
リボソームタンパク質抽出物(RPE)及び場合によってTh1促進アジュバントを含むワクチン
本発明は、寄生虫疾患を治療又は予防するための医薬品を調製するための、RPE及び場合によってTh1促進アジュバントを含む組成物及びその使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、寄生虫疾患を治療又は予防するための医薬品を調製するための、リボソームタンパク質抽出物(RPE)及び場合によってTh1促進アジュバントを含む組成物及びその使用に関する。
【背景技術】
【0002】
リーシュマニア症は、ヒト及びイヌを含めたさまざまな哺乳動物のマクロファージに主に感染する、リーシュマニア属(Leishmania)に属する細胞内寄生原虫によって引き起こされるいくつかの疾患を含む。寄生虫の種及びヒト宿主の免疫能力の状態に主として応じて、疾患のスペクトルは、自然治癒する皮膚リーシュマニア症(CL)から致命的な内臓リーシュマニア症(VL)又はカラアザールに及ぶ(18)。幼児リーシュマニア(Leishmania infantum)及びシャーガスリーシュマニア(L.chagasi)によって引き起こされるイヌ・内臓皮膚リーシュマニア症(VCL)は、地中海沿岸地方周辺、中東及びラテンアメリカの諸国で発見された重要な新興の動物原性感染症であり(16)、イヌがこれらの寄生虫の主要な保有宿主であり、サシチョウバエによるヒトへの伝染において中心的な役割を果たす(47)。感染の結果は、宿主の免疫系と種々の寄生種との相互作用によって決まり、今までのところリーシュマニア症の発症機序は不明のままであり、ヒト及びイヌにおける、リーシュマニアに対する免疫応答に関わるメカニズムについての知見は依然として限られている。大抵の場合、防御免疫は、T細胞由来のサイトカインによるマクロファージの活性化を誘導する古典的な細胞性免疫応答に関連付けられ、一方非治癒性の疾患は、強力な体液性応答の生成に関連付けられる(15、26)。寄生虫粗画分又は定義済みの寄生虫抗原をベースとする第2世代のワクチンを開発するための研究において取り組まれたのは、多種多様なアジュバントを用いたいくつかの実験モデルにおいてワクチン候補として試験された種々の寄生虫表面分子又は分泌分子の同定であった(1、17、22、46、48、49、52、54)。感染した動物又はヒト由来の血清を用いた発現ライブラリーのスクリーニングによっても、候補ワクチンとして数個の抗原を選択することができた((9)に総説がある)。その中で、感染したマウス又はヒト患者の細胞において、その細胞の位置に無関係に、主としてTh1型の免疫応答を引き出す抗原が、種々の動物モデルにおいて防御応答の生成に関連している(51、55、56)。一方、単離された抗原の一部は、VLに罹患しているヒト又はイヌにおける体液性応答、又は実験的に感染させたマウスにおけるTh2媒介体液性応答を優勢的に刺激する、細胞内に保存されているタンパク質である(3、36、38、40、42)。リーシュマニア症に罹患しているイヌにおいて、それらの抗原に対して誘導される不十分な体液性応答により、主に免疫複合体が有害作用を有するため、免疫病変、特にブドウ膜炎(13)、中枢神経系における損傷(14)又は腎炎(23、24、33、34)が生じると考えられている。また、VLを生じたヒトにおいて、IgG免疫複合体が存在することが、感染を消散できないことと相関があることが最近示され、感染した宿主に対して免疫複合体が悪影響を及ぼし得ることを実証している(30)。
【0003】
最初は良好なワクチン候補であるとみなされなかったにもかかわらず、感染プロセスの間に高度な体液性応答を誘導するタンパク質が、防御応答の誘導に関連している。例えば、寄生虫のチューブリン及びヒストンH2Bは、免疫ドナー由来のT細胞クローンによって認識され(39)、rK39は、免疫マウス由来のT細胞によって増殖及びIFN−γ産生を引き起こす(25)。寄生虫のH2B遺伝子、H3遺伝子及びH4遺伝子を用いた遺伝子による免疫によって、マウスの内臓リーシュマニア症モデルにおいて防御が誘導されることも示されている(27)。また、Th1促進アジュバントと共に投与される活性化Cキナーゼ(LACK)(32)、一部の寄生虫システインプロテイナーゼ(38、41)又は寄生虫ヌクレオソーム形成ヒストン(11、20)に対する受容体を免疫することにより、マウスモデルにおいて皮膚リーシュマニア症に対する防御に相関する免疫応答が生成する。
【0004】
進化的に保存されたリーシュマニアの抗原の中で、いくつかの証拠により、リボソームタンパク質が、リーシュマニア感染の間、免疫学的に関連する分子であることが示唆されている。一部の例では、リボソームの構成要素は、感染の間に細胞活性及びサイトカインの放出を調節するその能力を通して宿主の免疫系の機能不全に寄与し得る。したがって、大形リーシュマニア(L.major)のリボソームタンパク質S3aをBALB/cマウスに注射することにより、B細胞クローンのポリクローナル増大が誘導され、T細胞の増殖が阻害された(10)。また、推定60Sリボソームタンパク質L31をコードするDNAワクチンを用いた遺伝子による免疫により、マウスモデルにおいて、IL−10及びTh2サイトカインが誘導されることによって疾患が悪化する(44、53)。さらに、寄生虫の酸性Pタンパク質などの一部の寄生虫のリボソームタンパク質は、リーシュマニア症に罹患しているイヌ及びヒトにおける強力な体液性応答の生成に関連している((42)に総説がある)。
【発明の概要】
【発明が解決しようとする課題】
【0005】
今まで試みられているにも関わらず、依然としてリーシュマニア症などの寄生虫疾患に対して有益なワクチンはない。したがって、そのようなワクチンが依然として必要とされている。
【課題を解決するための手段】
【0006】
本研究において、我々は、RPE、特にリーシュマニアRPE(LRPE)が、幼児リーシュマニアに自然感染したイヌにおいて、及び大形リーシュマニアに実験的に感染させたマウスにおいて免疫応答の標的であることを示している。我々はさらに、LRPEをCpGオリゴデオキシヌクレオチド(ODN)などのTh1促進アジュバントと同時投与した際に、強力なTh1防御免疫応答が誘導されることを実証している。そのような組成物(Th1促進アジュバントと組み合わせたLRPE)は、ワクチンとして使用することが非常に魅力的である。本発明を以下さらに説明する。
【0007】
使用
本発明の第1の態様において、対象において寄生虫疾患を治療又は予防するための医薬品を調製するための、RPE及び場合によってTh1促進アジュバントの使用が提供される。
【0008】
好ましい実施形態において、RPEは、対象内に存在すると寄生虫疾患を引き起こす寄生虫細胞を用いて、
a.寄生虫細胞を溶解緩衝液と混合するステップと、
b.得られた混合物を遠心分離してサイトゾル抽出物を得るステップと、
c.得られたサイトゾル抽出物からRPEを調製するステップと
を行うことにより得られる。
【0009】
ステップaにおいて、寄生虫は原生動物を指すことが好ましい。好ましい寄生虫は本明細書において後で定義する。原生動物のステージは前鞭毛型であることがより好ましい。所望の量のRPEを調製するためにおよそ必要な寄生虫細胞の量は当業者には公知であろう。一般には、およそ500マイクログラムのRPEを調製するために、およそ3.109個の寄生虫細胞を使用する。溶解緩衝液は、寄生虫細胞の少なくとも一部を壊す緩衝液である。好ましい溶解緩衝液は、非イオン界面活性剤を含む。非イオン界面活性剤としてNonidet P40(NP40)を用いると良い結果が得られた。しかし、他の非イオン界面活性剤を使用してもよい。下記の溶解緩衝液を使用することが好ましい(緩衝液A):10mMのトリスHCl、pH8.0、150mMのNaCl、1.5mMのMgCl2及び0.5%のNP40(Roche)、並びにPMSF 1mM、Leupeptin 8μg/ml、Aprotinin 4μg/ml及びPentatin 8μg/mlなどのプロテアーゼ阻害剤を補充することが好ましい。適当量の寄生虫細胞(およそ細胞109個/緩衝液A1ml)を、一般には、エッペンドルフピペットを用いて溶解緩衝液と穏やかに混合する。
【0010】
ステップbにおいて、ステップaで得られた混合物に対し、遠心分離を4℃で少なくとも1ステップ行う。通常、第1の遠心分離ステップはおよそ3,000gでおよそ2分行う。得られた上澄みを、4℃で、およそ13,000gでおよそ15分、1回又は2回再度遠心分離することが好ましい。
【0011】
ステップcにおいて、得られた上澄みを(45)に記載の通りRPEを調製するために使用する。簡単に述べると、得られた上澄みを、Beckman TL100.3ローターを用いて、4℃、およそ90,000rpmでおよそ30分の高速遠心分離にかける。得られたペレットは粗製のリボソームペレットであり、緩衝液B(20mMのトリスHCl、pH7.4、500mMのAcNH4、100mMのMgCL2、5mMのβ−メルカプトエタエノール)に再懸濁させ、緩衝液A中、不連続なショ糖勾配(20/40%)で、TL100.3ローターを用いて、4℃、およそ90,000rpmで遠心分離する。得られたペレットはリボソームを含む。このペレットを、PBS(リン酸緩衝食塩水)に溶解させ、超音波処理し、−70℃で保存することが好ましい。
【0012】
リボソームタンパク質は、よく保存されたサイトゾルタンパク質である。したがって、本明細書で定義するRPEは、植物であろうと動物であろうと、哺乳動物、爬虫類、魚類、昆虫類であろうと原生生物などの染色体を持つ他の任意の生物であろうと、任意の真核生物から調製することができる。RPEは、疾患に近い生物、好ましくは系統樹内の寄生虫疾患を引き起こす生物から得ることが好ましい。したがって、寄生虫疾患の治療に使用されるRPE源として特に興味深いものは、原生動物、例えばプラスモジウム属(Plasmodium)、特にトリパノソーマ科のメンバー、より特定するとトリパノソーマ科の原生生物リーシュマニアの種々の種である。リーシュマニアには20を超える公知の種があり、大形リーシュマニアを含めた大形リーシュマニアコンプレックス、シャーガスリーシュマニア、ドノバンリーシュマニア(L.donovani)及び幼児リーシュマニアを含めたドノバンリーシュマニアコンプレックス、アマゾンリーシュマニア(L.amazonesis)及びメキシコリーシュマニア(L.mexicana)を含めたメキシコリーシュマニアコンプレックスを含むリーシュマニア亜属の種、並びにブラジルリーシュマニア(L.braziliensis)及びペルーリーシュマニア(L.peruviana)を含めたブラジルリーシュマニアコンプレックス及びガイアナリーシュマニア(L.guyanensis)及びパナマリーシュマニア(L.panamensis)を含めたガイアナリーシュマニアコンプレックスを含むビアンニア(Viannia)亜種が含まれる。特に興味深いプラスモジウム属の種は、熱帯熱マラリア原虫(Plasmodium falciparum)及び三日熱マラリア原虫(Plasmodium vivax)である。好ましい実施形態において、RPEは、リーシュマニア属の各種から得られ、大形リーシュマニア及び/又は幼児リーシュマニアから得ることが好ましい。他の好ましい実施形態において、RPEは、プラスモジウム属の各種から得られる。RPEは、本明細書で確認したいくつかの異なる生物由来のRPEを混合することにより調製してもよいことは当業者に理解されよう。所定のタンパク質を使用する代わりにRPEをワクチンに使用することは、RPEが異なる抗原を数多く含有するので、非常に魅力的である。これらの抗原はそれぞれ、治療される対象において免疫防御応答を潜在的に誘導し得る。さらに、抗原Aには応答するが、抗原Bには応答しない対象、及びその逆の対象がある。したがって、本明細書で定義するワクチンは、異なる抗原を数多く含有するので、広範囲の対象集団に対して使用することを企図している。好ましい実施形態において、RPEは、少なくとも1種のリボソームタンパク質及び/又は少なくとも1種のリボソームタンパク質抗原及び/又は少なくとも1種のリボソームタンパク質のタンパク質断片を含む。より好ましい実施形態において、RPEは、少なくとも2種のリボソームタンパク質及び/又は少なくとも2種のリボソームタンパク質抗原及び/又は少なくとも2種のリボソームタンパク質断片を含む。本明細書で定義するタンパク質断片は、少なくとも2、3、5、7、10、15、20、25、30個又はそれ以上の、リボソームタンパク質に対応する隣接アミノ酸を含むことが好ましい。一実施形態において、本明細書で定義するRPEは、幼児リーシュマニアの酸性リボソームタンパク質P0及び/又はブラジルリーシュマニア由来のリボソーム抗原LbeF4Aを含まない、又はそれらからなるものではない。
【0013】
Th1促進アジュバント(例えばCpG ODNモチーフを含むアジュバント)は文献(Liu N.,et al.,(2003),Nature Immunology,687−693)において、所定の抗原(ここではRPE)と一緒に用いた際に所与の抗原に対するTh1免疫応答を促進又は誘発することができ、その抗原と一緒に培養した際に処理した対象の脾細胞の上澄みから検出されるアジュバントとして定義されている。対照として、抗原及びアジュバントで処理していない、又は抗原でのみで処理した、同一対象の脾細胞集団において、Th1免疫応答の促進又は誘発について評価する。Th1免疫応答の誘発又は促進は、抗原を用いて処理した対象の脾細胞を培養すること、及び/又は抗原特異的IgG2a免疫グロブリンの産生を誘導することによって検出されるIFNγの誘導によって定義することが好ましい。このサイトカインの誘導についての評価は、実施例に記載の通り、脾細胞についてのELISAによって行うことが好ましい。IgG2aの誘導についての評価は、実施例に記載の通り、ELISA又はウェスタンブロットによって行うことが好ましい。脾細胞をRPE及びアジュバントで刺激するとIFNγ及び/又はIgG2aが誘導されることにより、アジュバントがTh1促進アジュバントであると認定されたことが示されることが好ましい。
【0014】
上記のTh1免疫応答の誘発又は促進についての最初の定義の代わりに、又はそれと組み合わせて、Th1免疫応答の誘発又は促進はさらに、Th2免疫応答の欠如(又は誘導の欠如)によって定義することができる。Th2免疫応答は、非処理の脾細胞と比べて、IL−4、IL−10の誘導が検出可能に増加すること、及び/又は検出可能なIgG1免疫グロブリンが産生されることを特徴とする。IL−4及び/又はIL−10の誘導についての評価は、実施例に記載の通り、脾細胞についてのELISAによって行うことが好ましい。IgG1の誘導についての評価は、実施例に記載の通り、ELISA又はウェスタンブロットによって行うことが好ましい。
【0015】
上記のTh1免疫応答の誘発又は促進についての最初の2つの定義の代わりに、又はそれと組み合わせて、Th1免疫応答の誘発又は促進はさらに、定義済みの抗原、この場合はRPEに対するINFγ/IL−10比及び/又はINFγ/IL−4比の増加、及び/又はIgG1/IgG2a比の減少が生じることによって定義することができる。好ましい実施形態において、これらの3つ以上の比のいずれかが変化する(上記のように増加又は減少する)ことにより、アジュバントがTh1の性質を有することが示される。言及した各サイトカインの誘導についての評価は、実施例に記載の通り、脾細胞についてのELISAによって行うことが好ましい。免疫グロブリンIgG1又はIgG2aの誘導についての評価は、実施例に記載の通り、ELISA又はウェスタンブロットによって行うことが好ましい。
【0016】
好ましい実施形態において、Th1促進アジュバントは、オリゴデオキシヌクレオチドである、又はオリゴデオキシヌクレオチドを含む、又はオリゴデオキシヌクレオチドからなる。オリゴデオキシヌクレオチド(ODN)は、Cがメチル化されていないCpGを含む、又はCがメチル化されていないCpGからなることがより好ましい(CpG ODN):3’プリン−CpG−5’ピリミジン。好ましいオリゴデオキシヌクレオチドは、ホスホロチオエート修飾されたODN配列を含む、又はホスホロチオエート修飾されたODN配列からなる。そのような修飾されたオリゴデオキシヌクレオチドを使用することは、それゆえに使用されるオリゴデオキシヌクレオチドの方が、修飾されていないオリゴヌクレオチドよりも安定しており、血液系に入っても容易に分解されないので、有利である。好ましいTh1促進アジュバントは、少なくとも1つ、少なくとも2つ又は少なくとも3つのCpGモチーフからなる、又はそれを含む。免疫刺激ODNの好ましい配列(5’→3’)は、TCAACGTTGA及びGCTAGCGTTAGCGTであった。当業者は、本明細書に明示した配列に制限されない。当業者は、他の配列を設計し、引き続いて、その配列について、本明細書で前に定義したTh1を促進する性質について試験することができる。この好ましい同定アジュバントCpG ODNは、実施例において、LRPEをこのTh1促進アジュバントと一緒に接種することにより、BALB/c系マウス及びC57BL/6系マウスの両方で、大形リーシュマニア寄生虫によるチャレンジに対する防御が誘導されることが実証されたので、非常に魅力的である。どちらのモデルでも、防御はIFN−γの特異的な産生に相関している。BALB/cでは、IL−4及びIL−10の産生に対する制限も検出された。
【0017】
