
ヤケヒョウヒダニ由来の低アレルゲン性DerP1およびDerP3タンパク質
本発明は、低アレルゲン活性を有する、組換えDer p 1/ProDer p 1/PreProDer p 1アレルゲン誘導体、ProDer p 3、または組換えProDer p 3/Der p 3/PreProDer p 3アレルゲン誘導体を提供することからなる、アレルギーの治療を提供する。また、アレルギー個体またはナイーブ個体においてTh1型免疫応答を刺激する前記変異アレルゲンを含んでなる医薬組成物も提供する。この医薬組成物は、野生型アレルゲンと接触した際のアレルギー反応の可能性を減少させるのに有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規の予防用および治療用製剤に関するものであり、この製剤は特定のアレルゲンに対するアレルギー反応の予防および/または軽減に有効である。さらに、本発明はヤケヒョウヒダニ(Dermatophagoides pteronyssinus)由来の主要なアレルゲンタンパク質である、アレルゲンDer p 1およびその前駆体型ProDer p 1の低アレルゲン性組換え誘導体に関する。特に、本発明の誘導体は、物理的に改変されたDer p 1もしくはProDer p 1、例えば熱処理されたタンパク質;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えDer p 1;組換えProDer p 1;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えProDer p 1;組換えPreProDer p 1;あるいは、ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えPreProDer p 1を包含する。また、Der p 1、ProDer p 1およびPreProDer p 1誘導体を発現させて精製する方法、ならびに免疫原性組成物およびワクチンを製剤化する方法も記載する。
【0002】
さらに本発明は、ヤケヒョウヒダニ由来のもう一つのアレルゲンタンパク質である、アレルゲンDer p 3ならびにその前駆体型ProDer p 3およびPreProDer p 3の低アレルゲン性組換え誘導体に関する。特に、本発明の誘導体には、物理的に改変されたDer p 3もしくはProDer p 3、例えば熱処理されたタンパク質;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えDer p 3;組換えProDer p 3;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えProDer p 3;組換えPreProDer p 3;あるいはジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えPreProDer p 3が包含される。また、Der p 3、ProDer p 3およびPreProDer p 3誘導体を発現させて精製する方法、ならびに免疫原性組成物およびワクチンを製剤化する方法も記載する。
【背景技術】
【0003】
アレルギー反応はヒトではよく見られるが、さまざまなアレルゲンが誘因となる可能性がある。アレルギーの個体はアレルゲンに対して敏感になり、血清中に高レベルのアレルゲン特異的IgEが存在し、Th2型サイトカイン(IL-4、IL-5およびIL-13)を産生するアレルゲン特異的T細胞集団を有することを特徴とする。アレルゲンの存在下における、マスト細胞および好塩基球の表面に存在するFcεRI受容体へのIgEの結合は、細胞の急速な脱顆粒をもたらし、さらにそれに続いてヒスタミン、ならびに他の、生成済み、および新たに生成されたメディエータの放出をもたらす。これに加えて、T細胞既往性応答の刺激は、結果として、IL-4およびIL-13の産生をもたらし、それとともにB細胞応答をさらにアレルゲン特異的IgEを産生するようにし向けて切り換えさせる。即時型および遅延型アレルギー反応の発生に関する詳細については、Joost Van Neeven ら, 1996, Immunology Today, 17, 526を参照されたい。アレルギーでない個体では、同じ抗原に対する免疫応答はIFN-γのようなTh1型サイトカインをさらに包含する可能性がある。このようなサイトカインは、高レベルのアレルゲン特異的IgEが含まれる高レベルのTh2型免疫応答を阻害することによって、アレルギー反応の発症を防止することができる。この点に関して重要なことは、IgE合成が、B細胞上のCD23(FcεRII)受容体に対するIgE/アレルゲン複合体の結合を介した阻害的フィードバックメカニズムによって、制御可能であるという事実である(Luo ら, J.Immunol., 1991, 146(7), 2122-9; Yu ら, 1994, Nature, 369(6483):753-6)。細胞結合型CD23を欠いた系では、IgE合成のこのような阻害は起こらない。
【0004】
世界の人口の20%を越える人々が、気管支喘息、アトピー性皮膚炎および通年性鼻炎といった、アレルゲンに対するIgEを介したI型アレルギー疾患に罹っている。こうしたアレルギー反応の現在の治療方法としては、抗ヒスタミン剤治療および/または抗炎症性コルチコステロイドの局所投与によって、ヒスタミン放出という症候的な影響を抑える方法がある。開発中の他の方法としては、宿主の免疫系を用いてマスト細胞の脱顆粒を防ぐ方法がある(Stanworth ら, EP 0 477 231 B1)。その他の免疫療法も報告されている(Hoyne ら, J.Exp.Med., 1993, 178, 1783-1788; Holt ら, Lancet, 1994, 344, 456-458)。
【0005】
遅延症状のみならず即時症状も薬理学的治療によって改善することができるが、I型アレルギーに対する唯一の治癒的なアプローチはアレルゲン特異的免疫療法である。しかしながら、この方法に関するいくつかの問題は未解決のままである。第1に、免疫療法は現在のところ、バッチごとに不均質である可能性のある全アレルゲン抽出物を用いて実施されている。さらに、これらのアレルゲン混合物は個々の患者の特性に向けて設計されていないので、好ましくない毒性タンパク質を含有しうる。第2に、天然アレルゲンを高用量で投与することは、激しいアナフィラキシー反応を引き起こす可能性があり、したがって、アレルゲンの用量を免疫療法の成功のために最適効率の高用量で使用できない場合が多い。最初の問題は、アレルゲン抽出物と比べて、より良い特性を有し、より高い生産性の組換えアレルゲンを用いた代替ワクチン接種によって対処された。第2の問題、すなわちアレルゲン抽出物の注射を繰り返すことによって引き起こされるアナフィラキシー反応のリスクは、IgE反応性を欠失もしくは変異導入によって変化させた組換え「低アレルゲン」の使用によって最小化することができる(Akdis, CA and Blaser, K, Regulation of specific immune responses by chemical and structural modifications of allergens, Int. Arch. Allergy Immunol., 2000, 121, 261-269)。
【0006】
アレルギーの治療および予防のための製剤が報告されているが、その製剤は、Th2型からTh1型への反応のシフトによって、アレルゲンに対する細胞性反応を改善するのみならず、IgEの産生をダウンレギュレートする方法を提供するものである(IL-4とIFN-γ産生Der p 1特異的T細胞との比の減少、またはその代わりにIL-5とIFN-γと比の減少によって判断される)。これは、たとえばWO 99/25823 に記載の、酵素活性の低下した組換えDer p 1のような組換えアレルゲンを使用することによって達成することができる。しかしながら、こうした組換えアレルゲンの免疫原性は、IgE合成誘導の点では野生型ProDer p 1の免疫原性と類似していると思われる。
【0007】
IgE結合活性の低下した、非アナフィラキシー型のアレルゲンが報告されている。アレルゲンエンジニアリングによって、アミノ酸残基の部位特異的変異導入もしくは特定のアミノ酸配列の欠失によりアレルゲンタンパク質のIgE結合能力を低下させることが可能となった。同時に、T細胞エピトープは維持されるのでT細胞活性化能力は依然として保たれる。このことは、結果はさまざまであるが異なるアレルゲンに関するいくつかのアプローチによって示されている。オオアワガエリ(timothy grass)花粉アレルゲンPhl p 5b(Schramm G ら, 1999, J Immunol.,162, 2406-14)、チリダニ類(ヒョウヒダニ)主要アレルゲンDerf2(Takai ら 2000, Eur. J. Biochem., 267, 6650-6656)、DerP2(Smith & Chapman 1996, Mol. Immunol. 33, 399-405)、およびDerf1(Takahashi K ら 2001, Int Arch Allergy Immunol.124, 454-60)に関する例が公表されている。ある研究は、ジスルフィド結合に関わるシステイン残基レベルでの点突然変異の導入による、低アレルゲンDerf1の生成を報告している(Takahashi K Int Arch Allergy Immunol. 2001;124(4):454-60., Takai T, Yasuhara T, Yokota T, Okumura Y)。しかしながら、野生型ProDerf1は酵母Pichia pastorisによってうまく分泌されはしたが、それとは対照的に、分子内ジスルフィド結合に関わるシステイン変異体は、分泌されなかった。
【0008】
ハウスダストのダニであるヤケヒョウヒダニ由来のアレルゲンは、アレルギー性過敏反応に関わる主要な原因因子の1つである。
【0009】
Der p 1
ヤケヒョウヒダニのグループ1アレルゲンであるDer p 1は主要アレルゲンであり、チリダニアレルギーの血清の80〜100%においてIgEと結合している(Chapman, M.D., ら (1983). J. Allergy Clin. Immunol., 72: 27-33; Krillis, S., ら (1984). J. Allergy Clin. Immunol., 74: 132-41)。このタンパク質はハウスダスト中に高濃度(100〜10000 ng/gダスト)で見出されることが多い(Platts-Mills and Chapman (1987). J.Allergy Clin. Immunol., 80: 755-75; Wahn, U., ら (1997). J. Allergy Clin. Immunol., 99: 763-69)が、Der p 1は糞便物とのみ関連するわけではなく、ある範囲の粒子と関連していると考えられている(DeLuca, ら (1999). J. Allergy Clin. Immunol., 103: 174-75)。100ngのレベルで感作に関わり、量の増加に伴い危険性は増大する。
【0010】
Der p 1をコードするcDNAはクローニングされ、配列決定されている(Chua, K., ら (1988). J. Exp. Med., 167: 175-82; Thomas, ら (1988). Int. Arch. Allergy Appl. Immunol., 85: 127-29; Chua, K., ら (1993). Int. Arch. Allergy Immunol., 101: 364-8):このアレルゲンは222個のアミノ酸残基からなるタンパク質で、計算上の分子量は25 KDaである。このアレルゲンは18アミノ酸シグナルペプチドおよび80アミノ酸N末端プロ配列を含めて、320個のアミノ酸残基からなる前駆体型として合成される。ProDer p 1の成熟プロセスは依然として不明であるが、タンパク質分解によるプロ領域の除去によって、または自己触媒的プロセシングによって、酵素が活性化されると考えられている。
【0011】
Der p 1配列は、システインプロテイナーゼの原型であるパパインの配列と30%の相同性を示した(Robinson, C., ら (1997). Clin. Exp. Allergy, 27 (1): 10-21)。パパインのタンパク質分解活性に関与する残基の大半は、活性部位のシステインおよびヒスチジン残基を含めて、Der p 1において保存されていた。Der p 1は入手しにくいため、このアレルゲンについての放射線結晶学データは得られていない。それにもかかわらずDer p 1の立体構造は、パパインおよびアクチニジンの放射線結晶学的構造に基づいて明らかにされている。Der p 1の構造は、基本的な構造的および機構的特徴が他のパパイン様システインプロテイナーゼと共通している。下記はDer p 1の立体構造モデルである(Topham, C.M, ら (1994) Protein engineering, 7 (7): 869-894)。Der p 1はN末端側配列およびC末端側配列がそれぞれ独立に形作る2つの球形ドメインを与える:基質結合性および触媒性残基はこれらのドメインの間の裂け目にあり、ドメインは可動性の外側のループで結び付けられている。
【0012】

【0013】
Der p 1のシステインプロテアーゼ活性は一般に認められているが、研究から、Der p 1は、活性部位が1つしかないにもかかわらず、独特の混合したシステイン/セリンプロテアーゼ活性を示すことが明らかになった(Hewitt, C.R.A., ら (1997). Clin. Exp. Allergy, 27: 201-207)。優先的な切断部位はシステインプロテアーゼ活性についてはグルタミン酸であり、セリンプロテアーゼ活性についてはアルギニンである。
【0014】
Der p 1は、特に気管支組織を保護するプロテアーゼインヒビターであるα1-アンチトリプシンを分解することによって(Kalsheker, ら (1996). Biochem. Biophys. Res. Comm., 221: 59-61)、および、気道上皮における細胞の密着結合をゆるめることによって(Wan, H., ら (2000). Clin. Exp. Allergy, 30:685-98)、気管支粘膜の透過性を増加させ、その結果、抗原提示細胞へのアクセスを容易にする。下記の模式図に示すように、Der p 1はタンパク質「オクルディン(occludin)」を切断することによって密着結合をゆるめ、樹状細胞による吸着を促進し、アレルギー反応を誘発する。
【0015】
【0016】
Der p 1は、IgE合成の制御に関わる低親和性IgE受容体であるCD23(FcεR II)を切断し(Hewitt. C., ら (1995). J. Exp. Med., 182: 1537-1544; Schulz, O., ら (1997). Eur. J. Immunol., 27: 584-588)、それによってIgE産生を促すことが明らかになった。他方では、Der p 1はIL-2受容体のαサブユニットであるCD25を切断する(Schulz, O., ら (1998). J. Exp. Med., 187: 271-275)。IL-2はTh1免疫応答の伝播に関わるサイトカインなので、その受容体が消化分解されると結果としてTh2応答に向かって傾く。Der p 1のタンパク質分解活性はヒトT細胞からのTh2サイトカイン放出を強め(Ghaemmaghami, A.M., ら (2001). Eur. J. Immunol., 31: 1211-1216)、バイスタンダー(bystander)アレルゲンにアジュバント活性を与える(Ghough L., ら (2001). Clin. Exp Allergy, 31: 1594-1598)ことも明らかになった。
【0017】
Der p 3
Der p 3はヤケヒョウヒダニのグループ3アレルゲンである。一般に主要抗原と考えられるが、Der p 3 IgE結合の推定値はかなり変動し、16%程度と低いこともあり(Heymann, P.W., ら (1989). J Allergy Clin Immunol., 83: 1055-1067)、グループ1および2アレルゲンと同様の結合力を有し100%と高いこともある(Stewart, G.A., ら (1992). Immunology, 75: 29-35)。
【0018】
Der p 3をコードするcDNAはクローニングされ、配列決定されている(Smith W.A., ら (1994). Clin. Exp Allergy, 24: 220-228):Der p 3は232アミノ酸残基からなるタンパク質であって、計算上の分子量は25 KDaである。このタンパク質は、18アミノ酸シグナルペプチドおよび11アミノ酸N末端プロ配列を有する、不活性なPrePro Der p 3前駆体として合成される。Der p 3はセリンプロテイナーゼの原型であるトリプシンに高い相同性を示すセリンプロテイナーゼであり(Stewart, G.A., ら (1992). Immunology, 75: 29-35)、活性部位に関わる残基を包含する(図4.9)。優先的な切断部位はアルギニンおよびリジンである。
【0019】
Der p 3とウシ トリプシンの間の配列相同性を下記に示す。*印で示す残基は触媒部位に関係している。
【0020】
【0021】
Der p 3は、肺上皮細胞上のプロテアーゼ活性化受容体2の活性化により、炎症性サイトカインGM-CSFおよびエオタキシンに対するシグナリング経路を始動させることが明らかになっている(Sum, G., ら (2001). J. Immunol., 167: 1014-1021)。実際、Der p 3は、膜貫通タンパク質オクルディン(occludin)を切断することによって、気道上皮における密着結合をさらにゆるめることができる(Wan, H. ら (2000). Clin. Exp. Allergy, 31: 279-294)。この特徴はDer p 1についても認められ、抗原提示細胞への優先的アクセスを提供する。
【0022】
本発明は、野生型アレルゲンと比較して低下したアレルゲン性を有する、ヤケヒョウヒダニDer p 1アレルゲン、またはその前駆体型であるProDer p 1/PreProDer p 1(以後“Der p 1/ProDer p 1/PreProDer p 1”と称する)の組換え誘導体の提供および使用に関する。アジュバントを添加した組換えタンパク質、またはNAVACに適したDer p 1/ProDer p 1/PreProDer p 1をコードするプラスミドという形態での本発明の組換え型Der p 1誘導体は、強力な予防的Th1を生じ、あるいはTh2免疫応答をTh1にシフトさせるための、予防用または治療用ワクチンとして使用される。低アレルゲン性誘導体を組換え発現系において良好に生成することができるが、これもまた本発明の一態様である。
【0023】
本発明はさらに、野生型アレルゲンと比較して低下したアレルゲン性を有する、ヤケヒョウヒダニDer p 3アレルゲン、またはその前駆体型であるProDer p 3/PreProDer p 3(以後“Der p 3/ProDer p 3/PreProDer p 3”と称する)の組換え誘導体の提供および使用に関する。アジュバントを添加した組換えタンパク質、またはNAVACに適したDer p 3/ProDer p 3/PreProDer p 3をコードするプラスミドという形態での本発明の組換え型Der p 3誘導体は、強力な予防的Th1を生じ、あるいはTh2免疫応答をTh1にシフトさせるための、予防用または治療用ワクチンとして使用される。低アレルゲン性誘導体を組換え発現系において良好に生成することができるが、これもまた本発明の一態様である。
【0024】
本発明はさらに、本明細書に記載の1つもしくは2つ以上のタンパク質アレルゲン、または組換え誘導体を組み合わせて提供し、使用することに関する。ある態様において、本発明は、本明細書に記載のDer p 1またはその誘導体を、本明細書に記載のDer p 3、ProDer p 3もしくはProDer p 1、またはその誘導体と組み合わせて、含んでなるか、またはそれらから構成される。好ましくは、本発明はDer p 1をProDer p 3と組み合わせて、またはそれと融合して提供する。さらに別の態様において、本発明は、本明細書に記載のDer p 3もしくはその誘導体を、本明細書に記載のDer p 1、ProDer p 3、PreProDer p 3もしくはProDer p 1、もしくはPreProDer p 1、またはその誘導体と組み合わせて、またはそれと融合して、含んでなるか、またはそれらから構成される。他の態様において、本発明は、Der p 1と融合したProDer p 3を含んでなる融合タンパク質を提供する。本明細書に記載のあらゆる融合タンパク質は、連続したヒスチジン、好ましくは6ヒスチジンを追加して含んでなることができる。ある態様において、融合タンパク質は、(His)6-ProDer p 3-Der p 1を含んでなるか、またはそれからなる。
【0025】
Der p 1は30 KDaタンパク質であり、クローニングおよび配列決定がなされている(Chua ら, 1988, J.Exp.Med., 167, 175-182)。成熟タンパク質は222アミノ酸残基を有することが知られている。Der p 1の配列は、パパインと31%の相同性を有し、酵素活性領域にではそれ以上の顕著な相同性を共有しているが、最も顕著なのはCys34-His170イオン対である(Topham ら, 上記)。Der p 1はダニの中腸で産生されるが、中腸におけるその役割はおそらく食物の消化に関係しているであろう。個々の糞便粒(それぞれ直径10〜40μm程度)には0.2 ng以下のタンパク質分解活性のあるDer p 1が混合されており、したがってDer p 1は容易にヒトの気道内に吸い込まれる。精製Der p 1標品を室温にて一晩保存すると、結果として、自己タンパク質分解により酵素活性はほとんど完全に失われる(Machado ら, 1996, Eur.J.Immunol. 26, 2972-2980)。Der p 1をコードするcDNA配列は、多くの哺乳動物および植物プロテイナーゼと同様にDer p 1が、320アミノ酸残基からなる不活性プレプロ酵素として合成され、後に222アミノ酸の成熟型にプロセシングされることが明らかにされた(Chua ら, 1988, J.Exp.Med., 167, 175-182; Chua ら, 1993, Int. Arch Allergy Immunol 101, 364-368)。ProDer p 1の成熟は現在まで知られていないが、このアレルゲンは80残基のプロ領域の切断によってプロセシングされると考えられている。
【発明の開示】
【0026】
本発明は、野生型アレルゲンと比較して著しく低下したアレルゲン活性を有する、組換えヤケヒョウヒダニDer p 1/ProDer p 1/PreProDer p 1タンパク質アレルゲン誘導体を提供する。タンパク質分子内のジスルフィド架橋を開裂させることによりタンパク質の立体構造を不安定化させること、またはタンパク質のある領域、例えばProDer p 1のアミノ酸227〜240(Der p 1配列の147〜160)を欠失させることにより、タンパク質形態の立体高次構造を損なうことをねらったいくつかの方法によって、アレルゲン活性を弱めることができる。前記アレルゲン誘導体は改変されていない野生型アレルゲンと比べて、以下のような、有利な点を有する:1)野生型アレルゲンによって刺激されるのと比較して、Th1型の免疫応答(たとえば、高いIgG2a)を増加させ、それによってワクチン接種を受けたホストの潜在的アレルギー性の抑制をもたらす、2)T細胞反応性は依然として保持しているがアレルゲン性が低下しており、そのため高用量の免疫原を全身投与するのにいっそう適している、3)天然Der p 1の結合についてIgEと競合するDer p 1特異的IgGを誘導する、4)エアロゾル化されたアレルゲン抽出物に曝露された後であっても、気道の好酸球増加を効果的に防御する。このような誘導体は治療用および予防用ワクチン製剤に使用するのに適しており、こうした製剤は医薬用に好適で、さらにはアレルギー反応の治療もしくは予防に適している。
【0027】
第1の態様によると本発明は、アレルゲン活性が好ましくは還元剤の存在下でタンパク質の熱処理といった物理的手段によって著しく低下している、たとえばほとんど、または完全になくなっている、組換えDer p 1/ProDer p 1/PreProDer p 1(すなわちDer p 1、ProDer p 1またはPreProDer p 1)アレルゲン誘導体を提供する。典型的には、Der p 1/ProDer p 1/PreProDer p 1タンパク質を、還元剤の存在下で約100℃にて数分間処理する。還元剤はβ-メルカプトエタノールもしくはDTTが好ましい。さらに好ましくは、タンパク質を50 mM β-メルカプトエタノールの存在下で約100℃にて5分間処理する。この処理はIgE結合エピトープのタンパク質高次構造の安定性に有害な影響を及ぼす。タンパク質はProDer p 1またはPreProDer p 1であることが好ましい。
【0028】
第2の態様において、本発明は、たとえば、コードしているcDNAまたはゲノムDNAに特定の変異を導入することによってアレルゲン活性が遺伝学的に損なわれた、組換えDer p 1/ProDer p 1/PreProDer p 1タンパク質誘導体を提供する。したがって、本発明の態様は、それ自体、遺伝的に変異した組換えDer p 1/ProDer p 1/PreProDer p 1を提供する。低アレルゲン性変異体を組換え的に生産する前に、天然配列内に変異を導入することによって、Der p 1/ProDer p 1/PreProDer p 1のアレルゲン性を弱めることができる。これは、置換、欠失または付加を導入することによって、またはタンパク質の立体構造を変化させることによって達成でき、タンパク質の立体構造は失われる。こうしたことは、中でも、タンパク質を断片として発現させることによって、またはジスルフィド架橋形成に関わるシステイン残基を欠失させることによって、またはタンパク質の三次構造が実質的に変化するような残基を欠失もしくは付加することによって達成されうる。好ましくは、2つのシステイン残基間の相互作用を変化させる効果のある変異を生じさせることができるが、典型的には天然型成熟Der p 1の4、31、65、71、103および117位(それぞれProDer p 1の84、111、145、151、183および197位に対応する)での単一変異である。本発明の変異タンパク質は、たとえば4および31、4および65、4および71、4および103、31および65、もしくは4および31および65位, または71および103、71および117、103および117、31および117、65および117、もしくは71および103および117位といった、2つ以上(3、4、5または全6個)のシステイン変異を含んでもよく、それによってさまざまなジスルフィド架橋に影響を及ぼす。誘導体は、上記のいずれかの位置に単一の変異を含んでなることが好ましい。もっとも好ましい変異はCys4(またはその代わりに、もしくはそれに加えて、Cys4のジスルフィド結合パートナーと考えられるCys117)に関する。このシステイン変異は欠失でもよいが、他の19の天然アミノ酸のいずれかへの置換が好ましい。好ましい置換は正電荷を有するアミノ酸残基を導入するものであり、その結果得られるタンパク質の立体構造をいっそう不安定化させる。たとえば、好ましい置換にはシステイン→アルギニン(またはリジン)置換がある。
【0029】
本発明のある態様において、誘導体は、システイン残基71、103および117がいずれもアラニンへと変異している三重変異を含んでなる。
【0030】
別の態様において、本発明はProDer p 1配列のアミノ酸227〜240が欠失した、ProDer p 1の一形態を提供する。これらのアミノ酸はDer p 1配列の147〜160に相当する。
【0031】
したがって本発明は、本明細書において、低アレルゲン性Der p 1/ProDer p 1/PreProDer p 1誘導体の例として与えられる6個の特定の変異、ならびにProDer p 1のアミノ酸227〜240(Der p 1の147〜160)が欠失しているさらなる変異によって例証されるが、それに限定されない。第1に、ProDer p 1のアレルゲン活性は、Der p 1タンパク質配列のCys4の位置でシステイン残基をアルギニン残基に置き換えることによって、実質的に減少し、好ましくは完全に消失するが、その配列を配列番号3に示す。第2に、ProDer p 1のアレルゲン活性は、(成熟Der p 1の配列と照合して計算された)下記の任意の位置でシステイン残基をアルギニン残基に置換することによって実質的に消失する:Der p 1タンパク質配列のCys31(配列番号5)、Cys65(配列番号7)、Cys71(配列番号9)、Cys103(配列番号11)、Cys117(配列番号13)。
【0032】
さらに、ProDer p 1のアレルゲン活性は、ProDer p 1のアミノ酸227〜240(Der p 1の147〜160)の欠失により、実質的に減少し、好ましくは完全に消失する(配列番号15)。
【0033】
Der p 1/ProDer p 1/PreProDer p 1の変異型は、Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするcDNAの、従来法による部位特異的変異導入(G. Winterら, Nature 1982, 299, 756 758 もしくはZollerおよびSmith, 1982; Nucl. Acids Res., 10, 6487 6500に記載)、または欠失変異導入(Chan および Smith, Nucl. Acids Res., 1984, 12, 2407 2419 もしくはG. Winter ら, Biochem. Soc. Trans., 1984, 12, 224-225に記載)によって調製することができる。
【0034】
低アレルゲン性ProDer p 1、PreProDer p 1、Der p 3、ProDer p 3もしくはPreProDer p 3をも本発明によって提供する。
【0035】
本発明は具体的に開示された配列に限定されず、変異して、IgE結合反応性および/またはヒスタミン放出活性が減少し、またはなくなっているにもかかわらず、T細胞反応性を保持し、および/または野生型アレルゲンに対する免疫応答を刺激する能力を保持するあらゆる低アレルゲン性アレルゲンをも包含する。変異アレルゲンのアレルゲン活性、および結果としてアレルゲン活性の減少は、下記の実施例の項に詳述する方法にしたがった、ヒスタミン放出活性またはIgE結合反応性の方法のいずれによっても野生型と比較することができる。
【0036】
「実質的に減少したアレルゲン活性」は、残存IgE結合活性によって測定されるアレルゲン活性が、天然-非修飾もしくは非変異-タンパク質の活性の最大50%、好ましくは最大20%、さらに好ましくは最大10%、さらにいっそう好ましくは最大5%、さらにその上好ましくは5%未満にまで、減少していることを意味する。あるいはまた、「実質的に減少したアレルゲン活性」を、変異体のヒスタミン放出を測定することによって評価することもできる。活性の実質的な減少は、天然タンパク質に比べて少なくとも100分の1に減少する場合であるが、好ましくは1000分の1、さらに好ましくは10000分の1である。
【0037】
