
リシル酸化酵素及びLOXL2由来の触媒ドメイン

【課題】 リシル酸化酵素及びリシル酸化酵素様酵素は、コラーゲンを架橋結合することによって、細胞外マトリクスの形成において重要な役割を果たすため、これらは重要な治療的目標である。それ故コラーゲン架橋結合酵素の阻害剤を選択する方法、これら酵素に結合する分子を同定する方法、及び様々な治療的使用(形態)(例えば、創傷治癒)のためのコラーゲン架橋結合活性の材料、の全てが所望される。
【解決手段】 本発明により、ヒト及びマウス由来のLOX及びLOXL2の単離された触媒ドメインのアミノ酸配列及びそれをエンコードするヌクレオチド配列が提供される。これらの単離された触媒ドメインの調製及び使用の方法も提供される。
【解決手段】 本発明により、ヒト及びマウス由来のLOX及びLOXL2の単離された触媒ドメインのアミノ酸配列及びそれをエンコードするヌクレオチド配列が提供される。これらの単離された触媒ドメインの調製及び使用の方法も提供される。
【発明の詳細な説明】
【関連出願の相互参照】
【0001】
この出願は、2009年8月21日に出願された米国仮出願番号61/235,776の利益を主張し、この開示は、全ての目的のために、その全てが本願に引用によって取り込まれる。
【0002】
[連邦政府の支援に関する陳述]
支援は、適用されない。
【技術分野】
【0003】
本出願は、酵素学及び分子生物学の分野に含まれる。
【背景技術】
【0004】
結合組織は、身体にある細胞及び器官に存在する骨格を提供する。脊椎動物における結合組織の一次構成要素は、細胞外マトリクスである。細胞外マトリクスの2つの主な構造的成分は、ゲル様の基質を形成するポリサッカライドであり、基質の中に埋め込まれている繊維性タンパク質である。これら繊維性タンパク質のうち、2つのもっとも一般的なものは、コラーゲンとエラスチンである。コラーゲン繊維は、コラーゲン原繊維の自己会合により形成され、三重らせんコラーゲン分子の架橋結合により、これら自身が会合する。この架橋結合は、リシル酸化酵素(LOX)及びそれに関連する酵素(「リシル酸化酵素様」 lysyl oxidase-like又は「LOXL」)によって触媒され、これらの全ては、リシン及びヒドロキシリシン残基のε‐アミノ基を脱アミノ化する能力を有する触媒ドメインを含み、かくて、ペプチジルリシンのペプチジル‐α‐アミノアジピック‐δ‐セミアルデヒド(アリシン)への変換をもたらす。アリシン残基は、自発的に、お互いに凝集することができ、かくて、コラーゲン分子の架橋結合をもたらす。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】国際公開WO 01/83702号
【特許文献2】国際公開WO 2004/047720号
【特許文献3】国際公開WO 2007/126457号
【特許文献4】米国特許出願公開US 2006/0127402号
【特許文献5】米国特許出願公開US 2007/0225242号
【特許文献6】米国特許出願公開US 2009/0053224号
【特許文献7】米国特許出願公開US 2009/0104201号
【非特許文献】
【0006】
【非特許文献1】Csiszar (2001) Prog. Nucleic Acid Res. and Molec. Biol. 70:1-32
【非特許文献2】Kirschmann et al. (2002) Cancer Research 62:4478-4483
【発明の概要】
【発明が解決しようとする課題】
【0007】
例えば、繊維症及び転移を含む様々な病状における細胞外マトリクスの関与は、次第に明らかになってきている。例えば、WO 01/83702(2001年11月8日)、WO 2004/047720(2004年6月10日)、WO 2007/126457(2007年8月11日)、US 2006/0127402(2006年6月15日)、US 2007/0225242(2007年9月27日)、US 2009/0053224(2009年2月26日)、US 2009/0104201(2009年4月23日)、Csiszar (2001) Prog. Nucleic Acid Res. and Molec. Biol. 70:1-32、Kirschmann et al. (2002) Cancer Research 62:4478-4483を参照する。リシル酸化酵素及びリシル酸化酵素様酵素は、コラーゲンを架橋結合することによって、細胞外マトリクスの形成において重要な役割を果たすので、これらは重要な治療的目標である。それ故、これらコラーゲン架橋結合酵素の阻害剤を選択する方法、これら酵素に結合する分子を同定する方法、及び様々な治療的使用(形態)(例えば、創傷治癒)のためのコラーゲン架橋結合活性の材料、の全てが所望される。
【課題を解決するための手段】
【0008】
本願開示は、ヒト及びマウス由来のリシル酸化酵素(LOX)及びリシル酸化酵素様2酵素(LOXL2)の単離された触媒ドメインを(これら触媒ドメインをエンコードする核酸と共に)提供する。かくて、次のペプチド及びこれらポリペプチドをエンコードする核酸が提供される(ヒトLOX触媒ドメイン、ヒトLOXL2触媒ドメイン、マウスLOX触媒ドメイン及びマウスLOXL2触媒ドメイン)。従って、本願開示は以下の形態を提供する。
【0009】
(形態1)ヒトリシル酸化酵素(LOX)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:1)。
【0010】
(形態2)ヒトリシル酸化酵素(LOX)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:2)。
【0011】
(形態3)ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:3)。
【0012】
(形態4)ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:4)。
【0013】
(形態5)マウス(murine)リシル酸化酵素(LOX)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:5)。
【0014】
(形態6)マウスリシル酸化酵素(LOX)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:6)。
【0015】
(形態7)マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:7)。
【0016】
(形態8)マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:8)。
【0017】
以下の形態もまた提供される。
【0018】
(形態1a)本願に記載の、ヒトリシル酸化酵素(LOX)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:1)、その触媒ドメインの外側のヒトLOXの配列を含まないポリペプチド。
【0019】
(形態2a)本願に記載の、ヒトリシル酸化酵素(LOX)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:2)、その触媒ドメインの外側のヒトLOXの配列をエンコードしないポリヌクレオチド。
【0020】
(形態3a)本願に記載の、ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:3)、その触媒ドメインの外側のヒトLOXL2の配列を含まないポリペプチド。
【0021】
(形態4a)本願に記載の、ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:4)、その触媒ドメインの外側のヒトLOXL2の配列をエンコードしないポリヌクレオチド。
【0022】
(形態5a)本願に記載の、マウスリシル酸化酵素(LOX)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:5)、その触媒ドメインの外側のマウスLOXの配列を含まないポリペプチド。
【0023】
(形態6a)本願に記載の、マウスリシル酸化酵素(LOX)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:6)、その触媒ドメインの外側のマウスLOXの配列をエンコードしないポリヌクレオチド。
【0024】
(形態7a)本願に記載の、マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:7)、その触媒ドメインの外側のマウスLOXL2の配列を含まないポリペプチド。
【0025】
(形態8a)本願に記載の、マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:8)、その触媒ドメインの外側のマウスLOXL2の配列をエンコードしないポリヌクレオチド。
【0026】
また、前述の核酸及び/又はポリヌクレオチドを含む発現ベクターも提供される。そのような発現ベクターは、任意に、プロモーター(例えば、T7プロモーター、T3プロモーター、SP6プロモーター、E.coli RNAポリメラーゼプロモーター、CMVプロモーター、SV40プロモーター、PGKプロモーター、EF‐1アルファプロモーター)、転写終結シグナル(例えば、SV40終結シグナル)、スプライス部位(例えば、SV40スプライス部位、ベータ‐グロブリンスプライス部位)、リボゾーム結合部位、シグナル配列(例えば、免疫グロブリンカッパシグナル配列)、エピトープタグ(例えば、myc)、精製タグ(例えば、His6)、複製起点及び薬剤選択マーカーを含む。リンカーアミノ酸をエンコードし、及び/又は制限酵素認識部位を含むリンカー配列又は、他のあらゆる種類のリンカー配列もまた、本願で開示される発現ベクターに存在し得る。
【0027】
従って、本願の開示はまた、以下の形態も提供する。
【0028】
(形態9)触媒ドメインの外側にあるヒトLOXのアミノ酸配列を含まず、ヒトLOXの触媒ドメインの全て又は一部のアミノ酸配列を含み、更に、1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、ヒトLOXの触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:9)。
【0029】
(形態10)形態9によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、ヒトLOXの触媒ドメイン、mycエピトープタグ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:10)。
【0030】
(形態11)形態10のポリヌクレオチドを含む発現ベクター。
【0031】
(形態12)形態11の発現ベクターであって、更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む発現ベクター。
【0032】
(形態13)触媒ドメインの外側にあるヒトLOXL2のアミノ酸配列を含まず、ヒトLOXL2の触媒ドメインの全て又は一部のアミノ酸配列を含み、更に、1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、ヒトLOXL2の触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:11)。
【0033】
(形態14)形態13によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、ヒトLOXL2の触媒ドメイン、mycエピトープタグ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:12)。
【0034】
(形態15)形態14のポリヌクレオチドを含む発現ベクター。
【0035】
(形態16)更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む形態15の発現ベクター。
【0036】
(形態17)触媒ドメインの外側にあるマウスLOXアミノ酸配列を含まず、マウスLOXの触媒ドメインの全て又は一部のアミノ酸配列を含み、更に1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、マウスLOXの触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:13)。
【0037】
(形態18)形態17によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、マウスLOXの触媒ドメイン、mycエピトープ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:14)。
【0038】
(形態19)形態18のポリヌクレオチドを含む発現ベクター。
【0039】
(形態20)更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む形態19の発現ベクター。
【0040】
(形態21)触媒ドメインの外側にあるマウスLOXL2のアミノ酸配列を含まず、マウスLOXL2の触媒ドメインの全て又は一部のアミノ酸配列を含み、更に1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、マウスLOXL2の触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:15)。
【0041】
(形態22)形態21によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、マウスLOXL2の触媒ドメイン、mycエピトープ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:16)。
【0042】
(形態23)形態22のポリヌクレオチドを含む発現ベクター。
【0043】
(形態24)更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む形態23の発現ベクター。
【図面の簡単な説明】
【0044】
【図1】図1は、phLOXMCD発現ベクターを導入(トランスフェクト)されたHEK293細胞による、ヒトリシル酸化酵素(hLOX)タンパク質の触媒ドメインの発現を表す。6つの安定にトランスフェクトされた細胞系統(番号23から28)の免疫ブロットによる解析を表す。左のパネルは、各6つの細胞系統からの無希釈増殖培地(neat CM)の試料(15μL)を含む、ポリアクリルアミドゲルの免疫ブロットを表す。中間のパネルは、可溶性細胞内物質の解析を表し、右のパネルは、不溶性細胞内物質の解析を表す。これらそれぞれのパネルにおいて、各レーンは、およそ105個細胞からの物質を含む。全てのブロットに、一次マウス抗His5抗体をプローブし、続いて、二次HRPコンジュゲートロバ抗マウス抗体と反応させた。HRP活性を、Chemi−Glow(登録商標)reagent(Alpha Innotech, San Leandro, CA)を使用して、可視化した。プロセスされた触媒ドメインは、26.9kDの予測分子量を有する。
【0045】
各パネルにおいて、「+」で標識されたレーンは、His6配列を含むタンパク質を含み、抗His5抗体の陽性対照として機能した。
【0046】
【図2】図2は、phLOXL2MCD発現ベクターをトランスフェクトされたHEK293細胞による、ヒトリシル酸化酵素様2(hLOXL2)タンパク質の触媒ドメインの発現を表す。左のパネルは、いくつかの安定にトランスフェクトされた細胞系統(各レーンの頂点に記載される数字により示される)からの全細胞溶解物の免疫ブロットを表し、各レーンは、約1×105個細胞からの物質を含む。右のパネルは、各6つの細胞系統(番号で示される)からの無希釈増殖培地(15μL Neat CM)の試料を含む、ポリアクリルアミドゲルの免疫ブロットを表す。全てのブロットに、一次マウス抗His5抗体をプローブし、続いて、二次HRPコンジュゲートロバ抗マウス抗体と反応させた。HRP活性を、Chemi−Glow(登録商標)reagent(Alpha Innotech, San Leandro, CA)を使用して、可視化した。プロセスされた触媒ドメインは、31kDの予測分子量を有する。
【0047】
左のパネルにおいて、「+」で標識されたレーンは、His6配列を含むタンパク質を含み、抗His5抗体の陽性対照として機能した。
【0048】
【図3】図3は、pmLOXMCD発現ベクターをトランスフェクトされたHEK293細胞による、マウスリシル酸化酵素(mLOX)タンパク質の触媒ドメインの発現を表す。5つの安定にトランスフェクトされた細胞系統(4、9、12、18及び19の番号が付けられている)の免疫ブロットによる解析を表す。左のパネルは、各6つの細胞系統からの無希釈増殖培地(neat CM)の試料(15μL)を含む、ポリアクリルアミドゲルの免疫ブロットを表す。右のパネルは、これら6つの系統のうち5つからの可溶性及び不溶性細胞内分画の解析を表し、各レーンは、およそ105個細胞からの物質を含む。全てのブロットに、一次マウス抗His5抗体をプローブし、続いて、二次HRPコンジュゲートロバ抗マウス抗体と反応させた。HRP活性を、Chemi−Glow(登録商標)reagent(Alpha Innotech, San Leandro, CA)を使用して、可視化した。プロセスされた触媒ドメインは、27kDの予測分子量を有する。
【0049】
【図4】図4は、ヒトとマウスのLOX触媒ドメインの分泌及びアミノ酸配列の違いを表す。詳細については実施例10を参照する。
【0050】
【図5】図5は、単離されたヒトLOXL2触媒ドメインのAmplex(登録商標)Red分析における、レゾルフィン生成の経時的解析を表す(590nmの吸光度により測定される)。詳細については実施例12を参照する。
【0051】
【図6】図6は、ヒト由来のポリペプチド(hLOX、hLOXL1、hLOXL2など)及びマウス由来のポリペプチド(mLOX及びmLOXL2)を含む、本願の開示における様々なポリペプチドのアミノ酸配列のアライメントを表す。アミノ酸残基の番号付けは、表示される最もN末端にある残基を「1」として開始し、当該技術分野における慣例に必ずしも従うものでは無い。European Bioinformatics Institute(EMBL-EBI)によって提供されたClustalW2プログラムに従って、アスタリスクは、表示された全てのポリペプチド間で共通した位置におけるアミノ酸の相同性を示し、一方で、コロンは、保存的な(conservative)アミノ酸置換を示し、単一の点は、半保存的な(semi-conservative)アミノ酸置換を示す。
【0052】
【図7】図7は、ヒト由来のポリペプチドのみを含む本願の開示における様々なポリペプチドのアミノ酸配列のアライメントを表す。番号付け及び記号の凡例は、図6において使用されたものと同一である。
【0053】
本願の開示の実施は、特段の指示のない限り、細胞生物学、毒物学、分子生物学、分子化学、細胞培養、免疫学、腫瘍学、組み換えDNAの分野及び、当該技術分野範囲内の関係する分野における標準の方法及び従来技術を採用する。そのような技術は、文献に記載され、それ故、当業者が利用できる。例えば、Alberts, B. et al., "Molecular Biology of the Cell," 5th edition, Garland Science, New York, NY, 2008、Voet, D. et al. "Fundamentals of Biochemistry: Life at the Molecular Level," 3rd edition,John Wiley & Sons, Hoboken, NJ, 2008、Sambrook, J. et al., "Molecular Cloning: A Laboratory Manual," 3rd edition, Cold Spring Harbor Laboratory Press, 2001、Ausubel, F. et al., "Current Protocols in Molecular Biology," John Wiley & Sons, New York, 1987及び定期的な更新、Freshney, R.I., "Culture of Animal Cells: A Manual of Basic Technique," 4th edition, John Wiley & Sons, Somerset, NJ, 2000、及び、シリーズ"Methods in Enzymology," Academic Press, San Diego, CAを参照する。
【0054】
[リシル酸化酵素型酵素]
本願で使用される場合、用語「リシル酸化酵素型酵素」又は「リシル酸化酵素」は、リシン及びヒドロキシリシン残基のε‐アミノ基の酸化的脱アミノ化を触媒する触媒ドメインを含むタンパク質のファミリーのメンバーであることを意味し、かくて、ペプチジルリシンのペプチジル‐α‐アミノアジピック‐δ−セミアルデヒド(アリシン)への変換をもたらし、アンモニア及び過酸化水素の化学量論的量を放出する。

【0055】
この反応は、ほとんどの場合、細胞外のコラーゲン及びエラスチンのリシン残基に生じる。アリシンのアルデヒド残基は、活性であり、同時に他のアリシン及びリシン残基と凝集することができ、かくて、コラーゲン分子の架橋結合をもたらし、コラーゲン原繊維を形成する。
【0056】
リシル酸化酵素は、ニワトリ、ラット、マウス、ウシ及びヒトから精製されている。すべてのリシル酸化酵素は、長さ約205アミノ酸であり、タンパク質のカルボキシ末端部分に位置し、酵素の活性部位を含む、共通の触媒ドメインを含む。活性部位は、Cu(II)原子に配位する4つのヒスチジン残基を含む保存アミノ酸配列を含む銅結合部位を含む。活性部位はまた、リシン残基とチロシン残基の間の分子内共有結合により形成されるリシルチロシルキノン(LTQ)補因子も含む(ラットリシル酸化酵素では、314番目のlys(lys314)及び349番目のtyr(tyr349)に相当し、ヒトリシル酸化酵素では、320番目のlys(lys320)及び355番目のtyr(tyr355)に相当する)。LTQ補因子を形成するチロシン残基を取り巻く配列はまた、リシル酸化酵素の間でも保存される。触媒ドメインはまた、5つのジスルフィド結合の形成に関与する10つの保存システイン残基をも含む。触媒ドメインはまた、フィブロネクチン結合ドメインをも含む。最終的に、成長因子及びサイトカインレセプタードメインに類似し、4つのシステイン残基を含むアミノ酸配列が、触媒ドメインに存在する。
【0057】
これらの保存領域が存在するにも関わらず、異なるリシル酸化型酵素は、これら触媒ドメインの内部及び外側の両方における、ヌクレオチド配列及びアミノ酸配列の相違する領域のお陰(virtue)で、他の異なるリシル酸化型酵素とお互いに区別され得る。
【0058】
単離され、特徴付けられた、この酵素のファミリーの最初のメンバーは、リシル酸化酵素(EC 1.4.3.13)であった。これはまた、タンパク質‐リシン6‐酸化酵素、タンパク質‐L‐リシン:酸素‐6‐酸化還元酵素(脱アミノ化)又はLOXとしても知られる。例えば、Harris et al., Biochim. Biophys. Acta (1974) 341:332-344、Rayton et al. (1979) J. Biol. Chem. 254:621-626、Stassen (1976) Biophys. Acta 438:49-60を参照する。
【0059】
更なるリシル酸化酵素型酵素が、続いて発見された。これらのタンパク質は、「LOX様」(LOX-like)又は「LOXL」と称されている。これらの全ては、上記の共通の触媒ドメインを含み、類似の酵素的活性を有する。これまでのところ、5つの異なったリシル酸化酵素が、ヒトとマウスの両方に存在することが知られている。すなわち、LOXタンパク質及び4つのLOXに関連するないしはLOX様のタンパク質であるLOXL1(これらはまた、「リシル酸化酵素様」「LOXL」又は「LOL」とも示される)、LOXL2(これは、「LOR−I」とも示される)、LOXL3及びLOXL4である。5つの異なるリシル酸化酵素型酵素をエンコードする各遺伝子は、異なった染色体上に存在する。例えば、Molnar et al. (2003) Biochim Biophys Acta. 1647:220- 24、Csiszar (2001) Prog. Nucl. Acid Res. 70:1-32、2001年11月8日公開されたWO 01/83702、及び 米国特許番号6,300,092を参照する(これらの全ては本願に引用として組み込まれる)。LOXL4といくつかの類似性を有するが、異なる発現パターンを有する、LOXCと呼ばれるLOX様タンパク質は、マウスEC細胞系統から単離されている(Ito et al. (2001) J. Biol. Chem. 276:24023-24029)。DmLOXL−1とDmLOXL−2の2つのリシル酸化酵素は、ショウジョウバエから単離されている。
【0060】
全てのリシル酸化酵素型酵素が、共通の触媒ドメインを共有しているにも関わらず、これらはまた、特にそのアミノ末端領域内において、お互いに異なる。LOXと比較して、4つのLOXLタンパク質は、アミノ末端伸張を有する。従って、ヒトプレプロLOX(例えば、シグナル配列の切断の前の一次翻訳産物、以下参照)は、417アミノ酸残基を含み、LOXL1は574(アミノ酸残基)を含み、LOXL2は638(アミノ酸残基)を含み、LOXL3は753(アミノ酸残基)を含み及びLOXL4は756(アミノ酸残基)を含む。当該技術分野における全長リシル酸化酵素型酵素の番号付けの慣例は、最もN末端のアミノ酸残基を「1」として開始し、シグナル配列から始まる。
【0061】
これらのアミノ末端領域の範囲内において、LOXL2、LOXL3及びLOXL4は、4つのスカベンジャーレセプターのシステインリッチ(SRCR)ドメインの繰り返しを含む。これらのドメインは、LOX又はLOXL1には存在しない。SRCRドメインは、分泌性のタンパク質、膜貫通型タンパク質又は細胞外マトリクスタンパク質に見られ、いくつかの分泌性の及びレセプタータンパク質におけるリガンド結合を媒介することが知られている(Hoheneste et al. (1999) Nat. Struct. Biol. 6:228-232、Sasaki et al. (1998) EMBO J. 17:1606-1613)。このSRCRドメインに加えて、LOXL3は、核局在シグナルをそのアミノ末端領域に含む。プロリンリッチドメインは、LOXL1に特有であると思われる(Molnar et al. (2003) Biochim. Biophys. Acta 1647:220-224)。様々なリシル酸化酵素型酵素はまた、それらのグリコシル化パターンの点でも異なる。
【0062】
組織分布もまた、リシル酸化酵素型酵素の間で異なる。ヒトLOXmRNAは、心臓、胎盤、睾丸、肺、腎臓及び子宮において高発現するが、脳及び肝臓においては、僅かに発現する。ヒトLOXL1のmRNAは、胎盤、腎臓、筋肉、心臓、肺及び膵臓において発現し、LOXと同様に、脳及び肝臓においては、(これらと比較して)はるかに少ないレベルで発現する(Kim et al. (1995) J. Biol. Chem. 270:7176-7182)。子宮、胎盤及び他の器官において、LOXL2mRNAは高いレベルで発現するが、LOX及びLOXLと同様に、脳及び肝臓においては、低いレベルで発現する(Jourdan Le-Saux et al.(1999) J. Biol. Chem. 274:12939:12944)。LOXL3mRNAが、睾丸、脾臓及び前立腺において高発現し、胎盤においては、中程度に(moderately)発現し、肝臓では発現しない一方で、LOXL4mRNAの高レベル(の発現)が肝臓において観察される(Huang et al. (2001) Matrix Biol. 20:153-157、Maki and Kivirikko (2001) Biochem. J. 355:381-387、Jourdan Le-Saux et al. (2001) Genomics 74:211-218、及びAsuncion et al. (2001) Matrix Biol. 20:487-491)。
【0063】
疾病における異なったリシル酸化酵素の発現及び/又は関与も変化し得る。例えば、Kagan (1994) Pathol. Res. Pract. 190:910-919、Murawaki et al. (1991) Hepatology 14:1167-1173、Siegel et al. (1978) Proc. Natl. Acad. Sci. USA 75:2945-2949、Jourdan Le-Saux et al. (1994) Biochem. Biophys. Res. Comm. 199:587-592、及び Kim et al. (1999) J. Cell Biochem. 72:181-188を参照する。リシル酸化酵素型酵素もまた、頭部及び首部の癌、膀胱癌、大腸(colon)癌、食道癌及び乳癌を含むいくつかの癌にも関係しているとされている。例えば、Wu et al. (2007) Cancer Res. 67:4123-4129、Gorough et al. (2007) J. Pathol. 212:74-82、Csiszar (2001) Prog. Nucl. Acid Res. 70:1-32及び、Kirschmann et al. (2002) Cancer Res. 62:4478-4483を参照する。
【0064】
従って、リシル酸化酵素型酵素は、構造及び機能においていくつかの重なりを示すが、その上、それぞれが異なった機能を有すると思われる。構造に関して、例えば、ヒトLOXタンパク質の触媒ドメインに対して挙げられる(raised)、ある抗体は、ヒトLOXL2に結合しない。機能に関して、標的とされるLOXの欠失は、マウスにおいて、出産の際に致死であると思われるが、一方、LOXL1の欠損は、深刻な発達表現型を引き起こさないことが報告されている(Hornstra et al. (2003) J. Biol. Chem. 278:14387-14393、Bronson et al. (2005) Neurosci. Lett. 390:118-122)。
【0065】
しかしながら、もっとも広く報告されているリシル酸化酵素型酵素の活性は、細胞外のコラーゲン及びエラスチンにおける特定のリシン残基の酸化であるが、リシル酸化酵素型酵素がまた、いくつかの細胞内プロセスにも関与することの証拠がある。例えば、いくつかのリシル酸化型酵素は、遺伝子発現を制御するという報告がある(Li et al. (1997) Proc. Natl. Acad. Sci. USA 94:12817-12822、Giampuzzi et al. (2000) J. Biol. Chem. 275:36341-36349)。加えて、LOXは、ヒストンH1におけるリシン残基を酸化することが報告されている。更なるLOXの細胞外の活性は、単核球、繊維芽細胞及び平滑筋細胞の走化性の誘導を含む(Lazarus et al. (1995) Matrix Biol. 14:727-731、Nelson et al. (1988) Proc. Soc. Exp. Biol. Med. 188:346-352)。LOX自体の発現は、TGF‐β、TNF‐α及びインターフェロンのような、いくつかの増殖因子及びステロイドにより誘導される(Csiszar (2001) Prog. Nucl. Acid Res. 70:1-32)。近年の研究により、LOXには、発生調節、腫瘍抑制、細胞運動性及び、細胞老化のような、異なった生物学的機能における他の役割があると考えられている。
【0066】
様々な材料由来のリシル酸化酵素(LOX)タンパク質の例は、次の配列の1つから発現し又は翻訳されるポリペプチドに実質的に相同であるアミノ酸配列を有する酵素を含む(EMBL/GenBank受託: M94054;AAA59525.1−−mRNA;S45875;AAB23549.1−mRNA;S78694;AAB21243.1−mRNA;AF039291;AAD02130.1−mRNA;BC074820;AAH74820.1−mRNA;BC074872;AAH74872.1−mRNA;M84150;AAA59541.1――ゲノムDNA。LOXの1つの実施形態は、ヒトリシル酸化酵素(hLOX)プレプロタンパク質である。
【0067】
リシル酸化酵素様酵素をエンコードする配列の例示的な開示は、次の通りである。LOXL1は、GenBank/EMBL BC015090;AAH15090.1で寄託されるmRNAによってエンコードされる。LOXL2は、GenBank/EMBL U89942で寄託されるmRNAによってエンコードされ、LOXL3は、GenBank/EMBL AF282619;AAK51671.1で寄託されるmRNAによってエンコードされ、及び、LOXL4は、GenBank/EMBL AF338441;AAK71934.1で寄託されるmRNAによってエンコードされる。
【0068】
プレプロペプチドとして知られるLOXタンパク質の一次翻訳産物は、1−21番目のアミノ酸に亘るシグナル配列を含む。このシグナル配列は、マウス及びヒトLOXの両方において、21番目のCys(Cys21)と22番目のAla(Ala22)の間の切断によって細胞内に放出され、LOXの形態の46−48kDaのポリペプチドを生じ、これはまた、本願において、全長形態と称される。プロポリペプチドは、ゴルジ体を通過する間に、N−グリコシル化され、50kDaタンパク質を生じ、続いて、細胞外環境に分泌される。この段階において、タンパク質は、触媒活性的に、不活性である。マウスLOXにおける168番目のGly(Gly168)と169番目のAsp(Asp169)の間での更なる切断(cleavage)、及びヒトLOXにおける174番目のGly(Gly174)と175番目のAsp(Asp175)の間での更なる切断によって、成熟した、酵素活性的に活性な、30−32kDAの酵素が生じ、18kDaのプロペプチドを放出する。この最終的な切断の事象は、骨形態発生タンパク質1(BMP−1)としても知られる、メタロエンドプロテアーゼのプロコラーゲンC−プロティナーゼによって触媒される。興味深いことに、この酵素はまた、LOXの基質であるコラーゲンのプロセシングにおいても機能する。N−グリコシル化ユニットは、その後に取り除かれる。
【0069】
潜在的なシグナルペプチド切断部位は、LOXL1、LOXL2、LOXL3及びLOXL4のアミノ末端に存在することが予測されている。予測シグナル切断部位は、LOXL1の25番目のGly(Gly25)と26番目のGln(Gln26)の間、LOXL2の25番目のAla(Ala25)と26番目のGln(Gln26)の間、LOXL3の25番目のGly(Gly25)と26番目のSer(Ser26)の間及び、LOXL4の23番目のArg(Arg23)と24番目のPro(Pro24)の間に存在する。
【0070】
LOXL1タンパク質におけるBMP−1切断部位は、354番目のSer(Ser354)と355番目のAsp(Asp355)の間に同定されている(Borel et al. (2001) J. Biol. Chem.276:48944-48949)。他のリシル酸化酵素型酵素における潜在的なBMP−1切断部位が、プロコラーゲン及びpro‐LOXにおいて、Ala/Gly‐Aspの配列(これには、しばしば、酸性又は荷電残基が続く)にある、BMP−1切断のコンセンサス配列に基づいて、予測されている。LOXL3における予測BMP−1切断部位は、447番目のGly(Gly447)と448番目のAsp(Asp448)の間に位置し、この部位でのプロセシングによって、成熟LOXと類似する大きさの成熟ペプチドを生じ得る。BMP−1の潜在的な切断部位はまた、LOXL4内でも、残基569番目のAla(Ala569)と570番目のAsp(Asp570)の間にも同定された(Kim et al. (2003) J. Biol. Chem. 278:52071-52074)。LOXL2もまた、LOXLファミリーの他のメンバーに類似の形でタンパク質分解的に(proteolytically)切断され、分泌され得る(Akiri et al.(2003) Cancer Res. 63:1657-1666)。
【0071】
リシル酸化型酵素における共通の触媒ドメインの存在に基づいて、活性部位が位置する、プロ酵素のC末端30kDa領域の特定のアミノ酸残基が、高度に保存される。より穏やかな程度の保存(約60−70%)が、プロペプチドドメインにおいて観察される。
【0072】
本願の開示の目的において、用語「リシル酸化型酵素」は、上述の5つのリシン酸化酵素(即ち、LOX、LOXL1(これはLOXLとも表示される)、LOXL2、LOXL3及びLOXL4)の全てを含み、LOX,LOXL1、LOXL2、LOXL3及びLOXL4の機能的なフラグメント及び/又は誘導体(これらは実質的に酵素的活性(例えば、リシル残基の脱アミノ化を触媒する能力)を保持する)もまた含む。アミノ酸配列又はポリヌクレオチド配列のコンテクスト(例えば、LOXL2「由来の」アミノ酸配列)において、「フラグメント及び/又は誘導体」とは、ポリペプチド又は核酸が、自然に生じるリシル酸化酵素型酵素タンパク質又はそれをエンコードする核酸の全て又は一部分の隣接(ないし連続)配列(contiguous sequence)を含むことを示すことを意味し、そして、タンパク質又は核酸が作成される材料又は方法について限定することを意味するものではない。
【0073】
典型的には、機能的なフラグメント又は誘導体は、少なくとも、50%のリシン酸化活性を保持する。いくつかの実施形態において、機能的なフラグメント又は誘導体は、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%又は、100%のリシン酸化活性を保持する。
【0074】
