
ライシルtRNA合成酵素の細胞内水準を調節して癌転移又は癌細胞の移動を調節する方法
本発明はライシルt-RNA合成酵素が原形質膜に転座(translocation)され、67LRと相互作用することにより、腫瘍(又は癌)細胞の移動を促進して癌の転移(metastasis)に影響を及ぼすとのライシルt-RNA合成酵素の新規な機能に関するものにして、より詳細には前記新規な機能を活用して、ライシルt-RNA合成酵素の細胞内水準を調節して、癌転移又は癌細胞の移動を調節する方法、癌の予防又は治療の為に、KRSの発現を抑制する構成を有する発現ベクターの用途、癌の予防又は治療の為に、KRSの活性を阻害する製剤の用途、癌転移又は癌細胞移動調節製剤をスクリーニングする方法及びKRS及び67LRの相互作用を阻害する製剤をスクリーニングする方法に関するものである。従って、本発明のKRSを利用して癌転移又は癌細胞移動を調節することができ、さらには、原形質膜のラミニン収容体(67LR)と関係した細胞内代謝を調節できる。本発明で究明したKRSとラミニン収容体の関係はこれと関係された多様な疾患又は疾病の治療、予防及び/又は診断に極めて有用に使用できる。

【発明の詳細な説明】
【技術分野】
【0001】
本発明はライシルtRNA合成酵素の新規な機能に関するものにして、より詳細には前記新規な機能を活用してライシルtRNA合成酵素の細胞内水準を調節して、癌転移又は癌細胞の移動を調節する方法、癌の予防又は治療の為に、KRSの発現を抑制する構成を有する発現べクターの用途、癌の予防又は治療の為に、KRS活性を阻害する製剤の用途、癌転移又は癌細胞移動調節製剤をスクリーニングする方法、及びKRS及び67LRの相互作用を阻害する製剤をスクリーニングする方法に関する。
【背景技術】
【0002】
癌(又は腫瘍)は、非正常的で非制御性で無秩序な細胞増殖の産物である。特に、破壊的な成長性、浸透性及び転移性があれば悪性に分類される。浸透性とは、周囲の組織を浸透又は破壊する性質にして、一般的に組織の境界をなす基底層を破壊させ腫瘍が局部的に伝播されることを意味し、たまたま体内の循環系にも流入される。転移とは、一般的にリンパ管(lymphotic)又は血管により、原発位置とは別の所に腫瘍細胞が広がることを意味する。転移はさらに、腸液性体腔又は他の空間を通じて直接伸長して腫瘍細胞を移動させることを意味する。
【0003】
現在、癌は主に3種の治療法、つまり、外科的な手術、放射線照射及び化学療法の内1種、又はこれらの組合せを通じて治療されている。手術は疾病組織を大部分除去することを含む。このような外科的手術は特定部位、例えば、乳房、結腸及び皮膚に位置した腫瘍を除去するには効果的ではあるものの、脊椎のように一部区域にある腫瘍を治療若しくは、白血病のような分散性腫瘍疾患を治療するのには使用できない。
【0004】
化学療法は細胞複製又は細胞代謝を崩壊させ、俗に、乳房、肺及び精巣の癌治療に多く使用されるものの、腫瘍疾病の治療に使用される全身性化学療法の副作用は癌治療を受ける患者等に最も問題となる。このような副作用の内で、目眩と嘔吐は最も一般的であり、深刻な副作用である。化学療法による副作用は患者の生命に大きな影響を及ぼし、治療に対する患者の順応性を急激に変化させ得る。さらに、化学治療剤と関連した副作用には一般的にこのような薬物投与の際、注意すべき容量制限毒性(DLT,Dose Limiting Toxicity)がある。例えば、粘膜炎は多様な抗癌剤、例えば、抗代謝物質細胞毒素剤である5−フルオロウラシル、メソトレキセート及び抗腫瘍抗生剤(例、ドキソルビシン)等に対する容量制限毒性がある。このような化学療法由来の副作用の内、大部分は甚だしい場合、入院を要するか若しくは痛症を治療する為に、鎮痛剤を必要とする場合もあり得る。このように化学治療剤及び放射線治療による副作用等は癌患者の臨床的処置の際、主要問題となっている。
【0005】
遺伝子治療とは、DNA組換え方法を利用して治療用遺伝子を患者の細胞内に導入させ、遺伝子結合を矯正又は細胞に新たな機能を追加させ、人体細胞の遺伝子変形を通じて各種遺伝疾患、癌、心血管疾患、感染性疾病、ウイルス細胞膜自家免疫疾患等のような疾患を治療又は予防する方法である。つまり、治療遺伝子を体内の望む臓器に伝達して細胞内で治療用或いは正常蛋白質が発現するようにして、疾病を治療することを遺伝子治療と言う。遺伝子治療は一般的な薬物による治療に比べて、優れた選択性を有することができ、他の治療法では調節し難い疾病の治療率及び治療速度を改善して、長期間適用することができる。遺伝子治療は単に疾病の症状を治療することに止どまることなく、疾病の原因を治療し、除去することである。このような遺伝子治療を効果的になす為には、治療遺伝子を望む標的細胞に伝達して、高い発現効率を得られるようにする遺伝子伝達技術が必要である。
【0006】
遺伝子伝達体は望む治療遺伝子を対象細胞に導入する為に、必要な媒介体として理想的な遺伝子伝達体は人体に無害で大量生産が容易で効率的に遺伝子を伝達することができ、持続的に遺伝子を発現できなければならない。遺伝子伝達体技術は遺伝子治療技術の核心要素にして、現在遺伝子治療に多く利用される代表的遺伝子伝達体には、アデノウィルス、アデノ付属ウイルス、レトロウイルスのようなウイルス性伝達体とリポソム、ポリエチレンイミンのような非ウイルス性伝達体がある。
【0007】
遺伝子治療戦略の内、腫瘍細胞を制御する戦略には腫瘍抑制遺伝子を利用する方法、腫瘍選択的殺傷ウイルスを利用する方法、自殺遺伝子を利用する方法、免疫調節遺伝子を導入する方法等がある。腫瘍抑制遺伝子を利用する方法は、多くの癌患者において遺伝子が欠損又は変形されているp53のような腫瘍抑制遺伝子を、原形で人体に伝達して癌を治療しようとする方法であって、腫瘍選択的殺傷ウイルスを利用する方法は、癌組織で変形されている腫瘍抑制遺伝子の活性を利用して、腫瘍細胞でのみ選択的に増殖できるウイルス遺伝子伝達体を人体に導入して、治療効果を挙げようとする方法にして、全て腫瘍細胞を直接殺傷させる戦略である。自殺遺伝子を利用する方法も、HSV-TKのような感受性遺伝子を導入して、腫瘍細胞の自殺を誘導する方法もこのような範疇に属する。反面、免疫調節遺伝子を導入する方法は、抗腫瘍免疫反応を増強せしめるインターロイキン12、インターロイキン4、インターロイキン7、ガンマインターフェロン、腫瘍壊死因子等の遺伝子を人体に伝達して、T細胞に腫瘍を認識するように誘発するか、又は腫瘍誘発蛋白質を遮断して細胞自殺を誘導して、間接的に疾病を治療する戦略である。アンジオスタチン、エンドスタチンのような血管生成抑制因子を発現させ、腫瘍への栄養供給を遮断して壊死させる方法もこのような間接的疾病治療戦略として広く認識されている。
【0008】
腫瘍の転移は、癌の生存率において決定的な要素である。67kDaのラミニン収容体(67LR)は、原形質膜に包埋された(embedded)非−インテグリン型収容体にして、癌の浸湿(湿潤)及び転移と関連がある。(Nelson, J. et al. The 67 kDa laminin receptor: structure, function and role in disease. Biosci. Rep. 28, 33-48(2008))。67LRはこれらの37kDa前球体(37LRP)の重合(dimerization)により生成されるものの、このような変化過程に対する分子的な細部機作はよく知られていない。37LRPはポリソーム形成に関与するリポソームサブユニットp40と同一である(Auth, D. & Brawerman, G. A 33-kDa polypeptide with homology to the laminin receptor: component of translation machinery. Proc. Natl. Acad. Sci. USA 89, 4368-4372(1992))。67LRはたまたま多様な種類の癌において、高濃度で観察される。(Nelson, J. et al. The 67 kDa laminin receptor: structure, function and role in disease. Biosci. Rep. 28, 33-48(2008); Menard, S., Castronovo, V., Tagliabue, E. & Sobel, M. E.New insights into the metastasis-associated 67 kD laminin receptor. J. Cell. Biochem. 67, 155-165(1997))。しかしながら、67LRの膜内存在における調節者や分子的機作は、未だに究明されていない。
【0009】
KRSは蛋白質合成の為、同族の(cognate)アミノ酸及び tRNAを接合させるaminoacyl-t-RNA synthetases(ARSs)に属する。これら原始酵素等は触媒的活性の他にも多面的(pleiotropic)特徴を有する(Park, S. G., Ewalt, K. L. & Kim, S. Functional expansion of aminoacyl-tRNA synthetases and their interacting factors: new perspectives on housekeepers. Trends Biochem. Sci . 30 , 569-574(2005))。反面、KRSを含む幾つかの哺乳類の ARSは構成蛋白質の多様な機能を調節する為に、分子貯蔵所(molecular reservoir)として作用する(Ray, P. S., Arif, A. & Fox, P. Macromolecular complexes as depots for releasable regulatory proteins. Trends Biochem. Sci. 32, 158-164(2007))巨大分子複合体を形成する(Lee, S. W., Cho, B. H.,Park, S. G. & Kim, S. Aminoacyl-tRNA synthetase complexes: beyond translation. J. Cell. Sci. 117, 3725-3734(2004); Han, J. M., Kim, J. Y. & Kim, S. Molecular network and functional implications of macromolecular tRNA synthetase complex. Biochem. Biophys. Res. Commun. 303, 985-993(2003))。ヒトKRSはRNAとその他蛋白質間の相互作用に関与する固有のN末端延長部位を含有している(Rho, S. B. et al. Genetic dissection of protein-protein interactions in multi-tRNA synthetase complex. Proc. Natl. Sci. Acad. USA 96, 4488-4493(1999); Francin, M., Kaminska, M., Kerjan,P. &Mirande. M. The N-terminal domain of mammalian Lysyl-tRNA synthetase is a functional tRNA-binding domain. J. Biol. Chem. 277, 1762-1769(2002))。
【発明の概要】
【発明が解決しようとする課題】
【0010】
ヒトKRSの機能的多様性と関連して、このペプチドの有意性を確認する為、本発明者等はヒトKRSのN末端116個アミノ酸ペプチドを分離して、イーストツーハイブリッドを利用した、HeLa細胞株 cQDNAライブラリからヒトKRSに結合する蛋白質をスクリーニングする探針(bait:エサ)用に使用した。前記スクリーニングから、本発明者等は本発明で結合可能性がある蛋白質の一つとして、37LRP/p40を確認し、KRSとラミニン収容体間の相互作用における機能的連関性を調査した。
【0011】
その結果、ライシルt-RNA合成酵素(lysyl-t-RNA synthetase,KRS)が原形質膜にある67LRを安定化させることにより、細胞移動及び癌転移を促進する等、KRSが原形質膜のラミニン収容体を通じて癌転移又は癌細胞移動に影響を及ぼすことを究明して本発明を完成した。
【0012】
本発明の目的は癌転移又は癌細胞移動に関して、ライシルt-RNA合成酵素(lysyl-t-RNA synthetase,KRS)の新たな用途を提供することである。
【0013】
前記のような目的を達成する為に、本発明はライシルt-RNA合成酵素の細胞内水準を調節して癌転移を調節する方法を提供する。
【0014】
本発明の他の目的を達成する為に、本発明はライシルt-RNA合成酵素の細胞内水準を調節して癌細胞株の移動(cancer cell migration)を調節する方法を提供する。
【0015】
本発明のさらに、他の目的を達成する為に、本発明はプロモータ及びこれと作動可能に連結されたKRSを、符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクターを有効成分として含む癌予防及び治療用組成物を提供する。
【0016】
本発明のさらに他の目的を達成する為に、本発明はプロモータ及びこれと作動可能に連結されたKRSを、符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクターを、これを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法を提供する。
【0017】
本発明のさらに他の目的を達成する為に、本発明は癌治療剤を製造する為のプロモータ及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを、符号化するポリヌクレオチドを含む発現ベクターの用途を提供する。
【0018】
本発明のさらに他の目的を達成する為に、本発明はKRSの活性を阻害する製剤を有効成分として含む癌予防及び治療用組成物を提供する。
【0019】
本発明のさらに他の目的を達成する為に、本発明はKRSの活性を阻害する製剤をこれを必要とする個体に有効量を投与することを含む癌の予防及び治療方法を提供する。
【0020】
本発明のさらに他の目的を達成する為に、本発明は癌の治療剤を製造する為の、KRSの活性を阻害する製剤の用途提供する。
本発明のさらに他の目的を達成する為に、本発明は、
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;及び
(c)選別された製剤が癌転移又は細胞移動を調節するか、否かをテストする段階を含む癌転移又は癌細胞移動調節製剤をスクリーニングする方法を提供する。
【0021】
本発明のさらに他の目的を達成する為に、本発明は、
(a)試験製剤の存在下でKRS、ラミニン収容体(67LR)及び試験製剤を接触させる段階;及び
(b)前記試験製剤がKRSとラミニン収容体の相互作用を調節するか、否かをテストする段階を含むKRSと、67LRK相互作用を阻害する製剤をスクリーニングする方法を提供する。
本発明のさらに他の目的を達成する為に、本発明は、
(a)試料において67LRの過発現の可否を分析する段階;及び
(b)67LRが過発現された試料においてKRSの過発現の可否を分析する段階を含む肺癌又は乳房癌を診断する方法を提供する。
【課題を解決するための手段】
【0022】
以下、本発明を詳細に説明する。
本発明はKRSが癌転移又は癌細胞移動に及ぼす影響を初めて究明した。つまり、本発明はKRSが原形質膜のラミニン収容体を通じて癌転移又は癌細胞移動に影響を及ぼすことを究明した。
【0023】
定 義
他の定義がない限り、本発明で使用される全ての技術的及び化学的用語は当業者等により、通常的に理解される同一な意味を有する。次の参考文献は、本発明の明細書に使用された多くの用語等の一般的な定義を有する技術(skill)の一つを提供する。(Singleton et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOTY(2nd ed. 1994); THE CAMBRIDGE DICTIONARY OF SCIENCE AND TECHNOLOGY(Walker ed., 1988); 及び Hale & Marham, THE HARPER COLLINS DICTIONARY OF BIOLOGY)。さらに、次の定義は本発明の実施の為に読者(reader)に提供される。
【0024】
本発明で“発現(expression)”とは、細胞で蛋白質又は核酸の生成を意味する。
【0025】
本発明で“宿主細胞(host cell)”とは、任意の手段(例:電気衝撃法、カルシウムホスフアターゼ沈澱法、微細注入法、形質転換法、ウイルス感染等)により、細胞内に導入された異種性DNAを含む原核又は眞核細胞を言う。
【0026】
本発明で“分離された(isolated)”とは、それの起源環境(例:若し自然的に生成されるものであれば自然環境)から除去された物質を意味する。例えば、自然的に生成される核酸、ポリペプチド、又は生きている動物に存在する細胞は、分離されたものではないものの、同一なポリヌクレオチド、ポリペプチド又は共同存在する物質の一部又は全部から分離された(separated)細胞は、以降、それが自然システムで再導入されるとしても、分離されたものである。前記核酸はベクターの一部の場合もあり得て、及び/又は、前記核酸又はポリペプチドの一部でも有り得て、前記ベクターや組成物は自然環境の一部でない分離されたものである。
【0027】
本発明でKRSの生体活性に対して“調節する(modulate)”とは、KRSの細胞内水準の変化を言う。KRS活性の調節は上向調節(up-regulation)(つまり、活性化又は促進)又は下向調節(down-regulation)(つまり、阻害又は抑制)の場合もあり得る。調節はKRSの細胞内水準、蛋白質の安定性、KRSの酵素的修飾(例:リン酸化)、結合特性(例:標的転写調節因子に対する結合)、又はKRSの他の生物学的、機能的及び免疫学的特性の変化を引起こすこともあり得る。KRSの生体活性の変化は、例えば、KRS遺伝子発現の増加又は減少、KRS蛋白質を符号化するmRNAの安定性、翻訳効率から招かれることもあり得る。KRS調節者(modulation)の作用方式(mode)は例えば、KRS蛋白質又はこれを符号化する遺伝子に対する結合を通じて直接的なものの場合もあり得る。さらに、前記変化はKRSを調節する他の物質(例:KRSを特異的にリン酸化させるカイネース)に対する結合及び/又は修飾(例:酵素学的に)を通じた間接的なものでもあり得る。
【0028】
本発明で“ポリペプチド(polypeptide)”は“ポリペプチド等(polypeptides)”又は“蛋白質(等)”と互換性を以て使用でき、例えば、自然状態の蛋白質において一般的に発見される通りアミノ酸残基の重合体を言う。
【0029】
本発明で“KRSポリペプチド”はライシルt-RNA合成酵素として知られているポリペプチドを言う。前記KRSポリペプチドは配列番号1で表示されるアミノ酸配列を有するポリペプチドでもあり得る(Genbank Accession No.NP-005539.1)。さらに、本発明のKRSはこれの機能的同等物を含む。
【0030】
前記機能的同等物とは、配列番号1で表示されるアミノ酸配列と少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上の配列相同性(つまり、同一性)を有するポリペプチドを言う。例えば、 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 100%の配列相同性を有するポリペプチドを含むものにして、配列番号1で表示されるポリペプチドと、実質的に同質の生理活性を呈するポリペプチドを言う。ここで、“実質的に同質の生理活性”とは、原形質膜のラミニン収容体と相互作用し、癌転移又は細胞移動を調節することを意味する。前記機能的同等物は配列番号1のアミノ酸配列の内、一部が付加、置換又は欠失の結果、生成されるものでもあり得る。前記にてアミノ酸置換は好ましくは、保存的置換である。天然に存在するアミノ酸の保存的置換の例は下記の通りである;脂肪族アミノ酸(Gly,Ala,Pro)、疎水性アミノ酸(Ile,Leu,Val)、芳香族アミノ酸(Phe,Tyr,Trp)、酸性アミノ酸(Asp,Glu)塩基性アミノ酸(His,Lys,Arg,Gln,Asn)及び硫黄含有アミノ酸(Gys,Met)、さらに、前記機能的同等物には本発明のKRSポリペプチドのアミノ酸配列上、アミノ酸の一部が欠失された変形体も含まれる。前記アミノ酸の欠失又は置換は好ましくは、本発明のポリペプチドの生理活性に直接的に関連されない域に位置している。さらに、アミノ酸の欠失は好ましくは、KRSポリペプチドの生理活性に直接関与しない部分に位置する。さらに、前記KRSポリペプチドのアミノ酸配列の両末端又は配列内に幾つかのアミノ酸が付加された変形体も含まれる。さらに、本発明の機能的同等物の範囲には、本発明に伴うポリペプチドの基本骨格及びこれの生理活性を維持しながらポリペプチドの一部化学構造が変形されたポリペプチド誘導体も含まれる。例えば、本発明のポリペプチドの安定性、貯蔵性、揮発性又は溶解度等を変形させる為の構造変更がこれに含まれる。
【0031】
本明細書で配列相同性及び同質性はKRSのアミノ酸配列(配列番号1)と候補配列を整列してギャップ(gaps)を導入した後、KRSのアミノ酸配列に対する候補配列のアミノ酸残基の百分率で定義される。必要な場合、最大百分率配列同質性を収得する為に、配列同質性の部分として保存的置換は考慮しない。さらに、KRSのアミノ酸配列のN−末端、又はC−末端又は内部伸長、欠損又は挿入は配列同質性又は相同性に影響を与える配列として解析されない。さらに、前記配列同質性は二つのポリペプチドのアミノ酸配列の類似した部分を比較する為に、使用される一般的な標準方法により決定できる。BLAST又はFASTAのようなコンピュータプログラムは、二つのポリペプチドをそれぞれのアミノ酸が最適にマッチングするように整列する(一つ又は二つの配列の全長配列に従い又は一つ又は二つの配列の予測された部分に従い)。前記プログラムはデフォルトオプニングペナルティ(gefault opening penalty)及び、デフォルトギャップペナルティ(default gap penalty)を提供してコンピュータプログラムと共に連係されて使用し得るPAM250(標準スコリングマトリックス; Dayhoff et al., in Atlas of Protein Sequence and Structure, vol 5, supp 3, 1978)のようなスコリングマトリックスを提供する。例えば、百分率同質性は次のように計算できる。一致する配列(identical matches)の総数に100を掛け、対応されるスパン(matched span)内のより長い配列の長さと二つの配列を整列する為に、より長い配列内に導入されたギャップ(gaps)の和で分ける。
【0032】
本発明に伴うKRSポリペプチドは天然で抽出するか、又は遺伝工学的方法により作製でき得る。例えば、通常的な方法により前記KRS又はこの機能的同等物を符号化する核酸(例:配列番号2(Genbank Accession No.D32053))を作製する。前記核酸は適切なプライマーを使用してPCR増幅することにより、作製し得る。他の方法で当業界に公知の標準方法により、例えば、自動DNA合成器(Biosearch又はApplied Biosystems社で販売するもの)を使用してDNA配列を合成することもできる。作製された核酸はこれに作動可能に連結され(operatively linked)、核酸の発現を調節する一つ以上の発現調節配列(expression control sequence)(例:プロモーター、インハンサー等)を含むベクターに挿入させ、これより形成された組換え発現ベクターで宿主細胞を形質転換させる。生成された形質転換体を前記核酸が発現されるに適切な培地及び条件下で培養して、培養物から前記核酸により発現された実質的に純粋なポリペプチドを回収する。前記回収は当業界に公知の方法(例えば、クロマトグラフィー)を利用して行え得る。前記にて“実質的に純粋なポリペプチド(substantially pure polypeptide)”とは、本発明に伴うポリペプチドが宿主細胞から由来した如何なる他の蛋白質も実質的に含まないことを意味する。本発明のポリペプチド合成の為の、遺伝工学的方法は次の文献を参考にし得る: Maniatis et al., Molecular Cloning; A laboratory Manual, Cold Spring Harbor laboratory, 1982; Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, N.Y., Second(1998) and Third(2000) Editions; Gene Expression Technology, Method in Enzymology, Genetics and Molecular Biology, Method in Enzymology, Guthrie & Fink(eds.), Academic Press, San Diego, Calif, 1991; Hitzeman et al., J. Biol. Chem., 255:12073-12080, 1990。
【0033】
さらに、本発明のポリペプチドは当業界に公知の化学的合成(Creighton, Proteins; Structures and Molecular Principles, W. H. Freeman and Co., NY, 1983)により容易に製造できる。代表的な方法としてこれらに限定されるものではないものの、液体又は固体状合成、断片凝縮、F-MOC又はT-BOC化合法が含むまれる。(Chemical Approaches to the Synthesis of Peptides and Proteins, Williams et al., Eds., CRC Press, Boca Raton Florida, 1997; A Practical Approach, Athert on & Sheppard, Eds., IRL Press, Oxford, England, 1989)。
【0034】
本発明で67kDaのラミニン収容体(67LR)は原形質膜に包埋された(embedded)非−インテグリーン型収容体にして、例えば、Genbank Accession No. NM_002295, S37431, AF284768, S37431, AF284768, J03799, XP_370865, XP_001083023.に記載された塩基配列又はアミノ酸配列を有し得る。
【0035】
本発明で“核酸”、“DNA配列”又は“ポリヌクレオチド”は単一-又は二重-鎖の形態でなるデオキシリボヌクレオチド又はリボヌクレオチドを言う。他の制限がない限り、自然的に生成されるヌクレオチドと類似した方法で核酸に混成化される自然的ヌクレオチドの公知のアナログも含まれる。
【0036】
本発明で“前記KRSを符号化するポリヌクレオチド”は配列番号1で表示されるアミノ酸配列、又はこれと少なくとも70%以上の配列相同性を有するアミノ酸配列を符号化する塩基配列を有し得る。前記核酸はDNA、cDNA及びRNA配列を全て含む。つまり、前記ポリヌクレオチドは配列番号1のアミノ酸配列又はこれと少なくとも70%以上の相同性を有するアミノ酸配列を符号化する塩基配列を有するか、若しくは、前記塩基配列に相補的な塩基配列を有し得る。好ましくは、配列番号2で表示される塩基配列を有する。前記核酸は天然で分離されたか、若しくは、上述した通り、遺伝工学的な方法により製造され得る。
【0037】
本発明で“アナログ(analog)”とは、標準物質(reference molecule)と構造的に類似するものの、標準物質の特異的置換基が置換により代替されることにより、標的や調節方法が変形された物質を言う。標準物質と比較するに、アナログは当業者により予想され得る通り、同一又は類似するか若しくは、向上した有用性(utility)を有する。向上した性質(例:標的物質に対するより高い結合親和力)を有する公知の化合物変異体を究明する為のアナログの合成及びスクリーニングは薬理化学的分野に公知の方法である。
【0038】
本発明で“相同体(homologues)”とは、蛋白質及び/又は蛋白質配列を言及する場合には、共通の祖先蛋白質(common ancestral protein)又は蛋白質配列で自然的に又は人為的に由来したことを示す。類似して核酸及び/又は核酸配列はそれらが共通の祖先核酸又は核酸配列から自然的に又は人為的に由来したときに相同する。
【0039】
本発明で“有効量(effective amount)”とは、細胞又は組織内でKRSの生体活性(例:細胞内水準等)を正常細胞や組織とは別に変更させる効果、又は癌転移又は癌細胞移動様態を対照群に比べて意味ある変化を示すことを言う。
【0040】
本発明で“接触(contacting)”とは、これの一般的な意味(normal meaning)であって、2個以上の製剤(例:2個のポリペプチド)を結合(combine)させるか、若しくは、製剤と細胞(例:蛋白質と細胞)を結合させることを言う。接触は試験管内(in vitro)で起こり得る。例えば、試験管又は他のコンテナー(container)で2個以上の製剤を結合させるか、若しくは、試験製剤と細胞又は細胞溶解物と試験製剤を結合させることである。さらに、接触は細胞又はインシツー(in situ)で起こることもあり得る。例えば、2個のポリペプチドを符号化する組換えポリヌクレオチドを、細胞内で共同発現(coexpression)させることにより、細胞又は細胞溶解物において2個のポリペプチドを接触させることである。
【0041】
本発明で“製剤(agent)”又は“試験製剤(test agent)”とは、任意の物質(substance)、分子(molecule)、元素(element)、化合物(compound)、実在物(entity)又はこれらの組合せを含む。例えば、これに限定はされないものの、蛋白質、ポリペプチド、小遺棄物質(small organic molecule)、多糖類(polysaccharide)、ポリヌクレオチド等を含む。さらに、自然産物(natural product)、合成化合物又は化学化合物又は2個以上の物質の組合せでもあり得る。別に指定されない限り、製剤、物質及び化合物は互換性あるように(interchangeably)使用できる。
【0042】
より具体的に本発明のスクリーニング方法でスクリーニングされ得る試験製剤は、ポリペプチド、ベーターターンミメチック(beta-turn mimetics)、多糖類、リン脂質、ホルモン、フロスタグランジン、ステロイド、芳香族化合物、ヘテロサイクリック化合物、ベンゾジアゼピン(benzodiazepines)、オリゴメリックN−置換グリシン(oligomeric N-substituted glycines)、オリゴカルバメート(oligocarbamates)、糖類(saccharides)、脂肪酸、プリン、ピリミジン又はこれらの誘導体、構造的アナログ又は組合せを含む。或る試験製剤は合成物質でもあり得る、他の試験製剤は天然物質でもあり得る。前記試験製剤は合成又は自然化合物のライブラリを含む広範囲で多様な出処から得られ得る。組合せ(combinatorial)ライブラリはステップ−バイ−ステップ方式で合成され得る多様な種類の化合物で生産され得る。多数の組合せライブラリの化合物等はESL(encoded synthetic libraries)方法(WO 95/12608,WO 93/06121,WO 94/08051,WO 95/395503及びWO 95/30642)により製造でき得る。ペプチドライブラリはファージデイスプレー方法(WO91/18980)により製造し得る。バクテリア、黴、植物及び動物抽出物形態の自然化合物のライブラリは商業的な出処から得るか、又はフィールド(field)で収集することができる。公知の薬理学的製剤が構造的アナログを製造する為に、アシル化、アルキル化、エステル化反応(esterification)、アミド化反応(amidification)のような指示又は無作為な化学的修飾に適用され得る。
【0043】
