
リグニン分解細菌
【課題】バイオマス中の難分解性物質であるリグニンの分解能力を持つ細菌の提供。また、その細菌を用いたリグニンの処理方法の提供。
【解決手段】リグニン分解酵素活性を有するラルストニア・ピッケティ(Ralstonia pickettii)、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)またはキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属することを特徴とする細菌。
木質系または草本系バイオマスの分解処理において、該バイオマスと上述の細菌とを接触させることを特徴とするバイオマスの処理方法。上述の細菌を用いてリグニンを低分子化することを特徴とするリグニンの処理方法。上述の細菌を含むことを特徴とするバイオマスの処理剤。
【解決手段】リグニン分解酵素活性を有するラルストニア・ピッケティ(Ralstonia pickettii)、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)またはキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属することを特徴とする細菌。
木質系または草本系バイオマスの分解処理において、該バイオマスと上述の細菌とを接触させることを特徴とするバイオマスの処理方法。上述の細菌を用いてリグニンを低分子化することを特徴とするリグニンの処理方法。上述の細菌を含むことを特徴とするバイオマスの処理剤。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、バイオマスの活用に関し、詳しくは、木質系及び草本系バイオマスの活用に関し、より詳しくはリグニンの分解能力を有する細菌およびそれを用いるリグニンの処理方法に関する。
【背景技術】
【0002】
生物由来の資源であるバイオマスを利用するバイオ燃料は、非化石燃料であり、これを消費して二酸化炭素を排出しても、大気中の二酸化炭素量は増減せず、二酸化炭素の排出はないものとして取り扱われる(カーボンニュートラル)。このため、バイオ燃料は、従来使用されてきた化石燃料に代わる新たなエネルギー源として注目を集めている。
【0003】
トウモロコシやサトウキビを原料とするバイオエタノールなどのバイオ燃料は、世界的に利用が進んでおり、増産が続いている。しかし、トウモロコシやサトウキビは、食糧としても利用される作物であり(第1世代バイオマス)、食糧との競合により食糧価格の高騰を招くという弊害も生じている。
そこで、近年では食糧とは競合しない木質系・草本系バイオマス(第2世代バイオマス)をバイオ燃料として利用するための研究、開発が進められている。
【0004】
木質系・草本系バイオマスは、主に、リグニン、セルロースおよびキシランにより構成されている。リグニン、セルロースおよびキシランはいずれもポリマーであり、バイオ燃料として利用するためには、低分子化が必要である。これらの構成要素の中でもリグニンは、難分解性成分として知られている。また、リグニンを希硫酸やアルカリ処理した際に生成するリグニン低分子化可溶化物はアルコール発酵の障害となるほか、セルラーゼやキシラナーゼなどの酵素を吸着し、さらに、廃液の発生原因ともなり、木質系・草本系バイオマス活用の大きな障害となっている。
【0005】
現在、木質系・草本系バイオマスの処理過程におけるリグニンの除去は、希硫酸やアルカリを用いる化学的処理、または熱処理が一般的である。しかし、化学的処理は前述のアルコール発酵障害と酵素吸着という問題に加え廃液の発生による2次汚染の問題があり、熱処理はエネルギー消費量が多く、新エネルギー源の開発手段として適当とは言い難く、さらに高コストとなるという問題点がある。
これらの問題を解消し、環境問題の要請にもこたえる方法として、リグニンを生物学的に処理する方法が考えられる。例えば、特許文献1には、糸状菌を用いてリグニンを分解処理する方法が開示されている。
しかし、糸状菌によるリグニンの分解は長時間を要するため、より効率的で簡便なリグニンの分解方法が求められている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2007−325604号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
従って、本発明の目的は、バイオマスに含まれるリグニンを、迅速に効率よく、かつ安価に処理しうる方法を提供することである。
【課題を解決するための手段】
【0008】
本発明者らは、糸状菌の代わりに細菌を用いることを想到し、リグニン分解酵素活性を有する細菌について鋭意研究および探索を行った。この結果、ラルストニア(Ralstonia)属、ステノトロホモナス(Stenotrophomonas)属、ストレプトマイセス(Streptmyces)属またはキラトコッカス(Chelatococcus)属に属する細菌がすぐれたリグニン分解酵素活性を有することを見いだし、上記目的を達成しうる本発明の完成に至った。
すなわち、本発明は、リグニン分解酵素活性を有するラルストニア・ピッケティ(Ralstonia pickettii)、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)またはキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属することを特徴とする細菌を提供する。
【0009】
本発明においては、前記リグニン分解酵素活性を有する細菌が、ストレプトマイセス・シュードグリセオラスKS−3株(受領番号NITE AP−918)であることが好ましい。
【0010】
また、本発明は、木質系または草本系バイオマスの分解処理において、該バイオマスと上述の細菌とを接触させることを特徴とするバイオマスの処理方法;上述の細菌を用いてリグニンを低分子化することを特徴とするリグニンの処理方法;および、上述の細菌を含むことを特徴とするバイオマスの処理剤を提供する。
【発明の効果】
【0011】
本発明によれば、バイオマスに含まれるリグニンを、迅速に効率よく、かつ安価に処理することができる。
【図面の簡単な説明】
【0012】
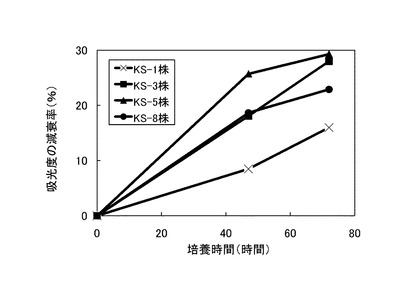
【図1】275nmの吸光度の減衰率を示すグラフである。
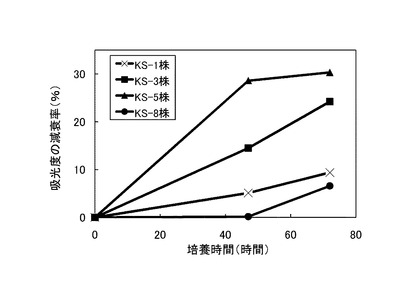
【図2】480nmの吸光度の減衰率を示すグラフである。
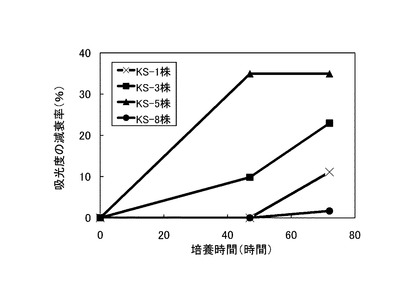
【図3】600nmの吸光度の減衰率を示すグラフである。
【発明を実施するための形態】
【0013】
次に、本発明を実施するための最良の形態を説明する。なお、本発明において「細菌」とは、原核生物のことをいう。
ラルストニア(Ralstonia)属は、分類学上、バークホルデリア科(Burkholderiaceae)ラルストニア属に属する。ラルストニア属に属する細菌は、寒天培地上では白色で直径1mm程度のコロニーを形成する。
