
リグニンペルオキシダーゼ変異体
【課題】耐熱性及び耐過酸化水素性に優れたリグニンペルオキシダーゼ変異体を提供する。
【解決手段】リグニンペルオキシダーゼ変異体は、白色腐朽菌由来の特定のアミノ酸配列、又は該アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなる。リグニンペルオキシダーゼ変異体は、前記のアミノ酸配列からなる組換えタンパク質である。
【解決手段】リグニンペルオキシダーゼ変異体は、白色腐朽菌由来の特定のアミノ酸配列、又は該アミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなる。リグニンペルオキシダーゼ変異体は、前記のアミノ酸配列からなる組換えタンパク質である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、白色腐朽菌に由来するリグニンペルオキシダーゼ変異体に関する。
【背景技術】
【0002】
近年、自動車用燃料として、ガソリン−エタノール混合燃料を用いることが検討されている。前記エタノールとしては、植物性物質の発酵、蒸留により得たバイオエタノールを用いると、土壌管理を厳密に行うことにより所謂カーボンニュートラル効果を得ることができ、二酸化炭素の排出量を低減して地球温暖化の防止に寄与できるものと考えられている。
【0003】
しかし、前記植物性物質として、例えばサトウキビ、トウモロコシ等の農作物を用いると、該農作物がエタノールの原料として大量に消費されることにより、食料としての供給量が減少するという問題がある。そこで、前記植物性物質として、食用ではないリグノセルロース系バイオマスを用いてエタノールを製造する技術が検討されている。前記リグノセルロース系バイオマスは、セルロースを含んでおり、該セルロースを酵素糖化によりグルコースに分解し、得られたグルコースを発酵させてバイオエタノールを得ることができる。
【0004】
ところが、前記リグノセルロースは、セルロースの他にヘミセルロース及びリグニンを主な構成成分としており、通常該セルロース及び該ヘミセルロースは、該リグニンにより強固に結合されている。従って、前記リグノセルロースはそのままでは前記酵素糖化反応が阻害されることとなり、予め前記リグニンを除去しておくことが望ましい。
【0005】
前記リグニンを除去する方法として、微生物または微生物が産生する酵素を用いることが考えられる。前記リグニンを分解する微生物としては白色腐朽菌が知られており、白色腐朽菌はリグニン分解酵素であるリグニンペルオキシダーゼを産生することが知られている(例えば、特許文献1参照)。
【0006】
前記リグニンペルオキシダーゼは、前記糖化前処理としてのリグニン除去の他、製紙工業におけるパルプ漂白の効率化、染料の分解、ダイオキシン等の難分解性環境汚染物質の浄化等のように幅広い産業に応用できる可能性を備えている(例えば、特許文献2,3参照)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2002−142772号公報
【特許文献2】特開平9−95885号公報
【特許文献3】特開2006−333758号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
一般に、酵素を工業的に利用する場合には、室温以上の高温で作用させることが好ましい。しかしながら、前記リグニンペルオキシダーゼは、耐熱性が低く、室温以上の高温、例えば37℃程度の温度で失活するという不都合がある。
【0009】
また、前記リグニンペルオキシダーゼは、過酸化水素存在下で前記リグニンを分解する酵素であるにも拘わらず、過酸化水素に対する耐性が低いという不都合がある。
【0010】
そこで、本発明は、かかる不都合を解消して、耐熱性及び耐過酸化水素性に優れたリグニンペルオキシダーゼ変異体を提供することを目的とする。
【課題を解決するための手段】
【0011】
かかる目的を達成するために、本発明のリグニンペルオキシダーゼ変異体は、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とする。
【0012】
前記第1のアミノ酸配列は、配列番号3で表されるアミノ酸配列からなる野生型のリグニンペルオキシダーゼにおいて、240番目のHをFに、241番目のTをLに、242番目のIをLに、それぞれアミノ酸置換したものである。この結果、本発明のリグニンペルオキシダーゼ変異体は、優れた耐熱性及び耐過酸化水素性を得ることができる。
【0013】
本発明のリグニンペルオキシダーゼ変異体は、前記第1のアミノ酸配列又は第2のアミノ酸配列からなる組換えタンパク質である。
【0014】
また、本発明は、前記組換えタンパク質をコードすることを特徴とする遺伝子にもある。本発明の遺伝子は、配列番号2で表される塩基配列からなる遺伝子であることが好ましい。
【0015】
本発明のリグニンペルオキシダーゼ変異体は、前記遺伝子を含有する組換えベクターを形質転換体に導入することにより発現させることができる。
【図面の簡単な説明】
【0016】
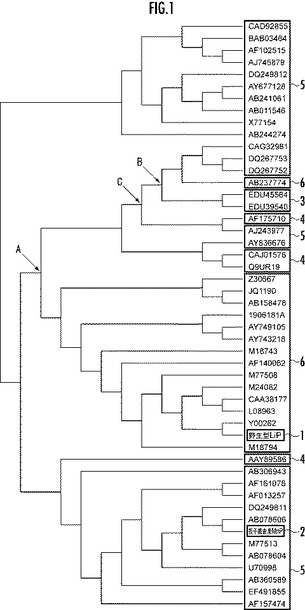
【図1】野生型のリグニンペルオキシダーゼと相同なアミノ酸配列からなるタンパク質48個の系統樹を示す図。
【図2】野生型のリグニンペルオキシダーゼのアミノ酸配列と、推定された先祖型のアミノ酸配列との多重配列を示す図。
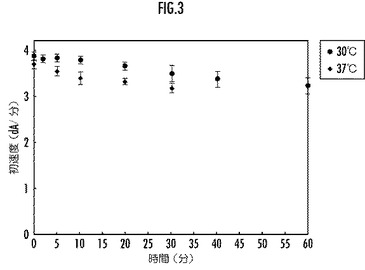
【図3】本発明のリグニンペルオキシダーゼ変異体の30℃と37℃とにおける酵素活性の経時変化を示すグラフ。
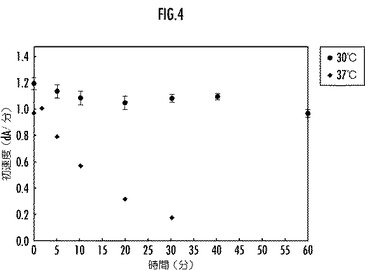
【図4】参考例としての野生型のリグニンペルオキシダーゼの30℃と37℃とにおける酵素活性の経時変化を示すグラフ。
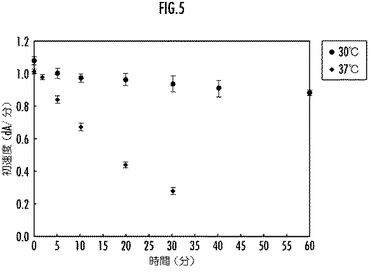
【図5】比較例としてのリグニンペルオキシダーゼ変異体の30℃と37℃とにおける酵素活性の経時変化を示すグラフ。
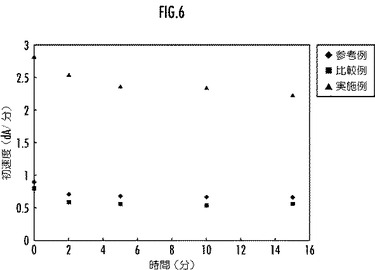