本発明の一つの利点は、広範なスペクトルの寄生虫疾患を治療するための医薬品、すなわち、異種間に特異的な医薬品の調製を可能にすることである。多くの寄生虫疾患において、特定の種に対して作製されたワクチンは、その特定の種に対してしか働かない。その一事例である寄生虫疾患は、リーシュマニア症である。現在のところ、この疾患は薬物により制御されているが、薬物治療では疾患の伝播を予防できず、強い効果がない場合が多い。好ましい実施形態において、寄生虫疾患はリーシュマニア症又はマラリアである。寄生虫疾患が、リーシュマニア属の各種によって、又はプラスモジウム属の各種によって引き起こされるものであることがより好ましい。別の好ましい実施形態において、寄生虫疾患は、RPEを得る種とは異なる種により引き起こされる。特に、リーシュマニア属の一種により引き起こされるリーシュマニア症は、リーシュマニア属の他の種由来のRPEをベースとする組成物を用いて治療することができる。一実施形態において、大形リーシュマニアにより引き起こされるリーシュマニア症は、幼児リーシュマニア由来のRPEを含む組成物で首尾よく治療される。或いは、マラリアなどの他の寄生虫疾患は、他種のRPEをベースとする組成物、例えば、幼児リーシュマニアのRPEをベースとする組成物で首尾よく治療され得る。
【0018】
本発明に照らして、対象はヒト又は動物を意味する。本発明の範囲に包含される動物としては、哺乳動物、特にイヌが挙げられる。
【0019】
好ましい実施形態において、本明細書で定義した医薬品は、感染及び/又は疾患、より好ましくは寄生虫による感染及び/又は寄生虫疾患と戦うための、ヒト又は動物の免疫系の能力を高めるために使用される。特に、ヒト又は動物の対象に投与するために使用され得る。本明細書で定義した医薬品は好ましくは、非経口的に、例えば、静脈内、皮下、腹腔内、筋肉内、動脈内又は病巣内という経路での注射又は点滴によって投与される。好ましい投与形態は、皮下である。医薬品は、当技術分野で公知の従来技法によって、薬学的に許容される培地又は送達ビヒクルと組み合わせることができる。例えば、RPE及び場合によってTh1促進アジュバントをリン酸緩衝食塩水(PBS)に溶解させることができる。非経口的に投与可能な組成物を調製する方法は、当技術分野で周知であり、例えば、Remington’s Pharmaceutical Sciences,Ed.AR Gennaro,20th edition,2000,Williams&Wilkins,PA,USAを含めたさまざまなソースに、より詳細が記載されている。医薬品は、治療有効用量、すなわち、本明細書で定義した感染及び/又は疾患と戦うために、ヒト又は動物の免疫系の能力を高める用量を投与することが好ましい。治療有効用量の本発明の医薬品により、真皮損傷の発生が予防及び/若しくは遅延され、かつ/又は耳及び/若しくは流入領域リンパ節(DLN)における寄生虫負荷量の有意な縮小が誘導されることが好ましい。真皮損傷の存在についての評価は、図6の説明文に記載している。寄生虫負荷量についての評価は、実施例に記載している。治療有効用量の本発明の医薬品により、最初に本発明の組成物を用いた1回のワクチン接種、続いて逐次的な1回の寄生虫による感染、及びおよそ±6週間の待機時間を含む期間の後に、真皮損傷の発生が予防されることが好ましく、及び/又は耳において寄生虫負荷量のおよそ3桁の縮小及び/又はDLNにおいて寄生虫負荷量のおよそ同様の桁の縮小が誘導されることが好ましい。好ましい実施形態において、本明細書で定義した医薬品はワクチンである。より好ましい実施形態において、少なくとも12μgのRPEがワクチンに使用される。さらに好ましい実施形態において、免疫応答をもたらすために少なくとも12〜20μgのRPEを、場合によって少なくとも50μgの、例えばCpG ODNなどのTh1促進アジュバントと組み合わせて使用する必要がある。本明細書で定義したワクチンは、予防ワクチン又は治療ワクチンであってよい。溶解させることができるRPE及び場合によってTh1促進アジュバントの体積は100〜500マイクロリットルで変動してよい。
【0020】
組成物
別の態様において、RPE及び場合によってTh1促進アジュバントを含む組成物が提供される。RPE及びTh1促進アジュバントは、本明細書ですでに定義した。好ましい実施形態において、組成物は、RPE及びTh1促進アジュバントからなる。好ましいTh1促進アジュバントはCpG ODNである。好ましい組成物は、PBSに溶解させたRPE及び場合によってTh1促進アジュバントを含む、又はそれらからなる。別の好ましい実施形態において、RPE及びTh1促進アジュバントを逐次的に投与することも本発明に包含される。したがって、その両方が対象に投与される限り、両成分が物理的に1つの単一組成物中に存在する必要はない。
【0021】
そのような組成物は、薬学的に許容できるアジュバント及び/又は担体をさらに含む。
【0022】
そのような組成物は、医薬品用であることが好ましい。その医薬品は、ワクチンであることが好ましい。医薬品及びワクチンは、本明細書ですでに広範に定義した。
【0023】
方法
他の態様において、本発明は、全て本明細書で定義した、寄生虫疾患を予防及び/若しくは治療し、かつ/又は寄生虫疾患の進行を遅延させ、かつ/又は真皮損傷の発生を予防及び/又は遅延させ、かつ/又は耳及び/若しくは流入領域リンパ節(DLN)において寄生虫負荷量の有意な縮小を誘導する方法を提供する。本方法において、本発明のワクチンは治療ワクチンとして機能する。一般には、感染と疾患発症の間には期間がある。この場合、ワクチンは、宿主において感染の病理学的影響を中和する免疫応答を引き出すことにより、疾患を予防及び/若しくは治療し、かつ/又は疾患の進行を遅延させる薬理学的免疫産物として作用する。治療ワクチンは、すでに感染又は疾患発症している患者において防御を誘導するという点において、予防ワクチンとは異なる。
【0024】
本明細書及び特許請求の範囲において、「含む」という動詞及びその活用は、その言葉に続く項目が含まれるが、特に言及していない項目が除外されるのではないことを指す非限定的な意味で使用している。さらに「からなる」という動詞は、「から本質的になる」と置き換えることができ、本明細書で定義した産物又は組成物又は保存混合物が、特に言及した成分と違う追加の成分(単数又は複数)を含んでよく、前記追加の成分(単数又は複数)は、本発明の独特な特性を変化させないことを意味する。
【0025】
さらに、不定冠詞「a(1つの)」又は「an(1つの)」により要素を参照することは、前後関係からその要素が唯一であることが明らかに必要とされているのでなければ、その要素が2つ以上存在する可能性を除外しない。したがって、不定冠詞「a(1つの)」又は「an(1つの)」は通常「at least one(少なくとも1つの)」を意味する。
【0026】
本明細書に引用した全ての特許文献及び非特許文献は、参照により全体として本明細書に完全に組み込まれている。
【0027】
本発明を以下の実施例によりさらに例証し、それにより本発明の範囲が限定されると解釈されるべきではない。
【図面の簡単な説明】
【0028】
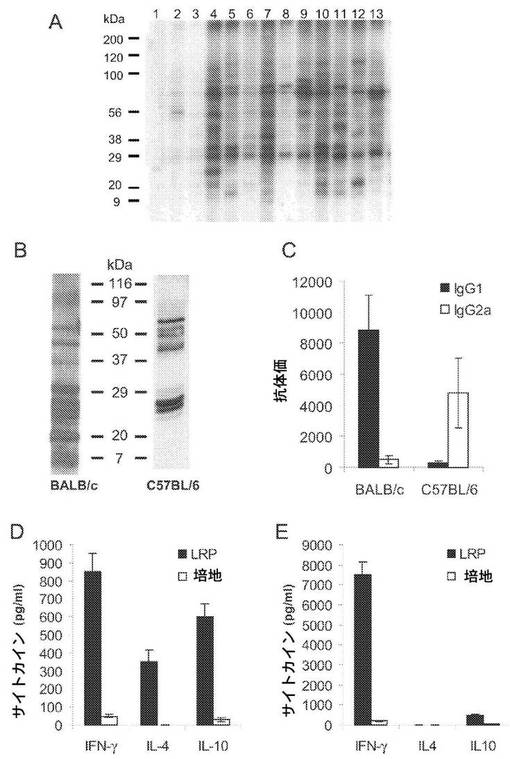
【図1】(A)幼児リーシュマニアのリボソームタンパク質を、直線的な10〜14%の勾配のSDS−PAGEゲルで電気泳動し、ニトロセルロースブロットに転写し、健康なイヌの血清(レーン1〜3)及びVCLに罹患しているイヌ由来の血清(レーン4〜13)と一緒にインキュベートした。血清は200倍希釈で利用した。二次試薬として、西洋ワサビペルオキシダーゼをコンジュゲートした抗イヌIgG抗体を使用した(B〜E)。BALB/cマウス4匹について、大形リーシュマニアの静止期前鞭毛型5×104個を左足蹠において皮下感染させ、チャレンジの8週間後に血清を得た。C57BL/6マウス4匹について、大形リーシュマニアの発育終末前鞭毛型300個を耳真皮において皮内感染させ、チャレンジの14週間後に血清を得た。両系統の感染前の結成についても、寄生虫によるチャレンジの前に得た。(B)大形リーシュマニアのリボソームタンパク質を、直線的な10〜14%の勾配のSDS−PAGEゲルで電気泳動し、ニトロセルロースブロットに転写し、感染させたBALB/cマウス又はC57BL/6マウス由来の保存血清と一緒にインキュベートした。血清は200倍希釈で利用した。感染前の血清でLRPに対する反応性を示したものはなかった(示さず)(C)両系統のマウスにおけるIgG1抗体及びIgG2a抗体のLRPに対する抗体価をELISAによって個々に決定した。(D)感染の8週間後にBALB/cマウスを安楽死させ、その膝窩部DLN細胞を、in vitroで、大形リーシュマニアLRP12μg/mlの存在下で又は培地のみで48時間培養した。培養物の上澄みについてELISAを行い、IFN−γ、IL−4及びIL−10のレベルを評価した。(E)感染の14週間後にC57BL/6マウスを安楽死させ、その上顎後方のDLNをDと同様に処理した。
【図2】(A)BALB/cマウスにおいて誘導される特異的な体液性応答の分析。BALB/cマウス(1群当たり6匹)について、大形リーシュマニアのリボソームタンパク質単独(LRP)、又はそれにCpG ODNアジュバントを加えたもの(LRP+CpG)を用いて、CpG ODNアジュバント単独(CpG)を用いて、又はPBS(生理食塩水)を用いて、右足蹠において3回皮下免疫した。3回目の免疫の14週間後、マウスから血を抜き、血清についてELISAを行い、IgG1アイソタイプ(黒い棒線)及びIgG2aアイソタイプ(白い棒線)の両方の特異的な抗LRP抗体応答について評価した。免疫前の血清で反応性を示したものはなかった。(B〜D)ワクチン接種の4週間後、マウスを安楽死させ、その脾臓を得、in vitro、LRPの存在下で(灰色の棒線)又は培地のみで(黒い棒線)48時間培養した。培養物の上澄みについてELISAを行い、IFN−γ(B)、IL−4(C)及びIL−10(D)のレベルを評価した。
【図3】(A)ワクチン接種したBALB/cマウスにおける大形リーシュマニア感染の経過。マウス(1群当たり6匹)を、図2に示したように皮下免疫した。最後の免疫の1カ月後、動物について、大形リーシュマニアの静止期前鞭毛型5×104個を左後部足蹠において感染させた。足蹠の腫大は、感染した足蹠と感染していない反対側の足蹠との厚さの違いとして得られる。結果は、2つの独立した実験についての平均及び標準偏差を表す。* チャレンジの8週間後の、LRP+CpG ODNをワクチン接種したマウス群における炎症対CpG ODNをワクチン接種したマウス群における炎症のP<0.001の有意差。(B)感染の8週間後、感染させた脚の膝窩部DLN及び脾臓における生存可能な寄生虫の数を、個々に限界希釈法により決定した。結果は、2つの独立した実験についての平均及び標準偏差を表す。* チャレンジの8週間後の、LRP+CpG ODNをワクチン接種したマウス群における寄生虫の体内総量対CpG ODNをワクチン接種したマウス群における寄生虫の体内総量のP<0.001の有意差。(C〜D)ワクチン接種したマウス及び感染させたマウスにおけるサイトカイン産生を、大形リーシュマニアによるチャレンジの8週間後に決定した。感染した脚の膝窩部DLNを得、in vitro、SLAの存在下で(白い棒線)、LRPの存在下で(灰色の棒線)又は培地単独で(黒い棒線)48時間培養した。培養物の上澄みについてELISAを行い、IL−4(C)、IL−10(D)、及びIFNγ(E)のレベルを試験した。この実験では同様の結果が繰り返された。(F)LRP+CpG ODNを用いたワクチン接種によって与えられた防御に付随するIFNγ産生におけるIL−12及びT細胞の関与の分析。大形リーシュマニアの静止期前鞭毛型5×104個でチャレンジした8週間後、CpG ODNを用いてワクチン接種したマウス(黒い棒線)及びLRP+CpG ODNを用いてワクチン接種したマウス(白い棒線)の膝窩部LNを得、抗IL−12抗体、抗CD4抗体又は抗CD8抗体のいずれか、及び対照モノクローナル抗体の存在下で、LRPを用いて培養して刺激した。78時間インキュベーションした後、ELISAによってIFNγのレベルを評価した。抗CD8モノクローナル抗体を用いた処理と対照抗体を用いた処理とにおけるIFNγ産生の差は、統計学的に有意であった(*P<0.05)。データは、同様の結果であった2つの独立した実験の代表的な実験に対応する。
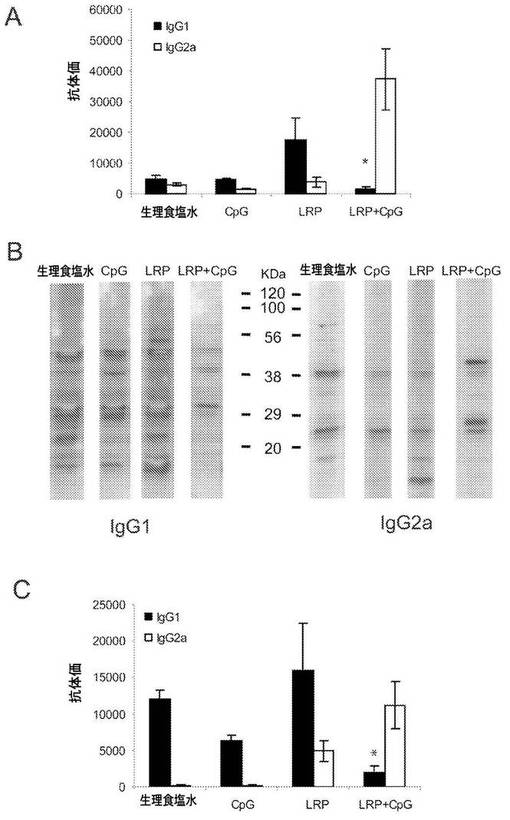
【図4】IgG1/IgG2a極性化の分析。(A)チャレンジの8週間後、血清試料を得、LRPに対するIgG1抗体及びIgG2a抗体の抗体価を、個々にELISAにより決定した。LRP+CpG ODNを用いてワクチン接種したマウスと他の3群とにおけるIgG1抗体価の差は、統計学的に有意であった(*P<0.01)。(B)大形リーシュマニアのLRPを、直線的な10〜14%の勾配のSDS−PAGEゲルで分離し、ニトロセルロースブロットに転写し、表示したマウス群由来の200倍希釈した保存血清と一緒にインキュベートした。IgG1アイソタイプ及びIgG2aアイソタイプ両方の抗体応答を示す。(C)同じ血清を利用して、IgG1及びIgG2aのSLAに対する抗体価を決定した。LRP+CpG ODNを用いてワクチン接種したマウスと他の3群とにおけるIgG1抗体価の差は、統計学的に有意であった(*P<0.02)。
【図5】BALB/cマウス6匹について、図3に示したように左足蹠においてLRP+CpG ODNを用いてワクチン接種し、感染させた。最初の寄生虫によるチャレンジの18週間後、マウスについて、大形リーシュマニアの発育終末前鞭毛型300個を用いて耳の皮内に再感染させた。対照として、無処置のマウス6匹についても耳の皮内にチャレンジした。(A)防御させ、再感染させたBALB/cマウスにおける大形リーシュマニアの感染経過。値は、平均損傷直径+標準偏差(SD)を表す。* チャレンジの7週間後の、再感染させたマウスにおける炎症対対照の感染させたマウスにおける炎症のP<0.0001の有意差。(B)再感染の7週間後、マウスを安楽死させ、耳真皮、脾臓及び局所DLNにおける寄生虫の生体内総量を、個々に定量化した。結果は、12匹分の耳及びDLNの平均±SDとして表す。* 再感染させたマウス対感染させた対照マウスのP<0.001の有意な減少。(C)安楽死させた後、対照マウス由来(黒い棒線)及び再感染させたマウス由来(白い棒線)の上顎後方のDLNを得、in vitro、SLAの存在下で、LRPの存在下で又は培地単独で48時間培養した。培養液の上澄みについてELISAを行うことにより、IFNγ、IL−4、及びIL−10のレベルを評価した。(D〜E)再チャレンジした7週間後、血清試料を得、LRP(D)及びSLA(E)に対するIgG1抗体及びIgG2a抗体の抗体価を、個々にELISAにより決定した。
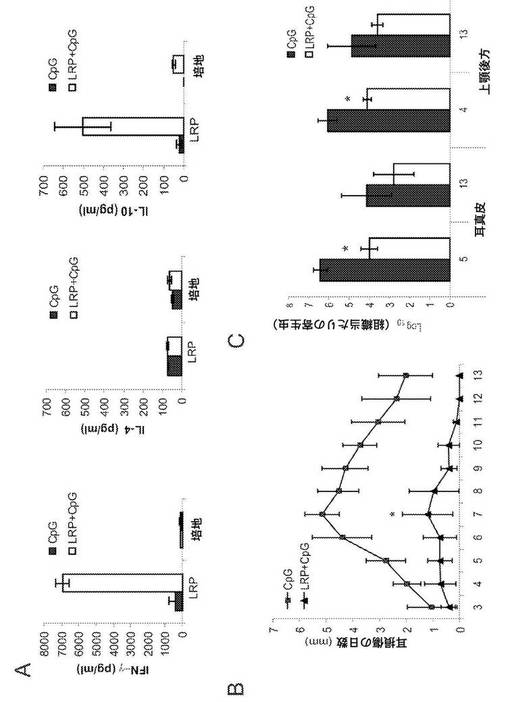
【図6】C57BL/6マウスにおける大形リーシュマニアの感染に対する防御。マウス(1群当たり6匹)について、LRP+CpG ODNを3回用いて、及びCpG ODNを単独で用いて、右足蹠において皮下免疫した。(A)ワクチン接種したC57BL76マウスの脾細胞によるINF−γ、IL−4及びIL−10の産生。CpG ODN(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種した4週間後、マウスを安楽死させ、その脾臓を得てin vitro、LRPの存在下又は培地単独で48時間培養した。培養物の上澄みについてELISAを行うことにより、サイトカインのレベルを評価した。(B)ワクチン接種したC57BL/6マウスにおける大形リーシュマニアの感染経過。1群当たり12匹のマウスを上記の通り免疫し、最後の接種の4週間後、マウスについて、大形リーシュマニアの発育終末前鞭毛型300個を耳の皮内に接種することにより感染させた。値は、平均損傷直径+標準偏差(SD)を表す。* 2つのマウス群間の炎症のP<0.001の有意な減少。(C)感染の5週間後(1群当たり6匹)及び感染の13週間後(1群当たり6匹)に定量した、CpG ODN(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種したマウス由来の耳真皮及び局所DLNにおける、寄生虫の生体内総量。結果は、12匹分の耳及びDLNの平均±SDを表す。* 両マウス群間のP<0.01の有意な減少。(D〜E)CpG(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種したマウスにおけるIFN−γの産生。感染させた5週間後、上顎後方の細胞を得、SLAの存在下で、LRPの存在下で及び培地単独(D)で培養し刺激した、又は抗IL−12抗体、抗CD4抗体又は抗CD8抗体及び対照のモノクローナル抗体の存在下でLRPを用いて刺激した。78時間インキュベーションした後、ELISAによりIFN−γのレベルを評価した。抗CD8モノクローナル抗体を用いた処理と対照抗体を用いた処理とにおけるIFN−γ産生の差は、統計学的に有意であった(*P<0.01)。(F)チャレンジした5週間後及び13週間後に血清試料を得、LRPに対するIgG1抗体及びIgG2a抗体の抗体価を、個々にELISAにより決定した。(G)CpG ODN(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種したマウスにおけるIL−10産生。感染させた5週間後、上顎後方の細胞を得、SLAの存在下で、LRPの存在下で又は培地単独で培養し刺激した。78時間インキュベーションした後、IL−10のレベルをELISAにより評価した。
【実施例】
【0029】
材料と方法
マウス系及び寄生虫
