
ラフォラ病遺伝子
【課題】ラフォラ病に関与する新規の遺伝子、EPM2B;その遺伝子によってコードされるタンパク質、malin;並びにラフォラ病を診断および治療する方法の提供。
【解決手段】ラフォラ病と関連する、RINGフィンガー・ドメインおよび6つのNHLモチーフを有するタンパク質をコードする、単離された核酸分子。当該核酸配列における突然変異を検出する段階を含む、ラフォラ病を検出する方法。
【解決手段】ラフォラ病と関連する、RINGフィンガー・ドメインおよび6つのNHLモチーフを有するタンパク質をコードする、単離された核酸分子。当該核酸配列における突然変異を検出する段階を含む、ラフォラ病を検出する方法。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、ラフォラ病に関与する新規の遺伝子、EPM2B;その遺伝子によってコードされるタンパク質、malin;並びにラフォラ病を診断および治療する方法に関する。
【背景技術】
【0002】
発明の背景
ラフォラ病(LD、OMIM 254780)は、青年期に発症する進行性てんかんで最も一般的かつ重篤な形態である。増加性発作は、認知症に向かって知らない間に進行する認知減退、および通常発症の10年以内の死が平行して生じる(1、2)(非特許文献1、2)。細胞レベルで、LDは、ポリグルコサン(またはラフォラ体)と呼ばれるデンプン様グルコース重合体(4)(非特許文献4)の小胞体(ER)随伴蓄積(reticulum-associated accumulation)(3)(非特許文献3)によって特徴付けられる。遺伝的形質は、遺伝的異質性をもった常染色体劣性であるが、臨床症状は同質である(5)(非特許文献5)。本発明者らは、炭水化物結合ドメインをもつ二重特異性ホスファターゼ(Laforinと名づけた)をコードする染色体6q24上のEPM2A遺伝子における突然変異が、LDを生じさせることを以前に発見した(6、7(非特許文献6、7)および国際公開公報第00/05405号(特許文献1))。
【0003】
当技術分野において、ラフォラ病の診断および治療を援助するためには、ラフォラ病に関与するその他の遺伝子を同定する必要がある。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】国際公開公報第00/05405号
【非特許文献】
【0005】
【非特許文献1】Lafora, G.R., Gluck, B. Beitrag zur histopathologie der myoklonischen epilepsie. Z Ges Neurol Psychiat 6, 1-14 (1911)
【非特許文献2】Minassian, B.A. Progressive myoclonus epilepsy with polyglucosan bodies: Lafora disease. Adv Neurol 89, 199-210 (2002)
【非特許文献3】Collins, G.H., Cowden, R.R. & Nevis, A.H. Myoclonus epilepsy with Lafora bodies. An ultrastructural and cytochemical study. Arch Pathol 86, 239-54(1968)
【非特許文献4】Sakai, M., Austin, J., Witmer, F. & Trueb, L. Studies in myoclonus epilepsy (Lafora body form). II. Polyglucosans in the systemic deposits of myoclonus epilepsy and in corpora amylacea. Neurology 20, 160-76 (1970)
【非特許文献5】Minassian, B.A. et al. Genetic locus heterogeneity in Lafora's progressive myoclonus epilepsy. Ann Neurol 45, 262-5 (1999)
【非特許文献6】Minassian, B.A. et al. Mutations in a gene encoding a novel protein tyrosine phosphatase cause progressive myoclonus epilepsy. Nat Genet 20, 171-4 (1998)
【非特許文献7】Minassian, B.A. et al. Mutation spectrum and predicted function of laforin in Lafora's progressive myoclonus epilepsy. Neurology 55, 341-6 (2000)
【発明の概要】
【0006】
発明の要約
本発明者らは、染色体6p22.3上に新規遺伝子EPM2Bを位置的にクローン化した。これは、それぞれユビキチンを媒介した特異的基質の制御(8、9)およびタンパク質-タンパク質相互作用(10-13)のためにデザインされる複合体の特徴である、推定上のRINGフィンガー・ドメインと6つのNHLモチーフをもつタンパク質とをコードする。malinと名付けられたタンパク質産物に対して有害作用を生じさせることが予測されるEPM2Bの21個の異なったDNA配列バリエーションは、LDと同時分離されることが39ファミリーにおいて見いだされた。laforinおよびmalinは、両方ともERに局在し、これらがニューロンのポリグルコサン蓄積およびてんかんを妨げるのに関連した経路で作動することを示唆する。また、本発明者らは、イヌにおけるEPM2Bを単離し、およびシーケンスし、LDをもつイヌにおけるEPM2Bの突然変異を示した。
【0007】
したがって、本発明は、ラフォラ病と関連し、かつ配列番号:1(図6A)(ヒトEPM2B)に示した配列を有する単離された核酸分子を提供する。また、本発明は、ラフォラ病と関連し、配列番号:3(図7A)(イヌEPM2B)に示した配列を有する単離された核酸分子を提供する。
【0008】
好ましくは、精製され、および単離された核酸分子は、以下を含む:
(a)配列番号:1(図6A)および配列番号:3(図7A)に示したとおりの、TがUであってもよい核酸配列;
(b)(a)に対して相補的な核酸配列;
(c)(a)もしくは(b)の核酸配列に対して実質的に配列相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;または、
(e)ストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする核酸配列。
【0009】
また、本発明は、ラフォラ病と関連した、RING型のZnフィンガーと6つのNHL繰り返しドメインとを含む、単離されたタンパク質を含む。本発明の好ましい態様において、タンパク質は、配列番号:2(図6B)(ヒトEPM2B)に示したとおりのアミノ酸配列を有する。もう一つの態様において、タンパク質は、配列番号:4(図7B)(イヌEPM2B)に示したアミノ酸配列を有する。
【0010】
表1に示すように、本発明者らは、ラフォラ病と関連するヒトEPM2B遺伝子において21個の異なる突然変異を見いだした。したがって、本発明は、哺乳類、好ましくはヒト由来の試料においてEPM2B遺伝子の突然変異を検出することを含む、ラフォラ病を検出する方法を提供する。好ましい態様において、突然変異は、表1の一覧表に記載されたものである。
【0011】
また、本発明者らは、ラフォラ病をもつイヌにおいてEPM2B遺伝子の突然変異を発見した。特に、研究した全ての病気に冒されたイヌは、Dと呼ばれ、かつ配列番号:3に示したイヌEPM2B配列のヌクレオチド番号1001で始まる配列

を有する12塩基反復の2対立遺伝子の伸長を有した。したがって、本発明は、EPM2B(配列番号:3)のイヌ配列のヌクレオチド番号1001で始まる配列
の繰り返しを検出することを含む、犬科の動物においてラフォラ病を検出する方法をさらに提供する。
【0012】
その他の本発明の特徴および効果は、以下の詳細な説明から明らかであろう。しかし、詳細な説明および具体的な実施例は、本発明の好ましい態様を示す一方で、本発明の範囲内の種々の変更および修飾がこの詳細な説明から当業者には明らかとなると考えられるので、例示目的のみによって与えられることが理解されるべきである。
【図面の簡単な説明】
【0013】
ここで図面に関して記載する。
【図1】6p22.3上のEPM2B領域。以前の2.2Mbの臨界区間(critical interval)は、マイクロサテライト・マーカーD6S1567およびBV12568(15)によって区切られた。BV012568境界は、F-Cファミリー由来のLD個体におけるホモ接合性の減少に基づいていた(LD6;図2を参照されたい)。D6S1567テロメア境界は、第2の血縁F-Cファミリー(15)(LD27)の病気に冒されていない同種におけるBV012563とD6S1567との間に起こる組換えによって定義した。LD38324ファミリーにおけるマーカーのホモ接合性鎖の破壊により、動原体境界をD6S1688にさらに再定義することができた(図2)。本領域は、7つの以前に注釈をつけた遺伝子および単一の1188bpをコードするエキソンを含む新たに発見されたEPM2B(図6A)を含む。代表的なヒトおよびマウスcDNA配列を、同様に推定上のATG開始およびAATAAAポリアデニル化シグナルを示してある。ATG開始は、インフレームのストップ(位置-60において)に続き、対応するAUGは、予測されるORFの初めに存在する。開始(CGCGCCAUGG)を囲むヌクレオチド配列は、真核生物の翻訳開始部位のコンセンサス配列(GCCA/GCCAUGG)の提唱された特徴を有する(29)。EPM2Bは、検出可能な亜鉛結合RINGフィンガーおよび6つのNHL繰り返しドメイン(図6B)を含む42.3kDaの(395aa)タンパク質(malin)をコードすることが予測される。RINGおよびNHLの頭字語は、これらを有することが同定された最初のタンパク質の記述、すなわち、それぞれヒト(30)のReally Interesting New Gene 1およびNCL-1(11)/HT2A(10)/LIN-41(13)遺伝子から生じる。RING-および/またはNHLドメインは、細胞質もしくは核局在化(8、9、12、18)の一方または両方を有することができる種々のタンパク質に存在することに留意すべきである。Malinは、RINGおよびNHLモチーフのみを有することが記載された今までに唯一のタンパク質である(この組み合わせをもつその他のタンパク質があるものの、これらは、RING-B-Box-コイルドコイルドメインなどのその他の関連するモチーフも有する)。共通のC332T(P111L)多型部位は、星印:(*)によって示してある(表1を参照されたい)。Malinは、予測されるラット(419aa)、マウス(401aa)、およびイヌ(402aa)タンパク質とそれぞれ79%、80%、および85%の相同性を共有する。可変アミノ酸は、主にタンパク質のカルボキシ末端およびアミノ末端に位置し、RINGフィンガーまたはNHLドメイン以外に位置した。BVで始まるマイクロサテライト・マーカーは、この研究において作製した。
【図2】LDファミリーにおけるハプロタイプ解析によるEPM2Bの臨界区間の絞り込みおよびEPM2B遺伝子における突然変異。a、動原体境界は、ファミリーLD38324の両発端者におけるホモ接合性の減少に基づいて、D6S1688まで狭くなり、840kbまで重要領域を絞り込んだ。b、EPM2Bの配列解析により、ファミリーLD6における(並びにLD7、LD27、およびLD28F-Cファミリーにおける、表1)ホモ接合性の76T>A変化を同定する。ファミリーLD38324における病気に冒された個体は、第5NHLドメインにフレームシフト突然変異を生じるジヌクレオチド欠失(1048-1049delGA)に関してホモ接合性であることが見いだされた。
【図3】人体組織におけるEPM2BのRNAハイブリダイゼーション発現解析。a、複数の組織ブロット(Clontech)をEPM2Bのコード領域の557bp断片とハイブリダイズさせた。2.4kb および1.5kbのサイズの2つの転写物が、全ての組織において同定された。b、同じ大きさの転写物が、試験した脳の全ての領域由来の組織において見いだされた。
【図4】患者のLD32817(EPM2B突然変異98T>C)由来の脳生検材料の電子顕微鏡写真。A:軸索(多数の正常な神経伝達物質小胞に留意されたい)。S:シナプス。LB:完全に樹状突起を占めているポリグルコサンフィラメント(PG)の密度の高い蓄積で構成されるラフォラ体(大きな丸い構造)。バーは、500nmに相当する。
【図5】malinおよびlaforinタンパク質の細胞局在。a、Myc-タグを付けたmalin(構築物pcDNA3mycEPM2B)は、核周辺で、並びに核内で明瞭な網状パターンを形成する。GRP94小胞体(ER)特異的マーカーに対する抗体での同時染色により、malinとの共局在化が明らかになる。b、ER特異的マーカーGRP94とのlaforin(構築物pcDNA3mycEPM2A(24))の細胞質アイソフォームの共局在化。
【図6A】図6A(配列番号:1)は、EPM2Bのヒト核酸配列およびアミノ酸配列を提供する。
【図6B】図6B(配列番号:2)は、EPM2Bのヒト核酸配列およびアミノ酸配列を提供する。
【図7A】図7A(配列番号:3)は、EPM2Bのイヌ核酸配列およびアミノ酸配列を提供する。
【図7B】図7B(配列番号:4)は、EPM2Bのイヌ核酸配列およびアミノ酸配列を提供する。
【図8】ヒトLDと同一の特徴および細胞分布パターンをもつ病気に冒されたMWHD組織において形成する大量のデンプン様ポリグルコサン蓄積(ラフォラ体;LB)を示す(1,29,30)。病気に冒されたMWHD組織は、9つの筋肉および2つの肝臓生検、並びに剖検で得られた皮膚試料および4つの全脳を含んだ。対照組織は、LBを示さず(2匹の正常MWHDおよび3匹の非LDてんかんイヌを含む7匹の年齢が一致した動物)、示していない。組織化学的スライドは、ジアスターゼ処理に続いて過ヨウ素酸シッフで染色し、これにより、ポリグルコサンの特異的検出が可能になる(1)。無数のLBは、ヒトLD(1)と同様に、病気に冒されたMWHDの脳の全体にわたって存在し、ニューロンに位置し、グリアに位置せず、かつニューロンの核周部および樹状突起に位置し、軸索に位置しなかった。a:大脳皮質の分子層におけるLB(矢印)(×100拡大)。b:樹状LBの例(星印)。この樹状突起と共にシナプスを形成する複数の軸索(A)は、LBを含まない(バー=500nm)。c:肝臓において、LB(矢印)は、ヒトと同様に、糖新生(門脈周囲)肝細胞にのみ存在した(29)。(PV:門脈。D:門脈小導管。×300)。d、e:皮膚において、LB(矢印)は、ヒトと同様に(30)、汗腺に、特にアポクリン汗腺を囲んでいる筋上皮細胞に(d、×100)、およびメロクリン汗腺の胆管細胞に(e:フットパッド(foot pad)皮膚組織、×250)に存在した。f:骨格筋において、LBは、再びヒトLD(29)と同様に、膜(矢じり)によって囲まれた空胞内に存在した(バー=500nm)。
【図9】MWHD LDが、病気に冒されたイヌにおける伸長突然変異を含むEPM2B遺伝子にリンクすることを示す。a:系統およびハプロタイプ。黒の数は、下行順に染色体CFA35マイクロサテライトREN256I01、REN282I22、D02001、REN157J09、REN214H22、REN94K23、REN166C14、REN172L08における遺伝子型である。赤においては、EPM2B遺伝子の位置およびそれをコードする多形12塩基反復におけるD繰り返しの数である。注:伸長突然変異は、減数分裂時に不安定である(たとえば、イヌ29からイヌ32への伸長の伝達を参照されたい)。E:正確な繰り返し数をもつ伸長対立遺伝子は、決定されなかった。ボックス:突然変異に隣接する遺伝子型のホモ接合性。b、マルチポイントLODスコア解析。
【図10】食肉目のイヌ科ファミリーでのEPM2B12塩基多型の進化、非MWHDてんかんのイヌにおける伸長突然変異の存在、およびEPM2B mRNAに対する伸長の不活性化効果を示す。a:イヌEPM2Bの12塩基反復配列およびオルソログ。D:正確な繰り返し。T:不完全な繰り返し(星印)。赤いヌクレオチド:繰り返し領域におけるイヌ配列との相違。b:イヌにおいてミオクローヌス痙攣と関連するEPM2B伸長突然変異。W:野生型。M:病気に冒されたMWHD。B:病気に冒されたバセット犬(Basset Hound)。C:病気に冒されていないいくつかのキャリアMWHD。矢じり:正常な対立遺伝子。他の全てのバンド:伸長変異体対立遺伝子。脱アミノ化が正常配列の存在下においてさえも、伸長された対立遺伝子のPCR検出をどれほどよく改善するかに留意されたい。c:リアルタイム定量的RT-PCRを使用した、病気に冒されたMWHDおよび対照に由来する骨格筋におけるEPM2B量(Gapdhに対して規準化した)(SYBR Green検出;補充法)。d:12塩基多型の進化。D配列は、ネコ種に存在しない。これは、全てのイヌに存在する。アルクト科において単一コピーとして、および犬科の動物において1または2つの多形状態として;両対立遺伝子は、ヘテロ接合性個体(2つの配列の列)で示してある。示した全ての現存する肉食動物は、1,000万年前(Ma)の後に現れた。
【発明を実施するための形態】
【0014】
発明の詳細な説明
本発明者らによる以前の研究は、LD患者の〜70%が染色体6q24(6,7,14)上のEPM2A遺伝子に劣性突然変異を有することを示唆した。残りの患者において原因となる遺伝子を同定するために、本発明者らは、初めに、フレンチ-カナディアン(French-Canadian:F-C)単離体(isolate)(15)に由来する非EPM2A LDファミリー(LD6、LD7、LD27、LD28)のサブセットに対して連鎖およびホモ接合性マッピングを行った(表1)。このアプローチにより、染色体6p22.3上の2.2Mbの領域に第2のLD遺伝子座(EPM2B)が局在することを導いた(図1)。これらのF-Cファミリー由来の病気に冒された全ての個体は、全ての臨界区間にわたって珍しいハプロタイプに関してホモ接合性であることが見いだされた(図2a)。さらに遺伝子座を絞り込むために、病気に冒された複数の個体を有する5つのさらなるLDファミリーを、臨界領域を包含するDNA配列から開発することができたあらゆるマイクロサテライト・マーカーを使用して調べた。4つのファミリーにおいて、病気に冒された個体が、臨界区間にわたって全てのマーカーについてホモ接合性であった。しかし、1つのファミリー(LD38324)では、病気に冒された2つの個体のホモ接合性の鎖が、臨界領域に部分的に及び、D6S1688とD6S1567との間の840kbにEPM2B遺伝子座を減少することができた(図1および2a)。
【0015】
I. 本発明の核酸分子
先に述べたとおり、本発明は、ラフォラ病に関与する単離された核酸分子に関する。用語「単離された」は、組換えDNA技術によって産生したときに、細胞形質成分もしくは培地が実質的にない核酸、または化学物質前駆体、または化学的に合成したときのその他の化学物質をいう。用語「核酸」は、DNAおよびRNAを含むことが意図され、二本鎖または一本鎖のいずれであってもよい。
【0016】
上述したとおり、本発明は、ラフォラ病と関連した、N末端部分にRING型のZnフィンガーとC末端部分に6つのNHL繰り返しドメインとを含むタンパク質をコードする単離された核酸分子を提供する。単離された核酸分子は、好ましくはラフォラ病と関連したEPM2B遺伝子である。本発明の態様において、単離された核酸分子は、配列番号:1(図6A)または配列番号:3(図7A)に示すとおりの配列を有する。
【0017】
配列番号:1および3に示した核酸配列(または、それぞれ図6Aおよび7A)、並びに表1において、または実施例3において特定した変異配列は、本明細書において、ひとまとめにして「本発明の核酸分子」ということができる。配列番号:2および4(または、それぞれ図6Bおよび7B)に示したアミノ酸配列、並びに表1において特定した変異配列は、本明細書において、ひとまとめにして「本発明のタンパク質」ということができる。
【0018】
好ましくは、精製および単離された核酸分子は、以下を含む:
(a)配列番号:1(図6A)もしくは配列番号:3(図7A)に示すとおりの核酸配列であって、TがUであってもよい核酸配列;
(b)(a)に対して相補的な核酸配列;
(c)(a)もしくは(b)の核酸配列に対して実質的に配列相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;または、
(e)ストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする核酸配列。
【0019】
用語「実質的に配列相同性を有する配列」は、(a)もしくは(b)の配列から、わずかな、または重要でない配列バリエーションを有する、すなわち配列が実質的に同じ方法で機能し、かつラフォラ病を検出し、調査し、または治療するために使用することができる核酸配列を意味する。バリエーションは、局部的な突然変異または構造修飾に起因していてもよい。実質的に相同性を有する核酸配列は、配列番号:1または配列番号:3に示したとおりの核酸配列と少なくとも65%、より好ましくは少なくとも85%、および最も好ましくは90〜95%の同一性を有する核酸配列を含む。
【0020】
「配列同一性」は、当技術分野において公知の方法に従って算出することができる。配列同一性は、最も好ましくはBLASTバージョン2.1アドバンスド・サーチのアルゴリズムによって評価される。BLASTは、http://www.ncbi.nlm.nih.gov/BLASTでオンラインで利用できる一連のプログラムである。アドバンスドblastサーチ(http://www.ncbi.nlm.nih.gov/blast/blast.cgi?Jform=1)は、デフォルト・パラメーターにセットされている(すなわち、Matrix BLOSUM62;ギャップ・存在コスト(Gap existence cost)11;残基あたりのギャップ・コスト(gap cost)1;ラムダ比0.85デフォルト)。BLAST検索に対する参照は、以下のとおりである:
。
【0021】
用語「ハイブリダイズする配列」は、ストリンジェントなハイブリダイゼーション条件下で、(a)、(b)、(c)、または(d)の配列にハイブリダイズすることができる核酸配列を意味する。DNAハイブリダイゼーションを促進する適切な「ストリンジェントなハイブリダイゼーション条件」は、当業者に公知であるか、またはMolecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6に見いだされ得る。本明細書に使用される用語「ストリンジェントなハイブリダイゼーション条件」は、溶液中の2つの相補的核酸分子間の選択的なハイブリダイゼーションを促進する条件が選択されることを意味する。ハイブリダイゼーションは、全てまたは一部の核酸配列分子に対して生じてもよい。ハイブリダイズされる部分は、ポリペプチドをコードするポリヌクレオチド配列のうちの1つに関して少なくとも50%の長さである。この点に関して、核酸二重鎖またはハイブリッドの安定性は、ナトリウム含有緩衝液において、ナトリウムイオン濃度、標識された核酸のG/C含量、核酸プローブの長さ(I)、および温度の関数であるTm(Tm = 81.5℃ - 16.6(Log10 [Na+]) + 0.41(%(G+C) - 600/I)によって決定される。したがって、ハイブリッド安定性を決定する洗浄条件のパラメーターは、ナトリウムイオン濃度および温度である。公知の核酸分子と類似するが、同一ではない分子を同定するためには、1%のミスマッチにより、Tmの約1℃の減少を生じると想定してもよく、たとえば、95%を超える同一性を有する核酸分子が調査される場合には、最終洗浄が5℃まで減少されると考えられる。これらの考慮に基づいて、ストリンジェントなハイブリダイゼーション条件は、以下のように定義することができる:5×塩化ナトリウム/クエン酸ナトリウム(SSC)/5×デンハート液/1.0%のSDSにおけるTm(上記の方程式に基づく)-5℃におけるハイブリダイゼーション、続く0.2×SSC/0.1%のSDSの60℃における洗浄。
【0022】
用語「類似体である核酸配列」は、(a)、(b)、または(c)の配列と比較して、修飾された核酸配列であって、修飾が本明細書に記載されている配列の有用性を変更しない核酸配列を意味する。修飾された配列または類似体は、(a)、(b)、または(c)に示した配列にわたって改善された性質を有していてもよい。類似体を調製する修飾の一つの例は、配列番号:1または3に示した配列の天然に存在する塩基(すなわち、アデニン、グアニン、シトシン、またはチミジン)のうちの1つを、以下のものなどの修飾塩基と置換することである:キサンチン、ヒポキサンチン、2-アミノアデニン、6-メチル、2-プロピル、およびその他のアルキルアデニン、5-ハロウラシル、5-ハロシトシン、6-アザウラシル、6-アザシトシン、および6-アザチミン、プソイドウラシル、4-チオウラシル、8-ハロアデニン、8-アミノアデニン、8-チオールアデニン、8-チオールアルキルアデニン、8-ヒドロキシルアデニン、およびその他の8位置換されたアデニン、8-ハログアニン、8アミノグアニン、8-チオールグアニン、8-チオールアルキルグアニン、8-ヒドロキシルグアニン、およびその他の8位置換されたグアニン、その他のアザおよびデアザウラシル、チミジン、シトシン、アデニン、またはグアニン、5-トリフルオロメチルウラシル、並びに5-トリフルオロシトシン。
【0023】
修飾のもう一つの例は、配列番号:1または配列番号:3に示した核酸分子において、リン酸骨格における修飾された亜リン酸もしくは酸素へテロ原子、短鎖アルキルもしくはシクロアルキルの糖間結合、または短鎖ヘテロ原子もしくは複素環の糖間結合を含む。たとえば、核酸配列は、ホスホロチオアート、リン酸トリエステル、メチルホスホナート、およびホスホロジチオアートを含んでもよい。
【0024】
本発明の核酸分子の類似体のさらなる例は、DNA(またはRNA)のデオキシリボース(またはリボース)リン酸骨格が、ペプチドにおいて見られるものと同様のポリアミド骨格で置換されたペプチド核酸(PNA)である(P.E. Nielsen et al., Science 1991, 254, 1497)。PNA類似体により、酵素による分解に耐性であること、並びにインビボおよびインビトロでの生存が延長されることが示されている。また、PNA鎖とDNA鎖の間には電荷相反がないため、PNAは、相補的なDNA配列に対してより強く結合する。その他の核酸類似体は、重合体骨格、環状骨格、または非環式骨格を含むヌクレオチドを含んでもよい。たとえば、ヌクレオチドは、モルホリノ骨格構造(米国特許第5,034,506号)を有していてもよい。また、類似体は、レポーター基、核酸配列の薬物動態学的または薬力学的な性質を改善するための基などの基を含んでいてもよい。
【0025】
本発明は、後述するように、本発明のタンパク質の切り詰め、並びに本発明のタンパク質の類似体および相同体、並びにこれらの切り詰めをコードする核酸分子を含むことが認識される。本発明のcDNAに対応するmRNAの選択的スプライシングによって生じる本発明の核酸分子の異型も、本発明によって包含されることがさらに認識される。
【0026】
また、遺伝暗号の縮重により、本発明の核酸配列とは異なる配列を有する単離されおよび精製された核酸分子も、本発明の範囲内である。このような核酸は、機能的に同等のタンパク質をコードするが、遺伝暗号の縮重のために上述した配列とは配列が異なる。
【0027】
また、本発明は、配列番号:1(図6A)または配列番号:3(図7A)に示した核酸分子と比較して突然変異を有し、該突然変異がラフォラ病と関連する単離された核分子を含む。好ましい態様において、突然変異は、表1に示した突然変異または実施例3に記載したイヌ突然変異のうちの1つから選択される。
【0028】
EPM2B遺伝子由来の核酸分子またはこれらの変異形態は、配列番号:1(図6A)および配列番号:3(図7A)に示すとおりの核酸配列、または表1に示した変異された配列の全部または一部に基づいて標識された核酸プローブを調製すること、並びにこの標識された核酸プローブを使用して、適切なDNAライブラリー(たとえば、cDNAまたはゲノムDNAライブラリー)をスクリーニングすることによって単離することができる。cDNAまたはゲノムDNAライブラリーのスクリーニングによって単離された核酸は、標準的な技術によってシーケンスすることができる。
【0029】
また、本発明の核酸分子は、ポリメラーゼ連鎖反応法(PCR)方法およびcDNAまたはゲノムDNAを使用して、核酸を選択的に増幅することによって単離することができる。PCRに使用するために、配列番号:1(図6A)および配列番号:3(図7A)に示すとおりの核酸分子、または表1に示した変異された配列から合成オリゴヌクレオチドプライマーをデザインすることができる。核酸は、これらのオリゴヌクレオチドプライマーおよび標準的なPCR増幅技術を使用して、cDNAまたはゲノムDNAから増幅することができる。こうして増幅された核酸は、適切なベクターにクローン化することができ、DNA配列解析によって特徴付けることができる。cDNAは、種々の技術によって、たとえば、Chirgwin et al., Biochemistry, 18, 5294-5299 (1979)のチオシアン酸グアニジニウム(guanidinium-thiocyanate)抽出法を使用することによって、全細胞mRNAを単離することにより、mRNAから調製してもよいことが認識される。次いで、逆転写酵素(たとえば、Gibco/BRL, Bethesda, MDから入手可能なモロニーMLV逆転写酵素、またはSeikagaku America, Inc., St. Petersburg, FLから入手可能なAMV逆転写酵素)を使用して、mRNAからcDNAが合成される。
【0030】
RNAである本発明の単離された核酸分子は、適切なベクターに本発明の新規のタンパク質をコードするcDNAをクローン化することによって単離することができ、これにより、cDNAを転写してmalinタンパク質をコードするRNA分子を産生することができる。たとえば、cDNAは、ベクターのバクテリオファージプロモーター(たとえば、T7プロモーター)の下流にクローン化することができ、cDNAをインビトロにおいてT7ポリメラーゼで転写させ、生じるRNAを標準的な技術によって単離することができる。
【0031】
また、本発明の核酸分子は、標準的な技術を使用して化学的に合成されてもよい。ポリデオキシヌクレオチドを化学的に合成する種々の方法が公知であり、ペプチド合成のように、市販されているDNA合成装置での完全に自動化された固相合成を含む(たとえば、Itakuraら、米国特許第4,598,049号;Caruthersら、米国特許第4,458,066号;並びにItakura米国特許第4,401,796号および同第4,373,071号を参照されたい)。
【0032】
本発明の核酸分子の開始コドンおよび非翻訳配列は、PC/Gene(IntelliGenetics Inc., Calif.)などの本目的のためにデザインされた現在利用できるコンピュータ・ソフトウェアを使用して決定してもよい。調節エレメントは、従来の技術を使用して同定することができる。エレメントの機能は、エレメントに機能的に結合されたリポーター遺伝子を発現するようにこれらのエレメントを使用して確認することができる。これらの構築物は、標準的な手順を使用して培養細胞に導入してもよい。DNA中の調節エレメントを同定することに加えて、このような構築物はまた、当技術分野において公知の技術を使用して、エレメントと相互作用するタンパク質を同定するために使用してもよい。
【0033】
アンチセンス核酸分子を産生するため転写のための通常の提示に対して、本発明の核酸分子の配列を逆にしてもよい。好ましくは、アンチセンス配列は、開始コドンまたは保存されていない領域の前の領域を逆にすることによって構築される。アンチセンス核酸分子を産生するため転写のための通常の提示に対して、特に、本発明の核酸分子もしくはこれらの断片、好ましくは配列番号:1(図6A)および配列番号:3(図7A)に示した核酸配列、または表1に示した変異された配列に含まれる核酸配列を逆にしてもよい。
【0034】
本発明もしくはこれらの断片のアンチセンス核酸分子は、天然に存在するヌクレオチドを使用して化学的に合成してもよく、または分子の生物学的安定性を増大するように、もしくはmRNAまたは天然の遺伝子と形成される二重鎖の物理安定度を増大するようにデザインされた多様に修飾されたヌクレオチド、たとえばホスホロチオアート誘導体およびアクリジン置換されたヌクレオチドでもよい。アンチセンス配列は、アンチセンス配列が、高効率制御領域(その活性は、ベクターが導入される細胞型によって決定してもよい)の制御下で産生される組換えプラスミド、ファージミド、または弱毒ウイルスの形態で細胞に導入された発現ベクターを使用して生物学的に産生してもよい。
【0035】
また、本発明は、本発明の新規タンパク質、および選択タンパク質、または選択可能なマーカータンパク質を含む融合タンパク質をコードする核酸を提供する(下記を参照されたい)。
【0036】
II. 本発明の新規のタンパク質
本発明は、本発明の核酸分子によってコードされる単離されたタンパク質をさらに含む。本発明の状況の範囲内で、本発明のタンパク質は、生物活性を保持する、種々のタンパク質一次構造形態を含んでいてもよい。
【0037】
