
リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を生産するための代謝改変細胞
【課題】リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を生産する細胞を提供する。
【解決手段】L−フェニルアラニンアンモニアリアーゼがフェニルアラニンからトランス−ケイヒ酸を作り、シンナメート4−ヒドロキシラーゼ(C4H)が該トランスケイヒ酸から4−クマル酸を作り、4−クマレート−CoAリガーゼ(4CL)が該4−クマル酸から4−クマロイル−CoAを作り、リスベラトロールシンターゼ(VST)が該4−クマロイル−CoAからリスベラトロールを作る経路によるか、又はL−フェニルアラニン−又はチロシン−アンモニアリアーゼ(PAL/TAL)が4−クマル酸を作り、4−クマレート−CoAリガーゼ(4CL)が該4−クマル酸から4−クマロイル−CoAを作り、リスベラトロールシンターゼ(VST)が該4−クマロイル−CoAからリスベラトロールを作る経路によりリスベラトロールを産生する組換え微生物。
【解決手段】L−フェニルアラニンアンモニアリアーゼがフェニルアラニンからトランス−ケイヒ酸を作り、シンナメート4−ヒドロキシラーゼ(C4H)が該トランスケイヒ酸から4−クマル酸を作り、4−クマレート−CoAリガーゼ(4CL)が該4−クマル酸から4−クマロイル−CoAを作り、リスベラトロールシンターゼ(VST)が該4−クマロイル−CoAからリスベラトロールを作る経路によるか、又はL−フェニルアラニン−又はチロシン−アンモニアリアーゼ(PAL/TAL)が4−クマル酸を作り、4−クマレート−CoAリガーゼ(4CL)が該4−クマル酸から4−クマロイル−CoAを作り、リスベラトロールシンターゼ(VST)が該4−クマロイル−CoAからリスベラトロールを作る経路によりリスベラトロールを産生する組換え微生物。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般的に、微生物細胞を用いたポリフェノールリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体、例えば、そのβ−グルコシドピセイドの生産に関する。さらに、本発明は、リスベラトロール又はそのような誘導体を作る天然微生物又は組換え微生物の、食物、餌及び飲料の生産のための使用に関する。
【背景技術】
【0002】
薬品の微生物からの生産は、バイオテクノロジーの重要な応用となっている。典型的には、そのようなバイオ生産法の開発の工程として、1)適当な微生物宿主の選択、2)副産物を導く代謝経路の排除、3)酵素活性レベルと転写レベルの両方で望ましい経路の調節解除、及び4)望ましい経路での適当な酵素の過剰発現が挙げられるだろう。好ましい態様において、本発明は、リスベラトロールの望ましい生合成のために必要な前駆体を供給する植物のフェニルプロパノイド経路の酵素により、フェニルアラニン又はチロシンからの炭素の流れを再指示する上記工程の組合せを使用している。
【0003】
リスベラトロール(すなわち、3,4,5−トリヒロドキシスチルベン)は、感染又は他のストレスに関連する現象に応答して植物中に活動的な防御機構を構成する低分子量の二次的代謝産物であるスチルベンフィトアレキシン類の群に属するフィトフェノールである。スチルベンフィトアレキシン類はスチルベン骨格(トランス−1,2−ジフェニルエチレン)をそれらの共通する基本構造として含み、この構造は他の基の付加により補われてもよい(非特許文献1及び2)。スチルベン類は一定の木(被子植物、裸子植物)に発見されてきたが、一部の多年草(フトモモ科、ブドウ科及びマメ科の種)にも発見されている。該化合物は、有害生物、特に菌類、細菌及び昆虫への毒性がある。ほんのわずかな植物がスチルベン類を合成する能力があるか、又は有害生物に対して十分な耐性を植物に与える量でスチルベン類を生産する能力を有する。
【0004】
スチルベンシンターゼによる基本的なスチルベン骨格の合成が探究されている。今までに、ピノシルビンシンターゼとリスベラトロールシンターゼという2種類の酵素がスチルベンシンターゼとして示された。現在までに、落花生(Arachis hypogaea)リスベラトロールシンターゼが最も詳細に特性付けられているので、その性質のほとんどが知られている(非特許文献3)。スチルベンシンターゼにより用いられる基質は、マロニル−CoA、シンナモイル−CoA又はクマロイル−CoAである。これらの基質は、フラボノイド、花の色素及び脂質等の他の重要な植物構成物の生合成にも同じく用いられるので、すべての植物に存在する。
【0005】
リスベラトロール(図1:トランス形)は2個の密に結合したフェノール環からなるのでポリフェノール類に属する。ユーカリ、エゾマツ及びユリ等の他の植物やクワの実及びピーナッツ等の他の食物に存在する一方で、リスベラトロールの最も豊富な天然源は、ワイン製造に使用されるヴィティス・ヴィニヘラ、ヴィティス・ラプラスカ及びヴィティス・マスカジン(ロタンディフォリア)のブドウである。この化合物は、ブドウの木、根、種及び茎に存在するが、その最大濃度は、50〜100μg/g(非特許文献4)を含む皮にある(非特許文献5)。赤ワインの醸造中、ブドウの皮は白ワインの醸造とは対照的にブドウの液汁に含まれるので、リスベラトロールは赤ワインのみに少量見られる。リスベラトロールは、その抗菌性に加えて、その心臓保護活性と発癌予防活性が認識されてきており、それは、血小板凝集の阻害剤であるフィトエストロゲン(非特許文献6、7及び8)や抗酸化物(非特許文献4及び9)として作用する。これらの性質は、所謂、フレンチパラドックス(すわなち、ワインを飲むフランス人は低運動で高脂肪食事にもかかわらず冠動脈心疾患の低い発生率)を説明する。最近、リスベラトロールは、共に寿命の延長に重要な役割を果たす酵母のSIR2遺伝子及び類似のヒト遺伝子SIRT1を活性化することもできることが示された。それ以来、リスベラトロールの寿命延長効果が非常に注目されている(非特許文献10及び11)。
【0006】
ライフエクステンションファンデーション等のアメリカの健康協同組合は、この医薬の非常に高い薬効を促進しつつあり、それによって、成功した商品化のために理想的な状態を推進しつつある。現在の生産工程は、主に、ブドウ果皮かタデのいずれかからのリスベラトロールの抽出に依存する。これは、労働集約的な過程であり、低い利益を生むため、新規で、さらに効率的で多収量の生産方法の開発に対する動機を促している。
【0007】
植物において、フェニルプロパノイド経路は、リグニン類、サリチレート類、クマリン類、ヒドロキシケイヒ酸アミド類、色素、フラボノイド類及びフィトアレキシン類等の多様な二次代謝化合物の合成に関与する。実際、植物におけるリスベラトロールの形成はフェニルプロパノイド経路を経る。アミノ酸のL−フェニルアラニンは、L−フェニルアラニンアンモニアリアーゼ(PAL)により非酸化脱アミノ化によりトランスケイヒ酸に変換される(図2)。次に、トランスケイヒ酸は、NADPH:チトクロームP450レダクターゼ(CPR)と共同するチトクロームP450モノオキシゲナーゼ酵素であるシンナメート−4−ヒドロキシラーゼ(C4H)によりパラ位でヒドロキシル化されて4−クマル酸(4−ヒドロキシケイヒ酸)となる。引き続いて、この4−クマル酸は4−クマレート−CoAリガーゼ(4CL)の作用により4−クマロイル−CoAに活性化される。最後に、リスベラトロールシンターゼ(VST)が4−クマロイル−CoAのフェニルプロパン単位のマロニルCoAへの縮合を触媒して、リスベラトロールの形成をもたらす。
【0008】
最近、ブドウ液汁に少量見られる4−クマル酸からリスベラトロールを作ることのできる酵母が開示された(非特許文献12)。S.セレヴィシエの実験室の菌株における4−クマロイル−CoAとそれに付随するリスベラトロールの生産は、雑種ポプラに由来する異種コエンザイム−Aリガーゼ遺伝子をブドウリスベラトロールシンターゼ遺伝子(vst1)とともに共発現させることにより達成された。リスベラトロールシンターゼの他の基質であるマノニル−CoAは酵母ですでに内因的に産生されており、新規な脂肪酸生合成にかかわる。研究により、S.セレヴィシエの細胞は、4−クマル酸を補った合成培地で培養した場合、微量のリスベラトロールを遊離形又はグルコシド結合形のいずれかで産生することができることが分かった。
【0009】
しかし、該酵母は低いリスベラトロール収率に弱点があり、ごくわずかな産業用培地のみにしか存在しない4−クマル酸の添加を必要とするために、商業的利用には適さないだろう。したがって、医薬と栄養補助食品の両方としてのリスベラトロールの利用を容易にし、広げるために、4−クマル酸の添加なしに、グルコースからリスベラトロールを直接産生することのできる酵母を得ることは非常に望ましい。
【0010】
最近の研究(非特許文献13)には、ポプラに由来するPAL、C4H及びCPRを導入することによる、S.セレヴィシエにおけるフェニルプロパノイド経路のエントリポイントの再構成が記載されている。この目的は、PALとC4Hを含む多酵素複合体(MEC)が、フェニルプロパノイド代謝へのエントリポイントにおいて機能的に重要かどうかを評価することにあった。組換え酵母に[3H]−フェニルアラニンを供給することにより、代謝された[3H]−フェニルアラニンの大多数が4−[3H]−クマル酸に取り込まれること、及びフェニルアラニン代謝はC4H活性を阻害することにより非常に減少することがわかった。さらに、PLAのみのエクスプレッサーが微小のフェニルアラニンをケイヒ酸に代謝した。[3H]−フェニルアラニンと[14C]−トランス−ケイヒ酸を同時にトリプルエクスプレッサーに供給した場合、内因的に合成された[3H]−トランス−ケイヒ酸の4−クマル酸へのチャネリングについていかなる証拠も見られなかった。したがって、PALとCH4により触媒される反応によりフェニルアラニンから4−クマル酸への効率的な炭素の流れは酵母におけるMECによるチャネリングを必要とするようには思えず、PALとC4Hの真の生化学的カップリングが炭素の流れをフェニルプロパノイド経路に引き込むのに十分であるように思える。さらに別の研究(非特許文献14)において、大腸菌による植物特異的なフラバノン類の生産は多様な異種起源のフェニルプロパノイド経路の3遺伝子(酵母Rhodotorula rubraに由来するPAL、放線菌ストレプトミセス coelicolorに由来する4CL及び甘草植物Glycyrrhiza echinataに由来するカルコンシンターゼ(CHS))を含む人工的な遺伝子クラスターの発現により達成された。細菌の4CL酵素はコエンザイムAをトランス−ケイヒ酸と4−クマル酸の両方に連結させたので、これらの経路はC4Hをバイパスした。さらに、Rhodotorula rubra由来のPALはフェニルアラニンとチロシンの両方を基質として利用する。したがって、該遺伝子クラスターを含有し、かつグルコースで増殖した大腸菌細胞は、少量の2種類のフラバノン(フェニルアラニンからピノセムブリン(0.29g/l)及びチロシンからナリンゲニン(0.17g/l))を産生した。さらに、大量のそれらの前駆体である4−クマル酸とトランス−ケイヒ酸(それぞれ0.47及び1.23mg/リットル)が蓄積された。さらに、これらの化合物の収率はフェニルアラニンとチロシンの添加により増加させることができた。
【0011】
双子葉植物に由来する酵素がフェニルアラニンのみを効率的に利用する一方で、幾つかの研究によって、単子葉植物及び一部の微生物に由来するPALがチロシンを同じく利用することが示された(非特許文献15)。そのような反応において、チロシン活性物はチロシンアンモニアリアーゼと命名されている(TAL、図3)。チロシンのTALによる変換は、C4HとCPRの仲介なしに4−クマル酸の直接の形成をもたらす。主に、両活性は同じポリペプチドに存在し、Kmとターンオーバー数の大きな違いにかかわらず、非常に類似の触媒効率を有する。しかし、植物に由来するほとんどのPAL/TAL酵素はチロシンよりもフェニルアラニンを好む。TAL活性のレベルはほとんどPAL活性よりも低いが、この差の程度は広範囲にわたって変化する。例えば、パセリ酵素はフェニルアラニンに対して15〜25μM及びチロシンに対して2.0〜8.0mMのKmと、それぞれ22秒−1と0.3秒−1のターンオーバー数を有する。対照的に、トウモロコシ酵素は、フェニルアラニンのみに対してチロシンよりも15倍高いKm値と約10倍高いターンオーバー数を有する。さらに、赤酵母であるRhodotorula glutinis(Rhodosporidium toruloides)と−rubraにおいて、TAL触媒活性はPAL触媒活性に近く、TAL/PAL比が約0.58である。これらの酵母のPAL酵素は異化機能としてフェニルアラニンを分解すると考えられ、形成されるトランス−ケイヒ酸はベンゾエートと他の細胞材料に変換される一方で、植物においては、それは、リグニン、イソフラボノイド類及び他のフェニルプロパノイド類の生合成において調節酵素にすぎないと考えられている。
【0012】
最近、光活性黄色タンパク質の発色団の生合成にかかわる仮説上の生合成チロシンアンモニアリアーゼ(TAL)をコードする細菌ロドバクテリア capsulatusで読み取り枠が発見された(非特許文献16)。これは、PAL相同遺伝子が細菌で発見された最初である。TAL遺伝子は単離され、大腸菌で過剰生産させた。チロシンの4−クマル酸への変換に関するKmとkcat値はそれぞれ15.6μMと27.7秒−1であり、L−フェニルアラニンのトランス−ケイヒ酸への変換に関してはそれぞれ1277μMと15.1秒−1であった。その小さいKmとわずかに高いkcatの結果として、酵素はL−フェニルアラニンよりもチロシンに対する強い好みを示し、フェニルアラニンに対するよりも約150倍高いチロシンに対する触媒効率(Km/kcat)を有する。動力学的研究によると、L−フェニルアラニンではなく、チロシンが生理学的条件下で酵素の天然の基質であることが立証されている。ごく最近、ある研究で、大腸菌でのフラボノイド類の産生に関してフェニルアラニンアンモニアリアーゼ、シンナメート−4−ヒドロキシラーゼ、4−クマレート−Coa−リガーゼ及びカルコンシンターゼの異種共発現が述べられている(非特許文献17)。しかし、すべての4遺伝子における同時発現は、非機能性のシンナメート−4−ヒドロキシラーゼのために成功しなかった。しかし、ロドバクテリア sphaeroidesからクローニングされた新しいチロシンアンモニアリアーゼをフェニルアラニンアンモニアリアーゼとシンナメート−4−ヒドロキシラーゼと置き換えることでその問題が解決され、フラボノンナリンゲニンの高水準生産が導かれた。さらに、ロドバクテリア sphaeroidesに由来する該チロシンアンモニアリガーゼは、大腸菌における4−クマル酸(すなわち、パラ−ヒドロキシケイヒ酸)の異種産生のためにも用いられている(特許文献1)。さらに、グルコースを4−クマル酸に変換する生体触媒のさらなる開発方法が記載されている。特許文献2には、大腸菌とサッカロマイセス・セレヴィシエ(出芽酵母)におけるクマル酸の産生のためにTrichosporon cutaneum酵母に由来するチロシンアンモニアリアーゼが開示されている。特許文献3には、チロシンを4−クマル酸へ直接変換する野生型PALの能力を明確に示すために、Rhodotorula glutinis酵母に由来する野生型PALの大腸菌への取り込みが記載されている。さらに、植物C4HとCPRに加えて、Rhodotorula glutinis酵母に由来する野生型PALの大腸菌とS.セレヴィシエとへの取り込みの例示も存在する。増強TAL活性を有する野生型酵母PAL Rhodotorula glutinisの突然誘発による生体触媒の開発も記載されている(特許文献4)。上記特許のいずれもリスベラトロールの産生のために4CL及びVSTの取り込みを請求するものではない。
【0013】
最近、糸状菌の麹菌(コウジカビ)が、植物中のナリンゲニン等のフラボノイド類の生合成に通常関与するカルコンシンターゼ(CHS)酵素を含むとの証拠が示された(非特許文献18)。実際、麹菌がフェニルプロパノイド−フラボノイド代謝に関与する主要な一組の遺伝子(すなわち、PAL、C4H及び4CL)を含むことも示した。しかし、麹菌がリスベラトロールシンターゼ等のスチルベンシンターゼを含むとの証拠はない。
【先行技術文献】
【特許文献】
【0014】
【特許文献1】US−A−2004059103
【特許文献2】US−A−2004023357
【特許文献3】US−A−2001053847
【特許文献4】US−A−6521748
【非特許文献】
【0015】
【非特許文献1】ハート及びシュリンプトン(Hart and Shrimpton)、1979年
【非特許文献2】ハート(Hart)、1981年
【非特許文献3】ショップナー及びキンドル(Schoppner and Kindl)、1984年
【非特許文献4】ジャングら(Jang et al.)、1997年
【非特許文献5】セロッティら(Celotti et al.)、1996年
【非特許文献6】コップら(Kopp et al.)、1998年
【非特許文献7】ゲームら(Gehm et al.)、1997年
【非特許文献8】ロボら(Lobo et al.)、1995年
【非特許文献9】ファング(Huang)、1997年
【非特許文献10】ハル(Hall)、2003年
【非特許文献11】コウジン(Couzin)、2004年
【非特許文献12】ベッカーら(Becker et al.)、2003年
【非特許文献13】ロー及びダグラス(Ro and Douglas)、2004年
【非特許文献14】Hwangら、2003年
【非特許文献15】Roslerら、1997年
【非特許文献16】キンドら(Kyndt et al.)、2002年
【非特許文献17】Wattsら、2004年
【非特許文献18】Seshimeら、2005年
【発明の概要】
【課題を解決するための手段】
【0016】
本発明は、
(1)4−クマル酸を生産し、さらに当該4−クマル酸からリスベラトロール又は当該リスベラトロールのオリゴマー誘導体又はグリコシド結合誘導体を生産する操作的な代謝経路を有する微生物において、
前記代謝経路において、前記4−クマル酸は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ及びシンナメート4−ヒドロキシラーゼの作用によってL−フェニルアラニンから、又は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼの作用によってチロシンから生成し、
4−クマロイル−CoAが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現する4−クマレート−CoAリガーゼによる酵素反応で前記4−クマル酸から生成し、
そして前記リスベラトロールが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するリスベラトロールシンターゼによって内在性マロニル−CoAを基質とする反応で前記4−クマロイル−CoAから生成することを特徴とする微生物、
(2)天然のNADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物で過剰発現している、又は、NADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物に組換えで導入されていることを特徴とする上記(1)に記載の微生物、
(3)チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする上記(1)又は(2)に記載の微生物、
(4)フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする上記(1)乃至(3)のいずれか一項に記載の微生物、
(5)真菌又は細菌であることを特徴とする上記(1)乃至(4)のいずれか一項に記載の微生物、
(6)発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有することを特徴とする上記(1)に記載の微生物、
(7)シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する上記(1)に記載の微生物、
(8)4−クマル酸の外部源の実質的な非存在下において、上記(1)乃至(7)のいずれか一項に記載の微生物を炭素基質に接触させる工程を含むことを特徴とする、リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法、
(9)前記炭素基質が、単糖類、小糖類及び多糖類からなる発酵性炭素基質の群より選択されることを特徴とする上記(8)に記載の方法、
(10)前記炭素基質が、エタノール、アセテート、グリセロール、ラクテート、及びアミノ酸で構成される非発酵性炭素基質からなる群より選択されることを特徴とする上記(8)に記載の方法、
(11)前記非発酵性炭素基質が、フェニルアラニン及びチロシンからなる群より選択されることを特徴とする上記(10)に記載の方法、
(12)さらに、生産されたリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を乳製品又は飲み物中の栄養補助食品として使用することを含む上記(8)乃至(11)のいずれか一項に記載の方法、
である。
【図面の簡単な説明】
【0017】
【図1】図1は、トランス−リスベラトロールの化学構造を示す。
【図2】図2は、L−ファニルアラニンに作用するフェニルアラニンアンモニアリアーゼを利用するフェニルプロパノイド経路を示す。
【図3】図3は、L−チロシンに作用するフェニルアラニンアンモニアリアーゼを利用する別の経路を示す。
【図4】図4は、100g/lのガラクトースで成長させたS.セレヴィシエ菌株であるFSSC−PALC4H4CLVST及びFSSC−TAL4CLVSTの抽出物のHPLCクロマトグラムを示す。60ナノグラムの純粋なリスベラトロールのクロマトグラムが含まれる。
【図5】図5は、純粋なトランス−リスベラトロール及び100g/lのガラクトースで成長させたS.セレヴィシエ菌株であるFSSC−PALC4H4CLVSTにより作られたトランス−リスベラトロールのUV吸収スペクトルを示す。
【図6】図6は、50g/lのグルコースで成長させた大腸菌株であるFSEC−TAL4CLVST及びFSECコントロールからの抽出物のHPLCクロマトグラムを示す。
【図7】図7は、20mg/lのクマル酸を添加した50g/lのグルコースで成長させた大腸菌株であるFSEC−TAL4CLVST及びFSECコントロールからの抽出物のHPLCクロマトグラムを示す。FSEC−TAL4CLVST菌株で作られたトランス−リスベラトロールのUV吸収スペクトルが含まれる。
【発明を実施するための形態】
【0018】
本発明は、少なくとも1種類の活性を含む操作的な代謝経路を有する微生物において、該経路が4−クマル酸を産生し、かつそれからリスベラトロール又はそのオリゴマー誘導体かグリコシド結合誘導体を産生する微生物を本願において提供する。そのような微生物は自然に存在してよく、適切なスクリーニング手順により単離してもよく、さらに好ましくは遺伝子組み換えされている。
【0019】
好ましくは、該リスベラトロール又は誘導体は、内在性のマロニル−CoAが基質である酵素により触媒される反応で作られ、好ましくは該リスベラトロールは4−クマロイル−CoAから生成される。
【0020】
好ましくは、該リスベラトロール又は誘導体は、微生物において、好ましくは該微生物に固有ではない該酵素をコードする核酸から発現するリスベラトロールシンターゼにより4−クマロイル−CoAから生成される。
【0021】
ここでは一般的に、文脈上他の意味に解すべき場合を除き、リスベラトロールへの言及は、特にピセイドを含む、そのオリゴマー誘導体かグリコシド結合誘導体への言及を含む。
【0022】
例えば、ある好ましい実施態様において、前記リスベラトロールシンターゼは、Arachis属に属する植物、例えば、A.glabatra、A.hypogaea、Rheum属に属する植物、例えば、R.tataricum、ヴィティス属に属する植物、例えば、V.ラプラスカ、V.riparaia、V.ヴィニヘラ、又はパイナス属、Piceea属、ユリ属、ユーカリ属、Parthenocissus属、ジッサス属、カロコルタス属、タデ属、Gnetum属、Artocarpus属、Nothofagus属、フェニックス属、ウシノケグサ属、スゲ属、シュロソウ属、ハカマカズラ又はPterolobium属のいずれかに由来するリスベラトロールシンターゼ(EC2.3.1.95)である。
【0023】
前記4−クマル酸は、好適には、酸素が基質であり、NADH又はNADPHが補因子であり、NAD+又はNADP+が産物である酵素により触媒される反応で酵素によりトランス−ケイヒ酸から生成されることが好ましい。
【0024】
例えば、該4−クマル酸はトランス−ケイヒ酸からシンナメート4−ヒドロキシラーゼにより作られてよく、該4−クマル酸は、好ましくは、微生物において、該微生物に固有ではない該酵素をコードする核酸から発現する。
【0025】
上記段落で言及されたものを含むある実施態様において、該シンナメート−4−ヒドロキシラーゼは植物又は微生物に由来するシンナメート−4−ヒドロキシラーゼ(EC1.14.13.11)である。植物は、シロイヌナズナ属、例えば、シロイヌナズナ、Citrusに属する植物、例えば、C.シネンシス、C.x paradisi、Phaseolus属する植物、例えば、P.ヴァガリス、パイナス属する植物、例えば、P.taeda、Populus属する植物、例えば、P.deltoides、P.tremuloides、P.trichocarpa、ナス属する植物、例えば、S.tuberosum、ヴィティス属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属する植物、例えば、Z.メイス又は他の植物属、例えば、Ammi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属に属してよい。微生物は、アスペルギルス属に属する真菌類、例えば麹菌であってよい。
【0026】
好ましくは、前記4−クマル酸は、アンモニアが生成される酵素触媒反応でチロシンから作られ、好適には、該4−クマル酸は、L−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼ、例えば酵母又は細菌に由来するチロシンアンモニアリアーゼ(EC4.3.1.5)によりチロシンから生成される。好適には、該チロシンアンモニアリアーゼは酵母Rhodotorula rubra又は細菌ロドバクテリア capsulatusに由来する。
【0027】
任意に、該チロシンアンモニアリアーゼは、微生物において、該微生物に固有ではない該酵素をコードする核酸から発現させる。
【0028】
もしくは、前記トランス−ケイヒ酸は、アンモニアが生成される酵素触媒反応でL−フェニルアラニンから作られてよく、好適には、該トランス−ケイヒ酸は、L−フェニルアラニンからフェニルアラニンアンモニアリアーゼにより形成する。
【0029】
ある好ましい実施態様において、該L−フェニルアラニンアンモニアリアーゼは、植物又は微生物に由来するL−フェニルアラニンアンモニアリアーゼ(EC4.3.1.5)である。植物は、シロイヌナズナ属に属する植物、例えば、シロイヌナズナ、Brassica属に属する植物、例えば、B.napus、B.rapa、シトラス属に属する植物、例えば、C.reticulata、C.clementinus、C.リモン、Phaseolus属に属する植物、例えば、P.coccineus、P.ヴァリガリス、パイナス属に属する植物、例えば、P.banksiana、P.monticola、P.パイナスター、P.sylvestris、P.taeda、Populus属に属する植物、例えば、P.balsamifera、P.deltoides、P.Canadensis、P.kitakamiensis、P.tremuloides、ナス属に属する植物、例えば、S.tuberosum、プラナス属に属する植物、例えば、P.avium、P.persica、ヴィティス属に属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属に属する植物、例えば、Z.メイス又は他の植物属、例えば、Agastache属、Ananas属、Asparagus属、Bromheadia属、Bambusa属、Beta属、Betula属、Cucumis属、カメリア属、Capsicum属、Cassia属、Catharanthus属、Cicer属、Citrullus属、Coffea属、Cucurbita属、Cynodon属、Daucus属、Dendrobium属、Dianthus属、Digitalis属、Dioscorea属、ユーカリ属、Gallus属、Ginkgo属、グリシン属、Hordeum属、Helianthus属、Ipomoea属、Lactuca属、リソスパーマム・オフィシナーレ属、ハス属、Lycopersicon属、Medicago属、マリュス属、Manihot属、Medicago属、マツバギク属、タバコ属、Olea属、オリザ属、Pisum属、Persea属、ペトロセリウム属、Phalaenopsis属、Phyllostachys属、Physcomitrella属、Picea属、Pyrus属、Quercus属、Raphanus属、Rehmannia属、キイチゴ属、Sorghum属、Sphenostylis属、Stellaria属、Stylosanthes属、コムギ属、Trifolium属、コムギ属、Vaccinium属、Vigna属、Zinnia属に属してよい。微生物は、Agaricus属に属する真菌類、例えば、A.bisporus、アスペルギルス属に属する真菌類、例えば、麹菌、A.nidulans、A.フミガーツス、Ustilago属に属する真菌類、例えば、U.maydis、ロドバクテリア属に属する細菌、例えば、R.capsulatus、Rhodotorula属に属する酵母、例えば、R.rubraであってよい。
【0030】
好適には、該L−フェニルアラニンアンモニアリアーゼは、微生物において、該微生物に固有ではない該酵素をコードする核酸から発現させる。
【0031】
好ましくは、4−クマロイル−CoAは、ATPとCoAが基質であり、かつADPが産物である酵素触媒反応で形成され、好適には、4−クマロイル−CoAは4−クマレート−CoAリガーゼにより触媒される反応で形成される。
【0032】
