると、16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片が提供される。ヒトパピローマウイルスは、子宮頸がん等の発がんの原因ウイルスであり、現在、100種類以上が確認されている。ヒトパピローマウイルスは、その感染部位により、上皮型(HPV1,5,8,14,20,21,25,47型等)、粘膜型(HPV6,11,16,18,31,33,35,39,41,45,51,52,56,58,59,68,70型等)に分類され、また、その発癌性により、低リスク群(6,11,41,42,43,44型等)、高リスク群(16,18,31,33,35,39,45,51,52,56,58,59,68,70型等)に分類される。
【0012】
HPVと子宮頸癌との関係は完全には明らかにされていないが、HPVのE6とE7遺伝子がその責任遺伝子であることが明らかとなっている。すなわち、発がん過程においてE6タンパク質がp53の不活化の他テロメラーゼの活性化や細胞トランスフォーメションを引き起こすなど、HPVのE6,E7タンパク質が癌タンパク質の性質を備え、細胞の不死化に関与していることが報告されている。具体的には、HPV16 E6およびE7ならびにHPV18 E6は、それぞれ配列番号1〜3に示すアミノ酸配列を有する。
【0013】
本発明者は、従来存在する抗体よりも感度および特異性に優れ、また、免疫ブロッティングや免疫沈降などの免疫化学的手法に適した、HPV16 E6タンパク質に対するモノクローナル抗体を分泌するハイブリドーマを作製した。
【0014】
従来市販の抗体C1P1は、HPV18 E6に対する抗体として作成されたものであるが(非特許文献1: Banks L et al. (1987)参照)、HPV18および16双方のE6タンパク質を認識するものとして市販されている。一方で、本発明にかかるモノクローナル抗体は非常に特異性が高く、HPV16 E6とは結合するが、HPV18 E6および/またはHPV16 E7とは実質的に結合しない。
【0015】
また、好ましくは、本発明にかかるモノクローナル抗体は、感度が非常に高く、通常の免疫ブロッティングにより30pgレベルのHPV16 E6を検出できる。一方で、市販の抗体C1P1のHPV16 E6に対する感度はおおよそ1μgである。子宮頸がんでは、約5pg/μg総タンパク質のHPV16 E6が発現しているが、本発明にかかるモノクローナル抗体によると、従来はできなかった子宮頸がんで発現しているレベルのE6タンパク質を容易に検出できる。
【0016】
上記モノクローナル抗体を産生するハイブリドーマの作製にあたっては、通常のハイブリドーマ作製方法を用いることができる(非特許文献4:Ed Harlow et al.等参照)。一般に、抗原として天然または合成のHPV18 E6またはその断片で動物を免疫し、得られた抗体産生細胞を骨髄腫細胞などと融合させる方法が用いられる。
【0017】
免疫動物として、例えば哺乳動物(例、マウス,ラット,モルモット,ネコ,イヌ,サル,ウサギ,ヒツジ,ヤギ,ハムスターなど),鳥類(例、ニワトリ,ガチョウ,アヒルなど)などを用いることができる。特に、マウス,ラット,ウサギなどが好ましい。免疫動物に誘導される抗体産生細胞としては、抗体を産生する細胞であれば脾臓,リンパ節,末梢血リンパ球などいずれ由来のものでもよいが、特に脾臓細胞が好適に用いられる。免疫方法は、例えばマウスを免疫する場合、皮下、腹腔内、静脈内、筋肉内、皮内等のいずれのルートからでも可能であるが、主として皮下、腹腔内、静脈内に(とりわけ皮下)投与することが好ましい。また、免疫間隔、免疫量等は適宜設定することができるが、例えば2週間隔で約2〜6回免疫し、最終免疫後、約1〜5回、好ましくは約2〜4日後に脾臓細胞を摘出する方法がよく用いられる。免疫量は1回にペプチド量として、マウス当り約0.1μg以上、好ましくは約10μg〜300μg用いることが望ましい。また、脾臓を摘出する前に、部分採血を行い、血中の抗体価の上昇を確認した上で、脾臓細胞を用いる融合実験を行うことが望ましい。
【0018】
