
リグナンメチル化酵素をコードする遺伝子
【課題】 メチル化リグナンを供給するために、リグナンにメチル基転移活性を有する酵素を提供する。
【解決手段】 リグナンにメチル基を転移する活性を有する酵素を同定し、当該酵素ポリペプチドのアミノ酸配列および当該ポリペプチドをコードするポリヌクレオチドの塩基配列を同定し、これらの配列情報に基づいて、メチル化リグナンを産生する形質転換体を作製すること。
【解決手段】 リグナンにメチル基を転移する活性を有する酵素を同定し、当該酵素ポリペプチドのアミノ酸配列および当該ポリペプチドをコードするポリヌクレオチドの塩基配列を同定し、これらの配列情報に基づいて、メチル化リグナンを産生する形質転換体を作製すること。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はリグナンにメチル基を転移する活性を有する酵素の遺伝子、当該メチル化酵素を利用してリグナン組成が変換された植物などに関する。更に詳しくは、本発明は、メチル化リグナンを合成する活性を有する酵素遺伝子、好ましくはゴマ由来のメチル化リグナンを合成する活性を有する酵素遺伝子、及びその利用に関する。
【背景技術】
【0002】
ゴマ属ゴマ(Sesamum indicum)は、ゴマ科(Pedaliaceae)に属する1年生の草本植物である。ゴマの原産地は、中央アフリカといわれている。ゴマは、約6000年の歴史を有する最古の栽培油糧植物とされ、世界中で栽培されてきた。ゴマは、古来から貴重な食品であり、健康に良い食品の代表として知られている。特に、ゴマ種子、ゴマ種子から得た油、ゴマ種子からの抽出物が利用されている(例えば、ゴマその科学と機能性、並木満夫編:丸善プラネット社(1998)参照)。ゴマ種子に含まれる成分は、その約50%が脂質であり、約20%が蛋白質である。ゴマに含まれる脂質の主要成分は、オレイン酸およびリノール酸を主体とするトリグリセリドである。ゴマはまた、ビタミンB1、B2、Eなどをさらに含む。リグナンと総称される植物の二次代謝物(例えば、セサミンおよびセサモリンなど)が、上記の成分以外にゴマに含まれており、これらは強い抗酸化性を有している(例えば、Biochemical Systematics and Ecology 13,133−139(1985)参照)。
【0003】
リグナンの生合成については、例えば、Lignans:biosynthesis and function,Comprehensive natural products chemistry,1:640−713(1999); Phytochemistry Rev. 2257−288 (2003)などに記載されている。
例えば、Phytochemistry Rev. 2257−288 (2003)には、コニフェリルアルコールが重合して合成されるピノレジノールが生合成上の最初のリグナンであること、そしてピノレジノールから個々の植物種において特有な生合成経路を経由して多様なリグナンが合成されることが示されている。このピノレジノールの合成に寄与するデリジェントタンパク質がレンギョウなどにおいて報告されている(例えば、Science 275, 362−366 (1997)等参照)。さらに、レンギョウのピノレジノール−ラリシレジノール還元酵素遺伝子(例えば、J.Biol.Chem.,271:29473(1996)、特表2001−507931号公報等参照)、Thuja plicataのピノレジノール−ラリシレジノール還元酵素遺伝子(例えば、J.Biol.Chem.,274:618(1999)等参照)ならびに組換えセコイソラリシレシノールデヒドロゲナーゼおよびその使用方法(例えば、J.Biol.Chem.,276(16):12614−23(2001)、特表2002−512790号公報等参照)が報告されている。さらに、Larrea tridentataから、ラレアトリシン水酸化酵素遺伝子がクローニングされている(例えば、Proc.Nat.Acad.Sci.USA,100:10641(2003)等参照)。
【0004】
ゴマリグナンの生合成についてはピノレジノールにピペリトール合成酵素が作用することによってピペリトールが合成され、次いで、このピペリトールにセサミン合成酵素が作用することによってセサミンが合成されるとされていた。しかしながら、ゴマからクローニングされたチトクロームP450タンパク質であるCYP81Q1は単独でピノレジノールからピペリトールを経て、セサミンを合成することが明らかにされた(国際公開公報WO2005/030944;図1参照)。
【0005】
近年、リグナンだけでなくリグナンの配糖体もまた注目されている。上記したリグナン分子のうちのいくつかは、配糖体として植物中に存在することが知られている。例えば、ゴマ種子中には、セサミノール配糖体(セサミノール2’−O−β−D−グルコピラノシド;セサミノール2’−O−β−D−グルコピラノシル(1−2)−O−β−D−グルコピラノシド;およびセサミノール2’−O−β−D−グルコピラノシル(1−2)−O−(−β−D−グルコピラノシル(1−6))−β−D−グルコピラノシド))、およびピノレジノール配糖体(ピノレジノール4’−O−β−D−グルコピラノシル(1−6)−β−D−グルコピラノシド;ピノレジノール4’−O−β−D−グルコピラノシル(1−2)−β−D−グルコピラノシド;ピノレジノール4’−O−β−D−グルコピラノシル(1−6)−O−(β−D−グルコピラノシル(1−6))β−D−グルコピラノシド;およびピノレジノールジ−O−β−D−グルコピラノシド))などが存在し、レンギョウには、(+)−ピノレジノール−4’−O−β−Dグルコシドおよび(−)−マタイレジノール−4−O−グルコシドが、アマには、セコラリシレジノールジグルコシドおよびピノレジノールジグルコシドなどが存在する(例えば、生薬学雑誌、第32巻、第194頁(1978); Tetrahedron,14:649(2003);Phytochemistry,58:587(2001)等参照)。
【0006】
ゴマに含まれるピノレジノール配糖体およびセサミノール配糖体(例えば、Katsuzaki,H.ら、Biosci.Biotech.Biochem.56,2087−2088(1992)等参照)は、水溶性領域で強い抗酸化性を示すので、脂溶性抗酸化剤(例えば、トコフェロール)とは異なる応用が期待されている。また、リグナン配糖体について以下のような作用機構が提唱されている:リグナン配糖体は、抗酸化性を示す官能基であるフェノール性水酸基が自身の有する糖によって保護されているが、体内へ摂取された後に、腸内細菌の有するβ−グルコシダーゼの作用によって加水分解され、アグリコン部分である脂溶性のリグナンが生成される。このアグリコンが腸内に吸収されて血液を経由して各種臓器に運ばれて、臓器の生体膜などにおける酸化障害を防ぐ。このような作用機構に基づいて、リグナン配糖体は動脈硬化予防食品としての応用が期待されている(例えば、T.Osawa:Anticarcinogenesis and radiation protection 2:p.327,Plenum Press,New Yo参照)。
【0007】
リグナン配糖体以外のリグナン派生体としてメチル化リグナンが知られている。メチル化リグナンもリグナン配糖体同様に抗酸化性を示す官能基であるフェノール性水酸基がメチル基によって保護されており、それによりメトキシ構造を有するリグナンである。フロフラン型のゴマリグナンの中にはピペリトールの4位の水酸基がメチル化されたコブシンおよびセサミノールの2‘位の水酸基がメチル化されたセサンゴリンが報告されている(非特許文献1:Pytochemistry 47, 583−591 (1998);および非特許文献2:J.Org.Chem. 27, 3232−3235 (1962)参照)(図1)。しかしながらこれらの合成反応を触媒する酵素は不明であり、フロフラン型リグナンをメチル化する酵素およびそれをコードする遺伝子はこれまでに精製されたり単離された報告はない。
なお、メチル基転移反応を触媒するという特定の機能を有するタンパク質は、異なる植物種においてもそのアミノ酸配列が類似しているということが知られている(例えば、非特許文献3:Plant Cell 14, 505−519 (2002)参照)。
【非特許文献1】Pytochemistry 47, 583−591 (1998)
【非特許文献2】J.Org.Chem. 27, 3232−3235 (1962)
【非特許文献3】Plant Cell 14, 505−519 (2002)
【発明の開示】
【発明が解決しようとする課題】
【0008】
植物の二次代謝物などの生合成経路を改変することによって、有用な物質の生成および/または有用な植物の育種が行われている。このような技術は、代謝工学と呼ばれている。このような技術を用いれば、任意の化合物を大量生産すること、および/または不要な物質の生産を抑制することが可能になる。従って、リグナン代謝経路に関与する遺伝子を用いてリグナンおよびその代謝物の合成を遺伝子工学的に行うことは、上述したようなこれらの物質の有用性に鑑みて産業上有用である。しかし、リグナン、特にゴマリグナンに代表されるフロフラン型リグナンの生合成に関与する遺伝子に関する知見は、前述のように限られており、さらに、メチル化フロフラン型リグナンの生成を触媒するメチル化酵素は見出されていないので、さらなる遺伝子の取得が望まれていた。
【課題を解決するための手段】
【0009】
本発明は、上記状況に鑑みてなされたもので、下記に示す、リグナンメチル基転移活性を有する酵素、それをコードするポリヌクレオチド、それを含有するベクター・細胞・形質転換体などを提供する。
【0010】
(1)以下の(a)〜(d)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチド;及び
(d)配列番号:2又は4のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
(2)配列番号:2又は4のアミノ酸配列、または当該アミノ酸配列に対して1又は数個のアミノ酸の付加、欠失及び/又は他のアミノ酸による置換によって修飾されているアミノ酸配列を有し、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする上記(1)に記載のポリヌクレオチド。
(3)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする上記(1)に記載のポリヌクレオチド。
(4)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドに対して、5xSSC、50℃の条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする上記(1)に記載のポリヌクレオチド。
(5)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(6)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(7)DNAである、上記(1)〜(6)のいずれかに記載のポリヌクレオチド。
(8)フロフラン型リグナンに対してメチル基を転移する活性を有するタンパク質をコードする上記(1)〜(7)のいずれかに記載のポリヌクレオチド。
(9)ピノレジノール及び/又はピペリトールに対してメチル基を転移する活性を有するタンパク質をコードする上記(8に記載のポリヌクレオチド。
(10)上記(1)〜(9)のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
(11)上記(1)〜(9)のいずれかに記載のポリヌクレオチドを含有するベクター。
(12)上記(11)に記載のベクターにより形質転換された宿主細胞。
(13)上記(12に記載の宿主細胞を培養し又は生育させ、当該宿主細胞からリグナンにメチル基を転移する活性を有するタンパク質を採取することを特徴とする該タンパク質の製造方法。
(14)上記(1)〜(9)のいずれかに記載のポリヌクレオチドが導入された植物体(例えば、ゴマ、レンギョウ又はアマ)もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体、又はそれら植物体の組織。
(15)上記(1)〜(9)のいずれかに記載のポリヌクレオチドを用いてリグナンにメチル基を転移する方法。
(16)上記(1)〜(9)のいずれかに記載のポリヌクレオチドを植物体に導入し、発現させることによって、産生するリグナン組成が改変された当該植物体もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体。
(17)上記(1)〜(9)に記載のポリヌクレオチドのフラグメントまたはその相補配列からなるポリヌクレオチド。
【発明の効果】
【0011】
本発明のポリペプチド(リグナンメチル化酵素)を利用すれば、例えば、生物(特に、植物)においてリグナンおよびメチル化リグナンの量を人為的に調節することができるという効果を奏する。また、これらの組換え酵素を用いてリグナンをメチル化することによって、インビトロおよびインビボにおいて物性(溶解度、動物の吸収効率など)を改変することもできる。また、本発明のポリヌクレオチドを利用することによって、これまでに自然界において未同定であったメチル化リグナンを人工的に生産することも可能となる。また、本発明のポリペプチドを利用して合成されるメチル化リグナンは、新規生理機能物質の出発物質又は中間体として有効に利用することができる。
【0012】
本発明のリグナンメチル化酵素を遺伝子組換え技術を用いて所望の生物に発現させることによって、ピノレジノールからモノメチル化ピノレジノールを、および/またはピペリトールからコブシンを人工的に生産することが可能となる。また、本発明のリグナンメチル化酵素を遺伝子組換え技術を用いて所望の生物に発現させることによって、リグナンとメチル化リグナンとの量を人為的に制御した植物および/または微生物を作出することができる。
また、コブシンまたはメチル化ピノレジノールを生産する植物において、本発明のリグナンメチル化酵素の発現を抑制することによって、アグリコンを遊離させて、リグナン(特に、ピペリトールおよび/またはピノレジノール)の量を増加させることもできる。
さらに、本発明のリグナンメチル化酵素を用いれば、新規メチル化リグナンであるモノメチル化ピノレジノールをピノレジノールから人工的に生産することができる。
【発明を実施するための最良の形式】
【0013】
本発明者らは、リグナン、特に、ピノレジノールおよび/またはピペリトールを主な基質にする新規メチル化酵素を見出すとともに、そのメチル化酵素がピノレジノールをメチル化することを見出した。これまでに、ジメチル化ピノレジノールであるユデスミンは同定されているがモノメチル化ピノレジノールは見出されていない。
本発明者らは、ゴマ種子由来の5000クローンからなるESTデータベースから相同性検索によりゴマメチル化酵素様遺伝子の部分配列を探索し、その結果2種類のメチル化酵素様遺伝子(以下SiOMT1およびSiOMT2)を得た。これらのSiOMT遺伝子は種子において強く発現していた。これらの完全長塩基配列をRACE法で取得し、大腸菌において発現させた。得られた組換えタンパク質をセサミノールまたはピノレジノールと反応させた後、HPLC分析、LC−MS分析、およびTOF−MS/MS分析を用いて酵素活性を測定した。その結果、SiOMT1がピペリトールをメチル化しコブシンを生成する反応を触媒する活性を有することが明らかとなった。さらにSiOMT1はピノレジノールをメチル化しモノメチル化ピノレジノールを生成する反応を触媒する活性を有することが明らかとなった。このメチル化リグナンはピノレジノール派生体であるユデスミンの中間体であると考えられる。アフリカゴマであるSesamum radiatumからPCRによりSiOMT1ホモログであるSrOMT1遺伝子をクローニングし、SrOMT1はSiOMT1と同様のメチル化活性を有することを確認した。
【0014】
なお、「リグナン」は、C6C3骨格を有するフェニルプロパノイド化合物2分子が、主としてこれらの8−8’位を介して重合(8,8’−結合)した化合物である。リグナンは、植物における生体防御機構に寄与していると考えられている(Phytochemistry Rev. 2, 371−390 (2,003)参照)。
代表的なリグナンとしては、ゴマに含まれる(+)−セサミン、(+)−セサミノール、(+)−セサモリン、(+)−ピノレジノール、(+)−ピペリトールおよび(+)−セサモリノール;レンギョウ(Forsythia intermedia)に含まれる(+)−ピノレジノール、(−)−アルクチゲニンおよび(−)−マタイレジノール;ソシマ(Daphne tangutica)に含まれる(−)−ピノレジノールおよび(−)−ラリシレジノール;アマ(Linum usitatissimum)に含まれる(+)−セコイソラリシレジノールなどが挙げられる。これらのリグナンの分子構造は多様である(Wood Research 90, 27−110 (2,003)等参照)。(+)−ピノレジノールに代表されるゴマリグナン類は最も幅広い植物種で同定されているフロフラン型のリグナン類に分類される。ゴマリグナンの1つであるセサミンは、多彩な生理活性作用を有しており、コレステロール代謝、肝機能および免疫機能の改善に有効である(例えば、ゴマその科学と機能性、並木満夫編:丸善プラネット社(1998)参照)。セサミンをゴマ種子またはゴマの絞り粕から分離精製する方法がすでに実用化されており(例えば、特開2001−139579公報、特開平10−7676号公報等)、セサミンを主成分とするアルコール分解促進作用を有する肝機能改善/増強剤が市販されている(商品名:セサミン、発売元:サントリー株式会社)。またセサミン以外の他のリグナン(例えば、セサミノール、セサモリンなど)もまた生理活性作用を有することが報告されている(例えば、日本農芸化学会誌、76:805−813(2002)参照)。このように、リグナン又はその誘導体は、種々の生理活性を有する生理活性物質あるいはその中間体として有用である。
【0015】
以下、本発明のリグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチド、当該ポリヌクレオチドにコードされるタンパク質、およびこれらの利用について詳述する。
【0016】
(1)ポリヌクレオチド
まず、本発明は、以下の(a)〜(d)のいずれかに記載のポリヌクレオチドを提供する。
(a)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチド;及び
(d)配列番号:2又は4のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【0017】
本明細書中、用語「ポリヌクレオチド」は、「遺伝子」、「核酸」または「核酸分子」と交換可能に使用され、ヌクレオチドの重合体が意図される。本明細書中、用語「塩基配列」は、「核酸配列」または「ヌクレオチド配列」と交換可能に使用され、デオキシリボヌクレオチド(A、G、CおよびTと省略される)の配列として示される。
本発明のポリヌクレオチドは、RNA(例えば、mRNA)の形態、またはDNAの形態(例えば、cDNAまたはゲノムDNA)で存在し得る。DNAは、二本鎖または一本鎖であり得る。一本鎖DNAまたはRNAは、コード鎖(センス鎖としても知られる)であり得るか、またはそれは、非コード鎖(アンチセンス鎖としても知られる)であり得る。
【0018】
本明細書中、用語「オリゴヌクレオチド」は、ヌクレオチドが数個ないし数十個(例えば、2〜60個)結合したものが意図され、「ポリヌクレオチド」と交換可能に使用される。オリゴヌクレオチドは、短いものはジヌクレオチド(二量体)、トリヌクレオチド(三量体)といわれ、長いものは30マーまたは100マーというように重合しているヌクレオチドの数で表される。オリゴヌクレオチドは、より長いポリヌクレオチドのフラグメントとして生成されても、化合合成されてもよい。
本明細書中、用語「ポリヌクレオチドの一部」は、そのポリヌクレオチドのフラグメントを意味し、例えば、少なくとも12nt(ヌクレオチド)、好ましくは約15nt、そしてより好ましくは少なくとも約20nt、なおより好ましくは少なくとも約30nt、そしてさらにより好ましくは少なくとも約40ntの長さのフラグメントが意図される。「少なくとも12ntの長さのフラグメント」は、例えば、配列番号1に示される塩基配列からの12以上の連続した塩基を含むフラグメントが意図される。本明細書を参照すれば配列番号1に示される塩基配列が提供されるので、当業者は,配列番号1に基づくDNAフラグメントを容易に作製することができる。例えば、制限エンドヌクレアーゼ切断または超音波による剪断は、種々のサイズのフラグメントを作製するために容易に使用され得る。あるいは、このようなフラグメントは、合成的に作製され得る。適切なフラグメント(オリゴヌクレオチド)が、Applied Biosystems Incorporated(ABI,850 Lincoln Center Dr.,Foster City,CA 94404)392型シンセサイザーなどによって合成される。
【0019】
本発明のポリヌクレオチドは、リグナンメチル化活性を有するポリペプチドをコードする。そのようなポリヌクレオチドは、典型的には、配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;あるいは配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドである。本発明の好ましいポリヌクレオチドは、配列番号:1又は3の塩基配列を有するポリヌクレオチド、あるいは配列番号:2又は4のアミノ酸配列を有するポリペプチドをコードするポリヌクレオチドである。
本発明のポリヌクレオチドは、それがコードするポリペプチドがリグナンメチル化活性を有する限り、配列番号:1又は3の塩基配列において1または複数個(例えば、1〜30個、1〜20個、1〜10個、1〜数個(例えば、6個)、1〜5個、1〜3個、1〜2個など)の塩基が欠失、挿入、置換、及び/または付加された変異体であってもよい。変異体は、コードもしくは非コード領域、またはその両方において変異され得る。コード領域における変異は、保存的または非保存的なアミノ酸の欠失、挿入、置換、または付加を生成し得る。
【0020】
本発明のポリヌクレオチドは、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドであって、配列番号:1または3の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドを含む。
【0021】
ハイブリダイゼーションは、Sambrookら、Molecular Cloning,A Laboratory Manual,2d Ed.,Cold Spring Harbor Laboratory(1989)に記載されている方法のような周知の方法で行うことができる。通常、温度が高いほど、塩濃度が低いほどストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同なポリヌクレオチドを取得することができる。適切なハイブリダイゼーション温度は、塩基配列やその塩基配列の長さによって異なり、例えば、アミノ酸6個をコードする18塩基からなるDNAフラグメントをプローブとして用いる場合、50℃以下の温度が好ましい。
【0022】
本明細書中、用語「ストリンジェントなハイブリダイゼーション条件」は、ハイブリダイゼーション溶液(50%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNAを含む)中にて42℃で一晩インキュベーションした後、約65℃にて0.1×SSC中でフィルターを洗浄することが意図される。ポリヌクレオチドの「一部」にハイブリダイズするポリヌクレオチドによって、参照ポリヌクレオチドの少なくとも約15ヌクレオチド(nt)、そしてより好ましくは少なくとも約20nt、さらにより好ましくは少なくとも約30nt、そしてさらにより好ましくは約30〜70ntにハイブリダイズするポリヌクレオチド(DNAまたはRNAのいずれか)が意図される。