【図6】本発明及び比較例としてのリグニンペルオキシダーゼ変異体と、参考例としての野生型のリグニンペルオキシダーゼとの過酸化水素存在下における酵素活性の経時変化を示すグラフ。
【発明を実施するための形態】
【0017】
次に、添付の図面を参照しながら本発明の実施の形態についてさらに詳しく説明する。
【0018】
本実施形態のリグニンペルオキシダーゼ変異体は、白色腐朽菌Phanerochaete chrysosporium UAMH 3641由来の野生型のリグニンペルオキシダーゼの変異体であり、配列番号1で表されるアミノ酸配列からなる。前記野生型のリグニンペルオキシダーゼは、配列番号3で表されるアミノ酸配列からなる。
【0019】
本実施形態のリグニンペルオキシダーゼ変異体の前記配列番号1で表されるアミノ酸配列は、配列番号3で表されるアミノ酸配列において、240番目のHをFに、241番目のTをLに、242番目のIをLに、それぞれアミノ酸置換したものである。また、本実施形態のリグニンペルオキシダーゼ変異体は、配列番号2で表される塩基配列からなる遺伝子によりコードされ、前記野生型のリグニンペルオキシダーゼは、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0020】
前記野生型のリグニンペルオキシダーゼは、耐熱性及び耐過酸化水素性を備えていない。しかし、ウーズ(Woese)らによって示された16srRNAによる系統樹を見ると、80℃以上で至適に生育する生物が該系統樹の根元に多いことが示されている。この事実から、真正細菌、真核生物、古細菌の共通の祖先は超好熱菌と推定され、先祖型のアミノ酸配列を有するか又はそれに近い配列を有するタンパク質は耐熱性を備えているものと考えられる。このような知見は、特開2002−247991号公報に記載されている。
【0021】
そこで、本実施形態では、リグニンを分解する酵素を含み、前記野生型のリグニンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質に関する系統樹を作成し、該系統樹における先祖型のアミノ酸配列について検討した。
【0022】
前記系統樹の作成に当たって、まず、リグニンを分解する酵素を含み、前記野生型のリグニンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質48個をDDBJ内のアミノ酸配列データベースからソフトウェアBLASTPを用いて抽出した。抽出したタンパク質は、リグニンを分解する酵素として、前記野生型のリグニンペルオキシダーゼの他、子のう菌由来のリグニンペルオキシダーゼ、担子菌由来のマンガンペルオキシダーゼ、リグニンペルオキシダーゼとマンガンペルオキシダーゼとのハイブリッド型酵素であるヴァーサタイルタイルペルオキシダーゼを含んでいる。尚、本明細書では、リグニンペルオキシダーゼを「LiP」、マンガンペルオキシダーゼを「MnP」、ヴァ−サタイルペルオキシダーゼ「VP」と略記することがある。
【0023】
次に、前記野生型LiP及び抽出したタンパク質の合わせて49個のタンパク質について、進化の系統を示す系統樹を作成した。前記系統樹は、ソフトウェアTreefinderとPhyMLとを用いて候補系統樹群を作成し、該候補系統樹群の各候補系統樹について、ソフトウェアPMALのCODEMLを用いて尤度を計算した。そして、最尤度を備える候補系統樹を前記49個のタンパク質の進化の系統を示す系統樹として選択した。選択した系統樹を図1に示す。
【0024】
図1において、符号1は野生型LiP、符号2は担子菌由来MnP、符号3は子のう菌由来LiP、符号4はVPを示す。また、符号5はMnPを示し、符号6はLiPを示す。抽出したタンパク質のうち、野生型LiP1及び担子菌由来MnP2を除く47個のタンパク質は論文等に発表されており、図1に示すDDBJ accession numbersにより特定することができる。
【0025】
また、前記担子菌由来MnPは、配列番号5で表されるアミノ酸配列からなり、配列番号6で表される塩基配列からなる遺伝子によりコードされる。
【0026】
図1において、分岐点Aは前記野生型LiPと他のタンパク質との分岐点であり、酵素の機能に関する分岐点である。また、分岐点Bは子のう菌と担子菌との分岐点であり、生物の種に関する分岐点である。
【0027】
そこで、次に、各分岐点A,Bのそれぞれに対応するタンパク質と、分岐点A,Bの中間の分岐点Cに対応するタンパク質とのアミノ酸配列をソフトウェアPMALのCODEMLにより推定し、前記野生型LiPのアミノ酸配列と、推定された3種のアミノ酸配列とをマルチプルアライメント(多重配列)した。前記マルチプルアライメントの結果を図2に示す。尚、前記野生型LiPは前記配列番号3で表されるアミノ酸配列からなり、分岐点A,B,Cに対応するタンパク質は、それぞれ配列番号7,8,9で表されるアミノ酸配列からなる。
【0028】
図2から、前記野生型LiPのアミノ酸配列の240〜242番目がH,T,Iであるのに対し、分岐点A,B,Cに対応するタンパク質のアミノ酸配列では、いずれもF,L,Lとなっていることが明らかである。そこで、前記野生型LiPのアミノ酸配列(配列番号3)中、240番目のHをFに、241番目のTをLに、242番目のIをLに、それぞれアミノ酸置換したアミノ酸配列を本実施形態の先祖型LiP変異体のアミノ酸配列(配列番号1)とした。
【0029】
次に、前記のようにして決定した先祖型LiP変異体のアミノ酸配列に基づいて、該アミノ酸配列をコードする遺伝子の構築を行った。
【0030】
前記遺伝子の構築のために、まず、前記野生型のリグニンペルオキシダーゼのcDNAを、Gateway system(商品名、Invitrogen社)を用いて、プラスミドであるpPDB−GBに導入し、ベクターとした。前記ベクターを、本明細書ではpPDB−LiPと記載する。尚、前記野生型LiPのcDNAは、白色腐朽菌Phanerochaete chrysosporium UAMH 3641由来の遺伝子であり、麻布大学環境生命科学部教授其木茂則氏から分譲を受けた。
【0031】
