
リグノセルロース系バイオマスの糖化方法
【課題】本発明は、広く利用されている糸状菌由来のセルラーゼによる、リグノセルロース系バイオマスの糖化を、効率よく行う方法を開発しようとするものである。
【解決手段】自然界では一種類の微生物ではなく、様々な微生物が生産する酵素が融合して作用することにより、バイオマス分解が行われている点に着目した。市販セルラーゼ製剤への放線菌由来糖化酵素の添加により、市販製剤の糖化能を高めることができた。本発明は、多様なリグノセルロース系バイオマスの糖化に対応できる糖化方法を提供するものである。
【解決手段】自然界では一種類の微生物ではなく、様々な微生物が生産する酵素が融合して作用することにより、バイオマス分解が行われている点に着目した。市販セルラーゼ製剤への放線菌由来糖化酵素の添加により、市販製剤の糖化能を高めることができた。本発明は、多様なリグノセルロース系バイオマスの糖化に対応できる糖化方法を提供するものである。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、リグノセルロース系バイオマスの糖化方法、より詳しくは、ストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素、好ましくはストレプトマイセス属に属する微生物以外の生物由来のリグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化するリグノセルロース系バイオマスの糖化方法等に関する。
【背景技術】
【0002】
バイオマスの利用は、新しい資源循環型の社会システムの構築の観点から重要である。バイオ燃料への世界的ニーズの高まりに対応して、糖質系バイオマス由来のバイオエタノール製造技術開発競争が世界的規模で繰り広げられている。特に、食料資源と競合しないリグノセルロース系バイオマスの利用技術開発が、欧米のみならず我が国においても最も重要なブレイクスルーとなりうると考えられている。現在セルロース系バイオマスの効率のよい糖化のため、酵素糖化による方法の研究開発が進められている。
【0003】
リグノセルロースは地球上で最も多量に存在する有機物で、構造性多糖のセルロース及びヘミセルロース、芳香族化合物の重合体のリグニンから構成される。セルロースはグルコースがβ−1,4−グリコシド結合により重合した直鎖状の多糖類であり、ヘミセルロースはセルロースと共に植物細胞壁を構成するセルロース以外の多糖類であって、ホモ多糖類及びヘテロ多糖類の総称をいう。ヘミセルロースとしては、例えば、マンナン、グルコマンナン、ガラクタン、キシラン、アラビナン、アラビノキシラン、アラビノガラクタン、ガラクトグルコマンナン、グルクロノキシラン、キシログルカンなどが挙げられる。また、リグニンは、高等植物の木化に関与する高分子のフェノール性化合物で、光合成(一次代謝)により同化された炭素化合物が更なる代謝(二次代謝)を受けることで合成されるフェニルプロパノイドのうち、p−クマリルアルコール・コニフェニルアルコール・シナピルアルコールという3種類のリグニンモノマーが、酵素(ラッカーゼ・ペルオキシダーゼ)の触媒の元で一電子酸化されフェノキシラジカルとなり、これがランダムなラジカルカップリングで高度に重合することにより三次元網目構造を形成した、巨大な生体高分子である。その構造は複雑で、いまだにはっきりとはわかっていない。リグニンは木材中の20〜30%を占めており、高等植物では生育に伴い、道管・仮道管・繊維などの組織でリグニンが生産される。生産されたリグニンはヘミセルロースと同じくセルロースミクロフィブリルに付着していく。まず細胞間層で堆積が始まり、徐々に一次壁、二次壁へと沈着する。同時にヘミセルロースも堆積し、木化してゆく。構造はランダムでアモルファスである。木部の組織はリグニン構造だけ残存して殆どの細胞は死細胞となり、通導・樹体支持を担う。腐朽・食害への抵抗性を有する。生体高分子としては反応性が乏しく、リグニンを分解することが可能な生物は白色腐朽菌のみであるが、白色腐朽菌などにより低分子化されたリグニンはバクテリアにより分解され、無機化することが知られている。
【0004】
リグノセルロース系バイオマスの糖化には、酸糖化と酵素糖化の2つの方法があり、技術改善が進められている(例えば、特許文献1や2)。酵素糖化法には酸糖化と比べ、低温で糖化できることや過分解が起こらず収率が高いなどの利点がある。
【0005】
酵素糖化に用いられる市販の糖化酵素セルラーゼ製剤のほとんどは糸状菌Trichoderma reesei由来であり、これは他の微生物に比べてセルラーゼ生産量が極めて多く、安価に大量の酵素製剤を製造することが可能であるためである。リグノセルロース系バイオマスの主要成分は、セルロース・ヘミセルロース・リグニンであるが、ヘミセルロースに着目すると、その種類や含有量はリグノセルロース系バイオマスの種類によって異なる。放線菌は糸状菌とは生産する糖化酵素の種類が異なっており、ヘミセルロース分解酵素が豊富な微生物である。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2009−189291号公報
【特許文献2】特開2008−266228号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の課題は、リグノセルロース系バイオマス糖化酵素、好ましくは広く利用されている糸状菌由来のセルラーゼ等、放線菌以外の生物に由来するリグノセルロース系バイオマス糖化酵素による、リグノセルロース系バイオマスの糖化を、効率よく行う方法を提供することにある。
【課題を解決するための手段】
【0008】
本発明は、自然界では一種類の微生物ではなく様々な微生物が生産する酵素が複合して作用することにより、バイオマス分解が行われていることから、放線菌が生産する酵素に着目し、放線菌由来糖化酵素を市販セルラーゼ製剤に添加したところ、リグノセルロース系バイオマスを効率よく糖化することができることを見いだし、本発明を完成するに至った。
【0009】
すなわち本発明は、(1)配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(2)配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(3)配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(4)配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(5)リグノセルロース系バイオマス糖化酵素が、ストレプトマイセス属に属する微生物以外の生物由来のリグノセルロース系バイオマス糖化酵素であることを特徴とする上記(1)〜(4)のいずれか記載のリグノセルロース系バイオマスの糖化方法に関する。
【0010】
また、本発明は、(6)配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法や、(7)配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法や、(8)配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法や、(9)配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法に関する。
【発明の効果】
【0011】
市販セルラーゼ製剤への放線菌由来糖化酵素の添加により、市販製剤の糖化能を高めることができた。本発明によると、多様なリグノセルロース系バイオマスの糖化に対応できる糖化方法を提供することができる。
【図面の簡単な説明】
【0012】
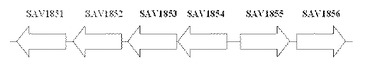
【図1】放線菌S. avermitilis ゲノム上の各酵素遺伝子の位置関係を示す図である。
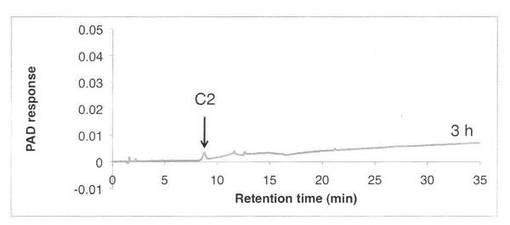
【図2】SAV1853によるCM−セルロースの分解プロダクトパターン(反応3時間)を示す図である。C2はセロビオースを示す。
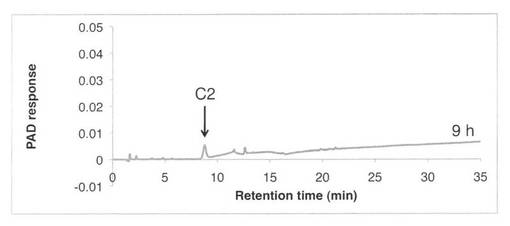
【図3】SAV1853によるCM−セルロースの分解プロダクトパターン(反応9時間)を示す図である。C2はセロビオースを示す。
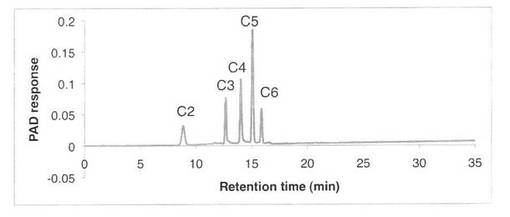
【図4】スタンダード(セロオリゴ糖)の解析パターンを示す図である。C2はセロビオース、C3はセロトリオース、C4はセロテトラオース、C5はセロペンタオース、C6はセロヘキサオースを示す。
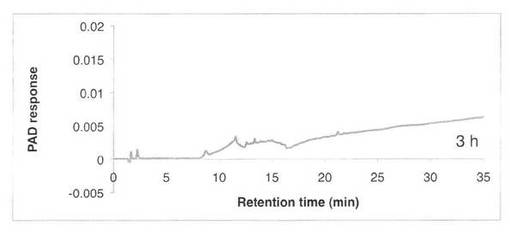
【図5】SAV1855によるCM−セルロースの分解プロダクトパターン(反応3時間)を示す図である。
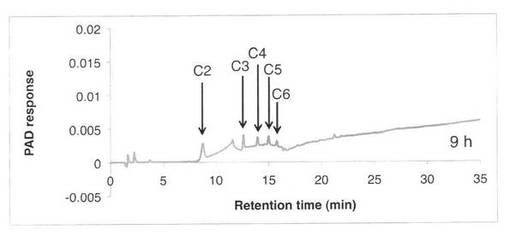
【図6】SAV1855によるCM−セルロースの分解プロダクトパターン(反応9時間)を示す図である。C2はセロビオース、C3はセロトリオース、C4はセロテトラオース、C5はセロペンタオース、C6はセロヘキサオースを示す。
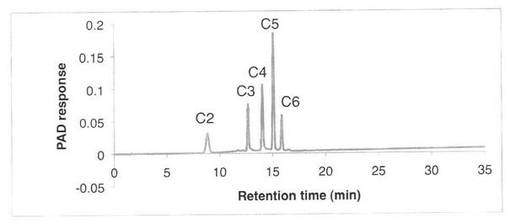
【図7】スタンダード(セロオリゴ糖)の解析パターンを示す図である。C2はセロビオース、C3はセロトリオース、C4はセロテトラオース、C5はセロペンタオース、C6はセロヘキサオースを示す。
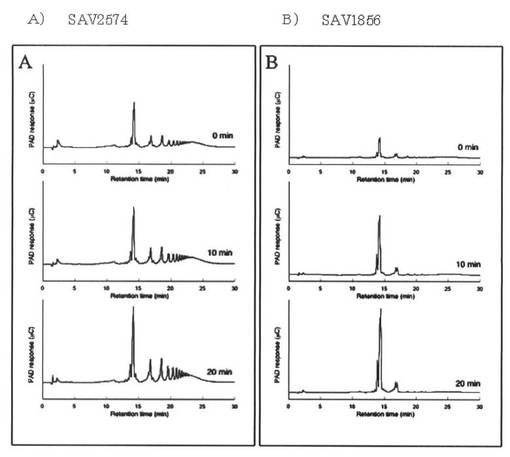
【図8】A)SAV2574、B)SAV1856によるTamarind xyloglucanの分解プロダクトパターンをそれぞれ示す図である。
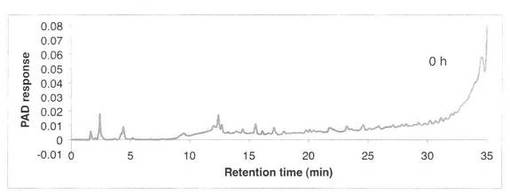
【図9】SAV1764による、ラミナリンの分解プロダクトパターン(反応0時間)を示す図である。
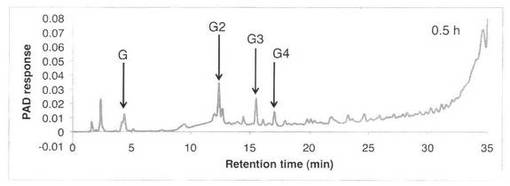
【図10】SAV1764による、ラミナリンの分解プロダクトパターン(反応0.5時間)を示す図である。Gはグルコース、G2はラミナリビオース、G3はラミナリトリオース、G4はラミナリテトラオースを示す。
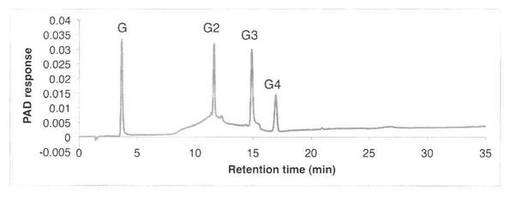
【図11】スタンダード(グルコース・ラミナリオリゴ糖)の解析パターンを示す図である。Gはグルコース、G2はラミナリビオース、G3はラミナリトリオース、G4はラミナリテトラオースを示す。
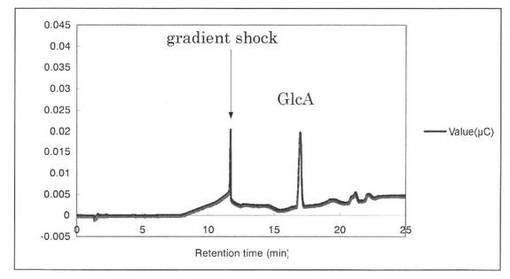
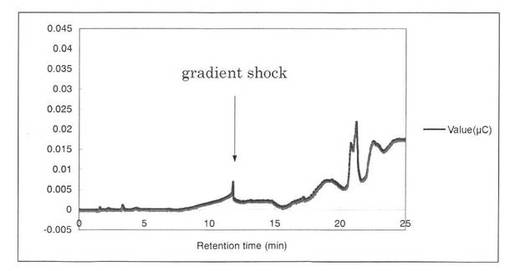
【図12】スタンダード(0.0005%グルクロン酸)の解析パターンを示す図である。グルクロン酸(GlcA)のピークの前に、gradient shockのピークが見られた。
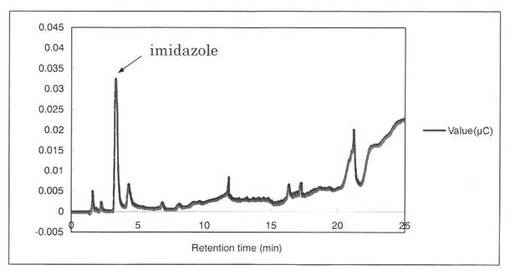
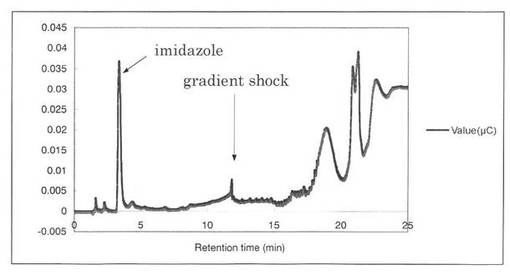
【図13】コントロールとして、SpGlcA115Aの解析パターンを示す図である。3.3分にイミダゾールのピークが見られた。
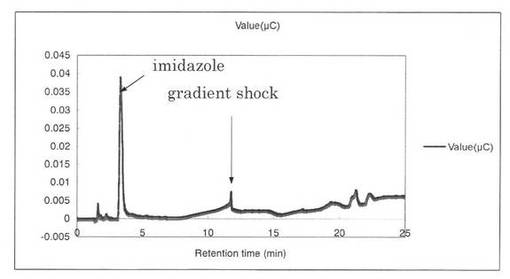
【図14】SpGlcA115Aによる、Beech wood xylanの分解プロダクトパターン(反応0時間)を示す図である。
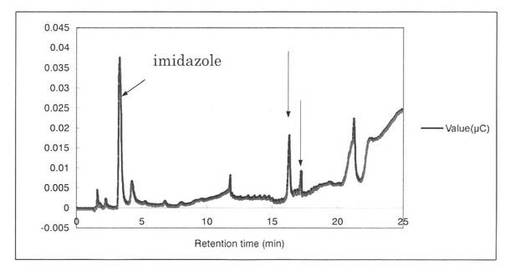
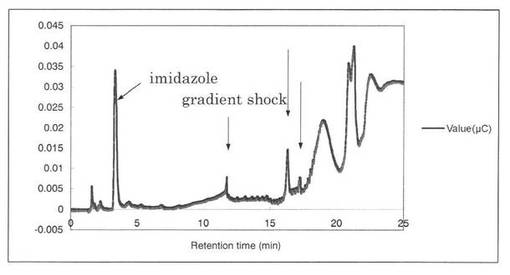
【図15】SpGlcA115Aによる、Beech wood xylanの分解プロダクトパターン(反応7時間)を示す図である。16.3分、17.2分の増加したピークを矢印で示す。
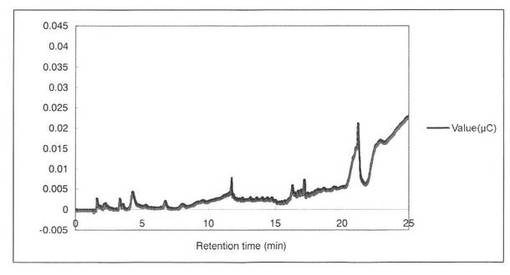
【図16】コントロールとして、Beech wood xylanのみの解析パターンを示す図である。
【図17】SpGlcA115Aによる、Birch wood xylanの分解プロダクトパターン(反応0時間)を示す図である。
【図18】SpGlcA115Aによる、Birch wood xylanの分解プロダクトパターン(反応7時間)を示す図である。
【図19】コントロールとして、Birch wood xylanのみの解析パターンを示す図である。
【図20】SpGlcA115Aによる、Birch wood xylanの分解のTLC解析結果を示す図である。
【図21】セルクラスト1.5Lによる、バイオマス(水熱処理イナワラ:160℃、10分間処理)の糖化率を示す図である。