さらに、本発明は、配列番号:1または3の塩基配列と少なくとも80%同一、より好ましくは少なくとも85%、90%、92%、95%、96%、97%、98%または99%同一である塩基配列からなるポリヌクレオチドを提供する。
【0023】
任意の特定の核酸分子が、例えば、配列番号:1または3に示される塩基配列に対して、少なくとも80%、85%、90%、92%、95%、96%、97%、98%、または99%同一であるか否かは、公知のコンピュータープログラム(例えば、Bestfit program(Wisconsin Sequence Analysis Package,Version 8 for Unix(登録商標),Genetics Computer Group,University Research Park,575 Science Drive,Madison,WI 53711)を使用して決定され得る。Bestfitは、SmithおよびWatermanの局所的相同性アルゴリズムを用いて、2つの配列間の最も良好な相同性セグメントを見出す(Advances in Applied Mathematics 2:482〜489(1981))。Bestfitまたは任意の他の配列整列プログラムを用いて、特定の配列が、本発明に従う参照配列に対して、例えば、95%同一であるか否かを決定する場合は、同一性のパーセントが参照塩基配列の全長にわたって計算され、そして参照配列におけるヌクレオチド数全体の5%までの相同性におけるギャップが許容されるように、パラメーターが設定される。
【0024】
特定の実施形態では、参照(QUERY)配列(本発明に係る配列)と対象配列との間の同一性(全体的な配列整列ともいわれる)は、Brutlagらのアルゴリズム(Comp.App.Biosci.6:237〜245(1990))に基づくFASTDBコンピュータープログラムを使用して決定される。%同一性を計算するために、DNA配列のFASTDB整列において使用される好ましいパラメーターは:Matrix=Unitary、k−tuple=4、Mismatch Penalty=1、Joining Penalty=30、Randomization Group Length=0、Cutoff Score=1、Gap Penalty=5、Gap Size Penalty=0.05、Window Size=500または対象塩基配列の長さ(どちらかより短い方)である。
【0025】
また、本発明は、上記ポリヌクレオチドのフラグメントまたはその相補配列からなるオリゴヌクレオチドを包含する。本発明のオリゴヌクレオチドがリグナンメチル化ポリペプチドをコードしない場合でさえ、当業者は、本発明のポリヌクレオチドが、ポリメラーゼ連鎖反応(PCR)のプライマーとして本発明のポリペプチドを作製するために使用され得ることを容易に理解する。リグナンメチル化ポリペプチドをコードしない本発明のオリゴヌクレオチドの他の用途としては、以下が挙げられる:(1)cDNAライブラリー中のリグナンメチル化酵素遺伝子またはその対立遺伝子もしくはスプライシング改変体の単離;(2)リグナンメチル化酵素遺伝子の正確な染色体位置を提供するための、分裂中期染色体スプレッドへのインサイチュハイブリダイゼーション(例えば、「FISH」)(Vermaら,Human Chromosomes:A Manual of Basic Techniques,Pergamon Press,New York(1988)に記載される);および(3)特定の組織におけるリグナンメチル化酵素mRNA発現を検出するためのノーザンブロット分析。
【0026】
本発明のポリヌクレオチドまたはオリゴヌクレオチドは、アンチセンスRNAメカニズムによる遺伝子発現操作のためのツールとして使用することができる。アンチセンスRNA技術によって、内因性遺伝子に由来する遺伝子産物の減少が観察される。本発明のオリゴヌクレオチドを導入することによって、リグナンメチル化活性を有するポリペプチドの含量を低下させ得、その結果、植物中のメチル化リグナン含量または含量比を制御(増加または減少)させることができる。本発明のポリヌクレオチドまたはオリゴヌクレオチドは、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配列を含む)などの配列を含むものであってもよい。
【0027】
本発明のポリヌクレオチドまたはオリゴヌクレオチドを取得する方法としては、本発明のポリヌクレオチドまたはオリゴヌクレオチドを含むDNA断片を単離する種々の公知の方法が挙げられる。例えば、本発明のポリヌクレオチドの塩基配列の一部と特異的にハイブリダイズするプローブを調製して、ゲノムDNAライブラリーまたはcDNAライブラリーをスクリーニングすれば、本発明のポリヌクレオチドまたはオリゴヌクレオチドを取得することができる。このようなプローブとしては、本発明のポリヌクレオチドの塩基配列またはその相補配列の少なくとも一部に特異的にハイブリダイズするポリヌクレオチド(オリゴヌクレオチド)であればよい。
【0028】
このようなハイブリダイゼーションによって選択されるポリヌクレオチドとしては、天然のポリヌクレオチド(例えば、ゴマ科植物やコケ科植物などの植物由来のポリヌクレオチド)が挙げられるが、植物以外に由来するポリヌクレオチドであってもよい。
本発明のポリヌクレオチドを取得する別の方法として、PCRを用いる方法が挙げられる。このPCR増幅方法は、例えば、本発明のポリヌクレオチドのcDNAの5’側および/または3’側の配列(またはその相補配列)を利用してプライマーを調製する工程、これらのプライマーを用いてゲノムDNA(またはcDNA)等をテンプレートにしてPCR増幅する工程を包含することを特徴としており、本方法を使用すれば、本発明のポリヌクレオチドを含むDNA断片を大量に取得することができる。
【0029】
本発明のポリヌクレオチドを取得するための供給源としては、特に限定されないが、ピペリトールまたはピノレジノールを含む生物材料であることが好ましい。本明細書中、用語「生物材料」は、生物学的サンプル(生物体から得られた組織サンプルまたは細胞サンプル)が意図される。後述する実施例においては、ゴマを用いているが、これに限定されない。
【0030】
本発明のポリヌクレオチドを使用すれば、形質転換体または細胞においてリグナンメチル化活性を有するポリペプチドを合成することができる。本発明のポリヌクレオチドを用いれば、ハイブリダイズするポリヌクレオチドを検出することによって、リグナンメチル化活性を有するポリペプチドを発現する生物を容易に検出することができる。
【0031】
本発明のオリゴヌクレオチドは、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドを検出するハイブリダイゼーションプローブまたは当該ポリヌクレオチドを増幅するためのプライマーとして利用することによって、リグナンメチル化活性を有するポリペプチドを発現する生物または組織を容易に検出することができる。なおさらに、上記オリゴヌクレオチドをアンチセンスオリゴヌクレオチドとして使用して、上記生物体またはその組織もしくは細胞におけるリグナンメチル化活性を有するポリペプチドの発現を抑制することができる。
【0032】
(2)ポリペプチド
本発明は、上記した本発明のポリヌクレオチドにコードされるタンパク質(ポリペプチド)をも提供する。そのようなポリペプチドは、典型的には、配列番号:2又は4のアミノ酸配列からなるタンパク質である。
本明細書中、用語「ポリペプチド」は、「ペプチド」または「タンパク質」と交換可能に使用される。また、ポリペプチドの「フラグメント」は、当該ポリペプチドの部分断片が意図される。本発明のポリペプチドはまた、天然供給源より単離されても、化学合成されてもよい。
【0033】
本発明のポリペプチドは、天然の精製産物、化学合成手順の産物、および原核生物宿主または真核生物宿主(例えば、細菌細胞、酵母細胞、高等植物細胞、昆虫細胞、および哺乳動物細胞を含む)から組換え技術によって産生された産物を含む。組換え産生手順において用いられる宿主に依存して、本発明のポリペプチドは、グリコシル化され得るか、または非グリコシル化され得る。さらに、本発明のポリペプチドはまた、いくつかの場合、宿主媒介プロセスの結果として、開始の改変メチオニン残基を含み得る。
また、本発明は、リグナンメチル化活性を有するポリペプチドを提供する。本明細書中、「リグナンメチル化活性」は、リグナンをメチル化する活性、すなわち、リグナンにメチル基を転移する活性が意図される。すなわち、本明細書中、「メチル化酵素」と「メチル基転移酵素」とは、相互交換可能に使用される。「リグナンメチル化活性」は、メチル基供与体であるSAMと基質であるリグナンとをリグナンメチル化酵素と反応させ、その反応生成物をHPLCまたはLC-MS分析することで測定又は確認することができる。一般的なメチル化酵素活性測定法は公知文献に記載のとおりである(公知文献:Toquin, V., et al. (2003) Plant Mol. Biol. 52, 495-509.,Gang, D. R., et al. (2002) Plant Cell 14, 505-519.)。
【0034】
また、本発明のポリペプチドは、配列番号:2または4のアミノ酸配列からなるポリペプチドの変異体であって、かつリグナンメチル化活性を有するポリペプチドを含む。
このような変異体は、配列番号:2又は4のアミノ酸配列において、1もしくは複数個(例えば、1〜30個、1〜20個、1〜10個、1〜数個(6個)、1〜3個、1〜2個など)のアミノ酸(アミノ酸残基)が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質を含む。「欠失、置換、挿入及び/又は付加」には、逆転、反復、およびタイプ置換(例えば、親水性の残基の別の残基への置換、しかし通常は強く親水性の残基を強く疎水性の残基には置換しない)が含まれる。特に、ポリペプチドにおける「中性」アミノ酸置換は、一般的にそのポリペプチドの活性にほとんど影響しない。
【0035】
ポリペプチドのアミノ酸配列中のいくつかのアミノ酸が、このポリペプチドの構造または機能に有意に影響することなく容易に改変され得ることは、当該分野において周知である。さらに、人為的に改変させるだけではく、天然のタンパク質において、当該タンパク質の構造または機能を有意に変化させない変異体が存在することもまた周知である。
当業者は、周知技術を使用してポリペプチドのアミノ酸配列において1または数個のアミノ酸を容易に変異させることができる。例えば、公知の点変異導入法に従えば、ポリペプチドをコードするポリヌクレオチドの任意の塩基を変異させることができる。また、ポリペプチドをコードするポリヌクレオチドの任意の部位に対応するプライマーを設計して欠失変異体または付加変異体を作製することができる。さらに、本明細書中に記載される方法を用いれば、作製した変異体が所望の活性を有するか否かを容易に決定し得る。
好ましい変異体は、保存性もしくは非保存性アミノ酸置換、欠失、または添加を有する。好ましくは、サイレント置換、添加、および欠失であり、特に好ましくは、保存性置換である。これらは、本発明のポリペプチド活性を変化させない。
【0036】
代表的に保存性置換と見られるのは、脂肪族アミノ酸Ala、Val、Leu、およびIleの中での1つのアミノ酸の別のアミノ酸への置換;ヒドロキシル残基SerおよびThrの交換、酸性残基AspおよびGluの交換、アミド残基AsnおよびGlnの間の置換、塩基性残基LysおよびArgの交換、ならびに芳香族残基Phe、Tyrの間の置換である。
上記に詳細に示されるように、どのアミノ酸の変化が表現型的にサイレントでありそうか(すなわち、機能に対して有意に有害な効果を有しそうにないか)に関するさらなるガイダンスは、Bowie, J.U.ら「Deciphering the Message in Protein Sequences: Tolerance to Amino Acid Substitutions」,Science 247:1306−1310 (1990)に記載されている。
【0037】
このような変異ポリペプチドは、上述したように、公知の変異ポリペプチド作製法により人為的に導入された変異を有するポリペプチドに限定されるものではなく、天然に存在するポリペプチドを単離精製したものであってもよい。
本発明のポリペプチドは、アミノ酸がペプチド結合しているポリペプチドであればよいが、これに限定されるものではなく、ポリペプチド以外の構造を含む複合ポリペプチドであってもよい。本明細書中で使用される場合、「ポリペプチド以外の構造」としては、糖鎖やイソプレノイド基等を挙げることができるが、特に限定されない。
また、本発明のポリペプチドは、付加的なポリペプチドを含むものであってもよい。付加的なポリペプチドとしては、例えば、HisやMyc、Flag等のエピトープ標識ポリペプチドが挙げられる。
【0038】
また、本発明のポリペプチドは、本発明のポリペプチドをコードするポリヌクレオチドを宿主細胞に導入して、そのポリペプチドを細胞内発現させた状態であってもよいし、細胞、組織などから単離精製されてもよい。
本発明のポリペプチドは、下記で詳述されるように組換え生成されても、化学合成されてもよい(例えば、Houghten,R.A.,Proc.Natl.Acad.Sci.USA 82:5131−5135(1985);米国特許第4,631,211号等参照)。
【0039】
本発明のポリペプチドは、リグナン(特に、ピノレジノールまたはピペリトール)のメチル化反応を触媒することができる。
【0040】
(3)本発明のポリペプチドまたはポリヌクレオチドの利用
本発明はさらに、本発明のポリペプチドまたはポリヌクレオチドを用いることによって生物(好ましくは、植物)中のリグナンおよびメチル化リグナンの量を制御(増加または減少)するための方法および当該制御された生物(好ましくは、植物)を提供する。
【0041】
(A)ベクター
本発明は、リグナンメチル化活性を有するポリペプチドを生成するために使用されるベクターを提供する。本発明のベクターは、インビトロ翻訳に用いるベクターであっても組換え発現に用いるベクターであってもよい。
本発明のベクターは、上述した本発明のポリヌクレオチドを含むものであれば、特に限定されない。例えば、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドのcDNAが挿入された組換え発現ベクターなどが挙げられる。組換え発現ベクターの作製方法としては、プラスミド、ファージ、またはコスミドなどを用いる方法が挙げられるが特に限定されない。
【0042】
ベクターの具体的な種類は特に限定されず、宿主細胞中で発現可能なベクターが適宜選択され得る。すなわち、宿主細胞の種類に応じて、確実に本発明のポリヌクレオチドを発現させるために適宜プロモーター配列を選択し、これと本発明のポリヌクレオチドを各種プラスミド等に組み込んだベクターを発現ベクターとして用いればよい。
本発明の発現ベクターは、導入されるべき宿主の種類に依存して、発現制御領域(例えば、プロモーター、ターミネーター、および/または複製起点等)を含有する。細菌用発現ベクターのプロモーターとしては、慣用的なプロモーター(例えば、trcプロモーター、tacプロモーター、lacプロモーター等)が使用され、酵母用プロモーターとしては、例えば、グリセルアルデヒド3リン酸デヒドロゲナーゼプロモーター、PH05プロモーター等が挙げられ、糸状菌用プロモーターとしては、例えば、アミラーゼ、trpC等が挙げられる。また動物細胞宿主用プロモーターとしては、ウイルス性プロモーター(例えば、SV40初期プロモーター、SV40後期プロモーター等)が挙げられる。植物体の形質転換に用いられる組換え発現ベクターは、当該植物内で本発明のポリヌクレオチドを発現させることが可能なベクターであれば特に限定されない。このようなベクターとしては、例えば、植物細胞内でポリヌクレオチドを構成的に発現させるプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクター、または外的な刺激によって誘導性に活性化されるプロモーターを有するベクターが挙げられる。
【0043】
発現ベクターの作製は、制限酵素および/またはリガーゼ等を用いる慣用的な手法に従って行うことができる。発現ベクターによる宿主の形質転換もまた、慣用的な手法に従って行うことができる。
上記発現ベクターを用いて形質転換された宿主を、培養、栽培または飼育した後、培養物等から慣用的な手法(例えば、濾過、遠心分離、細胞の破砕、ゲル濾過クロマトグラフィー、イオン交換クロマトグラフィー等)に従えば、目的タンパク質を回収、精製することができる。
【0044】
発現ベクターは、少なくとも1つの選択マーカーを含むことが好ましい。このようなマーカーとしては、真核生物細胞培養についてはジヒドロ葉酸レダクターゼ遺伝子またはネオマイシン耐性遺伝子、およびE.coliおよび他の細菌における培養についてはテトラサイクリン耐性遺伝子またはアンピシリン耐性遺伝子が挙げられる。
上記選択マーカーを用いれば、本発明のポリヌクレオチドが宿主細胞に導入されたか否か、さらには宿主細胞中で確実に発現しているか否かを確認することができる。あるいは、本発明のポリペプチドを融合ポリペプチド(例えば、GFPとの融合ポリペプチド)として発現させ、GFPの蛍光をマーカーとして用いてもよい。
【0045】
(B)形質転換体または細胞
本発明は、上述したリグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドが導入された形質転換体または細胞を提供する。本明細書中、用語「形質転換体」は、組織または器官だけでなく、生物個体を含むことが意図される。
形質転換体または細胞の作製方法(生産方法)は特に限定されないが、例えば、上述した組換えベクターを宿主に導入して形質転換する方法が挙げられる。ここで用いられる宿主細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いることができる。具体的には、例えば、大腸菌(Escherichia coli)等の細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母Schizosaccharomyces pombe)、線虫(Caenorhabditis elegans)、アフリカツメガエル(Xenopus laevis)の卵母細胞等を挙げることができる。上記の宿主細胞のための適切な培養培地および条件は当分野で周知である。また、形質転換の対象となる生物も特に限定されるものではなく、上記宿主細胞で例示した各種微生物、植物または動物が挙げられる。
【0046】
本発明の形質転換体または細胞は、これらの形質転換体または細胞が天然に有するリグナンおよび/またはメチル化リグナンの組成が改変されていることを特徴とする。本発明の形質転換体または細胞は、植物もしくはその子孫、またはこれら由来の組織であることが好ましく、ゴマ、レンギョウまたはアマであることが特に好ましい。このような形質転換体または細胞は、本発明のメチル化リグナン含有量を制御する方法を用いることによって、リグナンを産生する生物中のメチル化リグナンの含有量を増加または減少させることができる。
【0047】
本発明の形質転換体は、植物形質転換体であり得る。本実施形態の植物形質転換体は、本発明のポリヌクレオチドを含む組換えベクターを、当該ポリヌクレオチドによってコードされるポリペプチドが発現され得るように植物中に導入することによって取得される。
【0048】
組換え発現ベクターを用いる場合、植物体の形質転換に用いられる組換え発現ベクターは、当該植物内で本発明のポリヌクレオチドを発現させることが可能なベクターであれば特に限定されない。このようなベクターとしては、例えば、植物細胞内でポリヌクレオチドを構成的に発現させるプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクター、または外的な刺激によって誘導性に活性化されるプロモーターを有するベクターが挙げられる。
【0049】
本発明において形質転換の対象となる植物は、植物体全体、植物器官(例えば葉、花弁、茎、根、種子など)、植物組織(例えば表皮、師部、柔組織、木部、維管束、柵状組織、海綿状組織など)または植物培養細胞、あるいは種々の形態の植物細胞(例えば、懸濁培養細胞)、プロトプラスト、葉の切片、カルスなどのいずれをも意味する。形質転換に用いられる植物としては、特に限定されず、単子葉植物綱または双子葉植物綱に属する植物のいずれでもよい。
【0050】
植物への遺伝子の導入には、当業者に公知の形質転換方法(例えば、アグロバクテリウム法、遺伝子銃、PEG法、エレクトロポレーション法など)が用いられる。例えば、アグロバクテリウムを介する方法と直接植物細胞に導入する方法が周知である。アグロバクテリウム法を用いる場合は、構築した植物用発現ベクターを適当なアグロバクテリウム(例えば、アグロバクテリウム・チュメファシエンス(Agrobacterium tumefaciens))に導入し、この株をリーフディスク法(内宮博文著、植物遺伝子操作マニュアル(1990)27〜31頁、講談社サイエンティフィック、東京)などに従って無菌培養葉片に感染させ、形質転換植物を得ることができる。また、Nagelらの方法(Micribiol.Lett.、67、325(1990))が用いられ得る。この方法は、まず、例えば発現ベクターをアグロバクテリウムに導入し、次いで、形質転換されたアグロバクテリウムをPlantMolecular Biology Manual(S.B.Gelvinら、Academic Press Publishers)に記載の方法で植物細胞または植物組織に導入する方法である。ここで、「植物組織」とは、植物細胞の培養によって得られるカルスを含む。アグロバクテリウム法を用いて形質転換を行う場合には、バイナリーベクター(pBI121またはpPZP202など)を使用することができる。
【0051】
また、遺伝子を直接植物細胞または植物組織に導入する方法としては、エレクトロポレーション法、遺伝子銃法が知られている。遺伝子銃を用いる場合は、植物体、植物器官、植物組織自体をそのまま使用してもよく、切片を調製した後に使用してもよく、プロトプラストを調製して使用してもよい。このように調製した試料を遺伝子導入装置(例えばPDS−1000(BIO−RAD社)など)を用いて処理することができる。処理条件は植物または試料によって異なるが、通常は450〜2000psi程度の圧力、4〜12cm程度の距離で行う。
【0052】
遺伝子が導入された細胞または植物組織は、まずハイグロマイシン耐性などの薬剤耐性で選択され、次いで定法によって植物体に再生される。形質転換細胞から植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。
植物培養細胞を宿主として用いる場合は、形質転換は、組換えベクターを遺伝子銃、エレクトロポレーション法などで培養細胞に導入する。形質転換の結果得られるカルスやシュート、毛状根などは、そのまま細胞培養、組織培養または器官培養に用いることが可能であり、また従来知られている植物組織培養法を用い、適当な濃度の植物ホルモン(オーキシン、サイトカイニン、ジベレリン、アブシジン酸、エチレン、ブラシノライドなど)の投与などによって植物体に再生させることができる。
【0053】
遺伝子が植物に導入されたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法などによって行うことができる。例えば、形質転換植物からDNAを調製し、DNA特異的プライマーを設計してPCRを行う。
【0054】
本発明のポリヌクレオチドがゲノム内に組み込まれた形質転換植物体が一旦取得されれば、当該植物体の有性生殖または無性生殖によって子孫を得ることができる。また、当該植物体またはその子孫、あるいはこれらのクローンから、例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラストなどを得て、それらを基に当該植物体を量産することができる。したがって、本発明には、本発明のポリヌクレオチドが発現可能に導入された植物体、もしくは、当該植物体と同一の性質を有する当該植物体の子孫、またはこれら由来の組織も含まれる。