広く明言したとおり、本発明は、ラフォラ病と関連した、N末端側半分におけるRING型のZnフィンガーとC末端の方向に6つのNHL繰り返しドメインを含む単離されたタンパク質を提供する。好ましくは、亜鉛結合RINGフィンガー・モチーフ(C-X2-C-X16-C-X1-H-X2-C-X2-C-X14-C-X2-C)は、図6Bに示したmalinタンパク質の残基26〜71の間に位置する。RINGフィンガーの存在は、E3ユビキチン・リガーゼ機能を予測する。したがって、好ましい態様において、タンパク質は、ユビキチン・リガーゼ機能を有する。
【0038】
本発明の特定の態様において、タンパク質は、配列番号:2(図6B)(ヒトEPM2B)に示すとおりのアミノ酸配列を有する。もう一つの態様において、タンパク質は、配列番号:4(図7B)(イヌEPM2B)に示したアミノ酸配列を有する。
【0039】
全長アミノ酸配列に加えて、本発明のタンパク質は、本明細書に記載したとおりの、タンパク質の切り詰め、並びにタンパク質の類似体および相同体、並びにこれらの切り詰めを含む。切断されたタンパク質は、少なくとも15アミノ酸残基のペプチドを含んでいてもよい。
【0040】
本明細書に記載されている、配列番号:2(図6B)もしくは配列番号:4(図7B)に示したアミノ酸配列を有するタンパク質の類似体、および/またはこれらの切り詰めは、1つもしくは複数のアミノ酸置換、挿入、および/または欠失を含むアミノ酸配列を含んでもよいが、これらに限定されない。アミノ酸置換は、保存的または非保存的な性質であってもよい。保存的アミノ酸置換は、同様の電荷、大きさ、および/または疎水性の特徴のアミノ酸で、本発明のタンパク質の1つもしくは複数のアミノ酸を置換することを含む。保存的置換だけが作製されるとき、生じる類似体は、機能的に同等物であるはずである。非保存的置換は、異なる電荷、大きさ、および/または疎水性の特徴を有する1つもしくは複数のアミノ酸でアミノ酸配列の1つもしくは複数のアミノ酸を置換することを含む。
【0041】
1つまたは複数のアミノ酸挿入が、配列番号:2(図6B)または配列番号:4(図7B)に示したアミノ酸配列に導入されてもよい。アミノ酸挿入は、単一のアミノ酸残基または長さが2〜15アミノ酸の範囲の一連のアミノ酸からなってもよい。たとえば、アミノ酸の挿入は、タンパク質がもはや活性ではないように、標的配列を破壊するために使用されてもよい。この手順は、インビボで本発明のタンパク質の活性を阻害するために使用されてもよい。
【0042】
欠失は、配列番号:2(図6B)または配列番号:4(図7B)に示したアミノ酸配列からの1つもしくは複数のアミノ酸または離れた部分の除去からなっていてもよい。欠失されるアミノ酸は、隣接していても、または隣接していなくてもよい。欠失突然変異で生じる類似体の下限の長さは、約10アミノ酸、好ましくは100アミノ酸である。
【0043】
本発明のタンパク質の類似体は、タンパク質をコードするヌクレオチド配列に突然変異を導入することによって調製してもよい。本発明のタンパク質の類似体の発現のために構築されるヌクレオチド配列の突然変異は、コード配列のリーディングフレームを保存しなければならない。さらにまた、突然変異は、好ましくは、受容体mRNAの翻訳に悪影響を与え得る、ハイブリダイズして、ループまたはヘアピンなどの二次mRNA構造を生じさせ得る相補領域を生じない。
【0044】
突然変異は、制限部位に隣接して変異配列を含むオリゴヌクレオチドを合成して、天然の配列の断片に対するライゲーションを可能にすることによって、特定の遺伝子座に導入されてもよい。ライゲーションに続いて、生じる再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する類似体をコードする。
【0045】
または、必要とされる置換、欠失、または挿入にしたがって変更された特定のコドンを有する変更された遺伝子を提供するために、オリゴヌクレオチド部位特異的突然変異法を使用してもよい。また、本発明のタンパク質の欠失または切り詰めは、所望の欠失に隣接する制限エンドヌクレアーゼ部位を利用することによって構築されてもよい。制限後に、オーバーハングを埋めて、DNAを結合してもよい。上述した変更を作製する典型的な方法は、Sambrook et al(Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, 1989)によって開示されている。
【0046】
また、本発明のタンパク質は、本明細書に記載されているように、配列番号:2(図6B)もしくは配列番号:4(図7B)に示したアミノ酸配列の相同体、および/またはこれらの切り詰めを含む。このような相同体は、本発明のタンパク質を得るために使用されるプローブとストリンジェントなハイブリダイゼーション条件(本明細書のストリンジェントなハイブリダイゼーション条件の考察を参照されたい)下でハイブリダイズされるアミノ酸配列から構成されるアミノ酸配列のタンパク質である。好ましくは、本発明のタンパク質の相同体は、タンパク質の特徴であるチロシンホスファターゼ領域を有する。
【0047】
相同的タンパク質は、配列番号:2(図6B)または配列番号:4(図7B)に示すとおりのアミノ酸配列と少なくとも70%、好ましくは80〜90%の同一性を有するアミノ酸配列を有すタンパク質を含む。配列同一性は、本明細書に定義したとおりである。
【0048】
また、本発明は、本発明のタンパク質のアイソフォームを想定する。アイソフォームは、本発明のタンパク質と同じアミノ酸の数および種を含むが、異なる分子構造を有する。本発明によって想定されるアイソフォームは、本明細書に記載されているように、本発明のタンパク質と同じ性質を有するものである。
【0049】
本発明は、配列番号:2(図6B)または配列番号:4(図7B)に示したアミノ酸配列と比較して、突然変異を有し、該突然変異は、ラフォラ病と関連する単離されたタンパク質を含む。好ましい態様において、突然変異は、表1に示した突然変異または実施例3に記載されているイヌ突然変異のうちの1つから選択される。
【0050】
また、本発明は、融合タンパク質を産生するために、選択されたタンパク質、または選択可能なマーカータンパク質(下記を参照されたい)と結合した本発明のタンパク質を含む。加えて、本発明のタンパク質の免疫原性部分も本発明の範囲内である。
【0051】
本発明のタンパク質(切り詰め、類似体、その他を含む)は、組換えDNA法を使用して調製してもよい。したがって、本発明のタンパク質をコードする配列を有する本発明の核酸分子を、タンパク質の良好な発現を保証する適切な発現ベクターに公知の方法で組み込んでもよい。可能な発現ベクターは、ベクターが使用される宿主細胞と適合性を持つ限り、コスミド、プラスミド、または修飾されたウイルス(たとえば、複製欠損のレトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)を含むが、これらに限定されない。「宿主細胞のトランスフォーメーションのために適切な」発現ベクターは、発現ベクターが、本発明の核酸分子および制御配列を含み、発現のために使用される宿主細胞を基礎として選択され、これらが機能的に核酸分子に結合されていることを意味する。「機能的に結合される」とは、核酸が核酸を発現できる方法で制御配列に結合されことを意味することが意図される。
【0052】
したがって、本発明は、本発明の核酸分子、またはこれらの断片、並びに挿入されたタンパク質配列の転写および翻訳に必要な制御配列を含む本発明の組換え発現ベクターを想定する。適切な制御配列は、細菌、真菌、またはウイルスの遺伝子を含む種々の供与源に由来してもよい(たとえば、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)に記載されている制御配列を参照されたい)。適切な制御配列の選択は、選択される宿主細胞に依存的であり、当技術分野の当業者によって容易に達成されるであろう。このような制御配列の例は、転写プロモーターおよびエンハンサーまたはRNAポリメラーゼ結合配列、翻訳開始シグナルを含むリボソーム結合配列を含む。その上、選択される宿主細胞および使用されるベクターに応じて、複製開始点、さらなるDNA制限部位、エンハンサー、および転写誘導能を与える配列などのその他の配列を発現ベクターに組み込んでもよい。また、必要な制御配列が、天然のタンパク質および/またはその隣接領域によって供給されてもよいことが認識されるであろう。
【0053】
本発明は、アンチセンス方向で発現ベクターにクローン化された本発明のDNA核酸分子を含む組換え発現ベクターをさらに提供する。すなわち、DNA分子は、DNA分子の転写によって、配列番号:1または配列番号:3に示すとおりのヌクレオチドを含むヌクレオチド配列に対してアンチセンスであるRNA分子を発現することができる方法で、制御配列に機能的に結合される。アンチセンス核酸に機能的に結合された、アンチセンスRNA分子を持続的に発現させる制御配列を選択することができる。
【0054】
また、本発明の組換え発現ベクターは、本発明の組換え分子を形質転換またはトランスフェクトした宿主細胞の選択を容易にする選択可能なマーカー遺伝子を含んでもよい。選択可能なマーカー遺伝子の例は、特定の薬物に対する耐性を与える、G418およびハイグロマイシンなどのタンパク質、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼをコードする遺伝子である。選択可能なマーカー遺伝子の転写は、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼなどの選択可能なマーカータンパク質の濃度の変化によってモニターされる。選択可能なマーカー遺伝子が、ネオマイシン耐性などの抗生物質耐性を与えるタンパク質をコードする場合、形質転換体細胞は、G418によって選択することができる。選択可能なマーカー遺伝子を組み込んだ細胞は、生存すると考えられるが、その他の細胞は死滅する。これにより、本発明の組換え発現ベクターの発現の視覚化およびアッセイを可能にし、特に発現および表現型に対する突然変異の効果を決定することが可能となる。選択可能なマーカーは、関心対象の核酸から分離したベクターに導入することができることが認識される。
【0055】
また、組換え発現ベクターは、組換えタンパク質の発現の増大;組換えタンパク質の溶解性の増大;およびアフィニティー精製においてリガンドとして作用することによって標的組換えタンパク質の精製の援助をもたらす融合部分をコードする遺伝子を含んでいてもよい。たとえば、タンパク質分解切断部位を標的組換えタンパク質に付加して、融合タンパク質の精製後に融合部分から組換えタンパク質を分離させてもよい。
【0056】
組換え発現ベクターを宿主細胞に導入して、形質転換された宿主細胞を作製することができる。用語「形質転換された宿主細胞」は、本発明の組換え発現ベクターで形質転換またはトランスフェクトされた原核細胞および真核細胞を含むことが意図される。用語「で形質転換され」、「をトランスフェクトされ」「形質転換」、および「トランスフェクション」は、当技術分野において公知の多くの可能な技術のうちの1つによって、細胞に核酸(たとえば、ベクター)を導入することを含むことが意図される。たとえば、原核細胞は、電気穿孔法または塩化カルシウムを媒介したトランスフォーメーションによって核酸で形質転換することができる。核酸は、リン酸カルシウムもしくは塩化カルシウム共沈、DEAE-デキストランを媒介したトランスフェクション、リポフェクチン、電気穿孔法、または微量注入などの従来の技術を介して哺乳類細胞に導入することができる。宿主細胞を形質転換し、トランスフェクトするための適切な方法は、Sambrook et al.(Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press (1989))、およびその他の研究教科書において見いだすことができる。
【0057】
適切な宿主細胞は、多種多様な原核生物および真核生物の宿主細胞を含む。たとえば、本発明のタンパク質は、大腸菌(E. coli)などの細菌細胞、昆虫細胞(バキュロウイルスを使用する)、酵母細胞、または哺乳類細胞に発現されてもよい。その他の適切な宿主細胞は、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1991)において見いだすことができる。
【0058】
また、本発明のタンパク質は、固相合成などのタンパク質の化学において周知の技術を使用して、化学合成(Merrifield, 1964, J. Am. Chem. Assoc. 85:2149-2154)、または同種溶液における合成(Houbenweyl, 1987, Methods of Organic Chemistry, ed. E. Wansch, Vol.15 I and II, Thieme, Stuttgart)によって調製してもよい。
【0059】
III. 適用
本発明は、診断および治療的適用における、EPM2B核酸配列およびタンパク質およびこれらのモジュレーターの使用と同様に、抗体およびアンチセンス・オリゴヌクレオチドの調製、EPM2Bおよびこれらの変異された形態を研究するための実験系の調製、EPM26発現および/または活性を調整する物質の単離を含む(しかし、これらに限定されない)、本発明の核酸分子およびタンパク質の全ての使用を含む。いくつかの使用をさらに以下に記載してある。
【0060】
A.診断適用
前述のように、本発明者らは、遺伝子EPM2Bが、ラフォラ病の人々および犬科の動物において変異されていると断定した。結果として、本発明は、EPM2B遺伝子またはタンパク質の突然変異を検出することによってラフォラ病を検出する方法も含む。
【0061】
用語「突然変異」は、野生型配列と比較して、EPM2Bの核酸またはタンパク質配列における任意の変化または相違を意味する。突然変異は、ナンセンス突然変異、ミスセンス突然変異、フレームシフト突然変異、再編成突然変異、挿入突然変異、および欠失突然変異を含むが、これらに限定されない。
【0062】
試料は、体液(血液、尿、脳脊髄液、涙、唾液など)、組織、組織抽出物、細胞、および細胞抽出物を含むが、これらに限定されないEPM2B遺伝子またはタンパク質を含む任意の試料であることができる。
【0063】
(i)核酸配列における突然変異の検出
一つの態様において、本発明は、動物、好ましくは哺乳類、より好ましくは、ヒトまたは犬科の動物から得られる試料中のEPM2B遺伝子における突然変異を検出することを含む、ラフォラ病を検出するための方法を提供する。
【0064】
表1は、ラフォラ病患者のヒトEPM2Bにおいて見いだされるいくつかの突然変異を要約する。今日まで、フレームシフトを生じる8つの欠失および1つの挿入、8つのミスセンス、並びに1つのナンセンス変化を含む39ファミリーにおいてEPM2Bについて、21個の異なるDNA配列変化が見いだされた。各々の突然変異のために、スクリーニングアッセイ法を開発することができる。
【0065】
7つのファミリーにおいて同定された最も一般的な突然変異は、RINGフィンガー・ドメインにおいてプロリンからアラニンへの変化を生じるホモ接合性の205C→G移行である。したがって、一つの態様において、本発明は、EPM2B遺伝子(配列番号:1)の位置205におけるC→G突然変異を検出することを含む、ラフォラ病を検出する方法を提供する。
【0066】
本来の連鎖研究において使用した4血縁のF-Cファミリーで観察されるもう一つの突然変異は、全て、RINGフィンガー・ドメインの亜鉛結合能力に重要である7つの保存されたシステイン残基のうちの1つにシステインからセリンへの変化を生じるホモ接合性76T→A変化を有した。したがって、もう一つの態様において、本発明は、EPM2B遺伝子(配列番号:1)の位置76におけるT→A突然変異を検出することを含む、ラフォラ病を検出する方法を提供する。
【0067】
もう一つの突然変異は、LD38324ファミリーにおいて観察され、これはEPM2B遺伝子座(図2a)を絞り込む際に重要であって、第5NHL-ドメイン(図2b)においてフレームシフト突然変異を生じるホモ接合性2-bp欠失(1048-1049delGA)であった。したがって、さらなる態様において、本発明は、EPM2B遺伝子(配列番号:1)の位置1048および1049におけるGAの欠失を検出することを含むラフォラ病を検出する方法を提供する。
【0068】
実施例3に記載したように、発明者は、ラフォラ病のイヌにおけるEPM2B遺伝子の突然変異も発見した。特に、研究された病気に冒された全てのイヌは、Dと呼ばれ、かつ配列番号:3に示したイヌEPM2B配列のヌクレオチド番号1001で始まる配列
を有する12塩基反復の2対立遺伝子の伸長を有した。本発明者らは、この12ヌクレオチド繰り返しが、イヌ、オオカミ、キツネ、コヨーテ、およびジャッカルを含む犬科の動物スーパーファミリーに特異的であることを示し、この繰り返しは、イヌを大量の配列伸長にかかりやすくする素因になり、これがEPM2B遺伝子に有害であり、ラフォラ病を生じさせることを示した。本発明者らは、したがって、イヌを含む犬科の動物が、ラフォラてんかんにかかりやすいということを発見した。したがって、本発明は、EPM2B(配列番号:3)のイヌ配列におけるヌクレオチド番号1001で始まる配列
の繰り返しを検出することを含む、犬科の動物においてラフォラ病を検出する方法をさらに提供する。一つの態様において、本方法は、配列番号:5において、少なくとも3つの繰り返し、好ましくは少なくとも10の繰り返し、より好ましくは約14〜約26の繰り返しを検出することを含む。
【0069】
当業者であれば、実施例において論議したものに加え、および含む、多くの適切な方法をEPM2B遺伝子の突然変異を検出するために使用することができることを認識するであろう。使用することができる検出方法は、ポリメラーゼ連鎖反応法(PCR)、逆転写酵素PCR、直接シーケンシング電気泳動移動度、核酸ハイブリダイゼーション、蛍光インサイチュー・ハイブリダイゼーション、変性高速液体クロマトグラフィー、DNAチップ技術、および質量分析を含むが、これらに限定されない。一つの例において、試料中のラフォラ病遺伝子由来の核酸を単離するために、本発明の核酸配列からヌクレオチド・プローブを調製することができる。加えて、本明細書に記載されている核酸プローブを使用することもできる。ヌクレオチド・プローブは、32P、3H、14Cなどの適切なシグナルを提供し、十分な半減期を有する放射性の標識などの、検出可能な物質で標識されてもよい。使用してもよいその他の検出可能なマーカーは、特異的に標識された抗体、蛍光化合物、酵素、標識された抗原に特異的な抗体、および化学発光化合物によって認識される抗原を含む。適切な標識は、検出されるヌクレオチドに対するプローブのハイブリダイゼーションおよび結合の割合、並びにハイブリダイゼーションに利用できるヌクレオチドの量を考慮して選択してもよい。
【0070】
したがって、本発明は、試料を、核酸分子にハイブリダイズし、かつ検出可能なマーカーで標識された1つまたは複数のヌクレオチド・プローブと、ハイブリダイゼーション条件下で接触させる工程、および試料中の核酸分子とヌクレオチド・プローブとの間のハイブリダイゼーションの程度を決定する工程を含む、試料中のEPM2B遺伝子由来の核酸分子の存在を検出する方法にも関する。
【0071】
本発明の方法に使用してもよいハイブリダイゼーション条件は、当技術分野において公知であり、たとえばSambrook J, Fritch EF, Maniatis T. In: Molecular Cloning, A Laboratory Manual,1989. (Nolan C, Ed.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYに記載されている。ハイブリダイゼーション産物は、当技術分野において公知の技術を使用してアッセイしてもよい。ヌクレオチド・プローブは、本明細書に記載したように、検出可能なマーカーで標識してもよく、ハイブリダイゼーション産物は、検出可能なマーカーによって生じる検出可能なマーカーまたは検出可能な変化を検出することによってアッセイしてもよい。
【0072】
DNAプローブと試料をハイブリダイズさせる前に、試料中の核酸配列を増幅するために、EPM2B遺伝子に隣接するプライマーで試料を処理することができる。使用するプライマーは、本出願に記載されているものであってもよい。加えて、また本明細書において提供されるEPM2B遺伝子の配列により、新たなヌクレオチド配列の同定および単離、または合成が可能となり、たとえば下記にさらに詳細に論議したポリメラーゼ連鎖反応法(PCR)において本発明の核酸分子を増幅するためのプライマーとして使用してもよい。プライマーは、その他の種のゲノムDNAを増幅するために使用してもよい。PCR増幅された配列を、様々な種の遺伝子間の関係を決定するために調べることができる。
【0073】
PCRに使用するためのプライマーの長さおよび塩基は、これらが所望の配列の異なる鎖に対してハイブリダイズするように、および配列に沿った相対的位置で、テンプレートから分離するとき1つのプライマーから合成された伸長産物が、定義された長さの核酸へのその他のプライマーの伸長のテンプレートとして役立つことができるように選択される。本発明に使用してもよいプライマーは、オリゴヌクレオチド、すなわち本発明の核酸分子の2つ以上のデオキシリボヌクレオチドを含む分子であり、これらは、精製された制限エンドヌクレアーゼ消化物として天然に存在するか、またはたとえば、リン酸トリエステルおよびリン酸ジエステル法(Good et al Nucl. Acid Res 4:2157, 1977を参照されたい)または自動技術(たとえば、Conolly, B. A. Nucleic Acids Res. 15: 15 (7): 3131, 1987を参照されたい)などの当技術分野において公知の技術を使用して合成的に産生される。プライマーは、本発明のDNA配列に対して相補的であるプライマー伸長産物の合成を可能にする条件下に、すなわちヌクレオチド基質、DNAポリメラーゼなどの重合のための薬剤の存在下において、並びに適切な温度およびpHに配置されたときに合成の開始位置として作用することができる。好ましくは、プライマーは、プライマーのその他のコピーと塩基対形成することによって二次構造を形成しない配列か、またはヘアピン立体配置を形成する配列である。プライマーは、好ましくは約7〜25の間のヌクレオチドを含む。
【0074】
プライマーは、増幅産物の検出を可能にする検出可能なマーカーで標識してもよい。適切な検出可能なマーカーは、32P、35S、125I、および3Hなどの放射性のマーカー、化学発光マーカー、好ましくはルミノール、並びに蛍光マーカー、好ましくは塩化ダンシル、フルオルセイン-5-イソチオシアナート、および4-フルオロ-7-ニトロベンズ-2-axa-1,3ジアゾールなどの発光のマーカー、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β-ガラクトシダーゼ、アセチルコリンエステラーゼ、またはビオチンなどの酵素マーカーである。
【0075】
プライマーは、プライマーの十分な量が本発明の核酸分子またはこれらのオリゴヌクレオチド断片に対して相補的であり、増幅される配列を含むことを条件として、非相補的な配列を含んでもよいことが認識される。また、制限部位リンカーをプライマーに組み入れて、適切な制限酵素での増幅産物の消化を可能にして、増幅産物のクローニングおよびシーケンシングを容易にしてもよい。
【0076】
本発明のある態様において、増幅配列を形成することができる条件下において、ポリメラーゼ連鎖反応法において核酸分子または予め定められたこれらのオリゴヌクレオチド断片を増幅することができるプライマーで試料を処理して増幅配列を形成する工程、および増幅配列をアッセイする工程を含む、本発明の核酸分子の存在を決定する方法が提供される。
【0077】
ポリメラーゼ連鎖反応法は、一般にInnis et al, Academic Press, 1990 in Mullis et al., 米国特許第4,863,195号およびMullis, 米国特許第4,683,202号(これらは、参照として本明細書に組み入れられる)に記載されている、標的核酸配列を増幅するためのプロセスをいう。核酸テンプレートを増幅するための条件はM.A. Innis and D.H. Gelfand, PCR Protocols, A Guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989(これらも参照として本明細書に組み入れられる)に記載されている。
【0078】
増幅産物は、当技術分野において公知の技術を使用して、単離し、これらのそれぞれのサイズに基づいて区別することができる。たとえば、増幅後に、DNA試料をアガロースゲル上で分離して、臭化エチジウムで染色後に紫外(UV)光下で視覚化することができる。DNAは、所望のレベルに増幅してもよく、さらなる伸長反応を行って、放射性標識されたか、またはビオチン標識されたヌクレオシド三リン酸などの検出可能なマーカーを有するヌクレオチド誘導体を組み入れてもよい。また、プライマーは、上で考察したように検出可能なマーカーで標識してもよい。検出可能なマーカーは、制限および電気泳動分離、または当技術分野において公知のその他の技術によって解析してもよい。
【0079】
PCRを使用する本発明の方法において使用してもよい条件は、試料中のDNAおよび適切な相補的ハイブリダイゼーション・プライマーの存在下において、ハイブリダイゼーションおよび増幅反応を進行することができるものである。ポリメラーゼ連鎖反応法のために適した条件は、一般に当技術分野において公知である。たとえば、M.A. Innis and D.H. Gelfand, PCR Protocols, A guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989を参照されたい(これらは、参照として本明細書に組み入れられる)。好ましくは、PCRは、好熱性細菌サーマス・アクアティクス(Thermus aquatics)(Taqポリメラーゼ、GeneAmpキット、Perkin Elmer Cetus)から得られるポリメラーゼを利用し、またはその他の耐熱性ポリメラーゼを、DNA鋳型鎖を増幅するために使用してもよい。
【0080】
リガーゼ連鎖反応法(LCR)およびNASBAなどのその他の技術を本発明の核酸分子を増幅するために用いてもよいことが認識される(Barney in "PCR Methods and Applications", August 1991, Vol.1(1), page 5、および1989年6月14日に公開された欧州特許出願第0320308号、並びにMalekの米国特許出願第5,130,238号)。
【0081】
本発明のいくつかの態様において、本発明の配列における突然変異は、核酸分子の直接のシーケンシングによって検出してもよい。DNAを直接シーケンシングするための技術は、当技術分野において周知である。本発明の一つの態様において、突然変異は、サイクル・シーケンシングで検出されてもよく、これは、耐熱性ポリメラーゼ酵素、シーケンシング・プライマー、dNTP、および限定量の鎖を終結する蛍光性または放射性に標識されたddNTPの使用を含んでもよい。ポリアクリルアミドゲル電気泳動またはキャピラリー電気泳動法などのもう一つの技術を使用してシーケンシング反応の産物を分離し、続いて蛍光性または放射性の標識を検出してもよい。本発明の本態様の一つの例において、EMP2Bにおける突然変異は、Applied Biosystems 3700 DNA Analyzerまたは3730xl DNA Analyzer(商標)での自動シーケンシングを使用して決定することもできる。突然変異は、患者の配列を野生型の個体のものと、または公共データベースにおいて見いだされる参照配列と比較することによって同定してもよい。
【0082】
また、EMP2Bにおける突然変異は、米国特許第5,888,780号に開示され、かつMadison, WisconsinのThird Wave(商標) Technologiesから入手可能なCleavase(商標)Fragment Length Polymorphism(CFLP)アッセイ法を使用するInvader(商標)遺伝子タイピング系を使用して検出してもよい。
【0083】
本発明のその他の態様は、EMP2B遺伝子における突然変異の検出のためDNAチップ技術の使用を想定する。その他の適用の中で、DNAチップ技術により、予め定められた配列の高密度に空間的にアドレス指定可能なマイクロアレイ上に対する核酸試料のハイブリダイゼーション・パターンを解析することを介して目的の配列における突然変異を同定することができる。本発明の配列である突然変異の同定のために適したDNAチップ技術の一つの例は、米国特許第6,045,996号に開示されたAffymatrix GeneChip(商標)系である。この系は、何十万ものオリゴヌクレオチドプローブを含む高密度アレイを作製するために、フォトリソグラフィーおよび固相化学を使用する。Genechips(商標)は、特定の配列の再シーケンシングを容易にして、特定の突然変異を同定することができるようにデザインしてもよい。ハイブリダイゼーション・パターンのコンピュータに基づいた画像分析により、自動塩基呼び出し、および解析内に品質制御措置を組み入れることが可能になる。
【0084】
当業者であれば、本発明の配列内もしくは近くにおける病原性の欠失または再編成を検出するためにサザンブロッティング法またはノーザンブロッティング法を使用してもよいことを認識しているであろう。同様に、蛍光インサイチュー・ハイブリダイゼーション(FISH)、fiber-FISH、またはその他の高解像度の細胞遺伝学的方法を、本発明の配列を分裂させる再編成または欠失を検出するために使用してもよい。
【0085】
突然変異の検出のためのもう一つの技術は、変性HPLC解析である。したがって、本発明の一つの態様は、本発明の配列内におけるヘテロ接合性の突然変異または多型の同定のために、核酸のdHPLC解析のためのTransgenomic Wave(商標)機の使用を含む。
【0086】
また、本発明は、突然変異の遺伝子タイピングのための質量分析の使用も想定する。変異体および野生型の核酸分子は、野生型と変異体配列での異なる組成のために、質量が異なっており、異なるヌクレオチド配列の分子質量に基づいて突然変異を同定することができる。突然変異の遺伝子タイピングのための質量分析、特にマトリクス支援レーザー脱離イオン化飛行時間型(Matrix Assisted Laser Desorption Ionisation Time of Flight:MALDI-TOF)質量分析の使用は、関連した技術における当業者に周知である。たとえば、米国特許第6,043,031号は、特定の核酸配列を検出するための迅速かつ非常に正確な質量分析計に基づいたプロセスを記載する。SEQUENOM(商標)からのMass ARRAY(商標)プラットフォームは、一塩基多型を遺伝子タイピングすることができ、本発明に記載されているような突然変異を検出することができる、市販されている系の例である。
【0087】
(ii)Malinタンパク質の検出
もう一つの態様において、本発明は、ラフォラ病を有することが疑われる哺乳類、好ましくはヒトまたはイヌ由来の試料において、malinタンパク質が存在するか、または変異されているかどうかを決定することを含む、ラフォラ病を検出するための方法を提供する。
【0088】
本発明のmalinタンパク質は、後述する種々の免疫アッセイ法を使用してmalinに特異的である抗体を使用して、生体試料中で検出してもよい。変異されたmalinとのみ反応する抗体は、ラフォラ病を検出するための診断薬として有用である。したがって、タンパク質の変異された領域だけに結合する抗体が調製されることとなる。
【0089】
抗体を調製するために、従来法を使用することができる。たとえば、本発明のmalinタンパク質由来のペプチドを使用することにより、ポリクローナル抗血清またはモノクローナル抗体を標準的な方法を使用して作製することができる。哺乳類(たとえば、マウス、ハムスター、またはウサギ)に、哺乳類の抗体反応を引き出すペプチドの免疫原形態で免疫することができる。ペプチドに免疫原性を与えるための技術は、キャリアとの抱合または当技術分野において周知のその他の技術を含む。たとえば、ペプチドをアジュバントの存在下において投与することができる。免疫の過程は、プラズマまたは血清中の抗体力価の検出によってモニターすることができる。抗体のレベルを評価するために、標準的なELISAまたはその他の免疫アッセイ手順では、抗原として免疫原を使用することができる。免疫化に続いて、抗血清を得ることができ、必要であれば、ポリクローナル抗体を血清から単離することができる。
【0090】
モノクローナル抗体を産生するためには、抗体産生細胞(リンパ球)を免疫した動物から収集し、標準的な体細胞融合手順によって骨髄腫細胞と融合させることによって、これらの細胞を不死化し、ハイブリドーマ細胞を産生することができる。このような技術は、当技術分野において周知である(たとえば、当初Kohlerおよび Milstein (Nature 256, 495-497 (1975))によって開発されたハイブリドーマ技術、並びにヒトB-細胞ハイブリドーマ技術(Kozbor et al., Immunol. Today 4, 72 (1983))、ヒトモノクローナル抗体を産生するためのEBVハイブリドーマ技術(Cole et al., Monoclonal Antibodies in Cancer Therapy (1985) Allen R. Bliss, Inc., pages 77-96)、およびコンビナトリアル抗体ライブラリーのスクリーニング(Huse et al., Science 246, 1275 (1989))などのその他の技術)。ペプチドと特異的に反応する抗体の産生について、ハイブリドーマ細胞を免疫化学的にスクリーニングすることができ、モノクローナル抗体を単離することができる。したがって、本発明はまた、本発明のタンパク質に対する特異性を有するモノクローナル抗体を分泌するハイブリドーマ細胞を想定する。