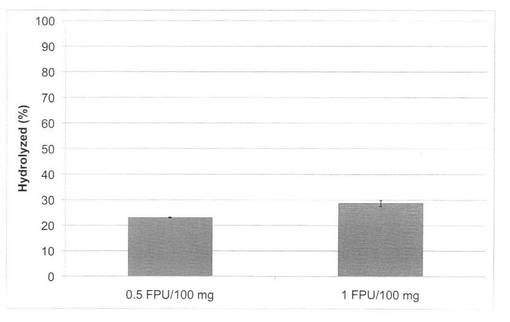
【図22】セルクラスト1.5Lによる、バイオマス(水熱処理イナワラ:180℃、10分間処理)の糖化率を示す図である。
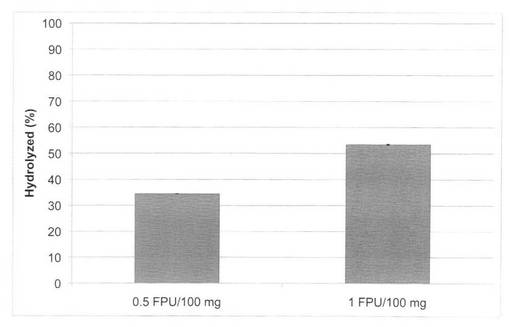
【図23】放線菌酵素(SAV1853)添加による、セルクラスト1.5Lの水熱処理イナワラ(160℃、10分間処理)の糖化率向上効果を示す図である。
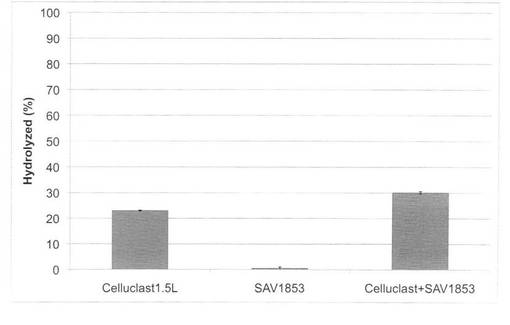
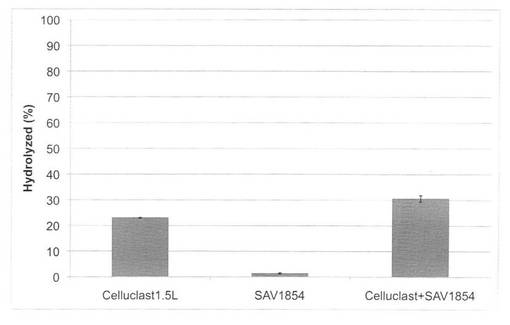
【図24】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理イナワラ(160℃、10分間処理)の糖化率向上効果を示す図である。
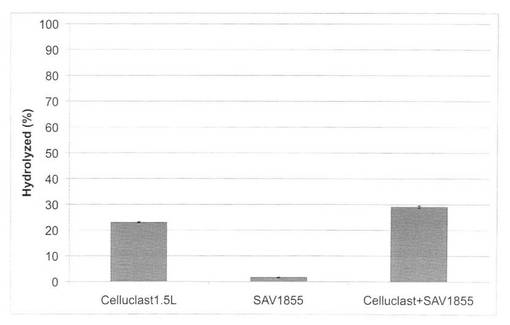
【図25】放線菌酵素(SAV1855)添加による、セルクラスト1.5Lの水熱処理イナワラ(160℃、10分間処理)の糖化率向上効果を示す図である。
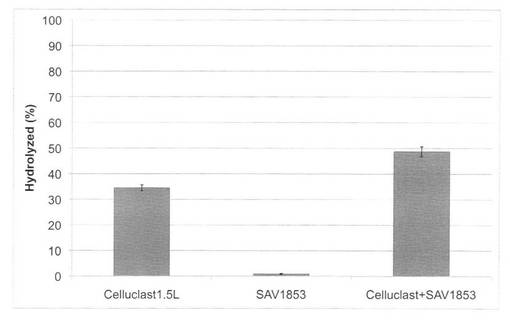
【図26】放線菌酵素(SAV1853)添加による、セルクラスト1.5Lの水熱処理イナワラ(180℃、10分間処理)の糖化率向上効果を示す図である。
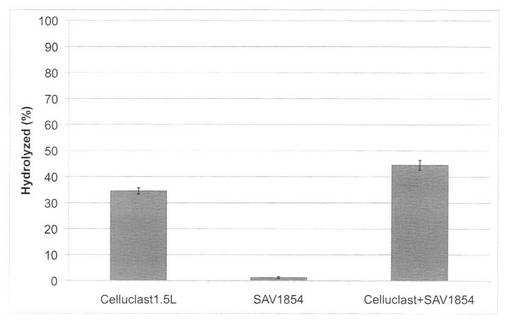
【図27】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理イナワラ(180℃、10分間処理)の糖化率向上効果を示す図である。
【図28】放線菌酵素(SAV1855)添加による、セルクラスト1.5Lの水熱処理イナワラ(180℃、10分間処理)の糖化率向上効果を示す図である。
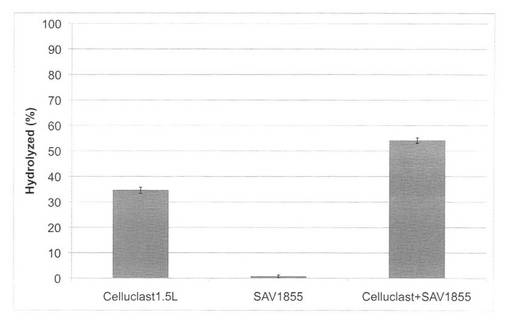
【図29】放線菌由来酵素(GH115;SpGlcA115A)添加による、キシラナーゼのキシランの糖化率向上効果を示す図である。
【図30】放線菌酵素(SAV1853)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図31】放線菌酵素(SAV1855)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図32】放線菌酵素(SAV1764)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図33】放線菌酵素(SAV2574)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図34】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの未処理イナワラの糖化率向上効果を示す図である。
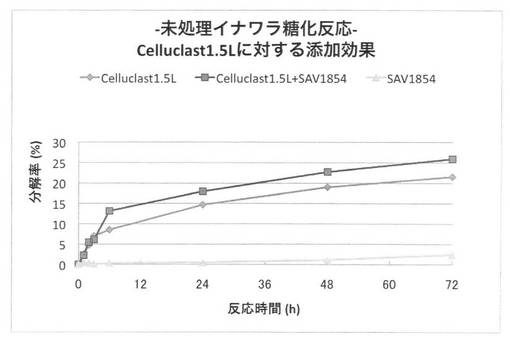
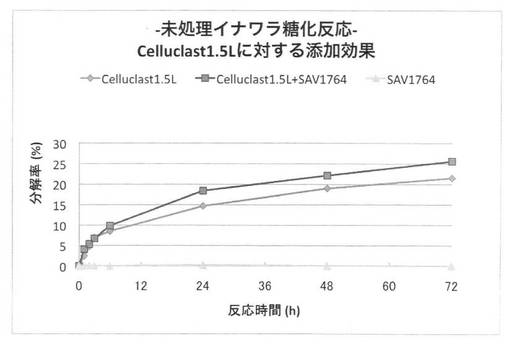
【図35】放線菌酵素(SAV1764)添加による、セルクラスト1.5Lの未処理イナワラの糖化率向上効果を示す図である。
【図36】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理イナワラの糖化率向上効果を示す図である。
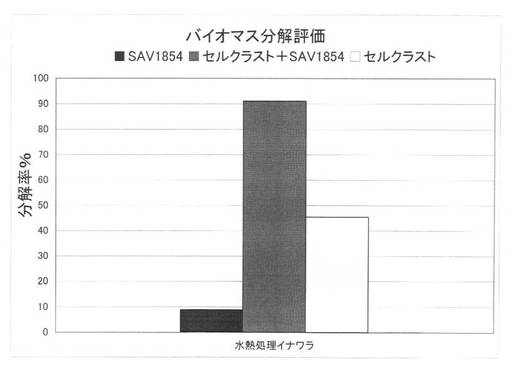
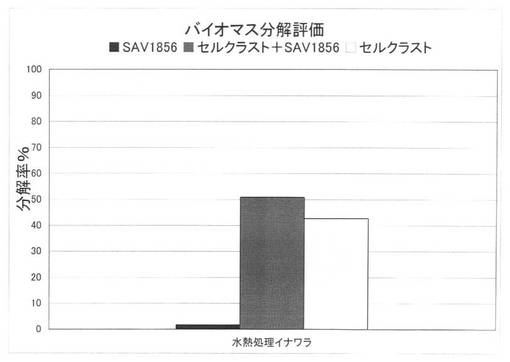
【図37】放線菌酵素(SAV1856)添加による、セルクラスト1.5Lの水熱処理イナワラの糖化率向上効果を示す図である。
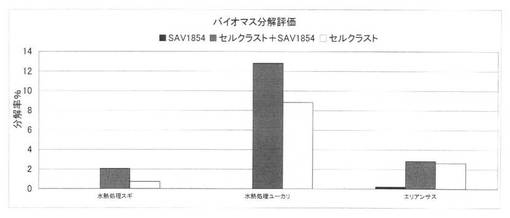
【図38】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理スギ、水熱処理ユーカリ、エリアンサスの糖化率向上効果を示す図である。
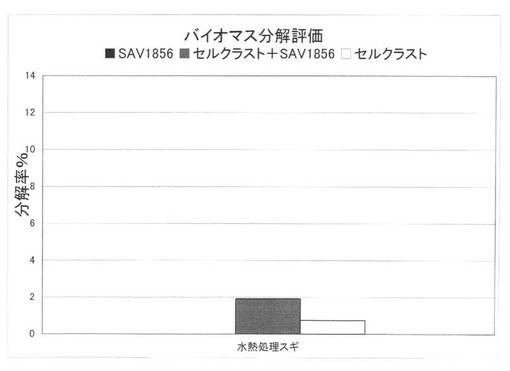
【図39】放線菌酵素(SAV1856)添加による、セルクラスト1.5Lの水熱処理スギの糖化率向上効果を示す図である。
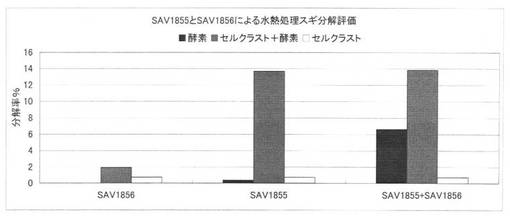
【図40】放線菌酵素(SAV1855、SAV1856)併用による水熱処理スギの糖化率向上効果を示す図である。
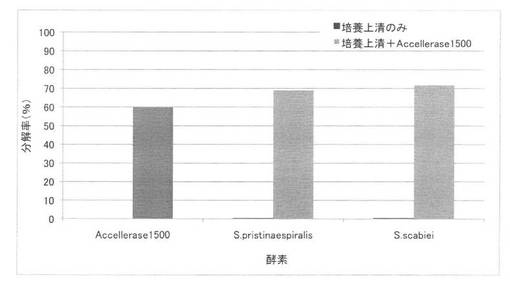
【図41】放線菌Streptomyces pristinaespiralis又はS. scabieiのイナワラ培養上清の添加では、アクセルレース1500のアルカリ処理イナワラ糖化率向上効果が得られないことを示す図である。
【発明を実施するための形態】
【0013】
本発明におけるリグノセルロース系バイオマスは、セルロース、ヘミセルロース、及びリグニンから構成されるリグノセルロースを主成分とするバイオマスを意味し、具体的には、木本植物・間伐材・廃材・おがくず等を含む木質バイオマス、竹・ケナフ・バガス・稲や麦わら・バナナ等の非木質バイオマス、葦・エレファントグラス等の草本植物、新聞・雑誌・台帳・段ボール等の回収古紙などを挙げることができる。
【0014】
上記のリグノセルロース系バイオマスに含まれる多糖類を加水分解することによって得られる単糖類としては、グルコース、ガラクトース、マンノース、フルクトース、キシロース、ラムノース、フコース及びアラビノースなどの単糖類、ガラクツロン酸、グルクロン酸などのウロン酸、水酸基がエステル化されたエステル誘導体、水酸基がメチル基などで誘導体化されたアルキル誘導体を例示することができる。
【0015】
本発明において、ストレプトマイセス属に属する微生物由来の組換えタンパク質としては、配列番号1、3、5、7、9、11又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトミセス・エバーミチルス(Streptomyces avermitilis)由来の組換えタンパク質や、配列番号2、4、6、8、10、12又は16で表されるアミノ酸配列からなるストレプトミセス・エバーミチルス由来の組換えタンパク質や、配列番号13で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス・プリスチナエスピラリス(Streptomyces pristinaespiralis)由来の組換えタンパク質や、配列番号14で表されるアミノ酸配列からなるストレプトマイセス・プリスチナエスピラリス由来の組換えタンパク質を挙げることができる。上記ストレプトマイセス・エバーミチルスとして、例えば、MA−4680株がある。このMA−4680株は、独立行政法人製品評価技術基盤機構 生物遺伝資源部門(NBRC)に、ストレプトマイセス・エバーミチルスMA−4680(NBRC番号:14893)として寄託されている。また、ストレプトマイセス・プリスチナエスピラリスとしては、独立行政法人製品評価技術基盤機構 生物遺伝資源部門(NBRC)に、ストレプトマイセス・プリスチナエスピラリス(NBRC番号:13074)として寄託されている菌株を例示することができる。
【0016】
上記組換えタンパク質は、そのアミノ酸配列に対応する公知のDNAの配列情報を基に、遺伝子工学的手法を用いて常法により調製することができる。例えば、ストレプトミセス・エバーミチルスやストレプトミセス・プリスチナエスピラリスより、常法に従ってゲノムDNAを調製し、次いで、このゲノムDNAから、配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなる遺伝子DNAに特有の適当なプローブ又はプライマーセット、好ましくは制限酵素の認識サイトを備えたプライマーセットを用いて所望クローンを選抜することにより、目的とする遺伝子DNAを取得することができる。かかる所定の遺伝子DNAを放線菌又は大腸菌用発現ベクターに適切にインテグレイトすることにより組換えベクターを構築し、この組換えベクターで放線菌又は大腸菌宿主を形質転換し、この形質転換体を培養することにより、目的とする組換えタンパク質を得ることができる。かかる組換えタンパク質は、マーカータンパク質及び/又はペプチドタグとが結合している融合タンパク質とすることもできる。このようにして得られる組換えタンパク質は通常の方法に従って、例えばイオン交換樹脂、分配クロマトグラフィー、ゲルクロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)、向流分配法等のペプチド化学の分野で汎用されている方法に従って、適宜その精製を行うことができる。
【0017】
本発明において、配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性、好ましくはリグノセルロース系バイオマス糖化酵素活性を有する組換えタンパク質(組換え変異タンパク質)における「1又は数個のアミノ酸残基が置換、欠失、挿入又は付加されたアミノ酸配列」とは、例えば1〜10個、好ましくは1〜5個、より好ましくは1〜3個、さらに好ましくは1〜2個の任意の数のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を意味する。かかる組換えタンパク質は、例えば配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的な手法等を用いて、これらDNAに変異を導入することにより、変異DNAを取得し調製することができる。上記遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989.(以後 "モレキュラークローニング第2版" と略す)、Current Protocols in Molecular Biology, Supplement 1〜38,John Wiley & Sons (1987-1997)等に記載の方法に準じて行うことができる。この変異DNAを適切な発現系を用いて発現させることにより、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質を得ることができる。
【0018】
本発明において、配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性、好ましくはリグノセルロース系バイオマス糖化酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質(組換え変異タンパク質)における「ストリジェントな条件下」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいい、より具体的には、70%以上、より好ましくは85%以上、さらに好ましくは95%以上の相同性を有するポリヌクレオチド同士がハイブリダイズし、それより相同性が低いポリヌクレオチド同士がハイブリダイズしない条件であり、例として、通常のサザンハイブリダイゼーションの洗浄条件である65℃、1×SSC、0.1%SDS、又は0.1×SSC、0.1%SDSに相当する塩濃度でハイブリダイズする条件を挙げることができる。また、上記「ストリンジェントな条件下でハイブリダイズするヌクレオチド」とは、DNA又はRNAなどの核酸をプローブとして使用し、コロニー・ハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるヌクレオチドを意味し、具体的には、コロニーあるいはプラーク由来のポリヌクレオチドの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2倍程度のSSC溶液(1倍濃度のSSC溶液の組成は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるヌクレオチドをあげることができる。ハイブリダイゼーションは、例えば、モレキュラー・クローニング、ア・ラボラトリーマニュアル(Molecular cloning, A laboratory manual)、第3版、第6章に記載の方法で行うことができる。
【0019】