ステノトロホモナス(Stenotrophomonas)属は、分類学上、キサントモナス科(Xanthomonadaceae)ステノトロホモナス属に属する。ステノトロホモナス属に属する細菌は、寒天培地上では白色でほぼ透明な直径0.2mm程度のコロニーを形成する。
ストレプトマイセス(Streptmyces)属は、分類学上、ストレプトマイセス科(Streptomycetaceae)ストレプトマイセス属に属する。ストレプトマイセスに属する細菌は、放線菌で寒天培地上では白〜灰色である直径0.3mm程度のコロニーを形成する。
キラトコッカス(Chelatococcus)属は、分類学上、ベイジェリンキー科(Beijerinckiaceae)キラトコッカス属に属する。キラトコッカスに属する細菌は、寒天培地上ではクリーム色で直径0.5mm程度のコロニーを形成する。
【0014】
上記ラルストニア・ピッケティに属する菌株の例としては、本発明者らが単離したラルストニア・ピッケティ(Ralstonia pickettii)KS−1株およびKS−9株を挙げることができる。
また、上記ステノトロホモナス・マルトフィリアに属する菌株の例としては、本発明者らが単離したステノトロホモナス・マルトフィリアKS−8株を挙げることができる。
また、上記ストレプトマイセス・シュードグリセオラスに属する菌株の例としては、本発明者らが単離したストレプトマイセス・シュードグリセオラスKS−3株を挙げることができる。このKS−3株は、本発明者らにより、受領日:2010年3月23日に受領番号「NITE AP−918」として、独立行政法人製品評価技術基盤機構 特許微生物寄託センターに寄託された。
また、上記キラトコッカス・アサッカロボランスに属する菌株の例としては、本発明者らが単離したキラトコッカス・アサッカロボランスKS−5株を挙げることができる。
【0015】
本発明の細菌のリグニン分解酵素活性は、少なくとも2種の酵素、リグニンパーオキシダーゼおよびマンガンペルオキシダーゼに由来する。
リグニンパーオキシダーゼは、過酸化水素の存在下でリグニンやそのモデル化合物(例えば、ベラトリルアルコール)の酸化分解を触媒する酵素である。リグニンパーオキシダーゼの働きによりベラトリルアルコールが酸化されベラトルムアルデヒドとなると、310nm付近の吸光度が増加する。この性質を利用して、糸状菌や担子菌類については、過酸化水素の存在下でベラトリルアルコール溶液に試験サンプルを添加し、ベラトリルアルコールからベラトルムアルデヒドへの酸化を吸光度測定により検出して、リグニンパーオキシダーゼ活性を測定する方法が確立されている。本発明者らは、この測定方法を細菌に応用して適用した。
【0016】
一方、マンガンペルオキシダーゼは、分子量4.6万の糖ヘムタンパク質であり、過酸化水素の存在下でMn2+からMn3+への酸化を触媒する酵素である。このMn3+が、リグニンやジメトキシフェノールなどのフェノール性基質を酸化分解する。2,6−ジメトキシフェノールがマンガンペルオキシダーゼにより酸化されると、470nm付近の吸光度が増加する。この性質を利用して、糸状菌や担子菌類については、過酸化水素の存在下で2,6−ジメトキシフェノール溶液に試験サンプルを添加し、2,6−ジメトキシフェノールの酸化を吸光度測定により検出して、マンガンペルオキシダーゼ活性を測定する方法が確立されている。本発明者らは、この測定方法を細菌に応用して適用した。
【0017】
また、細菌のリグニン分解能力を測定する方法としては、細菌をリグニン含有液体培地中で培養し、480nm付近の吸光度からリグニンの脱色を測定する方法や、275nm付近の吸光度からリグニンの芳香環の分解を測定する方法が知られている。
【0018】
本発明のリグニン分解酵素活性を有する細菌は、好気的条件下で培養し増殖することができる。培養液のpHは4〜9が好ましく、5〜8であることがより好ましい。
培養温度は10〜40℃であることが好ましく、より好ましくは20〜30℃である。
本発明のリグニン分解酵素活性を有する細菌の培地組成の一例として、表1に記載の成分を滅菌水で1Lにメスアップして調製した液体培地を挙げることができる。
【0019】

【0020】
培地中の炭素源としては、スギ微粉末などの木質系炭素源や、ケナフ微粉末、トマト茎微粉末、稲・麦ワラ微粉末などの草本系炭素源を挙げることができる。炭素源として、これらの特定の炭素源を唯一の炭素源として用いて培養することにより、難分解物であるリグニンを分解しうる菌体に対して栄養要求性に基づいて選択圧をかけることができるという利点がある。
窒素源の例としては、一般に酵母エキスが使用される。しかし、本発明者らにより、酵母エキス以外の窒素源を用いることにより、本発明のリグニン分解細菌培養液のリグニン分解酵素活性をさらに向上しうることが判明した。より高いリグニン分解酵素活性を得るために好適な窒素源は、菌株ごとに異なり、一概には言えないが、ラルストニア・ピッケティKS−1株では硝酸アンモニウムを用いることが好ましく、ストレプトマイセス・シュードグリセオラスKS−3株では硫酸アンモニウムまたは硝酸アンモニウムが好ましく、キラトコッカス・アサッカロボランスKS−5株では硝酸アンモニウムが好ましく、ステノトロホモナス・マルトフィリアKS−8株では硫酸アンモニウムが好ましい。これらの窒素源は酵母エキスに比較してはるかに安価であるため産業応用に好適である。
【0021】
本発明の細菌が産生するリグニンパーオキシダーゼおよびマンガンペルオキシダーゼは、上述の液体培地で培養した場合、菌外に分泌され、培養液がリグニン分解酵素活性を発揮する。したがって、菌体が木材などに直接接触して、そこで局所的にリグニンを分解する糸状菌とは異なり、本発明の細菌は、液体培養をすることにより、リグニン分解酵素を大量に入手することができるという利点がある。
本発明の応用としては、木質系バイオマスまたは草本系バイオマスを分解処理する際に、当該バイオマスを本発明の細菌を含む培養槽に投入して当該細菌が分泌するリグニン分解酵素を作用させ、難分解性のリグニンを選択的に低分子化し、分解することが考えられる。この生物学的リグニン分解処理により、バイオマスのこれまでの酸やアルカリによる分解処理の過程で発酵障害や酵素吸着を起こし、分解処理を妨げるリグニンを除去し、バイオマスに含まれるセルロースやキシランを分解しやすい状態にできる利点がある。
【0022】
また、本発明の細菌を微生物製剤として応用することが考えられる。本発明の細菌を微生物製剤とする場合、各細菌の酵素活性(例えば、リグニンパーオキシダーゼ活性、マンガンペルオキシダーゼ活性、キシラナーゼ活性、セルラーゼ活性)を把握した上で、使用目的に応じて各細菌を組み合わせることが好ましい。
【0023】
上記、処理対象のバイオマスの例として、建築廃材、稲わら、麦わら、古紙などのバイオマス資源を挙げることができる。また、これらのバイオマス資源を低分子化し、バイオエタノールの製造原料に利用することが考えられる。
さらに、処理対象のバイオマスの例として、木材パルプの製造工程で発生する黒液を挙げることができる。黒液は、従来、燃焼して処理されてきたが、本発明のリグニンの処理方法を黒液に適用して、バイオエタノールの原料として用いることにより、簡便でかつ環境にも配慮した形でリサイクルをすることができる。
【実施例】
【0024】
次に実施例を挙げて本発明をさらに具体的に説明するが、これらの実施例は本発明の単なる例示であって、本発明の限定を意図するものではない。
