
ランプブラシ染色体の生産
【課題】転写活性を有する染色体(即ち、ランプブラシ染色体)を、凝集クロマチンまたは核から誘導する方法を開示する。
【解決手段】凝集染色体を胚胞内容物と接触させる。好ましくは、核のエンベロープが破壊されるか除去される。異なる種の生体に由来する、異種の染色体及び胚胞系が好ましい。これはランプブラシ染色体を持たない、または他の技術で操作できない生体を分析することができるからである。このようなランプブラシ染色体は物質に結合させることができ、そして種々の分子的及び細胞学的技術、例えば染色体タンパク質の抗体染色、オートラジオグラフィー、電子及び光学顕微鏡、組織化学、イメージ分析、免疫蛍光、核酸のイン・サイツ(in situ)ハイブリダイゼーション、形態学などによって分析される。
【解決手段】凝集染色体を胚胞内容物と接触させる。好ましくは、核のエンベロープが破壊されるか除去される。異なる種の生体に由来する、異種の染色体及び胚胞系が好ましい。これはランプブラシ染色体を持たない、または他の技術で操作できない生体を分析することができるからである。このようなランプブラシ染色体は物質に結合させることができ、そして種々の分子的及び細胞学的技術、例えば染色体タンパク質の抗体染色、オートラジオグラフィー、電子及び光学顕微鏡、組織化学、イメージ分析、免疫蛍光、核酸のイン・サイツ(in situ)ハイブリダイゼーション、形態学などによって分析される。
【発明の詳細な説明】
【技術分野】
【0001】
1.発明の分野
本願発明は、転写活性を有する染色体(即ち、“ランプブラシ”染色体)を凝集したクロマチンまたは核から誘導する方法に関する。
【背景技術】
【0002】
2.関連技術の説明
100年以上も前に、Flemmingはサンショウウオ,アンビストーマ・メキシカナム(Ambystoma mexicanum)の卵母細胞の核や胚胞(GV)内の巨大な染色体について記載している。彼の初期の簡単な観察に続いてすぐ、脊椎でも無脊椎でも、多くの動物の胚胞内の巨大染色体の存在を確立する詳細な研究が行われた。Ruckertは、その染色体が燈油ランプの煙突部を掃除するために使用されるブラシに似ているという考えから、それをランプブラシ染色体(LBC)と名付けた。
【0003】
両生類の卵母細胞のランプブラシ染色体は、既知の中では最大級の染色体であり、匹敵するものとしてはドロソフィア(Drosophila)及び他のハエの巨大な多糸染色体のみである。それらの特徴は、染色体の主軸から横方向にループ状に突き出た、何百、何千という転写活性を有する領域の存在である。各ループは1個もしくは数個の活性な遺伝子からなる。形態及び/または、個々のループ及び主軸に沿った他の特徴的な部分(例えば、動原体、末端染色小粒、軸顆粒など)の分子組成における際立った変化によって、形態学、抗体結合及びイン・サイツ(in situ)の核酸ハイブリダイゼーションを組合せて用いて、慣用の光学顕微鏡によるこれらの染色体の遺伝子レベルでのマッピングが可能となる。
【0004】
ランプブラシ染色体は種々の生体内で見い出されるが、他の多くではよくわかるようには存在しておらず、幾つかの生体では胚胞の大きさが小さいために、検出できない場合もある。例えば、典型的なランプブラシ染色体がヒトの卵母細胞も含めた哺乳類の卵母細胞で生じるのかどうかは未だ議論の余地があり、例えランプブラシ染色体が生じるにしても、胚胞が非常に小さいので、このような染色体を従来技術によって分析することはできない。
【0005】
ランプブラシ染色体の全体的な構成及び機能上の特徴が、広範な実験作業と推論の的になっている(Callan,1986;Davidson,1986に概説されている)。LBCsは成長期の卵母細胞内に生じるため、相同染色体が1以上の交叉を伴って起きる減数分裂Iの静止した複糸期相中にある。各相同物はDAPI陽性の染色体軸をもち、姉妹染色体が密接に会合する転写不活性な領域に対応する。多くのループ対が染色小粒の軸の横方向へ広がる。これらは姉妹染色体が1つ1つ完全に分離している、転写活性を有する領域からなる。光学顕微鏡レベルの分析においてさえ、ループは均一な太さではなく、1以上の“細−太”領域を有しており、これが転写ユニットに対応する。電子顕微鏡によって、“細−太”形態は、転写ユニットが転写方向に沿っている発生期のRNP繊維の増大する長さを反映していることが示される。LBCsのこの構造はそれが転写活性状態にあることを反映している。
【0006】
ランプブラシ染色体上の詳細な形態的及び分子的情報が豊富であるにもかかわらず、その構造及び特にその高い転写活性という特徴についての根本的な問題は残ったままである。一時期、その構造は異常なものであると見られたが、体細胞の核でのループ化したクロマチンドメインの発見(PaulsonとLaemmli,1977;SaitohとLaemmli,1993;Yokotaら,1995)によって、ランプブラシ状態が通常の染色体としての良好な形であることが示されている。しかしながら、凝集した染色体をランプブラシ状態にするシス(cis)及びトランス(trans)の因子は同定されていない。
【0007】
LBCsについてのこれら及びその他の問題は、ランプブラシ状態を人工的に誘導し、操作できる系において、より容易に取りかかれるものになる。本出願において、我々は、異種の胚胞に注入された凝集クロマチンが数時間のうちに特有の転写活性を有するLBCsになることを開示する。この系は、LBCsの形態的及び分子的特性を確立する際のシス及びトランスの作働性因子の関連重要性を評価するのに有用であろう。それはまた、卵母細胞が現在の技術で取り扱うことができない、または典型的なランプブラシ段階を経ない生体からの転写活性を有する染色体の分析を可能とするものである。さらに、分裂間期の核からのLBCsは、蛍光イン・サイツハイブリダイゼーションによって染色される、分裂間期の染色体よりも高解像度である細胞遺伝的な分析に有用なループの様式を呈示する。このようなループの様式はマッピングを、本質的に単一遺伝子レベルで可能とする。
【発明の開示】
【課題を解決するための手段】
【0008】
本願発明の1つの態様において、精子または他の核からの染色体を卵母細胞の胚胞と接触させて、転写活性を有する染色体(即ち、ランプブラシ染色体)を生産する方法を提供する。このような例として、無傷の胚胞へのマイクロインジェクション、胚胞や細胞が存在しない系で分画された卵母細胞の抽出物への接触、及び胚胞内容物の分離分画とのインキュベーションがある。好ましくは、染色体及び胚胞が異種の生体由来である(即ち、異種間の方法)。
【0009】
染色体を含有する核は、精子、他の全能細胞、分裂期間細胞、体細胞または分化細胞、造血細胞、良性または転移性細胞、正常または形質転換された固形組織、非増殖性細胞及び非複製性細胞由来であってもよい。核の原料は、脊椎動物、例えば魚、哺乳動物、ハムスター、マウス、ラットまたはヒトであってもよい。
【0010】
胚胞及び卵母細胞は、好ましくは両生類(例えばカエル、イモリ、サンショウウオ、ヒキガエル)からのものであるが、好ましい染色体の原料ではない。異種間の方法において、生産された染色体は、胚胞を起源とするランプブラシ染色体に似た形態を有する。
【0011】
染色体を取り囲む膜、特に核エンベロープの破壊または除去は、胚胞内容物との接触に先立って行われる。界面活性剤,酵素,機械的な破壊,またはそれらの組合せを、膜の除去に使用してもよい。
【0012】
本願発明のもう1つの態様は、固相物質上の本願発明によって生産された転写活性を有する染色体(即ち、ランプブラシ染色体)を提供する。好ましくは、生産された染色体は、顕微鏡のガラススライドまたは電子顕微鏡のグリッドに結合させる。
【0013】
本願発明の利点としては、ランプブラシ染色体が生産される条件を操作し、特有のループの様式を有する分裂間期の核から染色体を提供し、このようなループ化された染色体を、本来ランプブラシ染色体を含んでいない生体または組織から生産する能力がある。
【発明を実施するための最良の形態】
【0014】
本願発明は、高度に凝集したクロマチン(例えば、精子頭部で見出されるようなもの)を、種々の分子的技術、例えばタンパク質への抗体結合及び核酸に対するイン・サイツハイブリダイゼーションなどによって細胞学的にマッピングし、分析することができる、転写活性を有する巨大なランプブラシ染色体に変換する方法を提供する。
【0015】
その方法は、形態的、オートラジオグラフィー、組織化学的染色、免疫蛍光及び他の細胞学的または分子的技術によって分析できるように、顕微鏡スライド上または他の物質上に染色体を生成する。
【0016】
1つの基本となる方法は、精子頭部を界面活性剤(リゾレシチンなど)で処理し、核のエンベロープを含む、覆っている膜を破壊するか除去することからなる。そして精子頭部は、生きている両生類の卵母細胞の巨大な核や胚胞(GV)にマイクロインジェクションされる。24−48時間の間に、精子頭部の高度に凝集した、不活性の染色体は、GVの核質に放出され、転写活性を有する巨大な“ランプブラシ”染色体に形質転換する。
【0017】
本願の目的は、顕微鏡で同定でき、かつ/または遺伝子的にマッピング可能な巨大な染色体を、精子頭部または、通常染色体が見えないかもしくは細胞学的に分析できない他の核から生成することである。考え得る使用としては、
a)染色体異常や不妊症の可能性となる原因の存在を評価するための、精子または別の全能細胞の分析。
b)不妊症の問題または遺伝的変異に連関する可能性がある染色体数または染色体異常の決定のための、ヒト精子を含む、生体からの精子細胞の分析。
c)染色体上の遺伝子位置、遺伝子構造、クロマチン構造、機能的遺伝学、または他の染色体成分(動原体,末端小粒,非遺伝的(non-genic)配列,転写制御要素など)についての情報を得るための、ヒトを含む、生体からの正常染色体の分析。
d)癌または他の疾患に関連する可能性がある染色体異常の存在を評価するための、体細胞または分化した細胞からの核の分析。
e)その他の可能な方法(ゲノムマッピング、核型付け(karyotyping)など)より高解像度での細胞遺伝的分析。
f)通常は巨大な染色体をもたない生体(ヒトを含む全ての哺乳動物)からの染色体の詳細なマッピング。
【0018】
精子頭部または他の核を取り囲む膜は、部分的または完全に、化学的に(界面活性剤、酵素またはそれらの組合せ)除去され、そして精子頭部は卵母細胞の核または胚胞(GV)にマイクロインジェクトされてもよい。数時間後、精子頭部または他の核は自然に、個々に認識される染色体(転写活性を有する“ランプブラシ”染色体)を生じる。
【0019】
本願発明を実施するための別の方法は、精子または核からの凝集した染色体をGV抽出物または分画された卵母細胞中でインキュベートすることです。このような細胞のない系は、DNA複製または細胞周期を再構築するために確立され(LohkaとMasui,1983及び1984)、更なる分画によって、LBCsを生成するのに関係するトランスの因子が同定されるだろう。
【0020】
ランプブラシ染色体が誘導する活性を富豊化または欠乏させた非細胞性の分画を調製するために、抽出物を非細胞性の分画法(密度浮遊または遠心沈降など)を用いて調製することができる。抽出物または分画は、沈殿法(硫安、熱変性、ポリエチレングリコールなど)、クロマトグラフィー(ゲルろ過、イオン交換、逆相、疎水相互作用、アフィニティーなど)及び電気泳動(クロマトフォーカシング、ネイティブまたは変性ポリアクリルアミドゲル移動)を含むタンパク質精製技術によってさらに分画してもよい。
【0021】
本願発明は、精子頭部からの染色体ついてと同じように、染色体を分裂間期の核からLBCsへ変換する。核を様々な組織から分離する方法が知られている。このような分離した核を、界面活性剤、酵素、または機械的な破壊で処理し、核エンベロープを除去する。そしてそれらは両生類の卵母細胞のGVへ注入され、巨大なランプブラシ染色体を生じるようになる。一般的に、分裂間期の核は精子頭部より凝集しておらず、より有効な広がりさえ有している。
【0022】
分裂間期の核を用いる利点は、正常及び腫瘍組織の両方を含めて、検査できる組織の数がより多いことである。腫瘍の染色体の構成を決定するためには、中期糸を得るためにその細胞を数日または数週間培養する必要がある。多くの場合において、増殖するための形質転換細胞を、もしくはイン・ビボの培養に適用される際の更なる染色体再配列を受けるゲノムを誘導することは不可能である。ゆえに、本願発明は、他では処理しにくい組織の染色体分析に対して提供される。
【0023】
細胞は、表現型(化学的または薬剤感受性、サイトカインまたはホルモンの応答性など)で選択されたり、検出可能なマーカー(抗体や受容体の相互作用を基礎とする分離またはソーティングなど)に従って分けられ、そして選択された細胞のLBCsの特定の遺伝子型に連関して分析される。
【0024】
両生類の胚胞抽出物も、本願発明において、生きている卵母細胞中の無傷の胚胞の代わりに使用されてもよい。精子または他の核からのランプブラシ染色体を組み立てるためのイン・ビトロ法は、マイクロインジェクションの必要性を省き、それによってその手順を大幅に簡略化し、その有用性を広げる。さらに、イン・ビトロ系は染色体の組み立て(assembly)に要求される因子の、ずっと深い分析を可能とする。例えば、多くの天然の、または遺伝子工学的に操作したタンパク質及び/または核酸は、イン・ビトロ系に容易に加えることができるが、生細胞に加えることはしばしば困難であったり、不可能であったりする。同様に、様々な構成分がイン・ビトロ抽出物から、抗体による阻害や免疫沈降、RNAの破壊を目的とするアンチセンスまたはリボザイム、化学的な阻害(ATPまたはGTPの水解、キナーゼまたはホスファターゼ阻害剤)、分画法、及び生細胞からよりずっと容易な他の技術によって除去できる。
【0025】
それゆえ、本願発明の最終目的は、転写活性を有するランプブラシ染色体を、いかなるタイプの核や他の染色体物質からでも、組成物を意のままに操作できるイン・ビトロ抽出物中において生成することである。
【0026】
本願明細書で引用される全ての書物、文献及び特許が、参照文献として全て、組み入れられている。
【実施例】
【0027】
以下の例は本願発明を例示するためのものである。しかしながら、本願発明の実施は、それらの例によって一切、限定されたり、制限されるたりするものではない。
【0028】
我々は、卵母細胞の胚胞(GV)に注入されるときの、脱膜化した精子頭部の挙動を調べた。ゼノパスのGVに注入されたゼノパスの精子頭部は直ちに膨張し、最初の数時間の内に、RNAポリメラーゼIIに対する抗体で染色し始めた。そのうちに、各精子頭部が染色体様の糸の緩んだ塊になり、24−48時間までに個々に識別できるランプブラシ染色体(LBCs)に解ける。精子由来のLBCsは、非複製性の単一の染色分体であったが、その形態及び免疫蛍光染色の性質は内因性のランプブラシ二価体と驚くほど良く似ていた。それは、部位特異的な“標識構造”と同様に、凝集した染色小粒の軸から広がった、転写活性を有するループを呈示した。3H−GTP及びアクチノマイシンの使用によって、転写は、精子頭部の初期の膨張及びRNAポリメラーゼIIの取得には必要ではないが、ランプブラシループの維持に対しては必要であることが実証された。スプライシングは、精子のLBCs形成の間のどの段階においても必要ではなかった。ゼノパスの精子頭部がイモリ,ノトフタルムスのGVに注入されたとき、生じた精子LBCsは、内因性のイモリLBCsと同様に、突出したRNAポリメラーゼII軸をもつ非常に長いループを呈示した。それはイモリ特異的なタンパク質に対する抗体で染色された。ゼノパスのGVへのカエル,ラナ・ピピエンス及びゼブラフィッシュ,ダニオ・レリオの精子頭部を含む、他の異種間の注入によって、LBCsが分類学的に遠い生体から誘導できることが実証された。GV系は凝集したクロマチンの転写活性を有するLBCsへの変換に必要な、シス及びトランス作働性因子を同定する。その系はまた、卵母細胞が典型的なランプブラシ相を経ない、もしくは現在の技術では操作できない生体からの細胞学的に分析可能な染色体を生成するのに有用である。