リシル酸化酵素型酵素の機能的なフラグメントは、本質的に、触媒活性を変更させない(天然のポリペプチド配列に関する)保存的なアミノ酸置換(複数)を含むことができることもまた意図される。用語「保存的なアミノ酸置換」は、或る共通の構造及び/又は特性を基にした、アミノ酸の群分け(grouping)を意味する。共通の構造に関して、アミノ酸は次のとおり、群分けできる。即ち、非極性側鎖を有するアミノ酸(グリシン、アラニン、バリン、ロイシン、イソロイシン、メチオニン、プロリン、フェニルアラニン及びトリプトファン)、非荷電極性側鎖を有するアミノ酸(セリン、スレオニン、アスパラギン、グルタミン、チロシン及びシステイン)、及び、荷電極性側鎖を有するアミノ酸(リシン、アルギニン、アスパラギン酸、グルタミン酸及びヒスチジン)に群分けされ得る。芳香族側鎖を含むアミノ酸の群は、フェニルアラニン、トリプトファン及びチロシンを含む。複素環の側鎖は、プロリン、トリプトファン及びヒスチジンに存在する。非極性側鎖を含むアミノ酸の群の中で、短い炭化水素側鎖を有するアミノ酸(グリシン、アラニン、バリン、ロイシン、イソロイシン)は、より長い非炭化水素側鎖を有するアミノ酸(メチオニン、プロリン、フェニルアラニン、トリプトファン)から区別され得る。荷電極性側鎖を有するアミノ酸の群の中で、酸性アミノ酸(アスパラギン酸、グルタミン酸)は、塩基性側鎖を有するアミノ酸(リシン、アルギニン及びヒスチジン)から区別され得る。
【0075】
個々のアミノ酸の共通の特性を定義するための機能的な方法は、相同的な生物の対応するタンパク質の間のアミノ酸変化の正規化された頻度を解析することである(Schulz, G. E. and R. H. Schirmer, Principles of Protein Structure, Springer-Verlag, 1979)。そのような解析によって、1つの群内のアミノ酸が、相同的なタンパク質において、相互に優先的に置換されることが定義され、そしてそれ故、タンパク質構造の全体において類似の影響を有し得る(Schulz, G. E. and R. H. Schirmer, Principles of Protein Structure, Springer-Verlag, 1979)。この種類の解析において、「保存的なアミノ酸置換」とは、あるアミノ酸残基の、アミノ酸側鎖の化学的及び物理的な特性(例えば、電荷、大きさ、疎水性/親水性)を共有する他の(アミノ酸残基)への置換を意味する。以下は、特定の化学的及び/又は物理的特性を共有するアミノ酸残基の例である。
【0076】
(1)Glu、Asp、Lys、Arg及びHisからなる、荷電基を含むアミノ酸。
【0077】
(2)Lys、Arg及びHisからなる、正荷電基を含むアミノ酸。
【0078】
(3)Glu及びAspからなる、負荷電基を含むアミノ酸。
【0079】
(4)Phe、Tyr及びTrpからなる、芳香族基を含むアミノ酸。
【0080】
(5)His及びTrpからなる、窒素環基を含むアミノ酸。
【0081】
(6)Val、Leu及びIleからなる、大きな脂肪族非極性基を含むアミノ酸。
【0082】
(7)Met及びCysからなる、僅かに極性な基を含むアミノ酸。
【0083】
(8)Ser、Thr、Asp、Asn、Gly、Ala、Glu、Gln及びProからなる、小さい残基の基を含むアミノ酸。
【0084】
(9)Val、Leu、Ile、Met及びCysからなる、脂肪族基を含むアミノ酸。
【0085】
(10)Ser及びThrからなる、ヒドロキシル基を含むアミノ酸。
【0086】
ある「保存的な置換」は、以下のアミノ酸残基の群(groups)内での置換を含み得る:gly,ala;val,ile,leu;asp,glu;asn,gln;ser,thr;lys,arg;及びphe,tyr。
【0087】
従って、上で例示された、アミノ酸の保存的な置換は、当業者に知られ、(置換の)結果の分子の生物学的活性を変更することなく一般的に行なわれ得る。この技術分野の当業者はまた、一般的に、ポリペプチドの本質的でない領域における単一のアミノ酸置換は、実質的に、生物学的活性を変更しないことも認識する。例えば、Watson, et al., "Molecular Biology of the Gene," 4th Edition, 1987, The Benjamin/Cummings Pub. Co., Menlo Park, CA, p.224を参照する。
【0088】
リシル酸化酵素に関する更なる情報については、例えば、Rucker et al. (1998) Am. J. Clin. Nutr. 67:996S-1002S及びKagan et al. (2003) J. Cell. Biochem 88:660-672を参照する。また、同一出願人による米国特許出願(公開)US 2009/0053224(2009年2月26日)及びUS 2009/0104201(2009年4月23日)を参照する。各々の開示は、リシル酸化酵素及びリシル酸化酵素様酵素、これら酵素のモジュレーター及び、これらの酵素のモジュレーターを同定する方法を記載する目的で、本願に引用によって取り込まれる。また、WO 2004/47720(2004年6月10日)、WO 2007/126457(2007年8月11日)、WO 2009/010974(2009年1月22日)、US 2006/0127402(2006年6月15日)、US 2007/0021365(2007年1月25日)及び、US 2007/0225242(2007年9月27日)を参照する。これらの全ては、リシル酸化酵素及びリシル酸化酵素様酵素、これらの酵素のモジュレーター及びこれら酵素のモジュレーターを同定する方法を記載する目的で、本願に引用によって取り込まれる。
【0089】
[リシル酸化酵素又はリシル酸化酵素型酵素の触媒ドメインを有するポリペプチド]
本願の開示は、上記のように、生化学及び分子生物学国際連合(International Union of Biochemistry and Molecular Biology)の命名に従うEC1.4.3.13に属するポリペプチドのような、リシル酸化酵素型酵素の活性を有するポリペプチドを提供する。全長のタンパク質は、シグナル配列及び/又はポリペプチド含む(この両方は、生体内で、タンパク質が、完全な触媒活性を獲得する前に切断され得る)。全長のリシル酸化酵素又はリシル酸化酵素様酵素は、(それぞれ)100、200、300、400、500、700より大で、754まで、又はそれより大のアミノ酸残基の長さであり得る。シグナル配列及び/又はプロペプチドを有さない成熟全長タンパク質は、(それぞれ)100、200、240、480、720、730より大で、740まで、又はそれより大のアミノ酸残基の長さであり得る。
【0090】
本願開示のポリペプチドは、成熟全長LOX又はLOXLタンパク質(例えば、シグナルペプチド及び/又はプロペプチドを有さない)由来であり得る。例示のポリペプチドは、図6及び7に記載のアミノ酸配列のいずれかを有することができる。
【0091】
図6は、ヒトLOX、ヒトLOXL1、ヒトLOXL2、ヒトLOXL3、ヒトLOXL4、マウスLOX及びマウスLOXL2由来のポリペプチドのアミノ酸配列のアライメントを表す。図7は、ヒトLOX/LOXLタンパク質のみから由来のポリペプチドの配列のアライメントを表す。便宜上、アミノ酸残基の番号付けは、図6及び7に表される、最もN末端の残基を「1」として開始する。本願開示のポリペプチドは、かくて、図6及び7に記載される配列に使用される番号付けシステム又は、上記のような当該技術分野で使用される番号付けシステムを参照して記載され得る。或いは、ポリペプチドは、本願で割り当てられた配列識別番号を参照して記載され得る。European Bioinformatics Institute(EMBL-EBI)によって提供されたClustalW2プログラムに使用される凡例に従って、図6及び7のアスタリスクは、表示される全てのポリペプチドの間で共通した位置における相同なアミノ酸を示し、一方で、コロンは、保存的なアミノ酸置換を示し、単一の点は、半保存的なアミノ酸置換を示す。
【0092】
本願開示のポリペプチドは、少なくとも約50、少なくとも約75、少なくとも約100、少なくとも約120、少なくとも約140、少なくとも約150、少なくとも約170、少なくとも約180、少なくとも約200、少なくとも約220、少なくとも約240まで、又はそれより大のリシル酸化酵素型酵素のC末端部分のアミノ酸を有するポリペプチドを含み得る。例えば、本願開示のポリペプチドは、図6のSEQ ID NO:1、SEQ ID NO:5又はSEQ ID NO:17によって表されるC末端の207アミノ酸残基を含み得る。本願開示のポリペプチドはまた、天然に生じた全長のリシル酸化酵素型酵素のアミノ酸配列に満たない(長さの)アミノ酸配列を有するポリペプチドも含む。ポリペプチドは、例えば、図6及び7に示された配列によって例示されるような、LOX又はLOXLタンパク質のC末端部位において、約25まで、約50まで、約75まで、約100まで、約120まで、約140まで、約160まで、約180まで、約200まで、約220まで、約230まで、又はそれより大のアミノ酸残基を含む。ポリペプチドは、シグナル配列、プロペプチド及び/又は、図6及び7に表される配列に対するN末端であるLOX又はLOXLタンパク質の任意の部分を除外し得る。
【0093】
本願開示のポリペプチドはまた、上記の当該技術分野において使用される番号付けシステムを参照して記載され得、この場合、アミノ酸番号「1」は、シグナル配列を含む全長のLOX又は全長のLOXLタンパク質の最もN末端のアミノ酸残基を示す。そのような番号付けに従って、ポリペプチドは、LOXにおける約190と240の間、約200と230の間、又は約210と220の間の残基番号(例えば、SEQ ID NO:1又はSEQ ID NO:5)で開始するアミノ酸配列を含むポリペプチドとして記載され得る。ポリペプチドは、LOXL1における約350と360の間、約340と370の間、又は約330と380の間の残基番号で開始するアミノ酸配列を含み得る。或いは、ポリペプチドは、LOXL2における約540と550の間、約530と560の間、又は約520と570の間の残基番号(例えば、SEQ ID NO:3又はSEQ ID NO:7)で開始するアミノ酸配列を含み得る。ポリペプチドはまた、LOXL3における約520と530の間、約510と540の間、又は約500と550の間の残基番号で開始するアミノ酸配列も含み得る。ポリペプチドはまた、LOXL4における約530と540の間、約540と550の間、又は約530と560の間の残基番号で開始するアミノ酸配列も含み得る。
【0094】
本願開示のポリペプチドは、1、2、3、4、5、6、7、8、9または10、又はそれより大のアミノ酸置換を有するポリペプチドを含む(例えば、図6及び7に示されるアミノ酸配列と比較される保存的なアミノ酸置換)。アミノ酸置換のガイダンスは、図6及び7で提供されるアライメントから得ることができる。置換され得るアミノ酸残基は、高度に保存されていない残基位置(たとえば、アスタリスク、コロン又は点によってマークされていない残基位置)に位置し得る。当業者は、表示されるアライメントを基にして、或る(特定の)残基位置のアミノ酸残基が、他の残基位置と比較して、ポリペプチド全体の物理的及び化学的特性を変更すること無しに、置換、欠損及び/又は挿入を許容し得ることを理解するだろう。
【0095】
本願開示のポリペプチドは、図6に表される何れかのポリペプチドと少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%、又は100%相同であるアミノ酸配列を有するポリペプチドを含む。例えば、図6に表されるマウスLOX触媒ドメインポリペプチドのアミノ酸配列は、ヒトLOX触媒ドメインポリペプチドのアミノ酸配列と98%のアミノ酸配列の相同性を有する。
【0096】
上記に従って、本願開示のポリペプチドから導き出せるタンパクは、LOX(GenBank受託番号NP_002308)、LOXL1(NP_005567)、LOXL2(NP_002309)、LOXL3(NP_115882)、及びLOXL4(NP_115587)を含む(これら前述のGenBank受託番号は、ヒトの配列を示す)。これら、ヒトにおいて発見されたタンパク質に加えて、これらLOX又はLOXLタンパク質はまた、マウス又は他の哺乳類において発見されたタンパク質をも含むことができる。例えば、LOX、LOXL1又はLOXL2タンパク質は、マウスを起源とすることができる(それぞれGenBank受託番号NP_034858、NP_034859、NP_201582)。
【0097】
対象のポリペプチドは、リシル酸化酵素型酵素(例えば、LOX又はLOXL2)由来のアミノ酸配列を含むことができ、更に、異種の(heterologous)アミノ酸配列を含む(例えば、LOX又はLOXL2タンパク質の一部分でないポリペプチドに結合したLOX又はLOXL2配列)。そのようなポリペプチドは、エピトープタグ、精製タグ、及び/又は検出可能な標識を含む融合タンパク質のような融合タンパク質であり得る。融合タンパク質は、任意に(optionally)、異種の配列とリシル酸化酵素型酵素由来のアミノ酸配列の間にリンカー配列を含むことができる。核酸配列が提供される場合、目的の融合タンパク質を生成する方法は、当該技術分野でよく知られている。他の異種の要素(elements)及び、例となる融合タンパク質は、以下により詳細に記載される。
【0098】
異種の要素を含む例となるポリペプチドは、myc及び/又はHis6タグを含み得、SEQ ID NO:9、SEQ ID NO:11、SEQ ID NO:13又はSEQ ID NO:15に記載されるような、隣接(flanking)リンカー配列を任意に含み得る。
【0099】
本願開示のポリペプチドは、レポーターポリペプチド(例えば、蛍光タンパク質、及び/又は分子にコンジュゲート(conjugate)するもの)に結合されるポリペプチドを更に含む。ポリペプチドにコンジュゲートされる分子は、担体分子又は、動物体内にあるとき、免疫反応を引き起こすことが知られる免疫原(例えばアジュバンド)であり得る。他のコンジュゲートとしては、輸送を促進し、及び/又は対象のポリペプチドの半減期を延長させるコンジュゲートを含み得る。
【0100】
本願開示のポリペプチドはまた、1つ以上のポリ(エチレングリコール)(PEG)部分(モイエティ moieties)をも含み得る。そのようなポリペプチドは、「PEG化された(PEGylated)ポリペプチド」と称される。ポリペプチドのペグ化に適した方法及び薬剤が当該技術分野でよく知られる。一般的に、ポリペプチドにコンジュゲーションするのに適したPEGは、概して、室温で水溶性あり、一般式R(O−CH2−CH2)nO−Rを有する。この式において、Rは、水素又は、アルキル又はアルカノール基のような保護基であり、さらにnは、1から1000の整数である。この式において、Rが、保護基の場合、これは、一般的に1から8つの炭素原子を含む。
【0101】
PEGは、ポリペプチド中のアミノ基(例えば、リシン残基のイプシロン(ε‐)アミノ基又は、ポリペプチドのN末端の自由アミノ基)又はカルボキシル基と反応する官能基を生じるように修飾された、すくなくとも1つのヒドロキシル基を有し得る。
【0102】
PEGの更なる誘導体は、アミノ基と選択的に反応しアミド誘導体を生じる末端のチオカルボン酸基(−COSH)を含む。他の実施形態において、PEGは、N‐ヒドロキシスクシンイミダートのような反応性エステルをPEG鎖の末端に含む。そのようなN‐ヒドロキシスクシンイミダート含有PEG分子は、(数値が)6.5−7.5近くの中性pHのような、特別のpH状態で、選択アミノ基と反応する。
【0103】
PEGは、ポリペプチドのアミノ酸残基と直接に、又はリンカーを介してコンジュゲートされ得る。いくつかの実施形態において、リンカーがポリペプチドに加えられ、リンカー修飾されたポリペプチドを形成する。そのようなリンカーは、様々な機能性(例えば、スルフヒドリル基、アミノ基又はカルボキシル基のような反応性の基)を提供し、PEG剤をリンカー修飾されたポリペプチドに連結する。
【0104】
ポリペプチドにコンジュゲートされるPEGは、直鎖状又は分岐鎖状であり得る。分岐鎖状のPEG誘導体は、米国特許番号5,643,575に記載のもの、「star−PGEs」、及び、Shearwater Polymers社カタログ「Polyethylene Glycol Derivatives 1997-1998」に記載されたような、複数の腕を有するPEGを含むがこれらに限定されない。StarPGEsは、例えば米国特許番号6,046,305に、既知の技術として記載されている。
【0105】
[リシル酸化酵素型酵素の触媒ドメインを有するポリペプチドをエンコードするポリヌクレオチド]
本願の開示は、リシル酸化酵素型酵素の活性を有する前記のポリペプチドをエンコードする核酸配列を含むポリヌクレオチドを考えている。本願で意図される核酸は、上記のポリペプチドの何れかをエンコードする核酸の連続(contiguous)配列と、少なくとも70%(例えば、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%又は100%)相同であるヌクレオチド配列を有する。パーセンテージ相同性は、比較される配列のより短い方を基に示される。初期設定のパラメーターを使用し、フィルターを使用しない、BLASTN(2.0.8) (Altschul et al. (1997) Nucl. Acids. Res. 25:3389-3402)のような良く知られたプログラムが、配列比較を作成するために採用され得る。核酸配列の相同性(例えば、相同なアミノ酸配列をエンコードする2つの異なったポリヌクレオチド間の核酸配列の相同性)は、遺伝コードの縮重に起因して、アミノ酸配列の相同性のパーセントと比較してより低いことがあり得る。
【0106】
本願開示のポリペプチドをエンコードするポリヌクレオチドの核酸配列の例は、SEQ ID NO:2(ヒトLOX由来)、SEQ ID NO:4(ヒトLOXL2由来)、SEQ ID NO:6(マウスLOX由来)、及びSEQ ID NO:8(マウスLOXL2由来)を含む。これらの核酸配列はまた、発現ベクターにも提供され得る。
【0107】
本願において提供される発現ベクターは、前述の核酸及び/又はポリヌクレオチドを含む。そのような発現ベクターは、プロモーター(例えば、T7プロモーター、T3プロモーター、SP6プロモーター、E.coliRNAポリメラーゼプロモーター、CMVプロモーター、SV40プロモーター、PGKプロモーター、EF‐1アルファプロモーター)、転写終結シグナル(例えば、SV40終結シグナル)、スプライス部位(例えば、SV40スプライス部位、ベータ‐グロブリンスプライス部位)、リボゾーム結合部位、シグナル配列(例えば、免疫グロブリンカッパシグナル配列)、エピトープタグ(例えば、myc、FLAG)、精製タグ(例えば、His6)、複製起点及び薬剤選択マーカーを含むことができる。リンカーアミノ酸をエンコードし、及び/又は制限酵素認識部位を含むリンカー配列又は、他の種類のリンカー配列はまた、本願で開示されるベクターにある対象ポリペプチドをエンコードする核酸に、機能可能(operably)に、結合され得る。
【0108】
[本願開示のポリペプチドを作成する方法]
本願開示のポリペプチドは、組み換え方法及び非組み換えの方法(例えば、化学合成)を含む任意の適切な方法によって製造され得る。対象のポリペプチドは、Merrifield (1963) J. Am. Chem. Soc. 85:2149及びMethods in Molecular Biology, Vol 35: Peptide Synthesis Protocolsに記載されるような、当業者によく知られた固相合成法(例えば、Fmoc‐又はt‐Boc化学)によって生成され得る。
【0109】
ポリペプチドが、組み換え技術を使用して生成される場合、方法は、任意の適切な構成及び任意の適切な宿主細胞を含むことができ、これは原核細胞又は真核細胞(例えば、微生物宿主細胞、酵母宿主細胞又は培養哺乳類宿主細胞)であり得る。遺伝子物質を宿主細胞に導入する方法は、例えば、形質転換法、電気穿孔法、リポフェクション法、コンジュゲーション法、リン酸カルシウム法などを含む。転移の方法は、導入された核酸エンコードポリペプチドの安定な発現をもたらすように選択され得る。核酸をエンコードするポリペプチドは、遺伝性エピソーム要素(inheritable episomal element)(例えば、プラスミド)として提供され得、又は、ゲノム的に組み込まれ得る。
【0110】
核酸をエンコードするポリペプチドを転移するための適切なベクターは、組成的に、変更することができる。組み込みのベクターは、条件的複製(conditionally replicative)であり得、又は、自殺型プラスミド、バクテリオファージなどであり得る。構成物(ないしはコンストラクト:construct)は、例えば、プロモーター、選択可能な遺伝子マーカー(例えば、抗生物質に対する抵抗性を与える遺伝子(例えば、ネオマイシン、G418、メトトレキサート、アンピシリン、カナマイシン、エリスロマイシン、クロラムフェニコール又はゲンタマイシン))、(宿主細胞、例えば、細菌性宿主細胞、における複製を促進するための)複製起点などを含む様々な要素を含むことができる。ベクターの選択は、増殖が所望される細胞の種類及び、増殖の目的のような様々な因子に応じてなされるであろう。特定のベクターは、所望のDNA配列を大量に増幅し及び作成するために役立つ。他のベクターは、培養中の細胞におけるタンパク質の発現に適する。更に、他のベクターは、他の動物における細胞での転移及び発現に適する。適切なベクターの選択は、当該技術分野の技術でよくなされる。多くのそのようなベクターは、市販されている。
【0111】
使用されるベクターは、選択可能な薬剤耐性マーカー及び異なる宿主細胞における自立した複製を提供する要素を含むエピソームのプラスミドを基にした発現ベクターであり得る。ベクターは、例えばShort Protocols in Molecular Biology, (1999) F. Ausubel, et al., eds., Wiley & Sonsを含む当業者に良く知られる多数の出版物に十分に記載されている。ベクターは、対象のポリペプチドをエンコードする核酸の発現をもたらし得、(又は、)対象の核酸の増殖をもたらし得、又はその両方をもたらし得る。
【0112】
構成物(constructs)は、例えば、目的のポリヌクレオチドを構成物(ないしはコンストラクト)の骨格に挿入することによって調製され得る(典型的には、ベクター内の切断制限酵素部位へのDNAリガーゼ付着によって、調製され得る)。或いは、所望のヌクレオチド配列は、相同組み換え又は部位特異的組み換え、又は1つ以上の増幅方法(例えばPCR)によって挿入され得る。典型的に、相同組み換えは、所望のヌクレオチド配列の側部にある相同領域をベクターに取り付けることにより達成され、一方で、部位特異的組み換えは、部位特異的組み換えを促進する配列の使用を介して達成され得る(例えば、cre−lox、att部位など)。そのような配列を含む核酸は、例えば、オリゴヌクレオチドのライゲーションによって付加され、又は、相同な領域及び所望のヌクレオチド配列の部分の両方を含むプライマーを使用するポリメラーゼ連鎖反応によって付加される。
【0113】
目的のポリペプチドの発現に関して、発現カセットが採用され得る。かくて、本願の開示は、対象の核酸を含む組み換え発現ベクターを提供する。発現ベクターは、転写調節配列及び翻訳調節配列を提供でき、また、誘導可能な又は構成的な発現も提供でき、そのコーディング領域は、転写開始領域(例えば、プロモーター又はエンハンサー)及び転写停止領域及び翻訳停止領域の転写制御下に、機能(作用)可能に、配置される。これらの制御領域は、対象のポリペプチドが由来するリシル酸化酵素型酵素に元から存在するものであり得、又は、外生的な材料由来であり得る。そのようなものとして、外生的な材料由来の制御領域は、対象のポリペプチドをエンコードする核酸に機能(作用)可能に結合する異種の要素を考慮し得る。一般的に、転写調節配列及び翻訳調節配列は、プロモーター配列、リボゾーム結合部位、転写開始配列及び転写停止配列、翻訳開始配列及び翻訳停止配列及び、エンハンサー配列又はアクチベータ配列を含むが、これらに限定されない。プロモーターは構成的か又は誘導可能のどちらかであり得、強力な構成的プロモーターであり得る(例えばT7など)。
【0114】
発現ベクターは、目的のタンパク質をエンコードする核酸配列の挿入をもたらすために、プロモーター配列の近くに位置する適当な制限部位を一般的に有する。発現宿主において作動する選択可能なマーカーは、ベクターを含む細胞の選択を容易にするために存在し得る。加えて、発現構成物は、更なる要素を含むことができる。例えば、発現ベクターは、1つ又は2つの複製システムを有し得、かくて、これは、例えば、発現のための哺乳類又は昆虫細胞において維持され、及びクローニング及び増幅のための原核生物の宿主において維持される。加えて、発現構成物は、形質転換宿主細胞の選択を可能にするための選択可能なマーカー遺伝子を含むことができる。選択遺伝子が、当該技術分野において良く知られており、使用される宿主細胞によって異なるだろう。
【0115】
本願開示のポリペプチドはまた、例えば、放射性標識、蛍光標識、ビオチン標識、免疫学的に検出可能な標識(例えば、ヘマグルチニン(HA)タグ、ポリヒスチジンタグ)などの検出可能な標識のような更なる要素をも含むことができる。更なる要素が、(例えば、融合ポリペプチドの形態で)、様々な方法を介して、開示のポリペプチドの発現を促進するために提供され得(例えば、宿主細胞内で、発現を促進するためのN末端メチオニン及び/又は異種のシグナル配列)、及び/又は単離を促進するために提供され得る(例えば、ビオチンタグ、免疫学的に検出可能なタグ)。ポリペプチドはまた、共有結合又は非共有結合を介して任意に支持体上に固定化され得る。対象のポリペプチドをエンコードする例となる核酸は、SEQ ID NO:10、SEQ ID NO:12、SEQ ID NO:14又は、SEQ ID NO:16を含むことができる。核酸は、mycタグ、His6タグ及び/又はリンカー配列を有する、ヒト又はマウスリシル酸化酵素型酵素由来のポリペプチドをエンコードすることができる。
【0116】
対象のポリペプチドの単離及び精製は、当該技術分野において知られる方法に従って達成され得る。用語「単離された」は、化合物(例えば、ポリペプチド又はポリヌクレオチド)が、自然界において、それに付いている全ての又はいくつかの化合物から分離されたことを意味することが意図される。用語「単離された」はまた、化合物が、製造の間(例えば、化学合成、組み換え発現、培養培地など)に、それに付いている全ての又はいくつかの成分から分離されたことをも意味する。
【0117】
例えば、本願の開示によるポリペプチドは、対象のポリペプチドを発現するために遺伝子操作された細胞の溶解物から単離され得、又は、細胞培養培地の上清から単離され得、又は、合成反応混合物から単離され得る。単離は、更に、免疫親和性精製によって達成され、これは、一般的に、試料が、ポリペプチドのエピトープ(例えば、リシル酸化酵素型酵素由来のアミノ酸配列に融合された異種のエピトープ)に対する抗体(これは、任意に固定化される)と接触すること、洗浄し、非特異的な結合材料を取り除くこと、及び特異的に結合するポリペプチドを溶出することを含む。単離されたポリペプチドは、更に、透析及び、タンパク質精製に通常採用される他の方法(例えば、金属キレートクロマトグラフィー、イオン交換及びサイズ排除)によって精製され得る。
【0118】
本願の開示はさらに、1つ以上の本願開示のポリペプチドをエンコードする外生的なポリヌクレオチドを含む組み換え宿主細胞を考えている。
【0119】
[スクリーニング方法]
リシル酸化酵素触媒活性を有する本願開示のポリペプチドは、リシル酸化酵素型酵素の活性のモジュレーターの選別をする方法に役立つ。前記の対象のポリペプチドの十分量の発現は、以下に記載されるスクリーニング分析用に基質を供給するために使用され得る。
【0120】
リシル酸化酵素型酵素の活性のモジュレーター(及び、モジュレーター活性の試験をされる候補化合物)は、例えばタンパク質(例えば、抗体、転写因子)及び核酸(例えば、三重鎖形成性オリゴヌクレオチド、アンチセンスオリゴヌクレオチド、リボゾーム、siRNA、shRNA)及び、合成有機法及び/又は組み合わせ化学(combinatorial chemistry)により得られるようなもの、小有機分子などのような高分子を含むことができる。
【0121】
リシル酸化酵素型酵素の活性のモジュレーターは、アクチベータ(アゴニスト)及び阻害剤(アンタゴニスト)の両方を含み、様々なスクリーニング分析を使用することにより選択され得る。1つの実施形態において、モジュレーターは、化合物が、本願で開示される触媒ドメインポリペプチドに結合するかを確認することにより同定され得、結合が生じた場合、当該化合物は、候補モジュレーターである。任意に、追加の試験が、そのような候補モジュレーターについて行なわれ得る。かわりとして、候補化合物を、本願で開示されるポリペプチドと接触させることができ、及び、ポリペプチドの生物学的活性が分析される。ポリペプチドの生物学的活性を変える化合物は、リシル酸化酵素型酵素のモジュレーターである。一般的に、ポリペプチドの生物学的活性を減少させる化合物は、酵素の阻害剤である。
【0122】
リシル酸化酵素型酵素の活性のモジュレーターを同定する他の方法は、細胞が1つ以上の開示のポリペプチドを発現している細胞培養中で、候補化合物をインキュベートすること、及び、細胞の、1つ以上の生物学的活性又は特性を分析することを含む。培地において、細胞の生物学的活性又は特性を変更する化合物は、リシル酸化酵素型酵素の活性の潜在的なモジュレーターである。分析され得る生物学的活性は、例えば、リシン酸化、過酸化物生成、アンモニア生成、リシル酸化酵素型酵素のレベル、リシル酸化型酵素をエンコードするmRNAのレベル、開示のポリペプチドのレベル、開示のポリペプチドをエンコードするmRNAのレベル及び/又は、リシル酸化酵素型酵素に特異的な1つ以上の機能を含む。
【0123】
前述の分析の更なる実施形態において、候補化合物との接触のない状態において、1つ以上の生物学的活性又は細胞特性が、1つ以上のリシル酸化酵素型酵素のレベル又は活性と関連付けられる。例えば、生物学活性は、遊走、走化性、上皮間葉転換(EMT)又は、間葉上皮転換(MET)のような細胞性機能であり得、及び、その変化は、1つ以上の、対照試料又は参考試料との比較によって検出される。
【0124】
対照もまた、使用され得る。例として、陰性対照試料は、候補化合物が加えられ、開示のポリペプチドの減少したレベルのものを有する培養物(又は開示のポリペプチドを発現しない培養物)又は、試験培養物として、候補化合物の添加が無いが、開示のポリペプチドと等量のものを有する培養物を含むことができる。対照には、模擬的(mock)候補化合物又は、リシル酸化酵素活性を活性化又は阻害することが知られる薬剤の追加を含むことができる。いくつかの実施形態において、開示のポリペプチドの異なったレベルのものを含む別の(separate)培養物が、候補化合物と接触される。もし生物学的活性における変化が観察された場合、及び、より高いレベルのポリペプチドを有する培養物において変化が大きい場合、当該化合物は、リシル酸化型酵素の活性のモジュレーターと確定される。化合物が、リシル酸化酵素型酵素の活性剤であるか又は阻害剤であるかの決定は、当該化合物によって誘導された表現型から明らかであり得、又は、1つ以上のリシル酸化酵素型酵素の酵素的活性における、当該化合物の効果の試験のような、更なる分析を要求し得る。
【0125】
上記のような、リシル酸化酵素型酵素の活性のモジュレーターを確定するための細胞培養及び酵素的分析の方法と同じく、生化学的に又は組み換え的にリシル酸化酵素型酵素を得る方法が当該技術分野において知られる。
【0126】
リシル酸化酵素型酵素の酵素的活性は、いくつかの異なった方法によって分析され得る。例えば、リシル酸化酵素の酵素的活性は、過酸化水素、アンモニウムイオン及び/又はアルデヒド生成の検出及び/又は定量によって評価され得、リシン酸化及び/又はコラーゲン架橋結合の分析によっても評価され得、又は、細胞性浸潤能、細胞接着、細胞増殖又は転移成長を測定することによっても評価され得る。例えば、Trackman et al. (1981) Anal. Biochem. 113:336-342、Kagan et al. (1982) Meth. Enzymol. 82A:637-649、Palamakumbura et al. (2002) Anal. Biochem. 300:245-251、Albini et al. (1987) Cancer Res. 47:3239-3245、Kamath et al. (2001) Cancer Res. 61:5933-5940、米国特許番号4,997,854及び米国特許出願公開番号2004/0248871を参照する。
【0127】
試験化合物は、例えば、小有機化合物(例えば、約50から約2500Daの間の分子量を有する有機分子)、核酸又はタンパク質を含むがこれらに限定されない。(単一の)化合物又は複数の化合物は、化学的に合成され、又は微生物学的に生成され、及び/又は、例えば、試料(例えば、植物、動物又は微生物などからの細胞抽出物)に含まれ得る。リシル酸化酵素型酵素のモジュレーターの分析をするための反応混合物は、無細胞の抽出物であり得、又は、細胞培養物又は組織培養物を含むことができる。複数の化合物は、例えば、反応混合物に添加され、培養培地に加えられ、細胞に注入され、又は遺伝子組み換え動物に、投与され得る。分析に採用される細胞又は組織は、例えば、微生物細胞、真菌細胞、昆虫細胞、脊椎動物細胞、哺乳類細胞、霊長類細胞、ヒト細胞であり得、又は、(人間を除く)遺伝子組み換え動物を含むことができ、又はこの動物から得られ得る。
【0128】
(単一の)化合物又は複数の化合物を含む試料が、開示の方法において同定された場合、続いて、これは、対象のポリペプチドを阻害し又は活性化する能力を有する化合物を含むとして同定された元の試料から化合物を単離することができ、又は、例えば、当該化合物が異なった複数の化合物からなる場合、各試料毎に異なった物質の数を減らし、元の試料の更なる分割をともなう方法を繰り返すために、元の試料を更に分割(subdivide)することもできる。試料の複雑さに応じて、上記のステップは、開示の方法に従って、単離された試料が限られた数の物質のみを含み又は、ただ1つの物質を含むまで、数回実施され得る(例えば、限界希釈によってなされる)。いくつかの実施形態において、試料は、類似の化学的及び/又は物理的特性を有する物質を含み、そして、いくつかの実施形態において、当該物質は、相同である。
【0129】
更に、上述の方法は、リシル酸化酵素型酵素由来の結合エピトープの構成に使用され得る。類似の手法(approach)は、抗p24(HIV−1)モノクローナル抗体のペプチド抗原に関して良く記載されている(Kramer (1997) Cell 97:799-809を参照する)。クラスタ化されたアミノ酸ペプチドライブラリを使用する、ペプチド‐抗体相互作用のフィンガープリント解析のための一般的手順は、Kramer (1995) Mol. Immunol. 32:459-465に記載されている。更に、リシル酸化酵素型酵素のアンタゴニストは、Doring (1994) Mol. Immunol. 31:1059-1067に記載されるような方法に従って、開示のポリペプチドと特異的に反応するモノクローナル抗体由来であり得、及びこれから同定され得る。
【0130】
リシル酸化酵素型酵素又は、本開示のポリペプチドのような目標物に対して特異的な親和性を有する化合物を同定するための大きなライブラリを作成し及びスクリーニングするためのいくつかの方法が当業者に知られる。これらの方法は、ファージ表示法を含み、そのランダム化されたペプチドは、ファージから表示され、固定化されたレセプターを使用した親和性クロマトグラフィーによりスクリーニングされる。例えば、WO 91/17271、WO 92/01047及び、米国特許番号5,223,409を参照する。他の手法において、固相支持体上に固定化されたポリマーの組み合わせライブラリ(combinatorial libraries)(例えば、「チップ」)は、フォトリソグラフィーを使用して合成される。例えば、米国特許番号5,143,854、WO 90/15070及びWO 92/10092を参照。固定化されたポリマーを、標識されたポリペプチド(例えば、リシル酸化酵素型酵素又は本願で開示されるようなポリペプチド)と接触させ、支持体をスキャンし、ラベルの位置を決定し、それにより、当該ポリペプチドに結合するポリマーを同定する。
【0131】
目的のポリペプチド(例えば、リシル酸化酵素型酵素又は開示のポリペプチド)の結合リガンド(例えば、活性剤又は阻害剤)を同定するために使用され得る、連続的なセルロース膜支持体でのペプチドライブラリの合成及びスクリーニングは、例えば、Kramer (1998) Methods Mol. Biol. 87: 25-39において記載される。そのような分析によって同定されるリガンドは、目的のタンパク質の候補モジュレーターであり、更なる試験のために選択され得る。この方法はまた、例えば、目的のタンパク質における結合部位及び認識モチーフを決定するためにも使用され得る。例えば、Rudiger (1997) EMBO J. 16:1501-1507及びWeiergraber (1996) FEBS Lett. 379:122-126を参照する。
【0132】
WO 98/25146は、所望の特性(例えば、ポリペプチド又はその結合相手をアゴナイズ(agonize)し、ポリペプチド又はその結合相手に結合し、又は、ポリペプチド又はその結合相手をアンタゴナイズ(antagonize)する能力)を有する化合物の複合体のスクリーニングライブラリの更なる方法を記載する。それらのライブラリにおける複合体は、供試化合物、化合物の合成における少なくとも1つのステップを証明するタグ及び、レポーター分子による修飾の影響を受けやすいテザーを含む。テザーの修飾は、複合体が、所望の特性を有する化合物を含むことを示すために使用される。タグは、そのような化合物の合成における少なくとも1つのステップを示すためにデコード(decode)され得る。リシル酸化酵素型酵素(又は、開示のポリペプチド)と相互作用する化合物を同定するための他の方法は、例えば、ファージ表示システムを使用したインビトロスクリーニング、フィルター結合分析及び、例えば、BIAcore装置(Pharmacia)を使用した相互作用の、「リアルタイムの」測定である。