【0091】
本明細書に使用される用語「抗体」は、本発明のタンパク質、またはこれらのペプチドと特異的に反応するこれらの断片も含むことが意図される。上記のものと同じ方法で利用するために、従来技術を使用して抗体を断片化し、断片をスクリーニングすることができる。たとえば、F(ab')2断片は、ペプシンで抗体を処理して作製することができる。生じるF(ab')2断片を処理してジスルフィド架橋を還元し、Fab'断片を産生することができる。
【0092】
キメラ抗体誘導体、すなわちヒト以外の動物の可変領域とヒトの定常領域を組み合わせた抗体分子も、本発明の範囲内において想定される。キメラ抗体分子は、たとえば、ヒト定常領域を有する、マウス、ラット、またはその他の種の抗体に由来する抗原結合ドメインを含むことができる。malinタンパク質を認識する免疫グロブリン可変領域を含むキメラ抗体を作製するために、従来法を使用してもよい(たとえば
を参照されたい)。
【0093】
本明細書に記載されている本発明のタンパク質と特異的に反応するモノクローナルまたはキメラ抗体は、可変領域の一部、特に抗原結合ドメインの保存されたフレームワーク領域が、ヒト起源であり、かつ高頻度可変領域だけが非ヒト起源であるヒト定常領域キメラを産生することによって、さらにヒト化することができる。このような免疫グロブリン分子は、当技術分野において公知の技術によって作製されてもよい(たとえばTeng et al., Proc. Natl. Acad. Sci. U.S.A., 80, 7308-7312 (1983); Kozbor et al., Immunology Today, 4, 7279 (1983); Olsson et al., Meth. Enzymol., 92, 3-16 (1982))、および国際公開公報第92/06193号または欧州特許第0239400号)。また、ヒト化抗体は、商業的に産生することができる(Scotgen Limited, 2 Holly Road, Twickenham, Middlesex, Great Britain)。
【0094】
本発明のタンパク質に対して反応性の、特異的な抗体、または抗体断片は、本発明の核酸分子から産生されるペプチドを用いて、細菌で発現した免疫グロブリン遺伝子、またはこれらの部分をコードする発現ライブラリーをスクリーニングすることによって作製されてもよい。たとえば、完全Fab断片、VH領域、およびFV領域は、ファージ発現ライブラリーを使用して、細菌で発現させることができる(Ward et al., Nature 341, 544-546: (1989); Huse et al., Science 246, 1275-1281 (1989);および McCafferty et al., Nature 348, 552-554 (1990)を参照されたい)。
【0095】
また、抗体は、DNA免疫を使用して調製してもよい。たとえば、本発明の(上記のとおりの)核酸を含む発現ベクターをマウスなどの適切な動物に注射してもよい。したがって、本発明のタンパク質がインビボで発現されて、抗体が誘導されると考えられる。抗体は、単離することができ、タンパク質免疫化のために上記のとおりに調製することができる。
【0096】
抗体は、種々の酵素、蛍光物質、発光材料、および放射性物質を含む検出可能なマーカーで標識してもよい。適切な酵素の例は、西洋ワサビペルオキシダーゼ、ビオチン、アルカリホスファターゼ、β-ガラクトシダーゼ、またはアセチルコリンエステラーゼを含み;適切な蛍光物質の例は、ウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリド、またはフィコエリトリンを含み;発光材料の例は、ルミノールを含み;および適切な放射性物質の例は、S-35、Cu-64、Ga-67、Zr-89、Ru-97、Tc-99m、Rh-105、Pd-109、In-111、I-123、I-125、I-131、Re-186、Au-198、Au-199、Pb-203、At-211、Pb-212、およびBi-212を含む。また、抗体は、リガンド結合対の一方のパートナーに対して標識するか、または抱合してもよい。代表例は、アビジン-ビオチンおよびリボフラビン-リボフラビン結合タンパク質を含む。上記した代表的標識をもつ、上で論議した抗体を抱合、または標識するための方法は、従来技術を使用して容易に達成し得る。
【0097】
本発明のタンパク質(たとえば、酵素抱合体または標識された誘導体)に対する抗体反応性は、種々の試料中における本発明のタンパク質を検出するために使用してもよく、たとえば、これらは、本発明のタンパク質の抗原決定基と抗体との間の結合相互作用に依存する任意の公知の免疫アッセイ法に使用してもよい。このようなアッセイ法の例は、放射免疫アッセイ法、酵素免疫測定法(たとえば、ELISA)、免疫蛍光、免疫沈降、ラテックス凝集反応法、赤血球凝集法、および組織化学的試験である。したがって、ラフォラ病の存在を診断するために、試料中の本発明のタンパク質の量を同定、または定量化するため抗体を使用してもよい。
【0098】
本発明の方法では、試料または濃縮された試料の予め定められた量が、抗体または標識抗体と混合される。本方法に使用される抗体の量は、選択される標識剤に依存的である。生じる、抗体または標識された抗体に結合したタンパク質は、従来の単離技術、たとえばば塩析、クロマトグラフィー、電気泳動法、ゲル濾過、分画、吸収、ポリアクリルアミドゲル電気泳動、凝集、またはこれらの組み合わせによって単離してもよい。
【0099】
試料または抗体は、不溶化してもよく、たとえば、試料または抗体は、公知の方法を使用して適切なキャリアと反応させることができる。適切なキャリアの例は、セファロースまたはアガロースビーズである。不溶化された試料または抗体が使用されるときは、抗体または反応していない抗体に結合したタンパク質は、洗浄することによって単離される。たとえば、試料がニトロセルロース膜にブロットされるときは、本発明のタンパク質に結合した抗体は、緩衝液、たとえばウシ血清アルブミン(BSA)を含むリン酸緩衝食塩水(PBS)で洗浄することにより、反応していない抗体から分離される。
【0100】
標識された抗体が使用されるときは、malinの存在は、試料中の本発明のタンパク質に結合した標識された抗体の量または反応していない標識された抗体の量を測定することによって決定することができる。標識された物質を測定する適切な方法は、標識剤に依存する。
【0101】
標識されていない抗体を本発明の方法に使用するときは、malinの存在は、凝集または沈澱を生じさせるために、抗体と特異的に相互作用する物質を使用してタンパク質に結合した抗体の量を測定することによって決定することができる。特に、本発明のタンパク質に特異的な抗体に対する標識された抗体を、反応混合物に添加することができる。本発明のタンパク質の存在は、標識剤のタイプに応じて、すでに記載されている技術の中から適切な方法によって決定することができる。本発明のタンパク質に特異的な抗体に対する抗体は、本明細書に記載した当技術分野において公知の従来法によって調製し、および標識することができる。本発明のタンパク質に特異的な抗体に対する抗体は、種特異的な抗免疫グロブリン抗体またはモノクローナル抗体であってもよく、たとえば、ヤギ抗ウサギ抗体を本発明のタンパク質に特異的なウサギ抗体を検出するために使用してもよい。
【0102】
(iii)キット
本発明の方法を実施するために適した試薬は、適切な容器にパックされて、必要な材料を提供する便利なキットにパックされてもよい。このようなキットは、本明細書に記載されている方法、および任意に本発明の方法を行う際に有用な適切な支援によって、試料中の本発明の核酸分子またはタンパク質を検出するために必要とされる全ての試薬を含んでもよい。
【0103】
本発明の一つの態様において、キットは、本発明の核酸分子またはこれらの予め定められたオリゴヌクレオチド断片を増幅することができるプライマー、ポリメラーゼ連鎖反応法において増幅された核酸分子またはこれらの予め定められた断片を産生するために必要とされる全ての試薬、および増幅された配列をアッセイするための手段を含む。また、キットは、PCR産物を消化するための制限酵素も含んでいてもよい。本発明のもう一つの態様において、キットは、本発明の核酸分子とハイブリダイズするヌクレオチド・プローブ、核酸分子とヌクレオチド・プローブのハイブリダイゼーションのために必要とされる試薬、およびその使用説明書を含む。本発明のさらなる態様において、キットは、本発明の抗体、および試料中において本発明のタンパク質に対して抗体を結合するために必要とされる試薬を含む。
【0104】
本発明の方法およびキットは、ラフォラ病を検出するために使用してもよい。試験してもよい試料は、血液、尿、血清、涙、唾液、糞便、組織、細胞などの身体材料を含む。ヒト試料に加えて、試料は、イヌなどの非ヒト霊長類および犬科の動物などの哺乳類に由来してもよい。
【0105】
本明細書に記載されている方法に従って試料を試験する前に、試料は、遠心分離および濾過などの当技術分野において公知の技術を使用して濃縮してもよい。本明細書に記載されているハイブリダイゼーションおよび/またはPCRに基づいた方法については、核酸は、当技術分野において公知の技術を使用して試験試料の細胞抽出物から抽出してもよい。
【0106】
B.EPM2B/Malinモジュレーター
上記した抗体およびアンチセンス・オリゴヌクレオチドに加えて、EPM2B発現または活性を調整するその他の物質、並びにmalinの変異した形態を調整する物質を同定してもよい。
【0107】
(i)Malinに結合する物質
malin活性に影響を及ぼす物質は、これらがmalinおよび/または変異したmalinと結合する能力に基づいて同定することができる。
【0108】
本発明のmalinと結合することができる物質は、潜在的にmalinに結合する物質とmalinを反応させることと、遊離の物質について、または複合体を形成していないmalinについて、またはmalinの活性について、複合体をアッセイすることによって同定され得る。特に、malinと相互作用するタンパク質を同定するために、酵母ツー・ハイブリッド・アッセイ系を使用してもよい(Fields, S. and Song, O., 1989, Nature, 340:245-247)。また、使用してもよい解析系には、ELISAを含む。
【0109】
したがって、本発明は、以下の工程をmalinと結合することができる物質を同定する方法であって:
(a)反応するmalinと試験物質を、malinと試験物質との間で複合体を形成することができる条件下で反応させる工程、および
(b)遊離の物質について、または複合体を形成していないmalinについて、malinと試験物質の複合体をアッセイする工程を含み、複合体の存在は、試験物質がmalinに結合できることを示す方法を提供する。
【0110】
アッセイ法に使用されるmalinタンパク質は、配列番号:2(図6B)もしくは配列番号:4(図7B)に示したアミノ酸配列を有してもよく、または本明細書に記載されている、LDと関連する変異されたタンパク質であってもよく、または本明細書に記載したような、これらの断片、類似体、誘導体、相同体、もしくはミメティックであってもよい。
【0111】
物質とmalinの複合体の形成を可能にする条件は、物質およびタンパク質の性質並びに量などの因子を考慮して選択してもよい。
【0112】
物質-タンパク質複合体、遊離の物質、または複合体を形成しないタンパク質は、従来の単離技術、たとえば、塩析、クロマトグラフィー、電気泳動法、ゲル濾過、分画、吸収、ポリアクリルアミドゲル電気泳動、凝集、またはこれらの組み合わせによって単離してもよい。malinもしくは物質に対する抗体、または標識されたmalin、または標識された物質を、成分のアッセイ法を容易にするために利用してもよい。抗体、タンパク質、または物質は、上記のような検出可能な物質で標識してもよい。
【0113】
本発明の方法に使用されるmalinまたは物質は、不溶化させてもよい。たとえば、malinまたは物質は、適切なキャリアに結合されてもよい。適切なキャリアの例は、アガロース、セルロース、デキストラン、セファデックス、セファロース、カルボキシメチルセルロースポリスチレン、濾紙、イオン交換樹脂、プラスチック・フィルム、プラスチック・チューブ、ガラスビーズ、ポリアミン-メチルビニル-エーテル-マレイン酸共重合体、アミノ酸共重合体、エチレン-マレイン酸共重合体、ナイロン、絹、その他である。キャリアは、たとえば、チューブ、試験プレート、ビーズ、盤、球、その他の形態であってもよい。
【0114】
不溶化されたタンパク質または物質は、公知の化学的または物理的な方法、たとえば、臭化シアンカップリングを使用して、適切な不溶性キャリアと材料を反応することによって調製してもよい。
【0115】
また、タンパク質または物質は、本明細書に記載されている方法を使用して細胞の表面上に発現させてもよい。
【0116】
また、本発明は、malinの作用のアンタゴニストまたはアゴニストのためのアッセイを想定する。
【0117】
本発明の方法を使用してアッセイすることができるアゴニストおよびアンタゴニストは、アゴニスト結合部位、競合的アンタゴニスト結合部位、非競合的アンタゴニスト結合部位、またはアロステリック部位を含む、1つもしくは複数のタンパク質または物質の結合部位に対して作用してもよいことが理解されるであろう。
【0118】
また、本発明は、malinのアゴニストの効果を阻害するアンタゴニストをスクリーニングすることを可能にする。したがって、本発明は、malinの同じ結合部位に対して競合する物質をアッセイするために使用してもよい。
【0119】
(ii)ペプチドミメティック
また、本発明は、本発明のmalinのペプチドミメティックおよび変異されたmalinタンパク質を含む。たとえば、malinの変異されたドメインに由来するペプチドは、変異されたタンパク質の天然の結合を模倣するような方法で関連する分子と直接的または間接的に相互作用する。このようなペプチドは、競合阻害剤、エンハンサー、ペプチドミメティック等を含んでもよい。実質的にこれらのペプチド類、並びにこれらのペプチドに対して相同的、相補的、またはさもなければ機能的もしくは構造的に同等な分子の全てを、本発明の目的を達成するために使用してもよい。
【0120】
「ペプチドミメティック」とは、分子間の相互作用においてペプチドの代用として役立つ構造である(総説については、Morgan et al (1989), Ann. Reports Med. Chem. 24:243-252を参照されたい)。ペプチドミメティックは、アミノ酸および/またはペプチド結合を含んでも、または含まなくてもよいが、ペプチド、または本発明のエンハンサーもしくは阻害剤の構造上および機能的な特徴を保持する合成構造を含む。また、ペプチドミメティックは、ペプトイド、オリゴペプトイド(Simon et al (1972) Proc. Natl. Acad, Sci USA 89:9367);および本発明のペプチドに対応するアミノ酸の全ての可能性のある配列を表す長さにデザインされたペプチドを含むペプチドライブラリーを含む。
【0121】
ペプチドミメティックは、D-アミノ酸によるL-アミノ酸の系統的置換、異なる電子的性質を有する基と側鎖の置換によって、およびアミド結合の置換によるペプチド結合の系統的置換によって得られる情報に基づいてデザインされてもよい。また、候補となるペプチドミメティックの活性についての高次構造上の要求を決定するために、局部的な高次構造上の制約を導入することができる。ミメティックは、逆向ターンの高次構造を安定化もしくは促進し、および分子の安定化を助けるために、等配電子アミド結合、またはD-アミノ酸を含んでもよい。特定の高次構造の状態にアミノ酸残基を拘束するために、環状アミノ酸類似体を使用してもよい。また、ミメティックには、ペプチドの二次構造の阻害剤のミメティックを含むことができる。これらの構造は、アミノ酸残基の三次元配向をタンパク質の公知の二次高次構造にモデル化することができる。また、N置換されたアミノ酸のオリゴマーであるペプトイドを使用してもよく、新規分子の化学的に多様なライブラリーを作製するためのモチーフとして使用することができる。
【0122】
また、本発明のペプチドは、薬剤開発のためのリード化合物を同定するために使用してもよい。本明細書に記載されたペプチドの構造は、NMRおよびX線結晶学などの多くの方法によって容易に決定することができる。配列が類似するが、これらが標的分子に誘発する生物活性が異なるペプチドの構造を比較すると、標的の構造-活性相関についての情報を提供することができる。構造-活性相関の調査から得られる情報は、修飾されたペプチド、または標的分子に関して予測される性質を試験することができるその他の小分子もしくはリード化合物のいずれかをデザインするために使用することができる。リード化合物の活性は、本明細書に記載されているものと同様のアッセイ法を使用して評価することができる。
【0123】
また、構造-活性相関についての情報は、共同結晶化研究から得てもよい。これらの研究では、所望の活性を有するペプチドを標的分子と結合させて結晶化し、複合体のX線構造を決定する。次いで、構造をその天然の状態の標的分子の構造と比較することができ、このような比較からの情報を、有することが予想される化合物をデザインするために使用してもよい。
【0124】
(iii)薬物のスクリーニング方法
一つの態様に従って、本発明は、変異したmalinタンパク質の活性を増減する能力について候補化合物をスクリーニングするための方法を可能にする。本方法は、malin活性をアッセイするため、候補もしくは試験化合物の有無における活性をアッセイするため、および化合物がmalin活性の増減をもたらすかどうかを決定するためのアッセイ系を提供することを含む。このような化合物は、ラフォラ病を治療する際に有用であり得る。
【0125】
したがって、本発明は、変異されたmalinタンパク質活性または発現に影響を及ぼす化合物を同定するための方法であって:
(a)malinタンパク質またはmalinタンパク質をコードする核酸と共に試験化合物をインキュベートする工程;および、
(b)malinタンパク質の活性または発現の量を決定し、対照(すなわち、試験物質の非存在下において)と比較する工程を含み、対照と比較したmalinタンパク質活性または発現の変化は、試験化合物がmalinタンパク質の活性または発現に対して効果を有することを示す方法を提供する。
【0126】
さらなる態様に従って、本発明によって、malinタンパク質の発現を増減する能力の候補化合物をスクリーニングする方法が可能となる。本方法は、候補化合物と共に細胞(本細胞は、リポーター遺伝子コード領域に操作可能な状態で連結されたmalin遺伝子の制御領域を含む)を置く工程、およびリポーター遺伝子の発現の変化を検出する工程を含む。
【0127】
一つの態様において、本発明により、変異されたmalin遺伝子を発現する株化細胞を候補化合物と共にインキュベートして、変異されたmalin発現に対するこれらの効果を試験する培養系が可能になる。このような培養系は、その他のタンパク質との相互作用を介して、malin発現もしくはその機能をアップレギュレートするか、またはダウンレギュレートする化合物を同定するために使用することができる。
【0128】
このような化合物は、培地に添加されるタンパク質化合物、化学薬品、および種々の薬物から選択することができる。選択された試験化合物(類)の存在下でのインキュベーション後、変異されたmalinの発現を、本明細書に含まれる実施例において説明したように、標準的なノーザンブロット法手順を使用してmalin mRNAのレベルを定量化することによって調べて、試験化合物の結果として発現の任意の変化を決定することができる。また、タンパク質発現を修飾するために開発された化合物の機能を試験するために、malinを発現する構築物をトランスフェクトされた細胞株を使用することができる。
【0129】
C.治療的使用
前述のように、本発明のEPM2B遺伝子およびmalinは、ラフォラ病に関与する可能性が高い。したがって、本発明は、ラフォラ病を治療する方法であって、その必要がある細胞または動物に対し、EPM2B/malin発現および/または活性を調整する薬剤の有効な量を投与することを含む方法を提供する。また、本発明はラフォラ病を治療するための、もしくはラフォラ病を治療するための薬物を調製するための、EPM2B/malin発現および/または活性を調整する薬剤の使用を提供する。
【0130】
用語「EPM2B/malin発現および/または活性を調整する薬剤」は、野生型EPM2B/malinと一致する動物において見いだされる変異されたEPM2B/malinの発現および/または活性を変化させることができる任意の物質を意味する。投与に使用してもよい薬剤の例は:野生型EPM2Bをコードする核酸分子;野生型malinタンパク質、並びにこれらの断片、類似体、誘導体、または相同体;抗体;アンチセンス核酸;ペプチドミメティック;および本明細書に記載されているスクリーニング法を使用して単離された物質であって、変異を症状のない人と同様のEPM2B/malinレベルおよび/または機能に修正することができるものを含む。
【0131】
本明細書に使用される用語「有効な量」は、所望の結果を達成するために必要な用量で、および期間で有効な量を意味する。
【0132】
本明細書に使用される用語「動物」は、ヒトおよびイヌを含む動物界の全ての一員を含む。
【0133】
一つの態様において、本発明は、変異されたmalinタンパク質の発現または生物活性を調整する薬剤の有効な量を、細胞または動物に投与することによってラフォラ病を治療する方法を提供する。また、本発明は、ラフォラ病を治療するために、またはラフォラ病を治療するための薬物を調製するために、変異されたmalinタンパク質の発現または生物活性を調整する薬剤の有効な量の使用を提供する。変異されたmalinの活性を阻害する物質は、ペプチドミメティック、malinアンタゴニスト、およびmalinに対する一定の抗体を含む。変異されたEPM2B遺伝子の発現を阻害する物質は、変異されたEPM2B核酸配列に対するアンチセンス・オリゴヌクレオチドを含む。
【0134】
もう一つの態様に従って、本発明は、ラフォラに対する潜在的な治療手段としての遺伝子治療であって、病気に冒されたいくつかの異なる細胞型において正常なmalinタンパク質をうまくコードするように、EPM2B遺伝子の正常なコピーを患者に導入する治療を可能にする。
【0135】
レトロウイルスのベクターは、これらが高い効率で感染および安定な組込みおよび発現をするので、特に体細胞遺伝子治療に使用することができる。しかし、ターゲットされる細胞は、分裂することができなければならず、正常タンパク質の発現レベルが高いはずである。全長正常EPM2B遺伝子は、レトロウイルスのベクターにクローン化することができ、その内因性のプロモーターからもしくはレトロウイルスの末端反復配列から、または関心対象の標的細胞タイプに特異的なプロモーターから駆動させることができる。使用することができるその他のウイルスのベクターは、アデノ随伴ウイルス、ワクシニアウイルス、ウシパピローマウイルス、またはエプスタイン・バーウイルスなどのヘルペスウイルスを含む。また、遺伝子導入は、インビトロでの感染に必要とされる非ウイルス手段を使用して達成することもできる。これは、リン酸カルシウム法、DEAEデキストラン法、電気穿孔法、陽イオン性または陰イオン性の脂質製剤(リポソーム)、およびプロトプラスト融合法を含む。これらの方法が利用できるものの、これらの多くは、より低効率である。
【0136】
変異されたEPM2B遺伝子機能を阻害するために、および治療薬のデザインの基礎として、アンチセンスに基づいたストラテジーを使用することができる。原理は、mRNAと相補アンチセンス種との間の細胞内ハイブリダイゼーションによって、遺伝子発現の配列特異的な抑制を達成することができるという仮説に基づく。RNAまたはDNAの有意鎖に高い特異性で結合するアンチセンス鎖ヌクレオチドを合成することが可能である。ハイブリッドRNA二重鎖の形成は、標的mRNAのプロセシング/輸送/翻訳、および/または安定性を妨げ得る。
【0137】
ハイブリダイゼーションは、アンチセンス効果を生じさせるために必要とされる。アンチセンス効果は、アンチセンス・オリゴヌクレオチドの使用、アンチセンスRNA、DNAの注入、およびアンチセンスRNA発現ベクターのトランスフェクションを含む種々のアプローチを使用して記載されている。
【0138】
治療的アンチセンス・ヌクレオチドは、オリゴヌクレオチドまたは発現されたヌクレオチドとして作製することができる。オリゴヌクレオチドは、通常15〜20核酸塩基の長さであるDNAの短い一本鎖である。発現されたヌクレオチドは、アデノウイルス、レトロウイルス、またはプラスミドベクターなどの、発現ベクターを使用して作製される。ベクターは、培養の細胞に、または患者に投与され、次いで、その細胞がアンチセンス・ヌクレオチドを作製する。発現ベクターは、アンチセンスRNAを産生するようにデザインすることができ、これは数ダースの塩基〜数千までで長さを変化させることができる。
【0139】
アンチセンス効果は、制御(センス)配列によって誘導することができる。表現型の変化の範囲は、非常に変わりやすい。アンチセンスによって誘導される表現型の効果は、生物学的なエンドポイント、タンパク質レベル、タンパク質活性化の測定、および標的mRNAレベルなどの基準の変化に基づく。
【0140】
D.薬学的組成物
EPM2Bおよび変異されたEPM2Bをコードする核酸、malinおよび変異されたmalinタンパク質、抗体、およびアンチセンス・オリゴヌクレオチド、並びにEPM2B/malinおよび/または変異されたEPM2B/malinを調整するその他の薬剤を含む上述した物質は、インビボでの投与に適した生物学的に適合性を有する形態で、被検者に投与するための薬学的組成物中に処方してもよい。「インビボでの投与に適した生物学的に適合性を有する形態」とは、治療的な効果が任意の有毒作用を上回って投与されるような物質の形態を意味する。物質は、ヒト、および動物を含む、生きている生物に投与してもよい。
【0141】
本発明の薬学的組成物の治療的に有効な量の投与は、所望の結果を達成するために必要な用量で、および期間で有効な量として定義される。たとえば、物質の治療的に有効な量は、個々の疾病状態、年齢、性、および重量、並びに個々に所望の反応を引き出すための物質の能力などの因子によって変更してもよい。投与法は、最適な治療反応を提供するように調整してもよい。たとえば、いくつかの分割用量を毎日投与してもよく、または用量は、治療状況の緊急性によって指示されて、比例して減らしていてもよい。
【0142】
活性物質は、注射(皮下の、静脈内の、その他)、経口投与、吸入、経皮の適用、または直腸の投与などによって、便利な方法で投与してもよい。投与の経路に応じて、活性物質は、酵素、酸、および化合物を不活性化するであろうその他の天然の条件の作用から化合物を保護する材料中に被覆されてもよい。活性物質が、たとえば、修飾されたEPM2B遺伝子をコードする核酸である場合、当技術分野において公知の技術を使用して送達してもよい。
【0143】
本明細書に記載されている組成物は、薬学的に許容される組成物の調製のためのそれ自体公知の方法によって調製することができ、これを、活性物質の有効な量が薬学的に許容される媒体との混合物中に組み合わされるように被検者に投与することができる。適切な媒体は、たとえばばRemington’s Pharmaceutical Sciences (Remington’s Pharmaceutical Sciences, Mack Publishing Company, Easton, Pa., USA 1985)またはHandbook of Pharmaceutical Additives (compiled by Michael and Irene Ash, Gower Publishing Limited, Aldershot, England (1995))に記載されている。これに基づいて、組成物は、排他的ではないものの、1つもしくは複数の薬学的に許容される媒体または希釈液と合わせた物質の溶液を含み、適切なpHを有する緩衝液に含まれてもよく、および/または生理液によって等浸透圧にしてもよい。この点に関しては、米国特許第5,843,456号を参照することができる。また、当業者に認識されるように、本明細書に記載されている物質の投与は、不活性ウイルスキャリアによるものであってもよい。
【0144】
E.実験モデル
また、本発明は、EPM2B遺伝子およびmalinタンパク質の機能を研究するための方法、並びに実験モデルを含む。EPM2B遺伝子を欠いた、もしくはmalin発現を部分的に欠いた細胞、組織、および非ヒト動物は、EPM2B遺伝子に特異的な欠失または変異を有する組換え発現ベクターを使用して開発してもよい。組換え発現ベクターを、相同組換えによってEPM2B遺伝子を不活性化または変化させるために使用して、これにより、EPM2B欠損細胞、組織、または動物を作製してもよい。
【0145】
ヌル対立遺伝子は、欠失突然変異によって胚幹性細胞などの細胞に作製してもよい。また、組換えEPM2B遺伝子を、EPM2Bを不活性化する挿入突然変異を含むように操作してもよい。次いで、このような構築物を、トランスフェクション、電気穿孔法、注入等の技術によって、胚幹性細胞などの細胞に導入してもよい。次いで、たとえばサザンブロッティング、ノーザンブロッティングによって、または本明細書に記載されている方法を使用してEPM2Bをアッセイすることによって、無傷のEPM2B遺伝子を欠いている細胞を同定してもよい。次いで、このような細胞を胚幹細胞に融合して、EPM2Bが欠損したトランスジェニック非ヒト動物を作製してもよい。突然変異の生殖系列伝達は、たとえば、インビトロで、8細胞胚などの初期胚で胚幹細胞を凝集すること;生じる胚盤胞をレシピエント女性へ移すこと、および;生じる凝集キメラの生殖系列伝達を発生させることによって達成してもよい。このような変異体動物を使用して、通常はEPM2B発現に依存的な、特異的な細胞群、発達パターン、およびインビボ・プロセスを定義してもよい。また、本発明は、EPM2B遺伝子の組織特異的ノックアウトの調製を含む。
【0146】
以下の非限定の実施例は、本発明の例示である:
【実施例】
【0147】
実施例1
本発明者らは、全ての利用できるデータベースを調べ、染色体6p22.3(図1)上の新たに定義された臨界領域において7つの注釈がついた遺伝子を見いだした。これらの予測される機能的特徴に基づいて、EPM2Bハプロタイプを有するLD患者のDNAシーケンシングを介した突然変異スクリーニングのため、それぞれの遺伝子を優先させたが、病原性変異体は同定されなかった。同時に、本発明者らの解析により、その他のより高等脊椎動物(ラット、マウス、およびイヌと、それぞれ75%、78%、87%のヌクレオチド同一性)におけるオーソロガスなユニット(タンパク質コード能力に相当)と広範な配列同一性を共有する、以前に特徴付けられていない、見かけ上単一エキソン(1188 bp)遺伝子を発見するに至った。また、EPM2Bと命名されたヒト遺伝子は、その5'末端に真核生物の翻訳開始部位のコンセンサス配列の提唱された特徴の全てを、およびその3'-末端に2つの推定ポリアデニル化シグナルを有した(図1)。さらに、ヒトおよびマウスにおける発現配列タグ(EST)およびcDNAデータは、単一エキソン・ユニットが本物の遺伝子であることを指示した。
【0148】
EPM2Bは、N末端の半分においてRING型のZnフィンガーとC末端方向に6つのNHL繰り返しドメインとを含む、発明者がmalin(フランス語の発作に関する疾患)と名付けた395アミノ酸(aa)タンパク質をコードすることが予測された(図1)。特に、RING-HC型のサイン配列(C-X2-C-X9-39-C-X1-3-H-X2-3-H-X2-C-X4-48-C-X2-C)(16、17)に対応した、亜鉛結合RINGフィンガー・モチーフ(C-X2-C-X16-C-X1-H-X2-C-X2-C-X14-C-X2-C)が、malinの残基26〜71間で同定された(0.0067のE値)。RINGフィンガーの存在は、E3ユビキチン・リガーゼ機能を予測させる(8、9、18)。E3ライゲーションは、E2から直接またはアダプタータンパク質を介して特異的基質にユビキチンを移行して、プロテアソーム系によってその除去を開始するユビキチン経路の最終的かつ特異的な工程である(8)。6つのNHLドメイン(10〜13)が、グリシンがリッチな約44残基モチーフおよび荷電残基のクラスターと共に発生源となった疎水性アミノ酸の存在に基づいて予測された(Pfamにより、0.011〜3.5の範囲のE値をもつNHLドメインについて6つの信頼される候補を検出した)(図1)。
【0149】
ノーザンブロット解析は、EPM2Bが脳の特定小領域を含む調べた全ての組織に(1.5kbおよび2.4kbのサイズの少なくとも2つの転写物として)存在することを示した(図3)。観察された転写物サイズは、予測されたATG開始部位と2つの異なるポリアデニル化シグナルとの間で予想される長さにほぼ一致する(図1)。さらに、発現プロフィールは、EPM2Aについて観察されるものと同様であり、両方とも、ラフォラ体が観察された全ての組織に存在した(2、19)。
【0150】