前記試験製剤は自然的に生成される蛋白質又はその断片でもあり得る。このような試験製剤は自然出処(natural source)、例えば、細胞又は組織溶解物から収得できる。ポリペプチド製剤のライブラリは例えば、通常的な方法により生成されるか、又は商業的に入手し得るcDNAライブラリから収得され得る。前記試験製剤はペプチド、例えば、約5〜30個、好ましくは、約5〜20個、より好ましくは約7〜15個のアミノ酸を有するペプチドでもあり得る。前記ペプチドは自然的に生成される蛋白質、ランダムペプチド又は“バイアス(biased)”ランダムペプチドの伝達物でもあり得る。
【0044】
らに、前記試験製剤は“核酸”でもあり得る。核酸試験製剤は自然的に生成される核酸でもあり得る。例えば、原核又は眞核ゲノムの伝達物の為に記載したのと類似するように使用できる。
【0045】
さらに、前記試験製剤は小分子(例:約1,000以下の分子量を有する分子)でもあり得る。小分子の調節製剤をスクリーニングする為の方法には、好ましくは、高速分析アッセイ(high throughput assay)が適用され得る。多くのアッセイが前記スクリーニングに有用である(Shultz, Bioorg. Med. Chem. Lett., 8:2409-2414, 1998; Weller, Mol. Drivers., 3:61-70, 1997; Fernandes, Curr. Opin. Chem. Biol., 2:597-603, 1998; and Sittampalam, Curr. Opin. Chem. Biol., 1:384-91, 1997)。
【0046】
本発明の方法にスクリーニングされる試験製剤のライブラリはKRS又はこれの断片若しくはアナログに対する構造研究を基に製造され得る。このような研究はKRSに結合する可能性がある試験製剤の究明を可能にする。KRSの3次元的構造は多様な方法、例えば、結晶学構造及び分子的モデリング(crystal structure and molecular modeling)で研究され得る。X-RAY結晶学を利用する蛋白質構造研究方法が文献に広く知られている。(Physical Bio-Chemistry, Van Holde, K. E.(Prentice-Hall, New Jersey 1971), pp.221-239, and Physical Chemistry with Applications to the Life Sciences, D. Eisengerg & D. C. Crothers(Benjamin Cummings, Menlo Park 1979)。KRSの構造に対するコンピューターモデリングはスクリーニングする為に、試験製剤のデザインの為の他の手段を提供する。分子的モデリング方法は文献に開示されている: U.S. Pat. No. 5, 612,894 and U.S. Pat. No. 5,583,973. さらに、蛋白質構造は中性子回折(neutron diffraction)及びNMR(nuclear magnetic resonance)により、決定し得る: Physical Chemistry, 4th Ed. Moore, W. J.(Prentice-Hall, New Jersey 1972)and NMR of Proteins and Nucleic Acids, K. Wuthrich(Wiley-Interscience, New York 1986)。
【0047】
以下、本発明を詳細に説明する。
【0048】
本発明者等は本発明のKRSは原形質膜で転座(translocation)され、67LRと相互作用することにより、腫瘍(又は癌)細胞の移動を促進して癌の転移(metastasis)に影響を及ぼすことを究明した。さらに、マウスを利用したin vivo実験を通じてKRSの過発現又は発現抑制が腫瘍(又は癌)細胞の転移を調節できることを究明した。
【0049】
従って、本発明はライシルt-RNA合成酵素(Lysyl t-RNA synthetase,KRS)の細胞内水準を調節(modulating)して癌転移を調節する方法を提供する。
【0050】
これをより詳細に説明すれば、本発明のライシルt-RNA合成酵素の細胞内水準を減少させる場合、癌転移を抑制することができ、ライシルt-RNA合成酵素の細胞内水準を増加させる場合癌転移を増進することもあり得る。
【0051】
細胞内水準の減少又は増加は前記にて記載したことと共に、当業者に公知の多様な方法で調節され得る。例えば、これに限定はされないものの、細胞内水準は転写段階における調節を通じて調節でき得る。転写段階における調節は当業者に公知の遺伝子の発現を増進させる為の方法、例えば、プロモーターにKRS又はこれに対する機能的同等物を符号化するポリヌクレオチドを連結した組換え発現ベクターを調節して前記遺伝子の発現を増進させる方法、又はKRS又はこれに対する機能的同等物を符号化する遺伝子周辺に前記遺伝子の発現が促進されるようにする発現調節配列を挿入する方法、又は遺伝子の発現を抑制する為の方法、例えば、プロモーター又は遺伝子部位の突然変異を誘導してプロモーター活性又は蛋白質の機能を阻害する方法、アンチセンス(antisense)遺伝子を発現させる方法、siRNA又はマイクロRNA(microRNA)方法等により実施し得る。
【0052】
転写後の段階における調節は、当業者に公知の蛋白質発現を増進又は阻害させる為の方法、例えば、KRS又はこれに対する機能的同等物を符号化する遺伝子を鋳型に、転写されたmRNAの安定性を増進又は阻害する方法、蛋白質又はポリペプチドの安定性を増進又は阻害する方法、又は蛋白質又はポリペプチドの活性を増進又は阻害する方法により実施し得る。
【0053】
前記方法のより具体的な例として1群イントロンタイプ、M1 RNAタイプ、ハンマーヘッド(hammerhead)型、又はヘアピン(hairpin)型又はマイクロRNA型等の転写されたmRNAに作用するRNAを符号化するDNA配列に形質転換するか、又は標的遺伝子配列と同一又は類似した配列を有するDNAの形質転換を通じた同時抑制(cosuppression)を誘導することができる。
【0054】
好ましくは、本発明でKRS又はこれに対する機能的同等物の細胞内水準を調節することは、前記ポリヌクレオチドを符号化するポリヌクレオチドの発現を増加又は減少させる方法により実施し得る。このような増加又は減少させる方法は、それぞれ当業者に公知の方法を使用し得るものの、例えば、プロモーターにKRS又はこれに対する機能的同等物を符号化するポリヌクレオチドを連結した組換え発現ベクターを製造して、その発現を増進又は、プロモーターに前記ポリヌクレオチドに対するアンチセンス又はsiRNAポリヌクレオチドを連結した組換え発現ベクターを製造してその発現を減少し得る。この際、前記KRS又はこれに対する機能的同等物を符号化するポリヌクレオチドは好ましくも、配列番号2で表示される塩基配列を有し得る。
【0055】
従って、本発明はライシルt-RNA合成酵素の細胞内水準を調節して癌細胞の移動(cancer migration)を調節する方法を提供する。細胞内水準の調節等に対しては前記にて記載した通りである。
【0056】
さらに、本発明のKRSはその発現を抑制させる場合、腫瘍(又は癌)の転移が抑制されるので、本発明はプロモーター及びこれと作動可能に連結されたKRSの発現を抑制する構造遺伝子を含む発現ベクター又はKRSに対する抗体を有効成分として含む癌の予防及び治療用組成物を提供する。前記にてKRSの発現を抑制する構造遺伝子はKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAでもあり得る。
【0057】
本発明の組成物が適用できる疾患は癌もあり得る。前記癌はこれに限定はされないものの、大腸癌、肺癌、肝臓癌、胃癌、食道癌、膵臓癌、胆嚢癌、腎臓癌、膀胱癌、前立腺癌、睾丸癌、子宮頸部癌、子宮内膜癌、絨毛癌、卵巣癌、乳癌、甲状腺癌、脳癌、頭頚部癌、悪性黒色腫、リンパ腫、再生不良性貧血等を含む。
【0058】
前記“プロモーター”とは、特定した宿主細胞で作動可能に連結された核酸配列の発現を調節するDNA配列を意味し、“作動可能に連結される(operably linked)”と言うことは、一つの核酸断片が他の核酸断片と結合され、その機能又は発現が他の核酸断片により影響を受けることを言う。従って、転写を調節する為の任意のオペレータ配列、適合したmRNAリボソーム結合部位をコーティングする配列及び転写及び解読の終結を調節する配列を追加して含め得る。前記プロモーターには全ての時間帯に常時的に目的遺伝子の発現を誘導するプロモーター又は特定した位置、時期に目的遺伝子の発現を誘導するプロモーターを使用することができ、その例としてはSV40プロモーター、CMVプロモーター、CAGプロモーター(Hitoshi Niwa et al., Gene, 108:193-199, 1991; Monahan et al., Gene Therapy, 7:24-30, 2000)35Sプロモーター(Odell et al., Nature 313:810-812, 1985)Rsyn7プロモーター(米国特許出願第08/991,601号)、ライスアクチン(rice actin)プロモーター(McElroy et al., Plant Cell 2:163-171, 1990)、ユビキチンプロモーター(Christensen et al., Plant Mol. Biol. 12:619-632, 1989)、ALSプロモーター(米国特許出願第08/409,297号)等がある。この他にも米国特許第5,608,149;第5,608,144号、第5,604,121号、第5,569,597号、第5,466,785号、第5,399,680号、第5,268,463号及び第5,608,142号等に開示されたプロモーターを全て使用し得る。
【0059】
一方、本発明はプロモーター及びこれと作動可能に連結されたKRSの発現を抑制する構造遺伝子を含む発現ベクター又はKRSに対する抗体を、これを必要とする個体に有効な量で投与する癌の予防及び治療方法を提供する。この際、構造遺伝子は前記記載の通りであるので、本発明はプロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体をこれを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法を提供する。
【0060】
本発明で“有効な量”とは、本発明の発現ベクターが投与対象である個体内で癌の予防又は治療する効果を表す量を言う。前記“個体”とは、好ましくは、哺乳動物、特にヒトを含む動物の場合も有り得て、動物より由来した細胞、組織、器官等でもあり得る。前記個体は治療が必要な患者(patient)の場合もあり得る。
【0061】
さらに、本発明は癌治療剤を製造する為の、プロモーター及び作動可能に連結されたKRSの発現を抑制する構造遺伝子を含む発現ベクター、又はKRSに対する抗体の用途を提供する。より詳細には、本発明は癌治療剤を製造する為のプロモーター及びこれと作動可能に連結されたKRSを、符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター、又はKRSに対する抗体の用途を提供する。前記にてプロモーター、KRS、発現ベクター、適用される癌に対しては前記記載の通りである。
【0062】
前記にてKRSに対する抗体は、KRSの抗原性部位に対して指示される特異的な蛋白質分子を意味する。本発明の目的上、前記抗体はKRS蛋白質に対して特異的に結合する抗体を意味し、タンクロン抗体、タンクロン抗体及び組換え抗体を全て含む。
【0063】
前記のようなKRSに対する抗体を生成することは、当業界に広く知られた技術を利用して容易に製造できる。タンクロン抗体は前記KRS蛋白質抗原を動物に注射し、動物から採血して抗体を含む血清を収得する当業界に広く知られた方法により生産することができる。このようなタンクロン抗体は羊、兎、山羊、猿、馬、豚、牛、犬等の任意の動物種の宿主から製造可能である。
【0064】
タンクロン抗体は当業界に広く知られたハイブリッドマ方法(hybridoma method) (hybridoma method) (Kohler and Milstein(1976)European Jounral of Immunology 6:511-519参照)、又はファージ抗体ライブラリ(Clackson et al, Nature, 352:624-628, 1991; Marks et al, J. Mol. Biol., 222:58, 1-597, 1991)技術を利用して製造され得る。
【0065】
ハイブリッドマ方法は肺癌診断マーカー蛋白質抗原を注射したマウスのような免疫学的に適合した宿主動物からの細胞を利用し、残りの一つの集団では癌又は骨髄腫細胞株を利用する。このような二つの集団の細胞等をポリエチレングリコールのような当業界に広く知られた方法により、融合させた後、抗体−生産細胞を標準的な組織培養方法により増殖させる。限界希釈法(limited dilution technique)によるサブクローニングにより、均一な細胞集団を収得した後、KRS蛋白質に対して特異的な抗体を生産できるハイブリッドマを標準技術により、試験管内で又は生体内で大量で培養する。前記のハイブリッドマが生産するタンクロン抗体は、精製せずに使用することもできるものの、最良の結果を得る為には、当業界に広く知られた方法により、高純度に精製して使用することが好ましい。ファージ抗体ライブラリ方法は、細胞内に存在する多様な肺癌マーカーに対する抗体遺伝子(Single chain fragmentvariable, scFv形態)を獲得してこれをファージの表面に融合蛋白質形態で発現することにより、抗体ライブラリを試験管内で製作し、このライブラリから肺癌蛋白質と結合するモノクロナル抗体を分離、製作する方法である。前記方法で製造された抗体はゲル電気泳動、透析、塩沈殿、イオン交換クロマトグラフィー、親和性クロマトグラフィー等の方法を利用して分離する。
【0066】
さらに、本発明の抗体は2個の全長の軽鎖及び2個の全長の重鎖を有する完全な形態のみならず、抗体分子の機能的な断片を含む。抗体分子の機能的な断片とは、少なくとも抗原結合機能を保有している断片を意味し、Feb, F(ab'), F(ab')2及びFv等がある。
【0067】
従って、本発明では、KRSの細胞内水準を減少させる場合、癌の転移を抑制して癌の予防及び治療に利用できることを究明したことから、本発明はKRSの活性を阻害する製剤を有効成分として含む癌の予防及び治療用組成物を提供する。従って、本発明はKRSの活性を阻害する製剤をこれを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法を提供し、癌治療剤を製造する為のKRSの活性を阻害する製剤の用途も提供する。癌、個体、有効な量等に対しては前記記載の通りである。
【0068】
KRSの活性を阻害する製剤KRSの発現を阻害、つまり、mRNA又は蛋白質水準における発現を阻害する製剤、例えば、KRSに対するantisense RNA又はsiRNAでも有り得て、発現されたKRSの活性を阻害する競争的阻害剤(competitive inhibitor)又は非競争的阻害剤、例えば、KRSに対する抗体でもあり得るものの、これに制限はされない。
【0069】
KRSの細胞内水準を減少させる場合、癌の転移を抑制して癌の予防及び治療に利用できるので、組成物、方法及び用途はそれ自体のみならず、従来公知の多様な癌の予防及び治療方法や抗癌剤と結合して利用され得る。つまり、本発明の組成物、方法等は癌の転移を抑制できるので、従来抗癌剤又は癌の予防及び治療方法と結び付けて治療に利用すれば癌が抑制され、主な腫瘍部位の治療を通じた癌の完治に効果的でもあり得る。
【0070】
本発明のポリペプチドに連結され得る抗癌剤又は癌の予防及び治療方法は、従来癌治療に使用されるものであれば制限なく使用できる。例えば、抗癌剤としてはパクリタキセル、トキソルビシン、ビンクリスチン、ダウノルビシン(daunorubicin)、ビンブラスチン(vinblastine)、アクチノマイシン-D(ACTINOMYCIN-D)、ドセタキセル(docetaxel)、エトポサイド(etoposide)、テニポサイド(teniposide)、ビサントレン(bisantrene)、ホモハリングトン(homoharringtonine)、グリーベック(Gleevec;STI-571)、シスプライン(cisplain)、5-フルオラシル(5-fluouracil)、アドリアマイシン(adriamycin)、メトソトレキサート(methotrexate)、ブスルフアン(busulfan)、クロラムブシル(chlorambucil)、シクロホスファミド(cyclophosphamide)、メルファラン(melphalan)、ニトロゲンムスタード(nitrogen mustard)、ニトロソウレア(nitrosourea)等がある。前記製剤と本発明の組成物又は予防及び治療方法の結合は、抗癌剤の種類及び量によって当業界に公知の技術により適切に行え得る。
【0071】
本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤は、経口的に又は非経口的に投与でき得る。経口投与は舌下適用も含む。非経口的投与は皮下注射、筋肉内注射及び静脈注射のような注射法及び点適法を含む。前記発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤は、薬学的に許容される担体と混合して各種薬学的剤形の形態で製造できる。前記“薬学的に許容される”とは、生理学的に許容されヒトに投与されるとき、通常的に胃腸障碍、眩気症等のようなアレルギー反応又はこれと類似した反応を起こさないことを言う。薬学的に許容される担体は経口投与には、結合剤、滑沢剤、崩解剤、賦形剤、可溶化剤、分散剤、安定化剤、懸濁化剤、色素及び香料を使用することができ、注射剤の場合には緩衝材、保存剤、無痛化剤、等張剤及び安定化剤を混合して使用することができ、局所投与用製剤の場合には、基剤、賦形剤、潤滑剤及び保存剤を使用し得る。このように本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤を含む薬学組成物の剤形は上述した通りの薬学的に許容される担体と混合して多様に製造し得る。例えば、経口投与時には錠剤、トポキ、カプセル、エリキシル、サスペンション、シロップ、ウェファー(wafer)等の形態で製造することができ、注射剤の場合には、単位投薬アンプル又は多数回投薬包含剤形態で製造できる。
【0072】
本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤の総有効量は単一投与量で投与することができ、多重投与量が長期間投与される分割治療方法により投与できる。本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤を含む組成物は疾患の程度及び/又は目的により、有効成分の含量を異にすることができるものの、通常的に1回投与時0.1μg乃至100mg、好ましくは1μg乃至10mgの有効容量で1日数回繰返し投与し得る。しかしながら、前記発現ベクター又はKRSの活性を阻害する製剤の濃度は、薬の投与経路及び治療回数のみならず、患者の年齢、体重、健康状態、性別、疾患の重症度、食餌及び排泄率等、多様な要因等を考慮して患者に対する有効投与量が決定されるものである為、このような点を考慮するに、当分野の通常的な知識を有する者であれば、適切な有効投与量を決定できるであろう。本発明に伴う発現ベクター又はKRSの活性を阻害する製剤を含有する組成物は本発明による効果を表す限り、その剤形、投与経路及び投与方法に特に制限はされない。
【0073】
一方、前記発現ベクターは感染(infection)、形質感染(transfection)又は形質導入(transduction)等の当業界に公知の方法により、発現型で標的細胞内に導入させることができる。
【0074】
プラスミド発現ベクターを利用した遺伝子伝達方法は、哺乳動物細胞に直接的にプラスミドDNAを伝達する方法にして、FDAから承認されたヒトに使用できる方法である(Nabel, E. G., et al., Science, 249:1285-1288, 1990)。プラスミドDNAはウイルスベクターとは異なり、均質に精製できる長所を有する。本発明で使用できるプラスミド発現ベクターには、当業界に公知の哺乳動物発現プラスミドを使用できる。例えば、これに限定はされないものの、pRK5(ヨーロッパ特許第307,247号)、pSV16B(国際特許公開第91/08291号)及びpVL1392(PharMingen)等が代表的である。前記プラスミド発現ベクター(plasmid expression vector)は当業界に公知の方法、例えば、これに限定はされないものの、形質感染、微細注射、形質導入、細胞融合、カルシウムホスフェート沈澱法、リポソーム媒介された形質感染(liposome-mediated transfection)、DEAEデキストランー媒介された形質感染(DEAE Dextran-mediated transfection)、ポリブレンー媒介された形質感染、電気穿孔法(electroporation)、遺伝子銃(gene gun)及び細胞内に遺伝子を流入させる為の他の公知方法により、標的細胞内に導入できる(Wu et al., J. Bio. Chem., 267:963-967, 1992; Wu and Wu, J. Bio. Chem., 263:14621-14624, 1988)。
【0075】
さらに、前記核酸を含むウイルス発現ベクターはこれに限定はされないものの、レトロウイルス(retrovirus)、アデノウイルス(adenovirus)、ヘペスウイルス(herpesvirus)及びアビポックスウイルス(avipox virus)、レンチウイルス(Lenti-virus)等が含まれる。前記レトロウイルスベクターはウイルス遺伝子が全て除去されているか、又は変更されて非−ウイルス蛋白質のウイルスベクターにより、感染された細胞内で造れるように作製されたものである。遺伝子療法の為の、レトロウイルスベクターの主要長所は、多量の遺伝子を複製細胞内に伝達し、細胞DNA内に伝達された遺伝子を正確に統合し、遺伝子形質感染後連続的な感染が誘発されないことである(Miller, A.D., Nature, 357:455-460,1992)。FDAより認証されたレトロ内ベクターPA317癌ホトロフィックレトロウイルスパッケージ細胞を利用して製造したものである(Miller, A.D. and Buttimore, C., Molec. Cell Biol., 6:2895-2902,1986)。非−レトロウイルスベクターは、前記にて言及した通りのアデノウイルスがある(Rosenfeld et al., Cell, 68:143-155, 1992; Jaffe et al., Nature Genetics, 1:372-378, 1992; Lemarchand et al., Proc. Natl. Acad. Sci. USA, 89:6482-6486, 1992)。アデノウイルスの主要長所は多量のDNA断片(36kbゲノム)を運搬し、極めて高い力価で非−複製細胞を感染させ得る能力があると言うことである。さらに、ハピスウイルスもヒト遺伝子療法の為、有用に使用できる(Wolfe, J.H., Nature Genetics, 1:379-384,1992)。以外にも、公知の適切なうウイルスベクターが本発明に使用できる。
【0076】
さらに、前記KRSの発現を抑制する構造遺伝子は他の方法、例えば、局所的に、非経口、経口、鼻腔、静脈、筋肉内、皮下又は他の適切な手段により投与できる。特に、前記ベクターは標的組織の腫瘍細胞を治療する為の、有効量で標的癌又は腫瘍細胞内に直接注射し得る。特に、目、胃腸管、泌尿生殖器官、肺及び気管支システムのような膵臓(body cavity)にある癌又は腫瘍の場合には、本発明の構造遺伝子(又は構造遺伝子を含む発現ベクター)を含有する薬学組成物を癌又は腫瘍により、影響を受ける有腔器官(hollow organ)内に針、導管(catheter)又は他の種類の輸送チューブを利用して直接注入できる。この時、X−ray、ソノグラム(sonogram)、の光繊維システム(fiberoptic visualization system)のような映像装置が標的組織の位置確認と針又は導管の注入に使用される。その他に直接的に到達できないか、又は分析学的に分離できない腫瘍又は癌の場合には、本発明の組成物を血液循環系内に投与できる。
【0077】
さらに、本発明は
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;及び
(c)選別された製剤が癌転移又は癌細胞移動を調節するか否かをテストする段階を含む癌転移又は癌細胞移動調節製剤をスクリーニングする方法を提供する。
【0078】
前記スクリーニング方法は、当業界に公知の多様な生化学的及び分子生物学的技術が前記方法を実施することに利用され得る。前記技術は下記の文献に開示されている:(Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, N.Y., Second(1998) and Third(2000) Editions; and Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York(1987-1999)。
【0079】
好ましくは、前記試験製剤が先にKRSの生物学的活性を調節する能力があるかをアッセイする(1次アッセイ段階)。具体的に1次段階では試験製剤の存在下で分離されKRSの生物学的活性をアッセイ(assay)して前記ポリペプチドの生物学的活性を調節する調整製剤(modulating agent)を究明(identify)する。より好ましくは、次の段階を含め得る。
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;
【0080】
1次アッセイ段階では多くの生物学的活性に対する調節がアッセイできる。例えば、試験製剤はKRSの発現水準、例えば、転写又は翻訳を調節する活性があるか、否かに付いてアッセイでき得る。さらに、前記試験製剤はKRSの細胞内水準又は安定性、例えば、翻訳後修飾(post-translational modification)又は加水分解を調節する活性があるか否かに付いてアッセイされ得る。
【0081】
以降、前記1次アッセイ段階を通じてKRSの生物学的活性を増加させる調節製剤を究明した後、前記試験製剤がラミニン収容体(67LR)、さらには、癌転移又は癌細胞移動を調節する能力があるかをKRSの存在下でさらに、テストする。(2次試験段階)、例えば、前記試験製剤は、癌転移又は癌細胞移動を調節する活性があるか否に付いてさらにテストする。
【0082】
上記にて言及した通り、本発明の方法により究明されたKRS−調節製剤は癌転移、又は癌細胞移動を調節できる。若し、前記1次アッセイ段階で究明された試験製剤がKRS−調節製剤の細胞内水準(例:転写活性の変更により)を調節すれば、それは癌転移又は癌細胞移動を調節できる。
【0083】
一方、若し、試験製剤KRSの細胞内水準でない他の活性を調節すれば、KRSに対する前記試験製剤の治療効果が癌転移又は癌細胞移動を実際に調節するか、否かを確認することが必要である。例えば、KRSのリン酸化活性を調節する試験製剤はKRSのリン酸化活性の調節が癌転移又は癌細胞移動を調節できるか、否かを確認する為にテストを重ねる必要がある。
【0084】
前記1次及び2次段階全て変わらない(intact)KRS及びこれの断片、アナログ又は機能的同等物を使用できる。これらアッセイに使用できる断片は一般的に、KRSの生物学的活性を一つ以上保有する。好ましくは、KRSの断片は配列番号1の1-72のアミノ酸残基を含め得る。さらに、前記断片又はアナログを含む融合蛋白質が試験製剤の為のスクリーニングに使用され得る。KRSの機能的同等物はアミノ酸欠失及び/又は挿入及び/又は置換を有するものの、KRSと同一な生体活性を保有することにより、本発明のスクリーニング方法を実施することに使用できる。
【0085】
当業界に通常的に実施されている多様なアッセイ等がKRSを調節する製剤を究明に使用できる。好ましくは、前記製剤は細胞基盤アッセイシステム(cell based assay system)でスクリーニングされ得る。例えば、スクリーニングする為の、典型的な細胞基盤アッセイにおいて(つまり、2次試験段階で)、試験製剤の存在下でリポーター遺伝子活性(例:酵素活性)を測定し、これを試験製剤の不在下におけるリポーター遺伝子の活性と比較する。前記リポーター遺伝子は当業界に公知の任意の検出可能なポリペプチド(反応又はリポーターポリペプチド)、例えば、蛍光又は燐光(phosphorescence)により検出可能なポリペプチドやそれが保有している酵素活性により、検出可能なポリペプチドを符号化し得る。検出可能な反応ポリペプチド(detectable response polypeptide)、例えば、ルシファーラゼ、α−グルクロニターゼ(glucuronidase)、α−カラクトシターゼ、クロラムペニコールアセチル転移酵素、緑色蛍光蛋白質、強化された緑色蛍光蛋白質及びヒトに分泌されたアルカリンリン酸化酵素でもあり得る。
【0086】
細胞−基盤アッセイにおいて、試験製剤(例:ペプチド又はポリペプチド)は宿主細胞内に存在する他のベクターにより、発現され得る。或る方法では、試験製剤のライブラリが前記ベクターライブラリ(例:cDNAライブラリ)により符号化される。前記ライブラリは当業界に公知の方法を利用して製造することができ(Sambrook et al. and Ausubel et al., supra)、又は多様な産業的出処から得られ得る。
【0087】
前記記載した細胞−基盤アッセイ以外に、非−細胞基盤方法によつてもスクリーニングされ得る。前記方法は、例えば、モビリティーシフトDNA結合アッセイ(mobility shift DNA-binding assays)、メチル化及びウラシル干渉アッセイ(methylation and uracil interference assays)、DNase 及びヒドロキシーラジカルフットプリンティング分析(DNase and hydroxyl radical footprinting analysis)、蛍光偏光(fluorescence polaryzation)及びUV交差結合(crosslinking)又は化学的架橋剤(cross-linkers)を含む。一般的な概要はAusubel等の文献に開示されている(Ausubel et al., supra, chapter 12, DNA-Protein Interaction)。核酸及びDNA/RNA結合蛋白質を含む共同−結合された蛋白質(co-associating protein)を分離する技術は、切断可能な架橋剤ジチオビス(dithiobis)(succinimidylpropionate)及び3,3′−ジチオビス(sulfosuccinimidyl-propionate)を含むUV交差結合又は化学的架橋剤を含む(McLaughlin, Am. J. Hum. Genet., 59:561-569, 1996; Tang, Biochemistry, 35:8216-8225, 1996; Lingner, Proc. Natl. Acad. Sci. U.S.A., 93:10712, 1996; and Chodosh, Mol. Cell. Biol., 6:4723-4733, 1986)。
【0088】
1次アッセイ段階:KRSを調節する製剤のスクリーニング
多くのアッセイシステムがKRSの調節因子の為の、試験製剤のスクリーニングに適用できる。上記にて言及した通り、前記スクリーニングは試験管内アッセイシステム又は細胞−基盤アッセイシステムを利用できる。このスクリーニング段階で、試験製剤はKRSとの結合、KRSの細胞内水準変更、又はKRSの他の生物学的活性の調節に対してスクリーニングされ得る。
【0089】
1)KRSに結合する製剤のスクリーニング
1次スクリーニング段階でKRSと試験製剤の結合が測定できる。KRSに対する試験製剤の結合は、例えば、標識された試験管内蛋白質−蛋白質結合アッセイ、(electrophoretic mobility shift assays)、蛋白質結合を検出する為の免疫アッセイ、機能的アッセイ(リン酸化アッセイ等)等のような多様な方法でアッセイできる(U.S. Pat. Nos. 4,366,241:4,376,110; 4,517,288 and 4,837,168; and Bevan et al., Trends in Biotechnology, 13:115-122, 1995; Ecker et al., Bio/Technology, 13:351-360, 1995; and Hodgson, Bio/Technology, 10:973-980, 1992)。試験製剤はKRSとの直接的な結合、例えば、KRSに対する抗体によるKRSポリペプチドとの共同−沈殿(co-immunoprecipitation)を検出することにより、確認できる。さらに、試験製剤はKRSと試験製剤の結合を表し得るシグナル、例えば、蛍光キンチング(quenching)を検出することにより確認できる。