次に、前記pPDB−LiPを鋳型として、次のプライマーと、DNAポリメラーゼ(TaKaRa Ex Taq(登録商標、タカラバイオ株式会社))とを用いてPCRを行い、該pPDB−LiPを増幅させた。
〔プライマー〕
pLiPf-S(配列番号12):5’−GGAGATATACATATGGCCACCTGTTCCAACGGCAA−3’
pLiPr(配列番号13):GTGCGGCCGCAAGCTTAAGCACCCGGAGGCGGAGGG−3’
前記PCRは、まず、前記pPDB−LiPと、前記プライマーと、前記DNAポリメラーゼとの混合物を、94℃の温度に3分間保持した後、さらに94℃の温度に30秒保持して熱変性処理した。次に、55℃に1分間保持してアニーリング反応を行った後、72℃の温度に2分間保持して伸長反応を行った。そして、前記熱変性処理と、アニーリング反応と、伸長反応とを1サイクルとして、このサイクルを25回繰り返した。
【0032】
次に、前記PCRにより増幅させた前記pPDB−LiPを制限酵素Nde IとHind IIIとで処理し、前記野生型LiPのcDNAに対応する遺伝子からなるDNA断片を得た。次に、前記DNA断片を、プラスミドであるpET21a(+)に導入し、ベクターとした。前記ベクターを、本明細書ではpET−LiPと記載する。そして、前記ベクターをEscherichia coli JM109を用いて増幅した。
【0033】
次に、前記増幅により得られたpET−LiPを鋳型として、次のプライマーと、QuickChange Lightning Site-Directed Mutagenesis Kit(商品名、Stratagene社))とを用いてPCRを行った。この結果、先祖型リグニンペルオキシダーゼ変異体のアミノ酸配列(配列番号1)をコードする遺伝子(配列番号2)を含むプラスミドが得られた。
〔プライマー〕
forward primer(配列番号14):5’−CGCATCCAGTCCGACTTCCTGCTGCCCGCGACTCACGC−3’
reverse primer(配列番号15):5’−GCGTGAGTCGCGGGCCAGCAGGAAGTCGGACTGGATGCG−3’
尚、前記プライマーにおいて、下線部が前記アミノ酸置換に関わる部分である。
【0034】
また、図2から、LiPのアミノ酸配列の117,118番目がA,Lであるのに対し、分岐点A,B,Cに対応するタンパク質のアミノ酸配列では、いずれもG,Vとなっていることが明らかである。そこで、LiPのアミノ酸配列中、117番目のAをGに、118番目のLをVに、それぞれアミノ酸置換したアミノ酸配列を比較のための先祖型LiP変異体のアミノ酸配列(配列番号10)とした。前記比較のための先祖型LiP変異体は、配列番号11で表される塩基配列からなる遺伝子によりコードされる。
【0035】
次に、前記のようにして決定した先祖型LiP変異体のアミノ酸配列に基づいて、該アミノ酸配列をコードする遺伝子の構築を行った。前記遺伝子の構築は、pET−LiPを鋳型とするPCRにおいて、次のプライマーを用いた以外は、本実施形態の先祖型LiP変異体のアミノ酸配列(配列番号1)をコードする遺伝子の構築と全く同一にして行った。この結果、比較のための先祖型LiP変異体のアミノ酸配列(配列番号10)をコードする遺伝子を含むプラスミドが得られた。
〔プライマー〕
forward primer(配列番号16):5’−TTCGCTGGTGCTGTCGGTGTTAGCAACTGCCCTGGT−3’
reverse primer(配列番号17):5’−ACCAGGGCAGTTGCTAACACCGACAGCACCAGGGAA−3’
尚、前記プライマーにおいて、下線部が前記アミノ酸置換に関わる部分である。
【0036】
次に、本実施形態及び比較のための先祖型LiP変異体のアミノ酸配列をコードする遺伝子を含むプラスミドをそれぞれベクターとすると共に、前記野生型LiPのcDNAに対応する遺伝子を導入したベクターとしてのpET−LiPを用い、各ベクターに含まれる遺伝子を発現させた。前記各遺伝子の発現は、前記各ベクターをそれぞれEscherichia coli BL21(DE3)に導入することにより次のようにして行った。
【0037】
まず、前記各ベクターを含むEscherichia coli BL21をアンピシリン0.1mg/mLを含むTerrific Btoth培地中、37℃の温度に保持して12時間培養した。前記Terrific Btoth培地は、1リットル当たりポリペプトン12g、酵母エキストラクト24g、グリセロール4ml、KH2PO42.31g、K2HPO412.5gを含む。
【0038】
前記培養後、前記培地にIPTGを添加して該培地中のIPTGの濃度が0.5mMになるようにして、タンパク質の発現を誘導し、さらに4時間培養した。このようにすることにより、前記Escherichia coli BL21の細胞内に、前記野生型LiP又はその変異体が産生される。
【0039】
その後、前記Escherichia coli BL21の菌体を回収し、第1の緩衝液中で超音波破砕した。前記第1の緩衝液は、10mM−Tris-HCl(pH8)、1mM−EDTA、5mM−DTT、50μM−フッ化フェニルメチルスルフォニル、1容量%−非イオン性界面活性剤(TritonX−100(商品名))を含む。
【0040】
前記野生型LiP又はその変異体は、いずれも封入体として発現されるので、前記第1の緩衝液中の不溶画分を回収し、該不溶画分1gに対して第2の緩衝液10mlで3回洗浄した。前記第2の緩衝液は、10mM−Tris-HCl(pH8)、1mM−EDTA、5mM−DTT、1容量%−非イオン性界面活性剤(TritonX−100(商品名))を含む。
【0041】
次に、前記不溶画分を、8Mの尿素と2mMのDTTとを含む水溶液中に室温で1時間保持し、前記野生型LiP又はその変異体を変性還元状態にした。次に、前記水溶液1容に、10mM−CaCl2、0.7mM−酸化グルタチオン、30μM−ヘミン、50mM−Tris-HCl(pH8)の混合物を3容の体積割合で加え、4倍に希釈し、攪拌しながら暗所に室温で12時間保持することにより、前記野生型LiP又はその変異体を活性型にした。