【0055】
また、種々の植物に対する形質転換方法が既に報告されている。本発明の形質転換体植物としては、例えば、ゴマ、イネ、タバコ、オオムギ、コムギ、ナタネ、ポテト、トマト、ポプラ、バナナ、ユーカリ、サツマイモ、タイズ、アルファルファ、ルーピン、トウモロコシ、カリフラワー、バラ、キク、カーネーション、キンギョソウ、シクラメン、ラン、トルコギキョウ、フリージア、ガーベラ、グラジオラス、カスミソウ、カランコエ、ユリ、ペラルゴニウム、ゼラニウム、ペチュニア、トレニア、チューリップ、レンギョウ、シロイヌナズナ、アマ、およびミヤコグサなどが挙げられるがこれらに限定されない。
好ましい実施形態において、ゴマを使用して、本発明の形質転換体を作製することができる。ゴマの形質転換体の作製方法としては、例えば、T.Asamizu:Transformation of sesame plants using MAT vector system:introduction of fatty acid desaturase genes.Sesame Newsletter 16:22〜25(2002)に記載されるような公知の方法が挙げられる。
このようにして得た形質転換ゴマを用いれば、当該ゴマ内でメチル化リグナンを生産するため、低コストかつ環境に低負荷な生産プロセスでメチル化リグナン(ピペリトール、および/またはピノレジノール)を生産することができる。
【0056】
他の好ましい実施形態において、本発明の形質転換体として、タバコを好適に用いることができる。タバコは、ペチュニアなどとともに、形質転換が容易な代表的植物であり、細胞壁を取り除いた細胞の1個(プロトプラスト)から、植物1個体への再生が可能である。この再生された植物1個体は、多細胞に由来する1個体とは異なりキメラ状に陥らないため、形質転換体の作製を効率よく行うことができる。
タバコの形質転換に好ましい方法としては、リーフディスク法が挙げられる。この方法は、操作が容易であり、かつ1枚の葉切片からいくつもの独立した形質転換体を得ることができる方法である。形質転換の方法はたとえば「新生物化学実験の手引き3 核酸の分離・分析と遺伝子実験法 化学同人 1996年」に記載されている。
このようにして得た形質転換タバコを用いれば、低コストかつ環境に低負荷な生産プロセスでリグナンメチル化酵素を生産することができる。
別の好ましい実施形態において、本発明の形質転換体として、イネを好適に用いることもできる。形質転換イネを用いれば、当該イネ内で低コストかつ環境に低負荷な生産プロセスでリグナンメチル化酵素を生産することができる。
【0057】
本発明の形質転換体は、リグナン(特に、ピノレジノールまたはピペリトール)を含む生物であれば、生物種を問わず、上記ポリヌクレオチドを導入することで当該メチル化リグナンを生産することができる。
本発明のポリペプチドをコードするポリヌクレオチドを含む組換え発現ベクターが導入された形質転換体を用いれば、植物などの生物に内在するリグナンをメチル化する反応を触媒することができるので、低コストかつ環境に対して低負荷な生産プロセスでメチル化リグナンの大量調製が可能になる。さらに本発明は、メチル化リグナンを大量調製することによって安価な食品または工業製品を提供することができる。
本発明の形質転換体を用いれば、リグナンメチル化反応を触媒するポリペプチドを低コストでありかつ環境に低負荷な条件下で提供することができる。
一実施形態において、本発明に係る細胞は、種々の細菌宿主であり得る。本実施形態に係る細胞は、本発明に係るポリヌクレオチドを含む組換えベクターを、当該ポリヌクレオチドによってコードされるポリペプチドが発現され得るように細胞中に導入することによって取得される。
【0058】
当業者は、本明細書中の記載に従えば、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドを含む組換え発現ベクターが導入されれば、細菌から高等植物までの広範な生物にリグナンメチル化能を付与することができるということを容易に理解する。
本発明の細胞は、リグナン(特に、ピノレジノールまたはピペリトール)を含む生物であれば、生物種を問わず、上記ポリヌクレオチドを導入することで当該メチル化リグナンを生産することができる。
【0059】
本発明のポリペプチドをコードするポリヌクレオチドを含む組換え発現ベクターが導入された細胞を用いれば、当該細胞内でリグナンメチル化反応を触媒することができるので、低コストかつ環境に対して低負荷な生産プロセスでメチル化リグナンの大量調製が可能になる。さらに本発明は、メチル化リグナンを大量調製することによって安価な食品または工業製品を提供することができる。
本発明に係る細胞を用いれば、リグナンメチル化反応を触媒するポリペプチドを低コストでありかつ環境に低負荷な条件下で提供することができる。
【0060】
(C)ポリペプチドの生産方法
本発明は、本発明のポリペプチドを生産する方法を提供する。本発明のポリペプチドの生産方法を用いれば、リグナンメチル化反応を触媒するポリペプチドを低コストでありかつ環境に低負荷な条件下で提供することが可能となる。また、本発明のポリペプチドの生産方法を用いれば、リグナンメチル化反応を触媒するポリペプチドを容易に生産することが可能となる。
【0061】
本発明のポリペプチドの生産方法では、本発明のポリペプチドをコードするポリヌクレオチドを含むベクターを用いることができる。
本発明のポリペプチドの生産方法は、上記ベクターを無細胞タンパク質合成系に用いることが好ましい。無細胞タンパク質合成系を用いる場合、種々の市販のキットを用いればよい。好ましくは、本実施形態のポリペプチドの生産方法は、上記ベクターと無細胞タンパク質合成液とをインキュベートする工程を包含する。
【0062】
また、本実施形態のポリペプチドの生産方法は、組換え発現系を用いることもできる。組換え発現系を用いる場合、本発明のポリヌクレオチドを組換え発現ベクターに組み込んだ後、公知の方法により発現可能に宿主に導入し、宿主内で翻訳されて得られる上記ポリペプチドを精製するという方法などを採用することができる。組換え発現ベクターは、プラスミドであってもなくてもよく、宿主に目的ポリヌクレオチドを導入することができればよい。好ましくは、本実施形態のポリペプチドの生産方法は、上記ベクターを宿主に導入する工程を包含する。
【0063】
宿主としては、原核生物または真核生物を用いることができる。原核生物宿主としては、例えば、Escherichia属に属する細菌(例えば、大腸菌(Escherichia coli)など)、例えば、Bacillus属に属する細菌(例えば、Bacillus subtilisなどを用いることができる。真核生物宿主としては、下等真核生物(例えば、酵母または糸状菌などの真核生物微生物)を用いることができる。酵母としては、Saccharomyces属微生物(例えば、Saccharomyces cerevisiaeなど)が挙げられ、糸状菌としては、Aspergillus属微生物(例えば、Aspergillus oryzae、Aspergillus nigerなど)、Penicillium属微生物が挙げられる。また、動物細胞または植物細胞が宿主として使用され得る。動物細胞としては、マウス、ハムスター、サル、ヒト等の細胞が挙げられる。さらに、昆虫細胞(例えば、カイコ細胞、またはカイコの成虫)もまた宿主として使用され得る。
上記の宿主細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いることができる。具体的には、例えば、大腸菌(Escherichia coli)等の細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母Schizosaccharomyces pombe)、線虫(Caenorhabditis elegans)、アフリカツメガエル(Xenopus laevis)の卵母細胞等を挙げることができるが、特に限定されない。上記の宿主細胞のための適切な培養培地および条件は当分野で周知である。
【0064】
このように宿主に外来ポリヌクレオチドを導入する場合、発現ベクターは、外来ポリヌクレオチドを発現するように宿主内で機能するプロモーターを組み込んであることが好ましい。組換え的に産生されたポリペプチドを精製する方法は、用いた宿主、ポリペプチドの性質によって異なるが、タグの利用等によって比較的容易に目的のポリペプチドを精製することが可能である。
【0065】
本実施形態に係るポリペプチドの生産方法は、本発明のポリペプチドを含む細胞または組織の抽出液から当該ポリペプチドを精製する工程をさらに包含することが好ましい。ポリペプチドを精製する工程は、周知の方法(例えば、細胞または組織を破壊した後に遠心分離して可溶性画分を回収する方法)で細胞や組織から細胞抽出液を調製した後、この細胞抽出液から周知の方法(例えば、硫安沈殿またはエタノール沈殿、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィー)によって精製する工程が好ましいが、これらに限定されない。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)が精製のために用いられる。
【0066】
本発明のポリペプチドは、本発明のポリペプチドを天然に発現する細胞または組織から当該ポリペプチドを精製することもできるし、化学合成によって調製することもできる。
【0067】
変異ポリペプチドを作製する方法についても、特に限定されない。例えば、部位特異的変異誘発法(例えば、Hashimoto−Gotoh,Gene 152,271−275(1995)参照)、PCR法を利用して塩基配列に点変異を導入し変異ポリペプチドを作製する方法、またはトランスポゾンの挿入による突然変異株作製法などの周知の変異ポリペプチド作製法を用いることによって、変異ポリペプチドを作製することができる。変異ポリペプチドの作製には市販のキットを利用してもよい。
このように、本発明のポリペプチドの生産方法は、少なくとも、リグナンメチル化活性を有するポリペプチドのアミノ酸配列、またはリグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドの塩基配列に基づいて公知慣用技術を用いればよいといえる。
【0068】
(D)メチル化リグナン生産方法
これまで、リグナンおよびメチル化リグナンの生産はゴマからの抽出により行われているので、大量生産ができない等の問題点を有していたが、本発明に従えば、リグナンおよびメチル化リグナンを低コストで大量生産できる。
本発明は、本発明のポリペプチドを発現する生物体または細胞を用いてメチル化リグナンを生産する方法を提供する。上記生物体は、天然の未改変生物体であっても組換え発現系を用いた形質転換体であってもよい。本発明のメチル化リグナン生産方法は、リグナン(特に、ピノレジノールまたはピペリトール)を効率よく生産することができる。
一実施形態において、本発明のメチル化リグナン生産方法は、本発明のポリペプチドをコードするポリヌクレオチドで形質転換された生物体またはその組織を用いてメチル化リグナンを生産することを特徴とする。好ましくは、上記生物体は、上述した形質転換体植物または細菌であり、特に好ましくは、大腸菌、ゴマ、レンギョウまたはアマである。
【0069】
本発明のメチル化リグナン生産方法は、本発明のポリペプチドをコードするポリヌクレオチドを上記生物体に導入する工程を包含する。ポリヌクレオチドを上記生物体に導入する工程としては、上述した種々の遺伝子導入方法を用いればよい。本実施形態のこの局面において、上記生物体は形質転換される前に産生したメチル化リグナンと形質転換後に産生するメチル化リグナンとの間でその組成が異なる。具体的には、上記生物体から得られるリグナンおよびメチル化リグナンは、その含有率が増加する。本実施形態のこの局面に係るメチル化リグナン生産方法は、上記生物体からメチル化リグナンを抽出する工程をさらに包含することが好ましい。
【0070】
また、本発明のメチル化リグナン生産方法は、本発明のポリペプチドを天然に発現する生物体に、本発明のオリゴヌクレオチドをアンチセンスオリゴヌクレオチドとして導入する工程を包含する。オリゴヌクレオチドを上記生物体に導入する工程としては、上述したアンチセンスRNA技術を用いればよい。本実施形態に係るメチル化リグナン生産方法は、上述した抗体またはオリゴヌクレオチドを用いて本発明のポリペプチドを天然に発現する生物体を同定する工程をさらに包含することが好ましい。本実施形態のこの局面に係るメチル化リグナン生産方法は、上記生物体からメチル化リグナンを抽出する工程をさらに包含することが好ましい。
本実施形態において、上記生物体は上記オリゴヌクレオチドを導入される前に産生したメチル化リグナンと導入後に産生するメチル化リグナンとの間でその組成が異なる。具体的には、上記生物体から得られるリグナンおよびメチル化リグナンは、その含有率が減少する。
【0071】
(E)食品および工業製品
本発明は、上述のメチル化リグナン生産方法により得られたメチル化リグナンを用いて製造される食品および工業製品を提供する。本項で記載する食品は、上述した形質転換植物体の種子、果実、切穂、塊茎、および/または塊根であっても、上述した形質転換植物体から抽出されたメチル化リグナンを用いて製造された食品(例えば、ゴマ、レンギョウまたはアマ、あるいはこれらの加工食品)であってもよい。本発明に係る食品または工業製品は、所望する量のリグナン(特に、ピノレジノールまたはピペリトール)を含有することができる。
例えば、上述のようにメチル化リグナンの含量が増加した本発明の形質転換植物から抽出されたメチル化リグナン抽出液は、メチル化リグナンの含量が高い食品として提供される。また、抽出したメチル化リグナンに限らず、上記形質転換植物体の種子、果実、切穂、塊茎、および/または塊根等もまた、メチル化リグナンを多く含む食品として提供される。メチル化リグナン組成を改変する対象は特に限定されるものではなく、植物以外にも動物、細菌、または酵母等のあらゆる生物を対象とすることが可能である。
また、リグナンおよびメチル化リグナンの独特な物性に基づいて、本発明のポリペプチドまたはポリヌクレオチドは、工業製品(例えば、フィルム、生分解性プラスティック、機能性繊維、潤滑油、または洗剤のような工業製品)の原料に利用され得る。
【0072】
本明細書において、ゴマのリグナンメチル化ポリペプチドを一例として記載されているが、本発明は、ゴマ由来のポリペプチドまたはポリヌクレオチドにのみ限定されるべきではなく、リグナンメチル化活性を有する全てのポリペプチド、およびその利用に関するものであることが、当業者には明白である。リグナンメチル化酵素は、植物、動物または微生物のいずれ由来でもよく、リグナンメチル化酵素活性を有していればリグナン量を制御することができる。さらに本発明は、リグナンメチル化酵素をコードするポリヌクレオチドを導入することによって作製されたリグナン量が調節された植物、その子孫またはこれらの組織に関し、その形態は切り花であってもよい。本発明のリグナンメチル化ポリペプチドを用いれば、メチル化リグナンの生成を促進または抑制することができる。また、慣用的な手法を用いれば、植物に上記ポリヌクレオチドを導入して当該ポリヌクレオチドを構成的あるいは組織特異的に発現させて、目的のポリペプチドの発現を増加させること、ならびに、アンチセンス法、同時サプレッション法およびRNAi法などを用いることによって目的のポリペプチドの発現を抑制することができるということを、当業者は容易に理解する。
本発明は、以下の実施例によってさらに詳細に説明されるが、これに限定されるべきではない。
【実施例】
【0073】
実施例において用いる分子生物学的手法は、他で詳述しない限り、国際公開WO2004/018682号公報(PCT/JP03/10500)またはMolecular Cloning(Sambrookら、Cold Spring Harbour Laboratory Press,1989)に記載の方法に従った。
【0074】
〔実施例1:ゴマ種子のEST解析〕
公知文献(国際公開公報WO2005/030944)に記載のゴマ種子由来のcDNAファージライブラリーをRapid Excisionキット(Stratagene社)を用いて製造業者が推奨する方法に従って大腸菌内にpBK-CMV(Stratagene社)に切り出した。このゴマ種子由来ESTを含む大腸菌コロニーをランダムに5000クローンをピックアップし、M13RVプライマー(配列番号5)およびM4(-20)プライマー(配列番号6)を用いて以下の条件でコロニーPCRを行った。
1×Ex−Taq buffer(TakaRa)、0.2mM dNTPs、プライマー各0.2pmol/μl、Ex−Taq polymerase 1.25Uからなる混合液に大腸菌コロニーをけん濁し、94℃で5分間の後、94℃で1分間、55℃で1分間、72℃で2分間の反応を30サイクル行ってPCR増幅した後、72℃で7分間保持した。
配列番号5:M13RV:5'-CAGGAAACAGCATTGAC
配列番号6:M4-20:5'-GTAAAACGACGGCCAGT
【0075】
これらのPCR産物をアガロースゲル電気泳動に供し、pBK−CMVに含まれるESTが特異的に増幅されていることをエチジウムブロマイド染色により確認した。これら5000種のPCR産物4μlを10unitのExonucleaseI(USB社)と2unitのShrimp Alkaline phosphatase(USB社)と混合し、37℃で30分保持し、80℃で20分保持し酵素反応を終了させた。この酵素反応液1μlを用いて以下の条件でダイレクトシークエンス反応を行った。シークエンス反応液は酵素反応液1μl、5xBigDyeSequencingBuffer ver.3.1(Applied Biosystems)3μl、BigDyeSequencingRR ver.3.1(Applied Biosystems) 2μl、1.6pmol/μlM13RVプライマー1μl、滅菌水13μlからなる。シークエンス反応は96℃1分間に続いて96℃10秒間、50℃5秒間、60℃4分間の反応を25サイクル行った。シークエンス反応液はDyeEx96(QIAGEN社)を用いて業者が推奨する方法で生成し、Sequencer model 3100(Applied Biosystems社)を用いて塩基配列を決定した。
【0076】
得られた5000種の塩基配列はBlastxを用いて相同検索を行い、その中からメチル化酵素遺伝子と相同性を有する2種類のゴマメチル化酵素(Sesamum indicum O-methyltransferase; 以下SiOMT)の部分配列を同定した。Blastx解析の条件は以下のとおりである。プログラム:Blastx ver.2.2.9、データベース:nr、遺伝コード:standard(1)、フィルタ:LOW complexity、Expect:10、Word size:3、マトリクス:BLOSUM62、Gap Costs:Existence 11、Extension 1。
【0077】
〔実施例2:ゴマメチル化酵素遺伝子の発現解析〕
EST解析により得られた二種類のSiOMT1およびSiOMT2の遺伝子発現パターンを解析するために、公知文献(国際公開公報WO2005/030944)に記載のゴマの器官別cDNAに対して以下の条件でRT−PCRを行った。
cDNA 1μl、1×Ex−Taq buffer(TakaRa)、0.2mM dNTPs、プライマー各0.2pmol/μl、Ex−Taq polymerase 1.25Uからなる。94℃で5分間の後、94℃で1分間、55℃で1分間、72℃で2分間の反応を32サイクル行ってPCR増幅した後、72℃で5分間保持した。SiOMT1およびSiOMT2特異的プライマーとしてSiOMT1-FW(配列番号7)とSiOMT1-RV(配列番号8)およびSiOMT2-FW(配列番号9)とSiOMT2-RV(配列番号10)を用いた。SiOMT遺伝子の発現量と内在性遺伝子の発現量とを比較するために、内部標準となる遺伝子のPCR反応を同時に行った。具体的には、Si18SrRNA−Fプライマー(配列番号11)およびSi18SrRNA−Rプライマ−(配列番号12)を用いてゴマの18S ribosomal RNA遺伝子(アクセッション番号:AF169853)のPCR反応を行った。
配列番号7:SiOMT1-FW:5‘-TTGCCCCATGTCATTCAAGAT
配列番号8:SiOMT1-RV:5‘-AAAATTCAGACTTATAACGATACCAAA
配列番号9:SiOMT2-FW:5‘-TTAGAAAAACTCAATTCGTCTAAT
配列番号10:SiOMT2-RV:5‘-CCTACATCCACGACGGAATCCAAA
配列番号11:Si18SrRNA−FW:5’−tatgcttgtctcaaagattaa
配列番号12:Si18SrRNA?RV:5’−aacatctaagggcatcacaga
【0078】
PCR産物を電気泳動し、エチジウムブロマイド染色を行った結果、両SiOMT遺伝子ともに種子において強い発現が確認された(図2の2A)。すなわち、両SiOMT遺伝子発現領域とメチル化リグナンが蓄積する領域が一致することを確認した。
【0079】
〔実施例3:ゴマメチル化酵素遺伝子のクローニング〕
両ESTクローンが推定ORFの5’領域を含んでいなかったため、5’Rapid Amplification of cDNA End(以下、5’RACE)法を使用して、それぞれのSiOMT遺伝子の5’領域を増幅した。具体的には、Gene Racer kit(Invitrogen社)を製造業者の推奨する方法に従って使用して、以下のプライマー(配列番号13〜16)を用いて、各々の5’領域を増幅した。
配列番号13:GR-SiOMT1-RV:5'-ccggcccactgttcgggtcctaacgggaaa
配列番号14:SiOMT1-NEST−RV:5'-gcaaatccacttcataaaaat
配列番号15:GR-SiOMT2-RV:5'-cctcgggttccgctctttctgctcccagaa
配列番号16:SiOMT2-NEST−RV:5'-atcaatttgggaaattacaaa
【0080】
SiOMT2に関してはESTクローンが推定ORFの3‘末端を含んでいなかったため、配列番号17および18のプライマーを用いて3’RACEを行った。
配列番号17:GR-SiOMT2-FW:5'-gaagatcgccccatgagcatgaaacccttt
配列番号18:SiOMT2-NEST−FW:5'-aacgtcgttctgggagcagaaaga
【0081】
RACE法によって得られた増幅断片の塩基配列を、プライマーウォーキング法によって決定し、SiOMT1およびSiOMT2遺伝子の完全長オープンリーディングフレームを含む塩基配列情報を得た(配列番号1および配列番号19)。
SiOMT1とSiOMT2はお互いにアミノ酸配列で29%の配列同一性を示した。配列同一性について、Clustal W alignmentプログラム(Mac Vector ver.7.2.2(Symanteccorporation))をデフォルト設定で使用した。
【0082】
得られたSiOMT遺伝子の完全長cDNA配列をBlastx解析(http://www.ncbi.nlm.nih.gov/BLAST/)によって相同性を有する既知タンパク質を検索した。Blastx解析の条件は以下のとおりである。
プログラム:Blastx ver.2.2.9、データベース:nr、遺伝コード:standard(1)、フィルタ:LOW complexity、Expect:10、Word size:3、マトリクス:BLOSUM62、Gap Costs:Existence 11、Extension 1。