【0090】
競争アッセイ(competition assays)はKRSに特異的に結合する試験製剤究明に適当なフォーマット(format)を提供する。前記フォーマットで、試験製剤はKRSと結合するものとして既に公知の化合物との競争を通じてスクリーニングされる。公知の結合化合物は合成化合物でもあり得る。さらに、それはKRSを特異的に認識する抗体、例えば、KRSに対する単一クロン抗体でもあり得る。若し、試験製剤が公知のKRSと公知の化合物の結合を阻害すれば、前記試験製剤はKRSにも結合する。
【0091】
多様な種類の競争アッセイらが当業界に知られている。例えば、固体状直接又は間接的酵素放射免疫測定法(solid phase direct or indirect radioimmunoassay, RIA)、固体状直接又は間接的酵素放射免疫測定法(EIA)、サンドイッチ競争アッセイ(Stahli et al., Methods in Enzymology, 9:242-2453, 1983);固体状直接バイオチンーアビチンEIA(Kirkland et al., J. Immunol., 137:3614-3619, 1986);固体状直接標識化されたアッセイ、固体状直接標識化されたサンドイッチアッセイ(Harlow and Lane, Antibodies, A laboratory Manual, Cold Spring Harbor Press, 1988);125 Iを利用した固体状直接標識RIA(Morel et al., Mol. Immuno., 25(1):7-15, 1988;固体状直接バイオチンーアビチンEIA(Cheung et al., Virology, 176:546-552, 1990);及び直接標識化されたRIA(Moldenhauer et al., Sacnd. J. Immunol., 32:77-82, 1990)、一般的に、前記アッセイ等は非標識化された試験製剤及び標識化された対照化合物を含めている細胞、又は個体表面に結合されている精製されたポリペプチドの利用を含む。競争的阻害は試験製剤の存在下で個体表面又は細胞に結合された標識の量を結合することにより測定される。競争アッセイにより究明された調節製剤は対照化合物と同一なエピトープに結合する製剤及び立体阻害(steric hindrance)が起こるようにする為、対照化合物により結合されたエピトープに十分に近接した隣接エピトープと結合する製剤を含む。通常的に競争阻害が過度に存在する時、一般的な標的ポリペプチドに対する対照化合物の特異的な結合が少なくとも50又は75%阻害されるであろう。
【0092】
前記スクリーニングアッセイは不溶性又は溶解性フォーマットでもあり得る。不溶性アッセイの一例はKRS又はこれの断片を固体状マトリックスで固定化したものである。以降、試験製剤が結合するに十分な時間の間、固体状マトリックスを試験製剤と接触させておく。以降、固体状マトリックスから結合されない物質は、洗滌して除去後、固体状に結合された製剤の存在を確認する。前記方法は、固体状マトリックスから結合された製剤を溶離して製剤を分離する段階をさらに含め得る。両者択一的に、KRSを固定化させる他の方法は製剤を固体状マトリックスに結合させた後、KRSを添加することである。
【0093】
溶解性アッセイは前記記載された幾つかの結合ライブラリスクリーニング方法を含む。溶解性アッセイフォーマットにおいて、試験製剤やKRSは全て固体支持台に結合しない。試験製剤に対するKRS又はこれの断片の結合は、例えば、KRS及び/又は試験製剤の蛍光により測定できる。蛍光は内在的か(intrinsic)若しくは蛍光物質(fluorophor)を有する成分の標識により与え得る。
幾つかの結合アッセイから、KRS、試験製剤又は第3の物質(例:KRSに結合する抗体)は、与えられた条件で前記ポリペプチドの確認、検出及び定量を容易にする為に、標識された状態で提供できる。つまり、共有的に結合又は検出可能な標識やグループ、又は架橋可能なグループにリンクされて提供できる。これら検出可能なグループ等は検出可能なポリペプチドグループ、例えば、アッセイ可能な酵素又は抗体エピトープを含む。両者択一的に前記検出可能なグループは放射線同位元素(例:125 I,32P,35S)又は化学発光性又は蛍光性グループのような他の検出可能なグループ又はラベルから選択できる。類似して、前記検出可能なグループは基質(substrate)、補助因子(cofactor)、阻害剤又は親和リガンドでもあり得る。
【0094】
2)KRSの他の生体活性を調節する製剤のスクリーニング
KRSの試験製剤の結合は、試験製剤がKRSの調節者であることを示す。さらに、これは前記製剤が癌転移又は癌細胞移動を調節する為に、ラミニン収容体の生体活性を調節し得ることを提示する。従って、KRSと結合する試験製剤はラミニン収容体活性を調節する能力を有しているかをさらにテストすべきである。
【0095】
両者択一的に、KRSと結合する試験製剤はKRSに対するその活性を測定する為に、さらに、調査され得る。そのような活性の存在、性質又は範囲は活性アッセイにより、テストされ得る。前記活性アッセイはKRSに試験製剤の結合が実際にKRSに対する調節活性を有することを確認できる。極めて広く前記調節アッセイ等はKRSの活性を調節する試験製剤を究明する為に、独立的に使用し得る。(つまり、KRSに結合する能力をアッセイする初段階なしに)、一般的に、前記方法等はKRSの生物学的活性をテストするに必要とする他の物質又は試薬の存在又は不在下でKRSを含むサンプルに試験製剤を添加し、KRSの生物学的活性の変更を測定することを含む。KRSの酵素又は他の生物学的活性を調節する製剤化をスクリーニングするアッセイに加えて、前記活性アッセイはKRSの発現又は細胞内水準の変化の為の試験管内スクリーニング及び生体内スクリーニングを含む。
【0096】
2次試験段階:癌転移又は癌細胞移動を調節する製剤のスクリーニング
一旦調節製剤がKRSと結合するか、及び/又はKRSの生物学的活性(細胞内水準を含む)を調節するものとして究明されると、前記製剤は癌転移又は癌細胞移動を調節する能力を有しているか、否かに付いて、さらにテストされ得る。前記調節製剤による癌転移又は癌細胞移動の調節は一般的に、KRSの存在下でテストされる。若し、細胞−基盤スクリーニングシステムを利用する場合、KRSは宿主細胞に導入された発現ベクターから発現され得る。両者択一的にKRSはスクリーニングシステムで宿主細胞により内生的に供給されることもあり得る。
【0097】
さらに、本発明は
(a)試験製剤の存在下でKRS、ラミニン収容体(67LR)及び試験製剤を接触させる段階;及び
(b)前記試験製剤がKRSとラミニン収容体の相互作用(interaction)を調節するかをテストする段階を含むKRSと67LRの相互作用を阻害する製剤をスクリーニングする方法を提供する。
【0098】
前記製剤はKRSとラミニン収容体(67LR)の相互作用を促進又は強化するものでも有り得て、逆に前記相互作用を抑制又は弱化させるものでもあり得る。
【0099】
前記(b)段階のテストは前記調節製剤が接触されない細胞又はこれの溶解物における67LRとKRSの相互作用水準に対する前記調節製剤が接触された細胞又はこれの溶解物における67LRとKRSの相互作用水準の相対的な変化を測定する段階を含め得る。
【0100】
前記スクリーニング方法は、前記にて言及したような標識された試験管内蛋白質−蛋白質結合アッセイ(試験管内プールーダーウンアッセイ)、EMSA(electrophoretic mobility shift assays)、蛋白質結合の為の、免疫アッセイ、機能的アッセイ(リン酸化アッセイ等)、酵母−2ハイブリッドアッセイ、非免疫沈澱アッセイ、免疫沈澱ウェスタンブロットアッセイ、免疫−共同−位置化アッセイ(immuno-co-localization)等当業界に公知の多様な方法で実施し得る。
【0101】
例えば、バクテリアリプレクサーLexA、又は酵母GAL4のDNA-結合ドメイン及び酵母GAL4蛋白質のトランスアクチベイション(transactivation)、ドメインにそれぞれ融合された、KRSと67LR、又はこれら蛋白質の一部又は相同体(homologues)を発現する酵母を利用して酵母−2ハイブリッドアッセイを行い得る(KIM, M. J. et al., Nat. Gent., 34:330-336, 2003)。KRSと67LRの相互作用はLexA蛋白質又はGAL4のDNA-結合ドメインに結合された調節配列を有しているプロモーターの統制下で、リポータ遺伝子の発現を誘導するトランスアクチベータを再構成するようになる。
【0102】
前記リポータ遺伝子では前記記載の通り、当業界に公知の任意の検出可能なポリペプチドを符号化する遺伝子(例:CAT(chloramphenicol acetyltransferase)、ルシファラーゼ、ベータカラクトシターゼ、ベーターグルコシターゼ、アルカリンホスファターゼ及びGFP(green fluorescent protein)等を使用できる。若し、KRSと67LR、又はこれら蛋白質の一部又は相同体の相互作用は試験製剤により、促進又は強化される場合、前記リポータ遺伝子の発現は正常条件に比べてさらに増加される。逆に前記相互作用が試験製剤により、阻害されるか又は弱化される場合、前記リポータ遺伝子は発現されないか、又は正常条件に比べて少なく発現される。
【0103】
さらに、リポータ遺伝子では酵母の成長を可能にする蛋白質(つまり、前記リポータ遺伝子が発現されない時、酵母の成長が抑制される)を符号化するものとして選択され得る。例えば、アミノ酸又は窒素塩基の為の生合成過程に関係ある酵素を符号化する栄養要求性(auxotropic)遺伝子(例:ADE3,HIS3等の酵母遺伝子又は他の種由来の同等遺伝子)でもあり得る。このシステムで発現されたKRSと67LR、又はこれら蛋白質の一部又は相同体の相互作用は試験製剤により、抑制又は弱化される場合、リポータ遺伝子は発現されないか、又は少なく発現する。従って、前記条件下で酵母の成長は停止又は遅くなる。このようなリポータ遺伝子の発現による効果は、肉眼や装置(例:顕微鏡)を通じて観察でき得る。 従って、本明細書では、肺癌又は乳癌患者を対象として67LR及びKRSの過発現可否を確認した結果、67LRが過発現された場合、KRSの過発現と肺癌又は乳癌との関連性が高いことが分った。(表1参照)。従って、本発明は
(a)試料で67LRの過発現可否を分析する段階;及び
(b)67LRが過発現された試料からKRSの過発現可否を分析する段階を含む肺癌又は乳癌を診断する方法を提供する。
【0104】
診断の為の、試料の採取及び処理と67LR及びKRSの過発現可否の分析は、通常的な分子生物学的技法を使用することができ、これは前記にて記載した通りである。
【0105】
本発明に記載された図面に対して下記の通り説明する
【0106】
図1乃至図6はヒトKRSとラミニン収容体の特異的相互作用を確認したものである。図1で全長のヒトKRSと37LRP/p40間の相互作用はイーストツーハイブリッド分析を通じて実施された。多重−ARS複合体の二つの酵素要素であるAIMP1及びAIMP2はそれぞれ陽性及び陰性対照群として使用された。陽性相互作用はx-galを含有した酵母培地で青色コロニーの形成により標識された。図2で、37LRPは35Sメチオニンの存在下でin vitro翻訳により合成され、これらをGST-KRS又はGSTと反応した(pull-down)、GST-KRSと共沈澱(co-precipitated)37LRPは自家放射撮影法を利用して測定した。図3で、KRS及び37LPRの存在に連関されたペプチド部位はイーストツーハイブリッド分析を通じて測定された。N−末端の細胞内部位(54乃至113番目アミノ酸)及びC末端の細胞外部位(137乃至210番目アミノ酸)を有している296個アミノ酸を有する37LRPは膜間ドメイン(113 乃至137番目アミノ酸)により区分される。ヒトKRS(597個のアミノ酸)のN−末端特異的拡張(約70個のアミノ酸)部位の次にOB-foldアンチコドン−結合性ドメイン(約70乃至214アミノ酸)と触媒ドメイン(約220乃至574アミノ酸)が位置している。図4で、Myc-KRSに感染されたA549細胞を溶解して抗−Myc及び抗−ラミニン収容体抗体で免疫ブロットを実施した。Myc-KRSを抗−Myc抗体と免疫沈澱させ、67LR及び37LRPで共沈澱させて免疫ブロットを行った。37LRP及び67LRの特異的ブロットの為、多重抗体であるH-141及びF-18(Santacruz)がそれぞれ使用された。(WCL:whole cell lysate)図5で、Myc-KRSで感染されたA549細胞の溶解物を指示した抗体にして、ウェスタンブロットを施行した。細胞は細胞質(C)及び膜分画(M)で分離され抗−Myc抗体で免疫沈澱され、37LRP及び67LR共沈澱されたのをウェスタンブロットで調査した。対照群としてIeGを使用した。図6で、ラミニン(10μg/ml,1h)処理した場合、67LRとKRS間の結合が増えたことを確認した。これを確認する為に、67LRを認識するantibody(abcam.cat# ab2508)で免疫沈澱(immunoprecipitation)し、左側のIgGラベルは兎から得た全体IgG(total IgG)として陰性対照群として使用された。0%SDS PAGEを遂行した後、PVDF膜に移してKRSと67LRを認識する抗体でそれぞれ免疫ブロットをした。
【0107】
図7乃至図12はラミニン−誘導性膜転移(translocation)及びKRSのリン酸化に対しては確認したものである。図7で、A549細胞をラミニン(10μg/ml)で処理してウェスタンブロットを利用して67LR、37LRP及びKRSの量(level)を時間帯別に観察した。Hsp90及びカドヘリン(Cad)をそれぞれ細胞質及び膜のマーカーとして使用した。図8で、A549細胞を1時間かけてラミニン処理、又は無処理して抗−67LR(MLuC5,Santacruz,sc-59732)(洗滌)及びKRS抗体(緑色)を使用して免疫蛍光染色を実施した。図9で、A549細胞をPCL-ガンマ、PKC及びPI3Kをそれぞれ抑制するU73122(U)、スタウロスポリン(staurosporin,ST)及びLY294002(LY)で3時間処理してラミニンで1時間処理後、これらリン酸化酵素抑制剤が67LR及びKRSの細胞質及び膜にどのような影響を及ぼすかを確認した。図10でA549細胞をMyc-KRSで感染させ、24時間培養した。以降、表示された(indicated)薬物で処理して前記の通りラミニンで処理した。Myc-KRSで免疫沈澱し、抗-p-Thr,-Ser,及び-Tyr抗体で免疫ブロットを実施した。図11で、A549細胞をMyc-KRSで感染させ、24時間培養した。感染されたLY294002で3時間処理し、ラミニンで1時間処理した。Myc-KRSは免疫沈澱され、67LRの共沈澱はウェスタンブロットで調査した。免疫沈澱法の対照群としてIgGを使用した。図12で、前記の通り、A549細胞をラミニン及びLY294002K存在又は不在下で培養した。EPRS(glutamyl-prolyl-tRNA sybthetase)はこれに特異的な抗体(AbCam)と免疫沈澱され、KRSの共沈澱はウェスタンブロットで確認した(上)。免疫−消盡上層液(immune-depleted supernatant:ID)で抗KRS及びEPRS抗体とウェスタンブロットを行った。
【0108】
図13乃至図17は、KRSは膜−結合性67LRを安定化させる点を確認したものである。図13で、A549細胞をsi-対照群(si-cont)又はsi-KRSで感染させ、ラミニンの存在又は不存在下で培養した。以降、細胞等を細胞質と膜分画で分離し、それぞれの分画における67LR及びKRSの量をウェスタンブロットで確認した。カドヘリン(赤色)及びhsp90をそれぞれ細胞質及び膜のマーカーとして使用した。図14で、A549細胞において膜結合性67LRの量は抗−LR抗体(MluC5)を利用して遊細胞分析器で観察された。空ベクター又はKRSプラスミドで感染された細胞等を24時間培養した。(上)67LRの量において、KRS抑制の効果を確認する為に、細胞をsi-KRS又はsi-対照群で感染させ、48時間培養した。(下)図15で、EV(空ベクター)又はKRSで感染させたA549細胞をG418で1週間選別して、抗−LR抗体を利用した免疫蛍光染色を利用して67LRの細胞内分布を確認した。膜に分布したLRは白色矢印で強調した。図16で、A549細胞をシクロヘキサミドで処理して新たな(de novo)蛋白質合成を抑制して細胞膜と細胞質内の67LR量において、KRS量の効果をウェスタンブロットで調査した。図17で67LRの細胞的安定性において、KRSの重要性はパルス−チェース実験を通じて調査した。293細胞をsi-KRS又はsi-対照群で感染させ、放射線メチオニンを1時間反応させた。67LRを67LRに特異的に反応する抗体(F-18,Santacruz)と免疫沈澱させ、SDS-PAGEにより分離させ後、自家放射撮影法を実施した。特異性siRNAによるKRSの抑制はウェスタンブロットにより行われ、ローデイング対照群としてチューブリンを使用した。
【0109】
図18乃至図22はKRSは67LRを通じて細胞移動(cell migration)及び癌転移を促進することを確認した。図18でA549細胞を表示されたプラスミドで感染させ、ラミニンの不存在又は存在下で培養し、これらの細胞移動に対する効果はトランスウェルチャンバーで移動した細胞を測定して確認した。膜を通過した細胞の数を計数して各パネルに表示した。実験は3回繰返した。図19で、前記の通り処理した細胞等をMMP-2活性及び量測定のためにそれぞれザイモグラフィー及びウェスタンブロット実施に使用した。図20で、乳癌細胞株である4T-1細胞を表示された(indicated)siRNAで感染させ、Balb/Cマウスの背に皮下注射した。21日経過後、マウスの肺を摘出して径1mm以上の腫瘍結節を計数した。図21で、外因性KRS(KRS-1,及びKRS-2)を発現する2種の互いに異なる4T-1細胞を前記の通り、接種し、30日経過後計数した。空ベクターを感染させた細胞を対照群として使用した。図22で、肺癌(上)及び乳癌(下)組織におけるKRS及び67LRの発現量をこれらそれぞれの抗体を利用した免疫組織化学染色法を使用して比較した。39個の肺癌及び40個の乳癌組織を利用して抗-KRS及び抗−67LR抗体で免疫組織化学染色を実施し、これらの発現量は正常組織のそれと比較された(各組織当り9個の試料)。ここに見える同一患者の代表的な対らはKRSと67LRの過発現を表す。KRSと67LRの統計学的相関関係は表1に表している。
【0110】
図23で、67LRの膜における量は発現に依存する点を示している。図23で、表示されたプラスミドで感染された293細胞を細胞質及び細胞膜分画で分離し、それぞれの分画で67LR、37LRP及びKRSの量を常用する抗体を利用したウェスタンブロットを行い確認した。
【0111】
図24乃至図27は、細胞移動、蛋白質合成及び細胞周期において、細胞内及び細胞のKRSの効果を確認したものである。図24で、ラミニン不存在下で培養されたA549細胞の移動は、トランスウエルチャンバーより移動した細胞を測定して確認した。図25で、KRSの化学走性(chemotactic)活性を見る為、表示された濃度でKRSを含有した無血清培地トランスウエルチャンバーの下のチャンバーに入れて、A549細胞を上のチャンバーに入れて培養した。培養6時間後、移動した細胞を計数した。図26で、A549細胞でKRSの量はsiRNAと外因性KRSの導入により、下落調節又は上昇調節された(図26及び図27パネル)。感染された細胞はそれぞれ48時間及び24時間培養され、無ーメチオニン培地で1時間培養して栄養欠乏状態を造り、放射線標識がなされたメチオニンで2時間標識した。洗滌後、細胞を4時間培養して0.5%トリトンX-100溶解液で溶解させて、放射線活性を液状閃光計数機(liquid scintillation counting)で測定した。図27で、A549細胞を表示された通りに感染させ、固定させてプロピジウムアイオダイド(Propidium iodide)で染色して流細胞分析器で分析した。
【0112】
図28乃至図30は癌転移において、KRS抑制の効果を確認したものである。
【0113】
図28で、標的蛋白質の発現において、si-KRS及びsi-DRSの効果はウェスタンブロットにより調節された。ローディング対照群としてはチューブリンを使用した。図29で、si-RNAが感染された細胞(1×106 )を前記の方法のように注射し、21日経過後腫瘍の大きさ及び嵩を測定することにより、1次腫瘍増殖においてKRS及びDRS抑制の効果を調査した。それぞれのグループは5匹のマウスを使用した。図30で、前記のマウスより摘出された肺は10%ホルマリン溶液に固定された。転移性腫瘍結節及び数は見られる通りである。
【0114】
図31乃至図33は癌転移において、KRS過発現の効果を確認したものである。図31で、KRS-1及びKRS-2細胞株の過発現はウェスタンブロットにより観察された。図32で、1次腫瘍増殖におけるKRS過発現の効果は互いに比較された。図33で、腫瘍転移におけるKRS過発現の効果は接種後30日目に調査された。それぞれのグループは4匹のマウスを使用した。
【図面の簡単な説明】
【0115】
【図1】図1はヒトKRSと37LRP/p40間の相互作用をイーストツーハイブリッド分析を通じて確認したものである。
【図2】図2はヒトKRSと37LRPの相互作用をプルダウン分析を通じて確認したものである。
【図3】図3はヒトKRSと37LRPの相互作用部位を確認したものである。
【図4】図4はKRSの67LR及び37LRPとの結合を確認する為に、Myc−KRSに感染されたA549細胞に対して抗−Myc及び抗−ラミニン収容体抗体を利用して、免疫ブロット分析をした結果である。
【図5】図5はKRSの67LR及び37LRPとの結合を確認する為に、Myc−KRS で感染されたA549細胞の溶解物に対するウェスタンブロット分析をした結果である。
【図6】図6は、ラミニン処理に伴うKRS及び67LRの結合を免疫沈澱で確認したものである。
【図7】図7は、A549細胞にラミニン処理の際、67LR,37LRP及びKRSの量(level)ウェスタンブロットで確認したものである。
【図8】図8は、A549細胞をラミニン処理又は無処理の際、67LR及びKRSの発現を免疫蛍光染色で確認したものである。
【図9】図9は、リン酸化酵素抑制剤が67LR及びKRSの細胞質及び膜における発現に及ぼす影響を確認したものである。
【図10】図10は、KRSを発現するA549細胞でラミニン及びリン酸化酵素抑制剤処理の際、リン酸化程度をp−Thr,−Ser,及び−Tyr抗体で免疫ブロットを実施して確認したものである。
【図11】図11は、KRSを発現するA549細胞でリン酸化されたKRSの67LRとの結合可否をウェスタンブロットで確認したものである。
【図12】図12は、KRSとEPRSの結合にラミニンが及ぼす影響をウェスタンブロット確認したものである。
【図13】図13は、si−対照群又はsi−KRSの形質感染の際、67LR及びKRSの量をウェスタンブロットで確認したものである。
【図14】図14は、A549細胞において、膜−結合性67LRの量を流細胞分析器で確認したものである。
【図15】図15は、EV(空ベクター)又はKRSで感染させたA549細胞に対して67LRの細胞内分布を免疫蛍光染色で確認したものである。
【図16】図16は、新たな蛋白質合成が抑制されたA549細胞で細胞膜と細胞質内の67LR量において、KRS量の効果をウェスタンブロットで確認したものである。
【図17】図17は、67LRの細胞的安定性にKRSが及ぼす影響をパルス−チェース実験で確認したものである。
【図18】図18は、KRS及び/又は67LRの発現抑制時、細胞移動に及ぼす影響を確認したものである。
【図19】図19は、KRS及び/又は67LRの発現抑制時、MMP−2活性及び量をザイモグラフィー及びウェスタンブロットで確認したものである。
【図20】図20は、4T−1細胞株が移植されたマウスでKRS発現抑制の際、腫瘍結節の数を表したものである。
【図21】図21は、4T−1細胞株が移植されたマウスでKRS発現増進の際、腫瘍結節の数を表したものである。
【図22】図22は、肺癌及び乳癌組織におけるKRS及び67LRの発現量を免疫組織化学染色法で確認したものである。
【図23】図23は、67LRの膜における量にKRS発現が及ぼす影響をウェスタンブロットで確認したものである。
【図24】図24は、ラミニン不存在下で培養されたA549細胞の移動を測定したものである。
【図25】図25は、細胞移動に対するKRSの化学走性(chemotactic)活性を測定したものである。
【図26】図26は、A549細胞でsiRNAと外因性KRSの導入に伴うKRSの量及び細胞内全体蛋白質合成量を確認したものである。
【図27】図27は、A549細胞でsiRNAと外因性KRSの導入に伴うKRSの量及び細胞周期を確認したものである。
【図28】図28は、標的蛋白質の発現において、si−KRS及びsi−DRSの効果をウェスタンブロットで確認したものである。
【図29】図29は、腫瘍細胞移植に伴う1次腫瘍増殖において、KRS及びDRS抑制の効果を確認したものである。
【図30】図30は、腫瘍細胞移植に伴う転移性腫瘍結節及び数を確認したものである。
【図31】図31は、KRS−1及びKRS−2細胞株のKRSの過発現をウェスタンブロットで確認したものである。
【図32】図32は、腫瘍細胞移植に伴う1次腫瘍増殖において、KRSの過発現の効果を確認したものである。
【図33】図33は、腫瘍細胞移植に伴う転移性腫瘍結節及び数を確認したものである。
【発明を実施するための形態】
【0116】
以下、本発明を実施例により詳細に説明する。
ただし、下記実施例は本発明を例示するもので、本発明の内容がこれに限定されるものではないものの、
【0117】
<実験方法>
1.細胞培養及び材料等
A549及びHEK293細胞は、ATCCから購入した。マウス乳癌(mammary carcinoma)4T-1細胞株は金ソンチン博士(カチョン医科大学)より得た。10%牛胎児血清(Fetal bovine serum, FBS)及び1%抗体を含む。RPMI(A549細胞及び4T-1細胞に対する)及びDMEM(Dulbecco's Modified Eagle Medium,他の細胞に対する)培地を細胞培養に使用した。37LRPを符号化するpcDNA3.1ベクターはタチバナヒロフミ博士(九州大学)から得た。Myc-が付着された(Myc-tagged)ヒトKRS及びDRSはpcDNA3ベクターのEcoRI/XhoI制限酵素部位にクローニングした。齧歯類KRS cDNAはRT-PCRで確保し、pcDNA3.1ベクターのHindIII/XhoI制限酵素部位にクローニングした。齧歯類及びヒトKRS及びDRSにターゲッティング(targeting)するsiRNAはInvitrogen社より購入した。siRNAの配列は要請すれば提供できるであろう。遺伝子ポーター(Gene porter, GTS)及びリポペクタミン2000(invitrogen)は形質感染試薬として使用された。LY294002及びスタウロスポリン(stauroaporin)はCalbiochemから購入し、サイクロヘキシミド(cyclohexiumide)及びラミニン(laminin, Engelbreth-Holm-Swarm murine sarcoma)はSigmaから購入した。
【0118】
2.免疫沈降及びウェスタンブロット
細胞を150mM NaCl、0.5%トリトンX-100, 0.1%SDS及び蛋白質分解酵素阻害剤を含む20mM Tris-HClバッファ(pH7.4,溶解バッファ)で溶解した。蛋白質抽出物を正常IgG及び蛋白質Gアガロスと2時間培養した後(incubation)、非特異的にIgGに結合した蛋白質を除去する為に遠心分離した。上澄液を精製された67LR抗体(F-18, Santacruz)と混合した後、揺さぶりながら4℃で2時間培養し、蛋白質Aアガロスを混合した。氷で冷たく冷却した溶解バッファを利用して3回洗滌し、沈澱物をSDSサンプルバッファに溶解させ、SDS-PAGEで分離した。互いに異なる分画でKRS及びLRの結合を確認する為に、pcDNA3.1-Myc-KRSで形質感染し、proteoextractキット(Calbiochem)を利用して製造社の指針に従って、プラスマメンブレーンとサイトプラズム分画を分離したあと、前記の通り、共同−免疫沈降を行った。蛋白質水準を分析する為に、蛋白質を細胞から抽出した後、10%SDS-PAGEで分離した。別に言及しない限り抗-LR抗体(Abcam, ab2508)を37LRP及び67LRの同時免疫ブロッティングに使用した。hsp90及びパンーカドヘリン(Pan-cadherin)に対する抗体はSantacruzから購入した。
【0119】
3.フローサイトメトリー
細胞サイクルをアドレス(address)する為に、培養された細胞を表示されたベクター又は化合物で形質感染又は処理して、70%エタノール4℃で1時間固定した後、氷で冷却したPBSで2回洗滌した。その後細胞をpropidium iodide(50μg/ml)、0.1% sodium citrate,0.3%NP40及びRNaseA(50μg/ml)で40分間染色した後、プローサイトメトリ(FACS Calibur, Beckton-Dickinson)を行った。それぞれの染色に対して、Cell Quest Proソフトウェアーを使用して20000細胞を分析した。細胞表面の67kDLRの量を分析する為に、1×106細胞をIgG又は67LRの細胞外ドメインを認識する抗−LR抗体(MLuC5,1ug)と培養した後、FITC2次抗体と培養した。PBSで洗滌してサンプルをFACSでスキャンした。
【0120】
4.免疫蛍光染色又は免疫組織化学染色
9mmカバースリップ(cover slip)上のA549細胞を70%メチルアルコールで固定した後、冷たいPBSで簡単に洗滌した。1%CAS,3%BSA及び0.5%triton X-100を含むブロッキングバッファで30分間処理した後、細胞をKRSに対する抗体(Abcam)、及びMLuC-5に対する抗体(Santacruz)で1時間培養した。アレキサ488及び568(invitrogen)を添加した後、室温で30分間処理した。冷たいPBSで30分間洗滌し、標本をレーザースキャニングマクロスコーピで観察した。乳癌及び肺癌に対する組織配列スライド(tissue array slide)をSuper-Biochip(韓国)から購入し、67LR及びKRSの発現水準を確認する為に、文献(Park, S. G. et al. Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. U S A 102, 6356-6361(2005))言及された相応する抗体と免疫組織化学染色を行った。67LR及びKRSの発現間の相関関係を計算する為に、ピアソンχ2テスト及びスチューデントtテストを使用して統計的分析を行った。P値が<0.05であれば意味あるものと見做した。全ての統計的分析はSPSS v11.5ソフトウェアー(SPSS,Chicago,Ill)を使用して行った。
【0121】
5.パルス−チェース実験(Pulse-chase experiment)
293細胞をリポベクターミン2000を利用して si-KRS又はsi対照群で形質感染させた。これをメチオニンが含まれていない培地で1時間培養後、[35S]メチオニン(50μCi/ml)を添加して1時間培養した。新鮮な培地で放射線メチオニンを洗い落とし、67LRをこれに特異的な抗体で免疫沈降させ、12%SDS-PAGEで分離し、BAS(FLA-3000,Fujifilm)を利用して露出させた(autoradiography)。67LRの量はMulti-gaugeプログラム(V3.0,Fujifilm)を利用して測定した。
【0122】
6.イーストツーハイブリッド(yeast two hybrid)分析