【0029】
(材料及び方法)
背景として有用で、一般的な参考文献はXenopus laevis: Practical Uses in Cell and Molecular Biology (Methods in Cell Biology, vol. 36, B.K. Kay and H.B. Peng, eds., Academic Press, San Diego, 1991)である。
【0030】
卵母細胞:雌ゼノパス・ラエビスは、0.1%アミノ安息香酸エチルエステルのメタンスルホン酸塩(トリカインメタンスルホン酸またはMS222,Sigma社製#A5040)を用いて麻酔した。卵巣サンプルを外科的に除去し、OR2食塩水の入った小さいペトリ皿中に18−20℃で保持した(Wallaceら,1973)。幾つかの実験においては、卵巣サンプルをクロストリジウム・ヒストリティカム(Clostridium histolyticum)からの粗精製コラゲナーゼで処理し、卵胞細胞の外皮層を取り除き、個々の卵母細胞に分離させた。Ca++のないOR2で0.2%としたコラゲナーゼ(タイプII,Sigma社製#C6885)中で室温、2−6時間の処理を行った。他の場合には、卵胞層が無傷の個々の卵母細胞または卵母細胞の小さい塊を宝石商摂子で分離した。
【0031】
GVの膨張:ゼノパスのランプブラシ染色体を研究するための詳細な説明が早くから刊行されたが(Gallら,1991)、現在における数種の決定的な修飾によって、GVの膨張の質が劇的に改善される(Gall,1998)。注入された卵母細胞を一時的に分離培地(83mMKCl,17mMNaCl,6.5mMNa2HPO4,3.5mMKH2PO4,1mMMgCl2,1mMジチオスレイトールpH7.0)に移した。GVを摂子で取り外し、20秒以内で膨張用培地(21mMKCl,4mMNaCl,1.6mMNa2HPO4,0.9mMKH2PO4,1mMMgCl2,10mMCaCl2,1mMジチオスレイトール,0.1%パラホルムアルデヒド,pH7.0)に移した。核エンベロープを宝石商摂子で除去し、核のゲルをピペットで膨張用チャンバーに移した。操作が速やかに行われたならば、核のゲルの分散が数分のうちに起こり、直ちに遠心することができる。スライドを特殊なホルダーに入れ、HS4ローター(Sorvall社製)で5000rpm(4800 g),4℃で30分間遠心した。GVの膨張をPBS中2%パラホルムアルデヒドでの1時間以上の処理で固定した。
【0032】
免疫蛍光:GVの膨張をリン酸緩衝性食塩水(PBS)中10%馬血清での15−30分間の処理でブロックし、そして一次抗体と1時間反応させた。3%馬血清での簡単な洗浄後、二次抗体でさらに1時間処理した。ウサギ血清を10%馬血清で1:200に希釈し、mAbsを希釈しない培地上清として使用した。二次抗体はCy3−またはフルオレセイン−結合ヤギ抗ウサギ血清(Cappel/Organon Teknika,Durham,NC)であった。調製物をフェニレンジアミンの1mg/ml(pH9)及び1mg/ml 4’,6−ジアミノ−2−フェニルインドール(DAPI)を含有する50%グリセリンにマウントした。
【0033】
精子頭部:精子頭部を、Gurdon(1976)に記載されるように、リゾレシチンで脱膜化した。ゼノパス・ラエビス及びシルラナ(ゼノパス)トロピカリスの精子頭部を記載するように(NewmeyerとWilson,1991)精巣から調製した。マウス,マス・マスカラス及びゼブラフィッシュ,ダニオ・レリオの精子を同様に調製した。マウスの場合には、成熟精子を精巣上体から取得し、ゼブラフィッシュの成熟精子を雄成体から圧搾した。コオロギ,アケタ・ドメスティカス(Acheta domesticus)の精子を、成虫からの6−8対の精巣を蒸留水中でホモジナイズすることによって取得し、成熟及びほとんど成熟した無傷の精子のままで、ほとんどの細胞を効果的に破壊した。精子を遠心で濃縮し、そして他の種についてと同様にリゾレシチンで処理した。各々、精子数を標準的な血球計で算出した。
【0034】
卵母細胞注入:幾つかの卵母細胞を、注入前にコラゲナーゼで部分的に脱胞化し、他の幾つかは酵素処理なしに使用した。脱胞化した卵母細胞は注射針で容易に刺し貫くことができるが、この利点は脱胞化に要する時間と処理した卵母細胞が幾分壊れやすいことで相殺される。注入前に、卵母細胞を1800 gで20−30分間遠心してGVを表面に浮かせるが、その位置は脱色領域として検出できる。ガラス針をキャピラリーチューブ(0.5mm i.d.,1.2mm o.d.)からバーティカルピペットピューラー(David Kopf Instruments,Tujunga,CA)を用いて作成した。使用前に、針はシリコン処理し、先端を摂子で掻いて内径を約10μmから20μmにした。針は後ろから鉱油を満たし、そして注入すべき溶液を先端から吸い上げる。4.8μlまたは9.6μlの注入をNANOJECTマイクロインジェクション装置(Drummond Scientific Co.,Broomall,PA)を付けた解剖用顕微鏡下で行い、プランジャーを利用して針の中の液体を押し出した。我々は、針を精子懸濁液で満たした後すぐに注入を行うことが良いことを見出した。顕微鏡スライドへスポットする試験は、放出された精子数が、満たした後の時間に連れて減少することを示したが、精子頭部の沈殿と針内部への付着によるものと推定された。
【0035】
転写及びスプライシング:転写を3Hラベル化GTPの卵母細胞の細胞質への注入によってモニターした。50%エタノール中の250μCiの3H−GTP(5.7Ci/mM,Amersham社製)を蒸留乾固し、1.25μlの水に再溶解した。約1μCi(5nl)を各卵母細胞に注入した。注入後の多くの時点で、GVの膨張を起こし、通常通り、2%パラホルムアルデヒドで固定化した。それらをエタノールで繰り返し脱水化し、アセトンから風乾させた。編入されなかったラベルを5%トリクロロ酢酸の4℃,5分間の処理で除去した。もう一度脱水化してアセトンから乾燥させた後、スライドをNTB2オートラジオグラフ用エマルジョン(Eastman Kodak社製)に浸した。オートラジオグラフの暴露を1.5dとし、続いてD19ディベロッパーで2分間現像した。GV内容物を、エマルジョンのクーマシーブルー処理によって染色した(Gallら,1991)。スプライシングを防ぐため、U2 snRNAに相補性のデオキシオリゴヌクレオチド10ngを精子を用いてGVに注入した(PanとPrives,1988;Tsvetkovら,1992)。そのオリゴはGV内の内因性U2 snRNAとハイブリダイズし、そしてRnase H活性によって1−4時間で完全に破壊される。注入24時間後に行ったLBCsの調製によって、全ての検出可能なU2 snRNAを欠失した異常に大きいループが示される(Tsvetkovら,1992)。
【0036】
精子頭部の注入
初期の実験は、脱膜化したゼノパス・ラエビスの精子頭部を、IV−V段階の卵母細胞(直径1.0mmから1.2mm)のX.ラエビスのGVへの注入に関するものであった。多くの実験において、注入体積は5−10nlで、表面上は精子頭部1−5個/nlを含有するが、注入された頭部はしばしば計算よりも低いものとなり、おそらく頭部の針内側への付着によるものと考えられる。注入後多くの時点で、GVを卵母細胞から手動で切断し、その内容物を特別に調製したウェル(well)スライドに分散することができ、GV内容物をガラス基質に確実に固着させるために、そのスライドを遠心する。我々は現在、各々が20個から60個の卵母細胞を含む数ダースの注入実験を行っている。典型的な実験として、注入された卵母細胞の約90%から約95%が24時間後で正常であり、これらの約60%から約90%がGV中に精子頭部を受けている。残りはおそらく、針がGVを刺し損ねたかもしくは精子頭部が針から噴出されない場合を表しているものと考えられる。
【0037】
精子頭部の初期膨張
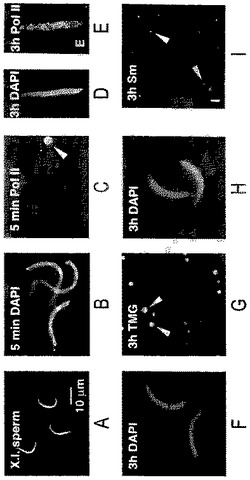
注入に用いられる、脱膜化したゼノパスの精子頭部は、約20μmの長さの線虫様構造をしており、中央付近に約1μmの最大径をもち、両端に向かってだんだん細くなっている(図1A)。GVへの注入後、それは、その形態とDNA特異的な染料であるDAPIによる強力な染色によって容易に識別できた。精子頭部における変化の最初の兆しは膨張であり(図1B−C)、それは数分のうちに、フェイズコントラストまたは示差干渉(DIC)顕微鏡によって検出可能な膜やエンベロープを形成することなしに起こった。その後の事象の起点は実験毎に変化したが、おそらく異なる雌からの卵母細胞による違いに関連するものと考えられる。多くの場合、膨張した精子頭部は最初の3−6時間、ほとんど変化しないままであったが(図1D−I)、別の実験では著しい脱凝集とLBC構造の最初の兆候が認められた(図2)。
【0038】
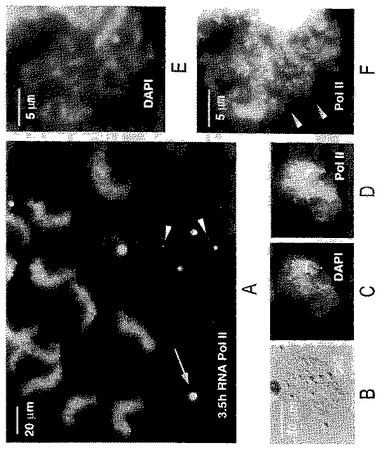
精子頭部の組成における変化をモニターするために、我々は、転写と核のスプライシング構成分を検出する3つの抗体:RNAポリメラーゼII(Pol II)に対するmAb H14(Bregmanら,1995),スプライシングsnRNAsと会合したSmタンパク質に対するmAb Y12(Lernerら,1981),snRNAsのトリメチルグアノシンキャップに対するmAb K121(Krainer,1988)。mAb H14は、リボヌクレオプロテイン(RNP)マトリックスは染色しないままで、横向きのループ軸のみを染色するため、LBCsの研究には特に重要である。このように、個々のループの経過を、例えかなりのオーバーラップがある領域においてさえ追うことが可能である。GV中の染色体外のオルガネラの中では、mAb H14は、本質的には他の全てを染色しないままで、コイル部(球体)に対して高い特異性を示す(図1C及び2A)。コイル部の染色がRNAポリメラーゼIIによるものか、交差反応性のエピトープによるものなのかは不明である。mAb Y12及びK121はsnRNPsの同定を可能とする。両方ともループのマトリックスを染色し(図3)、GV中のsnRNP含有部位がB−スナーポソームとして参照される(Wuら,1991)(図1G及びI)。
【0039】
精子頭部は注入後、最初の数時間のうちにmAb H14で染色され始めるが(図1D−E)、おそらくRNAポリメラーゼIIの、GV中に蓄えられたものからの取り込みによるものと推定される。この取り込みは転写に伴うものではなく、転写がアクチノマイシンによって阻害されるときでさえ起きる。初め、スプライシング因子は、Y12及びK121染色がないことによって示されるように(図1F−I)、取り込まれない。精子頭部が膨らみ続けるにつれ、RNAポリメラーゼIIに対して次第に強く染色される(図2A)。DNAに対するDAPI染色は、この膨張した頭部の内部に個々の染色分体が初めて示されたことを明らかにする。この初期段階においてさえ、注意深く観察すれば、DNAから広がったRNAポリメラーゼII反応性ループの存在が明らかになる(図2F,矢じり)。これらのループはRNA合成の始まりと発生期の転写物の集積の合図である。
【0040】
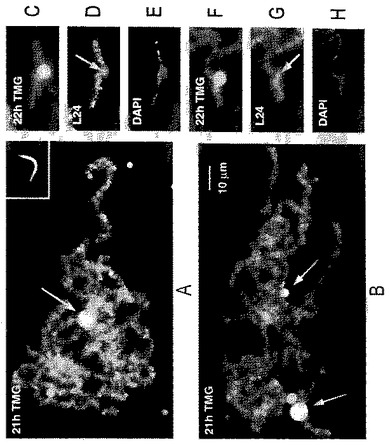
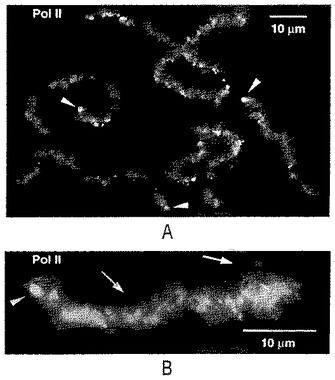
完全なランプブラシ構造の外観
精子頭部は最終的に、内因性のLBCsによく似た綿毛状の糸の緩い房に解けた(図3)。時間に連れて、これらの房は個々の染色体または染色体の小さな集まりへと分離し、我々は“精子LBCs”として称した。精子LBCsは、対になってはおらず、同じGV中の18個の内因性の二価体よりもわずかに短いため、容易に識別できた(図4)。精子のLBCsは、GVの体積中に満遍なく拡散するのではなく、一緒になったままであり、注入部位からほんのゆっくりとしか移動しないと推定された。多くの実験において、1個から約20個の精子までが単一のGVに注入された。この制限内では、精子のLBCsが、1つの精子頭部に由来しようと多くの精子頭部に由来しようと、形態及び染色性において類似しているという意味において、その系は飽和していなかった。
【0041】
全体的な構造において、精子LBCsは、同じ核内の内因性の染色体によく似ている。それらの各々は、DAPI陽性の染色小粒の軸を有しているが(図3E及びH)、内因性のLBCsよりも強くは染色されず、単一の染色分体と、2つの密に対となった姉妹染色分体との差を反映していると推定される。精子LBCs上のループは、xnf7に対するL24(Reddyら,1991)などの、発生期の転写物に会合する種々のタンパク質に対する抗体で染色され(図3D及びG)、それらはまた、Y12及びK121での染色によって実証されるように、スプライシング因子を含有していた(図3A−C及びF)。各々のループの軸は、RNAポリメラーゼIIに対するmAb H14で明確に輪郭が描かれた(図5B)。
【0042】
精子のLBCsは完全な染色体
X.ラエビスの18個のランプブラシ二価体のうち最も長いものは、最も短いもののちょうど2倍の長さであり、残りの染色体は、少しずつ大きくなる順次列を形成している(Muller,1974;Callan,1987)。精子LBCsは、特に長いまたは短いものもなく、大きさは同じ位である(図4及び5)。それゆえ見かけ上、それらは、あたかも精子クロマチンの不定のフラグメントに由来するように見える。しかしながら、より綿密な検査及び内因性の二価体との比較によって、そのほとんどが完全な無傷の染色体であることが示される。
【0043】
18個の二価体のうち、15個は長腕の端に目立つ末端顆粒を保有し、初期の文献においては末端小粒として照会されていた(Callan,1987)。この顆粒が、RNAポリメラーゼIIを含有するという直接的な証拠はないが、5S rRNAをコードする遺伝子を含有し、mAb H14で強く染色される。この顆粒は、お互いに、様々な組合せで融合する傾向がある。ほとんど全てのGVが、2つの相同体の末端顆粒が単一の顆粒に融合している少なくとも1個の二価体を有し(図6A)、非相同性の染色体の末端の間の融合は同じ頻度である(図2A,矢じり)。精子LBCsの原因を表わす調査によって、その多くがまた、mAb H14で染色される末端顆粒でキャップされる一端を有することが示される(図5A−B)。精子LBCs間の非相同性の融合は特に頻繁なものであり、見かけ上、分枝した染色体のように見える、多触手の配置を生じる(図6C)。
【0044】
精子LBCsが完全な染色体であるという最も強力な証拠は、特定の染色体の同定から得られる。中でも最も特徴的なLBCsとして同定されるのは14番である。この染色体の中央付近には、mAb Y12及びK121で強く染色される、卵型または球形の塊が付着する短い領域がある。このような塊は、精子LBCsの各々の房における顕著な特徴であり(図3A−B)、多くの場合、14番染色体が個々に認識される(図3C−H)。
【0045】