Blastx解析の結果、SiOMT1は、Catharanthsus roseus Caffeic acid-O-methyltransferase(COMT)(AAK20170)と74%の配列同一性を示し、SiOMT2は、Rosa hybrida(AAM23004)のOrcinol-O-methyltransferase(OOMT)と57%の配列同一性を示した。このように、両SiOMTと既知のメチル化酵素との配列同一性はいずれも高いとは言えない。従って、得られた両SiOMT遺伝子の機能を推定することはできなかった。すなわち、得られた両SiOMT遺伝子は、これまでに単離されていないリグナンメチル化酵素である可能性が非常に高い。
【0083】
〔実施例4:大腸菌発現ベクターの構築〕
SiOMT1に対して配列番号20および21のプライマー対を使用して、cDNAの開始メチオニンコドン(ATG)の上流にBglII部位を、終始コドンの下流にSalI部位を有するフラグメントをPCR増幅した。
一方、SiOMT2に対して配列番号22および23のプライマー対を使用して、cDNAの開始メチオニンコドン(ATG)の上流にBamHI部位を、終始コドンの下流にXhoI部位を有するフラグメントをPCR増幅した。PCRに用いたプライマーを以下に示す。
配列番号20:Bgl2-SiOMT1-FW:5'-aaaacatgtatggcggatcagtccgaggaagaagaggcttt
配列番号21:SalI-SiOMT1-RV:5'-attgtcgacttatgaaattccatgatccaaatatt
配列番号22:BamHI-SiOMT2-FW:5'-aaaggatccatggcgatggttaaccaaaagcaaaatctt
配列番号23:XhoI-SiOMT2-RV:5'-aaactcgagttaaggatatatctcgatgatagatctcaa
【0084】
PCR反応液(25μl)は、テンプレートとしてのゴマ種子cDNA、各プライマー0.2pmol/μl、1×KOD plus buffer (TOYOBO)、0.2mM dNTPs、1mM MgSO4、1U KOD plus polymeraseからなる。94℃で5分間反応させた後、94℃1分間、55℃1分間、72℃2分間の反応を30サイクル行ってPCR増幅した後、72℃で3分間保持した。得られた各PCR産物を、製造者が推奨する方法に従ってpCR4 Blunt−TOPO vector(Invitrogen)のマルチクローニング部位に挿入して、SiOMT1/pCR4 Blunt−TOPO(pSPB2678と称する)、SiOMT2/pCR4 Blunt−TOPO(pSPB2679と称する)を得た。
pSPB2678および、pSPB2679に含まれるSiOMT塩基配列を解析して、正しくPCRが行われたことを確認した。これらのプラスミドをPCRプライマーに付加した制限酵素サイトで消化して得られる完全長SiOMTを含む約1.1kbのDNA断片を、大腸菌発現ベクターであるpQE30ベクター(QIAGEN)のBamHI/SalIサイトに挿入して、SiOMT1/pQE30(pSPB2690と称する)、SiOMT2/pQE30(pSPB2686と称する)を得た。
【0085】
〔実施例5:組換えタンパク質の調製〕
上記において作製した大腸菌発現ベクターpSPB2690およびpSPB2686を用いて大腸菌株JM109(TOYOBO)を形質転換し、最終濃度20μg/mlのアンピシリンを含むLB培地中にて37℃で一晩前培養した。前培養液の一部をアンピシリン50μg/ml、カザミノ酸0.5%を含むM9培地(10ml)に添加して、A600=0.6〜1.0に達するまで振盪培養した。次いで、培養液に最終濃度0.5mMのIPTG(Isopropyl−β−D−thiogalactopyranoside)を加え、さらに30℃で一晩振盪培養した後、3000rpmにて4℃で10分間遠心分離を行って集菌した。菌体を10mlの緩衝液(30mM Tris−HCl(pH7.5)、2mM MgCl2、0.5mM EDTA,50μM APSMF)に懸濁した後に超音波処理を行って大腸菌を破砕し、次いで、15,000rpmにて4℃で10分間遠心分離を行い、得られた上清を粗酵素液として以下の活性測定に用いた。
【0086】
〔実施例6:ゴマリグナンメチル化酵素による生成物の分析〕
酵素反応の基質に用いるゴマリグナンは少量のDMSOに溶解した後に70%エタノールに溶解して基質溶液(1mg/ml)を調製した。ゴマリグナンは、例えば、公知の方法(日本農芸化学会誌 67:1693(1993))に従ってゴマから抽出および精製して得ることができる。基質溶液10μl、大腸菌で発現させたSiOMTの上記粗酵素液200μlおよび10mM S−Adenosyl-L-Mathianine(SAM) 10μlを反応チューブ中で混合した後、30℃で2時間反応させた。
0.1%トリフルオロ酢酸(TFA)を含む100%アセトニトリル(250μl)を反応チューブに添加することによって、酵素反応を停止させた。反応チューブをボルテックスミキサーで激しく攪拌した後、15,000rpmにて4℃で5分間遠心分離し、得られた上清をフィルタ(ポアサイズ0.45mm、4mm Millex−LH、Millipore)を用いて清浄化した後、液体高速クロマトグラフィー(以下HPLC)を用いてこの上清を分析した。リグナンおよびそのメチル化リグナンの分析条件は以下の通りである。
【0087】
C−30カラム(野村化学C30−UG−5、4.6mm×150mm)を用いて液体クロマトグラフィー(Lc−2010c(島津製作所))を行った。移動相には、A液として、0.1%TFA、B液として、0.1%TFAを含む90%アセトニトリルを用いた。A液80%:B液20%の混合液を用いてカラムを平衡化した(20分間)後、直線濃度勾配(A液80%:B液20%→A液10%:B液90%)を用いて20分間にわたって溶出(流速0.6ml/分)し、さらにA液10%:B液90%を用いて7分間溶出した。280nmの吸収を測定して、サンプル中に含有される化合物を検出した。化合物の各ピークを、SPD−10AV(島津製作所)を用いて190nm〜400nmのスペクトルを測定し、リグナンに特徴的な2つの吸収極大(230nmおよび280nm)を有する物質を探索した。この条件下で、ピノレジノール標準品は約11.8分、ピペリトール標準品は約15.5分、セサミノール標準品は約16.8分に検出される。
SiOMT1組換えタンパク質とピノレジノールとの反応液中において、リグナンのスペクトルを有する保持時間14.0分のピークAが新たに得られた(図3の3A)。また、SiOMT1組換えタンパク質とピペリトールとの反応液中において、リグナンのスペクトルを有する保持時間約17.7分のピークBが新たに得られた(図3の3B)。
【0088】
SiOMT2組換えタンパク質とピノレジノールまたはピペリトールとの反応液中において、新たな生成物は認められなかった(図4の3C、3D)。また、SiOMT1およびSiOMT2のセサミノールに対するリグナンメチル化活性は認められなかった。
【0089】
次に、LC−MS分析によって、SiOMT1により生成した新規リグナンの分子量を決定した(液体クロマトグラフィー(LC):Waters 2795(ウォーターズ社)、質量分析器:QUATRO micro(マイクロマス社))。ダイアイオンHP−20樹脂(三菱化学)を1ml充填したカラムを50%アセトン5mlで洗浄した後、水10mlを用いて平衡化した。SiOMT1により生成したメチル化リグナンを含む酵素反応液をカラムにロードし、5mlの水を用いて不純物を洗浄した後、80%アセトン2mlを用いて溶出した。エバポレーターを用いて溶出液を乾固させた後、1%ギ酸を含む90%アセトニトリル(100μl)中に溶解させてLC−MS分析試料とした。LC条件を以下に示す。
【0090】
Develosil C30−UG−3カラム(野村化学、3.0×150mm)を用い、移動相は、A液として水を、B液として100%アセトニトリルを、C液として1%ギ酸を、D液として100mM酢酸アンモニウム水溶液を用いた。10分間にわたる直線濃度勾配(A液60%:B液30%:C液5%:D液5%→A液10%:B液80%:C液5%:D液5%)を用いて溶出し、その後、A液10%:B液80%:C液5%:D液5%を用いて5分間溶出した(流速:常時0.2ml/分)。アンモニウム付加イオンとしてシグナルを検出した。
この条件下で、ピークAは約8.3分、ピークBは約11.3分に検出される(図3〜5)。イオンモードES+、Cone voltage 15V、Collision 2eVのMS条件を用いて、100〜800(m/z,30分)の範囲でMSスキャンし、210〜400nm(30分)の範囲のPDAを測定した。
上記条件下でのLC−MS分析の結果、ピークAはアンモニウムイオン付加分子量373(m/z)、ピークBはアンモニウムイオン付加分子量371(m/z)であった(図6の4A、図7の4B)。従って、これらのピークは、それぞれピノレジノール(アンモニウムイオン付加分子量359)のモノメチル化されたもの、およびピペリトール(アンモニウムイオン付加分子量374)のモノメチル化されたものであると同定した。
以上の結果からSiOMT1はゴマリグナンであるピノレジノールおよびピペリトールに対してモノメチル化する活性を有する酵素であることが明らかになった。
【0091】
〔実施例7:ゴマリグナンメチル化酵素ホモログ遺伝子の単離〕
リグナンメチル化活性を有する酵素遺伝子がゴマ種間において機能的および構造的に保存されているかどうかを明らかにするために、栽培ゴマ品種であるSesamum indicumとは細胞遺伝学的に異なるアフリカゴマ(Sesamum radiatum)から、SiOMT1のカウンターパート遺伝子(SrOMT1)を単離することを試みた。
S.radiatumはアフリカに現存するゴマ植物であり、細胞遺伝学解析によれば染色体数は2n=64とされており、栽培ゴマ品種であるS.indicum(2n=26)とは遺伝的に異なる系統である(参考文献、並木満夫、小林貞作「ゴマの科学」朝倉書店)。しかし、S.radiatum種子においても多様なリグナンが蓄積していることが知られている(参考文献、Bedigian,D.ら、Biochemical Systematics and Ecology 13:133−139(1985))。従って、S.radiatumのゲノム中にはS.indicumのSiOMT1に対応する遺伝子が含まれていることが期待される。
【0092】
S.radiatum種子のcDNAを用いてBgl2-SiOMT1-FW(配列番号20および)SalI-SiOMT1−RV(配列番号21)のプライマー対を用いて、PCRによりSrOMT1の増幅を試みた。
S.radiatum種子のcDNA 1μl、1×Ex−Taq buffer(TakaRa)、0.2mM dNTPs、プライマー各0.2pmol/μl、Ex−Taq polymerase 1.25Uからなる。94℃で5分間の後、94℃で1分間、55℃で1分間、72℃で2分間の反応を30サイクル行ってPCR増幅した後、72℃で5分間保持した。得られた約1.1kbのPCR産物を、製造者が推奨する方法に従ってpCR2−TOPO vector(Invitrogen社)のマルチクローニングサイトに挿入し、SrSiOMT1/pCR2−TOPO(pSPB2910)を得た。
プライマーウォーキング法によって、pSPB2910に含まれるSrOMT1の塩基配列(およびアミノ酸配列)を決定した(配列番号3および4)。SrOMT1は、SiOMT1に対してアミノ酸配列で89%の配列同一性を示した。配列同一性について、Clustal W alignmentプログラム(Mac Vector ver.7.2.2(Symanteccorporation))をデフォルト設定で使用した。この高い配列同一性はSrOMT1がSiOMT1のカウンターパート遺伝子であることを強く支持する。
【0093】
実施例2と同様の方法でRT−PCRを行い、SrOMT1の種子に特異的な遺伝子発現を確認した(図1)。
実施例4と同様にSrOMT1の大腸菌発現ベクターであるSrOMT1/pQE30(pSPB2911)を構築し、実施例5と同様に組換えタンパク質を調整し、実施例6と同様にその活性および生成物の分析を行った。
SrOMT1はSiOMT1と同様に、ピノレジノールおよびピペリトールに対してメチル化活性を示した(図2)。また、SiOMT1はセサミノールに対してメチル化活性を示さなかった。以上の結果より、SrOMT1はSiOMT1のカウンターパート遺伝子であり、本酵素遺伝子がゴマ種間で保存されていることが示された。
【0094】
〔実施例8:SiOMT1のゲノミックサザン解析〕
ゴマゲノム内におけるSiOMT1遺伝子のコピー数を明らかにするために、ゲノミックサザン解析を実施した。Nucleon Phytopure for Plant Extraction Kit(アマシャム社)を製造業者が推奨する方法に従って用いて、S.indicum(真瀬金品種)の葉からゲノムDNAを抽出した。抽出したゲノムDNA20μgを、EcoRI、HindIII,XhoIまたはXbaIで消化した後、アガロースゲルを用いた電気泳動にて分離した。このアガロースゲルを0.25M HClを用いて15分間加水分解した後、1.5M NaCl/0.5M NaOHの溶液を用いて変性させ(30分間)、1.5M NaClを含むTris−HCl(pH7.5)を添加することによって変性溶液で中和した(20分間)。次いで、アガロースゲル内のゲノムDNAを、20×SSC溶液中にてメンブレン(Hybribond−N、アマシャム社)に転写した。メンブレンに転写したゲノムDNAをUV照射によりメンブレンに結合させ、7%SDS、50%ホルムアミド、5×SSC、2%ブロッキング試薬、0.1%ラウロイルサルコシン、50mMリン酸ナトリウム緩衝液(pH7.0)からなるハイブリダイゼーション緩衝液(高SDS緩衝液:ロシュ社)を用いて、42℃で1時間プレハイブリダイゼーションを行った。
【0095】
pSPB2678をテンプレートとして、それぞれ配列番号20(Bgl2-SiOMT1−FW)および配列番号21(SalI−SiOMT1−RV)のプライマー対を用いるPCRによって、DIG標識化ハイブリダイゼーションプローブを作製した。PCR反応液は、pSPB2678プラスミド1ng、1×PCR緩衝液(タカラバイオ社)、2.5mM DIG−dNTP mixture(PCR DIG labeling Mix、ロシュ社)、0.2pmolの各プライマー、1U rTaq polymerase(タカラバイオ社)からなる。PCR反応を、95℃で30秒間、53℃で30秒間、72℃で2分間の反応を30サイクル行った。Sephadex G−50カラム−Fine(ベーリンガー社)を用いて精製したPCR産物をハイブリダイゼーションプローブとして用いた。このプローブを熱変性した後、プレハイブリダイゼーション液に15μl添加して、42℃で一晩インキュベートした。
【0096】
ハイブリダイゼーション後、高ストリンジェントな条件(0.2×SSCおよび0.1%SDSを含む溶液を用いて65℃で30分間を2回)によってメンブレンを洗浄した。DIGラベリング&デテクションキット(ロシュ社)を製造業者の指示書に従って用いて、ハイブリダイゼーションシグナルを検出した。
サザン解析の結果、いずれの制限酵素消化画分においても二本以上のバンドが検出されたので、SiOMT1と極めて相同性の高い遺伝子が少なくとも1つ以上ゴマゲノム内にコードされていることが示された(図8)。さらに言えば、ゴマゲノム中にはSiOMT1と機能的に重複もしくは類似した酵素遺伝子が複数存在することが期待される。
【0097】
〔実施例9:ゴマリグナンメチル化酵素のCOMT活性の測定〕
実施例3に示すようにデータベース検索によりSiOMT1およびSrOMT1は既知のタンパク質の中ではCOMTと最も高い配列同一性を示す。このことはゴマメチル化酵素がCOMT活性をも有していることを示唆する。
実施例5および実施例7で発現させたSiOMT1およびSrOMT1のカフェ酸に対するメチル化活性を検討した。0.4mg/mlカフェ酸10μl、大腸菌で発現させたSiOMTの粗酵素液200μlおよび10mM SAM 10μlを反応チューブ中で混合した後、30℃で1時間反応させた後、実施例6と同様の方法でHPLC分析に供した。
その結果、SiOMT1およびSrOMT1共にカフェ酸との反応液中に標準品フェルラー酸と保持時間が一致するピークC(保持時間約7.3分)が生成した(図9の6A〜6C))。本HPLC分析条件下ではカフェ酸は保持時間約3.9分に溶出される。
実施例6と同様の条件でLC−MS分析を行ったところピークCはアンモニウムイオン付加分子量195(m/z)であった(図10の6D)。従って、ピークCは、カフェ酸(アンモニウムイオン付加分子量181(m/z))がモノメチル化されて生成するフェルラー酸であることが確認された。これにより二種のリグナンメチル化酵素は共にCOMT活性をも有していることが明らかとなった。
【産業上の利用可能性】
【0098】
以上のように、本発明のポリペプチドおよびポリヌクレオチドは、メチル化リグナンの生産に有用である。また、本発明のポリヌクレオチドを発現可能に導入した形質転換体または細胞は、食品分野、各種工業分野において、メチル化リグナン、またはこれを用いる製品を生産する上で、極めて有用である。上記形質転換体が植物体である場合、植物体自体を食品として用いることができるので農業分野等において非常に有用である。
さらに、本発明のポリペプチドおよびポリヌクレオチドを、本発明者らが見出した他の酵素(ピペリトールおよびセサミンを合成する酵素であるSiP189)と組み合わせることによって、ゴマに限ることなく、特定のリグナン分子種の生産系の確立が可能となり、その結果、特定のリグナンおよびメチル化リグナンの生産量を拡大することができる。従って、本発明は、農業、食品産業、医薬品産業およびこれらの関連産業にわたる広範な利用が可能である。
【図面の簡単な説明】
【0099】
【図1】図1はゴマリグナンの構造と代謝経路図である。
【図2】2Aは、SiOMT1およびSiOMT2のゴマ(S.indicum cv. 真瀬金)器官別のRT−PCRによる遺伝子発現解析の結果である。2Bは、SrOMT1のゴマ(S.radiatum)器官別のRT−PCRによる遺伝子発現解析の結果である。
【図3】3Aは、大腸菌で発現させたSiOMT1とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。3Bは、大腸菌で発現させたSiOMT1とピペリトールとの酵素反応液のA280nmのクロマトグラムである。
【図4】3Cは、大腸菌で発現させたSiOMT2とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。3Dは、大腸菌で発現させたSiOMT2とピペリトールとの酵素反応液のA280nmのクロマトグラムである。
【図5】3Eは、大腸菌で発現させたSrOMT1とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。3Fは、大腸菌で発現させたSrOMT1とピペリトールとの酵素反応液のA280nmのクロマトグラムである。
【図6】4AはSiOMT1により生成されたメチル化ピノレジノールのLC−MSのクロマトグラムである。
【図7】4BはSiOMT1により生成されたメチル化ピペリトール(コブシン)のLC−MSのクロマトグラムである。
【図8】図8はSiOMT1遺伝子の完全長プローブを用いたゲノミックサザンハイブリダイゼーションの結果である。
【図9】6Aは、標準品のカフェ酸のA280nmのクロマトグラムである。6Bは、大腸菌で発現させたSiOMT1とカフェ酸との酵素反応液のA280nmのクロマトグラムである。6Cは、大腸菌で発現させたSrOMT1とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。
【図10】6DはSiOMT1により生成されたメチル化カフェ酸(フェルラー酸)のLC−MSのクロマトグラムである。
【技術分野】
【0001】
本発明はリグナンにメチル基を転移する活性を有する酵素の遺伝子、当該メチル化酵素を利用してリグナン組成が変換された植物などに関する。更に詳しくは、本発明は、メチル化リグナンを合成する活性を有する酵素遺伝子、好ましくはゴマ由来のメチル化リグナンを合成する活性を有する酵素遺伝子、及びその利用に関する。
【背景技術】
【0002】
ゴマ属ゴマ(Sesamum indicum)は、ゴマ科(Pedaliaceae)に属する1年生の草本植物である。ゴマの原産地は、中央アフリカといわれている。ゴマは、約6000年の歴史を有する最古の栽培油糧植物とされ、世界中で栽培されてきた。ゴマは、古来から貴重な食品であり、健康に良い食品の代表として知られている。特に、ゴマ種子、ゴマ種子から得た油、ゴマ種子からの抽出物が利用されている(例えば、ゴマその科学と機能性、並木満夫編:丸善プラネット社(1998)参照)。ゴマ種子に含まれる成分は、その約50%が脂質であり、約20%が蛋白質である。ゴマに含まれる脂質の主要成分は、オレイン酸およびリノール酸を主体とするトリグリセリドである。ゴマはまた、ビタミンB1、B2、Eなどをさらに含む。リグナンと総称される植物の二次代謝物(例えば、セサミンおよびセサモリンなど)が、上記の成分以外にゴマに含まれており、これらは強い抗酸化性を有している(例えば、Biochemical Systematics and Ecology 13,133−139(1985)参照)。
【0003】
リグナンの生合成については、例えば、Lignans:biosynthesis and function,Comprehensive natural products chemistry,1:640−713(1999); Phytochemistry Rev. 2257−288 (2003)などに記載されている。
例えば、Phytochemistry Rev. 2257−288 (2003)には、コニフェリルアルコールが重合して合成されるピノレジノールが生合成上の最初のリグナンであること、そしてピノレジノールから個々の植物種において特有な生合成経路を経由して多様なリグナンが合成されることが示されている。このピノレジノールの合成に寄与するデリジェントタンパク質がレンギョウなどにおいて報告されている(例えば、Science 275, 362−366 (1997)等参照)。さらに、レンギョウのピノレジノール−ラリシレジノール還元酵素遺伝子(例えば、J.Biol.Chem.,271:29473(1996)、特表2001−507931号公報等参照)、Thuja plicataのピノレジノール−ラリシレジノール還元酵素遺伝子(例えば、J.Biol.Chem.,274:618(1999)等参照)ならびに組換えセコイソラリシレシノールデヒドロゲナーゼおよびその使用方法(例えば、J.Biol.Chem.,276(16):12614−23(2001)、特表2002−512790号公報等参照)が報告されている。さらに、Larrea tridentataから、ラレアトリシン水酸化酵素遺伝子がクローニングされている(例えば、Proc.Nat.Acad.Sci.USA,100:10641(2003)等参照)。
【0004】
ゴマリグナンの生合成についてはピノレジノールにピペリトール合成酵素が作用することによってピペリトールが合成され、次いで、このピペリトールにセサミン合成酵素が作用することによってセサミンが合成されるとされていた。しかしながら、ゴマからクローニングされたチトクロームP450タンパク質であるCYP81Q1は単独でピノレジノールからピペリトールを経て、セサミンを合成することが明らかにされた(国際公開公報WO2005/030944;図1参照)。
【0005】