【0133】
これら方法の全ては、本願の開示に従って、リシル酸化酵素型酵素又はそれに関係するポリペプチドの活性剤/アゴニスト及び阻害剤/アンタゴニストを同定するために使用され得る。
【0134】
そのような活性化剤(アクチベータ)又は阻害剤の基礎的な構成のための様々な材料が採用され得、これは、例えば、開示のポリペプチドの模倣(模擬 mimetic)アナログを含む。開示のポリペプチドの模倣アナログ又は、その生物学的に活性なフラグメントは、例えば、生物学的活性に必須であると期待されるアミノ酸を置換することによって生じ得る(例えば、ステレオイソマー、即ちD−アミノ酸;例えば、Tsukida (1997) J. Med. Chem. 40:3534-3541を参照する)。更に、ポリペプチドフラグメントが、生物学的に活性なアナログをデザインするために使用される場合、プロ‐模倣化構成物(pro-mimetic components)が、少なくともいくつかの立体的形態特性(conformational properties、これは、元のポリペプチドの部分を取り除く際に失われていることもありうる)を再構築するために、ペプチドに組み込まれ得る(例えば、Nachman (1995) Regul. Pept. 57:359-370を参照する)。
【0135】
そのような天然アミノ酸配列の擬ペプチド(pseudopeptide)類似体は、とても効率的に、親タンパク質を模倣できる(Benkirane (1996) J. Biol. Chem. 271:33218-33224)。例えば、簡単に入手できるアキラルo−アミノ酸残基の開示のポリペプチド又は、そのフラグメントへの組み込みが、脂肪族化合物鎖のポリメチレンユニットによるアミド結合の置換をもたらし、それによって、ペプチド模倣物(peptide mimetic)を構成するための便利な戦略を提供する(Banerjee (1996) Biopolymers 39:769-777)。他のシステムにおける小ペプチドホルモンの超活性なペプチド模倣アナログもまた、記載されている(Zhang (1996) Biochem. Biophys. Res. Commun. 224:327-331)。
【0136】
ペプチド模倣物はまた、一連のアミドアルキル化を介して、ペプチド模倣組み合わせライブラリの合成によっても同定され得、続いて結果する化合物の試験(例えば、それらの結合及び免疫学的特性のための試験)を行なう。ペプチド模倣組み合わせライブラリの発生及び使用の方法が、記載されている。例えば、Ostresh, (1996) Methods in Enzymology 267:220-234及びDorner (1996) Bioorg. Med. Chem. 4:709-715を参照する。更に、開示のポリペプチドの3次元構造及び/又は結晶構造は、ペプチド模倣物をデザインするために使用され得る(Rose (1996) Biochemistry 35:12933-12944、Rutenber (1996) Bioorg. Med. Chem. 4:1545-1558)。
【0137】
天然の生物学的ポリペプチドの活性を模倣した低分子量合成分子(low-molecular-weight synthetic molecules)の、構造を元にしたデザイン及び合成が、例えば、Dowd (1998) Nature Biotechnol. 16:190-195、Kieber-Emmons (1997) Current Opinion Biotechnol. 8:435-441、Moore (1997) Proc. West Pharmacol. Soc. 40:115-119、Mathews (1997) Proc. West Pharmacol. Soc. 40:121-125及びMukhija (1998) European J. Biochem. 254:433-438に更に記載されている。
【0138】
また、例えば、リシル酸化酵素型酵素又は開示のポリペプチドの基質又はリガンドとして機能できる小有機化合物の模倣物をデザインし、合成し、評価することが可能であることも当業者によく知られている。例えば、細胞毒性において多剤耐性補助関連タンパク質(multidrug resistance assistance-associated protein)をアンタゴナイズ(antagonize)する際に、ハパロシンのD−グルコース模倣物がハパロシンと類似の性能を示したことが記載されている(Dinh (1998) J. Med. Chem. 41:981-987)。
【0139】
リシル酸化酵素型酵素及び本願で開示されるポリペプチドの構造は、例えば、小分子、ペプチド、ペプチド模倣物及び抗体のようなモジュレーターの選択を導くために研究され得る。リシル酸化酵素型酵素及び開示のポリペプチドの構造的特性は、リシル酸化酵素型酵素に結合し、又は、そのリガンド、基質、結合相手又はレセプターとして機能する、天然の又は合成の分子を同定するのに役立つ。例えば、Engleman (1997) J. Clin. Invest. 99:2284-2292を参照する。例えば、リシル酸化酵素型酵素の構造的モチーフの折りたたみシミュレーション及びコンピューター再設計は、適切なコンピュータープログラムを使用して行われ得る(Olszewski (1996) Proteins 25:286-299、Hoffman (1995) Comput. Appl. Biosci. 11:675-679)。タンパク質の折りたたみのコンピュータモデリングは、詳細なペプチド及びタンパク質構造の立体配座解析及びエネルギー解析に使用され得る(Monge (1995) J. Mol. Biol. 247:995-1012、Renouf (1995) Adv. Exp. Med. Biol. 376:37-45)。適切なプログラムが、相補的なペプチド配列のコンピューターを使用した調査を使用して、リシル酸化酵素型酵素及び/又は開示のポリペプチド上にあり、リガンド及び結合相手と相互作用する部位の同定に使用され得る(Fassina (1994) Immunomethods 5:114-120)。タンパク質及びペプチドのデザインのための更なるシステムが、当該技術分野において、例えば、Berry (1994) Biochem. Soc. Trans. 22:1033-1036、Wodak (1987), Ann. N.Y. Acad. Sci. 501:1-13、及びPabo (1986) Biochemistry 25:5987-5991に記載される。上記の構造解析から得られた結果は、例えば、有機分子、ペプチド及び、1つ以上のリシル酸化酵素型酵素の活性のモジュレーターとして機能するペプチド模倣物の作成に使用され得、そして、本開示のポリペプチドの模倣物の作成に使用され得る。
【0140】
抗体のような本願でスクリーニングされる阻害剤(inhibitors)は、競合阻害剤、不競合(uncompetitive)阻害剤、混合阻害剤又は非競合阻害剤(non-competitive)であり得る。競合阻害剤は、しばしば、基質と構造的な類似性を持ち、通常、活性部位に結合し、一般的に、より低い基質濃度において、より効率的である。競合阻害剤存在下では、見かけのKMが増加する。不競合阻害剤は、一般的に、酵素‐基質複合体又は、基質が活性部位に結合した後に結合可能になる部位に結合し、活性部位を変形させ得る。見かけのKM及びVmaxの両方が、不競合阻害剤の存在下で減少し、基質濃度は、阻害において少しの効果を有するか又は効果を有さない。従って、不競合阻害剤による阻害は、高基質濃度において、しばしば最も顕著である。混合阻害剤は、自由な(free)酵素及び、酵素‐基質複合体の両方に結合可能であり、かくて、基質結合及び触媒活性の両方に影響する。非競合阻害とは、阻害剤が、酵素及び酵素‐基質複合体に、同等の親和性を有して結合する、混合阻害の特別な例であり、当該阻害は、基質濃度に影響されない。非競合阻害剤は、一般的に、活性部位の外の領域で酵素と結合する。酵素阻害の更なる詳細については、例えば、上記のVoet et al. (2008)を参照する。リシル酸化酵素型酵素のような酵素に関して、その天然の基質(例えば、コラーゲン、エラスチン)は、通常、(インビボにおいて達成され得る任意の阻害剤の濃度と比較して)インビボにおいて非常に過度に存在し、阻害が基質濃度と無関係であるので、非競合阻害剤は、有益である。
【0141】
[抗体生成]
本願の開示はまた、開示のポリペプチドに特異的な抗体を生成する方法を提供する。抗体(例えば、抗体が自然に生じる環境以外の環境にある抗体)が単離され得る。対象のポリペプチドに特異的な、適切な抗体は、任意のアイソタイプの抗体、単鎖のFv、Fab、Fab、Fv、F(ab’)2、人工的な抗体、ヒト化抗体、これらのフラグメントなどを含む。
【0142】
適切な抗体は、宿主動物に、対象のポリペプチドの全部又は一部を含むペプチドを免疫することによって得られ得る。動物を免疫する場合、本願の開示は、対象のポリペプチドを含む免疫原性組成物(immunogenic composition)を提供する。対象の免疫原性組成物は、対象のポリペプチド及びアジュバンドを含み得る。アジュバンドは、当該技術分野において知られ、ヒト又は他の哺乳類に使用するのに適切なアジュバンドを含み得る。
【0143】
免疫するのに適切な宿主動物は、マウス、ラット、ヒツジ、ヤギ、ハムスター、ウサギ、ロバなどを含む。使用されるアジュバンド、追加免疫スケジュール、注入部位、適切な動物などを含む動物を免疫する方法は、当該技術分野において良く理解されており(例えば、Harlow et al., Antibodies: A Laboratory Manual, First Edition (1988) Cold Spring Harbor, N.Y.)、動物への生きている細胞の投与は、いくつかの哺乳類及び鳥類に関して記載されている(例えば、McKenzie et al. (1989) Oncogene 4:543-548、Scuderi et al. (1985) Med. Oncol. Tumor Pharmacother 2:233-242、Roth et al. (1984) Surgery 96:264-272及びDrebin et al. (1984) Nature 312:545-548)。免疫付与の後、抗体生成細胞の集団が生じる。細胞の集団は、当該技術分野における技術の1つとして良く知られるハイブリドーマ法を使用して生成され得る(例えば、前記のHarlowを参照する)。抗体を発現する細胞は、骨髄腫細胞又は形質転換細胞のような不死化細胞に融合され、これらは細胞培養において無制限に複製する能力を有し、それによって不死の、免疫グロブリン分泌細胞系統を生成する)。利用される不死化細胞系統は、特定の栄養素を利用するのに必要な酵素が欠損するように選択され得る。骨髄腫細胞のような多くのそのような系統は、当業者によく知られており、例えば、チミジンキナーゼ(TK)欠損細胞系統又は、ヒポキサンチン‐グアニンホスホリボキシル転移酵素(HGPRT)欠損細胞系統を含む。これらの欠損体は、例えば、ヒポキサンチン/アミノプテリン/チミジン(HAT)培地で増殖する能力によって融合細胞を選択することを可能にする。代わりの実施形態において、モノクローナル抗体を発現する細胞の集団は、ファージ表示法を使用して作成され得る。
【0144】
対象のポリペプチドに対する抗体はまた、遺伝子工学的技術によっても提供され得る。この技術において、標準のハイブリドーマ法と同様に、抗体生成細胞は、所望の抗原又は免疫原に感作される。免疫脾臓細胞又はハイブリドーマから単離されたメッセンジャーRNAは、PCR増幅を使用するcDNAを作成するための鋳型として使用される。それぞれ、最初の抗原特異性を保持する1つの重鎖遺伝子及び1つの軽鎖遺伝子を含むベクターのライブラリは、増幅された免疫グロブリンcDNAの適切な領域ないし部位(section)の、発現ベクターへの挿入により生成される。組み合わせライブラリは、重鎖遺伝子ライブラリを軽鎖遺伝子ライブラリと結合することによって構成され得る。これは、重鎖及び軽鎖(これらは、抗体分子のFabフラグメント又は抗原結合フラグメントに似る)を共発現するクローンのライブラリをもたらす。これらの遺伝子を有するベクターは、宿主(例えば、細菌、昆虫細胞、哺乳類細胞又は、他の適切なタンパク質生成宿主細胞)に共トランスフェクトされる。抗体遺伝子合成が、トランスフェクト宿主において誘導される場合、重鎖及び軽鎖タンパク質は、自己集合(self-assemble)し、抗原又は免疫原を使用するスクリーニングによって検出され得る活性な抗体を生成する。
【0145】
抗体生成の他の方法は、ファージ表示(法)を含む。ファージ表示(法)は、タンパク質相互作用の高‐スループットスクリーニング(high-throughput screening)に使用される。この方法において、ファージは、レパートリー又は組み合わせ抗体ライブラリ(例えば、ヒト又はマウス)から発現される抗原結合ドメインを表示するのに利用される。目的の、そして対象のポリペプチドを結合する抗原結合ドメインを発現するファージが、ポリペプチド目標を使用して、(例えば、標識されたポリペプチド又は、固体表面又はビーズに結合又は取り込まれるポリペプチドを使用して)選択又は同定される。これらの方法において使用されるファージは、典型的に、fd及びM13を含む線状ファージである。結合ドメインは、Fab、Fv(軽鎖又は重鎖由来の個々のFv領域)又は、ファージ遺伝子IIIタンパク質又は遺伝子VIIIタンパク質のどちらかに、組み換え的に結合されたジスルフィド安定化されたFv抗体ドメインを含むことができる。例示の方法は、例えば、EP 368684B1、米国特許番号5,969,108、Hoogenboom, H. R. and Chames (2000) Immunol. Today 21:371、Nagy et al. (2002) Nat. Med. 8:801、Huie et al. (2001) Proc. Natl. Acad. Sci. USA 98:2682、Lui et al. (2002) J. Mol. Biol. 315:1063(これら各々は、本願に引用として組み込まれる)に示される。いくつかの出版物(例えば、Marks et al. (1992) Bio/Technology 10:779-783)は、大きなファージライブラリを構成するための戦略としての組み合わせ感染(combinatorial infection)及びインビボ組み換えと同じく、鎖シャッフリング(chain shuffling)による高親和性ヒト抗体の生成を記載している。
【0146】
関係する実施形態において、リボゾーム表示は、表示基盤(display platform)として使用される。例えば、Hanes et al. (2000) Nat. Biotechnol. 18:1287、Wilson et al. (2001) Proc. Natl. Acad. Sci. USA 98:3750及び、Irving et al. (2001) J. Immunol. Methods 248:31を参照する。
【0147】
細胞表面ライブラリは、抗体のためにスクリーニングされ得る(Boder et al. (2000) Proc. Natl. Acad. Sci. USA 97:10701、Daugherty et al. (2000) J. Immunol. Methods 243:211)。そのような手段は、モノクローナル抗体の単離及びそれに続くクローニングのための伝統的なハイブリドーマ技術に、代わる方法を提供する。
【0148】
ファージ表示法において、機能的な抗体ドメインは、このドメインをエンコードするポリヌクレオチド配列を有するファージ粒子の表面に表示される。例えば、重鎖可変領域(VH)及び軽鎖可変領域(VL)をエンコードするDNA配列は増幅され、又はさもなければ、動物cDNAライブラリ(例えば、リンパ組織のヒト又はマウスcDNAライブラリ)又は合成cDNAライブラリから単離される。VH及びVL領域をエンコードするDNAは、scFvリンカーによってお互いに結合され得(例えば、PCRによって結合され得る)、ファージミドベクター(例えば、pCANTAB 6又はpComb 3HSS)にクローン化され得る。ベクターは、E.coliにエレクトロポレーションされ、そしてE.coliに補助ファージ(helper phage)を感染させる。VH又はVL領域をエンコードする配列は、通常は、ファージの遺伝子III配列又は遺伝子VIII配列のどちらかに組み換え的に融合される。目的の抗原(即ち、対象のポリペプチド)に結合する抗原結合ドメインを発現するファージは、抗原を使用して(例えば、標識された抗原又は、固体表面又はビーズに結合又は取り込まれた抗原を使用して)選択され又は同定され得る。
【0149】
ファージ表示法の更なる実施例は、PCT出願番号PCT/GB91/01134、国際公開WO 90/02809、WO 91/10737、WO 92/01047、WO 92/18619、WO 93/11236、WO 95/15982、WO 95/20401、及び米国特許番号5,698,426、5,223,409、5,403,484、5,580,717、5,427,908、5,750,753、5,821,047、5,571,698、5,427,908、5,516,637、5,780,225、5,658,727、5,733,743及び5,969,108(各々、本願に引用によってその全てが取り込まれる)に開示された方法を含む。
【0150】
[製剤、キット及び投与経路]
本願に開示されるような触媒ドメインを含む治療用組成物もまた提供される。そのような組成物は典型的に、触媒ドメイン(又は触媒ドメインをエンコードする核酸)、医薬的に許容可能な担体を含む。補助的な活性組成物もまた、当該構成物に組み込まれ得る。
【0151】
本願で使用される場合、用語「治療的に効果のある量」又は「効果的な量」は、単独で又は他の治療用剤と組み合わせて、細胞、組織又は対象(例えば、ヒト又は、霊長類、げっ歯類、ウシ、ウマ、ブタ、ヤギなどのヒトを除く動物のような哺乳類)に投与される場合、疾病の状態又は疾病の進行を予防し(妨げ)、又は改善するのに効果的な量である治療用剤の量を意味する。治療的に効果のある用量(dose)は更に、症状の完全な又は部分的な改善(例えば、関連する病状の処置、治癒、予防又は改善)、又はそのような状態の処置、治癒、予防又は改善の速度の増進をもたらすのに十分な化合物の量を意味する。薬剤の効果的な量は、投与したときに、1つ以上の用量(dose)において、個体内のリシル酸化酵素型酵素に関連する1つ以上の症状の重症度おいて(当該薬剤で処置しない場合の1つ以上の症状の重症度と比較して、)少なくとも約5%、少なくとも約10%、少なくとも約15%、少なくとも約20%、少なくとも約25%、少なくとも約30%、少なくとも約40%、少なくとも約50%、少なくとも約60%、少なくとも約70%、少なくとも約80%、少なくとも約90%又は90%超の減少をもたらす量である。そのような重症度における減少は、例えば、改善された創傷治癒又は促進された血管形成によって、明らかにされ得る。
【0152】
リシル酸化酵素型酵素の触媒ドメイン(例えばLOX又はLOXL2触媒ドメイン)の治療的に効果のある量は、疾病又は障害の種類、疾病又は障害の広さ、及び疾病又は障害を被る哺乳類の大きさによって異なる。
【0153】
様々な医薬組成物および、それらの調製及び使用の技術が、本願の開示を踏まえて、当業者に知られる。適切な医薬組成物及び、その投与のための技術の詳細なリストに関して、Remington's Pharmaceutical Sciences, 17th ed. 1985、Brunton et al., "Goodman and Gilman's The Pharmacological Basis of Therapeutics," McGraw-Hill, 2005、University of the Sciences in Philadelphia (eds.),"Remington: The Science and Practice of Pharmacy," Lippincott Williams & Wilkins, 2005、及び University of the Sciences in Philadelphia (eds.), "Remington: The Principles of Pharmacy Practice," Lippincott Williams & Wilkins, 2008のような文献を参照し得る。
【0154】
開示される治療用組成物は更に、医薬的に許容可能な物質、組成物又は、液体又は固形フィラー剤(solid filler)、希釈剤、添加剤、溶剤又は封入材料(即ち担体)のような賦形剤(vehicle)を含む。これらの担体には、対象の組成物を1つの器官又は身体の領域から他の器官又は身体の領域に輸送することに関与する。各担体は、製剤の他の材料と適合するという意味で「許容可能」であるべきであり、患者に無害であるべきである。医薬的に許容可能な担体として役立つことができる材料のいくつかの例は、糖(ラクトース、グルコース及びスクロースなど)、澱粉(トウモロコシ澱粉、ジャガイモ澱粉など)、セルロース及びその誘導体(カルボキシメチルセルロースナトリウム、エチルセルロース及び酢酸セルロースなど)、粉末トラガカント、麦芽、ゼラチン、タルク、添加剤(ココアバター及び座剤ワックス(suppository wax)など)、油(落花生油、綿の実油、ヒマワリ油、ごま油、オリーブ油、トウモロコシ油及び大豆油など)、グリコール(プロピレングリコールなど)、ポリオール(グリセリン、ソルビトール、マンニトール及びポリエチレングリコールなど)、エステル(エチルオレアート及びエチルラウラートなど)、寒天、緩衝剤(水酸化マグネシウム及び水酸化アルミニウムなど)、アルギン酸、発熱性物質除去水(pyrogen-free water)、等張食塩水、リンガー溶液、エチルアルコール、リン酸緩衝溶液、及び、製剤処方に採用される他の非毒性適合物質を含む。着色剤、放出剤、コーティング剤、甘味料、香味料、芳香剤、保存料及び酸化防止剤と同様に、湿潤剤、乳化剤及びラウリル硫酸ナトリウム及びステアリン酸マグネシウムのような潤滑剤もまた、当該組成物に存在し得る。
【0155】
本願の開示の他の視点は、本願に開示されるような、リシル酸化酵素型酵素の触媒ドメイン又は、そのような触媒ドメインをエンコードする核酸の、対象物への投与を行うためのキットに関する。1つの実施形態において、キットは、医薬的な担体に製剤化されるリシル酸化酵素型酵素の触媒ドメイン(又は、リシル酸化酵素型酵素の触媒ドメインをエンコードする核酸)を含む。
【0156】
製剤化及び輸送の方法は、処置される障害の種類及び部位に応じて一般的に適合される。例となる製剤は、例えば、静脈投与、動脈内投与、眼球内投与、筋肉内投与、又は皮下投与のような、非経口投与に適したものを含むがこれらに限定されず、ミセル、リポソーム又は薬剤開放カプセルにカプセル化された製剤(即ち、遅延放出用にデザインされた生体適合性のコーティング内に組み込まれた活性剤);摂取可能な製剤;クリーム、軟膏及びゲルのような局所使用の製剤;及び、吸入剤、エアロゾル及びスプレーのような他の製剤を含む。開示の化合物の用量は、処置を必要とする広さ及び重症度、投与される組成物の活性、対象物の全体的な健康及び、当業者に良く知られる他の検討事項に応じて、変化するであろう。
【0157】
更なる実施形態において、本願に記載される組成物は、局所的に輸送(ないし送達)される。局所的輸送は、例えば、創傷領域又は繊維化領域への非全身的な組成物の輸送を可能にし、全身的な輸送と比較した当該組成物の身体負荷量を減少させる。そのような局所輸送は、例えば、ステント及びカテーテルを含むがこれに限定されない様々な医学的に埋め込まれる装置を介して実現され得、又は、注射又は手術によって実現され得る。コーティング、インプランティング、埋めこみのための方法及び、さもなければ、所望の薬剤の、ステント及びカテーテルのような医学的な装置への取り付け方法が、当該技術分野において確立され、本願において考えられる。
【0158】
インプラントされたステントが、血栓溶解剤のような医学的な薬剤を移送するために使用されている。米国特許番号5,163,952は、ステント自体の材料の中に医学的な薬剤を有するように構成される(formulated)熱的形状記憶性拡張プラスチックステント装置(thermal memoried expanding plastic stent device)を開示する。米国特許番号5,092,877は、化合物の輸送に関連するコーティングを有することができるポリマー材料のステントを開示する。バイオ分解可能なポリマー及びバイオ吸収可能なポリマーを利用する種類の装置に向けられる他の特許は、米国特許番号4,916,193及び米国特許番号4,994,071を含む。例として、米国特許番号5,304,121は、ヒドロゲルポリマー及び細胞増殖阻害剤又はヘパリンのような予選択された化合物からなるステントに適用されるコーティングを開示する。治療用材料を有する、コートされた血管内ステントの作成方法が、米国特許番号5,464,650に記載され、ここでは、ポリマーコーティング材料は、溶媒に溶解され、治療用材料が、溶媒に分散している。溶媒は、続いて、ステントと接触の後蒸発される。
【0159】
米国特許番号6,120,536は、ステントを含む広範囲な種類の人工装具で使用する、更なるコーティングの種類を記載する。本願で記載される組成物の投与に役立つ更なる医学的な装置又は人工の装置の例は、血液交換装置、血管アクセスポート、中心静脈カテーテル、心血管カテーテル、体外循環回路、代用血管、ポンプ、心臓弁、及び心血管縫合糸を含むがこれに限定されない。
【0160】
ステント及びカテーテルを含む本願で記載される組成物でコートされた装置の使用は、組成物の、特異的な部位又は局在される部位への輸送を可能にする。そのような部位特異的輸送は、組成物の使用(及び/又はその用量)のための手段を提供し得、これは、そうでなければ溶解性、全身性の毒性の懸念又は他の問題をもたらす全身性の輸送の影響を受けにくい。例として、β‐アミノプロピオニトリル(BAPN)が、リシル酸化酵素型酵素の阻害剤として役立つことが知られているが、この化合物は、高い毒性があり、全身的に投与される場合、その効果的な使用に関して問題がある。活性薬剤又は、BAPNのような化合物の輸送のためのステント、カテーテル又は他の医学的な装置の使用は、目標とされる態様又は局所的な態様における効果的な用量の化合物の使用を許容し、かくて、そのような化合物と関連する全身性毒性効果を減少させる。
【0161】
[使用(形態)]
本願で開示されるポリペプチドは、いくつかの使用(形態ないし用途)を有する。例えば、これらは、リシル酸化酵素の酵素的活性(例えば、コラーゲン架橋結合)を分析するための標準品(例えば、陽性対照)として使用され得る。そのような分析の1つの使用(形態)としては、例えばLOX又はLOXL2タンパク質及び/又はその遺伝子の突然変異解析がある。リシル酸化酵素活性を有する触媒ドメインを含むポリペプチドはまた、リシル酸化酵素型酵素に、新しい抗体及びすでに存在する抗体を特徴付けるためにも使用され得る。
【0162】
本願で開示されるポリペプチドはまた、例えばLOX及び/又はLOXL2触媒ドメイン、小有機分子又は高分子(例えばポリペプチド)に結合する分子を同定するためにも使用され得る。これらの結合分子の或る特定のものは、触媒活性化のモジュレーター(即ち、活性化剤又は阻害剤)として機能し得る。
【0163】
本願で開示される対象のポリペプチドはまた、ヒト及びマウスLOX及びLOXL2タンパク質及び他のリシル酸化酵素型酵素の触媒的な活性領域に特異的に向けられる抗体の作成にも役立つ。そのような抗体は、例えば、リシル酸化酵素活性の病的に増加するレベルによって特徴付けられる疾病又は障害に関する医学的及び/又は治療的な応用に使用され得る。
【0164】
加えて、対象のポリペプチドは、コラーゲンの架橋結合を要求する医学的及び/又は治療的な応用に役立つ(例えば、創傷治癒及び糖尿病性潰瘍の処置)。(全長のタンパク質と比較して)より小さい大きさの単離された触媒ドメインは、例えば、改良された安定性及びより迅速な吸収によって、治療剤として、その使用を促進し得る。
【実施例】
【0165】
[実施例1:ヒトLOX触媒ドメイン]
ヒトLOXタンパク質の触媒ドメインを含むポリペプチドは、207アミノ酸残基を含み、次のアミノ酸配列を有する。
GLPDLVADPYYIQASTYVQKMSMYNLRCAAEENCLASTAYRADVRDYDHRVLLRFPQRVKNQGTSDFLPSRPRYSWEWHSCHQHYHSMDEFSHYDLLDANTQRRVAEGHKASFCLEDTSCDYGYHRRFACTAHTQGLSPGCYDTYGADIDCQWIDITDVKPGNYILKVSVNPSYLVPESDYTNNVVRCDIRYTGHHAYASGCTISPY
(SEQ ID NO:1)
【0166】
ある実施形態において、ヒトLOX触媒ドメインを含むポリペプチドは、上に表された配列における最初のグリシン及びロイシン残基(”GL”)を含まない。
【0167】
LOX触媒ドメインを含む対象のポリペプチドをエンコードするヒトDNA配列は、以下の通りである。
GGTCTCCCAGACCTGGTGGCCGACCCCTACTACATCCAGGCGTCCACGTACGTGCAGAAGATGTCCATGTACAACCTGAGATGCGCGGCGGAGGAAAACTGTCTGGCCAGTACAGCATACAGGGCAGATGTCAGAGATTATGATCACAGGGTGCTGCTCAGATTTCCCCAAAGAGTGAAAAACCAAGGGACATCAGATTTCTTACCCAGCCGACCAAGATATTCCTGGGAATGGCACAGTTGTCATCAACATTACCACAGTATGGATGAGTTTAGCCACTATGACCTGCTTGATGCCAACACCCAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGTTTCTGTCTTGAAGACACATCCTGTGACTATGGCTACCACAGGCGATTTGCATGTACTGCACACACACAGGGATTGAGTCCTGGCTGTTATGATACCTATGGTGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTAAAACCTGGAAACTATATCCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTTCCTGAATCTGACTATACCAACAATGTTGTGCGCTGTGACATTCGCTACACAGGACATCATGCGTATGCCTCAGGCTGCACAATTTCACCGTATTAG
(SEQ ID NO:2)
【0168】
[実施例2:ヒトLOXL2触媒ドメイン]
ヒトLOXL2タンパク質の触媒ドメインを含むポリペプチドは、次のアミノ酸配列を有する。
TAPDLVLNAEMVQQTTYLEDRPMFMLQCAMEENCLSASAAQTDPTTGYRRLLRFSSQIHNNGQSDFRPKNGRHAWIWHDCHRHYHSMEVFTHYDLLNLNGTKVAEGHKASFCLEDTECEGDIQKNYECANFGDQGITMGCWDMYRHDIDCQWVDITDVPPGDYLFQVVINPNFEVAESDYSNNIMKCRSRYDGHRIWMYNCHIGGSFSEETEKKFEHFSGLLNNQLSPQ
(SEQ ID NO:3)
【0169】
ある実施形態において、ヒトLOXL2触媒ドメインを含むポリペプチドは、上に表された配列における最初のスレオニン及びアラニン残基(”TA”)を含まない。
【0170】
LOXL2触媒ドメインをエンコードするヒトDNA配列は、以下の通りである。
ACCGCCCCTGACCTGGTCCTCAATGCGGAGATGGTGCAGCAGACCACCTACCTGGAGGACCGGCCCATGTTCATGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCGGCCTCAGCCGCGCAGACCGACCCCACCACGGGCTACCGCCGGCTCCTGCGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCCGACTTCCGGCCCAAGAACGGCCGCCACGCGTGGATCTGGCACGACTGTCACAGGCACTACCACAGCATGGAGGTGTTCACCCACTATGACCTGCTGAACCTCAATGGCACCAAGGTGGCAGAGGGCCACAAGGCCAGCTTCTGCTTGGAGGACACAGAATGTGAAGGAGACATCCAGAAGAATTACGAGTGTGCCAACTTCGGCGATCAGGGCATCACCATGGGCTGCTGGGACATGTACCGCCATGACATCGACTGCCAGTGGGTTGACATCACTGACGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTTATTAACCCCAACTTCGAGGTTGCAGAATCCGATTACTCCAACAACATCATGAAATGCAGGAGCCGCTATGACGGCCACCGCATCTGGATGTACAACTGCCACATAGGTGGTTCCTTCAGCGAAGAGACGGAAAAAAAGTTTGAGCACTTCAGCGGGCTCTTAAACAACCAGCTGTCCCCGCAGTAA
(SEQ ID NO:4)
【0171】
[実施例3:マウスLOX触媒ドメイン]
マウスLOXタンパク質の触媒ドメインを含むポリペプチドは、次のアミノ酸配列を有する。
GLPDLVPDPYYIQASTYVQKMSMYNLRCAAEENCLASSAYRADVRDYDHRVLLRFPQRVKNQGTSDFLPSRPRYSWEWHSCHQHYHSMDEFSHYDLLDANTQRRVAEGHKASFCLEDTSCDYGYHRRFACTAHTQGLSPGCYDTYAADIDCQWIDITDVQPGNYILKVSVNPSYLVPESDYTNNVVRCDIRYTGHHAYASGCTISPY
(SEQ ID NO:5)
【0172】
ある実施形態において、マウスLOX触媒ドメインを含むポリペプチドは、上に表された配列における最初のグリシン及びロイシン残基(”GL”)を含まない。
【0173】
LOX触媒ドメインを含むポリペプチドをエンコードするマウスDNA配列は、以下の通りである。
GGTCTCCCGGACCTGGTGCCCGACCCCTACTACATCCAGGCTTCCACGTACGTCCAGAAGATGTCTATGTACAACCTGAGATGCGCTGCGGAAGAAAACTGCCTGGCCAGTTCAGCATATAGGGCGGATGTCAGAGACTATGACCACAGGGTACTGCTACGATTTCCGCAAAGAGTGAAGAACCAAGGGACATCGGACTTCTTACCAAGCCGCCCTCGGTACTCCTGGGAGTGGCACAGCTGTCACCAACATTACCACAGCATGGACGAATTCAGCCACTATGACCTGCTTGATGCCAACACACAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGCTTCTGTCTGGAGGACACGTCCTGTGACTATGGGTACCACAGGCGCTTTGCGTGCACTGCACACACACAGGGATTGAGTCCTGGATGTTATGACACCTATGCGGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTACAACCTGGAAACTACATTCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTGCCTGAATCAGACTACACTAACAATGTTGTACGCTGTGACATTCGCTACACAGGACATCATGCCTATGCCTCAGGCTGCACAATTTCACCGTATTAG
(SEQ ID NO:6)
【0174】
[実施例4:マウスLOXL2触媒ドメイン]
マウスLOXL2タンパク質の触媒ドメインを含むポリペプチドは、次のアミノ酸配列を有する。
TAPDLVLNAEIVQQTAYLEDRPMSLLQCAMEENCLSASAVHTDPTRGHRRLLRFSSQIHNNGQSDFRPKNGRHAWIWHDCHRHYHSMEVFTYYDLLSLNGTKVAEGHKASFCLEDTECEGDIQKSYECANFGEQGITMGCWDMYRHDIDCQWIDITDVPPGDYLFQVVINPNYEVPESDFSNNIMKCRSRYDGYRIWMYNCHVGGAFSEETEQKFEHFSGLLNNQLSVQ
(SEQ ID NO:7)
【0175】
ある実施形態において、マウスLOXL2触媒ドメインを含むポリペプチドは、上に表された配列における最初のスレオニン及びアラニン残基(”TA”)を含まない。
【0176】
LOXL2触媒ドメインを含むポリペプチドをエンコードするマウスDNA配列は以下の通りである。
ACTGCACCTGACCTGGTGCTTAATGCTGAGATTGTCCAGCAGACTGCCTACCTGGAGGACAGGCCCATGTCCTTGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCCGCCTCCGCTGTGCACACCGACCCCACCAGAGGCCACCGGCGCCTTTTACGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCTGACTTCCGCCCCAAGAATGGCCGCCATGCGTGGATTTGGCACGACTGCCACAGGCACTACCACAGCATGGAAGTCTTCACTTACTATGACCTGCTGAGCCTCAACGGCACCAAGGTGGCTGAGGGCCACAAGGCCAGCTTCTGCCTGGAGGACACTGAGTGTGAGGGAGACATTCAGAAGAGTTACGAGTGTGCCAACTTTGGAGAACAAGGCATCACCATGGGCTGCTGGGACATGTACCGTCATGACATTGACTGCCAGTGGATAGACATCACCGATGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTCATTAACCCCAACTATGAAGTGCCAGAATCAGATTTCTCTAACAACATCATGAAGTGCAGGAGCCGCTATGATGGCTACCGCATCTGGATGTACAACTGTCACGTAGGTGGAGCCTTCAGTGAGGAGACAGAACAGAAGTTCGAACACTTCAGTGGACTTCTAAATAACCAGCTCTCTGTACAGTAA