ヒトKRSの多くの断片を符号化するcDNAを相応するプライマーを利用してPCRで得た。KRSに対するPCR産物をEcoRI及びXhoIで切断し、pEG202ベクター(LexA-融合蛋白質の製造の為)及びpJG4-5ベクター(B42-融合蛋白質の製造の為)の相応する位置に連結した。37LRP断片を符号化するcDNAはBarbara J,Ballermann博士(アルバータ大学)から得て、これをpJG4-5ベクターのEcoRI及びXhoI部位に挿入した。二つの融合蛋白質間の相互作用はX-gal-含有酵母培地上で青色コロニーの合成可否で分析した。
【0123】
7.In vitro結合分析
GST-KRS又はGSTを大腸菌ロゼタ(DE3)ストレンで発現させ、前記蛋白質抽出物を1%Triton X-100及び0.5% N-ラウリルサコシンが含有されたPBSバッフアで4℃で2時間グルタチオンーセパロスと混合した。ヒト37LRPはTNT Quick coupled Transcription/Translation(Promega)を使用し、pcDNA3-37LRPを鋳型に使用し、[35S]メチオニン存在下でin vitro translationにより合成した。合成された37LRPは前記 GST蛋白質混合物に添加し、1% Triton X-100, 0.5% N-ラウリルサコシン、1mM DTT, 2mM EDTA及び 300μMペニルメチルスルホニルフルオライドが含有されたPBSバッファで撹拌しながら4℃で4時間培養した後、0.5% Triton X-100を含有する同じバッファで6回洗滌した。その後セパロスビッドに結合された蛋白質を SDSサンプルバッフアで溶出し、SDS-PAGEで分離して放射線を測定した(autoradiograph)。
【0124】
8.細胞移動(cell migration)分析
細胞移動を先行文献(Park, S. G. et al. Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. USA 102, 6356-6361(2005))に記載した通り、ポリカボネート膜(8.0μm 空隙サイズ、Coster)を有する 24-ウェルトランスウェルチャンバで測定した。A549細胞を無血清培地に懸濁した後、各ウェル当り1X105 細胞の濃度で上位チャンバに入れた。表示された濃度の精製されたヒトKRS、ラミニン(10μg/ml)又はゼラチン(10μg/ml)を下位ウェルに入れ、細胞がCO2 培養器で37℃で6時間移動するようにした。細胞は70%メチルアルコールを含むPBSで30分間固定し、PBSで3回洗滌した。細胞はヘマトシリン(Sigma)で10分間染色した後、蒸留水で洗滌した。綿棒で膜の上の部分で移動しない細胞を除去した。膜をチャンバから分離した後、Gel Mount(Biomeda 米国)に乗せておいた(mount)。移動した細胞(膜の下位面に付着されたもの等)を顕微鏡下で(x20)4ヶ所を任意に選別して計測する方式で測定した。
【0125】
9.ザイモグラフィー(zymography)
表示されたsiRNA及び組換えKRS(又はDRS)を符号化するプラスミドで形質感染されたA549細胞をそれぞれ48時間及び24時間培養した後、10%FBSを含有する、RPMI培地に接触した(1x105 細胞/ウェル)、細胞を無血清RPMI培地で2時間飢餓処理(starving)した後、ラミニンを添加して10μg/mlで24時間培養した。20μlの培養培地を5xFODバッフア(4% SDS, 20%グリセロール及び0.01%ブロモフェノールブルーを含有する0.125M Tris-HCl, pH6.8)と混合した後、1mg/mlのゼラチンを含有する10%SDS-PAGEを行った。ゲルを2.5%トリトンx-100でそれぞれ20分間洗滌し、蒸留水でそれぞれ20分ずつ洗滌して反応バッフア(10mM CaCl2, 150mM NaCl, 1μM ZnCl2, 1% Triton X-100, 0.002% sodium azideを含有する50mM Tris-HCl, pH 7.5)と37℃で24時間培養した。ゲルを蒸留水で洗滌してクマシブルーR250で染色して35%メタノールで脱染(destain)した。
【0126】
10.生体内で癌転移実験
マウス乳癌4T-1細胞をsi KRS、si DRS又はsi−対照群で形質感染させ、24時間培養した。細胞(1x106)を6週令雌Balb/cマウスの背に皮下注射の方法で注入した。ターゲットに対するsiRNAの効果を形質感染後、48時以降に残った細胞で測定した。さらに、注入後3日から10日まで2日間隔で1次腫瘍(primary tumor)でターゲットに対するsiRNAの効果を相応する抗体を使用したウェスタンブロット分析で測定した。腫瘍の成長は週当り3回ずつ腫瘍の大きさをモニタリングした。これと同時に全体の体重も測定した。注入後21日目にマウスを殺し、1次腫瘍と肺をマウスから切除した。肺は10%ホルマリンで24時間固定した。肺に転移された腫瘍結節の数と大きさを測定し、径が1mmより大きい腫瘍結節に対しても別途記録した。1次腫瘍の質量も測定した。KRS過発現が腫瘍転移に及ぼす効果を確認する為に、齧歯類KRSベクター及び空ベクター(empty vector)を4T-1細胞に形質感染させ、安定的な形質感染体をG418の存在下で3週間培養して選別した。多くの単一コロニーを選びだして、ウェスタンブロットでKRS発現水準を比較した。対照群細胞より高い水準でKRSを発現する二つの異なるコロニー(KRS-1,KRS-2)を選択し、注入に使用した。注入後30日目にマウスを殺したことを除いては、前記方法と同一に以降実験を行った。
【0127】
<実験結果及び考察>
全長のKRSと37LRPとの特異的相互作用はイーストツーハイブリッド分析により確認された。LexA-KRSはKRSのパートナとして知られた(Kim, J.Y. et al. p38 is essential for the assembly and stability of macromolecular tRNA synthetase complex: Implications for its physiological significance, Proc. Natl. Acad. Sci. USA 99, 7912-7916(2002))AIMP2ばかりでなく、B42-37LRPと結合して青色コロニーを形成し、AIMP1に対してはそうではなかった(図1)。インビトロバインディングアッセイに対しては、[35S]メチオニンで表示された37LRPをGST-KRS又はGSTと混合し、グルタチオンーセファロースで沈澱させ、自家放射線撮影をした。37LRPはGST-KRSと共に沈澱したものの、GSTではなかった(図2)。イーストツーハイブリッド分析による消失地図(deletion mapping)でヒトKRSのN−末端延長部位とLRの C−末端細胞外ドメインがそれらの結合に関与することを確認した(図3)。
【0128】
細胞質内37LRPは膜−包埋性67LRで転換されるので、本発明者等はKRSが37LRP及び67LRに互いに別に結合するかを確認した。Myc-KRSを肺癌細胞株A549細胞に導入して、抗−Myc抗体と免疫沈澱させた。細胞溶解物のウェスタンブロット結果、67LRは37LRPより少ない量で存在した(図右側)。しかし、Myc-KRSは37LRPより67LRにより優先的に結合した(図左側)。本発明者等は以降、A549細胞質と原形質膜分画で分離してMyc-KRSは37LRP及び67LRの相互作用を確認した。37LRPは細胞質で67LRは原形質膜でそれぞれ優勢して観察された(図5右側)。KRSは細胞質においてより多くの量が観察されたものの、二つの分画全てにおいて存在した。二つの分画全てを抗−Myc-抗体と免疫沈澱させた時、例え、細胞質内少量の37LRPも沈澱されたものの、膜に存在する67LRが主にKRSと共に沈澱されたところ(図5左側)、これは膜に存在する67LR及びKRS間の信号的相互作用を示唆した。本発明者等は以降、KRSの細胞内分布がA549細胞のラミニン処理により、変化するかを細胞分画と免疫蛍光染色法を通じて調査した。ラミニン処理後、KRS及び67LRの膜内相互作用量は細胞質内のKRS及び37LRP量やこれらの発現には、殆ど変化なく漸次的に増加した(図7及び結果未図示)。免疫蛍光染色におても、さらに、67LR及びKRSがラミニン処理により膜側に移動することを説明してくれた(図8、それぞれ赤色及び緑色)。以降KRSの膜転座(translocation)が翻訳後変形(post-transcriptional modification)に関与するかを調査した。ホスホイノシチド3-OHキナーゼ(phosphoinositide 3-OH kinase, PI3K)(Shaw, L. M., Rabinovitz, I., Wang, H. H., Toker, A. & Mericurio. A.M. Activation of phosphoinositide 3-OH kinase by the alpha6beta4 integrin promotes carcinoma invasion. Cell 91, 949-960(1997))、蛋白質リン酸化酵素C(Protein kinase C, PKC)(Li, Y. Q. et al. Protein kinase C mediates the signal for interferon-gamma mRNA expression in cytotoxic T cells after their adhesion to laminin. Immunology 93, 455-461(1998))及びホスホリパーゼC−ガンマ(phospholipase C-gamma, PLC-gamma)(Vossmeyer, D., Hofmann, W., Loster, K., Reutter, W. & Danker, K. Phospholipase C-gamma binds alpha1beta1 integrin and modulates alpha1beta1 integrin-specific adhesion. J. Biol. Chem. 277, 4636-4643(2002); Kanner, S. B., Grosmaire, L. S., Ledbetter, J. A. & Damle, N. K. Beta 2-integrin LFA-1 signaling through phospholipase C-gamma 1 activation. Proc .Natl. Acad. Sci. USA 90, 7099-7103(1993))のような幾つかの互いに異なるリン酸化酵素等がラミニンにより、活性化されると知られている。このようなリン酸化酵素等の内、どのよなものがKRSのラミニン依存性膜転座に関与するかを確認する為に、本発明者等はそれぞれのキナーゼに特異的な抑制剤でそれぞれのリン酸化酵素を遮断し、この処理が如何にKRSのラミニン依存性膜転座に影響を及ぼすか、否かを確認した。膜分画でKRS及び67LRのラミニン依存的増加はPI3K抑制剤であるLY294002の存在下で抑制されるに反し、U73122又はスタウロスポリンで処理した細胞等は、対照群細胞と同様に67LRSのラミニン依存的増加を示した(図9上及び結果未図示)。これらの結果は、PI3KがKPSのラミニン誘導性リン酸化に連関されたものであることを暗示する。事実、チロシンでないトレオニンとセリンでリン酸化されたKRSはラミニン処理により、増加された反面、LY294002の存在下では抑制され、スタウロスポリンは如何なら影響も及ぼせなかった(図10)。本発明者等はさらに、KRSのラミニン誘導性リン酸化が67LRとの相互作用に必要であるか否かを調査した。LY294002処理はKRSと67LRのラミニン誘導性疾患を抑制した(図11)。細胞質KRSが多重−ARS複合体に固定されているので、本発明者等はさらに、KRSのラミニン依存的リン酸化がKRSと複合体の他の酵素合成成分であるグルタミルーフロリルーtRNA合成酵素(glutamyl-prolyl-tRNA synthetase, EPRS)と、共ー免疫沈澱することにより、多重−ARS複合体の結合に影響を及ぼすかを調査した。LY化合物の不在下で。ラミニン処理はKRSとEPRSの結合を減少させ、同時に免疫−消盡された溶解性分画(immune-depleted soluble fraction)のKRSを増加させた(図12上、下パネルの左側レーン)。これに反し、EPRSに結合したKRSは細胞がLY294002で前処理された時、ラミニン処理に影響が及ばなかったものの(図12上、下パネルの右側レーン)これはKRSのリン酸化がKRSと複合体のラミニン依存的結合に必要であることを示唆する。
【0129】
本発明者等は以降、KRSがA549細胞において、67LRの膜内存在量に影響を及ぼすかを確認した。67LRの量はラミニンにより増加するが、ラミニン効果はKRSがこれに対する特異的siRNAにより、抑制される時には消えるものの(図13左側)、これは67LRのラミニン依存的促進において、KRSの重要性を示唆する。本発明者等はさらに、遊細胞分析器を通じて膜に存在する67LRを調査した。膜に存在する67LRの量は細胞をKRSで形質感染させるか、又はsi-KRSで感染させた時、それぞれ増加及び減少した(図14)。ラミニン収容体の細胞内分布は空ベクター(EV)又はKRSに形質感染されたA549細胞間で、免疫蛍光染色を通じて比較された。ラミニン収容体は対照群と比較してKRS過発現細胞の膜部位に強く染色された(図15)。以降、KRS及び67LR間の陽性的相関関係(positive correlation)は、多様なKRS量に伴う膜及び細胞質内の67LR存在量を測定することにより確認された(図23)。
【0130】
本発明者等は、KRSがどのように67LRの膜内に存在量を増加させるかを調査した。KRSは37LRPから転写又は転換(conversion)を通じて67LRを促進することができる。しかしながら、KRSの感染はLR転写調節の暫定的役割を除いてはLR転写を増加させなかった(結果、未図示)。さらに、細胞質内で37LRPとよく結合しないので(図4、図5)、結果、が37LRPを67LRに転換する過程を促進するようである。本発明者等は、さらに、37LRPの変更が37LRPが67LRに転換されるにおいて、先行要件として知られているので(Landowski, T. H., Dratz, E.,A. & Starkey, J. R. Studies of the structure of the metastasis-associated 67 kDa laminin binding protein: fatty acid acylation and evidence supporting dimerization of the 32 kDa gene product to form the mature protein. Biochemistry 34, 11276-11287(1995); Buto, S. et al. Formation of the 67-kDa laminin receptor by acylation of the precursor. J. Cell. Biochem. 69, 244-251(1998))、KRSが37LRPの脂肪族アシル化(fatty acylation)を媒介するかを調査した。本発明において、KRSは37LRPの脂肪族アシル化に全く影響を及ぼさなかった(結果未図示)。結果、は膜に存在する67LRの細胞内安定性を増進できるので、本発明者等はKRSが膜に存在する67LRの食細胞作用(endocytosis)を妨害できるかを調査した。前記可能性を確認する為、本発明者等は、シクロヘキサミド(cyclohexamide)を利用して根源的に(de novo)蛋白質合成を阻止して、KRSが細胞膜と細胞質における67LRの存在量に影響を及ぼすかを実験した。KRSの発現がこれに特異的なsiRNAにより抑制される時には、67LRの膜内存在量が減少しながら同時に細胞質分画におけ量は増加した(図16左側)。これとは逆にKRSの過発現は前記の通り、膜内67LRの存在量を増加させた(図16右側)。このような結果を基に、KRSは67LRに細胞質内に再び入ることを抑制することにより、膜内67LRの存在量を増加させるようである。本発明者等は、さらに、パルス−チェース(pulse-chase)実験を通じて67LRの代謝転換(turn over)において、KRSの効果を調査した。初期蛋白質活性は放射線メチオニンで表示され、以降、シクロヘキサミドで抑制された。以降、67LRの消滅(disappearance)は時間的間隔をおいて、自家放射撮影により観察された。67LRはKRSがsiRNAにより抑制される時、急激に減少されたものの、この数値はsi対照群においては、時間間隔によって維持されたものと対照的である(図17)。従って、67LRの分解過程に対して追加的な研究が必要なものの、KRSは原形質膜で67LRと結合することにより、67LRの半減期を増加させ67LRの食細胞作用を抑制するものと見られる。
【0131】
本研究者等は以降、KRSの発現程度がラミニン依存的A549細胞移動に影響を及ぼすかをトランウェルメンブレーン分析を通じて調査した。対照群細胞の移動はラミニン処理により、平均6倍程促進された(図24及び図18)。しかしながら、ラミニン依存的細胞移動はKRSがsiRNAにより抑制された時減少した(図18,si-対照群及びsi-KRS)。これとは逆にKRS過発現はラミニン処理により、促進された細胞移動を増加させた(図18,EV及びKRS)。しかしながら、細胞移動においてKRSの効果は、ラミニン収容体がsiRNAにより抑制されると消えた(図18,si-LR,下パネル)。KRSは幾つかの癌細胞でサイトカインで分離されるので(Park, S. G. et al. Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. U S A 102, 6356-6361(2005))、本発明者等は細胞外因性KRSが細胞移動に影響を及ぼすかを調査した。A549細胞を互いに異なる濃度の精製されたKRSで処理した時、この分析でKRSの細胞外因性効果を除けば、細胞移動は殆ど影響を受けなかった(図25)。反面、細胞蛋白質の合成及び細胞周期は実験する間、KRSの抑制及び過発現に影響はされなかったものの、これはKRSの依存的細胞移動が前記過程の効果に起因しないことを示唆した(図26及び図27)。ラミニン処理がMMP-2(matrix metllo-proteinase-2)の活性化を招来するので(Givant-Horwitz, V., Davidson, B. & Reich, R. Laminin-induced signaling in tumor cells the role of the M(r) 67,000 laminin receptor. Cancer Res. 64, 3572-3579(2004))、 MMP-2のラミニン依存的活性化において、KRSの役割をin vitroザイモグラフィー分析を通じて調査した。MMO-2活性はラミニンにより活性化され、これはsi-KRSの存在下で抑制された(図19右側)。MMO-2の発現量はKRSに影響されなかった(図19下)。
【0132】
KRSが癌転移と関連された67LRを通じて細胞移動を促進できるので、肺に転移がよくなされるマウス乳癌4T-1細胞を使用してKRSの発現水準により、癌転移が影響されるのかを調査した。こめため、KRS又は多重-ARS複合体の他の構成要素であるDRS(aspartyl-tRNA synthetase)をそれらに特異的なsiRNAで発現を抑制し、如何に、KRS及びDRSの発現減少が癌転移に影響を及ぼすかを比較した。si-KRSの抑制効果をウェスタンブロットで確認した後(図28)、これら細胞それぞれ及びsi-対照群で処理した細胞をBalb/cマウスの背皮膚に皮下注射で注入した。注入された3種全て類似した質量及び嵩の腫瘍を発生させ、(図29)、KRSの水準が1次腫瘍の成長には影響しないことを知らせてくれた。肺を注入後21日目に分離し、転移性腫瘍結節(径1mmより大きいもの)の数を3グループ間で比較した。転移性結節の数は対照群及びDRSが抑制された細胞で得たのと比べてKRSの抑制により大きく減少した(図20及び図30)。逆に、前記方法によりKRSの過発現が癌転移を増進するかを調査した。我々はKRSを符号化するプラスミドの形質感染及びG418スクリーニングにより、KRSを安定的に過発現する4T-1細胞株を初めて構築した。構築された細胞株でKRS過発現はウェスタンブロットで確認し、空ベクターで形質感染された細胞等に比べて多量にKRSを発現する2個の互いに異なる細胞(KRS-1及びKRS-2)を選別した(図31)。これら細胞も類似した質量と大きさの1次腫瘍を生成した(図32)。これら細胞を注入した後、30日目に肺を検査した時、KRS過発現細胞二つとも対照群細胞に比べてより多くの結節を生成した(図21及び図33)。このような結果はKRSが生体内で癌転移を誘導することを暗示する。
【0133】
ラミニン収容体の癌特異的過発現は頻繁に観察されるので(Fontanini, G. et al.67-Kilodalton laminin receptor expression correlates with worse prognostic indicators in non-small cell lung carcinomas. Clin. Cancer Res. 3, 227-231(1997); Viacava, P. et al.The spectrum of 67-kD laminin receptor expression in breast carcinoma progression. J. Pathol. 182, 36-44(1997))、67LRの過発現が、さらにKRSの過発現と連関されているかを肺癌及び乳癌を例に67LR及びKRSの免疫組織化学染色により分析した。39個の検査した肺癌組織の中で67LR過発現は21事例(54%)で観察され、そのなかでKRS水準は19事例(約90%)で増加されていた(表1及び図22上位)。同様に40個の検査した乳癌患者の内、21事例で67LR過発現が観察された。これらの内、21事例全てKRSの水準が増加された(表1及び図22下位)。それらの同時−発現が転移において実際に連関されているかを確認すべきであるにも拘らず、二つの場合全て二つの蛋白質の発現間に密接な連関があらわれた。
【0134】
【表1】
【0135】
この際、前記表1に対しては下記の記載を参照できる:表1は腫瘍組織において67LRとKRS発現の相関関係を表したものである。表1では67LRの発現量がKRSの発現量と関係があるか否かを調べる為に、肺癌及び乳癌患者の組織マイクロアッセイ等を利用してそれぞれの抗体に対する免疫感染を実施し、前記二つの蛋白質等の相対的発現量を測定した。67LRの標識の為に、MluC5抗体が使用された。発現量は試料の染色濃度を通じて測定され、4個のグループに分類された(0,1,2,及び3点)。最終評価で試料等は正常グループ(0点又は1点)と過発現グループ(2点又は3点)に分類された。67LRとKRS発現の相関関係を評価する為の統計学的分析は、ピアソン χ2 テスト及びStudent tテストを通じて行った。この際、P値が0.05未満であれば(P<0.05)有効な数値として見做された。全ての統計学的分析はSPSS v11.5ソフトウェアー(SPSS, Chicago, Ill)を通じて行った。
【0136】
リポソム構成因子を含む多くの翻訳因子等が多面的であり(Wool, I. G. Extraribosomal functions of ribosomal proteins Trends Biochem. Sci. 21, 164-165(1996))、種々の腫瘍生成過程に関与されている(Lee, S. W., Kang, Y. S. & Kim, S.Multi-functional proteins in tumorigenesis: Aminoacyl-tRNA synthetases and translational components. Curr. Proteomics 3, 233-247(2006))。ここで我々は二つの翻訳因子であるKRS及びp40/37LRPが生体内で細胞移動及び癌転移に共に作用することを示した(図18乃至図22)。現時点ではこの二つの蛋白質の潜在的な連関進化的偶然か、若しくは以降究明される必要がある蛋白質合成において、生理学的理由を有しているかは知り兼ねない。ARS-複合体の構成要素の内、KRSは最も安定的な蛋白質であり、他の構成要素の安定性の為に必要であるものの(Han, J. M. et al. Hierarchical Network between the components of the multi-tRNA synthetase complex: Implications for complex formation. J. Biol. Chem. 281, 38663-38667(2006))、これは他の関係される蛋白質を安定化させるKRSの潜在能力を暗示するものである。ここで、我々はKRSの能力が67LRにおける細胞安定性にも拡大されることを示した(図17)。
【0137】
KRSの67LRとの連関は他の機能的暗示を有し得る。生理的条件下で、細胞質KRSの一部分がリン酸化され、多くの成長刺戟や生存信号により、ラミニン信号を媒介する67LRと結合する為に、原形質膜に移動される。癌細胞では過発現又はPI3Kのような過ー活性化された上位リン酸化酵素に因る結果にして、構造的な(constitutive)膜転移により、KRSの膜水準が非正常的に増加される。多分、このような過量のKRSが原形質膜を67LRを集めるか若しくは、排出することができる。さらに、PI3Kの活性化を減少させるのが頻繁に腫瘍成長及び転移と連関される点(Wymann, M. P. & Marone, R. Phosphoinositide 3-kinase in disease: timing, location, and scaffolding. Curr. Opin. Cell Biol. 17, 141-149(2005))及びラミPI3Kを通じて癌浸潤を増進する点(Baba, Y. et al. Laminin-332 promotes the invasion of oesophageal squamous cell carcinoma via PI3K activation . Br. J. Cancer 98, 974-980(2008))を念頭に置く価値がある。PI3Kの構造的な活性化は膜に移動させるKRSのリン酸化をリードすることができる。このよう条件等の一部又は全部が原形質膜での67LRの増加に寄与することができ、癌転移に対するラミニン信号伝達を増幅させ得る。癌の転移性拡張を調節する多くの研究が進行されてきた。このような点で、癌転移において67LRを通じたKRSの合成は癌診断及び治療の為の、従前には開拓されていない新たな窓を提供し得る。
【産業上の利用可能性】
【0138】
以上説明した通り、本発明者等はKRSが原形質膜に転座(translocation)され、67LRと相互作用することにより、腫瘍(又は癌)細胞の移動を促進して癌の転移に影響を及ぼすことを見い出した。さらに、マウスを利用したin vivo実験を通じてKRSの過発現又は発現抑制が癌の転移を調節し得ることを究明した。従って、本発明のKRSを利用して癌転移又は癌細胞移動を調節することができ、さらには、原形質膜のラミニン収容体(67LR)と関係した細胞内代謝を調節できる。本発明で究明したKRSとラミニン収容体の関係はこれと関係した多様な疾患又は疾病の治療、予防及び/又は診断に極めて有用に使用できる。
【技術分野】
【0001】
本発明はライシルtRNA合成酵素の新規な機能に関するものにして、より詳細には前記新規な機能を活用してライシルtRNA合成酵素の細胞内水準を調節して、癌転移又は癌細胞の移動を調節する方法、癌の予防又は治療の為に、KRSの発現を抑制する構成を有する発現べクターの用途、癌の予防又は治療の為に、KRS活性を阻害する製剤の用途、癌転移又は癌細胞移動調節製剤をスクリーニングする方法、及びKRS及び67LRの相互作用を阻害する製剤をスクリーニングする方法に関する。
【背景技術】
【0002】
癌(又は腫瘍)は、非正常的で非制御性で無秩序な細胞増殖の産物である。特に、破壊的な成長性、浸透性及び転移性があれば悪性に分類される。浸透性とは、周囲の組織を浸透又は破壊する性質にして、一般的に組織の境界をなす基底層を破壊させ腫瘍が局部的に伝播されることを意味し、たまたま体内の循環系にも流入される。転移とは、一般的にリンパ管(lymphotic)又は血管により、原発位置とは別の所に腫瘍細胞が広がることを意味する。転移はさらに、腸液性体腔又は他の空間を通じて直接伸長して腫瘍細胞を移動させることを意味する。
【0003】
現在、癌は主に3種の治療法、つまり、外科的な手術、放射線照射及び化学療法の内1種、又はこれらの組合せを通じて治療されている。手術は疾病組織を大部分除去することを含む。このような外科的手術は特定部位、例えば、乳房、結腸及び皮膚に位置した腫瘍を除去するには効果的ではあるものの、脊椎のように一部区域にある腫瘍を治療若しくは、白血病のような分散性腫瘍疾患を治療するのには使用できない。
【0004】
化学療法は細胞複製又は細胞代謝を崩壊させ、俗に、乳房、肺及び精巣の癌治療に多く使用されるものの、腫瘍疾病の治療に使用される全身性化学療法の副作用は癌治療を受ける患者等に最も問題となる。このような副作用の内で、目眩と嘔吐は最も一般的であり、深刻な副作用である。化学療法による副作用は患者の生命に大きな影響を及ぼし、治療に対する患者の順応性を急激に変化させ得る。さらに、化学治療剤と関連した副作用には一般的にこのような薬物投与の際、注意すべき容量制限毒性(DLT,Dose Limiting Toxicity)がある。例えば、粘膜炎は多様な抗癌剤、例えば、抗代謝物質細胞毒素剤である5−フルオロウラシル、メソトレキセート及び抗腫瘍抗生剤(例、ドキソルビシン)等に対する容量制限毒性がある。このような化学療法由来の副作用の内、大部分は甚だしい場合、入院を要するか若しくは痛症を治療する為に、鎮痛剤を必要とする場合もあり得る。このように化学治療剤及び放射線治療による副作用等は癌患者の臨床的処置の際、主要問題となっている。
【0005】
遺伝子治療とは、DNA組換え方法を利用して治療用遺伝子を患者の細胞内に導入させ、遺伝子結合を矯正又は細胞に新たな機能を追加させ、人体細胞の遺伝子変形を通じて各種遺伝疾患、癌、心血管疾患、感染性疾病、ウイルス細胞膜自家免疫疾患等のような疾患を治療又は予防する方法である。つまり、治療遺伝子を体内の望む臓器に伝達して細胞内で治療用或いは正常蛋白質が発現するようにして、疾病を治療することを遺伝子治療と言う。遺伝子治療は一般的な薬物による治療に比べて、優れた選択性を有することができ、他の治療法では調節し難い疾病の治療率及び治療速度を改善して、長期間適用することができる。遺伝子治療は単に疾病の症状を治療することに止どまることなく、疾病の原因を治療し、除去することである。このような遺伝子治療を効果的になす為には、治療遺伝子を望む標的細胞に伝達して、高い発現効率を得られるようにする遺伝子伝達技術が必要である。
【0006】
遺伝子伝達体は望む治療遺伝子を対象細胞に導入する為に、必要な媒介体として理想的な遺伝子伝達体は人体に無害で大量生産が容易で効率的に遺伝子を伝達することができ、持続的に遺伝子を発現できなければならない。遺伝子伝達体技術は遺伝子治療技術の核心要素にして、現在遺伝子治療に多く利用される代表的遺伝子伝達体には、アデノウィルス、アデノ付属ウイルス、レトロウイルスのようなウイルス性伝達体とリポソム、ポリエチレンイミンのような非ウイルス性伝達体がある。
【0007】
遺伝子治療戦略の内、腫瘍細胞を制御する戦略には腫瘍抑制遺伝子を利用する方法、腫瘍選択的殺傷ウイルスを利用する方法、自殺遺伝子を利用する方法、免疫調節遺伝子を導入する方法等がある。腫瘍抑制遺伝子を利用する方法は、多くの癌患者において遺伝子が欠損又は変形されているp53のような腫瘍抑制遺伝子を、原形で人体に伝達して癌を治療しようとする方法であって、腫瘍選択的殺傷ウイルスを利用する方法は、癌組織で変形されている腫瘍抑制遺伝子の活性を利用して、腫瘍細胞でのみ選択的に増殖できるウイルス遺伝子伝達体を人体に導入して、治療効果を挙げようとする方法にして、全て腫瘍細胞を直接殺傷させる戦略である。自殺遺伝子を利用する方法も、HSV-TKのような感受性遺伝子を導入して、腫瘍細胞の自殺を誘導する方法もこのような範疇に属する。反面、免疫調節遺伝子を導入する方法は、抗腫瘍免疫反応を増強せしめるインターロイキン12、インターロイキン4、インターロイキン7、ガンマインターフェロン、腫瘍壊死因子等の遺伝子を人体に伝達して、T細胞に腫瘍を認識するように誘発するか、又は腫瘍誘発蛋白質を遮断して細胞自殺を誘導して、間接的に疾病を治療する戦略である。アンジオスタチン、エンドスタチンのような血管生成抑制因子を発現させ、腫瘍への栄養供給を遮断して壊死させる方法もこのような間接的疾病治療戦略として広く認識されている。