上記組換え変異タンパク質における加水分解酵素活性、好ましくはリグノセルロース系バイオマス糖化酵素活性としては、塩基配列が配列番号1、アミノ酸配列が配列番号2の組換え変異タンパク質の場合はセロビオヒドロラーゼ活性を;塩基配列が配列番号3、アミノ酸配列が配列番号4の組換え変異タンパク質の場合はグルコマンナナーゼ活性を;塩基配列が配列番号5、アミノ酸配列が配列番号6の組換え変異タンパク質の場合はエンドグルカナーゼ活性を;塩基配列が配列番号7、アミノ酸配列が配列番号8の組換え変異タンパク質の場合はエキソ型キシログルカナーゼ活性を;塩基配列が配列番号9、アミノ酸配列が配列番号10の組換え変異タンパク質の場合はエンド型キシログルカナーゼ活性を;塩基配列が配列番号11、アミノ酸配列が配列番号12の組換え変異タンパク質の場合はエンド−β−1,3−グルカナーゼ活性を;塩基配列が配列番号13、アミノ酸配列が配列番号14の組換え変異タンパク質の場合はα−グルクロニダーゼ活性を;塩基配列が配列番号15、アミノ酸配列が配列番号16の組換え変異タンパク質の場合はキシラナーゼ活性を、それぞれ好適に例示することができる。
【0020】
本発明において、前記のストレプトマイセス属に属する微生物由来の組換えタンパク質や、上記組換え変異タンパク質と併用される、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来のリグノセルロース系バイオマス糖化酵素としては、セルロースの加水分解酵素(セルラーゼ)、ヘミセルロースの加水分解酵素(ヘミセルラーゼ)、リグニン分解酵素(リグニナーゼ)の他、ペクチン質を分解する酵素(ペクチナーゼ)も含まれる。これらのリグノセルロース系バイオマス糖化酵素は、市販酵素製剤を用いても、あるいは公知の方法で調製した酵素製剤を用いてもよい。また、セルラーゼ、ヘミセルラーゼ、リグニナーゼ、ペクチナーゼの2種以上の混合物を用いることもできる。
【0021】
上記セルラーゼとしては、エンド−β−1,4−グルカナーゼ、エキソ−β−1,4ーグルカナーゼ、β−グルコシダーゼ、カルボキシメチルセルラーゼ等を挙げることができ、その由来としては、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来であれば特に制限されず、高等植物、細菌、糸状菌、木材腐朽菌、シロアリの共生原生動物などに由来する天然のセルラーゼを用いてもよいし、遺伝子工学的方法で人工的に産生したものを用いてもよい。セルラーゼの由来として具体的には、トリコデルマ属(Trichoderma)菌、アスペルギルス属(Aspergillus)菌、フミコーラ属(Humicola)菌、スタフィロトリクム属(Staphylotrichum)菌、リゾプス属(Rhizopus)菌、ムコール属(Mucor)菌、アクレモニウム属(Acremonium)菌、カエトミウム属(Chaetomium)菌、アシドサーマス属(Acidothermus)菌、セルロモナス属(Cellulomonas)菌等を好適に例示することができる。
【0022】
上記ヘミセルラーゼとしては、β−キシラナーゼ、β−キシロシダーゼ、α−アラビノフラノシダーゼ、アセチルキシランエステラーゼ、グルクロニダーゼ、マンノシダーゼ、マンナナーゼ、アラビナナーゼ、ガラクタナーゼ等を挙げることができ、その由来としては、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来であれば特に制限されず、高等植物、細菌、糸状菌などに由来する天然のヘミセルラーゼを用いてもよいし、遺伝子工学的方法で人工的に産生したものを用いてもよい。ヘミセルラーゼの由来として具体的には、アスペルギルス属菌、ペニシリウム属(Penicillium)菌、フサリウム属(Fusarium)菌、トリコデルマ属菌、フミコーラ属菌、アシドサーマス属(Acidothermus)菌、アガリクス属(Agaricus)菌、バチルス属(Bacillus)菌、セルロリティカス属(Cellulolyticus)菌、サーモマイセス属(Thermomyces)菌、クロストリジウム属(Clostridium)菌、サーモトガ属(Thermotoga)菌、サーモアスクス属(Thermoascus)菌、カルドセラム属(Caldocellum)菌、サーモモノスポラ属(Thermomonospora)菌等を好適に例示することができる。
【0023】
上記リグニナーゼとしては、ラッカーゼ、リグニンペルオキシダーゼ、マンガンペルオキシダーゼ、フェノールオキシダーゼ等を挙げることができ、その由来としては、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来であれば特に制限されず、木材腐朽菌、細菌、糸状菌、木材腐朽菌などに由来する天然のリグニナーゼを用いてもよいし、遺伝子工学的方法で人工的に産生したものを用いてもよい。リグニナーゼの由来として具体的には、ファネロケーテ属(Phanerochaete)菌、プレイロータス属(Pleurotus)菌、トラメテス属(Trametes)菌、ガノデルマ属(Ganoderma)菌、レンチナス属(Lentinus)菌、ポリポラス属(Polyporus)菌、ステレウム属(Stereum)菌、フレビア属(Phlebia)菌等を好適に例示することができる。
【0024】
前記市販の酵素製剤としては、セルクラスト1.5L(ノボザイムズ社製)、アクセルレース1500(ジェネンコア協和社製)、TP−60(明治製菓社製)、ウルトラフロL(ノボザイムズ社製)、メイセラーゼ(明治製菓社製)、アクレモニウムセルラーゼ(明治製菓製)、マセロチーム(ヤクルト本社製)、セルロシンME(阪急バイオインダストリー製)、スミチームC(新日本化学工業社製)、スミチームX(新日本化学工業社製)、ビスコチーム(ノボザイムズ社製)、スクラーゼX(三菱化学フーズ社製)、ヘミセルラーゼ「アマノ」90(天野エンザイム社製)、グリンドアミルH(ダニスコ ジャパン社製)、ペントパン(ノボザイムズ社製)、及びアミログルコシダーゼ(メガザイム社製)などを挙げることができる。
【0025】
本発明おけるリグノセルロース系バイオマスの糖化工程は、従来公知の方法を適宜採用して行うことができる。まず、上記のリグノセルロース系バイオマス、好ましくはその粉砕処理物を水性媒体に懸濁して懸濁液を調製するが、水性媒体としては水、緩衝液、酸性水溶液、アルカリ性水溶液を挙げることができる。糖化(加水分解)反応は、リグノセルロース系バイオマスの粉砕物の懸濁液中に加水分解酵素を添加し、該懸濁液をインキュベート、好ましくは通気・撹拌することによって行うことができる。懸濁液中に含まれるリグノセルロース系バイオマスの濃度としては、バイオマスの種類、使用するリグノセルロース系バイオマス糖化酵素の種類によって適宜選択することができるが、水性媒体1Lに対して、通常、乾燥(無水)状態のバイオマス5〜300g、好ましくは50〜200gである。また、糖化反応条件として、pHは使用する酵素の至適pHが好ましいが、通常pH3〜9、好ましくはpH4〜7であり、また、温度は通常20〜70℃、好ましくは40〜50℃であり、反応時間は通常12〜144時間、好ましくは24〜72時間である。用いられるリグノセルロース系バイオマス糖化酵素量は、使用する酵素の種類によって適宜選択することができるが、懸濁するリグノセルロース系バイオマスの質量(乾物換算)に対し、好ましくは0.001〜15質量%、好ましくは0.01〜5質量%である。また、リグノセルロース系バイオマス糖化酵素量に対する組換えタンパク質の量は、使用するリグノセルロース系バイオマス糖化酵素及び組換えタンパク質の種類によって適宜選択することができるが、リグノセルロース系バイオマス糖化酵素の質量に対し、好ましくは0.05〜30質量%、好ましくは0.5〜15質量%である。
【0026】
また、リグノセルロース系バイオマス、好ましくはその粉砕物や爆砕処理物は、反応効率向上の観点から、加水分解反応に供する前に蒸煮処理を行うことが好ましい。さらに、加水分解反応の前に適宜酸又はアルカリ処理を行ってもよい。粉砕処理、爆砕処理、蒸煮処理、酸処理、アルカリ処理等は、従来公知の方法を用いることができる。
【0027】
本発明を実施例によりさらに具体的に説明するが、本発明の技術的範囲は実施例によって限定されるものではない。以下の実施例においては、まず、配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス属に属する微生物由来の組換えタンパク質(酵素)の基質特異性を調べた。次いで、かかる組換えタンパク質(酵素)と市販のセルラーゼ製剤とを併用し、市販のセルラーゼ製剤によるリグノセルロース系バイオマスの糖化効率が相乗的に向上することを確認した。
【実施例1】
【0028】
<Streptomyces avermitilis由来酵素について>
表1に、実施例で使用した放線菌Streptomyces avermitilis由来の酵素について示す。
SAV○○○○はStreptomyces avermitilisの遺伝子番号であり、以下その遺伝子がコードするタンパクを表している。
【0029】
【表1】

【0030】
<放線菌S. avermitilis ゲノム上の各遺伝子の位置関係>
SAV1853、SAV1854、SAV1855、SAV1856はゲノム上では図1で示される位置関係にある。
【0031】
<放線菌酵素の調製>
フォワードプライマーの5’側にNdeIサイト、リバースプライマーの5’側にHindIIIサイトをそれぞれ導入した配列番号17〜32のプライマーを用い、分泌シグナル配列を含む、配列番号1、3、5、7、9、11又は15で表される各放線菌酵素のORF全長のDNA配列をPCRで増幅した。増幅産物をpGEM-T Easy vector(プロメガ社製)にTAクローニングした。該ベクターに挿入されたPCR産物のDNA配列をシークエンスし、確認後、S.avermitilis由来7酵素は放線菌発現ベクター(特開2009−153516、Hatanaka, T., Onaka, H., Arima, J., Uraji, M., Uesugi, Y., Usuki, H., Nishimoto, Y., Iwabuchi, M., 2008. Protein Expr. Purif. 62, 244-248に記載のベクター)にNdeI-HindIIIサイトで導入した。作製した放線菌発現ベクターを放線菌Streptomyces lividans 1326に導入し、形質転換体を得た。形質転換体を液体培養することにより、組換えタンパク質は培養上清に分泌された。該培養液から菌体を除去し、得られた組換えタンパク質を含む培養上清を、粗酵素液とした。S.pristinaespiralis由来のSpGlcA115Aは、配列番号13で表される、分泌シグナル配列を含まない放線菌酵素のDNA配列をPCRで増幅し、増幅産物を大腸菌発現ベクターpET30にNdeI-HindIIIサイトで導入した。作製した大腸菌発現ベクターで形質転換した大腸菌を液体培養することにより、分泌シグナル配列を含まず、C末側にヒスチジンタグが融合した組換えタンパク質が大腸菌内に生産された。該培養液から回収した菌体を破砕した後、遠心操作を行い、得られた上清を粗酵素液とした。
【0032】
以下に各放線菌酵素の性質を示す。
<SAV1853>
SAV1853は分泌シグナル配列+CBM2+GH6のモジュラー構造からなり、配列番号1で示される塩基配列からなるDNAを発現させることにより得られる、配列番号2で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表2に示す。
【0033】
【表2】
【0034】
CM−セルロースの分解パターンを図2、3に、スタンダードとしてセロオリゴ糖の解析結果を図4に示す。反応液およびスタンダード溶液をHPAEC-PAD system(DX-500;Dionex社製)により解析した。CM−セルロースの分解物としてC2で示すセロビオースのみが生産された。すなわち、SAV1853はセロビオヒドロラーゼである。
【0035】
<SAV1854>
SAV1854は分泌シグナル配列+CBM2+GH5のモジュラー構造からなり、配列番号3で示される塩基配列からなるDNAを発現させることにより得られる、配列番号4で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表3に示す。
【0036】
【表3】
【0037】
SAV1854はKonjac glucomannanに対し高い活性を示した。すなわち、SAV1854はグルコマンナナーゼである。
【0038】
<SAV1855>
SAV1855は分泌シグナル配列+CBM2+GH48のモジュラー構造からなり、配列番号5で示される塩基配列からなるDNAを発現させることにより得られる、配列番号6で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表4に示す。
【0039】
【表4】
【0040】
CM−セルロースの分解パターンを図5、6に、スタンダードとしてセロオリゴの解析結果を図7に示す。反応液およびスタンダード溶液をHPAEC-PAD system(DX-500;Dionex社製)により解析した。CM−セルロースの分解物としてセロオリゴ糖が生産された。すなわち、SAV1855はエンドグルカナーゼである。