(SEQ ID NO:8)
【0177】
[実施例5:哺乳類細胞におけるヒトLOX触媒ドメインの発現]
ヒトLOX触媒ドメインを含むポリペプチドの発現に使用する発現カセットを、ヒトLOX cDNAを鋳型として使用してPCR増幅により構成した(GenBank NM_002317、Genecopoeia, Germantown, MDから入手した)。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro B ベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−hLOX MCD(以下、phLOXMCDとも称する)プラスミドを作成した。T7RNAポリメラーゼを使用したこのクローンの転写により、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、ヒトLOX触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列を有する。
(SEQ ID NO:9)
シグナルペプチド及びmycタグの配列には、下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQP及びGP)及びリンカー配列(NSAVD)の配列でエンコードされるアミノ酸配列を表す。
【0178】
このポリペプチド(これは、シグナル配列及びmycタグをエンコードするヌクレオチド示す下線が引かれた配列、及び小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccGGTCTCCCAGACCTGGTGGCCGACCCCTACTACATCCAGGCGTCCACGTACGTGCAGAAGATGTCCATGTACAACCTGAGATGCGCGGCGGAGGAAAACTGTCTGGCCAGTACAGCATACAGGGCAGATGTCAGAGATTATGATCACAGGGTGCTGCTCAGATTTCCCCAAAGAGTGAAAAACCAAGGGACATCAGATTTCTTACCCAGCCGACCAAGATATTCCTGGGAATGGCACAGTTGTCATCAACATTACCACAGTATGGATGAGTTTAGCCACTATGACCTGCTTGATGCCAACACCCAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGTTTCTGTCTTGAAGACACATCCTGTGACTATGGCTACCACAGGCGATTTGCATGTACTGCACACACACAGGGATTGAGTCCTGGCTGTTATGATACCTATGGTGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTAAAACCTGGAAACTATATCCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTTCCTGAATCTGACTATACCAACAATGTTGTGCGCTGTGACATTCGCTACACAGGACATCATGCGTATGCCTCAGGCTGCACAATTTCACCGTATgggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:10)
【0179】
phLOXMCD発現ベクターを、次のように、HEK293細胞にトランスフェクトした。7×105個の細胞を、6ウェル培養皿の1つのウェル(穴)にプレート(plate)し、37℃、5%CO2の環境下で、完全DMEM(Dulbecco's Modified Eagle's Medium+10%ウシ胎児血清+2mMのL−グルタミン)で増殖させた。終夜増殖の後、細胞に、Lipofectamine 2000(Invitrogen, Carlsbad, CA)を使用して、製造者の指示に従い、phLOXMCDをトランスフェクトした。4時間後、培地を吸引し、2mLの完全DMEMで交換し、トランスフェクト細胞を終夜で増殖させた。安定な細胞系統を、完全DMEM(cDMEM)+0.8mg/mLのHygromycin B(hygro)で、限界希釈(limiting dilution)により選択した。選択に残った個々のクローンを拡張し、hLOX触媒ドメインの発現に関するスクリーニングを次のように行った。拡張を、6ウェル培養皿中で細胞クローンを75%コンフルエンスな(confluence)状態まで増殖させることにより達成した。この時点で、培地を吸引し、新鮮なcDMEM+hygroを添加した。5日後、培地をハーベスト(harvest)し、以下に記載されるように、LOX触媒ドメインの存在を直接的に解析した。PBS+5mMのEDTAを培養皿に添加することにより細胞を培養皿の表面から取り外した。分離した細胞液(cell solution)を遠心分離に供した。約5×105個のペレット化した細胞(総計〜1×106個の細胞から得た)を0.1mLのM‐PER mammalian protein extraction reagent(Pierce, Rockford, IL)に溶解した。氷上での20分間のインキュベーションの後、細胞溶解物を、卓上遠心器で15000rpm、10分間の条件で遠心分離に供した。可溶性分画(上清)を回収し、以下に記載されるように、LOX触媒ドメインの存在を解析した。不溶性分画(ペレット)を、2回、M‐PERで洗浄し、M‐PERに再懸濁し、以下に記載されるように、LOX触媒ドメインの存在を解析した。
【0180】
[実施例6:タンパク質免疫ブロッティングによるヒトLOX触媒ドメインの検出]
6つの異なったクローンからの培地、可溶性分画及び不溶性分画の試料を、phLOXMCD発現ベクターにより発現されたLOX触媒ドメインの存在を(確認するために)免疫ブロッティングにより分析した。培地の分析に関して、150μLの馴化培地(conditioned medium)を50μLのNuPAGE(登録商標)4×LDS試料緩衝液(Invitrogen, Carlsbad, CA)と混合し、その結果得られた20μLの混合液をゲルの上にロードした(以下を参照)。可溶性及び不溶性の細胞内分画に関して、各試料は、1×105個の細胞に相当する。細胞内分画の試料を、NuPAGE(登録商標)、4×LDS試料緩衝液(Invitrogen, Carlsbad, CA)とNuPAGE(登録商標)10×還元剤とを混合し、これら両薬剤の終濃度が1倍(1×)の濃度になるようにし、5分間煮沸し、そして、NuPAGE(登録商標)Novex Bis−Trisゲル(4%‐12%アクリルアミド)を使用して、電気泳動に供した。ランニング緩衝液は、NuPAGE(登録商標)MOPS緩衝液(Invitrogen, Carlsbad, CA)であった。電気泳動の終了時に、タンパク質をゲルから、iBlot装置(Invitrogen, Carlsbad, CA)を使用してPVDF上に転写した。
【0181】
検出に関して、ブロットを、3%ウシ胎児血清(BSA)入りPBS+0.01% Tweenを使用して、3時間、室温でブロックした。続いて、ブロットをマウス抗His5抗体(Qiagen, Valencia, CA)と共に、1時間、室温で、インキュベートし、次にセイヨウワサビペルオキシダーゼ(HRP)コンジュゲートロバ抗マウス抗体(Jackson ImmunoResearch, West Grove, PA)と共に、1時間、室温でインキュベートし、Chemi−Glow(登録商標)(Alpha Innotech, San Leandro, CA)を使用して、発色させた(develop)。
【0182】
6つのクローン(23から28の番号で標識される)に関する結果を、図1に表す。増殖培地の分析を表した最も左のパネルは、クローン23における少量の分泌産物(矢印)を除いて、触媒ドメインの分泌が皆無か、それに近いことを示す。分泌タンパク質は、プロセスされていないポリペプチドの特徴である、明瞭な29kDの分子量を表す。これは、シグナルペプチドの切断の欠如(又は不完全な切断)に起因しているかも知れず、又は、分泌ポリペプチドの翻訳後修飾に起因しているかも知れない。
【0183】
phLOXMCD発現ベクターが一時的に(transiently)トランスフェクトされているHEK293細胞からの増殖培地のトリクロロ酢酸沈殿は、増殖培地における不溶性LOX触媒ドメインの存在を明らかにした。
【0184】
触媒ドメインの細胞内発現が、6つのクローン中、5つのクローンにおいて、可溶性及び不溶性分画の両方で検出された(図1、中央及び右のパネル)。加えて、高分子量の集合体(aggregates)が、不溶性細胞内分画において検出された。
【0185】
可溶性(であること)は、正しく折りたたまれ、プロセスされたポリペプチドであることの指標でありうる以上、可溶性細胞内ポリペプチド産物の存在は、適切に折りたたまれ、酵素的に活性な触媒ドメインが、トランスフェクト細胞内で生成されている可能性を示す。
【0186】
HEK293細胞に加えて、LOX触媒ドメインの発現が、更なる細胞種において観察された。CHOk1細胞におけるphLOXMCD発現ベクターの一時的な発現は、触媒ドメインの細胞内発現(これは、細胞溶解物全体の中に検出される)をもたらしたが、触媒ドメインの分泌は、検出されなかった。SW620細胞の一時的なトランスフェクションもまた、検出可能な分泌なしでの、細胞内発現をもたらした。
【0187】
[実施例7:哺乳類細胞におけるヒトLOXL2触媒ドメインの発現]
ヒトLOXL2触媒ドメインを含むポリペプチドの発現に使用する発現カセット(Genecopoeia, Germantown, MD)を、ヒトLOXL2 cDNAを鋳型として使用してPCR増幅により構成した。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro Bベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−hLOXL2 MCD(以下、phLOXL2MCDとも称する)発現ベクターを作成した。T7RNAポリメラーゼを使用したこのクローンの転写は、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、ヒトLOXL2触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列を有する。
(SEQ ID NO:11)
【0188】
シグナルペプチド及びmycタグの配列は下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQPARRARRTKL及びSRGGP)及びリンカー配列(NSAVD)でエンコードされるアミノ酸配列を表す。
【0189】
このポリペプチド(これは、リーダー(leader)及び、mycタグをエンコードする配列を示す下線が引かれた配列及び、小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccaggcgcgcgcgccgtacgaagcttACCGCCCCTGACCTGGTCCTCAATGCGGAGATGGTGCAGCAGACCACCTACCTGGAGGACCGGCCCATGTTCATGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCGGCCTCAGCCGCGCAGACCGACCCCACCACGGGCTACCGCCGGCTCCTGCGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCCGACTTCCGGCCCAAGAACGGCCGCCACGCGTGGATCTGGCACGACTGTCACAGGCACTACCACAGCATGGAGGTGTTCACCCACTATGACCTGCTGAACCTCAATGGCACCAAGGTGGCAGAGGGCCACAAGGCCAGCTTCTGCTTGGAGGACACAGAATGTGAAGGAGACATCCAGAAGAATTACGAGTGTGCCAACTTCGGCGATCAGGGCATCACCATGGGCTGCTGGGACATGTACCGCCATGACATCGACTGCCAGTGGGTTGACATCACTGACGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTTATTAACCCCAACTTCGAGGTTGCAGAATCCGATTACTCCAACAACATCATGAAATGCAGGAGCCGCTATGACGGCCACCGCATCTGGATGTACAACTGCCACATAGGTGGTTCCTTCAGCGAAGAGACGGAAAAAAAGTTTGAGCACTTCAGCGGGCTCTTAAACAACCAGCTGTCCCCGCAGtctcgaggagggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:12)
【0190】
phLOXL2MCD発現ベクターを、HEK293細胞にトランスフェクトし、安定な細胞系統を、限界希釈により選択し、そしてクローンを、phLOXMCDベクターについての実施例5に記載された方法と同じ方法を使用して拡張した。培地を分析のために回収し、細胞を分離し、実施例5に記載されたようにペレット化した。
【0191】
[実施例8:タンパク質免疫ブロッティングによるヒトLOXL2触媒ドメインの検出]
約5×105個の細胞を含むペレット化した細胞の試料(実施例7に記載されるようにして得たもの)を免疫ブロッティングにより解析した。約5×105個の細胞を含む細胞懸濁液を、6000rpm、10分間の条件で遠心分離に供し、上清を吸引した。ペレットを100μLのM−PER溶解緩衝液に再懸濁し、次に、氷上で20分間インキュベーションした。不溶性細胞残渣を、15000rpm、20分間の遠心分離により取り出し、75μLの上清(澄んだ細胞溶解物)を取り出し、30μLのNuPAGE(登録商標)4×LDS試料緩衝液(Invitrogen, Carlsbad, CA)と10μLのNuPAGE(登録商標)10×還元剤に混合した。混合物を5分間煮沸し、続いて、ゲルに適用するまで氷上に置いた。典型的に、20μLの各試料(〜1×105個の細胞に相当する)をNuPAGE(登録商標)Novex Bis−Trisゲル(4%−12%アクリルアミド)で電気泳動に供した。ランニング緩衝液は、NuPAGE(登録商標)MOPS緩衝液(Invitrogen, Carlsbad, CA)であった。電気泳動の終了時に、タンパク質をゲルから、iBlot装置(Invitrogen, Carlsbad, CA)を使用してPVDF上に転写した。
【0192】
検出に関して、ブロットを、phLOXMCD発現産物についての実施例6に記載されるように処理(プロセス)した。
【0193】
図2の左のパネルに表されるように、結果は、9つの異なったクローンにおけるLOXL2触媒ドメインの細胞内発現を示す。タンパク質の単量体の形態及び集合体(aggregated)の形態の両方が、検出された(単量体の形態が支配的であった(predominating))。
【0194】
ヒトLOXL2触媒ドメインが分泌されたかどうかを確認するために、phLOXMCD−トランスフェクト細胞についての実施例7に記載されるように、細胞クローンを得て、cDMEM+hygroで90分間増殖させ、この時点で、15μLの培地を取り除き、5μLの4×LDS試料緩衝液+還元剤(NuPAGE(登録商標)、Invitrogen, Carlsbad, CA)に混合し、5分間煮沸し、ポリアクリルアミドゲル電気泳動に供し、全細胞溶解物についての上記の方法により解析した。図2の右のパネルに表される結果は、ヒトLOX触媒ドメインと対照的に、ヒトLOXL2触媒ドメインは、無濃縮増殖培地で検出可能なほど大量にHEK293細胞により分泌されたことを表す。
【0195】
[実施例9:哺乳類細胞におけるマウスLOX触媒ドメインの発現]
マウスLOX触媒ドメインを含むポリペプチドの発現に使用する発現カセットを、マウスLOX cDNAを鋳型として使用してPCR増幅により構成した(GenBank BC018439、Invitrogen, Carlsbad, CAから入手した)。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro B ベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−mLOX MCD(以下、pmLOXMCDとも称する)発現ベクターを作成した。T7RNAポリメラーゼを使用したこのクローンの転写は、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、マウスLOX触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列を有する。
(SEQ ID NO:13)
シグナルペプチド及びmycタグの配列は下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQP及びGP)及びリンカー配列(NSAVD)の配列でエンコードされるアミノ酸配列を表す。
【0196】
このポリペプチド(これは、リーダー及びmycタグをエンコードする配列を示す下線が引かれた配列、及び小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccGGTCTCCCGGACCTGGTGCCCGACCCCTACTACATCCAGGCTTCCACGTACGTCCAGAAGATGTCTATGTACAACCTGAGATGCGCTGCGGAAGAAAACTGCCTGGCCAGTTCAGCATATAGGGCGGATGTCAGAGACTATGACCACAGGGTACTGCTACGATTTCCGCAAAGAGTGAAGAACCAAGGGACATCGGACTTCTTACCAAGCCGCCCTCGGTACTCCTGGGAGTGGCACAGCTGTCACCAACATTACCACAGCATGGACGAATTCAGCCACTATGACCTGCTTGATGCCAACACACAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGCTTCTGTCTGGAGGACACGTCCTGTGACTATGGGTACCACAGGCGCTTTGCGTGCACTGCACACACACAGGGATTGAGTCCTGGATGTTATGACACCTATGCGGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTACAACCTGGAAACTACATTCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTGCCTGAATCAGACTACACTAACAATGTTGTACGCTGTGACATTCGCTACACAGGACATCATGCCTATGCCTCAGGCTGCACAATTTCACCGTATgggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:14)
【0197】
pmLOXMCD発現ベクターを、HEK293細胞にトランスフェクトし、安定な細胞系統を、限界希釈により選択し、そしてクローンを、phLOXMCDベクターについての実施例5に記載された方法と同じ方法を使用して拡張した。培地を解析のために回収し、細胞を取り出し、実施例5に記載されるようにペレット化し、及び可溶性分各及び不溶性分画に分画した。
【0198】
マウスLOX触媒ドメインの発現を、ヒト触媒ドメインの解析についての実施例6に記載された方法と同じ方法を使用して、免疫ブロッティングにより分析した。図3に表される結果は、マウスLOX触媒ドメインが、ヒトLOX触媒ドメインとは対照的に、検出可能な量で分泌されることを示す(左のパネル)。mLOX触媒ドメインの細胞内発現プロファイルは、hLOX触媒ドメインのそれと、類似しており、可溶性及び不溶性分画の両方に単量体タンパク質を有し、不溶性分画において、集合体が存在する。
【0199】
[実施例10:哺乳類細胞からのヒトLOX触媒ドメインの分泌]
図4は、ヒトLOX触媒ドメインのアミノ酸配列(上)及びマウスLOX触媒ドメインのアミノ酸配列(下)の違いを表す模式図を含む。図に示すように、2つの配列は、4つの位置のみで異なる。上で示されるように、及び、図4の左のパネルで確認されるように、10倍濃縮された馴化増殖培地(10×CM)を使用すると、マウスLOX触媒ドメイン(M)をエンコードするベクターをトランスフェクトした細胞から、マウスLOX触媒ドメイン(M)が分泌されたが、一方で、ヒトLOX触媒ドメイン(H)は、分泌されなかった。それ故、ヒトLOX触媒ドメインの分泌を望むのであれば、1つの方法として、7、38、146又は160番目の残基の何れかを、ヒト配列からマウス配列へ変更することが可能であり(番号は、本願で開示されるphLOXMCD及びpmLOXMCD発現ベクターにおいて発現する触媒ドメインのアミノ酸配列における位置を意味する)、又は、これらの残基の任意の組み合わせ又はこれら残基の4つ全てをヒト配列からマウス配列へ変更することも可能である。例えば、次のアミノ酸配列の任意のもの又はその全て、又は任意の組み合わせ、の変更が可能である(A7P、T38S、G146A、K160Q)。
【0200】
反対に、マウスLOX触媒ドメインの分泌を減少させることを望み、かくて、このポリペプチドを細胞内により多く留めるのであれば、7、38、146及び160番目の残基をマウス配列からヒト配列に変更することが可能である。従って、例えば、次のアミノ酸配列の任意のもの又はその全て、又は任意の組み合わせ、の変更が可能である(P7A、S38T、A146G、Q160K)。
【0201】
[実施例11:哺乳類細胞におけるマウスLOXL2触媒ドメインの発現]
マウスLOXL2触媒ドメインを含むポリペプチドの発現に使用する発現カセットを、マウスLOXL2 cDNAを鋳型として使用してPCR増幅により構成する(GenBank NM033325を元にして商業的に合成される)。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro B ベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−mLOXL2 MCD(以下、pmLOXL2MCDとも称する)発現ベクターを作成する。T7RNAポリメラーゼを使用したこのクローンの転写は、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、マウスLOXL2触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列(これは、下線が引かれたリーダー配列及びmycタグを有する)を有する。
(SEQ ID NO:15)
【0202】
シグナルペプチド及びmycタグの配列は下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQP及びGP)及びリンカー配列(NSAVD)の配列でエンコードされるアミノ酸配列を表す。
【0203】
このポリペプチド(これは、リーダー及び、mycタグをエンコードする配列を示す下線が引かれた配列及び、小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccACTGCACCTGACCTGGTGCTTAATGCTGAGATTGTCCAGCAGACTGCCTACCTGGAGGACAGGCCCATGTCCTTGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCCGCCTCCGCTGTGCACACCGACCCCACCAGAGGCCACCGGCGCCTTTTACGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCTGACTTCCGCCCCAAGAATGGCCGCCATGCGTGGATTTGGCACGACTGCCACAGGCACTACCACAGCATGGAAGTCTTCACTTACTATGACCTGCTGAGCCTCAACGGCACCAAGGTGGCTGAGGGCCACAAGGCCAGCTTCTGCCTGGAGGACACTGAGTGTGAGGGAGACATTCAGAAGAGTTACGAGTGTGCCAACTTTGGAGAACAAGGCATCACCATGGGCTGCTGGGACATGTACCGTCATGACATTGACTGCCAGTGGATAGACATCACCGATGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTCATTAACCCCAACTATGAAGTGCCAGAATCAGATTTCTCTAACAACATCATGAAGTGCAGGAGCCGCTATGATGGCTACCGCATCTGGATGTACAACTGTCACGTAGGTGGAGCCTTCAGTGAGGAGACAGAACAGAAGTTCGAACACTTCAGTGGACTTCTAAATAACCAGCTCTCTGTACAGgggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:16)
【0204】
pmLOXL2MCD発現ベクターを、HEK293細胞にトランスフェクトし、安定な細胞系統を、限界希釈により選択し、そしてクローンを、phLOXMCDベクターについての実施例5に記載される方法と同じ方法を使用して拡張した。培地を解析のために回収し、細胞を取り出し、実施例5に記載されるように、ペレット化し及び可溶性分画及び不溶性分画に分画した。
【0205】
マウスLOXL2−由来のポリペプチドの発現を、ヒト触媒ドメインの解析についての実施例6に記載される方法と同じ方法を使用して、免疫ブロッティングにより分析した。
【0206】
[実施例12:一時的にトランスフェクトした細胞からの、酵素的に活性なヒトLOXL2触媒ドメインの発生]
293F細胞(これは、懸濁培養中で増殖するように適合されたHEK293細胞のサブクローンである、Invitrogen, Carlsbad, CA)を、製造者のプロトコルに従って、スピナーフラスコ中で増殖させた。一旦、培養液(の密度)を0.8−1.2×106個細胞/mLの密度まで到達させ、細胞培養液をCB22バッグ(WAVE, GE Health, Piscataway, NJ)に移した。293fectin reagent(Invitrogen, Carlsbad, CA)を使用して8リッターの細胞培養液に8mgのphLOXL2ベクターをトランスフェクトした。トランスフェクト細胞を、3日間、CB22バッグ内で増殖させ、その後、細胞培養液を、Allegra6Rベンチトップ遠心器(Beckman Coulter)で、スイングバケットローターを使用し、3000rpmで10分間、遠心分離に供した。上清を回収し、0.22mmのPES膜を介してフィルターろ過した。
【0207】
LOXL2由来ポリペプチドを、Ni−セファロース樹脂(GE Healthcare, Piscataway, NJ)によって、細胞培養液上清から精製した。樹脂を0.1MのTris‐Cl(pH8.0)を使用して平衡化した。馴化培地(即ち、細胞培養上清)を平衡化した樹脂にロードし、続いて、カラムを、10カラム容積の0.1MのTris‐Cl(pH8.0)、0.25Mの塩化ナトリウム、0.02Mのイミダゾールを使用して洗浄した。Hisタグ化された触媒ドメインを、5カラム容積の0.1MのTris‐Cl(pH8.0)、0.15Mの塩化ナトリウム、0.3Mのイミダゾールを使用して溶出した。溶出液は、Amicon Ultra 15 10kD MWCL(Millipore, Billerica, MA)で濃縮し、その濃縮された材料を0.05Mのホウ酸塩に対して、pH8.0で、4℃で終夜で透析した。試料を、SDSポリアクリルアミドゲル(4−12%勾配、Bis−Trisゲル、Invitrogen, Carlsbad, CA)で、還元条件で解析した。
【0208】
[実施例13:ヒトLOXL2の触媒ドメインの酵素的活性]
ヒトLOXL2の触媒ドメインを含むポリペプチドを、上記の実施例12に記載されたように生成した。酵素的活性は、過酸化水素の生成(1,5−ジアミノペンタンの脱アミノ化の後、LOXL2によって遊離させた過酸化水素)を、Amplex(登録商標)redの蛍光産物(レゾルフィン)へのHRP−触媒変換(HRP-catalyzed conversion)と組み合わせた生化学的な分析を使用して解析した(Palamakumbura et al. (2002) Anal. Biochem. 300:245-251)。反応プレートは、Corning社から入手した。Amplex(登録商標)Red reagentは、Invitrogen社(Carlsbad, CA)から入手した。セイヨウワサビペルオキシダーゼ(HRP)、1,5−ジアミノペンタン及び消泡剤は、Sigma社(St. Louis, MO)から入手した。他の薬剤の全ては、入手できる最高の品質のものであった。
【0209】
酵素混合物は、10μLのプールされたピーク分画(実施例12参照)を40μLの分析溶液(62.5mMのホウ酸ナトリウム(pH8.0)、5ユニット/mLのHRP、10ppmの消泡剤)に加えることによって、作成した。基質溶液は、50mMのホウ酸ナトリウム、100μMのAmplex(登録商標)red reagent、20mMの1,5−ジアミノペンタン、10ppmの消泡剤を含んでいた。反応を、50μLの酵素混合物を50μLの基質溶液と混合することによって開始した。1つの反応物はまた、2mMのβAPN(ベータ‐アミノプロプリオニトリル、これは、リシル酸化酵素及びLOXL2の触媒活性の阻害剤である)を含んでいた。反応混合物を、37℃で、1時間、蛍光(ex=544nm、em=590nm)を測定するように作られたMolecularDevices M5プレートリーダーを使用して、反応速度モード(kinetics mode)で、インキュベートした。データを、時間に対する蛍光応答の勾配として記録した。
【0210】
図5は、レゾルフィン生成の時間経過解析の形で、相対蛍光ユニット(RFU)で表される、この分析の結果を表す。ヒトLOXL2由来ポリペプチドは、測定可能な酵素的活性を示し、そしてこの活性は、βAPNによって阻害された。従って、ポリペプチドは、LOXL2の酵素的に活性な触媒ドメインを含む。
【関連出願の相互参照】
【0001】
この出願は、2009年8月21日に出願された米国仮出願番号61/235,776の利益を主張し、この開示は、全ての目的のために、その全てが本願に引用によって取り込まれる。
【0002】
[連邦政府の支援に関する陳述]
支援は、適用されない。
【技術分野】
【0003】
本出願は、酵素学及び分子生物学の分野に含まれる。
【背景技術】
【0004】
結合組織は、身体にある細胞及び器官に存在する骨格を提供する。脊椎動物における結合組織の一次構成要素は、細胞外マトリクスである。細胞外マトリクスの2つの主な構造的成分は、ゲル様の基質を形成するポリサッカライドであり、基質の中に埋め込まれている繊維性タンパク質である。これら繊維性タンパク質のうち、2つのもっとも一般的なものは、コラーゲンとエラスチンである。コラーゲン繊維は、コラーゲン原繊維の自己会合により形成され、三重らせんコラーゲン分子の架橋結合により、これら自身が会合する。この架橋結合は、リシル酸化酵素(LOX)及びそれに関連する酵素(「リシル酸化酵素様」 lysyl oxidase-like又は「LOXL」)によって触媒され、これらの全ては、リシン及びヒドロキシリシン残基のε‐アミノ基を脱アミノ化する能力を有する触媒ドメインを含み、かくて、ペプチジルリシンのペプチジル‐α‐アミノアジピック‐δ‐セミアルデヒド(アリシン)への変換をもたらす。アリシン残基は、自発的に、お互いに凝集することができ、かくて、コラーゲン分子の架橋結合をもたらす。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】国際公開WO 01/83702号
【特許文献2】国際公開WO 2004/047720号
【特許文献3】国際公開WO 2007/126457号
【特許文献4】米国特許出願公開US 2006/0127402号
【特許文献5】米国特許出願公開US 2007/0225242号
【特許文献6】米国特許出願公開US 2009/0053224号
【特許文献7】米国特許出願公開US 2009/0104201号
【非特許文献】
【0006】
【非特許文献1】Csiszar (2001) Prog. Nucleic Acid Res. and Molec. Biol. 70:1-32
【非特許文献2】Kirschmann et al. (2002) Cancer Research 62:4478-4483
【発明の概要】
【発明が解決しようとする課題】
【0007】
例えば、繊維症及び転移を含む様々な病状における細胞外マトリクスの関与は、次第に明らかになってきている。例えば、WO 01/83702(2001年11月8日)、WO 2004/047720(2004年6月10日)、WO 2007/126457(2007年8月11日)、US 2006/0127402(2006年6月15日)、US 2007/0225242(2007年9月27日)、US 2009/0053224(2009年2月26日)、US 2009/0104201(2009年4月23日)、Csiszar (2001) Prog. Nucleic Acid Res. and Molec. Biol. 70:1-32、Kirschmann et al. (2002) Cancer Research 62:4478-4483を参照する。リシル酸化酵素及びリシル酸化酵素様酵素は、コラーゲンを架橋結合することによって、細胞外マトリクスの形成において重要な役割を果たすので、これらは重要な治療的目標である。それ故、これらコラーゲン架橋結合酵素の阻害剤を選択する方法、これら酵素に結合する分子を同定する方法、及び様々な治療的使用(形態)(例えば、創傷治癒)のためのコラーゲン架橋結合活性の材料、の全てが所望される。
【課題を解決するための手段】
【0008】
本願開示は、ヒト及びマウス由来のリシル酸化酵素(LOX)及びリシル酸化酵素様2酵素(LOXL2)の単離された触媒ドメインを(これら触媒ドメインをエンコードする核酸と共に)提供する。かくて、次のペプチド及びこれらポリペプチドをエンコードする核酸が提供される(ヒトLOX触媒ドメイン、ヒトLOXL2触媒ドメイン、マウスLOX触媒ドメイン及びマウスLOXL2触媒ドメイン)。従って、本願開示は以下の形態を提供する。
【0009】
(形態1)ヒトリシル酸化酵素(LOX)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:1)。
【0010】
(形態2)ヒトリシル酸化酵素(LOX)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:2)。
【0011】
(形態3)ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:3)。
【0012】
(形態4)ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:4)。
【0013】
(形態5)マウス(murine)リシル酸化酵素(LOX)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:5)。
【0014】
(形態6)マウスリシル酸化酵素(LOX)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:6)。
【0015】
(形態7)マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインのアミノ酸配列を含むポリペプチド(SEQ ID NO:7)。
【0016】
(形態8)マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインをエンコードするヌクレオチド配列を含むポリヌクレオチド(SEQ ID NO:8)。
【0017】
以下の形態もまた提供される。
【0018】
(形態1a)本願に記載の、ヒトリシル酸化酵素(LOX)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:1)、その触媒ドメインの外側のヒトLOXの配列を含まないポリペプチド。
【0019】
(形態2a)本願に記載の、ヒトリシル酸化酵素(LOX)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:2)、その触媒ドメインの外側のヒトLOXの配列をエンコードしないポリヌクレオチド。
【0020】
(形態3a)本願に記載の、ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:3)、その触媒ドメインの外側のヒトLOXL2の配列を含まないポリペプチド。
【0021】
(形態4a)本願に記載の、ヒトリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:4)、その触媒ドメインの外側のヒトLOXL2の配列をエンコードしないポリヌクレオチド。
【0022】
(形態5a)本願に記載の、マウスリシル酸化酵素(LOX)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:5)、その触媒ドメインの外側のマウスLOXの配列を含まないポリペプチド。
【0023】
(形態6a)本願に記載の、マウスリシル酸化酵素(LOX)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:6)、その触媒ドメインの外側のマウスLOXの配列をエンコードしないポリヌクレオチド。
【0024】
(形態7a)本願に記載の、マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部のアミノ酸配列を含むポリペプチドであって(SEQ ID NO:7)、その触媒ドメインの外側のマウスLOXL2の配列を含まないポリペプチド。