雌のBALB/cマウスは実験開始時に6〜8週齢であり、Harlan Interfauna Iberica S.A.(Barcelona,Spain)から購入した。大形リーシュマニア寄生虫(クローンWHOM/IR/−173)及びクローンV1(MHOM/IL/80(Friedlin)を、BALB/cマウスで継代することにより毒性の状態に保った。大形リーシュマニアの無鞭毛型を膝窩部流入領域リンパ節(DLN)から得、20%ウシ胎児血清(FCS)を補充したシュナイダー培地(Schneider’s medium)(Gibco,BRL)中、26℃で、それらが後期静止期に達するまで培養することにより、前鞭毛型に変換させた。両方のクローンの前鞭毛型を、20%ウシ胎児血清(FCS)を補充したシュナイダー培地(Schneider’s medium)(Gibco,BRL)中、26℃で培養した。大形リーシュマニア(クローンV1)の感染性ステージの前鞭毛型(発育終末)を、ピーナッツ凝集素(Vector Laboratories,Burlingame,CA.)を用いたネガティブ選択によって静置培地から単離した。幼児リーシュマニア(MCAN/ES/96/BCN/150,MON−1)の前鞭毛型を、10%FCSを補充したRPMI培地(Gibco,BRL)中、26℃で培養した。
【0030】
CpG ODN及びリーシュマニア抗原
リーシュマニアのリボソームタンパク質抽出物(LRP)を調製するために、大形リーシュマニア及び幼児リーシュマニアの前鞭毛型を採取し、予備冷却したPBSで2回洗浄し、NP40(Roche Diagnostics,GmbH,Manheim Germany,cat.N.11332473001)1mlを含む溶解緩衝液(10mMのトリスClH pH8.0、150mMのNaCl、1.5mMのMgCl2及び0.5%NP40、PMSF 1mM、Leopeptin 8μg/ml、Aprotinin 4μg/ml及びPentatin 8μg/ml)に再懸濁させ、10回上下にピペッティングした。溶解後、試料を4℃、3,000×gで2分微量遠心し、核をペレット化した。上澄みを2回、4℃、13,000×gで15分微量遠心し、(45)に記載の通りサイトゾル上澄みからリボソームを調製した。簡単に述べると、サイトゾルを、Beckman TL100.3ローターを用いて、4℃、90,000rpmで30分の高速遠心分離にかけた。粗製のリボソームペレットを緩衝液A(20mMのトリスHCl、pH7.4、500mMのAcNH4、100mMのMgCL2、5mMのβ−メルカプトエタエノール)に再懸濁させ、緩衝液A中、不連続なショ糖勾配(20/40%)で、TL100.3ローターを用いて、4℃、90,000rpmで遠心分離した。洗浄したリボソームのペレットをPBSに溶解させ、超音波処理し、−70℃で保管した。
【0031】
大形リーシュマニアの全タンパク質(可溶性リーシュマニア抗原[SLA])を、PBSに懸濁させた大形リーシュマニアの静止期前鞭毛型を冷凍し解凍するサイクルを3回行うことにより調製した。細胞の溶解後、微量遠心を用いて12,000gで15分遠心分離することにより、可溶性抗原を不溶性画分から分離し、−70℃で保管した。CpGモチーフを含むホスホロチオエート修飾されたODN配列(CpG ODN)を、Isogen(The Netherlands)により合成した。免疫賦活性ODNの配列(5’→3’)は、TCAACGTTGA及びGCTAGCGTTAGCGTであった。
【0032】
免疫化及び寄生虫チャレンジ
BALB/cマウスについて、大形リーシュマニア12μgを単独で、又は大形リーシュマニア12μgとCpG ODN50μg(各免疫賦活性ODNを25μgずつ)を、CpG ODN(50μg)アジュバントを単独で、又はリン酸緩衝食塩水(PBS)を、右足蹠において皮下(s.c.)接種した。各群について、同じレジメンを用いて2週間後及び4週間後に追加免疫した。最初の寄生虫によるチャレンジは、最後の接種の4週間後に、大形リーシュマニアの静止期前鞭毛型(クローンWHOM/IR/−173)5×104個を、左の(未処置の)足蹠に皮下接種することにより行った。感染の進行を、決定用カリパスを用いて厚さを測定することにより追跡した。各動物の反対側の足蹠が対照値を表し、腫大を以下の通り計算した:左足蹠の厚さ−右足蹠の厚さ。損傷が壊死性になった時に動物を安楽死させた。再感染させるために、BALB/cマウス6匹について、上記の通りワクチン接種し感染させた。18週間後、大形リーシュマニアの発育終末前鞭毛型(クローンV1)300個を各マウスの両耳真皮に注射した。感染の進展を、決定用カリパスを用いて耳の損傷の硬結の直径を測定することによりモニターした。対照として、無処置のBALB/cマウス6匹の群についても、耳真皮において感染させた。
【0033】
C57BL/6マウスについて、大形リーシュマニアLRP12μg+CpG ODN50μg(各免疫賦活性ODNを25μgずつ)、及びCpG ODN(50μg)アジュバント50μgを単独で、足蹠に皮下注射した。これらのマウスについて、同じ免疫化レジメンを用いて2週間後及び4週間後に追加免疫した。最後の接種の4週間後に、大形リーシュマニアの発育終末前鞭毛型(クローンV1)300個を、マウスの両耳真皮に皮内(i.d.)接種することによって感染を行った。感染の進展を、決定用カリパスを用いて耳の損傷の硬結の直径を測定することによりモニターした。
【0034】
寄生虫の量子化
耳において、限界希釈アッセイ(6)により寄生虫の数を決定した。簡単に述べると、感染させたマウスから耳を回収した。感染させた耳の腹側及び背側のシートを分離した。耳のシートを、Liberase CI酵素ブレンド(50μg/ml)含有ダルベッコ改変イーグル培地(Dulbecco’s modified Eagle medium)に沈着させた。37℃で2時間インキュベートした後、組織を小片に切断し、ホモジナイズし、細胞ろ過器(孔サイズ70μm)を用いてろ過した。ホモジナイズした組織を、20%FCSを加えたシュナイダー培地(Schneider’s medium)を含有する96ウェルの平底マイクロタイタープレート中に連続的に希釈した。生存可能な寄生虫の数を、前鞭毛型が26℃でインキュベートして最大7日まで成長でき得る最大希釈から決定した。寄生虫の数は、感染させた耳(上顎後方)及び足蹠(膝窩部)の局所流入領域リンパ節(DLN)において、及び脾臓においても決定した。器官を回収し、機械的に切り離し、次いで上記の通り連続的に希釈した。寄生虫の負荷量は、全器官における寄生虫の数として表される。
【0035】
上澄み中のサイトカインの測定
脾臓及び対応する局所DLNを無菌的に取り出し、機械的に切り離し、完全なRPMI培地(10%FCS、2mMのグルタミン、及び10mMの2−メルカプトエタノールを補充したRPMI1640)に播種した。LRP(12μg/ml)又はSLA(12μg/ml)の存在下、37℃で48時間の間に、48ウェルプレートに細胞5×106個/mlを播種した。市販のELISAキット(Diaclone,Besancon,France)により、脾細胞及びDLN細胞の上澄みについて、IFN−γ、IL−10及びIL−4の放出を測定した。一部の例では、LRP12μ/mlで刺激したDLN細胞を、マウスCD4(GK1.5)、マウスIL−12(C17.8)、マウスCD8(53−6.7)のいずれかに対するモノクローナル抗体(mAb)10μg/mlの存在下でインキュベートした。適切なアイソタイプ整合対照についてもこのアッセイで分析した。抗体(no azide/low endotoxin(商標))は、BD)(PharMingen)から購入した。
【0036】
体液性応答の分析
ELISA又はウェスタンブロットにより、LRP又はSLAに対する特異的な抗体について血清試料を分析した。簡単に述べると、標準のELISAプレートを、100μlのLRP(PBS中5μg/ml)又はSLA(PBS中2μg/ml)を用いて室温で一晩コーティングした。抗体価を、血清を連続的に希釈することにより決定し、吸光度>0.2を生じる最大の血清希釈力価の逆数として定義した。アイソタイプ特異的な分析を、以下の西洋ワサビペルオキシダーゼをコンジュゲートした抗マウス免疫グロブリン(Nordic Immunological Laboratories,Tilburg,The Netherlands):抗IgG1(1/1000)及び抗IgG2a(1/500)を用いて行った。オルトフェニレンジアミンジヒドロクロライド−OPD−(Dako,A/S,Glostrup,Denmark)をELISAアッセイのペルオキシダーゼの基質として使用した。15分後、1MのH2SO4100μlを加えることにより反応を停止させ、吸光度を450nmで読み取った。
【0037】
ウェスタンブロット分析のために、幼児リーシュマニア及び大形リーシュマニアのリボソームタンパク質を得、Laemmli緩衝液に再懸濁させ、SDS−PAGEで分離し、ニトロセルロース膜(Amersham,Aylesbury,UK)に転写した。対照のイヌ、VCLリーシュマニア症に罹患しているイヌの血清、又は本研究において示した希釈で利用した種々のマウス群由来の血清を用いてブロットを探索した。二次抗体として、Nordic Immunological Laboratories (Tilburg,The Netherlands)から購入した、西洋ワサビペルオキシダーゼをコンジュゲートした、抗マウス抗IgG(1/1000)、抗IgG1(1/1000)、抗IgG2a(1/500)免疫グロブリン、及び抗イヌ抗IgG(1/2000)を使用した。
【0038】
統計分析
統計分析はスチューデントt検定により行った。P<0.05である場合に有意差があるとみなした。
【0039】
結果
感染の間のLRPの抗原性
リーシュマニアのリボソームタンパク質(LRP)の抗原性を分析するために、幼児リーシュマニアに自然感染したイヌ由来の血清の幼児リーシュマニアLRPに対する応答性を、ウェスタンブロットによりアッセイした。活動性疾患に罹患しているイヌ由来の血清が、LRP抽出物中の多数のタンパク質バンドを認識したことが観察された(図1A)。大形リーシュマニアに実験的に感染させたC57BL/6マウス及びBALB/cマウス由来の血清も、大形リーシュマニアLRP中の多くのタンパク質バンドを認識し、BALB/c感受性マウス由来の血清に存在するIgG抗体によって認識されるリボソームタンパク質の数の方が、C57BL/6耐性マウスの血清中のIgG抗体によって認識されるタンパク質の数よりも多い(図1B)。
【0040】
IgG1及びIgG2a抗体の誘導は、Th2型免疫応答及びTh1型免疫応答のマーカーとして使用することができるので(8)、我々は、大形リーシュマニアに感染させたマウスにおいてLRPに対するIgG1/IgG2a極性化の分析を行った。BALB/cマウスでは、抗LRP応答はIgG1アイソタイプが優勢であったのに対し、C57BL/6マウスでは、抗LRP応答はIgG2aアイソタイプが優勢であった(図1C)。両方のマウス系統において、in vitroでLRPを用いてDLN細胞を刺激した後のIFN−γ、IL−4及びIL−10の産生についても決定した。CLに罹患しているBALB/cマウスにおいて、LRP特異的なIFN−γの産生が検出されたが、IL−4及びIL−10の産生も強力に刺激され、INF−γ/IL−4比は約2.4、INF−γ/IL−10比は約1.4になっている(図1D)。一方、治癒したC57BL/6マウス由来のDLN細胞をLRPで刺激した場合、IL−4は検出されず(<7.5pg/ml)、高IFN−γ/IL−10比(約15)が得られた(図1E)。
【0041】
BALB/cマウスにおけるLRPの免疫原性
BALB/cマウスにおいて、リボソームタンパク質をCpG ODNの非存在下及び存在下で投与した後のLRPに対する免疫応答を調べた。LRPE+CpG ODNを用いたワクチン接種後、抗LRPE体液性応答はIgG2aアイソタイプが優勢であったのに対し、LRPE単独で免疫したマウス由来の血清では検出されたIgG1アイソタイプの抗体の抗体価は低かった(図2A)。LRPE+GpG ODNで免疫したマウス由来の脾臓細胞を、in vitroでLRPEを用いて刺激した後、その細胞から分泌されたIFN−γのレベルは、対照マウス由来の脾臓細胞及びLRPE単独で免疫したマウス由来の脾臓細胞から分泌されたものよりも高かった(図2B)。いずれの群においても、LRPEによる刺激後にIL−4産生の増加は見られなかった(図2C)。著しいことに、LRPE+CpG ODNを用いてワクチン接種した脾臓から確立した培養物の上澄みにおいて、特異的なIL−10が検出され、IFN−γ/IL−10比は約40であった(図2D)。全体として、これらの結果は、BALB/cマウスにおいて、アジュバント無しで投与したLRPEからは、これらの抗体に対してほんの弱いIgG1体液性応答しか誘導されないが、CpG ODNと同時投与すると、これらの抗原に対してTh1様応答が増強されることを実証している。
【0042】
LRPE+CpG ODNを用いたワクチン接種は大形リーシュマニアのチャレンジからBALB/cマウスを防御する
抗原に関連した疾患に誘導されるTh2応答をTh1応答に転送することは、リーシュマニアに対するワクチンを開発するための有望なアプローチ法であると考えられているので(7)、我々は、LRPE+CpG ODNを用いたワクチン接種によって大形リーシュマニア感染に対する防御を誘導することができるかどうかを分析した。図3Aは、LRPE+CpG ONDをワクチン接種したマウスの足蹠の腫大が、対照マウス及びLRPE単独でワクチン接種したマウスの足蹠の腫大(平均値約5.5mm)と比較して縮小されているので(8週目の平均値0.7mm)、LRPE+CpG ONDによって効果的な防御が誘導されることを示している。我々は次に、マウスの4群について、膝窩部DLN及び脾臓における寄生虫の負荷量を分析した。LRPE−CpGで免疫したマウス由来のDLNでは、他の群と比べて、約3logの寄生虫の生体内総量の縮小が見られた。さらに、対照マウス及びLRPE単独で免疫したマウスの脾臓では同様の寄生虫の負荷量が見られた一方、LRPE+CpG ODNをワクチン接種したマウスの脾臓では寄生虫が検出できなかった(図3B)。
【0043】
LRPE+CpG ODNに誘導される防御に関連する免疫学的パラメーターを決定するために、SLA主導又はLRPE主導のIL−4、IL−10及びIFN−γの産生をアッセイした。SLA特異的又はLRPE特異的に誘導されたIL−4及びIL−10の産生は、対照マウス(生理食塩水及びCpG)由来のDLN細胞及びLRPE単独で免疫したマウス由来のDLN細胞において検出された(図3C〜D)。一方、LRPE+CpG ODNを用いて免疫したマウス由来のDLN細胞が産生したIFN−γの量は、他の3群で検出されたIFN−γの量よりも多かった(図3E)。LRPEに特異的なIFN−γ産生に対するCD4+T細胞及びCD8+T細胞の寄与及びIL−12への依存についても分析した。図3Fに示されるように、IFN−γの産生は、抗IL−12モノクローナル抗体又は抗CD4モノクローナル抗体により完全に阻害された。DLN細胞培養物への抗CD8抗体の添加により、上澄み中のサイトカインの量が一部だけ縮小した。
【0044】
BALB/cマウスにおいて、疾患の進行に付随してIL−4に依存した高抗体価の抗体が産生されることから、我々は、感染の8週間後に、LRPEに対して引き出される体液性応答をELISAによって分析した。LRPE+CpG ODNを用いて免疫したマウスにおいて寄生虫のチャレンジによって引き出された、LRPEに対する抗体は、主にIgG2aアイソタイプであった。また、防御マウスの血清に存在するIgG1アイソタイプの抗LRPE抗体の抗体価は、LRPE単独で免疫したマウス及び2つの対照群と比較して低かった(図4A)。LRPEに対するIgG1抗体価の縮小は、LRPE+CpG ODNをワクチン接種したマウスの血清由来のIgG1抗体に認識されるリボソームタンパク質バンドの数の減少と相関した。図4Bに示されるように、LRPEウェスタンブロットにおいて、生理食塩水、CpG又はLRPE単独で免疫したマウスの血清由来のIgG1アイソタイプ抗体によって多くの数のタンパク質バンドが認識されたのに対して、防御マウスのIgG1抗体によって認識されたバンドはわずかであった。ウェスタンブロット分析により、LRPE+CpG ODNをワクチン接種したマウス由来の血清中のIgG2a抗体によって認識されるタンパク質バンドの数は、他の3群と比較して増加しなかったことも示された(図4B)。LRPE+CpG ODNを用いたワクチン接種によって、大形リーシュマニアの感染によって誘導される広域の抗リーシュマニア体液性応答も調整された。LRPE+CpG ODNを用いて免疫したマウスにおいて寄生虫のチャレンジによって引き出されたSLAに対する抗体は、主にIgG2aアイソタイプであり、IgG1アイソタイプの抗体の抗SLA抗体価は、他の3群において検出されたものより有意に低かった(図4C)。
【0045】
LRPE+CpG ODNをワクチン接種し、感染させたマウスは耳真皮における大形リーシュマニアの再感染に対して耐性である