前記4−クマレート−CoAリガーゼは、植物、微生物又は線虫に由来する4−クマレート−CoAリガーゼ(EC6.2.1.12)であってよい。植物は、Abies属、例えば、A.beshanzuensis、B.firma、B.holophylla、シロイヌナズナ属に属する植物、例えば、シロイヌナズナ、Brassica属に属する植物、例えば、B.napus、B.rapa、B.oleracea、シトラス属に属する植物、例えば、C.シネンシス、Larix属に属する植物、例えば、L.decidua、L.gmelinii、L.griffithiana、L.himalaica、L.kaempferi、L.laricina、L.mastersiana、L.occidentalis、L.potaninii、L.sibirica、L.speciosa、Phaseolus属に属する植物、例えば、P.acutifolius、P.coccineus、パイナス属に属する植物、例えば、P.armandii、P.banksiana、P.パイナスター、Populus属に属する植物、例えば、P.balsamifera、P.tomentosa、P.tremuloides、ナス属に属する植物、例えば、S.tuberosum、ヴィティス属に属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属に属する植物、例えば、Z.メイス又は他の植物属、例えば、Agastache属、アモルファ属、Cathaya属、Cedrus属、クロッカス属、ウシノケグサ属、グリシン属、Juglans属、Keteleeria属、リソスパーマム・オフィシナーレ属、Lolium属、ハス属、Lycopersicon属、マリュス属、Medicago属、マツバギク属、タバコ属、Nothotsuga属、オリザ属、ペラルゴニウム属、ペトロセリウム属、Physcomitrella属、Picea属、プラナス属、Pseudolarix属、Pseudotsuga属、ローザ属、キイチゴ属、Ryza属、Saccharum属、Suaeda属、Thellungiella属、コムギ属、Tsuga属に属してよい。微生物は、アスペルギルス属に属する糸状菌、例えば、A.flavus、A.nidulans、麹菌、A.フミガーツス、アカパンカビ属に属する糸状菌、例えば、アカパンカビ、Yarrowia属に属する真菌類、例えば、Y.lipolytica、Mycosphaerella属に属する真菌類、例えば、M.graminicola、マイコバクテリウム属に属する細菌、例えば、ウシ型結核菌、ハンセン菌、ヒト型結核菌、ナイセリア属に属する細菌、例えば、髄膜炎菌、ストレプトミセス属に属する細菌、例えば、S.coelicolor、ロドバクテリア属に属する細菌、例えば、R.capsulatus、鉤虫属に属する線虫類、例えば、A.ceylanicum、シノラブディス属に属する線虫類、例えば、C.エレガンス、捻転胃虫属に属する線虫類、例えば、H.contortus、Lumbricus属に属する線虫類、例えば、L.rubellus、Meilodogyne属に属する線虫類、例えば、M.hapla、Strongyloidus属に属する線虫類、例えば、S.rattii、糞線虫、Pristionchus属に属する線虫類、例えば、P.pacificusであってよい。
【0033】
任意に、NADPH:チトクロームP450レダクターゼ(CPR)は組換えで微生物に導入されている。これは非植物微生物に導入された植物CPRであってよい。あるいは、天然のNADPH:チトクロームP450レダクターゼ(CPR)が該微生物で過剰発現している。
【0034】
上記段落で言及されたものを含む好ましい実施態様において、前記NADPH:チトクロームP450レダクターゼは、シロイヌナズナ属に属する植物、例えば、シロイヌナズナ、シトラス属に属する植物、例えば、C.シネンシス、C.x paradisi、Phaseolus属に属する植物、例えば、P.ヴァガリス、パイナス属に属する植物、例えば、P.taeda、Populus属に属する植物、例えば、P.deltoides、P.tremuloides、P.trichocarpa、ナス属に属する植物、例えば、S.tuberosum、ヴィティス属に属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属に属する植物、例えば、Z.メイス又は他の植物属、例えば、Ammi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属に由来するNADPH:チトクロームP450レダクターゼ(EC1.6.2.4)である。
【0035】
微生物が天然に存在するものであってよい一方で、該代謝経路の各酵素をコードする少なくとも1つの遺伝子配列の少なくとも1つのコピーが該微生物に組換えで導入されているのが好ましい。
【0036】
該酵素をコードするコード配列の導入に加えて、又はその代わりに、該生物体中の該コード配列に天然では関連しないプロモーター配列等の1つ以上の発現シグナルを提供してよい。したがって、任意に、チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、該生物中の該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結され、及び/又はL−フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、該生物中の該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0037】
任意に、シンナメート4−ヒドロキシラーゼをコードする遺伝子配列の少なくとも1つのコピーが、天然かどうかにかかわらず、該生物において該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0038】
任意に、4−クマレート−CoAリガーゼをコードする遺伝子配列の少なくとも1つのコピーが、天然かどうかにかかわらず、該生物において該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0039】
任意に、リスベラトロールシンターゼをコードする遺伝子配列の少なくとも1つのコピーが、天然かどうかにかかわらず、該生物において該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0040】
発現シグナルとして、コード配列の上流(5’−非コード配列)、その内部又はその下流(3’−非コード配列)に配置され、かつ関連するコード配列の転写、RNAのプロセシング又は安定性、又は翻訳に影響を与えるヌクレオチド配列が挙げられる。そのような配列として、プロモーター、翻訳リーダー配列、イントロン及びポリアデニル化認識配列が挙げられよう。
【0041】
ある態様において、本発明は、第1酵素により触媒され、かつその反応工程がアンモニアを作る反応により第1代謝産物が第2代謝産物に変換され、酸素が基質であり、NADPH又はNADHが補因子であり、NADP+又はNAD+が生成物である第2酵素により触媒される反応で該第2代謝産物が第3代謝物に変換され、ATP及びCoAが基質であり、ADPが生成物である第3酵素により触媒される反応で第3代謝産物が第4代謝産物に変換され、内在性マロニル−CoAが基質である第4酵素により触媒される反応で第4代謝産物が第5代謝産物に変換される作動性代謝経路を有する代謝改変微生物を提供する。
【0042】
さらに、本発明は、第1酵素により触媒され、かつその反応工程が第2酵素の関与なしにアンモニアを作る反応により、第1代謝産物が第3代謝産物に変換され、ATP及びCoAが基質であり、ADPが生成物である第3酵素により触媒される反応で第3代謝産物が第4代謝産物に変換され、内在性マロニル−CoAが基質である第4酵素により触媒される反応で第4代謝産物が第5代謝産物に変換される作動性代謝経路を有する代謝改変微生物を提供する。
【0043】
上記微生物として、発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する微生物が挙げられる。
【0044】
それら微生物として、シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する微生物も挙げられる。
【0045】
現在の文脈において、「微生物」は、細菌等の微生物、酵母等の微細菌に関する。
【0046】
さらに具体的には、微生物は真菌であってよく、さらに具体的には、アスペルギルス属に属する糸状菌、例えば、黒色アスペルギルス、A.awamori、麹菌、A.nidulans、Saccharomyces属に属する酵母、例えば、S.セレヴィシエ、S.kluyveri、S.bayanus、S.exiguus、S.sevazzi、S.uvarum、Kluyveromyces属に属する酵母、例えば、K.lactis、K.marxianus var.marxianus、K.thermotolerans、カンジダ属に属する酵母、例えば、C.utilis、C.tropicalis、C.albicans、C.lipolytica、C.versatilis、ピチア属に属する酵母、例えば、P.stipidis、P.pastoris、P.sorbitophila、又は他の酵母属、例えば、クリプトコッカス属、Debaromyces属、Hansenula属、ピチア属、Yarrowia属、Zygosaccharomyces属又はシゾサッカロミセス属であってよい。他の微生物に関して、ペニシリウム属、クモノスカビ属、フサリウム属、Fusidium属、Gibberella属、ケカビ属、モルティエラ属、トリコデルマ属に属する種等の好適な糸状菌の非網羅的なリストが提供される。
【0047】
細菌に関して、好適な細菌の非網羅的なリストは以下の通り与えられる:バシラス属に属する種、エシェリキア属に属する種、乳酸菌属に属する種、Lactococcus属に属する種、コリネバクテリア属に属する種、アセトバクター属に属する種、アシネトバクター属に属する種、シュードモナス属に属する種等。
【0048】
本発明の好ましい微生物は、S.セレヴィシエ、黒色アスペルギルス、麹菌、大腸菌、乳酸連鎖球菌又は枯草菌であってよい。
【0049】
構築され、改変された微生物は、ケモスタット培養、バッチ培養、流加バッチ培養等の一般に知られている方法を用いて培養することができる。
【0050】
例えば、本発明は、非植物細胞を炭素基質に4−クマル酸の外部源の実質的な非存在下で接触させる工程を含むリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法であって、該細胞が該条件下でリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を作る能力を有し、該微生物が真菌及び細菌、特に酵母からなる群より選択することができる。
【0051】
前記炭素基質は、単糖類、小糖類及び多糖類、例えば、グルコース、フルクトース、ガラクトース、キシロース、アラビノース、マンノース、スクロース、ラクトース、エリトロース、スレオース及び/又はリボースからなる発酵性炭素基質の群より任意に選択される。該炭素基質は、さらに又は代わりに、エタノール、アセテート、グリセロール及び/又はラクテート等の非発酵性炭素基質からなる群より選択してもよい。該非発酵性炭素基質は、さらに又は代わりに、アミノ酸からなる群より選択してよく、フェニルアラニン及び/又はチロシンであってよい。
【0052】
別の態様において、本発明は、フェニルアラニンアンモニアリアーゼ、シンナメート4−ヒドロキシラーゼ、4−クマレート−CoA−リガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法、さらにチロシンアンモニアリアーゼ、4−クマレート−CoAリガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロールの製造方法を含む。
【0053】
そのように生成されるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体は乳製品又はビール等の飲み物中の栄養補助食品として用いることができる。
【0054】
本発明にしたがって生成されるリスベラトロールはシス−リスベラトロール又はトランス−リスベラトロールであってよいが、トランス型が通常は優勢であると予想できる。
【0055】
本発明は更に以下の形態を含む。
[1]少なくとも1種類の酵素活性を含む操作的な代謝経路を有する微生物において、前記経路が4−クマル酸を生産し、かつそれからリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を生産することを特徴とする微生物。
[2]前記リスベラトロールが、内在性のマロニル−CoAが基質である酵素により触媒される反応で生成されることを特徴とする上記[1]に記載の微生物。
[3]前記リスベラトロールが4−クマロイル−CoAから生成されることを特徴とする上記[1]又は[2]に記載の微生物。
[4]前記リスベラトロールが、4−クマロイル−CoAからリスベラトロールシンターゼにより生成されることを特徴とする上記[3]に記載の微生物。
[5]前記リスベラトロールシンターゼが、前記微生物中において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[4]に記載の微生物。
【0056】
[6]前記リスベラトロールシンターゼが、Arachis属に属する植物、Rheum属に属する植物、ヴィティス属に属する植物、又はパイナス属、Piceea属、ユリ属、ユーカリ属、Parthenocissus属、ジッサス属、カロコルタス属、タデ属、Gnetum属、Artocarpus属、Nothofagus属、フェニックス属、ウシノケグサ属、スゲ属、シュロソウ属、ハカマカズラ又はPterolobium属のいずれか1種類に由来するリスベラトロールシンターゼ(EC2.3.1.95)であることを特徴とする上記[5]に記載の微生物。
[7]前記4−クマル酸が、トランス−ケイヒ酸から生成されることを特徴とする前記いずれかの一項に記載の微生物。
[8]前記4−クマル酸が、酸素が基質であり、NADH又はNADPHが補因子であり、NAD又はNADPが産物である酵素により触媒される反応で前記酵素によりトランス−ケイヒ酸から生成されることを特徴とする上記[7]に記載の微生物。
[9]前記4−クマル酸が、トランス−ケイヒ酸からシンナメート4−ヒドロキシラーゼにより生成されることを特徴とする上記[8]に記載の微生物。
[10]前記シンナメート4−ヒドロキシラーゼが、微生物において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[9]に記載の微生物。
【0057】
[11]前記シンナメート−4−ヒドロキシラーゼが、シロイヌナズナ属に属する植物、シトラス属に属する植物、Phaseolus属に属する植物、パイナス属に属する植物、Populus属に属する植物、ナス属に属する植物、ヴィティス属に属する植物、ズィー属に属する植物、又はAmmi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属のいずれか1種類、及びアスペルギルス属に属する真菌類に由来するシンナメート−4−ヒドロキシラーゼ(EC1.14.13.11)であることを特徴とする上記[10]に記載の微生物。
[12]前記4−クマル酸が、アンモニアが生成される酵素触媒反応でチロシンから生成されることを特徴とする上記[1]〜[6]のいずれかに記載の微生物。
[13]前記4−クマル酸が、L−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼによりチロシンから生成されることを特徴とする上記[1]〜[4]のいずれかに記載の微生物。
[14]前記4−クマル酸が、酵母又は細菌に由来するチロシンアンモニアリアーゼ(EC4.3.1.5)により生成されることを特徴とする上記[13]に記載の微生物。
[15]前記チロシンアンモニアリアーゼが、酵母Rhodotorula rubra又は細菌ロドバクテリア capsulatusに由来することを特徴とする上記[14]に記載の微生物。
[16]前記チロシンアンモニアリアーゼが、微生物において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[12]〜[15]のいずれかに記載の微生物。
[17]前記トランス−ケイヒ酸が、アンモニアが生成される酵素触媒反応でL−フェニルアラニンから生成されることを特徴とする上記[9]〜[11]のいずれかに記載の微生物。
[18]前記トランス−ケイヒ酸が、L−フェニルアラニンからL−フェニルアラニンアンモニアリアーゼにより形成されることを特徴とする上記[17]に記載の微生物。
[19]前記トランス−ケイヒ酸が、シロイヌナズナ属に属する植物、Brassica属に属する植物、シトラス属に属する植物、Phaseolus属に属する植物、パイナス属に属する植物、Populus属に属する植物、ナス属に属する植物、プラナス属に属する植物、ヴィティス属に属する植物、ズィー属に属する植物、又はAgastache属、Ananas属、Asparagus属、Bromheadia属、Bambusa属、Beta属、Betula属、Cucumis属、カメリア属、Capsicum属、Cassia属、Catharanthus属、Cicer属、Citrullus属、Coffea属、Cucurbita属、Cynodon属、Daucus属、Dendrobium属、Dianthus属、Digitalis属、Dioscorea属、ユーカリ属、Gallus属、Ginkgo属、グリシン属、Hordeum属、Helianthus属、Ipomoea属、Lactuca属、リソスパーマム・オフィシナーレ属、ハス属、Lycopersicon属、Medicago属、マリュス属、Manihot属、Medicago属、マツバギク属、タバコ属、Olea属、オリザ属、Pisum属、Persea属、ペトロセリウム属、Phalaenopsis属、Phyllostachys属、Physcomitrella属、Picea属、Pyrus属、Quercus属、Raphanus属、Rehmannia属、キイチゴ属、Sorghum属、Sphenostylis属、Stellaria属、Stylosanthes属、コムギ属、Trifolium属、コムギ属、Vaccinium属、Vigna属又はZinnia属のいずれかに属する植物、又はアスペルギルス属に属する真菌類に由来するL−フェニルアラニンアンモニアリアーゼ(EC4.3.1.5)によりL−フェニルアラニンから形成されることを特徴とする上記[18]に記載の微生物。
[20]前記L−フェニルアラニンアンモニアリアーゼが、微生物において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[18]又は[19]に記載の微生物。
【0058】
[21]4−クマロイル−CoAが、ATPとCoAが基質であり、ADPが産物である酵素により触媒される反応で形成されることを特徴とする前記いずれかの一項に記載の微生物。
[22]4−クマロイル−CoAが、4−クマレート−CoAリガーゼにより触媒される反応で形成されることを特徴とする前記いずれかの一項に記載の微生物。
[23]前記4−クマレート−CoAリガーゼが、Abies属に属する植物、シロイヌナズナ属に属する植物、Brassica属に属する植物、シトラス属に属する植物、Larix属に属する植物、Phaseolus属に属する植物、パイナス属に属する植物、Populus属に属する植物、ナス属に属する植物、ヴィティス属に属する植物、ズィー属に属する植物、又はAgastache属、アモルファ属、Cathaya属、Cedrus属、クロッカス属、ウシノケグサ属、グリシン属、Juglans属、Keteleeria属、リソスパーマム・オフィシナーレ属、Lolium属、ハス属、Lycopersicon属、マリュス属、Medicago属、マツバギク属、タバコ属、Nothotsuga属、オリザ属、ペラルゴニウム属、ペトロセリウム属、Physcomitrella属、Picea属、プラナス属、Pseudolarix属、Pseudotsuga属、ローザ属、キイチゴ属、Ryza属、Saccharum属、Suaeda属、Thellungiella属、コムギ属又はTsuga属のいずれかに属する植物、アスペルギルス属に属する糸状菌、アカパンカビ属に属する糸状菌、Yarrowia属に属する真菌類、Mycosphaerella属に属する真菌類、マイコバクテリウム属に属する細菌、ナイセリア属に属する細菌、ストレプトミセス属に属する細菌、ロドバクテリア属に属する細菌、鉤虫属に属する線虫類、シノラブディス属に属する線虫類、捻転胃虫属に属する線虫類、Lumbricus属に属する線虫類、Meilodogyne属に属する線虫類、Strongyloidus属に属する線虫類、又はPristionchus属に属する線虫類に由来する4−クマレート−CoAリガーゼ(EC6.2.1.12)であることを特徴とする上記[22]に記載の微生物。
[24]前記代謝経路の各酵素をコードする少なくとも1つの遺伝子配列の少なくとも1つのコピーが、前記微生物に組換えで導入されていることを特徴とする前記いずれかの一項に記載の微生物。
[25]天然のNADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物で過剰発現していることを特徴とする上記[24]に記載の微生物。
【0059】
[26]NADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物に組換えで導入されていることを特徴とする上記[24]に記載の微生物。
[27]前記NADPH:チトクロームP450レダクターゼが、シロイヌナズナを含むArabidopsisに属する植物、C.シネンシス、C.x paradisiを含むCitrusに属する植物、P.ヴァガリスを含むPhaseolusに属する植物、P.taedaを含むPinusに属する植物、P.deltoides、P.tremuloides、P.trichocarpaを含むPopulusに属する植物、S.tuberosumを含むSolanumに属する植物、ヴィティス ヴィニヘラを含むヴィティスに属する植物、Z.メイスを含むZeaに属する植物、又はAmmi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属を含む他の植物属に由来するNADPH:チトクロームP450レダクターゼ(EC1.6.2.4)であることを特徴とする上記[26]に記載の微生物。
[28]チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[29]フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[30]シンナメート4−ヒドロキシラーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物において前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していること特徴とする前記いずれかの一項に記載の微生物。
【0060】
[31]4−クマレート−CoAリガーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物において前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[32]リスベラトロールシンターゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物において前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[33]真菌であることを特徴とする前記いずれかの一項に記載の微生物。
[34]糸状菌であることを特徴とする上記[33]に記載の微生物。
[35]アスペルギルス属に属する微生物であることを特徴とする上記[34]に記載の微生物。
【0061】
[36]菌株アスペルギルス niger又は麹菌であることを特徴とする上記[35]に記載の微生物。
[37]酵母であることを特徴とする上記[33]に記載の微生物。
[38]Saccharomyces属、Kluyveromyces属、カンジダ属、ピチア属、Debaromyces属、Hansenula属、Yarrowia属、Zygosaccharomyces属又はシゾサッカロミセス属に属する微生物であることを特徴とする上記[37]に記載の微生物。
[39]サッカロマイセス・セレヴィシエ、S.kluyveri、S.bayanus、S.exiguus、S.sevazzi、S.uvarum、Klyuveromyces lactis、K.marxianus var.marxianus、K.thermotolerans、Candida utilis、C.tropicalis、Pichia stipidis、P.pastoris、P.sorbitophila、Debaromyces hansenii、Hansenula polymorpha、Yarrowia lipolytica、Zygosaccharomyces rouxii又はシゾサッカロミセス・ポンベの菌株であることを特徴とする上記[38]に記載の微生物。
[40]細菌であることを特徴とする[1]〜[32]に記載の微生物。
【0062】
[41]エシェリキア属又はLactococcus属に属する微生物であることを特徴とする上記[40]に記載の微生物。
[42]大腸菌又は乳酸連鎖球菌の菌株であることを特徴とする上記[41]に記載の微生物。
[43]発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有することを特徴とする上記[1]に記載の微生物。
[44]シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する上記[1]に記載の微生物。
[45]適当なリスベラトロール産生代謝経路を有する非植物細胞を炭素基質に4−クマル酸の外部源の実質的な非存在下において接触させる工程を含むことを特徴とする、リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
【0063】
[46]前記非植物細胞が、真菌及び細菌からなる群より選択されることを特徴とする上記[45]に記載の方法。
[47]前記非植物細胞が、酵母からなる群より選択される真菌であることを特徴とする上記[46]に記載の方法。
[48]前記酵母がSaccharomyes種から選択されることを特徴とする上記[47]に記載の方法。
[49]前記非植物細胞が、上記[1]〜[44]に記載の微生物の細胞であることを特徴とする上記[45]〜[48]に記載の方法。
[50]前記炭素基質が、単糖類、小糖類及び多糖類からなる発酵性炭素基質の群より選択されることを特徴とする上記[45]〜[49]のいずれかに記載の方法。
【0064】
[51]前記発酵性炭素基質が、グルコース、フルクトース、ガラクトース、キシロース、アラビノース、マンノース、スクロース、ラクトース、エリトロース、スレオース、リボースであることを特徴とする上記[50]に記載の方法。
[52]前記炭素基質が、非発酵性炭素基質からなる群より選択されることを特徴とする上記[45]〜[49]のいずれかに記載の方法。
[53]前記非発酵性炭素基質が、エタノール、アセテート、グリセロール、ラクテートであることを特徴とする上記[52]に記載の方法。
[54]前記非発酵性炭素基質が、アミノ酸からなる群より選択されることを特徴とする上記[52]に記載の方法。
[55]前記非発酵性炭素基質が、フェニルアラニン及びチロシンからなる群より選択されることを特徴とする上記[54]に記載の方法。
【0065】
[56]フェニルアラニンアンモニアリアーゼ、シンナメート4−ヒドロキシラーゼ、4−クマレート−CoA−リガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
[57]チロシンアンモニアリアーゼ、4−クマレート−CoAリガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
[58]さらに、生産されたリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を乳製品又は飲み物中の栄養補助食品として使用することを含む上記[45]〜[57]のいずれかに記載の方法。
[59]前記リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体をビール中の栄養補助食品として使用することを特徴とする上記[58]に記載の方法。
【0066】
本発明の上記説明の容易な理解を助けるために、添付の図面を参照されたい。
【0067】
以下の限定されない実施例により本発明をさらに説明し、例示する。
【0068】
(実施例1)
PAL、TAL、C4H、CPR、4CL及びVSTをコードする遺伝子の単離
フェニルアラニンアンモニアリアーゼ(PAL2)(Cochraneら、2004年;配列番号1、配列番号2)、シンナメート4−ヒドロキシラーゼ(C4H)(Mizutaniら、1997年;配列番号3、配列番号4)及び4−クマレート:コエンザイムAリガーゼ(4CL1)(ハンバーガー及びハルブロック(Hamberger and Hahlbrock)、2004年;エルチングら(Ehlting et al.)、1999年;配列番号5、配列番号6)を、PCRにより、表1のプライマーを用いて、シロイヌナズナのcDNA(バイオキャット社(BioCat)、ハイデルベルグ、ドイツ)からPCRにより単離した。コハク酸とクマロイル−CoAのそれぞれに対する好ましい速度パラメーターのために、幾つかのシロイヌナズナ相同体のうち、PAL2と4CL1を選択した(コクレンら(Cochrane et al.)、2004年;ハンバーガー及びハルブロック(Hamberger and Hahlbrock)、2004年;エルチングら(Ehlting et al.)、1999年)。
【0069】