【0025】
<木質系または草本系バイオマスを炭素源とするリグニン分解微生物の探索>
木質系炭素源としてスギ微粉末を唯一の炭素源として生育する細菌を探索した。同様に、草本系炭素源としてケナフ微粉末を唯一の炭素源として生育する細菌およびトマト茎微粉末を唯一の炭素源として生育する細菌を探索した。
この結果、リグニン分解能力を有する細菌として、5種の細菌が分離された(KS−1株、KS−3株、KS−5株、KS−8株およびKS−9株)。
【0026】
これらの菌株について、16S rRNA系統解析を行った結果、KS−1株とKS−9株は、ラルストニア・ピッケティ(Ralstonia pickettii)に属する細菌であること、KS−8株は、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)に属する細菌であること、KS−3株は、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)に属する細菌であること、KS−5株は、キラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属する細菌であることが明らかとなった。なお、16SrRNAの相同性はKS−1株が100%、KS−3株が100%、KS−5株が97%、KS−8株が100%であった。
【0027】
ストレプトマイセス・シュードグリセオラスKS−3株は、本発明者らにより、受領番号:NITE AP−918として、受領日:2010年3月23日に独立行政法人製品評価技術基盤機構 特許微生物寄託センターに寄託された。
【0028】
<リグニン分解能力の測定>
[実施例1]
本発明のリグニン分解細菌であるラルストニア・ピッケティKS−1株によるリグニンの分解を、特開平9−67785号公報に記載された方法で測定した。具体的には、まず前培養として、表2に記載の組成の液体培地3mLにラルストニア・ピッケティKS−1株を1白金耳入れ、3日間、150rpmで回転振とう培養した。
【0029】
【0030】
次いで、表3に記載の組成の培地100mLを500mLの三角フラスコに注ぎ、これに前培養した菌体3mLを添加して、175rpmで回転振とう培養した。培養開始直後、培養開始から47時間後および72時間後に275nm、480nmおよび600nmの吸光度を測定した。275nmでは、リグニンの芳香環に由来する吸光度を測定し、480nmではリグニンの脱色、600nmでは菌体の増殖度を測定することにより、リグニンの分解率を算出した。なお、吸光度の測定には、培養液を10,000rpmで20分間遠心分離して得られた上清を用いた。
【0031】
【0032】
培養開始直後の吸光度に対する吸光度の減衰率を275nmについては図1に、480nmについては図2に、600nmについては図3に、それぞれ×印で示す。測定の結果、72時間後の吸光度は、実験開始時と比べて275nmで約16%、480nmで約9.4%、600nmでは約11.1%減衰した。これらの結果から、本発明のリグニン分解細菌であるラルストニア・ピッケティKS−1株がリグニン分解能力を持つことが示された。
【0033】
[実施例2〜4]
ラルストニア・ピッケティKS−1株の代わりに、それぞれストレプトマイセス・シュードグリセオラスKS−3株、キラトコッカス・アサッカロボランスKS−5株、ステノトロホモナス・マルトフィリアKS−8株を使用して、実施例1と同様の測定を行った。結果を図1〜3に、KS−3株については■で、KS−5株については▲で、KS−8株については●で示す。
これらの結果から、KS−3株、KS−5株、KS−8株のいずれもがリグニン分解能力を持つこと、ならびに、これらの中でもKS−3株とKS−5株は吸光度の減衰率が大きく、優れたリグニン分解能力を持つことが示された。
【0034】
<リグニン分解細菌ラルストニア・ピッケティKS−1株の菌学的性質>
[実施例5]
(1)リグニンパーオキシダーゼ活性測定法によるリグニン分解活性の分析
ラルストニア・ピッケティKS−1株のリグニン分解活性をリグニンパーオキシダーゼ(LiP)活性測定法により分析した。
【0035】
具体的には、以下の表4に示す1)から3)の3種の溶液を使用した。
【0036】
本実施例において、サンプル溶液は、KS−1株を表5に示す組成の液体培地で7日間、30℃で振とう培養した後の培養液を、10000rpmで20分間遠心分離して得られた上清を使用した。なお、炭素源としては、スギ微粉末、ケナフ微粉末またはトマト茎微粉末を使用した。
【0037】
基質溶液800μLとサンプル溶液100μLを1mL容のセル中で混合し、37℃で10分間静置した。その後、過酸化水素溶液100μLを加えて反応を開始した。反応直後の吸光度とブランクの吸光度(測定波長:310nm)とを比較し、反応直後の吸光度がブランクの吸光度の2倍以上である場合、酵素活性があると判断した(表7に+で示す)。実験の結果、反応直後の吸光度はブランクの吸光度の約2.66倍であり、KS−1株がリグニンパーオキシダーゼ活性を有することが明らかとなった。なお、ブランクとしては、過酸化水素溶液の代わりに水を使用したものを用いた。
【0038】
(2)マンガンペルオキシダーゼ活性測定法によるリグニン分解活性の分析
KS−1株のリグニン分解活性をマンガンペルオキシダーゼ(MnP)活性測定法により分析した。
【0039】
具体的には、以下の表6に示す1)から5)の5種の溶液を使用した。
【0040】
基質溶液50μL、20mM硫酸マンガン溶液50μL、マロン酸緩衝液700μLおよびサンプル溶液100μLを1mL容のセル中で混合し、37℃で10分間静置した。その後、過酸化水素溶液100μLを加えて反応を開始した。反応直後の吸光度とブランクの吸光度(測定波長:469nm)を比較し、反応直後の吸光度がブランクの吸光度にの2倍以上である場合、酵素活性があると判断した(表7に+で示す)。実験の結果、反応直後の吸光度はブランクの約1.65倍であり、KS−1株がマンガンペルオキシダーゼ活性を有することが明らかとなった。なお、ブランクとしては、過酸化水素溶液の代わりに水を使用したものを用いた。
【0041】
上記リグニン分解活性に加え、KS−1株のキシラン分解活性及びセルロース分解活性ならびに菌学的性質を検討した。結果を表7に示す。
なお、キシラン分解活性はキシラナーゼ活性を、セルロース分解活性はセルラーゼ活性を周知の方法により測定し、分析した。
【0042】
[実施例6]
ラルストニア・ピッケティに属するKS−9株の菌学的性質を実施例5と同様に分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度に対して約2.46倍であり、KS−9株がリグニンパーオキシダーゼ活性を有することが明らかとなった。一方、マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約1.30倍であった。分析結果を表7に示す。
【0043】
【0044】
<リグニン分解細菌ステノトロホモナス・マルトフィリアKS−8株の菌学的性質>
[実施例7]