LRPE+CpG ODNをワクチン接種し、感染させたマウスが、2回目の寄生虫によるチャレンジを制御できるかどうかを決定するために、BALB/cマウス6匹について、上記の通り足蹠においてワクチン接種し、感染させた。これらのマウス群の足蹠の腫大は、18週間の間<0.7mmであった(データは示さず)。次に、これらの防御マウスについて、大形リーシュマニアの発育終末前鞭毛型300個を用いて、耳真皮において再感染させた。無処置のマウス6匹の対照群についても感染させた。LRPE+CpG ODNをワクチン接種し、再感染させたマウスでは病変が見られなかったのに対し、対照マウスでは、7週目に開存性の耳損傷が発生したので、LRPE+CpG ODNをワクチン接種し、再感染させたマウスは真皮損傷の発生を防御することが観察された(図5A)。耳真皮及び上顎後方のDLNにおける寄生虫の負荷量も、2群間で有意差があった(図5B)。ワクチン接種し、再感染させたマウスの耳及びDLNにおける寄生虫の負荷量の低さは、脾臓に寄生虫が存在しないことと相関する。再感染後に起こる細胞性応答について知るために、LRPE又はSLAを用いたin vitroでの刺激後の上顎後方のDLN細胞によるIFN−γ、IL−4、及びIL−10の分泌について分析した(図5C)。対照マウスでは、マウスを足蹠において感染させた際にも起こるのと同様(3C〜E)、IFN−γの特異的産生が検出されたが、IL−4及びIL−10の産生も強力に刺激された。一方、ワクチン接種し、再感染させたマウスのDLN細胞は、特異的IFN−γを多量に産生し、IL−4及びIL−10はほとんど検出されなかった(図5C)。これに一致して、LRPE(図5D)及びSLA(図5E)に対するIgG体液性応答は、IgG2aアイソタイプによるものであった。
【0046】
LRPE+CpG ODNを用いたワクチン接種は、C57BL/6マウスにおける大形リーシュマニアのチャレンジに起因する真皮の病変に対する防御を付与する
BALB/cマウスでは、LRPE+CpG ODNを用いてワクチン接種すると、大形リーシュマニアの感染に対し、LRPEに対するTh2免疫応答がTh1免疫応答へ転送されることにより防御が起こることから、我々は、このワクチンを、リーシュマニア抗原に対して自然にTh1応答を生じるモデルであるC57BL/6マウスに投与した効果を分析した。C57BL/6マウスの群を、LRPE+CpG ODNで3回免疫し、対照マウスはCpG ODNアジュバントのみを受けさせた。接種により、LRPEを用いて刺激した脾臓細胞培養物の上澄みにおけるin vitroでのIFN−γ産生によって実証されたTh1応答が誘導された。LRPE+CpG ODNをワクチン接種したマウスから確立した脾臓細胞培養物の上澄みにおいて、特異的なIL−10の存在も検出されたのに対し、刺激後に特異的なIL−4産生は観察されなかった(図6A)。LRPE+CpG ODNをワクチン接種したマウスは、病変がほとんど又は全く観察されなかったので、真皮損傷の発生を防御した(図6B)。CpG ODNで免疫したマウスでは損傷が発生し、7週目でピークに達し、13週目にほとんど完全に治癒した。このモデルでは、感染部位における寄生虫の数が、損傷の発生の直前にピークに達するので(5)、我々は、5週目に耳及び局所DLN(上顎後方)における寄生虫の負荷量を決定した。ワクチン接種したマウスの耳真皮における寄生虫の数は、約300分の1に(LRPE+CpG ODNマウスでは寄生虫1.0×104個、CpG−ODNで免疫したマウスでは寄生虫3.0×106個)及びDLNマウスでは約40分の1に(LRPE+CpG ODNマウスでは寄生虫5.0×104個、CpG ODNマウスでは寄生虫2.0×104個)縮小した。治癒後、対照マウスでは(チャレンジの13週間後)、全ての群において寄生虫の数の縮小が観察された。
【0047】
防御に関連する免疫学的パラメーターを決定するために、抗原主導のIL−4、IL−10及びIFN−γの産生をアッセイした。チャレンジの5週間後、DLN細胞の培養物を確立し、LRPE又はSLAを用いて刺激した。図6Dに示される通り、ワクチン接種したマウス由来の細胞の方が、対照マウス由来の細胞よりも多くのSLA特異的及びLRPE特異的なIFN−γを産生した。LRPEに特異的なIFN−γ産生に対するCD4+T細胞及びCD8+T細胞の寄与及びIL−12への依存についても分析した。IFN−γの分泌は、抗IL−12モノクローナル抗体又は抗CD4モノクローナル抗体により完全に阻害された。抗CD8抗体での処理では、このサイトカインの量は一部だけ縮小した。これらのデータにより、C57BL/6において、LRPE+CpG ODNを用いたワクチン接種により、LRPEに対する早期の特異的Th1応答の誘導と相関する、皮膚及び局所DLNにおける病変及び寄生虫の生体内総量の縮小が誘導されることが実証された。これに一致して、対照マウスに対してワクチン接種したマウスにおいてIgG2a特異的な抗LRPE抗体が早く、高抗体価で検出された(図6F)。我々は、SLA抗原特異的又はLRPE抗原特異的なIL−10産生も検出し、それはワクチン接種したマウスと対照マウスとの間に統計的な差はなかった。
【0048】
考察
本研究において、我々は、多くの寄生虫リボソームタンパク質と反応する抗体が、VCLを生じたイヌ由来の血清、及び大形リーシュマニアに感染させたマウスにおいて観察されることを示し、そのことは、リーシュマニアの自然感染及び実験的感染の間、寄生虫のリボソームタンパク質が強力な抗原であることを示している。抗LRPE抗体応答はIgG1アイソタイプによるものが優勢であったので、感染させたBALB/cマウスのLRPEに対する応答はTh2型のものであった。この知見は、LRPEを用いて感染させたマウス由来のDLN細胞をin vitroで刺激した後に、培養物の上澄み中で多量のIL−4が検出されたという事実により補強される。LRPEは、INF−γのin vitro産生にも関係したが、匹敵するレベルの、寄生虫を破壊するための活性化シグナルに対して、感染したマクロファージを無反応性にする多面的な抗炎症サイトカインIL−10が検出された(31)。さらに、BALB/cマウスにおける疾患の促進におけるIL−4及びIL−10の影響は、相加的だと思われることから((37)に総説がある)、これらのサイトカインのLRPEによる刺激は、リボソーム抗原に対する宿主の応答が、BALB/cマウスにおいて寄生虫が拡大及び持続するのに好都合であることを示しているとみなすことができる。リーシュマニアによる感染後早期に、宿主の免疫系が寄生虫の細胞溶解によって放出される豊富なLRPEにより感作され、その後で、寄生虫の増殖の結果、追加免疫され得ると考えることができる。したがって、LRPEの強力な免疫原性及びその病原抗原的な役割は、おそらくその豊富さ及び抗原特異性(その進化的に保存された特性にもかかわらず)に依存している。一方、LRPEに対して応答するC57BL/6マウスはTh1型のものであり、IgG2a特異的抗体の生成及びin vitroでの高レベルの特異的INF−γ産生を伴い、IL−4レベルは検出不可能であった。また、我々は、感染の14週間後に、LRPEに特異的なIL−10産生を見出した。IFN−γ/IL−10比が、SLAを用いて細胞を刺激した場合(データは示さず)に得られたものと同様であったことから、我々は、これらの抗原によって誘導されるIL−10産生は、このモデルにおいて見られる、寄生虫が持続するのに好都合である調節性の応答に関連する可能性があることを示唆する(4)。我々のデータは、LRPEに対する応答が、感受性宿主の病変に関連したTh2に媒介される体液性応答及び耐性宿主におけるTh1に関連する防御応答の誘導に向かうバイアスに一致することを示している。
【0049】
いくつかのリーシュマニアのエピトープに対して誘導されるTh2応答のTh1応答への転送は、大形リーシュマニアの感染に対する防御を誘導するための有望な戦略になる可能性が高いので(7)、我々はまず、BALB/cマウスにおいて、異なるリーシュマニア抗原を用いて免疫した際のTh1に関連する長期免疫及び防御を与え(43)、さらにマウスにおいていくつかの寄生虫特異的Th2応答を抑制し得る(12、57)アジュバントであるCpG ODNと同時投与したLRPEの免疫原性について分析することを決めた。図2に示されるように、BALB/cマウスにおいて発生する免疫応答は、LRPEを用いてin vitroで刺激した後、免疫したマウスがIgG2aアイソタイプの抗LRPE抗体を発生し、ワクチン接種したマウス由来の脾細胞が多量のIFN−γを産生し、IL−4は産生しないので、Th1型であることが分かった。同様の特異的免疫応答が、C57BL/6マウスにおいて、LRPE+CpG ODNを用いて免疫した場合に生じた(図6A)。どちらのマウス系統においても、免疫した後にLRPE主導のIL−10産生も観察された。
【0050】
L31をコードする遺伝子ワクチンを用いたワクチン接種が、遺伝子ワクチン接種後の特異的IL−10産生に関係し得ることが報告されているにもかかわらず(44)、我々は、IL−10産生は、LRPE+CpG ODNの投与後に起こるTh1応答のホメオスタティックコントロールの方に関連している可能性があると考えている。事実、INF−γを産生するTh1細胞は、フィードバック制御の機構としてIL−10の産生にも関与している可能性があることが最近報告されている(2)。さらに、得られた高INF−γ/IL−10比の値により、ワクチンの結果について良好な予測が得られ得る(44)。
【0051】
ここで示したデータは、LRPE+CpG ODNの同時投与により、実験的な皮膚リーシュマニア症の2つの異なるモデル、BALB/cマウス(皮膚リーシュマニア症ワクチンのアッセイに広く用いられている)の足蹠における高接種量モデル、及びC57BL/6マウス(経路及び感染用量に関して、ヒト疾患をより厳密に再現するモデル)の耳における低接種量モデルにおいて防御が誘導されることを示している。ワクチン接種したBALB/cマウスでは膝窩部DLNにおける寄生虫の生体内総量が縮小しており、脾臓への寄生虫転移はなかった。また、非常に弱い炎症が感染した足蹠において検出された。ワクチン接種したC57BL/6マウスでは、耳真皮の損傷が防御され、皮膚及びDLNにおいて寄生虫の生体内総量が縮小したことが示された。この防御は、熱殺菌されたリーシュマニア抗原とCpG ODNを用いてワクチン接種したC57BL/6マウスによって示された防御に匹敵し、また、このモデルにおいて、LACK、LmSTI1及びTSAを含む多成分ワクチンについても試験した(28、29、43)。
【0052】
重要なことに、どちらの系統における防御も、LRPEに対するTh1特異的免疫応答の生成に相関している。細胞性応答についてのin vitro分析は、BALB/cマウスでは感染の8週間後、及びC57BL/6マウスでは5週間後(寄生虫の負荷量のピークに一致する(4))に測定した。LRPE+CpG ODNを用いてワクチン接種した両マウス系統由来のDLN細胞は、LRPEを用いて刺激した際、それぞれの対応する対照群よりも高いレベルのIFN−γを分泌した。IFN−γ応答は、IL−12依存性であり、CD4+T細胞により産生され、CD8+T細胞に対する寄与は少ないことが分かった。予想通り、両系統の防御マウスについて、SLA特異的IFN−γ産生は高度であった。BALB/cマウスにおいて、防御マウスにおけるTh1応答の生成は、LRPEに対する優勢なIgG2a特異的抗体の生成と相関する(図4A)。IL−10のin vitro産生において、系統間で、多少の違いが観察された。我々のデータは、LRPE及びSLAの両方を用いてin vitroで刺激した後、防御BALB/cマウスでは、対照マウスよりも有意に低いレベルのIL−10が産生されたことを示し、これは、このサイトカインがこのモデルの感受性に関与していることと一致している(35)。一方、対照マウス及びワクチン接種したC57BL/6マウスのどちらのLNCにおいても、LRPE又はSLAを用いてin vitroで刺激した後、感染の急性期の間IL−10が産生された(図6F)。IFN−γについて起こったように、IL−10レベルは、細胞を、SLAを用いて刺激した場合よりも、LRPEを用いて刺激した場合の方が高かった。IFN−γ及びIL−10の産生は同様のプロファイルに従ったという事実は、ワクチン接種後のIL−10産生が、宿主において強力なTh1によって引き起こされる有害な影響を制御するホメオスタティック機構を反映しているとみなすことができる(4)。驚くべきことに、我々は、LRPE+CpG ODNを用いてワクチン接種した後にBALB/cマウスにおいて見られる防御が、LRPE又はSLAを用いてin vitroで刺激した後のIL−4主導の抗原産生が有意に縮小したことにも関連していることを見出した。これらの細胞性応答は、in vivoで、リボソームタンパク質に対するTh2に媒介される抗体応答の転換に相関する。したがって、防御BALB/cマウス由来の血清は、抗体価、特にLRPE特異的なIgG1抗体によって認識される抗原の数について有意な減少を示した。さらに、LRPE+CpG ODNを用いたBALB/cマウスの免疫も、大形リーシュマニアの感染によってマウスにおいて誘発される広範な体液性応答に対して明らかに影響した(図5C)。したがって、ワクチン接種したマウスを感染させると、IgG1抗リーシュマニア特異的抗体の誘導が制限されるのに対して、他のアッセイした対照マウス群では、体液性応答が高度に誘導され、Th2型の抗体(すなわち、IgG1アイソタイプ)が優勢であった。全体として、どちらの系統のマウス由来のワクチン接種したマウスにおいて観察される防御も、LRPEに対するTh1応答の生成に相関し、またその結果、BALB/cマウスにおけるTh2主導のIL−4及びSLAに対するIL−10応答が下向き調節される。
【0053】
我々のデータは、防御BALB/cマウスが、追加的な感染に抵抗できる能力を彼らに付与する免疫学的状況(寄生虫への再曝露が極めて頻繁に起こり得る流行地域において利用され得るワクチンに魅力的な特徴)を獲得したことを示している。耳真皮における再チャレンジの後、これらのマウスは、大形リーシュマニアの感染に対して強固な防御を示した。真皮損傷の発生は非常に少なく(一部の例では、真皮損傷が完全にないことが検出された)、感染した耳及びDLNにおいて寄生虫数に相当な縮小がみられた。防御マウス由来のDLN細胞は、対照よりも大量のIFN−γを産生しないが(感染の7週間後に測定)、LRPE及びSLAに対するIgG2a抗体の抗体価は、これらのマウスが二次的なチャレンジ後に特異的なTh1防御応答を高めたとみなすことができる。驚くべきことに、これらのマウスにおける疾患関連サイトカインであるIL−4及びIL−10の特異的な産生は、非常に少なかった。これらのデータは、最初の寄生虫によるチャレンジの後に生成した免疫状態が非常に有力であり、再感染した部位からの寄生虫の迅速かつ効率的な排除を導くことを示している。
【0054】
ここで示したデータは、LRPE+CpG ODNを用いたワクチン接種が、耐性マウスと感受性マウスの両方において、リーシュマニアによる感染時に、免疫系の決定に直接影響を及ぼすことを実証している。我々の考えでは、リーシュマニアのような複雑な寄生虫に対するワクチンの生成は、ワクチン製剤中に種々の標的抗原を組み込み、これらの抗原が必要な免疫性を導く(主にCD4+及びCD8+IFN−γ媒介応答)ことを利用し、病変をもたらす病原抗原主導の免疫応答(IL−4 Th2主導及びIL−10不活性化応答)をTh1バイアスに転送することにより最適化され得る。それにもかかわらず、大形リーシュマニアのメタ1抗原で起こるように、これらの抗原の一部に対するTh2応答は、通常のTh1誘導剤によって転送されない場合があることを考慮しなければならない(50)。
(参考文献)

【技術分野】
【0001】
本発明は、寄生虫疾患を治療又は予防するための医薬品を調製するための、リボソームタンパク質抽出物(RPE)及び場合によってTh1促進アジュバントを含む組成物及びその使用に関する。
【背景技術】
【0002】
リーシュマニア症は、ヒト及びイヌを含めたさまざまな哺乳動物のマクロファージに主に感染する、リーシュマニア属(Leishmania)に属する細胞内寄生原虫によって引き起こされるいくつかの疾患を含む。寄生虫の種及びヒト宿主の免疫能力の状態に主として応じて、疾患のスペクトルは、自然治癒する皮膚リーシュマニア症(CL)から致命的な内臓リーシュマニア症(VL)又はカラアザールに及ぶ(18)。幼児リーシュマニア(Leishmania infantum)及びシャーガスリーシュマニア(L.chagasi)によって引き起こされるイヌ・内臓皮膚リーシュマニア症(VCL)は、地中海沿岸地方周辺、中東及びラテンアメリカの諸国で発見された重要な新興の動物原性感染症であり(16)、イヌがこれらの寄生虫の主要な保有宿主であり、サシチョウバエによるヒトへの伝染において中心的な役割を果たす(47)。感染の結果は、宿主の免疫系と種々の寄生種との相互作用によって決まり、今までのところリーシュマニア症の発症機序は不明のままであり、ヒト及びイヌにおける、リーシュマニアに対する免疫応答に関わるメカニズムについての知見は依然として限られている。大抵の場合、防御免疫は、T細胞由来のサイトカインによるマクロファージの活性化を誘導する古典的な細胞性免疫応答に関連付けられ、一方非治癒性の疾患は、強力な体液性応答の生成に関連付けられる(15、26)。寄生虫粗画分又は定義済みの寄生虫抗原をベースとする第2世代のワクチンを開発するための研究において取り組まれたのは、多種多様なアジュバントを用いたいくつかの実験モデルにおいてワクチン候補として試験された種々の寄生虫表面分子又は分泌分子の同定であった(1、17、22、46、48、49、52、54)。感染した動物又はヒト由来の血清を用いた発現ライブラリーのスクリーニングによっても、候補ワクチンとして数個の抗原を選択することができた((9)に総説がある)。その中で、感染したマウス又はヒト患者の細胞において、その細胞の位置に無関係に、主としてTh1型の免疫応答を引き出す抗原が、種々の動物モデルにおいて防御応答の生成に関連している(51、55、56)。一方、単離された抗原の一部は、VLに罹患しているヒト又はイヌにおける体液性応答、又は実験的に感染させたマウスにおけるTh2媒介体液性応答を優勢的に刺激する、細胞内に保存されているタンパク質である(3、36、38、40、42)。リーシュマニア症に罹患しているイヌにおいて、それらの抗原に対して誘導される不十分な体液性応答により、主に免疫複合体が有害作用を有するため、免疫病変、特にブドウ膜炎(13)、中枢神経系における損傷(14)又は腎炎(23、24、33、34)が生じると考えられている。また、VLを生じたヒトにおいて、IgG免疫複合体が存在することが、感染を消散できないことと相関があることが最近示され、感染した宿主に対して免疫複合体が悪影響を及ぼし得ることを実証している(30)。