11番染色体もまた、容易に同定される。突出した末端顆粒に加えて、mAb H14で明るく染色される、2つの軸をなす顆粒を有する(図6A)。長腕の端に最も近い軸性顆粒は、U2 snRNA遺伝子の配座であり、しばしば突出したループ対と会合する。もう1つの軸性顆粒は動原体のすぐ近くにある。11番染色体特有の特徴は、ほとんど認識されない染色小粒をもつ短腕の緩い構造と頻繁なダブルループブリッジである。図6B及びCは11番染色体の特徴を呈する精子LBCsの2例を示す。図6Bにおいて、今述べられた全ての特徴が認識され、図6Cにおいて、U2配座での軸性顆粒は明確ではないが、その染色体は11番として同定される。同定可能な染色体の追加例は、異種の精子頭部の注入を扱うセクションで指摘される。
【0046】
精子LBCsは単一の非複製性の染色分体を含有する
各々の精子は、半数体(haploid)数の非複製性染色分体を、X.ラエビスの場合では18個を含有する。結果として、各々の精子LBCは、DNA合成が精子の注入後に起こらなければ、非複製性の染色分体から構成されることになる。LohkaとMasui(1983)に示されているように、精子頭部がゼノパスの卵抽出物中でインキュベートされると、S期に入り、DNA合成を開始する。しかしながら、卵母細胞のS期は、減数分裂の開始前の分裂間期で起こり、ランプブラシ段階でのGV内のDNA合成についての確証はない。しかしながら、rDNAの異常な増幅が、両生類の卵母細胞内の太糸期で起こることから、GVのDNA合成能の評価にいくらか注意を払う必要がある(BrownとDawid,1968;Gall,1968)。
【0047】
ランプブラシ二価体の各相同体内の2つの染色分体の存在が、横方向のループの対になる性質から初めて推定された(Callan,1986中で論じられている)。精子LBCsが非複製性の染色分体からなるものならば、対にならないループを有するはずである。精子LBCのループの大多数が非常に短く、外観における評価では類似しているが、それが対にならない条件が幾つかの環境下で明らかにされている。第一に、特定のループが大きさや異常な形態のために識別されるとき、それは典型的な二価体であるように見えるが、いつも単一であり、決して対にならない(図7A−D)。第二に、幾つかのループは抗体で特異的に染色される。また、これらのループは精子LBCs上では単一である(図8E−F)。第三に、慣用のLBCsを引き伸すと、“ダブルループブリッジ”として知られる現象を呈する(Callan,1955;CallanとLloyd,1960)。ダブルループブリッジは、ループ対の塩基が伸長下で離れるときに起こる。本来対である姉妹ループであったものが、染色体の軸に沿って引き伸ばされた平行な糸のようになる。このようなダブルブリッジは、精子LBCsの場合には見られないが、代わりに“シングルループブリッジ”が比較的頻繁に認められる(図8G−H)。これらのブリッジは、精子LBCが単一の染色分体からなることを示している。
【0048】
転写及びスプライシング
ランプブラシループが実際にRNA合成に関与し、アクチノマイシンやアルファ−アマニチンでの転写阻害は、ループのRNPマトリックスの損失とループの収縮を導くことが何度も実証されている(Izawaら,1963;Schultzら,1981;Callan,1986)(図9G)。転写及び精子LBCsの形成における初期の事象とのありうる関係を調べるために、我々は、卵母細胞の細胞質へ3H−GTPを注入し、その後すぐに精子をGVに注入した。幾つかの卵母細胞をアクチノマイシン中で約1時間前もってプレインキュベートし、注入後アクチノマイシンに戻した。両セットの卵母細胞において、精子頭部は初期の膨張を経て、1時間までにはRNAポリメラーゼIIに対するmAb H14で強く染色された(図9A−B)。この最初の時点では、スプライシング構成分は精子頭部中に現れなかった(図9C−F)。アクチノマイシン中でさらにインキュベーションした後でも精子頭部に変化は認められず、最終的に形成された典型的な精子LBCsをOR2食塩水中に保った。精子及び3H−GTPを注入7時間後、OR2食塩水中に保存した卵母細胞において、核小体及び内因性染色体が強くラベル化され、膨張した精子頭部はラベル化されなかった(図10A)。3H−GTPはアクチノマイシン処理した卵母細胞においてはいかなる核構造にも編入されなかった(図10C)。ゆえに精子の膨張とRNAポリメラーゼIIの集積の初期段階が転写なしに起こったのである。1日後に検査した卵母細胞は、よく形成された精子LBCsを呈し、このときまでは活発に転写を行っていた(図10B)。
【0049】
アンチセンスオリゴデオキシヌクレオチドを用いる初期の実験によって、LBCsにおける転写はU2 snRNAが存在しなくても起こり得るものであり、ゆえにそれに伴うスプライシングを必要としないことが示された(Tsvetkovら,1992)。我々は、スプライシング機構が破壊された卵母細胞中の精子頭部の組み立てを実証する同様な実験を行った。図10D及びEは、2d前に精子を注入した卵母細胞からの、及び、2d前にU2 snRNAに対するオリゴデオキシヌクレオチドを注入した卵母細胞からの精子LBCを示す。この精子LBCはU2枯渇のGVに特有の2つの特徴、そのループが異常に突出していることと、mAb K121での染色が弱いことを呈する。標的としたU2 snRNAが注入後の最初の4時間弱のうちに破壊されるため(Tsvetkovら,1992)、この実験によって、精子LBCsがスプライシングなしで形成し得ることが実証される。
【0050】
まとめとして、精子膨張の初期段階及びRNAポリメラーゼIIの集積が通常、転写なしに起こり、転写がアクチノマイシンによって阻害されているときでさえ起こり得る。転写が、精子頭部からの細胞学的に識別される染色体の発生に必要かどうかは決定されていないが、典型的なランプブラシループの維持は転写に依存する。精子LBCsの形成にも維持にもそれに伴うスプライシングは必要とされない。
【0051】
異種間の注入
我々は、ある1つの種の脱膜化した精子頭部を別の種のGVに注入する幾つかの実験を行った。このような実験によって、LBCsの組み立てにおけるトランスの因子からシスの因子を区別すること、言い換えれば、精子クロマチン、タンパク質、及び宿主GVに供給される他の因子の相対的な寄与を評価することが可能になり始めている。
【0052】
シルラナ(=ゼノパス)・トロピカリスの精子をX.ラエビスのGVに注入した。S.トロピカリス由来の精子LBCsは、同種のX.ラエビス注入からのものと全く同じに見える。S.トロピカリスは半数体セットとして10個しか染色体を持たないから、特定の染色体を同定することは容易であり、精子LBCsが完全な染色体であるという付加的な証拠を提供する。ヒョウガエル,ラナ・ピピエンスもまたX.ラエビスのGVに注入された。また、生じた精子LBCsは、長さと一般的な形態において内因性のLBCsによく似ており、特定の染色体が認識できた(図7A−D)。DAPI染色で調べたところ(図7E)、精子LBCsの染色体軸は、比較が単一と対となった染色分体との間のものであるにもかかわらず、内因性のX.ラエビスの二価体のものより突出していた。強いDAPI染色はおそらく、R.ピピエンスがX.ラエビスよりも少ない染色体を有するが(n=13対n=18)より大きいゲノムを有する(6.8×109bp対2.8×109bp)結果、R.ピピエンスの“平均的な”染色分体は、X.ラエビスからの“平均的な”染色分体の約3倍のDNAを含有するという事実を反映していると考えられる。
【0053】
R.ピピエンスの精子LBCs上のループと内因性の二価体との類似は、予想されたものではなかった。R.ピピエンスのそれ自身の卵母細胞内のLBCsは、より大きいループを呈し、我々は精子LBCs上のより突出したループを見ることを予想していた。即ち、我々は、精子クロマチンの起源が、生じるLBCsの形態により大きい影響を与えることを期待していたのである。
【0054】
ゼブラフィッシュ,ダニオ・レリオの精子がX.ラエビスのGVに注入されたとき、生じたLBCsは、ずっと短いけれども、一般的な形態においては内因性のX.ラエビスの二価体に似ていた。図7F中のダニオ染色体は、観察された最も長いものである。ダニオは25個の半数体数及び1.7×109bpのゲノムDNA含量を有するため、“平均的な”ゼブラフィッシュの染色分体は、“平均的な”ゼノパスの染色分体の約0.44倍 のDNAを有する。
【0055】
我々が、X.ラエビスの精子をイモリ,ノトフタルムス・ビリデセンスのGVに注入したとき、予想し得ない結果が得られた。形態におけるゼノパス及びイモリLBCsの違いはかなりのもので、イモリのランプブラシループは既知のものの中で最も大きい部類であった(図8A−B)。N.ビリデセンスのGV中のX.ラエビスの精子LBCsが、全体の長さがずっと短いことを除いて、内因性のイモリの染色体と著しいほど似ていたことを見出して驚いた。この類似はループの3つの特徴、RNPマトリックスの長さ,形態、及びポリメラーゼII軸の突出に及んでいた。最も明らかな類似はループの長さにおいてであり、X.ラエビスの染色体で普通に見られるものより大きい(図8C−Hと図5−6を参照)。多くのループが、平均で内因性のイモリ染色体上のものほど長くはないが、長さ30μm−40μmであった。精子LBCループのRNPマトリックスは、イモリのLBCsのものと同じように形態において豊満なものであり、種類に富んでいる(図8D及びF)。ループ形態での同様な変化が、内因性のX.ラエビスのLBCs上で起こるが、あまり目立つものではない。目立つ特徴は、ループ軸のRNAポリメラーゼIIに対するmAb H14での染色である。正常なイモリLBCs及びイモリGV中のX.ラエビスの精子LBCsにおいて、この軸は普通、突出しているが(図8A及びC)、正常なX.ラエビスのLBCsにおいてはあまり目立たない(図6A)。
【0056】
我々は、イモリのGV中のX.ラエビスの精子LBCsに会合するタンパク質を調べるために、2つのイモリ−特異的な抗体を用いた。mAb A33はプロイロデレス・ウォルツル(Pleurodeles waltl)から最初に記載された亜鉛フィンガータンパク質(A33)を検出し(Lacroixら,1985;Belliniら,1995)、mAb SE5は、N.ビリデセンスからの非関連タンパク質(SE5)を検出する(RothとGall,1987)。両方の抗体とも、N.ビリデセンスのほとんどのRNPマトリックスを染色するが、LBCs上のループは染色せず、X.ラエビスの正常なLBCsとも反応しない。しかしながら、注入実験においては、両方ともX.ラエビスの精子LBCsを、同じGV中の内因性のN.ビリデセンスの染色体と区別がつかないほど染色した。この観察は、イモリのタンパク質がイモリのGV中のX.ラエビスの精子LBCsの組み立てに使用されることを実証する。
【0057】
他の異種間の注入実験が実施され、ある程度の成功を納めている。マウス,マス・マスカラス及びコオロギ,アケタ・ドメスティカスからのリゾレクチン処理した精子頭部がX.ラエビスのGVに注入された。両種の精子頭部が初期の膨張を経て、mAB H14で染色し始めた。しかしながら、どちらの場合においても、数個の短いループがマウスの核から広がっていたが、個々の染色体が見えるようにはならなかった。同じ精子頭部のサンプルは、ゼノパスの卵抽出物中で30−60分間のインキュベーション後、典型的なイン・ビトロの核を形成し、後者のアッセイにおける反応能がGV系での反応能に対しての十分な規準ではないことを示唆した。
【0058】
我々は、転写活性を有する典型的なランプブラシ染色体が、脱膜化した精子頭部が無傷の卵母細胞のGVに注入されると、精子のクロマチンから組み立てられることを示した。15年前、LohkaとMasuiは、このような精子頭部が、ゼノパスの卵の抽出物中でインキュベートされると、本質的に正常な前核に形質転換し(LohkaとMasui,1983及び1984)、この卵抽出物系が、DNA複製及び細胞周期の多くの側面を研究するのに使用されることを示した。GV及び卵抽出物系は基本的には違うものである。卵抽出物系においては、脱膜化した精子クロマチンが核エンベロープで取り囲まれるようになり、その後にDNA複製を経て有糸分裂に入る転写不活性な核となる。対照的に、GVの周辺環境に自由に曝されたままの同じ脱膜化した精子クロマチンは、複製を経ず、分離した、転写活性を有する巨大な染色体に形質転換する。
【0059】
両生類のGVは、初期胚となる運命の、転写及びスプライシング因子を含む大量の蓄積物質を含有することが実証されている。これらの因子はGVが注入されたDNAテンプレートを転写し、生じたRNAをプロセスできるようにする。GV抽出物は、Brownとその共同研究者の5S RNAについての広範な研究においてのように、イン・ビトロの転写をサポートするために使用されている。水性のGV抽出物は、GVが油下で分離され、そのため可溶性のコファクターやサポートに有効なU1 snRNAテンプレートからの転写物を保持するが、GVポリメラーゼII転写の研究に対しては制限的にしか使用されない。Gurdonによる初期の実験において、多くのタイプの体細胞性の核がゼノパスの卵母細胞の細胞質及びGVに注入され、その後の形態的及び転写に関わる事象が、かなり詳細に調べられた。GVに注入された核は膨張し、転写において活性となるが、明らかに無傷の核エンベロープを保持していた(Gurdon,1976)。ランプブラシ染色体はこれらの核の中に検出された。
【0060】
本願発明は、最大限凝集されたクロマチンが、比較的成熟した卵母細胞(デューモント(Dumont)段階IV−V)のGV中で1日以内に形質転換できることを示す。精子頭部は注入後、RNAポリメラーゼIIをほとんど瞬時に取得する。初期の膨張後、転写が始まり、付加因子が、snRNPを含む発生期の転写物に補給される。数倍のクロマチンがGV中で通常存在するように変換され、その方法では宿主のタンパク質が使用される。これらの特徴はGV中に蓄積されていることが知られている因子、RNAポリメラーゼII,hnRNP,snRNP,ヒストン及びヌクレオプラスミンなどの重要性を強調する。他の事柄の中で、これらの蓄積されている因子は、初期の胚形成の間のDNAからクロマチンへの変換において、及び中胞胚(mid-blastula)の転移後の転写の開始において使用される。正常なランプブラシ染色体は、転写機構がGV中では機能的であることを示し、精子の注入実験は、不活性のクロマチンが転写のために活性化され得ることを示す。
【0061】
精子クロマチン由来のLBCsの最終的な形は、形質転換が起こるGV内の因子によるのと同様に、クロマチン自身の特徴によって影響されるに違いない。X.ラエビスのGV内のX.ラエビスの精子クロマチンが、内因性のLBCsから区別できないLBCsを生じると予想される。他方、前記された異種間のプロセスの結果は期待できない。サンショウウオやある種のカエルのような、ある種の生体は非常に長いループをもつ巨大なLBCsを有し、また、ゼノパスのような他の種では並みのループをもつ中間的な大きさの染色体を有し、またさらに、わずかに検出できるループをもつ非常に短い染色体を有するものもあることが知られていた。これらの違いは多くの生体のゲノムDNA含量またはC値と論理的によく相関する。
【0062】