ステノトロホモナス・マルトフィリアKS−8株の菌学的性質を実施例5と同様にして分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.49倍であり、KS−8株がリグニンパーオキシダーゼ活性を有することが明らかとなった。マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約1.50倍であり、KS−8株がマンガンペルオキシダーゼ活性を有することが明らかとなった。分析結果を表8に示す。なお、表中の記号の意味は、表7と同様である。
【0045】
【0046】
<リグニン分解細菌ストレプトマイセス・シュードグリセオラスKS−3株の菌学的性質>
[実施例8]
ストレプトマイセス・シュードグリセオラスKS−3株の菌学的性質を実施例5と同様にして分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.28倍であり、KS−3株がリグニンパーオキシダーゼ活性を有することが明らかとなった。マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.0倍であり、KS−3株がマンガンペルオキシダーゼ活性を有することが明らかとなった。分析結果を表9に示す。なお、表中の記号の意味は、表7と同様である。
【0047】
【0048】
<リグニン分解細菌キラトコッカス・アサッカロボランスKS−5株の菌学的性質>
[実施例9]
キラトコッカス・アサッカロボランスKS−5株の菌学的性質を実施例5と同様にして分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.58倍であり、KS−5株がリグニンパーオキシダーゼ活性を有することが確認された。マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約1.57倍であり、KS−5株のマンガンペルオキシダーゼ活性が確認された。分析結果を表10に示す。なお、表中の記号の意味は、表7と同様である。
【0049】
【0050】
<リグニン分解酵素活性を増強する培養液の探索>
[実施例10]
表11に記載の液体培地5mLを試験管に注ぎ、これに本発明のリグニン分解細菌であるラルストニア・ピッケティKS−1株を1白金耳植菌し、ローラー回転式培養装置にセットして、7日間、30rpmで培養した。
培養終了後、培養液を10,000rpmで20分間遠心分離して得られた上清をサンプル溶液とした。
【0051】
【0052】
1mL容の吸光度測定用セルに、サンプル溶液100μLと表12に記載の基質溶液800μLを加えて混合し、37℃で10分間インキュベートした。その後表12に記載の過酸化水素溶液100μLを加え、よく混合して酵素反応を開始させ、反応開始から1分後に吸光度を測定した。
この吸光度の値を、ブランクの吸光度で割りリグニンパーオキシダーゼ活性値として2.20が得られた。なお、ブランクの吸光度は、過酸化水素溶液の代わりに水を用いたこと以外は同様の測定を行って得た。
【0053】
【0054】
[実施例11]
液体培地の窒素源として酵母エキスの代わりに硫酸アンモニウムを用いたこと以外は実施例10と同様にして吸光度測定を行い、リグニンパーオキシダーゼ活性値1.683を得た。
【0055】
[実施例12]
液体培地の窒素源として酵母エキスの代わりに硝酸アンモニウムを用いたこと以外は実施例10と同様にして吸光度測定を行い、リグニンパーオキシダーゼ活性値3.413を得た。結果のまとめを表13に示す。
実施例10〜12の結果から、ラルストニア・ピッケティKS−1株は、窒素源として硝酸アンモニウムを使用した場合に、優れたリグニンパーオキシダーゼ活性を示すことが明らかとなった。
【0056】
[実施例13〜15]
リグニン分解細菌として、ラルストニア・ピッケティKS−1株に代えてストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)KS−3株を用いたこと以外は、それぞれ実施例10〜12と同様にして吸光度測定を行った。窒素源として酵母エキスを用いた実施例13のリグニンパーオキシダーゼ活性は2.163、硫酸アンモニウムを用いた実施例14では4.314、硝酸アンモニウムを用いた実施例15では4.070であった。結果のまとめを表13に示す。
これらの結果から、ストレプトマイセス・シュードグリセオラスKS−3株は、窒素源として硫酸アンモニウムまたは硝酸アンモニウムを用いることにより、従来汎用されている酵母エキスを用いる場合と比べて2倍程度の酵素活性を得ることができる。
【0057】
[実施例16〜18]
リグニン分解細菌として、ラルストニア・ピッケティKS−1株に代えてキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)KS−5株を用いたこと以外は、それぞれ実施例10〜12と同様にして吸光度測定を行った。窒素源として酵母エキスを用いた実施例16のリグニンパーオキシダーゼ活性は2.478、硫酸アンモニウムを用いた実施例17では1.882、硝酸アンモニウムを用いた実施例18では3.188であった。結果のまとめを表13に示す。
これらの結果から、キラトコッカス・アサッカロボランスKS−5株は、窒素源として硝酸アンモニウムを用いることにより、優れたリグニンパーオキシダーゼ活性を示すことが明らかとなった。
【0058】
[実施例19〜21]
リグニン分解細菌として、ラルストニア・ピッケティKS−1株に代えてステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)KS−8株を用いたこと以外は、それぞれ実施例10〜12と同様にして吸光度測定を行った。窒素源として酵母エキスを用いた実施例19のリグニンパーオキシダーゼ活性は2.457、硫酸アンモニウムを用いた実施例20では3.921、硝酸アンモニウムを用いた実施例21では2.889であった。結果のまとめを表13に示す。
これらの結果から、ステノトロホモナス・マルトフィリアKS−8株は、窒素源として硫酸アンモニウムを用いることにより、優れたリグニンパーオキシダーゼ活性を示すことが明らかとなった。
【0059】
【産業上の利用可能性】
【0060】
本発明によれば、バイオマスに含まれるリグニンを、迅速に効率よく、かつ安価に処理することができるリグニン分解細菌が提供される。また、本発明のリグニン分解細菌を利用することにより、バイオマスに含まれるリグニンを迅速に効率よく、かつ安価に処理することができる。
【技術分野】
【0001】
本発明は、バイオマスの活用に関し、詳しくは、木質系及び草本系バイオマスの活用に関し、より詳しくはリグニンの分解能力を有する細菌およびそれを用いるリグニンの処理方法に関する。
【背景技術】
【0002】
生物由来の資源であるバイオマスを利用するバイオ燃料は、非化石燃料であり、これを消費して二酸化炭素を排出しても、大気中の二酸化炭素量は増減せず、二酸化炭素の排出はないものとして取り扱われる(カーボンニュートラル)。このため、バイオ燃料は、従来使用されてきた化石燃料に代わる新たなエネルギー源として注目を集めている。
【0003】
トウモロコシやサトウキビを原料とするバイオエタノールなどのバイオ燃料は、世界的に利用が進んでおり、増産が続いている。しかし、トウモロコシやサトウキビは、食糧としても利用される作物であり(第1世代バイオマス)、食糧との競合により食糧価格の高騰を招くという弊害も生じている。