【0008】
腫瘍の転移は、癌の生存率において決定的な要素である。67kDaのラミニン収容体(67LR)は、原形質膜に包埋された(embedded)非−インテグリン型収容体にして、癌の浸湿(湿潤)及び転移と関連がある。(Nelson, J. et al. The 67 kDa laminin receptor: structure, function and role in disease. Biosci. Rep. 28, 33-48(2008))。67LRはこれらの37kDa前球体(37LRP)の重合(dimerization)により生成されるものの、このような変化過程に対する分子的な細部機作はよく知られていない。37LRPはポリソーム形成に関与するリポソームサブユニットp40と同一である(Auth, D. & Brawerman, G. A 33-kDa polypeptide with homology to the laminin receptor: component of translation machinery. Proc. Natl. Acad. Sci. USA 89, 4368-4372(1992))。67LRはたまたま多様な種類の癌において、高濃度で観察される。(Nelson, J. et al. The 67 kDa laminin receptor: structure, function and role in disease. Biosci. Rep. 28, 33-48(2008); Menard, S., Castronovo, V., Tagliabue, E. & Sobel, M. E.New insights into the metastasis-associated 67 kD laminin receptor. J. Cell. Biochem. 67, 155-165(1997))。しかしながら、67LRの膜内存在における調節者や分子的機作は、未だに究明されていない。
【0009】
KRSは蛋白質合成の為、同族の(cognate)アミノ酸及び tRNAを接合させるaminoacyl-t-RNA synthetases(ARSs)に属する。これら原始酵素等は触媒的活性の他にも多面的(pleiotropic)特徴を有する(Park, S. G., Ewalt, K. L. & Kim, S. Functional expansion of aminoacyl-tRNA synthetases and their interacting factors: new perspectives on housekeepers. Trends Biochem. Sci . 30 , 569-574(2005))。反面、KRSを含む幾つかの哺乳類の ARSは構成蛋白質の多様な機能を調節する為に、分子貯蔵所(molecular reservoir)として作用する(Ray, P. S., Arif, A. & Fox, P. Macromolecular complexes as depots for releasable regulatory proteins. Trends Biochem. Sci. 32, 158-164(2007))巨大分子複合体を形成する(Lee, S. W., Cho, B. H.,Park, S. G. & Kim, S. Aminoacyl-tRNA synthetase complexes: beyond translation. J. Cell. Sci. 117, 3725-3734(2004); Han, J. M., Kim, J. Y. & Kim, S. Molecular network and functional implications of macromolecular tRNA synthetase complex. Biochem. Biophys. Res. Commun. 303, 985-993(2003))。ヒトKRSはRNAとその他蛋白質間の相互作用に関与する固有のN末端延長部位を含有している(Rho, S. B. et al. Genetic dissection of protein-protein interactions in multi-tRNA synthetase complex. Proc. Natl. Sci. Acad. USA 96, 4488-4493(1999); Francin, M., Kaminska, M., Kerjan,P. &Mirande. M. The N-terminal domain of mammalian Lysyl-tRNA synthetase is a functional tRNA-binding domain. J. Biol. Chem. 277, 1762-1769(2002))。
【発明の概要】
【発明が解決しようとする課題】
【0010】
ヒトKRSの機能的多様性と関連して、このペプチドの有意性を確認する為、本発明者等はヒトKRSのN末端116個アミノ酸ペプチドを分離して、イーストツーハイブリッドを利用した、HeLa細胞株 cQDNAライブラリからヒトKRSに結合する蛋白質をスクリーニングする探針(bait:エサ)用に使用した。前記スクリーニングから、本発明者等は本発明で結合可能性がある蛋白質の一つとして、37LRP/p40を確認し、KRSとラミニン収容体間の相互作用における機能的連関性を調査した。
【0011】
その結果、ライシルt-RNA合成酵素(lysyl-t-RNA synthetase,KRS)が原形質膜にある67LRを安定化させることにより、細胞移動及び癌転移を促進する等、KRSが原形質膜のラミニン収容体を通じて癌転移又は癌細胞移動に影響を及ぼすことを究明して本発明を完成した。
【0012】
本発明の目的は癌転移又は癌細胞移動に関して、ライシルt-RNA合成酵素(lysyl-t-RNA synthetase,KRS)の新たな用途を提供することである。
【0013】
前記のような目的を達成する為に、本発明はライシルt-RNA合成酵素の細胞内水準を調節して癌転移を調節する方法を提供する。
【0014】
本発明の他の目的を達成する為に、本発明はライシルt-RNA合成酵素の細胞内水準を調節して癌細胞株の移動(cancer cell migration)を調節する方法を提供する。
【0015】
本発明のさらに、他の目的を達成する為に、本発明はプロモータ及びこれと作動可能に連結されたKRSを、符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクターを有効成分として含む癌予防及び治療用組成物を提供する。
【0016】
本発明のさらに他の目的を達成する為に、本発明はプロモータ及びこれと作動可能に連結されたKRSを、符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクターを、これを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法を提供する。
【0017】
本発明のさらに他の目的を達成する為に、本発明は癌治療剤を製造する為のプロモータ及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを、符号化するポリヌクレオチドを含む発現ベクターの用途を提供する。
【0018】
本発明のさらに他の目的を達成する為に、本発明はKRSの活性を阻害する製剤を有効成分として含む癌予防及び治療用組成物を提供する。
【0019】
本発明のさらに他の目的を達成する為に、本発明はKRSの活性を阻害する製剤をこれを必要とする個体に有効量を投与することを含む癌の予防及び治療方法を提供する。
【0020】
本発明のさらに他の目的を達成する為に、本発明は癌の治療剤を製造する為の、KRSの活性を阻害する製剤の用途提供する。
本発明のさらに他の目的を達成する為に、本発明は、
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;及び
(c)選別された製剤が癌転移又は細胞移動を調節するか、否かをテストする段階を含む癌転移又は癌細胞移動調節製剤をスクリーニングする方法を提供する。
【0021】
本発明のさらに他の目的を達成する為に、本発明は、
(a)試験製剤の存在下でKRS、ラミニン収容体(67LR)及び試験製剤を接触させる段階;及び
(b)前記試験製剤がKRSとラミニン収容体の相互作用を調節するか、否かをテストする段階を含むKRSと、67LRK相互作用を阻害する製剤をスクリーニングする方法を提供する。
本発明のさらに他の目的を達成する為に、本発明は、
(a)試料において67LRの過発現の可否を分析する段階;及び
(b)67LRが過発現された試料においてKRSの過発現の可否を分析する段階を含む肺癌又は乳房癌を診断する方法を提供する。
【課題を解決するための手段】
【0022】
以下、本発明を詳細に説明する。
本発明はKRSが癌転移又は癌細胞移動に及ぼす影響を初めて究明した。つまり、本発明はKRSが原形質膜のラミニン収容体を通じて癌転移又は癌細胞移動に影響を及ぼすことを究明した。
【0023】
定 義
他の定義がない限り、本発明で使用される全ての技術的及び化学的用語は当業者等により、通常的に理解される同一な意味を有する。次の参考文献は、本発明の明細書に使用された多くの用語等の一般的な定義を有する技術(skill)の一つを提供する。(Singleton et al., DICTIONARY OF MICROBIOLOGY AND MOLECULAR BIOLOTY(2nd ed. 1994); THE CAMBRIDGE DICTIONARY OF SCIENCE AND TECHNOLOGY(Walker ed., 1988); 及び Hale & Marham, THE HARPER COLLINS DICTIONARY OF BIOLOGY)。さらに、次の定義は本発明の実施の為に読者(reader)に提供される。
【0024】
本発明で“発現(expression)”とは、細胞で蛋白質又は核酸の生成を意味する。
【0025】
本発明で“宿主細胞(host cell)”とは、任意の手段(例:電気衝撃法、カルシウムホスフアターゼ沈澱法、微細注入法、形質転換法、ウイルス感染等)により、細胞内に導入された異種性DNAを含む原核又は眞核細胞を言う。
【0026】
本発明で“分離された(isolated)”とは、それの起源環境(例:若し自然的に生成されるものであれば自然環境)から除去された物質を意味する。例えば、自然的に生成される核酸、ポリペプチド、又は生きている動物に存在する細胞は、分離されたものではないものの、同一なポリヌクレオチド、ポリペプチド又は共同存在する物質の一部又は全部から分離された(separated)細胞は、以降、それが自然システムで再導入されるとしても、分離されたものである。前記核酸はベクターの一部の場合もあり得て、及び/又は、前記核酸又はポリペプチドの一部でも有り得て、前記ベクターや組成物は自然環境の一部でない分離されたものである。
【0027】
本発明でKRSの生体活性に対して“調節する(modulate)”とは、KRSの細胞内水準の変化を言う。KRS活性の調節は上向調節(up-regulation)(つまり、活性化又は促進)又は下向調節(down-regulation)(つまり、阻害又は抑制)の場合もあり得る。調節はKRSの細胞内水準、蛋白質の安定性、KRSの酵素的修飾(例:リン酸化)、結合特性(例:標的転写調節因子に対する結合)、又はKRSの他の生物学的、機能的及び免疫学的特性の変化を引起こすこともあり得る。KRSの生体活性の変化は、例えば、KRS遺伝子発現の増加又は減少、KRS蛋白質を符号化するmRNAの安定性、翻訳効率から招かれることもあり得る。KRS調節者(modulation)の作用方式(mode)は例えば、KRS蛋白質又はこれを符号化する遺伝子に対する結合を通じて直接的なものの場合もあり得る。さらに、前記変化はKRSを調節する他の物質(例:KRSを特異的にリン酸化させるカイネース)に対する結合及び/又は修飾(例:酵素学的に)を通じた間接的なものでもあり得る。
【0028】
本発明で“ポリペプチド(polypeptide)”は“ポリペプチド等(polypeptides)”又は“蛋白質(等)”と互換性を以て使用でき、例えば、自然状態の蛋白質において一般的に発見される通りアミノ酸残基の重合体を言う。
【0029】
本発明で“KRSポリペプチド”はライシルt-RNA合成酵素として知られているポリペプチドを言う。前記KRSポリペプチドは配列番号1で表示されるアミノ酸配列を有するポリペプチドでもあり得る(Genbank Accession No.NP-005539.1)。さらに、本発明のKRSはこれの機能的同等物を含む。
【0030】
前記機能的同等物とは、配列番号1で表示されるアミノ酸配列と少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上の配列相同性(つまり、同一性)を有するポリペプチドを言う。例えば、 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99%, 100%の配列相同性を有するポリペプチドを含むものにして、配列番号1で表示されるポリペプチドと、実質的に同質の生理活性を呈するポリペプチドを言う。ここで、“実質的に同質の生理活性”とは、原形質膜のラミニン収容体と相互作用し、癌転移又は細胞移動を調節することを意味する。前記機能的同等物は配列番号1のアミノ酸配列の内、一部が付加、置換又は欠失の結果、生成されるものでもあり得る。前記にてアミノ酸置換は好ましくは、保存的置換である。天然に存在するアミノ酸の保存的置換の例は下記の通りである;脂肪族アミノ酸(Gly,Ala,Pro)、疎水性アミノ酸(Ile,Leu,Val)、芳香族アミノ酸(Phe,Tyr,Trp)、酸性アミノ酸(Asp,Glu)塩基性アミノ酸(His,Lys,Arg,Gln,Asn)及び硫黄含有アミノ酸(Gys,Met)、さらに、前記機能的同等物には本発明のKRSポリペプチドのアミノ酸配列上、アミノ酸の一部が欠失された変形体も含まれる。前記アミノ酸の欠失又は置換は好ましくは、本発明のポリペプチドの生理活性に直接的に関連されない域に位置している。さらに、アミノ酸の欠失は好ましくは、KRSポリペプチドの生理活性に直接関与しない部分に位置する。さらに、前記KRSポリペプチドのアミノ酸配列の両末端又は配列内に幾つかのアミノ酸が付加された変形体も含まれる。さらに、本発明の機能的同等物の範囲には、本発明に伴うポリペプチドの基本骨格及びこれの生理活性を維持しながらポリペプチドの一部化学構造が変形されたポリペプチド誘導体も含まれる。例えば、本発明のポリペプチドの安定性、貯蔵性、揮発性又は溶解度等を変形させる為の構造変更がこれに含まれる。
【0031】
本明細書で配列相同性及び同質性はKRSのアミノ酸配列(配列番号1)と候補配列を整列してギャップ(gaps)を導入した後、KRSのアミノ酸配列に対する候補配列のアミノ酸残基の百分率で定義される。必要な場合、最大百分率配列同質性を収得する為に、配列同質性の部分として保存的置換は考慮しない。さらに、KRSのアミノ酸配列のN−末端、又はC−末端又は内部伸長、欠損又は挿入は配列同質性又は相同性に影響を与える配列として解析されない。さらに、前記配列同質性は二つのポリペプチドのアミノ酸配列の類似した部分を比較する為に、使用される一般的な標準方法により決定できる。BLAST又はFASTAのようなコンピュータプログラムは、二つのポリペプチドをそれぞれのアミノ酸が最適にマッチングするように整列する(一つ又は二つの配列の全長配列に従い又は一つ又は二つの配列の予測された部分に従い)。前記プログラムはデフォルトオプニングペナルティ(gefault opening penalty)及び、デフォルトギャップペナルティ(default gap penalty)を提供してコンピュータプログラムと共に連係されて使用し得るPAM250(標準スコリングマトリックス; Dayhoff et al., in Atlas of Protein Sequence and Structure, vol 5, supp 3, 1978)のようなスコリングマトリックスを提供する。例えば、百分率同質性は次のように計算できる。一致する配列(identical matches)の総数に100を掛け、対応されるスパン(matched span)内のより長い配列の長さと二つの配列を整列する為に、より長い配列内に導入されたギャップ(gaps)の和で分ける。
【0032】
本発明に伴うKRSポリペプチドは天然で抽出するか、又は遺伝工学的方法により作製でき得る。例えば、通常的な方法により前記KRS又はこの機能的同等物を符号化する核酸(例:配列番号2(Genbank Accession No.D32053))を作製する。前記核酸は適切なプライマーを使用してPCR増幅することにより、作製し得る。他の方法で当業界に公知の標準方法により、例えば、自動DNA合成器(Biosearch又はApplied Biosystems社で販売するもの)を使用してDNA配列を合成することもできる。作製された核酸はこれに作動可能に連結され(operatively linked)、核酸の発現を調節する一つ以上の発現調節配列(expression control sequence)(例:プロモーター、インハンサー等)を含むベクターに挿入させ、これより形成された組換え発現ベクターで宿主細胞を形質転換させる。生成された形質転換体を前記核酸が発現されるに適切な培地及び条件下で培養して、培養物から前記核酸により発現された実質的に純粋なポリペプチドを回収する。前記回収は当業界に公知の方法(例えば、クロマトグラフィー)を利用して行え得る。前記にて“実質的に純粋なポリペプチド(substantially pure polypeptide)”とは、本発明に伴うポリペプチドが宿主細胞から由来した如何なる他の蛋白質も実質的に含まないことを意味する。本発明のポリペプチド合成の為の、遺伝工学的方法は次の文献を参考にし得る: Maniatis et al., Molecular Cloning; A laboratory Manual, Cold Spring Harbor laboratory, 1982; Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, N.Y., Second(1998) and Third(2000) Editions; Gene Expression Technology, Method in Enzymology, Genetics and Molecular Biology, Method in Enzymology, Guthrie & Fink(eds.), Academic Press, San Diego, Calif, 1991; Hitzeman et al., J. Biol. Chem., 255:12073-12080, 1990。
【0033】
さらに、本発明のポリペプチドは当業界に公知の化学的合成(Creighton, Proteins; Structures and Molecular Principles, W. H. Freeman and Co., NY, 1983)により容易に製造できる。代表的な方法としてこれらに限定されるものではないものの、液体又は固体状合成、断片凝縮、F-MOC又はT-BOC化合法が含むまれる。(Chemical Approaches to the Synthesis of Peptides and Proteins, Williams et al., Eds., CRC Press, Boca Raton Florida, 1997; A Practical Approach, Athert on & Sheppard, Eds., IRL Press, Oxford, England, 1989)。
【0034】
本発明で67kDaのラミニン収容体(67LR)は原形質膜に包埋された(embedded)非−インテグリーン型収容体にして、例えば、Genbank Accession No. NM_002295, S37431, AF284768, S37431, AF284768, J03799, XP_370865, XP_001083023.に記載された塩基配列又はアミノ酸配列を有し得る。
【0035】
本発明で“核酸”、“DNA配列”又は“ポリヌクレオチド”は単一-又は二重-鎖の形態でなるデオキシリボヌクレオチド又はリボヌクレオチドを言う。他の制限がない限り、自然的に生成されるヌクレオチドと類似した方法で核酸に混成化される自然的ヌクレオチドの公知のアナログも含まれる。
【0036】
本発明で“前記KRSを符号化するポリヌクレオチド”は配列番号1で表示されるアミノ酸配列、又はこれと少なくとも70%以上の配列相同性を有するアミノ酸配列を符号化する塩基配列を有し得る。前記核酸はDNA、cDNA及びRNA配列を全て含む。つまり、前記ポリヌクレオチドは配列番号1のアミノ酸配列又はこれと少なくとも70%以上の相同性を有するアミノ酸配列を符号化する塩基配列を有するか、若しくは、前記塩基配列に相補的な塩基配列を有し得る。好ましくは、配列番号2で表示される塩基配列を有する。前記核酸は天然で分離されたか、若しくは、上述した通り、遺伝工学的な方法により製造され得る。
【0037】
本発明で“アナログ(analog)”とは、標準物質(reference molecule)と構造的に類似するものの、標準物質の特異的置換基が置換により代替されることにより、標的や調節方法が変形された物質を言う。標準物質と比較するに、アナログは当業者により予想され得る通り、同一又は類似するか若しくは、向上した有用性(utility)を有する。向上した性質(例:標的物質に対するより高い結合親和力)を有する公知の化合物変異体を究明する為のアナログの合成及びスクリーニングは薬理化学的分野に公知の方法である。
【0038】
本発明で“相同体(homologues)”とは、蛋白質及び/又は蛋白質配列を言及する場合には、共通の祖先蛋白質(common ancestral protein)又は蛋白質配列で自然的に又は人為的に由来したことを示す。類似して核酸及び/又は核酸配列はそれらが共通の祖先核酸又は核酸配列から自然的に又は人為的に由来したときに相同する。
【0039】
本発明で“有効量(effective amount)”とは、細胞又は組織内でKRSの生体活性(例:細胞内水準等)を正常細胞や組織とは別に変更させる効果、又は癌転移又は癌細胞移動様態を対照群に比べて意味ある変化を示すことを言う。
【0040】
本発明で“接触(contacting)”とは、これの一般的な意味(normal meaning)であって、2個以上の製剤(例:2個のポリペプチド)を結合(combine)させるか、若しくは、製剤と細胞(例:蛋白質と細胞)を結合させることを言う。接触は試験管内(in vitro)で起こり得る。例えば、試験管又は他のコンテナー(container)で2個以上の製剤を結合させるか、若しくは、試験製剤と細胞又は細胞溶解物と試験製剤を結合させることである。さらに、接触は細胞又はインシツー(in situ)で起こることもあり得る。例えば、2個のポリペプチドを符号化する組換えポリヌクレオチドを、細胞内で共同発現(coexpression)させることにより、細胞又は細胞溶解物において2個のポリペプチドを接触させることである。
【0041】
本発明で“製剤(agent)”又は“試験製剤(test agent)”とは、任意の物質(substance)、分子(molecule)、元素(element)、化合物(compound)、実在物(entity)又はこれらの組合せを含む。例えば、これに限定はされないものの、蛋白質、ポリペプチド、小遺棄物質(small organic molecule)、多糖類(polysaccharide)、ポリヌクレオチド等を含む。さらに、自然産物(natural product)、合成化合物又は化学化合物又は2個以上の物質の組合せでもあり得る。別に指定されない限り、製剤、物質及び化合物は互換性あるように(interchangeably)使用できる。
【0042】
より具体的に本発明のスクリーニング方法でスクリーニングされ得る試験製剤は、ポリペプチド、ベーターターンミメチック(beta-turn mimetics)、多糖類、リン脂質、ホルモン、フロスタグランジン、ステロイド、芳香族化合物、ヘテロサイクリック化合物、ベンゾジアゼピン(benzodiazepines)、オリゴメリックN−置換グリシン(oligomeric N-substituted glycines)、オリゴカルバメート(oligocarbamates)、糖類(saccharides)、脂肪酸、プリン、ピリミジン又はこれらの誘導体、構造的アナログ又は組合せを含む。或る試験製剤は合成物質でもあり得る、他の試験製剤は天然物質でもあり得る。前記試験製剤は合成又は自然化合物のライブラリを含む広範囲で多様な出処から得られ得る。組合せ(combinatorial)ライブラリはステップ−バイ−ステップ方式で合成され得る多様な種類の化合物で生産され得る。多数の組合せライブラリの化合物等はESL(encoded synthetic libraries)方法(WO 95/12608,WO 93/06121,WO 94/08051,WO 95/395503及びWO 95/30642)により製造でき得る。ペプチドライブラリはファージデイスプレー方法(WO91/18980)により製造し得る。バクテリア、黴、植物及び動物抽出物形態の自然化合物のライブラリは商業的な出処から得るか、又はフィールド(field)で収集することができる。公知の薬理学的製剤が構造的アナログを製造する為に、アシル化、アルキル化、エステル化反応(esterification)、アミド化反応(amidification)のような指示又は無作為な化学的修飾に適用され得る。
【0043】
前記試験製剤は自然的に生成される蛋白質又はその断片でもあり得る。このような試験製剤は自然出処(natural source)、例えば、細胞又は組織溶解物から収得できる。ポリペプチド製剤のライブラリは例えば、通常的な方法により生成されるか、又は商業的に入手し得るcDNAライブラリから収得され得る。前記試験製剤はペプチド、例えば、約5〜30個、好ましくは、約5〜20個、より好ましくは約7〜15個のアミノ酸を有するペプチドでもあり得る。前記ペプチドは自然的に生成される蛋白質、ランダムペプチド又は“バイアス(biased)”ランダムペプチドの伝達物でもあり得る。
【0044】
らに、前記試験製剤は“核酸”でもあり得る。核酸試験製剤は自然的に生成される核酸でもあり得る。例えば、原核又は眞核ゲノムの伝達物の為に記載したのと類似するように使用できる。
【0045】
さらに、前記試験製剤は小分子(例:約1,000以下の分子量を有する分子)でもあり得る。小分子の調節製剤をスクリーニングする為の方法には、好ましくは、高速分析アッセイ(high throughput assay)が適用され得る。多くのアッセイが前記スクリーニングに有用である(Shultz, Bioorg. Med. Chem. Lett., 8:2409-2414, 1998; Weller, Mol. Drivers., 3:61-70, 1997; Fernandes, Curr. Opin. Chem. Biol., 2:597-603, 1998; and Sittampalam, Curr. Opin. Chem. Biol., 1:384-91, 1997)。
【0046】
本発明の方法にスクリーニングされる試験製剤のライブラリはKRS又はこれの断片若しくはアナログに対する構造研究を基に製造され得る。このような研究はKRSに結合する可能性がある試験製剤の究明を可能にする。KRSの3次元的構造は多様な方法、例えば、結晶学構造及び分子的モデリング(crystal structure and molecular modeling)で研究され得る。X-RAY結晶学を利用する蛋白質構造研究方法が文献に広く知られている。(Physical Bio-Chemistry, Van Holde, K. E.(Prentice-Hall, New Jersey 1971), pp.221-239, and Physical Chemistry with Applications to the Life Sciences, D. Eisengerg & D. C. Crothers(Benjamin Cummings, Menlo Park 1979)。KRSの構造に対するコンピューターモデリングはスクリーニングする為に、試験製剤のデザインの為の他の手段を提供する。分子的モデリング方法は文献に開示されている: U.S. Pat. No. 5, 612,894 and U.S. Pat. No. 5,583,973. さらに、蛋白質構造は中性子回折(neutron diffraction)及びNMR(nuclear magnetic resonance)により、決定し得る: Physical Chemistry, 4th Ed. Moore, W. J.(Prentice-Hall, New Jersey 1972)and NMR of Proteins and Nucleic Acids, K. Wuthrich(Wiley-Interscience, New York 1986)。
【0047】
以下、本発明を詳細に説明する。
【0048】
本発明者等は本発明のKRSは原形質膜で転座(translocation)され、67LRと相互作用することにより、腫瘍(又は癌)細胞の移動を促進して癌の転移(metastasis)に影響を及ぼすことを究明した。さらに、マウスを利用したin vivo実験を通じてKRSの過発現又は発現抑制が腫瘍(又は癌)細胞の転移を調節できることを究明した。
【0049】
従って、本発明はライシルt-RNA合成酵素(Lysyl t-RNA synthetase,KRS)の細胞内水準を調節(modulating)して癌転移を調節する方法を提供する。
【0050】
これをより詳細に説明すれば、本発明のライシルt-RNA合成酵素の細胞内水準を減少させる場合、癌転移を抑制することができ、ライシルt-RNA合成酵素の細胞内水準を増加させる場合癌転移を増進することもあり得る。
【0051】
細胞内水準の減少又は増加は前記にて記載したことと共に、当業者に公知の多様な方法で調節され得る。例えば、これに限定はされないものの、細胞内水準は転写段階における調節を通じて調節でき得る。転写段階における調節は当業者に公知の遺伝子の発現を増進させる為の方法、例えば、プロモーターにKRS又はこれに対する機能的同等物を符号化するポリヌクレオチドを連結した組換え発現ベクターを調節して前記遺伝子の発現を増進させる方法、又はKRS又はこれに対する機能的同等物を符号化する遺伝子周辺に前記遺伝子の発現が促進されるようにする発現調節配列を挿入する方法、又は遺伝子の発現を抑制する為の方法、例えば、プロモーター又は遺伝子部位の突然変異を誘導してプロモーター活性又は蛋白質の機能を阻害する方法、アンチセンス(antisense)遺伝子を発現させる方法、siRNA又はマイクロRNA(microRNA)方法等により実施し得る。
【0052】
転写後の段階における調節は、当業者に公知の蛋白質発現を増進又は阻害させる為の方法、例えば、KRS又はこれに対する機能的同等物を符号化する遺伝子を鋳型に、転写されたmRNAの安定性を増進又は阻害する方法、蛋白質又はポリペプチドの安定性を増進又は阻害する方法、又は蛋白質又はポリペプチドの活性を増進又は阻害する方法により実施し得る。
【0053】
前記方法のより具体的な例として1群イントロンタイプ、M1 RNAタイプ、ハンマーヘッド(hammerhead)型、又はヘアピン(hairpin)型又はマイクロRNA型等の転写されたmRNAに作用するRNAを符号化するDNA配列に形質転換するか、又は標的遺伝子配列と同一又は類似した配列を有するDNAの形質転換を通じた同時抑制(cosuppression)を誘導することができる。
【0054】