【0042】
次に、10mM−酢酸ナトリウム緩衝液(pH6)、5mM−CaCl2を含む溶液を用いて、前記野生型LiP又はその変異体を含む水溶液を、4℃の温度で12時間透析した。次に、前記透析により得られた溶液を、60000×gで20分間遠心分離し、上清を濾過膜(amicon YM-10(商品名、Millipore社))により濾過して濃縮した。
【0043】
次に、前記濃縮された溶液を、陰イオン交換カラム(DEAE Sepharose Fast Flow anion-exchange column(商品名、GE Healthcare Bioscience社)で分離し、前記野生型LiP又はその変異体を精製した。移動相は、10mM−酢酸ナトリウム緩衝液(pH6)、0.6M−NaCl、1mM−CaCl2を用いた。
【0044】
次に、前記陰イオン交換カラムで分離、精製された前記野生型LiP又はその変異体を含む液を、10mM−酢酸ナトリウム緩衝液(pH6)、1mM−CaCl2で3倍に希釈後、さらに、陰イオン交換カラム(HiTrapQ(商品名、GE Healthcare Bioscience社)で分離した。移動相は、10mM−酢酸ナトリウム緩衝液(pH6)、0.01〜0.6M−NaCl、1mM−CaCl2を用いた。この結果、活性を有する前記野生型LiP又はその変異体を得た。前記野生型LiP又はその変異体は、配列番号1で表されるアミノ酸配列からなる本実施形態のLiP変異体(実施例)、配列番号9で表されるアミノ酸配列からなる比較のためのLiP変異体(比較例)、配列番号3で表されるアミノ酸配列からなる野生型LiP(参考例)である。
【0045】
次に、前記野生型LiP又はその変異体の耐熱性を評価した。前記評価は、酒石酸の終濃度が50mMであると共に、前記野生型LiP又はその変異体の終濃度が200nMである溶液3mlを調製し、該溶液を30℃又は37℃の温度に保持したときのリグニン分解酵素活性の経時変化を測定することにより行った。前記リグニン分解酵素活性の経時変化は、所定時間後の残存活性を示す。
【0046】
前記リグニン分解酵素活性の残存活性の測定には、ベラトリルアルコールの酸化によるベラトリルアルデヒドの生成反応を用いた。具体的には、酒石酸の終濃度が50mM、前記野生型LiP又はその変異体の終濃度が20nMであると共に、ベラトリアルアルコールの終濃度が2mMである溶液0.5mlを調製し、終濃度が0.5mMになるようにH2O2を滴下して反応を開始した。ベラトリルアルデヒドの濃度は、310nmの波長の光線による吸光度で測定し、吸光係数9300M−1cm−1として算出した。前記条件で、1分当たり1μモルのベラトリルアルデヒドを生成する酵素活性を1enzyme unit と定義した。
【0047】
前記リグニンペルオキシダーゼ又はその変異体のリグニン分解酵素活性の経時変化を、図3(実施例)、図4(比較例)、図5(参考例)にそれぞれ示す。
【0048】
図3乃至図5から、本実施例のLiP変異体は、30℃及び37℃の温度におけるリグニン分解酵素活性について、所定時間後の残存活性が参考例の野生型LiPよりも大であり、優れた耐熱性を備えていることが明らかである。一方、配列番号3で表されるアミノ酸配列の他の部分をアミノ酸置換した比較例のLiP変異体は、30℃の温度におけるリグニン分解酵素活性については、所定時間後の残存活性が参考例の野生型LiPより大であるものの、37℃の温度では参考例の野生型LiPよりも小であり、十分な耐熱性を備えていないことが明らかである。
【0049】
次に、前記野生型LiP又はその変異体の耐過酸化水素性を評価した。前記評価は、酒石酸の終濃度が50mM、前記野生型LiP又はその変異体の終濃度が200nMであると共に、H2O2の終濃度が0.01mMである溶液3mlを調製し、該溶液を25℃の温度に保持したときのリグニン分解酵素活性の経時変化を測定することにより行った。
【0050】
前記リグニン分解酵素活性の残存活性の測定は、前記耐熱性評価の場合と全く同一にして行った。前記野生型LiP又はその変異体のリグニン分解酵素活性の経時変化を、図6に示す。
【0051】
図6から、本実施例のLiP変異体は、0.01mM−H2O2の存在下におけるリグニン分解酵素活性について、所定時間後の残存活性が参考例の野生型LiPよりも大であり、優れた耐過酸化水素性を備えていることが明らかである。一方、配列番号3で表されるアミノ酸配列の他の部分をアミノ酸置換した比較例のLiP変異体は、0.01mM−H2O2の存在下におけるリグニン分解酵素活性について、所定時間後の残存活性が参考例の野生型LiPよりも小であり、耐過酸化水素性が劣っていることが明らかである。
【0052】
尚、本実施形態では、本実施例のLiP変異体を配列番号1のアミノ酸配列で表されるものとしているが、配列番号1のアミノ酸配列において1又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列で表されるものであってもよい。
【技術分野】
【0001】
本発明は、白色腐朽菌に由来するリグニンペルオキシダーゼ変異体に関する。
【背景技術】
【0002】
近年、自動車用燃料として、ガソリン−エタノール混合燃料を用いることが検討されている。前記エタノールとしては、植物性物質の発酵、蒸留により得たバイオエタノールを用いると、土壌管理を厳密に行うことにより所謂カーボンニュートラル効果を得ることができ、二酸化炭素の排出量を低減して地球温暖化の防止に寄与できるものと考えられている。
【0003】
しかし、前記植物性物質として、例えばサトウキビ、トウモロコシ等の農作物を用いると、該農作物がエタノールの原料として大量に消費されることにより、食料としての供給量が減少するという問題がある。そこで、前記植物性物質として、食用ではないリグノセルロース系バイオマスを用いてエタノールを製造する技術が検討されている。前記リグノセルロース系バイオマスは、セルロースを含んでおり、該セルロースを酵素糖化によりグルコースに分解し、得られたグルコースを発酵させてバイオエタノールを得ることができる。
【0004】