近年、リグナンだけでなくリグナンの配糖体もまた注目されている。上記したリグナン分子のうちのいくつかは、配糖体として植物中に存在することが知られている。例えば、ゴマ種子中には、セサミノール配糖体(セサミノール2’−O−β−D−グルコピラノシド;セサミノール2’−O−β−D−グルコピラノシル(1−2)−O−β−D−グルコピラノシド;およびセサミノール2’−O−β−D−グルコピラノシル(1−2)−O−(−β−D−グルコピラノシル(1−6))−β−D−グルコピラノシド))、およびピノレジノール配糖体(ピノレジノール4’−O−β−D−グルコピラノシル(1−6)−β−D−グルコピラノシド;ピノレジノール4’−O−β−D−グルコピラノシル(1−2)−β−D−グルコピラノシド;ピノレジノール4’−O−β−D−グルコピラノシル(1−6)−O−(β−D−グルコピラノシル(1−6))β−D−グルコピラノシド;およびピノレジノールジ−O−β−D−グルコピラノシド))などが存在し、レンギョウには、(+)−ピノレジノール−4’−O−β−Dグルコシドおよび(−)−マタイレジノール−4−O−グルコシドが、アマには、セコラリシレジノールジグルコシドおよびピノレジノールジグルコシドなどが存在する(例えば、生薬学雑誌、第32巻、第194頁(1978); Tetrahedron,14:649(2003);Phytochemistry,58:587(2001)等参照)。
【0006】
ゴマに含まれるピノレジノール配糖体およびセサミノール配糖体(例えば、Katsuzaki,H.ら、Biosci.Biotech.Biochem.56,2087−2088(1992)等参照)は、水溶性領域で強い抗酸化性を示すので、脂溶性抗酸化剤(例えば、トコフェロール)とは異なる応用が期待されている。また、リグナン配糖体について以下のような作用機構が提唱されている:リグナン配糖体は、抗酸化性を示す官能基であるフェノール性水酸基が自身の有する糖によって保護されているが、体内へ摂取された後に、腸内細菌の有するβ−グルコシダーゼの作用によって加水分解され、アグリコン部分である脂溶性のリグナンが生成される。このアグリコンが腸内に吸収されて血液を経由して各種臓器に運ばれて、臓器の生体膜などにおける酸化障害を防ぐ。このような作用機構に基づいて、リグナン配糖体は動脈硬化予防食品としての応用が期待されている(例えば、T.Osawa:Anticarcinogenesis and radiation protection 2:p.327,Plenum Press,New Yo参照)。
【0007】
リグナン配糖体以外のリグナン派生体としてメチル化リグナンが知られている。メチル化リグナンもリグナン配糖体同様に抗酸化性を示す官能基であるフェノール性水酸基がメチル基によって保護されており、それによりメトキシ構造を有するリグナンである。フロフラン型のゴマリグナンの中にはピペリトールの4位の水酸基がメチル化されたコブシンおよびセサミノールの2‘位の水酸基がメチル化されたセサンゴリンが報告されている(非特許文献1:Pytochemistry 47, 583−591 (1998);および非特許文献2:J.Org.Chem. 27, 3232−3235 (1962)参照)(図1)。しかしながらこれらの合成反応を触媒する酵素は不明であり、フロフラン型リグナンをメチル化する酵素およびそれをコードする遺伝子はこれまでに精製されたり単離された報告はない。
なお、メチル基転移反応を触媒するという特定の機能を有するタンパク質は、異なる植物種においてもそのアミノ酸配列が類似しているということが知られている(例えば、非特許文献3:Plant Cell 14, 505−519 (2002)参照)。
【非特許文献1】Pytochemistry 47, 583−591 (1998)
【非特許文献2】J.Org.Chem. 27, 3232−3235 (1962)
【非特許文献3】Plant Cell 14, 505−519 (2002)
【発明の開示】
【発明が解決しようとする課題】
【0008】
植物の二次代謝物などの生合成経路を改変することによって、有用な物質の生成および/または有用な植物の育種が行われている。このような技術は、代謝工学と呼ばれている。このような技術を用いれば、任意の化合物を大量生産すること、および/または不要な物質の生産を抑制することが可能になる。従って、リグナン代謝経路に関与する遺伝子を用いてリグナンおよびその代謝物の合成を遺伝子工学的に行うことは、上述したようなこれらの物質の有用性に鑑みて産業上有用である。しかし、リグナン、特にゴマリグナンに代表されるフロフラン型リグナンの生合成に関与する遺伝子に関する知見は、前述のように限られており、さらに、メチル化フロフラン型リグナンの生成を触媒するメチル化酵素は見出されていないので、さらなる遺伝子の取得が望まれていた。
【課題を解決するための手段】
【0009】
本発明は、上記状況に鑑みてなされたもので、下記に示す、リグナンメチル基転移活性を有する酵素、それをコードするポリヌクレオチド、それを含有するベクター・細胞・形質転換体などを提供する。
【0010】
(1)以下の(a)〜(d)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチド;及び
(d)配列番号:2又は4のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
(2)配列番号:2又は4のアミノ酸配列、または当該アミノ酸配列に対して1又は数個のアミノ酸の付加、欠失及び/又は他のアミノ酸による置換によって修飾されているアミノ酸配列を有し、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする上記(1)に記載のポリヌクレオチド。
(3)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする上記(1)に記載のポリヌクレオチド。
(4)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドに対して、5xSSC、50℃の条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする上記(1)に記載のポリヌクレオチド。
(5)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(6)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する上記(1)に記載のポリヌクレオチド。
(7)DNAである、上記(1)〜(6)のいずれかに記載のポリヌクレオチド。
(8)フロフラン型リグナンに対してメチル基を転移する活性を有するタンパク質をコードする上記(1)〜(7)のいずれかに記載のポリヌクレオチド。
(9)ピノレジノール及び/又はピペリトールに対してメチル基を転移する活性を有するタンパク質をコードする上記(8に記載のポリヌクレオチド。
(10)上記(1)〜(9)のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
(11)上記(1)〜(9)のいずれかに記載のポリヌクレオチドを含有するベクター。
(12)上記(11)に記載のベクターにより形質転換された宿主細胞。
(13)上記(12に記載の宿主細胞を培養し又は生育させ、当該宿主細胞からリグナンにメチル基を転移する活性を有するタンパク質を採取することを特徴とする該タンパク質の製造方法。
(14)上記(1)〜(9)のいずれかに記載のポリヌクレオチドが導入された植物体(例えば、ゴマ、レンギョウ又はアマ)もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体、又はそれら植物体の組織。
(15)上記(1)〜(9)のいずれかに記載のポリヌクレオチドを用いてリグナンにメチル基を転移する方法。
(16)上記(1)〜(9)のいずれかに記載のポリヌクレオチドを植物体に導入し、発現させることによって、産生するリグナン組成が改変された当該植物体もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体。
(17)上記(1)〜(9)に記載のポリヌクレオチドのフラグメントまたはその相補配列からなるポリヌクレオチド。
【発明の効果】
【0011】
本発明のポリペプチド(リグナンメチル化酵素)を利用すれば、例えば、生物(特に、植物)においてリグナンおよびメチル化リグナンの量を人為的に調節することができるという効果を奏する。また、これらの組換え酵素を用いてリグナンをメチル化することによって、インビトロおよびインビボにおいて物性(溶解度、動物の吸収効率など)を改変することもできる。また、本発明のポリヌクレオチドを利用することによって、これまでに自然界において未同定であったメチル化リグナンを人工的に生産することも可能となる。また、本発明のポリペプチドを利用して合成されるメチル化リグナンは、新規生理機能物質の出発物質又は中間体として有効に利用することができる。
【0012】
本発明のリグナンメチル化酵素を遺伝子組換え技術を用いて所望の生物に発現させることによって、ピノレジノールからモノメチル化ピノレジノールを、および/またはピペリトールからコブシンを人工的に生産することが可能となる。また、本発明のリグナンメチル化酵素を遺伝子組換え技術を用いて所望の生物に発現させることによって、リグナンとメチル化リグナンとの量を人為的に制御した植物および/または微生物を作出することができる。
また、コブシンまたはメチル化ピノレジノールを生産する植物において、本発明のリグナンメチル化酵素の発現を抑制することによって、アグリコンを遊離させて、リグナン(特に、ピペリトールおよび/またはピノレジノール)の量を増加させることもできる。
さらに、本発明のリグナンメチル化酵素を用いれば、新規メチル化リグナンであるモノメチル化ピノレジノールをピノレジノールから人工的に生産することができる。
【発明を実施するための最良の形式】
【0013】
本発明者らは、リグナン、特に、ピノレジノールおよび/またはピペリトールを主な基質にする新規メチル化酵素を見出すとともに、そのメチル化酵素がピノレジノールをメチル化することを見出した。これまでに、ジメチル化ピノレジノールであるユデスミンは同定されているがモノメチル化ピノレジノールは見出されていない。
本発明者らは、ゴマ種子由来の5000クローンからなるESTデータベースから相同性検索によりゴマメチル化酵素様遺伝子の部分配列を探索し、その結果2種類のメチル化酵素様遺伝子(以下SiOMT1およびSiOMT2)を得た。これらのSiOMT遺伝子は種子において強く発現していた。これらの完全長塩基配列をRACE法で取得し、大腸菌において発現させた。得られた組換えタンパク質をセサミノールまたはピノレジノールと反応させた後、HPLC分析、LC−MS分析、およびTOF−MS/MS分析を用いて酵素活性を測定した。その結果、SiOMT1がピペリトールをメチル化しコブシンを生成する反応を触媒する活性を有することが明らかとなった。さらにSiOMT1はピノレジノールをメチル化しモノメチル化ピノレジノールを生成する反応を触媒する活性を有することが明らかとなった。このメチル化リグナンはピノレジノール派生体であるユデスミンの中間体であると考えられる。アフリカゴマであるSesamum radiatumからPCRによりSiOMT1ホモログであるSrOMT1遺伝子をクローニングし、SrOMT1はSiOMT1と同様のメチル化活性を有することを確認した。
【0014】
なお、「リグナン」は、C6C3骨格を有するフェニルプロパノイド化合物2分子が、主としてこれらの8−8’位を介して重合(8,8’−結合)した化合物である。リグナンは、植物における生体防御機構に寄与していると考えられている(Phytochemistry Rev. 2, 371−390 (2,003)参照)。
代表的なリグナンとしては、ゴマに含まれる(+)−セサミン、(+)−セサミノール、(+)−セサモリン、(+)−ピノレジノール、(+)−ピペリトールおよび(+)−セサモリノール;レンギョウ(Forsythia intermedia)に含まれる(+)−ピノレジノール、(−)−アルクチゲニンおよび(−)−マタイレジノール;ソシマ(Daphne tangutica)に含まれる(−)−ピノレジノールおよび(−)−ラリシレジノール;アマ(Linum usitatissimum)に含まれる(+)−セコイソラリシレジノールなどが挙げられる。これらのリグナンの分子構造は多様である(Wood Research 90, 27−110 (2,003)等参照)。(+)−ピノレジノールに代表されるゴマリグナン類は最も幅広い植物種で同定されているフロフラン型のリグナン類に分類される。ゴマリグナンの1つであるセサミンは、多彩な生理活性作用を有しており、コレステロール代謝、肝機能および免疫機能の改善に有効である(例えば、ゴマその科学と機能性、並木満夫編:丸善プラネット社(1998)参照)。セサミンをゴマ種子またはゴマの絞り粕から分離精製する方法がすでに実用化されており(例えば、特開2001−139579公報、特開平10−7676号公報等)、セサミンを主成分とするアルコール分解促進作用を有する肝機能改善/増強剤が市販されている(商品名:セサミン、発売元:サントリー株式会社)。またセサミン以外の他のリグナン(例えば、セサミノール、セサモリンなど)もまた生理活性作用を有することが報告されている(例えば、日本農芸化学会誌、76:805−813(2002)参照)。このように、リグナン又はその誘導体は、種々の生理活性を有する生理活性物質あるいはその中間体として有用である。
【0015】
以下、本発明のリグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチド、当該ポリヌクレオチドにコードされるタンパク質、およびこれらの利用について詳述する。
【0016】
(1)ポリヌクレオチド
まず、本発明は、以下の(a)〜(d)のいずれかに記載のポリヌクレオチドを提供する。
(a)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチド;及び
(d)配列番号:2又は4のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【0017】
本明細書中、用語「ポリヌクレオチド」は、「遺伝子」、「核酸」または「核酸分子」と交換可能に使用され、ヌクレオチドの重合体が意図される。本明細書中、用語「塩基配列」は、「核酸配列」または「ヌクレオチド配列」と交換可能に使用され、デオキシリボヌクレオチド(A、G、CおよびTと省略される)の配列として示される。
本発明のポリヌクレオチドは、RNA(例えば、mRNA)の形態、またはDNAの形態(例えば、cDNAまたはゲノムDNA)で存在し得る。DNAは、二本鎖または一本鎖であり得る。一本鎖DNAまたはRNAは、コード鎖(センス鎖としても知られる)であり得るか、またはそれは、非コード鎖(アンチセンス鎖としても知られる)であり得る。
【0018】
本明細書中、用語「オリゴヌクレオチド」は、ヌクレオチドが数個ないし数十個(例えば、2〜60個)結合したものが意図され、「ポリヌクレオチド」と交換可能に使用される。オリゴヌクレオチドは、短いものはジヌクレオチド(二量体)、トリヌクレオチド(三量体)といわれ、長いものは30マーまたは100マーというように重合しているヌクレオチドの数で表される。オリゴヌクレオチドは、より長いポリヌクレオチドのフラグメントとして生成されても、化合合成されてもよい。
本明細書中、用語「ポリヌクレオチドの一部」は、そのポリヌクレオチドのフラグメントを意味し、例えば、少なくとも12nt(ヌクレオチド)、好ましくは約15nt、そしてより好ましくは少なくとも約20nt、なおより好ましくは少なくとも約30nt、そしてさらにより好ましくは少なくとも約40ntの長さのフラグメントが意図される。「少なくとも12ntの長さのフラグメント」は、例えば、配列番号1に示される塩基配列からの12以上の連続した塩基を含むフラグメントが意図される。本明細書を参照すれば配列番号1に示される塩基配列が提供されるので、当業者は,配列番号1に基づくDNAフラグメントを容易に作製することができる。例えば、制限エンドヌクレアーゼ切断または超音波による剪断は、種々のサイズのフラグメントを作製するために容易に使用され得る。あるいは、このようなフラグメントは、合成的に作製され得る。適切なフラグメント(オリゴヌクレオチド)が、Applied Biosystems Incorporated(ABI,850 Lincoln Center Dr.,Foster City,CA 94404)392型シンセサイザーなどによって合成される。
【0019】
本発明のポリヌクレオチドは、リグナンメチル化活性を有するポリペプチドをコードする。そのようなポリヌクレオチドは、典型的には、配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;あるいは配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチドである。本発明の好ましいポリヌクレオチドは、配列番号:1又は3の塩基配列を有するポリヌクレオチド、あるいは配列番号:2又は4のアミノ酸配列を有するポリペプチドをコードするポリヌクレオチドである。
本発明のポリヌクレオチドは、それがコードするポリペプチドがリグナンメチル化活性を有する限り、配列番号:1又は3の塩基配列において1または複数個(例えば、1〜30個、1〜20個、1〜10個、1〜数個(例えば、6個)、1〜5個、1〜3個、1〜2個など)の塩基が欠失、挿入、置換、及び/または付加された変異体であってもよい。変異体は、コードもしくは非コード領域、またはその両方において変異され得る。コード領域における変異は、保存的または非保存的なアミノ酸の欠失、挿入、置換、または付加を生成し得る。
【0020】
本発明のポリヌクレオチドは、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドであって、配列番号:1または3の塩基配列と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズするポリヌクレオチドを含む。
【0021】
ハイブリダイゼーションは、Sambrookら、Molecular Cloning,A Laboratory Manual,2d Ed.,Cold Spring Harbor Laboratory(1989)に記載されている方法のような周知の方法で行うことができる。通常、温度が高いほど、塩濃度が低いほどストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同なポリヌクレオチドを取得することができる。適切なハイブリダイゼーション温度は、塩基配列やその塩基配列の長さによって異なり、例えば、アミノ酸6個をコードする18塩基からなるDNAフラグメントをプローブとして用いる場合、50℃以下の温度が好ましい。
【0022】
本明細書中、用語「ストリンジェントなハイブリダイゼーション条件」は、ハイブリダイゼーション溶液(50%ホルムアミド、5×SSC(150mMのNaCl、15mMのクエン酸三ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハート液、10%硫酸デキストラン、および20μg/mlの変性剪断サケ精子DNAを含む)中にて42℃で一晩インキュベーションした後、約65℃にて0.1×SSC中でフィルターを洗浄することが意図される。ポリヌクレオチドの「一部」にハイブリダイズするポリヌクレオチドによって、参照ポリヌクレオチドの少なくとも約15ヌクレオチド(nt)、そしてより好ましくは少なくとも約20nt、さらにより好ましくは少なくとも約30nt、そしてさらにより好ましくは約30〜70ntにハイブリダイズするポリヌクレオチド(DNAまたはRNAのいずれか)が意図される。
さらに、本発明は、配列番号:1または3の塩基配列と少なくとも80%同一、より好ましくは少なくとも85%、90%、92%、95%、96%、97%、98%または99%同一である塩基配列からなるポリヌクレオチドを提供する。
【0023】
任意の特定の核酸分子が、例えば、配列番号:1または3に示される塩基配列に対して、少なくとも80%、85%、90%、92%、95%、96%、97%、98%、または99%同一であるか否かは、公知のコンピュータープログラム(例えば、Bestfit program(Wisconsin Sequence Analysis Package,Version 8 for Unix(登録商標),Genetics Computer Group,University Research Park,575 Science Drive,Madison,WI 53711)を使用して決定され得る。Bestfitは、SmithおよびWatermanの局所的相同性アルゴリズムを用いて、2つの配列間の最も良好な相同性セグメントを見出す(Advances in Applied Mathematics 2:482〜489(1981))。Bestfitまたは任意の他の配列整列プログラムを用いて、特定の配列が、本発明に従う参照配列に対して、例えば、95%同一であるか否かを決定する場合は、同一性のパーセントが参照塩基配列の全長にわたって計算され、そして参照配列におけるヌクレオチド数全体の5%までの相同性におけるギャップが許容されるように、パラメーターが設定される。
【0024】
特定の実施形態では、参照(QUERY)配列(本発明に係る配列)と対象配列との間の同一性(全体的な配列整列ともいわれる)は、Brutlagらのアルゴリズム(Comp.App.Biosci.6:237〜245(1990))に基づくFASTDBコンピュータープログラムを使用して決定される。%同一性を計算するために、DNA配列のFASTDB整列において使用される好ましいパラメーターは:Matrix=Unitary、k−tuple=4、Mismatch Penalty=1、Joining Penalty=30、Randomization Group Length=0、Cutoff Score=1、Gap Penalty=5、Gap Size Penalty=0.05、Window Size=500または対象塩基配列の長さ(どちらかより短い方)である。
【0025】
また、本発明は、上記ポリヌクレオチドのフラグメントまたはその相補配列からなるオリゴヌクレオチドを包含する。本発明のオリゴヌクレオチドがリグナンメチル化ポリペプチドをコードしない場合でさえ、当業者は、本発明のポリヌクレオチドが、ポリメラーゼ連鎖反応(PCR)のプライマーとして本発明のポリペプチドを作製するために使用され得ることを容易に理解する。リグナンメチル化ポリペプチドをコードしない本発明のオリゴヌクレオチドの他の用途としては、以下が挙げられる:(1)cDNAライブラリー中のリグナンメチル化酵素遺伝子またはその対立遺伝子もしくはスプライシング改変体の単離;(2)リグナンメチル化酵素遺伝子の正確な染色体位置を提供するための、分裂中期染色体スプレッドへのインサイチュハイブリダイゼーション(例えば、「FISH」)(Vermaら,Human Chromosomes:A Manual of Basic Techniques,Pergamon Press,New York(1988)に記載される);および(3)特定の組織におけるリグナンメチル化酵素mRNA発現を検出するためのノーザンブロット分析。
【0026】
本発明のポリヌクレオチドまたはオリゴヌクレオチドは、アンチセンスRNAメカニズムによる遺伝子発現操作のためのツールとして使用することができる。アンチセンスRNA技術によって、内因性遺伝子に由来する遺伝子産物の減少が観察される。本発明のオリゴヌクレオチドを導入することによって、リグナンメチル化活性を有するポリペプチドの含量を低下させ得、その結果、植物中のメチル化リグナン含量または含量比を制御(増加または減少)させることができる。本発明のポリヌクレオチドまたはオリゴヌクレオチドは、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配列を含む)などの配列を含むものであってもよい。
【0027】