EPM2Bの完全なコード領域を、以前にEPM2Aに突然変異を有さないことが示された34人のLD発端者のコホートにおいてシーケンスした。この実施例において、フレームシフトを生じる8つの欠失および1つの挿入、7つのミスセンス、並びに1つのナンセンス変化を含む26ファミリーのEPM2Bにおいて、17個の異なるDNA配列変化が記載されている(表1)。これらの突然変異は、ホモ接合性(18)かつ複合ヘテロ接合性(8)の劣性状態の両方のファミリーにおいて見いだされた。最初の連鎖研究に使用した4血縁のF-Cファミリーは、全て、RINGフィンガー・ドメイン(図2b)の亜鉛結合能力に重要である7つの保存されたシステイン残基のうちの1つにシステインからセリンへの変化を生じるホモ接合性76T→A変化を有した。同定された最も共通する突然変異(7ファミリー)は、RINGフィンガー・ドメインにおいてプロリンからアラニンへの変化を生じるホモ接合性205C→G移行であった。EPM2B遺伝子座を絞り込む際に重要であったLD38324ファミリー(図2a)は、第5 NHLドメインにフレームシフト突然変異を生じるホモ接合性の2bp欠失(1048-1049delGA)を有した(図2b)。表1を参照し、ファミリーLD51において、発端者に由来するDNAは、入手不可能であったが、親は、両方とも468-469delAGのヘテロ接合性キャリアであり、これにより、LDの子供にホモ接合性のフレームシフト突然変異を生じることが予測される。検出される突然変異の全ては、推定RINGもしくはNHLモチーフに影響を及ぼすか、またはフレームシフトを生じるか、またはタンパク質の急激な構造変化(LD483は、260T→Cヌクレオチド変化を有し、ロイシンからプロリンへの変化を生じると考えられる)を生じさせることが予測される。4つのサイレントDNA配列をコードする変異体が同定された。これらの中で、3つのT312C(H104H)、G372C(G124G)、およびT1020C(G340G)は、それぞれ100個の対照染色体のうちの5つ、2つ、および1つに存在した。検出された最も共通する多型、C332T(P111L)(図1)は、100個の対照染色体のうちの42個で観察された。
【0151】
全体で、88%またはLDファミリーは、現在EPM2A(48%)およびEPM2B(40%)の突然変異によって説明することができる。これらの遺伝子のいずれにも検出可能な突然変異がない8ファミリーの観察は、さらなるLD遺伝子座位が存在し得ることを示唆する。
【0152】
成人ポリグルコサン小体疾患(APBD)を含む(20)、ポリグルコサン蓄積のあるその他の症状の中で、LDは、図4に示したように、軸索ではなくニューロン樹状突起における封入体の細胞内部位に関して独特である(21)。また、ERと形成性ポリグルコサン原繊維の物理的関連もLDに特異的である(3)。グリコーゲン分枝酵素の突然変異によって生じるAPBDにおいて(20、22、23)、ポリグルコサンは、サイズ、組成物、および数においてラフォラ体と見分けがつかないが、これらは、細胞体および軸索にのみ位置する(20)。APBDではなく、LDにおける発作表現型の存在は、LDのてんかんにおけるERに関連したポリグルコサンの樹状蓄積を暗示する。
【0153】
malinの細胞局在を調べるために、本発明者らは、エピトープタグを付けたEPM2B構築物をトランスフェクトし、これが実際にERに局在化し、培養細胞の核内においてわずかであることを見いだした(図5)。これらの結果は、それぞれ細胞質中のER(24-26)(図4b)および核において見いだされた、laforinのアイソフォームをコードするEPM2Aの2つの他の転写物(AおよびB)について観察される細胞局在と同じであった(27)。本発明者らのごく最近のデータは、転写物A特異的な突然変異の同定に基づいて、LDにおけるlaforinの細胞質型の機能喪失を暗示する(L.I. et al., 原稿投稿)。さらに、マウスのEPM2Aノックアウト(28)およびLD患者(1、2)の研究は、ER関連ポリグルコサン小体は、それぞれてんかんの発生に先行するか、または付随することを示した。
【0154】
したがって、簡単に説明すると、LDは、樹状突起においてポリグルコサンの不適当な排除(clearance)、およびその後の蓄積がニューロンのシナプス機能を妨げて、てんかん発生を生じることにより生じる。本発明者らは、今回、トランスジェニックLDマウスにおいて、laforinがポリグルコサン(およびグリコーゲンではない)と接触して、疾患遺伝子産物とLD病態との間の最初の物理的リンクを提供することを示した(E.M.C. et al., 準備中)。Laforinの唯一のその他の実験的に確証された機能は、二重特異性ホスファターゼのものであり(24、25)、これは、少なくとも1つのリンタンパク質中間体があり、中間体を介してこれが作用することを予測させる。可能性のある候補は、新たに発見されたEPM2A/P1 laforin相互作用タンパク質(26)、またはその他のさらにクローン化されたLD遺伝子、または任意のこれらの相互作用タンパク質の可能性がある。Malinは、そのNHLドメインを介した特異的なタンパク質-タンパク質相互作用、続くユビキチンを媒介した調節性標的の除去を経て関与して、laforinがラフォラ体およびてんかんに対してニューロンを保護するための重要な役割に寄与する可能性が高い。
【0155】
方法
試料
この研究に記載されている全ての患者は、形式的に、皮膚、骨格筋、肝臓、または脳の生検において特徴的なラフォラ体が存在することに基づいて、青年期発生進行性ミオクローヌスてんかんであると診断された。ホモ接合性マッピング、突然変異スクリーニング、または両方によって6q24におけるEPM2A遺伝子座の関与について、それぞれのLD個体およびこれらのファミリー・メンバー(入手できるならば)を調べた。
【0156】
遺伝子タイピングおよび突然変異スクリーニング
確立されたもの(D6S274、D6S285、D6S966、D6S1567、D6S1678、D6S1688、D6S1959)および新たなもの(BV012563、BV012730、BV012565、BV012566、BV012568)についての情報および増幅条件は、UniSTSおよびEntrez Nucleotidesデータベース(http://www.ncbi.nlm.nih.gov/)において見いだされる。DNA配列バリエーションは、PCR産物をシーケンシングすることによって検出した。EPM2Bをスクリーニングするために、重複断片を増幅する2セットのプライマー対を使用したEPM2B-1F:
とEPM2B-1R:
;EPM2B-2F:
とEPM2B-2R:
(図1を参照されたい)。PCRは、緩衝液[75mMのトリス-HCl(pH 8.8)、20mMの(NH4)2SO4、0.01% Tween 20、1.5mM MgCl2、1M Betaine、0.2mM dNTP、0.2μMのそれぞれのプライマー、2.5 UnitsのTaq Polymerase(MBI Fermentas)]中で50ngのDNAで行った。サイクル条件は、以下のとおりであった:94℃で3分間の最初の変性、続く94℃で30秒の変性、60℃で30秒のアニーリング、および72℃で30秒の伸長を35サイクル、72℃における10分間の最終伸長。PCR産物は、microCLEAN(Microzone Ltd.)を使用して精製した。3μl(100ng/μl)の精製したPCR産物をシーケンシングテンプレートとして使用した。全ての反応について、1μl(5pmol)のプライマー、1.5μlの5×シーケンシング緩衝液(Applied Biosystems)、1μlのBigDye Terminator v3.1、および7.5μlのH2Oを14μlの反応体積で使用した。Thermocycling(MJ Research, Inc.)条件は、96℃で30秒の変性;50℃で20秒のアニーリング;および60℃で4分の伸長;35サイクルであった。その後に全ての反応をマルチスクリーン-HVフィルタープレート(Millipore)を使用して精製し、ABI-3700を使用して解析した。LD患者において検出された全ての配列変異体は、50(100染色体)のランダムに選択したDNA試料のコレクションで調べた。
【0157】
遺伝子同定およびノーザンブロット
遺伝子注釈データおよびEST配列は、主にUniversity of California Santa Cruz(UCSC)ゲノム・ブラウザ(http://genome.ucsc.edu/)およびCelera Genomics(http://www.celera.com/)から得た。さらなる推定遺伝子は、Genescriptアルゴリズム(http://tcag.bioinfo.sickkids.on.ca/genescript/)およびヒトとマウス配列との間の多種VISTA(http://www-gsd.lbl.gov/vista/)整列を使用して注釈をつけた。RINGフィンガー・ドメインは、Pfam、Prosite、InterProScan、SMART、およびMotifScanによって予測した。NHLドメインは、PfamおよびInterProScanを使用して同定した。EPM2Bオルソログは、GenBank非重複性データベースに対するBLASTNおよびBLASTP解析を使用して同定した。配列分析によるmalinの細胞内局在性の予測は、PSORT IIを使用して行った。有意なシグナルペプチド配列(ER、ゴルジ複合体、リソソーム、および統合された必須原形質膜タンパク質の認識用)、ミトコンドリアのターゲティング配列、核局在化シグナル、およびペルオキシソームのターゲティング・シグナルは同定されなかった。human multiple-tissue blot Iおよびhuman brain blot II(Clontech)を、プライマー
および
を使用してEPM2Bのコード領域の557bp断片を増幅して作製された[32P ] dCTP標識されたプローブでプローブした。ハイブリダイゼーションおよび洗浄条件は、製造業者の説明書に従って行った。
【0158】
細胞内局在性および電子顕微鏡法
laforinの細胞質アイソフォームをコードするmycタグを付けたEPM2A転写物A発現構築物(pcDNA3mycEPM2A)が、記載されている(24)。mycタグを付けたEPM2B構築物は、同じ一般的なプロトコルを使用して作製した(pcDNA3mycEPM2B)。全長EPM2Bは、BamHI制限部位(下線を引いてある)と開始コドン(大文字)を含む(フォワード)プライマー
およびNotI部位(下線を引いてある)を含む(リバース)プライマー
を使用して、ゲノムDNAからPCRによって増幅した。この産物を哺乳類発現ベクターpcDNA3(Invitrogen)の対応する部位にクローン化した。次いで、5'KpnI-タグを付け、および3'BamHIタグを付けたプライマーで以前のmycを含むベクターから増幅した後に、インフレームでMycを導入した。pcDNA3mycEPM2AおよびpcDNA3mycEPM2B(2mg)をLipofectamine-Plus(Invitrogen)を使用してCos-7細胞にトランスフェクトし、5時間DMEM(Sigma-Aldrich)中で脂質-DNA複合体に曝露した。形質移入48時間後に、培養をPBS中で2回リンスし、アセトン:メタノール(1:1)混合物中で15分間-20℃で固定した。次いで、これらをmyc-laforinおよびERマーカーGRP94に対する抗体で染色した。培養を1時間(10%のBSA/PBS)ブロックし、室温で45分間抗Mycおよび抗ERと共にインキュベートした。スライドをPBSで洗浄し、ブロッキング溶液中の二次抗体(FITC標識されたヤギ抗マウス(1:400、緑色フィルターを介して検出可能);テキサスレッド標識されたロバ抗ヤギ(1:400、赤色フィルターを介して検出可能);Jackson Immuno Research Laboratories)と共にインキュベートした。マウンティング(Dako Anti-Fade)に続いて、これらを免疫蛍光光学顕微鏡によって解析した。電子顕微鏡検査のために、生検材料をLD患者から得られた冷却した汎用固定液に入れた。次いで、標準的なプロトコルを使用して、これを超微細構造的レベルで解析した。
【0159】
実施例2
実施例1に記載されている突然変異に加えて、本発明者らは、ラフォラ病と関連するEPM2Bにおいてさらなる4つの突然変異を見いだした。これらの突然変異は、実施例1と同じ様式で記載されていた。DNAは、実施例1に記載されているものと臨床的に類似したラフォラ病患者由来の血液から得た。EPM2Bにおける4つの突然変異は、以下のとおりである:1)位置606におけるヌクレオチドTの欠失;2)ヌクレオチド923におけるAからTへの変化;3)ヌクレオチド580におけるGからTへの変化;および4)ヌクレオチド199におけるGからTへの変化。後の突然変異は、スーダンおよびイタリアのファミリーにおいて見いだされた。さらなる突然変異は、表1の下段に一覧を記載してある。
【0160】
実施例3
ラフォラ病は、驚くべき頻度でイヌにおいても発見された。進行性ミオクローヌスてんかん(PME)は、英国において繁殖された人気のミニチュア・ワイアーヘアード・ダックスフンド(MWHD)において一般的である(〜5%)。本発明者らは、これらのイヌの臨床病理学的表現型を特徴づけ、これらがLDを有することを示した。本発明者らは、基礎になる疾患遺伝子座のマップを作製し、特異的疾患関連突然変異(EPM2B遺伝子において):いずれかの種における第1コーディング12塩基反復の伸長と、ヒトを越えた第1の病原性タンデム型繰り返し伸長とを同定した。本発明者らは、伸長突然変異が再発性であり、MWHD以外のてんかんのイヌにも影響を及ぼすことを示し、これがイヌ(およびその他の犬科の動物)に特有の配列変異により生じて、その種がLDにかかりやすいことを証明した。最後に、本発明者らは、キャリアおよび発症前動物において伸長対立遺伝子を検出し、MWHDおよび将来発症するイヌ集団に由来する疾患を根絶させるための試験を発明した。
【0161】
発病年令が6〜9歳である14匹の病気に冒されたMWHDを最初に評価した。ミオクローヌスは、頚部および肢筋の対称形の瞬間的な収縮からなり、イヌの後方突進を生じさせ;下顎が鳴るか、またはパチンと閉じられて、眼瞼が迅速にまばたきする。ミオクローヌスは、自発的に生じたが、突然の音、またはイヌの方へ転がるボールもしくは車に乗っている間の樹木を介してのストロボの光などの視覚刺激によって誘発することができる。ミオクローヌスは、ウサギを追っている間などのイヌが集中しているときはあまり可能性がなく、給餌時間で、もしくは訪問客があるなどの興奮または神経過敏であるとより可能性が高い。動物が急に座るかまたは倒れるアトニー性発作、並びに硬直、パドリング、発声、口から泡を吹くこと、排尿、および無意識を伴う全身性の発作は、共通していた。より老いたイヌは、失調症で、盲目で、これらの発作がより頻繁で、薬物に感受性が鈍かった。最終的には(年齢9〜12)、ミオクローヌスおよびドロップアタックにより、常にやむにやまれれずに安楽死させた。この臨床像は、基礎病理学と同様に(図8)、正確にヒトLD(2)を反復している。
【0162】
イヌがLDを有することを確認して、本発明者らは、最初にイヌEPM2A遺伝子をクローン化し、その関与を除外した。本発明者らは、次に疾患遺伝子座の局在を突き止めるために、ゲノム全体の走査を行った。本発明者らは、14匹の病気に冒されたイヌおよび4匹の冒されていない類縁において、全てのイヌ・ゲノムにわたって241個のイヌ特異的なマイクロサテライト・マーカー(31)の遺伝形質を決めた。冒されたイヌで最も多数がホモ接合性であり、かつ冒されていないイヌで最も多数がヘテロ接合性であった2つのマーカーは、REN94K23およびREN01G01であり、これらは小さな38Mbの染色体35(CFA35)に位置する。本発明者らは、図9aに示した、より広い血統に由来する多くのイヌを確かめて、CFA35上のさらなるマーカーの遺伝形質を決めた。組換え率θ=0.00における2.65の最大2ポイントLODスコア(two-point LOD score)がマーカーREN157J09について得られた。多点連鎖解析により、REN157J09を含む5つのマーカーを超えて3.38の最大LODスコアを生じ、イヌ連鎖研究についての3.2の有意な閾値を凌ぎ(32)(図9B)、これによりCFA35のこの領域には、MWHD LDが連鎖する。
【0163】
実施例1において論議したように、本発明者らは、ヒトEPM2B遺伝子座を染色体バンド6p22(15)にマップした。イヌCFA35は、その全体が、ヒト6p21.33-6p25.2(31)にシンテニーしており、したがって、ヒトEPM2B患者およびMWHD LDの場合において同じ遺伝子が変異される可能性があった。本発明者らは、最初にヒトEPM2Bをクローン化し(33)、その単一のエキソンを使用してRPCI-81イヌ細菌人工染色体(BAC)ライブラリーを探索するために、BAC 328A12を単離し、ヒトEPM2Bプライマーを使用するシーケンシングにより、イヌEPM2B(AY560905)の完全なオープンリーディングフレームを明らかにした。放射ハイブリッド・マッピングにより、EPM2BがマーカーREN157J09(LODスコア19.3)(MWHD LDが最大LODスコアで連鎖したものと同じマーカー(上記)(図9b))に連鎖した。全体として、これらのデータにより、EPM2BがMWHD疾患に対して生存できる候補であることを立証した。
【0164】
イヌEPM2B遺伝子のPCR増幅は、最初に病気に冒されたものでは失敗したが、病気に冒されていないMWHDにおいて遺伝子の5'の半分の領域全体ではうまくいった。この領域における正常な配列の調査により、2つの連続した同一の12塩基、および単一のヌクレオチドだけ異なった第3のコピーが明らかとなった(参照を容易にするために、完全な繰り返しコピーは、Dと名付け、および第3の不完全なコピーをTと名付けた)(図10a)。その他のシーケンスした種の対応する領域(図10a)は、繰り返しがなく、イヌ配列は、12ヌクレオチド、1回のDの繰り返しの長さだけ長い(図10a)。病気に冒された動物におけるこの領域全体を超えた増幅およびシーケンスのためのPCR条件の協奏的な改善は、最終的に成功し、MWHD LD突然変異を明らかにした:病気に冒された全ての動物は、それぞれの対立遺伝子にD配列の19〜26コピーをもつ12塩基反復の2対立遺伝子の伸長を有した(図9a、10b)。伸長は、インフレームであり、異常であるが、これらを含むmRNAが安定な場合には、タンパク質を作製することも可能かもしれない。
【0165】
本発明者らは、次に、定量的RT-PCR(補充方法)を使用して3匹の病気に冒されたイヌおよび2匹の対照からの骨格筋におけるEPM2B mRNAの量を比較することによって、mRNA安定性に対する伸長突然変異の効果を試験した。EPM2B mRNAレベルは、病気に冒された動物において900倍以上減少していた(図10c)。
【0166】
イヌEPM2Bを特徴付ける余分のD配列は、最初に、ドーベルマン・ピンシェル(Doberman Pinscher)起源のBACにおいて、および正常なMWHDにおいて検出された。本発明者らは、これが、いくつかの種類のイヌだけの特徴であり、少なくともMWHDの場合に、伸長およびLDにこれらがかかりやすいかどうかを考えた。本発明者らは、各々の128個の異なる種類から2匹の正常で無関係なイヌに由来するEPM2Bをシーケンスした。60パーセントの染色体は、3つの繰り返しを有し、40%は、2つの繰り返しを有した。3つの繰り返しをもつ対立遺伝子は、常に2つのDおよび1つのTから成っており、これらは1つのDと1つのTの2つの繰り返しをもつ。ほとんど全ての品種は、ホモまたはヘテロ接合性の状態で、両方の変異体の例を有した。異なる種類のうちの3種の4世代ファミリーにおいて試験した遺伝は、メンデルの法則によった。
【0167】
12塩基反復がイヌ種全体に存在することが示されたので、本発明者らは、その伸長が、その他の種類のイヌにおいても筋痙攣を生じさせるかどうかを考えた。本発明者らは、次の非MWHD PME症例を試験して臨床に提出し(バセット犬)、D繰り返しのホモ接合性の7倍の伸長(14コピー)を見いだした(図10b)。
【0168】
12塩基伸長突然変異の配列は、GおよびCヌクレオチドだけで構成される。正常な対立遺伝子の存在下では、伸長対立遺伝子のPCRが不可能であり、それゆえ、キャリアを決定することができなかった。キャリアを検出するために、本発明者らは、DNAを脱アミノし、メチル化されていないシトシンをウラシルに変換し、次いでPCR増幅し、これにより、変異誘発遺伝子の増幅、およびキャリアMWHD(図3b)の信頼できる検出が可能となった。また、脱アミノ化により、病気に冒された動物のPCR増幅および診断が非常に容易になった。
【0169】
上で使用した脱アミノ化法は、ヒト・シスタチンB遺伝子プロモーターに対して特異的な2〜3コピーの12塩基反復配列から生じ、かつシスタチンB発現(35)を妨げることによってEPM1(PMEのもう一つの形態)を生じさせる、現在まで唯一無二の12塩基反復の伸長突然変異の増幅のために開発された(34)。ヒトでは、2つのLD遺伝子(EPM2AおよびEPM2B)における60以上の突然変異は、PMEの最も一般的な原因ではない(http://projects.tcag.ca/lafora/)。その代わりに、単一の、何度も繰り返されるシスタチンBプロモーター伸長突然変異が、大部分のヒトPMEを引き起こす。本発明者らの知識の限りでは、イヌは、シスタチンB12の塩基反復配列を有しない可能性が高かったので、EPM1は、イヌにおいてこれまで記載されていなかった。LDは、一方ではイヌにおいて定期的に報告されている(バセット犬(36、37、38)、ミニアチュール(Miniature)(36)、およびスタンダード(39)プードル、ポインタ(40)、コルギス(Corgis)(41)、ビーグルズ(42、43、44)、およびMWHD(45、46))。これは、家畜化されたイヌに非常に多い集中的同系交配と相まって、EPM2B反復の反復性伸長事象によると思われる。その他の家畜化された動物では、LDは非常に珍しい:ウシ海綿状脳症(臨床的にLDに似ている)のための集団試験の間に同定された2頭のウシ(47)および1匹のネコ(48)。
【0170】
イヌ品種の障壁を越えた12塩基反復の存在(49)は、その起源がイヌに先んじていること、および、したがって、これが近縁種に存在するかもしれないことを示唆する。本発明者らは、食肉目における繰り返しの分布を決定し、これがイヌ科ファミリー(オオカミ、イヌ、キツネ、コヨーテ、およびジャッカル)に特異的であることを示した(図10d)。また、本発明者らは、2工程:D配列の出現、次いでその複製により生じる繰り返しの進化を識別することができた。肉食動物は、曉新世の〜6,000万年前(Ma)にネコ科およびイヌ上科に分かれた(50、51)。D配列は、現存するイヌ上科全体にわたって存在するが、ネコ科には存在しない(図10d)。したがって、これは、ネコ科(feline)-イヌ上科(canoid)がわかれた後に、その後にアルクト科(Arctoidae)とイヌ科に分岐する前の〜50Ma(図10d)に最初に現れた。アルクト科は、ユーラシア大陸上で全盛をきわめ、ここで、これらは、現存するクマ、スカンク、アライグマ、カワウソ、その他に進化した(50)。これらの種は、D配列の単一コピーを有する(図10d)。犬科の動物は、北アメリカ大陸で同等の生態学的地位を占有し(首尾よく、これは4,00万年間アルクト科(arctoid)侵略から守られる)(50)、〜10Ma〜今日までに、オオカミ、イヌ、キツネ、コヨーテ、およびジャッカル(51)に進化して、その全てが複製されたD配列(図10d)を有する。したがって、D配列の複製により、50Ma〜10Maのどこかで犬科の動物進化系統が生じた。生じるmalinの中間部分の可変性の4-アミノ酸伸長が(図10a)犬科の動物に有利なこのE3連鎖に固有性(property)を与えるかどうかは、分からないままである。
【0171】
てんかんは、イヌで最も一般的な神経疾患である。その有病率は、ヒトよりも少なくとも5倍高い(52)。本明細書において、本発明者らは、最初にイヌてんかん突然変異を発見した。これがどの程度イヌてんかんのその他の形態に寄与するかは、決定されないままである。
【0172】
英国では、現在、LDは、MWHD(毎年〜800匹のイヌが登録される品種)において広範囲にわたる。人気のキャリア雄親および繰り返し繁殖されたキャリアチャンピオンを介して、本疾患は、南アフリカおよび北アメリカまで広がった(データ示さず)。米国では、異なるダックスフンド・コート型が、同じ種類の変種と考えられ、異種交配されており、これが、ダックスフンドの全体にわたって疾患を蔓延させるかもしれない。その代わりに、本明細書に記載した研究から生じる突然変異検出試験により、繁殖を制御することによってこれを根絶できるはずである。一方、これらの動物は、愛されるペットであり、神経治療(neurologic care)を受けている。これらは、どの医療センターにおいても、ヒトLD患者よりもはるかに多い。これにより、本発明者らに対して、本疾病のための最高の治療を用いた重要な経験が与えられ、本発明者らは、これをヒト患者に適用している。
【0173】
本発明は、好ましい実施例であると現在みなされるものに関して記載したが、本発明は、開示された実施例に限定されないことは理解されるはずである。対照的に、本発明は、添付の特許請求の範囲の精神および範囲内に含まれる種々の改変および均等の処理を包含することが意図される。
【0174】
全ての刊行物、特許、および特許出願は、あたかも個々の刊行物、特許、または特許出願が特異的に、および個々に、その全体が参照として組み入れられることを示したのと同じ範囲で、これらの全体が本明細書に参照として組み入れられる。
【0175】
(表1)LD患者において同定されるEPM2B突然変異。
【0176】
明細書において言及した参照のための完全な引用
【技術分野】
【0001】
発明の分野
本発明は、ラフォラ病に関与する新規の遺伝子、EPM2B;その遺伝子によってコードされるタンパク質、malin;並びにラフォラ病を診断および治療する方法に関する。
【背景技術】
【0002】
発明の背景
ラフォラ病(LD、OMIM 254780)は、青年期に発症する進行性てんかんで最も一般的かつ重篤な形態である。増加性発作は、認知症に向かって知らない間に進行する認知減退、および通常発症の10年以内の死が平行して生じる(1、2)(非特許文献1、2)。細胞レベルで、LDは、ポリグルコサン(またはラフォラ体)と呼ばれるデンプン様グルコース重合体(4)(非特許文献4)の小胞体(ER)随伴蓄積(reticulum-associated accumulation)(3)(非特許文献3)によって特徴付けられる。遺伝的形質は、遺伝的異質性をもった常染色体劣性であるが、臨床症状は同質である(5)(非特許文献5)。本発明者らは、炭水化物結合ドメインをもつ二重特異性ホスファターゼ(Laforinと名づけた)をコードする染色体6q24上のEPM2A遺伝子における突然変異が、LDを生じさせることを以前に発見した(6、7(非特許文献6、7)および国際公開公報第00/05405号(特許文献1))。
【0003】
当技術分野において、ラフォラ病の診断および治療を援助するためには、ラフォラ病に関与するその他の遺伝子を同定する必要がある。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】国際公開公報第00/05405号
【非特許文献】
【0005】
【非特許文献1】Lafora, G.R., Gluck, B. Beitrag zur histopathologie der myoklonischen epilepsie. Z Ges Neurol Psychiat 6, 1-14 (1911)
【非特許文献2】Minassian, B.A. Progressive myoclonus epilepsy with polyglucosan bodies: Lafora disease. Adv Neurol 89, 199-210 (2002)
【非特許文献3】Collins, G.H., Cowden, R.R. & Nevis, A.H. Myoclonus epilepsy with Lafora bodies. An ultrastructural and cytochemical study. Arch Pathol 86, 239-54(1968)
【非特許文献4】Sakai, M., Austin, J., Witmer, F. & Trueb, L. Studies in myoclonus epilepsy (Lafora body form). II. Polyglucosans in the systemic deposits of myoclonus epilepsy and in corpora amylacea. Neurology 20, 160-76 (1970)
【非特許文献5】Minassian, B.A. et al. Genetic locus heterogeneity in Lafora's progressive myoclonus epilepsy. Ann Neurol 45, 262-5 (1999)
【非特許文献6】Minassian, B.A. et al. Mutations in a gene encoding a novel protein tyrosine phosphatase cause progressive myoclonus epilepsy. Nat Genet 20, 171-4 (1998)
【非特許文献7】Minassian, B.A. et al. Mutation spectrum and predicted function of laforin in Lafora's progressive myoclonus epilepsy. Neurology 55, 341-6 (2000)
【発明の概要】
【0006】
発明の要約
本発明者らは、染色体6p22.3上に新規遺伝子EPM2Bを位置的にクローン化した。これは、それぞれユビキチンを媒介した特異的基質の制御(8、9)およびタンパク質-タンパク質相互作用(10-13)のためにデザインされる複合体の特徴である、推定上のRINGフィンガー・ドメインと6つのNHLモチーフをもつタンパク質とをコードする。malinと名付けられたタンパク質産物に対して有害作用を生じさせることが予測されるEPM2Bの21個の異なったDNA配列バリエーションは、LDと同時分離されることが39ファミリーにおいて見いだされた。laforinおよびmalinは、両方ともERに局在し、これらがニューロンのポリグルコサン蓄積およびてんかんを妨げるのに関連した経路で作動することを示唆する。また、本発明者らは、イヌにおけるEPM2Bを単離し、およびシーケンスし、LDをもつイヌにおけるEPM2Bの突然変異を示した。
【0007】
したがって、本発明は、ラフォラ病と関連し、かつ配列番号:1(図6A)(ヒトEPM2B)に示した配列を有する単離された核酸分子を提供する。また、本発明は、ラフォラ病と関連し、配列番号:3(図7A)(イヌEPM2B)に示した配列を有する単離された核酸分子を提供する。
【0008】
好ましくは、精製され、および単離された核酸分子は、以下を含む:
(a)配列番号:1(図6A)および配列番号:3(図7A)に示したとおりの、TがUであってもよい核酸配列;
(b)(a)に対して相補的な核酸配列;
(c)(a)もしくは(b)の核酸配列に対して実質的に配列相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;または、
(e)ストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする核酸配列。
【0009】
また、本発明は、ラフォラ病と関連した、RING型のZnフィンガーと6つのNHL繰り返しドメインとを含む、単離されたタンパク質を含む。本発明の好ましい態様において、タンパク質は、配列番号:2(図6B)(ヒトEPM2B)に示したとおりのアミノ酸配列を有する。もう一つの態様において、タンパク質は、配列番号:4(図7B)(イヌEPM2B)に示したアミノ酸配列を有する。
【0010】
表1に示すように、本発明者らは、ラフォラ病と関連するヒトEPM2B遺伝子において21個の異なる突然変異を見いだした。したがって、本発明は、哺乳類、好ましくはヒト由来の試料においてEPM2B遺伝子の突然変異を検出することを含む、ラフォラ病を検出する方法を提供する。好ましい態様において、突然変異は、表1の一覧表に記載されたものである。
【0011】
また、本発明者らは、ラフォラ病をもつイヌにおいてEPM2B遺伝子の突然変異を発見した。特に、研究した全ての病気に冒されたイヌは、Dと呼ばれ、かつ配列番号:3に示したイヌEPM2B配列のヌクレオチド番号1001で始まる配列
を有する12塩基反復の2対立遺伝子の伸長を有した。したがって、本発明は、EPM2B(配列番号:3)のイヌ配列のヌクレオチド番号1001で始まる配列
の繰り返しを検出することを含む、犬科の動物においてラフォラ病を検出する方法をさらに提供する。
【0012】
その他の本発明の特徴および効果は、以下の詳細な説明から明らかであろう。しかし、詳細な説明および具体的な実施例は、本発明の好ましい態様を示す一方で、本発明の範囲内の種々の変更および修飾がこの詳細な説明から当業者には明らかとなると考えられるので、例示目的のみによって与えられることが理解されるべきである。
【図面の簡単な説明】
【0013】
ここで図面に関して記載する。