さまざまな免疫学的アッセイによって、変異アレルゲンの免疫原性を野生型アレルゲンと比較することができる。変異アレルゲンもしくは野生型アレルゲンのいずれかを用いたワクチン接種の後、in vitro T細胞アッセイによって、変異アレルゲンおよび野生型アレルゲンの交差反応性をアッセイすることができる。簡単に述べると、ワクチン接種を受けた動物から分離された脾臓T細胞をin vitroにて変異アレルゲンもしくは野生型アレルゲンのいずれか一方で再刺激した後、市販のELISAアッセイを用いてサイトカイン産生を測定するか、またはアレルゲン特異的T細胞の増殖をトリチウム標識したチミジンの取り込みによって、経時的にアッセイすることができる。また、免疫原性をELISAアッセイによって測定することも可能であり、当業者はその詳細を容易に決定することができよう。簡単に述べると、2種類のELISAアッセイが考えられる。1つ目は、野生型Der p 1で免疫したマウスの血清による変異型Der p 1の認識を評価することであり;2つ目は変異アレルゲンで免疫した動物の血清による野生型Der p 1の認識を評価することである。手短には、各ウェルを500 ngの精製された野生型もしくは変異型Der p 1で4℃にて一晩コーティングする。ブロッキング溶液(TBS-Tween 0.1%、1% BSA含有)とともにインキュベートした後、血清の連続希釈物を37℃にて1時間インキュベートする。ウェルを5回洗浄し、アルカリホスファターゼと結合した抗IgG抗体とともにインキュベートすることによって全IgGを明らかにする。変異型Der p 3の免疫原性も、上記のDer p 1に関する記載のように野生型Der p 3と比較することができる。
【0038】
本発明のもう一つの態様は、本明細書に開示のDer p 1/ProDer p 1/PreProDer p 1アレルゲンの変異型をコードする、単離された核酸を提供する。この核酸配列はDNA配列であることが好ましく、標準的なDNA合成技術、たとえば酵素による連結(D.M. Roberts ら, Biochemistry 1985, 24, 5090 5098)、化学合成、in vitro酵素重合、または上記技術を組み合わせることによって合成することができる。核酸配列は、想定される発現宿主において用いられるコドン使用頻度を模倣するように、最適化されたコドン使用頻度パターンを有することが好ましいが、さらに好ましくは高発現の哺乳動物(たとえばヒト)遺伝子のコドン使用頻度に類似する。好ましいDNA配列はコドンを最適化した配列であり、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号15および配列番号17に示す。
【0039】
本発明の別の態様は、本明細書に記載のDer p 3/ProDer p 3/PreProDer p 3アレルゲンの変異型をコードする、単離された核酸を提供する。この核酸配列はDNA配列であることが好ましく、標準的なDNA合成技術、たとえば酵素による連結(D.M. Roberts ら, Biochemistry 1985, 24, 5090 5098)、化学合成、in vitro酵素合成、または上記技術を組み合わせることによって合成することができる。核酸配列は、想定される発現宿主において用いられるコドン使用頻度を模倣するように、最適化されたコドン使用頻度パターンを有することが好ましいが、さらに好ましくは高発現の哺乳動物(たとえばヒト)遺伝子のコドン使用頻度に類似する。好ましいDNA配列を、配列番号20および21に示す。
【0040】
DNAの酵素的重合は、一般的には50 ml以下の容量で、10〜37℃にて、必要なヌクレオシド三リン酸、dATP, dCTP, dGTP およびdTTPを含有する適当なバッファー中で、DNAポリメラーゼI(Klenowフラグメント)のようなDNAポリメラーゼを用いて、in vitroで行うことができる。DNA断片の酵素による連結は、一般的には50ml以下の容量で、4℃から室温までの温度にて0.05M Tris (pH 7,4)、0.01M MgCl2、0.01Mジチオスレイトール、1mMスペルミジン、1mM ATPおよび0.1mg/mlウシ血清アルブミンといった適当なバッファー中でT4 DNAリガーゼのようなDNAリガーゼを用いて行うことができる。DNAポリマーもしくは断片の化学合成は、以下に記載の固相法を用いて、従来のリン酸トリエステル、亜リン酸エステルもしくはホスホルアミダイトの化学反応によって行うことができる('Chemical and Enzymatic Synthesis of Gene Fragments A Laboratory Manual' (H.G. GassenおよびA. Lang編), Verlag Chemie, Weinheim (1982)、または他の科学文献、たとえば、M.J. Gait, H.W.D. Matthes, M. Singh, B.S. Sproat, and R.C. Titmas, Nucleic Acids Research, 1982, 10, 6243; B.S. Sproat and W. Bannwarth, Tetrahedron Letters, 1983, 24, 5771; M.D. Matteucci and M.H Caruthers, Tetrahedron Letters, 1980, 21, 719; M.D. Matteucci and M.H. Caruthers, Journal of the American Chemical Society, 1981, 103, 3185; S.P. Adams ら, Journal of the American Chemical Society,1983, 105, 661; N.D. Sinha, J. Biernat, J. McMannus, and H. Koester, Nucleic Acids Research, 1984, 12, 4539; and H.W.D. Matthes ら, EMBO Journal, 1984, 3, 801)。
【0041】
あるいはまた、既知の技法(たとえば、相補的cDNA鎖を作製するためのmRNAの逆転写)および市販のcDNAキットを用いて、Der p 1/ProDer p 1/PreProDer p 1のmRNAからコード配列を導出することができる。Der p 3/ProDer p 3/PreProDer p 3のコード配列も上記のように導くことができる; PreProDer p 3ヌクレオチド配列のコドン使用頻度パターンは高発現細菌遺伝子に典型的なものである。
【0042】
驚くべきことに、ProDer p 3はDer p 3と比べて非常にアレルゲン性が低いことが明らかになった。
【0043】
ヌクレオチド配列のコドン使用頻度は、高発現ヒト遺伝子に典型的なものであることが望ましい。したがって、本発明の特定の態様において、変異したDer p 1/ProDer p 1/PreProDer p 1タンパク質をコードする複数のコドンを一緒に含んでなるヌクレオチド配列を提供するが、ここで、組換えダニタンパク質のアミノ酸配列をコードするために用いることのできるコドンの選択は、最適化された哺乳動物のコドン使用頻度をそっくり模するように変更され、その結果得られた遺伝子配列におけるコドンの使用頻度は、同一タンパク質をコードすると考えられる哺乳動物遺伝子と実質的に同一である。ヒトを含めて、哺乳動物のコドン使用頻度パターンは、文献に見出すことができる(たとえば、Nakamura ら 1996, Nucleic Acids Res. 24, 214-215を参照されたい)。
【0044】
DNA遺伝暗号には4つの文字があり(A、T、CおよびG)、これらを使用して3文字の「コドン」を綴り、このコドンは生物の遺伝子にコードされるタンパク質のアミノ酸を表す。DNA分子に沿ったコドンの一次配列は、遺伝子によってコードされるタンパク質におけるアミノ酸の一次配列に翻訳される。遺伝暗号は高度に縮重しており、61のコドンが20種の天然アミノ酸をコードし、3種のコドンが「終止」シグナルを表す。したがって、大半のアミノ酸は2以上のコドンによってコードされ、実際にいくつかのアミノ酸は4個以上の異なるコドンによってコードされる。
【0045】
所定のアミノ酸をコードするために2以上のコドンが使用できる場合、コドン使用頻度パターンはほとんどランダムではないことが認められている。異なる生物種は、コドンの選択において異なった偏りを示し、さらに高レベルで発現される遺伝子と低レベル発現遺伝子との間で、1つの種においてもコドンの使用は著しく異なることがある。こうした偏りはウイルス、植物、細菌、昆虫および哺乳動物細胞で相違しており、一部の種は、他の種より、ランダムなコドンとはかけ離れた強い偏りを示す。たとえば、ヒトおよび他の哺乳動物は特定の細菌もしくはウイルスほど偏りは強くない。こうした理由から、大腸菌(E. coli)において発現される哺乳動物遺伝子、または哺乳動物細胞において発現されるウイルス遺伝子が、効率よく発現するには不適当なコドンの配分を有する可能性はかなりある。しかしながら、大腸菌に適したコドン使用頻度パターンを有する遺伝子は、ヒトにおいても効率よく発現される可能性がある。宿主にはほとんど見られないコドンのクラスターが発現させるべき異種DNA配列中に存在することは、その宿主において低レベルの異種発現を予測するものであると考えられる。
【0046】
宿主でほとんど見られないコドンを宿主に好ましいコドンに変更すること(「コドン最適化」)が異種発現レベルを高めた例は、いくつか存在する。たとえば、BPV(ウシパピローマウイルス)後期遺伝子L1およびL2は哺乳動物のコドン使用頻度パターンにコドン最適化されたが、こうしたことが哺乳動物(Cos-1)細胞培養において野生型HPV配列より高い発現レベルを与えることが示された(Zhou et. al. J. Virol 1999. 73, 4972-4982)。この研究において、哺乳動物よりもBPVにおいて2倍より高い頻度で(使用頻度の比>2)出現するすべてのBPVコドン、および使用頻度の比が>1.5である大半のコドンが、優先的に使用される哺乳動物コドンに保存的に置き換えられた。WO97/31115, WO97/48370 およびWO98/34640 (Merck & Co., Inc.)において、HIV遺伝子もしくはそのセグメントのコドン最適化は、コドン最適化した配列を最適化の目的とした宿主動物においてDNAワクチンとして使用したとき、結果としてタンパク質発現の増加および免疫原性の向上をもたらすことが示された。
【0047】
本研究において、ヤケヒョウヒダニの各コドンは哺乳動物宿主に最適なコドンで保存的に置き換えられているため、その配列は全体として、好ましくは最適化されたコドンからなる(このことが望ましくない制限酵素切断部位、イントロンスプライス部位などを導入することになる場合を除く)。驚くべきことに、こうした最適化ProDer p 1/Der p 1配列はまた、酵母ではコドン使用頻度が異なっているにもかかわらず、酵母においても良好に発現する。
【0048】
本発明のさらに別の態様は、変異したDer p 1/ProDer p 1/PreProDer p 1タンパク質を調製する方法を提供するが、この方法は、組換え宿主細胞において当該タンパク質をコードするDNAを、コドンが最適化されている、いないにかかわらず、発現させること、および生成物を回収することを含んでなる;上記の方法はDer p 3/ProDer p 3/PreProDer p 3にも適用される。
【0049】
Der p 1は、その酵素活性、アレルゲン性および遺伝子クローニングについては十分に性質が明らかにされているものの、Der p 1の異種発現は問題があると報告されており(Chapman and Platts-Mills, J Immunol 1980;125:587-592)、これはおそらくこのシステインプロテイナーゼがPreProDer p 1前駆体として合成されるためである。タンパク質の高次構造に関わるシステイン残基が変異したDer p 1/ProDer p 1/PreProDer p 1配列の発現はさらにいっそう問題がある。したがって、本発明はさらに、こうした障害をすべて克服することによって変異タンパク質の製造を可能とし、ダニアレルギーに対する治療用および予防用ワクチンの産業的開発を可能にする方法を提供する。
【0050】
Der p 3/ProDer p 3/PreProDer p 3の変異タンパク質または組換えタンパク質の製造方法もまた提供される。
【0051】
変異の有無にかかわらずDer p 1/ProDer p 1/PreProDer p 1をマルトース結合タンパク質(MBP)融合タンパク質として発現させたとき、大腸菌においてタンパク質発現の実質的な改善が達成された。したがって、変異したProDer p/Der p 1タンパク質を大腸菌においてMBP融合タンパク質として発現させる方法を提供する。さらに、ジスルフィド結合がPichia pastorisにおける分泌に必須であると言われているにもかかわらず、驚くべきことに酵母におけるタンパク質発現の実質的な改善が、変異タンパク質について達成された(Takai ら 2001, Int. Arch. Allergy Immunol. 124, 454-460)。このことは、ダニの変異ProDer p/Der p 1タンパク質をコードするポリヌクレオチド配列を、高発現ヒト遺伝子に見られるコドン使用頻度に適合するように再度改変することによって達成され、それによって組換え抗原が天然ProDer p/Der p 1ダニアレルゲンと同じ高次構造および免疫学的特性を有することも可能となった。これまではPichia pastorisにおける発現はうまくいかないと報告されていた(Takai ら 2001, Int. Arch. Allergy Immunol. 124, 454-460)にもかかわらず、驚くべきことに、哺乳動物細胞での発現にコドンが最適化された変異型ProDer p 1のクローニングおよび発現を、Pichia pastorisにおいて達成することができた(一定の割合での分泌を伴う)。
【0052】
Maniatis et. al., Molecular Cloning - A Laboratory Manual; Cold Spring Harbor, 1982 1989に記載の従来の組換え技術によって本発明の方法を実施することができる。
【0053】
具体的には、本発明の方法は次のステップを含んでもよい:
1. 前記Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするヌクレオチド配列を含んでなるDNAポリマーを発現することができる、宿主細胞において複製可能な、または組み込み型の発現ベクターを調製すること;
2. 部位特異的変異導入によって、ジスルフィド結合に関わるシステイン残基を別の残基、好ましくはアルギニン残基に置き換えることにより、結果として得られるタンパク質のIgE結合活性を変化させること;
3. 宿主細胞を前記ベクターで形質転換すること;
4. DNAポリマーの発現を可能にする条件下で形質転換された宿主細胞を培養してタンパク質を産生させること;
5.タンパク質を回収すること。
【0054】
上記方法はDer p 3/ProDer p 3/PreProDer p 3にも適用可能である。
【0055】
「形質転換する」という用語は、本明細書において、たとえばGenetic Engineering; Eds. S.M. Kingsman and A.J. Kingsman; Blackwell Scientific Publications; Oxford, England, 1988に記載された従来の技術を利用して、適当なプラスミドもしくはウイルスベクターを用いて、形質転換、トランスフェクション、または感染によって外来DNAを宿主細胞内に導入することを表すために用いる。「形質転換された」または「形質転換体」という用語は、以後、当該の外来遺伝子を含有し発現する、結果として得られる宿主細胞に適用されるものとする。
【0056】
発現ベクターもまた新規であり、本発明の一部を成す。本発明のある特定の態様は、本発明のシステイン変異型Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするポリヌクレオチド配列を含んでなり、この配列の発現を指令する能力を有する発現ベクターを提供する。本発明の別の特定の態様は、システイン変異型Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするポリヌクレオチド配列を含んでなり、その配列の発現を指示する能力を有する発現ベクターを提供するが、ここで、このポリヌクレオチド配列のコドン使用頻度パターンは、高度に発現される哺乳動物遺伝子、好ましくは高度に発現されるヒト遺伝子に典型的なパターンである。ベクターは、細菌、昆虫、酵母、もしくは哺乳動物細胞、特にヒト細胞において異種DNAの発現を推進するのに適していると考えられる。
【0057】
本発明にしたがって、宿主細胞と適合するベクターを切断して無傷のレプリコンを有する線状DNAセグメントを準備し、前記線状セグメントを、その線状セグメントとともに所望の生成物をコードする1以上のDNA分子、たとえば、Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするDNAポリマーと、連結条件下で結合させることにより、複製可能な発現ベクターを調製することができる。
【0058】
上記ベクターは、本発明の変異したDer p 3/ProDer p 3/PreProDer p 3にも適用することができる。
【0059】
このようにしてDNAポリマーを、所望により、前もって形成する、またはベクターの構築時に形成することができる。
【0060】
ベクターの選択は、ある程度は宿主細胞によって決まるが、宿主細胞は原核生物または真核生物であってよい。適当なベクターにはプラスミド、バクテリオファージ、コスミドおよび組換えウイルスが含まれる。
【0061】
複製可能な発現ベクターの調製は、たとえば、Maniatis ら、上記引用、に記載の手順によって、DNAを制限切断、重合および連結する適当な酵素を用いて従来の方法で行うことができる。
【0062】
組換え宿主細胞は、本発明にしたがって、形質転換条件下で、宿主細胞を本発明の複製可能な発現ベクターで形質転換することにより調製される。適当な形質転換条件は通常のものであって、たとえば、上記のManiatis ら、または「DNA Cloning」 Vol. II, D.M. Glover ed., IRL Press Ltd, 1985に記載されている。
【0063】
形質転換条件の選択は宿主細胞によって決まる。したがって、大腸菌のような細菌宿主は、CaCl2溶液、またはRbCl、MnCl2、酢酸カリウムおよびグリセロールの混合物を含んでなる溶液で処理し、次いで3-[N-モルホリノ]-プロパンスルホン酸、RbClおよびグリセロールで処理してもよい。哺乳動物培養細胞は、細胞へのベクターDNAのカルシウム共沈、リポフェクション、またはエレクトロポレーションによって形質転換することができる。酵母に適合するベクターはまた、栄養要求性変異株に原栄養性を付与すること、または野生株に重金属耐性を付与することによって、形質転換に成功した株を選択できるようにするマーカーを保有する。酵母ベクターの制御配列には、解糖酵素のプロモーター(Hess ら, J. Adv. Enzyme Reg. 1968, 7, 149)、酸ホスファターゼをコードするPHO5遺伝子、CUP1遺伝子、ARG3遺伝子、GAL遺伝子プロモーター、および合成プロモーター配列が含まれる。酵母の発現に有用な他の調節因子は、ターミネーターおよびリーダー配列である。細胞からのタンパク質の分泌を指令するリーダー配列は、一般的には疎水性アミノ酸からなるシグナルペプチドをコードしているので、特に有用である。適当なシグナル配列をコードしている可能性があるのは、分泌される酵母タンパク質の遺伝子、たとえば酵母インベルターゼ遺伝子およびαファクター遺伝子、酸ホスファターゼ、キラートキシン、α接合因子遺伝子、および最近では酵母Kluyveromyces marxianusのINU1A遺伝子に由来する異種イヌリナーゼシグナル配列である。Pichia pastorisや出芽酵母Saccharomyces cerevisiaeにおける発現に適したベクターが開発されている。
【0064】
さまざまな誘導もしくは構成的プロモーターに基づく、さまざまなP. pastoris発現ベクターが利用可能である(Cereghino and Cregg, FEMS Microbiol. Rev. 2000,24:45-66)。細胞質内タンパク質および分泌タンパク質の産生のために、もっともよく利用されるP. pastorisベクターは、非常に強力で厳密に制御されたアルコールオキシダーゼ(AOX1)プロモーターを含有する。ベクターはまた、his4宿主における選択のためにP. pastorisヒスチジノールデヒドロゲナーゼ(HIS4)遺伝子をも含有する。外来タンパク質の分泌はシグナル配列の存在を必要とし、S. cerevisiaeプレプロα接合因子リーダー配列が、Pichia発現系において広く良好に使用されている。発現ベクターは、発現菌株の安定性を最大にするように、P. pastorisゲノムに組み込まれる。S. cerevisiaeの場合と同様に、P. pastoris発現ベクターの、宿主ゲノムと共通する配列(AOX1もしくはHIS4)内での切断は、そのゲノムの遺伝子座へのベクターの組み込みを効果的に標的とする相同組換えイベントを刺激する。一般に、組み込みコピーの発現カセットを複数含有する組換え菌株は、単一コピー菌株より多くの異種タンパク質を生成することができる。高コピー数の形質転換体を得るために最も有効な方法は、Pichia受容株をスフェロプラスト法で形質転換することを必要とする(Cregg ら, 1985, Mol.Cell.Biol. 5: 3376-3385)。
【0065】
本発明はまた、本発明の複製可能な発現ベクターで形質転換された宿主細胞にも及ぶ。
【0066】
形質転換された宿主細胞の、DNAポリマーの発現を可能にする条件下での培養は、従来の方法によって実施され、たとえば、Maniatis ら および「DNA Cloning」、上記に記載の通りである。従って、好ましくは細胞には栄養分を供給し、45℃以下の温度で培養する。
【0067】
生成物は、宿主細胞に応じて、従来の方法によって回収する。したがって、宿主細胞が大腸菌のような細菌である場合、その細胞を物理的、化学的もしくは酵素的に溶解し、その結果得られる溶菌液からタンパク質生成物を分離することができる。宿主細胞が哺乳動物細胞である場合、生成物は一般に、栄養培地から、または無細胞抽出液から分離することができる。従来のタンパク質分離技術には、選択的沈澱、吸着クロマトグラフィー、およびモノクローナル抗体アフィニティカラムを含めたアフィニティクロマトグラフィーがあげられる。
【0068】
あるいはまた、バキュロウイルスのような適当なベクターを用いて昆虫細胞において、形質転換されたショウジョウバエ細胞において、または哺乳動物CHO細胞においても発現させることができる。本発明の新規タンパク質はまた、EP-A-0 278 941の中でCSタンパク質について記載されているように、酵母細胞において発現させることもできる。
【0069】
本発明の低アレルゲン性Der p 1/ProDer p 1/PreProDer p 1誘導体を含んでなる、またはコドン最適化の如何に関わらず前記タンパク質をコードするポリヌクレオチド配列を含んでなる、医薬組成物、免疫原性組成物およびワクチン組成物も提供される。低アレルゲン性Der p 3/ProDer p 3/PreProDer p 3を含んでなる同様の組成物も提供される。好ましい実施形態において、DNA組成物は複数の粒子、好ましくは金粒子を含んでなり、この粒子は、ヤケヒョウヒダニアミノ酸配列をコードするポリヌクレオチド配列をコードするベクターを含んでなるDNAでコーティングされており、ここでこのポリヌクレオチド配列のコドン使用頻度パターンは高発現哺乳動物遺伝子、とりわけヒト遺伝子に特有のものである。
【0070】
本発明のポリヌクレオチド、およびコードされるポリペプチドは、治療薬もしくは予防薬として用途を見出すことができる。特に、DNAを、ワクチン接種すべき哺乳動物、たとえばヒトに投与する場合、本発明のポリヌクレオチド(天然ProDer p 1のポリヌクレオチド配列が含まれ、好ましくはコドン最適化されている)を、DNAワクチン接種(NAVAC)に使用することができる。RNAもしくはDNAのような核酸、好ましくはDNAは、上記のようなベクターの形で提供されるが、このベクターは哺乳動物の細胞において発現されうる。ポリヌクレオチドは利用可能なあらゆる方法によって投与することができる。たとえば、核酸を、注射によって、好ましくは皮内、皮下もしくは筋肉内に注入することができる。あるいはまた、粒子を介するDNAデリバリー(PMDD)のような核酸デリバリーデバイスを用いて、核酸を直接皮膚内に送達することができる。この方法では、不活性粒子(たとえば金ビーズ)を核酸でコーティングし、その粒子を、たとえば発射装置から高圧で放出することによって、受容側の表面(たとえば皮膚)を貫通可能とするのに十分な速度に加速する(本発明の核酸分子でコーティングされた粒子は本発明の範囲に含まれ、そうした粒子を充填したデリバリーデバイスも同様である)。
【0071】
裸のポリヌクレオチドもしくはベクターを患者に導入するのに適した技術には、適当なビヒクルを用いた局所適用が含まれる。核酸を局所的に、皮膚または粘膜表面に、たとえば、鼻内、経口、膣内または直腸内投与によって、投与することができる。裸のポリヌクレオチドもしくはベクターは、製薬上許容される添加剤、たとえばリン酸緩衝生理食塩水(PBS)とともに存在してもよい。DNAの取り込みは、ブピバカインといった促進剤を、DNA製剤に含めて、または別々に使用することによって、さらに促進することができる。核酸を直接、受容者に投与する別の方法には、超音波、電気刺激、エレクトロポレーションおよび米国特許第5,697,901号に記載のマイクロシード法(microseeding)がある。一般的に核酸は1pg〜1mgの範囲で投与され、粒子による遺伝子デリバリーでは好ましくは1pg〜1μg核酸、他の経路では10μg〜1mgの量で投与される。
【0072】
本発明の核酸配列は、遺伝子治療に有用な特化された運搬ベクターを用いて投与することもできる。遺伝子治療アプローチは、たとえばVerme ら, Nature 1997, 389:239-242によって論じられている。ウイルスベクター系も非ウイルスベクター系もいずれも利用することができる。ウイルスに基づく系には、レトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、カナリア痘およびワクシニアウイルスに基づく系が含まれる。非ウイルス性の系には、核酸の直接投与、マイクロスフェア封入技術(ラクチドとグリコリドの共重合体)およびリポソームに基づく系が含まれる。初回ワクチン接種後に追加免疫の注射を与えることが望ましい場合には、ウイルスおよび非ウイルスデリバリーシステムを組み合わせることができる。たとえば、プラスミドのような非ウイルスベクターを用いた初回の「プライム」DNAワクチン接種後、1回または複数回の「ブースト」ワクチン接種を、ウイルスベクターまたは非ウイルス性の系を用いて行う。
【0073】
このようにして、本発明者らは、ProDer p 1をコードするDNA(好ましくは哺乳動物用にコドン最適化されている)によるワクチン接種が、マウスモデルにおいてTh1応答を誘導し(特異的IgG2a抗体の力価は高く、特異的IgG1抗体の力価は低い)、特記すべきことに、抗ProDer p 1 IgEの不在をもたらすことを見出した。
【0074】
本発明の医薬組成物には、アジュバント化合物、またはタンパク質によって誘導される免疫応答を増強するのに役立つと考えられる他の物質が含まれうる。
【0075】
本発明のワクチン組成物は、免疫保護的な量の変異型もしくは組換え型Der p 1/ProDer p 1/PreProDer p 1低アレルゲン性タンパク質、または変異型もしくは組換え型Der p 3/ProDer p 3/PreProDer p 3低アレルゲン性タンパク質を含んでなる。「免疫保護的」という用語は、アレルギー疾患を回避または緩和するように、次のアレルギー誘発に対して免疫応答を誘導するために必要な量を表す。本発明のワクチンにおいて、タンパク質の水溶液をそのまま使用することができる。あるいはまた、タンパク質を、事前の凍結乾燥の有無にかかわらず、あらゆるさまざまな既知のアジュバントと混合、吸着、または共有結合させることができる。
【0076】
適当なアジュバントが市販されており、たとえば、フロイント不完全アジュバントおよび完全アジュバント(Difco Laboratories, Detroit, MI);Merck Adjuvant 65 (Merck and Company, Inc., Rahway, NJ);AS-2 (SmithKline Beecham, Philadelphia, PA);水酸化アルミニウムゲル(ミョウバン)もしくはリン酸アルミニウムといったアルミニウム塩;カルシウム、鉄もしくは亜鉛の塩;アシル化チロシンの不溶性懸濁物;アシル化糖類;陽イオンもしくは陰イオン誘導多糖類;ポリホスファゼン類;生分解性マイクロスフェア;モノホスホリルリピドAおよびquil Aといったものがある。GM-CSFまたはインターロイキン-2、-7、もしくは-12といったサイトカイン、およびケモカインもアジュバントとして使用することができる。
【0077】
本発明の製剤において、アジュバント組成物は主としてTh1型の免疫応答を誘導することが好ましい。高レベルのTh1型サイトカイン(たとえば、IFN-γ、TNFα、IL-2およびIL-12)は、投与された抗原に対する細胞性免疫応答の誘導に有利に働く傾向がある。応答の大部分がTh1型である好ましい実施形態において、Th1型サイトカインのレベルはTh2型サイトカインのレベルよりも非常に高まる。こうしたサイトカインのレベルは、標準的なアッセイによって容易に評価することができる。サイトカインファミリーに関する総説については、Mosmann and Coffman, Ann. Rev. Immunol. 7:145-173, 1989を参照されたい。
【0078】
したがって、主にTh1型応答の誘導に用いるのに適したアジュバントには、たとえば、モノホスホリルリピドA、好ましくは3-O-脱アシル化モノホスホリルリピドA(3D-MPL)のアルミニウム塩との組み合わせがある。他の既知のアジュバントで、優先的にTH1型免疫応答を誘導するものは、CpG含有オリゴヌクレオチドを包含する。このオリゴヌクレオチドは、CpGジヌクレオチドがメチル化されていないことを特徴とする。こうしたオリゴヌクレオチドはよく知られており、たとえば、国際公開WO 96/02555に記載されている。免疫賦活性DNA配列もまた、たとえば、Sato ら, Science 273:352, 1996によって記載されている。CpG含有オリゴヌクレオチドは、単独でも、他のアジュバントと組み合わせても使用することができる。たとえば、増強されたシステムは、CpG含有オリゴヌクレオチドとサポニン誘導体の組み合わせ、とりわけ国際公開WO 00/09159およびWO 00/62800に記載のようにCpGとQS21の組み合わせを伴う。製剤はそれに加えて、水中油型エマルジョンおよび/またはトコフェロールを含んでなることが好ましい。
【0079】
別の好ましいアジュバントはサポニン、好ましくはQS21(Aquila Biopharmaceuticals Inc., Framingham, MA)であり、これは単独でも他のアジュバントと組み合わせても使用することができる。たとえば、増強されたシステムは、国際公開WO 94/00153に記載の、QS21と3D-MPLの組み合わせのような、モノホスホリルリピドAとサポニン誘導体の組み合わせ、または国際公開WO 96/33739に記載の、QS21がコレステロールでクェンチされた、反応性の弱い組成物を伴う。他の好ましい製剤は、水中油型エマルジョンおよびトコフェロールを含んでなる。水中油型エマルジョン中にQS21、3D-MPLおよびトコフェロールを含む特に強力なアジュバント製剤が国際公開WO 95/17210に記載されている。
【0080】
水中油型エマルジョン中にQS21、3D-MPLおよびトコフェロールを含む特に強力なアジュバント製剤が国際公開WO 95/17210に記載されているが、これは好ましい製剤である。
【0081】
他の好ましいアジュバントには、Montanide ISA 720 (Seppic, France), SAF (Chiron, California, United States), ISCOMS (CSL), MF-59 (Chiron), Detox (Ribi, Hamilton, MT), RC-529 (Corixa, Hamilton, MT)および他のアミノアルキルグルコサミニド4-リン酸 (AGPs)がある。
【0082】