本発明のポリヌクレオチドまたはオリゴヌクレオチドを取得する方法としては、本発明のポリヌクレオチドまたはオリゴヌクレオチドを含むDNA断片を単離する種々の公知の方法が挙げられる。例えば、本発明のポリヌクレオチドの塩基配列の一部と特異的にハイブリダイズするプローブを調製して、ゲノムDNAライブラリーまたはcDNAライブラリーをスクリーニングすれば、本発明のポリヌクレオチドまたはオリゴヌクレオチドを取得することができる。このようなプローブとしては、本発明のポリヌクレオチドの塩基配列またはその相補配列の少なくとも一部に特異的にハイブリダイズするポリヌクレオチド(オリゴヌクレオチド)であればよい。
【0028】
このようなハイブリダイゼーションによって選択されるポリヌクレオチドとしては、天然のポリヌクレオチド(例えば、ゴマ科植物やコケ科植物などの植物由来のポリヌクレオチド)が挙げられるが、植物以外に由来するポリヌクレオチドであってもよい。
本発明のポリヌクレオチドを取得する別の方法として、PCRを用いる方法が挙げられる。このPCR増幅方法は、例えば、本発明のポリヌクレオチドのcDNAの5’側および/または3’側の配列(またはその相補配列)を利用してプライマーを調製する工程、これらのプライマーを用いてゲノムDNA(またはcDNA)等をテンプレートにしてPCR増幅する工程を包含することを特徴としており、本方法を使用すれば、本発明のポリヌクレオチドを含むDNA断片を大量に取得することができる。
【0029】
本発明のポリヌクレオチドを取得するための供給源としては、特に限定されないが、ピペリトールまたはピノレジノールを含む生物材料であることが好ましい。本明細書中、用語「生物材料」は、生物学的サンプル(生物体から得られた組織サンプルまたは細胞サンプル)が意図される。後述する実施例においては、ゴマを用いているが、これに限定されない。
【0030】
本発明のポリヌクレオチドを使用すれば、形質転換体または細胞においてリグナンメチル化活性を有するポリペプチドを合成することができる。本発明のポリヌクレオチドを用いれば、ハイブリダイズするポリヌクレオチドを検出することによって、リグナンメチル化活性を有するポリペプチドを発現する生物を容易に検出することができる。
【0031】
本発明のオリゴヌクレオチドは、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドを検出するハイブリダイゼーションプローブまたは当該ポリヌクレオチドを増幅するためのプライマーとして利用することによって、リグナンメチル化活性を有するポリペプチドを発現する生物または組織を容易に検出することができる。なおさらに、上記オリゴヌクレオチドをアンチセンスオリゴヌクレオチドとして使用して、上記生物体またはその組織もしくは細胞におけるリグナンメチル化活性を有するポリペプチドの発現を抑制することができる。
【0032】
(2)ポリペプチド
本発明は、上記した本発明のポリヌクレオチドにコードされるタンパク質(ポリペプチド)をも提供する。そのようなポリペプチドは、典型的には、配列番号:2又は4のアミノ酸配列からなるタンパク質である。
本明細書中、用語「ポリペプチド」は、「ペプチド」または「タンパク質」と交換可能に使用される。また、ポリペプチドの「フラグメント」は、当該ポリペプチドの部分断片が意図される。本発明のポリペプチドはまた、天然供給源より単離されても、化学合成されてもよい。
【0033】
本発明のポリペプチドは、天然の精製産物、化学合成手順の産物、および原核生物宿主または真核生物宿主(例えば、細菌細胞、酵母細胞、高等植物細胞、昆虫細胞、および哺乳動物細胞を含む)から組換え技術によって産生された産物を含む。組換え産生手順において用いられる宿主に依存して、本発明のポリペプチドは、グリコシル化され得るか、または非グリコシル化され得る。さらに、本発明のポリペプチドはまた、いくつかの場合、宿主媒介プロセスの結果として、開始の改変メチオニン残基を含み得る。
また、本発明は、リグナンメチル化活性を有するポリペプチドを提供する。本明細書中、「リグナンメチル化活性」は、リグナンをメチル化する活性、すなわち、リグナンにメチル基を転移する活性が意図される。すなわち、本明細書中、「メチル化酵素」と「メチル基転移酵素」とは、相互交換可能に使用される。「リグナンメチル化活性」は、メチル基供与体であるSAMと基質であるリグナンとをリグナンメチル化酵素と反応させ、その反応生成物をHPLCまたはLC-MS分析することで測定又は確認することができる。一般的なメチル化酵素活性測定法は公知文献に記載のとおりである(公知文献:Toquin, V., et al. (2003) Plant Mol. Biol. 52, 495-509.,Gang, D. R., et al. (2002) Plant Cell 14, 505-519.)。
【0034】
また、本発明のポリペプチドは、配列番号:2または4のアミノ酸配列からなるポリペプチドの変異体であって、かつリグナンメチル化活性を有するポリペプチドを含む。
このような変異体は、配列番号:2又は4のアミノ酸配列において、1もしくは複数個(例えば、1〜30個、1〜20個、1〜10個、1〜数個(6個)、1〜3個、1〜2個など)のアミノ酸(アミノ酸残基)が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質を含む。「欠失、置換、挿入及び/又は付加」には、逆転、反復、およびタイプ置換(例えば、親水性の残基の別の残基への置換、しかし通常は強く親水性の残基を強く疎水性の残基には置換しない)が含まれる。特に、ポリペプチドにおける「中性」アミノ酸置換は、一般的にそのポリペプチドの活性にほとんど影響しない。
【0035】
ポリペプチドのアミノ酸配列中のいくつかのアミノ酸が、このポリペプチドの構造または機能に有意に影響することなく容易に改変され得ることは、当該分野において周知である。さらに、人為的に改変させるだけではく、天然のタンパク質において、当該タンパク質の構造または機能を有意に変化させない変異体が存在することもまた周知である。
当業者は、周知技術を使用してポリペプチドのアミノ酸配列において1または数個のアミノ酸を容易に変異させることができる。例えば、公知の点変異導入法に従えば、ポリペプチドをコードするポリヌクレオチドの任意の塩基を変異させることができる。また、ポリペプチドをコードするポリヌクレオチドの任意の部位に対応するプライマーを設計して欠失変異体または付加変異体を作製することができる。さらに、本明細書中に記載される方法を用いれば、作製した変異体が所望の活性を有するか否かを容易に決定し得る。
好ましい変異体は、保存性もしくは非保存性アミノ酸置換、欠失、または添加を有する。好ましくは、サイレント置換、添加、および欠失であり、特に好ましくは、保存性置換である。これらは、本発明のポリペプチド活性を変化させない。
【0036】
代表的に保存性置換と見られるのは、脂肪族アミノ酸Ala、Val、Leu、およびIleの中での1つのアミノ酸の別のアミノ酸への置換;ヒドロキシル残基SerおよびThrの交換、酸性残基AspおよびGluの交換、アミド残基AsnおよびGlnの間の置換、塩基性残基LysおよびArgの交換、ならびに芳香族残基Phe、Tyrの間の置換である。
上記に詳細に示されるように、どのアミノ酸の変化が表現型的にサイレントでありそうか(すなわち、機能に対して有意に有害な効果を有しそうにないか)に関するさらなるガイダンスは、Bowie, J.U.ら「Deciphering the Message in Protein Sequences: Tolerance to Amino Acid Substitutions」,Science 247:1306−1310 (1990)に記載されている。
【0037】
このような変異ポリペプチドは、上述したように、公知の変異ポリペプチド作製法により人為的に導入された変異を有するポリペプチドに限定されるものではなく、天然に存在するポリペプチドを単離精製したものであってもよい。
本発明のポリペプチドは、アミノ酸がペプチド結合しているポリペプチドであればよいが、これに限定されるものではなく、ポリペプチド以外の構造を含む複合ポリペプチドであってもよい。本明細書中で使用される場合、「ポリペプチド以外の構造」としては、糖鎖やイソプレノイド基等を挙げることができるが、特に限定されない。
また、本発明のポリペプチドは、付加的なポリペプチドを含むものであってもよい。付加的なポリペプチドとしては、例えば、HisやMyc、Flag等のエピトープ標識ポリペプチドが挙げられる。
【0038】
また、本発明のポリペプチドは、本発明のポリペプチドをコードするポリヌクレオチドを宿主細胞に導入して、そのポリペプチドを細胞内発現させた状態であってもよいし、細胞、組織などから単離精製されてもよい。
本発明のポリペプチドは、下記で詳述されるように組換え生成されても、化学合成されてもよい(例えば、Houghten,R.A.,Proc.Natl.Acad.Sci.USA 82:5131−5135(1985);米国特許第4,631,211号等参照)。
【0039】
本発明のポリペプチドは、リグナン(特に、ピノレジノールまたはピペリトール)のメチル化反応を触媒することができる。
【0040】
(3)本発明のポリペプチドまたはポリヌクレオチドの利用
本発明はさらに、本発明のポリペプチドまたはポリヌクレオチドを用いることによって生物(好ましくは、植物)中のリグナンおよびメチル化リグナンの量を制御(増加または減少)するための方法および当該制御された生物(好ましくは、植物)を提供する。
【0041】
(A)ベクター
本発明は、リグナンメチル化活性を有するポリペプチドを生成するために使用されるベクターを提供する。本発明のベクターは、インビトロ翻訳に用いるベクターであっても組換え発現に用いるベクターであってもよい。
本発明のベクターは、上述した本発明のポリヌクレオチドを含むものであれば、特に限定されない。例えば、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドのcDNAが挿入された組換え発現ベクターなどが挙げられる。組換え発現ベクターの作製方法としては、プラスミド、ファージ、またはコスミドなどを用いる方法が挙げられるが特に限定されない。
【0042】
ベクターの具体的な種類は特に限定されず、宿主細胞中で発現可能なベクターが適宜選択され得る。すなわち、宿主細胞の種類に応じて、確実に本発明のポリヌクレオチドを発現させるために適宜プロモーター配列を選択し、これと本発明のポリヌクレオチドを各種プラスミド等に組み込んだベクターを発現ベクターとして用いればよい。
本発明の発現ベクターは、導入されるべき宿主の種類に依存して、発現制御領域(例えば、プロモーター、ターミネーター、および/または複製起点等)を含有する。細菌用発現ベクターのプロモーターとしては、慣用的なプロモーター(例えば、trcプロモーター、tacプロモーター、lacプロモーター等)が使用され、酵母用プロモーターとしては、例えば、グリセルアルデヒド3リン酸デヒドロゲナーゼプロモーター、PH05プロモーター等が挙げられ、糸状菌用プロモーターとしては、例えば、アミラーゼ、trpC等が挙げられる。また動物細胞宿主用プロモーターとしては、ウイルス性プロモーター(例えば、SV40初期プロモーター、SV40後期プロモーター等)が挙げられる。植物体の形質転換に用いられる組換え発現ベクターは、当該植物内で本発明のポリヌクレオチドを発現させることが可能なベクターであれば特に限定されない。このようなベクターとしては、例えば、植物細胞内でポリヌクレオチドを構成的に発現させるプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクター、または外的な刺激によって誘導性に活性化されるプロモーターを有するベクターが挙げられる。
【0043】
発現ベクターの作製は、制限酵素および/またはリガーゼ等を用いる慣用的な手法に従って行うことができる。発現ベクターによる宿主の形質転換もまた、慣用的な手法に従って行うことができる。
上記発現ベクターを用いて形質転換された宿主を、培養、栽培または飼育した後、培養物等から慣用的な手法(例えば、濾過、遠心分離、細胞の破砕、ゲル濾過クロマトグラフィー、イオン交換クロマトグラフィー等)に従えば、目的タンパク質を回収、精製することができる。
【0044】
発現ベクターは、少なくとも1つの選択マーカーを含むことが好ましい。このようなマーカーとしては、真核生物細胞培養についてはジヒドロ葉酸レダクターゼ遺伝子またはネオマイシン耐性遺伝子、およびE.coliおよび他の細菌における培養についてはテトラサイクリン耐性遺伝子またはアンピシリン耐性遺伝子が挙げられる。
上記選択マーカーを用いれば、本発明のポリヌクレオチドが宿主細胞に導入されたか否か、さらには宿主細胞中で確実に発現しているか否かを確認することができる。あるいは、本発明のポリペプチドを融合ポリペプチド(例えば、GFPとの融合ポリペプチド)として発現させ、GFPの蛍光をマーカーとして用いてもよい。
【0045】
(B)形質転換体または細胞
本発明は、上述したリグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドが導入された形質転換体または細胞を提供する。本明細書中、用語「形質転換体」は、組織または器官だけでなく、生物個体を含むことが意図される。
形質転換体または細胞の作製方法(生産方法)は特に限定されないが、例えば、上述した組換えベクターを宿主に導入して形質転換する方法が挙げられる。ここで用いられる宿主細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いることができる。具体的には、例えば、大腸菌(Escherichia coli)等の細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母Schizosaccharomyces pombe)、線虫(Caenorhabditis elegans)、アフリカツメガエル(Xenopus laevis)の卵母細胞等を挙げることができる。上記の宿主細胞のための適切な培養培地および条件は当分野で周知である。また、形質転換の対象となる生物も特に限定されるものではなく、上記宿主細胞で例示した各種微生物、植物または動物が挙げられる。
【0046】
本発明の形質転換体または細胞は、これらの形質転換体または細胞が天然に有するリグナンおよび/またはメチル化リグナンの組成が改変されていることを特徴とする。本発明の形質転換体または細胞は、植物もしくはその子孫、またはこれら由来の組織であることが好ましく、ゴマ、レンギョウまたはアマであることが特に好ましい。このような形質転換体または細胞は、本発明のメチル化リグナン含有量を制御する方法を用いることによって、リグナンを産生する生物中のメチル化リグナンの含有量を増加または減少させることができる。
【0047】
本発明の形質転換体は、植物形質転換体であり得る。本実施形態の植物形質転換体は、本発明のポリヌクレオチドを含む組換えベクターを、当該ポリヌクレオチドによってコードされるポリペプチドが発現され得るように植物中に導入することによって取得される。
【0048】
組換え発現ベクターを用いる場合、植物体の形質転換に用いられる組換え発現ベクターは、当該植物内で本発明のポリヌクレオチドを発現させることが可能なベクターであれば特に限定されない。このようなベクターとしては、例えば、植物細胞内でポリヌクレオチドを構成的に発現させるプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモーター)を有するベクター、または外的な刺激によって誘導性に活性化されるプロモーターを有するベクターが挙げられる。
【0049】
本発明において形質転換の対象となる植物は、植物体全体、植物器官(例えば葉、花弁、茎、根、種子など)、植物組織(例えば表皮、師部、柔組織、木部、維管束、柵状組織、海綿状組織など)または植物培養細胞、あるいは種々の形態の植物細胞(例えば、懸濁培養細胞)、プロトプラスト、葉の切片、カルスなどのいずれをも意味する。形質転換に用いられる植物としては、特に限定されず、単子葉植物綱または双子葉植物綱に属する植物のいずれでもよい。
【0050】
植物への遺伝子の導入には、当業者に公知の形質転換方法(例えば、アグロバクテリウム法、遺伝子銃、PEG法、エレクトロポレーション法など)が用いられる。例えば、アグロバクテリウムを介する方法と直接植物細胞に導入する方法が周知である。アグロバクテリウム法を用いる場合は、構築した植物用発現ベクターを適当なアグロバクテリウム(例えば、アグロバクテリウム・チュメファシエンス(Agrobacterium tumefaciens))に導入し、この株をリーフディスク法(内宮博文著、植物遺伝子操作マニュアル(1990)27〜31頁、講談社サイエンティフィック、東京)などに従って無菌培養葉片に感染させ、形質転換植物を得ることができる。また、Nagelらの方法(Micribiol.Lett.、67、325(1990))が用いられ得る。この方法は、まず、例えば発現ベクターをアグロバクテリウムに導入し、次いで、形質転換されたアグロバクテリウムをPlantMolecular Biology Manual(S.B.Gelvinら、Academic Press Publishers)に記載の方法で植物細胞または植物組織に導入する方法である。ここで、「植物組織」とは、植物細胞の培養によって得られるカルスを含む。アグロバクテリウム法を用いて形質転換を行う場合には、バイナリーベクター(pBI121またはpPZP202など)を使用することができる。
【0051】
また、遺伝子を直接植物細胞または植物組織に導入する方法としては、エレクトロポレーション法、遺伝子銃法が知られている。遺伝子銃を用いる場合は、植物体、植物器官、植物組織自体をそのまま使用してもよく、切片を調製した後に使用してもよく、プロトプラストを調製して使用してもよい。このように調製した試料を遺伝子導入装置(例えばPDS−1000(BIO−RAD社)など)を用いて処理することができる。処理条件は植物または試料によって異なるが、通常は450〜2000psi程度の圧力、4〜12cm程度の距離で行う。
【0052】
遺伝子が導入された細胞または植物組織は、まずハイグロマイシン耐性などの薬剤耐性で選択され、次いで定法によって植物体に再生される。形質転換細胞から植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。
植物培養細胞を宿主として用いる場合は、形質転換は、組換えベクターを遺伝子銃、エレクトロポレーション法などで培養細胞に導入する。形質転換の結果得られるカルスやシュート、毛状根などは、そのまま細胞培養、組織培養または器官培養に用いることが可能であり、また従来知られている植物組織培養法を用い、適当な濃度の植物ホルモン(オーキシン、サイトカイニン、ジベレリン、アブシジン酸、エチレン、ブラシノライドなど)の投与などによって植物体に再生させることができる。
【0053】
遺伝子が植物に導入されたか否かの確認は、PCR法、サザンハイブリダイゼーション法、ノーザンハイブリダイゼーション法などによって行うことができる。例えば、形質転換植物からDNAを調製し、DNA特異的プライマーを設計してPCRを行う。
【0054】
本発明のポリヌクレオチドがゲノム内に組み込まれた形質転換植物体が一旦取得されれば、当該植物体の有性生殖または無性生殖によって子孫を得ることができる。また、当該植物体またはその子孫、あるいはこれらのクローンから、例えば、種子、果実、切穂、塊茎、塊根、株、カルス、プロトプラストなどを得て、それらを基に当該植物体を量産することができる。したがって、本発明には、本発明のポリヌクレオチドが発現可能に導入された植物体、もしくは、当該植物体と同一の性質を有する当該植物体の子孫、またはこれら由来の組織も含まれる。
【0055】
また、種々の植物に対する形質転換方法が既に報告されている。本発明の形質転換体植物としては、例えば、ゴマ、イネ、タバコ、オオムギ、コムギ、ナタネ、ポテト、トマト、ポプラ、バナナ、ユーカリ、サツマイモ、タイズ、アルファルファ、ルーピン、トウモロコシ、カリフラワー、バラ、キク、カーネーション、キンギョソウ、シクラメン、ラン、トルコギキョウ、フリージア、ガーベラ、グラジオラス、カスミソウ、カランコエ、ユリ、ペラルゴニウム、ゼラニウム、ペチュニア、トレニア、チューリップ、レンギョウ、シロイヌナズナ、アマ、およびミヤコグサなどが挙げられるがこれらに限定されない。
好ましい実施形態において、ゴマを使用して、本発明の形質転換体を作製することができる。ゴマの形質転換体の作製方法としては、例えば、T.Asamizu:Transformation of sesame plants using MAT vector system:introduction of fatty acid desaturase genes.Sesame Newsletter 16:22〜25(2002)に記載されるような公知の方法が挙げられる。
このようにして得た形質転換ゴマを用いれば、当該ゴマ内でメチル化リグナンを生産するため、低コストかつ環境に低負荷な生産プロセスでメチル化リグナン(ピペリトール、および/またはピノレジノール)を生産することができる。
【0056】
他の好ましい実施形態において、本発明の形質転換体として、タバコを好適に用いることができる。タバコは、ペチュニアなどとともに、形質転換が容易な代表的植物であり、細胞壁を取り除いた細胞の1個(プロトプラスト)から、植物1個体への再生が可能である。この再生された植物1個体は、多細胞に由来する1個体とは異なりキメラ状に陥らないため、形質転換体の作製を効率よく行うことができる。
タバコの形質転換に好ましい方法としては、リーフディスク法が挙げられる。この方法は、操作が容易であり、かつ1枚の葉切片からいくつもの独立した形質転換体を得ることができる方法である。形質転換の方法はたとえば「新生物化学実験の手引き3 核酸の分離・分析と遺伝子実験法 化学同人 1996年」に記載されている。
このようにして得た形質転換タバコを用いれば、低コストかつ環境に低負荷な生産プロセスでリグナンメチル化酵素を生産することができる。
別の好ましい実施形態において、本発明の形質転換体として、イネを好適に用いることもできる。形質転換イネを用いれば、当該イネ内で低コストかつ環境に低負荷な生産プロセスでリグナンメチル化酵素を生産することができる。
【0057】
本発明の形質転換体は、リグナン(特に、ピノレジノールまたはピペリトール)を含む生物であれば、生物種を問わず、上記ポリヌクレオチドを導入することで当該メチル化リグナンを生産することができる。
本発明のポリペプチドをコードするポリヌクレオチドを含む組換え発現ベクターが導入された形質転換体を用いれば、植物などの生物に内在するリグナンをメチル化する反応を触媒することができるので、低コストかつ環境に対して低負荷な生産プロセスでメチル化リグナンの大量調製が可能になる。さらに本発明は、メチル化リグナンを大量調製することによって安価な食品または工業製品を提供することができる。
本発明の形質転換体を用いれば、リグナンメチル化反応を触媒するポリペプチドを低コストでありかつ環境に低負荷な条件下で提供することができる。
一実施形態において、本発明に係る細胞は、種々の細菌宿主であり得る。本実施形態に係る細胞は、本発明に係るポリヌクレオチドを含む組換えベクターを、当該ポリヌクレオチドによってコードされるポリペプチドが発現され得るように細胞中に導入することによって取得される。
【0058】
当業者は、本明細書中の記載に従えば、リグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドを含む組換え発現ベクターが導入されれば、細菌から高等植物までの広範な生物にリグナンメチル化能を付与することができるということを容易に理解する。
本発明の細胞は、リグナン(特に、ピノレジノールまたはピペリトール)を含む生物であれば、生物種を問わず、上記ポリヌクレオチドを導入することで当該メチル化リグナンを生産することができる。
【0059】