ところが、前記リグノセルロースは、セルロースの他にヘミセルロース及びリグニンを主な構成成分としており、通常該セルロース及び該ヘミセルロースは、該リグニンにより強固に結合されている。従って、前記リグノセルロースはそのままでは前記酵素糖化反応が阻害されることとなり、予め前記リグニンを除去しておくことが望ましい。
【0005】
前記リグニンを除去する方法として、微生物または微生物が産生する酵素を用いることが考えられる。前記リグニンを分解する微生物としては白色腐朽菌が知られており、白色腐朽菌はリグニン分解酵素であるリグニンペルオキシダーゼを産生することが知られている(例えば、特許文献1参照)。
【0006】
前記リグニンペルオキシダーゼは、前記糖化前処理としてのリグニン除去の他、製紙工業におけるパルプ漂白の効率化、染料の分解、ダイオキシン等の難分解性環境汚染物質の浄化等のように幅広い産業に応用できる可能性を備えている(例えば、特許文献2,3参照)。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】特開2002−142772号公報
【特許文献2】特開平9−95885号公報
【特許文献3】特開2006−333758号公報
【発明の概要】
【発明が解決しようとする課題】
【0008】
一般に、酵素を工業的に利用する場合には、室温以上の高温で作用させることが好ましい。しかしながら、前記リグニンペルオキシダーゼは、耐熱性が低く、室温以上の高温、例えば37℃程度の温度で失活するという不都合がある。
【0009】
また、前記リグニンペルオキシダーゼは、過酸化水素存在下で前記リグニンを分解する酵素であるにも拘わらず、過酸化水素に対する耐性が低いという不都合がある。
【0010】
そこで、本発明は、かかる不都合を解消して、耐熱性及び耐過酸化水素性に優れたリグニンペルオキシダーゼ変異体を提供することを目的とする。
【課題を解決するための手段】
【0011】
かかる目的を達成するために、本発明のリグニンペルオキシダーゼ変異体は、配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とする。
【0012】
前記第1のアミノ酸配列は、配列番号3で表されるアミノ酸配列からなる野生型のリグニンペルオキシダーゼにおいて、240番目のHをFに、241番目のTをLに、242番目のIをLに、それぞれアミノ酸置換したものである。この結果、本発明のリグニンペルオキシダーゼ変異体は、優れた耐熱性及び耐過酸化水素性を得ることができる。
【0013】
本発明のリグニンペルオキシダーゼ変異体は、前記第1のアミノ酸配列又は第2のアミノ酸配列からなる組換えタンパク質である。
【0014】
また、本発明は、前記組換えタンパク質をコードすることを特徴とする遺伝子にもある。本発明の遺伝子は、配列番号2で表される塩基配列からなる遺伝子であることが好ましい。
【0015】
本発明のリグニンペルオキシダーゼ変異体は、前記遺伝子を含有する組換えベクターを形質転換体に導入することにより発現させることができる。
【図面の簡単な説明】
【0016】
【図1】野生型のリグニンペルオキシダーゼと相同なアミノ酸配列からなるタンパク質48個の系統樹を示す図。
【図2】野生型のリグニンペルオキシダーゼのアミノ酸配列と、推定された先祖型のアミノ酸配列との多重配列を示す図。
【図3】本発明のリグニンペルオキシダーゼ変異体の30℃と37℃とにおける酵素活性の経時変化を示すグラフ。
【図4】参考例としての野生型のリグニンペルオキシダーゼの30℃と37℃とにおける酵素活性の経時変化を示すグラフ。
【図5】比較例としてのリグニンペルオキシダーゼ変異体の30℃と37℃とにおける酵素活性の経時変化を示すグラフ。
【図6】本発明及び比較例としてのリグニンペルオキシダーゼ変異体と、参考例としての野生型のリグニンペルオキシダーゼとの過酸化水素存在下における酵素活性の経時変化を示すグラフ。
【発明を実施するための形態】
【0017】
次に、添付の図面を参照しながら本発明の実施の形態についてさらに詳しく説明する。
【0018】
本実施形態のリグニンペルオキシダーゼ変異体は、白色腐朽菌Phanerochaete chrysosporium UAMH 3641由来の野生型のリグニンペルオキシダーゼの変異体であり、配列番号1で表されるアミノ酸配列からなる。前記野生型のリグニンペルオキシダーゼは、配列番号3で表されるアミノ酸配列からなる。
【0019】
本実施形態のリグニンペルオキシダーゼ変異体の前記配列番号1で表されるアミノ酸配列は、配列番号3で表されるアミノ酸配列において、240番目のHをFに、241番目のTをLに、242番目のIをLに、それぞれアミノ酸置換したものである。また、本実施形態のリグニンペルオキシダーゼ変異体は、配列番号2で表される塩基配列からなる遺伝子によりコードされ、前記野生型のリグニンペルオキシダーゼは、配列番号4で表される塩基配列からなる遺伝子によりコードされる。
【0020】
前記野生型のリグニンペルオキシダーゼは、耐熱性及び耐過酸化水素性を備えていない。しかし、ウーズ(Woese)らによって示された16srRNAによる系統樹を見ると、80℃以上で至適に生育する生物が該系統樹の根元に多いことが示されている。この事実から、真正細菌、真核生物、古細菌の共通の祖先は超好熱菌と推定され、先祖型のアミノ酸配列を有するか又はそれに近い配列を有するタンパク質は耐熱性を備えているものと考えられる。このような知見は、特開2002−247991号公報に記載されている。
【0021】
そこで、本実施形態では、リグニンを分解する酵素を含み、前記野生型のリグニンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質に関する系統樹を作成し、該系統樹における先祖型のアミノ酸配列について検討した。
【0022】