そこで、近年では食糧とは競合しない木質系・草本系バイオマス(第2世代バイオマス)をバイオ燃料として利用するための研究、開発が進められている。
【0004】
木質系・草本系バイオマスは、主に、リグニン、セルロースおよびキシランにより構成されている。リグニン、セルロースおよびキシランはいずれもポリマーであり、バイオ燃料として利用するためには、低分子化が必要である。これらの構成要素の中でもリグニンは、難分解性成分として知られている。また、リグニンを希硫酸やアルカリ処理した際に生成するリグニン低分子化可溶化物はアルコール発酵の障害となるほか、セルラーゼやキシラナーゼなどの酵素を吸着し、さらに、廃液の発生原因ともなり、木質系・草本系バイオマス活用の大きな障害となっている。
【0005】
現在、木質系・草本系バイオマスの処理過程におけるリグニンの除去は、希硫酸やアルカリを用いる化学的処理、または熱処理が一般的である。しかし、化学的処理は前述のアルコール発酵障害と酵素吸着という問題に加え廃液の発生による2次汚染の問題があり、熱処理はエネルギー消費量が多く、新エネルギー源の開発手段として適当とは言い難く、さらに高コストとなるという問題点がある。
これらの問題を解消し、環境問題の要請にもこたえる方法として、リグニンを生物学的に処理する方法が考えられる。例えば、特許文献1には、糸状菌を用いてリグニンを分解処理する方法が開示されている。
しかし、糸状菌によるリグニンの分解は長時間を要するため、より効率的で簡便なリグニンの分解方法が求められている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2007−325604号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
従って、本発明の目的は、バイオマスに含まれるリグニンを、迅速に効率よく、かつ安価に処理しうる方法を提供することである。
【課題を解決するための手段】
【0008】
本発明者らは、糸状菌の代わりに細菌を用いることを想到し、リグニン分解酵素活性を有する細菌について鋭意研究および探索を行った。この結果、ラルストニア(Ralstonia)属、ステノトロホモナス(Stenotrophomonas)属、ストレプトマイセス(Streptmyces)属またはキラトコッカス(Chelatococcus)属に属する細菌がすぐれたリグニン分解酵素活性を有することを見いだし、上記目的を達成しうる本発明の完成に至った。
すなわち、本発明は、リグニン分解酵素活性を有するラルストニア・ピッケティ(Ralstonia pickettii)、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)またはキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属することを特徴とする細菌を提供する。
【0009】
本発明においては、前記リグニン分解酵素活性を有する細菌が、ストレプトマイセス・シュードグリセオラスKS−3株(受領番号NITE AP−918)であることが好ましい。
【0010】
また、本発明は、木質系または草本系バイオマスの分解処理において、該バイオマスと上述の細菌とを接触させることを特徴とするバイオマスの処理方法;上述の細菌を用いてリグニンを低分子化することを特徴とするリグニンの処理方法;および、上述の細菌を含むことを特徴とするバイオマスの処理剤を提供する。
【発明の効果】
【0011】
本発明によれば、バイオマスに含まれるリグニンを、迅速に効率よく、かつ安価に処理することができる。
【図面の簡単な説明】
【0012】
【図1】275nmの吸光度の減衰率を示すグラフである。
【図2】480nmの吸光度の減衰率を示すグラフである。
【図3】600nmの吸光度の減衰率を示すグラフである。
【発明を実施するための形態】
【0013】
次に、本発明を実施するための最良の形態を説明する。なお、本発明において「細菌」とは、原核生物のことをいう。
ラルストニア(Ralstonia)属は、分類学上、バークホルデリア科(Burkholderiaceae)ラルストニア属に属する。ラルストニア属に属する細菌は、寒天培地上では白色で直径1mm程度のコロニーを形成する。
ステノトロホモナス(Stenotrophomonas)属は、分類学上、キサントモナス科(Xanthomonadaceae)ステノトロホモナス属に属する。ステノトロホモナス属に属する細菌は、寒天培地上では白色でほぼ透明な直径0.2mm程度のコロニーを形成する。
ストレプトマイセス(Streptmyces)属は、分類学上、ストレプトマイセス科(Streptomycetaceae)ストレプトマイセス属に属する。ストレプトマイセスに属する細菌は、放線菌で寒天培地上では白〜灰色である直径0.3mm程度のコロニーを形成する。
キラトコッカス(Chelatococcus)属は、分類学上、ベイジェリンキー科(Beijerinckiaceae)キラトコッカス属に属する。キラトコッカスに属する細菌は、寒天培地上ではクリーム色で直径0.5mm程度のコロニーを形成する。
【0014】
上記ラルストニア・ピッケティに属する菌株の例としては、本発明者らが単離したラルストニア・ピッケティ(Ralstonia pickettii)KS−1株およびKS−9株を挙げることができる。
また、上記ステノトロホモナス・マルトフィリアに属する菌株の例としては、本発明者らが単離したステノトロホモナス・マルトフィリアKS−8株を挙げることができる。
また、上記ストレプトマイセス・シュードグリセオラスに属する菌株の例としては、本発明者らが単離したストレプトマイセス・シュードグリセオラスKS−3株を挙げることができる。このKS−3株は、本発明者らにより、受領日:2010年3月23日に受領番号「NITE AP−918」として、独立行政法人製品評価技術基盤機構 特許微生物寄託センターに寄託された。
また、上記キラトコッカス・アサッカロボランスに属する菌株の例としては、本発明者らが単離したキラトコッカス・アサッカロボランスKS−5株を挙げることができる。
【0015】
本発明の細菌のリグニン分解酵素活性は、少なくとも2種の酵素、リグニンパーオキシダーゼおよびマンガンペルオキシダーゼに由来する。
リグニンパーオキシダーゼは、過酸化水素の存在下でリグニンやそのモデル化合物(例えば、ベラトリルアルコール)の酸化分解を触媒する酵素である。リグニンパーオキシダーゼの働きによりベラトリルアルコールが酸化されベラトルムアルデヒドとなると、310nm付近の吸光度が増加する。この性質を利用して、糸状菌や担子菌類については、過酸化水素の存在下でベラトリルアルコール溶液に試験サンプルを添加し、ベラトリルアルコールからベラトルムアルデヒドへの酸化を吸光度測定により検出して、リグニンパーオキシダーゼ活性を測定する方法が確立されている。本発明者らは、この測定方法を細菌に応用して適用した。
【0016】