好ましくは、本発明でKRS又はこれに対する機能的同等物の細胞内水準を調節することは、前記ポリヌクレオチドを符号化するポリヌクレオチドの発現を増加又は減少させる方法により実施し得る。このような増加又は減少させる方法は、それぞれ当業者に公知の方法を使用し得るものの、例えば、プロモーターにKRS又はこれに対する機能的同等物を符号化するポリヌクレオチドを連結した組換え発現ベクターを製造して、その発現を増進又は、プロモーターに前記ポリヌクレオチドに対するアンチセンス又はsiRNAポリヌクレオチドを連結した組換え発現ベクターを製造してその発現を減少し得る。この際、前記KRS又はこれに対する機能的同等物を符号化するポリヌクレオチドは好ましくも、配列番号2で表示される塩基配列を有し得る。
【0055】
従って、本発明はライシルt-RNA合成酵素の細胞内水準を調節して癌細胞の移動(cancer migration)を調節する方法を提供する。細胞内水準の調節等に対しては前記にて記載した通りである。
【0056】
さらに、本発明のKRSはその発現を抑制させる場合、腫瘍(又は癌)の転移が抑制されるので、本発明はプロモーター及びこれと作動可能に連結されたKRSの発現を抑制する構造遺伝子を含む発現ベクター又はKRSに対する抗体を有効成分として含む癌の予防及び治療用組成物を提供する。前記にてKRSの発現を抑制する構造遺伝子はKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAでもあり得る。
【0057】
本発明の組成物が適用できる疾患は癌もあり得る。前記癌はこれに限定はされないものの、大腸癌、肺癌、肝臓癌、胃癌、食道癌、膵臓癌、胆嚢癌、腎臓癌、膀胱癌、前立腺癌、睾丸癌、子宮頸部癌、子宮内膜癌、絨毛癌、卵巣癌、乳癌、甲状腺癌、脳癌、頭頚部癌、悪性黒色腫、リンパ腫、再生不良性貧血等を含む。
【0058】
前記“プロモーター”とは、特定した宿主細胞で作動可能に連結された核酸配列の発現を調節するDNA配列を意味し、“作動可能に連結される(operably linked)”と言うことは、一つの核酸断片が他の核酸断片と結合され、その機能又は発現が他の核酸断片により影響を受けることを言う。従って、転写を調節する為の任意のオペレータ配列、適合したmRNAリボソーム結合部位をコーティングする配列及び転写及び解読の終結を調節する配列を追加して含め得る。前記プロモーターには全ての時間帯に常時的に目的遺伝子の発現を誘導するプロモーター又は特定した位置、時期に目的遺伝子の発現を誘導するプロモーターを使用することができ、その例としてはSV40プロモーター、CMVプロモーター、CAGプロモーター(Hitoshi Niwa et al., Gene, 108:193-199, 1991; Monahan et al., Gene Therapy, 7:24-30, 2000)35Sプロモーター(Odell et al., Nature 313:810-812, 1985)Rsyn7プロモーター(米国特許出願第08/991,601号)、ライスアクチン(rice actin)プロモーター(McElroy et al., Plant Cell 2:163-171, 1990)、ユビキチンプロモーター(Christensen et al., Plant Mol. Biol. 12:619-632, 1989)、ALSプロモーター(米国特許出願第08/409,297号)等がある。この他にも米国特許第5,608,149;第5,608,144号、第5,604,121号、第5,569,597号、第5,466,785号、第5,399,680号、第5,268,463号及び第5,608,142号等に開示されたプロモーターを全て使用し得る。
【0059】
一方、本発明はプロモーター及びこれと作動可能に連結されたKRSの発現を抑制する構造遺伝子を含む発現ベクター又はKRSに対する抗体を、これを必要とする個体に有効な量で投与する癌の予防及び治療方法を提供する。この際、構造遺伝子は前記記載の通りであるので、本発明はプロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体をこれを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法を提供する。
【0060】
本発明で“有効な量”とは、本発明の発現ベクターが投与対象である個体内で癌の予防又は治療する効果を表す量を言う。前記“個体”とは、好ましくは、哺乳動物、特にヒトを含む動物の場合も有り得て、動物より由来した細胞、組織、器官等でもあり得る。前記個体は治療が必要な患者(patient)の場合もあり得る。
【0061】
さらに、本発明は癌治療剤を製造する為の、プロモーター及び作動可能に連結されたKRSの発現を抑制する構造遺伝子を含む発現ベクター、又はKRSに対する抗体の用途を提供する。より詳細には、本発明は癌治療剤を製造する為のプロモーター及びこれと作動可能に連結されたKRSを、符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター、又はKRSに対する抗体の用途を提供する。前記にてプロモーター、KRS、発現ベクター、適用される癌に対しては前記記載の通りである。
【0062】
前記にてKRSに対する抗体は、KRSの抗原性部位に対して指示される特異的な蛋白質分子を意味する。本発明の目的上、前記抗体はKRS蛋白質に対して特異的に結合する抗体を意味し、タンクロン抗体、タンクロン抗体及び組換え抗体を全て含む。
【0063】
前記のようなKRSに対する抗体を生成することは、当業界に広く知られた技術を利用して容易に製造できる。タンクロン抗体は前記KRS蛋白質抗原を動物に注射し、動物から採血して抗体を含む血清を収得する当業界に広く知られた方法により生産することができる。このようなタンクロン抗体は羊、兎、山羊、猿、馬、豚、牛、犬等の任意の動物種の宿主から製造可能である。
【0064】
タンクロン抗体は当業界に広く知られたハイブリッドマ方法(hybridoma method) (hybridoma method) (Kohler and Milstein(1976)European Jounral of Immunology 6:511-519参照)、又はファージ抗体ライブラリ(Clackson et al, Nature, 352:624-628, 1991; Marks et al, J. Mol. Biol., 222:58, 1-597, 1991)技術を利用して製造され得る。
【0065】
ハイブリッドマ方法は肺癌診断マーカー蛋白質抗原を注射したマウスのような免疫学的に適合した宿主動物からの細胞を利用し、残りの一つの集団では癌又は骨髄腫細胞株を利用する。このような二つの集団の細胞等をポリエチレングリコールのような当業界に広く知られた方法により、融合させた後、抗体−生産細胞を標準的な組織培養方法により増殖させる。限界希釈法(limited dilution technique)によるサブクローニングにより、均一な細胞集団を収得した後、KRS蛋白質に対して特異的な抗体を生産できるハイブリッドマを標準技術により、試験管内で又は生体内で大量で培養する。前記のハイブリッドマが生産するタンクロン抗体は、精製せずに使用することもできるものの、最良の結果を得る為には、当業界に広く知られた方法により、高純度に精製して使用することが好ましい。ファージ抗体ライブラリ方法は、細胞内に存在する多様な肺癌マーカーに対する抗体遺伝子(Single chain fragmentvariable, scFv形態)を獲得してこれをファージの表面に融合蛋白質形態で発現することにより、抗体ライブラリを試験管内で製作し、このライブラリから肺癌蛋白質と結合するモノクロナル抗体を分離、製作する方法である。前記方法で製造された抗体はゲル電気泳動、透析、塩沈殿、イオン交換クロマトグラフィー、親和性クロマトグラフィー等の方法を利用して分離する。
【0066】
さらに、本発明の抗体は2個の全長の軽鎖及び2個の全長の重鎖を有する完全な形態のみならず、抗体分子の機能的な断片を含む。抗体分子の機能的な断片とは、少なくとも抗原結合機能を保有している断片を意味し、Feb, F(ab'), F(ab')2及びFv等がある。
【0067】
従って、本発明では、KRSの細胞内水準を減少させる場合、癌の転移を抑制して癌の予防及び治療に利用できることを究明したことから、本発明はKRSの活性を阻害する製剤を有効成分として含む癌の予防及び治療用組成物を提供する。従って、本発明はKRSの活性を阻害する製剤をこれを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法を提供し、癌治療剤を製造する為のKRSの活性を阻害する製剤の用途も提供する。癌、個体、有効な量等に対しては前記記載の通りである。
【0068】
KRSの活性を阻害する製剤KRSの発現を阻害、つまり、mRNA又は蛋白質水準における発現を阻害する製剤、例えば、KRSに対するantisense RNA又はsiRNAでも有り得て、発現されたKRSの活性を阻害する競争的阻害剤(competitive inhibitor)又は非競争的阻害剤、例えば、KRSに対する抗体でもあり得るものの、これに制限はされない。
【0069】
KRSの細胞内水準を減少させる場合、癌の転移を抑制して癌の予防及び治療に利用できるので、組成物、方法及び用途はそれ自体のみならず、従来公知の多様な癌の予防及び治療方法や抗癌剤と結合して利用され得る。つまり、本発明の組成物、方法等は癌の転移を抑制できるので、従来抗癌剤又は癌の予防及び治療方法と結び付けて治療に利用すれば癌が抑制され、主な腫瘍部位の治療を通じた癌の完治に効果的でもあり得る。
【0070】
本発明のポリペプチドに連結され得る抗癌剤又は癌の予防及び治療方法は、従来癌治療に使用されるものであれば制限なく使用できる。例えば、抗癌剤としてはパクリタキセル、トキソルビシン、ビンクリスチン、ダウノルビシン(daunorubicin)、ビンブラスチン(vinblastine)、アクチノマイシン-D(ACTINOMYCIN-D)、ドセタキセル(docetaxel)、エトポサイド(etoposide)、テニポサイド(teniposide)、ビサントレン(bisantrene)、ホモハリングトン(homoharringtonine)、グリーベック(Gleevec;STI-571)、シスプライン(cisplain)、5-フルオラシル(5-fluouracil)、アドリアマイシン(adriamycin)、メトソトレキサート(methotrexate)、ブスルフアン(busulfan)、クロラムブシル(chlorambucil)、シクロホスファミド(cyclophosphamide)、メルファラン(melphalan)、ニトロゲンムスタード(nitrogen mustard)、ニトロソウレア(nitrosourea)等がある。前記製剤と本発明の組成物又は予防及び治療方法の結合は、抗癌剤の種類及び量によって当業界に公知の技術により適切に行え得る。
【0071】
本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤は、経口的に又は非経口的に投与でき得る。経口投与は舌下適用も含む。非経口的投与は皮下注射、筋肉内注射及び静脈注射のような注射法及び点適法を含む。前記発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤は、薬学的に許容される担体と混合して各種薬学的剤形の形態で製造できる。前記“薬学的に許容される”とは、生理学的に許容されヒトに投与されるとき、通常的に胃腸障碍、眩気症等のようなアレルギー反応又はこれと類似した反応を起こさないことを言う。薬学的に許容される担体は経口投与には、結合剤、滑沢剤、崩解剤、賦形剤、可溶化剤、分散剤、安定化剤、懸濁化剤、色素及び香料を使用することができ、注射剤の場合には緩衝材、保存剤、無痛化剤、等張剤及び安定化剤を混合して使用することができ、局所投与用製剤の場合には、基剤、賦形剤、潤滑剤及び保存剤を使用し得る。このように本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤を含む薬学組成物の剤形は上述した通りの薬学的に許容される担体と混合して多様に製造し得る。例えば、経口投与時には錠剤、トポキ、カプセル、エリキシル、サスペンション、シロップ、ウェファー(wafer)等の形態で製造することができ、注射剤の場合には、単位投薬アンプル又は多数回投薬包含剤形態で製造できる。
【0072】
本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤の総有効量は単一投与量で投与することができ、多重投与量が長期間投与される分割治療方法により投与できる。本発明の発現ベクター、KRSに対する抗体又はKRSの活性を阻害する製剤を含む組成物は疾患の程度及び/又は目的により、有効成分の含量を異にすることができるものの、通常的に1回投与時0.1μg乃至100mg、好ましくは1μg乃至10mgの有効容量で1日数回繰返し投与し得る。しかしながら、前記発現ベクター又はKRSの活性を阻害する製剤の濃度は、薬の投与経路及び治療回数のみならず、患者の年齢、体重、健康状態、性別、疾患の重症度、食餌及び排泄率等、多様な要因等を考慮して患者に対する有効投与量が決定されるものである為、このような点を考慮するに、当分野の通常的な知識を有する者であれば、適切な有効投与量を決定できるであろう。本発明に伴う発現ベクター又はKRSの活性を阻害する製剤を含有する組成物は本発明による効果を表す限り、その剤形、投与経路及び投与方法に特に制限はされない。
【0073】
一方、前記発現ベクターは感染(infection)、形質感染(transfection)又は形質導入(transduction)等の当業界に公知の方法により、発現型で標的細胞内に導入させることができる。
【0074】
プラスミド発現ベクターを利用した遺伝子伝達方法は、哺乳動物細胞に直接的にプラスミドDNAを伝達する方法にして、FDAから承認されたヒトに使用できる方法である(Nabel, E. G., et al., Science, 249:1285-1288, 1990)。プラスミドDNAはウイルスベクターとは異なり、均質に精製できる長所を有する。本発明で使用できるプラスミド発現ベクターには、当業界に公知の哺乳動物発現プラスミドを使用できる。例えば、これに限定はされないものの、pRK5(ヨーロッパ特許第307,247号)、pSV16B(国際特許公開第91/08291号)及びpVL1392(PharMingen)等が代表的である。前記プラスミド発現ベクター(plasmid expression vector)は当業界に公知の方法、例えば、これに限定はされないものの、形質感染、微細注射、形質導入、細胞融合、カルシウムホスフェート沈澱法、リポソーム媒介された形質感染(liposome-mediated transfection)、DEAEデキストランー媒介された形質感染(DEAE Dextran-mediated transfection)、ポリブレンー媒介された形質感染、電気穿孔法(electroporation)、遺伝子銃(gene gun)及び細胞内に遺伝子を流入させる為の他の公知方法により、標的細胞内に導入できる(Wu et al., J. Bio. Chem., 267:963-967, 1992; Wu and Wu, J. Bio. Chem., 263:14621-14624, 1988)。
【0075】
さらに、前記核酸を含むウイルス発現ベクターはこれに限定はされないものの、レトロウイルス(retrovirus)、アデノウイルス(adenovirus)、ヘペスウイルス(herpesvirus)及びアビポックスウイルス(avipox virus)、レンチウイルス(Lenti-virus)等が含まれる。前記レトロウイルスベクターはウイルス遺伝子が全て除去されているか、又は変更されて非−ウイルス蛋白質のウイルスベクターにより、感染された細胞内で造れるように作製されたものである。遺伝子療法の為の、レトロウイルスベクターの主要長所は、多量の遺伝子を複製細胞内に伝達し、細胞DNA内に伝達された遺伝子を正確に統合し、遺伝子形質感染後連続的な感染が誘発されないことである(Miller, A.D., Nature, 357:455-460,1992)。FDAより認証されたレトロ内ベクターPA317癌ホトロフィックレトロウイルスパッケージ細胞を利用して製造したものである(Miller, A.D. and Buttimore, C., Molec. Cell Biol., 6:2895-2902,1986)。非−レトロウイルスベクターは、前記にて言及した通りのアデノウイルスがある(Rosenfeld et al., Cell, 68:143-155, 1992; Jaffe et al., Nature Genetics, 1:372-378, 1992; Lemarchand et al., Proc. Natl. Acad. Sci. USA, 89:6482-6486, 1992)。アデノウイルスの主要長所は多量のDNA断片(36kbゲノム)を運搬し、極めて高い力価で非−複製細胞を感染させ得る能力があると言うことである。さらに、ハピスウイルスもヒト遺伝子療法の為、有用に使用できる(Wolfe, J.H., Nature Genetics, 1:379-384,1992)。以外にも、公知の適切なうウイルスベクターが本発明に使用できる。
【0076】
さらに、前記KRSの発現を抑制する構造遺伝子は他の方法、例えば、局所的に、非経口、経口、鼻腔、静脈、筋肉内、皮下又は他の適切な手段により投与できる。特に、前記ベクターは標的組織の腫瘍細胞を治療する為の、有効量で標的癌又は腫瘍細胞内に直接注射し得る。特に、目、胃腸管、泌尿生殖器官、肺及び気管支システムのような膵臓(body cavity)にある癌又は腫瘍の場合には、本発明の構造遺伝子(又は構造遺伝子を含む発現ベクター)を含有する薬学組成物を癌又は腫瘍により、影響を受ける有腔器官(hollow organ)内に針、導管(catheter)又は他の種類の輸送チューブを利用して直接注入できる。この時、X−ray、ソノグラム(sonogram)、の光繊維システム(fiberoptic visualization system)のような映像装置が標的組織の位置確認と針又は導管の注入に使用される。その他に直接的に到達できないか、又は分析学的に分離できない腫瘍又は癌の場合には、本発明の組成物を血液循環系内に投与できる。
【0077】
さらに、本発明は
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;及び
(c)選別された製剤が癌転移又は癌細胞移動を調節するか否かをテストする段階を含む癌転移又は癌細胞移動調節製剤をスクリーニングする方法を提供する。
【0078】
前記スクリーニング方法は、当業界に公知の多様な生化学的及び分子生物学的技術が前記方法を実施することに利用され得る。前記技術は下記の文献に開示されている:(Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press, N.Y., Second(1998) and Third(2000) Editions; and Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., New York(1987-1999)。
【0079】
好ましくは、前記試験製剤が先にKRSの生物学的活性を調節する能力があるかをアッセイする(1次アッセイ段階)。具体的に1次段階では試験製剤の存在下で分離されKRSの生物学的活性をアッセイ(assay)して前記ポリペプチドの生物学的活性を調節する調整製剤(modulating agent)を究明(identify)する。より好ましくは、次の段階を含め得る。
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;
【0080】
1次アッセイ段階では多くの生物学的活性に対する調節がアッセイできる。例えば、試験製剤はKRSの発現水準、例えば、転写又は翻訳を調節する活性があるか、否かに付いてアッセイでき得る。さらに、前記試験製剤はKRSの細胞内水準又は安定性、例えば、翻訳後修飾(post-translational modification)又は加水分解を調節する活性があるか否かに付いてアッセイされ得る。
【0081】
以降、前記1次アッセイ段階を通じてKRSの生物学的活性を増加させる調節製剤を究明した後、前記試験製剤がラミニン収容体(67LR)、さらには、癌転移又は癌細胞移動を調節する能力があるかをKRSの存在下でさらに、テストする。(2次試験段階)、例えば、前記試験製剤は、癌転移又は癌細胞移動を調節する活性があるか否に付いてさらにテストする。
【0082】
上記にて言及した通り、本発明の方法により究明されたKRS−調節製剤は癌転移、又は癌細胞移動を調節できる。若し、前記1次アッセイ段階で究明された試験製剤がKRS−調節製剤の細胞内水準(例:転写活性の変更により)を調節すれば、それは癌転移又は癌細胞移動を調節できる。
【0083】
一方、若し、試験製剤KRSの細胞内水準でない他の活性を調節すれば、KRSに対する前記試験製剤の治療効果が癌転移又は癌細胞移動を実際に調節するか、否かを確認することが必要である。例えば、KRSのリン酸化活性を調節する試験製剤はKRSのリン酸化活性の調節が癌転移又は癌細胞移動を調節できるか、否かを確認する為にテストを重ねる必要がある。
【0084】
前記1次及び2次段階全て変わらない(intact)KRS及びこれの断片、アナログ又は機能的同等物を使用できる。これらアッセイに使用できる断片は一般的に、KRSの生物学的活性を一つ以上保有する。好ましくは、KRSの断片は配列番号1の1-72のアミノ酸残基を含め得る。さらに、前記断片又はアナログを含む融合蛋白質が試験製剤の為のスクリーニングに使用され得る。KRSの機能的同等物はアミノ酸欠失及び/又は挿入及び/又は置換を有するものの、KRSと同一な生体活性を保有することにより、本発明のスクリーニング方法を実施することに使用できる。
【0085】
当業界に通常的に実施されている多様なアッセイ等がKRSを調節する製剤を究明に使用できる。好ましくは、前記製剤は細胞基盤アッセイシステム(cell based assay system)でスクリーニングされ得る。例えば、スクリーニングする為の、典型的な細胞基盤アッセイにおいて(つまり、2次試験段階で)、試験製剤の存在下でリポーター遺伝子活性(例:酵素活性)を測定し、これを試験製剤の不在下におけるリポーター遺伝子の活性と比較する。前記リポーター遺伝子は当業界に公知の任意の検出可能なポリペプチド(反応又はリポーターポリペプチド)、例えば、蛍光又は燐光(phosphorescence)により検出可能なポリペプチドやそれが保有している酵素活性により、検出可能なポリペプチドを符号化し得る。検出可能な反応ポリペプチド(detectable response polypeptide)、例えば、ルシファーラゼ、α−グルクロニターゼ(glucuronidase)、α−カラクトシターゼ、クロラムペニコールアセチル転移酵素、緑色蛍光蛋白質、強化された緑色蛍光蛋白質及びヒトに分泌されたアルカリンリン酸化酵素でもあり得る。
【0086】
細胞−基盤アッセイにおいて、試験製剤(例:ペプチド又はポリペプチド)は宿主細胞内に存在する他のベクターにより、発現され得る。或る方法では、試験製剤のライブラリが前記ベクターライブラリ(例:cDNAライブラリ)により符号化される。前記ライブラリは当業界に公知の方法を利用して製造することができ(Sambrook et al. and Ausubel et al., supra)、又は多様な産業的出処から得られ得る。
【0087】
前記記載した細胞−基盤アッセイ以外に、非−細胞基盤方法によつてもスクリーニングされ得る。前記方法は、例えば、モビリティーシフトDNA結合アッセイ(mobility shift DNA-binding assays)、メチル化及びウラシル干渉アッセイ(methylation and uracil interference assays)、DNase 及びヒドロキシーラジカルフットプリンティング分析(DNase and hydroxyl radical footprinting analysis)、蛍光偏光(fluorescence polaryzation)及びUV交差結合(crosslinking)又は化学的架橋剤(cross-linkers)を含む。一般的な概要はAusubel等の文献に開示されている(Ausubel et al., supra, chapter 12, DNA-Protein Interaction)。核酸及びDNA/RNA結合蛋白質を含む共同−結合された蛋白質(co-associating protein)を分離する技術は、切断可能な架橋剤ジチオビス(dithiobis)(succinimidylpropionate)及び3,3′−ジチオビス(sulfosuccinimidyl-propionate)を含むUV交差結合又は化学的架橋剤を含む(McLaughlin, Am. J. Hum. Genet., 59:561-569, 1996; Tang, Biochemistry, 35:8216-8225, 1996; Lingner, Proc. Natl. Acad. Sci. U.S.A., 93:10712, 1996; and Chodosh, Mol. Cell. Biol., 6:4723-4733, 1986)。
【0088】
1次アッセイ段階:KRSを調節する製剤のスクリーニング
多くのアッセイシステムがKRSの調節因子の為の、試験製剤のスクリーニングに適用できる。上記にて言及した通り、前記スクリーニングは試験管内アッセイシステム又は細胞−基盤アッセイシステムを利用できる。このスクリーニング段階で、試験製剤はKRSとの結合、KRSの細胞内水準変更、又はKRSの他の生物学的活性の調節に対してスクリーニングされ得る。
【0089】
1)KRSに結合する製剤のスクリーニング
1次スクリーニング段階でKRSと試験製剤の結合が測定できる。KRSに対する試験製剤の結合は、例えば、標識された試験管内蛋白質−蛋白質結合アッセイ、(electrophoretic mobility shift assays)、蛋白質結合を検出する為の免疫アッセイ、機能的アッセイ(リン酸化アッセイ等)等のような多様な方法でアッセイできる(U.S. Pat. Nos. 4,366,241:4,376,110; 4,517,288 and 4,837,168; and Bevan et al., Trends in Biotechnology, 13:115-122, 1995; Ecker et al., Bio/Technology, 13:351-360, 1995; and Hodgson, Bio/Technology, 10:973-980, 1992)。試験製剤はKRSとの直接的な結合、例えば、KRSに対する抗体によるKRSポリペプチドとの共同−沈殿(co-immunoprecipitation)を検出することにより、確認できる。さらに、試験製剤はKRSと試験製剤の結合を表し得るシグナル、例えば、蛍光キンチング(quenching)を検出することにより確認できる。
【0090】
競争アッセイ(competition assays)はKRSに特異的に結合する試験製剤究明に適当なフォーマット(format)を提供する。前記フォーマットで、試験製剤はKRSと結合するものとして既に公知の化合物との競争を通じてスクリーニングされる。公知の結合化合物は合成化合物でもあり得る。さらに、それはKRSを特異的に認識する抗体、例えば、KRSに対する単一クロン抗体でもあり得る。若し、試験製剤が公知のKRSと公知の化合物の結合を阻害すれば、前記試験製剤はKRSにも結合する。
【0091】
多様な種類の競争アッセイらが当業界に知られている。例えば、固体状直接又は間接的酵素放射免疫測定法(solid phase direct or indirect radioimmunoassay, RIA)、固体状直接又は間接的酵素放射免疫測定法(EIA)、サンドイッチ競争アッセイ(Stahli et al., Methods in Enzymology, 9:242-2453, 1983);固体状直接バイオチンーアビチンEIA(Kirkland et al., J. Immunol., 137:3614-3619, 1986);固体状直接標識化されたアッセイ、固体状直接標識化されたサンドイッチアッセイ(Harlow and Lane, Antibodies, A laboratory Manual, Cold Spring Harbor Press, 1988);125 Iを利用した固体状直接標識RIA(Morel et al., Mol. Immuno., 25(1):7-15, 1988;固体状直接バイオチンーアビチンEIA(Cheung et al., Virology, 176:546-552, 1990);及び直接標識化されたRIA(Moldenhauer et al., Sacnd. J. Immunol., 32:77-82, 1990)、一般的に、前記アッセイ等は非標識化された試験製剤及び標識化された対照化合物を含めている細胞、又は個体表面に結合されている精製されたポリペプチドの利用を含む。競争的阻害は試験製剤の存在下で個体表面又は細胞に結合された標識の量を結合することにより測定される。競争アッセイにより究明された調節製剤は対照化合物と同一なエピトープに結合する製剤及び立体阻害(steric hindrance)が起こるようにする為、対照化合物により結合されたエピトープに十分に近接した隣接エピトープと結合する製剤を含む。通常的に競争阻害が過度に存在する時、一般的な標的ポリペプチドに対する対照化合物の特異的な結合が少なくとも50又は75%阻害されるであろう。
【0092】
前記スクリーニングアッセイは不溶性又は溶解性フォーマットでもあり得る。不溶性アッセイの一例はKRS又はこれの断片を固体状マトリックスで固定化したものである。以降、試験製剤が結合するに十分な時間の間、固体状マトリックスを試験製剤と接触させておく。以降、固体状マトリックスから結合されない物質は、洗滌して除去後、固体状に結合された製剤の存在を確認する。前記方法は、固体状マトリックスから結合された製剤を溶離して製剤を分離する段階をさらに含め得る。両者択一的に、KRSを固定化させる他の方法は製剤を固体状マトリックスに結合させた後、KRSを添加することである。
【0093】
溶解性アッセイは前記記載された幾つかの結合ライブラリスクリーニング方法を含む。溶解性アッセイフォーマットにおいて、試験製剤やKRSは全て固体支持台に結合しない。試験製剤に対するKRS又はこれの断片の結合は、例えば、KRS及び/又は試験製剤の蛍光により測定できる。蛍光は内在的か(intrinsic)若しくは蛍光物質(fluorophor)を有する成分の標識により与え得る。
幾つかの結合アッセイから、KRS、試験製剤又は第3の物質(例:KRSに結合する抗体)は、与えられた条件で前記ポリペプチドの確認、検出及び定量を容易にする為に、標識された状態で提供できる。つまり、共有的に結合又は検出可能な標識やグループ、又は架橋可能なグループにリンクされて提供できる。これら検出可能なグループ等は検出可能なポリペプチドグループ、例えば、アッセイ可能な酵素又は抗体エピトープを含む。両者択一的に前記検出可能なグループは放射線同位元素(例:125 I,32P,35S)又は化学発光性又は蛍光性グループのような他の検出可能なグループ又はラベルから選択できる。類似して、前記検出可能なグループは基質(substrate)、補助因子(cofactor)、阻害剤又は親和リガンドでもあり得る。
【0094】
2)KRSの他の生体活性を調節する製剤のスクリーニング
KRSの試験製剤の結合は、試験製剤がKRSの調節者であることを示す。さらに、これは前記製剤が癌転移又は癌細胞移動を調節する為に、ラミニン収容体の生体活性を調節し得ることを提示する。従って、KRSと結合する試験製剤はラミニン収容体活性を調節する能力を有しているかをさらにテストすべきである。
【0095】
両者択一的に、KRSと結合する試験製剤はKRSに対するその活性を測定する為に、さらに、調査され得る。そのような活性の存在、性質又は範囲は活性アッセイにより、テストされ得る。前記活性アッセイはKRSに試験製剤の結合が実際にKRSに対する調節活性を有することを確認できる。極めて広く前記調節アッセイ等はKRSの活性を調節する試験製剤を究明する為に、独立的に使用し得る。(つまり、KRSに結合する能力をアッセイする初段階なしに)、一般的に、前記方法等はKRSの生物学的活性をテストするに必要とする他の物質又は試薬の存在又は不在下でKRSを含むサンプルに試験製剤を添加し、KRSの生物学的活性の変更を測定することを含む。KRSの酵素又は他の生物学的活性を調節する製剤化をスクリーニングするアッセイに加えて、前記活性アッセイはKRSの発現又は細胞内水準の変化の為の試験管内スクリーニング及び生体内スクリーニングを含む。