Rheum tataricumに由来するリスベラトロールシンターゼ(VST)のコード配列(サマピトら(Samappito et al)、2003年;配列番号7、配列番号8)及びロドバクテリア capsulatusに由来するチロシンアンモニアリアーゼ(TAL)(キンドら(Kyndt et al.)、2002年;配列番号11、配列番号12)は、S.セレヴィシエでの発現のために、www.entelechon.comのオンラインサービスバックトランスレーションツールを用いてコドン最適化を行って、それぞれ配列番号9と10及び配列番号13と14を得た。合成遺伝子組立てのためのオリゴはMWGバイオテック社(MWG Biotech)で構築され、合成遺伝子は、以下に説明するマーチンら(Martin et al.)(2003年)の若干改良された方法プロトコールを用いて、PCRにより組み立てられた。
【0070】
【表1】

【0071】
合成遺伝子の組立てのためにMWG社のプライマーをミリQ水に100pmole/μlの濃度に溶解した。各プライマーの5μlのアリコートをトータルミックス中で一緒にし、次にミリQ水で10倍希釈した。遺伝子は、融合DNAポリメラーゼ(Finnzymes)のために50μlあたり鋳型として5μlの希釈トータルミックスを用いたPCRにより組み立てた。PCRプログラムは以下の通りである。最初に98℃で30秒間、次に、1000塩基対あたり、98℃で10秒間、40℃で1分間及び72℃で1分間の30サイクル、及び最後に72℃で5分間とした。得られたPCR反応物から、20μl
を1%アガロースゲルで精製した。その結果はPCRスメアであり、欲する大きさの周囲の領域をアガロースゲルから切り取り、QiaQuick Gel Extraction Kit(キアゲン社(Qiagen))を用いて精製した。表1の(TAL及びVST用の)外側のプライマーによる最終的なPCRは、要求されたTAL遺伝子とVST遺伝子を与えた。Quickchange部位特異的変異誘発IIキット(ストラタジン社(Stratagene)、ラホーヤ、カリフォルニア州)を用いるか、又は幾つかの異なる大腸菌サブクローンに由来するオーバーラップエラーフリーDNAストレッチからPCRを用いて点突然変異を補正した。
【0072】
シロイヌナズナ(AR2)に由来するNADPH:チトクロームP450レダクターゼ(CPR)(ミズタニ及びオータ(Mizutani and Ohta)、1998年;配列番号17、配列番号18)及びS.セレヴィシエ(CPR1)に由来するNADPH:チトクロームP450レダクターゼ(CPR1)(アオヤマら(Aoyama et al.)、1978年;配列番号15、配列番号16)は、表1のプライマーを用いて、シロイヌナズナのcDNA(バイオキャット社(BioCat)、ハイデルベルグ、ドイツ)及びS.セレヴィシエのゲノムDNAからそれぞれ単離した。
【0073】
(実施例2)
PAL発現用の酵母ベクターの構築
実施例1に記載のように単離されたPALをコードする遺伝子を、EcoR1とSpe1の制限部位(表1)を含む5’突出を有するフォワードプライマーとリバースプライマーを用いるPCRにより再増幅した。増幅されたPAL PCR産物をEcoR1/Spe1により消化し、EcoR1/Spe1で消化されたpESC−URAベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−URA−PALを得た。遺伝子の配列は2つの異なるクローンの配列決定により検証した。
【0074】
(実施例3)
PAL及びC4H発現用の酵母ベクターの構築
実施例1に記載のように単離されたC4Hをコードする遺伝子を、Xho1とKpn1の制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いるPCRにより増幅した。増幅されたC4H PCR産物をXho1/Kpn1により消化し、同様に消化されたpESC−URA−PALベクターに連結した。得られたプラスミドpESC−URA−PAL−C4Hは、分岐GAL1/GAL10プロモーターの制御下にPALとC4Hをコードする遺伝子を含有した。C4Hをコードする遺伝子の配列は2つの異なるクローンの配列決定により検証した。
【0075】
(実施例4)
4CL発現用酵母ベクターの構築
4CLをコードする遺伝子を実施例1に記載のように単離した。増幅された4CL PCR産物をXba1/BamH1により消化し、Spe1/BglIIで消化されたpESC−TRPベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−TRP−4CLを得た。pESC−TRP−4CLの2つの異なるクローンの配列を決定して、クローニングされた遺伝子の配列を検証した。
【0076】
(実施例5)
4CL及びVST発現用の酵母ベクターの構築
VSTをコードする遺伝子を実施例1に記載のように単離した。増幅された合成VST遺伝子をBamH1/Xho1により消化し、BamH1/Xho1で消化されたpESC−TRP−4CL(実施例4)に連結した。得られたプラスミドpESC−TRP−4CL−VSTは、分岐GAL1/GAL10プロモーターの制御下に4CLとVSTをコードする遺伝子を含んでいた。VSTをコードする遺伝子の配列はpESC−TRP−4CL−VSTの2つの異なるクローンの配列決定により検証した。
【0077】
(実施例6)
TAL発現用酵母ベクターの構築
TALをコードする遺伝子を実施例1に記載のように単離した。増幅された合成TAL遺伝子をEcoR1/Spe1により消化し、EcoR1/Spe1で消化されたpESC−URAベクターに連結した。得られたプラスミドpESC−URA−TALは、分岐GAL1/GAL10プロモーターの制御下にTALをコードする遺伝子を含有した。その配列はpESC−URA−TALの2つの異なるクローンの配列決定により検証した。
【0078】
(実施例7)
S.セレヴィシエ内在性CPRの過剰発現用酵母ベクターの構築
S.セレヴィシエに由来するCPR(CPR1)をコードする遺伝子を実施例1に記載のように単離した。増幅されたCPR1遺伝子をXho1/HindIIIで消化し、Xho1/HindIIIで消化されたpESC−LEUベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−LEU−CPR1を得た。その配列はpESC−LEU−CPR1の2つの異なるクローンの配列決定により検証した。
【0079】
(実施例8)
シロイヌナズナ CPR(AR2)の過剰発現用酵母ベクターの構築
シロイヌナズナに由来するCPR(AR2)をコードする遺伝子を実施例1に記載のように単離した。増幅されたAR2遺伝子をBamH1/Xho1で消化し、BamH1/Xho1で消化されたpESC−LEUベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−LEU−AR2を得た。その配列はpESC−LEU−AR2の2つの異なるクローンの配列決定により検証した。
【0080】
(実施例9)
酵母S.セレヴィシエにおいてPAL、C4H、4CL及びVSTを用いてリスベラトロールを導く経路の発現
適当な遺伝子マーカーを含む酵母菌株を実施例2、実施例3、実施例4、実施例5、実施例6、実施例7及び実施例8で記載したベクターにより別々に、又は組合せて形質転換した。酵母細胞の形質転換は、当分野で公知の方法にしたがって、例えば、コンピテント細胞又は電気穿孔法を用いて実施した(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0081】
S.セレヴィシエ菌株CEN.PK113−5D(MATa ura3)をベクターpESC−URA−PAL(実施例2)で形質転換して、FSSC−PAL菌株を得て、pESC−URA−PAL−C4H(実施例3)で形質転換して、FSSC−PALC4H菌株を得た。S.セレヴィシエ菌株FS01267(MATa trp1 ura3)はpESC−URA−PAL−C4H及びpESC−TRP−4CL(実施例4)により同時形質転換し、形質転換された菌株をFSSC−PALC4H4CLと命名した。同菌株は、pESC−URA−PAL−C4H及びpESC−TRP−4CL−VST(実施例5)によっても同時形質転換して、FSSC−PALC4H4CLVST菌株を得た。
【0082】
(実施例10)
S.セレヴィシエにおいてTAL、4CL及びVSTを用いてリスベラトロールを導く経路の発現
S.セレヴィシエ菌株CEN.PK113−5D(MATa ura3)をベクターpESC−URA−TAL(実施例6)で別に形質転換して、FSSC−TAL菌株を得た。S.セレヴィシエ菌株FS01267(MATa trp1 ura3)をpESC−URA−TAL(実施例6)及びpESC−TRP−4CL(実施例4)で同時形質転換し、形質転換された菌株をFSSC−TAL4CLと命名した。同菌株は、pESC−URA−TALとpESC−TRP−4CL−VST(実施例5)とによっても同時形質転換して、FSSC−TAL4CLVST菌株を得た。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0083】
(実施例11)
S.セレヴィシエにおける過剰発現内在性CPRによるリスベラトロールを導く経路の発現
S.セレヴィシエ菌株FS01277(MATa ura3 leu2 trp1)を、ベクターpESC−URA−PAL−C4H(実施例3)、ベクターpESC−TRP−4CL(実施例4)及びベクターpESC−LEU−CPR1(実施例7)により同時形質転換した。形質転換された菌株をFSSC−PALC4H4CLVSTCPRと命名した。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0084】
(実施例12)
S.セレヴィシエにおける過剰発現シロイヌナズナ CPR(AR2)によるリスベラトロールを導く経路の発現
S.セレヴィシエ菌株FS01277(MATa ura3 leu2 trp1)を、ベクターpESC−URA−PAL−C4H(実施例3)、ベクターpESC−TRP−4CL(実施例4)及びベクターpESC−LEU−AR2(実施例8)により同時形質転換した。形質転換された菌株をFSSC−PALC4H4CLVSTAR2と命名した。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0085】
(実施例13)
振盪フラスコによる組換え酵母菌株による発酵
前記組換え酵母菌株を滅菌した接種用ループにより寒天プレートに接種し、ビタミン類、微量元素、5g/lのグルコース及び40g/l又は100g/lのガラクトースを含有する200mlの合成無機培地(ベルダインら(Verduyn et al.)、1992年)で増殖させた。500mlのコック付き振盪フラスコを3日間、30℃、160rpmでインキュベートした。
【0086】
(実施例14)
リスベラトロールの抽出
5000gで5分間の遠心により細胞を回収した。50mlの上清アリコートを20mlの酢酸エチルで一回抽出した。この酢酸エチルを凍結乾燥し、乾燥生成物を0.7mlメタノールで再溶解し、ろ過してHPLCバイアルに入れた。
【0087】
200ml培地から得られた細胞ペレットを1〜2mlの水に溶解し、3本の迅速調製チューブに分割し、ガラス玉で砕いた。これらの3チューブからの粗製抽出物を、50mlの縫工チューブ中の10mlの100%メタノール中にプールし、4℃の冷暗部屋でロータリーチャンバーにより48時間抽出した。48時間後、細胞デブリスを遠心により5分間、5000gで除去し、該メタノールを一晩の凍結乾燥により除去した。乾燥残留物を1mlのリン酸−クエン酸緩衝液(pH5.4)に再溶解し、アーモンドに由来する10単位のベータ−グルコシダーゼ(シグマ社(Sigma)製)を加えて、推定上グルコシド結合とされる構造物からリスベラトロールを遊離させた。混合物を3時間、37℃でインキュベートし、次に、1mlの酢酸エチルで2回抽出した。一緒にした酢酸エチルを凍結乾燥し、乾燥残留物を0.7mlのメタノールに再溶解し、ろ過してHPLCバイアルに入れた。
【0088】
(実施例15)
リスベラトロールの分析
(薄層クロマトグラフィー)
同一のTLCプレートによるケイヒ酸、クマル酸及びリスベラトロールの迅速な分離を可能とする薄層クロマトグラフィーに基づく方法を迅速なスクリーニング分析のために開発した。細胞と上清の両方を含む1mlの培養物アリコートを500マイクロリットルの酢酸エチルで抽出し、微小遠心機により13000rpmで30秒間遠心した。該酢酸エチルを乾燥し、メタノールに再溶解した。抽出物を、蛍光指示薬を含むシリカGプレート(0.2mm Alugram SIL G/UV254、マシュレ・ナーゲル社(Macherey-Nagel)製)により分析した。移動相はクロロホルム、酢酸エチル及び蟻酸の混合物(25:10:1)であった。
【0089】
(HPLC)
ケイヒ酸、クマル酸及びリスベラトロールの定量的分析のために、試料を、λ=306nmのUVダイオードアレイ検出前に高速液体クロマトグラフィー(HPLC)アジレント(Agilent)シリーズ1100システム(ヒューレットパッカード社(Hewlett Packard)製)による分離に供した。フェノメネックス社(Phenomenex)(トランス、カリフォルニア州、アメリカ合衆国)製Luna3マイクロメートルC18(100×2.00mm)カラムを40℃で用いた。移動相として、アセトニトリルとミリQ水(両方とも50ppmのトリフルオロ酢酸を含む)のグラジエントを0.4ml/分の流速で用いた。グラジエントのプロフィールは20分間にわたる15%アセトニトリルから100%アセトニトリルへのリニアであった。溶出時間はクマル酸で約3.4分、遊離トランス−リスベラトロールで5.5分、ケイヒ酸で6.8分であった。
【0090】
純粋なリスベラトロールスタンダードはケーマンケミカル社(Cayman chemical company)から購入し、純粋なクマル酸スタンダードとケイヒ酸スタンダードはシグマ社(Sigma)から購入した。
【0091】
(結果)
FSSC−PALC4H4CLVST菌株とFSSC−TAL4CLVST菌株を実施例13に記載のように100g/lのガラクトースで培養し、実施例14と実施例15にしたがって細胞内リスベラトロールの含量を分析した。さらに、空のベクターpESC−URA及びpESC−TRPのみを含むコントロール菌株のFSSCコントロールを含めた。HPLC分析は、FSSC−PALC4H4CLVST菌株とFSSC−TAL4CLVST菌株がトランス−リスベラトロールと同じ5.5分の保持時間を有する成分を含むことを示した(図4)。この結果は、約306nmのλmaxを有する純粋なトランス−リスベラトロールの吸収スペクトルに類似するUV吸収スペクトル(図5)によっても確認された。
【0092】
したがって、これらの結果は、トランス−リスベラトロールのインビボ生産を導くS.セレヴィシエの活性フェニル−プロパノイド経路の存在を示した。明確な増殖条件下にバッチ培養及び連続培養で菌株を培養し、及び/又は個々の酵素の発現/活性を最適化することによってリスベラトロールの生産が最も高い可能性で向上できるだろう。
【0093】
(実施例16)
大腸菌におけるTAL発現用細菌ベクターの構築
実施例1に記載したように単離されたTALをコードする遺伝子を、フォワードプライマー5’−CCG CTCGAG CGG ATG ACC CTG CAA TCT CAA ACA GCT AAA G−3’(配列番号33)とリバースプライマー5’−GC GGATCC TTA AGC AGG TGG ATC GGC AGC T−3’(配列番号34)(制限酵素部位XhoIとBamHIをそれぞれ有する5’突出を有する)を用いて、プラスミドpESC−URA−TAL(実施例6)から、PCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物のXhoIとBamHIとで消化したpET16bベクター(ノバゲン社(Novagen)製)への連結が可能となり、pET16b−TALが得られた。pET16bベクターはアンピシリン耐性遺伝子とT7プロモーターの両方を含んでいた。よって、上記手順は、T7プロモーターの制御下にTALをコードする遺伝子を含み、抗生物質選択マーカーを有するベクターをもたらした。TALをコードする遺伝子の配列はpET16b−TALの1つのクローンの配列決定により検証した。
【0094】
(実施例17)
大腸菌における4CLとVSTの発現用細菌ベクターの構築
実施例1に記載したように単離されたVSTをコードする遺伝子を、制限酵素BamHIとXhoIとにより4CLとVSTをコードする遺伝子を含む消化プラスミドpESC−TRP−4CL−VST(実施例5)から切り出した。カナマイシン耐性遺伝子を含有し、BamHIとSalIで消化したpET26bベクター(ノバゲン社(Novagen)製)にこのVST遺伝子を連結してpET26b−VSTを得た。制限酵素XhoIとSalIは適合性末端を有し、適当な連結を可能とした。pET26bベクターはカナマイシン耐性遺伝子とT7プロモーターの両方を含んでいた。よって、上記手順は、T7プロモーターの制御下にVSTをコードする遺伝子を含み、抗生物質選択マーカーを有するベクターをもたらした。
【0095】
実施例1に記載したように単離された4CLをコードする遺伝子を、フォワードプライマー5’−TG CCATGG CA ATGGCGCCAC AAGAACAAGC AGTTT−3’(配列番号35)とリバースプライマー5’−GC GGATCC CCT TCA CAA TCC ATT TGC TAG TTT TGCC−3’(配列番号36)(制限酵素部位NcoIとBamHIをそれぞれ有する5’突出を有する)を用いて、プラスミドpESC−URA−4CL−VST(実施例5)から、PCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物のNcoIとBamHIとで消化したpET16bベクター(ノバゲン社(Novagen)製)への連結を可能とした。得られたプラスミドpETl6b−4CLは、T7プロモーターの制御下に4CLをコードする遺伝子を含んでいた。T7プロモーターと4CLをコードする遺伝子との両方を、フォワードプライマー5’−TT GCGGCCGC AAA TCT CGA TCC CGC GAA ATT AAT ACG−3’(配列番号37)とリバースプライマー5’−CG CTCGAG CCT TCA CAA TCC ATT TGC TAG TTT TGCC−3’(配列番号38)(制限酵素部位NotIとXhoIをそれぞれ有する5’突出を有する)を用いて、プラスミドpETl6b−4CLから、PCRにより1つの断片として再増幅した。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、連結前にNotIXhoIとで消化したプラスミドpET26b−VSTへの制限PCR産物の連結を可能とした。得られたプラスミドpET26b−VST−4CLは、それぞれが個々のT7プロモーターの制御下にある2つの遺伝子(4CL及びVST)を含んでいた。
【0096】
(実施例18)
大腸菌において、TAL、4CL及びVSTを用いてリスベラトロールを導く経路の発現
細菌細胞の形質転換を当分野で公知の方法にしたがって、例えば、コンピテント細胞又は電気穿孔法を用いて実施した(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。大腸菌株BL21(DE3)(ノバゲン社(Novagen)製)を2種類のベクターpET16b−TAL(実施例16)とpET26b−VST−4CL(実施例17)とにより同時形質転換して、FSEC−TAL4CLVST菌株を得た。さらに、大腸菌株BL21(DE3)を、2種類の空のベクターpET16b(ノバゲン社(Novagen)製)とpET26b(ノバゲン社(Novagen)製)とで同時形質転換して、コントロール菌株として用いられるFSECコントロール菌株を得た。形質転換体は、100μg/mlのアンピシリンと60μg/mlのカナマイシンを含むルリア−ベルターニ(LB)培地により選択した。
【0097】
(実施例19)
振盪フラスコでの組換え大腸菌株による発酵
大腸菌BL21(DE3)の前培養物を、100μg/mlのアンピシリンと60μg/mlのカナマイシンを含む7mlのLB培地で、160rpm、37℃にてガラス管中で増殖させた。指数関数的に成長する前培養物を、50g/lのグルコース、5g/lのK2HPO4、80μg/mlのアンピシリン及び50μg/mlのカナマイシンを追加した200mlのLB培地を含む500mlバッフル付き振盪フラスコの接種に用いて、160rpm、37℃でインキュベートした。5時間後、3遺伝子(TAL、4CL及びVST)のそれぞれの上流に存在するT7プロモーターの誘導因子として、イソプロピルβ−チオガラクトピラノシド(IPTG)を1mMの最終濃度で加えた。37℃での48時間のインキュベーション時間の後、細胞を採取し、抽出手順に供し、産生したリスベラトロールの有無を分析した。
【0098】
(実施例20)
大腸菌のリスベラトロールの抽出と分析
抽出及び分析は、実施例14と実施例15に記載の方法を用いて実施した。
【0099】
(結果)
菌株FSEC−TAL4CLVST及びFSECコントロールを、実施例19に記載のように50g/lグルコースで培養し、実施例14と実施例15にしたがって細胞内リスベラトロールの含量を分析した。HPLC分析は、FSEC−TAL4CLVST菌株が、クマル酸と同じ3.4分の保持時間を有するかなりの量の成分を含むことを示した(図6)。しかし、その抽出物はトランス−リスベラトロールと同じ時間で溶出する成分を含んでいなかった。したがって、この結果は、チロシンアンモニアリアーゼ(TAL)は実際に活性はあるが、検出可能な量のリスベラトロールの生産を導くことはないことを示した。しかし、リスベラトロール形成の欠如は、i)非機能性クマレート−CoAリガーゼ(4CL)、ii)非機能性リスベラトロールシンターゼ(VST)、iii)非最適培養条件又はTALの非最適発現/活性により引き起こされるか、又はクマル酸の他の産物への分岐により引き起こされる非常に低いレベルのクマル酸との結果でありうるだろう。この仮説を評価するために、実施例19に記載の類似の培地を用いたが、ここでは20mg/lのクマル酸の存在下で菌株を増殖させた。FSEC−TAL4CLVSTの抽出物の引き続くHPLC分析は、トランス−リスベラトロールと同じ保持時間あたりに一群のピークを実際に示し、これはFSコントロールの抽出物には観察されなかった(図6)。実際、5.5分の保持時間を有するピークのUV吸収スペクトルは純粋なリスベラトロールのスペクトルに類似したが(図7)、そのようなスペクトルはコントロール菌株でのピークでは得られなかった。したがって、これらの結果は、リスベラトロールの生産を導きうる大腸菌の活性フェニルプロパノイド経路の存在を強く示唆する。明確な増殖条件下にバッチ培養及び連続培養で菌株を培養し、及び/又は個々の酵素の発現/活性を最適化することによって、クマル酸添加なしのリスベラトロールの生産が最も高い可能性で達成できるだろう。
【0100】
(実施例21)
乳酸連鎖球菌におけるPALとC4Hの発現用細菌ベクターの構築
下記実施例に使用されるプラスミドpSH71とその誘導体は、大腸菌と乳酸連鎖球菌に由来する複数の複製開始点を有する二機能性シャトルベクターである。それによって、宿主域特異性は大腸菌及び他の種の乳酸菌にわたる。したがって、乳酸連鎖球菌の形質転換は通常は問題なく進むが、他の種の乳酸菌の推定上の困難な形質転換は組換えプラスミドの構築のための中間ホストとして大腸菌を用いることにより克服することができる。プラスミドは1つ以上のマーカー遺伝子を持っており、それらを持つ微生物を、それらを持たない微生物から選択することができる。乳酸連鎖球菌に用いられる選択システムは、優性マーカー、例えば、エリスロマイシンとクロラムフェニコールに対する耐性に基づくが、糖質代謝に関与する遺伝子、ペプチダーゼ及び食品用マーカーに基づくシステムも記載されてきた。さらに、プラスミドは、組換え遺伝子の発現を可能とするプロモーター配列とターミネーター配列を含む。好適なプロモーターは、乳酸連鎖球菌の遺伝子、例えば、lacAから取られる。さらに、プラスミドはDNA断片のクローニングとそれに引き続く組換え体の同定を容易にする好適な独自の制限部位を含む。
【0101】
下記の実施例において、プラスミドは、pSH71−ERYrと呼ばれるエリスロマイシン耐性遺伝子又はpSH71−CMrと呼ばれるクロラムフェニコール耐性遺伝子のいずれかを含む。
【0102】
実施例1に記載のように単離されたPALをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を乳酸連鎖球菌に由来するlacAプロモーターを含む消化pSH71−ERYrベクターに連結することができる。得られたプラスミドpSH71−ERYr−PALは、乳酸連鎖球菌由来のlacAプロモーターの制御下にPALをコードする遺伝子を含有する。
【0103】
実施例1に記載のように単離されたC4Hをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−CMrベクターに連結して、pSH71−CMr−C4Hを得ることができる。lacAプロモーターとC4Hをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpSH71−CMr−C4HからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpSH71−ERYr−PALに連結することができる。得られたプラスミドpSH71−ERYr−PAL−C4Hは、個々のlacAプロモーターの制御下にPALとC4Hをそれぞれコードする遺伝子を含む。PALとC4Hをコードする遺伝子の配列を、pSH71−ERYr−PAL−C4Hの2つの異なるクローンの配列決定により検証する。
【0104】
(実施例22)
乳酸連鎖球菌におけるTAL発現用細菌ベクターの構築
実施例1に記載のように単離されたTALをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いてプラスミドpESC−URA−TAL(実施例6)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−ERYrベクターに連結することができる。得られたプラスミドpSH71−ERYr−TALは、乳酸連鎖球菌に由来するlacAプロモーターの制御下にTALをコードする遺伝子を含む。TALをコードする遺伝子の配列を、pSH71−ERYr−TALの2つの異なるクローンの配列決定により検証する。
【0105】
(実施例23)
乳酸連鎖球菌における4CLとVSTの発現用細菌ベクターの構築
実施例1に記載のように単離された4CLをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いてプラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−CMrベクターに連結することができる。得られたプラスミドpSH71−CMr−4CLは、乳酸桿菌(Lactobacillus lactis)由来のlacAプロモーターの制御下に4CLをコードする遺伝子を含有する。
【0106】
実施例1に記載のように単離されたVSTをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−ERYrベクターに連結することができる。得られたプラスミドpSH71−ERYr−VSTは、乳酸連鎖球菌に由来するlacAプロモーターの制御下にVSTをコードする遺伝子を含む。lacAプロモーターと、VSTをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpSH71−ERYr−VSTからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpSH71−CMr−4CLに連結することができる。得られたプラスミドpSH71−CMr−4CL−VSTは、個々のlacAプロモーターの制御下に4CL及びVSTをそれぞれコードする遺伝子を含む。4CLとVSTをコードする遺伝子の配列を、pSH71−CMr−4CL−VSTの2つの異なるクローンの配列決定により検証する。
【0107】
(実施例24)
乳酸連鎖球菌においてリスベラトロールを導く経路の発現
乳酸連鎖球菌株を、実施例21、実施例22及び実施例23に記載されたベクターで別々に、又は組合せて形質転換する。細菌細胞の形質転換は当分野で公知の方法にしたがって、例えば、コンピテント細胞又は電気穿孔法を用いて実施した(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体は、抗生物質のエリスロマイシンとクロラムフェニコールとを含有する培地上で選択し、画線を同培地上で精製した。
【0108】
別に、乳酸連鎖球菌株MG1363をベクターpSH71−ERYr−TAL(実施例22)により形質転換して、菌株FSLL−TALを得て、pSH71−ERYr−PAL−C4H(実施例21)により菌株FSLL−PALC4Hを得て、pSH71−CMr−4CL−VST(実施例23)により菌株FSLL−4CLVSTを得る。さらに、乳酸連鎖球菌株MG1363を、pSH71−ERYr−TAL(実施例22)及びpSH71−CMr−4CL−VST(実施例23)により同時形質転換し、この形質転換された菌株をFSLL−TAL4CLVSTと命名する。同菌株をpSH71−ERYr−PAL−C4H(実施例21)とpSH71−CMr−4CL−VST(実施例23)によっても同時形質転換して、菌株FSLL−PALC4H4CLVSTを得る。
【0109】
(実施例25)
発酵槽での組換え乳酸連鎖球菌株による発酵
組換え酵母菌株は、バッチ培養、流加バッチ培養又はケモスタット培養として操作される発酵槽で増殖させることができる。
【0110】
(バッチ培養と流加バッチ培養)
微生物を、1.5リットルの有効容積のバッフル付きバイオリアクターで、嫌気的、好気的又は微好気的条件で増殖させる。すべての培養物を30℃で、350rpmにてインキュベートする。pH6.6で一定になるように10MのKOHの自動添加により維持する。細胞を、以下の成分を補充した合成MS10培地中でラクトースにより増殖させて好気的条件下の増殖を可能とする:MnSO4(1.25×10−5g/l)、チアミン(1mg/l)及びDL−6,8−チオクト酸(2.5mg/l)。ラクトース濃度は、例えば、50g/lである。3倍高い濃度のK2HPO4とKH2PO4で緩衝化した上記培地により、振盪フラスコ内、30℃で増殖させた前培養物の細胞をバイオリアクターに接種する。接種前にN2(純度99.998%)を培地に流すことにより、かつ培養中にバイオリアクターの上部空間にN250ml/分の定速を維持することにより好気的条件を確保する。微好気的条件及び好気的培養のために用いられるバイオリアクターは、空気(DOT、100%)及びN2(DOT、0%)で較正されたポーラログラフィー酸素センサーを備える。好気的条件はバイオリアクターに空気を1vvmの速度で拡散させてDOTが80%を超えることを確保することで得られる。微好気的実験中、N2と大気の空気の混合物からなる気体を0.25vvmの速度でリアクターに拡散することによりDOTが5%の一定に保たれる。