前記系統樹の作成に当たって、まず、リグニンを分解する酵素を含み、前記野生型のリグニンペルオキシダーゼと相同のアミノ酸配列を備えるタンパク質48個をDDBJ内のアミノ酸配列データベースからソフトウェアBLASTPを用いて抽出した。抽出したタンパク質は、リグニンを分解する酵素として、前記野生型のリグニンペルオキシダーゼの他、子のう菌由来のリグニンペルオキシダーゼ、担子菌由来のマンガンペルオキシダーゼ、リグニンペルオキシダーゼとマンガンペルオキシダーゼとのハイブリッド型酵素であるヴァーサタイルタイルペルオキシダーゼを含んでいる。尚、本明細書では、リグニンペルオキシダーゼを「LiP」、マンガンペルオキシダーゼを「MnP」、ヴァ−サタイルペルオキシダーゼ「VP」と略記することがある。
【0023】
次に、前記野生型LiP及び抽出したタンパク質の合わせて49個のタンパク質について、進化の系統を示す系統樹を作成した。前記系統樹は、ソフトウェアTreefinderとPhyMLとを用いて候補系統樹群を作成し、該候補系統樹群の各候補系統樹について、ソフトウェアPMALのCODEMLを用いて尤度を計算した。そして、最尤度を備える候補系統樹を前記49個のタンパク質の進化の系統を示す系統樹として選択した。選択した系統樹を図1に示す。
【0024】
図1において、符号1は野生型LiP、符号2は担子菌由来MnP、符号3は子のう菌由来LiP、符号4はVPを示す。また、符号5はMnPを示し、符号6はLiPを示す。抽出したタンパク質のうち、野生型LiP1及び担子菌由来MnP2を除く47個のタンパク質は論文等に発表されており、図1に示すDDBJ accession numbersにより特定することができる。
【0025】
また、前記担子菌由来MnPは、配列番号5で表されるアミノ酸配列からなり、配列番号6で表される塩基配列からなる遺伝子によりコードされる。
【0026】
図1において、分岐点Aは前記野生型LiPと他のタンパク質との分岐点であり、酵素の機能に関する分岐点である。また、分岐点Bは子のう菌と担子菌との分岐点であり、生物の種に関する分岐点である。
【0027】
そこで、次に、各分岐点A,Bのそれぞれに対応するタンパク質と、分岐点A,Bの中間の分岐点Cに対応するタンパク質とのアミノ酸配列をソフトウェアPMALのCODEMLにより推定し、前記野生型LiPのアミノ酸配列と、推定された3種のアミノ酸配列とをマルチプルアライメント(多重配列)した。前記マルチプルアライメントの結果を図2に示す。尚、前記野生型LiPは前記配列番号3で表されるアミノ酸配列からなり、分岐点A,B,Cに対応するタンパク質は、それぞれ配列番号7,8,9で表されるアミノ酸配列からなる。
【0028】
図2から、前記野生型LiPのアミノ酸配列の240〜242番目がH,T,Iであるのに対し、分岐点A,B,Cに対応するタンパク質のアミノ酸配列では、いずれもF,L,Lとなっていることが明らかである。そこで、前記野生型LiPのアミノ酸配列(配列番号3)中、240番目のHをFに、241番目のTをLに、242番目のIをLに、それぞれアミノ酸置換したアミノ酸配列を本実施形態の先祖型LiP変異体のアミノ酸配列(配列番号1)とした。
【0029】
次に、前記のようにして決定した先祖型LiP変異体のアミノ酸配列に基づいて、該アミノ酸配列をコードする遺伝子の構築を行った。
【0030】
前記遺伝子の構築のために、まず、前記野生型のリグニンペルオキシダーゼのcDNAを、Gateway system(商品名、Invitrogen社)を用いて、プラスミドであるpPDB−GBに導入し、ベクターとした。前記ベクターを、本明細書ではpPDB−LiPと記載する。尚、前記野生型LiPのcDNAは、白色腐朽菌Phanerochaete chrysosporium UAMH 3641由来の遺伝子であり、麻布大学環境生命科学部教授其木茂則氏から分譲を受けた。
【0031】
次に、前記pPDB−LiPを鋳型として、次のプライマーと、DNAポリメラーゼ(TaKaRa Ex Taq(登録商標、タカラバイオ株式会社))とを用いてPCRを行い、該pPDB−LiPを増幅させた。
〔プライマー〕
pLiPf-S(配列番号12):5’−GGAGATATACATATGGCCACCTGTTCCAACGGCAA−3’
pLiPr(配列番号13):GTGCGGCCGCAAGCTTAAGCACCCGGAGGCGGAGGG−3’
前記PCRは、まず、前記pPDB−LiPと、前記プライマーと、前記DNAポリメラーゼとの混合物を、94℃の温度に3分間保持した後、さらに94℃の温度に30秒保持して熱変性処理した。次に、55℃に1分間保持してアニーリング反応を行った後、72℃の温度に2分間保持して伸長反応を行った。そして、前記熱変性処理と、アニーリング反応と、伸長反応とを1サイクルとして、このサイクルを25回繰り返した。
【0032】
次に、前記PCRにより増幅させた前記pPDB−LiPを制限酵素Nde IとHind IIIとで処理し、前記野生型LiPのcDNAに対応する遺伝子からなるDNA断片を得た。次に、前記DNA断片を、プラスミドであるpET21a(+)に導入し、ベクターとした。前記ベクターを、本明細書ではpET−LiPと記載する。そして、前記ベクターをEscherichia coli JM109を用いて増幅した。
【0033】
次に、前記増幅により得られたpET−LiPを鋳型として、次のプライマーと、QuickChange Lightning Site-Directed Mutagenesis Kit(商品名、Stratagene社))とを用いてPCRを行った。この結果、先祖型リグニンペルオキシダーゼ変異体のアミノ酸配列(配列番号1)をコードする遺伝子(配列番号2)を含むプラスミドが得られた。
〔プライマー〕
forward primer(配列番号14):5’−CGCATCCAGTCCGACTTCCTGCTGCCCGCGACTCACGC−3’
reverse primer(配列番号15):5’−GCGTGAGTCGCGGGCCAGCAGGAAGTCGGACTGGATGCG−3’
尚、前記プライマーにおいて、下線部が前記アミノ酸置換に関わる部分である。
【0034】
また、図2から、LiPのアミノ酸配列の117,118番目がA,Lであるのに対し、分岐点A,B,Cに対応するタンパク質のアミノ酸配列では、いずれもG,Vとなっていることが明らかである。そこで、LiPのアミノ酸配列中、117番目のAをGに、118番目のLをVに、それぞれアミノ酸置換したアミノ酸配列を比較のための先祖型LiP変異体のアミノ酸配列(配列番号10)とした。前記比較のための先祖型LiP変異体は、配列番号11で表される塩基配列からなる遺伝子によりコードされる。