【0096】
2次試験段階:癌転移又は癌細胞移動を調節する製剤のスクリーニング
一旦調節製剤がKRSと結合するか、及び/又はKRSの生物学的活性(細胞内水準を含む)を調節するものとして究明されると、前記製剤は癌転移又は癌細胞移動を調節する能力を有しているか、否かに付いて、さらにテストされ得る。前記調節製剤による癌転移又は癌細胞移動の調節は一般的に、KRSの存在下でテストされる。若し、細胞−基盤スクリーニングシステムを利用する場合、KRSは宿主細胞に導入された発現ベクターから発現され得る。両者択一的にKRSはスクリーニングシステムで宿主細胞により内生的に供給されることもあり得る。
【0097】
さらに、本発明は
(a)試験製剤の存在下でKRS、ラミニン収容体(67LR)及び試験製剤を接触させる段階;及び
(b)前記試験製剤がKRSとラミニン収容体の相互作用(interaction)を調節するかをテストする段階を含むKRSと67LRの相互作用を阻害する製剤をスクリーニングする方法を提供する。
【0098】
前記製剤はKRSとラミニン収容体(67LR)の相互作用を促進又は強化するものでも有り得て、逆に前記相互作用を抑制又は弱化させるものでもあり得る。
【0099】
前記(b)段階のテストは前記調節製剤が接触されない細胞又はこれの溶解物における67LRとKRSの相互作用水準に対する前記調節製剤が接触された細胞又はこれの溶解物における67LRとKRSの相互作用水準の相対的な変化を測定する段階を含め得る。
【0100】
前記スクリーニング方法は、前記にて言及したような標識された試験管内蛋白質−蛋白質結合アッセイ(試験管内プールーダーウンアッセイ)、EMSA(electrophoretic mobility shift assays)、蛋白質結合の為の、免疫アッセイ、機能的アッセイ(リン酸化アッセイ等)、酵母−2ハイブリッドアッセイ、非免疫沈澱アッセイ、免疫沈澱ウェスタンブロットアッセイ、免疫−共同−位置化アッセイ(immuno-co-localization)等当業界に公知の多様な方法で実施し得る。
【0101】
例えば、バクテリアリプレクサーLexA、又は酵母GAL4のDNA-結合ドメイン及び酵母GAL4蛋白質のトランスアクチベイション(transactivation)、ドメインにそれぞれ融合された、KRSと67LR、又はこれら蛋白質の一部又は相同体(homologues)を発現する酵母を利用して酵母−2ハイブリッドアッセイを行い得る(KIM, M. J. et al., Nat. Gent., 34:330-336, 2003)。KRSと67LRの相互作用はLexA蛋白質又はGAL4のDNA-結合ドメインに結合された調節配列を有しているプロモーターの統制下で、リポータ遺伝子の発現を誘導するトランスアクチベータを再構成するようになる。
【0102】
前記リポータ遺伝子では前記記載の通り、当業界に公知の任意の検出可能なポリペプチドを符号化する遺伝子(例:CAT(chloramphenicol acetyltransferase)、ルシファラーゼ、ベータカラクトシターゼ、ベーターグルコシターゼ、アルカリンホスファターゼ及びGFP(green fluorescent protein)等を使用できる。若し、KRSと67LR、又はこれら蛋白質の一部又は相同体の相互作用は試験製剤により、促進又は強化される場合、前記リポータ遺伝子の発現は正常条件に比べてさらに増加される。逆に前記相互作用が試験製剤により、阻害されるか又は弱化される場合、前記リポータ遺伝子は発現されないか、又は正常条件に比べて少なく発現される。
【0103】
さらに、リポータ遺伝子では酵母の成長を可能にする蛋白質(つまり、前記リポータ遺伝子が発現されない時、酵母の成長が抑制される)を符号化するものとして選択され得る。例えば、アミノ酸又は窒素塩基の為の生合成過程に関係ある酵素を符号化する栄養要求性(auxotropic)遺伝子(例:ADE3,HIS3等の酵母遺伝子又は他の種由来の同等遺伝子)でもあり得る。このシステムで発現されたKRSと67LR、又はこれら蛋白質の一部又は相同体の相互作用は試験製剤により、抑制又は弱化される場合、リポータ遺伝子は発現されないか、又は少なく発現する。従って、前記条件下で酵母の成長は停止又は遅くなる。このようなリポータ遺伝子の発現による効果は、肉眼や装置(例:顕微鏡)を通じて観察でき得る。 従って、本明細書では、肺癌又は乳癌患者を対象として67LR及びKRSの過発現可否を確認した結果、67LRが過発現された場合、KRSの過発現と肺癌又は乳癌との関連性が高いことが分った。(表1参照)。従って、本発明は
(a)試料で67LRの過発現可否を分析する段階;及び
(b)67LRが過発現された試料からKRSの過発現可否を分析する段階を含む肺癌又は乳癌を診断する方法を提供する。
【0104】
診断の為の、試料の採取及び処理と67LR及びKRSの過発現可否の分析は、通常的な分子生物学的技法を使用することができ、これは前記にて記載した通りである。
【0105】
本発明に記載された図面に対して下記の通り説明する
【0106】
図1乃至図6はヒトKRSとラミニン収容体の特異的相互作用を確認したものである。図1で全長のヒトKRSと37LRP/p40間の相互作用はイーストツーハイブリッド分析を通じて実施された。多重−ARS複合体の二つの酵素要素であるAIMP1及びAIMP2はそれぞれ陽性及び陰性対照群として使用された。陽性相互作用はx-galを含有した酵母培地で青色コロニーの形成により標識された。図2で、37LRPは35Sメチオニンの存在下でin vitro翻訳により合成され、これらをGST-KRS又はGSTと反応した(pull-down)、GST-KRSと共沈澱(co-precipitated)37LRPは自家放射撮影法を利用して測定した。図3で、KRS及び37LPRの存在に連関されたペプチド部位はイーストツーハイブリッド分析を通じて測定された。N−末端の細胞内部位(54乃至113番目アミノ酸)及びC末端の細胞外部位(137乃至210番目アミノ酸)を有している296個アミノ酸を有する37LRPは膜間ドメイン(113 乃至137番目アミノ酸)により区分される。ヒトKRS(597個のアミノ酸)のN−末端特異的拡張(約70個のアミノ酸)部位の次にOB-foldアンチコドン−結合性ドメイン(約70乃至214アミノ酸)と触媒ドメイン(約220乃至574アミノ酸)が位置している。図4で、Myc-KRSに感染されたA549細胞を溶解して抗−Myc及び抗−ラミニン収容体抗体で免疫ブロットを実施した。Myc-KRSを抗−Myc抗体と免疫沈澱させ、67LR及び37LRPで共沈澱させて免疫ブロットを行った。37LRP及び67LRの特異的ブロットの為、多重抗体であるH-141及びF-18(Santacruz)がそれぞれ使用された。(WCL:whole cell lysate)図5で、Myc-KRSで感染されたA549細胞の溶解物を指示した抗体にして、ウェスタンブロットを施行した。細胞は細胞質(C)及び膜分画(M)で分離され抗−Myc抗体で免疫沈澱され、37LRP及び67LR共沈澱されたのをウェスタンブロットで調査した。対照群としてIeGを使用した。図6で、ラミニン(10μg/ml,1h)処理した場合、67LRとKRS間の結合が増えたことを確認した。これを確認する為に、67LRを認識するantibody(abcam.cat# ab2508)で免疫沈澱(immunoprecipitation)し、左側のIgGラベルは兎から得た全体IgG(total IgG)として陰性対照群として使用された。0%SDS PAGEを遂行した後、PVDF膜に移してKRSと67LRを認識する抗体でそれぞれ免疫ブロットをした。
【0107】
図7乃至図12はラミニン−誘導性膜転移(translocation)及びKRSのリン酸化に対しては確認したものである。図7で、A549細胞をラミニン(10μg/ml)で処理してウェスタンブロットを利用して67LR、37LRP及びKRSの量(level)を時間帯別に観察した。Hsp90及びカドヘリン(Cad)をそれぞれ細胞質及び膜のマーカーとして使用した。図8で、A549細胞を1時間かけてラミニン処理、又は無処理して抗−67LR(MLuC5,Santacruz,sc-59732)(洗滌)及びKRS抗体(緑色)を使用して免疫蛍光染色を実施した。図9で、A549細胞をPCL-ガンマ、PKC及びPI3Kをそれぞれ抑制するU73122(U)、スタウロスポリン(staurosporin,ST)及びLY294002(LY)で3時間処理してラミニンで1時間処理後、これらリン酸化酵素抑制剤が67LR及びKRSの細胞質及び膜にどのような影響を及ぼすかを確認した。図10でA549細胞をMyc-KRSで感染させ、24時間培養した。以降、表示された(indicated)薬物で処理して前記の通りラミニンで処理した。Myc-KRSで免疫沈澱し、抗-p-Thr,-Ser,及び-Tyr抗体で免疫ブロットを実施した。図11で、A549細胞をMyc-KRSで感染させ、24時間培養した。感染されたLY294002で3時間処理し、ラミニンで1時間処理した。Myc-KRSは免疫沈澱され、67LRの共沈澱はウェスタンブロットで調査した。免疫沈澱法の対照群としてIgGを使用した。図12で、前記の通り、A549細胞をラミニン及びLY294002K存在又は不在下で培養した。EPRS(glutamyl-prolyl-tRNA sybthetase)はこれに特異的な抗体(AbCam)と免疫沈澱され、KRSの共沈澱はウェスタンブロットで確認した(上)。免疫−消盡上層液(immune-depleted supernatant:ID)で抗KRS及びEPRS抗体とウェスタンブロットを行った。
【0108】
図13乃至図17は、KRSは膜−結合性67LRを安定化させる点を確認したものである。図13で、A549細胞をsi-対照群(si-cont)又はsi-KRSで感染させ、ラミニンの存在又は不存在下で培養した。以降、細胞等を細胞質と膜分画で分離し、それぞれの分画における67LR及びKRSの量をウェスタンブロットで確認した。カドヘリン(赤色)及びhsp90をそれぞれ細胞質及び膜のマーカーとして使用した。図14で、A549細胞において膜結合性67LRの量は抗−LR抗体(MluC5)を利用して遊細胞分析器で観察された。空ベクター又はKRSプラスミドで感染された細胞等を24時間培養した。(上)67LRの量において、KRS抑制の効果を確認する為に、細胞をsi-KRS又はsi-対照群で感染させ、48時間培養した。(下)図15で、EV(空ベクター)又はKRSで感染させたA549細胞をG418で1週間選別して、抗−LR抗体を利用した免疫蛍光染色を利用して67LRの細胞内分布を確認した。膜に分布したLRは白色矢印で強調した。図16で、A549細胞をシクロヘキサミドで処理して新たな(de novo)蛋白質合成を抑制して細胞膜と細胞質内の67LR量において、KRS量の効果をウェスタンブロットで調査した。図17で67LRの細胞的安定性において、KRSの重要性はパルス−チェース実験を通じて調査した。293細胞をsi-KRS又はsi-対照群で感染させ、放射線メチオニンを1時間反応させた。67LRを67LRに特異的に反応する抗体(F-18,Santacruz)と免疫沈澱させ、SDS-PAGEにより分離させ後、自家放射撮影法を実施した。特異性siRNAによるKRSの抑制はウェスタンブロットにより行われ、ローデイング対照群としてチューブリンを使用した。
【0109】
図18乃至図22はKRSは67LRを通じて細胞移動(cell migration)及び癌転移を促進することを確認した。図18でA549細胞を表示されたプラスミドで感染させ、ラミニンの不存在又は存在下で培養し、これらの細胞移動に対する効果はトランスウェルチャンバーで移動した細胞を測定して確認した。膜を通過した細胞の数を計数して各パネルに表示した。実験は3回繰返した。図19で、前記の通り処理した細胞等をMMP-2活性及び量測定のためにそれぞれザイモグラフィー及びウェスタンブロット実施に使用した。図20で、乳癌細胞株である4T-1細胞を表示された(indicated)siRNAで感染させ、Balb/Cマウスの背に皮下注射した。21日経過後、マウスの肺を摘出して径1mm以上の腫瘍結節を計数した。図21で、外因性KRS(KRS-1,及びKRS-2)を発現する2種の互いに異なる4T-1細胞を前記の通り、接種し、30日経過後計数した。空ベクターを感染させた細胞を対照群として使用した。図22で、肺癌(上)及び乳癌(下)組織におけるKRS及び67LRの発現量をこれらそれぞれの抗体を利用した免疫組織化学染色法を使用して比較した。39個の肺癌及び40個の乳癌組織を利用して抗-KRS及び抗−67LR抗体で免疫組織化学染色を実施し、これらの発現量は正常組織のそれと比較された(各組織当り9個の試料)。ここに見える同一患者の代表的な対らはKRSと67LRの過発現を表す。KRSと67LRの統計学的相関関係は表1に表している。
【0110】
図23で、67LRの膜における量は発現に依存する点を示している。図23で、表示されたプラスミドで感染された293細胞を細胞質及び細胞膜分画で分離し、それぞれの分画で67LR、37LRP及びKRSの量を常用する抗体を利用したウェスタンブロットを行い確認した。
【0111】
図24乃至図27は、細胞移動、蛋白質合成及び細胞周期において、細胞内及び細胞のKRSの効果を確認したものである。図24で、ラミニン不存在下で培養されたA549細胞の移動は、トランスウエルチャンバーより移動した細胞を測定して確認した。図25で、KRSの化学走性(chemotactic)活性を見る為、表示された濃度でKRSを含有した無血清培地トランスウエルチャンバーの下のチャンバーに入れて、A549細胞を上のチャンバーに入れて培養した。培養6時間後、移動した細胞を計数した。図26で、A549細胞でKRSの量はsiRNAと外因性KRSの導入により、下落調節又は上昇調節された(図26及び図27パネル)。感染された細胞はそれぞれ48時間及び24時間培養され、無ーメチオニン培地で1時間培養して栄養欠乏状態を造り、放射線標識がなされたメチオニンで2時間標識した。洗滌後、細胞を4時間培養して0.5%トリトンX-100溶解液で溶解させて、放射線活性を液状閃光計数機(liquid scintillation counting)で測定した。図27で、A549細胞を表示された通りに感染させ、固定させてプロピジウムアイオダイド(Propidium iodide)で染色して流細胞分析器で分析した。
【0112】
図28乃至図30は癌転移において、KRS抑制の効果を確認したものである。
【0113】
図28で、標的蛋白質の発現において、si-KRS及びsi-DRSの効果はウェスタンブロットにより調節された。ローディング対照群としてはチューブリンを使用した。図29で、si-RNAが感染された細胞(1×106 )を前記の方法のように注射し、21日経過後腫瘍の大きさ及び嵩を測定することにより、1次腫瘍増殖においてKRS及びDRS抑制の効果を調査した。それぞれのグループは5匹のマウスを使用した。図30で、前記のマウスより摘出された肺は10%ホルマリン溶液に固定された。転移性腫瘍結節及び数は見られる通りである。
【0114】
図31乃至図33は癌転移において、KRS過発現の効果を確認したものである。図31で、KRS-1及びKRS-2細胞株の過発現はウェスタンブロットにより観察された。図32で、1次腫瘍増殖におけるKRS過発現の効果は互いに比較された。図33で、腫瘍転移におけるKRS過発現の効果は接種後30日目に調査された。それぞれのグループは4匹のマウスを使用した。
【図面の簡単な説明】
【0115】
【図1】図1はヒトKRSと37LRP/p40間の相互作用をイーストツーハイブリッド分析を通じて確認したものである。
【図2】図2はヒトKRSと37LRPの相互作用をプルダウン分析を通じて確認したものである。
【図3】図3はヒトKRSと37LRPの相互作用部位を確認したものである。
【図4】図4はKRSの67LR及び37LRPとの結合を確認する為に、Myc−KRSに感染されたA549細胞に対して抗−Myc及び抗−ラミニン収容体抗体を利用して、免疫ブロット分析をした結果である。
【図5】図5はKRSの67LR及び37LRPとの結合を確認する為に、Myc−KRS で感染されたA549細胞の溶解物に対するウェスタンブロット分析をした結果である。
【図6】図6は、ラミニン処理に伴うKRS及び67LRの結合を免疫沈澱で確認したものである。
【図7】図7は、A549細胞にラミニン処理の際、67LR,37LRP及びKRSの量(level)ウェスタンブロットで確認したものである。
【図8】図8は、A549細胞をラミニン処理又は無処理の際、67LR及びKRSの発現を免疫蛍光染色で確認したものである。
【図9】図9は、リン酸化酵素抑制剤が67LR及びKRSの細胞質及び膜における発現に及ぼす影響を確認したものである。
【図10】図10は、KRSを発現するA549細胞でラミニン及びリン酸化酵素抑制剤処理の際、リン酸化程度をp−Thr,−Ser,及び−Tyr抗体で免疫ブロットを実施して確認したものである。
【図11】図11は、KRSを発現するA549細胞でリン酸化されたKRSの67LRとの結合可否をウェスタンブロットで確認したものである。
【図12】図12は、KRSとEPRSの結合にラミニンが及ぼす影響をウェスタンブロット確認したものである。
【図13】図13は、si−対照群又はsi−KRSの形質感染の際、67LR及びKRSの量をウェスタンブロットで確認したものである。
【図14】図14は、A549細胞において、膜−結合性67LRの量を流細胞分析器で確認したものである。
【図15】図15は、EV(空ベクター)又はKRSで感染させたA549細胞に対して67LRの細胞内分布を免疫蛍光染色で確認したものである。
【図16】図16は、新たな蛋白質合成が抑制されたA549細胞で細胞膜と細胞質内の67LR量において、KRS量の効果をウェスタンブロットで確認したものである。
【図17】図17は、67LRの細胞的安定性にKRSが及ぼす影響をパルス−チェース実験で確認したものである。
【図18】図18は、KRS及び/又は67LRの発現抑制時、細胞移動に及ぼす影響を確認したものである。
【図19】図19は、KRS及び/又は67LRの発現抑制時、MMP−2活性及び量をザイモグラフィー及びウェスタンブロットで確認したものである。
【図20】図20は、4T−1細胞株が移植されたマウスでKRS発現抑制の際、腫瘍結節の数を表したものである。
【図21】図21は、4T−1細胞株が移植されたマウスでKRS発現増進の際、腫瘍結節の数を表したものである。
【図22】図22は、肺癌及び乳癌組織におけるKRS及び67LRの発現量を免疫組織化学染色法で確認したものである。
【図23】図23は、67LRの膜における量にKRS発現が及ぼす影響をウェスタンブロットで確認したものである。
【図24】図24は、ラミニン不存在下で培養されたA549細胞の移動を測定したものである。
【図25】図25は、細胞移動に対するKRSの化学走性(chemotactic)活性を測定したものである。
【図26】図26は、A549細胞でsiRNAと外因性KRSの導入に伴うKRSの量及び細胞内全体蛋白質合成量を確認したものである。
【図27】図27は、A549細胞でsiRNAと外因性KRSの導入に伴うKRSの量及び細胞周期を確認したものである。
【図28】図28は、標的蛋白質の発現において、si−KRS及びsi−DRSの効果をウェスタンブロットで確認したものである。
【図29】図29は、腫瘍細胞移植に伴う1次腫瘍増殖において、KRS及びDRS抑制の効果を確認したものである。
【図30】図30は、腫瘍細胞移植に伴う転移性腫瘍結節及び数を確認したものである。
【図31】図31は、KRS−1及びKRS−2細胞株のKRSの過発現をウェスタンブロットで確認したものである。
【図32】図32は、腫瘍細胞移植に伴う1次腫瘍増殖において、KRSの過発現の効果を確認したものである。
【図33】図33は、腫瘍細胞移植に伴う転移性腫瘍結節及び数を確認したものである。
【発明を実施するための形態】
【0116】
以下、本発明を実施例により詳細に説明する。
ただし、下記実施例は本発明を例示するもので、本発明の内容がこれに限定されるものではないものの、
【0117】
<実験方法>
1.細胞培養及び材料等
A549及びHEK293細胞は、ATCCから購入した。マウス乳癌(mammary carcinoma)4T-1細胞株は金ソンチン博士(カチョン医科大学)より得た。10%牛胎児血清(Fetal bovine serum, FBS)及び1%抗体を含む。RPMI(A549細胞及び4T-1細胞に対する)及びDMEM(Dulbecco's Modified Eagle Medium,他の細胞に対する)培地を細胞培養に使用した。37LRPを符号化するpcDNA3.1ベクターはタチバナヒロフミ博士(九州大学)から得た。Myc-が付着された(Myc-tagged)ヒトKRS及びDRSはpcDNA3ベクターのEcoRI/XhoI制限酵素部位にクローニングした。齧歯類KRS cDNAはRT-PCRで確保し、pcDNA3.1ベクターのHindIII/XhoI制限酵素部位にクローニングした。齧歯類及びヒトKRS及びDRSにターゲッティング(targeting)するsiRNAはInvitrogen社より購入した。siRNAの配列は要請すれば提供できるであろう。遺伝子ポーター(Gene porter, GTS)及びリポペクタミン2000(invitrogen)は形質感染試薬として使用された。LY294002及びスタウロスポリン(stauroaporin)はCalbiochemから購入し、サイクロヘキシミド(cyclohexiumide)及びラミニン(laminin, Engelbreth-Holm-Swarm murine sarcoma)はSigmaから購入した。
【0118】
2.免疫沈降及びウェスタンブロット
細胞を150mM NaCl、0.5%トリトンX-100, 0.1%SDS及び蛋白質分解酵素阻害剤を含む20mM Tris-HClバッファ(pH7.4,溶解バッファ)で溶解した。蛋白質抽出物を正常IgG及び蛋白質Gアガロスと2時間培養した後(incubation)、非特異的にIgGに結合した蛋白質を除去する為に遠心分離した。上澄液を精製された67LR抗体(F-18, Santacruz)と混合した後、揺さぶりながら4℃で2時間培養し、蛋白質Aアガロスを混合した。氷で冷たく冷却した溶解バッファを利用して3回洗滌し、沈澱物をSDSサンプルバッファに溶解させ、SDS-PAGEで分離した。互いに異なる分画でKRS及びLRの結合を確認する為に、pcDNA3.1-Myc-KRSで形質感染し、proteoextractキット(Calbiochem)を利用して製造社の指針に従って、プラスマメンブレーンとサイトプラズム分画を分離したあと、前記の通り、共同−免疫沈降を行った。蛋白質水準を分析する為に、蛋白質を細胞から抽出した後、10%SDS-PAGEで分離した。別に言及しない限り抗-LR抗体(Abcam, ab2508)を37LRP及び67LRの同時免疫ブロッティングに使用した。hsp90及びパンーカドヘリン(Pan-cadherin)に対する抗体はSantacruzから購入した。
【0119】
3.フローサイトメトリー
細胞サイクルをアドレス(address)する為に、培養された細胞を表示されたベクター又は化合物で形質感染又は処理して、70%エタノール4℃で1時間固定した後、氷で冷却したPBSで2回洗滌した。その後細胞をpropidium iodide(50μg/ml)、0.1% sodium citrate,0.3%NP40及びRNaseA(50μg/ml)で40分間染色した後、プローサイトメトリ(FACS Calibur, Beckton-Dickinson)を行った。それぞれの染色に対して、Cell Quest Proソフトウェアーを使用して20000細胞を分析した。細胞表面の67kDLRの量を分析する為に、1×106細胞をIgG又は67LRの細胞外ドメインを認識する抗−LR抗体(MLuC5,1ug)と培養した後、FITC2次抗体と培養した。PBSで洗滌してサンプルをFACSでスキャンした。
【0120】
4.免疫蛍光染色又は免疫組織化学染色
9mmカバースリップ(cover slip)上のA549細胞を70%メチルアルコールで固定した後、冷たいPBSで簡単に洗滌した。1%CAS,3%BSA及び0.5%triton X-100を含むブロッキングバッファで30分間処理した後、細胞をKRSに対する抗体(Abcam)、及びMLuC-5に対する抗体(Santacruz)で1時間培養した。アレキサ488及び568(invitrogen)を添加した後、室温で30分間処理した。冷たいPBSで30分間洗滌し、標本をレーザースキャニングマクロスコーピで観察した。乳癌及び肺癌に対する組織配列スライド(tissue array slide)をSuper-Biochip(韓国)から購入し、67LR及びKRSの発現水準を確認する為に、文献(Park, S. G. et al. Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. U S A 102, 6356-6361(2005))言及された相応する抗体と免疫組織化学染色を行った。67LR及びKRSの発現間の相関関係を計算する為に、ピアソンχ2テスト及びスチューデントtテストを使用して統計的分析を行った。P値が<0.05であれば意味あるものと見做した。全ての統計的分析はSPSS v11.5ソフトウェアー(SPSS,Chicago,Ill)を使用して行った。
【0121】
5.パルス−チェース実験(Pulse-chase experiment)
293細胞をリポベクターミン2000を利用して si-KRS又はsi対照群で形質感染させた。これをメチオニンが含まれていない培地で1時間培養後、[35S]メチオニン(50μCi/ml)を添加して1時間培養した。新鮮な培地で放射線メチオニンを洗い落とし、67LRをこれに特異的な抗体で免疫沈降させ、12%SDS-PAGEで分離し、BAS(FLA-3000,Fujifilm)を利用して露出させた(autoradiography)。67LRの量はMulti-gaugeプログラム(V3.0,Fujifilm)を利用して測定した。
【0122】
6.イーストツーハイブリッド(yeast two hybrid)分析
ヒトKRSの多くの断片を符号化するcDNAを相応するプライマーを利用してPCRで得た。KRSに対するPCR産物をEcoRI及びXhoIで切断し、pEG202ベクター(LexA-融合蛋白質の製造の為)及びpJG4-5ベクター(B42-融合蛋白質の製造の為)の相応する位置に連結した。37LRP断片を符号化するcDNAはBarbara J,Ballermann博士(アルバータ大学)から得て、これをpJG4-5ベクターのEcoRI及びXhoI部位に挿入した。二つの融合蛋白質間の相互作用はX-gal-含有酵母培地上で青色コロニーの合成可否で分析した。
【0123】
7.In vitro結合分析
GST-KRS又はGSTを大腸菌ロゼタ(DE3)ストレンで発現させ、前記蛋白質抽出物を1%Triton X-100及び0.5% N-ラウリルサコシンが含有されたPBSバッフアで4℃で2時間グルタチオンーセパロスと混合した。ヒト37LRPはTNT Quick coupled Transcription/Translation(Promega)を使用し、pcDNA3-37LRPを鋳型に使用し、[35S]メチオニン存在下でin vitro translationにより合成した。合成された37LRPは前記 GST蛋白質混合物に添加し、1% Triton X-100, 0.5% N-ラウリルサコシン、1mM DTT, 2mM EDTA及び 300μMペニルメチルスルホニルフルオライドが含有されたPBSバッファで撹拌しながら4℃で4時間培養した後、0.5% Triton X-100を含有する同じバッファで6回洗滌した。その後セパロスビッドに結合された蛋白質を SDSサンプルバッフアで溶出し、SDS-PAGEで分離して放射線を測定した(autoradiograph)。
【0124】
8.細胞移動(cell migration)分析
細胞移動を先行文献(Park, S. G. et al. Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. USA 102, 6356-6361(2005))に記載した通り、ポリカボネート膜(8.0μm 空隙サイズ、Coster)を有する 24-ウェルトランスウェルチャンバで測定した。A549細胞を無血清培地に懸濁した後、各ウェル当り1X105 細胞の濃度で上位チャンバに入れた。表示された濃度の精製されたヒトKRS、ラミニン(10μg/ml)又はゼラチン(10μg/ml)を下位ウェルに入れ、細胞がCO2 培養器で37℃で6時間移動するようにした。細胞は70%メチルアルコールを含むPBSで30分間固定し、PBSで3回洗滌した。細胞はヘマトシリン(Sigma)で10分間染色した後、蒸留水で洗滌した。綿棒で膜の上の部分で移動しない細胞を除去した。膜をチャンバから分離した後、Gel Mount(Biomeda 米国)に乗せておいた(mount)。移動した細胞(膜の下位面に付着されたもの等)を顕微鏡下で(x20)4ヶ所を任意に選別して計測する方式で測定した。
【0125】
9.ザイモグラフィー(zymography)
表示されたsiRNA及び組換えKRS(又はDRS)を符号化するプラスミドで形質感染されたA549細胞をそれぞれ48時間及び24時間培養した後、10%FBSを含有する、RPMI培地に接触した(1x105 細胞/ウェル)、細胞を無血清RPMI培地で2時間飢餓処理(starving)した後、ラミニンを添加して10μg/mlで24時間培養した。20μlの培養培地を5xFODバッフア(4% SDS, 20%グリセロール及び0.01%ブロモフェノールブルーを含有する0.125M Tris-HCl, pH6.8)と混合した後、1mg/mlのゼラチンを含有する10%SDS-PAGEを行った。ゲルを2.5%トリトンx-100でそれぞれ20分間洗滌し、蒸留水でそれぞれ20分ずつ洗滌して反応バッフア(10mM CaCl2, 150mM NaCl, 1μM ZnCl2, 1% Triton X-100, 0.002% sodium azideを含有する50mM Tris-HCl, pH 7.5)と37℃で24時間培養した。ゲルを蒸留水で洗滌してクマシブルーR250で染色して35%メタノールで脱染(destain)した。
【0126】
10.生体内で癌転移実験
マウス乳癌4T-1細胞をsi KRS、si DRS又はsi−対照群で形質感染させ、24時間培養した。細胞(1x106)を6週令雌Balb/cマウスの背に皮下注射の方法で注入した。ターゲットに対するsiRNAの効果を形質感染後、48時以降に残った細胞で測定した。さらに、注入後3日から10日まで2日間隔で1次腫瘍(primary tumor)でターゲットに対するsiRNAの効果を相応する抗体を使用したウェスタンブロット分析で測定した。腫瘍の成長は週当り3回ずつ腫瘍の大きさをモニタリングした。これと同時に全体の体重も測定した。注入後21日目にマウスを殺し、1次腫瘍と肺をマウスから切除した。肺は10%ホルマリンで24時間固定した。肺に転移された腫瘍結節の数と大きさを測定し、径が1mmより大きい腫瘍結節に対しても別途記録した。1次腫瘍の質量も測定した。KRS過発現が腫瘍転移に及ぼす効果を確認する為に、齧歯類KRSベクター及び空ベクター(empty vector)を4T-1細胞に形質感染させ、安定的な形質感染体をG418の存在下で3週間培養して選別した。多くの単一コロニーを選びだして、ウェスタンブロットでKRS発現水準を比較した。対照群細胞より高い水準でKRSを発現する二つの異なるコロニー(KRS-1,KRS-2)を選択し、注入に使用した。注入後30日目にマウスを殺したことを除いては、前記方法と同一に以降実験を行った。
【0127】
<実験結果及び考察>