しかしながら、全体的な染色体の長さがC値と相関するならば、ループの長さはなぜ、ループが単に、1個もしくは数個の転写ユニットに対応するというふうにならないのか?通常与えられる説明は、ランプブラシ段階での読み込み転写の存在に基づいている(Gallら,1983)。その最も単純な形において、転写ユニットが遺伝子間の全てのスペーサーを含み、平均してスペーサーの長さがC値とともに増加し、それゆえ転写ユニットは、高いC値をもつ生体では長くなる、という理由付けがある。このモデルはシスの要素にほとんど全ての重点を置き、与えられた生体の精子は、それが置かれるGVにかかわらず、類似のLBCsを形成することを予測させる。しかしながら、異種間の注入 −特にノトフタルムスのGV内のX.ラエビスの精子、この場合はイモリの染色体に著しく似た染色体を生じるのだが− に関する我々の結果は、トランスの因子が目覚しい役割を果たすことを論じるものである。この結論は、全体的なDNA含量に基づくLBCの形態の比較的単純なモデルの修飾を必要とするものである。
【0063】
LBCsの組み立てにおいて重要な因子を調べるためには、GV含量からの構成分を足したり引いたりする必要があるだろう。アンチセンスオリゴヌクレオチドは、GV内のRNAを分解するのに効果的に用いることができる(PrivesとFoukal,1991)。初期において我々は、LBC転写がそれに伴うスプライシングを必要としないこと(Tsvetkovら,1992)を示すために、U2 snRNAのノックアウトを用いたが、同じことが精子クロマチンのLBCへの形質転換ついても有効である。タンパク質の枯渇または付加は、抗体のGVへの注入が、ある程度うまく用いられているが(Bonaら,1981;Scheerら,1984)、問題は多い。分裂間期の核の組み立てのための卵抽出物に匹敵する、LBCの組み立てに対するイン・ビトロ系を持つことが高く望まれる。油下で分離されたGV(Paineら,1992)で我々の実験を繰り返すことにまず努力が払われ、これがうまくいけば精子のLBCへの形質転換をサポートするGVまたは卵母細胞からの細胞のない抽出物が調製される。
【0064】
本願発明は、染色体及び胚胞の種々の原料での使用を記載するものである。ここで例示される胚胞原料にはカエル及びイモリが含まれる。染色体を含有する核は、例えばカエル,マウス,ゼブラフィッシュ及びコオロギの組織から取得された。本願発明は、胚胞及び染色体含有の核の様々な他の原料での使用を意図している。特に、ヒト胚胞及び/またはヒト染色体原料が転写活性を有するランプブラシ染色体を、本願による、凝集したクロマチンまたは核から生成することを期待させる。
【0065】
本願発明が、実際の、及び好ましい態様であると現在考えられているものと関連して記載されている一方、本願発明は、開示された態様に制限され、限定されるものではないが、また一方では、添付された請求項の意味と範囲内に含まれる多くの修飾及び同意義の改作を網羅するように意図されている。
【0066】
ゆえに記載された発明での改変は、当業者にとって本願発明の新規な側面から外れることなく自明なものであり、このような改変は本願発明の範囲内に入ると解釈されるべきである。
【0067】
参考文献

【図面の簡単な説明】
【0068】
【図1】(A)注入に先立って脱膜化したゼノパス・ラエビス(Xenopus laevis)の精子頭部(DNAは4’, 6−ジアミジノ−2−フェニルインドールまたはDAPIで染色した)。(B)ゼノパスのGVに注入5分後の膨張した精子頭部(DAPI染色)。(C)パネルBと同じ領域をRNAポリメラーゼIIに対するmAb H14で染色:精子頭部は陰性だが、コイル部(球体)は明るく染色される(矢じり)。(D)GVに注入3時間後の精子頭部(DAPI染色)。(E)同じ精子頭部がRNAポリメラーゼIIに対するmAb H14での強い染色を示す。(F)GVに注入3時間後の精子頭部(DAPI染色)。(G)パネルFと同じ領域をsnRNAsのトリメチルグアノシンキャップ(trimethylguanosine cap)に対するmAb K121で染色:精子頭部は陰性だが、Bスナーポソーム(snurposome)は染色される(矢じり)。(H)GVに注入3時間後の精子頭部(DAPI染色)。(I)パネルHと同じ領域をsnRNAsのSmエピトープに対するmAb Y12で染色:精子頭部は陰性だが、Bスナーポソームは染色される(矢じり)。
【図2】(A)ゼノパスのGVへの注入3.5時間後のゼノパスの精子頭部の1群であり、広範囲に膨張し、RNAポリメラーゼIIに対するmAb H14で強く染色される。内因性ランプブラシ染色体は、その末端顆粒で2つの他の染色体に融合する(矢じり)。幾つかの明るく染色されるコイル部(球体)も表わされている(矢印)。(B)同じGVからの単一の精子頭部のDICイメージ。(C)DAPI染色は精子頭部内の個々の染色分体を明らかにする。(D)RNAポリメラーゼIIに対するmAb H14による染色は精子頭部で強い反応を示す。(E)DAPI−陽性の糸を示すパネルC部の高倍率像。(F)個々のポリメラーゼII−反応性ループ(矢じり)がパネルDからの同じ領域のこの拡大で見ることができる。
【図3】(A)及び(B)ゼノパスのGVに注入21時間後の2つのゼノパスの精子頭部。その各々が、内因性LBCsの特徴の大部分をもつ、染色体の緩い房に解けている。スプライシングsnRNAsのトリメチルグアノシンキャップに対するmAb K121での染色。精子LBCsのループはsnRNAsに対して強く染色する。染色体の各々の房はまた、1つの非常に大きいK121−反応性の塊及び幾つかのより小さい塊(矢印)を呈する。これらはほとんど確実に、二価体(bivalent)♯6,♯8及び♯14で生じる同様な塊に対応する。(C)中心近くに付くK121−反応性の大きな塊をもつ単一の精子LBC。その塊の位置及びこの染色体の相対的な大きさがNo.14として同定される。(D)同じ染色体がタンパク質xnf7に対する血清L24で二重染色された(Reddyら,1991)。この抗体は、内因性LBCsにおいてと同じように、ほとんどのランプブラシ染色体と同様にその塊中の小さい封入体(矢印)を染色する。(E)同じ染色体のDAPI染色は凝集した染色体の軸を示す。(F)−(H)一端でもう1つの染色体に付いている、精子LBC No.14と思われるもののもう1つの例のK121,L24及びDAPIの染色。パネルGの矢印はその塊の内側のL24−反応性顆粒を指す。
【図4】ゼノパスの精子頭部の注入21時間後の単一のゼノパスのGV内容物の全体像。精子頭部はLBCs特有の特徴を有する、染色分体の緩い房もしくは単一の染色分体に変換されている。このGVは15−20個の精子頭部を受け入れていると思われる。18個の内因性LBCs及びコイル部(球体)の大部分がこの領域の左上部のエリアを占める(点線部)。RNAポリメラーゼIIに対するmAb H14での染色。
【図5】(A)図4で示されたGVからの数個の精子LBCsの高解像度像で、RNAポリメラーゼIIに対するmAb H14での染色。染色体の全体に亘るファジーな外観は、横方向のループの軸の染色によるものである。加えて、mAb H14は、18個のゼノパスLBCsの15の長腕の端で認められる末端顆粒(矢じり)を染色する。(B)さらに高解像度の単一の精子LBCsで、mAb H14で染色。矢じりは末端顆粒を指し、矢印はループの軸(RNAポリメラーゼIIで変換されたDNA軸)を指す。
【図6】(A)長腕の端で末端顆粒によって同定される二価体♯11の(矢じり)、及び0.16及び0.80の位置での軸の顆粒(矢印)。この例では、0.80の位置に単一の融合顆粒がある。動原体がその融合顆粒のすぐ向うに位置するが、この染色では見えない。その短腕は特徴的な拡散構造を有する。RNAポリメラーゼIIに対するmAb H14での染色。(B)精子由来の染色体No.11。末端顆粒(矢じり)、2つの軸性顆粒(矢印)及び拡散短腕が認識される。(C)精子由来の染色体No.11のもう1つの例。0.16での軸顆粒はこのイメージでは明らかではないが、染色体は0.80での軸性顆粒(矢印)、拡散短腕、及び末端顆粒(矢じり)によって確かに同定され、ここで他の2つの染色体の末端顆粒に融合して三つ組み形となる。
【図7】(A)−(D)2日前にゼノパスのGVに注入されたラナ・ピピエンス(Rana pipiens)の精子頭部に由来する同じ精子LBCの4例のフェイズコントラストイメージ(phase contrast image)。この染色体の一端の近くに、その根元に突出した顆粒をもつ異常に大きなループがある(矢じり)。そのループは、非複製性染色分体で予想されるように、単一であることに注目。(E)パネルDと同領域の、突出したDNA軸を示すDAPI染色。(F)ゼブラフィッシュ,ダニオ・レリオ(Danio rerio)の精子頭部をゼノパスのGVに注入2日後の精子LBC。トリメチルグアノシンに対するmAb K121での染色。矢じりは異常に巨大なマトリックスをもつ2つのループを指す。
【図8】(A)イモリ,ノトフタルムス・ビリデセンス(Notophthalmus viridescens)のGVからの典型的なLBCの小部分で、RNAポリメラーゼIIに対するmAb H14で染色。(B)DICによる同じ領域の像。ゼノパスのLBCと比較して、イモリのLBCはより長いループ、より重厚に折り重なって突出したループマトリックス、及び異常で特有なRNAポリメラーゼII−反応性のループ軸を有する。加えて、イモリLBCsはゼノパスLBCsより長い。(C)2日前にイモリGVに注入したゼノパスの精子頭部に由来するLBC。RNAポリメラーゼIIに対する染色。(D)DICによる同じ領域の像。このゼノパスLBCが長さ以外、イモリLBCによく似ていることに注目。(E)ゼノパスの精子をイモリGVに注入2日後のゼノパス精子のLBC。xnf7に対する血清L24で染色(Reddyら,1991)。ほんの少しのループがこの抗体に強く反応する。染色体の端近くの大きなループ(矢印)が単一であり,卵母細胞のLBCでのループのように対ではないことに注目。このループは、2つのタンデムの転写ユニットからなり、そのうちの1つを染色。(F)フェイズコントラストによる同じ領域の像。(G)イモリGVへの注入2日後のゼノパス精子のLBC。mAb SE5での染色(RothとGall,1987)で、通常イモリのLBCループで染色し、ゼノパスのものは染色しない。染色は、イモリのタンパク質がゼノパスLBCを組み立てるのに使用されたことを実証する。矢印は下記の“単一ループブリッジ”を示す。(H)染色体の染色小粒の軸を示す同じ領域の像(DAPI染色)。
【図9】(A)ゼノパスのGVへの注入1時間後のゼノパスの精子頭部。卵母細胞を注入前に1時間、アクチノマイシンD(20mg/ml)中でインキュベートし、注入後薬剤に戻した(DAPI染色)。(B)同じ精子頭部をmAb H14で染色したものであり、例え転写が阻害されていても、精子によってRNAポリメラーゼIIが取り入れられることを示す。(C)アクチノマイシン処理した卵母細胞に注入1.5時間後の精子頭部(DAPI染色)。(D)同じ精子頭部をトリメチルグアノシンに対するMab K121での染色像であり、スプライシングsnRNAsが欠如していることを示す。(E)アクチノマイシン処理した卵母細胞に注入1.5時間後の精子頭部(DAPI染色)。(F)同じ精子頭部の、トリメチルグアノシンに対するmAB Y12での染色像であり、Smタンパク質が欠如していることを示す。矢じりは、Y12反応性のB−スナーポソームを指す。(G)パネルA及びBで示された精子頭部を含有する同じGVからの高コントラストの染色体(DAPI染色)。(H)同じエリアのmAB H14での染色像であり、末端顆粒を除いた染色体の反応の欠如を示す(矢じり)。コイル部(球体)は典型的な染色を示す(矢印)。
【図10】(A)3H−GTPをゼノパスの卵母細胞の細胞質に注入し、1時間後にゼノパスの精子頭部をそのGVに注入した。7時間後に行った、GV内容物のオートラジオグラフでは、精子頭部にラベルが認められないが(矢印)、3つの核小体では強いラベルが認められ(矢じり)、rRNA転写を示す(1.5d暴露)。(B)3H−GTP注入31時間後のラベル化した精子LBCsのオートラジオグラフ及びパネルAにあるような精子頭部。活発な転写が精子LBCsのループ上で起こっている。核小体(矢じり)は銀粒子によって黒く表わされる(1.5d暴露)。(C)卵母細胞はアクチノマイシンD(20mg/ml)中で1時間、プレインキュベートし、パネルAでのように3H−GTP及び精子頭部を注入し、薬剤に戻す。7時間後に行ったGV内容物のオートラジオグラフは、精子頭部(矢印)でも内因性核小体(矢じり)でもラベルは認められない(1.5d暴露)。(D)U2 snRNAについて欠乏させた卵母細胞からの精子LBCで、xnf7に対する血清L24での染色(Reddy,1991)。ループは良く染色され、大きな“塊”内部の顆粒は染色体に付いている(矢じり)。(E)トリメチルグアノシンに対するmAb K121で染色された、パネルDでの染色体。ループは無処理の卵母細胞に比べて(図3を比較)著しく減少した染色を示したが、K121反応性の塊は影響されない。
【技術分野】
【0001】
1.発明の分野
本願発明は、転写活性を有する染色体(即ち、“ランプブラシ”染色体)を凝集したクロマチンまたは核から誘導する方法に関する。
【背景技術】
【0002】
2.関連技術の説明
100年以上も前に、Flemmingはサンショウウオ,アンビストーマ・メキシカナム(Ambystoma mexicanum)の卵母細胞の核や胚胞(GV)内の巨大な染色体について記載している。彼の初期の簡単な観察に続いてすぐ、脊椎でも無脊椎でも、多くの動物の胚胞内の巨大染色体の存在を確立する詳細な研究が行われた。Ruckertは、その染色体が燈油ランプの煙突部を掃除するために使用されるブラシに似ているという考えから、それをランプブラシ染色体(LBC)と名付けた。
【0003】
両生類の卵母細胞のランプブラシ染色体は、既知の中では最大級の染色体であり、匹敵するものとしてはドロソフィア(Drosophila)及び他のハエの巨大な多糸染色体のみである。それらの特徴は、染色体の主軸から横方向にループ状に突き出た、何百、何千という転写活性を有する領域の存在である。各ループは1個もしくは数個の活性な遺伝子からなる。形態及び/または、個々のループ及び主軸に沿った他の特徴的な部分(例えば、動原体、末端染色小粒、軸顆粒など)の分子組成における際立った変化によって、形態学、抗体結合及びイン・サイツ(in situ)の核酸ハイブリダイゼーションを組合せて用いて、慣用の光学顕微鏡によるこれらの染色体の遺伝子レベルでのマッピングが可能となる。
【0004】
ランプブラシ染色体は種々の生体内で見い出されるが、他の多くではよくわかるようには存在しておらず、幾つかの生体では胚胞の大きさが小さいために、検出できない場合もある。例えば、典型的なランプブラシ染色体がヒトの卵母細胞も含めた哺乳類の卵母細胞で生じるのかどうかは未だ議論の余地があり、例えランプブラシ染色体が生じるにしても、胚胞が非常に小さいので、このような染色体を従来技術によって分析することはできない。
【0005】
ランプブラシ染色体の全体的な構成及び機能上の特徴が、広範な実験作業と推論の的になっている(Callan,1986;Davidson,1986に概説されている)。LBCsは成長期の卵母細胞内に生じるため、相同染色体が1以上の交叉を伴って起きる減数分裂Iの静止した複糸期相中にある。各相同物はDAPI陽性の染色体軸をもち、姉妹染色体が密接に会合する転写不活性な領域に対応する。多くのループ対が染色小粒の軸の横方向へ広がる。これらは姉妹染色体が1つ1つ完全に分離している、転写活性を有する領域からなる。光学顕微鏡レベルの分析においてさえ、ループは均一な太さではなく、1以上の“細−太”領域を有しており、これが転写ユニットに対応する。電子顕微鏡によって、“細−太”形態は、転写ユニットが転写方向に沿っている発生期のRNP繊維の増大する長さを反映していることが示される。LBCsのこの構造はそれが転写活性状態にあることを反映している。
【0006】