得られた抗体産生細胞を増殖性の細胞と融合してハイブリドーマとすることにより、安定的に抗体を産生することが可能になる。融合相手の細胞としては特に骨髄腫細胞(例えば、NS−1,P3−X63−AgUI,X45,SP2,X68−Ag8)が好適に用いられる。特にMOPC21(BALB/cマウス)由来の骨髄腫細胞、X68−Ag8およびその改良株が好ましく用いられる。これら改良株あるいは他の骨髄腫由来の細胞株は8−アザグアニン耐性であり、ヒポキサンチン・グアニンホスホリボシルトランスフェラーゼを欠くため、HAT培地(ヒポキサンチン−アミノプテリン−チミジン含有培地)では成育できず、融合後のハイブリドーマの選択に有利である。
【0019】
細胞融合は公知の方法に従って実施することができる。融合剤として、例えばポリエチレングリコール(PEG),センダイウイルスなどを用いることができる。特にPEGが好適に用いられる。PEGは、平均分子量1000〜6000のものが好ましく、特に好ましくはPEG4000が用いられる。PEGは好ましくは10〜80%さらに好ましくは40〜50%の濃度範囲で用いる。細胞融合は以下のように行うことができる。すなわち、抗体産生細胞と骨髄腫細胞をRPMI1640等の培地で洗い、これらを通常2:1〜10:1の比率で混合し、室温で700×g,5分間遠心して細胞ペレットを得る。細胞ペレットを37℃の恒温槽で温めながらほぐし、予め温めておいたPEG溶液を徐々に加えながら良く混ぜる。通常、PEG量は108個細胞当たり1mL加えられる。ついで、予め温めておいた培地を徐々に滴下、混和し、PEG濃度を下げる。通常は10分位の時間をかけ1〜30mLの培地を加える。室温で遠心して細胞を集め、10%ウシ胎児血清(以下、FCSと略記することがある)を含む培地で脾細胞として1〜2×106/mLの細胞濃度で96穴プレートに100μLずつ分注する。一夜放置後、HATおよび10%FCSを含む培地(HAT培地)100μLを加える。この操作を省略するため細胞融合後、細胞をHAT培地に懸濁してまいても良い。2〜3週間の間に数回の培地交換を行う。培地交換の方法としては通常プレートの各穴から培地100〜200μLを除き、新鮮なHAT培地100〜200μLを加える。この間、ハイブリドーマの出現が認められれば、出来るだけ早くスクリーニングを行い、目的とする抗体を産生しているハイブリドーマの成育しているウェルを検出する。得られたハイブリドーマは直ちにクローニングを行う。
【0020】
このような抗体産生ハイブリドーマのスクリーニングには、ELISA法など公知の種々の方法が使用できる。抗体活性陽性のハイブリドーマは直ちにクローニングに供されるが、通常これは限界希釈法などで容易に実施される。クローン化されたハイブリドーマの培養上清については、上記の方法でその抗体価を測定し、安定的に力価の高い抗体を産生するハイブリドーマを選択することができる。さらに、交差することが望まれないタンパク質(HPV18 E6等)への結合活性に基づいて抗体のスクリーニングをさらに行うことができる。
【0021】
上記モノクローナル抗体を産生するハイブリドーマとして、ハイブリドーマC−26,N−20G8,N−46A4またはN−47C5(FERM AP−20921,FERM AP−20922,FERM AP−20923またはFERM AP−20924)が、独立行政法人産業技術総合研究所特許生物寄託センターに寄託されている。
【0022】
本発明にかかるモノクローナル抗体の生成および蓄積は、本発明にかかるハイブリドーマを通常液体培地、好ましくは無血清培地中、またはヒト以外の温血動物(通常はマウス)の腹腔内で培養し実施することができる。特に高純度の抗体が必要な場合、無血清培地を使用することが望ましい。具体的には、液体培地として、例えば、抗生物質(ゲンタマイシン等)およびIL−6を添加したGIT培地(日本製薬株式会社)などを用いることができる。培養は通常約3〜60日間、好ましくは約3〜10日間、約30〜38℃、好ましくは約37℃で実施される。また、マウス腹腔内への移植については、1匹当り約2×105〜5×107個,好ましくは約1〜5×106個の抗体産生ハイブリドーマを腹腔内に移入し、約10〜35日間の飼育で十分量の抗体含有腹水液3〜10mlが得られる。
【0023】