【0041】
<SAV1856>
SAV1856は分泌シグナル配列+CBM2+GH74のモジュラー構造からなり、配列番号7で示される塩基配列からなるDNAを発現させることにより得られる、配列番号8で示されるアミノ酸配列からなる酵素である。
<SAV2574>
SAV2574は分泌シグナル配列+GH74のモジュラー構造からなり、配列番号9で示される塩基配列からなるDNAを発現させることにより得られる、配列番号10で示されるアミノ酸配列からなる酵素である。
各基質に対するSAV1856、SAV2574の活性(基質特異性)を表5に示す。
【0042】
【表5】
【0043】
SAV1856、SAV2574はTamarind xyloglucanに対して高い活性を示した。すなわち、SAV1856、SAV2574は共にキシログルカナーゼである。
【0044】
Tamarind xyloglucanの分解反応液をHPAEC-PAD system(DX-500;Dionex社製)により解析したパターンを図8に示す。SAV2574により様々な長さのキシログルカンオリゴ糖が遊離していることから、SAV2574はエンド型キシログルカナーゼであることが分かる。一方、SAV1856は、ある決まった長さのオリゴ糖が反応初期から生産され、反応時間を経て蓄積されている。この結果から、SAV1856はエキソ型キシログルカナーゼであると考えられる。
【0045】
<SAV1764>
SAV1764は分泌シグナル配列+GH16+CBM6のモジュラー構造からなり、配列番号11で示される塩基配列からなるDNAを発現させることにより得られる、配列番号12で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表6に示す。
【0046】
【表6】
【0047】
ラミナリンの分解パターンを図9、10に、スタンダードとしてグルコース・ラミナリオリゴ等の解析結果を図11に示す。反応液およびスタンダード溶液をHPAEC-PAD system(DX-500;Dionex社製)により解析した。ラミナリンの分解物としてグルコースGとラミナリオリゴ糖G2、G3、G4が生産された。すなわち、SAV1764はエンド−β−1,3−グルカナーゼである。
【0048】
<SAV2096>
SAV2096は分泌シグナル配列+GH10からなり、配列番号15で示される塩基配列からなるDNAを発現させることにより得られる、配列番号16で示されるアミノ酸配列からなる酵素で、キシラナーゼである。
【0049】
<Streptomyces pristinaespiralis由来酵素について>
表7に、以下で使用した放線菌Streptomyces pristinaespiralis由来の酵素SpGlcA115Aについて示す。
【0050】
【表7】
【0051】
<SpGlcA115A>
SpGlcA115Aは分泌シグナル配列+GH115からなる酵素であり、配列番号13で示される塩基配列からなるDNAを発現させることにより得られる、配列番号14で示されるアミノ酸配列からなる酵素である。
SpGlcA115Aによるキシラン分解活性を調べた。
【0052】
反応液
2%基質 (Beech wood xylan又はBirch wood xylan)
50mM acetate buffer、pH5.0
リコンビナントSpGlcA115A(10mg)
40℃、0−7時間
反応液をHPAEC-PAD system(DX-500;Dionex社製)により解析した結果を図12〜19に示す。スタンダードとしてグルクロン酸を解析した結果、グルクロン酸のピークの前に、gradient shockのピークが見られた(図12)。また、コントロールとしてSpGlcA115Aのみを解析した結果、3.3分にイミダゾールのピークが見られた(図13)。SpGlcA115AとBeech wood xylanを0時間(図14)又は7時間(図15)反応させた反応液の解析では、反応7時間において16.3分、17.2分のピークが増加した。スタンダードとして、基質Beech woodのみの解析結果を図16に示す。SpGlcA115AとBirch wood xylanを0時間又は7時間反応させた反応液の解析結果をそれぞれ図17、図18に示す。スタンダードとして、基質Birch woodのみの解析結果を図19に示す。
【0053】
反応液を以下に示す。
2% Birch wood xylan
50mM 酢酸バッファー(pH5.0)
40℃、0−48時間
酢酸エチル:酢酸:1−プロパノール:ギ酸: 水=25:10:5:1:15
スポッティング量
スタンダード(キシロース、グルクロン酸);1%反応液1μl
Sample; 反応液4μl
N-(1-naphtyl)ethylenediamine dihydrochloride1.8g/200mlエタノール/20ml conc.H2SO4
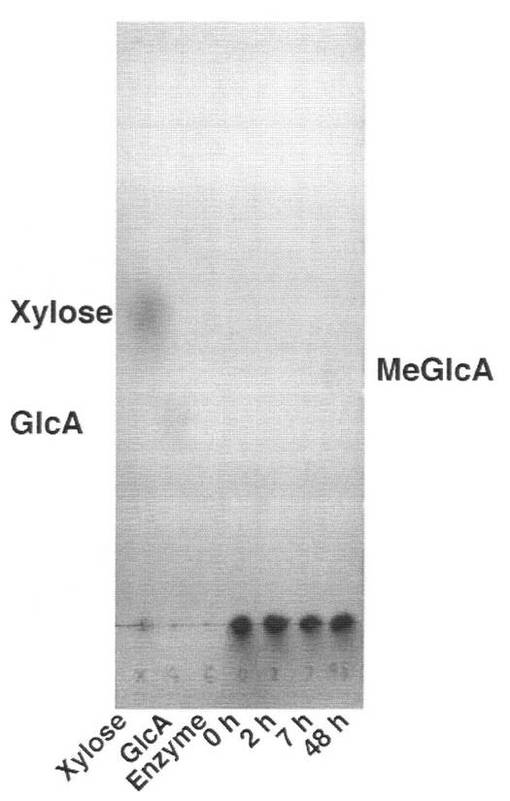
反応産物を、TLCにより検出した結果を図20に示す。MeGlcAに対応する弱いシグナルのスポットが検出された。すなわち、SpGlcA115Aはα−グルクロニダーゼである。
【実施例2】
【0054】
放線菌Streptomyces avermitilis NBRC14893由来酵素群を利用したイナワラの糖化方法
;セルクラスト1.5L(ノボザイムズ社製)と放線菌酵素による糖化方法
反応液の組成を表8に示す。
【0055】
【表8】
【0056】
反応:50℃、1000rpmで振とうしながら1日間(n=2又は3)
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上記反応系において使用した酵素タンパク質の量は(BCA法により測定)、
セルクラスト1.5L:4.5mg
放線菌酵素:約35μg
である。
【0057】
上の反応系において、セルクラスト1.5L(ノボザイムズ社製)をそれぞれ0.5FPU/100mg biomass又は1FPU/100mg biomassになるように加え、それぞれ160℃又は180℃で10分間処理した場合の糖化率は、図21、22のようになった。
【0058】
セルクラスト1.5L(ノボザイムズ社製)0.5FPU/100mg biomassの条件で、放線菌酵素(SAV1853、SAV1854、SAV1855)を添加し、160℃又は180℃で10分間処理した場合の糖化率を図23〜28に示す。微量の放線菌酵素を添加することにより、セルクラストを2倍量使用した糖化率とほぼ同等の糖化率を得ることができた。
【実施例3】
【0059】
放線菌由来酵素のイナワラ糖化への応用
;Streptomyces avermitilis由来GH10酵素(キシラナーゼ)
;Streptomyces pristinaespiralis由来GH115酵素(α−グルクロニダーゼ)SpGlcA115A
反応液の組成を表9に示す。
【0060】
【表9】
【0061】
反応:50℃、1000rpmで振とうしながら2日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
【0062】
上の反応系における、キシラン糖化率を図29に示す。微量のSpGlcA115A(GH115酵素)をキシラナーゼ(GH10酵素)に添加することにより、キシラン分解率を上昇させることができた。
【実施例4】
【0063】
放線菌Streptomyces avermitilis NBRC14893由来酵素群を利用したイナワラの糖化方法
;アクセルレース1500(ジェネンコア協和)と放線菌酵素による糖化方法
反応液の組成を表10に示す。
【0064】
【表10】
【0065】
反応:50℃、1000rpmで振とうしながら1日間(n=2又は3)
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上記反応系に使用した酵素タンパク質の量は(BCA法により測定)、
アクセルレース1500:350μg
放線菌酵素:約35μg
である。
【0066】
上の反応系において、放線菌酵素にSAV1853、SAV1855、SAV1764、SAV2574をそれぞれ添加した場合のアルカリ処理イナワラのキシラン糖化率は図30〜33となる。セルラーゼ製剤に各放線菌酵素を添加することにより、糖化率を上昇させることができた。
【実施例5】
【0067】
放線菌Streptomyces avermitilis NBRC14893由来酵素群を利用したイナワラの糖化方法
;セルクラスト1.5L(ノボザイムズ社製)と放線菌酵素による糖化方法
反応液の組成を表11に示す。
【0068】
【表11】
【0069】
反応:50℃、1000rpmで振とうしながら3日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上記反応系において使用した粗酵素タンパク量(Bradford法により測定)は、
X=60μg(SAV1854)、65μg(SAV1764)である。
上の反応系において、放線菌酵素にSAV1854又はSAV1764を使用した場合の未処理イナワラ粉末の糖化率を、それぞれ図34、35に示す。前処理を施していないイナワラの糖化において、セルラーゼ製剤に放線菌酵素を添加することにより、糖化率を上昇させることができた。
【実施例6】
【0070】
放線菌Streptomyces avermitilis NBRC14893由来酵素を利用したバイオマスの糖化方法
;セルクラスト1.5L(ノボザイムズ社製)と放線菌酵素による糖化方法
反応液の組成を表12に示す。
【0071】
【表12】
【0072】
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上の反応系において、放線菌酵素にSAV1854又はSAV1856を使用した場合の水熱処理イナワラの糖化率を、それぞれ図36、図37に示す。水熱処理を施したイナワラ、すなわちバイオマスの糖化において、セルラーゼ製剤に放線菌酵素を添加することにより、糖化率を上昇させることができた。
【0073】
反応液の組成を表13及び表14に示す。
【0074】
【表13】
【0075】
【表14】
【0076】
反応:45℃、1000rpmで振とうしながら1日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上の反応系において、SAV1854を添加した場合のセルクラスト1.5Lによる水熱処理スギ、水熱処理ユーカリ、未処理エリアンサス分解評価の結果を図38に、SAV1856を添加した場合のセルクラスト1.5Lによる水熱処理スギ分解評価の結果を図39に示す。分解率は小さいが、放線菌酵素を添加することにより、スギ、ユーカリのように分解の困難な基質でも糖化率を上昇させることができた。
【0077】
上の反応系において、SAV1855及びSAV1856を添加した場合のセルクラスト1.5Lによる水熱処理スギの分解評価の結果を図40に示す。放線菌酵素を併用した場合には、その組み合わせによりシナジー効果が得られ、放線菌酵素のみでの糖化率が上昇した。その結果、反応系の糖化率を高めることが可能である。
【実施例7】
【0078】
<放線菌のイナワラ培養上清をセルラーゼ製剤に添加した場合の糖化率>
【0079】
【表15】
【0080】
反応:50℃、1000rpmで振とうしながら1日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
アルカリ処理イナワラを基質とし、セルラーゼ製剤に加えて放線菌のイナワラ培養上清を添加した場合の糖化反応の結果を図41に示す。アクセルレース1500のみによる糖化率を一番左に、放線菌Streptomyces pristinaespiralis又はS. scabieiのイナワラ培養上清のみによる糖化率をそれぞれ左から2番目、4番目に、アクセルレース1500に各培養上清を添加した場合の糖化率をそれぞれ左から3番目、5番目のカラムで示した。アクセルレース1500に単に放線菌の培養上清を加えただけでは、糖化率は十分に上昇しなかった。
【技術分野】
【0001】
本発明は、リグノセルロース系バイオマスの糖化方法、より詳しくは、ストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素、好ましくはストレプトマイセス属に属する微生物以外の生物由来のリグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化するリグノセルロース系バイオマスの糖化方法等に関する。
【背景技術】
【0002】