一方、マンガンペルオキシダーゼは、分子量4.6万の糖ヘムタンパク質であり、過酸化水素の存在下でMn2+からMn3+への酸化を触媒する酵素である。このMn3+が、リグニンやジメトキシフェノールなどのフェノール性基質を酸化分解する。2,6−ジメトキシフェノールがマンガンペルオキシダーゼにより酸化されると、470nm付近の吸光度が増加する。この性質を利用して、糸状菌や担子菌類については、過酸化水素の存在下で2,6−ジメトキシフェノール溶液に試験サンプルを添加し、2,6−ジメトキシフェノールの酸化を吸光度測定により検出して、マンガンペルオキシダーゼ活性を測定する方法が確立されている。本発明者らは、この測定方法を細菌に応用して適用した。
【0017】
また、細菌のリグニン分解能力を測定する方法としては、細菌をリグニン含有液体培地中で培養し、480nm付近の吸光度からリグニンの脱色を測定する方法や、275nm付近の吸光度からリグニンの芳香環の分解を測定する方法が知られている。
【0018】
本発明のリグニン分解酵素活性を有する細菌は、好気的条件下で培養し増殖することができる。培養液のpHは4〜9が好ましく、5〜8であることがより好ましい。
培養温度は10〜40℃であることが好ましく、より好ましくは20〜30℃である。
本発明のリグニン分解酵素活性を有する細菌の培地組成の一例として、表1に記載の成分を滅菌水で1Lにメスアップして調製した液体培地を挙げることができる。
【0019】
【0020】
培地中の炭素源としては、スギ微粉末などの木質系炭素源や、ケナフ微粉末、トマト茎微粉末、稲・麦ワラ微粉末などの草本系炭素源を挙げることができる。炭素源として、これらの特定の炭素源を唯一の炭素源として用いて培養することにより、難分解物であるリグニンを分解しうる菌体に対して栄養要求性に基づいて選択圧をかけることができるという利点がある。
窒素源の例としては、一般に酵母エキスが使用される。しかし、本発明者らにより、酵母エキス以外の窒素源を用いることにより、本発明のリグニン分解細菌培養液のリグニン分解酵素活性をさらに向上しうることが判明した。より高いリグニン分解酵素活性を得るために好適な窒素源は、菌株ごとに異なり、一概には言えないが、ラルストニア・ピッケティKS−1株では硝酸アンモニウムを用いることが好ましく、ストレプトマイセス・シュードグリセオラスKS−3株では硫酸アンモニウムまたは硝酸アンモニウムが好ましく、キラトコッカス・アサッカロボランスKS−5株では硝酸アンモニウムが好ましく、ステノトロホモナス・マルトフィリアKS−8株では硫酸アンモニウムが好ましい。これらの窒素源は酵母エキスに比較してはるかに安価であるため産業応用に好適である。
【0021】
本発明の細菌が産生するリグニンパーオキシダーゼおよびマンガンペルオキシダーゼは、上述の液体培地で培養した場合、菌外に分泌され、培養液がリグニン分解酵素活性を発揮する。したがって、菌体が木材などに直接接触して、そこで局所的にリグニンを分解する糸状菌とは異なり、本発明の細菌は、液体培養をすることにより、リグニン分解酵素を大量に入手することができるという利点がある。
本発明の応用としては、木質系バイオマスまたは草本系バイオマスを分解処理する際に、当該バイオマスを本発明の細菌を含む培養槽に投入して当該細菌が分泌するリグニン分解酵素を作用させ、難分解性のリグニンを選択的に低分子化し、分解することが考えられる。この生物学的リグニン分解処理により、バイオマスのこれまでの酸やアルカリによる分解処理の過程で発酵障害や酵素吸着を起こし、分解処理を妨げるリグニンを除去し、バイオマスに含まれるセルロースやキシランを分解しやすい状態にできる利点がある。
【0022】
また、本発明の細菌を微生物製剤として応用することが考えられる。本発明の細菌を微生物製剤とする場合、各細菌の酵素活性(例えば、リグニンパーオキシダーゼ活性、マンガンペルオキシダーゼ活性、キシラナーゼ活性、セルラーゼ活性)を把握した上で、使用目的に応じて各細菌を組み合わせることが好ましい。
【0023】
上記、処理対象のバイオマスの例として、建築廃材、稲わら、麦わら、古紙などのバイオマス資源を挙げることができる。また、これらのバイオマス資源を低分子化し、バイオエタノールの製造原料に利用することが考えられる。
さらに、処理対象のバイオマスの例として、木材パルプの製造工程で発生する黒液を挙げることができる。黒液は、従来、燃焼して処理されてきたが、本発明のリグニンの処理方法を黒液に適用して、バイオエタノールの原料として用いることにより、簡便でかつ環境にも配慮した形でリサイクルをすることができる。
【実施例】
【0024】
次に実施例を挙げて本発明をさらに具体的に説明するが、これらの実施例は本発明の単なる例示であって、本発明の限定を意図するものではない。
【0025】
<木質系または草本系バイオマスを炭素源とするリグニン分解微生物の探索>
木質系炭素源としてスギ微粉末を唯一の炭素源として生育する細菌を探索した。同様に、草本系炭素源としてケナフ微粉末を唯一の炭素源として生育する細菌およびトマト茎微粉末を唯一の炭素源として生育する細菌を探索した。
この結果、リグニン分解能力を有する細菌として、5種の細菌が分離された(KS−1株、KS−3株、KS−5株、KS−8株およびKS−9株)。
【0026】
これらの菌株について、16S rRNA系統解析を行った結果、KS−1株とKS−9株は、ラルストニア・ピッケティ(Ralstonia pickettii)に属する細菌であること、KS−8株は、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)に属する細菌であること、KS−3株は、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)に属する細菌であること、KS−5株は、キラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属する細菌であることが明らかとなった。なお、16SrRNAの相同性はKS−1株が100%、KS−3株が100%、KS−5株が97%、KS−8株が100%であった。
【0027】
ストレプトマイセス・シュードグリセオラスKS−3株は、本発明者らにより、受領番号:NITE AP−918として、受領日:2010年3月23日に独立行政法人製品評価技術基盤機構 特許微生物寄託センターに寄託された。
【0028】
<リグニン分解能力の測定>
[実施例1]
本発明のリグニン分解細菌であるラルストニア・ピッケティKS−1株によるリグニンの分解を、特開平9−67785号公報に記載された方法で測定した。具体的には、まず前培養として、表2に記載の組成の液体培地3mLにラルストニア・ピッケティKS−1株を1白金耳入れ、3日間、150rpmで回転振とう培養した。
【0029】
【0030】
次いで、表3に記載の組成の培地100mLを500mLの三角フラスコに注ぎ、これに前培養した菌体3mLを添加して、175rpmで回転振とう培養した。培養開始直後、培養開始から47時間後および72時間後に275nm、480nmおよび600nmの吸光度を測定した。275nmでは、リグニンの芳香環に由来する吸光度を測定し、480nmではリグニンの脱色、600nmでは菌体の増殖度を測定することにより、リグニンの分解率を算出した。なお、吸光度の測定には、培養液を10,000rpmで20分間遠心分離して得られた上清を用いた。
【0031】