【0111】
(ケモスタット培養物)
ケモスタット培養物において、細胞は例えば1Lの有効容積のアプリコン(Applikon)実験室用発酵槽で、30℃、350rpmで増殖させることができる。希釈率(D)は、異なった値、例えば、0.050h−1、0.10h−1、0.15h−1又は0.20h−1に設定することができる。消泡剤(50μl/l)を追加した上記成長培地を用いて、5MのKOHの自動追加によりpHを一定、例えば6.6に保つ。ラクトースの濃度を異なる値、例えば、3.0g/l、6.0g/l、12.0g/l、15.0g/l又は18.0g/lに設定することができる。バイオリアクターに1mg/lの初期バイオマス濃度で接種し、指数増殖期の終わりにフィードポンプを作動させる。
【0112】
嫌気的定常状態は50ml/分のN2(純度99.998%)をバイオリアクターの上部空間に導入することにより得る。異なる無酸素性定常状態は、N2(純度99.998%)と大気の空気とから多様な比率で構成される250ml/分のガスをリアクターに拡散させることにより得ることができる。バイオリアクターに空気(100%DOT)とN2(0%DOT)を拡散させることにより酸素電極を較正する。
【0113】
すべての条件で、ガスはバイオリアクターに導入する前に滅菌ろ過する。オフガスは、−8℃未満に冷却した冷却器を経て、アコースティックガスアナライザーによりそのCO2とO2の体積含量が分析される。
【0114】
少なくとも5滞留時間後、及びバイオマスと発酵最終生成物の濃度が最後の少なくとも2滞留時間にわたって変化しない(5%相対偏差未満)の場合、培養は定常状態にあると考えられる。
【0115】
(実施例26)
乳酸連鎖球菌におけるリスベラトロールの抽出と分析
抽出と分析は実施例14と実施例15に記載された方法を用いて実施する。
【0116】
(実施例27)
アスペルギルス属に属する種におけるPALとC4Hの発現用真菌ベクターの構築
下記実施例で使用されるプラスミドは、黒色アスペルギルスと麹菌で自己プラスミド複製も維持する、アスペルギルス nidulans由来のAMA1開始複製配列を含有するpARp1から得られる(ゲムスら(Gems et al.)、1991年)。さらに、プラスミドは大腸菌の複製配列を含むシャトルベクターであるので、黒色アスペルギルス及び麹菌での固有の困難な形質転換が、組換えプラスミドの構築のために中間ホストとして大腸菌を用いることにより克服することができる。プラスミドは1つ以上のマーカー遺伝子を持っており、それらを持つ微生物を、それらを持たない微生物から選択することを可能とする。選択システムは、優性マーカー、例えば、ハイグロマイシンB、フレオマイシン及びブレオマイシンに対する耐性、又は異種マーカー、例えば、アミノ酸及びpyrG遺伝子に基づくことができる。さらに、プラスミドは組換え遺伝子の発現を可能とするプロモーター配列とターミネーター配列を含む。好適なプロモーターは、アスペルギルス nidulansの遺伝子、例えば、alcA、glaA、amy、niaD及びgpdAから得られる。さらに、プラスミドはDNA断片のクローニング及びその後の組換え体の同定を容易にする好適な独自の制限部位を含む。
【0117】
下記の実施例で用いられるプラスミドは強力な構成的gpdAプロモーターと栄養要求性マーカーを含み、これらすべてがアスペルギルス nidulansを起源とする。メチオニン生合成に関与する遺伝子methGを含むプラスミドをpAMA1−METと命名する。ヒスチジン生合成に関与する遺伝子hisAを含むプラスミドをpAMA1−HISと命名する。
【0118】
実施例1に記載のように単離されたPALをコードする遺伝子を、好適な制限部位を含
む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物をアスペルギルス nidulansに由来するgpdAプロモーターを含む消化pAMA1−METベクターに連結することができる。得られたプラスミドpAMA1−MET−PALは、アスペルギルス nidulans由来のgpdAプロモーターの制御下にPALをコードする遺伝子を含有する。
【0119】
実施例1に記載のように単離されたC4Hをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pAMA1−HISベクターに連結してpAMA1−HIS−C4Hを得ることができる。gpdAプロモーターと、C4Hをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpAMA1−HIS−C4HからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpAMA1−MET−PALに連結することができる。得られたプラスミドpAMA1−MET−PAL−C4Hは、アスペルギルス nidulansに由来する個々のpgdAプロモーターの制御下にPALとC4Hをそれぞれコードする遺伝子を含む。PALとC4Hをコードする遺伝子の配列をpAMA1−MET−PAL−C4Hの2つの異なるクローンの配列決定により検証する。
【0120】
(実施例28)
アスペルギルス属に属する種におけるTALの発現用真菌ベクターの構築
実施例1に記載のように単離されたTALをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−TAL(実施例6)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pAMA1−METベクターに連結することができる。得られたプラスミドpAMA1−MET−TALは、アスペルギルス nidulansに由来するgpdAプロモーターの制御下にTALをコードする遺伝子を含む。TALをコードする遺伝子の配列をpAMA1−MET−TALの2つの異なるクローンの配列決定により検証する。
【0121】
(実施例29)
アスペルギルス属に属する種における4CLとVSTの発現用真菌ベクターの構築
実施例1に記載のように単離された4CLをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を、アスペルギルス nidulans由来のgpdAプロモーターを含む消化pAMA1−HISベクターに連結することができる。得られたプラスミドpAMA1−HIS−4CLは、アスペルギルス nidulansに由来するgpdAプロモーターの制御下に4CLをコードする遺伝子を含む。
【0122】
実施例1に記載のように単離されたVSTをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pAMA1−METベクターに連結してpAMA1−MET−VSTを得ることができる。gpdAプロモーターと、VSTをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpAMA1−MET−VSTからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpAMA1−HIS−4CLに連結することができる。得られたプラスミドpAMA1−HIS−4CL−VSTは、アスペルギルス nidulansに由来する個々のpgdAプロモーターの制御下にC4LとVSTをそれぞれコードする遺伝子を含む。4CLとVSTをコードする遺伝子の配列をpAMA1−HIS−4CL−VSTの2つの異なるクローンの配列決定により検証する。
【0123】
(実施例30)
黒色アスペルギルスにおいてリスベラトロールを導く経路の発現
黒色アスペルギルス菌株を、実施例27、実施例28及び実施例29に記載されたベクターで、別々に、又は組合せて形質転換する。細菌細胞の形質転換は当分野で公知の方法にしたがって、例えば、電気穿孔法又は接合を用いて実施する(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体は、メチオニン及び/又はヒスチジンを欠く最小培地により選択する。
【0124】
ヒスチジンとメチオニンに対して栄養要求性のあるアスペルギルス nigerの菌株、例えば、別々にFGSC A919菌株(http://www.fgsc.netを参照)をベクターpAMA1−MET−TAL(実施例28)により形質転換して菌株FSAN−TALを得て、pAMA1−MET−PAL−C4H(実施例27)により菌株FSAN−PALC4Hを得て、pAMA1−HIS−4CL−VST(実施例29)により菌株FSAN−4CLVSTを得る。さらに、黒色アスペルギルス菌株FGSC A919をpAMA1−MET−TAL(実施例28)とpAMA1−HIS−4CL−VST(実施例29)とで同時形質転換して、形質転換菌株をFSAN−TAL4CLVSTと名付ける。また、同菌株をpAMA1−MET−PAL−C4H(実施例27)及びpAMA1−HIS−4CL−VST(実施例29)によっても同時形質転換して菌株FSAN−PALC4H4CLVSTを得る。
【0125】
(実施例31)
麹菌においてリスベラトロールを導く経路の発現
PAL、C4H及び4CLをコードする天然の遺伝子組を含有する麹菌の菌株(非特許文献18)であって、メチオニンに対して栄養要求性のある菌株をベクターpAMA1−MET−VST(実施例29)により形質転換して、菌株FSAO−VSTを得る。真菌細胞の形質転換は当分野で公知の方法にしたがって、例えば、電気穿孔法又は接合を用いて実施する(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体はメチオニンを欠く培地により選択する。
【0126】
(実施例32)
培養槽における黒色アスペルギルス及び麹菌の組換え菌株による発酵
組換え酵母菌株は、バッチ培養、流加バッチ培養又はケモスタット培養として操作される発酵槽で増殖させることができる。
【0127】
(バッチ培養及び流加バッチ培養)
微生物は、好気的条件下で、1.5リットルの有効容積のバッフル付きバイオリアクターで増殖させる。すべての培養物を30℃で、500rpmにてインキュベートする。pH6.0で一定となるように10MのKOHの自動添加により維持し、好気的条件は空気を1vvmの速度でバイオリアクターに拡散させてDOTが80%を超えることを確保することで得られる。以下の成分からなる合成培地中で細胞をグルコースで増殖させてバッチ培養で増殖させる:7.3g/lの(NH4)2SO4、1.5g/lのKH2PO4、1.0g/lのMgSO4・7H2O、1.0g/lのNaCl、0.1g/lのCaCl2・2H2O、0.1ml/lのシグマ社製の消泡剤、7.2mg/lのZnSO4・7H2O、1.3mg/lのCuSO4・5H2O、0.3mg/lのNiCl2・6H2O、3.5mg/lのMnCl2・4H2O及び6.9mg/lのFeSO4・7H2O。グルコース濃度は、例えば、10g/l、20g/l、30g/l、40g/l又は50g/lである。流加バッチ培養で細胞を増殖させるために、培地は、バッチ相中の7.3g/lの(NH4)2SO4、4.0g/lのKH2PO4、1.9g/lのMgSO4・7H2O、1.3g/lのNaCl、0.10g/lのCaCl2・2H2O、0.1ml/lのシグマ社製の消泡剤、7.2mg/lのZnSO4・7H2O、1.3mg/lのCuSO4・5H2O、0.3mg/lのNiCl2・6H2O、3.5mg/lのMnCl2・4H2O及び6.9mg/lのFeSO4・H2Oからなる。次に、リアクターに、例えば、285g/kgのグルコースと42g/kg(NH4)2SO4を供給する。
【0128】
前バッチからの遊離した菌糸体を用いて、バッチ培養物及び流加バッチ培養物に接種する。2.109/l濃度の胞子を用いて、pH2.5の前バッチ培養物に接種する。以下の成分が添加された29gの米により凍結乾燥胞子を増殖させて胞子を得る:6mlの15g/lのスクロース、2.3g/lの(NH4)2SO4、1.0g/lのKH2PO4、0.5g/lのMgSO4・7H2O、0.50g/lのNaCl、14.3mg/lのZnSO4・7H2O、2.5mg/CuSO4・5H2O、0.50mg/lのNiCl2・6H2O及び13.8mg/lのFeSO4・7H2O。これらの胞子は、30℃で、7〜14日間増殖して米粒上に胞子の黒い層を生じ、滅菌水中の100mlの0.1%Tween20を加えることにより胞子を回収する。すべての条件に関して、ガスはバイオリアクターに導入する前に滅菌ろ過する。オフガスは−8℃未満に冷却した冷却器を経て、アコースティックガスアナライザーによりそのCO2とO2の体積含量を分析する。
【0129】
ケモスタット培養
ケモスタット培養では、例えば1.5リットルの有効容積のバッフル付きバイオスタットB実験室用発酵槽で、30℃、500rpmで細胞を増殖させることができる。pH6.6で一定になるように10MのKOHの自動添加により維持し、好気的条件は、空気を1vvmの速度でバイオリアクターに拡散して、確実にDOTが80%を超えさせることにより得られる。希釈率(D)は、異なる値、例えば、0.050h−1、0.10h−1、0.15h−1又は0.20h−1に設定できる。以下の成分を有する最小成長培地を用いて、10Mの自動追加によりpHを一定、例えば、6.6に保持する:2.5g/lの(NH4)2SO4、0.75g/lのKH2PO4、1.0g/lのMgSO4・7H2O、1.0g/lのNaCl、0.1g/lのCaCl2・2H2O、0.1ml/lのシグマ社製消泡剤、7.2mg/lのZnSO4・7H2O、1.3mg/lのCuSO4・5H2O、0.3mg/lのNiCl2・6H2O、3.5mg/lのMnCl2・4H2O及び6.9mg/lのFeSO4・7H2O。グルコースの濃度は異なる値、例えば、3.0g/l、6.0g/l、12.0g/l、15.0g/l又は18.0g/lに設定することができる。バイオリアクターには上記のように前バッチ培養物からの遊離菌糸体が接種され、指数増殖期の終わりにフィードポンプを作動させる。
【0130】
すべての条件について、ガスはバイオリアクターに導入する前に滅菌ろ過する。オフガスは−8℃未満に冷却した冷却器を経て、アコースティックガスアナライザーによりそのCO2とO2の体積含量を分析する。
【0131】
少なくとも5滞留時間後、及びバイオマスグルコースとオフガスの組成が最後の2滞留時間にわたって変化しない(5%相対偏差未満)の場合、培養は定常状態にあると考えられる。
【0132】
(実施例33)
黒色アスペルギルス及び麹菌におけるリスベラトロールの抽出と分析
抽出と分析は実施例14と実施例15に記載された方法を用いて実施する。
【0133】
参考文献
米国特許出願公開第6521748号
米国特許出願公開第2001053847号
米国特許出願公開第2004059103号
米国特許出願公開第2004023357号
【0134】
Allina,S.M.,Pri−Hadash,A.,Theilmann,D.A.,Ellis,B.E.and Douglas,C.J.(1998)4−coumarate:Coenzyme A ligase in hybrid poplar.Properties of enzymes,cDNA cloning,and analysis of recombinant clones.Plant Physiol.116,743−754.
Aoyama,Y.,Yoshida,Y.,Kubota,S.,Kumaoka,H.and Furumichi,A.(1978).NADPH−cytochrome P−450 reductase of yeast microsomes.Arch.Biochem.Biophys.185,362−369.
Becker JV,Armstrong GO,van der Merwe MJ,Lambrechts MG,Vivier MA,Pretorius IS.(2003).Metabolic engineering of Saccharomyces cerevisiae for the synthesis of the
wine− related antioxidant resveratrol.FEMS Yeast Res.4,79−85.
【0135】
Blanquet,S.,Meunier,J.P.,Minekus,M.,Marol−Bonnin,S.,and Alric,M.(2003).Recombinant Saccharomyces cerevisiae Expressing P450 in Artificial Digestive Systems:a Model for Biodetoxication in the Human Digestive Environment.Appl.Environ.Microbiol.69,2884−2892.
Celotti E and others.(1996).Resveratrol content of some wines obtained from dried Valpolicella grapes:Recioto and Amarone.Journal of Chromatography A 730,47−52.
Cochrane,F.C,Davin,L.B.and Lewis N.G.(2004).The Arabidopsis phenylalanine ammonia lyase gene family:kinetic characterization of the four PAL isoforms.Phytochemistry 65,1557−1564.
Couzin,J.(2004)Aging Research’s Family Feud.Science 303,1276−1279.
【0136】
Ehlting,J.,Buttner,D.,Wang,Q.,Douglas,C.J.,Somssich,I.E.and Kombrink,E.(1999).Three 4−coumarate:coenzyme A ligases in Arabidopsis thaliana represents two evolutionary divergent classes in angiosperms.The plant journal.19,9−20.
Filpula,D.,Vaslet,CA.,Levy,A.,Sykes,A.and Strausberg,R.L.Nucleotide sequence of gene for phenylalanine ammonia−lyase from Rhodotorula rubra.(1988).Nucleic Acids Res.16,11381.
【0137】
Gehm,B.D.,McAndrews,J.M.,Chien,P.Y.and Jameson,J.L.(1997).Resveratrol,a polyphenolic compound found in grapes and wine,is an agonist for the estrogen receptor.Proc.Natl.Acad.Sci.USA 94,14138−14143.
Gems,D.,Johnstone,I.L.and Clutterbuck,A.J.(1991).An autonomously replicating plasmid transforms Aspergillus nidulans at high frequency.Gene 98,61−67.
Hain,R.,Reif,H.J.,Krause,E.,Langebartels,R.,Kindl,H.,Vornam,B.,Wiese,W.,Schmelzer,E.,Schreier,P.H.,Stocker,R.H.and Stenzel,K.(1993).Disease resistance results from foreign phytoalexin expression in a novel plant.Nature 361,153−156.
【0138】
Hwang EI,Kaneko M,Ohnishi Y,Horinouchi S.(2003).Production of plant−specific flavanones by Escherichia coli containing an artificial gene cluster.Appl.Environ.Microbiol.69,2699−706.
Huang,M−T.(1997).Diet for cancer prevention.Food Sci.24,713−727
Hart,J.H.(1981)Annu.Rev.Phytopathology 19,437−458.Hart,J.H.,Shrimpton,D.M.(1979)Phytopathology 69,1138−1143.
Hall,S.S.(2003)In Vino Vitalis? Compounds Activate Life− Extending Genes.Science 301,1165.
Hamberger,B.and Hahlbrock,K.(2004).The 4−coumarate:CoA ligase gene family in Arabidopsis thaliana comprises one rare,sinapate−activating and three commonly occurring isoenzymes.Proc.Natl.Acad.Sci .USA.101,2209−2214.
【0139】
Jang,M.,Cai,L.,Udeani,GO.,Slowing,KV.,Thomas,CF.,Beecher,CWW.,Fong,HHS.,Farnsworth,NR.,Kinghorn,AD.,Mehta,RG.,Moon,RC,Pezzuto,JM.(1997).Cancer Chemopreventive Activity of Resveratrol,a Natural Product Derived from Grapes.Science 275,218−220.
Jeandet,P.Bessis,R.,Maume,B.F.,Meunier,P.,Peyron,D.and Trollat,P.(1995).Effect of Enological Practices on the Resveratrol Isomer Content of Wine J.Agric.Food Chem.43,316−319.
Jeandet,P.Bessis,R.,Sbaghi,M.and Meunier,P.(1994).Occurence of a resveratrol β−D−glucoside in wine:Preliminary studies.Vitis 33,183−184.
【0140】
Koopmann,E.,Logemann,E.and Hahlbrock,K.(1999).Regulation and Functional Expression of Cinnamate 4−Hydroxylase from Parsley.Plant Physiol.119,49−55.
Kopp,P.(1998).Resveratrol,a phytooestrogen found in red wine.A possible explanation for the conundrum of the “French Paradox”? Eur.J.Endocrinol.138,619−620.
Kyndt JA,Meyer TE,Cusanovich MA,Van Beeumen JJ.(2002).Characterization of a bacterial tyrosine ammonia lyase,a biosynthetic enzyme for the photoactive yellow protein.FEBS Lett.512,240−244.
【0141】
LaGrange,D.C,Pretorius,I.S.and Van ZyI,W.H.(1997).Cloning of the Bacillus pumilus beta−xylosidase gene(xynB)and its expression in Saccharomyces cerevisiae.Appl.Microbiol.Biotechnol.47,262−266.
Lin X.,Kaul S.,Rounsley S.D.,Shea T.P.,Benito M.−I.,Town CD.,Fujii CY.,Mason T.M.,Bowman CL.,Barnstead M.E.,Feldblyum T.V.,Buell CR.,Ketchum K.A.,Lee J.J.,Ronning CM.,Koo H.L.,Moffat K.S.,Cronin L.A.,Shen M.,Pai G.,Van Aken S.,Umayam L.,Tallon L.J.,Gill J.E.,Adams M.D.,Carrera A.J.,Creasy T.H.,Goodman H.M.,Somerville CR.,Copenhaver G.P.,Preuss D.,Nierman W.C,White O.,Eisen J.A.,Salzberg S.L.,Fraser CM.,Venter J.C.(1999).Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana.Nature 402,761−768.
【0142】
Lobo,R.A.(1995).Benefits and risks of estrogen replacement therapy.Am.J.Obstet.Gynecol.173,982−989.
Martin,V.J.J.,Pitera,D.J.,Withers,S.T.,Newman,J.D.and Keasling,J.D.(2003).Engineering a mevalonate pathway in Escherichia coli for production of terpenoids.Nature biotechnology 21,796−802.
Mizutani,M.,Ohta,D.and Sato,R.(1997).Is
olation of a cDNA and a genomic clone encoding cinnamate 4−hydroxylase from Arabidopsis and its expression manner in planta.Plant Physiol.113,755−763.
【0143】
Ro,D.K.,Mah,N.,Ellis,B.E.and Douglas,CJ.(2001).Functional Characterization and Subcellular Localization of Poplar(Populus trichocarpa x Populus deltoides)Cinnamate 4−Hydroxylase.Plant Physiol.126,317−329.
Ro D.K.,Douglas CJ.(2004).Reconstitution of the entry point of plant phenylpropanoid metabolism in yeast(Saccharomyces cerevisiae):implications for control of metabolic flux into the phenylpropanoid pathway.J.Biol.Chem.279,2600−2607.
【0144】
Rosier J,Krekel F,Amrhein N,Schmid J.(1997).Maize phenylalanine ammonia−lyase has tyrosine ammonia−lyase activity.Plant
Physiol.113,175−179.activity.Plant physiol.113,175−179.
Samappito,S.,Page,J.E.,Schmidt,J.,De−Eknamkul,W.and Kutchan,T.M.(2003).Aromatic and pyrone polyketides synthesized by a stilbene synthase from Rheum tataricum.Phytochemistry 62,313−323.
Sambrook,J.,Fritsch,E.F.and Maniatis,T.(1989).Molecular Cloning,A Laboratory Manual,2d edition,Cold Spring Harbor,N.Y.
【0145】
Schoppner,A.;Kindl,H.(1984)Purification and properties of a stilbene synthase from induced cell suspension cultures of peanut.J.Biol.Chem.259,6806−6811.
Seshime,Y.,Juvvadi,P.R.,Fujii,I.and Kitamoto,K.(2005).Genomic evidences for the existence of a phenylpropanoid metabolic pathway in Aspergillus oryzae.Biochem Biophys Res Commun.337,747−51.
Urban P.,Mignotte,C,Kazmaier M.,Delorme F.And Pompon D.(1997).Cloning,Yeast Expression,and Characterization of the Coupling of Two Distantly Related Arabidopsis thaliana NADPH−Cytochrome 450 Reductases with P450 CYP73A5.J.Biol.Chem.272,191
76−19186.
【0146】
Watts,K.T.,Lee,P.C.and Schmidt−Dannert,C.(2004).Exploring recombinant flavonoid biosynthesis in metabolically engineered Escherichia coli.Chembiochem 5,500−507.