ランプブラシ染色体上の詳細な形態的及び分子的情報が豊富であるにもかかわらず、その構造及び特にその高い転写活性という特徴についての根本的な問題は残ったままである。一時期、その構造は異常なものであると見られたが、体細胞の核でのループ化したクロマチンドメインの発見(PaulsonとLaemmli,1977;SaitohとLaemmli,1993;Yokotaら,1995)によって、ランプブラシ状態が通常の染色体としての良好な形であることが示されている。しかしながら、凝集した染色体をランプブラシ状態にするシス(cis)及びトランス(trans)の因子は同定されていない。
【0007】
LBCsについてのこれら及びその他の問題は、ランプブラシ状態を人工的に誘導し、操作できる系において、より容易に取りかかれるものになる。本出願において、我々は、異種の胚胞に注入された凝集クロマチンが数時間のうちに特有の転写活性を有するLBCsになることを開示する。この系は、LBCsの形態的及び分子的特性を確立する際のシス及びトランスの作働性因子の関連重要性を評価するのに有用であろう。それはまた、卵母細胞が現在の技術で取り扱うことができない、または典型的なランプブラシ段階を経ない生体からの転写活性を有する染色体の分析を可能とするものである。さらに、分裂間期の核からのLBCsは、蛍光イン・サイツハイブリダイゼーションによって染色される、分裂間期の染色体よりも高解像度である細胞遺伝的な分析に有用なループの様式を呈示する。このようなループの様式はマッピングを、本質的に単一遺伝子レベルで可能とする。
【発明の開示】
【課題を解決するための手段】
【0008】
本願発明の1つの態様において、精子または他の核からの染色体を卵母細胞の胚胞と接触させて、転写活性を有する染色体(即ち、ランプブラシ染色体)を生産する方法を提供する。このような例として、無傷の胚胞へのマイクロインジェクション、胚胞や細胞が存在しない系で分画された卵母細胞の抽出物への接触、及び胚胞内容物の分離分画とのインキュベーションがある。好ましくは、染色体及び胚胞が異種の生体由来である(即ち、異種間の方法)。
【0009】
染色体を含有する核は、精子、他の全能細胞、分裂期間細胞、体細胞または分化細胞、造血細胞、良性または転移性細胞、正常または形質転換された固形組織、非増殖性細胞及び非複製性細胞由来であってもよい。核の原料は、脊椎動物、例えば魚、哺乳動物、ハムスター、マウス、ラットまたはヒトであってもよい。
【0010】
胚胞及び卵母細胞は、好ましくは両生類(例えばカエル、イモリ、サンショウウオ、ヒキガエル)からのものであるが、好ましい染色体の原料ではない。異種間の方法において、生産された染色体は、胚胞を起源とするランプブラシ染色体に似た形態を有する。
【0011】
染色体を取り囲む膜、特に核エンベロープの破壊または除去は、胚胞内容物との接触に先立って行われる。界面活性剤,酵素,機械的な破壊,またはそれらの組合せを、膜の除去に使用してもよい。
【0012】
本願発明のもう1つの態様は、固相物質上の本願発明によって生産された転写活性を有する染色体(即ち、ランプブラシ染色体)を提供する。好ましくは、生産された染色体は、顕微鏡のガラススライドまたは電子顕微鏡のグリッドに結合させる。
【0013】
本願発明の利点としては、ランプブラシ染色体が生産される条件を操作し、特有のループの様式を有する分裂間期の核から染色体を提供し、このようなループ化された染色体を、本来ランプブラシ染色体を含んでいない生体または組織から生産する能力がある。
【発明を実施するための最良の形態】
【0014】
本願発明は、高度に凝集したクロマチン(例えば、精子頭部で見出されるようなもの)を、種々の分子的技術、例えばタンパク質への抗体結合及び核酸に対するイン・サイツハイブリダイゼーションなどによって細胞学的にマッピングし、分析することができる、転写活性を有する巨大なランプブラシ染色体に変換する方法を提供する。
【0015】
その方法は、形態的、オートラジオグラフィー、組織化学的染色、免疫蛍光及び他の細胞学的または分子的技術によって分析できるように、顕微鏡スライド上または他の物質上に染色体を生成する。
【0016】
1つの基本となる方法は、精子頭部を界面活性剤(リゾレシチンなど)で処理し、核のエンベロープを含む、覆っている膜を破壊するか除去することからなる。そして精子頭部は、生きている両生類の卵母細胞の巨大な核や胚胞(GV)にマイクロインジェクションされる。24−48時間の間に、精子頭部の高度に凝集した、不活性の染色体は、GVの核質に放出され、転写活性を有する巨大な“ランプブラシ”染色体に形質転換する。
【0017】
本願の目的は、顕微鏡で同定でき、かつ/または遺伝子的にマッピング可能な巨大な染色体を、精子頭部または、通常染色体が見えないかもしくは細胞学的に分析できない他の核から生成することである。考え得る使用としては、
a)染色体異常や不妊症の可能性となる原因の存在を評価するための、精子または別の全能細胞の分析。
b)不妊症の問題または遺伝的変異に連関する可能性がある染色体数または染色体異常の決定のための、ヒト精子を含む、生体からの精子細胞の分析。
c)染色体上の遺伝子位置、遺伝子構造、クロマチン構造、機能的遺伝学、または他の染色体成分(動原体,末端小粒,非遺伝的(non-genic)配列,転写制御要素など)についての情報を得るための、ヒトを含む、生体からの正常染色体の分析。
d)癌または他の疾患に関連する可能性がある染色体異常の存在を評価するための、体細胞または分化した細胞からの核の分析。
e)その他の可能な方法(ゲノムマッピング、核型付け(karyotyping)など)より高解像度での細胞遺伝的分析。
f)通常は巨大な染色体をもたない生体(ヒトを含む全ての哺乳動物)からの染色体の詳細なマッピング。
【0018】
精子頭部または他の核を取り囲む膜は、部分的または完全に、化学的に(界面活性剤、酵素またはそれらの組合せ)除去され、そして精子頭部は卵母細胞の核または胚胞(GV)にマイクロインジェクトされてもよい。数時間後、精子頭部または他の核は自然に、個々に認識される染色体(転写活性を有する“ランプブラシ”染色体)を生じる。
【0019】
本願発明を実施するための別の方法は、精子または核からの凝集した染色体をGV抽出物または分画された卵母細胞中でインキュベートすることです。このような細胞のない系は、DNA複製または細胞周期を再構築するために確立され(LohkaとMasui,1983及び1984)、更なる分画によって、LBCsを生成するのに関係するトランスの因子が同定されるだろう。
【0020】
ランプブラシ染色体が誘導する活性を富豊化または欠乏させた非細胞性の分画を調製するために、抽出物を非細胞性の分画法(密度浮遊または遠心沈降など)を用いて調製することができる。抽出物または分画は、沈殿法(硫安、熱変性、ポリエチレングリコールなど)、クロマトグラフィー(ゲルろ過、イオン交換、逆相、疎水相互作用、アフィニティーなど)及び電気泳動(クロマトフォーカシング、ネイティブまたは変性ポリアクリルアミドゲル移動)を含むタンパク質精製技術によってさらに分画してもよい。
【0021】
本願発明は、精子頭部からの染色体ついてと同じように、染色体を分裂間期の核からLBCsへ変換する。核を様々な組織から分離する方法が知られている。このような分離した核を、界面活性剤、酵素、または機械的な破壊で処理し、核エンベロープを除去する。そしてそれらは両生類の卵母細胞のGVへ注入され、巨大なランプブラシ染色体を生じるようになる。一般的に、分裂間期の核は精子頭部より凝集しておらず、より有効な広がりさえ有している。
【0022】
分裂間期の核を用いる利点は、正常及び腫瘍組織の両方を含めて、検査できる組織の数がより多いことである。腫瘍の染色体の構成を決定するためには、中期糸を得るためにその細胞を数日または数週間培養する必要がある。多くの場合において、増殖するための形質転換細胞を、もしくはイン・ビボの培養に適用される際の更なる染色体再配列を受けるゲノムを誘導することは不可能である。ゆえに、本願発明は、他では処理しにくい組織の染色体分析に対して提供される。
【0023】
細胞は、表現型(化学的または薬剤感受性、サイトカインまたはホルモンの応答性など)で選択されたり、検出可能なマーカー(抗体や受容体の相互作用を基礎とする分離またはソーティングなど)に従って分けられ、そして選択された細胞のLBCsの特定の遺伝子型に連関して分析される。
【0024】
両生類の胚胞抽出物も、本願発明において、生きている卵母細胞中の無傷の胚胞の代わりに使用されてもよい。精子または他の核からのランプブラシ染色体を組み立てるためのイン・ビトロ法は、マイクロインジェクションの必要性を省き、それによってその手順を大幅に簡略化し、その有用性を広げる。さらに、イン・ビトロ系は染色体の組み立て(assembly)に要求される因子の、ずっと深い分析を可能とする。例えば、多くの天然の、または遺伝子工学的に操作したタンパク質及び/または核酸は、イン・ビトロ系に容易に加えることができるが、生細胞に加えることはしばしば困難であったり、不可能であったりする。同様に、様々な構成分がイン・ビトロ抽出物から、抗体による阻害や免疫沈降、RNAの破壊を目的とするアンチセンスまたはリボザイム、化学的な阻害(ATPまたはGTPの水解、キナーゼまたはホスファターゼ阻害剤)、分画法、及び生細胞からよりずっと容易な他の技術によって除去できる。
【0025】
それゆえ、本願発明の最終目的は、転写活性を有するランプブラシ染色体を、いかなるタイプの核や他の染色体物質からでも、組成物を意のままに操作できるイン・ビトロ抽出物中において生成することである。
【0026】
本願明細書で引用される全ての書物、文献及び特許が、参照文献として全て、組み入れられている。
【実施例】
【0027】
以下の例は本願発明を例示するためのものである。しかしながら、本願発明の実施は、それらの例によって一切、限定されたり、制限されるたりするものではない。
【0028】
我々は、卵母細胞の胚胞(GV)に注入されるときの、脱膜化した精子頭部の挙動を調べた。ゼノパスのGVに注入されたゼノパスの精子頭部は直ちに膨張し、最初の数時間の内に、RNAポリメラーゼIIに対する抗体で染色し始めた。そのうちに、各精子頭部が染色体様の糸の緩んだ塊になり、24−48時間までに個々に識別できるランプブラシ染色体(LBCs)に解ける。精子由来のLBCsは、非複製性の単一の染色分体であったが、その形態及び免疫蛍光染色の性質は内因性のランプブラシ二価体と驚くほど良く似ていた。それは、部位特異的な“標識構造”と同様に、凝集した染色小粒の軸から広がった、転写活性を有するループを呈示した。3H−GTP及びアクチノマイシンの使用によって、転写は、精子頭部の初期の膨張及びRNAポリメラーゼIIの取得には必要ではないが、ランプブラシループの維持に対しては必要であることが実証された。スプライシングは、精子のLBCs形成の間のどの段階においても必要ではなかった。ゼノパスの精子頭部がイモリ,ノトフタルムスのGVに注入されたとき、生じた精子LBCsは、内因性のイモリLBCsと同様に、突出したRNAポリメラーゼII軸をもつ非常に長いループを呈示した。それはイモリ特異的なタンパク質に対する抗体で染色された。ゼノパスのGVへのカエル,ラナ・ピピエンス及びゼブラフィッシュ,ダニオ・レリオの精子頭部を含む、他の異種間の注入によって、LBCsが分類学的に遠い生体から誘導できることが実証された。GV系は凝集したクロマチンの転写活性を有するLBCsへの変換に必要な、シス及びトランス作働性因子を同定する。その系はまた、卵母細胞が典型的なランプブラシ相を経ない、もしくは現在の技術では操作できない生体からの細胞学的に分析可能な染色体を生成するのに有用である。
【0029】
(材料及び方法)
背景として有用で、一般的な参考文献はXenopus laevis: Practical Uses in Cell and Molecular Biology (Methods in Cell Biology, vol. 36, B.K. Kay and H.B. Peng, eds., Academic Press, San Diego, 1991)である。
【0030】
卵母細胞:雌ゼノパス・ラエビスは、0.1%アミノ安息香酸エチルエステルのメタンスルホン酸塩(トリカインメタンスルホン酸またはMS222,Sigma社製#A5040)を用いて麻酔した。卵巣サンプルを外科的に除去し、OR2食塩水の入った小さいペトリ皿中に18−20℃で保持した(Wallaceら,1973)。幾つかの実験においては、卵巣サンプルをクロストリジウム・ヒストリティカム(Clostridium histolyticum)からの粗精製コラゲナーゼで処理し、卵胞細胞の外皮層を取り除き、個々の卵母細胞に分離させた。Ca++のないOR2で0.2%としたコラゲナーゼ(タイプII,Sigma社製#C6885)中で室温、2−6時間の処理を行った。他の場合には、卵胞層が無傷の個々の卵母細胞または卵母細胞の小さい塊を宝石商摂子で分離した。
【0031】
GVの膨張:ゼノパスのランプブラシ染色体を研究するための詳細な説明が早くから刊行されたが(Gallら,1991)、現在における数種の決定的な修飾によって、GVの膨張の質が劇的に改善される(Gall,1998)。注入された卵母細胞を一時的に分離培地(83mMKCl,17mMNaCl,6.5mMNa2HPO4,3.5mMKH2PO4,1mMMgCl2,1mMジチオスレイトールpH7.0)に移した。GVを摂子で取り外し、20秒以内で膨張用培地(21mMKCl,4mMNaCl,1.6mMNa2HPO4,0.9mMKH2PO4,1mMMgCl2,10mMCaCl2,1mMジチオスレイトール,0.1%パラホルムアルデヒド,pH7.0)に移した。核エンベロープを宝石商摂子で除去し、核のゲルをピペットで膨張用チャンバーに移した。操作が速やかに行われたならば、核のゲルの分散が数分のうちに起こり、直ちに遠心することができる。スライドを特殊なホルダーに入れ、HS4ローター(Sorvall社製)で5000rpm(4800 g),4℃で30分間遠心した。GVの膨張をPBS中2%パラホルムアルデヒドでの1時間以上の処理で固定した。
【0032】
免疫蛍光:GVの膨張をリン酸緩衝性食塩水(PBS)中10%馬血清での15−30分間の処理でブロックし、そして一次抗体と1時間反応させた。3%馬血清での簡単な洗浄後、二次抗体でさらに1時間処理した。ウサギ血清を10%馬血清で1:200に希釈し、mAbsを希釈しない培地上清として使用した。二次抗体はCy3−またはフルオレセイン−結合ヤギ抗ウサギ血清(Cappel/Organon Teknika,Durham,NC)であった。調製物をフェニレンジアミンの1mg/ml(pH9)及び1mg/ml 4’,6−ジアミノ−2−フェニルインドール(DAPI)を含有する50%グリセリンにマウントした。
【0033】
精子頭部:精子頭部を、Gurdon(1976)に記載されるように、リゾレシチンで脱膜化した。ゼノパス・ラエビス及びシルラナ(ゼノパス)トロピカリスの精子頭部を記載するように(NewmeyerとWilson,1991)精巣から調製した。マウス,マス・マスカラス及びゼブラフィッシュ,ダニオ・レリオの精子を同様に調製した。マウスの場合には、成熟精子を精巣上体から取得し、ゼブラフィッシュの成熟精子を雄成体から圧搾した。コオロギ,アケタ・ドメスティカス(Acheta domesticus)の精子を、成虫からの6−8対の精巣を蒸留水中でホモジナイズすることによって取得し、成熟及びほとんど成熟した無傷の精子のままで、ほとんどの細胞を効果的に破壊した。精子を遠心で濃縮し、そして他の種についてと同様にリゾレシチンで処理した。各々、精子数を標準的な血球計で算出した。