したがって、本明細書に記載のDer p 1/ProDer p 1/PreProDer p 1低アレルゲン性誘導体、およびアジュバントを含んでなる免疫原性組成物が提供されるが、このアジュバントは3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルもしくはエステルのうち1つまたは複数、またはこれらのアジュバントのうち2つ以上の組み合わせを含んでなる。免疫原性組成物中のDer p 1/ProDer p 1/PreProDer p 1低アレルゲン性誘導体は、水中油型もしくは油中水型エマルジョンビヒクル中で提供されることが好ましい。
【0083】
さらに、本明細書に記載のDer p 3/ProDer p 3/PreProDer p 3低アレルゲン性誘導体およびアジュバントを含んでなる免疫原性組成物が提供されるが、このアジュバントは、3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルもしくはエステルのうち1つまたは複数、またはこれらのアジュバントのうち2つ以上の組み合わせを含んでなる。免疫原性組成物中のDer p 3/ProDer p 3/PreProDer p 3低アレルゲン性誘導体は、水中油型もしくは油中水型エマルジョンビヒクル中で提供されることが好ましい。
【0084】
もう一つの態様において、本発明は医薬組成物を作製する方法を提供するが、その方法は、ジスルフィド架橋の形成に関わるDer p 1/ProDer p 1/PreProDer p 1/Der p 3/ProDer p 3/PreProDer p 3の1つまたは複数のシステイン残基を変異させるステップ、たとえばDer p 1の下記の残基:Cys4, Cys31, Cys65, Cys71, Cys103またはCys117を変異させることを包含する。さらに別の態様において、本発明は、ProDer p 1のアミノ酸残基227〜240(Der p 1の147〜160)を欠失させるステップを包含する、医薬組成物を作製する方法を提供する。
【0085】
本発明の方法はさらに、野生型Der p 1/ProDer p 1/PreProDer p 1ヌクレオチド配列のコドン使用頻度パターンを変更するステップ、またはポリヌクレオチド配列を合成によって作製するステップを含んでなり、高発現哺乳動物遺伝子に特有のコドン使用パターンを有し、かつコドンが最適化されたシステイン変異型ProDer p 1/ Der p 1アミノ酸配列、もしくは本発明にしたがって選択された残基が欠失しているProDer p 1/ Der p 1アミノ酸配列をコードする配列を生じる。ワクチン製剤は総じてVaccine Design(”The subunit and adjuvant approach” (eds. Powell M.F. & Newman M.J). (1995) Plenum Press New York)に記載されている。リポソーム内への封入はFullertonにより米国特許第4,235,877号に記載されている。タンパク質の巨大分子へのコンジュゲーションは、たとえば、Likhite、米国特許第4,372,945号およびArmorら、米国特許第4,474,757号によって開示されている。
【0086】
それぞれのワクチン用量に含まれる本発明のタンパク質の量は、典型的なワクチンにおいて重大な副作用なしに免疫保護応答を誘導する量として選択される。このような量は、どの特異的免疫原を用いるかによって、およびワクチンがアジュバントで増強されているかどうかによって変わってくる。一般に、各用量は1〜1000μgのタンパク質を含んでなると見込まれるが、1〜200μgが好ましい。具体的なワクチンの最適量は、被験体における抗体価の観察および他の反応に関わる標準的な調査によって確認することができる。本発明のワクチンは、成人または乳幼児に投与可能であるが、出生直後で実質的なTh2型免疫記憶反応が確立する前に、個体にワクチン接種することが好ましい。初回接種後、被験体は約4週間でブーストを受け、その後アレルギー反応のリスクが存在する限り、6ヶ月ごとに追加接種を繰り返すことが好ましい。
【0087】
ワクチンおよび医薬組成物は、密封アンプルもしくはバイアルといった、1回量または複数回量容器に入れて提供されることが考えられる。こうした容器は、使用するまで製剤の無菌性を保つように、密閉されていることが好ましい。一般に、製剤は油性もしくは水性ビヒクル中で懸濁液、溶液または乳濁液として保存してもよい。あるいはまた、ワクチンもしくは医薬組成物は、使用直前に無菌の液体担体を添加するだけでよい、凍結乾燥された状態で保存してもよい。
【0088】
本発明はワクチンを製造する方法も提供するが、この方法は、本発明のDer p 1/ProDer p 1/PreProDer p 1誘導体もしくはDer p 3/ProDer p 3/PreProDer p 3誘導体、またはそれらの誘導体を、本明細書に記載のプロセスによって精製するステップ、およびその結果得られたタンパク質を適当なアジュバント、希釈剤もしくは他の製薬上許容される添加剤と混合するステップを含んでなる。
【0089】
本発明はまた、本発明のタンパク質を製薬上許容される添加剤と混合することを含んでなる、ワクチン製剤を製造する方法を提供する。
【0090】
本発明の別の態様は、アレルギーを罹患しやすいまたはアレルギーに罹患した患者を免疫療法で治療するためのワクチンを製造するための、本出願で請求するタンパク質もしくはポリヌクレオチドの使用である。アレルギーを罹患しやすい、またはアレルギーに罹患した患者に製薬上活性な量の本明細書に記載の免疫原性組成物を投与することを含んでなる前記患者を治療する方法も、本発明により考慮される。
【0091】
本発明のさらなる態様は、ヒトにおいてアレルギー疾患(特にハウスダストダニアレルギー)を予防または緩和する方法を提供するものであるが、その方法は、免疫原として有効な量の本発明の変異型アレルゲンまたは本発明によるワクチンを、それを必要とする被験体に投与することを含んでなる。
【0092】
下記の実施例は、本発明を例証するものであるが本発明を限定するものではない。制限酵素および他の試薬は実質的にはメーカーの使用説明書にしたがって使用した。
【実施例】
【0093】
実施例1
一般的手法
1. SDS-PAGEおよびウェスタンブロット分析
タンパク質は、SDS-PAGEによって12.5%ポリアクリルアミドゲル上で分析した。電気泳動後、セミドライトランスブロットシステム(Bio-Rad)を用いて、タンパク質をニトロセルロース膜上に転写した。膜は、TBS-T(50mM Tris HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)中の0.5% Instagel(PB Gelatins)で30分間飽和させ、変性済みまたは天然型ProDer p 1に対して生じさせたマウスポリクローナル血清(ブロッキング溶液で希釈(1:5000))とともにインキュベートした。アルカリホスファターゼ結合ヤギ抗マウス抗体(Promega, 1:7500)および5-ブロモ-4-クロロ-3-インドリルリン酸(BCIP, Boehringer)/ニトロブルーテトラゾリウム(NBT, Sigma)を基質として用いて、免疫反応性物質を検出した。
【0094】
2. 糖鎖分析
Glycan Differenciation Kit (Boehringer)により、下記のレクチンを用いて糖鎖分析を行った:スノードロップ(Galanthus nivalis)アグルチニン(GNA)、セイヨウニワトコ(Sambucus nigra)アグルチニン(SNA)、イヌエンジュ(Maackia amurensis)アグルチニン(MAA)、ピーナッツアグルチニン(PNA)およびシロバナヨウシュチョウセンアサガオ(Datura stramonium)アグルチニン(DSA)。概略としては、精製タンパク質をSDS-PAGEからニトロセルロース膜上に転写した。膜を、ジゴキシゲニンと結合した異なるレクチンとともにインキュベートした。複合体はアルカリホスファターゼと結合した抗ジゴキシゲニン抗体を用いて検出した。
【0095】
3. 酵素アッセイ
1mM EDTAおよび20mM L-システインを含有する50 mM Tris-HCl pH 7中で、総容量1mlとして25℃にて酵素アッセイを行った。Cbz-Phe-Arg-7-アミノ-4-メチルクマリン(Cbz-Phe-Arg-AMC)およびBoc-Gln-Ala-Arg-7-アミノ-4-メチルクマリン(Boc-Gln-Ala-Arg-AMC)(Sigma)(いずれの基質も終濃度100μMとした)の加水分解を、SLM 8000分光蛍光光度計を用いてλex = 380nmおよびλem = 460nmでモニターした。システイン活性化アレルゲンを終濃度100 nMとなるように添加することによってアッセイを開始した。どのようなアッセイの前にも、精製Der p 1もしくはProDer p 1を、アプロチニン-およびp-アミノベンズアミジン-アガロース樹脂(Sigma)の混合物とともにインキュベートして、ありうる微量のセリンプロテアーゼ活性を除去した。
【0096】
4. タンパク質定量
ウシ血清アルブミンを標準として、ビシンコニン酸法(MicroBCA, Pierce)によって総タンパク質濃度を測定した。
【0097】
5. Der p 1 ELISA
Der p 1特異的モノクローナル抗体5H8および4C1(Indoor Biotechnologies)を用いてELISAキットによってDer p 1または組換えProDer p 1を検出した。アッセイに使用したDer p 1標準(UVA 93/03)の濃度は0.5μg/mlであった。
【0098】
6. IgE結合活性
イムノプレートを、Der p 1またはProDer p 1(1ウェル当たり500ng)で4℃にて一晩コーティングした。次にプレートを、ウェル当たり100μlのTBS-Tweenバッファー(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSAを添加した同じバッファー150μlで37℃にて1時間飽和させた。ヤケヒョウヒダニに対するアレルギー患者から採取した血清を1/8に希釈した後、37℃にて1時間インキュベートした。実験に使用した95の血清のうち、16の血清は、その特異的抗ヤケヒョウヒダニIgE値(RASTアッセイ)が58.1kU/Lから99kU/Lの範囲にあり、79の血清は、上限カットオフ値である100kU/Lを越えた。プレートをTBS-Tweenバッファーで5回洗浄し、マウス抗ヒトIgE抗体(Southern Biotechnology Associates)およびアルカリホスファターゼ結合ヤギ抗マウスIgG抗体(TBS-Tweenバッファーで1/7500に希釈、Promega)とともにインキュベート後、アレルゲン-IgE複合体を検出した。酵素活性は、ジエタノールアミンバッファー(pH 9.8)中に溶解したp-ニトロフェニルリン酸基質(Sigma)を用いて測定した。OD410nmを、Biorad Novapath ELISAリーダーで測定した。
【0099】
IgE阻害アッセイについては、プレートを同一濃度(0.12μM)のDer p 1もしくはProDer p 1でコーティングした。20のアレルギー患者由来ヒト血清のプール(RAST値>100kU/L)を、さまざまな濃度(3.6-0.002μM)の阻害剤としてのDer p 1 または組換えProDer p 1とともに4℃にて一晩プレインキュベートし、ELISAプレートに加えた。IgE結合は上記のように検出した。
【0100】
7. ヒスタミン放出
ヒスタミン放出はアレルギーの供血者の、ヘパリン処理末梢血由来の白血球を用いて、ヒスタミン-ELISAキット(Immunotech)により、アッセイした。好塩基球を、連続希釈した組換えProDer p 1またはDer p 1とともに37℃にて30分間インキュベートした。好塩基球中のヒスタミンの総量は、界面活性剤IGEPAL CA-630(Sigma)を用いた細胞破砕後に定量した。
【0101】
8. ProDer p 1の変性
100℃にて5分間、50mMβ-メルカプトエタノールの存在下で、組換えProDer p 1を変性させた。
【0102】
9. 免疫化
10匹のCBA/Jマウス(6週齢)群を、5μgの種々のタンパク質または100μgの種々のプラスミドDNAで1週間ごとに4回、免疫した。アジュバントとしてミョウバンの存在下で精製アレルゲンを注射した。対照として、マウス群をミョウバンまたはpJW4304 DNAベクターで免疫処置した。第7、14、21、28日にマウスの眼窩静脈叢から採血し、血清を採取した。
【0103】
10. 気管支吸入誘発
免疫後72時間以内にすべてのマウスをプレキシグラス(アクリル樹脂)容器(13 x 19 x 37.5 cm)に入れ、エアロゾル化したヤケヒョウヒダニ粗抽出物に7日間連続で20分間にわたり曝露した。ダニの粗抽出物の濃度は300μg/mlとした。エアロゾルは超音波ネブライザー(Syst’ AM)によって発生させた。ネブライザーの出力は0.5ml/分であり、エアロゾルの平均粒径は1〜5μmであった。対照としてはマウスにPBSを噴霧した。
【0104】
11. Der p 1特異的IgG、IgG1およびIgG2aの測定
血清の抗Der p 1 IgG、IgG1およびIgG2a抗体をELISAによってアッセイした。イムノプレートを、4℃にて16時間ProDer p 1(1ウェル当たり500ng)でコーティングした。プレートをTBS-Tween(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSAを添加した同じバッファー150μlで37℃にて1時間飽和させた。血清を飽和バッファーで連続希釈して、37℃で1時間インキュベートした。プレートをTBS-Tweenバッファーで5回洗浄し、抗原に結合した抗体を、アルカリホスファターゼと結合した二次抗体(ヤギ抗マウスIgG、Promega、USA)(TBS-Tweenバッファーで1/7500に希釈)によって検出した。酵素活性は、ジエタノールアミンバッファー(pH 9.8)に溶解したp-ニトロフェニルリン酸基質(Sigma)を用いて測定した。OD415nmはBiorad Novapath ELISAリーダーで測定した。
【0105】
マウス抗体のサブクラスは、上記のようにコーティングしたプレート、および二次抗体としてIgG1-もしくはIgG2a-特異的ビオチン標識モノクローナル抗体(ラット抗マウス、TBS-Tweenバッファーおよび1% BSAで1/7000希釈、Biosource)を用いて判別した。アルカリホスファターゼ結合ストレプトアビジン(1/1000希釈、Amersham)を各ウェルに添加した。酵素活性のアッセイは上記のように行った。
【0106】
すべての場合において、ELISAの力価は、測定したOD415の最大値の50%に相当するシグナルを与える希釈率の逆数とした。
【0107】
12. Der p 1特異的IgEの測定
イムノプレートを、4℃にて16時間ラット抗マウスIgE(1ウェル当たり10ng)でコーティングした。プレートをTBS-Tween(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSAを添加した同じバッファー150μlで37℃にて1時間飽和させた。血清を飽和バッファーで連続希釈して、37℃で1時間インキュベートした。その後ProDer p 1を飽和バッファーに添加して500ng/mlとした。結合したProDer p 1を、ビオチン標識抗Der p 1モノクローナル抗体4C1(Indoor Biotechnologies)の添加によって検出した。プレートをTBS-Tweenバッファーで5回洗浄し、抗原に結合した抗体を、アルカリホスファターゼと結合したストレプトアビジン(TBS-Tweenバッファーで1/7500に希釈)の添加によって検出した。酵素活性はジエタノールアミンバッファー(pH 9.8)に溶解したp-ニトロフェニルリン酸基質(Sigma)を用いて測定した。OD415nmはBiorad Novapath ELISAリーダーで測定した。
【0108】
13. 増殖アッセイ
Der p 1特異的T細胞増殖応答を測定するために、気管支吸入誘発の前後に免疫したマウスを犠牲にした。リンパ球を脾臓から分離した。15 mM HEPESおよび30μMβ-メルカプトエタノールを含有する10% FCSを添加したRPMI 1640で培養した細胞(1ウェル当たり4 x 105を3通り)を、96ウェルプレートにおいて連続希釈したダニ粗抽出物もしくはProDer p 1で刺激した(25μg/mlの濃度から始めて、10個の2倍希釈していった、抗原の希釈物をテストした)。対照としては、細胞をRPMI培地のみとともにインキュベートした。4日後に、細胞に1ウェル当たり1μCiの[3H]チミジン(Amersham)を加えて16時間パルス標識した。細胞を集め、3H-チミジンの取り込みをシンチレーション計数により測定した。増殖反応は4ウェルの平均として算出し、刺激指数(stimulation index, SI)として表した。2より大きいSIを陽性とした。
【0109】
14. サイトカインアッセイ
リンパ球培養上清におけるIFNγおよびIL-5のレベルをELISAアッセイで測定した。1μg/mlの抗マウスIL-5モノクローナル(PharMingen)または抗マウスIFNγ(Biosource)ポリクローナル抗体でプレートをコーティングした。プレートをTBS-Tweenバッファーで5回洗浄し、150μl TBS-Tween-BSAで37℃にて1時間飽和させた。脾細胞培養上清を連続希釈して添加し、37℃にて90分間インキュベートした。ビオチン標識抗マウスIL-5(PharMingen, 1μg/ml)または抗マウスIFNγ(Biosource, 0.2μg/ml)抗体を、37℃にて1時間、プレートに作用させた。抗原抗体複合体は、西洋ワサビペルオキシダーゼと結合したストレプトアビジン(1/10000希釈、Amersham)とともにインキュベートすることによって検出した。酵素活性は、基質としてテトラメチルベンジジン(TMB、Sigma)を用いて測定した。460nmでの吸光度をBiorad Novapath ELISAリーダーで測定した。サイトカイン濃度は、精製マウスIL-5またはIFNγを用いて作成した標準曲線に測定値を内挿することによって決定した。
【0110】
15. 気管支肺胞洗浄
最後のエアロゾル曝露の3日後、マウスを採血して犠牲にした。ただちに気管カニューレを通して、1mlハンクス平衡塩類溶液(HBSS)で肺を洗浄したが、この溶液は注入して3回の吸引によりそっと回収した。洗浄液を4℃にて400×gで10分間遠心した。細胞ペレットを300μlハンクス平衡塩類溶液(HBSS)に再懸濁し、トーマ(Thoma)血球計算盤で細胞を計数した。50μlのアリコートからのサイトスピン(Cytospin)標本を、示差的細胞計数のためにMay-Grunwald Giemsa染色によって染色した。
【0111】
実施例2
大腸菌におけるMBP-ProDer p 1の発現
1. MBP-ProDer p 1発現ベクターの構築
真核生物発現プラスミドpNIV 4846(ヒト化ProDer p 1 をコードするカセットを保有するpEE 14由来発現プラスミド(M.Massaer ら, International Archives of Allergy and Immunology, 2001, 125:32-43))をEag IおよびXba Iで切断後、ProDer p 1をコードする完全な合成cDNA(1-302 aa)(配列番号1)を単離した。Xba I制限切断の前に、ラージフラグメントDNAポリメラーゼ(Klenow)を用いてDNAを平滑末端化した。921 bp断片をMBP遺伝子の下流のpMAL-c2E(New England Biolabs)のAsp 718(平滑末端)-Xba I部位に挿入して、pNIV4854を導出した。配列番号1のcDNAによってコードされたProDer p 1のアミノ酸配列を、図2(配列番号2)に示す。
【0112】
2. 部位特異的変異導入
4、31または65位のDer p 1システイン残基(成熟ProDer p 1の数であり、ProDer p 1の84、111または145位に相当する)の変異導入は、プラスミドpNIV4854において、3個のシステインコドンのうちの1つを有するDNA断片を、変異を含む合成オリゴヌクレオチドで置換することにより行った。下記のオリゴヌクレオチドを使用した:
システイン残基4をアルギニンに変異させるための110bp Afl II-Sph I断片を作製(C4R)するためには、5’TTAAGACCCAGTTTGATCTCAACGCGGAGACCAACGCCCGTATCAACGGCAATGCCCCCGCTGAGATTGATCTGCGCCAGATGAGGACCGTGACTCCCATCCGCATGC3’(フォワード)および5’CGGATGGGAGTCACGGTCCTCATCTG GCGCAGATCAATCTCAGCGGGGGCATTGCCGTTGATACTACGGGCGTTGGTCTCCGCGTTGAGATCGAAACTGGGTC3’ (リバース)を使用、システイン残基31をアルギニンに変異させるための98bp Sph I-Blp I断片を作製(C31R)するためには、5’CAAGGCGGCCGTGGGTCTTGTTGGGCCTTTTCAGGCGTGGCCGCGACAG AGTCGGCATACCTCGCGTATCGGAATCAGAGCCTGGACCTCGC3’ (フォワード)および5’TCAGCGAGGTCCAGG CTCTGATTCCGATACGCGAGGTATGCCGACTCTGTCGCGGCCACGCCTGAAAAGGCCCAACAAGACCCACGGCCGCCTTGCATG3’ (リバース)を使用、システイン残基65をアルギニンに変異させるための82bp Afl II-Sph I断片を作製(C65R)するためには5’TGAGCAGGAGCTCGTTGACCGTGCCTCCCAACACGGATGTCATGGGGATACGATTCCCAGAGGTATCGAATACATCCAGCATA3’ (フォワード)および5’CTGGATGTATTCGATACCTCTGGGAATCGTATCCCCCATGACATCCGTGTTGGGAGGCACGGTCAACGCGCTCCTGC3’(リバース)を使用。
【0113】
結果として得られたプラスミドは、MBP遺伝子の下流にProDer p 1カセットを含有し、それぞれC4R、C31RおよびC65Rの変異を有しており、それぞれpNIV4870、pNIV4871およびpNIV4872と命名した。3つの変異はいずれもDNA配列決定によって検証した。C4R、C31RおよびC65R変異をそれぞれ保有する、変異したProDer p 1のアミノ酸配列を、それぞれ配列番号3、配列番号5および配列番号7に示す。これをコードする、対応する核酸配列は、配列番号4(C4R変異)、配列番号6(C31R変異)および配列番号8(C65R変異)に示す。
【0114】
3. 野生型および変異型MBP-ProDer p 1の発現および精製
異なる組換え発現ベクターを含有する大腸菌を、100μg/mlアンピシリンを含む869培地(A.Jacquet ら, Prot. Exp. Purif. 1999, 17, 392-400)中で37℃にて一晩培養した。その後細胞を1:100に希釈し、600nmでの光学密度が0.4〜0.6となるまで37℃にて増殖させた。イソプロピルβ-D-チオガラクトシド(IPTG)を終濃度0.3mMとなるように添加した。2時間インキュベートした後、細胞を、10000rpmで15分間の遠心によって回収した。
【0115】
1リットルの培養物から得られた細菌細胞ペレットを、1mMアプロチニンおよびAEBSFを含有する20mM Tris-HCl pH 7.5に再懸濁し、細胞破砕装置(Constant Systems Ltd, Warwick, UK)を用いて1800バールの圧力をかけて破砕した。溶菌液を150,000gにて60分間、超遠心した。超遠心の結果得られたペレットを20mM Tris-HCl pH 7.5で洗浄した。不溶性タンパク質は、6M尿素含有20mM Tris-HCl pH 7.5を用いて4℃にて一晩抽出した。懸濁液を150,000gで60分間、超遠心した。上清をそのまま、20mM Tris-HCl pH 7.5, 200mM NaCl, 1mM EDTAに対して一晩透析した。溶液を遠心して沈澱したタンパク質を除去し、そのまま、同じバッファーで平衡化されたアミロース樹脂(1×15cm)に加えた。A280nmがベースラインに達するまで、カラムを開始バッファーで洗浄した。カラムバッファーに10mMマルトースを添加することによってタンパク質を溶出させた。融合タンパク質を含有する画分をプールして濃縮した。精製タンパク質は−20℃で保存した。
【0116】
実施例3
CHO細胞における3種の異なるProDer p 1変異体の発現
1. 部位特異的変異導入
4、31または65位のDer p 1システイン残基の変異(成熟Der p 1の番号付けであり、ProDer p 1の84、111または145位に相当する)をプラスミドpNIV4846に導入した。プラスミドpNIV4870、pNIV4871およびpNIV4872はDer p 1カセットをMBP(実施例IIを参照されたい)遺伝子の下流に含有し、それぞれC4R、C31RおよびC65R変異を保有しているが、これらのプラスミドをそれぞれSfuI-XhoIで制限切断して、714bp断片を単離した。精製したDNA断片を、同じ制限酵素であらかじめ切断しておいたプラスミドpNIV4846に挿入した。その結果得られた、Der p 1変異体C4R、C31RおよびC65Rを含有するプラスミドをpNIV4873、pNIV4875およびpNIV4874と命名した。
【0117】
2. ProDer p 1を生成する安定なCHO-K1細胞株の一過性トランスフェクションおよび選択
プラスミドpNIV4873、pNIV4875およびpNIV4874によるDer p 1の産生を測定するために、COS細胞をリポフェクションによって一過的にトランスフェクトした。安定したDer p 1の発現のために、CHO-K1細胞をリポフェクションによって異なるプラスミドでトランスフェクトした。3週間の25μMメチオニルスルホキシミン(MSX)選択後、100μM MSX共存の下で遺伝子増幅を1ラウンド行った。
【0118】
実施例4
変性ProDer p 1はアレルギー血清に対してIgG結合反応性を示すがIgE結合反応性は示さない
ProDer p 1の変性型を低アレルゲン性ワクチンとして使用できるかを判定するために、変性(50mM βメルカプトエタノール存在下、100℃にて5分)ProDer p 1のIgGおよびIgE結合反応性をELISAテストでアッセイした。図1に示すように、変性ProDer p 1は天然型ProDer p 1に存在するIgGエピトープの大部分を保存していた。他方、この変性アレルゲンはそのIgE結合反応性の多くを失っていた。本発明者らのデータは、変性ProDer p 1がProDer p 1の低アレルゲン性変異体となりうることを示唆する。
【0119】
実施例5
MBP-ProDer p 1のIgE反応性
本実験の目的は、MBP-ProDer p 1と天然Der p 1のIgE反応性を比較することである。MBP-ProDer p 1の、アレルギー患者血清由来の特異的IgEとの反応性を、イムノプレートをDer p 1もしくはMBP-ProDer p 1で直接コーティングした直接ELISA法によって評価した。図2は、Der p 1およびMBP-ProDer p 1へのIgE結合の強い相関を示す。
【0120】
実施例6
MBP-ProDer p 1変異体のIgE結合反応性
MBP-ProDer p 1変異体のIgE結合能を、さまざまな形態のMBP-ProDer p 1でイムノプレートを直接コーティングする直接ELISAアッセイで測定した。RAST値>100 kU/Lである20人の別個のヤケヒョウヒダニ-アレルギー患者の血清から作った血清プールをアッセイに使用した。図3に示すように、変異体C31RおよびC65RのIgE結合反応性は、野生型MBP-ProDer p 1と比較して5%にまで大幅に減少していた。特筆すべきは、4位のシステイン残基をアルギニンに変異させたとき、MBP-ProDer p 1に対するIgEの反応性が認められなかった(0%残存)ことである。MBPまたは無関係なタンパク質と融合したMBPの、IgEを介した免疫認識は存在しなかったことから、IgE反応性はProDer p 1部分について特異的であった。20人の患者から得られた別の血清プールについても同様の結果が得られた。
【0121】
実施例7
さまざまな形態のProDer p 1のヒスタミン放出活性
天然Der p 1のアレルゲン活性をProDer p 1の組換え変異型誘導体の活性と比較するために、あるアレルギー患者由来の好塩基球をin vitroでさまざまな濃度のアレルゲンに曝露し、放出されるヒスタミンを測定した。図4に示すように、天然Der p 1は1ng/mlの濃度でさえも好塩基球からのヒスタミン放出を誘導することができた。対照的に、ProDer p 1の組換え変異型はこれよりも1000〜10000倍の高濃度ではじめてヒスタミンを放出させることができた。これらの結果は、ProDer p 1変異体が天然Der p 1より低いIgE結合反応性を示すことを明白に示す。
【0122】
実施例8
さまざまな形態のProDer p 1を用いた免疫原性実験
1. ハウスダストダニアレルギーの動物モデル
ハウスダストダニアレルギーの動物モデルを開発した。CBA/Jマウスに、ミョウバンをアジュバントとする精製Der p 1を注射した。1週間間隔で4回注射後、動物をヤケヒョウヒダニ抽出物による一連の気管支吸入誘発に供した(図5)。このモデルを用いて、種々の形態のDer p 1組換え体、ならびに種々のDNAをハウスダストダニアレルギーに対する予防ワクチンとしてテストした。
【0123】
実施例9
欠失型ProDer p 1の発現
1)PCRと、欠失させるべき断片の下流および上流の配列を含んでなる合成オリゴヌクレオチドを用いることによって、欠失を生じさせた。プラスミドpNIV4878(pNIV4878がProDer p 1のcDNA(ヒト化cDNA)を含有するpPIC9Kプラスミド(Invitrogen)であり、P. pastorisにおけるアレルゲンの発現に有利であることを想起されたい)のSnab I/Avr II断片を、増幅し欠失させた断片によって置き換えた。P. pastoris酵母をこの組換えプラスミドで形質転換し、ジェネティシン (G418)で選択後、0.25mg/ml G418に耐性のクローンを単離した。ProDer p 1カセットがシグナル配列の下流に位置しているものとして、メタノールで24または48時間誘導した後分泌されるProDer p 1Δ227〜240体の発現を調べた。
【0124】
2)PCRによる欠失の導入
プライマー1:5’-GCTATTACCGATACGTAGCTAGGG-3’
このプライマーは欠失すべき領域の下流のSnabI制限酵素切断部位を含む。
【0125】
プライマー2:5’-CCGTTGTCGCGATCCTTGATTCCGATGATGACAGCG-3’
このプライマーはしたがって、ProDer p 1配列のうち、欠失すべき領域の下流および上流にある部分に相同である。
【0126】
プライマー3:5’-CGGAATCAAGGATCGCGACAACGGGTATCAGCCAAACTACC-3’
この配列もまたProDer p 1配列の一部と相同であり、42bpの欠失を可能にする。加えて、これは、EcoRV切断部位の改変を可能にする点突然変異を含有する。
【0127】
プライマー4:5’-TAGGGGAGCTCAGATCTGATCCACTGAC-3’
【0128】
3)配列決定による欠失の確認
スフェロプラスト法を用いて、Pichia pastoris酵母を、あらかじめBglIIで線状とした組換えベクターによって形質転換した。形質転換体は、ヒスチジノールデヒドロゲナーゼ(His+)原栄養性について選別した。His+形質転換体のG418耐性に関するスクリーニングは、ジェネティシン(G418)(濃度を増加させていく)を含有する寒天上にクローンを播くことによって行った(0.25、1、2および4mg/ml)。30℃にて数日間インキュベートした後、数個の耐性株を得たが、それは0.25mg/ml G418の濃度についてのみであった。
【0129】