【0147】
以下は本明細書に現れるヌクレオチド配列とアミノ酸配列の概要である。
配列番号1は、フェニルアラニンアンモニアリアーゼ(PAL2)をコードするシロイヌナズナ由来のヌクレオチド配列である。
配列番号2は、配列番号1によりコードされるアミノ酸配列である。
配列番号3は、シンナメート4−ヒドロキシラーゼ(C4H)をコードするシロイヌナズナ由来のヌクレオチド配列である。
配列番号4は、配列番号3によりコードされるアミノ酸配列である。
配列番号5は、4−クマレート:コエンザイムAリガーゼ(4CL1)をコードするシロイヌナズナ由来のヌクレオチド配列である。
配列番号6は、配列番号5によりコードされるアミノ酸配列である。
配列番号7は、リスベラトロールシンターゼ(VST)をコードするRheum tataricum由来のヌクレオチド配列である。
配列番号8は、配列番号7によりコードされるアミノ酸配列である。
【0148】
配列番号9は、S.セレヴィシエでの発現のためにコドン最適化されたリスベラトロールシンターゼ(VST)をコードするRheum tataricum由来のヌクレオチド配列である。
配列番号10は、配列番号9によりコードされるアミノ酸配列である。
配列番号11は、チロシンアンモニアリアーゼ(TAL)をコードするロドバクテリア capsulatus由来のヌクレオチド配列である。
配列番号12は、配列番号11によりコードされるアミノ酸配列である。
配列番号13は、S.セレヴィシエでの発現のためにコドン最適化されたチロシンアンモニアリアーゼ(TAL)をコードするロドバクテリア capsulatus由来のヌクレオチド配列である。
配列番号14は、配列番号13によりコードされるアミノ酸配列である。
【0149】
配列番号15は、NADPH:チトクロームP450レダクターゼ(CPR1)をコードするS.セレヴィシエ由来のヌクレオチド配列である。
配列番号16は、配列番号15によりコードされるアミノ酸配列である。
配列番号17は、NADPH:チトクロームP450レダクターゼ(AR2)をコードするArabidopsis thalianus由来のヌクレオチド配列である。
配列番号18は、配列番号17によりコードされるアミノ酸配列である。
配列番号19〜32は、表1に現れる実施例1のプライマー配列である。
配列番号33〜34は、実施例16に現れるプライマー配列である。
配列番号35〜38は、実施例17に現れるプライマー配列である。
【技術分野】
【0001】
本発明は、一般的に、微生物細胞を用いたポリフェノールリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体、例えば、そのβ−グルコシドピセイドの生産に関する。さらに、本発明は、リスベラトロール又はそのような誘導体を作る天然微生物又は組換え微生物の、食物、餌及び飲料の生産のための使用に関する。
【背景技術】
【0002】
薬品の微生物からの生産は、バイオテクノロジーの重要な応用となっている。典型的には、そのようなバイオ生産法の開発の工程として、1)適当な微生物宿主の選択、2)副産物を導く代謝経路の排除、3)酵素活性レベルと転写レベルの両方で望ましい経路の調節解除、及び4)望ましい経路での適当な酵素の過剰発現が挙げられるだろう。好ましい態様において、本発明は、リスベラトロールの望ましい生合成のために必要な前駆体を供給する植物のフェニルプロパノイド経路の酵素により、フェニルアラニン又はチロシンからの炭素の流れを再指示する上記工程の組合せを使用している。
【0003】
リスベラトロール(すなわち、3,4,5−トリヒロドキシスチルベン)は、感染又は他のストレスに関連する現象に応答して植物中に活動的な防御機構を構成する低分子量の二次的代謝産物であるスチルベンフィトアレキシン類の群に属するフィトフェノールである。スチルベンフィトアレキシン類はスチルベン骨格(トランス−1,2−ジフェニルエチレン)をそれらの共通する基本構造として含み、この構造は他の基の付加により補われてもよい(非特許文献1及び2)。スチルベン類は一定の木(被子植物、裸子植物)に発見されてきたが、一部の多年草(フトモモ科、ブドウ科及びマメ科の種)にも発見されている。該化合物は、有害生物、特に菌類、細菌及び昆虫への毒性がある。ほんのわずかな植物がスチルベン類を合成する能力があるか、又は有害生物に対して十分な耐性を植物に与える量でスチルベン類を生産する能力を有する。
【0004】
スチルベンシンターゼによる基本的なスチルベン骨格の合成が探究されている。今までに、ピノシルビンシンターゼとリスベラトロールシンターゼという2種類の酵素がスチルベンシンターゼとして示された。現在までに、落花生(Arachis hypogaea)リスベラトロールシンターゼが最も詳細に特性付けられているので、その性質のほとんどが知られている(非特許文献3)。スチルベンシンターゼにより用いられる基質は、マロニル−CoA、シンナモイル−CoA又はクマロイル−CoAである。これらの基質は、フラボノイド、花の色素及び脂質等の他の重要な植物構成物の生合成にも同じく用いられるので、すべての植物に存在する。
【0005】
リスベラトロール(図1:トランス形)は2個の密に結合したフェノール環からなるのでポリフェノール類に属する。ユーカリ、エゾマツ及びユリ等の他の植物やクワの実及びピーナッツ等の他の食物に存在する一方で、リスベラトロールの最も豊富な天然源は、ワイン製造に使用されるヴィティス・ヴィニヘラ、ヴィティス・ラプラスカ及びヴィティス・マスカジン(ロタンディフォリア)のブドウである。この化合物は、ブドウの木、根、種及び茎に存在するが、その最大濃度は、50〜100μg/g(非特許文献4)を含む皮にある(非特許文献5)。赤ワインの醸造中、ブドウの皮は白ワインの醸造とは対照的にブドウの液汁に含まれるので、リスベラトロールは赤ワインのみに少量見られる。リスベラトロールは、その抗菌性に加えて、その心臓保護活性と発癌予防活性が認識されてきており、それは、血小板凝集の阻害剤であるフィトエストロゲン(非特許文献6、7及び8)や抗酸化物(非特許文献4及び9)として作用する。これらの性質は、所謂、フレンチパラドックス(すわなち、ワインを飲むフランス人は低運動で高脂肪食事にもかかわらず冠動脈心疾患の低い発生率)を説明する。最近、リスベラトロールは、共に寿命の延長に重要な役割を果たす酵母のSIR2遺伝子及び類似のヒト遺伝子SIRT1を活性化することもできることが示された。それ以来、リスベラトロールの寿命延長効果が非常に注目されている(非特許文献10及び11)。
【0006】
ライフエクステンションファンデーション等のアメリカの健康協同組合は、この医薬の非常に高い薬効を促進しつつあり、それによって、成功した商品化のために理想的な状態を推進しつつある。現在の生産工程は、主に、ブドウ果皮かタデのいずれかからのリスベラトロールの抽出に依存する。これは、労働集約的な過程であり、低い利益を生むため、新規で、さらに効率的で多収量の生産方法の開発に対する動機を促している。
【0007】
植物において、フェニルプロパノイド経路は、リグニン類、サリチレート類、クマリン類、ヒドロキシケイヒ酸アミド類、色素、フラボノイド類及びフィトアレキシン類等の多様な二次代謝化合物の合成に関与する。実際、植物におけるリスベラトロールの形成はフェニルプロパノイド経路を経る。アミノ酸のL−フェニルアラニンは、L−フェニルアラニンアンモニアリアーゼ(PAL)により非酸化脱アミノ化によりトランスケイヒ酸に変換される(図2)。次に、トランスケイヒ酸は、NADPH:チトクロームP450レダクターゼ(CPR)と共同するチトクロームP450モノオキシゲナーゼ酵素であるシンナメート−4−ヒドロキシラーゼ(C4H)によりパラ位でヒドロキシル化されて4−クマル酸(4−ヒドロキシケイヒ酸)となる。引き続いて、この4−クマル酸は4−クマレート−CoAリガーゼ(4CL)の作用により4−クマロイル−CoAに活性化される。最後に、リスベラトロールシンターゼ(VST)が4−クマロイル−CoAのフェニルプロパン単位のマロニルCoAへの縮合を触媒して、リスベラトロールの形成をもたらす。
【0008】
最近、ブドウ液汁に少量見られる4−クマル酸からリスベラトロールを作ることのできる酵母が開示された(非特許文献12)。S.セレヴィシエの実験室の菌株における4−クマロイル−CoAとそれに付随するリスベラトロールの生産は、雑種ポプラに由来する異種コエンザイム−Aリガーゼ遺伝子をブドウリスベラトロールシンターゼ遺伝子(vst1)とともに共発現させることにより達成された。リスベラトロールシンターゼの他の基質であるマノニル−CoAは酵母ですでに内因的に産生されており、新規な脂肪酸生合成にかかわる。研究により、S.セレヴィシエの細胞は、4−クマル酸を補った合成培地で培養した場合、微量のリスベラトロールを遊離形又はグルコシド結合形のいずれかで産生することができることが分かった。
【0009】
しかし、該酵母は低いリスベラトロール収率に弱点があり、ごくわずかな産業用培地のみにしか存在しない4−クマル酸の添加を必要とするために、商業的利用には適さないだろう。したがって、医薬と栄養補助食品の両方としてのリスベラトロールの利用を容易にし、広げるために、4−クマル酸の添加なしに、グルコースからリスベラトロールを直接産生することのできる酵母を得ることは非常に望ましい。
【0010】
最近の研究(非特許文献13)には、ポプラに由来するPAL、C4H及びCPRを導入することによる、S.セレヴィシエにおけるフェニルプロパノイド経路のエントリポイントの再構成が記載されている。この目的は、PALとC4Hを含む多酵素複合体(MEC)が、フェニルプロパノイド代謝へのエントリポイントにおいて機能的に重要かどうかを評価することにあった。組換え酵母に[3H]−フェニルアラニンを供給することにより、代謝された[3H]−フェニルアラニンの大多数が4−[3H]−クマル酸に取り込まれること、及びフェニルアラニン代謝はC4H活性を阻害することにより非常に減少することがわかった。さらに、PLAのみのエクスプレッサーが微小のフェニルアラニンをケイヒ酸に代謝した。[3H]−フェニルアラニンと[14C]−トランス−ケイヒ酸を同時にトリプルエクスプレッサーに供給した場合、内因的に合成された[3H]−トランス−ケイヒ酸の4−クマル酸へのチャネリングについていかなる証拠も見られなかった。したがって、PALとCH4により触媒される反応によりフェニルアラニンから4−クマル酸への効率的な炭素の流れは酵母におけるMECによるチャネリングを必要とするようには思えず、PALとC4Hの真の生化学的カップリングが炭素の流れをフェニルプロパノイド経路に引き込むのに十分であるように思える。さらに別の研究(非特許文献14)において、大腸菌による植物特異的なフラバノン類の生産は多様な異種起源のフェニルプロパノイド経路の3遺伝子(酵母Rhodotorula rubraに由来するPAL、放線菌ストレプトミセス coelicolorに由来する4CL及び甘草植物Glycyrrhiza echinataに由来するカルコンシンターゼ(CHS))を含む人工的な遺伝子クラスターの発現により達成された。細菌の4CL酵素はコエンザイムAをトランス−ケイヒ酸と4−クマル酸の両方に連結させたので、これらの経路はC4Hをバイパスした。さらに、Rhodotorula rubra由来のPALはフェニルアラニンとチロシンの両方を基質として利用する。したがって、該遺伝子クラスターを含有し、かつグルコースで増殖した大腸菌細胞は、少量の2種類のフラバノン(フェニルアラニンからピノセムブリン(0.29g/l)及びチロシンからナリンゲニン(0.17g/l))を産生した。さらに、大量のそれらの前駆体である4−クマル酸とトランス−ケイヒ酸(それぞれ0.47及び1.23mg/リットル)が蓄積された。さらに、これらの化合物の収率はフェニルアラニンとチロシンの添加により増加させることができた。
【0011】
双子葉植物に由来する酵素がフェニルアラニンのみを効率的に利用する一方で、幾つかの研究によって、単子葉植物及び一部の微生物に由来するPALがチロシンを同じく利用することが示された(非特許文献15)。そのような反応において、チロシン活性物はチロシンアンモニアリアーゼと命名されている(TAL、図3)。チロシンのTALによる変換は、C4HとCPRの仲介なしに4−クマル酸の直接の形成をもたらす。主に、両活性は同じポリペプチドに存在し、Kmとターンオーバー数の大きな違いにかかわらず、非常に類似の触媒効率を有する。しかし、植物に由来するほとんどのPAL/TAL酵素はチロシンよりもフェニルアラニンを好む。TAL活性のレベルはほとんどPAL活性よりも低いが、この差の程度は広範囲にわたって変化する。例えば、パセリ酵素はフェニルアラニンに対して15〜25μM及びチロシンに対して2.0〜8.0mMのKmと、それぞれ22秒−1と0.3秒−1のターンオーバー数を有する。対照的に、トウモロコシ酵素は、フェニルアラニンのみに対してチロシンよりも15倍高いKm値と約10倍高いターンオーバー数を有する。さらに、赤酵母であるRhodotorula glutinis(Rhodosporidium toruloides)と−rubraにおいて、TAL触媒活性はPAL触媒活性に近く、TAL/PAL比が約0.58である。これらの酵母のPAL酵素は異化機能としてフェニルアラニンを分解すると考えられ、形成されるトランス−ケイヒ酸はベンゾエートと他の細胞材料に変換される一方で、植物においては、それは、リグニン、イソフラボノイド類及び他のフェニルプロパノイド類の生合成において調節酵素にすぎないと考えられている。
【0012】
最近、光活性黄色タンパク質の発色団の生合成にかかわる仮説上の生合成チロシンアンモニアリアーゼ(TAL)をコードする細菌ロドバクテリア capsulatusで読み取り枠が発見された(非特許文献16)。これは、PAL相同遺伝子が細菌で発見された最初である。TAL遺伝子は単離され、大腸菌で過剰生産させた。チロシンの4−クマル酸への変換に関するKmとkcat値はそれぞれ15.6μMと27.7秒−1であり、L−フェニルアラニンのトランス−ケイヒ酸への変換に関してはそれぞれ1277μMと15.1秒−1であった。その小さいKmとわずかに高いkcatの結果として、酵素はL−フェニルアラニンよりもチロシンに対する強い好みを示し、フェニルアラニンに対するよりも約150倍高いチロシンに対する触媒効率(Km/kcat)を有する。動力学的研究によると、L−フェニルアラニンではなく、チロシンが生理学的条件下で酵素の天然の基質であることが立証されている。ごく最近、ある研究で、大腸菌でのフラボノイド類の産生に関してフェニルアラニンアンモニアリアーゼ、シンナメート−4−ヒドロキシラーゼ、4−クマレート−Coa−リガーゼ及びカルコンシンターゼの異種共発現が述べられている(非特許文献17)。しかし、すべての4遺伝子における同時発現は、非機能性のシンナメート−4−ヒドロキシラーゼのために成功しなかった。しかし、ロドバクテリア sphaeroidesからクローニングされた新しいチロシンアンモニアリアーゼをフェニルアラニンアンモニアリアーゼとシンナメート−4−ヒドロキシラーゼと置き換えることでその問題が解決され、フラボノンナリンゲニンの高水準生産が導かれた。さらに、ロドバクテリア sphaeroidesに由来する該チロシンアンモニアリガーゼは、大腸菌における4−クマル酸(すなわち、パラ−ヒドロキシケイヒ酸)の異種産生のためにも用いられている(特許文献1)。さらに、グルコースを4−クマル酸に変換する生体触媒のさらなる開発方法が記載されている。特許文献2には、大腸菌とサッカロマイセス・セレヴィシエ(出芽酵母)におけるクマル酸の産生のためにTrichosporon cutaneum酵母に由来するチロシンアンモニアリアーゼが開示されている。特許文献3には、チロシンを4−クマル酸へ直接変換する野生型PALの能力を明確に示すために、Rhodotorula glutinis酵母に由来する野生型PALの大腸菌への取り込みが記載されている。さらに、植物C4HとCPRに加えて、Rhodotorula glutinis酵母に由来する野生型PALの大腸菌とS.セレヴィシエとへの取り込みの例示も存在する。増強TAL活性を有する野生型酵母PAL Rhodotorula glutinisの突然誘発による生体触媒の開発も記載されている(特許文献4)。上記特許のいずれもリスベラトロールの産生のために4CL及びVSTの取り込みを請求するものではない。
【0013】
最近、糸状菌の麹菌(コウジカビ)が、植物中のナリンゲニン等のフラボノイド類の生合成に通常関与するカルコンシンターゼ(CHS)酵素を含むとの証拠が示された(非特許文献18)。実際、麹菌がフェニルプロパノイド−フラボノイド代謝に関与する主要な一組の遺伝子(すなわち、PAL、C4H及び4CL)を含むことも示した。しかし、麹菌がリスベラトロールシンターゼ等のスチルベンシンターゼを含むとの証拠はない。
【先行技術文献】
【特許文献】
【0014】
【特許文献1】US−A−2004059103
【特許文献2】US−A−2004023357
【特許文献3】US−A−2001053847
【特許文献4】US−A−6521748
【非特許文献】
【0015】
【非特許文献1】ハート及びシュリンプトン(Hart and Shrimpton)、1979年
【非特許文献2】ハート(Hart)、1981年
【非特許文献3】ショップナー及びキンドル(Schoppner and Kindl)、1984年
【非特許文献4】ジャングら(Jang et al.)、1997年
【非特許文献5】セロッティら(Celotti et al.)、1996年
【非特許文献6】コップら(Kopp et al.)、1998年
【非特許文献7】ゲームら(Gehm et al.)、1997年
【非特許文献8】ロボら(Lobo et al.)、1995年
【非特許文献9】ファング(Huang)、1997年
【非特許文献10】ハル(Hall)、2003年
【非特許文献11】コウジン(Couzin)、2004年
【非特許文献12】ベッカーら(Becker et al.)、2003年
【非特許文献13】ロー及びダグラス(Ro and Douglas)、2004年
【非特許文献14】Hwangら、2003年
【非特許文献15】Roslerら、1997年
【非特許文献16】キンドら(Kyndt et al.)、2002年
【非特許文献17】Wattsら、2004年
【非特許文献18】Seshimeら、2005年
【発明の概要】
【課題を解決するための手段】
【0016】
本発明は、
(1)4−クマル酸を生産し、さらに当該4−クマル酸からリスベラトロール又は当該リスベラトロールのオリゴマー誘導体又はグリコシド結合誘導体を生産する操作的な代謝経路を有する微生物において、
前記代謝経路において、前記4−クマル酸は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ及びシンナメート4−ヒドロキシラーゼの作用によってL−フェニルアラニンから、又は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼの作用によってチロシンから生成し、
4−クマロイル−CoAが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現する4−クマレート−CoAリガーゼによる酵素反応で前記4−クマル酸から生成し、
そして前記リスベラトロールが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するリスベラトロールシンターゼによって内在性マロニル−CoAを基質とする反応で前記4−クマロイル−CoAから生成することを特徴とする微生物、
(2)天然のNADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物で過剰発現している、又は、NADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物に組換えで導入されていることを特徴とする上記(1)に記載の微生物、
(3)チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする上記(1)又は(2)に記載の微生物、
(4)フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする上記(1)乃至(3)のいずれか一項に記載の微生物、
(5)真菌又は細菌であることを特徴とする上記(1)乃至(4)のいずれか一項に記載の微生物、
(6)発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有することを特徴とする上記(1)に記載の微生物、
(7)シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する上記(1)に記載の微生物、
(8)4−クマル酸の外部源の実質的な非存在下において、上記(1)乃至(7)のいずれか一項に記載の微生物を炭素基質に接触させる工程を含むことを特徴とする、リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法、
(9)前記炭素基質が、単糖類、小糖類及び多糖類からなる発酵性炭素基質の群より選択されることを特徴とする上記(8)に記載の方法、
(10)前記炭素基質が、エタノール、アセテート、グリセロール、ラクテート、及びアミノ酸で構成される非発酵性炭素基質からなる群より選択されることを特徴とする上記(8)に記載の方法、
(11)前記非発酵性炭素基質が、フェニルアラニン及びチロシンからなる群より選択されることを特徴とする上記(10)に記載の方法、
(12)さらに、生産されたリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を乳製品又は飲み物中の栄養補助食品として使用することを含む上記(8)乃至(11)のいずれか一項に記載の方法、
である。
【図面の簡単な説明】
【0017】
【図1】図1は、トランス−リスベラトロールの化学構造を示す。
【図2】図2は、L−ファニルアラニンに作用するフェニルアラニンアンモニアリアーゼを利用するフェニルプロパノイド経路を示す。
【図3】図3は、L−チロシンに作用するフェニルアラニンアンモニアリアーゼを利用する別の経路を示す。
【図4】図4は、100g/lのガラクトースで成長させたS.セレヴィシエ菌株であるFSSC−PALC4H4CLVST及びFSSC−TAL4CLVSTの抽出物のHPLCクロマトグラムを示す。60ナノグラムの純粋なリスベラトロールのクロマトグラムが含まれる。
【図5】図5は、純粋なトランス−リスベラトロール及び100g/lのガラクトースで成長させたS.セレヴィシエ菌株であるFSSC−PALC4H4CLVSTにより作られたトランス−リスベラトロールのUV吸収スペクトルを示す。
【図6】図6は、50g/lのグルコースで成長させた大腸菌株であるFSEC−TAL4CLVST及びFSECコントロールからの抽出物のHPLCクロマトグラムを示す。
【図7】図7は、20mg/lのクマル酸を添加した50g/lのグルコースで成長させた大腸菌株であるFSEC−TAL4CLVST及びFSECコントロールからの抽出物のHPLCクロマトグラムを示す。FSEC−TAL4CLVST菌株で作られたトランス−リスベラトロールのUV吸収スペクトルが含まれる。
【発明を実施するための形態】
【0018】
本発明は、少なくとも1種類の活性を含む操作的な代謝経路を有する微生物において、該経路が4−クマル酸を産生し、かつそれからリスベラトロール又はそのオリゴマー誘導体かグリコシド結合誘導体を産生する微生物を本願において提供する。そのような微生物は自然に存在してよく、適切なスクリーニング手順により単離してもよく、さらに好ましくは遺伝子組み換えされている。
【0019】
好ましくは、該リスベラトロール又は誘導体は、内在性のマロニル−CoAが基質である酵素により触媒される反応で作られ、好ましくは該リスベラトロールは4−クマロイル−CoAから生成される。
【0020】
好ましくは、該リスベラトロール又は誘導体は、微生物において、好ましくは該微生物に固有ではない該酵素をコードする核酸から発現するリスベラトロールシンターゼにより4−クマロイル−CoAから生成される。
【0021】
ここでは一般的に、文脈上他の意味に解すべき場合を除き、リスベラトロールへの言及は、特にピセイドを含む、そのオリゴマー誘導体かグリコシド結合誘導体への言及を含む。
【0022】
例えば、ある好ましい実施態様において、前記リスベラトロールシンターゼは、Arachis属に属する植物、例えば、A.glabatra、A.hypogaea、Rheum属に属する植物、例えば、R.tataricum、ヴィティス属に属する植物、例えば、V.ラプラスカ、V.riparaia、V.ヴィニヘラ、又はパイナス属、Piceea属、ユリ属、ユーカリ属、Parthenocissus属、ジッサス属、カロコルタス属、タデ属、Gnetum属、Artocarpus属、Nothofagus属、フェニックス属、ウシノケグサ属、スゲ属、シュロソウ属、ハカマカズラ又はPterolobium属のいずれかに由来するリスベラトロールシンターゼ(EC2.3.1.95)である。
【0023】
前記4−クマル酸は、好適には、酸素が基質であり、NADH又はNADPHが補因子であり、NAD+又はNADP+が産物である酵素により触媒される反応で酵素によりトランス−ケイヒ酸から生成されることが好ましい。
【0024】
例えば、該4−クマル酸はトランス−ケイヒ酸からシンナメート4−ヒドロキシラーゼにより作られてよく、該4−クマル酸は、好ましくは、微生物において、該微生物に固有ではない該酵素をコードする核酸から発現する。
【0025】
上記段落で言及されたものを含むある実施態様において、該シンナメート−4−ヒドロキシラーゼは植物又は微生物に由来するシンナメート−4−ヒドロキシラーゼ(EC1.14.13.11)である。植物は、シロイヌナズナ属、例えば、シロイヌナズナ、Citrusに属する植物、例えば、C.シネンシス、C.x paradisi、Phaseolus属する植物、例えば、P.ヴァガリス、パイナス属する植物、例えば、P.taeda、Populus属する植物、例えば、P.deltoides、P.tremuloides、P.trichocarpa、ナス属する植物、例えば、S.tuberosum、ヴィティス属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属する植物、例えば、Z.メイス又は他の植物属、例えば、Ammi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属に属してよい。微生物は、アスペルギルス属に属する真菌類、例えば麹菌であってよい。
【0026】
好ましくは、前記4−クマル酸は、アンモニアが生成される酵素触媒反応でチロシンから作られ、好適には、該4−クマル酸は、L−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼ、例えば酵母又は細菌に由来するチロシンアンモニアリアーゼ(EC4.3.1.5)によりチロシンから生成される。好適には、該チロシンアンモニアリアーゼは酵母Rhodotorula rubra又は細菌ロドバクテリア capsulatusに由来する。
【0027】
任意に、該チロシンアンモニアリアーゼは、微生物において、該微生物に固有ではない該酵素をコードする核酸から発現させる。
【0028】
もしくは、前記トランス−ケイヒ酸は、アンモニアが生成される酵素触媒反応でL−フェニルアラニンから作られてよく、好適には、該トランス−ケイヒ酸は、L−フェニルアラニンからフェニルアラニンアンモニアリアーゼにより形成する。
【0029】
ある好ましい実施態様において、該L−フェニルアラニンアンモニアリアーゼは、植物又は微生物に由来するL−フェニルアラニンアンモニアリアーゼ(EC4.3.1.5)である。植物は、シロイヌナズナ属に属する植物、例えば、シロイヌナズナ、Brassica属に属する植物、例えば、B.napus、B.rapa、シトラス属に属する植物、例えば、C.reticulata、C.clementinus、C.リモン、Phaseolus属に属する植物、例えば、P.coccineus、P.ヴァリガリス、パイナス属に属する植物、例えば、P.banksiana、P.monticola、P.パイナスター、P.sylvestris、P.taeda、Populus属に属する植物、例えば、P.balsamifera、P.deltoides、P.Canadensis、P.kitakamiensis、P.tremuloides、ナス属に属する植物、例えば、S.tuberosum、プラナス属に属する植物、例えば、P.avium、P.persica、ヴィティス属に属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属に属する植物、例えば、Z.メイス又は他の植物属、例えば、Agastache属、Ananas属、Asparagus属、Bromheadia属、Bambusa属、Beta属、Betula属、Cucumis属、カメリア属、Capsicum属、Cassia属、Catharanthus属、Cicer属、Citrullus属、Coffea属、Cucurbita属、Cynodon属、Daucus属、Dendrobium属、Dianthus属、Digitalis属、Dioscorea属、ユーカリ属、Gallus属、Ginkgo属、グリシン属、Hordeum属、Helianthus属、Ipomoea属、Lactuca属、リソスパーマム・オフィシナーレ属、ハス属、Lycopersicon属、Medicago属、マリュス属、Manihot属、Medicago属、マツバギク属、タバコ属、Olea属、オリザ属、Pisum属、Persea属、ペトロセリウム属、Phalaenopsis属、Phyllostachys属、Physcomitrella属、Picea属、Pyrus属、Quercus属、Raphanus属、Rehmannia属、キイチゴ属、Sorghum属、Sphenostylis属、Stellaria属、Stylosanthes属、コムギ属、Trifolium属、コムギ属、Vaccinium属、Vigna属、Zinnia属に属してよい。微生物は、Agaricus属に属する真菌類、例えば、A.bisporus、アスペルギルス属に属する真菌類、例えば、麹菌、A.nidulans、A.フミガーツス、Ustilago属に属する真菌類、例えば、U.maydis、ロドバクテリア属に属する細菌、例えば、R.capsulatus、Rhodotorula属に属する酵母、例えば、R.rubraであってよい。
【0030】
好適には、該L−フェニルアラニンアンモニアリアーゼは、微生物において、該微生物に固有ではない該酵素をコードする核酸から発現させる。
【0031】
好ましくは、4−クマロイル−CoAは、ATPとCoAが基質であり、かつADPが産物である酵素触媒反応で形成され、好適には、4−クマロイル−CoAは4−クマレート−CoAリガーゼにより触媒される反応で形成される。
【0032】
前記4−クマレート−CoAリガーゼは、植物、微生物又は線虫に由来する4−クマレート−CoAリガーゼ(EC6.2.1.12)であってよい。植物は、Abies属、例えば、A.beshanzuensis、B.firma、B.holophylla、シロイヌナズナ属に属する植物、例えば、シロイヌナズナ、Brassica属に属する植物、例えば、B.napus、B.rapa、B.oleracea、シトラス属に属する植物、例えば、C.シネンシス、Larix属に属する植物、例えば、L.decidua、L.gmelinii、L.griffithiana、L.himalaica、L.kaempferi、L.laricina、L.mastersiana、L.occidentalis、L.potaninii、L.sibirica、L.speciosa、Phaseolus属に属する植物、例えば、P.acutifolius、P.coccineus、パイナス属に属する植物、例えば、P.armandii、P.banksiana、P.パイナスター、Populus属に属する植物、例えば、P.balsamifera、P.tomentosa、P.tremuloides、ナス属に属する植物、例えば、S.tuberosum、ヴィティス属に属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属に属する植物、例えば、Z.メイス又は他の植物属、例えば、Agastache属、アモルファ属、Cathaya属、Cedrus属、クロッカス属、ウシノケグサ属、グリシン属、Juglans属、Keteleeria属、リソスパーマム・オフィシナーレ属、Lolium属、ハス属、Lycopersicon属、マリュス属、Medicago属、マツバギク属、タバコ属、Nothotsuga属、オリザ属、ペラルゴニウム属、ペトロセリウム属、Physcomitrella属、Picea属、プラナス属、Pseudolarix属、Pseudotsuga属、ローザ属、キイチゴ属、Ryza属、Saccharum属、Suaeda属、Thellungiella属、コムギ属、Tsuga属に属してよい。微生物は、アスペルギルス属に属する糸状菌、例えば、A.flavus、A.nidulans、麹菌、A.フミガーツス、アカパンカビ属に属する糸状菌、例えば、アカパンカビ、Yarrowia属に属する真菌類、例えば、Y.lipolytica、Mycosphaerella属に属する真菌類、例えば、M.graminicola、マイコバクテリウム属に属する細菌、例えば、ウシ型結核菌、ハンセン菌、ヒト型結核菌、ナイセリア属に属する細菌、例えば、髄膜炎菌、ストレプトミセス属に属する細菌、例えば、S.coelicolor、ロドバクテリア属に属する細菌、例えば、R.capsulatus、鉤虫属に属する線虫類、例えば、A.ceylanicum、シノラブディス属に属する線虫類、例えば、C.エレガンス、捻転胃虫属に属する線虫類、例えば、H.contortus、Lumbricus属に属する線虫類、例えば、L.rubellus、Meilodogyne属に属する線虫類、例えば、M.hapla、Strongyloidus属に属する線虫類、例えば、S.rattii、糞線虫、Pristionchus属に属する線虫類、例えば、P.pacificusであってよい。
【0033】
任意に、NADPH:チトクロームP450レダクターゼ(CPR)は組換えで微生物に導入されている。これは非植物微生物に導入された植物CPRであってよい。あるいは、天然のNADPH:チトクロームP450レダクターゼ(CPR)が該微生物で過剰発現している。
【0034】
上記段落で言及されたものを含む好ましい実施態様において、前記NADPH:チトクロームP450レダクターゼは、シロイヌナズナ属に属する植物、例えば、シロイヌナズナ、シトラス属に属する植物、例えば、C.シネンシス、C.x paradisi、Phaseolus属に属する植物、例えば、P.ヴァガリス、パイナス属に属する植物、例えば、P.taeda、Populus属に属する植物、例えば、P.deltoides、P.tremuloides、P.trichocarpa、ナス属に属する植物、例えば、S.tuberosum、ヴィティス属に属する植物、例えば、ヴィティス ヴィニヘラ、ズィー属に属する植物、例えば、Z.メイス又は他の植物属、例えば、Ammi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属に由来するNADPH:チトクロームP450レダクターゼ(EC1.6.2.4)である。
【0035】
微生物が天然に存在するものであってよい一方で、該代謝経路の各酵素をコードする少なくとも1つの遺伝子配列の少なくとも1つのコピーが該微生物に組換えで導入されているのが好ましい。
【0036】