【0034】
卵母細胞注入:幾つかの卵母細胞を、注入前にコラゲナーゼで部分的に脱胞化し、他の幾つかは酵素処理なしに使用した。脱胞化した卵母細胞は注射針で容易に刺し貫くことができるが、この利点は脱胞化に要する時間と処理した卵母細胞が幾分壊れやすいことで相殺される。注入前に、卵母細胞を1800 gで20−30分間遠心してGVを表面に浮かせるが、その位置は脱色領域として検出できる。ガラス針をキャピラリーチューブ(0.5mm i.d.,1.2mm o.d.)からバーティカルピペットピューラー(David Kopf Instruments,Tujunga,CA)を用いて作成した。使用前に、針はシリコン処理し、先端を摂子で掻いて内径を約10μmから20μmにした。針は後ろから鉱油を満たし、そして注入すべき溶液を先端から吸い上げる。4.8μlまたは9.6μlの注入をNANOJECTマイクロインジェクション装置(Drummond Scientific Co.,Broomall,PA)を付けた解剖用顕微鏡下で行い、プランジャーを利用して針の中の液体を押し出した。我々は、針を精子懸濁液で満たした後すぐに注入を行うことが良いことを見出した。顕微鏡スライドへスポットする試験は、放出された精子数が、満たした後の時間に連れて減少することを示したが、精子頭部の沈殿と針内部への付着によるものと推定された。
【0035】
転写及びスプライシング:転写を3Hラベル化GTPの卵母細胞の細胞質への注入によってモニターした。50%エタノール中の250μCiの3H−GTP(5.7Ci/mM,Amersham社製)を蒸留乾固し、1.25μlの水に再溶解した。約1μCi(5nl)を各卵母細胞に注入した。注入後の多くの時点で、GVの膨張を起こし、通常通り、2%パラホルムアルデヒドで固定化した。それらをエタノールで繰り返し脱水化し、アセトンから風乾させた。編入されなかったラベルを5%トリクロロ酢酸の4℃,5分間の処理で除去した。もう一度脱水化してアセトンから乾燥させた後、スライドをNTB2オートラジオグラフ用エマルジョン(Eastman Kodak社製)に浸した。オートラジオグラフの暴露を1.5dとし、続いてD19ディベロッパーで2分間現像した。GV内容物を、エマルジョンのクーマシーブルー処理によって染色した(Gallら,1991)。スプライシングを防ぐため、U2 snRNAに相補性のデオキシオリゴヌクレオチド10ngを精子を用いてGVに注入した(PanとPrives,1988;Tsvetkovら,1992)。そのオリゴはGV内の内因性U2 snRNAとハイブリダイズし、そしてRnase H活性によって1−4時間で完全に破壊される。注入24時間後に行ったLBCsの調製によって、全ての検出可能なU2 snRNAを欠失した異常に大きいループが示される(Tsvetkovら,1992)。
【0036】
精子頭部の注入
初期の実験は、脱膜化したゼノパス・ラエビスの精子頭部を、IV−V段階の卵母細胞(直径1.0mmから1.2mm)のX.ラエビスのGVへの注入に関するものであった。多くの実験において、注入体積は5−10nlで、表面上は精子頭部1−5個/nlを含有するが、注入された頭部はしばしば計算よりも低いものとなり、おそらく頭部の針内側への付着によるものと考えられる。注入後多くの時点で、GVを卵母細胞から手動で切断し、その内容物を特別に調製したウェル(well)スライドに分散することができ、GV内容物をガラス基質に確実に固着させるために、そのスライドを遠心する。我々は現在、各々が20個から60個の卵母細胞を含む数ダースの注入実験を行っている。典型的な実験として、注入された卵母細胞の約90%から約95%が24時間後で正常であり、これらの約60%から約90%がGV中に精子頭部を受けている。残りはおそらく、針がGVを刺し損ねたかもしくは精子頭部が針から噴出されない場合を表しているものと考えられる。
【0037】
精子頭部の初期膨張
注入に用いられる、脱膜化したゼノパスの精子頭部は、約20μmの長さの線虫様構造をしており、中央付近に約1μmの最大径をもち、両端に向かってだんだん細くなっている(図1A)。GVへの注入後、それは、その形態とDNA特異的な染料であるDAPIによる強力な染色によって容易に識別できた。精子頭部における変化の最初の兆しは膨張であり(図1B−C)、それは数分のうちに、フェイズコントラストまたは示差干渉(DIC)顕微鏡によって検出可能な膜やエンベロープを形成することなしに起こった。その後の事象の起点は実験毎に変化したが、おそらく異なる雌からの卵母細胞による違いに関連するものと考えられる。多くの場合、膨張した精子頭部は最初の3−6時間、ほとんど変化しないままであったが(図1D−I)、別の実験では著しい脱凝集とLBC構造の最初の兆候が認められた(図2)。
【0038】
精子頭部の組成における変化をモニターするために、我々は、転写と核のスプライシング構成分を検出する3つの抗体:RNAポリメラーゼII(Pol II)に対するmAb H14(Bregmanら,1995),スプライシングsnRNAsと会合したSmタンパク質に対するmAb Y12(Lernerら,1981),snRNAsのトリメチルグアノシンキャップに対するmAb K121(Krainer,1988)。mAb H14は、リボヌクレオプロテイン(RNP)マトリックスは染色しないままで、横向きのループ軸のみを染色するため、LBCsの研究には特に重要である。このように、個々のループの経過を、例えかなりのオーバーラップがある領域においてさえ追うことが可能である。GV中の染色体外のオルガネラの中では、mAb H14は、本質的には他の全てを染色しないままで、コイル部(球体)に対して高い特異性を示す(図1C及び2A)。コイル部の染色がRNAポリメラーゼIIによるものか、交差反応性のエピトープによるものなのかは不明である。mAb Y12及びK121はsnRNPsの同定を可能とする。両方ともループのマトリックスを染色し(図3)、GV中のsnRNP含有部位がB−スナーポソームとして参照される(Wuら,1991)(図1G及びI)。
【0039】
精子頭部は注入後、最初の数時間のうちにmAb H14で染色され始めるが(図1D−E)、おそらくRNAポリメラーゼIIの、GV中に蓄えられたものからの取り込みによるものと推定される。この取り込みは転写に伴うものではなく、転写がアクチノマイシンによって阻害されるときでさえ起きる。初め、スプライシング因子は、Y12及びK121染色がないことによって示されるように(図1F−I)、取り込まれない。精子頭部が膨らみ続けるにつれ、RNAポリメラーゼIIに対して次第に強く染色される(図2A)。DNAに対するDAPI染色は、この膨張した頭部の内部に個々の染色分体が初めて示されたことを明らかにする。この初期段階においてさえ、注意深く観察すれば、DNAから広がったRNAポリメラーゼII反応性ループの存在が明らかになる(図2F,矢じり)。これらのループはRNA合成の始まりと発生期の転写物の集積の合図である。
【0040】
完全なランプブラシ構造の外観
精子頭部は最終的に、内因性のLBCsによく似た綿毛状の糸の緩い房に解けた(図3)。時間に連れて、これらの房は個々の染色体または染色体の小さな集まりへと分離し、我々は“精子LBCs”として称した。精子LBCsは、対になってはおらず、同じGV中の18個の内因性の二価体よりもわずかに短いため、容易に識別できた(図4)。精子のLBCsは、GVの体積中に満遍なく拡散するのではなく、一緒になったままであり、注入部位からほんのゆっくりとしか移動しないと推定された。多くの実験において、1個から約20個の精子までが単一のGVに注入された。この制限内では、精子のLBCsが、1つの精子頭部に由来しようと多くの精子頭部に由来しようと、形態及び染色性において類似しているという意味において、その系は飽和していなかった。
【0041】
全体的な構造において、精子LBCsは、同じ核内の内因性の染色体によく似ている。それらの各々は、DAPI陽性の染色小粒の軸を有しているが(図3E及びH)、内因性のLBCsよりも強くは染色されず、単一の染色分体と、2つの密に対となった姉妹染色分体との差を反映していると推定される。精子LBCs上のループは、xnf7に対するL24(Reddyら,1991)などの、発生期の転写物に会合する種々のタンパク質に対する抗体で染色され(図3D及びG)、それらはまた、Y12及びK121での染色によって実証されるように、スプライシング因子を含有していた(図3A−C及びF)。各々のループの軸は、RNAポリメラーゼIIに対するmAb H14で明確に輪郭が描かれた(図5B)。
【0042】
精子のLBCsは完全な染色体
X.ラエビスの18個のランプブラシ二価体のうち最も長いものは、最も短いもののちょうど2倍の長さであり、残りの染色体は、少しずつ大きくなる順次列を形成している(Muller,1974;Callan,1987)。精子LBCsは、特に長いまたは短いものもなく、大きさは同じ位である(図4及び5)。それゆえ見かけ上、それらは、あたかも精子クロマチンの不定のフラグメントに由来するように見える。しかしながら、より綿密な検査及び内因性の二価体との比較によって、そのほとんどが完全な無傷の染色体であることが示される。
【0043】
18個の二価体のうち、15個は長腕の端に目立つ末端顆粒を保有し、初期の文献においては末端小粒として照会されていた(Callan,1987)。この顆粒が、RNAポリメラーゼIIを含有するという直接的な証拠はないが、5S rRNAをコードする遺伝子を含有し、mAb H14で強く染色される。この顆粒は、お互いに、様々な組合せで融合する傾向がある。ほとんど全てのGVが、2つの相同体の末端顆粒が単一の顆粒に融合している少なくとも1個の二価体を有し(図6A)、非相同性の染色体の末端の間の融合は同じ頻度である(図2A,矢じり)。精子LBCsの原因を表わす調査によって、その多くがまた、mAb H14で染色される末端顆粒でキャップされる一端を有することが示される(図5A−B)。精子LBCs間の非相同性の融合は特に頻繁なものであり、見かけ上、分枝した染色体のように見える、多触手の配置を生じる(図6C)。
【0044】
精子LBCsが完全な染色体であるという最も強力な証拠は、特定の染色体の同定から得られる。中でも最も特徴的なLBCsとして同定されるのは14番である。この染色体の中央付近には、mAb Y12及びK121で強く染色される、卵型または球形の塊が付着する短い領域がある。このような塊は、精子LBCsの各々の房における顕著な特徴であり(図3A−B)、多くの場合、14番染色体が個々に認識される(図3C−H)。
【0045】
11番染色体もまた、容易に同定される。突出した末端顆粒に加えて、mAb H14で明るく染色される、2つの軸をなす顆粒を有する(図6A)。長腕の端に最も近い軸性顆粒は、U2 snRNA遺伝子の配座であり、しばしば突出したループ対と会合する。もう1つの軸性顆粒は動原体のすぐ近くにある。11番染色体特有の特徴は、ほとんど認識されない染色小粒をもつ短腕の緩い構造と頻繁なダブルループブリッジである。図6B及びCは11番染色体の特徴を呈する精子LBCsの2例を示す。図6Bにおいて、今述べられた全ての特徴が認識され、図6Cにおいて、U2配座での軸性顆粒は明確ではないが、その染色体は11番として同定される。同定可能な染色体の追加例は、異種の精子頭部の注入を扱うセクションで指摘される。
【0046】
精子LBCsは単一の非複製性の染色分体を含有する
各々の精子は、半数体(haploid)数の非複製性染色分体を、X.ラエビスの場合では18個を含有する。結果として、各々の精子LBCは、DNA合成が精子の注入後に起こらなければ、非複製性の染色分体から構成されることになる。LohkaとMasui(1983)に示されているように、精子頭部がゼノパスの卵抽出物中でインキュベートされると、S期に入り、DNA合成を開始する。しかしながら、卵母細胞のS期は、減数分裂の開始前の分裂間期で起こり、ランプブラシ段階でのGV内のDNA合成についての確証はない。しかしながら、rDNAの異常な増幅が、両生類の卵母細胞内の太糸期で起こることから、GVのDNA合成能の評価にいくらか注意を払う必要がある(BrownとDawid,1968;Gall,1968)。
【0047】
ランプブラシ二価体の各相同体内の2つの染色分体の存在が、横方向のループの対になる性質から初めて推定された(Callan,1986中で論じられている)。精子LBCsが非複製性の染色分体からなるものならば、対にならないループを有するはずである。精子LBCのループの大多数が非常に短く、外観における評価では類似しているが、それが対にならない条件が幾つかの環境下で明らかにされている。第一に、特定のループが大きさや異常な形態のために識別されるとき、それは典型的な二価体であるように見えるが、いつも単一であり、決して対にならない(図7A−D)。第二に、幾つかのループは抗体で特異的に染色される。また、これらのループは精子LBCs上では単一である(図8E−F)。第三に、慣用のLBCsを引き伸すと、“ダブルループブリッジ”として知られる現象を呈する(Callan,1955;CallanとLloyd,1960)。ダブルループブリッジは、ループ対の塩基が伸長下で離れるときに起こる。本来対である姉妹ループであったものが、染色体の軸に沿って引き伸ばされた平行な糸のようになる。このようなダブルブリッジは、精子LBCsの場合には見られないが、代わりに“シングルループブリッジ”が比較的頻繁に認められる(図8G−H)。これらのブリッジは、精子LBCが単一の染色分体からなることを示している。
【0048】
転写及びスプライシング
ランプブラシループが実際にRNA合成に関与し、アクチノマイシンやアルファ−アマニチンでの転写阻害は、ループのRNPマトリックスの損失とループの収縮を導くことが何度も実証されている(Izawaら,1963;Schultzら,1981;Callan,1986)(図9G)。転写及び精子LBCsの形成における初期の事象とのありうる関係を調べるために、我々は、卵母細胞の細胞質へ3H−GTPを注入し、その後すぐに精子をGVに注入した。幾つかの卵母細胞をアクチノマイシン中で約1時間前もってプレインキュベートし、注入後アクチノマイシンに戻した。両セットの卵母細胞において、精子頭部は初期の膨張を経て、1時間までにはRNAポリメラーゼIIに対するmAb H14で強く染色された(図9A−B)。この最初の時点では、スプライシング構成分は精子頭部中に現れなかった(図9C−F)。アクチノマイシン中でさらにインキュベーションした後でも精子頭部に変化は認められず、最終的に形成された典型的な精子LBCsをOR2食塩水中に保った。精子及び3H−GTPを注入7時間後、OR2食塩水中に保存した卵母細胞において、核小体及び内因性染色体が強くラベル化され、膨張した精子頭部はラベル化されなかった(図10A)。3H−GTPはアクチノマイシン処理した卵母細胞においてはいかなる核構造にも編入されなかった(図10C)。ゆえに精子の膨張とRNAポリメラーゼIIの集積の初期段階が転写なしに起こったのである。1日後に検査した卵母細胞は、よく形成された精子LBCsを呈し、このときまでは活発に転写を行っていた(図10B)。
【0049】