【図1】6p22.3上のEPM2B領域。以前の2.2Mbの臨界区間(critical interval)は、マイクロサテライト・マーカーD6S1567およびBV12568(15)によって区切られた。BV012568境界は、F-Cファミリー由来のLD個体におけるホモ接合性の減少に基づいていた(LD6;図2を参照されたい)。D6S1567テロメア境界は、第2の血縁F-Cファミリー(15)(LD27)の病気に冒されていない同種におけるBV012563とD6S1567との間に起こる組換えによって定義した。LD38324ファミリーにおけるマーカーのホモ接合性鎖の破壊により、動原体境界をD6S1688にさらに再定義することができた(図2)。本領域は、7つの以前に注釈をつけた遺伝子および単一の1188bpをコードするエキソンを含む新たに発見されたEPM2B(図6A)を含む。代表的なヒトおよびマウスcDNA配列を、同様に推定上のATG開始およびAATAAAポリアデニル化シグナルを示してある。ATG開始は、インフレームのストップ(位置-60において)に続き、対応するAUGは、予測されるORFの初めに存在する。開始(CGCGCCAUGG)を囲むヌクレオチド配列は、真核生物の翻訳開始部位のコンセンサス配列(GCCA/GCCAUGG)の提唱された特徴を有する(29)。EPM2Bは、検出可能な亜鉛結合RINGフィンガーおよび6つのNHL繰り返しドメイン(図6B)を含む42.3kDaの(395aa)タンパク質(malin)をコードすることが予測される。RINGおよびNHLの頭字語は、これらを有することが同定された最初のタンパク質の記述、すなわち、それぞれヒト(30)のReally Interesting New Gene 1およびNCL-1(11)/HT2A(10)/LIN-41(13)遺伝子から生じる。RING-および/またはNHLドメインは、細胞質もしくは核局在化(8、9、12、18)の一方または両方を有することができる種々のタンパク質に存在することに留意すべきである。Malinは、RINGおよびNHLモチーフのみを有することが記載された今までに唯一のタンパク質である(この組み合わせをもつその他のタンパク質があるものの、これらは、RING-B-Box-コイルドコイルドメインなどのその他の関連するモチーフも有する)。共通のC332T(P111L)多型部位は、星印:(*)によって示してある(表1を参照されたい)。Malinは、予測されるラット(419aa)、マウス(401aa)、およびイヌ(402aa)タンパク質とそれぞれ79%、80%、および85%の相同性を共有する。可変アミノ酸は、主にタンパク質のカルボキシ末端およびアミノ末端に位置し、RINGフィンガーまたはNHLドメイン以外に位置した。BVで始まるマイクロサテライト・マーカーは、この研究において作製した。
【図2】LDファミリーにおけるハプロタイプ解析によるEPM2Bの臨界区間の絞り込みおよびEPM2B遺伝子における突然変異。a、動原体境界は、ファミリーLD38324の両発端者におけるホモ接合性の減少に基づいて、D6S1688まで狭くなり、840kbまで重要領域を絞り込んだ。b、EPM2Bの配列解析により、ファミリーLD6における(並びにLD7、LD27、およびLD28F-Cファミリーにおける、表1)ホモ接合性の76T>A変化を同定する。ファミリーLD38324における病気に冒された個体は、第5NHLドメインにフレームシフト突然変異を生じるジヌクレオチド欠失(1048-1049delGA)に関してホモ接合性であることが見いだされた。
【図3】人体組織におけるEPM2BのRNAハイブリダイゼーション発現解析。a、複数の組織ブロット(Clontech)をEPM2Bのコード領域の557bp断片とハイブリダイズさせた。2.4kb および1.5kbのサイズの2つの転写物が、全ての組織において同定された。b、同じ大きさの転写物が、試験した脳の全ての領域由来の組織において見いだされた。
【図4】患者のLD32817(EPM2B突然変異98T>C)由来の脳生検材料の電子顕微鏡写真。A:軸索(多数の正常な神経伝達物質小胞に留意されたい)。S:シナプス。LB:完全に樹状突起を占めているポリグルコサンフィラメント(PG)の密度の高い蓄積で構成されるラフォラ体(大きな丸い構造)。バーは、500nmに相当する。
【図5】malinおよびlaforinタンパク質の細胞局在。a、Myc-タグを付けたmalin(構築物pcDNA3mycEPM2B)は、核周辺で、並びに核内で明瞭な網状パターンを形成する。GRP94小胞体(ER)特異的マーカーに対する抗体での同時染色により、malinとの共局在化が明らかになる。b、ER特異的マーカーGRP94とのlaforin(構築物pcDNA3mycEPM2A(24))の細胞質アイソフォームの共局在化。
【図6A】図6A(配列番号:1)は、EPM2Bのヒト核酸配列およびアミノ酸配列を提供する。
【図6B】図6B(配列番号:2)は、EPM2Bのヒト核酸配列およびアミノ酸配列を提供する。
【図7A】図7A(配列番号:3)は、EPM2Bのイヌ核酸配列およびアミノ酸配列を提供する。
【図7B】図7B(配列番号:4)は、EPM2Bのイヌ核酸配列およびアミノ酸配列を提供する。
【図8】ヒトLDと同一の特徴および細胞分布パターンをもつ病気に冒されたMWHD組織において形成する大量のデンプン様ポリグルコサン蓄積(ラフォラ体;LB)を示す(1,29,30)。病気に冒されたMWHD組織は、9つの筋肉および2つの肝臓生検、並びに剖検で得られた皮膚試料および4つの全脳を含んだ。対照組織は、LBを示さず(2匹の正常MWHDおよび3匹の非LDてんかんイヌを含む7匹の年齢が一致した動物)、示していない。組織化学的スライドは、ジアスターゼ処理に続いて過ヨウ素酸シッフで染色し、これにより、ポリグルコサンの特異的検出が可能になる(1)。無数のLBは、ヒトLD(1)と同様に、病気に冒されたMWHDの脳の全体にわたって存在し、ニューロンに位置し、グリアに位置せず、かつニューロンの核周部および樹状突起に位置し、軸索に位置しなかった。a:大脳皮質の分子層におけるLB(矢印)(×100拡大)。b:樹状LBの例(星印)。この樹状突起と共にシナプスを形成する複数の軸索(A)は、LBを含まない(バー=500nm)。c:肝臓において、LB(矢印)は、ヒトと同様に、糖新生(門脈周囲)肝細胞にのみ存在した(29)。(PV:門脈。D:門脈小導管。×300)。d、e:皮膚において、LB(矢印)は、ヒトと同様に(30)、汗腺に、特にアポクリン汗腺を囲んでいる筋上皮細胞に(d、×100)、およびメロクリン汗腺の胆管細胞に(e:フットパッド(foot pad)皮膚組織、×250)に存在した。f:骨格筋において、LBは、再びヒトLD(29)と同様に、膜(矢じり)によって囲まれた空胞内に存在した(バー=500nm)。
【図9】MWHD LDが、病気に冒されたイヌにおける伸長突然変異を含むEPM2B遺伝子にリンクすることを示す。a:系統およびハプロタイプ。黒の数は、下行順に染色体CFA35マイクロサテライトREN256I01、REN282I22、D02001、REN157J09、REN214H22、REN94K23、REN166C14、REN172L08における遺伝子型である。赤においては、EPM2B遺伝子の位置およびそれをコードする多形12塩基反復におけるD繰り返しの数である。注:伸長突然変異は、減数分裂時に不安定である(たとえば、イヌ29からイヌ32への伸長の伝達を参照されたい)。E:正確な繰り返し数をもつ伸長対立遺伝子は、決定されなかった。ボックス:突然変異に隣接する遺伝子型のホモ接合性。b、マルチポイントLODスコア解析。
【図10】食肉目のイヌ科ファミリーでのEPM2B12塩基多型の進化、非MWHDてんかんのイヌにおける伸長突然変異の存在、およびEPM2B mRNAに対する伸長の不活性化効果を示す。a:イヌEPM2Bの12塩基反復配列およびオルソログ。D:正確な繰り返し。T:不完全な繰り返し(星印)。赤いヌクレオチド:繰り返し領域におけるイヌ配列との相違。b:イヌにおいてミオクローヌス痙攣と関連するEPM2B伸長突然変異。W:野生型。M:病気に冒されたMWHD。B:病気に冒されたバセット犬(Basset Hound)。C:病気に冒されていないいくつかのキャリアMWHD。矢じり:正常な対立遺伝子。他の全てのバンド:伸長変異体対立遺伝子。脱アミノ化が正常配列の存在下においてさえも、伸長された対立遺伝子のPCR検出をどれほどよく改善するかに留意されたい。c:リアルタイム定量的RT-PCRを使用した、病気に冒されたMWHDおよび対照に由来する骨格筋におけるEPM2B量(Gapdhに対して規準化した)(SYBR Green検出;補充法)。d:12塩基多型の進化。D配列は、ネコ種に存在しない。これは、全てのイヌに存在する。アルクト科において単一コピーとして、および犬科の動物において1または2つの多形状態として;両対立遺伝子は、ヘテロ接合性個体(2つの配列の列)で示してある。示した全ての現存する肉食動物は、1,000万年前(Ma)の後に現れた。
【発明を実施するための形態】
【0014】
発明の詳細な説明
本発明者らによる以前の研究は、LD患者の〜70%が染色体6q24(6,7,14)上のEPM2A遺伝子に劣性突然変異を有することを示唆した。残りの患者において原因となる遺伝子を同定するために、本発明者らは、初めに、フレンチ-カナディアン(French-Canadian:F-C)単離体(isolate)(15)に由来する非EPM2A LDファミリー(LD6、LD7、LD27、LD28)のサブセットに対して連鎖およびホモ接合性マッピングを行った(表1)。このアプローチにより、染色体6p22.3上の2.2Mbの領域に第2のLD遺伝子座(EPM2B)が局在することを導いた(図1)。これらのF-Cファミリー由来の病気に冒された全ての個体は、全ての臨界区間にわたって珍しいハプロタイプに関してホモ接合性であることが見いだされた(図2a)。さらに遺伝子座を絞り込むために、病気に冒された複数の個体を有する5つのさらなるLDファミリーを、臨界領域を包含するDNA配列から開発することができたあらゆるマイクロサテライト・マーカーを使用して調べた。4つのファミリーにおいて、病気に冒された個体が、臨界区間にわたって全てのマーカーについてホモ接合性であった。しかし、1つのファミリー(LD38324)では、病気に冒された2つの個体のホモ接合性の鎖が、臨界領域に部分的に及び、D6S1688とD6S1567との間の840kbにEPM2B遺伝子座を減少することができた(図1および2a)。
【0015】
I. 本発明の核酸分子
先に述べたとおり、本発明は、ラフォラ病に関与する単離された核酸分子に関する。用語「単離された」は、組換えDNA技術によって産生したときに、細胞形質成分もしくは培地が実質的にない核酸、または化学物質前駆体、または化学的に合成したときのその他の化学物質をいう。用語「核酸」は、DNAおよびRNAを含むことが意図され、二本鎖または一本鎖のいずれであってもよい。
【0016】
上述したとおり、本発明は、ラフォラ病と関連した、N末端部分にRING型のZnフィンガーとC末端部分に6つのNHL繰り返しドメインとを含むタンパク質をコードする単離された核酸分子を提供する。単離された核酸分子は、好ましくはラフォラ病と関連したEPM2B遺伝子である。本発明の態様において、単離された核酸分子は、配列番号:1(図6A)または配列番号:3(図7A)に示すとおりの配列を有する。
【0017】
配列番号:1および3に示した核酸配列(または、それぞれ図6Aおよび7A)、並びに表1において、または実施例3において特定した変異配列は、本明細書において、ひとまとめにして「本発明の核酸分子」ということができる。配列番号:2および4(または、それぞれ図6Bおよび7B)に示したアミノ酸配列、並びに表1において特定した変異配列は、本明細書において、ひとまとめにして「本発明のタンパク質」ということができる。
【0018】
好ましくは、精製および単離された核酸分子は、以下を含む:
(a)配列番号:1(図6A)もしくは配列番号:3(図7A)に示すとおりの核酸配列であって、TがUであってもよい核酸配列;
(b)(a)に対して相補的な核酸配列;
(c)(a)もしくは(b)の核酸配列に対して実質的に配列相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;または、
(e)ストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする核酸配列。
【0019】
用語「実質的に配列相同性を有する配列」は、(a)もしくは(b)の配列から、わずかな、または重要でない配列バリエーションを有する、すなわち配列が実質的に同じ方法で機能し、かつラフォラ病を検出し、調査し、または治療するために使用することができる核酸配列を意味する。バリエーションは、局部的な突然変異または構造修飾に起因していてもよい。実質的に相同性を有する核酸配列は、配列番号:1または配列番号:3に示したとおりの核酸配列と少なくとも65%、より好ましくは少なくとも85%、および最も好ましくは90〜95%の同一性を有する核酸配列を含む。
【0020】
「配列同一性」は、当技術分野において公知の方法に従って算出することができる。配列同一性は、最も好ましくはBLASTバージョン2.1アドバンスド・サーチのアルゴリズムによって評価される。BLASTは、http://www.ncbi.nlm.nih.gov/BLASTでオンラインで利用できる一連のプログラムである。アドバンスドblastサーチ(http://www.ncbi.nlm.nih.gov/blast/blast.cgi?Jform=1)は、デフォルト・パラメーターにセットされている(すなわち、Matrix BLOSUM62;ギャップ・存在コスト(Gap existence cost)11;残基あたりのギャップ・コスト(gap cost)1;ラムダ比0.85デフォルト)。BLAST検索に対する参照は、以下のとおりである:
。
【0021】
用語「ハイブリダイズする配列」は、ストリンジェントなハイブリダイゼーション条件下で、(a)、(b)、(c)、または(d)の配列にハイブリダイズすることができる核酸配列を意味する。DNAハイブリダイゼーションを促進する適切な「ストリンジェントなハイブリダイゼーション条件」は、当業者に公知であるか、またはMolecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6に見いだされ得る。本明細書に使用される用語「ストリンジェントなハイブリダイゼーション条件」は、溶液中の2つの相補的核酸分子間の選択的なハイブリダイゼーションを促進する条件が選択されることを意味する。ハイブリダイゼーションは、全てまたは一部の核酸配列分子に対して生じてもよい。ハイブリダイズされる部分は、ポリペプチドをコードするポリヌクレオチド配列のうちの1つに関して少なくとも50%の長さである。この点に関して、核酸二重鎖またはハイブリッドの安定性は、ナトリウム含有緩衝液において、ナトリウムイオン濃度、標識された核酸のG/C含量、核酸プローブの長さ(I)、および温度の関数であるTm(Tm = 81.5℃ - 16.6(Log10 [Na+]) + 0.41(%(G+C) - 600/I)によって決定される。したがって、ハイブリッド安定性を決定する洗浄条件のパラメーターは、ナトリウムイオン濃度および温度である。公知の核酸分子と類似するが、同一ではない分子を同定するためには、1%のミスマッチにより、Tmの約1℃の減少を生じると想定してもよく、たとえば、95%を超える同一性を有する核酸分子が調査される場合には、最終洗浄が5℃まで減少されると考えられる。これらの考慮に基づいて、ストリンジェントなハイブリダイゼーション条件は、以下のように定義することができる:5×塩化ナトリウム/クエン酸ナトリウム(SSC)/5×デンハート液/1.0%のSDSにおけるTm(上記の方程式に基づく)-5℃におけるハイブリダイゼーション、続く0.2×SSC/0.1%のSDSの60℃における洗浄。
【0022】
用語「類似体である核酸配列」は、(a)、(b)、または(c)の配列と比較して、修飾された核酸配列であって、修飾が本明細書に記載されている配列の有用性を変更しない核酸配列を意味する。修飾された配列または類似体は、(a)、(b)、または(c)に示した配列にわたって改善された性質を有していてもよい。類似体を調製する修飾の一つの例は、配列番号:1または3に示した配列の天然に存在する塩基(すなわち、アデニン、グアニン、シトシン、またはチミジン)のうちの1つを、以下のものなどの修飾塩基と置換することである:キサンチン、ヒポキサンチン、2-アミノアデニン、6-メチル、2-プロピル、およびその他のアルキルアデニン、5-ハロウラシル、5-ハロシトシン、6-アザウラシル、6-アザシトシン、および6-アザチミン、プソイドウラシル、4-チオウラシル、8-ハロアデニン、8-アミノアデニン、8-チオールアデニン、8-チオールアルキルアデニン、8-ヒドロキシルアデニン、およびその他の8位置換されたアデニン、8-ハログアニン、8アミノグアニン、8-チオールグアニン、8-チオールアルキルグアニン、8-ヒドロキシルグアニン、およびその他の8位置換されたグアニン、その他のアザおよびデアザウラシル、チミジン、シトシン、アデニン、またはグアニン、5-トリフルオロメチルウラシル、並びに5-トリフルオロシトシン。
【0023】
修飾のもう一つの例は、配列番号:1または配列番号:3に示した核酸分子において、リン酸骨格における修飾された亜リン酸もしくは酸素へテロ原子、短鎖アルキルもしくはシクロアルキルの糖間結合、または短鎖ヘテロ原子もしくは複素環の糖間結合を含む。たとえば、核酸配列は、ホスホロチオアート、リン酸トリエステル、メチルホスホナート、およびホスホロジチオアートを含んでもよい。
【0024】
本発明の核酸分子の類似体のさらなる例は、DNA(またはRNA)のデオキシリボース(またはリボース)リン酸骨格が、ペプチドにおいて見られるものと同様のポリアミド骨格で置換されたペプチド核酸(PNA)である(P.E. Nielsen et al., Science 1991, 254, 1497)。PNA類似体により、酵素による分解に耐性であること、並びにインビボおよびインビトロでの生存が延長されることが示されている。また、PNA鎖とDNA鎖の間には電荷相反がないため、PNAは、相補的なDNA配列に対してより強く結合する。その他の核酸類似体は、重合体骨格、環状骨格、または非環式骨格を含むヌクレオチドを含んでもよい。たとえば、ヌクレオチドは、モルホリノ骨格構造(米国特許第5,034,506号)を有していてもよい。また、類似体は、レポーター基、核酸配列の薬物動態学的または薬力学的な性質を改善するための基などの基を含んでいてもよい。
【0025】
本発明は、後述するように、本発明のタンパク質の切り詰め、並びに本発明のタンパク質の類似体および相同体、並びにこれらの切り詰めをコードする核酸分子を含むことが認識される。本発明のcDNAに対応するmRNAの選択的スプライシングによって生じる本発明の核酸分子の異型も、本発明によって包含されることがさらに認識される。
【0026】
また、遺伝暗号の縮重により、本発明の核酸配列とは異なる配列を有する単離されおよび精製された核酸分子も、本発明の範囲内である。このような核酸は、機能的に同等のタンパク質をコードするが、遺伝暗号の縮重のために上述した配列とは配列が異なる。
【0027】
また、本発明は、配列番号:1(図6A)または配列番号:3(図7A)に示した核酸分子と比較して突然変異を有し、該突然変異がラフォラ病と関連する単離された核分子を含む。好ましい態様において、突然変異は、表1に示した突然変異または実施例3に記載したイヌ突然変異のうちの1つから選択される。
【0028】
EPM2B遺伝子由来の核酸分子またはこれらの変異形態は、配列番号:1(図6A)および配列番号:3(図7A)に示すとおりの核酸配列、または表1に示した変異された配列の全部または一部に基づいて標識された核酸プローブを調製すること、並びにこの標識された核酸プローブを使用して、適切なDNAライブラリー(たとえば、cDNAまたはゲノムDNAライブラリー)をスクリーニングすることによって単離することができる。cDNAまたはゲノムDNAライブラリーのスクリーニングによって単離された核酸は、標準的な技術によってシーケンスすることができる。
【0029】
また、本発明の核酸分子は、ポリメラーゼ連鎖反応法(PCR)方法およびcDNAまたはゲノムDNAを使用して、核酸を選択的に増幅することによって単離することができる。PCRに使用するために、配列番号:1(図6A)および配列番号:3(図7A)に示すとおりの核酸分子、または表1に示した変異された配列から合成オリゴヌクレオチドプライマーをデザインすることができる。核酸は、これらのオリゴヌクレオチドプライマーおよび標準的なPCR増幅技術を使用して、cDNAまたはゲノムDNAから増幅することができる。こうして増幅された核酸は、適切なベクターにクローン化することができ、DNA配列解析によって特徴付けることができる。cDNAは、種々の技術によって、たとえば、Chirgwin et al., Biochemistry, 18, 5294-5299 (1979)のチオシアン酸グアニジニウム(guanidinium-thiocyanate)抽出法を使用することによって、全細胞mRNAを単離することにより、mRNAから調製してもよいことが認識される。次いで、逆転写酵素(たとえば、Gibco/BRL, Bethesda, MDから入手可能なモロニーMLV逆転写酵素、またはSeikagaku America, Inc., St. Petersburg, FLから入手可能なAMV逆転写酵素)を使用して、mRNAからcDNAが合成される。
【0030】
RNAである本発明の単離された核酸分子は、適切なベクターに本発明の新規のタンパク質をコードするcDNAをクローン化することによって単離することができ、これにより、cDNAを転写してmalinタンパク質をコードするRNA分子を産生することができる。たとえば、cDNAは、ベクターのバクテリオファージプロモーター(たとえば、T7プロモーター)の下流にクローン化することができ、cDNAをインビトロにおいてT7ポリメラーゼで転写させ、生じるRNAを標準的な技術によって単離することができる。
【0031】
また、本発明の核酸分子は、標準的な技術を使用して化学的に合成されてもよい。ポリデオキシヌクレオチドを化学的に合成する種々の方法が公知であり、ペプチド合成のように、市販されているDNA合成装置での完全に自動化された固相合成を含む(たとえば、Itakuraら、米国特許第4,598,049号;Caruthersら、米国特許第4,458,066号;並びにItakura米国特許第4,401,796号および同第4,373,071号を参照されたい)。
【0032】
本発明の核酸分子の開始コドンおよび非翻訳配列は、PC/Gene(IntelliGenetics Inc., Calif.)などの本目的のためにデザインされた現在利用できるコンピュータ・ソフトウェアを使用して決定してもよい。調節エレメントは、従来の技術を使用して同定することができる。エレメントの機能は、エレメントに機能的に結合されたリポーター遺伝子を発現するようにこれらのエレメントを使用して確認することができる。これらの構築物は、標準的な手順を使用して培養細胞に導入してもよい。DNA中の調節エレメントを同定することに加えて、このような構築物はまた、当技術分野において公知の技術を使用して、エレメントと相互作用するタンパク質を同定するために使用してもよい。
【0033】
アンチセンス核酸分子を産生するため転写のための通常の提示に対して、本発明の核酸分子の配列を逆にしてもよい。好ましくは、アンチセンス配列は、開始コドンまたは保存されていない領域の前の領域を逆にすることによって構築される。アンチセンス核酸分子を産生するため転写のための通常の提示に対して、特に、本発明の核酸分子もしくはこれらの断片、好ましくは配列番号:1(図6A)および配列番号:3(図7A)に示した核酸配列、または表1に示した変異された配列に含まれる核酸配列を逆にしてもよい。
【0034】
本発明もしくはこれらの断片のアンチセンス核酸分子は、天然に存在するヌクレオチドを使用して化学的に合成してもよく、または分子の生物学的安定性を増大するように、もしくはmRNAまたは天然の遺伝子と形成される二重鎖の物理安定度を増大するようにデザインされた多様に修飾されたヌクレオチド、たとえばホスホロチオアート誘導体およびアクリジン置換されたヌクレオチドでもよい。アンチセンス配列は、アンチセンス配列が、高効率制御領域(その活性は、ベクターが導入される細胞型によって決定してもよい)の制御下で産生される組換えプラスミド、ファージミド、または弱毒ウイルスの形態で細胞に導入された発現ベクターを使用して生物学的に産生してもよい。
【0035】
また、本発明は、本発明の新規タンパク質、および選択タンパク質、または選択可能なマーカータンパク質を含む融合タンパク質をコードする核酸を提供する(下記を参照されたい)。
【0036】
II. 本発明の新規のタンパク質
本発明は、本発明の核酸分子によってコードされる単離されたタンパク質をさらに含む。本発明の状況の範囲内で、本発明のタンパク質は、生物活性を保持する、種々のタンパク質一次構造形態を含んでいてもよい。
【0037】
広く明言したとおり、本発明は、ラフォラ病と関連した、N末端側半分におけるRING型のZnフィンガーとC末端の方向に6つのNHL繰り返しドメインを含む単離されたタンパク質を提供する。好ましくは、亜鉛結合RINGフィンガー・モチーフ(C-X2-C-X16-C-X1-H-X2-C-X2-C-X14-C-X2-C)は、図6Bに示したmalinタンパク質の残基26〜71の間に位置する。RINGフィンガーの存在は、E3ユビキチン・リガーゼ機能を予測する。したがって、好ましい態様において、タンパク質は、ユビキチン・リガーゼ機能を有する。
【0038】
本発明の特定の態様において、タンパク質は、配列番号:2(図6B)(ヒトEPM2B)に示すとおりのアミノ酸配列を有する。もう一つの態様において、タンパク質は、配列番号:4(図7B)(イヌEPM2B)に示したアミノ酸配列を有する。
【0039】
全長アミノ酸配列に加えて、本発明のタンパク質は、本明細書に記載したとおりの、タンパク質の切り詰め、並びにタンパク質の類似体および相同体、並びにこれらの切り詰めを含む。切断されたタンパク質は、少なくとも15アミノ酸残基のペプチドを含んでいてもよい。
【0040】
本明細書に記載されている、配列番号:2(図6B)もしくは配列番号:4(図7B)に示したアミノ酸配列を有するタンパク質の類似体、および/またはこれらの切り詰めは、1つもしくは複数のアミノ酸置換、挿入、および/または欠失を含むアミノ酸配列を含んでもよいが、これらに限定されない。アミノ酸置換は、保存的または非保存的な性質であってもよい。保存的アミノ酸置換は、同様の電荷、大きさ、および/または疎水性の特徴のアミノ酸で、本発明のタンパク質の1つもしくは複数のアミノ酸を置換することを含む。保存的置換だけが作製されるとき、生じる類似体は、機能的に同等物であるはずである。非保存的置換は、異なる電荷、大きさ、および/または疎水性の特徴を有する1つもしくは複数のアミノ酸でアミノ酸配列の1つもしくは複数のアミノ酸を置換することを含む。
【0041】
1つまたは複数のアミノ酸挿入が、配列番号:2(図6B)または配列番号:4(図7B)に示したアミノ酸配列に導入されてもよい。アミノ酸挿入は、単一のアミノ酸残基または長さが2〜15アミノ酸の範囲の一連のアミノ酸からなってもよい。たとえば、アミノ酸の挿入は、タンパク質がもはや活性ではないように、標的配列を破壊するために使用されてもよい。この手順は、インビボで本発明のタンパク質の活性を阻害するために使用されてもよい。
【0042】
欠失は、配列番号:2(図6B)または配列番号:4(図7B)に示したアミノ酸配列からの1つもしくは複数のアミノ酸または離れた部分の除去からなっていてもよい。欠失されるアミノ酸は、隣接していても、または隣接していなくてもよい。欠失突然変異で生じる類似体の下限の長さは、約10アミノ酸、好ましくは100アミノ酸である。
【0043】
本発明のタンパク質の類似体は、タンパク質をコードするヌクレオチド配列に突然変異を導入することによって調製してもよい。本発明のタンパク質の類似体の発現のために構築されるヌクレオチド配列の突然変異は、コード配列のリーディングフレームを保存しなければならない。さらにまた、突然変異は、好ましくは、受容体mRNAの翻訳に悪影響を与え得る、ハイブリダイズして、ループまたはヘアピンなどの二次mRNA構造を生じさせ得る相補領域を生じない。
【0044】
突然変異は、制限部位に隣接して変異配列を含むオリゴヌクレオチドを合成して、天然の配列の断片に対するライゲーションを可能にすることによって、特定の遺伝子座に導入されてもよい。ライゲーションに続いて、生じる再構築された配列は、所望のアミノ酸の挿入、置換、または欠失を有する類似体をコードする。
【0045】
または、必要とされる置換、欠失、または挿入にしたがって変更された特定のコドンを有する変更された遺伝子を提供するために、オリゴヌクレオチド部位特異的突然変異法を使用してもよい。また、本発明のタンパク質の欠失または切り詰めは、所望の欠失に隣接する制限エンドヌクレアーゼ部位を利用することによって構築されてもよい。制限後に、オーバーハングを埋めて、DNAを結合してもよい。上述した変更を作製する典型的な方法は、Sambrook et al(Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, 1989)によって開示されている。
【0046】
また、本発明のタンパク質は、本明細書に記載されているように、配列番号:2(図6B)もしくは配列番号:4(図7B)に示したアミノ酸配列の相同体、および/またはこれらの切り詰めを含む。このような相同体は、本発明のタンパク質を得るために使用されるプローブとストリンジェントなハイブリダイゼーション条件(本明細書のストリンジェントなハイブリダイゼーション条件の考察を参照されたい)下でハイブリダイズされるアミノ酸配列から構成されるアミノ酸配列のタンパク質である。好ましくは、本発明のタンパク質の相同体は、タンパク質の特徴であるチロシンホスファターゼ領域を有する。
【0047】
相同的タンパク質は、配列番号:2(図6B)または配列番号:4(図7B)に示すとおりのアミノ酸配列と少なくとも70%、好ましくは80〜90%の同一性を有するアミノ酸配列を有すタンパク質を含む。配列同一性は、本明細書に定義したとおりである。
【0048】
また、本発明は、本発明のタンパク質のアイソフォームを想定する。アイソフォームは、本発明のタンパク質と同じアミノ酸の数および種を含むが、異なる分子構造を有する。本発明によって想定されるアイソフォームは、本明細書に記載されているように、本発明のタンパク質と同じ性質を有するものである。
【0049】
本発明は、配列番号:2(図6B)または配列番号:4(図7B)に示したアミノ酸配列と比較して、突然変異を有し、該突然変異は、ラフォラ病と関連する単離されたタンパク質を含む。好ましい態様において、突然変異は、表1に示した突然変異または実施例3に記載されているイヌ突然変異のうちの1つから選択される。
【0050】
また、本発明は、融合タンパク質を産生するために、選択されたタンパク質、または選択可能なマーカータンパク質(下記を参照されたい)と結合した本発明のタンパク質を含む。加えて、本発明のタンパク質の免疫原性部分も本発明の範囲内である。
【0051】
本発明のタンパク質(切り詰め、類似体、その他を含む)は、組換えDNA法を使用して調製してもよい。したがって、本発明のタンパク質をコードする配列を有する本発明の核酸分子を、タンパク質の良好な発現を保証する適切な発現ベクターに公知の方法で組み込んでもよい。可能な発現ベクターは、ベクターが使用される宿主細胞と適合性を持つ限り、コスミド、プラスミド、または修飾されたウイルス(たとえば、複製欠損のレトロウイルス、アデノウイルス、およびアデノ随伴ウイルス)を含むが、これらに限定されない。「宿主細胞のトランスフォーメーションのために適切な」発現ベクターは、発現ベクターが、本発明の核酸分子および制御配列を含み、発現のために使用される宿主細胞を基礎として選択され、これらが機能的に核酸分子に結合されていることを意味する。「機能的に結合される」とは、核酸が核酸を発現できる方法で制御配列に結合されことを意味することが意図される。
【0052】
したがって、本発明は、本発明の核酸分子、またはこれらの断片、並びに挿入されたタンパク質配列の転写および翻訳に必要な制御配列を含む本発明の組換え発現ベクターを想定する。適切な制御配列は、細菌、真菌、またはウイルスの遺伝子を含む種々の供与源に由来してもよい(たとえば、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990)に記載されている制御配列を参照されたい)。適切な制御配列の選択は、選択される宿主細胞に依存的であり、当技術分野の当業者によって容易に達成されるであろう。このような制御配列の例は、転写プロモーターおよびエンハンサーまたはRNAポリメラーゼ結合配列、翻訳開始シグナルを含むリボソーム結合配列を含む。その上、選択される宿主細胞および使用されるベクターに応じて、複製開始点、さらなるDNA制限部位、エンハンサー、および転写誘導能を与える配列などのその他の配列を発現ベクターに組み込んでもよい。また、必要な制御配列が、天然のタンパク質および/またはその隣接領域によって供給されてもよいことが認識されるであろう。
【0053】
本発明は、アンチセンス方向で発現ベクターにクローン化された本発明のDNA核酸分子を含む組換え発現ベクターをさらに提供する。すなわち、DNA分子は、DNA分子の転写によって、配列番号:1または配列番号:3に示すとおりのヌクレオチドを含むヌクレオチド配列に対してアンチセンスであるRNA分子を発現することができる方法で、制御配列に機能的に結合される。アンチセンス核酸に機能的に結合された、アンチセンスRNA分子を持続的に発現させる制御配列を選択することができる。
【0054】