【0035】
次に、前記のようにして決定した先祖型LiP変異体のアミノ酸配列に基づいて、該アミノ酸配列をコードする遺伝子の構築を行った。前記遺伝子の構築は、pET−LiPを鋳型とするPCRにおいて、次のプライマーを用いた以外は、本実施形態の先祖型LiP変異体のアミノ酸配列(配列番号1)をコードする遺伝子の構築と全く同一にして行った。この結果、比較のための先祖型LiP変異体のアミノ酸配列(配列番号10)をコードする遺伝子を含むプラスミドが得られた。
〔プライマー〕
forward primer(配列番号16):5’−TTCGCTGGTGCTGTCGGTGTTAGCAACTGCCCTGGT−3’
reverse primer(配列番号17):5’−ACCAGGGCAGTTGCTAACACCGACAGCACCAGGGAA−3’
尚、前記プライマーにおいて、下線部が前記アミノ酸置換に関わる部分である。
【0036】
次に、本実施形態及び比較のための先祖型LiP変異体のアミノ酸配列をコードする遺伝子を含むプラスミドをそれぞれベクターとすると共に、前記野生型LiPのcDNAに対応する遺伝子を導入したベクターとしてのpET−LiPを用い、各ベクターに含まれる遺伝子を発現させた。前記各遺伝子の発現は、前記各ベクターをそれぞれEscherichia coli BL21(DE3)に導入することにより次のようにして行った。
【0037】
まず、前記各ベクターを含むEscherichia coli BL21をアンピシリン0.1mg/mLを含むTerrific Btoth培地中、37℃の温度に保持して12時間培養した。前記Terrific Btoth培地は、1リットル当たりポリペプトン12g、酵母エキストラクト24g、グリセロール4ml、KH2PO42.31g、K2HPO412.5gを含む。
【0038】
前記培養後、前記培地にIPTGを添加して該培地中のIPTGの濃度が0.5mMになるようにして、タンパク質の発現を誘導し、さらに4時間培養した。このようにすることにより、前記Escherichia coli BL21の細胞内に、前記野生型LiP又はその変異体が産生される。
【0039】
その後、前記Escherichia coli BL21の菌体を回収し、第1の緩衝液中で超音波破砕した。前記第1の緩衝液は、10mM−Tris-HCl(pH8)、1mM−EDTA、5mM−DTT、50μM−フッ化フェニルメチルスルフォニル、1容量%−非イオン性界面活性剤(TritonX−100(商品名))を含む。
【0040】
前記野生型LiP又はその変異体は、いずれも封入体として発現されるので、前記第1の緩衝液中の不溶画分を回収し、該不溶画分1gに対して第2の緩衝液10mlで3回洗浄した。前記第2の緩衝液は、10mM−Tris-HCl(pH8)、1mM−EDTA、5mM−DTT、1容量%−非イオン性界面活性剤(TritonX−100(商品名))を含む。
【0041】
次に、前記不溶画分を、8Mの尿素と2mMのDTTとを含む水溶液中に室温で1時間保持し、前記野生型LiP又はその変異体を変性還元状態にした。次に、前記水溶液1容に、10mM−CaCl2、0.7mM−酸化グルタチオン、30μM−ヘミン、50mM−Tris-HCl(pH8)の混合物を3容の体積割合で加え、4倍に希釈し、攪拌しながら暗所に室温で12時間保持することにより、前記野生型LiP又はその変異体を活性型にした。
【0042】
次に、10mM−酢酸ナトリウム緩衝液(pH6)、5mM−CaCl2を含む溶液を用いて、前記野生型LiP又はその変異体を含む水溶液を、4℃の温度で12時間透析した。次に、前記透析により得られた溶液を、60000×gで20分間遠心分離し、上清を濾過膜(amicon YM-10(商品名、Millipore社))により濾過して濃縮した。
【0043】
次に、前記濃縮された溶液を、陰イオン交換カラム(DEAE Sepharose Fast Flow anion-exchange column(商品名、GE Healthcare Bioscience社)で分離し、前記野生型LiP又はその変異体を精製した。移動相は、10mM−酢酸ナトリウム緩衝液(pH6)、0.6M−NaCl、1mM−CaCl2を用いた。
【0044】
次に、前記陰イオン交換カラムで分離、精製された前記野生型LiP又はその変異体を含む液を、10mM−酢酸ナトリウム緩衝液(pH6)、1mM−CaCl2で3倍に希釈後、さらに、陰イオン交換カラム(HiTrapQ(商品名、GE Healthcare Bioscience社)で分離した。移動相は、10mM−酢酸ナトリウム緩衝液(pH6)、0.01〜0.6M−NaCl、1mM−CaCl2を用いた。この結果、活性を有する前記野生型LiP又はその変異体を得た。前記野生型LiP又はその変異体は、配列番号1で表されるアミノ酸配列からなる本実施形態のLiP変異体(実施例)、配列番号9で表されるアミノ酸配列からなる比較のためのLiP変異体(比較例)、配列番号3で表されるアミノ酸配列からなる野生型LiP(参考例)である。
【0045】
次に、前記野生型LiP又はその変異体の耐熱性を評価した。前記評価は、酒石酸の終濃度が50mMであると共に、前記野生型LiP又はその変異体の終濃度が200nMである溶液3mlを調製し、該溶液を30℃又は37℃の温度に保持したときのリグニン分解酵素活性の経時変化を測定することにより行った。前記リグニン分解酵素活性の経時変化は、所定時間後の残存活性を示す。
【0046】
前記リグニン分解酵素活性の残存活性の測定には、ベラトリルアルコールの酸化によるベラトリルアルデヒドの生成反応を用いた。具体的には、酒石酸の終濃度が50mM、前記野生型LiP又はその変異体の終濃度が20nMであると共に、ベラトリアルアルコールの終濃度が2mMである溶液0.5mlを調製し、終濃度が0.5mMになるようにH2O2を滴下して反応を開始した。ベラトリルアルデヒドの濃度は、310nmの波長の光線による吸光度で測定し、吸光係数9300M−1cm−1として算出した。前記条件で、1分当たり1μモルのベラトリルアルデヒドを生成する酵素活性を1enzyme unit と定義した。