発現は、毎日、培地に0.5%メタノールを添加することによって誘導する。毎日、上清を回収するために培地を1ml採取する。ProDer p 1の発現はブロットにより示す。
【0130】
実施例10
三重変異型の形態のProDer p 1の発現
1)この実施例において、Der p 1の71、103および117位のシステイン残基をアラニンに変異させてある。Cys→Ala変異はジスルフィド架橋を壊すものの、Der p 1の構造に正電荷を導入することはない。この、より穏やかな構造破壊は、ProDer p 1の発現および分泌には影響を及ぼさないかもしれない。3つの変異を含んでなるオリゴヌクレオチドによって、これら3つの変異をPCRで導入した。
【0131】
実施例11
PreProDer p 3アレルゲンの構築
1. PreProDer p 3合成cDNAの構築
部分的に重複した10個のオリゴヌクレオチドのセットを用いて、PreProDer p 3 cDNAを合成した。これらのプライマーは、高発現の大腸菌細菌遺伝子のコドン選択に基づいて設計し、394 DNA/RNA Applied Biosystem Synthetizerによって作製した。縮重してコードされるアミノ酸は、もっとも一般的なコドンによってコードされるのではなく、個別のコドンの頻度を考慮した。たとえば、AAGまたはAAAは、高発現大腸菌細菌遺伝子においてそれぞれ21.45%および78.55%の頻度でリジン残基をコードする。このため、合成PreProDer p 3の各リジン残基にAAAコドンのみを選択するのではなく、同じコドン頻度になるように試みた。オリゴヌクレオチドは下記の通りである:
5’TCATGATCATCTACAACATTCTGATCGTACTCCTGCTGGCCATTAACACTTTGGCTAATCCGATCCTGCCGGCATCCCCGAACGCGACCATCGTTGGC 3’ (オリゴ1、コード)
5’CACCACAGAAGTGGCTACTAGACTGCAGGGAGATCTGATATGGGCACTCACCAGCCAGTGCTTTTTCGCCGCCAACGATGGTCGCG 3’ (オリゴ2、非コード)
5’GTAGCCACTTCTGTGGTGGTACTATTCTTGACGAATACTGGATCCTGACCGCGGCACACTGCGTGGCCGGCCAAACAGCGAGCAAACTCTCC 3’ (オリゴ3、コード)
5’GTCGATCTGGTAGCTATCATATTTTTCATGTGCGAAAATTTTAGCAACAGAGATCTTTTCGCCACCCAGTGAGTGTTTCAGGCTGTTGTAACGAATGGAGAGTTTGCTCGCTG 3’ (オリゴ4、非コード)
5’GATAGCTACCAGATCGACAATGACATTGCGCTGATCAAGCTGAAATCCCCTATGAAGCTGAACCAGAAAAACGCCAAAGCTGTGGGCCTGC 3’ (オリゴ5、コード)
5’CAGACGGCAGGGAGTAGCTGCCCTCTTCCAGATAACCCCAGCCAGAGACACGCACCTGGTCACCAACTTTTACATCCGAGCCTTTCGCCGGCAGGCCCACAGCTTTG 3’ (オリゴ6、非コード)
5’CTACTCCCTGCCGTCTGAATTACGCCGTGTTGATATCGCTGTGGTATCTCGCAAAGAATGTAACGAGCTGTACTCGAAAGCGAACGCTGAAGTCAC 3’ (オリゴ7、コード)
5’CCACCAGAATCGCCTTGACAAGAGTCCTTACCGCCGTTCGCAACATCACCACCGCAGATCATATTGTCGGTGACTTCAGCGTTCGC 3’ (オリゴ8、非コード)
5’CAAGGCGATTCTGGTGGGCCGGTGGTCGACGTTAAAAACAACCAGGTTGTAGGTATCGTTTCCTGGGGCTACGGTTGCGCACGTAAAGGC3’ (オリゴ9、コード)
5’AAGCTTTCAGTGGTGGTGGTGGTGGTGCTGGCTACGTTTAGATTCAATCCAATCGATAAAGTTACCAACGCGCGTGTACACACCCGGATAGCCTTTACGTGCGCAAC 3’ (オリゴ10、非コード)
【0132】
合成組換えPreProDer p 3 DNAの増幅のために、PCR反応において上記オリゴヌクレオチドを一緒にインキュベートした。Expand High Fidelity PCR System (Roche Diagnostics)を用いて下記の条件に従ってPCRを行った:30サイクル、94℃にて30秒間変性、52℃にて30秒間アニーリング、および72℃にて30秒間伸長。3’および5’末端プライマー(オリゴ1およびオリゴ10)を用いて、同じ条件で、生成産物を増幅した。その結果得られた812bp断片をpCRII-TOPOクローニングベクター(Invitrogen)にクローニングした。
【0133】
BamHIによる切断は、クローン1、3および9が正しく挿入されていることを示した。
【0134】
組換えPreProDer p 3の内部に相当するオリゴヌクレオチドを用いてインサートの配列を決定した:
5’AAGCTGAAATCCCCTATGAAGC3’ (コード)
5’CTCTTCCAGATAACCCCAGCC3’ (非コード)
【0135】
クローン3のみが正しいことが判明したが、5’コード末端の最初の6塩基は欠けていた。欠失塩基の付加は、2つの新たなオリゴヌクレオチドの使用によって達成した:
5’TTTTATTCATGATCATCTACAACATTCTGATCC3’ (オリゴ11、コード)
5’GATGCATGCTCGAGCGGC3’ (オリゴ12、非コード)
【0136】
オリゴヌクレオチドを、不完全なPreProDer p 3配列を保有するクローン3のDNAとともにインキュベートした。合成遺伝子の増幅は、Expand High Fidelity PCR Systemを用いたPCR反応(25サイクル、94℃にて30秒間変性、53℃にて30秒間アニーリング、および72℃にて30秒間伸長)によって達成した。その結果得られた断片を、pCRII-TOPOにクローニングした。PreProDer p 3 cDNAは、二重Rca I-XhoI制限切断後に単離し、NcoIおよびXhoIで切断されたpET15b発現ベクターにクローニングした。コンピテントAD494(DE3)pLys大腸菌細胞を、上記の結果得られたプラスミドによって形質転換し、ProDer p 3の発現を検出するために終濃度1mMのイソプロピル-チオガラクトシド(IPTG)(Duchefa)を培地に添加した。
【0137】
推定Der p 3シグナルペプチドの欠失は、PCRによって、2つの新たなプライマー5’CATATGAATCCGATCCTGCCGGCATCCCC3’ (オリゴ13、コード) および5’GGATCCTCACTGGCTACGTTTAGATTCAATCC3’ (オリゴ14、非コード)を使用して行った。ProDer p 3 cDNAの増幅は、Taqポリメラーゼ(Roche Diagnostics)を用いて15サイクルのPCRで行った:97℃にて30秒間変性、65℃にて30秒間アニーリング、および72℃にて1分間伸長。得られた750bp断片をpCRII-TOPOクローニングベクター(Invitrogen)にクローニングした。得られたプラスミドによって最もコンピテントな10個の大腸菌を形質転換した。9個のクローンは陽性に挿入されているようであった;EcoRIによる切断はクローン1、4、8が正しく挿入されていることを証明したが、一方、配列決定は、クローン4のみが正しい配列を有することを明らかにした。NdeIおよびXhoIによる切断後、ProDer p 3 cDNAを単離して、同じ酵素で切断したpET15bにクローニングした。その結果得られたプラスミドでBL21およびBL21 Star大腸菌(Invitrogen)株を形質転換した。培地へのIPTGの添加は、(His)6タグをN末端に保有する組換えProDer p 3の発現を誘導した。
【0138】
2. 大腸菌における組換えアレルゲンの発現
最も生産性の高い組換え大腸菌BL21 STARクローンを、100μg/mlアンピシリン(Pentrexyl)を含有する2リットルの液体869培地で培養した(37℃/250rpm)。培養物の620nmでの吸光度が0.5に達したとき、終濃度1mMのイソプロピル-チオガラクトシド(Duchefa)を添加することによって3時間、発現を誘導した。次いで、培養物を回収して11000gで遠心し−20℃で保存した。細菌ペレットを回収し、アプロチニン1/1000(Sigma)およびAEBSF 1/500(ICN)を添加した40mlの20mM Trisバッファー、pH7.5中に再懸濁した。続いて1500バールで細胞を破砕して−20℃で保存した。
【0139】
3. 破砕した大腸菌からの組換えProDer p 3の精製
回収した大腸菌細胞を20mM Trisバッファー、pH7.5、1mM アプロチニンおよび1mM AEBSFに再懸濁し、これを、1800キロバールの圧力下に細胞破砕装置(Cell D)で溶菌させた。溶菌液を149000gで1時間超遠心した。上清を除去し、組換えProDer p 3を含むペレットを次に、6M塩酸グアニジンを含有する50mM Tris-HClバッファー、pH7.5、40mlで4℃にて一晩抽出した。超遠心(45分間、149000g)後、抽出物の上清を3ml/分の流速で、抽出バッファーで平衡化済みのNickel-NTA Superflow カラム(1.6x5cm, Qiagen)に供した。カラムを0.5M PBS NaCl, pH7.5を用いて1.5ml/分で洗浄し、結合したタンパク質を復元させた。組換えProDer p 3はコンディショニングバッファーに200mMのイミダゾールを添加することによって4ml/分の流速で、溶出させた。組換えProDer p 3を含有する画分をプールし、限外濾過(Amicon-Millipore 再生セルロース限外濾過膜、NMWL 10kDa)によって濃縮した。このステップの間にバッファーをPBS, pH7.3で交換した。精製したタンパク質は−20℃で保存した。
【0140】
4. 天然ダニ全体抽出物からの天然Der p 3の精製
ヤケヒョウヒダニ抽出物を、最終濃度60%飽和硫酸アンモニウム沈澱に供した。超遠心(45分、149000g)後、上清を、2ml/分で、50mM Trisバッファー、0.5M NaCl、pH7.4で平衡化されたBenzamidine Sepharose 4 fast flow カラム(1.6x5cm, Pharmacia)に供した。Der p 3を、50mMグリシン-HClバッファー、pH2.5によってカラムから溶出させ、1mlの各フラクションを、75μl 1M Tris、pH9.5の添加によりただちに中和した。Der p 3を含有する画分をプールし濃縮して(Amicon-Millipore 再生セルロース限外濾過膜、NMWL 10kDa)、0.5ml/分にて、PBS pH7.3で平衡化済みのSuperdex 75 ゲル濾過クロマトグラフィーカラム (Pharmacia)に供した。精製Der p 3を濃縮し、−20℃にて保存した。
【0141】
5. SDS-PAGEおよびウェスタンブロット分析
タンパク質を12.5%ポリアクリルアミドゲルでのSDS-PAGEによって分析した。電気泳動後、タンパク質をセミドライ転写システム(Sigma-Aldrich)を用いてニトロセルロース膜に転写した。膜を、TBS-T(50mM Tris HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)中0.5% Instagel(PB Gelatins)で30分間飽和させ、ProDer p 3に対して生じたマウスポリクローナル血清をブロッキング溶液中に1:2500希釈したものとともにインキュベートした。アルカリホスファターゼ結合ヤギ抗マウス抗体(Promega、1:7500)および5-ブロモ-4-クロロ-3-インドリルリン酸(BCIP, Boehringer)/ニトロブルーテトラゾリウム(NBT, Sigma)を基質として用いて、免疫反応性物質を検出した。
【0142】
6. タンパク質定量
総タンパク質濃度は、標準タンパク質としてウシ血清アルブミンを用いてビシンコニン酸法(MicroBCA, Pierce)によって測定した。
【0143】
7. IgE結合活性
イムノプレートをDer p 3または組換えProDer p 3(1ウェル当たり500ng)で一晩4℃にてコーティングした。その後1ウェル当たり100μlのTBS-Tweenバッファー(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSA(Sigma)を添加した150μl同じバッファーで37℃にて1時間飽和させた。ヤケヒョウヒダニに対するアレルギー患者から得られた血清を1/8に希釈した後、37℃で1時間インキュベートした。実験に使用した47血清のうち、5血清はその特異的抗ヤケヒョウヒダニIgE値(RASTアッセイ)が0.7kU/L〜28.9kU/L、8血清は68.3kU/L〜94.1kU/Lであり、34血清は上限カットオフ値の100kU/Lを越えていた。アレルゲン-IgE複合体は、プレートをTBS-Tweenバッファーで5回洗浄し、ビオチン結合マウス抗ヒトIgE抗体(TBS-Tバッファーで1/2000に希釈、Southern Biotechnology Associates)およびストレプトアビジン-西洋ワサビペルオキシダーゼ(TBS-Tweenバッファーで1/1000に希釈、Amersham Life Science)とともにインキュベートした後検出した。酵素活性は、3,3’,5,5’-テトラメチルベンジジン(TMB) (Sigma)を用いて測定した。OD450nmはBiorad Novapath ELISAリーダーで測定した。
【0144】
8. 酵素アッセイ
総容量200μlで25℃にて、50mM Tris-HCl, pH 8において酵素アッセイを行った。N-α-ベンゾイル-L-アルギン-p-ニトロアニリド(Sigma)(終濃度1mM)の加水分解を、Biorad Novapath ELISAリーダーで405mにて測定した。アッセイは終濃度6.25μg/mlのアレルゲンの添加によって開始した。
【0145】
9. Der p 3のクローニングおよび発現
大腸菌における発現
Der p 3の未成熟型である、ProDer p 3を細菌で発現させた。12の合成オリゴヌクレオチドを用いて、かつ細菌における発現に向けて最適化したコドンを使用することにより、PreProDer p 3のcDNAを完全に合成した(図2)。ProDer p 3をコードするカセットを、発現ベクターpET-15bにおいてポリヒスチジンテールをコードする配列の下流にクローニングした。このベクターは、培地へのIPTGの添加によって誘導されるT7プロモーターを含有している。まず、ProDer p 3の発現条件の最適化を試みた。これを行うために、組換え細菌の培養を30℃および37℃で、2通りの誘導試薬濃度(IPTG 0.5および1 mM)で、1、2、3および16時間、行った。それぞれの細菌ペレットをフレンチプレスで溶菌した。溶菌液を20000 rpmで20分間遠心した。上清(細胞質画分)および/またはペレット(細胞片および不溶性生成物)中のProDer p 3の存在を、組換えProDer p 3のポリヒスチジンテールに対するマウス抗体を用いたウェスタンブロットによって明らかにした(図3)。
【0146】
発現試験は、温度およびIPTG濃度が、ProDer p 3の生成にごくわずかしか影響を及ぼさないことを示す。ProDer p 3は本質的に不溶性の形で発現され、その発現は、誘導時間2〜3時間が最適である。16時間誘導については可溶性型のProDer p 3が存在しないことに留意すべきである。クーマシーブルー染色後に検出されるので、ProDer p 3は発現されている。SDS-PAGEにおいて、このタンパク質は±32kDaのバンドとなっている(図4)。
【0147】
10. ProDer p 3の精製
組換えProDer p 3の精製プロトコールを開発した。ProDer p 3が不溶性の状態で発現されるならば、その組換えアレルゲンは変性条件(20mM Tris-HCl、6M塩酸グアニジン、pH7.5)でペレットを抽出することによって可溶化される。抽出の収率は80〜90%程度である。抽出物を、抽出バッファーで充填された約10mlのNi2+キレーティングセファロース樹脂カラム(直径1.6cm、高さ5cm)にかける。抽出バッファーでカラムを洗浄した後、付着したタンパク質は、PBSバッファー0.5M NaCl pH7.5を用いた直線再生濃度勾配によってカラム上で直接再生させる。濃度勾配をかける液量は200mlである。
【0148】
その後、溶出は、再生バッファー中のイミダゾール濃度を増加させて(20、50、100および200mM)バッファーを供することにより行われる。SDS-PAGEによる分析は、当該タンパク質が流出液中に存在せず、夾雑物がイミダゾール濃度20 mMで溶出するが、ProDer p 3はイミダゾール60mMから100mMの間で溶出することを示している。ProDer p 3はすべてクロマトグラフィー担体から解離させた(図10を参照されたい)。
【0149】
マイクロシークエンシングによってProDer p 3のN末端配列の分析を行った。その配列はヒスチジンテールと一致した。ベクターpET-15bにおいて、ProDer p 3のクローニング部位はヒスチジンテールをコードする配列の下流にあることに留意されたい。ProDer p 3とヒスチジンテールはトロンビン切断部位により分離されている。ProDer p 3の確実性を検証する目的で、ヒスチジンテールを取り除くために精製タンパク質をトロンビンで処理した。切断されたProDer p 3をもう一度マイクロシークエンシングすることによって、Der p 3のプロペプチドのN末端配列を明らかにした(N P I L P A S P N A T)。
【0150】
酵素活性:ProDer p 3は、天然Der p 3 が切断できる基質BAPNAに対して不活性である。
【0151】
11. IgE反応性
アレルゲンの直接コーティング
この予備的な結果は、ダニアレルギー患者のIgEについて、ProDer p 3の反応性がDer p 3と比べて低いことを示しているようである(図11)。
【0152】
2. ワクチン製剤
【表1】
【0153】
3. 抗体応答-結果
天然Der p 1を4回注射することによって免疫したマウスは、高力価のIgGおよびIgG1、低力価のIgG2ならびに大量のIgE抗体を産生したが、これは、天然Der p 1が強力なTh2免疫応答を誘導することを示唆する(表2および4)。
【0154】
抗Der p 1 IgGおよびIgG1抗体応答は天然型または変性型ProDer p 1を注射したマウスでも強力であった。天然型ProDer p 1を注射後、IgG2a力価はDer p 1を用いて得られた力価よりわずかに高くなったが、IgE力価はDer p 1で得られた力価と同程度かそれよりわずかに低くなった。天然型ProDer p 1で免疫したマウスとは対照的に、変性型ProDer p 1を注射した動物は、高いIgG2a力価を産生し、IgE抗体の産生は非常に低かった。予想通り、ミョウバンなしでのProDer p 1による免疫化は弱い免疫応答しか誘導しなかった(表4)。
【0155】
MBP-ProDer p 1野生型(WT)、C4R、C31RおよびC65Rで感作されたマウスは、特異的IgGおよびIgG1抗体の同様の産生を示した(表3)。最も高いIgG2a力価はMBP-ProDer p 1 WTおよびC31Rで免疫されたグループで観察された。特異的IgE力価は、MBP-ProDer p 1のどの変異体を注射してた場合でも低かった。ProDer p 1をコードするプラスミドでマウスを免疫した後にも同様の結果が得られた。
【表2】
【0156】
【表3】
【0157】
【表4】
【0158】
4. T細胞増殖応答-結果
エアロゾル投与の前(対照)および後に、免疫したマウスから分離された脾細胞の、ProDer p 1またはヤケヒョウヒダニ抽出物での刺激によるT細胞増殖応答を調べた。結果を表5(刺激指数)および表6(サイトカイン)に示す。
【0159】
さまざまな組換えProDer p 1変異体で免疫されたマウスにおいてアレルゲン特異的T細胞応答が検出された。脾細胞をProDer p 1で再刺激した場合に最も強い応答が観察された。T細胞反応性は投与とは無関係であると思われた。
【0160】
表5のこうした結果は、さまざまな形態のProDer p 1が共通のT細胞エピトープをDer p 1と共有していることを示した。さらに、熱変性または部位特異的変異導入によるProDer p 1の構造破壊は、ProDer p 1のT細胞反応性を変化させなかったので、こうした形態の変異体が、T細胞応答を刺激しうるIgE結合反応性の非常に弱い低アレルゲンとなることが確認された。
【表5】
【0161】
再刺激された脾細胞の培養上清におけるサイトカインIL-5およびIFNγの存在は、ELISAで測定した(表6)。[IFNγ]/[IL-5]比を比較したならば、ミョウバンをアジュバントとする天然Der p 1またはProDer p 1によるワクチン接種はIFNγよりもIL-5の産生をより強く誘導すると結論付けることができる。変性型ProDer p 1のみならずさまざまな形態のMBP- ProDer p 1(変異体および野生型)も、同程度のレベルの2つのサイトカインを誘導した。
【表6】
【0162】
5. 気管支肺胞洗浄−結果
天然Der p 1による感作とそれに続くエアロゾル化したハウスダストダニ抽出物への曝露は、非常に多数の気管支肺胞細胞を生じた(表7)。エアロゾル化したハウスダストダニ抽出物への7回の曝露は、Der p 1によるワクチン接種を受けた動物においてのみ、気道の好酸球増加を引き起こすことが示された。この群において、Der p 1に感作された動物がハウスダストダニ抽出物の噴霧を受けないか、またはエアロゾル化したPBSに曝露された場合は、気道の好酸球増加は認められなかった。
【0163】
さまざまな組換え型のProDer p 1によるワクチン接種は、エアロゾル化したハウスダストダニ(HDM)抽出物に曝露した後も、気道の好酸球増加を防止した。
【表7】
【0164】
実施例12
核酸ワクチン接種(NAVAC)用発現プラスミド
1. 核酸ワクチン接種のためのProDer p 1をコードするプラスミドの構築
ProDer p 1をコードするカセット(1-302aa)をプラスミドpNIV4846(上記参照)から切り出し、HindIIIおよびBglIIで制限切断して、あらかじめHindIIIおよびBglIIで切断しておいたプラスミドpJW4304に挿入した。その結果得られたプラスミドをpNIV4868と命名し、DNA配列決定によって確認した。
【0165】
2. 部位特異的変異導入
4、31または65位(これは成熟Der p 1の番号付けであり、ProDer p 1における84、111または145位に相当する)のProDer p 1システイン残基の変異を、プラスミドpNIV4868に導入した。プラスミドpNIV4870、pNIV4871およびpNIV4872は、ProDer p 1カセットをMBP遺伝子の下流に含有し、それぞれ変異C4R、C31RおよびC65Rを保有するものであるが、各々をAflII-BamHIで制限切断して699bpの断片を単離した。pNIV4868をAflII-HpaIで切断して480bpの断片を単離した。2つの精製DNA断片を、あらかじめHpaI-BamHIで切断したプラスミドpJW4304に挿入した。その結果得られたプラスミドはProDer p 1変異体C4R、C31RおよびC65Rを有しており、pNIV4879、pNIV4880およびpNIV4881と命名した。
【0166】
実施例13
Pichia pastorisにおけるProDer p 1の発現
1. ProDer p 1発現ベクターの構築
pNIV4868に由来するProDer p 1をコードするカセット(最適化された哺乳動物コドン使用頻度を有する全長1-302aa ProDer p 1 cDNA、)を以下のプライマーを用いてPCRによって増幅した:5’ACTGACAGGCCTCGGCCGAGCTCCATTAA3’ (下線はStuI 制限部位、フォワード)および5’CAGTCACCTAGGTCTAGACTC GAGGGGAT3’ (下線はAvrII 制限部位、リバース)。増幅された断片をpCR2.1 TOPOクローニングベクターにクローニングした。DNA配列決定によって正しいProDer p 1カセットを確認した。組換えTOPOベクターをStuI-AvrIIで切断し918bp断片を生じたが、これをSnaBI-AvrIIで制限切断したpPIC9K発現ベクターに導入した。得られたプラスミド、pNIV4878はProDer p 1カセットをS. cerevisiae α因子の下流に含有する。
【0167】
2. 部位特異的変異導入
非グリコシル化ProDer p 1(成熟Der p 1の数でN52Q)生産のための発現プラスミドを、4つのプライマーのセットを用いたオーバーラップ伸長(overlap extension)PCRによってpNIV4878から導出した。ProDer p 1アミノ酸配列71〜176をコードする317bp断片を増幅するために、以下のプライマー:5’GGCTTTCGAACACCTTAAGACCCAG3’ (プライマー1, 下線はAflII制限部位、フォワード)および5’GCTCCCTAGCTACGTA TCGGTAATAGC3’ (プライマー2, 下線はSnaBI制限部位、リバース)を用いた。
【0168】
ProDer p 1配列に変異N52Qを導入するために、以下のプライマー、5’CCTCGCGTATCGGCAACAGAGCCTGGACC3’ (プライマー3, 下線は変異N52Q、フォワード)および5’GGTCCAGGCTCTGTTGCC GATACGCGAGG3’ (プライマー4, 下線は変異N52Q、リバース)を用いた。
【0169】
変異した317bp AflII-SnaBI断片を、3段工程によって作製した。第1のPCRにおいて、プライマー1および4をpNIV4878と混合して〜200bpの断片を生成した。第2のPCRにおいて、プライマー2および3をpNIV4878と混合して〜140bp断片を生成した。2つのPCR産物をアガロースゲルで精製して、鋳型として3回目のPCRに使用し、〜340bp断片を得た。この精製断片をpCR2.1 TOPOクローニングベクター(Invitrogen)にクローニングした。変異はDNA配列決定によって確認した。組換えTOPOベクターをAflII-SnaBIで切断して317bp断片を生成し、これを同様に切断消化したpNIV4878に連結した。得られたプラスミド、pNIV4883はS. cerevisiae α因子の下流にProDer p 1 N52Qを含有する。
【0170】
Der p 1の4、31または65位(成熟Der p 1での番号付け)のシステイン残基の変異を有するProDer p 1の非グリコシル化変異体を得るために、プラスミドpNIV4873、pNIV4875およびpNIV4874についても同じプライマーのセットを用いてオーバーラップ伸長PCRを行った。その結果得られたプラスミドpNIV4884、4885および4886はそれぞれ、ProDer p 1 N52Q C4R、N52Q C31RおよびN52Q C65Rをコードする。
【0171】
3. P. pastorisの形質転換
スフェロプラスト形質転換法を用いてプラスミドpNIV4878をP. pastorisに導入した。形質転換体は、ヒスチジノールデヒドロゲナーゼ(His+)原栄養性で選択した。geneticin(G418)耐性についてのHis+形質転換体のスクリーニングは、含有するG418の濃度を増加させた寒天上にクローンを播くことによって、実施した。
【0172】
ProDer p 1 N52Q、ProDer p 1 N52Q C4R、N52Q C31RおよびN52Q C65Rをコードするプラスミドによる形質転換は、同様の方法によって行った。
【0173】
4. 組換え酵母によるProDer p 1の産生
G418耐性クローンをBMG培地中で30℃にてOD600nmが2〜6となるまで増殖させた。細胞を遠心によって回収し、OD600nmが1となるように100mlのBMG培地中に再懸濁した。6日間毎日0.5%メタノールを添加することによりProDer p 1の発現を誘導した。上清を遠心によって回収し、精製するまで−20℃で保存した。
【0174】
5. 酵母培養上清からのProDer p 1の精製
上清は水で10倍に希釈し、pHを9に調整した後、直接20mM Tris-HCl, pH9で平衡化済みのQ sepharoseカラムにかけた。カラムを開始バッファーで洗浄した。バッファー中のNaCl濃度を段階的に増加させることによってタンパク質の溶出を進行させた。ProDer p 1で富化された画分をプールし、Filtron膜(Omega Serie, カットオフ:10kD)による限外濾過によって濃縮した。ProDer p 1の精製は、PBS pH7.3で平衡化済みのsuperdex-75カラム(1 x 30cm, Pharmacia)でのゲル濾過クロマトグラフィーによって達成した。精製されたProDer p 1は濃縮し、−20℃で保存した。
【0175】
配列情報
【図面の簡単な説明】
【0176】
【図1】CHO細胞において発現された変性ProDer p 1のIgG-およびIgE-結合反応性。イムノプレートを500ng/ウェルの精製された天然もしくは変性ProDer p 1でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験(radioallergosorbent)に陽性の血清(1:8に希釈)とともにインキュベートした。結合したIgEもしくはIgGは、マウス抗ヒトIgEもしくはIgGおよびアルカリホスファターゼ標識抗マウスIgG抗体とともにインキュベートした後、酵素的にアッセイすることによって定量した。結果をOD410nm値として表示する。
【図2】MBP-ProDer p 1および天然Der pのIgE反応性の相関。イムノプレートを1ウェル当たり500ngの精製済みDer pもしくはMBP-ProDer p 1でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験に陽性の95血清(1:8に希釈)とともにインキュベートした。結合したIgEは、マウス抗ヒトIgEおよびアルカリホスファターゼ標識抗マウスIg抗体とともにインキュベートした後、酵素的にアッセイすることによって定量した。結果をOD410nm値として表示する。
【図3】C4R、C31RおよびC65Rの変異を有するMBP-ProDer p 1変異体のIgE結合反応性。イムノプレートを500ng/ウェルの野生型もしくは変異体MBP-ProDer p 1でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験に陽性の20血清(1:8に希釈)のプールとともにインキュベートした。結合したIgEは、マウス抗ヒトIgEおよびアルカリホスファターゼ標識抗マウスIgG抗体とともにインキュベートした後、酵素的にアッセイすることによって定量した。結果をOD410nm値として表示する。
【図4】アレルゲンのヒスタミン放出活性。アレルギーのある提供者の末梢血から分離された好塩基球を、種々のアレルゲンの連続希釈物で刺激した。細胞からのヒスタミンの放出はELISAによって測定した。好塩基球におけるヒスタミンの総量は、界面活性剤IGEPAL CA-630を用いて細胞を破壊した後、定量した。結果を、アレルゲンにより放出されたヒスタミンと、総ヒスタミンとの比として示す。
【図5】ハウスダストダニアレルギーの動物モデルの模式図。
【図6】メタノールで24および48時間誘導した後のP. pastorisにおけるProDer p 1Δ227〜240体の発現。組換えクローンの培養上清をSDS PAGEによって分析した。ブロットは、CHOにおいて発現されたProDer p 1に対するポリクローナルマウス血清を用いて明らかにした。レーン1,2: 無関係なタンパク質、レーン3: 誘導されていない酵母、レーン4: 精製ProDer p 1、レーン5: 24時間誘導後のクローン1、レーン6: 48時間誘導後のクローン1、レーン7: 24時間誘導後のクローン2、レーン8: 48時間誘導後のクローン2.