全長のKRSと37LRPとの特異的相互作用はイーストツーハイブリッド分析により確認された。LexA-KRSはKRSのパートナとして知られた(Kim, J.Y. et al. p38 is essential for the assembly and stability of macromolecular tRNA synthetase complex: Implications for its physiological significance, Proc. Natl. Acad. Sci. USA 99, 7912-7916(2002))AIMP2ばかりでなく、B42-37LRPと結合して青色コロニーを形成し、AIMP1に対してはそうではなかった(図1)。インビトロバインディングアッセイに対しては、[35S]メチオニンで表示された37LRPをGST-KRS又はGSTと混合し、グルタチオンーセファロースで沈澱させ、自家放射線撮影をした。37LRPはGST-KRSと共に沈澱したものの、GSTではなかった(図2)。イーストツーハイブリッド分析による消失地図(deletion mapping)でヒトKRSのN−末端延長部位とLRの C−末端細胞外ドメインがそれらの結合に関与することを確認した(図3)。
【0128】
細胞質内37LRPは膜−包埋性67LRで転換されるので、本発明者等はKRSが37LRP及び67LRに互いに別に結合するかを確認した。Myc-KRSを肺癌細胞株A549細胞に導入して、抗−Myc抗体と免疫沈澱させた。細胞溶解物のウェスタンブロット結果、67LRは37LRPより少ない量で存在した(図右側)。しかし、Myc-KRSは37LRPより67LRにより優先的に結合した(図左側)。本発明者等は以降、A549細胞質と原形質膜分画で分離してMyc-KRSは37LRP及び67LRの相互作用を確認した。37LRPは細胞質で67LRは原形質膜でそれぞれ優勢して観察された(図5右側)。KRSは細胞質においてより多くの量が観察されたものの、二つの分画全てにおいて存在した。二つの分画全てを抗−Myc-抗体と免疫沈澱させた時、例え、細胞質内少量の37LRPも沈澱されたものの、膜に存在する67LRが主にKRSと共に沈澱されたところ(図5左側)、これは膜に存在する67LR及びKRS間の信号的相互作用を示唆した。本発明者等は以降、KRSの細胞内分布がA549細胞のラミニン処理により、変化するかを細胞分画と免疫蛍光染色法を通じて調査した。ラミニン処理後、KRS及び67LRの膜内相互作用量は細胞質内のKRS及び37LRP量やこれらの発現には、殆ど変化なく漸次的に増加した(図7及び結果未図示)。免疫蛍光染色におても、さらに、67LR及びKRSがラミニン処理により膜側に移動することを説明してくれた(図8、それぞれ赤色及び緑色)。以降KRSの膜転座(translocation)が翻訳後変形(post-transcriptional modification)に関与するかを調査した。ホスホイノシチド3-OHキナーゼ(phosphoinositide 3-OH kinase, PI3K)(Shaw, L. M., Rabinovitz, I., Wang, H. H., Toker, A. & Mericurio. A.M. Activation of phosphoinositide 3-OH kinase by the alpha6beta4 integrin promotes carcinoma invasion. Cell 91, 949-960(1997))、蛋白質リン酸化酵素C(Protein kinase C, PKC)(Li, Y. Q. et al. Protein kinase C mediates the signal for interferon-gamma mRNA expression in cytotoxic T cells after their adhesion to laminin. Immunology 93, 455-461(1998))及びホスホリパーゼC−ガンマ(phospholipase C-gamma, PLC-gamma)(Vossmeyer, D., Hofmann, W., Loster, K., Reutter, W. & Danker, K. Phospholipase C-gamma binds alpha1beta1 integrin and modulates alpha1beta1 integrin-specific adhesion. J. Biol. Chem. 277, 4636-4643(2002); Kanner, S. B., Grosmaire, L. S., Ledbetter, J. A. & Damle, N. K. Beta 2-integrin LFA-1 signaling through phospholipase C-gamma 1 activation. Proc .Natl. Acad. Sci. USA 90, 7099-7103(1993))のような幾つかの互いに異なるリン酸化酵素等がラミニンにより、活性化されると知られている。このようなリン酸化酵素等の内、どのよなものがKRSのラミニン依存性膜転座に関与するかを確認する為に、本発明者等はそれぞれのキナーゼに特異的な抑制剤でそれぞれのリン酸化酵素を遮断し、この処理が如何にKRSのラミニン依存性膜転座に影響を及ぼすか、否かを確認した。膜分画でKRS及び67LRのラミニン依存的増加はPI3K抑制剤であるLY294002の存在下で抑制されるに反し、U73122又はスタウロスポリンで処理した細胞等は、対照群細胞と同様に67LRSのラミニン依存的増加を示した(図9上及び結果未図示)。これらの結果は、PI3KがKPSのラミニン誘導性リン酸化に連関されたものであることを暗示する。事実、チロシンでないトレオニンとセリンでリン酸化されたKRSはラミニン処理により、増加された反面、LY294002の存在下では抑制され、スタウロスポリンは如何なら影響も及ぼせなかった(図10)。本発明者等はさらに、KRSのラミニン誘導性リン酸化が67LRとの相互作用に必要であるか否かを調査した。LY294002処理はKRSと67LRのラミニン誘導性疾患を抑制した(図11)。細胞質KRSが多重−ARS複合体に固定されているので、本発明者等はさらに、KRSのラミニン依存的リン酸化がKRSと複合体の他の酵素合成成分であるグルタミルーフロリルーtRNA合成酵素(glutamyl-prolyl-tRNA synthetase, EPRS)と、共ー免疫沈澱することにより、多重−ARS複合体の結合に影響を及ぼすかを調査した。LY化合物の不在下で。ラミニン処理はKRSとEPRSの結合を減少させ、同時に免疫−消盡された溶解性分画(immune-depleted soluble fraction)のKRSを増加させた(図12上、下パネルの左側レーン)。これに反し、EPRSに結合したKRSは細胞がLY294002で前処理された時、ラミニン処理に影響が及ばなかったものの(図12上、下パネルの右側レーン)これはKRSのリン酸化がKRSと複合体のラミニン依存的結合に必要であることを示唆する。
【0129】
本発明者等は以降、KRSがA549細胞において、67LRの膜内存在量に影響を及ぼすかを確認した。67LRの量はラミニンにより増加するが、ラミニン効果はKRSがこれに対する特異的siRNAにより、抑制される時には消えるものの(図13左側)、これは67LRのラミニン依存的促進において、KRSの重要性を示唆する。本発明者等はさらに、遊細胞分析器を通じて膜に存在する67LRを調査した。膜に存在する67LRの量は細胞をKRSで形質感染させるか、又はsi-KRSで感染させた時、それぞれ増加及び減少した(図14)。ラミニン収容体の細胞内分布は空ベクター(EV)又はKRSに形質感染されたA549細胞間で、免疫蛍光染色を通じて比較された。ラミニン収容体は対照群と比較してKRS過発現細胞の膜部位に強く染色された(図15)。以降、KRS及び67LR間の陽性的相関関係(positive correlation)は、多様なKRS量に伴う膜及び細胞質内の67LR存在量を測定することにより確認された(図23)。
【0130】
本発明者等は、KRSがどのように67LRの膜内に存在量を増加させるかを調査した。KRSは37LRPから転写又は転換(conversion)を通じて67LRを促進することができる。しかしながら、KRSの感染はLR転写調節の暫定的役割を除いてはLR転写を増加させなかった(結果、未図示)。さらに、細胞質内で37LRPとよく結合しないので(図4、図5)、結果、が37LRPを67LRに転換する過程を促進するようである。本発明者等は、さらに、37LRPの変更が37LRPが67LRに転換されるにおいて、先行要件として知られているので(Landowski, T. H., Dratz, E.,A. & Starkey, J. R. Studies of the structure of the metastasis-associated 67 kDa laminin binding protein: fatty acid acylation and evidence supporting dimerization of the 32 kDa gene product to form the mature protein. Biochemistry 34, 11276-11287(1995); Buto, S. et al. Formation of the 67-kDa laminin receptor by acylation of the precursor. J. Cell. Biochem. 69, 244-251(1998))、KRSが37LRPの脂肪族アシル化(fatty acylation)を媒介するかを調査した。本発明において、KRSは37LRPの脂肪族アシル化に全く影響を及ぼさなかった(結果未図示)。結果、は膜に存在する67LRの細胞内安定性を増進できるので、本発明者等はKRSが膜に存在する67LRの食細胞作用(endocytosis)を妨害できるかを調査した。前記可能性を確認する為、本発明者等は、シクロヘキサミド(cyclohexamide)を利用して根源的に(de novo)蛋白質合成を阻止して、KRSが細胞膜と細胞質における67LRの存在量に影響を及ぼすかを実験した。KRSの発現がこれに特異的なsiRNAにより抑制される時には、67LRの膜内存在量が減少しながら同時に細胞質分画におけ量は増加した(図16左側)。これとは逆にKRSの過発現は前記の通り、膜内67LRの存在量を増加させた(図16右側)。このような結果を基に、KRSは67LRに細胞質内に再び入ることを抑制することにより、膜内67LRの存在量を増加させるようである。本発明者等は、さらに、パルス−チェース(pulse-chase)実験を通じて67LRの代謝転換(turn over)において、KRSの効果を調査した。初期蛋白質活性は放射線メチオニンで表示され、以降、シクロヘキサミドで抑制された。以降、67LRの消滅(disappearance)は時間的間隔をおいて、自家放射撮影により観察された。67LRはKRSがsiRNAにより抑制される時、急激に減少されたものの、この数値はsi対照群においては、時間間隔によって維持されたものと対照的である(図17)。従って、67LRの分解過程に対して追加的な研究が必要なものの、KRSは原形質膜で67LRと結合することにより、67LRの半減期を増加させ67LRの食細胞作用を抑制するものと見られる。
【0131】
本研究者等は以降、KRSの発現程度がラミニン依存的A549細胞移動に影響を及ぼすかをトランウェルメンブレーン分析を通じて調査した。対照群細胞の移動はラミニン処理により、平均6倍程促進された(図24及び図18)。しかしながら、ラミニン依存的細胞移動はKRSがsiRNAにより抑制された時減少した(図18,si-対照群及びsi-KRS)。これとは逆にKRS過発現はラミニン処理により、促進された細胞移動を増加させた(図18,EV及びKRS)。しかしながら、細胞移動においてKRSの効果は、ラミニン収容体がsiRNAにより抑制されると消えた(図18,si-LR,下パネル)。KRSは幾つかの癌細胞でサイトカインで分離されるので(Park, S. G. et al. Human lysyl-tRNA synthetase is secreted to trigger pro-inflammatory response, Proc. Natl. Acad. Sci. U S A 102, 6356-6361(2005))、本発明者等は細胞外因性KRSが細胞移動に影響を及ぼすかを調査した。A549細胞を互いに異なる濃度の精製されたKRSで処理した時、この分析でKRSの細胞外因性効果を除けば、細胞移動は殆ど影響を受けなかった(図25)。反面、細胞蛋白質の合成及び細胞周期は実験する間、KRSの抑制及び過発現に影響はされなかったものの、これはKRSの依存的細胞移動が前記過程の効果に起因しないことを示唆した(図26及び図27)。ラミニン処理がMMP-2(matrix metllo-proteinase-2)の活性化を招来するので(Givant-Horwitz, V., Davidson, B. & Reich, R. Laminin-induced signaling in tumor cells the role of the M(r) 67,000 laminin receptor. Cancer Res. 64, 3572-3579(2004))、 MMP-2のラミニン依存的活性化において、KRSの役割をin vitroザイモグラフィー分析を通じて調査した。MMO-2活性はラミニンにより活性化され、これはsi-KRSの存在下で抑制された(図19右側)。MMO-2の発現量はKRSに影響されなかった(図19下)。
【0132】
KRSが癌転移と関連された67LRを通じて細胞移動を促進できるので、肺に転移がよくなされるマウス乳癌4T-1細胞を使用してKRSの発現水準により、癌転移が影響されるのかを調査した。こめため、KRS又は多重-ARS複合体の他の構成要素であるDRS(aspartyl-tRNA synthetase)をそれらに特異的なsiRNAで発現を抑制し、如何に、KRS及びDRSの発現減少が癌転移に影響を及ぼすかを比較した。si-KRSの抑制効果をウェスタンブロットで確認した後(図28)、これら細胞それぞれ及びsi-対照群で処理した細胞をBalb/cマウスの背皮膚に皮下注射で注入した。注入された3種全て類似した質量及び嵩の腫瘍を発生させ、(図29)、KRSの水準が1次腫瘍の成長には影響しないことを知らせてくれた。肺を注入後21日目に分離し、転移性腫瘍結節(径1mmより大きいもの)の数を3グループ間で比較した。転移性結節の数は対照群及びDRSが抑制された細胞で得たのと比べてKRSの抑制により大きく減少した(図20及び図30)。逆に、前記方法によりKRSの過発現が癌転移を増進するかを調査した。我々はKRSを符号化するプラスミドの形質感染及びG418スクリーニングにより、KRSを安定的に過発現する4T-1細胞株を初めて構築した。構築された細胞株でKRS過発現はウェスタンブロットで確認し、空ベクターで形質感染された細胞等に比べて多量にKRSを発現する2個の互いに異なる細胞(KRS-1及びKRS-2)を選別した(図31)。これら細胞も類似した質量と大きさの1次腫瘍を生成した(図32)。これら細胞を注入した後、30日目に肺を検査した時、KRS過発現細胞二つとも対照群細胞に比べてより多くの結節を生成した(図21及び図33)。このような結果はKRSが生体内で癌転移を誘導することを暗示する。
【0133】
ラミニン収容体の癌特異的過発現は頻繁に観察されるので(Fontanini, G. et al.67-Kilodalton laminin receptor expression correlates with worse prognostic indicators in non-small cell lung carcinomas. Clin. Cancer Res. 3, 227-231(1997); Viacava, P. et al.The spectrum of 67-kD laminin receptor expression in breast carcinoma progression. J. Pathol. 182, 36-44(1997))、67LRの過発現が、さらにKRSの過発現と連関されているかを肺癌及び乳癌を例に67LR及びKRSの免疫組織化学染色により分析した。39個の検査した肺癌組織の中で67LR過発現は21事例(54%)で観察され、そのなかでKRS水準は19事例(約90%)で増加されていた(表1及び図22上位)。同様に40個の検査した乳癌患者の内、21事例で67LR過発現が観察された。これらの内、21事例全てKRSの水準が増加された(表1及び図22下位)。それらの同時−発現が転移において実際に連関されているかを確認すべきであるにも拘らず、二つの場合全て二つの蛋白質の発現間に密接な連関があらわれた。
【0134】
【表1】
【0135】
この際、前記表1に対しては下記の記載を参照できる:表1は腫瘍組織において67LRとKRS発現の相関関係を表したものである。表1では67LRの発現量がKRSの発現量と関係があるか否かを調べる為に、肺癌及び乳癌患者の組織マイクロアッセイ等を利用してそれぞれの抗体に対する免疫感染を実施し、前記二つの蛋白質等の相対的発現量を測定した。67LRの標識の為に、MluC5抗体が使用された。発現量は試料の染色濃度を通じて測定され、4個のグループに分類された(0,1,2,及び3点)。最終評価で試料等は正常グループ(0点又は1点)と過発現グループ(2点又は3点)に分類された。67LRとKRS発現の相関関係を評価する為の統計学的分析は、ピアソン χ2 テスト及びStudent tテストを通じて行った。この際、P値が0.05未満であれば(P<0.05)有効な数値として見做された。全ての統計学的分析はSPSS v11.5ソフトウェアー(SPSS, Chicago, Ill)を通じて行った。
【0136】
リポソム構成因子を含む多くの翻訳因子等が多面的であり(Wool, I. G. Extraribosomal functions of ribosomal proteins Trends Biochem. Sci. 21, 164-165(1996))、種々の腫瘍生成過程に関与されている(Lee, S. W., Kang, Y. S. & Kim, S.Multi-functional proteins in tumorigenesis: Aminoacyl-tRNA synthetases and translational components. Curr. Proteomics 3, 233-247(2006))。ここで我々は二つの翻訳因子であるKRS及びp40/37LRPが生体内で細胞移動及び癌転移に共に作用することを示した(図18乃至図22)。現時点ではこの二つの蛋白質の潜在的な連関進化的偶然か、若しくは以降究明される必要がある蛋白質合成において、生理学的理由を有しているかは知り兼ねない。ARS-複合体の構成要素の内、KRSは最も安定的な蛋白質であり、他の構成要素の安定性の為に必要であるものの(Han, J. M. et al. Hierarchical Network between the components of the multi-tRNA synthetase complex: Implications for complex formation. J. Biol. Chem. 281, 38663-38667(2006))、これは他の関係される蛋白質を安定化させるKRSの潜在能力を暗示するものである。ここで、我々はKRSの能力が67LRにおける細胞安定性にも拡大されることを示した(図17)。
【0137】
KRSの67LRとの連関は他の機能的暗示を有し得る。生理的条件下で、細胞質KRSの一部分がリン酸化され、多くの成長刺戟や生存信号により、ラミニン信号を媒介する67LRと結合する為に、原形質膜に移動される。癌細胞では過発現又はPI3Kのような過ー活性化された上位リン酸化酵素に因る結果にして、構造的な(constitutive)膜転移により、KRSの膜水準が非正常的に増加される。多分、このような過量のKRSが原形質膜を67LRを集めるか若しくは、排出することができる。さらに、PI3Kの活性化を減少させるのが頻繁に腫瘍成長及び転移と連関される点(Wymann, M. P. & Marone, R. Phosphoinositide 3-kinase in disease: timing, location, and scaffolding. Curr. Opin. Cell Biol. 17, 141-149(2005))及びラミPI3Kを通じて癌浸潤を増進する点(Baba, Y. et al. Laminin-332 promotes the invasion of oesophageal squamous cell carcinoma via PI3K activation . Br. J. Cancer 98, 974-980(2008))を念頭に置く価値がある。PI3Kの構造的な活性化は膜に移動させるKRSのリン酸化をリードすることができる。このよう条件等の一部又は全部が原形質膜での67LRの増加に寄与することができ、癌転移に対するラミニン信号伝達を増幅させ得る。癌の転移性拡張を調節する多くの研究が進行されてきた。このような点で、癌転移において67LRを通じたKRSの合成は癌診断及び治療の為の、従前には開拓されていない新たな窓を提供し得る。
【産業上の利用可能性】
【0138】
以上説明した通り、本発明者等はKRSが原形質膜に転座(translocation)され、67LRと相互作用することにより、腫瘍(又は癌)細胞の移動を促進して癌の転移に影響を及ぼすことを見い出した。さらに、マウスを利用したin vivo実験を通じてKRSの過発現又は発現抑制が癌の転移を調節し得ることを究明した。従って、本発明のKRSを利用して癌転移又は癌細胞移動を調節することができ、さらには、原形質膜のラミニン収容体(67LR)と関係した細胞内代謝を調節できる。本発明で究明したKRSとラミニン収容体の関係はこれと関係した多様な疾患又は疾病の治療、予防及び/又は診断に極めて有用に使用できる。
【特許請求の範囲】
【請求項1】
ライシルt-RNA合成酵素(Lysyl t-RNA synthetase,KRS)の細胞内水準を調節して癌転移(cancer metastasis)を調節する方法。
【請求項2】
ライシルt-RNA合成酵素の細胞内水準を減少させて癌転移を抑制することを特徴とする請求項1に記載の癌転移を調節する方法。
【請求項3】
ライシルt-RNA合成酵素の細胞内水準を増加させて癌転移を増進することを特徴とする請求項1に記載の癌転移を調節する方法。
【請求項4】
前記ライシルt-RNA合成酵素は配列番号1で表示されるアミノ酸配列からなることを特徴とする請求項1に記載の方法。
【請求項5】
ライシルt-RNA合成酵素の細胞内水準を調節して癌細胞の移動(cancer cell migration)を調節する方法。
【請求項6】
ライシルt-RNA合成酵素の細胞内水準を減少させ、癌細胞の移動を抑制することを特徴とする請求項5に記載の癌細胞の移動を調節する方法。
【請求項7】
ライシルt-RNA合成酵素の細胞内水準を増加させ、癌細胞の移動を促進することを特徴とする請求項5に記載の癌細胞の移動を調節する方法。
【請求項8】
前記ライシルt-RNA合成酵素は配列番号1で表示されるアミノ酸配列からなることを特徴とする請求項5に記載の方法。
【請求項9】
プロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体を有効成分として含む癌の予防及び治療用の組成物。
【請求項10】
前記癌は大腸癌、肺癌、肝臓癌、胃腸癌、食道癌、膵臓癌、胆嚢癌、腎臓癌、膀胱癌、前立腺癌、睾丸癌、子宮頸部癌、子宮内膜癌、絨毛癌、卵巣癌、乳癌、甲状腺癌、脳癌、頭頚部癌、悪性黒色腫、リンパ腫、再生不良性貧血からなる群より選ばれたことを特徴とする請求項9に記載の組成物。
【請求項11】
プロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体をこれを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法。
【請求項12】
癌治療剤を製造するためのプロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体の用途。
【請求項13】
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;
及び
(c)選別された製剤が癌転移又は細胞移動を調節するか否かをテストする段階を含む癌転移又は癌細胞移動調節をスクリーニングする方法。
【請求項14】
(a)試験製剤の存在下でKRS、ラミニン収容体(67LR)及び試験製剤を接触させる段階;及び
(b)前記試験製剤がKRSとラミニン収容体の相互作用(interaction)を調節するか否かをテストする段階を含むKRSと67LRの相互作用を阻害する製剤をスクリーニングする方法。
【請求項15】
(a)試料で67LRの過発現可否を分析する段階;及び
(b)67LRが過発現された試料からKRSの過発現可否を分析する段階を含む、肺癌又は乳癌を診断する方法。
【請求項1】
ライシルt-RNA合成酵素(Lysyl t-RNA synthetase,KRS)の細胞内水準を調節して癌転移(cancer metastasis)を調節する方法。
【請求項2】
ライシルt-RNA合成酵素の細胞内水準を減少させて癌転移を抑制することを特徴とする請求項1に記載の癌転移を調節する方法。
【請求項3】
ライシルt-RNA合成酵素の細胞内水準を増加させて癌転移を増進することを特徴とする請求項1に記載の癌転移を調節する方法。
【請求項4】
前記ライシルt-RNA合成酵素は配列番号1で表示されるアミノ酸配列からなることを特徴とする請求項1に記載の方法。
【請求項5】
ライシルt-RNA合成酵素の細胞内水準を調節して癌細胞の移動(cancer cell migration)を調節する方法。
【請求項6】
ライシルt-RNA合成酵素の細胞内水準を減少させ、癌細胞の移動を抑制することを特徴とする請求項5に記載の癌細胞の移動を調節する方法。
【請求項7】
ライシルt-RNA合成酵素の細胞内水準を増加させ、癌細胞の移動を促進することを特徴とする請求項5に記載の癌細胞の移動を調節する方法。
【請求項8】
前記ライシルt-RNA合成酵素は配列番号1で表示されるアミノ酸配列からなることを特徴とする請求項5に記載の方法。
【請求項9】
プロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体を有効成分として含む癌の予防及び治療用の組成物。
【請求項10】
前記癌は大腸癌、肺癌、肝臓癌、胃腸癌、食道癌、膵臓癌、胆嚢癌、腎臓癌、膀胱癌、前立腺癌、睾丸癌、子宮頸部癌、子宮内膜癌、絨毛癌、卵巣癌、乳癌、甲状腺癌、脳癌、頭頚部癌、悪性黒色腫、リンパ腫、再生不良性貧血からなる群より選ばれたことを特徴とする請求項9に記載の組成物。
【請求項11】
プロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体をこれを必要とする個体に有効な量で投与することを含む癌の予防及び治療方法。
【請求項12】
癌治療剤を製造するためのプロモーター及びこれと作動可能に連結されたKRSを符号化するポリヌクレオチドに対するアンチセンスRNA(antisense RNA)又はsiRNAを符号化するポリヌクレオチドを含む発現ベクター又はKRSに対する抗体の用途。
【請求項13】
(a)試験製剤の存在下でKRSと試験製剤を接触させる段階;
(b)KRSの活性を測定してKRSの活性を変化させる試験製剤を選別する段階;
及び
(c)選別された製剤が癌転移又は細胞移動を調節するか否かをテストする段階を含む癌転移又は癌細胞移動調節をスクリーニングする方法。
【請求項14】
(a)試験製剤の存在下でKRS、ラミニン収容体(67LR)及び試験製剤を接触させる段階;及び
(b)前記試験製剤がKRSとラミニン収容体の相互作用(interaction)を調節するか否かをテストする段階を含むKRSと67LRの相互作用を阻害する製剤をスクリーニングする方法。
【請求項15】
(a)試料で67LRの過発現可否を分析する段階;及び
(b)67LRが過発現された試料からKRSの過発現可否を分析する段階を含む、肺癌又は乳癌を診断する方法。
【図1】

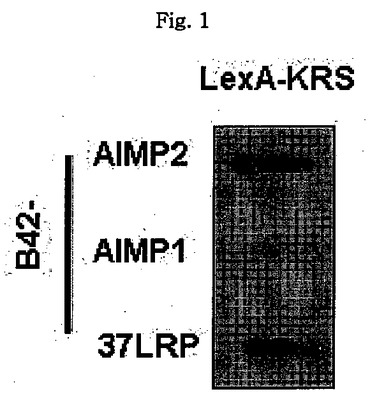
【図2】
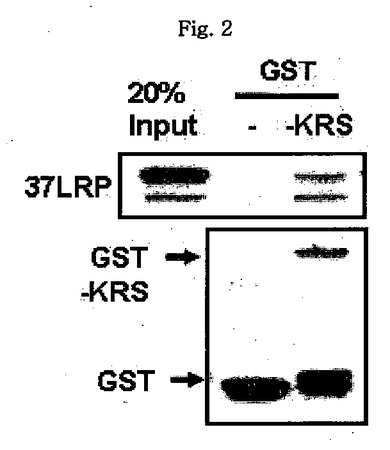
【図3】

【図4】
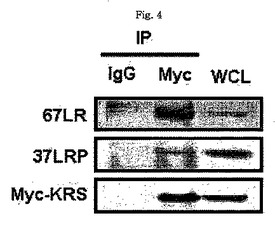
【図5】
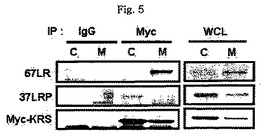
【図14】
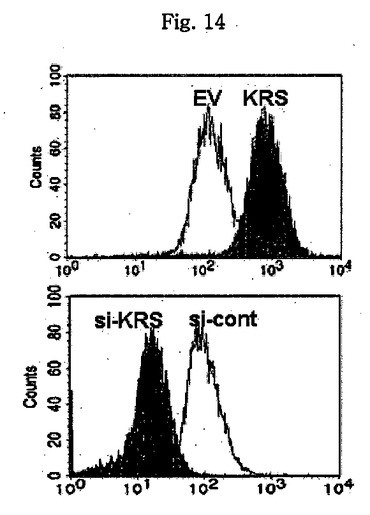
【図15】
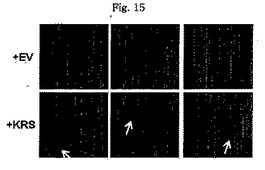
【図18】
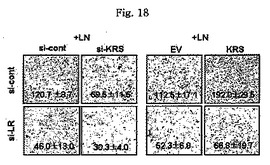
【図21】
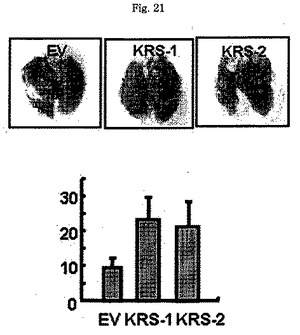
【図23】
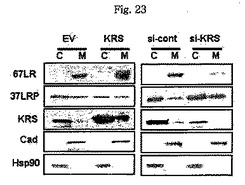
【図24】
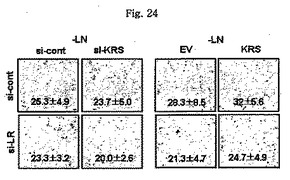
【図30】
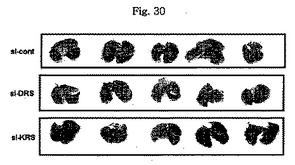
【図33】
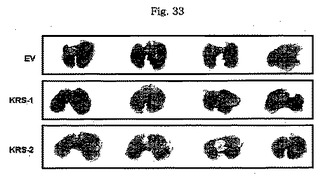
【図6】
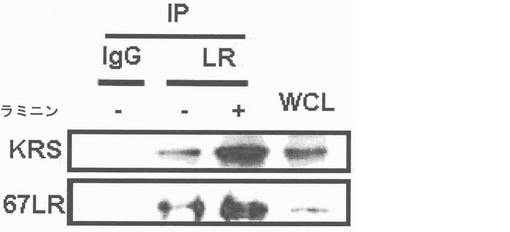
【図7】
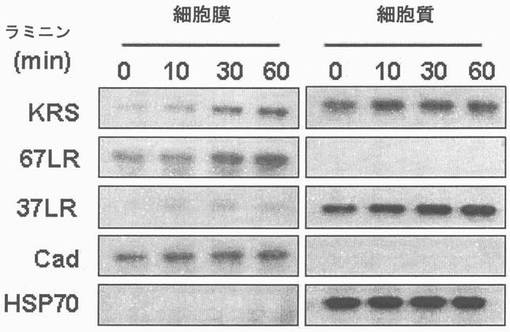
【図8】
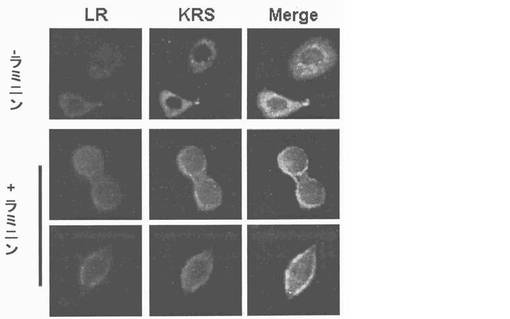
【図9】
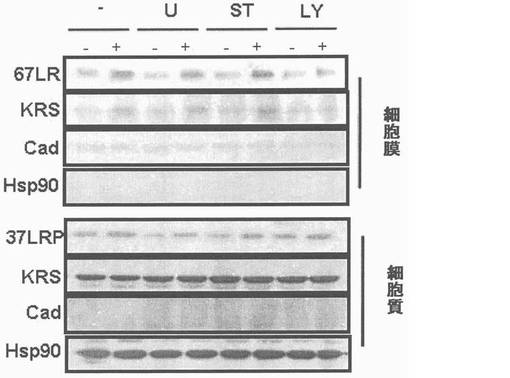
【図10】
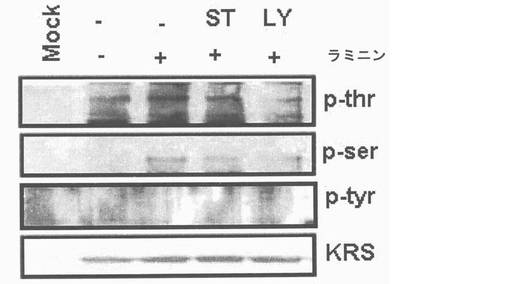
【図11】
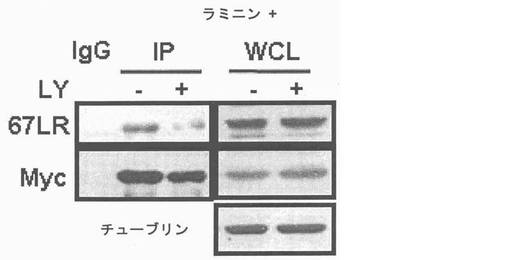
【図12】
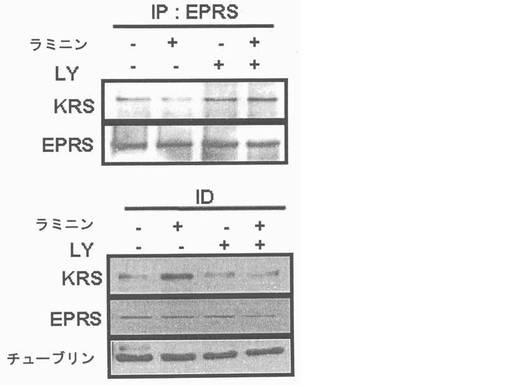
【図13】
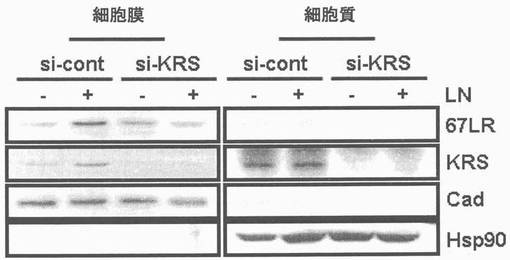
【図16】
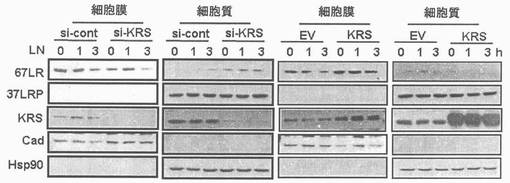
【図17】
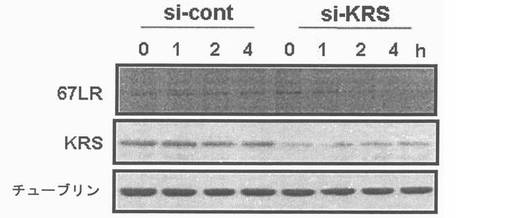
【図19】
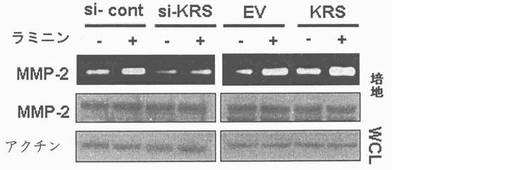
【図20】

【図22】
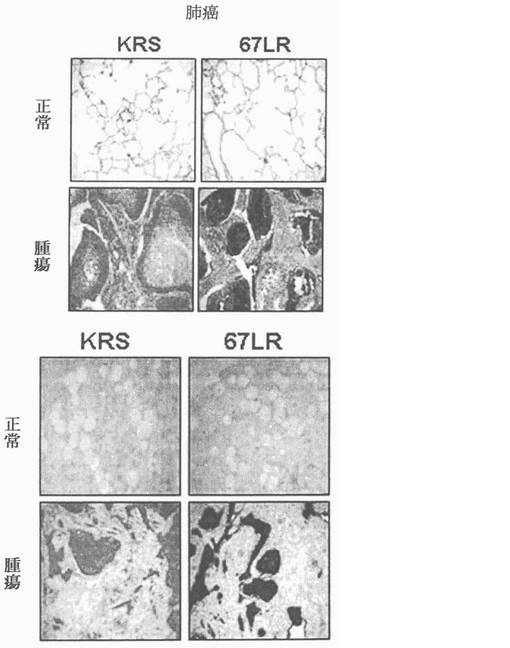
【図25】

【図26】
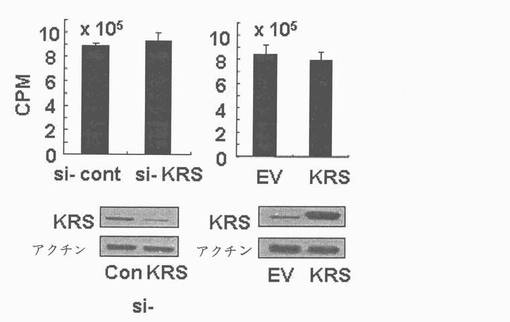
【図27】
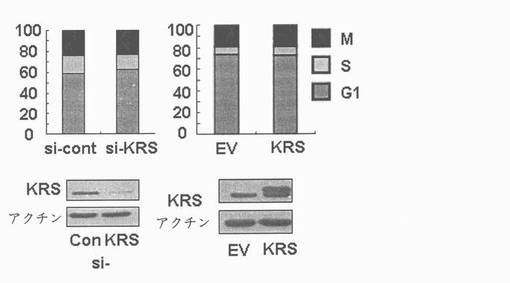
【図28】
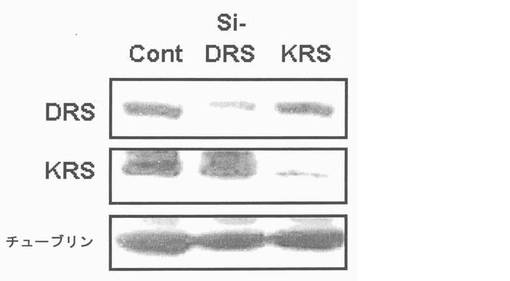
【図29】
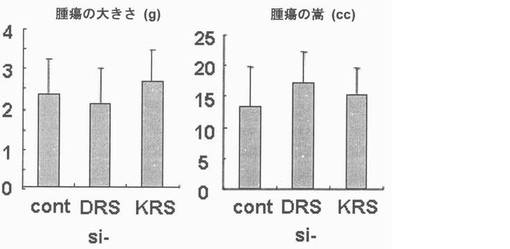
【図31】
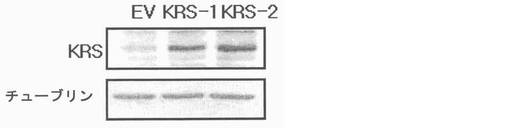
【図32】
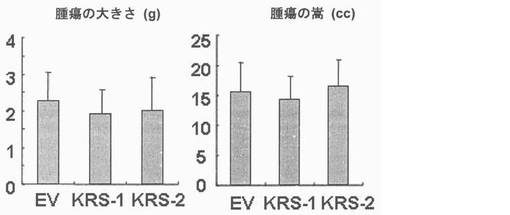
【図2】
【図3】
【図4】
【図5】
【図14】
【図15】
【図18】
【図21】
【図23】
【図24】
【図30】
【図33】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図16】
【図17】
【図19】
【図20】
【図22】
【図25】
【図26】
【図27】
【図28】
【図29】
【図31】
【図32】
【公表番号】特表2012−500256(P2012−500256A)
【公表日】平成24年1月5日(2012.1.5)
【国際特許分類】
【出願番号】特願2011−523721(P2011−523721)
【出願日】平成20年8月18日(2008.8.18)
【国際出願番号】PCT/KR2008/004785
【国際公開番号】WO2010/021415
【国際公開日】平成22年2月25日(2010.2.25)
【出願人】(503317485)ソウル ナショナル ユニバーシティー インダストリー ファウンデーション (25)
【Fターム(参考)】
【公表日】平成24年1月5日(2012.1.5)
【国際特許分類】
【出願日】平成20年8月18日(2008.8.18)
【国際出願番号】PCT/KR2008/004785
【国際公開番号】WO2010/021415
【国際公開日】平成22年2月25日(2010.2.25)
【出願人】(503317485)ソウル ナショナル ユニバーシティー インダストリー ファウンデーション (25)
【Fターム(参考)】
[ Back to top ]