アンチセンスオリゴデオキシヌクレオチドを用いる初期の実験によって、LBCsにおける転写はU2 snRNAが存在しなくても起こり得るものであり、ゆえにそれに伴うスプライシングを必要としないことが示された(Tsvetkovら,1992)。我々は、スプライシング機構が破壊された卵母細胞中の精子頭部の組み立てを実証する同様な実験を行った。図10D及びEは、2d前に精子を注入した卵母細胞からの、及び、2d前にU2 snRNAに対するオリゴデオキシヌクレオチドを注入した卵母細胞からの精子LBCを示す。この精子LBCはU2枯渇のGVに特有の2つの特徴、そのループが異常に突出していることと、mAb K121での染色が弱いことを呈する。標的としたU2 snRNAが注入後の最初の4時間弱のうちに破壊されるため(Tsvetkovら,1992)、この実験によって、精子LBCsがスプライシングなしで形成し得ることが実証される。
【0050】
まとめとして、精子膨張の初期段階及びRNAポリメラーゼIIの集積が通常、転写なしに起こり、転写がアクチノマイシンによって阻害されているときでさえ起こり得る。転写が、精子頭部からの細胞学的に識別される染色体の発生に必要かどうかは決定されていないが、典型的なランプブラシループの維持は転写に依存する。精子LBCsの形成にも維持にもそれに伴うスプライシングは必要とされない。
【0051】
異種間の注入
我々は、ある1つの種の脱膜化した精子頭部を別の種のGVに注入する幾つかの実験を行った。このような実験によって、LBCsの組み立てにおけるトランスの因子からシスの因子を区別すること、言い換えれば、精子クロマチン、タンパク質、及び宿主GVに供給される他の因子の相対的な寄与を評価することが可能になり始めている。
【0052】
シルラナ(=ゼノパス)・トロピカリスの精子をX.ラエビスのGVに注入した。S.トロピカリス由来の精子LBCsは、同種のX.ラエビス注入からのものと全く同じに見える。S.トロピカリスは半数体セットとして10個しか染色体を持たないから、特定の染色体を同定することは容易であり、精子LBCsが完全な染色体であるという付加的な証拠を提供する。ヒョウガエル,ラナ・ピピエンスもまたX.ラエビスのGVに注入された。また、生じた精子LBCsは、長さと一般的な形態において内因性のLBCsによく似ており、特定の染色体が認識できた(図7A−D)。DAPI染色で調べたところ(図7E)、精子LBCsの染色体軸は、比較が単一と対となった染色分体との間のものであるにもかかわらず、内因性のX.ラエビスの二価体のものより突出していた。強いDAPI染色はおそらく、R.ピピエンスがX.ラエビスよりも少ない染色体を有するが(n=13対n=18)より大きいゲノムを有する(6.8×109bp対2.8×109bp)結果、R.ピピエンスの“平均的な”染色分体は、X.ラエビスからの“平均的な”染色分体の約3倍のDNAを含有するという事実を反映していると考えられる。
【0053】
R.ピピエンスの精子LBCs上のループと内因性の二価体との類似は、予想されたものではなかった。R.ピピエンスのそれ自身の卵母細胞内のLBCsは、より大きいループを呈し、我々は精子LBCs上のより突出したループを見ることを予想していた。即ち、我々は、精子クロマチンの起源が、生じるLBCsの形態により大きい影響を与えることを期待していたのである。
【0054】
ゼブラフィッシュ,ダニオ・レリオの精子がX.ラエビスのGVに注入されたとき、生じたLBCsは、ずっと短いけれども、一般的な形態においては内因性のX.ラエビスの二価体に似ていた。図7F中のダニオ染色体は、観察された最も長いものである。ダニオは25個の半数体数及び1.7×109bpのゲノムDNA含量を有するため、“平均的な”ゼブラフィッシュの染色分体は、“平均的な”ゼノパスの染色分体の約0.44倍 のDNAを有する。
【0055】
我々が、X.ラエビスの精子をイモリ,ノトフタルムス・ビリデセンスのGVに注入したとき、予想し得ない結果が得られた。形態におけるゼノパス及びイモリLBCsの違いはかなりのもので、イモリのランプブラシループは既知のものの中で最も大きい部類であった(図8A−B)。N.ビリデセンスのGV中のX.ラエビスの精子LBCsが、全体の長さがずっと短いことを除いて、内因性のイモリの染色体と著しいほど似ていたことを見出して驚いた。この類似はループの3つの特徴、RNPマトリックスの長さ,形態、及びポリメラーゼII軸の突出に及んでいた。最も明らかな類似はループの長さにおいてであり、X.ラエビスの染色体で普通に見られるものより大きい(図8C−Hと図5−6を参照)。多くのループが、平均で内因性のイモリ染色体上のものほど長くはないが、長さ30μm−40μmであった。精子LBCループのRNPマトリックスは、イモリのLBCsのものと同じように形態において豊満なものであり、種類に富んでいる(図8D及びF)。ループ形態での同様な変化が、内因性のX.ラエビスのLBCs上で起こるが、あまり目立つものではない。目立つ特徴は、ループ軸のRNAポリメラーゼIIに対するmAb H14での染色である。正常なイモリLBCs及びイモリGV中のX.ラエビスの精子LBCsにおいて、この軸は普通、突出しているが(図8A及びC)、正常なX.ラエビスのLBCsにおいてはあまり目立たない(図6A)。
【0056】
我々は、イモリのGV中のX.ラエビスの精子LBCsに会合するタンパク質を調べるために、2つのイモリ−特異的な抗体を用いた。mAb A33はプロイロデレス・ウォルツル(Pleurodeles waltl)から最初に記載された亜鉛フィンガータンパク質(A33)を検出し(Lacroixら,1985;Belliniら,1995)、mAb SE5は、N.ビリデセンスからの非関連タンパク質(SE5)を検出する(RothとGall,1987)。両方の抗体とも、N.ビリデセンスのほとんどのRNPマトリックスを染色するが、LBCs上のループは染色せず、X.ラエビスの正常なLBCsとも反応しない。しかしながら、注入実験においては、両方ともX.ラエビスの精子LBCsを、同じGV中の内因性のN.ビリデセンスの染色体と区別がつかないほど染色した。この観察は、イモリのタンパク質がイモリのGV中のX.ラエビスの精子LBCsの組み立てに使用されることを実証する。
【0057】
他の異種間の注入実験が実施され、ある程度の成功を納めている。マウス,マス・マスカラス及びコオロギ,アケタ・ドメスティカスからのリゾレクチン処理した精子頭部がX.ラエビスのGVに注入された。両種の精子頭部が初期の膨張を経て、mAB H14で染色し始めた。しかしながら、どちらの場合においても、数個の短いループがマウスの核から広がっていたが、個々の染色体が見えるようにはならなかった。同じ精子頭部のサンプルは、ゼノパスの卵抽出物中で30−60分間のインキュベーション後、典型的なイン・ビトロの核を形成し、後者のアッセイにおける反応能がGV系での反応能に対しての十分な規準ではないことを示唆した。
【0058】
我々は、転写活性を有する典型的なランプブラシ染色体が、脱膜化した精子頭部が無傷の卵母細胞のGVに注入されると、精子のクロマチンから組み立てられることを示した。15年前、LohkaとMasuiは、このような精子頭部が、ゼノパスの卵の抽出物中でインキュベートされると、本質的に正常な前核に形質転換し(LohkaとMasui,1983及び1984)、この卵抽出物系が、DNA複製及び細胞周期の多くの側面を研究するのに使用されることを示した。GV及び卵抽出物系は基本的には違うものである。卵抽出物系においては、脱膜化した精子クロマチンが核エンベロープで取り囲まれるようになり、その後にDNA複製を経て有糸分裂に入る転写不活性な核となる。対照的に、GVの周辺環境に自由に曝されたままの同じ脱膜化した精子クロマチンは、複製を経ず、分離した、転写活性を有する巨大な染色体に形質転換する。
【0059】
両生類のGVは、初期胚となる運命の、転写及びスプライシング因子を含む大量の蓄積物質を含有することが実証されている。これらの因子はGVが注入されたDNAテンプレートを転写し、生じたRNAをプロセスできるようにする。GV抽出物は、Brownとその共同研究者の5S RNAについての広範な研究においてのように、イン・ビトロの転写をサポートするために使用されている。水性のGV抽出物は、GVが油下で分離され、そのため可溶性のコファクターやサポートに有効なU1 snRNAテンプレートからの転写物を保持するが、GVポリメラーゼII転写の研究に対しては制限的にしか使用されない。Gurdonによる初期の実験において、多くのタイプの体細胞性の核がゼノパスの卵母細胞の細胞質及びGVに注入され、その後の形態的及び転写に関わる事象が、かなり詳細に調べられた。GVに注入された核は膨張し、転写において活性となるが、明らかに無傷の核エンベロープを保持していた(Gurdon,1976)。ランプブラシ染色体はこれらの核の中に検出された。
【0060】
本願発明は、最大限凝集されたクロマチンが、比較的成熟した卵母細胞(デューモント(Dumont)段階IV−V)のGV中で1日以内に形質転換できることを示す。精子頭部は注入後、RNAポリメラーゼIIをほとんど瞬時に取得する。初期の膨張後、転写が始まり、付加因子が、snRNPを含む発生期の転写物に補給される。数倍のクロマチンがGV中で通常存在するように変換され、その方法では宿主のタンパク質が使用される。これらの特徴はGV中に蓄積されていることが知られている因子、RNAポリメラーゼII,hnRNP,snRNP,ヒストン及びヌクレオプラスミンなどの重要性を強調する。他の事柄の中で、これらの蓄積されている因子は、初期の胚形成の間のDNAからクロマチンへの変換において、及び中胞胚(mid-blastula)の転移後の転写の開始において使用される。正常なランプブラシ染色体は、転写機構がGV中では機能的であることを示し、精子の注入実験は、不活性のクロマチンが転写のために活性化され得ることを示す。
【0061】
精子クロマチン由来のLBCsの最終的な形は、形質転換が起こるGV内の因子によるのと同様に、クロマチン自身の特徴によって影響されるに違いない。X.ラエビスのGV内のX.ラエビスの精子クロマチンが、内因性のLBCsから区別できないLBCsを生じると予想される。他方、前記された異種間のプロセスの結果は期待できない。サンショウウオやある種のカエルのような、ある種の生体は非常に長いループをもつ巨大なLBCsを有し、また、ゼノパスのような他の種では並みのループをもつ中間的な大きさの染色体を有し、またさらに、わずかに検出できるループをもつ非常に短い染色体を有するものもあることが知られていた。これらの違いは多くの生体のゲノムDNA含量またはC値と論理的によく相関する。
【0062】
しかしながら、全体的な染色体の長さがC値と相関するならば、ループの長さはなぜ、ループが単に、1個もしくは数個の転写ユニットに対応するというふうにならないのか?通常与えられる説明は、ランプブラシ段階での読み込み転写の存在に基づいている(Gallら,1983)。その最も単純な形において、転写ユニットが遺伝子間の全てのスペーサーを含み、平均してスペーサーの長さがC値とともに増加し、それゆえ転写ユニットは、高いC値をもつ生体では長くなる、という理由付けがある。このモデルはシスの要素にほとんど全ての重点を置き、与えられた生体の精子は、それが置かれるGVにかかわらず、類似のLBCsを形成することを予測させる。しかしながら、異種間の注入 −特にノトフタルムスのGV内のX.ラエビスの精子、この場合はイモリの染色体に著しく似た染色体を生じるのだが− に関する我々の結果は、トランスの因子が目覚しい役割を果たすことを論じるものである。この結論は、全体的なDNA含量に基づくLBCの形態の比較的単純なモデルの修飾を必要とするものである。
【0063】
LBCsの組み立てにおいて重要な因子を調べるためには、GV含量からの構成分を足したり引いたりする必要があるだろう。アンチセンスオリゴヌクレオチドは、GV内のRNAを分解するのに効果的に用いることができる(PrivesとFoukal,1991)。初期において我々は、LBC転写がそれに伴うスプライシングを必要としないこと(Tsvetkovら,1992)を示すために、U2 snRNAのノックアウトを用いたが、同じことが精子クロマチンのLBCへの形質転換ついても有効である。タンパク質の枯渇または付加は、抗体のGVへの注入が、ある程度うまく用いられているが(Bonaら,1981;Scheerら,1984)、問題は多い。分裂間期の核の組み立てのための卵抽出物に匹敵する、LBCの組み立てに対するイン・ビトロ系を持つことが高く望まれる。油下で分離されたGV(Paineら,1992)で我々の実験を繰り返すことにまず努力が払われ、これがうまくいけば精子のLBCへの形質転換をサポートするGVまたは卵母細胞からの細胞のない抽出物が調製される。
【0064】
本願発明は、染色体及び胚胞の種々の原料での使用を記載するものである。ここで例示される胚胞原料にはカエル及びイモリが含まれる。染色体を含有する核は、例えばカエル,マウス,ゼブラフィッシュ及びコオロギの組織から取得された。本願発明は、胚胞及び染色体含有の核の様々な他の原料での使用を意図している。特に、ヒト胚胞及び/またはヒト染色体原料が転写活性を有するランプブラシ染色体を、本願による、凝集したクロマチンまたは核から生成することを期待させる。
【0065】
本願発明が、実際の、及び好ましい態様であると現在考えられているものと関連して記載されている一方、本願発明は、開示された態様に制限され、限定されるものではないが、また一方では、添付された請求項の意味と範囲内に含まれる多くの修飾及び同意義の改作を網羅するように意図されている。
【0066】
ゆえに記載された発明での改変は、当業者にとって本願発明の新規な側面から外れることなく自明なものであり、このような改変は本願発明の範囲内に入ると解釈されるべきである。
【0067】
参考文献
【図面の簡単な説明】
【0068】
【図1】(A)注入に先立って脱膜化したゼノパス・ラエビス(Xenopus laevis)の精子頭部(DNAは4’, 6−ジアミジノ−2−フェニルインドールまたはDAPIで染色した)。(B)ゼノパスのGVに注入5分後の膨張した精子頭部(DAPI染色)。(C)パネルBと同じ領域をRNAポリメラーゼIIに対するmAb H14で染色:精子頭部は陰性だが、コイル部(球体)は明るく染色される(矢じり)。(D)GVに注入3時間後の精子頭部(DAPI染色)。(E)同じ精子頭部がRNAポリメラーゼIIに対するmAb H14での強い染色を示す。(F)GVに注入3時間後の精子頭部(DAPI染色)。(G)パネルFと同じ領域をsnRNAsのトリメチルグアノシンキャップ(trimethylguanosine cap)に対するmAb K121で染色:精子頭部は陰性だが、Bスナーポソーム(snurposome)は染色される(矢じり)。(H)GVに注入3時間後の精子頭部(DAPI染色)。(I)パネルHと同じ領域をsnRNAsのSmエピトープに対するmAb Y12で染色:精子頭部は陰性だが、Bスナーポソームは染色される(矢じり)。
【図2】(A)ゼノパスのGVへの注入3.5時間後のゼノパスの精子頭部の1群であり、広範囲に膨張し、RNAポリメラーゼIIに対するmAb H14で強く染色される。内因性ランプブラシ染色体は、その末端顆粒で2つの他の染色体に融合する(矢じり)。幾つかの明るく染色されるコイル部(球体)も表わされている(矢印)。(B)同じGVからの単一の精子頭部のDICイメージ。(C)DAPI染色は精子頭部内の個々の染色分体を明らかにする。(D)RNAポリメラーゼIIに対するmAb H14による染色は精子頭部で強い反応を示す。(E)DAPI−陽性の糸を示すパネルC部の高倍率像。(F)個々のポリメラーゼII−反応性ループ(矢じり)がパネルDからの同じ領域のこの拡大で見ることができる。