【0025】
(形態8a)本願に記載の、マウスリシル酸化酵素様2(酵素)(LOXL2)の触媒ドメインの全て又は一部をエンコードするヌクレオチド配列含むポリヌクレオチドであって(SEQ ID NO:8)、その触媒ドメインの外側のマウスLOXL2の配列をエンコードしないポリヌクレオチド。
【0026】
また、前述の核酸及び/又はポリヌクレオチドを含む発現ベクターも提供される。そのような発現ベクターは、任意に、プロモーター(例えば、T7プロモーター、T3プロモーター、SP6プロモーター、E.coli RNAポリメラーゼプロモーター、CMVプロモーター、SV40プロモーター、PGKプロモーター、EF‐1アルファプロモーター)、転写終結シグナル(例えば、SV40終結シグナル)、スプライス部位(例えば、SV40スプライス部位、ベータ‐グロブリンスプライス部位)、リボゾーム結合部位、シグナル配列(例えば、免疫グロブリンカッパシグナル配列)、エピトープタグ(例えば、myc)、精製タグ(例えば、His6)、複製起点及び薬剤選択マーカーを含む。リンカーアミノ酸をエンコードし、及び/又は制限酵素認識部位を含むリンカー配列又は、他のあらゆる種類のリンカー配列もまた、本願で開示される発現ベクターに存在し得る。
【0027】
従って、本願の開示はまた、以下の形態も提供する。
【0028】
(形態9)触媒ドメインの外側にあるヒトLOXのアミノ酸配列を含まず、ヒトLOXの触媒ドメインの全て又は一部のアミノ酸配列を含み、更に、1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、ヒトLOXの触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:9)。
【0029】
(形態10)形態9によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、ヒトLOXの触媒ドメイン、mycエピトープタグ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:10)。
【0030】
(形態11)形態10のポリヌクレオチドを含む発現ベクター。
【0031】
(形態12)形態11の発現ベクターであって、更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む発現ベクター。
【0032】
(形態13)触媒ドメインの外側にあるヒトLOXL2のアミノ酸配列を含まず、ヒトLOXL2の触媒ドメインの全て又は一部のアミノ酸配列を含み、更に、1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、ヒトLOXL2の触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:11)。
【0033】
(形態14)形態13によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、ヒトLOXL2の触媒ドメイン、mycエピトープタグ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:12)。
【0034】
(形態15)形態14のポリヌクレオチドを含む発現ベクター。
【0035】
(形態16)更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む形態15の発現ベクター。
【0036】
(形態17)触媒ドメインの外側にあるマウスLOXアミノ酸配列を含まず、マウスLOXの触媒ドメインの全て又は一部のアミノ酸配列を含み、更に1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、マウスLOXの触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:13)。
【0037】
(形態18)形態17によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、マウスLOXの触媒ドメイン、mycエピトープ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:14)。
【0038】
(形態19)形態18のポリヌクレオチドを含む発現ベクター。
【0039】
(形態20)更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む形態19の発現ベクター。
【0040】
(形態21)触媒ドメインの外側にあるマウスLOXL2のアミノ酸配列を含まず、マウスLOXL2の触媒ドメインの全て又は一部のアミノ酸配列を含み、更に1つ以上のシグナル配列、エピトープタグ及びHis6精製タグを含むポリペプチドであって、例えば、シグナル配列、マウスLOXL2の触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチド(例えば、SEQ ID NO:15)。
【0041】
(形態22)形態21によるポリペプチドをエンコードするポリヌクレオチドであって、例えば、シグナル配列、マウスLOXL2の触媒ドメイン、mycエピトープ及びHis6精製タグをエンコードする配列を含むポリヌクレオチド(例えば、SEQ ID NO:16)。
【0042】
(形態23)形態22のポリヌクレオチドを含む発現ベクター。
【0043】
(形態24)更に、任意の1つのプロモーター、任意の組み合わせのプロモーター又は、全てのプロモーター、薬剤選択マーカー及び複製起点を含む形態23の発現ベクター。
【図面の簡単な説明】
【0044】
【図1】図1は、phLOXMCD発現ベクターを導入(トランスフェクト)されたHEK293細胞による、ヒトリシル酸化酵素(hLOX)タンパク質の触媒ドメインの発現を表す。6つの安定にトランスフェクトされた細胞系統(番号23から28)の免疫ブロットによる解析を表す。左のパネルは、各6つの細胞系統からの無希釈増殖培地(neat CM)の試料(15μL)を含む、ポリアクリルアミドゲルの免疫ブロットを表す。中間のパネルは、可溶性細胞内物質の解析を表し、右のパネルは、不溶性細胞内物質の解析を表す。これらそれぞれのパネルにおいて、各レーンは、およそ105個細胞からの物質を含む。全てのブロットに、一次マウス抗His5抗体をプローブし、続いて、二次HRPコンジュゲートロバ抗マウス抗体と反応させた。HRP活性を、Chemi−Glow(登録商標)reagent(Alpha Innotech, San Leandro, CA)を使用して、可視化した。プロセスされた触媒ドメインは、26.9kDの予測分子量を有する。
【0045】
各パネルにおいて、「+」で標識されたレーンは、His6配列を含むタンパク質を含み、抗His5抗体の陽性対照として機能した。
【0046】
【図2】図2は、phLOXL2MCD発現ベクターをトランスフェクトされたHEK293細胞による、ヒトリシル酸化酵素様2(hLOXL2)タンパク質の触媒ドメインの発現を表す。左のパネルは、いくつかの安定にトランスフェクトされた細胞系統(各レーンの頂点に記載される数字により示される)からの全細胞溶解物の免疫ブロットを表し、各レーンは、約1×105個細胞からの物質を含む。右のパネルは、各6つの細胞系統(番号で示される)からの無希釈増殖培地(15μL Neat CM)の試料を含む、ポリアクリルアミドゲルの免疫ブロットを表す。全てのブロットに、一次マウス抗His5抗体をプローブし、続いて、二次HRPコンジュゲートロバ抗マウス抗体と反応させた。HRP活性を、Chemi−Glow(登録商標)reagent(Alpha Innotech, San Leandro, CA)を使用して、可視化した。プロセスされた触媒ドメインは、31kDの予測分子量を有する。
【0047】
左のパネルにおいて、「+」で標識されたレーンは、His6配列を含むタンパク質を含み、抗His5抗体の陽性対照として機能した。
【0048】
【図3】図3は、pmLOXMCD発現ベクターをトランスフェクトされたHEK293細胞による、マウスリシル酸化酵素(mLOX)タンパク質の触媒ドメインの発現を表す。5つの安定にトランスフェクトされた細胞系統(4、9、12、18及び19の番号が付けられている)の免疫ブロットによる解析を表す。左のパネルは、各6つの細胞系統からの無希釈増殖培地(neat CM)の試料(15μL)を含む、ポリアクリルアミドゲルの免疫ブロットを表す。右のパネルは、これら6つの系統のうち5つからの可溶性及び不溶性細胞内分画の解析を表し、各レーンは、およそ105個細胞からの物質を含む。全てのブロットに、一次マウス抗His5抗体をプローブし、続いて、二次HRPコンジュゲートロバ抗マウス抗体と反応させた。HRP活性を、Chemi−Glow(登録商標)reagent(Alpha Innotech, San Leandro, CA)を使用して、可視化した。プロセスされた触媒ドメインは、27kDの予測分子量を有する。
【0049】
【図4】図4は、ヒトとマウスのLOX触媒ドメインの分泌及びアミノ酸配列の違いを表す。詳細については実施例10を参照する。
【0050】
【図5】図5は、単離されたヒトLOXL2触媒ドメインのAmplex(登録商標)Red分析における、レゾルフィン生成の経時的解析を表す(590nmの吸光度により測定される)。詳細については実施例12を参照する。
【0051】
【図6】図6は、ヒト由来のポリペプチド(hLOX、hLOXL1、hLOXL2など)及びマウス由来のポリペプチド(mLOX及びmLOXL2)を含む、本願の開示における様々なポリペプチドのアミノ酸配列のアライメントを表す。アミノ酸残基の番号付けは、表示される最もN末端にある残基を「1」として開始し、当該技術分野における慣例に必ずしも従うものでは無い。European Bioinformatics Institute(EMBL-EBI)によって提供されたClustalW2プログラムに従って、アスタリスクは、表示された全てのポリペプチド間で共通した位置におけるアミノ酸の相同性を示し、一方で、コロンは、保存的な(conservative)アミノ酸置換を示し、単一の点は、半保存的な(semi-conservative)アミノ酸置換を示す。
【0052】
【図7】図7は、ヒト由来のポリペプチドのみを含む本願の開示における様々なポリペプチドのアミノ酸配列のアライメントを表す。番号付け及び記号の凡例は、図6において使用されたものと同一である。
【0053】
本願の開示の実施は、特段の指示のない限り、細胞生物学、毒物学、分子生物学、分子化学、細胞培養、免疫学、腫瘍学、組み換えDNAの分野及び、当該技術分野範囲内の関係する分野における標準の方法及び従来技術を採用する。そのような技術は、文献に記載され、それ故、当業者が利用できる。例えば、Alberts, B. et al., "Molecular Biology of the Cell," 5th edition, Garland Science, New York, NY, 2008、Voet, D. et al. "Fundamentals of Biochemistry: Life at the Molecular Level," 3rd edition,John Wiley & Sons, Hoboken, NJ, 2008、Sambrook, J. et al., "Molecular Cloning: A Laboratory Manual," 3rd edition, Cold Spring Harbor Laboratory Press, 2001、Ausubel, F. et al., "Current Protocols in Molecular Biology," John Wiley & Sons, New York, 1987及び定期的な更新、Freshney, R.I., "Culture of Animal Cells: A Manual of Basic Technique," 4th edition, John Wiley & Sons, Somerset, NJ, 2000、及び、シリーズ"Methods in Enzymology," Academic Press, San Diego, CAを参照する。
【0054】
[リシル酸化酵素型酵素]
本願で使用される場合、用語「リシル酸化酵素型酵素」又は「リシル酸化酵素」は、リシン及びヒドロキシリシン残基のε‐アミノ基の酸化的脱アミノ化を触媒する触媒ドメインを含むタンパク質のファミリーのメンバーであることを意味し、かくて、ペプチジルリシンのペプチジル‐α‐アミノアジピック‐δ−セミアルデヒド(アリシン)への変換をもたらし、アンモニア及び過酸化水素の化学量論的量を放出する。
【0055】
この反応は、ほとんどの場合、細胞外のコラーゲン及びエラスチンのリシン残基に生じる。アリシンのアルデヒド残基は、活性であり、同時に他のアリシン及びリシン残基と凝集することができ、かくて、コラーゲン分子の架橋結合をもたらし、コラーゲン原繊維を形成する。
【0056】
リシル酸化酵素は、ニワトリ、ラット、マウス、ウシ及びヒトから精製されている。すべてのリシル酸化酵素は、長さ約205アミノ酸であり、タンパク質のカルボキシ末端部分に位置し、酵素の活性部位を含む、共通の触媒ドメインを含む。活性部位は、Cu(II)原子に配位する4つのヒスチジン残基を含む保存アミノ酸配列を含む銅結合部位を含む。活性部位はまた、リシン残基とチロシン残基の間の分子内共有結合により形成されるリシルチロシルキノン(LTQ)補因子も含む(ラットリシル酸化酵素では、314番目のlys(lys314)及び349番目のtyr(tyr349)に相当し、ヒトリシル酸化酵素では、320番目のlys(lys320)及び355番目のtyr(tyr355)に相当する)。LTQ補因子を形成するチロシン残基を取り巻く配列はまた、リシル酸化酵素の間でも保存される。触媒ドメインはまた、5つのジスルフィド結合の形成に関与する10つの保存システイン残基をも含む。触媒ドメインはまた、フィブロネクチン結合ドメインをも含む。最終的に、成長因子及びサイトカインレセプタードメインに類似し、4つのシステイン残基を含むアミノ酸配列が、触媒ドメインに存在する。
【0057】
これらの保存領域が存在するにも関わらず、異なるリシル酸化型酵素は、これら触媒ドメインの内部及び外側の両方における、ヌクレオチド配列及びアミノ酸配列の相違する領域のお陰(virtue)で、他の異なるリシル酸化型酵素とお互いに区別され得る。
【0058】
単離され、特徴付けられた、この酵素のファミリーの最初のメンバーは、リシル酸化酵素(EC 1.4.3.13)であった。これはまた、タンパク質‐リシン6‐酸化酵素、タンパク質‐L‐リシン:酸素‐6‐酸化還元酵素(脱アミノ化)又はLOXとしても知られる。例えば、Harris et al., Biochim. Biophys. Acta (1974) 341:332-344、Rayton et al. (1979) J. Biol. Chem. 254:621-626、Stassen (1976) Biophys. Acta 438:49-60を参照する。
【0059】
更なるリシル酸化酵素型酵素が、続いて発見された。これらのタンパク質は、「LOX様」(LOX-like)又は「LOXL」と称されている。これらの全ては、上記の共通の触媒ドメインを含み、類似の酵素的活性を有する。これまでのところ、5つの異なったリシル酸化酵素が、ヒトとマウスの両方に存在することが知られている。すなわち、LOXタンパク質及び4つのLOXに関連するないしはLOX様のタンパク質であるLOXL1(これらはまた、「リシル酸化酵素様」「LOXL」又は「LOL」とも示される)、LOXL2(これは、「LOR−I」とも示される)、LOXL3及びLOXL4である。5つの異なるリシル酸化酵素型酵素をエンコードする各遺伝子は、異なった染色体上に存在する。例えば、Molnar et al. (2003) Biochim Biophys Acta. 1647:220- 24、Csiszar (2001) Prog. Nucl. Acid Res. 70:1-32、2001年11月8日公開されたWO 01/83702、及び 米国特許番号6,300,092を参照する(これらの全ては本願に引用として組み込まれる)。LOXL4といくつかの類似性を有するが、異なる発現パターンを有する、LOXCと呼ばれるLOX様タンパク質は、マウスEC細胞系統から単離されている(Ito et al. (2001) J. Biol. Chem. 276:24023-24029)。DmLOXL−1とDmLOXL−2の2つのリシル酸化酵素は、ショウジョウバエから単離されている。
【0060】
全てのリシル酸化酵素型酵素が、共通の触媒ドメインを共有しているにも関わらず、これらはまた、特にそのアミノ末端領域内において、お互いに異なる。LOXと比較して、4つのLOXLタンパク質は、アミノ末端伸張を有する。従って、ヒトプレプロLOX(例えば、シグナル配列の切断の前の一次翻訳産物、以下参照)は、417アミノ酸残基を含み、LOXL1は574(アミノ酸残基)を含み、LOXL2は638(アミノ酸残基)を含み、LOXL3は753(アミノ酸残基)を含み及びLOXL4は756(アミノ酸残基)を含む。当該技術分野における全長リシル酸化酵素型酵素の番号付けの慣例は、最もN末端のアミノ酸残基を「1」として開始し、シグナル配列から始まる。
【0061】
これらのアミノ末端領域の範囲内において、LOXL2、LOXL3及びLOXL4は、4つのスカベンジャーレセプターのシステインリッチ(SRCR)ドメインの繰り返しを含む。これらのドメインは、LOX又はLOXL1には存在しない。SRCRドメインは、分泌性のタンパク質、膜貫通型タンパク質又は細胞外マトリクスタンパク質に見られ、いくつかの分泌性の及びレセプタータンパク質におけるリガンド結合を媒介することが知られている(Hoheneste et al. (1999) Nat. Struct. Biol. 6:228-232、Sasaki et al. (1998) EMBO J. 17:1606-1613)。このSRCRドメインに加えて、LOXL3は、核局在シグナルをそのアミノ末端領域に含む。プロリンリッチドメインは、LOXL1に特有であると思われる(Molnar et al. (2003) Biochim. Biophys. Acta 1647:220-224)。様々なリシル酸化酵素型酵素はまた、それらのグリコシル化パターンの点でも異なる。
【0062】
組織分布もまた、リシル酸化酵素型酵素の間で異なる。ヒトLOXmRNAは、心臓、胎盤、睾丸、肺、腎臓及び子宮において高発現するが、脳及び肝臓においては、僅かに発現する。ヒトLOXL1のmRNAは、胎盤、腎臓、筋肉、心臓、肺及び膵臓において発現し、LOXと同様に、脳及び肝臓においては、(これらと比較して)はるかに少ないレベルで発現する(Kim et al. (1995) J. Biol. Chem. 270:7176-7182)。子宮、胎盤及び他の器官において、LOXL2mRNAは高いレベルで発現するが、LOX及びLOXLと同様に、脳及び肝臓においては、低いレベルで発現する(Jourdan Le-Saux et al.(1999) J. Biol. Chem. 274:12939:12944)。LOXL3mRNAが、睾丸、脾臓及び前立腺において高発現し、胎盤においては、中程度に(moderately)発現し、肝臓では発現しない一方で、LOXL4mRNAの高レベル(の発現)が肝臓において観察される(Huang et al. (2001) Matrix Biol. 20:153-157、Maki and Kivirikko (2001) Biochem. J. 355:381-387、Jourdan Le-Saux et al. (2001) Genomics 74:211-218、及びAsuncion et al. (2001) Matrix Biol. 20:487-491)。
【0063】
疾病における異なったリシル酸化酵素の発現及び/又は関与も変化し得る。例えば、Kagan (1994) Pathol. Res. Pract. 190:910-919、Murawaki et al. (1991) Hepatology 14:1167-1173、Siegel et al. (1978) Proc. Natl. Acad. Sci. USA 75:2945-2949、Jourdan Le-Saux et al. (1994) Biochem. Biophys. Res. Comm. 199:587-592、及び Kim et al. (1999) J. Cell Biochem. 72:181-188を参照する。リシル酸化酵素型酵素もまた、頭部及び首部の癌、膀胱癌、大腸(colon)癌、食道癌及び乳癌を含むいくつかの癌にも関係しているとされている。例えば、Wu et al. (2007) Cancer Res. 67:4123-4129、Gorough et al. (2007) J. Pathol. 212:74-82、Csiszar (2001) Prog. Nucl. Acid Res. 70:1-32及び、Kirschmann et al. (2002) Cancer Res. 62:4478-4483を参照する。
【0064】
従って、リシル酸化酵素型酵素は、構造及び機能においていくつかの重なりを示すが、その上、それぞれが異なった機能を有すると思われる。構造に関して、例えば、ヒトLOXタンパク質の触媒ドメインに対して挙げられる(raised)、ある抗体は、ヒトLOXL2に結合しない。機能に関して、標的とされるLOXの欠失は、マウスにおいて、出産の際に致死であると思われるが、一方、LOXL1の欠損は、深刻な発達表現型を引き起こさないことが報告されている(Hornstra et al. (2003) J. Biol. Chem. 278:14387-14393、Bronson et al. (2005) Neurosci. Lett. 390:118-122)。
【0065】
しかしながら、もっとも広く報告されているリシル酸化酵素型酵素の活性は、細胞外のコラーゲン及びエラスチンにおける特定のリシン残基の酸化であるが、リシル酸化酵素型酵素がまた、いくつかの細胞内プロセスにも関与することの証拠がある。例えば、いくつかのリシル酸化型酵素は、遺伝子発現を制御するという報告がある(Li et al. (1997) Proc. Natl. Acad. Sci. USA 94:12817-12822、Giampuzzi et al. (2000) J. Biol. Chem. 275:36341-36349)。加えて、LOXは、ヒストンH1におけるリシン残基を酸化することが報告されている。更なるLOXの細胞外の活性は、単核球、繊維芽細胞及び平滑筋細胞の走化性の誘導を含む(Lazarus et al. (1995) Matrix Biol. 14:727-731、Nelson et al. (1988) Proc. Soc. Exp. Biol. Med. 188:346-352)。LOX自体の発現は、TGF‐β、TNF‐α及びインターフェロンのような、いくつかの増殖因子及びステロイドにより誘導される(Csiszar (2001) Prog. Nucl. Acid Res. 70:1-32)。近年の研究により、LOXには、発生調節、腫瘍抑制、細胞運動性及び、細胞老化のような、異なった生物学的機能における他の役割があると考えられている。
【0066】
様々な材料由来のリシル酸化酵素(LOX)タンパク質の例は、次の配列の1つから発現し又は翻訳されるポリペプチドに実質的に相同であるアミノ酸配列を有する酵素を含む(EMBL/GenBank受託: M94054;AAA59525.1−−mRNA;S45875;AAB23549.1−mRNA;S78694;AAB21243.1−mRNA;AF039291;AAD02130.1−mRNA;BC074820;AAH74820.1−mRNA;BC074872;AAH74872.1−mRNA;M84150;AAA59541.1――ゲノムDNA。LOXの1つの実施形態は、ヒトリシル酸化酵素(hLOX)プレプロタンパク質である。
【0067】
リシル酸化酵素様酵素をエンコードする配列の例示的な開示は、次の通りである。LOXL1は、GenBank/EMBL BC015090;AAH15090.1で寄託されるmRNAによってエンコードされる。LOXL2は、GenBank/EMBL U89942で寄託されるmRNAによってエンコードされ、LOXL3は、GenBank/EMBL AF282619;AAK51671.1で寄託されるmRNAによってエンコードされ、及び、LOXL4は、GenBank/EMBL AF338441;AAK71934.1で寄託されるmRNAによってエンコードされる。
【0068】
プレプロペプチドとして知られるLOXタンパク質の一次翻訳産物は、1−21番目のアミノ酸に亘るシグナル配列を含む。このシグナル配列は、マウス及びヒトLOXの両方において、21番目のCys(Cys21)と22番目のAla(Ala22)の間の切断によって細胞内に放出され、LOXの形態の46−48kDaのポリペプチドを生じ、これはまた、本願において、全長形態と称される。プロポリペプチドは、ゴルジ体を通過する間に、N−グリコシル化され、50kDaタンパク質を生じ、続いて、細胞外環境に分泌される。この段階において、タンパク質は、触媒活性的に、不活性である。マウスLOXにおける168番目のGly(Gly168)と169番目のAsp(Asp169)の間での更なる切断(cleavage)、及びヒトLOXにおける174番目のGly(Gly174)と175番目のAsp(Asp175)の間での更なる切断によって、成熟した、酵素活性的に活性な、30−32kDAの酵素が生じ、18kDaのプロペプチドを放出する。この最終的な切断の事象は、骨形態発生タンパク質1(BMP−1)としても知られる、メタロエンドプロテアーゼのプロコラーゲンC−プロティナーゼによって触媒される。興味深いことに、この酵素はまた、LOXの基質であるコラーゲンのプロセシングにおいても機能する。N−グリコシル化ユニットは、その後に取り除かれる。
【0069】
潜在的なシグナルペプチド切断部位は、LOXL1、LOXL2、LOXL3及びLOXL4のアミノ末端に存在することが予測されている。予測シグナル切断部位は、LOXL1の25番目のGly(Gly25)と26番目のGln(Gln26)の間、LOXL2の25番目のAla(Ala25)と26番目のGln(Gln26)の間、LOXL3の25番目のGly(Gly25)と26番目のSer(Ser26)の間及び、LOXL4の23番目のArg(Arg23)と24番目のPro(Pro24)の間に存在する。
【0070】
LOXL1タンパク質におけるBMP−1切断部位は、354番目のSer(Ser354)と355番目のAsp(Asp355)の間に同定されている(Borel et al. (2001) J. Biol. Chem.276:48944-48949)。他のリシル酸化酵素型酵素における潜在的なBMP−1切断部位が、プロコラーゲン及びpro‐LOXにおいて、Ala/Gly‐Aspの配列(これには、しばしば、酸性又は荷電残基が続く)にある、BMP−1切断のコンセンサス配列に基づいて、予測されている。LOXL3における予測BMP−1切断部位は、447番目のGly(Gly447)と448番目のAsp(Asp448)の間に位置し、この部位でのプロセシングによって、成熟LOXと類似する大きさの成熟ペプチドを生じ得る。BMP−1の潜在的な切断部位はまた、LOXL4内でも、残基569番目のAla(Ala569)と570番目のAsp(Asp570)の間にも同定された(Kim et al. (2003) J. Biol. Chem. 278:52071-52074)。LOXL2もまた、LOXLファミリーの他のメンバーに類似の形でタンパク質分解的に(proteolytically)切断され、分泌され得る(Akiri et al.(2003) Cancer Res. 63:1657-1666)。
【0071】
リシル酸化型酵素における共通の触媒ドメインの存在に基づいて、活性部位が位置する、プロ酵素のC末端30kDa領域の特定のアミノ酸残基が、高度に保存される。より穏やかな程度の保存(約60−70%)が、プロペプチドドメインにおいて観察される。
【0072】
本願の開示の目的において、用語「リシル酸化型酵素」は、上述の5つのリシン酸化酵素(即ち、LOX、LOXL1(これはLOXLとも表示される)、LOXL2、LOXL3及びLOXL4)の全てを含み、LOX,LOXL1、LOXL2、LOXL3及びLOXL4の機能的なフラグメント及び/又は誘導体(これらは実質的に酵素的活性(例えば、リシル残基の脱アミノ化を触媒する能力)を保持する)もまた含む。アミノ酸配列又はポリヌクレオチド配列のコンテクスト(例えば、LOXL2「由来の」アミノ酸配列)において、「フラグメント及び/又は誘導体」とは、ポリペプチド又は核酸が、自然に生じるリシル酸化酵素型酵素タンパク質又はそれをエンコードする核酸の全て又は一部分の隣接(ないし連続)配列(contiguous sequence)を含むことを示すことを意味し、そして、タンパク質又は核酸が作成される材料又は方法について限定することを意味するものではない。
【0073】
典型的には、機能的なフラグメント又は誘導体は、少なくとも、50%のリシン酸化活性を保持する。いくつかの実施形態において、機能的なフラグメント又は誘導体は、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%又は、100%のリシン酸化活性を保持する。
【0074】
リシル酸化酵素型酵素の機能的なフラグメントは、本質的に、触媒活性を変更させない(天然のポリペプチド配列に関する)保存的なアミノ酸置換(複数)を含むことができることもまた意図される。用語「保存的なアミノ酸置換」は、或る共通の構造及び/又は特性を基にした、アミノ酸の群分け(grouping)を意味する。共通の構造に関して、アミノ酸は次のとおり、群分けできる。即ち、非極性側鎖を有するアミノ酸(グリシン、アラニン、バリン、ロイシン、イソロイシン、メチオニン、プロリン、フェニルアラニン及びトリプトファン)、非荷電極性側鎖を有するアミノ酸(セリン、スレオニン、アスパラギン、グルタミン、チロシン及びシステイン)、及び、荷電極性側鎖を有するアミノ酸(リシン、アルギニン、アスパラギン酸、グルタミン酸及びヒスチジン)に群分けされ得る。芳香族側鎖を含むアミノ酸の群は、フェニルアラニン、トリプトファン及びチロシンを含む。複素環の側鎖は、プロリン、トリプトファン及びヒスチジンに存在する。非極性側鎖を含むアミノ酸の群の中で、短い炭化水素側鎖を有するアミノ酸(グリシン、アラニン、バリン、ロイシン、イソロイシン)は、より長い非炭化水素側鎖を有するアミノ酸(メチオニン、プロリン、フェニルアラニン、トリプトファン)から区別され得る。荷電極性側鎖を有するアミノ酸の群の中で、酸性アミノ酸(アスパラギン酸、グルタミン酸)は、塩基性側鎖を有するアミノ酸(リシン、アルギニン及びヒスチジン)から区別され得る。
【0075】
個々のアミノ酸の共通の特性を定義するための機能的な方法は、相同的な生物の対応するタンパク質の間のアミノ酸変化の正規化された頻度を解析することである(Schulz, G. E. and R. H. Schirmer, Principles of Protein Structure, Springer-Verlag, 1979)。そのような解析によって、1つの群内のアミノ酸が、相同的なタンパク質において、相互に優先的に置換されることが定義され、そしてそれ故、タンパク質構造の全体において類似の影響を有し得る(Schulz, G. E. and R. H. Schirmer, Principles of Protein Structure, Springer-Verlag, 1979)。この種類の解析において、「保存的なアミノ酸置換」とは、あるアミノ酸残基の、アミノ酸側鎖の化学的及び物理的な特性(例えば、電荷、大きさ、疎水性/親水性)を共有する他の(アミノ酸残基)への置換を意味する。以下は、特定の化学的及び/又は物理的特性を共有するアミノ酸残基の例である。
【0076】
(1)Glu、Asp、Lys、Arg及びHisからなる、荷電基を含むアミノ酸。
【0077】
(2)Lys、Arg及びHisからなる、正荷電基を含むアミノ酸。
【0078】
(3)Glu及びAspからなる、負荷電基を含むアミノ酸。
【0079】
(4)Phe、Tyr及びTrpからなる、芳香族基を含むアミノ酸。
【0080】
(5)His及びTrpからなる、窒素環基を含むアミノ酸。
【0081】
(6)Val、Leu及びIleからなる、大きな脂肪族非極性基を含むアミノ酸。
【0082】
(7)Met及びCysからなる、僅かに極性な基を含むアミノ酸。
【0083】
(8)Ser、Thr、Asp、Asn、Gly、Ala、Glu、Gln及びProからなる、小さい残基の基を含むアミノ酸。
【0084】
(9)Val、Leu、Ile、Met及びCysからなる、脂肪族基を含むアミノ酸。
【0085】
(10)Ser及びThrからなる、ヒドロキシル基を含むアミノ酸。
【0086】
ある「保存的な置換」は、以下のアミノ酸残基の群(groups)内での置換を含み得る:gly,ala;val,ile,leu;asp,glu;asn,gln;ser,thr;lys,arg;及びphe,tyr。
【0087】
従って、上で例示された、アミノ酸の保存的な置換は、当業者に知られ、(置換の)結果の分子の生物学的活性を変更することなく一般的に行なわれ得る。この技術分野の当業者はまた、一般的に、ポリペプチドの本質的でない領域における単一のアミノ酸置換は、実質的に、生物学的活性を変更しないことも認識する。例えば、Watson, et al., "Molecular Biology of the Gene," 4th Edition, 1987, The Benjamin/Cummings Pub. Co., Menlo Park, CA, p.224を参照する。
【0088】
リシル酸化酵素に関する更なる情報については、例えば、Rucker et al. (1998) Am. J. Clin. Nutr. 67:996S-1002S及びKagan et al. (2003) J. Cell. Biochem 88:660-672を参照する。また、同一出願人による米国特許出願(公開)US 2009/0053224(2009年2月26日)及びUS 2009/0104201(2009年4月23日)を参照する。各々の開示は、リシル酸化酵素及びリシル酸化酵素様酵素、これら酵素のモジュレーター及び、これらの酵素のモジュレーターを同定する方法を記載する目的で、本願に引用によって取り込まれる。また、WO 2004/47720(2004年6月10日)、WO 2007/126457(2007年8月11日)、WO 2009/010974(2009年1月22日)、US 2006/0127402(2006年6月15日)、US 2007/0021365(2007年1月25日)及び、US 2007/0225242(2007年9月27日)を参照する。これらの全ては、リシル酸化酵素及びリシル酸化酵素様酵素、これらの酵素のモジュレーター及びこれら酵素のモジュレーターを同定する方法を記載する目的で、本願に引用によって取り込まれる。
【0089】
[リシル酸化酵素又はリシル酸化酵素型酵素の触媒ドメインを有するポリペプチド]
本願の開示は、上記のように、生化学及び分子生物学国際連合(International Union of Biochemistry and Molecular Biology)の命名に従うEC1.4.3.13に属するポリペプチドのような、リシル酸化酵素型酵素の活性を有するポリペプチドを提供する。全長のタンパク質は、シグナル配列及び/又はポリペプチド含む(この両方は、生体内で、タンパク質が、完全な触媒活性を獲得する前に切断され得る)。全長のリシル酸化酵素又はリシル酸化酵素様酵素は、(それぞれ)100、200、300、400、500、700より大で、754まで、又はそれより大のアミノ酸残基の長さであり得る。シグナル配列及び/又はプロペプチドを有さない成熟全長タンパク質は、(それぞれ)100、200、240、480、720、730より大で、740まで、又はそれより大のアミノ酸残基の長さであり得る。
【0090】
本願開示のポリペプチドは、成熟全長LOX又はLOXLタンパク質(例えば、シグナルペプチド及び/又はプロペプチドを有さない)由来であり得る。例示のポリペプチドは、図6及び7に記載のアミノ酸配列のいずれかを有することができる。
【0091】
図6は、ヒトLOX、ヒトLOXL1、ヒトLOXL2、ヒトLOXL3、ヒトLOXL4、マウスLOX及びマウスLOXL2由来のポリペプチドのアミノ酸配列のアライメントを表す。図7は、ヒトLOX/LOXLタンパク質のみから由来のポリペプチドの配列のアライメントを表す。便宜上、アミノ酸残基の番号付けは、図6及び7に表される、最もN末端の残基を「1」として開始する。本願開示のポリペプチドは、かくて、図6及び7に記載される配列に使用される番号付けシステム又は、上記のような当該技術分野で使用される番号付けシステムを参照して記載され得る。或いは、ポリペプチドは、本願で割り当てられた配列識別番号を参照して記載され得る。European Bioinformatics Institute(EMBL-EBI)によって提供されたClustalW2プログラムに使用される凡例に従って、図6及び7のアスタリスクは、表示される全てのポリペプチドの間で共通した位置における相同なアミノ酸を示し、一方で、コロンは、保存的なアミノ酸置換を示し、単一の点は、半保存的なアミノ酸置換を示す。
【0092】
本願開示のポリペプチドは、少なくとも約50、少なくとも約75、少なくとも約100、少なくとも約120、少なくとも約140、少なくとも約150、少なくとも約170、少なくとも約180、少なくとも約200、少なくとも約220、少なくとも約240まで、又はそれより大のリシル酸化酵素型酵素のC末端部分のアミノ酸を有するポリペプチドを含み得る。例えば、本願開示のポリペプチドは、図6のSEQ ID NO:1、SEQ ID NO:5又はSEQ ID NO:17によって表されるC末端の207アミノ酸残基を含み得る。本願開示のポリペプチドはまた、天然に生じた全長のリシル酸化酵素型酵素のアミノ酸配列に満たない(長さの)アミノ酸配列を有するポリペプチドも含む。ポリペプチドは、例えば、図6及び7に示された配列によって例示されるような、LOX又はLOXLタンパク質のC末端部位において、約25まで、約50まで、約75まで、約100まで、約120まで、約140まで、約160まで、約180まで、約200まで、約220まで、約230まで、又はそれより大のアミノ酸残基を含む。ポリペプチドは、シグナル配列、プロペプチド及び/又は、図6及び7に表される配列に対するN末端であるLOX又はLOXLタンパク質の任意の部分を除外し得る。