バイオマスの利用は、新しい資源循環型の社会システムの構築の観点から重要である。バイオ燃料への世界的ニーズの高まりに対応して、糖質系バイオマス由来のバイオエタノール製造技術開発競争が世界的規模で繰り広げられている。特に、食料資源と競合しないリグノセルロース系バイオマスの利用技術開発が、欧米のみならず我が国においても最も重要なブレイクスルーとなりうると考えられている。現在セルロース系バイオマスの効率のよい糖化のため、酵素糖化による方法の研究開発が進められている。
【0003】
リグノセルロースは地球上で最も多量に存在する有機物で、構造性多糖のセルロース及びヘミセルロース、芳香族化合物の重合体のリグニンから構成される。セルロースはグルコースがβ−1,4−グリコシド結合により重合した直鎖状の多糖類であり、ヘミセルロースはセルロースと共に植物細胞壁を構成するセルロース以外の多糖類であって、ホモ多糖類及びヘテロ多糖類の総称をいう。ヘミセルロースとしては、例えば、マンナン、グルコマンナン、ガラクタン、キシラン、アラビナン、アラビノキシラン、アラビノガラクタン、ガラクトグルコマンナン、グルクロノキシラン、キシログルカンなどが挙げられる。また、リグニンは、高等植物の木化に関与する高分子のフェノール性化合物で、光合成(一次代謝)により同化された炭素化合物が更なる代謝(二次代謝)を受けることで合成されるフェニルプロパノイドのうち、p−クマリルアルコール・コニフェニルアルコール・シナピルアルコールという3種類のリグニンモノマーが、酵素(ラッカーゼ・ペルオキシダーゼ)の触媒の元で一電子酸化されフェノキシラジカルとなり、これがランダムなラジカルカップリングで高度に重合することにより三次元網目構造を形成した、巨大な生体高分子である。その構造は複雑で、いまだにはっきりとはわかっていない。リグニンは木材中の20〜30%を占めており、高等植物では生育に伴い、道管・仮道管・繊維などの組織でリグニンが生産される。生産されたリグニンはヘミセルロースと同じくセルロースミクロフィブリルに付着していく。まず細胞間層で堆積が始まり、徐々に一次壁、二次壁へと沈着する。同時にヘミセルロースも堆積し、木化してゆく。構造はランダムでアモルファスである。木部の組織はリグニン構造だけ残存して殆どの細胞は死細胞となり、通導・樹体支持を担う。腐朽・食害への抵抗性を有する。生体高分子としては反応性が乏しく、リグニンを分解することが可能な生物は白色腐朽菌のみであるが、白色腐朽菌などにより低分子化されたリグニンはバクテリアにより分解され、無機化することが知られている。
【0004】
リグノセルロース系バイオマスの糖化には、酸糖化と酵素糖化の2つの方法があり、技術改善が進められている(例えば、特許文献1や2)。酵素糖化法には酸糖化と比べ、低温で糖化できることや過分解が起こらず収率が高いなどの利点がある。
【0005】
酵素糖化に用いられる市販の糖化酵素セルラーゼ製剤のほとんどは糸状菌Trichoderma reesei由来であり、これは他の微生物に比べてセルラーゼ生産量が極めて多く、安価に大量の酵素製剤を製造することが可能であるためである。リグノセルロース系バイオマスの主要成分は、セルロース・ヘミセルロース・リグニンであるが、ヘミセルロースに着目すると、その種類や含有量はリグノセルロース系バイオマスの種類によって異なる。放線菌は糸状菌とは生産する糖化酵素の種類が異なっており、ヘミセルロース分解酵素が豊富な微生物である。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開2009−189291号公報
【特許文献2】特開2008−266228号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の課題は、リグノセルロース系バイオマス糖化酵素、好ましくは広く利用されている糸状菌由来のセルラーゼ等、放線菌以外の生物に由来するリグノセルロース系バイオマス糖化酵素による、リグノセルロース系バイオマスの糖化を、効率よく行う方法を提供することにある。
【課題を解決するための手段】
【0008】
本発明は、自然界では一種類の微生物ではなく様々な微生物が生産する酵素が複合して作用することにより、バイオマス分解が行われていることから、放線菌が生産する酵素に着目し、放線菌由来糖化酵素を市販セルラーゼ製剤に添加したところ、リグノセルロース系バイオマスを効率よく糖化することができることを見いだし、本発明を完成するに至った。
【0009】
すなわち本発明は、(1)配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(2)配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(3)配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(4)配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法や、(5)リグノセルロース系バイオマス糖化酵素が、ストレプトマイセス属に属する微生物以外の生物由来のリグノセルロース系バイオマス糖化酵素であることを特徴とする上記(1)〜(4)のいずれか記載のリグノセルロース系バイオマスの糖化方法に関する。
【0010】
また、本発明は、(6)配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法や、(7)配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法や、(8)配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法や、(9)配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法に関する。
【発明の効果】
【0011】
市販セルラーゼ製剤への放線菌由来糖化酵素の添加により、市販製剤の糖化能を高めることができた。本発明によると、多様なリグノセルロース系バイオマスの糖化に対応できる糖化方法を提供することができる。
【図面の簡単な説明】
【0012】
【図1】放線菌S. avermitilis ゲノム上の各酵素遺伝子の位置関係を示す図である。
【図2】SAV1853によるCM−セルロースの分解プロダクトパターン(反応3時間)を示す図である。C2はセロビオースを示す。
【図3】SAV1853によるCM−セルロースの分解プロダクトパターン(反応9時間)を示す図である。C2はセロビオースを示す。
【図4】スタンダード(セロオリゴ糖)の解析パターンを示す図である。C2はセロビオース、C3はセロトリオース、C4はセロテトラオース、C5はセロペンタオース、C6はセロヘキサオースを示す。
【図5】SAV1855によるCM−セルロースの分解プロダクトパターン(反応3時間)を示す図である。
【図6】SAV1855によるCM−セルロースの分解プロダクトパターン(反応9時間)を示す図である。C2はセロビオース、C3はセロトリオース、C4はセロテトラオース、C5はセロペンタオース、C6はセロヘキサオースを示す。
【図7】スタンダード(セロオリゴ糖)の解析パターンを示す図である。C2はセロビオース、C3はセロトリオース、C4はセロテトラオース、C5はセロペンタオース、C6はセロヘキサオースを示す。
【図8】A)SAV2574、B)SAV1856によるTamarind xyloglucanの分解プロダクトパターンをそれぞれ示す図である。
【図9】SAV1764による、ラミナリンの分解プロダクトパターン(反応0時間)を示す図である。
【図10】SAV1764による、ラミナリンの分解プロダクトパターン(反応0.5時間)を示す図である。Gはグルコース、G2はラミナリビオース、G3はラミナリトリオース、G4はラミナリテトラオースを示す。
【図11】スタンダード(グルコース・ラミナリオリゴ糖)の解析パターンを示す図である。Gはグルコース、G2はラミナリビオース、G3はラミナリトリオース、G4はラミナリテトラオースを示す。
【図12】スタンダード(0.0005%グルクロン酸)の解析パターンを示す図である。グルクロン酸(GlcA)のピークの前に、gradient shockのピークが見られた。
【図13】コントロールとして、SpGlcA115Aの解析パターンを示す図である。3.3分にイミダゾールのピークが見られた。
【図14】SpGlcA115Aによる、Beech wood xylanの分解プロダクトパターン(反応0時間)を示す図である。
【図15】SpGlcA115Aによる、Beech wood xylanの分解プロダクトパターン(反応7時間)を示す図である。16.3分、17.2分の増加したピークを矢印で示す。
【図16】コントロールとして、Beech wood xylanのみの解析パターンを示す図である。
【図17】SpGlcA115Aによる、Birch wood xylanの分解プロダクトパターン(反応0時間)を示す図である。
【図18】SpGlcA115Aによる、Birch wood xylanの分解プロダクトパターン(反応7時間)を示す図である。
【図19】コントロールとして、Birch wood xylanのみの解析パターンを示す図である。
【図20】SpGlcA115Aによる、Birch wood xylanの分解のTLC解析結果を示す図である。
【図21】セルクラスト1.5Lによる、バイオマス(水熱処理イナワラ:160℃、10分間処理)の糖化率を示す図である。
【図22】セルクラスト1.5Lによる、バイオマス(水熱処理イナワラ:180℃、10分間処理)の糖化率を示す図である。
【図23】放線菌酵素(SAV1853)添加による、セルクラスト1.5Lの水熱処理イナワラ(160℃、10分間処理)の糖化率向上効果を示す図である。
【図24】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理イナワラ(160℃、10分間処理)の糖化率向上効果を示す図である。
【図25】放線菌酵素(SAV1855)添加による、セルクラスト1.5Lの水熱処理イナワラ(160℃、10分間処理)の糖化率向上効果を示す図である。
【図26】放線菌酵素(SAV1853)添加による、セルクラスト1.5Lの水熱処理イナワラ(180℃、10分間処理)の糖化率向上効果を示す図である。
【図27】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理イナワラ(180℃、10分間処理)の糖化率向上効果を示す図である。
【図28】放線菌酵素(SAV1855)添加による、セルクラスト1.5Lの水熱処理イナワラ(180℃、10分間処理)の糖化率向上効果を示す図である。
【図29】放線菌由来酵素(GH115;SpGlcA115A)添加による、キシラナーゼのキシランの糖化率向上効果を示す図である。
【図30】放線菌酵素(SAV1853)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図31】放線菌酵素(SAV1855)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図32】放線菌酵素(SAV1764)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図33】放線菌酵素(SAV2574)添加による、アクセルレース1500のアルカリ処理イナワラの糖化率向上効果を示す図である。
【図34】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの未処理イナワラの糖化率向上効果を示す図である。
【図35】放線菌酵素(SAV1764)添加による、セルクラスト1.5Lの未処理イナワラの糖化率向上効果を示す図である。
【図36】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理イナワラの糖化率向上効果を示す図である。
【図37】放線菌酵素(SAV1856)添加による、セルクラスト1.5Lの水熱処理イナワラの糖化率向上効果を示す図である。
【図38】放線菌酵素(SAV1854)添加による、セルクラスト1.5Lの水熱処理スギ、水熱処理ユーカリ、エリアンサスの糖化率向上効果を示す図である。
【図39】放線菌酵素(SAV1856)添加による、セルクラスト1.5Lの水熱処理スギの糖化率向上効果を示す図である。
【図40】放線菌酵素(SAV1855、SAV1856)併用による水熱処理スギの糖化率向上効果を示す図である。
【図41】放線菌Streptomyces pristinaespiralis又はS. scabieiのイナワラ培養上清の添加では、アクセルレース1500のアルカリ処理イナワラ糖化率向上効果が得られないことを示す図である。
【発明を実施するための形態】
【0013】