【図3】(A)及び(B)ゼノパスのGVに注入21時間後の2つのゼノパスの精子頭部。その各々が、内因性LBCsの特徴の大部分をもつ、染色体の緩い房に解けている。スプライシングsnRNAsのトリメチルグアノシンキャップに対するmAb K121での染色。精子LBCsのループはsnRNAsに対して強く染色する。染色体の各々の房はまた、1つの非常に大きいK121−反応性の塊及び幾つかのより小さい塊(矢印)を呈する。これらはほとんど確実に、二価体(bivalent)♯6,♯8及び♯14で生じる同様な塊に対応する。(C)中心近くに付くK121−反応性の大きな塊をもつ単一の精子LBC。その塊の位置及びこの染色体の相対的な大きさがNo.14として同定される。(D)同じ染色体がタンパク質xnf7に対する血清L24で二重染色された(Reddyら,1991)。この抗体は、内因性LBCsにおいてと同じように、ほとんどのランプブラシ染色体と同様にその塊中の小さい封入体(矢印)を染色する。(E)同じ染色体のDAPI染色は凝集した染色体の軸を示す。(F)−(H)一端でもう1つの染色体に付いている、精子LBC No.14と思われるもののもう1つの例のK121,L24及びDAPIの染色。パネルGの矢印はその塊の内側のL24−反応性顆粒を指す。
【図4】ゼノパスの精子頭部の注入21時間後の単一のゼノパスのGV内容物の全体像。精子頭部はLBCs特有の特徴を有する、染色分体の緩い房もしくは単一の染色分体に変換されている。このGVは15−20個の精子頭部を受け入れていると思われる。18個の内因性LBCs及びコイル部(球体)の大部分がこの領域の左上部のエリアを占める(点線部)。RNAポリメラーゼIIに対するmAb H14での染色。
【図5】(A)図4で示されたGVからの数個の精子LBCsの高解像度像で、RNAポリメラーゼIIに対するmAb H14での染色。染色体の全体に亘るファジーな外観は、横方向のループの軸の染色によるものである。加えて、mAb H14は、18個のゼノパスLBCsの15の長腕の端で認められる末端顆粒(矢じり)を染色する。(B)さらに高解像度の単一の精子LBCsで、mAb H14で染色。矢じりは末端顆粒を指し、矢印はループの軸(RNAポリメラーゼIIで変換されたDNA軸)を指す。
【図6】(A)長腕の端で末端顆粒によって同定される二価体♯11の(矢じり)、及び0.16及び0.80の位置での軸の顆粒(矢印)。この例では、0.80の位置に単一の融合顆粒がある。動原体がその融合顆粒のすぐ向うに位置するが、この染色では見えない。その短腕は特徴的な拡散構造を有する。RNAポリメラーゼIIに対するmAb H14での染色。(B)精子由来の染色体No.11。末端顆粒(矢じり)、2つの軸性顆粒(矢印)及び拡散短腕が認識される。(C)精子由来の染色体No.11のもう1つの例。0.16での軸顆粒はこのイメージでは明らかではないが、染色体は0.80での軸性顆粒(矢印)、拡散短腕、及び末端顆粒(矢じり)によって確かに同定され、ここで他の2つの染色体の末端顆粒に融合して三つ組み形となる。
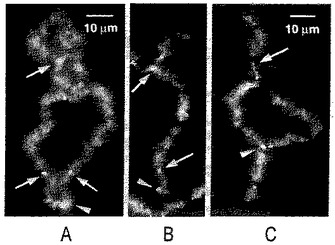
【図7】(A)−(D)2日前にゼノパスのGVに注入されたラナ・ピピエンス(Rana pipiens)の精子頭部に由来する同じ精子LBCの4例のフェイズコントラストイメージ(phase contrast image)。この染色体の一端の近くに、その根元に突出した顆粒をもつ異常に大きなループがある(矢じり)。そのループは、非複製性染色分体で予想されるように、単一であることに注目。(E)パネルDと同領域の、突出したDNA軸を示すDAPI染色。(F)ゼブラフィッシュ,ダニオ・レリオ(Danio rerio)の精子頭部をゼノパスのGVに注入2日後の精子LBC。トリメチルグアノシンに対するmAb K121での染色。矢じりは異常に巨大なマトリックスをもつ2つのループを指す。
【図8】(A)イモリ,ノトフタルムス・ビリデセンス(Notophthalmus viridescens)のGVからの典型的なLBCの小部分で、RNAポリメラーゼIIに対するmAb H14で染色。(B)DICによる同じ領域の像。ゼノパスのLBCと比較して、イモリのLBCはより長いループ、より重厚に折り重なって突出したループマトリックス、及び異常で特有なRNAポリメラーゼII−反応性のループ軸を有する。加えて、イモリLBCsはゼノパスLBCsより長い。(C)2日前にイモリGVに注入したゼノパスの精子頭部に由来するLBC。RNAポリメラーゼIIに対する染色。(D)DICによる同じ領域の像。このゼノパスLBCが長さ以外、イモリLBCによく似ていることに注目。(E)ゼノパスの精子をイモリGVに注入2日後のゼノパス精子のLBC。xnf7に対する血清L24で染色(Reddyら,1991)。ほんの少しのループがこの抗体に強く反応する。染色体の端近くの大きなループ(矢印)が単一であり,卵母細胞のLBCでのループのように対ではないことに注目。このループは、2つのタンデムの転写ユニットからなり、そのうちの1つを染色。(F)フェイズコントラストによる同じ領域の像。(G)イモリGVへの注入2日後のゼノパス精子のLBC。mAb SE5での染色(RothとGall,1987)で、通常イモリのLBCループで染色し、ゼノパスのものは染色しない。染色は、イモリのタンパク質がゼノパスLBCを組み立てるのに使用されたことを実証する。矢印は下記の“単一ループブリッジ”を示す。(H)染色体の染色小粒の軸を示す同じ領域の像(DAPI染色)。
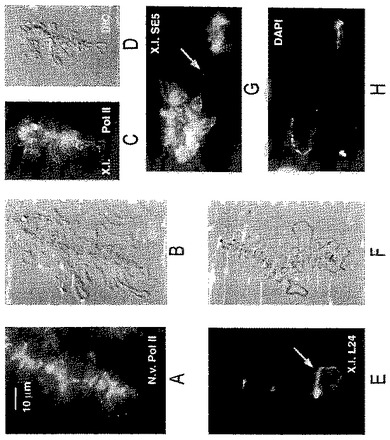
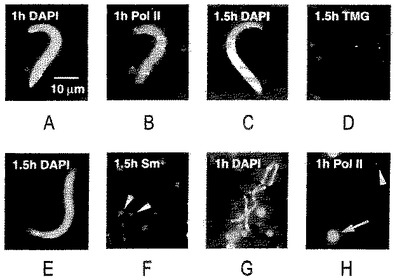
【図9】(A)ゼノパスのGVへの注入1時間後のゼノパスの精子頭部。卵母細胞を注入前に1時間、アクチノマイシンD(20mg/ml)中でインキュベートし、注入後薬剤に戻した(DAPI染色)。(B)同じ精子頭部をmAb H14で染色したものであり、例え転写が阻害されていても、精子によってRNAポリメラーゼIIが取り入れられることを示す。(C)アクチノマイシン処理した卵母細胞に注入1.5時間後の精子頭部(DAPI染色)。(D)同じ精子頭部をトリメチルグアノシンに対するMab K121での染色像であり、スプライシングsnRNAsが欠如していることを示す。(E)アクチノマイシン処理した卵母細胞に注入1.5時間後の精子頭部(DAPI染色)。(F)同じ精子頭部の、トリメチルグアノシンに対するmAB Y12での染色像であり、Smタンパク質が欠如していることを示す。矢じりは、Y12反応性のB−スナーポソームを指す。(G)パネルA及びBで示された精子頭部を含有する同じGVからの高コントラストの染色体(DAPI染色)。(H)同じエリアのmAB H14での染色像であり、末端顆粒を除いた染色体の反応の欠如を示す(矢じり)。コイル部(球体)は典型的な染色を示す(矢印)。
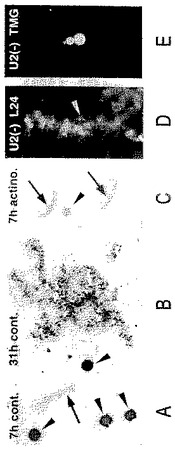
【図10】(A)3H−GTPをゼノパスの卵母細胞の細胞質に注入し、1時間後にゼノパスの精子頭部をそのGVに注入した。7時間後に行った、GV内容物のオートラジオグラフでは、精子頭部にラベルが認められないが(矢印)、3つの核小体では強いラベルが認められ(矢じり)、rRNA転写を示す(1.5d暴露)。(B)3H−GTP注入31時間後のラベル化した精子LBCsのオートラジオグラフ及びパネルAにあるような精子頭部。活発な転写が精子LBCsのループ上で起こっている。核小体(矢じり)は銀粒子によって黒く表わされる(1.5d暴露)。(C)卵母細胞はアクチノマイシンD(20mg/ml)中で1時間、プレインキュベートし、パネルAでのように3H−GTP及び精子頭部を注入し、薬剤に戻す。7時間後に行ったGV内容物のオートラジオグラフは、精子頭部(矢印)でも内因性核小体(矢じり)でもラベルは認められない(1.5d暴露)。(D)U2 snRNAについて欠乏させた卵母細胞からの精子LBCで、xnf7に対する血清L24での染色(Reddy,1991)。ループは良く染色され、大きな“塊”内部の顆粒は染色体に付いている(矢じり)。(E)トリメチルグアノシンに対するmAb K121で染色された、パネルDでの染色体。ループは無処理の卵母細胞に比べて(図3を比較)著しく減少した染色を示したが、K121反応性の塊は影響されない。
【特許請求の範囲】
【請求項1】
転写活性を有する巨大なランプブラシ(lampbrush)染色体を生産する方法であって、
高度に凝集して不活性な染色体の原料を、部分的または完全に周囲の膜を除去するために処理し;
脱膜化した不活性な染色体を卵母細胞の胚胞の内容物に接触させ;そして
不活性な染色体及び胚胞内容物を、転写活性を有する、個々に識別できるランプブラシ染色体を生産するのに十分な時間、インキュベートすることからなる上記方法。
【請求項2】
高度に凝集して不活性な染色体の原料が、精子の頭部または核からなる、請求項1記載の方法。
【請求項3】
核が分裂間期(interphase)の核であるところの、請求項2記載の方法。
【請求項4】
不活性な染色体の原料を取り囲む膜が、界面活性剤,酵素,またはそれらの組合せによって化学的に除去されることを特徴とする、請求項1記載の方法。
【請求項5】
不活性な染色体の原料を取り囲む膜が機械的に除去されることを特徴とする、請求項1記載の方法。
【請求項6】
請求項1記載の方法によって生産されるランプブラシ染色体を形成することに関連するトランス(trans)の因子を同定する方法であり、不活性の染色体を胚胞または分画された卵母細胞のイン・ビトロの、細胞のない抽出物中でインキュベートされることを特徴とする上記方法。
【請求項7】
方法が異種間のものであり、高度に凝集した不活性の染色体が、卵母細胞の胚胞とは異なる生体からのものであることを特徴とする、請求項1記載の方法。
【請求項8】
異種間の方法が、脱膜化したある種の精子頭部を別の種の胚胞に注入することからなることを特徴とする、請求項7記載の方法。
【請求項9】
該方法が、精子クロマチン及び胚胞によって供給される因子の相対的な寄与を評価するために染色体を染色することをさらに含むことを特徴とする、請求項8記載の方法。
【請求項10】
不活性な染色体の原料がゼノパス・ラエビス(Xenopus laevis),シルラナ・トロピカリス(Silurana tropicalis),ラナ・ピピエンス(Rana pipiens),マス・マスカラス(Mus musculus),アケタ・ドメスティクス(Acheta domestics)及びダニオ・レリオ(Danio rerio)からなる群から選択される生体であるところの、請求項7記載の方法。
【請求項11】
胚胞がゼノパス・ラエビス及びノトフタルムス・ビリデセンス(Notophthalmus viridescens)からなる群から選択される生体からのものであるところの、請求項7記載の方法。
【請求項12】
染色体の異常について分析する方法であって、
不活性な染色体を卵母細胞の胚胞の内容物と接触させ;
不活性な染色体と胚胞内容物を、転写活性を有する、個々に識別されるランプブラシ染色体を生産するのに十分な時間、インキュベートし;そして
転写活性を有する、個々に認識されるランプブラシ染色体を細胞遺伝的な分析に当てることからなる上記方法。
【請求項13】
細胞遺伝的な分析がゲノムマッピング,核型付け(karyotyping),遺伝子構築及び機能的遺伝学からなる群から選択されるところの、請求項12記載の方法。
【請求項1】
転写活性を有する巨大なランプブラシ(lampbrush)染色体を生産する方法であって、
高度に凝集して不活性な染色体の原料を、部分的または完全に周囲の膜を除去するために処理し;
脱膜化した不活性な染色体を卵母細胞の胚胞の内容物に接触させ;そして
不活性な染色体及び胚胞内容物を、転写活性を有する、個々に識別できるランプブラシ染色体を生産するのに十分な時間、インキュベートすることからなる上記方法。
【請求項2】
高度に凝集して不活性な染色体の原料が、精子の頭部または核からなる、請求項1記載の方法。
【請求項3】
核が分裂間期(interphase)の核であるところの、請求項2記載の方法。
【請求項4】
不活性な染色体の原料を取り囲む膜が、界面活性剤,酵素,またはそれらの組合せによって化学的に除去されることを特徴とする、請求項1記載の方法。
【請求項5】
不活性な染色体の原料を取り囲む膜が機械的に除去されることを特徴とする、請求項1記載の方法。
【請求項6】
請求項1記載の方法によって生産されるランプブラシ染色体を形成することに関連するトランス(trans)の因子を同定する方法であり、不活性の染色体を胚胞または分画された卵母細胞のイン・ビトロの、細胞のない抽出物中でインキュベートされることを特徴とする上記方法。
【請求項7】
方法が異種間のものであり、高度に凝集した不活性の染色体が、卵母細胞の胚胞とは異なる生体からのものであることを特徴とする、請求項1記載の方法。
【請求項8】
異種間の方法が、脱膜化したある種の精子頭部を別の種の胚胞に注入することからなることを特徴とする、請求項7記載の方法。
【請求項9】
該方法が、精子クロマチン及び胚胞によって供給される因子の相対的な寄与を評価するために染色体を染色することをさらに含むことを特徴とする、請求項8記載の方法。
【請求項10】
不活性な染色体の原料がゼノパス・ラエビス(Xenopus laevis),シルラナ・トロピカリス(Silurana tropicalis),ラナ・ピピエンス(Rana pipiens),マス・マスカラス(Mus musculus),アケタ・ドメスティクス(Acheta domestics)及びダニオ・レリオ(Danio rerio)からなる群から選択される生体であるところの、請求項7記載の方法。
【請求項11】
胚胞がゼノパス・ラエビス及びノトフタルムス・ビリデセンス(Notophthalmus viridescens)からなる群から選択される生体からのものであるところの、請求項7記載の方法。
【請求項12】
染色体の異常について分析する方法であって、
不活性な染色体を卵母細胞の胚胞の内容物と接触させ;
不活性な染色体と胚胞内容物を、転写活性を有する、個々に識別されるランプブラシ染色体を生産するのに十分な時間、インキュベートし;そして
転写活性を有する、個々に認識されるランプブラシ染色体を細胞遺伝的な分析に当てることからなる上記方法。
【請求項13】
細胞遺伝的な分析がゲノムマッピング,核型付け(karyotyping),遺伝子構築及び機能的遺伝学からなる群から選択されるところの、請求項12記載の方法。
【図1】

【図2】
【図3】
【図4】

【図5】
【図6】
【図7】

【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2009−100749(P2009−100749A)
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願番号】特願2008−300377(P2008−300377)
【出願日】平成20年11月26日(2008.11.26)
【分割の表示】特願平11−528942の分割
【原出願日】平成10年11月16日(1998.11.16)
【出願人】(500026234)カーネギー インスチチューション オブ ワシントン (25)
【Fターム(参考)】
【公開日】平成21年5月14日(2009.5.14)
【国際特許分類】
【出願日】平成20年11月26日(2008.11.26)
【分割の表示】特願平11−528942の分割
【原出願日】平成10年11月16日(1998.11.16)
【出願人】(500026234)カーネギー インスチチューション オブ ワシントン (25)
【Fターム(参考)】
[ Back to top ]