また、本発明の組換え発現ベクターは、本発明の組換え分子を形質転換またはトランスフェクトした宿主細胞の選択を容易にする選択可能なマーカー遺伝子を含んでもよい。選択可能なマーカー遺伝子の例は、特定の薬物に対する耐性を与える、G418およびハイグロマイシンなどのタンパク質、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼをコードする遺伝子である。選択可能なマーカー遺伝子の転写は、β-ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、またはホタルルシフェラーゼなどの選択可能なマーカータンパク質の濃度の変化によってモニターされる。選択可能なマーカー遺伝子が、ネオマイシン耐性などの抗生物質耐性を与えるタンパク質をコードする場合、形質転換体細胞は、G418によって選択することができる。選択可能なマーカー遺伝子を組み込んだ細胞は、生存すると考えられるが、その他の細胞は死滅する。これにより、本発明の組換え発現ベクターの発現の視覚化およびアッセイを可能にし、特に発現および表現型に対する突然変異の効果を決定することが可能となる。選択可能なマーカーは、関心対象の核酸から分離したベクターに導入することができることが認識される。
【0055】
また、組換え発現ベクターは、組換えタンパク質の発現の増大;組換えタンパク質の溶解性の増大;およびアフィニティー精製においてリガンドとして作用することによって標的組換えタンパク質の精製の援助をもたらす融合部分をコードする遺伝子を含んでいてもよい。たとえば、タンパク質分解切断部位を標的組換えタンパク質に付加して、融合タンパク質の精製後に融合部分から組換えタンパク質を分離させてもよい。
【0056】
組換え発現ベクターを宿主細胞に導入して、形質転換された宿主細胞を作製することができる。用語「形質転換された宿主細胞」は、本発明の組換え発現ベクターで形質転換またはトランスフェクトされた原核細胞および真核細胞を含むことが意図される。用語「で形質転換され」、「をトランスフェクトされ」「形質転換」、および「トランスフェクション」は、当技術分野において公知の多くの可能な技術のうちの1つによって、細胞に核酸(たとえば、ベクター)を導入することを含むことが意図される。たとえば、原核細胞は、電気穿孔法または塩化カルシウムを媒介したトランスフォーメーションによって核酸で形質転換することができる。核酸は、リン酸カルシウムもしくは塩化カルシウム共沈、DEAE-デキストランを媒介したトランスフェクション、リポフェクチン、電気穿孔法、または微量注入などの従来の技術を介して哺乳類細胞に導入することができる。宿主細胞を形質転換し、トランスフェクトするための適切な方法は、Sambrook et al.(Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory press (1989))、およびその他の研究教科書において見いだすことができる。
【0057】
適切な宿主細胞は、多種多様な原核生物および真核生物の宿主細胞を含む。たとえば、本発明のタンパク質は、大腸菌(E. coli)などの細菌細胞、昆虫細胞(バキュロウイルスを使用する)、酵母細胞、または哺乳類細胞に発現されてもよい。その他の適切な宿主細胞は、Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1991)において見いだすことができる。
【0058】
また、本発明のタンパク質は、固相合成などのタンパク質の化学において周知の技術を使用して、化学合成(Merrifield, 1964, J. Am. Chem. Assoc. 85:2149-2154)、または同種溶液における合成(Houbenweyl, 1987, Methods of Organic Chemistry, ed. E. Wansch, Vol.15 I and II, Thieme, Stuttgart)によって調製してもよい。
【0059】
III. 適用
本発明は、診断および治療的適用における、EPM2B核酸配列およびタンパク質およびこれらのモジュレーターの使用と同様に、抗体およびアンチセンス・オリゴヌクレオチドの調製、EPM2Bおよびこれらの変異された形態を研究するための実験系の調製、EPM26発現および/または活性を調整する物質の単離を含む(しかし、これらに限定されない)、本発明の核酸分子およびタンパク質の全ての使用を含む。いくつかの使用をさらに以下に記載してある。
【0060】
A.診断適用
前述のように、本発明者らは、遺伝子EPM2Bが、ラフォラ病の人々および犬科の動物において変異されていると断定した。結果として、本発明は、EPM2B遺伝子またはタンパク質の突然変異を検出することによってラフォラ病を検出する方法も含む。
【0061】
用語「突然変異」は、野生型配列と比較して、EPM2Bの核酸またはタンパク質配列における任意の変化または相違を意味する。突然変異は、ナンセンス突然変異、ミスセンス突然変異、フレームシフト突然変異、再編成突然変異、挿入突然変異、および欠失突然変異を含むが、これらに限定されない。
【0062】
試料は、体液(血液、尿、脳脊髄液、涙、唾液など)、組織、組織抽出物、細胞、および細胞抽出物を含むが、これらに限定されないEPM2B遺伝子またはタンパク質を含む任意の試料であることができる。
【0063】
(i)核酸配列における突然変異の検出
一つの態様において、本発明は、動物、好ましくは哺乳類、より好ましくは、ヒトまたは犬科の動物から得られる試料中のEPM2B遺伝子における突然変異を検出することを含む、ラフォラ病を検出するための方法を提供する。
【0064】
表1は、ラフォラ病患者のヒトEPM2Bにおいて見いだされるいくつかの突然変異を要約する。今日まで、フレームシフトを生じる8つの欠失および1つの挿入、8つのミスセンス、並びに1つのナンセンス変化を含む39ファミリーにおいてEPM2Bについて、21個の異なるDNA配列変化が見いだされた。各々の突然変異のために、スクリーニングアッセイ法を開発することができる。
【0065】
7つのファミリーにおいて同定された最も一般的な突然変異は、RINGフィンガー・ドメインにおいてプロリンからアラニンへの変化を生じるホモ接合性の205C→G移行である。したがって、一つの態様において、本発明は、EPM2B遺伝子(配列番号:1)の位置205におけるC→G突然変異を検出することを含む、ラフォラ病を検出する方法を提供する。
【0066】
本来の連鎖研究において使用した4血縁のF-Cファミリーで観察されるもう一つの突然変異は、全て、RINGフィンガー・ドメインの亜鉛結合能力に重要である7つの保存されたシステイン残基のうちの1つにシステインからセリンへの変化を生じるホモ接合性76T→A変化を有した。したがって、もう一つの態様において、本発明は、EPM2B遺伝子(配列番号:1)の位置76におけるT→A突然変異を検出することを含む、ラフォラ病を検出する方法を提供する。
【0067】
もう一つの突然変異は、LD38324ファミリーにおいて観察され、これはEPM2B遺伝子座(図2a)を絞り込む際に重要であって、第5NHL-ドメイン(図2b)においてフレームシフト突然変異を生じるホモ接合性2-bp欠失(1048-1049delGA)であった。したがって、さらなる態様において、本発明は、EPM2B遺伝子(配列番号:1)の位置1048および1049におけるGAの欠失を検出することを含むラフォラ病を検出する方法を提供する。
【0068】
実施例3に記載したように、発明者は、ラフォラ病のイヌにおけるEPM2B遺伝子の突然変異も発見した。特に、研究された病気に冒された全てのイヌは、Dと呼ばれ、かつ配列番号:3に示したイヌEPM2B配列のヌクレオチド番号1001で始まる配列
を有する12塩基反復の2対立遺伝子の伸長を有した。本発明者らは、この12ヌクレオチド繰り返しが、イヌ、オオカミ、キツネ、コヨーテ、およびジャッカルを含む犬科の動物スーパーファミリーに特異的であることを示し、この繰り返しは、イヌを大量の配列伸長にかかりやすくする素因になり、これがEPM2B遺伝子に有害であり、ラフォラ病を生じさせることを示した。本発明者らは、したがって、イヌを含む犬科の動物が、ラフォラてんかんにかかりやすいということを発見した。したがって、本発明は、EPM2B(配列番号:3)のイヌ配列におけるヌクレオチド番号1001で始まる配列
の繰り返しを検出することを含む、犬科の動物においてラフォラ病を検出する方法をさらに提供する。一つの態様において、本方法は、配列番号:5において、少なくとも3つの繰り返し、好ましくは少なくとも10の繰り返し、より好ましくは約14〜約26の繰り返しを検出することを含む。
【0069】
当業者であれば、実施例において論議したものに加え、および含む、多くの適切な方法をEPM2B遺伝子の突然変異を検出するために使用することができることを認識するであろう。使用することができる検出方法は、ポリメラーゼ連鎖反応法(PCR)、逆転写酵素PCR、直接シーケンシング電気泳動移動度、核酸ハイブリダイゼーション、蛍光インサイチュー・ハイブリダイゼーション、変性高速液体クロマトグラフィー、DNAチップ技術、および質量分析を含むが、これらに限定されない。一つの例において、試料中のラフォラ病遺伝子由来の核酸を単離するために、本発明の核酸配列からヌクレオチド・プローブを調製することができる。加えて、本明細書に記載されている核酸プローブを使用することもできる。ヌクレオチド・プローブは、32P、3H、14Cなどの適切なシグナルを提供し、十分な半減期を有する放射性の標識などの、検出可能な物質で標識されてもよい。使用してもよいその他の検出可能なマーカーは、特異的に標識された抗体、蛍光化合物、酵素、標識された抗原に特異的な抗体、および化学発光化合物によって認識される抗原を含む。適切な標識は、検出されるヌクレオチドに対するプローブのハイブリダイゼーションおよび結合の割合、並びにハイブリダイゼーションに利用できるヌクレオチドの量を考慮して選択してもよい。
【0070】
したがって、本発明は、試料を、核酸分子にハイブリダイズし、かつ検出可能なマーカーで標識された1つまたは複数のヌクレオチド・プローブと、ハイブリダイゼーション条件下で接触させる工程、および試料中の核酸分子とヌクレオチド・プローブとの間のハイブリダイゼーションの程度を決定する工程を含む、試料中のEPM2B遺伝子由来の核酸分子の存在を検出する方法にも関する。
【0071】
本発明の方法に使用してもよいハイブリダイゼーション条件は、当技術分野において公知であり、たとえばSambrook J, Fritch EF, Maniatis T. In: Molecular Cloning, A Laboratory Manual,1989. (Nolan C, Ed.), Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYに記載されている。ハイブリダイゼーション産物は、当技術分野において公知の技術を使用してアッセイしてもよい。ヌクレオチド・プローブは、本明細書に記載したように、検出可能なマーカーで標識してもよく、ハイブリダイゼーション産物は、検出可能なマーカーによって生じる検出可能なマーカーまたは検出可能な変化を検出することによってアッセイしてもよい。
【0072】
DNAプローブと試料をハイブリダイズさせる前に、試料中の核酸配列を増幅するために、EPM2B遺伝子に隣接するプライマーで試料を処理することができる。使用するプライマーは、本出願に記載されているものであってもよい。加えて、また本明細書において提供されるEPM2B遺伝子の配列により、新たなヌクレオチド配列の同定および単離、または合成が可能となり、たとえば下記にさらに詳細に論議したポリメラーゼ連鎖反応法(PCR)において本発明の核酸分子を増幅するためのプライマーとして使用してもよい。プライマーは、その他の種のゲノムDNAを増幅するために使用してもよい。PCR増幅された配列を、様々な種の遺伝子間の関係を決定するために調べることができる。
【0073】
PCRに使用するためのプライマーの長さおよび塩基は、これらが所望の配列の異なる鎖に対してハイブリダイズするように、および配列に沿った相対的位置で、テンプレートから分離するとき1つのプライマーから合成された伸長産物が、定義された長さの核酸へのその他のプライマーの伸長のテンプレートとして役立つことができるように選択される。本発明に使用してもよいプライマーは、オリゴヌクレオチド、すなわち本発明の核酸分子の2つ以上のデオキシリボヌクレオチドを含む分子であり、これらは、精製された制限エンドヌクレアーゼ消化物として天然に存在するか、またはたとえば、リン酸トリエステルおよびリン酸ジエステル法(Good et al Nucl. Acid Res 4:2157, 1977を参照されたい)または自動技術(たとえば、Conolly, B. A. Nucleic Acids Res. 15: 15 (7): 3131, 1987を参照されたい)などの当技術分野において公知の技術を使用して合成的に産生される。プライマーは、本発明のDNA配列に対して相補的であるプライマー伸長産物の合成を可能にする条件下に、すなわちヌクレオチド基質、DNAポリメラーゼなどの重合のための薬剤の存在下において、並びに適切な温度およびpHに配置されたときに合成の開始位置として作用することができる。好ましくは、プライマーは、プライマーのその他のコピーと塩基対形成することによって二次構造を形成しない配列か、またはヘアピン立体配置を形成する配列である。プライマーは、好ましくは約7〜25の間のヌクレオチドを含む。
【0074】
プライマーは、増幅産物の検出を可能にする検出可能なマーカーで標識してもよい。適切な検出可能なマーカーは、32P、35S、125I、および3Hなどの放射性のマーカー、化学発光マーカー、好ましくはルミノール、並びに蛍光マーカー、好ましくは塩化ダンシル、フルオルセイン-5-イソチオシアナート、および4-フルオロ-7-ニトロベンズ-2-axa-1,3ジアゾールなどの発光のマーカー、西洋ワサビペルオキシダーゼ、アルカリホスファターゼ、β-ガラクトシダーゼ、アセチルコリンエステラーゼ、またはビオチンなどの酵素マーカーである。
【0075】
プライマーは、プライマーの十分な量が本発明の核酸分子またはこれらのオリゴヌクレオチド断片に対して相補的であり、増幅される配列を含むことを条件として、非相補的な配列を含んでもよいことが認識される。また、制限部位リンカーをプライマーに組み入れて、適切な制限酵素での増幅産物の消化を可能にして、増幅産物のクローニングおよびシーケンシングを容易にしてもよい。
【0076】
本発明のある態様において、増幅配列を形成することができる条件下において、ポリメラーゼ連鎖反応法において核酸分子または予め定められたこれらのオリゴヌクレオチド断片を増幅することができるプライマーで試料を処理して増幅配列を形成する工程、および増幅配列をアッセイする工程を含む、本発明の核酸分子の存在を決定する方法が提供される。
【0077】
ポリメラーゼ連鎖反応法は、一般にInnis et al, Academic Press, 1990 in Mullis et al., 米国特許第4,863,195号およびMullis, 米国特許第4,683,202号(これらは、参照として本明細書に組み入れられる)に記載されている、標的核酸配列を増幅するためのプロセスをいう。核酸テンプレートを増幅するための条件はM.A. Innis and D.H. Gelfand, PCR Protocols, A Guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989(これらも参照として本明細書に組み入れられる)に記載されている。
【0078】
増幅産物は、当技術分野において公知の技術を使用して、単離し、これらのそれぞれのサイズに基づいて区別することができる。たとえば、増幅後に、DNA試料をアガロースゲル上で分離して、臭化エチジウムで染色後に紫外(UV)光下で視覚化することができる。DNAは、所望のレベルに増幅してもよく、さらなる伸長反応を行って、放射性標識されたか、またはビオチン標識されたヌクレオシド三リン酸などの検出可能なマーカーを有するヌクレオチド誘導体を組み入れてもよい。また、プライマーは、上で考察したように検出可能なマーカーで標識してもよい。検出可能なマーカーは、制限および電気泳動分離、または当技術分野において公知のその他の技術によって解析してもよい。
【0079】
PCRを使用する本発明の方法において使用してもよい条件は、試料中のDNAおよび適切な相補的ハイブリダイゼーション・プライマーの存在下において、ハイブリダイゼーションおよび増幅反応を進行することができるものである。ポリメラーゼ連鎖反応法のために適した条件は、一般に当技術分野において公知である。たとえば、M.A. Innis and D.H. Gelfand, PCR Protocols, A guide to Methods and Applications M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White eds, pp3-12, Academic Press 1989を参照されたい(これらは、参照として本明細書に組み入れられる)。好ましくは、PCRは、好熱性細菌サーマス・アクアティクス(Thermus aquatics)(Taqポリメラーゼ、GeneAmpキット、Perkin Elmer Cetus)から得られるポリメラーゼを利用し、またはその他の耐熱性ポリメラーゼを、DNA鋳型鎖を増幅するために使用してもよい。
【0080】
リガーゼ連鎖反応法(LCR)およびNASBAなどのその他の技術を本発明の核酸分子を増幅するために用いてもよいことが認識される(Barney in "PCR Methods and Applications", August 1991, Vol.1(1), page 5、および1989年6月14日に公開された欧州特許出願第0320308号、並びにMalekの米国特許出願第5,130,238号)。
【0081】
本発明のいくつかの態様において、本発明の配列における突然変異は、核酸分子の直接のシーケンシングによって検出してもよい。DNAを直接シーケンシングするための技術は、当技術分野において周知である。本発明の一つの態様において、突然変異は、サイクル・シーケンシングで検出されてもよく、これは、耐熱性ポリメラーゼ酵素、シーケンシング・プライマー、dNTP、および限定量の鎖を終結する蛍光性または放射性に標識されたddNTPの使用を含んでもよい。ポリアクリルアミドゲル電気泳動またはキャピラリー電気泳動法などのもう一つの技術を使用してシーケンシング反応の産物を分離し、続いて蛍光性または放射性の標識を検出してもよい。本発明の本態様の一つの例において、EMP2Bにおける突然変異は、Applied Biosystems 3700 DNA Analyzerまたは3730xl DNA Analyzer(商標)での自動シーケンシングを使用して決定することもできる。突然変異は、患者の配列を野生型の個体のものと、または公共データベースにおいて見いだされる参照配列と比較することによって同定してもよい。
【0082】
また、EMP2Bにおける突然変異は、米国特許第5,888,780号に開示され、かつMadison, WisconsinのThird Wave(商標) Technologiesから入手可能なCleavase(商標)Fragment Length Polymorphism(CFLP)アッセイ法を使用するInvader(商標)遺伝子タイピング系を使用して検出してもよい。
【0083】
本発明のその他の態様は、EMP2B遺伝子における突然変異の検出のためDNAチップ技術の使用を想定する。その他の適用の中で、DNAチップ技術により、予め定められた配列の高密度に空間的にアドレス指定可能なマイクロアレイ上に対する核酸試料のハイブリダイゼーション・パターンを解析することを介して目的の配列における突然変異を同定することができる。本発明の配列である突然変異の同定のために適したDNAチップ技術の一つの例は、米国特許第6,045,996号に開示されたAffymatrix GeneChip(商標)系である。この系は、何十万ものオリゴヌクレオチドプローブを含む高密度アレイを作製するために、フォトリソグラフィーおよび固相化学を使用する。Genechips(商標)は、特定の配列の再シーケンシングを容易にして、特定の突然変異を同定することができるようにデザインしてもよい。ハイブリダイゼーション・パターンのコンピュータに基づいた画像分析により、自動塩基呼び出し、および解析内に品質制御措置を組み入れることが可能になる。
【0084】
当業者であれば、本発明の配列内もしくは近くにおける病原性の欠失または再編成を検出するためにサザンブロッティング法またはノーザンブロッティング法を使用してもよいことを認識しているであろう。同様に、蛍光インサイチュー・ハイブリダイゼーション(FISH)、fiber-FISH、またはその他の高解像度の細胞遺伝学的方法を、本発明の配列を分裂させる再編成または欠失を検出するために使用してもよい。
【0085】
突然変異の検出のためのもう一つの技術は、変性HPLC解析である。したがって、本発明の一つの態様は、本発明の配列内におけるヘテロ接合性の突然変異または多型の同定のために、核酸のdHPLC解析のためのTransgenomic Wave(商標)機の使用を含む。
【0086】
また、本発明は、突然変異の遺伝子タイピングのための質量分析の使用も想定する。変異体および野生型の核酸分子は、野生型と変異体配列での異なる組成のために、質量が異なっており、異なるヌクレオチド配列の分子質量に基づいて突然変異を同定することができる。突然変異の遺伝子タイピングのための質量分析、特にマトリクス支援レーザー脱離イオン化飛行時間型(Matrix Assisted Laser Desorption Ionisation Time of Flight:MALDI-TOF)質量分析の使用は、関連した技術における当業者に周知である。たとえば、米国特許第6,043,031号は、特定の核酸配列を検出するための迅速かつ非常に正確な質量分析計に基づいたプロセスを記載する。SEQUENOM(商標)からのMass ARRAY(商標)プラットフォームは、一塩基多型を遺伝子タイピングすることができ、本発明に記載されているような突然変異を検出することができる、市販されている系の例である。
【0087】
(ii)Malinタンパク質の検出
もう一つの態様において、本発明は、ラフォラ病を有することが疑われる哺乳類、好ましくはヒトまたはイヌ由来の試料において、malinタンパク質が存在するか、または変異されているかどうかを決定することを含む、ラフォラ病を検出するための方法を提供する。
【0088】
本発明のmalinタンパク質は、後述する種々の免疫アッセイ法を使用してmalinに特異的である抗体を使用して、生体試料中で検出してもよい。変異されたmalinとのみ反応する抗体は、ラフォラ病を検出するための診断薬として有用である。したがって、タンパク質の変異された領域だけに結合する抗体が調製されることとなる。
【0089】
抗体を調製するために、従来法を使用することができる。たとえば、本発明のmalinタンパク質由来のペプチドを使用することにより、ポリクローナル抗血清またはモノクローナル抗体を標準的な方法を使用して作製することができる。哺乳類(たとえば、マウス、ハムスター、またはウサギ)に、哺乳類の抗体反応を引き出すペプチドの免疫原形態で免疫することができる。ペプチドに免疫原性を与えるための技術は、キャリアとの抱合または当技術分野において周知のその他の技術を含む。たとえば、ペプチドをアジュバントの存在下において投与することができる。免疫の過程は、プラズマまたは血清中の抗体力価の検出によってモニターすることができる。抗体のレベルを評価するために、標準的なELISAまたはその他の免疫アッセイ手順では、抗原として免疫原を使用することができる。免疫化に続いて、抗血清を得ることができ、必要であれば、ポリクローナル抗体を血清から単離することができる。
【0090】
モノクローナル抗体を産生するためには、抗体産生細胞(リンパ球)を免疫した動物から収集し、標準的な体細胞融合手順によって骨髄腫細胞と融合させることによって、これらの細胞を不死化し、ハイブリドーマ細胞を産生することができる。このような技術は、当技術分野において周知である(たとえば、当初Kohlerおよび Milstein (Nature 256, 495-497 (1975))によって開発されたハイブリドーマ技術、並びにヒトB-細胞ハイブリドーマ技術(Kozbor et al., Immunol. Today 4, 72 (1983))、ヒトモノクローナル抗体を産生するためのEBVハイブリドーマ技術(Cole et al., Monoclonal Antibodies in Cancer Therapy (1985) Allen R. Bliss, Inc., pages 77-96)、およびコンビナトリアル抗体ライブラリーのスクリーニング(Huse et al., Science 246, 1275 (1989))などのその他の技術)。ペプチドと特異的に反応する抗体の産生について、ハイブリドーマ細胞を免疫化学的にスクリーニングすることができ、モノクローナル抗体を単離することができる。したがって、本発明はまた、本発明のタンパク質に対する特異性を有するモノクローナル抗体を分泌するハイブリドーマ細胞を想定する。
【0091】
本明細書に使用される用語「抗体」は、本発明のタンパク質、またはこれらのペプチドと特異的に反応するこれらの断片も含むことが意図される。上記のものと同じ方法で利用するために、従来技術を使用して抗体を断片化し、断片をスクリーニングすることができる。たとえば、F(ab')2断片は、ペプシンで抗体を処理して作製することができる。生じるF(ab')2断片を処理してジスルフィド架橋を還元し、Fab'断片を産生することができる。
【0092】
キメラ抗体誘導体、すなわちヒト以外の動物の可変領域とヒトの定常領域を組み合わせた抗体分子も、本発明の範囲内において想定される。キメラ抗体分子は、たとえば、ヒト定常領域を有する、マウス、ラット、またはその他の種の抗体に由来する抗原結合ドメインを含むことができる。malinタンパク質を認識する免疫グロブリン可変領域を含むキメラ抗体を作製するために、従来法を使用してもよい(たとえば
を参照されたい)。
【0093】
本明細書に記載されている本発明のタンパク質と特異的に反応するモノクローナルまたはキメラ抗体は、可変領域の一部、特に抗原結合ドメインの保存されたフレームワーク領域が、ヒト起源であり、かつ高頻度可変領域だけが非ヒト起源であるヒト定常領域キメラを産生することによって、さらにヒト化することができる。このような免疫グロブリン分子は、当技術分野において公知の技術によって作製されてもよい(たとえばTeng et al., Proc. Natl. Acad. Sci. U.S.A., 80, 7308-7312 (1983); Kozbor et al., Immunology Today, 4, 7279 (1983); Olsson et al., Meth. Enzymol., 92, 3-16 (1982))、および国際公開公報第92/06193号または欧州特許第0239400号)。また、ヒト化抗体は、商業的に産生することができる(Scotgen Limited, 2 Holly Road, Twickenham, Middlesex, Great Britain)。
【0094】
本発明のタンパク質に対して反応性の、特異的な抗体、または抗体断片は、本発明の核酸分子から産生されるペプチドを用いて、細菌で発現した免疫グロブリン遺伝子、またはこれらの部分をコードする発現ライブラリーをスクリーニングすることによって作製されてもよい。たとえば、完全Fab断片、VH領域、およびFV領域は、ファージ発現ライブラリーを使用して、細菌で発現させることができる(Ward et al., Nature 341, 544-546: (1989); Huse et al., Science 246, 1275-1281 (1989);および McCafferty et al., Nature 348, 552-554 (1990)を参照されたい)。
【0095】
また、抗体は、DNA免疫を使用して調製してもよい。たとえば、本発明の(上記のとおりの)核酸を含む発現ベクターをマウスなどの適切な動物に注射してもよい。したがって、本発明のタンパク質がインビボで発現されて、抗体が誘導されると考えられる。抗体は、単離することができ、タンパク質免疫化のために上記のとおりに調製することができる。
【0096】
抗体は、種々の酵素、蛍光物質、発光材料、および放射性物質を含む検出可能なマーカーで標識してもよい。適切な酵素の例は、西洋ワサビペルオキシダーゼ、ビオチン、アルカリホスファターゼ、β-ガラクトシダーゼ、またはアセチルコリンエステラーゼを含み;適切な蛍光物質の例は、ウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリド、またはフィコエリトリンを含み;発光材料の例は、ルミノールを含み;および適切な放射性物質の例は、S-35、Cu-64、Ga-67、Zr-89、Ru-97、Tc-99m、Rh-105、Pd-109、In-111、I-123、I-125、I-131、Re-186、Au-198、Au-199、Pb-203、At-211、Pb-212、およびBi-212を含む。また、抗体は、リガンド結合対の一方のパートナーに対して標識するか、または抱合してもよい。代表例は、アビジン-ビオチンおよびリボフラビン-リボフラビン結合タンパク質を含む。上記した代表的標識をもつ、上で論議した抗体を抱合、または標識するための方法は、従来技術を使用して容易に達成し得る。
【0097】
本発明のタンパク質(たとえば、酵素抱合体または標識された誘導体)に対する抗体反応性は、種々の試料中における本発明のタンパク質を検出するために使用してもよく、たとえば、これらは、本発明のタンパク質の抗原決定基と抗体との間の結合相互作用に依存する任意の公知の免疫アッセイ法に使用してもよい。このようなアッセイ法の例は、放射免疫アッセイ法、酵素免疫測定法(たとえば、ELISA)、免疫蛍光、免疫沈降、ラテックス凝集反応法、赤血球凝集法、および組織化学的試験である。したがって、ラフォラ病の存在を診断するために、試料中の本発明のタンパク質の量を同定、または定量化するため抗体を使用してもよい。
【0098】
本発明の方法では、試料または濃縮された試料の予め定められた量が、抗体または標識抗体と混合される。本方法に使用される抗体の量は、選択される標識剤に依存的である。生じる、抗体または標識された抗体に結合したタンパク質は、従来の単離技術、たとえばば塩析、クロマトグラフィー、電気泳動法、ゲル濾過、分画、吸収、ポリアクリルアミドゲル電気泳動、凝集、またはこれらの組み合わせによって単離してもよい。
【0099】
試料または抗体は、不溶化してもよく、たとえば、試料または抗体は、公知の方法を使用して適切なキャリアと反応させることができる。適切なキャリアの例は、セファロースまたはアガロースビーズである。不溶化された試料または抗体が使用されるときは、抗体または反応していない抗体に結合したタンパク質は、洗浄することによって単離される。たとえば、試料がニトロセルロース膜にブロットされるときは、本発明のタンパク質に結合した抗体は、緩衝液、たとえばウシ血清アルブミン(BSA)を含むリン酸緩衝食塩水(PBS)で洗浄することにより、反応していない抗体から分離される。
【0100】
標識された抗体が使用されるときは、malinの存在は、試料中の本発明のタンパク質に結合した標識された抗体の量または反応していない標識された抗体の量を測定することによって決定することができる。標識された物質を測定する適切な方法は、標識剤に依存する。
【0101】
標識されていない抗体を本発明の方法に使用するときは、malinの存在は、凝集または沈澱を生じさせるために、抗体と特異的に相互作用する物質を使用してタンパク質に結合した抗体の量を測定することによって決定することができる。特に、本発明のタンパク質に特異的な抗体に対する標識された抗体を、反応混合物に添加することができる。本発明のタンパク質の存在は、標識剤のタイプに応じて、すでに記載されている技術の中から適切な方法によって決定することができる。本発明のタンパク質に特異的な抗体に対する抗体は、本明細書に記載した当技術分野において公知の従来法によって調製し、および標識することができる。本発明のタンパク質に特異的な抗体に対する抗体は、種特異的な抗免疫グロブリン抗体またはモノクローナル抗体であってもよく、たとえば、ヤギ抗ウサギ抗体を本発明のタンパク質に特異的なウサギ抗体を検出するために使用してもよい。
【0102】
(iii)キット
本発明の方法を実施するために適した試薬は、適切な容器にパックされて、必要な材料を提供する便利なキットにパックされてもよい。このようなキットは、本明細書に記載されている方法、および任意に本発明の方法を行う際に有用な適切な支援によって、試料中の本発明の核酸分子またはタンパク質を検出するために必要とされる全ての試薬を含んでもよい。
【0103】
本発明の一つの態様において、キットは、本発明の核酸分子またはこれらの予め定められたオリゴヌクレオチド断片を増幅することができるプライマー、ポリメラーゼ連鎖反応法において増幅された核酸分子またはこれらの予め定められた断片を産生するために必要とされる全ての試薬、および増幅された配列をアッセイするための手段を含む。また、キットは、PCR産物を消化するための制限酵素も含んでいてもよい。本発明のもう一つの態様において、キットは、本発明の核酸分子とハイブリダイズするヌクレオチド・プローブ、核酸分子とヌクレオチド・プローブのハイブリダイゼーションのために必要とされる試薬、およびその使用説明書を含む。本発明のさらなる態様において、キットは、本発明の抗体、および試料中において本発明のタンパク質に対して抗体を結合するために必要とされる試薬を含む。
【0104】
本発明の方法およびキットは、ラフォラ病を検出するために使用してもよい。試験してもよい試料は、血液、尿、血清、涙、唾液、糞便、組織、細胞などの身体材料を含む。ヒト試料に加えて、試料は、イヌなどの非ヒト霊長類および犬科の動物などの哺乳類に由来してもよい。