本発明におけるリグノセルロース系バイオマスは、セルロース、ヘミセルロース、及びリグニンから構成されるリグノセルロースを主成分とするバイオマスを意味し、具体的には、木本植物・間伐材・廃材・おがくず等を含む木質バイオマス、竹・ケナフ・バガス・稲や麦わら・バナナ等の非木質バイオマス、葦・エレファントグラス等の草本植物、新聞・雑誌・台帳・段ボール等の回収古紙などを挙げることができる。
【0014】
上記のリグノセルロース系バイオマスに含まれる多糖類を加水分解することによって得られる単糖類としては、グルコース、ガラクトース、マンノース、フルクトース、キシロース、ラムノース、フコース及びアラビノースなどの単糖類、ガラクツロン酸、グルクロン酸などのウロン酸、水酸基がエステル化されたエステル誘導体、水酸基がメチル基などで誘導体化されたアルキル誘導体を例示することができる。
【0015】
本発明において、ストレプトマイセス属に属する微生物由来の組換えタンパク質としては、配列番号1、3、5、7、9、11又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトミセス・エバーミチルス(Streptomyces avermitilis)由来の組換えタンパク質や、配列番号2、4、6、8、10、12又は16で表されるアミノ酸配列からなるストレプトミセス・エバーミチルス由来の組換えタンパク質や、配列番号13で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス・プリスチナエスピラリス(Streptomyces pristinaespiralis)由来の組換えタンパク質や、配列番号14で表されるアミノ酸配列からなるストレプトマイセス・プリスチナエスピラリス由来の組換えタンパク質を挙げることができる。上記ストレプトマイセス・エバーミチルスとして、例えば、MA−4680株がある。このMA−4680株は、独立行政法人製品評価技術基盤機構 生物遺伝資源部門(NBRC)に、ストレプトマイセス・エバーミチルスMA−4680(NBRC番号:14893)として寄託されている。また、ストレプトマイセス・プリスチナエスピラリスとしては、独立行政法人製品評価技術基盤機構 生物遺伝資源部門(NBRC)に、ストレプトマイセス・プリスチナエスピラリス(NBRC番号:13074)として寄託されている菌株を例示することができる。
【0016】
上記組換えタンパク質は、そのアミノ酸配列に対応する公知のDNAの配列情報を基に、遺伝子工学的手法を用いて常法により調製することができる。例えば、ストレプトミセス・エバーミチルスやストレプトミセス・プリスチナエスピラリスより、常法に従ってゲノムDNAを調製し、次いで、このゲノムDNAから、配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなる遺伝子DNAに特有の適当なプローブ又はプライマーセット、好ましくは制限酵素の認識サイトを備えたプライマーセットを用いて所望クローンを選抜することにより、目的とする遺伝子DNAを取得することができる。かかる所定の遺伝子DNAを放線菌又は大腸菌用発現ベクターに適切にインテグレイトすることにより組換えベクターを構築し、この組換えベクターで放線菌又は大腸菌宿主を形質転換し、この形質転換体を培養することにより、目的とする組換えタンパク質を得ることができる。かかる組換えタンパク質は、マーカータンパク質及び/又はペプチドタグとが結合している融合タンパク質とすることもできる。このようにして得られる組換えタンパク質は通常の方法に従って、例えばイオン交換樹脂、分配クロマトグラフィー、ゲルクロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)、向流分配法等のペプチド化学の分野で汎用されている方法に従って、適宜その精製を行うことができる。
【0017】
本発明において、配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性、好ましくはリグノセルロース系バイオマス糖化酵素活性を有する組換えタンパク質(組換え変異タンパク質)における「1又は数個のアミノ酸残基が置換、欠失、挿入又は付加されたアミノ酸配列」とは、例えば1〜10個、好ましくは1〜5個、より好ましくは1〜3個、さらに好ましくは1〜2個の任意の数のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列を意味する。かかる組換えタンパク質は、例えば配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAに対し、変異原となる薬剤と接触作用させる方法、紫外線を照射する方法、遺伝子工学的な手法等を用いて、これらDNAに変異を導入することにより、変異DNAを取得し調製することができる。上記遺伝子工学的手法の一つである部位特異的変異誘発法は特定の位置に特定の変異を導入できる手法であることから有用であり、Molecular Cloning: A laboratory Mannual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.,1989.(以後 "モレキュラークローニング第2版" と略す)、Current Protocols in Molecular Biology, Supplement 1〜38,John Wiley & Sons (1987-1997)等に記載の方法に準じて行うことができる。この変異DNAを適切な発現系を用いて発現させることにより、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるタンパク質を得ることができる。
【0018】
本発明において、配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性、好ましくはリグノセルロース系バイオマス糖化酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質(組換え変異タンパク質)における「ストリジェントな条件下」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいい、より具体的には、70%以上、より好ましくは85%以上、さらに好ましくは95%以上の相同性を有するポリヌクレオチド同士がハイブリダイズし、それより相同性が低いポリヌクレオチド同士がハイブリダイズしない条件であり、例として、通常のサザンハイブリダイゼーションの洗浄条件である65℃、1×SSC、0.1%SDS、又は0.1×SSC、0.1%SDSに相当する塩濃度でハイブリダイズする条件を挙げることができる。また、上記「ストリンジェントな条件下でハイブリダイズするヌクレオチド」とは、DNA又はRNAなどの核酸をプローブとして使用し、コロニー・ハイブリダイゼーション法、プラークハイブリダイゼーション法、あるいはサザンブロットハイブリダイゼーション法等を用いることにより得られるヌクレオチドを意味し、具体的には、コロニーあるいはプラーク由来のポリヌクレオチドの断片を固定化したフィルターを用いて、0.7〜1.0MのNaCl存在下、65℃でハイブリダイゼーションを行った後、0.1〜2倍程度のSSC溶液(1倍濃度のSSC溶液の組成は、150mM塩化ナトリウム、15mMクエン酸ナトリウム)を用い、65℃条件下でフィルターを洗浄することにより同定できるヌクレオチドをあげることができる。ハイブリダイゼーションは、例えば、モレキュラー・クローニング、ア・ラボラトリーマニュアル(Molecular cloning, A laboratory manual)、第3版、第6章に記載の方法で行うことができる。
【0019】
上記組換え変異タンパク質における加水分解酵素活性、好ましくはリグノセルロース系バイオマス糖化酵素活性としては、塩基配列が配列番号1、アミノ酸配列が配列番号2の組換え変異タンパク質の場合はセロビオヒドロラーゼ活性を;塩基配列が配列番号3、アミノ酸配列が配列番号4の組換え変異タンパク質の場合はグルコマンナナーゼ活性を;塩基配列が配列番号5、アミノ酸配列が配列番号6の組換え変異タンパク質の場合はエンドグルカナーゼ活性を;塩基配列が配列番号7、アミノ酸配列が配列番号8の組換え変異タンパク質の場合はエキソ型キシログルカナーゼ活性を;塩基配列が配列番号9、アミノ酸配列が配列番号10の組換え変異タンパク質の場合はエンド型キシログルカナーゼ活性を;塩基配列が配列番号11、アミノ酸配列が配列番号12の組換え変異タンパク質の場合はエンド−β−1,3−グルカナーゼ活性を;塩基配列が配列番号13、アミノ酸配列が配列番号14の組換え変異タンパク質の場合はα−グルクロニダーゼ活性を;塩基配列が配列番号15、アミノ酸配列が配列番号16の組換え変異タンパク質の場合はキシラナーゼ活性を、それぞれ好適に例示することができる。
【0020】
本発明において、前記のストレプトマイセス属に属する微生物由来の組換えタンパク質や、上記組換え変異タンパク質と併用される、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来のリグノセルロース系バイオマス糖化酵素としては、セルロースの加水分解酵素(セルラーゼ)、ヘミセルロースの加水分解酵素(ヘミセルラーゼ)、リグニン分解酵素(リグニナーゼ)の他、ペクチン質を分解する酵素(ペクチナーゼ)も含まれる。これらのリグノセルロース系バイオマス糖化酵素は、市販酵素製剤を用いても、あるいは公知の方法で調製した酵素製剤を用いてもよい。また、セルラーゼ、ヘミセルラーゼ、リグニナーゼ、ペクチナーゼの2種以上の混合物を用いることもできる。
【0021】
上記セルラーゼとしては、エンド−β−1,4−グルカナーゼ、エキソ−β−1,4ーグルカナーゼ、β−グルコシダーゼ、カルボキシメチルセルラーゼ等を挙げることができ、その由来としては、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来であれば特に制限されず、高等植物、細菌、糸状菌、木材腐朽菌、シロアリの共生原生動物などに由来する天然のセルラーゼを用いてもよいし、遺伝子工学的方法で人工的に産生したものを用いてもよい。セルラーゼの由来として具体的には、トリコデルマ属(Trichoderma)菌、アスペルギルス属(Aspergillus)菌、フミコーラ属(Humicola)菌、スタフィロトリクム属(Staphylotrichum)菌、リゾプス属(Rhizopus)菌、ムコール属(Mucor)菌、アクレモニウム属(Acremonium)菌、カエトミウム属(Chaetomium)菌、アシドサーマス属(Acidothermus)菌、セルロモナス属(Cellulomonas)菌等を好適に例示することができる。
【0022】
上記ヘミセルラーゼとしては、β−キシラナーゼ、β−キシロシダーゼ、α−アラビノフラノシダーゼ、アセチルキシランエステラーゼ、グルクロニダーゼ、マンノシダーゼ、マンナナーゼ、アラビナナーゼ、ガラクタナーゼ等を挙げることができ、その由来としては、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来であれば特に制限されず、高等植物、細菌、糸状菌などに由来する天然のヘミセルラーゼを用いてもよいし、遺伝子工学的方法で人工的に産生したものを用いてもよい。ヘミセルラーゼの由来として具体的には、アスペルギルス属菌、ペニシリウム属(Penicillium)菌、フサリウム属(Fusarium)菌、トリコデルマ属菌、フミコーラ属菌、アシドサーマス属(Acidothermus)菌、アガリクス属(Agaricus)菌、バチルス属(Bacillus)菌、セルロリティカス属(Cellulolyticus)菌、サーモマイセス属(Thermomyces)菌、クロストリジウム属(Clostridium)菌、サーモトガ属(Thermotoga)菌、サーモアスクス属(Thermoascus)菌、カルドセラム属(Caldocellum)菌、サーモモノスポラ属(Thermomonospora)菌等を好適に例示することができる。
【0023】
上記リグニナーゼとしては、ラッカーゼ、リグニンペルオキシダーゼ、マンガンペルオキシダーゼ、フェノールオキシダーゼ等を挙げることができ、その由来としては、ストレプトマイセス属に属する微生物以外、好ましくは放線菌以外の生物由来であれば特に制限されず、木材腐朽菌、細菌、糸状菌、木材腐朽菌などに由来する天然のリグニナーゼを用いてもよいし、遺伝子工学的方法で人工的に産生したものを用いてもよい。リグニナーゼの由来として具体的には、ファネロケーテ属(Phanerochaete)菌、プレイロータス属(Pleurotus)菌、トラメテス属(Trametes)菌、ガノデルマ属(Ganoderma)菌、レンチナス属(Lentinus)菌、ポリポラス属(Polyporus)菌、ステレウム属(Stereum)菌、フレビア属(Phlebia)菌等を好適に例示することができる。