液体培地あるいは腹水液中の抗体の精製は、公知の生化学的手法を組み合わせることにより実施できる。例えば、抗体含有液を遠心分離後、硫酸アンモニウムや硫酸ナトリウムを用いて上清液を塩析する。得られたタンパク質沈殿物を適当な緩衝液に溶解し透析後カラムクロマトグラフィー(DEAEイオン交換カラム,ヒドロキシアパタイトカラム,ゲルろ過カラム,プロテインAカラム,プロテインGカラムなど),イムノアフィニティークロマトグラフィーなどに供し、目的とする抗体を分離し、精製することができる。以上のような分離および精製操作により、例えば500mLの液体培地からタンパク質重量比で90%以上の純度の抗体を約5〜30mg得ることができる。また50mLの腹水液からは同様の抗体が10〜200mg得られる。特に、プロテインAあるいはGカラムを用いたクロマトグラフィーにゲルろ過カラム担体、DEAEイオン交換カラム担体、ヒドロキシアパタイトなどを用いたクロマトグラフィーを組み合わせることによりさらに高純度の抗体を得ることができる。
【0024】
以上のようにして得られた抗体は、タンパク質分解酵素(パパイン,ペプシンなど)処理ならびに還元剤処理などにより、結合能を保持したままFab,Fab’あるいはF(ab’)2断片を得ることができ、本発明にかかる抗体と同様の目的で用いることができる。また、抗体の抗原認識部位を含む可変領域をコードするDNAを取得し、これに遺伝子操作技術(非特許文献5:Steplewski Z et al.)を用いてヒトIgGの定常領域コードする遺伝子を結合させ、マウス−ヒトキメラ抗体を作製することもできる。
【0025】
上記したように、本発明にかかるモノクローナル抗体には、より高い感度および特異性で、16型ヒトパピローマウイルスE6タンパク質を検出または定量することができ、ひいては、子宮頸がんの素因、前がん病変および/またはがん病変の診断に好適に適用することができる。被検試料として、血漿、血清、尿、脳脊髄液、腹水、胸水、羊水等の体液や、痰、便などを用いることができる。これらの試料は、そのまま、または各種緩衝液で希釈または抽出後濃縮し、免疫学的測定の試料とすることができる。また、本発明にかかるモノクローナル抗体によると、これらのためのキットを提供することができる。キットは、免疫化学的測定に用いられるその他の試薬をさらに含むことができる。
【実施例】
【0026】
以下に、本発明の実施例を、添付図面を参照しながら説明する。もっとも、本発明は、以下に説明する実施例によって限定されるものではない。
【0027】
[実験例1:免疫感作]
〔材料〕
・抗原:抗体作成のために、HPV16 E6タンパク質のアミノ末端16アミノ酸残基またはカルボキシ末端12アミノ酸残基からなる合成ペプチド1、2を免疫原として使用した(図1参照)。
ペプチド1:MFQDPQERPRKLPQLC(配列番号4)
ペプチド2:CRSSRTRRETQL(配列番号5)
・ミエローマ細胞株:ハイブリドーマ作成のために、マウス骨髄腫細胞株(P3-X63-Ag8.653)を使用した(非特許文献6:Kearney JF et al.参照)。
・培地:ハイブリドーマの培養のために、ゲンタマイシンを終濃度0.1mg/mlで、rHuman IL-6(Invitrogen Diaclone #IM-14)を終濃度1ng/mlで含む無血清培地GIT(日本製薬株式会社)を使用した(非特許文献7:S.Sasai et al.、特許文献1:特開昭60-145088号参照)。
【0028】
〔プロトコール〕
HPV16 E6に対する抗体を産生するハイブリドーマを、非特許文献4(Ed Harlow et al.)に記載の手順に準じて作製した。前記免疫原をフロイントコンプリートアジュバンド(FCA)とともに投与することで、マウスを免疫感作した。このマウスから採取した脾臓細胞とマウス骨髄腫細胞株(P3-X63-Ag8.653)とをPEGを用いて細胞融合させることで、ハイブリドーマを作製した。得られたハイブリドーマを、抗原として用いた上記ペプチド、および全長E6タンパク質を用いてスクリーニングおよびクローニングした。ペプチド1を抗原としたものからはハイブリドーマN−20G8,N−46A4およびN−47C5が、ペプチド2を抗原としたものからはハイブリドーマC−26が得られた。
【0029】
[実験例2:抗体の特徴づけ]
〔実験例2−1:免疫ブロッティング〕