本発明のポリペプチドをコードするポリヌクレオチドを含む組換え発現ベクターが導入された細胞を用いれば、当該細胞内でリグナンメチル化反応を触媒することができるので、低コストかつ環境に対して低負荷な生産プロセスでメチル化リグナンの大量調製が可能になる。さらに本発明は、メチル化リグナンを大量調製することによって安価な食品または工業製品を提供することができる。
本発明に係る細胞を用いれば、リグナンメチル化反応を触媒するポリペプチドを低コストでありかつ環境に低負荷な条件下で提供することができる。
【0060】
(C)ポリペプチドの生産方法
本発明は、本発明のポリペプチドを生産する方法を提供する。本発明のポリペプチドの生産方法を用いれば、リグナンメチル化反応を触媒するポリペプチドを低コストでありかつ環境に低負荷な条件下で提供することが可能となる。また、本発明のポリペプチドの生産方法を用いれば、リグナンメチル化反応を触媒するポリペプチドを容易に生産することが可能となる。
【0061】
本発明のポリペプチドの生産方法では、本発明のポリペプチドをコードするポリヌクレオチドを含むベクターを用いることができる。
本発明のポリペプチドの生産方法は、上記ベクターを無細胞タンパク質合成系に用いることが好ましい。無細胞タンパク質合成系を用いる場合、種々の市販のキットを用いればよい。好ましくは、本実施形態のポリペプチドの生産方法は、上記ベクターと無細胞タンパク質合成液とをインキュベートする工程を包含する。
【0062】
また、本実施形態のポリペプチドの生産方法は、組換え発現系を用いることもできる。組換え発現系を用いる場合、本発明のポリヌクレオチドを組換え発現ベクターに組み込んだ後、公知の方法により発現可能に宿主に導入し、宿主内で翻訳されて得られる上記ポリペプチドを精製するという方法などを採用することができる。組換え発現ベクターは、プラスミドであってもなくてもよく、宿主に目的ポリヌクレオチドを導入することができればよい。好ましくは、本実施形態のポリペプチドの生産方法は、上記ベクターを宿主に導入する工程を包含する。
【0063】
宿主としては、原核生物または真核生物を用いることができる。原核生物宿主としては、例えば、Escherichia属に属する細菌(例えば、大腸菌(Escherichia coli)など)、例えば、Bacillus属に属する細菌(例えば、Bacillus subtilisなどを用いることができる。真核生物宿主としては、下等真核生物(例えば、酵母または糸状菌などの真核生物微生物)を用いることができる。酵母としては、Saccharomyces属微生物(例えば、Saccharomyces cerevisiaeなど)が挙げられ、糸状菌としては、Aspergillus属微生物(例えば、Aspergillus oryzae、Aspergillus nigerなど)、Penicillium属微生物が挙げられる。また、動物細胞または植物細胞が宿主として使用され得る。動物細胞としては、マウス、ハムスター、サル、ヒト等の細胞が挙げられる。さらに、昆虫細胞(例えば、カイコ細胞、またはカイコの成虫)もまた宿主として使用され得る。
上記の宿主細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いることができる。具体的には、例えば、大腸菌(Escherichia coli)等の細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母Schizosaccharomyces pombe)、線虫(Caenorhabditis elegans)、アフリカツメガエル(Xenopus laevis)の卵母細胞等を挙げることができるが、特に限定されない。上記の宿主細胞のための適切な培養培地および条件は当分野で周知である。
【0064】
このように宿主に外来ポリヌクレオチドを導入する場合、発現ベクターは、外来ポリヌクレオチドを発現するように宿主内で機能するプロモーターを組み込んであることが好ましい。組換え的に産生されたポリペプチドを精製する方法は、用いた宿主、ポリペプチドの性質によって異なるが、タグの利用等によって比較的容易に目的のポリペプチドを精製することが可能である。
【0065】
本実施形態に係るポリペプチドの生産方法は、本発明のポリペプチドを含む細胞または組織の抽出液から当該ポリペプチドを精製する工程をさらに包含することが好ましい。ポリペプチドを精製する工程は、周知の方法(例えば、細胞または組織を破壊した後に遠心分離して可溶性画分を回収する方法)で細胞や組織から細胞抽出液を調製した後、この細胞抽出液から周知の方法(例えば、硫安沈殿またはエタノール沈殿、酸抽出、陰イオンまたは陽イオン交換クロマトグラフィー、ホスホセルロースクロマトグラフィー、疎水性相互作用クロマトグラフィー、アフィニティークロマトグラフィー、ヒドロキシアパタイトクロマトグラフィー、およびレクチンクロマトグラフィー)によって精製する工程が好ましいが、これらに限定されない。最も好ましくは、高速液体クロマトグラフィー(「HPLC」)が精製のために用いられる。
【0066】
本発明のポリペプチドは、本発明のポリペプチドを天然に発現する細胞または組織から当該ポリペプチドを精製することもできるし、化学合成によって調製することもできる。
【0067】
変異ポリペプチドを作製する方法についても、特に限定されない。例えば、部位特異的変異誘発法(例えば、Hashimoto−Gotoh,Gene 152,271−275(1995)参照)、PCR法を利用して塩基配列に点変異を導入し変異ポリペプチドを作製する方法、またはトランスポゾンの挿入による突然変異株作製法などの周知の変異ポリペプチド作製法を用いることによって、変異ポリペプチドを作製することができる。変異ポリペプチドの作製には市販のキットを利用してもよい。
このように、本発明のポリペプチドの生産方法は、少なくとも、リグナンメチル化活性を有するポリペプチドのアミノ酸配列、またはリグナンメチル化活性を有するポリペプチドをコードするポリヌクレオチドの塩基配列に基づいて公知慣用技術を用いればよいといえる。
【0068】
(D)メチル化リグナン生産方法
これまで、リグナンおよびメチル化リグナンの生産はゴマからの抽出により行われているので、大量生産ができない等の問題点を有していたが、本発明に従えば、リグナンおよびメチル化リグナンを低コストで大量生産できる。
本発明は、本発明のポリペプチドを発現する生物体または細胞を用いてメチル化リグナンを生産する方法を提供する。上記生物体は、天然の未改変生物体であっても組換え発現系を用いた形質転換体であってもよい。本発明のメチル化リグナン生産方法は、リグナン(特に、ピノレジノールまたはピペリトール)を効率よく生産することができる。
一実施形態において、本発明のメチル化リグナン生産方法は、本発明のポリペプチドをコードするポリヌクレオチドで形質転換された生物体またはその組織を用いてメチル化リグナンを生産することを特徴とする。好ましくは、上記生物体は、上述した形質転換体植物または細菌であり、特に好ましくは、大腸菌、ゴマ、レンギョウまたはアマである。
【0069】
本発明のメチル化リグナン生産方法は、本発明のポリペプチドをコードするポリヌクレオチドを上記生物体に導入する工程を包含する。ポリヌクレオチドを上記生物体に導入する工程としては、上述した種々の遺伝子導入方法を用いればよい。本実施形態のこの局面において、上記生物体は形質転換される前に産生したメチル化リグナンと形質転換後に産生するメチル化リグナンとの間でその組成が異なる。具体的には、上記生物体から得られるリグナンおよびメチル化リグナンは、その含有率が増加する。本実施形態のこの局面に係るメチル化リグナン生産方法は、上記生物体からメチル化リグナンを抽出する工程をさらに包含することが好ましい。
【0070】
また、本発明のメチル化リグナン生産方法は、本発明のポリペプチドを天然に発現する生物体に、本発明のオリゴヌクレオチドをアンチセンスオリゴヌクレオチドとして導入する工程を包含する。オリゴヌクレオチドを上記生物体に導入する工程としては、上述したアンチセンスRNA技術を用いればよい。本実施形態に係るメチル化リグナン生産方法は、上述した抗体またはオリゴヌクレオチドを用いて本発明のポリペプチドを天然に発現する生物体を同定する工程をさらに包含することが好ましい。本実施形態のこの局面に係るメチル化リグナン生産方法は、上記生物体からメチル化リグナンを抽出する工程をさらに包含することが好ましい。
本実施形態において、上記生物体は上記オリゴヌクレオチドを導入される前に産生したメチル化リグナンと導入後に産生するメチル化リグナンとの間でその組成が異なる。具体的には、上記生物体から得られるリグナンおよびメチル化リグナンは、その含有率が減少する。
【0071】
(E)食品および工業製品
本発明は、上述のメチル化リグナン生産方法により得られたメチル化リグナンを用いて製造される食品および工業製品を提供する。本項で記載する食品は、上述した形質転換植物体の種子、果実、切穂、塊茎、および/または塊根であっても、上述した形質転換植物体から抽出されたメチル化リグナンを用いて製造された食品(例えば、ゴマ、レンギョウまたはアマ、あるいはこれらの加工食品)であってもよい。本発明に係る食品または工業製品は、所望する量のリグナン(特に、ピノレジノールまたはピペリトール)を含有することができる。
例えば、上述のようにメチル化リグナンの含量が増加した本発明の形質転換植物から抽出されたメチル化リグナン抽出液は、メチル化リグナンの含量が高い食品として提供される。また、抽出したメチル化リグナンに限らず、上記形質転換植物体の種子、果実、切穂、塊茎、および/または塊根等もまた、メチル化リグナンを多く含む食品として提供される。メチル化リグナン組成を改変する対象は特に限定されるものではなく、植物以外にも動物、細菌、または酵母等のあらゆる生物を対象とすることが可能である。
また、リグナンおよびメチル化リグナンの独特な物性に基づいて、本発明のポリペプチドまたはポリヌクレオチドは、工業製品(例えば、フィルム、生分解性プラスティック、機能性繊維、潤滑油、または洗剤のような工業製品)の原料に利用され得る。
【0072】
本明細書において、ゴマのリグナンメチル化ポリペプチドを一例として記載されているが、本発明は、ゴマ由来のポリペプチドまたはポリヌクレオチドにのみ限定されるべきではなく、リグナンメチル化活性を有する全てのポリペプチド、およびその利用に関するものであることが、当業者には明白である。リグナンメチル化酵素は、植物、動物または微生物のいずれ由来でもよく、リグナンメチル化酵素活性を有していればリグナン量を制御することができる。さらに本発明は、リグナンメチル化酵素をコードするポリヌクレオチドを導入することによって作製されたリグナン量が調節された植物、その子孫またはこれらの組織に関し、その形態は切り花であってもよい。本発明のリグナンメチル化ポリペプチドを用いれば、メチル化リグナンの生成を促進または抑制することができる。また、慣用的な手法を用いれば、植物に上記ポリヌクレオチドを導入して当該ポリヌクレオチドを構成的あるいは組織特異的に発現させて、目的のポリペプチドの発現を増加させること、ならびに、アンチセンス法、同時サプレッション法およびRNAi法などを用いることによって目的のポリペプチドの発現を抑制することができるということを、当業者は容易に理解する。
本発明は、以下の実施例によってさらに詳細に説明されるが、これに限定されるべきではない。
【実施例】
【0073】
実施例において用いる分子生物学的手法は、他で詳述しない限り、国際公開WO2004/018682号公報(PCT/JP03/10500)またはMolecular Cloning(Sambrookら、Cold Spring Harbour Laboratory Press,1989)に記載の方法に従った。
【0074】
〔実施例1:ゴマ種子のEST解析〕
公知文献(国際公開公報WO2005/030944)に記載のゴマ種子由来のcDNAファージライブラリーをRapid Excisionキット(Stratagene社)を用いて製造業者が推奨する方法に従って大腸菌内にpBK-CMV(Stratagene社)に切り出した。このゴマ種子由来ESTを含む大腸菌コロニーをランダムに5000クローンをピックアップし、M13RVプライマー(配列番号5)およびM4(-20)プライマー(配列番号6)を用いて以下の条件でコロニーPCRを行った。
1×Ex−Taq buffer(TakaRa)、0.2mM dNTPs、プライマー各0.2pmol/μl、Ex−Taq polymerase 1.25Uからなる混合液に大腸菌コロニーをけん濁し、94℃で5分間の後、94℃で1分間、55℃で1分間、72℃で2分間の反応を30サイクル行ってPCR増幅した後、72℃で7分間保持した。
配列番号5:M13RV:5'-CAGGAAACAGCATTGAC
配列番号6:M4-20:5'-GTAAAACGACGGCCAGT
【0075】
これらのPCR産物をアガロースゲル電気泳動に供し、pBK−CMVに含まれるESTが特異的に増幅されていることをエチジウムブロマイド染色により確認した。これら5000種のPCR産物4μlを10unitのExonucleaseI(USB社)と2unitのShrimp Alkaline phosphatase(USB社)と混合し、37℃で30分保持し、80℃で20分保持し酵素反応を終了させた。この酵素反応液1μlを用いて以下の条件でダイレクトシークエンス反応を行った。シークエンス反応液は酵素反応液1μl、5xBigDyeSequencingBuffer ver.3.1(Applied Biosystems)3μl、BigDyeSequencingRR ver.3.1(Applied Biosystems) 2μl、1.6pmol/μlM13RVプライマー1μl、滅菌水13μlからなる。シークエンス反応は96℃1分間に続いて96℃10秒間、50℃5秒間、60℃4分間の反応を25サイクル行った。シークエンス反応液はDyeEx96(QIAGEN社)を用いて業者が推奨する方法で生成し、Sequencer model 3100(Applied Biosystems社)を用いて塩基配列を決定した。
【0076】
得られた5000種の塩基配列はBlastxを用いて相同検索を行い、その中からメチル化酵素遺伝子と相同性を有する2種類のゴマメチル化酵素(Sesamum indicum O-methyltransferase; 以下SiOMT)の部分配列を同定した。Blastx解析の条件は以下のとおりである。プログラム:Blastx ver.2.2.9、データベース:nr、遺伝コード:standard(1)、フィルタ:LOW complexity、Expect:10、Word size:3、マトリクス:BLOSUM62、Gap Costs:Existence 11、Extension 1。
【0077】
〔実施例2:ゴマメチル化酵素遺伝子の発現解析〕
EST解析により得られた二種類のSiOMT1およびSiOMT2の遺伝子発現パターンを解析するために、公知文献(国際公開公報WO2005/030944)に記載のゴマの器官別cDNAに対して以下の条件でRT−PCRを行った。
cDNA 1μl、1×Ex−Taq buffer(TakaRa)、0.2mM dNTPs、プライマー各0.2pmol/μl、Ex−Taq polymerase 1.25Uからなる。94℃で5分間の後、94℃で1分間、55℃で1分間、72℃で2分間の反応を32サイクル行ってPCR増幅した後、72℃で5分間保持した。SiOMT1およびSiOMT2特異的プライマーとしてSiOMT1-FW(配列番号7)とSiOMT1-RV(配列番号8)およびSiOMT2-FW(配列番号9)とSiOMT2-RV(配列番号10)を用いた。SiOMT遺伝子の発現量と内在性遺伝子の発現量とを比較するために、内部標準となる遺伝子のPCR反応を同時に行った。具体的には、Si18SrRNA−Fプライマー(配列番号11)およびSi18SrRNA−Rプライマ−(配列番号12)を用いてゴマの18S ribosomal RNA遺伝子(アクセッション番号:AF169853)のPCR反応を行った。
配列番号7:SiOMT1-FW:5‘-TTGCCCCATGTCATTCAAGAT
配列番号8:SiOMT1-RV:5‘-AAAATTCAGACTTATAACGATACCAAA
配列番号9:SiOMT2-FW:5‘-TTAGAAAAACTCAATTCGTCTAAT
配列番号10:SiOMT2-RV:5‘-CCTACATCCACGACGGAATCCAAA
配列番号11:Si18SrRNA−FW:5’−tatgcttgtctcaaagattaa
配列番号12:Si18SrRNA?RV:5’−aacatctaagggcatcacaga
【0078】
PCR産物を電気泳動し、エチジウムブロマイド染色を行った結果、両SiOMT遺伝子ともに種子において強い発現が確認された(図2の2A)。すなわち、両SiOMT遺伝子発現領域とメチル化リグナンが蓄積する領域が一致することを確認した。
【0079】
〔実施例3:ゴマメチル化酵素遺伝子のクローニング〕
両ESTクローンが推定ORFの5’領域を含んでいなかったため、5’Rapid Amplification of cDNA End(以下、5’RACE)法を使用して、それぞれのSiOMT遺伝子の5’領域を増幅した。具体的には、Gene Racer kit(Invitrogen社)を製造業者の推奨する方法に従って使用して、以下のプライマー(配列番号13〜16)を用いて、各々の5’領域を増幅した。
配列番号13:GR-SiOMT1-RV:5'-ccggcccactgttcgggtcctaacgggaaa
配列番号14:SiOMT1-NEST−RV:5'-gcaaatccacttcataaaaat
配列番号15:GR-SiOMT2-RV:5'-cctcgggttccgctctttctgctcccagaa
配列番号16:SiOMT2-NEST−RV:5'-atcaatttgggaaattacaaa
【0080】
SiOMT2に関してはESTクローンが推定ORFの3‘末端を含んでいなかったため、配列番号17および18のプライマーを用いて3’RACEを行った。
配列番号17:GR-SiOMT2-FW:5'-gaagatcgccccatgagcatgaaacccttt
配列番号18:SiOMT2-NEST−FW:5'-aacgtcgttctgggagcagaaaga
【0081】
RACE法によって得られた増幅断片の塩基配列を、プライマーウォーキング法によって決定し、SiOMT1およびSiOMT2遺伝子の完全長オープンリーディングフレームを含む塩基配列情報を得た(配列番号1および配列番号19)。
SiOMT1とSiOMT2はお互いにアミノ酸配列で29%の配列同一性を示した。配列同一性について、Clustal W alignmentプログラム(Mac Vector ver.7.2.2(Symanteccorporation))をデフォルト設定で使用した。
【0082】
得られたSiOMT遺伝子の完全長cDNA配列をBlastx解析(http://www.ncbi.nlm.nih.gov/BLAST/)によって相同性を有する既知タンパク質を検索した。Blastx解析の条件は以下のとおりである。
プログラム:Blastx ver.2.2.9、データベース:nr、遺伝コード:standard(1)、フィルタ:LOW complexity、Expect:10、Word size:3、マトリクス:BLOSUM62、Gap Costs:Existence 11、Extension 1。
Blastx解析の結果、SiOMT1は、Catharanthsus roseus Caffeic acid-O-methyltransferase(COMT)(AAK20170)と74%の配列同一性を示し、SiOMT2は、Rosa hybrida(AAM23004)のOrcinol-O-methyltransferase(OOMT)と57%の配列同一性を示した。このように、両SiOMTと既知のメチル化酵素との配列同一性はいずれも高いとは言えない。従って、得られた両SiOMT遺伝子の機能を推定することはできなかった。すなわち、得られた両SiOMT遺伝子は、これまでに単離されていないリグナンメチル化酵素である可能性が非常に高い。
【0083】
〔実施例4:大腸菌発現ベクターの構築〕
SiOMT1に対して配列番号20および21のプライマー対を使用して、cDNAの開始メチオニンコドン(ATG)の上流にBglII部位を、終始コドンの下流にSalI部位を有するフラグメントをPCR増幅した。
一方、SiOMT2に対して配列番号22および23のプライマー対を使用して、cDNAの開始メチオニンコドン(ATG)の上流にBamHI部位を、終始コドンの下流にXhoI部位を有するフラグメントをPCR増幅した。PCRに用いたプライマーを以下に示す。
配列番号20:Bgl2-SiOMT1-FW:5'-aaaacatgtatggcggatcagtccgaggaagaagaggcttt
配列番号21:SalI-SiOMT1-RV:5'-attgtcgacttatgaaattccatgatccaaatatt
配列番号22:BamHI-SiOMT2-FW:5'-aaaggatccatggcgatggttaaccaaaagcaaaatctt
配列番号23:XhoI-SiOMT2-RV:5'-aaactcgagttaaggatatatctcgatgatagatctcaa
【0084】
PCR反応液(25μl)は、テンプレートとしてのゴマ種子cDNA、各プライマー0.2pmol/μl、1×KOD plus buffer (TOYOBO)、0.2mM dNTPs、1mM MgSO4、1U KOD plus polymeraseからなる。94℃で5分間反応させた後、94℃1分間、55℃1分間、72℃2分間の反応を30サイクル行ってPCR増幅した後、72℃で3分間保持した。得られた各PCR産物を、製造者が推奨する方法に従ってpCR4 Blunt−TOPO vector(Invitrogen)のマルチクローニング部位に挿入して、SiOMT1/pCR4 Blunt−TOPO(pSPB2678と称する)、SiOMT2/pCR4 Blunt−TOPO(pSPB2679と称する)を得た。
pSPB2678および、pSPB2679に含まれるSiOMT塩基配列を解析して、正しくPCRが行われたことを確認した。これらのプラスミドをPCRプライマーに付加した制限酵素サイトで消化して得られる完全長SiOMTを含む約1.1kbのDNA断片を、大腸菌発現ベクターであるpQE30ベクター(QIAGEN)のBamHI/SalIサイトに挿入して、SiOMT1/pQE30(pSPB2690と称する)、SiOMT2/pQE30(pSPB2686と称する)を得た。
【0085】
〔実施例5:組換えタンパク質の調製〕
上記において作製した大腸菌発現ベクターpSPB2690およびpSPB2686を用いて大腸菌株JM109(TOYOBO)を形質転換し、最終濃度20μg/mlのアンピシリンを含むLB培地中にて37℃で一晩前培養した。前培養液の一部をアンピシリン50μg/ml、カザミノ酸0.5%を含むM9培地(10ml)に添加して、A600=0.6〜1.0に達するまで振盪培養した。次いで、培養液に最終濃度0.5mMのIPTG(Isopropyl−β−D−thiogalactopyranoside)を加え、さらに30℃で一晩振盪培養した後、3000rpmにて4℃で10分間遠心分離を行って集菌した。菌体を10mlの緩衝液(30mM Tris−HCl(pH7.5)、2mM MgCl2、0.5mM EDTA,50μM APSMF)に懸濁した後に超音波処理を行って大腸菌を破砕し、次いで、15,000rpmにて4℃で10分間遠心分離を行い、得られた上清を粗酵素液として以下の活性測定に用いた。
【0086】
〔実施例6:ゴマリグナンメチル化酵素による生成物の分析〕
酵素反応の基質に用いるゴマリグナンは少量のDMSOに溶解した後に70%エタノールに溶解して基質溶液(1mg/ml)を調製した。ゴマリグナンは、例えば、公知の方法(日本農芸化学会誌 67:1693(1993))に従ってゴマから抽出および精製して得ることができる。基質溶液10μl、大腸菌で発現させたSiOMTの上記粗酵素液200μlおよび10mM S−Adenosyl-L-Mathianine(SAM) 10μlを反応チューブ中で混合した後、30℃で2時間反応させた。