【0003】
最初は良好なワクチン候補であるとみなされなかったにもかかわらず、感染プロセスの間に高度な体液性応答を誘導するタンパク質が、防御応答の誘導に関連している。例えば、寄生虫のチューブリン及びヒストンH2Bは、免疫ドナー由来のT細胞クローンによって認識され(39)、rK39は、免疫マウス由来のT細胞によって増殖及びIFN−γ産生を引き起こす(25)。寄生虫のH2B遺伝子、H3遺伝子及びH4遺伝子を用いた遺伝子による免疫によって、マウスの内臓リーシュマニア症モデルにおいて防御が誘導されることも示されている(27)。また、Th1促進アジュバントと共に投与される活性化Cキナーゼ(LACK)(32)、一部の寄生虫システインプロテイナーゼ(38、41)又は寄生虫ヌクレオソーム形成ヒストン(11、20)に対する受容体を免疫することにより、マウスモデルにおいて皮膚リーシュマニア症に対する防御に相関する免疫応答が生成する。
【0004】
進化的に保存されたリーシュマニアの抗原の中で、いくつかの証拠により、リボソームタンパク質が、リーシュマニア感染の間、免疫学的に関連する分子であることが示唆されている。一部の例では、リボソームの構成要素は、感染の間に細胞活性及びサイトカインの放出を調節するその能力を通して宿主の免疫系の機能不全に寄与し得る。したがって、大形リーシュマニア(L.major)のリボソームタンパク質S3aをBALB/cマウスに注射することにより、B細胞クローンのポリクローナル増大が誘導され、T細胞の増殖が阻害された(10)。また、推定60Sリボソームタンパク質L31をコードするDNAワクチンを用いた遺伝子による免疫により、マウスモデルにおいて、IL−10及びTh2サイトカインが誘導されることによって疾患が悪化する(44、53)。さらに、寄生虫の酸性Pタンパク質などの一部の寄生虫のリボソームタンパク質は、リーシュマニア症に罹患しているイヌ及びヒトにおける強力な体液性応答の生成に関連している((42)に総説がある)。
【発明の概要】
【発明が解決しようとする課題】
【0005】
今まで試みられているにも関わらず、依然としてリーシュマニア症などの寄生虫疾患に対して有益なワクチンはない。したがって、そのようなワクチンが依然として必要とされている。
【課題を解決するための手段】
【0006】
本研究において、我々は、RPE、特にリーシュマニアRPE(LRPE)が、幼児リーシュマニアに自然感染したイヌにおいて、及び大形リーシュマニアに実験的に感染させたマウスにおいて免疫応答の標的であることを示している。我々はさらに、LRPEをCpGオリゴデオキシヌクレオチド(ODN)などのTh1促進アジュバントと同時投与した際に、強力なTh1防御免疫応答が誘導されることを実証している。そのような組成物(Th1促進アジュバントと組み合わせたLRPE)は、ワクチンとして使用することが非常に魅力的である。本発明を以下さらに説明する。
【0007】
使用
本発明の第1の態様において、対象において寄生虫疾患を治療又は予防するための医薬品を調製するための、RPE及び場合によってTh1促進アジュバントの使用が提供される。
【0008】
好ましい実施形態において、RPEは、対象内に存在すると寄生虫疾患を引き起こす寄生虫細胞を用いて、
a.寄生虫細胞を溶解緩衝液と混合するステップと、
b.得られた混合物を遠心分離してサイトゾル抽出物を得るステップと、
c.得られたサイトゾル抽出物からRPEを調製するステップと
を行うことにより得られる。
【0009】
ステップaにおいて、寄生虫は原生動物を指すことが好ましい。好ましい寄生虫は本明細書において後で定義する。原生動物のステージは前鞭毛型であることがより好ましい。所望の量のRPEを調製するためにおよそ必要な寄生虫細胞の量は当業者には公知であろう。一般には、およそ500マイクログラムのRPEを調製するために、およそ3.109個の寄生虫細胞を使用する。溶解緩衝液は、寄生虫細胞の少なくとも一部を壊す緩衝液である。好ましい溶解緩衝液は、非イオン界面活性剤を含む。非イオン界面活性剤としてNonidet P40(NP40)を用いると良い結果が得られた。しかし、他の非イオン界面活性剤を使用してもよい。下記の溶解緩衝液を使用することが好ましい(緩衝液A):10mMのトリスHCl、pH8.0、150mMのNaCl、1.5mMのMgCl2及び0.5%のNP40(Roche)、並びにPMSF 1mM、Leupeptin 8μg/ml、Aprotinin 4μg/ml及びPentatin 8μg/mlなどのプロテアーゼ阻害剤を補充することが好ましい。適当量の寄生虫細胞(およそ細胞109個/緩衝液A1ml)を、一般には、エッペンドルフピペットを用いて溶解緩衝液と穏やかに混合する。
【0010】
ステップbにおいて、ステップaで得られた混合物に対し、遠心分離を4℃で少なくとも1ステップ行う。通常、第1の遠心分離ステップはおよそ3,000gでおよそ2分行う。得られた上澄みを、4℃で、およそ13,000gでおよそ15分、1回又は2回再度遠心分離することが好ましい。
【0011】
ステップcにおいて、得られた上澄みを(45)に記載の通りRPEを調製するために使用する。簡単に述べると、得られた上澄みを、Beckman TL100.3ローターを用いて、4℃、およそ90,000rpmでおよそ30分の高速遠心分離にかける。得られたペレットは粗製のリボソームペレットであり、緩衝液B(20mMのトリスHCl、pH7.4、500mMのAcNH4、100mMのMgCL2、5mMのβ−メルカプトエタエノール)に再懸濁させ、緩衝液A中、不連続なショ糖勾配(20/40%)で、TL100.3ローターを用いて、4℃、およそ90,000rpmで遠心分離する。得られたペレットはリボソームを含む。このペレットを、PBS(リン酸緩衝食塩水)に溶解させ、超音波処理し、−70℃で保存することが好ましい。
【0012】
リボソームタンパク質は、よく保存されたサイトゾルタンパク質である。したがって、本明細書で定義するRPEは、植物であろうと動物であろうと、哺乳動物、爬虫類、魚類、昆虫類であろうと原生生物などの染色体を持つ他の任意の生物であろうと、任意の真核生物から調製することができる。RPEは、疾患に近い生物、好ましくは系統樹内の寄生虫疾患を引き起こす生物から得ることが好ましい。したがって、寄生虫疾患の治療に使用されるRPE源として特に興味深いものは、原生動物、例えばプラスモジウム属(Plasmodium)、特にトリパノソーマ科のメンバー、より特定するとトリパノソーマ科の原生生物リーシュマニアの種々の種である。リーシュマニアには20を超える公知の種があり、大形リーシュマニアを含めた大形リーシュマニアコンプレックス、シャーガスリーシュマニア、ドノバンリーシュマニア(L.donovani)及び幼児リーシュマニアを含めたドノバンリーシュマニアコンプレックス、アマゾンリーシュマニア(L.amazonesis)及びメキシコリーシュマニア(L.mexicana)を含めたメキシコリーシュマニアコンプレックスを含むリーシュマニア亜属の種、並びにブラジルリーシュマニア(L.braziliensis)及びペルーリーシュマニア(L.peruviana)を含めたブラジルリーシュマニアコンプレックス及びガイアナリーシュマニア(L.guyanensis)及びパナマリーシュマニア(L.panamensis)を含めたガイアナリーシュマニアコンプレックスを含むビアンニア(Viannia)亜種が含まれる。特に興味深いプラスモジウム属の種は、熱帯熱マラリア原虫(Plasmodium falciparum)及び三日熱マラリア原虫(Plasmodium vivax)である。好ましい実施形態において、RPEは、リーシュマニア属の各種から得られ、大形リーシュマニア及び/又は幼児リーシュマニアから得ることが好ましい。他の好ましい実施形態において、RPEは、プラスモジウム属の各種から得られる。RPEは、本明細書で確認したいくつかの異なる生物由来のRPEを混合することにより調製してもよいことは当業者に理解されよう。所定のタンパク質を使用する代わりにRPEをワクチンに使用することは、RPEが異なる抗原を数多く含有するので、非常に魅力的である。これらの抗原はそれぞれ、治療される対象において免疫防御応答を潜在的に誘導し得る。さらに、抗原Aには応答するが、抗原Bには応答しない対象、及びその逆の対象がある。したがって、本明細書で定義するワクチンは、異なる抗原を数多く含有するので、広範囲の対象集団に対して使用することを企図している。好ましい実施形態において、RPEは、少なくとも1種のリボソームタンパク質及び/又は少なくとも1種のリボソームタンパク質抗原及び/又は少なくとも1種のリボソームタンパク質のタンパク質断片を含む。より好ましい実施形態において、RPEは、少なくとも2種のリボソームタンパク質及び/又は少なくとも2種のリボソームタンパク質抗原及び/又は少なくとも2種のリボソームタンパク質断片を含む。本明細書で定義するタンパク質断片は、少なくとも2、3、5、7、10、15、20、25、30個又はそれ以上の、リボソームタンパク質に対応する隣接アミノ酸を含むことが好ましい。一実施形態において、本明細書で定義するRPEは、幼児リーシュマニアの酸性リボソームタンパク質P0及び/又はブラジルリーシュマニア由来のリボソーム抗原LbeF4Aを含まない、又はそれらからなるものではない。
【0013】
Th1促進アジュバント(例えばCpG ODNモチーフを含むアジュバント)は文献(Liu N.,et al.,(2003),Nature Immunology,687−693)において、所定の抗原(ここではRPE)と一緒に用いた際に所与の抗原に対するTh1免疫応答を促進又は誘発することができ、その抗原と一緒に培養した際に処理した対象の脾細胞の上澄みから検出されるアジュバントとして定義されている。対照として、抗原及びアジュバントで処理していない、又は抗原でのみで処理した、同一対象の脾細胞集団において、Th1免疫応答の促進又は誘発について評価する。Th1免疫応答の誘発又は促進は、抗原を用いて処理した対象の脾細胞を培養すること、及び/又は抗原特異的IgG2a免疫グロブリンの産生を誘導することによって検出されるIFNγの誘導によって定義することが好ましい。このサイトカインの誘導についての評価は、実施例に記載の通り、脾細胞についてのELISAによって行うことが好ましい。IgG2aの誘導についての評価は、実施例に記載の通り、ELISA又はウェスタンブロットによって行うことが好ましい。脾細胞をRPE及びアジュバントで刺激するとIFNγ及び/又はIgG2aが誘導されることにより、アジュバントがTh1促進アジュバントであると認定されたことが示されることが好ましい。
【0014】
上記のTh1免疫応答の誘発又は促進についての最初の定義の代わりに、又はそれと組み合わせて、Th1免疫応答の誘発又は促進はさらに、Th2免疫応答の欠如(又は誘導の欠如)によって定義することができる。Th2免疫応答は、非処理の脾細胞と比べて、IL−4、IL−10の誘導が検出可能に増加すること、及び/又は検出可能なIgG1免疫グロブリンが産生されることを特徴とする。IL−4及び/又はIL−10の誘導についての評価は、実施例に記載の通り、脾細胞についてのELISAによって行うことが好ましい。IgG1の誘導についての評価は、実施例に記載の通り、ELISA又はウェスタンブロットによって行うことが好ましい。
【0015】
上記のTh1免疫応答の誘発又は促進についての最初の2つの定義の代わりに、又はそれと組み合わせて、Th1免疫応答の誘発又は促進はさらに、定義済みの抗原、この場合はRPEに対するINFγ/IL−10比及び/又はINFγ/IL−4比の増加、及び/又はIgG1/IgG2a比の減少が生じることによって定義することができる。好ましい実施形態において、これらの3つ以上の比のいずれかが変化する(上記のように増加又は減少する)ことにより、アジュバントがTh1の性質を有することが示される。言及した各サイトカインの誘導についての評価は、実施例に記載の通り、脾細胞についてのELISAによって行うことが好ましい。免疫グロブリンIgG1又はIgG2aの誘導についての評価は、実施例に記載の通り、ELISA又はウェスタンブロットによって行うことが好ましい。
【0016】
好ましい実施形態において、Th1促進アジュバントは、オリゴデオキシヌクレオチドである、又はオリゴデオキシヌクレオチドを含む、又はオリゴデオキシヌクレオチドからなる。オリゴデオキシヌクレオチド(ODN)は、Cがメチル化されていないCpGを含む、又はCがメチル化されていないCpGからなることがより好ましい(CpG ODN):3’プリン−CpG−5’ピリミジン。好ましいオリゴデオキシヌクレオチドは、ホスホロチオエート修飾されたODN配列を含む、又はホスホロチオエート修飾されたODN配列からなる。そのような修飾されたオリゴデオキシヌクレオチドを使用することは、それゆえに使用されるオリゴデオキシヌクレオチドの方が、修飾されていないオリゴヌクレオチドよりも安定しており、血液系に入っても容易に分解されないので、有利である。好ましいTh1促進アジュバントは、少なくとも1つ、少なくとも2つ又は少なくとも3つのCpGモチーフからなる、又はそれを含む。免疫刺激ODNの好ましい配列(5’→3’)は、TCAACGTTGA及びGCTAGCGTTAGCGTであった。当業者は、本明細書に明示した配列に制限されない。当業者は、他の配列を設計し、引き続いて、その配列について、本明細書で前に定義したTh1を促進する性質について試験することができる。この好ましい同定アジュバントCpG ODNは、実施例において、LRPEをこのTh1促進アジュバントと一緒に接種することにより、BALB/c系マウス及びC57BL/6系マウスの両方で、大形リーシュマニア寄生虫によるチャレンジに対する防御が誘導されることが実証されたので、非常に魅力的である。どちらのモデルでも、防御はIFN−γの特異的な産生に相関している。BALB/cでは、IL−4及びIL−10の産生に対する制限も検出された。
【0017】
本発明の一つの利点は、広範なスペクトルの寄生虫疾患を治療するための医薬品、すなわち、異種間に特異的な医薬品の調製を可能にすることである。多くの寄生虫疾患において、特定の種に対して作製されたワクチンは、その特定の種に対してしか働かない。その一事例である寄生虫疾患は、リーシュマニア症である。現在のところ、この疾患は薬物により制御されているが、薬物治療では疾患の伝播を予防できず、強い効果がない場合が多い。好ましい実施形態において、寄生虫疾患はリーシュマニア症又はマラリアである。寄生虫疾患が、リーシュマニア属の各種によって、又はプラスモジウム属の各種によって引き起こされるものであることがより好ましい。別の好ましい実施形態において、寄生虫疾患は、RPEを得る種とは異なる種により引き起こされる。特に、リーシュマニア属の一種により引き起こされるリーシュマニア症は、リーシュマニア属の他の種由来のRPEをベースとする組成物を用いて治療することができる。一実施形態において、大形リーシュマニアにより引き起こされるリーシュマニア症は、幼児リーシュマニア由来のRPEを含む組成物で首尾よく治療される。或いは、マラリアなどの他の寄生虫疾患は、他種のRPEをベースとする組成物、例えば、幼児リーシュマニアのRPEをベースとする組成物で首尾よく治療され得る。
【0018】
本発明に照らして、対象はヒト又は動物を意味する。本発明の範囲に包含される動物としては、哺乳動物、特にイヌが挙げられる。
【0019】
好ましい実施形態において、本明細書で定義した医薬品は、感染及び/又は疾患、より好ましくは寄生虫による感染及び/又は寄生虫疾患と戦うための、ヒト又は動物の免疫系の能力を高めるために使用される。特に、ヒト又は動物の対象に投与するために使用され得る。本明細書で定義した医薬品は好ましくは、非経口的に、例えば、静脈内、皮下、腹腔内、筋肉内、動脈内又は病巣内という経路での注射又は点滴によって投与される。好ましい投与形態は、皮下である。医薬品は、当技術分野で公知の従来技法によって、薬学的に許容される培地又は送達ビヒクルと組み合わせることができる。例えば、RPE及び場合によってTh1促進アジュバントをリン酸緩衝食塩水(PBS)に溶解させることができる。非経口的に投与可能な組成物を調製する方法は、当技術分野で周知であり、例えば、Remington’s Pharmaceutical Sciences,Ed.AR Gennaro,20th edition,2000,Williams&Wilkins,PA,USAを含めたさまざまなソースに、より詳細が記載されている。医薬品は、治療有効用量、すなわち、本明細書で定義した感染及び/又は疾患と戦うために、ヒト又は動物の免疫系の能力を高める用量を投与することが好ましい。治療有効用量の本発明の医薬品により、真皮損傷の発生が予防及び/若しくは遅延され、かつ/又は耳及び/若しくは流入領域リンパ節(DLN)における寄生虫負荷量の有意な縮小が誘導されることが好ましい。真皮損傷の存在についての評価は、図6の説明文に記載している。寄生虫負荷量についての評価は、実施例に記載している。治療有効用量の本発明の医薬品により、最初に本発明の組成物を用いた1回のワクチン接種、続いて逐次的な1回の寄生虫による感染、及びおよそ±6週間の待機時間を含む期間の後に、真皮損傷の発生が予防されることが好ましく、及び/又は耳において寄生虫負荷量のおよそ3桁の縮小及び/又はDLNにおいて寄生虫負荷量のおよそ同様の桁の縮小が誘導されることが好ましい。好ましい実施形態において、本明細書で定義した医薬品はワクチンである。より好ましい実施形態において、少なくとも12μgのRPEがワクチンに使用される。さらに好ましい実施形態において、免疫応答をもたらすために少なくとも12〜20μgのRPEを、場合によって少なくとも50μgの、例えばCpG ODNなどのTh1促進アジュバントと組み合わせて使用する必要がある。本明細書で定義したワクチンは、予防ワクチン又は治療ワクチンであってよい。溶解させることができるRPE及び場合によってTh1促進アジュバントの体積は100〜500マイクロリットルで変動してよい。