該酵素をコードするコード配列の導入に加えて、又はその代わりに、該生物体中の該コード配列に天然では関連しないプロモーター配列等の1つ以上の発現シグナルを提供してよい。したがって、任意に、チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、該生物中の該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結され、及び/又はL−フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、該生物中の該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0037】
任意に、シンナメート4−ヒドロキシラーゼをコードする遺伝子配列の少なくとも1つのコピーが、天然かどうかにかかわらず、該生物において該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0038】
任意に、4−クマレート−CoAリガーゼをコードする遺伝子配列の少なくとも1つのコピーが、天然かどうかにかかわらず、該生物において該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0039】
任意に、リスベラトロールシンターゼをコードする遺伝子配列の少なくとも1つのコピーが、天然かどうかにかかわらず、該生物において該遺伝子配列に天然では関連しない発現シグナルに作動可能に連結される。
【0040】
発現シグナルとして、コード配列の上流(5’−非コード配列)、その内部又はその下流(3’−非コード配列)に配置され、かつ関連するコード配列の転写、RNAのプロセシング又は安定性、又は翻訳に影響を与えるヌクレオチド配列が挙げられる。そのような配列として、プロモーター、翻訳リーダー配列、イントロン及びポリアデニル化認識配列が挙げられよう。
【0041】
ある態様において、本発明は、第1酵素により触媒され、かつその反応工程がアンモニアを作る反応により第1代謝産物が第2代謝産物に変換され、酸素が基質であり、NADPH又はNADHが補因子であり、NADP+又はNAD+が生成物である第2酵素により触媒される反応で該第2代謝産物が第3代謝物に変換され、ATP及びCoAが基質であり、ADPが生成物である第3酵素により触媒される反応で第3代謝産物が第4代謝産物に変換され、内在性マロニル−CoAが基質である第4酵素により触媒される反応で第4代謝産物が第5代謝産物に変換される作動性代謝経路を有する代謝改変微生物を提供する。
【0042】
さらに、本発明は、第1酵素により触媒され、かつその反応工程が第2酵素の関与なしにアンモニアを作る反応により、第1代謝産物が第3代謝産物に変換され、ATP及びCoAが基質であり、ADPが生成物である第3酵素により触媒される反応で第3代謝産物が第4代謝産物に変換され、内在性マロニル−CoAが基質である第4酵素により触媒される反応で第4代謝産物が第5代謝産物に変換される作動性代謝経路を有する代謝改変微生物を提供する。
【0043】
上記微生物として、発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する微生物が挙げられる。
【0044】
それら微生物として、シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する微生物も挙げられる。
【0045】
現在の文脈において、「微生物」は、細菌等の微生物、酵母等の微細菌に関する。
【0046】
さらに具体的には、微生物は真菌であってよく、さらに具体的には、アスペルギルス属に属する糸状菌、例えば、黒色アスペルギルス、A.awamori、麹菌、A.nidulans、Saccharomyces属に属する酵母、例えば、S.セレヴィシエ、S.kluyveri、S.bayanus、S.exiguus、S.sevazzi、S.uvarum、Kluyveromyces属に属する酵母、例えば、K.lactis、K.marxianus var.marxianus、K.thermotolerans、カンジダ属に属する酵母、例えば、C.utilis、C.tropicalis、C.albicans、C.lipolytica、C.versatilis、ピチア属に属する酵母、例えば、P.stipidis、P.pastoris、P.sorbitophila、又は他の酵母属、例えば、クリプトコッカス属、Debaromyces属、Hansenula属、ピチア属、Yarrowia属、Zygosaccharomyces属又はシゾサッカロミセス属であってよい。他の微生物に関して、ペニシリウム属、クモノスカビ属、フサリウム属、Fusidium属、Gibberella属、ケカビ属、モルティエラ属、トリコデルマ属に属する種等の好適な糸状菌の非網羅的なリストが提供される。
【0047】
細菌に関して、好適な細菌の非網羅的なリストは以下の通り与えられる:バシラス属に属する種、エシェリキア属に属する種、乳酸菌属に属する種、Lactococcus属に属する種、コリネバクテリア属に属する種、アセトバクター属に属する種、アシネトバクター属に属する種、シュードモナス属に属する種等。
【0048】
本発明の好ましい微生物は、S.セレヴィシエ、黒色アスペルギルス、麹菌、大腸菌、乳酸連鎖球菌又は枯草菌であってよい。
【0049】
構築され、改変された微生物は、ケモスタット培養、バッチ培養、流加バッチ培養等の一般に知られている方法を用いて培養することができる。
【0050】
例えば、本発明は、非植物細胞を炭素基質に4−クマル酸の外部源の実質的な非存在下で接触させる工程を含むリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法であって、該細胞が該条件下でリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を作る能力を有し、該微生物が真菌及び細菌、特に酵母からなる群より選択することができる。
【0051】
前記炭素基質は、単糖類、小糖類及び多糖類、例えば、グルコース、フルクトース、ガラクトース、キシロース、アラビノース、マンノース、スクロース、ラクトース、エリトロース、スレオース及び/又はリボースからなる発酵性炭素基質の群より任意に選択される。該炭素基質は、さらに又は代わりに、エタノール、アセテート、グリセロール及び/又はラクテート等の非発酵性炭素基質からなる群より選択してもよい。該非発酵性炭素基質は、さらに又は代わりに、アミノ酸からなる群より選択してよく、フェニルアラニン及び/又はチロシンであってよい。
【0052】
別の態様において、本発明は、フェニルアラニンアンモニアリアーゼ、シンナメート4−ヒドロキシラーゼ、4−クマレート−CoA−リガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法、さらにチロシンアンモニアリアーゼ、4−クマレート−CoAリガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロールの製造方法を含む。
【0053】
そのように生成されるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体は乳製品又はビール等の飲み物中の栄養補助食品として用いることができる。
【0054】
本発明にしたがって生成されるリスベラトロールはシス−リスベラトロール又はトランス−リスベラトロールであってよいが、トランス型が通常は優勢であると予想できる。
【0055】
本発明は更に以下の形態を含む。
[1]少なくとも1種類の酵素活性を含む操作的な代謝経路を有する微生物において、前記経路が4−クマル酸を生産し、かつそれからリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を生産することを特徴とする微生物。
[2]前記リスベラトロールが、内在性のマロニル−CoAが基質である酵素により触媒される反応で生成されることを特徴とする上記[1]に記載の微生物。
[3]前記リスベラトロールが4−クマロイル−CoAから生成されることを特徴とする上記[1]又は[2]に記載の微生物。
[4]前記リスベラトロールが、4−クマロイル−CoAからリスベラトロールシンターゼにより生成されることを特徴とする上記[3]に記載の微生物。
[5]前記リスベラトロールシンターゼが、前記微生物中において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[4]に記載の微生物。
【0056】
[6]前記リスベラトロールシンターゼが、Arachis属に属する植物、Rheum属に属する植物、ヴィティス属に属する植物、又はパイナス属、Piceea属、ユリ属、ユーカリ属、Parthenocissus属、ジッサス属、カロコルタス属、タデ属、Gnetum属、Artocarpus属、Nothofagus属、フェニックス属、ウシノケグサ属、スゲ属、シュロソウ属、ハカマカズラ又はPterolobium属のいずれか1種類に由来するリスベラトロールシンターゼ(EC2.3.1.95)であることを特徴とする上記[5]に記載の微生物。
[7]前記4−クマル酸が、トランス−ケイヒ酸から生成されることを特徴とする前記いずれかの一項に記載の微生物。
[8]前記4−クマル酸が、酸素が基質であり、NADH又はNADPHが補因子であり、NAD又はNADPが産物である酵素により触媒される反応で前記酵素によりトランス−ケイヒ酸から生成されることを特徴とする上記[7]に記載の微生物。
[9]前記4−クマル酸が、トランス−ケイヒ酸からシンナメート4−ヒドロキシラーゼにより生成されることを特徴とする上記[8]に記載の微生物。
[10]前記シンナメート4−ヒドロキシラーゼが、微生物において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[9]に記載の微生物。
【0057】
[11]前記シンナメート−4−ヒドロキシラーゼが、シロイヌナズナ属に属する植物、シトラス属に属する植物、Phaseolus属に属する植物、パイナス属に属する植物、Populus属に属する植物、ナス属に属する植物、ヴィティス属に属する植物、ズィー属に属する植物、又はAmmi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属のいずれか1種類、及びアスペルギルス属に属する真菌類に由来するシンナメート−4−ヒドロキシラーゼ(EC1.14.13.11)であることを特徴とする上記[10]に記載の微生物。
[12]前記4−クマル酸が、アンモニアが生成される酵素触媒反応でチロシンから生成されることを特徴とする上記[1]〜[6]のいずれかに記載の微生物。
[13]前記4−クマル酸が、L−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼによりチロシンから生成されることを特徴とする上記[1]〜[4]のいずれかに記載の微生物。
[14]前記4−クマル酸が、酵母又は細菌に由来するチロシンアンモニアリアーゼ(EC4.3.1.5)により生成されることを特徴とする上記[13]に記載の微生物。
[15]前記チロシンアンモニアリアーゼが、酵母Rhodotorula rubra又は細菌ロドバクテリア capsulatusに由来することを特徴とする上記[14]に記載の微生物。
[16]前記チロシンアンモニアリアーゼが、微生物において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[12]〜[15]のいずれかに記載の微生物。
[17]前記トランス−ケイヒ酸が、アンモニアが生成される酵素触媒反応でL−フェニルアラニンから生成されることを特徴とする上記[9]〜[11]のいずれかに記載の微生物。
[18]前記トランス−ケイヒ酸が、L−フェニルアラニンからL−フェニルアラニンアンモニアリアーゼにより形成されることを特徴とする上記[17]に記載の微生物。
[19]前記トランス−ケイヒ酸が、シロイヌナズナ属に属する植物、Brassica属に属する植物、シトラス属に属する植物、Phaseolus属に属する植物、パイナス属に属する植物、Populus属に属する植物、ナス属に属する植物、プラナス属に属する植物、ヴィティス属に属する植物、ズィー属に属する植物、又はAgastache属、Ananas属、Asparagus属、Bromheadia属、Bambusa属、Beta属、Betula属、Cucumis属、カメリア属、Capsicum属、Cassia属、Catharanthus属、Cicer属、Citrullus属、Coffea属、Cucurbita属、Cynodon属、Daucus属、Dendrobium属、Dianthus属、Digitalis属、Dioscorea属、ユーカリ属、Gallus属、Ginkgo属、グリシン属、Hordeum属、Helianthus属、Ipomoea属、Lactuca属、リソスパーマム・オフィシナーレ属、ハス属、Lycopersicon属、Medicago属、マリュス属、Manihot属、Medicago属、マツバギク属、タバコ属、Olea属、オリザ属、Pisum属、Persea属、ペトロセリウム属、Phalaenopsis属、Phyllostachys属、Physcomitrella属、Picea属、Pyrus属、Quercus属、Raphanus属、Rehmannia属、キイチゴ属、Sorghum属、Sphenostylis属、Stellaria属、Stylosanthes属、コムギ属、Trifolium属、コムギ属、Vaccinium属、Vigna属又はZinnia属のいずれかに属する植物、又はアスペルギルス属に属する真菌類に由来するL−フェニルアラニンアンモニアリアーゼ(EC4.3.1.5)によりL−フェニルアラニンから形成されることを特徴とする上記[18]に記載の微生物。
[20]前記L−フェニルアラニンアンモニアリアーゼが、微生物において、前記微生物に固有ではない前記酵素をコードする核酸から発現することを特徴とする上記[18]又は[19]に記載の微生物。
【0058】
[21]4−クマロイル−CoAが、ATPとCoAが基質であり、ADPが産物である酵素により触媒される反応で形成されることを特徴とする前記いずれかの一項に記載の微生物。
[22]4−クマロイル−CoAが、4−クマレート−CoAリガーゼにより触媒される反応で形成されることを特徴とする前記いずれかの一項に記載の微生物。
[23]前記4−クマレート−CoAリガーゼが、Abies属に属する植物、シロイヌナズナ属に属する植物、Brassica属に属する植物、シトラス属に属する植物、Larix属に属する植物、Phaseolus属に属する植物、パイナス属に属する植物、Populus属に属する植物、ナス属に属する植物、ヴィティス属に属する植物、ズィー属に属する植物、又はAgastache属、アモルファ属、Cathaya属、Cedrus属、クロッカス属、ウシノケグサ属、グリシン属、Juglans属、Keteleeria属、リソスパーマム・オフィシナーレ属、Lolium属、ハス属、Lycopersicon属、マリュス属、Medicago属、マツバギク属、タバコ属、Nothotsuga属、オリザ属、ペラルゴニウム属、ペトロセリウム属、Physcomitrella属、Picea属、プラナス属、Pseudolarix属、Pseudotsuga属、ローザ属、キイチゴ属、Ryza属、Saccharum属、Suaeda属、Thellungiella属、コムギ属又はTsuga属のいずれかに属する植物、アスペルギルス属に属する糸状菌、アカパンカビ属に属する糸状菌、Yarrowia属に属する真菌類、Mycosphaerella属に属する真菌類、マイコバクテリウム属に属する細菌、ナイセリア属に属する細菌、ストレプトミセス属に属する細菌、ロドバクテリア属に属する細菌、鉤虫属に属する線虫類、シノラブディス属に属する線虫類、捻転胃虫属に属する線虫類、Lumbricus属に属する線虫類、Meilodogyne属に属する線虫類、Strongyloidus属に属する線虫類、又はPristionchus属に属する線虫類に由来する4−クマレート−CoAリガーゼ(EC6.2.1.12)であることを特徴とする上記[22]に記載の微生物。
[24]前記代謝経路の各酵素をコードする少なくとも1つの遺伝子配列の少なくとも1つのコピーが、前記微生物に組換えで導入されていることを特徴とする前記いずれかの一項に記載の微生物。
[25]天然のNADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物で過剰発現していることを特徴とする上記[24]に記載の微生物。
【0059】
[26]NADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物に組換えで導入されていることを特徴とする上記[24]に記載の微生物。
[27]前記NADPH:チトクロームP450レダクターゼが、シロイヌナズナを含むArabidopsisに属する植物、C.シネンシス、C.x paradisiを含むCitrusに属する植物、P.ヴァガリスを含むPhaseolusに属する植物、P.taedaを含むPinusに属する植物、P.deltoides、P.tremuloides、P.trichocarpaを含むPopulusに属する植物、S.tuberosumを含むSolanumに属する植物、ヴィティス ヴィニヘラを含むヴィティスに属する植物、Z.メイスを含むZeaに属する植物、又はAmmi属、Avicennia属、カメリア属、Camptotheca属、Catharanthus属、グリシン属、Helianthus属、ハス属、マツバギク属、Physcomitrella属、Ruta属、Saccharum属、Vigna属を含む他の植物属に由来するNADPH:チトクロームP450レダクターゼ(EC1.6.2.4)であることを特徴とする上記[26]に記載の微生物。
[28]チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[29]フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[30]シンナメート4−ヒドロキシラーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物において前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していること特徴とする前記いずれかの一項に記載の微生物。
【0060】
[31]4−クマレート−CoAリガーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物において前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[32]リスベラトロールシンターゼをコードする遺伝子配列の少なくとも1つのコピーが、前記生物において前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする前記いずれかの一項に記載の微生物。
[33]真菌であることを特徴とする前記いずれかの一項に記載の微生物。
[34]糸状菌であることを特徴とする上記[33]に記載の微生物。
[35]アスペルギルス属に属する微生物であることを特徴とする上記[34]に記載の微生物。
【0061】
[36]菌株アスペルギルス niger又は麹菌であることを特徴とする上記[35]に記載の微生物。
[37]酵母であることを特徴とする上記[33]に記載の微生物。
[38]Saccharomyces属、Kluyveromyces属、カンジダ属、ピチア属、Debaromyces属、Hansenula属、Yarrowia属、Zygosaccharomyces属又はシゾサッカロミセス属に属する微生物であることを特徴とする上記[37]に記載の微生物。
[39]サッカロマイセス・セレヴィシエ、S.kluyveri、S.bayanus、S.exiguus、S.sevazzi、S.uvarum、Klyuveromyces lactis、K.marxianus var.marxianus、K.thermotolerans、Candida utilis、C.tropicalis、Pichia stipidis、P.pastoris、P.sorbitophila、Debaromyces hansenii、Hansenula polymorpha、Yarrowia lipolytica、Zygosaccharomyces rouxii又はシゾサッカロミセス・ポンベの菌株であることを特徴とする上記[38]に記載の微生物。
[40]細菌であることを特徴とする[1]〜[32]に記載の微生物。
【0062】
[41]エシェリキア属又はLactococcus属に属する微生物であることを特徴とする上記[40]に記載の微生物。
[42]大腸菌又は乳酸連鎖球菌の菌株であることを特徴とする上記[41]に記載の微生物。
[43]発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有することを特徴とする上記[1]に記載の微生物。
[44]シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する上記[1]に記載の微生物。
[45]適当なリスベラトロール産生代謝経路を有する非植物細胞を炭素基質に4−クマル酸の外部源の実質的な非存在下において接触させる工程を含むことを特徴とする、リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
【0063】
[46]前記非植物細胞が、真菌及び細菌からなる群より選択されることを特徴とする上記[45]に記載の方法。
[47]前記非植物細胞が、酵母からなる群より選択される真菌であることを特徴とする上記[46]に記載の方法。
[48]前記酵母がSaccharomyes種から選択されることを特徴とする上記[47]に記載の方法。
[49]前記非植物細胞が、上記[1]〜[44]に記載の微生物の細胞であることを特徴とする上記[45]〜[48]に記載の方法。
[50]前記炭素基質が、単糖類、小糖類及び多糖類からなる発酵性炭素基質の群より選択されることを特徴とする上記[45]〜[49]のいずれかに記載の方法。
【0064】
[51]前記発酵性炭素基質が、グルコース、フルクトース、ガラクトース、キシロース、アラビノース、マンノース、スクロース、ラクトース、エリトロース、スレオース、リボースであることを特徴とする上記[50]に記載の方法。
[52]前記炭素基質が、非発酵性炭素基質からなる群より選択されることを特徴とする上記[45]〜[49]のいずれかに記載の方法。
[53]前記非発酵性炭素基質が、エタノール、アセテート、グリセロール、ラクテートであることを特徴とする上記[52]に記載の方法。
[54]前記非発酵性炭素基質が、アミノ酸からなる群より選択されることを特徴とする上記[52]に記載の方法。
[55]前記非発酵性炭素基質が、フェニルアラニン及びチロシンからなる群より選択されることを特徴とする上記[54]に記載の方法。
【0065】
[56]フェニルアラニンアンモニアリアーゼ、シンナメート4−ヒドロキシラーゼ、4−クマレート−CoA−リガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
[57]チロシンアンモニアリアーゼ、4−クマレート−CoAリガーゼ及びリスベラトロールシンターゼをコードするヌクレオチド配列の異種発現によるリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
[58]さらに、生産されたリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を乳製品又は飲み物中の栄養補助食品として使用することを含む上記[45]〜[57]のいずれかに記載の方法。
[59]前記リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体をビール中の栄養補助食品として使用することを特徴とする上記[58]に記載の方法。
【0066】
本発明の上記説明の容易な理解を助けるために、添付の図面を参照されたい。
【0067】
以下の限定されない実施例により本発明をさらに説明し、例示する。
【0068】
(実施例1)
PAL、TAL、C4H、CPR、4CL及びVSTをコードする遺伝子の単離
フェニルアラニンアンモニアリアーゼ(PAL2)(Cochraneら、2004年;配列番号1、配列番号2)、シンナメート4−ヒドロキシラーゼ(C4H)(Mizutaniら、1997年;配列番号3、配列番号4)及び4−クマレート:コエンザイムAリガーゼ(4CL1)(ハンバーガー及びハルブロック(Hamberger and Hahlbrock)、2004年;エルチングら(Ehlting et al.)、1999年;配列番号5、配列番号6)を、PCRにより、表1のプライマーを用いて、シロイヌナズナのcDNA(バイオキャット社(BioCat)、ハイデルベルグ、ドイツ)からPCRにより単離した。コハク酸とクマロイル−CoAのそれぞれに対する好ましい速度パラメーターのために、幾つかのシロイヌナズナ相同体のうち、PAL2と4CL1を選択した(コクレンら(Cochrane et al.)、2004年;ハンバーガー及びハルブロック(Hamberger and Hahlbrock)、2004年;エルチングら(Ehlting et al.)、1999年)。
【0069】
Rheum tataricumに由来するリスベラトロールシンターゼ(VST)のコード配列(サマピトら(Samappito et al)、2003年;配列番号7、配列番号8)及びロドバクテリア capsulatusに由来するチロシンアンモニアリアーゼ(TAL)(キンドら(Kyndt et al.)、2002年;配列番号11、配列番号12)は、S.セレヴィシエでの発現のために、www.entelechon.comのオンラインサービスバックトランスレーションツールを用いてコドン最適化を行って、それぞれ配列番号9と10及び配列番号13と14を得た。合成遺伝子組立てのためのオリゴはMWGバイオテック社(MWG Biotech)で構築され、合成遺伝子は、以下に説明するマーチンら(Martin et al.)(2003年)の若干改良された方法プロトコールを用いて、PCRにより組み立てられた。
【0070】
【表1】
【0071】
合成遺伝子の組立てのためにMWG社のプライマーをミリQ水に100pmole/μlの濃度に溶解した。各プライマーの5μlのアリコートをトータルミックス中で一緒にし、次にミリQ水で10倍希釈した。遺伝子は、融合DNAポリメラーゼ(Finnzymes)のために50μlあたり鋳型として5μlの希釈トータルミックスを用いたPCRにより組み立てた。PCRプログラムは以下の通りである。最初に98℃で30秒間、次に、1000塩基対あたり、98℃で10秒間、40℃で1分間及び72℃で1分間の30サイクル、及び最後に72℃で5分間とした。得られたPCR反応物から、20μl
を1%アガロースゲルで精製した。その結果はPCRスメアであり、欲する大きさの周囲の領域をアガロースゲルから切り取り、QiaQuick Gel Extraction Kit(キアゲン社(Qiagen))を用いて精製した。表1の(TAL及びVST用の)外側のプライマーによる最終的なPCRは、要求されたTAL遺伝子とVST遺伝子を与えた。Quickchange部位特異的変異誘発IIキット(ストラタジン社(Stratagene)、ラホーヤ、カリフォルニア州)を用いるか、又は幾つかの異なる大腸菌サブクローンに由来するオーバーラップエラーフリーDNAストレッチからPCRを用いて点突然変異を補正した。
【0072】
シロイヌナズナ(AR2)に由来するNADPH:チトクロームP450レダクターゼ(CPR)(ミズタニ及びオータ(Mizutani and Ohta)、1998年;配列番号17、配列番号18)及びS.セレヴィシエ(CPR1)に由来するNADPH:チトクロームP450レダクターゼ(CPR1)(アオヤマら(Aoyama et al.)、1978年;配列番号15、配列番号16)は、表1のプライマーを用いて、シロイヌナズナのcDNA(バイオキャット社(BioCat)、ハイデルベルグ、ドイツ)及びS.セレヴィシエのゲノムDNAからそれぞれ単離した。
【0073】
(実施例2)
PAL発現用の酵母ベクターの構築
実施例1に記載のように単離されたPALをコードする遺伝子を、EcoR1とSpe1の制限部位(表1)を含む5’突出を有するフォワードプライマーとリバースプライマーを用いるPCRにより再増幅した。増幅されたPAL PCR産物をEcoR1/Spe1により消化し、EcoR1/Spe1で消化されたpESC−URAベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−URA−PALを得た。遺伝子の配列は2つの異なるクローンの配列決定により検証した。
【0074】
(実施例3)
PAL及びC4H発現用の酵母ベクターの構築
実施例1に記載のように単離されたC4Hをコードする遺伝子を、Xho1とKpn1の制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いるPCRにより増幅した。増幅されたC4H PCR産物をXho1/Kpn1により消化し、同様に消化されたpESC−URA−PALベクターに連結した。得られたプラスミドpESC−URA−PAL−C4Hは、分岐GAL1/GAL10プロモーターの制御下にPALとC4Hをコードする遺伝子を含有した。C4Hをコードする遺伝子の配列は2つの異なるクローンの配列決定により検証した。
【0075】
(実施例4)
4CL発現用酵母ベクターの構築
4CLをコードする遺伝子を実施例1に記載のように単離した。増幅された4CL PCR産物をXba1/BamH1により消化し、Spe1/BglIIで消化されたpESC−TRPベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−TRP−4CLを得た。pESC−TRP−4CLの2つの異なるクローンの配列を決定して、クローニングされた遺伝子の配列を検証した。
【0076】
(実施例5)
4CL及びVST発現用の酵母ベクターの構築
VSTをコードする遺伝子を実施例1に記載のように単離した。増幅された合成VST遺伝子をBamH1/Xho1により消化し、BamH1/Xho1で消化されたpESC−TRP−4CL(実施例4)に連結した。得られたプラスミドpESC−TRP−4CL−VSTは、分岐GAL1/GAL10プロモーターの制御下に4CLとVSTをコードする遺伝子を含んでいた。VSTをコードする遺伝子の配列はpESC−TRP−4CL−VSTの2つの異なるクローンの配列決定により検証した。
【0077】
(実施例6)
TAL発現用酵母ベクターの構築
TALをコードする遺伝子を実施例1に記載のように単離した。増幅された合成TAL遺伝子をEcoR1/Spe1により消化し、EcoR1/Spe1で消化されたpESC−URAベクターに連結した。得られたプラスミドpESC−URA−TALは、分岐GAL1/GAL10プロモーターの制御下にTALをコードする遺伝子を含有した。その配列はpESC−URA−TALの2つの異なるクローンの配列決定により検証した。
【0078】
(実施例7)
S.セレヴィシエ内在性CPRの過剰発現用酵母ベクターの構築
S.セレヴィシエに由来するCPR(CPR1)をコードする遺伝子を実施例1に記載のように単離した。増幅されたCPR1遺伝子をXho1/HindIIIで消化し、Xho1/HindIIIで消化されたpESC−LEUベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−LEU−CPR1を得た。その配列はpESC−LEU−CPR1の2つの異なるクローンの配列決定により検証した。
【0079】
(実施例8)
シロイヌナズナ CPR(AR2)の過剰発現用酵母ベクターの構築
シロイヌナズナに由来するCPR(AR2)をコードする遺伝子を実施例1に記載のように単離した。増幅されたAR2遺伝子をBamH1/Xho1で消化し、BamH1/Xho1で消化されたpESC−LEUベクター(ストラタジン社(Stratagene))に連結して、ベクターpESC−LEU−AR2を得た。その配列はpESC−LEU−AR2の2つの異なるクローンの配列決定により検証した。
【0080】
(実施例9)
酵母S.セレヴィシエにおいてPAL、C4H、4CL及びVSTを用いてリスベラトロールを導く経路の発現
適当な遺伝子マーカーを含む酵母菌株を実施例2、実施例3、実施例4、実施例5、実施例6、実施例7及び実施例8で記載したベクターにより別々に、又は組合せて形質転換した。酵母細胞の形質転換は、当分野で公知の方法にしたがって、例えば、コンピテント細胞又は電気穿孔法を用いて実施した(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0081】
S.セレヴィシエ菌株CEN.PK113−5D(MATa ura3)をベクターpESC−URA−PAL(実施例2)で形質転換して、FSSC−PAL菌株を得て、pESC−URA−PAL−C4H(実施例3)で形質転換して、FSSC−PALC4H菌株を得た。S.セレヴィシエ菌株FS01267(MATa trp1 ura3)はpESC−URA−PAL−C4H及びpESC−TRP−4CL(実施例4)により同時形質転換し、形質転換された菌株をFSSC−PALC4H4CLと命名した。同菌株は、pESC−URA−PAL−C4H及びpESC−TRP−4CL−VST(実施例5)によっても同時形質転換して、FSSC−PALC4H4CLVST菌株を得た。
【0082】
(実施例10)
S.セレヴィシエにおいてTAL、4CL及びVSTを用いてリスベラトロールを導く経路の発現
S.セレヴィシエ菌株CEN.PK113−5D(MATa ura3)をベクターpESC−URA−TAL(実施例6)で別に形質転換して、FSSC−TAL菌株を得た。S.セレヴィシエ菌株FS01267(MATa trp1 ura3)をpESC−URA−TAL(実施例6)及びpESC−TRP−4CL(実施例4)で同時形質転換し、形質転換された菌株をFSSC−TAL4CLと命名した。同菌株は、pESC−URA−TALとpESC−TRP−4CL−VST(実施例5)とによっても同時形質転換して、FSSC−TAL4CLVST菌株を得た。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0083】
(実施例11)
S.セレヴィシエにおける過剰発現内在性CPRによるリスベラトロールを導く経路の発現
S.セレヴィシエ菌株FS01277(MATa ura3 leu2 trp1)を、ベクターpESC−URA−PAL−C4H(実施例3)、ベクターpESC−TRP−4CL(実施例4)及びベクターpESC−LEU−CPR1(実施例7)により同時形質転換した。形質転換された菌株をFSSC−PALC4H4CLVSTCPRと命名した。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0084】
(実施例12)
S.セレヴィシエにおける過剰発現シロイヌナズナ CPR(AR2)によるリスベラトロールを導く経路の発現
S.セレヴィシエ菌株FS01277(MATa ura3 leu2 trp1)を、ベクターpESC−URA−PAL−C4H(実施例3)、ベクターpESC−TRP−4CL(実施例4)及びベクターpESC−LEU−AR2(実施例8)により同時形質転換した。形質転換された菌株をFSSC−PALC4H4CLVSTAR2と命名した。形質転換体は、ウラシル及び/又はトリプトファンを欠如する培地上で選択し、画線を同培地上で精製した。
【0085】
(実施例13)
振盪フラスコによる組換え酵母菌株による発酵
前記組換え酵母菌株を滅菌した接種用ループにより寒天プレートに接種し、ビタミン類、微量元素、5g/lのグルコース及び40g/l又は100g/lのガラクトースを含有する200mlの合成無機培地(ベルダインら(Verduyn et al.)、1992年)で増殖させた。500mlのコック付き振盪フラスコを3日間、30℃、160rpmでインキュベートした。
【0086】
(実施例14)
リスベラトロールの抽出