【0047】
前記リグニンペルオキシダーゼ又はその変異体のリグニン分解酵素活性の経時変化を、図3(実施例)、図4(比較例)、図5(参考例)にそれぞれ示す。
【0048】
図3乃至図5から、本実施例のLiP変異体は、30℃及び37℃の温度におけるリグニン分解酵素活性について、所定時間後の残存活性が参考例の野生型LiPよりも大であり、優れた耐熱性を備えていることが明らかである。一方、配列番号3で表されるアミノ酸配列の他の部分をアミノ酸置換した比較例のLiP変異体は、30℃の温度におけるリグニン分解酵素活性については、所定時間後の残存活性が参考例の野生型LiPより大であるものの、37℃の温度では参考例の野生型LiPよりも小であり、十分な耐熱性を備えていないことが明らかである。
【0049】
次に、前記野生型LiP又はその変異体の耐過酸化水素性を評価した。前記評価は、酒石酸の終濃度が50mM、前記野生型LiP又はその変異体の終濃度が200nMであると共に、H2O2の終濃度が0.01mMである溶液3mlを調製し、該溶液を25℃の温度に保持したときのリグニン分解酵素活性の経時変化を測定することにより行った。
【0050】
前記リグニン分解酵素活性の残存活性の測定は、前記耐熱性評価の場合と全く同一にして行った。前記野生型LiP又はその変異体のリグニン分解酵素活性の経時変化を、図6に示す。
【0051】
図6から、本実施例のLiP変異体は、0.01mM−H2O2の存在下におけるリグニン分解酵素活性について、所定時間後の残存活性が参考例の野生型LiPよりも大であり、優れた耐過酸化水素性を備えていることが明らかである。一方、配列番号3で表されるアミノ酸配列の他の部分をアミノ酸置換した比較例のLiP変異体は、0.01mM−H2O2の存在下におけるリグニン分解酵素活性について、所定時間後の残存活性が参考例の野生型LiPよりも小であり、耐過酸化水素性が劣っていることが明らかである。
【0052】
尚、本実施形態では、本実施例のLiP変異体を配列番号1のアミノ酸配列で表されるものとしているが、配列番号1のアミノ酸配列において1又は数個のアミノ酸が欠失、置換又は付加されたアミノ酸配列で表されるものであってもよい。
【特許請求の範囲】
【請求項1】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とするリグニンペルオキシダーゼ変異体。
【請求項2】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とする組換えタンパク質。
【請求項3】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードすることを特徴とする遺伝子。
【請求項4】
請求項3記載の遺伝子において、配列番号2で表される塩基配列からなることを特徴とする遺伝子。
【請求項5】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有することを特徴とする組換えベクター。
【請求項6】
請求項5記載の組換えベクターにおいて、配列番号2で表される塩基配列からなる遺伝子を含有することを特徴とする組換えベクター。
【請求項7】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【請求項8】
請求項7記載の形質転換体において、配列番号2で表される塩基配列からなる遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【請求項1】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とするリグニンペルオキシダーゼ変異体。
【請求項2】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなることを特徴とする組換えタンパク質。
【請求項3】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードすることを特徴とする遺伝子。
【請求項4】
請求項3記載の遺伝子において、配列番号2で表される塩基配列からなることを特徴とする遺伝子。
【請求項5】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有することを特徴とする組換えベクター。
【請求項6】
請求項5記載の組換えベクターにおいて、配列番号2で表される塩基配列からなる遺伝子を含有することを特徴とする組換えベクター。
【請求項7】
配列番号1で表される第1のアミノ酸配列、又は第1のアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加された第2のアミノ酸配列からなる組換えタンパク質をコードする遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【請求項8】
請求項7記載の形質転換体において、配列番号2で表される塩基配列からなる遺伝子を含有する組換えベクターを含むことを特徴とする形質転換体。
【図1】


【図2】

【図3】
【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2011−188771(P2011−188771A)
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願番号】特願2010−56389(P2010−56389)
【出願日】平成22年3月12日(2010.3.12)
【出願人】(592068200)学校法人東京薬科大学 (32)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願日】平成22年3月12日(2010.3.12)
【出願人】(592068200)学校法人東京薬科大学 (32)
【出願人】(000005326)本田技研工業株式会社 (23,863)
【Fターム(参考)】
[ Back to top ]