得られたハイブリドーマにより産生されるモノクローナル抗体の検出感度および特異性を試験した。プラスミドまたはレトロウイルスベクターを用いてHPV16 E6を発現させた細胞、および子宮頸がん細胞株を用いた。免疫ブロッティングは、非特許文献8(Terai et al.)に記載された方法に準じて行った。5%(v/v)プロテアーゼインヒビターカクテル(ナカライテスク株式会社)を添加したリシスバッファー(0.5% NP4O,1mM DTT,50mM NaCl,25mM Tris-HCl,pH8.0,0.02% NaN3)を用いて全細胞タンパク質を抽出した。細胞タンパク質をSDS-PAGEにより分離し、セミドライトランスファーシステム(アトー株式会社)によりJmmobilon-Pメンブレン(Millipore,Bedford,MA)にブロットした。上記ハイブリドーマ細胞株の調整培地、および市販の抗E6モノクローナル抗体(C1P5, Carbiochem, Ab-1)を、それぞれ1/2および1/100の希釈率でプローブとして用いた。二次抗体として、ホースラディッシュペルオキシダーゼで標識した抗マウス免疫グロブリンを用いた。ローディングコントロールとして、1/1000の希釈率の抗βアクチンヤギポリクローナル抗体(Santa Cruz, sc-1616)および抗ヤギ免疫グロブリンでβアクチンをプローブした。また、ローディングコントロールとして、抗HA抗体(クローン16B12)でHAタグをプローブした。Lumi-light plusウェスタンブロッティング基質(Roche)により視覚化したタンパク質を、LAS3000 CCDイメージングシステム(富士写真フイルム株式会社)により検出および定量化した。
【0030】
図2に示すように、ハイブリドーマ細胞株C−26から産生された抗体は、プラスミドを用いて発現させたE6タンパク質を、最もよく使われている市販抗体(C1P5)よりも約10倍の感度で検出できた。しかしながら、C−26から産生された抗体は、レトロウイルスベクターを用いて発現させたE6タンパク質や子宮頸がん細胞株で発現するE6タンパク質を検出することはできなかった(図示せず)。
【0031】
また、図3に示すように、図2においてC1P5(市販抗体)がかろうじて検出できた量の1/1000量(10ng)のE6タンパク質をN−46A4等は余裕を持って検出できた。その他のクローンも同様にE6タンパク質を高い感度で検出可能であった。しかし、得られた抗体はN末端に変異を導入したE6タンパク質(N末端から8番目(Arg)、9番目(Pro)、10番目(Arg)のアミノ酸残基をそれぞれSer,Ala,Thrに置換したもの)は認識しなかった。ここから、これらの抗体はE6タンパク質のN末端を特異的に認識していることが確認される。
【0032】
また、図4に示すように、ハイブリドーマN−46A4から産生された抗体は、HPV16陽性である全ての子宮頸がん細胞株(CaSki,SiHa,QG−U,Qg−H,SKGIIIb)で発現するE6タンパク質を10μgの細胞タンパク質を用いて検出できた。しかし、ハイブリドーマN−46A4から産生された抗体は、HPV18陽性のHeLa細胞のE6は認識しなかった。
【0033】
同様に、大腸菌で発現させ、すでに定量されたMBP融合タンパク質を用いて、免疫ブロッティング法により抗体の感度を定量した。図5に示すように、ハイブリドーマN−46A4から産生された抗体は、MBP−16E6 100pg(E6タンパク質の重量に換算すると約30pg)を2分の露出で検出できた。しかしながら、ハイブリドーマN−46A4から産生された抗体は、その100倍量(10ng)のMBP−18E6、MBP−16E7およびMBP−lacZタンパク質とは2分の露出でも全く反応しなかった。市販の抗体C1P5は、10ngのMBP−18E6を1分の露出で検出できたが(図示せず)、10ngのMBP−16E6を60分の露出によっても全く検出できなかった。
【0034】
なお、大腸菌での合成に用いたMBP−E6の遺伝子にはE6の開始コドンが残っており、非融合のE6タンパク質も一部産生されたと思われる。クマシーブリリアントブルー(CBB)染色の結果から(図示せず)、非融合16E6タンパク質の量はMBP−16E6の約10%程度であった。図5の結果と併せると、上記抗体N−46A4による非融合16E6タンパク質の検出感度は約30pgであり、融合タンパク質から推定される感度と大きな開きはなかった。