5000gで5分間の遠心により細胞を回収した。50mlの上清アリコートを20mlの酢酸エチルで一回抽出した。この酢酸エチルを凍結乾燥し、乾燥生成物を0.7mlメタノールで再溶解し、ろ過してHPLCバイアルに入れた。
【0087】
200ml培地から得られた細胞ペレットを1〜2mlの水に溶解し、3本の迅速調製チューブに分割し、ガラス玉で砕いた。これらの3チューブからの粗製抽出物を、50mlの縫工チューブ中の10mlの100%メタノール中にプールし、4℃の冷暗部屋でロータリーチャンバーにより48時間抽出した。48時間後、細胞デブリスを遠心により5分間、5000gで除去し、該メタノールを一晩の凍結乾燥により除去した。乾燥残留物を1mlのリン酸−クエン酸緩衝液(pH5.4)に再溶解し、アーモンドに由来する10単位のベータ−グルコシダーゼ(シグマ社(Sigma)製)を加えて、推定上グルコシド結合とされる構造物からリスベラトロールを遊離させた。混合物を3時間、37℃でインキュベートし、次に、1mlの酢酸エチルで2回抽出した。一緒にした酢酸エチルを凍結乾燥し、乾燥残留物を0.7mlのメタノールに再溶解し、ろ過してHPLCバイアルに入れた。
【0088】
(実施例15)
リスベラトロールの分析
(薄層クロマトグラフィー)
同一のTLCプレートによるケイヒ酸、クマル酸及びリスベラトロールの迅速な分離を可能とする薄層クロマトグラフィーに基づく方法を迅速なスクリーニング分析のために開発した。細胞と上清の両方を含む1mlの培養物アリコートを500マイクロリットルの酢酸エチルで抽出し、微小遠心機により13000rpmで30秒間遠心した。該酢酸エチルを乾燥し、メタノールに再溶解した。抽出物を、蛍光指示薬を含むシリカGプレート(0.2mm Alugram SIL G/UV254、マシュレ・ナーゲル社(Macherey-Nagel)製)により分析した。移動相はクロロホルム、酢酸エチル及び蟻酸の混合物(25:10:1)であった。
【0089】
(HPLC)
ケイヒ酸、クマル酸及びリスベラトロールの定量的分析のために、試料を、λ=306nmのUVダイオードアレイ検出前に高速液体クロマトグラフィー(HPLC)アジレント(Agilent)シリーズ1100システム(ヒューレットパッカード社(Hewlett Packard)製)による分離に供した。フェノメネックス社(Phenomenex)(トランス、カリフォルニア州、アメリカ合衆国)製Luna3マイクロメートルC18(100×2.00mm)カラムを40℃で用いた。移動相として、アセトニトリルとミリQ水(両方とも50ppmのトリフルオロ酢酸を含む)のグラジエントを0.4ml/分の流速で用いた。グラジエントのプロフィールは20分間にわたる15%アセトニトリルから100%アセトニトリルへのリニアであった。溶出時間はクマル酸で約3.4分、遊離トランス−リスベラトロールで5.5分、ケイヒ酸で6.8分であった。
【0090】
純粋なリスベラトロールスタンダードはケーマンケミカル社(Cayman chemical company)から購入し、純粋なクマル酸スタンダードとケイヒ酸スタンダードはシグマ社(Sigma)から購入した。
【0091】
(結果)
FSSC−PALC4H4CLVST菌株とFSSC−TAL4CLVST菌株を実施例13に記載のように100g/lのガラクトースで培養し、実施例14と実施例15にしたがって細胞内リスベラトロールの含量を分析した。さらに、空のベクターpESC−URA及びpESC−TRPのみを含むコントロール菌株のFSSCコントロールを含めた。HPLC分析は、FSSC−PALC4H4CLVST菌株とFSSC−TAL4CLVST菌株がトランス−リスベラトロールと同じ5.5分の保持時間を有する成分を含むことを示した(図4)。この結果は、約306nmのλmaxを有する純粋なトランス−リスベラトロールの吸収スペクトルに類似するUV吸収スペクトル(図5)によっても確認された。
【0092】
したがって、これらの結果は、トランス−リスベラトロールのインビボ生産を導くS.セレヴィシエの活性フェニル−プロパノイド経路の存在を示した。明確な増殖条件下にバッチ培養及び連続培養で菌株を培養し、及び/又は個々の酵素の発現/活性を最適化することによってリスベラトロールの生産が最も高い可能性で向上できるだろう。
【0093】
(実施例16)
大腸菌におけるTAL発現用細菌ベクターの構築
実施例1に記載したように単離されたTALをコードする遺伝子を、フォワードプライマー5’−CCG CTCGAG CGG ATG ACC CTG CAA TCT CAA ACA GCT AAA G−3’(配列番号33)とリバースプライマー5’−GC GGATCC TTA AGC AGG TGG ATC GGC AGC T−3’(配列番号34)(制限酵素部位XhoIとBamHIをそれぞれ有する5’突出を有する)を用いて、プラスミドpESC−URA−TAL(実施例6)から、PCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物のXhoIとBamHIとで消化したpET16bベクター(ノバゲン社(Novagen)製)への連結が可能となり、pET16b−TALが得られた。pET16bベクターはアンピシリン耐性遺伝子とT7プロモーターの両方を含んでいた。よって、上記手順は、T7プロモーターの制御下にTALをコードする遺伝子を含み、抗生物質選択マーカーを有するベクターをもたらした。TALをコードする遺伝子の配列はpET16b−TALの1つのクローンの配列決定により検証した。
【0094】
(実施例17)
大腸菌における4CLとVSTの発現用細菌ベクターの構築
実施例1に記載したように単離されたVSTをコードする遺伝子を、制限酵素BamHIとXhoIとにより4CLとVSTをコードする遺伝子を含む消化プラスミドpESC−TRP−4CL−VST(実施例5)から切り出した。カナマイシン耐性遺伝子を含有し、BamHIとSalIで消化したpET26bベクター(ノバゲン社(Novagen)製)にこのVST遺伝子を連結してpET26b−VSTを得た。制限酵素XhoIとSalIは適合性末端を有し、適当な連結を可能とした。pET26bベクターはカナマイシン耐性遺伝子とT7プロモーターの両方を含んでいた。よって、上記手順は、T7プロモーターの制御下にVSTをコードする遺伝子を含み、抗生物質選択マーカーを有するベクターをもたらした。
【0095】
実施例1に記載したように単離された4CLをコードする遺伝子を、フォワードプライマー5’−TG CCATGG CA ATGGCGCCAC AAGAACAAGC AGTTT−3’(配列番号35)とリバースプライマー5’−GC GGATCC CCT TCA CAA TCC ATT TGC TAG TTT TGCC−3’(配列番号36)(制限酵素部位NcoIとBamHIをそれぞれ有する5’突出を有する)を用いて、プラスミドpESC−URA−4CL−VST(実施例5)から、PCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物のNcoIとBamHIとで消化したpET16bベクター(ノバゲン社(Novagen)製)への連結を可能とした。得られたプラスミドpETl6b−4CLは、T7プロモーターの制御下に4CLをコードする遺伝子を含んでいた。T7プロモーターと4CLをコードする遺伝子との両方を、フォワードプライマー5’−TT GCGGCCGC AAA TCT CGA TCC CGC GAA ATT AAT ACG−3’(配列番号37)とリバースプライマー5’−CG CTCGAG CCT TCA CAA TCC ATT TGC TAG TTT TGCC−3’(配列番号38)(制限酵素部位NotIとXhoIをそれぞれ有する5’突出を有する)を用いて、プラスミドpETl6b−4CLから、PCRにより1つの断片として再増幅した。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、連結前にNotIXhoIとで消化したプラスミドpET26b−VSTへの制限PCR産物の連結を可能とした。得られたプラスミドpET26b−VST−4CLは、それぞれが個々のT7プロモーターの制御下にある2つの遺伝子(4CL及びVST)を含んでいた。
【0096】
(実施例18)
大腸菌において、TAL、4CL及びVSTを用いてリスベラトロールを導く経路の発現
細菌細胞の形質転換を当分野で公知の方法にしたがって、例えば、コンピテント細胞又は電気穿孔法を用いて実施した(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。大腸菌株BL21(DE3)(ノバゲン社(Novagen)製)を2種類のベクターpET16b−TAL(実施例16)とpET26b−VST−4CL(実施例17)とにより同時形質転換して、FSEC−TAL4CLVST菌株を得た。さらに、大腸菌株BL21(DE3)を、2種類の空のベクターpET16b(ノバゲン社(Novagen)製)とpET26b(ノバゲン社(Novagen)製)とで同時形質転換して、コントロール菌株として用いられるFSECコントロール菌株を得た。形質転換体は、100μg/mlのアンピシリンと60μg/mlのカナマイシンを含むルリア−ベルターニ(LB)培地により選択した。
【0097】
(実施例19)
振盪フラスコでの組換え大腸菌株による発酵
大腸菌BL21(DE3)の前培養物を、100μg/mlのアンピシリンと60μg/mlのカナマイシンを含む7mlのLB培地で、160rpm、37℃にてガラス管中で増殖させた。指数関数的に成長する前培養物を、50g/lのグルコース、5g/lのK2HPO4、80μg/mlのアンピシリン及び50μg/mlのカナマイシンを追加した200mlのLB培地を含む500mlバッフル付き振盪フラスコの接種に用いて、160rpm、37℃でインキュベートした。5時間後、3遺伝子(TAL、4CL及びVST)のそれぞれの上流に存在するT7プロモーターの誘導因子として、イソプロピルβ−チオガラクトピラノシド(IPTG)を1mMの最終濃度で加えた。37℃での48時間のインキュベーション時間の後、細胞を採取し、抽出手順に供し、産生したリスベラトロールの有無を分析した。
【0098】
(実施例20)
大腸菌のリスベラトロールの抽出と分析
抽出及び分析は、実施例14と実施例15に記載の方法を用いて実施した。
【0099】
(結果)
菌株FSEC−TAL4CLVST及びFSECコントロールを、実施例19に記載のように50g/lグルコースで培養し、実施例14と実施例15にしたがって細胞内リスベラトロールの含量を分析した。HPLC分析は、FSEC−TAL4CLVST菌株が、クマル酸と同じ3.4分の保持時間を有するかなりの量の成分を含むことを示した(図6)。しかし、その抽出物はトランス−リスベラトロールと同じ時間で溶出する成分を含んでいなかった。したがって、この結果は、チロシンアンモニアリアーゼ(TAL)は実際に活性はあるが、検出可能な量のリスベラトロールの生産を導くことはないことを示した。しかし、リスベラトロール形成の欠如は、i)非機能性クマレート−CoAリガーゼ(4CL)、ii)非機能性リスベラトロールシンターゼ(VST)、iii)非最適培養条件又はTALの非最適発現/活性により引き起こされるか、又はクマル酸の他の産物への分岐により引き起こされる非常に低いレベルのクマル酸との結果でありうるだろう。この仮説を評価するために、実施例19に記載の類似の培地を用いたが、ここでは20mg/lのクマル酸の存在下で菌株を増殖させた。FSEC−TAL4CLVSTの抽出物の引き続くHPLC分析は、トランス−リスベラトロールと同じ保持時間あたりに一群のピークを実際に示し、これはFSコントロールの抽出物には観察されなかった(図6)。実際、5.5分の保持時間を有するピークのUV吸収スペクトルは純粋なリスベラトロールのスペクトルに類似したが(図7)、そのようなスペクトルはコントロール菌株でのピークでは得られなかった。したがって、これらの結果は、リスベラトロールの生産を導きうる大腸菌の活性フェニルプロパノイド経路の存在を強く示唆する。明確な増殖条件下にバッチ培養及び連続培養で菌株を培養し、及び/又は個々の酵素の発現/活性を最適化することによって、クマル酸添加なしのリスベラトロールの生産が最も高い可能性で達成できるだろう。
【0100】
(実施例21)
乳酸連鎖球菌におけるPALとC4Hの発現用細菌ベクターの構築
下記実施例に使用されるプラスミドpSH71とその誘導体は、大腸菌と乳酸連鎖球菌に由来する複数の複製開始点を有する二機能性シャトルベクターである。それによって、宿主域特異性は大腸菌及び他の種の乳酸菌にわたる。したがって、乳酸連鎖球菌の形質転換は通常は問題なく進むが、他の種の乳酸菌の推定上の困難な形質転換は組換えプラスミドの構築のための中間ホストとして大腸菌を用いることにより克服することができる。プラスミドは1つ以上のマーカー遺伝子を持っており、それらを持つ微生物を、それらを持たない微生物から選択することができる。乳酸連鎖球菌に用いられる選択システムは、優性マーカー、例えば、エリスロマイシンとクロラムフェニコールに対する耐性に基づくが、糖質代謝に関与する遺伝子、ペプチダーゼ及び食品用マーカーに基づくシステムも記載されてきた。さらに、プラスミドは、組換え遺伝子の発現を可能とするプロモーター配列とターミネーター配列を含む。好適なプロモーターは、乳酸連鎖球菌の遺伝子、例えば、lacAから取られる。さらに、プラスミドはDNA断片のクローニングとそれに引き続く組換え体の同定を容易にする好適な独自の制限部位を含む。
【0101】
下記の実施例において、プラスミドは、pSH71−ERYrと呼ばれるエリスロマイシン耐性遺伝子又はpSH71−CMrと呼ばれるクロラムフェニコール耐性遺伝子のいずれかを含む。
【0102】
実施例1に記載のように単離されたPALをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を乳酸連鎖球菌に由来するlacAプロモーターを含む消化pSH71−ERYrベクターに連結することができる。得られたプラスミドpSH71−ERYr−PALは、乳酸連鎖球菌由来のlacAプロモーターの制御下にPALをコードする遺伝子を含有する。
【0103】
実施例1に記載のように単離されたC4Hをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−CMrベクターに連結して、pSH71−CMr−C4Hを得ることができる。lacAプロモーターとC4Hをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpSH71−CMr−C4HからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpSH71−ERYr−PALに連結することができる。得られたプラスミドpSH71−ERYr−PAL−C4Hは、個々のlacAプロモーターの制御下にPALとC4Hをそれぞれコードする遺伝子を含む。PALとC4Hをコードする遺伝子の配列を、pSH71−ERYr−PAL−C4Hの2つの異なるクローンの配列決定により検証する。
【0104】
(実施例22)
乳酸連鎖球菌におけるTAL発現用細菌ベクターの構築
実施例1に記載のように単離されたTALをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いてプラスミドpESC−URA−TAL(実施例6)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−ERYrベクターに連結することができる。得られたプラスミドpSH71−ERYr−TALは、乳酸連鎖球菌に由来するlacAプロモーターの制御下にTALをコードする遺伝子を含む。TALをコードする遺伝子の配列を、pSH71−ERYr−TALの2つの異なるクローンの配列決定により検証する。
【0105】
(実施例23)
乳酸連鎖球菌における4CLとVSTの発現用細菌ベクターの構築
実施例1に記載のように単離された4CLをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いてプラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−CMrベクターに連結することができる。得られたプラスミドpSH71−CMr−4CLは、乳酸桿菌(Lactobacillus lactis)由来のlacAプロモーターの制御下に4CLをコードする遺伝子を含有する。
【0106】
実施例1に記載のように単離されたVSTをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅した。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pSH71−ERYrベクターに連結することができる。得られたプラスミドpSH71−ERYr−VSTは、乳酸連鎖球菌に由来するlacAプロモーターの制御下にVSTをコードする遺伝子を含む。lacAプロモーターと、VSTをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpSH71−ERYr−VSTからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpSH71−CMr−4CLに連結することができる。得られたプラスミドpSH71−CMr−4CL−VSTは、個々のlacAプロモーターの制御下に4CL及びVSTをそれぞれコードする遺伝子を含む。4CLとVSTをコードする遺伝子の配列を、pSH71−CMr−4CL−VSTの2つの異なるクローンの配列決定により検証する。
【0107】
(実施例24)
乳酸連鎖球菌においてリスベラトロールを導く経路の発現
乳酸連鎖球菌株を、実施例21、実施例22及び実施例23に記載されたベクターで別々に、又は組合せて形質転換する。細菌細胞の形質転換は当分野で公知の方法にしたがって、例えば、コンピテント細胞又は電気穿孔法を用いて実施した(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体は、抗生物質のエリスロマイシンとクロラムフェニコールとを含有する培地上で選択し、画線を同培地上で精製した。
【0108】
別に、乳酸連鎖球菌株MG1363をベクターpSH71−ERYr−TAL(実施例22)により形質転換して、菌株FSLL−TALを得て、pSH71−ERYr−PAL−C4H(実施例21)により菌株FSLL−PALC4Hを得て、pSH71−CMr−4CL−VST(実施例23)により菌株FSLL−4CLVSTを得る。さらに、乳酸連鎖球菌株MG1363を、pSH71−ERYr−TAL(実施例22)及びpSH71−CMr−4CL−VST(実施例23)により同時形質転換し、この形質転換された菌株をFSLL−TAL4CLVSTと命名する。同菌株をpSH71−ERYr−PAL−C4H(実施例21)とpSH71−CMr−4CL−VST(実施例23)によっても同時形質転換して、菌株FSLL−PALC4H4CLVSTを得る。
【0109】
(実施例25)
発酵槽での組換え乳酸連鎖球菌株による発酵
組換え酵母菌株は、バッチ培養、流加バッチ培養又はケモスタット培養として操作される発酵槽で増殖させることができる。
【0110】
(バッチ培養と流加バッチ培養)
微生物を、1.5リットルの有効容積のバッフル付きバイオリアクターで、嫌気的、好気的又は微好気的条件で増殖させる。すべての培養物を30℃で、350rpmにてインキュベートする。pH6.6で一定になるように10MのKOHの自動添加により維持する。細胞を、以下の成分を補充した合成MS10培地中でラクトースにより増殖させて好気的条件下の増殖を可能とする:MnSO4(1.25×10−5g/l)、チアミン(1mg/l)及びDL−6,8−チオクト酸(2.5mg/l)。ラクトース濃度は、例えば、50g/lである。3倍高い濃度のK2HPO4とKH2PO4で緩衝化した上記培地により、振盪フラスコ内、30℃で増殖させた前培養物の細胞をバイオリアクターに接種する。接種前にN2(純度99.998%)を培地に流すことにより、かつ培養中にバイオリアクターの上部空間にN250ml/分の定速を維持することにより好気的条件を確保する。微好気的条件及び好気的培養のために用いられるバイオリアクターは、空気(DOT、100%)及びN2(DOT、0%)で較正されたポーラログラフィー酸素センサーを備える。好気的条件はバイオリアクターに空気を1vvmの速度で拡散させてDOTが80%を超えることを確保することで得られる。微好気的実験中、N2と大気の空気の混合物からなる気体を0.25vvmの速度でリアクターに拡散することによりDOTが5%の一定に保たれる。
【0111】
(ケモスタット培養物)
ケモスタット培養物において、細胞は例えば1Lの有効容積のアプリコン(Applikon)実験室用発酵槽で、30℃、350rpmで増殖させることができる。希釈率(D)は、異なった値、例えば、0.050h−1、0.10h−1、0.15h−1又は0.20h−1に設定することができる。消泡剤(50μl/l)を追加した上記成長培地を用いて、5MのKOHの自動追加によりpHを一定、例えば6.6に保つ。ラクトースの濃度を異なる値、例えば、3.0g/l、6.0g/l、12.0g/l、15.0g/l又は18.0g/lに設定することができる。バイオリアクターに1mg/lの初期バイオマス濃度で接種し、指数増殖期の終わりにフィードポンプを作動させる。
【0112】
嫌気的定常状態は50ml/分のN2(純度99.998%)をバイオリアクターの上部空間に導入することにより得る。異なる無酸素性定常状態は、N2(純度99.998%)と大気の空気とから多様な比率で構成される250ml/分のガスをリアクターに拡散させることにより得ることができる。バイオリアクターに空気(100%DOT)とN2(0%DOT)を拡散させることにより酸素電極を較正する。
【0113】
すべての条件で、ガスはバイオリアクターに導入する前に滅菌ろ過する。オフガスは、−8℃未満に冷却した冷却器を経て、アコースティックガスアナライザーによりそのCO2とO2の体積含量が分析される。
【0114】
少なくとも5滞留時間後、及びバイオマスと発酵最終生成物の濃度が最後の少なくとも2滞留時間にわたって変化しない(5%相対偏差未満)の場合、培養は定常状態にあると考えられる。
【0115】
(実施例26)
乳酸連鎖球菌におけるリスベラトロールの抽出と分析
抽出と分析は実施例14と実施例15に記載された方法を用いて実施する。
【0116】
(実施例27)
アスペルギルス属に属する種におけるPALとC4Hの発現用真菌ベクターの構築
下記実施例で使用されるプラスミドは、黒色アスペルギルスと麹菌で自己プラスミド複製も維持する、アスペルギルス nidulans由来のAMA1開始複製配列を含有するpARp1から得られる(ゲムスら(Gems et al.)、1991年)。さらに、プラスミドは大腸菌の複製配列を含むシャトルベクターであるので、黒色アスペルギルス及び麹菌での固有の困難な形質転換が、組換えプラスミドの構築のために中間ホストとして大腸菌を用いることにより克服することができる。プラスミドは1つ以上のマーカー遺伝子を持っており、それらを持つ微生物を、それらを持たない微生物から選択することを可能とする。選択システムは、優性マーカー、例えば、ハイグロマイシンB、フレオマイシン及びブレオマイシンに対する耐性、又は異種マーカー、例えば、アミノ酸及びpyrG遺伝子に基づくことができる。さらに、プラスミドは組換え遺伝子の発現を可能とするプロモーター配列とターミネーター配列を含む。好適なプロモーターは、アスペルギルス nidulansの遺伝子、例えば、alcA、glaA、amy、niaD及びgpdAから得られる。さらに、プラスミドはDNA断片のクローニング及びその後の組換え体の同定を容易にする好適な独自の制限部位を含む。
【0117】
下記の実施例で用いられるプラスミドは強力な構成的gpdAプロモーターと栄養要求性マーカーを含み、これらすべてがアスペルギルス nidulansを起源とする。メチオニン生合成に関与する遺伝子methGを含むプラスミドをpAMA1−METと命名する。ヒスチジン生合成に関与する遺伝子hisAを含むプラスミドをpAMA1−HISと命名する。
【0118】
実施例1に記載のように単離されたPALをコードする遺伝子を、好適な制限部位を含
む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物をアスペルギルス nidulansに由来するgpdAプロモーターを含む消化pAMA1−METベクターに連結することができる。得られたプラスミドpAMA1−MET−PALは、アスペルギルス nidulans由来のgpdAプロモーターの制御下にPALをコードする遺伝子を含有する。
【0119】
実施例1に記載のように単離されたC4Hをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−PAL−C4H(実施例3)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pAMA1−HISベクターに連結してpAMA1−HIS−C4Hを得ることができる。gpdAプロモーターと、C4Hをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpAMA1−HIS−C4HからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpAMA1−MET−PALに連結することができる。得られたプラスミドpAMA1−MET−PAL−C4Hは、アスペルギルス nidulansに由来する個々のpgdAプロモーターの制御下にPALとC4Hをそれぞれコードする遺伝子を含む。PALとC4Hをコードする遺伝子の配列をpAMA1−MET−PAL−C4Hの2つの異なるクローンの配列決定により検証する。
【0120】
(実施例28)
アスペルギルス属に属する種におけるTALの発現用真菌ベクターの構築
実施例1に記載のように単離されたTALをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−URA−TAL(実施例6)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pAMA1−METベクターに連結することができる。得られたプラスミドpAMA1−MET−TALは、アスペルギルス nidulansに由来するgpdAプロモーターの制御下にTALをコードする遺伝子を含む。TALをコードする遺伝子の配列をpAMA1−MET−TALの2つの異なるクローンの配列決定により検証する。
【0121】
(実施例29)
アスペルギルス属に属する種における4CLとVSTの発現用真菌ベクターの構築
実施例1に記載のように単離された4CLをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を、アスペルギルス nidulans由来のgpdAプロモーターを含む消化pAMA1−HISベクターに連結することができる。得られたプラスミドpAMA1−HIS−4CLは、アスペルギルス nidulansに由来するgpdAプロモーターの制御下に4CLをコードする遺伝子を含む。
【0122】
実施例1に記載のように単離されたVSTをコードする遺伝子を、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpESC−TRP−4CL−VST(実施例5)からPCRにより再増幅する。遺伝子の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化pAMA1−METベクターに連結してpAMA1−MET−VSTを得ることができる。gpdAプロモーターと、VSTをコードする遺伝子とを、好適な制限部位を含む5’突出を有するフォワードプライマーとリバースプライマーを用いて、プラスミドpAMA1−MET−VSTからPCRにより1つの断片として再増幅する。DNA断片の5’末端と3’末端における該制限酵素部位の導入により、制限PCR産物を消化プラスミドpAMA1−HIS−4CLに連結することができる。得られたプラスミドpAMA1−HIS−4CL−VSTは、アスペルギルス nidulansに由来する個々のpgdAプロモーターの制御下にC4LとVSTをそれぞれコードする遺伝子を含む。4CLとVSTをコードする遺伝子の配列をpAMA1−HIS−4CL−VSTの2つの異なるクローンの配列決定により検証する。
【0123】
(実施例30)
黒色アスペルギルスにおいてリスベラトロールを導く経路の発現
黒色アスペルギルス菌株を、実施例27、実施例28及び実施例29に記載されたベクターで、別々に、又は組合せて形質転換する。細菌細胞の形質転換は当分野で公知の方法にしたがって、例えば、電気穿孔法又は接合を用いて実施する(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体は、メチオニン及び/又はヒスチジンを欠く最小培地により選択する。
【0124】
ヒスチジンとメチオニンに対して栄養要求性のあるアスペルギルス nigerの菌株、例えば、別々にFGSC A919菌株(http://www.fgsc.netを参照)をベクターpAMA1−MET−TAL(実施例28)により形質転換して菌株FSAN−TALを得て、pAMA1−MET−PAL−C4H(実施例27)により菌株FSAN−PALC4Hを得て、pAMA1−HIS−4CL−VST(実施例29)により菌株FSAN−4CLVSTを得る。さらに、黒色アスペルギルス菌株FGSC A919をpAMA1−MET−TAL(実施例28)とpAMA1−HIS−4CL−VST(実施例29)とで同時形質転換して、形質転換菌株をFSAN−TAL4CLVSTと名付ける。また、同菌株をpAMA1−MET−PAL−C4H(実施例27)及びpAMA1−HIS−4CL−VST(実施例29)によっても同時形質転換して菌株FSAN−PALC4H4CLVSTを得る。
【0125】
(実施例31)
麹菌においてリスベラトロールを導く経路の発現
PAL、C4H及び4CLをコードする天然の遺伝子組を含有する麹菌の菌株(非特許文献18)であって、メチオニンに対して栄養要求性のある菌株をベクターpAMA1−MET−VST(実施例29)により形質転換して、菌株FSAO−VSTを得る。真菌細胞の形質転換は当分野で公知の方法にしたがって、例えば、電気穿孔法又は接合を用いて実施する(例えば、サムブルックら(Sambrook et al.)(1989年)を参照)。形質転換体はメチオニンを欠く培地により選択する。
【0126】
(実施例32)
培養槽における黒色アスペルギルス及び麹菌の組換え菌株による発酵
組換え酵母菌株は、バッチ培養、流加バッチ培養又はケモスタット培養として操作される発酵槽で増殖させることができる。
【0127】
(バッチ培養及び流加バッチ培養)
微生物は、好気的条件下で、1.5リットルの有効容積のバッフル付きバイオリアクターで増殖させる。すべての培養物を30℃で、500rpmにてインキュベートする。pH6.0で一定となるように10MのKOHの自動添加により維持し、好気的条件は空気を1vvmの速度でバイオリアクターに拡散させてDOTが80%を超えることを確保することで得られる。以下の成分からなる合成培地中で細胞をグルコースで増殖させてバッチ培養で増殖させる:7.3g/lの(NH4)2SO4、1.5g/lのKH2PO4、1.0g/lのMgSO4・7H2O、1.0g/lのNaCl、0.1g/lのCaCl2・2H2O、0.1ml/lのシグマ社製の消泡剤、7.2mg/lのZnSO4・7H2O、1.3mg/lのCuSO4・5H2O、0.3mg/lのNiCl2・6H2O、3.5mg/lのMnCl2・4H2O及び6.9mg/lのFeSO4・7H2O。グルコース濃度は、例えば、10g/l、20g/l、30g/l、40g/l又は50g/lである。流加バッチ培養で細胞を増殖させるために、培地は、バッチ相中の7.3g/lの(NH4)2SO4、4.0g/lのKH2PO4、1.9g/lのMgSO4・7H2O、1.3g/lのNaCl、0.10g/lのCaCl2・2H2O、0.1ml/lのシグマ社製の消泡剤、7.2mg/lのZnSO4・7H2O、1.3mg/lのCuSO4・5H2O、0.3mg/lのNiCl2・6H2O、3.5mg/lのMnCl2・4H2O及び6.9mg/lのFeSO4・H2Oからなる。次に、リアクターに、例えば、285g/kgのグルコースと42g/kg(NH4)2SO4を供給する。
【0128】
前バッチからの遊離した菌糸体を用いて、バッチ培養物及び流加バッチ培養物に接種する。2.109/l濃度の胞子を用いて、pH2.5の前バッチ培養物に接種する。以下の成分が添加された29gの米により凍結乾燥胞子を増殖させて胞子を得る:6mlの15g/lのスクロース、2.3g/lの(NH4)2SO4、1.0g/lのKH2PO4、0.5g/lのMgSO4・7H2O、0.50g/lのNaCl、14.3mg/lのZnSO4・7H2O、2.5mg/CuSO4・5H2O、0.50mg/lのNiCl2・6H2O及び13.8mg/lのFeSO4・7H2O。これらの胞子は、30℃で、7〜14日間増殖して米粒上に胞子の黒い層を生じ、滅菌水中の100mlの0.1%Tween20を加えることにより胞子を回収する。すべての条件に関して、ガスはバイオリアクターに導入する前に滅菌ろ過する。オフガスは−8℃未満に冷却した冷却器を経て、アコースティックガスアナライザーによりそのCO2とO2の体積含量を分析する。
【0129】
ケモスタット培養
ケモスタット培養では、例えば1.5リットルの有効容積のバッフル付きバイオスタットB実験室用発酵槽で、30℃、500rpmで細胞を増殖させることができる。pH6.6で一定になるように10MのKOHの自動添加により維持し、好気的条件は、空気を1vvmの速度でバイオリアクターに拡散して、確実にDOTが80%を超えさせることにより得られる。希釈率(D)は、異なる値、例えば、0.050h−1、0.10h−1、0.15h−1又は0.20h−1に設定できる。以下の成分を有する最小成長培地を用いて、10Mの自動追加によりpHを一定、例えば、6.6に保持する:2.5g/lの(NH4)2SO4、0.75g/lのKH2PO4、1.0g/lのMgSO4・7H2O、1.0g/lのNaCl、0.1g/lのCaCl2・2H2O、0.1ml/lのシグマ社製消泡剤、7.2mg/lのZnSO4・7H2O、1.3mg/lのCuSO4・5H2O、0.3mg/lのNiCl2・6H2O、3.5mg/lのMnCl2・4H2O及び6.9mg/lのFeSO4・7H2O。グルコースの濃度は異なる値、例えば、3.0g/l、6.0g/l、12.0g/l、15.0g/l又は18.0g/lに設定することができる。バイオリアクターには上記のように前バッチ培養物からの遊離菌糸体が接種され、指数増殖期の終わりにフィードポンプを作動させる。
【0130】
すべての条件について、ガスはバイオリアクターに導入する前に滅菌ろ過する。オフガスは−8℃未満に冷却した冷却器を経て、アコースティックガスアナライザーによりそのCO2とO2の体積含量を分析する。
【0131】
少なくとも5滞留時間後、及びバイオマスグルコースとオフガスの組成が最後の2滞留時間にわたって変化しない(5%相対偏差未満)の場合、培養は定常状態にあると考えられる。
【0132】
(実施例33)
黒色アスペルギルス及び麹菌におけるリスベラトロールの抽出と分析
抽出と分析は実施例14と実施例15に記載された方法を用いて実施する。
【0133】
参考文献
米国特許出願公開第6521748号
米国特許出願公開第2001053847号
米国特許出願公開第2004059103号
米国特許出願公開第2004023357号
【0134】
Allina,S.M.,Pri−Hadash,A.,Theilmann,D.A.,Ellis,B.E.and Douglas,C.J.(1998)4−coumarate:Coenzyme A ligase in hybrid poplar.Properties of enzymes,cDNA cloning,and analysis of recombinant clones.Plant Physiol.116,743−754.