【0105】
本明細書に記載されている方法に従って試料を試験する前に、試料は、遠心分離および濾過などの当技術分野において公知の技術を使用して濃縮してもよい。本明細書に記載されているハイブリダイゼーションおよび/またはPCRに基づいた方法については、核酸は、当技術分野において公知の技術を使用して試験試料の細胞抽出物から抽出してもよい。
【0106】
B.EPM2B/Malinモジュレーター
上記した抗体およびアンチセンス・オリゴヌクレオチドに加えて、EPM2B発現または活性を調整するその他の物質、並びにmalinの変異した形態を調整する物質を同定してもよい。
【0107】
(i)Malinに結合する物質
malin活性に影響を及ぼす物質は、これらがmalinおよび/または変異したmalinと結合する能力に基づいて同定することができる。
【0108】
本発明のmalinと結合することができる物質は、潜在的にmalinに結合する物質とmalinを反応させることと、遊離の物質について、または複合体を形成していないmalinについて、またはmalinの活性について、複合体をアッセイすることによって同定され得る。特に、malinと相互作用するタンパク質を同定するために、酵母ツー・ハイブリッド・アッセイ系を使用してもよい(Fields, S. and Song, O., 1989, Nature, 340:245-247)。また、使用してもよい解析系には、ELISAを含む。
【0109】
したがって、本発明は、以下の工程をmalinと結合することができる物質を同定する方法であって:
(a)反応するmalinと試験物質を、malinと試験物質との間で複合体を形成することができる条件下で反応させる工程、および
(b)遊離の物質について、または複合体を形成していないmalinについて、malinと試験物質の複合体をアッセイする工程を含み、複合体の存在は、試験物質がmalinに結合できることを示す方法を提供する。
【0110】
アッセイ法に使用されるmalinタンパク質は、配列番号:2(図6B)もしくは配列番号:4(図7B)に示したアミノ酸配列を有してもよく、または本明細書に記載されている、LDと関連する変異されたタンパク質であってもよく、または本明細書に記載したような、これらの断片、類似体、誘導体、相同体、もしくはミメティックであってもよい。
【0111】
物質とmalinの複合体の形成を可能にする条件は、物質およびタンパク質の性質並びに量などの因子を考慮して選択してもよい。
【0112】
物質-タンパク質複合体、遊離の物質、または複合体を形成しないタンパク質は、従来の単離技術、たとえば、塩析、クロマトグラフィー、電気泳動法、ゲル濾過、分画、吸収、ポリアクリルアミドゲル電気泳動、凝集、またはこれらの組み合わせによって単離してもよい。malinもしくは物質に対する抗体、または標識されたmalin、または標識された物質を、成分のアッセイ法を容易にするために利用してもよい。抗体、タンパク質、または物質は、上記のような検出可能な物質で標識してもよい。
【0113】
本発明の方法に使用されるmalinまたは物質は、不溶化させてもよい。たとえば、malinまたは物質は、適切なキャリアに結合されてもよい。適切なキャリアの例は、アガロース、セルロース、デキストラン、セファデックス、セファロース、カルボキシメチルセルロースポリスチレン、濾紙、イオン交換樹脂、プラスチック・フィルム、プラスチック・チューブ、ガラスビーズ、ポリアミン-メチルビニル-エーテル-マレイン酸共重合体、アミノ酸共重合体、エチレン-マレイン酸共重合体、ナイロン、絹、その他である。キャリアは、たとえば、チューブ、試験プレート、ビーズ、盤、球、その他の形態であってもよい。
【0114】
不溶化されたタンパク質または物質は、公知の化学的または物理的な方法、たとえば、臭化シアンカップリングを使用して、適切な不溶性キャリアと材料を反応することによって調製してもよい。
【0115】
また、タンパク質または物質は、本明細書に記載されている方法を使用して細胞の表面上に発現させてもよい。
【0116】
また、本発明は、malinの作用のアンタゴニストまたはアゴニストのためのアッセイを想定する。
【0117】
本発明の方法を使用してアッセイすることができるアゴニストおよびアンタゴニストは、アゴニスト結合部位、競合的アンタゴニスト結合部位、非競合的アンタゴニスト結合部位、またはアロステリック部位を含む、1つもしくは複数のタンパク質または物質の結合部位に対して作用してもよいことが理解されるであろう。
【0118】
また、本発明は、malinのアゴニストの効果を阻害するアンタゴニストをスクリーニングすることを可能にする。したがって、本発明は、malinの同じ結合部位に対して競合する物質をアッセイするために使用してもよい。
【0119】
(ii)ペプチドミメティック
また、本発明は、本発明のmalinのペプチドミメティックおよび変異されたmalinタンパク質を含む。たとえば、malinの変異されたドメインに由来するペプチドは、変異されたタンパク質の天然の結合を模倣するような方法で関連する分子と直接的または間接的に相互作用する。このようなペプチドは、競合阻害剤、エンハンサー、ペプチドミメティック等を含んでもよい。実質的にこれらのペプチド類、並びにこれらのペプチドに対して相同的、相補的、またはさもなければ機能的もしくは構造的に同等な分子の全てを、本発明の目的を達成するために使用してもよい。
【0120】
「ペプチドミメティック」とは、分子間の相互作用においてペプチドの代用として役立つ構造である(総説については、Morgan et al (1989), Ann. Reports Med. Chem. 24:243-252を参照されたい)。ペプチドミメティックは、アミノ酸および/またはペプチド結合を含んでも、または含まなくてもよいが、ペプチド、または本発明のエンハンサーもしくは阻害剤の構造上および機能的な特徴を保持する合成構造を含む。また、ペプチドミメティックは、ペプトイド、オリゴペプトイド(Simon et al (1972) Proc. Natl. Acad, Sci USA 89:9367);および本発明のペプチドに対応するアミノ酸の全ての可能性のある配列を表す長さにデザインされたペプチドを含むペプチドライブラリーを含む。
【0121】
ペプチドミメティックは、D-アミノ酸によるL-アミノ酸の系統的置換、異なる電子的性質を有する基と側鎖の置換によって、およびアミド結合の置換によるペプチド結合の系統的置換によって得られる情報に基づいてデザインされてもよい。また、候補となるペプチドミメティックの活性についての高次構造上の要求を決定するために、局部的な高次構造上の制約を導入することができる。ミメティックは、逆向ターンの高次構造を安定化もしくは促進し、および分子の安定化を助けるために、等配電子アミド結合、またはD-アミノ酸を含んでもよい。特定の高次構造の状態にアミノ酸残基を拘束するために、環状アミノ酸類似体を使用してもよい。また、ミメティックには、ペプチドの二次構造の阻害剤のミメティックを含むことができる。これらの構造は、アミノ酸残基の三次元配向をタンパク質の公知の二次高次構造にモデル化することができる。また、N置換されたアミノ酸のオリゴマーであるペプトイドを使用してもよく、新規分子の化学的に多様なライブラリーを作製するためのモチーフとして使用することができる。
【0122】
また、本発明のペプチドは、薬剤開発のためのリード化合物を同定するために使用してもよい。本明細書に記載されたペプチドの構造は、NMRおよびX線結晶学などの多くの方法によって容易に決定することができる。配列が類似するが、これらが標的分子に誘発する生物活性が異なるペプチドの構造を比較すると、標的の構造-活性相関についての情報を提供することができる。構造-活性相関の調査から得られる情報は、修飾されたペプチド、または標的分子に関して予測される性質を試験することができるその他の小分子もしくはリード化合物のいずれかをデザインするために使用することができる。リード化合物の活性は、本明細書に記載されているものと同様のアッセイ法を使用して評価することができる。
【0123】
また、構造-活性相関についての情報は、共同結晶化研究から得てもよい。これらの研究では、所望の活性を有するペプチドを標的分子と結合させて結晶化し、複合体のX線構造を決定する。次いで、構造をその天然の状態の標的分子の構造と比較することができ、このような比較からの情報を、有することが予想される化合物をデザインするために使用してもよい。
【0124】
(iii)薬物のスクリーニング方法
一つの態様に従って、本発明は、変異したmalinタンパク質の活性を増減する能力について候補化合物をスクリーニングするための方法を可能にする。本方法は、malin活性をアッセイするため、候補もしくは試験化合物の有無における活性をアッセイするため、および化合物がmalin活性の増減をもたらすかどうかを決定するためのアッセイ系を提供することを含む。このような化合物は、ラフォラ病を治療する際に有用であり得る。
【0125】
したがって、本発明は、変異されたmalinタンパク質活性または発現に影響を及ぼす化合物を同定するための方法であって:
(a)malinタンパク質またはmalinタンパク質をコードする核酸と共に試験化合物をインキュベートする工程;および、
(b)malinタンパク質の活性または発現の量を決定し、対照(すなわち、試験物質の非存在下において)と比較する工程を含み、対照と比較したmalinタンパク質活性または発現の変化は、試験化合物がmalinタンパク質の活性または発現に対して効果を有することを示す方法を提供する。
【0126】
さらなる態様に従って、本発明によって、malinタンパク質の発現を増減する能力の候補化合物をスクリーニングする方法が可能となる。本方法は、候補化合物と共に細胞(本細胞は、リポーター遺伝子コード領域に操作可能な状態で連結されたmalin遺伝子の制御領域を含む)を置く工程、およびリポーター遺伝子の発現の変化を検出する工程を含む。
【0127】
一つの態様において、本発明により、変異されたmalin遺伝子を発現する株化細胞を候補化合物と共にインキュベートして、変異されたmalin発現に対するこれらの効果を試験する培養系が可能になる。このような培養系は、その他のタンパク質との相互作用を介して、malin発現もしくはその機能をアップレギュレートするか、またはダウンレギュレートする化合物を同定するために使用することができる。
【0128】
このような化合物は、培地に添加されるタンパク質化合物、化学薬品、および種々の薬物から選択することができる。選択された試験化合物(類)の存在下でのインキュベーション後、変異されたmalinの発現を、本明細書に含まれる実施例において説明したように、標準的なノーザンブロット法手順を使用してmalin mRNAのレベルを定量化することによって調べて、試験化合物の結果として発現の任意の変化を決定することができる。また、タンパク質発現を修飾するために開発された化合物の機能を試験するために、malinを発現する構築物をトランスフェクトされた細胞株を使用することができる。
【0129】
C.治療的使用
前述のように、本発明のEPM2B遺伝子およびmalinは、ラフォラ病に関与する可能性が高い。したがって、本発明は、ラフォラ病を治療する方法であって、その必要がある細胞または動物に対し、EPM2B/malin発現および/または活性を調整する薬剤の有効な量を投与することを含む方法を提供する。また、本発明はラフォラ病を治療するための、もしくはラフォラ病を治療するための薬物を調製するための、EPM2B/malin発現および/または活性を調整する薬剤の使用を提供する。
【0130】
用語「EPM2B/malin発現および/または活性を調整する薬剤」は、野生型EPM2B/malinと一致する動物において見いだされる変異されたEPM2B/malinの発現および/または活性を変化させることができる任意の物質を意味する。投与に使用してもよい薬剤の例は:野生型EPM2Bをコードする核酸分子;野生型malinタンパク質、並びにこれらの断片、類似体、誘導体、または相同体;抗体;アンチセンス核酸;ペプチドミメティック;および本明細書に記載されているスクリーニング法を使用して単離された物質であって、変異を症状のない人と同様のEPM2B/malinレベルおよび/または機能に修正することができるものを含む。
【0131】
本明細書に使用される用語「有効な量」は、所望の結果を達成するために必要な用量で、および期間で有効な量を意味する。
【0132】
本明細書に使用される用語「動物」は、ヒトおよびイヌを含む動物界の全ての一員を含む。
【0133】
一つの態様において、本発明は、変異されたmalinタンパク質の発現または生物活性を調整する薬剤の有効な量を、細胞または動物に投与することによってラフォラ病を治療する方法を提供する。また、本発明は、ラフォラ病を治療するために、またはラフォラ病を治療するための薬物を調製するために、変異されたmalinタンパク質の発現または生物活性を調整する薬剤の有効な量の使用を提供する。変異されたmalinの活性を阻害する物質は、ペプチドミメティック、malinアンタゴニスト、およびmalinに対する一定の抗体を含む。変異されたEPM2B遺伝子の発現を阻害する物質は、変異されたEPM2B核酸配列に対するアンチセンス・オリゴヌクレオチドを含む。
【0134】
もう一つの態様に従って、本発明は、ラフォラに対する潜在的な治療手段としての遺伝子治療であって、病気に冒されたいくつかの異なる細胞型において正常なmalinタンパク質をうまくコードするように、EPM2B遺伝子の正常なコピーを患者に導入する治療を可能にする。
【0135】
レトロウイルスのベクターは、これらが高い効率で感染および安定な組込みおよび発現をするので、特に体細胞遺伝子治療に使用することができる。しかし、ターゲットされる細胞は、分裂することができなければならず、正常タンパク質の発現レベルが高いはずである。全長正常EPM2B遺伝子は、レトロウイルスのベクターにクローン化することができ、その内因性のプロモーターからもしくはレトロウイルスの末端反復配列から、または関心対象の標的細胞タイプに特異的なプロモーターから駆動させることができる。使用することができるその他のウイルスのベクターは、アデノ随伴ウイルス、ワクシニアウイルス、ウシパピローマウイルス、またはエプスタイン・バーウイルスなどのヘルペスウイルスを含む。また、遺伝子導入は、インビトロでの感染に必要とされる非ウイルス手段を使用して達成することもできる。これは、リン酸カルシウム法、DEAEデキストラン法、電気穿孔法、陽イオン性または陰イオン性の脂質製剤(リポソーム)、およびプロトプラスト融合法を含む。これらの方法が利用できるものの、これらの多くは、より低効率である。
【0136】
変異されたEPM2B遺伝子機能を阻害するために、および治療薬のデザインの基礎として、アンチセンスに基づいたストラテジーを使用することができる。原理は、mRNAと相補アンチセンス種との間の細胞内ハイブリダイゼーションによって、遺伝子発現の配列特異的な抑制を達成することができるという仮説に基づく。RNAまたはDNAの有意鎖に高い特異性で結合するアンチセンス鎖ヌクレオチドを合成することが可能である。ハイブリッドRNA二重鎖の形成は、標的mRNAのプロセシング/輸送/翻訳、および/または安定性を妨げ得る。
【0137】
ハイブリダイゼーションは、アンチセンス効果を生じさせるために必要とされる。アンチセンス効果は、アンチセンス・オリゴヌクレオチドの使用、アンチセンスRNA、DNAの注入、およびアンチセンスRNA発現ベクターのトランスフェクションを含む種々のアプローチを使用して記載されている。
【0138】
治療的アンチセンス・ヌクレオチドは、オリゴヌクレオチドまたは発現されたヌクレオチドとして作製することができる。オリゴヌクレオチドは、通常15〜20核酸塩基の長さであるDNAの短い一本鎖である。発現されたヌクレオチドは、アデノウイルス、レトロウイルス、またはプラスミドベクターなどの、発現ベクターを使用して作製される。ベクターは、培養の細胞に、または患者に投与され、次いで、その細胞がアンチセンス・ヌクレオチドを作製する。発現ベクターは、アンチセンスRNAを産生するようにデザインすることができ、これは数ダースの塩基〜数千までで長さを変化させることができる。
【0139】
アンチセンス効果は、制御(センス)配列によって誘導することができる。表現型の変化の範囲は、非常に変わりやすい。アンチセンスによって誘導される表現型の効果は、生物学的なエンドポイント、タンパク質レベル、タンパク質活性化の測定、および標的mRNAレベルなどの基準の変化に基づく。
【0140】
D.薬学的組成物
EPM2Bおよび変異されたEPM2Bをコードする核酸、malinおよび変異されたmalinタンパク質、抗体、およびアンチセンス・オリゴヌクレオチド、並びにEPM2B/malinおよび/または変異されたEPM2B/malinを調整するその他の薬剤を含む上述した物質は、インビボでの投与に適した生物学的に適合性を有する形態で、被検者に投与するための薬学的組成物中に処方してもよい。「インビボでの投与に適した生物学的に適合性を有する形態」とは、治療的な効果が任意の有毒作用を上回って投与されるような物質の形態を意味する。物質は、ヒト、および動物を含む、生きている生物に投与してもよい。
【0141】
本発明の薬学的組成物の治療的に有効な量の投与は、所望の結果を達成するために必要な用量で、および期間で有効な量として定義される。たとえば、物質の治療的に有効な量は、個々の疾病状態、年齢、性、および重量、並びに個々に所望の反応を引き出すための物質の能力などの因子によって変更してもよい。投与法は、最適な治療反応を提供するように調整してもよい。たとえば、いくつかの分割用量を毎日投与してもよく、または用量は、治療状況の緊急性によって指示されて、比例して減らしていてもよい。
【0142】
活性物質は、注射(皮下の、静脈内の、その他)、経口投与、吸入、経皮の適用、または直腸の投与などによって、便利な方法で投与してもよい。投与の経路に応じて、活性物質は、酵素、酸、および化合物を不活性化するであろうその他の天然の条件の作用から化合物を保護する材料中に被覆されてもよい。活性物質が、たとえば、修飾されたEPM2B遺伝子をコードする核酸である場合、当技術分野において公知の技術を使用して送達してもよい。
【0143】
本明細書に記載されている組成物は、薬学的に許容される組成物の調製のためのそれ自体公知の方法によって調製することができ、これを、活性物質の有効な量が薬学的に許容される媒体との混合物中に組み合わされるように被検者に投与することができる。適切な媒体は、たとえばばRemington’s Pharmaceutical Sciences (Remington’s Pharmaceutical Sciences, Mack Publishing Company, Easton, Pa., USA 1985)またはHandbook of Pharmaceutical Additives (compiled by Michael and Irene Ash, Gower Publishing Limited, Aldershot, England (1995))に記載されている。これに基づいて、組成物は、排他的ではないものの、1つもしくは複数の薬学的に許容される媒体または希釈液と合わせた物質の溶液を含み、適切なpHを有する緩衝液に含まれてもよく、および/または生理液によって等浸透圧にしてもよい。この点に関しては、米国特許第5,843,456号を参照することができる。また、当業者に認識されるように、本明細書に記載されている物質の投与は、不活性ウイルスキャリアによるものであってもよい。
【0144】
E.実験モデル
また、本発明は、EPM2B遺伝子およびmalinタンパク質の機能を研究するための方法、並びに実験モデルを含む。EPM2B遺伝子を欠いた、もしくはmalin発現を部分的に欠いた細胞、組織、および非ヒト動物は、EPM2B遺伝子に特異的な欠失または変異を有する組換え発現ベクターを使用して開発してもよい。組換え発現ベクターを、相同組換えによってEPM2B遺伝子を不活性化または変化させるために使用して、これにより、EPM2B欠損細胞、組織、または動物を作製してもよい。
【0145】
ヌル対立遺伝子は、欠失突然変異によって胚幹性細胞などの細胞に作製してもよい。また、組換えEPM2B遺伝子を、EPM2Bを不活性化する挿入突然変異を含むように操作してもよい。次いで、このような構築物を、トランスフェクション、電気穿孔法、注入等の技術によって、胚幹性細胞などの細胞に導入してもよい。次いで、たとえばサザンブロッティング、ノーザンブロッティングによって、または本明細書に記載されている方法を使用してEPM2Bをアッセイすることによって、無傷のEPM2B遺伝子を欠いている細胞を同定してもよい。次いで、このような細胞を胚幹細胞に融合して、EPM2Bが欠損したトランスジェニック非ヒト動物を作製してもよい。突然変異の生殖系列伝達は、たとえば、インビトロで、8細胞胚などの初期胚で胚幹細胞を凝集すること;生じる胚盤胞をレシピエント女性へ移すこと、および;生じる凝集キメラの生殖系列伝達を発生させることによって達成してもよい。このような変異体動物を使用して、通常はEPM2B発現に依存的な、特異的な細胞群、発達パターン、およびインビボ・プロセスを定義してもよい。また、本発明は、EPM2B遺伝子の組織特異的ノックアウトの調製を含む。
【0146】
以下の非限定の実施例は、本発明の例示である:
【実施例】
【0147】
実施例1
本発明者らは、全ての利用できるデータベースを調べ、染色体6p22.3(図1)上の新たに定義された臨界領域において7つの注釈がついた遺伝子を見いだした。これらの予測される機能的特徴に基づいて、EPM2Bハプロタイプを有するLD患者のDNAシーケンシングを介した突然変異スクリーニングのため、それぞれの遺伝子を優先させたが、病原性変異体は同定されなかった。同時に、本発明者らの解析により、その他のより高等脊椎動物(ラット、マウス、およびイヌと、それぞれ75%、78%、87%のヌクレオチド同一性)におけるオーソロガスなユニット(タンパク質コード能力に相当)と広範な配列同一性を共有する、以前に特徴付けられていない、見かけ上単一エキソン(1188 bp)遺伝子を発見するに至った。また、EPM2Bと命名されたヒト遺伝子は、その5'末端に真核生物の翻訳開始部位のコンセンサス配列の提唱された特徴の全てを、およびその3'-末端に2つの推定ポリアデニル化シグナルを有した(図1)。さらに、ヒトおよびマウスにおける発現配列タグ(EST)およびcDNAデータは、単一エキソン・ユニットが本物の遺伝子であることを指示した。
【0148】
EPM2Bは、N末端の半分においてRING型のZnフィンガーとC末端方向に6つのNHL繰り返しドメインとを含む、発明者がmalin(フランス語の発作に関する疾患)と名付けた395アミノ酸(aa)タンパク質をコードすることが予測された(図1)。特に、RING-HC型のサイン配列(C-X2-C-X9-39-C-X1-3-H-X2-3-H-X2-C-X4-48-C-X2-C)(16、17)に対応した、亜鉛結合RINGフィンガー・モチーフ(C-X2-C-X16-C-X1-H-X2-C-X2-C-X14-C-X2-C)が、malinの残基26〜71間で同定された(0.0067のE値)。RINGフィンガーの存在は、E3ユビキチン・リガーゼ機能を予測させる(8、9、18)。E3ライゲーションは、E2から直接またはアダプタータンパク質を介して特異的基質にユビキチンを移行して、プロテアソーム系によってその除去を開始するユビキチン経路の最終的かつ特異的な工程である(8)。6つのNHLドメイン(10〜13)が、グリシンがリッチな約44残基モチーフおよび荷電残基のクラスターと共に発生源となった疎水性アミノ酸の存在に基づいて予測された(Pfamにより、0.011〜3.5の範囲のE値をもつNHLドメインについて6つの信頼される候補を検出した)(図1)。
【0149】
ノーザンブロット解析は、EPM2Bが脳の特定小領域を含む調べた全ての組織に(1.5kbおよび2.4kbのサイズの少なくとも2つの転写物として)存在することを示した(図3)。観察された転写物サイズは、予測されたATG開始部位と2つの異なるポリアデニル化シグナルとの間で予想される長さにほぼ一致する(図1)。さらに、発現プロフィールは、EPM2Aについて観察されるものと同様であり、両方とも、ラフォラ体が観察された全ての組織に存在した(2、19)。
【0150】
EPM2Bの完全なコード領域を、以前にEPM2Aに突然変異を有さないことが示された34人のLD発端者のコホートにおいてシーケンスした。この実施例において、フレームシフトを生じる8つの欠失および1つの挿入、7つのミスセンス、並びに1つのナンセンス変化を含む26ファミリーのEPM2Bにおいて、17個の異なるDNA配列変化が記載されている(表1)。これらの突然変異は、ホモ接合性(18)かつ複合ヘテロ接合性(8)の劣性状態の両方のファミリーにおいて見いだされた。最初の連鎖研究に使用した4血縁のF-Cファミリーは、全て、RINGフィンガー・ドメイン(図2b)の亜鉛結合能力に重要である7つの保存されたシステイン残基のうちの1つにシステインからセリンへの変化を生じるホモ接合性76T→A変化を有した。同定された最も共通する突然変異(7ファミリー)は、RINGフィンガー・ドメインにおいてプロリンからアラニンへの変化を生じるホモ接合性205C→G移行であった。EPM2B遺伝子座を絞り込む際に重要であったLD38324ファミリー(図2a)は、第5 NHLドメインにフレームシフト突然変異を生じるホモ接合性の2bp欠失(1048-1049delGA)を有した(図2b)。表1を参照し、ファミリーLD51において、発端者に由来するDNAは、入手不可能であったが、親は、両方とも468-469delAGのヘテロ接合性キャリアであり、これにより、LDの子供にホモ接合性のフレームシフト突然変異を生じることが予測される。検出される突然変異の全ては、推定RINGもしくはNHLモチーフに影響を及ぼすか、またはフレームシフトを生じるか、またはタンパク質の急激な構造変化(LD483は、260T→Cヌクレオチド変化を有し、ロイシンからプロリンへの変化を生じると考えられる)を生じさせることが予測される。4つのサイレントDNA配列をコードする変異体が同定された。これらの中で、3つのT312C(H104H)、G372C(G124G)、およびT1020C(G340G)は、それぞれ100個の対照染色体のうちの5つ、2つ、および1つに存在した。検出された最も共通する多型、C332T(P111L)(図1)は、100個の対照染色体のうちの42個で観察された。
【0151】
全体で、88%またはLDファミリーは、現在EPM2A(48%)およびEPM2B(40%)の突然変異によって説明することができる。これらの遺伝子のいずれにも検出可能な突然変異がない8ファミリーの観察は、さらなるLD遺伝子座位が存在し得ることを示唆する。
【0152】
成人ポリグルコサン小体疾患(APBD)を含む(20)、ポリグルコサン蓄積のあるその他の症状の中で、LDは、図4に示したように、軸索ではなくニューロン樹状突起における封入体の細胞内部位に関して独特である(21)。また、ERと形成性ポリグルコサン原繊維の物理的関連もLDに特異的である(3)。グリコーゲン分枝酵素の突然変異によって生じるAPBDにおいて(20、22、23)、ポリグルコサンは、サイズ、組成物、および数においてラフォラ体と見分けがつかないが、これらは、細胞体および軸索にのみ位置する(20)。APBDではなく、LDにおける発作表現型の存在は、LDのてんかんにおけるERに関連したポリグルコサンの樹状蓄積を暗示する。
【0153】
malinの細胞局在を調べるために、本発明者らは、エピトープタグを付けたEPM2B構築物をトランスフェクトし、これが実際にERに局在化し、培養細胞の核内においてわずかであることを見いだした(図5)。これらの結果は、それぞれ細胞質中のER(24-26)(図4b)および核において見いだされた、laforinのアイソフォームをコードするEPM2Aの2つの他の転写物(AおよびB)について観察される細胞局在と同じであった(27)。本発明者らのごく最近のデータは、転写物A特異的な突然変異の同定に基づいて、LDにおけるlaforinの細胞質型の機能喪失を暗示する(L.I. et al., 原稿投稿)。さらに、マウスのEPM2Aノックアウト(28)およびLD患者(1、2)の研究は、ER関連ポリグルコサン小体は、それぞれてんかんの発生に先行するか、または付随することを示した。
【0154】
したがって、簡単に説明すると、LDは、樹状突起においてポリグルコサンの不適当な排除(clearance)、およびその後の蓄積がニューロンのシナプス機能を妨げて、てんかん発生を生じることにより生じる。本発明者らは、今回、トランスジェニックLDマウスにおいて、laforinがポリグルコサン(およびグリコーゲンではない)と接触して、疾患遺伝子産物とLD病態との間の最初の物理的リンクを提供することを示した(E.M.C. et al., 準備中)。Laforinの唯一のその他の実験的に確証された機能は、二重特異性ホスファターゼのものであり(24、25)、これは、少なくとも1つのリンタンパク質中間体があり、中間体を介してこれが作用することを予測させる。可能性のある候補は、新たに発見されたEPM2A/P1 laforin相互作用タンパク質(26)、またはその他のさらにクローン化されたLD遺伝子、または任意のこれらの相互作用タンパク質の可能性がある。Malinは、そのNHLドメインを介した特異的なタンパク質-タンパク質相互作用、続くユビキチンを媒介した調節性標的の除去を経て関与して、laforinがラフォラ体およびてんかんに対してニューロンを保護するための重要な役割に寄与する可能性が高い。
【0155】
方法
試料
この研究に記載されている全ての患者は、形式的に、皮膚、骨格筋、肝臓、または脳の生検において特徴的なラフォラ体が存在することに基づいて、青年期発生進行性ミオクローヌスてんかんであると診断された。ホモ接合性マッピング、突然変異スクリーニング、または両方によって6q24におけるEPM2A遺伝子座の関与について、それぞれのLD個体およびこれらのファミリー・メンバー(入手できるならば)を調べた。
【0156】
遺伝子タイピングおよび突然変異スクリーニング
確立されたもの(D6S274、D6S285、D6S966、D6S1567、D6S1678、D6S1688、D6S1959)および新たなもの(BV012563、BV012730、BV012565、BV012566、BV012568)についての情報および増幅条件は、UniSTSおよびEntrez Nucleotidesデータベース(http://www.ncbi.nlm.nih.gov/)において見いだされる。DNA配列バリエーションは、PCR産物をシーケンシングすることによって検出した。EPM2Bをスクリーニングするために、重複断片を増幅する2セットのプライマー対を使用したEPM2B-1F:
とEPM2B-1R:
;EPM2B-2F:
とEPM2B-2R:
(図1を参照されたい)。PCRは、緩衝液[75mMのトリス-HCl(pH 8.8)、20mMの(NH4)2SO4、0.01% Tween 20、1.5mM MgCl2、1M Betaine、0.2mM dNTP、0.2μMのそれぞれのプライマー、2.5 UnitsのTaq Polymerase(MBI Fermentas)]中で50ngのDNAで行った。サイクル条件は、以下のとおりであった:94℃で3分間の最初の変性、続く94℃で30秒の変性、60℃で30秒のアニーリング、および72℃で30秒の伸長を35サイクル、72℃における10分間の最終伸長。PCR産物は、microCLEAN(Microzone Ltd.)を使用して精製した。3μl(100ng/μl)の精製したPCR産物をシーケンシングテンプレートとして使用した。全ての反応について、1μl(5pmol)のプライマー、1.5μlの5×シーケンシング緩衝液(Applied Biosystems)、1μlのBigDye Terminator v3.1、および7.5μlのH2Oを14μlの反応体積で使用した。Thermocycling(MJ Research, Inc.)条件は、96℃で30秒の変性;50℃で20秒のアニーリング;および60℃で4分の伸長;35サイクルであった。その後に全ての反応をマルチスクリーン-HVフィルタープレート(Millipore)を使用して精製し、ABI-3700を使用して解析した。LD患者において検出された全ての配列変異体は、50(100染色体)のランダムに選択したDNA試料のコレクションで調べた。
【0157】
遺伝子同定およびノーザンブロット
遺伝子注釈データおよびEST配列は、主にUniversity of California Santa Cruz(UCSC)ゲノム・ブラウザ(http://genome.ucsc.edu/)およびCelera Genomics(http://www.celera.com/)から得た。さらなる推定遺伝子は、Genescriptアルゴリズム(http://tcag.bioinfo.sickkids.on.ca/genescript/)およびヒトとマウス配列との間の多種VISTA(http://www-gsd.lbl.gov/vista/)整列を使用して注釈をつけた。RINGフィンガー・ドメインは、Pfam、Prosite、InterProScan、SMART、およびMotifScanによって予測した。NHLドメインは、PfamおよびInterProScanを使用して同定した。EPM2Bオルソログは、GenBank非重複性データベースに対するBLASTNおよびBLASTP解析を使用して同定した。配列分析によるmalinの細胞内局在性の予測は、PSORT IIを使用して行った。有意なシグナルペプチド配列(ER、ゴルジ複合体、リソソーム、および統合された必須原形質膜タンパク質の認識用)、ミトコンドリアのターゲティング配列、核局在化シグナル、およびペルオキシソームのターゲティング・シグナルは同定されなかった。human multiple-tissue blot Iおよびhuman brain blot II(Clontech)を、プライマー