【0035】
このように、上記抗体は約30pgのHPV16 E6タンパク質を検出できる一方で、その100倍量のHPV18 E6およびHPV16 E7であっても免疫ブロッティングで検出しない。すなわち、HPV18 E6およびHPV16 E7と比べて100倍以上の感度で、上記抗体はHPV16 E6を検出できることがわかる。
【0036】
これらの結果を基にSiHa細胞タンパク質20μg中の16E6タンパク質量を半定量すると約100pgとなった。なお、MBP融合タンパク質の定量は、総タンパク質量とMBP−E6の位置のバンドの濃さから判定した。
【0037】
なお、C1P5は18E6とβガラクトシダーゼとの融合タンパク質を抗原として取られた抗体であり、16E6にも反応すると報告されている(実際図2でも検出できている)。また、非特許文献1(Banks L et al. (1987))においては[35S]システイン標識したHeLa細胞で発現しているHPV18 E6タンパク質を免疫沈降で検出できることが示されている。C1P5はHPV18 E6タンパク質に対してはかなり良い抗体であることが分かったが、HPV16 E6タンパク質に対しては免疫ブロッティングでも免疫沈降法でも非常に感度の悪い(ほぼ使い物にならない)抗体であることが分かった。具体的には、C1P5は10ngのMBP−18E6タンパク質を1分以内の露光で楽々検出できた、おそらく1ngも検出できると推定されるのに対し、MBP−16E6は10ngを60分の露光でも全く検出できなかった。図2,3の結果と併せると、C1P5の16E6タンパク質の検出感度は約300ng(MBP−16E6の重量で約1μg)と推定される。
【0038】
〔実験例2−2:免疫沈降〕
得られたハイブリドーマにより産生されるモノクローナル抗体の免疫沈降の効率を試験した。レトロウイルスベクターを用いてHPV16 E6タンパク質を発現させた細胞、および子宮頸がん細胞株を用いた。免疫沈降は、非特許文献9(Kiyono T)に記載された方法に準じて行った。2%(v/v)プロテアーゼインヒビターカクテル(ナカライテスク株式会社)を添加したリシスバッファー(10mM Tris-HCl(pH7.8),1% NP-40,150mM NaCl,1mM EDTA)を用いて全細胞タンパク質を抽出した。得られた抽出液と、上記ハイブリドーマ細胞株の調整培地1mLまたは市販の抗Myc抗体(9E10.2)もしくは抗HA抗体(16B12)との混合物にプロテインGセファロース4B(Pharmacia)を加え4℃で1時間インキュベートした。その後、プロテインGセファロース4Bを洗浄し、SDS-PAGEおよび抗HA抗体(16B12)による免疫ブロッティングにより免疫沈降の効率を確認した。
【0039】
図6に示すように、各抗体はHAタグを3個もつE6タンパク質を効率よく免疫沈降した。抗HA抗体の方が免疫沈降効率が高いが、これはエピトープを3個持つため相乗的に効率が上がっているためと考えられる。各抗体も約10〜14%のE6タンパク質を免疫沈降しており、一般的にその効率は高いと考えられる
【0040】
図6とほぼ同じ条件でハイブリドーマC−26から産生された抗体と市販の抗体C1P5の免疫沈降の効率を比較した。図7に示すように、C−26から産生された抗体は、HAタグを3個もつE6タンパク質を約5%免疫沈降した。これに対し、C1P5は抗原をほとんど(1%以下)免疫沈降できなかった。図5の結果と併せると、少なくとも我々の実験条件ではC1P5はHPV18 E6タンパク質に特異的な抗体ではあるが、免疫ブロッティングでも免疫沈降でもHPV16 E6タンパク質の検出感度は著しく低いことが分かる。対照と して市販の抗HA抗体(16B12)を用いた。図6と同様に抗HA抗体の方が免疫沈降効率が高いが、これはエピトープを3個持つため相乗的に効率が上がっているためと考えられる。なお、C−26から産生された抗体は免疫沈降にも使えるが、その効率は、図6のN−20G8,N−46A4,N−47C5から産生された抗体の方が優れていた。