【図7】野生型(AcaNucSeq)および細菌コドン最適化(EcoNucSeq)PreProDer p 3の配列比較。各コドンの下に示す推定アミノ酸配列は、一文字記号で表記した。リーダーペプチドおよびプロペプチド配列は、それぞれイタリックおよび下線によって示す。
【図8】ProDer p 3の発現。0.5もしくは1 mM IPTGを用いて1、2、3および16時間誘導した後、細菌を破砕した。細胞質画分(S)および不溶性画分(C)を、ProDer p 3の存在についてウェスタンブロットで解析した。
【図9】SDS-PAGEにおけるProDer p 3のクーマシーブルー染色後の検出。レーン1:細胞質画分レーン2:不溶性画分の洗液レーン3:不溶性画分矢印はProDer p 3の位置を示す。
【図10】Ni2+-NTA クロマトグラフィーによるProDer p 3の精製。開始バッファー中の濃度を次第に増加させたイミダゾールを添加することによって、結合したタンパク質を溶出した。画分をSDS-PAGE分析したが、左図はクーマシーブルー染色後の分析であり、右図は抗His抗体を用いたウェスタンブロットによる分析である。FTはフロースルー(flow-through;通り抜け)の略である。20、40、45、60、100はイミダゾール濃度(mM)を表す。Rは精製後のNi2+NTA樹脂を表す。
【図11】組換えProDer p 3および天然Der p 3のIgE反応性の相関。イムノプレートを1ウェル当たり500ngの精製済み天然Der p 3もしくは組換えProDer p 3でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験に陽性の血清(1:8に希釈)とともにインキュベートした。結合したIgEは、マウスビオチン標識抗ヒトIgEおよびアルカリホスファターゼ標識ストレプトアビジンとともにインキュベートした後、酵素的にアッセイすることによって検出した。結果をOD410nm値として表示する。
【技術分野】
【0001】
本発明は、新規の予防用および治療用製剤に関するものであり、この製剤は特定のアレルゲンに対するアレルギー反応の予防および/または軽減に有効である。さらに、本発明はヤケヒョウヒダニ(Dermatophagoides pteronyssinus)由来の主要なアレルゲンタンパク質である、アレルゲンDer p 1およびその前駆体型ProDer p 1の低アレルゲン性組換え誘導体に関する。特に、本発明の誘導体は、物理的に改変されたDer p 1もしくはProDer p 1、例えば熱処理されたタンパク質;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えDer p 1;組換えProDer p 1;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えProDer p 1;組換えPreProDer p 1;あるいは、ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えPreProDer p 1を包含する。また、Der p 1、ProDer p 1およびPreProDer p 1誘導体を発現させて精製する方法、ならびに免疫原性組成物およびワクチンを製剤化する方法も記載する。
【0002】
さらに本発明は、ヤケヒョウヒダニ由来のもう一つのアレルゲンタンパク質である、アレルゲンDer p 3ならびにその前駆体型ProDer p 3およびPreProDer p 3の低アレルゲン性組換え誘導体に関する。特に、本発明の誘導体には、物理的に改変されたDer p 3もしくはProDer p 3、例えば熱処理されたタンパク質;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えDer p 3;組換えProDer p 3;ジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えProDer p 3;組換えPreProDer p 3;あるいはジスルフィド結合形成に関わる1つまたは複数のシステイン残基を変異させた遺伝子改変組換えPreProDer p 3が包含される。また、Der p 3、ProDer p 3およびPreProDer p 3誘導体を発現させて精製する方法、ならびに免疫原性組成物およびワクチンを製剤化する方法も記載する。
【背景技術】
【0003】
アレルギー反応はヒトではよく見られるが、さまざまなアレルゲンが誘因となる可能性がある。アレルギーの個体はアレルゲンに対して敏感になり、血清中に高レベルのアレルゲン特異的IgEが存在し、Th2型サイトカイン(IL-4、IL-5およびIL-13)を産生するアレルゲン特異的T細胞集団を有することを特徴とする。アレルゲンの存在下における、マスト細胞および好塩基球の表面に存在するFcεRI受容体へのIgEの結合は、細胞の急速な脱顆粒をもたらし、さらにそれに続いてヒスタミン、ならびに他の、生成済み、および新たに生成されたメディエータの放出をもたらす。これに加えて、T細胞既往性応答の刺激は、結果として、IL-4およびIL-13の産生をもたらし、それとともにB細胞応答をさらにアレルゲン特異的IgEを産生するようにし向けて切り換えさせる。即時型および遅延型アレルギー反応の発生に関する詳細については、Joost Van Neeven ら, 1996, Immunology Today, 17, 526を参照されたい。アレルギーでない個体では、同じ抗原に対する免疫応答はIFN-γのようなTh1型サイトカインをさらに包含する可能性がある。このようなサイトカインは、高レベルのアレルゲン特異的IgEが含まれる高レベルのTh2型免疫応答を阻害することによって、アレルギー反応の発症を防止することができる。この点に関して重要なことは、IgE合成が、B細胞上のCD23(FcεRII)受容体に対するIgE/アレルゲン複合体の結合を介した阻害的フィードバックメカニズムによって、制御可能であるという事実である(Luo ら, J.Immunol., 1991, 146(7), 2122-9; Yu ら, 1994, Nature, 369(6483):753-6)。細胞結合型CD23を欠いた系では、IgE合成のこのような阻害は起こらない。
【0004】
世界の人口の20%を越える人々が、気管支喘息、アトピー性皮膚炎および通年性鼻炎といった、アレルゲンに対するIgEを介したI型アレルギー疾患に罹っている。こうしたアレルギー反応の現在の治療方法としては、抗ヒスタミン剤治療および/または抗炎症性コルチコステロイドの局所投与によって、ヒスタミン放出という症候的な影響を抑える方法がある。開発中の他の方法としては、宿主の免疫系を用いてマスト細胞の脱顆粒を防ぐ方法がある(Stanworth ら, EP 0 477 231 B1)。その他の免疫療法も報告されている(Hoyne ら, J.Exp.Med., 1993, 178, 1783-1788; Holt ら, Lancet, 1994, 344, 456-458)。
【0005】
遅延症状のみならず即時症状も薬理学的治療によって改善することができるが、I型アレルギーに対する唯一の治癒的なアプローチはアレルゲン特異的免疫療法である。しかしながら、この方法に関するいくつかの問題は未解決のままである。第1に、免疫療法は現在のところ、バッチごとに不均質である可能性のある全アレルゲン抽出物を用いて実施されている。さらに、これらのアレルゲン混合物は個々の患者の特性に向けて設計されていないので、好ましくない毒性タンパク質を含有しうる。第2に、天然アレルゲンを高用量で投与することは、激しいアナフィラキシー反応を引き起こす可能性があり、したがって、アレルゲンの用量を免疫療法の成功のために最適効率の高用量で使用できない場合が多い。最初の問題は、アレルゲン抽出物と比べて、より良い特性を有し、より高い生産性の組換えアレルゲンを用いた代替ワクチン接種によって対処された。第2の問題、すなわちアレルゲン抽出物の注射を繰り返すことによって引き起こされるアナフィラキシー反応のリスクは、IgE反応性を欠失もしくは変異導入によって変化させた組換え「低アレルゲン」の使用によって最小化することができる(Akdis, CA and Blaser, K, Regulation of specific immune responses by chemical and structural modifications of allergens, Int. Arch. Allergy Immunol., 2000, 121, 261-269)。
【0006】
アレルギーの治療および予防のための製剤が報告されているが、その製剤は、Th2型からTh1型への反応のシフトによって、アレルゲンに対する細胞性反応を改善するのみならず、IgEの産生をダウンレギュレートする方法を提供するものである(IL-4とIFN-γ産生Der p 1特異的T細胞との比の減少、またはその代わりにIL-5とIFN-γと比の減少によって判断される)。これは、たとえばWO 99/25823 に記載の、酵素活性の低下した組換えDer p 1のような組換えアレルゲンを使用することによって達成することができる。しかしながら、こうした組換えアレルゲンの免疫原性は、IgE合成誘導の点では野生型ProDer p 1の免疫原性と類似していると思われる。
【0007】
IgE結合活性の低下した、非アナフィラキシー型のアレルゲンが報告されている。アレルゲンエンジニアリングによって、アミノ酸残基の部位特異的変異導入もしくは特定のアミノ酸配列の欠失によりアレルゲンタンパク質のIgE結合能力を低下させることが可能となった。同時に、T細胞エピトープは維持されるのでT細胞活性化能力は依然として保たれる。このことは、結果はさまざまであるが異なるアレルゲンに関するいくつかのアプローチによって示されている。オオアワガエリ(timothy grass)花粉アレルゲンPhl p 5b(Schramm G ら, 1999, J Immunol.,162, 2406-14)、チリダニ類(ヒョウヒダニ)主要アレルゲンDerf2(Takai ら 2000, Eur. J. Biochem., 267, 6650-6656)、DerP2(Smith & Chapman 1996, Mol. Immunol. 33, 399-405)、およびDerf1(Takahashi K ら 2001, Int Arch Allergy Immunol.124, 454-60)に関する例が公表されている。ある研究は、ジスルフィド結合に関わるシステイン残基レベルでの点突然変異の導入による、低アレルゲンDerf1の生成を報告している(Takahashi K Int Arch Allergy Immunol. 2001;124(4):454-60., Takai T, Yasuhara T, Yokota T, Okumura Y)。しかしながら、野生型ProDerf1は酵母Pichia pastorisによってうまく分泌されはしたが、それとは対照的に、分子内ジスルフィド結合に関わるシステイン変異体は、分泌されなかった。
【0008】
ハウスダストのダニであるヤケヒョウヒダニ由来のアレルゲンは、アレルギー性過敏反応に関わる主要な原因因子の1つである。
【0009】
Der p 1
ヤケヒョウヒダニのグループ1アレルゲンであるDer p 1は主要アレルゲンであり、チリダニアレルギーの血清の80〜100%においてIgEと結合している(Chapman, M.D., ら (1983). J. Allergy Clin. Immunol., 72: 27-33; Krillis, S., ら (1984). J. Allergy Clin. Immunol., 74: 132-41)。このタンパク質はハウスダスト中に高濃度(100〜10000 ng/gダスト)で見出されることが多い(Platts-Mills and Chapman (1987). J.Allergy Clin. Immunol., 80: 755-75; Wahn, U., ら (1997). J. Allergy Clin. Immunol., 99: 763-69)が、Der p 1は糞便物とのみ関連するわけではなく、ある範囲の粒子と関連していると考えられている(DeLuca, ら (1999). J. Allergy Clin. Immunol., 103: 174-75)。100ngのレベルで感作に関わり、量の増加に伴い危険性は増大する。
【0010】
Der p 1をコードするcDNAはクローニングされ、配列決定されている(Chua, K., ら (1988). J. Exp. Med., 167: 175-82; Thomas, ら (1988). Int. Arch. Allergy Appl. Immunol., 85: 127-29; Chua, K., ら (1993). Int. Arch. Allergy Immunol., 101: 364-8):このアレルゲンは222個のアミノ酸残基からなるタンパク質で、計算上の分子量は25 KDaである。このアレルゲンは18アミノ酸シグナルペプチドおよび80アミノ酸N末端プロ配列を含めて、320個のアミノ酸残基からなる前駆体型として合成される。ProDer p 1の成熟プロセスは依然として不明であるが、タンパク質分解によるプロ領域の除去によって、または自己触媒的プロセシングによって、酵素が活性化されると考えられている。
【0011】
Der p 1配列は、システインプロテイナーゼの原型であるパパインの配列と30%の相同性を示した(Robinson, C., ら (1997). Clin. Exp. Allergy, 27 (1): 10-21)。パパインのタンパク質分解活性に関与する残基の大半は、活性部位のシステインおよびヒスチジン残基を含めて、Der p 1において保存されていた。Der p 1は入手しにくいため、このアレルゲンについての放射線結晶学データは得られていない。それにもかかわらずDer p 1の立体構造は、パパインおよびアクチニジンの放射線結晶学的構造に基づいて明らかにされている。Der p 1の構造は、基本的な構造的および機構的特徴が他のパパイン様システインプロテイナーゼと共通している。下記はDer p 1の立体構造モデルである(Topham, C.M, ら (1994) Protein engineering, 7 (7): 869-894)。Der p 1はN末端側配列およびC末端側配列がそれぞれ独立に形作る2つの球形ドメインを与える:基質結合性および触媒性残基はこれらのドメインの間の裂け目にあり、ドメインは可動性の外側のループで結び付けられている。
【0012】
【0013】
Der p 1のシステインプロテアーゼ活性は一般に認められているが、研究から、Der p 1は、活性部位が1つしかないにもかかわらず、独特の混合したシステイン/セリンプロテアーゼ活性を示すことが明らかになった(Hewitt, C.R.A., ら (1997). Clin. Exp. Allergy, 27: 201-207)。優先的な切断部位はシステインプロテアーゼ活性についてはグルタミン酸であり、セリンプロテアーゼ活性についてはアルギニンである。
【0014】
Der p 1は、特に気管支組織を保護するプロテアーゼインヒビターであるα1-アンチトリプシンを分解することによって(Kalsheker, ら (1996). Biochem. Biophys. Res. Comm., 221: 59-61)、および、気道上皮における細胞の密着結合をゆるめることによって(Wan, H., ら (2000). Clin. Exp. Allergy, 30:685-98)、気管支粘膜の透過性を増加させ、その結果、抗原提示細胞へのアクセスを容易にする。下記の模式図に示すように、Der p 1はタンパク質「オクルディン(occludin)」を切断することによって密着結合をゆるめ、樹状細胞による吸着を促進し、アレルギー反応を誘発する。
【0015】
【0016】
Der p 1は、IgE合成の制御に関わる低親和性IgE受容体であるCD23(FcεR II)を切断し(Hewitt. C., ら (1995). J. Exp. Med., 182: 1537-1544; Schulz, O., ら (1997). Eur. J. Immunol., 27: 584-588)、それによってIgE産生を促すことが明らかになった。他方では、Der p 1はIL-2受容体のαサブユニットであるCD25を切断する(Schulz, O., ら (1998). J. Exp. Med., 187: 271-275)。IL-2はTh1免疫応答の伝播に関わるサイトカインなので、その受容体が消化分解されると結果としてTh2応答に向かって傾く。Der p 1のタンパク質分解活性はヒトT細胞からのTh2サイトカイン放出を強め(Ghaemmaghami, A.M., ら (2001). Eur. J. Immunol., 31: 1211-1216)、バイスタンダー(bystander)アレルゲンにアジュバント活性を与える(Ghough L., ら (2001). Clin. Exp Allergy, 31: 1594-1598)ことも明らかになった。
【0017】
Der p 3
Der p 3はヤケヒョウヒダニのグループ3アレルゲンである。一般に主要抗原と考えられるが、Der p 3 IgE結合の推定値はかなり変動し、16%程度と低いこともあり(Heymann, P.W., ら (1989). J Allergy Clin Immunol., 83: 1055-1067)、グループ1および2アレルゲンと同様の結合力を有し100%と高いこともある(Stewart, G.A., ら (1992). Immunology, 75: 29-35)。
【0018】
Der p 3をコードするcDNAはクローニングされ、配列決定されている(Smith W.A., ら (1994). Clin. Exp Allergy, 24: 220-228):Der p 3は232アミノ酸残基からなるタンパク質であって、計算上の分子量は25 KDaである。このタンパク質は、18アミノ酸シグナルペプチドおよび11アミノ酸N末端プロ配列を有する、不活性なPrePro Der p 3前駆体として合成される。Der p 3はセリンプロテイナーゼの原型であるトリプシンに高い相同性を示すセリンプロテイナーゼであり(Stewart, G.A., ら (1992). Immunology, 75: 29-35)、活性部位に関わる残基を包含する(図4.9)。優先的な切断部位はアルギニンおよびリジンである。
【0019】
Der p 3とウシ トリプシンの間の配列相同性を下記に示す。*印で示す残基は触媒部位に関係している。
【0020】
【0021】
Der p 3は、肺上皮細胞上のプロテアーゼ活性化受容体2の活性化により、炎症性サイトカインGM-CSFおよびエオタキシンに対するシグナリング経路を始動させることが明らかになっている(Sum, G., ら (2001). J. Immunol., 167: 1014-1021)。実際、Der p 3は、膜貫通タンパク質オクルディン(occludin)を切断することによって、気道上皮における密着結合をさらにゆるめることができる(Wan, H. ら (2000). Clin. Exp. Allergy, 31: 279-294)。この特徴はDer p 1についても認められ、抗原提示細胞への優先的アクセスを提供する。
【0022】
本発明は、野生型アレルゲンと比較して低下したアレルゲン性を有する、ヤケヒョウヒダニDer p 1アレルゲン、またはその前駆体型であるProDer p 1/PreProDer p 1(以後“Der p 1/ProDer p 1/PreProDer p 1”と称する)の組換え誘導体の提供および使用に関する。アジュバントを添加した組換えタンパク質、またはNAVACに適したDer p 1/ProDer p 1/PreProDer p 1をコードするプラスミドという形態での本発明の組換え型Der p 1誘導体は、強力な予防的Th1を生じ、あるいはTh2免疫応答をTh1にシフトさせるための、予防用または治療用ワクチンとして使用される。低アレルゲン性誘導体を組換え発現系において良好に生成することができるが、これもまた本発明の一態様である。
【0023】
本発明はさらに、野生型アレルゲンと比較して低下したアレルゲン性を有する、ヤケヒョウヒダニDer p 3アレルゲン、またはその前駆体型であるProDer p 3/PreProDer p 3(以後“Der p 3/ProDer p 3/PreProDer p 3”と称する)の組換え誘導体の提供および使用に関する。アジュバントを添加した組換えタンパク質、またはNAVACに適したDer p 3/ProDer p 3/PreProDer p 3をコードするプラスミドという形態での本発明の組換え型Der p 3誘導体は、強力な予防的Th1を生じ、あるいはTh2免疫応答をTh1にシフトさせるための、予防用または治療用ワクチンとして使用される。低アレルゲン性誘導体を組換え発現系において良好に生成することができるが、これもまた本発明の一態様である。
【0024】
本発明はさらに、本明細書に記載の1つもしくは2つ以上のタンパク質アレルゲン、または組換え誘導体を組み合わせて提供し、使用することに関する。ある態様において、本発明は、本明細書に記載のDer p 1またはその誘導体を、本明細書に記載のDer p 3、ProDer p 3もしくはProDer p 1、またはその誘導体と組み合わせて、含んでなるか、またはそれらから構成される。好ましくは、本発明はDer p 1をProDer p 3と組み合わせて、またはそれと融合して提供する。さらに別の態様において、本発明は、本明細書に記載のDer p 3もしくはその誘導体を、本明細書に記載のDer p 1、ProDer p 3、PreProDer p 3もしくはProDer p 1、もしくはPreProDer p 1、またはその誘導体と組み合わせて、またはそれと融合して、含んでなるか、またはそれらから構成される。他の態様において、本発明は、Der p 1と融合したProDer p 3を含んでなる融合タンパク質を提供する。本明細書に記載のあらゆる融合タンパク質は、連続したヒスチジン、好ましくは6ヒスチジンを追加して含んでなることができる。ある態様において、融合タンパク質は、(His)6-ProDer p 3-Der p 1を含んでなるか、またはそれからなる。
【0025】
Der p 1は30 KDaタンパク質であり、クローニングおよび配列決定がなされている(Chua ら, 1988, J.Exp.Med., 167, 175-182)。成熟タンパク質は222アミノ酸残基を有することが知られている。Der p 1の配列は、パパインと31%の相同性を有し、酵素活性領域にではそれ以上の顕著な相同性を共有しているが、最も顕著なのはCys34-His170イオン対である(Topham ら, 上記)。Der p 1はダニの中腸で産生されるが、中腸におけるその役割はおそらく食物の消化に関係しているであろう。個々の糞便粒(それぞれ直径10〜40μm程度)には0.2 ng以下のタンパク質分解活性のあるDer p 1が混合されており、したがってDer p 1は容易にヒトの気道内に吸い込まれる。精製Der p 1標品を室温にて一晩保存すると、結果として、自己タンパク質分解により酵素活性はほとんど完全に失われる(Machado ら, 1996, Eur.J.Immunol. 26, 2972-2980)。Der p 1をコードするcDNA配列は、多くの哺乳動物および植物プロテイナーゼと同様にDer p 1が、320アミノ酸残基からなる不活性プレプロ酵素として合成され、後に222アミノ酸の成熟型にプロセシングされることが明らかにされた(Chua ら, 1988, J.Exp.Med., 167, 175-182; Chua ら, 1993, Int. Arch Allergy Immunol 101, 364-368)。ProDer p 1の成熟は現在まで知られていないが、このアレルゲンは80残基のプロ領域の切断によってプロセシングされると考えられている。
【発明の開示】
【0026】
本発明は、野生型アレルゲンと比較して著しく低下したアレルゲン活性を有する、組換えヤケヒョウヒダニDer p 1/ProDer p 1/PreProDer p 1タンパク質アレルゲン誘導体を提供する。タンパク質分子内のジスルフィド架橋を開裂させることによりタンパク質の立体構造を不安定化させること、またはタンパク質のある領域、例えばProDer p 1のアミノ酸227〜240(Der p 1配列の147〜160)を欠失させることにより、タンパク質形態の立体高次構造を損なうことをねらったいくつかの方法によって、アレルゲン活性を弱めることができる。前記アレルゲン誘導体は改変されていない野生型アレルゲンと比べて、以下のような、有利な点を有する:1)野生型アレルゲンによって刺激されるのと比較して、Th1型の免疫応答(たとえば、高いIgG2a)を増加させ、それによってワクチン接種を受けたホストの潜在的アレルギー性の抑制をもたらす、2)T細胞反応性は依然として保持しているがアレルゲン性が低下しており、そのため高用量の免疫原を全身投与するのにいっそう適している、3)天然Der p 1の結合についてIgEと競合するDer p 1特異的IgGを誘導する、4)エアロゾル化されたアレルゲン抽出物に曝露された後であっても、気道の好酸球増加を効果的に防御する。このような誘導体は治療用および予防用ワクチン製剤に使用するのに適しており、こうした製剤は医薬用に好適で、さらにはアレルギー反応の治療もしくは予防に適している。
【0027】
第1の態様によると本発明は、アレルゲン活性が好ましくは還元剤の存在下でタンパク質の熱処理といった物理的手段によって著しく低下している、たとえばほとんど、または完全になくなっている、組換えDer p 1/ProDer p 1/PreProDer p 1(すなわちDer p 1、ProDer p 1またはPreProDer p 1)アレルゲン誘導体を提供する。典型的には、Der p 1/ProDer p 1/PreProDer p 1タンパク質を、還元剤の存在下で約100℃にて数分間処理する。還元剤はβ-メルカプトエタノールもしくはDTTが好ましい。さらに好ましくは、タンパク質を50 mM β-メルカプトエタノールの存在下で約100℃にて5分間処理する。この処理はIgE結合エピトープのタンパク質高次構造の安定性に有害な影響を及ぼす。タンパク質はProDer p 1またはPreProDer p 1であることが好ましい。
【0028】
第2の態様において、本発明は、たとえば、コードしているcDNAまたはゲノムDNAに特定の変異を導入することによってアレルゲン活性が遺伝学的に損なわれた、組換えDer p 1/ProDer p 1/PreProDer p 1タンパク質誘導体を提供する。したがって、本発明の態様は、それ自体、遺伝的に変異した組換えDer p 1/ProDer p 1/PreProDer p 1を提供する。低アレルゲン性変異体を組換え的に生産する前に、天然配列内に変異を導入することによって、Der p 1/ProDer p 1/PreProDer p 1のアレルゲン性を弱めることができる。これは、置換、欠失または付加を導入することによって、またはタンパク質の立体構造を変化させることによって達成でき、タンパク質の立体構造は失われる。こうしたことは、中でも、タンパク質を断片として発現させることによって、またはジスルフィド架橋形成に関わるシステイン残基を欠失させることによって、またはタンパク質の三次構造が実質的に変化するような残基を欠失もしくは付加することによって達成されうる。好ましくは、2つのシステイン残基間の相互作用を変化させる効果のある変異を生じさせることができるが、典型的には天然型成熟Der p 1の4、31、65、71、103および117位(それぞれProDer p 1の84、111、145、151、183および197位に対応する)での単一変異である。本発明の変異タンパク質は、たとえば4および31、4および65、4および71、4および103、31および65、もしくは4および31および65位, または71および103、71および117、103および117、31および117、65および117、もしくは71および103および117位といった、2つ以上(3、4、5または全6個)のシステイン変異を含んでもよく、それによってさまざまなジスルフィド架橋に影響を及ぼす。誘導体は、上記のいずれかの位置に単一の変異を含んでなることが好ましい。もっとも好ましい変異はCys4(またはその代わりに、もしくはそれに加えて、Cys4のジスルフィド結合パートナーと考えられるCys117)に関する。このシステイン変異は欠失でもよいが、他の19の天然アミノ酸のいずれかへの置換が好ましい。好ましい置換は正電荷を有するアミノ酸残基を導入するものであり、その結果得られるタンパク質の立体構造をいっそう不安定化させる。たとえば、好ましい置換にはシステイン→アルギニン(またはリジン)置換がある。
【0029】
本発明のある態様において、誘導体は、システイン残基71、103および117がいずれもアラニンへと変異している三重変異を含んでなる。
【0030】
別の態様において、本発明はProDer p 1配列のアミノ酸227〜240が欠失した、ProDer p 1の一形態を提供する。これらのアミノ酸はDer p 1配列の147〜160に相当する。
【0031】
したがって本発明は、本明細書において、低アレルゲン性Der p 1/ProDer p 1/PreProDer p 1誘導体の例として与えられる6個の特定の変異、ならびにProDer p 1のアミノ酸227〜240(Der p 1の147〜160)が欠失しているさらなる変異によって例証されるが、それに限定されない。第1に、ProDer p 1のアレルゲン活性は、Der p 1タンパク質配列のCys4の位置でシステイン残基をアルギニン残基に置き換えることによって、実質的に減少し、好ましくは完全に消失するが、その配列を配列番号3に示す。第2に、ProDer p 1のアレルゲン活性は、(成熟Der p 1の配列と照合して計算された)下記の任意の位置でシステイン残基をアルギニン残基に置換することによって実質的に消失する:Der p 1タンパク質配列のCys31(配列番号5)、Cys65(配列番号7)、Cys71(配列番号9)、Cys103(配列番号11)、Cys117(配列番号13)。
【0032】
さらに、ProDer p 1のアレルゲン活性は、ProDer p 1のアミノ酸227〜240(Der p 1の147〜160)の欠失により、実質的に減少し、好ましくは完全に消失する(配列番号15)。
【0033】
Der p 1/ProDer p 1/PreProDer p 1の変異型は、Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするcDNAの、従来法による部位特異的変異導入(G. Winterら, Nature 1982, 299, 756 758 もしくはZollerおよびSmith, 1982; Nucl. Acids Res., 10, 6487 6500に記載)、または欠失変異導入(Chan および Smith, Nucl. Acids Res., 1984, 12, 2407 2419 もしくはG. Winter ら, Biochem. Soc. Trans., 1984, 12, 224-225に記載)によって調製することができる。
【0034】
低アレルゲン性ProDer p 1、PreProDer p 1、Der p 3、ProDer p 3もしくはPreProDer p 3をも本発明によって提供する。
【0035】
本発明は具体的に開示された配列に限定されず、変異して、IgE結合反応性および/またはヒスタミン放出活性が減少し、またはなくなっているにもかかわらず、T細胞反応性を保持し、および/または野生型アレルゲンに対する免疫応答を刺激する能力を保持するあらゆる低アレルゲン性アレルゲンをも包含する。変異アレルゲンのアレルゲン活性、および結果としてアレルゲン活性の減少は、下記の実施例の項に詳述する方法にしたがった、ヒスタミン放出活性またはIgE結合反応性の方法のいずれによっても野生型と比較することができる。
【0036】
「実質的に減少したアレルゲン活性」は、残存IgE結合活性によって測定されるアレルゲン活性が、天然-非修飾もしくは非変異-タンパク質の活性の最大50%、好ましくは最大20%、さらに好ましくは最大10%、さらにいっそう好ましくは最大5%、さらにその上好ましくは5%未満にまで、減少していることを意味する。あるいはまた、「実質的に減少したアレルゲン活性」を、変異体のヒスタミン放出を測定することによって評価することもできる。活性の実質的な減少は、天然タンパク質に比べて少なくとも100分の1に減少する場合であるが、好ましくは1000分の1、さらに好ましくは10000分の1である。
【0037】
さまざまな免疫学的アッセイによって、変異アレルゲンの免疫原性を野生型アレルゲンと比較することができる。変異アレルゲンもしくは野生型アレルゲンのいずれかを用いたワクチン接種の後、in vitro T細胞アッセイによって、変異アレルゲンおよび野生型アレルゲンの交差反応性をアッセイすることができる。簡単に述べると、ワクチン接種を受けた動物から分離された脾臓T細胞をin vitroにて変異アレルゲンもしくは野生型アレルゲンのいずれか一方で再刺激した後、市販のELISAアッセイを用いてサイトカイン産生を測定するか、またはアレルゲン特異的T細胞の増殖をトリチウム標識したチミジンの取り込みによって、経時的にアッセイすることができる。また、免疫原性をELISAアッセイによって測定することも可能であり、当業者はその詳細を容易に決定することができよう。簡単に述べると、2種類のELISAアッセイが考えられる。1つ目は、野生型Der p 1で免疫したマウスの血清による変異型Der p 1の認識を評価することであり;2つ目は変異アレルゲンで免疫した動物の血清による野生型Der p 1の認識を評価することである。手短には、各ウェルを500 ngの精製された野生型もしくは変異型Der p 1で4℃にて一晩コーティングする。ブロッキング溶液(TBS-Tween 0.1%、1% BSA含有)とともにインキュベートした後、血清の連続希釈物を37℃にて1時間インキュベートする。ウェルを5回洗浄し、アルカリホスファターゼと結合した抗IgG抗体とともにインキュベートすることによって全IgGを明らかにする。変異型Der p 3の免疫原性も、上記のDer p 1に関する記載のように野生型Der p 3と比較することができる。
【0038】
本発明のもう一つの態様は、本明細書に開示のDer p 1/ProDer p 1/PreProDer p 1アレルゲンの変異型をコードする、単離された核酸を提供する。この核酸配列はDNA配列であることが好ましく、標準的なDNA合成技術、たとえば酵素による連結(D.M. Roberts ら, Biochemistry 1985, 24, 5090 5098)、化学合成、in vitro酵素重合、または上記技術を組み合わせることによって合成することができる。核酸配列は、想定される発現宿主において用いられるコドン使用頻度を模倣するように、最適化されたコドン使用頻度パターンを有することが好ましいが、さらに好ましくは高発現の哺乳動物(たとえばヒト)遺伝子のコドン使用頻度に類似する。好ましいDNA配列はコドンを最適化した配列であり、配列番号4、配列番号6、配列番号8、配列番号10、配列番号12、配列番号14、配列番号15および配列番号17に示す。
【0039】
本発明の別の態様は、本明細書に記載のDer p 3/ProDer p 3/PreProDer p 3アレルゲンの変異型をコードする、単離された核酸を提供する。この核酸配列はDNA配列であることが好ましく、標準的なDNA合成技術、たとえば酵素による連結(D.M. Roberts ら, Biochemistry 1985, 24, 5090 5098)、化学合成、in vitro酵素合成、または上記技術を組み合わせることによって合成することができる。核酸配列は、想定される発現宿主において用いられるコドン使用頻度を模倣するように、最適化されたコドン使用頻度パターンを有することが好ましいが、さらに好ましくは高発現の哺乳動物(たとえばヒト)遺伝子のコドン使用頻度に類似する。好ましいDNA配列を、配列番号20および21に示す。
【0040】