および
を使用してEPM2Bのコード領域の557bp断片を増幅して作製された[32P ] dCTP標識されたプローブでプローブした。ハイブリダイゼーションおよび洗浄条件は、製造業者の説明書に従って行った。
【0158】
細胞内局在性および電子顕微鏡法
laforinの細胞質アイソフォームをコードするmycタグを付けたEPM2A転写物A発現構築物(pcDNA3mycEPM2A)が、記載されている(24)。mycタグを付けたEPM2B構築物は、同じ一般的なプロトコルを使用して作製した(pcDNA3mycEPM2B)。全長EPM2Bは、BamHI制限部位(下線を引いてある)と開始コドン(大文字)を含む(フォワード)プライマー
およびNotI部位(下線を引いてある)を含む(リバース)プライマー
を使用して、ゲノムDNAからPCRによって増幅した。この産物を哺乳類発現ベクターpcDNA3(Invitrogen)の対応する部位にクローン化した。次いで、5'KpnI-タグを付け、および3'BamHIタグを付けたプライマーで以前のmycを含むベクターから増幅した後に、インフレームでMycを導入した。pcDNA3mycEPM2AおよびpcDNA3mycEPM2B(2mg)をLipofectamine-Plus(Invitrogen)を使用してCos-7細胞にトランスフェクトし、5時間DMEM(Sigma-Aldrich)中で脂質-DNA複合体に曝露した。形質移入48時間後に、培養をPBS中で2回リンスし、アセトン:メタノール(1:1)混合物中で15分間-20℃で固定した。次いで、これらをmyc-laforinおよびERマーカーGRP94に対する抗体で染色した。培養を1時間(10%のBSA/PBS)ブロックし、室温で45分間抗Mycおよび抗ERと共にインキュベートした。スライドをPBSで洗浄し、ブロッキング溶液中の二次抗体(FITC標識されたヤギ抗マウス(1:400、緑色フィルターを介して検出可能);テキサスレッド標識されたロバ抗ヤギ(1:400、赤色フィルターを介して検出可能);Jackson Immuno Research Laboratories)と共にインキュベートした。マウンティング(Dako Anti-Fade)に続いて、これらを免疫蛍光光学顕微鏡によって解析した。電子顕微鏡検査のために、生検材料をLD患者から得られた冷却した汎用固定液に入れた。次いで、標準的なプロトコルを使用して、これを超微細構造的レベルで解析した。
【0159】
実施例2
実施例1に記載されている突然変異に加えて、本発明者らは、ラフォラ病と関連するEPM2Bにおいてさらなる4つの突然変異を見いだした。これらの突然変異は、実施例1と同じ様式で記載されていた。DNAは、実施例1に記載されているものと臨床的に類似したラフォラ病患者由来の血液から得た。EPM2Bにおける4つの突然変異は、以下のとおりである:1)位置606におけるヌクレオチドTの欠失;2)ヌクレオチド923におけるAからTへの変化;3)ヌクレオチド580におけるGからTへの変化;および4)ヌクレオチド199におけるGからTへの変化。後の突然変異は、スーダンおよびイタリアのファミリーにおいて見いだされた。さらなる突然変異は、表1の下段に一覧を記載してある。
【0160】
実施例3
ラフォラ病は、驚くべき頻度でイヌにおいても発見された。進行性ミオクローヌスてんかん(PME)は、英国において繁殖された人気のミニチュア・ワイアーヘアード・ダックスフンド(MWHD)において一般的である(〜5%)。本発明者らは、これらのイヌの臨床病理学的表現型を特徴づけ、これらがLDを有することを示した。本発明者らは、基礎になる疾患遺伝子座のマップを作製し、特異的疾患関連突然変異(EPM2B遺伝子において):いずれかの種における第1コーディング12塩基反復の伸長と、ヒトを越えた第1の病原性タンデム型繰り返し伸長とを同定した。本発明者らは、伸長突然変異が再発性であり、MWHD以外のてんかんのイヌにも影響を及ぼすことを示し、これがイヌ(およびその他の犬科の動物)に特有の配列変異により生じて、その種がLDにかかりやすいことを証明した。最後に、本発明者らは、キャリアおよび発症前動物において伸長対立遺伝子を検出し、MWHDおよび将来発症するイヌ集団に由来する疾患を根絶させるための試験を発明した。
【0161】
発病年令が6〜9歳である14匹の病気に冒されたMWHDを最初に評価した。ミオクローヌスは、頚部および肢筋の対称形の瞬間的な収縮からなり、イヌの後方突進を生じさせ;下顎が鳴るか、またはパチンと閉じられて、眼瞼が迅速にまばたきする。ミオクローヌスは、自発的に生じたが、突然の音、またはイヌの方へ転がるボールもしくは車に乗っている間の樹木を介してのストロボの光などの視覚刺激によって誘発することができる。ミオクローヌスは、ウサギを追っている間などのイヌが集中しているときはあまり可能性がなく、給餌時間で、もしくは訪問客があるなどの興奮または神経過敏であるとより可能性が高い。動物が急に座るかまたは倒れるアトニー性発作、並びに硬直、パドリング、発声、口から泡を吹くこと、排尿、および無意識を伴う全身性の発作は、共通していた。より老いたイヌは、失調症で、盲目で、これらの発作がより頻繁で、薬物に感受性が鈍かった。最終的には(年齢9〜12)、ミオクローヌスおよびドロップアタックにより、常にやむにやまれれずに安楽死させた。この臨床像は、基礎病理学と同様に(図8)、正確にヒトLD(2)を反復している。
【0162】
イヌがLDを有することを確認して、本発明者らは、最初にイヌEPM2A遺伝子をクローン化し、その関与を除外した。本発明者らは、次に疾患遺伝子座の局在を突き止めるために、ゲノム全体の走査を行った。本発明者らは、14匹の病気に冒されたイヌおよび4匹の冒されていない類縁において、全てのイヌ・ゲノムにわたって241個のイヌ特異的なマイクロサテライト・マーカー(31)の遺伝形質を決めた。冒されたイヌで最も多数がホモ接合性であり、かつ冒されていないイヌで最も多数がヘテロ接合性であった2つのマーカーは、REN94K23およびREN01G01であり、これらは小さな38Mbの染色体35(CFA35)に位置する。本発明者らは、図9aに示した、より広い血統に由来する多くのイヌを確かめて、CFA35上のさらなるマーカーの遺伝形質を決めた。組換え率θ=0.00における2.65の最大2ポイントLODスコア(two-point LOD score)がマーカーREN157J09について得られた。多点連鎖解析により、REN157J09を含む5つのマーカーを超えて3.38の最大LODスコアを生じ、イヌ連鎖研究についての3.2の有意な閾値を凌ぎ(32)(図9B)、これによりCFA35のこの領域には、MWHD LDが連鎖する。
【0163】
実施例1において論議したように、本発明者らは、ヒトEPM2B遺伝子座を染色体バンド6p22(15)にマップした。イヌCFA35は、その全体が、ヒト6p21.33-6p25.2(31)にシンテニーしており、したがって、ヒトEPM2B患者およびMWHD LDの場合において同じ遺伝子が変異される可能性があった。本発明者らは、最初にヒトEPM2Bをクローン化し(33)、その単一のエキソンを使用してRPCI-81イヌ細菌人工染色体(BAC)ライブラリーを探索するために、BAC 328A12を単離し、ヒトEPM2Bプライマーを使用するシーケンシングにより、イヌEPM2B(AY560905)の完全なオープンリーディングフレームを明らかにした。放射ハイブリッド・マッピングにより、EPM2BがマーカーREN157J09(LODスコア19.3)(MWHD LDが最大LODスコアで連鎖したものと同じマーカー(上記)(図9b))に連鎖した。全体として、これらのデータにより、EPM2BがMWHD疾患に対して生存できる候補であることを立証した。
【0164】
イヌEPM2B遺伝子のPCR増幅は、最初に病気に冒されたものでは失敗したが、病気に冒されていないMWHDにおいて遺伝子の5'の半分の領域全体ではうまくいった。この領域における正常な配列の調査により、2つの連続した同一の12塩基、および単一のヌクレオチドだけ異なった第3のコピーが明らかとなった(参照を容易にするために、完全な繰り返しコピーは、Dと名付け、および第3の不完全なコピーをTと名付けた)(図10a)。その他のシーケンスした種の対応する領域(図10a)は、繰り返しがなく、イヌ配列は、12ヌクレオチド、1回のDの繰り返しの長さだけ長い(図10a)。病気に冒された動物におけるこの領域全体を超えた増幅およびシーケンスのためのPCR条件の協奏的な改善は、最終的に成功し、MWHD LD突然変異を明らかにした:病気に冒された全ての動物は、それぞれの対立遺伝子にD配列の19〜26コピーをもつ12塩基反復の2対立遺伝子の伸長を有した(図9a、10b)。伸長は、インフレームであり、異常であるが、これらを含むmRNAが安定な場合には、タンパク質を作製することも可能かもしれない。
【0165】
本発明者らは、次に、定量的RT-PCR(補充方法)を使用して3匹の病気に冒されたイヌおよび2匹の対照からの骨格筋におけるEPM2B mRNAの量を比較することによって、mRNA安定性に対する伸長突然変異の効果を試験した。EPM2B mRNAレベルは、病気に冒された動物において900倍以上減少していた(図10c)。
【0166】
イヌEPM2Bを特徴付ける余分のD配列は、最初に、ドーベルマン・ピンシェル(Doberman Pinscher)起源のBACにおいて、および正常なMWHDにおいて検出された。本発明者らは、これが、いくつかの種類のイヌだけの特徴であり、少なくともMWHDの場合に、伸長およびLDにこれらがかかりやすいかどうかを考えた。本発明者らは、各々の128個の異なる種類から2匹の正常で無関係なイヌに由来するEPM2Bをシーケンスした。60パーセントの染色体は、3つの繰り返しを有し、40%は、2つの繰り返しを有した。3つの繰り返しをもつ対立遺伝子は、常に2つのDおよび1つのTから成っており、これらは1つのDと1つのTの2つの繰り返しをもつ。ほとんど全ての品種は、ホモまたはヘテロ接合性の状態で、両方の変異体の例を有した。異なる種類のうちの3種の4世代ファミリーにおいて試験した遺伝は、メンデルの法則によった。
【0167】
12塩基反復がイヌ種全体に存在することが示されたので、本発明者らは、その伸長が、その他の種類のイヌにおいても筋痙攣を生じさせるかどうかを考えた。本発明者らは、次の非MWHD PME症例を試験して臨床に提出し(バセット犬)、D繰り返しのホモ接合性の7倍の伸長(14コピー)を見いだした(図10b)。
【0168】
12塩基伸長突然変異の配列は、GおよびCヌクレオチドだけで構成される。正常な対立遺伝子の存在下では、伸長対立遺伝子のPCRが不可能であり、それゆえ、キャリアを決定することができなかった。キャリアを検出するために、本発明者らは、DNAを脱アミノし、メチル化されていないシトシンをウラシルに変換し、次いでPCR増幅し、これにより、変異誘発遺伝子の増幅、およびキャリアMWHD(図3b)の信頼できる検出が可能となった。また、脱アミノ化により、病気に冒された動物のPCR増幅および診断が非常に容易になった。
【0169】
上で使用した脱アミノ化法は、ヒト・シスタチンB遺伝子プロモーターに対して特異的な2〜3コピーの12塩基反復配列から生じ、かつシスタチンB発現(35)を妨げることによってEPM1(PMEのもう一つの形態)を生じさせる、現在まで唯一無二の12塩基反復の伸長突然変異の増幅のために開発された(34)。ヒトでは、2つのLD遺伝子(EPM2AおよびEPM2B)における60以上の突然変異は、PMEの最も一般的な原因ではない(http://projects.tcag.ca/lafora/)。その代わりに、単一の、何度も繰り返されるシスタチンBプロモーター伸長突然変異が、大部分のヒトPMEを引き起こす。本発明者らの知識の限りでは、イヌは、シスタチンB12の塩基反復配列を有しない可能性が高かったので、EPM1は、イヌにおいてこれまで記載されていなかった。LDは、一方ではイヌにおいて定期的に報告されている(バセット犬(36、37、38)、ミニアチュール(Miniature)(36)、およびスタンダード(39)プードル、ポインタ(40)、コルギス(Corgis)(41)、ビーグルズ(42、43、44)、およびMWHD(45、46))。これは、家畜化されたイヌに非常に多い集中的同系交配と相まって、EPM2B反復の反復性伸長事象によると思われる。その他の家畜化された動物では、LDは非常に珍しい:ウシ海綿状脳症(臨床的にLDに似ている)のための集団試験の間に同定された2頭のウシ(47)および1匹のネコ(48)。
【0170】
イヌ品種の障壁を越えた12塩基反復の存在(49)は、その起源がイヌに先んじていること、および、したがって、これが近縁種に存在するかもしれないことを示唆する。本発明者らは、食肉目における繰り返しの分布を決定し、これがイヌ科ファミリー(オオカミ、イヌ、キツネ、コヨーテ、およびジャッカル)に特異的であることを示した(図10d)。また、本発明者らは、2工程:D配列の出現、次いでその複製により生じる繰り返しの進化を識別することができた。肉食動物は、曉新世の〜6,000万年前(Ma)にネコ科およびイヌ上科に分かれた(50、51)。D配列は、現存するイヌ上科全体にわたって存在するが、ネコ科には存在しない(図10d)。したがって、これは、ネコ科(feline)-イヌ上科(canoid)がわかれた後に、その後にアルクト科(Arctoidae)とイヌ科に分岐する前の〜50Ma(図10d)に最初に現れた。アルクト科は、ユーラシア大陸上で全盛をきわめ、ここで、これらは、現存するクマ、スカンク、アライグマ、カワウソ、その他に進化した(50)。これらの種は、D配列の単一コピーを有する(図10d)。犬科の動物は、北アメリカ大陸で同等の生態学的地位を占有し(首尾よく、これは4,00万年間アルクト科(arctoid)侵略から守られる)(50)、〜10Ma〜今日までに、オオカミ、イヌ、キツネ、コヨーテ、およびジャッカル(51)に進化して、その全てが複製されたD配列(図10d)を有する。したがって、D配列の複製により、50Ma〜10Maのどこかで犬科の動物進化系統が生じた。生じるmalinの中間部分の可変性の4-アミノ酸伸長が(図10a)犬科の動物に有利なこのE3連鎖に固有性(property)を与えるかどうかは、分からないままである。
【0171】
てんかんは、イヌで最も一般的な神経疾患である。その有病率は、ヒトよりも少なくとも5倍高い(52)。本明細書において、本発明者らは、最初にイヌてんかん突然変異を発見した。これがどの程度イヌてんかんのその他の形態に寄与するかは、決定されないままである。
【0172】
英国では、現在、LDは、MWHD(毎年〜800匹のイヌが登録される品種)において広範囲にわたる。人気のキャリア雄親および繰り返し繁殖されたキャリアチャンピオンを介して、本疾患は、南アフリカおよび北アメリカまで広がった(データ示さず)。米国では、異なるダックスフンド・コート型が、同じ種類の変種と考えられ、異種交配されており、これが、ダックスフンドの全体にわたって疾患を蔓延させるかもしれない。その代わりに、本明細書に記載した研究から生じる突然変異検出試験により、繁殖を制御することによってこれを根絶できるはずである。一方、これらの動物は、愛されるペットであり、神経治療(neurologic care)を受けている。これらは、どの医療センターにおいても、ヒトLD患者よりもはるかに多い。これにより、本発明者らに対して、本疾病のための最高の治療を用いた重要な経験が与えられ、本発明者らは、これをヒト患者に適用している。
【0173】
本発明は、好ましい実施例であると現在みなされるものに関して記載したが、本発明は、開示された実施例に限定されないことは理解されるはずである。対照的に、本発明は、添付の特許請求の範囲の精神および範囲内に含まれる種々の改変および均等の処理を包含することが意図される。
【0174】
全ての刊行物、特許、および特許出願は、あたかも個々の刊行物、特許、または特許出願が特異的に、および個々に、その全体が参照として組み入れられることを示したのと同じ範囲で、これらの全体が本明細書に参照として組み入れられる。
【0175】
(表1)LD患者において同定されるEPM2B突然変異。
【0176】
明細書において言及した参照のための完全な引用
【特許請求の範囲】
【請求項1】
ラフォラ病と関連する、RINGフィンガー・ドメインおよび6つのNHLモチーフを有するタンパク質をコードする、単離された核酸分子。
【請求項2】
配列番号:1(図6A)または配列番号:3(図7)に示すとおりの配列を有する、請求項1記載の核酸。
【請求項3】
以下を含む、請求項1記載の単離された核酸分子:
(a)配列番号:1(図6A)もしくは配列番号:3(図7A)に示すとおりの核酸配列であって、TがUであってもよい核酸配列;
(b)(a)の相補的な核酸配列;
(c)(a)もしくは(b)の核酸配列に対して実質的に配列相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;または、
(e)ストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする核酸配列。
【請求項4】
動物由来の試料において、請求項1〜3のいずれか一項記載の核酸配列における突然変異を検出する段階を含む、ラフォラ病を検出する方法。
【請求項5】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号205におけるCからGへの変化を検出する段階を含む、請求項4記載の方法。
【請求項6】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号76におけるTからAへの変化を検出する段階を含む、請求項4記載の方法。
【請求項7】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置1048および1049におけるヌクレオチドGAの欠失を検出する段階を含む、請求項4記載の方法。
【請求項8】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置468および469におけるヌクレオチドAGの欠失を検出する段階を含む、請求項4記載の方法。
【請求項9】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号992におけるヌクレオチドGの欠失を検出する段階を含む、請求項4記載の方法。
【請求項10】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置373〜382における10bpの欠失を検出する段階を含む、請求項4記載の方法。
【請求項11】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置661〜692における32bpの欠失を検出する段階を含む、請求項4記載の方法。
【請求項12】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号260におけるTからCへの変化を検出する段階を含む、請求項4記載の方法。
【請求項13】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号905におけるAからCへの変化を検出する段階を含む、請求項4記載の方法。
【請求項14】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号98におけるTからCへの変化を検出する段階を含む、請求項4記載の方法。
【請求項15】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号892における2つのTの挿入を検出する段階を含む、請求項4記載の方法。
【請求項16】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号436におけるGからAへの変化を検出する段階を含む、請求項4記載の方法。
【請求項17】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号1100におけるヌクレオチドTの欠失を検出する段階を含む、請求項4記載の方法。
【請求項18】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置606におけるヌクレオチドTの欠失を検出する段階を含む、請求項4記載の方法。
【請求項19】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号923におけるAからTへの変化を検出する段階を含む、請求項4記載の方法。
【請求項20】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号580におけるGからTへの変化を検出する段階を、含む請求項4記載の方法。
【請求項21】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号199におけるGからTへの変化を検出する段階を含む、請求項4記載の方法。
【請求項22】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号838におけるGからAへの変化を検出する段階を含む、請求項4記載の方法。
【請求項23】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号676におけるCからTへの変化を検出する段階を含む、請求項4記載の方法。
【請求項24】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置468におけるヌクレオチドAの欠失を検出する段階を含む、請求項4記載の方法
【請求項25】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置204におけるヌクレオチドCの欠失を検出する段階を含む、請求項4記載の方法。
【請求項26】
表1に示したとおりEPM2B遺伝子における1つまたは複数の突然変異を検出する段階を含む、請求項4記載の方法。
【請求項27】
配列番号:3に示したEPM2Bのイヌ配列のヌクレオチド位置1001における配列
の繰り返しを検出する段階を含む、請求項4記載の方法。
【請求項28】
配列番号:5の少なくとも3つの繰り返しを検出する段階を含む、請求項22記載の方法。
【請求項29】
配列番号:5の少なくとも10の繰り返しを検出する段階を含む、請求項22記載の方法。
【請求項30】
配列番号:5の約14〜約26の繰り返しを検出する段階を含む、請求項22記載の方法。
【請求項31】
動物が、ヒトである、請求項4〜26のいずれか一項記載の方法。
【請求項32】
動物が、犬科の動物である、請求項4および26〜30のいずれか一項記載の方法。
【請求項33】
動物がイヌである、請求項4および26〜30のいずれか一項記載の方法。
【請求項34】
ラフォラ病に関連した、RINGフィンガー・ドメインおよび6つのNHLドメインを有する単離されたタンパク質。
【請求項35】
配列番号:2(図6B)または配列番号:4(図7)に示すとおりのアミノ酸配列を有する、請求項34記載のタンパク質。
【請求項36】
請求項34または35記載のタンパク質の突然変異を検出する段階を含む、ラフォラ病を検出するための方法。
【請求項37】
表1に示すとおりのEPM2Bタンパク質の突然変異を検出する段階を含む、請求項36記載の方法。
【請求項38】
配列番号:1または配列番号:3に示すとおりの核酸配列の突然変異の検出のための試薬を含む、請求項4〜33または37のいずれか一項記載の方法を実施するためのキット。
【請求項39】
配列番号:2または配列番号:5に示したとおりのタンパク質配列の突然変異の検出のための試薬を含む、請求項36または37記載の方法を実施するためのキット。
【請求項1】
ラフォラ病と関連する、RINGフィンガー・ドメインおよび6つのNHLモチーフを有するタンパク質をコードする、単離された核酸分子。
【請求項2】
配列番号:1(図6A)または配列番号:3(図7)に示すとおりの配列を有する、請求項1記載の核酸。
【請求項3】
以下を含む、請求項1記載の単離された核酸分子:
(a)配列番号:1(図6A)もしくは配列番号:3(図7A)に示すとおりの核酸配列であって、TがUであってもよい核酸配列;
(b)(a)の相補的な核酸配列;
(c)(a)もしくは(b)の核酸配列に対して実質的に配列相同性を有する核酸配列;
(d)(a)、(b)、もしくは(c)の核酸配列の類似体である核酸配列;または、
(e)ストリンジェントなハイブリダイゼーション条件下で(a)、(b)、(c)、もしくは(d)の核酸配列に対してハイブリダイズする核酸配列。
【請求項4】
動物由来の試料において、請求項1〜3のいずれか一項記載の核酸配列における突然変異を検出する段階を含む、ラフォラ病を検出する方法。
【請求項5】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号205におけるCからGへの変化を検出する段階を含む、請求項4記載の方法。
【請求項6】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号76におけるTからAへの変化を検出する段階を含む、請求項4記載の方法。
【請求項7】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置1048および1049におけるヌクレオチドGAの欠失を検出する段階を含む、請求項4記載の方法。
【請求項8】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置468および469におけるヌクレオチドAGの欠失を検出する段階を含む、請求項4記載の方法。
【請求項9】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号992におけるヌクレオチドGの欠失を検出する段階を含む、請求項4記載の方法。
【請求項10】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置373〜382における10bpの欠失を検出する段階を含む、請求項4記載の方法。
【請求項11】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置661〜692における32bpの欠失を検出する段階を含む、請求項4記載の方法。
【請求項12】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号260におけるTからCへの変化を検出する段階を含む、請求項4記載の方法。
【請求項13】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号905におけるAからCへの変化を検出する段階を含む、請求項4記載の方法。
【請求項14】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号98におけるTからCへの変化を検出する段階を含む、請求項4記載の方法。
【請求項15】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号892における2つのTの挿入を検出する段階を含む、請求項4記載の方法。
【請求項16】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号436におけるGからAへの変化を検出する段階を含む、請求項4記載の方法。
【請求項17】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号1100におけるヌクレオチドTの欠失を検出する段階を含む、請求項4記載の方法。
【請求項18】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置606におけるヌクレオチドTの欠失を検出する段階を含む、請求項4記載の方法。
【請求項19】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号923におけるAからTへの変化を検出する段階を含む、請求項4記載の方法。
【請求項20】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号580におけるGからTへの変化を検出する段階を、含む請求項4記載の方法。
【請求項21】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号199におけるGからTへの変化を検出する段階を含む、請求項4記載の方法。
【請求項22】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号838におけるGからAへの変化を検出する段階を含む、請求項4記載の方法。
【請求項23】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド番号676におけるCからTへの変化を検出する段階を含む、請求項4記載の方法。
【請求項24】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置468におけるヌクレオチドAの欠失を検出する段階を含む、請求項4記載の方法
【請求項25】
配列番号:1に示したEPM2B遺伝子配列のヌクレオチド位置204におけるヌクレオチドCの欠失を検出する段階を含む、請求項4記載の方法。
【請求項26】
表1に示したとおりEPM2B遺伝子における1つまたは複数の突然変異を検出する段階を含む、請求項4記載の方法。
【請求項27】
配列番号:3に示したEPM2Bのイヌ配列のヌクレオチド位置1001における配列
の繰り返しを検出する段階を含む、請求項4記載の方法。
【請求項28】
配列番号:5の少なくとも3つの繰り返しを検出する段階を含む、請求項22記載の方法。
【請求項29】
配列番号:5の少なくとも10の繰り返しを検出する段階を含む、請求項22記載の方法。
【請求項30】
配列番号:5の約14〜約26の繰り返しを検出する段階を含む、請求項22記載の方法。
【請求項31】
動物が、ヒトである、請求項4〜26のいずれか一項記載の方法。
【請求項32】
動物が、犬科の動物である、請求項4および26〜30のいずれか一項記載の方法。
【請求項33】
動物がイヌである、請求項4および26〜30のいずれか一項記載の方法。
【請求項34】
ラフォラ病に関連した、RINGフィンガー・ドメインおよび6つのNHLドメインを有する単離されたタンパク質。
【請求項35】
配列番号:2(図6B)または配列番号:4(図7)に示すとおりのアミノ酸配列を有する、請求項34記載のタンパク質。
【請求項36】
請求項34または35記載のタンパク質の突然変異を検出する段階を含む、ラフォラ病を検出するための方法。
【請求項37】
表1に示すとおりのEPM2Bタンパク質の突然変異を検出する段階を含む、請求項36記載の方法。
【請求項38】
配列番号:1または配列番号:3に示すとおりの核酸配列の突然変異の検出のための試薬を含む、請求項4〜33または37のいずれか一項記載の方法を実施するためのキット。
【請求項39】
配列番号:2または配列番号:5に示したとおりのタンパク質配列の突然変異の検出のための試薬を含む、請求項36または37記載の方法を実施するためのキット。
【図1】

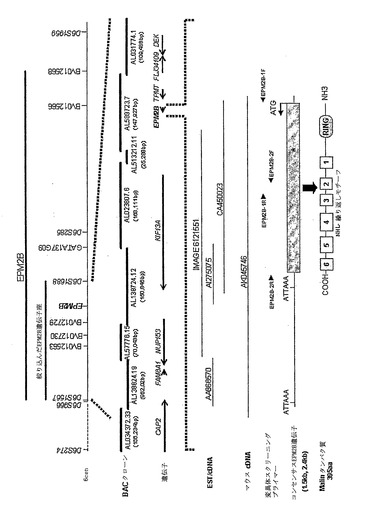
【図2】
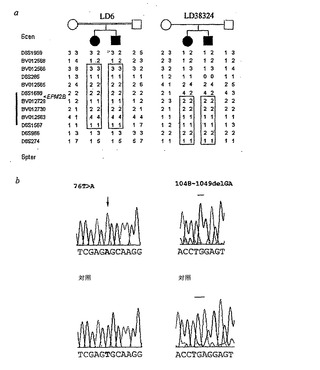
【図3】
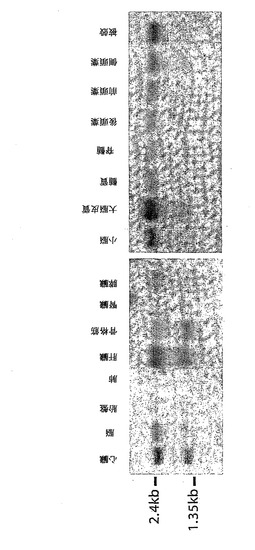
【図4】
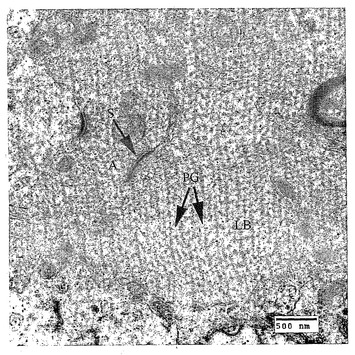
【図5】
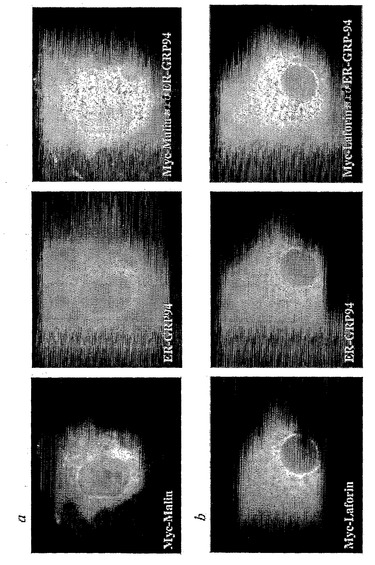
【図6A】

【図6B】

【図7A】

【図7B】

【図8】
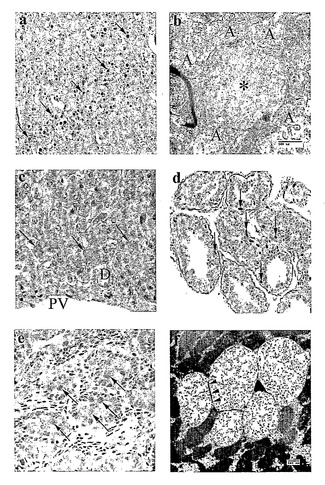
【図9】
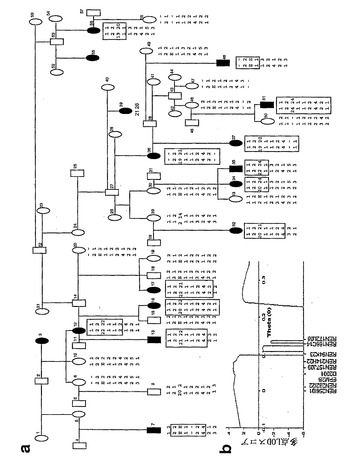
【図10】
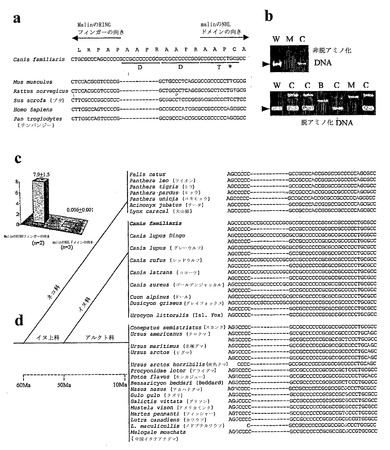
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7A】
【図7B】
【図8】
【図9】
【図10】
【公開番号】特開2012−95651(P2012−95651A)
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願番号】特願2011−261115(P2011−261115)
【出願日】平成23年11月30日(2011.11.30)
【分割の表示】特願2006−522189(P2006−522189)の分割
【原出願日】平成16年7月30日(2004.7.30)
【出願人】(501181075)
【氏名又は名称原語表記】THE HOSPITAL FOR SICK CHILDREN
【Fターム(参考)】
【公開日】平成24年5月24日(2012.5.24)
【国際特許分類】
【出願日】平成23年11月30日(2011.11.30)
【分割の表示】特願2006−522189(P2006−522189)の分割
【原出願日】平成16年7月30日(2004.7.30)
【出願人】(501181075)
【氏名又は名称原語表記】THE HOSPITAL FOR SICK CHILDREN
【Fターム(参考)】
[ Back to top ]