【0041】
表1に以上の結果の概要を示す。
【表1】
【図面の簡単な説明】
【0042】
【図1】HPV16 E6のORFがコードするタンパク質と抗原に用いたペプチドのアミノ酸配列を示す。ペプチド1:MFQDPQERPRKLPQLC(配列番号1),ペプチド2:CRSSRTRRETQL(配列番号2)。
【図2】本発明にかかるモノクローナル抗体(C−26)による免疫ブロッティングの結果を示す。比較として従来市販の抗体C1P5によるものも示す。レーン1:293FT/p2816:pEF-MYC16E6SD(293FT細胞にMYCタグ融合HPV16 E6の発現ベクターを導入したもの。);レーン2:293FT/p5735:pEF6/16E6SD(293FT細胞にHPV16 E6の発現ベクターを導入したもの。)。各々、総細胞タンパク質10μg。露光時間:C−26=60分;C1P5=164分。
【図3】本発明にかかるモノクローナル抗体(N−20G8,N−46A4,N−47C5)による免疫ブロッティングの結果を示す。レーン1:293FT/p5735:pEF6/16E6SD;レーン2:HMEC4/3HA-16E6(HMEC4細胞に3HAタグ融合HPV16 E6発現ベクターを導入したもの。);レーン3:HMEC4/3HA-16E6-SAT(HPV16 E6のN末端変異体を発現する以外はレーン2と同じ);レーン4:HMEC4/mock(HMEC4細胞に発現ベクターを導入していないもの。)。総細胞タンパク質:レーン1 10ng,レーン2〜3 20μg。
【図4】本発明にかかるモノクローナル抗体(N−46A4)による免疫ブロッティングの結果を示す。293FT/p5735/16E6(293FT細胞にHPV16 E6の発現ベクターを導入したもの。);0:mock(293FT細胞に発現ベクターを導入していないもの。);HMEC4/3HA-16E6(HMEC4細胞に3HA-16E6の発現ベクターを導入したもの。);3Y1/PG10/3HA-16E6(3Y1/PG10細胞に3HA-16E6の発現ベクターを導入したもの。)。総細胞タンパク質:293FT/p5735/16E6 1/100, 1/1000は各々100ng,10ng,その他は10μg。
【図5】本発明にかかるモノクローナル抗体(N−46A4)による免疫ブロッティングの結果を示す。比較として従来市販の抗体C1P5によるものも示す。総細胞タンパク質:HelaおよびSiHa 20μg,その他図示の通り。
【図6】本発明にかかるモノクローナル抗体(N−20G8,N−46A4,N−47C5)による免疫沈降の結果を示す。293FT/3HA-16E6SD(293FT細胞に一過性遺伝子導入により3HA-16E6を高発現させた細胞)の細胞抽出液を用いた。免疫沈降のネガティブコントロールとして抗Myc抗体(9E10.2)を、ポジティブコントロールとして抗HA抗体(16B12)を用いた。定量のために、用いた細胞抽出液の2容量%をSDS-PAGEおよび免疫ブロッティングに供した。
【図7】本発明にかかるモノクローナル抗体(C−26)による免疫沈降の結果を示す。免疫沈降のネガティブコントロールとして抗Myc抗体(9E10.2)を、ポジティブコントロールとして抗HA抗体(16B12)を用いた。定量のために、用いた細胞抽出液の2、10容量%をSDS-PAGEおよび免疫ブロッティングに供した。
【特許請求の範囲】
【請求項1】
16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片であって、
16型ヒトパピローマウイルスE6タンパク質と結合し、
18型ヒトパピローマウイルスE6タンパク質および/または16型ヒトパピローマウイルスE7タンパク質と実質的に結合しない、
モノクローナル抗体またはその結合活性断片。
【請求項2】
ハイブリドーマC−26,N−20G8,N−46A4またはN−47C5(FERM AP−20921,FERM AP−20922,FERM AP−20923またはFERM AP−20924)から産生され得る、請求項1に記載のモノクローナル抗体またはその結合活性断片。
【請求項3】
請求項1に記載の抗体を産生するハイブリドーマ。
【請求項4】