【0020】
組成物
別の態様において、RPE及び場合によってTh1促進アジュバントを含む組成物が提供される。RPE及びTh1促進アジュバントは、本明細書ですでに定義した。好ましい実施形態において、組成物は、RPE及びTh1促進アジュバントからなる。好ましいTh1促進アジュバントはCpG ODNである。好ましい組成物は、PBSに溶解させたRPE及び場合によってTh1促進アジュバントを含む、又はそれらからなる。別の好ましい実施形態において、RPE及びTh1促進アジュバントを逐次的に投与することも本発明に包含される。したがって、その両方が対象に投与される限り、両成分が物理的に1つの単一組成物中に存在する必要はない。
【0021】
そのような組成物は、薬学的に許容できるアジュバント及び/又は担体をさらに含む。
【0022】
そのような組成物は、医薬品用であることが好ましい。その医薬品は、ワクチンであることが好ましい。医薬品及びワクチンは、本明細書ですでに広範に定義した。
【0023】
方法
他の態様において、本発明は、全て本明細書で定義した、寄生虫疾患を予防及び/若しくは治療し、かつ/又は寄生虫疾患の進行を遅延させ、かつ/又は真皮損傷の発生を予防及び/又は遅延させ、かつ/又は耳及び/若しくは流入領域リンパ節(DLN)において寄生虫負荷量の有意な縮小を誘導する方法を提供する。本方法において、本発明のワクチンは治療ワクチンとして機能する。一般には、感染と疾患発症の間には期間がある。この場合、ワクチンは、宿主において感染の病理学的影響を中和する免疫応答を引き出すことにより、疾患を予防及び/若しくは治療し、かつ/又は疾患の進行を遅延させる薬理学的免疫産物として作用する。治療ワクチンは、すでに感染又は疾患発症している患者において防御を誘導するという点において、予防ワクチンとは異なる。
【0024】
本明細書及び特許請求の範囲において、「含む」という動詞及びその活用は、その言葉に続く項目が含まれるが、特に言及していない項目が除外されるのではないことを指す非限定的な意味で使用している。さらに「からなる」という動詞は、「から本質的になる」と置き換えることができ、本明細書で定義した産物又は組成物又は保存混合物が、特に言及した成分と違う追加の成分(単数又は複数)を含んでよく、前記追加の成分(単数又は複数)は、本発明の独特な特性を変化させないことを意味する。
【0025】
さらに、不定冠詞「a(1つの)」又は「an(1つの)」により要素を参照することは、前後関係からその要素が唯一であることが明らかに必要とされているのでなければ、その要素が2つ以上存在する可能性を除外しない。したがって、不定冠詞「a(1つの)」又は「an(1つの)」は通常「at least one(少なくとも1つの)」を意味する。
【0026】
本明細書に引用した全ての特許文献及び非特許文献は、参照により全体として本明細書に完全に組み込まれている。
【0027】
本発明を以下の実施例によりさらに例証し、それにより本発明の範囲が限定されると解釈されるべきではない。
【図面の簡単な説明】
【0028】
【図1】(A)幼児リーシュマニアのリボソームタンパク質を、直線的な10〜14%の勾配のSDS−PAGEゲルで電気泳動し、ニトロセルロースブロットに転写し、健康なイヌの血清(レーン1〜3)及びVCLに罹患しているイヌ由来の血清(レーン4〜13)と一緒にインキュベートした。血清は200倍希釈で利用した。二次試薬として、西洋ワサビペルオキシダーゼをコンジュゲートした抗イヌIgG抗体を使用した(B〜E)。BALB/cマウス4匹について、大形リーシュマニアの静止期前鞭毛型5×104個を左足蹠において皮下感染させ、チャレンジの8週間後に血清を得た。C57BL/6マウス4匹について、大形リーシュマニアの発育終末前鞭毛型300個を耳真皮において皮内感染させ、チャレンジの14週間後に血清を得た。両系統の感染前の結成についても、寄生虫によるチャレンジの前に得た。(B)大形リーシュマニアのリボソームタンパク質を、直線的な10〜14%の勾配のSDS−PAGEゲルで電気泳動し、ニトロセルロースブロットに転写し、感染させたBALB/cマウス又はC57BL/6マウス由来の保存血清と一緒にインキュベートした。血清は200倍希釈で利用した。感染前の血清でLRPに対する反応性を示したものはなかった(示さず)(C)両系統のマウスにおけるIgG1抗体及びIgG2a抗体のLRPに対する抗体価をELISAによって個々に決定した。(D)感染の8週間後にBALB/cマウスを安楽死させ、その膝窩部DLN細胞を、in vitroで、大形リーシュマニアLRP12μg/mlの存在下で又は培地のみで48時間培養した。培養物の上澄みについてELISAを行い、IFN−γ、IL−4及びIL−10のレベルを評価した。(E)感染の14週間後にC57BL/6マウスを安楽死させ、その上顎後方のDLNをDと同様に処理した。
【図2】(A)BALB/cマウスにおいて誘導される特異的な体液性応答の分析。BALB/cマウス(1群当たり6匹)について、大形リーシュマニアのリボソームタンパク質単独(LRP)、又はそれにCpG ODNアジュバントを加えたもの(LRP+CpG)を用いて、CpG ODNアジュバント単独(CpG)を用いて、又はPBS(生理食塩水)を用いて、右足蹠において3回皮下免疫した。3回目の免疫の14週間後、マウスから血を抜き、血清についてELISAを行い、IgG1アイソタイプ(黒い棒線)及びIgG2aアイソタイプ(白い棒線)の両方の特異的な抗LRP抗体応答について評価した。免疫前の血清で反応性を示したものはなかった。(B〜D)ワクチン接種の4週間後、マウスを安楽死させ、その脾臓を得、in vitro、LRPの存在下で(灰色の棒線)又は培地のみで(黒い棒線)48時間培養した。培養物の上澄みについてELISAを行い、IFN−γ(B)、IL−4(C)及びIL−10(D)のレベルを評価した。
【図3】(A)ワクチン接種したBALB/cマウスにおける大形リーシュマニア感染の経過。マウス(1群当たり6匹)を、図2に示したように皮下免疫した。最後の免疫の1カ月後、動物について、大形リーシュマニアの静止期前鞭毛型5×104個を左後部足蹠において感染させた。足蹠の腫大は、感染した足蹠と感染していない反対側の足蹠との厚さの違いとして得られる。結果は、2つの独立した実験についての平均及び標準偏差を表す。* チャレンジの8週間後の、LRP+CpG ODNをワクチン接種したマウス群における炎症対CpG ODNをワクチン接種したマウス群における炎症のP<0.001の有意差。(B)感染の8週間後、感染させた脚の膝窩部DLN及び脾臓における生存可能な寄生虫の数を、個々に限界希釈法により決定した。結果は、2つの独立した実験についての平均及び標準偏差を表す。* チャレンジの8週間後の、LRP+CpG ODNをワクチン接種したマウス群における寄生虫の体内総量対CpG ODNをワクチン接種したマウス群における寄生虫の体内総量のP<0.001の有意差。(C〜D)ワクチン接種したマウス及び感染させたマウスにおけるサイトカイン産生を、大形リーシュマニアによるチャレンジの8週間後に決定した。感染した脚の膝窩部DLNを得、in vitro、SLAの存在下で(白い棒線)、LRPの存在下で(灰色の棒線)又は培地単独で(黒い棒線)48時間培養した。培養物の上澄みについてELISAを行い、IL−4(C)、IL−10(D)、及びIFNγ(E)のレベルを試験した。この実験では同様の結果が繰り返された。(F)LRP+CpG ODNを用いたワクチン接種によって与えられた防御に付随するIFNγ産生におけるIL−12及びT細胞の関与の分析。大形リーシュマニアの静止期前鞭毛型5×104個でチャレンジした8週間後、CpG ODNを用いてワクチン接種したマウス(黒い棒線)及びLRP+CpG ODNを用いてワクチン接種したマウス(白い棒線)の膝窩部LNを得、抗IL−12抗体、抗CD4抗体又は抗CD8抗体のいずれか、及び対照モノクローナル抗体の存在下で、LRPを用いて培養して刺激した。78時間インキュベーションした後、ELISAによってIFNγのレベルを評価した。抗CD8モノクローナル抗体を用いた処理と対照抗体を用いた処理とにおけるIFNγ産生の差は、統計学的に有意であった(*P<0.05)。データは、同様の結果であった2つの独立した実験の代表的な実験に対応する。
【図4】IgG1/IgG2a極性化の分析。(A)チャレンジの8週間後、血清試料を得、LRPに対するIgG1抗体及びIgG2a抗体の抗体価を、個々にELISAにより決定した。LRP+CpG ODNを用いてワクチン接種したマウスと他の3群とにおけるIgG1抗体価の差は、統計学的に有意であった(*P<0.01)。(B)大形リーシュマニアのLRPを、直線的な10〜14%の勾配のSDS−PAGEゲルで分離し、ニトロセルロースブロットに転写し、表示したマウス群由来の200倍希釈した保存血清と一緒にインキュベートした。IgG1アイソタイプ及びIgG2aアイソタイプ両方の抗体応答を示す。(C)同じ血清を利用して、IgG1及びIgG2aのSLAに対する抗体価を決定した。LRP+CpG ODNを用いてワクチン接種したマウスと他の3群とにおけるIgG1抗体価の差は、統計学的に有意であった(*P<0.02)。
【図5】BALB/cマウス6匹について、図3に示したように左足蹠においてLRP+CpG ODNを用いてワクチン接種し、感染させた。最初の寄生虫によるチャレンジの18週間後、マウスについて、大形リーシュマニアの発育終末前鞭毛型300個を用いて耳の皮内に再感染させた。対照として、無処置のマウス6匹についても耳の皮内にチャレンジした。(A)防御させ、再感染させたBALB/cマウスにおける大形リーシュマニアの感染経過。値は、平均損傷直径+標準偏差(SD)を表す。* チャレンジの7週間後の、再感染させたマウスにおける炎症対対照の感染させたマウスにおける炎症のP<0.0001の有意差。(B)再感染の7週間後、マウスを安楽死させ、耳真皮、脾臓及び局所DLNにおける寄生虫の生体内総量を、個々に定量化した。結果は、12匹分の耳及びDLNの平均±SDとして表す。* 再感染させたマウス対感染させた対照マウスのP<0.001の有意な減少。(C)安楽死させた後、対照マウス由来(黒い棒線)及び再感染させたマウス由来(白い棒線)の上顎後方のDLNを得、in vitro、SLAの存在下で、LRPの存在下で又は培地単独で48時間培養した。培養液の上澄みについてELISAを行うことにより、IFNγ、IL−4、及びIL−10のレベルを評価した。(D〜E)再チャレンジした7週間後、血清試料を得、LRP(D)及びSLA(E)に対するIgG1抗体及びIgG2a抗体の抗体価を、個々にELISAにより決定した。
【図6】C57BL/6マウスにおける大形リーシュマニアの感染に対する防御。マウス(1群当たり6匹)について、LRP+CpG ODNを3回用いて、及びCpG ODNを単独で用いて、右足蹠において皮下免疫した。(A)ワクチン接種したC57BL76マウスの脾細胞によるINF−γ、IL−4及びIL−10の産生。CpG ODN(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種した4週間後、マウスを安楽死させ、その脾臓を得てin vitro、LRPの存在下又は培地単独で48時間培養した。培養物の上澄みについてELISAを行うことにより、サイトカインのレベルを評価した。(B)ワクチン接種したC57BL/6マウスにおける大形リーシュマニアの感染経過。1群当たり12匹のマウスを上記の通り免疫し、最後の接種の4週間後、マウスについて、大形リーシュマニアの発育終末前鞭毛型300個を耳の皮内に接種することにより感染させた。値は、平均損傷直径+標準偏差(SD)を表す。* 2つのマウス群間の炎症のP<0.001の有意な減少。(C)感染の5週間後(1群当たり6匹)及び感染の13週間後(1群当たり6匹)に定量した、CpG ODN(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種したマウス由来の耳真皮及び局所DLNにおける、寄生虫の生体内総量。結果は、12匹分の耳及びDLNの平均±SDを表す。* 両マウス群間のP<0.01の有意な減少。(D〜E)CpG(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種したマウスにおけるIFN−γの産生。感染させた5週間後、上顎後方の細胞を得、SLAの存在下で、LRPの存在下で及び培地単独(D)で培養し刺激した、又は抗IL−12抗体、抗CD4抗体又は抗CD8抗体及び対照のモノクローナル抗体の存在下でLRPを用いて刺激した。78時間インキュベーションした後、ELISAによりIFN−γのレベルを評価した。抗CD8モノクローナル抗体を用いた処理と対照抗体を用いた処理とにおけるIFN−γ産生の差は、統計学的に有意であった(*P<0.01)。(F)チャレンジした5週間後及び13週間後に血清試料を得、LRPに対するIgG1抗体及びIgG2a抗体の抗体価を、個々にELISAにより決定した。(G)CpG ODN(黒い棒線)又はLRP+CpG ODN(白い棒線)を用いてワクチン接種したマウスにおけるIL−10産生。感染させた5週間後、上顎後方の細胞を得、SLAの存在下で、LRPの存在下で又は培地単独で培養し刺激した。78時間インキュベーションした後、IL−10のレベルをELISAにより評価した。
【実施例】
【0029】
材料と方法
マウス系及び寄生虫
雌のBALB/cマウスは実験開始時に6〜8週齢であり、Harlan Interfauna Iberica S.A.(Barcelona,Spain)から購入した。大形リーシュマニア寄生虫(クローンWHOM/IR/−173)及びクローンV1(MHOM/IL/80(Friedlin)を、BALB/cマウスで継代することにより毒性の状態に保った。大形リーシュマニアの無鞭毛型を膝窩部流入領域リンパ節(DLN)から得、20%ウシ胎児血清(FCS)を補充したシュナイダー培地(Schneider’s medium)(Gibco,BRL)中、26℃で、それらが後期静止期に達するまで培養することにより、前鞭毛型に変換させた。両方のクローンの前鞭毛型を、20%ウシ胎児血清(FCS)を補充したシュナイダー培地(Schneider’s medium)(Gibco,BRL)中、26℃で培養した。大形リーシュマニア(クローンV1)の感染性ステージの前鞭毛型(発育終末)を、ピーナッツ凝集素(Vector Laboratories,Burlingame,CA.)を用いたネガティブ選択によって静置培地から単離した。幼児リーシュマニア(MCAN/ES/96/BCN/150,MON−1)の前鞭毛型を、10%FCSを補充したRPMI培地(Gibco,BRL)中、26℃で培養した。
【0030】
CpG ODN及びリーシュマニア抗原
リーシュマニアのリボソームタンパク質抽出物(LRP)を調製するために、大形リーシュマニア及び幼児リーシュマニアの前鞭毛型を採取し、予備冷却したPBSで2回洗浄し、NP40(Roche Diagnostics,GmbH,Manheim Germany,cat.N.11332473001)1mlを含む溶解緩衝液(10mMのトリスClH pH8.0、150mMのNaCl、1.5mMのMgCl2及び0.5%NP40、PMSF 1mM、Leopeptin 8μg/ml、Aprotinin 4μg/ml及びPentatin 8μg/ml)に再懸濁させ、10回上下にピペッティングした。溶解後、試料を4℃、3,000×gで2分微量遠心し、核をペレット化した。上澄みを2回、4℃、13,000×gで15分微量遠心し、(45)に記載の通りサイトゾル上澄みからリボソームを調製した。簡単に述べると、サイトゾルを、Beckman TL100.3ローターを用いて、4℃、90,000rpmで30分の高速遠心分離にかけた。粗製のリボソームペレットを緩衝液A(20mMのトリスHCl、pH7.4、500mMのAcNH4、100mMのMgCL2、5mMのβ−メルカプトエタエノール)に再懸濁させ、緩衝液A中、不連続なショ糖勾配(20/40%)で、TL100.3ローターを用いて、4℃、90,000rpmで遠心分離した。洗浄したリボソームのペレットをPBSに溶解させ、超音波処理し、−70℃で保管した。
【0031】
大形リーシュマニアの全タンパク質(可溶性リーシュマニア抗原[SLA])を、PBSに懸濁させた大形リーシュマニアの静止期前鞭毛型を冷凍し解凍するサイクルを3回行うことにより調製した。細胞の溶解後、微量遠心を用いて12,000gで15分遠心分離することにより、可溶性抗原を不溶性画分から分離し、−70℃で保管した。CpGモチーフを含むホスホロチオエート修飾されたODN配列(CpG ODN)を、Isogen(The Netherlands)により合成した。免疫賦活性ODNの配列(5’→3’)は、TCAACGTTGA及びGCTAGCGTTAGCGTであった。