【0093】
本願開示のポリペプチドはまた、上記の当該技術分野において使用される番号付けシステムを参照して記載され得、この場合、アミノ酸番号「1」は、シグナル配列を含む全長のLOX又は全長のLOXLタンパク質の最もN末端のアミノ酸残基を示す。そのような番号付けに従って、ポリペプチドは、LOXにおける約190と240の間、約200と230の間、又は約210と220の間の残基番号(例えば、SEQ ID NO:1又はSEQ ID NO:5)で開始するアミノ酸配列を含むポリペプチドとして記載され得る。ポリペプチドは、LOXL1における約350と360の間、約340と370の間、又は約330と380の間の残基番号で開始するアミノ酸配列を含み得る。或いは、ポリペプチドは、LOXL2における約540と550の間、約530と560の間、又は約520と570の間の残基番号(例えば、SEQ ID NO:3又はSEQ ID NO:7)で開始するアミノ酸配列を含み得る。ポリペプチドはまた、LOXL3における約520と530の間、約510と540の間、又は約500と550の間の残基番号で開始するアミノ酸配列も含み得る。ポリペプチドはまた、LOXL4における約530と540の間、約540と550の間、又は約530と560の間の残基番号で開始するアミノ酸配列も含み得る。
【0094】
本願開示のポリペプチドは、1、2、3、4、5、6、7、8、9または10、又はそれより大のアミノ酸置換を有するポリペプチドを含む(例えば、図6及び7に示されるアミノ酸配列と比較される保存的なアミノ酸置換)。アミノ酸置換のガイダンスは、図6及び7で提供されるアライメントから得ることができる。置換され得るアミノ酸残基は、高度に保存されていない残基位置(たとえば、アスタリスク、コロン又は点によってマークされていない残基位置)に位置し得る。当業者は、表示されるアライメントを基にして、或る(特定の)残基位置のアミノ酸残基が、他の残基位置と比較して、ポリペプチド全体の物理的及び化学的特性を変更すること無しに、置換、欠損及び/又は挿入を許容し得ることを理解するだろう。
【0095】
本願開示のポリペプチドは、図6に表される何れかのポリペプチドと少なくとも80%、少なくとも90%、少なくとも95%、少なくとも99%、又は100%相同であるアミノ酸配列を有するポリペプチドを含む。例えば、図6に表されるマウスLOX触媒ドメインポリペプチドのアミノ酸配列は、ヒトLOX触媒ドメインポリペプチドのアミノ酸配列と98%のアミノ酸配列の相同性を有する。
【0096】
上記に従って、本願開示のポリペプチドから導き出せるタンパクは、LOX(GenBank受託番号NP_002308)、LOXL1(NP_005567)、LOXL2(NP_002309)、LOXL3(NP_115882)、及びLOXL4(NP_115587)を含む(これら前述のGenBank受託番号は、ヒトの配列を示す)。これら、ヒトにおいて発見されたタンパク質に加えて、これらLOX又はLOXLタンパク質はまた、マウス又は他の哺乳類において発見されたタンパク質をも含むことができる。例えば、LOX、LOXL1又はLOXL2タンパク質は、マウスを起源とすることができる(それぞれGenBank受託番号NP_034858、NP_034859、NP_201582)。
【0097】
対象のポリペプチドは、リシル酸化酵素型酵素(例えば、LOX又はLOXL2)由来のアミノ酸配列を含むことができ、更に、異種の(heterologous)アミノ酸配列を含む(例えば、LOX又はLOXL2タンパク質の一部分でないポリペプチドに結合したLOX又はLOXL2配列)。そのようなポリペプチドは、エピトープタグ、精製タグ、及び/又は検出可能な標識を含む融合タンパク質のような融合タンパク質であり得る。融合タンパク質は、任意に(optionally)、異種の配列とリシル酸化酵素型酵素由来のアミノ酸配列の間にリンカー配列を含むことができる。核酸配列が提供される場合、目的の融合タンパク質を生成する方法は、当該技術分野でよく知られている。他の異種の要素(elements)及び、例となる融合タンパク質は、以下により詳細に記載される。
【0098】
異種の要素を含む例となるポリペプチドは、myc及び/又はHis6タグを含み得、SEQ ID NO:9、SEQ ID NO:11、SEQ ID NO:13又はSEQ ID NO:15に記載されるような、隣接(flanking)リンカー配列を任意に含み得る。
【0099】
本願開示のポリペプチドは、レポーターポリペプチド(例えば、蛍光タンパク質、及び/又は分子にコンジュゲート(conjugate)するもの)に結合されるポリペプチドを更に含む。ポリペプチドにコンジュゲートされる分子は、担体分子又は、動物体内にあるとき、免疫反応を引き起こすことが知られる免疫原(例えばアジュバンド)であり得る。他のコンジュゲートとしては、輸送を促進し、及び/又は対象のポリペプチドの半減期を延長させるコンジュゲートを含み得る。
【0100】
本願開示のポリペプチドはまた、1つ以上のポリ(エチレングリコール)(PEG)部分(モイエティ moieties)をも含み得る。そのようなポリペプチドは、「PEG化された(PEGylated)ポリペプチド」と称される。ポリペプチドのペグ化に適した方法及び薬剤が当該技術分野でよく知られる。一般的に、ポリペプチドにコンジュゲーションするのに適したPEGは、概して、室温で水溶性あり、一般式R(O−CH2−CH2)nO−Rを有する。この式において、Rは、水素又は、アルキル又はアルカノール基のような保護基であり、さらにnは、1から1000の整数である。この式において、Rが、保護基の場合、これは、一般的に1から8つの炭素原子を含む。
【0101】
PEGは、ポリペプチド中のアミノ基(例えば、リシン残基のイプシロン(ε‐)アミノ基又は、ポリペプチドのN末端の自由アミノ基)又はカルボキシル基と反応する官能基を生じるように修飾された、すくなくとも1つのヒドロキシル基を有し得る。
【0102】
PEGの更なる誘導体は、アミノ基と選択的に反応しアミド誘導体を生じる末端のチオカルボン酸基(−COSH)を含む。他の実施形態において、PEGは、N‐ヒドロキシスクシンイミダートのような反応性エステルをPEG鎖の末端に含む。そのようなN‐ヒドロキシスクシンイミダート含有PEG分子は、(数値が)6.5−7.5近くの中性pHのような、特別のpH状態で、選択アミノ基と反応する。
【0103】
PEGは、ポリペプチドのアミノ酸残基と直接に、又はリンカーを介してコンジュゲートされ得る。いくつかの実施形態において、リンカーがポリペプチドに加えられ、リンカー修飾されたポリペプチドを形成する。そのようなリンカーは、様々な機能性(例えば、スルフヒドリル基、アミノ基又はカルボキシル基のような反応性の基)を提供し、PEG剤をリンカー修飾されたポリペプチドに連結する。
【0104】
ポリペプチドにコンジュゲートされるPEGは、直鎖状又は分岐鎖状であり得る。分岐鎖状のPEG誘導体は、米国特許番号5,643,575に記載のもの、「star−PGEs」、及び、Shearwater Polymers社カタログ「Polyethylene Glycol Derivatives 1997-1998」に記載されたような、複数の腕を有するPEGを含むがこれらに限定されない。StarPGEsは、例えば米国特許番号6,046,305に、既知の技術として記載されている。
【0105】
[リシル酸化酵素型酵素の触媒ドメインを有するポリペプチドをエンコードするポリヌクレオチド]
本願の開示は、リシル酸化酵素型酵素の活性を有する前記のポリペプチドをエンコードする核酸配列を含むポリヌクレオチドを考えている。本願で意図される核酸は、上記のポリペプチドの何れかをエンコードする核酸の連続(contiguous)配列と、少なくとも70%(例えば、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも98%、少なくとも99%又は100%)相同であるヌクレオチド配列を有する。パーセンテージ相同性は、比較される配列のより短い方を基に示される。初期設定のパラメーターを使用し、フィルターを使用しない、BLASTN(2.0.8) (Altschul et al. (1997) Nucl. Acids. Res. 25:3389-3402)のような良く知られたプログラムが、配列比較を作成するために採用され得る。核酸配列の相同性(例えば、相同なアミノ酸配列をエンコードする2つの異なったポリヌクレオチド間の核酸配列の相同性)は、遺伝コードの縮重に起因して、アミノ酸配列の相同性のパーセントと比較してより低いことがあり得る。
【0106】
本願開示のポリペプチドをエンコードするポリヌクレオチドの核酸配列の例は、SEQ ID NO:2(ヒトLOX由来)、SEQ ID NO:4(ヒトLOXL2由来)、SEQ ID NO:6(マウスLOX由来)、及びSEQ ID NO:8(マウスLOXL2由来)を含む。これらの核酸配列はまた、発現ベクターにも提供され得る。
【0107】
本願において提供される発現ベクターは、前述の核酸及び/又はポリヌクレオチドを含む。そのような発現ベクターは、プロモーター(例えば、T7プロモーター、T3プロモーター、SP6プロモーター、E.coliRNAポリメラーゼプロモーター、CMVプロモーター、SV40プロモーター、PGKプロモーター、EF‐1アルファプロモーター)、転写終結シグナル(例えば、SV40終結シグナル)、スプライス部位(例えば、SV40スプライス部位、ベータ‐グロブリンスプライス部位)、リボゾーム結合部位、シグナル配列(例えば、免疫グロブリンカッパシグナル配列)、エピトープタグ(例えば、myc、FLAG)、精製タグ(例えば、His6)、複製起点及び薬剤選択マーカーを含むことができる。リンカーアミノ酸をエンコードし、及び/又は制限酵素認識部位を含むリンカー配列又は、他の種類のリンカー配列はまた、本願で開示されるベクターにある対象ポリペプチドをエンコードする核酸に、機能可能(operably)に、結合され得る。
【0108】
[本願開示のポリペプチドを作成する方法]
本願開示のポリペプチドは、組み換え方法及び非組み換えの方法(例えば、化学合成)を含む任意の適切な方法によって製造され得る。対象のポリペプチドは、Merrifield (1963) J. Am. Chem. Soc. 85:2149及びMethods in Molecular Biology, Vol 35: Peptide Synthesis Protocolsに記載されるような、当業者によく知られた固相合成法(例えば、Fmoc‐又はt‐Boc化学)によって生成され得る。
【0109】
ポリペプチドが、組み換え技術を使用して生成される場合、方法は、任意の適切な構成及び任意の適切な宿主細胞を含むことができ、これは原核細胞又は真核細胞(例えば、微生物宿主細胞、酵母宿主細胞又は培養哺乳類宿主細胞)であり得る。遺伝子物質を宿主細胞に導入する方法は、例えば、形質転換法、電気穿孔法、リポフェクション法、コンジュゲーション法、リン酸カルシウム法などを含む。転移の方法は、導入された核酸エンコードポリペプチドの安定な発現をもたらすように選択され得る。核酸をエンコードするポリペプチドは、遺伝性エピソーム要素(inheritable episomal element)(例えば、プラスミド)として提供され得、又は、ゲノム的に組み込まれ得る。
【0110】
核酸をエンコードするポリペプチドを転移するための適切なベクターは、組成的に、変更することができる。組み込みのベクターは、条件的複製(conditionally replicative)であり得、又は、自殺型プラスミド、バクテリオファージなどであり得る。構成物(ないしはコンストラクト:construct)は、例えば、プロモーター、選択可能な遺伝子マーカー(例えば、抗生物質に対する抵抗性を与える遺伝子(例えば、ネオマイシン、G418、メトトレキサート、アンピシリン、カナマイシン、エリスロマイシン、クロラムフェニコール又はゲンタマイシン))、(宿主細胞、例えば、細菌性宿主細胞、における複製を促進するための)複製起点などを含む様々な要素を含むことができる。ベクターの選択は、増殖が所望される細胞の種類及び、増殖の目的のような様々な因子に応じてなされるであろう。特定のベクターは、所望のDNA配列を大量に増幅し及び作成するために役立つ。他のベクターは、培養中の細胞におけるタンパク質の発現に適する。更に、他のベクターは、他の動物における細胞での転移及び発現に適する。適切なベクターの選択は、当該技術分野の技術でよくなされる。多くのそのようなベクターは、市販されている。
【0111】
使用されるベクターは、選択可能な薬剤耐性マーカー及び異なる宿主細胞における自立した複製を提供する要素を含むエピソームのプラスミドを基にした発現ベクターであり得る。ベクターは、例えばShort Protocols in Molecular Biology, (1999) F. Ausubel, et al., eds., Wiley & Sonsを含む当業者に良く知られる多数の出版物に十分に記載されている。ベクターは、対象のポリペプチドをエンコードする核酸の発現をもたらし得、(又は、)対象の核酸の増殖をもたらし得、又はその両方をもたらし得る。
【0112】
構成物(constructs)は、例えば、目的のポリヌクレオチドを構成物(ないしはコンストラクト)の骨格に挿入することによって調製され得る(典型的には、ベクター内の切断制限酵素部位へのDNAリガーゼ付着によって、調製され得る)。或いは、所望のヌクレオチド配列は、相同組み換え又は部位特異的組み換え、又は1つ以上の増幅方法(例えばPCR)によって挿入され得る。典型的に、相同組み換えは、所望のヌクレオチド配列の側部にある相同領域をベクターに取り付けることにより達成され、一方で、部位特異的組み換えは、部位特異的組み換えを促進する配列の使用を介して達成され得る(例えば、cre−lox、att部位など)。そのような配列を含む核酸は、例えば、オリゴヌクレオチドのライゲーションによって付加され、又は、相同な領域及び所望のヌクレオチド配列の部分の両方を含むプライマーを使用するポリメラーゼ連鎖反応によって付加される。
【0113】
目的のポリペプチドの発現に関して、発現カセットが採用され得る。かくて、本願の開示は、対象の核酸を含む組み換え発現ベクターを提供する。発現ベクターは、転写調節配列及び翻訳調節配列を提供でき、また、誘導可能な又は構成的な発現も提供でき、そのコーディング領域は、転写開始領域(例えば、プロモーター又はエンハンサー)及び転写停止領域及び翻訳停止領域の転写制御下に、機能(作用)可能に、配置される。これらの制御領域は、対象のポリペプチドが由来するリシル酸化酵素型酵素に元から存在するものであり得、又は、外生的な材料由来であり得る。そのようなものとして、外生的な材料由来の制御領域は、対象のポリペプチドをエンコードする核酸に機能(作用)可能に結合する異種の要素を考慮し得る。一般的に、転写調節配列及び翻訳調節配列は、プロモーター配列、リボゾーム結合部位、転写開始配列及び転写停止配列、翻訳開始配列及び翻訳停止配列及び、エンハンサー配列又はアクチベータ配列を含むが、これらに限定されない。プロモーターは構成的か又は誘導可能のどちらかであり得、強力な構成的プロモーターであり得る(例えばT7など)。
【0114】
発現ベクターは、目的のタンパク質をエンコードする核酸配列の挿入をもたらすために、プロモーター配列の近くに位置する適当な制限部位を一般的に有する。発現宿主において作動する選択可能なマーカーは、ベクターを含む細胞の選択を容易にするために存在し得る。加えて、発現構成物は、更なる要素を含むことができる。例えば、発現ベクターは、1つ又は2つの複製システムを有し得、かくて、これは、例えば、発現のための哺乳類又は昆虫細胞において維持され、及びクローニング及び増幅のための原核生物の宿主において維持される。加えて、発現構成物は、形質転換宿主細胞の選択を可能にするための選択可能なマーカー遺伝子を含むことができる。選択遺伝子が、当該技術分野において良く知られており、使用される宿主細胞によって異なるだろう。
【0115】
本願開示のポリペプチドはまた、例えば、放射性標識、蛍光標識、ビオチン標識、免疫学的に検出可能な標識(例えば、ヘマグルチニン(HA)タグ、ポリヒスチジンタグ)などの検出可能な標識のような更なる要素をも含むことができる。更なる要素が、(例えば、融合ポリペプチドの形態で)、様々な方法を介して、開示のポリペプチドの発現を促進するために提供され得(例えば、宿主細胞内で、発現を促進するためのN末端メチオニン及び/又は異種のシグナル配列)、及び/又は単離を促進するために提供され得る(例えば、ビオチンタグ、免疫学的に検出可能なタグ)。ポリペプチドはまた、共有結合又は非共有結合を介して任意に支持体上に固定化され得る。対象のポリペプチドをエンコードする例となる核酸は、SEQ ID NO:10、SEQ ID NO:12、SEQ ID NO:14又は、SEQ ID NO:16を含むことができる。核酸は、mycタグ、His6タグ及び/又はリンカー配列を有する、ヒト又はマウスリシル酸化酵素型酵素由来のポリペプチドをエンコードすることができる。
【0116】
対象のポリペプチドの単離及び精製は、当該技術分野において知られる方法に従って達成され得る。用語「単離された」は、化合物(例えば、ポリペプチド又はポリヌクレオチド)が、自然界において、それに付いている全ての又はいくつかの化合物から分離されたことを意味することが意図される。用語「単離された」はまた、化合物が、製造の間(例えば、化学合成、組み換え発現、培養培地など)に、それに付いている全ての又はいくつかの成分から分離されたことをも意味する。
【0117】
例えば、本願の開示によるポリペプチドは、対象のポリペプチドを発現するために遺伝子操作された細胞の溶解物から単離され得、又は、細胞培養培地の上清から単離され得、又は、合成反応混合物から単離され得る。単離は、更に、免疫親和性精製によって達成され、これは、一般的に、試料が、ポリペプチドのエピトープ(例えば、リシル酸化酵素型酵素由来のアミノ酸配列に融合された異種のエピトープ)に対する抗体(これは、任意に固定化される)と接触すること、洗浄し、非特異的な結合材料を取り除くこと、及び特異的に結合するポリペプチドを溶出することを含む。単離されたポリペプチドは、更に、透析及び、タンパク質精製に通常採用される他の方法(例えば、金属キレートクロマトグラフィー、イオン交換及びサイズ排除)によって精製され得る。
【0118】
本願の開示はさらに、1つ以上の本願開示のポリペプチドをエンコードする外生的なポリヌクレオチドを含む組み換え宿主細胞を考えている。
【0119】
[スクリーニング方法]
リシル酸化酵素触媒活性を有する本願開示のポリペプチドは、リシル酸化酵素型酵素の活性のモジュレーターの選別をする方法に役立つ。前記の対象のポリペプチドの十分量の発現は、以下に記載されるスクリーニング分析用に基質を供給するために使用され得る。
【0120】
リシル酸化酵素型酵素の活性のモジュレーター(及び、モジュレーター活性の試験をされる候補化合物)は、例えばタンパク質(例えば、抗体、転写因子)及び核酸(例えば、三重鎖形成性オリゴヌクレオチド、アンチセンスオリゴヌクレオチド、リボゾーム、siRNA、shRNA)及び、合成有機法及び/又は組み合わせ化学(combinatorial chemistry)により得られるようなもの、小有機分子などのような高分子を含むことができる。
【0121】
リシル酸化酵素型酵素の活性のモジュレーターは、アクチベータ(アゴニスト)及び阻害剤(アンタゴニスト)の両方を含み、様々なスクリーニング分析を使用することにより選択され得る。1つの実施形態において、モジュレーターは、化合物が、本願で開示される触媒ドメインポリペプチドに結合するかを確認することにより同定され得、結合が生じた場合、当該化合物は、候補モジュレーターである。任意に、追加の試験が、そのような候補モジュレーターについて行なわれ得る。かわりとして、候補化合物を、本願で開示されるポリペプチドと接触させることができ、及び、ポリペプチドの生物学的活性が分析される。ポリペプチドの生物学的活性を変える化合物は、リシル酸化酵素型酵素のモジュレーターである。一般的に、ポリペプチドの生物学的活性を減少させる化合物は、酵素の阻害剤である。
【0122】
リシル酸化酵素型酵素の活性のモジュレーターを同定する他の方法は、細胞が1つ以上の開示のポリペプチドを発現している細胞培養中で、候補化合物をインキュベートすること、及び、細胞の、1つ以上の生物学的活性又は特性を分析することを含む。培地において、細胞の生物学的活性又は特性を変更する化合物は、リシル酸化酵素型酵素の活性の潜在的なモジュレーターである。分析され得る生物学的活性は、例えば、リシン酸化、過酸化物生成、アンモニア生成、リシル酸化酵素型酵素のレベル、リシル酸化型酵素をエンコードするmRNAのレベル、開示のポリペプチドのレベル、開示のポリペプチドをエンコードするmRNAのレベル及び/又は、リシル酸化酵素型酵素に特異的な1つ以上の機能を含む。
【0123】
前述の分析の更なる実施形態において、候補化合物との接触のない状態において、1つ以上の生物学的活性又は細胞特性が、1つ以上のリシル酸化酵素型酵素のレベル又は活性と関連付けられる。例えば、生物学活性は、遊走、走化性、上皮間葉転換(EMT)又は、間葉上皮転換(MET)のような細胞性機能であり得、及び、その変化は、1つ以上の、対照試料又は参考試料との比較によって検出される。
【0124】
対照もまた、使用され得る。例として、陰性対照試料は、候補化合物が加えられ、開示のポリペプチドの減少したレベルのものを有する培養物(又は開示のポリペプチドを発現しない培養物)又は、試験培養物として、候補化合物の添加が無いが、開示のポリペプチドと等量のものを有する培養物を含むことができる。対照には、模擬的(mock)候補化合物又は、リシル酸化酵素活性を活性化又は阻害することが知られる薬剤の追加を含むことができる。いくつかの実施形態において、開示のポリペプチドの異なったレベルのものを含む別の(separate)培養物が、候補化合物と接触される。もし生物学的活性における変化が観察された場合、及び、より高いレベルのポリペプチドを有する培養物において変化が大きい場合、当該化合物は、リシル酸化型酵素の活性のモジュレーターと確定される。化合物が、リシル酸化酵素型酵素の活性剤であるか又は阻害剤であるかの決定は、当該化合物によって誘導された表現型から明らかであり得、又は、1つ以上のリシル酸化酵素型酵素の酵素的活性における、当該化合物の効果の試験のような、更なる分析を要求し得る。
【0125】
上記のような、リシル酸化酵素型酵素の活性のモジュレーターを確定するための細胞培養及び酵素的分析の方法と同じく、生化学的に又は組み換え的にリシル酸化酵素型酵素を得る方法が当該技術分野において知られる。
【0126】
リシル酸化酵素型酵素の酵素的活性は、いくつかの異なった方法によって分析され得る。例えば、リシル酸化酵素の酵素的活性は、過酸化水素、アンモニウムイオン及び/又はアルデヒド生成の検出及び/又は定量によって評価され得、リシン酸化及び/又はコラーゲン架橋結合の分析によっても評価され得、又は、細胞性浸潤能、細胞接着、細胞増殖又は転移成長を測定することによっても評価され得る。例えば、Trackman et al. (1981) Anal. Biochem. 113:336-342、Kagan et al. (1982) Meth. Enzymol. 82A:637-649、Palamakumbura et al. (2002) Anal. Biochem. 300:245-251、Albini et al. (1987) Cancer Res. 47:3239-3245、Kamath et al. (2001) Cancer Res. 61:5933-5940、米国特許番号4,997,854及び米国特許出願公開番号2004/0248871を参照する。
【0127】
試験化合物は、例えば、小有機化合物(例えば、約50から約2500Daの間の分子量を有する有機分子)、核酸又はタンパク質を含むがこれらに限定されない。(単一の)化合物又は複数の化合物は、化学的に合成され、又は微生物学的に生成され、及び/又は、例えば、試料(例えば、植物、動物又は微生物などからの細胞抽出物)に含まれ得る。リシル酸化酵素型酵素のモジュレーターの分析をするための反応混合物は、無細胞の抽出物であり得、又は、細胞培養物又は組織培養物を含むことができる。複数の化合物は、例えば、反応混合物に添加され、培養培地に加えられ、細胞に注入され、又は遺伝子組み換え動物に、投与され得る。分析に採用される細胞又は組織は、例えば、微生物細胞、真菌細胞、昆虫細胞、脊椎動物細胞、哺乳類細胞、霊長類細胞、ヒト細胞であり得、又は、(人間を除く)遺伝子組み換え動物を含むことができ、又はこの動物から得られ得る。
【0128】
(単一の)化合物又は複数の化合物を含む試料が、開示の方法において同定された場合、続いて、これは、対象のポリペプチドを阻害し又は活性化する能力を有する化合物を含むとして同定された元の試料から化合物を単離することができ、又は、例えば、当該化合物が異なった複数の化合物からなる場合、各試料毎に異なった物質の数を減らし、元の試料の更なる分割をともなう方法を繰り返すために、元の試料を更に分割(subdivide)することもできる。試料の複雑さに応じて、上記のステップは、開示の方法に従って、単離された試料が限られた数の物質のみを含み又は、ただ1つの物質を含むまで、数回実施され得る(例えば、限界希釈によってなされる)。いくつかの実施形態において、試料は、類似の化学的及び/又は物理的特性を有する物質を含み、そして、いくつかの実施形態において、当該物質は、相同である。
【0129】
更に、上述の方法は、リシル酸化酵素型酵素由来の結合エピトープの構成に使用され得る。類似の手法(approach)は、抗p24(HIV−1)モノクローナル抗体のペプチド抗原に関して良く記載されている(Kramer (1997) Cell 97:799-809を参照する)。クラスタ化されたアミノ酸ペプチドライブラリを使用する、ペプチド‐抗体相互作用のフィンガープリント解析のための一般的手順は、Kramer (1995) Mol. Immunol. 32:459-465に記載されている。更に、リシル酸化酵素型酵素のアンタゴニストは、Doring (1994) Mol. Immunol. 31:1059-1067に記載されるような方法に従って、開示のポリペプチドと特異的に反応するモノクローナル抗体由来であり得、及びこれから同定され得る。
【0130】
リシル酸化酵素型酵素又は、本開示のポリペプチドのような目標物に対して特異的な親和性を有する化合物を同定するための大きなライブラリを作成し及びスクリーニングするためのいくつかの方法が当業者に知られる。これらの方法は、ファージ表示法を含み、そのランダム化されたペプチドは、ファージから表示され、固定化されたレセプターを使用した親和性クロマトグラフィーによりスクリーニングされる。例えば、WO 91/17271、WO 92/01047及び、米国特許番号5,223,409を参照する。他の手法において、固相支持体上に固定化されたポリマーの組み合わせライブラリ(combinatorial libraries)(例えば、「チップ」)は、フォトリソグラフィーを使用して合成される。例えば、米国特許番号5,143,854、WO 90/15070及びWO 92/10092を参照。固定化されたポリマーを、標識されたポリペプチド(例えば、リシル酸化酵素型酵素又は本願で開示されるようなポリペプチド)と接触させ、支持体をスキャンし、ラベルの位置を決定し、それにより、当該ポリペプチドに結合するポリマーを同定する。
【0131】
目的のポリペプチド(例えば、リシル酸化酵素型酵素又は開示のポリペプチド)の結合リガンド(例えば、活性剤又は阻害剤)を同定するために使用され得る、連続的なセルロース膜支持体でのペプチドライブラリの合成及びスクリーニングは、例えば、Kramer (1998) Methods Mol. Biol. 87: 25-39において記載される。そのような分析によって同定されるリガンドは、目的のタンパク質の候補モジュレーターであり、更なる試験のために選択され得る。この方法はまた、例えば、目的のタンパク質における結合部位及び認識モチーフを決定するためにも使用され得る。例えば、Rudiger (1997) EMBO J. 16:1501-1507及びWeiergraber (1996) FEBS Lett. 379:122-126を参照する。
【0132】
WO 98/25146は、所望の特性(例えば、ポリペプチド又はその結合相手をアゴナイズ(agonize)し、ポリペプチド又はその結合相手に結合し、又は、ポリペプチド又はその結合相手をアンタゴナイズ(antagonize)する能力)を有する化合物の複合体のスクリーニングライブラリの更なる方法を記載する。それらのライブラリにおける複合体は、供試化合物、化合物の合成における少なくとも1つのステップを証明するタグ及び、レポーター分子による修飾の影響を受けやすいテザーを含む。テザーの修飾は、複合体が、所望の特性を有する化合物を含むことを示すために使用される。タグは、そのような化合物の合成における少なくとも1つのステップを示すためにデコード(decode)され得る。リシル酸化酵素型酵素(又は、開示のポリペプチド)と相互作用する化合物を同定するための他の方法は、例えば、ファージ表示システムを使用したインビトロスクリーニング、フィルター結合分析及び、例えば、BIAcore装置(Pharmacia)を使用した相互作用の、「リアルタイムの」測定である。
【0133】
これら方法の全ては、本願の開示に従って、リシル酸化酵素型酵素又はそれに関係するポリペプチドの活性剤/アゴニスト及び阻害剤/アンタゴニストを同定するために使用され得る。
【0134】
そのような活性化剤(アクチベータ)又は阻害剤の基礎的な構成のための様々な材料が採用され得、これは、例えば、開示のポリペプチドの模倣(模擬 mimetic)アナログを含む。開示のポリペプチドの模倣アナログ又は、その生物学的に活性なフラグメントは、例えば、生物学的活性に必須であると期待されるアミノ酸を置換することによって生じ得る(例えば、ステレオイソマー、即ちD−アミノ酸;例えば、Tsukida (1997) J. Med. Chem. 40:3534-3541を参照する)。更に、ポリペプチドフラグメントが、生物学的に活性なアナログをデザインするために使用される場合、プロ‐模倣化構成物(pro-mimetic components)が、少なくともいくつかの立体的形態特性(conformational properties、これは、元のポリペプチドの部分を取り除く際に失われていることもありうる)を再構築するために、ペプチドに組み込まれ得る(例えば、Nachman (1995) Regul. Pept. 57:359-370を参照する)。
【0135】
そのような天然アミノ酸配列の擬ペプチド(pseudopeptide)類似体は、とても効率的に、親タンパク質を模倣できる(Benkirane (1996) J. Biol. Chem. 271:33218-33224)。例えば、簡単に入手できるアキラルo−アミノ酸残基の開示のポリペプチド又は、そのフラグメントへの組み込みが、脂肪族化合物鎖のポリメチレンユニットによるアミド結合の置換をもたらし、それによって、ペプチド模倣物(peptide mimetic)を構成するための便利な戦略を提供する(Banerjee (1996) Biopolymers 39:769-777)。他のシステムにおける小ペプチドホルモンの超活性なペプチド模倣アナログもまた、記載されている(Zhang (1996) Biochem. Biophys. Res. Commun. 224:327-331)。
【0136】
ペプチド模倣物はまた、一連のアミドアルキル化を介して、ペプチド模倣組み合わせライブラリの合成によっても同定され得、続いて結果する化合物の試験(例えば、それらの結合及び免疫学的特性のための試験)を行なう。ペプチド模倣組み合わせライブラリの発生及び使用の方法が、記載されている。例えば、Ostresh, (1996) Methods in Enzymology 267:220-234及びDorner (1996) Bioorg. Med. Chem. 4:709-715を参照する。更に、開示のポリペプチドの3次元構造及び/又は結晶構造は、ペプチド模倣物をデザインするために使用され得る(Rose (1996) Biochemistry 35:12933-12944、Rutenber (1996) Bioorg. Med. Chem. 4:1545-1558)。
【0137】
天然の生物学的ポリペプチドの活性を模倣した低分子量合成分子(low-molecular-weight synthetic molecules)の、構造を元にしたデザイン及び合成が、例えば、Dowd (1998) Nature Biotechnol. 16:190-195、Kieber-Emmons (1997) Current Opinion Biotechnol. 8:435-441、Moore (1997) Proc. West Pharmacol. Soc. 40:115-119、Mathews (1997) Proc. West Pharmacol. Soc. 40:121-125及びMukhija (1998) European J. Biochem. 254:433-438に更に記載されている。
【0138】
また、例えば、リシル酸化酵素型酵素又は開示のポリペプチドの基質又はリガンドとして機能できる小有機化合物の模倣物をデザインし、合成し、評価することが可能であることも当業者によく知られている。例えば、細胞毒性において多剤耐性補助関連タンパク質(multidrug resistance assistance-associated protein)をアンタゴナイズ(antagonize)する際に、ハパロシンのD−グルコース模倣物がハパロシンと類似の性能を示したことが記載されている(Dinh (1998) J. Med. Chem. 41:981-987)。
【0139】
リシル酸化酵素型酵素及び本願で開示されるポリペプチドの構造は、例えば、小分子、ペプチド、ペプチド模倣物及び抗体のようなモジュレーターの選択を導くために研究され得る。リシル酸化酵素型酵素及び開示のポリペプチドの構造的特性は、リシル酸化酵素型酵素に結合し、又は、そのリガンド、基質、結合相手又はレセプターとして機能する、天然の又は合成の分子を同定するのに役立つ。例えば、Engleman (1997) J. Clin. Invest. 99:2284-2292を参照する。例えば、リシル酸化酵素型酵素の構造的モチーフの折りたたみシミュレーション及びコンピューター再設計は、適切なコンピュータープログラムを使用して行われ得る(Olszewski (1996) Proteins 25:286-299、Hoffman (1995) Comput. Appl. Biosci. 11:675-679)。タンパク質の折りたたみのコンピュータモデリングは、詳細なペプチド及びタンパク質構造の立体配座解析及びエネルギー解析に使用され得る(Monge (1995) J. Mol. Biol. 247:995-1012、Renouf (1995) Adv. Exp. Med. Biol. 376:37-45)。適切なプログラムが、相補的なペプチド配列のコンピューターを使用した調査を使用して、リシル酸化酵素型酵素及び/又は開示のポリペプチド上にあり、リガンド及び結合相手と相互作用する部位の同定に使用され得る(Fassina (1994) Immunomethods 5:114-120)。タンパク質及びペプチドのデザインのための更なるシステムが、当該技術分野において、例えば、Berry (1994) Biochem. Soc. Trans. 22:1033-1036、Wodak (1987), Ann. N.Y. Acad. Sci. 501:1-13、及びPabo (1986) Biochemistry 25:5987-5991に記載される。上記の構造解析から得られた結果は、例えば、有機分子、ペプチド及び、1つ以上のリシル酸化酵素型酵素の活性のモジュレーターとして機能するペプチド模倣物の作成に使用され得、そして、本開示のポリペプチドの模倣物の作成に使用され得る。
【0140】
抗体のような本願でスクリーニングされる阻害剤(inhibitors)は、競合阻害剤、不競合(uncompetitive)阻害剤、混合阻害剤又は非競合阻害剤(non-competitive)であり得る。競合阻害剤は、しばしば、基質と構造的な類似性を持ち、通常、活性部位に結合し、一般的に、より低い基質濃度において、より効率的である。競合阻害剤存在下では、見かけのKMが増加する。不競合阻害剤は、一般的に、酵素‐基質複合体又は、基質が活性部位に結合した後に結合可能になる部位に結合し、活性部位を変形させ得る。見かけのKM及びVmaxの両方が、不競合阻害剤の存在下で減少し、基質濃度は、阻害において少しの効果を有するか又は効果を有さない。従って、不競合阻害剤による阻害は、高基質濃度において、しばしば最も顕著である。混合阻害剤は、自由な(free)酵素及び、酵素‐基質複合体の両方に結合可能であり、かくて、基質結合及び触媒活性の両方に影響する。非競合阻害とは、阻害剤が、酵素及び酵素‐基質複合体に、同等の親和性を有して結合する、混合阻害の特別な例であり、当該阻害は、基質濃度に影響されない。非競合阻害剤は、一般的に、活性部位の外の領域で酵素と結合する。酵素阻害の更なる詳細については、例えば、上記のVoet et al. (2008)を参照する。リシル酸化酵素型酵素のような酵素に関して、その天然の基質(例えば、コラーゲン、エラスチン)は、通常、(インビボにおいて達成され得る任意の阻害剤の濃度と比較して)インビボにおいて非常に過度に存在し、阻害が基質濃度と無関係であるので、非競合阻害剤は、有益である。
【0141】
[抗体生成]