DNAの酵素的重合は、一般的には50 ml以下の容量で、10〜37℃にて、必要なヌクレオシド三リン酸、dATP, dCTP, dGTP およびdTTPを含有する適当なバッファー中で、DNAポリメラーゼI(Klenowフラグメント)のようなDNAポリメラーゼを用いて、in vitroで行うことができる。DNA断片の酵素による連結は、一般的には50ml以下の容量で、4℃から室温までの温度にて0.05M Tris (pH 7,4)、0.01M MgCl2、0.01Mジチオスレイトール、1mMスペルミジン、1mM ATPおよび0.1mg/mlウシ血清アルブミンといった適当なバッファー中でT4 DNAリガーゼのようなDNAリガーゼを用いて行うことができる。DNAポリマーもしくは断片の化学合成は、以下に記載の固相法を用いて、従来のリン酸トリエステル、亜リン酸エステルもしくはホスホルアミダイトの化学反応によって行うことができる('Chemical and Enzymatic Synthesis of Gene Fragments A Laboratory Manual' (H.G. GassenおよびA. Lang編), Verlag Chemie, Weinheim (1982)、または他の科学文献、たとえば、M.J. Gait, H.W.D. Matthes, M. Singh, B.S. Sproat, and R.C. Titmas, Nucleic Acids Research, 1982, 10, 6243; B.S. Sproat and W. Bannwarth, Tetrahedron Letters, 1983, 24, 5771; M.D. Matteucci and M.H Caruthers, Tetrahedron Letters, 1980, 21, 719; M.D. Matteucci and M.H. Caruthers, Journal of the American Chemical Society, 1981, 103, 3185; S.P. Adams ら, Journal of the American Chemical Society,1983, 105, 661; N.D. Sinha, J. Biernat, J. McMannus, and H. Koester, Nucleic Acids Research, 1984, 12, 4539; and H.W.D. Matthes ら, EMBO Journal, 1984, 3, 801)。
【0041】
あるいはまた、既知の技法(たとえば、相補的cDNA鎖を作製するためのmRNAの逆転写)および市販のcDNAキットを用いて、Der p 1/ProDer p 1/PreProDer p 1のmRNAからコード配列を導出することができる。Der p 3/ProDer p 3/PreProDer p 3のコード配列も上記のように導くことができる; PreProDer p 3ヌクレオチド配列のコドン使用頻度パターンは高発現細菌遺伝子に典型的なものである。
【0042】
驚くべきことに、ProDer p 3はDer p 3と比べて非常にアレルゲン性が低いことが明らかになった。
【0043】
ヌクレオチド配列のコドン使用頻度は、高発現ヒト遺伝子に典型的なものであることが望ましい。したがって、本発明の特定の態様において、変異したDer p 1/ProDer p 1/PreProDer p 1タンパク質をコードする複数のコドンを一緒に含んでなるヌクレオチド配列を提供するが、ここで、組換えダニタンパク質のアミノ酸配列をコードするために用いることのできるコドンの選択は、最適化された哺乳動物のコドン使用頻度をそっくり模するように変更され、その結果得られた遺伝子配列におけるコドンの使用頻度は、同一タンパク質をコードすると考えられる哺乳動物遺伝子と実質的に同一である。ヒトを含めて、哺乳動物のコドン使用頻度パターンは、文献に見出すことができる(たとえば、Nakamura ら 1996, Nucleic Acids Res. 24, 214-215を参照されたい)。
【0044】
DNA遺伝暗号には4つの文字があり(A、T、CおよびG)、これらを使用して3文字の「コドン」を綴り、このコドンは生物の遺伝子にコードされるタンパク質のアミノ酸を表す。DNA分子に沿ったコドンの一次配列は、遺伝子によってコードされるタンパク質におけるアミノ酸の一次配列に翻訳される。遺伝暗号は高度に縮重しており、61のコドンが20種の天然アミノ酸をコードし、3種のコドンが「終止」シグナルを表す。したがって、大半のアミノ酸は2以上のコドンによってコードされ、実際にいくつかのアミノ酸は4個以上の異なるコドンによってコードされる。
【0045】
所定のアミノ酸をコードするために2以上のコドンが使用できる場合、コドン使用頻度パターンはほとんどランダムではないことが認められている。異なる生物種は、コドンの選択において異なった偏りを示し、さらに高レベルで発現される遺伝子と低レベル発現遺伝子との間で、1つの種においてもコドンの使用は著しく異なることがある。こうした偏りはウイルス、植物、細菌、昆虫および哺乳動物細胞で相違しており、一部の種は、他の種より、ランダムなコドンとはかけ離れた強い偏りを示す。たとえば、ヒトおよび他の哺乳動物は特定の細菌もしくはウイルスほど偏りは強くない。こうした理由から、大腸菌(E. coli)において発現される哺乳動物遺伝子、または哺乳動物細胞において発現されるウイルス遺伝子が、効率よく発現するには不適当なコドンの配分を有する可能性はかなりある。しかしながら、大腸菌に適したコドン使用頻度パターンを有する遺伝子は、ヒトにおいても効率よく発現される可能性がある。宿主にはほとんど見られないコドンのクラスターが発現させるべき異種DNA配列中に存在することは、その宿主において低レベルの異種発現を予測するものであると考えられる。
【0046】
宿主でほとんど見られないコドンを宿主に好ましいコドンに変更すること(「コドン最適化」)が異種発現レベルを高めた例は、いくつか存在する。たとえば、BPV(ウシパピローマウイルス)後期遺伝子L1およびL2は哺乳動物のコドン使用頻度パターンにコドン最適化されたが、こうしたことが哺乳動物(Cos-1)細胞培養において野生型HPV配列より高い発現レベルを与えることが示された(Zhou et. al. J. Virol 1999. 73, 4972-4982)。この研究において、哺乳動物よりもBPVにおいて2倍より高い頻度で(使用頻度の比>2)出現するすべてのBPVコドン、および使用頻度の比が>1.5である大半のコドンが、優先的に使用される哺乳動物コドンに保存的に置き換えられた。WO97/31115, WO97/48370 およびWO98/34640 (Merck & Co., Inc.)において、HIV遺伝子もしくはそのセグメントのコドン最適化は、コドン最適化した配列を最適化の目的とした宿主動物においてDNAワクチンとして使用したとき、結果としてタンパク質発現の増加および免疫原性の向上をもたらすことが示された。
【0047】
本研究において、ヤケヒョウヒダニの各コドンは哺乳動物宿主に最適なコドンで保存的に置き換えられているため、その配列は全体として、好ましくは最適化されたコドンからなる(このことが望ましくない制限酵素切断部位、イントロンスプライス部位などを導入することになる場合を除く)。驚くべきことに、こうした最適化ProDer p 1/Der p 1配列はまた、酵母ではコドン使用頻度が異なっているにもかかわらず、酵母においても良好に発現する。
【0048】
本発明のさらに別の態様は、変異したDer p 1/ProDer p 1/PreProDer p 1タンパク質を調製する方法を提供するが、この方法は、組換え宿主細胞において当該タンパク質をコードするDNAを、コドンが最適化されている、いないにかかわらず、発現させること、および生成物を回収することを含んでなる;上記の方法はDer p 3/ProDer p 3/PreProDer p 3にも適用される。
【0049】
Der p 1は、その酵素活性、アレルゲン性および遺伝子クローニングについては十分に性質が明らかにされているものの、Der p 1の異種発現は問題があると報告されており(Chapman and Platts-Mills, J Immunol 1980;125:587-592)、これはおそらくこのシステインプロテイナーゼがPreProDer p 1前駆体として合成されるためである。タンパク質の高次構造に関わるシステイン残基が変異したDer p 1/ProDer p 1/PreProDer p 1配列の発現はさらにいっそう問題がある。したがって、本発明はさらに、こうした障害をすべて克服することによって変異タンパク質の製造を可能とし、ダニアレルギーに対する治療用および予防用ワクチンの産業的開発を可能にする方法を提供する。
【0050】
Der p 3/ProDer p 3/PreProDer p 3の変異タンパク質または組換えタンパク質の製造方法もまた提供される。
【0051】
変異の有無にかかわらずDer p 1/ProDer p 1/PreProDer p 1をマルトース結合タンパク質(MBP)融合タンパク質として発現させたとき、大腸菌においてタンパク質発現の実質的な改善が達成された。したがって、変異したProDer p/Der p 1タンパク質を大腸菌においてMBP融合タンパク質として発現させる方法を提供する。さらに、ジスルフィド結合がPichia pastorisにおける分泌に必須であると言われているにもかかわらず、驚くべきことに酵母におけるタンパク質発現の実質的な改善が、変異タンパク質について達成された(Takai ら 2001, Int. Arch. Allergy Immunol. 124, 454-460)。このことは、ダニの変異ProDer p/Der p 1タンパク質をコードするポリヌクレオチド配列を、高発現ヒト遺伝子に見られるコドン使用頻度に適合するように再度改変することによって達成され、それによって組換え抗原が天然ProDer p/Der p 1ダニアレルゲンと同じ高次構造および免疫学的特性を有することも可能となった。これまではPichia pastorisにおける発現はうまくいかないと報告されていた(Takai ら 2001, Int. Arch. Allergy Immunol. 124, 454-460)にもかかわらず、驚くべきことに、哺乳動物細胞での発現にコドンが最適化された変異型ProDer p 1のクローニングおよび発現を、Pichia pastorisにおいて達成することができた(一定の割合での分泌を伴う)。
【0052】
Maniatis et. al., Molecular Cloning - A Laboratory Manual; Cold Spring Harbor, 1982 1989に記載の従来の組換え技術によって本発明の方法を実施することができる。
【0053】
具体的には、本発明の方法は次のステップを含んでもよい:
1. 前記Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするヌクレオチド配列を含んでなるDNAポリマーを発現することができる、宿主細胞において複製可能な、または組み込み型の発現ベクターを調製すること;
2. 部位特異的変異導入によって、ジスルフィド結合に関わるシステイン残基を別の残基、好ましくはアルギニン残基に置き換えることにより、結果として得られるタンパク質のIgE結合活性を変化させること;
3. 宿主細胞を前記ベクターで形質転換すること;
4. DNAポリマーの発現を可能にする条件下で形質転換された宿主細胞を培養してタンパク質を産生させること;
5.タンパク質を回収すること。
【0054】
上記方法はDer p 3/ProDer p 3/PreProDer p 3にも適用可能である。
【0055】
「形質転換する」という用語は、本明細書において、たとえばGenetic Engineering; Eds. S.M. Kingsman and A.J. Kingsman; Blackwell Scientific Publications; Oxford, England, 1988に記載された従来の技術を利用して、適当なプラスミドもしくはウイルスベクターを用いて、形質転換、トランスフェクション、または感染によって外来DNAを宿主細胞内に導入することを表すために用いる。「形質転換された」または「形質転換体」という用語は、以後、当該の外来遺伝子を含有し発現する、結果として得られる宿主細胞に適用されるものとする。
【0056】
発現ベクターもまた新規であり、本発明の一部を成す。本発明のある特定の態様は、本発明のシステイン変異型Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするポリヌクレオチド配列を含んでなり、この配列の発現を指令する能力を有する発現ベクターを提供する。本発明の別の特定の態様は、システイン変異型Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするポリヌクレオチド配列を含んでなり、その配列の発現を指示する能力を有する発現ベクターを提供するが、ここで、このポリヌクレオチド配列のコドン使用頻度パターンは、高度に発現される哺乳動物遺伝子、好ましくは高度に発現されるヒト遺伝子に典型的なパターンである。ベクターは、細菌、昆虫、酵母、もしくは哺乳動物細胞、特にヒト細胞において異種DNAの発現を推進するのに適していると考えられる。
【0057】
本発明にしたがって、宿主細胞と適合するベクターを切断して無傷のレプリコンを有する線状DNAセグメントを準備し、前記線状セグメントを、その線状セグメントとともに所望の生成物をコードする1以上のDNA分子、たとえば、Der p 1/ProDer p 1/PreProDer p 1タンパク質をコードするDNAポリマーと、連結条件下で結合させることにより、複製可能な発現ベクターを調製することができる。
【0058】
上記ベクターは、本発明の変異したDer p 3/ProDer p 3/PreProDer p 3にも適用することができる。
【0059】
このようにしてDNAポリマーを、所望により、前もって形成する、またはベクターの構築時に形成することができる。
【0060】
ベクターの選択は、ある程度は宿主細胞によって決まるが、宿主細胞は原核生物または真核生物であってよい。適当なベクターにはプラスミド、バクテリオファージ、コスミドおよび組換えウイルスが含まれる。
【0061】
複製可能な発現ベクターの調製は、たとえば、Maniatis ら、上記引用、に記載の手順によって、DNAを制限切断、重合および連結する適当な酵素を用いて従来の方法で行うことができる。
【0062】
組換え宿主細胞は、本発明にしたがって、形質転換条件下で、宿主細胞を本発明の複製可能な発現ベクターで形質転換することにより調製される。適当な形質転換条件は通常のものであって、たとえば、上記のManiatis ら、または「DNA Cloning」 Vol. II, D.M. Glover ed., IRL Press Ltd, 1985に記載されている。
【0063】
形質転換条件の選択は宿主細胞によって決まる。したがって、大腸菌のような細菌宿主は、CaCl2溶液、またはRbCl、MnCl2、酢酸カリウムおよびグリセロールの混合物を含んでなる溶液で処理し、次いで3-[N-モルホリノ]-プロパンスルホン酸、RbClおよびグリセロールで処理してもよい。哺乳動物培養細胞は、細胞へのベクターDNAのカルシウム共沈、リポフェクション、またはエレクトロポレーションによって形質転換することができる。酵母に適合するベクターはまた、栄養要求性変異株に原栄養性を付与すること、または野生株に重金属耐性を付与することによって、形質転換に成功した株を選択できるようにするマーカーを保有する。酵母ベクターの制御配列には、解糖酵素のプロモーター(Hess ら, J. Adv. Enzyme Reg. 1968, 7, 149)、酸ホスファターゼをコードするPHO5遺伝子、CUP1遺伝子、ARG3遺伝子、GAL遺伝子プロモーター、および合成プロモーター配列が含まれる。酵母の発現に有用な他の調節因子は、ターミネーターおよびリーダー配列である。細胞からのタンパク質の分泌を指令するリーダー配列は、一般的には疎水性アミノ酸からなるシグナルペプチドをコードしているので、特に有用である。適当なシグナル配列をコードしている可能性があるのは、分泌される酵母タンパク質の遺伝子、たとえば酵母インベルターゼ遺伝子およびαファクター遺伝子、酸ホスファターゼ、キラートキシン、α接合因子遺伝子、および最近では酵母Kluyveromyces marxianusのINU1A遺伝子に由来する異種イヌリナーゼシグナル配列である。Pichia pastorisや出芽酵母Saccharomyces cerevisiaeにおける発現に適したベクターが開発されている。
【0064】
さまざまな誘導もしくは構成的プロモーターに基づく、さまざまなP. pastoris発現ベクターが利用可能である(Cereghino and Cregg, FEMS Microbiol. Rev. 2000,24:45-66)。細胞質内タンパク質および分泌タンパク質の産生のために、もっともよく利用されるP. pastorisベクターは、非常に強力で厳密に制御されたアルコールオキシダーゼ(AOX1)プロモーターを含有する。ベクターはまた、his4宿主における選択のためにP. pastorisヒスチジノールデヒドロゲナーゼ(HIS4)遺伝子をも含有する。外来タンパク質の分泌はシグナル配列の存在を必要とし、S. cerevisiaeプレプロα接合因子リーダー配列が、Pichia発現系において広く良好に使用されている。発現ベクターは、発現菌株の安定性を最大にするように、P. pastorisゲノムに組み込まれる。S. cerevisiaeの場合と同様に、P. pastoris発現ベクターの、宿主ゲノムと共通する配列(AOX1もしくはHIS4)内での切断は、そのゲノムの遺伝子座へのベクターの組み込みを効果的に標的とする相同組換えイベントを刺激する。一般に、組み込みコピーの発現カセットを複数含有する組換え菌株は、単一コピー菌株より多くの異種タンパク質を生成することができる。高コピー数の形質転換体を得るために最も有効な方法は、Pichia受容株をスフェロプラスト法で形質転換することを必要とする(Cregg ら, 1985, Mol.Cell.Biol. 5: 3376-3385)。
【0065】
本発明はまた、本発明の複製可能な発現ベクターで形質転換された宿主細胞にも及ぶ。
【0066】
形質転換された宿主細胞の、DNAポリマーの発現を可能にする条件下での培養は、従来の方法によって実施され、たとえば、Maniatis ら および「DNA Cloning」、上記に記載の通りである。従って、好ましくは細胞には栄養分を供給し、45℃以下の温度で培養する。
【0067】
生成物は、宿主細胞に応じて、従来の方法によって回収する。したがって、宿主細胞が大腸菌のような細菌である場合、その細胞を物理的、化学的もしくは酵素的に溶解し、その結果得られる溶菌液からタンパク質生成物を分離することができる。宿主細胞が哺乳動物細胞である場合、生成物は一般に、栄養培地から、または無細胞抽出液から分離することができる。従来のタンパク質分離技術には、選択的沈澱、吸着クロマトグラフィー、およびモノクローナル抗体アフィニティカラムを含めたアフィニティクロマトグラフィーがあげられる。
【0068】
あるいはまた、バキュロウイルスのような適当なベクターを用いて昆虫細胞において、形質転換されたショウジョウバエ細胞において、または哺乳動物CHO細胞においても発現させることができる。本発明の新規タンパク質はまた、EP-A-0 278 941の中でCSタンパク質について記載されているように、酵母細胞において発現させることもできる。
【0069】
本発明の低アレルゲン性Der p 1/ProDer p 1/PreProDer p 1誘導体を含んでなる、またはコドン最適化の如何に関わらず前記タンパク質をコードするポリヌクレオチド配列を含んでなる、医薬組成物、免疫原性組成物およびワクチン組成物も提供される。低アレルゲン性Der p 3/ProDer p 3/PreProDer p 3を含んでなる同様の組成物も提供される。好ましい実施形態において、DNA組成物は複数の粒子、好ましくは金粒子を含んでなり、この粒子は、ヤケヒョウヒダニアミノ酸配列をコードするポリヌクレオチド配列をコードするベクターを含んでなるDNAでコーティングされており、ここでこのポリヌクレオチド配列のコドン使用頻度パターンは高発現哺乳動物遺伝子、とりわけヒト遺伝子に特有のものである。
【0070】
本発明のポリヌクレオチド、およびコードされるポリペプチドは、治療薬もしくは予防薬として用途を見出すことができる。特に、DNAを、ワクチン接種すべき哺乳動物、たとえばヒトに投与する場合、本発明のポリヌクレオチド(天然ProDer p 1のポリヌクレオチド配列が含まれ、好ましくはコドン最適化されている)を、DNAワクチン接種(NAVAC)に使用することができる。RNAもしくはDNAのような核酸、好ましくはDNAは、上記のようなベクターの形で提供されるが、このベクターは哺乳動物の細胞において発現されうる。ポリヌクレオチドは利用可能なあらゆる方法によって投与することができる。たとえば、核酸を、注射によって、好ましくは皮内、皮下もしくは筋肉内に注入することができる。あるいはまた、粒子を介するDNAデリバリー(PMDD)のような核酸デリバリーデバイスを用いて、核酸を直接皮膚内に送達することができる。この方法では、不活性粒子(たとえば金ビーズ)を核酸でコーティングし、その粒子を、たとえば発射装置から高圧で放出することによって、受容側の表面(たとえば皮膚)を貫通可能とするのに十分な速度に加速する(本発明の核酸分子でコーティングされた粒子は本発明の範囲に含まれ、そうした粒子を充填したデリバリーデバイスも同様である)。
【0071】
裸のポリヌクレオチドもしくはベクターを患者に導入するのに適した技術には、適当なビヒクルを用いた局所適用が含まれる。核酸を局所的に、皮膚または粘膜表面に、たとえば、鼻内、経口、膣内または直腸内投与によって、投与することができる。裸のポリヌクレオチドもしくはベクターは、製薬上許容される添加剤、たとえばリン酸緩衝生理食塩水(PBS)とともに存在してもよい。DNAの取り込みは、ブピバカインといった促進剤を、DNA製剤に含めて、または別々に使用することによって、さらに促進することができる。核酸を直接、受容者に投与する別の方法には、超音波、電気刺激、エレクトロポレーションおよび米国特許第5,697,901号に記載のマイクロシード法(microseeding)がある。一般的に核酸は1pg〜1mgの範囲で投与され、粒子による遺伝子デリバリーでは好ましくは1pg〜1μg核酸、他の経路では10μg〜1mgの量で投与される。
【0072】
本発明の核酸配列は、遺伝子治療に有用な特化された運搬ベクターを用いて投与することもできる。遺伝子治療アプローチは、たとえばVerme ら, Nature 1997, 389:239-242によって論じられている。ウイルスベクター系も非ウイルスベクター系もいずれも利用することができる。ウイルスに基づく系には、レトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ヘルペスウイルス、カナリア痘およびワクシニアウイルスに基づく系が含まれる。非ウイルス性の系には、核酸の直接投与、マイクロスフェア封入技術(ラクチドとグリコリドの共重合体)およびリポソームに基づく系が含まれる。初回ワクチン接種後に追加免疫の注射を与えることが望ましい場合には、ウイルスおよび非ウイルスデリバリーシステムを組み合わせることができる。たとえば、プラスミドのような非ウイルスベクターを用いた初回の「プライム」DNAワクチン接種後、1回または複数回の「ブースト」ワクチン接種を、ウイルスベクターまたは非ウイルス性の系を用いて行う。
【0073】
このようにして、本発明者らは、ProDer p 1をコードするDNA(好ましくは哺乳動物用にコドン最適化されている)によるワクチン接種が、マウスモデルにおいてTh1応答を誘導し(特異的IgG2a抗体の力価は高く、特異的IgG1抗体の力価は低い)、特記すべきことに、抗ProDer p 1 IgEの不在をもたらすことを見出した。
【0074】
本発明の医薬組成物には、アジュバント化合物、またはタンパク質によって誘導される免疫応答を増強するのに役立つと考えられる他の物質が含まれうる。
【0075】
本発明のワクチン組成物は、免疫保護的な量の変異型もしくは組換え型Der p 1/ProDer p 1/PreProDer p 1低アレルゲン性タンパク質、または変異型もしくは組換え型Der p 3/ProDer p 3/PreProDer p 3低アレルゲン性タンパク質を含んでなる。「免疫保護的」という用語は、アレルギー疾患を回避または緩和するように、次のアレルギー誘発に対して免疫応答を誘導するために必要な量を表す。本発明のワクチンにおいて、タンパク質の水溶液をそのまま使用することができる。あるいはまた、タンパク質を、事前の凍結乾燥の有無にかかわらず、あらゆるさまざまな既知のアジュバントと混合、吸着、または共有結合させることができる。
【0076】
適当なアジュバントが市販されており、たとえば、フロイント不完全アジュバントおよび完全アジュバント(Difco Laboratories, Detroit, MI);Merck Adjuvant 65 (Merck and Company, Inc., Rahway, NJ);AS-2 (SmithKline Beecham, Philadelphia, PA);水酸化アルミニウムゲル(ミョウバン)もしくはリン酸アルミニウムといったアルミニウム塩;カルシウム、鉄もしくは亜鉛の塩;アシル化チロシンの不溶性懸濁物;アシル化糖類;陽イオンもしくは陰イオン誘導多糖類;ポリホスファゼン類;生分解性マイクロスフェア;モノホスホリルリピドAおよびquil Aといったものがある。GM-CSFまたはインターロイキン-2、-7、もしくは-12といったサイトカイン、およびケモカインもアジュバントとして使用することができる。
【0077】
本発明の製剤において、アジュバント組成物は主としてTh1型の免疫応答を誘導することが好ましい。高レベルのTh1型サイトカイン(たとえば、IFN-γ、TNFα、IL-2およびIL-12)は、投与された抗原に対する細胞性免疫応答の誘導に有利に働く傾向がある。応答の大部分がTh1型である好ましい実施形態において、Th1型サイトカインのレベルはTh2型サイトカインのレベルよりも非常に高まる。こうしたサイトカインのレベルは、標準的なアッセイによって容易に評価することができる。サイトカインファミリーに関する総説については、Mosmann and Coffman, Ann. Rev. Immunol. 7:145-173, 1989を参照されたい。
【0078】
したがって、主にTh1型応答の誘導に用いるのに適したアジュバントには、たとえば、モノホスホリルリピドA、好ましくは3-O-脱アシル化モノホスホリルリピドA(3D-MPL)のアルミニウム塩との組み合わせがある。他の既知のアジュバントで、優先的にTH1型免疫応答を誘導するものは、CpG含有オリゴヌクレオチドを包含する。このオリゴヌクレオチドは、CpGジヌクレオチドがメチル化されていないことを特徴とする。こうしたオリゴヌクレオチドはよく知られており、たとえば、国際公開WO 96/02555に記載されている。免疫賦活性DNA配列もまた、たとえば、Sato ら, Science 273:352, 1996によって記載されている。CpG含有オリゴヌクレオチドは、単独でも、他のアジュバントと組み合わせても使用することができる。たとえば、増強されたシステムは、CpG含有オリゴヌクレオチドとサポニン誘導体の組み合わせ、とりわけ国際公開WO 00/09159およびWO 00/62800に記載のようにCpGとQS21の組み合わせを伴う。製剤はそれに加えて、水中油型エマルジョンおよび/またはトコフェロールを含んでなることが好ましい。
【0079】
別の好ましいアジュバントはサポニン、好ましくはQS21(Aquila Biopharmaceuticals Inc., Framingham, MA)であり、これは単独でも他のアジュバントと組み合わせても使用することができる。たとえば、増強されたシステムは、国際公開WO 94/00153に記載の、QS21と3D-MPLの組み合わせのような、モノホスホリルリピドAとサポニン誘導体の組み合わせ、または国際公開WO 96/33739に記載の、QS21がコレステロールでクェンチされた、反応性の弱い組成物を伴う。他の好ましい製剤は、水中油型エマルジョンおよびトコフェロールを含んでなる。水中油型エマルジョン中にQS21、3D-MPLおよびトコフェロールを含む特に強力なアジュバント製剤が国際公開WO 95/17210に記載されている。
【0080】
水中油型エマルジョン中にQS21、3D-MPLおよびトコフェロールを含む特に強力なアジュバント製剤が国際公開WO 95/17210に記載されているが、これは好ましい製剤である。
【0081】
他の好ましいアジュバントには、Montanide ISA 720 (Seppic, France), SAF (Chiron, California, United States), ISCOMS (CSL), MF-59 (Chiron), Detox (Ribi, Hamilton, MT), RC-529 (Corixa, Hamilton, MT)および他のアミノアルキルグルコサミニド4-リン酸 (AGPs)がある。
【0082】
したがって、本明細書に記載のDer p 1/ProDer p 1/PreProDer p 1低アレルゲン性誘導体、およびアジュバントを含んでなる免疫原性組成物が提供されるが、このアジュバントは3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルもしくはエステルのうち1つまたは複数、またはこれらのアジュバントのうち2つ以上の組み合わせを含んでなる。免疫原性組成物中のDer p 1/ProDer p 1/PreProDer p 1低アレルゲン性誘導体は、水中油型もしくは油中水型エマルジョンビヒクル中で提供されることが好ましい。
【0083】
さらに、本明細書に記載のDer p 3/ProDer p 3/PreProDer p 3低アレルゲン性誘導体およびアジュバントを含んでなる免疫原性組成物が提供されるが、このアジュバントは、3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルもしくはエステルのうち1つまたは複数、またはこれらのアジュバントのうち2つ以上の組み合わせを含んでなる。免疫原性組成物中のDer p 3/ProDer p 3/PreProDer p 3低アレルゲン性誘導体は、水中油型もしくは油中水型エマルジョンビヒクル中で提供されることが好ましい。
【0084】
もう一つの態様において、本発明は医薬組成物を作製する方法を提供するが、その方法は、ジスルフィド架橋の形成に関わるDer p 1/ProDer p 1/PreProDer p 1/Der p 3/ProDer p 3/PreProDer p 3の1つまたは複数のシステイン残基を変異させるステップ、たとえばDer p 1の下記の残基:Cys4, Cys31, Cys65, Cys71, Cys103またはCys117を変異させることを包含する。さらに別の態様において、本発明は、ProDer p 1のアミノ酸残基227〜240(Der p 1の147〜160)を欠失させるステップを包含する、医薬組成物を作製する方法を提供する。
【0085】
本発明の方法はさらに、野生型Der p 1/ProDer p 1/PreProDer p 1ヌクレオチド配列のコドン使用頻度パターンを変更するステップ、またはポリヌクレオチド配列を合成によって作製するステップを含んでなり、高発現哺乳動物遺伝子に特有のコドン使用パターンを有し、かつコドンが最適化されたシステイン変異型ProDer p 1/ Der p 1アミノ酸配列、もしくは本発明にしたがって選択された残基が欠失しているProDer p 1/ Der p 1アミノ酸配列をコードする配列を生じる。ワクチン製剤は総じてVaccine Design(”The subunit and adjuvant approach” (eds. Powell M.F. & Newman M.J). (1995) Plenum Press New York)に記載されている。リポソーム内への封入はFullertonにより米国特許第4,235,877号に記載されている。タンパク質の巨大分子へのコンジュゲーションは、たとえば、Likhite、米国特許第4,372,945号およびArmorら、米国特許第4,474,757号によって開示されている。
【0086】
それぞれのワクチン用量に含まれる本発明のタンパク質の量は、典型的なワクチンにおいて重大な副作用なしに免疫保護応答を誘導する量として選択される。このような量は、どの特異的免疫原を用いるかによって、およびワクチンがアジュバントで増強されているかどうかによって変わってくる。一般に、各用量は1〜1000μgのタンパク質を含んでなると見込まれるが、1〜200μgが好ましい。具体的なワクチンの最適量は、被験体における抗体価の観察および他の反応に関わる標準的な調査によって確認することができる。本発明のワクチンは、成人または乳幼児に投与可能であるが、出生直後で実質的なTh2型免疫記憶反応が確立する前に、個体にワクチン接種することが好ましい。初回接種後、被験体は約4週間でブーストを受け、その後アレルギー反応のリスクが存在する限り、6ヶ月ごとに追加接種を繰り返すことが好ましい。
【0087】
ワクチンおよび医薬組成物は、密封アンプルもしくはバイアルといった、1回量または複数回量容器に入れて提供されることが考えられる。こうした容器は、使用するまで製剤の無菌性を保つように、密閉されていることが好ましい。一般に、製剤は油性もしくは水性ビヒクル中で懸濁液、溶液または乳濁液として保存してもよい。あるいはまた、ワクチンもしくは医薬組成物は、使用直前に無菌の液体担体を添加するだけでよい、凍結乾燥された状態で保存してもよい。
【0088】
本発明はワクチンを製造する方法も提供するが、この方法は、本発明のDer p 1/ProDer p 1/PreProDer p 1誘導体もしくはDer p 3/ProDer p 3/PreProDer p 3誘導体、またはそれらの誘導体を、本明細書に記載のプロセスによって精製するステップ、およびその結果得られたタンパク質を適当なアジュバント、希釈剤もしくは他の製薬上許容される添加剤と混合するステップを含んでなる。
【0089】
本発明はまた、本発明のタンパク質を製薬上許容される添加剤と混合することを含んでなる、ワクチン製剤を製造する方法を提供する。
【0090】
本発明の別の態様は、アレルギーを罹患しやすいまたはアレルギーに罹患した患者を免疫療法で治療するためのワクチンを製造するための、本出願で請求するタンパク質もしくはポリヌクレオチドの使用である。アレルギーを罹患しやすい、またはアレルギーに罹患した患者に製薬上活性な量の本明細書に記載の免疫原性組成物を投与することを含んでなる前記患者を治療する方法も、本発明により考慮される。
【0091】
本発明のさらなる態様は、ヒトにおいてアレルギー疾患(特にハウスダストダニアレルギー)を予防または緩和する方法を提供するものであるが、その方法は、免疫原として有効な量の本発明の変異型アレルゲンまたは本発明によるワクチンを、それを必要とする被験体に投与することを含んでなる。
【0092】
下記の実施例は、本発明を例証するものであるが本発明を限定するものではない。制限酵素および他の試薬は実質的にはメーカーの使用説明書にしたがって使用した。
【実施例】
【0093】
実施例1
一般的手法
1. SDS-PAGEおよびウェスタンブロット分析
タンパク質は、SDS-PAGEによって12.5%ポリアクリルアミドゲル上で分析した。電気泳動後、セミドライトランスブロットシステム(Bio-Rad)を用いて、タンパク質をニトロセルロース膜上に転写した。膜は、TBS-T(50mM Tris HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)中の0.5% Instagel(PB Gelatins)で30分間飽和させ、変性済みまたは天然型ProDer p 1に対して生じさせたマウスポリクローナル血清(ブロッキング溶液で希釈(1:5000))とともにインキュベートした。アルカリホスファターゼ結合ヤギ抗マウス抗体(Promega, 1:7500)および5-ブロモ-4-クロロ-3-インドリルリン酸(BCIP, Boehringer)/ニトロブルーテトラゾリウム(NBT, Sigma)を基質として用いて、免疫反応性物質を検出した。