【0032】
免疫化及び寄生虫チャレンジ
BALB/cマウスについて、大形リーシュマニア12μgを単独で、又は大形リーシュマニア12μgとCpG ODN50μg(各免疫賦活性ODNを25μgずつ)を、CpG ODN(50μg)アジュバントを単独で、又はリン酸緩衝食塩水(PBS)を、右足蹠において皮下(s.c.)接種した。各群について、同じレジメンを用いて2週間後及び4週間後に追加免疫した。最初の寄生虫によるチャレンジは、最後の接種の4週間後に、大形リーシュマニアの静止期前鞭毛型(クローンWHOM/IR/−173)5×104個を、左の(未処置の)足蹠に皮下接種することにより行った。感染の進行を、決定用カリパスを用いて厚さを測定することにより追跡した。各動物の反対側の足蹠が対照値を表し、腫大を以下の通り計算した:左足蹠の厚さ−右足蹠の厚さ。損傷が壊死性になった時に動物を安楽死させた。再感染させるために、BALB/cマウス6匹について、上記の通りワクチン接種し感染させた。18週間後、大形リーシュマニアの発育終末前鞭毛型(クローンV1)300個を各マウスの両耳真皮に注射した。感染の進展を、決定用カリパスを用いて耳の損傷の硬結の直径を測定することによりモニターした。対照として、無処置のBALB/cマウス6匹の群についても、耳真皮において感染させた。
【0033】
C57BL/6マウスについて、大形リーシュマニアLRP12μg+CpG ODN50μg(各免疫賦活性ODNを25μgずつ)、及びCpG ODN(50μg)アジュバント50μgを単独で、足蹠に皮下注射した。これらのマウスについて、同じ免疫化レジメンを用いて2週間後及び4週間後に追加免疫した。最後の接種の4週間後に、大形リーシュマニアの発育終末前鞭毛型(クローンV1)300個を、マウスの両耳真皮に皮内(i.d.)接種することによって感染を行った。感染の進展を、決定用カリパスを用いて耳の損傷の硬結の直径を測定することによりモニターした。
【0034】
寄生虫の量子化
耳において、限界希釈アッセイ(6)により寄生虫の数を決定した。簡単に述べると、感染させたマウスから耳を回収した。感染させた耳の腹側及び背側のシートを分離した。耳のシートを、Liberase CI酵素ブレンド(50μg/ml)含有ダルベッコ改変イーグル培地(Dulbecco’s modified Eagle medium)に沈着させた。37℃で2時間インキュベートした後、組織を小片に切断し、ホモジナイズし、細胞ろ過器(孔サイズ70μm)を用いてろ過した。ホモジナイズした組織を、20%FCSを加えたシュナイダー培地(Schneider’s medium)を含有する96ウェルの平底マイクロタイタープレート中に連続的に希釈した。生存可能な寄生虫の数を、前鞭毛型が26℃でインキュベートして最大7日まで成長でき得る最大希釈から決定した。寄生虫の数は、感染させた耳(上顎後方)及び足蹠(膝窩部)の局所流入領域リンパ節(DLN)において、及び脾臓においても決定した。器官を回収し、機械的に切り離し、次いで上記の通り連続的に希釈した。寄生虫の負荷量は、全器官における寄生虫の数として表される。
【0035】
上澄み中のサイトカインの測定
脾臓及び対応する局所DLNを無菌的に取り出し、機械的に切り離し、完全なRPMI培地(10%FCS、2mMのグルタミン、及び10mMの2−メルカプトエタノールを補充したRPMI1640)に播種した。LRP(12μg/ml)又はSLA(12μg/ml)の存在下、37℃で48時間の間に、48ウェルプレートに細胞5×106個/mlを播種した。市販のELISAキット(Diaclone,Besancon,France)により、脾細胞及びDLN細胞の上澄みについて、IFN−γ、IL−10及びIL−4の放出を測定した。一部の例では、LRP12μ/mlで刺激したDLN細胞を、マウスCD4(GK1.5)、マウスIL−12(C17.8)、マウスCD8(53−6.7)のいずれかに対するモノクローナル抗体(mAb)10μg/mlの存在下でインキュベートした。適切なアイソタイプ整合対照についてもこのアッセイで分析した。抗体(no azide/low endotoxin(商標))は、BD)(PharMingen)から購入した。
【0036】
体液性応答の分析
ELISA又はウェスタンブロットにより、LRP又はSLAに対する特異的な抗体について血清試料を分析した。簡単に述べると、標準のELISAプレートを、100μlのLRP(PBS中5μg/ml)又はSLA(PBS中2μg/ml)を用いて室温で一晩コーティングした。抗体価を、血清を連続的に希釈することにより決定し、吸光度>0.2を生じる最大の血清希釈力価の逆数として定義した。アイソタイプ特異的な分析を、以下の西洋ワサビペルオキシダーゼをコンジュゲートした抗マウス免疫グロブリン(Nordic Immunological Laboratories,Tilburg,The Netherlands):抗IgG1(1/1000)及び抗IgG2a(1/500)を用いて行った。オルトフェニレンジアミンジヒドロクロライド−OPD−(Dako,A/S,Glostrup,Denmark)をELISAアッセイのペルオキシダーゼの基質として使用した。15分後、1MのH2SO4100μlを加えることにより反応を停止させ、吸光度を450nmで読み取った。
【0037】
ウェスタンブロット分析のために、幼児リーシュマニア及び大形リーシュマニアのリボソームタンパク質を得、Laemmli緩衝液に再懸濁させ、SDS−PAGEで分離し、ニトロセルロース膜(Amersham,Aylesbury,UK)に転写した。対照のイヌ、VCLリーシュマニア症に罹患しているイヌの血清、又は本研究において示した希釈で利用した種々のマウス群由来の血清を用いてブロットを探索した。二次抗体として、Nordic Immunological Laboratories (Tilburg,The Netherlands)から購入した、西洋ワサビペルオキシダーゼをコンジュゲートした、抗マウス抗IgG(1/1000)、抗IgG1(1/1000)、抗IgG2a(1/500)免疫グロブリン、及び抗イヌ抗IgG(1/2000)を使用した。
【0038】
統計分析
統計分析はスチューデントt検定により行った。P<0.05である場合に有意差があるとみなした。
【0039】
結果
感染の間のLRPの抗原性
リーシュマニアのリボソームタンパク質(LRP)の抗原性を分析するために、幼児リーシュマニアに自然感染したイヌ由来の血清の幼児リーシュマニアLRPに対する応答性を、ウェスタンブロットによりアッセイした。活動性疾患に罹患しているイヌ由来の血清が、LRP抽出物中の多数のタンパク質バンドを認識したことが観察された(図1A)。大形リーシュマニアに実験的に感染させたC57BL/6マウス及びBALB/cマウス由来の血清も、大形リーシュマニアLRP中の多くのタンパク質バンドを認識し、BALB/c感受性マウス由来の血清に存在するIgG抗体によって認識されるリボソームタンパク質の数の方が、C57BL/6耐性マウスの血清中のIgG抗体によって認識されるタンパク質の数よりも多い(図1B)。
【0040】
IgG1及びIgG2a抗体の誘導は、Th2型免疫応答及びTh1型免疫応答のマーカーとして使用することができるので(8)、我々は、大形リーシュマニアに感染させたマウスにおいてLRPに対するIgG1/IgG2a極性化の分析を行った。BALB/cマウスでは、抗LRP応答はIgG1アイソタイプが優勢であったのに対し、C57BL/6マウスでは、抗LRP応答はIgG2aアイソタイプが優勢であった(図1C)。両方のマウス系統において、in vitroでLRPを用いてDLN細胞を刺激した後のIFN−γ、IL−4及びIL−10の産生についても決定した。CLに罹患しているBALB/cマウスにおいて、LRP特異的なIFN−γの産生が検出されたが、IL−4及びIL−10の産生も強力に刺激され、INF−γ/IL−4比は約2.4、INF−γ/IL−10比は約1.4になっている(図1D)。一方、治癒したC57BL/6マウス由来のDLN細胞をLRPで刺激した場合、IL−4は検出されず(<7.5pg/ml)、高IFN−γ/IL−10比(約15)が得られた(図1E)。
【0041】
BALB/cマウスにおけるLRPの免疫原性
BALB/cマウスにおいて、リボソームタンパク質をCpG ODNの非存在下及び存在下で投与した後のLRPに対する免疫応答を調べた。LRPE+CpG ODNを用いたワクチン接種後、抗LRPE体液性応答はIgG2aアイソタイプが優勢であったのに対し、LRPE単独で免疫したマウス由来の血清では検出されたIgG1アイソタイプの抗体の抗体価は低かった(図2A)。LRPE+GpG ODNで免疫したマウス由来の脾臓細胞を、in vitroでLRPEを用いて刺激した後、その細胞から分泌されたIFN−γのレベルは、対照マウス由来の脾臓細胞及びLRPE単独で免疫したマウス由来の脾臓細胞から分泌されたものよりも高かった(図2B)。いずれの群においても、LRPEによる刺激後にIL−4産生の増加は見られなかった(図2C)。著しいことに、LRPE+CpG ODNを用いてワクチン接種した脾臓から確立した培養物の上澄みにおいて、特異的なIL−10が検出され、IFN−γ/IL−10比は約40であった(図2D)。全体として、これらの結果は、BALB/cマウスにおいて、アジュバント無しで投与したLRPEからは、これらの抗体に対してほんの弱いIgG1体液性応答しか誘導されないが、CpG ODNと同時投与すると、これらの抗原に対してTh1様応答が増強されることを実証している。
【0042】
LRPE+CpG ODNを用いたワクチン接種は大形リーシュマニアのチャレンジからBALB/cマウスを防御する
抗原に関連した疾患に誘導されるTh2応答をTh1応答に転送することは、リーシュマニアに対するワクチンを開発するための有望なアプローチ法であると考えられているので(7)、我々は、LRPE+CpG ODNを用いたワクチン接種によって大形リーシュマニア感染に対する防御を誘導することができるかどうかを分析した。図3Aは、LRPE+CpG ONDをワクチン接種したマウスの足蹠の腫大が、対照マウス及びLRPE単独でワクチン接種したマウスの足蹠の腫大(平均値約5.5mm)と比較して縮小されているので(8週目の平均値0.7mm)、LRPE+CpG ONDによって効果的な防御が誘導されることを示している。我々は次に、マウスの4群について、膝窩部DLN及び脾臓における寄生虫の負荷量を分析した。LRPE−CpGで免疫したマウス由来のDLNでは、他の群と比べて、約3logの寄生虫の生体内総量の縮小が見られた。さらに、対照マウス及びLRPE単独で免疫したマウスの脾臓では同様の寄生虫の負荷量が見られた一方、LRPE+CpG ODNをワクチン接種したマウスの脾臓では寄生虫が検出できなかった(図3B)。
【0043】
LRPE+CpG ODNに誘導される防御に関連する免疫学的パラメーターを決定するために、SLA主導又はLRPE主導のIL−4、IL−10及びIFN−γの産生をアッセイした。SLA特異的又はLRPE特異的に誘導されたIL−4及びIL−10の産生は、対照マウス(生理食塩水及びCpG)由来のDLN細胞及びLRPE単独で免疫したマウス由来のDLN細胞において検出された(図3C〜D)。一方、LRPE+CpG ODNを用いて免疫したマウス由来のDLN細胞が産生したIFN−γの量は、他の3群で検出されたIFN−γの量よりも多かった(図3E)。LRPEに特異的なIFN−γ産生に対するCD4+T細胞及びCD8+T細胞の寄与及びIL−12への依存についても分析した。図3Fに示されるように、IFN−γの産生は、抗IL−12モノクローナル抗体又は抗CD4モノクローナル抗体により完全に阻害された。DLN細胞培養物への抗CD8抗体の添加により、上澄み中のサイトカインの量が一部だけ縮小した。
【0044】
BALB/cマウスにおいて、疾患の進行に付随してIL−4に依存した高抗体価の抗体が産生されることから、我々は、感染の8週間後に、LRPEに対して引き出される体液性応答をELISAによって分析した。LRPE+CpG ODNを用いて免疫したマウスにおいて寄生虫のチャレンジによって引き出された、LRPEに対する抗体は、主にIgG2aアイソタイプであった。また、防御マウスの血清に存在するIgG1アイソタイプの抗LRPE抗体の抗体価は、LRPE単独で免疫したマウス及び2つの対照群と比較して低かった(図4A)。LRPEに対するIgG1抗体価の縮小は、LRPE+CpG ODNをワクチン接種したマウスの血清由来のIgG1抗体に認識されるリボソームタンパク質バンドの数の減少と相関した。図4Bに示されるように、LRPEウェスタンブロットにおいて、生理食塩水、CpG又はLRPE単独で免疫したマウスの血清由来のIgG1アイソタイプ抗体によって多くの数のタンパク質バンドが認識されたのに対して、防御マウスのIgG1抗体によって認識されたバンドはわずかであった。ウェスタンブロット分析により、LRPE+CpG ODNをワクチン接種したマウス由来の血清中のIgG2a抗体によって認識されるタンパク質バンドの数は、他の3群と比較して増加しなかったことも示された(図4B)。LRPE+CpG ODNを用いたワクチン接種によって、大形リーシュマニアの感染によって誘導される広域の抗リーシュマニア体液性応答も調整された。LRPE+CpG ODNを用いて免疫したマウスにおいて寄生虫のチャレンジによって引き出されたSLAに対する抗体は、主にIgG2aアイソタイプであり、IgG1アイソタイプの抗体の抗SLA抗体価は、他の3群において検出されたものより有意に低かった(図4C)。
【0045】
LRPE+CpG ODNをワクチン接種し、感染させたマウスは耳真皮における大形リーシュマニアの再感染に対して耐性である
LRPE+CpG ODNをワクチン接種し、感染させたマウスが、2回目の寄生虫によるチャレンジを制御できるかどうかを決定するために、BALB/cマウス6匹について、上記の通り足蹠においてワクチン接種し、感染させた。これらのマウス群の足蹠の腫大は、18週間の間<0.7mmであった(データは示さず)。次に、これらの防御マウスについて、大形リーシュマニアの発育終末前鞭毛型300個を用いて、耳真皮において再感染させた。無処置のマウス6匹の対照群についても感染させた。LRPE+CpG ODNをワクチン接種し、再感染させたマウスでは病変が見られなかったのに対し、対照マウスでは、7週目に開存性の耳損傷が発生したので、LRPE+CpG ODNをワクチン接種し、再感染させたマウスは真皮損傷の発生を防御することが観察された(図5A)。耳真皮及び上顎後方のDLNにおける寄生虫の負荷量も、2群間で有意差があった(図5B)。ワクチン接種し、再感染させたマウスの耳及びDLNにおける寄生虫の負荷量の低さは、脾臓に寄生虫が存在しないことと相関する。再感染後に起こる細胞性応答について知るために、LRPE又はSLAを用いたin vitroでの刺激後の上顎後方のDLN細胞によるIFN−γ、IL−4、及びIL−10の分泌について分析した(図5C)。対照マウスでは、マウスを足蹠において感染させた際にも起こるのと同様(3C〜E)、IFN−γの特異的産生が検出されたが、IL−4及びIL−10の産生も強力に刺激された。一方、ワクチン接種し、再感染させたマウスのDLN細胞は、特異的IFN−γを多量に産生し、IL−4及びIL−10はほとんど検出されなかった(図5C)。これに一致して、LRPE(図5D)及びSLA(図5E)に対するIgG体液性応答は、IgG2aアイソタイプによるものであった。
【0046】
LRPE+CpG ODNを用いたワクチン接種は、C57BL/6マウスにおける大形リーシュマニアのチャレンジに起因する真皮の病変に対する防御を付与する
BALB/cマウスでは、LRPE+CpG ODNを用いてワクチン接種すると、大形リーシュマニアの感染に対し、LRPEに対するTh2免疫応答がTh1免疫応答へ転送されることにより防御が起こることから、我々は、このワクチンを、リーシュマニア抗原に対して自然にTh1応答を生じるモデルであるC57BL/6マウスに投与した効果を分析した。C57BL/6マウスの群を、LRPE+CpG ODNで3回免疫し、対照マウスはCpG ODNアジュバントのみを受けさせた。接種により、LRPEを用いて刺激した脾臓細胞培養物の上澄みにおけるin vitroでのIFN−γ産生によって実証されたTh1応答が誘導された。LRPE+CpG ODNをワクチン接種したマウスから確立した脾臓細胞培養物の上澄みにおいて、特異的なIL−10の存在も検出されたのに対し、刺激後に特異的なIL−4産生は観察されなかった(図6A)。LRPE+CpG ODNをワクチン接種したマウスは、病変がほとんど又は全く観察されなかったので、真皮損傷の発生を防御した(図6B)。CpG ODNで免疫したマウスでは損傷が発生し、7週目でピークに達し、13週目にほとんど完全に治癒した。このモデルでは、感染部位における寄生虫の数が、損傷の発生の直前にピークに達するので(5)、我々は、5週目に耳及び局所DLN(上顎後方)における寄生虫の負荷量を決定した。ワクチン接種したマウスの耳真皮における寄生虫の数は、約300分の1に(LRPE+CpG ODNマウスでは寄生虫1.0×104個、CpG−ODNで免疫したマウスでは寄生虫3.0×106個)及びDLNマウスでは約40分の1に(LRPE+CpG ODNマウスでは寄生虫5.0×104個、CpG ODNマウスでは寄生虫2.0×104個)縮小した。治癒後、対照マウスでは(チャレンジの13週間後)、全ての群において寄生虫の数の縮小が観察された。
【0047】