0.1%トリフルオロ酢酸(TFA)を含む100%アセトニトリル(250μl)を反応チューブに添加することによって、酵素反応を停止させた。反応チューブをボルテックスミキサーで激しく攪拌した後、15,000rpmにて4℃で5分間遠心分離し、得られた上清をフィルタ(ポアサイズ0.45mm、4mm Millex−LH、Millipore)を用いて清浄化した後、液体高速クロマトグラフィー(以下HPLC)を用いてこの上清を分析した。リグナンおよびそのメチル化リグナンの分析条件は以下の通りである。
【0087】
C−30カラム(野村化学C30−UG−5、4.6mm×150mm)を用いて液体クロマトグラフィー(Lc−2010c(島津製作所))を行った。移動相には、A液として、0.1%TFA、B液として、0.1%TFAを含む90%アセトニトリルを用いた。A液80%:B液20%の混合液を用いてカラムを平衡化した(20分間)後、直線濃度勾配(A液80%:B液20%→A液10%:B液90%)を用いて20分間にわたって溶出(流速0.6ml/分)し、さらにA液10%:B液90%を用いて7分間溶出した。280nmの吸収を測定して、サンプル中に含有される化合物を検出した。化合物の各ピークを、SPD−10AV(島津製作所)を用いて190nm〜400nmのスペクトルを測定し、リグナンに特徴的な2つの吸収極大(230nmおよび280nm)を有する物質を探索した。この条件下で、ピノレジノール標準品は約11.8分、ピペリトール標準品は約15.5分、セサミノール標準品は約16.8分に検出される。
SiOMT1組換えタンパク質とピノレジノールとの反応液中において、リグナンのスペクトルを有する保持時間14.0分のピークAが新たに得られた(図3の3A)。また、SiOMT1組換えタンパク質とピペリトールとの反応液中において、リグナンのスペクトルを有する保持時間約17.7分のピークBが新たに得られた(図3の3B)。
【0088】
SiOMT2組換えタンパク質とピノレジノールまたはピペリトールとの反応液中において、新たな生成物は認められなかった(図4の3C、3D)。また、SiOMT1およびSiOMT2のセサミノールに対するリグナンメチル化活性は認められなかった。
【0089】
次に、LC−MS分析によって、SiOMT1により生成した新規リグナンの分子量を決定した(液体クロマトグラフィー(LC):Waters 2795(ウォーターズ社)、質量分析器:QUATRO micro(マイクロマス社))。ダイアイオンHP−20樹脂(三菱化学)を1ml充填したカラムを50%アセトン5mlで洗浄した後、水10mlを用いて平衡化した。SiOMT1により生成したメチル化リグナンを含む酵素反応液をカラムにロードし、5mlの水を用いて不純物を洗浄した後、80%アセトン2mlを用いて溶出した。エバポレーターを用いて溶出液を乾固させた後、1%ギ酸を含む90%アセトニトリル(100μl)中に溶解させてLC−MS分析試料とした。LC条件を以下に示す。
【0090】
Develosil C30−UG−3カラム(野村化学、3.0×150mm)を用い、移動相は、A液として水を、B液として100%アセトニトリルを、C液として1%ギ酸を、D液として100mM酢酸アンモニウム水溶液を用いた。10分間にわたる直線濃度勾配(A液60%:B液30%:C液5%:D液5%→A液10%:B液80%:C液5%:D液5%)を用いて溶出し、その後、A液10%:B液80%:C液5%:D液5%を用いて5分間溶出した(流速:常時0.2ml/分)。アンモニウム付加イオンとしてシグナルを検出した。
この条件下で、ピークAは約8.3分、ピークBは約11.3分に検出される(図3〜5)。イオンモードES+、Cone voltage 15V、Collision 2eVのMS条件を用いて、100〜800(m/z,30分)の範囲でMSスキャンし、210〜400nm(30分)の範囲のPDAを測定した。
上記条件下でのLC−MS分析の結果、ピークAはアンモニウムイオン付加分子量373(m/z)、ピークBはアンモニウムイオン付加分子量371(m/z)であった(図6の4A、図7の4B)。従って、これらのピークは、それぞれピノレジノール(アンモニウムイオン付加分子量359)のモノメチル化されたもの、およびピペリトール(アンモニウムイオン付加分子量374)のモノメチル化されたものであると同定した。
以上の結果からSiOMT1はゴマリグナンであるピノレジノールおよびピペリトールに対してモノメチル化する活性を有する酵素であることが明らかになった。
【0091】
〔実施例7:ゴマリグナンメチル化酵素ホモログ遺伝子の単離〕
リグナンメチル化活性を有する酵素遺伝子がゴマ種間において機能的および構造的に保存されているかどうかを明らかにするために、栽培ゴマ品種であるSesamum indicumとは細胞遺伝学的に異なるアフリカゴマ(Sesamum radiatum)から、SiOMT1のカウンターパート遺伝子(SrOMT1)を単離することを試みた。
S.radiatumはアフリカに現存するゴマ植物であり、細胞遺伝学解析によれば染色体数は2n=64とされており、栽培ゴマ品種であるS.indicum(2n=26)とは遺伝的に異なる系統である(参考文献、並木満夫、小林貞作「ゴマの科学」朝倉書店)。しかし、S.radiatum種子においても多様なリグナンが蓄積していることが知られている(参考文献、Bedigian,D.ら、Biochemical Systematics and Ecology 13:133−139(1985))。従って、S.radiatumのゲノム中にはS.indicumのSiOMT1に対応する遺伝子が含まれていることが期待される。
【0092】
S.radiatum種子のcDNAを用いてBgl2-SiOMT1-FW(配列番号20および)SalI-SiOMT1−RV(配列番号21)のプライマー対を用いて、PCRによりSrOMT1の増幅を試みた。
S.radiatum種子のcDNA 1μl、1×Ex−Taq buffer(TakaRa)、0.2mM dNTPs、プライマー各0.2pmol/μl、Ex−Taq polymerase 1.25Uからなる。94℃で5分間の後、94℃で1分間、55℃で1分間、72℃で2分間の反応を30サイクル行ってPCR増幅した後、72℃で5分間保持した。得られた約1.1kbのPCR産物を、製造者が推奨する方法に従ってpCR2−TOPO vector(Invitrogen社)のマルチクローニングサイトに挿入し、SrSiOMT1/pCR2−TOPO(pSPB2910)を得た。
プライマーウォーキング法によって、pSPB2910に含まれるSrOMT1の塩基配列(およびアミノ酸配列)を決定した(配列番号3および4)。SrOMT1は、SiOMT1に対してアミノ酸配列で89%の配列同一性を示した。配列同一性について、Clustal W alignmentプログラム(Mac Vector ver.7.2.2(Symanteccorporation))をデフォルト設定で使用した。この高い配列同一性はSrOMT1がSiOMT1のカウンターパート遺伝子であることを強く支持する。
【0093】
実施例2と同様の方法でRT−PCRを行い、SrOMT1の種子に特異的な遺伝子発現を確認した(図1)。
実施例4と同様にSrOMT1の大腸菌発現ベクターであるSrOMT1/pQE30(pSPB2911)を構築し、実施例5と同様に組換えタンパク質を調整し、実施例6と同様にその活性および生成物の分析を行った。
SrOMT1はSiOMT1と同様に、ピノレジノールおよびピペリトールに対してメチル化活性を示した(図2)。また、SiOMT1はセサミノールに対してメチル化活性を示さなかった。以上の結果より、SrOMT1はSiOMT1のカウンターパート遺伝子であり、本酵素遺伝子がゴマ種間で保存されていることが示された。
【0094】
〔実施例8:SiOMT1のゲノミックサザン解析〕
ゴマゲノム内におけるSiOMT1遺伝子のコピー数を明らかにするために、ゲノミックサザン解析を実施した。Nucleon Phytopure for Plant Extraction Kit(アマシャム社)を製造業者が推奨する方法に従って用いて、S.indicum(真瀬金品種)の葉からゲノムDNAを抽出した。抽出したゲノムDNA20μgを、EcoRI、HindIII,XhoIまたはXbaIで消化した後、アガロースゲルを用いた電気泳動にて分離した。このアガロースゲルを0.25M HClを用いて15分間加水分解した後、1.5M NaCl/0.5M NaOHの溶液を用いて変性させ(30分間)、1.5M NaClを含むTris−HCl(pH7.5)を添加することによって変性溶液で中和した(20分間)。次いで、アガロースゲル内のゲノムDNAを、20×SSC溶液中にてメンブレン(Hybribond−N、アマシャム社)に転写した。メンブレンに転写したゲノムDNAをUV照射によりメンブレンに結合させ、7%SDS、50%ホルムアミド、5×SSC、2%ブロッキング試薬、0.1%ラウロイルサルコシン、50mMリン酸ナトリウム緩衝液(pH7.0)からなるハイブリダイゼーション緩衝液(高SDS緩衝液:ロシュ社)を用いて、42℃で1時間プレハイブリダイゼーションを行った。
【0095】
pSPB2678をテンプレートとして、それぞれ配列番号20(Bgl2-SiOMT1−FW)および配列番号21(SalI−SiOMT1−RV)のプライマー対を用いるPCRによって、DIG標識化ハイブリダイゼーションプローブを作製した。PCR反応液は、pSPB2678プラスミド1ng、1×PCR緩衝液(タカラバイオ社)、2.5mM DIG−dNTP mixture(PCR DIG labeling Mix、ロシュ社)、0.2pmolの各プライマー、1U rTaq polymerase(タカラバイオ社)からなる。PCR反応を、95℃で30秒間、53℃で30秒間、72℃で2分間の反応を30サイクル行った。Sephadex G−50カラム−Fine(ベーリンガー社)を用いて精製したPCR産物をハイブリダイゼーションプローブとして用いた。このプローブを熱変性した後、プレハイブリダイゼーション液に15μl添加して、42℃で一晩インキュベートした。
【0096】
ハイブリダイゼーション後、高ストリンジェントな条件(0.2×SSCおよび0.1%SDSを含む溶液を用いて65℃で30分間を2回)によってメンブレンを洗浄した。DIGラベリング&デテクションキット(ロシュ社)を製造業者の指示書に従って用いて、ハイブリダイゼーションシグナルを検出した。
サザン解析の結果、いずれの制限酵素消化画分においても二本以上のバンドが検出されたので、SiOMT1と極めて相同性の高い遺伝子が少なくとも1つ以上ゴマゲノム内にコードされていることが示された(図8)。さらに言えば、ゴマゲノム中にはSiOMT1と機能的に重複もしくは類似した酵素遺伝子が複数存在することが期待される。
【0097】
〔実施例9:ゴマリグナンメチル化酵素のCOMT活性の測定〕
実施例3に示すようにデータベース検索によりSiOMT1およびSrOMT1は既知のタンパク質の中ではCOMTと最も高い配列同一性を示す。このことはゴマメチル化酵素がCOMT活性をも有していることを示唆する。
実施例5および実施例7で発現させたSiOMT1およびSrOMT1のカフェ酸に対するメチル化活性を検討した。0.4mg/mlカフェ酸10μl、大腸菌で発現させたSiOMTの粗酵素液200μlおよび10mM SAM 10μlを反応チューブ中で混合した後、30℃で1時間反応させた後、実施例6と同様の方法でHPLC分析に供した。
その結果、SiOMT1およびSrOMT1共にカフェ酸との反応液中に標準品フェルラー酸と保持時間が一致するピークC(保持時間約7.3分)が生成した(図9の6A〜6C))。本HPLC分析条件下ではカフェ酸は保持時間約3.9分に溶出される。
実施例6と同様の条件でLC−MS分析を行ったところピークCはアンモニウムイオン付加分子量195(m/z)であった(図10の6D)。従って、ピークCは、カフェ酸(アンモニウムイオン付加分子量181(m/z))がモノメチル化されて生成するフェルラー酸であることが確認された。これにより二種のリグナンメチル化酵素は共にCOMT活性をも有していることが明らかとなった。
【産業上の利用可能性】
【0098】
以上のように、本発明のポリペプチドおよびポリヌクレオチドは、メチル化リグナンの生産に有用である。また、本発明のポリヌクレオチドを発現可能に導入した形質転換体または細胞は、食品分野、各種工業分野において、メチル化リグナン、またはこれを用いる製品を生産する上で、極めて有用である。上記形質転換体が植物体である場合、植物体自体を食品として用いることができるので農業分野等において非常に有用である。
さらに、本発明のポリペプチドおよびポリヌクレオチドを、本発明者らが見出した他の酵素(ピペリトールおよびセサミンを合成する酵素であるSiP189)と組み合わせることによって、ゴマに限ることなく、特定のリグナン分子種の生産系の確立が可能となり、その結果、特定のリグナンおよびメチル化リグナンの生産量を拡大することができる。従って、本発明は、農業、食品産業、医薬品産業およびこれらの関連産業にわたる広範な利用が可能である。
【図面の簡単な説明】
【0099】
【図1】図1はゴマリグナンの構造と代謝経路図である。
【図2】2Aは、SiOMT1およびSiOMT2のゴマ(S.indicum cv. 真瀬金)器官別のRT−PCRによる遺伝子発現解析の結果である。2Bは、SrOMT1のゴマ(S.radiatum)器官別のRT−PCRによる遺伝子発現解析の結果である。
【図3】3Aは、大腸菌で発現させたSiOMT1とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。3Bは、大腸菌で発現させたSiOMT1とピペリトールとの酵素反応液のA280nmのクロマトグラムである。
【図4】3Cは、大腸菌で発現させたSiOMT2とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。3Dは、大腸菌で発現させたSiOMT2とピペリトールとの酵素反応液のA280nmのクロマトグラムである。
【図5】3Eは、大腸菌で発現させたSrOMT1とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。3Fは、大腸菌で発現させたSrOMT1とピペリトールとの酵素反応液のA280nmのクロマトグラムである。
【図6】4AはSiOMT1により生成されたメチル化ピノレジノールのLC−MSのクロマトグラムである。
【図7】4BはSiOMT1により生成されたメチル化ピペリトール(コブシン)のLC−MSのクロマトグラムである。
【図8】図8はSiOMT1遺伝子の完全長プローブを用いたゲノミックサザンハイブリダイゼーションの結果である。
【図9】6Aは、標準品のカフェ酸のA280nmのクロマトグラムである。6Bは、大腸菌で発現させたSiOMT1とカフェ酸との酵素反応液のA280nmのクロマトグラムである。6Cは、大腸菌で発現させたSrOMT1とピノレジノールとの酵素反応液のA280nmのクロマトグラムである。
【図10】6DはSiOMT1により生成されたメチル化カフェ酸(フェルラー酸)のLC−MSのクロマトグラムである。
【特許請求の範囲】
【請求項1】
以下の(a)〜(d)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチド;及び
(d)配列番号:2又は4のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【請求項2】
配列番号:2又は4のアミノ酸配列、または当該アミノ酸配列に対して1又は数個のアミノ酸の付加、欠失及び/又は他のアミノ酸による置換によって修飾されているアミノ酸配列を有し、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする前記請求項1に記載のポリヌクレオチド。
【請求項3】
配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする請求項1に記載のポリヌクレオチド。
【請求項4】
配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドに対して、5xSSC、50℃の条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする前記請求項1に記載のポリヌクレオチド。
【請求項5】
配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有する前記請求項1に記載のポリヌクレオチド。
【請求項6】
配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する前記請求項1に記載のポリヌクレオチド。
【請求項7】
DNAである、前記請求項1〜6のいずれかに記載のポリヌクレオチド。
【請求項8】
フロフラン型リグナンに対してメチル基を転移する活性を有するタンパク質をコードする前記請求項1〜7のいずれかに記載のポリヌクレオチド。
【請求項9】
ピノレジノール及び/又はピペリトールに対してメチル基を転移する活性を有するタンパク質をコードする前記請求項8に記載のポリヌクレオチド。
【請求項10】
前記請求項1〜9のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
【請求項11】
前記請求項1〜9のいずれかに記載のポリヌクレオチドを含有するベクター。
【請求項12】
請求項11に記載のベクターにより形質転換された宿主細胞。
【請求項13】
請求項12に記載の宿主細胞を培養し又は生育させ、当該宿主細胞からリグナンにメチル基を転移する活性を有するタンパク質を採取することを特徴とする該タンパク質の製造方法。
【請求項14】
請求項1〜9のいずれかに記載のポリヌクレオチドが導入された植物体もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体、又はそれら植物体の組織。
【請求項15】
請求項1〜9のいずれかに記載のポリヌクレオチドを用いてリグナンにメチル基を転移する方法。
【請求項16】
請求項1〜9のいずれかに記載のポリヌクレオチドを植物体に導入し、発現させることによって、産生するリグナン組成が改変された当該植物体もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体。
【請求項1】
以下の(a)〜(d)のいずれかに記載のポリヌクレオチド:
(a)配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有するポリヌクレオチド;
(b)配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド;
(c)配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリンジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチド;及び
(d)配列番号:2又は4のアミノ酸配列において、1もしくは複数個のアミノ酸が欠失、置換、挿入及び/又は付加したアミノ酸配列からなり、かつリグナンにメチル基を転移する活性を有するタンパク質をコードするポリヌクレオチドを含有するポリヌクレオチド。
【請求項2】
配列番号:2又は4のアミノ酸配列、または当該アミノ酸配列に対して1又は数個のアミノ酸の付加、欠失及び/又は他のアミノ酸による置換によって修飾されているアミノ酸配列を有し、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする前記請求項1に記載のポリヌクレオチド。
【請求項3】
配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドとストリジェントな条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする請求項1に記載のポリヌクレオチド。
【請求項4】
配列番号:1又は3の塩基配列の一部又は全部と相補的な塩基配列からなるポリヌクレオチドに対して、5xSSC、50℃の条件下でハイブリダイズし、かつリグナンにメチル基を転移する活性を有するタンパク質をコードする前記請求項1に記載のポリヌクレオチド。
【請求項5】
配列番号:1又は3の塩基配列からなるポリヌクレオチドを含有する前記請求項1に記載のポリヌクレオチド。
【請求項6】
配列番号:2又は4のアミノ酸配列からなるタンパク質をコードするポリヌクレオチドを含有する前記請求項1に記載のポリヌクレオチド。
【請求項7】
DNAである、前記請求項1〜6のいずれかに記載のポリヌクレオチド。
【請求項8】
フロフラン型リグナンに対してメチル基を転移する活性を有するタンパク質をコードする前記請求項1〜7のいずれかに記載のポリヌクレオチド。
【請求項9】
ピノレジノール及び/又はピペリトールに対してメチル基を転移する活性を有するタンパク質をコードする前記請求項8に記載のポリヌクレオチド。
【請求項10】
前記請求項1〜9のいずれかに記載のポリヌクレオチドにコードされるタンパク質。
【請求項11】
前記請求項1〜9のいずれかに記載のポリヌクレオチドを含有するベクター。
【請求項12】
請求項11に記載のベクターにより形質転換された宿主細胞。
【請求項13】
請求項12に記載の宿主細胞を培養し又は生育させ、当該宿主細胞からリグナンにメチル基を転移する活性を有するタンパク質を採取することを特徴とする該タンパク質の製造方法。
【請求項14】
請求項1〜9のいずれかに記載のポリヌクレオチドが導入された植物体もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体、又はそれら植物体の組織。
【請求項15】
請求項1〜9のいずれかに記載のポリヌクレオチドを用いてリグナンにメチル基を転移する方法。
【請求項16】
請求項1〜9のいずれかに記載のポリヌクレオチドを植物体に導入し、発現させることによって、産生するリグナン組成が改変された当該植物体もしくは当該植物体と同一の性質を有する該植物体の子孫となる植物体。
【図1】


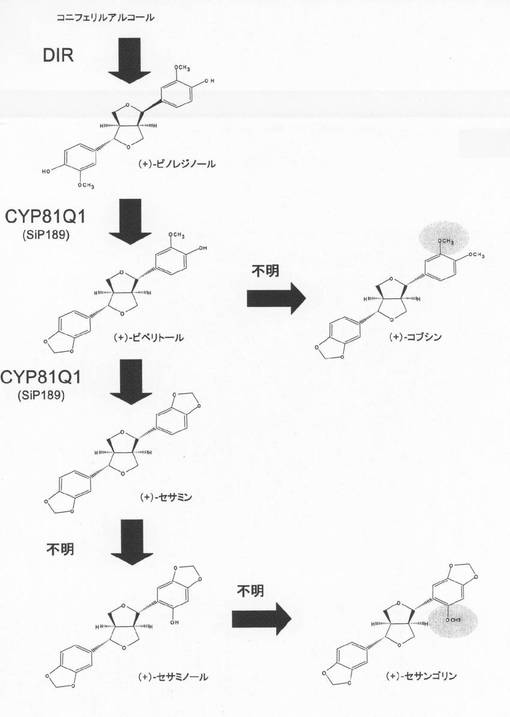
【図2】
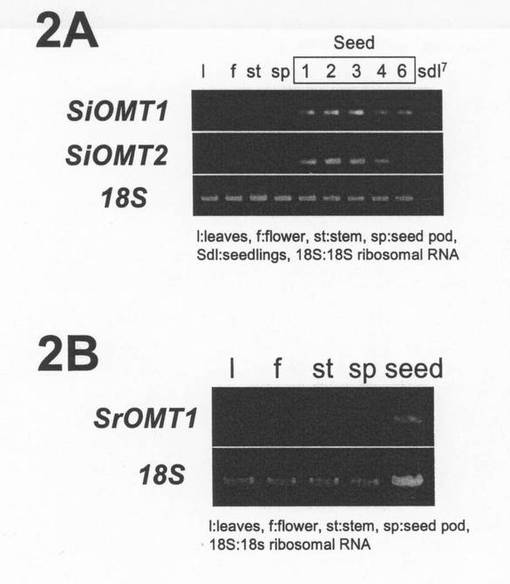
【図3】
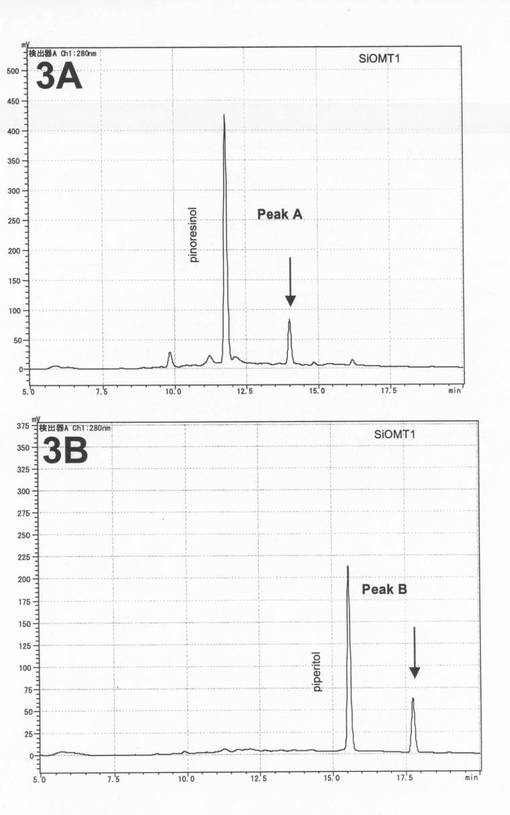
【図4】
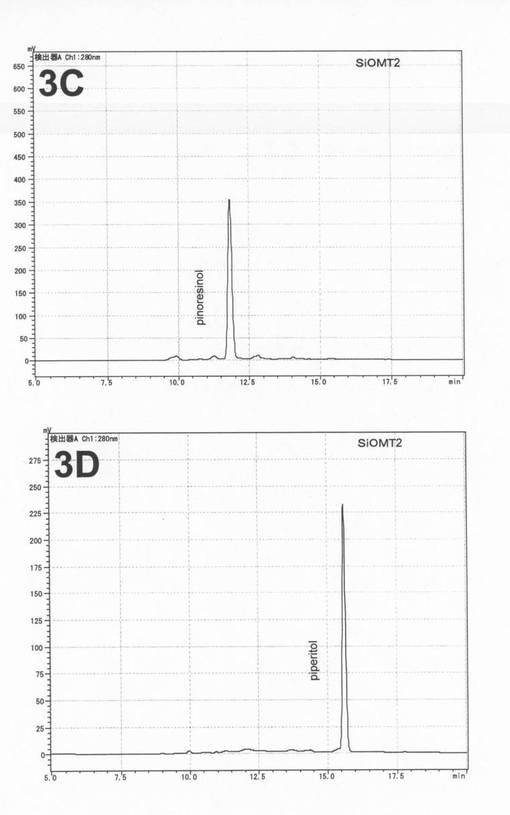
【図5】
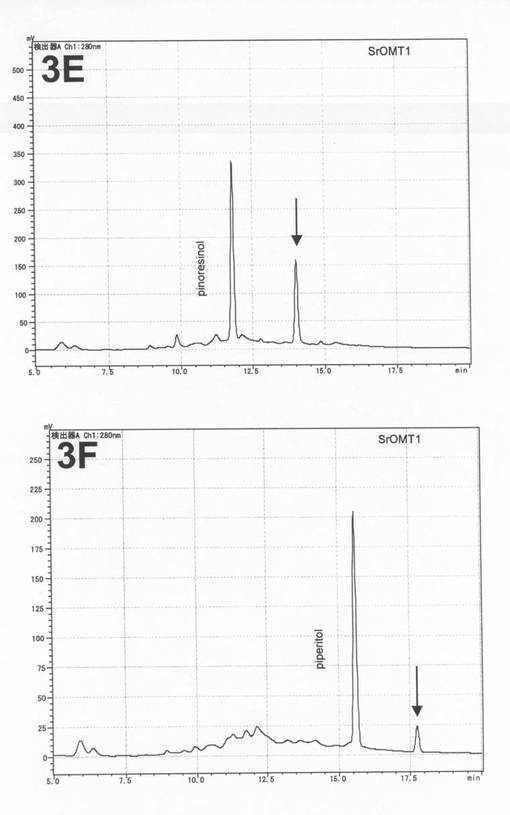
【図6】
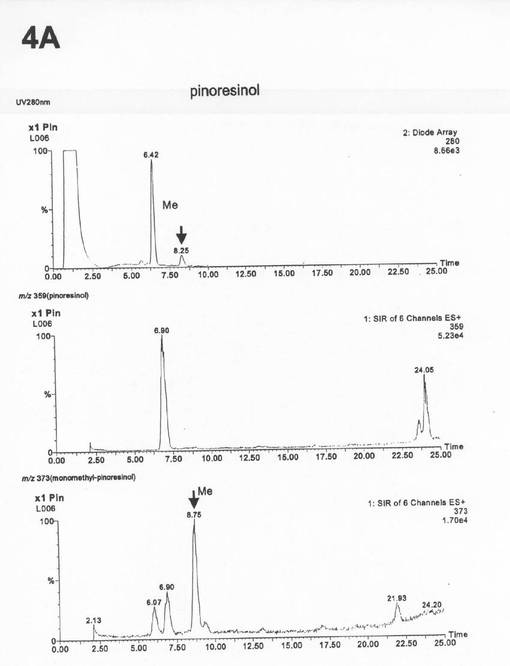
【図7】
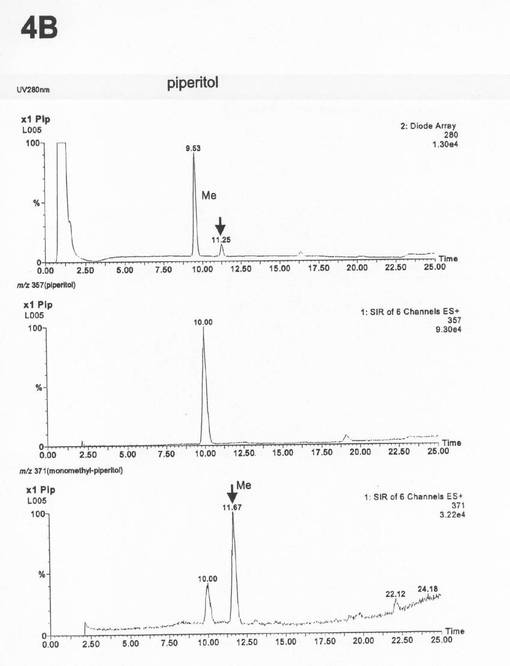
【図8】
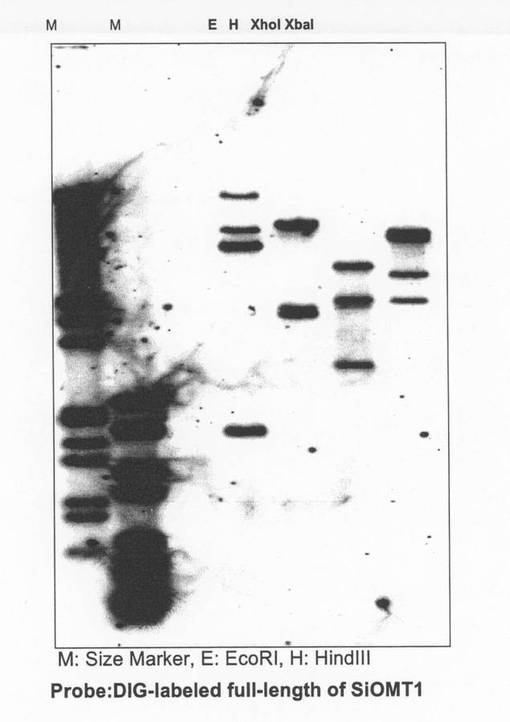
【図9】
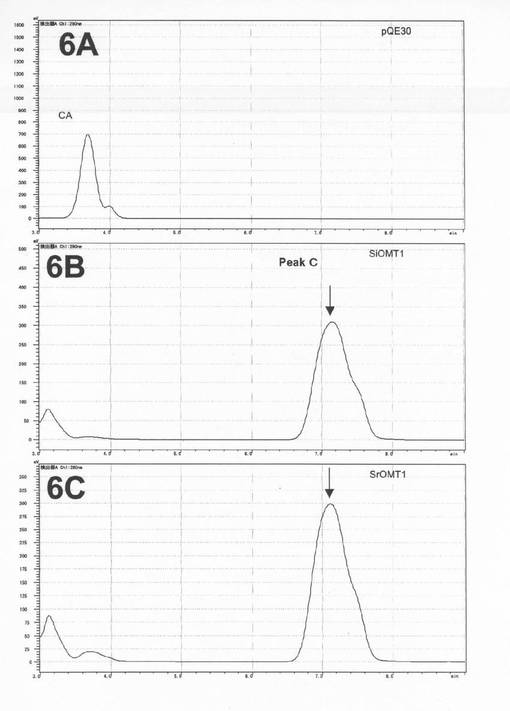
【図10】
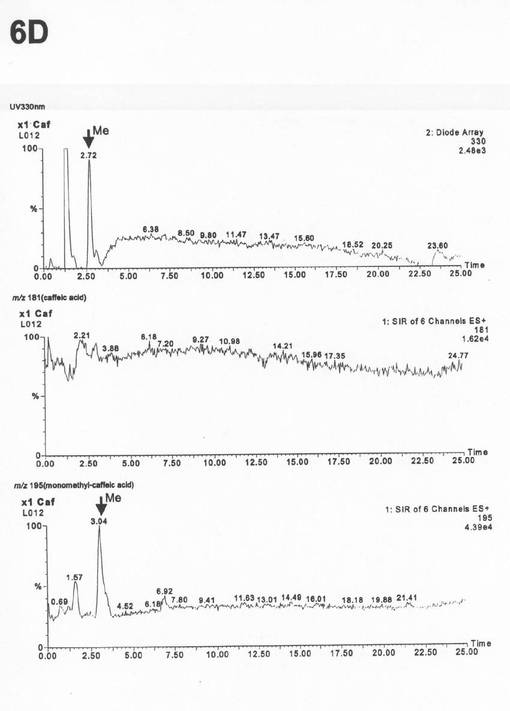
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2007−259789(P2007−259789A)
【公開日】平成19年10月11日(2007.10.11)
【国際特許分類】
【出願番号】特願2006−90877(P2006−90877)
【出願日】平成18年3月29日(2006.3.29)
【出願人】(000001904)サントリー株式会社 (319)
【Fターム(参考)】
【公開日】平成19年10月11日(2007.10.11)
【国際特許分類】
【出願日】平成18年3月29日(2006.3.29)
【出願人】(000001904)サントリー株式会社 (319)
【Fターム(参考)】
[ Back to top ]