ハイブリドーマC−26,N−20G8,N−46A4またはN−47C5(FERM AP−20921,FERM AP−20922,FERM AP−20923またはFERM AP−20924)である、請求項3に記載のハイブリドーマ。
【請求項5】
請求項3または4に記載のハイブリドーマを用いることを特徴とする、モノクローナル抗体またはその結合活性断片の調製方法。
【請求項6】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を含む、16型ヒトパピローマウイルスE6タンパク質を検出または定量するためのキット。
【請求項7】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を用いることを特徴とする、16型ヒトパピローマウイルスE6タンパク質を検出または定量する方法。
【請求項8】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を含む、子宮頸がんの素因、前がん病変および/またはがん病変を診断するためのキット。
【請求項9】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を用いることを特徴とする、子宮頸がんの素因、前がん病変および/またはがん病変を診断する方法。
【請求項1】
16型ヒトパピローマウイルスE6タンパク質に対するモノクローナル抗体またはその結合活性断片であって、
16型ヒトパピローマウイルスE6タンパク質と結合し、
18型ヒトパピローマウイルスE6タンパク質および/または16型ヒトパピローマウイルスE7タンパク質と実質的に結合しない、
モノクローナル抗体またはその結合活性断片。
【請求項2】
ハイブリドーマC−26,N−20G8,N−46A4またはN−47C5(FERM AP−20921,FERM AP−20922,FERM AP−20923またはFERM AP−20924)から産生され得る、請求項1に記載のモノクローナル抗体またはその結合活性断片。
【請求項3】
請求項1に記載の抗体を産生するハイブリドーマ。
【請求項4】
ハイブリドーマC−26,N−20G8,N−46A4またはN−47C5(FERM AP−20921,FERM AP−20922,FERM AP−20923またはFERM AP−20924)である、請求項3に記載のハイブリドーマ。
【請求項5】
請求項3または4に記載のハイブリドーマを用いることを特徴とする、モノクローナル抗体またはその結合活性断片の調製方法。
【請求項6】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を含む、16型ヒトパピローマウイルスE6タンパク質を検出または定量するためのキット。
【請求項7】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を用いることを特徴とする、16型ヒトパピローマウイルスE6タンパク質を検出または定量する方法。
【請求項8】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を含む、子宮頸がんの素因、前がん病変および/またはがん病変を診断するためのキット。
【請求項9】
請求項1または2に記載のモノクローナル抗体またはその結合活性断片を用いることを特徴とする、子宮頸がんの素因、前がん病変および/またはがん病変を診断する方法。
【図1】


【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2007−314476(P2007−314476A)
【公開日】平成19年12月6日(2007.12.6)
【国際特許分類】
【出願番号】特願2006−146261(P2006−146261)
【出願日】平成18年5月26日(2006.5.26)
【出願人】(803000056)財団法人ヒューマンサイエンス振興財団 (341)
【Fターム(参考)】
【公開日】平成19年12月6日(2007.12.6)
【国際特許分類】
【出願日】平成18年5月26日(2006.5.26)
【出願人】(803000056)財団法人ヒューマンサイエンス振興財団 (341)
【Fターム(参考)】
[ Back to top ]