防御に関連する免疫学的パラメーターを決定するために、抗原主導のIL−4、IL−10及びIFN−γの産生をアッセイした。チャレンジの5週間後、DLN細胞の培養物を確立し、LRPE又はSLAを用いて刺激した。図6Dに示される通り、ワクチン接種したマウス由来の細胞の方が、対照マウス由来の細胞よりも多くのSLA特異的及びLRPE特異的なIFN−γを産生した。LRPEに特異的なIFN−γ産生に対するCD4+T細胞及びCD8+T細胞の寄与及びIL−12への依存についても分析した。IFN−γの分泌は、抗IL−12モノクローナル抗体又は抗CD4モノクローナル抗体により完全に阻害された。抗CD8抗体での処理では、このサイトカインの量は一部だけ縮小した。これらのデータにより、C57BL/6において、LRPE+CpG ODNを用いたワクチン接種により、LRPEに対する早期の特異的Th1応答の誘導と相関する、皮膚及び局所DLNにおける病変及び寄生虫の生体内総量の縮小が誘導されることが実証された。これに一致して、対照マウスに対してワクチン接種したマウスにおいてIgG2a特異的な抗LRPE抗体が早く、高抗体価で検出された(図6F)。我々は、SLA抗原特異的又はLRPE抗原特異的なIL−10産生も検出し、それはワクチン接種したマウスと対照マウスとの間に統計的な差はなかった。
【0048】
考察
本研究において、我々は、多くの寄生虫リボソームタンパク質と反応する抗体が、VCLを生じたイヌ由来の血清、及び大形リーシュマニアに感染させたマウスにおいて観察されることを示し、そのことは、リーシュマニアの自然感染及び実験的感染の間、寄生虫のリボソームタンパク質が強力な抗原であることを示している。抗LRPE抗体応答はIgG1アイソタイプによるものが優勢であったので、感染させたBALB/cマウスのLRPEに対する応答はTh2型のものであった。この知見は、LRPEを用いて感染させたマウス由来のDLN細胞をin vitroで刺激した後に、培養物の上澄み中で多量のIL−4が検出されたという事実により補強される。LRPEは、INF−γのin vitro産生にも関係したが、匹敵するレベルの、寄生虫を破壊するための活性化シグナルに対して、感染したマクロファージを無反応性にする多面的な抗炎症サイトカインIL−10が検出された(31)。さらに、BALB/cマウスにおける疾患の促進におけるIL−4及びIL−10の影響は、相加的だと思われることから((37)に総説がある)、これらのサイトカインのLRPEによる刺激は、リボソーム抗原に対する宿主の応答が、BALB/cマウスにおいて寄生虫が拡大及び持続するのに好都合であることを示しているとみなすことができる。リーシュマニアによる感染後早期に、宿主の免疫系が寄生虫の細胞溶解によって放出される豊富なLRPEにより感作され、その後で、寄生虫の増殖の結果、追加免疫され得ると考えることができる。したがって、LRPEの強力な免疫原性及びその病原抗原的な役割は、おそらくその豊富さ及び抗原特異性(その進化的に保存された特性にもかかわらず)に依存している。一方、LRPEに対して応答するC57BL/6マウスはTh1型のものであり、IgG2a特異的抗体の生成及びin vitroでの高レベルの特異的INF−γ産生を伴い、IL−4レベルは検出不可能であった。また、我々は、感染の14週間後に、LRPEに特異的なIL−10産生を見出した。IFN−γ/IL−10比が、SLAを用いて細胞を刺激した場合(データは示さず)に得られたものと同様であったことから、我々は、これらの抗原によって誘導されるIL−10産生は、このモデルにおいて見られる、寄生虫が持続するのに好都合である調節性の応答に関連する可能性があることを示唆する(4)。我々のデータは、LRPEに対する応答が、感受性宿主の病変に関連したTh2に媒介される体液性応答及び耐性宿主におけるTh1に関連する防御応答の誘導に向かうバイアスに一致することを示している。
【0049】
いくつかのリーシュマニアのエピトープに対して誘導されるTh2応答のTh1応答への転送は、大形リーシュマニアの感染に対する防御を誘導するための有望な戦略になる可能性が高いので(7)、我々はまず、BALB/cマウスにおいて、異なるリーシュマニア抗原を用いて免疫した際のTh1に関連する長期免疫及び防御を与え(43)、さらにマウスにおいていくつかの寄生虫特異的Th2応答を抑制し得る(12、57)アジュバントであるCpG ODNと同時投与したLRPEの免疫原性について分析することを決めた。図2に示されるように、BALB/cマウスにおいて発生する免疫応答は、LRPEを用いてin vitroで刺激した後、免疫したマウスがIgG2aアイソタイプの抗LRPE抗体を発生し、ワクチン接種したマウス由来の脾細胞が多量のIFN−γを産生し、IL−4は産生しないので、Th1型であることが分かった。同様の特異的免疫応答が、C57BL/6マウスにおいて、LRPE+CpG ODNを用いて免疫した場合に生じた(図6A)。どちらのマウス系統においても、免疫した後にLRPE主導のIL−10産生も観察された。
【0050】
L31をコードする遺伝子ワクチンを用いたワクチン接種が、遺伝子ワクチン接種後の特異的IL−10産生に関係し得ることが報告されているにもかかわらず(44)、我々は、IL−10産生は、LRPE+CpG ODNの投与後に起こるTh1応答のホメオスタティックコントロールの方に関連している可能性があると考えている。事実、INF−γを産生するTh1細胞は、フィードバック制御の機構としてIL−10の産生にも関与している可能性があることが最近報告されている(2)。さらに、得られた高INF−γ/IL−10比の値により、ワクチンの結果について良好な予測が得られ得る(44)。
【0051】
ここで示したデータは、LRPE+CpG ODNの同時投与により、実験的な皮膚リーシュマニア症の2つの異なるモデル、BALB/cマウス(皮膚リーシュマニア症ワクチンのアッセイに広く用いられている)の足蹠における高接種量モデル、及びC57BL/6マウス(経路及び感染用量に関して、ヒト疾患をより厳密に再現するモデル)の耳における低接種量モデルにおいて防御が誘導されることを示している。ワクチン接種したBALB/cマウスでは膝窩部DLNにおける寄生虫の生体内総量が縮小しており、脾臓への寄生虫転移はなかった。また、非常に弱い炎症が感染した足蹠において検出された。ワクチン接種したC57BL/6マウスでは、耳真皮の損傷が防御され、皮膚及びDLNにおいて寄生虫の生体内総量が縮小したことが示された。この防御は、熱殺菌されたリーシュマニア抗原とCpG ODNを用いてワクチン接種したC57BL/6マウスによって示された防御に匹敵し、また、このモデルにおいて、LACK、LmSTI1及びTSAを含む多成分ワクチンについても試験した(28、29、43)。
【0052】
重要なことに、どちらの系統における防御も、LRPEに対するTh1特異的免疫応答の生成に相関している。細胞性応答についてのin vitro分析は、BALB/cマウスでは感染の8週間後、及びC57BL/6マウスでは5週間後(寄生虫の負荷量のピークに一致する(4))に測定した。LRPE+CpG ODNを用いてワクチン接種した両マウス系統由来のDLN細胞は、LRPEを用いて刺激した際、それぞれの対応する対照群よりも高いレベルのIFN−γを分泌した。IFN−γ応答は、IL−12依存性であり、CD4+T細胞により産生され、CD8+T細胞に対する寄与は少ないことが分かった。予想通り、両系統の防御マウスについて、SLA特異的IFN−γ産生は高度であった。BALB/cマウスにおいて、防御マウスにおけるTh1応答の生成は、LRPEに対する優勢なIgG2a特異的抗体の生成と相関する(図4A)。IL−10のin vitro産生において、系統間で、多少の違いが観察された。我々のデータは、LRPE及びSLAの両方を用いてin vitroで刺激した後、防御BALB/cマウスでは、対照マウスよりも有意に低いレベルのIL−10が産生されたことを示し、これは、このサイトカインがこのモデルの感受性に関与していることと一致している(35)。一方、対照マウス及びワクチン接種したC57BL/6マウスのどちらのLNCにおいても、LRPE又はSLAを用いてin vitroで刺激した後、感染の急性期の間IL−10が産生された(図6F)。IFN−γについて起こったように、IL−10レベルは、細胞を、SLAを用いて刺激した場合よりも、LRPEを用いて刺激した場合の方が高かった。IFN−γ及びIL−10の産生は同様のプロファイルに従ったという事実は、ワクチン接種後のIL−10産生が、宿主において強力なTh1によって引き起こされる有害な影響を制御するホメオスタティック機構を反映しているとみなすことができる(4)。驚くべきことに、我々は、LRPE+CpG ODNを用いてワクチン接種した後にBALB/cマウスにおいて見られる防御が、LRPE又はSLAを用いてin vitroで刺激した後のIL−4主導の抗原産生が有意に縮小したことにも関連していることを見出した。これらの細胞性応答は、in vivoで、リボソームタンパク質に対するTh2に媒介される抗体応答の転換に相関する。したがって、防御BALB/cマウス由来の血清は、抗体価、特にLRPE特異的なIgG1抗体によって認識される抗原の数について有意な減少を示した。さらに、LRPE+CpG ODNを用いたBALB/cマウスの免疫も、大形リーシュマニアの感染によってマウスにおいて誘発される広範な体液性応答に対して明らかに影響した(図5C)。したがって、ワクチン接種したマウスを感染させると、IgG1抗リーシュマニア特異的抗体の誘導が制限されるのに対して、他のアッセイした対照マウス群では、体液性応答が高度に誘導され、Th2型の抗体(すなわち、IgG1アイソタイプ)が優勢であった。全体として、どちらの系統のマウス由来のワクチン接種したマウスにおいて観察される防御も、LRPEに対するTh1応答の生成に相関し、またその結果、BALB/cマウスにおけるTh2主導のIL−4及びSLAに対するIL−10応答が下向き調節される。
【0053】
我々のデータは、防御BALB/cマウスが、追加的な感染に抵抗できる能力を彼らに付与する免疫学的状況(寄生虫への再曝露が極めて頻繁に起こり得る流行地域において利用され得るワクチンに魅力的な特徴)を獲得したことを示している。耳真皮における再チャレンジの後、これらのマウスは、大形リーシュマニアの感染に対して強固な防御を示した。真皮損傷の発生は非常に少なく(一部の例では、真皮損傷が完全にないことが検出された)、感染した耳及びDLNにおいて寄生虫数に相当な縮小がみられた。防御マウス由来のDLN細胞は、対照よりも大量のIFN−γを産生しないが(感染の7週間後に測定)、LRPE及びSLAに対するIgG2a抗体の抗体価は、これらのマウスが二次的なチャレンジ後に特異的なTh1防御応答を高めたとみなすことができる。驚くべきことに、これらのマウスにおける疾患関連サイトカインであるIL−4及びIL−10の特異的な産生は、非常に少なかった。これらのデータは、最初の寄生虫によるチャレンジの後に生成した免疫状態が非常に有力であり、再感染した部位からの寄生虫の迅速かつ効率的な排除を導くことを示している。
【0054】
ここで示したデータは、LRPE+CpG ODNを用いたワクチン接種が、耐性マウスと感受性マウスの両方において、リーシュマニアによる感染時に、免疫系の決定に直接影響を及ぼすことを実証している。我々の考えでは、リーシュマニアのような複雑な寄生虫に対するワクチンの生成は、ワクチン製剤中に種々の標的抗原を組み込み、これらの抗原が必要な免疫性を導く(主にCD4+及びCD8+IFN−γ媒介応答)ことを利用し、病変をもたらす病原抗原主導の免疫応答(IL−4 Th2主導及びIL−10不活性化応答)をTh1バイアスに転送することにより最適化され得る。それにもかかわらず、大形リーシュマニアのメタ1抗原で起こるように、これらの抗原の一部に対するTh2応答は、通常のTh1誘導剤によって転送されない場合があることを考慮しなければならない(50)。
(参考文献)
【特許請求の範囲】
【請求項1】
対象において寄生虫疾患を治療又は予防するための医薬品を調製するための、リボソームタンパク質抽出物及び場合によってTh1促進アジュバントの使用。
【請求項2】
前記医薬品がワクチンである、請求項1に記載の使用。
【請求項3】
前記リボソームタンパク質抽出物が、対象内に存在すると寄生虫疾患を引き起こす寄生虫細胞を用いて、
a.寄生虫細胞を溶解緩衝液と混合するステップと、
b.得られた混合物を遠心分離してサイトゾル抽出物を得るステップと、
c.得られたサイトゾル抽出物からリボソームタンパク質抽出物を調製するステップと
を行うことにより得られる、請求項1又は2に記載の使用。
【請求項4】
前記リボソームタンパク質抽出物が、リーシュマニア属の各種(Leishmania species)、好ましくは大形リーシュマニア(Leishmania major)から得られる、請求項1から3までのいずれか一項に記載の使用。
【請求項5】
前記Th1促進アジュバントがCpG ODNである、請求項1から4までのいずれか一項に記載の使用。
【請求項6】
前記寄生虫疾患がリーシュマニア症又はマラリアである、請求項1から5までのいずれか一項に記載の使用。
【請求項7】
前記寄生虫疾患が、リーシュマニア属の各種(Leishmania species)又はプラスモディウム属の各種(Plasmodium species)によって引き起こされる、請求項1から6までのいずれか一項に記載の使用。
【請求項8】
前記寄生虫疾患が、リボソームタンパク質抽出物を得る種とは異なる種によって引き起こされる、請求項1から7までのいずれか一項に記載の使用。
【請求項9】
リボソームタンパク質抽出物及び場合によってTh1促進アジュバントを含む組成物。
【請求項10】
リボソームタンパク質抽出物及びTh1促進アジュバントからなる、請求項9に記載の組成物。
【請求項11】
前記Th1促進アジュバントがCpG ODNである、請求項9又は10に記載の組成物。
【請求項12】
薬学的に許容されるアジュバント及び/又は担体をさらに含む、請求項9から11までのいずれか一項に記載の組成物。
【請求項13】
医薬品として使用するための、請求項9から12までのいずれか一項に記載の組成物。
【請求項14】
前記医薬品がワクチンである、請求項12に記載の組成物。
【請求項1】
対象において寄生虫疾患を治療又は予防するための医薬品を調製するための、リボソームタンパク質抽出物及び場合によってTh1促進アジュバントの使用。
【請求項2】
前記医薬品がワクチンである、請求項1に記載の使用。
【請求項3】
前記リボソームタンパク質抽出物が、対象内に存在すると寄生虫疾患を引き起こす寄生虫細胞を用いて、
a.寄生虫細胞を溶解緩衝液と混合するステップと、
b.得られた混合物を遠心分離してサイトゾル抽出物を得るステップと、
c.得られたサイトゾル抽出物からリボソームタンパク質抽出物を調製するステップと
を行うことにより得られる、請求項1又は2に記載の使用。
【請求項4】
前記リボソームタンパク質抽出物が、リーシュマニア属の各種(Leishmania species)、好ましくは大形リーシュマニア(Leishmania major)から得られる、請求項1から3までのいずれか一項に記載の使用。
【請求項5】
前記Th1促進アジュバントがCpG ODNである、請求項1から4までのいずれか一項に記載の使用。
【請求項6】
前記寄生虫疾患がリーシュマニア症又はマラリアである、請求項1から5までのいずれか一項に記載の使用。
【請求項7】
前記寄生虫疾患が、リーシュマニア属の各種(Leishmania species)又はプラスモディウム属の各種(Plasmodium species)によって引き起こされる、請求項1から6までのいずれか一項に記載の使用。
【請求項8】
前記寄生虫疾患が、リボソームタンパク質抽出物を得る種とは異なる種によって引き起こされる、請求項1から7までのいずれか一項に記載の使用。
【請求項9】
リボソームタンパク質抽出物及び場合によってTh1促進アジュバントを含む組成物。
【請求項10】
リボソームタンパク質抽出物及びTh1促進アジュバントからなる、請求項9に記載の組成物。
【請求項11】
前記Th1促進アジュバントがCpG ODNである、請求項9又は10に記載の組成物。
【請求項12】
薬学的に許容されるアジュバント及び/又は担体をさらに含む、請求項9から11までのいずれか一項に記載の組成物。
【請求項13】
医薬品として使用するための、請求項9から12までのいずれか一項に記載の組成物。
【請求項14】
前記医薬品がワクチンである、請求項12に記載の組成物。
【図1】

【図2】

【図3】

【図4】
【図5】

【図6−1】
【図6−2】

【図2】
【図3】
【図4】
【図5】
【図6−1】
【図6−2】
【公表番号】特表2011−509968(P2011−509968A)
【公表日】平成23年3月31日(2011.3.31)
【国際特許分類】
【出願番号】特願2010−542611(P2010−542611)
【出願日】平成21年1月14日(2009.1.14)
【国際出願番号】PCT/EP2009/050334
【国際公開番号】WO2009/090175
【国際公開日】平成21年7月23日(2009.7.23)
【出願人】(510197070)
【Fターム(参考)】
【公表日】平成23年3月31日(2011.3.31)
【国際特許分類】
【出願日】平成21年1月14日(2009.1.14)
【国際出願番号】PCT/EP2009/050334
【国際公開番号】WO2009/090175
【国際公開日】平成21年7月23日(2009.7.23)
【出願人】(510197070)
【Fターム(参考)】
[ Back to top ]