本願の開示はまた、開示のポリペプチドに特異的な抗体を生成する方法を提供する。抗体(例えば、抗体が自然に生じる環境以外の環境にある抗体)が単離され得る。対象のポリペプチドに特異的な、適切な抗体は、任意のアイソタイプの抗体、単鎖のFv、Fab、Fab、Fv、F(ab’)2、人工的な抗体、ヒト化抗体、これらのフラグメントなどを含む。
【0142】
適切な抗体は、宿主動物に、対象のポリペプチドの全部又は一部を含むペプチドを免疫することによって得られ得る。動物を免疫する場合、本願の開示は、対象のポリペプチドを含む免疫原性組成物(immunogenic composition)を提供する。対象の免疫原性組成物は、対象のポリペプチド及びアジュバンドを含み得る。アジュバンドは、当該技術分野において知られ、ヒト又は他の哺乳類に使用するのに適切なアジュバンドを含み得る。
【0143】
免疫するのに適切な宿主動物は、マウス、ラット、ヒツジ、ヤギ、ハムスター、ウサギ、ロバなどを含む。使用されるアジュバンド、追加免疫スケジュール、注入部位、適切な動物などを含む動物を免疫する方法は、当該技術分野において良く理解されており(例えば、Harlow et al., Antibodies: A Laboratory Manual, First Edition (1988) Cold Spring Harbor, N.Y.)、動物への生きている細胞の投与は、いくつかの哺乳類及び鳥類に関して記載されている(例えば、McKenzie et al. (1989) Oncogene 4:543-548、Scuderi et al. (1985) Med. Oncol. Tumor Pharmacother 2:233-242、Roth et al. (1984) Surgery 96:264-272及びDrebin et al. (1984) Nature 312:545-548)。免疫付与の後、抗体生成細胞の集団が生じる。細胞の集団は、当該技術分野における技術の1つとして良く知られるハイブリドーマ法を使用して生成され得る(例えば、前記のHarlowを参照する)。抗体を発現する細胞は、骨髄腫細胞又は形質転換細胞のような不死化細胞に融合され、これらは細胞培養において無制限に複製する能力を有し、それによって不死の、免疫グロブリン分泌細胞系統を生成する)。利用される不死化細胞系統は、特定の栄養素を利用するのに必要な酵素が欠損するように選択され得る。骨髄腫細胞のような多くのそのような系統は、当業者によく知られており、例えば、チミジンキナーゼ(TK)欠損細胞系統又は、ヒポキサンチン‐グアニンホスホリボキシル転移酵素(HGPRT)欠損細胞系統を含む。これらの欠損体は、例えば、ヒポキサンチン/アミノプテリン/チミジン(HAT)培地で増殖する能力によって融合細胞を選択することを可能にする。代わりの実施形態において、モノクローナル抗体を発現する細胞の集団は、ファージ表示法を使用して作成され得る。
【0144】
対象のポリペプチドに対する抗体はまた、遺伝子工学的技術によっても提供され得る。この技術において、標準のハイブリドーマ法と同様に、抗体生成細胞は、所望の抗原又は免疫原に感作される。免疫脾臓細胞又はハイブリドーマから単離されたメッセンジャーRNAは、PCR増幅を使用するcDNAを作成するための鋳型として使用される。それぞれ、最初の抗原特異性を保持する1つの重鎖遺伝子及び1つの軽鎖遺伝子を含むベクターのライブラリは、増幅された免疫グロブリンcDNAの適切な領域ないし部位(section)の、発現ベクターへの挿入により生成される。組み合わせライブラリは、重鎖遺伝子ライブラリを軽鎖遺伝子ライブラリと結合することによって構成され得る。これは、重鎖及び軽鎖(これらは、抗体分子のFabフラグメント又は抗原結合フラグメントに似る)を共発現するクローンのライブラリをもたらす。これらの遺伝子を有するベクターは、宿主(例えば、細菌、昆虫細胞、哺乳類細胞又は、他の適切なタンパク質生成宿主細胞)に共トランスフェクトされる。抗体遺伝子合成が、トランスフェクト宿主において誘導される場合、重鎖及び軽鎖タンパク質は、自己集合(self-assemble)し、抗原又は免疫原を使用するスクリーニングによって検出され得る活性な抗体を生成する。
【0145】
抗体生成の他の方法は、ファージ表示(法)を含む。ファージ表示(法)は、タンパク質相互作用の高‐スループットスクリーニング(high-throughput screening)に使用される。この方法において、ファージは、レパートリー又は組み合わせ抗体ライブラリ(例えば、ヒト又はマウス)から発現される抗原結合ドメインを表示するのに利用される。目的の、そして対象のポリペプチドを結合する抗原結合ドメインを発現するファージが、ポリペプチド目標を使用して、(例えば、標識されたポリペプチド又は、固体表面又はビーズに結合又は取り込まれるポリペプチドを使用して)選択又は同定される。これらの方法において使用されるファージは、典型的に、fd及びM13を含む線状ファージである。結合ドメインは、Fab、Fv(軽鎖又は重鎖由来の個々のFv領域)又は、ファージ遺伝子IIIタンパク質又は遺伝子VIIIタンパク質のどちらかに、組み換え的に結合されたジスルフィド安定化されたFv抗体ドメインを含むことができる。例示の方法は、例えば、EP 368684B1、米国特許番号5,969,108、Hoogenboom, H. R. and Chames (2000) Immunol. Today 21:371、Nagy et al. (2002) Nat. Med. 8:801、Huie et al. (2001) Proc. Natl. Acad. Sci. USA 98:2682、Lui et al. (2002) J. Mol. Biol. 315:1063(これら各々は、本願に引用として組み込まれる)に示される。いくつかの出版物(例えば、Marks et al. (1992) Bio/Technology 10:779-783)は、大きなファージライブラリを構成するための戦略としての組み合わせ感染(combinatorial infection)及びインビボ組み換えと同じく、鎖シャッフリング(chain shuffling)による高親和性ヒト抗体の生成を記載している。
【0146】
関係する実施形態において、リボゾーム表示は、表示基盤(display platform)として使用される。例えば、Hanes et al. (2000) Nat. Biotechnol. 18:1287、Wilson et al. (2001) Proc. Natl. Acad. Sci. USA 98:3750及び、Irving et al. (2001) J. Immunol. Methods 248:31を参照する。
【0147】
細胞表面ライブラリは、抗体のためにスクリーニングされ得る(Boder et al. (2000) Proc. Natl. Acad. Sci. USA 97:10701、Daugherty et al. (2000) J. Immunol. Methods 243:211)。そのような手段は、モノクローナル抗体の単離及びそれに続くクローニングのための伝統的なハイブリドーマ技術に、代わる方法を提供する。
【0148】
ファージ表示法において、機能的な抗体ドメインは、このドメインをエンコードするポリヌクレオチド配列を有するファージ粒子の表面に表示される。例えば、重鎖可変領域(VH)及び軽鎖可変領域(VL)をエンコードするDNA配列は増幅され、又はさもなければ、動物cDNAライブラリ(例えば、リンパ組織のヒト又はマウスcDNAライブラリ)又は合成cDNAライブラリから単離される。VH及びVL領域をエンコードするDNAは、scFvリンカーによってお互いに結合され得(例えば、PCRによって結合され得る)、ファージミドベクター(例えば、pCANTAB 6又はpComb 3HSS)にクローン化され得る。ベクターは、E.coliにエレクトロポレーションされ、そしてE.coliに補助ファージ(helper phage)を感染させる。VH又はVL領域をエンコードする配列は、通常は、ファージの遺伝子III配列又は遺伝子VIII配列のどちらかに組み換え的に融合される。目的の抗原(即ち、対象のポリペプチド)に結合する抗原結合ドメインを発現するファージは、抗原を使用して(例えば、標識された抗原又は、固体表面又はビーズに結合又は取り込まれた抗原を使用して)選択され又は同定され得る。
【0149】
ファージ表示法の更なる実施例は、PCT出願番号PCT/GB91/01134、国際公開WO 90/02809、WO 91/10737、WO 92/01047、WO 92/18619、WO 93/11236、WO 95/15982、WO 95/20401、及び米国特許番号5,698,426、5,223,409、5,403,484、5,580,717、5,427,908、5,750,753、5,821,047、5,571,698、5,427,908、5,516,637、5,780,225、5,658,727、5,733,743及び5,969,108(各々、本願に引用によってその全てが取り込まれる)に開示された方法を含む。
【0150】
[製剤、キット及び投与経路]
本願に開示されるような触媒ドメインを含む治療用組成物もまた提供される。そのような組成物は典型的に、触媒ドメイン(又は触媒ドメインをエンコードする核酸)、医薬的に許容可能な担体を含む。補助的な活性組成物もまた、当該構成物に組み込まれ得る。
【0151】
本願で使用される場合、用語「治療的に効果のある量」又は「効果的な量」は、単独で又は他の治療用剤と組み合わせて、細胞、組織又は対象(例えば、ヒト又は、霊長類、げっ歯類、ウシ、ウマ、ブタ、ヤギなどのヒトを除く動物のような哺乳類)に投与される場合、疾病の状態又は疾病の進行を予防し(妨げ)、又は改善するのに効果的な量である治療用剤の量を意味する。治療的に効果のある用量(dose)は更に、症状の完全な又は部分的な改善(例えば、関連する病状の処置、治癒、予防又は改善)、又はそのような状態の処置、治癒、予防又は改善の速度の増進をもたらすのに十分な化合物の量を意味する。薬剤の効果的な量は、投与したときに、1つ以上の用量(dose)において、個体内のリシル酸化酵素型酵素に関連する1つ以上の症状の重症度おいて(当該薬剤で処置しない場合の1つ以上の症状の重症度と比較して、)少なくとも約5%、少なくとも約10%、少なくとも約15%、少なくとも約20%、少なくとも約25%、少なくとも約30%、少なくとも約40%、少なくとも約50%、少なくとも約60%、少なくとも約70%、少なくとも約80%、少なくとも約90%又は90%超の減少をもたらす量である。そのような重症度における減少は、例えば、改善された創傷治癒又は促進された血管形成によって、明らかにされ得る。
【0152】
リシル酸化酵素型酵素の触媒ドメイン(例えばLOX又はLOXL2触媒ドメイン)の治療的に効果のある量は、疾病又は障害の種類、疾病又は障害の広さ、及び疾病又は障害を被る哺乳類の大きさによって異なる。
【0153】
様々な医薬組成物および、それらの調製及び使用の技術が、本願の開示を踏まえて、当業者に知られる。適切な医薬組成物及び、その投与のための技術の詳細なリストに関して、Remington's Pharmaceutical Sciences, 17th ed. 1985、Brunton et al., "Goodman and Gilman's The Pharmacological Basis of Therapeutics," McGraw-Hill, 2005、University of the Sciences in Philadelphia (eds.),"Remington: The Science and Practice of Pharmacy," Lippincott Williams & Wilkins, 2005、及び University of the Sciences in Philadelphia (eds.), "Remington: The Principles of Pharmacy Practice," Lippincott Williams & Wilkins, 2008のような文献を参照し得る。
【0154】
開示される治療用組成物は更に、医薬的に許容可能な物質、組成物又は、液体又は固形フィラー剤(solid filler)、希釈剤、添加剤、溶剤又は封入材料(即ち担体)のような賦形剤(vehicle)を含む。これらの担体には、対象の組成物を1つの器官又は身体の領域から他の器官又は身体の領域に輸送することに関与する。各担体は、製剤の他の材料と適合するという意味で「許容可能」であるべきであり、患者に無害であるべきである。医薬的に許容可能な担体として役立つことができる材料のいくつかの例は、糖(ラクトース、グルコース及びスクロースなど)、澱粉(トウモロコシ澱粉、ジャガイモ澱粉など)、セルロース及びその誘導体(カルボキシメチルセルロースナトリウム、エチルセルロース及び酢酸セルロースなど)、粉末トラガカント、麦芽、ゼラチン、タルク、添加剤(ココアバター及び座剤ワックス(suppository wax)など)、油(落花生油、綿の実油、ヒマワリ油、ごま油、オリーブ油、トウモロコシ油及び大豆油など)、グリコール(プロピレングリコールなど)、ポリオール(グリセリン、ソルビトール、マンニトール及びポリエチレングリコールなど)、エステル(エチルオレアート及びエチルラウラートなど)、寒天、緩衝剤(水酸化マグネシウム及び水酸化アルミニウムなど)、アルギン酸、発熱性物質除去水(pyrogen-free water)、等張食塩水、リンガー溶液、エチルアルコール、リン酸緩衝溶液、及び、製剤処方に採用される他の非毒性適合物質を含む。着色剤、放出剤、コーティング剤、甘味料、香味料、芳香剤、保存料及び酸化防止剤と同様に、湿潤剤、乳化剤及びラウリル硫酸ナトリウム及びステアリン酸マグネシウムのような潤滑剤もまた、当該組成物に存在し得る。
【0155】
本願の開示の他の視点は、本願に開示されるような、リシル酸化酵素型酵素の触媒ドメイン又は、そのような触媒ドメインをエンコードする核酸の、対象物への投与を行うためのキットに関する。1つの実施形態において、キットは、医薬的な担体に製剤化されるリシル酸化酵素型酵素の触媒ドメイン(又は、リシル酸化酵素型酵素の触媒ドメインをエンコードする核酸)を含む。
【0156】
製剤化及び輸送の方法は、処置される障害の種類及び部位に応じて一般的に適合される。例となる製剤は、例えば、静脈投与、動脈内投与、眼球内投与、筋肉内投与、又は皮下投与のような、非経口投与に適したものを含むがこれらに限定されず、ミセル、リポソーム又は薬剤開放カプセルにカプセル化された製剤(即ち、遅延放出用にデザインされた生体適合性のコーティング内に組み込まれた活性剤);摂取可能な製剤;クリーム、軟膏及びゲルのような局所使用の製剤;及び、吸入剤、エアロゾル及びスプレーのような他の製剤を含む。開示の化合物の用量は、処置を必要とする広さ及び重症度、投与される組成物の活性、対象物の全体的な健康及び、当業者に良く知られる他の検討事項に応じて、変化するであろう。
【0157】
更なる実施形態において、本願に記載される組成物は、局所的に輸送(ないし送達)される。局所的輸送は、例えば、創傷領域又は繊維化領域への非全身的な組成物の輸送を可能にし、全身的な輸送と比較した当該組成物の身体負荷量を減少させる。そのような局所輸送は、例えば、ステント及びカテーテルを含むがこれに限定されない様々な医学的に埋め込まれる装置を介して実現され得、又は、注射又は手術によって実現され得る。コーティング、インプランティング、埋めこみのための方法及び、さもなければ、所望の薬剤の、ステント及びカテーテルのような医学的な装置への取り付け方法が、当該技術分野において確立され、本願において考えられる。
【0158】
インプラントされたステントが、血栓溶解剤のような医学的な薬剤を移送するために使用されている。米国特許番号5,163,952は、ステント自体の材料の中に医学的な薬剤を有するように構成される(formulated)熱的形状記憶性拡張プラスチックステント装置(thermal memoried expanding plastic stent device)を開示する。米国特許番号5,092,877は、化合物の輸送に関連するコーティングを有することができるポリマー材料のステントを開示する。バイオ分解可能なポリマー及びバイオ吸収可能なポリマーを利用する種類の装置に向けられる他の特許は、米国特許番号4,916,193及び米国特許番号4,994,071を含む。例として、米国特許番号5,304,121は、ヒドロゲルポリマー及び細胞増殖阻害剤又はヘパリンのような予選択された化合物からなるステントに適用されるコーティングを開示する。治療用材料を有する、コートされた血管内ステントの作成方法が、米国特許番号5,464,650に記載され、ここでは、ポリマーコーティング材料は、溶媒に溶解され、治療用材料が、溶媒に分散している。溶媒は、続いて、ステントと接触の後蒸発される。
【0159】
米国特許番号6,120,536は、ステントを含む広範囲な種類の人工装具で使用する、更なるコーティングの種類を記載する。本願で記載される組成物の投与に役立つ更なる医学的な装置又は人工の装置の例は、血液交換装置、血管アクセスポート、中心静脈カテーテル、心血管カテーテル、体外循環回路、代用血管、ポンプ、心臓弁、及び心血管縫合糸を含むがこれに限定されない。
【0160】
ステント及びカテーテルを含む本願で記載される組成物でコートされた装置の使用は、組成物の、特異的な部位又は局在される部位への輸送を可能にする。そのような部位特異的輸送は、組成物の使用(及び/又はその用量)のための手段を提供し得、これは、そうでなければ溶解性、全身性の毒性の懸念又は他の問題をもたらす全身性の輸送の影響を受けにくい。例として、β‐アミノプロピオニトリル(BAPN)が、リシル酸化酵素型酵素の阻害剤として役立つことが知られているが、この化合物は、高い毒性があり、全身的に投与される場合、その効果的な使用に関して問題がある。活性薬剤又は、BAPNのような化合物の輸送のためのステント、カテーテル又は他の医学的な装置の使用は、目標とされる態様又は局所的な態様における効果的な用量の化合物の使用を許容し、かくて、そのような化合物と関連する全身性毒性効果を減少させる。
【0161】
[使用(形態)]
本願で開示されるポリペプチドは、いくつかの使用(形態ないし用途)を有する。例えば、これらは、リシル酸化酵素の酵素的活性(例えば、コラーゲン架橋結合)を分析するための標準品(例えば、陽性対照)として使用され得る。そのような分析の1つの使用(形態)としては、例えばLOX又はLOXL2タンパク質及び/又はその遺伝子の突然変異解析がある。リシル酸化酵素活性を有する触媒ドメインを含むポリペプチドはまた、リシル酸化酵素型酵素に、新しい抗体及びすでに存在する抗体を特徴付けるためにも使用され得る。
【0162】
本願で開示されるポリペプチドはまた、例えばLOX及び/又はLOXL2触媒ドメイン、小有機分子又は高分子(例えばポリペプチド)に結合する分子を同定するためにも使用され得る。これらの結合分子の或る特定のものは、触媒活性化のモジュレーター(即ち、活性化剤又は阻害剤)として機能し得る。
【0163】
本願で開示される対象のポリペプチドはまた、ヒト及びマウスLOX及びLOXL2タンパク質及び他のリシル酸化酵素型酵素の触媒的な活性領域に特異的に向けられる抗体の作成にも役立つ。そのような抗体は、例えば、リシル酸化酵素活性の病的に増加するレベルによって特徴付けられる疾病又は障害に関する医学的及び/又は治療的な応用に使用され得る。
【0164】
加えて、対象のポリペプチドは、コラーゲンの架橋結合を要求する医学的及び/又は治療的な応用に役立つ(例えば、創傷治癒及び糖尿病性潰瘍の処置)。(全長のタンパク質と比較して)より小さい大きさの単離された触媒ドメインは、例えば、改良された安定性及びより迅速な吸収によって、治療剤として、その使用を促進し得る。
【実施例】
【0165】
[実施例1:ヒトLOX触媒ドメイン]
ヒトLOXタンパク質の触媒ドメインを含むポリペプチドは、207アミノ酸残基を含み、次のアミノ酸配列を有する。
GLPDLVADPYYIQASTYVQKMSMYNLRCAAEENCLASTAYRADVRDYDHRVLLRFPQRVKNQGTSDFLPSRPRYSWEWHSCHQHYHSMDEFSHYDLLDANTQRRVAEGHKASFCLEDTSCDYGYHRRFACTAHTQGLSPGCYDTYGADIDCQWIDITDVKPGNYILKVSVNPSYLVPESDYTNNVVRCDIRYTGHHAYASGCTISPY
(SEQ ID NO:1)
【0166】
ある実施形態において、ヒトLOX触媒ドメインを含むポリペプチドは、上に表された配列における最初のグリシン及びロイシン残基(”GL”)を含まない。
【0167】
LOX触媒ドメインを含む対象のポリペプチドをエンコードするヒトDNA配列は、以下の通りである。
GGTCTCCCAGACCTGGTGGCCGACCCCTACTACATCCAGGCGTCCACGTACGTGCAGAAGATGTCCATGTACAACCTGAGATGCGCGGCGGAGGAAAACTGTCTGGCCAGTACAGCATACAGGGCAGATGTCAGAGATTATGATCACAGGGTGCTGCTCAGATTTCCCCAAAGAGTGAAAAACCAAGGGACATCAGATTTCTTACCCAGCCGACCAAGATATTCCTGGGAATGGCACAGTTGTCATCAACATTACCACAGTATGGATGAGTTTAGCCACTATGACCTGCTTGATGCCAACACCCAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGTTTCTGTCTTGAAGACACATCCTGTGACTATGGCTACCACAGGCGATTTGCATGTACTGCACACACACAGGGATTGAGTCCTGGCTGTTATGATACCTATGGTGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTAAAACCTGGAAACTATATCCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTTCCTGAATCTGACTATACCAACAATGTTGTGCGCTGTGACATTCGCTACACAGGACATCATGCGTATGCCTCAGGCTGCACAATTTCACCGTATTAG
(SEQ ID NO:2)
【0168】
[実施例2:ヒトLOXL2触媒ドメイン]
ヒトLOXL2タンパク質の触媒ドメインを含むポリペプチドは、次のアミノ酸配列を有する。
TAPDLVLNAEMVQQTTYLEDRPMFMLQCAMEENCLSASAAQTDPTTGYRRLLRFSSQIHNNGQSDFRPKNGRHAWIWHDCHRHYHSMEVFTHYDLLNLNGTKVAEGHKASFCLEDTECEGDIQKNYECANFGDQGITMGCWDMYRHDIDCQWVDITDVPPGDYLFQVVINPNFEVAESDYSNNIMKCRSRYDGHRIWMYNCHIGGSFSEETEKKFEHFSGLLNNQLSPQ
(SEQ ID NO:3)
【0169】
ある実施形態において、ヒトLOXL2触媒ドメインを含むポリペプチドは、上に表された配列における最初のスレオニン及びアラニン残基(”TA”)を含まない。
【0170】
LOXL2触媒ドメインをエンコードするヒトDNA配列は、以下の通りである。
ACCGCCCCTGACCTGGTCCTCAATGCGGAGATGGTGCAGCAGACCACCTACCTGGAGGACCGGCCCATGTTCATGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCGGCCTCAGCCGCGCAGACCGACCCCACCACGGGCTACCGCCGGCTCCTGCGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCCGACTTCCGGCCCAAGAACGGCCGCCACGCGTGGATCTGGCACGACTGTCACAGGCACTACCACAGCATGGAGGTGTTCACCCACTATGACCTGCTGAACCTCAATGGCACCAAGGTGGCAGAGGGCCACAAGGCCAGCTTCTGCTTGGAGGACACAGAATGTGAAGGAGACATCCAGAAGAATTACGAGTGTGCCAACTTCGGCGATCAGGGCATCACCATGGGCTGCTGGGACATGTACCGCCATGACATCGACTGCCAGTGGGTTGACATCACTGACGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTTATTAACCCCAACTTCGAGGTTGCAGAATCCGATTACTCCAACAACATCATGAAATGCAGGAGCCGCTATGACGGCCACCGCATCTGGATGTACAACTGCCACATAGGTGGTTCCTTCAGCGAAGAGACGGAAAAAAAGTTTGAGCACTTCAGCGGGCTCTTAAACAACCAGCTGTCCCCGCAGTAA
(SEQ ID NO:4)
【0171】
[実施例3:マウスLOX触媒ドメイン]
マウスLOXタンパク質の触媒ドメインを含むポリペプチドは、次のアミノ酸配列を有する。
GLPDLVPDPYYIQASTYVQKMSMYNLRCAAEENCLASSAYRADVRDYDHRVLLRFPQRVKNQGTSDFLPSRPRYSWEWHSCHQHYHSMDEFSHYDLLDANTQRRVAEGHKASFCLEDTSCDYGYHRRFACTAHTQGLSPGCYDTYAADIDCQWIDITDVQPGNYILKVSVNPSYLVPESDYTNNVVRCDIRYTGHHAYASGCTISPY
(SEQ ID NO:5)
【0172】
ある実施形態において、マウスLOX触媒ドメインを含むポリペプチドは、上に表された配列における最初のグリシン及びロイシン残基(”GL”)を含まない。
【0173】
LOX触媒ドメインを含むポリペプチドをエンコードするマウスDNA配列は、以下の通りである。
GGTCTCCCGGACCTGGTGCCCGACCCCTACTACATCCAGGCTTCCACGTACGTCCAGAAGATGTCTATGTACAACCTGAGATGCGCTGCGGAAGAAAACTGCCTGGCCAGTTCAGCATATAGGGCGGATGTCAGAGACTATGACCACAGGGTACTGCTACGATTTCCGCAAAGAGTGAAGAACCAAGGGACATCGGACTTCTTACCAAGCCGCCCTCGGTACTCCTGGGAGTGGCACAGCTGTCACCAACATTACCACAGCATGGACGAATTCAGCCACTATGACCTGCTTGATGCCAACACACAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGCTTCTGTCTGGAGGACACGTCCTGTGACTATGGGTACCACAGGCGCTTTGCGTGCACTGCACACACACAGGGATTGAGTCCTGGATGTTATGACACCTATGCGGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTACAACCTGGAAACTACATTCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTGCCTGAATCAGACTACACTAACAATGTTGTACGCTGTGACATTCGCTACACAGGACATCATGCCTATGCCTCAGGCTGCACAATTTCACCGTATTAG
(SEQ ID NO:6)
【0174】
[実施例4:マウスLOXL2触媒ドメイン]
マウスLOXL2タンパク質の触媒ドメインを含むポリペプチドは、次のアミノ酸配列を有する。
TAPDLVLNAEIVQQTAYLEDRPMSLLQCAMEENCLSASAVHTDPTRGHRRLLRFSSQIHNNGQSDFRPKNGRHAWIWHDCHRHYHSMEVFTYYDLLSLNGTKVAEGHKASFCLEDTECEGDIQKSYECANFGEQGITMGCWDMYRHDIDCQWIDITDVPPGDYLFQVVINPNYEVPESDFSNNIMKCRSRYDGYRIWMYNCHVGGAFSEETEQKFEHFSGLLNNQLSVQ
(SEQ ID NO:7)
【0175】
ある実施形態において、マウスLOXL2触媒ドメインを含むポリペプチドは、上に表された配列における最初のスレオニン及びアラニン残基(”TA”)を含まない。
【0176】
LOXL2触媒ドメインを含むポリペプチドをエンコードするマウスDNA配列は以下の通りである。
ACTGCACCTGACCTGGTGCTTAATGCTGAGATTGTCCAGCAGACTGCCTACCTGGAGGACAGGCCCATGTCCTTGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCCGCCTCCGCTGTGCACACCGACCCCACCAGAGGCCACCGGCGCCTTTTACGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCTGACTTCCGCCCCAAGAATGGCCGCCATGCGTGGATTTGGCACGACTGCCACAGGCACTACCACAGCATGGAAGTCTTCACTTACTATGACCTGCTGAGCCTCAACGGCACCAAGGTGGCTGAGGGCCACAAGGCCAGCTTCTGCCTGGAGGACACTGAGTGTGAGGGAGACATTCAGAAGAGTTACGAGTGTGCCAACTTTGGAGAACAAGGCATCACCATGGGCTGCTGGGACATGTACCGTCATGACATTGACTGCCAGTGGATAGACATCACCGATGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTCATTAACCCCAACTATGAAGTGCCAGAATCAGATTTCTCTAACAACATCATGAAGTGCAGGAGCCGCTATGATGGCTACCGCATCTGGATGTACAACTGTCACGTAGGTGGAGCCTTCAGTGAGGAGACAGAACAGAAGTTCGAACACTTCAGTGGACTTCTAAATAACCAGCTCTCTGTACAGTAA
(SEQ ID NO:8)
【0177】
[実施例5:哺乳類細胞におけるヒトLOX触媒ドメインの発現]
ヒトLOX触媒ドメインを含むポリペプチドの発現に使用する発現カセットを、ヒトLOX cDNAを鋳型として使用してPCR増幅により構成した(GenBank NM_002317、Genecopoeia, Germantown, MDから入手した)。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro B ベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−hLOX MCD(以下、phLOXMCDとも称する)プラスミドを作成した。T7RNAポリメラーゼを使用したこのクローンの転写により、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、ヒトLOX触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列を有する。
(SEQ ID NO:9)
シグナルペプチド及びmycタグの配列には、下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQP及びGP)及びリンカー配列(NSAVD)の配列でエンコードされるアミノ酸配列を表す。
【0178】
このポリペプチド(これは、シグナル配列及びmycタグをエンコードするヌクレオチド示す下線が引かれた配列、及び小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccGGTCTCCCAGACCTGGTGGCCGACCCCTACTACATCCAGGCGTCCACGTACGTGCAGAAGATGTCCATGTACAACCTGAGATGCGCGGCGGAGGAAAACTGTCTGGCCAGTACAGCATACAGGGCAGATGTCAGAGATTATGATCACAGGGTGCTGCTCAGATTTCCCCAAAGAGTGAAAAACCAAGGGACATCAGATTTCTTACCCAGCCGACCAAGATATTCCTGGGAATGGCACAGTTGTCATCAACATTACCACAGTATGGATGAGTTTAGCCACTATGACCTGCTTGATGCCAACACCCAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGTTTCTGTCTTGAAGACACATCCTGTGACTATGGCTACCACAGGCGATTTGCATGTACTGCACACACACAGGGATTGAGTCCTGGCTGTTATGATACCTATGGTGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTAAAACCTGGAAACTATATCCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTTCCTGAATCTGACTATACCAACAATGTTGTGCGCTGTGACATTCGCTACACAGGACATCATGCGTATGCCTCAGGCTGCACAATTTCACCGTATgggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:10)
【0179】
phLOXMCD発現ベクターを、次のように、HEK293細胞にトランスフェクトした。7×105個の細胞を、6ウェル培養皿の1つのウェル(穴)にプレート(plate)し、37℃、5%CO2の環境下で、完全DMEM(Dulbecco's Modified Eagle's Medium+10%ウシ胎児血清+2mMのL−グルタミン)で増殖させた。終夜増殖の後、細胞に、Lipofectamine 2000(Invitrogen, Carlsbad, CA)を使用して、製造者の指示に従い、phLOXMCDをトランスフェクトした。4時間後、培地を吸引し、2mLの完全DMEMで交換し、トランスフェクト細胞を終夜で増殖させた。安定な細胞系統を、完全DMEM(cDMEM)+0.8mg/mLのHygromycin B(hygro)で、限界希釈(limiting dilution)により選択した。選択に残った個々のクローンを拡張し、hLOX触媒ドメインの発現に関するスクリーニングを次のように行った。拡張を、6ウェル培養皿中で細胞クローンを75%コンフルエンスな(confluence)状態まで増殖させることにより達成した。この時点で、培地を吸引し、新鮮なcDMEM+hygroを添加した。5日後、培地をハーベスト(harvest)し、以下に記載されるように、LOX触媒ドメインの存在を直接的に解析した。PBS+5mMのEDTAを培養皿に添加することにより細胞を培養皿の表面から取り外した。分離した細胞液(cell solution)を遠心分離に供した。約5×105個のペレット化した細胞(総計〜1×106個の細胞から得た)を0.1mLのM‐PER mammalian protein extraction reagent(Pierce, Rockford, IL)に溶解した。氷上での20分間のインキュベーションの後、細胞溶解物を、卓上遠心器で15000rpm、10分間の条件で遠心分離に供した。可溶性分画(上清)を回収し、以下に記載されるように、LOX触媒ドメインの存在を解析した。不溶性分画(ペレット)を、2回、M‐PERで洗浄し、M‐PERに再懸濁し、以下に記載されるように、LOX触媒ドメインの存在を解析した。
【0180】
[実施例6:タンパク質免疫ブロッティングによるヒトLOX触媒ドメインの検出]
6つの異なったクローンからの培地、可溶性分画及び不溶性分画の試料を、phLOXMCD発現ベクターにより発現されたLOX触媒ドメインの存在を(確認するために)免疫ブロッティングにより分析した。培地の分析に関して、150μLの馴化培地(conditioned medium)を50μLのNuPAGE(登録商標)4×LDS試料緩衝液(Invitrogen, Carlsbad, CA)と混合し、その結果得られた20μLの混合液をゲルの上にロードした(以下を参照)。可溶性及び不溶性の細胞内分画に関して、各試料は、1×105個の細胞に相当する。細胞内分画の試料を、NuPAGE(登録商標)、4×LDS試料緩衝液(Invitrogen, Carlsbad, CA)とNuPAGE(登録商標)10×還元剤とを混合し、これら両薬剤の終濃度が1倍(1×)の濃度になるようにし、5分間煮沸し、そして、NuPAGE(登録商標)Novex Bis−Trisゲル(4%‐12%アクリルアミド)を使用して、電気泳動に供した。ランニング緩衝液は、NuPAGE(登録商標)MOPS緩衝液(Invitrogen, Carlsbad, CA)であった。電気泳動の終了時に、タンパク質をゲルから、iBlot装置(Invitrogen, Carlsbad, CA)を使用してPVDF上に転写した。
【0181】
検出に関して、ブロットを、3%ウシ胎児血清(BSA)入りPBS+0.01% Tweenを使用して、3時間、室温でブロックした。続いて、ブロットをマウス抗His5抗体(Qiagen, Valencia, CA)と共に、1時間、室温で、インキュベートし、次にセイヨウワサビペルオキシダーゼ(HRP)コンジュゲートロバ抗マウス抗体(Jackson ImmunoResearch, West Grove, PA)と共に、1時間、室温でインキュベートし、Chemi−Glow(登録商標)(Alpha Innotech, San Leandro, CA)を使用して、発色させた(develop)。
【0182】
6つのクローン(23から28の番号で標識される)に関する結果を、図1に表す。増殖培地の分析を表した最も左のパネルは、クローン23における少量の分泌産物(矢印)を除いて、触媒ドメインの分泌が皆無か、それに近いことを示す。分泌タンパク質は、プロセスされていないポリペプチドの特徴である、明瞭な29kDの分子量を表す。これは、シグナルペプチドの切断の欠如(又は不完全な切断)に起因しているかも知れず、又は、分泌ポリペプチドの翻訳後修飾に起因しているかも知れない。