【0094】
2. 糖鎖分析
Glycan Differenciation Kit (Boehringer)により、下記のレクチンを用いて糖鎖分析を行った:スノードロップ(Galanthus nivalis)アグルチニン(GNA)、セイヨウニワトコ(Sambucus nigra)アグルチニン(SNA)、イヌエンジュ(Maackia amurensis)アグルチニン(MAA)、ピーナッツアグルチニン(PNA)およびシロバナヨウシュチョウセンアサガオ(Datura stramonium)アグルチニン(DSA)。概略としては、精製タンパク質をSDS-PAGEからニトロセルロース膜上に転写した。膜を、ジゴキシゲニンと結合した異なるレクチンとともにインキュベートした。複合体はアルカリホスファターゼと結合した抗ジゴキシゲニン抗体を用いて検出した。
【0095】
3. 酵素アッセイ
1mM EDTAおよび20mM L-システインを含有する50 mM Tris-HCl pH 7中で、総容量1mlとして25℃にて酵素アッセイを行った。Cbz-Phe-Arg-7-アミノ-4-メチルクマリン(Cbz-Phe-Arg-AMC)およびBoc-Gln-Ala-Arg-7-アミノ-4-メチルクマリン(Boc-Gln-Ala-Arg-AMC)(Sigma)(いずれの基質も終濃度100μMとした)の加水分解を、SLM 8000分光蛍光光度計を用いてλex = 380nmおよびλem = 460nmでモニターした。システイン活性化アレルゲンを終濃度100 nMとなるように添加することによってアッセイを開始した。どのようなアッセイの前にも、精製Der p 1もしくはProDer p 1を、アプロチニン-およびp-アミノベンズアミジン-アガロース樹脂(Sigma)の混合物とともにインキュベートして、ありうる微量のセリンプロテアーゼ活性を除去した。
【0096】
4. タンパク質定量
ウシ血清アルブミンを標準として、ビシンコニン酸法(MicroBCA, Pierce)によって総タンパク質濃度を測定した。
【0097】
5. Der p 1 ELISA
Der p 1特異的モノクローナル抗体5H8および4C1(Indoor Biotechnologies)を用いてELISAキットによってDer p 1または組換えProDer p 1を検出した。アッセイに使用したDer p 1標準(UVA 93/03)の濃度は0.5μg/mlであった。
【0098】
6. IgE結合活性
イムノプレートを、Der p 1またはProDer p 1(1ウェル当たり500ng)で4℃にて一晩コーティングした。次にプレートを、ウェル当たり100μlのTBS-Tweenバッファー(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSAを添加した同じバッファー150μlで37℃にて1時間飽和させた。ヤケヒョウヒダニに対するアレルギー患者から採取した血清を1/8に希釈した後、37℃にて1時間インキュベートした。実験に使用した95の血清のうち、16の血清は、その特異的抗ヤケヒョウヒダニIgE値(RASTアッセイ)が58.1kU/Lから99kU/Lの範囲にあり、79の血清は、上限カットオフ値である100kU/Lを越えた。プレートをTBS-Tweenバッファーで5回洗浄し、マウス抗ヒトIgE抗体(Southern Biotechnology Associates)およびアルカリホスファターゼ結合ヤギ抗マウスIgG抗体(TBS-Tweenバッファーで1/7500に希釈、Promega)とともにインキュベート後、アレルゲン-IgE複合体を検出した。酵素活性は、ジエタノールアミンバッファー(pH 9.8)中に溶解したp-ニトロフェニルリン酸基質(Sigma)を用いて測定した。OD410nmを、Biorad Novapath ELISAリーダーで測定した。
【0099】
IgE阻害アッセイについては、プレートを同一濃度(0.12μM)のDer p 1もしくはProDer p 1でコーティングした。20のアレルギー患者由来ヒト血清のプール(RAST値>100kU/L)を、さまざまな濃度(3.6-0.002μM)の阻害剤としてのDer p 1 または組換えProDer p 1とともに4℃にて一晩プレインキュベートし、ELISAプレートに加えた。IgE結合は上記のように検出した。
【0100】
7. ヒスタミン放出
ヒスタミン放出はアレルギーの供血者の、ヘパリン処理末梢血由来の白血球を用いて、ヒスタミン-ELISAキット(Immunotech)により、アッセイした。好塩基球を、連続希釈した組換えProDer p 1またはDer p 1とともに37℃にて30分間インキュベートした。好塩基球中のヒスタミンの総量は、界面活性剤IGEPAL CA-630(Sigma)を用いた細胞破砕後に定量した。
【0101】
8. ProDer p 1の変性
100℃にて5分間、50mMβ-メルカプトエタノールの存在下で、組換えProDer p 1を変性させた。
【0102】
9. 免疫化
10匹のCBA/Jマウス(6週齢)群を、5μgの種々のタンパク質または100μgの種々のプラスミドDNAで1週間ごとに4回、免疫した。アジュバントとしてミョウバンの存在下で精製アレルゲンを注射した。対照として、マウス群をミョウバンまたはpJW4304 DNAベクターで免疫処置した。第7、14、21、28日にマウスの眼窩静脈叢から採血し、血清を採取した。
【0103】
10. 気管支吸入誘発
免疫後72時間以内にすべてのマウスをプレキシグラス(アクリル樹脂)容器(13 x 19 x 37.5 cm)に入れ、エアロゾル化したヤケヒョウヒダニ粗抽出物に7日間連続で20分間にわたり曝露した。ダニの粗抽出物の濃度は300μg/mlとした。エアロゾルは超音波ネブライザー(Syst’ AM)によって発生させた。ネブライザーの出力は0.5ml/分であり、エアロゾルの平均粒径は1〜5μmであった。対照としてはマウスにPBSを噴霧した。
【0104】
11. Der p 1特異的IgG、IgG1およびIgG2aの測定
血清の抗Der p 1 IgG、IgG1およびIgG2a抗体をELISAによってアッセイした。イムノプレートを、4℃にて16時間ProDer p 1(1ウェル当たり500ng)でコーティングした。プレートをTBS-Tween(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSAを添加した同じバッファー150μlで37℃にて1時間飽和させた。血清を飽和バッファーで連続希釈して、37℃で1時間インキュベートした。プレートをTBS-Tweenバッファーで5回洗浄し、抗原に結合した抗体を、アルカリホスファターゼと結合した二次抗体(ヤギ抗マウスIgG、Promega、USA)(TBS-Tweenバッファーで1/7500に希釈)によって検出した。酵素活性は、ジエタノールアミンバッファー(pH 9.8)に溶解したp-ニトロフェニルリン酸基質(Sigma)を用いて測定した。OD415nmはBiorad Novapath ELISAリーダーで測定した。
【0105】
マウス抗体のサブクラスは、上記のようにコーティングしたプレート、および二次抗体としてIgG1-もしくはIgG2a-特異的ビオチン標識モノクローナル抗体(ラット抗マウス、TBS-Tweenバッファーおよび1% BSAで1/7000希釈、Biosource)を用いて判別した。アルカリホスファターゼ結合ストレプトアビジン(1/1000希釈、Amersham)を各ウェルに添加した。酵素活性のアッセイは上記のように行った。
【0106】
すべての場合において、ELISAの力価は、測定したOD415の最大値の50%に相当するシグナルを与える希釈率の逆数とした。
【0107】
12. Der p 1特異的IgEの測定
イムノプレートを、4℃にて16時間ラット抗マウスIgE(1ウェル当たり10ng)でコーティングした。プレートをTBS-Tween(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSAを添加した同じバッファー150μlで37℃にて1時間飽和させた。血清を飽和バッファーで連続希釈して、37℃で1時間インキュベートした。その後ProDer p 1を飽和バッファーに添加して500ng/mlとした。結合したProDer p 1を、ビオチン標識抗Der p 1モノクローナル抗体4C1(Indoor Biotechnologies)の添加によって検出した。プレートをTBS-Tweenバッファーで5回洗浄し、抗原に結合した抗体を、アルカリホスファターゼと結合したストレプトアビジン(TBS-Tweenバッファーで1/7500に希釈)の添加によって検出した。酵素活性はジエタノールアミンバッファー(pH 9.8)に溶解したp-ニトロフェニルリン酸基質(Sigma)を用いて測定した。OD415nmはBiorad Novapath ELISAリーダーで測定した。
【0108】
13. 増殖アッセイ
Der p 1特異的T細胞増殖応答を測定するために、気管支吸入誘発の前後に免疫したマウスを犠牲にした。リンパ球を脾臓から分離した。15 mM HEPESおよび30μMβ-メルカプトエタノールを含有する10% FCSを添加したRPMI 1640で培養した細胞(1ウェル当たり4 x 105を3通り)を、96ウェルプレートにおいて連続希釈したダニ粗抽出物もしくはProDer p 1で刺激した(25μg/mlの濃度から始めて、10個の2倍希釈していった、抗原の希釈物をテストした)。対照としては、細胞をRPMI培地のみとともにインキュベートした。4日後に、細胞に1ウェル当たり1μCiの[3H]チミジン(Amersham)を加えて16時間パルス標識した。細胞を集め、3H-チミジンの取り込みをシンチレーション計数により測定した。増殖反応は4ウェルの平均として算出し、刺激指数(stimulation index, SI)として表した。2より大きいSIを陽性とした。
【0109】
14. サイトカインアッセイ
リンパ球培養上清におけるIFNγおよびIL-5のレベルをELISAアッセイで測定した。1μg/mlの抗マウスIL-5モノクローナル(PharMingen)または抗マウスIFNγ(Biosource)ポリクローナル抗体でプレートをコーティングした。プレートをTBS-Tweenバッファーで5回洗浄し、150μl TBS-Tween-BSAで37℃にて1時間飽和させた。脾細胞培養上清を連続希釈して添加し、37℃にて90分間インキュベートした。ビオチン標識抗マウスIL-5(PharMingen, 1μg/ml)または抗マウスIFNγ(Biosource, 0.2μg/ml)抗体を、37℃にて1時間、プレートに作用させた。抗原抗体複合体は、西洋ワサビペルオキシダーゼと結合したストレプトアビジン(1/10000希釈、Amersham)とともにインキュベートすることによって検出した。酵素活性は、基質としてテトラメチルベンジジン(TMB、Sigma)を用いて測定した。460nmでの吸光度をBiorad Novapath ELISAリーダーで測定した。サイトカイン濃度は、精製マウスIL-5またはIFNγを用いて作成した標準曲線に測定値を内挿することによって決定した。
【0110】
15. 気管支肺胞洗浄
最後のエアロゾル曝露の3日後、マウスを採血して犠牲にした。ただちに気管カニューレを通して、1mlハンクス平衡塩類溶液(HBSS)で肺を洗浄したが、この溶液は注入して3回の吸引によりそっと回収した。洗浄液を4℃にて400×gで10分間遠心した。細胞ペレットを300μlハンクス平衡塩類溶液(HBSS)に再懸濁し、トーマ(Thoma)血球計算盤で細胞を計数した。50μlのアリコートからのサイトスピン(Cytospin)標本を、示差的細胞計数のためにMay-Grunwald Giemsa染色によって染色した。
【0111】
実施例2
大腸菌におけるMBP-ProDer p 1の発現
1. MBP-ProDer p 1発現ベクターの構築
真核生物発現プラスミドpNIV 4846(ヒト化ProDer p 1 をコードするカセットを保有するpEE 14由来発現プラスミド(M.Massaer ら, International Archives of Allergy and Immunology, 2001, 125:32-43))をEag IおよびXba Iで切断後、ProDer p 1をコードする完全な合成cDNA(1-302 aa)(配列番号1)を単離した。Xba I制限切断の前に、ラージフラグメントDNAポリメラーゼ(Klenow)を用いてDNAを平滑末端化した。921 bp断片をMBP遺伝子の下流のpMAL-c2E(New England Biolabs)のAsp 718(平滑末端)-Xba I部位に挿入して、pNIV4854を導出した。配列番号1のcDNAによってコードされたProDer p 1のアミノ酸配列を、図2(配列番号2)に示す。
【0112】
2. 部位特異的変異導入
4、31または65位のDer p 1システイン残基(成熟ProDer p 1の数であり、ProDer p 1の84、111または145位に相当する)の変異導入は、プラスミドpNIV4854において、3個のシステインコドンのうちの1つを有するDNA断片を、変異を含む合成オリゴヌクレオチドで置換することにより行った。下記のオリゴヌクレオチドを使用した:
システイン残基4をアルギニンに変異させるための110bp Afl II-Sph I断片を作製(C4R)するためには、5’TTAAGACCCAGTTTGATCTCAACGCGGAGACCAACGCCCGTATCAACGGCAATGCCCCCGCTGAGATTGATCTGCGCCAGATGAGGACCGTGACTCCCATCCGCATGC3’(フォワード)および5’CGGATGGGAGTCACGGTCCTCATCTG GCGCAGATCAATCTCAGCGGGGGCATTGCCGTTGATACTACGGGCGTTGGTCTCCGCGTTGAGATCGAAACTGGGTC3’ (リバース)を使用、システイン残基31をアルギニンに変異させるための98bp Sph I-Blp I断片を作製(C31R)するためには、5’CAAGGCGGCCGTGGGTCTTGTTGGGCCTTTTCAGGCGTGGCCGCGACAG AGTCGGCATACCTCGCGTATCGGAATCAGAGCCTGGACCTCGC3’ (フォワード)および5’TCAGCGAGGTCCAGG CTCTGATTCCGATACGCGAGGTATGCCGACTCTGTCGCGGCCACGCCTGAAAAGGCCCAACAAGACCCACGGCCGCCTTGCATG3’ (リバース)を使用、システイン残基65をアルギニンに変異させるための82bp Afl II-Sph I断片を作製(C65R)するためには5’TGAGCAGGAGCTCGTTGACCGTGCCTCCCAACACGGATGTCATGGGGATACGATTCCCAGAGGTATCGAATACATCCAGCATA3’ (フォワード)および5’CTGGATGTATTCGATACCTCTGGGAATCGTATCCCCCATGACATCCGTGTTGGGAGGCACGGTCAACGCGCTCCTGC3’(リバース)を使用。
【0113】
結果として得られたプラスミドは、MBP遺伝子の下流にProDer p 1カセットを含有し、それぞれC4R、C31RおよびC65Rの変異を有しており、それぞれpNIV4870、pNIV4871およびpNIV4872と命名した。3つの変異はいずれもDNA配列決定によって検証した。C4R、C31RおよびC65R変異をそれぞれ保有する、変異したProDer p 1のアミノ酸配列を、それぞれ配列番号3、配列番号5および配列番号7に示す。これをコードする、対応する核酸配列は、配列番号4(C4R変異)、配列番号6(C31R変異)および配列番号8(C65R変異)に示す。
【0114】
3. 野生型および変異型MBP-ProDer p 1の発現および精製
異なる組換え発現ベクターを含有する大腸菌を、100μg/mlアンピシリンを含む869培地(A.Jacquet ら, Prot. Exp. Purif. 1999, 17, 392-400)中で37℃にて一晩培養した。その後細胞を1:100に希釈し、600nmでの光学密度が0.4〜0.6となるまで37℃にて増殖させた。イソプロピルβ-D-チオガラクトシド(IPTG)を終濃度0.3mMとなるように添加した。2時間インキュベートした後、細胞を、10000rpmで15分間の遠心によって回収した。
【0115】
1リットルの培養物から得られた細菌細胞ペレットを、1mMアプロチニンおよびAEBSFを含有する20mM Tris-HCl pH 7.5に再懸濁し、細胞破砕装置(Constant Systems Ltd, Warwick, UK)を用いて1800バールの圧力をかけて破砕した。溶菌液を150,000gにて60分間、超遠心した。超遠心の結果得られたペレットを20mM Tris-HCl pH 7.5で洗浄した。不溶性タンパク質は、6M尿素含有20mM Tris-HCl pH 7.5を用いて4℃にて一晩抽出した。懸濁液を150,000gで60分間、超遠心した。上清をそのまま、20mM Tris-HCl pH 7.5, 200mM NaCl, 1mM EDTAに対して一晩透析した。溶液を遠心して沈澱したタンパク質を除去し、そのまま、同じバッファーで平衡化されたアミロース樹脂(1×15cm)に加えた。A280nmがベースラインに達するまで、カラムを開始バッファーで洗浄した。カラムバッファーに10mMマルトースを添加することによってタンパク質を溶出させた。融合タンパク質を含有する画分をプールして濃縮した。精製タンパク質は−20℃で保存した。
【0116】
実施例3
CHO細胞における3種の異なるProDer p 1変異体の発現
1. 部位特異的変異導入
4、31または65位のDer p 1システイン残基の変異(成熟Der p 1の番号付けであり、ProDer p 1の84、111または145位に相当する)をプラスミドpNIV4846に導入した。プラスミドpNIV4870、pNIV4871およびpNIV4872はDer p 1カセットをMBP(実施例IIを参照されたい)遺伝子の下流に含有し、それぞれC4R、C31RおよびC65R変異を保有しているが、これらのプラスミドをそれぞれSfuI-XhoIで制限切断して、714bp断片を単離した。精製したDNA断片を、同じ制限酵素であらかじめ切断しておいたプラスミドpNIV4846に挿入した。その結果得られた、Der p 1変異体C4R、C31RおよびC65Rを含有するプラスミドをpNIV4873、pNIV4875およびpNIV4874と命名した。
【0117】
2. ProDer p 1を生成する安定なCHO-K1細胞株の一過性トランスフェクションおよび選択
プラスミドpNIV4873、pNIV4875およびpNIV4874によるDer p 1の産生を測定するために、COS細胞をリポフェクションによって一過的にトランスフェクトした。安定したDer p 1の発現のために、CHO-K1細胞をリポフェクションによって異なるプラスミドでトランスフェクトした。3週間の25μMメチオニルスルホキシミン(MSX)選択後、100μM MSX共存の下で遺伝子増幅を1ラウンド行った。
【0118】
実施例4
変性ProDer p 1はアレルギー血清に対してIgG結合反応性を示すがIgE結合反応性は示さない
ProDer p 1の変性型を低アレルゲン性ワクチンとして使用できるかを判定するために、変性(50mM βメルカプトエタノール存在下、100℃にて5分)ProDer p 1のIgGおよびIgE結合反応性をELISAテストでアッセイした。図1に示すように、変性ProDer p 1は天然型ProDer p 1に存在するIgGエピトープの大部分を保存していた。他方、この変性アレルゲンはそのIgE結合反応性の多くを失っていた。本発明者らのデータは、変性ProDer p 1がProDer p 1の低アレルゲン性変異体となりうることを示唆する。
【0119】
実施例5
MBP-ProDer p 1のIgE反応性
本実験の目的は、MBP-ProDer p 1と天然Der p 1のIgE反応性を比較することである。MBP-ProDer p 1の、アレルギー患者血清由来の特異的IgEとの反応性を、イムノプレートをDer p 1もしくはMBP-ProDer p 1で直接コーティングした直接ELISA法によって評価した。図2は、Der p 1およびMBP-ProDer p 1へのIgE結合の強い相関を示す。
【0120】
実施例6
MBP-ProDer p 1変異体のIgE結合反応性
MBP-ProDer p 1変異体のIgE結合能を、さまざまな形態のMBP-ProDer p 1でイムノプレートを直接コーティングする直接ELISAアッセイで測定した。RAST値>100 kU/Lである20人の別個のヤケヒョウヒダニ-アレルギー患者の血清から作った血清プールをアッセイに使用した。図3に示すように、変異体C31RおよびC65RのIgE結合反応性は、野生型MBP-ProDer p 1と比較して5%にまで大幅に減少していた。特筆すべきは、4位のシステイン残基をアルギニンに変異させたとき、MBP-ProDer p 1に対するIgEの反応性が認められなかった(0%残存)ことである。MBPまたは無関係なタンパク質と融合したMBPの、IgEを介した免疫認識は存在しなかったことから、IgE反応性はProDer p 1部分について特異的であった。20人の患者から得られた別の血清プールについても同様の結果が得られた。
【0121】
実施例7
さまざまな形態のProDer p 1のヒスタミン放出活性
天然Der p 1のアレルゲン活性をProDer p 1の組換え変異型誘導体の活性と比較するために、あるアレルギー患者由来の好塩基球をin vitroでさまざまな濃度のアレルゲンに曝露し、放出されるヒスタミンを測定した。図4に示すように、天然Der p 1は1ng/mlの濃度でさえも好塩基球からのヒスタミン放出を誘導することができた。対照的に、ProDer p 1の組換え変異型はこれよりも1000〜10000倍の高濃度ではじめてヒスタミンを放出させることができた。これらの結果は、ProDer p 1変異体が天然Der p 1より低いIgE結合反応性を示すことを明白に示す。
【0122】
実施例8
さまざまな形態のProDer p 1を用いた免疫原性実験
1. ハウスダストダニアレルギーの動物モデル
ハウスダストダニアレルギーの動物モデルを開発した。CBA/Jマウスに、ミョウバンをアジュバントとする精製Der p 1を注射した。1週間間隔で4回注射後、動物をヤケヒョウヒダニ抽出物による一連の気管支吸入誘発に供した(図5)。このモデルを用いて、種々の形態のDer p 1組換え体、ならびに種々のDNAをハウスダストダニアレルギーに対する予防ワクチンとしてテストした。
【0123】
実施例9
欠失型ProDer p 1の発現
1)PCRと、欠失させるべき断片の下流および上流の配列を含んでなる合成オリゴヌクレオチドを用いることによって、欠失を生じさせた。プラスミドpNIV4878(pNIV4878がProDer p 1のcDNA(ヒト化cDNA)を含有するpPIC9Kプラスミド(Invitrogen)であり、P. pastorisにおけるアレルゲンの発現に有利であることを想起されたい)のSnab I/Avr II断片を、増幅し欠失させた断片によって置き換えた。P. pastoris酵母をこの組換えプラスミドで形質転換し、ジェネティシン (G418)で選択後、0.25mg/ml G418に耐性のクローンを単離した。ProDer p 1カセットがシグナル配列の下流に位置しているものとして、メタノールで24または48時間誘導した後分泌されるProDer p 1Δ227〜240体の発現を調べた。
【0124】
2)PCRによる欠失の導入
プライマー1:5’-GCTATTACCGATACGTAGCTAGGG-3’
このプライマーは欠失すべき領域の下流のSnabI制限酵素切断部位を含む。
【0125】
プライマー2:5’-CCGTTGTCGCGATCCTTGATTCCGATGATGACAGCG-3’
このプライマーはしたがって、ProDer p 1配列のうち、欠失すべき領域の下流および上流にある部分に相同である。
【0126】
プライマー3:5’-CGGAATCAAGGATCGCGACAACGGGTATCAGCCAAACTACC-3’
この配列もまたProDer p 1配列の一部と相同であり、42bpの欠失を可能にする。加えて、これは、EcoRV切断部位の改変を可能にする点突然変異を含有する。
【0127】
プライマー4:5’-TAGGGGAGCTCAGATCTGATCCACTGAC-3’
【0128】
3)配列決定による欠失の確認
スフェロプラスト法を用いて、Pichia pastoris酵母を、あらかじめBglIIで線状とした組換えベクターによって形質転換した。形質転換体は、ヒスチジノールデヒドロゲナーゼ(His+)原栄養性について選別した。His+形質転換体のG418耐性に関するスクリーニングは、ジェネティシン(G418)(濃度を増加させていく)を含有する寒天上にクローンを播くことによって行った(0.25、1、2および4mg/ml)。30℃にて数日間インキュベートした後、数個の耐性株を得たが、それは0.25mg/ml G418の濃度についてのみであった。
【0129】
発現は、毎日、培地に0.5%メタノールを添加することによって誘導する。毎日、上清を回収するために培地を1ml採取する。ProDer p 1の発現はブロットにより示す。
【0130】
実施例10
三重変異型の形態のProDer p 1の発現
1)この実施例において、Der p 1の71、103および117位のシステイン残基をアラニンに変異させてある。Cys→Ala変異はジスルフィド架橋を壊すものの、Der p 1の構造に正電荷を導入することはない。この、より穏やかな構造破壊は、ProDer p 1の発現および分泌には影響を及ぼさないかもしれない。3つの変異を含んでなるオリゴヌクレオチドによって、これら3つの変異をPCRで導入した。
【0131】
実施例11
PreProDer p 3アレルゲンの構築
1. PreProDer p 3合成cDNAの構築
部分的に重複した10個のオリゴヌクレオチドのセットを用いて、PreProDer p 3 cDNAを合成した。これらのプライマーは、高発現の大腸菌細菌遺伝子のコドン選択に基づいて設計し、394 DNA/RNA Applied Biosystem Synthetizerによって作製した。縮重してコードされるアミノ酸は、もっとも一般的なコドンによってコードされるのではなく、個別のコドンの頻度を考慮した。たとえば、AAGまたはAAAは、高発現大腸菌細菌遺伝子においてそれぞれ21.45%および78.55%の頻度でリジン残基をコードする。このため、合成PreProDer p 3の各リジン残基にAAAコドンのみを選択するのではなく、同じコドン頻度になるように試みた。オリゴヌクレオチドは下記の通りである:
5’TCATGATCATCTACAACATTCTGATCGTACTCCTGCTGGCCATTAACACTTTGGCTAATCCGATCCTGCCGGCATCCCCGAACGCGACCATCGTTGGC 3’ (オリゴ1、コード)
5’CACCACAGAAGTGGCTACTAGACTGCAGGGAGATCTGATATGGGCACTCACCAGCCAGTGCTTTTTCGCCGCCAACGATGGTCGCG 3’ (オリゴ2、非コード)
5’GTAGCCACTTCTGTGGTGGTACTATTCTTGACGAATACTGGATCCTGACCGCGGCACACTGCGTGGCCGGCCAAACAGCGAGCAAACTCTCC 3’ (オリゴ3、コード)
5’GTCGATCTGGTAGCTATCATATTTTTCATGTGCGAAAATTTTAGCAACAGAGATCTTTTCGCCACCCAGTGAGTGTTTCAGGCTGTTGTAACGAATGGAGAGTTTGCTCGCTG 3’ (オリゴ4、非コード)
5’GATAGCTACCAGATCGACAATGACATTGCGCTGATCAAGCTGAAATCCCCTATGAAGCTGAACCAGAAAAACGCCAAAGCTGTGGGCCTGC 3’ (オリゴ5、コード)
5’CAGACGGCAGGGAGTAGCTGCCCTCTTCCAGATAACCCCAGCCAGAGACACGCACCTGGTCACCAACTTTTACATCCGAGCCTTTCGCCGGCAGGCCCACAGCTTTG 3’ (オリゴ6、非コード)
5’CTACTCCCTGCCGTCTGAATTACGCCGTGTTGATATCGCTGTGGTATCTCGCAAAGAATGTAACGAGCTGTACTCGAAAGCGAACGCTGAAGTCAC 3’ (オリゴ7、コード)
5’CCACCAGAATCGCCTTGACAAGAGTCCTTACCGCCGTTCGCAACATCACCACCGCAGATCATATTGTCGGTGACTTCAGCGTTCGC 3’ (オリゴ8、非コード)
5’CAAGGCGATTCTGGTGGGCCGGTGGTCGACGTTAAAAACAACCAGGTTGTAGGTATCGTTTCCTGGGGCTACGGTTGCGCACGTAAAGGC3’ (オリゴ9、コード)
5’AAGCTTTCAGTGGTGGTGGTGGTGGTGCTGGCTACGTTTAGATTCAATCCAATCGATAAAGTTACCAACGCGCGTGTACACACCCGGATAGCCTTTACGTGCGCAAC 3’ (オリゴ10、非コード)
【0132】
合成組換えPreProDer p 3 DNAの増幅のために、PCR反応において上記オリゴヌクレオチドを一緒にインキュベートした。Expand High Fidelity PCR System (Roche Diagnostics)を用いて下記の条件に従ってPCRを行った:30サイクル、94℃にて30秒間変性、52℃にて30秒間アニーリング、および72℃にて30秒間伸長。3’および5’末端プライマー(オリゴ1およびオリゴ10)を用いて、同じ条件で、生成産物を増幅した。その結果得られた812bp断片をpCRII-TOPOクローニングベクター(Invitrogen)にクローニングした。
【0133】
BamHIによる切断は、クローン1、3および9が正しく挿入されていることを示した。
【0134】
組換えPreProDer p 3の内部に相当するオリゴヌクレオチドを用いてインサートの配列を決定した:
5’AAGCTGAAATCCCCTATGAAGC3’ (コード)
5’CTCTTCCAGATAACCCCAGCC3’ (非コード)
【0135】
クローン3のみが正しいことが判明したが、5’コード末端の最初の6塩基は欠けていた。欠失塩基の付加は、2つの新たなオリゴヌクレオチドの使用によって達成した:
5’TTTTATTCATGATCATCTACAACATTCTGATCC3’ (オリゴ11、コード)
5’GATGCATGCTCGAGCGGC3’ (オリゴ12、非コード)
【0136】
オリゴヌクレオチドを、不完全なPreProDer p 3配列を保有するクローン3のDNAとともにインキュベートした。合成遺伝子の増幅は、Expand High Fidelity PCR Systemを用いたPCR反応(25サイクル、94℃にて30秒間変性、53℃にて30秒間アニーリング、および72℃にて30秒間伸長)によって達成した。その結果得られた断片を、pCRII-TOPOにクローニングした。PreProDer p 3 cDNAは、二重Rca I-XhoI制限切断後に単離し、NcoIおよびXhoIで切断されたpET15b発現ベクターにクローニングした。コンピテントAD494(DE3)pLys大腸菌細胞を、上記の結果得られたプラスミドによって形質転換し、ProDer p 3の発現を検出するために終濃度1mMのイソプロピル-チオガラクトシド(IPTG)(Duchefa)を培地に添加した。
【0137】
推定Der p 3シグナルペプチドの欠失は、PCRによって、2つの新たなプライマー5’CATATGAATCCGATCCTGCCGGCATCCCC3’ (オリゴ13、コード) および5’GGATCCTCACTGGCTACGTTTAGATTCAATCC3’ (オリゴ14、非コード)を使用して行った。ProDer p 3 cDNAの増幅は、Taqポリメラーゼ(Roche Diagnostics)を用いて15サイクルのPCRで行った:97℃にて30秒間変性、65℃にて30秒間アニーリング、および72℃にて1分間伸長。得られた750bp断片をpCRII-TOPOクローニングベクター(Invitrogen)にクローニングした。得られたプラスミドによって最もコンピテントな10個の大腸菌を形質転換した。9個のクローンは陽性に挿入されているようであった;EcoRIによる切断はクローン1、4、8が正しく挿入されていることを証明したが、一方、配列決定は、クローン4のみが正しい配列を有することを明らかにした。NdeIおよびXhoIによる切断後、ProDer p 3 cDNAを単離して、同じ酵素で切断したpET15bにクローニングした。その結果得られたプラスミドでBL21およびBL21 Star大腸菌(Invitrogen)株を形質転換した。培地へのIPTGの添加は、(His)6タグをN末端に保有する組換えProDer p 3の発現を誘導した。
【0138】
2. 大腸菌における組換えアレルゲンの発現
最も生産性の高い組換え大腸菌BL21 STARクローンを、100μg/mlアンピシリン(Pentrexyl)を含有する2リットルの液体869培地で培養した(37℃/250rpm)。培養物の620nmでの吸光度が0.5に達したとき、終濃度1mMのイソプロピル-チオガラクトシド(Duchefa)を添加することによって3時間、発現を誘導した。次いで、培養物を回収して11000gで遠心し−20℃で保存した。細菌ペレットを回収し、アプロチニン1/1000(Sigma)およびAEBSF 1/500(ICN)を添加した40mlの20mM Trisバッファー、pH7.5中に再懸濁した。続いて1500バールで細胞を破砕して−20℃で保存した。
【0139】
3. 破砕した大腸菌からの組換えProDer p 3の精製
回収した大腸菌細胞を20mM Trisバッファー、pH7.5、1mM アプロチニンおよび1mM AEBSFに再懸濁し、これを、1800キロバールの圧力下に細胞破砕装置(Cell D)で溶菌させた。溶菌液を149000gで1時間超遠心した。上清を除去し、組換えProDer p 3を含むペレットを次に、6M塩酸グアニジンを含有する50mM Tris-HClバッファー、pH7.5、40mlで4℃にて一晩抽出した。超遠心(45分間、149000g)後、抽出物の上清を3ml/分の流速で、抽出バッファーで平衡化済みのNickel-NTA Superflow カラム(1.6x5cm, Qiagen)に供した。カラムを0.5M PBS NaCl, pH7.5を用いて1.5ml/分で洗浄し、結合したタンパク質を復元させた。組換えProDer p 3はコンディショニングバッファーに200mMのイミダゾールを添加することによって4ml/分の流速で、溶出させた。組換えProDer p 3を含有する画分をプールし、限外濾過(Amicon-Millipore 再生セルロース限外濾過膜、NMWL 10kDa)によって濃縮した。このステップの間にバッファーをPBS, pH7.3で交換した。精製したタンパク質は−20℃で保存した。
【0140】
4. 天然ダニ全体抽出物からの天然Der p 3の精製
ヤケヒョウヒダニ抽出物を、最終濃度60%飽和硫酸アンモニウム沈澱に供した。超遠心(45分、149000g)後、上清を、2ml/分で、50mM Trisバッファー、0.5M NaCl、pH7.4で平衡化されたBenzamidine Sepharose 4 fast flow カラム(1.6x5cm, Pharmacia)に供した。Der p 3を、50mMグリシン-HClバッファー、pH2.5によってカラムから溶出させ、1mlの各フラクションを、75μl 1M Tris、pH9.5の添加によりただちに中和した。Der p 3を含有する画分をプールし濃縮して(Amicon-Millipore 再生セルロース限外濾過膜、NMWL 10kDa)、0.5ml/分にて、PBS pH7.3で平衡化済みのSuperdex 75 ゲル濾過クロマトグラフィーカラム (Pharmacia)に供した。精製Der p 3を濃縮し、−20℃にて保存した。
【0141】
5. SDS-PAGEおよびウェスタンブロット分析