【0024】
前記市販の酵素製剤としては、セルクラスト1.5L(ノボザイムズ社製)、アクセルレース1500(ジェネンコア協和社製)、TP−60(明治製菓社製)、ウルトラフロL(ノボザイムズ社製)、メイセラーゼ(明治製菓社製)、アクレモニウムセルラーゼ(明治製菓製)、マセロチーム(ヤクルト本社製)、セルロシンME(阪急バイオインダストリー製)、スミチームC(新日本化学工業社製)、スミチームX(新日本化学工業社製)、ビスコチーム(ノボザイムズ社製)、スクラーゼX(三菱化学フーズ社製)、ヘミセルラーゼ「アマノ」90(天野エンザイム社製)、グリンドアミルH(ダニスコ ジャパン社製)、ペントパン(ノボザイムズ社製)、及びアミログルコシダーゼ(メガザイム社製)などを挙げることができる。
【0025】
本発明おけるリグノセルロース系バイオマスの糖化工程は、従来公知の方法を適宜採用して行うことができる。まず、上記のリグノセルロース系バイオマス、好ましくはその粉砕処理物を水性媒体に懸濁して懸濁液を調製するが、水性媒体としては水、緩衝液、酸性水溶液、アルカリ性水溶液を挙げることができる。糖化(加水分解)反応は、リグノセルロース系バイオマスの粉砕物の懸濁液中に加水分解酵素を添加し、該懸濁液をインキュベート、好ましくは通気・撹拌することによって行うことができる。懸濁液中に含まれるリグノセルロース系バイオマスの濃度としては、バイオマスの種類、使用するリグノセルロース系バイオマス糖化酵素の種類によって適宜選択することができるが、水性媒体1Lに対して、通常、乾燥(無水)状態のバイオマス5〜300g、好ましくは50〜200gである。また、糖化反応条件として、pHは使用する酵素の至適pHが好ましいが、通常pH3〜9、好ましくはpH4〜7であり、また、温度は通常20〜70℃、好ましくは40〜50℃であり、反応時間は通常12〜144時間、好ましくは24〜72時間である。用いられるリグノセルロース系バイオマス糖化酵素量は、使用する酵素の種類によって適宜選択することができるが、懸濁するリグノセルロース系バイオマスの質量(乾物換算)に対し、好ましくは0.001〜15質量%、好ましくは0.01〜5質量%である。また、リグノセルロース系バイオマス糖化酵素量に対する組換えタンパク質の量は、使用するリグノセルロース系バイオマス糖化酵素及び組換えタンパク質の種類によって適宜選択することができるが、リグノセルロース系バイオマス糖化酵素の質量に対し、好ましくは0.05〜30質量%、好ましくは0.5〜15質量%である。
【0026】
また、リグノセルロース系バイオマス、好ましくはその粉砕物や爆砕処理物は、反応効率向上の観点から、加水分解反応に供する前に蒸煮処理を行うことが好ましい。さらに、加水分解反応の前に適宜酸又はアルカリ処理を行ってもよい。粉砕処理、爆砕処理、蒸煮処理、酸処理、アルカリ処理等は、従来公知の方法を用いることができる。
【0027】
本発明を実施例によりさらに具体的に説明するが、本発明の技術的範囲は実施例によって限定されるものではない。以下の実施例においては、まず、配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス属に属する微生物由来の組換えタンパク質(酵素)の基質特異性を調べた。次いで、かかる組換えタンパク質(酵素)と市販のセルラーゼ製剤とを併用し、市販のセルラーゼ製剤によるリグノセルロース系バイオマスの糖化効率が相乗的に向上することを確認した。
【実施例1】
【0028】
<Streptomyces avermitilis由来酵素について>
表1に、実施例で使用した放線菌Streptomyces avermitilis由来の酵素について示す。
SAV○○○○はStreptomyces avermitilisの遺伝子番号であり、以下その遺伝子がコードするタンパクを表している。
【0029】
【表1】
【0030】
<放線菌S. avermitilis ゲノム上の各遺伝子の位置関係>
SAV1853、SAV1854、SAV1855、SAV1856はゲノム上では図1で示される位置関係にある。
【0031】
<放線菌酵素の調製>
フォワードプライマーの5’側にNdeIサイト、リバースプライマーの5’側にHindIIIサイトをそれぞれ導入した配列番号17〜32のプライマーを用い、分泌シグナル配列を含む、配列番号1、3、5、7、9、11又は15で表される各放線菌酵素のORF全長のDNA配列をPCRで増幅した。増幅産物をpGEM-T Easy vector(プロメガ社製)にTAクローニングした。該ベクターに挿入されたPCR産物のDNA配列をシークエンスし、確認後、S.avermitilis由来7酵素は放線菌発現ベクター(特開2009−153516、Hatanaka, T., Onaka, H., Arima, J., Uraji, M., Uesugi, Y., Usuki, H., Nishimoto, Y., Iwabuchi, M., 2008. Protein Expr. Purif. 62, 244-248に記載のベクター)にNdeI-HindIIIサイトで導入した。作製した放線菌発現ベクターを放線菌Streptomyces lividans 1326に導入し、形質転換体を得た。形質転換体を液体培養することにより、組換えタンパク質は培養上清に分泌された。該培養液から菌体を除去し、得られた組換えタンパク質を含む培養上清を、粗酵素液とした。S.pristinaespiralis由来のSpGlcA115Aは、配列番号13で表される、分泌シグナル配列を含まない放線菌酵素のDNA配列をPCRで増幅し、増幅産物を大腸菌発現ベクターpET30にNdeI-HindIIIサイトで導入した。作製した大腸菌発現ベクターで形質転換した大腸菌を液体培養することにより、分泌シグナル配列を含まず、C末側にヒスチジンタグが融合した組換えタンパク質が大腸菌内に生産された。該培養液から回収した菌体を破砕した後、遠心操作を行い、得られた上清を粗酵素液とした。
【0032】
以下に各放線菌酵素の性質を示す。
<SAV1853>
SAV1853は分泌シグナル配列+CBM2+GH6のモジュラー構造からなり、配列番号1で示される塩基配列からなるDNAを発現させることにより得られる、配列番号2で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表2に示す。
【0033】
【表2】
【0034】
CM−セルロースの分解パターンを図2、3に、スタンダードとしてセロオリゴ糖の解析結果を図4に示す。反応液およびスタンダード溶液をHPAEC-PAD system(DX-500;Dionex社製)により解析した。CM−セルロースの分解物としてC2で示すセロビオースのみが生産された。すなわち、SAV1853はセロビオヒドロラーゼである。
【0035】
<SAV1854>
SAV1854は分泌シグナル配列+CBM2+GH5のモジュラー構造からなり、配列番号3で示される塩基配列からなるDNAを発現させることにより得られる、配列番号4で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表3に示す。
【0036】
【表3】
【0037】
SAV1854はKonjac glucomannanに対し高い活性を示した。すなわち、SAV1854はグルコマンナナーゼである。
【0038】
<SAV1855>
SAV1855は分泌シグナル配列+CBM2+GH48のモジュラー構造からなり、配列番号5で示される塩基配列からなるDNAを発現させることにより得られる、配列番号6で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表4に示す。
【0039】
【表4】
【0040】
CM−セルロースの分解パターンを図5、6に、スタンダードとしてセロオリゴの解析結果を図7に示す。反応液およびスタンダード溶液をHPAEC-PAD system(DX-500;Dionex社製)により解析した。CM−セルロースの分解物としてセロオリゴ糖が生産された。すなわち、SAV1855はエンドグルカナーゼである。
【0041】
<SAV1856>
SAV1856は分泌シグナル配列+CBM2+GH74のモジュラー構造からなり、配列番号7で示される塩基配列からなるDNAを発現させることにより得られる、配列番号8で示されるアミノ酸配列からなる酵素である。
<SAV2574>
SAV2574は分泌シグナル配列+GH74のモジュラー構造からなり、配列番号9で示される塩基配列からなるDNAを発現させることにより得られる、配列番号10で示されるアミノ酸配列からなる酵素である。
各基質に対するSAV1856、SAV2574の活性(基質特異性)を表5に示す。
【0042】
【表5】
【0043】
SAV1856、SAV2574はTamarind xyloglucanに対して高い活性を示した。すなわち、SAV1856、SAV2574は共にキシログルカナーゼである。
【0044】
Tamarind xyloglucanの分解反応液をHPAEC-PAD system(DX-500;Dionex社製)により解析したパターンを図8に示す。SAV2574により様々な長さのキシログルカンオリゴ糖が遊離していることから、SAV2574はエンド型キシログルカナーゼであることが分かる。一方、SAV1856は、ある決まった長さのオリゴ糖が反応初期から生産され、反応時間を経て蓄積されている。この結果から、SAV1856はエキソ型キシログルカナーゼであると考えられる。
【0045】
<SAV1764>
SAV1764は分泌シグナル配列+GH16+CBM6のモジュラー構造からなり、配列番号11で示される塩基配列からなるDNAを発現させることにより得られる、配列番号12で示されるアミノ酸配列からなる酵素である。
各基質に対する活性(基質特異性)を表6に示す。
【0046】
【表6】
【0047】
ラミナリンの分解パターンを図9、10に、スタンダードとしてグルコース・ラミナリオリゴ等の解析結果を図11に示す。反応液およびスタンダード溶液をHPAEC-PAD system(DX-500;Dionex社製)により解析した。ラミナリンの分解物としてグルコースGとラミナリオリゴ糖G2、G3、G4が生産された。すなわち、SAV1764はエンド−β−1,3−グルカナーゼである。
【0048】
<SAV2096>
SAV2096は分泌シグナル配列+GH10からなり、配列番号15で示される塩基配列からなるDNAを発現させることにより得られる、配列番号16で示されるアミノ酸配列からなる酵素で、キシラナーゼである。
【0049】
<Streptomyces pristinaespiralis由来酵素について>
表7に、以下で使用した放線菌Streptomyces pristinaespiralis由来の酵素SpGlcA115Aについて示す。
【0050】
【表7】
【0051】
<SpGlcA115A>
SpGlcA115Aは分泌シグナル配列+GH115からなる酵素であり、配列番号13で示される塩基配列からなるDNAを発現させることにより得られる、配列番号14で示されるアミノ酸配列からなる酵素である。
SpGlcA115Aによるキシラン分解活性を調べた。
【0052】
反応液
2%基質 (Beech wood xylan又はBirch wood xylan)
50mM acetate buffer、pH5.0
リコンビナントSpGlcA115A(10mg)
40℃、0−7時間
反応液をHPAEC-PAD system(DX-500;Dionex社製)により解析した結果を図12〜19に示す。スタンダードとしてグルクロン酸を解析した結果、グルクロン酸のピークの前に、gradient shockのピークが見られた(図12)。また、コントロールとしてSpGlcA115Aのみを解析した結果、3.3分にイミダゾールのピークが見られた(図13)。SpGlcA115AとBeech wood xylanを0時間(図14)又は7時間(図15)反応させた反応液の解析では、反応7時間において16.3分、17.2分のピークが増加した。スタンダードとして、基質Beech woodのみの解析結果を図16に示す。SpGlcA115AとBirch wood xylanを0時間又は7時間反応させた反応液の解析結果をそれぞれ図17、図18に示す。スタンダードとして、基質Birch woodのみの解析結果を図19に示す。
【0053】
反応液を以下に示す。
2% Birch wood xylan
50mM 酢酸バッファー(pH5.0)
40℃、0−48時間
酢酸エチル:酢酸:1−プロパノール:ギ酸: 水=25:10:5:1:15
スポッティング量
スタンダード(キシロース、グルクロン酸);1%反応液1μl
Sample; 反応液4μl