【0032】
培養開始直後の吸光度に対する吸光度の減衰率を275nmについては図1に、480nmについては図2に、600nmについては図3に、それぞれ×印で示す。測定の結果、72時間後の吸光度は、実験開始時と比べて275nmで約16%、480nmで約9.4%、600nmでは約11.1%減衰した。これらの結果から、本発明のリグニン分解細菌であるラルストニア・ピッケティKS−1株がリグニン分解能力を持つことが示された。
【0033】
[実施例2〜4]
ラルストニア・ピッケティKS−1株の代わりに、それぞれストレプトマイセス・シュードグリセオラスKS−3株、キラトコッカス・アサッカロボランスKS−5株、ステノトロホモナス・マルトフィリアKS−8株を使用して、実施例1と同様の測定を行った。結果を図1〜3に、KS−3株については■で、KS−5株については▲で、KS−8株については●で示す。
これらの結果から、KS−3株、KS−5株、KS−8株のいずれもがリグニン分解能力を持つこと、ならびに、これらの中でもKS−3株とKS−5株は吸光度の減衰率が大きく、優れたリグニン分解能力を持つことが示された。
【0034】
<リグニン分解細菌ラルストニア・ピッケティKS−1株の菌学的性質>
[実施例5]
(1)リグニンパーオキシダーゼ活性測定法によるリグニン分解活性の分析
ラルストニア・ピッケティKS−1株のリグニン分解活性をリグニンパーオキシダーゼ(LiP)活性測定法により分析した。
【0035】
具体的には、以下の表4に示す1)から3)の3種の溶液を使用した。
【0036】
本実施例において、サンプル溶液は、KS−1株を表5に示す組成の液体培地で7日間、30℃で振とう培養した後の培養液を、10000rpmで20分間遠心分離して得られた上清を使用した。なお、炭素源としては、スギ微粉末、ケナフ微粉末またはトマト茎微粉末を使用した。
【0037】
基質溶液800μLとサンプル溶液100μLを1mL容のセル中で混合し、37℃で10分間静置した。その後、過酸化水素溶液100μLを加えて反応を開始した。反応直後の吸光度とブランクの吸光度(測定波長:310nm)とを比較し、反応直後の吸光度がブランクの吸光度の2倍以上である場合、酵素活性があると判断した(表7に+で示す)。実験の結果、反応直後の吸光度はブランクの吸光度の約2.66倍であり、KS−1株がリグニンパーオキシダーゼ活性を有することが明らかとなった。なお、ブランクとしては、過酸化水素溶液の代わりに水を使用したものを用いた。
【0038】
(2)マンガンペルオキシダーゼ活性測定法によるリグニン分解活性の分析
KS−1株のリグニン分解活性をマンガンペルオキシダーゼ(MnP)活性測定法により分析した。
【0039】
具体的には、以下の表6に示す1)から5)の5種の溶液を使用した。
【0040】
基質溶液50μL、20mM硫酸マンガン溶液50μL、マロン酸緩衝液700μLおよびサンプル溶液100μLを1mL容のセル中で混合し、37℃で10分間静置した。その後、過酸化水素溶液100μLを加えて反応を開始した。反応直後の吸光度とブランクの吸光度(測定波長:469nm)を比較し、反応直後の吸光度がブランクの吸光度にの2倍以上である場合、酵素活性があると判断した(表7に+で示す)。実験の結果、反応直後の吸光度はブランクの約1.65倍であり、KS−1株がマンガンペルオキシダーゼ活性を有することが明らかとなった。なお、ブランクとしては、過酸化水素溶液の代わりに水を使用したものを用いた。
【0041】
上記リグニン分解活性に加え、KS−1株のキシラン分解活性及びセルロース分解活性ならびに菌学的性質を検討した。結果を表7に示す。
なお、キシラン分解活性はキシラナーゼ活性を、セルロース分解活性はセルラーゼ活性を周知の方法により測定し、分析した。
【0042】
[実施例6]
ラルストニア・ピッケティに属するKS−9株の菌学的性質を実施例5と同様に分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度に対して約2.46倍であり、KS−9株がリグニンパーオキシダーゼ活性を有することが明らかとなった。一方、マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約1.30倍であった。分析結果を表7に示す。
【0043】
【0044】
<リグニン分解細菌ステノトロホモナス・マルトフィリアKS−8株の菌学的性質>
[実施例7]
ステノトロホモナス・マルトフィリアKS−8株の菌学的性質を実施例5と同様にして分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.49倍であり、KS−8株がリグニンパーオキシダーゼ活性を有することが明らかとなった。マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約1.50倍であり、KS−8株がマンガンペルオキシダーゼ活性を有することが明らかとなった。分析結果を表8に示す。なお、表中の記号の意味は、表7と同様である。
【0045】
【0046】
<リグニン分解細菌ストレプトマイセス・シュードグリセオラスKS−3株の菌学的性質>
[実施例8]
ストレプトマイセス・シュードグリセオラスKS−3株の菌学的性質を実施例5と同様にして分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.28倍であり、KS−3株がリグニンパーオキシダーゼ活性を有することが明らかとなった。マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.0倍であり、KS−3株がマンガンペルオキシダーゼ活性を有することが明らかとなった。分析結果を表9に示す。なお、表中の記号の意味は、表7と同様である。
【0047】
【0048】
<リグニン分解細菌キラトコッカス・アサッカロボランスKS−5株の菌学的性質>
[実施例9]
キラトコッカス・アサッカロボランスKS−5株の菌学的性質を実施例5と同様にして分析した。
この結果、リグニンパーオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約2.58倍であり、KS−5株がリグニンパーオキシダーゼ活性を有することが確認された。マンガンペルオキシダーゼ活性測定では、反応直後の吸光度はブランクの吸光度の約1.57倍であり、KS−5株のマンガンペルオキシダーゼ活性が確認された。分析結果を表10に示す。なお、表中の記号の意味は、表7と同様である。
【0049】
【0050】
<リグニン分解酵素活性を増強する培養液の探索>
[実施例10]
表11に記載の液体培地5mLを試験管に注ぎ、これに本発明のリグニン分解細菌であるラルストニア・ピッケティKS−1株を1白金耳植菌し、ローラー回転式培養装置にセットして、7日間、30rpmで培養した。
培養終了後、培養液を10,000rpmで20分間遠心分離して得られた上清をサンプル溶液とした。
【0051】
【0052】
1mL容の吸光度測定用セルに、サンプル溶液100μLと表12に記載の基質溶液800μLを加えて混合し、37℃で10分間インキュベートした。その後表12に記載の過酸化水素溶液100μLを加え、よく混合して酵素反応を開始させ、反応開始から1分後に吸光度を測定した。