タンパク質を12.5%ポリアクリルアミドゲルでのSDS-PAGEによって分析した。電気泳動後、タンパク質をセミドライ転写システム(Sigma-Aldrich)を用いてニトロセルロース膜に転写した。膜を、TBS-T(50mM Tris HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)中0.5% Instagel(PB Gelatins)で30分間飽和させ、ProDer p 3に対して生じたマウスポリクローナル血清をブロッキング溶液中に1:2500希釈したものとともにインキュベートした。アルカリホスファターゼ結合ヤギ抗マウス抗体(Promega、1:7500)および5-ブロモ-4-クロロ-3-インドリルリン酸(BCIP, Boehringer)/ニトロブルーテトラゾリウム(NBT, Sigma)を基質として用いて、免疫反応性物質を検出した。
【0142】
6. タンパク質定量
総タンパク質濃度は、標準タンパク質としてウシ血清アルブミンを用いてビシンコニン酸法(MicroBCA, Pierce)によって測定した。
【0143】
7. IgE結合活性
イムノプレートをDer p 3または組換えProDer p 3(1ウェル当たり500ng)で一晩4℃にてコーティングした。その後1ウェル当たり100μlのTBS-Tweenバッファー(50mM Tris-HCl pH 7.5, 150mM NaCl, 0.1% Tween 80)で5回洗浄し、1% BSA(Sigma)を添加した150μl同じバッファーで37℃にて1時間飽和させた。ヤケヒョウヒダニに対するアレルギー患者から得られた血清を1/8に希釈した後、37℃で1時間インキュベートした。実験に使用した47血清のうち、5血清はその特異的抗ヤケヒョウヒダニIgE値(RASTアッセイ)が0.7kU/L〜28.9kU/L、8血清は68.3kU/L〜94.1kU/Lであり、34血清は上限カットオフ値の100kU/Lを越えていた。アレルゲン-IgE複合体は、プレートをTBS-Tweenバッファーで5回洗浄し、ビオチン結合マウス抗ヒトIgE抗体(TBS-Tバッファーで1/2000に希釈、Southern Biotechnology Associates)およびストレプトアビジン-西洋ワサビペルオキシダーゼ(TBS-Tweenバッファーで1/1000に希釈、Amersham Life Science)とともにインキュベートした後検出した。酵素活性は、3,3’,5,5’-テトラメチルベンジジン(TMB) (Sigma)を用いて測定した。OD450nmはBiorad Novapath ELISAリーダーで測定した。
【0144】
8. 酵素アッセイ
総容量200μlで25℃にて、50mM Tris-HCl, pH 8において酵素アッセイを行った。N-α-ベンゾイル-L-アルギン-p-ニトロアニリド(Sigma)(終濃度1mM)の加水分解を、Biorad Novapath ELISAリーダーで405mにて測定した。アッセイは終濃度6.25μg/mlのアレルゲンの添加によって開始した。
【0145】
9. Der p 3のクローニングおよび発現
大腸菌における発現
Der p 3の未成熟型である、ProDer p 3を細菌で発現させた。12の合成オリゴヌクレオチドを用いて、かつ細菌における発現に向けて最適化したコドンを使用することにより、PreProDer p 3のcDNAを完全に合成した(図2)。ProDer p 3をコードするカセットを、発現ベクターpET-15bにおいてポリヒスチジンテールをコードする配列の下流にクローニングした。このベクターは、培地へのIPTGの添加によって誘導されるT7プロモーターを含有している。まず、ProDer p 3の発現条件の最適化を試みた。これを行うために、組換え細菌の培養を30℃および37℃で、2通りの誘導試薬濃度(IPTG 0.5および1 mM)で、1、2、3および16時間、行った。それぞれの細菌ペレットをフレンチプレスで溶菌した。溶菌液を20000 rpmで20分間遠心した。上清(細胞質画分)および/またはペレット(細胞片および不溶性生成物)中のProDer p 3の存在を、組換えProDer p 3のポリヒスチジンテールに対するマウス抗体を用いたウェスタンブロットによって明らかにした(図3)。
【0146】
発現試験は、温度およびIPTG濃度が、ProDer p 3の生成にごくわずかしか影響を及ぼさないことを示す。ProDer p 3は本質的に不溶性の形で発現され、その発現は、誘導時間2〜3時間が最適である。16時間誘導については可溶性型のProDer p 3が存在しないことに留意すべきである。クーマシーブルー染色後に検出されるので、ProDer p 3は発現されている。SDS-PAGEにおいて、このタンパク質は±32kDaのバンドとなっている(図4)。
【0147】
10. ProDer p 3の精製
組換えProDer p 3の精製プロトコールを開発した。ProDer p 3が不溶性の状態で発現されるならば、その組換えアレルゲンは変性条件(20mM Tris-HCl、6M塩酸グアニジン、pH7.5)でペレットを抽出することによって可溶化される。抽出の収率は80〜90%程度である。抽出物を、抽出バッファーで充填された約10mlのNi2+キレーティングセファロース樹脂カラム(直径1.6cm、高さ5cm)にかける。抽出バッファーでカラムを洗浄した後、付着したタンパク質は、PBSバッファー0.5M NaCl pH7.5を用いた直線再生濃度勾配によってカラム上で直接再生させる。濃度勾配をかける液量は200mlである。
【0148】
その後、溶出は、再生バッファー中のイミダゾール濃度を増加させて(20、50、100および200mM)バッファーを供することにより行われる。SDS-PAGEによる分析は、当該タンパク質が流出液中に存在せず、夾雑物がイミダゾール濃度20 mMで溶出するが、ProDer p 3はイミダゾール60mMから100mMの間で溶出することを示している。ProDer p 3はすべてクロマトグラフィー担体から解離させた(図10を参照されたい)。
【0149】
マイクロシークエンシングによってProDer p 3のN末端配列の分析を行った。その配列はヒスチジンテールと一致した。ベクターpET-15bにおいて、ProDer p 3のクローニング部位はヒスチジンテールをコードする配列の下流にあることに留意されたい。ProDer p 3とヒスチジンテールはトロンビン切断部位により分離されている。ProDer p 3の確実性を検証する目的で、ヒスチジンテールを取り除くために精製タンパク質をトロンビンで処理した。切断されたProDer p 3をもう一度マイクロシークエンシングすることによって、Der p 3のプロペプチドのN末端配列を明らかにした(N P I L P A S P N A T)。
【0150】
酵素活性:ProDer p 3は、天然Der p 3 が切断できる基質BAPNAに対して不活性である。
【0151】
11. IgE反応性
アレルゲンの直接コーティング
この予備的な結果は、ダニアレルギー患者のIgEについて、ProDer p 3の反応性がDer p 3と比べて低いことを示しているようである(図11)。
【0152】
2. ワクチン製剤
【表1】
【0153】
3. 抗体応答-結果
天然Der p 1を4回注射することによって免疫したマウスは、高力価のIgGおよびIgG1、低力価のIgG2ならびに大量のIgE抗体を産生したが、これは、天然Der p 1が強力なTh2免疫応答を誘導することを示唆する(表2および4)。
【0154】
抗Der p 1 IgGおよびIgG1抗体応答は天然型または変性型ProDer p 1を注射したマウスでも強力であった。天然型ProDer p 1を注射後、IgG2a力価はDer p 1を用いて得られた力価よりわずかに高くなったが、IgE力価はDer p 1で得られた力価と同程度かそれよりわずかに低くなった。天然型ProDer p 1で免疫したマウスとは対照的に、変性型ProDer p 1を注射した動物は、高いIgG2a力価を産生し、IgE抗体の産生は非常に低かった。予想通り、ミョウバンなしでのProDer p 1による免疫化は弱い免疫応答しか誘導しなかった(表4)。
【0155】
MBP-ProDer p 1野生型(WT)、C4R、C31RおよびC65Rで感作されたマウスは、特異的IgGおよびIgG1抗体の同様の産生を示した(表3)。最も高いIgG2a力価はMBP-ProDer p 1 WTおよびC31Rで免疫されたグループで観察された。特異的IgE力価は、MBP-ProDer p 1のどの変異体を注射してた場合でも低かった。ProDer p 1をコードするプラスミドでマウスを免疫した後にも同様の結果が得られた。
【表2】
【0156】
【表3】
【0157】
【表4】
【0158】
4. T細胞増殖応答-結果
エアロゾル投与の前(対照)および後に、免疫したマウスから分離された脾細胞の、ProDer p 1またはヤケヒョウヒダニ抽出物での刺激によるT細胞増殖応答を調べた。結果を表5(刺激指数)および表6(サイトカイン)に示す。
【0159】
さまざまな組換えProDer p 1変異体で免疫されたマウスにおいてアレルゲン特異的T細胞応答が検出された。脾細胞をProDer p 1で再刺激した場合に最も強い応答が観察された。T細胞反応性は投与とは無関係であると思われた。
【0160】
表5のこうした結果は、さまざまな形態のProDer p 1が共通のT細胞エピトープをDer p 1と共有していることを示した。さらに、熱変性または部位特異的変異導入によるProDer p 1の構造破壊は、ProDer p 1のT細胞反応性を変化させなかったので、こうした形態の変異体が、T細胞応答を刺激しうるIgE結合反応性の非常に弱い低アレルゲンとなることが確認された。
【表5】
【0161】
再刺激された脾細胞の培養上清におけるサイトカインIL-5およびIFNγの存在は、ELISAで測定した(表6)。[IFNγ]/[IL-5]比を比較したならば、ミョウバンをアジュバントとする天然Der p 1またはProDer p 1によるワクチン接種はIFNγよりもIL-5の産生をより強く誘導すると結論付けることができる。変性型ProDer p 1のみならずさまざまな形態のMBP- ProDer p 1(変異体および野生型)も、同程度のレベルの2つのサイトカインを誘導した。
【表6】
【0162】
5. 気管支肺胞洗浄−結果
天然Der p 1による感作とそれに続くエアロゾル化したハウスダストダニ抽出物への曝露は、非常に多数の気管支肺胞細胞を生じた(表7)。エアロゾル化したハウスダストダニ抽出物への7回の曝露は、Der p 1によるワクチン接種を受けた動物においてのみ、気道の好酸球増加を引き起こすことが示された。この群において、Der p 1に感作された動物がハウスダストダニ抽出物の噴霧を受けないか、またはエアロゾル化したPBSに曝露された場合は、気道の好酸球増加は認められなかった。
【0163】
さまざまな組換え型のProDer p 1によるワクチン接種は、エアロゾル化したハウスダストダニ(HDM)抽出物に曝露した後も、気道の好酸球増加を防止した。
【表7】
【0164】
実施例12
核酸ワクチン接種(NAVAC)用発現プラスミド
1. 核酸ワクチン接種のためのProDer p 1をコードするプラスミドの構築
ProDer p 1をコードするカセット(1-302aa)をプラスミドpNIV4846(上記参照)から切り出し、HindIIIおよびBglIIで制限切断して、あらかじめHindIIIおよびBglIIで切断しておいたプラスミドpJW4304に挿入した。その結果得られたプラスミドをpNIV4868と命名し、DNA配列決定によって確認した。
【0165】
2. 部位特異的変異導入
4、31または65位(これは成熟Der p 1の番号付けであり、ProDer p 1における84、111または145位に相当する)のProDer p 1システイン残基の変異を、プラスミドpNIV4868に導入した。プラスミドpNIV4870、pNIV4871およびpNIV4872は、ProDer p 1カセットをMBP遺伝子の下流に含有し、それぞれ変異C4R、C31RおよびC65Rを保有するものであるが、各々をAflII-BamHIで制限切断して699bpの断片を単離した。pNIV4868をAflII-HpaIで切断して480bpの断片を単離した。2つの精製DNA断片を、あらかじめHpaI-BamHIで切断したプラスミドpJW4304に挿入した。その結果得られたプラスミドはProDer p 1変異体C4R、C31RおよびC65Rを有しており、pNIV4879、pNIV4880およびpNIV4881と命名した。
【0166】
実施例13
Pichia pastorisにおけるProDer p 1の発現
1. ProDer p 1発現ベクターの構築
pNIV4868に由来するProDer p 1をコードするカセット(最適化された哺乳動物コドン使用頻度を有する全長1-302aa ProDer p 1 cDNA、)を以下のプライマーを用いてPCRによって増幅した:5’ACTGACAGGCCTCGGCCGAGCTCCATTAA3’ (下線はStuI 制限部位、フォワード)および5’CAGTCACCTAGGTCTAGACTC GAGGGGAT3’ (下線はAvrII 制限部位、リバース)。増幅された断片をpCR2.1 TOPOクローニングベクターにクローニングした。DNA配列決定によって正しいProDer p 1カセットを確認した。組換えTOPOベクターをStuI-AvrIIで切断し918bp断片を生じたが、これをSnaBI-AvrIIで制限切断したpPIC9K発現ベクターに導入した。得られたプラスミド、pNIV4878はProDer p 1カセットをS. cerevisiae α因子の下流に含有する。
【0167】
2. 部位特異的変異導入
非グリコシル化ProDer p 1(成熟Der p 1の数でN52Q)生産のための発現プラスミドを、4つのプライマーのセットを用いたオーバーラップ伸長(overlap extension)PCRによってpNIV4878から導出した。ProDer p 1アミノ酸配列71〜176をコードする317bp断片を増幅するために、以下のプライマー:5’GGCTTTCGAACACCTTAAGACCCAG3’ (プライマー1, 下線はAflII制限部位、フォワード)および5’GCTCCCTAGCTACGTA TCGGTAATAGC3’ (プライマー2, 下線はSnaBI制限部位、リバース)を用いた。
【0168】
ProDer p 1配列に変異N52Qを導入するために、以下のプライマー、5’CCTCGCGTATCGGCAACAGAGCCTGGACC3’ (プライマー3, 下線は変異N52Q、フォワード)および5’GGTCCAGGCTCTGTTGCC GATACGCGAGG3’ (プライマー4, 下線は変異N52Q、リバース)を用いた。
【0169】
変異した317bp AflII-SnaBI断片を、3段工程によって作製した。第1のPCRにおいて、プライマー1および4をpNIV4878と混合して〜200bpの断片を生成した。第2のPCRにおいて、プライマー2および3をpNIV4878と混合して〜140bp断片を生成した。2つのPCR産物をアガロースゲルで精製して、鋳型として3回目のPCRに使用し、〜340bp断片を得た。この精製断片をpCR2.1 TOPOクローニングベクター(Invitrogen)にクローニングした。変異はDNA配列決定によって確認した。組換えTOPOベクターをAflII-SnaBIで切断して317bp断片を生成し、これを同様に切断消化したpNIV4878に連結した。得られたプラスミド、pNIV4883はS. cerevisiae α因子の下流にProDer p 1 N52Qを含有する。
【0170】
Der p 1の4、31または65位(成熟Der p 1での番号付け)のシステイン残基の変異を有するProDer p 1の非グリコシル化変異体を得るために、プラスミドpNIV4873、pNIV4875およびpNIV4874についても同じプライマーのセットを用いてオーバーラップ伸長PCRを行った。その結果得られたプラスミドpNIV4884、4885および4886はそれぞれ、ProDer p 1 N52Q C4R、N52Q C31RおよびN52Q C65Rをコードする。
【0171】
3. P. pastorisの形質転換
スフェロプラスト形質転換法を用いてプラスミドpNIV4878をP. pastorisに導入した。形質転換体は、ヒスチジノールデヒドロゲナーゼ(His+)原栄養性で選択した。geneticin(G418)耐性についてのHis+形質転換体のスクリーニングは、含有するG418の濃度を増加させた寒天上にクローンを播くことによって、実施した。
【0172】
ProDer p 1 N52Q、ProDer p 1 N52Q C4R、N52Q C31RおよびN52Q C65Rをコードするプラスミドによる形質転換は、同様の方法によって行った。
【0173】
4. 組換え酵母によるProDer p 1の産生
G418耐性クローンをBMG培地中で30℃にてOD600nmが2〜6となるまで増殖させた。細胞を遠心によって回収し、OD600nmが1となるように100mlのBMG培地中に再懸濁した。6日間毎日0.5%メタノールを添加することによりProDer p 1の発現を誘導した。上清を遠心によって回収し、精製するまで−20℃で保存した。
【0174】
5. 酵母培養上清からのProDer p 1の精製
上清は水で10倍に希釈し、pHを9に調整した後、直接20mM Tris-HCl, pH9で平衡化済みのQ sepharoseカラムにかけた。カラムを開始バッファーで洗浄した。バッファー中のNaCl濃度を段階的に増加させることによってタンパク質の溶出を進行させた。ProDer p 1で富化された画分をプールし、Filtron膜(Omega Serie, カットオフ:10kD)による限外濾過によって濃縮した。ProDer p 1の精製は、PBS pH7.3で平衡化済みのsuperdex-75カラム(1 x 30cm, Pharmacia)でのゲル濾過クロマトグラフィーによって達成した。精製されたProDer p 1は濃縮し、−20℃で保存した。
【0175】
配列情報
【図面の簡単な説明】
【0176】
【図1】CHO細胞において発現された変性ProDer p 1のIgG-およびIgE-結合反応性。イムノプレートを500ng/ウェルの精製された天然もしくは変性ProDer p 1でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験(radioallergosorbent)に陽性の血清(1:8に希釈)とともにインキュベートした。結合したIgEもしくはIgGは、マウス抗ヒトIgEもしくはIgGおよびアルカリホスファターゼ標識抗マウスIgG抗体とともにインキュベートした後、酵素的にアッセイすることによって定量した。結果をOD410nm値として表示する。
【図2】MBP-ProDer p 1および天然Der pのIgE反応性の相関。イムノプレートを1ウェル当たり500ngの精製済みDer pもしくはMBP-ProDer p 1でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験に陽性の95血清(1:8に希釈)とともにインキュベートした。結合したIgEは、マウス抗ヒトIgEおよびアルカリホスファターゼ標識抗マウスIg抗体とともにインキュベートした後、酵素的にアッセイすることによって定量した。結果をOD410nm値として表示する。
【図3】C4R、C31RおよびC65Rの変異を有するMBP-ProDer p 1変異体のIgE結合反応性。イムノプレートを500ng/ウェルの野生型もしくは変異体MBP-ProDer p 1でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験に陽性の20血清(1:8に希釈)のプールとともにインキュベートした。結合したIgEは、マウス抗ヒトIgEおよびアルカリホスファターゼ標識抗マウスIgG抗体とともにインキュベートした後、酵素的にアッセイすることによって定量した。結果をOD410nm値として表示する。
【図4】アレルゲンのヒスタミン放出活性。アレルギーのある提供者の末梢血から分離された好塩基球を、種々のアレルゲンの連続希釈物で刺激した。細胞からのヒスタミンの放出はELISAによって測定した。好塩基球におけるヒスタミンの総量は、界面活性剤IGEPAL CA-630を用いて細胞を破壊した後、定量した。結果を、アレルゲンにより放出されたヒスタミンと、総ヒスタミンとの比として示す。
【図5】ハウスダストダニアレルギーの動物モデルの模式図。
【図6】メタノールで24および48時間誘導した後のP. pastorisにおけるProDer p 1Δ227〜240体の発現。組換えクローンの培養上清をSDS PAGEによって分析した。ブロットは、CHOにおいて発現されたProDer p 1に対するポリクローナルマウス血清を用いて明らかにした。レーン1,2: 無関係なタンパク質、レーン3: 誘導されていない酵母、レーン4: 精製ProDer p 1、レーン5: 24時間誘導後のクローン1、レーン6: 48時間誘導後のクローン1、レーン7: 24時間誘導後のクローン2、レーン8: 48時間誘導後のクローン2.
【図7】野生型(AcaNucSeq)および細菌コドン最適化(EcoNucSeq)PreProDer p 3の配列比較。各コドンの下に示す推定アミノ酸配列は、一文字記号で表記した。リーダーペプチドおよびプロペプチド配列は、それぞれイタリックおよび下線によって示す。
【図8】ProDer p 3の発現。0.5もしくは1 mM IPTGを用いて1、2、3および16時間誘導した後、細菌を破砕した。細胞質画分(S)および不溶性画分(C)を、ProDer p 3の存在についてウェスタンブロットで解析した。
【図9】SDS-PAGEにおけるProDer p 3のクーマシーブルー染色後の検出。レーン1:細胞質画分レーン2:不溶性画分の洗液レーン3:不溶性画分矢印はProDer p 3の位置を示す。
【図10】Ni2+-NTA クロマトグラフィーによるProDer p 3の精製。開始バッファー中の濃度を次第に増加させたイミダゾールを添加することによって、結合したタンパク質を溶出した。画分をSDS-PAGE分析したが、左図はクーマシーブルー染色後の分析であり、右図は抗His抗体を用いたウェスタンブロットによる分析である。FTはフロースルー(flow-through;通り抜け)の略である。20、40、45、60、100はイミダゾール濃度(mM)を表す。Rは精製後のNi2+NTA樹脂を表す。
【図11】組換えProDer p 3および天然Der p 3のIgE反応性の相関。イムノプレートを1ウェル当たり500ngの精製済み天然Der p 3もしくは組換えProDer p 3でコーティングし、ヤケヒョウヒダニに対して放射性アレルゲン吸着試験に陽性の血清(1:8に希釈)とともにインキュベートした。結合したIgEは、マウスビオチン標識抗ヒトIgEおよびアルカリホスファターゼ標識ストレプトアビジンとともにインキュベートした後、酵素的にアッセイすることによって検出した。結果をOD410nm値として表示する。
【特許請求の範囲】
【請求項1】
組換えヤケヒョウヒダニ(Dermatophagoides pteronyssinus)Der p 1タンパク質アレルゲン誘導体において、そのアレルゲン誘導体が野生型アレルゲンに比べて著しく低下したアレルゲン活性を有し、その誘導体が遺伝的に変異しており、さらにその変異体が3つの次のような変異:システイン71残基の変異、システイン103残基の変異、およびシステイン117残基の変異を含んでなる、前記組換えヤケヒョウヒダニDer p 1タンパク質アレルゲン誘導体。
【請求項2】
組換えヤケヒョウヒダニ(Dermatophagoides pteronyssinus)Der p 1タンパク質アレルゲン誘導体において、そのアレルゲン誘導体が野生型アレルゲンに比べて著しく低下したアレルゲン活性を有し、その誘導体が遺伝的に変異しており、さらにその変異体がDer p 1のアミノ酸残基147〜160またはProDer p 1の残基227〜140の欠失を含んでなる、前記組換えヤケヒョウヒダニDer p 1タンパク質アレルゲン誘導体。
【請求項3】
配列番号15および配列番号17からなる群から選択される配列のいずれかを有する組換え変異型アレルゲン。
【請求項4】
請求項1〜3のいずれか1項に記載の変異型アレルゲンをコードする、単離された核酸分子。
【請求項5】
コドン使用頻度パターンが高発現哺乳動物遺伝子のパターンに類似している、請求項4に記載の核酸配列。
【請求項6】
請求項4または5に記載の核酸を含有する発現ベクター。
【請求項7】
請求項4もしくは5に記載の核酸配列によって、または請求項6に記載のベクターによって形質転換された宿主細胞。
【請求項8】
請求項1〜3のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲン、または請求項4〜7に記載のコードするポリヌクレオチド、および、場合によってアジュバントを含んでなる、免疫原性組成物。
【請求項9】
アジュバントがTh1型免疫応答を優先的に刺激する物質である、請求項8に記載の免疫原性組成物。
【請求項10】
アジュバントが、3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルまたはエステルの1種以上、あるいはこれらのアジュバントの2種以上の組み合わせを含んでなる、請求項8または9に記載の免疫原性組成物。
【請求項11】
アレルゲンが水中油型または油中水型エマルションビヒクル中で与えられる、請求項8〜10のいずれか1項に記載の免疫原性組成物。
【請求項12】
医薬用の、請求項8〜11のいずれか1項に記載の免疫原性組成物。
【請求項13】
請求項1〜3のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲンの、アレルギー治療用医薬の製造における使用。
【請求項14】
アレルギー反応に苦しむ患者を治療する、またはアレルギー反応を起こしやすい患者を予防する方法であって、請求項8〜12のいずれか1項に記載の免疫原性組成物を前記個体に投与することを含んでなる、前記方法。
【請求項15】
Der p 3と比較して著しく低下したアレルゲン活性を有する、ヤケヒョウヒダニProDer p 3もしくはPreProDer p 3タンパク質アレルゲン、またはその誘導体。
【請求項16】
前記アレルゲンもしくはその誘導体が熱処理されている、請求項15に記載のアレルゲンもしくはその誘導体。
【請求項17】
前記アレルゲンもしくはその誘導体が遺伝的に変異している、請求項15に記載のアレルゲンもしくはその誘導体。
【請求項18】
変異がシステイン残基の変異を含んでなる、請求項17に記載のアレルゲンもしくはその誘導体。
【請求項19】
配列番号19の配列を有する、組換えアレルゲン。
【請求項20】
請求項17〜19のいずれか1項に記載の変異型アレルゲンをコードする、単離された核酸分子。
【請求項21】
配列番号20または21のいずれかの配列を有する、単離された核酸分子。
【請求項22】
請求項20または21に記載の核酸を含有する、発現ベクター。
【請求項23】
請求項20もしくは21に記載の核酸配列によって、または請求項22に記載のベクターによって、形質転換された宿主細胞。
【請求項24】
請求項15〜19のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲン、または請求項20もしくは21に記載のコードするポリヌクレオチド、および、場合によって、アジュバントを含んでなる、免疫原性組成物。
【請求項25】
アジュバントがTh1型免疫応答を優先的に刺激する物質である、請求項24に記載の免疫原性組成物。
【請求項26】
アジュバントが、3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルもしくはエステルの1種以上、またはこれらのアジュバントの2種以上の組み合わせを含んでなる、請求項24または25に記載の免疫原性組成物。
【請求項27】
アレルゲンを水中油型または油中水型エマルションビヒクル中で提供する、請求項24〜26のいずれか1項に記載の免疫原性組成物。
【請求項28】
医薬用の、請求項24〜27のいずれか1項に記載の免疫原性組成物。
【請求項29】
請求項15〜19のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲンの、アレルギー治療用医薬の製造における使用。
【請求項30】
アレルギー反応に苦しむ患者を治療する、またはアレルギー反応を起こしやすい患者を予防する方法であって、請求項24〜28のいずれか1項に記載の免疫原性組成物を前記個体に投与することを含んでなる、前記方法。
【請求項1】
組換えヤケヒョウヒダニ(Dermatophagoides pteronyssinus)Der p 1タンパク質アレルゲン誘導体において、そのアレルゲン誘導体が野生型アレルゲンに比べて著しく低下したアレルゲン活性を有し、その誘導体が遺伝的に変異しており、さらにその変異体が3つの次のような変異:システイン71残基の変異、システイン103残基の変異、およびシステイン117残基の変異を含んでなる、前記組換えヤケヒョウヒダニDer p 1タンパク質アレルゲン誘導体。
【請求項2】
組換えヤケヒョウヒダニ(Dermatophagoides pteronyssinus)Der p 1タンパク質アレルゲン誘導体において、そのアレルゲン誘導体が野生型アレルゲンに比べて著しく低下したアレルゲン活性を有し、その誘導体が遺伝的に変異しており、さらにその変異体がDer p 1のアミノ酸残基147〜160またはProDer p 1の残基227〜140の欠失を含んでなる、前記組換えヤケヒョウヒダニDer p 1タンパク質アレルゲン誘導体。
【請求項3】
配列番号15および配列番号17からなる群から選択される配列のいずれかを有する組換え変異型アレルゲン。
【請求項4】
請求項1〜3のいずれか1項に記載の変異型アレルゲンをコードする、単離された核酸分子。
【請求項5】
コドン使用頻度パターンが高発現哺乳動物遺伝子のパターンに類似している、請求項4に記載の核酸配列。
【請求項6】
請求項4または5に記載の核酸を含有する発現ベクター。
【請求項7】
請求項4もしくは5に記載の核酸配列によって、または請求項6に記載のベクターによって形質転換された宿主細胞。
【請求項8】
請求項1〜3のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲン、または請求項4〜7に記載のコードするポリヌクレオチド、および、場合によってアジュバントを含んでなる、免疫原性組成物。
【請求項9】
アジュバントがTh1型免疫応答を優先的に刺激する物質である、請求項8に記載の免疫原性組成物。
【請求項10】
アジュバントが、3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルまたはエステルの1種以上、あるいはこれらのアジュバントの2種以上の組み合わせを含んでなる、請求項8または9に記載の免疫原性組成物。
【請求項11】
アレルゲンが水中油型または油中水型エマルションビヒクル中で与えられる、請求項8〜10のいずれか1項に記載の免疫原性組成物。
【請求項12】
医薬用の、請求項8〜11のいずれか1項に記載の免疫原性組成物。
【請求項13】
請求項1〜3のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲンの、アレルギー治療用医薬の製造における使用。
【請求項14】
アレルギー反応に苦しむ患者を治療する、またはアレルギー反応を起こしやすい患者を予防する方法であって、請求項8〜12のいずれか1項に記載の免疫原性組成物を前記個体に投与することを含んでなる、前記方法。
【請求項15】
Der p 3と比較して著しく低下したアレルゲン活性を有する、ヤケヒョウヒダニProDer p 3もしくはPreProDer p 3タンパク質アレルゲン、またはその誘導体。
【請求項16】
前記アレルゲンもしくはその誘導体が熱処理されている、請求項15に記載のアレルゲンもしくはその誘導体。
【請求項17】
前記アレルゲンもしくはその誘導体が遺伝的に変異している、請求項15に記載のアレルゲンもしくはその誘導体。
【請求項18】
変異がシステイン残基の変異を含んでなる、請求項17に記載のアレルゲンもしくはその誘導体。
【請求項19】
配列番号19の配列を有する、組換えアレルゲン。
【請求項20】
請求項17〜19のいずれか1項に記載の変異型アレルゲンをコードする、単離された核酸分子。
【請求項21】
配列番号20または21のいずれかの配列を有する、単離された核酸分子。
【請求項22】
請求項20または21に記載の核酸を含有する、発現ベクター。
【請求項23】
請求項20もしくは21に記載の核酸配列によって、または請求項22に記載のベクターによって、形質転換された宿主細胞。
【請求項24】
請求項15〜19のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲン、または請求項20もしくは21に記載のコードするポリヌクレオチド、および、場合によって、アジュバントを含んでなる、免疫原性組成物。
【請求項25】
アジュバントがTh1型免疫応答を優先的に刺激する物質である、請求項24に記載の免疫原性組成物。
【請求項26】
アジュバントが、3D-MPL、QS21、CpGオリゴヌクレオチド、ポリエチレンエーテルもしくはエステルの1種以上、またはこれらのアジュバントの2種以上の組み合わせを含んでなる、請求項24または25に記載の免疫原性組成物。
【請求項27】
アレルゲンを水中油型または油中水型エマルションビヒクル中で提供する、請求項24〜26のいずれか1項に記載の免疫原性組成物。
【請求項28】
医薬用の、請求項24〜27のいずれか1項に記載の免疫原性組成物。
【請求項29】
請求項15〜19のいずれか1項に記載の組換えタンパク質もしくは変異アレルゲンの、アレルギー治療用医薬の製造における使用。
【請求項30】
アレルギー反応に苦しむ患者を治療する、またはアレルギー反応を起こしやすい患者を予防する方法であって、請求項24〜28のいずれか1項に記載の免疫原性組成物を前記個体に投与することを含んでなる、前記方法。
【図1】

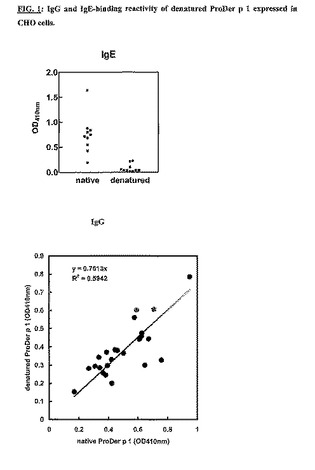
【図2】
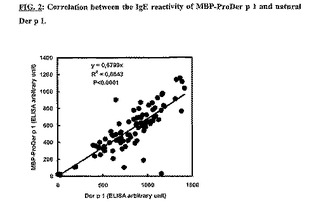
【図3】
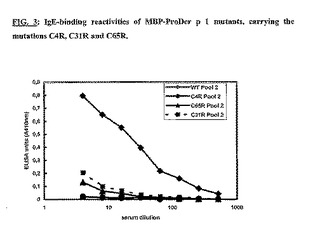
【図4】
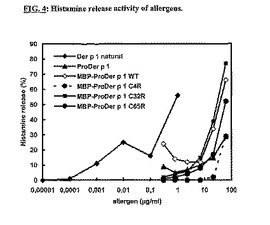
【図5】
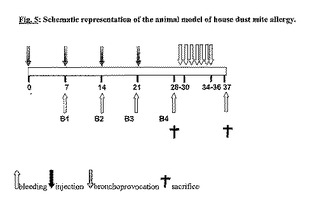
【図6】
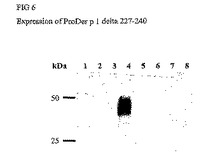
【図7】

【図8】
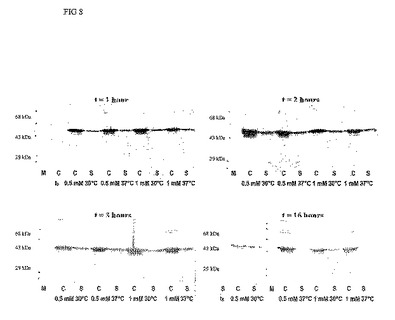
【図9】
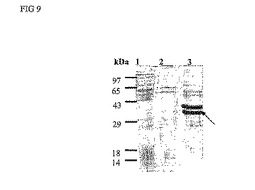
【図10】
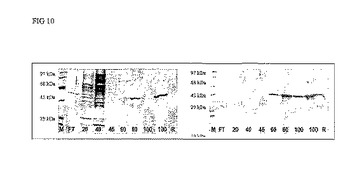
【図11】
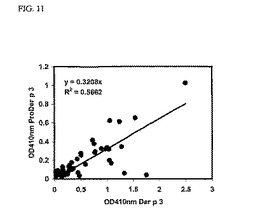
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2007−525150(P2007−525150A)
【公表日】平成19年9月6日(2007.9.6)
【国際特許分類】
【出願番号】特願2006−501950(P2006−501950)
【出願日】平成16年2月24日(2004.2.24)
【国際出願番号】PCT/EP2004/001850
【国際公開番号】WO2004/076481
【国際公開日】平成16年9月10日(2004.9.10)
【出願人】(397062700)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (37)
【Fターム(参考)】
【公表日】平成19年9月6日(2007.9.6)
【国際特許分類】
【出願日】平成16年2月24日(2004.2.24)
【国際出願番号】PCT/EP2004/001850
【国際公開番号】WO2004/076481
【国際公開日】平成16年9月10日(2004.9.10)
【出願人】(397062700)グラクソスミスクライン バイオロジカルズ ソシエテ アノニム (37)
【Fターム(参考)】
[ Back to top ]