N-(1-naphtyl)ethylenediamine dihydrochloride1.8g/200mlエタノール/20ml conc.H2SO4
反応産物を、TLCにより検出した結果を図20に示す。MeGlcAに対応する弱いシグナルのスポットが検出された。すなわち、SpGlcA115Aはα−グルクロニダーゼである。
【実施例2】
【0054】
放線菌Streptomyces avermitilis NBRC14893由来酵素群を利用したイナワラの糖化方法
;セルクラスト1.5L(ノボザイムズ社製)と放線菌酵素による糖化方法
反応液の組成を表8に示す。
【0055】
【表8】
【0056】
反応:50℃、1000rpmで振とうしながら1日間(n=2又は3)
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上記反応系において使用した酵素タンパク質の量は(BCA法により測定)、
セルクラスト1.5L:4.5mg
放線菌酵素:約35μg
である。
【0057】
上の反応系において、セルクラスト1.5L(ノボザイムズ社製)をそれぞれ0.5FPU/100mg biomass又は1FPU/100mg biomassになるように加え、それぞれ160℃又は180℃で10分間処理した場合の糖化率は、図21、22のようになった。
【0058】
セルクラスト1.5L(ノボザイムズ社製)0.5FPU/100mg biomassの条件で、放線菌酵素(SAV1853、SAV1854、SAV1855)を添加し、160℃又は180℃で10分間処理した場合の糖化率を図23〜28に示す。微量の放線菌酵素を添加することにより、セルクラストを2倍量使用した糖化率とほぼ同等の糖化率を得ることができた。
【実施例3】
【0059】
放線菌由来酵素のイナワラ糖化への応用
;Streptomyces avermitilis由来GH10酵素(キシラナーゼ)
;Streptomyces pristinaespiralis由来GH115酵素(α−グルクロニダーゼ)SpGlcA115A
反応液の組成を表9に示す。
【0060】
【表9】
【0061】
反応:50℃、1000rpmで振とうしながら2日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
【0062】
上の反応系における、キシラン糖化率を図29に示す。微量のSpGlcA115A(GH115酵素)をキシラナーゼ(GH10酵素)に添加することにより、キシラン分解率を上昇させることができた。
【実施例4】
【0063】
放線菌Streptomyces avermitilis NBRC14893由来酵素群を利用したイナワラの糖化方法
;アクセルレース1500(ジェネンコア協和)と放線菌酵素による糖化方法
反応液の組成を表10に示す。
【0064】
【表10】
【0065】
反応:50℃、1000rpmで振とうしながら1日間(n=2又は3)
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上記反応系に使用した酵素タンパク質の量は(BCA法により測定)、
アクセルレース1500:350μg
放線菌酵素:約35μg
である。
【0066】
上の反応系において、放線菌酵素にSAV1853、SAV1855、SAV1764、SAV2574をそれぞれ添加した場合のアルカリ処理イナワラのキシラン糖化率は図30〜33となる。セルラーゼ製剤に各放線菌酵素を添加することにより、糖化率を上昇させることができた。
【実施例5】
【0067】
放線菌Streptomyces avermitilis NBRC14893由来酵素群を利用したイナワラの糖化方法
;セルクラスト1.5L(ノボザイムズ社製)と放線菌酵素による糖化方法
反応液の組成を表11に示す。
【0068】
【表11】
【0069】
反応:50℃、1000rpmで振とうしながら3日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上記反応系において使用した粗酵素タンパク量(Bradford法により測定)は、
X=60μg(SAV1854)、65μg(SAV1764)である。
上の反応系において、放線菌酵素にSAV1854又はSAV1764を使用した場合の未処理イナワラ粉末の糖化率を、それぞれ図34、35に示す。前処理を施していないイナワラの糖化において、セルラーゼ製剤に放線菌酵素を添加することにより、糖化率を上昇させることができた。
【実施例6】
【0070】
放線菌Streptomyces avermitilis NBRC14893由来酵素を利用したバイオマスの糖化方法
;セルクラスト1.5L(ノボザイムズ社製)と放線菌酵素による糖化方法
反応液の組成を表12に示す。
【0071】
【表12】
【0072】
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上の反応系において、放線菌酵素にSAV1854又はSAV1856を使用した場合の水熱処理イナワラの糖化率を、それぞれ図36、図37に示す。水熱処理を施したイナワラ、すなわちバイオマスの糖化において、セルラーゼ製剤に放線菌酵素を添加することにより、糖化率を上昇させることができた。
【0073】
反応液の組成を表13及び表14に示す。
【0074】
【表13】
【0075】
【表14】
【0076】
反応:45℃、1000rpmで振とうしながら1日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
上の反応系において、SAV1854を添加した場合のセルクラスト1.5Lによる水熱処理スギ、水熱処理ユーカリ、未処理エリアンサス分解評価の結果を図38に、SAV1856を添加した場合のセルクラスト1.5Lによる水熱処理スギ分解評価の結果を図39に示す。分解率は小さいが、放線菌酵素を添加することにより、スギ、ユーカリのように分解の困難な基質でも糖化率を上昇させることができた。
【0077】
上の反応系において、SAV1855及びSAV1856を添加した場合のセルクラスト1.5Lによる水熱処理スギの分解評価の結果を図40に示す。放線菌酵素を併用した場合には、その組み合わせによりシナジー効果が得られ、放線菌酵素のみでの糖化率が上昇した。その結果、反応系の糖化率を高めることが可能である。
【実施例7】
【0078】
<放線菌のイナワラ培養上清をセルラーゼ製剤に添加した場合の糖化率>
【0079】
【表15】
【0080】
反応:50℃、1000rpmで振とうしながら1日間
反応停止:100℃、20分加熱処理
還元糖量測定法:ソモギー・ネルソン法
糖化率:還元糖量をグルコース換算とした。
アルカリ処理イナワラを基質とし、セルラーゼ製剤に加えて放線菌のイナワラ培養上清を添加した場合の糖化反応の結果を図41に示す。アクセルレース1500のみによる糖化率を一番左に、放線菌Streptomyces pristinaespiralis又はS. scabieiのイナワラ培養上清のみによる糖化率をそれぞれ左から2番目、4番目に、アクセルレース1500に各培養上清を添加した場合の糖化率をそれぞれ左から3番目、5番目のカラムで示した。アクセルレース1500に単に放線菌の培養上清を加えただけでは、糖化率は十分に上昇しなかった。
【特許請求の範囲】
【請求項1】
配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項2】
配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項3】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項4】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項5】
リグノセルロース系バイオマス糖化酵素が、ストレプトマイセス属に属する微生物以外の生物由来のリグノセルロース系バイオマス糖化酵素であることを特徴とする請求項1〜4のいずれか記載のリグノセルロース系バイオマスの糖化方法。
【請求項6】
配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【請求項7】
配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【請求項8】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【請求項9】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【請求項1】
配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項2】
配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項3】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項4】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質のいずれか1種又は2種以上を、リグノセルロース系バイオマス糖化酵素と併用して、リグノセルロース系バイオマスを糖化することを特徴とするリグノセルロース系バイオマスの糖化方法。
【請求項5】
リグノセルロース系バイオマス糖化酵素が、ストレプトマイセス属に属する微生物以外の生物由来のリグノセルロース系バイオマス糖化酵素であることを特徴とする請求項1〜4のいずれか記載のリグノセルロース系バイオマスの糖化方法。
【請求項6】
配列番号2、4、6、8、10、12、14又は16で表されるアミノ酸配列からなるストレプトマイセス(Streptomyces)属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【請求項7】
配列番号2、4、6、8、10、12、14又は16記載のアミノ酸配列において、1又は数個のアミノ酸残基が置換、欠失、挿入又は付加され、かつ加水分解酵素活性を有する組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【請求項8】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列からなるDNAを発現させることにより得られるストレプトマイセス属に属する微生物由来の組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【請求項9】
配列番号1、3、5、7、9、11、13又は15で表される塩基配列と相補的な配列とストリンジェントな条件でハイブリダイズし、かつ加水分解酵素活性を有するタンパク質をコードするポリヌクレオチドでコードされる組換えタンパク質をリグノセルロース系バイオマス糖化酵素として使用する方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】

【図30】
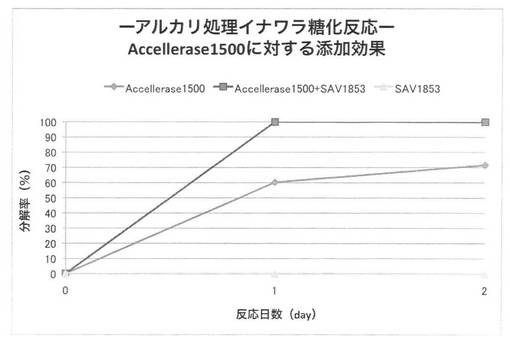
【図31】
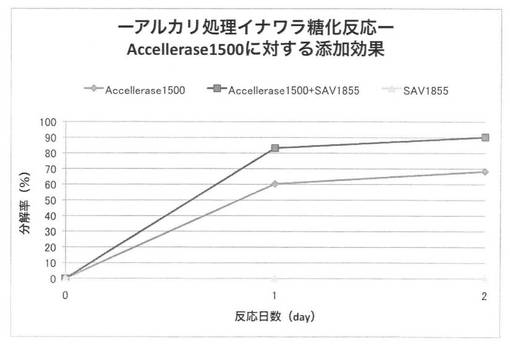
【図32】
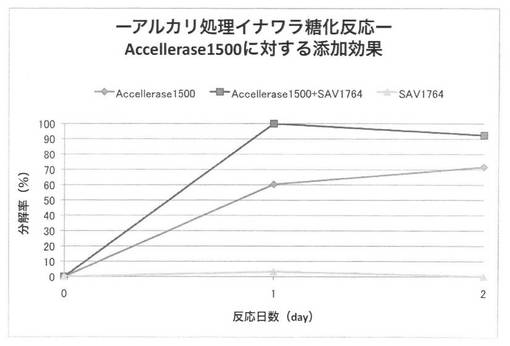
【図33】
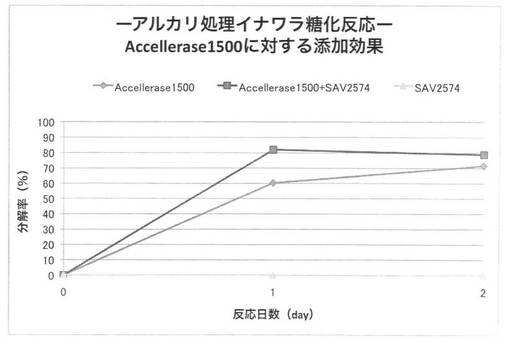
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38】
【図39】
【図40】
【図41】
【公開番号】特開2012−39895(P2012−39895A)
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願番号】特願2010−181548(P2010−181548)
【出願日】平成22年8月16日(2010.8.16)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、経済産業省、独立行政法人新エネルギー・産業技術開発機構「新エネルギー技術研究開発/バイオマスエネルギー等高効率転換技術開発(先導技術開発)」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願日】平成22年8月16日(2010.8.16)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、経済産業省、独立行政法人新エネルギー・産業技術開発機構「新エネルギー技術研究開発/バイオマスエネルギー等高効率転換技術開発(先導技術開発)」委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]