【0183】
phLOXMCD発現ベクターが一時的に(transiently)トランスフェクトされているHEK293細胞からの増殖培地のトリクロロ酢酸沈殿は、増殖培地における不溶性LOX触媒ドメインの存在を明らかにした。
【0184】
触媒ドメインの細胞内発現が、6つのクローン中、5つのクローンにおいて、可溶性及び不溶性分画の両方で検出された(図1、中央及び右のパネル)。加えて、高分子量の集合体(aggregates)が、不溶性細胞内分画において検出された。
【0185】
可溶性(であること)は、正しく折りたたまれ、プロセスされたポリペプチドであることの指標でありうる以上、可溶性細胞内ポリペプチド産物の存在は、適切に折りたたまれ、酵素的に活性な触媒ドメインが、トランスフェクト細胞内で生成されている可能性を示す。
【0186】
HEK293細胞に加えて、LOX触媒ドメインの発現が、更なる細胞種において観察された。CHOk1細胞におけるphLOXMCD発現ベクターの一時的な発現は、触媒ドメインの細胞内発現(これは、細胞溶解物全体の中に検出される)をもたらしたが、触媒ドメインの分泌は、検出されなかった。SW620細胞の一時的なトランスフェクションもまた、検出可能な分泌なしでの、細胞内発現をもたらした。
【0187】
[実施例7:哺乳類細胞におけるヒトLOXL2触媒ドメインの発現]
ヒトLOXL2触媒ドメインを含むポリペプチドの発現に使用する発現カセット(Genecopoeia, Germantown, MD)を、ヒトLOXL2 cDNAを鋳型として使用してPCR増幅により構成した。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro Bベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−hLOXL2 MCD(以下、phLOXL2MCDとも称する)発現ベクターを作成した。T7RNAポリメラーゼを使用したこのクローンの転写は、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、ヒトLOXL2触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列を有する。
(SEQ ID NO:11)
【0188】
シグナルペプチド及びmycタグの配列は下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQPARRARRTKL及びSRGGP)及びリンカー配列(NSAVD)でエンコードされるアミノ酸配列を表す。
【0189】
このポリペプチド(これは、リーダー(leader)及び、mycタグをエンコードする配列を示す下線が引かれた配列及び、小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccaggcgcgcgcgccgtacgaagcttACCGCCCCTGACCTGGTCCTCAATGCGGAGATGGTGCAGCAGACCACCTACCTGGAGGACCGGCCCATGTTCATGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCGGCCTCAGCCGCGCAGACCGACCCCACCACGGGCTACCGCCGGCTCCTGCGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCCGACTTCCGGCCCAAGAACGGCCGCCACGCGTGGATCTGGCACGACTGTCACAGGCACTACCACAGCATGGAGGTGTTCACCCACTATGACCTGCTGAACCTCAATGGCACCAAGGTGGCAGAGGGCCACAAGGCCAGCTTCTGCTTGGAGGACACAGAATGTGAAGGAGACATCCAGAAGAATTACGAGTGTGCCAACTTCGGCGATCAGGGCATCACCATGGGCTGCTGGGACATGTACCGCCATGACATCGACTGCCAGTGGGTTGACATCACTGACGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTTATTAACCCCAACTTCGAGGTTGCAGAATCCGATTACTCCAACAACATCATGAAATGCAGGAGCCGCTATGACGGCCACCGCATCTGGATGTACAACTGCCACATAGGTGGTTCCTTCAGCGAAGAGACGGAAAAAAAGTTTGAGCACTTCAGCGGGCTCTTAAACAACCAGCTGTCCCCGCAGtctcgaggagggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:12)
【0190】
phLOXL2MCD発現ベクターを、HEK293細胞にトランスフェクトし、安定な細胞系統を、限界希釈により選択し、そしてクローンを、phLOXMCDベクターについての実施例5に記載された方法と同じ方法を使用して拡張した。培地を分析のために回収し、細胞を分離し、実施例5に記載されたようにペレット化した。
【0191】
[実施例8:タンパク質免疫ブロッティングによるヒトLOXL2触媒ドメインの検出]
約5×105個の細胞を含むペレット化した細胞の試料(実施例7に記載されるようにして得たもの)を免疫ブロッティングにより解析した。約5×105個の細胞を含む細胞懸濁液を、6000rpm、10分間の条件で遠心分離に供し、上清を吸引した。ペレットを100μLのM−PER溶解緩衝液に再懸濁し、次に、氷上で20分間インキュベーションした。不溶性細胞残渣を、15000rpm、20分間の遠心分離により取り出し、75μLの上清(澄んだ細胞溶解物)を取り出し、30μLのNuPAGE(登録商標)4×LDS試料緩衝液(Invitrogen, Carlsbad, CA)と10μLのNuPAGE(登録商標)10×還元剤に混合した。混合物を5分間煮沸し、続いて、ゲルに適用するまで氷上に置いた。典型的に、20μLの各試料(〜1×105個の細胞に相当する)をNuPAGE(登録商標)Novex Bis−Trisゲル(4%−12%アクリルアミド)で電気泳動に供した。ランニング緩衝液は、NuPAGE(登録商標)MOPS緩衝液(Invitrogen, Carlsbad, CA)であった。電気泳動の終了時に、タンパク質をゲルから、iBlot装置(Invitrogen, Carlsbad, CA)を使用してPVDF上に転写した。
【0192】
検出に関して、ブロットを、phLOXMCD発現産物についての実施例6に記載されるように処理(プロセス)した。
【0193】
図2の左のパネルに表されるように、結果は、9つの異なったクローンにおけるLOXL2触媒ドメインの細胞内発現を示す。タンパク質の単量体の形態及び集合体(aggregated)の形態の両方が、検出された(単量体の形態が支配的であった(predominating))。
【0194】
ヒトLOXL2触媒ドメインが分泌されたかどうかを確認するために、phLOXMCD−トランスフェクト細胞についての実施例7に記載されるように、細胞クローンを得て、cDMEM+hygroで90分間増殖させ、この時点で、15μLの培地を取り除き、5μLの4×LDS試料緩衝液+還元剤(NuPAGE(登録商標)、Invitrogen, Carlsbad, CA)に混合し、5分間煮沸し、ポリアクリルアミドゲル電気泳動に供し、全細胞溶解物についての上記の方法により解析した。図2の右のパネルに表される結果は、ヒトLOX触媒ドメインと対照的に、ヒトLOXL2触媒ドメインは、無濃縮増殖培地で検出可能なほど大量にHEK293細胞により分泌されたことを表す。
【0195】
[実施例9:哺乳類細胞におけるマウスLOX触媒ドメインの発現]
マウスLOX触媒ドメインを含むポリペプチドの発現に使用する発現カセットを、マウスLOX cDNAを鋳型として使用してPCR増幅により構成した(GenBank BC018439、Invitrogen, Carlsbad, CAから入手した)。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro B ベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−mLOX MCD(以下、pmLOXMCDとも称する)発現ベクターを作成した。T7RNAポリメラーゼを使用したこのクローンの転写は、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、マウスLOX触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列を有する。
(SEQ ID NO:13)
シグナルペプチド及びmycタグの配列は下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQP及びGP)及びリンカー配列(NSAVD)の配列でエンコードされるアミノ酸配列を表す。
【0196】
このポリペプチド(これは、リーダー及びmycタグをエンコードする配列を示す下線が引かれた配列、及び小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccGGTCTCCCGGACCTGGTGCCCGACCCCTACTACATCCAGGCTTCCACGTACGTCCAGAAGATGTCTATGTACAACCTGAGATGCGCTGCGGAAGAAAACTGCCTGGCCAGTTCAGCATATAGGGCGGATGTCAGAGACTATGACCACAGGGTACTGCTACGATTTCCGCAAAGAGTGAAGAACCAAGGGACATCGGACTTCTTACCAAGCCGCCCTCGGTACTCCTGGGAGTGGCACAGCTGTCACCAACATTACCACAGCATGGACGAATTCAGCCACTATGACCTGCTTGATGCCAACACACAGAGGAGAGTGGCTGAAGGCCACAAAGCAAGCTTCTGTCTGGAGGACACGTCCTGTGACTATGGGTACCACAGGCGCTTTGCGTGCACTGCACACACACAGGGATTGAGTCCTGGATGTTATGACACCTATGCGGCAGACATAGACTGCCAGTGGATTGATATTACAGATGTACAACCTGGAAACTACATTCTAAAGGTCAGTGTAAACCCCAGCTACCTGGTGCCTGAATCAGACTACACTAACAATGTTGTACGCTGTGACATTCGCTACACAGGACATCATGCCTATGCCTCAGGCTGCACAATTTCACCGTATgggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:14)
【0197】
pmLOXMCD発現ベクターを、HEK293細胞にトランスフェクトし、安定な細胞系統を、限界希釈により選択し、そしてクローンを、phLOXMCDベクターについての実施例5に記載された方法と同じ方法を使用して拡張した。培地を解析のために回収し、細胞を取り出し、実施例5に記載されるようにペレット化し、及び可溶性分各及び不溶性分画に分画した。
【0198】
マウスLOX触媒ドメインの発現を、ヒト触媒ドメインの解析についての実施例6に記載された方法と同じ方法を使用して、免疫ブロッティングにより分析した。図3に表される結果は、マウスLOX触媒ドメインが、ヒトLOX触媒ドメインとは対照的に、検出可能な量で分泌されることを示す(左のパネル)。mLOX触媒ドメインの細胞内発現プロファイルは、hLOX触媒ドメインのそれと、類似しており、可溶性及び不溶性分画の両方に単量体タンパク質を有し、不溶性分画において、集合体が存在する。
【0199】
[実施例10:哺乳類細胞からのヒトLOX触媒ドメインの分泌]
図4は、ヒトLOX触媒ドメインのアミノ酸配列(上)及びマウスLOX触媒ドメインのアミノ酸配列(下)の違いを表す模式図を含む。図に示すように、2つの配列は、4つの位置のみで異なる。上で示されるように、及び、図4の左のパネルで確認されるように、10倍濃縮された馴化増殖培地(10×CM)を使用すると、マウスLOX触媒ドメイン(M)をエンコードするベクターをトランスフェクトした細胞から、マウスLOX触媒ドメイン(M)が分泌されたが、一方で、ヒトLOX触媒ドメイン(H)は、分泌されなかった。それ故、ヒトLOX触媒ドメインの分泌を望むのであれば、1つの方法として、7、38、146又は160番目の残基の何れかを、ヒト配列からマウス配列へ変更することが可能であり(番号は、本願で開示されるphLOXMCD及びpmLOXMCD発現ベクターにおいて発現する触媒ドメインのアミノ酸配列における位置を意味する)、又は、これらの残基の任意の組み合わせ又はこれら残基の4つ全てをヒト配列からマウス配列へ変更することも可能である。例えば、次のアミノ酸配列の任意のもの又はその全て、又は任意の組み合わせ、の変更が可能である(A7P、T38S、G146A、K160Q)。
【0200】
反対に、マウスLOX触媒ドメインの分泌を減少させることを望み、かくて、このポリペプチドを細胞内により多く留めるのであれば、7、38、146及び160番目の残基をマウス配列からヒト配列に変更することが可能である。従って、例えば、次のアミノ酸配列の任意のもの又はその全て、又は任意の組み合わせ、の変更が可能である(P7A、S38T、A146G、Q160K)。
【0201】
[実施例11:哺乳類細胞におけるマウスLOXL2触媒ドメインの発現]
マウスLOXL2触媒ドメインを含むポリペプチドの発現に使用する発現カセットを、マウスLOXL2 cDNAを鋳型として使用してPCR増幅により構成する(GenBank NM033325を元にして商業的に合成される)。触媒ドメインをエンコードする配列を含む増幅産物を、pSecTag2/Hygro B ベクター(Invitrogen, Carlsbad, CA)にクローン化し、pSecTag2hygro−mLOXL2 MCD(以下、pmLOXL2MCDとも称する)発現ベクターを作成する。T7RNAポリメラーゼを使用したこのクローンの転写は、(N末端側から、C末端側へ順に)免疫グロブリンカッパシグナル配列、マウスLOXL2触媒ドメイン、mycエピトープタグ及びHis6精製タグを含むポリペプチドをエンコードするmRNAを作成する。このポリペプチドは、次のアミノ酸配列(これは、下線が引かれたリーダー配列及びmycタグを有する)を有する。
(SEQ ID NO:15)
【0202】
シグナルペプチド及びmycタグの配列は下線が引かれている。イタリック体で表される配列は、ベクター配列における全部又は一部及びクローニングに使用する制限部位(AAQP及びGP)及びリンカー配列(NSAVD)の配列でエンコードされるアミノ酸配列を表す。
【0203】
このポリペプチド(これは、リーダー及び、mycタグをエンコードする配列を示す下線が引かれた配列及び、小文字形で示されたリンカー配列を有する)をエンコードするDNA配列は以下の通りである。
ATGGAGACAGACACACTCCTGCTATGGGTACTGCTGCTCTGGGTTCCAGGTTCCACTGGTGACgcggcccagccggccACTGCACCTGACCTGGTGCTTAATGCTGAGATTGTCCAGCAGACTGCCTACCTGGAGGACAGGCCCATGTCCTTGCTGCAGTGTGCCATGGAGGAGAACTGCCTCTCCGCCTCCGCTGTGCACACCGACCCCACCAGAGGCCACCGGCGCCTTTTACGCTTCTCCTCCCAGATCCACAACAATGGCCAGTCTGACTTCCGCCCCAAGAATGGCCGCCATGCGTGGATTTGGCACGACTGCCACAGGCACTACCACAGCATGGAAGTCTTCACTTACTATGACCTGCTGAGCCTCAACGGCACCAAGGTGGCTGAGGGCCACAAGGCCAGCTTCTGCCTGGAGGACACTGAGTGTGAGGGAGACATTCAGAAGAGTTACGAGTGTGCCAACTTTGGAGAACAAGGCATCACCATGGGCTGCTGGGACATGTACCGTCATGACATTGACTGCCAGTGGATAGACATCACCGATGTGCCCCCTGGAGACTACCTGTTCCAGGTTGTCATTAACCCCAACTATGAAGTGCCAGAATCAGATTTCTCTAACAACATCATGAAGTGCAGGAGCCGCTATGATGGCTACCGCATCTGGATGTACAACTGTCACGTAGGTGGAGCCTTCAGTGAGGAGACAGAACAGAAGTTCGAACACTTCAGTGGACTTCTAAATAACCAGCTCTCTGTACAGgggcccGAACAAAAACTCATCTCAGAAGAGGATCTGaatagcgccgtcgacCATCATCATCATCATCATTGA
(SEQ ID NO:16)
【0204】
pmLOXL2MCD発現ベクターを、HEK293細胞にトランスフェクトし、安定な細胞系統を、限界希釈により選択し、そしてクローンを、phLOXMCDベクターについての実施例5に記載される方法と同じ方法を使用して拡張した。培地を解析のために回収し、細胞を取り出し、実施例5に記載されるように、ペレット化し及び可溶性分画及び不溶性分画に分画した。
【0205】
マウスLOXL2−由来のポリペプチドの発現を、ヒト触媒ドメインの解析についての実施例6に記載される方法と同じ方法を使用して、免疫ブロッティングにより分析した。
【0206】
[実施例12:一時的にトランスフェクトした細胞からの、酵素的に活性なヒトLOXL2触媒ドメインの発生]
293F細胞(これは、懸濁培養中で増殖するように適合されたHEK293細胞のサブクローンである、Invitrogen, Carlsbad, CA)を、製造者のプロトコルに従って、スピナーフラスコ中で増殖させた。一旦、培養液(の密度)を0.8−1.2×106個細胞/mLの密度まで到達させ、細胞培養液をCB22バッグ(WAVE, GE Health, Piscataway, NJ)に移した。293fectin reagent(Invitrogen, Carlsbad, CA)を使用して8リッターの細胞培養液に8mgのphLOXL2ベクターをトランスフェクトした。トランスフェクト細胞を、3日間、CB22バッグ内で増殖させ、その後、細胞培養液を、Allegra6Rベンチトップ遠心器(Beckman Coulter)で、スイングバケットローターを使用し、3000rpmで10分間、遠心分離に供した。上清を回収し、0.22mmのPES膜を介してフィルターろ過した。
【0207】
LOXL2由来ポリペプチドを、Ni−セファロース樹脂(GE Healthcare, Piscataway, NJ)によって、細胞培養液上清から精製した。樹脂を0.1MのTris‐Cl(pH8.0)を使用して平衡化した。馴化培地(即ち、細胞培養上清)を平衡化した樹脂にロードし、続いて、カラムを、10カラム容積の0.1MのTris‐Cl(pH8.0)、0.25Mの塩化ナトリウム、0.02Mのイミダゾールを使用して洗浄した。Hisタグ化された触媒ドメインを、5カラム容積の0.1MのTris‐Cl(pH8.0)、0.15Mの塩化ナトリウム、0.3Mのイミダゾールを使用して溶出した。溶出液は、Amicon Ultra 15 10kD MWCL(Millipore, Billerica, MA)で濃縮し、その濃縮された材料を0.05Mのホウ酸塩に対して、pH8.0で、4℃で終夜で透析した。試料を、SDSポリアクリルアミドゲル(4−12%勾配、Bis−Trisゲル、Invitrogen, Carlsbad, CA)で、還元条件で解析した。
【0208】
[実施例13:ヒトLOXL2の触媒ドメインの酵素的活性]
ヒトLOXL2の触媒ドメインを含むポリペプチドを、上記の実施例12に記載されたように生成した。酵素的活性は、過酸化水素の生成(1,5−ジアミノペンタンの脱アミノ化の後、LOXL2によって遊離させた過酸化水素)を、Amplex(登録商標)redの蛍光産物(レゾルフィン)へのHRP−触媒変換(HRP-catalyzed conversion)と組み合わせた生化学的な分析を使用して解析した(Palamakumbura et al. (2002) Anal. Biochem. 300:245-251)。反応プレートは、Corning社から入手した。Amplex(登録商標)Red reagentは、Invitrogen社(Carlsbad, CA)から入手した。セイヨウワサビペルオキシダーゼ(HRP)、1,5−ジアミノペンタン及び消泡剤は、Sigma社(St. Louis, MO)から入手した。他の薬剤の全ては、入手できる最高の品質のものであった。
【0209】
酵素混合物は、10μLのプールされたピーク分画(実施例12参照)を40μLの分析溶液(62.5mMのホウ酸ナトリウム(pH8.0)、5ユニット/mLのHRP、10ppmの消泡剤)に加えることによって、作成した。基質溶液は、50mMのホウ酸ナトリウム、100μMのAmplex(登録商標)red reagent、20mMの1,5−ジアミノペンタン、10ppmの消泡剤を含んでいた。反応を、50μLの酵素混合物を50μLの基質溶液と混合することによって開始した。1つの反応物はまた、2mMのβAPN(ベータ‐アミノプロプリオニトリル、これは、リシル酸化酵素及びLOXL2の触媒活性の阻害剤である)を含んでいた。反応混合物を、37℃で、1時間、蛍光(ex=544nm、em=590nm)を測定するように作られたMolecularDevices M5プレートリーダーを使用して、反応速度モード(kinetics mode)で、インキュベートした。データを、時間に対する蛍光応答の勾配として記録した。
【0210】
図5は、レゾルフィン生成の時間経過解析の形で、相対蛍光ユニット(RFU)で表される、この分析の結果を表す。ヒトLOXL2由来ポリペプチドは、測定可能な酵素的活性を示し、そしてこの活性は、βAPNによって阻害された。従って、ポリペプチドは、LOXL2の酵素的に活性な触媒ドメインを含む。
【特許請求の範囲】
【請求項1】
ヒトLOX(SEQ ID NO:1)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、ヒトLOXの触媒ドメインの外側に位置するヒトLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項2】
請求項1のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項3】
SEQ ID NO:2において記載されるヌクレオチド配列を有する請求項2のポリヌクレオチド。
【請求項4】
ヒトLOXL2(SEQ ID NO:3)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、ヒトLOX2[LOXL2の誤記]の触媒ドメインの外側に位置するヒトLOX2由来の配列を含まないことを特徴とするポリペプチド。
【請求項5】
請求項4のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項6】
SEQ ID NO:4において記載されるヌクレオチド配列を有する請求項5のポリヌクレオチド。
【請求項7】
マウスLOX(SEQ ID NO:5)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、マウスLOXの触媒ドメインの外側に位置するマウスLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項8】
請求項7のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項9】
SEQ ID NO:6において記載されるヌクレオチド配列を有する請求項8のポリヌクレオチド。
【請求項10】
マウスLOXL2(SEQ ID NO:7)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、マウスLOXL2の触媒ドメインの外側に位置するマウスLOXL2由来の配列を含まないことを特徴とするポリペプチド。
【請求項11】
請求項10のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項12】
SEQ ID NO:8において記載されるヌクレオチド配列を有する請求項11のポリヌクレオチド。
【請求項13】
シグナル配列、ヒトLOXの触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、ヒトLOXの触媒ドメインの外側に位置するヒトLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項14】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項13のポリペプチド。
【請求項15】
前記エピトープタグがmycタグであることを特徴とする請求項13のポリペプチド。
【請求項16】
前記精製タグが、His6タグであることを特徴とする請求項13のポリペプチド。
【請求項17】
SEQ ID NO:9において記載されるアミノ酸配列を有する請求項13のポリペプチド。
【請求項18】
請求項13のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項19】
SEQ ID NO:10において記載されるヌクレオチド配列を有する請求項18のポリヌクレオチド。
【請求項20】
請求項18のポリヌクレオチドを含む発現ベクター。
【請求項21】
請求項19のポリヌクレオチドを含む発現ベクター。
【請求項22】
シグナル配列、ヒトLOXL2の触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、ヒトLOXL2の触媒ドメインの外側に位置するヒトLOXL2由来の配列を含まないことを特徴とするポリペプチド。
【請求項23】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項22のポリペプチド。
【請求項24】
前記エピトープタグがmycタグであることを特徴とする請求項22のポリペプチド。
【請求項25】
前記精製タグがHis6タグであることを特徴とする請求項22のポリペプチド。
【請求項26】
SEQ ID NO:11において記載されるアミノ酸配列を有する請求項22のポリペプチド。
【請求項27】
請求項22のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項28】
SEQ ID NO:12において記載されるヌクレオチド配列を有する請求項27のポリヌクレオチド。
【請求項29】
請求項27のポリヌクレオチドを含む発現ベクター。
【請求項30】
請求項28のポリヌクレオチドを含む発現ベクター。
【請求項31】
シグナル配列、マウスLOXの触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、マウスLOXの触媒ドメインの外側に位置するマウスLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項32】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項31のポリペプチド。
【請求項33】
前記エピトープタグがmycタグであることを特徴とする請求項31のポリペプチド。
【請求項34】
前記精製タグがHis6タグであることを特徴とする請求項31のポリペプチド。
【請求項35】
SEQ ID NO:13において記載されるアミノ酸配列を有する請求項31のポリペプチド。
【請求項36】
請求項31のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項37】
SEQ ID NO:14において記載されるヌクレオチド配列を有する請求項36のポリヌクレオチド。
【請求項38】
請求項36のポリヌクレオチドを含む発現ベクター。
【請求項39】
請求項37のポリヌクレオチドを含む発現ベクター。
【請求項40】
シグナル配列、マウスLOXL2の触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、マウスLOXL2の触媒ドメインの外側に位置するマウスLOXL2由来の配列を含まないことを特徴とするポリペプチド。
【請求項41】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項40のポリペプチド。
【請求項42】
前記エピトープタグがmycタグであることを特徴とする請求項40のポリペプチド。
【請求項43】
前記精製タグがHis6タグであることを特徴とする請求項40のポリペプチド。
【請求項44】
SEQ ID NO:15において記載されるアミノ酸配列を有する請求項40のポリペプチド。
【請求項45】
請求項40のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項46】
SEQ ID NO:16において記載されるヌクレオチド配列を有する請求項45のポリヌクレオチド。
【請求項47】
請求項45のポリヌクレオチドを含む発現ベクター。
【請求項48】
請求項46のポリヌクレオチドを含む発現ベクター。
【請求項1】
ヒトLOX(SEQ ID NO:1)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、ヒトLOXの触媒ドメインの外側に位置するヒトLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項2】
請求項1のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項3】
SEQ ID NO:2において記載されるヌクレオチド配列を有する請求項2のポリヌクレオチド。
【請求項4】
ヒトLOXL2(SEQ ID NO:3)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、ヒトLOX2[LOXL2の誤記]の触媒ドメインの外側に位置するヒトLOX2由来の配列を含まないことを特徴とするポリペプチド。
【請求項5】
請求項4のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項6】
SEQ ID NO:4において記載されるヌクレオチド配列を有する請求項5のポリヌクレオチド。
【請求項7】
マウスLOX(SEQ ID NO:5)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、マウスLOXの触媒ドメインの外側に位置するマウスLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項8】
請求項7のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項9】
SEQ ID NO:6において記載されるヌクレオチド配列を有する請求項8のポリヌクレオチド。
【請求項10】
マウスLOXL2(SEQ ID NO:7)の触媒ドメインのアミノ酸配列を含むポリペプチドであって、前記ポリペプチドは、マウスLOXL2の触媒ドメインの外側に位置するマウスLOXL2由来の配列を含まないことを特徴とするポリペプチド。
【請求項11】
請求項10のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項12】
SEQ ID NO:8において記載されるヌクレオチド配列を有する請求項11のポリヌクレオチド。
【請求項13】
シグナル配列、ヒトLOXの触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、ヒトLOXの触媒ドメインの外側に位置するヒトLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項14】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項13のポリペプチド。
【請求項15】
前記エピトープタグがmycタグであることを特徴とする請求項13のポリペプチド。
【請求項16】
前記精製タグが、His6タグであることを特徴とする請求項13のポリペプチド。
【請求項17】
SEQ ID NO:9において記載されるアミノ酸配列を有する請求項13のポリペプチド。
【請求項18】
請求項13のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項19】
SEQ ID NO:10において記載されるヌクレオチド配列を有する請求項18のポリヌクレオチド。
【請求項20】
請求項18のポリヌクレオチドを含む発現ベクター。
【請求項21】
請求項19のポリヌクレオチドを含む発現ベクター。
【請求項22】
シグナル配列、ヒトLOXL2の触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、ヒトLOXL2の触媒ドメインの外側に位置するヒトLOXL2由来の配列を含まないことを特徴とするポリペプチド。
【請求項23】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項22のポリペプチド。
【請求項24】
前記エピトープタグがmycタグであることを特徴とする請求項22のポリペプチド。
【請求項25】
前記精製タグがHis6タグであることを特徴とする請求項22のポリペプチド。
【請求項26】
SEQ ID NO:11において記載されるアミノ酸配列を有する請求項22のポリペプチド。
【請求項27】
請求項22のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項28】
SEQ ID NO:12において記載されるヌクレオチド配列を有する請求項27のポリヌクレオチド。
【請求項29】
請求項27のポリヌクレオチドを含む発現ベクター。
【請求項30】
請求項28のポリヌクレオチドを含む発現ベクター。
【請求項31】
シグナル配列、マウスLOXの触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、マウスLOXの触媒ドメインの外側に位置するマウスLOX由来の配列を含まないことを特徴とするポリペプチド。
【請求項32】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項31のポリペプチド。
【請求項33】
前記エピトープタグがmycタグであることを特徴とする請求項31のポリペプチド。
【請求項34】
前記精製タグがHis6タグであることを特徴とする請求項31のポリペプチド。
【請求項35】
SEQ ID NO:13において記載されるアミノ酸配列を有する請求項31のポリペプチド。
【請求項36】
請求項31のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項37】
SEQ ID NO:14において記載されるヌクレオチド配列を有する請求項36のポリヌクレオチド。
【請求項38】
請求項36のポリヌクレオチドを含む発現ベクター。
【請求項39】
請求項37のポリヌクレオチドを含む発現ベクター。
【請求項40】
シグナル配列、マウスLOXL2の触媒ドメイン、エピトープタグ及び精製タグを含むポリペプチドであって、前記ポリペプチドは、マウスLOXL2の触媒ドメインの外側に位置するマウスLOXL2由来の配列を含まないことを特徴とするポリペプチド。
【請求項41】
前記シグナル配列が免疫グロブリンカッパシグナル配列であることを特徴とする請求項40のポリペプチド。
【請求項42】
前記エピトープタグがmycタグであることを特徴とする請求項40のポリペプチド。
【請求項43】
前記精製タグがHis6タグであることを特徴とする請求項40のポリペプチド。
【請求項44】
SEQ ID NO:15において記載されるアミノ酸配列を有する請求項40のポリペプチド。
【請求項45】
請求項40のポリペプチドをエンコードするヌクレオチド配列を含むポリヌクレオチド。
【請求項46】
SEQ ID NO:16において記載されるヌクレオチド配列を有する請求項45のポリヌクレオチド。
【請求項47】
請求項45のポリヌクレオチドを含む発現ベクター。
【請求項48】
請求項46のポリヌクレオチドを含む発現ベクター。
【図6】


【図7】

【図1】
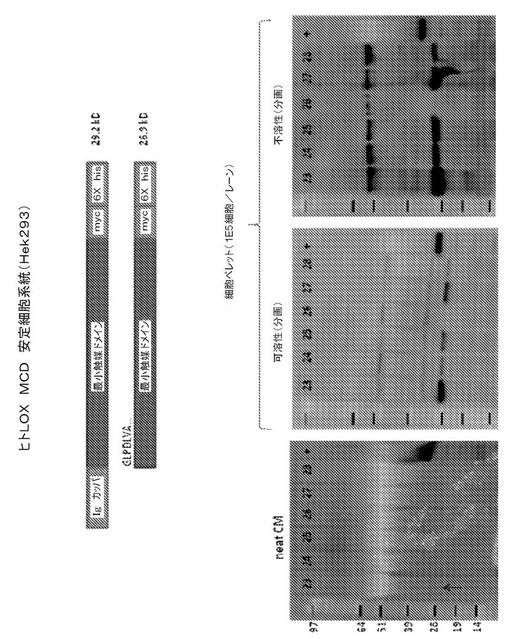
【図2】
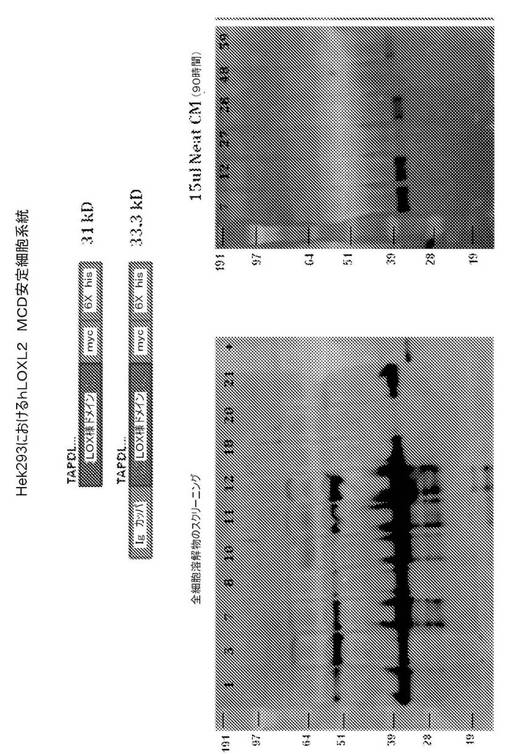
【図3】
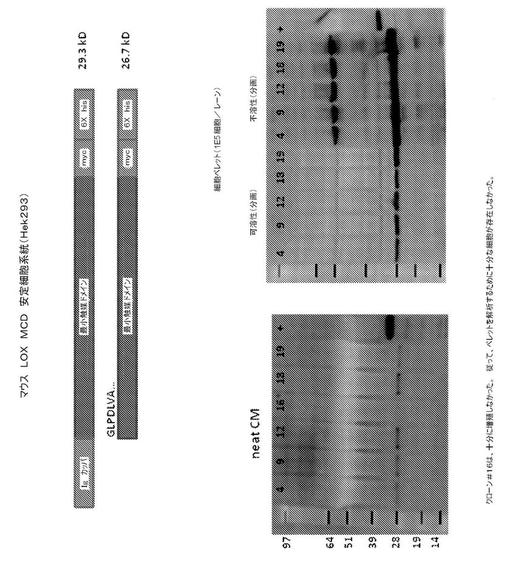
【図4】
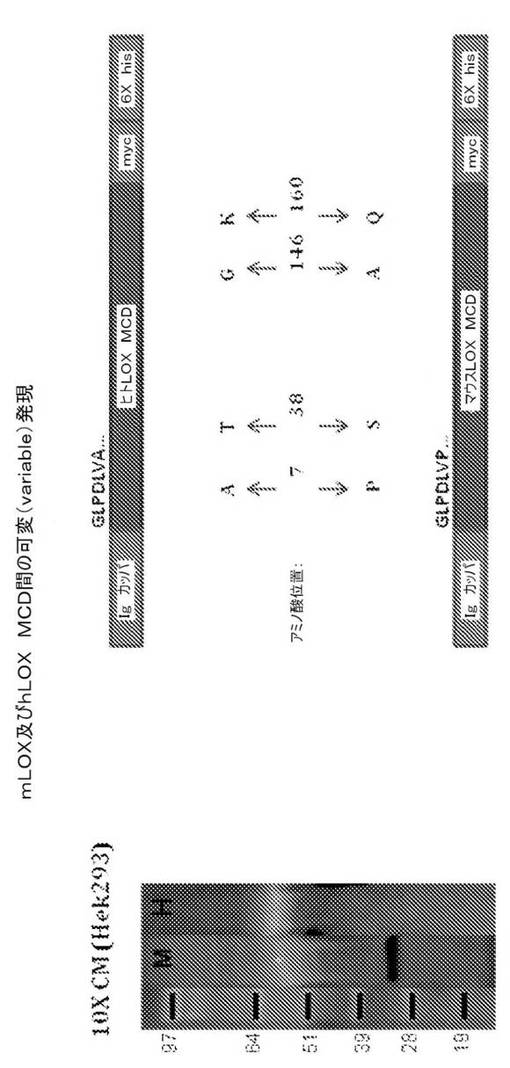
【図5】
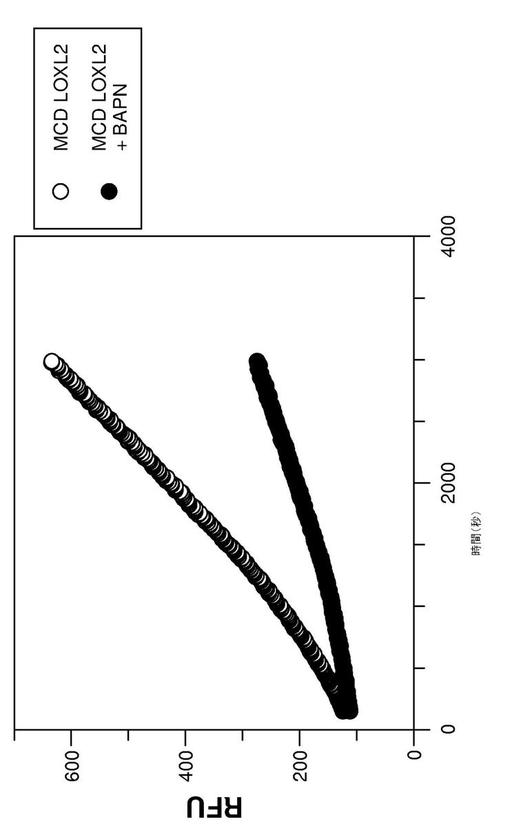
【図7】
【図1】
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2013−502226(P2013−502226A)
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願番号】特願2012−525734(P2012−525734)
【出願日】平成22年8月20日(2010.8.20)
【国際出願番号】PCT/US2010/046192
【国際公開番号】WO2011/022667
【国際公開日】平成23年2月24日(2011.2.24)
【出願人】(512043588)ギリアド バイオロジクス,インク. (7)
【氏名又は名称原語表記】GILEAD BIOLOGICS,INC.
【Fターム(参考)】
【公表日】平成25年1月24日(2013.1.24)
【国際特許分類】
【出願日】平成22年8月20日(2010.8.20)
【国際出願番号】PCT/US2010/046192
【国際公開番号】WO2011/022667
【国際公開日】平成23年2月24日(2011.2.24)
【出願人】(512043588)ギリアド バイオロジクス,インク. (7)
【氏名又は名称原語表記】GILEAD BIOLOGICS,INC.
【Fターム(参考)】
[ Back to top ]