Aoyama,Y.,Yoshida,Y.,Kubota,S.,Kumaoka,H.and Furumichi,A.(1978).NADPH−cytochrome P−450 reductase of yeast microsomes.Arch.Biochem.Biophys.185,362−369.
Becker JV,Armstrong GO,van der Merwe MJ,Lambrechts MG,Vivier MA,Pretorius IS.(2003).Metabolic engineering of Saccharomyces cerevisiae for the synthesis of the
wine− related antioxidant resveratrol.FEMS Yeast Res.4,79−85.
【0135】
Blanquet,S.,Meunier,J.P.,Minekus,M.,Marol−Bonnin,S.,and Alric,M.(2003).Recombinant Saccharomyces cerevisiae Expressing P450 in Artificial Digestive Systems:a Model for Biodetoxication in the Human Digestive Environment.Appl.Environ.Microbiol.69,2884−2892.
Celotti E and others.(1996).Resveratrol content of some wines obtained from dried Valpolicella grapes:Recioto and Amarone.Journal of Chromatography A 730,47−52.
Cochrane,F.C,Davin,L.B.and Lewis N.G.(2004).The Arabidopsis phenylalanine ammonia lyase gene family:kinetic characterization of the four PAL isoforms.Phytochemistry 65,1557−1564.
Couzin,J.(2004)Aging Research’s Family Feud.Science 303,1276−1279.
【0136】
Ehlting,J.,Buttner,D.,Wang,Q.,Douglas,C.J.,Somssich,I.E.and Kombrink,E.(1999).Three 4−coumarate:coenzyme A ligases in Arabidopsis thaliana represents two evolutionary divergent classes in angiosperms.The plant journal.19,9−20.
Filpula,D.,Vaslet,CA.,Levy,A.,Sykes,A.and Strausberg,R.L.Nucleotide sequence of gene for phenylalanine ammonia−lyase from Rhodotorula rubra.(1988).Nucleic Acids Res.16,11381.
【0137】
Gehm,B.D.,McAndrews,J.M.,Chien,P.Y.and Jameson,J.L.(1997).Resveratrol,a polyphenolic compound found in grapes and wine,is an agonist for the estrogen receptor.Proc.Natl.Acad.Sci.USA 94,14138−14143.
Gems,D.,Johnstone,I.L.and Clutterbuck,A.J.(1991).An autonomously replicating plasmid transforms Aspergillus nidulans at high frequency.Gene 98,61−67.
Hain,R.,Reif,H.J.,Krause,E.,Langebartels,R.,Kindl,H.,Vornam,B.,Wiese,W.,Schmelzer,E.,Schreier,P.H.,Stocker,R.H.and Stenzel,K.(1993).Disease resistance results from foreign phytoalexin expression in a novel plant.Nature 361,153−156.
【0138】
Hwang EI,Kaneko M,Ohnishi Y,Horinouchi S.(2003).Production of plant−specific flavanones by Escherichia coli containing an artificial gene cluster.Appl.Environ.Microbiol.69,2699−706.
Huang,M−T.(1997).Diet for cancer prevention.Food Sci.24,713−727
Hart,J.H.(1981)Annu.Rev.Phytopathology 19,437−458.Hart,J.H.,Shrimpton,D.M.(1979)Phytopathology 69,1138−1143.
Hall,S.S.(2003)In Vino Vitalis? Compounds Activate Life− Extending Genes.Science 301,1165.
Hamberger,B.and Hahlbrock,K.(2004).The 4−coumarate:CoA ligase gene family in Arabidopsis thaliana comprises one rare,sinapate−activating and three commonly occurring isoenzymes.Proc.Natl.Acad.Sci .USA.101,2209−2214.
【0139】
Jang,M.,Cai,L.,Udeani,GO.,Slowing,KV.,Thomas,CF.,Beecher,CWW.,Fong,HHS.,Farnsworth,NR.,Kinghorn,AD.,Mehta,RG.,Moon,RC,Pezzuto,JM.(1997).Cancer Chemopreventive Activity of Resveratrol,a Natural Product Derived from Grapes.Science 275,218−220.
Jeandet,P.Bessis,R.,Maume,B.F.,Meunier,P.,Peyron,D.and Trollat,P.(1995).Effect of Enological Practices on the Resveratrol Isomer Content of Wine J.Agric.Food Chem.43,316−319.
Jeandet,P.Bessis,R.,Sbaghi,M.and Meunier,P.(1994).Occurence of a resveratrol β−D−glucoside in wine:Preliminary studies.Vitis 33,183−184.
【0140】
Koopmann,E.,Logemann,E.and Hahlbrock,K.(1999).Regulation and Functional Expression of Cinnamate 4−Hydroxylase from Parsley.Plant Physiol.119,49−55.
Kopp,P.(1998).Resveratrol,a phytooestrogen found in red wine.A possible explanation for the conundrum of the “French Paradox”? Eur.J.Endocrinol.138,619−620.
Kyndt JA,Meyer TE,Cusanovich MA,Van Beeumen JJ.(2002).Characterization of a bacterial tyrosine ammonia lyase,a biosynthetic enzyme for the photoactive yellow protein.FEBS Lett.512,240−244.
【0141】
LaGrange,D.C,Pretorius,I.S.and Van ZyI,W.H.(1997).Cloning of the Bacillus pumilus beta−xylosidase gene(xynB)and its expression in Saccharomyces cerevisiae.Appl.Microbiol.Biotechnol.47,262−266.
Lin X.,Kaul S.,Rounsley S.D.,Shea T.P.,Benito M.−I.,Town CD.,Fujii CY.,Mason T.M.,Bowman CL.,Barnstead M.E.,Feldblyum T.V.,Buell CR.,Ketchum K.A.,Lee J.J.,Ronning CM.,Koo H.L.,Moffat K.S.,Cronin L.A.,Shen M.,Pai G.,Van Aken S.,Umayam L.,Tallon L.J.,Gill J.E.,Adams M.D.,Carrera A.J.,Creasy T.H.,Goodman H.M.,Somerville CR.,Copenhaver G.P.,Preuss D.,Nierman W.C,White O.,Eisen J.A.,Salzberg S.L.,Fraser CM.,Venter J.C.(1999).Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana.Nature 402,761−768.
【0142】
Lobo,R.A.(1995).Benefits and risks of estrogen replacement therapy.Am.J.Obstet.Gynecol.173,982−989.
Martin,V.J.J.,Pitera,D.J.,Withers,S.T.,Newman,J.D.and Keasling,J.D.(2003).Engineering a mevalonate pathway in Escherichia coli for production of terpenoids.Nature biotechnology 21,796−802.
Mizutani,M.,Ohta,D.and Sato,R.(1997).Is
olation of a cDNA and a genomic clone encoding cinnamate 4−hydroxylase from Arabidopsis and its expression manner in planta.Plant Physiol.113,755−763.
【0143】
Ro,D.K.,Mah,N.,Ellis,B.E.and Douglas,CJ.(2001).Functional Characterization and Subcellular Localization of Poplar(Populus trichocarpa x Populus deltoides)Cinnamate 4−Hydroxylase.Plant Physiol.126,317−329.
Ro D.K.,Douglas CJ.(2004).Reconstitution of the entry point of plant phenylpropanoid metabolism in yeast(Saccharomyces cerevisiae):implications for control of metabolic flux into the phenylpropanoid pathway.J.Biol.Chem.279,2600−2607.
【0144】
Rosier J,Krekel F,Amrhein N,Schmid J.(1997).Maize phenylalanine ammonia−lyase has tyrosine ammonia−lyase activity.Plant
Physiol.113,175−179.activity.Plant physiol.113,175−179.
Samappito,S.,Page,J.E.,Schmidt,J.,De−Eknamkul,W.and Kutchan,T.M.(2003).Aromatic and pyrone polyketides synthesized by a stilbene synthase from Rheum tataricum.Phytochemistry 62,313−323.
Sambrook,J.,Fritsch,E.F.and Maniatis,T.(1989).Molecular Cloning,A Laboratory Manual,2d edition,Cold Spring Harbor,N.Y.
【0145】
Schoppner,A.;Kindl,H.(1984)Purification and properties of a stilbene synthase from induced cell suspension cultures of peanut.J.Biol.Chem.259,6806−6811.
Seshime,Y.,Juvvadi,P.R.,Fujii,I.and Kitamoto,K.(2005).Genomic evidences for the existence of a phenylpropanoid metabolic pathway in Aspergillus oryzae.Biochem Biophys Res Commun.337,747−51.
Urban P.,Mignotte,C,Kazmaier M.,Delorme F.And Pompon D.(1997).Cloning,Yeast Expression,and Characterization of the Coupling of Two Distantly Related Arabidopsis thaliana NADPH−Cytochrome 450 Reductases with P450 CYP73A5.J.Biol.Chem.272,191
76−19186.
【0146】
Watts,K.T.,Lee,P.C.and Schmidt−Dannert,C.(2004).Exploring recombinant flavonoid biosynthesis in metabolically engineered Escherichia coli.Chembiochem 5,500−507.
【0147】
以下は本明細書に現れるヌクレオチド配列とアミノ酸配列の概要である。
配列番号1は、フェニルアラニンアンモニアリアーゼ(PAL2)をコードするシロイヌナズナ由来のヌクレオチド配列である。
配列番号2は、配列番号1によりコードされるアミノ酸配列である。
配列番号3は、シンナメート4−ヒドロキシラーゼ(C4H)をコードするシロイヌナズナ由来のヌクレオチド配列である。
配列番号4は、配列番号3によりコードされるアミノ酸配列である。
配列番号5は、4−クマレート:コエンザイムAリガーゼ(4CL1)をコードするシロイヌナズナ由来のヌクレオチド配列である。
配列番号6は、配列番号5によりコードされるアミノ酸配列である。
配列番号7は、リスベラトロールシンターゼ(VST)をコードするRheum tataricum由来のヌクレオチド配列である。
配列番号8は、配列番号7によりコードされるアミノ酸配列である。
【0148】
配列番号9は、S.セレヴィシエでの発現のためにコドン最適化されたリスベラトロールシンターゼ(VST)をコードするRheum tataricum由来のヌクレオチド配列である。
配列番号10は、配列番号9によりコードされるアミノ酸配列である。
配列番号11は、チロシンアンモニアリアーゼ(TAL)をコードするロドバクテリア capsulatus由来のヌクレオチド配列である。
配列番号12は、配列番号11によりコードされるアミノ酸配列である。
配列番号13は、S.セレヴィシエでの発現のためにコドン最適化されたチロシンアンモニアリアーゼ(TAL)をコードするロドバクテリア capsulatus由来のヌクレオチド配列である。
配列番号14は、配列番号13によりコードされるアミノ酸配列である。
【0149】
配列番号15は、NADPH:チトクロームP450レダクターゼ(CPR1)をコードするS.セレヴィシエ由来のヌクレオチド配列である。
配列番号16は、配列番号15によりコードされるアミノ酸配列である。
配列番号17は、NADPH:チトクロームP450レダクターゼ(AR2)をコードするArabidopsis thalianus由来のヌクレオチド配列である。
配列番号18は、配列番号17によりコードされるアミノ酸配列である。
配列番号19〜32は、表1に現れる実施例1のプライマー配列である。
配列番号33〜34は、実施例16に現れるプライマー配列である。
配列番号35〜38は、実施例17に現れるプライマー配列である。
【特許請求の範囲】
【請求項1】
4−クマル酸を生産し、さらに当該4−クマル酸からリスベラトロール又は当該リスベラトロールのオリゴマー誘導体又はグリコシド結合誘導体を生産する操作的な代謝経路を有する微生物において、
前記代謝経路において、前記4−クマル酸は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ及びシンナメート4−ヒドロキシラーゼの作用によってL−フェニルアラニンから、又は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼの作用によってチロシンから生成し、
4−クマロイル−CoAが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現する4−クマレート−CoAリガーゼによる酵素反応で前記4−クマル酸から生成し、
そして前記リスベラトロールが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するリスベラトロールシンターゼによって内在性マロニル−CoAを基質とする反応で前記4−クマロイル−CoAから生成することを特徴とする微生物。
【請求項2】
天然のNADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物で過剰発現している、又は、NADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物に組換えで導入されていることを特徴とする請求項1に記載の微生物。
【請求項3】
チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする請求項1又は2に記載の微生物。
【請求項4】
フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする請求項1乃至3のいずれか一項に記載の微生物。
【請求項5】
真菌又は細菌であることを特徴とする請求項1乃至4のいずれか一項に記載の微生物。
【請求項6】
発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有することを特徴とする請求項1に記載の微生物。
【請求項7】
シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する請求項1に記載の微生物。
【請求項8】
4−クマル酸の外部源の実質的な非存在下において、請求項1乃至7のいずれか一項に記載の微生物を炭素基質に接触させる工程を含むことを特徴とする、リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
【請求項9】
前記炭素基質が、単糖類、小糖類及び多糖類からなる発酵性炭素基質の群より選択されることを特徴とする請求項8に記載の方法。
【請求項10】
前記炭素基質が、エタノール、アセテート、グリセロール、ラクテート、及びアミノ酸で構成される非発酵性炭素基質からなる群より選択されることを特徴とする請求項8に記載の方法。
【請求項11】
前記非発酵性炭素基質が、フェニルアラニン及びチロシンからなる群より選択されることを特徴とする請求項10に記載の方法。
【請求項12】
さらに、生産されたリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を乳製品又は飲み物中の栄養補助食品として使用することを含む請求項8乃至11のいずれか一項に記載の方法。
【請求項1】
4−クマル酸を生産し、さらに当該4−クマル酸からリスベラトロール又は当該リスベラトロールのオリゴマー誘導体又はグリコシド結合誘導体を生産する操作的な代謝経路を有する微生物において、
前記代謝経路において、前記4−クマル酸は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ及びシンナメート4−ヒドロキシラーゼの作用によってL−フェニルアラニンから、又は、
当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するL−フェニルアラニンアンモニアリアーゼ又はチロシンアンモニアリアーゼの作用によってチロシンから生成し、
4−クマロイル−CoAが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現する4−クマレート−CoAリガーゼによる酵素反応で前記4−クマル酸から生成し、
そして前記リスベラトロールが、当該微生物に固有でない酵素をコードする核酸から、当該微生物中で発現するリスベラトロールシンターゼによって内在性マロニル−CoAを基質とする反応で前記4−クマロイル−CoAから生成することを特徴とする微生物。
【請求項2】
天然のNADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物で過剰発現している、又は、NADPH:チトクロームP450レダクターゼ(CPR)が、前記微生物に組換えで導入されていることを特徴とする請求項1に記載の微生物。
【請求項3】
チロシンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする請求項1又は2に記載の微生物。
【請求項4】
フェニルアラニンアンモニアリアーゼをコードする遺伝子配列の少なくとも1つのコピーが、前記微生物中の前記遺伝子配列に天然では関連しない発現シグナルに作動可能に連結していることを特徴とする請求項1乃至3のいずれか一項に記載の微生物。
【請求項5】
真菌又は細菌であることを特徴とする請求項1乃至4のいずれか一項に記載の微生物。
【請求項6】
発現シグナルに作動可能に連結されたフェニルアラニンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたシンナメート−4−ヒドロキシラーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有することを特徴とする請求項1に記載の微生物。
【請求項7】
シンナメート−4−ヒドロキシラーゼ活性を欠如し、発現シグナルに作動可能に連結されたチロシンアンモニアリアーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結された4−クマレート−CoA−リガーゼをコードする異種DNA配列の1つ以上のコピーを含有し、発現シグナルに作動可能に連結されたリスベラトロールシンターゼをコードする異種DNA配列の1つ以上のコピーを含有する請求項1に記載の微生物。
【請求項8】
4−クマル酸の外部源の実質的な非存在下において、請求項1乃至7のいずれか一項に記載の微生物を炭素基質に接触させる工程を含むことを特徴とする、リスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体の製造方法。
【請求項9】
前記炭素基質が、単糖類、小糖類及び多糖類からなる発酵性炭素基質の群より選択されることを特徴とする請求項8に記載の方法。
【請求項10】
前記炭素基質が、エタノール、アセテート、グリセロール、ラクテート、及びアミノ酸で構成される非発酵性炭素基質からなる群より選択されることを特徴とする請求項8に記載の方法。
【請求項11】
前記非発酵性炭素基質が、フェニルアラニン及びチロシンからなる群より選択されることを特徴とする請求項10に記載の方法。
【請求項12】
さらに、生産されたリスベラトロール又はそのオリゴマー誘導体又はグリコシド結合誘導体を乳製品又は飲み物中の栄養補助食品として使用することを含む請求項8乃至11のいずれか一項に記載の方法。
【図1】

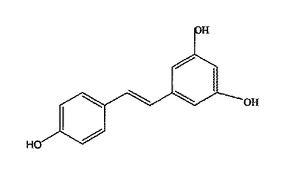
【図2】
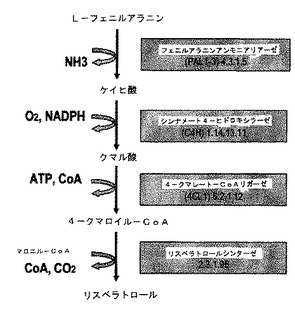
【図3】
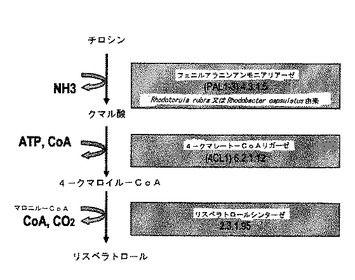
【図4】
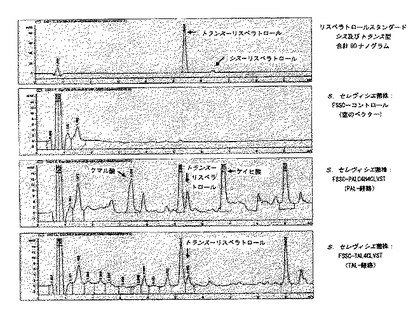
【図5】
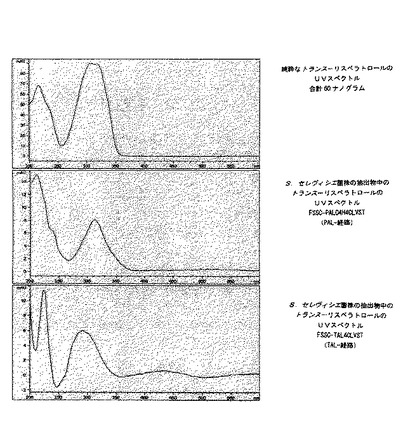
【図6】
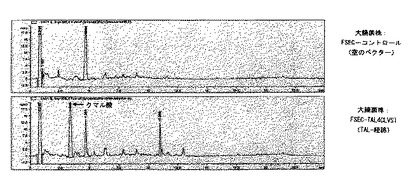
【図7】
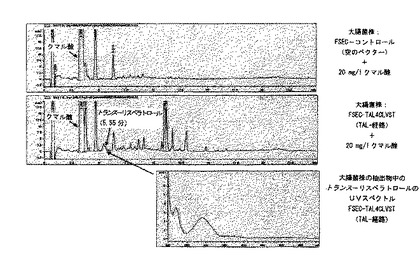
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2013−78337(P2013−78337A)
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2012−288206(P2012−288206)
【出願日】平成24年12月28日(2012.12.28)
【分割の表示】特願2007−555639(P2007−555639)の分割
【原出願日】平成18年2月21日(2006.2.21)
【出願人】(513000403)
【氏名又は名称原語表記】Evolva SA
【住所又は居所原語表記】Duggingerstrasse 23, CH−4123 Reinach, Switzerland
【Fターム(参考)】
【公開日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願日】平成24年12月28日(2012.12.28)
【分割の表示】特願2007−555639(P2007−555639)の分割
【原出願日】平成18年2月21日(2006.2.21)
【出願人】(513000403)
【氏名又は名称原語表記】Evolva SA
【住所又は居所原語表記】Duggingerstrasse 23, CH−4123 Reinach, Switzerland
【Fターム(参考)】
[ Back to top ]