この吸光度の値を、ブランクの吸光度で割りリグニンパーオキシダーゼ活性値として2.20が得られた。なお、ブランクの吸光度は、過酸化水素溶液の代わりに水を用いたこと以外は同様の測定を行って得た。
【0053】
【0054】
[実施例11]
液体培地の窒素源として酵母エキスの代わりに硫酸アンモニウムを用いたこと以外は実施例10と同様にして吸光度測定を行い、リグニンパーオキシダーゼ活性値1.683を得た。
【0055】
[実施例12]
液体培地の窒素源として酵母エキスの代わりに硝酸アンモニウムを用いたこと以外は実施例10と同様にして吸光度測定を行い、リグニンパーオキシダーゼ活性値3.413を得た。結果のまとめを表13に示す。
実施例10〜12の結果から、ラルストニア・ピッケティKS−1株は、窒素源として硝酸アンモニウムを使用した場合に、優れたリグニンパーオキシダーゼ活性を示すことが明らかとなった。
【0056】
[実施例13〜15]
リグニン分解細菌として、ラルストニア・ピッケティKS−1株に代えてストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)KS−3株を用いたこと以外は、それぞれ実施例10〜12と同様にして吸光度測定を行った。窒素源として酵母エキスを用いた実施例13のリグニンパーオキシダーゼ活性は2.163、硫酸アンモニウムを用いた実施例14では4.314、硝酸アンモニウムを用いた実施例15では4.070であった。結果のまとめを表13に示す。
これらの結果から、ストレプトマイセス・シュードグリセオラスKS−3株は、窒素源として硫酸アンモニウムまたは硝酸アンモニウムを用いることにより、従来汎用されている酵母エキスを用いる場合と比べて2倍程度の酵素活性を得ることができる。
【0057】
[実施例16〜18]
リグニン分解細菌として、ラルストニア・ピッケティKS−1株に代えてキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)KS−5株を用いたこと以外は、それぞれ実施例10〜12と同様にして吸光度測定を行った。窒素源として酵母エキスを用いた実施例16のリグニンパーオキシダーゼ活性は2.478、硫酸アンモニウムを用いた実施例17では1.882、硝酸アンモニウムを用いた実施例18では3.188であった。結果のまとめを表13に示す。
これらの結果から、キラトコッカス・アサッカロボランスKS−5株は、窒素源として硝酸アンモニウムを用いることにより、優れたリグニンパーオキシダーゼ活性を示すことが明らかとなった。
【0058】
[実施例19〜21]
リグニン分解細菌として、ラルストニア・ピッケティKS−1株に代えてステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)KS−8株を用いたこと以外は、それぞれ実施例10〜12と同様にして吸光度測定を行った。窒素源として酵母エキスを用いた実施例19のリグニンパーオキシダーゼ活性は2.457、硫酸アンモニウムを用いた実施例20では3.921、硝酸アンモニウムを用いた実施例21では2.889であった。結果のまとめを表13に示す。
これらの結果から、ステノトロホモナス・マルトフィリアKS−8株は、窒素源として硫酸アンモニウムを用いることにより、優れたリグニンパーオキシダーゼ活性を示すことが明らかとなった。
【0059】
【産業上の利用可能性】
【0060】
本発明によれば、バイオマスに含まれるリグニンを、迅速に効率よく、かつ安価に処理することができるリグニン分解細菌が提供される。また、本発明のリグニン分解細菌を利用することにより、バイオマスに含まれるリグニンを迅速に効率よく、かつ安価に処理することができる。
【特許請求の範囲】
【請求項1】
リグニン分解酵素活性を有するラルストニア・ピッケティ(Ralstonia pickettii)、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)またはキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属することを特徴とする細菌。
【請求項2】
ストレプトマイセス・シュードグリセオラスKS−3株(受領番号NITE AP−918)である請求項1に記載の細菌。
【請求項3】
木質系または草本系バイオマスの分解処理において、該バイオマスと請求項1または2に記載の細菌とを接触させることを特徴とするバイオマスの処理方法。
【請求項4】
請求項1または2に記載の細菌を用いてリグニンを低分子化することを特徴とするリグニンの処理方法。
【請求項5】
請求項1または2に記載の細菌を含むことを特徴とするバイオマスの処理剤。
【請求項1】
リグニン分解酵素活性を有するラルストニア・ピッケティ(Ralstonia pickettii)、ステノトロホモナス・マルトフィリア(Stenotrophomonas maltophilia)、ストレプトマイセス・シュードグリセオラス(Streptomyces pseudogriseolus)またはキラトコッカス・アサッカロボランス(Chelatococcus asaccharovorans)に属することを特徴とする細菌。
【請求項2】
ストレプトマイセス・シュードグリセオラスKS−3株(受領番号NITE AP−918)である請求項1に記載の細菌。
【請求項3】
木質系または草本系バイオマスの分解処理において、該バイオマスと請求項1または2に記載の細菌とを接触させることを特徴とするバイオマスの処理方法。
【請求項4】
請求項1または2に記載の細菌を用いてリグニンを低分子化することを特徴とするリグニンの処理方法。
【請求項5】
請求項1または2に記載の細菌を含むことを特徴とするバイオマスの処理剤。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2011−193848(P2011−193848A)
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願番号】特願2010−66846(P2010−66846)
【出願日】平成22年3月23日(2010.3.23)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 研究集会名;平成21年度 日本生物工学会大会、主催者名;社団法人 日本生物工学会、公開日;平成21年9月23日
【出願人】(500433225)学校法人中部大学 (105)
【出願人】(000156581)日鉄環境エンジニアリング株式会社 (67)
【Fターム(参考)】
【公開日】平成23年10月6日(2011.10.6)
【国際特許分類】
【出願日】平成22年3月23日(2010.3.23)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 研究集会名;平成21年度 日本生物工学会大会、主催者名;社団法人 日本生物工学会、公開日;平成21年9月23日
【出願人】(500433225)学校法人中部大学 (105)
【出願人】(000156581)日鉄環境エンジニアリング株式会社 (67)
【Fターム(参考)】
[ Back to top ]